Перечень последовательностей
[0000] Настоящая заявка содержит перечень последовательностей, который был подан в электронном виде в формате ASCII и включен в данный документ посредством ссылки во всей своей полноте. Указанная копия в формате ASCII, созданная 06 декабря 2019 года, имеет название PAT058359-WO-PCT_SL.txt и размер 285253 байт.
Область техники, к которой относится изобретение
[0001] Настоящее изобретение в целом относится к антителам к PMEL17 или их фрагментам, их конъюгатам, включая их конъюгаты с ингибитором GNAQ/GNA11, а также вариантам их применения для лечения или предупреждения рака.
Предпосылки изобретения
[0002] PMEL17 (также обозначаемый как gp100 и SILV) представляет собой однопролетный трансмембранный белок типа I, продуцируемый меланоцитами и принимающий участие в синтезе меланина. По мере созревания PMEL17 временно экспрессируется на поверхности клетки, прежде чем попасть в меланосомы, где PMEL17 подвергается разрушению на различные домены, которые мультимеризуются с образованием фибриллярных листов. Такая структура затем служит опорой для улавливания меланина. Экспрессия PMEL17 в меланосомах регулируется MITF, онкогеном линий дифференцировки, и было обнаружено, что она активируется во множестве первичных и метастатических подкожных и увеальных меланом. Временная экспрессия на поверхности клетки и последующая интернализация PMEL17 делает его подходящей мишенью для разработки конъюгата антитела и лекарственного средства (ADC) для лечения меланомы.
PMEL17 и рак
[0003] По мере созревания PMEL17 в значительной степени подвергается процессингу под воздействием пропротеиновых конвертаз. Белок расщепляется между V467/K468, образуя два субдомена, Mα с N-конца и Mβ с C-конца, предположительно поддерживаемых посредством дисульфидного мостика. После выхода из аппарата Гольджи некоторые молекулы PMEL17 временно экспрессируются на поверхности клетки. Большая часть PMEL17 затем перенаправляется в меланоциты для дальнейшего созревания, в то время как некоторое количество PMEL17, по-видимому, удаляется. После дополнительного ферментативного расщепления PMEL17 распадается на различные домены, которые реорганизуются и образуют фибриллярные листы, в которых происходит полимеризация меланина (Valencia JC, et al. Sorting of Pmel17 to melanosomes through the plasma membrane by AP1 and AP2: evidence for the polarized nature of melanocytes. J Cell Sci. 2006 Mar 15;119(Pt 6):1080-91; Theos AC, et al. The Silver locus product Pmel17/gp100/Silv/ME20: controversial in name and in function. Pigment Cell Res. 2005 Oct;18(5):322-36).
[0004] PMEL17 представляет собой потенциальную терапевтическую мишень для лечения меланомы. PMEL17 является прямой транскрипционной мишенью для MITF, онкогена линии дифференцировки в меланоме, как показали исследования экспрессии мРНК (Du J, et al. MLANA/MART1 and SILV/PMEL17/GP100 are transcriptionally regulated by MITF in melanocytes and melanoma. Am J Pathol. 2003 Jul;163(1):333-43). Экспрессия PMEL17 ограничена линиями дифференцировки меланоцитов, которые включают меланоциты кожи, меланоциты волосяной луковицы, пигментный эпителий сетчатки, пигментированный ресничный эпителий и, возможно, меланоциты сосудистой оболочки сетчатки. PMEL17 также экспрессируется на высоком уровне в опухолях меланоцитарного происхождения, таких как подкожная и увеальная меланома. В отличие от этого исследования мРНК продемонстрировали, что экспрессия PMEL17 ограничена в других типах опухолей и нормальных тканях (Wagner SN, Wagner C, Schultewolter T, Goos M. Analysis of Pmel17/gp100 expression in primary human tissue specimens: implications for melanoma immuno-and gene-therapy. Cancer Immunol Immunother. 1997 Jun;44(4):239-47). Кроме того, ранее было описано, что соединения на основе ADC и ImmTAC, целенаправленно воздействующие на PMEL17, вызывают специфическое уничтожение меланомы in vivo и in vitro, и в настоящее время они проходят клинические испытания (Chen Y, et al. The melanosomal protein PMEL17 as a target for antibody drug conjugate therapy in melanoma. J Biol Chem. 2012 Jul 13;287(29):24082-91. doi:10.1074/jbc.M112.361485. Epub 2012 May 21).
GNAQ/GNA11 и рак
[0005] Гены GNAQ и GNA11 кодируют альфа-субъединицу гетеротримерных G-белков Gq/11, которые практически повсеместно экспрессируются и действуют как бинарные молекулярные переключатели, которые циклически переключаются между активным, связанным с гуанозинтрифосфатом (GTP), и неактивным, связанным с гуанозиндифосфатом (GDP), состояниями. GTP-связанные Gαq и Gα11 активируют β-изоформы фосфолипазы C, которая запускает ряд путей передачи сигналов посредством генерации вторичных мессенджеров IP3 и DAG. Прекращение передачи сигналов обусловлено гидролизом GTP, опосредованным собственной GTPазной активностью этих Gα-белков. Было показано, что Gq и G11 участвуют в широком спектре физиологических функций, включая активацию тромбоцитов, гипертрофию миокарда и регуляцию тонуса гладких мышц.
[0006] Онкогенные мутации в GNAQ или GNA11 встречаются не более чем в 90% случаев увеальной меланомы (UM) и в ~ 2-3% меланомы кожи. Примерно 95% этих мутаций затрагивают кодоны 209 (Q209) в Ras-подобном домене, что приводит к полной или частичной потере активности GTPазы и, таким образом, блокирует GNAQ/11 в его активном состоянии. Q209 GNAQ/11 являются доминантно действующими онкогенами, которые трансформируют меланоциты, запуская активацию нескольких путей, включая PKC/MAPK, Rho/Rac, β-катенин и YAP. Хотя путь PKC/MAPK был показан как один из факторов, способствующих GNAQ-опосредованному онкогенезу, многочисленные доказательства свидетельствуют о том, что мутантный GNAQ/11 регулирует дополнительные пути, которые также, вероятно, играют роль в онкогенезе UM (т. е. YAP, β-катенин). Интересно, что еще одна соматическая активирующая мутация в GNAQ (R183Q) была недавно описана как причина синдрома Стерджа-Вебера (SWS), нейрокожного нарушения, характеризующегося капиллярной мальформацией (винные пятна), а также сосудистыми мальформациями хориоидальных и лептоменингеальных сосудов. Таким образом, GNAQ и GNA11 представляют собой потенциальные терапевтические мишени для лечения увеальной и кожной меланомы.
Конъюгаты антитела и лекарственного средства
[0007] Конъюгаты антитела и лекарственного средства ("ADC") нашли применение в местной доставке цитотоксических средств при лечении рака (см., например, Lambert, Curr. Opinion In Pharmacology 5:543-549, 2005). ADC обеспечивают возможность нацеленной доставки фрагмента, представляющего собой лекарственное средство, при которой может достигаться максимальная эффективность при минимальной токсичности. ADC включают антитело, выбранное по его способности связываться с клеткой, на которую нацелено терапевтическое вмешательство, соединенное с лекарственным средством, выбранным по его цитостатической или цитотоксической активности. Таким образом, связывание антитела с целевой клеткой обеспечивает доставку лекарственного средства в сайт, в котором требуется его терапевтический эффект.
[0008] Было раскрыто множество антител для применения в ADC, которые распознают и селективно связываются с целевыми клетками, например раковыми клетками. Несмотря на масштабную работу с ADC, связывания антитела с конкретной мишенью, представляющей интерес, недостаточно для прогнозирования успеха в применениях ADC. Примеры факторов, которые могут воздействовать на терапевтическую эффективность ADC (кроме особенностей, присущих мишени), включают различные аспекты, которые необходимо модифицировать путем тонкой настройки, такие как оптимальная аффинность антитела как баланс между мишень-опосредованным распределением (TMDD) и управляющим эффективностью воздействием, оценка Fc-опосредованных функций (антителозависимая клеточноопосредованная цитотоксичность, ADCC), способ конъюгации (сайт-специфическая или нет), соотношение молекул лекарственного средства/полезной нагрузки у такого конъюгата с каждым антителом ("DAR" или "соотношение лекарственное средство/антитело"), расщепляемость или стабильность линкера, стабильность ADC и склонность ADC к образованию агрегатов.
[0009] Остается потребность в антителах, способах прикрепления и цитотоксических полезных нагрузках с улучшенными свойствами для применения в качестве эффективных терапевтических композиций на основе ADC и способов, связанных с ними.
Краткое описание настоящего изобретения
[0010] В одном варианте осуществления настоящей заявки раскрыты антитело или его антигенсвязывающий фрагмент, которые связывают PMEL17, содержащие
вариабельную область тяжелой цепи, которая содержит CDR1 (определяющая комплементарность область 1) тяжелой цепи под SEQ ID NO:1, 4, 5 или 7, CDR2 (определяющая комплементарность область 2) тяжелой цепи под SEQ ID NO:2, 6 или 8 и CDR3 (определяющая комплементарность область 3) тяжелой цепи под SEQ ID NO:3 или 9; и вариабельную область легкой цепи, которая содержит CDR1 (определяющая комплементарность область 1) легкой цепи под SEQ ID NO:14, 17 или 20, CDR2 (определяющая комплементарность область 2) легкой цепи под SEQ ID NO:15 или 18 и CDR3 (определяющая комплементарность область 3) легкой цепи под SEQ ID NO:16 или 19;
вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO:33, 36, 37 или 39, CDR2 тяжелой цепи под SEQ ID NO:34, 38 или 40; CDR3 тяжелой цепи под SEQ ID NO:35 или 41; CDR1 легкой цепи под SEQ ID NO:46, 49 или 52; CDR2 легкой цепи под SEQ ID NO:47 или 50 и CDR3 легкой цепи под SEQ ID NO:48 или 51;
вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO:5, 7, 57 или 60, CDR2 тяжелой цепи под SEQ ID NO:58, 61 или 62; CDR3 тяжелой цепи под SEQ ID NO:59 или 63; CDR1 легкой цепи под SEQ ID NO:68, 71 или 74; CDR2 легкой цепи под SEQ ID NO:69 или 72 и CDR3 легкой цепи под SEQ ID NO:70 или 73;
вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO:79, 82, 83 или 85, CDR2 тяжелой цепи под SEQ ID NO:80, 84 или 86; CDR3 тяжелой цепи под SEQ ID NO:81 или 87; CDR1 легкой цепи под SEQ ID NO:92, 95 или 98; CDR2 легкой цепи под SEQ ID NO:93 или 96 и CDR3 легкой цепи под SEQ ID NO:94 или 97;
вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO:103, 106, 107 или 109, CDR2 тяжелой цепи под SEQ ID NO:104, 108 или 110; CDR3 тяжелой цепи под SEQ ID NO:105 или 111; CDR1 легкой цепи под SEQ ID NO:49, 52 или 116; CDR2 легкой цепи под SEQ ID NO:47 или 50 и CDR3 легкой цепи под SEQ ID NO:117 или 118;
вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO:123, 126, 127 или 129, CDR2 тяжелой цепи под SEQ ID NO:124, 128 или 130; CDR3 тяжелой цепи под SEQ ID NO:125 или 131; CDR1 легкой цепи под SEQ ID NO:136, 139 или 142; CDR2 легкой цепи под SEQ ID NO:137 или 140 и CDR3 легкой цепи под SEQ ID NO:138 или 141;
вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO:123, 126, 127 или 129, CDR2 тяжелой цепи под SEQ ID NO:124, 128 или 130; CDR3 тяжелой цепи под SEQ ID NO:147 или 148; CDR1 легкой цепи под SEQ ID NO:153, 156 или 158; CDR2 легкой цепи под SEQ ID NO:50 или 154 и CDR3 легкой цепи под SEQ ID NO:155 или 157;
вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO:103, 106, 107 или 109, CDR2 тяжелой цепи под SEQ ID NO:104, 108 или 110; CDR3 тяжелой цепи под SEQ ID NO:163 или 164; CDR1 легкой цепи под SEQ ID NO:49, 52 или 116; CDR2 легкой цепи под SEQ ID NO:47 или 50 и CDR3 легкой цепи под SEQ ID NO:169 или 170;
вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO:175, 178, 179 или 181, CDR2 тяжелой цепи под SEQ ID NO:176, 180 или 182; CDR3 тяжелой цепи под SEQ ID NO:177 или 183; CDR1 легкой цепи под SEQ ID NO:49, 52 или 116; CDR2 легкой цепи под SEQ ID NO:47 или 50 и CDR3 легкой цепи под SEQ ID NO:188 или 189;
вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 103, 106, 107 или 109, CDR2 тяжелой цепи под SEQ ID NO: 104, 108 или 110; CDR3 тяжелой цепи под SEQ ID NO:194 или 195; CDR1 легкой цепи под SEQ ID NO: 49, 52 или 116; CDR2 легкой цепи под SEQ ID NO: 47 или 50 и CDR3 легкой цепи под SEQ ID NO:200 или 201;
вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO:206, 209, 210 или 212, CDR2 тяжелой цепи под SEQ ID NO:207, 211 или 213; CDR3 тяжелой цепи под SEQ ID NO:208 или 214; CDR1 легкой цепи под SEQ ID NO:153, 156 или 158; CDR2 легкой цепи под SEQ ID NO:50 или 154 и CDR3 легкой цепи под SEQ ID NO:219 или 220;
вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 206, 209, 210 или 212, CDR2 тяжелой цепи под SEQ ID NO: 207, 211 или 213; CDR3 тяжелой цепи под SEQ ID NO:225 или 226; CDR1 легкой цепи под SEQ ID NO:136, 139 или 142; CDR2 легкой цепи под SEQ ID NO:137 или 140 и CDR3 легкой цепи под SEQ ID NO:231 или 232;
вариабельную область тяжелой цепи, которая содержит вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 206, 209, 210 или 212, HCDR2 под SEQ ID NO: 207, 211 или 213 и HCDR3 под SEQ ID NO:237 или 238; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:243, 245 или 247, LCDR2 под SEQ ID NO:47 или 50 и LCDR3 под SEQ ID NO:244 или 246;
вариабельную область тяжелой цепи, которая содержит вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 206, 209, 210 или 212, HCDR2 под SEQ ID NO: 207, 211 или 213 и HCDR3 под SEQ ID NO:252 или 253; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:153, 156 или 158, LCDR2 под SEQ ID NO:50 или 154 и LCDR3 под SEQ ID NO:258 или 259;
CDR1 тяжелой цепи под SEQ ID NO:1, CDR2 тяжелой цепи под SEQ ID NO:2, CDR3 тяжелой цепи под SEQ ID NO:3, CDR1 легкой цепи под SEQ ID NO:14, CDR2 легкой цепи под SEQ ID NO:15 и CDR3 легкой цепи под SEQ ID NO:16;
CDR1 тяжелой цепи под SEQ ID NO: 4, CDR2 тяжелой цепи под SEQ ID NO:2, CDR3 тяжелой цепи под SEQ ID NO:3, CDR1 легкой цепи под SEQ ID NO:14, CDR2 легкой цепи под SEQ ID NO:15 и CDR3 легкой цепи под SEQ ID NO:16;
CDR1 тяжелой цепи под SEQ ID NO:5, CDR2 тяжелой цепи под SEQ ID NO:6, CDR3 тяжелой цепи под SEQ ID NO:3, CDR1 легкой цепи под SEQ ID NO:17, CDR2 легкой цепи под SEQ ID NO: 18 и CDR3 легкой цепи под SEQ ID NO: 19;
CDR1 тяжелой цепи под SEQ ID NO:7, CDR2 тяжелой цепи под SEQ ID NO:8, CDR3 тяжелой цепи под SEQ ID NO:9, CDR1 легкой цепи под SEQ ID NO:20, CDR2 легкой цепи под SEQ ID NO:18 и CDR3 легкой цепи под SEQ ID NO:16;
CDR1 тяжелой цепи под SEQ ID NO:33, CDR2 тяжелой цепи под SEQ ID NO:34, CDR3 тяжелой цепи под SEQ ID NO:35, CDR1 легкой цепи под SEQ ID NO:46, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:48;
CDR1 тяжелой цепи под SEQ ID NO:36, CDR2 тяжелой цепи под SEQ ID NO:34, CDR3 тяжелой цепи под SEQ ID NO:35, CDR1 легкой цепи под SEQ ID NO:46, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:48;
CDR1 тяжелой цепи под SEQ ID NO:37, CDR2 тяжелой цепи под SEQ ID NO:38, CDR3 тяжелой цепи под SEQ ID NO:35, CDR1 легкой цепи под SEQ ID NO:49, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:51;
CDR1 тяжелой цепи под SEQ ID NO: 39, CDR2 тяжелой цепи под SEQ ID NO:40, CDR3 тяжелой цепи под SEQ ID NO:41, CDR1 легкой цепи под SEQ ID NO:52, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:48;
CDR1 тяжелой цепи под SEQ ID NO:57, CDR2 тяжелой цепи под SEQ ID NO:58, CDR3 тяжелой цепи под SEQ ID NO:59, CDR1 легкой цепи под SEQ ID NO:68, CDR2 легкой цепи под SEQ ID NO:69 и CDR3 легкой цепи под SEQ ID NO:70;
CDR1 тяжелой цепи под SEQ ID NO:60, CDR2 тяжелой цепи под SEQ ID NO:58, CDR3 тяжелой цепи под SEQ ID NO:59, CDR1 легкой цепи под SEQ ID NO:68, CDR2 легкой цепи под SEQ ID NO:69 и CDR3 легкой цепи под SEQ ID NO:70;
CDR1 тяжелой цепи под SEQ ID NO:5, CDR2 тяжелой цепи под SEQ ID NO:61, CDR3 тяжелой цепи под SEQ ID NO:59, CDR1 легкой цепи под SEQ ID NO:71, CDR2 легкой цепи под SEQ ID NO:72 и CDR3 легкой цепи под SEQ ID NO:73;
CDR1 тяжелой цепи под SEQ ID NO:7, CDR2 тяжелой цепи под SEQ ID NO:62, CDR3 тяжелой цепи под SEQ ID NO:63, CDR1 легкой цепи под SEQ ID NO:74, CDR2 легкой цепи под SEQ ID NO:72 и CDR3 легкой цепи под SEQ ID NO:70;
CDR1 тяжелой цепи под SEQ ID NO:79, CDR2 тяжелой цепи под SEQ ID NO:80, CDR3 тяжелой цепи под SEQ ID NO:81, CDR1 легкой цепи под SEQ ID NO:92, CDR2 легкой цепи под SEQ ID NO:93 и CDR3 легкой цепи под SEQ ID NO:94;
CDR1 тяжелой цепи под SEQ ID NO:82, CDR2 тяжелой цепи под SEQ ID NO:80, CDR3 тяжелой цепи под SEQ ID NO:81, CDR1 легкой цепи под SEQ ID NO:92, CDR2 легкой цепи под SEQ ID NO:93 и CDR3 легкой цепи под SEQ ID NO:94;
CDR1 тяжелой цепи под SEQ ID NO:83, CDR2 тяжелой цепи под SEQ ID NO:84, CDR3 тяжелой цепи под SEQ ID NO:81, CDR1 легкой цепи под SEQ ID NO:95, CDR2 легкой цепи под SEQ ID NO:96 и CDR3 легкой цепи под SEQ ID NO: 97;
CDR1 тяжелой цепи под SEQ ID NO: 85, CDR2 тяжелой цепи под SEQ ID NO:86, CDR3 тяжелой цепи под SEQ ID NO:87, CDR1 легкой цепи под SEQ ID NO:98, CDR2 легкой цепи под SEQ ID NO:96 и CDR3 легкой цепи под SEQ ID NO:94;
CDR1 тяжелой цепи под SEQ ID NO:103, CDR2 тяжелой цепи под SEQ ID NO:104, CDR3 тяжелой цепи под SEQ ID NO:105, CDR1 легкой цепи под SEQ ID NO: 116; CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:117;
CDR1 тяжелой цепи под SEQ ID NO:106, CDR2 тяжелой цепи под SEQ ID NO:104, CDR3 тяжелой цепи под SEQ ID NO:105, CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:117;
CDR1 тяжелой цепи под SEQ ID NO:107, CDR2 тяжелой цепи под SEQ ID NO:108, CDR3 тяжелой цепи под SEQ ID NO:105, CDR1 легкой цепи под SEQ ID NO:49, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:118;
CDR1 тяжелой цепи под SEQ ID NO:109, CDR2 тяжелой цепи под SEQ ID NO:110, CDR3 тяжелой цепи под SEQ ID NO:111, CDR1 легкой цепи под SEQ ID NO:52, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:117;
CDR1 тяжелой цепи под SEQ ID NO:123, CDR2 тяжелой цепи под SEQ ID NO:124, CDR3 тяжелой цепи под SEQ ID NO:125, CDR1 легкой цепи под SEQ ID NO:136, CDR2 легкой цепи под SEQ ID NO:137 и CDR3 легкой цепи под SEQ ID NO:138;
CDR1 тяжелой цепи под SEQ ID NO:126, CDR2 тяжелой цепи под SEQ ID NO:124, CDR3 тяжелой цепи под SEQ ID NO:125, CDR1 легкой цепи под SEQ ID NO:136, CDR2 легкой цепи под SEQ ID NO:137 и CDR3 легкой цепи под SEQ ID NO:138;
CDR1 тяжелой цепи под SEQ ID NO:127, CDR2 тяжелой цепи под SEQ ID NO:128, CDR3 тяжелой цепи под SEQ ID NO:125, CDR1 легкой цепи под SEQ ID NO:139, CDR2 легкой цепи под SEQ ID NO:140 и CDR3 легкой цепи под SEQ ID NO: 141;
CDR1 тяжелой цепи под SEQ ID NO: 129, CDR2 тяжелой цепи под SEQ ID NO:130, CDR3 тяжелой цепи под SEQ ID NO:131, CDR1 легкой цепи под SEQ ID NO:142, CDR2 легкой цепи под SEQ ID NO:140 и CDR3 легкой цепи под SEQ ID NO:138;
CDR1 тяжелой цепи под SEQ ID NO:123, CDR2 тяжелой цепи под SEQ ID NO:124, CDR3 тяжелой цепи под SEQ ID NO:147, CDR1 легкой цепи под SEQ ID NO:153, CDR2 легкой цепи под SEQ ID NO:154 и CDR3 легкой цепи под SEQ ID NO:155;
CDR1 тяжелой цепи под SEQ ID NO:126, CDR2 тяжелой цепи под SEQ ID NO:124, CDR3 тяжелой цепи под SEQ ID NO:147, CDR1 легкой цепи под SEQ ID NO:153, CDR2 легкой цепи под SEQ ID NO: 154 и CDR3 легкой цепи под SEQ ID NO:155;
CDR1 тяжелой цепи под SEQ ID NO:127, CDR2 тяжелой цепи под SEQ ID NO:128, CDR3 тяжелой цепи под SEQ ID NO:147, CDR1 легкой цепи под SEQ ID NO:156, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:157;
CDR1 тяжелой цепи под SEQ ID NO: 129, CDR2 тяжелой цепи под SEQ ID NO:130, CDR3 тяжелой цепи под SEQ ID NO:148, CDR1 легкой цепи под SEQ ID NO:158, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:155;
CDR1 тяжелой цепи под SEQ ID NO:103, CDR2 тяжелой цепи под SEQ ID NO:104, CDR3 тяжелой цепи под SEQ ID NO:163, CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:169;
CDR1 тяжелой цепи под SEQ ID NO:106, CDR2 тяжелой цепи под SEQ ID NO:104, CDR3 тяжелой цепи под SEQ ID NO:163, CDR1 легкой цепи под SEQ ID NO:116, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:169;
CDR1 тяжелой цепи под SEQ ID NO:107, CDR2 тяжелой цепи под SEQ ID NO:108, CDR3 тяжелой цепи под SEQ ID NO:163, CDR1 легкой цепи под SEQ ID NO:49, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:170;
CDR1 тяжелой цепи под SEQ ID NO: 109, CDR2 тяжелой цепи под SEQ ID NO:110, CDR3 тяжелой цепи под SEQ ID NO:164, CDR1 легкой цепи под SEQ ID NO:52, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:169;
CDR1 тяжелой цепи под SEQ ID NO:175, CDR2 тяжелой цепи под SEQ ID NO:176, CDR3 тяжелой цепи под SEQ ID NO:177, CDR1 легкой цепи под SEQ ID NO:116, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:188;
CDR1 тяжелой цепи под SEQ ID NO:178, CDR2 тяжелой цепи под SEQ ID NO:176, CDR3 тяжелой цепи под SEQ ID NO:177, CDR1 легкой цепи под SEQ ID NO:116, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:188;
CDR1 тяжелой цепи под SEQ ID NO:179, CDR2 тяжелой цепи под SEQ ID NO:180, CDR3 тяжелой цепи под SEQ ID NO:177, CDR1 легкой цепи под SEQ ID NO:49, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:189;
CDR1 тяжелой цепи под SEQ ID NO: 181, CDR2 тяжелой цепи под SEQ ID NO:182, CDR3 тяжелой цепи под SEQ ID NO:183, CDR1 легкой цепи под SEQ ID NO:52, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:188;
CDR1 тяжелой цепи под SEQ ID NO: 103, CDR2 тяжелой цепи под SEQ ID NO: 104, CDR3 тяжелой цепи под SEQ ID NO:194, CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO:200;
CDR1 тяжелой цепи под SEQ ID NO: 106, CDR2 тяжелой цепи под SEQ ID NO: 104, CDR3 тяжелой цепи под SEQ ID NO:194, CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO:200;
CDR1 тяжелой цепи под SEQ ID NO: 107, CDR2 тяжелой цепи под SEQ ID NO: 108, CDR3 тяжелой цепи под SEQ ID NO:194, CDR1 легкой цепи под SEQ ID NO: 49, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 201;
CDR1 тяжелой цепи под SEQ ID NO: 109, CDR2 тяжелой цепи под SEQ ID NO: 110, CDR3 тяжелой цепи под SEQ ID NO:195, CDR1 легкой цепи под SEQ ID NO: 52, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO:200;
CDR1 тяжелой цепи под SEQ ID NO:206, CDR2 тяжелой цепи под SEQ ID NO:207, CDR3 тяжелой цепи под SEQ ID NO:208, CDR1 легкой цепи под SEQ ID NO:153, CDR2 легкой цепи под SEQ ID NO:154 и CDR3 легкой цепи под SEQ ID NO:219;
CDR1 тяжелой цепи под SEQ ID NO:209, CDR2 тяжелой цепи под SEQ ID NO:207, CDR3 тяжелой цепи под SEQ ID NO:208, CDR1 легкой цепи под SEQ ID NO:153, CDR2 легкой цепи под SEQ ID NO: 154 и CDR3 легкой цепи под SEQ ID NO:219;
CDR1 тяжелой цепи под SEQ ID NO:210, CDR2 тяжелой цепи под SEQ ID NO:211, CDR3 тяжелой цепи под SEQ ID NO:208, CDR1 легкой цепи под SEQ ID NO:156, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:220;
CDR1 тяжелой цепи под SEQ ID NO: 212, CDR2 тяжелой цепи под SEQ ID NO:213, CDR3 тяжелой цепи под SEQ ID NO:214, CDR1 легкой цепи под SEQ ID NO:158, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:219;
CDR1 тяжелой цепи под SEQ ID NO: 206, CDR2 тяжелой цепи под SEQ ID NO: 207, CDR3 тяжелой цепи под SEQ ID NO:225, CDR1 легкой цепи под SEQ ID NO:136, CDR2 легкой цепи под SEQ ID NO:137 и CDR3 легкой цепи под SEQ ID NO:231;
CDR1 тяжелой цепи под SEQ ID NO: 209, CDR2 тяжелой цепи под SEQ ID NO: 207, CDR3 тяжелой цепи под SEQ ID NO:225, CDR1 легкой цепи под SEQ ID NO:136, CDR2 легкой цепи под SEQ ID NO:137 и CDR3 легкой цепи под SEQ ID NO:231;
CDR1 тяжелой цепи под SEQ ID NO: 210, CDR2 тяжелой цепи под SEQ ID NO: 211, CDR3 тяжелой цепи под SEQ ID NO:225, CDR1 легкой цепи под SEQ ID NO:139, CDR2 легкой цепи под SEQ ID NO:140 и CDR3 легкой цепи под SEQ ID NO: 232;
CDR1 тяжелой цепи под SEQ ID NO: 212, CDR2 тяжелой цепи под SEQ ID NO: 213, CDR3 тяжелой цепи под SEQ ID NO: 226, CDR1 легкой цепи под SEQ ID NO:142; CDR2 легкой цепи под SEQ ID NO: 140 и CDR3 легкой цепи под SEQ ID NO:231;
вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 206, HCDR2 под SEQ ID NO: 207 и HCDR3 под SEQ ID NO:237, и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:243, LCDR2 под SEQ ID NO:47 и LCDR3 под SEQ ID NO:244;
вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 209, HCDR2 под SEQ ID NO: 207 и HCDR3 под SEQ ID NO:237, и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:243, LCDR2 под SEQ ID NO:47 и LCDR3 под SEQ ID NO:244;
вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 210, HCDR2 под SEQ ID NO: 211 и HCDR3 под SEQ ID NO:237, и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:245, LCDR2 под SEQ ID NO:50 и LCDR3 под SEQ ID NO:246;
вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 212, HCDR2 под SEQ ID NO: 213 и HCDR3 под SEQ ID NO:238, и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:247, LCDR2 под SEQ ID NO: 50 и LCDR3 под SEQ ID NO:244;
вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 206, HCDR2 под SEQ ID NO: 207 и HCDR3 под SEQ ID NO:252, и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:153, LCDR2 под SEQ ID NO: 154 и LCDR3 под SEQ ID NO:258;
вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 209, HCDR2 под SEQ ID NO: 207 и HCDR3 под SEQ ID NO:252, и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:153, LCDR2 под SEQ ID NO:154 и LCDR3 под SEQ ID NO:258;
вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 210, HCDR2 под SEQ ID NO: 211 и HCDR3 под SEQ ID NO:252, и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:156, LCDR2 под SEQ ID NO:50 и LCDR3 под SEQ ID NO:259; или
вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 212, HCDR2 под SEQ ID NO: 213 и HCDR3 под SEQ ID NO: 253, и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:158, LCDR2 под SEQ ID NO:50 и LCDR3 под SEQ ID NO:258.
[0011] Антитело или его антигенсвязывающий фрагмент, которые связывают PMEL17 по настоящей заявке, также могут содержать:
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:10, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:21;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:10, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:25;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:10, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:29;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:42, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:53;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:64, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:75;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:88, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:99;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:112, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:119;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:132, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:143;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:149, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:159;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:165, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:171;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:184, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:190;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:196, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:202;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:215, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:221;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:227, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:233;
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:239, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:248; или
вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:254, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:260.
[0012] В другом варианте осуществления антитело или его антигенсвязывающий фрагмент, которые связывают PMEL17, содержат:
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:12, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:23;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:12, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:27;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:12, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:31;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:44, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:55;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:66, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:77;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:90, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:101;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:114, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:121;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:134, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:145;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:151, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:161;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:167, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:173;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:186, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:192;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:198, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:204;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:217, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:223;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:229, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:235;
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:241, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:250; или
тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:256, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:262.
[0013] Антитело или его антигенсвязывающий фрагмент, описанные в данном документе, могут содержать одну или несколько цистеиновых замен. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент содержат одну или несколько цистеиновых замен, выбранных из S152C, S375C или обеих S152C и S375C, в тяжелой цепи антитела или его антигенсвязывающего фрагмента, где нумерация положения соответствует системе EU. Антитело, раскрытое в данном документе, может представлять собой моноклональное антитело.
[0014] В одном аспекте конъюгат антитела и лекарственного средства по настоящему изобретению представляет собой конъюгат формулы (C):
Ab-(LA-(D)n)y (C),
где
D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11;
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
LA представляет собой линкер;
n равняется 1, 2, 3 или 4 и
y равняется 1, 2, 3 или 4,
где фрагмент, представляющий собой линкер-лекарственное средство -(LA-(D)n), ковалентно присоединен к антителу или его антигенсвязывающему фрагменту.
[0015] В другом аспекте конъюгатов антитела и лекарственного средства формулы (C) LA представляет собой расщепляемый линкер, содержащий один или несколько линкерных компонентов, выбранных из саморасщепляющегося спейсера, фосфатной группы, карбонатной группы и бивалентного пептидного линкера.
[0016] В другом аспекте конъюгат антитела и лекарственного средства формулы (C) представляет собой конъюгат формулы (C-1):
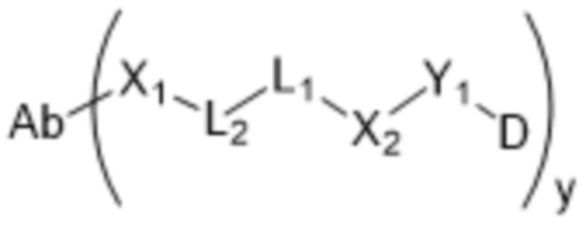 (C-1),
(C-1),
где
D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11;
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
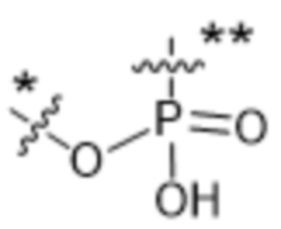
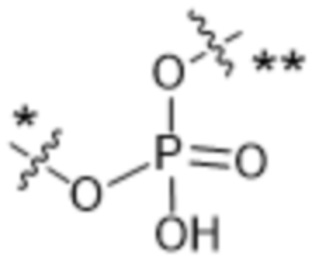
Y1 представляет собой 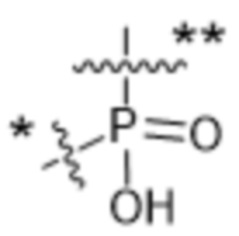 ,
,  ,
,  ,
, 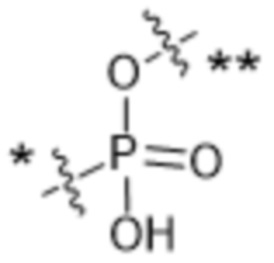 ,
, 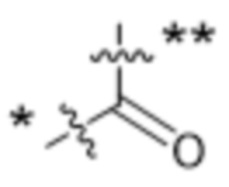 ,
, 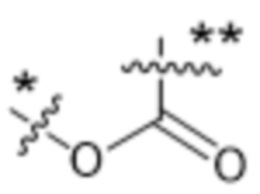 ,
, 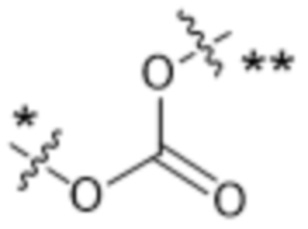 или
или 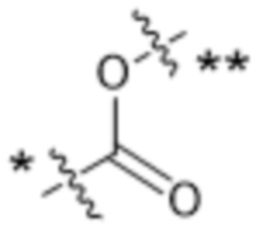 , где * в Y1 указывает на точку присоединения к X2, а ** в Y1 указывает на точку присоединения к D;
, где * в Y1 указывает на точку присоединения к X2, а ** в Y1 указывает на точку присоединения к D;
L1 представляет собой бивалентный пептидный линкер;
L2 представляет собой связь или линкер, и
y равняется 1, 2, 3 или 4.
[0017] В настоящей заявке также раскрыты фармацевтические композиции, содержащие антитела или их антигенсвязывающие фрагменты, раскрытые в данном документе, и фармацевтически приемлемый носитель. В настоящей заявке также раскрыты фармацевтические композиции, содержащие конъюгаты антитела и лекарственного средства, раскрытые в данном документе, и фармацевтически приемлемый носитель.
[0018] В настоящей заявке также раскрыты способы лечения или предупреждения рака нуждающегося в этом у пациента, предусматривающие введение указанному пациенту конъюгатов антитела и лекарственного средства или фармацевтических композиций, раскрытых в данном документе, где рак экспрессирует PMEL17, содержит мутацию гена GNAQ или GNA11, или рак экспрессирует PMEL17 и содержит мутацию GNAQ, GNA11 или обе.
[0019] В некоторых вариантах осуществления способов лечения или предупреждения рака конъюгат антитела и лекарственного средства или фармацевтическую композицию вводят пациенту в комбинации с одним или несколькими дополнительными терапевтическими соединениями. В одном варианте осуществления одно или несколько дополнительных терапевтических соединений выбраны из стандартного химиотерапевтического средства, ингибитора MDM2, ингибитора MRC2, ингибитора PKC, ингибитора MAPK, костимулирующей молекулы или ингибитора контрольных точек иммунного ответа. В одном варианте осуществления костимулирующая молекула выбрана из агониста лиганда OX40, CD2, CD27, CDS, ICAM-1, LFA-1 (CD11a/CD18), ICOS (CD278), 4-1BB (CD137), GITR, CD30, CD40, BAFFR, HVEM, CD7, LIGHT, NKG2C, SLAMF7, NKp80, CD160, B7-H3, STING, или CD83. В другом варианте осуществления ингибитор контрольных точек иммунного ответа выбран из ингибитора PD-1, PD-L1, PD-L2, CTLA4, TIM3, LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и/или TGFR бета.
[0020] В настоящей заявке также раскрыты конъюгаты антитела и лекарственного средства или фармацевтические композиции, раскрытые в данном документе, для применения в качестве лекарственного препарата. В одном варианте осуществления конъюгаты антитела и лекарственного средства или фармацевтические композиции, раскрытые в данном документе, предназначены для применения в лечении или предупреждении рака, экспрессирующего PMEL17, или рака, который содержит мутацию гена GNAQ или GNA11, у нуждающегося в этом пациента.
[0021] В одном варианте осуществления настоящей заявки раскрыто применение антител или их антигенсвязывающих фрагментов, конъюгатов антитела и лекарственного средства или фармацевтических композиций, раскрытых в данном документе, для лечения или предупреждения рака, экспрессирующего PMEL17, у нуждающегося в этом пациента.
[0022] В одном варианте осуществления настоящей заявки раскрыто применение антител или их антигенсвязывающих фрагментов, конъюгатов антитела и лекарственного средства или фармацевтических композиций, раскрытых в данном документе, для лечения или предупреждения рака, экспрессирующего PMEL17, или рака, который содержит мутацию гена GNAQ или GNA11, у нуждающегося в этом пациента. В одном варианте осуществления настоящей заявки раскрыто применение антител или их антигенсвязывающих фрагментов, конъюгатов антитела и лекарственного средства или фармацевтических композиций, раскрытых в данном документе, в изготовлении лекарственного препарата.
[0023] В одном варианте осуществления рак экспрессирует PMEL17 или содержит мутацию гена GNAQ или GNA11. В одном варианте осуществления рак представляет собой увеальную меланому, подкожную меланому, гепатоцеллюлярную карциному или метастатическую форму данных видов рака.
[0024] В настоящей заявке также раскрыты нуклеиновые кислоты, которые кодируют антитела или антигенсвязывающие фрагменты, раскрытые в данном документе. В одном варианте осуществления нуклеиновая кислота содержит нуклеотидную последовательность под SEQ ID NO: 13, 24, 28, 32, 45, 56, 67, 78, 91, 102, 115, 122, 135, 146, 152, 162, 168, 174, 187, 193, 199, 205, 218, 224, 230, 236, 242, 251, 257, или 26. В данной заявке также раскрыты векторы, содержащие нуклеиновые кислоты, и клетки-хозяева, содержащие векторы или нуклеиновые кислоты. В данной заявке также раскрыт способ получения антител или антигенсвязывающих фрагментов, раскрытых в данном документе, предусматривающий культивирование клетки-хозяина и извлечения антитела из культуры клеток. В одном варианте осуществления способ извлечения антитела из культуры клеток предусматривает следующие стадии:
a) удаление клеток и фильтрование культуры;
b) проведение очистки культуры посредством аффинной хроматографии;
c) инактивация любых вирусов в культуре путем доведения pH до 3,4-3,6, затем повторного доведения pH до 5,8-6,2 и фильтрования культуры;
d) проведение очистки культуры посредством катионообменной хроматографии и осуществления восстановления культуры на колонке;
e) осуществление анионообменной хроматографии в отношении культуры;
f) удаление вирусов посредством нанофильтрации;
g) фильтрование культуры, содержащей антитело, и
h) получение очищенного антитела.
[0025] В настоящей заявке также раскрыт способ получения конъюгата антитела к PMEL17 и лекарственного средства, предусматривающий:
(a) предварительное образование фрагмента, представляющего собой линкер-лекарственное средство, следующей формулы (B):
R8-LB-(D)n (B),
где
D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11;
R8 представляет собой реакционноспособную группу;
LB представляет собой расщепляемый или нерасщепляемый линкер и
n равняется 1, 2, 3 или 4;
(b) конъюгирование указанного фрагмента, представляющего собой линкер-лекарственное средство, с антителом, извлеченным из культуры клеток с применением способа получения антитела или антигенсвязывающего фрагмента, раскрытого в данном документе, с получением конъюгата антитела и лекарственного средства; и
(c) проведение очистки конъюгата антитела и лекарственного средства.
[0026] В настоящей заявке также раскрыт диагностический реагент, содержащий антитело или его антигенсвязывающий фрагмент, раскрытые в данном документе. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент являются меченными с помощью радиоактивной метки, флуорофора, хромофора, средства для визуализации или иона металла.
Краткое описание графических материалов
[0027] На фиг. 1A - 1B показаны иллюстративные данные об in vitro активности против UM ингибиторов GNAQ/11, соединения (A1) и соединения (A2).
[0028] На фиг. 2 показаны иллюстративные данные об активности ингибиторов GNAQ/11, соединения (A1) и соединения (A2), в отношении индуцирования апоптоза в клетках увеальной меланомы.
[0029] На фиг. 3A - 3C показаны иллюстративные данные об ингибировании GNAQ/11 соединением (A1) и соединением (A2). Соединение (A1) и соединение (A2) снижали уровни IP1 (фиг. 3A) и относительную пролиферацию (фиг. 3B) в клетках 92.1. Иммуноблоты с образцами клеток 92.1, обработанных соединением (A1) и соединением (A2), показали снижение передачи сигналов ERK (фиг. 3C).
[0030] На фиг. 4A - 4D показаны иллюстративные данные о метаболической стабильности и PK-показателям соединения (A1). Исчезновение соединения (A1) (фиг. 4A), а также появление формы с раскрытым кольцом соединения (A8) (фиг. 4B) контролировали в течение 24 ч. За исключением крыс суммирование % оставшегося соединения (A1) и % образовавшегося соединения (A8) демонстрирует стехиометрию в течение 24 часов (фиг. 4C). PK соединения (A1) после внутривенного введения мышам характеризуется очень высоким клиренсом и объемом распределения от умеренного до высокого (фиг. 4D).
[0031] На фиг. 5A - 5D показаны иллюстративные данные о метаболической стабильности и PK-показателям соединения (A1) и соединения (A2). In vitro стабильность соединения (A2) тестировали в плазме крови и в крови различных видов (фиг. 5A) Соединение (A2) показало приемлемую химическую стабильность в трех различных системах (фиг. 5B). PK соединения (A2) у самок мышей balb/c показала высокий клиренс и короткий период полувыведения (фиг. 5C). Соединение (A1) и соединение (A2) были стабильны в буфере при pH 5,6 и в лизосомах в течение 4 ч (фиг. 5D).
[0032] На фиг. 6A - 6B показаны иллюстративные данные об in vitro активности ADC к PMEL17-(B1) против увеальной меланомы. Данные представлены в виде среднего значения для 3 независимых повторов и относительно клеток, обработанных PBS (контроль).
[0033] На фиг. 7 показаны иллюстративные данные об ADC к PMEL17-(B1), индуцирующих апоптоз в клетках увеальной меланомы. Данные представлены в виде среднего значения для 3 независимых повторов.
[0034] На фиг. 8A - 8B показаны иллюстративные данные об in vitro активности ADC к PMEL17-(B2) и mAb к PMEL17 против увеальной меланомы. Данные представлены в виде среднего значения для 3 независимых повторов и относительно клеток, обработанных PBS (контроль).
[0035] На фиг. 9 показаны иллюстративные данные об ингибировании GNAQ/11 с помощью ADC к PMEL17-(B1) и к PMEL17-(B2) в клетках увеальной меланомы. Уровни IP1 (нM) представлены в виде среднего значения для 3 независимых повторов.
[0036] На фиг. 10A - 10D показаны иллюстративные данные об активности связывания антител к PMEL17 с интактными тромбоцитами и клетками увеальной меланомы.
[0037] На фиг. 11A - 11C показаны иллюстративные данные о влиянии соединения (A1) и ADC к PMEL17-(B1) на агрегацию тромбоцитов человека.
[0038] На фиг. 12A - 12E показаны иллюстративные данные о противоопухолевой активности in vivo для ADC к PMEL17-(B1). G1-(B1) ингибировал рост опухоли дозозависимым образом (фиг. 12A). Значения являются средними ± SEM; размер выборки (n=5-12 мышей в группе). Исходный объем опухоли в день 0 составлял примерно 200-250 мм3. В течение 14 дней после обработки потеря веса тела не наблюдалась (фиг. 12B). Значения являются средними ± SEM; размер выборки (n=4 мыши в группе). Обработка с использованием G1-(B1) приводила к ингибированию передачи сигналов GNAQ и ингибированию пролиферации опухолевых клеток, о чем свидетельствуют сниженные уровни pERK и Ki67 соответственно (фиг. 12C). Кроме того, G1-(B1) индуцировал апоптоз клеток по сравнению с получавшими среду-носитель и изотипический контроль 3207-(B1) мышами, что коррелировало с накоплением в опухолевых клетках ADC G1-(B1), обнаруживаемого при окрашивании с IgG (фиг. 12C) После ингибирования GNAQ изменений в уровнях MITF и PMEL17 не наблюдали (фиг. 12C). У мышей, получавших G1-(B1), не наблюдали ингибирования агрегации тромбоцитов в течение 7 дней (фиг. 12D и E).
[0039] На фиг. 13A - 13C показаны иллюстративные данные о влиянии ADC G1-(B1) на мышиной модели увеальной меланомы с метастазами в печень и легкие. Отдельные изображения от каждой мыши представлены в день 45 после в/в инъекции клеток 92.1 с люциферазой (непосредственно перед началом обработки) и через 12 дней после обработки (фиг. 13A и 13B); размер выборки (n=6 мышей в группе). Исходный BLI для метастазов в печень в день 0 составил приблизительно 2,8*109 фотонов/сек/см2. Опухоли в легких (сигнал биолюминесценции) на фиг. 13B обозначены черной стрелкой. Соответствующее изменение веса тела (% по сравнению с днем 15) оценивали 2-3 раза в неделю до и после обработки с использованием 20 мг/кг G1-(B1) (серые круги). Значения на фиг. 13C являются средними ± SEM; размер выборки (n=5-6 мышей в группе). Исходная масса тела в день 15 составляла примерно 21 г.
[0040] На фиг. 14A - 14E показаны иллюстративные данные о PK-показателях ADC G1-(B1). Фармакокинетический профиль (общие уровни IgG) G1-(B1) показал несколько непропорциональное увеличение воздействия при дозе от 7,5 до 30 мг/кг у «голых» мышей (фиг. 14A). У мышей с опухолью измеряли концентрации свободной полезной нагрузки после введения либо связывающего мишень G1-(B1), либо изотипического контроля 3207-(B1). Выраженное (> 4-кратное) увеличение доставки в опухоль полезной нагрузки соединения (A1) можно было наблюдать при использовании целенаправленно воздействующего ADC (фиг. 14B). Превращение соединения (A1) (незакрашенные круги) в его форму с раскрытым кольцом, соединение (A8), (закрашенные круги) при конъюгировании с антителом было показано in vivo на мышах (фиг. 14C). Виды воздействия в исследовании эффективности in vivo, сравнивающем два разных формата DAR2 с форматом DAR4 для G1-(B1) и с форматом Fc-молчащий DAR4, показали наиболее низкий клиренс для ADC DAR2 (E152C) и Fc-молчащего DAR4, тогда как воздействие DAR2 (S375C) уменьшается быстрее (фиг. 14D). На фиг. 14E показана концентрация конъюгатов 3207 (контрольное изотипическое антитело)-(B1) DAR4 (E152C, S375C) и 3207 (контрольное изотипическое антитело)-(B1) Fc-молчащий DAR4 с течением времени.
[0041] На фиг. 15A - 15C показаны иллюстративные данные об in vitro стабильности ADC к PMEL17-GNAQ/11i в буфере, плазме крови мыши, крысы и человека и in vivo стабильности ADC к PMEL17-GNAQ/11i у мышей.
[0042] На фиг. 16A - 16B показаны иллюстративные данные об in vivo эффективности G1-E152C-DAR2-(B1), G1-S375C-DAR2-(B1), Fc-молчащего G1-(B1) на модели ксенотрансплантата увеальной меланомы. Значения представлены в виде среднего ± SEM; размер выборки (n=5-6 мышей в группе). Исходный объем опухоли в день 0 составлял примерно 300-325 мм3.
[0043] На фиг. 17A - 17B показаны иллюстративные данные об in vitro активности ADC к PMEL17-(B1) против увеальной меланомы. Данные представлены в виде среднего значения для 3 независимых повторов и относительно клеток, обработанных PBS (контроль).
[0044] На фиг. 18 показаны иллюстративные данные о противоопухолевой активности in vivo ADC к PMEL17-(B1).
[0045] На фиг. 19A - 19B показаны иллюстративные данные иммуногистохимического анализа опухолевых биоптатов от пациентов с метастатической увеальной меланомой.
[0046] На фиг. 20A - 20C показаны иллюстративные данные сенсограммы по оценке эпитоп-специфической сортировки антител к PMEL. На фиг. 20A проиллюстрированы стадии связывания. На фиг. 20B показана сенсограмма, когда антитело G1 3J LC иммобилизуют первым, а 17A9 пропускают. На фиг. 20С показана сенсограмма, когда 17A9 иммобилизуют первым, а G1 3J LC пропускают. В обоих случаях наблюдается связывание при пропускании второго антитела, позволяя предположить, что G1 3J LC и 17A9 связываются с разными эпитопами PMEL человека.
Подробное описание изобретения
Определения
[0047] Если не указано иное, подразумевается, что следующие термины и фразы, используемые в данном документе, имеют следующие значения.
[0048] Термин "алкил" относится к одновалентной насыщенной углеводородной цепи, содержащей указанное число атомов углерода. Например, C1-C6алкил относится к алкильной группе, содержащей от 1 до 6 атомов углерода. Алкильная группа может быть прямой или разветвленной. Типичная разветвленная алкильная группа содержит одну, две или три ветви. Примеры алкильных групп включают без ограничения, метил, этил, пропил (н-пропил и изопропил), бутил (н-бутил, изобутил, втор-бутил и трет-бутил), пентил (н-пентил, изопентил и неопентил) и гексил.
[0049] Используемый в данном документе термин "расщепляемый" относится к связывающей группе или линкерному компоненту, которые соединяют два фрагмента за счет ковалентных связей, но разъединяются с разрывом ковалентной связи между фрагментами при физиологически соответствующих условиях; как правило, расщепляемая связывающая группа разрывается in vivo быстрее во внутриклеточной среде, чем при нахождении вне клетки, что вызывает предпочтительное высвобождение полезной нагрузки внутри целевой клетки. Расщепление может быть ферментативным или неферментативным, но обычно приводит к высвобождению полезной нагрузки из антитела без разрушения антитела. При расщеплении некоторая часть связывающей группы или линкерного компонента может оставаться присоединенной к полезной нагрузке или высвобождение полезной нагрузки может происходить без какого-либо остатка связывающей группы.
[0050] Используемый в данном документе термин "нерасщепляемый" относится к связывающей группе или линкерному компоненту, которые практически не подвергаются разъединению при физиологических условиях, например, они устойчивы по меньшей мере в такой же степени, как и часть конъюгата, представляющая собой антитело или антигенсвязывающий фрагмент. Такие связывающие группы иногда называют ‘устойчивыми', что означает, что они в достаточной степени устойчивы к разрушению, чтобы удержать полезную нагрузку, присоединенную к антителу или антигенсвязывающему фрагменту до тех пор, пока антитело или антигенсвязывающий фрагмент сами по себе по меньшей мере частично не разрушатся, т. е. разрушение антитела или антигенсвязывающего фрагмента предшествует расщеплению связывающей группы in vivo. При разрушении части, представляющей собой антитело, у ADC с устойчивой или нерасщепляемой связывающей группой часть или вся связывающая группа, например, одна или несколько аминокислотных групп из антитела, могут оставаться присоединенными к полезной нагрузке или фрагменту, представляющему собой лекарственное средство, которые доставляются in vivo.
[0051] Используемый в данном документе термин "антитело" относится к полипептиду из семейства иммуноглобулинов, который способен к связыванию соответствующего антигена нековалентным, обратимым и специфическим образом. Например, встречающееся в природе антитело IgG представляет собой тетрамер, содержащий по меньшей мере две тяжелые (H) цепи и две легкие (L) цепи, соединенные между собой посредством дисульфидных связей. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно обозначаемой в данном документе как VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно обозначаемой в данном документе как VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена, CL. Области VH и VL могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), которые чередуются с более консервативными областями, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, которые включают различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента.
[0052] Термин "антитело" включает без ограничения моноклональные антитела, человеческие антитела, гуманизированные антитела, химерные антитела и антиидиотипические (анти-Id) антитела (включая, например, анти-Id антитела к антителам по настоящему изобретению). Антитела могут относиться к любому изотипу/классу (например, IgG, IgE, IgM, IgD, IgA и IgY) или подклассу (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2).
[0053] "Определяющие комплементарность домены" или "определяющие комплементарность области" ("CDR") взаимозаменяемо относятся к гипервариабельным областям VL и VH. В цепях антитела CDR представляют собой сайт связывания белка-мишени, который обуславливает специфичность в отношении такого белка-мишени. В каждой человеческой VL или VH имеется по три CDR (CDR1-3, пронумерованные последовательно от N-конца), составляющие приблизительно 15-20% от вариабельных доменов. CDR являются структурно комплементарными эпитопу белка-мишени и, таким образом, непосредственно ответственны за специфичность связывания. Остальные отрезки VL или VH, так называемые каркасные области, проявляют меньшую изменчивость аминокислотной последовательности (Kuby, Immunology, 4th ed., Chapter 4. W.H. Freeman & Co., New York, 2000).
[0054] Положения CDR и каркасных областей могут быть определены с использованием различных определений, широко известных из уровня техники, например, определений по Кабату, Чотиа, международной базы данных ImMunoGeneTics (IMGT) (во всемирной сети по адресу www.imgt.org/) и AbM (см., например, Johnson et al., Nucleic Acids Res., 29:205-206 (2001); Chothia and Lesk, J. Mol. Biol., 196:901-917 (1987); Chothia et al., Nature, 342:877-883 (1989); Chothia et al., J. Mol. Biol., 227:799-817 (1992); Al-Lazikani et al., J.Mol.Biol., 273:927-748 (1997)). Определения антигенсвязывающих активных центров также описаны в следующих источниках: Ruiz et al., Nucleic Acids Res., 28:219-221 (2000); и Lefranc, M.P., Nucleic Acids Res., 29:207-209 (2001); MacCallum et al., J. Mol. Biol., 262:732-745 (1996); и Martin et al., Proc. Natl. Acad. Sci. USA, 86:9268-9272 (1989); Martin et al., Methods Enzymol., 203:121-153 (1991) и Rees et al., в Sternberg M.J.E. (ed.), Protein Structure Prediction, Oxford University Press, Oxford, 141-172 (1996).
[0055] Как легкая, так и тяжелая цепи подразделяются на области структурной и функциональной гомологии. Термины "константный" и "вариабельный" используются в функциональном смысле. В связи с этим следует понимать, что вариабельные домены из частей как легкой (VL), так и тяжелой (VH) цепей определяют распознавание антигена и специфичность. Напротив, константные домены легкой цепи (CL) и тяжелой цепи (CH1, CH2 или CH3) придают важные биологические свойства, такие как секреция, перемещение через плаценту, связывание с рецептором Fc, связывание с комплементом и т. п. Принято, что нумерация доменов константной области идет по возрастанию по мере их удаления от антигенсвязывающего участка или аминоконца антитела. N-конец представляет собой вариабельную область, а на C-конце находится константная область; домены CH3 и CL фактически содержат карбоксиконцевые домены тяжелой и легкой цепи соответственно.
[0056] Используемый в данном документе термин "антигенсвязывающий фрагмент" относится к одной или нескольким частям антитела, которые сохраняют способность специфически взаимодействовать (например, посредством связывания, стерического несоответствия, стабилизации/дестабилизации, пространственного распределения) с эпитопом антигена. Примеры связывающих фрагментов включают без ограничения одноцепочечные Fv (scFv), антитела верблюжьих, соединенные дисульфидными связями Fv (sdFv), Fab-фрагменты, F(ab')-фрагменты, одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; F(ab)2-фрагмент, двухвалентный фрагмент, содержащий два Fab-фрагмента, соединенные дисульфидным мостиком в шарнирной области; Fd-фрагмент, состоящий из доменов VH и CH1; Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела; dAb-фрагмент (Ward et al., Nature 341:544-546, 1989), который состоит из домена VH; а также выделенную определяющую комплементарность область (CDR) или другие эпитопсвязывающие фрагменты антитела.
[0057] Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, они могут быть соединены с применением рекомбинантных способов с помощью синтетического линкера, что обеспечивает их получение в виде одной белковой цепи, в которой области VL и VH соединяются в пару с образованием одновалентной молекулы (известной как одноцепочечный Fv ("scFv"); см., например, Bird et al., Science 242:423-426, 1988; и Huston et al., Proc. Natl. Acad. Sci. 85:5879-5883, 1988). Предусматривается, что такие одноцепочечные антитела также охватываются термином "антигенсвязывающий фрагмент". Такие антигенсвязывающие фрагменты получают с применением традиционных методик, известных специалистам в данной области техники, и фрагменты подвергают скринингу на применимость таким же способом, как и интактные антитела.
[0058] Антигенсвязывающие фрагменты также могут быть включены в состав однодоменных антител, максиантител, миниантител, однодоменных антител, интраантител, диантител, триантител, тетраантител, v-NAR и бис-scFv (см., например, Hollinger and Hudson, Nature Biotechnology 23:1126-1136, 2005). Антигенсвязывающие фрагменты можно прививать на остовы на основе полипептидов, таких как фибронектин типа III (Fn3) (см. патент США № 6703199, в котором описаны монотела на основе полипептида фибронектина).
[0059] Антигенсвязывающие фрагменты можно вводить в состав одноцепочечных молекул, содержащих пару тандемных сегментов Fv (VH-CH1-VH-CH1), которые вместе с комплементарными полипептидами легкой цепи образуют пару антигенсвязывающих областей (Zapata et al., Protein Eng. 8:1057-1062, 1995 и патент США № 5641870).
[0060] Используемый в данном документе термин "моноклональное антитело" или "композиция на основе моноклонального антитела" относится к полипептидам, включающим антитела и антигенсвязывающие фрагменты, которые характеризуются по сути идентичной аминокислотной последовательностью или происходят из одного генетического источника. Данный термин также охватывает препараты на основе молекул антител одного молекулярного состава. Композиция на основе моноклональных антител проявляет одну специфичность и аффинность связывания в отношении конкретного эпитопа.
[0061] Используемый в данном документе термин "человеческое антитело" охватывает антитела, имеющие вариабельные области, в которых как каркасные, так и CDR-области получены из последовательностей, происходящих от человека. Кроме того, если антитело содержит константную область, то константная область также происходит из таких человеческих последовательностей, например, последовательностей зародышевой линии человека, или мутантных вариантов последовательностей зародышевой линии человека, или антитела, содержащего консенсусные каркасные последовательности, полученные за счет анализа человеческих каркасных последовательностей, например, как описано в Knappik et al., J. Mol. Biol. 296:57-86, 2000). Также предусматриваются антитела, происходящие из человеческих последовательностей, где одна или несколько CDR были подвергнуты мутированию в процессе созревания аффинности или для целей изготовления/конъюгации с полезной нагрузкой. См. Kilpatrick et al., "Rapid development of affinity matured monoclonal antibodies using RIMMS", Hybridoma. 1997 Aug;16(4):381-9.
[0062] Человеческие антитела по настоящему изобретению могут включать аминокислотные остатки, которые не закодированы в человеческих последовательностях (например, мутации, введенные посредством случайного или сайт-специфического мутагенеза in vitro или за счет соматических мутаций in vivo, или консервативная замена, которая содействует стабильности или облегчает изготовление).
[0063] Используемый в данном документе термин "распознавать" относится к антителу или его антигенсвязывающему фрагменту, которые отыскивают свой эпитоп и взаимодействуют (например, связываются) с ним, независимо от того является ли эпитоп линейным или конформационным. Термин "эпитоп" относится к сайту на антигене, с которым специфически связываются антитело или антигенсвязывающий фрагмент по настоящему изобретению. Эпитопы могут быть образованы как смежными аминокислотами, так и несмежными аминокислотами, размещаемыми рядом за счет третичной укладки белка. Эпитопы, образуемые из смежных аминокислот, как правило, сохраняются при воздействии денатурирующих растворителей, в то время как эпитопы, образуемые за счет третичной укладки, как правило, утрачиваются при обработке денатурирующими растворителями. Эпитоп, как правило, содержит по меньшей мере 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 аминокислот в уникальной пространственной конформации. Способы определения пространственной конформации эпитопов включают методики из уровня техники, например, рентгеноструктурную кристаллографию и 2-мерный ядерный магнитный резонанс (см., например, Epitope Mapping Protocols в Methods in Molecular Biology, Vol. 66, G. E. Morris, Ed. (1996)).
[0064] Используемый в данном документе термин "аффинность" относится к силе взаимодействия между антителом и антигеном в отдельных антигенных сайтах. В пределах каждого антигенного сайта вариабельная область "плеча" антитела взаимодействует с антигеном посредством слабых нековалентных сил во многих сайтах; при этом чем больше взаимодействий, тем сильнее аффинность.
[0065] Термин "выделенное антитело" относится к антителу, которое по сути не содержит другие антитела с отличающейся антигенной специфичностью. Однако выделенное антитело, которое специфически связывается с одним антигеном, может характеризоваться перекрестной реактивностью в отношении других антигенов. Более того, выделенное антитело может по сути не содержать другие клеточные материалы и/или химические вещества.
[0066] Термин "соответствующая последовательность зародышевой линии человека" относится к последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность или подпоследовательность человеческой вариабельной области, которые обладают наиболее высокой установленной идентичностью аминокислотной последовательности с референтной аминокислотной последовательностью или подпоследовательностью вариабельной области в сравнении со всеми другими всеми другими известными аминокислотными последовательностями вариабельной области, закодированными в последовательностях вариабельной области иммуноглобулина зародышевой линии человека. Соответствующая последовательность зародышевой линии человека также может относиться к аминокислотной последовательности или подпоследовательности человеческой вариабельной области, характеризующимся наиболее высокой идентичностью аминокислотной последовательности с референтной аминокислотной последовательностью или подпоследовательностью вариабельной области в сравнении со всеми другими подвергнутыми оценке аминокислотными последовательностями вариабельной области. Соответствующей последовательностью зародышевой линии человека могут быть только каркасные области, только определяющие комплементарность области, каркасные и определяющие комплементарность области, вариабельный сегмент (как определено выше) или другие комбинации последовательностей или подпоследовательностей, которые составляют вариабельную область. Идентичность последовательности может быть определена с применением способов, описанных в данном документе, например, выравнивания двух последовательностей с применением BLAST, ALIGN или другого алгоритма выравнивания, известного из уровня техники. Соответствующая последовательность нуклеиновой кислоты или аминокислотная последовательность зародышевой линии человека может характеризоваться по меньшей мере приблизительно 90%, 91, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, или 100% идентичностью последовательности с референтной последовательностью нуклеиновой кислоты или аминокислотной последовательностью вариабельной области. Соответствующие последовательности зародышевой линии человека могут быть определены, например, с помощью международной базы данных ImMunoGeneTics (IMGT), находящейся в открытом доступе (во всемирной сети по адресу www.imgt.org/) и V-base (во всемирной сети по адресу vbase.mrc-cpe.cam.ac.uk).
[0067] Фраза "специфически связывает" или "селективно связывает", когда она применяется в контексте описания взаимодействия между антигеном (например, белком) и антителом, фрагментом антитела или связывающим средством, происходящим из антитела, относится к реакции связывания, которая является определяющей для установления присутствия антигена в неоднородной популяции белков и других биологических веществ, например, в биологическом образце, например, крови, сыворотке крови, плазме крови или образце ткани. Таким образом, при некоторых обозначенных условиях проведения иммунологического анализа антитела или связывающие средства, характеризующиеся конкретной специфичностью связывания, связываются с конкретным антигеном в по меньшей мере два раза сильнее, чем фоновый уровень, и практически не связываются в значительном количестве с другими антигенами, присутствующими в образце. В одном варианте осуществления при обозначенных условиях иммунологического анализа антитело или связывающее средство с конкретной специфичностью связывания связывается с конкретным антигеном в по меньшей мере десять (10) раз сильнее относительно фонового уровня, и практически не связывается в значительном количестве с другими антигенами, присутствующими в образце. Специфическое связывание с антителом или связывающим средством в таких условиях может предусматривать то, что антитело или средство должно отбираться по его специфичности в отношении конкретного белка. При желании или необходимости данный отбор можно проводить путем отбрасывания антител, которые вступают в перекрестные реакции с молекулами от другого вида (например, мыши или крысы) или других подтипов. В качестве альтернативы в некоторых вариантах осуществления отбирают антитела или фрагменты антител, которые вступают в перекрестные реакции с некоторыми требуемыми молекулами.
[0068] Целый ряд форматов иммунологического анализа может применяться для отбора антител, характеризующихся специфической иммунной реактивностью в отношении конкретного белка. Например, твердофазные иммунологические анализы ELISA традиционно применяются для отбора антител, характеризующихся специфической реактивностью в отношении белка (см., например, Harlow & Lane, Using Antibodies, A Laboratory Manual (1998), где описаны форматы и условия иммунологического анализа, которые можно применять для определения специфической иммунной реактивности). Как правило, реакция специфического или селективного связывания будет приводить к сигналу, в по меньшей мере два раза превышающему фоновый уровень, и, что более типично, в по меньшей мере 10-100 раз превышающему фоновый уровень.
[0069] Термин "равновесная константа диссоциации (KD [M])" относится к константе скорости диссоциации (kd [время-1]), поделенной на константу скорости ассоциации (ka [время-1, M-1]). Равновесные константы диссоциации могут быть измерены с применением любого способа, известного из уровня техники. Обычно антитела по настоящему изобретению будут характеризоваться равновесной константой диссоциации, составляющей менее приблизительно 10-7 или 10-8 M, например, менее приблизительно 10-9 M или 10-10 M, в некоторых вариантах осуществления менее приблизительно 10-11 M, 10-12 M или 10-13 M.
[0070] Термин "биодоступность" относится к системной доступности (т. е. уровням в крови/плазме крови) заданного количества лекарственного средства, вводимого пациенту. Биодоступность представляет собой абсолютный термин, обозначающий показатель как времени (скорости), в течение которого лекарственное средство достигает общего кровообращения из введенной лекарственной формы, так и общего количества (величины) лекарственного средства в нем.
[0071] Используемая в данном документе фраза "состоящий фактически из" относится к родам или видам активных фармацевтических средств, включенных в способ или композицию, а также любым вспомогательным веществам, не проявляющим активность в отношении намеченной цели применения способов или композиций. В некоторых вариантах осуществления фраза "состоящий фактически из" однозначно исключает включение одного или нескольких дополнительных активных средств, отличных от конъюгата антитела и лекарственного средства по настоящему изобретению. В некоторых вариантах осуществления фраза "состоящий фактически из" однозначно исключает включение одного или нескольких дополнительных активных средств, отличных от конъюгата антитела и лекарственного средства по настоящему изобретению и второго средства, вводимого совместно.
[0072] Термин "аминокислота" относится к встречающимся в природе, синтетическим и неприродным аминокислотам, а также аналогам аминокислот и миметикам аминокислот, которые функционируют подобно встречающимся в природе аминокислотам. Встречающиеся в природе аминокислоты представляют собой аминокислоты, закодированные в генетическом коде, а также такие аминокислоты, которые были впоследствии модифицированы, например, гидроксипролин, γ-карбоксиглутамат и O-фосфосерин. Аналоги аминокислот относятся к соединениям, которые характеризуются такой же основной химической структурой, что и встречающаяся в природе аминокислота, то есть имеют α-углерод, который связан с водородом, карбоксильной группой, аминогруппой и R-группой, например, к гомосерину, норлейцину, метионинсульфоксиду, метионинметилсульфонию. Такие аналоги имеют модифицированные R-группы (например, норлейцин) или модифицированные пептидные остовы, но сохраняют такую же основную химическую структуру, как и встречающаяся в природе аминокислота. Миметики аминокислот относятся к химическим соединениям, которые имеют структуру, отличающуюся от общей химической структуры аминокислоты, но которые функционируют подобно встречающейся в природе аминокислоте.
[0073] Термин "консервативно модифицированный вариант" применяется в отношении как аминокислотных последовательностей, так и последовательностей нуклеиновой кислоты. Применительно к конкретным последовательностям нуклеиновой кислоты, консервативно модифицированные варианты относятся к тем нуклеиновым кислотам, которые кодируют идентичные или фактически идентичные аминокислотные последовательности, или же, если нуклеиновая кислота не кодирует аминокислотную последовательность, к фактически идентичным последовательностям. Вследствие вырожденности генетического кода любой заданный белок кодируется большим количеством функционально идентичных нуклеиновых кислот. Например, все кодоны GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в каждом положении, в котором кодоном задан аланин, кодон может быть изменен на любой из соответствующих описанных кодонов без изменения кодируемого полипептида. Такие варианты нуклеиновой кислоты являются "молчащими вариациями", которые представляют собой одну разновидность вариаций с консервативными модификациями. В данном документе каждая последовательность нуклеиновой кислоты, которая кодирует полипептид, также описывает каждый возможный молчащий вариант нуклеиновой кислоты. Специалист в данной области техники будет осознавать, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который обычно является единственным кодоном для метионина, и TGG, который обычно является единственным кодоном для триптофана) может быть модифицирован с получением функционально идентичной молекулы. Соответственно, каждый молчащий вариант нуклеиновой кислоты, которая кодирует полипептид, неявно определен в каждой описанной последовательности.
[0074] В случае полипептидных последовательностей "консервативно модифицированные варианты" охватывают отдельные замены, делеции или добавления в полипептидной последовательности, которые приводят к замене аминокислоты на аналогичную по химическим свойствам аминокислоту. Таблицы консервативных замен, обеспечивающие функционально аналогичные аминокислоты, хорошо известны из уровня техники. Такие консервативно модифицированные варианты дополняют, а не исключают полиморфные варианты, межвидовые гомологи и аллели по настоящему изобретению. Следующие восемь групп содержат аминокислоты, которые являются консервативными заменами друг для друга: 1) аланин (A), глицин (G); 2) аспарагиновая кислота (D), глутаминовая кислота (E); 3) аспарагин (N), глутамин (Q); 4) аргинин (R), лизин (K); 5) изолейцин (I), лейцин (L), метионин (M), валин (V); 6) фенилаланин (F), тирозин (Y), триптофан (W); 7) серин (S), треонин (T) и 8) цистеин (C), метионин (M) (см., например, Creighton, Proteins (1984)). В некоторых вариантах осуществления термин "консервативные модификации последовательности" используется для обозначения аминокислотных модификаций, которые не оказывают значительного влияния на характеристики связывания антитела, содержащего аминокислотную последовательность, или не изменяют их.
[0075] Используемый в данном документе термин "оптимизированная" относится к нуклеотидной последовательности, кодирующей аминокислотную последовательность, которая была изменена с применением кодонов, предпочтительных в продуцирующих клетке или организме, обычно в эукариотической клетке, например, дрожжевой клетке, клетке Pichia, грибной клетке, клетке Trichoderma, клетке яичника китайского хомячка (CHO) или человеческой клетке. Оптимизированную нуклеотидную последовательность конструируют таким образом, чтобы полностью или насколько это возможно сохранить аминокислотную последовательность, изначально закодированную в исходной нуклеотидной последовательности, которая также известна как "родительская" последовательность.
[0076] Термины "процент идентичности" или "процентная идентичность" в контексте двух или более нуклеиновых кислот или полипептидных последовательностей относятся к степени, в которой две или более последовательности или подпоследовательности являются одинаковыми. Две последовательности являются "идентичными", если они имеют одинаковую последовательность из аминокислот или нуклеотидов на протяжении области, подлежащей сравнению. Две последовательности являются "по сути идентичными", если две последовательности имеют указанную процентную долю аминокислотных остатков или нуклеотидов, которые являются одинаковыми (т. е. 60% идентичность, необязательно 65%, 70%, 75%, 80%, 85%, 90%, 95% или 99% идентичность на протяжении указанной области или, если не указано, на протяжении всей последовательности), при сравнении и выравнивании для обеспечения максимального соответствия на протяжении окна сравнения или обозначенной области, что измерено с применением одного из следующих алгоритмов сравнения последовательности или посредством ручного выравнивания и визуального просмотра. Необязательно идентичность существует на протяжении области, длина которой составляет по меньшей мере приблизительно 30 нуклеотидов (или 10 аминокислот), или более предпочтительно на протяжении области, длина которой составляет 100-500 или 1000 или более нуклеотидов (или 20, 50, 200 или более аминокислот).
[0077] При сравнении последовательностей обычно одна последовательность выступает в качестве референтной последовательности, с которой сравнивают тестируемые последовательности. При использовании алгоритма сравнения последовательностей тестируемую и референтную последовательности вводят в компьютер, если это необходимо, устанавливают координаты подпоследовательностей и устанавливают программные параметры алгоритма для анализа последовательностей. Могут применяться программные параметры по умолчанию или можно устанавливать альтернативные параметры. На основании программных параметров алгоритм сравнения последовательностей затем рассчитывает значения процента идентичности последовательностей для тестируемых последовательностей относительно референтной последовательности.
[0078] Используемое в данном документе "окно сравнения" предусматривает ссылку на сегмент из любого количества смежных положений, выбранных из группы, состоящей из от 20 до 600, обычно от приблизительно 50 до приблизительно 200, чаще от приблизительно 100 до приблизительно 150, в котором последовательность можно сравнивать с референтной последовательностью с таким же количеством смежных положений, после того как две последовательности подвергли оптимальному выравниванию. Способы выравнивания последовательностей для проведения сравнения хорошо известны из уровня техники. Оптимальное выравнивание последовательностей для сравнения можно проводить, например, с помощью алгоритма поиска локальной гомологии Смита-Уотермана, Adv. Appl. Math. 2:482c (1970), с помощью алгоритма выравнивания областей гомологии Нидлмана-Вунша, J. Mol. Biol. 48:443 (1970), с помощью способа поиска сходства Пирсона-Липмана, Proc. Natl. Acad. Sci. USA 85:2444 (1988), с помощью компьютерных реализаций таких алгоритмов (GAP, BESTFIT, FASTA и TFASTA в составе пакета программного обеспечения Wisconsin Genetics, Genetics Computer Group, 575 Science Dr., Мэдисон, Висконсин) или с помощью ручного выравнивания и визуального просмотра (см., например, Brent et al., Current Protocols in Molecular Biology, 2003).
[0079] Два примера алгоритмов, которые подходят для определения процента идентичности последовательностей и сходства последовательностей, представляют собой алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul et al., Nuc. Acids Res. 25:3389-3402, 1977 и Altschul et al., J. Mol. Biol. 215:403-410, 1990 соответственно. Программное обеспечение для осуществления анализов BLAST доступно для общего пользования в Национальном центре биотехнологической информации. На первом этапе данный алгоритм предусматривает идентификацию пар последовательностей с высоким показателем сходства (HSP) за счет идентификации коротких слов длиной W в запрашиваемой последовательности, которые либо совпадают, либо удовлетворяют некоторому пороговому баллу T с положительным значением при выравнивании со словом такой же длины в последовательности из базы данных. T известен под названием пороговый балл соседних слов (Altschul et al., выше). Эти первоначальные совпадения соседних слов выступают в качестве "затравок" для начала поисков, предназначенных для нахождения более длинных HSP, содержащие их. Совпадения слов продлеваются в обоих направлениях вдоль каждой последовательности до тех пор, пока может увеличиваться суммарный балл выравнивания. В случае нуклеотидных последовательностей суммарные баллы рассчитывают с применением параметров M (вознаграждающий балл за пару совпадающих остатков; всегда >0) и N (штрафной балл за несовпадающие остатки; всегда <0). В случае аминокислотных последовательностей для подсчета суммарного балла применяют матрицу замен. Продление совпадений слов в каждом направлении останавливается, когда суммарный балл выравнивания уменьшается на величину X относительно своего максимального достигнутого значения; суммарный балл стремится к нулю или ниже вследствие накопления одного или нескольких выравниваний остатков с отрицательными баллами; или достигается конец любой из последовательностей. Параметры W, T и X алгоритма BLAST определяют чувствительность и скорость выравнивания. В программе BLASTN (для нуклеотидных последовательностей) в качестве параметров по умолчанию используется длина слова (W), составляющая 11, ожидание (E), составляющее 10, M=5, N= -4 и сравнение обеих нитей. В случае аминокислотных последовательностей в программе BLASTP в качестве параметров по умолчанию используется длина слова, составляющая 3, и ожидание (E), составляющее 10, и матрица замен BLOSUM62 (см. Henikoff and Henikoff (1989) Proc. Natl. Acad. Sci. USA 89:10915), выравнивания (B), составляющие 50, ожидание (E), составляющее 10, M=5, N= -4 и сравнение обеих нитей.
[0080] Алгоритм BLAST также осуществляет статистический анализ сходства между двумя последовательностями (см., например, Karlin and Altschul, Proc. Natl. Acad. Sci. USA 90:5873-5787, 1993). Одной мерой сходства, предусмотренной в алгоритме BLAST, является наименьшая суммарная вероятность (P (N)), которая указывает на вероятность, с которой совпадение между двумя нуклеотидными или аминокислотными последовательностями возникло случайно. Например, нуклеиновая кислота считается подобной референтной последовательности, если наименьшая суммарная вероятность при сравнении тестируемой нуклеиновой кислоты с референтной нуклеиновой кислотой составляет менее приблизительно 0,2, более предпочтительно менее приблизительно 0,01 и наиболее предпочтительно менее приблизительно 0,001.
[0081] Процентную идентичность между двумя аминокислотными последовательностями также можно определить с использованием алгоритма из E. Meyers и W. Miller Comput. Appl. Biosci. 4:11-17, (1988) который был включен в программу ALIGN (версия 2.0), с применением таблицы весов замен остатков PAM120, штрафа за продление гэпа, составляющего 12, и штрафа за открытие гэпа, составляющего 4. Кроме того, процентная идентичность между двумя аминокислотными последовательностями может быть определена с применением алгоритма Нидлмана-Вунша, J. Mol. Biol. 48:444-453, (1970), который был включен в программу GAP в составе пакета программного обеспечения GCG (доступного на www.gcg.com), с применением либо матрицы BLOSUM62, либо матрицы PAM250, и также штрафа за открытие гэпа, составляющего 16, 14, 12, 10, 8, 6 или 4, и штрафа за продление гэпа, составляющего 1, 2, 3, 4, 5 или 6.
[0082] Помимо процента идентичности последовательностей, упомянутого выше, еще одним показателем того, что две последовательности нуклеиновой кислоты или полипептида являются по сути идентичными, является то, что полипептид, кодируемый первой нуклеиновой кислотой, является иммунологически перекрестно реактивным с антителами, продуцирование которых индуцировано полипептидом, кодируемым второй нуклеиновой кислотой, как описано ниже. Таким образом, полипептид, как правило, является по сути идентичным второму полипептиду, например, если два пептида отличаются только консервативными заменами. Другим показателем того, что две последовательности нуклеиновой кислоты являются по сути идентичными, является то, что две молекулы или их комплементарные цепи гибридизируются друг с другом в жестких условиях, как описано ниже. Еще одним показателем того, что две последовательности нуклеиновой кислоты являются по сути идентичными, является то, что для амплификации последовательности могут применяться одни и те же праймеры.
[0083] Термин "нуклеиновая кислота" используется в данном документе взаимозаменяемо с термином "полинуклеотид" и относится к дезоксирибонуклеотидам или рибонуклеотидам и полимерам на их основе в одно- или двухнитевой форме. Термин охватывает нуклеиновые кислоты, содержащие известные аналоги нуклеотидов или модифицированные остатки или связи в остове, которые являются синтетическими, встречающимися в природе и не встречающимися в природе, которые характеризуются свойствами связывания, подобными референтной нуклеиновой кислоте, и которые метаболизируются подобно референтным нуклеотидам. Примеры таких аналогов включают без ограничения фосфоротиоаты, фосфорамидаты, метилфосфонаты, хиральные метилфосфонаты, 2-O-метилрибонуклеотиды, пептидонуклеиновые кислоты (PNA).
[0084] Если не указано иное, конкретная последовательность нуклеиновой кислоты также неочевидно охватывает ее консервативно модифицированные варианты (например, замены на основе вырожденных кодонов) и комплементарные последовательности, а также конкретно указанную последовательность. В частности, как подробно описано ниже, замены на основе вырожденных кодонов можно проводить за счет создания последовательностей, в которых третье положение в одном или нескольких выбранных (или всех) кодонах заменено на любой из канонических нуклеозидов и/или остаток дезоксиинозина (Batzer et al., Nucleic Acid Res. 19:5081, 1991; Ohtsuka et al., J. Biol. Chem. 260:2605-2608, 1985 и Rossolini et al., Mol. Cell. Probes 8:91-98, 1994).
[0085] В контексте нуклеиновых кислот термин "функционально связанный" относится к функциональной взаимосвязи двух или более сегментов полинуклеотида (например, ДНК). Как правило, он относится к функциональной взаимосвязи регулирующей транскрипцию последовательности с транскрибируемой последовательностью. Например, промоторная или энхансерная последовательность является функционально связанной с кодирующей последовательностью, если она стимулирует или модулирует транскрипцию кодирующей последовательности в соответствующей клетке-хозяине или другой системе экспрессии. Обычно регулирующие транскрипцию промоторные последовательности, которые являются функционально связанными с транскрибируемой последовательностью, являются физически смежными с транскрибируемой последовательностью, т. е. они функционируют в цис-положении. Однако некоторые регулирующие транскрипцию последовательности, такие как энхансеры, не обязательно должны быть физически смежными или располагаться в непосредственной близости от кодирующих последовательностей, транскрипцию которых они усиливают.
[0086] Термины "полипептид" и "белок" используются в данном документе взаимозаменяемо для обозначения полимера из аминокислотных остатков. Термины применимы к полимерам из аминокислот, в которых один или несколько аминокислотных остатков представляют собой искусственный химический миметик соответствующей встречающейся в природе аминокислоты, а также к полимерам из встречающихся в природе аминокислот и полимеру из не встречающихся в природе аминокислот. Если не указано иное, конкретная полипептидная последовательность также неочевидно охватывает ее консервативно модифицированные варианты.
[0087] Используемый в данном документе термин "конъюгат антитела и лекарственного средства" или "иммуноконъюгат" относится к связи антитела или его антигенсвязывающего фрагмента с другим средством, таким как химиотерапевтическое средство, токсин, иммунотерапевтическое средство, зонд для визуализации и т. п. Связь может представлять собой ковалентные связи или нековалентные взаимодействия, такие как посредством электростатических сил. Для образования конъюгата антитела и лекарственного средства могут использоваться различные линкеры, известные из уровня техники. Кроме того, конъюгат антитела и лекарственного средства может предусматриваться в форме слитого белка, который может экспрессироваться с полинуклеотида, кодирующего иммуноконъюгат. Используемый в данном документе "слитый белок" относится к белкам, созданным за счет соединения двух или более генов или фрагментов генов, которые изначально кодировали отдельные белки (в том числе пептиды и полипептиды). Трансляция слитого гена приводит к единому белку с функциональными свойствами, происходящими из каждого из исходных белков.
[0088] Термин "субъект" охватывает человека и отличных от человека животных. Отличные от человека животные охватывают всех позвоночных, например, млекопитающих и отличных от млекопитающих животных, как, например, отличных от человека приматов, овцу, собаку, корову, кур, амфибий и рептилий. За исключением случаев, когда это отмечается, термины "пациент" или "субъект" используются в данном документе взаимозаменяемо.
[0089] Используемый в данном документе термин "цитотоксин" или "цитотоксическое средство" относится к любому средству, которое является вредным для роста и пролиферации клеток и которое может функционировать с уменьшением, ингибированием или разрушением клетки или злокачественной опухоли.
[0090] Используемый в данном документе термин "противораковое средство" относится к любому средству, которое может использоваться для лечения или предупреждения нарушения пролиферации клеток, такого как рак, в том числе без ограничения к цитотоксическим средствам, химиотерапевтическим средствам, лучевой терапии и средствам для лучевой терапии, нацеливающим противораковым средствам и иммунотерапевтическим средствам.
[0091] Используемый в данном документе термин "фрагмент, представляющий собой лекарственное средство" или "полезная нагрузка" относится к химическому фрагменту, который конъюгирован с антителом или антигенсвязывающим фрагментом по настоящему изобретению, и может охватывать любое терапевтическое или диагностическое средство, например, противораковое, противовоспалительное, противоинфекционное (например, противогрибковое, антибактериальное, антипаразитарное, противовирусное) или анестезирующее средство. Например, фрагмент, представляющий собой лекарственное средство, может представлять собой противораковое средство, такое как цитотоксин. В определенных вариантах осуществления фрагмент, представляющий собой лекарственное средство, представляет собой соединение-ингибитор мишени. Кроме того, полезная нагрузка может представлять собой зонд для биофизического обнаружения, флуорофор, спиновую метку, зонд для инфракрасного обнаружения, зонд для обнаружения аффинности, хелатор, зонд для спектроскопического обнаружения, радиоактивный зонд, липидную молекулу, полиэтиленгликоль, полимер, спиновую метку, ДНК, РНК, белок, пептид, покрытие, антитело, фрагмент антитела, наночастицу, квантовую точку, липосому, частицу PLGA, сахарид или полисахарид.
[0092] В некоторых вариантах осуществления фрагмент, представляющий собой лекарственное средство, или полезная нагрузка представляют собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11 (ингибитор GNAQ/GNA11). В некоторых вариантах осуществления ингибитор GNAQ/11 представляет собой молекулу, которая ингибирует GNAQ/11-опосредованное продуцирование IP3 и/или проявляет дозозависимый антипролиферативный эффект в клетках, зависящих от передачи сигналов GNAQ/11 (т. е. GNAQ/11-мутантных клетках увеальной меланомы). В некоторых вариантах осуществления ингибитор GNAQ/11 представляет собой соединение, которое стабилизирует GNAQ/11 в неактивном GDP-связанном состоянии и предотвращает высвобождение GDP или связывается при активном GTP-связанном состоянии и предотвращает взаимодействие GNAQ/11 с нижестоящими эффекторами. В некоторых вариантах осуществления ингибитор GNAQ/11 действует путем ингибирования мутантного GNAQ и/или GNA11, такого как тот, который содержит мутацию Q209L/P. Способы присоединения таких фрагментов, представляющих собой лекарственное средство, к линкеру, совместимому с нацеливающим фрагментом, приведены в настоящем изобретении наряду со способами, известными из уровня техники. См., например, Singh et al., (2009) Therapeutic Antibodies: Methods and Protocols, vol. 525, 445-457.
[0093] GNAQ (альфа-субъединица гуаниннуклеотид-связывающего белка G(q), также известная как CMC1, G-ALPHA-q, GAQ, SWS и субъединица альфа-q G-белка) и GNA11 (субъединица альфа-11 гуаниннуклеотид-связывающего белка, также известная как FBH, FBH2, FHH2, GNA-11, HHC2, HYPOC2 и субъединица альфа 11 G-белка) являются тесно связанными GTPазами, которые составляют α-субъединицы гетеротримерных G-белков, действующих ниже рецепторов, связанных с G-белками (GPCR). α-Субъединицы действуют как переключатель для активации G-белков путем замены гуанозиндифосфата (GDP) на гуанозинтрифосфат (GTP), что приводит к активации отдельных нижестоящих эффекторов. Активация прекращается за счет собственной активности GTPазы, когда GTP гидролизуется до GDP (Van Raamsdonk et al., 2010, N Engl J Med.; 363(23):2191-9). Классическая активация каскада Gq-белков происходит посредством фосфолипазы C-β (PLC-β), которая гидролизует фосфолипид фосфатидилинозитол-4,5-бифосфат с высвобождением двух высокоактивных вторичных мессенджеров: D-миоинозитол-1,4,5-трифосфата (IP3) и диацилглицерина (DAG). После временного увеличения внутриклеточного Ca2+ IP3 быстро превращается в IP2, IP1 и миоинозитол. С другой стороны, DAG активирует протеинкиназу C (PKC), что приводит к каскаду фосфорилирования RAF, MEK и ERK, которые перемещаются в ядро для регулирования пролиферации и выживания клеток (Krantz et al., 2017, Clin Ophthalmol.; 11:279-289).
[0094] Последовательности нуклеиновых кислот и аминокислотные последовательности GNAQ человека были опубликованы в GenBank под следующими №№ доступа:
[0095] NP_002063 (SEQ ID NO:268)
1 mtlesimacc lseeakearr indeierqlr rdkrdarrel kllllgtges gkstfikqmr
61 iihgsgysde dkrgftklvy qniftamqam iramdtlkip ykyehnkaha qlvrevdvek
121 vsafenpyvd aikslwndpg iqecydrrre yqlsdstkyy lndldrvadp aylptqqdvl
181 rvrvpttgii eypfdlqsvi frmvdvggqr serrkwihcf envtsimflv alseydqvlv
241 esdnenrmee skalfrtiit ypwfqnssvi lflnkkdlle ekimyshlvd yfpeydgpqr
301 daqaarefil kmfvdlnpds dkiiyshftc atdtenirfv faavkdtilq lnlkeynlv
[0096] NM_002072 (SEQ ID NO:269)
1 agactatccg ctcccaccgc gcccccggcc cacctggtgg ccccggccct ggccgccgcc
61 cccgcggcgg ttcccggagc tcgtcccgga cgcgcgcccg ggcggcgggg gctcggcggc
121 caccgctgcc tcgggggagc gagggcggga gggtgtgtgt gcgcgctgtg agcagggggt
181 gccggcgggg ctgcagcgga ggcactttgg aagaatgact ctggagtcca tcatggcgtg
241 ctgcctgagc gaggaggcca aggaagcccg gcggatcaac gacgagatcg agcggcagct
301 ccgcagggac aagcgggacg cccgccggga gctcaagctg ctgctgctcg ggacaggaga
361 gagtggcaag agtacgttta tcaagcagat gagaatcatc catgggtcag gatactctga
421 tgaagataaa aggggcttca ccaagctggt gtatcagaac atcttcacgg ccatgcaggc
481 catgatcaga gccatggaca cactcaagat cccatacaag tatgagcaca ataaggctca
541 tgcacaatta gttcgagaag ttgatgtgga gaaggtgtct gcttttgaga atccatatgt
601 agatgcaata aagagtttat ggaatgatcc tggaatccag gaatgctatg atagacgacg
661 agaatatcaa ttatctgact ctaccaaata ctatcttaat gacttggacc gcgtagctga
721 ccctgcctac ctgcctacgc aacaagatgt gcttagagtt cgagtcccca ccacagggat
781 catcgaatac ccctttgact tacaaagtgt cattttcaga atggtcgatg tagggggcca
841 aaggtcagag agaagaaaat ggatacactg ctttgaaaat gtcacctcta tcatgtttct
901 agtagcgctt agtgaatatg atcaagttct cgtggagtca gacaatgaga accgaatgga
961 ggaaagcaag gctctcttta gaacaattat cacatacccc tggttccaga actcctcggt
1021 tattctgttc ttaaacaaga aagatcttct agaggagaaa atcatgtatt cccatctagt
1081 cgactacttc ccagaatatg atggacccca gagagatgcc caggcagccc gagaattcat
1141 tctgaagatg ttcgtggacc tgaacccaga cagtgacaaa attatctact cccacttcac
1201 gtgcgccaca gacaccgaga atatccgctt tgtctttgct gccgtcaagg acaccatcct
1261 ccagttgaac ctgaaggagt acaatctggt ctaattgtgc ctcctagaca cccgccctgc
1321 ccttccctgg tgggctattg aagatacaca agagggactg tatttctgtg gaaaacaatt
1381 tgcataatac taatttattg ccgtcctgga ctctgtgtga gcgtgtccac agagtttgta
1441 gtaaatatta tgattttatt taaactattc agaggaaaaa cagaggatgc tgaagtacag
1501 tcccagcaca tttcctctct atcttttttt taggcaaaac cttgtgactc agtgtatttt
1561 aaattctcag tcatgcactc acaaagataa gacttgtttc tttctgtctc tctctctttt
1621 tcttttctat ggagcaaaac aaagctgatt tccctttttt cttcccccgc taattcatac
1681 ctccctcctg atgtttttcc caggttacaa tggcctttat cctagttcca ttcttggtca
1741 agtttttctc tcaaatgata cagtcaggac acatcgttcg atttaagcca tcatcagctt
1801 aatttaagtt tgtagttttt gctgaaggat tatatgtatt aatacttacg gttttaaatg
1861 tgttgctttg gatacacaca tagtttcttt tttaatagaa tatactgtct tgtctcactt
1921 tggactggga cagtggatgc ccatctaaaa gttaagtgtc atttctttta gatgtttacc
1981 ttcagccata gcttgattgc tcagagaaat atgcagaagg caggatcaaa gacacacagg
2041 agtcctttct tttgaaatgc cacgtgccat tgtctttcct cccttctttg cttctttttc
2101 ttaccctctc tttcaattgc agatgccaaa aaagatgcca acagacacta cattacccta
2161 atggctgcta cccagaacct ttttataggt tgttcttaat ttttttgttg ttgttgttca
2221 agcttttcct ttcttttttt tcttggtgtt tgggccacga ttttaaaatg acttttatta
2281 tgggtatgtg ttgccaaagc tggctttttg tcaaataaaa tgaatacgaa cttaaaaaat
2341 aaaagctggt atcttaaaat gtaagagagt aagactgtga agcctaaaat gactggctga
2401 gaatgaacca gaaatgccat ttgccaaaca gttgtaacta gaaatttgat tctcacggtc
2461 cattcttttc tttgtcctta agatgacatt gttagtgttc acgtcccatg ttcagtgtcc
2521 aaaccggcaa tgtaaaaagt atcctgtgtg gtttaacagg aaatctgttt atgtctcttt
2581 atttgaaacc agttttactc tcagtggttc tttaagttca atgaagtctg ccaggaacat
2641 tggttggtag tattattccg acacctttaa tttccaaaat ctgaagttcc tgctagttta
2701 ccaccttcat gatcttcttg aactggtaac tgattaggtt gaacttatgg aagatttgtg
2761 gacttaactc aaaagtaacc tctcagtgtt ctatagaaca tgtatttgtg taactgaacc
2821 taccaggaga aatgtttgga attctatatg tgcaattttt caacaaatgc aaaaaaaata
2881 cagcacatgt attgacaagc ttctgtcaag cagcttgagt tgaaatttga tttaagaaaa
2941 taaatcatga ttgttcaaag ctgctgggac gttagaatta ggccatgata ctggtctcat
3001 tttaactaca gtggtatttg gcactagtgt aaacttccat ataaatcact cttttggaac
3061 aacaaagggg gagggagaaa aatcacggcc tgttaaatga gtaccaaagc cgcccaacag
3121 taatgagatg ttctcatcct tgattctccc agcctcaaac aacacagctt actttttttt
3181 tcccttgctc agaaagtacc tgtaatttaa caaacagact gcctgtaggt atagtgcaat
3241 tacaaatgct ctaatcattg tacatacatc tctcttgata ttgcagcatc catactggct
3301 ttgtaatcat taattttttg gcagattgaa tgtgctgtat tgatatgtat ctatgtaatt
3361 gtattgtatg tctatagcta attcacgttt tgaataatgt tattttattt acttttttaa
3421 gagaggagaa tgtaaatttg tcagtttatt tctgactagg gatattttct ttccatttag
3481 aaaagaagaa aaaaaaaaaa ccttactgtc atacagagcg gtactagcgt cgtgctgtat
3541 aaaatcattt gcacattcct gagtagaggt atactgatta taagacccaa aggtaatttc
3601 atagcaaaat acataaaatc agtcggagct tttatacaaa catggaaacc aactttgtag
3661 aacttttgcc atttgatcta ggattggaat atgagctttt atacaattca tattcttatt
3721 tggcaaatgc acagtttagt attacctctc tgatggcctt tactagaaag gcagttttag
3781 aagctattgt gatccactaa ggaaatgttt taacagctag agaccactgc ttgcctgaaa
3841 gggcgttctt aaatttggtg cagcaaaaaa aaaaaaaaaa aaaaaaaaaa ttaaacaaca
3901 acatttgaag gcctacagtg tgtatagaga aaacctcatc acaagatcat aagtgttaca
3961 gttttaggga atcaagatat tctatttaat agagctatag taaatgtagt caattaaacc
4021 tgatctcaaa gcttgaagaa gctgagcaaa acagggaaag attgttatat ttgtctttat
4081 gaaattggga tggaatttgc tatgcagaat tgaggtttgt ggcttcgctg ttcctgtagg
4141 gtgcatgaca agatcccttc tcttgagaaa ggaaaaaatt gatcacccta gcagcagtga
4201 tgcatagaaa cctaatttta gccacaccag tcaatcgaag ctaaaggatt ttcttttttg
4261 tttcttcggg gttttattga aggggctagg ggcgggacgg gattcttttc agttttgtat
4321 aaaaacaaag tttactcatg ctttatatta tattgtgatt gcaagcgtta taagcgtgtg
4381 ccactggcct cctattgttg atgcttaggt aatggaggcc tgtggtgagt tttatggtga
4441 cttgggcatg tcttattcaa aaacaaaaac ataaaacaca gaaacctttc ttcagcatac
4501 caaggcaagc agccatttca tgactcactt aacacattgc agtgtaccag tttacagatg
4561 atttttccct ttttgcgtga catggcagtt ctaaccccca gagaattcct tatttgtaaa
4621 ttggaagttt ctactatgcc ttacagagct taaattcaga agtttgtgcc tcatatctga
4681 aacaaaggga aataacacac ccattcaaaa gtaaataaat ctcctagaag tttttgtttt
4741 taacatttcc atataaagag ctctgttgaa tgtcatgaat agactggaaa aaaaaatttt
4801 aagaacctgc atatgttgtt tactagcaga tgacaactac aaaaggaatc tgaagaacac
4861 gtaaaacttg tatttttttt tttttggtag attaactagc aggcctattt taaaaaggta
4921 attcagctaa agggcaattt acttttttgt acttcagact atcttgattg tcaaagtgta
4981 cgaactgtaa ttttaaaatt tatactgcca catgattgta aattttagtt gtcttaagtt
5041 aggaattggt gaaaagctat ttatgctgga tttgggtcaa aatgacttat ttgcaaaaaa
5101 ataaataatg ggaagaaagg gctgtataat gaaatactgc aagactcaca tattggttgg
5161 aaatttccct caaatcacct accgattacc cttgatttcc ctttgttttc agtttctcaa
5221 aacgaatgaa atgaaatata gcagaatgtt aacccatata aaaataaagt gtacccaaat
5281 attgtaatgt atattgctgc tcttcttcaa attaaataag ggtttaaaac cacttaattg
5341 gtaatcaaca tctcaattga tacaaataag gtgtgcttgg tatacattaa tattttcttc
5401 caaagatata tctttggtta gaaacacaaa aaaataaaac tagtaatatt gtatgtttat
5461 ctatctctac atatttccag catatgtagc gttaatagat ctgtcctggt aactgtgtct
5521 ttgggatttc attttggttc catcaaatta ggaaaagaaa tggcttagtt gtatatgatt
5581 agctagagat ttttggagcc agacacctgc tgtttagtag ataacttagt acagacccta
5641 aacttgtcat ttgtttttct cacagaatag ccatttcctg ctgtcttccc aatgatcact
5701 gccctttcaa taacactctt gcctctagaa tcatatgttc aaagtatgaa tacacaccta
5761 gcacatagta ggtgctcaaa tattaatttc ctccttgcct tccttatcta ccctgtgtcc
5821 tccatttccc cgtatgattc caacccaata tagcaaatga catttacatg ttatgaaaac
5881 atctattggg taaaatcaga tcttggataa agaaattctg acttttatat aagcttttgg
5941 tagacagaaa aaacagaaag gtattcgttg gtagaacatt tttaagttca ggaaagaaag
6001 ctggaataat actacgtaac tttgtccagg ttactttgac tgaaacacgt ttttggtgga
6061 tttcttttcc tcaaagaact ctctaaatgc aactccttgc tggattcctc acccatcatc
6121 ctgttggaaa cccttactag acctatgtat ttagggagtt ttgtcagaaa acatttttaa
6181 cttgcagtat ttaaaagaat atttactgtt cctaaaatgt cattcaaatg catgtactgt
6241 ctattgtttg gggatgggaa ctagttttgc aaaaaacacc taatgttgta taataatgcc
6301 ccaatgatct tgctggttaa aaatacagta tttttggcca taa
[0097] Последовательность нуклеиновой кислоты и аминокислотная последовательность GNA11 человека была опубликована в GenBank под следующими №№ доступа:
[0098] NP_002058 (SEQ ID NO:270)
1 mtlesmmacc lsdevkeskr inaeiekqlr rdkrdarrel kllllgtges gkstfikqmr
61 iihgagysee dkrgftklvy qniftamqam irametlkil ykyeqnkana llirevdvek
121 vttfehqyvs aiktlwedpg iqecydrrre yqlsdsakyy ltdvdriatl gylptqqdvl
181 rvrvpttgii eypfdlenii frmvdvggqr serrkwihcf envtsimflv alseydqvlv
241 esdnenrmee skalfrtiit ypwfqnssvi lflnkkdlle dkilyshlvd yfpefdgpqr
301 daqaarefil kmfvdlnpds dkiiyshftc atdtenirfv faavkdtilq lnlkeynlv
[0099] NM_002067 (SEQ ID NO:271)
1 aggttgtccg gcgctgtcgc tcggttgcgg cggctgcggt tggcggtggc tgcggcggcg
61 gcgcgggctg agtgcggccg cgcgggagtc cgcggctggc gcggcccgag cggggacccg
121 gcggctcgcc aggcggcggc cgaggcgggg cgggccggcc cggggccgag ggccggtggc
181 cgaggccgga gggccgcggc gggcggcggc cgaggcggct ccggccaggg ccgggccggg
241 ggccgggggg cggcggcggg caggcggccg cgtcggccgg ggccgggacg atgactctgg
301 agtccatgat ggcgtgttgc ctgagcgatg aggtgaagga gtccaagcgg atcaacgccg
361 agatcgagaa gcagctgcgg cgggacaagc gcgacgcccg gcgcgagctc aagctgctgc
421 tgctcggcac gggcgagagc gggaagagca cgttcatcaa gcagatgcgc atcatccacg
481 gcgccggcta ctcggaggag gacaagcgcg gcttcaccaa gctcgtctac cagaacatct
541 tcaccgccat gcaggccatg atccgggcca tggagacgct caagatcctc tacaagtacg
601 agcagaacaa ggccaatgcg ctcctgatcc gggaggtgga cgtggagaag gtgaccacct
661 tcgagcatca gtacgtcagt gccatcaaga ccctgtggga ggacccgggc atccaggaat
721 gctacgaccg caggcgcgag taccagctct ccgactctgc caagtactac ctgaccgacg
781 ttgaccgcat cgccaccttg ggctacctgc ccacccagca ggacgtgctg cgggtccgcg
841 tgcccaccac cggcatcatc gagtaccctt tcgacctgga gaacatcatc ttccggatgg
901 tggatgtggg gggccagcgg tcggagcgga ggaagtggat ccactgcttt gagaacgtga
961 catccatcat gtttctcgtc gccctcagcg aatacgacca agtcctggtg gagtcggaca
1021 acgagaaccg gatggaggag agcaaagccc tgttccggac catcatcacc tacccctggt
1081 tccagaactc ctccgtcatc ctcttcctca acaagaagga cctgctggag gacaagatcc
1141 tgtactcgca cctggtggac tacttccccg agttcgatgg tccccagcgg gacgcccagg
1201 cggcgcggga gttcatcctg aagatgttcg tggacctgaa ccccgacagc gacaagatca
1261 tctactcaca cttcacgtgt gccaccgaca cggagaacat ccgcttcgtg ttcgcggccg
1321 tgaaggacac catcctgcag ctcaacctca aggagtacaa cctggtctga gcgcccaggc
1381 ccagggagac gggatggaga cacggggcag gaccttcctt ccacggagcc tgcggctgcc
1441 gggcgggtgg cgctgccgag tccgggccgg ggcctctgcc cgcgggagga gatttttttt
1501 tttcatattt ttaacaaatg gtttttattt cacagttatc aggggatgta catctctccc
1561 tccgtacact tcgcgcacct tctcaccttt tgtcaacggc aaaggcagcc tttttctggc
1621 cttgacttat ggctcgcttt tttctaaaaa aaaaaaaaaa agaaagaaag aaaaaaagca
1681 acgaaacata aaacacacaa gcgccccgtg cccccagtga ctctgggcct cacagagccc
1741 ccgccagcca gcatggggcc ccgccctgca gccagtcacg cgcccccaca ccgcagcccc
1801 ccgtggctgt ccttccaacc ccacgtgctt tttctttctc ctgcccgctt cttttcttca
1861 tcacaaaagg cgtggagact cggagacgga cgtttttccc cttttttaag ttattgacgc
1921 ccagcgcgcc tcgcctcttc acccatcaac gctgtgcttt gcccactgga ctcctgaaga
1981 gggggtgggg ggctccctcg gtcgcccacc ctgggaagtg cctaaccttt tattttattt
2041 tatttttttg aggaaaaaga acgcctgact cacaggttga agaaacaccc tgggccctct
2101 ctcatggccg ggttccccgt ccctctgcag aggctgggaa gggtccccgg gctggagcca
2161 cgggggcttc tctgggctgt gcctccgggg ccaacactgg ctgcttgggg ctgcccgggg
2221 actccagagg gctgcacggc caccctgccc tggctagagc gcaccccacc ggagcccacg
2281 tgggctgggc ggctggaggg atggtccccc ggtgacactg ggagaaaggc cacttggatg
2341 ggggcgtttc tgttttgttc cgctttgtga tgtcaccaat ttggaaacag cgagggtggg
2401 tggggacttt tacagaatat tctcaggtgt gtacccgaga ggcagagaga gggacgtggc
2461 cggcagctct gtgcgtggcc ttgtcccaag cacttgcgcc cgcccccgag cgccgccccc
2521 ggggagcggg aagccagcac tcgcactttg gccaggggcg cgtggaaggt ggtggcaggc
2581 accggcctgg gcagcttcca ggcctggctg gccacgacca cggcccgagg gggagcccgc
2641 caggccacgc cgcactgagc cacagccccg ggggccgcct cccggggccc cttgaggcac
2701 tgaggcaccg agactggttc tccccgagag actcggaagg tggggaacga ggggactgtg
2761 tttggggagg tggctttttc gtctgctgtt gactgaacac tacagcgccc tgtggttccg
2821 ggcttcgcac agctgtccca gggatggatc gcctgtgctg ccttcgcccg ccgccacacc
2881 gggaccctgc acggctgctt ctggcctcga cagatgacaa aagaaacagc cccaaaatac
2941 gaccactcca accagcagtt cccgcctgcc tgcccgccac tgtcaggcct gccctggcct
3001 cctcgtccgc agggctgtct gctggcttct gggggcagaa gagcggggag ccccgtggaa
3061 gggtcagggg agaccaggtc agggcagcta catttctggt gatcagcccc atggggagac
3121 ggggctggcg ggataccccc cccccggctt ccccacacca cttctgtctc acccggaagc
3181 gtcctttttt tgtgccaggt gtctacctaa gagggttggt gccagaagcc ccccatggcg
3241 agtgctgggg cccggcggtg ccctggggga gcagatgggg ccacccctgg cagggccgct
3301 acaacctttt ccagcagcgg agccctctgg ggggcctgtg cttgtggcat ctctgagggc
3361 ctagattgca caaggtgacc tggccgtggc ctgagggtgg agtcgcccag cacgcaggcc
3421 ggggcgctgc ggggctaagt attaggcctt cccagggagg gggcgtgcca agcatcccag
3481 agccgggctg ggaccgccaa aacgtcgtgg cctggatcct ctgggtctga gtgcctgatc
3541 ccctgccccc caaaaaagca gaggtaggtg ttgcaggccc agggcagggg tgcctgcccc
3601 aggagagtcc caggcagtgg ttctcgtgcc agtggcaccc aggggcaagg acagccaacc
3661 cccacccttg ccacgtgtgg ggccacgtgg gcatgtgggg tgtgtgtttt taccttggtg
3721 aatctcacct gccaacgatt tctcgtgagt gccgaccacc ttctccgacc atgttacgcc
3781 cgggcggcag cagcccccgg ccactgcaaa cccatgccct gggtcccccg gctcccccag
3841 ggaggcatcc ccgtgccaat gtcccccagt ggtggcagca gatcctgtgg ccggcctggc
3901 ggacgggacc cagtgatact tgtatattac acagtcctga tttcagacaa tttcaacctt
3961 aatctattta aaaaagaata ttctatacaa gctgttttta agccttttac catttgaaat
4021 gcatgtgttg tgcgcgttgg ggatgggagg aggggctgag gagcggctca gtgtcacctc
4081 ccacagccac cggccctgac ccttaatcca gacaccgatg gaagtcgact tttcatatct
4141 ttctcctgaa atgaactctg ttttaaattg gaataaattt tgttcctaaa
[00100] "Опухоль" относится к росту и пролиферации неопластических клеток, неважно злокачественных или доброкачественных, а также всех предраковых и раковых клеток и тканей.
