ОБЛАСТЬ ТЕХНИКИ
[01] Настоящее изобретение принадлежит к области иммунотерапии рака и относится к способу лечения рака, включающему способ лечения плотных (солидных) опухолей, причем указанный способ включает применение антитела к CD25.
УРОВЕНЬ ТЕХНИКИ
[02] Иммунотерапия рака включает применение собственной иммунной системы субъекта для лечения или предотвращения рака. Иммунотерапевтические средства основаны на том факте, что раковые клетки часто имеют незначительно отличающиеся молекулы на своей поверхности, которые могут детектироваться иммунной системой. Эти молекулы, или раковые антигены, чаще всего являются белками, но также включают такие молекулы как углеводы. Таким образом, иммунотерапия включает индукцию иммунной системы для атаки опухолевых клеток за счет этих антигенов-мишеней. Однако злокачественные опухоли, в частности, плотные опухоли, или разные виды гематологического рака могут избегать надзора иммунной системы с помощью различных механизмов, как присущих опухолевой клетке, так и опосредуемых компонентами микросреды опухоли. Среди последних в качестве критических факторов были предложены инфильтрация опухоли регуляторными Т-клетками (Трег-клетками или Трег) и, более конкретно, неблагоприятный баланс эффекторных Т-клеток (Тэфф) по отношению к Трег (т.е. низкое отношение Тэфф к Трег) (Smyth М. et al., 2014, Immunol Cell Biol. 92, 473-4). [03] С момента открытия Трег-клеток было обнаружено, что они играют ключевую роль в опосредовании иммунного гомеостаза, а также стимулируют развитие и поддержание периферической толерантности. Однако в случае рака их роль более сложна. Поскольку раковые клетки экспрессируют как собственные, так и связанные с опухолью антигены, присутствие Трег, которые стремятся ослабить ответ эффекторных клеток, может способствовать прогрессированию опухоли. Инфильтрация Трег в развившихся опухолях, соответственно, представляет собой одно из основных препятствий для эффективных противоопухолевых ответов и лечения разных видов рака в целом. Полагают, что механизмы подавления, используемые Трег, вносят значительный вклад в ограничение или даже неэффективность современных видов терапии, в частности, видов иммунотерапии, которые основаны на индукции или стимуляции противоопухолевых ответов (Onishi И et al, 2012 Anticanc. Res. 32, 997-1003).
[04] Истощение Трег в качестве терапевтического подхода к лечению рака представляет собой подход, который поддерживается исследованиями, показавшими вклад Трег в развитие и прогрессирование опухолей в моделях на мышах. Более того, инфильтрация опухоли Трег также была связана с худшим прогнозом при некоторых видах рака человека (Shang В et al., 2015, Sci Rep.5:15179). Было продемонстрировано, что Трег-клетки способствуют развитию и прогрессированию опухолей в моделях на мышах и что их отсутствие приводит к задержке прогрессирования опухоли (Elpek et al., 2007 J Immunol. 178(11):6840-8; Golgher et al., 2002; Eur J Immunol. 32(11):3267-75, Jones et al., 2002 Cancer Immun. 22;2:1; Onizuka et al., 1999 Cancer Res. 59(13):3128-33.; Shimizu et al., 1999, J Immunol. 163(10):5211-8). У человека сильная инфильтрация опухоли 1 per-клетками и, что более важно, низкое отношение эффекторных Т (Тэфф) клеток к Трег-клеткам связано с неблагоприятными исходами при многих видах рака человека (Shang et al., 2015). Напротив, высокое отношение Тэфф/Трег клеток связано с благоприятными ответами на иммунотерапию как у человека, так и у мышей (Hodi et al., 2008, Proc. Natl. Acad. Sci. USA, 105, 3005 3010; Quezada et al., 2006, J Clin Invest. 116(7): 1935-45). Однако истощение Трег в опухолях является сложным, и результаты исследований в данной области были противоречивыми.
[05] CD25 представляет собой одну из потенциальных молекулярных мишеней для достижения истощения Трег.CD25, также известный как альфа-цепь высокоаффинного рецептора интерлейкина-2 (IL-2Ra), конститутивно экспрессируется на высоких уровнях на Трег-клетках и отсутствует или экспрессируется на низких уровнях на эффекторных Т-клетках и поэтому является перспективной мишенью для истощения Трег. Взаимодействие IL-2/CD25 было объектом нескольких исследований в моделях на мышах, большинство из которых было связано с применением РС61, крысиного антитела к мышиному CD25 (Setiady Y et al., 2010. Eur J Immunol. 40:780-6). Связывание CD25 и разные виды функциональной активности этого антитела сравнивали с аналогичными параметрами для панели моноклональных антител, созданных разными авторами (Lowenthal J.W et al., 1985. J. Immunol., 135, 3988-3994; Moreau, J.-L et al., 1987. Eur. J. Immunol. 17, 929-935; Volk HD et al., 1989 Clin. exp. Immunol. 76, 121-5; Dantal J et al., 1991, Transplantation 52:110 5). Несмотря на то, что первоначальные исследования продемонстрировали профилактическую, но не терапевтическую активность РС61, недавнее исследование показало, что оптимизированный по Fc вариант этого антитела к CD25 привел к внутриопухолевому истощению Трег и обеспечивает значительную терапевтическую пользу в некоторых моделях опухолей на мышах (Vargas A et al., 2017, Immunity 48(6), 577-586). Доступные антитела к CD25, такие как РС61, блокируют или ингибируют связывание IL-2 с CD25, как и многие другие антитела к мышиному CD25, и большинство из них раскрыто как антитела к человеческому CD25; см., например, WO 2004/045512, WO 2006/108670, WO 1993/011238, WO 1990/007861 и WO 2017/174331. Например, базиликсимаб и даклизумаб представляют собой антитела к человеческому CD25, которые ингибируют связывание IL-2 с CD25, и были разработаны для снижения активации эффекторных Т-клеток. Базиликсимаб представляет собой химерное мышиное и человеческое антитело к CD25, одобренное в настоящее время для лечения заболеваний «трансплантат против хозяина», и даклизумаб представляет собой гуманизированное антитело к CD25, одобренное для лечения рассеянного склероза. Однако другие антитела к CD25 все еще допускают связывание IL-2 с CD25, такие как клон 7D4 (антитело к мышиному CD25), клон МА251 (антитело к человеческому CD25) или 7G7B6 (антитело к человеческому CD25) (Rubin et al,1985, Hybridoma 4(2) 91-102, Tanaka et al, 1986, Microbiol. Immunol 30(4), 373-388). 7G7B6 было использовано в качестве исследовательского антитела и предложено в качестве нацеливающего фрагмента для нацеливания радионуклида к СВ25-экспрессирующим лимфомам (Zhang et al, 2009, Cancer Biother RadiopHarm 24(3), 303-309).
[06] Например, 7D4 представляет собой крысиное антитело IgM к мышиному CD25, которое широко применяли для детектирования СВ25-положительных клеток в присутствии или после обработки РС61 или антителами, имеющими близкие связывающие свойства (Onizuka S et al., 1999. Cane Res. 59, 3128 3133). В очень небольшом количестве документов раскрыто какое-либо функциональное свойство антитела 7D4-IgM, по отдельности или в сравнении с РС61 (Kohm A et al., 2006, J Immunol. 176: 3301-5; Hallett W et al., 2008. Biol Blood Marrow Transplant 14:1088-1099; Fecci P et al., 2006 Clin Cancer Res. 12:4294-4305; McNeill A et al., 2007. Scand J Immunol 65: 63-9; Setiady Y et al., 2010. Eur. J. Immunol. 40: 780-6; Couper К et al., 2007. J Immunol. 178: 4136-4146). Действительно, в уровне техники отсутствуют сведения о возможности адаптации или модификации каким-либо образом изотипа или других структурных признаков 7D4 для того чтобы получить улучшенное антитело для применения в терапии рака.
[07] Однако способность 7D4-IgM (как такового или в виде модифицированного антитела) или любого антитела к человеческому CD25, сконструированного или охарактеризованного как обладающего признаками связывания CD25, сходными с таковыми у 7D4 для мышиного CD25, такого как 7G7B6 или М-А251, не оценивали детально в отношении оптимизированного истощения Трег-клеток внутри опухолей, по отдельности или в комбинации с другими антителами или другими противораковыми соединениями. Как обсуждалось выше, инфильтрация Трег-клеток в опухоли и, в частности, низкое отношение Тэфф-клеток к Трег-клеткам, может привести к неблагоприятному клиническому исходу. CD25 был выявлен как маркер Трег и, соответственно, может быть перспективной мишенью для терапевтических антител, предназначенных для истощения Трег. Важно отметить, что CD25 представляет собой альфа-субъединицу рецептора для IL-2, и IL-2 является ключевым цитокином для ответов Тэфф. Антитела к CD25, которые до настоящего времени проходили клинические исследования, при истощении 1рег~ клеток также блокируют передачу сигналов IL-2 за счет CD25. Авторы настоящего изобретения обнаружили, что такая блокада передачи сигналов IL-2 ограничивает ответы Тэфф и что антитело к CD25, которое не блокирует передачу сигналов IL-2, может эффективно истощать Трег-клетки, одновременно с этим позволяя IL-2 стимулировать Тэфф-клетки, обеспечивая антитела, которые проявляют сильный противораковый эффект. Таким образом, в данной области техники существует потребность в способе лечения рака, включающем истощение Трег, в частности, позволяющем при этом IL-2 стимулировать Тэфф-клетки, в частности, за счет применения соответствующих антител к CD25.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[08] Согласно настоящему изобретению предложены антитела к CD25 и варианты применения антител к CD25, которые характеризуются структурными элементами, которые обеспечивают как связывание CD25, без существенного блокирования связывания интерлейкина 2 (IL-2) с CD25 или передачи сигналов IL-2 за счет CD25, так и эффективное истощение Трег, в частности, внутри опухолей. Структурные и функциональные признаки 7D4-IgM (описанные в отношении мышиного CD25) были модифицированы для того чтобы обеспечить антитела, которые демонстрируют неожиданно улучшенные признаки в отношении применения для истощения Трег и эффективность против опухолей, по отдельности или в комбинации с другими противораковыми агентами. Также были охарактеризованы структурные и функциональные признаки дополнительных антител к CD25, которые не блокируют связывание интерлейкина 2 с CD25 (и не блокируют передачу сигналов IL-2 за счет CD25) и эффективно истощают Трег. Эти результаты можно применять для определения и создания дополнительных антител к человеческому CD25, которые обеспечивают сопоставимые эффекты против опухолей у субъектов-людей. В настоящем документе ссылки на «антитела к CD25» и тому подобное включают их антигенсвязывающие фрагменты, а также варианты (включая варианты с созревшей аффинностью), если из контекста не следует иное.
[09] Согласно основному аспекту настоящего изобретения предложен способ лечения субъекта-человека, имеющего рак, включающий этап введения субъекту антитела к CD25, причем указанный субъект имеет опухоль (предпочтительно плотную опухоль), причем указанное антитело не ингибирует связывание интерлейкина-2 (IL-2) с CD25.
[010] Указания на «не блокирует», «неблокирующее», «не блокирующее IL-2», «без блокирования» и эквивалентная терминология в настоящем документе (применительно к отсутствию блокирования связывания IL-2 с CD25 в присутствии антитела к CD25) включают варианты реализации, в которых антитело к CD25 не блокирует передачу сигналов IL-2 за счет CD25. Это означает, что антитело к CD25 согласно настоящему изобретению ингибирует менее 50% передачи сигналов IL-2 за счет CD25 по сравнению с передачей сигналов IL-2 в отсутствие антител. Предпочтительно антитело к CD25 ингибирует менее приблизительно 40%, 35%, 30%, предпочтительно менее приблизительно 25% передачи сигналов IL-2 по сравнению с передачей сигналов IL-2 в отсутствие антител.
[011] Согласно одному варианту реализации антитело к CD25 конкурирует с антителом 7G7B6 за связывание с человеческим CD25; и/или конкурирует с антителом МА251 за связывание с человеческим CD25.
[012] Согласно одному варианту реализации антитело к CD25 связывается с тем же эпитопом, который распознается антителом 7G7B6, и/или связывается с тем же эпитопом, который распознается антителом МА251.
[013] Согласно одному варианту реализации антитело к CD25 специфически связывается с эпитопом человеческого CD25, причем указанный эпитоп содержит один или более остатков аминокислот, содержащихся в одном или более аминокислотных участках, выбранных из аминокислот 150-163 из SEQ ID NO: 1 (YQCVQGYRALHRGP), аминокислот 166-186 из SEQ ID NO: 1 (SVCKMTHGKTRWTQPQLICTG), аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и аминокислот 70-88 из SEQ ID NO: 1 (NSSHSSWDNQCQCTSSATR). Предпочтительно эпитоп содержит по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять, по меньшей мере десять, по меньшей мере одиннадцать, по меньшей мере двенадцать, по меньшей мере тринадцать, по меньшей мере четырнадцать, по меньшей мере пятнадцать, по меньшей мере шестнадцать, по меньшей мере семнадцать, по меньшей мере восемнадцать или более остатков аминокислот, содержащихся в одном из нескольких аминокислотных участков, выбранных из аминокислот 150-163 из SEQ ID NO: 1 (YQCVQGYRALHRGP), аминокислот 166-186 из SEQ ID NO: 1 (SVCKMTHGKTRWTQPQLICTG), аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и/или аминокислот 70-88 из SEQ ID NO: 1 (NSSHSSWDNQCQCTSSATR).
[014] Согласно одному варианту реализации антитело к CD25 специфически связывается с эпитопом человеческого CD25, причем указанный эпитоп содержит по меньшей мере одну последовательность, выбранную из: аминокислот 150-158 из SEQ ID NO: 1 (YQCVQGYRA), аминокислот 176-180 из SEQ ID NO: 1 (RWTQP), аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и аминокислот 74-84 из SEQ ID NO: 1 (SSWDNQCQCTS).
[015] Согласно одному варианту реализации антитело к CD25 специфически связывается с эпитопом человеческого CD25, причем указанный эпитоп содержит по меньшей мере одну последовательность, выбранную из аминокислот, причем указанный эпитоп содержит по меньшей мере одну последовательность, выбранную из аминокислот 150-158 из SEQ ID NO: 1 (YQCVQGYRA), аминокислот 166-180 из SEQ ID NO: 1 (SVCKMTHGKTRWTQP), аминокислот 176-186 из SEQ ID NO: 1 (RWTQPQLICTG), аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и аминокислот 74-84 из SEQ ID NO: 1 (SSWDNQCQCTS).
[016] Согласно одному варианту реализации антитело к CD25 специфически связывается с эпитопом человеческого CD25, причем указанный эпитоп содержит по меньшей мере одну последовательность, выбранную из: аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR), аминокислот 70-84 из SEQ ID NO: 1 (NSSHSSWDNQCQCTS) и аминокислот 150-158 из SEQ ID NO: 1 (YQCVQGYRA).
[017] Согласно одному варианту реализации антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR). Согласно одному варианту реализации антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и аминокислот 150-160 из SEQ ID NO: 1 (YQCVQGYRALH). Согласно другому варианту реализации антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и аминокислот 74-88 из SEQ ID NO: 1 (SSWDNQCQCTSSATR). Согласно другому варианту реализации антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 150-163 из SEQ ID NO: 1 (YQCVQGYRALHRGP), аминокислот 166-180 из SEQ ID NO: 1 (SVCKMTHGKTRWTQP), аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и аминокислот 74-88 из SEQ ID NO: 1 (SSWDNQCQCTSSATR).
[018] Согласно одному варианту реализации антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 176-180 из SEQ ID NO: 1 (RWTQP). Согласно одному варианту реализации антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 166-180 из SEQ ID NO: 1 (SVCKMTHGKTRWTQP). Согласно одному варианту реализации антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 176-186 из SEQ ID NO: 1 (RWTQPQLICTG).
[019] Согласно одному варианту реализации антитело к CD25 специфически связывается с эпитопом, содержащим последовательность аминокислот 150-158 из SEQ ID NO: 1 (YQCVQGYRA) и аминокислот 176-180 из SEQ ID NO: 1 (RWTQP). Согласно одному варианту реализации антитело к CD25 специфически связывается с эпитопом, содержащим последовательность аминокислот 150-158 из SEQ ID NO: 1 (YQCVQGYRA) и аминокислот 176-186 из SEQ ID NO: 1 (RWTQPQLICTG). Согласно одному варианту реализации антитело к CD25 специфически связывается с эпитопом, содержащим последовательность аминокислот 150-163 из SEQ ID NO: 1 (YQCVQGYRALHRGP) и аминокислот 166-180 из SEQ ID NO: 1 (SVCKMTHGKTRWTQP).
[020] Согласно одному варианту реализации антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 74-84 из SEQ ID NO: 1 (SSWDNQCQCTSSATR). Согласно одному варианту реализации антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 70-84 из SEQ ID NO: 1 (NSSHSSWDNQCQCTS).
[021] Авторы настоящего изобретения неожиданно обнаружили, что антитела, которые связываются с конкретными эпитопами CD25, включая антитела, которые конкурируют с 7G7B6 и/или МА251 за связывание с CD25, можно применять в лечении рака, в частности, плотных опухолей. Такие антитела все еще позволяют передавать сигналы IL-2 за счет CD25, связанного антителом, и авторы настоящего изобретения впервые обнаружили, что помимо истощения Трег- клеток антитела, применяемые в настоящем изобретении, позволяют Тэфф-клеткам оптимально проявлять их противораковые эффекты, по меньшей мере частично, обеспечивая возможность связывания IL-2 и передачу сигналов за счет CD25, экспрессируемого на Тэфф-клетках.
[022] Такие антитела предпочтительно имеют константу диссоциации (Kd) для CD25 менее 10-7 М и/или константу диссоциации для по меньшей мере одного активирующего рецептора Fcγ менее приблизительно 10-6 М. Предпочтительно антитело имеет константу диссоциации (Kd) для CD25 в пределах 10-8 или 10-9, или 10-10, или 10-11, или 10-12, или 10-13, или ниже. Наиболее предпочтительно такое антитело представляет собой человеческое антитело IgG1, которое связывается по меньшей мере с одним активирующим рецептором Fcγ с высокой аффинностью и истощает инфильтрирующие опухоль регуляторные Т-клетки. Наиболее предпочтительно антитело к CD25 характеризуется другими признаками, связанными с рецепторами Fcγ, в частности:
(a) связывается с рецепторами Fcγ с соотношением активирующих и ингибирующему (А/I), превосходящим 1; и/или
(b) связывается с FcγRIIa с более высокой аффинностью, чем оно связывается с FcγRIIb.
[023] Учитывая применение антитела к CD25 в терапевтических способах, оно может проявлять дополнительные предпочтительные признаки. Антитело к CD25 предпочтительно представляет собой моноклональное антитело, в частности, человеческое, химерное или гуманизированное антитело. Антитело может представлять собой его вариант с созревшей аффинностью, необязательно гуманизированный вариант или вариант с созревшей аффинностью 7G7B6 или МА251. Более того, ввиду его взаимодействия с иммунными клетками и/или другими компонентами иммунной системы для проявления его активности, антитело к CD25 может дополнительно вызывать усиленный ответ КЗЦ, АЗКЦ и/или АЗКФ, предпочтительно повышенный ответ АЗКЦ и/или АЗКФ, более предпочтительно повышенный ответ АЗКЦ, по сравнению с существующими клиническими антителами к человеческому CD25, даклизумабом и базиликсимабом. Согласно некоторым вариантам реализации антитело к CD25 может вызывать сниженный ответ КЗЦ по сравнению с существующими клиническими антителами к человеческому CD25, даклизумабом и базиликсимабом, более предпочтительно антитело к CD25 не вызывает ответ КЗЦ.
[024] Антитело к CD25 согласно настоящему изобретению (как определено выше в общих чертах и более детально в подробном описании) можно применять в способах лечения субъекта-человека, причем указанное антитело к CD25 вводят субъекту. Согласно одному варианту реализации субъект имеет рак. Предпочтительно субъект имеет развившуюся плотную опухоль (предпочтительно в способе, дополнительно включающем этап выявления субъекта, имеющего плотную опухоль). Такие способы могут дополнительно включать введение дополнительного терапевтического агента указанному субъекту. Согласно одному варианту реализации дополнительный агент может представлять собой ингибитор контрольной точки иммунитета для указанного субъекта, например, в форме антитела, связывающего и ингибирующего белок контрольной точки иммунитета. Предпочтительный ингибитор контрольной точки иммунитета представляет собой антагонист PD-1, который может представлять собой антитело к PD-1 или антитело к PD-L1. В более общем случае антитело к CD25 можно применять в способах истощения регуляторных Т-клеток в плотной опухоли у субъекта, включающих этап введения указанного антитела к CD25 указанному субъекту.
[025] Согласно дополнительному аспекту антитело к CD25 согласно настоящему изобретению можно применять для изготовления лекарственного средства для лечения рака у субъекта-человека, причем предпочтительно указанный субъект имеет опухоль, предпочтительно плотную опухоль. Указанное антитело можно вводить в комбинации с дополнительным терапевтическим агентом, предпочтительно с дополнительным противораковым терапевтическим агентом, например, с ингибитором контрольной точки иммунитета, предпочтительно с антагонистом пути PD-1/PD-L1, противораковой вакциной и/или применять в комбинации с видами терапии, которые являются стандартом лечения, такими как химиотерапия или лучевая терапия.
[026] Согласно дополнительному аспекту настоящего изобретения предложена комбинация антитела к CD25, определенного выше, с другим противораковым соединением (предпочтительно ингибитором контрольной точки иммунитета или другими соединениями, как указано в подробном описании) для применения в лечении рака у субъекта-человека, причем предпочтительно указанный субъект имеет плотную опухоль, и противораковое соединение (например, ингибитор контрольной точки иммунитета, такой как антагонист PD-1, или цитокин, такой как интерлейкин 2), может быть введен одновременно, по отдельности или последовательно. В данном объеме настоящего изобретения также предложен набор для применения в лечении рака, содержащий антитело к CD25, определенное выше, и противораковое соединение (например, ингибитор контрольной точки иммунитета, такой как антагонист PD-1).
[027] Согласно дополнительному аспекту настоящего изобретения также предложена фармацевтическая композиция, содержащая антитело к CD25, определенное выше, в фармацевтически приемлемой среде. Такая композиция также может содержать противораковое соединение (например, ингибитор контрольной точки иммунитета, такой как антагонист PD-1).
[028] Согласно еще одному аспекту настоящего изобретения также предложено биспецифическое антитело, содержащее:
(a) первый антигенсвязывающий фрагмент, который связывается с CD25; и
(b) второй антигенсвязывающий фрагмент, который связывается с другим антигеном; причем указанное антитело к CD25 не ингибирует связывание интерлейкина-2 (IL-2) с CD25, и предпочтительно биспецифическое антитело представляет собой антитело IgG1, которое связывается с по меньшей мере одним активирующим рецептором Fcγ с высокой аффинностью и истощает инфильтрирующие опухоль регуляторные Т-клетки. Предпочтительно такой второй антигенсвязывающий фрагмент связывается с антигеном, выбранным из белка контрольной точки иммунитета или связанного с опухолью антигена, или может представлять собой или может быть основан на антителе к человеческому активирующему рецептору Fc (антитело к FcγRI, антитело к FcγRIIa, антитело к FcγRIII) или на антагонистическом антителе к человеческому FcγRIIb. Второй антигенсвязывающий фрагмент как таковой может связываться с FcRIIb. Согласно другому варианту он может связываться с FcγRI, FcγRIIa и/или FcγRIII с антагонистической активностью.
[029] Предпочтительно такое биспецифическое антитело содержит второй антигенсвязывающий фрагмент, который связывает белок контрольной точки иммунитета, который выбран из группы, состоящей из PD-1, CTLA-4, BTLA, KIR, LAG3, VISTA, TIGIT, TIM3, PD-L1, В7Н3, В7Н4, PD-L2, CD80, CD86, HVEM, LLT1, GAL9, GITR, ОХ40, CD137 и ICOS. Такой белок контрольной точки иммунитета предпочтительно экспрессируется на опухолевой клетке. Предпочтительно белок контрольной точки иммунитета выбран из PD-1, PD-L1 и CTLA-4. Второй антигенсвязывающий фрагмент, который связывается с белком контрольной точки иммунитета, может содержаться в коммерчески доступном антителе, которое действует как ингибитор контрольной точки иммунитета, например:
(a) в случае PD-1 антитело к PD-1 может представлять собой ниволумаб или пембролизумаб.
(b) в случае PD-L1 антитело к PD-L1 представляет собой атезолизумаб;
(c) в случае CTLA-4 антитело к CTLA-4 представляет собой ипилимумаб.
[030] Такое биспецифическое антитело может быть представлено в любом коммерчески доступном формате, включая DuoBody®, BiTE DART, CrossMab, «выступы-во-впадины», Triomab® или другой подходящий молекулярный формат биспецифического антитела и его фрагментов.
[031] Согласно другому варианту такое биспецифическое антитело содержит второй антигенсвязывающий фрагмент, который связывается с антигеном, связанным с опухолью. В этом альтернативном варианте такие антигены и соответствующие антитела включают, но не ограничиваются ими, CD22 (блинатумомаб), CD20 (ритуксимаб, тозитумомаб), CD56 (лорвотузумаб), CD66e/CEA (лабетузумаб), CD152/CTLA-4 (ипилимумаб), CD221/IGF1R (MK-0646), CD326/эпкам (эдреколомаб), CD340/HER2 (трастузумаб, пертузумаб) и EGFR (цетуксимаб, панитумумаб).
[032] Комбинацию антитела к CD25 согласно настоящему изобретению с другим противораковым соединением, а также биспецифическими антителами, определенными выше, можно применять в способе лечения рака, включающем этап введения указанной комбинации или указанного биспецифического антитела субъекту, в частности, если субъект имеет плотную опухоль, и можно применять в лечении рака у субъекта.
[033] Другие объекты настоящего изобретения, включая дополнительные определения антител к человеческому CD25 согласно настоящему изобретению и вариантов их применения в способах лечения рака, в фармацевтических композициях, в комбинации с другими противораковыми соединениями, в биспецифических антителах, представлены в подробном описании и в примерах.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[034] Согласно настоящему изобретению предложен способ лечения или предотвращения рака у субъекта, предпочтительно если субъект имеет плотную опухоль, включающий этап введения указанному субъекту антитела, которое связывается с CD25, причем указанные антитела к CD25 характеризуются структурными элементами, которые обеспечивают как связывание CD25, без нарушения связывания интерлейкина 2 или передачи сигналов за счет CD25, так и эффективное истощение Трег, в частности, внутри опухолей. Антитело, которое связывается с CD25, определенное в настоящем изобретении, можно применять в лечении или предотвращении рака, предпочтительно плотной опухоли. В другом варианте, согласно настоящему изобретению предложено применение антитела, которое связывается с CD25 и обеспечивает как связывание CD25, без нарушения связывания интерлейкина 2 с CD25, так и эффективное истощение Трег для изготовления лекарственного средства для лечения или предотвращения рака, предпочтительно плотной опухоли. Согласно настоящему изобретению также предложено применение антитела, которое связывает CD25 и которое обеспечивает как связывание CD25, без существенного нарушения связывания интерлейкина 2 с CD25, так и истощение Трег в лечении или предотвращении рака, предпочтительно плотной опухоли.
[035] Авторы настоящего изобретения обнаружили, что на CD25 можно нацелено воздействовать с применением антитела к CD25, которое не ингибирует (или по существу не ингибирует) связывание интерлейкина 2 с CD25 или передачу сигналов IL-2 за счет CD25, для истощения регуляторных Т-клеток в терапевтических условиях, например, в развившейся плотной опухоли. Авторы настоящего изобретения обнаружили, что антитела к CD25, не блокирующие IL-2, имеющие изотип, который усиливает их связывание с активирующими рецепторами Fc-гамма, приводят к эффективному истощению инфильтрирующих опухоль регуляторных Т-клеток, при этом обеспечивая оптимальный ответ Тэфф, терапевтический подход, который может быть, например, связан (в комбинации с биспецифическими антителами или в составе биспецифичных антител) с другими нацеленными на рак соединениями, такими как соединения, нацеленные на белок контрольной точки иммунитета, связанный с опухолью антиген или ингибирующий рецептор Fcγ. Эти результаты также позволяют комбинировать применение антитела к CD25 с интерлейкином-2 в соответствующих дозах для лечения рака.
[036] CD25 представляет собой альфа-цепь рецептора IL-2 и обнаруживается на активированных Т-клетках, регуляторных Т-клетках, активированных В-клетках, некоторых NK-T-клетках, некоторых тимоцитах, миелоидных предшественниках и олигодендроцитах. CD25 связывается с CD122 и CD132 с образованием гетеротримерного комплекса, который действует как высокоаффинный рецептор IL-2. Консенсусная последовательность человеческого CD25 представлена ниже в SEQ ID NO: 1 (номер доступа в Uniprot Р01589; внеклеточный домен зрелого человеческого CD25, соответствующий аминокислотам 22-240, подчеркнут и представлен как SEQ ID NO: 2):

[037] В настоящем документе термин «антитело, которое связывает CD25» относится к антителу, которое способно связываться с субъединицей CD25 рецептора IL-2. Эта субъединица также известна как альфа-субъединица рецептора IL-2. Такое антитело также называется в настоящем документе «антитело к CD25».
[038] Антитело к CD25 представляет собой антитело, способное специфически связываться с субъединицей CD25 (антигеном) рецептора IL-2. Подразумевается, что «специфическое связывание», «связываться специфически» и «специфически связываться» означает, что антитело имеет константу диссоциации (Kd) для антигена, представляющего интерес, менее приблизительно 10-6 М, 10-7 М, 10-8 М, 10-9 М, 10-10 М, 10-11 M, 10-12 M или 10-13 М. В предпочтительном варианте реализации константа диссоциации составляет менее 10-8 М, например, находится в пределах 10-9 М, 10-10 М, 10-11 М, 10-12 М или 10-13 М.
[039] В настоящем документе термин «антитело» относится как к интактным молекулам иммуноглобулинов, так и к их фрагментам, которые содержат антигенсвязывающий сайт, и включает поликлональные, моноклональные, генетически модифицированные и иным образом модифицированные формы антител, включая, но не ограничиваясь ими, химерные антитела, гуманизированные антитела, гетероконъюгатные и/или мультиспецифические антитела (например, биспецифические антитела, диатела, тритела и тетратела) и антигенсвязывающие фрагменты антител, включая, например, Fab', F(ab')2, Fab, Fv, rIgG, гибриды полипептид-Fc, одноцепочечные варианты (фрагменты scFv, VHH, транс-тела®, аффитела®, однодоменные антитела акул, одноцепочечные или тандемные диатела (TandAb®), VHH, антикалины®, нанотела®, минитела, BiTE®, бициклические пептиды и другие альтернативные каркасы иммуноглобулиновых белков). Согласно некоторым вариантам реализации антитело может не содержать ковалентную модификацию (например, присоединение гликана), которую оно имело бы при выработке в природных условиях. Согласно некоторым вариантам реализации антитело может содержать ковалентную модификацию (например, присоединение гликана, детектируемый фрагмент, терапевтический фрагмент, каталитический фрагмент или другую химическую группу, обеспечивающую улучшенную стабильность или введение антитела, такую как полиэтиленгликоль). Согласно некоторым вариантам реализации антитело может быть в форме замаскированного антитела (например, протела®). Замаскированное антитело может содержать блокирующий или «маскирующий» пептид, который специфически связывается с антигенсвязывающей поверхностью антитела и препятствует связыванию антигена этого антитела. Маскирующий пептид связан с антителом с помощью расщепляемого линкера (например, под действием протеазы). Селективное расщепление линкера в желаемой среде, т.е. в опухолевой среде, позволяет маскирующему/блокирующему пептиду диссоциировать, обеспечивая связывание антигена в опухоли, что ограничивает потенциальные проблемы токсичности. «Антитело» также может относиться к антителам верблюдовых (антитела, содержащие только тяжелую цепь) и антитело подобным молекулам, таким как антикалины (Skerra (2008) FEBS J 275, 2677-83). Согласно некоторым вариантам реализации антитело является поликлональным или олигоклональным, которое получают в виде панели антител, каждое из которых связано с одной последовательностью антитела и связывает более или менее различающиеся эпитопы в пределах антигена (такие как различные эпитопы в пределах внеклеточного домена человеческого CD25, которые связаны с различными эталонными антителами к человеческому CD25). Поликлональные или олигоклональные антитела могут быть обеспечены в одном препарате для разных видов медицинского применения, как описано в литературе (Kearns JD et al., 2015. Mol Cancer Ther. 14:1625-36). [040] Согласно одному аспекту настоящего изобретения антитело является моноклональным. Дополнительно или альтернативно антитело может быть гуманизированным или человеческим. Согласно дополнительному аспекту антитело является человеческим или, в любом случае, представляет собой антитело, которое имеет формат и признаки, обеспечивающие его применение и введение субъектам-людям. Согласно одному аспекту настоящего изобретения антитела могут представлять собой гуманизированные варианты 7G7B6 или МА251 с созревшей аффинностью. Антитело с созревшей аффинностью имеет по меньшей мере на 10% более высокую аффинность в отношении CD25, и/или имеет последовательности CDR, которые по меньшей мере на 80%, предпочтительно на 90% идентичны CDR исходной последовательности (для всех последовательностей). Антитела с созревшей аффинностью представляют собой антитело с одной или более измененными аминокислотами в одном или более CDR, что приводит к получению антитела с улучшенной аффинностью в отношении CD25 по сравнению с исходным видом, не имеющим измененных аминокислот.
[041] Антитела (Ab) и иммуноглобулины (Ig) представляют собой гликопротеины, имеющие одинаковые структурные характеристики. Иммуноглобулины могут относиться к любому классу, такому как IgA, IgD, IgG, IgE или IgM. Иммуноглобулины могут быть любого подкласса, такого как IgG1, IgG2, IgG3 или IgG4. Согласно предпочтительному аспекту настоящего изобретения антитело к CD25 относится к классу IgG, предпочтительно к подклассу IgG1. Согласно одному аспекту антитело к CD25 относится к подклассу человеческого IgG1. Согласно другому варианту в одном аспекте антитело к CD25 относится к подклассу человеческого IgG2.
[042] Область Fc антител IgG взаимодействует с несколькими клеточными рецепторами Fcγ (FcγR), чтобы стимулировать и регулировать последующие эффекторные механизмы. Существует пять активирующих рецепторов, а именно FcγRI (CD64), FcγRIIa (CD32a), FcγRIIc (CD32c), FcγRIIIa (CD16a) и FcγRIIIb (CD16b), и один ингибирующий рецептор FcγRIIb (CD32b). Взаимодействие антител IgG с иммунной системой контролируется и опосредуется FcγR, которые передают иммунной системе информацию, воспринимаемую и собираемую антителами, обеспечивая связь между врожденной и адаптивной иммунными системами, и, в частности, в случае биотерапевтических средств (Hayes J et al., 2016. J Mlamm Res 9: 209 219).
[043] Подклассы IgG различаются по своей способности связываться с FcγR, и такое различное связывание определяет их способность вызывать ряд функциональных ответов. Например, у человека FcγRIIIa является основным рецептором, вовлеченным в активацию антителозависимой клеточной цитотоксичности (АЗКЦ), и IgG3, за которым следует IgG1, проявляет наибольшие значения аффинности в отношении этого рецептора, что отражает их способность эффективно индуцировать АЗКЦ. Несмотря на то, что было показано, что IgG2 слабо связывается с этим рецептором, антитело к CD25, относящееся к человеческому изотипу IgG2, как было обнаружено, также эффективно истощает Трег.
[044] Согласно предпочтительному варианту реализации настоящего изобретения антитело связывает FcγR с высокой аффинностью, предпочтительно связывает активирующий рецептор с высокой аффинностью. Предпочтительно антитело связывает FcγRI и/или FcγRIIa, и/или FcγRIIIa с высокой аффинностью. Согласно конкретному варианту реализации антитело связывается по меньшей мере с одним активирующим рецептором Fcγ с константой диссоциации менее приблизительно 10-6 М, 10-7 М, 10-8 М, 10-9 М или 10-10 М.
[045] Согласно одному аспекту антитело представляет собой антитело IgG1, предпочтительно человеческое антитело IgG1, которое способно связываться с по меньшей мере одним активирующим рецептором Fc. Например, антитело может связываться с одним или более рецепторами, выбранными из FcγRI, FcγRIIa, FcγRIIc, FcγRIIIa и FcγRIIIb. Согласно одному аспекту антитело способно связываться с FcγRIIIa. Согласно одному аспекту антитело способно связываться с FcγRIIIa и FcγRIIa и необязательно с FcγRI. Согласно одному аспекту антитело способно связываться с этими рецепторами с высокой аффинностью, например, с константой диссоциации менее приблизительно 10-7 М, 10-8 М, 10-9 М или 10-10 М.
[046] Согласно одному аспекту антитело связывает ингибирующий рецептор FcγRIIb с низкой аффинностью. Согласно одному аспекту антитело связывает FcγRIIb с константой диссоциации выше приблизительно 10-7 М, выше приблизительно 10-6 М или выше приблизительно 10-5 М.
[047] Согласно предпочтительному варианту реализации настоящего изобретения антитело к CD25 относится к подклассу IgG1 и предпочтительно обладает активностью АЗКЦ и/или АЗКФ, как описано в настоящем документе, в частности, в отношении клеток человеческого происхождения. Как описано ранее (Nimmerjahn F et al., 2005. Science, 310:1510-2), изотип mIgG2a (который соответствует человеческому изотипу IgG1) связывается со всеми подтипами FcγR с высоким отношением активирующих к ингибирующим (А/I), которое по меньшей мере превосходит 1. Напротив, другие изотипы (такие как изотип rIgG1) связываются с сопоставимой аффинностью только с одним активирующим FcγR (FcγRIII), а также ингибирующим FcγRIIb, что приводит к низкому отношению А/I (<1). Такое более низкое отношение А/I может коррелировать с более низким внутриопухолевым истощением Трег и более низкой противоопухолевой терапевтической активностью изотипа. Несмотря на известный профиль связывания FcγR для антител человеческого изотипа IgG2, значительное истощение Трег также может быть достигнуто при применении человеческого изотипа IgG2 антитела к CD25. Следовательно, согласно одному варианту реализации антитело к CD25 относится к подклассу IgG2.
[048] Согласно предпочтительному варианту реализации антитело к CD25, описанное в настоящем документе, связывает человеческий CD25, предпочтительно с высокой аффинностью. Еще более предпочтительно антитело к CD25 связывается с внеклеточной областью человеческого CD25, как показано выше. Согласно одному аспекту настоящего изобретения предложено антитело к CD25, описанное в настоящем документе. В частности, в Примерах представлены экспериментальные данные, полученные при применении антитела, которое секретируется гибридомой 7D4. Как указано в уровне техники, это антитело является специфичным для мышиного CD25 и, согласно результатам сравнения панели моноклональных антител (включая РС61), связывается с одним из трех эпитопов в мышином CD25, который отличается от сайта связывания IL-2, и не блокирует связывание IL-2 с CD25. Например, было показано, что 7D4 связывает мышиный CD25 в эпитопе, содержащем аминокислоты 184-194 (REHHRFLASEE) в [последовательности Uniprot Р01590]). Анализы с применением 7D4 и мышиного CD25, описанные в литературе (например, Setiady Y et al., 2010. Eur. J. Immunol. 40: 780-6; McNeill A et al., 2007. Scand J Immunol. 65:63-9; Teege S et al., 2015, Sci Rep 5: 8959), вместе с теми, которые раскрыты в Примерах, включая рекомбинантные антитела, содержащие СВ25-связывающий домен 7D4 или антител к человеческому CD25, не блокирующих IL-2, называемых МА-251 и 7G7B6, могут быть адаптированы для характеристики тех человеческих антител, которые распознают человеческий CD25 и функциональные особенности которых аналогичны таковым 7D4, на уровне как взаимодействия с CD25 (в частности, отсутствие блокирования связывания IL-2), так и с рецепторами Fcγ (в частности, предпочтительное связывание одного или более человеческих активирующих рецепторов Fcγ и эффективное истощение Трег), при связи с соответствующим изотипом, как описано в примерах.
[049] Согласно одному аспекту настоящего изобретения антитела конкурируют с антителом 7G7B6 за связывание с человеческим CD25; и/или связываются с тем же эпитопом или эпитопами, которые распознаются антителом 7G7B6. 7G7B6 представляет собой моноклональное антитело, имеющее мышиный изотип IgG2a, который распознает человеческий CD25. 7G7B6 содержит вариабельную область тяжелой цепи, имеющую последовательность:

и вариабельную область легкой цепи, имеющую последовательность:

[050] Согласно одному варианту реализации антитело содержит тяжелую цепь, содержащую аминокислотную последовательность GFTLDSYGVS (SEQ ID NO: 7) в качестве CDR1 вариабельной тяжелой цепи; аминокислотную последовательность GVTSSGGSAYYADSV (SEQ ID NO: 8) в качестве CDR2 вариабельной тяжелой цепи, аминокислотную последовательность DRYVYTGGYLYHYGMDL (SEQ ID NO: 9) в качестве CDR3 вариабельной тяжелой цепи, и содержит легкую цепь, содержащую аминокислотную последовательность RASQSISDYLA (SEQ ID NO: 11) в качестве CDR1 вариабельной легкой цепи; аминокислотную последовательность YAASTLPF (SEQ ID NO: 12) в качестве CDR2 вариабельной легкой цепи, аминокислотную последовательность QGTYDSSDWYWA (SEQ ID NO: 13) в качестве CDR3 вариабельной легкой цепи. Антитело может конкурировать с 7G7B6 за связывание с человеческим CD25. Предпочтительно антитело содержит тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую последовательность:
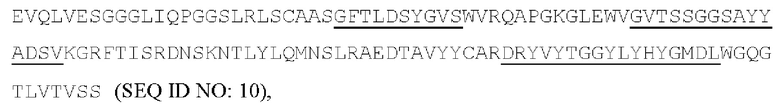
и легкую цепь, содержащую вариабельную область легкой цепи, содержащую последовательность:
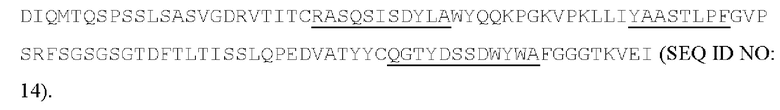
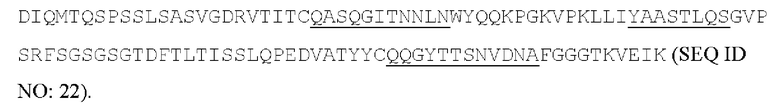
[051] Согласно другому варианту реализации антитело содержит тяжелую цепь, содержащую аминокислотную последовательность SGFSVDIYDMS (SEQ ID NO: 15) в качестве CDR1 вариабельной тяжелой цепи; аминокислотную последовательность YISSSLGATYYADSV (SEQ ID NO: 16) в качестве CDR2 вариабельной тяжелой цепи, аминокислотную последовательность ERIYSVYTLDYYAMDL (SEQ ID NO: 17) в качестве CDR3 вариабельной тяжелой цепи, и содержат легкую цепь, содержащую аминокислотную последовательность QASQGITNNLN (SEQ ID NO: 19) в качестве CDR1 вариабельной легкой цепи; аминокислотную последовательность YAASTLQS (SEQ ID NO: 20) в качестве CDR2 вариабельной легкой цепи, аминокислотную последовательность QQGYTTSNVDNA (SEQ ID NO: 21) в качестве CDR3 вариабельной легкой цепи. Антитело может конкурировать с 7G7B6 за связывание с человеческим CD25. Предпочтительно антитело содержит тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую последовательность:

и легкую цепь, содержащую вариабельную область легкой цепи, содержащую последовательность:
[052] Согласно одному варианту реализации антитело, которое может конкурировать с 7G7B6 за связывание с человеческим CD25, содержит тяжелую цепь, содержащую аминокислотную последовательность:

и содержит легкую цепь, содержащую аминокислотную последовательность:

[053] Согласно одному аспекту настоящего изобретения антитела конкурируют с антителом МА251 за связывание с человеческим CD25; и/или связываются с тем же эпитопом или эпитопами, которые распознаются антителом МА251. МА251 представляет собой моноклональное антитело, имеющее мышиный изотип, которое распознает человеческий CD25. МА251 содержит вариабельную область тяжелой цепи, имеющую последовательность:

и вариабельную область легкой цепи, имеющую последовательность:

[054] Согласно одному варианту реализации антитело, которое может конкурировать с МА251 за связывание с человеческим CD25, содержит тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность:
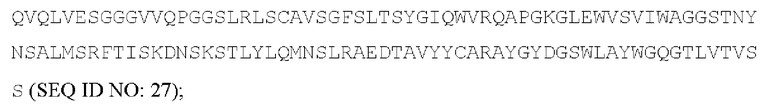
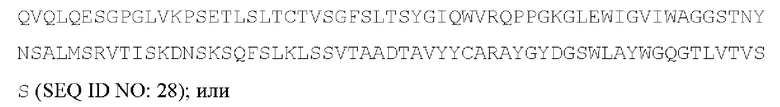
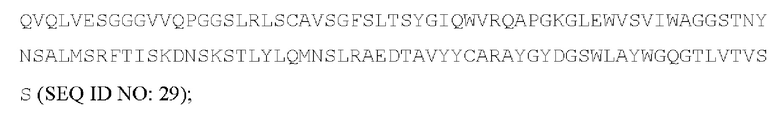
и содержит легкую цепь, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность:

