Изобретение относится к биологИи размножения человека и животных, точнее - к эндокринологии размножения, а также кле-- точной биологии и может быть использовано для получения клеток, являющихся удобной экспериментальной моделью для изучения молекулярно-клеточных механизмов действия гормонов и факторов роста (ФР) в процессах регуляции роста фолликулов яичников.
Целью данного изобретения является создание способа получения культуры клеток гранулезы, обладающей большей однородностью и большей чувствительностью к действию ФР по сравнению с прототипом.
Поставленная цель достигается отбором для культивирования малодифференцированных клеток яйценосного бугорка и преантральной гранулезы путем одноразового прокола фолликула, а также выбором в качестве источника материала неполовозрелых лабораторных животных чистых линий.
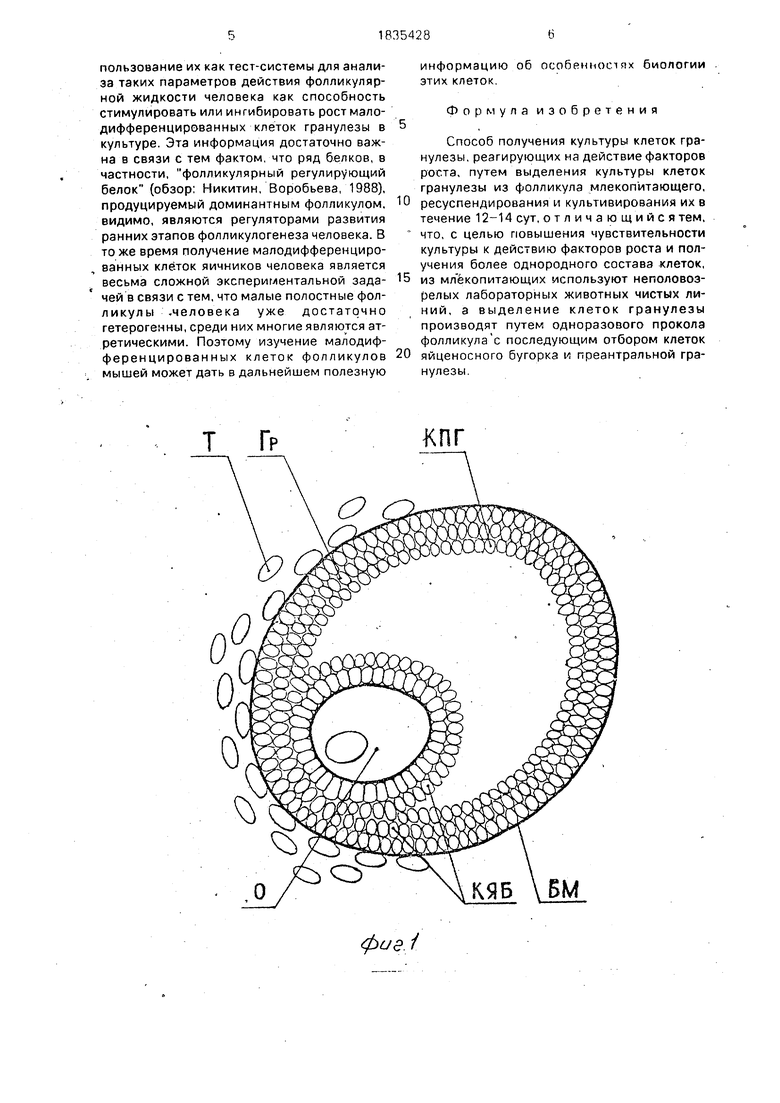
Предлагаемый способ иллюстрируется сопровождающими заявку рисунками. На рис. 1. представлена схема строения фолликула яичника. Принятые сокращения: Т. клетки теки; Гр. - клетки гранулезы; Б.М. - базальная мембрана; О. - ооцит; КЯБ. - клетки яйценосного бугорка; ПГ. - клетки преантральной гранулезы.