[00101] Термин "противоопухолевая активность" означает снижение скорости пролиферации, жизнеспособности или метастатической активности опухолевых клеток. Например, противоопухолевая активность может быть показана за счет падения скорости роста аномальных клеток, которое появляется во время терапии, или стабилизации или уменьшения размера опухоли, или более длительного выживания, обусловленного терапия, по сравнению с контролем без терапии. Такую активность можно оценить с применением принятых in vitro или in vivo моделей опухоли, в том числе без ограничения ксенотрансплантатных моделей, моделей с аллогенными трансплантатами, моделей MMTV и других известных моделей, известных из уровня техники для исследования противоопухолевой активности.
[00102] Термин "злокачественная опухоль" относится к недоброкачественной опухоли или раку. Используемый в данном документе термин "рак" охватывает злокачественную опухоль, которая характеризуется дерегулированным или неконтролируемым ростом клеток. Иллюстративные типы рака включают: карциномы, саркомы, лейкозы и лимфомы.
[00103] Термин "рак" охватывает первичные злокачественные опухоли (например, опухоли, клетки которых не мигрировали в места локализации в организме субъекта, отличные от места локализации исходной опухоли) и вторичные злокачественные опухоли (например, опухоли, развившиеся за счет метастазирования, миграции опухолевых клеток во вторичные места локализации, которые отличны от места локализации исходной опухоли).
[00104] Термин "PMEL17" (также называемый премеланосомным белком (PMEL), D12S53E, ME20, ME20-M, ME20M, P1, P100, gp100, SI, SIL и гомологом белка локуса silver (SILV)) относится к однопролетному трансмембранному белку типа I, продуцируемому меланоцитами и принимающему участие в синтезе меланина. Последовательность нуклеиновой кислоты и аминокислотная последовательность PMEL17 человек были опубликованы в GenBank под следующими №№ доступа: NP_008859, NP_001307050, NP_001307051, NP_001186982, NP_001186983 (аминокислотные последовательности) и NM_006928, NM_001200053, NM_001200054, NM_001320121, NM_001320122 (нуклеотидные последовательности). Используемый в данном документе термин "PMEL17" применяют для совместного обозначения всех встречающихся в природе изоформ белка PMEL17 или его варианта.
[00105] NP_008859 (SEQ ID NO:272)
1 mdlvlkrcll hlavigalla vgatkvprnq dwlgvsrqlr tkawnrqlyp ewteaqrldc
61 wrggqvslkv sndgptliga nasfsialnf pgsqkvlpdg qviwvnntii ngsqvwggqp
121 vypqetddac ifpdggpcps gswsqkrsfv yvwktwgqyw qvlggpvsgl sigtgramlg
181 thtmevtvyh rrgsrsyvpl ahsssaftit dqvpfsvsvs qlraldggnk hflrnqpltf
241 alqlhdpsgy laeadlsytw dfgdssgtli sralvvthty lepgpvtaqv vlqaaiplts
301 cgsspvpgtt dghrptaeap nttagqvptt evvgttpgqa ptaepsgtts vqvpttevis
361 tapvqmptae stgmtpekvp vsevmgttla emstpeatgm tpaevsivvl sgttaaqvtt
421 tewvettare lpipepegpd assimstesi tgslgplldg tatlrlvkrq vpldcvlyry
481 gsfsvtldiv qgiesaeilq avpsgegdaf eltvscqggl pkeacmeiss pgcqppaqrl
541 cqpvlpspac qlvlhqilkg gsgtyclnvs ladtnslavv stqlimpgqe aglgqvpliv
601 gillvlmavv lasliyrrrl mkqdfsvpql phssshwlrl prifcscpig enspllsgqq
661 v
[00106] NP_001307050 (SEQ ID NO:273)
1 mdlvlkrcll hlavigalla vgatkvprnq dwlgvsrqlr tkawnrqlyp ewteaqrldc
61 wrggqvslkv sndgptliga nasfsialnf pgsqkvlpdg qviwvnntii ngsqvwggqp
121 vypqetddac ifpdggpcps gswsqkrsfv yvwktwgqyw qvlggpvsgl sigtgramlg
181 thtmevtvyh rrgsrsyvpl ahsssaftit dqvpfsvsvs qlraldggnk hflrnqpltf
241 alqlhdpsgy laeadlsytw dfgdssgtli sralvvthty lepgpvtaqv vlqaaiplts
301 cgsspvpgtt dghrptaeap nttagqvptt evvgttpgqa ptaepsgtts vqvpttevis
361 tapvqmptae staaqvttte wvettarelp ipepegpdas simstesitg slgplldgta
421 tlrlvkrqvp ldcvlyrygs fsvtldivqg iesaeilqav psgegdafel tvscqgglpk
481 eacmeisspg cqppaqrlcq pvlpspacql vlhqilkggs gtyclnvsla dtnslavvst
541 qlimpvpgil ltgqeaglgq vplivgillv lmavvlasli yrrrlmkqdf svpqlphsss
601 hwlrlprifc scpigenspl lsgqqv
[00107] NP_001307051 (SEQ ID NO:274)
1 mdlvlkrcll hlavigalla vgatkvprnq dwlgvsrqlr tkawnrqlyp ewteaqrldc
61 wrggqvslkv sndgptliga nasfsialnf pgsqkvlpdg qviwvnntii ngsqvwggqp
121 vypqetddac ifpdggpcps gswsqkrsfv yvwktwgqyw qvlggpvsgl sigtgramlg
181 thtmevtvyh rrgsrsyvpl ahsssaftit dqvpfsvsvs qlraldggnk hflrnqpltf
241 alqlhdpsgy laeadlsytw dfgdssgtli sralvvthty lepgpvtaqv vlqaaiplts
301 cgsspvpgtt dghrptaeap nttagqvptt evvgttpgqa ptaepsgtts vqvpttevis
361 tapvqmptae staaqvttte wvettarelp ipepegpdas simstesitg slgplldgta
421 tlrlvkrqvp ldcvlyrygs fsvtldivqg iesaeilqav psgegdafel tvscqgglpk
481 eacmeisspg cqppaqrlcq pvlpspacql vlhqilkggs gtyclnvsla dtnslavvst
541 qlimpgqeag lgqvplivgi llvlmavvla sliyrrrlmk qdfsvpqlph ssshwlrlpr
601 ifcscpigen spllsgqqv
[00108] NP_001186982 (SEQ ID NO:275)
1 mdlvlkrcll hlavigalla vgatkgsqvw ggqpvypqet ddacifpdgg pcpsgswsqk
61 rsfvyvwktw gqywqvlggp vsglsigtgr amlgthtmev tvyhrrgsrs yvplahsssa
121 ftitdqvpfs vsvsqlrald ggnkhflrnq pltfalqlhd psgylaeadl sytwdfgdss
181 gtlisralvv thtylepgpv taqvvlqaai pltscgsspv pgttdghrpt aeapnttagq
241 vpttevvgtt pgqaptaeps gttsvqvptt evistapvqm ptaestgmtp ekvpvsevmg
301 ttlaemstpe atgmtpaevs ivvlsgttaa qvtttewvet tarelpipep egpdassims
361 tesitgslgp lldgtatlrl vkrqvpldcv lyrygsfsvt ldivqgiesa eilqavpsge
421 gdafeltvsc qgglpkeacm eisspgcqpp aqrlcqpvlp spacqlvlhq ilkggsgtyc
481 lnvsladtns lavvstqlim pgqeaglgqv plivgillvl mavvlasliy rrrlmkqdfs
541 vpqlphsssh wlrlprifcs cpigenspll sgqqv
[00109] NP_001186983 (SEQ ID NO:276)
1 mdlvlkrcll hlavigalla vgatkvprnq dwlgvsrqlr tkawnrqlyp ewteaqrldc
61 wrggqvslkv sndgptliga nasfsialnf pgsqkvlpdg qviwvnntii ngsqvwggqp
121 vypqetddac ifpdggpcps gswsqkrsfv yvwktwgqyw qvlggpvsgl sigtgramlg
181 thtmevtvyh rrgsrsyvpl ahsssaftit dqvpfsvsvs qlraldggnk hflrnqpltf
241 alqlhdpsgy laeadlsytw dfgdssgtli sralvvthty lepgpvtaqv vlqaaiplts
301 cgsspvpgtt dghrptaeap nttagqvptt evvgttpgqa ptaepsgtts vqvpttevis
361 tapvqmptae stgmtpekvp vsevmgttla emstpeatgm tpaevsivvl sgttaaqvtt
421 tewvettare lpipepegpd assimstesi tgslgplldg tatlrlvkrq vpldcvlyry
481 gsfsvtldiv qgiesaeilq avpsgegdaf eltvscqggl pkeacmeiss pgcqppaqrl
541 cqpvlpspac qlvlhqilkg gsgtyclnvs ladtnslavv stqlimpvpg illtgqeagl
601 gqvplivgil lvlmavvlas liyrrrlmkq dfsvpqlphs sshwlrlpri fcscpigens
661 pllsgqqv
[00110] NM_006928 (SEQ ID NO:277)
1 cccagcgctc ctccccgcaa atgatcccgc cccaggggcc tatcccagtc cccccagtgc
61 ctttggttgc tggagggaag aacacaatgg atctggtgct aaaaagatgc cttcttcatt
121 tggctgtgat aggtgctttg ctggctgtgg gggctacaaa agtacccaga aaccaggact
181 ggcttggtgt ctcaaggcaa ctcagaacca aagcctggaa caggcagctg tatccagagt
241 ggacagaagc ccagagactt gactgctgga gaggtggtca agtgtccctc aaggtcagta
301 atgatgggcc tacactgatt ggtgcaaatg cctccttctc tattgccttg aacttccctg
361 gaagccaaaa ggtattgcca gatgggcagg ttatctgggt caacaatacc atcatcaatg
421 ggagccaggt gtggggagga cagccagtgt atccccagga aactgacgat gcctgcatct
481 tccctgatgg tggaccttgc ccatctggct cttggtctca gaagagaagc tttgtttatg
541 tctggaagac ctggggccaa tactggcaag ttctaggggg cccagtgtct gggctgagca
601 ttgggacagg cagggcaatg ctgggcacac acaccatgga agtgactgtc taccatcgcc
661 ggggatcccg gagctatgtg cctcttgctc attccagctc agccttcacc attactgacc
721 aggtgccttt ctccgtgagc gtgtcccagt tgcgggcctt ggatggaggg aacaagcact
781 tcctgagaaa tcagcctctg acctttgccc tccagctcca tgaccccagt ggctatctgg
841 ctgaagctga cctctcctac acctgggact ttggagacag tagtggaacc ctgatctctc
901 gggcacttgt ggtcactcat acttacctgg agcctggccc agtcactgcc caggtggtcc
961 tgcaggctgc cattcctctc acctcctgtg gctcctcccc agttccaggc accacagatg
1021 ggcacaggcc aactgcagag gcccctaaca ccacagctgg ccaagtgcct actacagaag
1081 ttgtgggtac tacacctggt caggcgccaa ctgcagagcc ctctggaacc acatctgtgc
1141 aggtgccaac cactgaagtc ataagcactg cacctgtgca gatgccaact gcagagagca
1201 caggtatgac acctgagaag gtgccagttt cagaggtcat gggtaccaca ctggcagaga
1261 tgtcaactcc agaggctaca ggtatgacac ctgcagaggt atcaattgtg gtgctttctg
1321 gaaccacagc tgcacaggta acaactacag agtgggtgga gaccacagct agagagctac
1381 ctatccctga gcctgaaggt ccagatgcca gctcaatcat gtctacggaa agtattacag
1441 gttccctggg ccccctgctg gatggtacag ccaccttaag gctggtgaag agacaagtcc
1501 ccctggattg tgttctgtat cgatatggtt ccttttccgt caccctggac attgtccagg
1561 gtattgaaag tgccgagatc ctgcaggctg tgccgtccgg tgagggggat gcatttgagc
1621 tgactgtgtc ctgccaaggc gggctgccca aggaagcctg catggagatc tcatcgccag
1681 ggtgccagcc ccctgcccag cggctgtgcc agcctgtgct acccagccca gcctgccagc
1741 tggttctgca ccagatactg aagggtggct cggggacata ctgcctcaat gtgtctctgg
1801 ctgataccaa cagcctggca gtggtcagca cccagcttat catgcctggt caagaagcag
1861 gccttgggca ggttccgctg atcgtgggca tcttgctggt gttgatggct gtggtccttg
1921 catctctgat atataggcgc agacttatga agcaagactt ctccgtaccc cagttgccac
1981 atagcagcag tcactggctg cgtctacccc gcatcttctg ctcttgtccc attggtgaga
2041 acagccccct cctcagtggg cagcaggtct gagtactctc atatgatgct gtgattttcc
2101 tggagttgac agaaacacct atatttcccc cagtcttccc tgggagacta ctattaactg
2161 aaataaatac tcagagcctg aaaaaaaaaa aaaaa
[00111] NM_001200053 (SEQ ID NO:278)
1 gggcctatcc cagtcccccc agtgcctttg gttgctggag ggaagaacac aatggatctg
61 gtgctaaaaa gatgccttct tcatttggct gtgataggtg ctttgctggc tgtgggggct
121 acaaaaggga gccaggtgtg gggaggacag ccagtgtatc cccaggaaac tgacgatgcc
181 tgcatcttcc ctgatggtgg accttgccca tctggctctt ggtctcagaa gagaagcttt
241 gtttatgtct ggaagacctg gggccaatac tggcaagttc tagggggccc agtgtctggg
301 ctgagcattg ggacaggcag ggcaatgctg ggcacacaca ccatggaagt gactgtctac
361 catcgccggg gatcccggag ctatgtgcct cttgctcatt ccagctcagc cttcaccatt
421 actgaccagg tgcctttctc cgtgagcgtg tcccagttgc gggccttgga tggagggaac
481 aagcacttcc tgagaaatca gcctctgacc tttgccctcc agctccatga ccccagtggc
541 tatctggctg aagctgacct ctcctacacc tgggactttg gagacagtag tggaaccctg
601 atctctcggg cacttgtggt cactcatact tacctggagc ctggcccagt cactgcccag
661 gtggtcctgc aggctgccat tcctctcacc tcctgtggct cctccccagt tccaggcacc
721 acagatgggc acaggccaac tgcagaggcc cctaacacca cagctggcca agtgcctact
781 acagaagttg tgggtactac acctggtcag gcgccaactg cagagccctc tggaaccaca
841 tctgtgcagg tgccaaccac tgaagtcata agcactgcac ctgtgcagat gccaactgca
901 gagagcacag gtatgacacc tgagaaggtg ccagtttcag aggtcatggg taccacactg
961 gcagagatgt caactccaga ggctacaggt atgacacctg cagaggtatc aattgtggtg
1021 ctttctggaa ccacagctgc acaggtaaca actacagagt gggtggagac cacagctaga
1081 gagctaccta tccctgagcc tgaaggtcca gatgccagct caatcatgtc tacggaaagt
1141 attacaggtt ccctgggccc cctgctggat ggtacagcca ccttaaggct ggtgaagaga
1201 caagtccccc tggattgtgt tctgtatcga tatggttcct tttccgtcac cctggacatt
1261 gtccagggta ttgaaagtgc cgagatcctg caggctgtgc cgtccggtga gggggatgca
1321 tttgagctga ctgtgtcctg ccaaggcggg ctgcccaagg aagcctgcat ggagatctca
1381 tcgccagggt gccagccccc tgcccagcgg ctgtgccagc ctgtgctacc cagcccagcc
1441 tgccagctgg ttctgcacca gatactgaag ggtggctcgg ggacatactg cctcaatgtg
1501 tctctggctg ataccaacag cctggcagtg gtcagcaccc agcttatcat gcctggtcaa
1561 gaagcaggcc ttgggcaggt tccgctgatc gtgggcatct tgctggtgtt gatggctgtg
1621 gtccttgcat ctctgatata taggcgcaga cttatgaagc aagacttctc cgtaccccag
1681 ttgccacata gcagcagtca ctggctgcgt ctaccccgca tcttctgctc ttgtcccatt
1741 ggtgagaaca gccccctcct cagtgggcag caggtctgag tactctcata tgatgctgtg
1801 attttcctgg agttgacaga aacacctata tttcccccag tcttccctgg gagactacta
1861 ttaactgaaa taaatactca gagcctgaaa aaaaaaaaaa aa
[00112] NM_001200054 (SEQ ID NO:279)
1 gggcctatcc cagtcccccc agtgcctttg gttgctggag ggaagaacac aatggatctg
61 gtgctaaaaa gatgccttct tcatttggct gtgataggtg ctttgctggc tgtgggggct
121 acaaaagtac ccagaaacca ggactggctt ggtgtctcaa ggcaactcag aaccaaagcc
181 tggaacaggc agctgtatcc agagtggaca gaagcccaga gacttgactg ctggagaggt
241 ggtcaagtgt ccctcaaggt cagtaatgat gggcctacac tgattggtgc aaatgcctcc
301 ttctctattg ccttgaactt ccctggaagc caaaaggtat tgccagatgg gcaggttatc
361 tgggtcaaca ataccatcat caatgggagc caggtgtggg gaggacagcc agtgtatccc
421 caggaaactg acgatgcctg catcttccct gatggtggac cttgcccatc tggctcttgg
481 tctcagaaga gaagctttgt ttatgtctgg aagacctggg gccaatactg gcaagttcta
541 gggggcccag tgtctgggct gagcattggg acaggcaggg caatgctggg cacacacacc
601 atggaagtga ctgtctacca tcgccgggga tcccggagct atgtgcctct tgctcattcc
661 agctcagcct tcaccattac tgaccaggtg cctttctccg tgagcgtgtc ccagttgcgg
721 gccttggatg gagggaacaa gcacttcctg agaaatcagc ctctgacctt tgccctccag
781 ctccatgacc ccagtggcta tctggctgaa gctgacctct cctacacctg ggactttgga
841 gacagtagtg gaaccctgat ctctcgggca cttgtggtca ctcatactta cctggagcct
901 ggcccagtca ctgcccaggt ggtcctgcag gctgccattc ctctcacctc ctgtggctcc
961 tccccagttc caggcaccac agatgggcac aggccaactg cagaggcccc taacaccaca
1021 gctggccaag tgcctactac agaagttgtg ggtactacac ctggtcaggc gccaactgca
1081 gagccctctg gaaccacatc tgtgcaggtg ccaaccactg aagtcataag cactgcacct
1141 gtgcagatgc caactgcaga gagcacaggt atgacacctg agaaggtgcc agtttcagag
1201 gtcatgggta ccacactggc agagatgtca actccagagg ctacaggtat gacacctgca
1261 gaggtatcaa ttgtggtgct ttctggaacc acagctgcac aggtaacaac tacagagtgg
1321 gtggagacca cagctagaga gctacctatc cctgagcctg aaggtccaga tgccagctca
1381 atcatgtcta cggaaagtat tacaggttcc ctgggccccc tgctggatgg tacagccacc
1441 ttaaggctgg tgaagagaca agtccccctg gattgtgttc tgtatcgata tggttccttt
1501 tccgtcaccc tggacattgt ccagggtatt gaaagtgccg agatcctgca ggctgtgccg
1561 tccggtgagg gggatgcatt tgagctgact gtgtcctgcc aaggcgggct gcccaaggaa
1621 gcctgcatgg agatctcatc gccagggtgc cagccccctg cccagcggct gtgccagcct
1681 gtgctaccca gcccagcctg ccagctggtt ctgcaccaga tactgaaggg tggctcgggg
1741 acatactgcc tcaatgtgtc tctggctgat accaacagcc tggcagtggt cagcacccag
1801 cttatcatgc ctgtgcctgg gattcttctc acaggtcaag aagcaggcct tgggcaggtt
1861 ccgctgatcg tgggcatctt gctggtgttg atggctgtgg tccttgcatc tctgatatat
1921 aggcgcagac ttatgaagca agacttctcc gtaccccagt tgccacatag cagcagtcac
1981 tggctgcgtc taccccgcat cttctgctct tgtcccattg gtgagaacag ccccctcctc
2041 agtgggcagc aggtctgagt actctcatat gatgctgtga ttttcctgga gttgacagaa
2101 acacctatat ttcccccagt cttccctggg agactactat taactgaaat aaatactcag
2161 agcctgaaaa aaaaaaaaaa a
[00113] NM_001320121 (SEQ ID NO:280)
1 gggcctatcc cagtcccccc agtgcctttg gttgctggag ggaagaacac aatggatctg
61 gtgctaaaaa gatgccttct tcatttggct gtgataggtg ctttgctggc tgtgggggct
121 acaaaagtac ccagaaacca ggactggctt ggtgtctcaa ggcaactcag aaccaaagcc
181 tggaacaggc agctgtatcc agagtggaca gaagcccaga gacttgactg ctggagaggt
241 ggtcaagtgt ccctcaaggt cagtaatgat gggcctacac tgattggtgc aaatgcctcc
301 ttctctattg ccttgaactt ccctggaagc caaaaggtat tgccagatgg gcaggttatc
361 tgggtcaaca ataccatcat caatgggagc caggtgtggg gaggacagcc agtgtatccc
421 caggaaactg acgatgcctg catcttccct gatggtggac cttgcccatc tggctcttgg
481 tctcagaaga gaagctttgt ttatgtctgg aagacctggg gccaatactg gcaagttcta
541 gggggcccag tgtctgggct gagcattggg acaggcaggg caatgctggg cacacacacc
601 atggaagtga ctgtctacca tcgccgggga tcccggagct atgtgcctct tgctcattcc
661 agctcagcct tcaccattac tgaccaggtg cctttctccg tgagcgtgtc ccagttgcgg
721 gccttggatg gagggaacaa gcacttcctg agaaatcagc ctctgacctt tgccctccag
781 ctccatgacc ccagtggcta tctggctgaa gctgacctct cctacacctg ggactttgga
841 gacagtagtg gaaccctgat ctctcgggca cttgtggtca ctcatactta cctggagcct
901 ggcccagtca ctgcccaggt ggtcctgcag gctgccattc ctctcacctc ctgtggctcc
961 tccccagttc caggcaccac agatgggcac aggccaactg cagaggcccc taacaccaca
1021 gctggccaag tgcctactac agaagttgtg ggtactacac ctggtcaggc gccaactgca
1081 gagccctctg gaaccacatc tgtgcaggtg ccaaccactg aagtcataag cactgcacct
1141 gtgcagatgc caactgcaga gagcacagct gcacaggtaa caactacaga gtgggtggag
1201 accacagcta gagagctacc tatccctgag cctgaaggtc cagatgccag ctcaatcatg
1261 tctacggaaa gtattacagg ttccctgggc cccctgctgg atggtacagc caccttaagg
1321 ctggtgaaga gacaagtccc cctggattgt gttctgtatc gatatggttc cttttccgtc
1381 accctggaca ttgtccaggg tattgaaagt gccgagatcc tgcaggctgt gccgtccggt
1441 gagggggatg catttgagct gactgtgtcc tgccaaggcg ggctgcccaa ggaagcctgc
1501 atggagatct catcgccagg gtgccagccc cctgcccagc ggctgtgcca gcctgtgcta
1561 cccagcccag cctgccagct ggttctgcac cagatactga agggtggctc ggggacatac
1621 tgcctcaatg tgtctctggc tgataccaac agcctggcag tggtcagcac ccagcttatc
1681 atgcctgtgc ctgggattct tctcacaggt caagaagcag gccttgggca ggttccgctg
1741 atcgtgggca tcttgctggt gttgatggct gtggtccttg catctctgat atataggcgc
1801 agacttatga agcaagactt ctccgtaccc cagttgccac atagcagcag tcactggctg
1861 cgtctacccc gcatcttctg ctcttgtccc attggtgaga acagccccct cctcagtggg
1921 cagcaggtct gagtactctc atatgatgct gtgattttcc tggagttgac agaaacacct
1981 atatttcccc cagtcttccc tgggagacta ctattaactg aaataaatac tcagagcctg
2041 a
[00114] NM_001320122 (SEQ ID NO:281)
1 gggcctatcc cagtcccccc agtgcctttg gttgctggag ggaagaacac aatggatctg
61 gtgctaaaaa gatgccttct tcatttggct gtgataggtg ctttgctggc tgtgggggct
121 acaaaagtac ccagaaacca ggactggctt ggtgtctcaa ggcaactcag aaccaaagcc
181 tggaacaggc agctgtatcc agagtggaca gaagcccaga gacttgactg ctggagaggt
241 ggtcaagtgt ccctcaaggt cagtaatgat gggcctacac tgattggtgc aaatgcctcc
301 ttctctattg ccttgaactt ccctggaagc caaaaggtat tgccagatgg gcaggttatc
361 tgggtcaaca ataccatcat caatgggagc caggtgtggg gaggacagcc agtgtatccc
421 caggaaactg acgatgcctg catcttccct gatggtggac cttgcccatc tggctcttgg
481 tctcagaaga gaagctttgt ttatgtctgg aagacctggg gccaatactg gcaagttcta
541 gggggcccag tgtctgggct gagcattggg acaggcaggg caatgctggg cacacacacc
601 atggaagtga ctgtctacca tcgccgggga tcccggagct atgtgcctct tgctcattcc
661 agctcagcct tcaccattac tgaccaggtg cctttctccg tgagcgtgtc ccagttgcgg
721 gccttggatg gagggaacaa gcacttcctg agaaatcagc ctctgacctt tgccctccag
781 ctccatgacc ccagtggcta tctggctgaa gctgacctct cctacacctg ggactttgga
841 gacagtagtg gaaccctgat ctctcgggca cttgtggtca ctcatactta cctggagcct
901 ggcccagtca ctgcccaggt ggtcctgcag gctgccattc ctctcacctc ctgtggctcc
961 tccccagttc caggcaccac agatgggcac aggccaactg cagaggcccc taacaccaca
1021 gctggccaag tgcctactac agaagttgtg ggtactacac ctggtcaggc gccaactgca
1081 gagccctctg gaaccacatc tgtgcaggtg ccaaccactg aagtcataag cactgcacct
1141 gtgcagatgc caactgcaga gagcacagct gcacaggtaa caactacaga gtgggtggag
1201 accacagcta gagagctacc tatccctgag cctgaaggtc cagatgccag ctcaatcatg
1261 tctacggaaa gtattacagg ttccctgggc cccctgctgg atggtacagc caccttaagg
1321 ctggtgaaga gacaagtccc cctggattgt gttctgtatc gatatggttc cttttccgtc
1381 accctggaca ttgtccaggg tattgaaagt gccgagatcc tgcaggctgt gccgtccggt
1441 gagggggatg catttgagct gactgtgtcc tgccaaggcg ggctgcccaa ggaagcctgc
1501 atggagatct catcgccagg gtgccagccc cctgcccagc ggctgtgcca gcctgtgcta
1561 cccagcccag cctgccagct ggttctgcac cagatactga agggtggctc ggggacatac
1621 tgcctcaatg tgtctctggc tgataccaac agcctggcag tggtcagcac ccagcttatc
1681 atgcctggtc aagaagcagg ccttgggcag gttccgctga tcgtgggcat cttgctggtg
1741 ttgatggctg tggtccttgc atctctgata tataggcgca gacttatgaa gcaagacttc
1801 tccgtacccc agttgccaca tagcagcagt cactggctgc gtctaccccg catcttctgc
1861 tcttgtccca ttggtgagaa cagccccctc ctcagtgggc agcaggtctg agtactctca
1921 tatgatgctg tgattttcct ggagttgaca gaaacaccta tatttccccc agtcttccct
1981 gggagactac tattaactga aataaatact cagagcctga
[00115] Термин "вариант" относится к полипептиду, который имеет по сути идентичную аминокислотную последовательность с референтным полипептидом, или кодируется практически идентичной нуклеотидной последовательностью и может характеризоваться одной или несколькими активностями референтного полипептида. Например, вариант может характеризоваться приблизительно 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более высокой идентичностью последовательности с референтным полипептидом, при этом он сохраняет одну или несколько активностей референтного полипептида.
[00116] Используемые в данном документе термины "лечить", "осуществление лечения" или "лечение" любого заболевания или нарушения в одном варианте осуществления относятся к уменьшению тяжести заболевания или нарушения (т. е. к замедлению, или остановке, или уменьшению развития заболевания или по меньшей мере одного из его клинических симптомов). В другом варианте осуществления "лечить", "осуществление лечения" или "лечение" относятся к облегчению или уменьшению тяжести по меньшей мере одного физического параметра, в том числе таких, которые могут не ощущаться пациентом. В еще одном варианте осуществления "лечить", "осуществление лечения" или "лечение" означают модулирование заболевания или нарушения либо физически (например, стабилизацию явного симптома), либо физиологически (например, стабилизацию физического параметра), либо с помощью того и другого.
[00117] Используемый в данном документе термин "предупреждать", "осуществление предупреждения" или "предупреждение" любого заболевания или нарушения относится к профилактическому лечению заболевания или нарушения или замедлению начала или прогрессирования заболевания или нарушения.
[00118] Термин "терапевтически приемлемое количество" или "терапевтически эффективная доза" взаимозаменяемо относится к количеству, достаточному для произведения требуемого результата (т. е. уменьшения размера опухоли, ингибирования роста опухоли, предотвращения метастазирования, ингибирования или предупреждения вирусной, бактериальной, грибковой или паразитарной инфекции). В некоторых вариантах осуществления терапевтически приемлемое количество не индуцирует или не вызывает нежелательные побочные эффекты. В некоторых вариантах осуществления терапевтически приемлемое количество индуцирует или вызывает побочные эффекты, но только такие, которые являются допустимыми с точки зрения медицинских работников, учитывая состояние пациента. Терапевтически приемлемое количество можно определять, вначале вводя низкую дозу, а затем постепенно увеличивая данную дозу до тех пор, пока не будет достигнут требуемый эффект. "Профилактически эффективная доза" и "терапевтически эффективная доза" молекул по настоящему изобретению может предупреждать проявление или, соответственно, приводить к уменьшению тяжести симптомов заболевания, в том числе симптомов, ассоциированных с раком.
[00119] Термин "совместное введение" относится к присутствию двух активных средств в крови индивидуума. Активные средства, которые вводятся совместно, могут доставляться одновременно или последовательно.
[00120] В настоящем изобретении представлены антитела, фрагменты антител (например, антигенсвязывающие фрагменты) и их конъюгаты с лекарственным средством, т. е. конъюгаты антитела и лекарственного средства или ADC, которые связываются с PMEL17. В частности, в настоящем изобретении представлены антитела и фрагменты антител (например, антигенсвязывающие фрагменты), которые связываются с PMEL17 и подвергаются интернализации после такого связывания. Антитела и фрагменты антител (например, антигенсвязывающие фрагменты) по настоящему изобретению могут применяться для получения конъюгатов антитела и лекарственного средства. Кроме того, в настоящем изобретении представлены конъюгаты антитела и лекарственного средства, которые характеризуются требуемыми фармакокинетическими характеристиками и другими требуемыми свойствами, и, таким образом, они могут применяться для лечения или предупреждения рака, экспрессирующего PMEL17. В настоящем изобретении дополнительно представлены фармацевтические композиции, содержащие конъюгаты антитела и лекарственного средства по настоящему изобретению; а также способы изготовления и применения таких фармацевтических композиций для лечения или предупреждения рака.
Фрагмент, представляющий собой лекарственное средство (D)
[00121] В одном аспекте фрагмент, представляющий собой лекарственное средство (D), конъюгата антитела и лекарственного средства по настоящему изобретению представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11 (ингибитор GNAQ/GNA11).
[00122] В другом аспекте фрагмент, представляющий собой лекарственное средство (D), конъюгата антитела и лекарственного средства по настоящему изобретению представляет собой ингибитор GNAQ.
[00123] В другом аспекте фрагмент, представляющий собой лекарственное средство (D), конъюгата антитела и лекарственного средства по настоящему изобретению представляет собой ингибитор GNA11.
[00124] В другом аспекте фрагмент, представляющий собой лекарственное средство (D), конъюгата антитела и лекарственного средства по настоящему изобретению представляет собой ингибитор GNAQ и GNA11 (ингибитор GNAQ/GNA11).
[00125] В другом аспекте фрагмент, представляющий собой лекарственное средство, конъюгата антитела и лекарственного средства по настоящему изобретению представляет собой соединение, имеющее структуру формулы (A):
 (A),
(A),
где R0 представляет собой метил или этил, R1 представляет собой метил или изопропил и R2 представляет собой метил или этил.
[00126] В другом аспекте фрагмент, представляющий собой лекарственное средство, конъюгата антитела и лекарственного средства по настоящему изобретению представляет собой соединение (A1), имеющее следующую структуру:
 (A1).
(A1).
[00127] В другом аспекте фрагмент, представляющий собой лекарственное средство, конъюгата антитела и лекарственного средства по настоящему изобретению представляет собой соединение (A2), имеющее следующую структуру:
 (A2).
(A2).
[00128] В другом аспекте фрагмент, представляющий собой лекарственное средство, конъюгата антитела и лекарственного средства по настоящему изобретению представляет собой соединение (A3), имеющее следующую структуру:
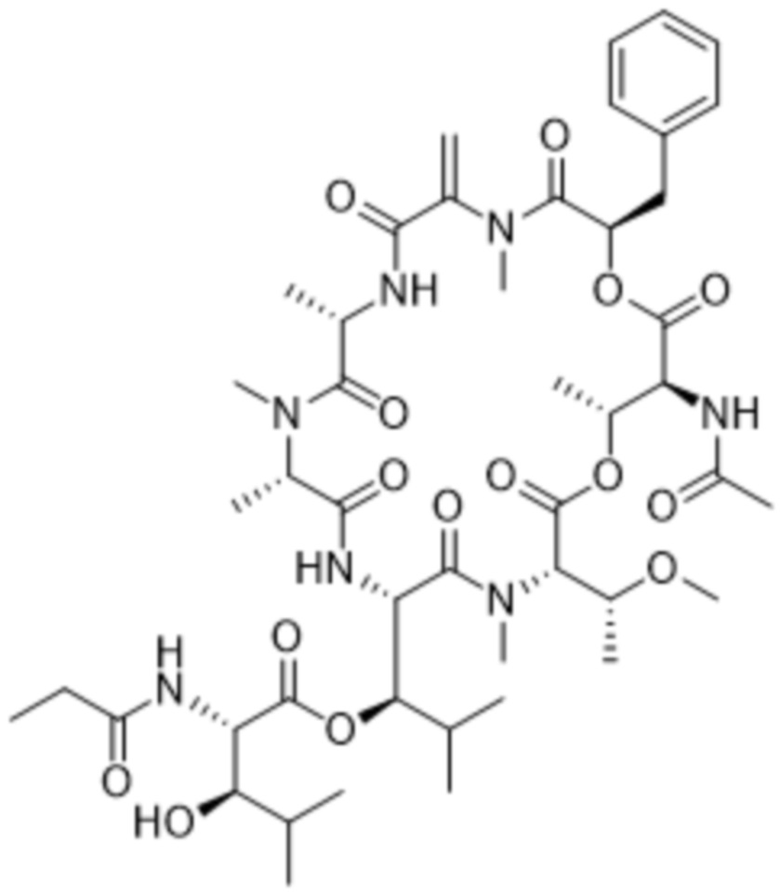 (A3).
(A3).
В таблице 1 представлена ингибирующая активность соединений (A1), (A2) и (A3), полученная с использованием анализа, описанного в примере 5.
Таблица 1
92.1GNAQ Q209L
Фрагмент, представляющий собой линкер-лекарственное средство (LA-(D)n)
[00129] Во втором аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению содержит один или несколько фрагментов, представляющих собой лекарственное средство, ковалентно присоединенных к линкеру (LA), где каждый из одного или нескольких фрагментов, представляющих собой лекарственное средство, независимо выбран из ингибитора GNAQ, ингибитора GNA11 или ингибитора GNAQ и GNA11 (ингибитора GNAQ/GNA11).
[00130] В другом аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению содержит один или несколько фрагментов, представляющих собой лекарственное средство, ковалентно присоединенных к линкеру (LA), где каждый из одного или нескольких фрагментов, представляющих собой лекарственное средство, независимо выбран из ингибитора GNAQ.
[00131] В другом аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению содержит один или несколько фрагментов, представляющих собой лекарственное средство, присоединенных к линкеру (LA), где каждый из одного или нескольких фрагментов, представляющих собой лекарственное средство, независимо выбран из ингибитора GNA11.
[00132] В другом аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению содержит один или несколько фрагментов, представляющих собой лекарственное средство, ковалентно присоединенных к линкеру (LA), где каждый из одного или нескольких фрагментов, представляющих собой лекарственное средство, независимо выбран из ингибитора GNAQ и GNA11 (ингибитора GNAQ/GNA11).
[00133] В другом аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению содержит один или несколько фрагментов, представляющих собой лекарственное средство, ковалентно присоединенных к линкеру (LA), где линкер (LA) представляет собой расщепляемый линкер и каждый из одного или нескольких фрагментов, представляющих собой лекарственное средство, независимо выбран из ингибитора GNAQ, ингибитора GNA11 или ингибитора GNAQ и GNA11 (ингибитора GNAQ/GNA11).
[00134] В другом аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению содержит один или несколько фрагментов, представляющих собой лекарственное средство, ковалентно присоединенных к линкеру (LA), где линкер (LA) представляет собой расщепляемый линкер и каждый из одного или нескольких фрагментов, представляющих собой лекарственное средство, независимо выбран из ингибитора GNAQ.
[00135] В другом аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению содержит один или несколько фрагментов, представляющих собой лекарственное средство, ковалентно присоединенных к линкеру (LA), где линкер (LA) представляет собой расщепляемый линкер и каждый из одного или нескольких фрагментов, представляющих собой лекарственное средство, независимо выбран из ингибитора GNA11.
[00136] В другом аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению содержит один или несколько фрагментов, представляющих собой лекарственное средство, ковалентно присоединенных к линкеру (LA), где линкер (LA) представляет собой расщепляемый линкер и каждый из одного или нескольких фрагментов, представляющих собой лекарственное средство, независимо выбран из ингибитора GNAQ и GNA11 (ингибитора GNAQ/GNA11).
[00137] В другом аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению содержит один или несколько фрагментов, представляющих собой лекарственное средство, ковалентно присоединенных к линкеру (LA), где линкер (LA) представляет собой нерасщепляемый линкер и каждый из одного или нескольких фрагментов, представляющих собой лекарственное средство, независимо выбран из ингибитора GNAQ, ингибитора GNA11 или ингибитора GNAQ и GNA11 (ингибитора GNAQ/GNA11).
[00138] В другом аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению содержит один или несколько фрагментов, представляющих собой лекарственное средство, ковалентно присоединенных к линкеру (LA), где линкер (LA) представляет собой нерасщепляемый линкер и каждый из одного или нескольких фрагментов, представляющих собой лекарственное средство, независимо выбран из ингибитора GNAQ.
[00139] В другом аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению содержит один или несколько фрагментов, представляющих собой лекарственное средство, ковалентно присоединенных к линкеру (LA), где линкер (LA) представляет собой нерасщепляемый линкер и каждый из одного или нескольких фрагментов, представляющих собой лекарственное средство, независимо выбран из ингибитора GNA11.
[00140] В другом аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению содержит один или несколько фрагментов, представляющих собой лекарственное средство, ковалентно присоединенных к линкеру (LA), где линкер (LA) представляет собой нерасщепляемый линкер и каждый из одного или нескольких фрагментов, представляющих собой лекарственное средство, независимо выбраны из ингибитора GNAQ и GNA11 (ингибитора GNAQ/GNA11).
[00141] В другом аспекте линкер (LA) фрагмента, представляющего собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению имеет следующую формулу:
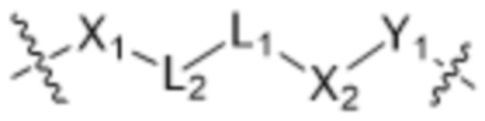
где
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
Y1 представляет собой , , , , , , или , где * в Y1 указывает на точку присоединения к X2, а ** в Y1 указывает на другую точку присоединения;
L1 представляет собой бивалентный пептидный линкер и
L2 представляет собой связь или линкер.
[00142] В другом аспекте фрагмент, представляющий собой линкер-лекарственное средство ((LA-(D)n)), конъюгата антитела и лекарственного средства по настоящему изобретению имеет следующую формулу:
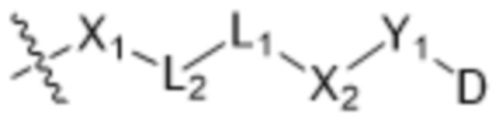
где
D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11;
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
Y1 представляет собой , , , , , , или , где * в Y1 указывает на точку присоединения к X2, а ** в Y1 указывает на точку присоединения к D;
L1 представляет собой бивалентный пептидный линкер и
L2 представляет собой связь или линкер.
Соединения линкер-лекарственное средство (LB-(D)n)
[00143] В одном аспекте линкер-лекарственное средство по настоящему изобретению представляют собой соединение, имеющее структуру формулы (B), или его стереоизомеры или фармацевтически приемлемые соли:
R8-LB-(D)n (B),
где
D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11;
R8 представляет собой реакционноспособную группу;
LB представляет собой расщепляемый линкер или нерасщепляемый линкер и
n равняется 1, 2, 3 или 4.
[00144] В одном аспекте линкер-лекарственное средство по настоящему изобретению имеет структуру формулы (B) или его стереоизомеры или фармацевтически приемлемые соли, где
D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11;
R8 представляет собой реакционноспособную группу;
LB представляет собой расщепляемый линкер, содержащий один или несколько линкерных компонентов, выбранных из саморасщепляющегося спейсера, фосфатной группы, карбонатной группы и бивалентного пептидного линкера, и
n равняется 1, 2, 3 или 4.
[00145] В одном аспекте линкер-лекарственное средство по настоящему изобретению представляют собой соединение, имеющее структуру формулы (B-1), или его стереоизомеры или фармацевтически приемлемые соли:
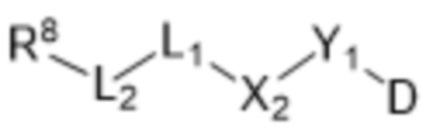 (B-1),
(B-1),
где
D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11;
R8 представляет собой реакционноспособную группу;
X2 представляет собой саморасщепляющийся спейсер;
Y1 представляет собой , , , , , , или , где * в Y1 указывает на точку присоединения к X2, а ** в Y1 указывает на точку присоединения к D;
L1 представляет собой бивалентный пептидный линкер и
L2 представляет собой связь или линкер.
[00146] Определенные аспекты и примеры соединений линкер-лекарственное средство по настоящему изобретению представлены в следующем перечне дополнительных пронумерованных вариантов осуществления. Следует понимать, что признаки, указанные в каждом варианте осуществления, можно объединять с другими указанными признаками с получением дополнительных вариантов осуществления настоящего изобретения.
[00147] Вариант осуществления 1. Соединение формулы (B) или формулы (B-1) или его стереоизомеры или фармацевтически приемлемая соль, где D представляет собой ингибитор GNAQ.
[00148] Вариант осуществления 2. Соединение формулы (B) или формулы (B-1) или его стереоизомеры или фармацевтически приемлемая соль, где D представляет собой ингибитор GNA11.
[00149] Вариант осуществления 3. Соединение формулы (B) или формулы (B-1) или его стереоизомеры или фармацевтически приемлемая соль, где D представляет собой ингибитор GNAQ и GNA11.
[00150] Вариант осуществления 4. Соединение формулы (B) или формулы (B-1) или его стереоизомеры или фармацевтически приемлемая соль, где D представляет собой
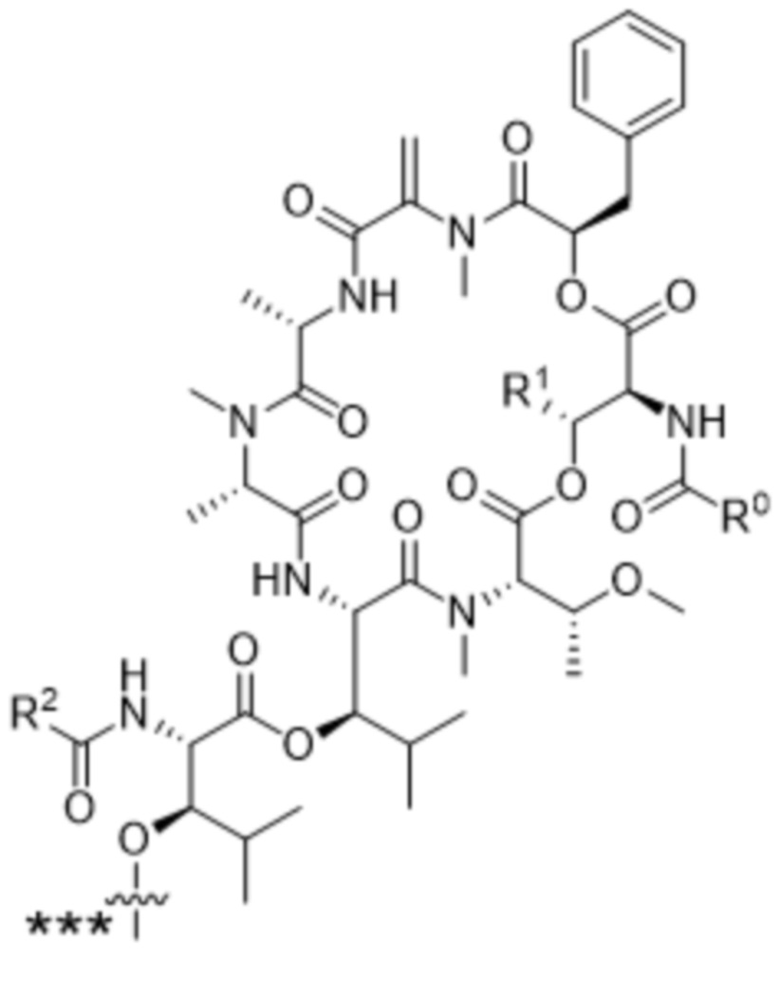 ,
,
где R0 представляет собой метил или этил, R1 представляет собой метил или изопропил, R2 представляет собой метил или этил и *** указывает на точку присоединения к LB или Y1.
[00151] Вариант осуществления 5. Соединение формулы (B) или формулы (B-1) или его стереоизомеры или фармацевтически приемлемая соль, где D представляет собой
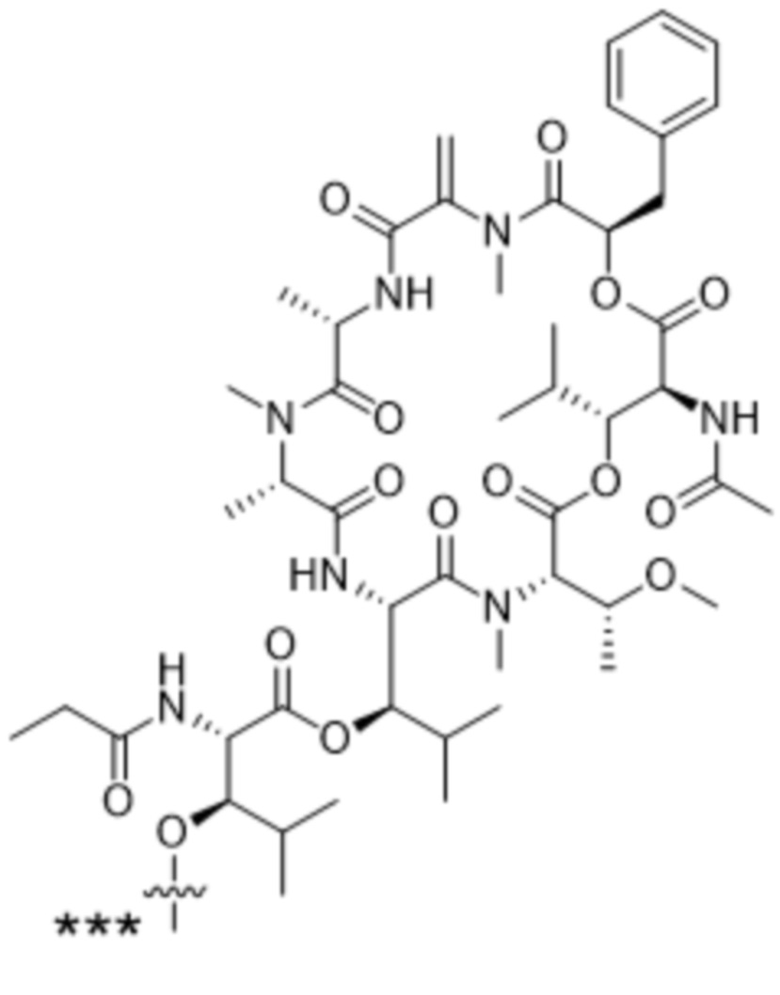 ,
,
где *** указывает на точку присоединения к LB или Y1.
[00152] Вариант осуществления 6. Соединение формулы (B) или формулы (B-1) или его стереоизомеры или фармацевтически приемлемая соль, где D представляет собой
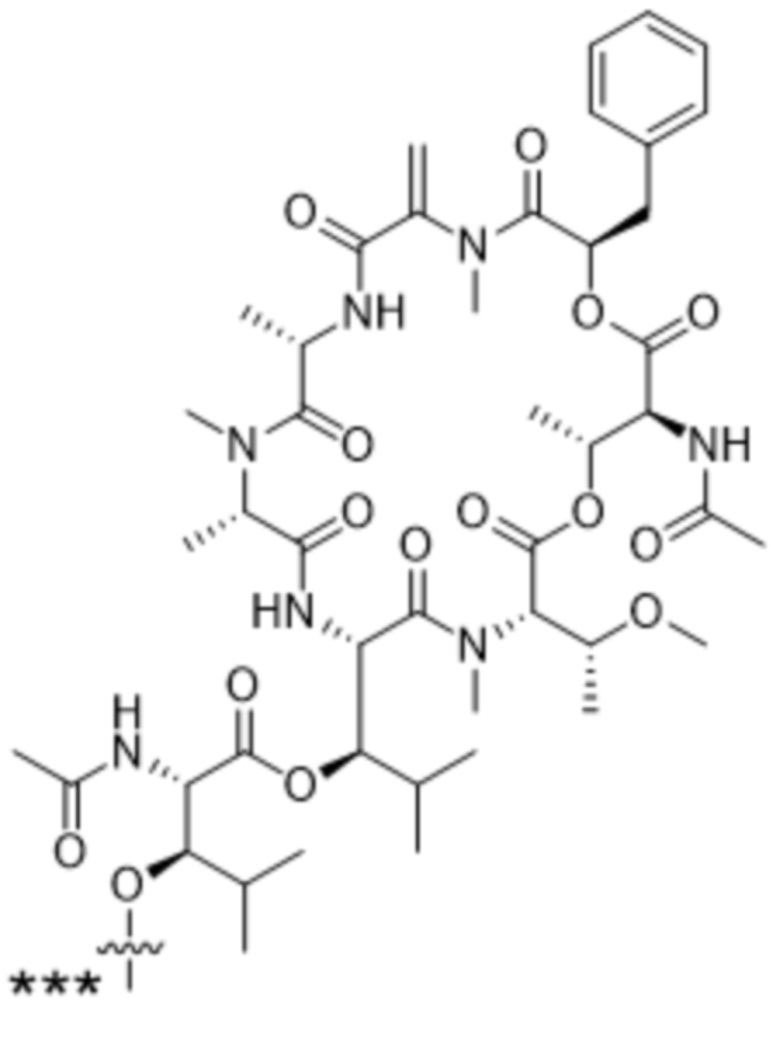 ,
,
где *** указывает на точку присоединения к LB или Y1.
[00153] Вариант осуществления 7. Соединение формулы (B) или формулы (B-1) или его стереоизомеры или фармацевтически приемлемая соль, где D представляет собой
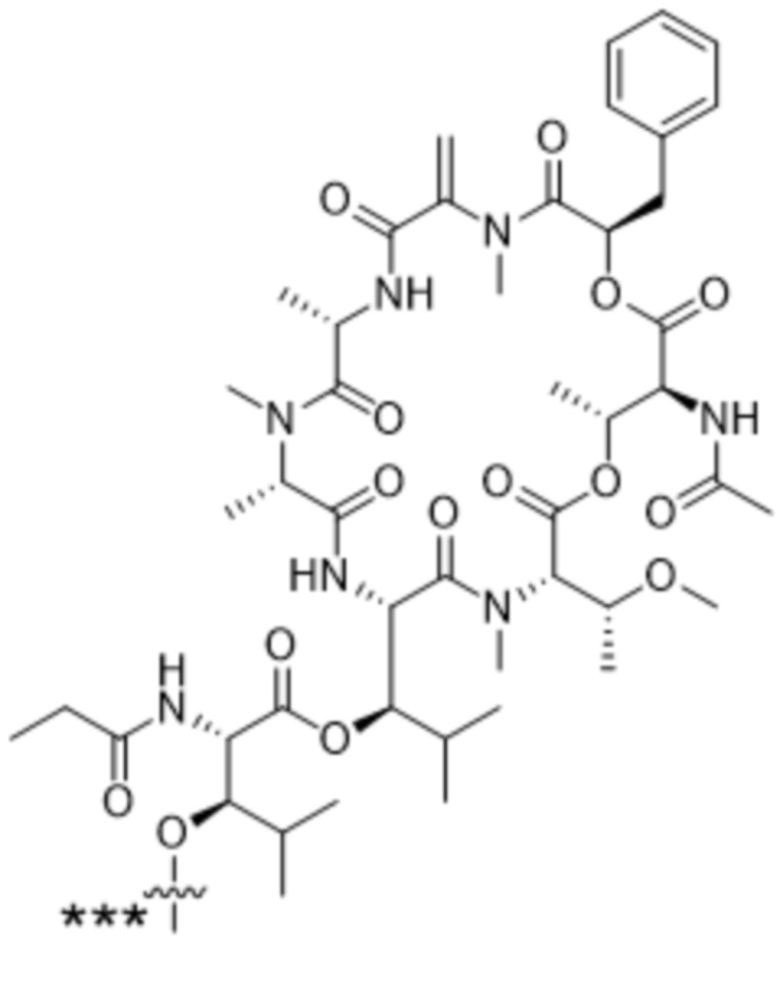 ,
,
где *** указывает на точку присоединения к LB или Y1.
[00154] Вариант осуществления 8. Соединение формулы (B) или формулы (B-1) или его стереоизомеры или фармацевтически приемлемая соль, имеющее структуру формулы (B-2) или его фармацевтически приемлемой соли:
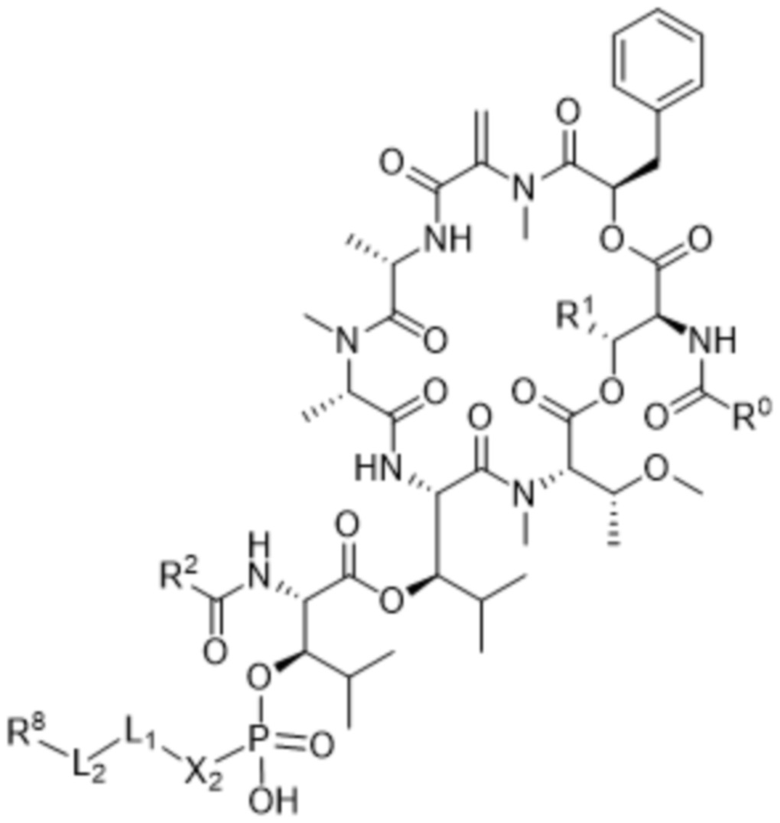 (B-2),
(B-2),
где
R0 представляет собой метил или этил;
R1 представляет собой метил или изопропил;
R2 представляет собой метил или этил и
X2, L1, L2 и R8 представляют собой такие, которые определены в соединениях формулы (B-1) выше.
[00155] Вариант осуществления 9. Соединение формулы (B), формулы (B-1) или формулы (B-2) или его стереоизомеры или фармацевтически приемлемая соль, имеющее структуру формулы (B-2a) или его фармацевтически приемлемой соли:
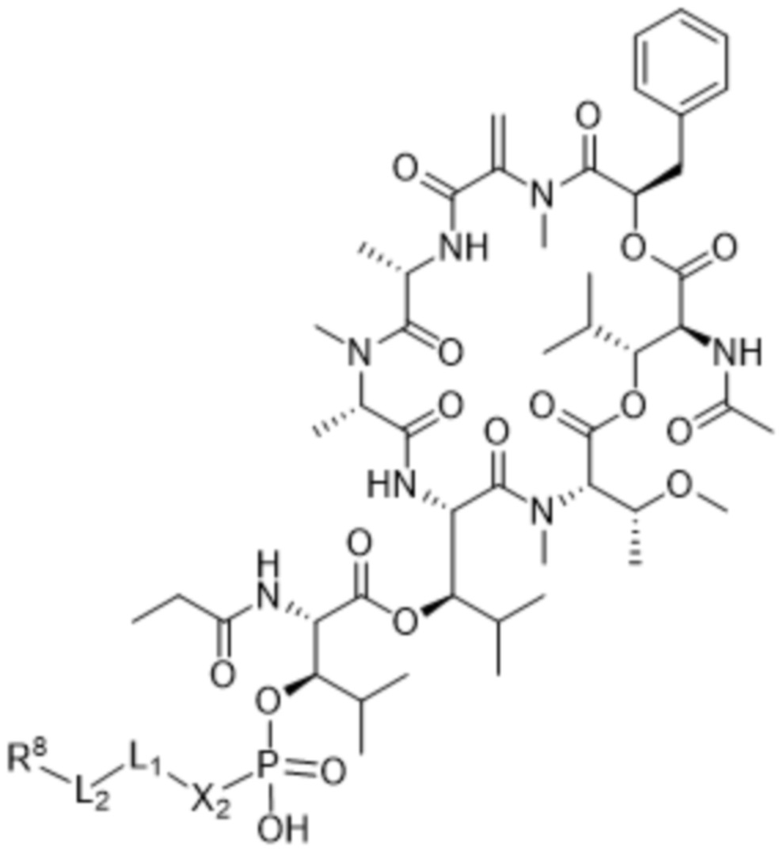 (B-2a),
(B-2a),
где
X2, L1, L2 и R8 представляют собой такие, которые определены в соединениях формулы (B-1) выше.
[00156] Вариант осуществления 10. Соединение формулы (B), формулы (B-1) или формулы (B-2) или его стереоизомеры или фармацевтически приемлемая соль, имеющее структуру формулы (B-2b) или его фармацевтически приемлемой соли:
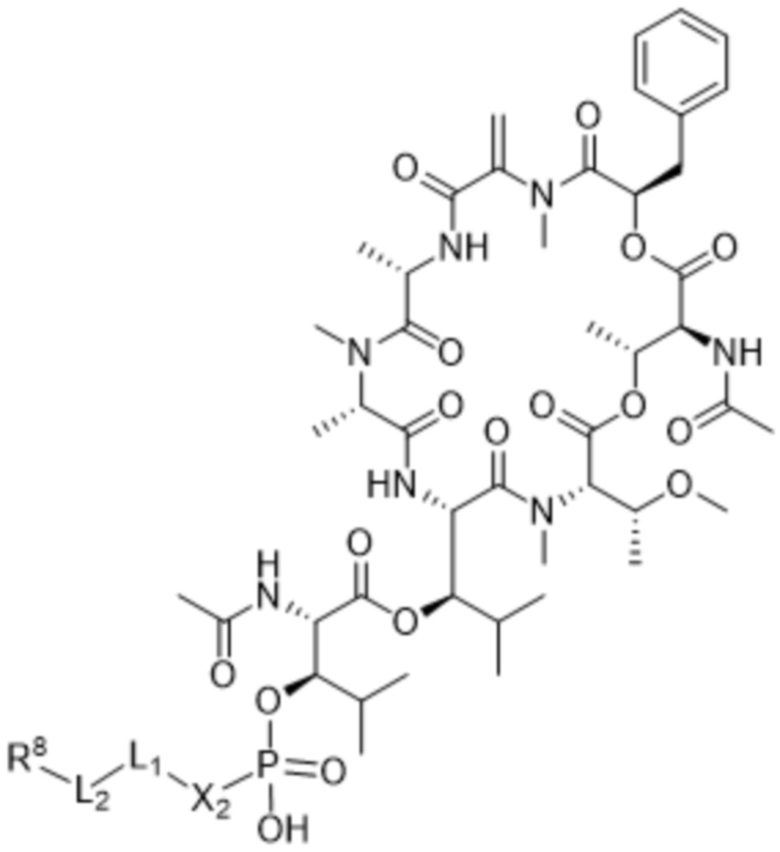 (B-2b),
(B-2b),
где
X2, L1, L2 и R8 представляют собой такие, которые определены в соединениях формулы (B-1) выше.
[00157] Вариант осуществления 11. Соединение формулы (B), формулы (B-1) или формулы (B-2) или его стереоизомеры или фармацевтически приемлемая соль, имеющее структуру формулы (B-2c) или его фармацевтически приемлемой соли:
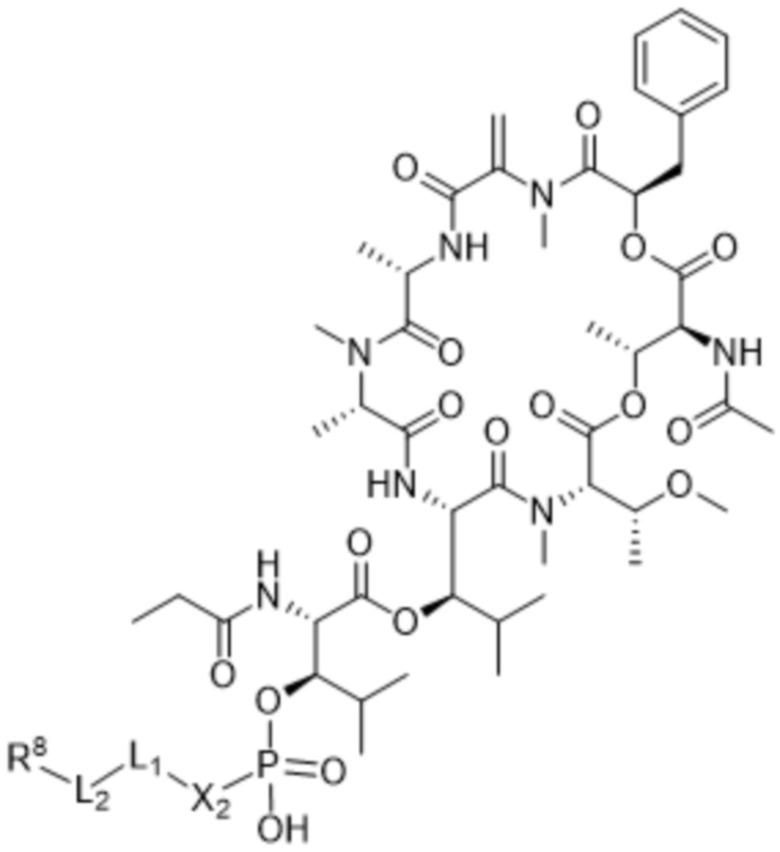 (B-2c),
(B-2c),
где
X2, L1, L2 и R8 представляют собой такие, которые определены в соединениях формулы (B-1) выше.
[00158] Вариант осуществления 12. Соединение формулы (B) или формулы (B-1) или его стереоизомеры или фармацевтически приемлемая соль, имеющее структуру формулы (B-3) или его фармацевтически приемлемой соли:
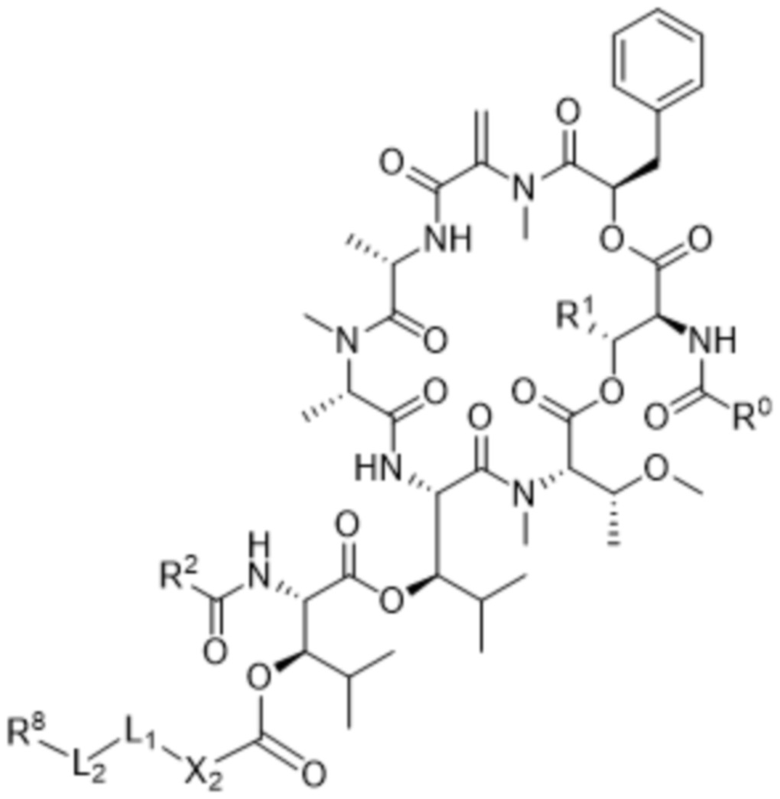 (B-3),
(B-3),
где
R0 представляет собой метил или этил;
R1 представляет собой метил или изопропил;
R2 представляет собой метил или этил и
X2, L1, L2 и R8 представляют собой такие, которые определены в соединениях формулы (B-1) выше.
[00159] Вариант осуществления 13. Соединение формулы (B), формулы (B-1) или формулы (B-3) или его стереоизомеры или фармацевтически приемлемая соль, имеющее структуру формулы (B-3a) или его фармацевтически приемлемой соли:
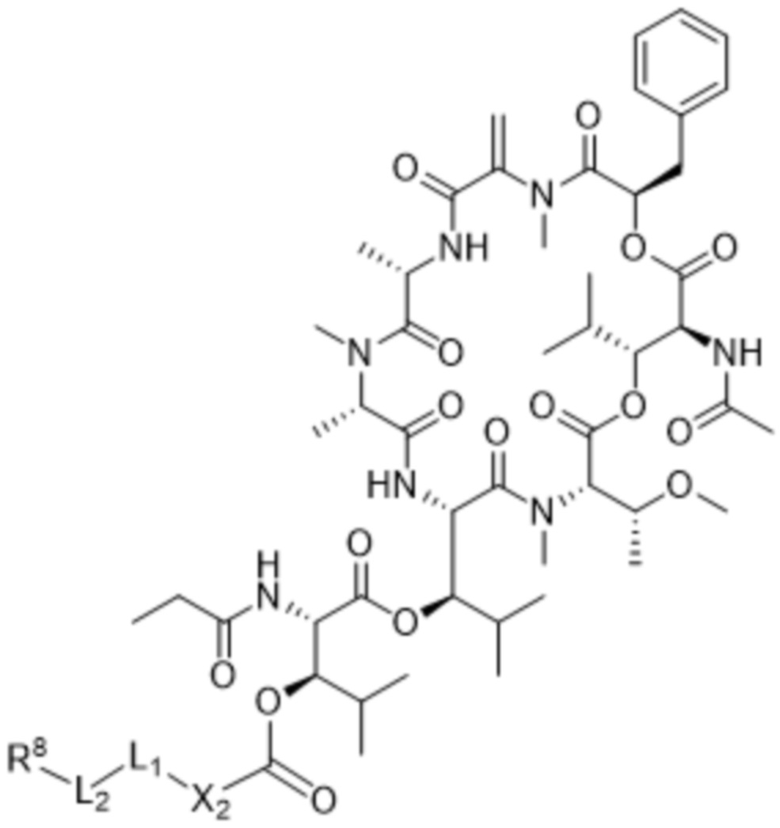 (B-3a),
(B-3a),
где
X2, L1, L2 и R8 представляют собой такие, которые определены в соединениях формулы (B-1) выше.
[00160] Вариант осуществления 14. Соединение формулы (B), формулы (B-1) или формулы (B-3) или его стереоизомеры или фармацевтически приемлемая соль, имеющее структуру формулы (B-3b) или его фармацевтически приемлемой соли:
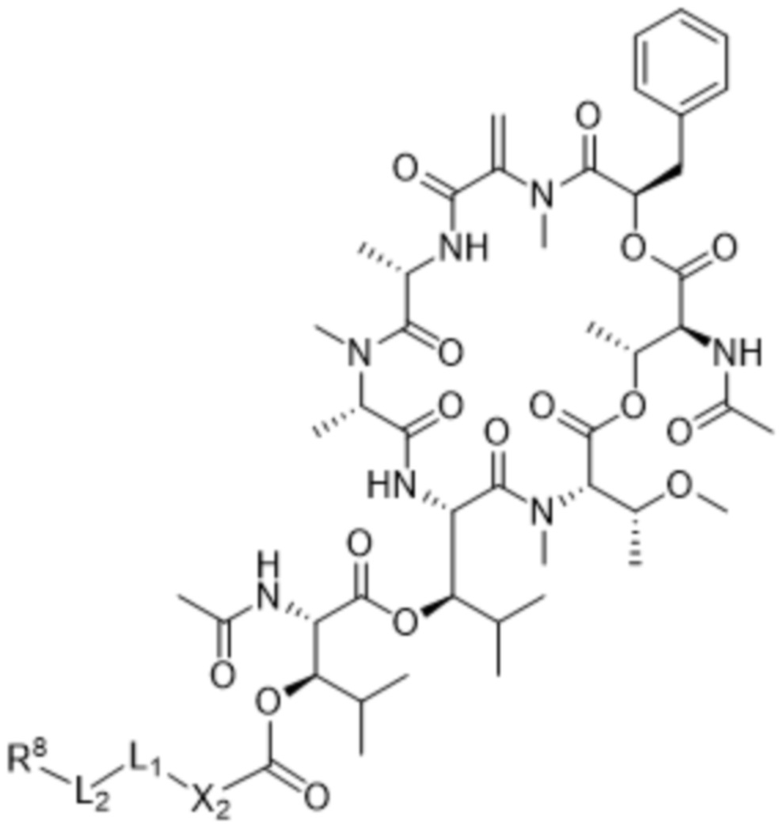 (B-3b),
(B-3b),
где
X2, L1, L2 и R8 представляют собой такие, которые определены в соединениях формулы (B-1) выше.
[00161] Вариант осуществления 15. Соединение формулы (B), формулы (B-1) или формулы (B-3) или его стереоизомеры или фармацевтически приемлемая соль, имеющее структуру формулы (B-3c) или его фармацевтически приемлемой соли:
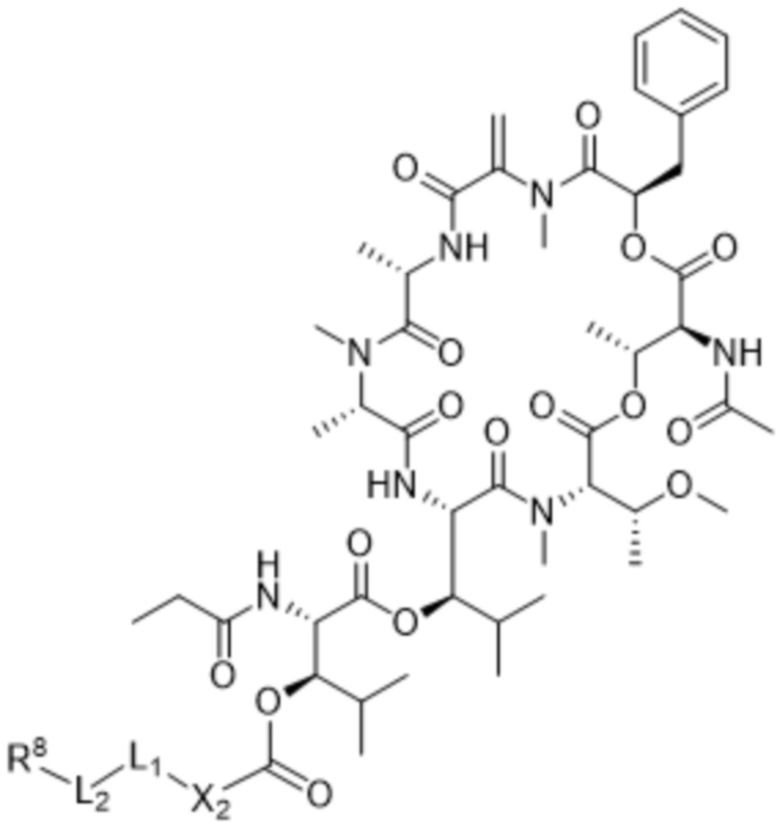 (B-3c),
(B-3c),
где
X2, L1, L2 и R8 представляют собой такие, которые определены в соединениях формулы (B-1) выше.
[00162] Вариант осуществления 16. Соединение формулы (B-2) согласно варианту осуществления 8, формулы (B-3) согласно варианту осуществления 12 или его фармацевтически приемлемая соль:
где
R0 представляет собой метил или этил;
R1 представляет собой метил или изопропил;
R2 представляет собой метил или этил;
X2 представляет собой саморасщепляющийся спейсер, выбранный из
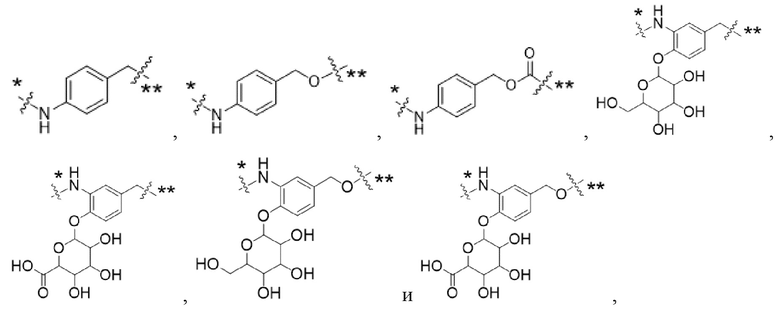
где * в X2 указывает на точку присоединения к L1 и ** в X2 указывает на точку присоединения к  группе или точку присоединения к
группе или точку присоединения к  группе;
группе;
L1 представляет собой бивалентный пептидный линкер, содержащий 2-4 аминокислотных остатка;
L2 представляет собой линкер,
R8 выбран из  , -N3, -ONH2, -NR4C(=O)CH=CH2, SH, -SSR13, -S(=O)2(CH=CH2), -NR4S(=O)2(CH=CH2), -NR4C(=O)CH2Br, -NR4C(=O)CH2I, -NHC(=O)CH2Br, -NHC(=O)CH2I, -C(=O)NHNH2,
, -N3, -ONH2, -NR4C(=O)CH=CH2, SH, -SSR13, -S(=O)2(CH=CH2), -NR4S(=O)2(CH=CH2), -NR4C(=O)CH2Br, -NR4C(=O)CH2I, -NHC(=O)CH2Br, -NHC(=O)CH2I, -C(=O)NHNH2, 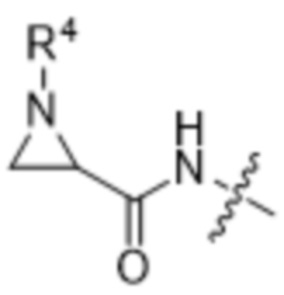 , -CO2H, -NH2,
, -CO2H, -NH2, 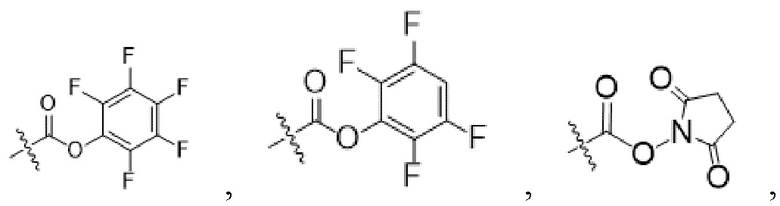
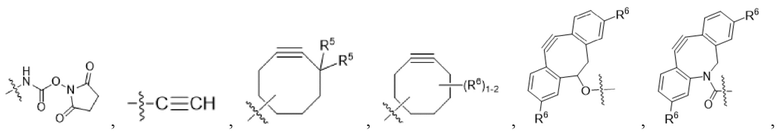
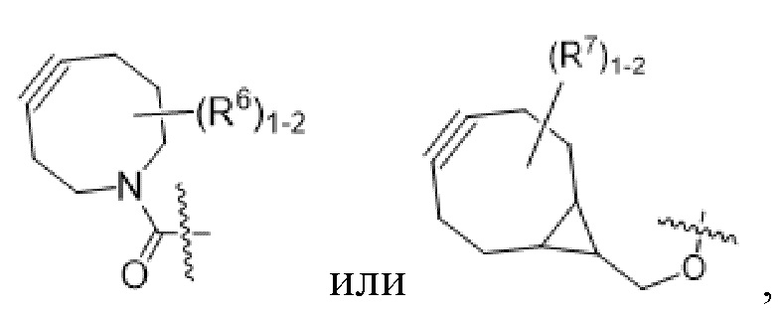
где каждый R4 независимо выбран из H и C1-C6алкила;
каждый R5 независимо выбран из H, C1-C6алкила, F, Cl и -OH;
каждый R6 независимо выбран из H, C1-C6алкила, F, Cl, -NH2, -OCH3, -OCH2CH3, -N(CH3)2, -CN, -NO2 и -OH
и
каждый R7 независимо выбран из H, C1-6алкила, фтора, бензилокси, замещенного -C(=O)OH, бензила, замещенного -C(=O)OH, C1-4алкокси, замещенного -C(=O)OH, и C1-4алкила, замещенного -C(=O)OH.
[00163] Вариант осуществления 17. Соединение согласно любому из вариантов осуществления 1-16, где X2 представляет собой саморасщепляющийся спейсер, выбранный из 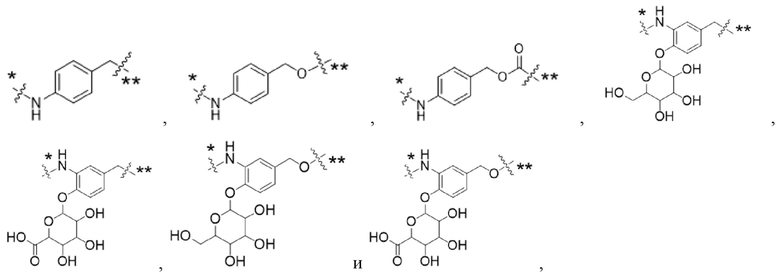
где * в X2 указывает на точку присоединения к L1 и ** в X2 указывает на точку присоединения к Y1, точку присоединения к группе или точку присоединения к группе.
[00164] Вариант осуществления 18. Соединение согласно любому из вариантов осуществления 1-17, где X2 представляет собой 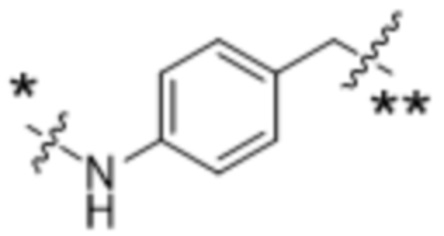 , где * в X2 указывает на точку присоединения к L1 и ** в X2 указывает на точку присоединения к Y1, точку присоединения к группе или точку присоединения к группе .
, где * в X2 указывает на точку присоединения к L1 и ** в X2 указывает на точку присоединения к Y1, точку присоединения к группе или точку присоединения к группе .
[00165] Вариант осуществления 19. Соединение согласно любому из вариантов осуществления 1-18, где L1 представляет собой бивалентный пептидный линкер, содержащий 2-4 аминокислотных остатка.
[00166] Вариант осуществления 20. Соединение согласно любому из вариантов осуществления 1-18, где L1 представляет собой бивалентный пептидный линкер, содержащий аминокислотный остаток, выбранный из валина, цитруллина, лизина, изолейцина, фенилаланина, метионина, аспарагина, пролина, аланина, лейцина, триптофана и тирозина.
[00167] Вариант осуществления 21. Соединение согласно любому из вариантов осуществления 1-18, где L1 представляет собой бивалентный пептидный линкер, содержащий по меньшей мере один остаток валина (Val) или цитруллина (Cit).
[00168] Вариант осуществления 22. Соединение согласно любому из вариантов осуществления 1-18, где L1 представляет собой бивалентный дипептидный линкер, выбранный из ValCit, PheLys, ValAla и ValLys.
[00169] Вариант осуществления 23. Соединение согласно любому из вариантов осуществления 1-18, где L1 представляет собой бивалентный дипептидный линкер, выбранный из
где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2.
[00170] Вариант осуществления 24. Соединение согласно любому из вариантов осуществления 1-18, где L1 представляет собой ValCit.
[00171] Вариант осуществления 25. Соединение согласно любому из вариантов осуществления 1-18, где
L1 представляет собой  (ValCit), где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2.
(ValCit), где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2.
[00172] Вариант осуществления 26. Соединение согласно любому из вариантов осуществления 1-25, где L2 представляет собой линкер.
[00173] Вариант осуществления 27. Соединение согласно любому из вариантов осуществления 1-25, где L2 представляет собой линкер, выбранный из
-*C(=O)((CH2)mO)p(CH2)m**-, -*C(=O)(CH2)m**-, -*C(=O)(CH2)nNHC(=O)(CH2)m**-, -*C(=O)(CH2)mNHC(=O)((CH2)mO)p(CH2)m**-, -* ((CH2)mO)p(CH2)m**-, -* ((CH2)mO)p(CH2)m**-, -(CH2)m-, -*(CH2)mNHC(=O)(CH2)m**-, -* (CH2)mNHC(=O)(CH2)mC(=O)NH(CH2)m**-, -* ((CH2)mO)p(CH2)mNHC(=O)(CH2)m**-, -* *((CH2)mO)pCH2)mC(=O)NH(CH2)m**-, -* (CH2)mC(R3)2**-, и -* (CH2)mC(R3)2SS(CH2)mNHC(=O)(CH2)m**-, где * в L2 указывает на точку присоединения к L1 и ** в L2 указывает на точку присоединения к R8;
и где
каждый R3 независимо выбран из H и C1-C6алкила;
каждый m независимо выбран из 1, 2, 3, 4, 5, 6, 7, 8, 9 и 10 и
каждый p независимо выбран из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 и 14.
[00174] Вариант осуществления 28. Соединение согласно любому из вариантов осуществления 1-25, где L2 представляет собой -*C(=O)((CH2)mO)p(CH2)m**- или -*C(=O)(CH2)m**-, где * в L2 указывает на точку присоединения к L1 и ** в L2 указывает на точку присоединения к R8,
и где каждый m независимо выбран из 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 и p равняется 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или 14.
[00175] Вариант осуществления 29. Соединение согласно любому из вариантов осуществления 1-25, где L2 представляет собой 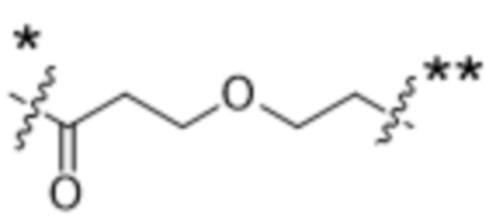 ,
, 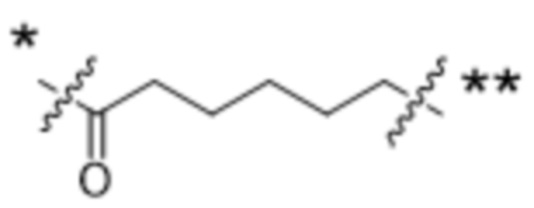 или
или 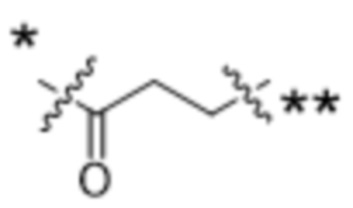 , где * в L2 указывает на точку присоединения к L1 и ** в L2 указывает на точку присоединения к R8.
, где * в L2 указывает на точку присоединения к L1 и ** в L2 указывает на точку присоединения к R8.
[00176] Вариант осуществления 30. Соединение согласно любому из вариантов осуществления 1-29, где R8 представляет собой 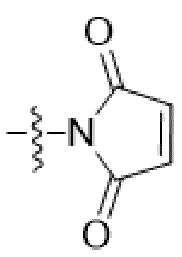 , -N3, -ONH2, -NR4C(=O)CH=CH2, SH, -SSR13, -S(=O)2(CH=CH2), -NR4S(=O)2(CH=CH2), -NR4C(=O)CH2Br, -NR4C(=O)CH2I, -NHC(=O)CH2Br, -NHC(=O)CH2I, -C(=O)NHNH2, , -CO2H, -NH2,
, -N3, -ONH2, -NR4C(=O)CH=CH2, SH, -SSR13, -S(=O)2(CH=CH2), -NR4S(=O)2(CH=CH2), -NR4C(=O)CH2Br, -NR4C(=O)CH2I, -NHC(=O)CH2Br, -NHC(=O)CH2I, -C(=O)NHNH2, , -CO2H, -NH2,
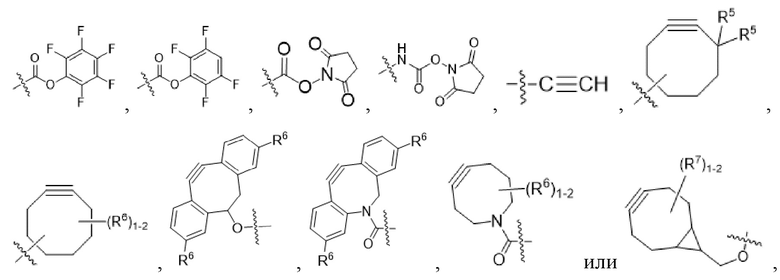
где каждый R4 независимо выбран из H и C1-C6алкила;
каждый R5 независимо выбран из H, C1-C6алкила, F, Cl и -OH;
каждый R6 независимо выбран из H, C1-C6алкила, F, Cl, -NH2, -OCH3, -OCH2CH3, -N(CH3)2, -CN, -NO2 и -OH
и
каждый R7 независимо выбран из H, C1-6алкила, фтора, бензилокси, замещенного -C(=O)OH, бензила, замещенного -C(=O)OH, C1-4алкокси, замещенного -C(=O)OH, и C1-4алкила, замещенного -C(=O)OH.
[00177] Вариант осуществления 31. Соединение согласно любому из вариантов осуществления 1-29, где R8 представляет собой , -ONH2, 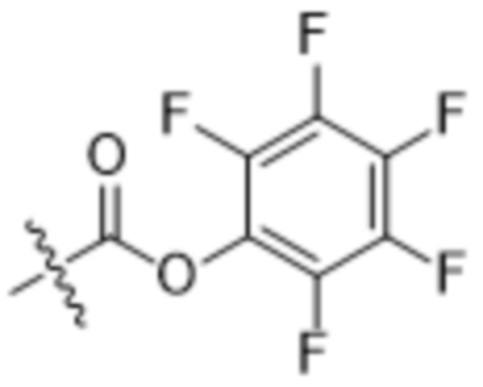 ,
, 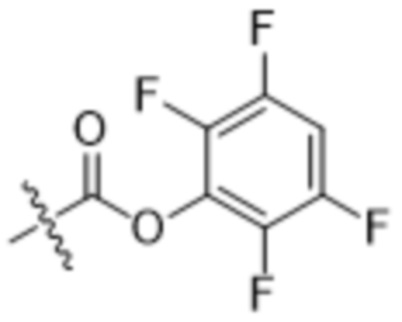 или
или 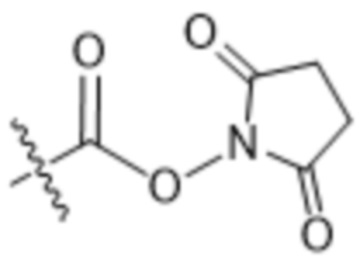 .
.
[00178] Вариант осуществления 32. Соединение согласно любому из вариантов осуществления 1-29, где R8 представляет собой .
[00179] Вариант осуществления 33. Соединение формулы (B-2) согласно варианту осуществления 8 или его фармацевтически приемлемая соль:
где
R0 представляет собой метил или этил;
R1 представляет собой метил или изопропил;
R2 представляет собой метил или этил;
X2 представляет собой 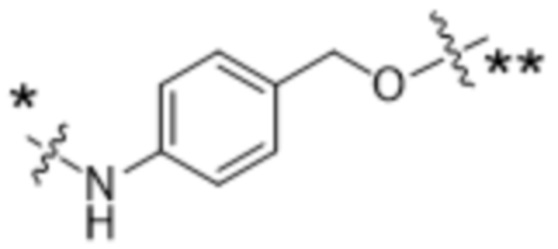 , где * в X2 указывает на точку присоединения к L1 и ** в X2 указывает на точку присоединения к группе ;
, где * в X2 указывает на точку присоединения к L1 и ** в X2 указывает на точку присоединения к группе ;
L1 представляет собой 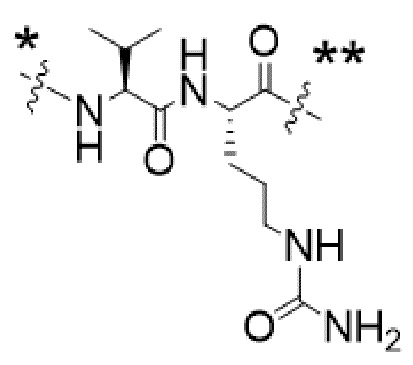 (ValCit), где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2;
(ValCit), где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2;
L2 представляет собой или , где * в L2 указывает на точку присоединения к L1 и ** в L2 указывает на точку присоединения к R8,
и
R8 представляет собой .
[00180] Вариант осуществления 34. Соединение формулы (B-3) согласно варианту осуществления 12 или его фармацевтически приемлемая соль:
где
R0 представляет собой метил или этил;
R1 представляет собой метил или изопропил;
R2 представляет собой метил или этил;
X2 представляет собой , где * в X2 указывает на точку присоединения к L1 и ** в X2 указывает на точку присоединения к группе 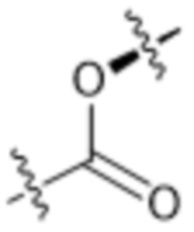 ;
;
L1 представляет собой 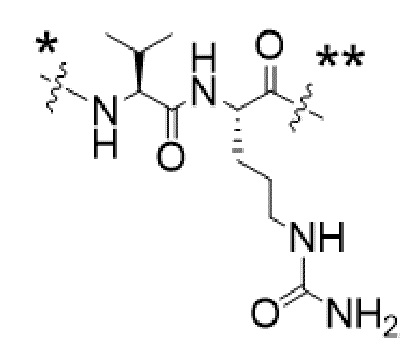 (ValCit), где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2;
(ValCit), где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2;
L2 представляет собой или , где * в L2 указывает на точку присоединения к L1 и ** в L2 указывает на точку присоединения к R8,
и
R8 представляет собой .
[00181] Вариант осуществления 35. Соединение формулы (B), формулы (B-1) или формулы (B-2), где соединение представляет собой
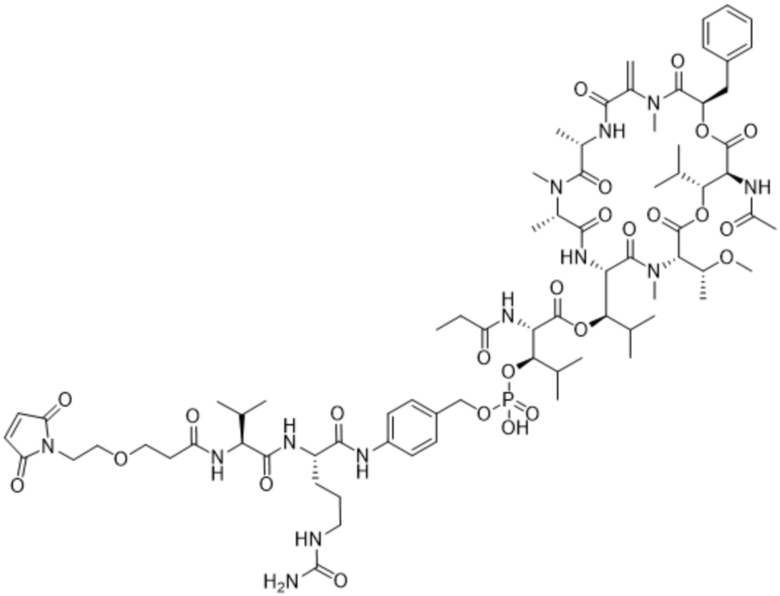 (B1).
(B1).