[055] Согласно одному варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 23, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 25. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 23, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 26. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 24, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 25. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 24, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 26. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 27, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 30. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 27, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 31. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 27, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 32. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 27, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 33. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 28, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 30. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 28, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 31. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 28, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 32. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 28, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 33. Согласно другому варианту реализации антитело содержит вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 29, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 30. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 29, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 31. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 29, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 32. Согласно другому варианту реализации антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 29, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 33.
[056] Согласно одному аспекту антитела конкурируют как с антителом 7G7B6, так и с антителом МА251 за связывание с человеческим CD25. Согласно одному аспекту антитела связываются с одним и тем же эпитопом или эпитопами, распознаваемыми 7G7B6 и распознаваемыми МА251.
[057] Конкуренция между антителом 7G7B6 или антителом МА251 и другим антителом может быть измерена, например, как обсуждается в Примерах и известно в данной области техники. Согласно некоторым вариантам реализации конкуренцию между двумя антителами, такими как 7G7B6 или МА251, и другим антителом определяют путем добавления другого антитела к анализу и измерения взаимодействия между антителом 7G7B6 или МА251 и человеческим CD25. Один такой анализ представляет собой анализ на основе Octet, в котором определяют одновременное связывание антитела 7G7B6 или МА251, другого антитела и рекомбинантного человеческого CD25. Если детектируют связывание двух антител с рекомбинантным человеческим CD25, то антитела не являются конкурирующими. Согласно другому варианту один такой анализ представляет собой твердофазный иммуноферментный анализ (ИФА), в котором детектируют связывание антител 7G7B6 или МА251 с рекомбинантным человеческим CD25. Если наблюдаемый сигнал уменьшается при добавлении другого антитела (например, уменьшается по меньшей мере на 75%), то последнее антитело является конкурентом антител 7G7B6 или МА251. Одновременное связывание антител 7G7B6 или МА251 и другого антитела с клетками, экспрессирующими человеческий CD25, также можно детектировать с помощью проточной питометрии.
[058] Согласно одному аспекту настоящего изобретения предложено антитело к CD25, которое специфически связывается с эпитопом человеческого CD25, причем указанный эпитоп содержит один или более остатков аминокислот из одного или более аминокислотных участков, выбранных из аминокислот 150-163 из SEQ ID NO: 1 (YQCVQGYRALHRGP), аминокислот 166-186 из SEQ ID NO: 1 (SVCKMTHGKTRWTQPQLICTG), аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и аминокислот 70-88 из SEQ ID NO: 1 (NSSHSSWDNQCQCTSSATR). Предпочтительно эпитоп содержит по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять или более остатков из выбранных аминокислотных участков. Более предпочтительно эпитоп содержит последовательность, выбранную из аминокислот 150-158 из SEQ ID NO: 1 (YQCVQGYRA), аминокислот 176-180 из SEQ ID NO: 1 (RWTQP), аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и аминокислот 74-84 из SEQ ID NO: 1 (SSWDNQCQCTS) и их комбинаций. Эти эпитопы отличаются от сайта связывания IL-2 в человеческом CD25, и антитела, связывающиеся с таким эпитопом, как описано в Примерах, не блокируют связывание IL-2 с CD25.
[059] Согласно предпочтительному варианту реализации способ лечения субъекта-человека, имеющего рак, включает этап введения субъекту антитела к CD25 согласно настоящему изобретению, причем указанный субъект предпочтительно имеет плотную опухоль, и при этом указанное антитело к CD25 предпочтительно представляет собой человеческое антитело IgG1, которое не ингибирует связывание интерлейкина 2 с CD25 и которое связывается с по меньшей мере одним активирующим рецептором Fcγ, выбранным из FcγRI (CD64), FcγRIIc (CD32c) и FcγRIIIa (CD16a), с высокой аффинностью, и истощает инфильтрирующие опухоль регуляторные Т-клетки. Предпочтительно антитело к CD25 имеет константу диссоциации (Kd) для CD25 менее 10-7 М, предпочтительно менее 10-8 М. Более предпочтительно антитело к CD25 связывает человеческий CD25, оказывая влияние на связывание IL-2 и истощение Трег, которое сходно с таковым 7D4 на мышиный CD25 или 7G7B6 и МА251 на человеческий CD25. Согласно другому варианту реализации антитело к CD25 связывается с рецепторами Fcγ с отношением активирующих к ингибирующим (А/I), превосходящим 1, и/или связывается с FcγRI (CD64), FcγRIIc (CD32c), FcγRIIIa (CD16a) и/или FcγRIIa (CD32a) с более высокой аффинностью, чем оно связывается с FcγRIIb (CD32b).
[060] CD25-связывающий домен антитела 7D4 был клонирован и экспрессирован в виде рекомбинантного белка, гибридизованного с соответствующей константной областью. Последовательность CD25-связывающего домена антитела 7D4, а также его специфичность в отношении различных эпитопов во внеклеточном домене CD25 и/или другие виды его функциональной активности можно применять для сравнения антител-кандидатов к CD25, которые получают и подвергают скринингу с помощью любой подходящей методики (например, путем выращивания панелей гибридом из CD25-иммунизированных грызунов или создания библиотек рекомбинантных антител и последующего скрининга этих репертуаров антител с фрагментами CD25 для получения функциональной характеристики, как описано в настоящем документе). Антитела к CD25, которые впоследствии идентифицируют, также могут быть получены в виде рекомбинантных антител, в частности, в виде полноразмерных антител или в виде фрагментов или вариантов, которые описаны в настоящем документе.
[061] Нативные антитела и иммуноглобулины обычно представляют собой гетеротетрамерные гликопротеины с массой приблизительно 150000 дальтон, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей. Каждая тяжелая цепь имеет на аминоконце вариабельный домен (VH), за которым расположен ряд константных доменов. Каждая легкая цепь имеет вариабельный домен на аминоконце (VL) и константный домен на карбоксиконце.
[062] Вариабельные области способны взаимодействовать со структурно комплементарной антигенной мишенью и характеризуются отличиями в аминокислотной последовательности от антител другой антигенной специфичности. Вариабельные области как Н, так и L цепей содержат аминокислотные последовательности, способные специфически связываться с антигенными мишенями. В пределах этих последовательностей имеются меньшие последовательности, называемые «гипервариабельными» из-за их сильной изменчивости между антителами различной специфичности. Такие гипервариабельные участки также называют «области, определяющие комплементарность» или «CDR».
[063] Эти участки CDR обеспечивают основную специфичность антитела в отношении конкретной антигенной детерминанта ой структуры. CDR представляют собой несмежные участки аминокислот в пределах вариабельных областей, но было обнаружено, что, независимо от вида, положения этих важнейших аминокислотных последовательностей в вариабельных областях тяжелой и легкой цепей являются сходными в пределах аминокислотных последовательностей вариабельных цепей. Каждая из вариабельной тяжелой и легкой цепей всех антител имеет 3 участка CDR, каждый из которых не является смежным с другими (называемые L1, L2, L3, H1, Н2, Н3), для соответствующей легкой (L) и тяжелой (Н) цепей. Общепринятые участки CDR были описаны ранее (Kabat et al., 1977. J Biol Chem 252, 6609-6616).
[064] Антитела согласно настоящему изобретению могут функционировать за счет комплементзависимой цитотоксичности (КЗЦ) и/или антителозависимой клеточной цитотоксичности (АЗКЦ) и/или антителозависимого клеточного фагоцитоза (АЗКФ), а также любого другого механизма, который позволяет оказывать нацеленное действие, блокировать пролиферацию и/или истощать Tpeг-клетки.
[065] «Комплементзависимая цитотоксичность» (КЗЦ) относится к лизису экспрессирующих антиген клеток антителом согласно настоящему изобретению в присутствии комплемента.
[066] «Антителозависимая клеточная цитотоксичность» (АЗКЦ) относится к опосредуемой клетками реакции, в которой неспецифические цитотоксические клетки, которые экспрессируют рецепторы Fc (FcR) (например, природные клетки-киллеры (NK), нейтрофилы и макрофаги), распознают связанное антитело на клетке-мишени и тем самым приводят к лизису клетки-мишени.
[067] «Антителозависимый клеточный фагоцитоз» (АЗКФ) относится к опосредуемой клетками реакции, в которой фагоциты (такие как макрофаги), которые экспрессируют рецепторы Fc (FcR), распознают связанное антитело на клетке-мишени и тем самым приводят к фагоцитозу клетки-мишени.
[068] КЗЦ, АЗКЦ и АЗКФ могут быть измерены с использованием анализов, которые известны и доступны в данной области техники (Clynes et al. (1998) Proc Natl Acad Sci USA 95, 652-6) и обсуждаются в примерах. Константная область антитела важна для способности антитела связывать комплемент и опосредовать зависимую от клеток цитотоксичность и фагоцитоз. Таким образом, как обсуждается в настоящем документе, изотип антитела может быть выбран на основе того, желательно ли для антитела опосредовать цитотоксичность/фагоцитоз.
[069] Как обсуждается в настоящем документе, в одном варианте реализации настоящего изобретения применяют антитело к CD25, которое не ингибирует связывание интерлейкина 2 и которое приводит к истощению Трег-клеток. Например, можно применять антитело к CD25, которое не ингибирует связывание интерлейкина 2 с CD25 и вызывает сильный ответ КЗЦ и/или сильный ответ АЗКЦ, и/или сильный ответ АЗКФ. Способы увеличения КЗЦ, АЗКЦ и/или АЗКФ известны в данной области техники. Например, ответ КЗЦ может быть увеличен с помощью мутаций в антителе, которые увеличивают аффинность связывания C1q (ldusogie et at. (2001) J Immunol 166, 2571-5).
[070] Ссылки в настоящем документе на «не ингибирует связывание интерлейкина-2 с CD25» могут быть выражены в другом случае как антитело к CD25 представляет собой антитело, не блокирующее IL-2, или «неблокирующее» антитело (применительно к отсутствию блокирования связывания IL-2 с CD25 в присутствии антитела к CD25), т.е. антитело не блокирует связывание интерлейкина-2 с CD25 и, в частности, не ингибирует передачу сигналов с участием интерлейкина-2 в клетках, экспрессирующих CD25. Ссылки на «неблокирующее», «не блокирующее IL-2», «не блокирует» или «без блокирования» и тому подобное (применительно к отсутствию блокирования связывания IL-2 с CD25 в присутствии антитела к CD25) включают варианты реализации, в которых антитело к CD25 согласно настоящему изобретению не блокирует передачу сигналов IL-2 за счет CD25. т.е. антитело к CD25 ингибирует менее 50% передачи сигналов IL-2 по сравнению с передачей сигналов IL-2 в отсутствие антител. Согласно конкретным вариантам реализации настоящего изобретения, как описано в настоящем документе, антитело к CD25 ингибирует менее приблизительно 40%, 35%, 30%, предпочтительно менее приблизительно 25% передачи сигналов IL-2 по сравнению с передачей сигналов IL-2 в отсутствие антител. Антитела к CD25, не блокирующие IL-2, обеспечивают связывание с CD25, не нарушая связывание IL-2 с CD25 или по существу не нарушая связывание IL-2 с CD25. Ссылки в настоящем документе на антитело, не блокирующее IL-2, могут быть выражены в другом случае как антитело к CD25, которое «не ингибирует связывание интерлейкина-2 с CD25», или как антитело к CD25, которое «не ингибирует передачу сигналов IL-2».
[071] Некоторые антитела к CD25 могут допускать связывание IL-2 с CD25, но при этом блокируют передачу сигналов через рецептор CD25. Такие антитела к CD25 не включены в объем настоящего изобретения. Вместо этого антитела к CD25, не блокирующие IL-2, допускают связывание IL-2 с CD25, чтобы стимулировать по меньшей мере 50% от уровня передачи сигналов через рецептор CD25 по сравнению с передачей сигналов в отсутствие антитела к CD25.
[072] Передача сигналов IL-2 за счет CD25 может быть измерена способами, обсуждаемыми в Примерах и известными в данной области техники. Сравнение передачи сигналов IL-2 в присутствии и в отсутствие антитела-агента к CD25 может происходить в одинаковых или по существу одинаковых условиях.
[073] Согласно некоторым вариантам реализации передача сигналов IL-2 может быть определена путем измерения уровней фосфорилированного белка STAT5 в клетках с использованием стандартного анализа фосфорилирования Stat-5. Например, анализ фосфорилирования Stat-5 для измерения передачи сигналов IL-2 может включать культивирование мононуклеарных клеток периферической крови (МКПК) в присутствии антитела к CD25 в концентрации 10 мкг/мл в течение 30 минут и затем добавление различных концентраций IL-2 (например, 10 Ед/мл или различных концентраций 0,25 Ед/мл, 0,74 Ед/мл, 2,22 Ед/мл, 6,66 Ед/мл или 20 Ед/мл) в течение 10 минут. Затем клетки могут быть пермеабилизированы, и уровни белка STAT5 могут быть затем измерены с помощью содержащего флуоресцентную метку антитела к фосфорилированному пептиду STAT5, проанализированного методом проточной цитометрии. Процент блокирования передачи сигналов IL-2 можно рассчитать следующим образом: % блокирования = 100×[(% Stat5+ клеток в группе без обработки антителом - % Stat5+ клеток в группе 10 мкг/мл антитела)/(% Stat5+ клеток в группе без обработки антителом).
[074] АЗКЦ можно повысить с помощью способов, которые позволяют удалить фукозный фрагмент из гликана антитела, например, путем выработки антитела в клеточной линии YB2/0 или путем введения специфических мутаций в часть Fc человеческого IgG1 (например, S298A/E333A/K334A, S239D/I332E/A330L, G236A/S239D/A330L/I332E) (Lazar et al. (2006) Proc Natl Acad Sci USA 103, 2005-2010, Smith et al. (2012) Proc Natl 25 Acad Sci USA 109, 6181-6). АЗКФ также можно повысить путем введения специфических мутаций в часть Fc человеческого IgG1 (Richards et al. (2008) Mol Cancer Ther 7, 2517-27).
[075] Согласно предпочтительному варианту реализации настоящего изобретения антитело оптимизировано, чтобы вызывать ответ АЗКЦ, т.е. ответ АЗКЦ усилен, повышен или улучшен по сравнению с другими антителами к CD25, включая те, которые не ингибируют связывание интерлейкина 2 с CD25, и, например, немодифицированными моноклональными антителами к CD25.
[076] Согласно предпочтительному варианту реализации настоящего изобретения антитело оптимизировано, чтобы вызывать ответ АЗКФ, т.е. ответ АЗКФ усилен, повышен или улучшен по сравнению с другими антителами к CD25, включая те, которые не ингибируют связывание интерлейкина 2 с CD25, и, например, немодифицированными моноклональными антителами к CD25.
[077] В настоящем документе термин «химерное антитело» может относиться к антителу, имеющему вариабельные последовательности, происходящие из иммуноглобулина из одного вида, такого как антитело крысы или мыши, и константные области иммуноглобулина из другого вида, такого как человеческое антитело. Согласно некоторым вариантам реализации химерное антитело может иметь константную область, которая улучшена для индукции АЗКЦ.
[078] Антитела согласно настоящему изобретению также могут быть частично или полностью синтетическими, причем по меньшей мере часть полипептидных цепей антител синтезирована и необязательно оптимизирована для связывания с их когнатным антигеном. Такие антитела могут представлять собой химерные или гуманизированные антитела и могут быть полностью тетрамерными по структуре или могут быть димерными и содержать только одну тяжелую цепь и одну легкую цепь.
[079] Антитела согласно настоящему изобретению также могут представлять собой моноклональные антитела. В настоящем документе термин «моноклональное антитело» не ограничивается антителами, полученными с помощью технологии гибридом. Термин «моноклональное антитело» относится к антителу, происходящему из одного клона, включая любой эукариотический, прокариотический или фаговый клон, а не к способу, с помощью которого оно получено.
[080] Антитела согласно настоящему изобретению также могут представлять собой человеческие антитела. В настоящем документе термин «человеческое антитело» относится к антителам, имеющим вариабельные области, в которых как каркасные участки, так и участки CDR происходят из последовательностей иммуноглобулинов человеческой зародышевой линии. Кроме того, если антитело содержит константную область, константная область также происходит из последовательностей иммуноглобулинов человеческой зародышевой линии. Человеческие антитела согласно настоящему изобретению могут содержать остатки аминокислот, не кодируемые последовательностями иммуноглобулинов человеческой зародышевой линии (например, мутации, введенные с помощью случайного или сайт-специфического мутагенеза в условиях in vitro или с помощью соматической мутации в условиях in vivo).
[081] Антитело к CD25, обладающее признаками, описанными в настоящем документе, представляет собой еще один объект настоящего изобретения. Антитело к CD25 можно применять в медицине. Согласно дополнительному варианту реализации настоящего изобретения предложен способ лечения заболевания у субъекта, включающий введение антитела к CD25, которое не ингибирует связывание интерлейкина-2 (IL-2) с CD25 или передачу сигналов IL-2 за счет CD25. Предпочтительно заболевание представляет собой рак, в частности, для лечения плотных опухолей.
[082] Согласно другому варианту реализации настоящего изобретения предложены молекулы нуклеиновых кислот, кодирующие антитела к CD25, определенные в настоящем документе. Согласно некоторым вариантам реализации такие предложенные молекулы нуклеиновых кислот могут содержать оптимизированные по кодонам последовательности нуклеиновых кислот и/или могут быть включены в кассеты для экспрессии в соответствующих векторах нуклеиновых кислот для экспрессии в клетках-хозяевах, таких как, например, клетки бактерий, дрожжей, насекомых, рыб, мышей, обезьян или человека. Согласно некоторым вариантам реализации настоящего изобретения предложены клетки-хозяева, содержащие гетерологичные молекулы нуклеиновых кислот (например, ДНК-векторы), которые экспрессируют желательное антитело.
[083] Согласно некоторым вариантам реализации настоящего изобретения предложены способы получения выделенного антитела к CD25, определенного выше. Согласно некоторым вариантам реализации такие способы могут включать культивирование клетки-хозяина, которая содержит нуклеиновые кислоты (например, гетерологичные нуклеиновые кислоты, которые могут содержать векторы и/или могут быть доставлены в клетку-хозяина с помощью векторов). Предпочтительно клетка-хозяин (и/или гетерологичные последовательности нуклеиновых кислот) приспособлена и сконструирована так, что антитело или его антигенсвязывающий фрагмент или его вариант секретируются из клетки-хозяина и выделяются из супернатантов клеточной культуры.
[084] Антитела согласно настоящему изобретению могут быть моноспецифическими, биспецифическими или мультиспецифическими. «Мультиспецифические антитела» могут быть специфическими в отношении разных эпитопов одного целевого антигена или полипептида или могут содержать антигенсвязывающие домены, специфические в отношении более чем одного целевого антигена или полипептида (Kufer et at. (2004) Trends Biotechnol 22, 238-44).
[085] Согласно одному аспекту настоящего изобретения антитело представляет собой моноспецифическое антитело. Как подробнее обсуждается ниже, согласно альтернативному аспекту антитело представляет собой биспецифическое антитело.
[086] В настоящем документе термин «биспецифическое антитело» относится к антителу, способному связываться с двумя различными эпитопами как на одном антигене или полипептиде, так и на двух разных антигенах или полипептидах.
[087] Биспецифические антитела согласно настоящему изобретению, обсуждаемые в настоящем документе, могут быть получены с помощью биологических способов, таких как соматическая гибридизация; или генетических способов, таких как экспрессия ненативной последовательности ДНК, кодирующей желательную структуру антитела, в клеточной линии или в организме; химических способов (например, с помощью химического связывания, генетической гибридизации, нековалентной связи или иного способа с одним или более молекулярными соединениями, такими как другое антитело или фрагмент антитела); или их комбинации.
[088] Технологии и продукты, позволяющие получать моноспецифичность или биспецифичность, известны в данной области техники, как подробно рассматривается в литературе, также в отношении альтернативных форматов, конъюгатов антитело-лекарственный препарат, способов конструирования антител, способов скрининга в условиях in vitro, константных областей, посттрансляционных и химических модификаций, улучшенного признака для запуска гибели раковых клеток, такого как модификация Fc (Tiller K and Tessier Р, 2015 Annu Rev Biomed Eng. 17: 191-216; Speiss С et al., 2015. Molecular Immunology 67 95-106; Weiner G, 2015. Nat Rev Cancer, 15: 361-370; Fan G et al., 2015. J Hematol Oncol 8:130). Такое биспецифическое антитело может быть обеспечено в любом коммерчески доступном формате, включая DuoBody®, BiTE DART, CrossMab, «выступы-во-впадины», Triomab® или другой подходящий молекулярный формат и его фрагменты.
[089] В настоящем документе термин «эпитоп» или «антигенная детерминанта» относится к сайту на антигене, с которым связывается антитело. Как хорошо известно в данной области техники, эпитопы могут быть образованы как из смежных аминокислот (линейный эпитоп), так и из несмежных аминокислот, расположенных рядом в результате третичного сворачивания белка (конформациоиные эпитопы). Эпитопы, образованные из смежных аминокислот, как правило, сохраняются при воздействии денатурирующих растворителей, тогда как эпитопы, образованные в результате третичного сворачивания, обычно исчезают при обработке денатурирующими растворителями. Эпитоп, как правило, содержит по меньшей мере 3 и более обычно по меньшей мере 5 или 8-10 аминокислот в уникальной пространственной конформации. Способы определения пространственной конформации эпитопов хорошо известны в данной области техники и включают, например, рентгеновскую кристаллографию и двумерный ядерный магнитный резонанс. См., например, Epitope Mapping Protocols в Methods in Molecular Biology, Vol. 66, Glenn E. Morris, Ed (1996). Например, антитело согласно настоящему изобретению может распознавать конформационный эпитоп, с которым связываются антитела 7G7B6 или МА251. Согласно одному варианту реализации конформационный эпитоп содержит по меньшей мере две последовательности, выбранные из аминокислот 150-158 из SEQ ID NO: 1 (YQCVQGYRA), аминокислот 176-180 из SEQ ID NO: 1 (RWTQP), аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и аминокислот 74-84 из SEQ ID NO: 1 (SSWDNQCQCTS).
[090] Согласно некоторым вариантам реализации антитело к CD25 может быть включено в агент, который дополнительно содержит конъюгированный полезный груз, такой как терапевтический или диагностический агент, в частности, для терапии или диагностики рака. Можно применять конъюгаты антител к CD25 с радионуклидами или токсинами. Примеры обычно применяемых радионуклидов включают, например, Y90, I131 и Cu67, помимо прочих, и примеры обычно применяемых токсинов включают доксорубицин и калихеамицин. Согласно дополнительному варианту реализации антитело к CD25 может быть модифицировано, чтобы иметь измененный период полужизни. Способы изменения периода полужизни известны в данной области техники. Согласно некоторым вариантам реализации антитело к CD25 не конъюгировано с другим терапевтическим или диагностическим агентом. В частности, согласно некоторым вариантам реализации антитело к CD25 не конъюгировано с радионуклидом, т.е. согласно некоторым вариантам реализации антитело к CD25 не содержит радиоактивную метку.
[091] Согласно предпочтительному варианту реализации настоящего изобретения субъектом любого из аспектов настоящего изобретения, описанных в настоящем документе, является млекопитающее, предпочтительно кошка, собака, лошадь, осел, овца, свинья, коза, корова, хомяк, мышь, крыса, кролик или морская свинка, но наиболее предпочтительно субъект представляет собой человека. Таким образом, во всех аспектах настоящего изобретения, описанных в настоящем документе, субъект предпочтительно представляет собой человека.
[092] В настоящем документе термины «рак», «раковый» или «злокачественный» относятся или описывают физиологическое состояние у млекопитающих, которое, как правило, характеризуется нерегулируемым размножением клеток.
[093] Примеры рака включают, но не ограничиваются ими, карциному, лимфому, лейкоз, бластому и саркому. Более конкретные примеры таких видов рака включают плоскоклеточную карциному, миелому, мелкоклеточный рак легких, немелкоклеточный рак легких, глиому, гепатоцеллюлярную карциному (ГЦК), лимфому Ходжкина, неходжкинскую лимфому, острый миелолейкоз (ОМЛ), множественную миелому, рак желудочно-кишечного тракта, рак почек, рак яичников, рак печени, лимфобластный лейкоз, лимфоцитарный лейкоз, колоректальный рак, рак эндометрия, рак почек, рак простаты, рак щитовидной железы, меланому, хондросаркому, нейробластому, рак поджелудочной железы, мультиформную глиобластому, рак шейки матки, рак головного мозга, рак желудка, рак мочевого пузыря, гепатому, рак молочной железы, карциному толстой кишки и рак головы и шеи.
[094] Согласно одному аспекту рак включает плотную опухоль. Примеры плотных опухолей включают саркомы (включая виды рака, возникающие из трансформированных клеток мезенхимного происхождения в тканях, таких как губчатая кость, хрящ, жир, мышца, сосудистая ткань, кроветворная ткань или фиброзная соединительная ткань), карциномы (включая опухоли, возникающие из эпителиальных клеток), мезотелиому, нейробластому, ретинобластому и др. Виды рака, вовлекающие плотные опухоли, включают, но не ограничиваются ими, рак головного мозга, рак легких, рак желудка, рак двенадцатиперстной кишки, рак пищевода, рак молочной железы, рак толстой и прямой кишки, рак почек, рак мочевого пузыря, рак почек, рак поджелудочной железы, рак простаты, рак яичников, меланому, рак ротовой полости, саркому, рак глаз, рак щитовидной железы, рак уретры, рак влагалища, рак шеи, лимфому и тому подобное.
[095] Согласно одному аспекту виды рака включают опухоли, экспрессирующие CD25, включая, но не ограничиваясь ими, лимфомы, такие как лимфомы Ходжкина, и лимфоцитарные лейкозы, такие как хронический лимфолейкоз (ХЛЛ).
[096] Согласно одному аспекту настоящего изобретения рак идентифицируют по наличию специфических, имеющих отношение к опухоли маркеров и антигенов, таких как CD20, HER2, PD-1, PD-L1, SLAM7F, CD47, CD137, CD134, TIM3, CD25, GITR, CD25, EGFR и т.д., или рак представляет собой рак, который был идентифицирован как имеющий биомаркер, называемый высокой нестабильностью микросателлита (MSI-H), или дефицит репарации ошибок репликации (dMMR). Кроме того, антитела можно применять, если идентификация специфического, имеющего отношение к опухоли маркера, антигена или биомаркеров была использована при определении предраковых, неинвазивных состояний вышеуказанных видов рака у пациента, таких как рак in situ, тлеющая миелома, моноклональная гаммапатия неопределенного значения, цервикальная интраэпителиальная неоплазия, MALTomas/GALTomes и различные лимфопролиферативные расстройства. Предпочтительно в некоторых вариантах реализации субъект, которого лечат, имеет плотную опухоль.
[097] Согласно одному аспекту настоящего изобретения рак выбран из меланомы, немелкоклеточного рака легких, рака почек, рака яичников, рака мочевого пузыря, саркомы и рака толстой кишки. Согласно предпочтительному аспекту настоящего изобретения рак выбран из меланомы, рака яичников, немелкоклеточного рака легких и рака почек. Согласно одному варианту реализации рак не является меланомой, раком яичников или раком молочной железы. Согласно предпочтительному аспекту рак представляет собой саркому, рак толстой кишки, меланому или колоректальный рак или в более общем случае любой рак человека, для которого клеточная линия 4Т1, МСА205, В16, СТ26 или МС38 может представлять собой доклинические модели для подтверждения соединений, которые можно применять для их терапевтического контроля.
[098] В настоящем документе термин «опухоль», применительно к субъекту, у которого диагностирован или у которого подозревают рак, относится к злокачественному или потенциально злокачественному новообразованию или массе ткани любого размера и включает первичные опухоли и вторичные новообразования. Термины «рак», «злокачественная опухоль», «новообразование», «опухоль» и «карцинома» также могут использоваться в настоящем документе взаимозаменяемо для обозначения опухолей и опухолевых клеток, которые проявляют относительно патологическое, неконтролируемое и/или автономное размножение, так, что они проявляют аберрантный фенотип размножения, характеризующийся значительной потерей контроля пролиферации клеток. В целом, клетки, представляющие интерес для детектирования или лечения, включают предраковые (например, доброкачественные), злокачественные, предметастатические, метастатические и неметастатические клетки. Идеи настоящего изобретения могут быть подходящими для любого и всех видов рака.
[099] В настоящем документе «плотные (солидные) опухоли» - это патологический рост или масса ткани, которая обычно не содержит кисты или области с жидким содержимым, в частности, опухоли и/или метастазы (где бы они ни находились), отличные от лейкоза или разных видов неплотного рака лимфатической системы. Плотные опухоли могут быть доброкачественными или злокачественными. Различные типы плотных опухолей названы по типу клеток, которые их образуют, и/или ткани или органа, в котором они расположены. Примеры плотных опухолей включают саркомы (включая виды рака, возникающие из трансформированных клеток мезенхимного происхождения в тканях, таких как губчатая кость, хрящ, жир, мышца, сосудистая ткань, кроветворная ткань или фиброзная соединительная ткань), карциномы (включая опухоли, возникающие из эпителиальных клеток), меланомы, лимфомы, мезотелиому, нейробластому и ретинобластому.
[0100] Особенно предпочтительные виды рака в соответствии с настоящим изобретением включают те, которые характеризуются наличием плотной опухоли, т.е. субъект не имеет опухоль, отличную от плотной. Во всех аспектах изобретения, обсуждаемых в настоящем документе, предпочтительно рак представляет собой плотную опухоль, т.е. субъект имеет плотную опухоль (и не имеет опухоль, отличную от плотной).
[0101] В настоящем документе ссылка на «лечить» или «лечение» рака определяет достижение по меньшей мере одного положительного терапевтического эффекта, такого как, например, уменьшение количества раковых клеток, уменьшение размера опухоли, уменьшение скорости инфильтрации раковых клеток в периферические органы или уменьшение скорости метастазирования опухоли или роста опухоли.
[0102] Положительные терапевтические эффекты при раке могут быть измерены несколькими способами (например, Weber (2009) J Nucl Med 50, 1S-10S). Например, в отношении ингибирования роста опухоли, согласно стандартам Национального института онкологии США (NCI), Т/С≤42% представляет минимальный уровень противоопухолевой активности. Т/С<10% считается высоким уровнем противоопухолевой активности, при этом Т/С (%)=медианный объем опухоли после лечения/медианный объем опухоли в контроле × 100. Согласно некоторым вариантам реализации лечение, достигнутое при применении терапевтически эффективного количества, представляет собой любое из выживаемости без прогрессирования заболевания (PFS), выживаемости без признаков заболевания (DFS) или общей выживаемости (OS). PFS, также называемая «время до прогрессирования опухоли», указывает период времени в ходе лечения и после него, когда рак не развивается, и включает время, в течение которого пациенты испытывали полный или частичный ответ, а также время, в течение которого пациенты имели стабильное заболевание. DFS относится к периоду времени в ходе лечения и после него, когда пациент не имеет признаков заболевания. OS относится к увеличению продолжительности жизни по сравнению с наивными индивидуумами или пациентами, не получавшими лечение.
[0103] В настоящем документе ссылка на «предотвращение» (или профилактику) относится к задержке или предотвращению возникновения симптомов рака. Предотвращение может быть абсолютным (так, что заболевание не возникает) или может быть эффективным только у некоторых индивидуумов или в течение ограниченного периода времени.
[0104] Согласно предпочтительному аспекту настоящего изобретения субъект имеет развившуюся опухоль, т.е. у субъекта уже есть опухоль, например, которая классифицирована как плотная опухоль. В этой связи изобретение, описанное в настоящем документе, можно применять, если субъект уже имеет опухоль, такую как плотная опухоль. В этой связи изобретение обеспечивает терапевтический вариант, который можно применять для лечения существующей опухоли. Согласно одному аспекту настоящего изобретения субъект имеет существующую плотную опухоль. Настоящее изобретение можно применять для предотвращения или предпочтительно для лечения у субъектов, уже имеющих плотную опухоль. Согласно одному аспекту настоящее изобретение не применяют для предотвращения или профилактики.
[0105] Согласно одному аспекту при применении настоящего изобретения регрессия опухоли может быть усилена, рост опухоли может быть нарушен или уменьшен, и/или время выживания может быть увеличено, как описано в настоящем документе, например, по сравнению с другими способами лечения рака (например, стандартными способами лечения конкретного рака).
[0106] Согласно одному аспекту настоящего изобретения способ лечения или предотвращения рака, описанный в настоящем документе, дополнительно включает этап выявления субъекта, имеющего рак, предпочтительно выявление субъекта, который имеет опухоль, такую как плотная опухоль. Согласно одному варианту реализации способ может включать выявление субъекта, имеющего гематологический рак.
[0107] Схема дозирования терапии, описанной в настоящем документе, которая эффективна для лечения пациента, имеющего рак, может изменяться в зависимости от таких факторов как патологическое состояние, возраст и масса пациента, а также способность терапии вызывать противораковый ответ у субъекта. Выбор подходящей дозировки будет находиться в пределах компетенции квалифицированного специалиста в данной области техники. Например, 0,01, 0,1, 0,3, 0,5, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40 или 50 мг/кг. Согласно некоторым вариантам реализации такое количество представляет собой стандартную лекарственную форму (или ее целую фракцию), подходящую для введения в соответствии со схемой дозирования, которая, как определено, коррелирует с желательным или благоприятным исходом при введении в соответствующую популяцию (т.е. с терапевтической схемой дозирования).
[0108] Антитело согласно любому аспекту настоящего изобретения, как описано в настоящем документе, может быть в форме фармацевтической композиции, которая дополнительно содержит фармацевтически приемлемый носитель, разбавитель или вспомогательное вещество. Эти композиции включают, например, жидкие, полутвердые и твердые лекарственные формы, такие как жидкие растворы (например, растворы для инъекций и инфузий), дисперсии или суспензии, таблетки, пилюли или липосомы. Согласно некоторым вариантам реализации предпочтительная форма может зависеть от предполагаемого способа введения и/или терапевтического применения. Фармацевтические композиции, содержащие антитело, можно вводить любым подходящим способом, известным в данной области техники, включая, но не ограничиваясь ими, пероральный, слизистый, ингаляционный, местный, буккальный, назальный, ректальный или парентеральный (например, внутривенный, инфузионный, внутриопухолевый, интранодальный, подкожный, внутрибрюшинный, внутримышечный, внутрикожный, трансдермальный или другие способы введения, включающие физическое нарушение целостности ткани субъекта и введение фармацевтической композиции через повреждение в ткани). Такой состав, например, может быть в форме раствора для инъекции или инфузии, который подходит для внутрикожного, внутриопухолевого или подкожного введения или для внутривенной инфузии. Введение может включать периодическое дозирование. Согласно другому варианту введение может включать непрерывное дозирование (например, перфузию) в течение по меньшей мере выбранного периода времени, одновременно или в период времени между введением других соединений.
[0109] Согласно некоторым вариантам реализации антитело может быть получено с носителями, которые защищают его от быстрого высвобождения и/или разрушения, такими как состав с контролируемым высвобождением, такой как имплантаты, трансдермальные пластыри, и микрокапсулированные системы доставки. Можно применять биоразлагаемые, биосовместимые полимеры.
[0110] Квалифицированные специалисты в данной области техники поймут, например, что путь доставки (например, пероральный в сравнении с внутривенным, в сравнении с подкожным, в сравнении с внутриопухолевым и т.д.) может влиять на количество дозы и/или необходимое количество дозы может влиять на путь доставки. Например, если интерес представляют особенно высокие концентрации агента в конкретном сайте или месте (например, в опухоли), желательной и/или подходящей может быть узконаправленная доставка (например, в этом примере, внутриопухолевая доставка). Другие факторы, которые следует учитывать при оптимизации путей и/или схемы дозирования для конкретной терапевтической схемы, могут включать, например, конкретный рак, который лечат (например, тип, стадия, расположение и т.д.), клиническое состояние субъекта (например, возраст, общее состояние здоровья и т.д.), наличие или отсутствие комбинированной терапии и другие факторы, известные врачам.
[0111] Фармацевтические композиции, как правило, должны быть стерильными и стабильными в условиях производства и хранения. Композиция может быть изготовлена в виде раствора, микроэмульсии, дисперсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного препарата. Стерильные растворы для инъекций могут быть получены путем встраивания антитела в необходимом количестве в подходящем растворителе с одним или комбинацией ингредиентов, перечисленных выше, по мере необходимости, с последующей стерилизацией путем фильтрации. Составы для парентерального введения включают, но не ограничиваются ими, суспензии, растворы, эмульсии в масляных или водных носителях, пасты и имплантируемые составы с замедленным высвобождением или биоразлагаемые составы, обсуждаемые в настоящем документе. Стерильные составы для инъекций могут быть получены с использованием нетоксичного разбавителя или растворителя, приемлемого для парентерального введения. Каждая фармацевтическая композиция для применения в соответствии с настоящим изобретением может включать фармацевтически приемлемые диспергирующие агенты, смачивающие агенты, суспендирующие агенты, изотонические агенты, покрытия, антибактериальные и противогрибковые агенты, носители, вспомогательные вещества, соли или стабилизаторы, которые нетоксичны для субъектов в применяемых дозировках и концентрациях. Предпочтительно такая композиция может дополнительно содержать фармацевтически приемлемый носитель или вспомогательное вещество для применения в лечении рака, которые совместимы с конкретным способом и/или местом введения, например, для парентерального (например, подкожной, внутрикожной или внутривенной инъекции), внутриопухолевого или околоопухолевого введения.
[0112] Несмотря на то, что вариант реализации способа лечения или композиций для применения в соответствии с настоящим изобретением может быть неэффективным в достижении положительного терапевтического эффекта у каждого субъекта, он должен его достичь при применении фармацевтических композиций и схем дозирования, которые соответствуют требованиям Надлежащей медицинской практики, и статистически значимого количества субъектов, определяемого с помощью любого статистического теста, известного в данной области техники, такого как t-критерий Стьюдента, χ2-критерий, U-критерий Манна-Уитни, критерий Крускала-Уоллиса (Н-критерий), тест Джонкира-Терпстры и тест Вилкоксона.
[0113] Если в настоящем документе выше и далее упоминается опухоль, опухолевое заболевание, карцинома или рак, также подразумевается метастазирование в исходном органе или ткани и/или в любом другом месте, альтернативно или дополнительно, независимо от местоположения опухоли и/или метастазов.
[0114] Как обсуждается в настоящем документе, настоящее изобретение относится к истощению регуляторных Т-клеток (Трег). Таким образом, согласно одному аспекту настоящего изобретения антитело к CD25, которое не ингибирует связывание интерлейкина 2 с CD25, также истощает или уменьшает количество инфильтрирующих опухоль регуляторных Т-клеток. Согласно одному аспекту истощение происходит за счет АЗКЦ. Согласно другому аспекту истощение происходит за счет АЗКФ.
[0115] В этой связи настоящее изобретение обеспечивает способ истощения регуляторных Т-клеток в опухоли у субъекта, включающий введение указанному субъекту антитела к CD25, которое не ингибирует связывание интерлейкина 2 с CD25. Согласно предпочтительному варианту реализации Трег истощают в плотной опухоли. Под «истощенны» подразумевается, что количество, соотношение или процент Трег уменьшается по сравнению с тем, когда не вводят антитело к CD25, которое не ингибирует связывание интерлейкина 2 с CD25. Согласно конкретным вариантам реализации настоящего изобретения, описанным в настоящем документе, истощается более приблизительно 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 99% инфильтрирующих опухоль регуляторных Т-клеток.
[0116] В настоящем документе «регуляторные Т-клетки» («Трег», «Трег-клетки» или «Трег») относятся к линии CD4+ Т-лимфоцитов, специализирующихся на контроле аутоиммунитета, аллергии и инфекции. Как правило, они регулируют активность популяций Т-клеток, но они также могут влиять на определенные типы клеток врожденной иммунной системы. Обычно Трег идентифицируют по экспрессии биомаркеров CD4, CD25 и Foxp3. Природные Трег-клетки обычно составляют приблизительно 5-10% периферических CD4+ Т-лимфоцитов. Однако в микросреде опухоли (т.е. инфильтрирующие опухоль Трег-клетки) они могут составлять до 20-30% от общей популяции CD4+ Т-лимфоцитов.
[0117] Активированные человеческие Трег-клетки могут непосредственно уничтожать клетки-мишени, такие как эффекторные Т-клетки и АПК, за счет перфорин- или гранзим В-зависимых путей; Трег-клетки, экспрессирующие связанный с цитотоксическими Т-лимфоцитами антиген 4 (CTLA4+), индуцируют экспрессию индоламин-2,3-диоксигеназы (IDO) АПК, а они, в свою очередь, подавляют активацию Т-клеток за счет снижения количества триптофана; Трег-клетки могут высвобождать интерлейкин-10 (ИЛ-10) и трансформирующий фактор роста (ТФР-β) в условиях in vivo и, таким образом, непосредственно ингибируют активацию Т-клеток и подавляют функцию АПК, ингибируя экспрессию молекул ГКГС, CD80, CD86 и ИЛ-12. Трег-клетки также могут подавлять иммунитет, экспрессируя высокие уровни CTLA4, который может связываться с CD80 и CD86 на антигенпрезентирующих клетках и предотвращать надлежащую активацию эффекторных Т-клеток.
[0118] Согласно предпочтительному варианту реализации настоящего изобретения отношение эффекторных Т-клеток к регуляторным Т-клеткам в плотной опухоли увеличивается. Согласно некоторым вариантам реализации отношение эффекторных Т-клеток к регуляторным Т-клеткам в плотной опухоли увеличивается до более чем 5, 10, 15, 20, 40 или 80.
[0119] Иммунная эффекторная клетка относится к иммунной клетке, которая участвует в эффекторной фазе иммунного ответа. Примерные иммунные клетки включают клетки миелоидного или лимфоидного происхождения, например, лимфоциты (например, В-клетки и Т-клетки, включая цитолитические Т-клетки (CTL)), клетки-киллеры, природные клетки-киллеры, макрофаги, моноциты, эозинофилы, нейтрофилы, полиморфноядерные клетки, гранулоциты, тучные клетки и базофилы.
[0120] Иммунные эффекторные клетки, участвующие в эффекторной фазе иммунного ответа, экспрессируют специфические рецепторы Fc и выполняют специфические иммунные функции. Эффекторная клетка может индуцировать антителозависимую клеточную цитотоксичность (АЗКЦ), например, нейтрофил, способный индуцировать АЗКЦ. Например, моноциты, макрофаги, нейтрофилы, эозинофилы и лимфоциты, которые экспрессируют FcαR, участвуют в специфическом уничтожении клеток-мишеней и презентации антигенов другим компонентам иммунной системы или связывании с клетками, которые презентируют антигены. Эффекторная клетка также может поглощать путем фагоцитоза антиген-мишень, клетку-мишень или микроорганизм. Как обсуждается в настоящем документе, антитела согласно настоящему изобретению могут быть оптимизированы по их способности индуцировать АЗКЦ.
[0121] Согласно некоторым вариантам реализации другой агент против рака можно вводить в комбинации с антителом с помощью одного и того же или разных путей доставки и/или в соответствии с разными схемами. Альтернативно или дополнительно, согласно некоторым вариантам реализации одну или более доз первого активного агента вводят по существу одновременно, и согласно некоторым вариантам реализации, с помощью общего пути и/или как часть одной композиции, с одним или более другими активными агентами. Квалифицированные специалисты в данной области техники также поймут, что некоторые варианты реализации видов комбинированной терапии, обеспеченных в соответствии с настоящим изобретением, достигают синергетических эффектов; в некоторых таких вариантах реализации доза одного или более агентов, применяемых в комбинации, может существенно отличаться (например, может быть ниже) и/или может быть доставлена альтернативным путем, отличным от стандартного, предпочтительного или необходимого, когда этот агент применяется в другой терапевтической схеме (например, в виде монотерапии и/или как часть другой комбинированной терапии).
[0122] Согласно некоторым вариантам реализации, в которых два или более активных агентов применяют в соответствии с настоящим изобретением, такие агенты могут быть введены одновременно или последовательно. Согласно некоторым вариантам реализации введение одного агента специально рассчитано по времени относительно введения другого агента. Например, согласно некоторым вариантам реализации первый агент вводят так, что наблюдается конкретный эффект (или который, как ожидается, будет наблюдаться, например, на основании популяционных исследований, показывающих корреляцию между конкретной схемой дозирования и конкретным эффектом, представляющим интерес). Согласно некоторым вариантам реализации желательные относительные схемы дозирования для агентов, вводимых в комбинации, можно оценить или определить эмпирически, например, с использованием моделей в условиях ex vivo, in vivo и/или in vitro; согласно некоторым вариантам реализации такую оценку или эмпирическое определение производят в условиях in vivo, в популяции пациентов (например, для установления корреляции) или альтернативно в отношении конкретного пациента, представляющего интерес.
[0123] Согласно другому аспекту настоящего изобретения антитело к CD25, которое не ингибирует связывание интерлейкина 2 с CD25, обладает улучшенными терапевтическими эффектами в комбинации с ингибитором контрольной точки иммунитета. Комбинированная терапия с применением антитела к CD25, которое не ингибирует связывание интерлейкина 2 с CD25, и ингибитора контрольной точки иммунитета может оказывать синергетический эффект в лечении развившихся опухолей. Данные в отношении PD-1/PD-L1 в Примерах в настоящем документе относятся к нарушению взаимодействия PD-1/PD-L1. Таким образом, взаимодействие между рецептором PD-1 и лигандом PD-L1 может быть блокировано, что приводит к «блокаде PD-1». Согласно одному аспекту комбинация может привести к усилению регрессии опухоли, усилению нарушения или уменьшения роста опухоли, и/или время выживания может быть увеличено при применении настоящего изобретения, как описано в настоящем документе, например, по сравнению либо с антителами к CD25, либо с блокадой PD-1/PD-L1 по отдельности (непосредственно, с применением антитела к PD1, или косвенно, с применением антитела к PD-L1). Если антитело к CD25 не ингибирует связывание интерлейкина 2 с CD25, комбинированная терапия с применением антитела к CD25 и ингибитора контрольной точки иммунитета также может дополнительно включать введение интерлейкина-2 в дозировке, подходящей для лечения рака.
[0124] В настоящем документе «контрольная точка иммунной системы» или «белок контрольной точки иммунитета» относятся к белкам, принадлежащим к ингибиторным путям в иммунной системе, в частности, для модуляции ответов Т-клеток. В нормальных физиологических условиях контрольные точки иммунной системы имеют решающее значение для предотвращения аутоиммунитета, в частности, во время ответа на патоген. Раковые клетки могут изменить регуляцию экспрессии белков контрольной точки иммунитета, чтобы избежать иммунного надзора.
[0125] Примеры белков контрольной точки иммунитета включают, но не ограничиваются ими, PD-1, CTLA-4, BTLA, KIR, LAG3, TIGIT, CD155, В7Н3, В7Н4, VISTA и TIM3, а также ОХ40, GITR, ICOS, 4-1ВВ и HVEM. Белки контрольной точки иммунитета также могут относиться к белкам, которые связываются с другими белками контрольной точки иммунитета. Такие белки включают PD-L1, PD-L2, CD80, CD86, HVEM, LLT1 и GAL9.
[0126] «Ингибиторы белка контрольной точки иммунитета» относятся к любому белку, который может нарушать передачу сигналов и/или белок-белковые взаимодействия, опосредуемые белком контрольной точки иммунитета. Согласно одному аспекту настоящего изобретения белок контрольной точки иммунитета представляет собой PD-1 или PD-L1. Согласно предпочтительному аспекту настоящего изобретения, как описано в настоящем документе, ингибитор контрольной точки иммунитета нарушает взаимодействия PD-1/PD-L1 за счет антител к PD-1 или PD-L1.
[0127] В этой связи согласно настоящему изобретению также предложен способ лечения рака, включающий введение субъекту антитела к CD25, которое не ингибирует связывание интерлейкина 2 с CD25, и дополнительного терапевтического агента, предпочтительно ингибитора контрольной точки иммунитета. Согласно настоящему изобретению также предложено антитело к CD25, которое не ингибирует связывание интерлейкина 2 с CD25, и дополнительный терапевтический агент, предпочтительно ингибитор контрольной точки иммунитета, для применения в лечении рака.
[0128] Согласно настоящему изобретению дополнительно предложено применение антитела к CD25, которое не ингибирует связывание интерлейкина-2 с CD25, и дополнительного терапевтического агента, предпочтительно ингибитора контрольной точки иммунитета, для изготовления лекарственного средства для лечения рака. Введение антитела к CD25, которое не ингибирует связывание интерлейкина-2 с CD25, и дополнительного терапевтического агента, такого как ингибитор контрольной точки иммунитета, может быть одновременным, по отдельности или последовательным.
[0129] Согласно настоящему изобретению предложена комбинация антитела к CD25, которое не ингибирует связывание интерлейкина-2 с CD25, и дополнительного терапевтического агента, предпочтительно ингибитора контрольной точки иммунитета, для применения в лечении рака у субъекта, причем указанное антитело к CD25, которое не ингибирует связывание интерлейкина-2 с CD25, и дополнительный терапевтический агент, такой как ингибитор контрольной точки иммунитета, вводят одновременно, по отдельности или последовательно. Такое антитело к человеческому CD25, которое не ингибирует связывание интерлейкина-2 с CD25 и представляет собой человеческий изотип IgG1, можно применять, в частности, в комбинации с антителами, нацеленными на контрольные точки иммунной системы, но не содержащими последовательности, которые обеспечивают АЗКЦ, АЗКФ и/или КЗЦ.
[0130] Согласно альтернативному аспекту настоящего изобретения предложено антитело к CD25, которое не ингибирует связывание интерлейкина 2 с CD25, для применения в лечении рака, причем указанное антитело предназначено для введения в комбинации с дополнительным терапевтическим агентом, предпочтительно с ингибитором контрольной точкой иммунной системы. Согласно настоящему изобретению также предложено применение антитела к CD25, которое не ингибирует связывание интерлейкина 2 с CD25, в изготовлении лекарственного средства для лечения рака, причем указанное лекарственное средство предназначено для введения в комбинации с дополнительным терапевтическим агентом, предпочтительно с ингибитором контрольной точки иммунитета.
[0131] Согласно настоящему изобретению предложена фармацевтическая композиция, содержащая антитело к CD25, которое не ингибирует связывание интерлейкина 2 с CD25, и необязательно дополнительный терапевтический агент, предпочтительно ингибитор контрольной точки иммунитета, в фармацевтически приемлемой среде. Как обсуждалось выше, ингибитор контрольной точки иммунитета может представлять собой ингибитор PD-1, т.е. антагонист PD-1.
[0132] PD-1 (белок запрограммированной гибели клеток 1), также известный как CD279, представляет собой рецептор клеточной поверхности, экспрессируемый на активированных Т-клетках и В-клетках. Было показано, что взаимодействие с его лигандами ослабляет ответы Т-клеток как в условиях in vitro, так и в условиях in vivo. PD-1 связывает два лиганда, PD-L1 и PD-L2. PD-1 принадлежит к суперсемейству иммуноглобулинов. Передача сигналов с участием PD-1 требует связывания с лигандом PD-1 в непосредственной близости от пептидного антигена, представленного главным комплексом гистосовместимости (ГКГС) (Freeman (2008) Proc Natl Acad Sci USA 105, 10275-6). Соответственно, белки, антитела или небольшие молекулы, которые предотвращают колигирование PD-1 и TCR на мембране Т-клеток, можно применять в качестве антагонистов PD-1.
[0133] Согласно одному варианту реализации антагонист рецептора PD-1 представляет собой антитело к PD-1 или его антигенсвязывающий фрагмент, который специфически связывается с PD-1 и блокирует связывание PD-L1 с PD-1. Антитело к PD-1 может представлять собой моноклональное антитело. Антитело к PD-1 может представлять собой человеческое или гуманизированное антитело. Антитело к PD-1 представляет собой антитело, способное специфически связываться с рецептором PD-1. Антитела к PD-1, известные в данной области техники, включают ниволумаб и пембролизумаб.
[0134] Антагонисты PD-1 согласно настоящему изобретению также включают соединения или агенты, которые либо связываются с лигандом PD-1, либо блокируют его, чтобы нарушить или ингибировать связывание лиганда с рецептором PD-1, либо непосредственно связываются и блокируют рецептор PD-1, не индуцируя передачу ингибирующих сигналов через рецептор PD-1. В частности, антагонисты PD-1 включают низкомолекулярные ингибиторы пути передачи сигналов с участием PD-1/PD-L1. Согласно другому варианту антагонист рецептора PD-1 может непосредственно связываться с рецептором PD-1, не запуская передачу ингибирующих сигналов, а также связывается с лигандом рецептора PD-1, чтобы уменьшить или ингибировать запуск передачи сигналов через рецептор PD-1 под действием лиганда. При уменьшении числа и/или количества лигандов, которые связываются с рецептором PD-1 и запускают передачу ингибирующего сигнала, меньшее количество клеток ослабляется отрицательным сигналом, который доставляется при передаче сигнала с участием PD-1, и может быть достигнут более устойчивый иммунный ответ.
[0135] Согласно одному варианту реализации антагонист рецептора PD-1 представляет собой антитело к PD-L1 или его антигенсвязывающий фрагмент, который специфически связывается с PD-L1 и блокирует связывание PD-L1 с PD-1. Антитело к PD-L1 может представлять собой моноклональное антитело. Антитело к PD-L1 может представлять собой человеческое или гуманизированное антитело, такое как атезолизумаб (MPDL3280A).
[0136] Согласно настоящему изобретению также предложен способ лечения рака, включающий введение субъекту антитела к CD25, которое не ингибирует связывание интерлейкина 2 с CD25, и антитела, которое является агонистом костимулирующего пути, активирующего Т-клетки. Антитела-агонисты к костимулирующему пути, активирующему Т-клетки, включают, но не ограничиваются ими, антитела-агонисты к ICOS, GITR, ОХ40, CD40, LIGHT и 4-1ВВ.
