(21)4200182/28-13
22)24.02.87
(46)07.01.89. Вюл. № 1
(71)Новосибирский медицинский институт
(72) А.Б.Пупьппев
(53) 663.15 (088.8)
(56) Покровский А.А., Тутельян В.А.
Лизосомы. - М.: Наука, 1976.
Bohley Р,, Kirscke H.,-Langner I., Ansorge S. Preparative gevinnung hochgereinigter lysosoraenenryme aus ratten leben. - FEBS Lett., 1969, 5. p.233.
(54) СПОСОБ ЭКСТРАКЦИИ ЛИЗОСОМАПЬНЫХ ФЕРМЕНТОВ
(57) Изобретение относится к области биохимии, а именно к способам очистки ферментов. Целью изобретения является упрощение процесса и увеличение выхода целевого продукта. Способ предусматривает, дифференциальную экстракцию лизосомальных- ферментов из грубо- очгаценного препарата митохондриально- мукомольной субклеточной фракции в присутствии дигитонина в концентрациях 0,3-0,4 мМ. 2 ил., 2 табл.

| название | год | авторы | номер документа |
|---|---|---|---|
| ПРИМЕНЕНИЕ HSP70 В КАЧЕСТВЕ РЕГУЛЯТОРА ФЕРМЕНТАТИВНОЙ АКТИВНОСТИ | 2009 |
|
RU2571946C2 |
| ПРИМЕНЕНИЕ HSP70 В КАЧЕСТВЕ РЕГУЛЯТОРА ФЕРМЕНТАТИВНОЙ АКТИВНОСТИ | 2009 |
|
RU2521672C2 |
| Конструкция генетическая для экспрессии в лизосомах клеток млекопитающих родопсина NsXeR из организма наногалоархеон Nanosalina, позволяющего повышать pH лизосом | 2023 |
|
RU2820973C1 |
| Способ моделирования функциональных сдвигов ультраструктур клетки | 1984 |
|
SU1343285A1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ЧУВСТВИТЕЛЬНОСТИ К ВНУТРИВЕННОЙ ЛАЗЕРОТЕРАПИИ БОЛЬНЫХ ИШЕМИЧЕСКОЙ БОЛЕЗНЬЮ СЕРДЦА | 1991 |
|
RU2012885C1 |
| Способ определения активности биопрепаратов | 1976 |
|
SU675649A1 |
| Способ определения реакции организма на воздействие стрессорных факторов | 1984 |
|
SU1293657A1 |
| Способ получения фаголизосомальных структур фагоцитирующих клеток | 1983 |
|
SU1141338A1 |
| Способ получения лизосомального полиферментного препарата для лечения инфицированнных очагов | 1975 |
|
SU577035A1 |
| СПОСОБ ФАРМАКОЛОГИЧЕСКОЙ ЗАЩИТЫ ДЕЦЕНТРАЛИЗОВАННОЙ ПОЧКИ | 1991 |
|
RU2008901C1 |

СП
00
Ферменты
10
Изобретение относится к биотехно- лйгии, а именно к способам очистки фйрментов.
Цель изобретения - ускорение и уйрощение способа и увеличение вы- целевого продукта.
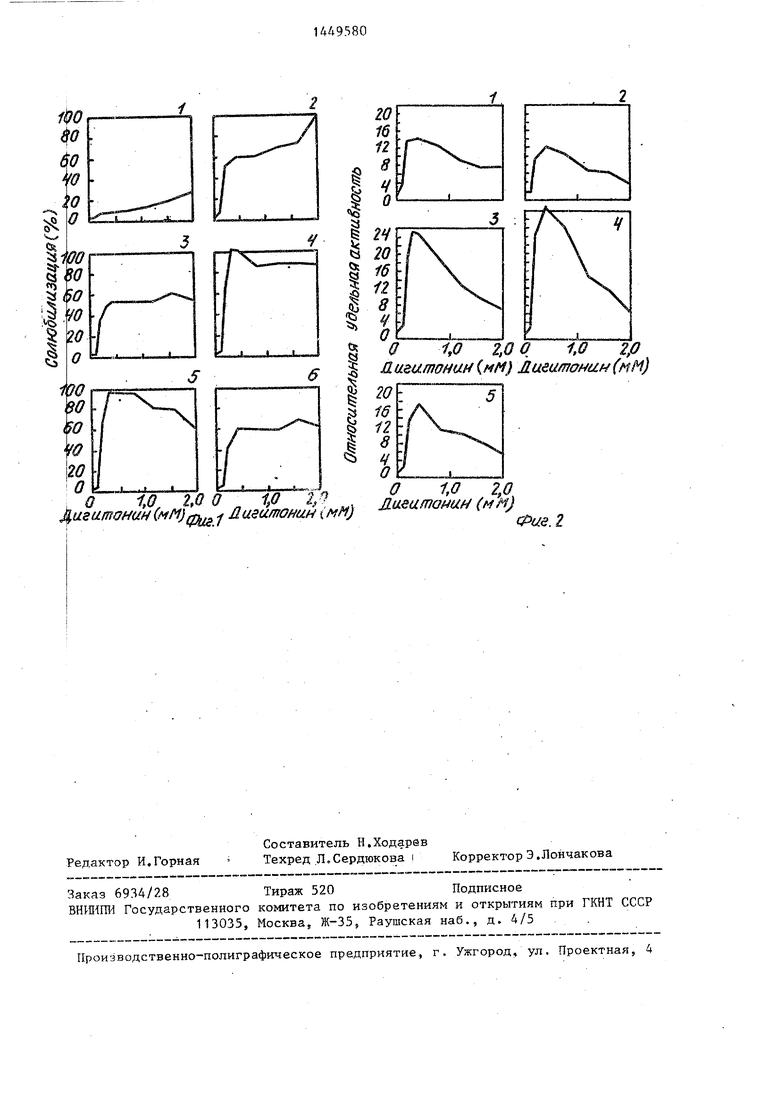
I На фиг.1 и 2 представлены графиЙ, П(ясняюр1ие предлагаемый способ.