[00182] Вариант осуществления 36. Соединение формулы (B), формулы (B-1) или формулы (B-2), где соединение представляет собой
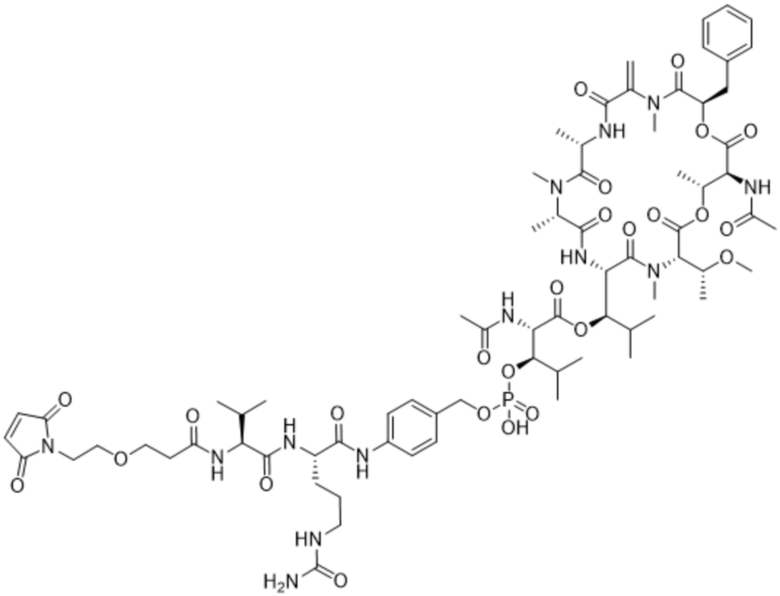 (B2).
(B2).
[00183] Вариант осуществления 37. Соединение формулы (B), формулы (B-1) или формулы (B-2), где соединение представляет собой
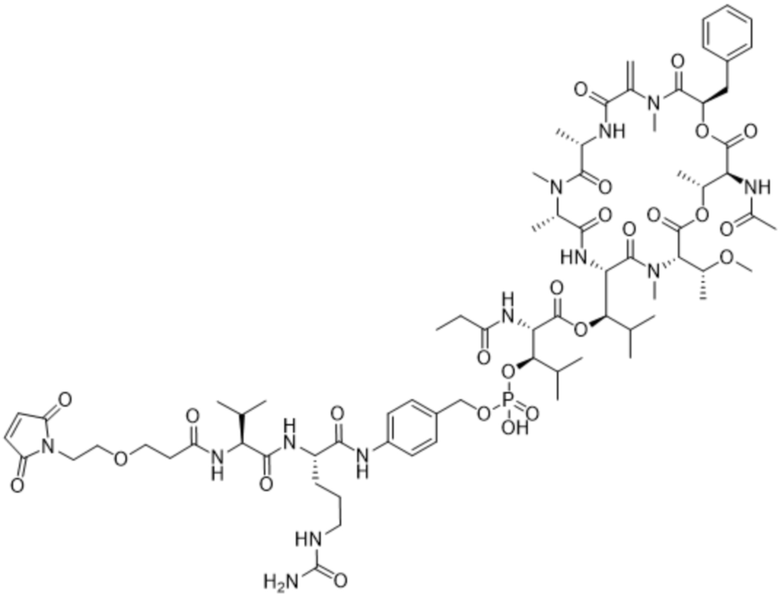 (B3).
(B3).
[00184] Вариант осуществления 38. Соединение формулы (B), формулы (B-1) или формулы (B-2), где соединение представляет собой
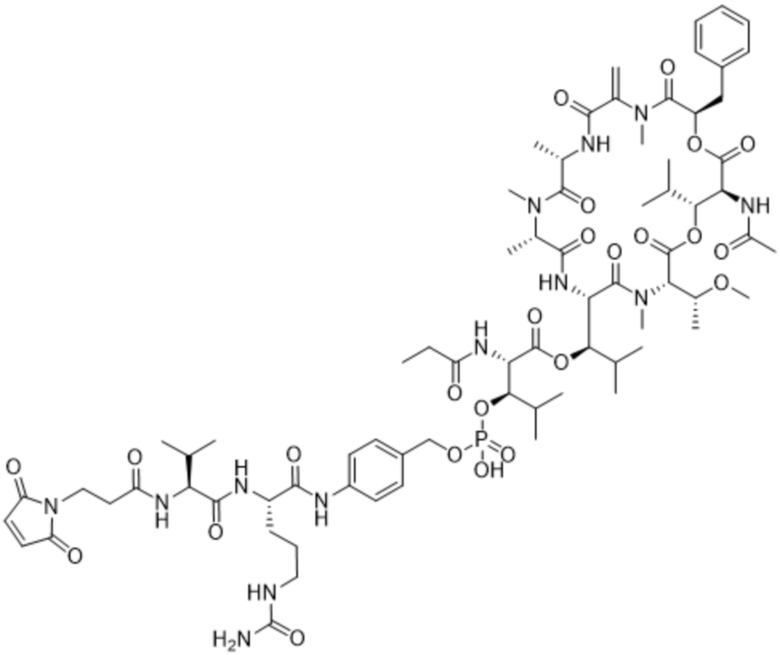 (B4).
(B4).
[00185] Вариант осуществления 39. Соединение формулы (B), формулы (B-1) или формулы (B-2), где соединение представляет собой
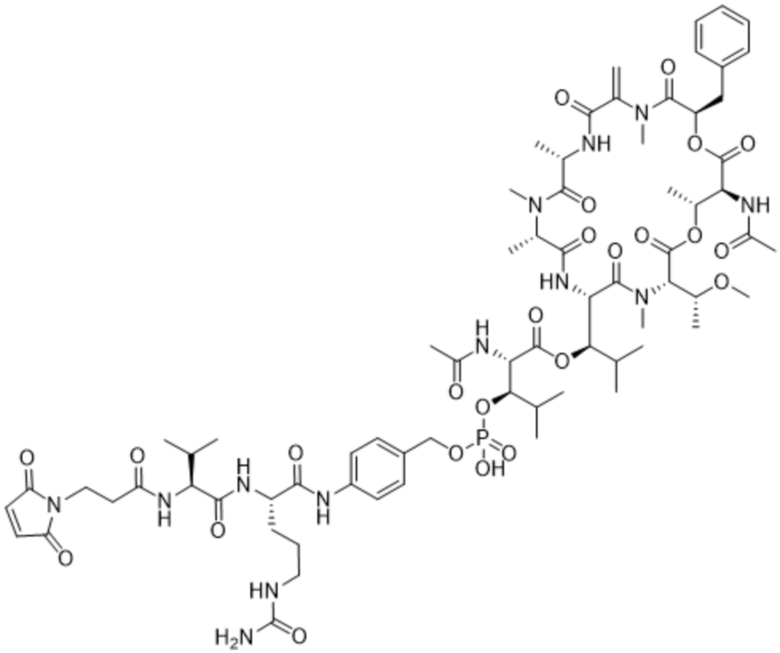 (B5).
(B5).
[00186] Вариант осуществления 40. Соединение формулы (B), формулы (B-1) или формулы (B-2), где соединение представляет собой
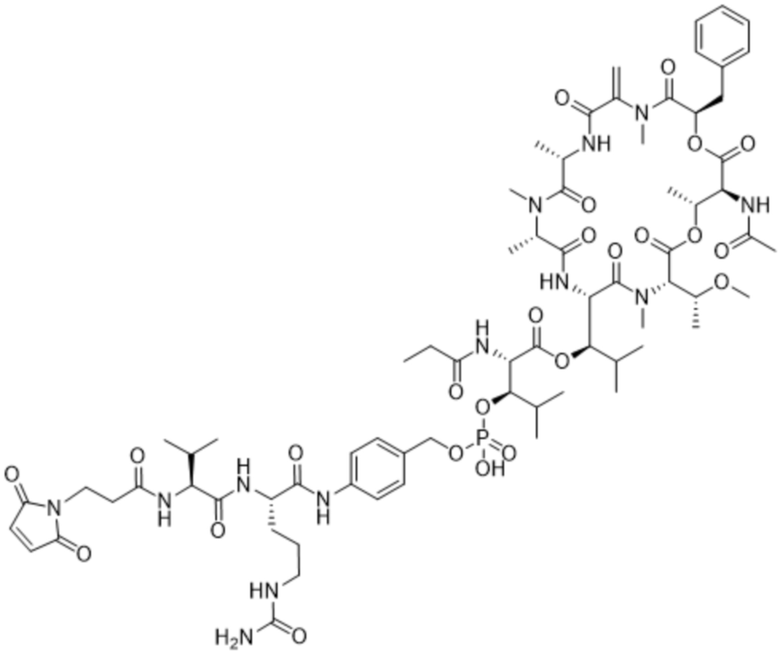 (B6).
(B6).
[00187] Вариант осуществления 41. Соединение формулы (B), формулы (B-1) или формулы (B-3), где соединение представляет собой
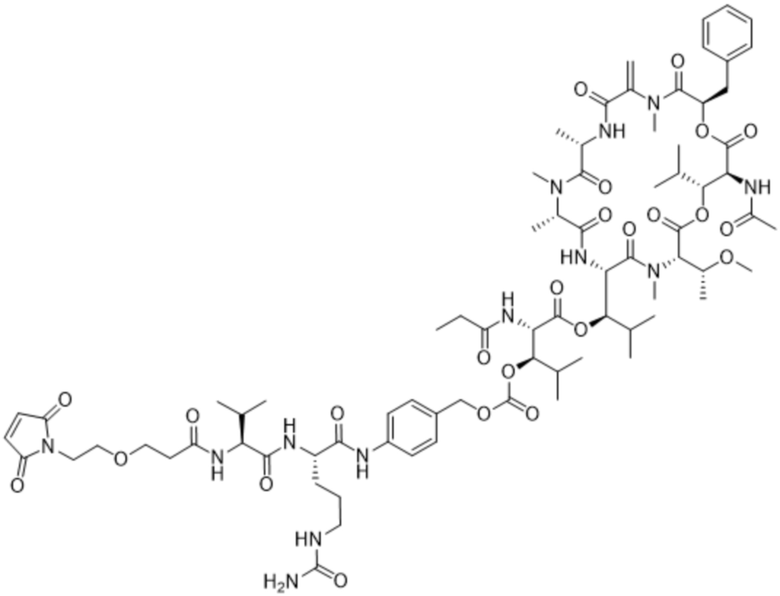 (B7).
(B7).
[00188] Вариант осуществления 42. Соединение формулы (B), формулы (B-1) или формулы (B-3), где соединение представляет собой
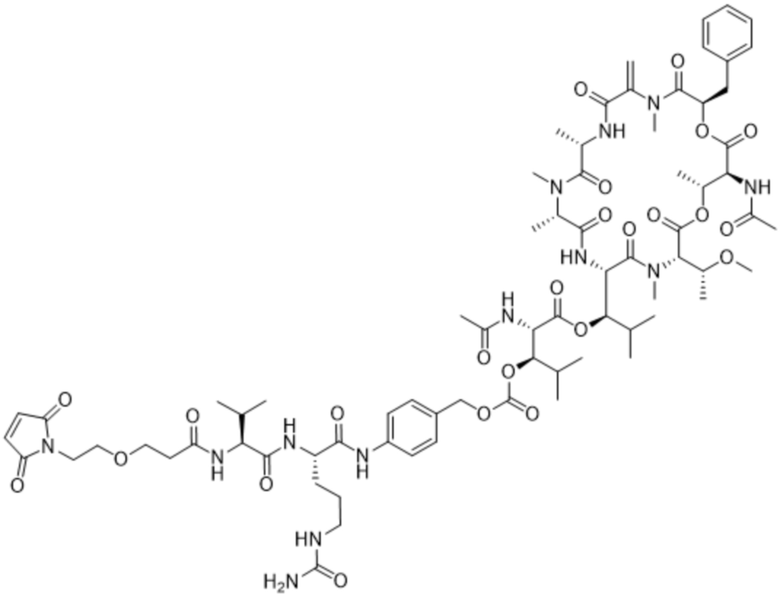 (B8).
(B8).
[00189] Вариант осуществления 43. Соединение формулы (B), формулы (B-1) или формулы (B-3), где соединение представляет собой
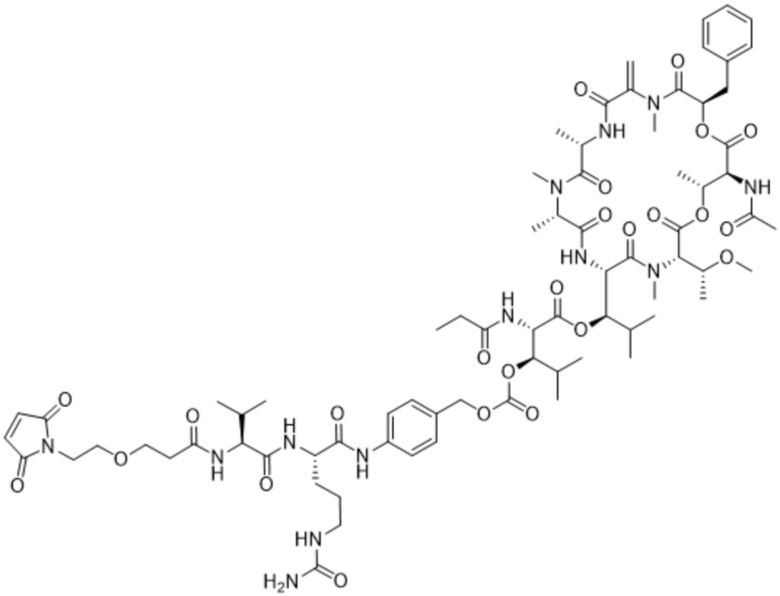 (B9).
(B9).
[00190] Вариант осуществления 44. Соединение формулы (B), формулы (B-1) или формулы (B-3), где соединение представляет собой
 (B10).
(B10).
[00191] Вариант осуществления 45. Соединение формулы (B), формулы (B-1) или формулы (B-3), где соединение представляет собой
 (B11).
(B11).
[00192] Вариант осуществления 46. Соединение формулы (B), формулы (B-1) или формулы (B-3), где соединение представляет собой
 (B12).
(B12).
Конъюгаты антитела и лекарственного средства
[00193] В одном аспекте конъюгат антитела и лекарственного средства по настоящему изобретению представляет собой конъюгат формулы (C):
Ab-(LA-(D)n)y (C),
где
D представляет собой фрагмент, представляющий собой лекарственное средство;
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
LA представляет собой линкер;
n равняется 1, 2, 3 или 4 и
y равняется 1, 2, 3 или 4,
где фрагмент, представляющий собой линкер-лекарственное средство (LA-(D)n), ковалентно присоединен к антителу или его антигенсвязывающему фрагменту.
[00194] В одном аспекте конъюгат антитела и лекарственного средства по настоящему изобретению имеет структуру формулы (C), где
D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11;
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
LA представляет собой расщепляемый линкер, содержащий один или несколько линкерных компонентов, выбранных из саморасщепляющегося спейсера, фосфатной группы, карбонатной группы и бивалентного пептидного линкера;
n равняется 1, 2, 3 или 4 и
y равняется 1, 2, 3 или 4,
где фрагмент, представляющий собой линкер-лекарственное средство (LA-(D)n), ковалентно присоединен к антителу или его антигенсвязывающему фрагменту.
[00195] В одном аспекте конъюгат антитела и лекарственного средства формулы (C) представляет собой конъюгат формулы (C-1):
(C-1),
где
D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11;
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
Y1 представляет собой , , , , , , или , где * в Y1 указывает на точку присоединения к X2, а ** в Y1 указывает на точку присоединения к D;
L1 представляет собой бивалентный пептидный линкер;
L2 представляет собой связь или линкер, и
y равняется 1, 2, 3 или 4.
[00196] В конъюгатах формулы (C) один или несколько фрагментов, представляющих собой линкер-лекарственное средство (LB-(D)n), могут быть ковалентно присоединены к антителу Ab или его антигенсвязывающему фрагменту, тем самым соединяя ковалентной связью один или несколько фрагментов, представляющих собой лекарственные средства D, с антителом Ab или его антигенсвязывающим фрагментом посредством линкера, LA. LA представляет собой любой химический фрагмент, который способен соединять антитело Ab или его антигенсвязывающий фрагмент с одним или несколькими фрагментами, представляющими собой лекарственные средства D. Конъюгаты формулы (C), где один или несколько фрагментов, представляющих собой лекарственные средства D, ковалентно связаны с антителом Ab или его антигенсвязывающим фрагментом, могут быть образованы с применением реагента на основе бифункционального или мультифункционального линкера, имеющего одну или несколько реакционноспособных функциональных групп, которые являются одинаковыми или разными. Одну из реакционноспособных функциональных групп реагента на основе бифункционального или мультифункционального линкера применяют для осуществления реакции с группой на антителе Ab или его антигенсвязывающем фрагменте, в качестве примера тиольной или аминной (например, на цистеине, N-конце или боковой цепи аминокислоты, такой как лизин), с образованием ковалентной связи с одним концом линкера LA. Такие реакционноспособные функциональные группы реагента на основе бифункционального или мультифункционального линкера включают без ограничения малеимидную, тиольную и NHS-сложноэфирную. Другую реакционноспособную функциональную группу или группы реагента на основе бифункционального или мультифункционального линкера применяют для ковалентного присоединения одного или нескольких фрагментов, представляющих собой лекарственные средства D, к линкеру LA.
[00197] В одном аспекте LA представляет собой расщепляемый линкер. В другом аспекте LA представляет собой нерасщепляемый линкер. В некоторых аспектах LA представляет собой кислотолабильный линкер, фотолабильный линкер, расщепляемый пептидазами линкер, расщепляемый эстеразами линкер, расщепляемый гликозидазами линкер, расщепляемый фосфодиэстеразами линкер, линкер с восстанавливаемой дисульфидной связью, гидрофильный линкер или линкер на основе дикарбоновой кислоты.
[00198] В одном аспекте LA представляет собой расщепляемый линкер, содержащий один или несколько линкерных компонентов, выбранных из саморасщепляющегося спейсера, фосфатной группы, карбонатной группы и бивалентного пептидного линкера.
[00199] В одном аспекте LA представляет собой расщепляемый линкер, содержащий один или несколько линкерных компонентов, выбранных из саморасщепляющегося спейсера, фосфатной группы, карбонатной группы, бивалентного пептидного линкера и бивалентной связывающей группы.
[00200] В одном аспекте LA представляет собой расщепляемый линкер, содержащий один или несколько линкерных компонентов, выбранных из саморасщепляющегося спейсера, фосфатной группы и бивалентного пептидного линкера.
[00201] В одном аспекте LA представляет собой расщепляемый линкер, содержащий один или несколько линкерных компонентов, выбранных из саморасщепляющегося спейсера, фосфатной группы, бивалентного пептидного линкера и бивалентной связывающей группы.
[00202] В другом аспекте линкер (LA) имеет следующую формулу:
где
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
Y1 представляет собой , , , , , , или , где * в Y1 указывает на точку присоединения к X2 и ** в Y1 указывает на другую точку присоединения;
L1 представляет собой бивалентный пептидный линкер и
L2 представляет собой связь или линкер.
[00203] В другом аспекте линкер (LA) имеет следующую формулу:
где
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
Y1 представляет собой , , или , где * в Y1 указывает на точку присоединения к X2;
L1 представляет собой бивалентный пептидный линкер и
L2 представляет собой связь или линкер.
[00204] В другом аспекте линкер (LA) имеет следующую формулу:
где
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
Y1 представляет собой , , или , где * в Y1 указывает на точку присоединения к X2;
L1 представляет собой бивалентный пептидный линкер и
L2 представляет собой связь или линкер.
[00205] Хотя в конкретной молекуле конъюгата соотношение лекарственного средства и антитела имеет точное целочисленное значение (например, произведение n и y в формуле (С), известно, что зачастую значение будет представлять собой среднее значение, когда его применяют для описания образца, содержащего множество молекул, вследствие некоторой степени гетерогенности, обычно сопряженной со стадией конъюгации. Средняя нагрузка для образца конъюгата обозначается в данном документе как соотношение лекарственного средства и антитела или «DAR». В некоторых аспектах DAR составляет от приблизительно 1 до приблизительно 5, но обычно составляет приблизительно 1, 2, 3 или 4. В некоторых аспектах по меньшей мере 50% образца по весу составляет соединение, характеризующееся средним DAR плюс или минус 2, и предпочтительно по меньшей мере 50% образца составляет конъюгат, который характеризуется средним DAR плюс или минус 1. Другие аспекты включают конъюгаты, в которых DAR составляет приблизительно 2. В некоторых аспектах DAR, составляющее ‘приблизительно y', означает, что измеренное значение DAR находится в пределах 20% от произведения n и y в формуле (I). В некоторых аспектах DAR, составляющее ‘приблизительно n', означает, что измеренное значение DAR находится в пределах 20% от n в формуле (II).
[00206] В одном аспекте среднее молярное соотношение лекарственного средства и антитела в конъюгатах формулы (C) (т. е. среднее значение произведения n и y, также известное как соотношение лекарственного средства и антитела (DAR)) составляет от приблизительно 1 до приблизительно 10, от приблизительно 1 до приблизительно 6 (например, 0,9, 1,0, 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4, 2,5, 2,6, 2,7, 2,8, 2,9, 3,0, 3,1, 3,2, 3,3, 3,4, 3,5, 3,6, 3,7, 3,8, 3,9, 4,0, 4,1, 4,2, 4,3, 4,4, 4,5, 4,6, 4,7, 4,8, 4,9, 5,0, 5,1, 5,2, 5,3, 5,4, 5,5, 5,6, 5,7, 5,8, 5,9, 6,0), от приблизительно 1 до приблизительно 5, от приблизительно 1,5 до приблизительно 4,5 или от приблизительно 2 до приблизительно 4.
[00207] В одном аспекте представленный в настоящем изобретении конъюгат характеризуется по сути высокой степью чистотой и имеет один или несколько из следующих признаков: (a) более приблизительно 90% (например, более или равно приблизительно 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%), предпочтительно более приблизительно 95% разновидностей конъюгата являются мономерными, (b) уровень неконъюгированного линкера в препарате на основе конъюгата составляет менее приблизительно 10% (например, менее или равно приблизительно 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1%, или 0%) (относительно общего количества линкера), (c) менее 10% молекул конъюгата являются сшитыми (например, менее или равно приблизительно 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1%, или 0%), (d) уровень свободного лекарственного средства (ADP-индуцируемого ингибитора агрегации тромбоцитов, например, ингибитора GNAQ, ингибитора GNA11 или ингибитора GNAQ и ингибитора GNA11) в препарате на основе конъюгата составляет менее приблизительно 2% (например, менее или равно приблизительно 1,5%, 1,4%, 1,3%, 1,2%, 1,1%, 1,0%, 0,9%, 0,8%, 0,7%, 0,6%, 0,5%, 0,4%, 0,3%, 0,2%, 0,1%, или 0%) (моль/моль относительно общего количества лекарственного средства).
[00208] Определенные аспекты и примеры конъюгатов антитела и лекарственного средства по настоящему изобретению представлены в следующем перечне дополнительных пронумерованных вариантов осуществления. Следует понимать, что признаки, указанные в каждом варианте осуществления, можно объединять с другими указанными признаками с получением дополнительных вариантов осуществления настоящего изобретения.
[00209] Вариант осуществления 47. Конъюгат формулы (C) или формулы (C-1), где D представляет собой ингибитор GNAQ.
[00210] Вариант осуществления 48. Конъюгат формулы (C) или формулы (C-1), где D представляет собой ингибитор GNA11.
[00211] Вариант осуществления 49. Конъюгат формулы (C) или формулы (C-1), где D представляет собой ингибитор GNAQ и GNA11.
[00212] Вариант осуществления 50. Конъюгат формулы (C) или формулы (C-1), где D представляет собой
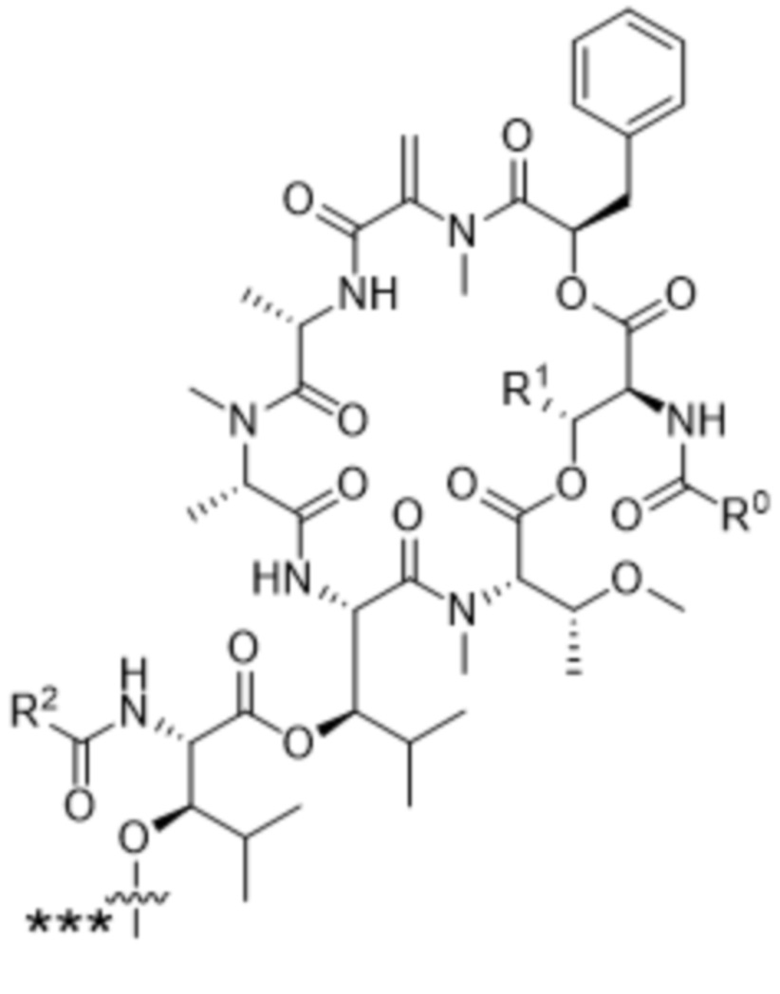 ,
,
где R0 представляет собой метил или этил, R1 представляет собой метил или изопропил, R2 представляет собой метил или этил и *** указывает на точку присоединения к LA или Y1.
[00213] Вариант осуществления 51. Конъюгат формулы (C) или формулы (C-1), где D представляет собой
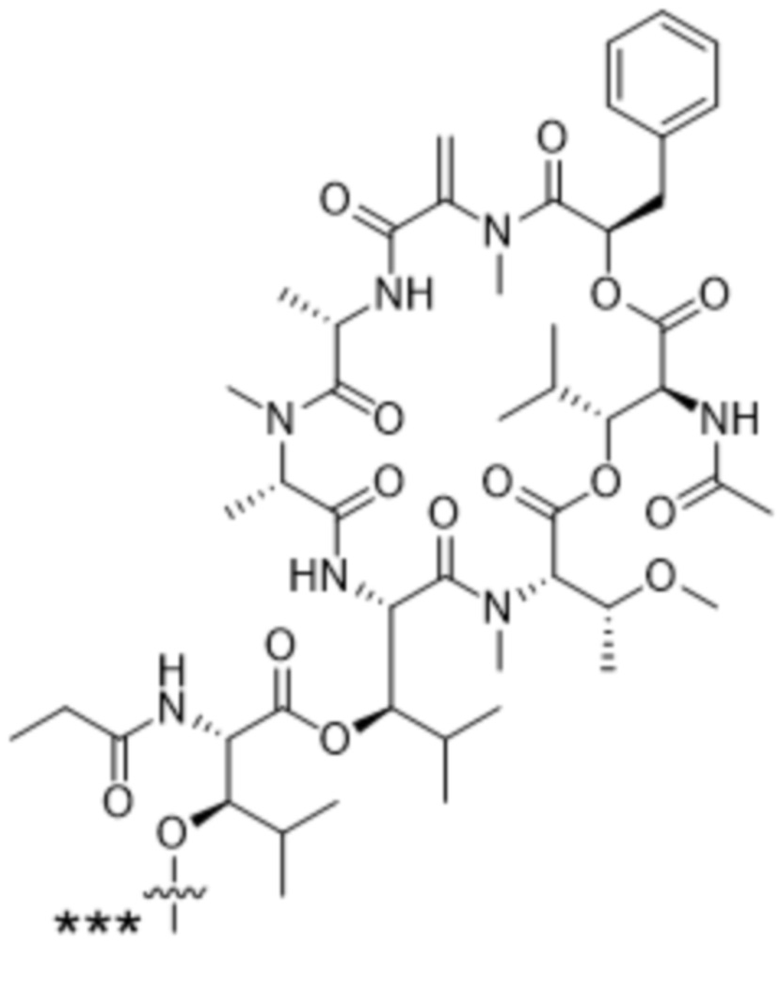 ,
,
где *** указывает на точку присоединения к LA или Y1.
[00214] Вариант осуществления 52. Конъюгат формулы (C) или формулы (C-1), где D представляет собой
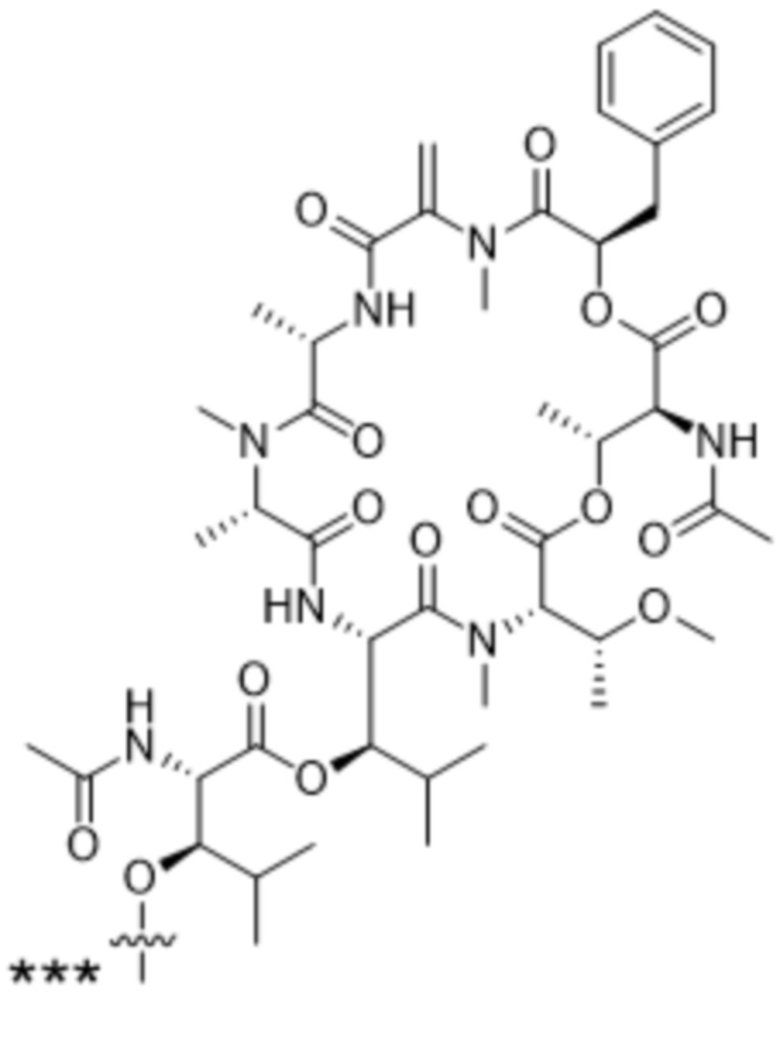 ,
,
где *** указывает на точку присоединения к LA или Y1.
[00215] Вариант осуществления 53. Конъюгат формулы (C) или формулы (C-1), где D представляет собой
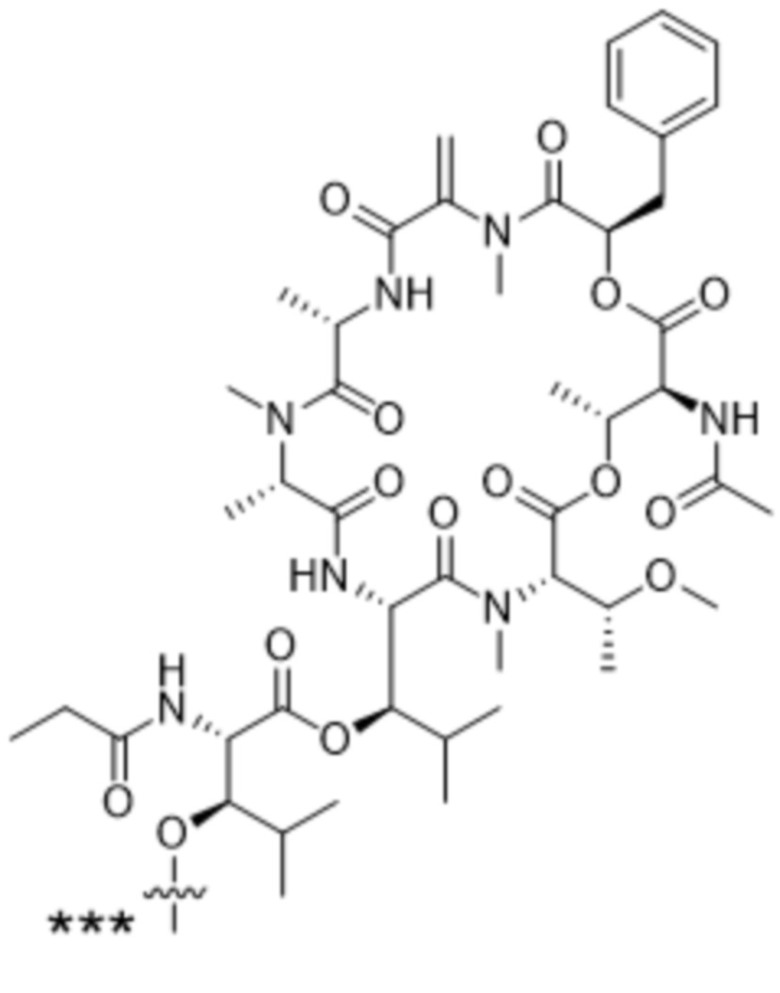 ,
,
где *** указывает на точку присоединения к LA или Y1.
[00216] Вариант осуществления 54. Конъюгат формулы (C) или формулы (C-1), имеющий структуру формулы (C-2):
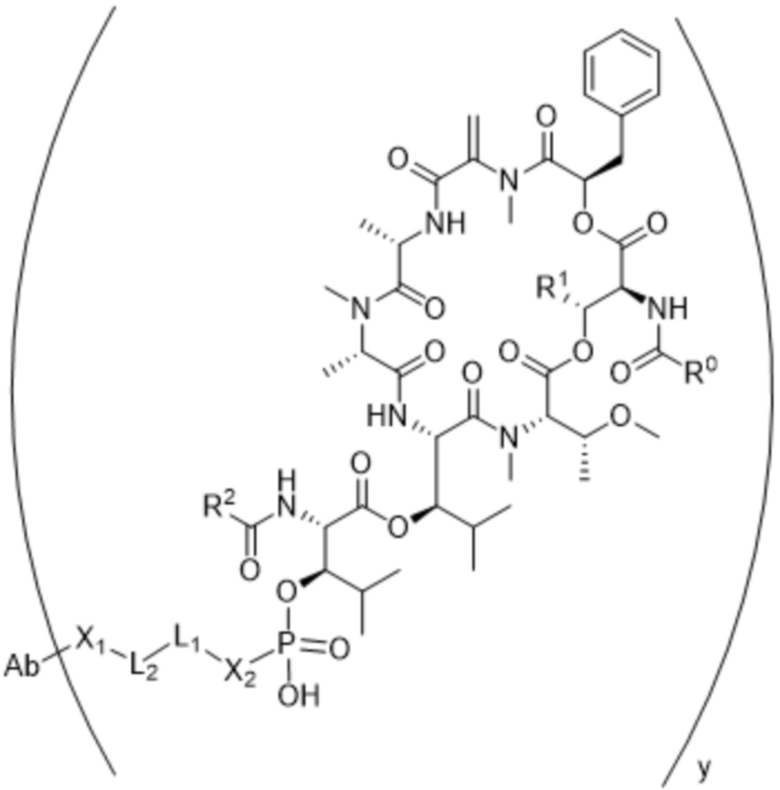 (C-2),
(C-2),
где
R0 представляет собой метил или этил;
R1 представляет собой метил или изопропил;
R2 представляет собой метил или этил;
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
L1 представляет собой бивалентный пептидный линкер;
L2 представляет собой связь или линкер, и
y равняется 1, 2, 3 или 4.
[00217] Вариант осуществления 55. Конъюгат формулы (C) или формулы (C-1), имеющий структуру формулы (C-2a):
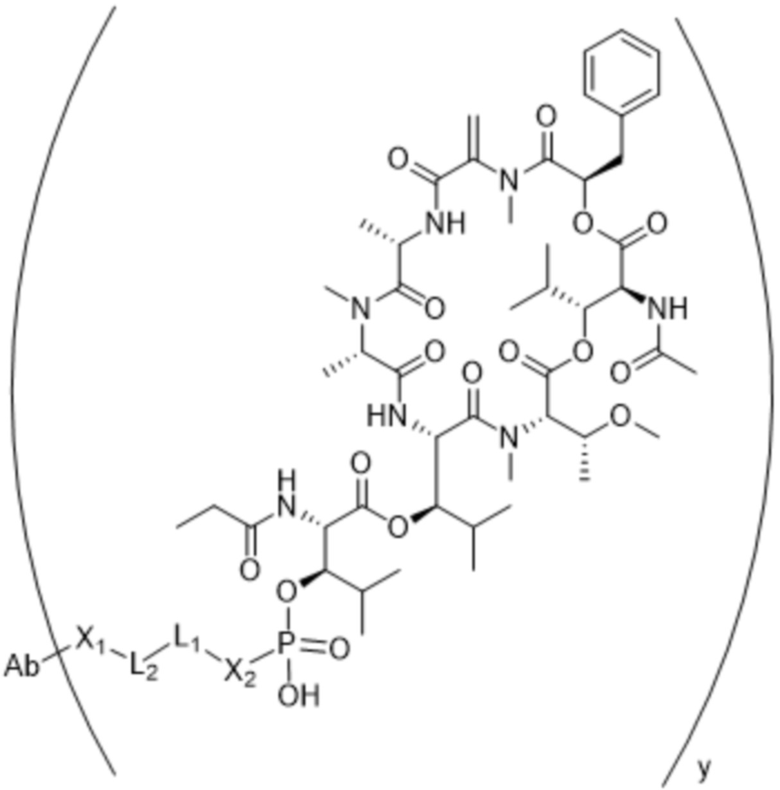 (C-2a),
(C-2a),
где
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
L1 представляет собой бивалентный пептидный линкер;
L2 представляет собой связь или линкер, и
y равняется 1, 2, 3 или 4.
[00218] Вариант осуществления 56. Конъюгат формулы (C) или формулы (C-1), имеющий структуру формулы (C-2b):
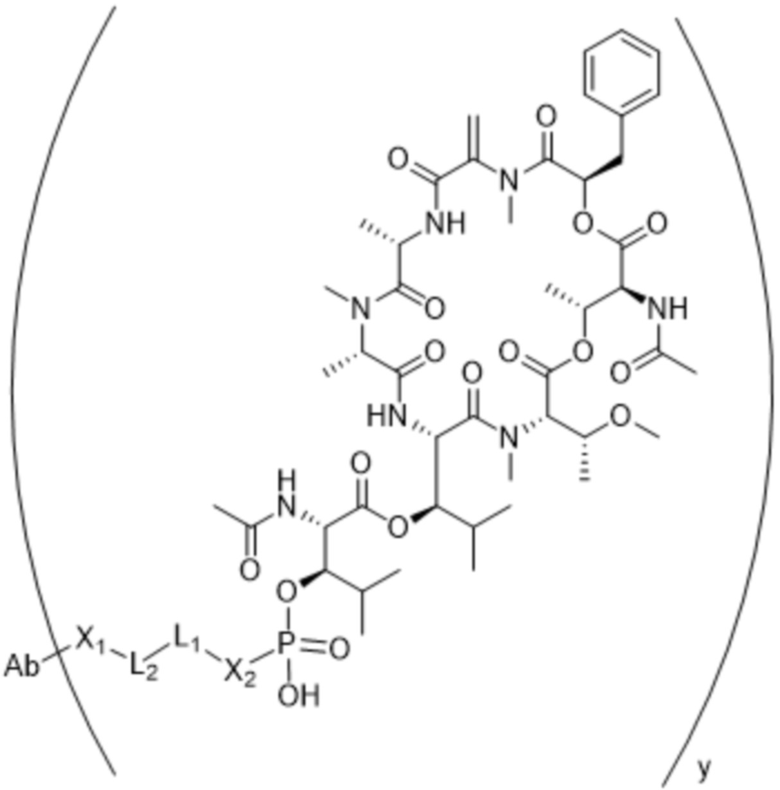 (C-2b),
(C-2b),
где
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
L1 представляет собой бивалентный пептидный линкер;
L2 представляет собой связь или линкер, и
y равняется 1, 2, 3 или 4.
[00219] Вариант осуществления 57. Конъюгат формулы (C) или формулы (C-1), имеющий структуру формулы (C-2c):
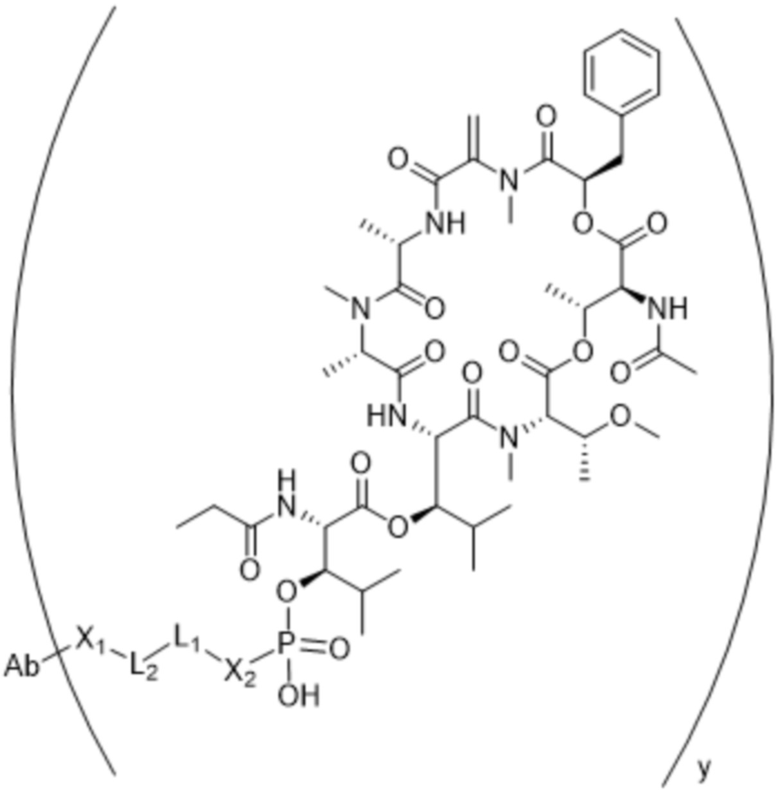 (C-2c),
(C-2c),
где
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
L1 представляет собой бивалентный пептидный линкер;
L2 представляет собой связь или линкер, и
y равняется 1, 2, 3 или 4.
[00220] Вариант осуществления 58. Конъюгат формулы (C) или формулы (C-1), имеющий структуру формулы (C-3):
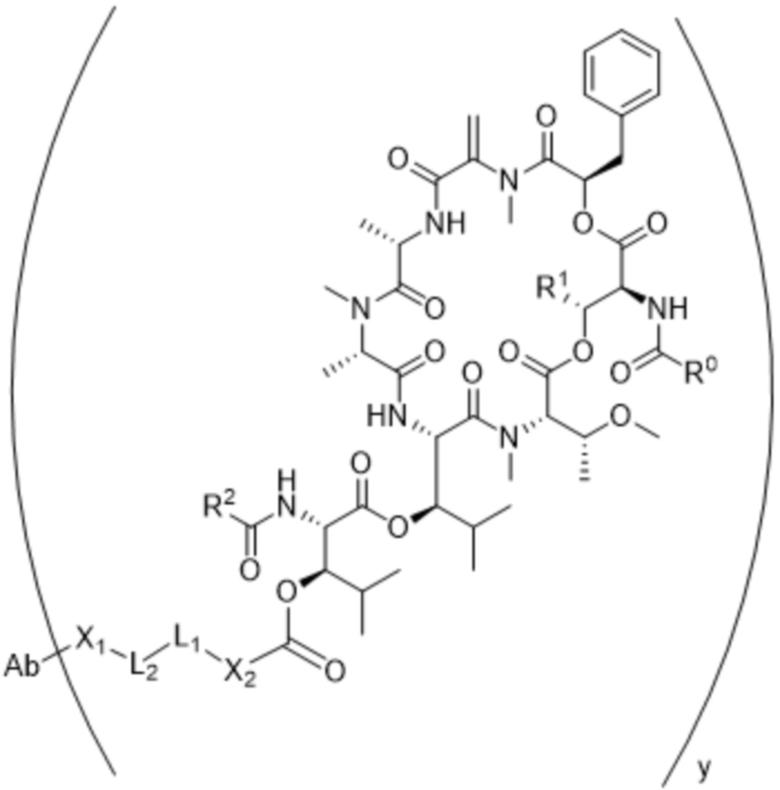 (C-3),
(C-3),
где
R0 представляет собой метил или этил;
R1 представляет собой метил или изопропил;
R2 представляет собой метил или этил;
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
L1 представляет собой бивалентный пептидный линкер;
L2 представляет собой связь или линкер, и
y равняется 1, 2, 3 или 4.
[00221] Вариант осуществления 59. Конъюгат формулы (C) или формулы (C-1), имеющий структуру формулы (C-3a):
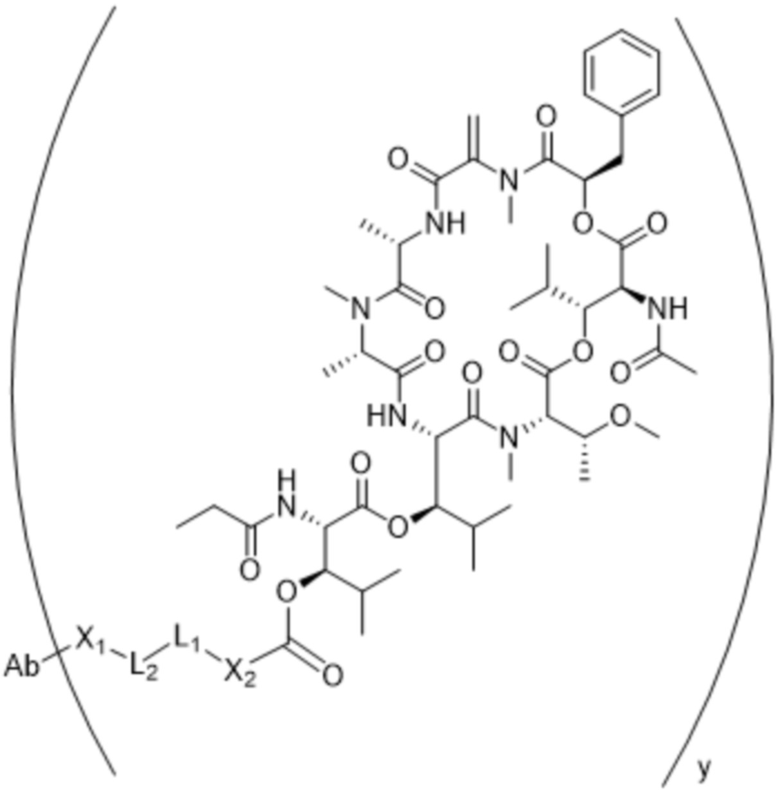 (C-3a),
(C-3a),
где
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
L1 представляет собой бивалентный пептидный линкер;
L2 представляет собой связь или линкер, и
y равняется 1, 2, 3 или 4.
[00222] Вариант осуществления 60. Конъюгат формулы (C) или формулы (C-1), имеющий структуру формулы (C-3b):
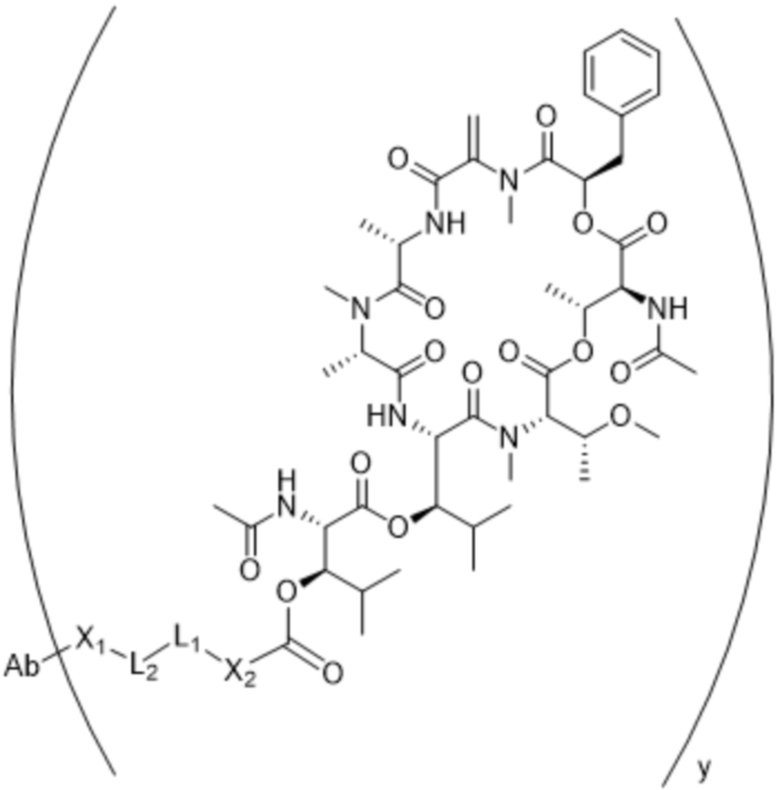 (C-3b),
(C-3b),
где
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
L1 представляет собой бивалентный пептидный линкер;
L2 представляет собой связь или линкер, и
y равняется 1, 2, 3 или 4.
[00223] Вариант осуществления 61. Конъюгат формулы (C) или формулы (C-1), имеющий структуру формулы (C-3c):
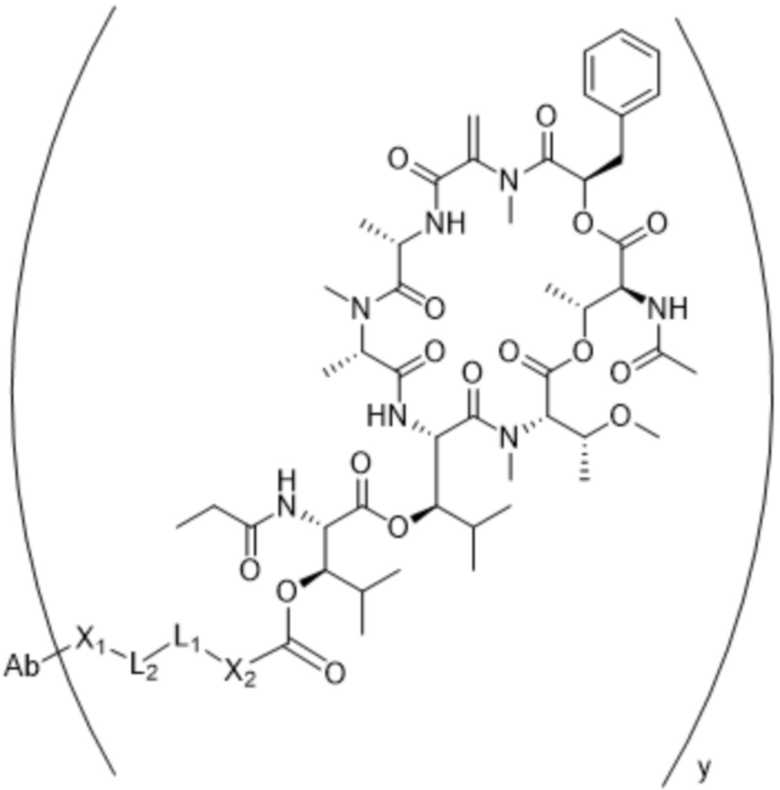 (C-3c),
(C-3c),
где
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
L1 представляет собой бивалентный пептидный линкер;
L2 представляет собой связь или линкер, и
y равняется 1, 2, 3 или 4.
[00224] Вариант осуществления 62. Конъюгат формулы (C-2) согласно варианту осуществления 54 или конъюгат формулы (C-3) согласно варианту осуществления 58:
где
R0 представляет собой метил или этил;
R1 представляет собой метил или изопропил;
R2 представляет собой метил или этил;
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
X2 представляет собой саморасщепляющийся спейсер, выбранный из
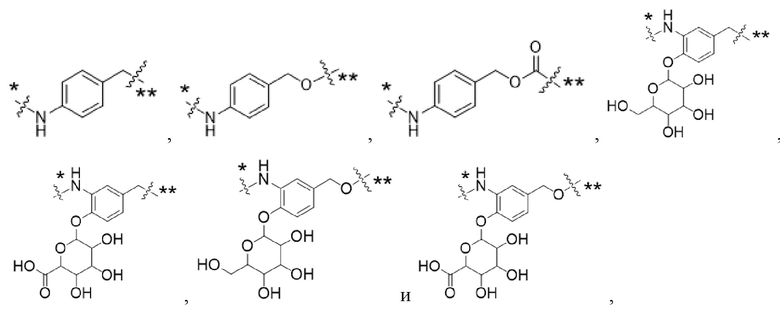
где * в X2 указывает на точку присоединения к L1 и ** в X2 указывает на точку присоединения к группе или точку присоединения к группе;
L1 представляет собой бивалентный пептидный линкер, содержащий 2-4 аминокислотных остатка;
L2 представляет собой линкер;
X1 представляет собой бивалентную связывающую группу, выбранную из
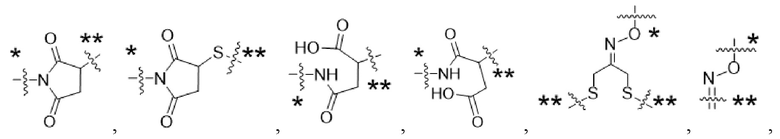
-*NR4C(=O)CH2**-, -*NHC(=O)CH2**-, -*S(=O)2CH2CH2**-, -* (CH2)2S(=O)2CH2CH2**-, -*NR4S(=O)2CH2CH2**-, -*NR4C(=O)CH2CH2**-, -NH-, -C(=O)-, -*NHC(=O) **-, -*CH2NHCH2CH2**-, -*NHCH2CH2**-, -S-,
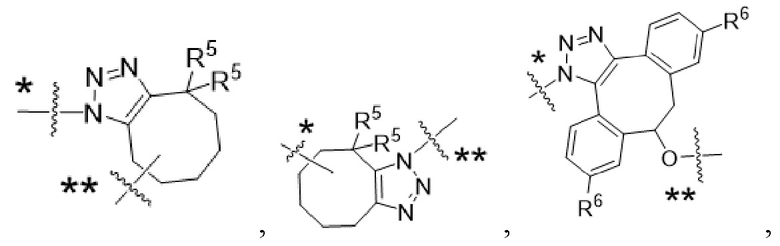
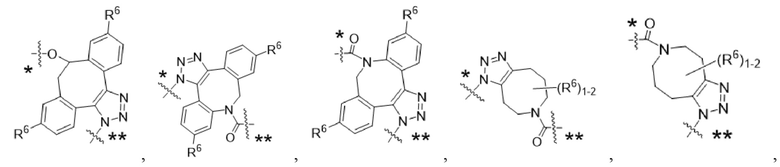
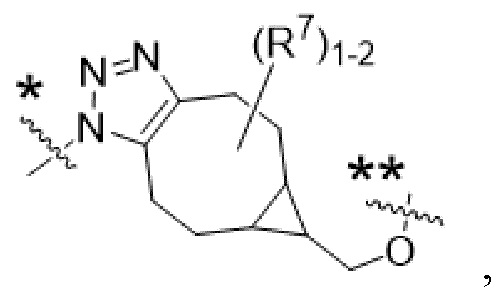 **-, где * в X1 указывает на точку присоединения к L2 и ** в X1 указывает на точку присоединения к Ab;
**-, где * в X1 указывает на точку присоединения к L2 и ** в X1 указывает на точку присоединения к Ab;
и где
каждый R4 независимо выбран из H и C1-C6алкила;
каждый R5 независимо выбран из H, C1-C6алкила, F, Cl и -OH;
каждый R6 независимо выбран из H, C1-C6алкила, F, Cl, -NH2, -OCH3, -OCH2CH3, -N(CH3)2, -CN, -NO2 и -OH
и
каждый R7 независимо выбран из H, C1-6алкила, фтора, бензилокси, замещенного -C(=O)OH, бензила, замещенного -C(=O)OH, C1-4алкокси, замещенного -C(=O)OH, и C1-4алкила, замещенного -C(=O)OH,
и
y равняется 1, 2, 3 или 4.
[00225] Вариант осуществления 63. Конъюгат согласно любому из вариантов осуществления 54-62, где X2 представляет собой саморасщепляющийся спейсер, выбранный из
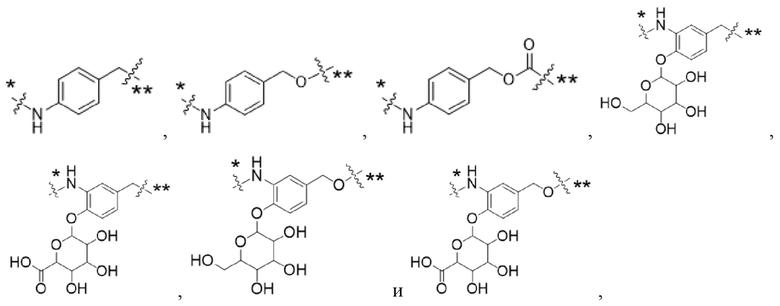
где * в X2 указывает на точку присоединения к L1 и ** в X2 указывает на точку присоединения к Y1, или точку присоединения к группе , или точку присоединения к группе .
[00226] Вариант осуществления 64. Конъюгат согласно любому из вариантов осуществления 54-63, где X2 представляет собой , где * в X2 указывает на точку присоединения к L1 и ** в X2 указывает на точку присоединения к Y1, или точку присоединения к группе , или точку присоединения к группе .
[00227] Вариант осуществления 65. Конъюгат согласно любому из вариантов осуществления 54-64, где L1 представляет собой бивалентный пептидный линкер, содержащий 2-4 аминокислотных остатка.
[00228] Вариант осуществления 66. Конъюгат согласно любому из вариантов осуществления 54-65, где L1 представляет собой бивалентный пептидный линкер, содержащий аминокислотный остаток, выбранный из валина, цитруллина, лизина, изолейцина, фенилаланина, метионина, аспарагина, пролина, аланина, лейцина, триптофана и тирозина.
[00229] Вариант осуществления 67. Конъюгат согласно любому из вариантов осуществления 54-64, где L1 представляет собой бивалентный пептидный линкер, содержащий по меньшей мере один остаток валина (Val) или цитруллина (Cit).
[00230] Вариант осуществления 68. Конъюгат согласно любому из вариантов осуществления 54-64, где L1 представляет собой бивалентный дипептидный линкер, выбранный из ValCit, PheLys, ValAla и ValLys.
[00231] Вариант осуществления 69. Конъюгат согласно любому из вариантов осуществления 54-64, где L1 представляет собой бивалентный дипептидный линкер, выбранный из
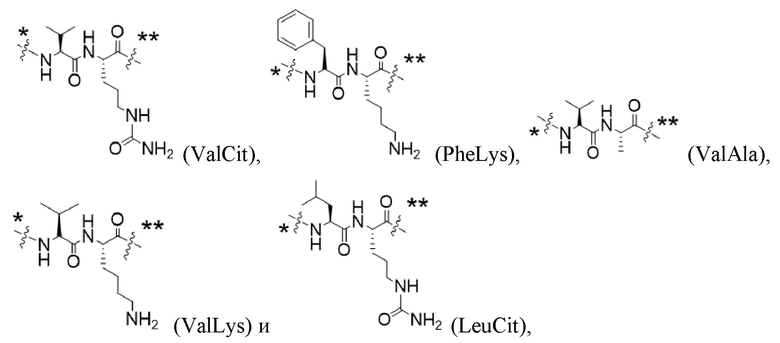
где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2.
[00232] Вариант осуществления 70. Конъюгат согласно любому из вариантов осуществления 54-64, где L1 представляет собой ValCit.
[00233] Вариант осуществления 71. Конъюгат согласно любому из вариантов осуществления 54-64, где
L1 представляет собой 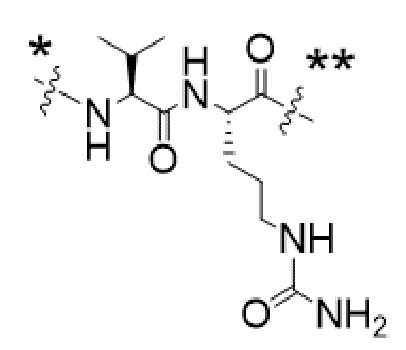 (ValCit), где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2.
(ValCit), где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2.
[00234] Вариант осуществления 72. Конъюгат согласно любому из вариантов осуществления 54-71, где L2 представляет собой линкер.
[00235] Вариант осуществления 73. Конъюгат согласно любому из вариантов осуществления 54-71, где L2 представляет собой линкер, выбранный из
-*C(=O)((CH2)mO)p(CH2)m**-, -*C(=O)(CH2)m**-, -*C(=O)(CH2)nNHC(=O)(CH2)m**-, -*C(=O)(CH2)mNHC(=O)((CH2)mO)p(CH2)m**-, -* ((CH2)mO)p(CH2)m**-, -* ((CH2)mO)p(CH2)m**-, -(CH2)m-, -*(CH2)mNHC(=O)(CH2)m**-, -* (CH2)mNHC(=O)(CH2)mC(=O)NH(CH2)m**-, -* ((CH2)mO)p(CH2)mNHC(=O)(CH2)m**-, -* *((CH2)mO)pCH2)mC(=O)NH(CH2)m**-, -* (CH2)mC(R3)2**-, и -* (CH2)mC(R3)2SS(CH2)mNHC(=O)(CH2)m**-, где * в L2 указывает на точку присоединения к L1 и ** в L2 указывает на точку присоединения к X1;
и где
каждый R3 независимо выбран из H и C1-C6алкила;
каждый m независимо выбран из 1, 2, 3, 4, 5, 6, 7, 8, 9 и 10 и
каждый p независимо выбран из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 и 14.
[00236] Вариант осуществления 74. Конъюгат согласно любому из вариантов осуществления 54-71, где L2 представляет собой -*C(=O)((CH2)mO)p(CH2)m**- или -*C(=O)(CH2)m**-, где * в L2 указывает на точку присоединения к L1 и ** в L2 указывает на точку присоединения к X1,
и где каждый m независимо выбран из 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 и p равняется 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или 14.
[00237] Вариант осуществления 75. Конъюгат согласно любому из вариантов осуществления 54-71, где L2 представляет собой , или , где * в L2 указывает на точку присоединения к L1 и ** в L2 указывает на точку присоединения к X1.
[00238] Вариант осуществления 76. Конъюгат согласно любому из вариантов осуществления 54-75,
где X1 представляет собой бивалентную связывающую группу, выбранную из
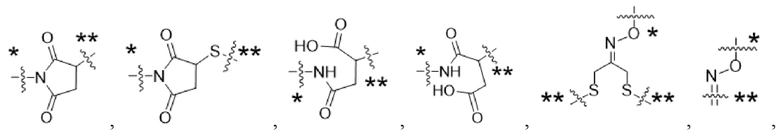
-*NR4C(=O)CH2**-, -*NHC(=O)CH2**-, -*S(=O)2CH2CH2**-, -* (CH2)2S(=O)2CH2CH2**-, -*NR4S(=O)2CH2CH2**-, -*NR4C(=O)CH2CH2**-, -NH-, -C(=O)-, -*NHC(=O) **-, -*CH2NHCH2CH2**-, -*NHCH2CH2**-, -S-,
**-, где * в X1 указывает на точку присоединения к L2 и ** в X1 указывает на точку присоединения к Ab;
и где
каждый R4 независимо выбран из H и C1-C6алкила;
каждый R5 независимо выбран из H, C1-C6алкила, F, Cl и -OH;
каждый R6 независимо выбран из H, C1-C6алкила, F, Cl, -NH2, -OCH3, -OCH2CH3, -N(CH3)2, -CN, -NO2 и -OH
и
каждый R7 независимо выбран из H, C1-6алкила, фтора, бензилокси, замещенного -C(=O)OH, бензила, замещенного -C(=O)OH, C1-4алкокси, замещенного -C(=O)OH, и C1-4алкила, замещенного -C(=O)OH.
[00239] Вариант осуществления 77. Конъюгат согласно любому из вариантов осуществления 54-75,
где X1 представляет собой бивалентную связывающую группу, выбранную из
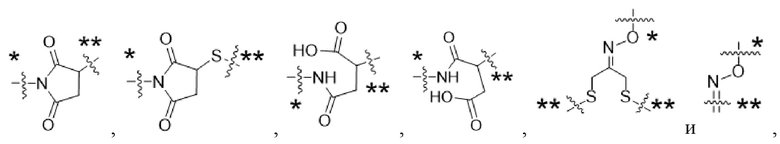
где * в X1 указывает на точку присоединения к L2 и ** в X1 указывает на точку присоединения к Ab.
[00240] Вариант осуществления 78. Конъюгат согласно любому из вариантов осуществления 54-75,
где X1 представляет собой бивалентную связывающую группу, выбранную из
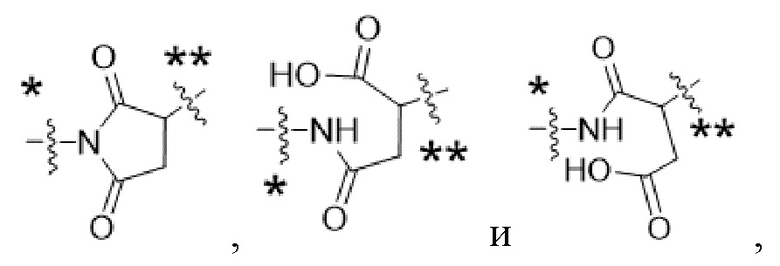
где * в X1 указывает на точку присоединения к L2 и ** в X1 указывает на точку присоединения к Ab.
[00241] Вариант осуществления 79. Конъюгат формулы (C-2) согласно варианту осуществления 54, где
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
R0 представляет собой метил или этил;
R1 представляет собой метил или изопропил;
R2 представляет собой метил или этил;
X1 представляет собой бивалентную связывающую группу, выбранную из
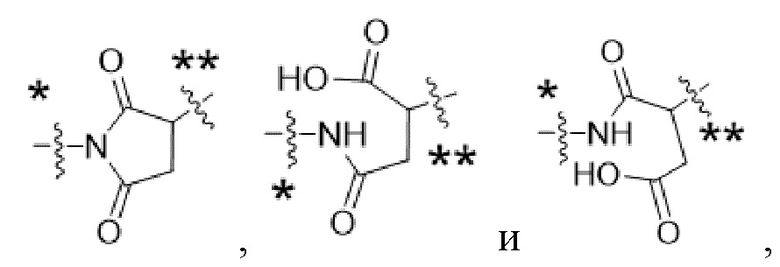
где * в X1 указывает на точку присоединения к L2 и ** в X1 указывает на точку присоединения к Ab,
X2 представляет собой , где * в X2 указывает на точку присоединения к L1 и ** в X2 указывает на точку присоединения к группе  ;
;
L1 представляет собой  (ValCit), где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2;
(ValCit), где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2;
L2 представляет собой или , где * в L2 указывает на точку присоединения к L1 и ** в L2 указывает на точку присоединения к R8,
и
y равняется 1, 2, 3 или 4.
[00242] Вариант осуществления 80. Конъюгат формулы (C-3) согласно варианту осуществления 58, где
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека;
R0 представляет собой метил или этил;
R1 представляет собой метил или изопропил;
R2 представляет собой метил или этил;
X1 представляет собой бивалентную связывающую группу, выбранную из
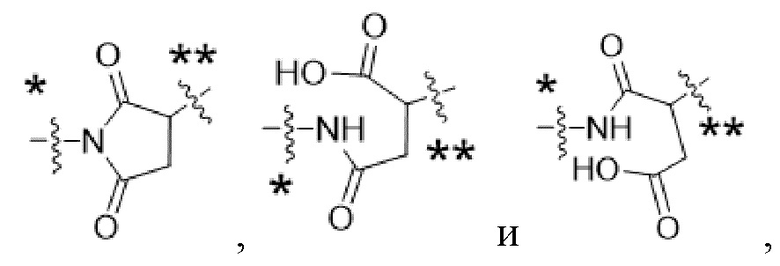
где * в X1 указывает на точку присоединения к L2 и ** в X1 указывает на точку присоединения к Ab,
X2 представляет собой , где * в X2 указывает на точку присоединения к L1 и ** в X2 указывает на точку присоединения к группе 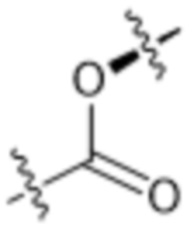 ;
;
L1 представляет собой 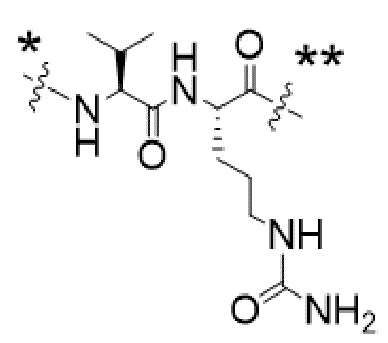 (ValCit), где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2;
(ValCit), где * в L1 указывает на точку присоединения к L2 и ** в L1 указывает на точку присоединения к X2;
L2 представляет собой или , где * в L2 указывает на точку присоединения к L1 и ** в L2 указывает на точку присоединения к R8,
и
y равняется 1, 2, 3 или 4.
[00243] Вариант осуществления 81. Конъюгат формулы (C), формулы (C-1) или формулы (C-2), имеющий структуру:
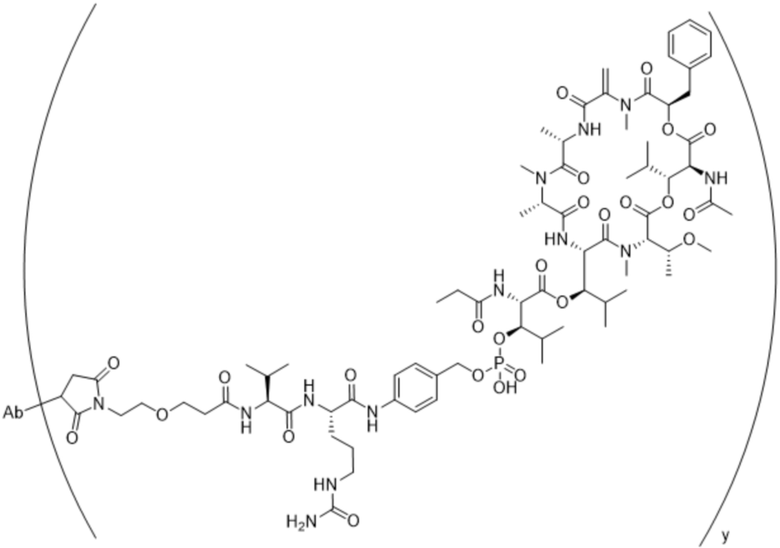
где
y равняется 2 или 4 и
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека.
[00244] Вариант осуществления 82. Конъюгат формулы (C), формулы (C-1) или формулы (C-2), имеющий структуру:
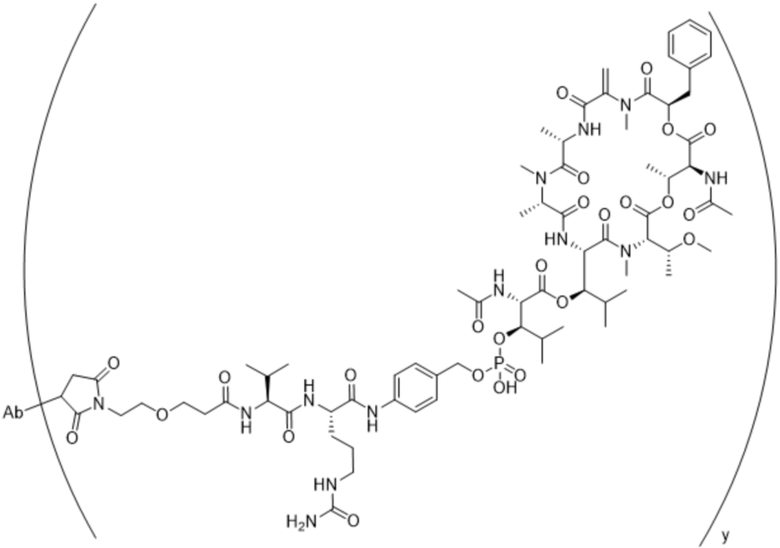
где
y равняется 2 или 4 и
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека.
[00245] Вариант осуществления 83. Конъюгат формулы (C), формулы (C-1) или формулы (C-2), имеющий структуру:
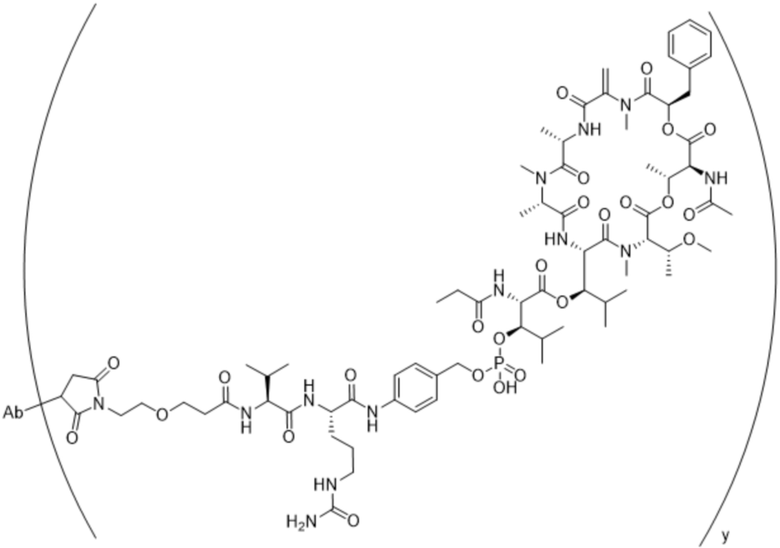
где
y равняется 2 или 4 и
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека.
[00246] Вариант осуществления 84. Конъюгат формулы (C), формулы (C-1) или формулы (C-2), имеющий структуру:
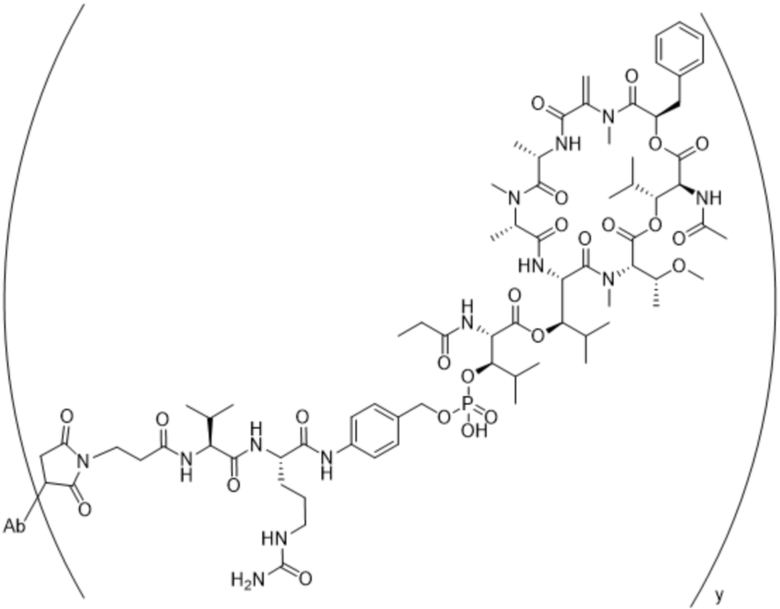
где
y равняется 2 или 4 и
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека.
[00247] Вариант осуществления 85. Конъюгат формулы (C), формулы (C-1) или формулы (C-2), имеющий структуру:
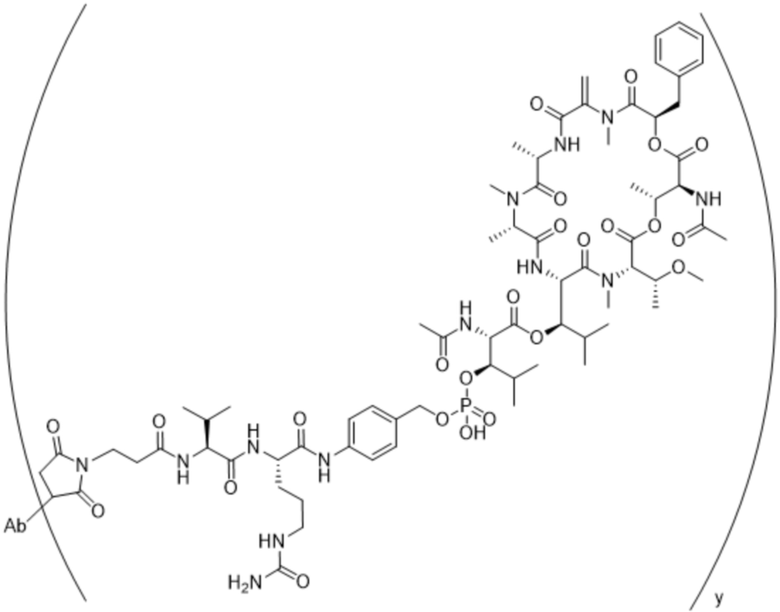
где
y равняется 2 или 4 и
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека.
[00248] Вариант осуществления 86. Конъюгат формулы (C), формулы (C-1) или формулы (C-2), имеющий структуру:
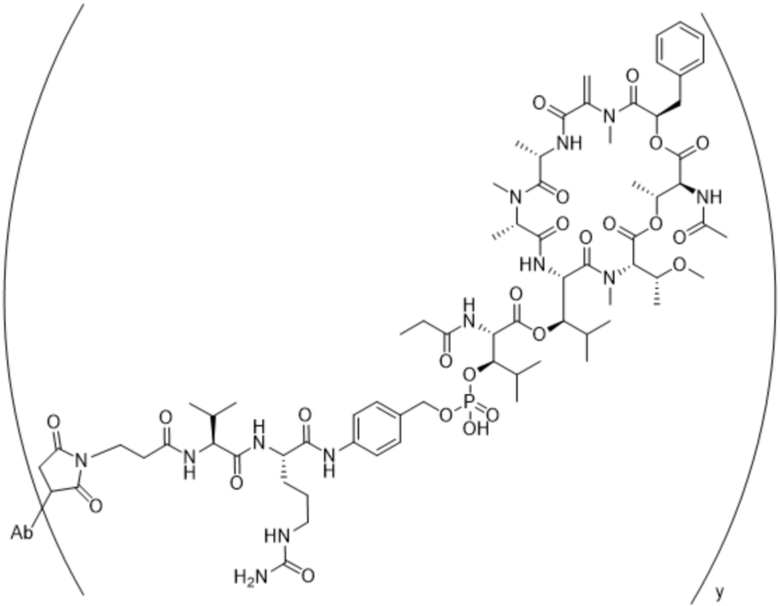
где
y равняется 2 или 4 и
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека.
[00249] Вариант осуществления 87. Конъюгат формулы (C), формулы (C-1) или формулы (C-3), имеющий структуру:
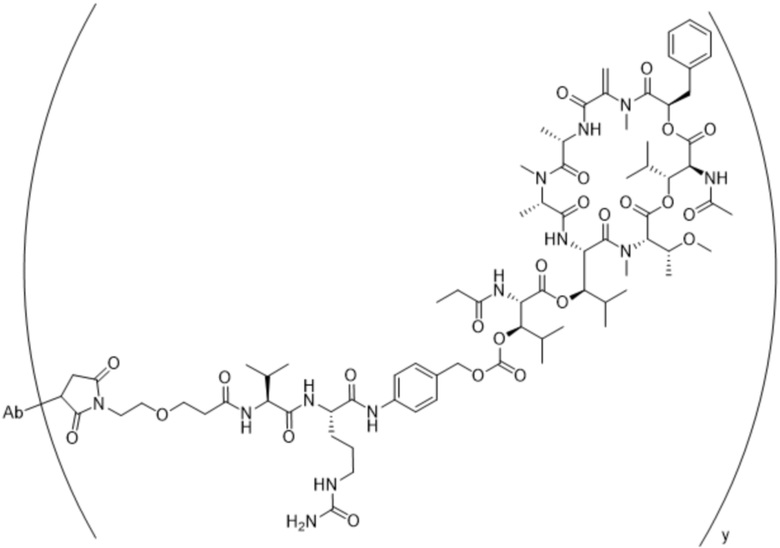
где
y равняется 2 или 4 и
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека.
[00250] Вариант осуществления 88. Конъюгат формулы (C), формулы (C-1) или формулы (C-3), имеющий структуру:
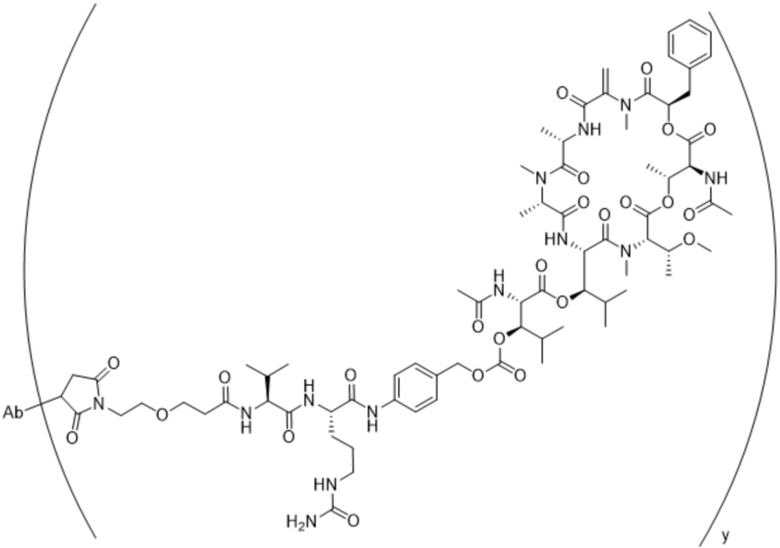
где
y равняется 2 или 4 и
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека.
[00251] Вариант осуществления 89. Конъюгат формулы (C), формулы (C-1) или формулы (C-3), имеющий структуру:
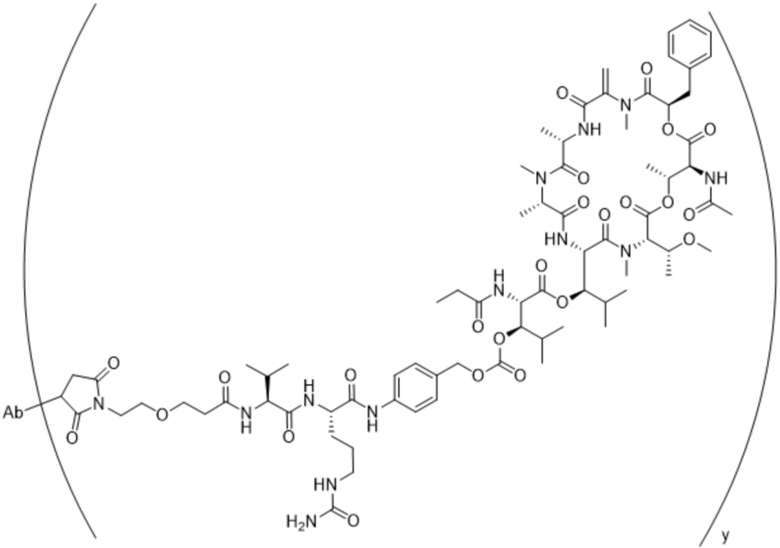
где
y равняется 2 или 4 и
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека.
[00252] Вариант осуществления 90. Конъюгат формулы (C), формулы (C-1) или формулы (C-3), имеющий структуру:
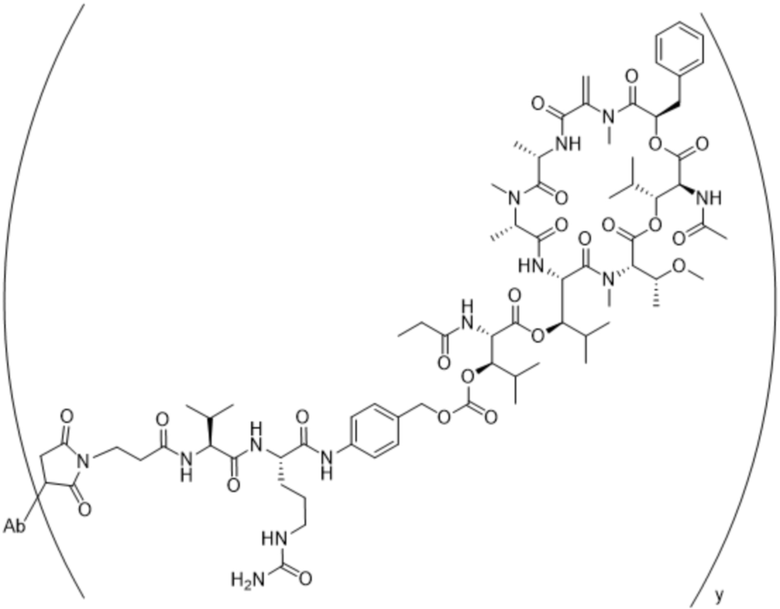
где
y равняется 2 или 4 и
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека.
[00253] Вариант осуществления 91. Конъюгат формулы (C), формулы (C-1) или формулы (C-3), имеющий структуру:
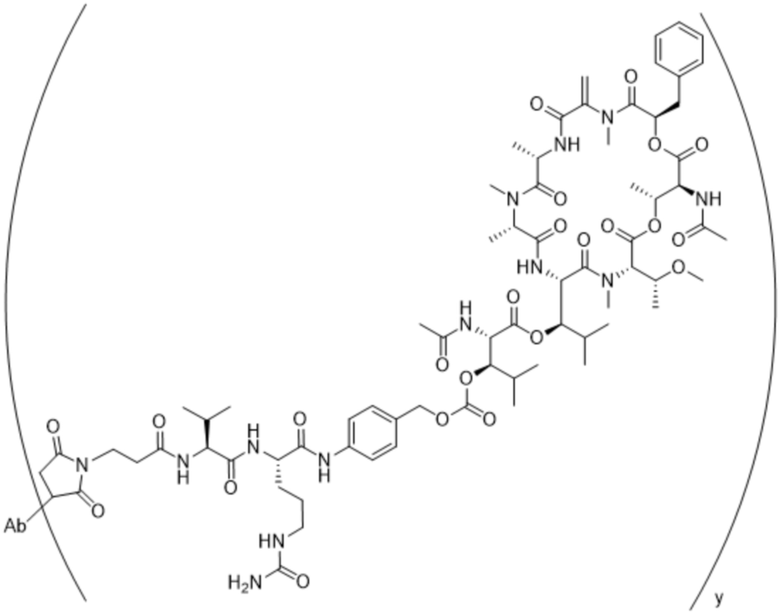
где
y равняется 2 или 4 и
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека.
[00254] Вариант осуществления 92. Конъюгат формулы (C), формулы (C-1) или формулы (C-3), имеющий структуру:
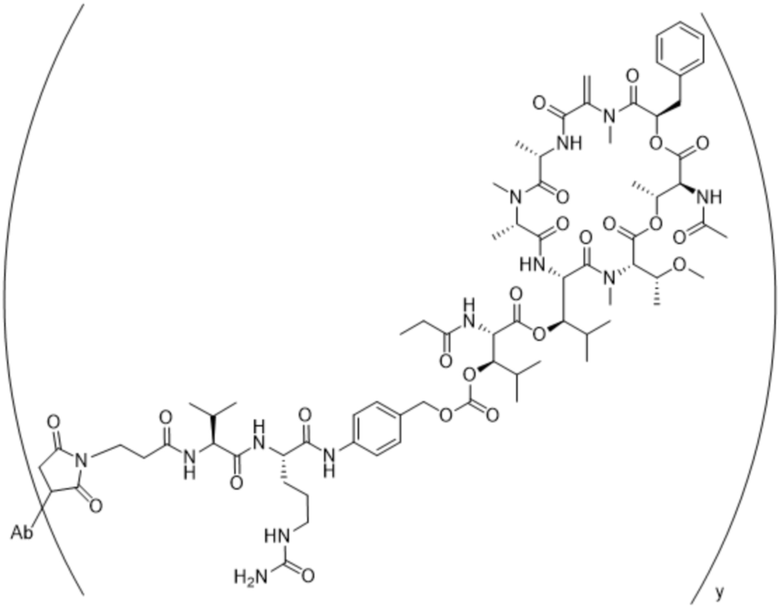
где
y равняется 2 или 4 и
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека.
[00255] Кроме того, антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению могут быть конъюгированы с фрагментом, представляющим собой лекарственное средство, модифицирующее заданный биологический ответ. Фрагменты, представляющие собой лекарственные средства, не следует рассматривать как ограниченные классическими химическими терапевтическими средствами. Например, фрагмент, представляющий собой лекарственное средство, может представлять собой белок, пептид или полипептид, обладающие требуемой биологической активностью. Такие белки могут включать, например, токсин, такой как абрин, рицин A, экзотоксин Pseudomonas, холерный токсин или дифтерийный токсин; белок, такой как фактор некроза опухоли, α-интерферон, β-интерферон, фактор роста нервов, фактор роста тромбоцитов, тканевой активатор плазминогена, цитокин, апоптотическое средство, антиангиогенное средство или модификатор биологического ответа, такой как, например, лимфокин.
[00256] В одном варианте осуществления антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению конъюгированы с фрагментом, представляющим собой лекарственное средство, таким как цитотоксин, лекарственное средство (например, иммуносупрессор) или радиотоксин. Примеры цитотоксинов включают без ограничения таксаны (см., например, международные (PCT) заявки на патент №№ WO 01/38318 и PCT/US03/02675), средства, алкилирующие ДНК (например, аналоги CC-1065), антрациклины, аналоги тубулизина, аналоги дуокармицина, ауристатин E, ауристатин F, майтанзиноиды и цитотоксические средства, содержащие фрагмент, представляющий собой реакционноспособный полиэтиленгликоль (см., например, Sasse et al., J. Antibiot. (Tokyo), 53, 879-85 (2000), Suzawa et al., Bioorg. Med. Chem., 8, 2175-84 (2000), Ichimura et al., J. Antibiot. (Tokyo), 44, 1045-53 (1991), Francisco et al., Blood (2003) (электронная публикация до печатной публикации), патенты США №№ 5475092, 6340701, 6372738 и 6436931, публикацию заявки на патент США № 2001/0036923 A1, находящиеся на рассмотрении заявки на патент США с порядковыми №№ 10/024290 и 10/116053 и международную (PCT) заявку на патент № WO 01/49698), таксон, цитохалазин B, грамицидин D, этидия бромид, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, t. колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин, а также их аналоги или гомологи. Терапевтические средства также включают, например, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил, декарбазин), разрушающие средства (например, мехлорэтамин, тиотепу, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин C и цис-дихлордиаминплатину (II) (DDP), цисплатин, антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (AMC)) и антимитотические средства (например, винкристин и винбластин). (См. например, US20090304721 от Seattle Genetics).
[00257] Другие примеры цитотоксинов, которые могут быть конъюгированы с антителами, фрагментами антител (антигенсвязывающими фрагментами) или функциональными эквивалентами по настоящему изобретению, включают дуокармицины, калихеамицины, майтанзины и ауристатины, а также их производные.
[00258] Различные типы цитотоксинов, линкеров и способы конъюгирования терапевтических средств с антителами известны из уровня техники, см., например, Saito et al., (2003) Adv. Drug Deliv. Rev. 55:199-215; Trail et al., (2003) Cancer Immunol. Immunother. 52:328-337; Payne, (2003) Cancer Cell 3:207-212; Allen, (2002) Nat. Rev. Cancer 2:750-763; Pastan and Kreitman, (2002) Curr. Opin. Investig. Drugs 3:1089-1091; Senter and Springer, (2001) Adv. Drug Deliv. Rev. 53:247-264.
[00259] Антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению также можно конъюгировать с радиоактивным изотопом для создания цитотоксических радиофармацевтических препаратов, называемых радиоиммуноконъюгаты. Примеры радиоактивных изотопов, которые можно конъюгировать с антителами для применения в диагностических или терапевтических целях, включают без ограничения йод-131, индий-111, иттрий-90 и лютеций-177. Способы получения радиоиммуноконъюгатов определены в уровне техники. Примеры коммерчески доступных радиоиммуноконъюгатов включают Zevalin™ (DEC Pharmaceuticals) и Bexxar™ (Corixa Pharmaceuticals), а также подобные подходы можно использовать для получения радиоиммуноконъюгатов с применением антител по настоящему изобретению. В некоторых вариантах осуществления макроциклический хелатор представляет собой 1,4,7,10-тетраазациклододекан-N, N',N'',N'''-тетрауксусную кислоту (DOTA), которая может быть прикреплена к антителу через линкерную молекулу. Такие линкерные молекулы широко известны из уровня техники и описаны в Denardo et al., (1998) Clin Cancer Res. 4(10):2483-90; Peterson et al., (1999) Bioconjug. Chem. 10(4):553-7 и Zimmerman et al., (1999) Nucl. Med. Biol. 26(8):943-50, каждый из которых включен посредством ссылки во всей своей полноте.
[00260] Антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению также можно конъюгировать с гетерологичным белком или полипептидом (или их фрагментом, предпочтительно с полипептидом из по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90 или по меньшей мере 100 аминокислот) для создания слитых белков. В частности, в настоящем изобретении представлены слитые белки, содержащие фрагмент антитела (например, антигенсвязывающий фрагмент), описанный в данном документе (например, Fab-фрагмент, Fd-фрагмент, Fv-фрагмент, F(ab)2-фрагмент, домен VH, CDR VH, домен VL или CDR VL), и гетерологичный белок, полипептид или пептид.
[00261] Дополнительные слитые белки можно получать посредством методик перетасовки генов, перетасовки мотивов, перетасовки экзонов и/или перетасовки кодонов (в совокупности называемых "перетасовкой ДНК"). Перетасовку ДНК можно использовать для изменения активности антител по настоящему изобретению или их фрагментов (например, получения антител или их фрагментов с более высокими уровнями аффинности и более низкими значениями скорости диссоциации). См. в общем патенты США №№ 5605793, 5811238, 5830721, 5834252 и 5837458; Patten et al., (1997) Curr. Opinion Biotechnol. 8:724-33; Harayama, (1998) Trends Biotechnol. 16(2):76-82; Hansson et al., (1999) J. Mol. Biol. 287:265-76 и Lorenzo and Blasco, (1998) Biotechniques 24(2):308-313 (каждый из данных патентов и публикаций тем самым включен посредством ссылки во всей своей полноте). Антитела или их фрагменты или кодируемые антитела или их фрагменты можно изменять путем воздействия на них случайного мутагенеза посредством ПЦР с внесением ошибок, случайной вставки нуклеотидов или других способов до проведения рекомбинации. Полинуклеотид, кодирующий антитело или его фрагмент, которые специфически связываются с антигеном, может быть подвергнут рекомбинации с одним или несколькими компонентами, мотивами, отрезками, частями, доменами, фрагментами и т. д. одной или нескольких гетерологичных молекул.
[00262] Более того, антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению могут быть конъюгированы с маркерными последовательностями, такими как пептид, для облегчения очистки. В предпочтительных вариантах осуществления маркерная аминокислотная последовательность представляет собой гексагистидиновый пептид (SEQ ID NO: 267), такой как, в частности, метка, представленная в векторе pQE (QIAGEN, Inc., 9259 Eton Avenue, Чатсворт, Калифорния, 91311), из которых многие являются коммерчески доступными. Как описано в Gentz et al., (1989) Proc. Natl. Acad. Sci. USA 86:821-824, например, гексагистидин (SEQ ID NO: 267) обеспечивает удобную очистку слитого белка. Другие пептидные метки, применимые для очистки, включают без ограничения гемагглютининовую ("HA") метку, которая соответствует эпитопу, полученному из белка гемагглютинина вируса гриппа (Wilson et al., (1984) Cell 37:767), и метку "FLAG" (A. Einhauer et al., J. Biochem. Biophys. Methods 49: 455-465, 2001). Согласно настоящему изобретению для повышения эффективности антител или антигенсвязывающих фрагментов их также можно конъюгировать с проникающими в опухоль пептидами.
[00263] В других вариантах осуществления антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению конъюгируют с диагностическим или детектируемым средством. Такие иммуноконъюгаты могут быть применимыми для контроля или прогнозирования проявления, развития, прогрессирования и/или степени тяжести заболевания или нарушения в рамках процедуры клинического тестирования, такой как определение эффективности конкретной терапии. Такие диагностика и детекция могут осуществляться путем связывания антитела с детектируемыми веществами, в том числе без ограничения с различными ферментами, такими как без ограничения пероксидаза хрена, щелочная фосфатаза, бета-галактозидаза или ацетилхолинэстераза; простетическими группами, такими как без ограничения стрептавидин/биотин и авидин/биотин; флуоресцентными соединениями, такими как без ограничения Alexa Fluor 350, Alexa Fluor 405, Alexa Fluor 430, Alexa Fluor 488, Alexa Fluor 500, Alexa Fluor 514, Alexa Fluor 532, Alexa Fluor 546, Alexa Fluor 555, Alexa Fluor 568, Alexa Fluor 594, Alexa Fluor 610, Alexa Fluor 633, Alexa Fluor 647, Alexa Fluor 660, Alexa Fluor 680, Alexa Fluor 700, Alexa Fluor 750, умбеллиферон, флуоресцеин, флуоресцеина изотиоцианат, родамин, дихлортриазиниламинфлуоресцеин, дансилхлорид или фикоэритрин; люминесцентными соединениями, такими как без ограничения люминол; биолюминесцентными соединениями, такими как без ограничения люцифераза, люциферин и экворин; радиоактивными соединениями, такими как без ограничения йод (131I, 125I, 123I и 121I), углерод (14C), сера (35S), тритий (3H), индий (115In, 113In, 112In и 111In), технеций (99Tc), таллий (201Ti), галлий (68Ga, 67Ga), палладий (103Pd), молибден (99Mo), ксенон (133Xe), фтор (18F), 153Sm, 177Lu, 159Gd, 149Pm, 140La, 175Yb, 166Ho, 90Y, 47Sc, 186Re, 188Re, 142Pr, 105Rh, 97Ru, 68Ge, 57Co, 65Zn, 85Sr, 32P, 153Gd, 169Yb, 51Cr, 54Mn, 75Se, 64Cu, 113Sn, и 117Sn; и позитронно-активными металлами с применением различных видов позитронной эмиссионной томографии, и нерадиоактивными ионами парамагнитных металлов.
[00264] Антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению также могут быть прикреплены к твердым подложкам, которые особенно применимы для иммунологических анализов или очистки целевого антигена. Такие твердые подложки включают без ограничения стекло, целлюлозу, полиакриламид, нейлон, полистирол, поливинилхлорид или полипропилен.
3. Конъюгация и получение ADC
Способы получения конъюгатов на основе антитела формулы (C), формулы (C-1) и формулы (C-2)
[00265] Общая схема реакции образования конъюгатов формулы (C) показана на схеме 1 ниже:
схема 1,
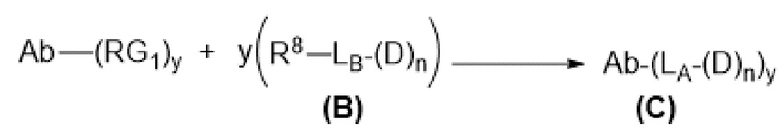
где RG1 представляет собой реакционноспособную группу на антителе Ab или его антигенсвязывающем фрагменте, исключительно в качестве примера тиольную, аминную или кетоновую, которая вступает в реакцию с совместимой реакционноспособной группой, R8, присоединенной к соединению линкер-лекарственное средство, обеспечивая тем самым ковалентную связь антитела Ab или его антигенсвязывающего фрагмента с одним или несколькими фрагментами, представляющими собой линкер-лекарственное средство. Неограничивающими примерами таких реакций с участием групп RG1 и R8 является малеимидная группа (R8), которая вступает в реакцию с тиольной группой (RG1) с получением сукцинимидного кольца, или гидроксиламинная группа (R8), которая вступает в реакцию с кетоновой группой (RG1) с получением оксима.
В одном варианте осуществления D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11 (ингибитор GNAQ/GNA11), La представляет собой линкер, дополнительно содержащий бивалентную связывающую группу, образуемую при вступлении RG1 и R8 в реакцию, n равняется 1, 2, 3 или 4 и y равняется 1, 2, 3 или 4.
[00266] Общая схема реакции образования конъюгатов формулы (C-1) показана на схеме 2 ниже:
схема 2,
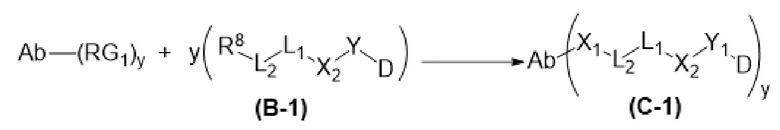
где RG1 представляет собой реакционноспособную группу на антителе Ab или его антигенсвязывающем фрагменте, исключительно в качестве примера тиольную, аминную или кетоновую, которая вступает в реакцию с совместимой реакционноспособной группой R8, присоединенной к фрагменту, представляющему собой линкер-лекарственное средство, обеспечивая тем самым ковалентную связь антитела Ab или его антигенсвязывающего фрагмента с одним или несколькими фрагментами, представляющими собой линкер-лекарственное средство. Неограничивающими примерами таких реакций с участием групп RG1 и R8 является малеимидная группа (R8), которая вступает в реакцию с тиольной группой (RG1) с получением сукцинимидного кольца, или гидроксиламинная группа (R8), которая вступает в реакцию с кетоновой группой (RG1) с получением оксима.