[0137] Другой способ лечения рака включает введение антитела к CD25, которое не ингибирует связывание интерлейкина 2 с CD25, и соединения, которое уменьшает, блокирует, ингибирует и/или оказывает антагонистическое действие на FcγRIIb (CD32b). Такой антагонист FcγRIIb может представлять собой небольшую молекулу, нарушающую FcγRIIb-индуцированную внутриклеточную передачу сигналов, модифицированные антитела, которые не связывают ингибирующий рецептор FcγRIIb, или антитело к человеческому FcγRIIb (антитело к CD32b). Например, были охарактеризованы противоопухолевые свойства антител-антагонистов к человеческому FcγRIIb (Roghanian А et al., 2015, Cancer Cell. 27, 473^88; Rozan С et al., 2013, Mol Cancer Ther. 12:1481-91; WO 2015173384; WO 2008002933).
[0138] Согласно другому аспекту настоящего изобретения предложено биспецифическое антитело, содержащее:
(a) первый антигенсвязывающий фрагмент, который связывается с CD25; и
(b) второй антигенсвязывающий фрагмент, который связывается с белком контрольной точки иммунитета, связанным с опухолью антигеном, антитело к человеческому активирующему рецептору Fc (FcγRI, FcγRIIa, FcγRIII) или антитело-антагонист к человеческому FcγRIIb;
причем указанное антитело к CD25 не ингибирует связывание интерлейкина-2 (IL-2) с CD25 и предпочтительно представляет собой биспецифическое антитело IgG1, которое связывается по меньшей мере с одним активирующим рецептором Fcγ с высокой аффинностью и истощает инфильтрирующие опухоль регуляторные Т-клетки. Согласно предпочтительному варианту реализации второй антигенсвязывающий фрагмент связывается с PD-L1.
[0139] В настоящем документе «связанный с опухолью антиген» относится к антигенам, экспрессируемым на опухолевых клетках, что делает их отличимыми от расположенных рядом с ними нераковых клеток, и включает, но не ограничивается ими, CD20, CD38, PD-L1, EGFR, EGFRV3, СЕА, TYRP1 и HER2. Были опубликованы различные обзорные статьи, в которых описаны значимые связанные с опухолью антигены и соответствующие терапевтически пригодные противоопухолевые антитела-агенты (см., например, Sliwkowski & Mellman (2013) Science 341, 192-8). Такие антигены и соответствующие антитела включают, но не ограничиваются ими, CD22 (блинатумомаб), CD20 (ритуксимаб, тозитумомаб), CD56 (лорвотузумаб), CD66e/CEA (лабетузумаб), CD152/CTLA-4 (ипилимумаб), CD221/IGF1R (MK-0646), CD326/Epcam (эдреколомаб), CD340/HER2 (трастузумаб, пертузумаб) и EGFR (цетуксимаб, панитумумаб).
[0140] Согласно одному аспекту биспецифическое антитело в соответствии с настоящим изобретением, описанное в настоящем документе, приводит к АЗКЦ или, согласно одному аспекту, к усиленной АЗКЦ.
[0141] Биспецифическое антитело может связываться со специфическим эпитопом на CD25, который не влияет на связывание IL-2 с CD25, и со специфическим эпитопом на белке контрольной точки иммунитета или связанном с опухолью антигене, как определено в настоящем документе. Согласно предпочтительному варианту реализации второй антигенсвязывающий фрагмент связывается с PD-L1. Согласно предпочтительному аспекту настоящего изобретения предложено биспецифическое антитело, содержащее:
(a) первый антигенсвязывающий фрагмент, который связывается с CD25 и не влияет на связывание IL-2 с CD25; и
(b) второй антигенсвязывающий фрагмент, который связывается с белком контрольной точки иммунитета, экспрессируемым на опухолевой клетке.
[0142] Согласно конкретному варианту реализации белок контрольной точки иммунитета, экспрессируемый на опухолевой клетке, представляет собой PD-L1, VISTA, GAL9, В7Н3 или В7Н4. Также предпочтительно антитело к CD25 представляет собой антитело IgG1, которое не влияет на связывание IL-2 с CD25 и которое связывается с по меньшей мере одним активирующим рецептором Fcγ с высокой аффинностью и истощает инфильтрирующие опухоль регуляторные Т-клетки. Согласно другому варианту антитело к CD25 представляет собой человеческое антитело IgG2, которое истощает инфильтрирующие опухоль регуляторные Т-клетки. Согласно одному конкретному варианту реализации антитело к CD25 представляет собой человеческое антитело IgG2, которое связывается с по меньшей мере одним активирующим рецептором Fcγ с высокой аффинностью, предпочтительно с FcγRIIa.
[0143] Квалифицированный специалист в данной области техники сможет получить биспецифическое антитело, используя известные способы. Биспецифическое антитело в соответствии с настоящим изобретением можно применять в любом из аспектов настоящего изобретения, как описано в настоящем документе. Предпочтительно второй антигенсвязывающий фрагмент в биспецифическом антителе в соответствии с настоящим изобретением связывается с человеческим PD-1, человеческим PD-L1 или человеческим CTLA-4.
[0144] Согласно одному аспекту биспецифическое антитело может связываться с CD25 и с иммуно модулирующими рецепторами, экспрессируемыми на высоких уровнях на инфильтрирующих опухоль Трег, например, CTLA4, ICOS, GITR, 4-1ВВ или ОХ40.
[0145] Согласно настоящему изобретению также предложен набор, который содержит антитело к CD25, описанное в настоящем документе, и дополнительный терапевтический агент, предпочтительно ингибитор контрольной точки иммунитета, предпочтительно антагонист PD-1 (непосредственно, с применением антитела к PD1, или косвенно, с применением антитела к PD-L1), как обсуждается в настоящем документе. Согласно одному аспекту ингибитор контрольной точки иммунитета представляет собой антитело к PD-L1. Согласно альтернативному варианту реализации набор содержит антитело к CD25, описанное в настоящем документе, и антитело, которое является агонистом костимулирующего пути, активирующего Т-клетки. Набор может содержать инструкции по применению.
[0146] Любой аспект настоящего изобретения, описанный в настоящем документе, может быть выполнен в комбинации с дополнительными терапевтическими агентами, в частности, с дополнительными видами терапии рака. В частности, антитело к CD25 и необязательно ингибитор контрольной точки иммунитета в соответствии с настоящим изобретением можно вводить в комбинации с костимулирующими антителами, химиотерапией и/или радиотерапией (путем применения внешнего облучения тела или путем введения радиоконъюгированных соединений), терапией на основе цитокинов, нацеленной терапией, терапией моноклональными антителами, вакциной или адъювантом или любой их комбинацией.
[0147] В настоящем документе химиотерапевтическое соединение относится к соединению, которое разрушает клетку, т.е. соединение уменьшает жизнеспособность клетки. Химиотерапевтическое соединение может представлять собой цитотоксический лекарственный препарат. Предусмотренный химиотерапевтический агент включает, но не ограничивается ими, алкилирующие агенты, антрациклины, эпотилоны, нитрозомочевины, этиленимины/метилмеламин, алкилсульфонаты, алкилирующие агенты, антиметаболиты, аналоги пиримидина, эпиподофилотоксины, ферменты, такие как L-аспарагиназа; модификаторы биологического ответа, такие как ИФН-α, ИФН-γ, IL-2, ИЛ-12, Г-КСФ и ГМ-КСФ; координационные комплексы платины, такие как цисплатин, оксалиплатин и карбоплатин, антрацендионы, замещенную мочевину, такую как гидроксимочевина, производные метилгидразина, включая N-метилгидразин (MIH) и прокарбазин, адренокортикальные супрессоры, такие как митотан (o,p'-DDD) и аминоглутетимид; гормоны и антагонисты, включая антагонисты адренокортикостероидов, такие как преднизон и его эквиваленты, дексаметазон и аминоглутетимид; прогестин, такой как капроат гидроксипрогестерона, ацетат медроксипрогестерона и ацетат мегестрола; эстроген, такой как диэтилстилбэстрол и эквиваленты этинилэстрадиола; антиэстроген, такой как тамоксифен; андрогены, включая пропионат тестостерона и флуоксиместерон/эквиваленты; антиандрогены, такие как флутамид, аналоги гонадотропин-рилизинг-гормона и лейпролид; и нестероидные антиандрогены, такие как флутамид.
[0148] Дополнительная терапия рака также может включать введение противораковой вакцины. В настоящем документе «противораковые вакцины» (cancer vaccines) относятся к терапевтическим противораковым вакцинам, вводимым пациентам с раком и предназначенным для уничтожения раковых клеток за счет усиления собственных иммунных ответов пациента. Противораковые вакцины включают вакцины опухолевых клеток (аутологичных и аллогенных), вакцины на основе дендритных клеток (полученных в условиях ex vivo и активированных пептидами), противораковые вакцины на основе белков/пептидов и генетические вакцины (вакцины на основе ДНК, РНК и вирусов). Соответственно, терапевтические противораковые вакцины, в принципе, можно применять для ингибирования дальнейшего роста распространенных видов рака и/или рецидивирующих опухолей, которые устойчивы к обычным видам терапии, таким как хирургическое вмешательство, лучевая терапия и химиотерапия. Вакцины на основе опухолевых клеток (аутологичных и аллогенных) включают вакцины, генетически модифицированные для секреции растворимых иммуностимулирующих агентов, таких как цитокины (IL-2, ИФН-γ, ИЛ-12, ГМ-КСФ, FLT3L), одноцепочечных Fv-антител к иммуномодулирующим рецепторам (PD-1, CTLA-4, GITR, ICOS, ОХ40, 4-1ВВ) и/или экспрессии на их мембране лиганда иммуностимулирующих рецепторов, такого как лиганд ICOS, лиганд 4-1ВВ, лиганд GITR и/или лиганд ОХ40, помимо прочих. Согласно одному варианту реализации противораковая вакцина может представлять собой противоопухолевую вакцину GVAX.
[0149] Дополнительная терапия рака может представлять собой другие антитела или низкомолекулярные реагенты, которые уменьшают иммунную регуляцию на периферии и в микросреде опухоли, например, молекулы, которые нацелены на пути TGF-бета, IDO (индоламиндезоксигеназу), аргиназу и/или CSF1R.
[0150] «В комбинации» может относиться к введению дополнительной терапии перед, одновременно или после введения любого аспекта в соответствии с настоящим изобретением.
[0151] Настоящее изобретение будет описано далее с помощью нижеследующих примеров, которые предназначены для облегчения реализации настоящего изобретения обычным специалистом в данной области техники и никоим образом не предназначены для ограничения объема настоящего изобретения, со ссылкой на чертежи.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[0152] Фигура 1: представлена характеристика антител к мышиному CD25, 7D4 и РС61, использованных в Примерах. Оценку влияния на связывание IL-2 выполняли с использованием анализа перекрестного блокирования в тандемном формате. Биотинилированный мышиный CD25 загружали на сенсоры SA. Затем сенсоры подвергали воздействию 100 нМ мышиного IL-2 и затем добавляли одно из антител к мышиному CD25 в момент времени 150 с. Дополнительное связывание антителом после ассоциации с IL-2 указывает на антитело к мышиному CD25, которое не блокирует связывание с IL-2, при этом отсутствие дополнительного связывания указывает на блокирование лиганда. РС-61 mIgG2a нарушало взаимодействие мышиного IL-2 с мышиным CD25, в отличие от 7D4 (А). Взаимодействие мышиного IL-2/мышиного CD25 в присутствии рекомбинантного антитела к мышиному CD25, 7D4 (mIgG1), оценивали с использованием стандартного анализа перекрестного блокирования в формате «сэндвич». 7D4 (mIgG1) загружали на сенсоры AHQ, и незанятые Fc-связывающие сайты на сенсоре блокировали нецелевым человеческим антителом IgG1. Затем сенсоры подвергали воздействию 100 нМ рекомбинантного мышиного CD25 (R&D Systems; каталожный номер 2438-RM-050), и затем рекомбинантного мышиного IL-2 (Peprotech; каталожный номер 212-12). Дополнительное связывание мышиным IL-2 после ассоциации 7D4 (mIgG1)-мышиный CD25 указывает на незанятый эпитоп, при этом как 7D4 (mIgG1), так и мышиный IL-2 не конкурируют за эпитоп в мышином CD25, поскольку связывание с мышиным CD25 происходит одновременно (В). Связывание с мышиным CD25 определяли с использованием клеток СНО, экспрессирующих мышиный CD25, для антител, связывающих человеческий CD25, даклизумаба (DAC), антитела РС61 (mIgG2a) (исходного антитела к CD25, полученного из клона РС-61 с мышиным IgG2a и константными κ-областями, которые связаны с АЗКЦ) и антитела a7D4 (mIgG1) (антитела к CD25, полученного из клона 7D4 с мышиным IgG1 и константными κ-областями). IgG к мышиному CD25 связывались с клеточным экспрессируемым mCD25. CHO-mCD25 вносили по аликвотам в 96-луночные аналитические планшеты (50000 клеток/лунку) и инкубировали с 0,1 мл раствора, содержащего антитело (антитело в концентрации 100 нМ в фосфатно-солевом буфере (ФСБ)+0,1% бычьего сывороточного альбумина), при 25°С в течение 15 минут. Клетки трижды промывали ледяным ФСБ+0,1% бычьего сывороточного альбумина и затем метили R-ФЭ-конъюгированным козьим антителом к человеческому IgG (специфическим для γ-цепи) (Southern Biotech, каталожный номер 2040-09) и анализировали методом проточной цитометрии (для распознавания мертвых клеток использовали йодид пропидия). Для даклизумаба (DAC) связывание не детектировали, в то время РС61 (mIgG2a) и 7D4 (mIgG1) проявляют явное связывание с такими клетками (С).
[0153] Фигура 2: показан эффект антител к мышиному CD25 на индукцию экспрессии гранзима В CD4 Т-клетками при стимуляции антителом к CD3 и антителом к CD28. Общую популяцию CD4-положительных Т-клеток выделяли с использованием CD4 микрогранул из лимфатических узлов и селезенки мыши и метили фиолетовыми красителями CellTrace™ (ThermoFisher), чтобы измерить пролиферацию клеток. Меченые Т-клетки (105) высевали с фидерными клетками (105; отрицательная фракция CD90.2, с использованием набора для общей популяции мышиных Т-клеток Dynabeads) в 96-луночный планшет. В лунки добавляли антитело к CD3 (клон 145-2С11, BioXcell, каталожный номер ВЕ0001-1; 1 мкг/мл) и антитело к CD28 (клон 37.51, каталожный номер BioXcell ВЕ0015-1; 0,5 мкг/мл) (за исключением контрольного образца не стимулированных меченых Т-клеток) для активации CD4 Т-клеток и индукции пролиферации и выработки гранзима В. Затем в лунки, содержащие меченые CD4 Т-клетки, антитело к CD3 и антитело к CD28, добавляли следующие антитела (в концентрации 25 мкг/мл) (лунки, содержащие только меченые Т-клетки и антитело к CD3 и антитело к CD28, использовали в качестве отрицательного контроля): PC61(mIgG2a), 7D4(mIgG1) или нейтрализующее антитело к мышиному IL-2 (клон Jes6-1A12, BioXcell 6032988564), используемое в качестве положительного контроля, чтобы показать, как влияет блокирование взаимодействия между IL-2 и его рецептором на активацию Т-клеток. Образцы меченых Т-клеток затем инкубировали в течение приблизительно 84 ч. Затем клетки фиксировали и пермеабилизировали перед окрашиванием антителом к мышиному гранзиму В (клон GB11, Invitrogen). После этого клетки анализировали методом проточной цитометрии для определения экспрессии гранзима В и разбавления фиолетовых красителей CellTrace (BV450). Процент меченых Т-клеток, которые пролиферируют и экспрессируют гранзим В (GnzB), показан в верхнем левом квадранте графика, соответствующего каждому виду лечения (Q9), тогда как процент меченых Т-клеток, которые пролиферируют, но не экспрессируют GnzB, показан в нижнем левом квадранте графика, соответствующего каждому виду лечения (Q12).
[0154] Фигура 3: показан эффект в условиях in vivo антител к мышиному CD25, имеющих одинаковый изотип (мышиный IgG2a), которые либо блокируют, либо не блокируют взаимодействие между CD25 и IL-2, при введении в присутствии антитела к мышиному PD1 или без него (aPD1; клон RMP1-14), на иммунных клетках. Шесть групп мышей получали подкожные инъекции опухолевых клеток МСА205 (5×105 15) на 0 день и получали лечение по отдельности, как указано на графиках. В четырех группах антитело к мышиному CD25 (либо аРС61 mIgG2a, либо a7D4 mIgG2a; 200 мкг) вводили внутрибрюшинно на 5 день. В трех группах aPD1 (100 мкг) вводили внутрибрюшинно на 6 день и 9 день. Опухоли и лимфатические узлы собирали на 12 день и затем обрабатывали для окрашивания клеток в соответствии с желаемым типом и анализировали методом проточной цитометрии, как указано на каждой панели, с использованием следующих антител: антитело к CD3 (клон 17А2, Biolegend), антитело к CD4 (клон RM4-5, BD biosciences), антитело к CD8 (клон 53-6.7, Biolegend) и антитело к FoxP3 (клон FJK-16s, eBiosciences). Внутриядерное окрашивание FoxP3 выполняли с использованием набора буферов для окрашивания фактора транскрипции FoxP3 (eBioscience). Представлен процент CD4-положительных/Рохр3-положительных регуляторных Т-клеток и CD4-положительных/FoxP3-отрицательных эффекторных CD4 Т-клеток (CD4 Тэфф) в лимфатических узлах и ИОЛ, а также соотношение эффекторных CD8-положительных Т-клеток/Трег-клеток и CD4 Тэфф/Трег-клеток. Анализ данных выполняли в Flowjo, версия 10.0.8 (Tree Star Inc.). Статистические анализы выполняли в Prism 6 (GraphPad Software, Inc.); Значения p рассчитывали с использованием дисперсионного анализа Краскела-Уоллиса и апостериорного критерия Данна (ns=p>0,05; ****=р<0,0001).
[0155] Фигура 4: показан эффект антител к мышиному CD25, имеющих одинаковый изотип (IgG2a), которые либо блокируют, либо не блокируют взаимодействие между CD25 и IL-2, в присутствии антитела к мышиному PD1 или без него (aPD1; клон RMP1-14) на выработку гранзима В пролиферирующими Т-клетками в условиях in vivo. Образец клеток получали с использованием модели на основе МСА205 в шести группах лечения, как показано на Фигуре 3. Опухолевые клетки окрашивали в соответствии с желаемым типом и анализировали методом проточной цитометрии, как указано на каждой панели, с использованием следующих антител: антитело к CD3 (РеСу7, клон 145-2С11, Ebioscience, 25003182), антитело к CD4 (V500, клон RM4-5, BD biosciences, 560782), антитело к CD8 (BV785, клон 53-6.7, Biolegend, 100750), антитело к гранзиму В (АРС, клон GB11; Invitrogen, grb05) и Ki67 (V450, клон SolA15; eBiosciences, 48569882). Внутриядерное окрашивание Ki67 и гранзима В выполняли с использованием набора буферов для окрашивания фактора транскрипции FoxP3 (eBioscience, 00-5523-00). Сравнивали процент GnzB-положительных, а также общее количество GnzB-положительных пролиферирующих (о чем свидетельствует положительное окрашивание Ki67) CD4-положительных или CD8-положительных Т-клеток. Статистические анализы выполняли как для Фигуры 3 (ns=p>0,05; *=р<0,05; **=р<0,01; ***=р<0,001; ****=р<0,0001).
[0156] Фигура 5: показан эффект антитела к мышиному CD25 (изотип IgG2a), которое вводят в комбинации с антителом к PD-1 (клон RMP1-14) или без него, на эрадикацию развившихся опухолей в модели СТ26 на мышах. Оба антитела, 7D4m2a и PC61m2a, представляют собой антитела к мышиному CD25, истощающие Трег, однако одно не блокирует IL-2 (7D4m2a), а другое блокирует IL-2 (PC61m2). Кривые роста отдельных мышей в зависимости от времени были установлены для каждой группы лечения. Число выживших без опухолей через 50 дней указано на каждом графике. Клетки СТ26, использованные для имплантации, собирали во время логарифмической фазы размножения и ресуспендировали в холодном ФСБ. На 1 день (D1) исследования каждой мыши вводили путем подкожной инъекции в правый бок 3×105 клеток (ОД мл клеточной суспензии). Антитело к мышиному CD25 вводили путем внутрибрюшинной инъекции (10 мг/кг) на 6 день (когда детектировали пальпируемые опухоли). Антитело к мышиному PD1 вводили путем внутрибрюшинной инъекции (100 мкг/инъекцию) на 7 день, 10 день, 14 день и 17 день. Опухоли измеряли штангенциркулем по двум измерениям два раза в неделю для контроля роста. Размер опухоли в мм3 рассчитывали по формуле: объем опухоли = (w2×l)/2, где w = ширина и l = длина опухоли в мм. Конечной точкой исследования был объем опухоли 4000 мм3 или 50 дней, в зависимости от того, что наступило раньше (точки данных, которые остановились на разных, более ранних днях, связаны со смертью мышей; количество животных, выживших в конце эксперимента, указано на каждой панели).
[0157] Фигура 6: показаны кривые роста опухоли СТ26 у отдельных мышей, которые не получали лечение (ФСБ, только носитель) или которых лечили с применением антител IgG2a к мышиному CD25, оба из которых истощают Трег, однако одно блокирует IL-2 (7D4m2a), а другое не блокирует IL-2 (PC61m2), и затем комбинировали с антителом к мышиному PD-L1, клон 10F.9G2 (aPDL1; клон 10F.9G2), либо не применяли его. Модель, схема и анализ данных аналогичны таковым для Фигуры 5.
[0158] Фигура 7: показан эффект антитела к мышиному CD25 (изотип IgG2a), которое вводили в комбинации с антителом к PD-1 (клон RMP1-14) или без него, на эрадикацию развившихся опухолей в модели МС38 на мышах. Испытанные антитела представляют собой антитела, описанные на Фигуре 5. Кривые роста отдельных мышей в зависимости от времени были установлены для каждой группы лечения. Количество выживших без опухолей через 35 дней указано на каждом графике. Клетки карциномы толстой кишки МС38, использованные для имплантации, собирали во время логарифмической фазы размножения и ресуспендировали в холодном ФСБ. Каждой мыши вводили путем подкожной инъекции в правый бок 5×105 опухолевых клеток (0,1 мл суспензии клеток). Опухоли контролировали по мере того, как их объемы достигали целевого диапазона от 100 до 150 мм3. Через двадцать два дня после имплантации опухоли, на 1 день исследования, животных с индивидуальными объемами опухоли, которые находились в пределах диапазона 75-126 мм3, разделяли на девять групп (n=10), при этом средние объемы опухолей составили приблизительно 106 мм3. Лечение начинали на 1 день у мышей с развившимися опухолями МС38. Эффекты каждого лечения сравнивали с контрольной группой, получавшей носитель, которая получала ФСБ внутрибрюшинно (в/б) на 1 день, 2 день, 5 день, 9 день и 12 день. Антитело к PD1 вводили внутрибрюшинно в дозе 100 мкг/животное, два раза в неделю в течение двух недель (2 р./неделю×2), начиная со 2 дня. 7D4m2a и PC61m2a вводили внугрибрюшинно один раз на 1 день в дозе 200 мкг/животное. Измерения опухоли проводили два раза в неделю. Конечной точкой исследования был объем опухоли 4000 мм3 или 35 дней, в зависимости от того, что наступило раньше (точки данных, которые остановились на разных, более ранних днях, связаны со смертью мышей; количество животных, выживших в конце эксперимента, указано на каждой панели).
[0159] Фигура 8: показаны кривые роста опухоли МС38 у отдельных мышей, которые не получали лечение (ФСБ, только носитель), которым вводили антитела IgG2a к мышиному CD25, оба из которых истощают Трег, но одно не блокирует IL-2 (7D4m2a), а другое блокирует IL-2 (PC61m2), и затем комбинировали с антителом к мышиному PD-L1, клон 10F.9G2 (aPDL1; клон 10F.9G2), или не применяли его. Модель, схема и анализ данных аналогичны таковым для Фигуры 7.
[0160] Фигура 9: оценка терапевтической активности 7D4 mIgG2a, не блокирующего IL-2, Трег-истощающего антитела к мышиному CD25, по отдельности (D) и в комбинации с IL-2-нейтрализующим антителом (ThermoFisher; JES6-1A12) (Е), или блокирующего IL-2, не истощающего антитела мышиного изотипа IgG1 к мышиному CD25 (РС61, мышиный IgG1) (F) у самок мышей BALB/c, несущих сингенные опухоли толстой кишки СТ26. Для сравнения испытывали активность контрольного мышиного IgG2 (A), IL-2-нейтрализующего антитела по отдельности (В) и блокирующего IL-2 антитела к CD25 по отдельности (С).
[0161] Фигура 10: показана консенсусная последовательность человеческого CD25 (код Uniprot Р01589), называемая в настоящем документе SEQ ID NO: 1. Подчеркнут внеклеточный домен зрелого CD25, соответствующий аминокислотам 22-240. Указано положение эпитопов из антител к CD25, не блокирующих ИЛ, выявленных заранее: эпитопы 1 (полные и короткие эпитопы), эпитоп 2 (полные и короткие эпитопы), эпитоп 3 и эпитоп 4 (полные и короткие эпитопы). Положение эпитопа базиликсимаба и даклизумаба (указан как DAC) также выявлено.
[0162] Фигура 11: характеристика связывания (A) 7D4, (В) РС61 и (С) 2Е4 с CD25, экспрессируемым на клетках СНО, экспрессирующих CD25, при повышении концентрации антител и сравнение с контролем мышиного изотипа IgG2a.
[0163] Фигура 12: анализ очищенных антител (mIgG2a) к rmCD25, несущему His-метку, с помощью SPR на Biacore 2000, (A) 7D4, (В) 2Е4.
[0164] Фигура 13: анализ конкуренции антител к mCD25 с rmCD25, несущим His-метку, на Octet96. Показано связывание вторым антителом после захвата 7D4 на этапах ассоциации с сенсором и антигеном. Конкурентное связывание с mCD25 наблюдается между 7D4 и 2Е4 (А), но не между 7D4 и РС61 (В).
[0165] Фигура 14: характеристика 7D4, РС61 и 2Е4 в сравнении с контролем мышиного изотипа IgG2a или в отсутствие первичного антитела в отношении блокирования передачи сигналов IL-2 в анализе фосфорилирования STAT5 с использованием Т-клеток, выделенных из спленоцитов C57BL/6. Клетки инкубировали с 50 мкг/мл антитела, и затем с 50 Ед/мл IL-2. Анализ был ограничен процентом Трег-клеток, фосфорилирующих STAT5.
[0166] Фигура 15: истощение Трег в условиях in vivo у мышей balb/c, несущих опухоль 4Т1, после введения мышиного антитела к мышиному CD25 (7D4). (А)-(С): процент (%) отрицательных по CD4, CD4+ и CD25+FoxP3+ клеток, соответственно, в цельной крови на 3 день после введения дозы. (D)-(F): % отрицательных по CD4, CD4+ и CD25+FoxP3+ клеток, соответственно, в опухоли на 3 день после введения дозы. (G)-(I): % отрицательных по CD4, CD4+ и CD25+FoxP3+ клеток, соответственно, в цельной крови на 9 день после введения дозы. (J)-(L): % отрицательных по CD4, CD4+ и CD25+FoxP3+ клеток, соответственно, в опухоли на 9 день после введения дозы.
[0167] Фигура 16: характеристика связывания мышиного (В) или химерного (А, С и D) клона 7G7B6 к человеческому CD25 с CD25, экспрессируемым на клетках Karpas 299 (А), дифференцированных в условиях in vitro человеческих Трег-клетках (В), клетках SU-DHL-1 (С) или клетках SR-786 (D), при повышении концентрации антител и сравнение с контролем человеческого изотипа IgG1.
[0168] Фигура 17: характеристика 7G7B6 в сравнении с контролем мышиного изотипа IgG2a, контролем человеческого изотипа IgG1, даклизумабом или в отсутствие первичного антитела в отношении блокирования передачи сигналов IL-2 в анализе фосфорилирования STAT5 с использованием МКПК человеческого происхождения. Клетки инкубировали с 10 мкг/мл антитела с последующим повышением концентрации IL-2 (как показано на Фигурах). Анализ был ограничен процентом CD3-положительных клеток, фосфорилирующих STAT5.
[0169] Фигура 18: функциональная характеристика химерного 7G7B6 в сравнении с контролем человеческого изотипа IgG1, даклизумабом или коммерчески доступным нейтрализующим мышиным антителом к человеческому IL-2 в качестве положительного контроля (клон: AB12-3G4) с использованием общей популяции Т-клеток. Клетки инкубировали с 10 мкг/мл антитела, затем активировали гранулами CD3/CD28 в течение 72 часов перед анализом методом проточной цитометрии. Результаты показывают процент гранзим В-положительных пролиферирующих CD4 Т-клеток.
[0170] Фигура 19: функциональная характеристика химерного 7G7B6 в сравнении с контролем человеческого изотипа IgG1 в отношении уничтожения CD25-положительных клеточных линий в анализе АЗКЦ. Клетки с высокой или низкой экспрессией CD25, клетки SU-DHL-1 (А) или SR-786 (В), соответственно, культивировали совместно с очищенными NK-клетками в присутствии различных концентраций антител (как показано на Фигурах). Лизис клеток-мишеней измеряли по высвобождению кальцеина в супернатант через четыре часа после добавления к NK-клеткам. Данные нормировали к обработанным сапонином контролям.
[0171] Фигура 20: функциональная характеристика химерного 7G7B6 в сравнении с контролем человеческого изотипа IgG1 в отношении фагоцитоза дифференцированных в условиях in vitro Трег-клеток в анализе АЗКФ. Трег культивировали совместно с дифференцированными под действием MCSF макрофагами в присутствии различных концентраций антител (как показано на Фигурах). Анализ методом двухцветной проточной цитометрии выполняли с окрашенными по CD14+ макрофагами и Трег, мечеными красителем eFluor450. Остаточные клетки-мишени были определены как клетки, которые были eFluor450-краситель+/СВ14-. Как полагали, клетки с двойной меткой (eFluor450-краситель+/CD14+) представляют фагоцитоз мишеней макрофагами. Фагоцитоз клеток-мишеней рассчитывали по следующему уравнению: % фагоцитоза = 100×[(процент двойных положительных мишеней)/(процент двойных положительных мишеней + процент остаточных мишеней)].
[0172] Фигура 21: характеристика связывания МА-251 с CD25, экспрессируемым на клетках Karpas 299, при повышении концентрации антител и сравнение с контролем мышиного изотипа IgG1.
[0173] Фигура 22: характеристика МА-251 в сравнении с контролем мышиного изотипа IgG1, контролем человеческого изотипа IgG1, даклизумабом или в отсутствие первичного антитела. Блокирование передачи сигналов IL-2 в анализе фосфорилирования STAT5 оценивали с использованием МКПК человеческого происхождения. Клетки инкубировали с 10 мкг/мл антитела и затем с повышающимися концентрациями IL-2 (как показано на Фигурах). Анализ был ограничен процентом CD3-положительных клеток, фосфорилирующих STAT5.
[0174] Фигура 23: характеристика связывания МА-251 и IL-2 с CD25. Нарушение связывания IL-2-лиганда с CD25 выполняли на системе Forte Bio Octet Red384 (Pall Forte Bio Corp., США) с использованием стандартного анализа биннинга в формате «сэндвич». Антитело МА251 загружали на сенсоры AHQ, и незанятые Fc-связывающие сайты на сенсоре блокировали нецелевым человеческим антителом IgG1. Сенсоры подвергали воздействию 100 нМ человеческого CD25 и затем 100 нМ человеческого IL-2. Данные обрабатывали с использованием программного обеспечения для анализа данных Forte Bio 7.0. Дополнительное связывание человеческим IL-2 после ассоциации антигена указывает на незанятый эпитоп (не является конкурентом), тогда как отсутствие связывания указывает на блокирование эпитопа (конкурент).
[0175] Фигура 24: анализ конкуренции антител к CD25 в Octet. Связывание первого антитела с иммобилизованным rhCD25 с последующим повторным внесением либо первого антитела (в качестве контроля), либо второго антитела. Моноклональные антитела (МАТ), которые не блокируют сигнал IL-2, конкурируют друг с другом или с 7G7B6 и МА251 и не конкурируют с исследовательским даклизумабом или исследовательским базиликсимабом (Фигуры 24 (A)-(N)). (А)-(С) анализ конкуренции 7G7B6; (D)-(F) анализ конкуренции МА251; (G)-(I) и (N) анализ конкуренции антитела 3; (J)-(M) анализ конкуренции антитела 1. МАТ, которые блокируют передачу сигналов IL-2 (TSK031), действительно конкурируют с исследовательским даклизумабом и исследовательским базиликсимабом и не конкурируют с 7G7B6 (Фигуры 24 (O)-(Q)).
[0176] Фигура 25: модель в условиях in vivo, показывающая подавление роста опухоли после введения дозы: носителя (А) и (С); или антитела 1 (В), (D) и (Е).
[0177] Фигура 26: определение аффинности очищенных антител (IgG1) к rhCD25, несущему His-метку, с помощью анализа на основе SPR на Biacore 2000. A) 7g7B6ch, В) MA251ch, С) антитело 1, D) антитело 3 и Е) даклизумаб (контроль), или с помощью интерферометрии биослоев на приборе Octet Red 96 (F).
[0178] Фигура 27: характеристика связывания антитела 1 с CD25, экспрессируемым на дифференцированных в условиях in vitro человеческих Трег-клетках (А), клетках SU-DHL-1 (В) или клетках SR-786 (С), при повышении концентрации антител и сравнение с контролем человеческого изотипа IgG1.
[0179] Фигура 28: характеристика связывания антитела 1 с CD25, экспрессируемым на активированных гранулами CD3/CD28 человеческих Т-клетках (А) и на общей популяции Т-клеток (В), затем селективно пропущенных в области CD4+ и CD8+ Т-клеток, при повышении концентрации антител и сравнение с контролем человеческого изотипа IgG1.
[0180] Фигура 29: показано неконкурентное связывание антитела 1 и IL-2 (А) и конкурентное связывание IL-2-конкурирующего антитела с IL-2 (В), измеренное с помощью интерферометрии биослоев на Octet Red384 с использованием стандартного анализа биннинга в формате «сэндвич». Антитело к человеческому CD25, антитело 1, загружали на сенсоры AHQ. Затем сенсоры подвергали воздействию 100 нМ человеческого CD25 и затем человеческого IL-2. Дополнительное связывание человеческим IL-2 после ассоциации антигена указывает на незанятый эпитоп (не является конкурентом), тогда как отсутствие связывания указывает на блокирование эпитопа (конкурент).
[0181] Фигура 30: показано неконкурентное связывание антитела 1 и даклизумаба с CD25, измеренное с помощью интерферометрии биослоев в системе Octet Red384 с использованием стандартного анализа биннинга в формате «сэндвич». Эталонное моноклональное антитело к человеческому CD25, даклизумаб, загружали на сенсоры AHQ. Затем сенсоры подвергали воздействию 100 нМ антигена человеческого CD25 и затем антитела к человеческому CD25 (антитело 1). Дополнительное связывание вторым антителом после ассоциации антигена указывает на незанятый эпитоп (не является конкурентом), тогда как отсутствие связывания указывает на блокирование эпитопа (конкурент).
[0182] Фигура 31: характеристика антитела 1 в сравнении с контролем человеческого изотипа IgG1, даклизумабом или в отсутствие первичного антитела в отношении блокирования передачи сигналов IL-2 в анализе фосфорилирования STAT5 с использованием МКПК человеческого происхождения. Клетки инкубировали с 10 мкг/мл антитела и затем с повышающимися концентрациями IL-2 (как показано на Фигурах). Анализ был ограничен процентом CD3-положительных клеток, фосфорилирующих STAT5.
[0183] Фигура 32: функциональная характеристика антитела 1 в сравнении с контролем человеческого изотипа IgG1, даклизумабом или коммерчески доступным нейтрализующим мышиным антителом к человеческому IL-2 в качестве положительного контроля (клон: AB12-3G4) с использованием общей популяции Т-клеток. Клетки инкубировали с 10 мкг/мл антитела, затем активировали гранулами CD3/CD28 в течение 72 часов перед анализом методом проточной цитометрии. Результаты показывают процент гранзим В-положительных пролиферирующих CD4 (А) или CD8 (В) Т-клеток.
[0184] Фигура 33: функциональная характеристика антитела 1 в сравнении с контролем человеческого изотипа IgG1 в отношении уничтожения CD25-положительных клеточных линий в анализе АЗКЦ. Клетки с высокой или низкой экспрессией CD25, клетки SU-DHL-1 (А) или клетки SR-786 (В), соответственно, культивировали совместно с очищенными NK-клетками в присутствии различных концентраций антител (как показано на Фигурах). Лизис клеток-мишеней измеряли по высвобождению кальцеина в супернатант через четыре часа после добавления к NK-клеткам. Данные были нормированы к обработанным сапонином контролям.
[0185] Фигура 34: функциональная характеристика антитела 1 в сравнении с контролем человеческого изотипа IgG1 в отношении фагоцитоза дифференцированных в условиях in vitro Трег-клеток в анализе АЗКФ. Трег культивировали совместно с дифференцированными под действием MCSF (колонне стимулирующе го макрофагального фактора) макрофагами в присутствии различных концентраций антител (как показано на Фигурах). Анализ методом двухцветной проточной цитометрии выполняли с окрашенными по CD14+ макрофагами и Трег, мечеными красителем eFluor450. Остаточные клетки-мишени определяли как клетки, которые представляли собой eFluor450-краситель+/CD14-. Как полагали, клетки с двойной меткой (eFluor50-краситель+/CD14+) представляют фагоцитоз мишеней макрофагами. Фагоцитоз клеток-мишеней рассчитывали по следующему уравнению: % фагоцитоза = 100×[(процент двойных положительных мишеней)/(процент двойных положительных мишеней + процент остаточных мишеней)].
[0186] Фигура 35: характеристика связывания антитела 3 с CD25, экспрессируемым на дифференцированных в условиях in vitro человеческих Трег-клетках (А), клетках SU-DHL-1 (В) или клетках SR-786 (С), при повышении концентрации антител и сравнение с контролем человеческого изотипа IgG1.
[0187] Фигура 36: характеристика связывания антитела 3 с CD25, экспрессируемым на активированных гранулами CD3/CD28 человеческих Т-клетках (А) и (В) или Т-клетках яванской макаки (С) и (D) общей популяции Т-клеток, затем селективно пропущенных в области CD4+ и CD8+ Т-клеток, при повышении концентрации антител и сравнение с контролем человеческого изотипа IgG1.
[0188] Фигура 37: показано неконкурентное связывание антитела 3 и IL-2, измеренное с помощью интерферометрии биослоев на Octet Red384 с использованием стандартного анализа биннинга в формате «сэндвич». Антитело к человеческому CD25, антитело 3, загружали на сенсоры AHQ. Затем сенсоры подвергали воздействию 100 нМ человеческого CD25 и затем человеческого IL-2. Дополнительное связывание человеческим IL-2 после ассоциации антигена указывает на незанятый эпитоп (не является конкурентом).
[0189] Фигура 38: показано неконкурентное связывание антитела 3 и даклизумаба с CD25, измеренное с помощью интерферометрии биослоев в системе Octet Red384 с использованием стандартного анализа биннинга в формате «сэндвич». Эталонное моноклональное антитело к человеческому CD25, даклизумаб, загружали на сенсоры AHQ. Затем сенсоры подвергали воздействию 100 нМ антигена человеческого CD25 и затем антитела к человеческому CD25 (антитело 3). Дополнительное связывание вторым антителом после ассоциации антигена указывает на незанятый эпитоп (не является конкурентом), тогда как отсутствие связывания указывает на блокирование эпитопа (конкурент).
[0190] Фигура 39: характеристика антитела 3 в сравнении с контролем человеческого изотипа IgG1, даклизумабом или в отсутствие первичного антитела в отношении блокирования передачи сигналов IL-2 в анализе фосфорилирования STAT5 с использованием МКПК человеческого происхождения. Клетки инкубировали с 10 мкг/мл антитела и затем с повышающимися концентрациями IL-2 (как показано на Фигурах). Анализ был ограничен процентом CD3-положительных клеток, фосфорилирующих STAT5.
[0191] Фигура 40: функциональная характеристика антитела 3 в сравнении с контролем человеческого изотипа IgG1, даклизумабом или коммерчески доступным нейтрализующим мышиным антителом к человеческому IL-2 в качестве положительного контроля (клон: AB12-3G4) с использованием общей популяции Т-клеток. Клетки инкубировали с 10 мкг/мл антитела, затем активировали гранулами CD3/CD28 в течение 72 часов перед анализом методом проточной цитометрии. Результаты показывают процент гранзим В-положительных пролиферирующих CD4 (А) или CD8 (В) Т-клеток.
[0192] Фигура 41: функциональная характеристика антитела 3 в сравнении с контролем человеческого изотипа IgG1 в отношении уничтожения CD25-положительных клеточных линий в анализе АЗКЦ. Клетки с высокой или низкой экспрессией CD25, клетки SU-DHL-1 (А) или клетки SR-786 (В), соответственно, культивировали совместно с очищенными NK-клетками в присутствии различных концентраций антител (как показано на Фигурах). Лизис клеток-мишеней измеряли по высвобождению кальцеина в супернатант через четыре часа после добавления к NK-клеткам. Данные были нормированы к обработанным сапонином контролям.
[0193] Фигура 42: функциональная характеристика антитела 3 в сравнении с контролем человеческого изотипа IgG1 в отношении фагоцитоза дифференцированных в условиях in vitro Трег-клеток в анализе АЗКФ. Трег культивировали совместно с дифференцированными под действием MCSF макрофагами в присутствии различных концентраций антител (как показано на Фигурах). Анализ методом двухцветной проточной цитометрии проводили с окрашенными по CD14+ макрофагами и Трег, мечеными красителем eFluor450. Остаточные клетки-мишени определяли как клетки, которые представляли собой eFluor450-краситель+/CD14-. Как полагали, клетки с двойной меткой (eFluor450-краситель+/CD14+) представляют фагоцитоз мишеней макрофагами. Фагоцитоз клеток-мишеней рассчитывали по следующему уравнению: % фагоцитоза = 100×[(процент двойных положительных мишеней)/(процент двойных положительных мишеней + процент остаточных мишеней)].
[0194] Фигура 43: характеристика связывания антитела 4 с CD25, экспрессируемым на дифференцированных в условиях in vitro человеческих Трег-клетках (А), клетках SU-DHL-1 (В) или клетках SR-786 (С), при повышении концентрации антител и сравнение с контролем человеческого изотипа IgG1.
[0195] Фигура 44: характеристика связывания антитела 4 с немодифицированными клетками CHO-S (отрицательный контроль) (А) или клетками cyno-CD25-CHO-S (В) при концентрации антитела 100 нМ и сравнение с контролем человеческого изотипа IgG1.
[0196] Фигура 45: показано неконкурентное связывание антитела 4 и IL-2, измеренное с помощью интерферометрии биослоев на Octet Red384 с использованием стандартного анализа биннинга в формате «сэндвич». Антитело к человеческому CD25, антитело 4, загружали на сенсоры AHQ. Затем сенсоры подвергали воздействию 100 нМ человеческого CD25 и затем человеческого IL-2. Дополнительное связывание человеческим IL-2 после ассоциации антигена указывает на незанятый эпитоп (не является конкурентом).
[0197] Фигура 46: показано неконкурентное связывание антитела 4 и даклизумаба с CD25, измеренное с помощью интерферометрии биослоев в системе Octet Red384 с использованием стандартного анализа биннинга в формате «сэндвич». Эталонное моноклональное антитело к человеческому CD25, даклизумаб, загружали на сенсоры AHQ. Затем сенсоры подвергали воздействию 100 нМ антигена человеческого CD25 и затем антитела к человеческому CD25 (антитело 4). Дополнительное связывание вторым антителом после ассоциации антигена указывает на незанятый эпитоп (не является конкурентом), тогда как отсутствие связывания указывает на блокирование эпитопа (конкурент).
[0198] Фигура 47: характеристика антитела 4 в сравнении с контролем человеческого изотипа IgG1, даклизумабом или в отсутствие первичного антитела в отношении блокирования передачи сигналов IL-2 в анализе фосфорилирования STAT5 с использованием МКПК человеческого происхождения. Клетки инкубировали с 10 мкг/мл антитела и затем с повышающимися концентрациями IL-2 (как показано на Фигурах). Анализ был ограничен процентом CD3-положительных клеток, фосфорилирующих STAT5.
[0199] Фигура 48: функциональная характеристика антитела 4 в сравнении с контролем человеческого изотипа IgG1, даклизумабом или коммерчески доступным нейтрализующим мышиным антителом к человеческому IL-2 в качестве положительного контроля (клон: AB12-3G4) с использованием общей популяции Т-клеток. Клетки инкубировали с 10 мкг/мл антитела, затем активировали гранулами CD3/CD28 в течение 72 часов перед анализом методом проточной цитометрии. Результаты показывают процент гранзим В-положительных пролиферирующих CD4 (А) или CD8 (В) Т-клеток.
[0200] Фигура 49: функциональная характеристика антитела 4 в сравнении с контролем человеческого изотипа IgG1 в отношении уничтожения СВ25-положительных клеточных линий в анализе АЗКЦ. Клетки с высокой или низкой экспрессией CD25, клетки SU-DHL-1 (А) или клетки SR-786 (В), соответственно, культивировали совместно с очищенными NK-клетками в присутствии различных концентраций антител (как показано на Фигурах). Лизис клеток-мишеней измеряли по высвобождению кальцеина в супернатант через четыре часа после добавления к NK-клеткам. Данные были нормированы к обработанным сапонином контролям.
[0201] Фигура 50: функциональная характеристика антитела 4 в сравнении с контролем человеческого изотипа IgG1 в отношении индукции АЗКФ в репортерном биоанализе. Клетки SU-DHL-1, экспрессирующие CD25, культивировали совместно с Т-клетками Jurkat, генетически модифицированными для экспрессии FcγRIIa и NFAT-элемента ответа, который запускает экспрессию люциферазы (NFAT-RE-luc2), в присутствии различных концентраций антител (как показано на Фигурах).
[0202] Фигура 51: характеристика связывания антитела 2 с CD25, экспрессируемым на дифференцированных в условиях in vitro человеческих Трег-клетках (А), клетках SU-DHL-1 (В) или клетках SR-786 (С), при повышении концентрации антител и сравнение с контролем человеческого изотипа IgG1.
[0203] Фигура 52: характеристика связывания антитела 2 с CD25, экспрессируемым на активированных гранулами CD3/CD28 человеческих Т-клетках (А) и (В) или Т-клетках яванской макаки (С) и (D) общей популяции Т-клеток, затем селективно пропущенных в области CD4+ и CD8+ Т-клеток, при повышении концентрации антител и сравнение с контролем человеческого изотипа IgG1.
[0204] Фигура 53: показано неконкурентное связывание антитела 2 и IL-2 (А), а также конкурентное связывание антитела, конкурирующего с IL-2, и IL-2 (В), измеренное с помощью интерферометрии биослоев на Octet Red384 с использованием стандартного анализа биннинга в формате «сэндвич». Антитело к человеческому CD25, антитело 2, загружали на сенсоры AHQ. Затем сенсоры подвергали воздействию 100 нМ человеческого CD25 и затем человеческого IL-2. Дополнительное связывание человеческим IL-2 после ассоциации антигена указывает на незанятый эпитоп (не является конкурентом).
[0205] Фигура 54: показано неконкурентное связывание антитела 2 и даклизумаба с CD25, измеренное с помощью интерферометрии биослоев в системе Octet Red384 с использованием стандартного анализа биннинга в формате «сэндвич». Эталонное моноклональное антитело к человеческому CD25, даклизумаб, загружали на сенсоры AHQ. Затем сенсоры подвергали воздействию 100 нМ антигена человеческого CD25 и затем антитела к человеческому CD25 (антитело 2). Дополнительное связывание вторым антителом после ассоциации антигена указывает на незанятый эпитоп (не является конкурентом), тогда как отсутствие связывания указывает на блокирование эпитопа (конкурент).
[0206] Фигура 55: характеристика антитела 2 в сравнении с контролем человеческого изотипа IgG1, даклизумабом или в отсутствие первичного антитела в отношении блокирования передачи сигналов IL-2 в анализе фосфорилирования STAT5 с использованием МКПК человеческого происхождения. Клетки инкубировали с 10 мкг/мл антитела и затем с повышающимися концентрациями IL-2 (как показано на Фигурах). Анализ был ограничен процентом CD3-положительных клеток, фосфорилирующих STAT5.
[0207] Фигура 56: функциональная характеристика антитела 2 в сравнении с контролем человеческого изотипа IgG1 в отношении уничтожения СВ25-положительных клеточных линий в анализе АЗКЦ. Клетки с высокой или низкой экспрессией CD25, клетки SU-DHL-1 (А) или клетки SR-786 (В), соответственно, культивировали совместно с очищенными NK-клетками в присутствии различных концентраций антител (как показано на Фигурах). Лизис клеток-мишеней измеряли по высвобождению кальцеина в супернатант через четыре часа после добавления к NK-клеткам. Данные были нормированы к обработанным сапонином контролям.
[0208] Фигура 57: функциональная характеристика антитела 2 в сравнении с контролем человеческого изотипа IgG1 в отношении фагоцитоза дифференцированных в условиях in vitro Трег-клеток в анализе АЗКФ. Трег культивировали совместно с дифференцированными под действием MCSF макрофагами в присутствии различных концентраций антител (как показано на Фигурах). Анализ методом двухцветной проточной цитометрии проводили с окрашенными по CD14+макрофагами и Трег, мечеными красителем eFluor450. Остаточные клетки-мишени определяли как клетки, которые представляли собой eFluor450-краситель+/СВ14-. Как полагали, клетки с двойной меткой (eFluor450-краситель+/СВ14+) представляют фагоцитоз мишеней макрофагами. Фагоцитоз клеток-мишеней рассчитывали по следующему уравнению: % фагоцитоза=100×[(процент двойных положительных мишеней)/(процент двойных положительных мишеней + процент остаточных мишеней)].
[0209] Фигура 58: характеристика связывания антитела 5 с CD25, экспрессируемым на клетках Karpas 299, при повышении концентрации антитела и сравнение с контролем человеческого изотипа IgG1.
[0210] Фигура 59: характеристика антитела 5 в отношении блокирования передачи сигналов IL-2 в анализе фосфорилирования STAT5 с использованием МКПК человеческого происхождения. Мышиное антитело к человеческому белку, МА-251, использовали в качестве неблокирующего контроля, тогда как клинический даклизумаб-высокопродуктивный процесс (DAC HYP) использовали в качестве блокирующего контроля в сравнении с контролем мышиного изотипа IgG1, контролем человеческого изотипа IgG1 или в отсутствие первичного антитела, соответственно. Клетки инкубировали с 10 мкг/мл антитела и затем 10 Ед/мл IL-2. Анализ был ограничен процентом CD3-положительных клеток, фосфорилирующих STAT5.
[0211] Фигура 60: анализы конкуренции в Octet. Связывание антитела 1 с иммобилизованным rhCD25 с последующим повторным внесением либо первого антитела (антитело 1 в качестве контроля), либо второго антитела, либо конкурента IL-2, например, исследовательского даклизумаба и базиликеимаба, либо не конкурента IL-2, например, 7G7B6. Антитело 1 не конкурирует с блокаторами сигнала IL-2 исследовательским базиликсимабом (А) и даклизумабом (В), в то время как оно конкурирует с 7G7B6 (не является блокатором IL-2) (С).
[0212] Фигура 61: характеристика антитела 5 в сравнении с Fc-подавленным контрольным антителом к человеческому CD25 в отношении индукции АЗКЦ в репортерном биоанализе. Клетки SR-786, экспрессирующие CD25, культивировали совместно с Т-клетками Jurkat, генетически модифицированными для экспрессии FcyRIIIa и NFAT-элемента ответа, который запускает экспрессию люциферазы (NFAT-RE-luc2), в присутствии различных концентраций антител (как показано на Фигурах).
[0213] Фигура 62: функциональная характеристика антитела 5 в сравнении с контролем человеческого изотипа IgG1 в отношении фагоцитоза дифференцированных в условиях in vitro Трег-клеток в анализе АЗКФ. Трег культивировали совместно с дифференцированными под действием MCSF макрофагами в присутствии различных концентраций антител (как показано на Фигурах). Анализ методом двухцветной проточной цитометрии выполняли с окрашенными по CD14+макрофагами и Трег, мечеными красителем eFluor450. Остаточные клетки-мишени определяли как клетки, которые представляли собой eFluor450-краситель+/СВ14-. Как полагали, клетки с двойной меткой (eFluor450-краситель+/СВ14+) представляют фагоцитоз мишеней макрофагами. Фагоцитоз клеток-мишеней рассчитывали по следующему уравнению: % фагоцитоза=100×[(процент двойных положительных мишеней)/(процент двойных положительных мишеней + процент остаточных мишеней)].
[0214] Фигура 63: характеристика связывания антитела 6, антитела 7, антитела 8 и антитела 9 с CD25, экспрессируемым на клетках Karpas 299, при повышении концентрации антител и сравнение с контролем человеческого изотипа IgG1.
[0215] Фигура 64: анализы конкуренции в Octet. Связывание первого антитела (антитела 7) с иммобилизованным rhCD25 с последующим повторным внесением либо первого антитела (в качестве контроля), либо второго антитела даклизумаба (А) или базиликсимаба (В).
[0216] Фигура 65: характеристика антитела 7 в сравнении с контролем человеческого изотипа IgG1, даклизумабом-Нур или в отсутствие первичного антитела в отношении блокирования передачи сигналов IL-2 в анализе фосфорилирования STAT5 с использованием МКПК человеческого происхождения. Клетки инкубировали с 10 мкг/мл антитела и затем 10 Ед/мл IL-2. Анализ был ограничен процентом CD3-положительных клеток, фосфорилирующих STAT5.
[0217] Фигура 66: функциональная характеристика антитела 7 в сравнении с Fc-подавленным контрольным антителом к человеческому CD25 в отношении индукции АЗКЦ в репортерном биоанализе. Клетки SR-786, экспрессирующие CD25, культивировали совместно с Т-клетками Jurkat, генетически модифицированными для экспрессии FcγRIIIa и NFAT-элемента ответа, который запускает экспрессию люциферазы (NFAT-RE-luc2), в присутствии различных концентраций антител (как показано на Фигурах).
[0218] Фигура 67: функциональная характеристика антитела 7 в сравнении с Fc-подавленным контрольным антителом к человеческому CD25 в отношении фагоцитоза дифференцированных в условиях in vitro Трег-клеток в анализе АЗКФ. 1рег культивировали совместно с дифференцированными под действием MCSF макрофагами в присутствии различных концентраций антител (как показано на Фигурах). Анализ методом двухцветной проточной цитометрии выполняли с окрашенными по CD14+ макрофагами и ТРег, мечеными красителем eFluor450. Остаточные клетки-мишени определяли как клетки, которые были eFluor450-краситель+/СВ14-. Как полагали, клетки с двойной меткой (eFluor450-краситель+/СВ14+) представляют фагоцитоз мишеней макрофагами. Фагоцитоз клеток-мишеней рассчитывали по следующему уравнению: % фагоцитоза=100×[(процент двойных положительных мишеней)/(процент двойных положительных мишеней + процент остаточных мишеней)].
[0219] Фигура 68: характеристика связывания антитела 10, антитела 11, антитела 12, антитела 12, антитела 13, антитела 14, антитела 15, антитела 16, антитела 17, антитела 18, антитела 19, антитела 20 и антитела 21 с CD25, экспрессируемым на клетках Karpas 299, при повышении концентрации антител и сравнение с контролем человеческого изотипа IgG1.
[0220] Фигура 69: анализы конкуренции в Octet. Связывание первого антитела (антитела 19) с иммобилизованным rhCD25 с последующим повторным внесением либо первого антитела (в качестве контроля), либо второго антитела даклизумаба (А) или базиликсимаба (В).
[0221] Фигура 70: характеристика антитела 10, антитела 11, антитела 12, антитела 12, антитела 13, антитела 14, антитела 15, антитела 16, антитела 17, антитела 18, антитела 19, антитела 20 и антитела 21 в сравнении с контролем человеческого изотипа IgG1, даклизумабом-Нур или в отсутствие первичного антитела в отношении блокирования передачи сигналов IL-2 в анализе фосфорилирования STAT5 с использованием МКПК человеческого происхождения. Клетки инкубировали с 10 мкг/мл антитела и затем 10 Ед/мл IL-2. Анализ был ограничен процентом CD3-положительных клеток, фосфорилирующих STAT5.
[0222] Фигура 71: функциональная характеристика антитела 19 в сравнении с Fc-подавленным контрольным антителом к человеческому CD25 в отношении индукции АЗКЦ в репортерном биоанализе. Клетки SR-786, экспрессирующие CD25, культивировали совместно с Т-клетками Jurkat, генетически модифицированными для экспрессии FcyRIIIa и NFAT-элемента ответа, который запускает экспрессию люциферазы (NFAT-RE-luc2), в присутствии различных концентраций антител (как показано на Фигурах).
[0223] Фигура 72: функциональная характеристика антитела 19 в сравнении с Fc-подавленным контрольным антителом к человеческому CD25 в отношении фагоцитоза дифференцированных в условиях in vitro Трег-клеток в анализе АЗКФ. 1рег культивировали совместно с дифференцированными под действием MCSF макрофагами в присутствии различных концентраций антител (как показано на Фигурах). Анализ методом двухцветной проточной цитометрии выполняли с окрашенными по CD14+ макрофагами и Трег, мечеными красителем eFluor450. Остаточные клетки-мишени определяли как клетки, которые представляли собой eFluor450-краситель+/CD14-. Как полагали, клетки с двойной меткой (eFluor450-краситель+/CD14+) представляют фагоцитоз мишеней макрофагами. Фагоцитоз клеток-мишеней рассчитывали по следующему уравнению: % фагоцитоза=100×[(процент двойных положительных мишеней)/(процент двойных положительных мишеней + процент остаточных мишеней)].
[0224] Фигура 73: функциональная характеристика антитела 19, антитела 12 и антитела 20 в сравнение с Fc-подавленным контрольным антителом к человеческому CD25 в отношении фагоцитоза дифференцированных в условиях in vitro Трег-клеток в анализе АЗКФ. Трег культивировали совместно с дифференцированными под действием MCSF макрофагами в присутствии различных концентраций антител (как показано на Фигурах). Анализ методом двухцветной проточной цитометрии выполняли с окрашенными по CD14+ макрофагами и Трег, мечеными красителем eFluor450. Остаточные клетки-мишени были определены как клетки, которые были eFluor450-краситель+/CD14-. Как полагали, клетки с двойной меткой (eFluor450-краситель/CD14+) представляют фагоцитоз мишеней макрофагами. Фагоцитоз клеток-мишеней рассчитывали по следующему уравнению: % фагоцитоза=100×[(процент двойных положительных мишеней)/(процент двойных положительных мишеней + процент остаточных мишеней)].
[0225] Фигура 74: терапевтическая активность не блокирующего IL-2 антитела к CD25, мышиного IgG2a 7D4, в комбинации с GVAX в модели устойчивой к иммунотерапии В16 В16. Отдельных мышей лечили Gvax по отдельности или в комбинации с 7D4.
[0226] Фигура 75: терапевтическая активность не блокирующих IL-2 антител к CD25 (7D4 и 2Е4) в сравнении с блокирующим IL-2 антителом (РС61) в модели опухоли СТ26 с использованием самок мышей BALB/c. Неблокирующие антитела к CD25, 7D4 и 2Е4, проявляют эффективную терапевтическую активность против плотных опухолей. Как 7D4, так и 2Е4 более эффективно, чем блокирующее IL-2 антитело, РС61.
[0227] Фигура 76: оценка терапевтической активности не блокирующего IL-2 антитела к CD25, 7D4 mIgG2a, в модели МСА205 при однократных и повторных инъекциях в комбинации с антителом к мышиному PD-L1. *указывает на то, что мыши были живы в конце эксперимента.
Примеры
[0228] Пример 1 - Характеристика в условиях in vitro и получение рекомбинантных антител к мышиному CD25, истощающих Трег, которые либо не блокируют IL-2, либо блокируют IL-2
Материалы и методы
[0229] Происхождение антител и их рекомбинантное получение
Последовательность вариабельных областей тяжелой и легкой цепей крысиного антитела к мышиному CD25, РС61, выделяли из гибридомы РС-61.5.3 (каталожный номер АТСС TIB-222) с помощью быстрой амплификации концов кДНК (RACE) и затем клонировали в константные области мышиного IgG2a и κ-цепи (или соответствующие последовательности мышиного IgG1, которые выделяли из коммерческих плазмид (Invivogen)).
[0230] Затем каждую цепь антител субклонировали в ретровирусный вектор, полученный из вируса лейкоза мышей (MLV). Для предварительных экспериментов антитело получали с использованием клеток К562, трансдуцированных векторами, кодирующими как тяжелые, так и легкие цепи. Антитело очищали из супернатантов с использованием колонки с белком G HiTrap MabSelect (GE Healthcare), подвергали диализу в фосфатно-солевом буфере (ФСБ), концентрировали и стерилизовали путем фильтрации.
[0231] Переклонированная последовательность ДНК вариабельной тяжелой цепи, направленной против мышиного CD25, из антитела РС-61.5.3 (мышиный IgG2a) кодирует следующую последовательность белка:

[0232] Переклонированная последовательность ДНК вариабельной легкой цепи, направленной против мышиного CD25, из антитела РС-61.5.3 (мышиный IgG2a) кодирует следующую последовательность белка:

[0233] Секвенирование 7D4-IgM выполняли на гибридоме 7D4 (ЕСАСС, 88111402). Общую РНК или мРНК экстрагировали и проводили обратную транскрипцию для получения кДНК для тяжелой и легкой цепей антитела. Вариабельные тяжелые и вариабельные легкие цепи амплифицировали с использованием вырожденных прямых праймеров, которые связывались либо в сигнальном пептиде, либо в каркасном участке 1, и обратного праймера, который связывался в константной области антитела. Амплифицированные гены клонировали и секвенировали в соответствии со стандартным подходом. кДНК получали с помощью обратной транскрипции и к 3'-концу кДНК добавляли го мо полимерный хвост.Затем гены вариабельного домена антитела амплифицировали с использованием геноспецифических праймеров с последующим стандартным подходом клонирования и секвенирования. ДНК секвенировали с помощью обычного секвенирования по Сэнгеру, и данные анализировали с использованием программного обеспечения DNASTAR Lasergene. Последовательности сигнального пептида и вариабельного домена идентифицировали путем сравнения с известными последовательностями в базе данных IMGT.
[0234] Гены, кодирующие вариабельные тяжелые и вариабельные легкие домены, были оптимизированы по кодонам для экспрессии в человеческой клеточной линии и синтезированы с сайтами рестрикции NheI и AvaI, расположенными в 5'- и 3'-направлении относительно гена. Чтобы вставить ген вариабельного тяжелого домена 7D4 в отдельные векторы экспрессии, содержащие константные домены мышиных IgG1 и IgG2a, проводили клонирование с использованием расщепления рестриктазами. Чтобы вставить ген вариабельного легкого домена 7F4 в вектор экспрессии, содержащий константный домен мышиной каппа-цепи, проводили клонирование с использованием расщепления рестриктазами. Суспензию клеток НЕК293, культивированных в бессывороточных средах, подвергали химической трансфекции вектором экспрессии, несущим тяжелую и легкую цепи, и культивировали в течение еще 6 дней при 37°С в среде с 5% СО2 и при встряхивании при 140 об./мин. Культуры собирали с помощью центрифугирования при 4000 об./мин и дополнительно очищали с помощью фильтрации через фильтр с размером пор 0,22 мкм. Супернатант загружали на колонку с белком А, предварительно уравновешенную ФСБ, рН=7,2, элюировали цитратом натрия, рН=3,5, и уравновешивали 10% (об./об.) 0,5 М Tris, рН=9,0. Буфер в растворе нейтрализованного антитела заменяли ФСБ, рН=7,2, с использованием обессоливающей колонки и концентрировали, при необходимости, с использованием центробежного концентратора с отсечением по молекулярной массе 30 кДа. Концентрацию белка определяли путем измерения поглощения при 280 нм, и чистоту определяли с помощью электрофореза в ДСН-ПААГ.
[0235] Переклонированная последовательность ДНК тяжелой цепи, направленной против мышиного CD25, из антитела 7D4 (мышиный IgG1) кодирует следующую последовательность белка:

[0236] Переклонированная последовательность ДНК вариабельной тяжелой цепи, направленной против мышиного CD25, из антитела 7D4 (мышиный IgG2a) кодирует следующую последовательность белка:

[0237] Переклонированная последовательность ДНК легкой каппа-цепи, направленной против мышиного CD25, как для антитела 7D4 (mIg1), так и для антитела 7D4 (mIg2a) (мышиный IgG2a) кодирует следующую последовательность белка:

[0238] 2Е4 получали из гибридомы 2Е4 (подарок д-ра Этана М. Шевах (Ethan М. Shevach), Национальный институт здоровья). Секвенирование гибридомы выполняли с помощью запатентованной технологии на основе секвенирования следующего поколения (NGS). Образцы РНК использовали для создания библиотеки кДНК. Библиотеки секвенировали на платформе Illumina. Сборку de novo использовали для реконструкции транскриптома образца из необработанных данных. Последовательности вариабельных доменов идентифицировали путем сравнения с известными последовательностями.
[0239] Последовательность белка вариабельного тяжелого домена для антитела к мышиному CD25, 2Е4 (мышиный IgG1), имеет следующую последовательность белка:
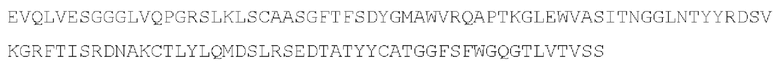
[0240] Последовательность белка вариабельного легкого домена для антитела к мышиному CD25, 2Е4 (mIg1), имеет следующую последовательность белка:
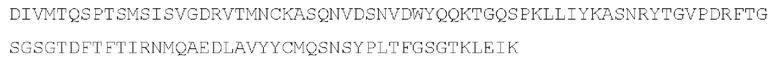
[0241] Оценка аффинности рекомбинантных антител в отношении мышиного CD25 Измерения аффинности с помощью ForteBio выполняли на Octet RED384, в целом, как описано ранее (см., например, Estep Р et al., 2013. Mabs. 5(2), 270-8). В общих чертах, измерения аффинности с помощью ForteBio выполняли, загружая IgG в режиме реального времени на сенсоры AHQ. Сенсоры уравновешивали в автономном режиме в буфере для анализа в течение 30 минут и затем контролировали в режиме реального времени в течение 60 секунд для установления исходного уровня. Сенсоры с нагруженными IgG подвергали воздействию 100 нМ антигена в течение 3 минут и затем переносили в буфер для анализа на 3 мин для измерения в автономном режиме. Все кинетики анализировали с использованием модели связывания в соотношении 1:1.
[0242] Результаты
Две мышиные гибридомы отбирали в качестве эталонного антитела для оценки связывания CD25 и Трег-истощающих свойств антител к мышиному CD25, которые либо не блокируют IL-2, либо блокируют IL-2: 7D4 (мышиный изотип IgM) и РС61 (мышиный изотип IgG1), соответственно. Свойства, относящиеся к связыванию IL-2, которые были описаны в литературе, предварительно подтверждали с использованием исходных нерекомбинантных антител и рекомбинантного мышиного IL-2 (Фиг. 1А). Рекомбинантные варианты таких антител получали для испытания антител, изотип которых является более активным и целесообразным для функциональных исследований (например, для истощения Трег или влияния на другие иммунные клетки). Кроме того, в случае 7D4, требовалось изменение изотипа, поскольку свойства агрегации антител IgM могут отрицательно влиять на результаты анализов. Рекомбинантный 7D4 (mIgG1), как и исходное нерекомбинантное антитело изотипа IgM, все еще допускает связывание мышиного IL-2 с мышиным CD25 (Фиг. 1B). 7D4 (mIgG1) также связывает мышиный CD25 на клеточной поверхности, аналогично рекомбинантному РС61 (IgG2a), тогда как эталонное антитело к человеческому CD25 не связывает его (Фиг. 1С).
[0243] Последовательность ДНК, кодирующую вариабельный домен тяжелой цепи 7D4 (а также РС61), также клонировали в вектор, который обеспечивает экспрессию домена, связывающего мышиный CD25, с мышиным изотипом IgG2a (функционально соответствующим человеческому IgG1). Таким способом можно сравнить два рекомбинантных антитела к мышиному CD25 с оптимизированной активностью АЗКЦ, которые могут эффективно истощать внутриопухолевые Трег, но проявляют различные свойства в отношении связывания мышиного IL-2 с мышиным CD25. Испытывали аффинность полученных рекомбинантных антител к мышиному CD25 в отношении CD25. Другой изотип (мышиный IgG2a или мышиный IgG1) не оказывает отрицательного влияния на это свойство, поскольку значения Kd сопоставимы между рекомбинантными антителами на основе 7D4 (приблизительно 1 нМ) и сопоставимы с Kd для РС61 (mIgG2a), которая, как измерено, составляет 4,6 нМ.
[0244] Функциональные свойства этих рекомбинантных антител также сравнивали в анализе в условиях in vitro для определения их влияния на выработку гранзима В в ответ на стимуляцию антителом к CD3 и антителом к CD28 (Фиг. 2). Гранзим В (GnzB) представляет собой сериновую протеиназу, экспрессируемую Т-клетками памяти и NK-клетками, а также активированными CD4 и CD8 Т-клетками, которые экспрессируют на высоких уровнях и секретируют GnzB во время иммунологических ответов. Этот фермент является важным посредником гибели клеток, патологии тканей и заболевания. С помощью цитокинов и антител можно оказывать отрицательное влияние на стимуляцию и пролиферацию Т-клеток в условиях in vitro под действием антитела к CD3 и антитела к CD28 (при этом >80% CD4 Т-клеток как пролиферируют, так и экспрессируют GnzB). Если эту стимуляцию выполняют в комбинации с нейтрализующим антителом к IL-2, ингибируется выработка гранзима В, но не пролиферация: частота клеток, которые как пролиферируют, так и вырабатывают GnzB, падает с >80% до<1%, а частота пролиферирующих клеток остается >90%. Это указывает на то, что выработка гранзима В, но не пролиферация клеток, зависит от передачи сигналов IL-2. Подобное снижение уровня Т-клеток, вырабатывающих гранзим В, наблюдается при добавлении РС61 (mIgG1) к стимулированным Т-клеткам. Однако 7D4 (mIgG1) в основном сохраняет способность CD4 Т-клеток отвечать на стимуляцию антителом к CD3 и антителом к CD28, вырабатывая GnzB (>65% клеток продолжают вырабатывать GnzB и пролиферировать). Эти результаты подтверждают, что антитела на основе РС61 блокируют передачу сигналов IL-2, тогда как 7D4 оказывает лишь незначительное влияние на эту передачу сигналов и, таким образом, может применяться в качестве суррогатного антитела для оценки терапевтического потенциала антитела к человеческому CD25, в частности, в отношении истощения Трег и специфических для опухоли свойств, которые будут влиять на передачу сигналов IL-2.
[0245] ПРИМЕР 2 - Трег-истощающие и противораковые свойства рекомбинантных антител к мышиному CD25, истощающих Трег, которые либо не блокируют IL-2, либо блокируют IL-2 Материалы и метод
[0246] Мыши
Исследования в условиях in vivo были выполнены Charles River Discovery Services North Carolina (CR Discovery Services). Самки мышей (BALB/c AnNcrl, Charles River) и самки мышей C57BL/6 (C57BL/6Ncrl, Charles River) были в возрасте от семи до девяти недель в начале исследования. CR Discovery Services, в частности, удовлетворяет рекомендациям Руководства по уходу и использованию лабораторных животных в отношении ограничений, условий содержания, хирургических процедур, нормативных требований к кормам и жидкостям, а также ветеринарной помощи. Программа по уходу за животными и их использованию в CR Discovery Services аккредитована Международной ассоциацией по оценке и аккредитации лабораторных исследований на животных, которая обеспечивает соответствие принятым стандартам по уходу и использованию лабораторных животных.
[0247] Клеточные линии и тканевая культура
Опухолевые клетки МСА205 (слабоиммуногенные клетки фибросаркомы, индуцированной 3-метилхолантреном; от Г. Креме (G. Kroemer), Институт исследований рака Густава Русси) культивировали в модифицированной Дульбекко среде Игла (DMEM, Sigma), дополненной 10% фетальной телячьей сыворотки (ФТС, Sigma), 100 Ед/мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ L-глутамина (все от Gibco). Клетки карциномы толстой кишки мыши МС38 (CR Discovery Services) выращивали до середины логарифмической фазы в модифицированной Дульбекко среде Игла (DMEM), содержащей 10% фетальной бычьей сыворотки, 2 мМ глутамина, 100 единиц/мл пенициллина G, 100 мкг/мл сульфата стрептомицина и 25 мкг/мл гентамицина. Клетки карциномы толстой кишки мыши СТ26 (CR Discovery Services) выращивали в среде RPMI-1640, содержащей 10% фетальной бычьей сыворотки, 2 мМ глутамина, 100 Ед/мл натриевой соли пенициллина G, 100 мкг/мл сульфата стрептомицина и 25 мкг/мл гентамицина. Все опухолевые клетки культивировали в колбах для тканевых культур в инкубаторе с увлажнением при 37°С в атмосфере 5% СО2 и 95% воздуха. Клетки К562, использованные для получения антител, культивировали в среде Дульбекко в модификации Искова (IMDM), не содержащей феноловый красный, с добавлением 10% ФТС, истощенной по IgG (Life Technologies).
[0248] Эксперименты с опухолями в условиях in vivo
Культивируемые опухолевые клетки обрабатывали трипсином (МСА205) или оставляли без обработки (МС38 и СТ26), промывали, ресуспендировали в ФСБ и инъецировали подкожно (п/к) в бок (5 × 105 клеток для моделей МСА205 и МС38 у мышей C57BL/6; 3 × 105 15 клеток для моделей СТ26 у мышей BALB/c). Антитела вводили путем инъекции внутрибрюшинно (в/б) в моменты времени, указанные в подписях к фигурам. Для функциональных экспериментов опухоли и дренирующие лимфатические узлы собирали через 12 дней после имплантации опухоли и обрабатывали для анализа методом проточной цитометрии, как описано (Simpson et al. (2013) J Exp Med 210, 1695- 710). Для терапевтических экспериментов опухоли измеряли два раза в неделю, и объемы рассчитывали как произведение трех ортогональных диаметров.
[0249] Проточная цитометрия
Регистрацию сигнала выполняли с помощью BD LSR II Fortessa (BD Biosciences). Использовали следующие антитела: антитело к CD3 (клон 145-2С11, ebioscience, 25003182), антитело к CD4 (клон RM4-5, BD biosciences, 560782), антитело к CD8 (клон 53-6.7, Biolegend, 100750), антитело к гранзиму В (клон GB11, Invitrogen), антитело к FoxP3 (клон FJK-16s, eBiosciences) и Ki67 (клон SolA15, eBiosciences, 48569882). Лимфатические узлы (паховые, подмышечные и плечевые) и опухоли мышей иссекали в бессывороточной RPMI. Лимфатические узлы диспергировали через фильтр с размером пор 70 мкм, тогда как опухоли механически разрушали с использованием gentleMACS (Miltenyl Biotech) и расщепляли с использованием смеси 0,33 мг/мл ДНКазы (Sigma-Aldrich) и 0,27 мг/мл либеразы TL (Roche) в бессывороточной RPMI в течение 30 мин при 37°С. Опухоли фильтровали через фильтр с размером пор 70 мкм, и полученные суспензии отдельных опухолевых клеток обогащали лейкоцитами путем пропускания через градиент Ficoll-paque (GE Healthcare). Опухоли и лимфатические узлы (LN) промывали в полной RPMI, повторно суспендировали в буфере для FACS (500 мл ФСБ, 2% ФТС, 2 мМ ЭДТА) и помещали в 96-луночные планшеты с круглым дном. Концентрированную реакционную смесь поверхностных антител готовили в рекомендованном производителем разведении: антитело к CD3 (клон 145-2С11, ebioscience, 25003182), антитело к CD4 (клон RM4-5, BD biosciences, 560782), антитело к CD8 (клон 53-6.7, Biolegend, 100750). В концентрированную реакционную смесь поверхностных антител также включали подходящий для фиксации краситель для оценки жизнеспособности (eFlour780, eBioscience). После пермеабилизации в течение 20 минут с использованием набора буферов для внутриклеточной фиксации и пермеабилизации (eBioscience) применяли панель для внутриклеточного окрашивания, состоящую из следующих антител, использованных в разведении, рекомендованном производителями: антитело к гранзиму В (клон GB11, Invitrogen), антитело к FoxP3 (клон FJK-16s, eBiosciences) и Ki67 (клон SolA15, eBiosciences, 48569882).
[0250] Результаты
Модель саркомы МСА205 на мышах позволяет получать мышей, у которых за короткое время можно оценить иммунологический ответ и общую эффективность против плотной опухоли для панели иммуномодулирующих соединений. В частности, рекомбинантные антитела к мышиному CD25 на основе мышиного IgG2a испытывали для оценки изменений в субпопуляциях Т-клеток, которые представлены как инфильтрирующие опухоль лимфоциты, или в периферических лимфатических узлах, а также роста опухоли и жизнеспособности мышей, подвергшихся воздействию МСА205. Дополнительное антитело (антитело к мышиному PD1) включали в исследование в качестве отрицательного контроля иммунологического влияния на Трег.
[0251] Иммунологический анализ показал, что антитело 7D4, при клонировании в остов мышиного IgG2a, проявляет способность, эквивалентную таковой РС61 (мышиный IgG2a), в отношении истощения Трег и последующего увеличения отношения Тэфф к Трег как в опухоли, так и на периферии, тогда как антитело к PD1 является неэффективным как по отдельности, так и в комбинации (Фиг. 3). Соответственно, любой дополнительный эффект, который измеряли с использованием 7D4 (mIgG2a) в качестве суррогатного антитела для антитела к человеческому CD25, не блокирующего IL-2, по-видимому, не связан с изменениями свойств, относящихся к истощению Трег.
[0252] Мыши, являющиеся моделью МСА205, которых лечили с применением 7D4, также имели более высокий процент GnzB-положительных клеток, таких как пролиферирующие СВ4-положительные и CDS-положительные Т-клетки, в отношении не только лечения антителом к PD1, но также IL-2-блокирующим антителом РС61 (mIg2a). У таких мышей, получавших лечение, 7D4 (mIg2a), как и РС61 (mIg2a), не только не оказывает отрицательного влияния на Тэфф клетки, но оно также увеличивает частоту клеток Тэфф по сравнению с РС61 (mIg2a), это указывает на еще более высокую противоопухолевую активность антитела к человеческому CD25, которое не блокирует взаимодействие IL-2/CD25 (Фиг. 4).
[0253] Применение антитела к человеческому CD25, функционально эквивалентного 7D4 (mIg2a) для иммунотерапии рака, в частности, для плотных опухолей, может быть испытано в модели МСА205 на мышах, но также в других моделях, таких как модели СТ26 и МС38 (карцинома толстой кишки) или В16 (меланома). Оба антитела IgG2a к мышиному CD25 проявляли терапевтическую активность против развившихся опухолей СТ26 при введении в комбинации с антителами к PD1. Интересно отметить, что, при применении в качестве монотерапии, антитело 7D4 (mIg2a), не блокирующее IL-2, проявляет явно более высокую терапевтическую активность, чем антитело на основе РС61, имеющее тот же изотип.В конце эксперимента все мыши, которых лечили 7D4 (mIg2a), проявляли контроль роста опухоли только до объема менее 50 мм3, в то время как ни у одной из мышей, которых лечили РС61 (mIg2a), не наблюдали опухоль менее 50 мм3, при этом опухоли у 8 из 10 мышей даже достигли конечной точки 2000 мм3. Это также иллюстрируется различием выживаемости, при этом все мыши, которых лечили 7D4 (mIg2a), были живы на 50 день по сравнению с всего 2 из 10 мышей, которых лечили РС61 (mIg2a). Действительно, если эффективность РС61 (mIg2a) значительно улучшается при комбинировании с антителом к PD1, эффективность 7D4 (mIg2a) дополнительно не улучшается, по меньшей мере при применении этого антитела в данной концентрации.
[0254] Поскольку эти антитела на основе 7D4 и РС61 показали сходную способность истощать Трег (см. Фиг. 3), такое различие в эффективности можно объяснить, по меньшей мере частично, меньшим влиянием 7D4 (mIg2a) на взаимодействие между IL-2 и его рецептором. Это свидетельствует о том, что отсутствие активности, направленной на блокирование IL-2/рецептор IL-2, не только не ухудшает терапевтическую активность, но оно также может обеспечить терапевтическое преимущество. Следовательно, эти данные подтверждают отбор антитела, нацеленного на CD25, не блокирующего IL-2/рецептор IL-2, для применения в терапии рака. Эти предпочтительные свойства антитела 7D4 (mIg2a) также были подтверждены при применении антитела к мышиному PD-L1 в той же модели СТ26 на мышах (Фиг. 6) или при применении модели МС38 на мышах с теми же комбинациями антител (Фиг. 7 и Фиг. 8).
[0255] Эти данные свидетельствуют о том, что свойства антител, направленные на истощение Трег и связывание CD25, на основе свойств 7D4 и с соответствующим изотипом, можно применять в комбинации с другими противораковыми соединениями, такими как антитела, нацеленные на белки контрольной точки иммунитета (например, против PD-1 и против PD-L1) или против других мишеней, имеющих отношение к раку. Этот подход можно осуществлять с помощью получения и введения двух продуктов в виде новой смеси моноспецифических антител или в качестве новых биспецифических антител. Этот подход, который включает конструирование биспецифических антител, сочетающих два антигенсвязывающих свойства и терапевтически целесообразный изотип (например, человеческий IgG1), может быть подтвержден с использованием технологии DuoBody®, которая позволяет эффективно связывать отдельные тяжелые и легкие цепи из двух различных моноспецифических антител, которые вырабатываются по отдельности и содержат единичные соответствующие точечные мутации в домене СНЗ, что обеспечивает обмен Fab в одном гетеромерном белке (Labrijn AF et al., Nat Protoc. 2014, 9:2450-63). Функциональные свойства таких продуктов DuoBody на основе 7D4 (например, включая антитело к PD1 или антитело к PD-L1) можно оценить с использованием моделей взаимодействия и истощения клеток, которые использовали для подтверждения антител на основе 7D4 и комбинаций антител, как описано выше.
[0256] Эти результаты также указывают на то, что связывающие свойства 7D4 в отношении мышиного CD25, без нарушения взаимодействия IL-2 с его рецептором и передачи сигналов IL-2 в клетках, экспрессирующих CD25, можно применять в антителе к человеческому CD25, в котором изотип выбран в соответствии с этим механизмом действия (например, человеческий IgG1). Действительно, некоторые другие свойства могут быть рассмотрены для скрининга антител-кандидатов к человеческому CD25 с дополнительными улучшенными свойствами в отношении их получения, применения и/или введения для лечения рака и, в частности, плотных опухолей.
[0257] Эти свойства также могут быть определены в отношении признаков известного антитела к человеческому CD25, такого как хумакс-ТАС, базиликсимаб или даклизумаб, все из которых имеют Kd для человеческого CD25 в наномолярном диапазоне, но все блокируют связывание человеческого IL-2 с человеческим CD25 (используя клон М-А251 в качестве потенциального эталонного антитела к человеческому CD25, не блокирующего IL-2, который должен быть включен в отбор антитела к человеческому CD25 согласно настоящему изобретению).
[0258] Эти признаки могут представлять собой одно или более из следующих:
- аффинность в отношении выделенного рекомбинантного мономерного человеческого CD25 с KD менее 25 нМ, предпочтительно менее 10 нМ и еще более предпочтительно менее 1 нМ (как установлено с использованием таких технологий как Octet, Kinexa, ИФА или др.);
- перекрестная реактивность в отношении выделенного рекомбинантного мономерного CD25 яванской макаки с KD менее 75 нМ, предпочтительно менее 30 нМ и еще более предпочтительно менее 3 нМ (как установлено с использованием таких технологий как Octet, Kinexa, ИФА или др.);
- аффинность в отношении рекомбинантного мономерного человеческого CD25 на поверхности клеток СНО или MJ с KD менее 100 нМ, предпочтительно менее 10 нМ и даже более предпочтительно менее 1 нМ (как установлено с использованием таких технологий как проточная цитометрия, ИФА на основе клеток или др.);
- аффинность в отношении рекомбинантного мономерного CD25 яванской макаки на поверхности клеток СНО с KD менее 300 нМ, предпочтительно менее 30 нМ и еще более предпочтительно менее 3 нМ (как установлено с использованием таких технологий как проточная цитометрия, ИФА на основе клеток или др.);
- связывание человеческих Трег-клеток с KD менее 100 нМ, предпочтительно менее 10 нМ и еще более предпочтительно менее 1 нМ (как установлено с использованием таких технологий как проточная цитометрия, ИФА на основе клеток или др.);
- связывание Трег-клеток яванской макаки с KD менее 300 нМ, предпочтительно менее 30 нМ и еще более предпочтительно менее 3 нМ (как установлено с использованием таких технологий как проточная цитометрия, ИФА на основе клеток или др);
- отсутствие ингибирования взаимодействия между рекомбинантным человеческим IL-2 и рекомбинантным человеческим CD25 в биохимическом анализе (менее 25% связывания IL-2 с CD25 блокируется при скрининге, описанном в Примере 1);
- отсутствие IL-2-индуцированной передачи сигналов в клеточном анализе, таком как фосфорилирование STAT5 в активированных CDS-положительных или CD4-положительных Т-клетках или СВ25-экспрессирующих клеточных линиях, или активация гранзима В после активации CD4-положительных Т-клеток в анализе (ингибируется менее 25% сигнала в исходных условиях, как описано в Примере 1); и/или
- соответствующая оценка эффективности в клеточных анализах, таких как анализ АЗКЦ, АЗКФ и/или КЗЦ в клеточных линиях, экспрессирующих человеческий CD25, или первичных Трег-клетках (с ЭК50 менее 10 нМ, предпочтительно менее 1 нМ и даже более предпочтительно менее 0,1 нМ).
[0259] Пример 3. Дополнительные эксперименты в модели на мышах в условиях in vivo с применением антител к мышиному CD25, не блокирующих IL-2
[0260] Материалы и методы
Терапевтическая активность антитела, не блокирующего IL-2. Самкам мышей BALB/c, полученным из Charles River, вводили путем инъекции 3 × 105 опухолевых клеток СТ26 в 0% матригеле подкожно в бок, n=15 на группу. Животных рандомизировали в группы лечения в зависимости от массы тела на 1 день. Лечение начинали на 6 день, и мыши получили одну инъекцию каждого антитела (мышиный изотип IgG2a, IL-2-нейтрализующее антитело, РС61 mIgG1, антитело мышиного изотипа IgG1 к мышиному CD25, блокирующее передачу сигналов IL-2, и 7D4 mIgG2a, антитело мышиного изотипа IgG2a к мышиному CD25, не блокирующее передачу сигналов IL-2) в дозе 200 мкг/животное. Животные получили либо монотерапию, с одной группой на антитело, либо комбинированную терапию 7D4 mIgG2a и нейтрализующего IL-2 антитела или 7D4 mIgG2a и антитела PCI mIgG1. Мышей умерщвляли, когда объем опухоли достигал 2000 мм3 или через 50 дней, в зависимости от того, что было достигнуто первым.
[0261] Терапевтическая активность антител, не блокирующих IL-2, по сравнению с блокирующими антителами
3 × 105 клеток СТ26 имплантировали подкожно в бок. Соотнесение пар выполняли на 0 день, когда опухоли достигли 30-60 мм3, и было начато лечение. На 1 день и каждые две недели после этого вводили внутрибрюшинно дозу 10 мг/кг. Группы лечили с применением IL-2-нейтрализующего антитела РС61-m2a, антитела 7D4, не блокирующего IL-2, антитела 2Е4, не блокирующего IL-2, или оставляли без лечения.
[0262] Терапевтическая активность антитела, не блокирующего IL-2, в комбинации с терапией aPDL1
Мышам вводили путем подкожной инъекции 50000 опухолевых клеток МСА205, n=10 на группу или n=5, как показано на фигурах. Животных рандомизировали в группы лечения. Животные получали либо монотерапию, либо 7D4 mIgG2a, либо aPD-L1 (клон 10F.9G2), комбинированное лечение 7D4 mIgG2a с aPD-L1 (клон 10F.9G2), либо лечение не применяли. Группы получали либо: a7D4 mIgG2a по отдельности 10 день (200 мкг), aPD-L1 rIgG2b (10F.9G2) - 6, 9 и 12 день (200 мкг), комбинацию aPD-L1 + a7D4 (aPDL-1 на 6, 9 и 12 день и a7D4 на 10 день) или комбинацию aPD-L1+a7D4 (aPDL-1 на 6, 9 и 12 день и a7D4 на 10 день), дополнительное введение a7D4 на 15 день+aPD-L1 на 18 день (только 5 мышей).
[0263] Результаты
Истощающее антитело mIgG2a 7D4 к CD25, которое не блокирует IL-2, индуцировало отторжение опухоли у получавших лечение мышей, тогда как другие антитела не оказывали влияния в качестве монотерапии по сравнению с контролем мышиного изотипа IgG2a. Комбинация с IL-2-блокирующими антителами, как с mIgG1 РС61, так и с нанотелом к IL-2, устраняла терапевтическую активность антитела 7D4 mIgG2a, не блокирующего IL-2 (Фигура 13). Это демонстрирует, что отсутствие способности 7D4 mIgG2a блокировать IL-2 является ключевым для терапевтической активности. Это также указывает на то, что терапевтическая активность этого антитела зависит от противоопухолевого иммунного ответа, опосредуемого эффекторными Т-клетками, которые зависят от передачи сигналов IL-2, для достижения оптимальной активности. Эти результаты показывают, что отсутствие блокирующей активности по отношению к IL-2/CD25 необходимо для оптимальной терапевтической активности антитела, нацеленного на CD25, и поддерживает применение антитела к CD25, не блокирующего IL-2, описанного в настоящем документе, в терапии рака.
[0264] Эти результаты дополнительно показывают, что отсутствие блокирующей активности в отношении IL-2/CD25 не оказывает вредного воздействия на терапевтическую активность антител и поддерживает применение антитела к CD25, не блокирующего IL-2, описанного в настоящем документе, в терапии рака.
[0265] Эти результаты дополнительно показали, что антитела 7D4 и 2Е4, не блокирующие IL-2, являются более эффективными, чем блокирующее IL-2 антитело, РС61. Неблокирующие антитела к CD25, 7D4 и 2Е4, проявляют эффективную терапевтическую активность против плотных опухолей (Фигура 75).
[0266] Результаты показали, что однократные или повторные инъекции антитела 7D4 к aCD25, не блокирующего IL-2, после начала терапии aPDL1 усиливают противоопухолевые ответы. Антитело к CD25 защищает и стимулирует Тэфф клетки, активированные после обработки aPDL1 (Фигура 76).
[0267] Пример 4 - характеристика эпитопов антител к CD25, не блокирующих IL-2
[0268] Биннинг эпитопов
Биннинг эпитопов антител выполняли на системе Forte Bio Octet Red384 (Pall Forte Bio Corporation, Менло-Парк, Калифорния, США) с использованием стандартного анализа биннинга в формате «сэндвич». Антитело к мышиному CD25, РС61, загружали на сенсоры АМС, и незанятые Fc-связывающие сайты на сенсоре блокировали нецелевым мышиным антителом IgG1. Затем сенсоры подвергали воздействию 15 нМ целевого антигена и затем антитела 7D4. Данные обрабатывали с использованием программного обеспечения для анализа данных ForteBio 7.0. Дополнительное связывание вторым антителом после ассоциации антигена указывает на незанятый эпитоп (не является конкурентом), в то время как отсутствие связывания указывает на блокирование эпитопа (конкурент).
[0269] Картирование эпитопов антител к CD25, не блокирующих IL-2
Различные наборы линейных пептидов, однопетлевых пептидов, пептидов, имитирующих β-виток, дисульфидные мостики, прерывистые дисульфидные мостики, прерывистые эпитопы, представляющих последовательность человеческого CD25 (запись в Uniprot № Р01589), синтезировали с использованием твердофазного Fmoc-синтеза (Pepscan BV, The Netherlands; Timmermann P et al., 2007 J. Mol. Recognit., 20, 283-99; Langedijk JP et al., 2011, Analytical Biochemistry. 417: 149-155). Связывание антител с каждым из синтезированных пептидов испытывали методом ИФА (Pepscan, Нидерланды). Пептидные массивы инкубировали с раствором первичного антитела (в течение ночи при 4°С). После промывки пептидные массивы инкубировали с соответствующим конъюгатом пероксидазы и антитела в разведении 1/1000 (2010-05; Southern Biotech) в течение одного часа при 25°С. После промывки добавляли субстрат пероксидазы сульфонат 2,2'-азино-ди-3-этилбензтиазолина (ABTS) и 20 мкл/мл 3% Н2О2. Через час измеряли развитие окрашивания. Развитие окрашивания определяли количественно с помощью камеры с зарядовой связью (CCD) и системы обработки изображений. Значения, полученные от CCD-камеры, варьировались от 0 до 3000 мЕОП, аналогично стандартному считывателю для 96-луночных планшетов для ИФА. Чтобы проверить качество синтезированных пептидов, отдельный набор положительных и отрицательных контрольных пептидов синтезировали в это же время и подвергали скринингу с нецелевыми контрольными антителами.
[0270] Результаты
Биннинг эпитопов выполняли для того чтобы определить, связываются ли антитела с эпитопами, перекрывающимися с эпитопами для коммерчески доступного мышиного антитела к человеческому CD25, не блокирующего IL-2, 7G7B6. Антитела дополнительно характеризовали для определения эпитопов для антител, не блокирующих IL-2. Эпитоп блокирующего антитела к мышиному CD25, РС61, определяли для сравнения в качестве контроля. Результаты картирования эпитопа представлены в таблице 1 для антител к человеческому CD25 и в таблице 2 для антител к мышиному CD25:
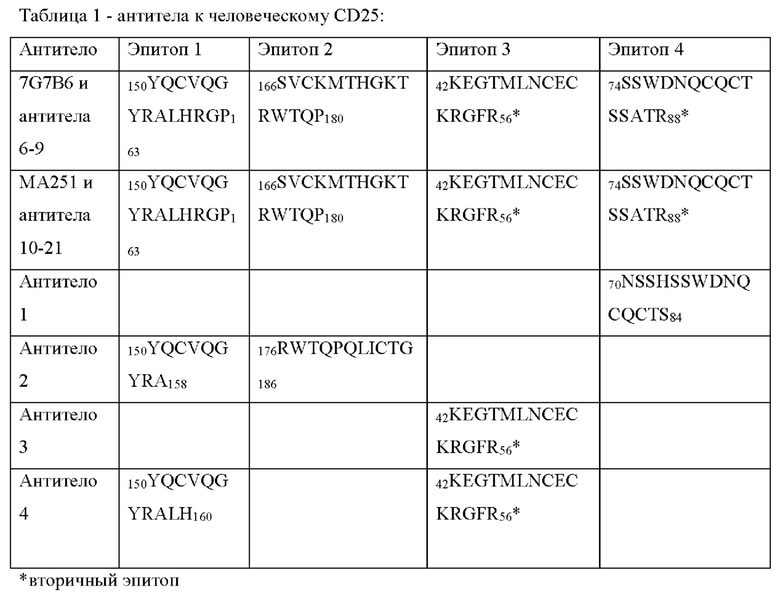
Нумерация аминокислотных (ак.) последовательностей основана на человеческом CD25, взятом из последовательности, опубликованной под номером доступа в Uniprot Р01589.

Нумерация аминокислотной (ак.) последовательности основана на мышином CD25, взятом из последовательности, опубликованной под номером доступа в Uniprot Р01590.
[0271] Исследование с картированием эпитопов, которое выполняли с использованием технологии Pepscan, свидетельствует о том, что антитела к человеческому белку связывают человеческий CD25 в эпитопе, который не перекрывается с сайтом связывания IL-2 на CD25. Антитела к человеческому белку связываются с другим эпитопом, нежели базиликсимаб и даклизумаб. Эпитоп для базиликсимаба и даклизумаба содержит остатки в области аминокислот 137-143 (SEQ ID NO: 1), которая перекрывается со стороной взаимодействия CD25 с IL-2 (Binder М et al, Cancer Res 2007 vol 67 (8): 3518-23). Неблокирующие антитела к мышиному CD25, 2Е4 и 7D4, распознают другой эпитоп, нежели РС61.
[0272] Пример 5: Характеристика мышиных антител к CD25
Связывание антител с клетками СНО, экспрессирующими мышиный CD25
Связывание с клетками СНО, экспрессирующими CD25, исследовали путем окрашивания испытываемых изделий (первичных антител к CD25, 7D1, РС61 и 2Е4) с применением 30 мг/мл антител с последующим полулогарифмическим последовательным разведением (7 точек) в течение 30 минут на льду. После этого окрашивали вторичным антителом (Alexa Fluor 647-AffiniPure козий фрагмент Fab к человеческому IgG (H+L) - (Jackson ImmunoResearch)) в концентрации 1 мг/мл в течение 30 минут на льду. Все образцы окрашивали с двумя повторами. Живые клетки селективно пропускали с использованием параметров FSC по отношению к SSC с помощью проточной цитометрии во время отбора образцов. Среднюю интенсивность флуоресценции (MFI) окрашенных клеток наносили на XY-диаграмму, на которой в графической форме представлена зависимость MFI от логарифма концентрации, и данные аппроксимировали с помощью кривой нелинейной регрессии, на основании которой рассчитывали ЭК50. Результаты, представленные на Фигуре 11, подтвердили, что антитела к мышиному CD25 связываются с клетками СНО, экспрессирующими мышиный CD25.
[0273] Измерения аффинности антител к mCD25
Аффинность антител к мышиному CD25, 7D4, РС61 и 2Е4, определяли путем измерения их KD с помощью SPR в Biacore 2000 с использованием сенсорного чипа СМ-5 при температуре окружающей среды 25°С во время эксперимента. Антитело к мышиному белку первоначально иммобилизовали во всех проточных ячейках в буфере для анализа (рН=7,4, 10 мМ HEPES, 150 мМ NaCl, 3 мМ ЭДТА, 0,05% Твин 20) до 16000-18000 единиц ответа (RU) в течение 10 минут. Лиганд (испытываемые изделия-антитела) последовательно загружали до уровня захвата 119-163 RU. Затем анализируемое вещество (His-меченый рекомбинантный мышиный CD25) связывали в буфере для анализа из 2-кратного разведения, начиная с 800 нМ, с самой низкой концентрацией 3,13 нМ, в течение 6 минут. Диссоциацию проводили в буфере для анализа в течение 10 минут. Этапы регенерации между концентрациями образца выполняли в 10 мМ глицине с рН=1,7 в течение 10 минут. Скорость потока 25 мкл/мин поддерживали на протяжении всего процесса. Кинетические данные аппроксимировали с использованием модели связывания бивалентных аналитов с помощью программного обеспечения для общего анализа данных, предоставленного Biacore, с вычитанием эталона. Результаты анализа на основе SPR представлены на Фигуре 12. Значения Kd, которые были установлены в этом анализе для антител к мышиному CD25, являются следующими: для 7D4, 2,6×109 М; для 2Е4, 114×10-9 М, и для РС61, 3,6×10-9 М (результат не представлен).
[0274] Конкуренция антител к мышиному белку в Octet
Исследование конкуренции антител выполняли на системе Forte Bio Octet Red96 (Pall Forte Bio Corp., США) с использованием стандартного анализа биннинга в формате «сэндвич». 10 нМ антитела к мышиному CD25 загружали на сенсоры АМС в течение 900 с, и незанятые Fc-связывающие сайты на сенсоре блокировали нецелевым мышиным антителом IgG2a. Сенсоры подвергали воздействию 15 нМ антигена-мишени (His-меченый CD25 мыши) в течение 600 с и затем второго антитела к CD25 (также 10 нМ). Данные обрабатывали с использованием программного обеспечения для анализа данных Forte Bio 9.0. Дополнительное связывание вторым антителом после ассоциации антигена указывает на незанятый эпитоп, тогда как отсутствие связывания указывает на блокирование эпитопа.
[0275] Конкурентное связывание с mCD25 наблюдали между 7D4 и 2Е4 (Фигура 13 (А)), но не между 7D4 и РС61 (Фигура 13 (В)).
[0276] Исследование передачи сигналов IL-2 в условиях in vitro с помощью анализа фосфорилирования STAT5:
Общую популяцию Т-клеток выделяли из спленоцитов с использованием набора Dynabeads® FlowComp™ для общей популяции мышиных Т-клеток (CD90.2) от Invitrogen (каталожный номер 11465D). 200000 клеток высевали и оставляли на 2 часа при 37°С. Антитела добавляли в концентрации 50 мкг/мл и инкубировали с клетками в течение 30 минут при 37°С, после чего клетки стимулировали IL-2 (50 Ед/мл) в течение 10 минут при 37°С.
Индуцированное IL-2 фосфорилирование STAT5 прекращалось, когда клетки фиксировали и пермеабилизировали с помощью набора буферов для окрашивания фактора транскрипции Foxp3 eBioscience™ (Invitrogen) и обрабатывали буфером BD Phosflow Perm Buffer III (BD Biosciences). Затем клетки одновременно окрашивали поверхностными и внутриклеточными антителами, мечеными флуорохромом (STAT5-Alexa Fluor 647, клон 47/stat5/pY694 BD Bioscience, CD3-PerCP-Cy5.5, клон 17A2 Biolegend, CD4-PE, клон RM4-5 Biolegend, FoxP3-AF488 клон FJK-16s Ebioscience), образцы отбирали на проточном цитометре Fortessa LSR Х20 (BD Bioscience) и анализировали с использованием программного обеспечения BD FACSDIVA. Дублеты исключали с использованием FCS-H по отношению к FCS-A, и лимфоциты определяли с использованием параметров SSC-A по отношению к FCS-A. CD3+ Т-клетки определяли с использованием графика CD3 PerCP-Су5.5-А по отношению к FCS-A, и для определения популяции Stat5+CD3+ Т-клеток на гистограмме была показана пропущенная популяция, показывающая количество клеток по сравнению с STAT5 Alexa Fluor 647-А. Процент блокирования передачи сигналов IL-2 рассчитывали следующим образом: % блокирования = 100×[(% Stat5+ клеток в группе без обработки антителом - % Stat5+ клеток в группе 50 мкг/мл антитела)/(% Stat5+ клеток в группе без обработки антителом)]. Дальнейший анализ фосфорилирования STAT5 по различными подгруппам Т-клеток (CD4+, CD8+, CD4+FoxP3-) также оценивали путем селективного пропускания соответствующих подгрупп и анализировали, как описано выше. Для построения графиков и проведения статистического анализа использовали GraphPad Prism v7 (результаты не представлены). Результаты представлены на Фигуре 14.
[0277] Результаты:
Дополнительно оценивали способность антител к мышиному белку, 7D4 и 2Е4, связывать CD25 и не нарушать передачу сигналов IL-2 клеток-мишеней, экспрессирующих CD25. 7D4 и 2Е4, не блокирующие IL-2, конкурируют за связывание с CD25, в то время как РС61 (блокатор передачи сигналов IL-2) не конкурирует с 2Е4 или 7D4 за связывание CD25 (Фигура 12).
[0278] Анализ STAT5 подтвердил, что 7D4 и 2Е4 не блокировали передачу сигналов IL-2, в то время как «блокирующее» антитело РС61 блокировало передачу сигналов IL-2 (Фигура 14).
[0279] Пример 6: истощение Трег в условиях in vivo
По 1×105 клеток 4Т1 в 200 мкл среды RPMI-1640 имплантировали в жировую ткань в области 2 грудного ребра мышей Balb/c. Когда опухоли достигали 50-100 мм3, мышей рандомизировали и вводили внутрибрюшинно однократную фиксированную дозу мышиного антитела к мышиному CD25 (7D4) 2 мкг, 20 мкг или 200 мкг на мышь. На 3 и 9 день опухолевые ткани и цельную кровь выделяли для иммунофенотипирования.
[0280] Результаты:
Антитело 7D4 проявляло активность, направленную на истощение Трег, как в цельной крови, так и в опухолевой ткани на основании анализа на 3 и 9 день методом иммунофенотипирования (Фигура 15).
[0281] Пример 7: Характеристика антитела к CD25, 7G76B
Связывание антител к CD25 с клетками, экспрессирующими человеческий CD25:
Оценивали связывание 7G76B с линиями клеток лимфомы человека, Karpas 299, SU-DHL-1 и SR-786, и дифференцированными в условиях in vitro Трег-клетками. Связывание с CD25-экспрессирующими человеческими клеточными линиями (SU-DHL-1 и SR-786) исследовали, сначала блокируя клетки с применением Trustain (Biolegend) перед инкубацией с антителами к CD25, протитрованными в полулогарифмических последовательных разведениях, начиная с самой высокой концентрации 20 мкг/мл, в течение 30 минут при 4°С, с последующей промывкой и инкубацией с ФЭ-конъюгированным антителом к Fc человеческого IgG (Biolegend). Клетки повторно промывали и ресуспендировали в буфере для FACS, содержащем DAPI, и отбирали образцы на Intellicyt iQue. Живые клетки селективно пропускали с использованием параметров FSC по отношению к SSC с помощью проточной цитометрии во время отбора образцов. Значения геометрического среднего интенсивности окрашенных клеток наносили на XY-диаграмму, на которой в графической форме представлена зависимость геометрического среднего интенсивности от логарифма концентрации, и данные аппроксимировали с использованием кривой нелинейной регрессии, на основании которой рассчитывали ЭК50.
[0282] Связывание с CD25-экспрессирующими клетками Karpas 299 и дифференцированными в условиях in vitro Трег исследовали с помощью окрашивания испытываемых изделий (первичных антител к CD25) с применением 30 мг/мл антител с последующими полулогарифмическими последовательными разведениями (7 точек) в течение 30 минут на льду. После этого окрашивали вторичным антителом (Alexa Fluor 647-AffiniPure козий фрагмент Fab к человеческому IgG (H+L) - (Jackson ImmunoResearch)) в концентрации 1 мг/мл в течение 30 минут на льду. Все образцы окрашивали с двумя повторами. Живые клетки подвергали селективному пропусканию с использованием параметров FSC по отношению к SSC с помощью проточной цитометрии во время отбора образцов. Среднюю интенсивность флуоресценции (MFI) окрашенных клеток наносили на XY-диаграмму, на которой в графической форме была представлена зависимость MFI от логарифма концентрации, и данные аппроксимировали с использованием кривой нелинейной регрессии, на основании которой рассчитывали ЭК50. Результаты, представленные на Фигуре 16 и Фигуре 21, подтвердили, что антитела к CD25 связываются с клетками, экспрессирующими CD25.
[0283] Исследование передачи сигналов IL-2 в условиях in vitro с помощью анализа фосфорилирования STAT5:
Блокирование IL-2 характеризовали с использованием анализа фосфорилирования STAT5, в котором исследовали передачу сигналов IL-2. Замороженные ранее МКПК (Stemcell Technologies) культивировали в 96-луночных планшетах с U-образным дном в присутствии 10 мкг/мл антител к CD25 в течение 30 минут перед добавлением IL-2 (Peprotech) в различных концентрациях 0,1 Ед/мл, 1 Ед/мл или 10 Ед/мл в течение 10 минут в RPMI-1640 (Life Technologies), содержащей 10% ФБС (Sigma), 2 мМ L-глутамина (Life Technologies) и 10000 Ед/мл пенициллина-стрептомицина (Sigma). Индуцированное IL-2 фосфорилирование STAT5 прекращалось, когда клетки фиксировали и пермеабилизировали с помощью набора буферов для окрашивания фактора транскрипции Foxp3 eBioscience™ (Invitrogen) и обрабатывали буфером BD Phosflow Perm Buffer III (BD Biosciences). Затем клетки одновременно окрашивали поверхностными и внутриклеточными антителами, мечеными флуорохромом (STAT5-Alexa Fluor 647 клон 47/stat5/pY694 BD Bioscience, CD3-PerCP-Cy5.5 клон UCHT1 Biolegend, CD4-BV510 клон SK3 BD Bioscience, CD8-Alexa Fluor 700 клон RPA-T8 Invitrogen, CD45RA-PE-Cy7 клон HI100 Invitrogen, FoxP3-Alexa Fluor 488 клон 236A/E7 Invitrogen), образцы отбирали на проточном цитометре Fortessa LSR Х20 (BD Bioscience) и анализировали с использованием программного обеспечения BD FACSDIVA. Дублеты исключали с использованием FCS-H по отношению к FCS-A, и лимфоциты определяли с использованием параметров SSC-A по отношению к FCS-A. CD3+ Т-клетки определяли с использованием графика CD3 PerCP-Су5.5-А по отношению к FCS-A, и для определения популяции Stat5+CD3+ Т-клеток на гистограмме была показана пропущенная популяция, показывающая количество клеток по сравнению с STAT5 Alexa Fluor 647-А. Процент блокирования передачи сигналов IL-2 рассчитывали следующим образом: % блокирования =100×[(% Stat5+ клеток без обработки антителом - % Stat5+ клеток в группе 10 мкг/мл антитела)/(% Stat5+ клеток без обработки антителом)]. Дальнейший анализ фосфорилирования STAT5 различными подгруппами Т-клеток (CD4+, CD8+, CD4+FoxP3+, наивными и Т-клетками памяти) также оценивали путем селективного пропускания соответствующих подгрупп и анализировали, как описано выше. Для построения графиков и проведения статистического анализа использовали GraphPad Prism v7 (результаты не представлены). Результаты представлены на Фигуре 17 и Фигуре 22.
[0284] Анализ активации Т-клеток в условиях in vitro:
Влияние передачи сигналов IL-2 на ответы Тэфф характеризовали в анализе активации Т-клеток, в котором исследовали активацию внутриклеточного гранзима В (GrB) и пролиферацию. Замороженную ранее общую популяцию первичных человеческих Т-клеток (Stemcell Technologies) метили красителем для оценки пролиферации клеток eFluor450 (Invitrogen) в соответствии с рекомендацией производителя и добавляли в лунки 96-луночного планшета с U-образным дном в количестве 1×105 клеток/лунку в RPMI-1640 (Life Technologies), содержащей 10% ФБС (Sigma), 2 мМ L-глутамина (Life Technologies) и 10000 Ед/мл пенициллина-стрептомицина (Sigma). Затем клетки обрабатывали 10 мкг/мл антител к CD25 или контрольных антител с последующим введением человеческого Т-активатора CD3/CD28 (соотношение клеток и гранул 20:1; Gibco) и инкубировали в течение 72 часов при 37°С и 5% СО2 в инкубаторе с увлажнением. Для оценки активации Т-клеток клетки окрашивали красителем eBioscience Fixable Viability Dye efluor780 (Invitrogen), затем мечеными флуорохромом антителами для поверхностных маркеров Т-клеток (CD3-PerCP-Cy5.5 клон UCHT1 Biolegend, CD4-BV510 клон SK3 BD Bioscience, CD8-Alexa Fluor 700 клон RPA-T8 Invitrogen, CD45RA-PE-Cy7 клон HI100 Invitrogen, CD25-BUV737 клон 2А3 BD Bioscience), после этого фиксировали и пермеабилизировали с помощью набора буферов для окрашивания фактора транскрипции Foxp3 eBioscience™ (Invitrogen) перед окрашиванием внутриклеточного GrB и внутриядерного FoxP3 (гранзим В-РЕ, клон GB11 BD Bioscience, FoxP3-APC клон 236А/Е7). Образцы отбирали на проточном цитометре Fortessa LSR Х20 (BD Bioscience) и анализировали с использованием программного обеспечения BD FACSDIVA. Дублеты исключали с использованием FCS-H по отношению к FCS-A, лимфоциты определяли с использованием параметров SSC-A по отношению к FCS-A. Подгруппы CD4+ и CD8+ Т-клеток, селективно пропущенные в области живых CD3+ лимфоцитов, оценивали с использованием графика GrB-PE-A по отношению к пролиферации на основании сигнала eFluor450-A. Результаты представлены в виде процента пролиферирующих GrB-положительных клеток от общей популяции CD4+ Т-клеток. Для построения графиков и статистического анализа использовали GraphPad Prism v7. Результаты представлены на Фигуре 18.
[0285] Анализ АЗКЦ в условиях in vitro:
Анализы антителозависимой клеточной цитотоксичности (анализы АЗКЦ) выполняли для характеристики антител к человеческому CD25 с использованием SU-DHL-1 или SR-786 (CD25-положительных) человеческих клеточных линий в качестве клеток-мишеней и человеческих NK-клеток в качестве источника эффекторных клеток. NK-клетки выделяли из МКПК здоровых доноров с использованием набора для выделения NK-клеток с отрицательной селекцией (Stemcell Technologies). NK-клетки культивировали в течение ночи в присутствии 2 нг/мл IL-2 (Peprotech). Клетки-мишени SU-DHL-1 или SR-786 загружали кальцеином-АМ (Thermofisher) и высевали с 4 повторами для каждого условия в присутствии антител к CD25 или изотипических антител в течение 30 минут при 37°С и 5% СО2. После инкубации NK-клетки добавляли в лунки в соотношении мишень:эффектор (Т:Е) 1:10 (10000 клеток-мишеней и 100000 эффекторных клеток) и инкубировали в течение 4 часов при 37°С и 5% CO2. Считывание флуоресценции кальцеина в супернатанте выполняли на считывателе для планшетов BMG Fluostar. Процент специфического лизиса рассчитывали относительно только клеток-мишеней (лизис 0%) и клеток-мишеней, обработанных 0,1% сапонином (лизис 100%). Графики исходных данных получали с использованием Graphpad Prism v7 для построения кривых зависимости от дозы. Процент лизиса клеток-мишеней наносили на XY-диаграмму, на которой в графической форме представлена зависимость нормированного процента высвобождения кальцеина-AM от логарифма концентрации, и данные аппроксимировали с использованием кривой нелинейной регрессии, на основании которой рассчитывали ЭК50. Результаты представлены на Фигуре 19.
[0286] Анализ АЗКФ в условиях in vitro:
Анализы антителозависимого клеточного фагоцитоза (АЗКФ) выполняли с использованием дифференцированных в условиях in vitro Трег в качестве клеток-мишенеи и макрофагов моноцитарного происхождения в качестве эффекторных клеток. МКПК выделяли из лейкоцитарных конусов с помощью центрифугирования в градиенте фиколла. Моноциты (CD14+ клетки) выделяли с использованием микрогранул CD14 (Miltenyi Biotec). Моноциты культивировали в течение 5 дней в присутствии 50 нг/мл М-КСФ в RPMI-1640 (Life Technologies), содержащей 10% ФБС (Sigma), 2 мМ L-глутамина (Life Technologies) и 10000 Ед/мл пенициллина-стрептомицина (Sigma), свежие среды, содержащие М-КСФ, добавляли через 3 дня. Регуляторные Т-клетки (Трег) выделяли с использованием набора для дифференцировки человеческих Трег-клеток (R&D Systems). Эти клетки инкубировали при 37°С и 5% СО2 в инкубаторе с увлажнением в течение 5 дней и метили красителем eFluor450 (Invitrogen) в соответствии с рекомендациями производителя. На 5 день макрофаги и меченые красителем eFluor450 Трег культивировали в течение 4 часов при соотношении эффектор/мишень 10:1 в присутствии антител к CD25 или контролей, как описано далее. Клетки-мишени (Трег) добавляли в количестве 1×104 клеток/лунку, тогда как эффекторные клетки (макрофаги) добавляли в количестве 1×105 клеток/лунку, чтобы получить соотношение эффекторов к мишеням 10:1. Затем добавляли антитела к CD25 в максимальной концентрации 1 мкг/мл с последующим логарифмическими последовательными разведениями (7 точек) с двумя повторами. Клетки и антитела инкубировали в течение 4 часов при 37°С и 5% СО2. Для оценки АЗКФ клетки помещали на лед, окрашивали маркером клеточной поверхности CD14 (CD14- PerCP-Су5.5 клон MfP9 BD Biosciences) и фиксировали буфером для фиксации eBioscience. Анализ методом двухцветной проточной цитометрии выполняли с использованием Fortessa LSR Х20. Остаточные клетки-мишени определяли как клетки, которые были eFluor450-краситель+/CD14-. Макрофаги определяли как CD14+. Содержащие двойную метку клетки (eFluor450-краситель+/CD14+) рассматривали как фагоцитоз мишеней макрофагами. Фагоцитоз клеток-мишеней рассчитывали по следующему уравнению: % фагоцитоза = 100×[(процент двойных положительных мишеней)/(процент двойных положительных мишеней + процент остаточных мишеней)]. Результаты представлены на Фигуре 20.
[0287] Статистика:
Программное обеспечение Prism (GraphPad) использовали для аппроксимации кривой, чтобы определить значения ЭК50 и максимальную активность.
[0288] Человеческие антитела не блокируют взаимодействие IL-2-CD25 Нарушение связывания IL-2-лиганда с CD25 выполняли на системе Forte Bio Octet Red384 (Pall Forte Bio Corp., США) с использованием стандартного анализа биннинга в формате «сэндвич». Антитело МА251 загружали на сенсоры AHQ, и незанятые Fc-связывающие сайты на сенсоре блокировали нецелевым человеческим антителом IgG1. Сенсоры подвергали воздействию 100 нМ человеческого CD25 и затем 100 нМ человеческого IL-2. Данные обрабатывали с использованием программного обеспечения для анализа данных Forte Bio 7.0. Дополнительное связывание IL-2 человека после ассоциации антигена указывает на незанятый эпитоп (не является конкурентом), тогда как отсутствие связывания указывает на блокирование эпитопа (конкурент).
[0289] Результаты:
Антитело 7G7B6 дополнительно оценивали в отношении его способности не нарушать передачу сигналов IL-2 и его способности уничтожать экспрессирующие CD25 клетки-мишени. В анализе STAT5 7G7B6 не блокировало передачу сигналов IL-2, независимо от испытанных концентраций IL-2, в то время как эталонное антитело, даклизумаб, полностью блокировало передачу сигналов IL-2 (Фигура 17). Даклизумаб, который, как было показано, блокирует взаимодействие CD25 с IL-2 через так называемый эпитоп «Тас» (Queen С et al, 1989, PNAS. 86 (24): 10029-10033, и Bielekova В, 2013, Neurotherapeutics, 10 (1): 55-67), связывается с другим эпитопом, нежели 7G7B6 (Фигура 10 и Фигура 24В), это может объяснить, почему даклизумаб блокирует передачу сигналов IL-2, a 7G7B6 не блокирует передачу сигналов IL-2 в анализе фосфорилирования STAT5 (Фигура 17). Помимо этого даклизумаб снижает эффекторные ответы активированных Т-клеток, вероятно, поскольку он блокирует передачу сигналов IL-2, в то время как 7G7B6, которое не блокирует передачу сигналов IL-2, не оказывает отрицательного влияния на ответы Т-клеток (Фигура 18). Наконец, химерное антитело 7G7B6 уничтожает CD25-экспрессирующие клетки, опухолевые клетки или регуляторные Т-клетки за счет АЗКЦ (Фигура 19) и АЗКФ (Фигура 20) по сравнению с антителом изотипа IgG1.
[0290] В заключение следует отметить, что 7G7B6, как химерное антитело, было охарактеризовано и демонстрирует эффективное уничтожение CD25-положительных клеток (Трег или раковых клеточных линий) и не нарушает передачу сигналов IL-2 и, следовательно, не ингибирует эффекторные ответы Т-клеток. Соответственно, 7G7B6 представляет собой антитело, истощающее Трег, которое можно применять для лечения рака в виде монотерапии или в комбинации.
[0291] Антитело МА-251 дополнительно оценивали в отношении его способности не нарушать передачу сигналов IL-2. Антитело МА251 оценивали в конкурентном анализе IL-2-CD25 с помощью Octet. Наблюдали одновременное связывание IL-2 и МА251 с CD25 (Фигура 23), это указывает на то, что МА251 связывается неконкурентным способом. В анализе STAT5 МА-251 не блокировало передачу сигналов IL-2, независимо от испытанных концентраций IL-2, в то время как эталонное антитело, даклизумаб, полностью блокировало передачу сигналов IL-2 (Фигура 22). Даклизумаб, который, как было показано, блокирует взаимодействие CD25 с IL-2 за счет так называемого эпитопа «Тас» (Queen С et al, 1989 и Bielekova В, 2013), связывается с другим эпитопом, нежели МА-251 (Фигура 10 и Фигура 24 (Е)), это может объяснить, почему даклизумаб блокирует передачу сигналов IL-2, а МА-251 не блокирует передачу сигналов IL-2 в анализе фосфорилирования STAT5 (Фигура 22).
[0292] Пример 8: конкурентный анализ антитела к человеческому CD25
Конкурентные анализы антител проводили на системе Forte Bio Octet Red96 (Pall Forte Bio Corp., США) с использованием стандартного анализа последовательного связывания. 26,8 нМ рекомбинантного человеческого CD25, несущего His-метку, загружали на биосенсоры Ni-NTA в течение 200 с. После этапа регистрации исходного уровня в кинетическом буфере сенсоры подвергали воздействию 66,6 нМ первого антитела в течение 600 с или 1800 с с последующим введением второго антитела к CD25 (также в концентрации 66,6 нМ в течение 600 с или 1800 с). Данные обрабатывали с использованием программного обеспечения для анализа данных Forte Bio 9.0. Дополнительное связывание вторым антителом указывает на незанятый эпитоп (нет конкуренции за эпитоп), тогда как отсутствие связывания указывает на блокирование эпитопа (конкуренция за эпитоп).
[0293] Результаты
Моноклональные антитела, не блокирующие передачу сигналов IL-2 (антитело 1 и антитело 3), конкурируют друг с другом или с 7G7B6 и МА251, однако они не конкурируют с исследовательским даклизумабом или исследовательским базиликсимабом (примеры (A)-(N), Фигура 24). Блокаторы передачи сигналов IL-2 (т.е. TSK031) действительно конкурируют с исследовательским даклизумабом или исследовательским базиликсимабом и не конкурируют с 7G7B6 (примеры (O)-(Q), Фигура 24).
[0294] Пример 9: Терапевтический анализ неблокирующего антитела На 0 день в правый бок имплантировали 1×107 клеток SU-DHL-1 в 200 мкл RPMI-1640. На 12 день мышей с пальпируемыми опухолями рандомизировали для получения либо носителя, либо антитела 1 в дозе 2 мг/кг два раза в неделю. На 15 день мышей с размером опухоли 100-200 мм3 рандомизировали и вводили им либо носитель, либо антитело 1 в дозе 2 мг/кг дважды в неделю, либо однократную дозу антитела 1 в дозе 10 мг/кг.
[0295] Результаты
Антитело 1 предотвращало рост опухоли у 9/10 мышей с пальпируемыми опухолями, которым вводили дозу 2 мг/кг два раза в неделю (Фигура 25 (А)-(В)). У мышей с размером опухоли 100-200 мм3 антитело 1 также предотвращало рост опухоли в дозах 2 мг/кг два раза в неделю и в однократной дозе 10 мг/кг (Фигура 25 (С)-(Е)).
[0296] Пример 10. Измерение аффинности антител к человеческому CD25 Аффинность антител к человеческому CD25 определяли путем измерения их KD с помощью SPR в Biacore 2000 с использованием сенсорного чипа СМ-5 при температуре окружающей среды 25°С во время эксперимента. Антитело к человеческому белку первоначально иммобилизовали во всех проточных ячейках в буфере для анализа (рН=7,4, 10 мМ HEPES, 150 мМ NaCl, 3 мМ ЭДТА, 0,05% Твин 20) до 12000-14000 RU в течение 10 минут. Лиганд (испытываемые изделия-антитела) последовательно загружали до уровня захвата 145-190 RU. Затем аналит (рекомбинантный человеческий CD25, несущий His-метку) связывали в буфере для анализа из 2-кратного разведения, начиная с 400 нМ, с самой низкой концентрацией 3,13 нМ, в течение 6 минут. Диссоциацию выполняли в буфере для анализа в течение 10 минут. Этапы регенерации между концентрациями образца выполняли в 10 мМ глицине с рН=1,7 в течение 10 минут. Скорость потока 25 мкл/мин поддерживали на протяжении всего процесса. Кинетические данные аппроксимировали с использованием программного обеспечения для общего анализа конформационных изменений в реакции с двумя состояниями, предоставленного Biacore, с вычитанием эталона для Фигуры 26 (С) и Фигуры 26(D). Модель Ленгмюра 1:1 с вычитанием эталона использовали для Фигуры 26 (А), Фигуры 26 (В) и Фигуры 26 (Е).
[0297] Измерения аффинности с помощью ForteBio выполняли на Octet RED384, в целом, как описано ранее (см., например, Estep Р et al., 2013. Mabs. 5 (2), 270-8).
[0298] Согласно другому варианту аффинность антител к человеческому CD25 определяли путем измерения их Kd методом интерферометрии биослоев на системе Octet Red 96 (Pall Forte Bio Corp., США). Сенсоры уравновешивали в автономном режиме в кинетическом буфере в течение 10 минут и затем контролировали в режиме реального времени в течение 60 секунд для установления исходного уровня. По 13,32 нМ антитела загружали в биосенсор АНС в течение 200 с и затем вносили различные концентрации His-меченого rhCD25 (последовательные разведения 1:3, от 50 нМ до 0,54 нМ) в течение 600 с и обеспечивали время для диссоциации в кинетическом буфере в течение 400 с. Кинетические данные аппроксимировали с использованием программного обеспечения для общего анализа аппроксимации в соотношении 1:1, предоставленного Pall Forte Bio, с вычитанием эталона. Результаты представлены на Фигуре 26 (F).
[0299] Результаты:
Результаты представлены на Фигуре 26. Значения Kd, которые были установлены в этом анализе для антител к CD25, являются следующими: для антитела 1, 3,2×10-9 М; для антитела 3, 3,8×10-9 М, и для даклизумаба 0,61×10-9 М.
[0300] Пример 11: Характеристика антител к CD25 - антитела с 1 по 21
Связывание антител к CD25 с CD25-экспрессирующими клетками:
Выявленные соединения-кандидаты оценивали по связыванию с линиями клеток лимфомы человека, такими как клетки Karpas 299, SU-DHL-1 и SR-786, дифференцированные в условиях in vitro Трег-клетки, активированные МКПК человека или яванской макаки, линия Т-клеток яванской макаки HSC-F и клетки СНО.
[0301] Связывание с С025-экспрессирующими клеточными линиями человека (SU-DHL-1 и SR-786) исследовали, сначала блокируя клетки с применением Trustain (Biolegend) перед инкубацией с антителами к CD25, протитрованными в полулогарифмических последовательных разведениях, начиная с самой высокой концентрации 20 мкг/мл, в течение 30 минут при 4°С, с последующей промывкой и инкубацией с ФЭ-конъюгированным антителом к Fc человеческого IgG (Biolegend). Клетки повторно промывали и ресуспендировали в буфере для FACS, содержащем DAPI, и образцы отбирали на Intellicyt iQue. Живые клетки подвергали селективному пропусканию с использованием параметров FSC по отношению к SSC с помощью проточной цитометрии во время отбора образцов. Значения геометрического среднего интенсивности окрашенных клеток наносили на XY-диаграмму, на которой в графической форме представлена зависимость геометрического среднего интенсивности от логарифма концентрации, и данные аппроксимировали с использованием кривой нелинейной регрессии, на основании которой рассчитывали ЭК50.
[0302] Связывание с CD25-экспрессирующими клетками Karpas 299 и дифференцированными в условиях in vitro Трег исследовали путем окрашивания испытываемых изделий (первичных антител к CD25) с применением 30 мг/мл антител с последующими полулогарифмическими последовательными разведениями (7 точек) в течение 30 минут на льду. После этого выполняли окрашивание вторичным антителом (Alexa Fluor 647-AffiniPure козий фрагмент Fab к человеческому IgG (H+L) или Alexa Fluor 647-AffiniPure кроличий фрагмент F(ab')2 к фрагменту Feγ человеческого IgG - (Jackson ImmunoResearch)) в концентрации 1 мг/мл в течение 30 минут на льду. Все образцы окрашивали с двумя повторами. Живые клетки подвергали селективному пропусканию с использованием параметров FSC по отношению к SSC с помощью проточной цитометрии во время отбора образцов. Среднюю интенсивность флуоресценции (MFI) окрашенных клеток наносили на XY-диаграмму, на которой в графической форме представлена зависимость MFI от логарифма концентрации, и данные аппроксимировали с использованием кривой нелинейной регрессии, на основании которой рассчитывали ЭК50.
[0303] Связывание с CD25-экспрессирующим активированными МКПК человека и яванской макаки исследовали путем окрашивания испытываемых изделий (первичных антител к CD25) с применением 20 мг/мл антител с последующими полулогарифмическими последовательными разведениями (7 точек) в течение 30 минут на льду. После этого выполняли окрашивание вторичным антителом (кроличий F(ab')2 к человеческому Fcγ - (Jackson ImmunoResearch)) в концентрации 5 мг/мл в течение 30 минут на льду. Все образцы окрашивали с тремя повторами. Чтобы свести к минимуму индуцированную перекрестным связыванием гибель клеток, опосредуемую связыванием вторичного антитела, клеточные линии исследовали в когортах окрашивания по 4 испытываемых изделия за один раз. Живые лимфоциты селективно пропускали с использованием параметров FSC по отношению к SSC с помощью проточной цитометрии во время отбора образцов. Среднюю интенсивность флуоресценции (MFI) селективно пропущенных подгрупп CD4+ и CD8+ Т-клеток наносили на XY-диаграмму, на которой в графической форме представлена зависимость MFI от логарифма концентрации, и данные аппроксимировали с использованием кривой нелинейной регрессии, на основании которой рассчитывали ЭК50.
[0304] Связывание с CD25-экспрессирующей линией Т-клеток яванской макаки HSC-F исследовали путем окрашивания испытываемых изделий (первичных антител к CD25) с применением 20 мг/мл антител с последующими полулогарифмическими последовательными разведениями (7 точек) в течение 30 минут на льду. После этого выполняли окрашивание вторичным антителом (кроличий F(ab')2 к человеческому Fcγ - (Jackson ImmunoResearch)) в концентрации 5 мг/мл в течение 30 минут на льду. Все образцы окрашивали с тремя повторами. Чтобы свести к минимуму индуцированную перекрестным связыванием гибель клеток, опосредуемую связыванием вторичного антитела, клеточные линии исследовали в когортах окрашивания по 4 испытываемых изделия за один раз. Живые лимфоциты селективно пропускали с использованием параметров FSC по отношению к SSC с помощью проточной цитометрии во время отбора образцов. Среднюю интенсивность флуоресценции (MFI) живых клеток наносили на XY-диаграмму, на которой в графической форме представлена зависимость MFI от логарифма концентрации, и данные аппроксимировали с использованием кривой нелинейной регрессии, на основании которой рассчитывали ЭК50.
[0305] Также исследовали связывание с CD25-экспрессирующими клетками СНО. Приблизительно 100000 клеток, сверхэкспрессирующих антиген, промывали промывочным буфером и инкубировали с 100 мкл 100 нМ IgG в течение 15 минут при комнатной температуре. Затем клетки дважды промывали промывочным буфером и инкубировали с 100 мкл Human-ФЭ в разведении 1:100 в течение 15 минут на льду. Затем клетки дважды промывали промывочным буфером и анализировали на анализаторе FACS Canto II (BD Biosciences). Немодифицированную клеточную линию СНО также использовали в качестве отрицательного контроля.
[0306] Исследование передачи сигналов IL-2 в условиях in vitro с помощью анализа фосфорилирования STAT5:
Блокирование IL-2 характеризовали с использованием анализа фосфорилирования STAT5, в котором исследовали передачу сигналов IL-2. Замороженные ранее МКПК (Stemcell Technologies) культивировали в 96-луночных планшетах с U-образным дном в присутствии 10 мкг/мл антител к CD25 в течение 30 минут перед добавлением IL-2 (Peprotech) в различных концентрациях 10 Ед/мл или 0,25 Ед/мл, 0,74 Ед/мл, 2,22 Ед/мл, 6,66 Ед/мл или 20 Ед/мл в течение 10 минут в RPMI-1640 (Life Technologies), содержащей 10% ФБС (Sigma), 2 мМ L-глутамина (Life Technologies) и 10000 Ед/мл пенициллина-стрептомицина (Sigma). Индуцированное IL-2 фосфорилирование STAT5 прекращалось, когда клетки фиксировали и пермеабилизировали с помощью набора буферов для окрашивания фактора транскрипции Foxp3 eBioscience™ (Invitrogen) и обрабатывали буфером BD Phosflow Perm Buffer III (BD Biosciences). Затем клетки одновременно окрашивали поверхностными и внутриклеточными антителами, мечеными флуорохромом (STAT5-Alexa Fluor 647 клон 47/stat5/pY694 BD Bioscience, CD3-PerCP-Cy5.5 клон UCHT1 Biolegend, CD4-BV510 клон SK3 BD Bioscience, CD8-Alexa Fluor 700 клон RPA-T8 Invitrogen, CD45RA-PE-Cy7 клон HI100 Invitrogen, FoxP3-Alexa Fluor 488 клон 236A/E7 Invitrogen), образцы отбирали на проточном цитометре Fortessa LSR Х20 (BD Bioscience) и анализировали с использованием программного обеспечения BD FACSDIVA. Дублеты исключали с использованием FCS-H по отношению к FCS-A, и лимфоциты определяли с использованием параметров SSC-A по отношению к FCS-A. CD3+ Т-клетки определяли с использованием графика CD3 PerCP-Су5.5-А по отношению к FCS-A, и для определения популяции Stat5+CD3+ Т-клеток на гистограмме была показана пропущенная популяция, показывающая количество клеток по сравнению с STAT5 Alexa Fluor 647-А. Процент блокирования передачи сигналов IL-2 рассчитывали следующим образом: % блокирования = 100×[(% Stat5+ клеток без обработки антителом - % Stat5+ клеток в группе 10 мкг/мл антитела)/(% Stat5+ клеток без обработки антителом)]. Дальнейший анализ фосфорилирования STAT5 различными подгруппами Т-клеток (CD4+, CD8+, CD4+ FoxP3+, наивными и Т-клетками памяти) также оценивали путем селективного пропускания соответствующих подгрупп и анализировали, как описано выше. Для построения графиков и проведения статистического анализа использовали GraphPad Prism v7.
[0307] Анализ активации Т-клеток в условиях in vitro:
Влияние передачи сигналов IL-2 на ответы Тэфф характеризовали в анализе активации Т-клеток, в котором исследовали активацию внутриклеточного гранзима В (GrB) и пролиферацию. Замороженную ранее общую популяцию первичных человеческих Т-клеток (Stemcell Technologies) метили красителем для оценки пролиферации клеток eFluor450 (Invitrogen) в соответствии с рекомендацией производителя и добавляли в лунки 96-луночного планшета с U-образным дном в количестве 1×105 клеток/лунку в RPMI-1640 (Life Technologies), содержащей 10% ФБС (Sigma), 2 мМ L-глутамина (Life Technologies) и 10000 Ед/мл пенициллина-стрептомицина (Sigma). Затем клетки обрабатывали 10 мкг/мл антител к CD25 или контрольных антител с последующим введением человеческого Т-активатора CD3/CD28 (соотношение клеток и гранул 20:1; Gibco) и инкубировали в течение 72 часов при 37°С и 5% СО2 в инкубаторе с увлажнением. Для оценки активации Т-клеток клетки окрашивали красителем eBioscience Fixable Viability Dye efluor780 (Invitrogen), затем мечеными флуорохромом антителами для поверхностных маркеров Т-клеток (CD3-PerCP-Cy5.5 клон UCHT1 Biolegend, CD4-BV510 клон SK3 BD Bioscience, CD8-Alexa Fluor 700 клон RPA-T8 Invitrogen, CD45RA-PE-Cy7 клон НИ 00 Invitrogen, CD25-BUV737 клон 2А3 BD Bioscience), после этого фиксировали и пермеабилизировали с помощью набора буферов для окрашивания фактора транскрипции Foxp3 eBioscience™ (Invitrogen) перед окрашиванием внутриклеточного GrB и внутриядерного FoxP3 (гранзим В-РЕ, клон GB11 BD Bioscience, FoxP3-APC клон 236А/Е7). Образцы отбирали на проточном цитометре Fortessa LSR Х20 (BD Bioscience) и анализировали с использованием программного обеспечения BD FACSDIVA. Дублеты исключали с использованием FCS-H по отношению к FCS-A, и лимфоциты определяли с использованием параметров SSC-A по отношению к FCS-A. Подгруппы CD4+ и CD8+ Т-клеток, селективно пропущенные в области живых CD3+ лимфоцитов, оценивали с использованием графика GrB-PE-A по отношению к пролиферации на основании сигнала eFluor450-A. Результаты представлены в виде процента пролиферирующих GrB-положительных клеток от общей популяции CD4+ или CD8+ Т-клеток. Для построения графиков и статистического анализа использовали GraphPad Prism v7.
[0308] Анализ АЗКЦ в условиях in vitro:
Анализы антителозависимой клеточной цитотоксичности (анализы АЗКЦ) выполняли для характеристики антител к человеческому CD25 с использованием человеческих клеточных линий SU-DHL-1 или SR-786 (СВ25-положительных) в качестве клеток-мишеней и человеческих NK-клеток в качестве источника эффекторных клеток. NK-клетки выделяли из МКПК здоровых доноров с использованием набора для выделения NK-клеток с отрицательной селекцией (Stemcell Technologies). NK-клетки культивировали в течение ночи в присутствии 2 нг/мл IL-2 (Peprotech). Клетки-мишени SU-DHL-1 или SR-786 загружали кальцеином-АМ (Thermofisher) и высевали с 4 повторами для каждого условия в присутствии антител к CD25 или изотопических антител на 30 минут при 37°С и 5% СО2. После инкубации NK-клетки добавляли в лунки в соотношении мишень : эффектор (Т:Е) 1:10 (10000 клеток-мишеней и 100000 эффекторных клеток) и инкубировали в течение 4 часов при 37°С и 5% СО2. Считывание флуоресценции кальцеина в супернатанте проводили на считывателе для планшетов BMG Fluostar. Процент специфического лизиса рассчитывали относительно только клеток-мишеней (лизис 0%) и клеток-мишеней, обработанных 0,1% сапонином (лизис 100%). Графики исходных данных получали с использованием Graphpad Prism v7 для построения кривых зависимости от дозы. Процент лизиса клеток-мишеней наносили на XY-диаграмму, на которой в графической форме представлена зависимость нормированного процента высвобождения кальцеина-AM от логарифма концентрации, и данные аппроксимировали с использованием кривой нелинейной регрессии, на основании которой рассчитывали ЭК50.
[0309] АЗКЦ также определяли в анализе с использованием репортерной системы на основе люциферазы. Клетки SR786, экспрессирующие CD25, называемые в настоящем документе клетками-мишенями (Т), инкубировали в течение 20 минут при 37°С с различными концентрациями МАТ к CD25 (или контрольным IgG) в среде с низким содержанием IgG, содержащей ФБС (4% ФБС в RPMI). Затем к смеси клетка-МАТ добавляли АЗКЦ-эффекторные (Е) клетки в соотношении Е:Т равном 1:1. Эффекторными клетками являлись клетки Jurkat, стабильно трансфецированные репортерной системой на основе люциферазы и сверхэкспрессирующие CD16/FcγRIIIA (Promega). После инкубации в течение ночи при 37°С клетки лизировали и измеряли активность люциферазы по испусканию люминесценции при гидролизе специфического субстрата люциферазы в соответствии с инструкцией производителя (протокол Promega Bio-Glow).
[0310] Анализ АЗКФ в условиях in vitro с использованием дифференцированных в условиях in vitro макрофагов и Трег-клеток.'
Анализы антителозависимого клеточного фагоцитоза (АЗКФ) выполняли с использованием дифференцированных в условиях in vitro Трег в качестве клеток-мишенеи и макрофагов моноцитарного происхождения в качестве эффекторных клеток. МКПК выделяли из лейкоцитарных конусов с помощью центрифугирования в градиенте фиколла. Моноциты (клетки CD14+) выделяли с использованием микрогранул CD14 (Miltenyi Biotec). Моноциты культивировали в течение 5 дней в присутствии 50 нг/мл М-КСФ в RPMI-1640 (Life Technologies), содержащей 10% ФБС (Sigma), 2 мМ L-глутамина (Life Technologies) и 10000 Ед/мл пенициллина-стрептомицина (Sigma), свежие среды, содержащие М-КСФ, добавляли через 3 дня. Регуляторные Т-клетки (Трег) выделяли с использованием набора для дифференцировки Трег-клеток человека (R&D Systems). Эти клетки инкубировали при 37°С и 5% СО2 в инкубаторе с увлажнением в течение 5 дней и метили красителем eFluor450 (Invitrogen) в соответствии с рекомендациями производителя. На 5 день макрофаги и Трег, меченые красителем eFluor450, совместно культивировали в течение 4 часов при соотношении эффектор/мишень 10:1 в присутствии антител к CD25 или контролей, как описано далее. Клетки-мишени (Трег) добавляли в количестве 1×104 клеток/лунку, тогда как эффекторные клетки (макрофаги) добавляли в количестве 1×105 клеток/лунку, чтобы получить соотношение эффекторов к мишени 10:1. Затем добавляли антитела к CD25 в максимальной концентрации 1 мкг/мл с последующими логарифмическими последовательными разведениями (7 точек) с двумя повторами. Клетки и антитела инкубировали в течение 4 часов при 37°С и 5% СО2. Для оценки АЗКФ клетки помещали на лед, окрашивали маркером клеточной поверхности CD14 (CD14-PerCP-Cy5.5 клон MfP9 BD Biosciences) и фиксировали буфером для фиксации eBioscience. Анализ методом двухцветной проточной цитометрии выполняли с использованием Fortessa LSR Х20. Остаточные клетки-мишени определяли как клетки, которые были eFluor450-краситель+/CD14-. Макрофаги определяли как CD14+. Как полагали, клетки с двойной меткой (eFluor450-краситель+/CD14+) представляют фагоцитоз мишеней макрофагами. Фагоцитоз клеток-мишеней рассчитывали по следующему уравнению: % фагоцитоза = 100×[(процент двойных положительных мишеней)/(процент двойных положительных мишеней + процент остаточных мишеней)].
[0311] Анализ АЗКФ в условиях in vitro с использованием репортерного анализа FcγRIIa-H:
Эффекторные клетки для биоанализа АЗКФ (FcγRIIa-H) получали от Promega (каталожный номер G9881/5; партия №0000261099). По 5000 клеток/лунку клеток-мишеней SU-DHL-1 высевали (25 мкл/лунку) с использованием 96-луночного белого полистирольного планшета (Costar; каталожный номер 3917). Испытываемые антитела последовательно разводили, используя 3-кратные разведения, и по 25 мкл добавляли к клеткам. По 50000 эффекторных клеток добавляли на лунку в объеме 25 мкл, чтобы получить соотношение эффекторных клеток и клеток-мишеней 10:1. Все клетки-мишени, антитела и эффекторные клетки высевали с использованием сред для культивирования клеток. Планшет инкубировали в течение 18 часов при 37°С. Затем планшеты извлекали из инкубатора и выдерживали при комнатной температуре в течение 20 минут. В каждую лунку добавляли по 60 мкл субстратного буфера для анализа люциферазы Bio-Glo с последующей 30 мин инкубацией, и измеряли люминесценцию с использованием системы GloMax Multi Detection System (Promega).
[0312] Статистика:
Программное обеспечение Prism (GraphPad) использовали для аппроксимации кривой, чтобы определить значения ЭК50 и максимальную активность.
[0313] Результаты
Оценивали способность антитела 1 не нарушать передачу сигналов IL-2 и его способность уничтожать клетки-мишени, экспрессирующие CD25. Результаты анализа связывания с rhCD25 и анализа конкурентного связывания IL-2 представлены на Фигурах 27 и 28. Анализ связывания лиганда с использованием Octet выявил, что антитело 1 не оказывало отрицательного влияния на связывание IL-2 с CD25 (Фигура 29). Это было подтверждено в анализе STAT5, в котором антитело 1 не блокировало передачу сигналов IL-2, независимо от испытываемых концентраций IL-2, в то время как эталонное антитело, даклизумаб, полностью блокировало передачу сигналов IL-2 (Фигура 31). Даклизумаб, который, как было показано, блокирует взаимодействие CD25 с IL-2 за счет так называемого эпитопа «Тас» (Queen С et al, 1989 и Bielekova В, 2013), связывается с другим эпитопом, чем антитело 1 (Фигура 30), это может объяснить, почему даклизумаб блокирует передачу сигналов IL-2, а антитело 1 не блокирует передачу сигналов IL-2 в анализе фосфорилирования STAT5 (Фигура 31). Помимо этого даклизумаб снижает эффекторные ответы активированных Т-клеток, вероятно, поскольку он блокирует передачу сигналов IL-2, в то время как антитело 1, которое не блокирует передачу сигналов IL-2, не оказывает отрицательного влияния на ответы Т-клеток (Фигура 32). Наконец, антитело 1 вызывает гибель СВ25-экспрессирующих клеток, опухолевых клеток или регуляторных Т-клеток за счет АЗКЦ (Фигура 33) и АЗКФ (Фигура 34) по сравнению с антителом изотипа IgG1.
[0314] В заключение следует отметить, что антитело 1 было охарактеризовано и демонстрирует эффективное уничтожение СВ25-положительных клеток (Трег или раковых клеточных линий) и не нарушает передачу сигналов IL-2 и, следовательно, не ингибирует эффекторные ответы Т-клеток. Соответственно, антитело 1 представляет собой антитело, истощающее Трег, которое можно применять для лечения рака в виде монотерапии или в комбинации.
[0315] Оценивали способность антитела 3 не нарушать передачу сигналов IL-2 и его способность уничтожать клетки-мишени, экспрессирующие CD25. Результаты анализа связывания с rhCD25 и анализа конкурентного связывания IL-2 представлены на Фигурах 35 и 36. Анализ связывания лиганда с использованием Octet выявил, что антитело 3 не оказывает отрицательного влияния на связывание IL-2 с CD25 (Фигура 37). Это было подтверждено в анализе STAT5, в котором антитело 3 не блокировало передачу сигналов IL-2, независимо от испытанных концентраций IL-2, в то время как эталонное антитело, даклизумаб, полностью блокировало передачу сигналов IL-2 (Фигура 39). Даклизумаб, который, как было показано, блокирует взаимодействие CD25 с IL-2 за счет так называемого эпитопа «Тас», связывается с другим эпитопом, чем антитело 3 (Фигура 38), это может объяснить, почему даклизумаб блокирует передачу сигналов IL-2 и антитело 3 не блокирует передачу сигналов IL-2 в анализе фосфорилирования STAT5 (Фигура 39). Помимо этого даклизумаб снижает эффекторные ответы активированных Т-клеток, вероятно, поскольку он блокирует передачу сигналов IL-2, в то время как антитело 3, которое не блокирует передачу сигналов IL-2, оказывает лишь минимальное влияние, если таковое имеется, на ответы Т-клеток по сравнению условием без антител или с изотопическим контролем (Фигура 40). Наконец, антитело 3 уничтожает CD25-экспрессирующие клетки, опухолевые клетки или регуляторные Т-клетки за счет АЗКЦ (Фигура 41) и АЗКФ (Фигура 42) по сравнению с изотипическим антителом IgGl.
[0316] В заключение следует отметить, что антитело 3 было охарактеризовано и демонстрирует эффективное уничтожение СВ25-положительных клеток (Трег или раковых клеточных линий) и не нарушает передачу сигналов IL-2 и, следовательно, не ингибирует эффекторные ответы Т-клеток. Соответственно, антитело 3 представляет собой антитело, истощающее Трег, которое можно применять для лечения рака в виде монотерапии или в комбинации.
[0317] Оценивали способность антитела 4 не нарушать передачу сигналов IL-2 и его способность уничтожать клетки-мишени, экспрессирующие CD25. Результаты анализа связывания с rhCD25 и анализа конкурентного связывания IL-2 представлены на Фигурах 43 и 44. Анализ связывания лиганда с использованием Octet выявил, что антитело 4 не оказывает отрицательного влияния на связывание IL-2 с CD25 (Фигура 45). Это было подтверждено в анализе STAT5, в котором антитело 4 не блокировало передачу сигналов IL-2, независимо от испытанных концентраций IL-2, в то время как эталонное антитело, даклизумаб, полностью блокировало передачу сигналов IL-2 (Фигура 47). Даклизумаб, который, как было показано, блокирует взаимодействие CD25 с IL-2 за счет так называемого эпитопа «Тас», связывается с другим эпитопом, чем антитело 4 (Фигура 46), это может объяснить, почему даклизумаб блокирует передачу сигналов IL-2, а антитело 4 не блокирует передачу сигналов IL-2 в анализе фосфорилирования STAT5 (Фигура 47). Помимо этого даклизумаб снижает эффекторные ответы активированных Т-клеток, вероятно, поскольку он блокирует передачу сигналов IL-2, в то время как антитело 4, которое не блокирует передачу сигналов IL-2, не оказывает отрицательного влияния на ответы Т-клеток (Фигура 48). Наконец, антитело 4 уничтожает СВ25-экспрессирующие клетки, опухолевые клетки или регуляторные Т-клетки за счет АЗКЦ (Фигура 49) и АЗКФ (Фигура 50) по сравнению с изотопическим антителом IgG1.
[0318] В заключение следует отметить, что антитело 4 было охарактеризовано и демонстрирует эффективное уничтожение СВ25-положительных клеток (Трег или раковых клеточных линий) и не нарушает передачу сигналов IL-2 и, следовательно, не ингибирует эффекторные ответы Т-клеток. Соответственно, антитело 4 представляет собой антитело, истощающее Трег, которое можно применять для лечения рака в виде монотерапии или в комбинации.
[0319] Оценивали способность антитела 2 не нарушать передачу сигналов IL-2 и его способность уничтожать клетки-мишени, экспрессирующие CD25. Результаты анализа связывания с rhCD25 и анализа конкурентного связывания IL-2 представлены на Фигурах 51 и 52. Анализ связывания лиганда с использованием Octet выявил, что антитело 2 не оказывает отрицательного влияния на связывание IL-2 с CD25 (Фигура 53). Это было подтверждено в анализе STAT5, в котором антитело 2 не блокировало передачу сигналов IL-2, независимо от испытанных концентраций IL-2, в то время как эталонное антитело, даклизумаб, полностью блокировало передачу сигналов IL-2 (Фигура 55). Даклизумаб, который, как было показано, блокирует взаимодействие CD25 с IL-2 за счет так называемого эпитопа «Тас», связывается с другим эпитопом, чем антитело 2 (Фигура 54), это может объяснить, почему даклизумаб блокирует передачу сигналов IL-2, а антитело 4 не блокирует передачу сигналов IL-2 в анализе фосфорилирования STAT5 (Фигура 55). Наконец, антитело 2 уничтожает СВ25-экспрессирующие клетки, опухолевые клетки или регуляторные Т-клетки за счет АЗКЦ (Фигура 56) и АЗКФ (Фигура 57) по сравнению с изотипическим антителом IgG1.
[0320] В заключение следует отметить, что антитело 2 было охарактеризовано и демонстрирует эффективное уничтожение СВ25-положительных клеток (Трег или раковых клеточных линий) и не нарушает передачу сигналов IL-2 и, следовательно, не ингибирует эффекторные ответы Т-клеток. Соответственно, антитело 2 представляет собой антитело, истощающее Трег, которое можно применять для лечения рака в виде монотерапии или в комбинации.
[0321] Антитело 5 характеризуется тем, что содержит вариабельную область тяжелой цепи, содержащую последовательность:
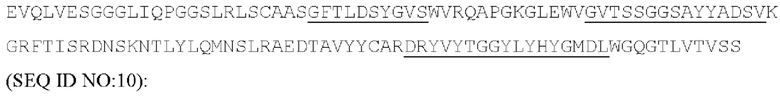
и вариабельную легкую цепь, содержащую последовательность:

[0322] Последовательности гипервариабельных участков (CDR; т.е. CDR1, CDR2 и CDR3), как указано выше, и каркасных участков (FR) определяли в соответствии со схемой нумерации Kabat.
[0323] Оценивали способность антитела 5 не нарушать передачу сигналов IL-2 и его способность уничтожать клетки-мишени, экспрессирующие CD25. Результаты анализа связывания с rhCD25 представлены на Фигурах 58. Анализ STAT5 показал, что антитело 5 не блокировало испытываемую передачу сигналов IL-2, в то время как антитело даклизумаб полностью блокировало передачу сигналов IL-2 (Фигура 59). Конкурентный анализ показал, что антитело 5 не конкурирует с блокаторами сигнала IL-2, даклизумабом или базиликсимабом, Фигуры 60 (А) и (В), однако оно действительно конкурирует с 7G7B6 (не блокирующим IL-2) (Фигура 60 (С)). Наконец, антитело 5 уничтожает CD25-экспрессирующие клетки, опухолевые клетки или регуляторные Т-клетки за счет АЗКЦ (Фигура 61) и АЗКФ (Фигура 61) по сравнению с контрольным Fc-подавленным антителом к человеческому CD25.
[0324] В заключение следует отметить, что антитело 5 было охарактеризовано и демонстрирует эффективное уничтожение СВ25-положительных клеток (Трег или раковых клеточных линий) и не нарушает передачу сигналов IL-2 и, следовательно, не ингибирует эффекторные ответы Т-клеток. Соответственно, антитело 5 представляет собой антитело, истощающее Трег, которое можно применять для лечения рака в виде монотерапии или в комбинации.
[0325] Антитело 6, антитело 7, антитело 8 и антитело 9 охарактеризованы как содержащие следующие последовательности:
[0326] Антитело 6 содержит вариабельную область тяжелой цепи, содержащую последовательность:

и вариабельную легкую цепь, содержащую последовательность:
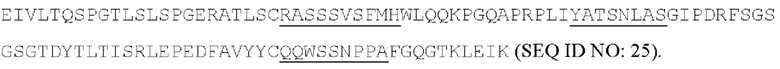
[0327] Антитело 7 содержит вариабельную область тяжелой цепи, содержащую последовательность:

и вариабельную легкую цепь, содержащую последовательность:
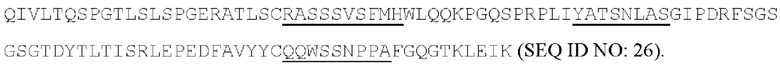
[0328] Антитело 8 содержит вариабельную область тяжелой цепи, содержащую последовательность:

и вариабельную легкую цепь, содержащую последовательность:
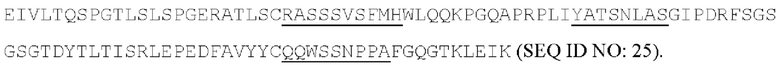
[0329] Антитело 9 содержит вариабельную область тяжелой цепи, содержащую последовательность:

и вариабельную легкую цепь, содержащую последовательность:
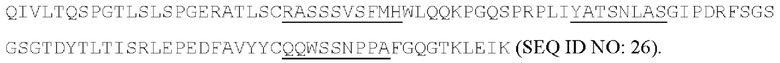
[0330] Последовательности гипервариабельных участков (CDR; т.е. CDR1, CDR2 и CDR3), как указано выше, и каркасных участков (FR) определяли в соответствии со схемой нумерации Kabat.
[0331] Результаты картирования эпитопов указывали на то, что антитело 6, антитело 7, антитело 8 и антитело 9 связывают человеческий CD25 в области аминокислот 150-163 (YQCVQGYRALHRGP) и аминокислот 166-180 (SVCKMTHGKTRWTQP) из SEQ ID NO: 1, и связывают внеклеточные белковые последовательности человеческого CD25 со значением Kd в диапазоне от 10-8 М до 10-10 М.
[0332] Оценивали способность антитела 6, антитела 7, антитела 8 и антитела 9 не нарушать передачу сигналов IL-2 и их способность уничтожать клетки-мишени, экспрессирующие CD25. Результаты связывания с rhCD25 представлены на Фигуре 63. Анализ STAT5 показал, что антитела не блокировали испытываемую передачу сигналов IL-2, в то время как антитело даклизумаб полностью блокировало передачу сигналов IL-2 (Фигура 65). Конкурентный анализ показал, что антитело 7 не конкурирует с блокаторами сигнала IL-2, даклизумабом или базиликеимабом, Фигура 64 (А) и (В). Наконец, антитело 7 уничтожает С025-экспрессирующие клетки, опухолевые клетки или регуляторные Т-клетки за счет АЗКЦ (Фигура 66) и АЗКФ (Фигура 67) по сравнению с контрольным Fc-подавленным антителом к человеческому CD25.
[0333] В заключение следует отметить, что антитело 6, антитело 7, антитело 8 и антитело 9 были охарактеризованы и демонстрируют эффективное уничтожение CD25-положительных клеток (Трег или раковых клеточных линий) и не нарушают передачу сигналов IL-2 и, следовательно, не ингибируют эффекторные ответы Т-клеток. Соответственно, антитела представляют собой антитела, истощающие iper? которые можно применять для лечения рака в виде монотерапии или в комбинации.
[0334] Антитело 10, антитело 11, антитело 12, антитело 12, антитело 13, антитело 14, антитело 15, антитело 16, антитело 17, антитело 18, антитело 19, антитело 20, антитело 21, антитела характеризуются тем, что содержат вариабельную область тяжелой цепи, содержащую последовательность:




[0335] Последовательности гипервариабельных участков (CDR; т.е. CDR1, CDR2 и CDR3), как указано выше, и каркасных участков (FR) определяли в соответствии со схемой нумерации Kabat.
[0336] Результаты картирования эпитопа указывают на то, что антитело 10, антитело 11, антитело 12, антитело 12, антитело 13, антитело 14, антитело 15, антитело 16, антитело 17, антитело 18, антитело 19, антитело 20, антитело 21 связывают человеческий CD25 в области аминокислот 150-163 (YQCVQGYRALHRGP) и аминокислот 166-180 (SVCKMTHGKTRWTQP) из SEQ ID NO: 1, и связывают внеклеточные белковые последовательности человеческого CD25 со значением Kd в диапазоне от 10-8 М до 10-10 М.
[0337] Оценивали способность антител не нарушать передачу сигналов IL-2 и их способность уничтожать клетки-мишени, экспрессирующие CD25. Результаты связывания с rhCD25 представлены на Фигуре 68. Анализ STAT5 показал, что антитела не блокировали испытываемую передачу сигналов IL-2, в то время как антитело даклизумаб полностью блокировало передачу сигналов IL-2 (Фигура 70). Конкурентный анализ показал, что антитело 19 не конкурирует с блокаторами сигналов IL-2, даклизумабом или базиликсимабом, Фигура 69 (А) и (В). Наконец, антитело 12, антитело 19 и антитело 20 уничтожали СВ25-экспрессирующие клетки, опухолевые клетки или регуляторные Т-клетки за счет АЗКЦ (Фигура 71) и АЗКФ (Фигура 72 и 73) по сравнению с контрольным Fc-подавленным антителом к человеческому CD25.
[0338] В заключение следует отметить, что антитело 10, антитело 11, антитело 12, антитело 12, антитело 13, антитело 14, антитело 15, антитело 16, антитело 17, антитело 18, антитело 19, антитело 20, антитело 21 были охарактеризованы и демонстрируют эффективное уничтожение CD25-положительных клеток (Трег или раковых клеточных линий) и не нарушают передачу сигналов IL-2 и, следовательно, не ингибируют эффекторные ответы Т-клеток. Соответственно, антитела представляют собой антитела, истощающие Трег, которые можно применять для лечения рака в виде монотерапии или в комбинации.
[0339] Пример 12: Терапевтический анализ в комбинации с противораковой вакциной
Определяли терапевтическую активность антитела к CD25, мышиного IgG2a 7D4, не блокирующего IL-2, в комбинации с GVAX в модели устойчивой к иммунотерапии В16 В16 на мышах. На 0 день внутрикожно имплантировали 50×103 клеток В16 В16. На 5 день внутрибрюшинно вводили 200 мкг антитела к CD25, не блокирующего IL-2, или не применяли его. На 6, 9 и 12 дни мышей обрабатывали 1×106 облученных (150 Гр) клеток В16 В16, к которым в качестве адъюванта был добавлен ГМ-КСФ (GVAX), либо не проводили обработку. Рост опухолей и выживаемость мышей контролировали до 33 дня. Результаты представлены на Фигуре 74.
[0340] В модели В16 В16 для комбинации GVAX и неблокирующего антитела к CD25, 7D4, наблюдали синергетический эффект. Следовательно, введение 7D4 с противораковой вакциной усиливало индуцированный вакциной противоопухолевый ответ. Эти результаты показывают, что не блокирующее IL-2 и истощающее антитело к CD25 можно применять в комбинации с противораковыми вакцинами для лечения рака у человека. Кроме того, эти данные показывают, что не блокирующее IL-2 и истощающее антитело способно усиливать индуцированные вакциной иммунные ответы с потенциально более широкими вариантами применения, чем рак.
[0341] Все публикации, упомянутые в приведенном выше описании, включены в него посредством ссылки. Различные модификации и варианты описанных способов и систем настоящего изобретения будут очевидны для специалистов в данной области техники без отступления от объема и сущности настоящего изобретения. Несмотря на то, что настоящее изобретение было описано в отношении конкретных предпочтительных вариантов реализации, следует понимать, что заявленное изобретение не должно ненадлежащим образом ограничиваться такими конкретными вариантами реализации. Действительно, различные модификации описанных способов реализации настоящего изобретения, которые очевидны для специалистов в области молекулярной биологии, клеточной иммунологии или в смежных областях, предназначены для включения в объем следующей формулы изобретения.
--->
Перечень последовательностей
<110> Tusk Therapeutics Ltd
<120> FC-ОПТИМИЗИРОВАННЫЕ АНТИТЕЛА К CD25 ДЛЯ ИСТОЩЕНИЯ ОПУХОЛЕСПЕЦИФИЧЕСКИХ
КЛЕТОК
<130> P115692WO
<160> 40
<170> PatentIn, версия 3.5
<210> 1
<211> 272
<212> PRT/БЕЛОК
<213> homosapien
<400> 1
Met Asp Ser Tyr Leu Leu Met Trp Gly Leu Leu Thr Phe Ile Met Val
1 5 10 15
Pro Gly Cys Gln Ala Glu Leu Cys Asp Asp Asp Pro Pro Glu Ile Pro
20 25 30
His Ala Thr Phe Lys Ala Met Ala Tyr Lys Glu Gly Thr Met Leu Asn
35 40 45
Cys Glu Cys Lys Arg Gly Phe Arg Arg Ile Lys Ser Gly Ser Leu Tyr
50 55 60
Met Leu Cys Thr Gly Asn Ser Ser His Ser Ser Trp Asp Asn Gln Cys
65 70 75 80
Gln Cys Thr Ser Ser Ala Thr Arg Asn Thr Thr Lys Gln Val Thr Pro
85 90 95
Gln Pro Glu Glu Gln Lys Glu Arg Lys Thr Thr Glu Met Gln Ser Pro
100 105 110
Met Gln Pro Val Asp Gln Ala Ser Leu Pro Gly His Cys Arg Glu Pro
115 120 125
Pro Pro Trp Glu Asn Glu Ala Thr Glu Arg Ile Tyr His Phe Val Val
130 135 140
Gly Gln Met Val Tyr Tyr Gln Cys Val Gln Gly Tyr Arg Ala Leu His
145 150 155 160
Arg Gly Pro Ala Glu Ser Val Cys Lys Met Thr His Gly Lys Thr Arg
165 170 175
Trp Thr Gln Pro Gln Leu Ile Cys Thr Gly Glu Met Glu Thr Ser Gln
180 185 190
Phe Pro Gly Glu Glu Lys Pro Gln Ala Ser Pro Glu Gly Arg Pro Glu
195 200 205
Ser Glu Thr Ser Cys Leu Val Thr Thr Thr Asp Phe Gln Ile Gln Thr
210 215 220
Glu Met Ala Ala Thr Met Glu Thr Ser Ile Phe Thr Thr Glu Tyr Gln
225 230 235 240
Val Ala Val Ala Gly Cys Val Phe Leu Leu Ile Ser Val Leu Leu Leu
245 250 255
Ser Gly Leu Thr Trp Gln Arg Arg Gln Arg Lys Ser Arg Arg Thr Ile
260 265 270
<210> 2
<211> 219
<212> PRT/БЕЛОК
<213> homosapien
<400> 2
Glu Leu Cys Asp Asp Asp Pro Pro Glu Ile Pro His Ala Thr Phe Lys
1 5 10 15
Ala Met Ala Tyr Lys Glu Gly Thr Met Leu Asn Cys Glu Cys Lys Arg
20 25 30
Gly Phe Arg Arg Ile Lys Ser Gly Ser Leu Tyr Met Leu Cys Thr Gly
35 40 45
Asn Ser Ser His Ser Ser Trp Asp Asn Gln Cys Gln Cys Thr Ser Ser
50 55 60
Ala Thr Arg Asn Thr Thr Lys Gln Val Thr Pro Gln Pro Glu Glu Gln
65 70 75 80
Lys Glu Arg Lys Thr Thr Glu Met Gln Ser Pro Met Gln Pro Val Asp
85 90 95
Gln Ala Ser Leu Pro Gly His Cys Arg Glu Pro Pro Pro Trp Glu Asn
100 105 110
Glu Ala Thr Glu Arg Ile Tyr His Phe Val Val Gly Gln Met Val Tyr
115 120 125
Tyr Gln Cys Val Gln Gly Tyr Arg Ala Leu His Arg Gly Pro Ala Glu
130 135 140
Ser Val Cys Lys Met Thr His Gly Lys Thr Arg Trp Thr Gln Pro Gln
145 150 155 160
Leu Ile Cys Thr Gly Glu Met Glu Thr Ser Gln Phe Pro Gly Glu Glu
165 170 175
Lys Pro Gln Ala Ser Pro Glu Gly Arg Pro Glu Ser Glu Thr Ser Cys
180 185 190
Leu Val Thr Thr Thr Asp Phe Gln Ile Gln Thr Glu Met Ala Ala Thr
195 200 205
Met Glu Thr Ser Ile Phe Thr Thr Glu Tyr Gln
210 215
<210> 3
<211> 123
<212> PRT/БЕЛОК
<213> Мышиная
<400> 3
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Gln Pro Arg Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu Glu Leu Val
35 40 45
Ala Thr Ile Asn Gly Tyr Gly Asp Thr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met Tyr Phe Cys
85 90 95
Ala Arg Asp Arg Asp Tyr Gly Asn Ser Tyr Tyr Tyr Ala Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 4
<211> 106
<212> PRT/БЕЛОК
<213> Мышиная
<400> 4
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Arg Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Phe Met
20 25 30
His Trp Leu Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Ser Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Thr Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Ala
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 5
<211> 119
<212> PRT/БЕЛОК
<213> Мышиная
<400> 5
Gln Val Gln Leu Lys Glu Ser Gly Pro Gly Leu Val Ala Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Tyr
20 25 30
Gly Ile Gln Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ala Gly Gly Ser Thr Asn Tyr Asn Ser Ala Leu Met
50 55 60
Ser Arg Leu Ser Ile Ser Lys Asp Asn Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Arg Ala Tyr Gly Tyr Asp Gly Ser Trp Leu Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 6
<211> 106
<212> PRT/БЕЛОК
<213> Мышиная
<400> 6
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Phe
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Asn Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Asp Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 7
<211> 10
<212> PRT/БЕЛОК
<213> Мышиная
<400> 7
Gly Phe Thr Leu Asp Ser Tyr Gly Val Ser
1 5 10
<210> 8
<211> 15
<212> PRT/БЕЛОК
<213> Мышиная
<400> 8
Gly Val Thr Ser Ser Gly Gly Ser Ala Tyr Tyr Ala Asp Ser Val
1 5 10 15
<210> 9
<211> 17
<212> PRT/БЕЛОК
<213> Мышиная
<400> 9
Asp Arg Tyr Val Tyr Thr Gly Gly Tyr Leu Tyr His Tyr Gly Met Asp
1 5 10 15
Leu
<210> 10
<211> 125
<212> PRT/БЕЛОК
<213> Мышиная
<400> 10
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Leu Asp Ser Tyr
20 25 30
Gly Val Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Thr Ser Ser Gly Gly Ser Ala Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Arg Tyr Val Tyr Thr Gly Gly Tyr Leu Tyr His Tyr Gly Met
100 105 110
Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 11
<211> 11
<212> PRT/БЕЛОК
<213> Мышиная
<400> 11
Arg Ala Ser Gln Ser Ile Ser Asp Tyr Leu Ala
1 5 10
<210> 12
<211> 8
<212> PRT/БЕЛОК
<213> Мышиная
<400> 12
Tyr Ala Ala Ser Thr Leu Pro Phe
1 5
<210> 13
<211> 12
<212> PRT/БЕЛОК
<213> Мышиная
<400> 13
Gln Gly Thr Tyr Asp Ser Ser Asp Trp Tyr Trp Ala
1 5 10
<210> 14
<211> 109
<212> PRT/БЕЛОК
<213> Мышиная
<400> 14
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Asp Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Pro Phe Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Gly Thr Tyr Asp Ser Ser Asp
85 90 95
Trp Tyr Trp Ala Phe Gly Gly Gly Thr Lys Val Glu Ile
100 105
<210> 15
<211> 11
<212> PRT/БЕЛОК
<213> Мышиная
<400> 15
Ser Gly Phe Ser Val Asp Ile Tyr Asp Met Ser
1 5 10
<210> 16
<211> 15
<212> PRT/БЕЛОК
<213> Мышиная
<400> 16
Tyr Ile Ser Ser Ser Leu Gly Ala Thr Tyr Tyr Ala Asp Ser Val
1 5 10 15
<210> 17
<211> 16
<212> PRT/БЕЛОК
<213> Мышиная
<400> 17
Glu Arg Ile Tyr Ser Val Tyr Thr Leu Asp Tyr Tyr Ala Met Asp Leu
1 5 10 15
<210> 18
<211> 125
<212> PRT/БЕЛОК
<213> Мышиная
<400> 18
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Val Asp Ile Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Ser Leu Gly Ala Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Arg Ile Tyr Ser Val Tyr Thr Leu Asp Tyr Tyr Ala Met
100 105 110
Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 19
<211> 11
<212> PRT/БЕЛОК
<213> Мышиная
<400> 19
Gln Ala Ser Gln Gly Ile Thr Asn Asn Leu Asn
1 5 10
<210> 20
<211> 8
<212> PRT/БЕЛОК
<213> Мышиная
<400> 20
Tyr Ala Ala Ser Thr Leu Gln Ser
1 5
<210> 21
<211> 12
<212> PRT/БЕЛОК
<213> Мышиная
<400> 21
Gln Gln Gly Tyr Thr Thr Ser Asn Val Asp Asn Ala
1 5 10
<210> 22
<211> 110
<212> PRT/БЕЛОК
<213> Мышиная
<400> 22
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Gly Ile Thr Asn Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Gln Gly Tyr Thr Thr Ser Asn
85 90 95
Val Asp Asn Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 23
<211> 123
<212> PRT/БЕЛОК
<213> Мышиная
<400> 23
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ser Thr Ile Asn Gly Tyr Gly Asp Thr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Asp Tyr Gly Asn Ser Tyr Tyr Tyr Ala Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 24
<211> 123
<212> PRT/БЕЛОК
<213> Мышиная
<400> 24
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ser Thr Ile Asn Gly Tyr Gly Asp Thr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Asp Arg Asp Tyr Gly Asn Ser Tyr Tyr Tyr Ala Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 25
<211> 106
<212> PRT/БЕЛОК
<213> Мышиная
<400> 25
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Ser Ser Val Ser Phe Met
20 25 30
His Trp Leu Gln Gln Lys Pro Gly Gln Ala Pro Arg Pro Leu Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Ala
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 26
<211> 106
<212> PRT/БЕЛОК
<213> Мышиная
<400> 26
Gln Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Ser Ser Val Ser Phe Met
20 25 30
His Trp Leu Gln Gln Lys Pro Gly Gln Ser Pro Arg Pro Leu Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Ala
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 27
<211> 119
<212> PRT/БЕЛОК
<213> homosapien
<400> 27
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Tyr
20 25 30
Gly Ile Gln Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Trp Ala Gly Gly Ser Thr Asn Tyr Asn Ser Ala Leu Met
50 55 60
Ser Arg Val Thr Ile Ser Lys Asp Asn Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Tyr Gly Tyr Asp Gly Ser Trp Leu Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 28
<211> 119
<212> PRT/БЕЛОК
<213> Мышиная
<400> 28
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Tyr
20 25 30
Gly Ile Gln Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Trp Ala Gly Gly Ser Thr Asn Tyr Asn Ser Ala Leu Met
50 55 60
Ser Arg Val Thr Ile Ser Lys Asp Asn Ser Lys Ser Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Tyr Gly Tyr Asp Gly Ser Trp Leu Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 29
<211> 119
<212> PRT/БЕЛОК
<213> Мышиная
<400> 29
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Ser Leu Thr Ser Tyr
20 25 30
Gly Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Trp Ala Gly Gly Ser Thr Asn Tyr Asn Ser Ala Leu Met
50 55 60
Ser Arg Phe Thr Ile Ser Lys Asp Asn Ser Lys Ser Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Tyr Gly Tyr Asp Gly Ser Trp Leu Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 30
<211> 106
<212> PRT/БЕЛОК
<213> Мышиная
<400> 30
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Pro Leu Ile Phe
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 31
<211> 106
<212> PRT/БЕЛОК
<213> Мышиная
<400> 31
Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Pro Leu Ile Phe
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 32
<211> 106
<212> PRT/БЕЛОК
<213> Мышиная
<400> 32
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Leu Ile Phe
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 33
<211> 106
<212> PRT/БЕЛОК
<213> Мышиная
<400> 33
Gln Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Pro Leu Ile Phe
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 34
<211> 139
<212> PRT/БЕЛОК
<213> Мышиная
<400> 34
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val
20 25 30
Arg Pro Gly Thr Ser Val Lys Leu Ser Cys Lys Val Ser Gly Asp Thr
35 40 45
Ile Thr Ala Tyr Tyr Ile His Phe Val Lys Gln Arg Pro Gly Gln Gly
50 55 60
Leu Glu Trp Ile Gly Arg Ile Asp Pro Glu Asp Asp Ser Thr Glu Tyr
65 70 75 80
Ala Glu Lys Phe Lys Asn Lys Ala Thr Ile Thr Ala Asn Thr Ser Ser
85 90 95
Asn Thr Ala His Leu Lys Tyr Ser Arg Leu Thr Ser Glu Asp Thr Ala
100 105 110
Thr Tyr Phe Cys Thr Thr Asp Asn Met Gly Ala Thr Glu Phe Val Tyr
115 120 125
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
130 135
<210> 35
<211> 129
<212> PRT/БЕЛОК
<213> Мышиная
<400> 35
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Gln Val Val Leu Thr Gln Pro Lys Ser Val Ser Ala
20 25 30
Ser Leu Glu Ser Thr Val Lys Leu Ser Cys Lys Leu Asn Ser Gly Asn
35 40 45
Ile Gly Ser Tyr Tyr Met His Trp Tyr Gln Gln Arg Glu Gly Arg Ser
50 55 60
Pro Thr Asn Leu Ile Tyr Arg Asp Asp Lys Arg Pro Asp Gly Ala Pro
65 70 75 80
Asp Arg Phe Ser Gly Ser Ile Asp Ile Ser Ser Asn Ser Ala Phe Leu
85 90 95
Thr Ile Asn Asn Val Gln Thr Glu Asp Glu Ala Met Tyr Phe Cys His
100 105 110
Ser Tyr Asp Gly Arg Met Tyr Ile Phe Gly Gly Gly Thr Lys Leu Thr
115 120 125
Val
<210> 36
<211> 438
<212> PRT/БЕЛОК
<213> Мышиная
<400> 36
Glu Val Gln Leu Gln Gln Ser Gly Ala Ala Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Ser Phe Pro Asp Ser
20 25 30
Trp Val Thr Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Phe Pro Asn Ser Gly Ala Thr Asn Phe Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Thr Ser Glu Asp Ser Ala Ile Tyr Tyr Cys
85 90 95
Thr Arg Leu Asp Tyr Gly Tyr Trp Gly Gln Gly Val Met Val Thr Val
100 105 110
Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly
115 120 125
Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys
130 135 140
Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu
145 150 155 160
Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr
165 170 175
Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Gln
180 185 190
Thr Val Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp
195 200 205
Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr
210 215 220
Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp
225 230 235 240
Val Leu Met Ile Ser Leu Thr Pro Lys Val Thr Cys Val Val Val Asp
245 250 255
Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp
260 265 270
Val Glu Val His Thr Ala Gln Thr Lys Pro Arg Glu Glu Gln Ile Asn
275 280 285
Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Leu His Gln Asp Trp
290 295 300
Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro
305 310 315 320
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala
325 330 335
Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp
340 345 350
Lys Val Ser Leu Thr Cys Met Ile Thr Asn Phe Phe Pro Glu Asp Ile
355 360 365
Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn
370 375 380
Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys
385 390 395 400
Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys
405 410 415
Ser Val Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu
420 425 430
Ser His Ser Pro Gly Lys
435
<210> 37
<211> 444
<212> PRT/БЕЛОК
<213> Мышиная
<400> 37
Glu Val Gln Leu Gln Gln Ser Gly Ala Ala Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Ser Phe Pro Asp Ser
20 25 30
Trp Val Thr Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Phe Pro Asn Ser Gly Ala Thr Asn Phe Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Thr Ser Glu Asp Ser Ala Ile Tyr Tyr Cys
85 90 95
Thr Arg Leu Asp Tyr Gly Tyr Trp Gly Gln Gly Val Met Val Thr Val
100 105 110
Ser Ser Ala Lys Thr Thr Ala Pro Ser Val Tyr Pro Leu Ala Pro Val
115 120 125
Cys Gly Asp Thr Thr Gly Ser Ser Val Thr Leu Gly Cys Leu Val Lys
130 135 140
Gly Tyr Phe Pro Glu Pro Val Thr Leu Thr Trp Asn Ser Gly Ser Leu
145 150 155 160
Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr
165 170 175
Thr Leu Ser Ser Ser Val Thr Val Thr Ser Ser Thr Trp Pro Ser Gln
180 185 190
Ser Ile Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp
195 200 205
Lys Lys Ile Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys
210 215 220
Lys Cys Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile
260 265 270
Ser Trp Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr
275 280 285
His Arg Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro
290 295 300
Ile Gln His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val
305 310 315 320
Asn Asn Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro
325 330 335
Lys Gly Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu
340 345 350
Glu Glu Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp
355 360 365
Phe Met Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr
370 375 380
Glu Leu Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Tyr Phe Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu
405 410 415
Arg Asn Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His
420 425 430
His Thr Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
435 440
<210> 38
<211> 219
<212> PRT/БЕЛОК
<213> Мышиная
<400> 38
Asp Val Val Leu Thr Gln Thr Pro Pro Thr Leu Ser Ala Thr Ile Gly
1 5 10 15
Gln Ser Val Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asn Trp Leu Leu Gln Arg Pro Gly Gln Pro
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Ala Ser Arg Leu Glu Ser Gly Val Pro
50 55 60
Asn Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Gly Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Val Gln Ser
85 90 95
Ser His Phe Pro Asn Thr Phe Gly Val Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
115 120 125
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
145 150 155 160
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
180 185 190
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
195 200 205
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 39
<211> 114
<212> PRT/БЕЛОК
<213> Мышиная
<400> 39
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met Ala Trp Val Arg Gln Ala Pro Thr Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Thr Asn Gly Gly Leu Asn Thr Tyr Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Cys Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Thr Gly Gly Phe Ser Phe Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser
<210> 40
<211> 107
<212> PRT/БЕЛОК
<213> Мышиная
<400> 40
Asp Ile Val Met Thr Gln Ser Pro Thr Ser Met Ser Ile Ser Val Gly
1 5 10 15
Asp Arg Val Thr Met Asn Cys Lys Ala Ser Gln Asn Val Asp Ser Asn
20 25 30
Val Asp Trp Tyr Gln Gln Lys Thr Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Arg Asn Met Gln Ala
65 70 75 80
Glu Asp Leu Ala Val Tyr Tyr Cys Met Gln Ser Asn Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<---
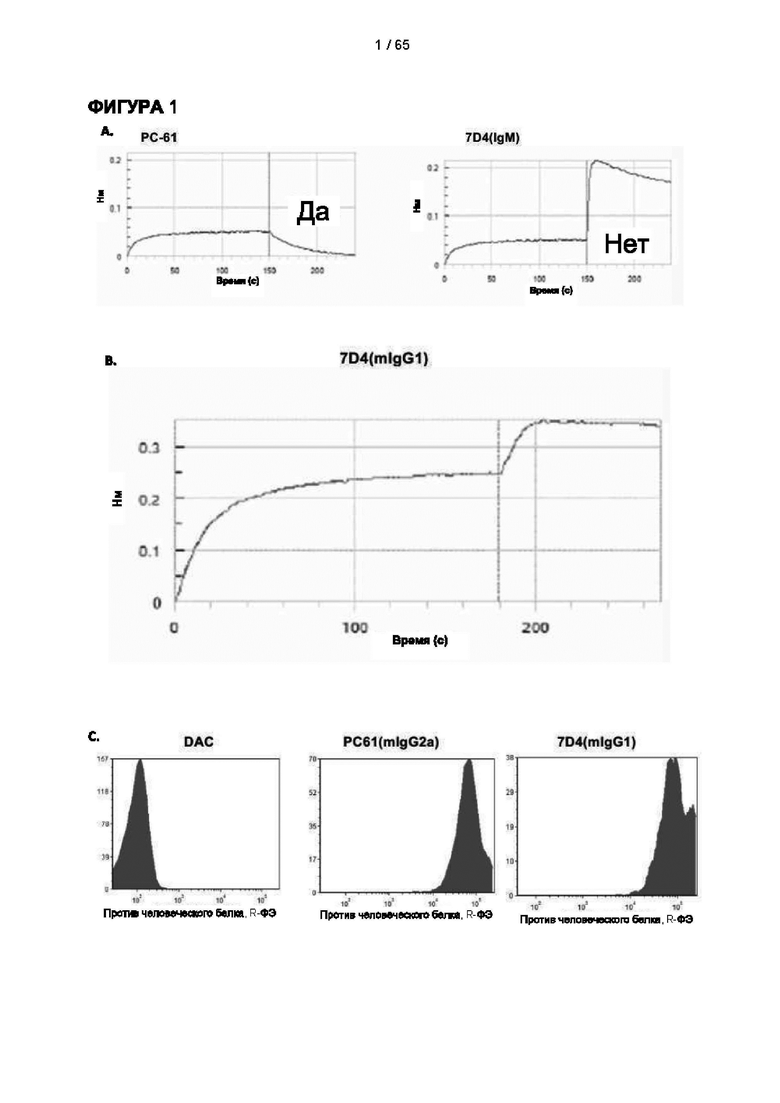
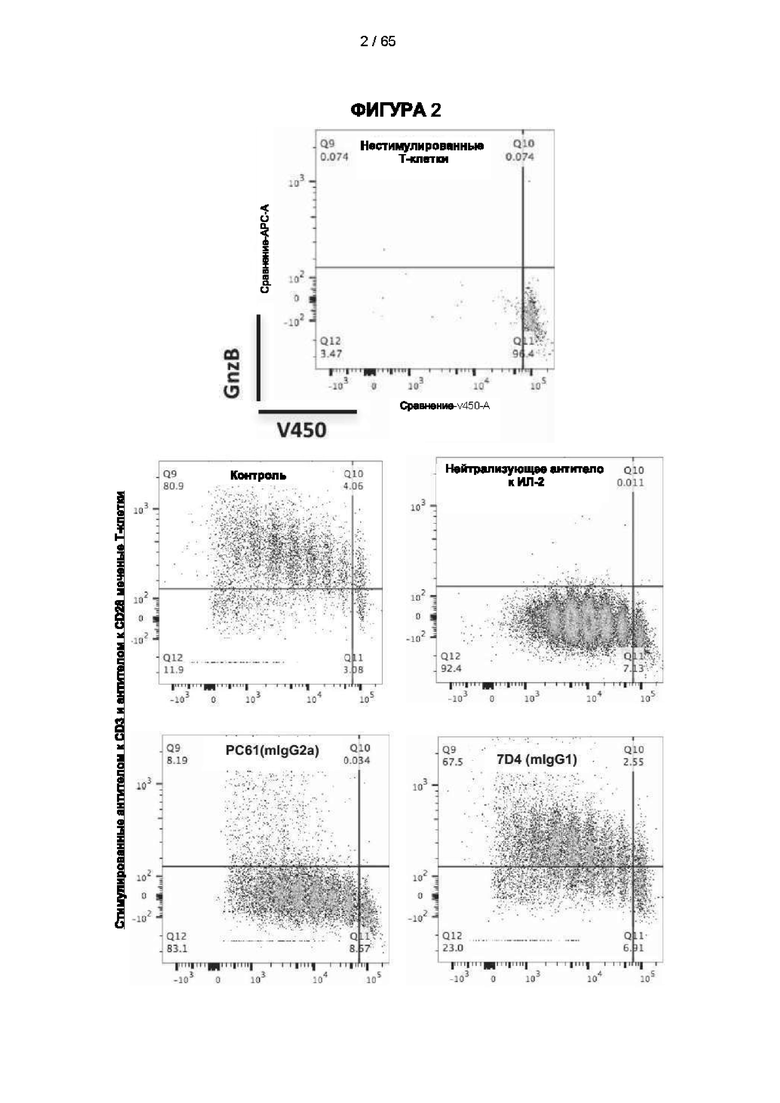
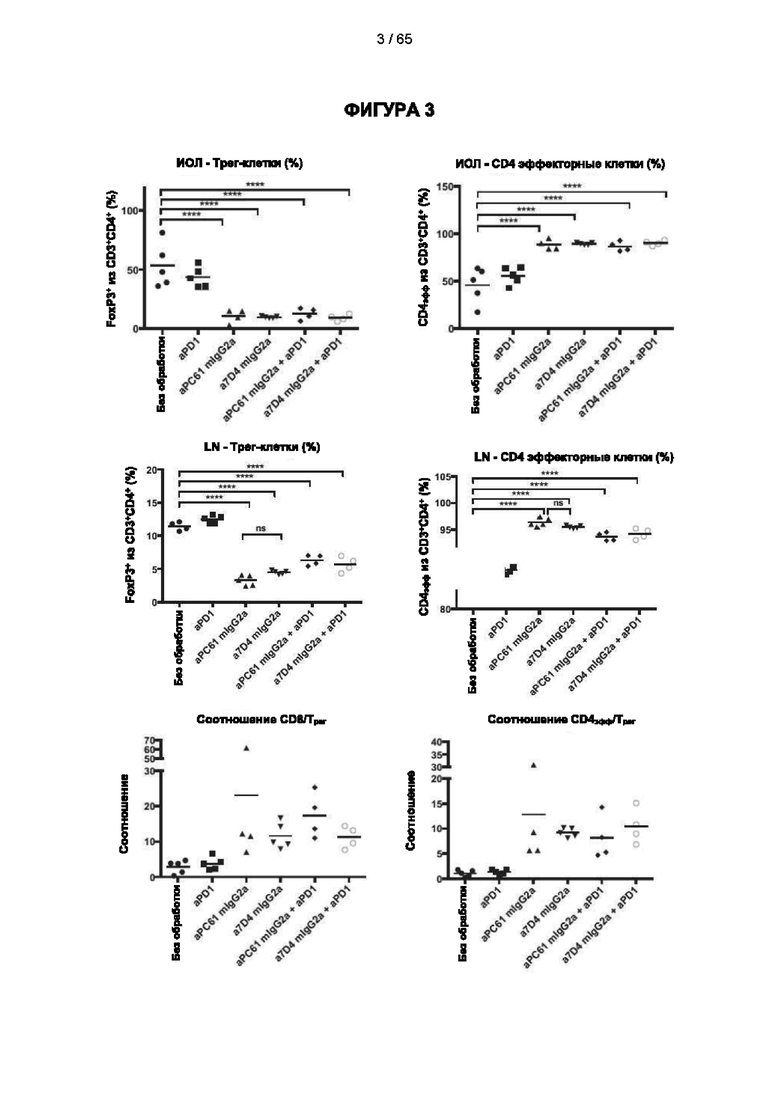
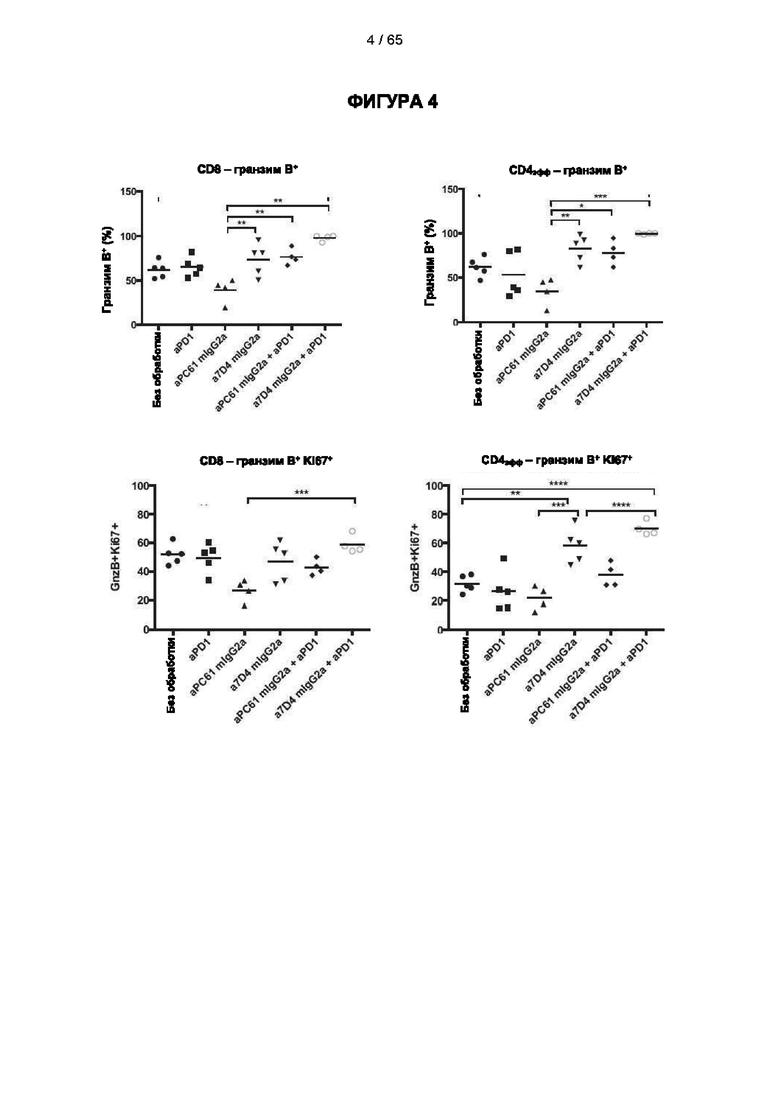
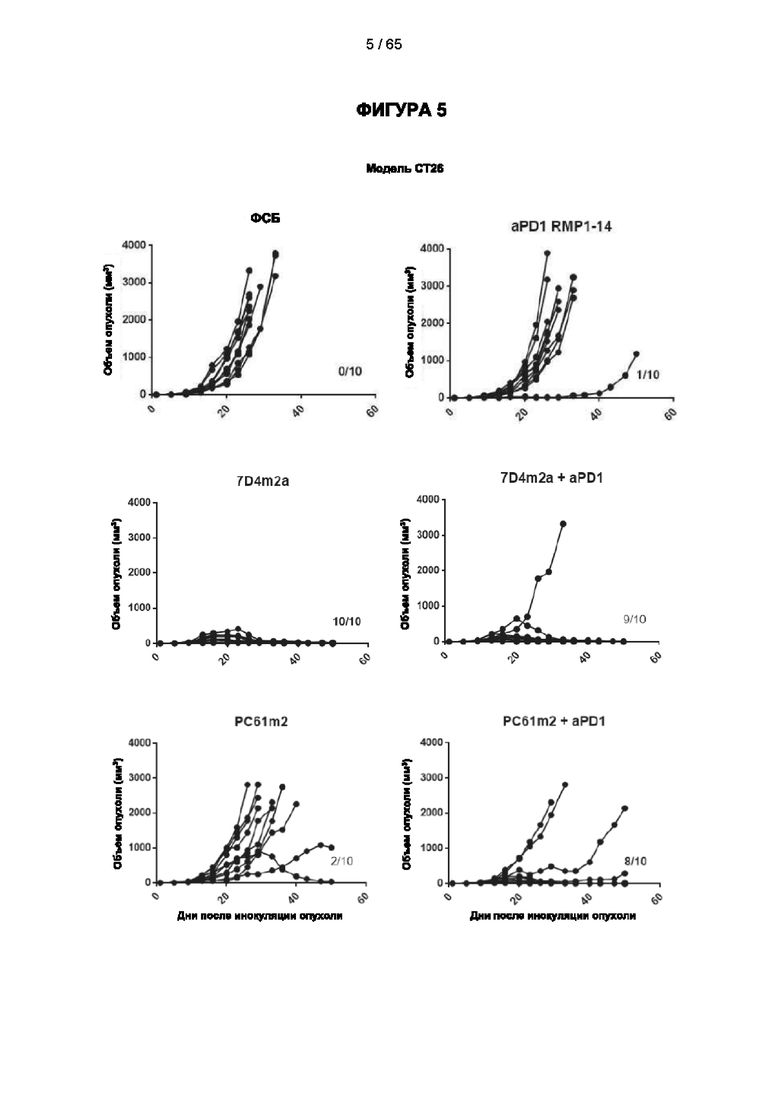
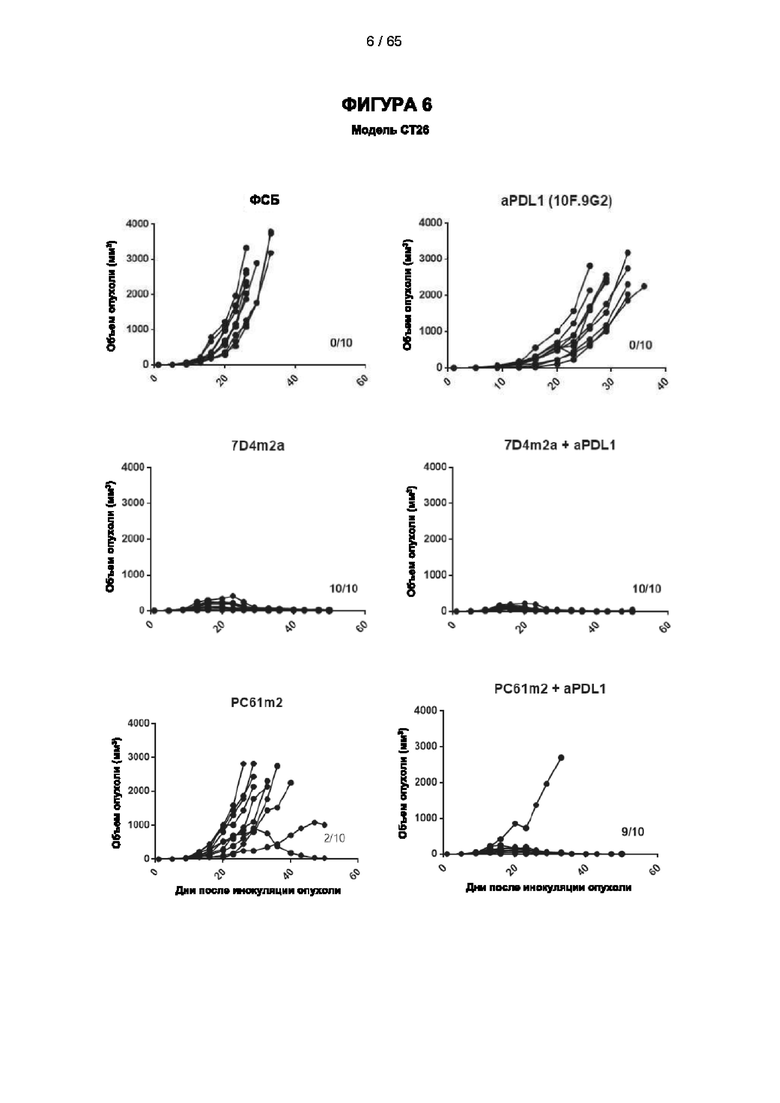
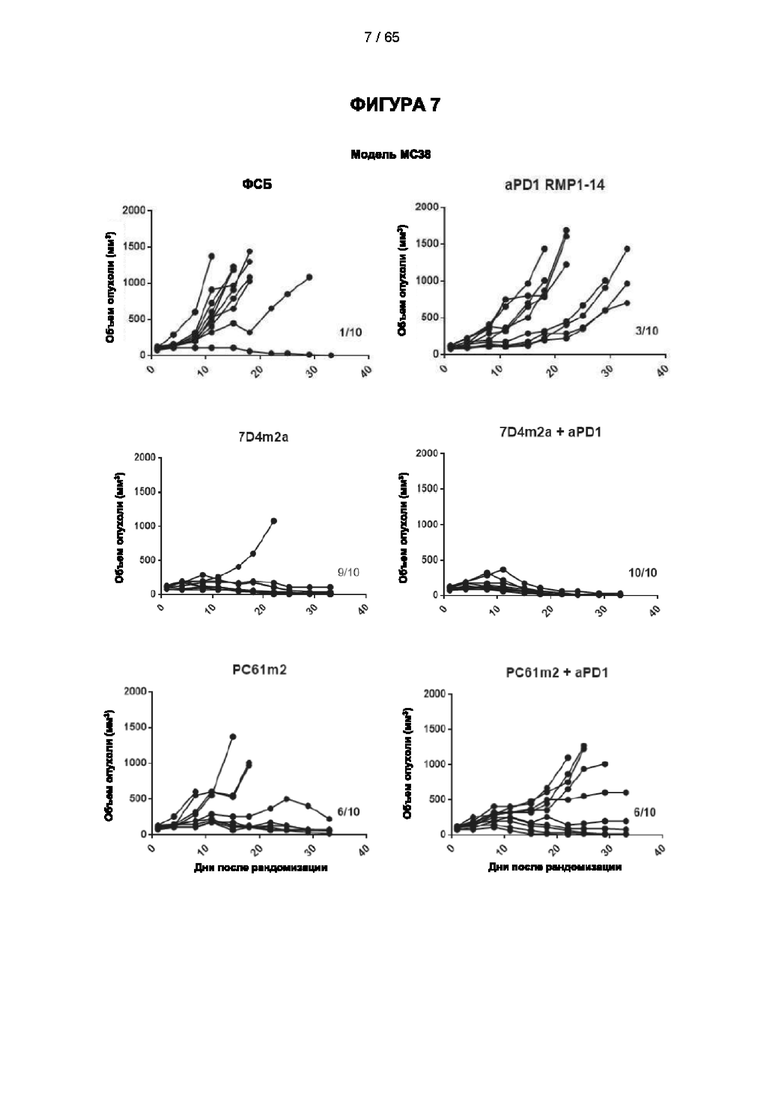
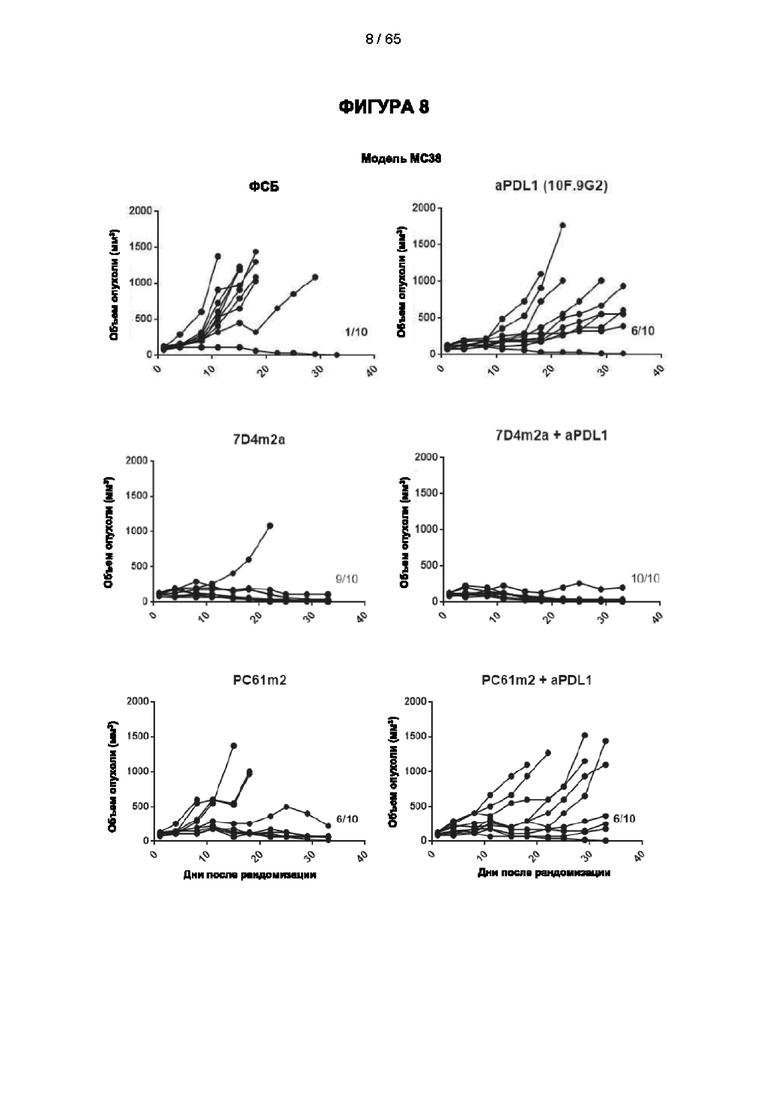
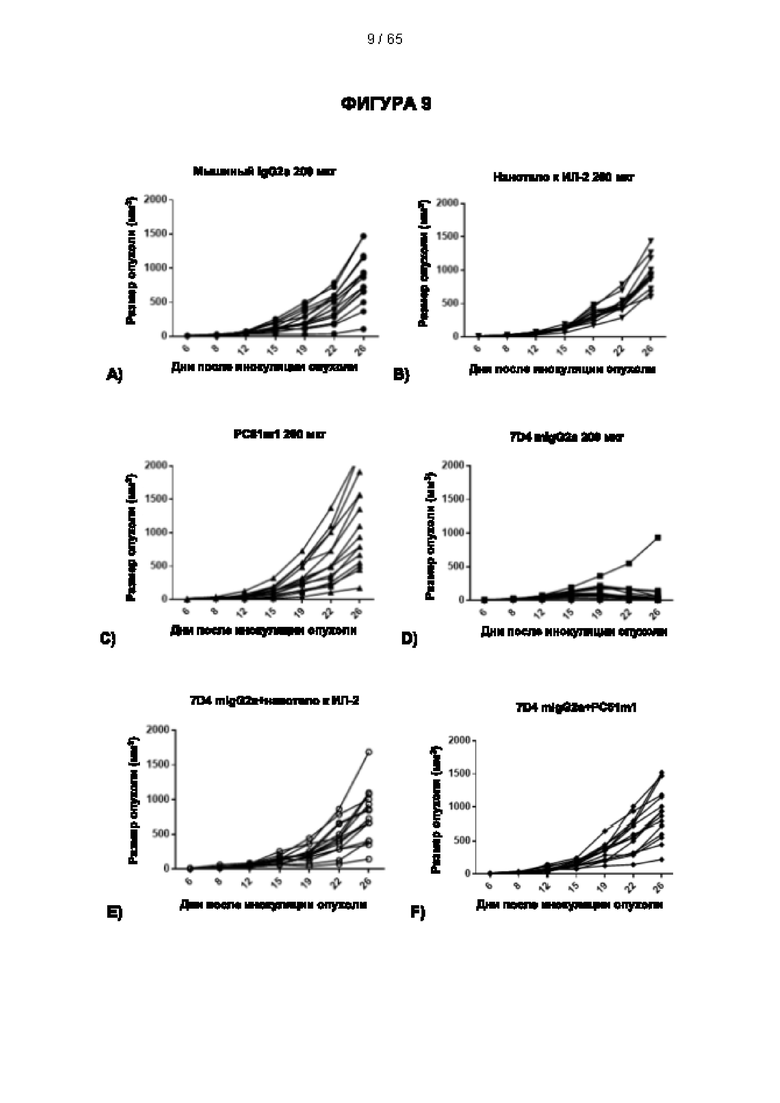