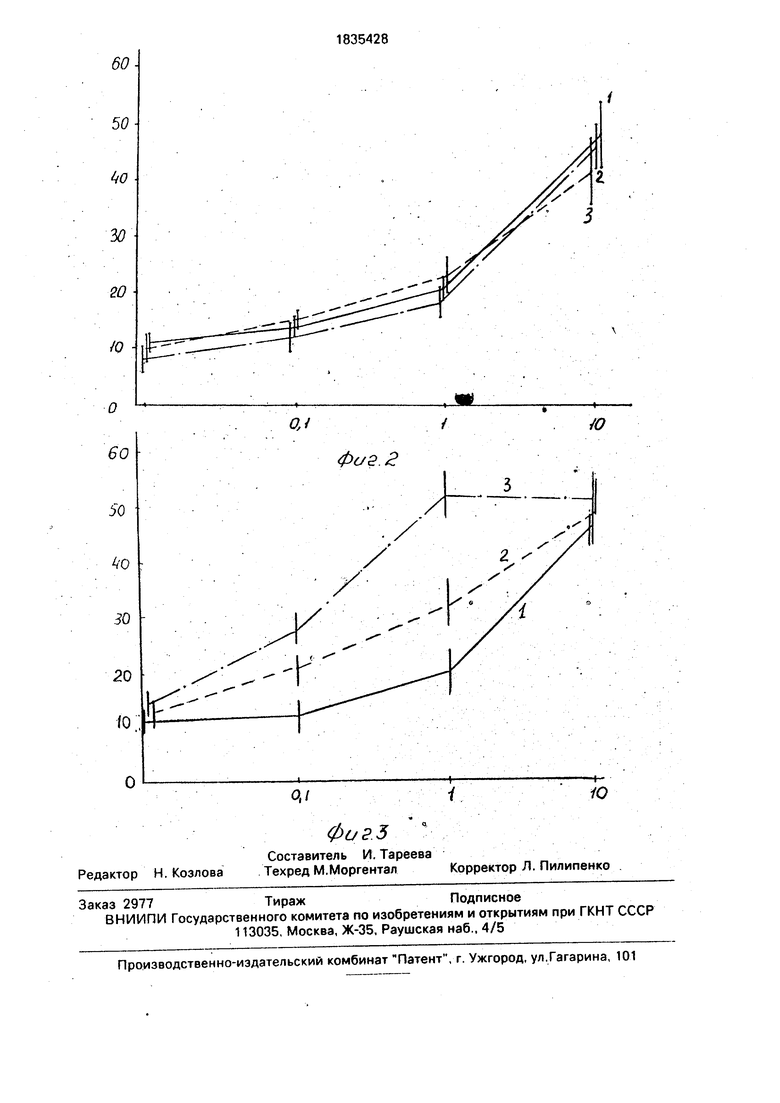
На рис. 2 представлены данные по влиянию ЭФР на эффективность клонообразо- вания культуры малодифференцированных клеток гранулезы, выделенных предложенным нами способом.
На рис. 3 представлены данные по влиянию ЭФР на эффективность клонообразо- вания культуры клеток гранулезы мышей, выделенных по способу, предложенному в прототипе. По оси абсцисс на этих графиках отмечена концентрация сыворотки в среде, По оси ординат - число выросших клонов.
(Л
С
со
СА) СЛ
4 Ю 00
Вертикальные отрезки на графиках - 95%- ные доверительные интервалы. На графиках кривая 1 соответствует эффективности кло- нообразования клеток в контроле. Кривая:2 - эффективность клонообразования клеток в среде, содержащей ЭФР в дозе 1 нг/мл; Кривая 3 - эффективность клонообразования клеток в среде, содержащей ЭФР в дозе 10 нг/мл.
Как видно из приведенных результатов . клетки гранулезы, выделенные предложенным нами способом, реагируют на ЭФР в среде с низким содержанием сыворотки стимуляцией клонообразования. Клетки, выделенные по способу, описанному в прототипе, не реагируют на ЭФР стимуляцией роста.
Предлагаемый способ осуществляется следующим образом. Клетки гранулезы получали из яичников неполовозрелых мышей гибридов FI CBA/C57 В (возраст мышей 20- 25 сут.). Яичники в стерильных условиях выделяли в капли питательной среды на чашки Петри, очищали от окружающих тканей и переносили в свежие капли среды. Тонкими вольфрамовыми иглами прокалывали большие антральные фолликулы и микропипеткой отбирали ооциты. окруженные клетками яйценосного бугорка, и небольшое количество клеток гранупезы, выделяющихся из фолликула в виде шлейфа за ооцитом, это, видимо, клетки преантральной области фолликулов. Полученные таким образом клетки высевали в клональной плотности (1000 клеток на 3 см чашку Петри) и культивировали в среде с различным содержанием сыворотки (0, 0,1 и 10%).и ЭФР (1 и 10 нг/мл). На 12-14 день культивирования клетки фиксировали непосредственно в чашках и анализировали эффективность клонообразования клеток.
Как видно из данных, представленных, на рис. 2, клетки, полученные описанным способом, реагируют стимуляцией роста на введение в среду культивирования ЭФР . Клетки, выделенные по способу, предложенному в работе Господаровича и Билецки (Gospodarowicz, Bialecki, 1979), а именно с помощью разрыва стенки фолликула, выделения его содержимого в среду, обработки ферментами и последующего клонирования не реагируют на наличие в среде культивирования ЭФР, как это видно из приведен- . ных данных (рис. 3).. В среде с низким содержанием сыворотки (0,1 и 1 %) чувствительность клеток к действию ЭФР была наибольшей. Так, число клонов, содержащих более 10 клеток1, увеличивалось в 4 раза, что соответствовало 10-кратному приросту числа клеток.о
При получении клеток по методу Госпа- даровича и Билецки (Gospodarowicz, Blaleckl, 1979) выделяется и значительное количество лютеинизирующихся клеток, которые не реагируют на ЭФР. Так, нами показано, что лютеинизирующиеся клетки фолликулов мышей, которые были получены через 56 час после введения животным сывороточного гонадотропина (СЖК), также не
реагируют на ЭФР в среде. При выделении клеток по методу прототипа велика вероятность загрязнения культуры клетками теки. В то же время было показано, что введение клеток теки в -культуру клеток гранулезы
5 приводит к утрате последними способности реагировать на введение в среду ЭФР.
Следует отметить, что примененные нами концентрации ЭФР (1 и 10 нг/мл) значительно ниже чем е прототипе (20 нг/мл) и
0 более близки к физиологическим. Максимальное стимулирующее влияние ФР на пролиферацию клеток гранулезы проявляется в среде с низким содержанием сыворотки. По-видимому, при высокой
5 концентрации сыворотки все рецепторы клеток оккупируются факторами роста, содержащимися в сыворотке, ли же в этом случае проявляется влияние компонентов сыворотки, ингибирующих действие ЭФР
0 (гонадотропинов, трансформирующего фактора роста типа /3-/ ТФР и др.). Так, недавно показано, что р ТФР в среде с высоким содержанием сыворотки препятствует стимуляции роста клеток гранулезы свиней под
5 влиянием ЭФР, а в среде с 0,1 и 0,25% сыворотки усиливает влияние ЭФР (обзор: Воробьева, 1989).
Знание механизмов регуляции пролиферации клеток фолликула имеет первостепен0 ное значение для биологии репродукции. В настоящее время вопрос о факторах, регулирующих рост клеток гранулезы в организме животных, пересматривается. Общепринято, что ФСГ, ЛГ и эстрадиол являются регулято5 рами роста фолликулов в яичнике. Однако, показано, что высокоочищенные препараты ЛГ и ФСГ не индуцируют рост клеток гранулезы в культуре. Более того, ФСГ, ЛГ и эстрадиол ингибируют реинициацию
0
репликации и деление клеток в этих условиях (обзор: Воробьева, 1989). Таким образом, создание клеточных моделей, позволяющих изучать вопросы регуляции роста .клеток гранулезы достаточно актуально. 5 из вышеизложенного следует, что мало- дифференцированные клетки гранулезы яичников мышей обладают такими же особенностями реагирования на ЭФР, как и клетки человека, что делает возможным использование их как тест-системы для анализа таких параметров действия фолликулярной жидкости человека как способность стимулировать или ингибировать рост малодифференцированных клеток гранулезы в культуре. Эта информация достаточно важна в связи с тем фактом, что ряд белков, в частности, фолликулярный регулирующий белок (обзор: Никитин, Воробьева, 1988), продуцируемый доминантным фолликулом. видимо, являются регуляторами развития ранних этапов фолликулогенеза человека. В то же время получение малодифференцированных клеток яичников человека является весьма сложной экспериментальной задачей в связи с тем, что малые полостные фол- ликулы .человека уже достаточно гетерогенны, среди них многие являются ат- ретическими. Поэтому изучение малодифференцированных клеток фолликулов мышей может дать в дальнейшем полезную
0
5
0
информацию об особенностях биологии этих клеток.