В одном варианте осуществления D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11 (ингибитор GNAQ/GNA11), X1 представляет собой бивалентную связывающую группу, образуемую при вступлении RG1 и R8 в реакцию (например сукцинимидное кольцо или оксим), Y1 представляет собой фосфатную группу, X2 представляет собой саморасщепляющийся спейсер, L1 представляет собой бивалентный пептидный линкер, L2 представляет собой связь или линкер, и y равняется 1, 2, 3 или 4.
[00267] Общая схема реакции образования конъюгатов формулы (C-2) показана на схеме 3 ниже:
схема 3,
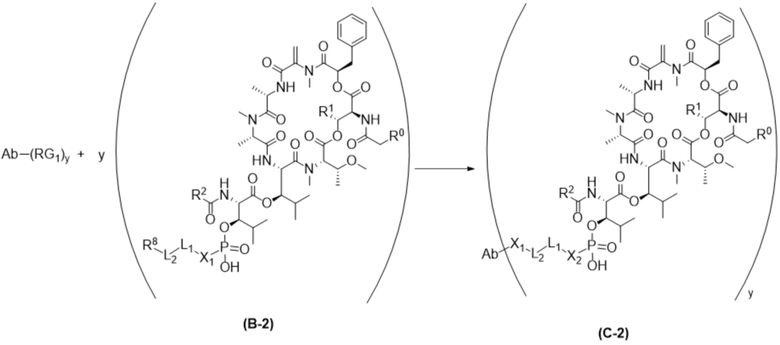
где RG1 представляет собой реакционноспособную группу на антителе Ab или его антигенсвязывающем фрагменте, исключительно в качестве примера тиольную, аминную или кетоновую, которая вступает в реакцию с совместимой реакционноспособной группой R8, присоединенной к фрагменту, представляющему собой линкер-лекарственное средство, обеспечивая тем самым ковалентную связь антитела Ab или его антигенсвязывающего фрагмента с одним или несколькими фрагментами, представляющими собой линкер-лекарственное средство. Неограничивающими примерами таких реакций с участием групп RG1 и R8 является малеимидная группа (R8), которая вступает в реакцию с тиольной группой (RG1) с получением сукцинимидного кольца, или гидроксиламинная группа (R8), которая вступает в реакцию с кетоновой группой (RG1) с получением оксима.
В данном случае R0 представляет собой метил или этил, R1 представляет собой метил или изопропил, R2 представляет собой метил или этил, X1 представляет собой бивалентную связывающую группу, образуемую при вступлении RG1 и R8 в реакцию (например сукцинимидное кольцо или оксим), X2 представляет собой саморасщепляющийся спейсер, L1 представляет собой бивалентный пептидный линкер, L2 представляет собой связь или линкер, и y равняется 1, 2, 3 или 4.
[00268] Общая схема реакции образования конъюгатов формулы (C-2) показана на схеме 4 ниже:
схема 4,
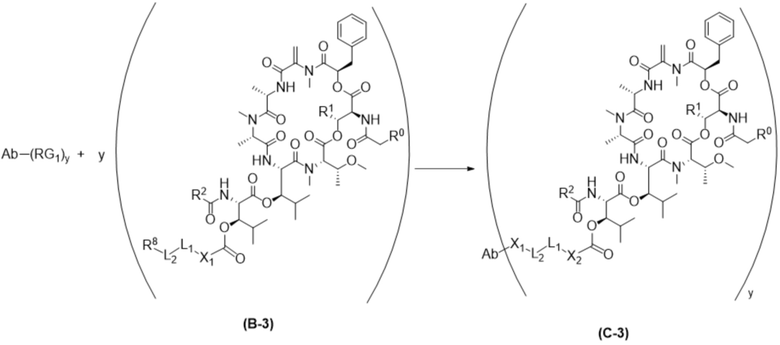
где RG1 представляет собой реакционноспособную группу на антителе Ab или его антигенсвязывающем фрагменте, исключительно в качестве примера тиольную, аминную или кетоновую, которая вступает в реакцию с совместимой реакционноспособной группой R8, присоединенной к фрагменту, представляющему собой линкер-лекарственное средство, обеспечивая тем самым ковалентную связь антитела Ab или его антигенсвязывающего фрагмента с одним или несколькими фрагментами, представляющими собой линкер-лекарственное средство. Неограничивающими примерами таких реакций с участием групп RG1 и R8 является малеимидная группа (R8), которая вступает в реакцию с тиольной группой (RG1) с получением сукцинимидного кольца, или гидроксиламинная группа (R8), которая вступает в реакцию с кетоновой группой (RG1) с получением оксима.
В данном случае R0 представляет собой метил или этил, R1 представляет собой метил или изопропил, R2 представляет собой метил или этил, X1 представляет собой бивалентную связывающую группу, образуемую при вступлении RG1 и R8 в реакцию (например сукцинимидное кольцо или оксим), X2 представляет собой саморасщепляющийся спейсер, L1 представляет собой бивалентный пептидный линкер, L2 представляет собой связь или линкер, и y равняется 1, 2, 3 или 4.
Способ конъюгации с введенными путем конструирования цистеиновыми остатками антитела
[00269] Конъюгаты по настоящему изобретению могут быть получены с применением цистеиновых остатков, введенных путем конструирования в антитело, например, с помощью сайт-направленного мутагенеза. Такие сайт-специфические конъюгаты являются гомогенными и характеризуются улучшенными свойствами (Junutula JR, Raab H, Clark S, Bhakta S, Leipold DD, Weir S, Chen Y, Simpson M, Tsai SP, Dennis MS, Lu Y, Meng YG, Ng C, Yang J, Lee CC, Duenas E, Gorrell J, Katta V, Kim A, McDorman K, Flagella K, Venook R, Ross S, Spencer SD, Lee Wong W, Lowman HB, Vandlen R, Sliwkowski MX, Scheller RH, Polakis P, Mallet W. (2008) Nature Biotechnology 26:925-932.)
[00270] Поскольку введенные путем конструирования цистеины в антителах, экспрессируемых в клетках млекопитающих, во время биосинтеза модифицируются за счет аддуктов (дисульфидов), таких как глутатион (GSH) и/или цистеин (Chen et al. 2009), изначально экспрессированные введенные путем конструирования цистеиновые остатки в продукте не реагируют с реагентами, вступающими в реакцию с тиолами, такими как малеимидная или бром- или йод-ацетамидная группы. Для осуществления конъюгации полезной нагрузки с введенным путем конструирования цистеином после экспрессии глутатионовый или цистеиновый аддукты должны быть удалены путем восстановления данных дисульфидных аддуктов, что обычно вызывает также восстановление нативных дисульфидов в экспрессированном белке. Удаление аддуктов у введенных путем конструирования цистеинов можно осуществлять, подвергая вначале антитело воздействию восстановителя, например, дитиотреитола (DTT), TCEP или восстановленного цистеина, с последующим проведением процедуры, которая обеспечивает возможность повторного окисления всех нативных дисульфидных связей антитела для восстановления и/или стабилизации функциональной структуры антитела.
[00271] Можно использовать несколько способов восстановления и повторного окисления антител с введенными путем конструирования остатками цистеина для получения конъюгатов антитела и лекарственного средства. Попытки следовать ранее описанным в литературе протоколам повторного окисления с применением высоких концентраций CuSO4 приводили к осаждению белка (Junutula JR, Raab H, Clark S, Bhakta S, Leipold DD, Weir S, Chen Y, Simpson M, Tsai SP, Dennis MS, Lu Y, Meng YG, Ng C, Yang J, Lee CC, Duenas E, Gorrell J, Katta V, Kim A, McDorman K, Flagella K, Venook R, Ross S, Spencer SD, Lee Wong W, Lowman HB, Vandlen R, Sliwkowski MX, Scheller RH, Polakis P, Mallet W. (2008) Nature Biotechnology 26:925). Авторы настоящего изобретения успешно подготовили и получили конъюгаты антитела и лекарственного средства с помощью нескольких различных способов восстановления и повторного окисления антитела.
[00272] Ниже приводится способ восстановления и повторного окисления антител с введенными путем конструирования остатками цистеина для получения конъюгатов антитела и лекарственного средства: свежеполученный DTT добавляют к очищенным антителам с Cys-мутацией до конечной концентрации, составляющей 10 мМ. После инкубации с DTT при комнатной температуре в течение 1 часа смесь диализировали против PBS при 4°C в течение трех дней с ежедневной заменой буфера для удаления DTT и повторного окисления нативных дисульфидных связей антитела. Альтернативный способ заключается в удалении восстанавливающих реагентов путем пропускания через обессоливающую колонку, такую как Sephadex G-25, уравновешенную с помощью PBS. После того как белок полностью восстановился к обессоленным образцам необязательно добавляют 1 мМ окисленного аскорбата (дигидроаскорбиновой кислоты) и инкубации для повторного окисления проводят в течение 20-24 часов.
[00273] В другом иллюстративном способе освобождение введенных путем конструирования Cys-остатков осуществляют путем добавления полностью восстановленного цистеина при концентрации 20 мМ к антителам, связанным со смолой на основе сефарозы с белком A. Восстановление аддуктов на Cys достигается путем инкубации в течение примерно 30-60 минут при комнатной температуре, затем восстановитель быстро удаляют путем промывания смолы с использованием 50 объемов слоя PBS. Повторное окисление восстановленного антитела достигается инкубацией промытой взвеси при комнатной температуре с добавлением или без добавления 50-2000 нМ CuCl2 в качестве катализатора. За исключением применения сульфата меди в примерах из данного документа применялся каждый из протоколов, описанных в данном документе, с получением аналогичных результатов. Повторное окисление восстанавливает внутрицепочечные дисульфиды, в то время как диализ, обессоливание или хроматография с белком A удаляет восстановитель, а также цистеины и глутатионы, изначально присоединенные к введенному(введенным) путем конструирования цистеину(цистеинам) антитела. Для отслеживания процесса повторного окисления обычно применяют HPLC-хроматографию с обращенной фазой: антитела загружают в колонку PLRP-S (4000 Å, 50 мм x 2,1 мм, Agilent), нагретую до 80°C и элюируют с использованием линейного градиента 30-45% CH3CN в воде, содержащей 0,1% TFA, при 1,5 мл/мин и пике обнаружения при 215, 254 и 280 нм.
[00274] После повторного окисления антитело конъюгируют с соединением линкер-лекарственное средство, например, с соединениями формулы (B), формулы (B-1), формулы (B-2) или формулы (B-3) (см. схемы 1-4). В качестве примера, соединение формулы (B), формулы (B-1), формулы (B-2) или формулы (B-3) добавляют к повторно окисленному Cys-мутантному антителу в количестве 5-10 молярных эквивалентов относительно антитела в буфере PBS (pH 7,2). Инкубации проводят в течение 1-2 часов. Процесс конъюгации отслеживают с помощью HPLC с обращенной фазой, которая может отделить конъюгированные антитела от неконъюгированных. Реакционные смеси в процессе конъюгации анализируют на колонке PRLP-S (4000 Å, 50 мм x 2,1 мм, Agilent), нагретой до 80°C, а элюирование колонки проводят с использованием линейного градиента 30-60% ацетонитрила в воде, содержащей 0,1% TFA при расходе 1,5 мл/мин. Элюирование белков из колонки отслеживают при 280 нм, 254 нм и 215 нм.
[00275] В качестве альтернативы для антител, связанных со смолой с белком А, после повторного окисления антитела смолу промывают 10 объемами колонки PBS, а затем смолу ресуспендируют в равном объеме PBS и добавляют в 8-кратном избытке соединение формулы (B), формулы (B-1), формулы (B-2) или формулы (B-3) (в DMSO) и инкубируют при комнатной температуре в течение 2 часов. Затем смолу промывают 50 объемами колонки PBS, и полученный конъюгат антитела и лекарственного средства элюируют из смолы с белком А, нейтрализуют 1/10 объема 1 М трис, pH 9,0, и осуществляют замену буфера на соответствующий буфер для проведения препаративной эксклюзионной хроматографии (если это необходимо).
[00276] Иммуноконъюгаты также характеризуют на предмет средней нагрузки фрагмента, представляющего собой лекарственное средство, на фрагмент, представляющий собой антитело связывания, обычно называемой соотношение лекарственного средства и антитела (DAR). Значение DAR экстраполируют, например, на основании данных LC-MS для восстановленных и дегликозилированных образцов. LC/MS обеспечивает возможность получения количественной оценки среднего числа молекул полезной нагрузки (фрагмент, представляющий собой лекарственное средство), прикрепленных к антителу в ADC. HPLC разделяет антитело на легкую и тяжелую цепи, а также разделяет тяжелую цепь (HC) и легкую цепь (LC) в соответствии с числом групп линкер-полезная нагрузка на цепь. Данные масс-спектров позволяют идентифицировать разновидности компонентов в смеси, например, LC, LC+1, LC+2, HC, HC+1, HC+2 и т. д. На основании средней нагрузки цепей LC и HC для ADC может быть рассчитано среднее DAR. DAR для данного образца иммуноконъюгата представляет собой среднее число молекул лекарственного средства (полезная нагрузка), прикрепленных к тетрамерному антителу, содержащему две легкие цепи и две тяжелые цепи.
[00277] На протяжении текста настоящей заявки может встречаться несоответствие между текстом описания и перечнем последовательностей, при этом текст описания имеет преимущественную силу.
Таблица 2. Примеры антител к PMEL17 по настоящему изобретению
[00278] Другие антитела по настоящему изобретению включают антитела, в которых аминокислоты или нуклеиновые кислоты, кодирующие аминокислоты, были подвергнуты мутации, но при этом они характеризуются по меньшей мере 60, 70, 80, 90 или 95 процентной идентичностью с последовательностями, описанными в таблице 2. В некоторых вариантах осуществления 1, 2, 3, 4 или 5 аминокислот были подвергнуты мутации в вариабельных областях по сравнению с вариабельными областями, показанными в последовательности, описанной в таблице 2, при этом они сохраняют практически такую же терапевтическую активность, как и антитела, перечисленные в таблице 2.
[00279] Поскольку каждое из данных антител может связываться с PMEL17, последовательности (аминокислотные последовательности и нуклеотидные последовательности, кодирующие аминокислотные последовательности) VH, VL, полноразмерной легкой цепи и полноразмерной тяжелой цепи можно подвергать "смешиванию и подбору" для создания других PMEL17-связывающих антител по настоящему изобретению. Такие подвергнутые "смешиванию и подбору" PMEL17-связывающие антитела можно тестировать с применением анализов связывания, известных из уровня техники (например, разновидностей ELISA и других анализов, описанных в разделе "Примеры"). Когда такие цепи подвергают смешиванию и подбору, последовательность VH из конкретной пары VH/VL следует заменить структурно подобной последовательностью VH. Аналогичным образом последовательность полноразмерной тяжелой цепи из конкретной пары полноразмерная тяжелая цепь/полноразмерная легкая цепь следует заменить структурно подобной последовательностью полноразмерной тяжелой цепи. Аналогичным образом последовательность VL из конкретной пары VH/VL следует заменить структурно подобной последовательностью VL. Аналогичным образом последовательность полноразмерной легкой цепи из конкретной пары полноразмерная тяжелая цепь/полноразмерная легкая цепь следует заменить структурно подобной последовательностью полноразмерной легкой цепи. Соответственно, в одном аспекте в настоящем изобретении представлены выделенное моноклональное антитело или его антигенсвязывающая область, имеющие вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 10, 42, 64, 88, 112, 132, 149, 165, 184, 196, 215, 227, 239 или 254; и вариабельную область легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 21, 25, 29, 53, 75, 99, 119, 143, 159, 171, 190, 202, 221, 233, 248 или 260; где антитело специфично связывается с PMEL17.
[00280] В другом аспекте в настоящем изобретении представлены (i) выделенное моноклональное антитело, имеющее полноразмерную тяжелую цепь, содержащую аминокислотную последовательность, оптимизированную для экспрессии в клетке системы экспрессии млекопитающего, выбранную из группы, состоящей из SEQ ID NOs: 12, 44, 66, 90, 114, 134, 151, 167, 186, 198, 217, 229, 241 или 256; и полноразмерную легкую цепь, содержащую аминокислотную последовательность, оптимизированную для экспрессии в клетке млекопитающего, выбранную из группы, состоящей из SEQ ID NOs: 23, 27, 31, 55, 77, 101, 121, 145, 161, 173, 192, 204, 223, 235, 250 или 262; или (ii) функциональный белок, содержащий его антигенсвязывающую часть.
[00281] В другом аспекте настоящего изобретения представлены PMEL17-связывающие антитела, которые содержат CDR1, CDR2 и CDR3 тяжелой цепи и легкой цепи, описанные в таблице 2, или их комбинации. Аминокислотные последовательности CDR1 VH антител показаны, например, под SEQ ID NOs: 1, 4, 5, 7, 33, 36, 37, 39, 57, 60, 79, 82, 83, 85, 103, 106, 107, 109, 123, 126, 127, 129, 175, 178, 179, 181, 206, 209, 210, и 212. Аминокислотные последовательности CDR2 VH антител показаны, например, под SEQ ID NOs: 2, 6, 8, 34, 38, 40, 58, 61, 62, 80, 84, 86, 104, 108, 110, 124, 128, 130, 176, 180, 182, 207, 211, и 213. Аминокислотные последовательности CDR3 VH антител показаны, например, под SEQ ID NOs: 3, 9, 35, 41, 59, 63, 81, 87, 105, 111, 125, 131, 147, 148, 163, 164, 177, 183, 194, 195, 208, 214, 225, 226, 237, 238, 252, и 253. Аминокислотные последовательности CDR1 VL антител показаны, например, под SEQ ID NOs: 14, 17, 20, 46, 49, 52, 68, 71, 74, 92, 95, 98, 116, 136, 139, 142, 153, 156, 158, 243, 245, и 247. Аминокислотные последовательности CDR2 VL антител показаны, например, под SEQ ID Nos: 15, 18, 47, 50, 69, 72, 93, 96, 137, 140, и 154. Аминокислотные последовательности CDR3 VL антител показаны, например, под SEQ ID NOs: 16, 19, 48, 51, 70, 73, 94, 97, 117, 118, 138, 141, 155, 157, 169, 170188, 189, 200, 201, 219, 220, 231, 232, 244, 246, 258, и 259.
[00282] С учетом того, что каждое из данных антител может связываться с PMEL17 и что специфичность связывания антигена в первую очередь обеспечивается областями CDR1, 2 и 3, последовательности CDR1, CDR2 и CDR3 VH и последовательности CDR1, CDR2 и CDR3 VL могут быть подвергнуты "смешиванию и подбору" (т. е. CDR из разных антител могут быть подвергнуты смешиванию и подбору. Такие подвергнутые "смешиванию и подбору" антитела, связывающие PMEL17, можно тестировать с применением анализов связывания, известных из уровня техники и описанных в разделе "Примеры" (например, вариантов ELISA). Когда последовательности CDR VH подвергают смешиванию и подбору, последовательность CDR1, CDR2 и/или CDR3 из конкретной последовательности VH следует заменить на структурно подобную(подобные) последовательность(последовательности) CDR. Аналогичным образом, когда последовательности CDR VL подвергают смешиванию и подбору, последовательность CDR1, CDR2 и/или CDR3 из конкретной последовательности VL следует заменить на структурно подобную(подобные) последовательность(последовательности) CDR. Рядовому специалисту в данной области техники будет совершенно очевидно, что новые последовательности VH и VL можно создавать путем замены одной или нескольких последовательностей областей CDR VH и/или VL структурно подобными последовательностями из последовательностей CDR, показанных в данном документе для моноклональных антител по настоящему изобретению.
[00283] Соответственно, в некоторых вариантах осуществления в настоящем изобретении представлены выделенное моноклональное антитело или его антигенсвязывающая область, содержащие CDR1 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 1, 4, 5, 7, 33, 36, 37, 39, 57, 60, 79, 82, 83, 85, 103, 106, 107, 109, 123, 126, 127, 129, 175, 178, 179, 181, 206, 209, 210, и 212; CDR2 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 2, 6, 8, 34, 38, 40, 58, 61, 62, 80, 84, 86, 104, 108, 110, 124, 128, 130, 176, 180, 182, 207, 211, и 213; CDR3 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 3, 9, 35, 41, 59, 63, 81, 87, 105, 111, 125, 131, 147, 148, 163, 164, 177, 183, 194, 195, 208, 214, 225, 226, 237, 238, 252, и 253; CDR1 легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 14, 17, 20, 46, 49, 52, 68, 71, 74, 92, 95, 98, 116, 136, 139, 142, 153, 156, 158, 243, 245, и 247; CDR2 легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 15, 18, 47, 50, 69, 72, 93, 96, 137, 140, и 154; и CDR3 легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 16, 19, 48, 51, 70, 73, 94, 97, 117, 118, 138, 141, 155, 157, 169, 170, 188, 189, 200, 201, 219, 220, 231, 232, 244, 246, 258, и 259; где антитело специфично связывает PMEL17.
[00284] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат CDR1 тяжелой цепи под SEQ ID NO:1, 4, 5 или 7, CDR2 тяжелой цепи под SEQ ID NO:2, 6 или 8; CDR3 тяжелой цепи под SEQ ID NO:3 или 9; CDR1 легкой цепи под SEQ ID NO:14, 17 или 20; CDR2 легкой цепи под SEQ ID NO:15 или 18 и CDR3 легкой цепи под SEQ ID NO:16 или 19.
[00285] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат CDR1 тяжелой цепи под SEQ ID NO:33, 36, 37 или 39, CDR2 тяжелой цепи под SEQ ID NO:34, 38 или 40; CDR3 тяжелой цепи под SEQ ID NO:35 или 41; CDR1 легкой цепи под SEQ ID NO:46, 49 или 52; CDR2 легкой цепи под SEQ ID NO:47 или 50 и CDR3 легкой цепи под SEQ ID NO:48 или 51.
[00286] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат CDR1 тяжелой цепи под SEQ ID NO:5, 7, 57 или 60, CDR2 тяжелой цепи под SEQ ID NO:58, 61 или 62; CDR3 тяжелой цепи под SEQ ID NO:59 или 63; CDR1 легкой цепи под SEQ ID NO:68, 71 или 74; CDR2 легкой цепи под SEQ ID NO:69 или 72 и CDR3 легкой цепи под SEQ ID NO:70 или 73.
[00287] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат CDR1 тяжелой цепи под SEQ ID NO:79, 82, 83 или 85, CDR2 тяжелой цепи под SEQ ID NO:80, 84 или 86; CDR3 тяжелой цепи под SEQ ID NO:81 или 87; CDR1 легкой цепи под SEQ ID NO:92, 95 или 98; CDR2 легкой цепи под SEQ ID NO:93 или 96 и CDR3 легкой цепи под SEQ ID NO:94 или 97.
[00288] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат CDR1 тяжелой цепи под SEQ ID NO:103, 106, 107 или 109, CDR2 тяжелой цепи под SEQ ID NO:104, 108 или 110; CDR3 тяжелой цепи под SEQ ID NO:105 или 111; CDR1 легкой цепи под SEQ ID NO:49, 52 или 116; CDR2 легкой цепи под SEQ ID NO:47 или 50 и CDR3 легкой цепи под SEQ ID NO:117 или 118.
[00289] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат CDR1 тяжелой цепи под SEQ ID NO:123, 126, 127 или 129, CDR2 тяжелой цепи под SEQ ID NO:124, 128 или 130; CDR3 тяжелой цепи под SEQ ID NO:125 или 131; CDR1 легкой цепи под SEQ ID NO:136, 139 или 142; CDR2 легкой цепи под SEQ ID NO:137 или 140 и CDR3 легкой цепи под SEQ ID NO:138 или 141.
[00290] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат CDR1 тяжелой цепи под SEQ ID NO:123, 126, 127 или 129, CDR2 тяжелой цепи под SEQ ID NO:124, 128 или 130; CDR3 тяжелой цепи под SEQ ID NO:147 или 148; CDR1 легкой цепи под SEQ ID NO:153, 156 или 158; CDR2 легкой цепи под SEQ ID NO:50 или 154 и CDR3 легкой цепи под SEQ ID NO:155 или 157.
[00291] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат CDR1 тяжелой цепи под SEQ ID NO:103, 106, 107 или 109, CDR2 тяжелой цепи под SEQ ID NO:104, 108 или 110; CDR3 тяжелой цепи под SEQ ID NO:163 или 164; CDR1 легкой цепи под SEQ ID NO:49, 52 или 116; CDR2 легкой цепи под SEQ ID NO:47 или 50 и CDR3 легкой цепи под SEQ ID NO:169 или 170.
[00292] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат CDR1 тяжелой цепи под SEQ ID NO:175, 178, 179 или 181, CDR2 тяжелой цепи под SEQ ID NO:176, 180 или 182; CDR3 тяжелой цепи под SEQ ID NO:177 или 183; CDR1 легкой цепи под SEQ ID NO:49, 52 или 116; CDR2 легкой цепи под SEQ ID NO:47 или 50 и CDR3 легкой цепи под SEQ ID NO:188 или 189.
[00293] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат CDR1 тяжелой цепи под SEQ ID NO: 103, 106, 107 или 109, CDR2 тяжелой цепи под SEQ ID NO: 104, 108 или 110; CDR3 тяжелой цепи под SEQ ID NO:194 или 195; CDR1 легкой цепи под SEQ ID NO: 49, 52 или 116; CDR2 легкой цепи под SEQ ID NO: 47 или 50 и CDR3 легкой цепи под SEQ ID NO:200 или 201.
[00294] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат CDR1 тяжелой цепи под SEQ ID NO:206, 209, 210 или 212, CDR2 тяжелой цепи под SEQ ID NO:207, 211 или 213; CDR3 тяжелой цепи под SEQ ID NO:208 или 214; CDR1 легкой цепи под SEQ ID NO:153, 156 или 158; CDR2 легкой цепи под SEQ ID NO:50 или 154 и CDR3 легкой цепи под SEQ ID NO:219 или 220.
[00295] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат CDR1 тяжелой цепи под SEQ ID NO: 206, 209, 210 или 212, CDR2 тяжелой цепи под SEQ ID NO: 207, 211 или 213; CDR3 тяжелой цепи под SEQ ID NO:225 или 226; CDR1 легкой цепи под SEQ ID NO:136, 139 или 142; CDR2 легкой цепи под SEQ ID NO:137 или 140 и CDR3 легкой цепи под SEQ ID NO:231 или 232.
[00296] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 206, 209, 210 или 212, HCDR2 под SEQ ID NO: 207, 211 или 213 и HCDR3 под SEQ ID NO:237 или 238; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:243, 245 или 247, LCDR2 под SEQ ID NO:47 или 50 и LCDR3 под SEQ ID NO:244 или 246.
[00297] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 206, 209, 210 или 212, HCDR2 под SEQ ID NO: 207, 211 или 213 и HCDR3 под SEQ ID NO:252 или 253; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:153, 156 или 158, LCDR2 под SEQ ID NO:50 или 154 и LCDR3 под SEQ ID NO:258 или 259.
[00298] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) CDR1 тяжелой цепи под SEQ ID NO:1, CDR2 тяжелой цепи под SEQ ID NO:2, CDR3 тяжелой цепи под SEQ ID NO:3, CDR1 легкой цепи под SEQ ID NO:14, CDR2 легкой цепи под SEQ ID NO:15 и CDR3 легкой цепи под SEQ ID NO:16;
b) CDR1 тяжелой цепи под SEQ ID NO: 4, CDR2 тяжелой цепи под SEQ ID NO:2, CDR3 тяжелой цепи под SEQ ID NO:3, CDR1 легкой цепи под SEQ ID NO:14, CDR2 легкой цепи под SEQ ID NO:15 и CDR3 легкой цепи под SEQ ID NO:16;
c) CDR1 тяжелой цепи под SEQ ID NO:5, CDR2 тяжелой цепи под SEQ ID NO:6, CDR3 тяжелой цепи под SEQ ID NO:3, CDR1 легкой цепи под SEQ ID NO:17, CDR2 легкой цепи под SEQ ID NO: 18 и CDR3 легкой цепи под SEQ ID NO: 19 или
d) CDR1 тяжелой цепи под SEQ ID NO:7, CDR2 тяжелой цепи под SEQ ID NO:8, CDR3 тяжелой цепи под SEQ ID NO:9, CDR1 легкой цепи под SEQ ID NO:20, CDR2 легкой цепи под SEQ ID NO:18 и CDR3 легкой цепи под SEQ ID NO:16.
[00299] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) CDR1 тяжелой цепи под SEQ ID NO:33, CDR2 тяжелой цепи под SEQ ID NO:34, CDR3 тяжелой цепи под SEQ ID NO:35, CDR1 легкой цепи под SEQ ID NO:46, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:48;
b) CDR1 тяжелой цепи под SEQ ID NO:36, CDR2 тяжелой цепи под SEQ ID NO:34, CDR3 тяжелой цепи под SEQ ID NO:35, CDR1 легкой цепи под SEQ ID NO:46, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:48;
c) CDR1 тяжелой цепи под SEQ ID NO:37, CDR2 тяжелой цепи под SEQ ID NO:38, CDR3 тяжелой цепи под SEQ ID NO:35, CDR1 легкой цепи под SEQ ID NO:49, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:51 или
d) CDR1 тяжелой цепи под SEQ ID NO: 39, CDR2 тяжелой цепи под SEQ ID NO:40, CDR3 тяжелой цепи под SEQ ID NO:41, CDR1 легкой цепи под SEQ ID NO:52, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:48.
[00300] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из a) CDR1 тяжелой цепи под SEQ ID NO:57, CDR2 тяжелой цепи под SEQ ID NO:58, CDR3 тяжелой цепи под SEQ ID NO:59, CDR1 легкой цепи под SEQ ID NO:68, CDR2 легкой цепи под SEQ ID NO:69 и CDR3 легкой цепи под SEQ ID NO:70; b) CDR1 тяжелой цепи под SEQ ID NO:60, CDR2 тяжелой цепи под SEQ ID NO:58, CDR3 тяжелой цепи под SEQ ID NO:59, CDR1 легкой цепи под SEQ ID NO:68, CDR2 легкой цепи под SEQ ID NO:69 и CDR3 легкой цепи под SEQ ID NO:70;
c) CDR1 тяжелой цепи под SEQ ID NO:5, CDR2 тяжелой цепи под SEQ ID NO:61, CDR3 тяжелой цепи под SEQ ID NO:59, CDR1 легкой цепи под SEQ ID NO:71, CDR2 легкой цепи под SEQ ID NO:72 и CDR3 легкой цепи под SEQ ID NO:73 или d) CDR1 тяжелой цепи под SEQ ID NO:7, CDR2 тяжелой цепи под SEQ ID NO:62, CDR3 тяжелой цепи под SEQ ID NO:63, CDR1 легкой цепи под SEQ ID NO:74, CDR2 легкой цепи под SEQ ID NO:72 и CDR3 легкой цепи под SEQ ID NO:70.
[00301] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) CDR1 тяжелой цепи под SEQ ID NO:79, CDR2 тяжелой цепи под SEQ ID NO:80, CDR3 тяжелой цепи под SEQ ID NO:81, CDR1 легкой цепи под SEQ ID NO:92, CDR2 легкой цепи под SEQ ID NO:93 и CDR3 легкой цепи под SEQ ID NO:94;
b) CDR1 тяжелой цепи под SEQ ID NO:82, CDR2 тяжелой цепи под SEQ ID NO:80, CDR3 тяжелой цепи под SEQ ID NO:81, CDR1 легкой цепи под SEQ ID NO:92, CDR2 легкой цепи под SEQ ID NO:93 и CDR3 легкой цепи под SEQ ID NO:94;
c) CDR1 тяжелой цепи под SEQ ID NO:83, CDR2 тяжелой цепи под SEQ ID NO:84, CDR3 тяжелой цепи под SEQ ID NO:81, CDR1 легкой цепи под SEQ ID NO:95, CDR2 легкой цепи под SEQ ID NO:96 и CDR3 легкой цепи под SEQ ID NO: 97 или
d) CDR1 тяжелой цепи под SEQ ID NO: 85, CDR2 тяжелой цепи под SEQ ID NO:86, CDR3 тяжелой цепи под SEQ ID NO:87, CDR1 легкой цепи под SEQ ID NO:98, CDR2 легкой цепи под SEQ ID NO:96 и CDR3 легкой цепи под SEQ ID NO:94.
[00302] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) CDR1 тяжелой цепи под SEQ ID NO:103, CDR2 тяжелой цепи под SEQ ID NO:104, CDR3 тяжелой цепи под SEQ ID NO:105, CDR1 легкой цепи под SEQ ID NO: 116; CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:117;
b) CDR1 тяжелой цепи под SEQ ID NO:106, CDR2 тяжелой цепи под SEQ ID NO:104, CDR3 тяжелой цепи под SEQ ID NO:105, CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:117;
c) CDR1 тяжелой цепи под SEQ ID NO:107, CDR2 тяжелой цепи под SEQ ID NO:108, CDR3 тяжелой цепи под SEQ ID NO:105, CDR1 легкой цепи под SEQ ID NO:49, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:118 или
d) CDR1 тяжелой цепи под SEQ ID NO:109, CDR2 тяжелой цепи под SEQ ID NO:110, CDR3 тяжелой цепи под SEQ ID NO:111, CDR1 легкой цепи под SEQ ID NO:52, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:117.
[00303] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) CDR1 тяжелой цепи под SEQ ID NO:123, CDR2 тяжелой цепи под SEQ ID NO:124, CDR3 тяжелой цепи под SEQ ID NO:125, CDR1 легкой цепи под SEQ ID NO:136, CDR2 легкой цепи под SEQ ID NO:137 и CDR3 легкой цепи под SEQ ID NO:138;
b) CDR1 тяжелой цепи под SEQ ID NO:126, CDR2 тяжелой цепи под SEQ ID NO:124, CDR3 тяжелой цепи под SEQ ID NO:125, CDR1 легкой цепи под SEQ ID NO:136, CDR2 легкой цепи под SEQ ID NO:137 и CDR3 легкой цепи под SEQ ID NO:138;
c) CDR1 тяжелой цепи под SEQ ID NO:127, CDR2 тяжелой цепи под SEQ ID NO:128, CDR3 тяжелой цепи под SEQ ID NO:125, CDR1 легкой цепи под SEQ ID NO:139, CDR2 легкой цепи под SEQ ID NO:140 и CDR3 легкой цепи под SEQ ID NO: 141 или
d) CDR1 тяжелой цепи под SEQ ID NO: 129, CDR2 тяжелой цепи под SEQ ID NO:130, CDR3 тяжелой цепи под SEQ ID NO:131, CDR1 легкой цепи под SEQ ID NO:142, CDR2 легкой цепи под SEQ ID NO:140 и CDR3 легкой цепи под SEQ ID NO:138.
[00304] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) CDR1 тяжелой цепи под SEQ ID NO:123, CDR2 тяжелой цепи под SEQ ID NO:124, CDR3 тяжелой цепи под SEQ ID NO:147, CDR1 легкой цепи под SEQ ID NO:153, CDR2 легкой цепи под SEQ ID NO:154 и CDR3 легкой цепи под SEQ ID NO:155;
b) CDR1 тяжелой цепи под SEQ ID NO:126, CDR2 тяжелой цепи под SEQ ID NO:124, CDR3 тяжелой цепи под SEQ ID NO:147, CDR1 легкой цепи под SEQ ID NO:153, CDR2 легкой цепи под SEQ ID NO: 154 и CDR3 легкой цепи под SEQ ID NO:155;
c) CDR1 тяжелой цепи под SEQ ID NO:127, CDR2 тяжелой цепи под SEQ ID NO:128, CDR3 тяжелой цепи под SEQ ID NO:147, CDR1 легкой цепи под SEQ ID NO:156, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:157 или
d) CDR1 тяжелой цепи под SEQ ID NO: 129, CDR2 тяжелой цепи под SEQ ID NO:130, CDR3 тяжелой цепи под SEQ ID NO:148, CDR1 легкой цепи под SEQ ID NO:158, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:155.
[00305] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) CDR1 тяжелой цепи под SEQ ID NO:103, CDR2 тяжелой цепи под SEQ ID NO:104, CDR3 тяжелой цепи под SEQ ID NO:163, CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:169;
b) CDR1 тяжелой цепи под SEQ ID NO:106, CDR2 тяжелой цепи под SEQ ID NO:104, CDR3 тяжелой цепи под SEQ ID NO:163, CDR1 легкой цепи под SEQ ID NO:116, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:169;
c) CDR1 тяжелой цепи под SEQ ID NO:107, CDR2 тяжелой цепи под SEQ ID NO:108, CDR3 тяжелой цепи под SEQ ID NO:163, CDR1 легкой цепи под SEQ ID NO:49, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:170 или
d) CDR1 тяжелой цепи под SEQ ID NO: 109, CDR2 тяжелой цепи под SEQ ID NO:110, CDR3 тяжелой цепи под SEQ ID NO:164, CDR1 легкой цепи под SEQ ID NO:52, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:169.
[00306] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) CDR1 тяжелой цепи под SEQ ID NO:175, CDR2 тяжелой цепи под SEQ ID NO:176, CDR3 тяжелой цепи под SEQ ID NO:177, CDR1 легкой цепи под SEQ ID NO:116, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:188;
b) CDR1 тяжелой цепи под SEQ ID NO:178, CDR2 тяжелой цепи под SEQ ID NO:176, CDR3 тяжелой цепи под SEQ ID NO:177, CDR1 легкой цепи под SEQ ID NO:116, CDR2 легкой цепи под SEQ ID NO:47 и CDR3 легкой цепи под SEQ ID NO:188;
c) CDR1 тяжелой цепи под SEQ ID NO:179, CDR2 тяжелой цепи под SEQ ID NO:180, CDR3 тяжелой цепи под SEQ ID NO:177, CDR1 легкой цепи под SEQ ID NO:49, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:189 или
d) CDR1 тяжелой цепи под SEQ ID NO: 181, CDR2 тяжелой цепи под SEQ ID NO:182, CDR3 тяжелой цепи под SEQ ID NO:183, CDR1 легкой цепи под SEQ ID NO:52, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:188.
[00307] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) CDR1 тяжелой цепи под SEQ ID NO: 103, CDR2 тяжелой цепи под SEQ ID NO: 104, CDR3 тяжелой цепи под SEQ ID NO:194, CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO:200;
b) CDR1 тяжелой цепи под SEQ ID NO: 106, CDR2 тяжелой цепи под SEQ ID NO: 104, CDR3 тяжелой цепи под SEQ ID NO:194, CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO:200;
c) CDR1 тяжелой цепи под SEQ ID NO: 107, CDR2 тяжелой цепи под SEQ ID NO: 108, CDR3 тяжелой цепи под SEQ ID NO:194, CDR1 легкой цепи под SEQ ID NO: 49, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 201 или
d) CDR1 тяжелой цепи под SEQ ID NO: 109, CDR2 тяжелой цепи под SEQ ID NO: 110, CDR3 тяжелой цепи под SEQ ID NO:195, CDR1 легкой цепи под SEQ ID NO: 52, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO:200.
[00308] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) CDR1 тяжелой цепи под SEQ ID NO:206, CDR2 тяжелой цепи под SEQ ID NO:207, CDR3 тяжелой цепи под SEQ ID NO:208, CDR1 легкой цепи под SEQ ID NO:153, CDR2 легкой цепи под SEQ ID NO:154 и CDR3 легкой цепи под SEQ ID NO:219;
b) CDR1 тяжелой цепи под SEQ ID NO:209, CDR2 тяжелой цепи под SEQ ID NO:207, CDR3 тяжелой цепи под SEQ ID NO:208, CDR1 легкой цепи под SEQ ID NO:153, CDR2 легкой цепи под SEQ ID NO: 154 и CDR3 легкой цепи под SEQ ID NO:219;
c) CDR1 тяжелой цепи под SEQ ID NO:210, CDR2 тяжелой цепи под SEQ ID NO:211, CDR3 тяжелой цепи под SEQ ID NO:208, CDR1 легкой цепи под SEQ ID NO:156, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:220 или
d) CDR1 тяжелой цепи под SEQ ID NO: 212, CDR2 тяжелой цепи под SEQ ID NO:213, CDR3 тяжелой цепи под SEQ ID NO:214, CDR1 легкой цепи под SEQ ID NO:158, CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:219.
[00309] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) CDR1 тяжелой цепи под SEQ ID NO: 206, CDR2 тяжелой цепи под SEQ ID NO: 207, CDR3 тяжелой цепи под SEQ ID NO:225, CDR1 легкой цепи под SEQ ID NO:136, CDR2 легкой цепи под SEQ ID NO:137 и CDR3 легкой цепи под SEQ ID NO:231;
b) CDR1 тяжелой цепи под SEQ ID NO: 209, CDR2 тяжелой цепи под SEQ ID NO: 207, CDR3 тяжелой цепи под SEQ ID NO:225, CDR1 легкой цепи под SEQ ID NO:136, CDR2 легкой цепи под SEQ ID NO:137 и CDR3 легкой цепи под SEQ ID NO:231;
c) CDR1 тяжелой цепи под SEQ ID NO: 210, CDR2 тяжелой цепи под SEQ ID NO: 211, CDR3 тяжелой цепи под SEQ ID NO:225, CDR1 легкой цепи под SEQ ID NO:139, CDR2 легкой цепи под SEQ ID NO:140 и CDR3 легкой цепи под SEQ ID NO: 232 или
d) CDR1 тяжелой цепи под SEQ ID NO: 212, CDR2 тяжелой цепи под SEQ ID NO: 213, CDR3 тяжелой цепи под SEQ ID NO: 226, CDR1 легкой цепи под SEQ ID NO:142; CDR2 легкой цепи под SEQ ID NO: 140 и CDR3 легкой цепи под SEQ ID NO:231.
[00310] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) вариабельной области тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 206, HCDR2 под SEQ ID NO: 207 и HCDR3 под SEQ ID NO:237, и вариабельной области легкой цепи, которая содержит LCDR1 под SEQ ID NO:243, LCDR2 под SEQ ID NO:47 и LCDR3 под SEQ ID NO:244;
b) вариабельной области тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 209, HCDR2 под SEQ ID NO: 207 и HCDR3 под SEQ ID NO:237, и вариабельной области легкой цепи, которая содержит LCDR1 под SEQ ID NO:243, LCDR2 под SEQ ID NO:47 и LCDR3 под SEQ ID NO:244;
c) вариабельной области тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 210, HCDR2 под SEQ ID NO: 211 и HCDR3 под SEQ ID NO:237, и вариабельной области легкой цепи, которая содержит LCDR1 под SEQ ID NO:245, LCDR2 под SEQ ID NO:50 и LCDR3 под SEQ ID NO:246; или
d) вариабельной области тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 212, HCDR2 под SEQ ID NO: 213 и HCDR3 под SEQ ID NO:238, и вариабельной области легкой цепи, которая содержит LCDR1 под SEQ ID NO:247, LCDR2 под SEQ ID NO: 50 и LCDR3 под SEQ ID NO:244.
[00311] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат последовательности CDR, выбранные из
a) вариабельной области тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 206, HCDR2 под SEQ ID NO: 207 и HCDR3 под SEQ ID NO:252, и вариабельной области легкой цепи, которая содержит LCDR1 под SEQ ID NO:153, LCDR2 под SEQ ID NO: 154 и LCDR3 под SEQ ID NO:258;
b) вариабельной области тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 209, HCDR2 под SEQ ID NO: 207 и HCDR3 под SEQ ID NO:252, и вариабельной области легкой цепи, которая содержит LCDR1 под SEQ ID NO:153, LCDR2 под SEQ ID NO:154 и LCDR3 под SEQ ID NO:258;
c) вариабельной области тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 210, HCDR2 под SEQ ID NO: 211 и HCDR3 под SEQ ID NO:252, и вариабельной области легкой цепи, которая содержит LCDR1 под SEQ ID NO:156, LCDR2 под SEQ ID NO:50 и LCDR3 под SEQ ID NO:259; или
d) вариабельной области тяжелой цепи, которая содержит HCDR1 под SEQ ID NO: 212, HCDR2 под SEQ ID NO: 213 и HCDR3 под SEQ ID NO: 253, и вариабельной области легкой цепи, которая содержит LCDR1 под SEQ ID NO:158, LCDR2 под SEQ ID NO:50 и LCDR3 под SEQ ID NO:258.
[00312] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:10, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:21.
[00313] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:10, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:25.
[00314] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:10, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:29.
[00315] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:42, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:53.
[00316] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:64, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:75.
[00317] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:88, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:99.
[00318] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:112, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:119.
[00319] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:132, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:143.
[00320] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:149, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:159.
[00321] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:165, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:171.
[00322] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:184, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:190.
[00323] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:196, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:202.
[00324] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:215, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:221.
[00325] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:227, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:233.
[00326] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:239, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:248.
[00327] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:254, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:260.
[00328] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:12, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:23.
[00329] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:12, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:27.
[00330] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:12, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:31.
[00331] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:44, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:55.
[00332] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:66, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:77.
[00333] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:90, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:101.
[00334] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:114, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:121.
[00335] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:134, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:145.
[00336] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:151, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:161.
[00337] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:167, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:173.
[00338] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:186, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:192.
[00339] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:198, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:204.
[00340] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:217, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:223.
[00341] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:229, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:235.
[00342] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:241, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:250.
[00343] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с PMEL17, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:256, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:262.
[00344] В определенных вариантах осуществления антитело, которое специфически связывается с PMEL17, представляет собой антитело или фрагмент антитела (например, антигенсвязывающий фрагмент), которые описаны в таблице 2.
1. Идентификация эпитопов и антител, которые связываются с одним и тем же эпитопом
[00345] В настоящем изобретении также представлены антитела и фрагменты антител (например, антигенсвязывающие фрагменты), которые специфически связываются с тем же эпитопом, что и антитела к PMEL17, описанные в таблице 2, или вступают в перекрестную конкуренцию с антителами, описанными в таблице 2. Следовательно, дополнительные антитела и фрагменты антител (например, антигенсвязывающие фрагменты) можно идентифицировать на основе их способности к перекрестной конкуренции (например, к конкурентному ингибированию связывания статистически значимым образом) с другими антителами по настоящему изобретению в анализах связывания PMEL17, например с помощью BIACORE, или анализов, известных специалистам в данной области техники для измерения связывания. Способность тестируемого антитела ингибировать связывание антител и фрагментов антител (например, антигенсвязывающих фрагментов) по настоящему изобретению с PMEL17 (например, PMEL17 человека) демонстрирует, что тестируемое антитело может конкурировать с таким антителом или фрагментом антитела (например, антигенсвязывающими фрагментами) за связывание с PMEL17; при этом согласно неограничивающей теории такое антитело может связываться с тем же или родственным (например, структурно подобным или пространственно близким или перекрывающимся) эпитопом на белке PMEL17, что и антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), с которым оно конкурирует. В определенных вариантах осуществления антитела, которые связываются с тем же эпитопом на PMEL17, что и антитела или фрагменты антител (например, антигенсвязывающие фрагменты), описанные в таблице 2, представляют собой человеческие или гуманизированные моноклональные антитела. Такие человеческие или гуманизированные моноклональные антитела могут быть получены и выделены, как описано в данном документе.
2. Дополнительное изменение каркасной области в области Fc
[00346] Иммуноконъюгаты по настоящему изобретению могут содержать модифицированные антитела или их антигенсвязывающие фрагменты, которые дополнительно содержат модификации каркасных остатков в пределах VH и/или VL, например, для улучшения свойств антитела. В некоторых вариантах осуществления модификации каркасной области осуществляют для снижения иммуногенности антитела. Например, один подход заключается в том, что один или несколько каркасных остатков "мутируют к первоначальному виду" в направлении соответствующей последовательности зародышевой линии. Более конкретно, антитело, которое подверглось соматической мутации, может содержать каркасные остатки, которые отличаются от последовательности зародышевой линии, из которой происходит антитело. Такие остатки могут быть идентифицированы посредством сравнения каркасных последовательностей антитела с последовательностями зародышевой линии, из которых происходит антитело. Чтобы вернуть последовательности каркасной области в их конфигурацию зародышевой линии, соматические мутации можно вводить для "мутирования к первоначальному виду" в последовательности зародышевой линии, например, с помощью сайт-направленного мутагенеза. Подразумевается, что такие подвергнутые "мутации к первоначальному виду" антитела также охватываются настоящим изобретением.
[00347] Другой тип модификации каркасной области предусматривает мутацию одного или нескольких остатков в пределах каркасной области или даже в пределах одной или нескольких CDR-областей для удаления T-клеточных эпитопов со снижением тем самым потенциальной иммуногенности антитела. Данный подход также называют "деиммунизацией", и он более подробно описан в публикации заявки на патент США № 20030153043 от Carr et al.
[00348] В качестве дополнения или альтернативы модификациям, осуществляемым в пределах каркасных областей или областей CDR, антитела по настоящему изобретению можно конструировать таким образом, чтобы они включали модификации в пределах области Fc, как правило, для изменения одного или нескольких функциональных свойств антитела, таких как период полужизни в сыворотке крови, фиксация комплемента, связывание с Fc-рецептором и/или антигензависимая клеточная цитотоксичность (ADCC). Кроме того, антитело по настоящему изобретению может быть модифицировано химически (например, к антителу можно присоединять один или несколько химических фрагментов), или оно может быть модифицировано для изменения характера его гликозилирования, чтобы, опять-таки, изменить одно или несколько функциональных свойств антитела. Каждый из данных вариантов осуществления более подробно описан ниже.
[00349] В одном варианте осуществления шарнирную область CH1 модифицируют таким образом, что меняется число цистеиновых остатков в шарнирной области, например, увеличивается или уменьшается. Данный подход дополнительно описан в патенте США № 5677425 от Bodmer et al. Число цистеиновых остатков в шарнирной области CH1 изменяют, чтобы, например, облегчить сборку легкой и тяжелой цепей или чтобы увеличить или уменьшить стабильность антитела.
[00350] В некоторых вариантах осуществления антитело или фрагмент антитела, раскрытые в данном документе, содержат модифицированные или введенные путем конструирования аминокислотные остатки, например один или несколько цистеиновых остатков, в качестве сайтов для конъюгации с фрагментом, представляющим собой лекарственное средство (Junutula JR, et al., Nat Biotechnol 2008, 26:925-932). В одном варианте осуществления настоящего изобретения представлены модифицированные антитело или фрагмент антитела, содержащие замену одной или нескольких аминокислот на цистеин в положениях, описанных в данном документе. Сайты для цистеиновой замены расположены в константных областях антитела или фрагмента антитела и, таким образом, применимы ко множеству антител или фрагментов антител, при этом сайты выбирают для обеспечения стабильных и однородных конъюгатов. Модифицированные антитело или фрагмент могут иметь одну, две или более цистеиновых замен, и такие замены могут применяться в комбинации с другими способами модификации и конъюгации, описанными в данном документе. Способы для вставки цистеина в специфические местоположения в антителе известны из уровня техники, см., например, Lyons et al, (1990) Protein Eng., 3:703-708, WO 2011/005481, WO2014/124316, WO 2015/138615. В определенных вариантах осуществления модифицированное антитело содержит замену одной или нескольких аминокислот на цистеин в своей константной области в положении, выбранном из положений 117, 119, 121, 124, 139, 152, 153, 155, 157, 164, 169, 171, 174, 189, 191, 195, 197, 205, 207, 246, 258, 269, 274, 286, 288, 290, 292, 293, 320, 322, 326, 333, 334, 335, 337, 344, 355, 360, 375, 382, 390, 392, 398, 400 и 422 тяжелой цепи антитела, и где нумерация положений соответствует системе EU. В некоторых вариантах осуществления модифицированные антитело или фрагмент антитела содержат замену одной или нескольких аминокислот на цистеин в своей константной области в положении, выбранном из положений 107, 108, 109, 114, 129, 142, 143, 145, 152, 154, 156, 159, 161, 165, 168, 169, 170, 182, 183, 197, 199, и 203 легкой цепи антитела или фрагмента антитела, где нумерация положений соответствует системе EU и где легкая цепь представляет собой человеческую легкую каппа-цепь. В некоторых вариантах осуществления модифицированные антитело или фрагмент такого антитела содержат комбинацию из замен двух или более аминокислот на цистеин в своих константных областях, где комбинации предусматривают замены в положениях 375 тяжелой цепи антитела, положении 152 тяжелой цепи антитела, положении 360 тяжелой цепи антитела или положении 107 легкой цепи антитела, при этом нумерация положений соответствует системе EU. В некоторых вариантах осуществления модифицированные антитело или фрагмент такого антитела содержат замену одной аминокислоты на цистеин в своих константных областях, где замена находится в положении 375 тяжелой цепи антитела, положении 152 тяжелой цепи антитела, положении 360 тяжелой цепи антитела, положении 107 легкой цепи антитела, положении 165 легкой цепи антитела или положении 159 легкой цепи антитела, при этом нумерация положений соответствует системе EU, и где легкая цепь представляет собой каппа-цепь. В конкретных вариантах осуществления модифицированные антитело или фрагмент такого антитела содержат комбинацию из замен двух аминокислот на цистеин в своих константных областях, где комбинации предусматривают замены в положениях 375 тяжелой цепи антитела и положении 152 тяжелой цепи антитела, где нумерация положений соответствует системе EU. В конкретных вариантах осуществления модифицированные антитело или фрагмент такого антитела содержат замену одной аминокислоты на цистеин в положении 360 тяжелой цепи антитела, где нумерация положений соответствует системе EU. В других конкретных вариантах осуществления модифицированные антитело или фрагмент такого антитела содержат замену одной аминокислоты на цистеин в положении 107 легкой цепи антитела, и где нумерация положений соответствует системе EU, и где легкая цепь представляет собой каппа-цепь.
[00351] В дополнительных вариантах осуществления антитела или фрагменты антител (например, антигенсвязывающий фрагмент), применимые в иммуноконъюгатах по настоящему изобретению, включают модифицированные или сконструированные антитела, такие как антитело, модифицированное для введения одной или нескольких других реакционноспособных аминокислот (отличных от цистеина), в том числе Pcl, пирролизина, пептидных меток (таких как метки S6, A1 и ybbR) и неприродных аминокислот, вместо по меньшей мере одной аминокислоты из нативной последовательности, обеспечивая за счет этого реакционноспособный сайт на антителе или антигенсвязывающем фрагменте для конъюгации с фрагментом, представляющим собой лекарственное средство, или фрагментом, представляющим собой линкер-лекарственное средство, с комплементарной реакционной группой. Например, антитела или фрагменты антител могут быть модифицированы для введения Pcl или пирролизина (W. Ou, et al., (2011) PNAS 108 (26), 10437-10442; WO2014124258) или неприродных аминокислот (J.Y. Axup, et al., Proc Natl Acad Sci U S A, 109 (2012), pp. 16101-16106; обзор см. в C.C. Liu and P.G. Schultz (2010) Annu Rev Biochem 79, 413-444; C.H. Kim, et al., (2013) Curr Opin Chem Biol. 17, 412-419) в качестве сайтов для конъюгации с лекарственным средством. Аналогичным образом в антитело могут быть введены пептидные метки для способов ферментативной конъюгации (Strop P., et al., Chem Biol. 2013, 20(2):161-7; Rabuka D., Curr Opin Chem Biol. 2010 Dec;14(6):790-6; Rabuka D, et al., Nat Protoc. 2012, 7(6):1052-67). Одним другим примером является применение 4'-фосфопантетеинилтрансфераз (PPTазы) для конъюгации аналогов кофермента A (WO2013184514) и (Grünewald et al., (2015) Bioconjugate Chem. 26 (12), 2554-62). Способы конъюгации таких модифицированных или сконструированных антител с полезными нагрузками или комбинациями линкер-полезная нагрузка известны из уровня техники.
[00352] В другом варианте осуществления для снижения биологического периода полужизни антитела подвергают мутации Fc-шарнирную область антитела. Более конкретно, одну или несколько аминокислотных мутаций вводят в пограничную область между доменами CH2-CH3 фрагмента Fc-шарнир, вследствие чего антитело характеризуется нарушенным связыванием с белком A стафилококка (SpA) по сравнению со связыванием нативного домена Fc-шарнир с SpA. Данный подход описан более подробно описан в патенте США № 6165745 от Ward et al.
[00353] В других вариантах осуществления область Fc изменяют путем замены по меньшей мере одного аминокислотного остатка на отличный аминокислотный остаток для изменения эффекторных функций антитела. Например, одну или несколько аминокислот можно заменить на отличный аминокислотный остаток, вследствие чего антитело характеризуется измененной аффинностью в отношении эффекторного лиганда, но сохраняет способность к связыванию антигена исходного антитела. Эффекторный лиганд, аффинность в отношении которого изменяют, может представлять собой, например, Fc-рецептор или компонент C1 комплемента. Данный подход описан, например, в патентах США №№ 5624821 и 5648260, оба от Winter et al.
[00354] В другом варианте осуществления одну или несколько аминокислот, выбранных из аминокислотных остатков, можно заменить на отличный аминокислотный остаток, вследствие чего антитело характеризуется измененным связыванием C1q и/или сниженной или устраненной комплементзависимой цитотоксичностью (CDC). Данный подход описан, например, в патенте США № 6194551 от Idusogie et al.
[00355] В другом варианте осуществления один или несколько аминокислотных остатков изменяют с тем, чтобы таким образом изменить способность антитела к фиксации комплемента. Данный подход описан, например, в PCT-публикации WO 94/29351 от Bodmer et al. Аллотипические аминокислотные остатки охватывают без ограничения константную область тяжелой цепи подклассов IgG1, IgG2 и IgG3, а также константную область легкой цепи изотипов каппа-цепи, как описано в Jefferis et al., MAbs. 1:332-338 (2009).
[00356] Комплексы на основе слитого белка с антителом, содержащим такие мутации, опосредуют сниженные антителозависимую клеточную цитотоксичность (ADCC) или комплементзависимую цитотоксичность (CDC) или их отсутствие. В некоторых вариантах осуществления аминокислотные остатки L234 и L235 в константной области IgG1 заменены на A234 и A235. В некоторых вариантах осуществления аминокислотный остаток N267 в константной области IgG1 заменен на A267. В некоторых вариантах осуществления аминокислотные остатки D265 и P329 в константной области IgG1 заменены на A265 и A329. Также можно использовать другие мутации, подавляющие Fc антитела.
[00357] В другом варианте осуществления один или несколько аминокислотных остатков изменяют с тем, чтобы таким образом изменить способность антитела к фиксации комплемента. Данный подход описан, например, в PCT-публикации WO 94/29351 от Bodmer et al. В конкретном варианте осуществления одна или несколько аминокислот антитела или его антигенсвязывающего фрагмента по настоящему изобретению заменены на один или несколько аллотипических аминокислотных остатков. Аллотипические аминокислотные остатки также включают без ограничения константную область тяжелой цепи подклассов IgG1, IgG2 и IgG3, а также константную область легкой цепи изотипа каппа-цепи, как это описано в Jefferis et al., MAbs. 1:332-338 (2009).
[00358] В еще одном варианте осуществления модифицируют характер гликозилирования антитела. Например, можно получать aгликозилированное антитело (т. е. антитело, у которого отсутствует гликозилирование). Характер гликозилирования может быть изменен, например, для повышения аффинности антитела в отношении "антигена". Такие модификации углеводов можно осуществлять, например, посредством изменения одного или нескольких сайтов гликозилирования в пределах последовательности антитела. Например, можно произвести одну или несколько аминокислотных замен, которые приводят к устранению одного или нескольких сайтов гликозилирования в каркасной области вариабельной области, тем самым устраняя гликозилирования в этом сайте. Такое агликозилирование может повышать аффинность антитела в отношении антигена. Такой подход описан, например, патентах США №№ 5714350 и 6350861 от Co et al.
[00359] В другом варианте осуществления антитело модифицируют для увеличения его биологического периода полужизни. Возможны различные подходы. Например, можно вводить одну или несколько из следующих мутаций: T252L, T254S, T256F, как описано в патенте США № 6277375 от Ward. В качестве альтернативы для увеличения биологического периода полужизни антитело можно изменять в пределах области CH1 или CL таким образом, чтобы оно содержало эпитоп для связывания с рецептором реутилизации, взятый из двух петель домена CH2 из области Fc IgG, как описано в патентах США №№ 5869046 и 6121022 от Presta et al.
3. Получение антител к PMEL17
[00360] Антитела к PMEL17 и фрагменты таких антител (например, антигенсвязывающие фрагменты) могут быть получены с помощью любых средств, известных из уровня техники, в том числе без ограничения посредством рекомбинантной экспрессии, химического синтеза и ферментативного расщепления тетрамеров антител, при этом полноразмерные моноклональные антитела можно получать, например, с использованием гибридом или посредством рекомбинантного продуцирования. Рекомбинантная экспрессия может осуществляться в любых подходящих клетках-хозяевах, известных из уровня техники, например, в клетках-хозяевах, представляющих собой клетки млекопитающих, бактериальных клетках-хозяевах, дрожжевых клетках-хозяевах, клетках-хозяевах, представляющих собой клетки насекомых, и т. д.
[00361] В настоящем изобретении дополнительно представлены полинуклеотиды, кодирующие антитела, описанные в данном документе, например, полинуклеотиды, кодирующие вариабельные области или сегменты тяжелой или легкой цепей, содержащие определяющие комплементарность области, описанные в данном документе. В некоторых вариантах осуществления полинуклеотид, кодирующий вариабельные области тяжелой цепи, характеризуется по меньшей мере 85%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, или 100% идентичностью последовательности нуклеиновой кислоты с полинуклеотидом, выбранным из группы, состоящей из SEQ ID NOs: 11, 43, 65, 89, 113, 133, 150, 166, 185, 197, 216, 228, 240, и 255. В некоторых вариантах осуществления полинуклеотид, кодирующий вариабельные области легкой цепи, характеризуется по меньшей мере 85%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, или 100% идентичностью последовательности нуклеиновой кислоты с полинуклеотидом, выбранным из группы, состоящей из SEQ ID NOs: 22, 26, 30, 54, 76, 100, 120, 144, 160, 172, 191, 203, 222, 234, 249, и 261.
[00362] В некоторых вариантах осуществления полинуклеотид, кодирующий тяжелую цепь, характеризуется по меньшей мере 85%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, или 100% идентичностью последовательности нуклеиновой кислоты с полинуклеотидом под SEQ ID NO: 13, 45, 67, 91, 115, 135, 152, 168, 187, 199, 218, 230, 242, и 257. В некоторых вариантах осуществления полинуклеотид, кодирующий легкую цепь, характеризуется по меньшей мере 85%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, или 100% идентичностью последовательности нуклеиновой кислоты с полинуклеотидом под SEQ ID NO: 24, 28, 32, 56, 78, 102, 122, 146, 162, 174, 193, 205, 224, 236, 251, и 263.
[00363] Полинуклеотиды по настоящему изобретению могут кодировать только последовательность вариабельной области антитела к PMEL17. Они также могут кодировать как вариабельную область, так и константную область антитела. Некоторые из полинуклеотидных последовательностей кодируют полипептид, который содержит вариабельные области как тяжелой цепи, так и легкой цепи одного из иллюстративных мышиных антител к PMEL17. Некоторые другие полинуклеотиды кодируют два полипептидных сегмента, которые в значительной степени идентичны вариабельным областям тяжелой цепи и легкой цепи, соответственно, одного из мышиных антител.
[00364] Полинуклеотидные последовательности можно получать посредством твердофазного синтеза ДНК de novo или посредством ПЦР-мутагенеза существующей последовательности (например, последовательностей, которые описаны в разделе "Примеры" ниже), кодирующей антитело к PMEL17 или его связывающий фрагмент. Прямой химический синтез нуклеиновых кислот можно осуществлять с помощью способов, известных из уровня техники, таких как фосфотриэфирный способ из Narang et al., Meth. Enzymol. 68:90, 1979; фосфодиэфирный способ из Brown et al., Meth. Enzymol. 68:109, 1979; диэтилфосфорамидитный способ из Beaucage et al., Tetra Lett., 22:1859, 1981 и способ с использованием твердой подложки из патента США № 4458066. Введение мутаций в полинуклеотидную последовательность посредством ПЦР можно осуществлять так, как это описано, например, в PCR Technology: Principles and Applications for DNA Amplification, H.A. Erlich (Ed.), Freeman Press, NY, NY, 1992; PCR Protocols: A Guide to Methods and Applications, Innis et al. (Ed.), Academic Press, San Diego, CA, 1990; Mattila et al., Nucleic Acids Res. 19:967, 1991 и Eckert et al., PCR Methods and Applications 1:17, 1991.
[00365] Также в настоящем изобретении представлены векторы экспрессии и клетки-хозяева для продуцирования описанных выше антител к PMEL17. Различные векторы экспрессии можно использовать для экспрессии полинуклеотидов, кодирующих цепи или связывающие фрагменты антитела к PMEL17. Как вирусные, так и невирусные векторы экспрессии можно применять для продуцирования антител в клетке-хозяине, представляющей собой клетку млекопитающего. Невирусные векторы и системы включают плазмиды, эписомные векторы, как правило, с кассетой экспрессии для экспрессии белка или РНК, а также искусственные человеческие хромосомы (см., например, Harrington et al., Nat Genet. 15:345, 1997). Например, невирусные векторы, применимые для экспрессии полинуклеотидов и полипептидов, связывающих PMEL17, в клетках млекопитающих (например, человеческих), включают pThioHis A, B и C, pcDNA™ 3.1/His, pEBVHis A, B и C (Invitrogen, Сан-Диего, Калифорния), векторы MPSV и ряд других векторов, известных из уровня техники экспрессии других белков. Применимые вирусные векторы включают векторы на основе ретровирусов, аденовирусов, аденоассоциированных вирусов, вирусов герпеса, векторы на основе SV40, папилломавируса, вируса Эпштейна-Барр HBP, векторы на основе вируса коровьей оспы и вируса леса Семлики (SFV). См. выше Brent et al.; Smith, Annu. Rev. Microbiol. 49:807, 1995 и Rosenfeld et al., Cell 68:143, 1992.
[00366] Выбор вектора экспрессии зависит от предполагаемых клеток-хозяев, в которых следует экспрессировать вектор. Как правило, векторы экспрессии содержат промотор и другие регуляторные последовательности (например, энхансеры), которые функционально связаны с полинуклеотидами, кодирующими цепь или фрагмент антитела к PMEL17. В некоторых вариантах осуществления индуцируемый промотор используют для предотвращения экспрессии встроенных последовательностей в условиях, отличающихся от индуцирующих. Индуцируемые промоторы включают, например, арабинозный, lacZ, металлотионеиновый промотор или промотор белка теплового шока. Культуры трансформированных организмов можно размножать в неиндуцирующих условиях без смещения популяции в направлении кодирующих последовательностей, продукты экспрессии которых лучше переносятся клетками-хозяевами. Для эффективной экспрессии цепи или фрагмента антитела к PMEL17 кроме промоторов также могут быть необходимы или потребоваться другие регуляторные элементы. Такие элементы, как правило, включают кодон инициации трансляции ATG и смежный сайт связывания рибосомы или другие последовательности. Кроме того, эффективность экспрессии можно повысить за счет включения энхансеров, соответствующих применяемой клеточной системе (см., например, Scharf et al., Results Probl. Cell Differ. 20:125, 1994 и Bittner et al., Meth. Enzymol., 153:516, 1987). Например, для повышения экспрессии в клетках-хозяевах млекопитающих можно использовать энхансер SV40 или энхансер CMV.
[00367] В векторах экспрессии также может быть предусмотрено положение сигнальной последовательности секреции для образования слитого белка с полипептидами, кодируемыми вставленными последовательностями антитела к PMEL17. Чаще вставляемые последовательности антитела к PMEL17 соединяют с сигнальными последовательностями до включения в вектор. Векторы, которые должны использоваться для получения последовательностей, кодирующих вариабельные домены легкой и тяжелой цепей антитела к PMEL17, иногда также кодируют константные области или их части. Такие векторы обеспечивают возможность экспрессии вариабельных областей в виде слитых белков с константными областями, в результате чего получают интактные антитела или их фрагменты. Как правило, такие константные области являются человеческими.
[00368] Клетки-хозяева, несущие и экспрессирующие цепи антитела к PMEL17, могут быть либо прокариотическими, либо эукариотическими. E. coli является одним прокариотическим хозяином, применимым для клонирования и экспрессии полинуклеотидов по настоящему изобретению. Другие подходящие для применения микробные хозяева включают бацилл, таких как Bacillus subtilis, и других энтеробактерий, таких как Salmonella, Serratia, и различные виды Pseudomonas. Для этих прокариотических хозяев также можно создать векторы экспрессии, которые, как правило, содержат последовательности для управления экспрессией, совместимые с клеткой-хозяином (например, точку начала репликации). Кроме того, будет присутствовать любое число разнообразных хорошо известных промоторов, как, например, лактозная промоторная система, триптофановая (trp) промоторная система, бета-лактамазная промоторная система или промоторная система из фага лямбда. Как правило, промоторы контролируют экспрессию, необязательно с помощью операторной последовательности, и имеют последовательности сайта связывания рибосомы и т. п. для инициации и завершения транскрипции и трансляции. Для экспрессии полипептидов, связывающих PMEL17, по настоящему изобретению также можно использовать другие микроорганизмы, такие как дрожжи. Также можно применять клетки насекомых в сочетании с бакуловирусными векторами.
[00369] В некоторых предпочтительных вариантах осуществления для экспрессии и продуцирования полипептидов, связывающих PMEL17, по настоящему изобретению используют клетки-хозяева, представляющие собой клетки млекопитающих. Например, они могут представлять собой либо линию клеток гибридомы, экспрессирующих эндогенные гены иммуноглобулинов (например, клоны гибридомы из клеток миеломы, как описано в разделе "Примеры"), либо линию клеток млекопитающих, несущих экзогенный вектор экспрессии (например, клетки миеломы SP2/0, приведенные в качестве примера ниже). Они охватывают любые нормальные мортальные или нормальные или аномальные иммортальные клетки животных или человека. Например, был разработан ряд подходящих линий клеток-хозяев, способных секретировать интактные иммуноглобулины, в том числе линии клеток CHO, различные линии клеток Cos, клетки HeLa, линии клеток миеломы, трансформированные B-клетки и гибридомы. Применение тканевой культуры клеток млекопитающих для экспрессии полипептидов в общих чертах обсуждается, например, в Winnacker, From Genes to Clones, VCH Publishers, N.Y., N.Y., 1987. Векторы экспрессии для клеток-хозяев, представляющих собой клетки млекопитающих, могут включать последовательности для управления экспрессией, такие как точка начала репликации, промотор и энхансер (см., например, Queen et al., Immunol. Rev. 89:49-68, 1986), и необходимые сайты, несущие информацию для процессинга, такие как сайты связывания рибосом, сайты сплайсинга РНК, сайты полиаденилирования и последовательности терминатора транскрипции. Такие векторы экспрессии обычно содержат промоторы, происходящие из генов млекопитающих или из вирусов млекопитающих. Подходящие промоторы могут быть конститутивными, специфическими в отношении типа клеток, специфическими в отношении стадии развития и/или модулируемыми или регулируемыми. Применимые промоторы включают без ограничения металлотионеиновый промотор, конститутивный большой поздний промотор аденовируса, дексаметазон-индуцируемый промотор MMTV, промотор SV40, промотор polIII MRP, конститутивный промотор MPSV, тетрациклин-индуцируемый промотор CMV (такой как немедленно-ранний промотор CMV человека), конститутивный промотор CMV и комбинации промотор-энхансер, известные из уровня техники.
[00370] Способы введения векторов экспрессии, содержащих представляющие интерес полинуклеотидные последовательности, варьируют в зависимости от типа клетки-хозяина. Например, трансфекцию с использованием хлорида кальция обычно используют для прокариотических клеток, тогда как обработку фосфатом кальция или электропорацию можно применять для других клеток-хозяев (см. выше в общих чертах Sambrook et al.). A Laboratory Manual, 4th ed.). Другие способы включают, например, электропорацию, обработку фосфатом кальция, трансформацию, опосредованную липосомами, инъекцию и микроинъекцию, баллистические способы, виросомы, иммунолипосомы, конъюгаты поликатион:нуклеиновая кислота, депротеинизированную ДНК, искусственные вирионы, слияние со структурным белком VP22 вируса герпеса (Elliot and O'Hare, Cell 88:223, 1997), усиленное средством поглощение ДНК и трансдукцию ex vivo. Чтобы обеспечить длительное получение рекомбинантных белков с высоким выходом, зачастую будет требоваться стабильная экспрессия. Например, линии клеток, которые стабильно экспрессируют цепи или связывающие фрагменты антитела к PMEL17, можно получить с использованием векторов экспрессии по настоящему изобретению, которые содержат вирусные точки начала репликации или эндогенные экспрессионные элементы и ген селектируемого маркера. После введения вектора клетки можно оставить расти в течение 1-2 дней в обогащенной среде перед их переносом на селективную среду. Задачей селектируемого маркера является придание устойчивости к факторам отбора, и его присутствие обеспечивает возможность роста клеток, которые успешно экспрессируют введенные последовательности, в селективных средах. Пролиферацию устойчивых стабильно трансфицированных клеток можно обеспечивать с применением методик тканевой культуры, соответствующих типу клеток.
Варианты применения в терапии
[00371] Антитела, фрагменты антител (например, антигенсвязывающие фрагменты) и конъюгаты антитела и лекарственного средства по настоящему изобретению применимы в виде разнообразных вариантов применения, в том числе без ограничения лечения или предупреждения рака, такого как типы солидного рака или гемобластозы. В некоторых вариантах осуществления антитела, фрагменты антител (например, антигенсвязывающие фрагменты) и конъюгаты антитела и лекарственного средства по настоящему изобретению применимы для ингибирования роста опухоли, индуцирования дифференцировки, уменьшения объема опухоли и/или снижения туморогенности опухоли. Способы применения могут представлять собой in vitro, ex vivo или in vivo способы.
[00372] В одном аспекте антитела, фрагменты антител (например, антигенсвязывающие фрагменты) и конъюгаты антитела и лекарственного средства по настоящему изобретению применимы для выявления присутствия PMEL17 в биологическом образце. Используемый в данном документе термин "осуществление обнаружения" охватывает количественное или качественное обнаружение. В некоторых вариантах осуществления биологический образец содержит клетку или ткань. В определенных вариантах осуществления такие ткани включают нормальные и/или раковые ткани, которые экспрессируют PMEL17 на более высоких уровнях по сравнению с другими тканями.
[00373] В одном аспекте настоящего изобретения представлен способ обнаружения присутствия PMEL17 в биологическом образце. В определенных вариантах осуществления способ предусматривает приведение биологического образца в контакт с антителом к PMEL17 в условиях, допускающих связывание антитела с антигеном, и выявление того, формируется ли комплекс между антителом и антигеном.
[00374] В одном аспекте настоящего изобретения представлен способ диагностики нарушения, ассоциированного с повышенной экспрессией PMEL17. В определенных вариантах осуществления способ предусматривает приведение тестируемой клетки в контакт с антителом к PMEL17; определение уровня экспрессии (или количественно, или качественно) PMEL17 на тестируемой клетке путем обнаружения связывания антитела к PMEL17 с антигеном PMEL17 и сравнение уровня экспрессии PMEL17 в тестируемой клетке с уровнем экспрессии PMEL17 на контрольной клетке (например, нормальной клетке, происходящей из той же ткани, что и тестируемая клетка, или на клетке, которая экспрессирует PMEL17 на уровнях, сравнимых с такими уровнями у нормальной клетки), где более высокий уровень экспрессии PMEL17 на тестируемой клетке по сравнению с контрольной клеткой указывает на присутствие нарушения, ассоциированного с повышенной экспрессией PMEL17. В определенных вариантах осуществления тестируемую клетку получают от индивидуума с подозрением на наличие нарушения, ассоциированного с повышенной экспрессией PMEL17. В некоторых вариантах осуществления нарушение представляет собой нарушение пролиферации клеток, такое как рак или опухоль. В определенных вариантах осуществления способ предусматривает измерение числа копий гена PMEL17 в тестируемой клетке.
[00375] В определенных вариантах осуществления способ диагностики или обнаружения, такой как описываемые выше, предусматривает обнаружение связывания антитела к PMEL17 с PMEL17, экспрессируемым на поверхности клетки или в препарате мембраны, полученном из клетки, экспрессирующей PMEL17 на своей поверхности. Иллюстративным анализом для обнаружения связывания антитела к PMEL17 с PMEL17, экспрессируемым на поверхности клетки, является "FACS-анализ".
[00376] Для обнаружения связывания антитела к PMEL17 с PMEL17 могут применяться некоторые другие способы. Такие способы включают без ограничения анализы связывания антигена, которые широко известны из уровня техники, такие как вестерн-блоттинг, радиоиммунологические анализы, ELISA (твердофазный иммуноферментный анализ), иммунологические "сэндвич"-анализы, анализы иммунопреципитации, флуоресцентные иммунологические анализы, иммунологические анализы с белком A и иммуногистохимическое исследование (IHC).
[00377] В определенных вариантах осуществления антитела к PMEL17 являются мечеными. Метки включают без ограничения метки или фрагменты, которые обнаруживают непосредственно (такие как флуоресцентные, хромофорные, электронноплотные, хемилюминесцентные и радиоактивные метки), а также фрагменты, такие как ферменты или лиганды, которые обнаруживают опосредованно, например, за счет ферментативной реакции или молекулярного взаимодействия.
[00378] В определенных вариантах осуществления антитела к PMEL17 являются иммобилизированными на нерастворимой матрице. Иммобилизация приводит к отделению антитела к PMEL17 от любого белка PMEL17, который остается свободным в растворе. Традиционно это осуществляют или посредством перевода антитела к PMEL17 в нерастворимую форму перед процедурой анализа в результате адсорбции на водонерастворимой матрице или поверхности (Bennich et al, патент США № 3720760), или соединения ковалентной связью (например, с использованием сшивания глутаральдегидом), или посредством перевода антитела к PMEL17 в нерастворимую форму после образования комплекса между антителом к PMEL17 и белком PMEL17, например, посредством иммунного осаждения.
[00379] Любой из вышеизложенных вариантов осуществления диагностики или обнаружения может проводиться с использованием иммуноконъюгата по настоящему изобретению вместо антитела к PMEL17 или в дополнение к нему.
[00380] В одном варианте осуществления настоящего изобретения представлен способ лечения или предупреждения заболевания, предусматривающий введение пациенту антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства по настоящему изобретению. В настоящем изобретении также представлено применение антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства по настоящему изобретению для лечения или предупреждения заболевания у пациента. В некоторых вариантах осуществления настоящего изобретения представлены антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или конъюгаты антитела и лекарственного средства по настоящему изобретению для применения в лечении или предупреждении заболевания у пациента. В дополнительных вариантах осуществления настоящего изобретения представлено применение антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства по настоящему изобретению в изготовлении лекарственного препарата, предназначенного для лечения или предупреждения заболевания у пациента.
[00381] В некоторых вариантах осуществления заболевание, которое лечат с использованием антител, фрагментов антител (например, антигенсвязывающих фрагментов) и конъюгатов антитела и лекарственного средства по настоящему изобретению, представляет собой рак. В определенных вариантах осуществления рак характеризуется наличием клеток, экспрессирующих PMEL17, с которыми связываются антитела, фрагменты антител (например, антигенсвязывающие фрагменты) и конъюгаты антитела и лекарственного средства по настоящему изобретению. В определенных вариантах осуществления рак характеризуется повышением экспрессии PMEL17 по сравнению с уровнем экспрессии у здорового пациента. В некоторых вариантах осуществления экспрессия PMEL17 может быть измерена по повышению РНК PMEL17. В других вариантах осуществления рак характеризуется повышением числа копий ДНК PMEL17. Специалистам в данной области техники известны другие способы измерения или определения уровней экспрессии PMEL17. В определенных вариантах осуществления рак характеризуется мутацией, например активирующей мутацией, затрагивающей Q209 или R183, в гене GNAQ и/или GNA11. Примеры заболеваний, которые можно лечить и/или предупреждать, включают без ограничения меланому, увеальную меланому, гепатоцеллюлярную карциному и метастатическую форму данных видов рака.
[00382] В настоящем изобретении представлены способы лечения или предупреждения рака, предусматривающие введение терапевтически эффективного количества антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства по настоящему изобретению. В некоторых вариантах осуществления рак представляет собой солидный рак, такой как меланома, увеальная меланома, гепатоцеллюлярная карцинома или метастатическая форма данных видов рака. В некоторых вариантах осуществления субъектом является человек. В некоторых вариантах осуществления рак представляет собой устойчивый рак и/или рецидивирующий рак.
[00383] В некоторых вариантах осуществления настоящего изобретения представлены способы ингибирования роста опухоли, предусматривающие введение субъекту терапевтически эффективного количества антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства по настоящему изобретению. В некоторых вариантах осуществления опухоль представляет собой солидный рак, такой как меланома, увеальная меланома, гепатоцеллюлярная карцинома или метастатическая форма данных видов рака. В некоторых вариантах осуществления субъектом является человек. В некоторых вариантах осуществления субъект имеет опухоль или у него была удалена опухоль.
[00384] В некоторых вариантах осуществления опухоль экспрессирует PMEL17, с которым связываются антитела к PMEL17. В некоторых вариантах осуществления опухоль сверхэкспрессирует PMEL17 человека. В некоторых вариантах осуществления опухоль характеризуется повышенным числом копий гена PMEL17. В некоторых вариантах осуществления опухоль характеризуется мутацией, например активирующей мутацией, затрагивающей Q209 или R183, в гене GNAQ и/или GNA11.
[00385] В настоящем изобретении также представлены способы отбора пациентов для лечения антителами, фрагментами антител (например, антигенсвязывающими фрагментами) или конъюгатами антитела и лекарственного средства по настоящему изобретению, предусматривающие введение терапевтически эффективного количества указанных антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства. В определенных аспектах настоящего изобретения способы предусматривают отбор пациента путем измерения экспрессии PMEL17. В определенных аспектах настоящего изобретения способы предусматривают отбор пациента путем идентификации мутации, например активирующей мутации, затрагивающей Q209 или R183, в гене GNAQ или GNA11. В некоторых вариантах осуществления способы предусматривают измерение уровня экспрессии PMEL17 у пациента, а также определение гена GNAQ и/или GNA11.
[00386] В случае лечения или предупреждения заболеваний подходящая доза антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства по настоящему изобретению зависит от различных факторов, таких как тип заболевания, подлежащего лечению, тяжесть и течение заболевания, ответная реакция заболевания, предыдущая терапия, клинический анамнез пациента и т. д. Антитело или средство могут вводиться один раз или на протяжении курса лечения, длящегося от нескольких дней до нескольких месяцев, или до осуществления излечения, или до достижения ослабления болезненного состояния (например, уменьшения размера опухоли). Оптимальные схемы приема можно рассчитать на основании измерений накопления лекарственного средства в организме пациента, и они будут варьироваться в зависимости от относительной активности индивидуальных антитела, фрагментов антитела (например, антигенсвязывающего фрагмента) или конъюгатов антитела и лекарственного средства. Лечащий врач может оценить частоту повторения введения дозы на основании измеренного времени удержания и концентраций лекарственного средства в физиологических жидкостях или тканях.
Комбинированная терапия
[00387] В определенных случаях антитело, фрагмент антитела (например, антигенсвязывающий фрагмент) или конъюгат антитела и лекарственного средства по настоящему изобретению объединяют с другими методами терапии, такими как хирургия и лучевая терапия, терапевтическими средствами, такими как другие противораковые средства, противоаллергические средства, средства против тошноты (или противорвотные средства), обезболивающие средства, цитопротекторные средства и их комбинации.
[00388] В одном варианте осуществления антитело, фрагмент антитела (например, антигенсвязывающий фрагмент) или конъюгат антитела и лекарственного средства по настоящему изобретению объединяют в фармацевтическом комбинированном составе со вторым соединением, характеризующимся противораковыми свойствами, или схема введения доз предусматривает комбинированную терапию с ним. Второе соединение фармацевтического комбинированного состава или схемы введения доз может характеризоваться видами активности, которые взаимно дополняют антитело или иммуноконъюгат из комбинации, вследствие чего они не оказывают неблагоприятного эффекта друг на друга. Например, антитело, фрагмент антитела (например, антигенсвязывающий фрагмент) или конъюгат антитела и лекарственного средства по настоящему изобретению могут вводиться в комбинации без ограничения с химиотерапевтическим средством, иммуномодулирующими средствами, ингибитором тирозинкиназы, ингибитором пути передачи нисходящего сигнала с участием GNAQ/GNA11, ингибиторами IAP, ингибиторами Bcl2, ингибиторами Mcl1 и другими ингибиторами GNAQ/GNA11.
[00389] Используемый в данном документе термин "фармацевтическая комбинация" относится либо к фиксированной комбинации в форме единицы дозирования, либо к нефиксированной комбинации или набору из частей для комбинированного введения, где два или более терапевтических средств могут вводиться независимо в одно и то же время или по отдельности в пределах временных интервалов, в частности, когда такие временные интервалы обеспечивают возможность демонстрации кооперативного, например синергетического, эффекта партнеров по комбинации.
[00390] Термин "комбинированная терапия" относится к введению двух или более терапевтических средств для лечения или предупреждения терапевтического состояния или нарушения, описанного в настоящем изобретении. Такое введение охватывает совместное введение этих терапевтических средств практически одновременно, например, в одной капсуле, имеющей фиксированное соотношение активных ингредиентов. В качестве альтернативы такое введение охватывает совместное введение в виде нескольких или отдельных контейнеров (например, капсулы, порошки и жидкости) для каждого активного ингредиента. Порошки и/или жидкости могут быть восстановлены или разбавлены до требуемой дозы перед введением. Кроме того, такое введение также охватывает применение каждого типа терапевтического средства последовательным образом, либо приблизительно в одно и то же время, либо в разное время. В любом случае схема лечения будет обеспечивать приносящие пользу эффекты комбинации лекарственных средств при лечении или предупреждении состояний или нарушений, описанных в данном документе.
[00391] Комбинированная терапия может обеспечивать "синергию" и оказаться "синергетической", т. e. эффект, достигаемый, когда активные ингредиенты применяются вместе, превышает сумму эффектов, которые являются результатом применения соединений по отдельности. Синергетический эффект может быть достигнут, когда активные ингредиенты: (1) составлены вместе и вводятся или доставляются одновременно в виде комбинированного состава с однократной дозой; (2) доставляются по очереди или параллельно как отдельные составы или (3) применяется какая-либо другая схема. При доставке в виде терапии с чередованием синергетический эффект может достигаться, когда соединения вводят или доставляют одно за другим, например, посредством различных инъекций в отдельных шприцах. В целом, во время терапии с чередованием эффективную дозу каждого активного ингредиента вводят одну за другой, т. е. сериями, в то время как при комбинированной терапии эффективную дозу двух или более активных ингредиентов вводят вместе.
[00392] Стандартные химиотерапевтические средства, рассматриваемые для применения в вариантах комбинированной терапии, включают анастрозол (Arimidex®), бикалутамид (Casodex®), блеомицина сульфат (Blenoxane®), бусульфан (Myleran®), инъекционный бусульфан (Busulfex®), капецитабин (Xeloda®), N4-пентоксикарбонил-5-дезокси-5-фторцитидин, карбоплатин (Paraplatin®), кармустин (BiCNU®), хлорамбуцил (Leukeran®), цисплатин (Platinol®), кладрибин (Leustatin®), циклофосфамид (Cytoxan® или Neosar®), цитарабин, цитозина арабинозид (Cytosar-U®), цитарабин в форме липосом для инъекций (DepoCyt®), дакарбазин (DTIC-Dome®), дактиномицин (актиномицин D, Cosmegan), даунорубицина гидрохлорид (Cerubidine®), даунорубицина цитрат в форме липосом для инъекций (DaunoXome®), дексаметазон, доцетаксел (Taxotere®), доксорубицина гидрохлорид (Adriamycin®, Rubex®), этопозид (Vepesid®), флударабина фосфат (Fludara®), 5-фторурацил (Adrucil®, Efudex®), флутамид (Eulexin®), тезацитибин, гемцитабин (дифтордезоксицитидин), гидроксимочевину (Hydrea®), идарубицин (Idamycin®), ифосфамид (IFEX®), иринотекан (Camptosar®), L-аспарагиназу (ELSPAR®), лейковорин кальция, мелфалан (Alkeran®), 6-меркаптопурин (Purinethol®), метотрексат (Folex®), митоксантрон (Novantrone®), милотарг, паклитаксел (Taxol®), феникс (иттрий 90/MX-DTPA), пентостатин, полифероспан 20 с имплантатом кармустина (Gliadel®), тамоксифена цитрат (Nolvadex®), тенипозид (Vumon®), 6-тиогуанин, тиотепу, тирапазамин (Tirazone®), топотекана гидрохлорид для инъекций (Hycamptin®), винбластин (Velban®), винкристин (Oncovin®) и винорелбин (Navelbine®), а также пеметрексед.
[00393] В одном аспекте настоящего изобретения представлен способ лечения или предупреждения рака путем введения нуждающемуся в этом субъекту конъюгата антитела и лекарственного средства по настоящему изобретению в комбинации с одним или несколькими ингибиторами MDM2, ингибиторами PKC, ингибиторами PRC2, ингибиторами MAPK, ингибиторами GPCR, ингибиторами тирозинкиназы, в том числе без ограничения ингибиторами BTK, ингибиторами EGFR, ингибиторами Her2, ингибиторами Her3, ингибиторами IGFR и ингибиторами Met.
[00394] Например, ингибиторы MDM2 включают без ограничения RG7112 (RO5045337); RG7388 (RO5503781, идазанутлин); MI-77301 (SAR405838); MK-8242 (SCH-900242); AMG232; CGM097; DS3032b; HDM201 и ALRN-6924.
[00395] Например, ингибиторы PKC включают без ограничения баланол; рилузол; стауроспорин; энзастаурин; δV1-1 (KAI-9803 или делкасертиб); εV1-2 (KAI-1678); апринокарсен; мидостаурин (PKC412); UCN-01 (7-гидрокси-стауроспорин); роттлерин (5,7-дигидрокси-2,2-диметил-6-(2,4,6-тригидрокси-3-метил-5-ацетилбензил)-8-циннамоил-1,2-хромен) и бриостатин 1.
[00396] Например, ингибиторы PRC2 включают без ограничения EI1; EPZ011989; EPZ005687; тетраметилпиперидинилбензамиды; UNC1999 и GSK126.
[00397] Например, ингибиторы MAPK включают без ограничения вемурафениб (Zelboraf); дабрафениб (Tafinlar); энкорафениб (Braftovi); траметиниб (Mekinist); кобиметиниб (Cotellic); биниметиниб (Mektovi) и уликсертиниб.
[00398] Например, ингибиторы тирозинкиназы включают без ограничения ибрутиниб (PCI-32765); эрлотиниба гидрохлорид (Tarceva®); линифаниб (N-[4-(3-амино-1H-индазол-4-ил)фенил]-N'-(2-фтор-5-метилфенил)мочевина, также известный как ABT 869, доступный от Genentech); сунитиниба малат (Sutent®); босутиниб (4-[(2,4-дихлор-5-метоксифенил)амино]-6-метокси-7-[3-(4-метилпиперазин-1-ил)пропокси]хинолин-3-карбонитрил, также известный как SKI-606 и описанный в патенте США № 6780996); дазатиниб (Sprycel®); пазопаниб (Votrient®); сорафениб (Nexavar®); зактима (ZD6474) и иматиниб или иматиниба мезилат (Gilvec® и Gleevec®).
[00399] Ингибиторы рецептора эпидермального фактора роста рецептор (EGFR) ингибиторы включают без ограничения, эрлотиниба гидрохлорид (Tarceva®), гефитиниб (Iressa®); N-[4-[(3-хлор-4-фторфенил)амино]-7-[[(3''S'')-тетрагидро-3-фуранил]окси]-6-хиназолинил]-4(диметиламино)-2-бутенамид, Tovok®); вандетаниб (Caprelsa®); лапатиниб (Tykerb®); (3R,4R)-4-амино-1-((4-((3-метоксифенил)амино)пирроло[2,1-f][1,2,4]триазин-5-ил)метил)пиперидин-3-ол (BMS690514); канертиниба дигидрохлорид (CI-1033); 6-[4-[(4-этил-1-пиперазинил)метил]фенил]-N-[(1R)-1-фенилэтил]- 7H-пирроло[2,3-d]пиримидин-4-амин (AEE788, CAS 497839-62-0); мубритиниб (TAK165); пелитиниб (EKB569); афатиниб (BIBW2992); нератиниб (HKI-272); N-[4-[[1-[(3-фторфенил)метил]-1H-индазол-5-ил]амино]-5-метилпирроло[2,1-f][1,2,4]триазин-6-ил]-карбаминовую кислоту, (3S)-3-морфолинилметиловый сложный эфир (BMS599626); N-(3,4-дихлор-2-фторфенил)-6-метокси-7-[[(3aα,5β,6aα)-октагидро-2-метилциклопента[c]пиррол-5-ил]метокси]-4-хиназолинамин (XL647, CAS 781613-23-8) и 4-[4-[[(1R)-1-фенилэтил]амино]-7H-пирроло[2,3-d]пиримидин-6-ил]-фенол (PKI166, CAS 187724-61-4).
[00400] Антитела, связывающие EGFR, включают без ограничения, цетуксимаб (Erbitux®); панитумумаб (Vectibix®); матузумаб (EMD-72000); нимотузумаб (hR3); залутумумаб; TheraCIM h-R3; MDX0447 (CAS 339151-96-1) и ch806 (mAb-806, CAS 946414-09-1).
[00401] Ингибиторы рецептора эпидермального фактора роста 2 (рецептора Her2) человека (также известного как Neu, ErbB-2, CD340 или p185) включают без ограничения трастузумаб (Herceptin®); пертузумаб (Omnitarg®); трастузумаб эмтанзин (Kadcyla®); нератиниб (HKI-272, (2E)-N-[4-[[3-хлор-4-[(пиридин-2-ил)метокси]фенил]амино]-3-циано-7-этоксихинолин-6-ил]-4-(диметиламино)бут-2-енамид и описанный в PCT-публикации № WO 05/028443); лапатиниб или лапатиниба дитозилат (Tykerb®); (3R,4R)-4-амино-1-((4-((3-метоксифенил)амино)пирроло[2,1-f][1,2,4]триазин-5-ил)метил)пиперидин-3-ол (BMS690514); (2E)-N-[4-[(3-хлор-4-фторфенил)амино]-7-[[(3S)-тетрагидро-3-фуранил]окси]-6-хиназолинил]-4-(диметиламино)-2-бутенамид (BIBW-2992, CAS 850140-72-6); N-[4-[[1-[(3-фторфенил)метил]-1H-индазол-5-ил]амино]-5-метилпирроло[2,1-f][1,2,4]триазин-6-ил]-карбаминовую кислоту, (3S)-3-морфолинилметиловый сложный эфир (BMS 599626, CAS 714971-09-2); канертиниба дигидрохлорид (PD183805 или CI-1033) и N-(3,4-дихлор-2-фторфенил)-6-метокси-7-[[(3aα,5β,6aα)-октагидро-2-метилциклопента[c]пиррол-5-ил]метокси]-4-хиназолинамин (XL647, CAS 781613-23-8).
[00402] Ингибиторы Her3 включают без ограничения LJM716, MM-121, AMG-888, RG7116, REGN-1400, AV-203, MP-RM-1, MM-111 и MEHD-7945A.
[00403] Ингибиторы Met включают без ограничения кабозатиниб (XL184, CAS 849217-68-1); форетиниб (GSK1363089, ранее XL880, CAS 849217-64-7); тивантиниб (ARQ197, CAS 1000873-98-2); 1-(2-гидрокси-2-метилпропил)-N-(5-(7-метоксихинолин-4-илокси)пиридин-2-ил)-5-метил-3-оксо-2-фенил-2,3-дигидро-1H-пиразол-4-карбоксамид (AMG 458); кризотиниб (Xalkori®, PF-02341066); (3Z)-5-(2,3-дигидро-1H-индол-1-илсульфонил)-3-({3,5-диметил-4-[(4-метилпиперазин-1-ил)карбонил]-1H-пиррол-2-ил}метилен)-1,3-дигидро-2H-индол-2-он (SU11271); (3Z)-N-(3-хлорфенил)-3-({3,5-диметил-4-[(4-метилпиперазин-1-ил)карбонил]-1H-пиррол-2-ил}метилен)-N-метил-2-оксоиндолин-5-сульфонамид (SU11274); (3Z)-N-(3-хлорфенил)-3-{[3,5-диметил-4-(3-морфолин-4-илпропил)-1H-пиррол-2-ил]метилен}-N-метил-2-оксоиндолин-5-сульфонамид (SU11606); 6-[дифтор[6-(1-метил-1H-пиразол-4-ил)-1,2,4-триазоло[4,3-b]пиридазин-3-ил]метил]-хинолин (JNJ38877605, CAS 943540-75-8); 2-[4-[1-(хинолин-6-илметил)-1H-[1,2,3]триазоло[4,5-b]пиразин-6-ил]-1H-пиразол-1-ил]этанол (PF04217903, CAS 956905-27-4); N-((2R)-1,4-диоксан-2-илметил)-N-метил-N'-[3-(1-метил-1H-пиразол-4-ил)-5-оксо-5H-бензо[4,5]циклогепта[1,2-b]пиридин-7-ил]сульфамид (MK2461, CAS 917879-39-1); 6-[[6-(1-метил-1H-пиразол-4-ил)-1,2,4-триазоло[4,3-b]пиридазин-3-ил]тио]-хинолин (SGX523, CAS 1022150-57-7) и (3Z)-5-[[(2,6-дихлорфенил)метил]сульфонил]-3-[[3,5-диметил-4-[[(2R)-2-(1-пирролидинилметил)-1-пирролидинил]карбонил]-1H-пиррол-2-ил]метилен]-1,3-дигидро-2H-индол-2-он (PHA665752, CAS 477575-56-7).
[00404] Ингибиторы IGF1R включают без ограничения, BMS-754807, XL-228, OSI-906, GSK0904529A, A-928605, AXL1717, KW-2450, MK0646, AMG479, IMCA12, MEDI-573 и BI836845. Обзор см., например, в Yee, JNCI, 104; 975 (2012).
[00405] В другом аспекте настоящего изобретения представлен способ лечения или предупреждения рака путем введения нуждающемуся в этом субъекту конъюгата антитела и лекарственного средства по настоящему изобретению в комбинации с одним или несколькими ингибиторами пути передачи нисходящего сигнала с участием GNAQ/GNA11, в том числе без ограничения ингибиторами β-аррестина, ингибиторами GRK, ингибиторами MAPK, ингибиторами PI3K, ингибиторами JAK и т. д.
[00406] Например, ингибиторы фосфоинозитид 3-киназы (PI3K) включают без ограничения иделалисиб (Zydelig, GS-1101, Cal-101), 4-[2-(1H-индазол-4-ил)-6-[[4-(метилсульфонил)пиперазин-1-ил]метил]тиено[3,2-d]пиримидин-4-ил]морфолин (также известный как GDC 0941 и описанный в публикациях согласно PCT №№ WO 09/036082 и WO 09/055730); 2-метил-2-[4-[3-метил-2-оксо-8-(хинолин-3-ил)-2,3-дигидроимидазо[4,5-c]хинолин-1-ил]фенил]пропионитрил (также известный как BEZ 235 или NVP-BEZ 235 и описанный в публикации согласно PCT № WO 06/122806); 4-(трифторметил)-5-(2,6-диморфолинoпиримидин-4-ил)пиридин-2-амин (также известный как BKM120 или NVP-BKM120 и описанный в публикации согласно PCT № WO2007/084786); тозасертиб (VX680 или MK-0457, CAS 639089-54-6); (5Z)-5-[[4-(4-пиридинил)-6-хинолинил]метилен]-2,4-тиазолидиндион (GSK1059615, CAS 958852-01-2); (1E,4S,4aR,5R,6aS,9aR)-5-(ацетилокси)-1-[(ди-2-пропениламино)метилен]-4,4a,5,6,6a,8,9,9a-октагидро-11-гидрокси-4-(метоксиметил)-4a,6a-диметил-циклопента[5,6]нафто[1,2-c]пиран-2,7,10(1H)-трион (PX866, CAS 502632-66-8) и 8-фенил-2-(морфолин-4-ил)-хромен-4-он (LY294002, CAS 154447-36-6).
[00407] В еще одном аспекте настоящего изобретения представлен способ лечения или предупреждения рака путем введения нуждающемуся в этом субъекту конъюгата антитела и лекарственного средства по настоящему изобретению в комбинации с одним или несколькими проапоптотическими средствами, в том числе без ограничения ингибиторами IAP, ингибиторами Bcl2, ингибиторами MCl1, средствами Trail, ингибиторами Chk.
[00408] Например, ингибиторы IAP включают без ограничения LCL161, GDC-0917, AEG-35156, AT406 и TL32711. Другие примеры ингибиторов IAP включают без ограничения раскрытые в WO04/005284, WO 04/007529, WO05/097791, WO 05/069894, WO 05/069888, WO 05/094818, US2006/0014700, US2006/0025347, WO 06/069063, WO 06/010118, WO 06/017295 и WO08/134679, все из которых включены в данный документ посредством ссылки.
[00409] Ингибиторы BCL-2 включают без ограничения венетоклакс (также известный как GDC-0199, ABT-199, RG7601); 4-[4-[[2-(4-хлорфенил)-5,5-диметил-1-циклогексен-1-ил]метил]-1-пиперазинил]-N-[[4-[[(1R)-3-(4-морфолинил)-1-[(фенилтио)метил]пропил]амино]-3-[(трифторметил)сульфонил]фенил]сульфонил]бензамид (также известный как ABT-263 и описанный в PCT-публикации № WO 09/155386); тетрокарцин A; антимицин; госсипол ((-)BL-193); обатоклакс; этил-2-амино-6-циклопентил-4-(1-циано-2-этокси-2-оксоэтил)-4H-хромон-3-карбоксилат (HA14-1); облимерсен (G3139, Genasense®); пептид Bak BH3; (-)-госсипол-уксусную кислоту (AT-101); 4-[4-[(4'-хлор[1,1'-бифенил]-2-ил)метил]-1-пиперазинил]-N-[[4-[[(1R)-3-(диметиламино)-1-[(фенилтио)метил]пропил]амино]-3-нитрофенил]сульфонил]-бензамид (ABT-737, CAS 852808-04-9) и навитоклакс (ABT-263, CAS 923564-51-6).
