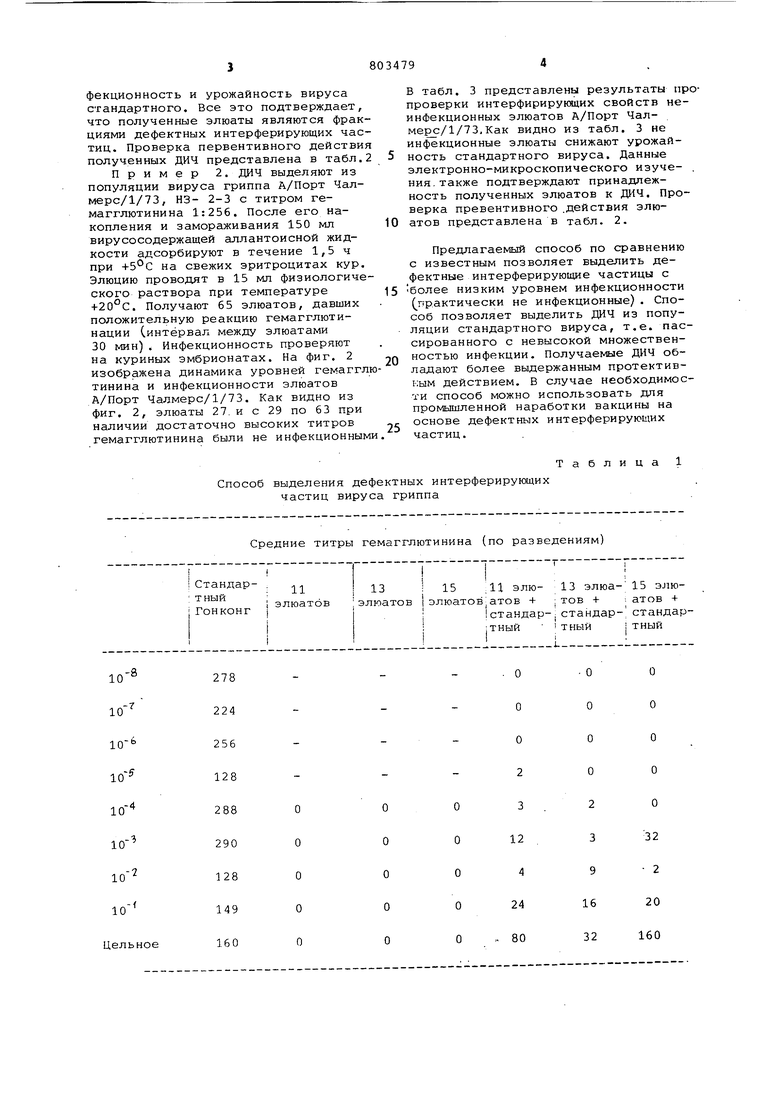
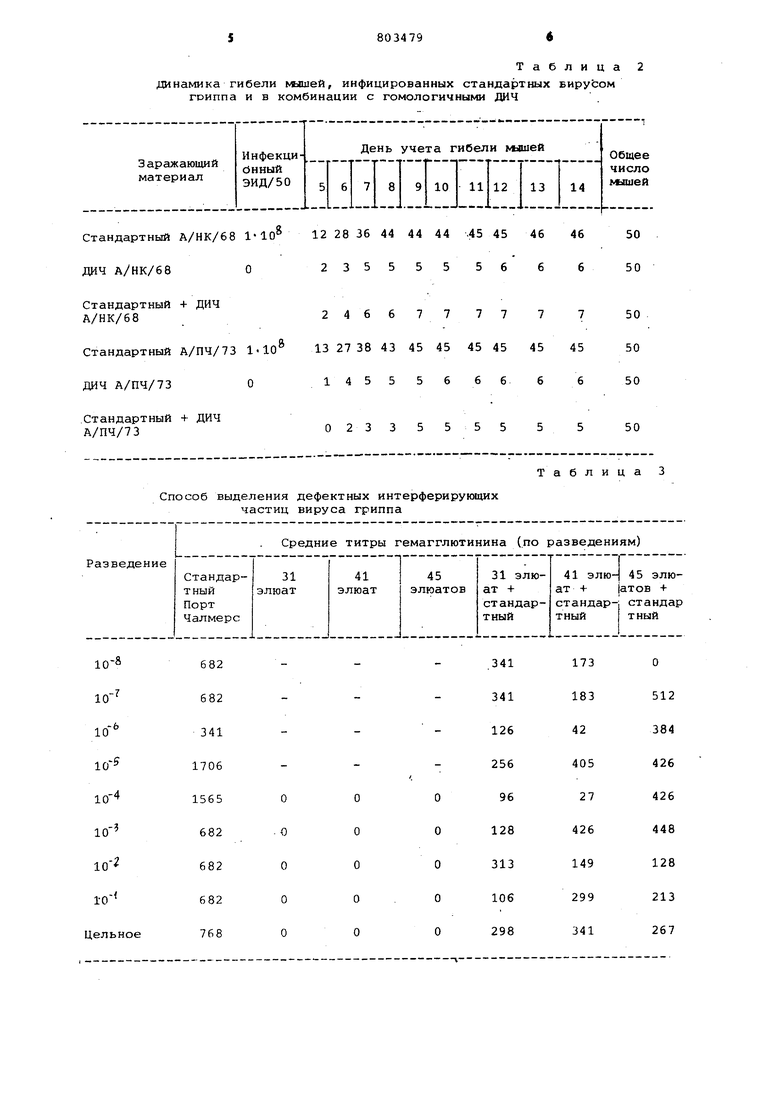
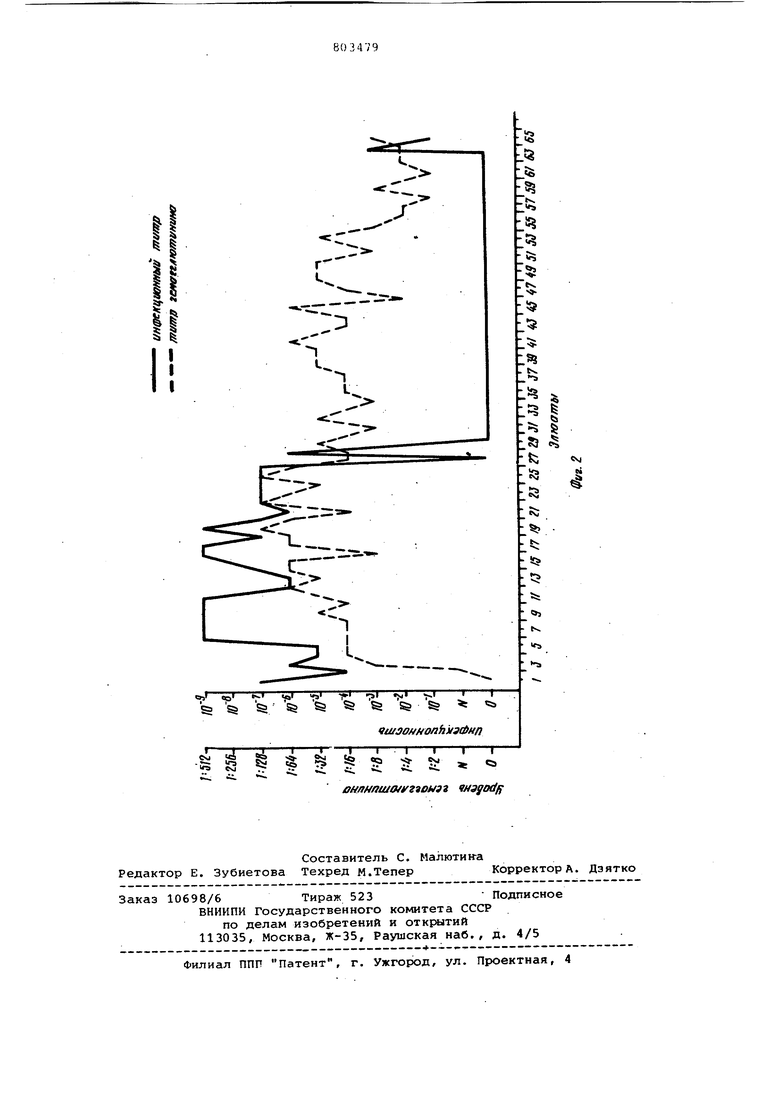
- Изобретение относится к области вирусологии и может быть использовано для разработки вакцин на основе дефектных интерферирующих частиц а также для различных вирусологических исследований с их применением Известен способ получения .дефект ных интерферирующих частиц вируса гриппа путем заражения вирусом грип па куриных эмбрионов с последующим отбором вирусосодержащей аллантоисной Жидкости и выделением целевого продукта l , Однако известный способ не позволяет полностью освободиться от примеси инфекционного вируса, получаемый целевой продукт содержит большое количество аллантоисных бел ков. Целью изобретения является вьщеление целевого продукта свободного от примесей инфекционного вируса и аллантоисных белков. Эта цель достигается тем, что СПОСО.6 получения дефектных интерферирующих частиц вируса гриппа осуществляют путем заражения вирусом гриппа .куриных эмбрионов с последующим отбором вирусосодержащей аллан тоисной жидкости и выделением.целевого продукта, при этом вирусосодержащую жидкость повергают замораживанию - оттаиванию и центрифу гированию, затем полученную надосадочную жидкость адсорбируют на куриные эритроциты и элюируют целевой продукт охлажденным физиологическим раствором при соотношении объемов физиологического раствора и эритроцитов 1:1,5-0,5. 1 Способ осуществляют следующим об разом. Получение вируса. Накопление вируса гриппа проводя на куриных эмбрионах. Вирус с титром гемагглютинина от 1:64 до 1:512 раститровывают до предельных разведений, после чего заражают куриные эмбрионы. Инкубация куриных эмбрионов проходит при температуре+35,5 в течение 72 ч, накопленный вирус выдерживают при температуре -20 С в течение 3-10 сут. После размораживания в пул берут материалы с положительной реакцией гемагглютинации. Пул центрифугируют при 2,5 тыс. об/мин для удаления белков и других примесей. Сбор фракций вируса. Фракционирование проводят на све жих или формалинизированных эритроцитах кур, предварительно отмытых стерильным физиологическим раствором. Вирус адсорбируют на эритроцитах в течение 1-1,5 ч при температу (+)4 -(, Соотношение вирусосодержащей жидкости и эритроцитов lU:i. После адсорбции смесь центрифугируют при 1,5 -тыс,об/мин в течение 5 мин. Снимают надосадочную жидкость. Для промывания к эритроцитам добавляют охлажденный физиологический раствор ( + ) в соотношении 2:1, эритроциты размешивают пастеровской пипеткой и центрифуги- . руют при 1,5 тыс.