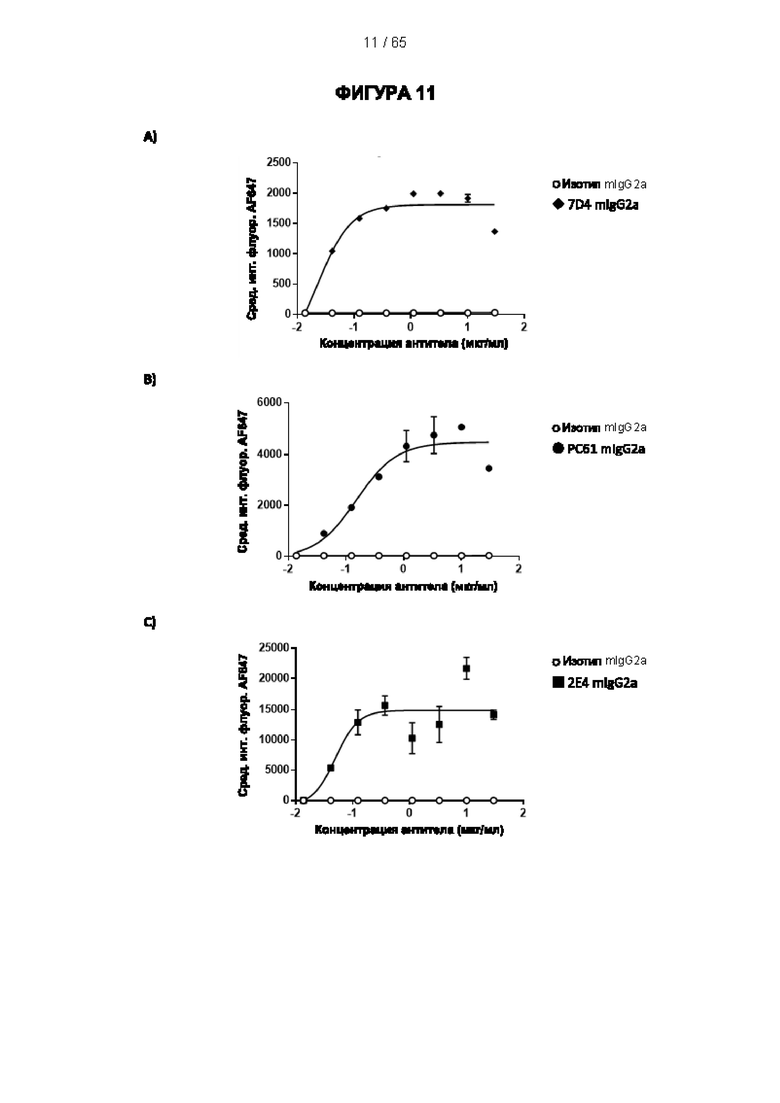
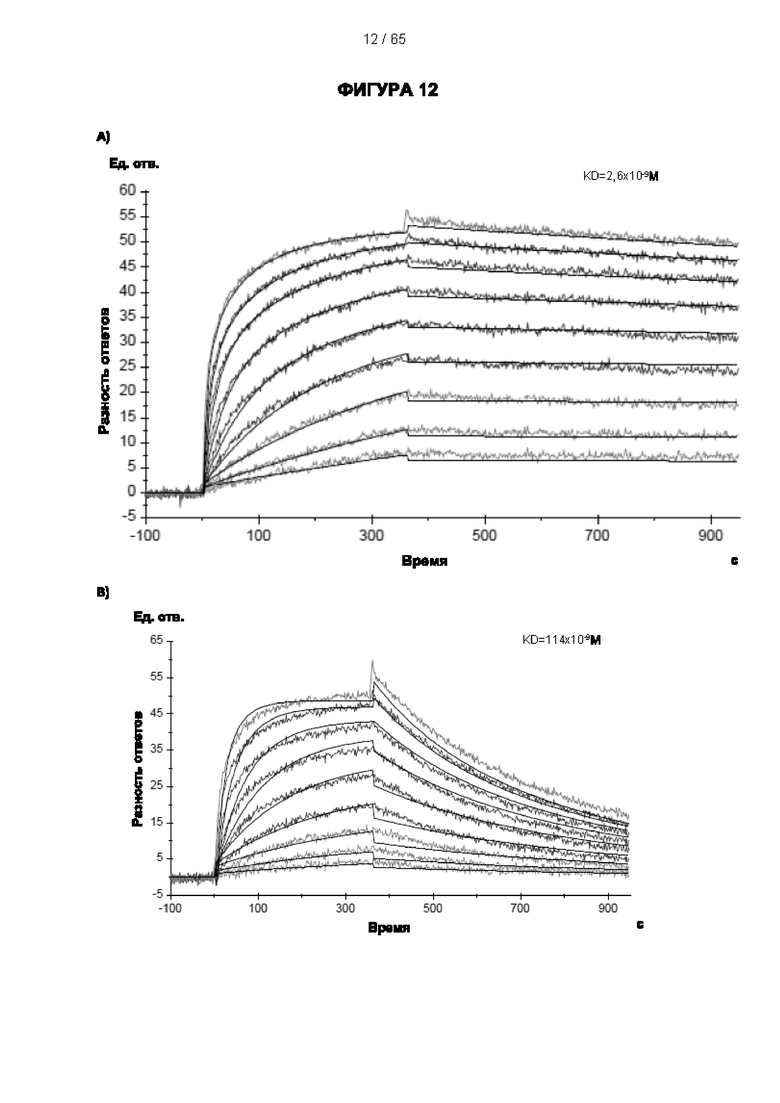
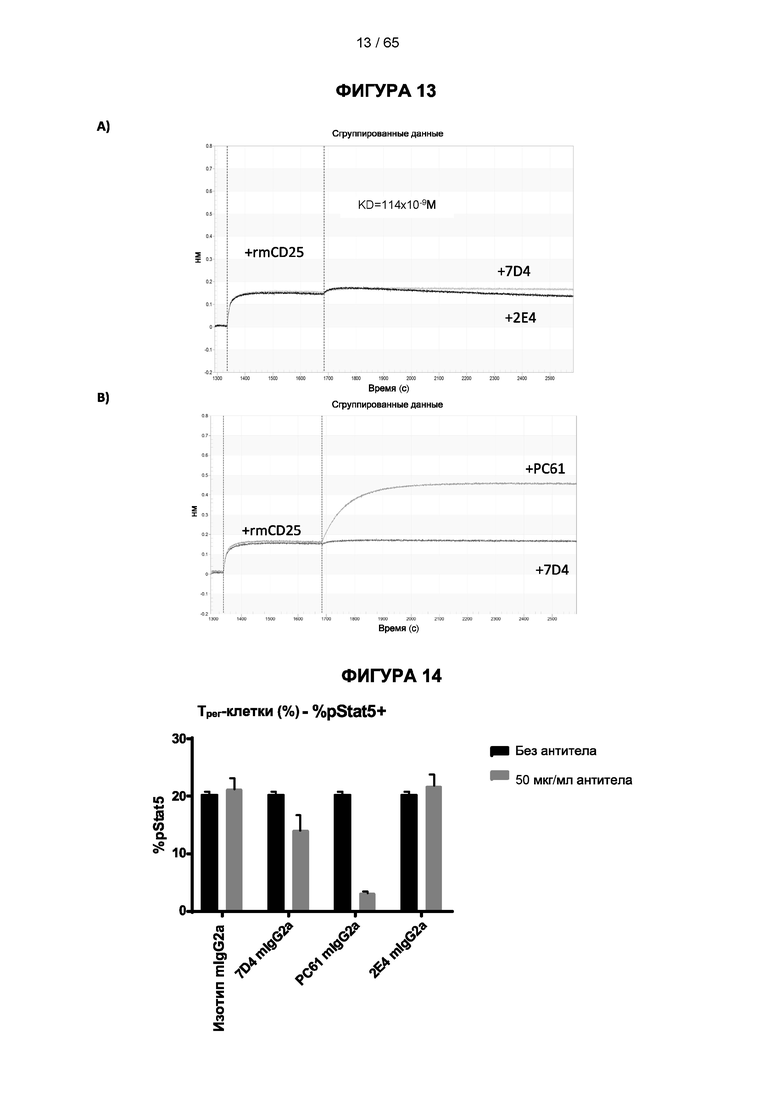
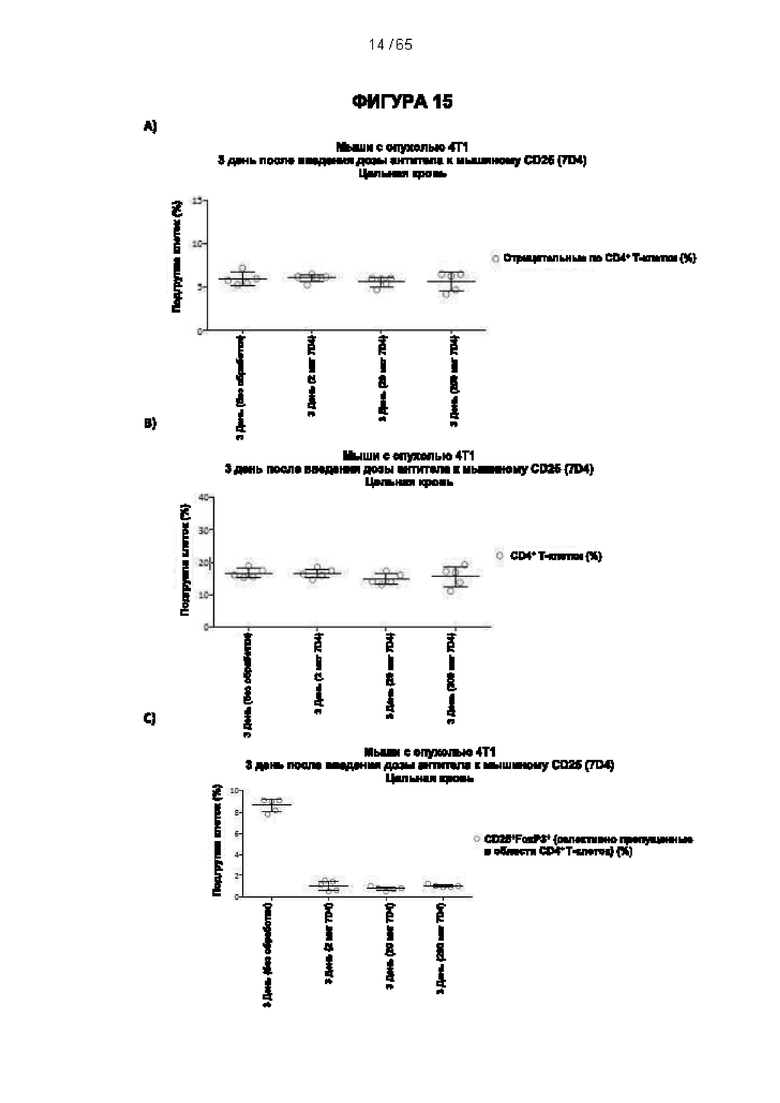
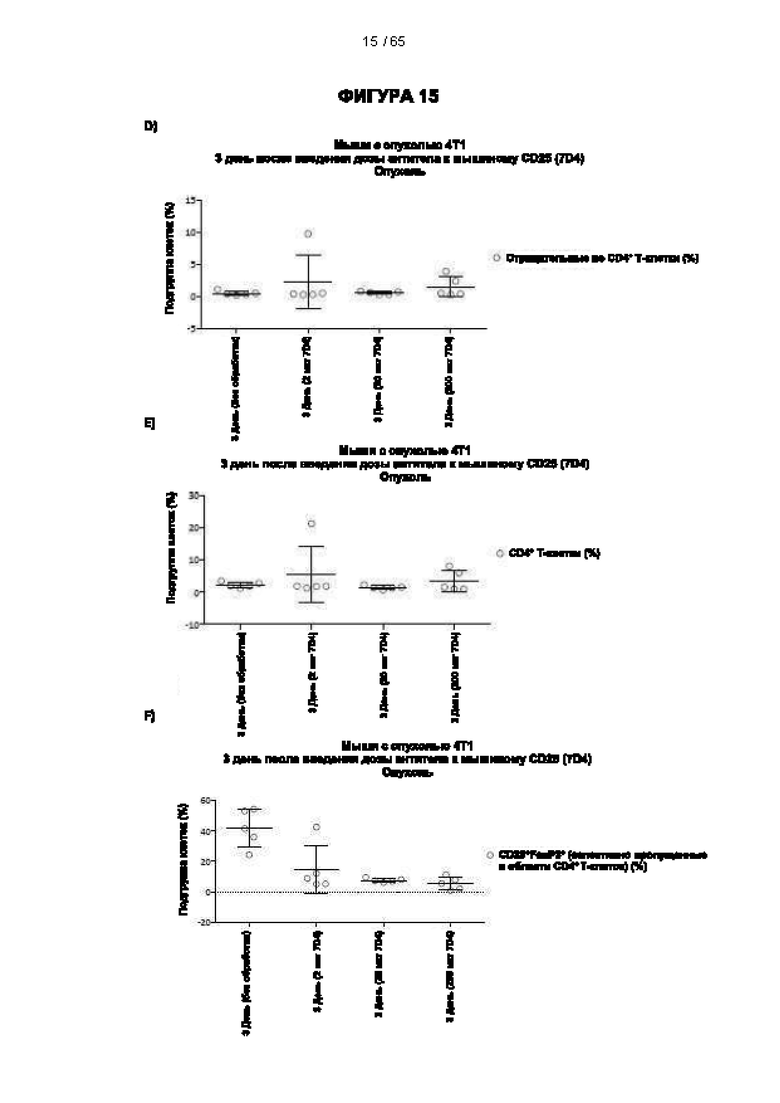
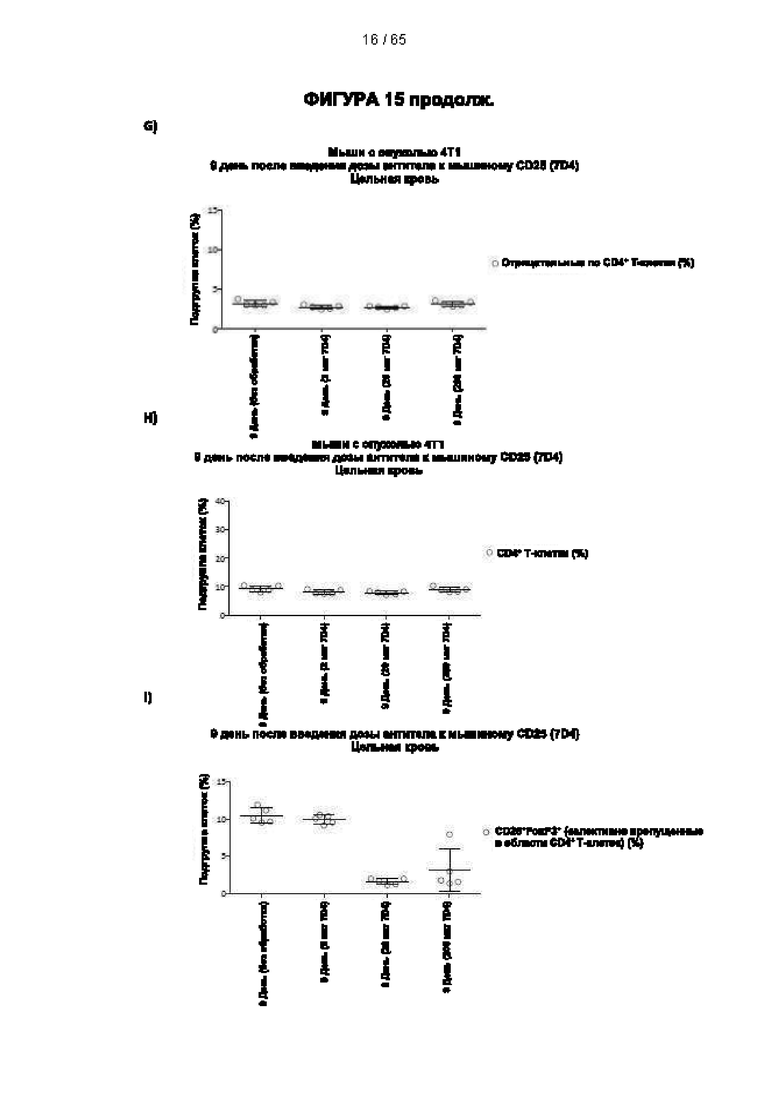
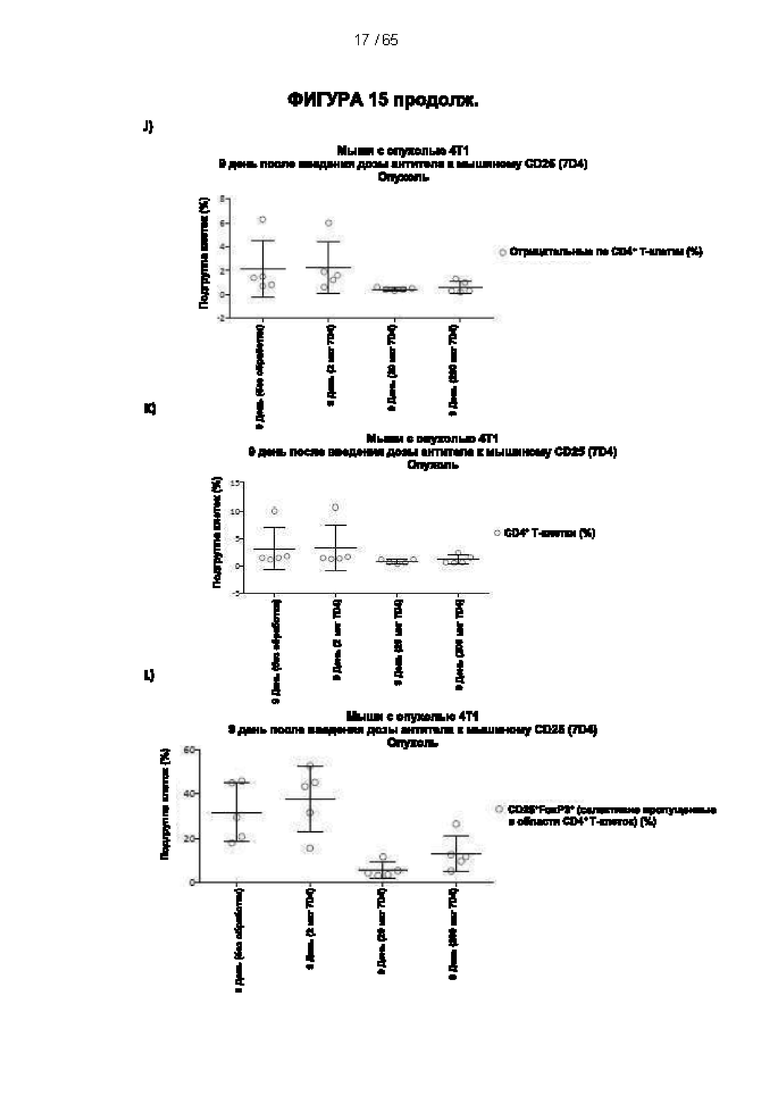
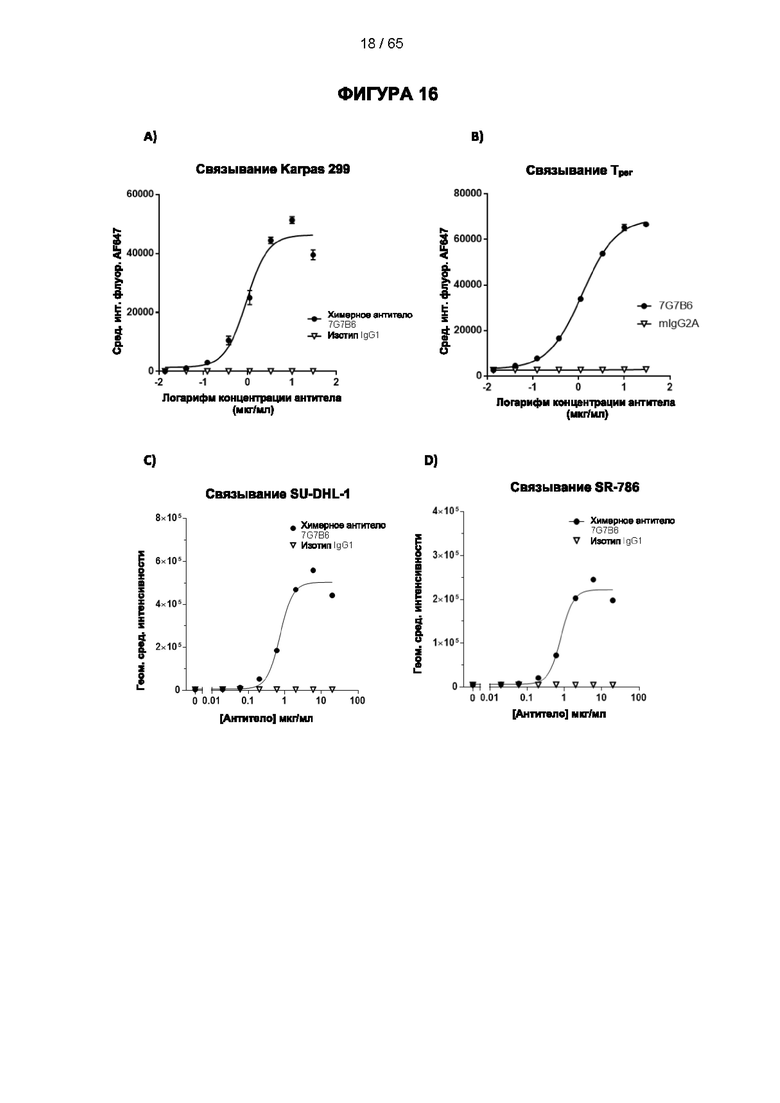
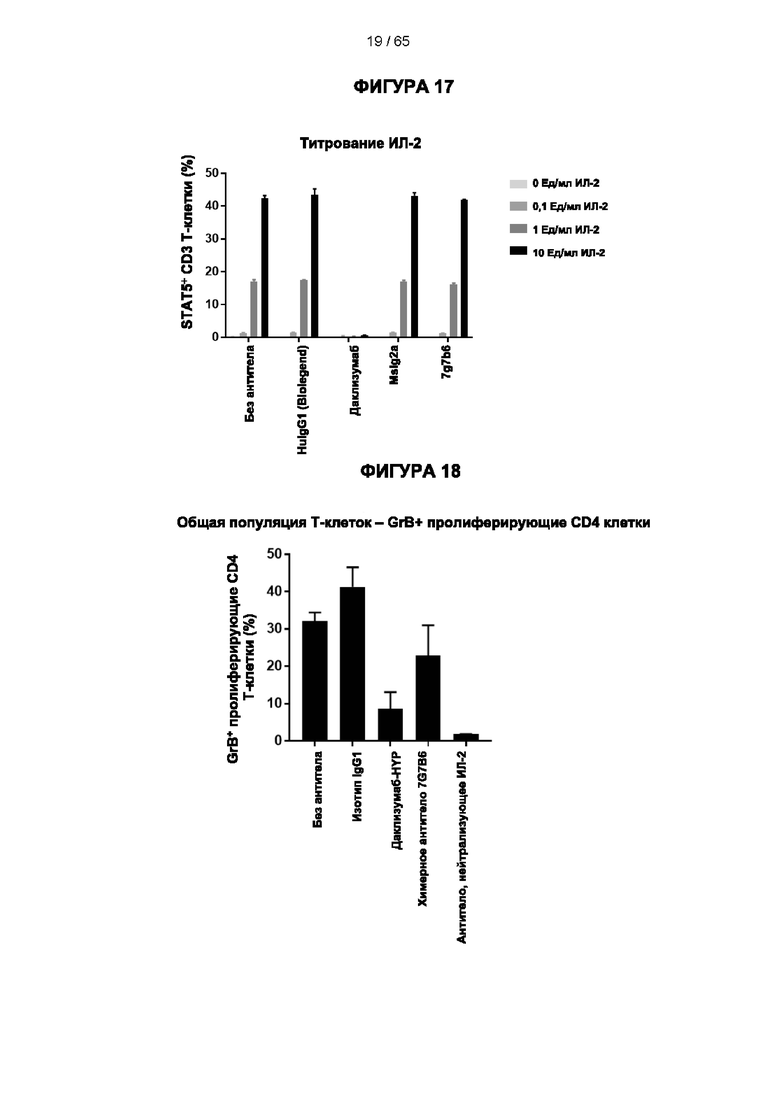
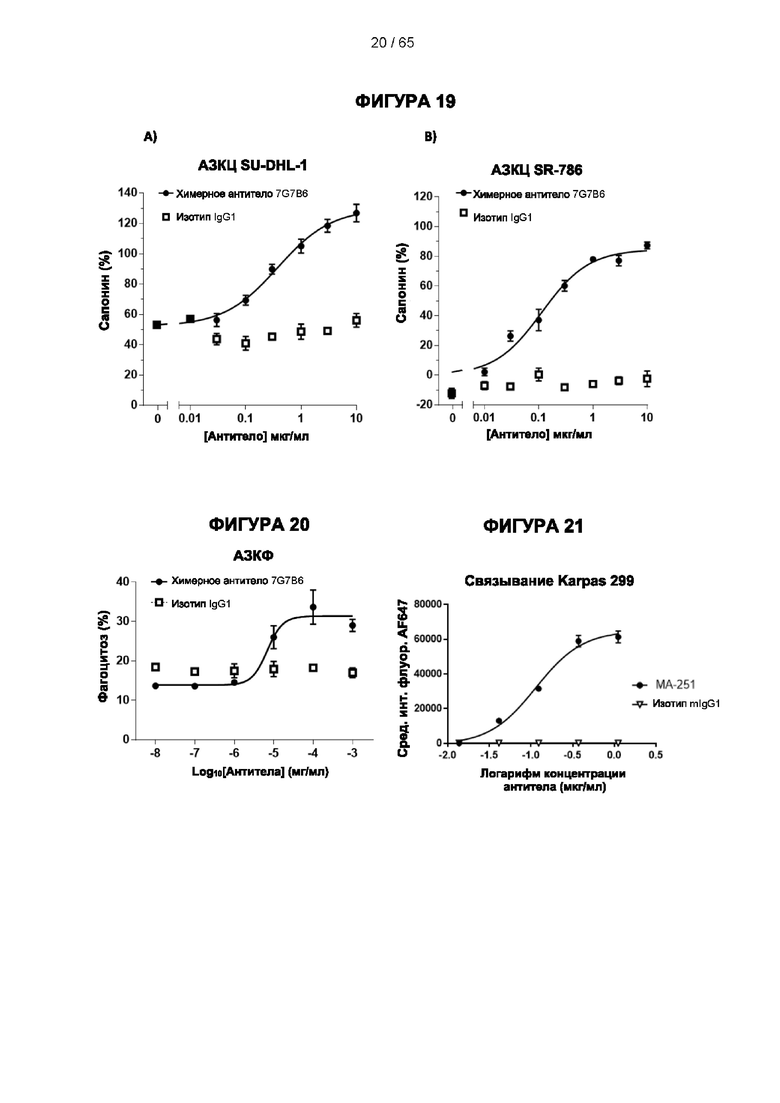
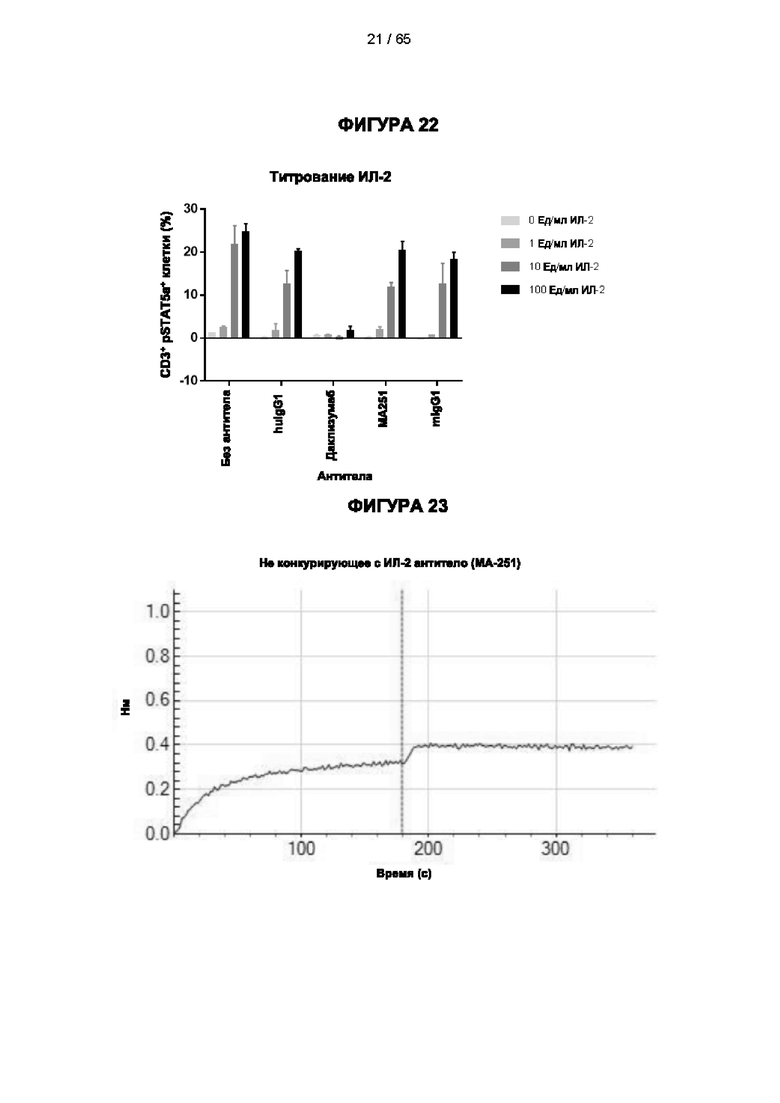
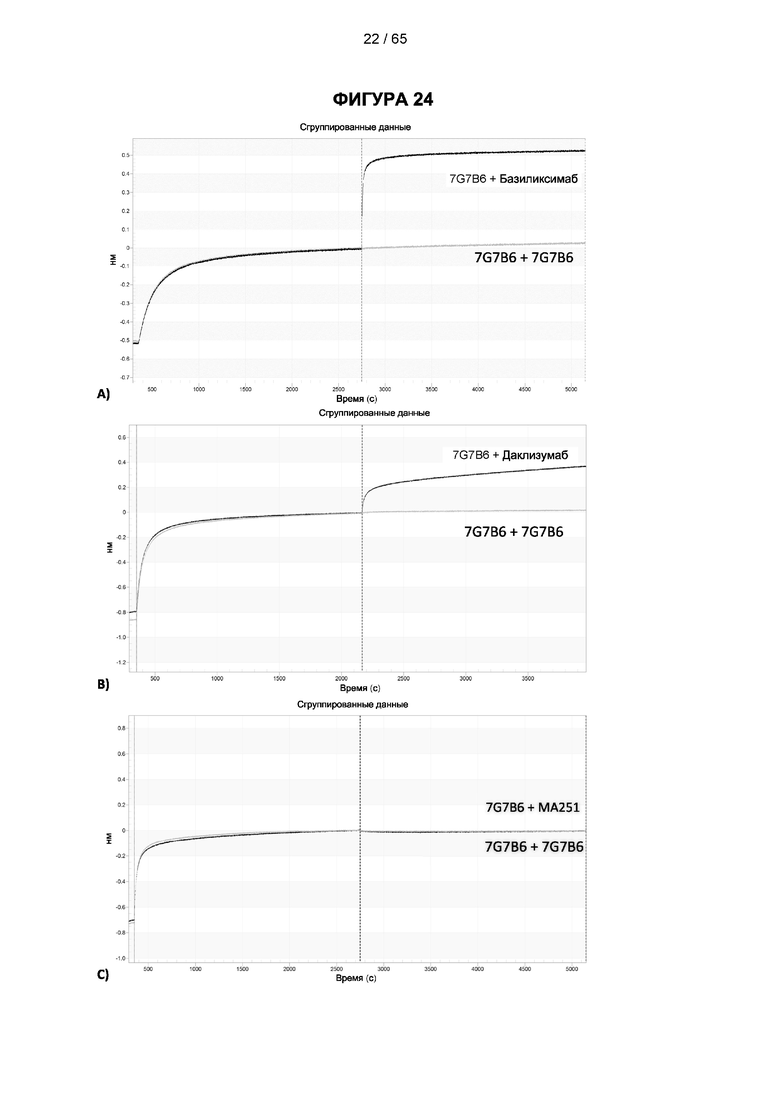
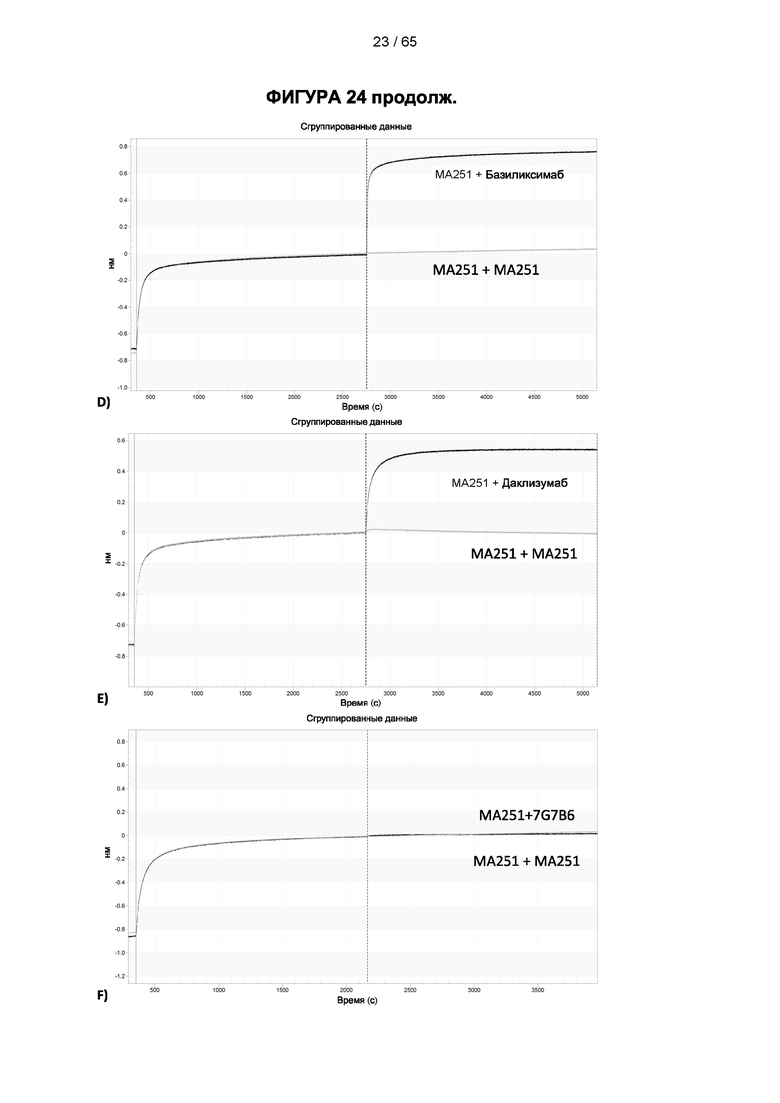
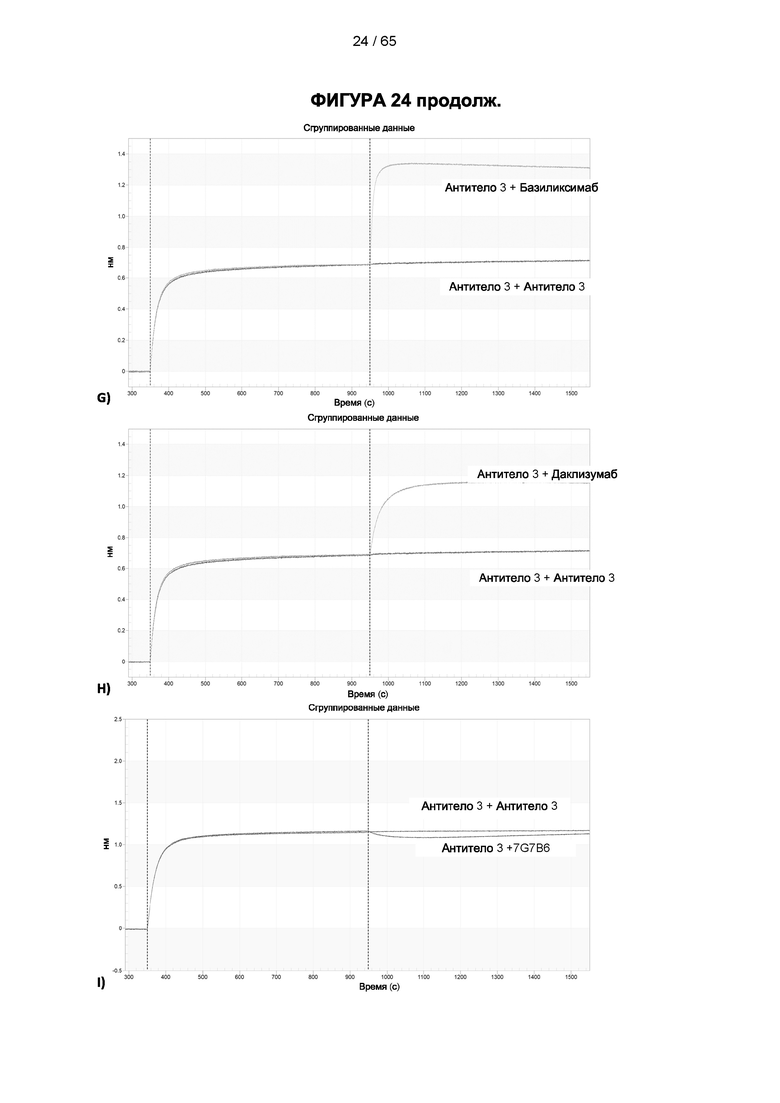
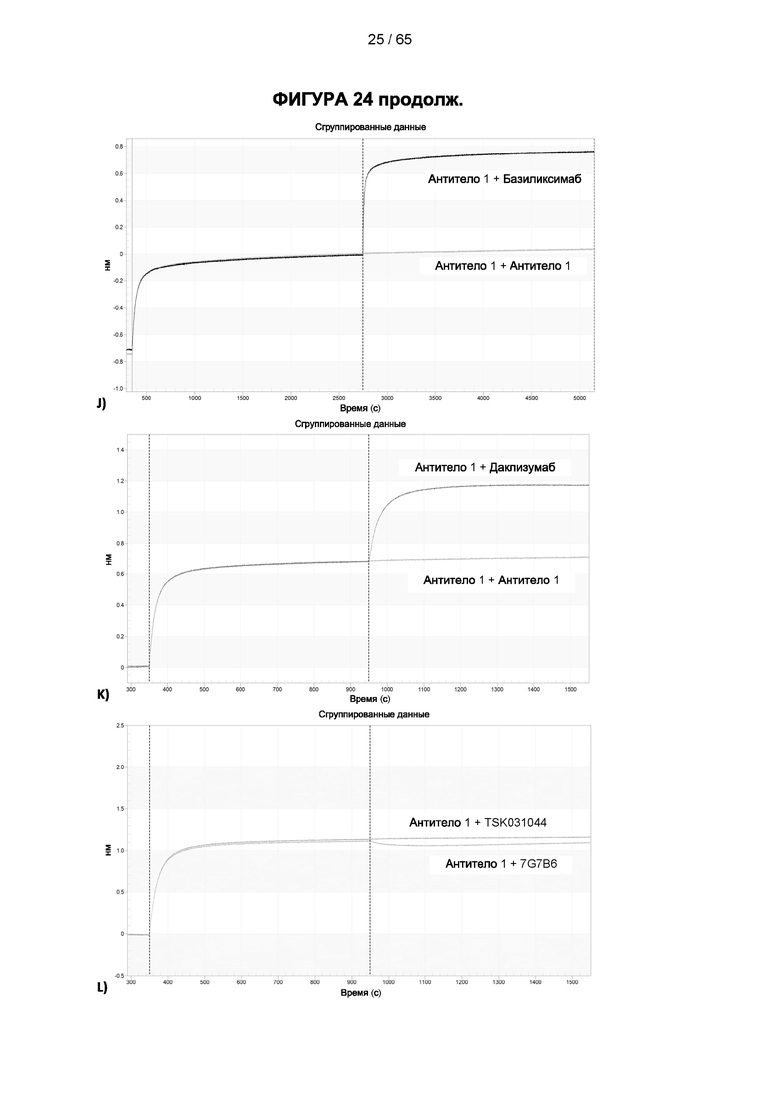
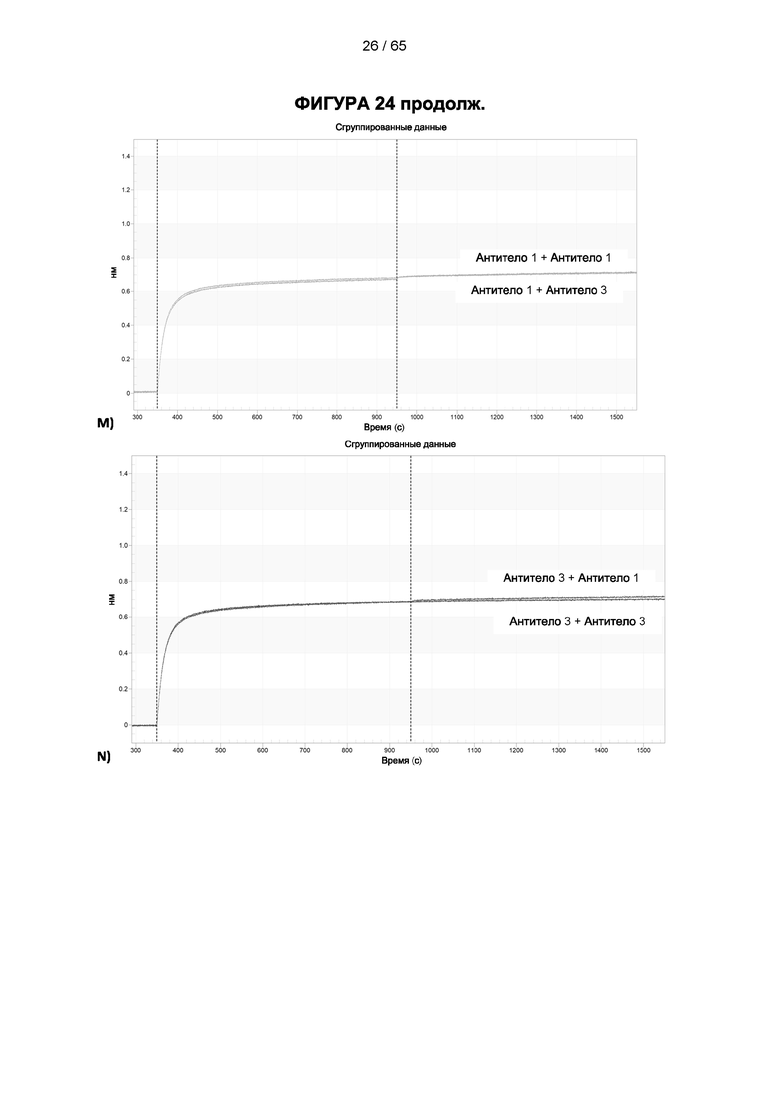
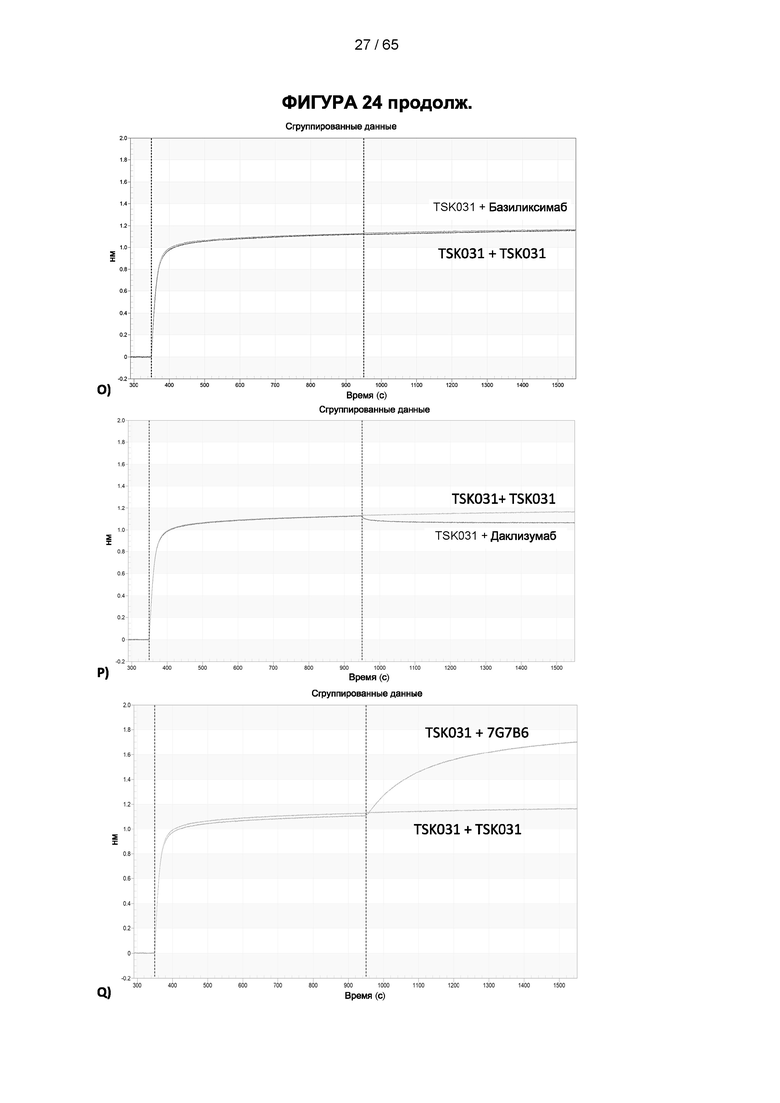
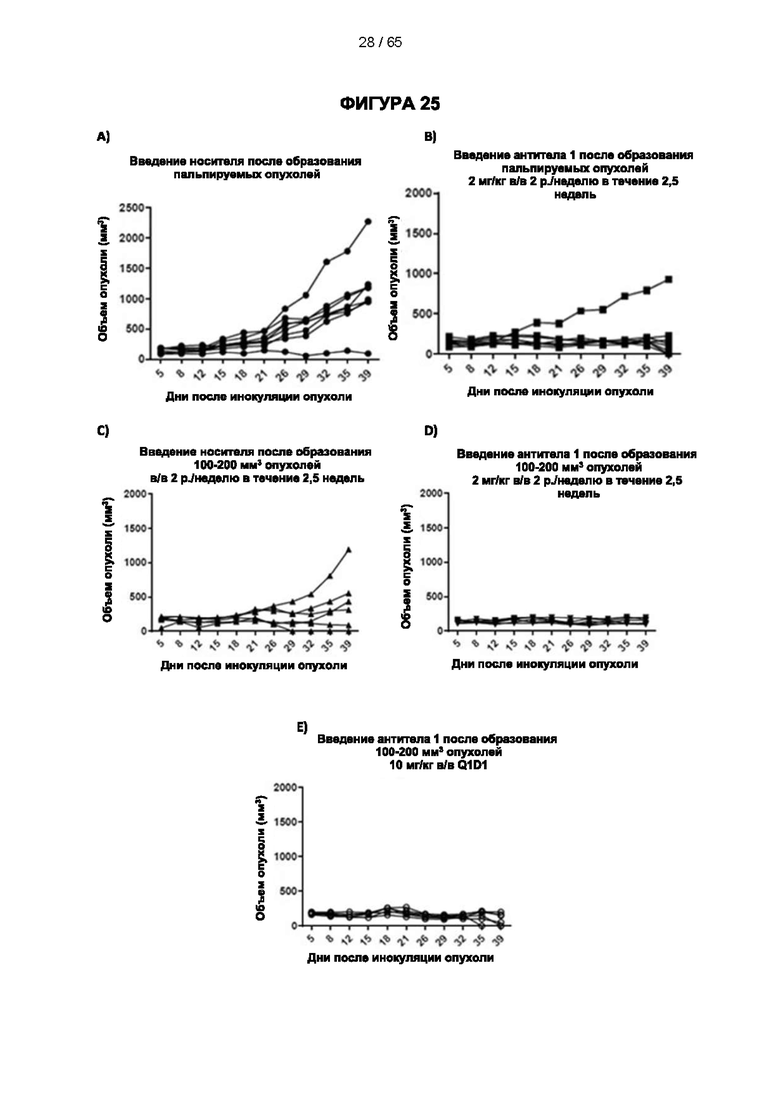
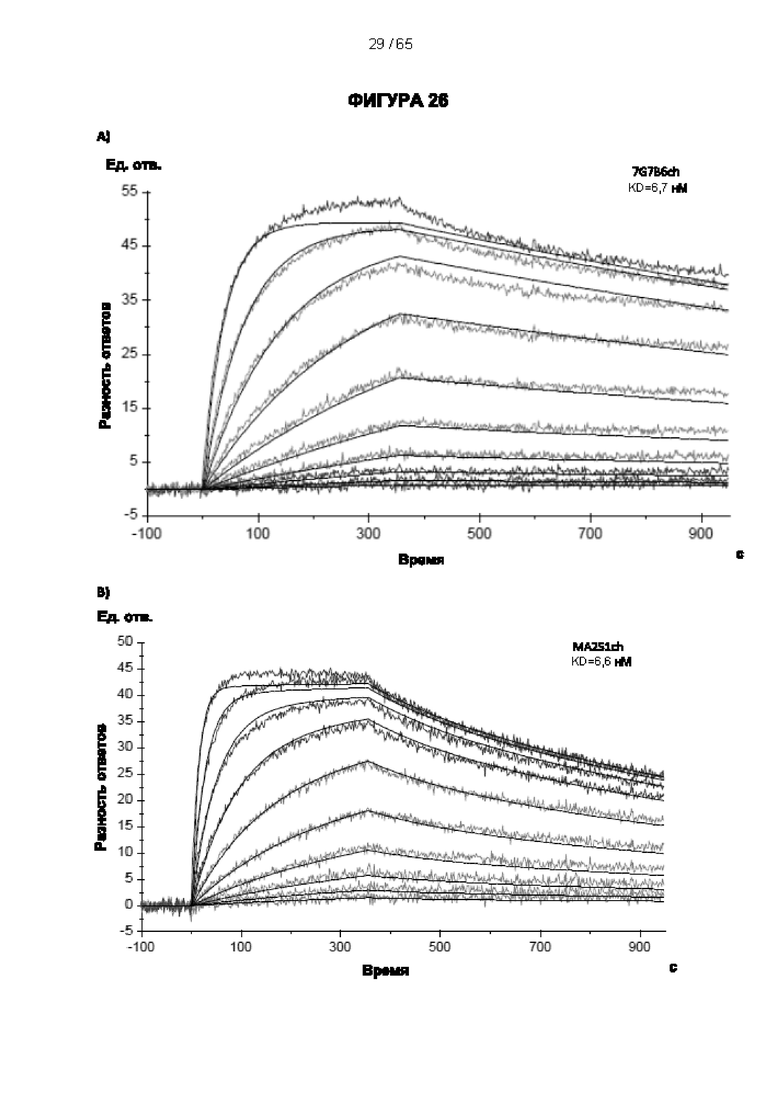
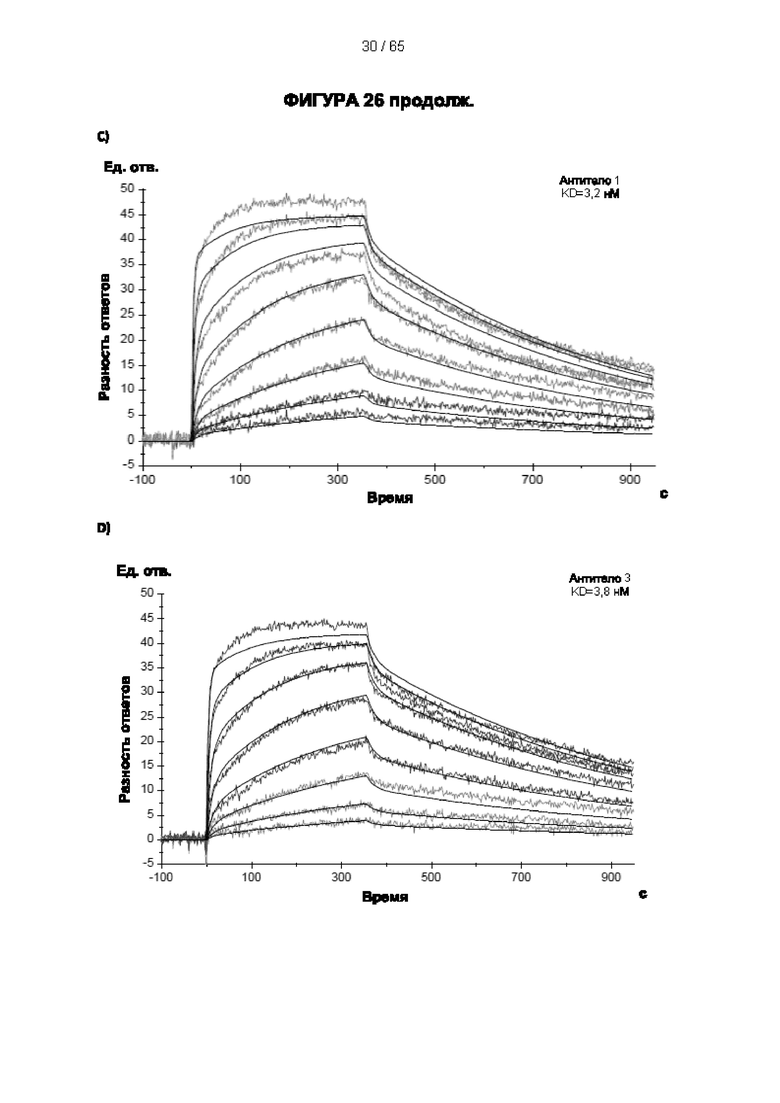
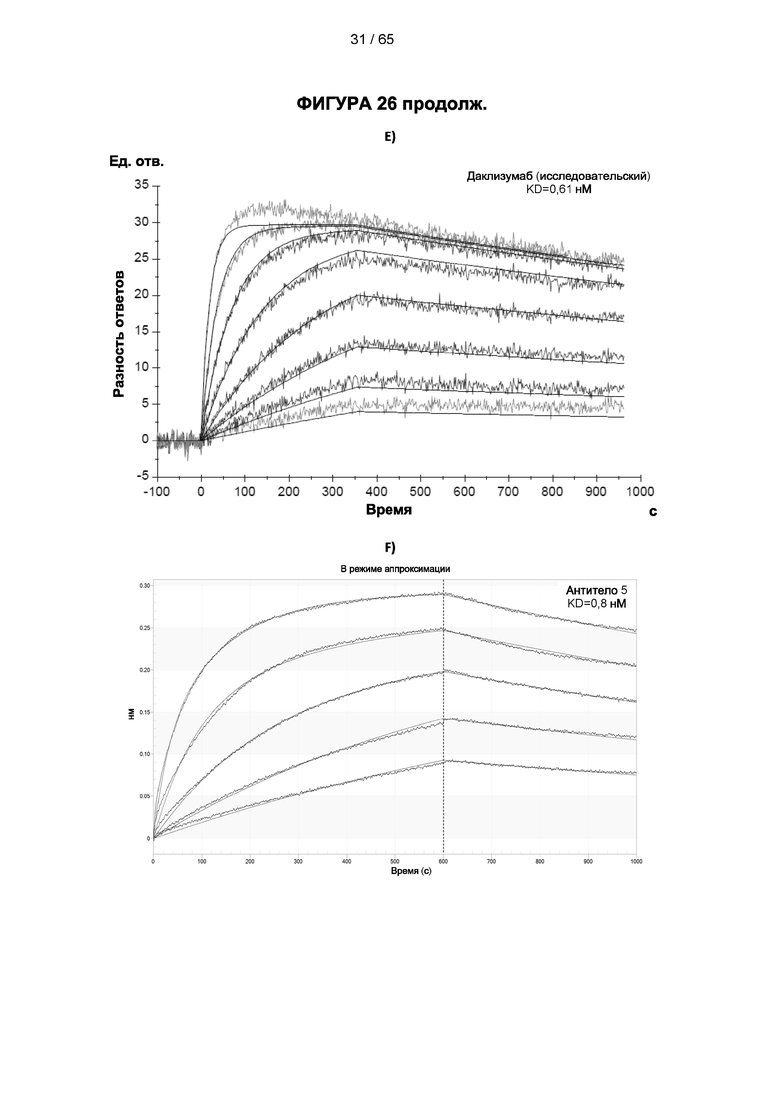
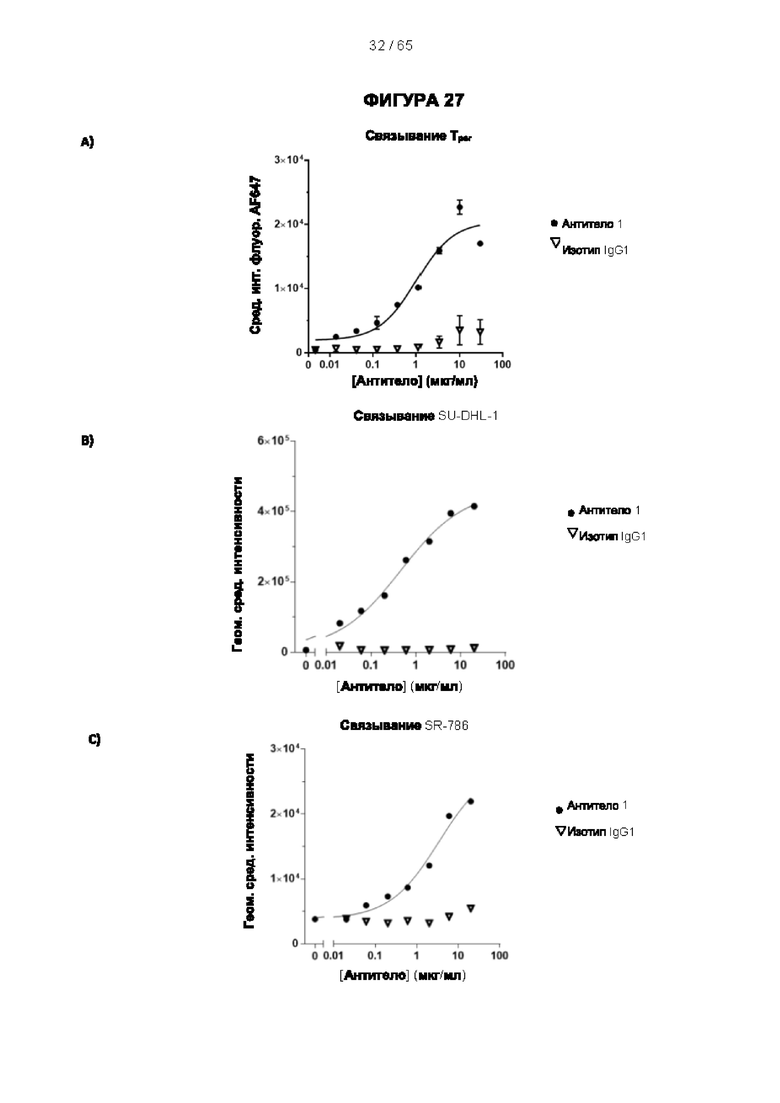
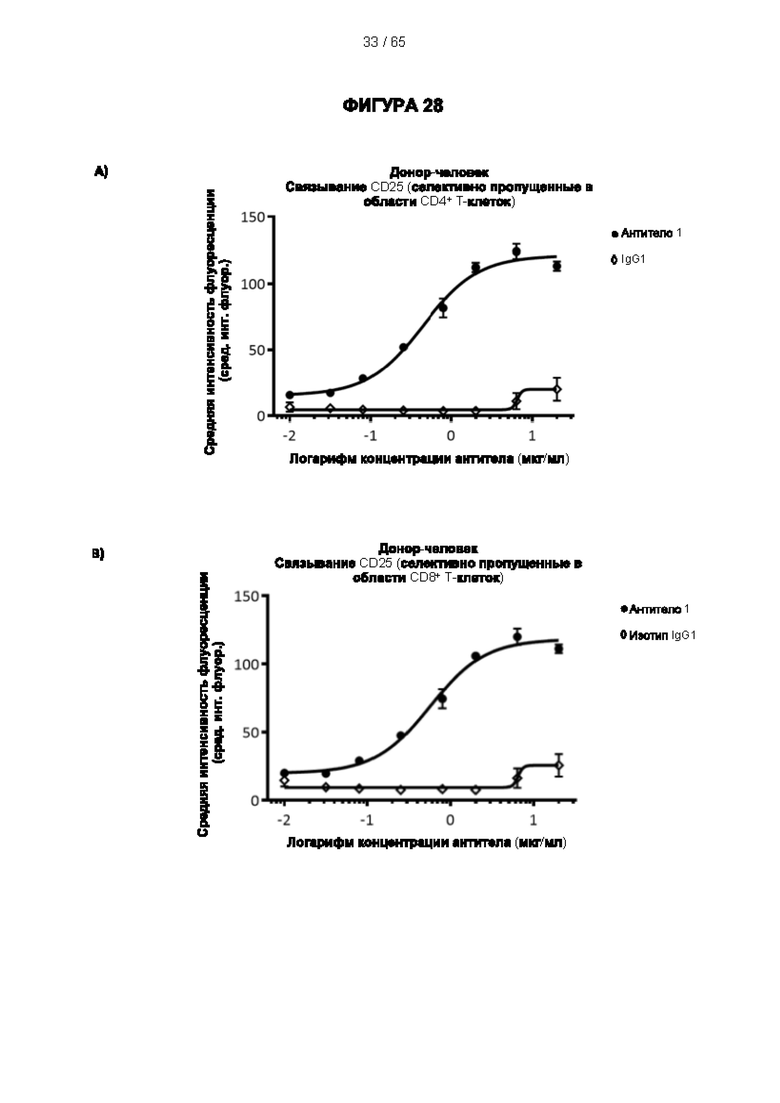
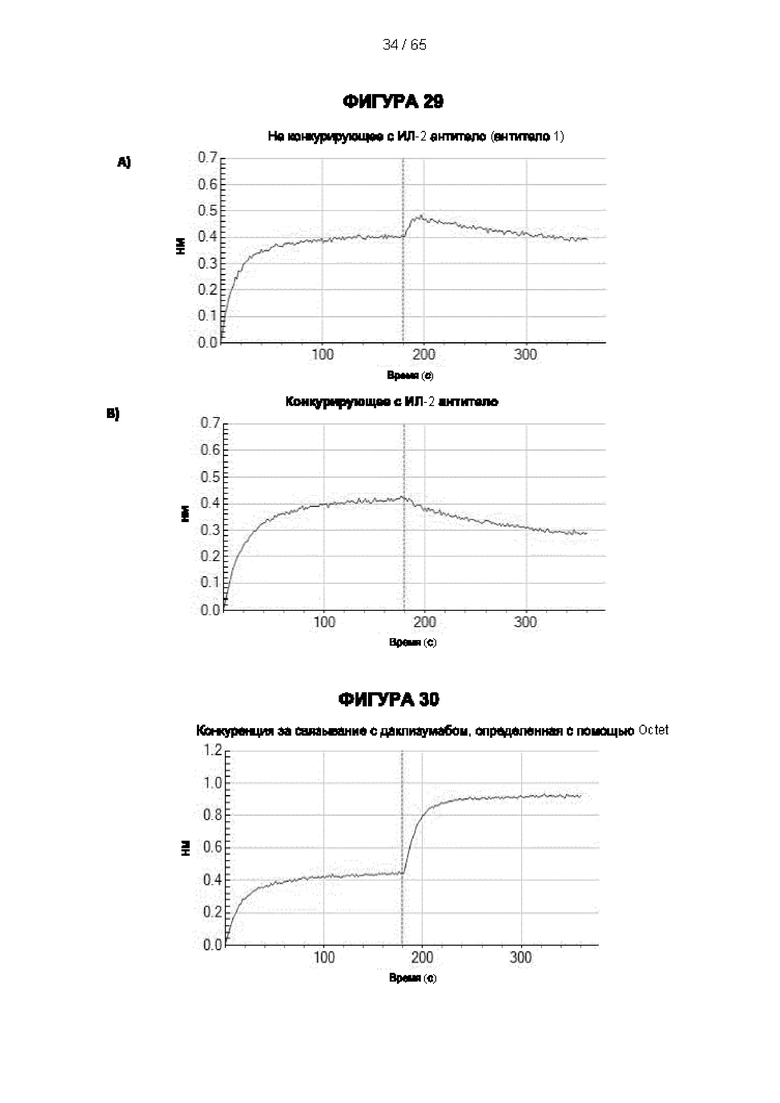
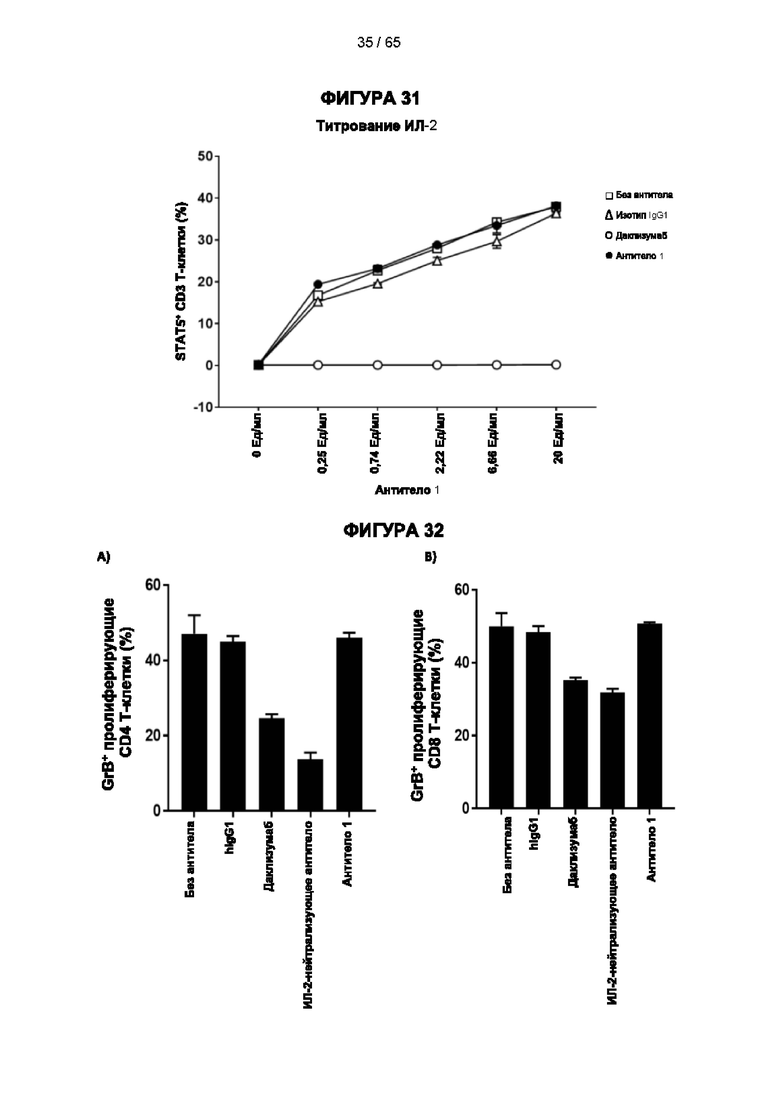
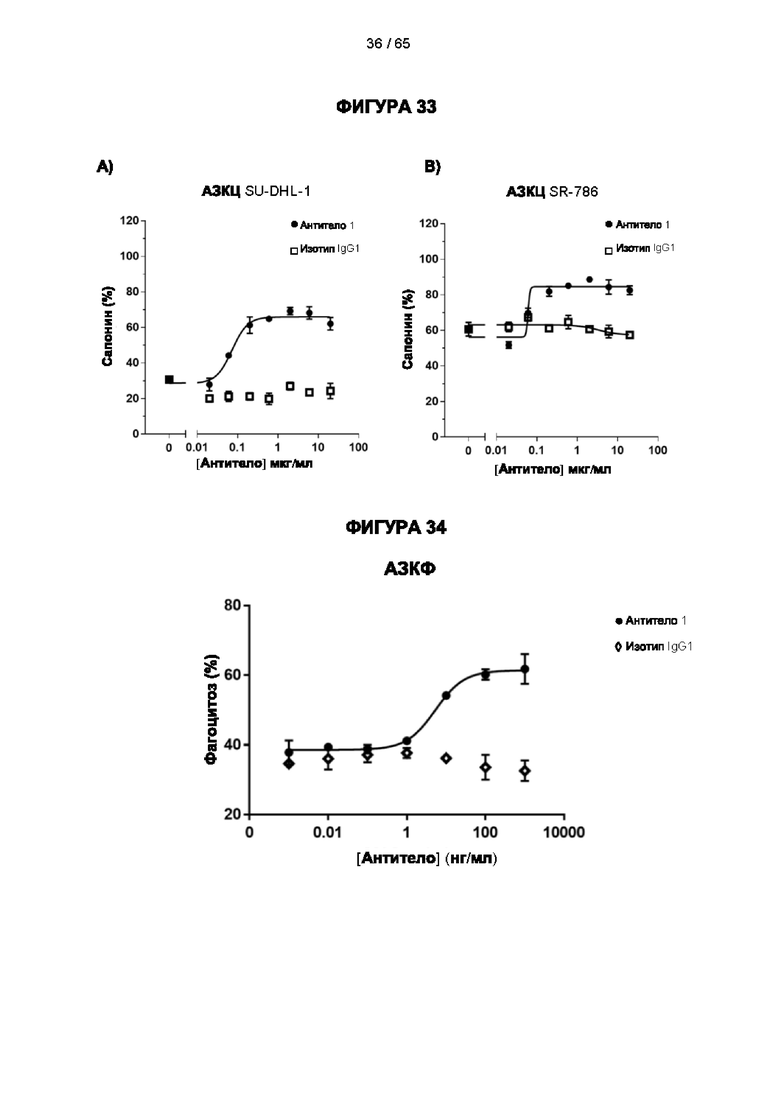
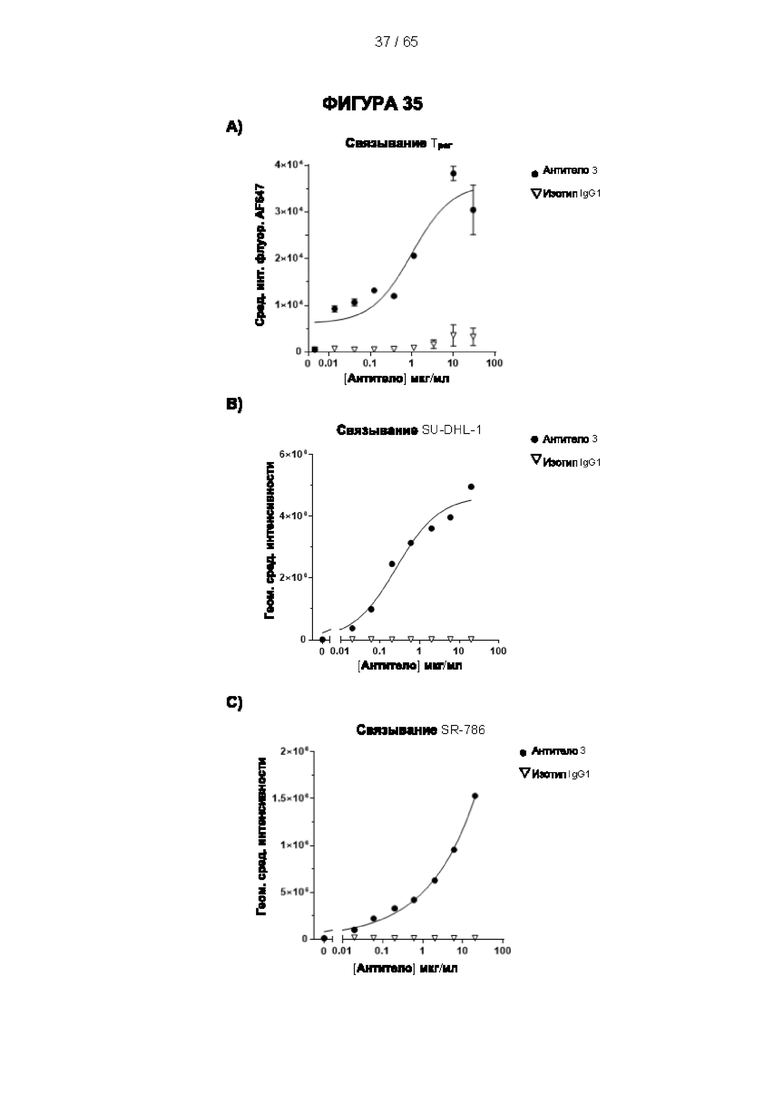
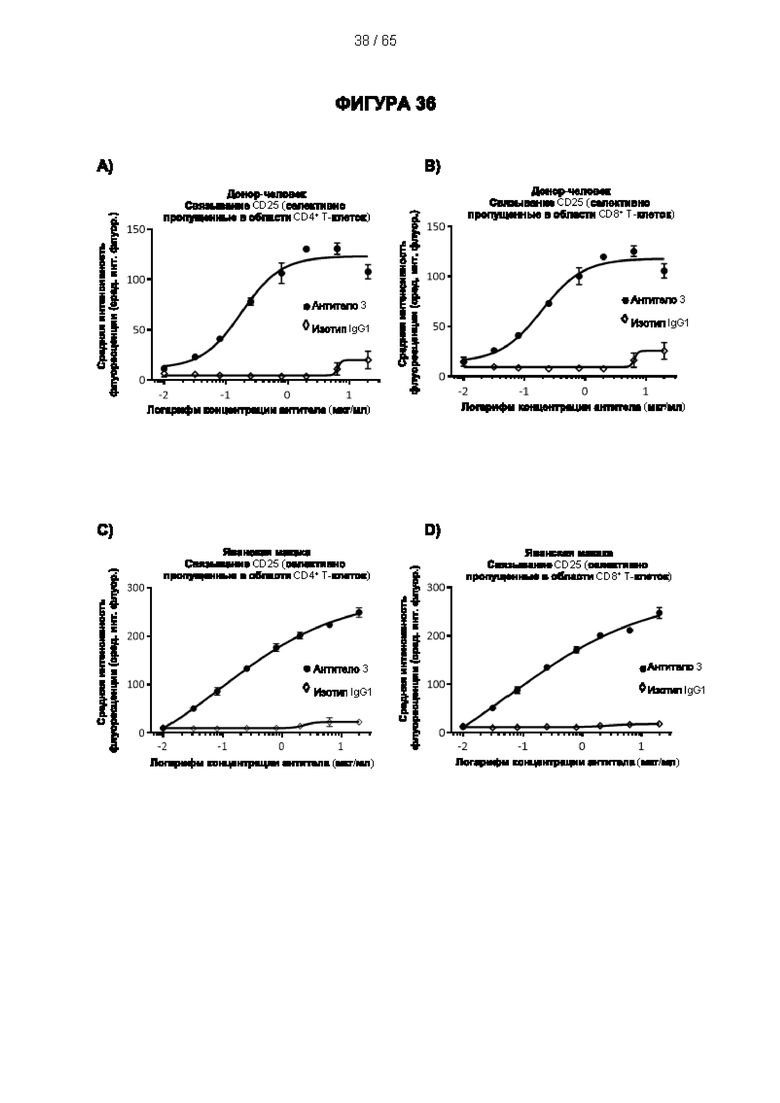
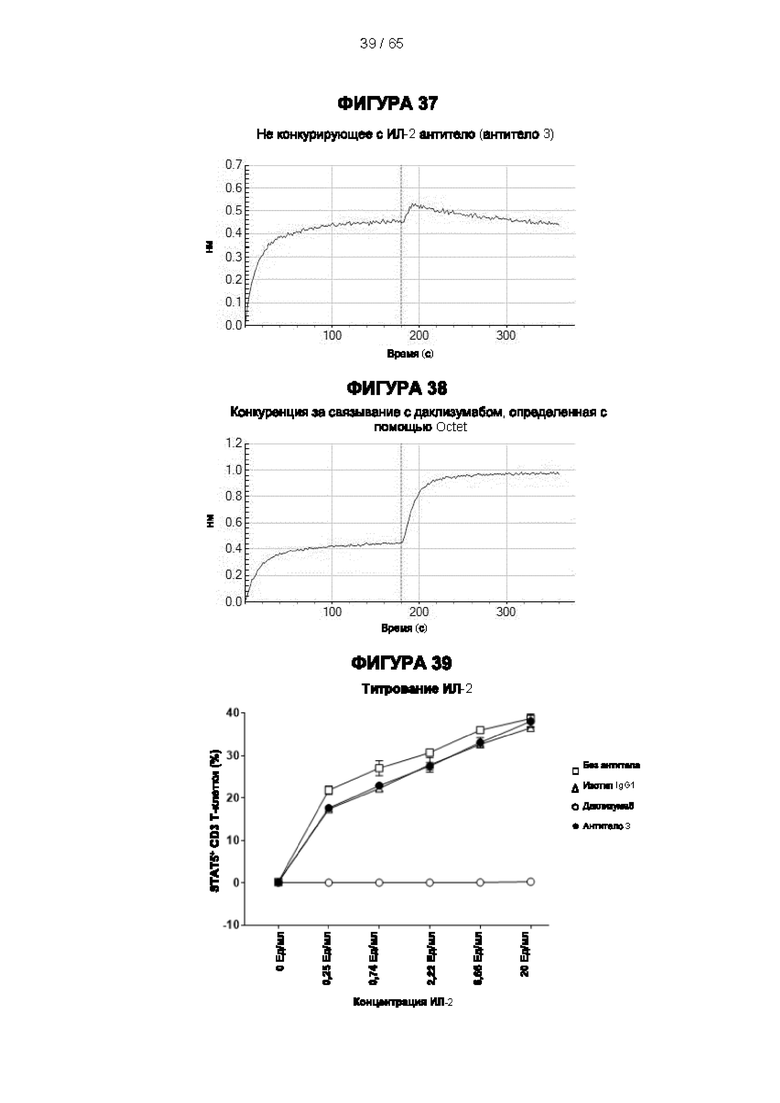
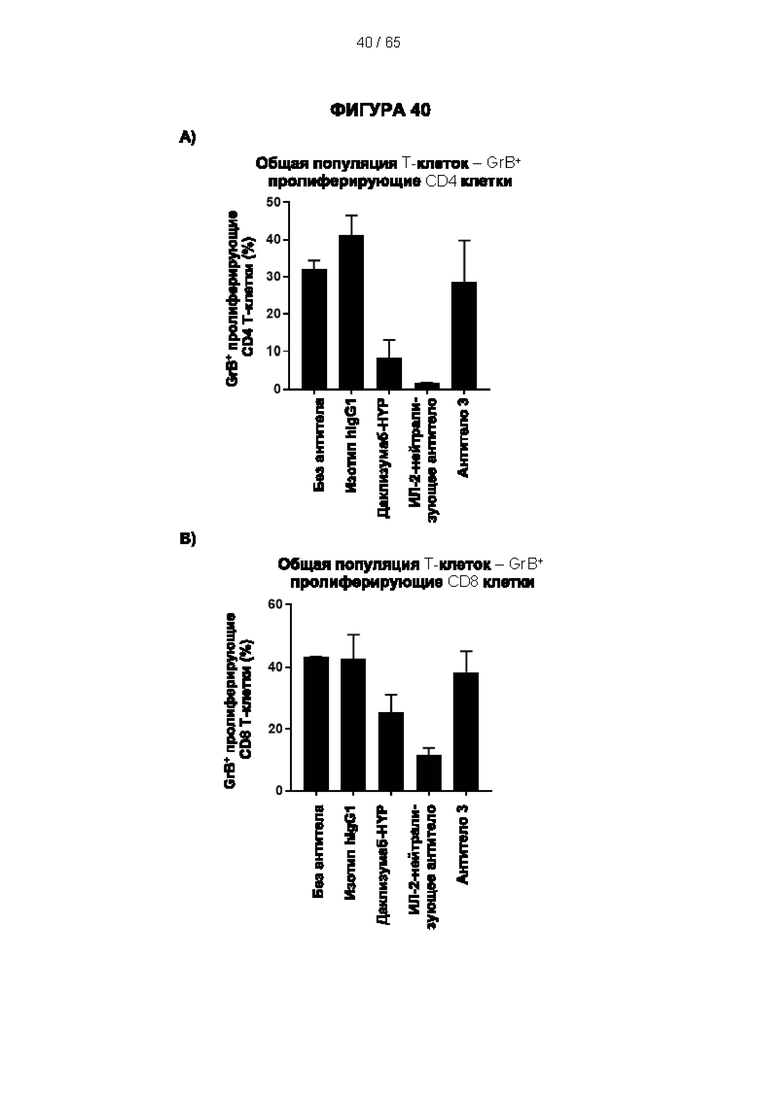
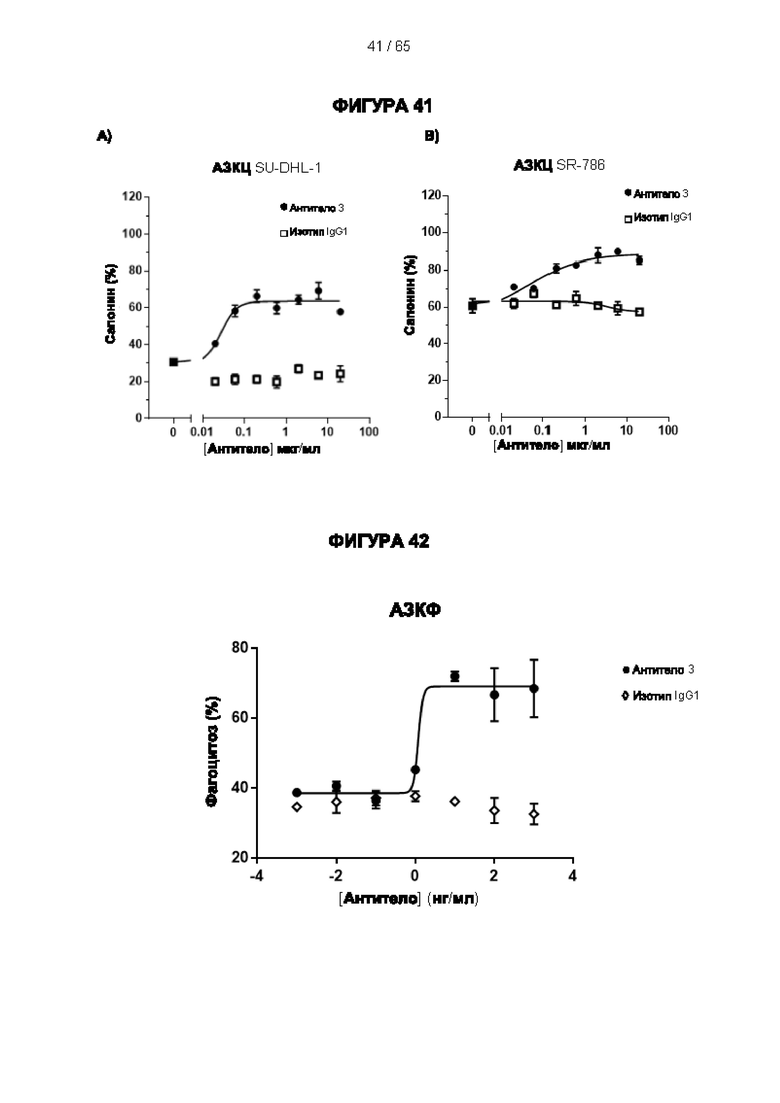
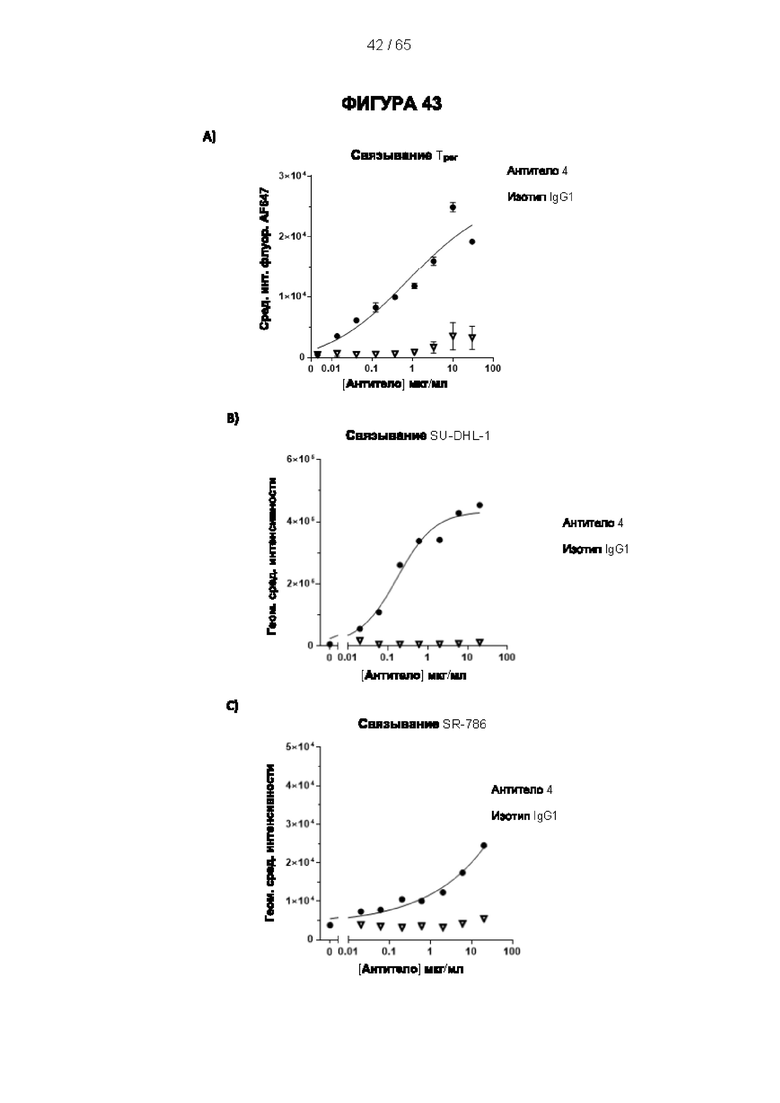
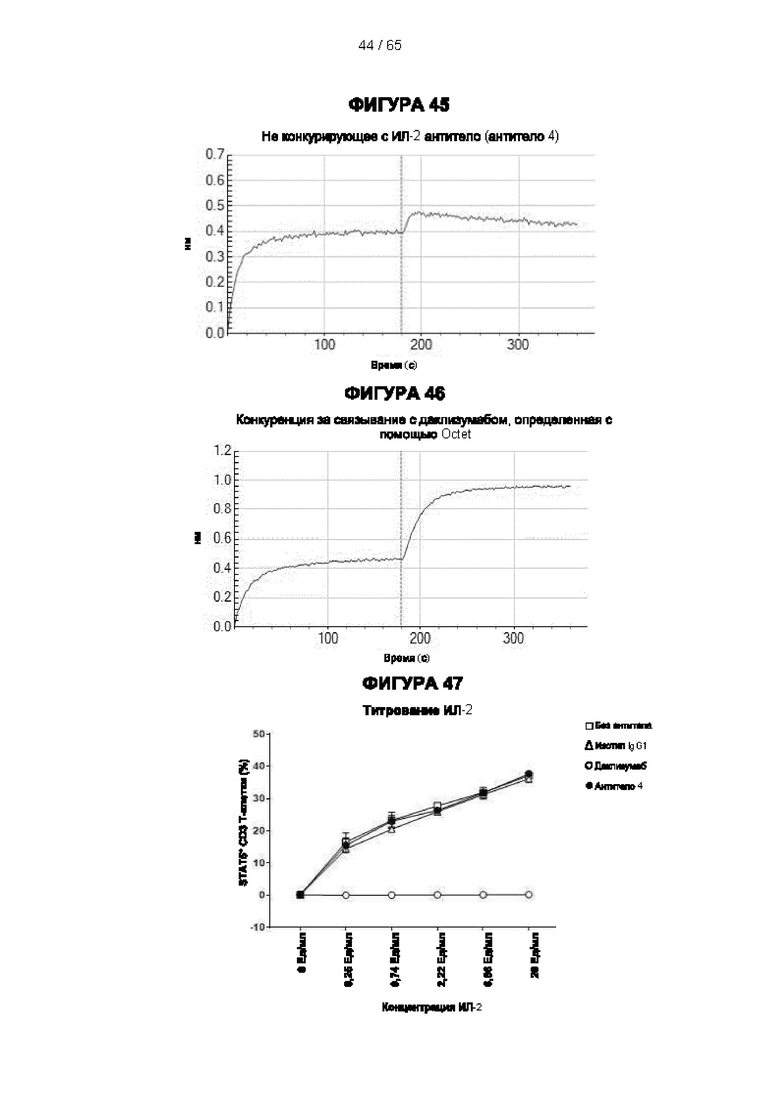
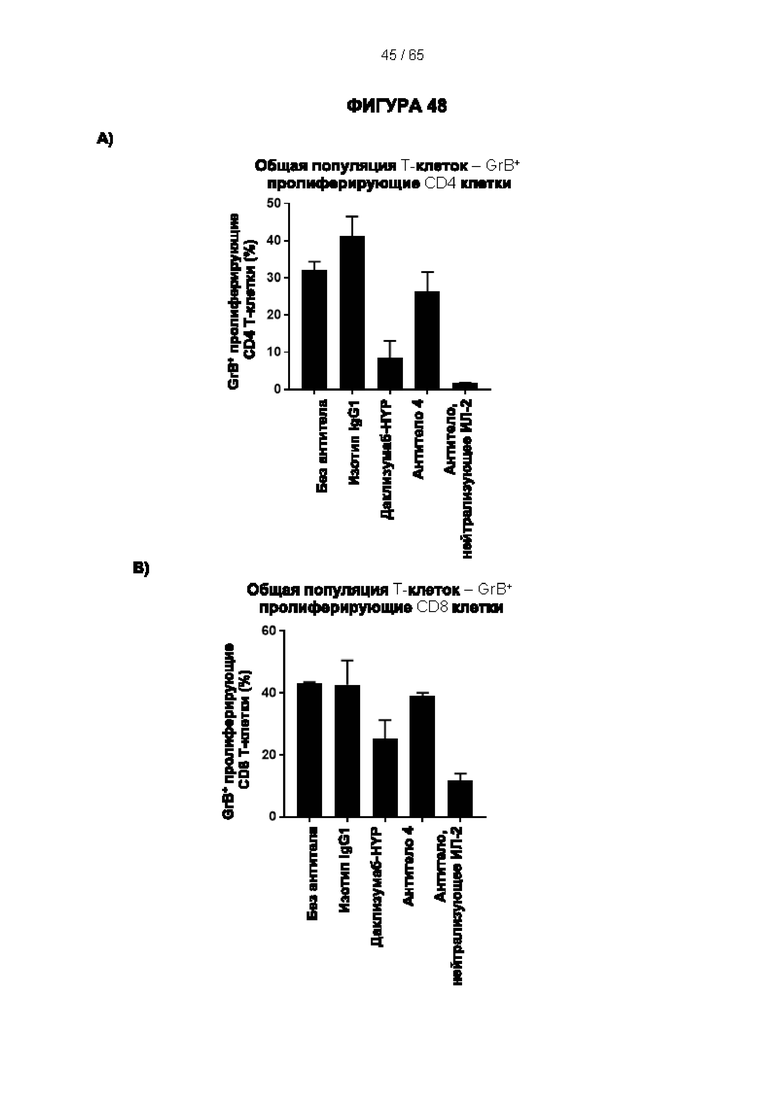
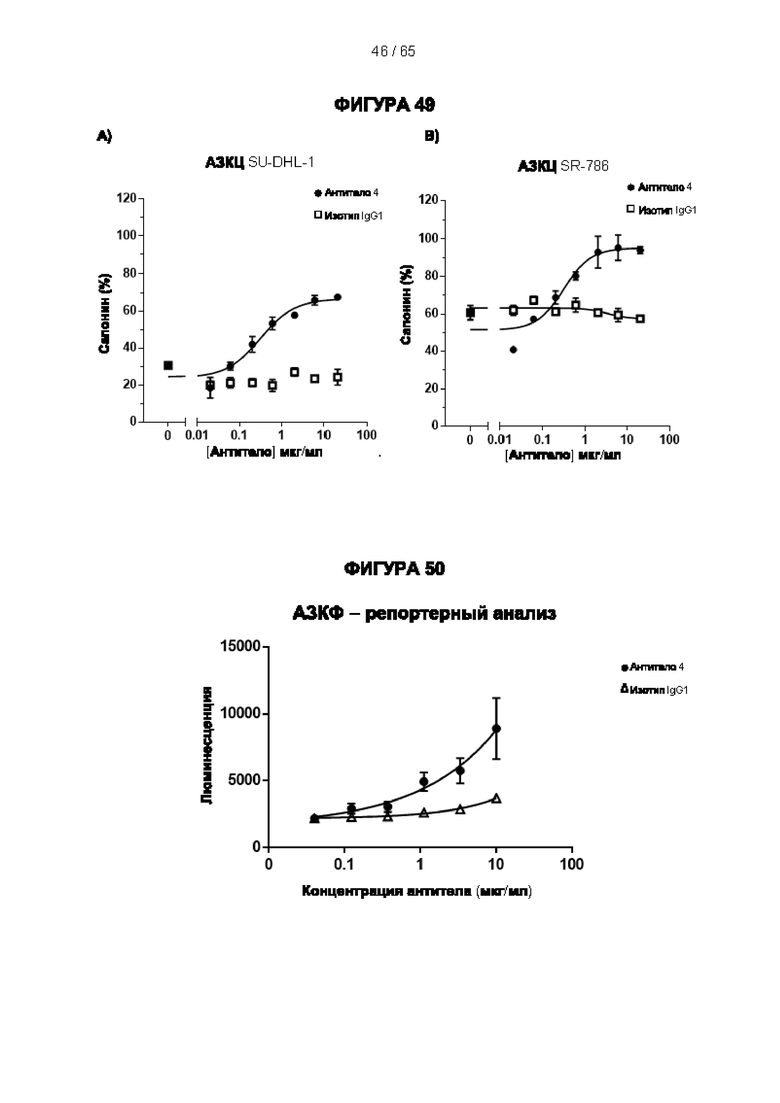
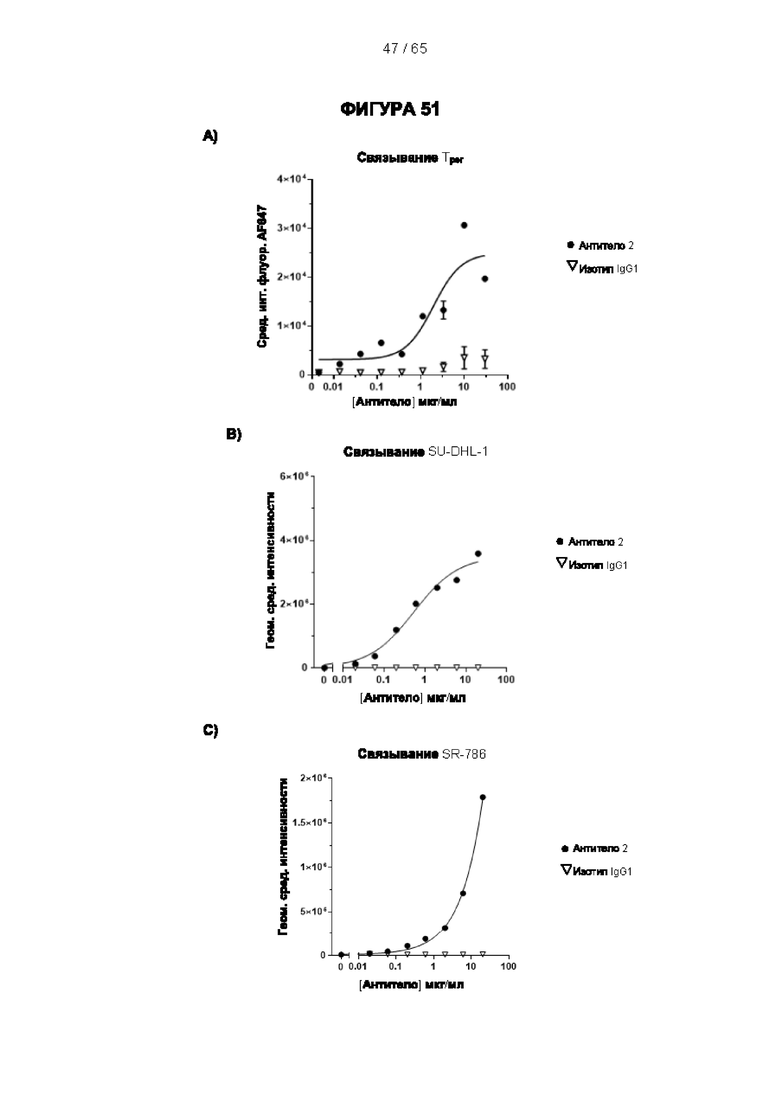
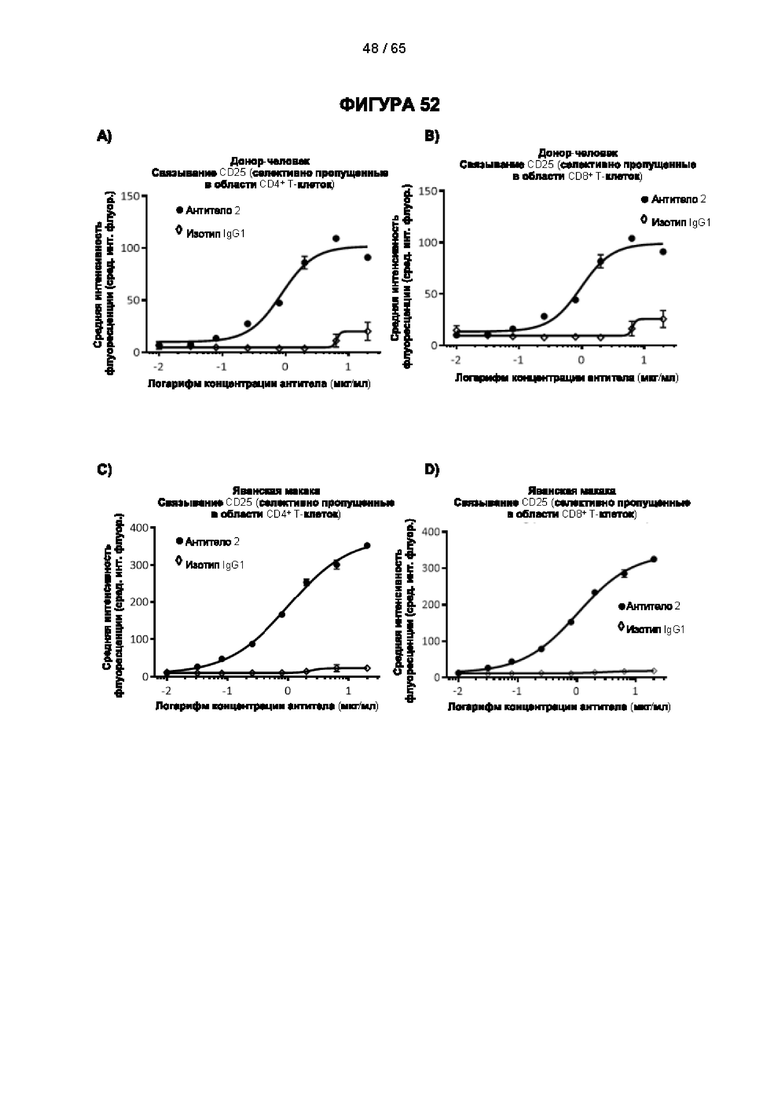
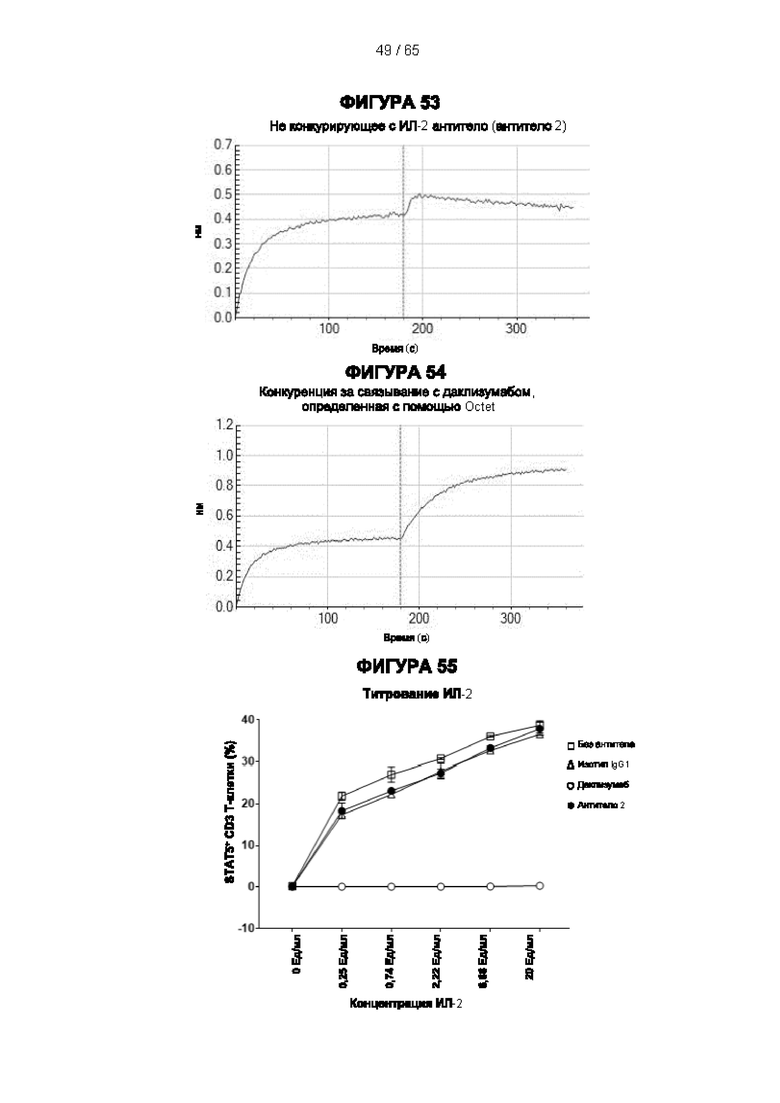
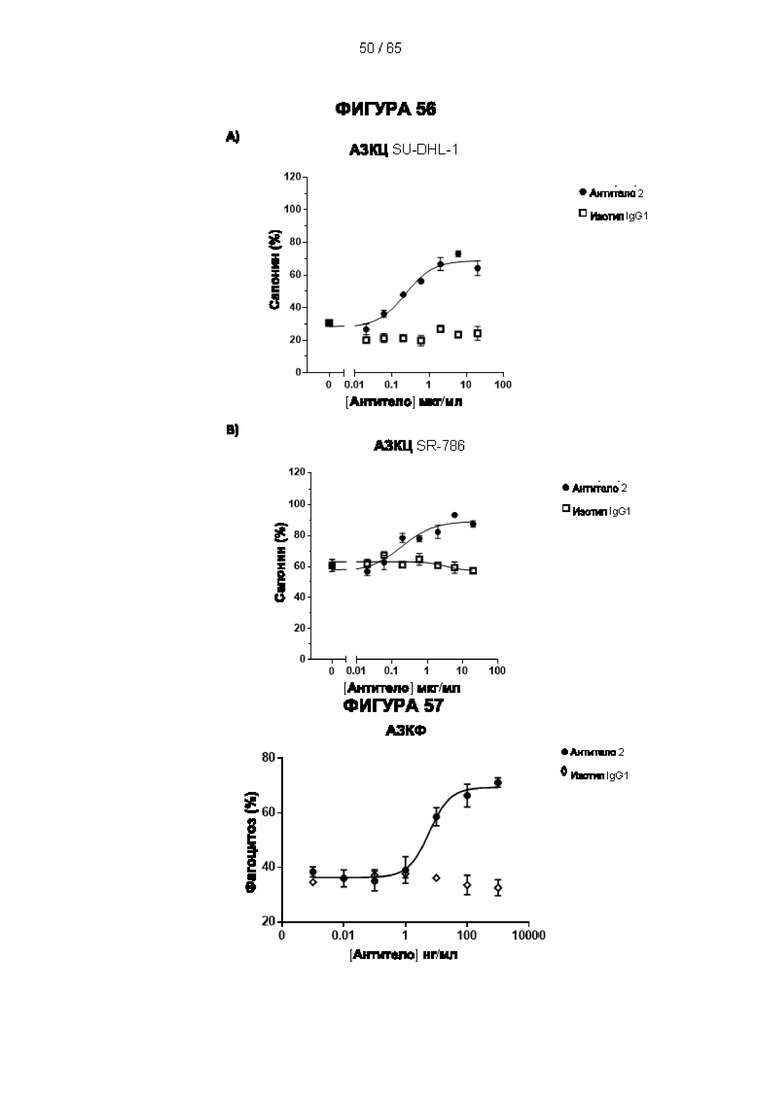
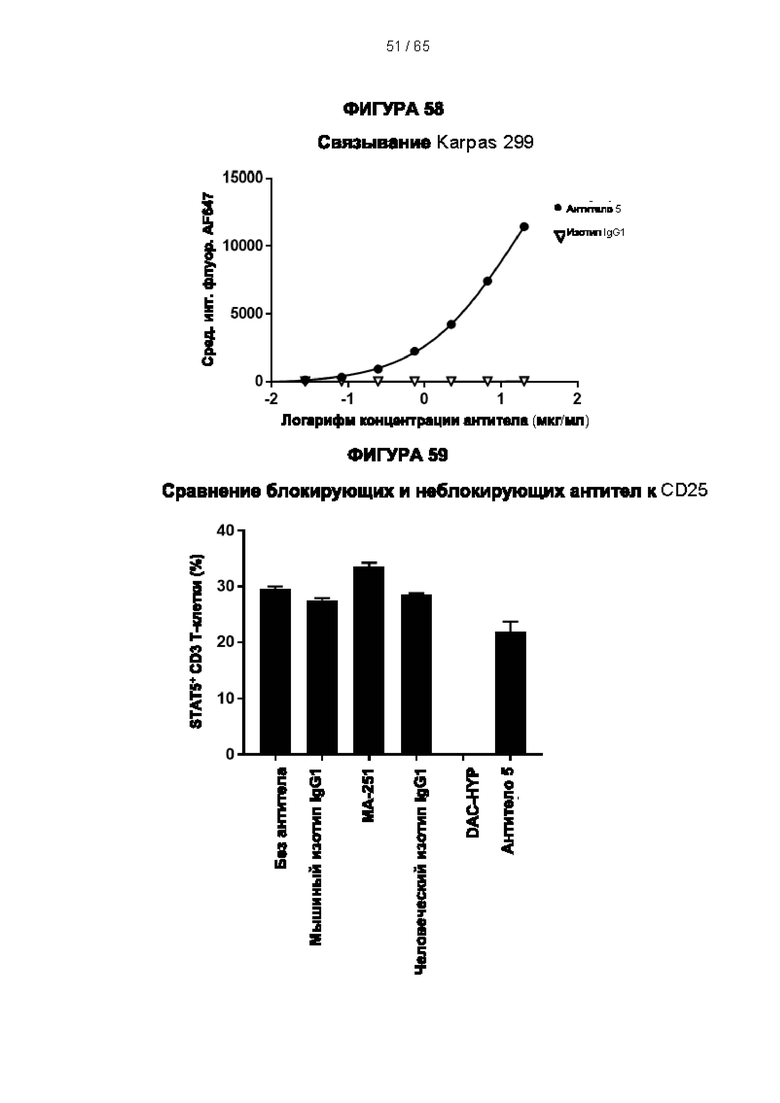
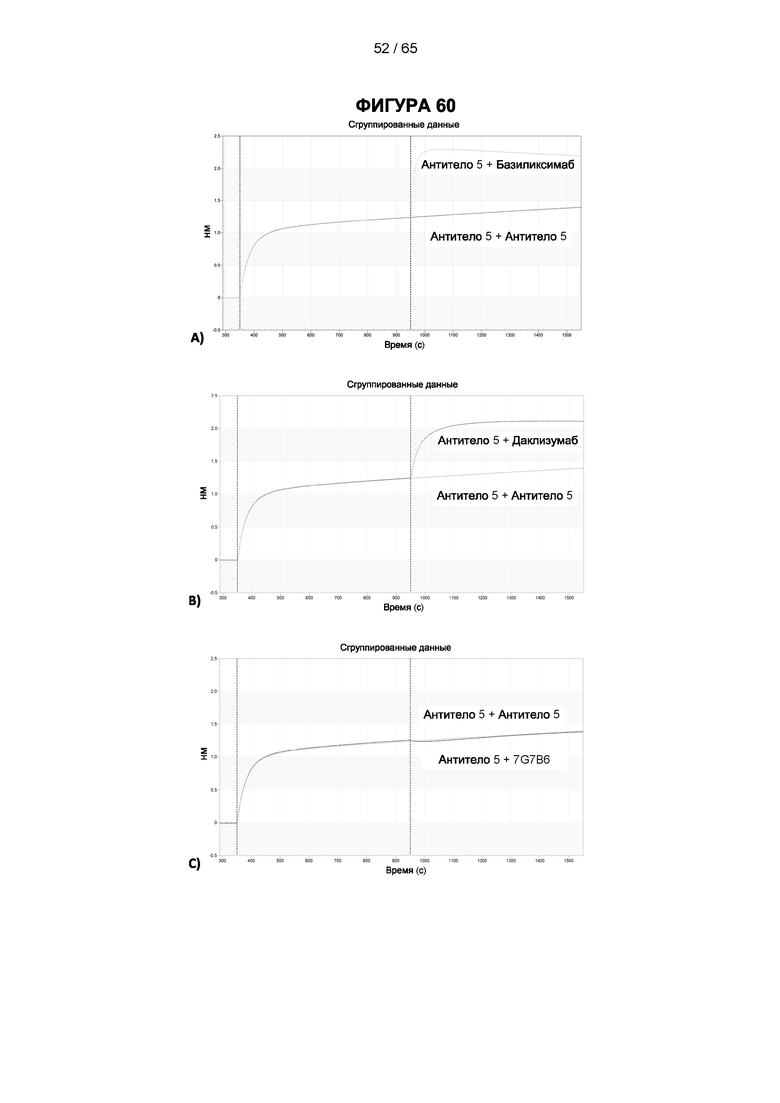
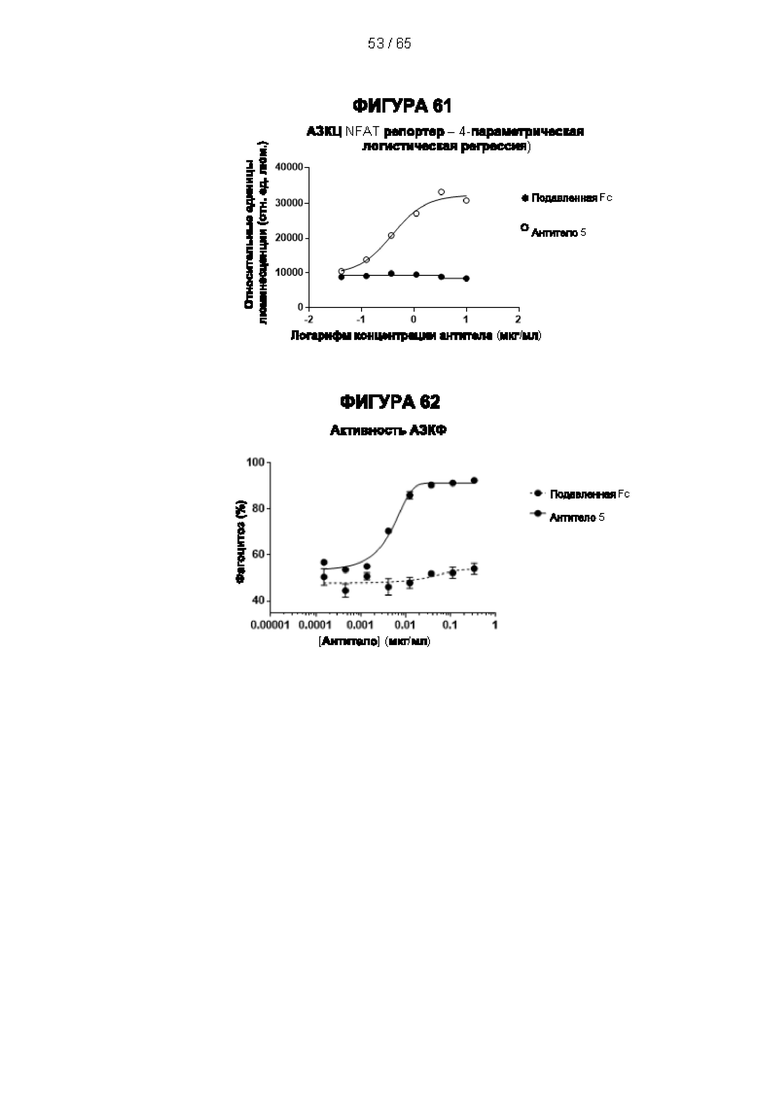
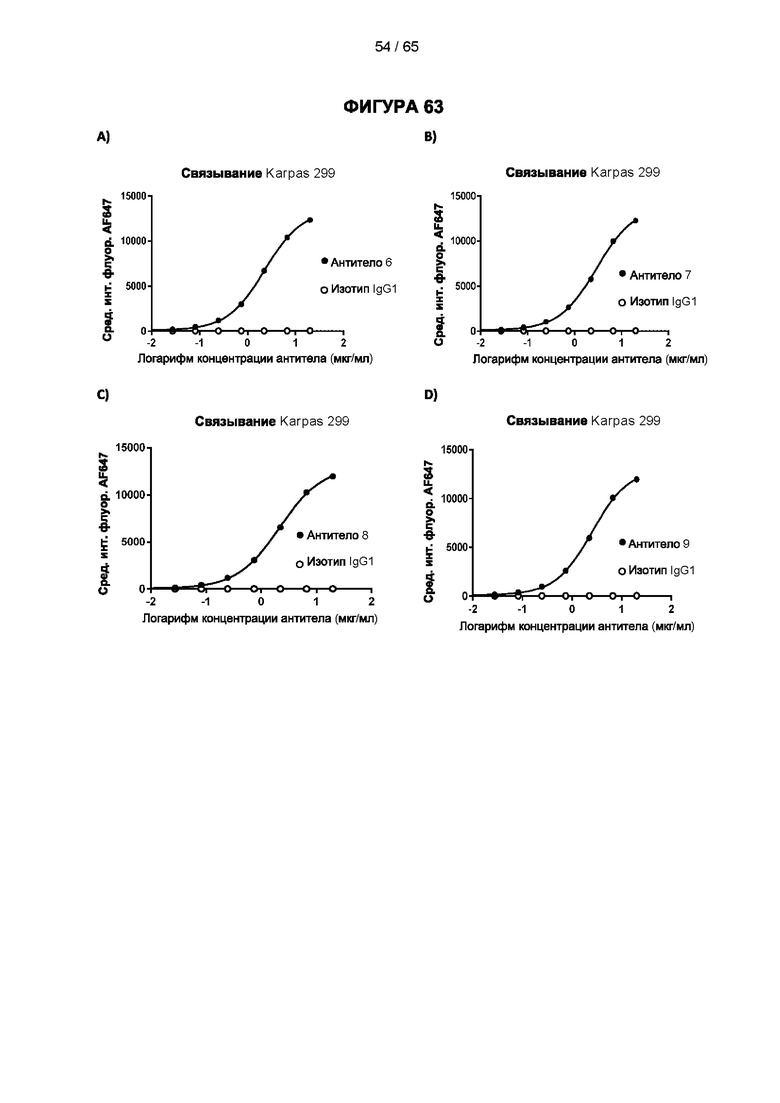
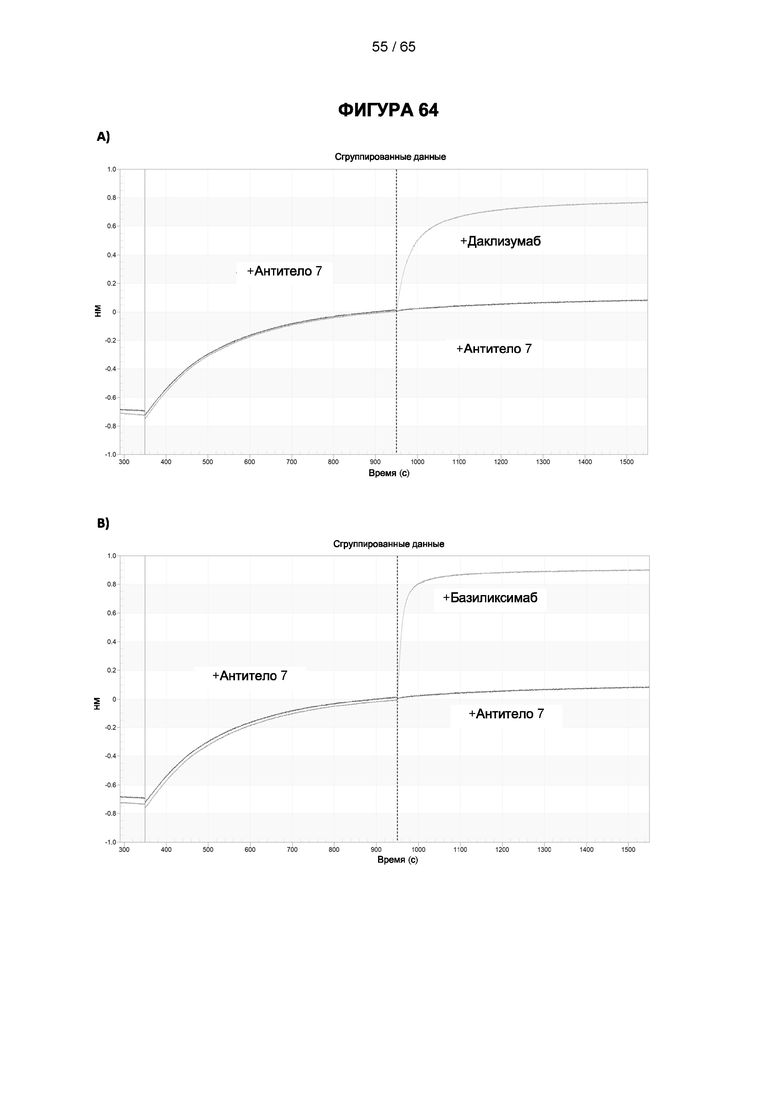
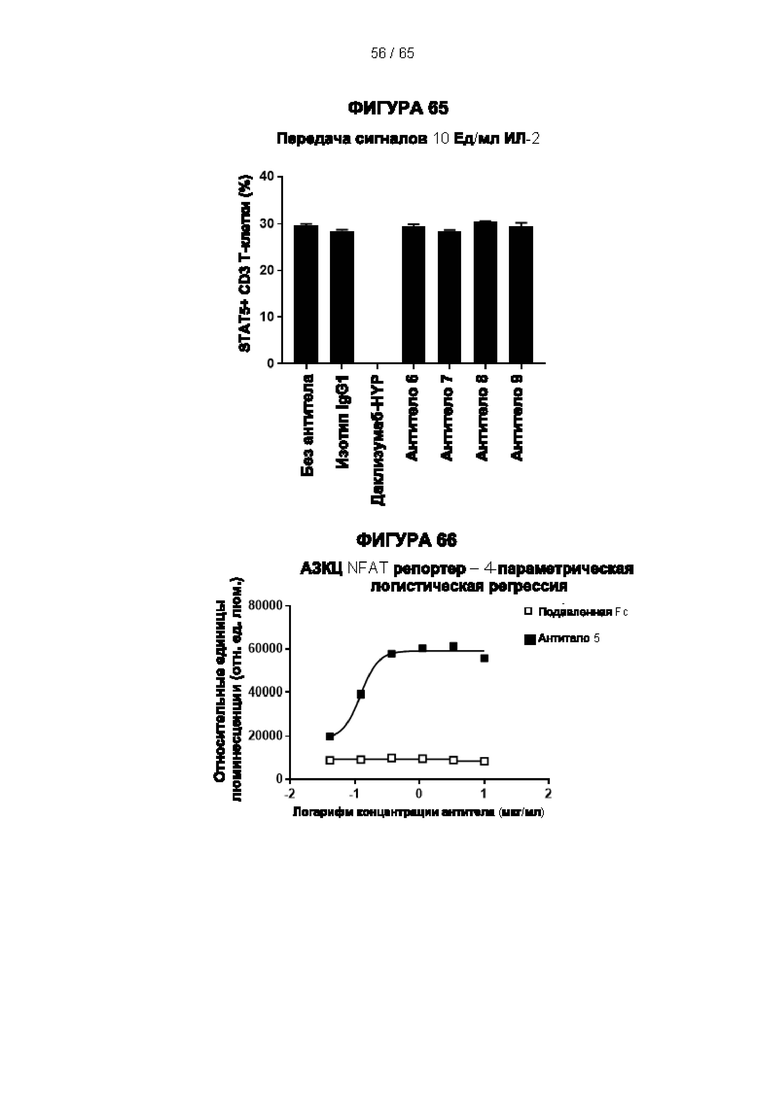
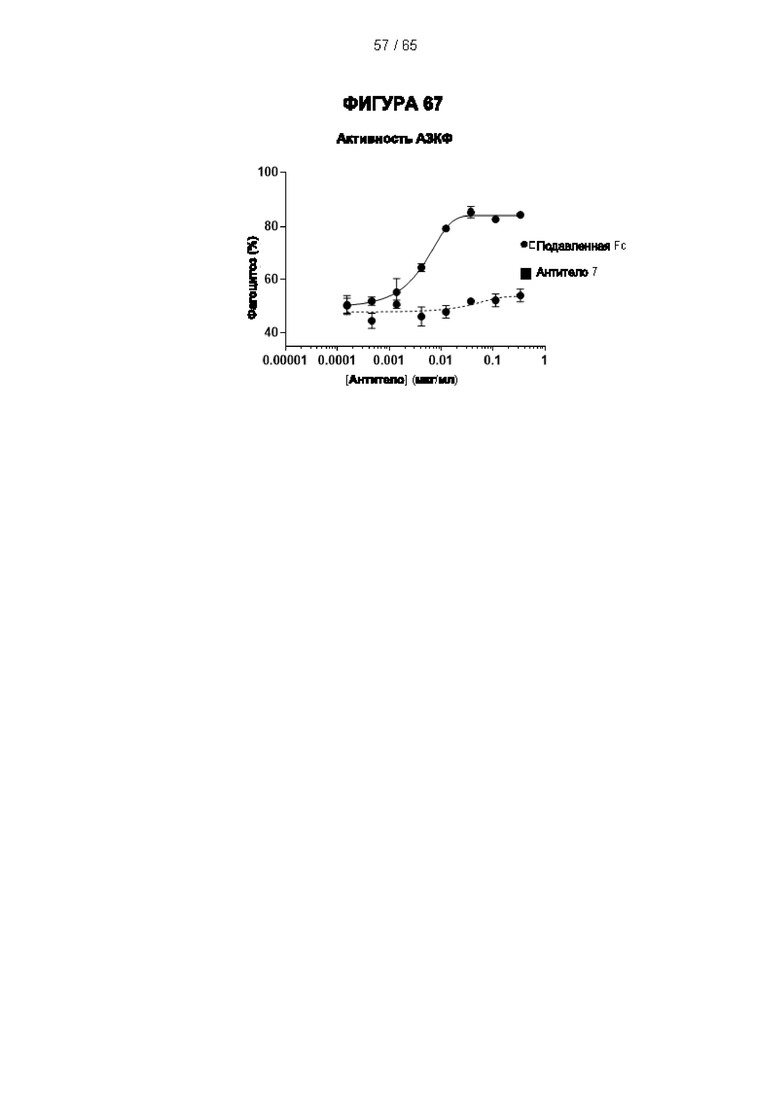
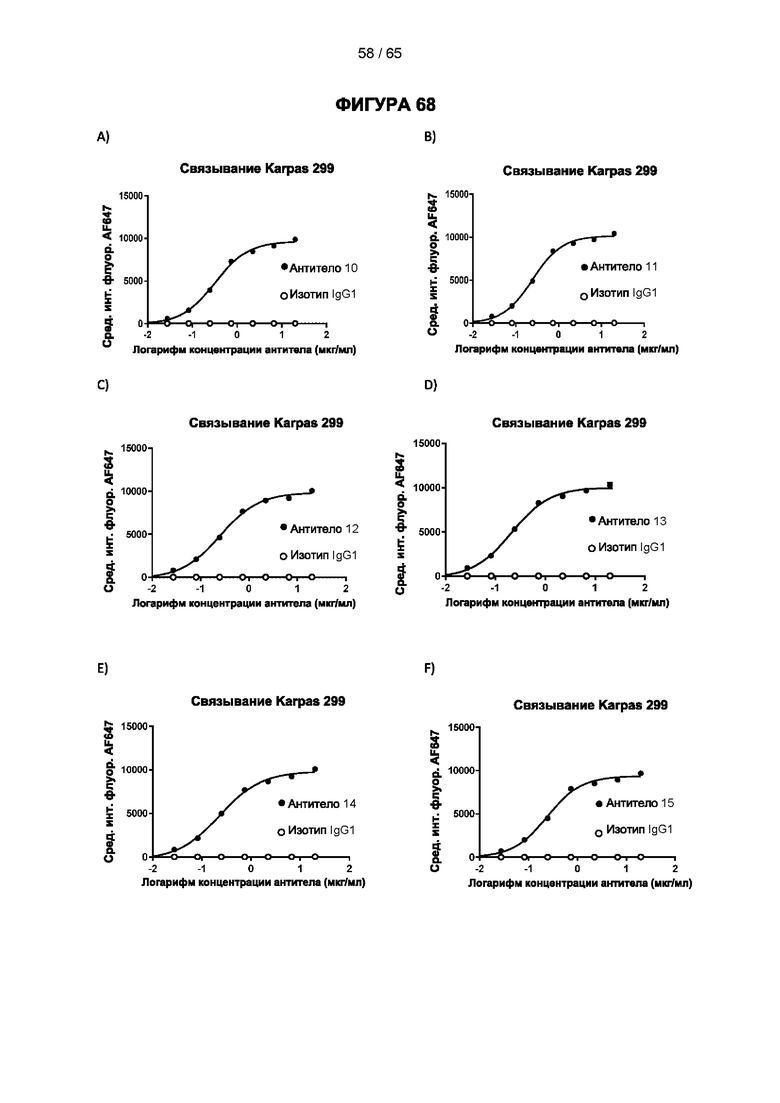
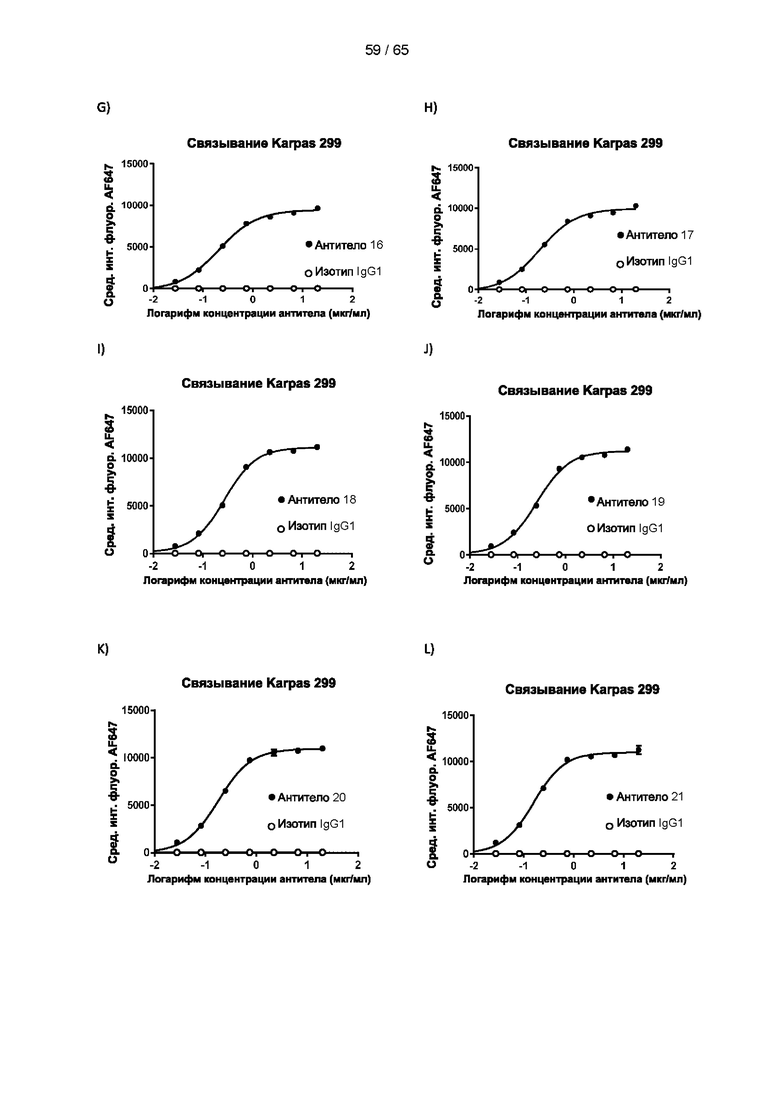
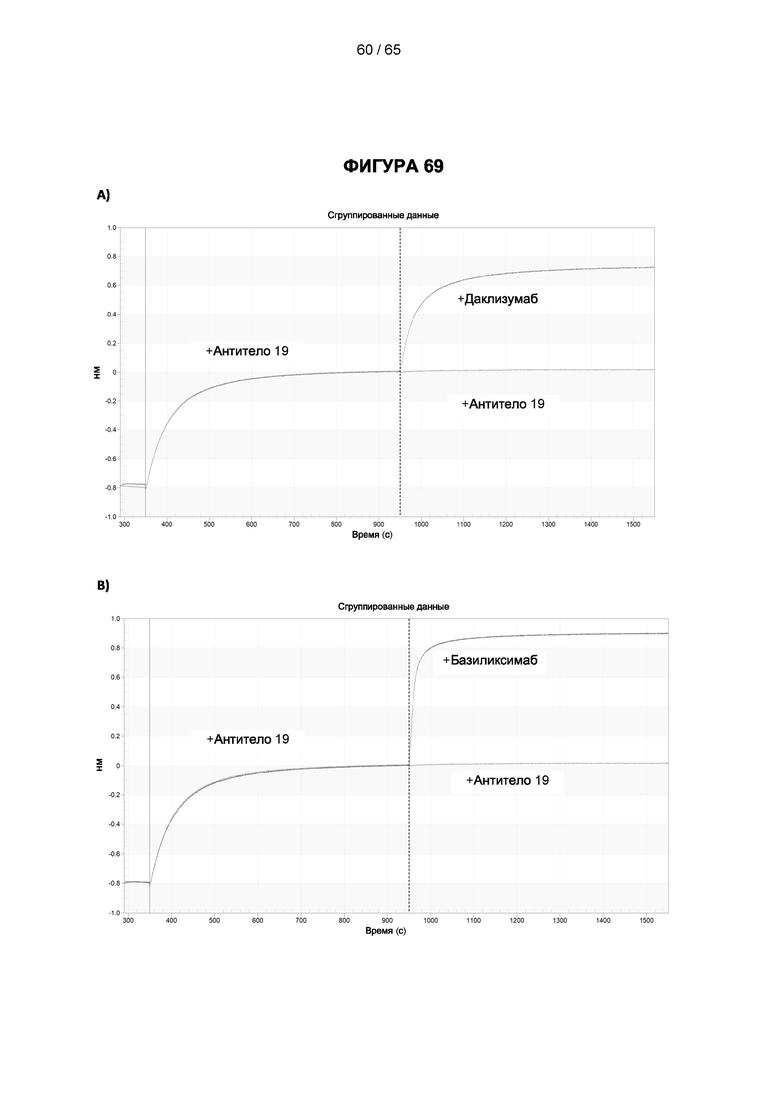
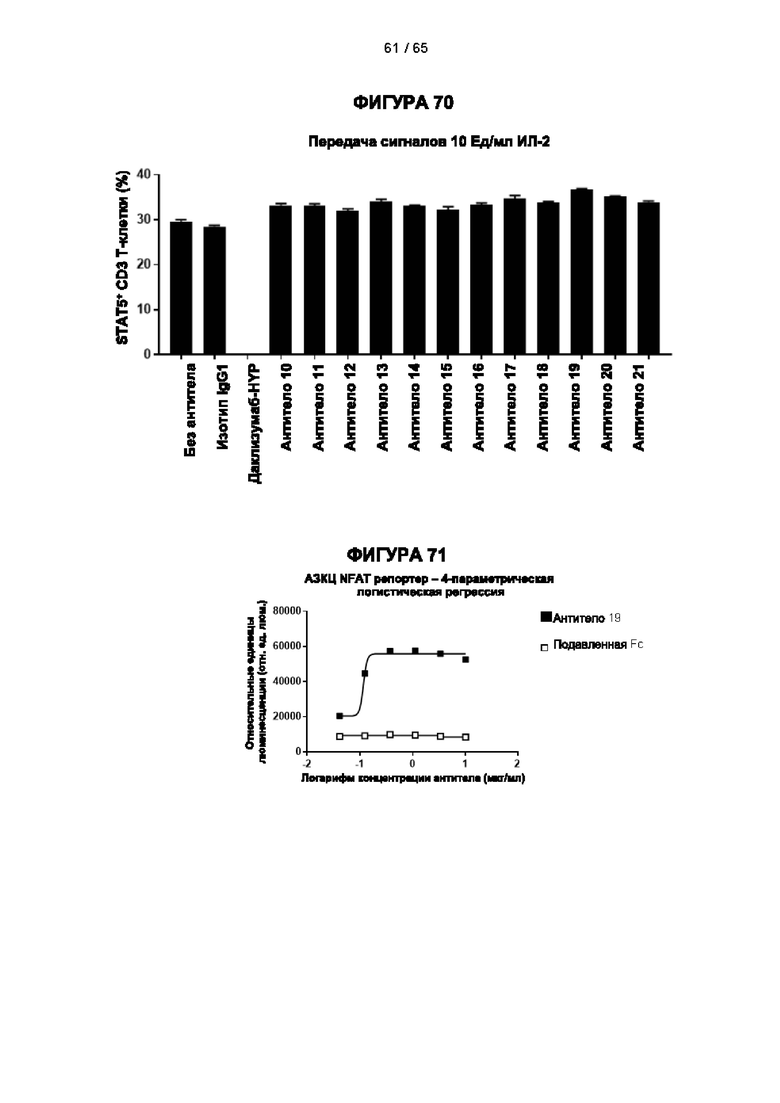
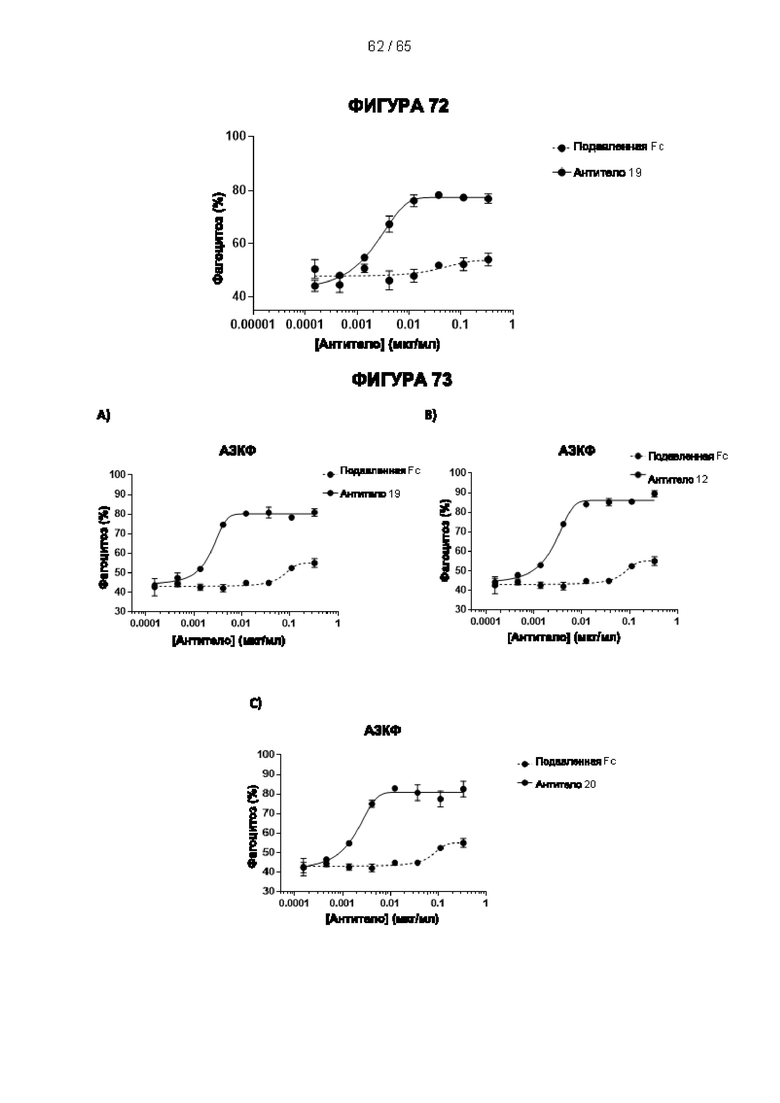
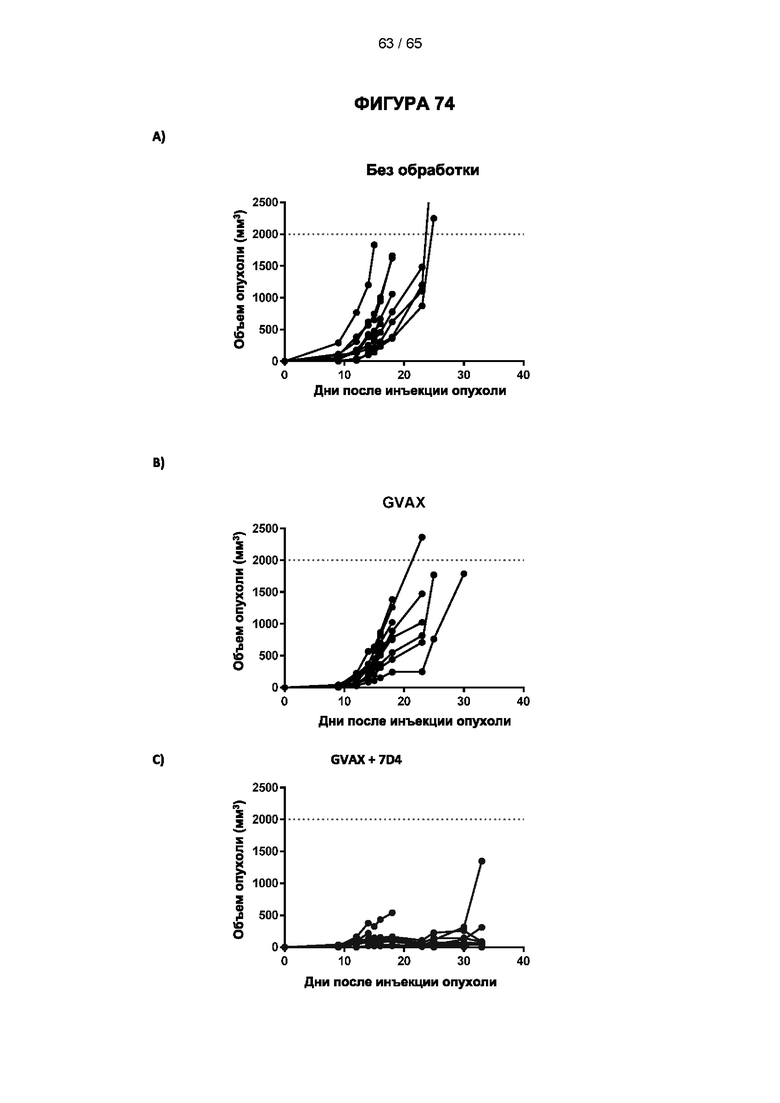
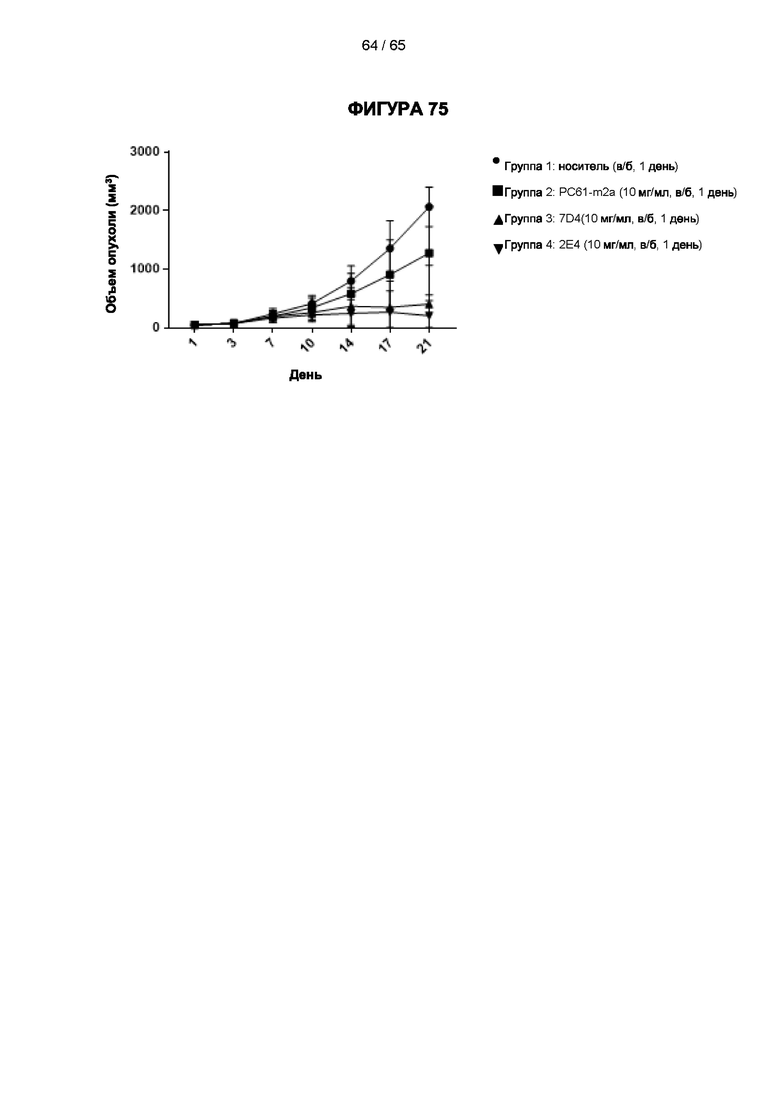
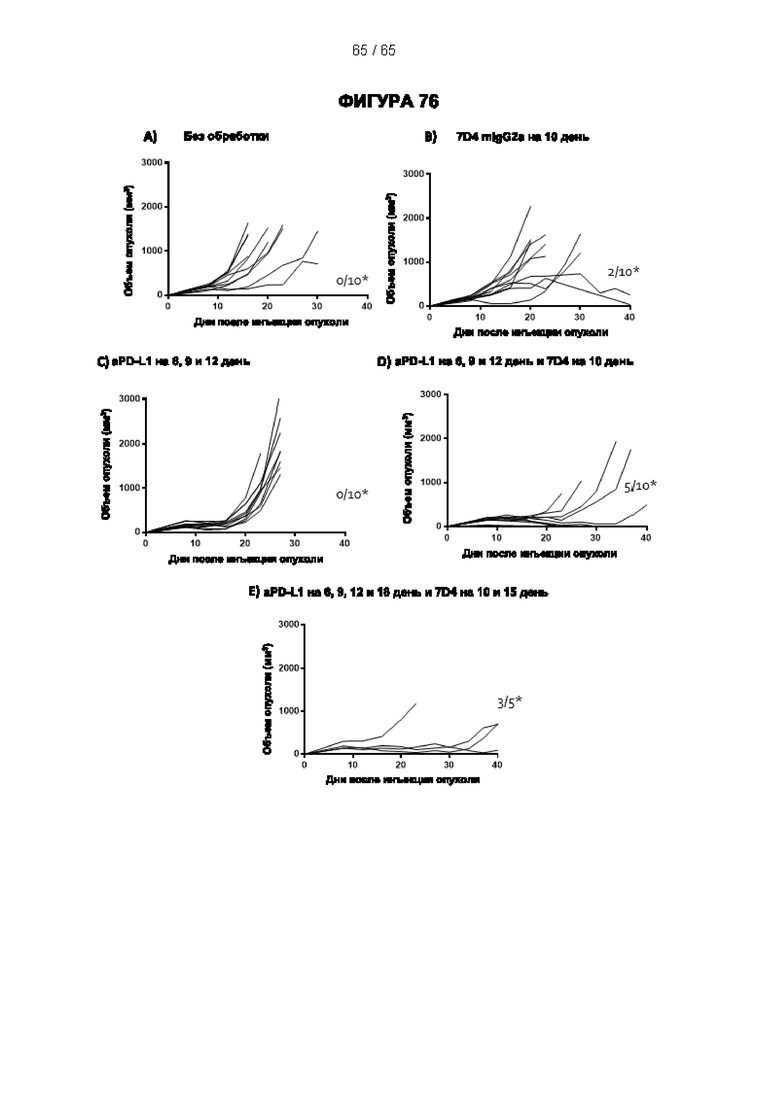
Изобретение относится к области биотехнологии, а именно к способу лечения субъекта-человека, имеющего рак, экспрессирующий CD25, включающему этап введения субъекту человеческого антитела IgG1 к CD25, причем указанный субъект имеет плотную опухоль, причем указанное антитело не ингибирует связывание интерлейкина-2 (IL-2) с CD25. Изобретение обеспечивает возможность лечения плотных (солидных) опухолей. 29 з.п. ф-лы, 76 ил., 3 табл.
1. Способ лечения субъекта-человека, имеющего рак, экспрессирующий CD25, включающий этап введения субъекту человеческого антитела IgG1 к CD25, причем указанный субъект имеет плотную опухоль, причем указанное антитело не ингибирует связывание интерлейкина-2 (IL-2) с CD25.
2. Способ по п. 1, характеризующийся тем, что указанное антитело к CD25 конкурирует с антителом 7G7B6 за связывание с человеческим CD25; и/или конкурирует с антителом MA251 за связывание с человеческим CD25.
3. Способ по п. 1 или 2, характеризующийся тем, что указанное антитело к CD25 связывается с тем же эпитопом, который распознается антителом 7G7B6, и/или эпитопом, который распознается антителом MA251.
4. Способ по любому из пп. 1-3, характеризующийся тем, что указанное антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR).
5. Способ по любому из пп. 1-3, характеризующийся тем, что указанное антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и аминокислот 150-160 из SEQ ID NO: 1 (YQCVQGYRALH).
6. Способ по любому из пп. 1-3, характеризующийся тем, что указанное антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и аминокислот 74-84 из SEQ ID NO: 1 (SSWDNQCQCTSSATR).
7. Способ по любому из пп. 1-3, характеризующийся тем, что указанное антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 150-163 из SEQ ID NO: 1 (YQCVQGYRALHRGP), аминокислот 166-180 из SEQ ID NO: 1 (SVCKMTHGKTRWTQP), аминокислот 42-56 из SEQ ID NO: 1 (KEGTMLNCECKRGFR) и аминокислот 74-84 из SEQ ID NO: 1 (SSWDNQCQCTSSATR).
8. Способ по любому из пп. 1-3, характеризующийся тем, что указанное антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 150-158 из SEQ ID NO: 1 (YQCVQGYRA) и аминокислот 176-180 из SEQ ID NO: 1 (RWTQP).
9. Способ по любому из пп. 1-3, характеризующийся тем, что указанное антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 150-158 из SEQ ID NO: 1 (YQCVQGYRA) и аминокислот 176-186 из SEQ ID NO: 1 (RWTQPQLICTG).
10. Способ по любому из пп. 1-3, характеризующийся тем, что указанное антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 150-163 из SEQ ID NO: 1 (YQCVQGYRALHRGP) и аминокислот 166-180 из SEQ ID NO: 1 (SVCKMTHGKTRWTQP).
11. Способ по любому из пп. 1-3, характеризующийся тем, что указанное антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 74-84 из SEQ ID NO: 1 (SSWDNQCQCTS).
12. Способ по любому из пп. 1-3, характеризующийся тем, что указанное антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 70-84 из SEQ ID NO: 1 (NSSHSSWDNQCQCTS).
13. Способ по любому из пп. 1-3, характеризующийся тем, что указанное антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 176-180 из SEQ ID NO: 1 (RWTQP).
14. Способ по любому из пп. 1-3, характеризующийся тем, что указанное антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 166-180 из SEQ ID NO: 1 (SVCKMTHGKTRWTQP).
