Изобретение относится к способам получения ДНК-переносчика, имекхцего нуклеотидную последовательность, кодирующую гормон роста.
Последовательность оснований ДНК (дезоксирибонуклеиновая кислота) хра нит генетическую информацию , и исполь зуется в качестве кода, в соответствии с которым клетка синтезирует последовательность аминокислот всех протеинов. Некоторые части этой последовательности являются регуляторами, контролирукнцими время и количество синтезируемых протеинов (природа этих контролирующих элементов исследована только частично). Кроме того последовательность оснований в каждой нити используется в качестве шаблона для репликации ДНК, которая осуществляется при делении клетки.
Процесс, при помощи которого несущая генетическую информацию последовательность оснований в ДНК используется для образования последовательности аминокислот протеинов,в самых общих чертах является универсальным для всех живых организмов. Установлено, что каждая аминокислота, содержащаяся в протеин.ах, определяется одним или несколькими трину- клеотидами или последовательностями триплетов. Для каждого протеина суще ствует соответствующий сегмент ДНК, содержащий последовательность триплетов, соответствующих протеиновой последовательности аминокислот. Генетический код представлен в таблице.
В биологическом процессе преобразования информации, содержащейся в последовательности нуклеотид,ов, в структуре последовательности а 1ино- кислот на первой стадии осуществляется транскрипция. На этой стадии локальный сегмент ДНК, содержащий последовательность, которая относит ся к определенному протеину, копируется на РНК (рибонуклеиновой кислоте). I РНК является полинуклеоти- дом, аналогичным ДНК, но вместо рибо вы здесь содержится дезоксирибоза, а вместо тимина используется урацил. Пары оснований в РНК могут быть взаимосвязаны также как и в ДНК. Таким образом, в результате транскрипции РНК последовательности ну- клеотидов из ДНК РНК становится
10
15 к
20
25
057772
комплементарной по отношению к после дующему копированию. Такая РНК назы вается матричной РНК ( мРНК) из-за ее промежуточного положения между
I) генетическим аппаратом и аппаратом синтеза протеина клетки.
Внутри клетки мРНК происходит сложный процесс, в котором участвуют различные ферменты и органеллы внутри клетки, в результате которого синтезируется определенная последовательность аминокислот. Этот процесс называется трансляцией мРНК. В результате дополнительной обработ-.- ки последовательность аминокислот, сичтезированная в течение процесса трансляции, превращается в функциональный протеин.
Многие протеины, имеющие важное значение в медицине, в частности гормон роста, протеины, участвующие в механизме регулирования кровя ного давления, а также большое количество других ферментов, обнаружены в клетках или вырабатываются; клетками высших организмов, например позвоночных. Однако получить их в необходимом количестве экстрагированием из организма, особенно
30 из организма человека, очень трудно. Поэтому возникает необходимость в получении протеинов из искусственно выращенных клеток. Применение известных; способов выращивания
35 биологической ткани нецелесообразно, поскольку клетки растут медленно, наблюдается их низкий выход, используется дорогостоящая среда для выращивания и условия выращи40 вания необходимо жестко контролиро-; вать. Кроме того, как правило трудно поддерживать такие условия выращивания клеток, чтобы добиться искомых свойств деления.
Мик;юорганизмы, например, бактерии, относительно просто выращивать в среде, обладакмцей определенными химическими свойствами. Современная технология процессов ферментации в достаточной степени развита и хорошо поддается контролю. Микроорганизмы быстро размножаются, что обеспечивает их высокий выход. Кроме того, некоторые микроорганизмы достаточно 55 хорошо исследованы с генетической точки зрения.
Таким образом, необходимо найти переносчика генетического кода про50
теина, представляющего медицинскую ценность, из организма, который вы- рабатывает протеин, в нормальных условиях в подходящий микроорганизм. В этом случае протеин синтезируется микроорганизмами при контролируемых условиях роста и в необходимых количествах. Предлагаемый способ позволяет значительно снизить общую стоимость получения искомого протеина и обеспечивает, возможность выделения и транспортирования генетической последовательности, которая программирует получение определенного протеина в микроорганизме, имеющего вполне определенную генетическута структуру, а также исследования механизма синтеза такого протеина в контролируемых условиях и механизма дальнейшей обработ ки протеина после стадии синтеза. Кроме того, выделенные генетические последовательности можно модифицировать с целью получения генетического кода для синтеза других протеинов, имеющих другие терапевтические или функциональные свойства.
Цель изобретения - получение вектора, имеющего нуклеотидную последовательность, кодирукмцую гормон ростэ, путем вь;целения молекулы ДНК спецкгльной последовательности нукле отида и ее транспортировки в микроорганизм а также обнаружения исходной последовательности нуклеотидов ДНК после репликации в указанном организме.
Предлагаемый способ состоит из четырех основных стадий .
I. Выделение искомой популяции ,клетки из высшего организма.
Имеется два потенциальных источ- .ника последовательности, несущей генетический код для синтеза протеина: сама ДНК организма-источника и РНК, полученная после транскрипции на матрице ДНК Мерами безопасности предусмотрено введение генов человека в рекомбинированную ДНК и затем в клетку бактерии только после их тщательного отбора при помощи специального оборудования, предназ- наченного для экспериментов, связан- -ных с большой степенью риска (Р4). I
В больщинстве тканей желез или других органов клетки связаны между собой слоистой сетью соединительных тканей, которые состоят в основ
205777
ном из коллагена, но могут содержать в зависимости от ткани и другие структурные протеины, полисахариды и минеральные отложения. При выделе5 НИИ клеток из соединительной ткани необходимы специальные средства.
Таким образом, выделение и очистка типа клеток заключает в.себе две основные стадии: высвобождение кле-т
10 ток из матрицы соединительной ткани и отделение клеток искомого типа от всех клеток других типов, которые содержатся в ткани.
Содержание искомой мРНК можно
15 увеличить используя способность
клетки реагировать на внешние возмущения, например обработку популяции клетки гормоном, определенную темпе- ратуру и использован ие специальных пи2Q тательных или других химических вещест в, Например,при отделении мРНК гормона роста крысы (ГРК) обработка выращиваемых клеток гипофиза крысы тироид- ным гормоном и глюкокортикоидами
25 значительно увеличивает количество мРНК гормона роста.
И. Вьщеление мРНК. Применение предлагаемого способа позволяет полностью выделить активную РНК
3Q из экстракта клеток.