[00410] Агонисты проапоптических рецепторов (PARA), в том числе DR4 (TRAILR1) и DR5 (TRAILR2), включают без ограничения дуланермин (AMG-951, RhApo2L/TRAIL); мапатумумаб (HRS-ETR1, CAS 658052-09-6); лексатумумаб (HGS-ETR2, CAS 845816-02-6); апомаб (Apomab®); конатумумаб (AMG655, CAS 896731-82-1) и тигатузумаб (CS1008, CAS 946415-34-5, доступен от Daiichi Sankyo).
[00411] Ингибиторы киназы контрольных точек (CHK) включают без ограничения 7-гидроксистауроспорин (UCN-01); 6-бром-3-(1-метил-1H-пиразол-4-ил)-5-(3R)-3-пиперидинил-пиразолo[1,5-a]пиримидин-7-амин (SCH900776, CAS 891494-63-6); 5-(3-фторфенил)-3-уреидотиофен-2-карбоновой кислоты N-[(S)-пиперидин-3-ил]амид (AZD7762, CAS 860352-01-8); 4-[((3S)-1-азабицикло[2.2.2]окт-3-ил)амино]-3-(1H-бензимидазол-2-ил)-6-хлорхинолин-2(1H)-он (CHIR 124, CAS 405168-58-3); 7-аминодактиномицин (7-AAD), изогранулатимид, дебромгимениалдизин; N-[5-бром-4-метил-2-[(2S)-2-морфолинилметокси]-фенил]-N'-(5-метил-2-пиразинил)мочевину (LY2603618, CAS 911222-45-2); сульфорафан (CAS 4478-93-7, 4-метилсульфинилбутилизотиоцианат); 9,10,11,12-тетрагидро- 9,12-эпокси-1H-дииндолo[1,2,3-fg:3',2',1'-kl]пирроло[3,4-i][1,6]бензодиазоцин-1,3(2H)-дион (SB-218078, CAS 135897-06-2) и TAT-S216A (YGRKKRRQRRRLYRSPAMPENL (SEQ ID NO: 282)) и CBP501 ((d-Bpa)sws(d-Phe-F5)(d-Cha)rrrqrr).
[00412] В дополнительном варианте осуществления настоящего изобретения представлен способ лечения или предупреждения рака путем введения субъекту, нуждающемуся в этом, конъюгата антитела и лекарственного средства по настоящему изобретению в комбинации с одним или несколькими иммуномодуляторами (например, одним или несколькими из активатора костимулирующей молекулы или ингибитора молекулы иммунных контрольных точек).
[00413] В некоторых вариантах осуществления иммуномодулятор представляет собой активатор костимулирующей молекулы. В одном варианте осуществления агонист костимулирующей молекулы выбран из агониста (например, агонистического антитела или его антигенсвязывающего фрагмента, или растворимого слитого белка) OX40, CD2, CD27, CDS, ICAM-1, LFA-1 (CD11a/CD18), ICOS (CD278), 4-1BB (CD137), GITR, CD30, CD40, BAFFR, HVEM, CD7, LIGHT, NKG2C, SLAMF7, NKp80, CD160, B7-H3, STING или лиганда CD83.
[00414] В некоторых вариантах осуществления иммуномодулятор представляет собой ингибитор молекулы контрольных точек иммунного ответа. В одном варианте осуществления иммуномодулятор представляет собой ингибитор PD-1, PD-L1, PD-L2, CTLA4, TIM3, LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и/или TGFR-бета. В одном варианте осуществления ингибитор молекулы иммунных контрольных точек ингибирует PD-1, PD-L1, LAG-3, TIM-3 или CTLA4 или любую их комбинацию. Термин "ингибирование" или "ингибитор" подразумевает снижение определенного параметра, например, активности данной молекулы, например, ингибитора контрольных точек иммунного ответа. Например, данный термин подразумевает ингибирование активности, например, активности PD-1 или PD-L1, на по меньшей мере 5%, 10%, 20%, 30%, 40%, 50% или больше. Таким образом, ингибирование не обязательно должно составлять 100%.
[00415] Ингибирование ингибирующей молекулы может происходить на уровне ДНК, РНК или белка. В некоторых вариантах осуществления для подавления экспрессии ингибирующей молекулы может применяться ингибирующая нуклеиновая кислота (например, dsRNA, siRNA или shRNA). В других вариантах осуществления ингибитор ингибирующего сигнала представляет собой полипептид, например, растворимый лиганд (например, Ig, связывающий PD-1, или Ig, связывающий CTLA-4) или антитело или его антигенсвязывающий фрагмент, которые связываются с ингибирующей молекулой; например, антитело или его фрагмент (также обозначаемые в данном документе как "молекула антитела"), которые связываются с PD-1, PD-L1, PD-L2, CTLA4, TIM3, LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и/или TGFR-бета или их комбинацией.
[00416] В одном варианте осуществления молекула антитела представляет собой полное антитело или его фрагмент (например, Fab, F(ab')2, Fv или одноцепочечный Fv-фрагмент (scFv)). В других вариантах осуществления молекула антитела имеет константную область тяжелой цепи (Fc), выбранную, например, из константных областей тяжелой цепи IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgD и IgE; в частности, выбранную, например, из константных областей тяжелой цепи IgG1, IgG2, IgG3 и IgG4, более конкретно, константной области тяжелой цепи IgG1 или IgG4 (например, IgG1 или IgG4 человека). В одном варианте осуществления константной область тяжелой цепи получена из IgG1 человека или IgG4 человека. В одном варианте осуществления константная область изменена, например, подвергнута мутации для модификации свойств молекулы антитела (например, повышения или снижения одного или нескольких из связывания с рецептором Fc, гликозилирования антител, числа цистеиновых остатков, функции эффекторных клеток или функции комплемента).
[00417] В некоторых вариантах осуществления молекула антитела находится в форме биспецифической или полиспецифической молекулы антитела. В одном варианте осуществления молекула биспецифического антитела характеризуется первой специфичностью связывания с PD-1 или PD-L1 и второй специфичностью связывания, например, второй специфичностью связывания с TIM-3, LAG-3 или PD-L2. В одном варианте осуществления молекула биспецифического антитела связывается с PD-1 или PD-L1 и TIM-3. В другом варианте осуществления молекула биспецифического антитела связывается с PD-1 или PD-L1 и LAG-3. В другом варианте осуществления молекула биспецифического антитела связывается с PD-1 и PD-L1. В еще одном варианте осуществления молекула биспецифического антитела связывается с PD-1 и PD-L2. В другом варианте осуществления молекула биспецифического антитела связывается с TIM-3 и LAG-3. Любая комбинация вышеуказанных молекул может быть составлена в молекулу полиспецифического антитела, например, триспецифическое антитело, которое предусматривает первую специфичность связывания с PD-1 или PD-1, а также вторую и третью специфичности связывания с двумя или более из TIM-3, LAG-3 или PD-L2.
[00418] В некоторых вариантах осуществления иммуномодулятор представляет собой ингибитор of PD-1, например PD-1 человека. В другом варианте осуществления иммуномодулятор представляет собой ингибитор PD-L1, например PD-L1 человека. В одном варианте осуществления ингибитор PD-1 или PD-L1 представляет собой молекулу антитела к PD-1 или PD-L1. Ингибитор PD-1 или PD-L1 может вводиться отдельно или в комбинации с другими иммуномодуляторами, например, в комбинации с ингибитором LAG-3, TIM-3 или CTLA4. В иллюстративном варианте осуществления ингибитор PD-1 или PD-L1, например, молекулу антитела к PD-1 или PD-L1, вводят в комбинации с ингибитором LAG-3, например, молекулой антитела к LAG-3. В другом варианте осуществления ингибитор PD-1 или PD-L1, например, молекулу антитела к PD-1 или PD-L1, вводят в комбинации ингибитором TIM-3, например, молекулой антитела к TIM-3. В других вариантах осуществления ингибитор PD-1 или PD-L1, например, молекулу антитела к PD-1, вводят в комбинации с ингибитором LAG-3, например, молекулой антитела к LAG-3, и ингибитором TIM-3, например, молекулой антитела к TIM-3. Другие комбинации иммуномодуляторов с ингибитором PD-1 (например, с одним или несколькими из PD-L2, CTLA4, TIM3, LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и/или TGFR) также предусмотрены в пределах настоящего изобретения. Любая из молекул антитела, известная из уровня техники или раскрытая в данном документе, может использоваться в вышеуказанной комбинации ингибиторов молекулы контрольных точек иммунного ответа.
[00419] В одном варианте осуществления ингибитор PD-1 представляет собой антитело к PD-1, выбранное из ниволумаба, пембролизумаба или пидилизумаба. В некоторых вариантах осуществления антитело к PD-1 представляет собой ниволумаб. К альтернативным названиям ниволумаба относятся MDX-1106, MDX-1106-04, ONO-4538 или BMS-936558. В некоторых вариантах осуществления антитело к PD-1 представляет собой ниволумаб (регистрационный номер CAS: 946414-94-4). Ниволумаб представляет собой полностью человеческое моноклональное антитело IgG4, которое специфически блокирует PD1. Ниволумаб (клон 5C4) и другие человеческие моноклональные антитела, которые специфически связываются с PD1, раскрыты в патенте США № 8008449 и PCT-публикации № WO2006/121168.
[00420] В других вариантах осуществления антитело к PD-1 представляет собой пембролизумаб. Пембролизумаб (торговое наименование KEYTRUDA, ранее ламбролизумаб, также известный как Merck 3745, MK-3475 или SCH-900475) представляет собой гуманизированное моноклональное антитело IgG4, которое связывается с PD1. Пембролизумаб раскрыт, например, в Hamid, O. et al. (2013) New England Journal of Medicine 369 (2): 134-44, PCT-публикации № WO2009/114335 и патенте США № 8354509.
[00421] В некоторых вариантах осуществления антитело к PD-1 представляет собой пидилизумаб. Пидилизумаб (CT-011; Cure Tech) представляет собой гуманизированное моноклональное антитело IgG1k, которое связывается с PD1. Пидилизумаб и другие гуманизированные моноклональные антитела к PD-1 раскрыты в PCT-публикации № WO2009/101611. Другие антитела к PD1 раскрыты в патенте США № 8609089, публикации заявки на патент США № 2010028330 и/или публикации заявки на патент США № 20120114649. Другие антитела к PD1 включают AMP 514 (Amplimmune).
[00422] В некоторых вариантах осуществления ингибитор PD-1 представляет собой PDR001, также известный как спартализумаб, или любое другое антитело к PD-1, раскрытое в WO2015/112900.
[00423] В некоторых вариантах осуществления ингибитор PD-1 представляет собой иммуноадгезин (например, иммуноадгезин, содержащий внеклеточную или PD-1-связывающую часть PD-L1 или PD-L2, слитую с константной областью (например, областью Fc последовательности иммуноглобулина). В некоторых вариантах осуществления ингибитор PD-1 представляет собой AMP-224.
[00424] В некоторых вариантах осуществления ингибитор PD-L1 представляет собой антитело к PD-L1. В некоторых вариантах осуществления ингибитор на основе антитела к PD-L1 выбран из YW243.55.S70, MPDL3280A, MEDI-4736 или MDX-1105MSB-0010718C (также называемого A09-246-2), раскрытого, например, WO 2013/0179174, и имеет последовательность, раскрытую в данном документе (или последовательность практически идентичную или сходную с ней, например, последовательность, которая характеризуется по меньшей мере 85%, 90%, 95% или более высокой идентичностью с указанной последовательностью).
[00425] В одном варианте осуществления ингибитор PD-L1 представляет собой MDX-1105. MDX-1105, также известный как BMS-936559, представляет собой антитело к PD-L1, описанное в PCT-публикации № WO2007/005874.
[00426] В одном варианте осуществления ингибитор PD-L1 представляет собой YW243.55.S70. Антитело YW243.55.S70 представляет собой антитело к PD-L1, описанное в PCT-публикации № WO 2010/077634 (последовательности вариабельных областей тяжелой и легкой цепей показаны под SEQ ID No: 20 и 21, соответственно).
[00427] В одном варианте осуществления ингибитор PD-L1 представляет собой MDPL3280A (Genentech/Roche). MDPL3280A представляет собой человеческое моноклональное антитело IgG1 с оптимизированной Fc, которое связывается с PD-L1. MDPL3280A и другие человеческие моноклональные антитела к PD-L1 раскрыты в патенте США № 7943743 и публикации заявки на патент США № 20120039906.
[00428] В других вариантах осуществления ингибитор PD-L2 представляет собой AMP-224. AMP-224 представляет собой растворимый рецептор на основе слитого белка PD-L2 и Fc, который блокирует взаимодействие между PD1 и B7-H1 (B7-DCIg; Amplimmune; например, раскрытый в PCT-публикациях №№ WO2010/027827 и WO2011/066342).
[00429] В одном варианте осуществления ингибитор LAG-3 представляет собой молекулу антитела к LAG-3. В одном варианте осуществления ингибитор LAG-3 представляет собой BMS-986016. В одном варианте осуществления ингибитор LAG-3 представляет собой LAG525 или любое другое антитело к LAG3, раскрытое в WO2015/138920.
[00430] В одном варианте осуществления ингибитор TIM-3 представляет собой молекулу антитела к TIM3. В одном варианте осуществления ингибитор TIM-3 представляет собой MBG453 или любое другое антитело к TIM3, раскрытое в WO2015/117002.
Фармацевтические композиции
[00431] Для получения фармацевтических или стерильных композиций, содержащих иммуноконъюгаты, иммуноконъюгаты по настоящему изобретению смешивают с фармацевтически приемлемым носителем или вспомогательным веществом. Композиции могут дополнительно содержать одно или несколько других терапевтических средств, которые подходят для лечения или предупреждения рака, экспрессирующего PMEL17 (включающего без ограничения подкожную меланому, увеальную меланому, гепатоцеллюлярную карциному и метастатическую форму данных видов рака).
[00432] Составы терапевтических и диагностических средств могут быть получены путем смешивания с физиологически приемлемыми носителями, вспомогательными веществами или стабилизаторами в форме, например, лиофилизированных порошков, взвесей, водных растворов, лосьонов или суспензий (см., например, Hardman et al., Goodman and Gilman's The Pharmacological Basis of Therapeutics, McGraw-Hill, New York, N.Y., 2001; Gennaro, Remington: The Science and Practice of Pharmacy, Lippincott, Williams, and Wilkins, New York, N.Y., 2000; Avis, et al. (eds.), Pharmaceutical Dosage Forms: Parenteral Medications, Marcel Dekker, NY, 1993; Lieberman, et al. (eds.), Pharmaceutical Dosage Forms: tablets, Marcel Dekker, NY, 1990; Lieberman, et al. (eds.) Pharmaceutical Dosage Forms: Disperse Systems, Marcel Dekker, NY, 1990; Weiner and Kotkoskie, Excipient Toxicity and Safety, Marcel Dekker, Inc., New York, N.Y., 2000).
[00433] Выбор схемы введения для терапевтического средства зависит от нескольких факторов, в том числе интенсивности метаболизма сыворотки или ткани для объекта, уровня симптомов, иммуногенности объекта и доступности клеток-мишеней в биологической матрице. В некоторых вариантах осуществления схема введения максимизирует количество терапевтического средства, доставляемого пациенту, в соответствии с приемлемым уровнем побочных эффектов. Соответственно, количество доставляемого биологического средства частично зависит от конкретного объекта и тяжести состояния, подлежащего лечению. Руководства по подбору подходящих доз антител, цитокинов и малых молекул являются доступными (см., например, Wawrzynczak, Antibody Therapy, Bios Scientific Pub. Ltd, Oxfordshire, UK, 1996; Kresina (ed.), Monoclonal Antibodies, Cytokines and Arthritis, Marcel Dekker, New York, N.Y., 1991; Bach (ed.), Monoclonal Antibodies and Peptide Therapy in Autoimmune Diseases, Marcel Dekker, New York, N.Y., 1993; Baert et al., New Engl. J. Med. 348:601-608, 2003; Milgrom et al., New Engl. J. Med. 341:1966-1973, 1999; Slamon et al., New Engl. J. Med. 344:783-792, 2001; Beniaminovitz et al., New Engl. J. Med. 342:613-619, 2000; Ghosh et al., New Engl. J. Med. 348:24-32, 2003; Lipsky et al., New Engl. J. Med. 343:1594-1602, 2000).
[00434] Определение подходящей дозы выполняет лечащий врач, например, с применением параметров или факторов, которые, как известно или ожидаемо из уровня техники, воздействуют на лечение или предупреждение или, как прогнозируют, воздействуют на лечение или предупреждение. Обычно начинают дозирование с количества, несколько меньшего, чем оптимальная доза, и после этого ее повышают небольшими шагами до тех пор, пока не достигается требуемый или оптимальный эффект по сравнению с любыми отрицательными побочными эффектами. Важные диагностические показатели включают, например, показатели, являющиеся симптомами воспаления, или уровень продуцируемых воспалительных цитокинов.
[00435] Фактические уровни дозы активных ингредиентов в фармацевтических композициях по настоящему изобретению могут варьироваться для того, чтобы получить количество активного ингредиента, которое является эффективным для достижения требуемого терапевтического ответа у конкретного пациента, эффективным в отношении композиции и способа введения, при этом не токсично для пациента. Выбранный уровень дозы будет зависеть от ряда фармакокинетических факторов, в том числе активности конкретных используемых композиций по настоящему изобретению или их сложного эфира, соли или амида, пути введения, времени введения, скорости выведения конкретного используемого соединения, длительности лечения, других лекарственных средств, соединений и/или материалов, применяемых в комбинации с конкретными используемыми композициями, возраста, пола, веса тела, состояния, общего состояния здоровья и анамнеза пациента, подлежащего лечению, и аналогичных факторов, известных в области техники медицины.
[00436] Композиции, содержащие антитела или их фрагменты по настоящему изобретению, могут вводиться в помощью непрерывной инфузии или с помощью доз с интервалами, составляющими, например, один день, одну неделю, или вводиться 1-7 раз в неделю, раз в две недели, раз в три недели, раз в четыре недели, раз в пять недель, раз в шесть недель, раз в семь недель или раз в восемь недель. Дозы могут вводиться внутривенно, подкожно, местно, перорально, назально, ректально, внутримышечно, внутрь мозга или посредством ингаляции. Специфический протокол введения доз является таким, который предусматривает максимальную дозу или частоту введения доз, позволяющие избежать значительных нежелательных побочных эффектов.
[00437] Для иммуноконъюгатов по настоящему изобретению доза, вводимая пациенту, может составлять от 0,0001 мг/кг до 100 мг/кг веса тела пациента. Доза может быть составлять от 0,0001 мг/кг до 30 мг/кг, от 0,0001 мг/кг до 20 мг/кг, от 0,0001 мг/кг и 10 мг/кг, от 0,0001 мг/кг до 5 мг/кг, от 0,0001 до 2 мг/кг, от 0,0001 до 1 мг/кг, от 0,0001 мг/кг до 0,75 мг/кг, от 0,0001 мг/кг до 0,5 мг/кг, от 0,0001 мг/кг до 0,25 мг/кг, от 0,0001 до 0,15 мг/кг, от 0,0001 до 0,10 мг/кг, от 0,001 до 0,5 мг/кг, от 0,01 до 0,25 мг/кг или от 0,01 до 0,10 мг/кг веса тела пациента. Доза антител или их фрагментов по настоящему изобретению может быть рассчитана с применением массы пациента в килограммах (кг), умноженной на подлежащую введению дозу в мг/кг.
[00438] Дозы иммуноконъюгатов по настоящему изобретению могут быть повторными, а введения могут быть разделены менее чем 1 днем, по меньшей мере 1 днем, 2 днями, 3 днями, 5 днями, 10 днями, 15 днями, 30 днями, 45 днями, 2 месяцами, 75 днями, 3 месяцами, 4 месяцами, 5 месяцами или по меньшей мере 6 месяцами. В некоторых вариантах осуществления иммуноконъюгаты по настоящему изобретению можно давать два раза в неделю, раз в неделю, раз в две недели, раз в три недели, раз в четыре недели или менее часто. В конкретном варианте осуществления дозы иммуноконъюгатов по настоящему изобретению вводятся повторно через каждые 2 недели.
[00439] Эффективное количество для конкретного пациента может варьироваться в зависимости от таких факторов, как подлежащее лечению состояние, общее состояние здоровья пациента, способ, путь и доза введения и тяжесть побочных эффектов (см., например, Maynard et al., A Handbook of SOPs for Good Clinical Practice, Interpharm Press, Boca Raton, Fla., 1996; Dent, Good Laboratory and Good Clinical Practice, Urch Publ., London, UK, 2001).
[00440] Путем введение может быть, например, местное нанесение или нанесение на кожу, инъекция или инфузия посредством подкожного, внутривенного, внутрибрюшинного, внутримозгового, внутримышечного, внутриглазного, внутриартериального, внутриспинномозгового, внутриочагового введения, или системы замедленного высвобождения или имплантат (см., например, Sidman et al., Biopolymers 22:547-556, 1983; Langer et al., J. Biomed. Mater. Res. 15:167-277, 1981; Langer, Chem. Tech. 12:98-105, 1982; Epstein et al., Proc. Natl. Acad. Sci. USA 82:3688-3692, 1985; Hwang et al., Proc. Natl. Acad. Sci. USA 77:4030-4034, 1980; патенты США №№ 6350466 и 6316024). При необходимости композиция может также содержать солюбилизирующее средство и местный анестетик, такой как лидокаин, для облегчения боли в месте инъекции, или их оба. Кроме того, также можно использовать ингаляционное введение, например, за счет применения ингалятора или распылителя и состава со средством в виде аэрозоля. См., например, патенты США №№ 6019968, 5985320, 5985309, 5934272, 5874064, 5855913, 5290540 и 4880078; и публикации согласно PCT №№ WO 92/19244, WO 97/32572, WO 97/44013, WO 98/31346 и WO 99/66903, каждый из которых включен в данный документ посредством ссылки во всей своей полноте.
[00441] Композицию по настоящему изобретению также вводят с помощью одного или нескольких путей введения с применением одного или нескольких из целого ряда способов, известных из уровня техники. Как будет понятно специалисту в данной области техники, путь и/или способ введения будут варьироваться в зависимости от необходимых результатов. Выбранные пути введения иммуноконъюгатов по настоящему изобретению включают внутривенный, внутримышечный, внутрикожный, внутрибрюшинный, подкожный пути введения, введение в спинной мозг или другие парентеральные пути введения, например, с помощью инъекции или инфузии. Парентеральное введение может представлять способы введения, отличные от энтерального и местного введения, обычно производимые с помощью инъекции, и оно охватывает без ограничения внутривенные, внутримышечные, внутриартериальные, интратекальные, внутрикапсульные, интраорбитальные, внутрисердечные, внутрикожные, внутрибрюшинные, транстрахеальные, подкожные, субкутикулярные, внутрисуставные, подкапсулярные, субарахноидальные, интраспинальные, эпидуральные и интрастернальные инъекцию и инфузию. В качестве альтернативы композицию по настоящему изобретению можно вводить посредством отличного от парентерального пути, такого как местный, эпидермальный пути введения или введение через слизистые оболочки, например, интраназально, перорально, вагинально, ректально, сублингвально или местно. В одном варианте осуществления иммуноконъюгаты по настоящему изобретению вводят с помощью инфузии. В другом варианте осуществления иммуноконъюгаты по настоящему изобретению вводят подкожно.
[00442] Если иммуноконъюгаты по настоящему изобретению вводятся в системе контролируемого высвобождения или замедленного высвобождения, для осуществления контролируемого или замедленного высвобождения может применяться насос (см. Langer, выше; Sefton, CRC Crit. Ref Biomed. Eng. 14:20, 1987; Buchwald et al., Surgery 88:507, 1980; Saudek et al., N. Engl. J. Med. 321:574, 1989). Для осуществления контролируемого или замедленного высвобождения видов терапии по настоящему изобретению могут применяться полимерные материалы (см., например, Medical Applications of Controlled Release, Langer and Wise (eds.), CRC Pres., Boca Raton, Fla., 1974; Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen and Ball (eds.), Wiley, New York, 1984; Ranger and Peppas, J. Macromol. Sci. Rev. Macromol. Chem. 23:61, 1983; см. также Levy et al., Science 228:190, 1985; During et al., Ann. Neurol. 25:351, 1989; Howard et al., J. Neurosurg. 7 1:105, 1989; патент США № 5679377; патент США № 5916597; патент США № 5912015; патент США № 5989463; патент США № 5128326; PCT-публикацию № WO 99/15154 и PCT-публикацию № WO 99/20253. Примеры полимеров, используемых в составах с замедленным высвобождением, включают без ограничения поли(2-гидроксиэтилметакрилат), поли(метилметакрилат), поли(акриловую кислоту), сополимер(этилена и винилацетата), поли(метакриловую кислоту), полигликолиды (PLG), полиангидриды, поли(N-винилпирролидон), поли(виниловый спирт), полиакриламид, поли(этиленгликоль), полилактиды (PLA), сополимеры(лактида и гликолидов) (PLGA) и сложные полиортоэфиры. В одном варианте осуществления полимер, используемый в составе с замедленным высвобождением, является инертным, не содержит выщелачиваемых примесей, стабильный при хранении, стерильный и биологически разлагаемый. Систему контролируемого или замедленного высвобождения можно поместить вблизи профилактической или терапевтической мишени, при этом требуется только часть системной дозы (см., например, Goodson, в Medical Applications of Controlled Release, выше, vol. 2, pp. 115-138, 1984).
[00443] Системы контролируемого высвобождения рассмотрены в обзоре Langer, Science 249:1527-1533, 1990). Любую методику, известную специалисту в данной области техники, можно применять для получения составов с замедленным высвобождением, содержащих один или несколько иммуноконъюгатов по настоящему изобретению. См., например, патент США № 4526938, PCT-публикацию WO 91/05548, PCT-публикацию WO 96/20698, Ning et al., Radiotherapy & Oncology 39:179-189, 1996; Song et al., PDA Journal of Pharmaceutical Science & Technology 50:372-397, 1995; Cleek et al., Pro. Int'l. Symp. Control. Rel. Bioact. Mater. 24:853-854, 1997; и Lam et al., Proc. Int'l. Symp. Control Rel. Bioact. Mater. 24:759-760, 1997, каждый из которых включен в данный документ посредством ссылки во всей своей полноте.
[00444] Если иммуноконъюгаты по настоящему изобретению вводятся местно, они могут быть составлены в форме мази, крема, трансдермального пластыря, лосьона, геля, спрея, аэрозоля, раствора, эмульсии или другой формы, широко известной специалисту в данной области техники. См., например, Remington's Pharmaceutical Sciences and Introduction to Pharmaceutical Dosage Forms, 19th ed., Mack Pub. Co., Easton, Pa. (1995). В случае нераспыляемых местных лекарственных форм обычно используются вязкие-полутвердые или твердые формы, содержащие носитель или одно или несколько вспомогательных веществ, совместимые с местным нанесением и, в некоторых случаях, характеризующиеся динамической вязкостью больше, чем у воды. Подходящие составы включают без ограничения растворы, суспензии, эмульсии, кремы, мази, порошки, линименты, бальзамы и т. п., которые при необходимости стерилизуют или смешивают со вспомогательными средствами (например, консервантами, стабилизаторами, смачивающими средствами, буферами или солями) для воздействия на различные свойства, такие как, например, осмотическое давление. Другие подходящие местные лекарственные формы охватывают распыляемые аэрозольные препараты, где активный ингредиент, в некоторых случаях в комбинации с твердым или жидким инертным носителем, упакован в смеси с летучим веществом под давлением (например, газообразным пропеллентом, таким как фреон) или в бутылке-пульверизаторе. При необходимости в фармацевтические композиции и лекарственные формы также можно добавлять увлажнители или увлажняющие средства. Примеры таких дополнительных ингредиентов широко известны из уровня техники.
[00445] Если композиции, содержащие иммуноконъюгаты вводят интраназально, они могут быть составлены в форме аэрозоля, спрея, тумана или в форме капель. В частности, в целях удобства профилактические или терапевтические средства для применения согласно настоящему изобретению можно доставлять в форме спрея-аэрозоля из пакетов под давлением или с помощью небулайзера с применением подходящего пропеллента (например, дихлордифторметана, трихлорфторметана, дихлортетрафторэтана, диоксида углерода или другого подходящего газа). В случае аэрозоля под давлением единица дозирования может определяться за счет обеспечения клапана для доставки отмеренного количества. Капсулы и картриджи (например, состоящие из желатина) для применения в ингаляторе или инсуффляторе можно составлять содержащими порошковую смесь соединения и подходящей порошковой основы, такой как лактоза или крахмал.
[00446] Способы совместного введения или лечения с применением второго терапевтического средства, например, цитокина, стероида, химиотерапевтического средства, антибиотика, или облучения, известны из уровня техники (см., например, Hardman et al., (eds.) (2001) Goodman and Gilman's The Pharmacological Basis of Therapeutics, 10th ed., McGraw-Hill, New York, N.Y.; Poole and Peterson (eds.) (2001) Pharmacotherapeutics for Advanced Practice:A Practical Approach, Lippincott, Williams & Wilkins, Phila., Pa.; Chabner and Longo (eds.) (2001) Cancer Chemotherapy and Biotherapy, Lippincott, Williams & Wilkins, Phila., Pa.). Эффективное количество терапевтического средства может снижать симптомы на по меньшей мере 10%; по меньшей мере 20%; по меньшей мере приблизительно 30%; по меньшей мере 40% или по меньшей мере 50%.
[00447] Дополнительные виды терапии (например, профилактические или терапевтические средства), которые можно применять в комбинации с иммуноконъюгатами по настоящему изобретению, можно применять с интервалом менее 5 минут, с интервалом менее 30 минут, с интервалом 1 час, с интервалом приблизительно 1 час, с интервалом от приблизительно 1 до приблизительно 2 часов, с интервалом от приблизительно 2 часов до приблизительно 3 часов, с интервалом от приблизительно 3 часов до приблизительно 4 часов, с интервалом от приблизительно 4 часов до приблизительно 5 часов, с интервалом от приблизительно 5 часов до приблизительно 6 часов, с интервалом от приблизительно 6 часов до приблизительно 7 часов, с интервалом от приблизительно 7 часов до приблизительно 8 часов, с интервалом от приблизительно 8 часов до приблизительно 9 часов, с интервалом от приблизительно 9 часов до приблизительно 10 часов, с интервалом от приблизительно 10 часов до приблизительно 11 часов, с интервалом от приблизительно 11 часов до приблизительно 12 часов, с интервалом от приблизительно 12 часов до 18 часов, с интервалом от 18 часов до 24 часов, с интервалом от 24 часов до 36 часов, с интервалом от 36 часов до 48 часов, с интервалом от 48 часов до 52 часов, с интервалом от 52 часов до 60 часов, с интервалом от 60 часов до 72 часов, с интервалом от 72 часов до 84 часов, с интервалом от 84 часов до 96 часов или с интервалом от 96 часов до 120 часов от введения иммуноконъюгатов по настоящему изобретению. Два или более видов терапии могут применяться во время одного и того же визита пациента.
[00448] В некоторых вариантах осуществления иммуноконъюгаты по настоящему изобретению могут составляться, чтобы гарантировать точное распределение in vivo. Например, гематоэнцефалический барьер (BBB) не пропускает множество высокогидрофильных соединений. Чтобы гарантировать то, что терапевтические соединения по настоящему изобретению пересекают BBB (при необходимости), их можно составлять, например, в липосомах. Способы изготовления липосомы, см., например, в патентах США №№ 4522811; 5374548 и 5399331. Липосомы могут содержать один или несколько фрагментов, которые селективно транспортируются в специфические клетки или органы, с улучшением тем самым нацеленной доставки лекарственного средства (см., например, Ranade, (1989) J. Clin. Pharmacol. 29:685). Иллюстративные нацеливающие фрагменты включают фолат или биотин (см., например, патент США № 5416016 от Low et al.); маннозиды (Umezawa et al., (1988) Biochem. Biophys. Res. Commun. 153:1038); антитела (Bloeman et al., (1995) FEBS Lett. 357:140; Owais et al., (1995) Antimicrob. Agents Chemother. 39:180); рецептор поверхностно-активного белка A (Briscoe et al., (1995) Am. J. Physiol. 1233:134); p 120 (Schreier et al., (1994) J. Biol. Chem. 269:9090); см. также K. Keinanen; M. L. Laukkanen (1994) FEBS Lett. 346:123; J. J. Killion; I. J. Fidler (1994) Immunomethods 4:273.
[00449] В настоящем изобретении предусмотрены протоколы для введения субъекту, нуждающемуся в этом, фармацевтической композиции, содержащей иммуноконъюгаты по настоящему изобретению, отдельно или в комбинации с другими видами терапии. Виды терапии (например, профилактические или терапевтические средства) из видов комбинированной терапии по настоящему изобретению могут применяться в отношении субъекта одновременно или последовательно. Терапия (например, профилактические или терапевтические средства) из видов комбинированной терапии по настоящему изобретению также можно вводить циклически. Циклическая терапия предусматривает введение первого средства терапии (например, первого профилактического или терапевтического средства) в течение некоторого периода времени, а затем введения второго средства терапии (например, второго профилактического или терапевтического средства) в течение некоторого периода времени и повторения такого последовательного введения, т. е. цикла, с целью снижения развития устойчивости к одному из видов терапии (например, средствам), для недопущения или снижения побочных эффектов одного из видов терапии (например, средств) и/или для улучшения эффективности видов терапии.
[00450] Виды терапии (например, профилактические или терапевтические средства) из видов комбинированной терапии по настоящему изобретению могут применяться в отношении субъекта одновременно.
[00451] Термин "одновременно" не ограничивается применением видов терапии (например, профилактических или терапевтических средств) точно в одно и то же время, а скорее означает, что фармацевтическую композицию, содержащую антитела или их фрагменты по настоящему изобретению, вводят субъекту последовательно и в течение временного интервала таким образом, чтобы конъюгаты антитела и лекарственного средства по настоящему изобретению могли действовать вместе с другим(-и) видом(-ами) терапии для обеспечения большей пользы, чем если бы их вводили иным способом. Например, каждый вид терапии может применяться в отношении субъекта в одно и то же время или последовательно в любом порядке в различные моменты времени; однако, если их не применяют в одно и то же время, их следует применять достаточно близко по времени, чтобы обеспечить требуемый терапевтический или профилактический эффект. Каждый вид терапии можно применять в отношении субъекта по отдельности, в любой подходящей форме и с помощью любого подходящего пути. В различных вариантах осуществления виды терапии (например, профилактические или терапевтические средства) применяют в отношении субъекта с интервалом менее 5 минут, с интервалом менее 15 минут, с интервалом менее 30 минут, с интервалом менее 1 часа, с интервалом приблизительно 1 час, с интервалом от приблизительно 1 часа до приблизительно 2 часов, с интервалом от приблизительно 2 часов до приблизительно 3 часов, с интервалом от приблизительно 3 часов до приблизительно 4 часов, с интервалом от приблизительно 4 часов до приблизительно 5 часов, с интервалом от приблизительно 5 часов до приблизительно 6 часов, с интервалом от приблизительно 6 часов до приблизительно 7 часов, с интервалом от приблизительно 7 часов до приблизительно 8 часов, с интервалом от приблизительно 8 часов до приблизительно 9 часов, с интервалом от приблизительно 9 часов до приблизительно 10 часов, с интервалом от приблизительно 10 часов до приблизительно 11 часов, с интервалом от приблизительно 11 часов до приблизительно 12 часов, с интервалом 24 часа, с интервалом 48 часов, с интервалом 72 часа или с интервалом 1 неделя. В других вариантах осуществления два или более видов терапии (например, профилактические или терапевтические средства) применяются во время одного и того же визита пациента.
[00452] Профилактические или терапевтические средства из видов комбинированной терапии могут вводиться субъекту в одной и той же фармацевтической композиции. В качестве альтернативы профилактические или терапевтические средства из видов комбинированной терапии могут вводиться субъекту одновременно в отдельных фармацевтических композициях. Профилактические или терапевтические средства могут вводиться субъекту с помощью одного и того же или различных путей введения. Профилактические или терапевтические средства из видов комбинированной терапии могут вводиться субъекту в одной и той же фармацевтической композиции. В качестве альтернативы профилактические или терапевтические средства из видов комбинированной терапии могут вводиться субъекту одновременно в отдельных фармацевтических композициях. Профилактические или терапевтические средства могут вводиться субъекту с помощью одного и того же или различных путей введения.
Примеры
Пример 1: cинтез иллюстративных соединений линкер-лекарственное средство
Пример 1-1: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-22-изопропил-3-((R)-1-метоксиэтил)-4,9,10,12,16-пентаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((((4-((S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)(гидрокси)фосфорил)окси)-4-метил-2-пропионамидопентаноата(B1)
Стадия 1: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-22-изопропил-3-((R)-1-метоксиэтил)-4,9,10,12,16-пентаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((гидроксигидрофосфорил)окси)-4-метил-2-пропионамидопентаноата (1-1):
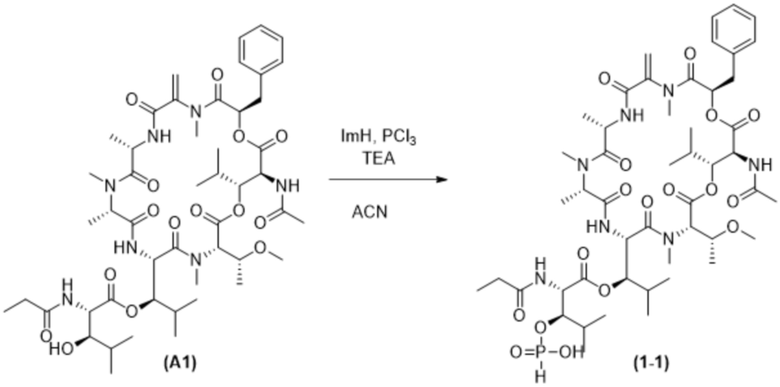
[00453] имидазол (102 мг, 1,49 ммоль, 15 экв.) растворяли в ацетонитриле (ACN) (1,4 мл) и охлаждали на ледяной бане (наблюдается дробление ImH, и смесь поднимали из ледяной бани для растворения ImH) пока она оставалась холодной. Затем по каплям добавляли трихлорид фосфора (1,0 М в ACN) (499 мкл, 0,499 ммоль, 5 экв.) (с получением в результате белой суспензии) и смесь перемешивали в течение 10 мин. Затем добавляли триэтиламин (250 мкл, 1,796 ммоль, 18 экв.) и смесь перемешивали в течение 40 мин, после чего получали (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-22-изопропил-3-((R)-1-метоксиэтил)-4,9,10,12,16-пентаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-гидрокси-4-метил-2-пропионамидопентаноат (A1) (100 мг, 0,100 ммоль, 1,0 экв., соединение (A1) посредством способа, описанного в примере 3-1) с добавлением ACN (1,7 мл). Желтовато-оранжевую гетерогенную смесь нагревали до комнатной температуры и перемешивали в течение в общей сложности 60 минут. Смесь обрабатывали водой (0,2 мл), и материал очищали посредством обращенно-фазовой флэш-хроматографии (0-100% ACN/вода, колонка C18 40 грам, нейтральная подвижная фаза), фракции продукта собирали и лиофилизировали с получением H-фосфоната (1-1) в виде желтовато-белого аморфного порошка. LCMS: MH+ = 1066,3, 0,78 мин (колонка Acquity UPLC BEH C18 1,7 мкм, 2-98%, 2 мин, прогонка с водой/MeCN +0,1% NH4OH, основный способ).
Стадия 2: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-22-изопропил-3-((R)-1-метоксиэтил)-4,9,10,12,16-пентаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((((4-((S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)(гидрокси)фосфорил)окси)-4-метил-2-пропионамидопентаноата (B1):
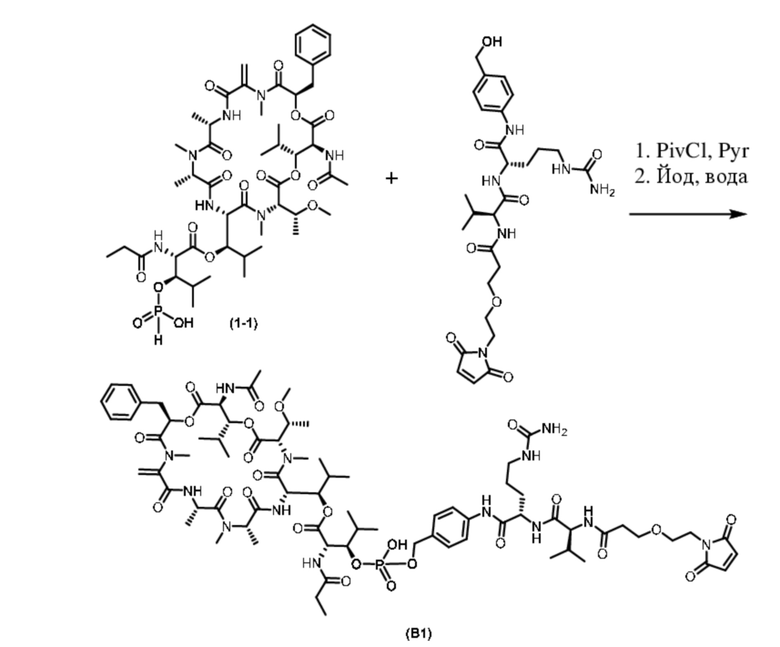
[00454] (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-22-изопропил-3-((R)-1-метоксиэтил)-4,9,10,12,16-пентаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((гидроксигидрофосфорил)окси)-4-метил-2-пропионамидопентаноат (1-1) (100 мг, 0,094 ммоль, 1,0 экв.) и (S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-N-(4-(гидроксиметил)фенил)-5-уреидопентанамид (108 мг, 0,188 ммоль, 2,0 экв., CAS № 2055041-37-5) (оба лиофилизированных порошка) переносили во флакон объемом 10 мл и растворяли в пиридине (4 мл). Затем по каплям добавляли пивалоилхлорид (0,058 мл, 0,469 ммоль, 5 экв.) с получением бледно-желтого раствора. Смесь перемешивали при комнатной температуре в течение 10 минут, а затем добавляли дополнительно 1,0 экв. пивалоилхлорида. Свежеприготовленный раствор йода (47,6 мг, 0,188 ммоль, 2,0 экв.) в смеси пиридин-вода (14:1, 750 мкл) добавляли с получением темно-коричневого прозрачного раствора. Смесь перемешивали в течение 25 минут и непосредственно очищали с помощью обращенно-фазовой флэш-хроматографии (колонка C-18 40 г, 0% Ac/MeCN 3 минуты, затем 0-60% ACN/вода в течение 15 минут, нейтральный способ) с получением (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-22-изопропил-3-((R)-1-метоксиэтил)-4,9,10,12,16-пентаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((((4-((S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)(гидрокси)фосфорил)окси)-4-метил-2-пропионамидопентаноата (B-1). HRMS; MH+ = 1638,7700, 2,84 мин.
Пример 1-2: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-2-ацетамидо-3-((((4-((S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)(гидрокси)фосфорил)окси)-4-метилпентаноата (B2)
Стадия 1: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-2-ацетамидо-3-((гидроксигидрофосфорил)окси)-4-метилпентаноата (1-2):
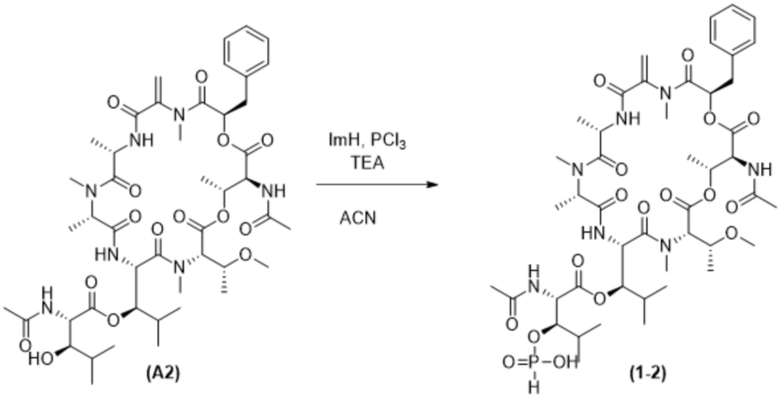
[00455] имидазол (85 мг, 1,25 ммоль, 15 экв.) растворяли в ацетонитриле (ACN) (2,5 мл) и охлаждали на ледяной бане (наблюдается дробление ImH, и смесь поднимали из ледяной бани для растворения ImH) пока она оставалась холодной. Затем по каплям добавляли трихлорид фосфора (36,4 мкл растворяли в 0,5 мл MeCN, 0,417 ммоль, 5 экв.) (с получением в результате белой суспензии) и смесь перемешивали в течение 10 мин. Затем добавляли триэтиламин (174 мкл, 1,25 ммоль, 15 экв.) и смесь перемешивали в течение 40 мин, после чего получали (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-2-ацетамидо-3-гидрокси-4-метилпентаноат (A2) (80 мг, 0,084 ммоль, 1,0 экв., соединение (A2) посредством способа, описанного в примере 3-2, с добавлением ACN (1,7 мл). Желтовато-оранжевую гетерогенную смесь нагревали до комнатной температуры и перемешивали в течение в общей сложности 30 минут. Смесь обрабатывали водой (1 мл), и материал очищали посредством обращенно-фазовой флэш-хроматографии (0-100% ACN/вода, колонка C18 40 грам, нейтральная подвижная фаза), фракции продукта собирали и лиофилизировали с получением H-фосфоната (1-2) в виде желтовато-белого аморфного порошка. LCMS: MH+ = 1024,3, 0,78 мин (колонка Acquity UPLC BEH C18 1,7 мкм, 2-98%, 2 мин, прогонка с водой/MeCN +0,1% NH4OH, основный способ).
Стадия 2: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-2-ацетамидо-3-((((4-((S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)(гидрокси)фосфорил)окси)-4-метилпентаноата:
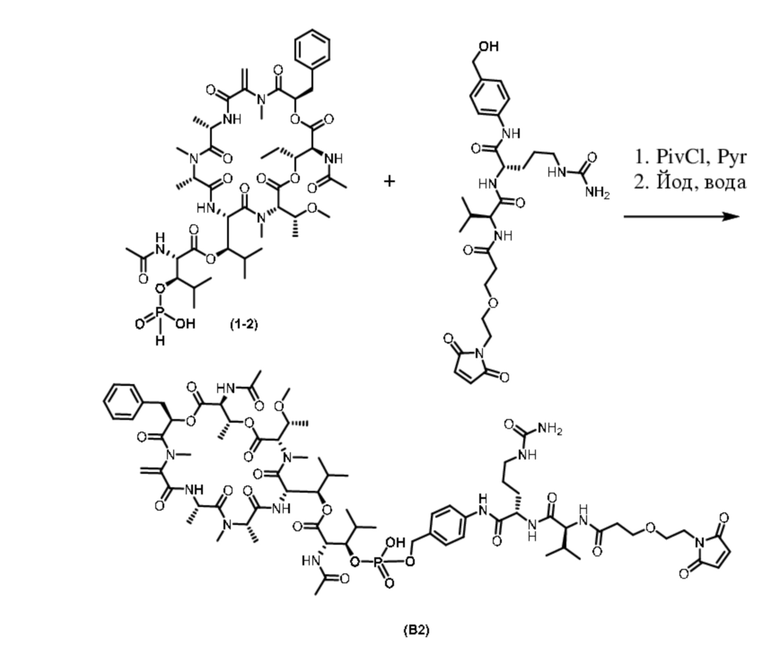
[00456] (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-2-ацетамидо-3-((гидроксигидрофосфорил)окси)-4-метилпентаноат (1-2) (50 мг, 0,049 ммоль, 1,0 экв.) и (S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-N-(4-(гидроксиметил)фенил)-5-уреидопентанамид (CAS № 2055041-37-5) (33,7 мг, 0,059 ммоль, 1,2 экв.) (оба лиофилизированных порошка) переносили во флакон объемом 10 мл и растворяли в пиридине (1 мл). Затем по каплям добавляли пивалоилхлорид (0,042 мл, 0,342 ммоль, 7 экв.) с получением бледно-желтого раствора. Смесь перемешивали при комнатной температуре в течение 30 минут. Свежеприготовленный раствор йода (49,6 мг, 0,195 ммоль, 4 экв.) в смеси пиридин-вода (20:1, 500 мкл) добавляли с получением темно-коричневого прозрачного раствора. Смесь перемешивали в течение 30 минут и непосредственно очищали с помощью обращенно-фазовой флэш-хроматографии (колонка C-18 40 г, 0% Ac/MeCN 3 минуты, затем 0-70% ACN/вода в течение 15 минут, нейтральный способ) с получением (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-22-изопропил-3-((R)-1-метоксиэтил)-4,9,10,12,16-пентаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((((4-((S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)(гидрокси)фосфорил)окси)-4-метил-2-пропионамидопентаноата (B2). HRMS; MH+ = 1595,7200, 2,25 мин.
Пример 1-3: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((((4-((S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)(гидрокси)фосфорил)окси)-4-метил-2-пропионамидопентаноата (B3)
(B3)
[00457] Соединение (B3) можно получить с использованием процедур, аналогичных способам, описанным в примере 1-1, за исключением стадии 1, где соединение (A3) (из примера 3-3) используют вместо соединения (A1).
Пример 1-4: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-22-изопропил-3-((R)-1-метоксиэтил)-4,9,10,12,16-пентаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((((4-((S)-2-((S)-2-(3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)(гидрокси)фосфорил)окси)-4-метил-2-пропионамидопентаноата (B4)
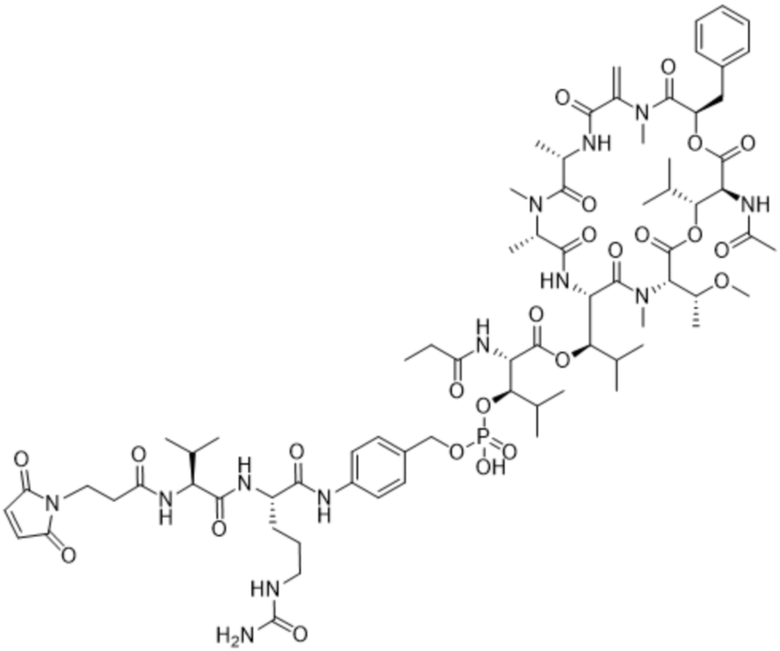 (B4)
(B4)
[00458] Соединение (B4) получали с использованием процедур, аналогичных способам, описанным в примере 1-1, за исключением стадии 2, где (S)-2-((S)-2-(3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропанамидо)-3-метилбутанамидо)-N-(4-(гидроксиметил)фенил)-5-уреидопентанамид (CAS № 1949793-46-7) использовали вместо (S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-N-(4-(гидроксиметил)фенил)-5-уреидопентанамида (CAS № 2055041-37-5). HRMS; MH+ = 1594,5400, 2,88 мин.
Пример 1-5: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-2-ацетамидо-3-((((4-((S)-2-((S)-2-(3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)(гидрокси)фосфорил)окси)-4-метилпентаноата (B5)
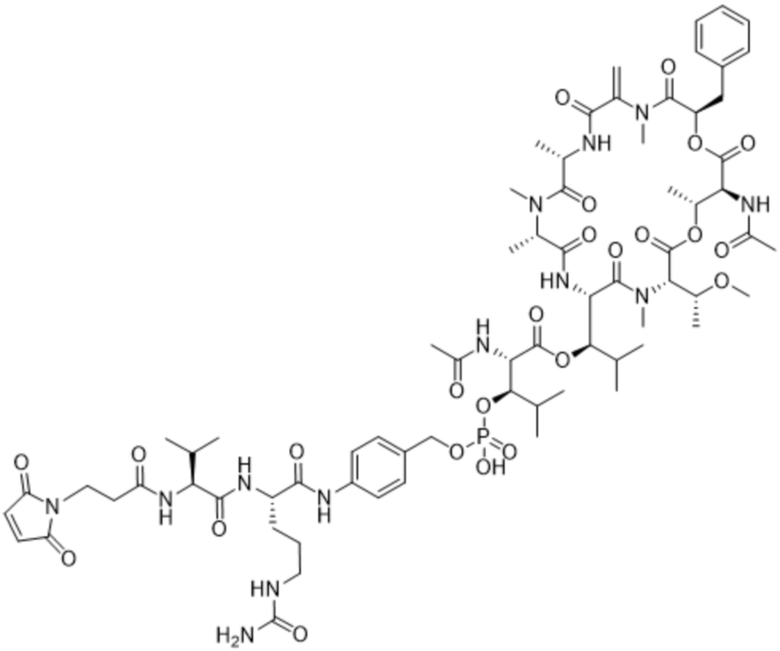 (B5)
(B5)
[00459] Соединение (B5) можно получить с использованием процедур, аналогичных способам, описанным в примере 1-1, за исключением стадии 1, где соединение (A2) (из примера 3-2) использовали вместо соединения (A1), и стадии 2, где (S)-2-((S)-2-(3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропанамидо)-3-метилбутанамидо)-N-(4-(гидроксиметил)фенил)-5-уреидопентанамид (CAS № 1949793-46-7) использовали вместо (S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-N-(4-(гидроксиметил)фенил)-5-уреидопентанамида (CAS № 2055041-37-5).
Пример 1-6: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((((4-((S)-2-((S)-2-(3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)(гидрокси)фосфорил)окси)-4-метил-2-пропионамидопентаноата (B6)
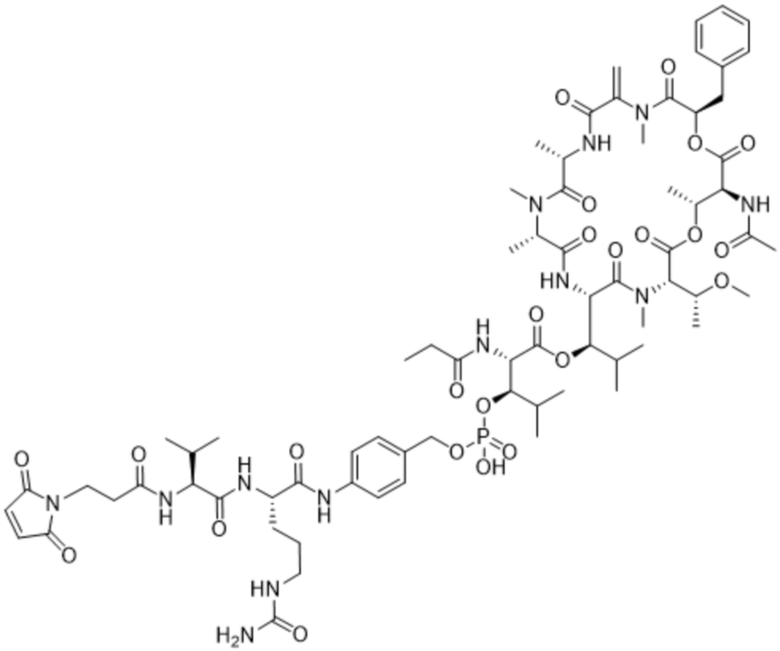 (B6)
(B6)
[00460] Соединение (B6) можно получить с использованием процедур, аналогичных способам, описанным в примере 1-1, за исключением стадии 1, где соединение (A3) (из примера 3-3) использовали вместо соединения (A1), и стадии 2, где (S)-2-((S)-2-(3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропанамидо)-3-метилбутанамидо)-N-(4-(гидроксиметил)фенил)-5-уреидопентанамид (CAS № 1949793-46-7) использовали вместо (S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-N-(4-(гидроксиметил)фенил)-5-уреидопентанамида (CAS № 2055041-37-5).
Пример 1-7: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-22-изопропил-3-((R)-1-метоксиэтил)-4,9,10,12,16-пентаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((((4-((S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)карбонил)окси)-4-метил-2-пропионамидопентаноата (B7)
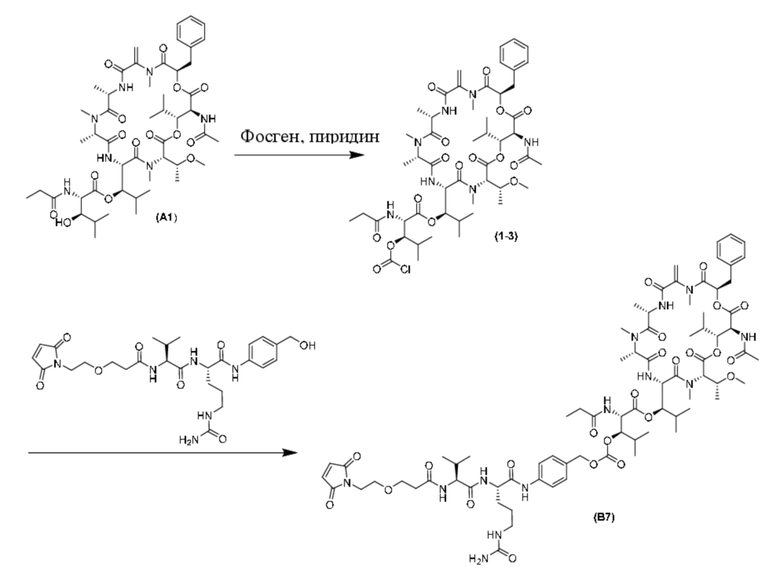
[00461] Соединение (B7) можно получить в результате взаимодействия хлорформиата (1-3) с (S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-N-(4-(гидроксиметил)фенил)-5-уреидопентанамидом (CAS № 2055041-37-5). Хлорформиат (1-3) можно получить в результате взаимодействия соединения (A1) с фосгеном.
Пример 1-8: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-2-ацетамидо-3-((((4-((S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)карбонил)окси)-4-метилпентаноата (B8)
(B8)
[00462] Соединение (B8) можно получить с использованием способа, описанного в примере 1-7, за исключением того, что соединение (A2) используют вместо соединения (A1).
Пример 1-9: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((((4-((S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)карбонил)окси)-4-метил-2-пропионамидопентаноата (B9)
(B9)
[00463] Соединение (B9) можно получить с использованием способа, описанного в примере 1-7, за исключением того, что соединение (A3) используют вместо соединения (A1).
Пример 1-10: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-22-изопропил-3-((R)-1-метоксиэтил)-4,9,10,12,16-пентаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((((4-((S)-2-((S)-2-(3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)карбонил)окси)-4-метил-2-пропионамидопентаноата (B10)
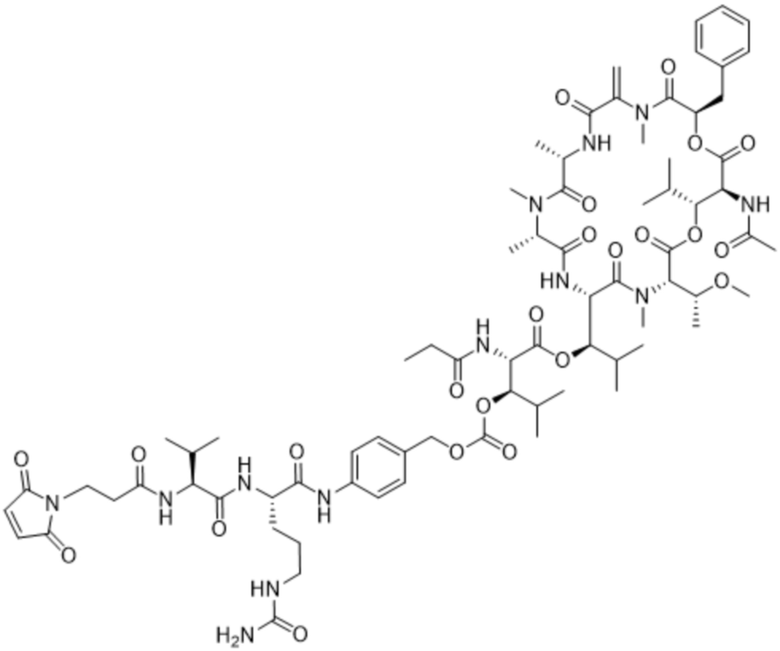 (B10)
(B10)
[00464] Соединение (B10) можно получить с использованием способа, описанного в примере 1-7, за исключением того, что (S)-2-((S)-2-(3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропанамидо)-3-метилбутанамидо)-N-(4-(гидроксиметил)фенил)-5-уреидопентанамид (CAS № 1949793-46-7) используют вместо (S)-2-((S)-2-(3-(2-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)этокси)пропанамидо)-3-метилбутанамидо)-N-(4-(гидроксиметил)фенил)-5-уреидопентанамида (CAS № 2055041-37-5).
Пример 1-11: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-2-ацетамидо-3-((((4-((S)-2-((S)-2-(3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)карбонил)окси)-4-метилпентаноата (B11)
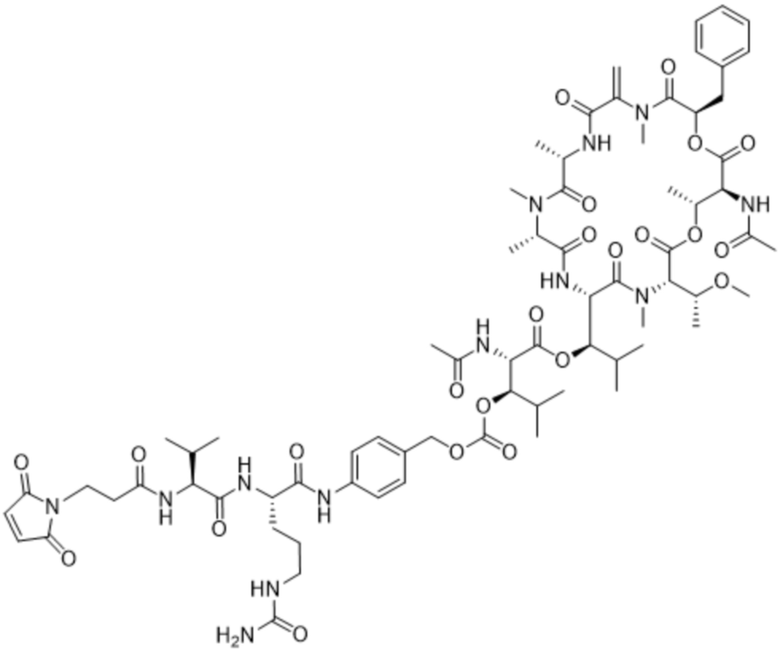 (B11)
(B11)
[00465] Соединение (B11) можно получить с использованием способа, описанного в примере 1-10, за исключением того, что соединение (A2) используют вместо соединения (A1).
Пример 1-12: синтез (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-((((4-((S)-2-((S)-2-(3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропанамидо)-3-метилбутанамидо)-5-уреидопентанамидо)бензил)окси)карбонил)окси)-4-метил-2-пропионамидопентаноата (B12)
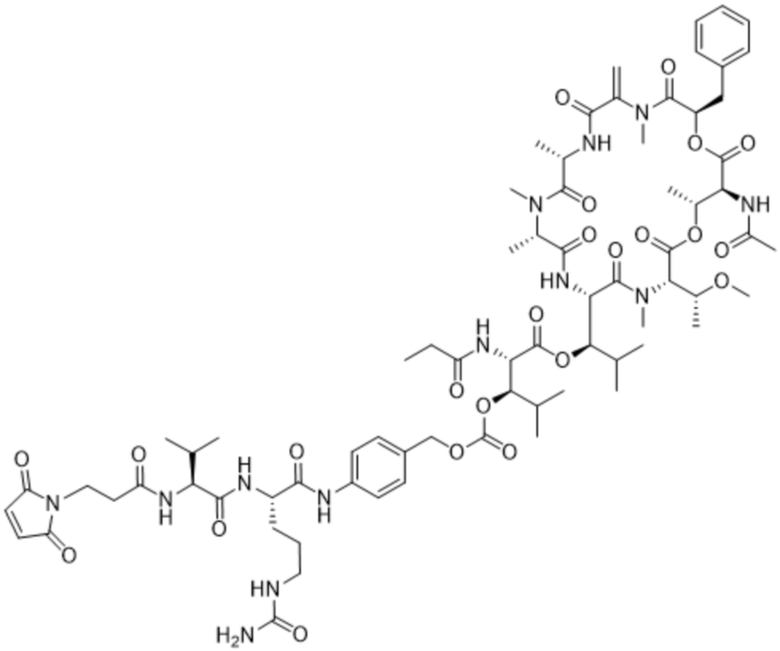 (B12)
(B12)
[00466] Соединение (B12) можно получить с использованием способа, описанного в примере 1-10, за исключением того, что соединение (A3) используют вместо соединения (A1).
Пример 2: получение антител к PMEL17
Пример 2-1: получение линий клеток, экспрессирующих PMEL17
[00467] Полноразмерные гены PMEL17 человека, макака-крабоеда и крысы синтезировали на основе аминокислотных последовательностей из баз данных GenBank или Uniprot . Все синтезированные фрагменты ДНК клонировали в подходящие векторы экспрессии.
[00468] Сконструированные линии клеток, стабильно экспрессирующие PMEL17, получали и культивировали в условиях отбора, подходящих для получения линий клеток, стабильно экспрессирующих PMEL17.
Пример 2-2: пэннинг целых клеток против PMEL17
[00469] Фагмидные библиотеки основаны на концепциях HuCAL PLATINUM® (Knappik et al., 2000) и Ylanthia (Tiller et al., 2013) и в них используют технологию CysDisplayTM для экспрессии Fab на поверхности фага (Lohning, 2001).
[00470] Для каждого пэннинга приблизительно 4×1013 HuCAL PLATINUM® фаг-антител или приблизительно 1×1014 Ylanthia® фаг-антител блокировали в PBS/5% FCS. Параллельно по 0,5-1,0×107 клеток-мишеней, экспрессирующих антиген PMEL17, и 0,5-1,0×107 адсорбционных клеток без экспрессии антигена PMEL17 на пул фагов ресуспендировали в 1 мл PBS/5% FCS для блокирования на льду. Блокированные целевые клетки центрифугировали, ресуспендировали в предварительно блокированных фаговых частицах и инкубировали в течение 2 ч при 4°С на встряхивателе. Комплексы фаг-клетка трижды промывали в PBS/5% FCS. Элюирование специфически связанного фага из клеток-мишеней проводили посредством 10-мин кислотного элюирования с 0,1 М глицин-HCl/0,5 М NaCl, pH 2,2. После центрифугирования супернатант (элюат) нейтрализовали добавлением 2 М незабуференного триса. Для удаления связывания фага с молекулами клеточной поверхности, отличными от целевого антигена, трижды проводили постадсорбцию с 0,5-1,0×107 адсорбционных клеток каждая. Конечный супернатант использовали для инфицирования 14 мл культуры E. coli TG1, выращенной до OD600 0,6-0,8. Культуру инкубировали 45 мин на водяной бане при 37°C для заражения фагами. Осадок бактерий ресуспендировали в 2x среде YT, высевали на чашки с агаром LB/Cam и инкубировали в течение ночи при 30°C. Колонии соскребали с чашек и использовали для высвобождения фага и амплификации фага. Амплифицированный фаг использовали для следующего цикла пэннинга. Второй и третий циклы пэннинга целых клеток проводили согласно протоколу для первого цикла.
Пример 2-3: субклонирование, экспрессия и скрининг Fab-фрагментов
[00471] Для обеспечения быстрой экспрессии растворимого Fab, кодирующие Fab вставки выбранного фага HuCAL PLATINUM® субклонировали из дисплейного вектора pMORPH®30 в вектор экспрессии pMORPH®x11_FH. Субклонирование выполняли посредством тройного расщепления с использованием EcoRI, XbaI и BmtI. После трансформации E. coli TG1-F- экспрессию отдельных клонов и получение периплазматических экстрактов, содержащих Fab-фрагменты HuCAL®, проводили так, как это описано ранее (Rauchenberger et al., 2003).
[00472] Кодирующие Fab вставки выбранного фага Ylanthia® субклонировали из дисплейного вектора pYPdis10 в вектор экспрессии pYBex10_Fab_FH. Субклонирование выполняли посредством тройного расщепления с использованием XbaI, EcoRI-HF и PstI-HF. После трансформации E. coli TG1-F- экспрессию отдельных клонов и получение периплазматических экстрактов, содержащих Fab-фрагменты Ylanthia®, проводили так, как это описано ранее (Rauchenberger et al., 2003).
[00473] Экспрессию Fab-фрагментов, кодируемых pMORPH®x11_Fab_FH и pYBex10_Fab_FH, в клетках E. coli TG1 F- осуществляли в культурах во встряхиваемых колбах с использованием 500 мл 2x среды YT, дополненной 0,1% глюкозой и 34 мкг/мл хлорамфеникола. Культуры встряхивали при 30°C до достижения значения OD600, составляющего 0,5. Экспрессию Fab индуцировали посредством добавления IPTG (изопропил-β-D-тиогалактопиранозид) в конечной концентрации 0,75 мM и дальнейшего культивирования в течение 20 ч при 30°C. Клетки собирали и разрушали с использованием лизоцима. His6-меченные (SEQ ID NO: 267) Fab-фрагменты выделяли с использованием IMAC (Bio-Rad, Германия) и элюировали с использованием имидазола. Замену буфера на 1x PBS Дульбекко (pH 7,2) проводили с использованием колонок PD10 (GE Healthcare, Германия). Образцы подвергали стерилизующей фильтрации (0,2 мкм). Концентрации белков определяли с помощью УФ-спектрофотометрии. Чистоту образцов анализировали в 15% геле для SDS-PAGE в денатурирующих восстанавливающих условиях. Гомогенность препаратов Fab определяли в нативном состоянии посредством эксклюзионной хроматографии (HP-SEC) с калибровочными стандартами.
[00474] При скрининге FACS очищенные Fab титровали на различных линиях клеток, экспрессирующих PMEL17, и не экспрессирующих PMEL17 (для контроля). Клетки собирали с использованием Accutase, доводили до ~ 4×106 клеток/мл в буфере для FACS (PBS, 3% FCS и 0,02% азид натрия) и хранили на льду, чтобы избежать интернализации. 15 мкл клеточной суспензии на лунку переносили в 384-луночные планшеты с V-образным дном (Greiner, кат. № 781280) и инкубировали с 15 мкл Fab в различных концентрациях (в основном от 200 до 3,5×10-3 нм) в течение 1 часа при 4°C, осторожно встряхивая. После инкубации клетки трижды промывали буфером для FACS. После каждой стадии промывки клетки центрифугировали (250 x g, 4 мин, 4°C) и осторожно ресуспендировали. Добавляли 15 мкл детектирующего антитела, конъюгированного с PE (PE-конъюгированное козье антитело к человеческому IgG, специфичное к F(ab`)2-фрагменту, 1:150 в буфере для FACS; Jackson Immuno Research, кат. № 109-116-097), или Alexa Fluor (Alexa Fluor-конъюгированное козье антитело к человеческому IgG, специфичное к F(ab`)2-фрагменту, 1:150 в буфере для FACS; Jackson Immuno Research, кат. № 109-606-097) и образцы инкубировали от 45 минут до 1 часа на льду в темноте, осторожно встряхивая. После 3 стадий промывки клетки ресуспендировали в 30 мкл буфера для FACS и образцы измеряли с использованием устройства IntelliCyt HTFC.
[00475] Для дополнительной проверки специфичности идентифицированных антител антиген иммунологически иммобилизовали из клеточного лизата и использовали в качестве материала покрытия для скрининга на основе ELISA. Клетки, экспрессирующие PMEL17 и не экспрессирующие PMEL17 (используемые в качестве отрицательного контроля), центрифугировали (250 g, 5 мин) и ресуспендировали в лизирующем буфере (трис-лизирующий буфер MSD, Meso Scale Discovery, R60TX-2), содержащем ингибиторы протеаз (полная смесь ингибиторов протеаз без EDTA, Roche, 11 873 580 001), от 1 х 107 до 1 х 108 клеток/мл и инкубировали 30 мин при 4°C. Лизат центрифугировали при 13000 об/мин в течение 5 минут для удаления остатков клеток и супернатант разделяли на аликвоты и хранили при -80°C. Предварительные испытания показали, что лизат можно замораживать и оттаивать 3 раза без значительной потери сигнала в ELISA.
[00476] Для проведения скрининга на основе ELISA IgG к His иммобилизовали в течение ночи в 384-луночном планшете maxisorb (R&D systems, MAB050) 10 мкг/мл, 20 мкл на лунку. Планшеты блокировали 3% BSA и 20 мкл очищенного Fab переносили в разных концентрациях (в основном от 400 до 0,2 нМ) в каждую лунку на 1 час. Планшет промывали 3 раза и добавляли 20 мкл лизата клеток, содержащего PMEL17, в концентрации 2×105 лизированных клеток/лунка в течение 1 часа. После 3 дополнительных промывок присутствие PMEL17 выявляли с использованием средства на основе биотинилированного антитела к PMEL17 (5 мкг/мл, 20 мкл на лунку) и стрептавидина-ECL (Dianova, 13MSA37, 1:1500, 20 мкл на лунку). В планшет добавляли 20 мкл на лунку 1X буфера для считывания MSD (Meso Scale Discovery, R92TC-2) и сигналы регистрировали с использованием MSD Sector Imager 6000.
Пример 2-4: превращение в IgG
[00477] Превращение выбранных Fab в IgG осуществляли посредством способа на основе ПЦР в 96-луночном формате. Векторы для бактериальной экспрессии Fab pMORPH®x11_FH и pYBex10_Fab_FH преобразовывали в векторы для экспрессии IgG у млекопитающих, pMORPH®4 и pYMex10 (для клонов HuCAL и Ylanthia соответственно).
[00478] ДНК плазмиды pMORPHx11_FH сперва амплифицировали посредством ПЦР с использованием биотинилированного праймера, специфичного для лидерной области phoA, и небиотинилированного праймера, специфичного для бактериального домена CL. Амплифицированный продукт захватывали на гранулах со стрептавидином, расщепляли с использованием BsiWI или HpaI (для клонов VLkappa или VLlambda соответственно), промывали и затем снова расщепляли с использованием MfeI. Данная процедура привела к высвобождению в супернатант очищенного каркаса вектора, в котором уже отсутствовала бактериальная константная область легкой цепи (CL) и лидерная последовательность тяжелой цепи phoA. Затем каппа- или лямбда-специфическую кассету экспрессии pIN у млекопитающего клонировали в каркас вектора, несущий CL млекопитающего, сайт polyA, промотор CMV и лидерную последовательность тяжелой цепи млекопитающих. На второй стадии ПЦР вновь созданную вставку Fab снова амплифицировали с использованием биотинилированного праймера, специфичного для области CH1, и небиотинилированного праймера, связывающегося внутри бактериальной лидерной последовательности ompA. ПЦР-продукт захватывали на гранулах со стрептавидином, расщепляли с использованием EcoRV, промывали и расщепляли с использованием BlpI, что приводило к высвобождению очищенной вставки в супернатант. И наконец, вставки клонировали в акцепторный вектор Fab_Cys для экспрессии в клетках млекопитающих.
[00479] ДНК плазмиды pYBex10_Fab_FH сперва амплифицировали посредством ПЦР с использованием биотинилированного праймера, специфичного для лидерной области phoA, и небиотинилированного праймера, специфичного для бактериального домена CL. Амплифицированный продукт захватывали на гранулах со стрептавидином, расщепляли с использованием NheI, промывали и затем снова расщепляли с использованием KpnI. Данная процедура привела к высвобождению в супернатант очищенного каркаса вектора, в котором уже отсутствовала бактериальная константная область легкой цепи (CL) и лидерная последовательность тяжелой цепи phoA. Затем каппа- или лямбда-специфическую кассету экспрессии pIN у млекопитающего клонировали в каркас вектора, несущий CL млекопитающего, сайт polyA, промотор CMV и лидерную последовательность тяжелой цепи млекопитающих. На второй стадии ПЦР вновь созданную вставку Fab снова амплифицировали с использованием биотинилированного праймера, специфичного для области CH1, и небиотинилированного праймера, связывающегося внутри бактериальной лидерной последовательности ompA. ПЦР-продукт захватывали на гранулах со стрептавидином, расщепляли с использованием XhoI, промывали и расщепляли с использованием NdeI, что приводило к высвобождению очищенной вставки в супернатант. И наконец, вставки клонировали в акцепторный вектор Fab_Cys для экспрессии в клетках млекопитающих.
[00480] После трансформации клеток E. coli XL-1 blue отдельные клоны контролировали посредством ПЦР для отбора колоний и секвенирования полной области вставки.
Пример 2-5: подтверждающий скрининг IgG человека
[00481] Препараты ДНК отдельных колоний получали с использованием соответствующего набора для подготовки ДНК в сочетании с устройством BioRobot®8000. Индивидуальные концентрации ДНК определяли посредством УФ-спектрофотометрии. Эукариотические клетки HEK293 c18 (ATCC № CRL-10852) использовали в 96-луночной системе экспрессии для создания кондиционированных супернатантов клеточных культур, содержащих полноразмерный IgG. Эукариотические клетки HEK293 c18 высевали в 96-луночный планшет с плоским дном до плотности ~4×104 клеток/50 мкл/лунка накануне и трансфицировали равными количествами ДНК-вектора экспрессии Ig. После инкубации в течение 40-50 часов при 37°C и 6% CO2 супернатанты культур переносили в 96-луночный планшет с U-образным дном и очищали центрифугированием. Полученные в результате супернатанты Ig тестировали посредством ELISA с захватом антителами к Fd для расчета концентрации Ig в соответствии с известными стандартами и хранили при -20°C для последующего использования в анализах специфичности и/или функционального скрининга.
[00482] ДНК представляющих интерес клонов подвергали секвенированию с праймером CMV_HC_for (CTC TAG CGC CAC CAT GAA ACA (SEQ ID NO: 264)) для домена VH и IgG_const_for (AGC CCA GCA ACA CCA AGG (SEQ ID NO: 265)) для Fc-домена с последующим секвенированием легкой цепи с праймером промотора T7 (TAA TAC GAC TCA CTA TAG GG (SEQ ID NO: 266)), чтобы получить полную информацию о последовательности IgG.
[00483] FACS-скрининг проводили в формате 384-луночного планшета с использованием платформы для скрининга HTFC от IntelliCyt. Клетки собирали с использованием Accutase, доводили до ~ 4×106 клеток/мл в буфере для FACS (PBS, 3% FCS и 0,02% азид натрия) и хранили на льду, чтобы избежать интернализации. 15 мкл клеточной суспензии на лунку переносили в 384-луночные планшеты с V-образным дном (Greiner, кат. № 781280) и инкубировали с 15 мкл супернатанта, содержащего IgG (или разбавленные очищенные контрольные антитела), в течение 1 часа при 4°C, осторожно встряхивая. После инкубации клетки трижды промывали буфером для FACS. После каждой стадии промывки клетки центрифугировали (250 x g, 4 мин, 4°C) и осторожно ресуспендировали. Добавляли 15 мкл детектирующего антитела, конъюгированного с PE (PE-конъюгированное козье антитело к человеческому IgG, специфичное к F(ab`)2-фрагменту, 1:150 в буфере для FACS; Jackson Immuno Research, кат. № 109-116-097), и образцы инкубировали от 45 минут до 1 часа на льду в темноте, осторожно встряхивая. После 3 стадий промывки клетки ресуспендировали в 30 мкл буфера для FACS и образцы измеряли с использованием устройства IntelliCyt HTFC.
Пример 2-6: получение IgG человека и Fab_Cys человека
[00484] Эукариотические клетки HKB11 трансфицировали ДНК-вектором экспрессии pMORPH®4 или pYMex10, кодирующим как тяжелую, так и легкую цепи IgG. Супернатант клеточной культуры собирали в день 3 или 7 после трансфекции. Супернатант клеточной культуры подвергали стандартной аффинной хроматографии с белком А (MabSelect SURE, GE Healthcare) и Capture Select IgG-CH1 (ВАС) для IgG человека и Fab_Cys человека соответственно. Если не указано иное, проводили замену буфера на 1x PBS Дульбекко (pH 7,2, Invitrogen) и образцы подвергали стерилизующей фильтрации (размер пор 0,2 мкм). Чистоту IgG анализировали в денатурирующих, восстанавливающих и невосстанавливающих условиях с использованием Labchip System (GXII, Perkin Elmer, США) или в SDS-PAGE. Концентрации белка определяли посредством УФ-спектрофотометрии и выполняли HP-SEC для анализа препаратов IgG в нативном состоянии.
Пример 2-7: получение антител G1 и G4 к PMEL17
[00485] Для первого цикла пэннинга приблизительно 4×1013 фага HuCAL PLATINUM® блокировали с использованием PBS/5% FBS. Параллельно по 1,0×107 клеток-мишеней, экспрессирующих антиген PMEL17, и 1,5-3,0×107 адсорбционных клеток без экспрессии антигена PMEL17 на пул фагов ресуспендировали в 1 мл PBS/5% FCS для блокирования на льду. Для предварительной очистки от неспецифического связывания фаг инкубировали с 0,5×107 адсорбционных клеток без экспрессии антигена PMEL17 на пул фага в течение 30 минут на льду. После инкубации клетки осаждали центрифугированием и супернатант добавляли к свежему образцу 0,5×107 адсорбционных клеток без экспрессии антигена PMEL17. Использовали те же условия инкубации, и предварительную абсорбцию снова использовали по отношению к свежему образцу 0,5×107 адсорбционных клеток без экспрессии антигена PMEL17, всего в течение трех циклов предварительной абсорбции. После окончательной предварительной абсорбции супернатант добавляли к 1,0×107 клеток-мишеней, экспрессирующих антиген PMEL17, и инкубировали на льду в течение 2 часов с периодическим перемешиванием. Комплексы фаг-клетка трижды промывали в PBS/5% FCS. Элюирование специфически связанного фага из клеток-мишеней проводили путем 10-минутного кислотного элюирования с 0,1 М глицин-HCl/0,5 М NaCl, pH 2,5. После центрифугирования супернатант (элюат) нейтрализовали добавлением 2 М незабуференного триса, и этот элюат использовали для инфицирования 14 мл культуры E. coli TG1 F', выращенной до OD600 0,6-0,8. Культуру инкубировали в течение 20 минут при 37°C, а затем еще 25 минут при 37°C со встряхиванием при 200 об./мин для инфицирования фагом. Осадки бактерий ресуспендировали в 2x среде YT, высевали на чашки с агаром LB/хлорамфеникол и инкубировали в течение ночи при 30°C. Колонии соскребали с чашек и использовали для высвобождения фага и амплификации фага. Амплифицированный фаг использовали для следующего цикла пэннинга. Второй и третий циклы пэннинга выполняли аналогично за исключением того, что для уменьшения фона и повышения эффективности предварительной очистки использовали только приблизительно 1×1012 фага HuCAL. После последнего цикла пэннинга плазмидную ДНК получали из каждого выхода фага и Fab-содержащие вставки клонировали в бактериальный вектор экспрессии. После лигирования и трансформации отдельные колонии собирали с посевом в 2XT/хлорамфеникол и культивировали. Экспрессию Fab индуцировали добавлением 0,25 мМ IPTG в культурах, выращенных в течение ночи в 96-луночных микротитровальных планшетах при 25°C. Осадки клеток лизировали 0,1% лизоцима в PBS и затем блокировали добавлением BSA до конечной концентрации 1%. Окрашивание FACS проводили на экспрессирующих PMEL17 и контрольных клетках.
Пример 2-8: кажущиеся аффинности антител к PMEL17
[00486] Очищенные IgG титровали на различных линиях клеток, экспрессирующих PMEL17 и не экспрессирующих PMEL17 (для контроля), для определения значений EC50. Клетки собирали с использованием Accutase, доводили до ~ 4×106 клеток/мл в буфере для FACS (PBS, 3% FCS и 0,02% азид натрия) и хранили на льду, чтобы избежать интернализации. 15 мкл клеточной суспензии на лунку переносили в 384-луночные планшеты с V-образным дном (Greiner, кат. № 781280) и инкубировали с 15 мкл IgG в различных концентрациях (в основном от 200 до 3,5×10-3 нм) в течение 1 часа при 4°C, осторожно встряхивая. После инкубации клетки трижды промывали буфером для FACS. После каждой стадии промывки клетки центрифугировали (250 х g, 5 мин, 4°C) и осторожно ресуспендировали. Добавляли 15 мкл детектирующего антитела, конъюгированного с PE (PE-конъюгированное козье антитело к человеческому IgG, специфичное к F(ab`)2-фрагменту, 1:150 в буфере для FACS; Jackson Immuno Research, кат. № 109-116-097), или Alexa Fluor (Alexa Fluor-конъюгированное козье антитело к человеческому IgG, специфичное к F(ab`)2-фрагменту, 1:150 в буфере для FACS; Jackson Immuno Research, кат. № 109-606-097) и образцы инкубировали от 45 минут до 1 часа на льду в темноте, осторожно встряхивая. После 3 стадий промывки клетки ресуспендировали в 30 мкл буфера для FACS и образцы измеряли с использованием устройства IntelliCyt HTFC. Значения EC50 рассчитывали с использованием программного обеспечения Graphpad Prism. Линия клеток меланомы G-361 экспрессирует PMEL17 человека на более низком уровне по сравнению с линией клеток HKB11, сверхэкспрессирующей PMEL17 человека, и, следовательно, позволяет более точно ранжировать клоны в зависимости от их кажущейся аффинности к клеткам. В таблице 3 показано, что были достигнуты различные значения EC50 в диапазоне от ~ 200 пМ до ~ 70 нМ.
Таблица 3. Кажущиеся аффинности (FACS, EC50 (нM)) и перекрестная реактивность антител к PMEL17.
(Alexa Fluor)
(Alexa Fluor)
(мышь)
(Alexa Fluor)
Пример 2-9: перекрестная реактивность антител к PMEL17
[00487] Перекрестную специфичность IgG оценивали в дозозависимом FACS G-361 (меланома человека), HKB11-PMEL17 яванского макака, HKB11-PMEL17 крысы и B16-F10 (меланома, экспрессирующая PMEL17 мыши) (таблица 3). Тестирование на G-361 проводили с использованием детектирующего антитела, меченного PE. В отличие от этого необходимо было использовать детектирующее антитело, меченное Alexa-Fluor, для тестирования кандидатов на других клеточных линиях с тем, чтобы достичь обнаруживаемых сигналов.
[00488] IgG классифицировали на несколько групп эпитопов в соответствии с их профилями перекрестной специфичности. Тем не менее, различные профили перекрестной реактивности IgG предполагают, что их мишенью являются по меньшей мере 4 эпитопа (человек/макак/крыса/мышь, человек/макак/крыса, человек/макак/мышь и человек/макак).
Пример 2-10: анализ ADC
[00489] IgG тестировали дозозависимо в отношении их способности к интернализации и индуцированию гибели различных клеток, экспрессирующих PMEL17, как в анализе ADC с полезной нагрузкой, так и после прямой конъюгации.
[00490] Для анализа ADC с полезной нагрузкой использовали соединение Fab-ZAP (козье антитело к человеческому mAb, соединенное с сапорином; ATS Biotechnology, кат. № IT-51), которое специфически связывается с Fc человека и соединено с цитотоксическим элементом сапорином. Клетки собирали с использованием Accutase и доводили до 5×104 клеток/мл. По 50 мкл суспензии клеток переносили на лунку в 96-луночный белый планшет с плоским прозрачным дном (Corning, 3610) и инкубировали в течение ночи при 37°C, 5% CO2. На следующий день антитела-кандидаты инкубировали при различных концентрациях (чаще всего от 44 до 7×10-3 нМ) с соединением Fab-ZAP при 8 нМ в течение 30 минут при 37°C. Затем к клеткам-мишеням добавляли по 50 мкл на лунку комплексов IgG-Fab-ZAP. Для контроля готовили лунки только с клетками, с клетками, инкубированными только с IgG-кандидатами (= контроль 100% жизнеспособности), и с клетками, инкубированными только с Fab-ZAP (для проверки неспецифического уничтожения вторичным реагентом). Конечная концентрация IgG составляла от 22 до 3,5×10-3 нМ и 4 нМ Fab-ZAP. Планшеты инкубировали 72 ч при 37°C и 5% CO2. Количество жизнеспособных клеток оценивали с использованием CellTiter-Glo (Promega, № G7571), а люминесценцию детектировали посредством Tecan Infinite 500. Затем жизнеспособность нормализовали относительно контроля клетки+только IgG.
[00491] Почти все IgG могли эффективно уничтожать клетки меланомы G-361 (максимальное уничтожение >80%) и демонстрировали ограниченное уничтожение клеток 293T, не экспрессирующих PMEL17 (максимальное уничтожение <30%). Уничтожение G-361 также достигалось в диапазоне EC50, что может быть связано с аффинностью или эпитопом. Перекрестная специфичность также может быть подтверждена для большинства IgG, реакционноспособных в отношении человека/макака/крысы/мыши, человека/макака/крысы и человека/макака/мыши, т. е. IgG, специфичные для PMEL17 мыши, могут также уничтожать B16-F10 (меланома мыши).
Пример 2-11: конструирование антител к PMEL17
[00492] Как правило, все процессы конструирования выполняли с использованием стратегий на основе ПЦР. Процессы конструирования включали следующие аспекты: приведение антител к зародышевой линии, удаление сайтов PTM, конструирование pI и оптимизация кодонов. После синтеза и сборки посредством ПЦР с перекрывающимися праймерами повторно сконструированные фрагменты VH и VL субклонировали в соответствующие каркасы вектора для последующей экспрессии IgG.
Пример 3: синтез соединений A1-A3
Пример 3-1: способ выделения (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-22-изопропил-3-((R)-1-метоксиэтил)-4,9,10,12,16-пентаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-гидрокси-4-метил-2-пропионамидопентаноата (A1) из сушеных листьев Ardisia crenata
(A1)
[00493] Соединение A1 выделяли посредством способов, описанных в публикации патента Японии № JP62283999. Стадия 1: экстрагирование: 25 кг сушеных листьев Ardisia crenata измельчали до мелкого порошка и экстрагировали с использованием 500 л метанола. После фильтрации экстракт выпаривали досуха. Остаток растворяли в 100 л этилацетата и пять раз экстрагировали с использованием 100 л деионизированной воды (добавляли хлорид натрия для улучшения разделения фаз). Органический слой выпаривали досуха. Остаток растворяли в 50 л смеси ацетонитрил/вода (9/1) и экстрагировали с использованием 50 л гептана. Слой ацетонитрил/вода выпаривали до тех пор, пока не оставалась только вода. Затем его экстрагировали с использованием 50 л этилацетата. Слой этилацетата упаривали досуха с получением 109 г неочищенного экстракта.
[00494] Стадия 2: обезжиривание: неочищенный экстракт растворяли в 25 л смеси ацетонитрил/вода (9/1) и трижды экстрагировали с использованием 25 л гептана. Слой ацетонитрил/вода выпаривали до тех пор, пока не оставалась только вода. Затем его экстрагировали с использованием 25 л этилацетата. Слой этилацетата упаривали досуха с получением 40 г неочищенного экстракта.
[00495] Стадия 3: флэш-хроматография: неочищенный экстракт растворяли в смеси ацетон/метанол (1/1), адсорбировали на 100 г Isolute (диатомитовая земля) и выпаривали досуха. Поводили флэш-хроматографию на силикагеле с тройной системой растворителей с циклогексаном (элюент A), этилацетатом (элюент B) и метанолом (элюент C) (подробности эксперимента: колонка: RediSep Rf 120 г; скорость потока: 85 мл/мин; градиент: 0 мин: 75% A, 25% B, 0% C; 3 мин: 75% A, 25% B, 0% C; 10 мин: 0% A, 100% B, 0% C; 17,5 мин: 0% A, 90% B 10% C; 25 мин: 0% A, 50% B, 50% C; 30 мин: 0% A, 50% B, 50% C; различные сегменты градиента линейно связаны во времени). Провели четыре цикла с 10 г экстракта в каждом. Использовали фракционирование по времени, и собранные фракции анализировали с использованием UPLC-UV-MS на присутствие соединения (A1). Фракции, содержащие соединение (A1), объединяли и выпаривали досуха, получая фракцию весом 25 г.
[00496] Стадия 4: эксклюзионная хроматография (SEC): фракцию весом 25 г растворяли в 400 мл метанола и дополнительно фракционировали посредством SEC на колонке (длина 25 см, диаметр 12,5 см), заполненной сефадексом LH20 и метанолом в качестве элюента. Собирали фракции по 200 мл каждая, и собранные фракции анализировали посредством UPLC-UV-MS на присутствие соединения (A1). Фракции, содержащие соединение (A1), объединяли и выпаривали досуха, получая обогащенную фракцию весом 6,2 г.
[00497] Стадия 5: 1-я препаративная HPLC: обогащенную фракцию весом 6,2 г растворяли в 12 мл смеси метанол/диметилсульфоксид (1/1) и дополнительно фракционировали посредством препаративной HPLC (подробности эксперимента: колонка: Sunfire C18, 30×150 мм, размер частиц 5 мкм; элюент A: деионизированная вода с 0,1% муравьиной кислотой, элюент B: метанол с 0,1% муравьиной кислотой; скорость потока 60 мл/мин; градиент: 0 мин: 35% A, 65% B; 0,5 мин: 35% A, 65% B; 17 мин: 15% A, 85% B; 15,0 мин: 0% A, 100% B; 19 мин: 0% A, 100% B; 19,1 мин: 35% A, 65% B; 20 мин: 35% A, 65% B; различные сегменты градиента линейно связаны во времени). Фракционирование инициировали масс-спектрометрией. Провели 15 циклов с обогащенной фракцией весом 410 мг каждый. Фракции, содержащие соединение (A1), объединяли и выпаривали досуха с получением полуочищенной фракции весом 488 мг с расчетным содержанием соединения (A1) 70%.
[00498] Стадия 6: 2-я препаративная HPLC: Полуочищенную фракцию весом 488 мг растворяли в 4 мл метанола и дополнительно фракционировали посредством препаративной HPLC (подробности эксперимента: колонка: X-Select PFP, 19×250 мм, размер частиц 5 мкм; элюент A: деионизированная вода с 0,1% муравьиной кислотой, элюент B: ацетонитрил с 0,1% муравьиной кислотой; скорость потока 30 мл/мин; градиент: 0 мин: 45% A, 55% B; 0,5 мин: 45% A, 55% B; 24 мин: 25% A, 75% B; 24,0 мин: 0% A, 100% B; 27,5 мин: 0% A, 100% B; 27,6 мин: 45% A, 55% B; 30,5 мин: 45% A, 55% B; различные сегменты градиента линейно связаны во времени). Фракционирование инициировали масс-спектрометрией. Провели пять циклов с полуочищенной фракцией весом 98 мг каждый. Фракции, содержащие соединение (A1), объединяли и выпаривали досуха с получением 287 мг соединения (A1) чистотой > 95%.
[00499] Соединение (A1): время удерживания: 4,73 мин, молекулярная формула [M+H]+: C49H76N7O15+, расчетная моноизотопная масса [M+H]+: 1002,5394 Да, фактическая масса: 1002,5391 Да. Подробности эксперимента: колонка: ACQUITY UPLC BEH C18, 2,1×100 мм, размер частиц 1,7 мкм; элюент A: деионизированная вода с 0,1% муравьиной кислотой, элюент B: ацетонитрил с 0,1% муравьиной кислотой; скорость потока 0,9 мл/мин; линейный градиент: 0 мин: 95% A, 5% B до 6,4 мин: 0% A, 100% B. 1H ЯМР (600 МГц, Ацетонитрил-d3) δ 8,36 (d, J=9,4 Гц, 1H), 7,50 (d, J=10,0 Гц, 1H), 7,38-7,22 (m, 6H), 6,94 (d, J=4,4 Гц, 1H), 6,77 (d, J=9,9 Гц, 1H), 5,39 (d, J=9,8 Гц, 1H), 5,34 (dd, J=8,8, 3,8 Гц, 1H), 5,31-5,25 (m, 2H), 5,21-5,15 (m, 2H), 5,11 (dd, J=9,9, 1,9 Гц, 1H), 4,90-4,84 (m, 1H), 4,70 (q, J=6,9 Гц, 1H), 4,39 (dd, J=7,7, 2,1 Гц, 1H), 4,10 (d, J=9,8 Гц, 1H), 3,82-3,74 (m, 1H), 3,60 (ddd, J=9,8, 4,4, 2,0 Гц, 1H), 3,38 (s, 3H), 3,23 (s, 3H), 3,16-3,11 (m, 1H), 2,94-2,85 (m, 1H), 2,84 (s, 3H), 2,66 (s, 3H), 2,55-2,43 (m, 2H), 2,16 (s, 3H), 1,96-1,90 (m, 1H), 1,89-1,74 (m, 2H), 1,37 (d, J=7,0 Гц, 3H), 1,32 (d, J=6,6 Гц, 3H), 1,19 (d, J=6,5 Гц, 3H), 1,17 (d, J=6,0 Гц, 3H), 1,12 (d, J=7,7 Гц, 3H), 1,06 (d, J=6,8 Гц, 3H), 0,96 (d, J=6,6 Гц, 3H), 0,88 (d, J=6,6 Гц, 3H), 0,85 (d, J=7,0 Гц, 3H), 0,77 (d, J=6,5 Гц, 3H).
Пример 3-2: способ получения (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-2-ацетамидо-3-гидрокси-4-метилпентаноата (A2)
(A2)
Соединение (A2) получали посредством способов, описанных в [M. Taniguchi et al., Tetrahedron 59 (2003) 4533-4538]. Выделение проводили посредством способов, описанных для соединения (A1) в примере 3-1. Затем материал характеризовали посредством UPLC-UV-HRMS, 1D-ЯМР- и 2D-ЯМР-экспериментов. Соединение (A2): время удерживания: 4,20 мин, молекулярная формула [M+H]+: C46H70N7O15+, расчетная моноизотопная масса [M+H]+: 960,4924 Да, фактическая масса: 960,4914 Да. 1H ЯМР (600 МГц, 1,4-диоксан-d8) δ 8,36 (d, J=9,2 Гц, 1H), 7,37 (d, J=10,0 Гц, 1H), 7,31-7,21 (m, 4H), 7,21-7,16 (m, 2H), 6,79-6,72 (m, 2H), 5,40-5,37 (m, 1H), 5,36-5,33 (m, 1H), 5,30-5,28 (m, 2H), 5,20 (dd, J=9,0, 3,7 Гц, 1H), 5,13 (d, J=1,9 Гц, 1H), 5,04 (dd, J=9,9, 1,4 Гц, 1H), 4,89-4,82 (m, 1H), 4,73 (q, J=6,9 Гц, 1H), 4,39 (dd, J=8,0, 2,0 Гц, 1H), 4,04 (d, J=9,9 Гц, 1H), 3,80-3,73 (m, 1H), 3,65 (ddd, J=9,8, 4,3, 1,9 Гц, 1H), 3,37 (s, 3H), 3,21 (s, 3H), 3,11 (dd, J=14,7, 3,7 Гц, 1H), 2,89 (dd, J=14,7, 8,9 Гц, 1H), 2,84 (s, 3H), 2,62 (s, 3H), 2,14 (s, 3H), 2,12 (s, 3H), 1,99-1,90 (m, 1H), 1,75-1,69 (m, 1H), 1,35 (d, J=6,8 Гц, 3H), 1,30 (d, J=6,5 Гц, 3H), 1,18-1,14 (m, 6H), 1,11 (d, J=6,5 Гц, 3H), 1,04 (d, J=6,7 Гц, 3H), 0,86 (d, J=6,8 Гц, 3H), 0,78 (d, J=6,6 Гц, 4H).
Пример 3-3: способ получения (R)-1-((3S,6S,9S,12S,18R,21S,22R)-21-ацетамидо-18-бензил-3-((R)-1-метоксиэтил)-4,9,10,12,16,22-гексаметил-15-метилен-2,5,8,11,14,17,20-гептаоксо-1,19-диокса-4,7,10,13,16-пентаазациклодокозан-6-ил)-2-метилпропил-(2S,3R)-3-гидрокси-4-метил-2-пропионамидопентаноата (A3)
(A3)
Соединение (A3) получали посредством способов, описанных в [M. Taniguchi et al. Tetrahedron 59 (2003) 4533-4538]. Выделение проводили посредством способов, описанных для соединения (A1) в примере 3-1. Затем материал характеризовали посредством UPLC-UV-HRMS, 1D-ЯМР- и 2D-ЯМР-экспериментов. Соединение (A3): время удерживания: 4,42 мин, молекулярная формула [M+H]+: C47H72N7O15+, расчетная моноизотопная масса [M+H]+: 974,5081 Да, фактическая масса: 974,5098 Да. 1H ЯМР (600 МГц, 1,4-диоксан-d8) δ 8,37 (d, J=9,1 Гц, 1H), 7,38 (d, J=9,9 Гц, 1H), 7,31-7,23 (m, 4H), 7,22-7,16 (m, 1H), 7,12 (d, J=7,7 Гц, 1H), 6,78-6,72 (m, 2H), 5,41-5,33 (m, 2H), 5,32-5,26 (m, 2H), 5,20 (dd, J=8,8, 3,7 Гц, 1H), 5,12 (d, J=2,2 Гц, 1H), 5,04 (dd, J=9,9, 1,7 Гц, 1H), 4,89-4,81 (m, 1H), 4,72 (q, J=7,0 Гц, 1H), 4,40 (dd, J=7,8, 2,1 Гц, 1H), 4,04 (d, J=9,8 Гц, 1H), 3,81-3,73 (m, 1H), 3,68-3,64 (m, 1H), 3,38 (s, 3H), 3,21 (s, 3H), 3,11 (dd, J=14,6, 3,7 Гц, 1H), 2,89 (dd, J=14,7, 8,9 Гц, 1H), 2,83 (s, 3H), 2,62 (s, 3H), 2,51-2,41 (m, 2H), 2,14 (s, 3H), 1,99-1,89 (m, 1H), 1,78-1,68 (m, 1H), 1,34 (d, J=6,9 Гц, 3H), 1,30 (d, J=6,5 Гц, 3H), 1,18-1,14 (m, 6H), 1,13-1,08 (m, 6H), 1,04 (d, J=6,7 Гц, 3H), 0,86 (d, J=6,8 Гц, 3H), 0,78 (d, J=6,6 Гц, 3H).