об/мин в течение 5 мин. . После снятия промывной жидкости к эритроцитам добавляют охлажденный физиологический раствор в соотношении 1:1 эритроциты размешивают пастеровской пипеткой и оставляют для элюции при комнатной температуре. Через 1 ч проводят центрифугирование (1,5 тыс.об/мин 5 мин) и отсасывание пастеровской пипеткой элюата. К эритроцитам доливают новую порцию охлажденного .физиологического раствора, Элюаты снимают с интервалом в 30 или 60 мин до значительного снижения их титров гемагглютинина (0-1:4, Для предотвращения прироста в используемый физиологический раствор и в пробирки для элюатов добавляют смесь антибиотиков. Проверка элюатов, У полученных элюатов кроме гемагглютинирующей активности проверяют инфекционность на куриных эмбрионах или на хорионаллантоиснык оболочках куриного эмбриона (хло). Инкубацию куриных эмбрионов и ХАО проводят при температуре 35,5°С в течение 48 ч. Пример 1, Для выделения дефектных интерферирующих частиц (дач) берут вирус А/Гонконг/1/68, НЗ 2-2 с титром гемагглютинина 1:128, После его накопления на куриных эмбрионах и замораживания 100 мл вирусосодержащей аллантоисной жидкости адсорбируют в течение 1 ч на свежих эритроцитах кур, Элюцию проводят в 10 мл физиологического, раствора при температуре +21°С. Получают 21 элюат, дающие положительную реакцию гемагглютинации. Инфекционность элюатов проверяют на куриных эмбрионах. На фиг. 1 изображена динамика уровней гемагглюуина- и инфекционности элюатов А/Гонконг/1/68, Как видно из фиг.1 элюаты с 11 по 18 при наличии высоких титров гемагглютинина оказались не инфекционными. При электронномикроскопическом исследовании элюатов выявлено различие в строении вирусных частиц из неинфекционных элюатов и высокоинфекционных. Для вирусных частиц из неинфекционных элюатов характерно отсутствие части генома. В табл, 1 представлены результаты проверки интерферирующих свойств элюатов. Как видно из табл.1 элюаты, содержащие ДИЧ, снижают ийфекционность и урожайность вируса стандартного. Все это подтверждает что полученные элюаты являются фрак циями дефектных интерферирующих час тиц. Проверка первентивного действи полученных ДИЧ представлена в табл Пример 2. ДИЧ выделяют из популяции вируса гриппа А/Порт Чалмерс/1/73, НЗ- 2-3 с титром гемагглютинина 1:256. После его накопления и замораживания 150 мл вирусосодержащей аллантоисной жидкости адсорбируют в течение 1,5 ч при на свежих эритроцитах кур Элюцию проводят в 15 мл физиологиче ского раствора при температуре +20°С. Получают 65 элюатов, давших положительную реакцию гемагглютинации Синтёрвал между элюатами 30 мин). Инфекционность проверяют на куриных эмбрионатах. На фиг. 2 изображена динамика уровней гемагг тинина и инфекционности элюатов А/Порт Чалмерс/1/73. Как видно из фиг. 2, элюаты 27. и с 29 по 63 при наличии достаточно высоких титров гемагглютинина были не инфекционны
Способ выделения дефектных интерферирующих частиц вируса гриппа
Таблица в табл. 3 представлены результаты пропроверки интерфирирующих свойств неинфекционных элюатов А/Порт Чалмерс/1/73,Как видно из табл. 3 не инфекционные элюаты снижают урожайность стандартного вируса. Данные электронно-микроскопического изучения, также подтверждают принадлежность полученных элюатов к ДИЧ. Проверка превентивного .действия элюатов представлена в табл. 2. Предлагаемый способ по сравнению с известным позволяет выделить дефектные интерферирующие частицы с более низким уровнем инфекционности (практически не инфекционные) . Способ позволяет выделить ДИЧ из популяции стандартного вируса, т.е. пассированного с невысокой множественностью инфекции. Получаемые ДИЧ обладают более выдержанным протективкым действием. В случае необходимости способ можно использовать для промышленной наработки вакцины на основе дефектных интерферирующих частиц.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ определения эпидемичности вируса гриппа | 1980 |
|
SU875847A1 |
| СПОСОБ ПРОИЗВОДСТВА ВАКЦИНЫ ПРОТИВ ГРИППА | 2009 |
|
RU2423995C1 |
| Способ культивирования вируса гриппа | 1988 |
|
SU1564185A1 |
| СПОСОБ ПОЛУЧЕНИЯ ЖИВОЙ ГРИППОЗНОЙ ВАКЦИНЫ | 1995 |
|
RU2080124C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЖИВОЙ ГРИППОЗНОЙ ВАКЦИНЫ | 1991 |
|
RU2033182C1 |
| Способ получения противогриппозных антител | 1985 |