Формула изобретения
Способ получения культуры клеток гранулезы, реагирующих на действие факторов роста, путем выделения культуры клеток гранулезы из фолликула млекопитающего, ресуспендирования и культивирования их в течение 12-14 сут, отличающийся тем, что, с целью повышения чувствительности культуры к действию факторов роста и получения более однородного состава клеток, из мл ёкопитающих используют неполовозрелых лабораторных животных чистых линий, а выделение клеток гранулезы производят путем одноразового прокола фолликула с последующим отбором клеток яйценосного бугорка и преантральной гранулезы.

| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОТБОРА ПАЦИЕНТОК С СИНДРОМОМ "СЛАБОГО" ОТВЕТА ЯИЧНИКОВ ДЛЯ ПРОВЕДЕНИЯ ПРОГРАММЫ "ДОНАЦИЯ ООЦИТОВ" | 2014 |
|
RU2565449C1 |
| Белково-пептидный комплекс, повышающий жизнеспособность фолликулов в яичниках млекопитающих | 2017 |
|
RU2660587C1 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ IN VITRO ОВАРИАЛЬНЫХ ФОЛЛИКУЛОВ | 2001 |
|
RU2286384C2 |
| НУКЛЕОТИДНЫЕ И АМИНОКИСЛОТНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ ФАКТОРОВ ЯЙЦЕКЛЕТОК ДЛЯ ИЗМЕНЕНИЯ РОСТА ЯИЧНИКОВЫХ ФОЛЛИКУЛОВ IN VIVO ИЛИ IN VITRO | 2001 |
|
RU2270862C2 |
| СПОСОБ ЭКСТРАКОРПОРАЛЬНОГО КУЛЬТИВИРОВАНИЯ ООЦИТОВ КОРОВ | 2015 |
|
RU2602448C1 |
| ПЛАЗМИДА ДЛЯ ЭКСПРЕССИИ В КЛЕТКАХ СНО РЕКОМБИНАНТНОГО ФОЛЛИКУЛОСТИМУЛИРУЮЩЕГО ГОРМОНА (ФСГ) ЧЕЛОВЕКА, ПЛАЗМИДА ДЛЯ ЭКСПРЕССИИ В КЛЕТКАХ СНО БЕТА-СУБЪЕДИНИЦЫ РЕКОМБИНАНТНОГО ФСГ ЧЕЛОВЕКА, КЛЕТКА СНО - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ФСГ ЧЕЛОВЕКА И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО ГОРМОНА | 2013 |
|
RU2560596C2 |
| РЕЦЕПТОР ФОЛЛИКУЛОСТИМУЛИРУЮЩЕГО ГОРМОНА (FSH) ЧЕЛОВЕКА, ДНК, ВЕКТОР, СПОСОБ ПОЛУЧЕНИЯ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 1992 |
|
RU2193599C2 |
| Средство для лечения женского бесплодия и бесплодия самок животных | 2017 |
|
RU2740922C2 |
| Питательная среда для культивирования ооцитов крупного рогатого скота in vitro | 2021 |
|
RU2778147C1 |
| СПОСОБ ЭКСТРАКОРПОРАЛЬНОГО КУЛЬТИВИРОВАНИЯ ООЦИТОВ КОРОВ | 2009 |
|
RU2410063C1 |
Использование: биология размножения человека и животных, точнее - эндокринология размножения, а также клеточная биология для получения клеток, являющихся удобной экспериментальной моделью для изучения молекулярно-клеточных механизмов действия гормонов и факторов роста в процессах регуляции роста фолликулов яичников. Сущность изобретения: для культивирования отбирают малодифференцированные клетки яйценосного бугорка и преантральной гранулезы путем одноразового прокола фолликула, а также в качестве источника материала выбирают неполовозрелых лабораторных животных чистых линий.
Т ГР
кпг
фие. 1
1835428
| Charming С.P., Ledwitz-Rlgby | |||
| Methods for assessing hormone mediated differentiation of ovarin cells in culture and in shortterm incubation // Methods Enzymol | |||
| Машина для изготовления проволочных гвоздей | 1922 |
|
SU39A1 |