Пример 4: способ получения конъюгатов антитела к PMEL17 и лекарственного средства
[00500] Антитело инкубировали со смолой с белком A RMP (GE) в соотношении 10 мг Ab и 1 мл смолы в PBS в течение 15 минут при перемешивании в одноразовой колонке подходящего размера. Цистеин-HCl добавляли до конечной концентрации 20 мМ и инкубировали при перемешивании в течение 30 мин при комнатной температуре, давая возможность реакционноспособным цистеинам деблокироваться. Смолу быстро промывали 50 объемами колонки PBS на вакуумном коллекторе. Затем смолу ресуспендировали в равном объеме PBS, содержащем 250 нМ CuCl2. Преобразование межцепочечных дисульфидов антител контролировали по временным точкам. В каждой временной точке извлекали 25 мкл суспензии смолы, добавляли 1 мкл 20 мМ соединения (B1) или соединения (B2) и пробиркой осторожно постукивали несколько раз. Смолу центрифугировали, супернатант удаляли, а затем элюировали с использованием 50 мкл элюирующего буфера для антител (Thermo). Смолу осаждали и супернатант анализировали посредством обращенно-фазовой хроматографии с использованием колонки Agilent PLRP-S 4000A, 5 мкм, 4,6×50 мм (буфер A представляет собой воду, 0,1% TFA, буфер B ацетонитрил, 0,1% TFA, колонку выдерживают при 80°C, скорость потока 1,5 мл/мин).
[00501] Как только определяли, что межцепочечные дисульфидные связи антитела преобразованы, смолу промывали 10 объемами колонки PBS, ресуспендировали в равном объеме PBS и добавляли 8 эквивалентов полезной нагрузки с линкером (20 мМ) в DMSO, а затем инкубировали при комнатной температуре 2 часа. Затем смолу промывали 50 объемами колонки PBS. ADC элюировали из смолы с белком А посредством буфера для элюирования антител и нейтрализовали 1/10 объема 1 М трис, pH 9,0. Затем в ADC проводили замену буфера на PBS или другой подходящий буфер, и при необходимости проводили препаративную эксклюзионную хроматографию для удаления агрегатов (S200 Increase; GE). Проводили следующие анализы: аналитическую SEC для определения процентного содержания мономера, масс-спектроскопию для определения DAR, тест LAL для определения нагрузки эндотоксином и определяли концентрацию белка с использованием коэффициента экстинкции при A280 и молекулярной массы антитела.
Пример 5: in vitro активность ингибиторов GNAQ/11, соединения (A1) и соединения (A2) в отношении увеальной меланомы
[00502] Клетки увеальной меланомы 92.1 и аденокарциномы протоков поджелудочной железы MIAPACA высевали с низкой плотностью клеток в 96-луночные планшеты и обрабатывали возрастающими концентрациями соединения (A1) или соединения (A2) согласно указанному. После обработки лекарственным средством в течение 96 или 120 часов жизнеспособность и пролиферацию клеток определяли с использованием анализа жизнеспособности на основе резазурина. Если вкратце, клетки инкубировали с раствором на основе резазурина и изменение цвета выявляли по оптической плотности посредством спектрофотометра и использовали для регистрации жизнеспособности клеток. Данные представлены в виде среднего значения для 3 независимых повторов и относительно клеток, обработанных PBS (контроль). В таблице 4 показана концентрация, вызывающая 50% подавление роста (GI50), для соединения (A2) и соединения (A1). Соединение (А2) и соединение (А1) проявляли сильную зависимую от мишени активность против UM (фиг. 1).
Таблица 4. Подавляющая рост активность соединения (A1) и соединения (A2) в клетках UM
GI50, нM
GI50, нM
Пример 6: соединение (A1) и соединение (A2) индуцируют апоптоз в клетках увеальной меланомы
[00503] Клетки 92.1 и MIAPACA высевали в 96-луночные планшеты (5000 клеток на лунку) и обрабатывали возрастающими концентрациями соединения (A1), соединения (A2) и фиксированной дозой стауроспорина (100 нМ, положительный контроль) согласно указанному. После обработки лекарственными средствами в течение 96 ч клетки анализировали посредством проточной цитометрии с активацией флуоресценции с использованием антитела к аннексину V, конъюгированного с флуоресцентным красителем. Данные представлены в виде среднего значения для 3 независимых повторов. Соединение (A2) и соединение (A1) индуцировали апоптоз в клетках UM GNAQ/11-зависимым образом (фиг. 2).
Пример 7: анализ ингибирования GNAQ/11 соединением (A1) и соединением (A2) в клетках увеальной меланомы
[00504] Увеальную меланому 92.1 обрабатывали возрастающими концентрациями соединения (А1) или соединения (А2) согласно указанному. После обработки лекарственными средствами в течение ночи клетки обрабатывали для определения уровней IP1 с использованием TR-FRET (резонансный перенос энергии флуоресценции с временным разрешением) или уровней белка посредством вестерн-блоттинга. На фиг. 3А показаны уровни IP1 (нМ) в клетках 92.1, обработанных DMSO (контроль), соединением (А2) или соединением (A1). На фиг. 3B показана корреляция между уровнями IP1 и относительной пролиферацией в обработанных соединением (A1) клетках 92.1. На фиг. 3С показаны иммуноблоты образцов клеток 92.1, обработанных соединением (А1) и соединением (А2), для определения влияния на передачу сигналов ERK.
Пример 8: метаболическая стабильность и PK-показатели соединения (A1)
[00505] Стабильность соединения (A1) в плазме крови исследовали в плазме крови мышей, крыс, обезьян и человека и сравнивали с буфером. Как исчезновение соединения (A1) (фиг. 4A), так и появление формы соединения (A1) с раскрытым кольцом, имеющей структуру соединения (A4) 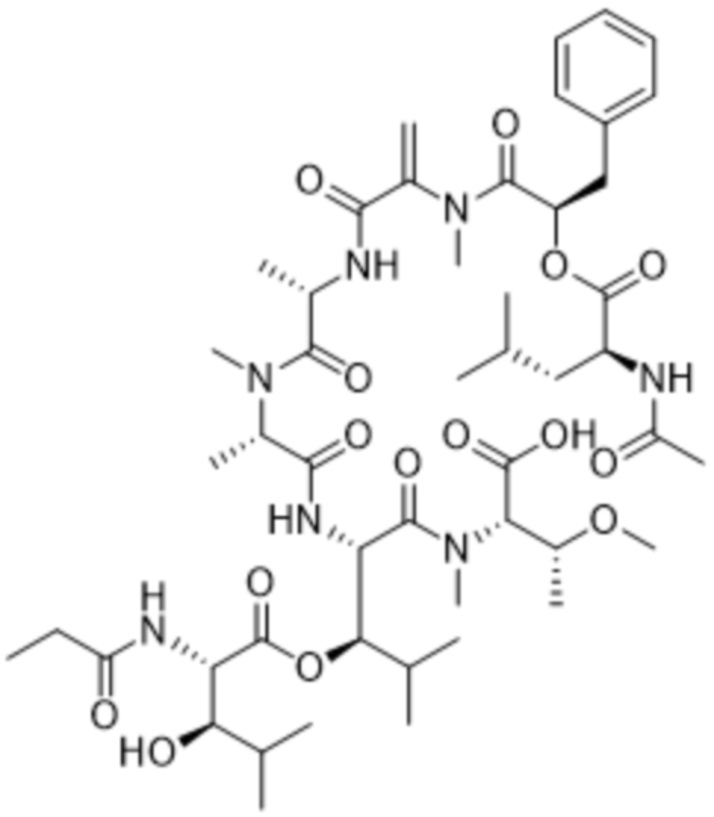 , соединения (A6)
, соединения (A6) 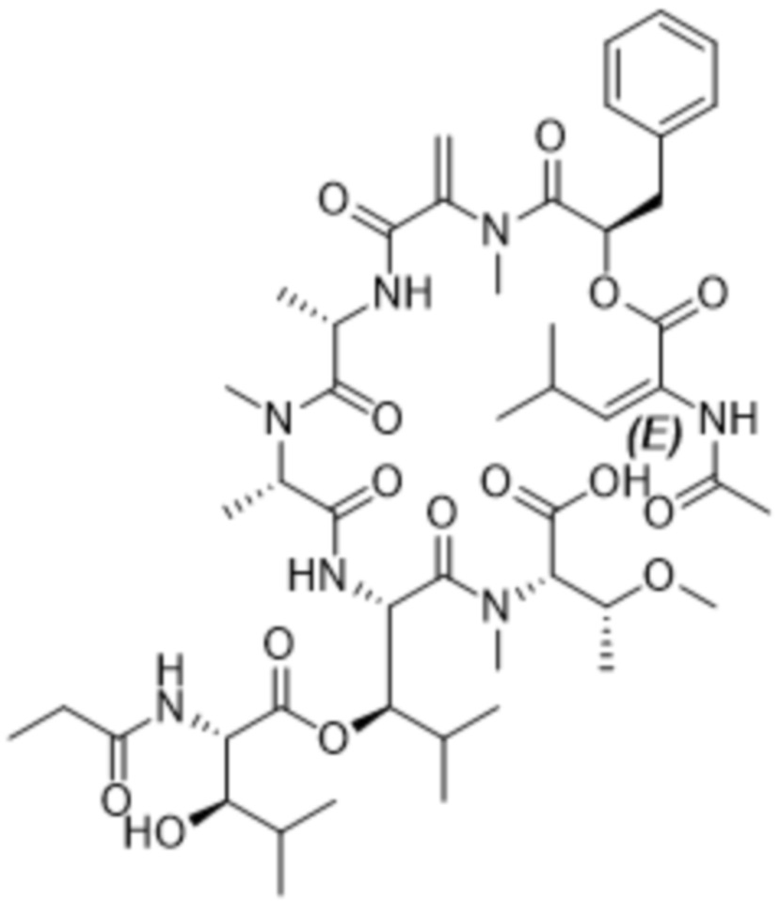 или соединения (A8)
или соединения (A8) 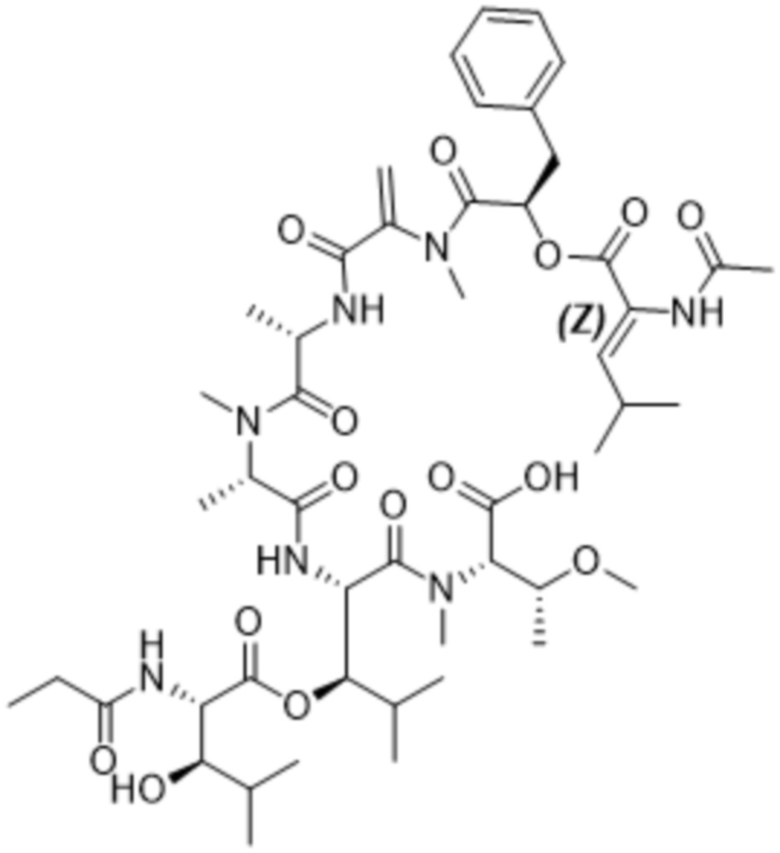 (фиг. 4B), контролировали в течение 24 ч. Очевидно, что трансформация в плазме крови человека происходит несколько быстрее, чем у других видов. За исключением крыс суммирование % оставшегося соединения (A1) и % образовавшегося соединения (A8) характеризуется стехиометрией в течение 24 ч, указывая на то, что какие-либо другие преобразования, по-видимому, не играют значительной роли (фиг. 4C).
(фиг. 4B), контролировали в течение 24 ч. Очевидно, что трансформация в плазме крови человека происходит несколько быстрее, чем у других видов. За исключением крыс суммирование % оставшегося соединения (A1) и % образовавшегося соединения (A8) характеризуется стехиометрией в течение 24 ч, указывая на то, что какие-либо другие преобразования, по-видимому, не играют значительной роли (фиг. 4C).
[00506] PK соединения (A1) после внутривенного введения мышам характеризовалась очень высоким клиренсом и объемом распределения от умеренного до высокого. Наблюдалось несколько непропорциональное увеличение воздействия доз в диапазоне 0,2-0,8 мг/кг (фиг. 4D). Ниже приведена таблица 5, в которой обобщены все показатели PK.
Таблица 5. PK-показатели соединения (A1)
Пример 9: метаболическая стабильность и PK-показатели соединения (A2)
[00507] In vitro стабильность соединения (A2) тестировали в плазме крови и в крови различных видов (фиг. 5A) Соединение (A2) показало приемлемую химическую стабильность в трех различных системах (фиг. 5B). PK соединения (A2) у самок мышей balb/c показала высокий клиренс и короткий период полувыведения (фиг. 5C). В таблице 6 обобщены некоторые показатели PK. Соединение (А2) показало высокий истинный клиренс в микросомах печени мыши, крысы и человека, но низкий клиренс в гепатоцитах, вероятно, из-за ограниченной проницаемости мембраны (таблица 7). Инкубация соединения (A2) с печеночной фракцией S9 и с плазмой крови показала аналогичные периоды полужизни. Соединение (A2) и соединение (A1) были стабильны в буфере при pH 5,6 и в лизосомах в течение 4 ч (фиг. 5D).
Таблица 6. PK-показатели соединения (A2) у самок мышей balb/c (1 мг/кг в/в)
Таблица 7. Стабильность соединения (A2)
Пример 10: in vitro активность ADC к PMEL17-(B1) в отношении увеальной меланомы
[00508] Родительские и не воздействующие целенаправленно контрольные (NT) или с трансдукцией PMEL17-shRNA клетки 92.1 высевали с низкой плотностью клеток в 96-луночные планшеты и обрабатывали возрастающими концентрациями 3207-(B1) (изотипический контроль), G4-(B1) и G1-(B1) в присутствии или в отсутствие доксициклина согласно указанному. После обработки лекарственным средством в течение 96 или 120 часов жизнеспособность и пролиферацию клеток определяли с использованием анализа жизнеспособности на основе резазурина. Если вкратце, клетки инкубировали с раствором на основе резазурина и изменение цвета выявляли по оптической плотности посредством спектрофотометра и использовали для регистрации жизнеспособности клеток. Данные представлены в виде среднего значения для 3 независимых повторов и относительно клеток, обработанных PBS (контроль) (фиг. 6). ADC к PMEL17-(B1) ингибируют пролиферацию клеток увеальной меланомы зависимым от PMEL17 и дозы образом.
Пример 11: ADC к PMEL17-(B1) индуцируют апоптоз в клетках увеальной меланомы
[00509] Клетки 92.1 высевали в 96-луночные планшеты (5000 клеток на лунку) и обрабатывали возрастающими концентрациями соединения (A1), 3207-(B1) (изотипический контроль), G4-(B1) и G1-(B1) согласно указанному. После обработки лекарственными средствами в течение 96 ч клетки анализировали посредством проточной цитометрии с активацией флуоресценции с использованием антитела к аннексину V, конъюгированного с флуоресцентным красителем. Данные представлены в виде среднего значения для 3 независимых повторов. Как G4-(B1), так и G1-(B1) индуцировали апоптоз в клетках 92.1 дозозависимым образом (фиг. 7).
Пример 12: in vitro активность ADC к PMEL17-(B2) и mAb к PMEL17 в отношении увеальной меланомы
[00510] Клетки увеальной меланомы 92.1 высевали с низкой плотностью клеток в 96-луночные планшеты и обрабатывали возрастающими концентрациями 3207-(B2) (изотипический контроль), G4-(B2) и G1-(B2) и mAb к PMEL17-G1 и -G4 согласно указанному. После обработки лекарственным средством в течение 96 или 120 часов жизнеспособность и пролиферацию клеток определяли с использованием анализа жизнеспособности на основе резазурина. Данные представлены в виде среднего значения для 3 независимых повторов и относительно клеток, обработанных PBS (контроль) (фиг. 8). ADC к PMEL17-(B2) и mAb к PMEL17 проявляли минимальный антипролиферативный эффект в клетках увеальной меланомы.
Пример 13: анализ ингибирования GNAQ/11 с использованием ADC к PMEL17-(B1) и к PMEL17-(B2) в клетках увеальной меланомы
[00511] Увеальную меланому 92.1 обрабатывали DMSO, соединением (A1) (8 нM) и возрастающими концентрациями ADC изотипического-(B1), к PMEL17-(B1) и к PMEL17-(B2) согласно указанному. После обработки лекарственными средствами в течение 2 дней клетки обрабатывали для определения уровней IP1 с использованием TR-FRET (резонансный перенос энергии флуоресценции с временным разрешением). Уровни IP1 (нM) представлены в виде среднего значения для 3 независимых повторов. Как G1-(B1), так и G1-(B2) ингибируют мутантный GNAQ и GNA11 дозозависимым образом, о чем свидетельствуют сниженные уровни IP1 (фиг. 9).
Пример 14: анализ связывания антител к PMEL17 с интактными тромбоцитами и клетками увеальной меланомы
[00512] Линию клеток увеальной меланомы 92.1 и тромбоциты, выделенные из плазмы крови человека, богатой тромбоцитами (PRP от 3 доноров-людей), анализировали посредством проточной цитометрии с активированной флуоресценцией с использованием антител к PMEL17 G1 (черная линия) и G4 (серая линия) и вторичного антитела к IgG человека, конъюгированного с флуоресцентным красителем, или только конъюгированного вторичного антитела (светло-серая линия) (фиг. 10). В отличие от клеток увеальной меланомы 92.1, тромбоциты человека не демонстрируют экспрессию PMEL17 на клеточной поверхности.
Пример 15: влияние соединения (A1) и ADC к PMEL17-(B1) на агрегацию тромбоцитов человека
[00513] Богатую тромбоцитами плазму крови человека (PRP) инкубировали с ADC 17A9-(B1), Ab к PMEL, конъюгированным с B1, или соединением (A1) (0,1, 1 и 150 нМ) в течение 4, 16 или 24 ч. Затем измеряли агрегацию тромбоцитов после обработки активирующим тромбиновый рецептор пептидом 6 (TRAP6) в концентрации 32 мкМ. В то время как соединение (A1) индуцировало агрегацию тромбоцитов, антитело к PMEL17-(B1) не индуцировало агрегацию тромбоцитов в течение 24 часов (фиг. 11).
Пример 16: ADC к PMEL17-(B1) ингибируют рост опухоли на мышиной модели
[00514] Самок "голых" мышей с подкожными ксенотрансплантатами 92.1 с люциферазой обрабатывали посредством однократного в/в введения 3207-(B1), G1-(B1) или среды-носителя в указанных дозах. Обработку (однократную в/в инъекцию) проводили через 17 дней после инокуляции опухоли. Значения являются средними ± SEM; размер выборки (n=5-12 мышей в группе). Исходный объем опухоли в день 0 составлял примерно 200-250 мм3. G1-(B1) ингибировал рост опухоли дозозависимым образом (фиг. 12A).
[00515] Изменение веса тела самок наивных "голых" мышей после обработки с G1-(B1) контролировали при указанных дозах. Обработку (однократную в/в инъекцию) проводили в день 0. Значения являются средними ± SEM; размер выборки (n=4 мыши в группе). Исходный вес тела в день 0 составлял примерно 24 г. Вес тела измеряли каждый день в течение 14 дней после обработки лекарственным средством. В течение 14 дней после обработки потеря веса тела не наблюдалась (фиг. 12B).
[00516] Проводили иммуногистохимический анализ опухолевой ткани из подкожных ксенотрансплантатов 92.1 с люциферазой, обработанных средой-носителем, 3207-(B1) (7,5 мг/кг веса тела) или G1-(B1) (7,5 мг/кг веса тела). Опухоли фиксировали в 10% (об./об.) формалине с нейтральным буфером в течение 24 ч при комнатной температуре, затем промывали PBS, обрабатывали для обезвоживания, очищали и заливали в парафин. Готовили срезы толщиной 3 мкм и обрабатывали для окрашивания гематоксилином и эозином (Г+Э) и для иммуногистохимии. Иммуногистохимическое окрашивание проводили на фиксированных формалином и залитых парафином срезах ткани с использованием полностью автоматизированной системы Bond-RX (Leica) для окрашивания антителами к hIgG (ThermoFischer) и антителами к gp100 (PMEL17) (клон EP4863; Abcam) и Discovery XT (Ventana Medical System), полностью автоматизированной системы для окрашивания антителами к Ki67 (клон Sp6; NeoMarkers), антителами к pERK1/2 (клон D13.14.4E; Cell Signaling), антителами к MITF (Sigma) и антителами к расщепленной каспазе-3 (Cell Signaling). Если вкратце, обработка с использованием G1-(B1) приводила к ингибированию передачи сигналов GNAQ и ингибированию пролиферации опухолевых клеток, о чем свидетельствуют сниженные уровни pERK и Ki67 соответственно (фиг. 12C). Кроме того, G1-(B1) индуцировал апоптоз клеток по сравнению с мышами, обработанными средой-носителем и 3207-(B1), что коррелировало с накоплением в опухолевых клетках ADC G1-(B1), обнаруживаемого при окрашивании IgG (фиг. 12C). После обработки с G1-(A1) изменений в уровнях MITF и PMEL17 не наблюдалось (фиг. 12C).
[00517] Тестировали агрегацию тромбоцитов цельной крови у наивных мышей, обработанных средой-носителем, G1-(B1) (20 мг/кг веса тела) или соединением (A1) (0,01, 0,03, 0,1 и 0,3 мг/кг веса тела). Животных умерщвляли через 5 мин после однократного в/в введения и кровь собирали через полую вену. Через 1 ч измеряли агрегацию в цельной крови после активации тромбоцитов посредством ADP в концентрации 6,5 мкМ (фиг. 12D). В отличие от соединения (A1) G1-(B1) не оказывал какого-либо значительного влияния на ADP-индуцированную агрегацию тромбоцитов in vivo.
[00518] Измеряли агрегацию тромбоцитов цельной крови у наивных мышей, обработанных средой-носителем и G1-(B1) (7,5 и 30 мг/кг веса тела), через 24 часа или 7 дн. после введения дозы. Мышей умерщвляли через 24 ч или 7 дн. после однократной в/в инъекции и кровь собирали через полую вену. Через 1 ч измеряли агрегацию в цельной крови после активации тромбоцитов посредством ADP в концентрации 6,5 мкМ. У мышей, получавших G1-(B1), не наблюдалось ингибирования агрегации тромбоцитов в течение 7 дней (фиг. 12E).
Пример 17: влияние G1-(B1) на мышиной модели увеальной меланомы с метастазами в печень и легкие
[00519] Самкам мышей NOD-Scid внутривенно вводили клетки 92.1 с люциферазой (примерно 2 миллиона клеток на животное) и измеряли биолюминесценцию дважды в неделю для оценки образования опухоли. Через 45 дней у всех мышей с инъекцией развились метастазы в печень и легкие, что было обнаружено по сигналу биолюминесценции (BLI) (фиг. 13A и 13B). Мышей с опухолями обрабатывали однократной в/в инъекцией G1-(B1) (20 мг/кг веса тела) и биолюминесценцию контролировали с течением времени в качестве показателя прогрессирования опухоли. Отдельные изображения от каждой мыши представлены в день 45 после в/в инъекции клеток 92.1 с люциферазой (непосредственно перед началом обработки) и через 12 дней после обработки (фиг. 13A и 13B); размер выборки (n=6 мышей в группе). Исходный BLI для метастазов в печень в день 0 составил приблизительно 2,8*109 фотонов/сек/см2. Опухоли в легких (сигнал биолюминесценции) на фиг. 13B обозначены черной стрелкой. Снижение биолюминесценции in vivo и ex vivo указывает, что G1-(B1) индуцировал регрессию метастазов в печени и легких после однократной в/в дозы 20 мг/кг.
[00520] Соответствующее изменение веса тела (% по сравнению с днем 15) оценивали 2-3 раза в неделю до и после обработки с использованием 20 мг/кг G1-(B1) (серые круги). Значения являются средними ± SEM; размер выборки (n=5-6 мышей в группе). Исходный вес тела в день 15 составлял примерно 21 г. Обработка с использованием G1-(B1) привела к восстановлению веса тела модели увеальной меланомы с метастазами в печень и легкие.
Пример 18: PK-показатели ADC G1-(B1).
[00521] Фармакокинетический профиль G1-(B1) показал несколько непропорциональное увеличение воздействия в дозе от 7,5 до 30 мг/кг у "голых" мышей (фиг. 14A). Клиренс, объем распределения и период полужизни находятся в типичном диапазоне для ADC. В таблице 8 обобщены некоторые показатели PK.
[00522] У мышей с опухолью измеряли концентрации свободной полезной нагрузки после введения либо связывающего мишень G1-(B1), либо изотипического контроля 3207-(B1). Выраженное (> 4-кратное) увеличение доставки в опухоль полезной нагрузки соединения (A1) можно было наблюдать при использовании целенаправленно воздействующего ADC (фиг. 14B). Превращение соединения (A1) (незакрашенные круги) в его форму с раскрытым кольцом соединения (A1), имеющую структуру соединения (A4) , соединения (A6) или соединения (A8) (закрашенные круги), при конъюгировании с антителом было показано in vivo на мышах (фиг. 14C). Виды воздействия в исследовании эффективности in vivo, сравнивающем два разных формата DAR2 с форматом DAR4 для G1-(B1) и с форматом Fc-молчащий DAR4, показали наиболее низкий клиренс для ADC DAR2 (E152C) и Fc-молчащего DAR4, тогда как воздействие DAR2 (S375C) уменьшается быстрее (фиг. 14D). Из сравнения PK не нацеленных 3207-ADC (фиг. 14E) можно сделать вывод, что "молчание" Fc оказывает существенное влияние на снижение клиренса формата DAR4.
Таблица 8. PK-показатели ADC G1-(B1)
Пример 19: стабильность in vitro ADC к PMEL17-GNAQ/11i в буфере, плазме крови мыши, крысы и человека и стабильность in vivo ADC к PMEL17-GNAQ/11i у мышей
[00523] Антитело к PMEL17G1_E152C_S375C_(B2) (G1-(B2)) добавляли в концентрации 100 мкг/мл в соответствующую матрицу и инкубировали при 37ºC. Образцы отбирали в 0, 1, 2 и 4 часа и мгновенно замораживали при -70ºC. Аликвоты каждого образца подвергали иммунопреципитации с использованием гранул Capture Select FcXL и инкубировали с буфером, содержащим папаин, для высвобождения полезной нагрузки из ADC, захваченного гранулами. Затем высвободившуюся полезную нагрузку анализировали посредством HPLC MS для определения относительных уровней соединения (A2)-PO4 (т. e. 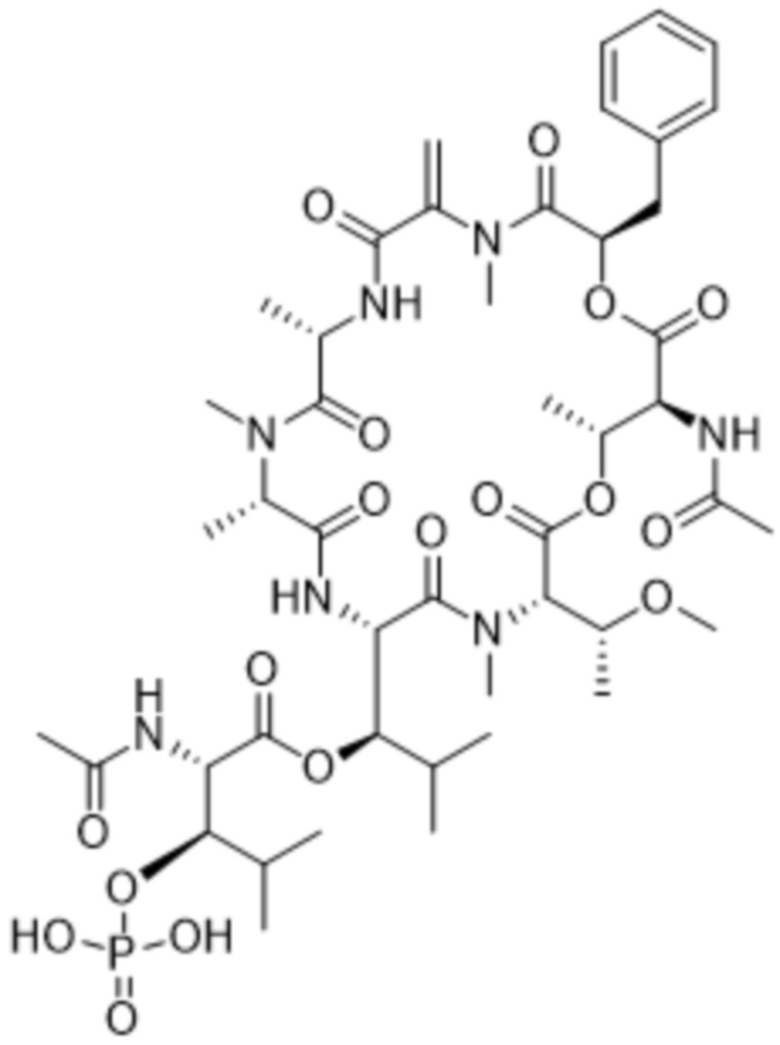 из
из 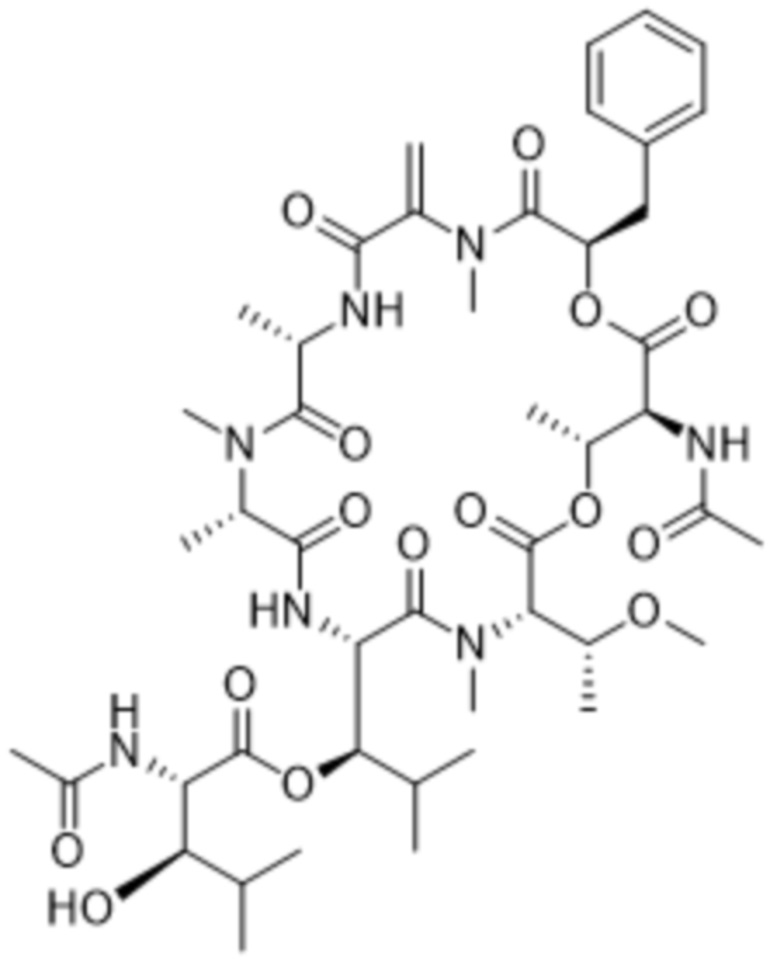 , см. пример 1.2) и соединения (A2)-PO4 с раскрытым кольцом, имеющего структуру соединения (A5)-PO4 (т. e. соединения (A5)-PO4
, см. пример 1.2) и соединения (A2)-PO4 с раскрытым кольцом, имеющего структуру соединения (A5)-PO4 (т. e. соединения (A5)-PO4 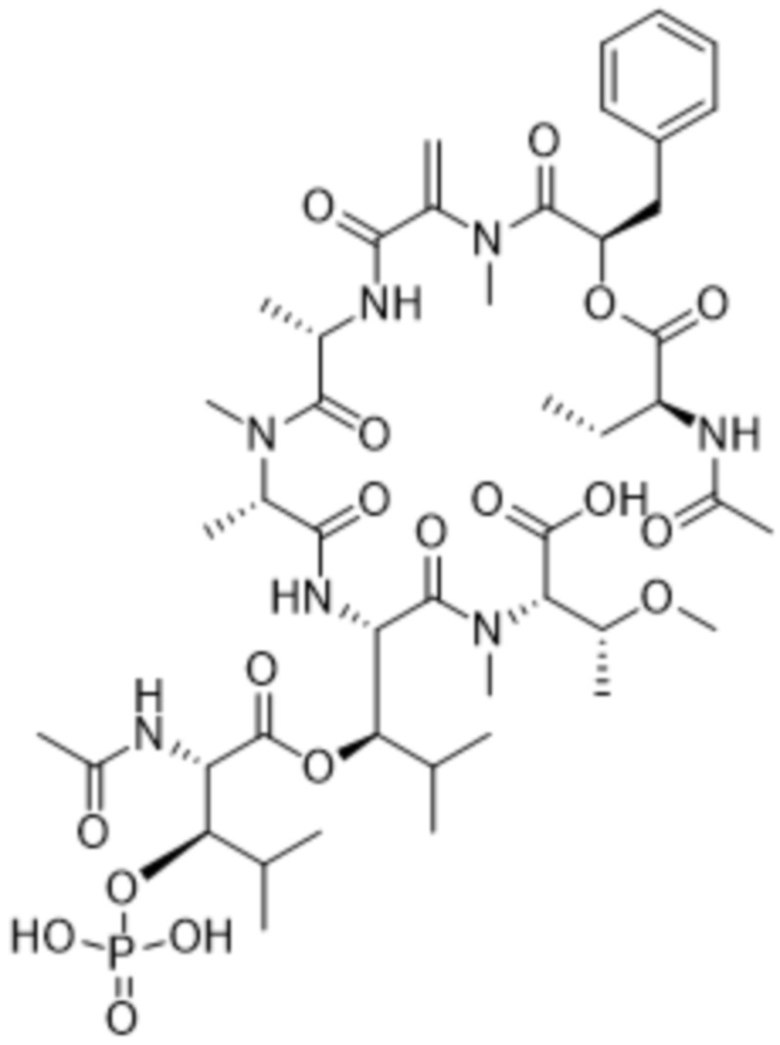 из
из 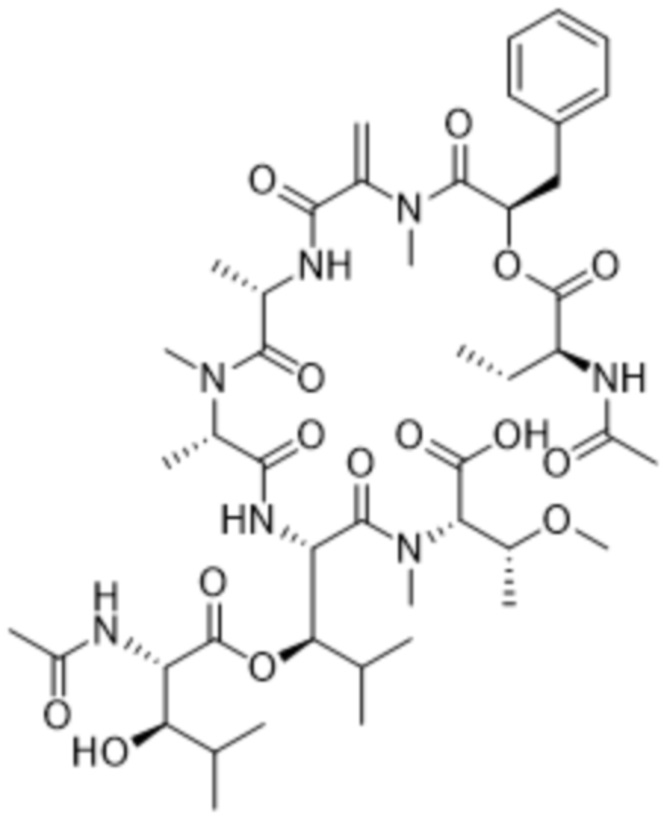 ), соединения (A7)-PO4
), соединения (A7)-PO4 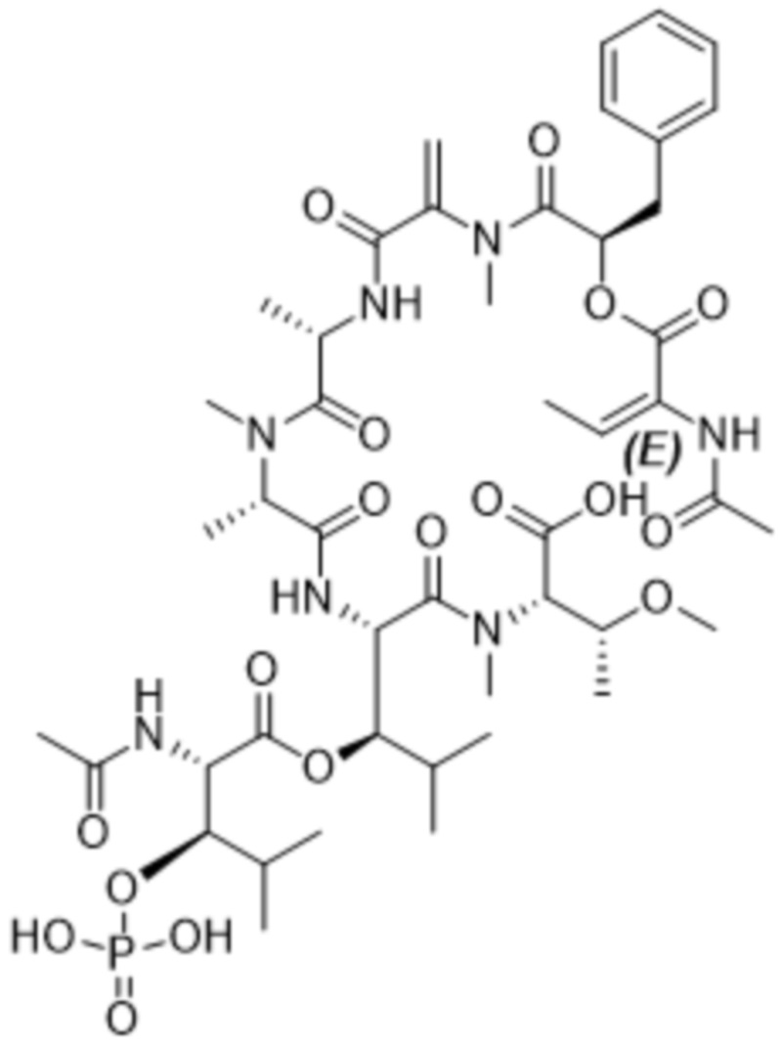 или соединения (A9)-PO4
или соединения (A9)-PO4 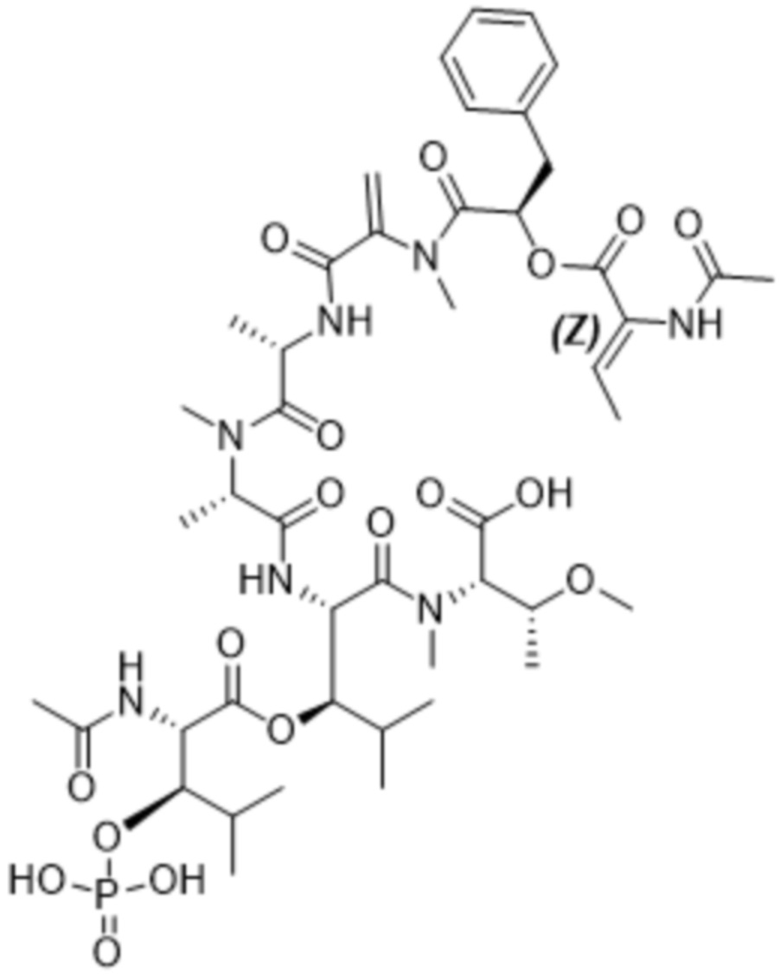 (где соединение (A2)-PO4 с раскрытым кольцом - неактивная полезная нагрузка). График отображает процент высвободившейся полезной нагрузки, которая присутствовала в виде соединения (A2)-PO4 на протяжении эксперимента (фиг. 15A).
(где соединение (A2)-PO4 с раскрытым кольцом - неактивная полезная нагрузка). График отображает процент высвободившейся полезной нагрузки, которая присутствовала в виде соединения (A2)-PO4 на протяжении эксперимента (фиг. 15A).
[00524] Антитело к PMEL17G1_E152C_S375C_(B1) (G1-(B1)), антитело к PMEL17G1_E152C_(B1) (G1-E152C-DAR2-(B1)), антитело к PMEL17G1_S375C_(B1) (G1-S375C-DAR2-(B1)) и Fc-молчащее антитело к PMEL17G1_E152C_S375C_(B1) (Fc-молчащий G1-(B1)) добавляли в концентрации 100 мкг/мл в соответствующую матрицу и инкубировали при 37ºC. Образцы собирали в 0, 2, 4 и 6 часов и мгновенно замораживали при -70ºC. Аликвоты каждого образца подвергали иммунопреципитации с использованием гранул Capture Select FcXL и инкубировали с буфером, содержащим папаин, для высвобождения полезной нагрузки из ADC, захваченного гранулами. Затем высвободившуюся полезную нагрузку анализировали посредством HPLC MS для определения относительных уровней соединения (A1)-PO4 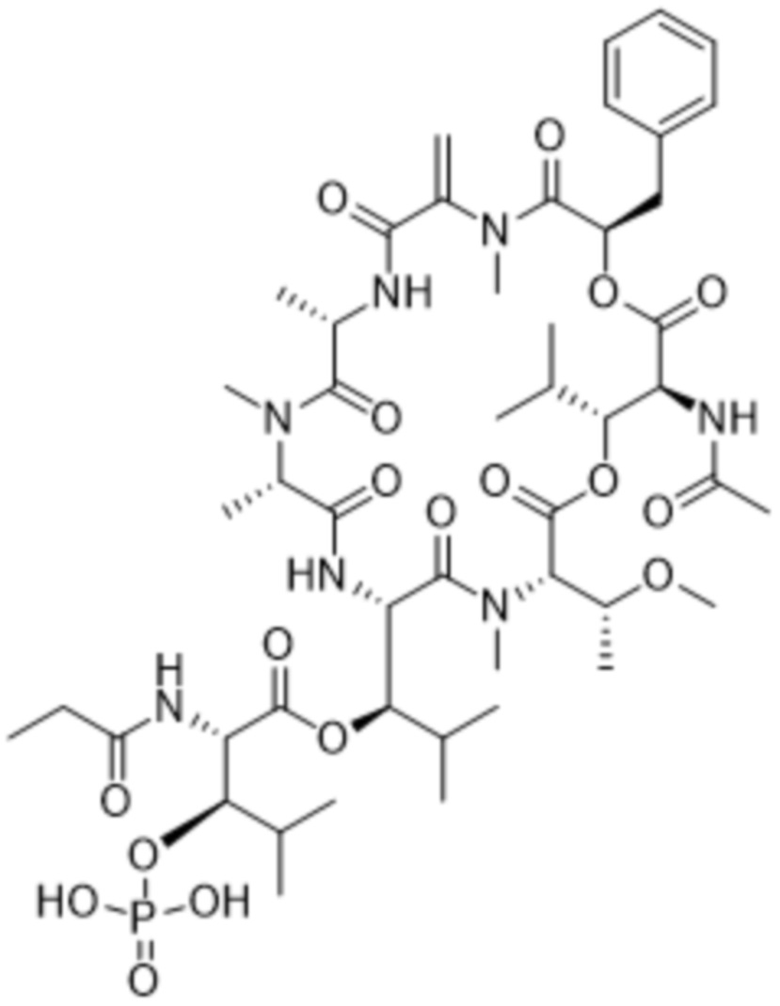 и соединения (A1)-PO4 с раскрытым кольцом, имеющего структуру соединения (A4)-PO4
и соединения (A1)-PO4 с раскрытым кольцом, имеющего структуру соединения (A4)-PO4 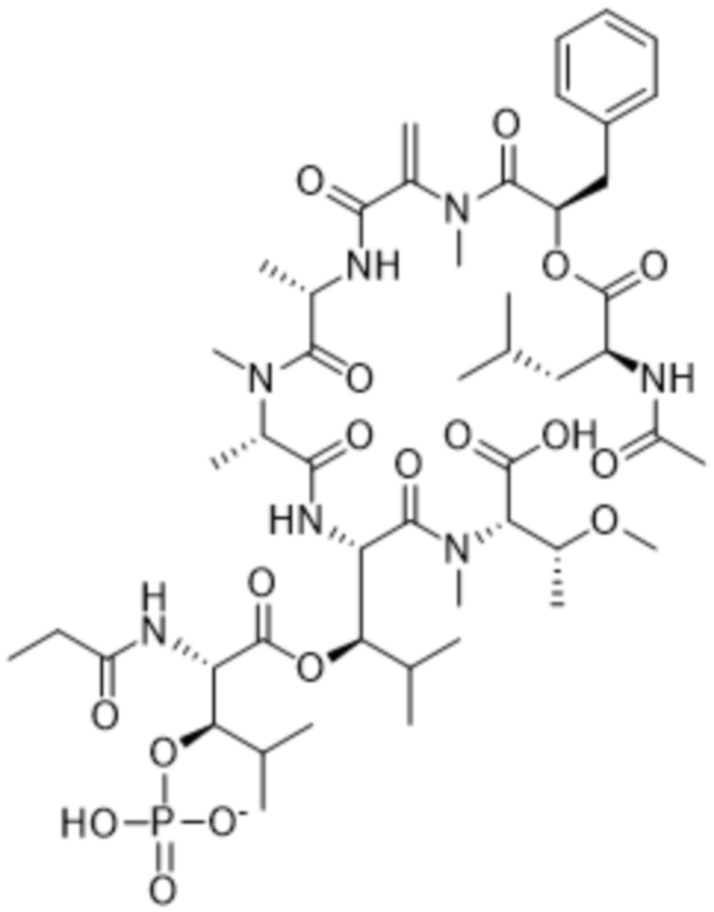 , соединения (A6)-PO4
, соединения (A6)-PO4 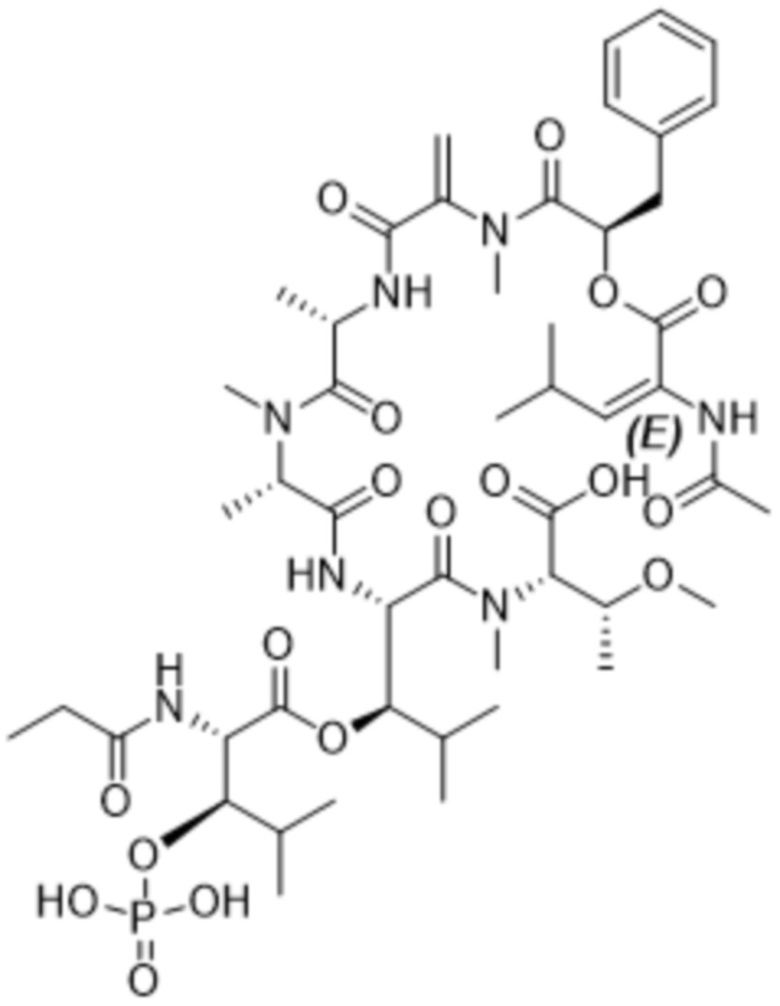 , или соединения (A8)-PO4
, или соединения (A8)-PO4 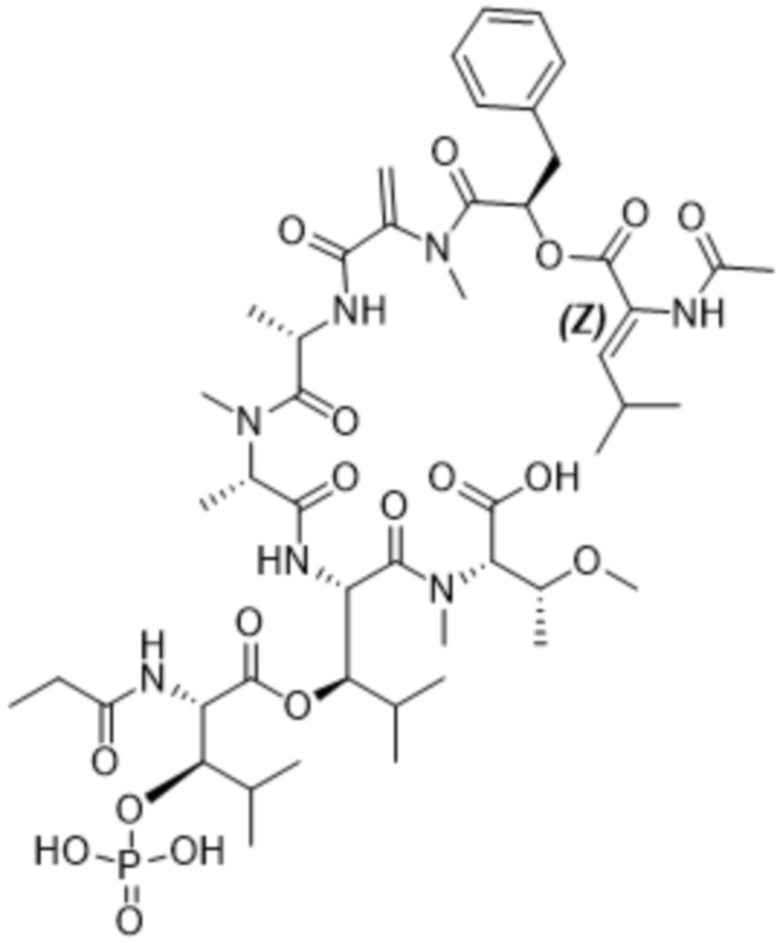 (где соединения (A1)-PO4 с раскрытым кольцом
(где соединения (A1)-PO4 с раскрытым кольцом 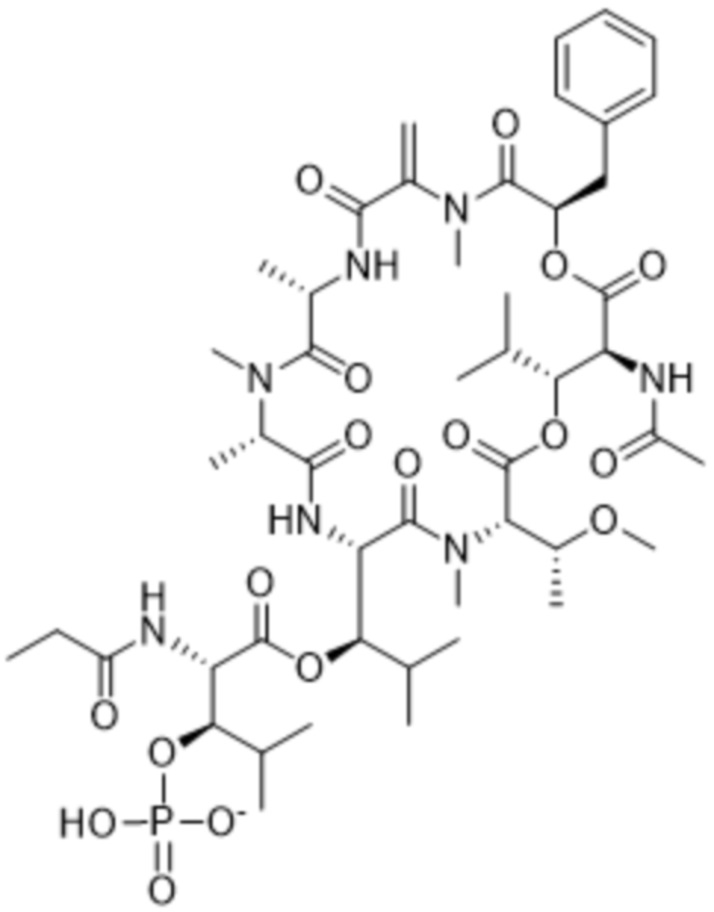 представляют собой неактивную полезную нагрузку). График отображает процент высвободившейся полезной нагрузки, которая присутствовала в виде соединения (A1)-PO4 на протяжении эксперимента для каждой из различных конструкций ADC (фиг. 15B).
представляют собой неактивную полезную нагрузку). График отображает процент высвободившейся полезной нагрузки, которая присутствовала в виде соединения (A1)-PO4 на протяжении эксперимента для каждой из различных конструкций ADC (фиг. 15B).
[00525] Каждое антитело к PMEL17G1_E152C_S375C_(B1) (G1-(B1)), антитело к PMEL17G1_E152C_(B1) (G1-E152C-DAR2-(B1)), антитело к PMEL17G1_S375C_(B1) (G1-S375C-DAR2-(B1)) и Fc-молчащее антитело к PMEL17G1_E152C_S375C_(B1) (Fc-молчащий G1-(B1)) вводили внутривенно "голым" мышам в дозе 20 мг/кг (веса тела). Девяти мышам вводили дозу каждой из описанных выше конструкций ADC. У трех животных на группу (конструкция на основе ADC) после умерщвления отбирали кровь для сбора образцов сыворотки крови для анализа через 24 ч, 7 и 14 дней после введения дозы. Аликвоты каждого образца подвергали иммунопреципитации с использованием гранул Capture Select FcXL и инкубировали с буфером, содержащим папаин, для высвобождения полезной нагрузки из ADC, захваченного гранулами. Затем высвободившуюся полезную нагрузку анализировали посредством HPLC MS для определения относительных уровней соединения (A1)-PO4 и соединения (A1)-PO4 с раскрытым кольцом, имеющего структуру соединения (A4)-PO4, соединения (A6)-PO4 или соединения (A8)-PO4 (где соединения (A1)-PO4 с раскрытым кольцом представляют собой неактивную полезную нагрузку). На графике показан процент полезной нагрузки, присутствующей на ADC в активной форме соединения (A1)-PO4 (при условии, что соединение (A1)-PO4+соединение (A8)-PO4=100%) (фиг. 15C). Исходные значения оценивали исходя из результатов экспериментов in vitro.
Пример 20: эффективность in vivo G1-E152C-DAR2-(B1), G1-S375C-DAR2-(B1), Fc-молчащего G1-(B1) в модели ксенотрансплантата увеальной меланомы
[00526] Самок "голых" мышей, несущих подкожные ксенотрансплантаты 92.1 с люциферазой, обрабатывали посредством 3207-E152C_S375C-DAR4-(B1), G1-E152C_S375C-DAR4-(B1), G1-E152C-DAR2-(B1), G1-S375C-DAR2-(B1), Fc-молчащий 3207-(B1) и Fc-молчащий G1-DAR4 в дозе 7,5 мг/кг. Обработки (однократную в/в инъекцию) проводили через 17 дней после инокуляции опухоли. Значения представлены в виде среднего ± SEM; размер выборки (n=5-6 мышей в группе). Исходный объем опухоли в день 0 составлял примерно 300-325 мм3 (фиг. 16). G1-E152C-DAR2-(B1), Fc-молчащий G1-DAR4 и G1-E152C_S375C-DAR4-(B1) проявляли сопоставимую противоопухолевую активность в модели ксенотрансплантата 92.1 увеальной меланомы, тогда как G1-S375C-DAR2-(B1) и не воздействующие целенаправленно 3207-ADC не оказывали влияния на рост опухоли.
Пример 21: in vitro активность G1-3J-DAR4-(B1), G1-3R-DAR4-(B1), G1-DAR4-(B1), G1-3J-DAR2 (E152C)-(B1), Fc-молчащего G1-3J -DAR2 (E152C)-(B1), 3207-DAR2 (E152C)-(B1) и G1-DAR2 (E152C)-(B1)
[00527] На фиг. 17A клетки 92.1 (левая панель) и MP41 (правая панель) высевали с низкой плотностью клеток в 96-луночные планшеты и обрабатывали возрастающими концентрациями G1-3J-DAR4-(B1), G1-3R-DAR4-(B1), G1-DAR4-(B1) согласно указанному. После обработки лекарственным средством в течение 96 или 120 часов жизнеспособность и пролиферацию клеток определяли с использованием анализа жизнеспособности на основе резазурина. Если вкратце, клетки инкубировали с раствором на основе резазурина и изменение цвета выявляли по оптической плотности посредством спектрофотометра и использовали для регистрации жизнеспособности клеток. Данные представлены в виде среднего значения для 3 независимых повторов и относительно клеток, обработанных PBS (контроль). Значение GI50 (концентрация для 50% максимального ингибирования пролиферации клеток) для каждого ADC и линии клеток показано в таблице 9. ADC к PMEL17-(B1) ингибируют пролиферацию клеток увеальной меланомы зависимым от PMEL17 и дозы образом.
Таблица 9. Подавляющая рост активность G1-3J-DAR4-(B1), G1-3R-DAR4-(B1), G1-DAR4-(B1) в клетках UM
GI50, нM
GI50, нM
GI50, нM
[00528] На фиг. 17B показаны результаты анализа пролиферации клеток в линии клеток 92.1 с использованием G1-3J-DAR2 (E152C)-(B1), Fc-молчащего G1-3J-DAR2 (E152C)-(B1), 3207-DAR2 (E152C)-(B1) и G1-DAR2 (E152C)-(B1), описанного на фиг. 17A. GI50 показана в таблице 10.
Таблица 10. Подавляющая рост активность G1-3J-DAR2 (E152C)-(B1), Fc-молчащего G1-3J -DAR2 (E152C)-(B1), 3207-DAR2 (E152C)-(B1) и G1-DAR2 (E152C)-(B1) в клетках UM
GI50, нM
GI50, нM
GI50, нM
GI50, нM
Пример 22: ADC к PMEL17-(B1) ингибируют рост опухоли в мышиной модели
[00529] Самок "голых" мышей с подкожными ксенотрансплантатами 92.1 с люциферазой обрабатывали посредством однократного в/в введения 3207-(B1), G1-3J(E152C), G1-3J-DAR2 (E152C)-(B1), Fc-молчащего G1-3J-DAR2 (E152C)-(B1), G1-DAR2 (E152C)-(B1) или среды-носителя в указанных дозах. Обработку (однократную в/в инъекцию) проводили через 14-17 дней после инокуляции опухоли. Значения являются средними ± SEM; размер выборки (n=5-12 мышей в группе). Исходный объем опухоли в день 0 составлял примерно 200-250 мм3. G1-3J-DAR2 (E152C)-(B1) и Fc-молчащий G1-3J-DAR2 (E152C)-(B1) ингибировали рост опухоли дозозависимым образом (фиг. 18).
Пример 23: иммуногистохимический анализ опухолевых биоптатов от пациентов с метастатической увеальной меланомой
[00530] Опухоли фиксировали в 10% (об./об.) формалине с нейтральным буфером в течение 24 ч при комнатной температуре, затем промывали PBS, обрабатывали для обезвоживания, очищали и заливали в парафин. Получали срезы толщиной 3 мкм, которые подвергали обработке для иммуногистохимии. Иммуногистохимическое окрашивание проводили на фиксированных формалином, залитых парафином срезах ткани с использованием полностью автоматизированной системы Bond-RX (Leica) с целью окрашивания антителами к gp100 (PMEL17) (мышиное моноклональное HMB45). Образцы метастатической увеальной меланомы демонстрируют высокую и относительно однородную экспрессию PMEL17 (фиг. 19A). Количественная оценка уровней экспрессии PMEL17 и доли PMEL17-положительных клеток в образцах пациентов с метастатической увеальной меланомой (фиг. 19B, фиг. 19C).
Пример 24: конкурентный анализ связывания
[00531] SPR использовали для эпитоп-специфичной сортировки антител к PMEL на приборе T200 Biacore (кат. № 28975001, GE Healthcare Life Sciences).
[00532] Поверхностный плазмонный резонанс (SPR) представляет собой метод измерения бимолекулярных взаимодействий в режиме реального времени в среде без меток. Молекулы иммобилизуют на поверхности сенсора, по которой пропускают раствор образца. Взаимодействие между иммобилизованной молекулой и пропускаемым образцом вызывает изменение показателя преломления. Изменение показателя преломления представляет собой изменение угла, с которым поляризованный свет пониженной интенсивности отражается от стеклянной плоскости, являющейся подложкой. Это изменение угла вызвано связыванием или диссоциацией молекул с поверхности сенсора, пропорционально массе связанного материала и регистрируется прибором на сенсограмме. Сенсограмма показывает возрастающий ответ при взаимодействии молекул, ответ остается постоянным, если взаимодействие достигает равновесия, и ответ уменьшается, когда образец заменяют буфером и партнеры по взаимодействию диссоциируют. Из ответов событий ассоциации и диссоциации определяют скорость ассоциации (ka), скорость диссоциации (kd) и общую аффинность (KD). Для эпитоп-специфичной сортировки KD не определены.
[00533] На первой стадии антитела G1 LC 3J и 17A9 непосредственно иммобилизовали на поверхности чипа CM5 посредством иммобилизации по амино-группе для достижения приблизительно 2000 RU соответственно на проточной ячейке 2 и проточной ячейке 3. Проточная ячейка 1 не имела иммобилизованного на ней антитела и служила референтной Fc. Иммобилизованные антитела на Fc2 и Fc3 являются основными антителами в данном эксперименте.
[00534] После иммобилизации PMEL человека пропускали на второй стадии при 20 нМ через все проточные кюветы в течение 60 секунд при 30 мкл/мин.
[00535] На третьей и последней стадии пропускают только антитело G1 LC 3J, или антитело G1 LC 3J с антителом 17A9 через Fc2, или только антитело 17A9, или антитело 17A9 с антителом G1 LC 3J через Fc3 (фиг. 20A). Антитела пропускали при общей концентрации 500 нМ через обе проточные кюветы в течение 120 секунд при 30 мкл/мин для ассоциации, после чего следовала фаза диссоциации продолжительностью 120 секунд при 30 мкл/мин с рабочим буфером.
[00536] Как и ожидалось, когда G1 LC 3J было основным антителом, не наблюдалось сигнала связывания при пропускании G13J. Связывание 100 RU наблюдали при протекании 17A9, позволяя предположить, что оно связывается с другим эпитопом, чем G1 LC 3J (фиг. 20B). Аналогичные тенденции наблюдались, когда 17A9 было основным антителом. На стадии 3 не наблюдается связывания с самим собой, но наблюдается связывание при пропускании G1 LC 3J (фиг. 20C). Это указывает на то, что G1 LC 3J и 17A9 связываются с разными эпитопами.
[00537] Регенерацию проводили в конце каждого цикла на всех проточных ячейках. Буфером для регенерации служил Glycine 2.0, который пропускали при 30 мкл/мин в течение 30 секунд.
[00538] Данные анализировали с использованием программного обеспечения Biacore T200 Evaluation версии 3.0.
[00539] Следует понимать, что примеры и варианты осуществления, описанные в данном документе, предназначены только для иллюстративных целей, и что различные модификации или изменения в их контексте будут предложены специалистам в данной области техники и они должны быть включены в сущность и область действия настоящей заявки и объем прилагаемой формулы изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> NOVARTIS AG
<120> АНТИТЕЛА К PMEL17 И КОНЪЮГАТЫ НА ИХ ОСНОВЕ
<130> PAT058359-WO-PCT
<140>
<141>
<150> 62/803,110
<151> 2019-02-08
<150> 62/783,565
<151> 2018-12-21
<160> 282
<170> PatentIn версия 3.5
<210> 1
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 1
Gly Gly Thr Phe Ser Asp Tyr Ala Ile Thr
1 5 10
<210> 2
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 2
Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 3
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 3
Glu Gly Gly Leu Leu Thr Asp Ile Ser Tyr Ser Arg Tyr Trp Phe Ala
1 5 10 15
Tyr
<210> 4
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 4
Asp Tyr Ala Ile Thr
1 5
<210> 5
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 5
Gly Gly Thr Phe Ser Asp Tyr
1 5
<210> 6
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 6
Ile Pro Ile Phe Gly Thr
1 5
<210> 7
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 7
Gly Gly Thr Phe Ser Asp Tyr Ala
1 5
<210> 8
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 8
Ile Ile Pro Ile Phe Gly Thr Ala
1 5
<210> 9
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 9
Ala Arg Glu Gly Gly Leu Leu Thr Asp Ile Ser Tyr Ser Arg Tyr Trp
1 5 10 15
Phe Ala Tyr
<210> 10
<211> 126
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 10
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Asp Tyr
20 25 30
Ala Ile Thr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Leu Leu Thr Asp Ile Ser Tyr Ser Arg Tyr Trp
100 105 110
Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 11
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 11
caggtgcaat tggtgcagag cggtgccgaa gtgaaaaaac cgggcagcag cgtgaaagtt 60
agctgcaaag catccggagg gacgttttct gactacgcta tcacttgggt gcgccaggcc 120
ccgggccagg gcctcgagtg gatgggcggt atcatcccga tcttcggcac tgcgaactac 180
gcccagaaat ttcagggccg ggtgaccatt accgccgatg aaagcaccag caccgcctat 240
atggaactga gcagcctgcg cagcgaagat acggccgtgt attattgcgc gcgtgaaggt 300
ggtctgctga ctgacatctc ttactctcgt tactggttcg cttactgggg ccaaggcacc 360
ctggtgactg ttagctca 378
<210> 12
<211> 456
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 12
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Asp Tyr
20 25 30
Ala Ile Thr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Leu Leu Thr Asp Ile Ser Tyr Ser Arg Tyr Trp
100 105 110
Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Cys Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 13
<211> 1368
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 13
caggtgcaat tggtgcagag cggtgccgaa gtgaaaaaac cgggcagcag cgtgaaagtt 60
agctgcaaag catccggagg gacgttttct gactacgcta tcacttgggt gcgccaggcc 120
ccgggccagg gcctcgagtg gatgggcggt atcatcccga tcttcggcac tgcgaactac 180
gcccagaaat ttcagggccg ggtgaccatt accgccgatg aaagcaccag caccgcctat 240
atggaactga gcagcctgcg cagcgaagat acggccgtgt attattgcgc gcgtgaaggt 300
ggtctgctga ctgacatctc ttactctcgt tactggttcg cttactgggg ccaaggcacc 360
ctggtgactg ttagctcagc tagcaccaag ggcccaagtg tgtttcccct ggcccccagc 420
agcaagtcta cttccggcgg aactgctgcc ctgggttgcc tggtgaagga ctacttcccc 480
tgtcccgtga cagtgtcctg gaactctggg gctctgactt ccggcgtgca caccttcccc 540
gccgtgctgc agagcagcgg cctgtacagc ctgagcagcg tggtgacagt gccctccagc 600
tctctgggaa cccagaccta tatctgcaac gtgaaccaca agcccagcaa caccaaggtg 660
gacaagagag tggagcccaa gagctgcgac aagacccaca cctgcccccc ctgcccagct 720
ccagaactgc tgggagggcc ttccgtgttc ctgttccccc ccaagcccaa ggacaccctg 780
atgatcagca ggacccccga ggtgacctgc gtggtggtgg acgtgtccca cgaggaccca 840
gaggtgaagt tcaactggta cgtggacggc gtggaggtgc acaacgccaa gaccaagccc 900
agagaggagc agtacaacag cacctacagg gtggtgtccg tgctgaccgt gctgcaccag 960
gactggctga acggcaaaga atacaagtgc aaagtctcca acaaggccct gccagcccca 1020
atcgaaaaga caatcagcaa ggccaagggc cagccacggg agccccaggt gtacaccctg 1080
ccccccagcc gggaggagat gaccaagaac caggtgtccc tgacctgtct ggtgaagggc 1140
ttctacccct gtgatatcgc cgtggagtgg gagagcaacg gccagcccga gaacaactac 1200
aagaccaccc ccccagtgct ggacagcgac ggcagcttct tcctgtacag caagctgacc 1260
gtggacaagt ccaggtggca gcagggcaac gtgttcagct gcagcgtgat gcacgaggcc 1320
ctgcacaacc actacaccca gaagtccctg agcctgagcc ccggcaag 1368
<210> 14
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 14
Ser Gly Asp Ala Leu Arg Asp Lys Phe Val Tyr
1 5 10
<210> 15
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 15
Asp Asp Asn Asn Arg Pro Ser
1 5
<210> 16
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 16
Gln Ser Trp Asp His Ser Tyr Ser Leu Val Val
1 5 10
<210> 17
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 17
Asp Ala Leu Arg Asp Lys Phe
1 5
<210> 18
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 18
Asp Asp Asn
1
<210> 19
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 19
Trp Asp His Ser Tyr Ser Leu Val
1 5
<210> 20
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 20
Ala Leu Arg Asp Lys Phe
1 5
<210> 21
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 21
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Ala Leu Arg Asp Lys Phe Val
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asn Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Asp His Ser Tyr Ser Leu
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 22
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 22
gatatcgaac tgacccagcc gccgagcgtg agcgtgagcc cgggccagac cgcgagcatt 60
acctgtagcg gcgatgctct gcgtgacaaa ttcgtttact ggtaccagca gaaaccgggc 120
caggcgccgg tgctggtgat ctacgacgac aacaaccgtc cgagcggcat cccggaacgt 180
tttagcggat ccaacagcgg caacaccgcg accctgacca ttagcggcac ccaggcggaa 240
gacgaagcgg attattactg ccagtcttgg gaccattctt actctctggt tgtgtttggc 300
ggcggcacga agttaactgt cctg 324
<210> 23
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 23
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Ala Leu Arg Asp Lys Phe Val
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asn Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Asp His Ser Tyr Ser Leu
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 24
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 24
gatatcgaac tgacccagcc gccgagcgtg agcgtgagcc cgggccagac cgcgagcatt 60
acctgtagcg gcgatgctct gcgtgacaaa ttcgtttact ggtaccagca gaaaccgggc 120
caggcgccgg tgctggtgat ctacgacgac aacaaccgtc cgagcggcat cccggaacgt 180
tttagcggat ccaacagcgg caacaccgcg accctgacca ttagcggcac ccaggcggaa 240
gacgaagcgg attattactg ccagtcttgg gaccattctt actctctggt tgtgtttggc 300
ggcggcacga agttaactgt cctgggacaa cctaaggccg ctccctccgt gaccctgttc 360
ccccccagct ccgaggaact gcaggccaac aaggccaccc tggtgtgcct gatcagcgac 420
ttctaccctg gcgccgtgac cgtggcctgg aaggccgaca gcagccccgt gaaggccggc 480
gtggagacaa ccacccccag caagcagagc aacaacaagt acgccgccag cagctacctg 540
agcctgaccc ccgagcagtg gaagagccac agaagctaca gctgccaggt cacccacgag 600
ggcagcaccg tggagaaaac cgtggccccc accgagtgca gc 642
<210> 25
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 25
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu Arg Asp Lys Phe Val
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asn Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Asp His Ser Tyr Ser Leu
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 26
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 26
agctacgagc tgacccagcc gctgtcggtg tcagtcgccc tgggacaaac tgccaggatc 60
acttgttccg gggacgcatt gcgggacaag ttcgtgtact ggtaccagca gaagccgggt 120
caagccccag tgctcgtgat ctacgacgac aacaaccggc cttccggtat ccccgaacgc 180
ttctccggat ccaatagcgg aaacaccgcc accctgacca tttcgagagc tcaggccggg 240
gatgaagcgg actactactg ccagtcatgg gatcactcgt actccctcgt cgtgtttgga 300
ggcggcacga agcttactgt gctg 324
<210> 27
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 27
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu Arg Asp Lys Phe Val
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asn Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Asp His Ser Tyr Ser Leu
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 28
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 28
agctacgagc tgacccagcc gctgtcggtg tcagtcgccc tgggacaaac tgccaggatc 60
acttgttccg gggacgcatt gcgggacaag ttcgtgtact ggtaccagca gaagccgggt 120
caagccccag tgctcgtgat ctacgacgac aacaaccggc cttccggtat ccccgaacgc 180
ttctccggat ccaatagcgg aaacaccgcc accctgacca tttcgagagc tcaggccggg 240
gatgaagcgg actactactg ccagtcatgg gatcactcgt actccctcgt cgtgtttgga 300
ggcggcacga agcttactgt gctgggccag cctaaggccg ctccctccgt gaccctgttc 360
ccccccagct ccgaggaact gcaggccaac aaggccaccc tggtgtgcct gatcagcgac 420
ttctaccctg gcgccgtgac cgtggcctgg aaggccgaca gcagccccgt gaaggccggc 480
gtggagacaa ccacccccag caagcagagc aacaacaagt acgccgccag cagctacctg 540
agcctgaccc ccgagcagtg gaagagccac agaagctaca gctgccaggt cacccacgag 600
ggcagcaccg tggagaaaac cgtggccccc accgagtgca gc 642
<210> 29
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 29
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Ala Leu Arg Asp Lys Phe Val
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asn Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Asp His Ser Tyr Ser Leu
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 30
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 30
tcgtacgagc tcactcaacc gccttctgtg tccgtgtcac ccgggcagac tgcctccatt 60
acctgttcgg gagatgccct gcgcgacaag tttgtgtact ggtaccagca gaagcccgga 120
cagtcgccag tgctcgtcat ctatgacgac aacaacagac cttccggtat cccggaacgg 180
ttcagcggaa gcaattccgg caacaccgct accctgacca ttagcggcac tcaggccatg 240
gacgaagcgg attactactg ccaatcctgg gaccactcat actcccttgt ggtgttcggt 300
ggcggaacga agctgaccgt cctg 324
<210> 31
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 31
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Ala Leu Arg Asp Lys Phe Val
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asn Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Asp His Ser Tyr Ser Leu
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 32
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 32
tcgtacgagc tcactcaacc gccttctgtg tccgtgtcac ccgggcagac tgcctccatt 60
acctgttcgg gagatgccct gcgcgacaag tttgtgtact ggtaccagca gaagcccgga 120
cagtcgccag tgctcgtcat ctatgacgac aacaacagac cttccggtat cccggaacgg 180
ttcagcggaa gcaattccgg caacaccgct accctgacca ttagcggcac tcaggccatg 240
gacgaagcgg attactactg ccaatcctgg gaccactcat actcccttgt ggtgttcggt 300
ggcggaacga agctgaccgt cctgggccag cctaaggccg ctccctccgt gaccctgttc 360
ccccccagct ccgaggaact gcaggccaac aaggccaccc tggtgtgcct gatcagcgac 420
ttctaccctg gcgccgtgac cgtggcctgg aaggccgaca gcagccccgt gaaggccggc 480
gtggagacaa ccacccccag caagcagagc aacaacaagt acgccgccag cagctacctg 540
agcctgaccc ccgagcagtg gaagagccac agaagctaca gctgccaggt cacccacgag 600
ggcagcaccg tggagaaaac cgtggccccc accgagtgca gc 642
<210> 33
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 33
Gly Gly Thr Phe Ser Thr Tyr Ala Ile Ser
1 5 10
<210> 34
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 34
Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 35
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 35
Glu Val Arg Met Ile Phe Asp Tyr
1 5
<210> 36
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 36
Thr Tyr Ala Ile Ser
1 5
<210> 37
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 37
Gly Gly Thr Phe Ser Thr Tyr
1 5
<210> 38
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 38
Ile Pro Ile Leu Gly Ile
1 5
<210> 39
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 39
Gly Gly Thr Phe Ser Thr Tyr Ala
1 5
<210> 40
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 40
Ile Ile Pro Ile Leu Gly Ile Ala
1 5
<210> 41
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 41
Ala Arg Glu Val Arg Met Ile Phe Asp Tyr
1 5 10
<210> 42
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 42
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Thr Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Val Arg Met Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 43
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 43
caggtgcaat tggtgcagag cggtgccgaa gtgaaaaaac cgggcagcag cgtgaaagtt 60
agctgcaaag catccggagg gacgttttct acttacgcta tctcttgggt gcgccaggcc 120
ccgggccagg gcctcgagtg gatgggccgt atcatcccga tcctgggcat cgcgaactac 180
gcccagaaat ttcagggccg ggtgaccatt accgccgatg aaagcaccag caccgcctat 240
atggaactga gcagcctgcg cagcgaagat acggccgtgt attattgcgc gcgtgaagtt 300
cgtatgatct tcgattactg gggccaaggc accctggtga ctgttagctc a 351
<210> 44
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 44
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Thr Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Val Arg Met Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 45
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 45
caggtgcaat tggtgcagag cggtgccgaa gtgaaaaaac cgggcagcag cgtgaaagtt 60
agctgcaaag catccggagg gacgttttct acttacgcta tctcttgggt gcgccaggcc 120
ccgggccagg gcctcgagtg gatgggccgt atcatcccga tcctgggcat cgcgaactac 180
gcccagaaat ttcagggccg ggtgaccatt accgccgatg aaagcaccag caccgcctat 240
atggaactga gcagcctgcg cagcgaagat acggccgtgt attattgcgc gcgtgaagtt 300
cgtatgatct tcgattactg gggccaaggc accctggtga ctgttagctc agcctctacg 360
aaaggcccaa gcgtatttcc cctggctcct tctagtaaat caacctcagg tggtacagca 420
gcccttggct gcctggtcaa agactatttc ccctgtccgg tgaccgtctc atggaactca 480
ggtgctttga catctggtgt gcatacattc ccagctgtgc tgcaaagtag tggactgtac 540
agcctttcca gcgtggtcac ggtgccaagt agctccttgg gtactcagac ttatatctgc 600
aatgtgaacc acaagccctc taacacgaag gtggacaagc gcgtggagcc caaatcttgc 660
gataagacgc atacttgtcc cccatgccct gctcctgagc tgttgggagg cccgtcagtg 720
ttcttgttcc ctccgaagcc taaggacact ttgatgataa gtaggacacc agaggtgact 780
tgcgtggtgg ttgatgtgtc ccatgaagat cccgaggtca aatttaattg gtacgtagat 840
ggtgtcgaag ttcacaatgc taagactaag ccaagggaag agcagtacaa cagtacatat 900
agggtagtct ccgtgctgac agtcctccac caggactggt tgaacggcaa ggaatacaaa 960
tgtaaggtgt caaacaaagc tctgcctgct cccattgaga aaacaatctc taaagccaaa 1020
ggccagccga gagagcccca agtctacact ttgcccccga gcagggagga aatgaccaag 1080
aatcaggtga gtctgacgtg cctcgtcaaa ggattttatc catgcgatat tgcagttgaa 1140
tgggagagca atggccagcc agagaacaac tataaaacca caccacccgt gctcgactct 1200
gatggcagct tcttcctcta tagcaagctg acagtcgata aatctcgctg gcagcaaggc 1260
aatgtgttct cctgctccgt catgcacgag gctttgcata accattatac tcaaaaatct 1320
ctgtccctgt cacctggtaa a 1341
<210> 46
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 46
Arg Ala Ser Gln Ser Ile Ser Ser Tyr Leu Ala
1 5 10
<210> 47
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 47
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 48
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 48
Gln Gln Ser Tyr Asp Tyr Tyr Thr
1 5
<210> 49
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 49
Ser Gln Ser Ile Ser Ser Tyr
1 5
<210> 50
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 50
Ala Ala Ser
1
<210> 51
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 51
Ser Tyr Asp Tyr Tyr
1 5
<210> 52
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 52
Gln Ser Ile Ser Ser Tyr
1 5
<210> 53
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 53
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Asp Tyr Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 54
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 54
gatatccaga tgacccagag cccgagcagc ctgagcgcca gcgtgggcga tcgcgtgacc 60
attacctgca gagccagcca gtctatttct tcttacctgg cttggtacca gcagaaaccg 120
ggcaaagcgc cgaaactatt aatctacgct gcttcttctc tgcaaagcgg cgtgccgagc 180
cgctttagcg gcagcggatc cggcaccgat ttcaccctga ccattagctc tctgcaaccg 240
gaagactttg cgacctatta ttgccagcag tcttacgact actacacctt tggccagggc 300
acgaaagttg aaattaaa 318
<210> 55
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 55
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Asp Tyr Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 56
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 56
gatatccaga tgacccagag cccgagcagc ctgagcgcca gcgtgggcga tcgcgtgacc 60
attacctgca gagccagcca gtctatttct tcttacctgg cttggtacca gcagaaaccg 120
ggcaaagcgc cgaaactatt aatctacgct gcttcttctc tgcaaagcgg cgtgccgagc 180
cgctttagcg gcagcggatc cggcaccgat ttcaccctga ccattagctc tctgcaaccg 240
gaagactttg cgacctatta ttgccagcag tcttacgact actacacctt tggccagggc 300
acgaaagttg aaattaaacg tacggtggca gctccgtctg ttttcatctt tccacctagc 360
gacgagcaac tcaaaagtgg tacagcatcc gtggtttgtc tgctgaacaa tttttacccc 420
agggaggcta aggtccagtg gaaagtcgat aacgctcttc agtctggcaa cagtcaggag 480
agcgtcacag agcaggactc taaggatagc acttatagtc tgtcctccac gctgacactg 540
tctaaagcgg attatgagaa gcacaaggtt tacgcctgtg aggtaacgca ccaaggactc 600
tcctccccag ttaccaaatc tttcaacaga ggagaatgt 639
<210> 57
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 57
Gly Gly Thr Phe Ser Asp Tyr Ala Ile Ser
1 5 10
<210> 58
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 58
Gly Ile Ile Pro Ile Phe Gly Asp Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 59
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 59
Glu Gly Ser Ser Tyr Phe Tyr Met Ala Tyr
1 5 10
<210> 60
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 60
Asp Tyr Ala Ile Ser
1 5
<210> 61
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 61
Ile Pro Ile Phe Gly Asp
1 5
<210> 62
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 62
Ile Ile Pro Ile Phe Gly Asp Ala
1 5
<210> 63
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 63
Ala Arg Glu Gly Ser Ser Tyr Phe Tyr Met Ala Tyr
1 5 10
<210> 64
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 64
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Asp Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Asp Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ser Ser Tyr Phe Tyr Met Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 65
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 65
caggttcagc tggtgcagtc tggcgccgaa gtgaagaaac ctggcagcag cgtgaaggtg 60
tcctgcaaag caagcggcgg caccttcagc gattacgcca tctcttgggt ccgacaggcc 120
cctggacaag gcttggaatg gatgggcggc atcatcccca tcttcggcga cgccaattac 180
gcccagaaat tccagggcag agtgaccatc accgccgacg agtctacaag caccgcctac 240
atggaactga gcagcctgag aagcgaggac accgccgtgt actactgtgc cagagagggc 300
agcagctact tctacatggc ctattggggc cagggcaccc tggtcacagt tagctct 357
<210> 66
<211> 449
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 66
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Asp Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Asp Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ser Ser Tyr Phe Tyr Met Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 67
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 67
caggttcagc tggtgcagtc tggcgccgaa gtgaagaaac ctggcagcag cgtgaaggtg 60
tcctgcaaag caagcggcgg caccttcagc gattacgcca tctcttgggt ccgacaggcc 120
cctggacaag gcttggaatg gatgggcggc atcatcccca tcttcggcga cgccaattac 180
gcccagaaat tccagggcag agtgaccatc accgccgacg agtctacaag caccgcctac 240
atggaactga gcagcctgag aagcgaggac accgccgtgt actactgtgc cagagagggc 300
agcagctact tctacatggc ctattggggc cagggcaccc tggtcacagt tagctctgct 360
agcaccaagg gcccaagtgt gtttcccctg gcccccagca gcaagtctac ttccggcgga 420
actgctgccc tgggttgcct ggtgaaggac tacttcccct gtcccgtgac agtgtcctgg 480
aactctgggg ctctgacttc cggcgtgcac accttccccg ccgtgctgca gagcagcggc 540
ctgtacagcc tgagcagcgt ggtgacagtg ccctccagct ctctgggaac ccagacctat 600
atctgcaacg tgaaccacaa gcccagcaac accaaggtgg acaagagagt ggagcccaag 660
agctgcgaca agacccacac ctgccccccc tgcccagctc cagaactgct gggagggcct 720
tccgtgttcc tgttcccccc caagcccaag gacaccctga tgatcagcag gacccccgag 780
gtgacctgcg tggtggtgga cgtgtcccac gaggacccag aggtgaagtt caactggtac 840
gtggacggcg tggaggtgca caacgccaag accaagccca gagaggagca gtacaacagc 900
acctacaggg tggtgtccgt gctgaccgtg ctgcaccagg actggctgaa cggcaaagaa 960
tacaagtgca aagtctccaa caaggccctg ccagccccaa tcgaaaagac aatcagcaag 1020
gccaagggcc agccacggga gccccaggtg tacaccctgc cccccagccg ggaggagatg 1080
accaagaacc aggtgtccct gacctgtctg gtgaagggct tctacccctg tgatatcgcc 1140
gtggagtggg agagcaacgg ccagcccgag aacaactaca agaccacccc cccagtgctg 1200
gacagcgacg gcagcttctt cctgtacagc aagctgaccg tggacaagtc caggtggcag 1260
cagggcaacg tgttcagctg cagcgtgatg cacgaggccc tgcacaacca ctacacccag 1320
aagtccctga gcctgagccc cggcaag 1347
<210> 68
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 68
Ser Gly Asp Asn Ile Gly Ser Lys Leu Ala Ser
1 5 10
<210> 69
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 69
Asp Asp Ser Asn Arg Pro Ser
1 5
<210> 70
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 70
Ala Ala Thr Ala Gly Asp Arg Trp Ala Tyr Val
1 5 10
<210> 71
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 71
Asp Asn Ile Gly Ser Lys Leu
1 5
<210> 72
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 72
Asp Asp Ser
1
<210> 73
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 73
Thr Ala Gly Asp Arg Trp Ala Tyr
1 5
<210> 74
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 74
Asn Ile Gly Ser Lys Leu
1 5
<210> 75
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 75
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Asn Ile Gly Ser Lys Leu Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Thr Ala Gly Asp Arg Trp Ala
85 90 95
Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 76
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 76
agctatgagc tgacacagcc tctgtccgtg tctgtggctc tgggacagac cgccagaatc 60
acctgtagcg gcgacaacat cggcagcaag ctggcctctt ggtatcagca gaagcctgga 120
caggcccctg tgctggtcat ctacgacgac agcaatagac ccagcggcat ccccgagaga 180
ttcagcggca gcaatagcgg caataccgcc acactgacca tcagcagagc acaggctggc 240
gacgaggccg attactattg tgctgccaca gccggcgaca gatgggccta tgtttttggc 300
ggcggaacaa agctgaccgt gctg 324
<210> 77
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 77
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Asn Ile Gly Ser Lys Leu Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Thr Ala Gly Asp Arg Trp Ala
85 90 95
Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 78
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 78
agctatgagc tgacacagcc tctgtccgtg tctgtggctc tgggacagac cgccagaatc 60
acctgtagcg gcgacaacat cggcagcaag ctggcctctt ggtatcagca gaagcctgga 120
caggcccctg tgctggtcat ctacgacgac agcaatagac ccagcggcat ccccgagaga 180
ttcagcggca gcaatagcgg caataccgcc acactgacca tcagcagagc acaggctggc 240
gacgaggccg attactattg tgctgccaca gccggcgaca gatgggccta tgtttttggc 300
ggcggaacaa agctgaccgt gctgggacag cctaaggccg ctccctccgt gaccctgttc 360
ccccccagct ccgaggaact gcaggccaac aaggccaccc tggtgtgcct gatcagcgac 420
ttctaccctg gcgccgtgac cgtggcctgg aaggccgaca gcagccccgt gaaggccggc 480
gtggagacaa ccacccccag caagcagagc aacaacaagt acgccgccag cagctacctg 540
agcctgaccc ccgagcagtg gaagagccac agaagctaca gctgccaggt cacccacgag 600
ggcagcaccg tggagaaaac cgtggccccc accgagtgca gc 642
<210> 79
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 79
Gly Phe Thr Phe Ser Ser Phe Gly Met Ser
1 5 10
<210> 80
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 80
Ala Ile Ser Tyr Ser Gly Ser Asp Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 81
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 81
Asp Val Gly Val Met Asp Tyr
1 5
<210> 82
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 82
Ser Phe Gly Met Ser
1 5
<210> 83
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 83
Gly Phe Thr Phe Ser Ser Phe
1 5
<210> 84
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 84
Ser Tyr Ser Gly Ser Asp
1 5
<210> 85
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 85
Gly Phe Thr Phe Ser Ser Phe Gly
1 5
<210> 86
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 86
Ile Ser Tyr Ser Gly Ser Asp Thr
1 5
<210> 87
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 87
Ala Arg Asp Val Gly Val Met Asp Tyr
1 5
<210> 88
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 88
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Ser Gly Ser Asp Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Val Gly Val Met Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 89
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 89
caggttcagc tgctggaatc tggcggagga ctggttcaac ctggcggctc tctgagactg 60
tcttgtgccg ccagcggctt caccttcagc agctttggca tgagctgggt ccgacaggcc 120
cctggcaaag gacttgaatg ggtgtccgcc atcagctaca gcggcagcga tacctactac 180
gccgacagcg tgaagggcag attcaccatc tccagagaca acagcaagaa caccctgtac 240
ctgcagatga acagcctgag agccgaggac accgccgtgt actactgtgc cagagatgtg 300
ggcgtgatgg actattgggg ccagggcaca ctggtcaccg ttagctct 348
<210> 90
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 90
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Ser Gly Ser Asp Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Val Gly Val Met Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 91
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 91
caggttcagc tgctggaatc tggcggagga ctggttcaac ctggcggctc tctgagactg 60
tcttgtgccg ccagcggctt caccttcagc agctttggca tgagctgggt ccgacaggcc 120
cctggcaaag gacttgaatg ggtgtccgcc atcagctaca gcggcagcga tacctactac 180
gccgacagcg tgaagggcag attcaccatc tccagagaca acagcaagaa caccctgtac 240
ctgcagatga acagcctgag agccgaggac accgccgtgt actactgtgc cagagatgtg 300
ggcgtgatgg actattgggg ccagggcaca ctggtcaccg ttagctctgc tagcaccaag 360
ggcccaagtg tgtttcccct ggcccccagc agcaagtcta cttccggcgg aactgctgcc 420
ctgggttgcc tggtgaagga ctacttcccc tgtcccgtga cagtgtcctg gaactctggg 480
gctctgactt ccggcgtgca caccttcccc gccgtgctgc agagcagcgg cctgtacagc 540
ctgagcagcg tggtgacagt gccctccagc tctctgggaa cccagaccta tatctgcaac 600
gtgaaccaca agcccagcaa caccaaggtg gacaagagag tggagcccaa gagctgcgac 660
aagacccaca cctgcccccc ctgcccagct ccagaactgc tgggagggcc ttccgtgttc 720
ctgttccccc ccaagcccaa ggacaccctg atgatcagca ggacccccga ggtgacctgc 780
gtggtggtgg acgtgtccca cgaggaccca gaggtgaagt tcaactggta cgtggacggc 840
gtggaggtgc acaacgccaa gaccaagccc agagaggagc agtacaacag cacctacagg 900
gtggtgtccg tgctgaccgt gctgcaccag gactggctga acggcaaaga atacaagtgc 960
aaagtctcca acaaggccct gccagcccca atcgaaaaga caatcagcaa ggccaagggc 1020
cagccacggg agccccaggt gtacaccctg ccccccagcc gggaggagat gaccaagaac 1080
caggtgtccc tgacctgtct ggtgaagggc ttctacccct gtgatatcgc cgtggagtgg 1140
gagagcaacg gccagcccga gaacaactac aagaccaccc ccccagtgct ggacagcgac 1200
ggcagcttct tcctgtacag caagctgacc gtggacaagt ccaggtggca gcagggcaac 1260
gtgttcagct gcagcgtgat gcacgaggcc ctgcacaacc actacaccca gaagtccctg 1320
agcctgagcc ccggcaag 1338
<210> 92
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 92
Ser Gly Asp Asn Leu Gly Thr Tyr Tyr Ala His
1 5 10
<210> 93
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 93
Ser Gln Ser His Arg Pro Ser
1 5
<210> 94
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 94
Gly Ala Trp Asp Ala Pro Ser Pro Glu Leu Val
1 5 10
<210> 95
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 95
Asp Asn Leu Gly Thr Tyr Tyr
1 5
<210> 96
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 96
Ser Gln Ser
1
<210> 97
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 97
Trp Asp Ala Pro Ser Pro Glu Leu
1 5
<210> 98
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 98
Asn Leu Gly Thr Tyr Tyr
1 5
<210> 99
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 99
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Asn Leu Gly Thr Tyr Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Ser Gln Ser His Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ala Trp Asp Ala Pro Ser Pro Glu
85 90 95
Leu Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 100
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 100
agctatgagc tgacacagcc tctgtccgtg tctgtggctc tgggacagac cgccagaatc 60
acctgtagcg gcgataacct gggcacctac tacgcccact ggtatcagca gaagcctgga 120
caggctcccg tgctggtcat ctacagccag tctcacagac ccagcggcat ccccgagaga 180
ttcagcggca gcaatagcgg caataccgcc acactgacca tcagcagagc acaggctggc 240
gacgaggccg attactattg tggcgcttgg gacgccccat ctcctgagct tgtttttggc 300
ggaggcacca agctgacagt gctg 324
<210> 101
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 101
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Asn Leu Gly Thr Tyr Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Ser Gln Ser His Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ala Trp Asp Ala Pro Ser Pro Glu
85 90 95
Leu Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 102
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 102
agctatgagc tgacacagcc tctgtccgtg tctgtggctc tgggacagac cgccagaatc 60
acctgtagcg gcgataacct gggcacctac tacgcccact ggtatcagca gaagcctgga 120
caggctcccg tgctggtcat ctacagccag tctcacagac ccagcggcat ccccgagaga 180
ttcagcggca gcaatagcgg caataccgcc acactgacca tcagcagagc acaggctggc 240
gacgaggccg attactattg tggcgcttgg gacgccccat ctcctgagct tgtttttggc 300
ggaggcacca agctgacagt gctgggacag cctaaggccg ctccctccgt gaccctgttc 360
ccccccagct ccgaggaact gcaggccaac aaggccaccc tggtgtgcct gatcagcgac 420
ttctaccctg gcgccgtgac cgtggcctgg aaggccgaca gcagccccgt gaaggccggc 480
gtggagacaa ccacccccag caagcagagc aacaacaagt acgccgccag cagctacctg 540
agcctgaccc ccgagcagtg gaagagccac agaagctaca gctgccaggt cacccacgag 600
ggcagcaccg tggagaaaac cgtggccccc accgagtgca gc 642
<210> 103
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 103
Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
1 5 10
<210> 104
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 104
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 105
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 105
Ala Phe Arg Leu Tyr Trp Leu Asp Val
1 5
<210> 106
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 106
Ser Tyr Ala Met Ser
1 5
<210> 107
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 107
Gly Phe Thr Phe Ser Ser Tyr
1 5
<210> 108
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 108
Ser Gly Ser Gly Gly Ser
1 5
<210> 109
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 109
Gly Phe Thr Phe Ser Ser Tyr Ala
1 5
<210> 110
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 110
Ile Ser Gly Ser Gly Gly Ser Thr
1 5
<210> 111
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 111
Ala Arg Ala Phe Arg Leu Tyr Trp Leu Asp Val
1 5 10
<210> 112
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 112
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Phe Arg Leu Tyr Trp Leu Asp Val Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 113
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 113
gaagttcagc tgctggaatc tggcggagga ctggttcaac ctggcggctc tctgagactg 60
tcttgtgccg ccagcggctt cacctttagc agctacgcca tgagctgggt ccgacaggct 120
cctggcaaag gccttgaatg ggtgtccgcc atctctggct ctggcggcag cacatattac 180
gccgactctg tgaagggcag attcaccatc agccgggaca acagcaagaa caccctgtac 240
ctgcagatga acagcctgag agccgaggac accgccgtgt actattgtgc cagagccttc 300
cggctgtact ggctggatgt ttggggacag ggcaccctgg tcacagtgtc atct 354
<210> 114
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 114
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Phe Arg Leu Tyr Trp Leu Asp Val Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 115
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 115
gaagttcagc tgctggaatc tggcggagga ctggttcaac ctggcggctc tctgagactg 60
tcttgtgccg ccagcggctt cacctttagc agctacgcca tgagctgggt ccgacaggct 120
cctggcaaag gccttgaatg ggtgtccgcc atctctggct ctggcggcag cacatattac 180
gccgactctg tgaagggcag attcaccatc agccgggaca acagcaagaa caccctgtac 240
ctgcagatga acagcctgag agccgaggac accgccgtgt actattgtgc cagagccttc 300
cggctgtact ggctggatgt ttggggacag ggcaccctgg tcacagtgtc atctgctagc 360
accaagggcc caagtgtgtt tcccctggcc cccagcagca agtctacttc cggcggaact 420
gctgccctgg gttgcctggt gaaggactac ttcccctgtc ccgtgacagt gtcctggaac 480
tctggggctc tgacttccgg cgtgcacacc ttccccgccg tgctgcagag cagcggcctg 540
tacagcctga gcagcgtggt gacagtgccc tccagctctc tgggaaccca gacctatatc 600
tgcaacgtga accacaagcc cagcaacacc aaggtggaca agagagtgga gcccaagagc 660
tgcgacaaga cccacacctg ccccccctgc ccagctccag aactgctggg agggccttcc 720
gtgttcctgt tcccccccaa gcccaaggac accctgatga tcagcaggac ccccgaggtg 780
acctgcgtgg tggtggacgt gtcccacgag gacccagagg tgaagttcaa ctggtacgtg 840