Выделяемая мРНК является одной полинуклеотидной нитью, не имеющей пары среди других комплементарных нитей. Следовательно,гидролитическое рассечение одной фосфо- диэфирной связи в последовательности может привести к образованию полной молекулы, которая непригодна для транспортировки полной генетической последовательности в микро- организм. Широко распространенный, очень активный и исключительно устойчивый фермент РНК-азы находится на поверхности, обнаруживается при помощи известных способов промывки и иногда загрязняет используемые химические материалы, что особенно нежелательно при работе с экстрактами клеток поджелудочной железы, так как поджелудочная железа является источником пищеварительных ферментов и поэтому богата РНК- азой. Однако проблема загрязнения РНК-азой актуальна и при работе с
другими тканями. 55
Предлагаемый способ подавления активности РНК-азы применим к любым тканям, однако наиболее эффективен
35
40
45
50
при выделении полной мРНК из отделенного участка ткани поджелудоч ной железы.
.Согласно предлагаемому способу используются хаостропный анион, хаостропный катион и разрывающий дисульфидные связи агент как при разрушении клетки, так и в течение всех операций, которые необходимы для вьщеления свободной РНК из протеина. Эффективность комбинированного действия указанных агентов была доказана при их использовании для выделения полной мРНК с хорошим выходом из участков Лангерханса поджелудочной железы крысы.
Выбор соответствукщих каостроп™ ных ионов зависит от их растворимости в водных средах. К хаостроп ным катионам относятся гуанидин, карбомоил-, гуанилгуанидин, литий и т.д.; к хаостропным анионам - иодид, перхлорат, тиоцианат, дийодо- салицинат и т.п. Относительная эффективность солей, образованных комбинированием таких катионов и анионов, частично определяется их растворимостью, , Например, дийодо- салшдинат лития обладает более сильным денатурирующим свойством, чем гуанидинтиоцианат, но его растворимость составляет всего 0,1 Ми, кроме того, он является относительно дорогостоящим материалом. Пред™ почтительно используют комбинацию катион-анион, так как она легкодоступна и хорошо растворяется в водны средах до 5 М.
Тиоловые соединения, такие как Э-меркаптоэтанол,дитиотрейтол,цйс-
теин, пропанолдимеркаптан, используются для разрыва внутримолекулярных дисульфидных связей в прот.еи нах при помощи тиолдисульфидной реакции обмена. Наиболее предпочти- тельно применение р-меркаптоэтано- ла, поскольку он легкодоступен и имеет невысокую стоимость растворятся в воде, так как для доведения обменной реакции до конца необходим чтобы :тиоловое соединение было в большом избытке по сравнению с. внутримолекулярными дисульфидами.
Эффективность хаос гропного агента непосредственно зависит и от его 5 онцентрации; предпочтительно исползуется максимально возможная концентрация .
Ю is
20 2.5 о
,,
р41
,|
f
5
На выход неповрежденной мРНК на стадии выделения влияет скорость, с которой денатурируется РНК-аза, а также масштаб процесса денатурации. Поэтому преимущественно используют ; гуанидинтиоцианат по сравнению с гидрохлоридом, несмотря на то, что активность гидрохлорида в качестве денатурирующего агента лишь незначительно отличается от активности тиоцианата. Эффективность денатурирующего агента определяется пороговой концентрацией, которая необходима для достижения полной денатура ции протеина, скорость-которой зависит от концентрации денатурирунлцего агента относительно порогового значения, изменяющегося от 5 до 10-и степени. Поэтому денатурирующее средство, незначительно более сильное, чем гуанидингидрохлорид, может денатурировать протеин в несколько раз быстрее при той же концентрации.
Анализ показал, что целесообразно использовать денатурирукхцее средство с низким пороговым значением денатурирующей концентрации и высокой растворимостью в водной , среде. Исходя из этого целесообразнее применять гуанидинтиоционат, а не дийодсалицилат лития, хотя последний обладает более сильным денатурирукяцим действием (растворимость гуанидинтиоционата намного Ъыше, поэтому его можно использовать в концентрациях, при которых гораздо быстрее подавляется активность РНК-азы). Кроме того, гуанидинтиоционат по сравнению с гидро- хлоридной солью, имеющей сравнимую степень растворимости, обладает несколько большей динатурирующей способностью.
Использование агента для разрыва дисульфидной связи в комбинации с денатурирующим средством увеличивает эффективность последнего ,1 обеспечивая получение полностью развернутой молекулы РНК- азы. Если некоторое количество РНК-азы остается в качестве примеси в процессе получения мРНК, она неактивна даже при отсутствии денатурирующего средства и тиола. Аген ты для разрыва дисульфидных связей содержащие тиоловые группы эффективны при любой концентрации, обес7
печивающей рассечение внутримолеку лярных дисульфидных связей. Однако верхний предел концентрации ограничен тем, что соединения при высоких концентрациях зловонны. Уста- 1новлено, что оптимальной концентрацией Р-меркаптоэтанола является О,05-1,0 М, а для выделения полной РНК из поджелудочной железы крысы концентрация составляет 0,2 М.
Величина рН среды в процессе экстрагирования мРЫК из клеток может составлять 5,0 - 8,0.
После стадии разрушения клетки РНК выделяется из основной массы протеина клетки и ДНК известными способами, например осаждением из этанола, в соответствии с которым РНК селективно осаждается в осадок В соответствии с предлагаемым способом эту стадию можно исключить, при этом гомогенат вводят в раство 5,7 М хлорида цезия, находящийся в камере центрифуги, что обеспечивает высокий выход РНК, свободной от ДНК и протеина.
В результате осуществления всех описанных выше стадий из гомогената клеток выделяется полна Т НК. Однако только часть этой РНК является искомой мРНК. Для извлече НЦЯ искомой мРНК используется способность мРНК в клетках высших организмов после транскрип1дии присоединять полиадениловую кислоту. Такая мРНК, содержащая последовательность полиА (аденин) которые присоединены к ней, может быть селективно выделена при помощ хроматографии на колоннах, содер- ,жащих целлюлозу с присоединением -опиготимидилата. Выполнение всех описанных выше стадий достаточно для получения из источников, богатых РНК-азой, существенно чистой, полной, пригодной для копирования (трансляции) мРНК. Очистка мРНК и .последующие лабораторные стадии могут быть осуществлены из клеток любого организма.
Б некоторых случаях, например когда в качестве источника мРНК используются клетки ткани с достаточно низким содержанием РНК-азы, для снижения активности РНК-азы могут применяться известные способы
Ш. Образование кДНК (комплементарной ДНК).