15. Способ по любому из пп. 1-3, характеризующийся тем, что указанное антитело к CD25 связывается с эпитопом, содержащим последовательность аминокислот 176-186 из SEQ ID NO: 1 (RWTQPQLICTG).
16. Способ по любому из пп. 1-15, характеризующийся тем, что указанное антитело к CD25 представляет собой антитело IgG1, которое связывается по меньшей мере с одним активирующим рецептором Fcγ, выбранным из FcγRI, FcγRIIc и/или FcγRIIIa, с высокой аффинностью и истощает инфильтрирующие опухоль регуляторные Т-клетки.
17. Способ по любому из пп. 1-16, характеризующийся тем, что указанное антитело выбрано из группы, состоящей из:
(a) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 3, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 4;
(b) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 5, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 6;
(c) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 10, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 14;
(d) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 18, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 22;
(e) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 23, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 25;
(f) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 23, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 26;
(g) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 24, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 25;
(h) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 24, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 26;
(i) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 27, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 30;
(j) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 27, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 31;
(k) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 27, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 32;
(l) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 27, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 33;
(m) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 28, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 30;
(n) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 28, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 31;
(o) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 28, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 32;
(p) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 28, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 33;
(q) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 29, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 30;
(r) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 29, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 31;
(s) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 29, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 32; и
(t) антитела, содержащего тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 29, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 33.
18. Способ по любому из пп. 1-17, характеризующийся тем, что указанное антитело к CD25 имеет константу диссоциации (Kd) для CD25 менее 10-7 М.
19. Способ по любому из пп. 1-18, характеризующийся тем, что указанное антитело к CD25 ингибирует передачу сигналов IL-2 менее чем на 50%.
20. Способ по любому из пп. 1-19, характеризующийся тем, что указанное антитело к CD25 представляет собой моноклональное антитело.
21. Способ по любому из пп. 1-20, характеризующийся тем, что указанное антитело к CD25 представляет собой человеческое, химерное или гуманизированное антитело.
22. Способ по любому из пп. 1-21, характеризующийся тем, что указанное антитело представляет собой его варианты с созревшей аффинностью.
23. Способ по любому из пп. 1-22, характеризующийся тем, что указанное антитело к CD25 вызывает усиленный ответ КЗЦ, АЗКЦ и/или АЗКФ, предпочтительно повышенный ответ АЗКЦ и/или АЗКФ, более предпочтительно повышенный ответ АЗКЦ.
24. Способ по любому из пп. 1-23, характеризующийся тем, что указанное антитело к CD25 вводят субъекту, который имеет развившуюся опухоль.
25. Способ по любому из пп. 1-24, характеризующийся тем, что указанный способ дополнительно включает этап выявления субъекта, который имеет плотную опухоль.
26. Способ по любому из пп. 1-25, характеризующийся тем, что указанный способ дополнительно включает введение указанному субъекту ингибитора контрольной точки иммунитета.
27. Способ по п. 26, характеризующийся тем, что указанный ингибитор контрольной точки иммунитета представляет собой антагонист PD-1.
28. Способ по п. 27, характеризующийся тем, что указанный антагонист PD-1 представляет собой антитело к PD-1 или антитело к PD-L1.
29. Способ по любому из пп. 1-28, характеризующийся тем, что указанный способ дополнительно включает введение противораковой вакцины.
30. Способ по п. 29, характеризующийся тем, что указанная противораковая вакцина представляет собой противораковую вакцину GVAX.
| СПОСОБЫ ИДЕНТИФИКАЦИИ, ВЫДЕЛЕНИЯ, ИСТОЩЕНИЯ И ОБОГАЩЕНИЯ ПОПУЛЯЦИЙ TR1 КЛЕТОК, ПОПУЛЯЦИИ TR1 КЛЕТОК, ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ, СПОСОБ МОНИТОРИНГА ЭФФЕКТА ТЕРАПИИ | 2011 |
|
RU2627445C2 |
| СПОСОБ ЛЕЧЕНИЯ АУТОИММУННОГО ЗАБОЛЕВАНИЯ (ВАРИАНТЫ) | 2009 |
|
RU2539110C2 |
| СРЕДСТВО, ПРЕДНАЗНАЧЕННОЕ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ АУТОИММУННОГО ЗАБОЛЕВАНИЯ И ДЛЯ ОБРАЗОВАНИЯ РЕГУЛЯТОРНЫХ Т-КЛЕТОК | 2009 |
|
RU2531936C2 |
| WO 2017174331 A1, 12.10.2017. | |||