gacggcgtgg aggtgcacaa cgccaagacc aagcccagag aggagcagta caacagcacc 900
tacagggtgg tgtccgtgct gaccgtgctg caccaggact ggctgaacgg caaagaatac 960
aagtgcaaag tctccaacaa ggccctgcca gccccaatcg aaaagacaat cagcaaggcc 1020
aagggccagc cacgggagcc ccaggtgtac accctgcccc ccagccggga ggagatgacc 1080
aagaaccagg tgtccctgac ctgtctggtg aagggcttct acccctgtga tatcgccgtg 1140
gagtgggaga gcaacggcca gcccgagaac aactacaaga ccaccccccc agtgctggac 1200
agcgacggca gcttcttcct gtacagcaag ctgaccgtgg acaagtccag gtggcagcag 1260
ggcaacgtgt tcagctgcag cgtgatgcac gaggccctgc acaaccacta cacccagaag 1320
tccctgagcc tgagccccgg caag 1344
<210> 116
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 116
Arg Ala Ser Gln Ser Ile Ser Ser Tyr Leu Asn
1 5 10
<210> 117
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 117
Gln Gln Val Tyr Ser Ala Pro Val Thr
1 5
<210> 118
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 118
Val Tyr Ser Ala Pro Val
1 5
<210> 119
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 119
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Val Tyr Ser Ala Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 120
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 120
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gagcatcagc agctacctga actggtatca gcagaagccc 120
ggcaaggccc ctaaactgct gatctatgcc gccagctctc tgcagtctgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgac ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ctgccagcag gtctacagcg cccctgtgac atttggccag 300
ggcaccaagg tggaaatcaa g 321
<210> 121
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 121
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Val Tyr Ser Ala Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 122
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 122
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gagcatcagc agctacctga actggtatca gcagaagccc 120
ggcaaggccc ctaaactgct gatctatgcc gccagctctc tgcagtctgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgac ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ctgccagcag gtctacagcg cccctgtgac atttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 123
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 123
Gly Phe Thr Phe Ser Asn Tyr Trp Ile Ser
1 5 10
<210> 124
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 124
Arg Ile Lys Ser Lys Thr Tyr Gly Gly Thr Thr Asp Tyr Ala Glu Pro
1 5 10 15
Val Lys Gly
<210> 125
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 125
Thr Ser Arg Arg Ser Tyr Ala Phe Asp Tyr
1 5 10
<210> 126
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 126
Asn Tyr Trp Ile Ser
1 5
<210> 127
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 127
Gly Phe Thr Phe Ser Asn Tyr
1 5
<210> 128
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 128
Lys Ser Lys Thr Tyr Gly Gly Thr
1 5
<210> 129
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 129
Gly Phe Thr Phe Ser Asn Tyr Trp
1 5
<210> 130
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 130
Ile Lys Ser Lys Thr Tyr Gly Gly Thr Thr
1 5 10
<210> 131
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 131
Ala Arg Thr Ser Arg Arg Ser Tyr Ala Phe Asp Tyr
1 5 10
<210> 132
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 132
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Tyr Gly Gly Thr Thr Asp Tyr Ala Glu
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Thr Ser Arg Arg Ser Tyr Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 133
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 133
gaagtgcagc tggtggaatc tggcggcgga cttgtgaaac ctggcggctc tctgagactg 60
agctgtgccg cttccggctt caccttcagc aactactgga tcagctgggt ccgacaggcc 120
cctggcaaag gacttgagtg ggtcggacgg atcaagagca agacctacgg cggcaccacc 180
gattatgccg agcctgtgaa gggcagattc accatcagcc gggacgacag caagaacacc 240
ctgtacctgc agatgaacag cctgaaaacc gaggacaccg ccgtgtacta ctgcgccaga 300
accagcagaa gaagctacgc cttcgactac tggggccagg gcacactggt taccgttagc 360
tct 363
<210> 134
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 134
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Tyr Gly Gly Thr Thr Asp Tyr Ala Glu
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Thr Ser Arg Arg Ser Tyr Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 135
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 135
gaagtgcagc tggtggaatc tggcggcgga cttgtgaaac ctggcggctc tctgagactg 60
agctgtgccg cttccggctt caccttcagc aactactgga tcagctgggt ccgacaggcc 120
cctggcaaag gacttgagtg ggtcggacgg atcaagagca agacctacgg cggcaccacc 180
gattatgccg agcctgtgaa gggcagattc accatcagcc gggacgacag caagaacacc 240
ctgtacctgc agatgaacag cctgaaaacc gaggacaccg ccgtgtacta ctgcgccaga 300
accagcagaa gaagctacgc cttcgactac tggggccagg gcacactggt taccgttagc 360
tctgctagca ccaagggccc aagtgtgttt cccctggccc ccagcagcaa gtctacttcc 420
ggcggaactg ctgccctggg ttgcctggtg aaggactact tcccctgtcc cgtgacagtg 480
tcctggaact ctggggctct gacttccggc gtgcacacct tccccgccgt gctgcagagc 540
agcggcctgt acagcctgag cagcgtggtg acagtgccct ccagctctct gggaacccag 600
acctatatct gcaacgtgaa ccacaagccc agcaacacca aggtggacaa gagagtggag 660
cccaagagct gcgacaagac ccacacctgc cccccctgcc cagctccaga actgctggga 720
gggccttccg tgttcctgtt cccccccaag cccaaggaca ccctgatgat cagcaggacc 780
cccgaggtga cctgcgtggt ggtggacgtg tcccacgagg acccagaggt gaagttcaac 840
tggtacgtgg acggcgtgga ggtgcacaac gccaagacca agcccagaga ggagcagtac 900
aacagcacct acagggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 960
aaagaataca agtgcaaagt ctccaacaag gccctgccag ccccaatcga aaagacaatc 1020
agcaaggcca agggccagcc acgggagccc caggtgtaca ccctgccccc cagccgggag 1080
gagatgacca agaaccaggt gtccctgacc tgtctggtga agggcttcta cccctgtgat 1140
atcgccgtgg agtgggagag caacggccag cccgagaaca actacaagac caccccccca 1200
gtgctggaca gcgacggcag cttcttcctg tacagcaagc tgaccgtgga caagtccagg 1260
tggcagcagg gcaacgtgtt cagctgcagc gtgatgcacg aggccctgca caaccactac 1320
acccagaagt ccctgagcct gagccccggc aag 1353
<210> 136
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 136
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala
1 5 10
<210> 137
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 137
Asp Ala Ser Ser Leu Glu Ser
1 5
<210> 138
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 138
Gln Gln Ile Thr Arg Tyr Pro Val Thr
1 5
<210> 139
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 139
Ser Gln Ser Ile Ser Ser Trp
1 5
<210> 140
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 140
Asp Ala Ser
1
<210> 141
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 141
Ile Thr Arg Tyr Pro Val
1 5
<210> 142
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 142
Gln Ser Ile Ser Ser Trp
1 5
<210> 143
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 143
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ile Thr Arg Tyr Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 144
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 144
gacatccaga tgacacagag ccccagcaca ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gagcatctcc tcttggctgg cctggtatca gcagaagcct 120
ggcaaggccc ctaagctgct gatctacgat gccagcagcc tggaaagcgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgag ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ctgccagcag atcacaagat accccgtgac ctttggccag 300
ggcaccaagg tggaaatcaa g 321
<210> 145
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 145
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ile Thr Arg Tyr Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 146
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 146
gacatccaga tgacacagag ccccagcaca ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gagcatctcc tcttggctgg cctggtatca gcagaagcct 120
ggcaaggccc ctaagctgct gatctacgat gccagcagcc tggaaagcgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgag ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ctgccagcag atcacaagat accccgtgac ctttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 147
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 147
Val Ser Gly Tyr Tyr Ser His Ser Gly Gly Phe Asp Val
1 5 10
<210> 148
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 148
Ala Arg Val Ser Gly Tyr Tyr Ser His Ser Gly Gly Phe Asp Val
1 5 10 15
<210> 149
<211> 124
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 149
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Tyr Gly Gly Thr Thr Asp Tyr Ala Glu
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Val Ser Gly Tyr Tyr Ser His Ser Gly Gly Phe Asp
100 105 110
Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 150
<211> 372
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 150
gaagtgcagc tggtggaatc tggcggcgga cttgtgaaac ctggcggctc tctgagactg 60
agctgtgccg cttccggctt caccttcagc aactactgga tcagctgggt ccgacaggcc 120
cctggcaaag gacttgagtg ggtcggacgg atcaagagca agacctacgg cggcaccacc 180
gattatgccg agcctgtgaa gggcagattc accatcagcc gggacgacag caagaacacc 240
ctgtacctgc agatgaacag cctgaaaacc gaggacaccg ccgtgtacta ctgcgccaga 300
gtgtctggct actactctca cagcggcggc tttgatgtgt ggggccaggg aacactggtc 360
accgttagtt ct 372
<210> 151
<211> 454
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 151
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Tyr Gly Gly Thr Thr Asp Tyr Ala Glu
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Val Ser Gly Tyr Tyr Ser His Ser Gly Gly Phe Asp
100 105 110
Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 152
<211> 1362
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 152
gaagtgcagc tggtggaatc tggcggcgga cttgtgaaac ctggcggctc tctgagactg 60
agctgtgccg cttccggctt caccttcagc aactactgga tcagctgggt ccgacaggcc 120
cctggcaaag gacttgagtg ggtcggacgg atcaagagca agacctacgg cggcaccacc 180
gattatgccg agcctgtgaa gggcagattc accatcagcc gggacgacag caagaacacc 240
ctgtacctgc agatgaacag cctgaaaacc gaggacaccg ccgtgtacta ctgcgccaga 300
gtgtctggct actactctca cagcggcggc tttgatgtgt ggggccaggg aacactggtc 360
accgttagtt ctgctagcac caagggccca agtgtgtttc ccctggcccc cagcagcaag 420
tctacttccg gcggaactgc tgccctgggt tgcctggtga aggactactt cccctgtccc 480
gtgacagtgt cctggaactc tggggctctg acttccggcg tgcacacctt ccccgccgtg 540
ctgcagagca gcggcctgta cagcctgagc agcgtggtga cagtgccctc cagctctctg 600
ggaacccaga cctatatctg caacgtgaac cacaagccca gcaacaccaa ggtggacaag 660
agagtggagc ccaagagctg cgacaagacc cacacctgcc ccccctgccc agctccagaa 720
ctgctgggag ggccttccgt gttcctgttc ccccccaagc ccaaggacac cctgatgatc 780
agcaggaccc ccgaggtgac ctgcgtggtg gtggacgtgt cccacgagga cccagaggtg 840
aagttcaact ggtacgtgga cggcgtggag gtgcacaacg ccaagaccaa gcccagagag 900
gagcagtaca acagcaccta cagggtggtg tccgtgctga ccgtgctgca ccaggactgg 960
ctgaacggca aagaatacaa gtgcaaagtc tccaacaagg ccctgccagc cccaatcgaa 1020
aagacaatca gcaaggccaa gggccagcca cgggagcccc aggtgtacac cctgcccccc 1080
agccgggagg agatgaccaa gaaccaggtg tccctgacct gtctggtgaa gggcttctac 1140
ccctgtgata tcgccgtgga gtgggagagc aacggccagc ccgagaacaa ctacaagacc 1200
acccccccag tgctggacag cgacggcagc ttcttcctgt acagcaagct gaccgtggac 1260
aagtccaggt ggcagcaggg caacgtgttc agctgcagcg tgatgcacga ggccctgcac 1320
aaccactaca cccagaagtc cctgagcctg agccccggca ag 1362
<210> 153
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 153
Arg Ala Ser Gln Gly Ile Ser Asn Tyr Leu Ala
1 5 10
<210> 154
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 154
Ala Ala Ser Thr Leu Gln Ser
1 5
<210> 155
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 155
Gln Lys Thr Trp Arg Thr Pro Gly Thr
1 5
<210> 156
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 156
Ser Gln Gly Ile Ser Asn Tyr
1 5
<210> 157
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 157
Thr Trp Arg Thr Pro Gly
1 5
<210> 158
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 158
Gln Gly Ile Ser Asn Tyr
1 5
<210> 159
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 159
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Lys Thr Trp Arg Thr Pro Gly
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 160
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 160
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gggcatcagc aactacctgg cctggtatca gcagaaaccc 120
ggcaaggtgc ccaagctgct gatctacgct gccagcacac tgcagagcgg agtgcctagc 180
agattttctg gcagcggctc cggcaccgat ttcaccctga ccatatctag cctgcagcca 240
gaggacgtgg ccacctacta ctgtcagaaa acctggcgga cccctggcac atttggccag 300
ggaacaaagg tggaaatcaa g 321
<210> 161
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 161
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Lys Thr Trp Arg Thr Pro Gly
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 162
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 162
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gggcatcagc aactacctgg cctggtatca gcagaaaccc 120
ggcaaggtgc ccaagctgct gatctacgct gccagcacac tgcagagcgg agtgcctagc 180
agattttctg gcagcggctc cggcaccgat ttcaccctga ccatatctag cctgcagcca 240
gaggacgtgg ccacctacta ctgtcagaaa acctggcgga cccctggcac atttggccag 300
ggaacaaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 163
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 163
Ser Arg Leu Ile Ala Pro Trp Leu Asp Tyr
1 5 10
<210> 164
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 164
Ala Arg Ser Arg Leu Ile Ala Pro Trp Leu Asp Tyr
1 5 10
<210> 165
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 165
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Leu Ile Ala Pro Trp Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 166
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 166
gaagttcagc tgctggaatc tggcggagga ctggttcaac ctggcggctc tctgagactg 60
tcttgtgccg ccagcggctt cacctttagc agctacgcca tgagctgggt ccgacaggct 120
cctggcaaag gccttgaatg ggtgtccgcc atctctggct ctggcggcag cacatattac 180
gccgactctg tgaagggcag attcaccatc agccgggaca acagcaagaa caccctgtac 240
ctgcagatga acagcctgag agccgaggac accgccgtgt actactgtgc cagaagcaga 300
ctgatcgccc cttggctgga ttattggggc cagggcacac tggtcaccgt gtcatct 357
<210> 167
<211> 449
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 167
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Arg Leu Ile Ala Pro Trp Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 168
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 168
gaagttcagc tgctggaatc tggcggagga ctggttcaac ctggcggctc tctgagactg 60
tcttgtgccg ccagcggctt cacctttagc agctacgcca tgagctgggt ccgacaggct 120
cctggcaaag gccttgaatg ggtgtccgcc atctctggct ctggcggcag cacatattac 180
gccgactctg tgaagggcag attcaccatc agccgggaca acagcaagaa caccctgtac 240
ctgcagatga acagcctgag agccgaggac accgccgtgt actactgtgc cagaagcaga 300
ctgatcgccc cttggctgga ttattggggc cagggcacac tggtcaccgt gtcatctgct 360
agcaccaagg gcccaagtgt gtttcccctg gcccccagca gcaagtctac ttccggcgga 420
actgctgccc tgggttgcct ggtgaaggac tacttcccct gtcccgtgac agtgtcctgg 480
aactctgggg ctctgacttc cggcgtgcac accttccccg ccgtgctgca gagcagcggc 540
ctgtacagcc tgagcagcgt ggtgacagtg ccctccagct ctctgggaac ccagacctat 600
atctgcaacg tgaaccacaa gcccagcaac accaaggtgg acaagagagt ggagcccaag 660
agctgcgaca agacccacac ctgccccccc tgcccagctc cagaactgct gggagggcct 720
tccgtgttcc tgttcccccc caagcccaag gacaccctga tgatcagcag gacccccgag 780
gtgacctgcg tggtggtgga cgtgtcccac gaggacccag aggtgaagtt caactggtac 840
gtggacggcg tggaggtgca caacgccaag accaagccca gagaggagca gtacaacagc 900
acctacaggg tggtgtccgt gctgaccgtg ctgcaccagg actggctgaa cggcaaagaa 960
tacaagtgca aagtctccaa caaggccctg ccagccccaa tcgaaaagac aatcagcaag 1020
gccaagggcc agccacggga gccccaggtg tacaccctgc cccccagccg ggaggagatg 1080
accaagaacc aggtgtccct gacctgtctg gtgaagggct tctacccctg tgatatcgcc 1140
gtggagtggg agagcaacgg ccagcccgag aacaactaca agaccacccc cccagtgctg 1200
gacagcgacg gcagcttctt cctgtacagc aagctgaccg tggacaagtc caggtggcag 1260
cagggcaacg tgttcagctg cagcgtgatg cacgaggccc tgcacaacca ctacacccag 1320
aagtccctga gcctgagccc cggcaag 1347
<210> 169
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 169
Gln Gln Val Tyr Gly Ser Pro Pro Thr
1 5
<210> 170
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 170
Val Tyr Gly Ser Pro Pro
1 5
<210> 171
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 171
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Val Tyr Gly Ser Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 172
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 172
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gagcatcagc agctacctga actggtatca gcagaagccc 120
ggcaaggccc ctaaactgct gatctatgcc gccagctctc tgcagtctgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgac ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ctgccagcag gtctacggca gccctcctac atttggccag 300
ggcaccaagg tggaaatcaa g 321
<210> 173
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 173
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Val Tyr Gly Ser Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 174
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 174
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gagcatcagc agctacctga actggtatca gcagaagccc 120
ggcaaggccc ctaaactgct gatctatgcc gccagctctc tgcagtctgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgac ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ctgccagcag gtctacggca gccctcctac atttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 175
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 175
Gly Tyr Ser Phe Thr Ser Tyr Trp Ile Gly
1 5 10
<210> 176
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 176
Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe Gln
1 5 10 15
Gly
<210> 177
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 177
Gly Ser Ser Ala Ala Ser Gly Leu Ser Gly Asp Leu
1 5 10
<210> 178
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 178
Ser Tyr Trp Ile Gly
1 5
<210> 179
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 179
Gly Tyr Ser Phe Thr Ser Tyr
1 5
<210> 180
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 180
Tyr Pro Gly Asp Ser Asp
1 5
<210> 181
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 181
Gly Tyr Ser Phe Thr Ser Tyr Trp
1 5
<210> 182
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 182
Ile Tyr Pro Gly Asp Ser Asp Thr
1 5
<210> 183
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 183
Ala Arg Gly Ser Ser Ala Ala Ser Gly Leu Ser Gly Asp Leu
1 5 10
<210> 184
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 184
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ser Ala Ala Ser Gly Leu Ser Gly Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 185
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 185
gaagttcagc tggtgcagtc tggcgccgaa gtgaagaagc ctggcgagag cctgaagatc 60
tcctgcaaag gcagcggcta cagcttcacc agctactgga tcggctgggt ccgacagatg 120
cctggcaaag gccttgagtg gatgggcatc atctaccccg gcgacagcga caccagatac 180
agccctagct ttcagggcca agtgaccatc agcgccgaca agagcatcag cacagcctac 240
ctgcagtggt ccagcctgaa ggcctctgac accgccatgt actactgtgc cagaggaagc 300
tctgccgcct ctggactgtc tggcgatctt tggggacagg gcacactggt cacagtgtct 360
agt 363
<210> 186
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 186
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ser Ala Ala Ser Gly Leu Ser Gly Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 187
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 187
gaagttcagc tggtgcagtc tggcgccgaa gtgaagaagc ctggcgagag cctgaagatc 60
tcctgcaaag gcagcggcta cagcttcacc agctactgga tcggctgggt ccgacagatg 120
cctggcaaag gccttgagtg gatgggcatc atctaccccg gcgacagcga caccagatac 180
agccctagct ttcagggcca agtgaccatc agcgccgaca agagcatcag cacagcctac 240
ctgcagtggt ccagcctgaa ggcctctgac accgccatgt actactgtgc cagaggaagc 300
tctgccgcct ctggactgtc tggcgatctt tggggacagg gcacactggt cacagtgtct 360
agtgctagca ccaagggccc aagtgtgttt cccctggccc ccagcagcaa gtctacttcc 420
ggcggaactg ctgccctggg ttgcctggtg aaggactact tcccctgtcc cgtgacagtg 480
tcctggaact ctggggctct gacttccggc gtgcacacct tccccgccgt gctgcagagc 540
agcggcctgt acagcctgag cagcgtggtg acagtgccct ccagctctct gggaacccag 600
acctatatct gcaacgtgaa ccacaagccc agcaacacca aggtggacaa gagagtggag 660
cccaagagct gcgacaagac ccacacctgc cccccctgcc cagctccaga actgctggga 720
gggccttccg tgttcctgtt cccccccaag cccaaggaca ccctgatgat cagcaggacc 780
cccgaggtga cctgcgtggt ggtggacgtg tcccacgagg acccagaggt gaagttcaac 840
tggtacgtgg acggcgtgga ggtgcacaac gccaagacca agcccagaga ggagcagtac 900
aacagcacct acagggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 960
aaagaataca agtgcaaagt ctccaacaag gccctgccag ccccaatcga aaagacaatc 1020
agcaaggcca agggccagcc acgggagccc caggtgtaca ccctgccccc cagccgggag 1080
gagatgacca agaaccaggt gtccctgacc tgtctggtga agggcttcta cccctgtgat 1140
atcgccgtgg agtgggagag caacggccag cccgagaaca actacaagac caccccccca 1200
gtgctggaca gcgacggcag cttcttcctg tacagcaagc tgaccgtgga caagtccagg 1260
tggcagcagg gcaacgtgtt cagctgcagc gtgatgcacg aggccctgca caaccactac 1320
acccagaagt ccctgagcct gagccccggc aag 1353
<210> 188
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 188
Gln Gln Asp Tyr Tyr Ser Pro Phe Thr
1 5
<210> 189
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 189
Asp Tyr Tyr Ser Pro Phe
1 5
<210> 190
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 190
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Asp Tyr Tyr Ser Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 191
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 191
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gagcatcagc agctacctga actggtatca gcagaagccc 120
ggcaaggccc ctaaactgct gatctatgcc gccagctctc tgcagtctgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgac ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ctgccagcag gactactaca gccccttcac ctttggccag 300
ggcaccaagg tggaaatcaa g 321
<210> 192
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 192
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Asp Tyr Tyr Ser Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 193
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 193
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gagcatcagc agctacctga actggtatca gcagaagccc 120
ggcaaggccc ctaaactgct gatctatgcc gccagctctc tgcagtctgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgac ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ctgccagcag gactactaca gccccttcac ctttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 194
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 194
Ala Tyr Lys Leu Ser Trp Leu Asp Leu
1 5
<210> 195
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 195
Ala Arg Ala Tyr Lys Leu Ser Trp Leu Asp Leu
1 5 10
<210> 196
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 196
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Tyr Lys Leu Ser Trp Leu Asp Leu Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 197
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 197
gaagttcagc tgctggaatc tggcggagga ctggttcaac ctggcggctc tctgagactg 60
tcttgtgccg ccagcggctt cacctttagc agctacgcca tgagctgggt ccgacaggct 120
cctggcaaag gccttgaatg ggtgtccgcc atctctggct ctggcggcag cacatattac 180
gccgactctg tgaagggcag attcaccatc agccgggaca acagcaagaa caccctgtac 240
ctgcagatga acagcctgag agccgaggac accgccgtgt actattgtgc cagagcctac 300
aagctgagct ggctggatct ttggggccag ggcacactgg tcacagtgtc atct 354
<210> 198
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 198
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Tyr Lys Leu Ser Trp Leu Asp Leu Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 199
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 199
gaagttcagc tgctggaatc tggcggagga ctggttcaac ctggcggctc tctgagactg 60
tcttgtgccg ccagcggctt cacctttagc agctacgcca tgagctgggt ccgacaggct 120
cctggcaaag gccttgaatg ggtgtccgcc atctctggct ctggcggcag cacatattac 180
gccgactctg tgaagggcag attcaccatc agccgggaca acagcaagaa caccctgtac 240
ctgcagatga acagcctgag agccgaggac accgccgtgt actattgtgc cagagcctac 300
aagctgagct ggctggatct ttggggccag ggcacactgg tcacagtgtc atctgctagc 360
accaagggcc caagtgtgtt tcccctggcc cccagcagca agtctacttc cggcggaact 420
gctgccctgg gttgcctggt gaaggactac ttcccctgtc ccgtgacagt gtcctggaac 480
tctggggctc tgacttccgg cgtgcacacc ttccccgccg tgctgcagag cagcggcctg 540
tacagcctga gcagcgtggt gacagtgccc tccagctctc tgggaaccca gacctatatc 600
tgcaacgtga accacaagcc cagcaacacc aaggtggaca agagagtgga gcccaagagc 660
tgcgacaaga cccacacctg ccccccctgc ccagctccag aactgctggg agggccttcc 720
gtgttcctgt tcccccccaa gcccaaggac accctgatga tcagcaggac ccccgaggtg 780
acctgcgtgg tggtggacgt gtcccacgag gacccagagg tgaagttcaa ctggtacgtg 840
gacggcgtgg aggtgcacaa cgccaagacc aagcccagag aggagcagta caacagcacc 900
tacagggtgg tgtccgtgct gaccgtgctg caccaggact ggctgaacgg caaagaatac 960
aagtgcaaag tctccaacaa ggccctgcca gccccaatcg aaaagacaat cagcaaggcc 1020
aagggccagc cacgggagcc ccaggtgtac accctgcccc ccagccggga ggagatgacc 1080
aagaaccagg tgtccctgac ctgtctggtg aagggcttct acccctgtga tatcgccgtg 1140
gagtgggaga gcaacggcca gcccgagaac aactacaaga ccaccccccc agtgctggac 1200
agcgacggca gcttcttcct gtacagcaag ctgaccgtgg acaagtccag gtggcagcag 1260
ggcaacgtgt tcagctgcag cgtgatgcac gaggccctgc acaaccacta cacccagaag 1320
tccctgagcc tgagccccgg caag 1344
<210> 200
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 200
Gln Gln Val Trp Tyr Ala Pro Val Thr
1 5
<210> 201
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 201
Val Trp Tyr Ala Pro Val
1 5
<210> 202
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 202
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Val Trp Tyr Ala Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 203
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 203
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gagcatcagc agctacctga actggtatca gcagaagccc 120
ggcaaggccc ctaaactgct gatctatgcc gccagctctc tgcagtctgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgac ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ctgccagcaa gtttggtacg cccctgtgac ctttggccag 300
ggcaccaagg tggaaatcaa g 321
<210> 204
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 204
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Val Trp Tyr Ala Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 205
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 205
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gagcatcagc agctacctga actggtatca gcagaagccc 120
ggcaaggccc ctaaactgct gatctatgcc gccagctctc tgcagtctgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgac ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ctgccagcaa gtttggtacg cccctgtgac ctttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 206
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 206
Gly Phe Thr Phe Ser Asn Ala Trp Met Ser
1 5 10
<210> 207
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 207
Arg Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala Pro
1 5 10 15
Val Lys Gly
<210> 208
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 208
Thr Ile Tyr Pro Ser Ala Pro Ser Ser Ser Leu Asp Tyr
1 5 10
<210> 209
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 209
Asn Ala Trp Met Ser
1 5
<210> 210
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 210
Gly Phe Thr Phe Ser Asn Ala
1 5
<210> 211
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 211
Lys Ser Lys Thr Asp Ala Gly Thr
1 5
<210> 212
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 212
Gly Phe Thr Phe Ser Asn Ala Trp
1 5
<210> 213
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 213
Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr
1 5 10
<210> 214
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 214
Ala Arg Thr Ile Tyr Pro Ser Ala Pro Ser Ser Ser Leu Asp Tyr
1 5 10 15
<210> 215
<211> 124
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 215
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Thr Ile Tyr Pro Ser Ala Pro Ser Ser Ser Leu Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 216
<211> 372
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 216
gaagtgcagc tggtggaatc tggcggcgga cttgtgaaac ctggcggctc tctgagactg 60
agctgtgccg cttccggctt caccttcagc aatgcctgga tgagctgggt ccgacaggcc 120
cctggaaaag gccttgagtg ggtcggacgg atcaagagca agaccgatgc cggcaccacc 180
gattatgctg cccctgtgaa gggcagattc accatcagca gggacgacag caagaacacc 240
ctgtacctgc agatgaacag cctgaaaacc gaggacaccg ccgtgtacta ctgcgccaga 300
acaatctacc ccagcgctcc tagcagcagc ctggattatt ggggccaggg cacactggtc 360
accgtgtcat ct 372
<210> 217
<211> 454
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 217
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Thr Ile Tyr Pro Ser Ala Pro Ser Ser Ser Leu Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 218
<211> 1362
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 218
gaagtgcagc tggtggaatc tggcggcgga cttgtgaaac ctggcggctc tctgagactg 60
agctgtgccg cttccggctt caccttcagc aatgcctgga tgagctgggt ccgacaggcc 120
cctggaaaag gccttgagtg ggtcggacgg atcaagagca agaccgatgc cggcaccacc 180
gattatgctg cccctgtgaa gggcagattc accatcagca gggacgacag caagaacacc 240
ctgtacctgc agatgaacag cctgaaaacc gaggacaccg ccgtgtacta ctgcgccaga 300
acaatctacc ccagcgctcc tagcagcagc ctggattatt ggggccaggg cacactggtc 360
accgtgtcat ctgctagcac caagggccca agtgtgtttc ccctggcccc cagcagcaag 420
tctacttccg gcggaactgc tgccctgggt tgcctggtga aggactactt cccctgtccc 480
gtgacagtgt cctggaactc tggggctctg acttccggcg tgcacacctt ccccgccgtg 540
ctgcagagca gcggcctgta cagcctgagc agcgtggtga cagtgccctc cagctctctg 600
ggaacccaga cctatatctg caacgtgaac cacaagccca gcaacaccaa ggtggacaag 660
agagtggagc ccaagagctg cgacaagacc cacacctgcc ccccctgccc agctccagaa 720
ctgctgggag ggccttccgt gttcctgttc ccccccaagc ccaaggacac cctgatgatc 780
agcaggaccc ccgaggtgac ctgcgtggtg gtggacgtgt cccacgagga cccagaggtg 840
aagttcaact ggtacgtgga cggcgtggag gtgcacaacg ccaagaccaa gcccagagag 900
gagcagtaca acagcaccta cagggtggtg tccgtgctga ccgtgctgca ccaggactgg 960
ctgaacggca aagaatacaa gtgcaaagtc tccaacaagg ccctgccagc cccaatcgaa 1020
aagacaatca gcaaggccaa gggccagcca cgggagcccc aggtgtacac cctgcccccc 1080
agccgggagg agatgaccaa gaaccaggtg tccctgacct gtctggtgaa gggcttctac 1140
ccctgtgata tcgccgtgga gtgggagagc aacggccagc ccgagaacaa ctacaagacc 1200
acccccccag tgctggacag cgacggcagc ttcttcctgt acagcaagct gaccgtggac 1260
aagtccaggt ggcagcaggg caacgtgttc agctgcagcg tgatgcacga ggccctgcac 1320
aaccactaca cccagaagtc cctgagcctg agccccggca ag 1362
<210> 219
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 219
Gln Gln Leu Ile Phe Phe Pro Leu Thr
1 5
<210> 220
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 220
Leu Ile Phe Phe Pro Leu
1 5
<210> 221
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 221
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Gln Leu Ile Phe Phe Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 222
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 222
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gggcatcagc aactacctgg cctggtatca gcagaaaccc 120
ggcaaggtgc ccaagctgct gatctacgct gccagcacac tgcagagcgg agtgcctagc 180
agattttctg gcagcggctc cggcaccgat ttcaccctga ccatatctag cctgcagcca 240
gaggacgtgg ccacctacta ttgccagcag ctgatcttct tccctctgac ctttggccag 300
ggcaccaagg tggaaatcaa g 321
<210> 223
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 223
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Gln Leu Ile Phe Phe Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 224
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 224
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gggcatcagc aactacctgg cctggtatca gcagaaaccc 120
ggcaaggtgc ccaagctgct gatctacgct gccagcacac tgcagagcgg agtgcctagc 180
agattttctg gcagcggctc cggcaccgat ttcaccctga ccatatctag cctgcagcca 240
gaggacgtgg ccacctacta ttgccagcag ctgatcttct tccctctgac ctttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 225
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 225
Ala Ser His Arg Leu His Ser Leu Phe Asp Val
1 5 10
<210> 226
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 226
Ala Arg Ala Ser His Arg Leu His Ser Leu Phe Asp Val
1 5 10
<210> 227
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 227
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Ala Ser His Arg Leu His Ser Leu Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 228
<211> 366
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 228
gaagtgcagc tggtggaatc tggcggcgga cttgtgaaac ctggcggctc tctgagactg 60
agctgtgccg cttccggctt caccttcagc aatgcctgga tgagctgggt ccgacaggcc 120
cctggaaaag gccttgagtg ggtcggacgg atcaagagca agaccgatgc cggcaccacc 180
gattatgctg cccctgtgaa gggcagattc accatcagca gggacgacag caagaacacc 240
ctgtacctgc agatgaacag cctgaaaacc gaggacaccg ccgtgtacta ctgtgccaga 300
gcctctcaca gactgcacag cctgtttgac gtgtggggcc agggaacact ggtcaccgtt 360
agttct 366
<210> 229
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 229
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Ala Ser His Arg Leu His Ser Leu Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 230
<211> 1356
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 230
gaagtgcagc tggtggaatc tggcggcgga cttgtgaaac ctggcggctc tctgagactg 60
agctgtgccg cttccggctt caccttcagc aatgcctgga tgagctgggt ccgacaggcc 120
cctggaaaag gccttgagtg ggtcggacgg atcaagagca agaccgatgc cggcaccacc 180
gattatgctg cccctgtgaa gggcagattc accatcagca gggacgacag caagaacacc 240
ctgtacctgc agatgaacag cctgaaaacc gaggacaccg ccgtgtacta ctgtgccaga 300
gcctctcaca gactgcacag cctgtttgac gtgtggggcc agggaacact ggtcaccgtt 360
agttctgcta gcaccaaggg cccaagtgtg tttcccctgg cccccagcag caagtctact 420
tccggcggaa ctgctgccct gggttgcctg gtgaaggact acttcccctg tcccgtgaca 480
gtgtcctgga actctggggc tctgacttcc ggcgtgcaca ccttccccgc cgtgctgcag 540
agcagcggcc tgtacagcct gagcagcgtg gtgacagtgc cctccagctc tctgggaacc 600
cagacctata tctgcaacgt gaaccacaag cccagcaaca ccaaggtgga caagagagtg 660
gagcccaaga gctgcgacaa gacccacacc tgccccccct gcccagctcc agaactgctg 720
ggagggcctt ccgtgttcct gttccccccc aagcccaagg acaccctgat gatcagcagg 780
acccccgagg tgacctgcgt ggtggtggac gtgtcccacg aggacccaga ggtgaagttc 840
aactggtacg tggacggcgt ggaggtgcac aacgccaaga ccaagcccag agaggagcag 900
tacaacagca cctacagggt ggtgtccgtg ctgaccgtgc tgcaccagga ctggctgaac 960
ggcaaagaat acaagtgcaa agtctccaac aaggccctgc cagccccaat cgaaaagaca 1020
atcagcaagg ccaagggcca gccacgggag ccccaggtgt acaccctgcc ccccagccgg 1080
gaggagatga ccaagaacca ggtgtccctg acctgtctgg tgaagggctt ctacccctgt 1140
gatatcgccg tggagtggga gagcaacggc cagcccgaga acaactacaa gaccaccccc 1200
ccagtgctgg acagcgacgg cagcttcttc ctgtacagca agctgaccgt ggacaagtcc 1260
aggtggcagc agggcaacgt gttcagctgc agcgtgatgc acgaggccct gcacaaccac 1320
tacacccaga agtccctgag cctgagcccc ggcaag 1356
<210> 231
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 231
Gln Gln Gly Leu Phe Tyr Pro His Thr
1 5
<210> 232
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 232
Gly Leu Phe Tyr Pro His
1 5
<210> 233
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 233
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Leu Phe Tyr Pro His
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 234
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 234
gacatccaga tgacacagag ccccagcaca ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gagcatctcc tcttggctgg cctggtatca gcagaagcct 120
ggcaaggccc ctaagctgct gatctacgat gccagcagcc tggaaagcgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgag ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ttgtcagcag ggcctgttct accctcacac ctttggccag 300
ggcaccaagg tggaaatcaa g 321
<210> 235
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 235
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Leu Phe Tyr Pro His
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 236
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 236
gacatccaga tgacacagag ccccagcaca ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gagcatctcc tcttggctgg cctggtatca gcagaagcct 120
ggcaaggccc ctaagctgct gatctacgat gccagcagcc tggaaagcgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgag ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ttgtcagcag ggcctgttct accctcacac ctttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 237
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 237
Asp Glu Tyr Pro Trp Gly Trp Phe Asp Val
1 5 10
<210> 238
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 238
Ala Arg Asp Glu Tyr Pro Trp Gly Trp Phe Asp Val
1 5 10
<210> 239
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 239
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Asp Glu Tyr Pro Trp Gly Trp Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 240
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 240
gaagtgcagc tggtggaatc tggcggcgga cttgtgaaac ctggcggctc tctgagactg 60
agctgtgccg cttccggctt caccttcagc aatgcctgga tgagctgggt ccgacaggcc 120
cctggaaaag gccttgagtg ggtcggacgg atcaagagca agaccgatgc cggcaccacc 180
gattatgctg cccctgtgaa gggcagattc accatcagca gggacgacag caagaacacc 240
ctgtacctgc agatgaacag cctgaaaacc gaggacaccg ccgtgtacta ctgcgccaga 300
gatgagtacc cctggggatg gttcgatgtg tggggacagg gaaccctggt caccgttagt 360
tct 363
<210> 241
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 241
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Asp Glu Tyr Pro Trp Gly Trp Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 242
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 242
gaagtgcagc tggtggaatc tggcggcgga cttgtgaaac ctggcggctc tctgagactg 60
agctgtgccg cttccggctt caccttcagc aatgcctgga tgagctgggt ccgacaggcc 120
cctggaaaag gccttgagtg ggtcggacgg atcaagagca agaccgatgc cggcaccacc 180
gattatgctg cccctgtgaa gggcagattc accatcagca gggacgacag caagaacacc 240
ctgtacctgc agatgaacag cctgaaaacc gaggacaccg ccgtgtacta ctgcgccaga 300
gatgagtacc cctggggatg gttcgatgtg tggggacagg gaaccctggt caccgttagt 360
tctgctagca ccaagggccc aagtgtgttt cccctggccc ccagcagcaa gtctacttcc 420
ggcggaactg ctgccctggg ttgcctggtg aaggactact tcccctgtcc cgtgacagtg 480
tcctggaact ctggggctct gacttccggc gtgcacacct tccccgccgt gctgcagagc 540
agcggcctgt acagcctgag cagcgtggtg acagtgccct ccagctctct gggaacccag 600
acctatatct gcaacgtgaa ccacaagccc agcaacacca aggtggacaa gagagtggag 660
cccaagagct gcgacaagac ccacacctgc cccccctgcc cagctccaga actgctggga 720
gggccttccg tgttcctgtt cccccccaag cccaaggaca ccctgatgat cagcaggacc 780
cccgaggtga cctgcgtggt ggtggacgtg tcccacgagg acccagaggt gaagttcaac 840
tggtacgtgg acggcgtgga ggtgcacaac gccaagacca agcccagaga ggagcagtac 900
aacagcacct acagggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 960
aaagaataca agtgcaaagt ctccaacaag gccctgccag ccccaatcga aaagacaatc 1020
agcaaggcca agggccagcc acgggagccc caggtgtaca ccctgccccc cagccgggag 1080
gagatgacca agaaccaggt gtccctgacc tgtctggtga agggcttcta cccctgtgat 1140
atcgccgtgg agtgggagag caacggccag cccgagaaca actacaagac caccccccca 1200
gtgctggaca gcgacggcag cttcttcctg tacagcaagc tgaccgtgga caagtccagg 1260
tggcagcagg gcaacgtgtt cagctgcagc gtgatgcacg aggccctgca caaccactac 1320
acccagaagt ccctgagcct gagccccggc aag 1353
<210> 243
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 243
Arg Ala Ser Gln Gly Ile Ser Ser Trp Leu Ala
1 5 10
<210> 244
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 244
Gln Gln Tyr Ile Phe Tyr Pro Leu Thr
1 5
<210> 245
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 245
Ser Gln Gly Ile Ser Ser Trp
1 5
<210> 246
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 246
Tyr Ile Phe Tyr Pro Leu
1 5
<210> 247
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 247
Gln Gly Ile Ser Ser Trp
1 5
<210> 248
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 248
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ile Phe Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 249
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 249
gacatccaga tgacacagag ccctagctcc gtgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gggcatctct tcttggctgg cctggtatca gcagaagcct 120
ggcaaggccc ctaagctgct gatctatgcc gcttccagtc tgcagagcgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgac ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ctgccagcag tacatcttct accctctgac cttcggccag 300
ggcaccaagg tggaaatcaa g 321
<210> 250
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 250
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ile Phe Tyr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 251
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 251
gacatccaga tgacacagag ccctagctcc gtgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gggcatctct tcttggctgg cctggtatca gcagaagcct 120
ggcaaggccc ctaagctgct gatctatgcc gcttccagtc tgcagagcgg cgtgccaagc 180
agattttctg gcagcggctc tggcaccgac ttcaccctga ccatatctag cctgcagcca 240
gaggacttcg ccacctacta ctgccagcag tacatcttct accctctgac cttcggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 252
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 252
Val Ala Ser Pro Ser Ala Pro Gly Gly Phe Asp Tyr
1 5 10
<210> 253
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 253
Ala Arg Val Ala Ser Pro Ser Ala Pro Gly Gly Phe Asp Tyr
1 5 10
<210> 254
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 254
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Val Ala Ser Pro Ser Ala Pro Gly Gly Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 255
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 255
gaagtgcagc tggtggaatc tggcggcgga cttgtgaaac ctggcggctc tctgagactg 60
agctgtgccg cttccggctt caccttcagc aatgcctgga tgagctgggt ccgacaggcc 120
cctggaaaag gccttgagtg ggtcggacgg atcaagagca agaccgatgc cggcaccacc 180
gattatgctg cccctgtgaa gggcagattc accatcagca gggacgacag caagaacacc 240
ctgtacctgc agatgaacag cctgaaaacc gaggacaccg ccgtgtacta ctgcgccaga 300
gtggcttctc cttctgctcc cggcggattc gattattggg gccagggaac actggtcacc 360
gtgtctagt 369
<210> 256
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 256
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Val Ala Ser Pro Ser Ala Pro Gly Gly Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 257
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 257
gaagtgcagc tggtggaatc tggcggcgga cttgtgaaac ctggcggctc tctgagactg 60
agctgtgccg cttccggctt caccttcagc aatgcctgga tgagctgggt ccgacaggcc 120
cctggaaaag gccttgagtg ggtcggacgg atcaagagca agaccgatgc cggcaccacc 180
gattatgctg cccctgtgaa gggcagattc accatcagca gggacgacag caagaacacc 240
ctgtacctgc agatgaacag cctgaaaacc gaggacaccg ccgtgtacta ctgcgccaga 300
gtggcttctc cttctgctcc cggcggattc gattattggg gccagggaac actggtcacc 360
gtgtctagtg ctagcaccaa gggcccaagt gtgtttcccc tggcccccag cagcaagtct 420
acttccggcg gaactgctgc cctgggttgc ctggtgaagg actacttccc ctgtcccgtg 480
acagtgtcct ggaactctgg ggctctgact tccggcgtgc acaccttccc cgccgtgctg 540
cagagcagcg gcctgtacag cctgagcagc gtggtgacag tgccctccag ctctctggga 600
acccagacct atatctgcaa cgtgaaccac aagcccagca acaccaaggt ggacaagaga 660
gtggagccca agagctgcga caagacccac acctgccccc cctgcccagc tccagaactg 720
ctgggagggc cttccgtgtt cctgttcccc cccaagccca aggacaccct gatgatcagc 780
aggacccccg aggtgacctg cgtggtggtg gacgtgtccc acgaggaccc agaggtgaag 840
ttcaactggt acgtggacgg cgtggaggtg cacaacgcca agaccaagcc cagagaggag 900
cagtacaaca gcacctacag ggtggtgtcc gtgctgaccg tgctgcacca ggactggctg 960
aacggcaaag aatacaagtg caaagtctcc aacaaggccc tgccagcccc aatcgaaaag 1020
acaatcagca aggccaaggg ccagccacgg gagccccagg tgtacaccct gccccccagc 1080
cgggaggaga tgaccaagaa ccaggtgtcc ctgacctgtc tggtgaaggg cttctacccc 1140
tgtgatatcg ccgtggagtg ggagagcaac ggccagcccg agaacaacta caagaccacc 1200
cccccagtgc tggacagcga cggcagcttc ttcctgtaca gcaagctgac cgtggacaag 1260
tccaggtggc agcagggcaa cgtgttcagc tgcagcgtga tgcacgaggc cctgcacaac 1320
cactacaccc agaagtccct gagcctgagc cccggcaag 1359
<210> 258
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 258
Gln Gln Ser Leu Phe Ala Pro Phe Thr
1 5
<210> 259
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 259
Ser Leu Phe Ala Pro Phe
1 5
<210> 260
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 260
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Gln Ser Leu Phe Ala Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 261
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 261
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gggcatcagc aactacctgg cctggtatca gcagaaaccc 120
ggcaaggtgc ccaagctgct gatctacgct gccagcacac tgcagagcgg agtgcctagc 180
agattttctg gcagcggctc cggcaccgat ttcaccctga ccatatctag cctgcagcca 240
gaggacgtgg ccacctacta ctgtcagcag agcctgttcg cccctttcac ctttggccag 300
ggcaccaagg tggaaatcaa g 321
<210> 262
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 262
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Gln Ser Leu Phe Ala Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 263
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 263
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gggcatcagc aactacctgg cctggtatca gcagaaaccc 120
ggcaaggtgc ccaagctgct gatctacgct gccagcacac tgcagagcgg agtgcctagc 180
agattttctg gcagcggctc cggcaccgat ttcaccctga ccatatctag cctgcagcca 240
gaggacgtgg ccacctacta ctgtcagcag agcctgttcg cccctttcac ctttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 264
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
Синтетический олигонуклеотид"
<400> 264
ctctagcgcc accatgaaac a 21
<210> 265
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
Синтетический олигонуклеотид"
<400> 265
agcccagcaa caccaagg 18
<210> 266
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
Синтетический олигонуклеотид"
<400> 266
taatacgact cactataggg 20
<210> 267
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
Синтетическая 6xHis метка"
<400> 267
His His His His His His
1 5
<210> 268
<211> 359
<212> БЕЛОК
<213> Homo sapiens
<400> 268
Met Thr Leu Glu Ser Ile Met Ala Cys Cys Leu Ser Glu Glu Ala Lys
1 5 10 15
Glu Ala Arg Arg Ile Asn Asp Glu Ile Glu Arg Gln Leu Arg Arg Asp
20 25 30
Lys Arg Asp Ala Arg Arg Glu Leu Lys Leu Leu Leu Leu Gly Thr Gly
35 40 45
Glu Ser Gly Lys Ser Thr Phe Ile Lys Gln Met Arg Ile Ile His Gly
50 55 60
Ser Gly Tyr Ser Asp Glu Asp Lys Arg Gly Phe Thr Lys Leu Val Tyr
65 70 75 80
Gln Asn Ile Phe Thr Ala Met Gln Ala Met Ile Arg Ala Met Asp Thr
85 90 95
Leu Lys Ile Pro Tyr Lys Tyr Glu His Asn Lys Ala His Ala Gln Leu
100 105 110
Val Arg Glu Val Asp Val Glu Lys Val Ser Ala Phe Glu Asn Pro Tyr
115 120 125
Val Asp Ala Ile Lys Ser Leu Trp Asn Asp Pro Gly Ile Gln Glu Cys
130 135 140
Tyr Asp Arg Arg Arg Glu Tyr Gln Leu Ser Asp Ser Thr Lys Tyr Tyr
145 150 155 160
Leu Asn Asp Leu Asp Arg Val Ala Asp Pro Ala Tyr Leu Pro Thr Gln
165 170 175
Gln Asp Val Leu Arg Val Arg Val Pro Thr Thr Gly Ile Ile Glu Tyr
180 185 190
Pro Phe Asp Leu Gln Ser Val Ile Phe Arg Met Val Asp Val Gly Gly
195 200 205
Gln Arg Ser Glu Arg Arg Lys Trp Ile His Cys Phe Glu Asn Val Thr
210 215 220
Ser Ile Met Phe Leu Val Ala Leu Ser Glu Tyr Asp Gln Val Leu Val
225 230 235 240
Glu Ser Asp Asn Glu Asn Arg Met Glu Glu Ser Lys Ala Leu Phe Arg
245 250 255
Thr Ile Ile Thr Tyr Pro Trp Phe Gln Asn Ser Ser Val Ile Leu Phe
260 265 270
Leu Asn Lys Lys Asp Leu Leu Glu Glu Lys Ile Met Tyr Ser His Leu
275 280 285
Val Asp Tyr Phe Pro Glu Tyr Asp Gly Pro Gln Arg Asp Ala Gln Ala
290 295 300
Ala Arg Glu Phe Ile Leu Lys Met Phe Val Asp Leu Asn Pro Asp Ser
305 310 315 320
Asp Lys Ile Ile Tyr Ser His Phe Thr Cys Ala Thr Asp Thr Glu Asn
325 330 335
Ile Arg Phe Val Phe Ala Ala Val Lys Asp Thr Ile Leu Gln Leu Asn
340 345 350
Leu Lys Glu Tyr Asn Leu Val
355
<210> 269
<211> 6343
<212> ДНК
<213> Homo sapiens
<400> 269
agactatccg ctcccaccgc gcccccggcc cacctggtgg ccccggccct ggccgccgcc 60
cccgcggcgg ttcccggagc tcgtcccgga cgcgcgcccg ggcggcgggg gctcggcggc 120
caccgctgcc tcgggggagc gagggcggga gggtgtgtgt gcgcgctgtg agcagggggt 180
gccggcgggg ctgcagcgga ggcactttgg aagaatgact ctggagtcca tcatggcgtg 240
ctgcctgagc gaggaggcca aggaagcccg gcggatcaac gacgagatcg agcggcagct 300
ccgcagggac aagcgggacg cccgccggga gctcaagctg ctgctgctcg ggacaggaga 360
gagtggcaag agtacgttta tcaagcagat gagaatcatc catgggtcag gatactctga 420
tgaagataaa aggggcttca ccaagctggt gtatcagaac atcttcacgg ccatgcaggc 480
catgatcaga gccatggaca cactcaagat cccatacaag tatgagcaca ataaggctca 540
tgcacaatta gttcgagaag ttgatgtgga gaaggtgtct gcttttgaga atccatatgt 600
agatgcaata aagagtttat ggaatgatcc tggaatccag gaatgctatg atagacgacg 660
agaatatcaa ttatctgact ctaccaaata ctatcttaat gacttggacc gcgtagctga 720
ccctgcctac ctgcctacgc aacaagatgt gcttagagtt cgagtcccca ccacagggat 780
catcgaatac ccctttgact tacaaagtgt cattttcaga atggtcgatg tagggggcca 840
aaggtcagag agaagaaaat ggatacactg ctttgaaaat gtcacctcta tcatgtttct 900
agtagcgctt agtgaatatg atcaagttct cgtggagtca gacaatgaga accgaatgga 960
ggaaagcaag gctctcttta gaacaattat cacatacccc tggttccaga actcctcggt 1020
tattctgttc ttaaacaaga aagatcttct agaggagaaa atcatgtatt cccatctagt 1080
cgactacttc ccagaatatg atggacccca gagagatgcc caggcagccc gagaattcat 1140
tctgaagatg ttcgtggacc tgaacccaga cagtgacaaa attatctact cccacttcac 1200
gtgcgccaca gacaccgaga atatccgctt tgtctttgct gccgtcaagg acaccatcct 1260
ccagttgaac ctgaaggagt acaatctggt ctaattgtgc ctcctagaca cccgccctgc 1320
ccttccctgg tgggctattg aagatacaca agagggactg tatttctgtg gaaaacaatt 1380
tgcataatac taatttattg ccgtcctgga ctctgtgtga gcgtgtccac agagtttgta 1440
gtaaatatta tgattttatt taaactattc agaggaaaaa cagaggatgc tgaagtacag 1500
tcccagcaca tttcctctct atcttttttt taggcaaaac cttgtgactc agtgtatttt 1560
aaattctcag tcatgcactc acaaagataa gacttgtttc tttctgtctc tctctctttt 1620
tcttttctat ggagcaaaac aaagctgatt tccctttttt cttcccccgc taattcatac 1680
ctccctcctg atgtttttcc caggttacaa tggcctttat cctagttcca ttcttggtca 1740
agtttttctc tcaaatgata cagtcaggac acatcgttcg atttaagcca tcatcagctt 1800
aatttaagtt tgtagttttt gctgaaggat tatatgtatt aatacttacg gttttaaatg 1860
tgttgctttg gatacacaca tagtttcttt tttaatagaa tatactgtct tgtctcactt 1920
tggactggga cagtggatgc ccatctaaaa gttaagtgtc atttctttta gatgtttacc 1980
ttcagccata gcttgattgc tcagagaaat atgcagaagg caggatcaaa gacacacagg 2040
agtcctttct tttgaaatgc cacgtgccat tgtctttcct cccttctttg cttctttttc 2100
ttaccctctc tttcaattgc agatgccaaa aaagatgcca acagacacta cattacccta 2160
atggctgcta cccagaacct ttttataggt tgttcttaat ttttttgttg ttgttgttca 2220
agcttttcct ttcttttttt tcttggtgtt tgggccacga ttttaaaatg acttttatta 2280
tgggtatgtg ttgccaaagc tggctttttg tcaaataaaa tgaatacgaa cttaaaaaat 2340
aaaagctggt atcttaaaat gtaagagagt aagactgtga agcctaaaat gactggctga 2400
gaatgaacca gaaatgccat ttgccaaaca gttgtaacta gaaatttgat tctcacggtc 2460
cattcttttc tttgtcctta agatgacatt gttagtgttc acgtcccatg ttcagtgtcc 2520
aaaccggcaa tgtaaaaagt atcctgtgtg gtttaacagg aaatctgttt atgtctcttt 2580
atttgaaacc agttttactc tcagtggttc tttaagttca atgaagtctg ccaggaacat 2640
tggttggtag tattattccg acacctttaa tttccaaaat ctgaagttcc tgctagttta 2700
ccaccttcat gatcttcttg aactggtaac tgattaggtt gaacttatgg aagatttgtg 2760
gacttaactc aaaagtaacc tctcagtgtt ctatagaaca tgtatttgtg taactgaacc 2820
taccaggaga aatgtttgga attctatatg tgcaattttt caacaaatgc aaaaaaaata 2880
cagcacatgt attgacaagc ttctgtcaag cagcttgagt tgaaatttga tttaagaaaa 2940
taaatcatga ttgttcaaag ctgctgggac gttagaatta ggccatgata ctggtctcat 3000
tttaactaca gtggtatttg gcactagtgt aaacttccat ataaatcact cttttggaac 3060
aacaaagggg gagggagaaa aatcacggcc tgttaaatga gtaccaaagc cgcccaacag 3120
taatgagatg ttctcatcct tgattctccc agcctcaaac aacacagctt actttttttt 3180
tcccttgctc agaaagtacc tgtaatttaa caaacagact gcctgtaggt atagtgcaat 3240
tacaaatgct ctaatcattg tacatacatc tctcttgata ttgcagcatc catactggct 3300
ttgtaatcat taattttttg gcagattgaa tgtgctgtat tgatatgtat ctatgtaatt 3360
gtattgtatg tctatagcta attcacgttt tgaataatgt tattttattt acttttttaa 3420
gagaggagaa tgtaaatttg tcagtttatt tctgactagg gatattttct ttccatttag 3480
aaaagaagaa aaaaaaaaaa ccttactgtc atacagagcg gtactagcgt cgtgctgtat 3540
aaaatcattt gcacattcct gagtagaggt atactgatta taagacccaa aggtaatttc 3600
atagcaaaat acataaaatc agtcggagct tttatacaaa catggaaacc aactttgtag 3660
aacttttgcc atttgatcta ggattggaat atgagctttt atacaattca tattcttatt 3720
tggcaaatgc acagtttagt attacctctc tgatggcctt tactagaaag gcagttttag 3780
aagctattgt gatccactaa ggaaatgttt taacagctag agaccactgc ttgcctgaaa 3840
gggcgttctt aaatttggtg cagcaaaaaa aaaaaaaaaa aaaaaaaaaa ttaaacaaca 3900
acatttgaag gcctacagtg tgtatagaga aaacctcatc acaagatcat aagtgttaca 3960
gttttaggga atcaagatat tctatttaat agagctatag taaatgtagt caattaaacc 4020
tgatctcaaa gcttgaagaa gctgagcaaa acagggaaag attgttatat ttgtctttat 4080
gaaattggga tggaatttgc tatgcagaat tgaggtttgt ggcttcgctg ttcctgtagg 4140
gtgcatgaca agatcccttc tcttgagaaa ggaaaaaatt gatcacccta gcagcagtga 4200
tgcatagaaa cctaatttta gccacaccag tcaatcgaag ctaaaggatt ttcttttttg 4260
tttcttcggg gttttattga aggggctagg ggcgggacgg gattcttttc agttttgtat 4320
aaaaacaaag tttactcatg ctttatatta tattgtgatt gcaagcgtta taagcgtgtg 4380
ccactggcct cctattgttg atgcttaggt aatggaggcc tgtggtgagt tttatggtga 4440
cttgggcatg tcttattcaa aaacaaaaac ataaaacaca gaaacctttc ttcagcatac 4500
caaggcaagc agccatttca tgactcactt aacacattgc agtgtaccag tttacagatg 4560
atttttccct ttttgcgtga catggcagtt ctaaccccca gagaattcct tatttgtaaa 4620
ttggaagttt ctactatgcc ttacagagct taaattcaga agtttgtgcc tcatatctga 4680
aacaaaggga aataacacac ccattcaaaa gtaaataaat ctcctagaag tttttgtttt 4740
taacatttcc atataaagag ctctgttgaa tgtcatgaat agactggaaa aaaaaatttt 4800
aagaacctgc atatgttgtt tactagcaga tgacaactac aaaaggaatc tgaagaacac 4860
gtaaaacttg tatttttttt tttttggtag attaactagc aggcctattt taaaaaggta 4920
attcagctaa agggcaattt acttttttgt acttcagact atcttgattg tcaaagtgta 4980
cgaactgtaa ttttaaaatt tatactgcca catgattgta aattttagtt gtcttaagtt 5040
aggaattggt gaaaagctat ttatgctgga tttgggtcaa aatgacttat ttgcaaaaaa 5100
ataaataatg ggaagaaagg gctgtataat gaaatactgc aagactcaca tattggttgg 5160
aaatttccct caaatcacct accgattacc cttgatttcc ctttgttttc agtttctcaa 5220
aacgaatgaa atgaaatata gcagaatgtt aacccatata aaaataaagt gtacccaaat 5280
attgtaatgt atattgctgc tcttcttcaa attaaataag ggtttaaaac cacttaattg 5340
gtaatcaaca tctcaattga tacaaataag gtgtgcttgg tatacattaa tattttcttc 5400
caaagatata tctttggtta gaaacacaaa aaaataaaac tagtaatatt gtatgtttat 5460
ctatctctac atatttccag catatgtagc gttaatagat ctgtcctggt aactgtgtct 5520
ttgggatttc attttggttc catcaaatta ggaaaagaaa tggcttagtt gtatatgatt 5580
agctagagat ttttggagcc agacacctgc tgtttagtag ataacttagt acagacccta 5640
aacttgtcat ttgtttttct cacagaatag ccatttcctg ctgtcttccc aatgatcact 5700
gccctttcaa taacactctt gcctctagaa tcatatgttc aaagtatgaa tacacaccta 5760
gcacatagta ggtgctcaaa tattaatttc ctccttgcct tccttatcta ccctgtgtcc 5820
tccatttccc cgtatgattc caacccaata tagcaaatga catttacatg ttatgaaaac 5880
atctattggg taaaatcaga tcttggataa agaaattctg acttttatat aagcttttgg 5940
tagacagaaa aaacagaaag gtattcgttg gtagaacatt tttaagttca ggaaagaaag 6000
ctggaataat actacgtaac tttgtccagg ttactttgac tgaaacacgt ttttggtgga 6060
tttcttttcc tcaaagaact ctctaaatgc aactccttgc tggattcctc acccatcatc 6120
ctgttggaaa cccttactag acctatgtat ttagggagtt ttgtcagaaa acatttttaa 6180
cttgcagtat ttaaaagaat atttactgtt cctaaaatgt cattcaaatg catgtactgt 6240
ctattgtttg gggatgggaa ctagttttgc aaaaaacacc taatgttgta taataatgcc 6300
ccaatgatct tgctggttaa aaatacagta tttttggcca taa 6343
<210> 270
<211> 359
<212> БЕЛОК
<213> Homo sapiens
<400> 270
Met Thr Leu Glu Ser Met Met Ala Cys Cys Leu Ser Asp Glu Val Lys
1 5 10 15
Glu Ser Lys Arg Ile Asn Ala Glu Ile Glu Lys Gln Leu Arg Arg Asp
20 25 30
Lys Arg Asp Ala Arg Arg Glu Leu Lys Leu Leu Leu Leu Gly Thr Gly
35 40 45
Glu Ser Gly Lys Ser Thr Phe Ile Lys Gln Met Arg Ile Ile His Gly
50 55 60
Ala Gly Tyr Ser Glu Glu Asp Lys Arg Gly Phe Thr Lys Leu Val Tyr
65 70 75 80
Gln Asn Ile Phe Thr Ala Met Gln Ala Met Ile Arg Ala Met Glu Thr
85 90 95
Leu Lys Ile Leu Tyr Lys Tyr Glu Gln Asn Lys Ala Asn Ala Leu Leu
100 105 110
Ile Arg Glu Val Asp Val Glu Lys Val Thr Thr Phe Glu His Gln Tyr
115 120 125
Val Ser Ala Ile Lys Thr Leu Trp Glu Asp Pro Gly Ile Gln Glu Cys
130 135 140
Tyr Asp Arg Arg Arg Glu Tyr Gln Leu Ser Asp Ser Ala Lys Tyr Tyr
145 150 155 160
Leu Thr Asp Val Asp Arg Ile Ala Thr Leu Gly Tyr Leu Pro Thr Gln
165 170 175
Gln Asp Val Leu Arg Val Arg Val Pro Thr Thr Gly Ile Ile Glu Tyr
180 185 190
Pro Phe Asp Leu Glu Asn Ile Ile Phe Arg Met Val Asp Val Gly Gly
195 200 205
Gln Arg Ser Glu Arg Arg Lys Trp Ile His Cys Phe Glu Asn Val Thr
210 215 220
Ser Ile Met Phe Leu Val Ala Leu Ser Glu Tyr Asp Gln Val Leu Val
225 230 235 240
Glu Ser Asp Asn Glu Asn Arg Met Glu Glu Ser Lys Ala Leu Phe Arg
245 250 255
Thr Ile Ile Thr Tyr Pro Trp Phe Gln Asn Ser Ser Val Ile Leu Phe
260 265 270
Leu Asn Lys Lys Asp Leu Leu Glu Asp Lys Ile Leu Tyr Ser His Leu
275 280 285
Val Asp Tyr Phe Pro Glu Phe Asp Gly Pro Gln Arg Asp Ala Gln Ala
290 295 300
Ala Arg Glu Phe Ile Leu Lys Met Phe Val Asp Leu Asn Pro Asp Ser
305 310 315 320
Asp Lys Ile Ile Tyr Ser His Phe Thr Cys Ala Thr Asp Thr Glu Asn
325 330 335
Ile Arg Phe Val Phe Ala Ala Val Lys Asp Thr Ile Leu Gln Leu Asn
340 345 350
Leu Lys Glu Tyr Asn Leu Val
355
<210> 271
<211> 4190
<212> ДНК
<213> Homo sapiens
<400> 271
aggttgtccg gcgctgtcgc tcggttgcgg cggctgcggt tggcggtggc tgcggcggcg 60
gcgcgggctg agtgcggccg cgcgggagtc cgcggctggc gcggcccgag cggggacccg 120
gcggctcgcc aggcggcggc cgaggcgggg cgggccggcc cggggccgag ggccggtggc 180
cgaggccgga gggccgcggc gggcggcggc cgaggcggct ccggccaggg ccgggccggg 240
ggccgggggg cggcggcggg caggcggccg cgtcggccgg ggccgggacg atgactctgg 300
agtccatgat ggcgtgttgc ctgagcgatg aggtgaagga gtccaagcgg atcaacgccg 360
agatcgagaa gcagctgcgg cgggacaagc gcgacgcccg gcgcgagctc aagctgctgc 420
tgctcggcac gggcgagagc gggaagagca cgttcatcaa gcagatgcgc atcatccacg 480
gcgccggcta ctcggaggag gacaagcgcg gcttcaccaa gctcgtctac cagaacatct 540
tcaccgccat gcaggccatg atccgggcca tggagacgct caagatcctc tacaagtacg 600
agcagaacaa ggccaatgcg ctcctgatcc gggaggtgga cgtggagaag gtgaccacct 660
tcgagcatca gtacgtcagt gccatcaaga ccctgtggga ggacccgggc atccaggaat 720
gctacgaccg caggcgcgag taccagctct ccgactctgc caagtactac ctgaccgacg 780
ttgaccgcat cgccaccttg ggctacctgc ccacccagca ggacgtgctg cgggtccgcg 840
tgcccaccac cggcatcatc gagtaccctt tcgacctgga gaacatcatc ttccggatgg 900
tggatgtggg gggccagcgg tcggagcgga ggaagtggat ccactgcttt gagaacgtga 960
catccatcat gtttctcgtc gccctcagcg aatacgacca agtcctggtg gagtcggaca 1020
acgagaaccg gatggaggag agcaaagccc tgttccggac catcatcacc tacccctggt 1080
tccagaactc ctccgtcatc ctcttcctca acaagaagga cctgctggag gacaagatcc 1140
tgtactcgca cctggtggac tacttccccg agttcgatgg tccccagcgg gacgcccagg 1200
cggcgcggga gttcatcctg aagatgttcg tggacctgaa ccccgacagc gacaagatca 1260
tctactcaca cttcacgtgt gccaccgaca cggagaacat ccgcttcgtg ttcgcggccg 1320
tgaaggacac catcctgcag ctcaacctca aggagtacaa cctggtctga gcgcccaggc 1380
ccagggagac gggatggaga cacggggcag gaccttcctt ccacggagcc tgcggctgcc 1440
gggcgggtgg cgctgccgag tccgggccgg ggcctctgcc cgcgggagga gatttttttt 1500
tttcatattt ttaacaaatg gtttttattt cacagttatc aggggatgta catctctccc 1560
tccgtacact tcgcgcacct tctcaccttt tgtcaacggc aaaggcagcc tttttctggc 1620
cttgacttat ggctcgcttt tttctaaaaa aaaaaaaaaa agaaagaaag aaaaaaagca 1680
acgaaacata aaacacacaa gcgccccgtg cccccagtga ctctgggcct cacagagccc 1740
ccgccagcca gcatggggcc ccgccctgca gccagtcacg cgcccccaca ccgcagcccc 1800
ccgtggctgt ccttccaacc ccacgtgctt tttctttctc ctgcccgctt cttttcttca 1860
tcacaaaagg cgtggagact cggagacgga cgtttttccc cttttttaag ttattgacgc 1920
ccagcgcgcc tcgcctcttc acccatcaac gctgtgcttt gcccactgga ctcctgaaga 1980
gggggtgggg ggctccctcg gtcgcccacc ctgggaagtg cctaaccttt tattttattt 2040
tatttttttg aggaaaaaga acgcctgact cacaggttga agaaacaccc tgggccctct 2100
ctcatggccg ggttccccgt ccctctgcag aggctgggaa gggtccccgg gctggagcca 2160
cgggggcttc tctgggctgt gcctccgggg ccaacactgg ctgcttgggg ctgcccgggg 2220
actccagagg gctgcacggc caccctgccc tggctagagc gcaccccacc ggagcccacg 2280
tgggctgggc ggctggaggg atggtccccc ggtgacactg ggagaaaggc cacttggatg 2340
ggggcgtttc tgttttgttc cgctttgtga tgtcaccaat ttggaaacag cgagggtggg 2400
tggggacttt tacagaatat tctcaggtgt gtacccgaga ggcagagaga gggacgtggc 2460
cggcagctct gtgcgtggcc ttgtcccaag cacttgcgcc cgcccccgag cgccgccccc 2520
ggggagcggg aagccagcac tcgcactttg gccaggggcg cgtggaaggt ggtggcaggc 2580
accggcctgg gcagcttcca ggcctggctg gccacgacca cggcccgagg gggagcccgc 2640
caggccacgc cgcactgagc cacagccccg ggggccgcct cccggggccc cttgaggcac 2700
tgaggcaccg agactggttc tccccgagag actcggaagg tggggaacga ggggactgtg 2760
tttggggagg tggctttttc gtctgctgtt gactgaacac tacagcgccc tgtggttccg 2820
ggcttcgcac agctgtccca gggatggatc gcctgtgctg ccttcgcccg ccgccacacc 2880
gggaccctgc acggctgctt ctggcctcga cagatgacaa aagaaacagc cccaaaatac 2940
gaccactcca accagcagtt cccgcctgcc tgcccgccac tgtcaggcct gccctggcct 3000
cctcgtccgc agggctgtct gctggcttct gggggcagaa gagcggggag ccccgtggaa 3060
gggtcagggg agaccaggtc agggcagcta catttctggt gatcagcccc atggggagac 3120
ggggctggcg ggataccccc cccccggctt ccccacacca cttctgtctc acccggaagc 3180
gtcctttttt tgtgccaggt gtctacctaa gagggttggt gccagaagcc ccccatggcg 3240
agtgctgggg cccggcggtg ccctggggga gcagatgggg ccacccctgg cagggccgct 3300
acaacctttt ccagcagcgg agccctctgg ggggcctgtg cttgtggcat ctctgagggc 3360
ctagattgca caaggtgacc tggccgtggc ctgagggtgg agtcgcccag cacgcaggcc 3420
ggggcgctgc ggggctaagt attaggcctt cccagggagg gggcgtgcca agcatcccag 3480
agccgggctg ggaccgccaa aacgtcgtgg cctggatcct ctgggtctga gtgcctgatc 3540
ccctgccccc caaaaaagca gaggtaggtg ttgcaggccc agggcagggg tgcctgcccc 3600
aggagagtcc caggcagtgg ttctcgtgcc agtggcaccc aggggcaagg acagccaacc 3660
cccacccttg ccacgtgtgg ggccacgtgg gcatgtgggg tgtgtgtttt taccttggtg 3720
aatctcacct gccaacgatt tctcgtgagt gccgaccacc ttctccgacc atgttacgcc 3780
cgggcggcag cagcccccgg ccactgcaaa cccatgccct gggtcccccg gctcccccag 3840
ggaggcatcc ccgtgccaat gtcccccagt ggtggcagca gatcctgtgg ccggcctggc 3900
ggacgggacc cagtgatact tgtatattac acagtcctga tttcagacaa tttcaacctt 3960
aatctattta aaaaagaata ttctatacaa gctgttttta agccttttac catttgaaat 4020
gcatgtgttg tgcgcgttgg ggatgggagg aggggctgag gagcggctca gtgtcacctc 4080
ccacagccac cggccctgac ccttaatcca gacaccgatg gaagtcgact tttcatatct 4140
ttctcctgaa atgaactctg ttttaaattg gaataaattt tgttcctaaa 4190
<210> 272
<211> 661
<212> БЕЛОК
<213> Homo sapiens
<400> 272
Met Asp Leu Val Leu Lys Arg Cys Leu Leu His Leu Ala Val Ile Gly
1 5 10 15
Ala Leu Leu Ala Val Gly Ala Thr Lys Val Pro Arg Asn Gln Asp Trp
20 25 30
Leu Gly Val Ser Arg Gln Leu Arg Thr Lys Ala Trp Asn Arg Gln Leu
35 40 45
Tyr Pro Glu Trp Thr Glu Ala Gln Arg Leu Asp Cys Trp Arg Gly Gly
50 55 60
Gln Val Ser Leu Lys Val Ser Asn Asp Gly Pro Thr Leu Ile Gly Ala
65 70 75 80
Asn Ala Ser Phe Ser Ile Ala Leu Asn Phe Pro Gly Ser Gln Lys Val
85 90 95
Leu Pro Asp Gly Gln Val Ile Trp Val Asn Asn Thr Ile Ile Asn Gly
100 105 110
Ser Gln Val Trp Gly Gly Gln Pro Val Tyr Pro Gln Glu Thr Asp Asp
115 120 125
Ala Cys Ile Phe Pro Asp Gly Gly Pro Cys Pro Ser Gly Ser Trp Ser
130 135 140
Gln Lys Arg Ser Phe Val Tyr Val Trp Lys Thr Trp Gly Gln Tyr Trp
145 150 155 160
Gln Val Leu Gly Gly Pro Val Ser Gly Leu Ser Ile Gly Thr Gly Arg
165 170 175
Ala Met Leu Gly Thr His Thr Met Glu Val Thr Val Tyr His Arg Arg
180 185 190
Gly Ser Arg Ser Tyr Val Pro Leu Ala His Ser Ser Ser Ala Phe Thr
195 200 205
Ile Thr Asp Gln Val Pro Phe Ser Val Ser Val Ser Gln Leu Arg Ala
210 215 220
Leu Asp Gly Gly Asn Lys His Phe Leu Arg Asn Gln Pro Leu Thr Phe
225 230 235 240
Ala Leu Gln Leu His Asp Pro Ser Gly Tyr Leu Ala Glu Ala Asp Leu
245 250 255
Ser Tyr Thr Trp Asp Phe Gly Asp Ser Ser Gly Thr Leu Ile Ser Arg
260 265 270
Ala Leu Val Val Thr His Thr Tyr Leu Glu Pro Gly Pro Val Thr Ala
275 280 285
Gln Val Val Leu Gln Ala Ala Ile Pro Leu Thr Ser Cys Gly Ser Ser
290 295 300
Pro Val Pro Gly Thr Thr Asp Gly His Arg Pro Thr Ala Glu Ala Pro
305 310 315 320
Asn Thr Thr Ala Gly Gln Val Pro Thr Thr Glu Val Val Gly Thr Thr
325 330 335
Pro Gly Gln Ala Pro Thr Ala Glu Pro Ser Gly Thr Thr Ser Val Gln
340 345 350
Val Pro Thr Thr Glu Val Ile Ser Thr Ala Pro Val Gln Met Pro Thr
355 360 365
Ala Glu Ser Thr Gly Met Thr Pro Glu Lys Val Pro Val Ser Glu Val
370 375 380
Met Gly Thr Thr Leu Ala Glu Met Ser Thr Pro Glu Ala Thr Gly Met
385 390 395 400
Thr Pro Ala Glu Val Ser Ile Val Val Leu Ser Gly Thr Thr Ala Ala
405 410 415
Gln Val Thr Thr Thr Glu Trp Val Glu Thr Thr Ala Arg Glu Leu Pro
420 425 430
Ile Pro Glu Pro Glu Gly Pro Asp Ala Ser Ser Ile Met Ser Thr Glu
435 440 445
Ser Ile Thr Gly Ser Leu Gly Pro Leu Leu Asp Gly Thr Ala Thr Leu
450 455 460
Arg Leu Val Lys Arg Gln Val Pro Leu Asp Cys Val Leu Tyr Arg Tyr
465 470 475 480
Gly Ser Phe Ser Val Thr Leu Asp Ile Val Gln Gly Ile Glu Ser Ala