205777
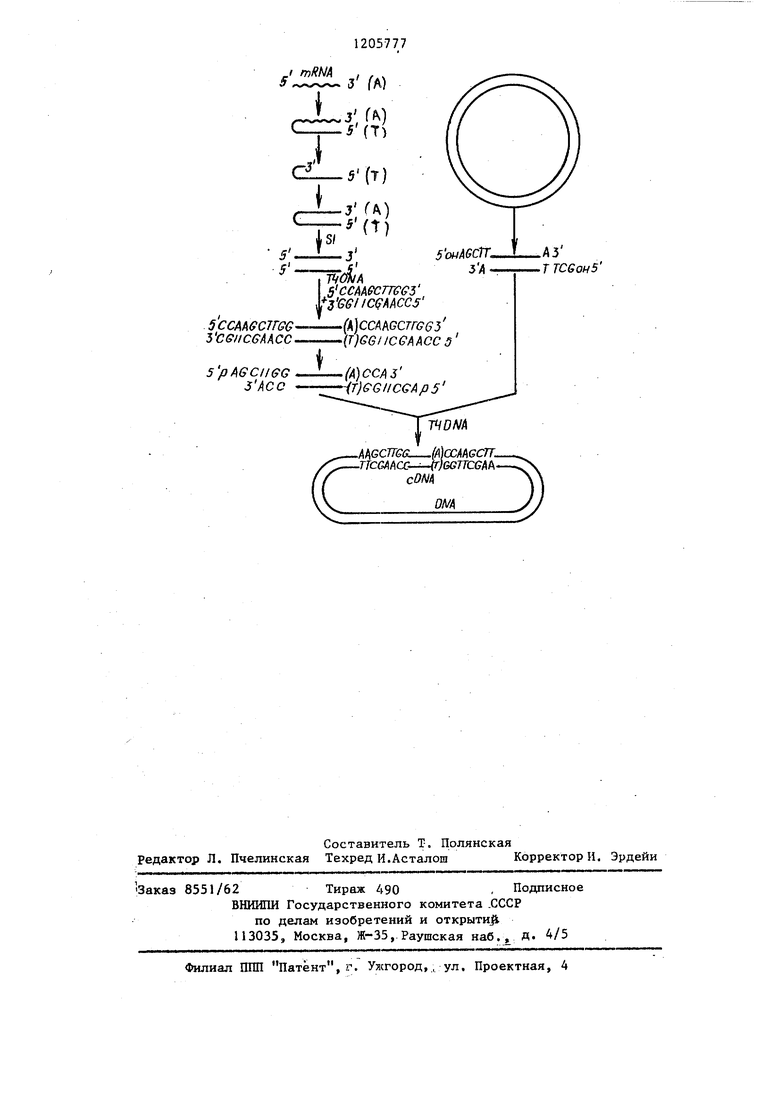
На чертеже схематически представлены оставшиеся стадии предлагаемого способа.
Первая стадия. Образование после- 5 довательности ДНК, комплементарной к выделенной мРНК.
В качестве фермента для данной .реакции выбрана обратная транскрип- тиза, хотя можно использовать любой 10 другой фермент, способный образовывать точную копию нити к ДНК, используя мРНК в качестве шаблона. Эта реакция протекает при известных условиях с использованием мРНК в 15 качестве .шаблона . и смеси четыре г. дезоксинуклеозидтрифосфатов в ка- честве предшествующих соединений для ДНК из одной нити. Для того, чтобы можно было следить за течением 20 реакции, один из дезоксинуклеозидов- трифосфатов метят при помощи в альфа-позиции; метку можно исполь зовать и в качестве ориентира для извлечения продукта после стадии 25 отделения, например, при хроматогра- фии и электрофорезе, а также при проведении количественных оценок на стадии извлечения.
Как показано на чертеже, продук- 3Q том реакции с обратной транскрип- тазо: 1 «является структура из двух нитб, имеющая форму приколки для волос, с нековалентной связью между нитью РНК и нитью ДНК.
Продукт реакции с обратной транс- криптазой получают из реакционной смеси путем, например. Экстрагирования из фенола, хроматографии на сефадексе G-100 и осаждения из этанола.
После синтеза кДНК в результате ферментной реакции шаблон РНК можно удалить. Селекторное разрушение РНК в присутствии ДНК, по предлагаемому способу осуществляют щелочным гидролизом с высокой избирательностью и возможностью регулирования, обеспечивания необходимое значение величины рН.
После щелочного гидролиза и последующей нейтрализации, если это необходимо, кДНК, меченная , может быть сконцентрирована путем осаждения этанолом.
55 Синтез имеющей форму приколки для волос молекулы кДНК из двух нитей достигается при помощи соответствующего фермента, такого как ДНК-
35
40
45
50
полимераза или обратная транскрипта за. Реакционные условия анг1логичны условиям, которые использовались ранее, включая использование меченного оС - Р нуклеозидфосфата. Обратную транскриптазу можно получить, например, из вируса avian mallob- lastosis.
После образования молекулы кДНК, имеющей форму приколки для волос, ДНК извлекают из реакционной смеси путем экстрагирования из фенола,, хроматографии на сефадексе и осаждения из этанола: при этом ДНК очищается от загрязняющего протеина.
Молекулу. ДНК в форме приколки для волос можно превратить в более предпочтительную структуру ДНК из двух нитей при помощи удаления петли из одной нити, соединяющей концы комплементарных нитей. Для этого проводят обработку ферментами, пригодными для гидролитического рассечения фрагментов ДНК, содержащих одну нить, например S1-нуклеазой, выделенной из Aspergillus. oryzae, в результате чего получают высокий выход молекул кДНК, концы которых соединены основанием. Затем осу ществляют экстрагирование, хроматографию и осаждение из этанола.
При необходимости доля молекул кДНК,имеющих срезанные концы,может быть увеличена при помощи обработки ДНК полимеразой- , которая выделяется из микроорганизмов E.coli в присутствии четырех дезоксинуклеозидтри- фосфатов. Комбинированное действие ферментов эндонуклеазы и полимеразы приводи т к удалению лк)бого выступающего 3 -конца и к заполнению любого выступающего 5 -конца. При этом обеспечивается участие максимальной доли молекул кДНК в последующих реакциях с лигазами,
Затем осуществляют обработку концов кДНК с тем, чтобы обеспечить соответствующие последовательности на каждом конце, содержащие места узнавания для эндонуклеазы ограничения. Выбор участка ДНК, который присоединяется к концам, определяется материалами, с которь и предстоит манипулировать. Последовательность, которую присоединяют к концам, выбирают в зависимости от выбранного фермента,эндонуклеазы ограничения, который,в свою очередь.
0577.710
зависит от переносчика ДНК, с которым кДНК рекомбинирует. Выбранный при этом плазмид должен иметь по крайней мере одно звено, по которо-
5 му происходит рассечение при обработке эндонуклеазой ограничения. Например, плазмид рМВЭ содержит одну точку ограничения для фермента Hind Ш. Фермент Hind Ш выделяют из бактерий
10 Hemophilus influenzae и очищают (Смит Х.О., Уилкоксом К,В. J. Мо1. Biol, 1970, 51/379). Фермент Нае Ш получают из Hemophilus aegyptipris, и очищают ( Миддлтон Дж.Х, , Эдге- ,
15 лем М.Н., Хатчисоном III, С.А. - J. virol, 1972, 10, 42/1. Фермент, полученный из Hemophilus suis, названный Hsu 1, катализирует гидролиз тех же мест узнавания, что и фермент
20 Hind Ш. Следовательно эти два фермента могут заменять друг друга.
С целью присоединения к концам двойной кДНК предпочтительно использовать химически синтезированный
25 декануклеотид из двух нитей, содержащий последовательность м ест узнавания для фермента Hind Ш. Известны различные синтетические последовательности мест узнавания, поэтому можно
.jg получить концы двойной ДНК, восприимчивые к действию любой из- эндону-f клеаз ограничения.
Присоединение последовательности мест узнавания к концам кДНК осуществляют известными метода.ми. Выбор осуп ествляют на основе реакции, которая завершается лигацией срезанных концов, которая катализируется ли- газой ДНК, о чищенной по. известному способу ( Панет А. и др. Biochemistry, 1973,, 12,5045;.
Продуктом реакции лигации срезанных концов между кДНК со срезанными .концами и большим молярным избытком декануклеотида из двух нитей, содержащего в качестве, эндонуклеазы мест узнавания фермент Hind Ш, является кДНК, содержащая на каждом конце последо вательность мест узнавания фермента Hind Ш.: Обработка продукта реакции эндоку клеазой Hind Ш приводит к рассечению молекулы в месте узнавания с образованием самокомплементарных молекул, состоящих из одной нити с концами 5 (см. чертеж).
1, Образование переносчика ре- комбинированной ДНК.
с целью образования рекомбинатов с кДНК можно использовать разные
35
40
45
50
55
11
жизнеспособные плазмиды ДНК,Основные требования заключаются в том, чтобы переносчик ДНК мог проникать в клетку-хозяина, подвергаться в ней репликации и содержать генетический детерминант, при помощи которого можно было бы выбрать те клетки, , которые получили указанный переносчик. В качестве соответствующих переносчиков предпочтительно используют, в Ч встности производные бактериофагов лямбда и производные плазмиды cot El.
Плазмиды, полученные из col El, имеют молекулы относительно небольшого размера, молекулярный вес по-; рядка нескольких миллионов и, кроме того, обладают тем свойством, что число копий плазмид ДНК на одну клетку-хозяина может быть увеличено от 20-40 при нормальных условиях до 1000 и более при обработке этих клеток хлорамфениколом. Способность плазмид распространять генетические факторы внутри клетки-хозяина обеспечивает возможность при соответ- ствукнцих условиях, контролируемых исследователем, заставить клетку- хозяина вырабатывать протеины, генетический код на синтез которых был i epeHeceH плазмидой. Таким образом, целесообразно, в качестве переносчика ДНК использовать производные , выделенные из со 1 Е1, в частности плазмиды рМВ-9, несущие ген, контролирующий устойчивость к тетрациклину, а также рБР-313, рВР-315, рВР-316, рВР-317 и рВР-322, которые несут ген, контролирующий устойчивость к амплицилину. Наличие генов, контролирующих устойчивость к лекарственным препаратам, позво-, ляет селекционировать клетки, кото- содержат плазмиды, так как популяции таких клеток выращивают в присутствии лекарственных препаратов. Клетки,которые не содержат плазмид, не имеют возможности расти или образовывать популяции. Согласно предлагаемому способу используют плазмиды, выделенные из col. Е1, содержащие, одно.место узнавания для фермента Hind Ш.
Рекомбинированные плазмиды образуются в результате смешения плазми- ды ДНК, обработанной эндонуклеазой ограничения, с кДНК, содержащей концевые группы послеаналогичной обработки. Для сведения к минимуму
0577712
;вероятнос ти образования комбинации сегментов кДНК друг с другом плазмиду ДНК добавляют в молярном избытке по сравнению с кДНК. Это относится к
5 большинству плазмид, которые исполь- . зуются без введенного фрагмента кДНК. Следовательно, трансформированные клетки не содержат рекомбинирован- ных плазмид кДНК, Поэтому процесс
10 селекции является очень трудоемким и продолжительным. Попытки синтезировать носители ДНК, содержащие места для действия эндонуклеазы ограничения, в середине соответствующего
15 гена, так, чтобы введение рекомбина- та способствовало разделению гена, привели к снижению функции гена как носителя информации, Предпочтитель- -но используют способ для снижения
20 числа колоний, к которым затем применяют рекомбинированную плазмиду, согласно которому отрезки плазмиды ДНК обрабатывают эндонуклеазой ограничения и щелочной фосфатазой,
25 вследствие, чего удаляются 5 -терминальные фосфаты, находящиеся на концах, образовавшихся в результате действия эндонуклеазы, и предотвращается самолигация плазмиды ДНК.
30 Следовательно, образованиецикла, а значит и трансформация, зависит от введения в молекулу фрагмента ДНК, содержащего фосфорированные 5 -терминалы. Этот способ снижает относительную частоту трансформации при отсутствии рекомбинации до менее Ю
Предлагаемый способ основан на реакции, катализатором которой служит лигаза ДНК, между 5 -концевой фосфатной и З -концевой гидро- ксильной группами ДНК. Если терминальный 5 - фосфат отсутствует, то реакция не осуществляется.
Соединение ДНК из двух нитей представлено в таблице (концевые группы отмечены гидроксилом (ОН) или фосфатом(ОРОг Н 2 j.
В первом случае 5 -фосфаты на- .
50 ходятся на обоих концах реагента, в результате чего между двумя Нитями установлена ковалентная связь. Во втором случае только одна из нитей, вступающих в связь, содержит
55 терминальный 5 - фосфат, в результате чего между одной парой нитей установлена ковалентная связь, тогда как на другой нити разрыв сохра40
13
ияется. Нить, не содержащая кова лентной сэязи, остается связанной со всей молекулой благодаря связывающему взаимодействию между атомами водорода комплементарных пар основа™ НИИ на противоположных нитях. В третьем случае ни один из концов не содержит 5 фосфата и реакция сое цинения отсутствует.
Таким образом, нежелательных реакций соединения можно избежать обработав соответствующие концы молекулы реагента с целью удаления 5 - юсфатных групп любым известным способом а tie разрушакмцим структуру ДНК, в частности гидролизом, катализируемым ферментом - щелочной фос фатазой.
Аналогичным способом выделяется кДНК, содержащая генетический код
Схема реакции между 5 -концевой фосфатной и З -концевой гидроксильной группами ДНК, катализируемой лигазой
Случай
Реагенты
ОН
-fHjOi
ОРОзН ,ОЫ
он он
.
он
он он
он он
05777
для синтеза ГРК, рекомбинируется с плазмидой и транспортируется в E.coli. После продолжительной репликации в E.coli вьщеляется после™
5 довательность нуклеотидов, содержащая приблизительно 800 нуклеотидов по длине. -Было установлено,, что она содержит полную последовательность j содержащую генетический
10 код для синтеза ГРК, а также некоторые части предшествугацих пептидов и часть 5 - области, которая не участвовала в трансляции.
Предлагаемый способ можно исполь15 зовать для вьщеления и очистки гв нов высшего организма, включая гены человека, а также тр анспортировки |ИХ в микроорганизм для репликаций. В таблице приведены конкретные
20 примеры вьшолнения предлагаемого способа.
Лигаза
-С - Р - ,,
О - Р - О
5 3
, Oh
о - р - о он
Нет реакции
15
Культивированные клетки гипофиза крысы (подклон семейства кл.еток GH-U использовали в качестве источника мРНК для синтеза ГКР. В э тих клетках, если рост происходит в нормальных условиях, мРНК гормона роста содержится в небольших количествах 1 - 3% от общего содержания РНК, имеющей полиА. Однако, содержание мРНК гормона роста значительно увеличивается в результате стимулирующего действия тироидных гормонов и глюкокортикоидов. РНК вьщеляется из 5x10 клеток, выращиваемых в суспензии, в которую с . целью получения гормона роста за 4 дня до сбора клеток добавляют 1 мМ дексаметазона и 10 мМ. L-три- йодтиронина. Полиаденилированная РНК вьщеляется из цитоплазменных разделякщих мембран культивируемых клеток известными способами. Поли- аденилированная РНК, выделенная из гипофизных клеток, содержит мРНК, кодирующую гормон роста. Клетки делают более вязкими обработкой трипсина и обрабатывают ледяным 25 мМ раствором фосфата калия и 0,1 М NaCi- при рН 7,4. Затем клетки инкубируют 10 мин при О с в 10 мМ NaCZ 10 мМ Грис -НСЕ , рН 8,5 и 3 мМ MgCij., 5 мл/г клеток. Далее. клетки разрушают в гомогенизаторе Доунса. Ядра удаляют дентрифугиро- ранием гомогената при 800 G в течение 5 мин. Супернатант изготовляют из 0,5% (.вес/об.) додедилсульфата натрия (НДС), 1 мМ этилендиаминтетр уксусной кислоты (ЭДТК) и 100 ьй NaCf. РНК экстрагируют фенолхлоро- формом и осаждают объемами этанола. Таблетку РНК, полученную центрифугированием осадка при 10000 G в течение 20 мин, растворяют в водном растворе, содержащем 0,5% (вес/об.; НДС, 5 мМ NaCf и 1 мМ ЭДТ после чего инкубируют и течение 30 мин при 37 с вместе с 100 мг/мл самопереваренной проназы. Из инкубированной смеси РНК повторно экстргируют фенолхлороформом и осаждают этанолом, после чего растворяют в воде до концентрации в 2 мг/мп и хранят при .
Получают 10 клеток - 10 мг цито плазмовой РНК, включая рибосомную РНК и РНК-1и;реносчик, в дополнение к полиаденилированной РНК.
1205777
16
Полиаденилированную ГНК выделяют при помощи хроматографического анализа всего препарата РНК на олиго-(.дТ)- целлюлозе. Выход 30 мг из 5x10
клеток.
Синтез двунитьевой кДНК (где одна нить имела нуклеотидную последовательность , комплементарную мРНК) завершают реакцией, катализируемой
обратной транскриптазой,содержащей смесь дезоксиаденозинтрифосфат(дАТФ), дезоксицитидинТрифосфат (дЦТФ) де- зоксигуанозинтрифосфат (дГТФ),дезок- ,ситимидинтрифосфат (дТТФ), выделеннум как описано выше мРНК, олиго- .з -целлюлозу Supra и обратную транскриптазу.
Обратную транскриптазу, выделенную из вируса avian macloblastosis, используют для того, чтобы скопировать полную молекулу полиаденилиро- ванной РНК, полученную из участков Лангерханса крысы, на кДНК. Реакции осуществляют в растворе 50 мМ трис- НСГ (рН 8,3) , 9 мМ MgCr, 30 мМ хлорида натрия, 20 мМ р-меркаптоэтанола, 1 мМ каждого из трех нерадиоактивных дезоксириОунуклеозидтрифосфатов,
250 мМ четвертого дезоксинуклеозидзгт
триофосфата, меченного Р в альфа- позиции, с удельной радиоактивностью 50-200 Ки/моль, 20 мг/мл олиго-дТ,..,, 100 мг/мл полиаденилированной РНК и 200 ед./мл обратной тpa cкpиптaзы. Смесь инкубируют при в течение 50 мин. После добавления 45 мМ . ЭДТК-Na, раствор экстрагируют насьш енным водой фенолом, водяной слой хроматографируют на сёфадексе
G-100 с колонкой диаметром 0,3 см и высотой 1 о см в смеси 10 мМ трис- НС Г. (рИ 9,0) , 100 мМ хлорида натрия, 2 мМ ЭДТК. Нуклеиновую кислоту, элюированную в пустой объем, осаж-
дают этанолом после добавления
ацетата аммония, до 0,25 М ( рН 6,OJ , Осадок собирают центрифугированием. Полученный осадок растворяют в 50 мл свежеприготс/вленного О, 1 М раство- .
ра гидрата окиси натрия и инкуби-. руют при в течение 20 мин с целью гидролиза РНК, Смесь нейтрализуют добавлением 1М раствора ацетата натрия (рИ 4,5), а меченную
р кДНК осаждают .этанолом, а затем вновь растворяют в воде« Образовавшийся промежуточный продукт представляет собой однонитевую кДНК,
17
которую используют для синтеза требуемого двунитевого ДНК-продукта. С целью идентификации однонитевую кДНК анализируют для выяснения возможности ее применения для синтеза гормона роста.
Отдельные порции кДНК, состоящей из одной нити, анализируют на натуральных полиакриламидных гелях. Гели сушат и ДНК, меченную Р, анализируют при помощи авторадиографии с. использованием пленки Kodak No- sereen-2T. При помощи диаграммы электрофореза установлено, что кДНК представляет собой гетероди- сперсоид/ После разделения при помощи электрофореза на геле, обнаружена слабая полоса поглощения, соответствующая ДНК с примерно 800 парами оснований, что характерно ЦIя кДНК, кодирующей гормон роста,
Пример 1. Полученную кДНК, состоящую из одной нити, обрабатывают обратной транскриптазой с целью синтеза комплементарной нити.
Реакционная смесь содержит 50 мИ (рН 8,3) ; 9 мМ MgCf, 10 iM дитиотрейтол; 50 мМ трех немеченных дезоксирибонуклеози;.-- триофосфатов; 1 мМ меченного P в альфа- позиции нуклеозидтрифосфата удельной радиоактивностью 1-10 Ки/мМ, 50 мг/мл кДНК и 220 ед. /мл обратной тра криптазы. Реакционного смесь инкубируют при АЗ С в течение 120 мин.
..Реакцию прекращают добавлением ЭДТК-Ма2 (до 25 мМ) . Смесь экстра гируют фенолом и подвергают хрома- тографическому анализу на сефадек™ се G-100, а затем осаждают этанолом. Порции продукта реакции с показателем от 500 до 100 отсчетов в 1 мин анализируют при помощи электрфреза на геле.
Полоса поглощения гетеродисперсо да сосредоточена вокруг 800 нуклео- тидов по длине,.
При обработке полной кДНК, на кторой скопирована структура мРНК клетки гипофиза крысы, э,ндонуклеа- зой Hhal образуется два основных фрагмента ДНК (установлено при помщи разделения 7 лектрофорезом), причем один содержит примерно 320 нукл отидов (фрагмент А), а второй.240 нуклеотидов фрагмент В).
205777 «
На основе известной последовательности аминокислот ГКР и других из- вестных данных и гормонах роста уста-
5
0
5
0
5
0
5
новлено, что эти фрагменты являются в действительности теми частями, на которых.содержится генетический код , контролирующий синтез ГРК.
При воздействии на кДНК из двух нитей,, содержащую 800 пар оснований и выделенную при помощи электрофореза эндонуклеазой Hhal, при анализе среди основных продуктов рассечения были обнаружены два фрагмента,соответствующие по длине фрагментам А и В.
Таким образом, двунитьевую кДНК, содержащую примерно 800 пар оснований, получили с помоЬчью реакции, катализируемой обратной транскрипта- ЗОЙ, содержащей в начале ДНК, имеюг- щую нуклеотидную последовательность, кодирующую ГРК.
Так как кДНК, контролирующую син. тез ГРК, содержащую приб аизительно 800 пар оснований, не очищают перед обработкой эндонуклеазой огр аниче- НИН, ДНК необходимо подвергнуть обра- ботке с тем, чтобы удалить все несое- диненные концы. Обработка с целью удаления таких непарных концов производится перед стадией разделения электрофорезом в смеси, содержащей 25 мл 60 мМ трис-НС (рН 7,5) , 8 мМ хлорида магния, 10 мМ (f3 -мер-, каптоэтанола, 1 мМ АТФ и 200 мМ каждого из дАТФ;дТТФ,дГТФ идЦТФ.Смесь инкубируют с полимеразой 1 ДНК { 1 ед.) , выделенной из штамма Е. со 11 при 10 С в течение 10 мин с тем, чтобы удалит } все выступающие З -концы и заполнить все 5 -выступающие концы.
кДНК, контролирующую синтез ГРК и содержащую приблизительно 800 пар оснований, обрабатьшают химически синтезированным линкером Hind Ш.
Полученный в результате реакции продукт, молекула которого состоит из двух нитей, с концентрацией 2-5 мг/мл обрабатывают 30 ед. S 1
нуклеазы имеющей активность 1200 ед/г-ш, в смеси, содержащей 0,03 М ацетата натрия, (рЕ 4,6) 0,3 М хлорида , 4,5 мМ хлорида цинка, при в течение 30 мин, а
затем инкубируют еще в течение
15 мин при . С целью прекращения ферментной реакции в смесь добавляют трис-основание с конечной
19
концентрацией 0,1 м ЭДТК до 25 мМ и тРНК, вьщеленную из E.coli,,до 40 мг/мл. После экстрагирования реакционной смеси этанолом и хрома- ографического анализа на сефадек- се G-100, меченную Р кДНК элюи- руют в пустой объем и осаждают при помощи этанола. В результате./ такой обработки обеспечивается высокий вьпсод молекул кДНК, концы которой связаны основанием. I Декамеры Hind Ш добавляют на каждый конец молекулы кДНК для обеспечения возможности соединения реакционноспособных концов гибридизацией или ковалентной связью кДНК, полученной, расщеплением вектора- переносчика ограничительным ферментом Hind Ш. Лигация декамера Hind Ш с кДНК осуществляется при помощи инкубации при 14°С в смеси 66 гМ трис-HCf (рН 7,6), 6,6 мМ хлорида магния, I гМ АТФ, 10 мМ дитиотрейтола, 3 мМ декамера Hind Ш с показателем 10 отсчетов в минуту и лигазы ДНК Т4, приблизительно 500 ед./мл в течение 1 ч. Для дезактивации лигазы реакционную смесь нагревают до и выдерживают в течение 5 мин. Предварительно к пищеварительным ферментам, содержащим 150 ед./мл Hsu I или Hind Ш добавляют KCf с конечной концентрцией 50 мМ, р - меркаптоэтанол с конечной концентрацией 1 мМ и ЭДТК с конечной концентрацией 0,1 мМ; смесь вьщерживают в течение 2 ч при . Эндонуклеазы Hind Ш и Нао Ш. Продукт реакции анализируют при помощи электрофореза на геле, при этом был обнаружен пик, соответствующий последовательности приблизительно 850 нукпеотидов наряду с отдельными фрагментами рассеченного декамера Hind Ш.
Плазмиду рВР-322, содержащую ген, контролирующий устойчивость к ампицилину, и единственное место узнавания для фермента Hind Ш, расположенное внутри указанного гена, рассекают в месте узнавания для Hind Ш эндонуклеазой Hsu I, затем обрабатывают щелочньм фосфатом типа PAPF. Фермент вводят в реакционную смесь с концентрацией О, 1 ДНК, Реакционну ) смесь инкубируют в 25 мМ трис-HCf при рН 8 в течение 30 мин при 65 С, затем удаляют фос05777 0
фатазу путем экстрагирования в феноле.
Полученный продукт представляет собой молекулу ДНК, приготовленную
5 расщеплением вектора-переносчика (в данном случае плазмида рВГ-322) ограничительным ферментом (в данном случае эндонуклеазой Hind Ш). Перед фосфатазной обработкой расщепленные
10 концы вектора-переносчика ДНК, генерированные в реакции расщепления Hind f l, способны соединяться друг с другом с помощью гибридизации или присоединяться к двунитевой
15 кДНК с образованием гибридной или ковалентной связи. После осаждения этанолом плазмидную ДНК, обработан-,, ную фосфатазой, добавляют в кДНК, содержащую фермент Hind Ш, соединяю20 щий концы молекулы, в молярном соотношении 3 моль плазмиды и 1 моль кДНК.
Судя по удельной радиоактивности, в реакции применено приблизительно
25 10-50 мг кДНК и 100 мг плазмидной ДНК. Смесь инкубируют в смеси 66 мМ трис-НСЕ ( рН 7,6), 6,6 мМ хлорида магния, 10 мМ дитиотрейтол.а и 1 мМ АТФ в течение 1 ч при 10 С в присут-
30 ствии 50 ед/мл лигазы ДНК Т4.
Смесь, в которой протекает лига- ция, добавляют непосредственно в суспензию штамма E.coli Х-1776, которую для трансформации приготавливают следующим образом.
I
Клетки выращивают до удельной
плотности примерно 2x10 клеток/мл в 50 мл среды, содержащей триптон 10 г/л, экстракт дрожжей 5 г/л, хлорид натрия 10 г/л, гидрат окиси натрия 2 мМ, диаминопимелиловая кислота 10 мг/мл и тимин 40 мг/мл при 37°С. Клетки собирают центрифугированием в течение 5 мин со скорое-
5 тью 6000 G при , вновь суспендируют в 20 мл холодного 10 мМ раствора хлорида натрия, центрифугируют, суспендируют в 20 мл буфера трансформации, сддержащем 75 мМ CaCl ,
50 хлорида натрия и 10 мМ
трис (рН 7,5) затем смесь выдерживают в течение 5 мин на льду. Клетки центрифугируют и вновь суспендируют в буфере трансформации 0,5 мл.
55 Трансформацию осуществляют путем смешивания 100 мл суспензии ,;слеток с 50 МП рекомбината ДНК (1 мг/мл). Смесь инкубируют при О С в течение
40
21
15 мин, затем в течение 4 мин при 25°С и в течение 30 мин при О С. Клетки переносят на питательный агар для роста в селективных условиях. Рекомбинированные ,колонии ; селекционируют вьфащиванием на питательной среде, содержащей ампи цилин, а если роста не обнаруживается, то на питательной среде содержащей тетрациклин с концентрацией 2 мкг/мл. Получено 10 таких колоний клеток, каждая из которых содержит плазмид с приблизительно 800 парами оснований, молекула кото рого разрывается под воздействием фермента Flind Ш. Такие вводимые рекомбииатные плазмиды обозначают рГРК. Для дальнейшего анализа выбрали одну колонию рекомбинированных бактерий, обозначенных E.coli X Х-1776/рГ К-1, Из культуры E.coli Х-1776/рГРК-1 приготовили плазмиду ДНК, обозначенную рГРК-1.
ДНК гормона роста крысы, содержащую 800 пар оснований, выделяют в препаративных количествах 5-30 мг из рекомбинированного клона рГРК-1 путем расщепления Hind Ш эндонук леазой и анализируют -последователь- кость нуклеотидов, Нуклеотидная последовательность содержит участки 5 областей, которые не участвовали в трансляции, а также последовательность 26 аминокислот, которые были обнаружены в протеине, предшествукядем гормону роста, перед стадией выделения.
Таким образом, приблизительно 800 пар оснований выделены из плазмиды рГРК-1, закодированной для гормона, роста и представляющей собой вектор-переносчик ДНК, имеющий нуклеотидную последовательность кодирующую гормон роста.
Пример 2. Вьщеление мРНК, контролирующей синтез ГРЧ, проводят аналогично примеру 1, однако биологическим источником материала яв-- ляется ткань гипофиза человека. Пять доброкачественных гипофизов человека, замороженных в жидком азоте после хирургического удаления вес которых 0,4 - 1,5 г , оттаивают и тонко измельчают в 4М растворе гуанидинтиоционата, содержащем Ш буферный раствор р -меркап трзтанола (рН 5,0 при 4°С. Гомоге- нат наслаивают на 1,2 мл 5,7 -М рас205777 . 22 .
вора хлорида цэзия, содержащего 100 мМ ЭДТК и центрифигируют в течение 18 ч со скоростью 37000 об/мин на ультра-центрифуге Бекман с рото5 ром типа SW 50.1. При этом РНК
собирается на дне пробирки. Далее ,проводится очистка с использованием колонны на олиго-дТ осаждение при помощи градиентного раствора цукро10, зы. Примерно 10% вьщеленной таким образом РНК содер;кит генетический . код для синтеза гормона роста, что установлено путем введения радиоактивного соединения, содержащего
15 аминокислоты в гормон, подавляюций ;рост, который выпадает в осадок в .- системе рслеток, в которых не происходила трансляция и полученных из зародьшзей пшеницы. кДНК гормона рос20 та человека получают аналогично примеру 1. кДНК гормона роста фракционируют при помощи электрофореза . на геле и материал с примерно 800 нуклеотидами по длине выбирают для
25 размножения. Выбранную фракцию обрабатывают полимеразой1 ДНК согласно примеру 1, затем обрабатывают линкером Hind III с целью сцепления концов. Далее кДНК рекомбинируется
30 с плазмидой рВР-322, обработанной щелочной фосфатазой в присутствии лигазы ДНК. Клетки штамма Е. coli Х-1776 трансформируют рекомбинантом ДНК, а затем штамм, содержащий ДНК гормона роста, извлекают. Штамм, содержащий ДНК гормона роста, выращивают в препаративных количествах, из него выделяют ДНК гормона роста, а затем определяют последовательность нуклеодидов. Было установлено, что вьщеленная ДНК гормона роста содержит нуклеотиды, содержащие информацию для синтеза полной последовательности аминокислот гормона роста человека. Первые 23 аминокислоты гормона роста имеют вид: HjN-Phe- -Pyo-Thr-Ile-Pro-Leu-Ser-Arg-Leu- Phe-Asp-Asn-Ala-Met-Leu-Arg-Ala-His- Arg Leu-His-Glu-Leu.