i Способ заключается в вьщелепии бфльшей части клеточных лизосом в ей став в митохо ндриально-лиз о сомал ьной ф акции гомогената печени, получаемой достаточно просто и быстро дом дифференциального центрифугирова-J5 Белок Н11Я, с последующим использованием особенностей химического состава ли- зосомальной мембраны, которая в сравнении с мембранами других органелл, в частности митохондрий, обогащена 20 холестерином. Способностью к специфи- ч гскому связыванию с холестерином (точнее с 3,|5-оксистероидами) с последующей- лабилизацией мембраны обла- .дает стероидный гликозид дигитонин. 25 Пэи этом чувствительность лизосом к д; гитонину максимальна среди всех основных цитоплазматических структур И: наиболее существенно отличается от Т&ковой митохондрий.
П р и м е рИ. Печень животных, умерщвленных декапитацией, перфузи- , рЬтот холодным раствором 0,25 М саха- , 1 мМ ЭДТА, рН 7,4 (стандартный pjacTBOp сахарозы - СРС), Гомогенат г| ечени (ткань: среда Г. 3) подвергают 1| трифугированию (700 g, 13 мин). Осажденную ядерную фракцию промывают Дважды ресуспендированием в 15 мл Холодного СРС с последующим переосаж- Дением частиц при 700 g, 11 мин. Объединенный супернатант центрифуги- 1|)уют на препаративной центрифуге К-70 (Янецки,. ГДР) со скоростной йасадкой при 13600 об/мин (13000 о)
Содержание активности лизосомаль- ных ферментов в митохондриально-ли- зосомальной фракции (Mtni) приведено в табл.1.
Таблица 1
Содержание активности, % от содержания в го- могенате печени
Относительная удельная активность
31,2t1,1
1
30
Относительная удельная активност лизосомальных ферментов находится в пределах от 1,93 для кислой фосфата зы до 2,73 для катепсина D.
П р и м е р 2. Дигитонин растворяют в диметилсульфоксиде и разводя СРС до концентрации 4 мМ. Суспензию органелл митохондриально-лизосомаль ной фракции смешивают в соотношении 1:1 с раствором дигитонина 0,8 мМ и вьщерживают во льду в течение 30 мин Затем смесь центрифугируют (18000 g, 15 мин). Надосадок, представляющий
40
t1 мин,, Осадок митохондриально-лизо- прозрачную желтоватую жидкость, ис- сомальной фракции промывают ресуспен- пользуют в дальнейшем как экстракт Жированием в 15 мл СРС и повторным
(рсаждением при 13600 об/мин, 11 мин. Конечный осадок суспендируют в 15 мл СРС (концентрация белка 32-42 мг/мл) . Все операции проводят при 0-4 С.
Выход активности лизосомальных (|)ерментов в объединенной митохондри- альной (митохондриалБНо-лизосомаль- Ной) фракции 62,0-84,5% при содержании белка 31,2% от содержания в гомо- генате пячени.
лизосом. Установлено, что растворитель диметилсульфоксид не вызывает повреждения лизосомальиой мембраны,
50 Действие различных концентраций дигитЪнина в изотоничных растворах сахарозы на солюбилизацию лизосомал ных ферментов представлено на фиг,1 До концентрации дигитонина 0,1 мМ
55 (включительно) заметного освобождения ферментов лизосом не происходит (фиг,1, график 1 - белок, 2- кисла фосфатаза, 3 - кислая РНКаза, 4 Содержание активности лизосомаль- ных ферментов в митохондриально-ли- зосомальной фракции (Mtni) приведено в табл.1.
Таблица 1
ты
Содержание активности, % от содержания в го- могенате печени
Относительная удельная активность
31,2t1,1
Относительная удельная активность лизосомальных ферментов находится в пределах от 1,93 для кислой фосфата- зы до 2,73 для катепсина D.
П р и м е р 2. Дигитонин растворяют в диметилсульфоксиде и разводят СРС до концентрации 4 мМ. Суспензию органелл митохондриально-лизосомаль- ной фракции смешивают в соотношении 1:1 с раствором дигитонина 0,8 мМ и вьщерживают во льду в течение 30 мин. Затем смесь центрифугируют (18000 g, 15 мин). Надосадок, представляющий
прозрачную желтоватую жидкость, ис- пользуют в дальнейшем как экстракт
прозрачную желтоватую жидкость, ис- пользуют в дальнейшем как экстракт
лизосом. Установлено, что растворитель диметилсульфоксид не вызывает повреждения лизосомальиой мембраны,
50 Действие различных концентраций дигитЪнина в изотоничных растворах сахарозы на солюбилизацию лизосомальных ферментов представлено на фиг,1. До концентрации дигитонина 0,1 мМ
55 (включительно) заметного освобождения ферментов лизосом не происходит (фиг,1, график 1 - белок, 2- кислая фосфатаза, 3 - кислая РНКаза, 4 - -галактоэидлза, 5 - катепсины В, L Н, 6 - катепсин D). В интервале 0,2- 0,4 мМ лигитонина наблюдается резкий прирост солюбилизируемой активности ферментов, заканчивающийся выходом на плато. Последующее увеличение концентрации дигитонина до 1,6 мМ слабо отражается на приросте активности ферментов, хотя в интервале 1,6-2,0 мМ отмечено разнонаправленное действие дигитонина на активност кислой фосфатазы и цистеиновых катеп синов (катепсинов В, L, Н).
Степень солюбилизации отдельных ферментов различна. При 0,4 мМ дигитонина полному освобождению из лизо- сом подвергаются активность /3-галак- озидазы и активность цистеиновых катепсинов. В этих же условиях кислая i
14
- -.-«ъ f ffftf л.-.хл. q 71
фосфатаза, кислая РНКаза и катепсин D солюбилизируются на 53-60%,
Выход активности лизосомальных ферментов (от их содержания в гомо- генате печени) при концентрации дигитонина 0,4 мМ от 32% для кислой РНКазы и 38% для кислой фосфатазы до 72% для /3-галактозидазы и 81% для цистеиновых катепсинов. При дальнейшем увеличении концентрации дигитони- на заметньй прирост вьпсода активности наблюдается только для кислой фосфатазы.
Освобождение белка органелл развивается медленно до концентрации
Концентрация дигитонина, мМ
0,2
0,4. 0,8
101,5.4,4 102,1.1,4 105,5.1,5
Формула изобретения
Способ экстракции лизосомальных ферментов, включающий вьщеление фракции лизосом из исходного сьфья методом дифференциального центрифугирования с последующим разрушением мембран лизосом и получением фракции матрикса органелл с использованием экстрагирующего агента, о т л и ч аL, 2- ий и ть п-
1449580
дигитонина 0,8 мМ (фиг.1, график 1)о и далее заметно возрастает,
В соответствии с данными по солю- g билизации ферментов относительная удельная активность (ОУА) ферментов лизосом резко возрастает при 0,2 мМ дигитонина и достигает максимальных значений при 0,4 мМ (фиг.2, график 10 1 - кислая фосфагаза, 2 - кислая РНКаза, 3 - /3-галактозидаза, 4 - катепсины В, L, Н, 5 - катепсин D). Наибольшие значения ОУА получены для цистеиновых катепсинов (30,7) и И - 5 галактозидазы (24,9), а наименьшие - для частично мембраносвязанных ферментов - кислой РНКазы (12,1) и кислой фосфатазы (14,1). При концентрации дигитонина 0,Я мМ и вьше наблюдается
I
0 резкое снижение ОУА лизосомальных
ферментов экстракта органелл в результате заметного прироста солюбшш- зации белка, что особенно отчетливо отражается на ОУА /3-галактозидазы 5 и катепсинов.
Дигитонин не оказывает действ ия на активность лизосойальных ферментов.
I
Влияние дигитонина на активность лизосомальных ферментов митохондри- ально-лизосомальной фракции, обработанной 0,1%-ным тритоном Х-100(М±т) показано в табл.2.
Таблица2
50
ю щ и и с я тем, что, с целью упрощения процесса и увеличения выхода целевого продукта, в качестве исходного сырья используют грубоочищенньй
55 препарат митохондриально-лизрсомаль- ной субклеточной фракции, гомогената печени, а в качестве экстрагирующего агента применяют дигитонин с концен. трацией 0,3-0,4 мМ.
0
ю о
N
iqo
30
|j ;|i
I
20
$0
(W
W
|2 0
6
1,0 2,00 1.0 2P I HuLQumoHUH (мМ) Лиеитоиан (нН)
I § 16 /
I q
0
0 1,0 2A
,ан(,) , (м н)
Фие.1
0 1,0 2A
, (м н)
Фие.1