|
SU1301409A1 |
| СПОСОБ ПОЛУЧЕНИЯ ТЕТРАВАЛЕНТНОЙ СУБЪЕДИНИЧНОЙ ПРОТИВОГРИППОЗНОЙ ВАКЦИНЫ | 2019 |
|
RU2740751C1 |
| Штамм вируса гриппа А/Киев/3304/84, используемый для приготовления гриппозного диагностикума | 1987 |
|
SU1463758A1 |
| СПОСОБ ПРОФИЛАКТИКИ БОЛЕЗНИ НЬЮКАСЛА | 2008 |
|
RU2403060C2 |
| Штамм В (Ленинград/104/84)20 вируса гриппа, используемый для приготовления инактивированной гриппозной вакцины | 1986 |
|
SU1389288A1 |
СПОСОБ ПОЛУЧЕНИЯ ДЕФЕКТНЫХ ИНТЕРФЕРИРУЮЩИХ ЧАСТИЦ ВИРУСА ГРИППА путем заражения вирусом гриппа куриных эмбрионов с последующим отбором вирусосодержащей аллантоисной жидкости и выделением целевого продукта , отличающийся тем, что, с целью выделения целевого продукта, свободного от примесей инфекционного вируса и аллантоисных белков, вирусосодержащую жидкость. подвергают замораживанию-оттаиванию и центрифугированию, затем пот лученную надосадочную жидкость адсорбируют на куриные эритроциты и элюируют целевой продукт охлажденным физиологическим раствором при' Соотношении объемов физиологического раствора и эритроцитов 1:1,5 - 0,5.Ф(Лсuufcfitaofftfv ттщг аптр •еятлюягнан»I г 3 1, S 6^ В S vnijijufsisnteiafotiЭяюоты Лп.'00Оее N^ ^ со
Средние титры гемагглютинина (по разведениям) Стандар- . : il3 :15 Т НЫИ. I ; 1 элюатов ; элюатов j элюат л-ОНКОНГ)i
-8
10
278
1022410 256
,- 128
10
г 4
10 288
-3
10 290
-2 128
10
10- 149 160
ельное I
. О
О
. О
О О О О
О О 2 3
О О О 2 3 9
32
12 4
- 2
20
16 32
24 ВО
160 ;Ц ЭЛЮ- 13 элюа- 15 элю. ,атов + ; тов + ; атов + I- 1 стандар-Iстандар- стандарj ТНЫЙ I ТНЫЙ 1ТНЫЙ -.1
динамика гибели мышей, инфицированных стандартных вирусом гриппа и в комбинации с гомологичными ДИЧ 1228 36 44 44 Стандартный А/НК/68 110° 23555 ДИЧ А/НК/68 Стандартный + ДИЧ 24667 А/НК/68 1327 38 43 45 Стандартный А/ПЧ/73 1-10 14555 ДИЧ А/ПЧ/73 О Стандартный + ДИЧ 02335 А/ПЧ/73
Способ выделения дефектных интерферирующих частиц вируса гриппа
Таблица 2
Таблица 3 44 .45 4546 4650 5566 650 7777 750 45 45 4545 4550 6666 650 5555550
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Вишниченко В.К | |||
| и Петере В.В | |||
| Плотностной анализ стандартных и дефектных вирионов, "Вопросы вирусологии", 1974, № 6, с | |||
| Колосниковая решетка с охлажденными водой колосниками | 1925 |
|
SU688A1 |