5Q Нуклеотидная последовательность одной нити ДНК гормона роста человека,а также последовательность ДНК,которая соответствуем последовательности мРНК, контролирующей синтез ГРЧ (исключение
,.j составляет то.,что вмРНК V заменяет Т),. показаны ниже.Числа относятся к последовательности кислот ГРЧ,начиная с конечной аминогруппы.
35
40
45
u И
9
г-t
о ы
iH
(9 X
и
М
н ж
3
о.
О)
ся fs
01 ч
Щ (Х|
(в , О
U
01
л
V
I-}
э
JS
X
r-l
о
Kl
м
п
в) (К
1
о
б
и
S
о .
о
i S
о
н н
CJ
g
и
и н о
S
о
о
S
g
н
S
и
U
о и о о
о и и (J
S
и
S у S S
U
S
б
ё
S
S
U
н
S
и
о. о
5
о о
b
I Е.
fnRNA , . 5 з fA)
.j ГА)
, Is/
-j;
S ccAi ecTrees 3 Gff//C fAACCS
(K}CCAf(GCTrffe5 {T)GG//CGAACC3
f
.{A}OCA3 i.r}(GIICGAp5
мА
мАбСГТ, 5 A
A3
Г ТСбонВ
ТЧОШ f(GCTreG-L(A CCAAGCrr.
TTccMa: -)GGTrcGAt
cDNK OM











| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения рекомбинантной плазмидной ДНК,кодирующей гормон роста крысы или человека | 1980 |
|
SU1436887A3 |
| Способ получения рекомбинантной ДНК,кодирующей нуклеотидную последовательность инсулина | 1978 |
|
SU1308199A3 |
| Способ конструирования рекомбинантной плазмидной ДНК, кодирующей бычий гормон роста | 1981 |
|
SU1471949A3 |
| ФАКТОРЫ, ДЕЙСТВУЮЩИЕ НА АКТИВНОСТЬ ФЕРМЕНТА, ВЫСВОБОЖДАЮЩЕГО РЕЦЕПТОР ФАКТОРА НЕКРОЗА ОПУХОЛЕЙ | 1999 |
|
RU2279478C2 |
| ТРАНСГЕННЫЕ РАСТЕНИЯ, ЭКСПРЕССИРУЮЩИЕ ГЕН АЦЕТИЛ-СОА-КАРБОКСИЛАЗЫ | 1996 |
|
RU2187555C2 |
| СПОСОБЫ И ВЕЩЕСТВА ДЛЯ ПРИДАНИЯ РАСТЕНИЯМ УСТОЙЧИВОСТИ К ЗАБОЛЕВАНИЯМ | 1996 |
|
RU2203320C2 |
| Способ получения фрагмента нуклеиновой кислоты, кодирующего проинсулин человека | 1980 |
|
SU1491345A3 |
| ФРАГМЕНТ РЕКОМБИНАНТНОЙ РНК (ВАРИАНТЫ), РИБОНУКЛЕОПРОТЕИНОВЫЙ КОМПЛЕКС (ВАРИАНТЫ), ПЛАЗМИДА | 1990 |
|
RU2132877C1 |
| ЭРИТРОФЕРРОН И ERFE-ПОЛИПЕПТИДЫ И СПОСОБЫ РЕГУЛЯЦИИ МЕТАБОЛИЗМА ЖЕЛЕЗА | 2013 |
|
RU2684216C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ, РАСПОЛОЖЕННАЯ ВЫШЕ ГЕНА CARP, ВЕКТОР, СОДЕРЖАЩИЙ ЭТУ ПОСЛЕДОВАТЕЛЬНОСТЬ, И ИХ ИСПОЛЬЗОВАНИЕ | 2001 |
|
RU2283865C2 |

Составитель Т, Цолянская Редактор Л. Пчелинская Техред И. Ас галош Корректор И. Эрдейи
Заказ 8551/62 Тираж 490, Подписное
ВНИИПИ Государственного комитета .СССР
по делам изобретений и oткpытиjl П3035, Москва, Ж-35, Раушская наб., Д. 4/5
Филиал ШШ Патент, г. Ужгород,, ул. Проектная, 4
| Ruogeon F et al | |||
| Insertion of a rabbit beta globin gene sequence into un E | |||
| coli plasmid - Nucleic acids Res, 1975, 2, 2365-2378 | |||
| Видоизменение прибора для получения стереоскопических впечатлений от двух изображений различного масштаба | 1919 |
|
SU54A1 |
| СПОСОБ ПОЛУЧЕНИЯ ВЕКТОРА, ИМЕЩЕГО НУКЛЕОТИДНУЮ-ПОСЛЕДОВАТЕЛЬНОСТЬ, КОДИРУЩУЮ ПРОТЕИН, включакяций вьщеление клеток животной ткани, экстракцию мРНК из клеток, кодирующей протеин в присутствии ингибитора РНК-азы, вьщеление мРНК, последующий синтез комплементарной ДНК и конструирование нукле- отидной последовательности, кодирующей протеин, отлича.ющий- с я тем, что, с целью получения вектора, имеющего нуклеотидную по следовательность, кодирующую гормон роста, в качестве клеточной ткани животного используют клетки гипофиза, в качестве ингибитора РНК-азы используют раствор 4М гуани- динзтиоцианата и 0,2 М бетамеркап- тоэтанола, а вектор синтезируют путем реакции комплементарной ДНК с ДНК-молекулой, имеющей реакционно- способные концы, способные соединяться мелщу собой или с комплементарной ДНК при условии предотвращения соединения реакционноспособных концов молекулы ДНК | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Способ по п | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| СО | |||