485 490 495
Glu Ile Leu Gln Ala Val Pro Ser Gly Glu Gly Asp Ala Phe Glu Leu
500 505 510
Thr Val Ser Cys Gln Gly Gly Leu Pro Lys Glu Ala Cys Met Glu Ile
515 520 525
Ser Ser Pro Gly Cys Gln Pro Pro Ala Gln Arg Leu Cys Gln Pro Val
530 535 540
Leu Pro Ser Pro Ala Cys Gln Leu Val Leu His Gln Ile Leu Lys Gly
545 550 555 560
Gly Ser Gly Thr Tyr Cys Leu Asn Val Ser Leu Ala Asp Thr Asn Ser
565 570 575
Leu Ala Val Val Ser Thr Gln Leu Ile Met Pro Gly Gln Glu Ala Gly
580 585 590
Leu Gly Gln Val Pro Leu Ile Val Gly Ile Leu Leu Val Leu Met Ala
595 600 605
Val Val Leu Ala Ser Leu Ile Tyr Arg Arg Arg Leu Met Lys Gln Asp
610 615 620
Phe Ser Val Pro Gln Leu Pro His Ser Ser Ser His Trp Leu Arg Leu
625 630 635 640
Pro Arg Ile Phe Cys Ser Cys Pro Ile Gly Glu Asn Ser Pro Leu Leu
645 650 655
Ser Gly Gln Gln Val
660
<210> 273
<211> 626
<212> БЕЛОК
<213> Homo sapiens
<400> 273
Met Asp Leu Val Leu Lys Arg Cys Leu Leu His Leu Ala Val Ile Gly
1 5 10 15
Ala Leu Leu Ala Val Gly Ala Thr Lys Val Pro Arg Asn Gln Asp Trp
20 25 30
Leu Gly Val Ser Arg Gln Leu Arg Thr Lys Ala Trp Asn Arg Gln Leu
35 40 45
Tyr Pro Glu Trp Thr Glu Ala Gln Arg Leu Asp Cys Trp Arg Gly Gly
50 55 60
Gln Val Ser Leu Lys Val Ser Asn Asp Gly Pro Thr Leu Ile Gly Ala
65 70 75 80
Asn Ala Ser Phe Ser Ile Ala Leu Asn Phe Pro Gly Ser Gln Lys Val
85 90 95
Leu Pro Asp Gly Gln Val Ile Trp Val Asn Asn Thr Ile Ile Asn Gly
100 105 110
Ser Gln Val Trp Gly Gly Gln Pro Val Tyr Pro Gln Glu Thr Asp Asp
115 120 125
Ala Cys Ile Phe Pro Asp Gly Gly Pro Cys Pro Ser Gly Ser Trp Ser
130 135 140
Gln Lys Arg Ser Phe Val Tyr Val Trp Lys Thr Trp Gly Gln Tyr Trp
145 150 155 160
Gln Val Leu Gly Gly Pro Val Ser Gly Leu Ser Ile Gly Thr Gly Arg
165 170 175
Ala Met Leu Gly Thr His Thr Met Glu Val Thr Val Tyr His Arg Arg
180 185 190
Gly Ser Arg Ser Tyr Val Pro Leu Ala His Ser Ser Ser Ala Phe Thr
195 200 205
Ile Thr Asp Gln Val Pro Phe Ser Val Ser Val Ser Gln Leu Arg Ala
210 215 220
Leu Asp Gly Gly Asn Lys His Phe Leu Arg Asn Gln Pro Leu Thr Phe
225 230 235 240
Ala Leu Gln Leu His Asp Pro Ser Gly Tyr Leu Ala Glu Ala Asp Leu
245 250 255
Ser Tyr Thr Trp Asp Phe Gly Asp Ser Ser Gly Thr Leu Ile Ser Arg
260 265 270
Ala Leu Val Val Thr His Thr Tyr Leu Glu Pro Gly Pro Val Thr Ala
275 280 285
Gln Val Val Leu Gln Ala Ala Ile Pro Leu Thr Ser Cys Gly Ser Ser
290 295 300
Pro Val Pro Gly Thr Thr Asp Gly His Arg Pro Thr Ala Glu Ala Pro
305 310 315 320
Asn Thr Thr Ala Gly Gln Val Pro Thr Thr Glu Val Val Gly Thr Thr
325 330 335
Pro Gly Gln Ala Pro Thr Ala Glu Pro Ser Gly Thr Thr Ser Val Gln
340 345 350
Val Pro Thr Thr Glu Val Ile Ser Thr Ala Pro Val Gln Met Pro Thr
355 360 365
Ala Glu Ser Thr Ala Ala Gln Val Thr Thr Thr Glu Trp Val Glu Thr
370 375 380
Thr Ala Arg Glu Leu Pro Ile Pro Glu Pro Glu Gly Pro Asp Ala Ser
385 390 395 400
Ser Ile Met Ser Thr Glu Ser Ile Thr Gly Ser Leu Gly Pro Leu Leu
405 410 415
Asp Gly Thr Ala Thr Leu Arg Leu Val Lys Arg Gln Val Pro Leu Asp
420 425 430
Cys Val Leu Tyr Arg Tyr Gly Ser Phe Ser Val Thr Leu Asp Ile Val
435 440 445
Gln Gly Ile Glu Ser Ala Glu Ile Leu Gln Ala Val Pro Ser Gly Glu
450 455 460
Gly Asp Ala Phe Glu Leu Thr Val Ser Cys Gln Gly Gly Leu Pro Lys
465 470 475 480
Glu Ala Cys Met Glu Ile Ser Ser Pro Gly Cys Gln Pro Pro Ala Gln
485 490 495
Arg Leu Cys Gln Pro Val Leu Pro Ser Pro Ala Cys Gln Leu Val Leu
500 505 510
His Gln Ile Leu Lys Gly Gly Ser Gly Thr Tyr Cys Leu Asn Val Ser
515 520 525
Leu Ala Asp Thr Asn Ser Leu Ala Val Val Ser Thr Gln Leu Ile Met
530 535 540
Pro Val Pro Gly Ile Leu Leu Thr Gly Gln Glu Ala Gly Leu Gly Gln
545 550 555 560
Val Pro Leu Ile Val Gly Ile Leu Leu Val Leu Met Ala Val Val Leu
565 570 575
Ala Ser Leu Ile Tyr Arg Arg Arg Leu Met Lys Gln Asp Phe Ser Val
580 585 590
Pro Gln Leu Pro His Ser Ser Ser His Trp Leu Arg Leu Pro Arg Ile
595 600 605
Phe Cys Ser Cys Pro Ile Gly Glu Asn Ser Pro Leu Leu Ser Gly Gln
610 615 620
Gln Val
625
<210> 274
<211> 619
<212> БЕЛОК
<213> Homo sapiens
<400> 274
Met Asp Leu Val Leu Lys Arg Cys Leu Leu His Leu Ala Val Ile Gly
1 5 10 15
Ala Leu Leu Ala Val Gly Ala Thr Lys Val Pro Arg Asn Gln Asp Trp
20 25 30
Leu Gly Val Ser Arg Gln Leu Arg Thr Lys Ala Trp Asn Arg Gln Leu
35 40 45
Tyr Pro Glu Trp Thr Glu Ala Gln Arg Leu Asp Cys Trp Arg Gly Gly
50 55 60
Gln Val Ser Leu Lys Val Ser Asn Asp Gly Pro Thr Leu Ile Gly Ala
65 70 75 80
Asn Ala Ser Phe Ser Ile Ala Leu Asn Phe Pro Gly Ser Gln Lys Val
85 90 95
Leu Pro Asp Gly Gln Val Ile Trp Val Asn Asn Thr Ile Ile Asn Gly
100 105 110
Ser Gln Val Trp Gly Gly Gln Pro Val Tyr Pro Gln Glu Thr Asp Asp
115 120 125
Ala Cys Ile Phe Pro Asp Gly Gly Pro Cys Pro Ser Gly Ser Trp Ser
130 135 140
Gln Lys Arg Ser Phe Val Tyr Val Trp Lys Thr Trp Gly Gln Tyr Trp
145 150 155 160
Gln Val Leu Gly Gly Pro Val Ser Gly Leu Ser Ile Gly Thr Gly Arg
165 170 175
Ala Met Leu Gly Thr His Thr Met Glu Val Thr Val Tyr His Arg Arg
180 185 190
Gly Ser Arg Ser Tyr Val Pro Leu Ala His Ser Ser Ser Ala Phe Thr
195 200 205
Ile Thr Asp Gln Val Pro Phe Ser Val Ser Val Ser Gln Leu Arg Ala
210 215 220
Leu Asp Gly Gly Asn Lys His Phe Leu Arg Asn Gln Pro Leu Thr Phe
225 230 235 240
Ala Leu Gln Leu His Asp Pro Ser Gly Tyr Leu Ala Glu Ala Asp Leu
245 250 255
Ser Tyr Thr Trp Asp Phe Gly Asp Ser Ser Gly Thr Leu Ile Ser Arg
260 265 270
Ala Leu Val Val Thr His Thr Tyr Leu Glu Pro Gly Pro Val Thr Ala
275 280 285
Gln Val Val Leu Gln Ala Ala Ile Pro Leu Thr Ser Cys Gly Ser Ser
290 295 300
Pro Val Pro Gly Thr Thr Asp Gly His Arg Pro Thr Ala Glu Ala Pro
305 310 315 320
Asn Thr Thr Ala Gly Gln Val Pro Thr Thr Glu Val Val Gly Thr Thr
325 330 335
Pro Gly Gln Ala Pro Thr Ala Glu Pro Ser Gly Thr Thr Ser Val Gln
340 345 350
Val Pro Thr Thr Glu Val Ile Ser Thr Ala Pro Val Gln Met Pro Thr
355 360 365
Ala Glu Ser Thr Ala Ala Gln Val Thr Thr Thr Glu Trp Val Glu Thr
370 375 380
Thr Ala Arg Glu Leu Pro Ile Pro Glu Pro Glu Gly Pro Asp Ala Ser
385 390 395 400
Ser Ile Met Ser Thr Glu Ser Ile Thr Gly Ser Leu Gly Pro Leu Leu
405 410 415
Asp Gly Thr Ala Thr Leu Arg Leu Val Lys Arg Gln Val Pro Leu Asp
420 425 430
Cys Val Leu Tyr Arg Tyr Gly Ser Phe Ser Val Thr Leu Asp Ile Val
435 440 445
Gln Gly Ile Glu Ser Ala Glu Ile Leu Gln Ala Val Pro Ser Gly Glu
450 455 460
Gly Asp Ala Phe Glu Leu Thr Val Ser Cys Gln Gly Gly Leu Pro Lys
465 470 475 480
Glu Ala Cys Met Glu Ile Ser Ser Pro Gly Cys Gln Pro Pro Ala Gln
485 490 495
Arg Leu Cys Gln Pro Val Leu Pro Ser Pro Ala Cys Gln Leu Val Leu
500 505 510
His Gln Ile Leu Lys Gly Gly Ser Gly Thr Tyr Cys Leu Asn Val Ser
515 520 525
Leu Ala Asp Thr Asn Ser Leu Ala Val Val Ser Thr Gln Leu Ile Met
530 535 540
Pro Gly Gln Glu Ala Gly Leu Gly Gln Val Pro Leu Ile Val Gly Ile
545 550 555 560
Leu Leu Val Leu Met Ala Val Val Leu Ala Ser Leu Ile Tyr Arg Arg
565 570 575
Arg Leu Met Lys Gln Asp Phe Ser Val Pro Gln Leu Pro His Ser Ser
580 585 590
Ser His Trp Leu Arg Leu Pro Arg Ile Phe Cys Ser Cys Pro Ile Gly
595 600 605
Glu Asn Ser Pro Leu Leu Ser Gly Gln Gln Val
610 615
<210> 275
<211> 575
<212> БЕЛОК
<213> Homo sapiens
<400> 275
Met Asp Leu Val Leu Lys Arg Cys Leu Leu His Leu Ala Val Ile Gly
1 5 10 15
Ala Leu Leu Ala Val Gly Ala Thr Lys Gly Ser Gln Val Trp Gly Gly
20 25 30
Gln Pro Val Tyr Pro Gln Glu Thr Asp Asp Ala Cys Ile Phe Pro Asp
35 40 45
Gly Gly Pro Cys Pro Ser Gly Ser Trp Ser Gln Lys Arg Ser Phe Val
50 55 60
Tyr Val Trp Lys Thr Trp Gly Gln Tyr Trp Gln Val Leu Gly Gly Pro
65 70 75 80
Val Ser Gly Leu Ser Ile Gly Thr Gly Arg Ala Met Leu Gly Thr His
85 90 95
Thr Met Glu Val Thr Val Tyr His Arg Arg Gly Ser Arg Ser Tyr Val
100 105 110
Pro Leu Ala His Ser Ser Ser Ala Phe Thr Ile Thr Asp Gln Val Pro
115 120 125
Phe Ser Val Ser Val Ser Gln Leu Arg Ala Leu Asp Gly Gly Asn Lys
130 135 140
His Phe Leu Arg Asn Gln Pro Leu Thr Phe Ala Leu Gln Leu His Asp
145 150 155 160
Pro Ser Gly Tyr Leu Ala Glu Ala Asp Leu Ser Tyr Thr Trp Asp Phe
165 170 175
Gly Asp Ser Ser Gly Thr Leu Ile Ser Arg Ala Leu Val Val Thr His
180 185 190
Thr Tyr Leu Glu Pro Gly Pro Val Thr Ala Gln Val Val Leu Gln Ala
195 200 205
Ala Ile Pro Leu Thr Ser Cys Gly Ser Ser Pro Val Pro Gly Thr Thr
210 215 220
Asp Gly His Arg Pro Thr Ala Glu Ala Pro Asn Thr Thr Ala Gly Gln
225 230 235 240
Val Pro Thr Thr Glu Val Val Gly Thr Thr Pro Gly Gln Ala Pro Thr
245 250 255
Ala Glu Pro Ser Gly Thr Thr Ser Val Gln Val Pro Thr Thr Glu Val
260 265 270
Ile Ser Thr Ala Pro Val Gln Met Pro Thr Ala Glu Ser Thr Gly Met
275 280 285
Thr Pro Glu Lys Val Pro Val Ser Glu Val Met Gly Thr Thr Leu Ala
290 295 300
Glu Met Ser Thr Pro Glu Ala Thr Gly Met Thr Pro Ala Glu Val Ser
305 310 315 320
Ile Val Val Leu Ser Gly Thr Thr Ala Ala Gln Val Thr Thr Thr Glu
325 330 335
Trp Val Glu Thr Thr Ala Arg Glu Leu Pro Ile Pro Glu Pro Glu Gly
340 345 350
Pro Asp Ala Ser Ser Ile Met Ser Thr Glu Ser Ile Thr Gly Ser Leu
355 360 365
Gly Pro Leu Leu Asp Gly Thr Ala Thr Leu Arg Leu Val Lys Arg Gln
370 375 380
Val Pro Leu Asp Cys Val Leu Tyr Arg Tyr Gly Ser Phe Ser Val Thr
385 390 395 400
Leu Asp Ile Val Gln Gly Ile Glu Ser Ala Glu Ile Leu Gln Ala Val
405 410 415
Pro Ser Gly Glu Gly Asp Ala Phe Glu Leu Thr Val Ser Cys Gln Gly
420 425 430
Gly Leu Pro Lys Glu Ala Cys Met Glu Ile Ser Ser Pro Gly Cys Gln
435 440 445
Pro Pro Ala Gln Arg Leu Cys Gln Pro Val Leu Pro Ser Pro Ala Cys
450 455 460
Gln Leu Val Leu His Gln Ile Leu Lys Gly Gly Ser Gly Thr Tyr Cys
465 470 475 480
Leu Asn Val Ser Leu Ala Asp Thr Asn Ser Leu Ala Val Val Ser Thr
485 490 495
Gln Leu Ile Met Pro Gly Gln Glu Ala Gly Leu Gly Gln Val Pro Leu
500 505 510
Ile Val Gly Ile Leu Leu Val Leu Met Ala Val Val Leu Ala Ser Leu
515 520 525
Ile Tyr Arg Arg Arg Leu Met Lys Gln Asp Phe Ser Val Pro Gln Leu
530 535 540
Pro His Ser Ser Ser His Trp Leu Arg Leu Pro Arg Ile Phe Cys Ser
545 550 555 560
Cys Pro Ile Gly Glu Asn Ser Pro Leu Leu Ser Gly Gln Gln Val
565 570 575
<210> 276
<211> 668
<212> БЕЛОК
<213> Homo sapiens
<400> 276
Met Asp Leu Val Leu Lys Arg Cys Leu Leu His Leu Ala Val Ile Gly
1 5 10 15
Ala Leu Leu Ala Val Gly Ala Thr Lys Val Pro Arg Asn Gln Asp Trp
20 25 30
Leu Gly Val Ser Arg Gln Leu Arg Thr Lys Ala Trp Asn Arg Gln Leu
35 40 45
Tyr Pro Glu Trp Thr Glu Ala Gln Arg Leu Asp Cys Trp Arg Gly Gly
50 55 60
Gln Val Ser Leu Lys Val Ser Asn Asp Gly Pro Thr Leu Ile Gly Ala
65 70 75 80
Asn Ala Ser Phe Ser Ile Ala Leu Asn Phe Pro Gly Ser Gln Lys Val
85 90 95
Leu Pro Asp Gly Gln Val Ile Trp Val Asn Asn Thr Ile Ile Asn Gly
100 105 110
Ser Gln Val Trp Gly Gly Gln Pro Val Tyr Pro Gln Glu Thr Asp Asp
115 120 125
Ala Cys Ile Phe Pro Asp Gly Gly Pro Cys Pro Ser Gly Ser Trp Ser
130 135 140
Gln Lys Arg Ser Phe Val Tyr Val Trp Lys Thr Trp Gly Gln Tyr Trp
145 150 155 160
Gln Val Leu Gly Gly Pro Val Ser Gly Leu Ser Ile Gly Thr Gly Arg
165 170 175
Ala Met Leu Gly Thr His Thr Met Glu Val Thr Val Tyr His Arg Arg
180 185 190
Gly Ser Arg Ser Tyr Val Pro Leu Ala His Ser Ser Ser Ala Phe Thr
195 200 205
Ile Thr Asp Gln Val Pro Phe Ser Val Ser Val Ser Gln Leu Arg Ala
210 215 220
Leu Asp Gly Gly Asn Lys His Phe Leu Arg Asn Gln Pro Leu Thr Phe
225 230 235 240
Ala Leu Gln Leu His Asp Pro Ser Gly Tyr Leu Ala Glu Ala Asp Leu
245 250 255
Ser Tyr Thr Trp Asp Phe Gly Asp Ser Ser Gly Thr Leu Ile Ser Arg
260 265 270
Ala Leu Val Val Thr His Thr Tyr Leu Glu Pro Gly Pro Val Thr Ala
275 280 285
Gln Val Val Leu Gln Ala Ala Ile Pro Leu Thr Ser Cys Gly Ser Ser
290 295 300
Pro Val Pro Gly Thr Thr Asp Gly His Arg Pro Thr Ala Glu Ala Pro
305 310 315 320
Asn Thr Thr Ala Gly Gln Val Pro Thr Thr Glu Val Val Gly Thr Thr
325 330 335
Pro Gly Gln Ala Pro Thr Ala Glu Pro Ser Gly Thr Thr Ser Val Gln
340 345 350
Val Pro Thr Thr Glu Val Ile Ser Thr Ala Pro Val Gln Met Pro Thr
355 360 365
Ala Glu Ser Thr Gly Met Thr Pro Glu Lys Val Pro Val Ser Glu Val
370 375 380
Met Gly Thr Thr Leu Ala Glu Met Ser Thr Pro Glu Ala Thr Gly Met
385 390 395 400
Thr Pro Ala Glu Val Ser Ile Val Val Leu Ser Gly Thr Thr Ala Ala
405 410 415
Gln Val Thr Thr Thr Glu Trp Val Glu Thr Thr Ala Arg Glu Leu Pro
420 425 430
Ile Pro Glu Pro Glu Gly Pro Asp Ala Ser Ser Ile Met Ser Thr Glu
435 440 445
Ser Ile Thr Gly Ser Leu Gly Pro Leu Leu Asp Gly Thr Ala Thr Leu
450 455 460
Arg Leu Val Lys Arg Gln Val Pro Leu Asp Cys Val Leu Tyr Arg Tyr
465 470 475 480
Gly Ser Phe Ser Val Thr Leu Asp Ile Val Gln Gly Ile Glu Ser Ala
485 490 495
Glu Ile Leu Gln Ala Val Pro Ser Gly Glu Gly Asp Ala Phe Glu Leu
500 505 510
Thr Val Ser Cys Gln Gly Gly Leu Pro Lys Glu Ala Cys Met Glu Ile
515 520 525
Ser Ser Pro Gly Cys Gln Pro Pro Ala Gln Arg Leu Cys Gln Pro Val
530 535 540
Leu Pro Ser Pro Ala Cys Gln Leu Val Leu His Gln Ile Leu Lys Gly
545 550 555 560
Gly Ser Gly Thr Tyr Cys Leu Asn Val Ser Leu Ala Asp Thr Asn Ser
565 570 575
Leu Ala Val Val Ser Thr Gln Leu Ile Met Pro Val Pro Gly Ile Leu
580 585 590
Leu Thr Gly Gln Glu Ala Gly Leu Gly Gln Val Pro Leu Ile Val Gly
595 600 605
Ile Leu Leu Val Leu Met Ala Val Val Leu Ala Ser Leu Ile Tyr Arg
610 615 620
Arg Arg Leu Met Lys Gln Asp Phe Ser Val Pro Gln Leu Pro His Ser
625 630 635 640
Ser Ser His Trp Leu Arg Leu Pro Arg Ile Phe Cys Ser Cys Pro Ile
645 650 655
Gly Glu Asn Ser Pro Leu Leu Ser Gly Gln Gln Val
660 665
<210> 277
<211> 2195
<212> ДНК
<213> Homo sapiens
<400> 277
cccagcgctc ctccccgcaa atgatcccgc cccaggggcc tatcccagtc cccccagtgc 60
ctttggttgc tggagggaag aacacaatgg atctggtgct aaaaagatgc cttcttcatt 120
tggctgtgat aggtgctttg ctggctgtgg gggctacaaa agtacccaga aaccaggact 180
ggcttggtgt ctcaaggcaa ctcagaacca aagcctggaa caggcagctg tatccagagt 240
ggacagaagc ccagagactt gactgctgga gaggtggtca agtgtccctc aaggtcagta 300
atgatgggcc tacactgatt ggtgcaaatg cctccttctc tattgccttg aacttccctg 360
gaagccaaaa ggtattgcca gatgggcagg ttatctgggt caacaatacc atcatcaatg 420
ggagccaggt gtggggagga cagccagtgt atccccagga aactgacgat gcctgcatct 480
tccctgatgg tggaccttgc ccatctggct cttggtctca gaagagaagc tttgtttatg 540
tctggaagac ctggggccaa tactggcaag ttctaggggg cccagtgtct gggctgagca 600
ttgggacagg cagggcaatg ctgggcacac acaccatgga agtgactgtc taccatcgcc 660
ggggatcccg gagctatgtg cctcttgctc attccagctc agccttcacc attactgacc 720
aggtgccttt ctccgtgagc gtgtcccagt tgcgggcctt ggatggaggg aacaagcact 780
tcctgagaaa tcagcctctg acctttgccc tccagctcca tgaccccagt ggctatctgg 840
ctgaagctga cctctcctac acctgggact ttggagacag tagtggaacc ctgatctctc 900
gggcacttgt ggtcactcat acttacctgg agcctggccc agtcactgcc caggtggtcc 960
tgcaggctgc cattcctctc acctcctgtg gctcctcccc agttccaggc accacagatg 1020
ggcacaggcc aactgcagag gcccctaaca ccacagctgg ccaagtgcct actacagaag 1080
ttgtgggtac tacacctggt caggcgccaa ctgcagagcc ctctggaacc acatctgtgc 1140
aggtgccaac cactgaagtc ataagcactg cacctgtgca gatgccaact gcagagagca 1200
caggtatgac acctgagaag gtgccagttt cagaggtcat gggtaccaca ctggcagaga 1260
tgtcaactcc agaggctaca ggtatgacac ctgcagaggt atcaattgtg gtgctttctg 1320
gaaccacagc tgcacaggta acaactacag agtgggtgga gaccacagct agagagctac 1380
ctatccctga gcctgaaggt ccagatgcca gctcaatcat gtctacggaa agtattacag 1440
gttccctggg ccccctgctg gatggtacag ccaccttaag gctggtgaag agacaagtcc 1500
ccctggattg tgttctgtat cgatatggtt ccttttccgt caccctggac attgtccagg 1560
gtattgaaag tgccgagatc ctgcaggctg tgccgtccgg tgagggggat gcatttgagc 1620
tgactgtgtc ctgccaaggc gggctgccca aggaagcctg catggagatc tcatcgccag 1680
ggtgccagcc ccctgcccag cggctgtgcc agcctgtgct acccagccca gcctgccagc 1740
tggttctgca ccagatactg aagggtggct cggggacata ctgcctcaat gtgtctctgg 1800
ctgataccaa cagcctggca gtggtcagca cccagcttat catgcctggt caagaagcag 1860
gccttgggca ggttccgctg atcgtgggca tcttgctggt gttgatggct gtggtccttg 1920
catctctgat atataggcgc agacttatga agcaagactt ctccgtaccc cagttgccac 1980
atagcagcag tcactggctg cgtctacccc gcatcttctg ctcttgtccc attggtgaga 2040
acagccccct cctcagtggg cagcaggtct gagtactctc atatgatgct gtgattttcc 2100
tggagttgac agaaacacct atatttcccc cagtcttccc tgggagacta ctattaactg 2160
aaataaatac tcagagcctg aaaaaaaaaa aaaaa 2195
<210> 278
<211> 1902
<212> ДНК
<213> Homo sapiens
<400> 278
gggcctatcc cagtcccccc agtgcctttg gttgctggag ggaagaacac aatggatctg 60
gtgctaaaaa gatgccttct tcatttggct gtgataggtg ctttgctggc tgtgggggct 120
acaaaaggga gccaggtgtg gggaggacag ccagtgtatc cccaggaaac tgacgatgcc 180
tgcatcttcc ctgatggtgg accttgccca tctggctctt ggtctcagaa gagaagcttt 240
gtttatgtct ggaagacctg gggccaatac tggcaagttc tagggggccc agtgtctggg 300
ctgagcattg ggacaggcag ggcaatgctg ggcacacaca ccatggaagt gactgtctac 360
catcgccggg gatcccggag ctatgtgcct cttgctcatt ccagctcagc cttcaccatt 420
actgaccagg tgcctttctc cgtgagcgtg tcccagttgc gggccttgga tggagggaac 480
aagcacttcc tgagaaatca gcctctgacc tttgccctcc agctccatga ccccagtggc 540
tatctggctg aagctgacct ctcctacacc tgggactttg gagacagtag tggaaccctg 600
atctctcggg cacttgtggt cactcatact tacctggagc ctggcccagt cactgcccag 660
gtggtcctgc aggctgccat tcctctcacc tcctgtggct cctccccagt tccaggcacc 720
acagatgggc acaggccaac tgcagaggcc cctaacacca cagctggcca agtgcctact 780
acagaagttg tgggtactac acctggtcag gcgccaactg cagagccctc tggaaccaca 840
tctgtgcagg tgccaaccac tgaagtcata agcactgcac ctgtgcagat gccaactgca 900
gagagcacag gtatgacacc tgagaaggtg ccagtttcag aggtcatggg taccacactg 960
gcagagatgt caactccaga ggctacaggt atgacacctg cagaggtatc aattgtggtg 1020
ctttctggaa ccacagctgc acaggtaaca actacagagt gggtggagac cacagctaga 1080
gagctaccta tccctgagcc tgaaggtcca gatgccagct caatcatgtc tacggaaagt 1140
attacaggtt ccctgggccc cctgctggat ggtacagcca ccttaaggct ggtgaagaga 1200
caagtccccc tggattgtgt tctgtatcga tatggttcct tttccgtcac cctggacatt 1260
gtccagggta ttgaaagtgc cgagatcctg caggctgtgc cgtccggtga gggggatgca 1320
tttgagctga ctgtgtcctg ccaaggcggg ctgcccaagg aagcctgcat ggagatctca 1380
tcgccagggt gccagccccc tgcccagcgg ctgtgccagc ctgtgctacc cagcccagcc 1440
tgccagctgg ttctgcacca gatactgaag ggtggctcgg ggacatactg cctcaatgtg 1500
tctctggctg ataccaacag cctggcagtg gtcagcaccc agcttatcat gcctggtcaa 1560
gaagcaggcc ttgggcaggt tccgctgatc gtgggcatct tgctggtgtt gatggctgtg 1620
gtccttgcat ctctgatata taggcgcaga cttatgaagc aagacttctc cgtaccccag 1680
ttgccacata gcagcagtca ctggctgcgt ctaccccgca tcttctgctc ttgtcccatt 1740
ggtgagaaca gccccctcct cagtgggcag caggtctgag tactctcata tgatgctgtg 1800
attttcctgg agttgacaga aacacctata tttcccccag tcttccctgg gagactacta 1860
ttaactgaaa taaatactca gagcctgaaa aaaaaaaaaa aa 1902
<210> 279
<211> 2181
<212> ДНК
<213> Homo sapiens
<400> 279
gggcctatcc cagtcccccc agtgcctttg gttgctggag ggaagaacac aatggatctg 60
gtgctaaaaa gatgccttct tcatttggct gtgataggtg ctttgctggc tgtgggggct 120
acaaaagtac ccagaaacca ggactggctt ggtgtctcaa ggcaactcag aaccaaagcc 180
tggaacaggc agctgtatcc agagtggaca gaagcccaga gacttgactg ctggagaggt 240
ggtcaagtgt ccctcaaggt cagtaatgat gggcctacac tgattggtgc aaatgcctcc 300
ttctctattg ccttgaactt ccctggaagc caaaaggtat tgccagatgg gcaggttatc 360
tgggtcaaca ataccatcat caatgggagc caggtgtggg gaggacagcc agtgtatccc 420
caggaaactg acgatgcctg catcttccct gatggtggac cttgcccatc tggctcttgg 480
tctcagaaga gaagctttgt ttatgtctgg aagacctggg gccaatactg gcaagttcta 540
gggggcccag tgtctgggct gagcattggg acaggcaggg caatgctggg cacacacacc 600
atggaagtga ctgtctacca tcgccgggga tcccggagct atgtgcctct tgctcattcc 660
agctcagcct tcaccattac tgaccaggtg cctttctccg tgagcgtgtc ccagttgcgg 720
gccttggatg gagggaacaa gcacttcctg agaaatcagc ctctgacctt tgccctccag 780
ctccatgacc ccagtggcta tctggctgaa gctgacctct cctacacctg ggactttgga 840
gacagtagtg gaaccctgat ctctcgggca cttgtggtca ctcatactta cctggagcct 900
ggcccagtca ctgcccaggt ggtcctgcag gctgccattc ctctcacctc ctgtggctcc 960
tccccagttc caggcaccac agatgggcac aggccaactg cagaggcccc taacaccaca 1020
gctggccaag tgcctactac agaagttgtg ggtactacac ctggtcaggc gccaactgca 1080
gagccctctg gaaccacatc tgtgcaggtg ccaaccactg aagtcataag cactgcacct 1140
gtgcagatgc caactgcaga gagcacaggt atgacacctg agaaggtgcc agtttcagag 1200
gtcatgggta ccacactggc agagatgtca actccagagg ctacaggtat gacacctgca 1260
gaggtatcaa ttgtggtgct ttctggaacc acagctgcac aggtaacaac tacagagtgg 1320
gtggagacca cagctagaga gctacctatc cctgagcctg aaggtccaga tgccagctca 1380
atcatgtcta cggaaagtat tacaggttcc ctgggccccc tgctggatgg tacagccacc 1440
ttaaggctgg tgaagagaca agtccccctg gattgtgttc tgtatcgata tggttccttt 1500
tccgtcaccc tggacattgt ccagggtatt gaaagtgccg agatcctgca ggctgtgccg 1560
tccggtgagg gggatgcatt tgagctgact gtgtcctgcc aaggcgggct gcccaaggaa 1620
gcctgcatgg agatctcatc gccagggtgc cagccccctg cccagcggct gtgccagcct 1680
gtgctaccca gcccagcctg ccagctggtt ctgcaccaga tactgaaggg tggctcgggg 1740
acatactgcc tcaatgtgtc tctggctgat accaacagcc tggcagtggt cagcacccag 1800
cttatcatgc ctgtgcctgg gattcttctc acaggtcaag aagcaggcct tgggcaggtt 1860
ccgctgatcg tgggcatctt gctggtgttg atggctgtgg tccttgcatc tctgatatat 1920
aggcgcagac ttatgaagca agacttctcc gtaccccagt tgccacatag cagcagtcac 1980
tggctgcgtc taccccgcat cttctgctct tgtcccattg gtgagaacag ccccctcctc 2040
agtgggcagc aggtctgagt actctcatat gatgctgtga ttttcctgga gttgacagaa 2100
acacctatat ttcccccagt cttccctggg agactactat taactgaaat aaatactcag 2160
agcctgaaaa aaaaaaaaaa a 2181
<210> 280
<211> 2041
<212> ДНК
<213> Homo sapiens
<400> 280
gggcctatcc cagtcccccc agtgcctttg gttgctggag ggaagaacac aatggatctg 60
gtgctaaaaa gatgccttct tcatttggct gtgataggtg ctttgctggc tgtgggggct 120
acaaaagtac ccagaaacca ggactggctt ggtgtctcaa ggcaactcag aaccaaagcc 180
tggaacaggc agctgtatcc agagtggaca gaagcccaga gacttgactg ctggagaggt 240
ggtcaagtgt ccctcaaggt cagtaatgat gggcctacac tgattggtgc aaatgcctcc 300
ttctctattg ccttgaactt ccctggaagc caaaaggtat tgccagatgg gcaggttatc 360
tgggtcaaca ataccatcat caatgggagc caggtgtggg gaggacagcc agtgtatccc 420
caggaaactg acgatgcctg catcttccct gatggtggac cttgcccatc tggctcttgg 480
tctcagaaga gaagctttgt ttatgtctgg aagacctggg gccaatactg gcaagttcta 540
gggggcccag tgtctgggct gagcattggg acaggcaggg caatgctggg cacacacacc 600
atggaagtga ctgtctacca tcgccgggga tcccggagct atgtgcctct tgctcattcc 660
agctcagcct tcaccattac tgaccaggtg cctttctccg tgagcgtgtc ccagttgcgg 720
gccttggatg gagggaacaa gcacttcctg agaaatcagc ctctgacctt tgccctccag 780
ctccatgacc ccagtggcta tctggctgaa gctgacctct cctacacctg ggactttgga 840
gacagtagtg gaaccctgat ctctcgggca cttgtggtca ctcatactta cctggagcct 900
ggcccagtca ctgcccaggt ggtcctgcag gctgccattc ctctcacctc ctgtggctcc 960
tccccagttc caggcaccac agatgggcac aggccaactg cagaggcccc taacaccaca 1020
gctggccaag tgcctactac agaagttgtg ggtactacac ctggtcaggc gccaactgca 1080
gagccctctg gaaccacatc tgtgcaggtg ccaaccactg aagtcataag cactgcacct 1140
gtgcagatgc caactgcaga gagcacagct gcacaggtaa caactacaga gtgggtggag 1200
accacagcta gagagctacc tatccctgag cctgaaggtc cagatgccag ctcaatcatg 1260
tctacggaaa gtattacagg ttccctgggc cccctgctgg atggtacagc caccttaagg 1320
ctggtgaaga gacaagtccc cctggattgt gttctgtatc gatatggttc cttttccgtc 1380
accctggaca ttgtccaggg tattgaaagt gccgagatcc tgcaggctgt gccgtccggt 1440
gagggggatg catttgagct gactgtgtcc tgccaaggcg ggctgcccaa ggaagcctgc 1500
atggagatct catcgccagg gtgccagccc cctgcccagc ggctgtgcca gcctgtgcta 1560
cccagcccag cctgccagct ggttctgcac cagatactga agggtggctc ggggacatac 1620
tgcctcaatg tgtctctggc tgataccaac agcctggcag tggtcagcac ccagcttatc 1680
atgcctgtgc ctgggattct tctcacaggt caagaagcag gccttgggca ggttccgctg 1740
atcgtgggca tcttgctggt gttgatggct gtggtccttg catctctgat atataggcgc 1800
agacttatga agcaagactt ctccgtaccc cagttgccac atagcagcag tcactggctg 1860
cgtctacccc gcatcttctg ctcttgtccc attggtgaga acagccccct cctcagtggg 1920
cagcaggtct gagtactctc atatgatgct gtgattttcc tggagttgac agaaacacct 1980
atatttcccc cagtcttccc tgggagacta ctattaactg aaataaatac tcagagcctg 2040
a 2041
<210> 281
<211> 2020
<212> ДНК
<213> Homo sapiens
<400> 281
gggcctatcc cagtcccccc agtgcctttg gttgctggag ggaagaacac aatggatctg 60
gtgctaaaaa gatgccttct tcatttggct gtgataggtg ctttgctggc tgtgggggct 120
acaaaagtac ccagaaacca ggactggctt ggtgtctcaa ggcaactcag aaccaaagcc 180
tggaacaggc agctgtatcc agagtggaca gaagcccaga gacttgactg ctggagaggt 240
ggtcaagtgt ccctcaaggt cagtaatgat gggcctacac tgattggtgc aaatgcctcc 300
ttctctattg ccttgaactt ccctggaagc caaaaggtat tgccagatgg gcaggttatc 360
tgggtcaaca ataccatcat caatgggagc caggtgtggg gaggacagcc agtgtatccc 420
caggaaactg acgatgcctg catcttccct gatggtggac cttgcccatc tggctcttgg 480
tctcagaaga gaagctttgt ttatgtctgg aagacctggg gccaatactg gcaagttcta 540
gggggcccag tgtctgggct gagcattggg acaggcaggg caatgctggg cacacacacc 600
atggaagtga ctgtctacca tcgccgggga tcccggagct atgtgcctct tgctcattcc 660
agctcagcct tcaccattac tgaccaggtg cctttctccg tgagcgtgtc ccagttgcgg 720
gccttggatg gagggaacaa gcacttcctg agaaatcagc ctctgacctt tgccctccag 780
ctccatgacc ccagtggcta tctggctgaa gctgacctct cctacacctg ggactttgga 840
gacagtagtg gaaccctgat ctctcgggca cttgtggtca ctcatactta cctggagcct 900
ggcccagtca ctgcccaggt ggtcctgcag gctgccattc ctctcacctc ctgtggctcc 960
tccccagttc caggcaccac agatgggcac aggccaactg cagaggcccc taacaccaca 1020
gctggccaag tgcctactac agaagttgtg ggtactacac ctggtcaggc gccaactgca 1080
gagccctctg gaaccacatc tgtgcaggtg ccaaccactg aagtcataag cactgcacct 1140
gtgcagatgc caactgcaga gagcacagct gcacaggtaa caactacaga gtgggtggag 1200
accacagcta gagagctacc tatccctgag cctgaaggtc cagatgccag ctcaatcatg 1260
tctacggaaa gtattacagg ttccctgggc cccctgctgg atggtacagc caccttaagg 1320
ctggtgaaga gacaagtccc cctggattgt gttctgtatc gatatggttc cttttccgtc 1380
accctggaca ttgtccaggg tattgaaagt gccgagatcc tgcaggctgt gccgtccggt 1440
gagggggatg catttgagct gactgtgtcc tgccaaggcg ggctgcccaa ggaagcctgc 1500
atggagatct catcgccagg gtgccagccc cctgcccagc ggctgtgcca gcctgtgcta 1560
cccagcccag cctgccagct ggttctgcac cagatactga agggtggctc ggggacatac 1620
tgcctcaatg tgtctctggc tgataccaac agcctggcag tggtcagcac ccagcttatc 1680
atgcctggtc aagaagcagg ccttgggcag gttccgctga tcgtgggcat cttgctggtg 1740
ttgatggctg tggtccttgc atctctgata tataggcgca gacttatgaa gcaagacttc 1800
tccgtacccc agttgccaca tagcagcagt cactggctgc gtctaccccg catcttctgc 1860
tcttgtccca ttggtgagaa cagccccctc ctcagtgggc agcaggtctg agtactctca 1920
tatgatgctg tgattttcct ggagttgaca gaaacaccta tatttccccc agtcttccct 1980
gggagactac tattaactga aataaatact cagagcctga 2020
<210> 282
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 282
Tyr Gly Arg Lys Lys Arg Arg Gln Arg Arg Arg Leu Tyr Arg Ser Pro
1 5 10 15
Ala Met Pro Glu Asn Leu
20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ БЕЛКА-1 ЗАПРОГРАММИРОВАННОЙ КЛЕТОЧНОЙ СМЕРТИ (PD-1) И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2725950C1 |
| C3-СВЯЗЫВАЮЩИЕ АГЕНТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2802307C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО К NAV1.7 | 2019 |
|
RU2776821C2 |
| Антитела против лиганда-1 запрограммированной смерти (PD-L1) и их применение | 2017 |
|
RU2721582C1 |
| УСОВЕРШЕНСТВОВАННЫЕ ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2019 |
|
RU2810748C2 |
| АНТИ-CEACAM1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2754876C2 |
| СПОСОБ ПОЛУЧЕНИЯ КОНЪЮГАТА АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО НА ОСНОВЕ ЭРИБУЛИНА | 2020 |
|
RU2840013C1 |
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С СОРТИЛИНОМ И ПОДАВЛЯЮТ СВЯЗЫВАНИЕ ПРОГРАНУЛИНА | 2016 |
|
RU2735639C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, КОТОРОЕ СПЕЦИФИЧЕСКИ СВЯЗЫВАЕТСЯ С IL-4Rα, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2808563C2 |
| БЕЛОК, СВЯЗЫВАЮЩИЙ АНТИГЕН ST2 | 2021 |
|
RU2841164C2 |
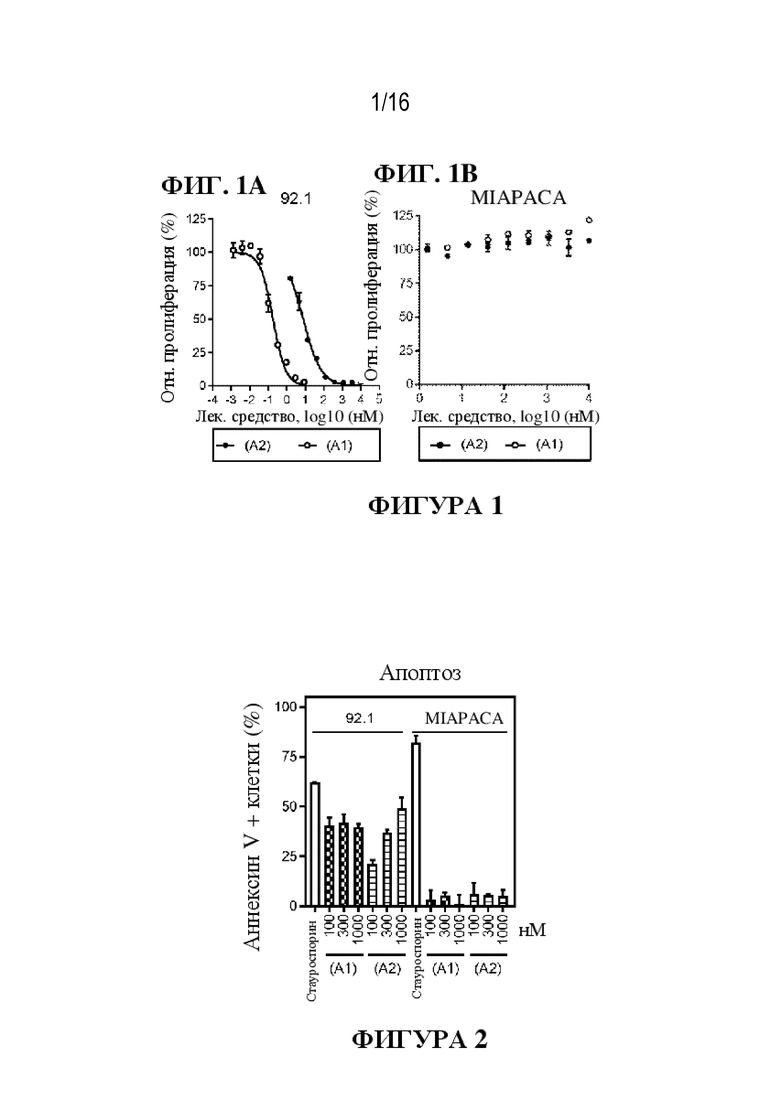
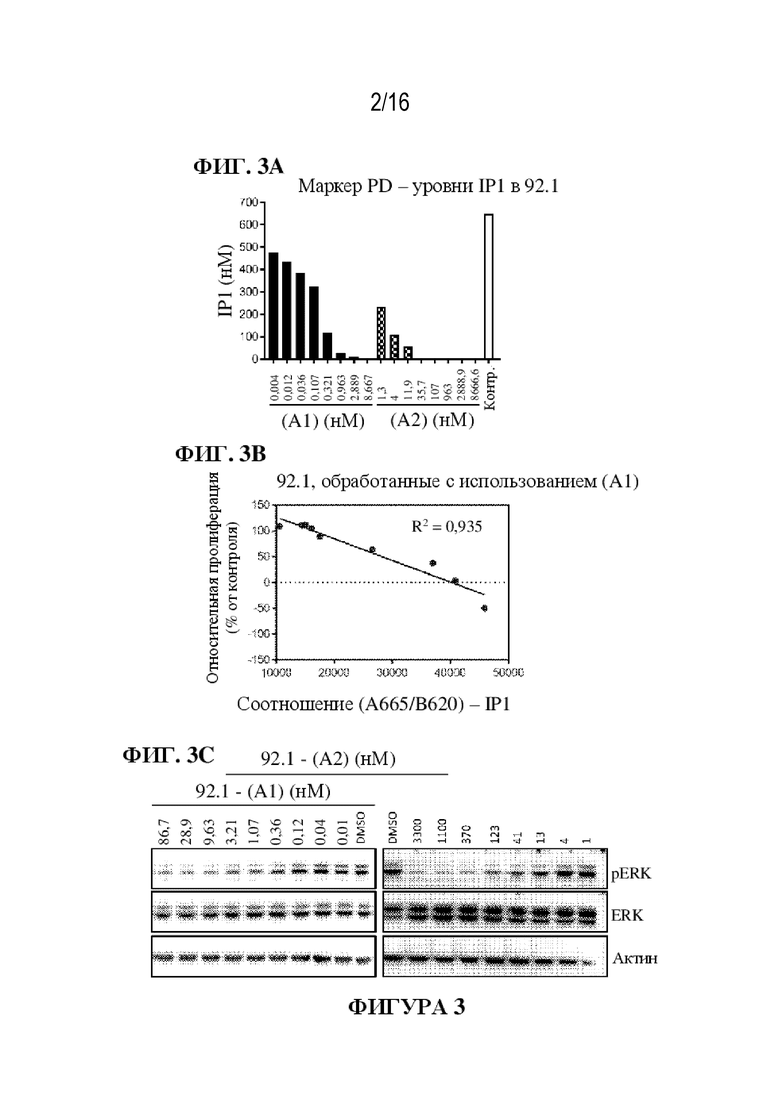
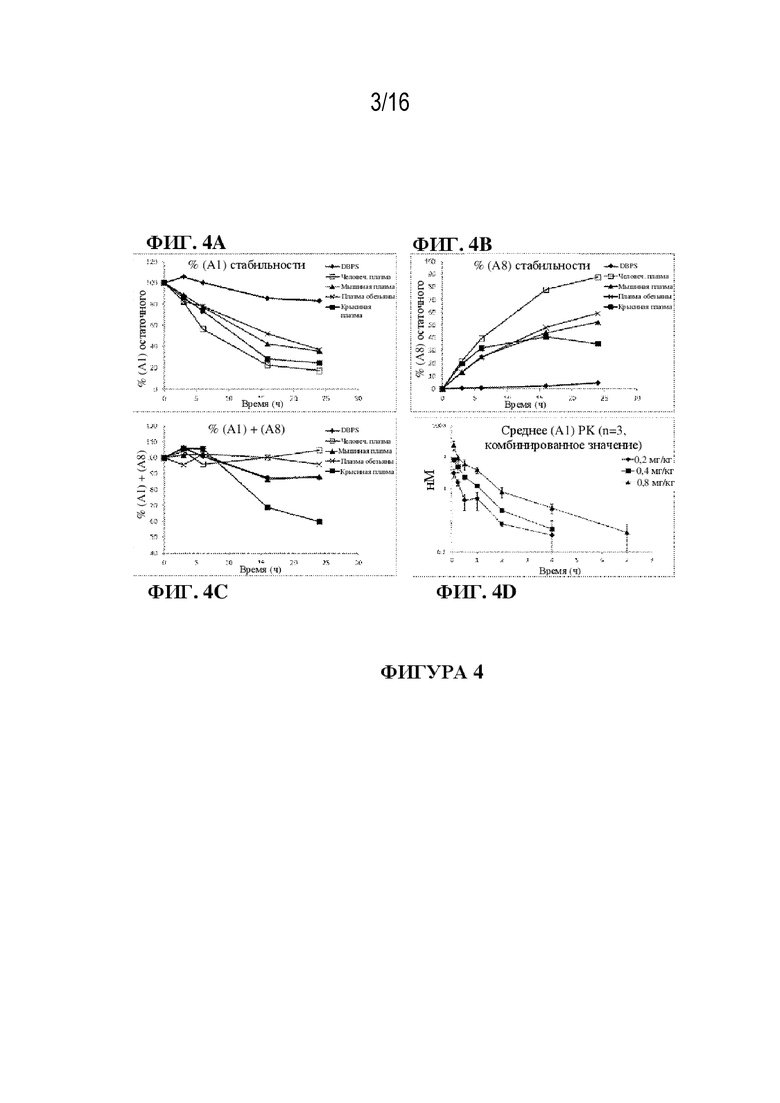
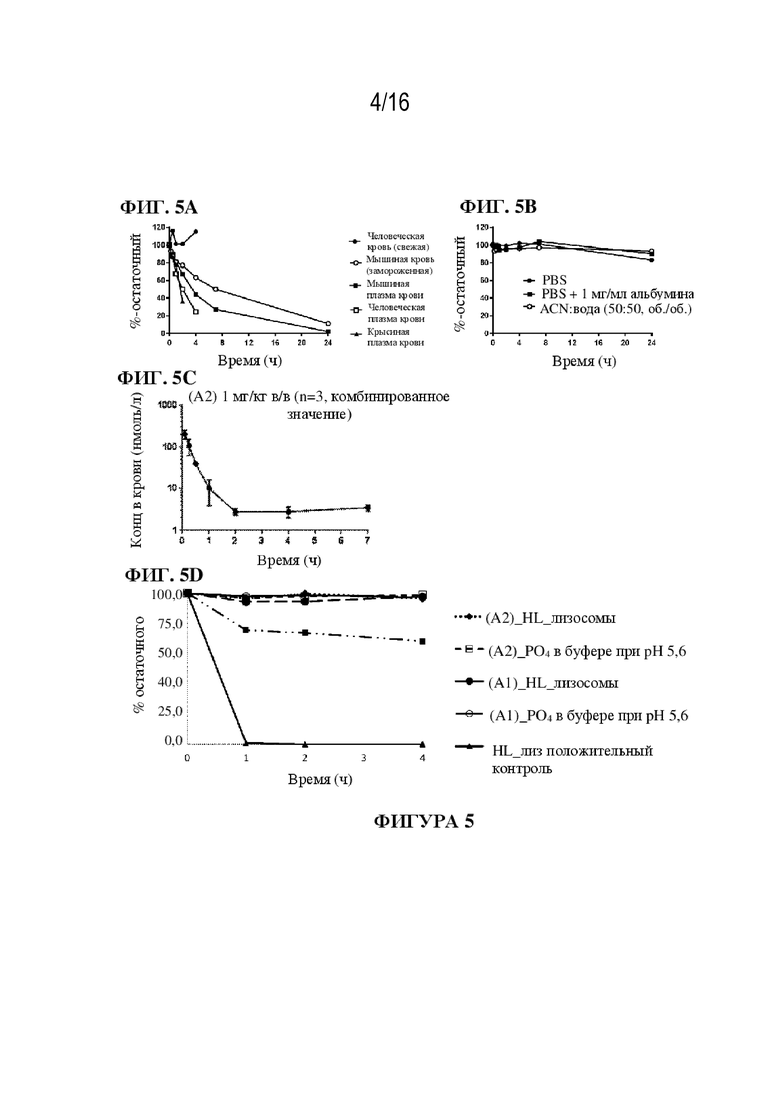
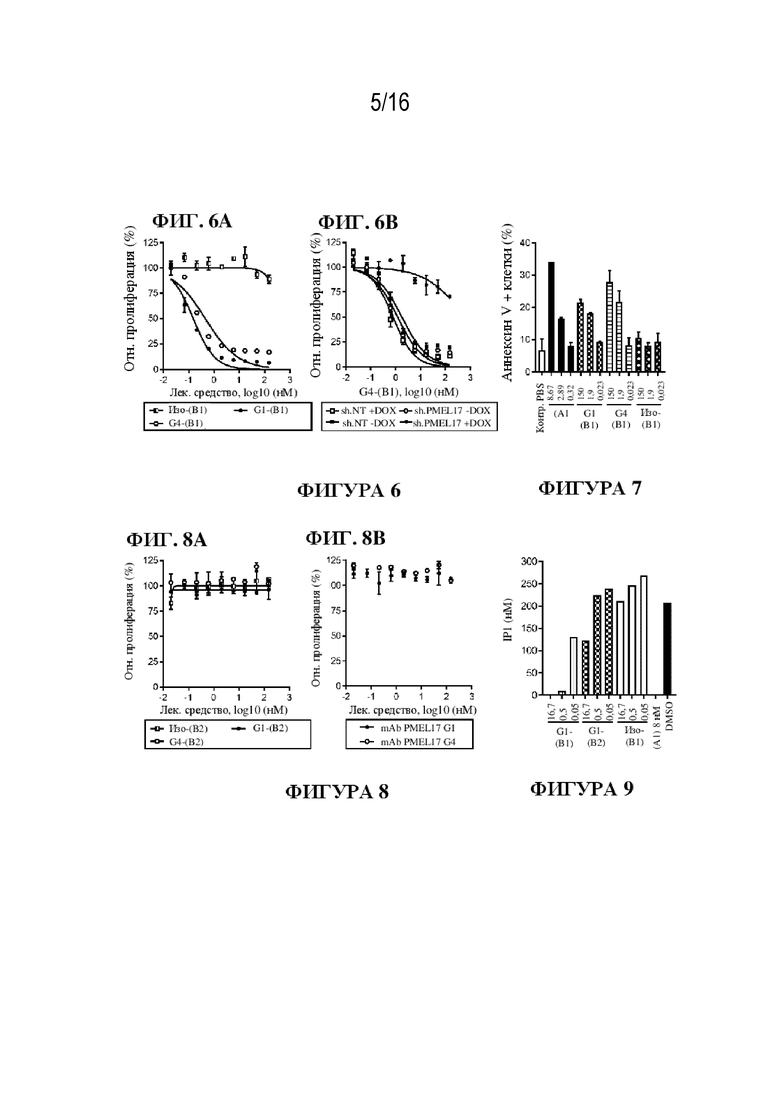
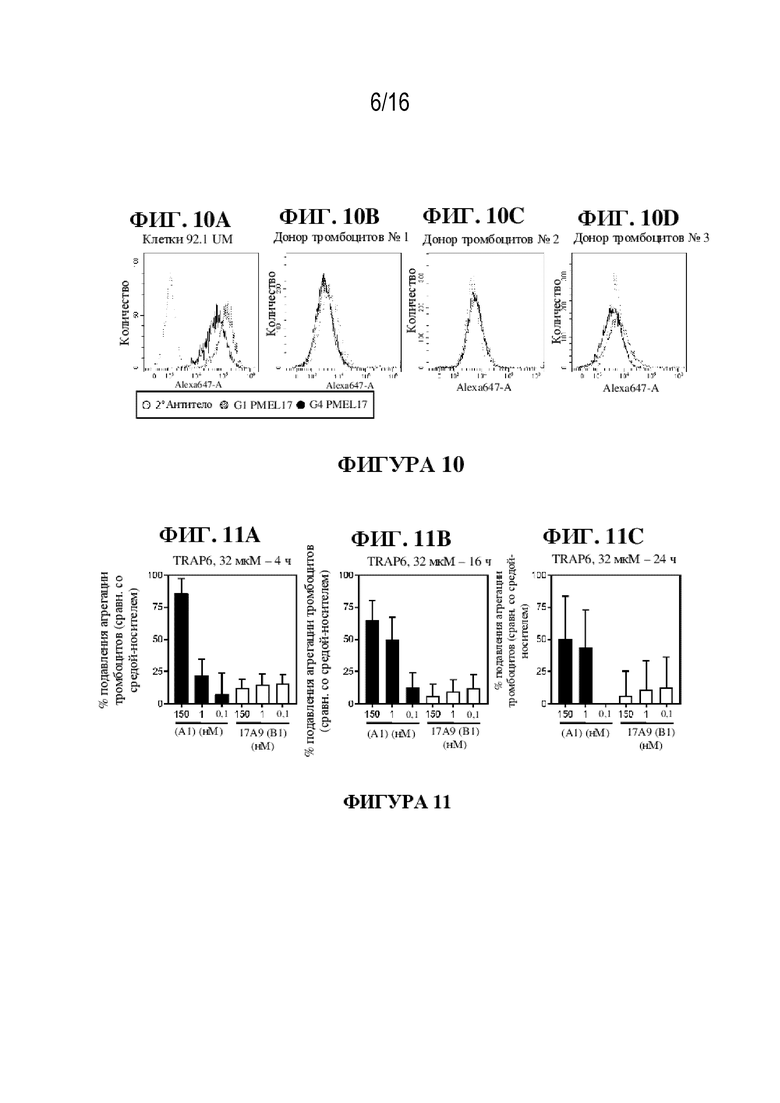
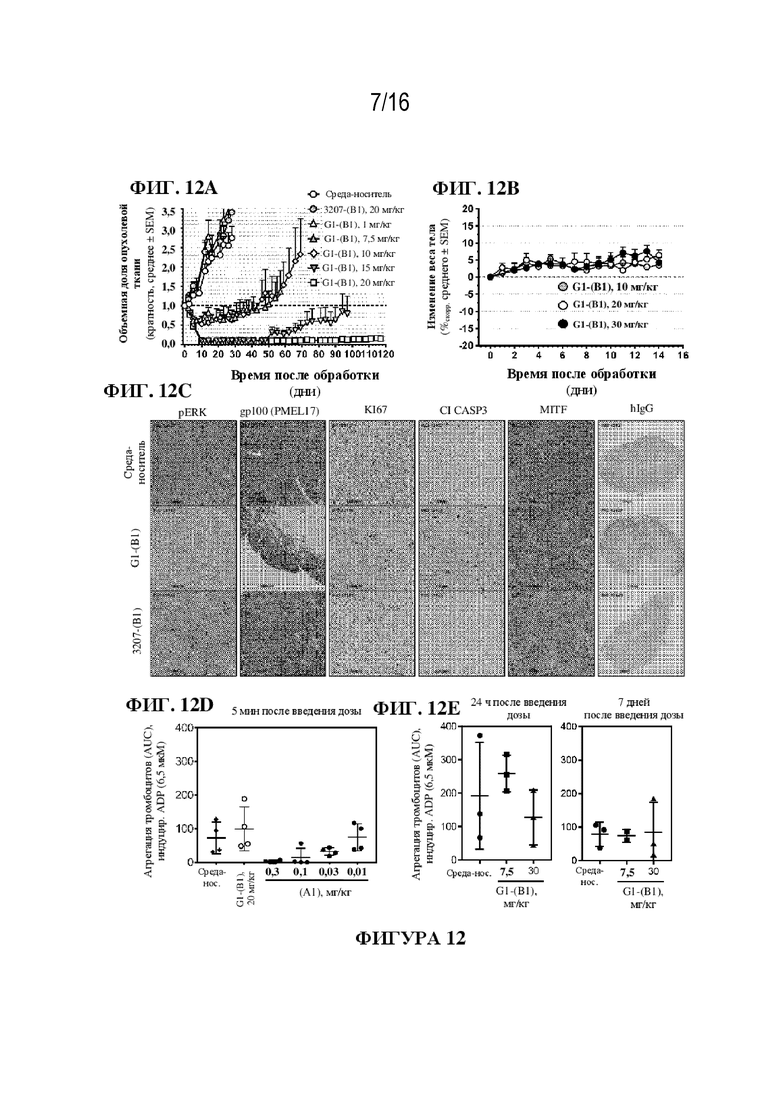
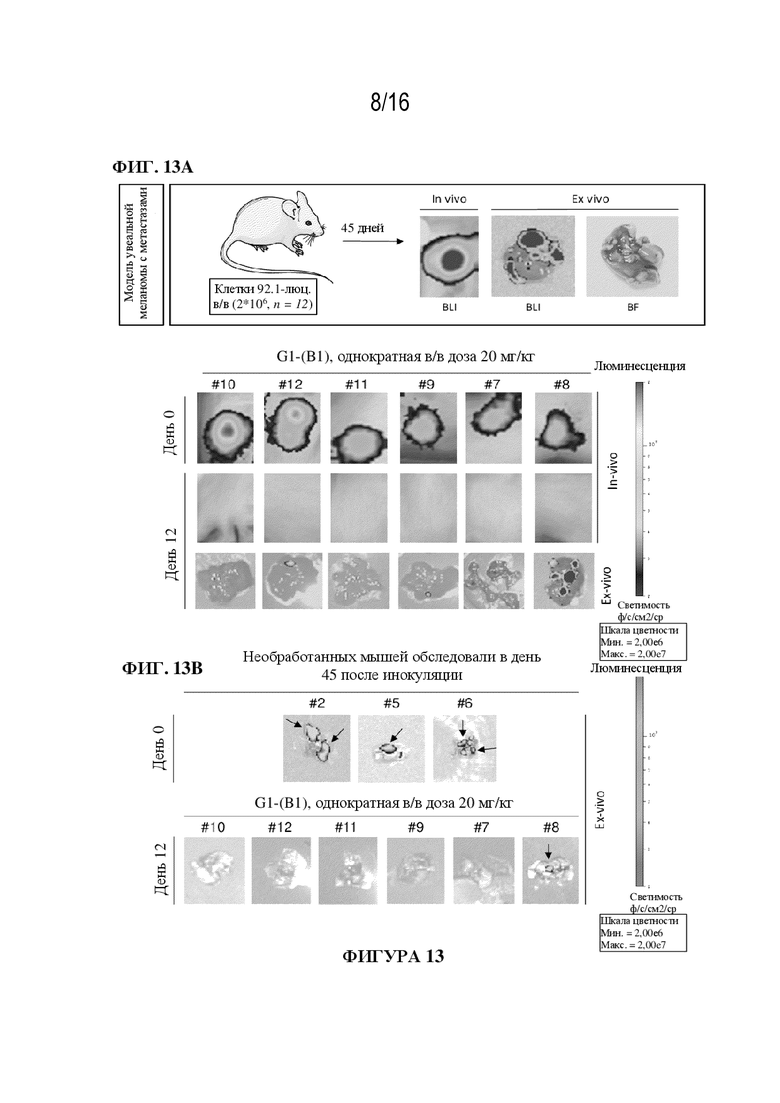
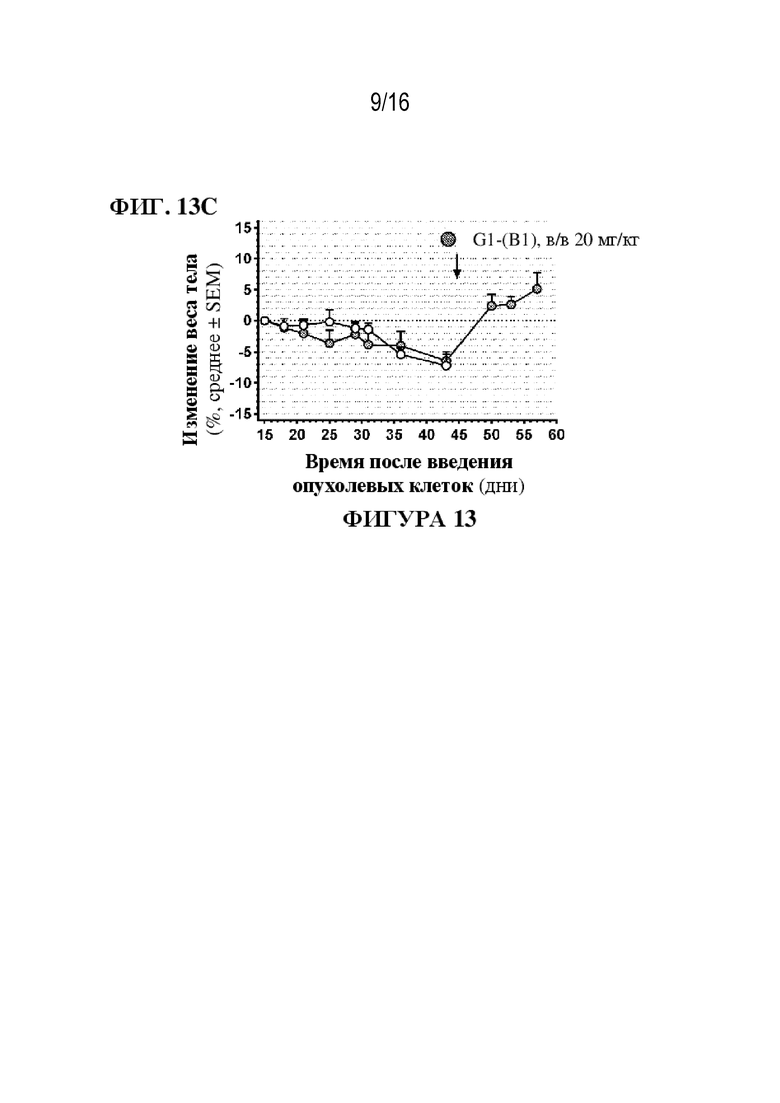
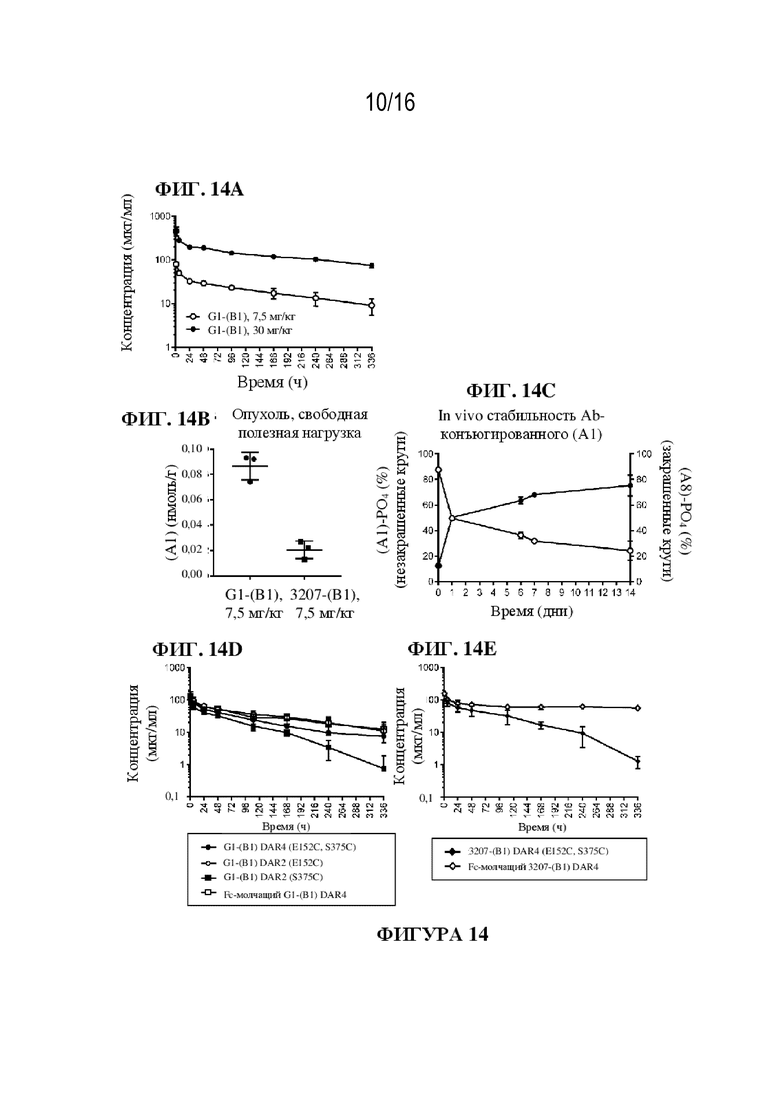
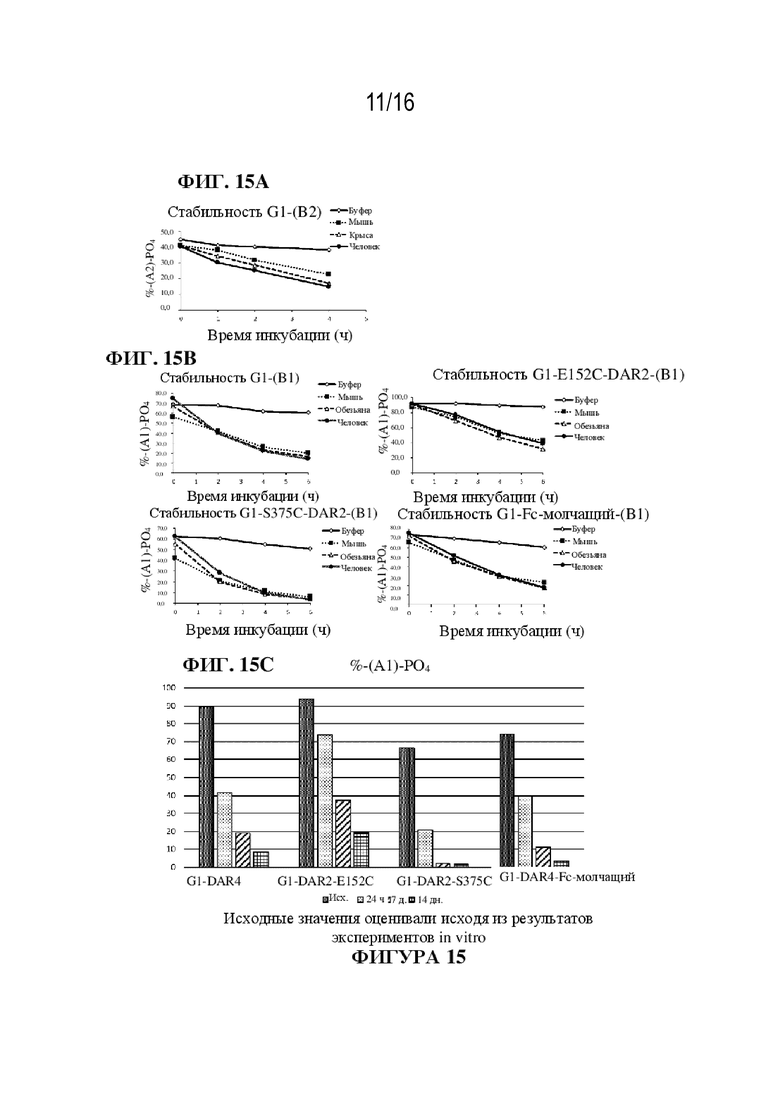
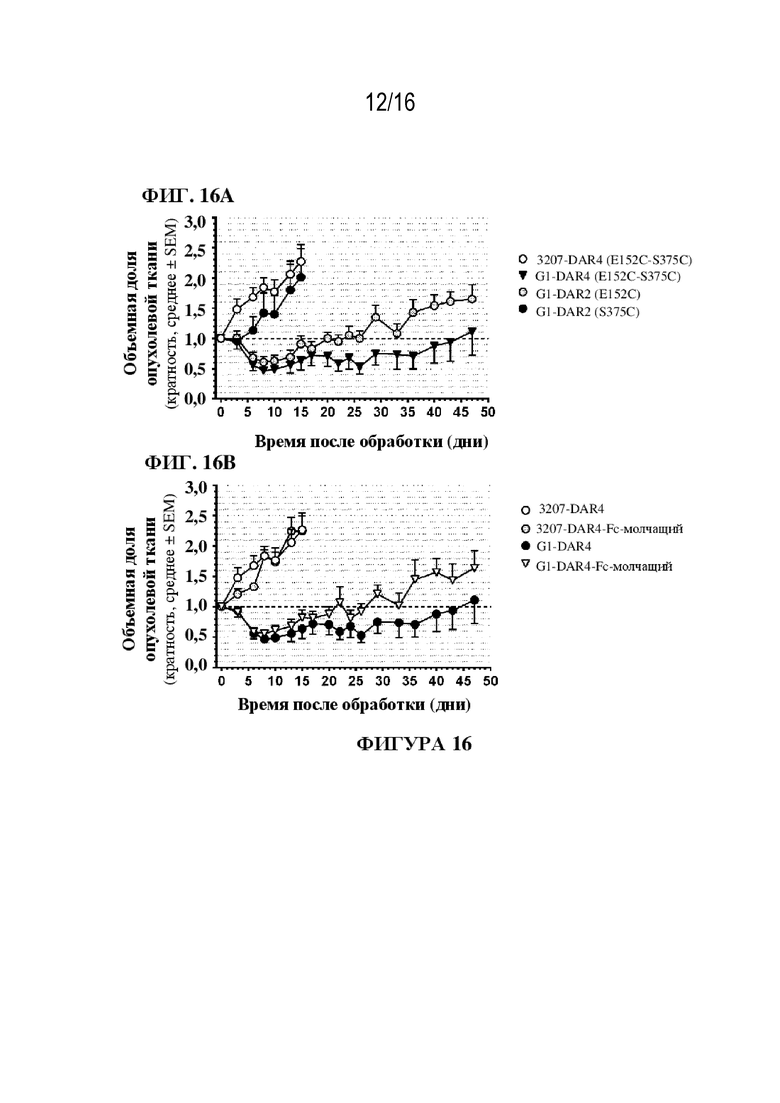
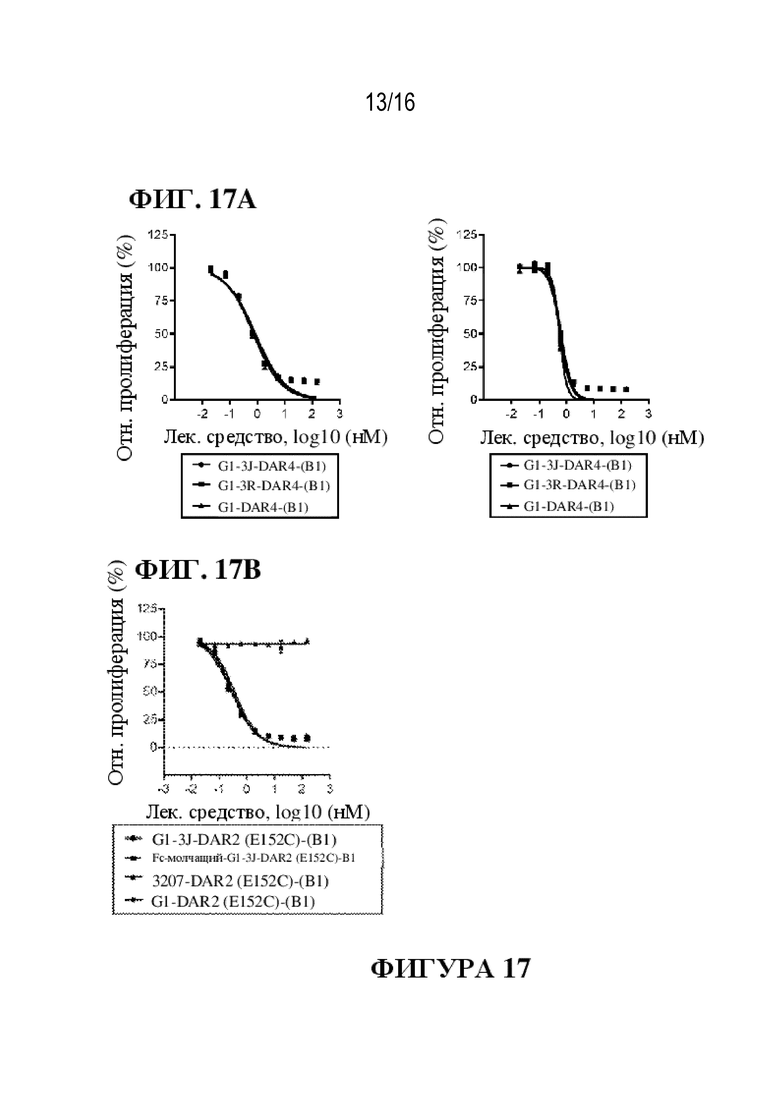
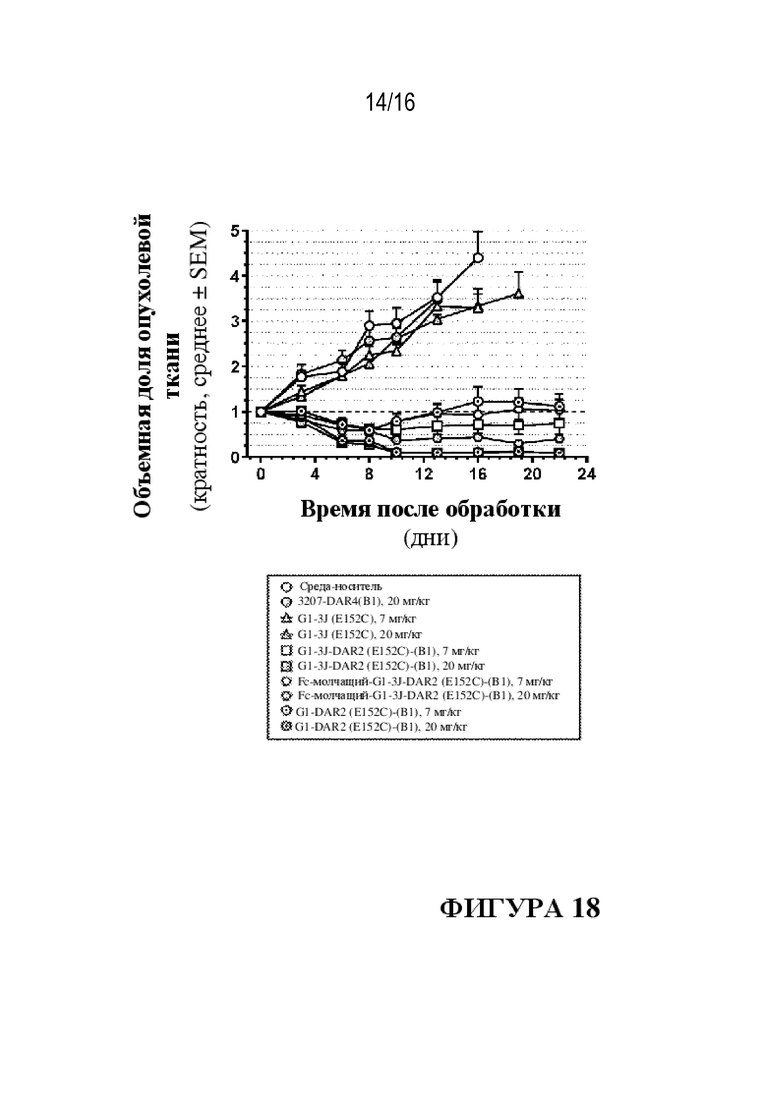
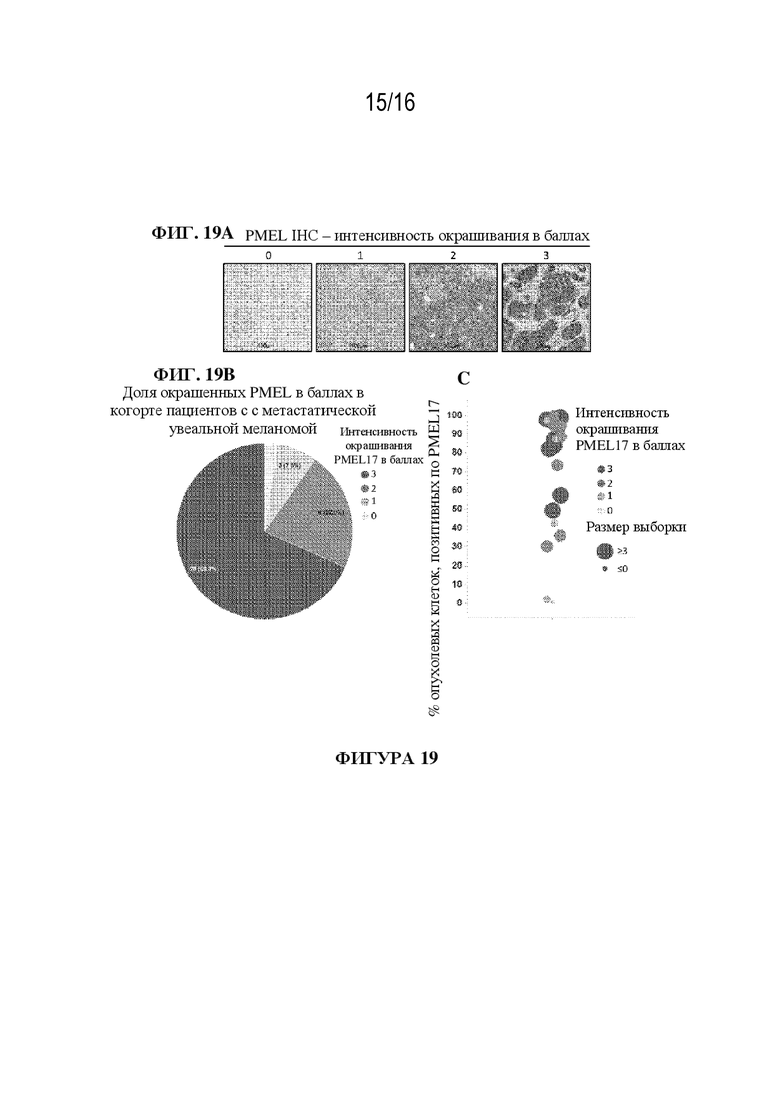
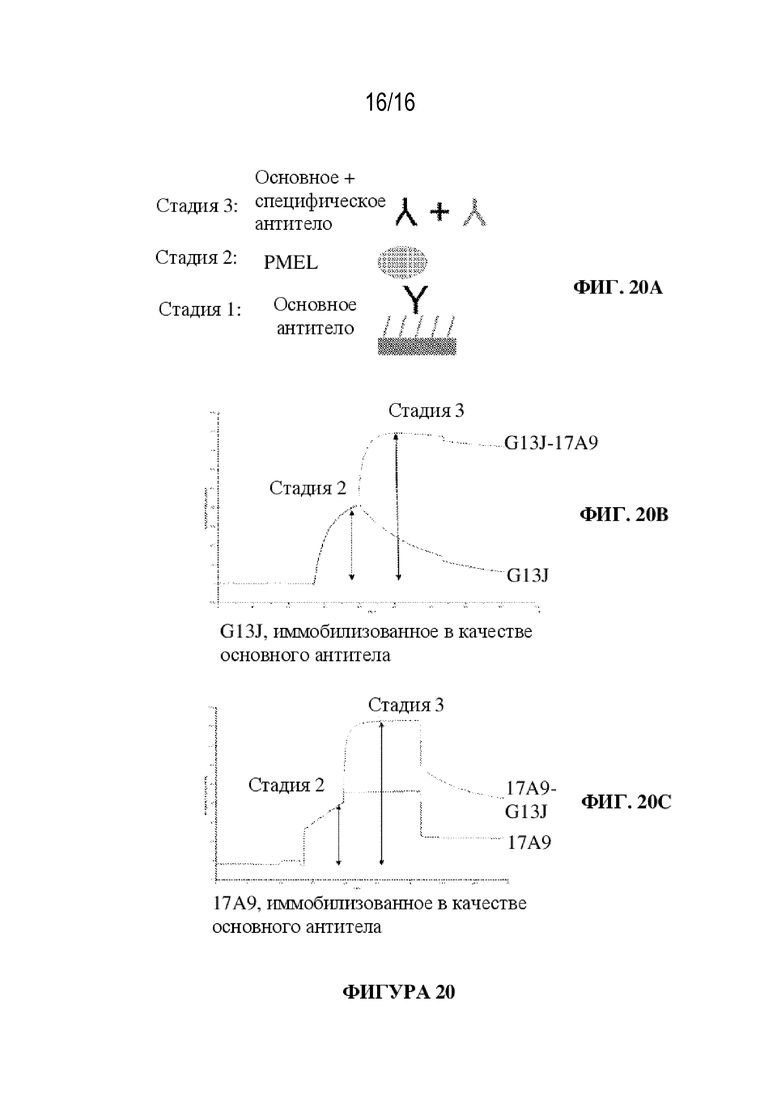
Группа изобретений относится к биотехнологии. Представлены: антитела или их антигенсвязывающие фрагменты, которые связываются с белком PMEL17 человека, которые содержат определенные аминокислотные последовательности, способ их получения и применения для лечения или предупреждения рака у нуждающегося в этом пациента, конъюгаты антитела и лекарственного средства, которые связываются с белком PMEL17 человека, содержащие определенную структуру, способ их получения и применение для лечения или предупреждения рака у нуждающегося в этом пациента, фармацевтические композиции для лечения или предупреждения рака у нуждающегося в этом пациента; способ лечения или предупреждения рака у нуждающегося в этом пациента. Также раскрыты: нуклеиновая кислота, которая кодирует антитело или антигенсвязывающий фрагмент, клетка-хозяин для получения антитела или антигенсвязывающего фрагмента, диагностический реагент для диагностики заболевания, связанного с экспрессией PMEL17. Изобретение применяется для лечения рака, где рак экспрессирует PMEL17, рак содержит мутацию гена GNAQ и/или гена GNA11 или рак экспрессирует PMEL17 и содержит мутацию гена GNAQ и/или гена GNA11. 25 н. и 22 з.п. ф-лы, 20 ил., 10 табл., 2 пр.
1. Антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека, содержащие:
a) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 1, CDR2 тяжелой цепи под SEQ ID NO: 2 и CDR3 тяжелой цепи под SEQ ID NO: 3; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 14, CDR2 легкой цепи под SEQ ID NO: 15 и CDR3 легкой цепи под SEQ ID NO: 16;
b) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 4, CDR2 тяжелой цепи под SEQ ID NO: 2 и CDR3 тяжелой цепи под SEQ ID NO: 3; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 14, CDR2 легкой цепи под SEQ ID NO: 15 и CDR3 легкой цепи под SEQ ID NO: 16;
c) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 5, CDR2 тяжелой цепи под SEQ ID NO: 6 и CDR3 тяжелой цепи под SEQ ID NO: 3; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 17, CDR2 легкой цепи под SEQ ID NO: 18 и CDR3 легкой цепи под SEQ ID NO: 19;
d) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 7, CDR2 тяжелой цепи под SEQ ID NO: 8 и CDR3 тяжелой цепи под SEQ ID NO: 9; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 20, CDR2 легкой цепи под SEQ ID NO: 18 и CDR3 легкой цепи под SEQ ID NO: 16;
e) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 33, CDR2 тяжелой цепи под SEQ ID NO: 34 и CDR3 тяжелой цепи под SEQ ID NO:35; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 46, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO: 48;
f) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 36, CDR2 тяжелой цепи под SEQ ID NO: 34 и CDR3 тяжелой цепи под SEQ ID NO:35; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 46, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO: 48;
g) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 37, CDR2 тяжелой цепи под SEQ ID NO: 38 и CDR3 тяжелой цепи под SEQ ID NO: 35; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 49, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 51;
h) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 39, CDR2 тяжелой цепи под SEQ ID NO: 40 и CDR3 тяжелой цепи под SEQ ID NO: 41; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 52, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 48;
i) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 57, CDR2 тяжелой цепи под SEQ ID NO: 58 и CDR3 тяжелой цепи под SEQ ID NO: 59; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 68, CDR2 легкой цепи под SEQ ID NO: 69 и CDR3 легкой цепи под SEQ ID NO: 70;
j) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 60, CDR2 тяжелой цепи под SEQ ID NO: 58 и CDR3 тяжелой цепи под SEQ ID NO: 59; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 68, CDR2 легкой цепи под SEQ ID NO: 69 и CDR3 легкой цепи под SEQ ID NO: 70;
k) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 5, CDR2 тяжелой цепи под SEQ ID NO: 61 и CDR3 тяжелой цепи под SEQ ID NO: 59; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 71, CDR2 легкой цепи под SEQ ID NO: 72 и CDR3 легкой цепи под SEQ ID NO: 73;
l) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 7, CDR2 тяжелой цепи под SEQ ID NO: 62 и CDR3 тяжелой цепи под SEQ ID NO: 63; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 74, CDR2 легкой цепи под SEQ ID NO: 72 и CDR3 легкой цепи под SEQ ID NO: 70;
m) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 79, CDR2 тяжелой цепи под SEQ ID NO: 80 и CDR3 тяжелой цепи под SEQ ID NO: 81; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 92, CDR2 легкой цепи под SEQ ID NO: 93 и CDR3 легкой цепи под SEQ ID NO: 94;
n) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 82, CDR2 тяжелой цепи под SEQ ID NO: 80 и CDR3 тяжелой цепи под SEQ ID NO: 81; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 92, CDR2 легкой цепи под SEQ ID NO: 93 и CDR3 легкой цепи под SEQ ID NO: 94;
o) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 83, CDR2 тяжелой цепи под SEQ ID NO: 84 и CDR3 тяжелой цепи под SEQ ID NO: 81; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 95, CDR2 легкой цепи под SEQ ID NO: 96 и CDR3 легкой цепи под SEQ ID NO: 97;
p) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 85, CDR2 тяжелой цепи под SEQ ID NO: 86 и CDR3 тяжелой цепи под SEQ ID NO: 87; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 98, CDR2 легкой цепи под SEQ ID NO: 96 и CDR3 легкой цепи под SEQ ID NO: 94;
q) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 103, CDR2 тяжелой цепи под SEQ ID NO: 104 и CDR3 тяжелой цепи под SEQ ID NO: 105; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO:117;
r) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 106, CDR2 тяжелой цепи под SEQ ID NO: 104 и CDR3 тяжелой цепи под SEQ ID NO: 105; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO: 117;
s) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 107, CDR2 тяжелой цепи под SEQ ID NO: 108 и CDR3 тяжелой цепи под SEQ ID NO: 105; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 49, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 118;
t) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 109, CDR2 тяжелой цепи под SEQ ID NO: 110 и CDR3 тяжелой цепи под SEQ ID NO: 111; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 52, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO:117;
u) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 123, CDR2 тяжелой цепи под SEQ ID NO: 124 и CDR3 тяжелой цепи под SEQ ID NO: 125; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 136, CDR2 легкой цепи под SEQ ID NO: 137 и CDR3 легкой цепи под SEQ ID NO: 138;
v) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 126, CDR2 тяжелой цепи под SEQ ID NO: 124 и CDR3 тяжелой цепи под SEQ ID NO: 125; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 136, CDR2 легкой цепи под SEQ ID NO: 137 и CDR3 легкой цепи под SEQ ID NO: 138;
w) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 127, CDR2 тяжелой цепи под SEQ ID NO: 128 и CDR3 тяжелой цепи под SEQ ID NO: 125; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 139, CDR2 легкой цепи под SEQ ID NO: 140 и CDR3 легкой цепи под SEQ ID NO: 141;
x) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 129, CDR2 тяжелой цепи под SEQ ID NO: 130 и CDR3 тяжелой цепи под SEQ ID NO: 131; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 142, CDR2 легкой цепи под SEQ ID NO: 140 и CDR3 легкой цепи под SEQ ID NO: 138;
y) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 123, CDR2 тяжелой цепи под SEQ ID NO: 124 и CDR3 тяжелой цепи под SEQ ID NO: 147; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 153, CDR2 легкой цепи под SEQ ID NO: 154 и CDR3 легкой цепи под SEQ ID NO: 155;
z) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 126, CDR2 тяжелой цепи под SEQ ID NO: 124 и CDR3 тяжелой цепи под SEQ ID NO: 147; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 153, CDR2 легкой цепи под SEQ ID NO: 154 и CDR3 легкой цепи под SEQ ID NO: 155;
aa) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 127, CDR2 тяжелой цепи под SEQ ID NO: 128 и CDR3 тяжелой цепи под SEQ ID NO: 147; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 156, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 157;
bb) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 129, CDR2 тяжелой цепи под SEQ ID NO: 130 и CDR3 тяжелой цепи под SEQ ID NO: 148; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 158, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 155;
cc) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 103, CDR2 тяжелой цепи под SEQ ID NO: 104 и CDR3 тяжелой цепи под SEQ ID NO: 163; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO: 169;
dd) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 106, CDR2 тяжелой цепи под SEQ ID NO: 104 и CDR3 тяжелой цепи под SEQ ID NO: 163; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO: 169;
ee) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 107, CDR2 тяжелой цепи под SEQ ID NO: 108 и CDR3 тяжелой цепи под SEQ ID NO: 163; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 49, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 170;
ff) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 109, CDR2 тяжелой цепи под SEQ ID NO: 110 и CDR3 тяжелой цепи под SEQ ID NO: 164; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 52, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 169;
gg) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 175, CDR2 тяжелой цепи под SEQ ID NO: 176 и CDR3 тяжелой цепи под SEQ ID NO: 177; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO: 188;
hh) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 178, CDR2 тяжелой цепи под SEQ ID NO: 176 и CDR3 тяжелой цепи под SEQ ID NO: 177; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO: 188;
ii) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 179, CDR2 тяжелой цепи под SEQ ID NO: 180 и CDR3 тяжелой цепи под SEQ ID NO: 177; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 49, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 189;
jj) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 181, CDR2 тяжелой цепи под SEQ ID NO: 182 и CDR3 тяжелой цепи под SEQ ID NO: 183; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 52, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 188;
kk) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 103, CDR2 тяжелой цепи под SEQ ID NO: 104 и CDR3 тяжелой цепи под SEQ ID NO: 194; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO: 200;
ll) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 106, CDR2 тяжелой цепи под SEQ ID NO: 104 и CDR3 тяжелой цепи под SEQ ID NO: 194; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 116, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO: 200;
mm) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 107, CDR2 тяжелой цепи под SEQ ID NO: 108 и CDR3 тяжелой цепи под SEQ ID NO: 194; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 49, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 201;
nn) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 109, CDR2 тяжелой цепи под SEQ ID NO: 110 и CDR3 тяжелой цепи под SEQ ID NO: 195; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 52, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 200;
oo) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 206, CDR2 тяжелой цепи под SEQ ID NO: 207 и CDR3 тяжелой цепи под SEQ ID NO: 208; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 153, CDR2 легкой цепи под SEQ ID NO: 154 и CDR3 легкой цепи под SEQ ID NO: 219;
pp) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 209, CDR2 тяжелой цепи под SEQ ID NO: 207 и CDR3 тяжелой цепи под SEQ ID NO: 208; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 153, CDR2 легкой цепи под SEQ ID NO: 154 и CDR3 легкой цепи под SEQ ID NO: 219;
qq) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 210, CDR2 тяжелой цепи под SEQ ID NO: 211 и CDR3 тяжелой цепи под SEQ ID NO: 208; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 156, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 220;
rr) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 212, CDR2 тяжелой цепи под SEQ ID NO: 213 и CDR3 тяжелой цепи под SEQ ID NO: 214; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 158, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 219;
ss) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 206, CDR2 тяжелой цепи под SEQ ID NO: 207 и CDR3 тяжелой цепи под SEQ ID NO: 225; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 136, CDR2 легкой цепи под SEQ ID NO: 137 и CDR3 легкой цепи под SEQ ID NO: 231;
tt) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 209, CDR2 тяжелой цепи под SEQ ID NO: 207 и CDR3 тяжелой цепи под SEQ ID NO: 225; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 136, CDR2 легкой цепи под SEQ ID NO: 137 и CDR3 легкой цепи под SEQ ID NO: 231;
uu) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 210, CDR2 тяжелой цепи под SEQ ID NO: 211 и CDR3 тяжелой цепи под SEQ ID NO:225; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 139, CDR2 легкой цепи под SEQ ID NO: 140 и CDR3 легкой цепи под SEQ ID NO: 232;
vv) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 212, CDR2 тяжелой цепи под SEQ ID NO: 213 и CDR3 тяжелой цепи под SEQ ID NO: 226; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 142; CDR2 легкой цепи под SEQ ID NO: 140 и CDR3 легкой цепи под SEQ ID NO: 231;
ww) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 206, CDR2 тяжелой цепи под SEQ ID NO: 207 и CDR3 тяжелой цепи под SEQ ID NO: 237; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 243, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO: 244;
xx) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 209, CDR2 тяжелой цепи под SEQ ID NO: 207 и CDR3 тяжелой цепи под SEQ ID NO: 237; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 243, CDR2 легкой цепи под SEQ ID NO: 47 и CDR3 легкой цепи под SEQ ID NO: 244;
yy) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 210, CDR2 тяжелой цепи под SEQ ID NO: 211 и CDR3 тяжелой цепи под SEQ ID NO: 237; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 245, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 246;
zz) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 212, CDR2 тяжелой цепи под SEQ ID NO: 213 и CDR3 тяжелой цепи под SEQ ID NO: 238; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 247, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 244;
aaa) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 206, CDR2 тяжелой цепи под SEQ ID NO: 207 и CDR3 тяжелой цепи под SEQ ID NO: 252; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 153, CDR2 легкой цепи под SEQ ID NO: 154 и CDR3 легкой цепи под SEQ ID NO: 258;
bbb) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 209, CDR2 тяжелой цепи под SEQ ID NO: 207 и CDR3 тяжелой цепи под SEQ ID NO: 252; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 153, CDR2 легкой цепи под SEQ ID NO: 154 и CDR3 легкой цепи под SEQ ID NO: 258;
ccc) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 210, CDR2 тяжелой цепи под SEQ ID NO: 211 и CDR3 тяжелой цепи под SEQ ID NO: 252; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 156, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 259; или
ddd) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 212, CDR2 тяжелой цепи под SEQ ID NO: 213 и CDR3 тяжелой цепи под SEQ ID NO: 253; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 158, CDR2 легкой цепи под SEQ ID NO: 50 и CDR3 легкой цепи под SEQ ID NO: 258.
2. Антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека, содержащие:
a) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 10, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 21;
b) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 10, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 25;
c) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 10, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 29;
d) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 42, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 53;
e) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 64, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 75;
f) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 88, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 99;
g) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 112, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 119;
h) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 132, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 143;
i) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 149, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 159;
j) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 165, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 171;
k) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 184, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 190;
l) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 196, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 202;
m) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 215, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 221;
n) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 227, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 233;
o) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 239, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 248; или
p) вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 254, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 260.
3. Антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека, содержащие:
a) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 12, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 23;
b) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 12, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 27;
c) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 12, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 31;
d) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 44, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 55;
e) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 66, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 77;
f) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 90, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 101;
g) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 114, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 121;
h) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 134, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 145;
i) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 151, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 161;
j) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 167, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 173;
k) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 186, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 192;
l) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 198, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 204;
m) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 217, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 223;
n) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 229, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 235;
o) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 241, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 250; или
p) тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 256, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 262.
4. Антитело по любому из пп. 1-3, где антитело или его антигенсвязывающий фрагмент содержат одну или несколько цистеиновых замен.
5. Антитело по п. 4, где одна или несколько цистеиновых замен выбраны из Е152С, S375C или как Е152С, так и S375C в тяжелой цепи антитела или его антигенсвязывающего фрагмента, где нумерация положения соответствует системе EU.
6. Антитело по любому из пп. 1-5, где указанное антитело представляет собой моноклональное антитело.
7. Конъюгат антитела и лекарственного средства, который связывается с белком PMEL17 человека, при этом конъюгат антитела и лекарственного средства содержит формулу (C):
Ab-(LA-(D)n)y (C),
где D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11;
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека, согласно любому из пп. 1-6;
LA представляет собой линкер;
n равняется 1, 2, 3 или 4 и
y равняется 1, 2, 3 или 4.
8. Конъюгат антитела и лекарственного средства по п. 7, где n равняется 1.
9. Конъюгат антитела и лекарственного средства по п. 7 или 8, где y составляет 2.
10. Конъюгат антитела и лекарственного средства по любому из пп. 7-9, где указанный линкер представляет собой расщепляемый линкер или нерасщепляемый линкер.
11. Конъюгат антитела и лекарственного средства по п. 10, где линкер предусматривает пептидный линкер ValCit.
12. Конъюгат антитела и лекарственного средства по любому из пп. 7-11, где D является ингибитором GNAQ и GNA11.
13. Конъюгат антитела и лекарственного средства по любому из пп. 7-11, где D представляет собой
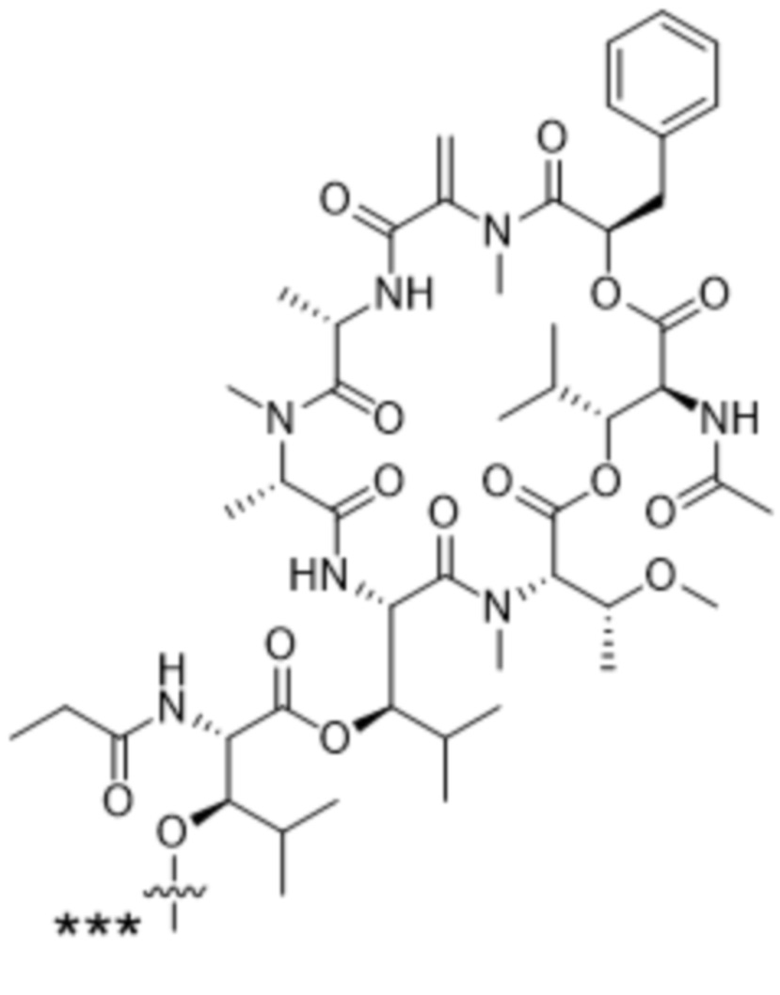 .
.
14. Конъюгат антитела и лекарственного средства по любому из пп. 7-11, где D представляет собой
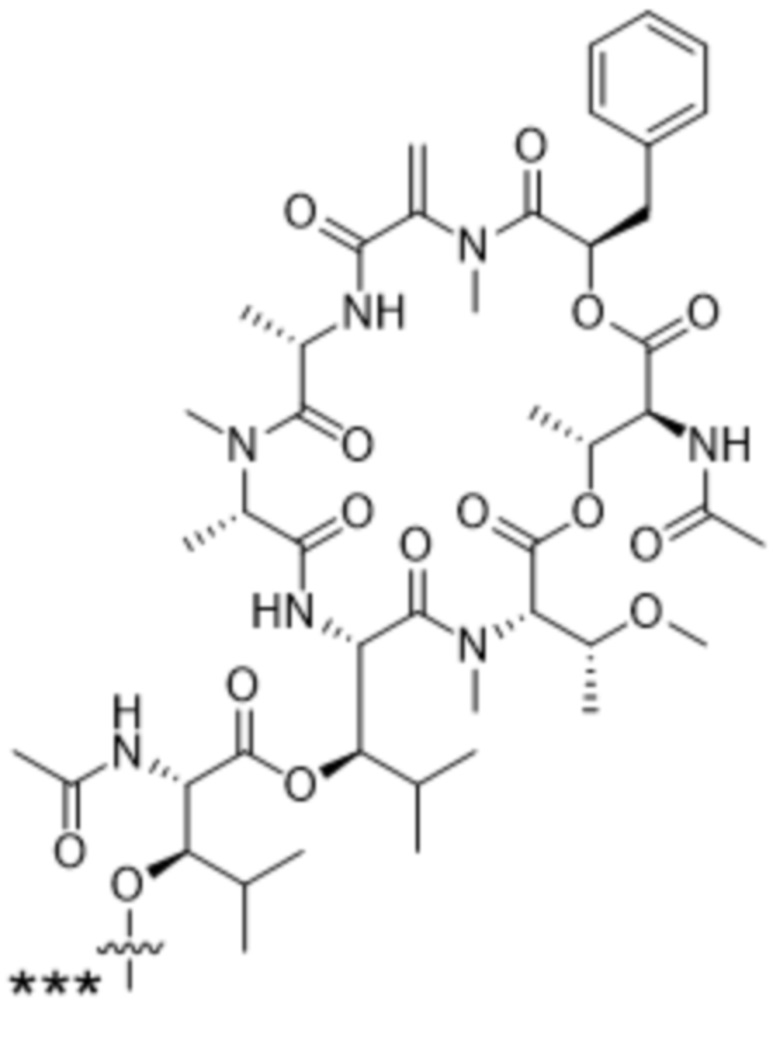 .
.
15. Конъюгат антитела и лекарственного средства по любому из пп. 7-13, содержащий следующую структуру:
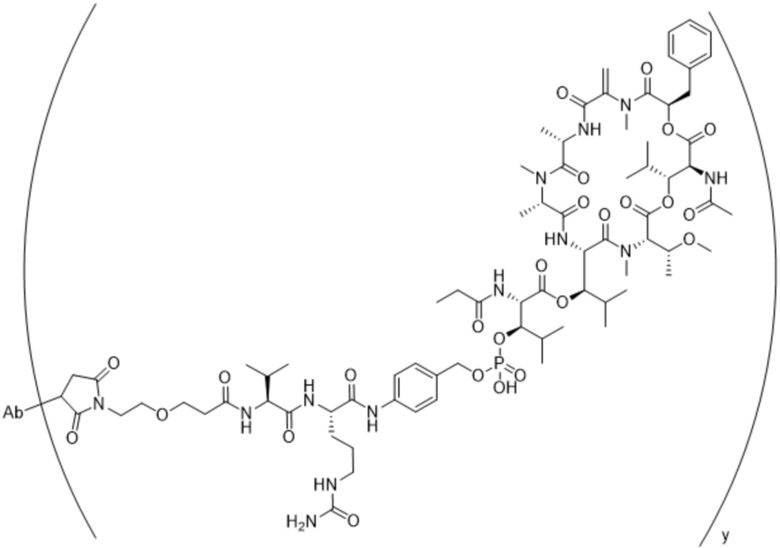 .
.
16. Конъюгат антитела и лекарственного средства по любому из пп. 7-12 или 14, содержащий следующую структуру:
 .
.
17. Конъюгат антитела и лекарственного средства, который связывается с белком PMEL17 человека, при этом конъюгат антитела и лекарственного средства содержит формулу (C-2)
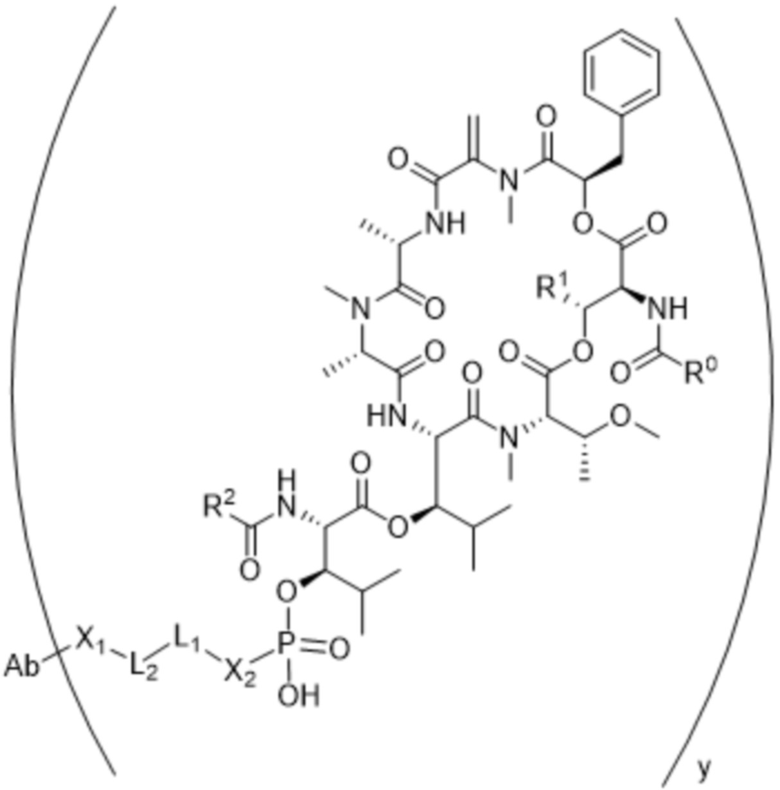 (C-2),
(C-2),
где R0 представляет собой метил или этил;
R1 представляет собой метил или изопропил;
R2 представляет собой метил или этил;
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека, согласно любому из пп. 1-6;
X1 представляет собой бивалентную связывающую группу;
X2 представляет собой саморасщепляющийся спейсер;
L1 представляет собой бивалентный пептидный линкер;
L2 представляет собой связь или линкер и
y равняется 1, 2, 3 или 4.
18. Конъюгат антитела и лекарственного средства, который связывается с белком PMEL17 человека, при этом конъюгат антитела и лекарственного средства содержит следующую структуру:
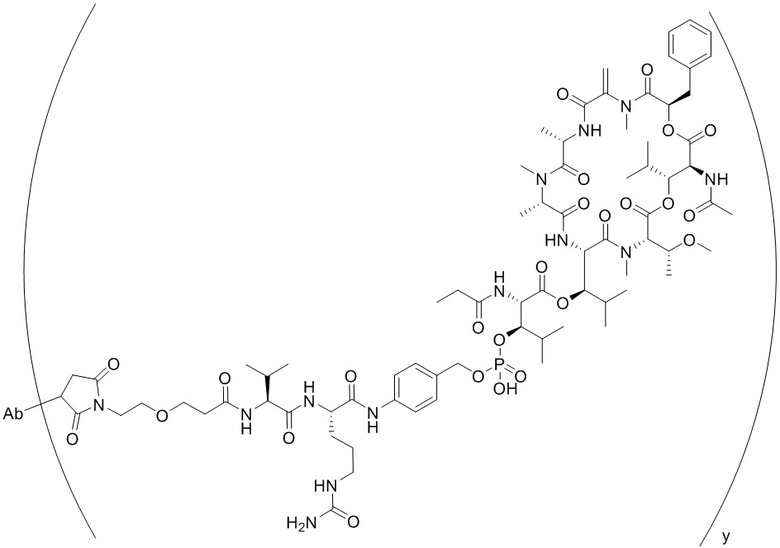 ,
,
где Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека, содержащие:
a) вариабельную область тяжелой цепи (VH), которая содержит аминокислотную последовательность SEQ ID NO: 10, и вариабельную область легкой цепи (VL), которая содержит аминокислотную последовательность SEQ ID NO: 21;
b) вариабельную область тяжелой цепи (VH), которая содержит аминокислотную последовательность SEQ ID NO: 10, и вариабельную область легкой цепи (VL), которая содержит аминокислотную последовательность SEQ ID NO: 25;
c) вариабельную область тяжелой цепи (VH), которая содержит аминокислотную последовательность SEQ ID NO: 10, и вариабельную область легкой цепи (VL), которая содержит аминокислотную последовательность SEQ ID NO: 29;
d) тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 12, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 23;
e) тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 12, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 27; или
f) тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 12, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 31; и
где y равняется 2.
19. Конъюгат антитела и лекарственного средства, который связывается с белком PMEL17 человека, при этом конъюгат антитела и лекарственного средства содержит следующую структуру:
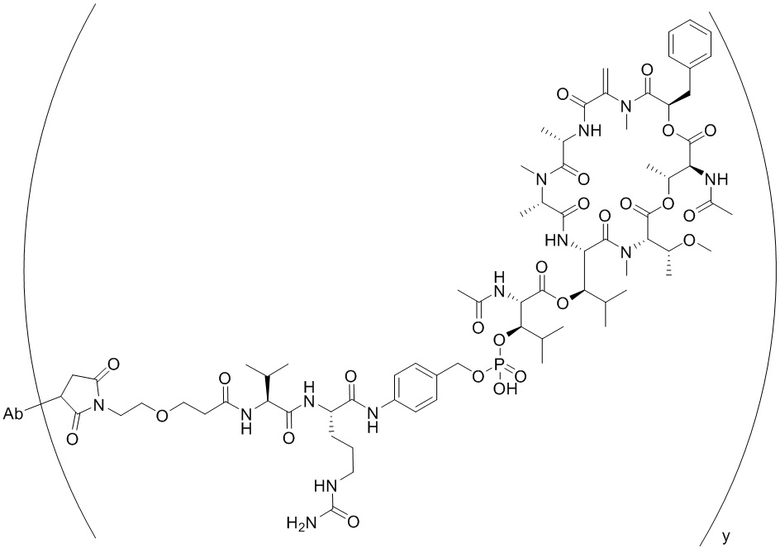 ,
,
где Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека, содержащие:
a) вариабельную область тяжелой цепи (VH), которая содержит аминокислотную последовательность SEQ ID NO: 10, и вариабельную область легкой цепи (VL), которая содержит аминокислотную последовательность SEQ ID NO: 21;
b) вариабельную область тяжелой цепи (VH), которая содержит аминокислотную последовательность SEQ ID NO: 10, и вариабельную область легкой цепи (VL), которая содержит аминокислотную последовательность SEQ ID NO: 25;
c) вариабельную область тяжелой цепи (VH), которая содержит аминокислотную последовательность SEQ ID NO: 10, и вариабельную область легкой цепи (VL), которая содержит аминокислотную последовательность SEQ ID NO: 29;
d) тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 12, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 23;
e) тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 12, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 27; или
f) тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 12, и легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 31; и
где y равняется 2.
20. Фармацевтическая композиция для лечения или предупреждения рака у нуждающегося в этом пациента, при этом композиция содержит терапевтически эффективное количество антитела или его антигенсвязывающего фрагмента по любому из пп. 1-6 и фармацевтически приемлемый носитель, где рак экспрессирует PMEL17, рак содержит мутацию гена GNAQ и/или гена GNA11 или рак экспрессирует PMEL17 и содержит мутацию гена GNAQ и/или гена GNA11.
21. Фармацевтическая композиция для лечения или предупреждения рака у нуждающегося в этом пациента, при этом композиция содержит терапевтически эффективное количество конъюгата антитела и лекарственного средства по любому из пп. 7-19 и фармацевтически приемлемый носитель, где рак экспрессирует PMEL17, рак содержит мутацию гена GNAQ и/или гена GNA11 или рак экспрессирует PMEL17 и содержит мутацию гена GNAQ и/или гена GNA11.
22. Способ лечения или предупреждения рака у нуждающегося в этом пациента, предусматривающий введение указанному пациенту конъюгата антитела и лекарственного средства по любому из пп. 7-19 или фармацевтической композиции по п. 20 или 21, где рак экспрессирует PMEL17, рак содержит мутацию гена GNAQ и/или гена GNA11 или рак экспрессирует PMEL17 и содержит мутацию гена GNAQ и/или гена GNA11.
23. Способ по п. 22, где конъюгат антитела и лекарственного средства или фармацевтическую композицию вводят пациенту в комбинации с одним или несколькими дополнительными терапевтическими соединениями.
24. Способ по п. 23, где одно или несколько дополнительных терапевтических соединений выбраны из стандартного химиотерапевтического средства, ингибитора MDM2, ингибитора MRC2, ингибитора PKC, ингибитора MAPK, костимулирующей молекулы или ингибитора контрольных точек иммунного ответа.
25. Способ по п. 24, где костимулирующая молекула выбрана из агониста OX40, CD2, CD27, ICAM-1, LFA-1 (CD11a/CD18), ICOS (CD278), 4-1BB (CD137), GITR, CD30, CD40, BAFFR, HVEM, CD7, LIGHT, NKG2C, SLAMF7, NKp80, CD160, B7-H3, STING или лиганда CD83.
26. Способ по п. 24, где ингибитор контрольных точек иммунного ответа выбран из ингибитора PD-1, PD-L1, PD-L2, CTLA4, TIM3, LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и/или TGFR-бета.
27. Конъюгат антитела и лекарственного средства по любому из пп. 7-19 или фармацевтическая композиция по п. 20 или 21 для применения в лечении или предупреждении рака у нуждающегося в этом пациента, при этом рак экспрессирует PMEL17, рак содержит мутацию гена GNAQ и/или гена GNA11 или рак экспрессирует PMEL17 и содержит мутацию гена GNAQ и/или гена GNA11.
28. Применение антитела или его антигенсвязывающего фрагмента по любому из пп. 1-6, конъюгата антитела и лекарственного средства по любому из пп. 7-19 или фармацевтической композиции по п. 20 или 21 для лечения или предупреждения рака у нуждающегося в этом пациента, при этом рак экспрессирует PMEL17, рак содержит мутацию гена GNAQ и/или гена GNA11 или рак экспрессирует PMEL17 и содержит мутацию гена GNAQ и/или гена GNA11.
29. Применение антитела или его антигенсвязывающего фрагмента по любому из пп. 1-6, конъюгата антитела и лекарственного средства по любому из пп. 7-19 или фармацевтической композиции по п. 20 или 21 в изготовлении лекарственного препарата для лечения или предупреждения рака у нуждающегося в этом пациента, где рак экспрессирует PMEL17, рак содержит мутацию гена GNAQ и/или гена GNA11 или рак экспрессирует PMEL17 и содержит мутацию гена GNAQ и/или гена GNA11.
30. Способ по любому из пп. 22-26, конъюгат антитела и лекарственного средства или фармацевтической композиции по п. 27, применение по п. 28 или 29, где рак представляет собой увеальную меланому, подкожную меланому, гепатоцеллюлярную карциному или метастатическую форму данных видов рака.
31. Нуклеиновая кислота, которая кодирует антитело или антигенсвязывающий фрагмент по любому из пп. 1-6.
32. Нуклеиновая кислота по п. 31, где нуклеиновая кислота содержит нуклеотидную последовательность под SEQ ID NO: 13, 24, 28, 32, 45, 56, 67, 78, 91, 102, 115, 122, 135, 146, 152, 162, 168, 174, 187, 193, 199, 205, 218, 224, 230, 236, 242, 251, 257 или 263.
33. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 31 или 32.
34. Клетка-хозяин для получения антитела или антигенсвязывающего фрагмента, которые связываются с белком PMEL17 человека, при этом указанная клетка-хозяин содержит нуклеиновую кислоту по п. 31 или 32, вектор экспрессии по п. 33.
35. Способ получения антитела или антигенсвязывающего фрагмента, предусматривающий культивирование клетки-хозяина по п. 34 и извлечение антитела или антигенсвязывающего фрагмента из культуры клеток.
36. Способ по п. 35, где извлечение антитела или антигенсвязывающего фрагмента из культуры клеток предусматривает следующие стадии:
a) удаление клеток и фильтрование культуры;
b) проведение очистки культуры посредством аффинной хроматографии;
c) инактивация любых вирусов в культуре путем доведения pH до 3,4-3,6, затем повторного доведения pH до 5,8-6,2 и фильтрования культуры;
d) проведение очистки культуры посредством катионообменной хроматографии и осуществления восстановления культуры на колонке;
e) осуществление анионообменной хроматографии в отношении культуры;
f) удаление вирусов посредством нанофильтрации;
g) фильтрование культуры, содержащей антитело или антигенсвязывающий фрагмент, и
h) получение очищенного антитела или антигенсвязывающего фрагмента.
37. Способ получения конъюгата антитела и лекарственного средства, который связывается с белком PMEL17 человека, согласно любому из пп. 7-19, предусматривающий:
(a) предварительное образование фрагмента, представляющего собой линкер-лекарственное средство, следующей формулы (B):
R8-LB-(D)n (B),
где D представляет собой ингибитор GNAQ, ингибитор GNA11 или ингибитор GNAQ и GNA11;
R8 представляет собой реакционноспособную группу;
LB представляет собой расщепляемый или нерасщепляемый линкер и
n равняется 1, 2, 3 или 4;
(b) конъюгирование указанного фрагмента, представляющего собой линкер-лекарственное средство, с антителом, извлеченным из культуры клеток по п. 35 или 36, с получением конъюгата антитела и лекарственного средства; и
(c) проведение очистки конъюгата антитела и лекарственного средства.
38. Диагностический реагент для диагностики заболевания, связанного с экспрессией PMEL17, при этом указанный диагностический реагент содержит антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6.
39. Диагностический реагент по п. 38, где антитело или его антигенсвязывающий фрагмент являются меченными посредством радиоактивной метки, флуорофора, хромофора, средства для визуализации или иона металла.
40. Конъюгат антитела и лекарственного средства, который связывается с белком PMEL17 человека, при этом конъюгат антитела и лекарственного средства содержит следующую структуру
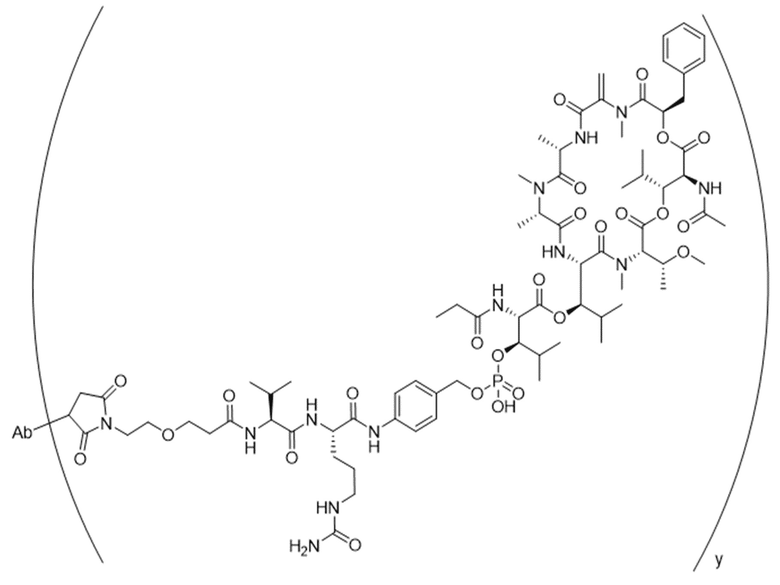
где Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека, содержащее:
a) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 1, CDR2 тяжелой цепи под SEQ ID NO: 2 и CDR3 тяжелой цепи под SEQ ID NO: 3; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 14, CDR2 легкой цепи под SEQ ID NO: 15 и CDR3 легкой цепи под SEQ ID NO: 16;
b) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 4, CDR2 тяжелой цепи под SEQ ID NO: 2 и CDR3 тяжелую цепи под SEQ ID NO: 3; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 14, CDR2 легкой цепи под SEQ ID NO: 15 и CDR3 легкой цепи под SEQ ID NO: 16;
c) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 5, CDR2 тяжелой цепи под SEQ ID NO: 6 и CDR3 тяжелой цепи под SEQ ID NO: 3; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 17, CDR2 легкой цепи под SEQ ID NO: 18 и CDR3 легкой цепи под SEQ ID NO: 19;
d) вариабельную область тяжелой цепи, которая содержит CDR1 тяжелой цепи под SEQ ID NO: 7, CDR2 тяжелой цепи под SEQ ID NO: 8 CDR3 тяжелой цепи под SEQ ID NO: 9; и вариабельную область легкой цепи, которая содержит CDR1 легкой цепи под SEQ ID NO: 20, CDR2 легкой цепи под SEQ ID NO: 18 и CDR3 легкой цепи под SEQ ID NO: 16; и
где у равняется 2.
41. Конъюгат антитела и лекарственного средства, который связывается с белком PMEL17 человека, при этом конъюгат антитела и лекарственного средства содержит следующую структуру:
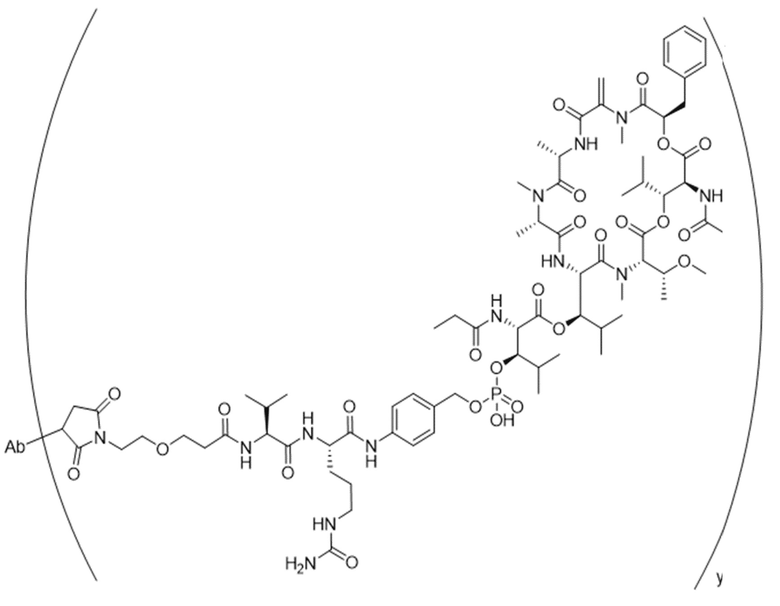
где Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека, содержащее вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO: 10, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO: 25; и
где у равняется 2.
42. Конъюгат антитела и лекарственного средства, который связывается с белком PMEL17 человека, при этом конъюгат антитела и лекарственного средства содержит следующую структуру:
где Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые связываются с белком PMEL17 человека, содержащее тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO: 12, за исключением того, что аминокислота в положении 375 представляет собой S, где положение пронумеровано в соответствии с системой EU, и легкую цепь, содержащая аминокислотную последовательность под SEQ ID NO: 27; и
где у равняется 2.
43. Фармацевтическая композиция для лечения или предупреждения рака у нуждающегося в этом пациента, при этом фармацевтическая композиция содержит терапевтически эффективное количество конъюгата антитела и лекарственного средства по любому из пп. 40-42 и фармацевтически приемлемый носитель, и где рак экспрессирует PMEL17, рак содержит мутацию гена GNAQ и/или гена GNA11 или рак экспрессирует PMEL17 и содержит мутацию гена GNAQ и/или гена GNA11.
44. Способ лечения или предупреждения рака у нуждающегося в этом пациента, предусматривающий введение указанному пациенту конъюгата антитела и лекарственного средства по любому из пп. 40-42 или фармацевтической композиции по п. 43, где рак экспрессирует PMEL17, рак содержит мутацию гена GNAQ и/или гена GNA11 или рак экспрессирует PMEL17 и содержит мутацию гена GNAQ и/или гена GNA11.
45. Конъюгат антитела и лекарственного средства по любому из пп. 40-42 или фармацевтическая композиция по п. 43 для применения в способе лечения или предупреждения рака у нуждающегося в этом пациента, где рак экспрессирует PMEL17, рак содержит мутацию гена GNAQ и/или гена GNA11 или рак экспрессирует PMEL17 и содержит мутацию гена GNAQ и/или гена GNA11.
46. Применение конъюгата антитела и лекарственного средства по любому из пп. 40-42 или фармацевтической композиции по п. 43 в способе лечения рака у нуждающегося в этом пациента, где рак экспрессирует PMEL17, рак содержит мутацию гена GNAQ и/или гена GNA11 или рак экспрессирует PMEL17 и содержит мутацию гена GNAQ и/или гена GNA11.
47. Применение конъюгата антитела и лекарственного средства по любому из пп. 40-42 или фармацевтической композиции по п. 43 в изготовлении лекарственного препарата для лечения или предупреждения рака у нуждающегося в этом пациента, где рак экспрессирует PMEL17, рак содержит мутацию гена GNAQ и/или гена GNA11 или рак экспрессирует PMEL17 и содержит мутацию гена GNAQ и/или гена GNA11.
| Y | |||
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| СПОСОБ ИЗГОТОВЛЕНИЯ ЧЕРТЕЖЕЙ ДЛЯ ОДНООБРАЗНОЙ РАСКРОЙКИ ПРЕДМЕТОВ ОДЕЖДЫ | 1919 |
|
SU287A1 |
| Солесос | 1922 |
|
SU29A1 |
| WO 2013165940 A1, 07.11.2013 | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |

