Изобретение относится к медицине, а именно к лепрологии, и может быть использовано для оптимизации существующих моделей лепры на животных.
Многие исследования по изучению иммунитета и, в частности, в лепрологии выполняются на животных с искусственно созданными дефектами иммунной системы. Из практики медицины известен ряд способов снижения иммунитета и, в частности, снижения функциональной способности клеток системы мононуклеарных фагоцитов (СМФ).
Известен способ снижения иммунитета подопытных животных (мыши) путем облучения их рентгеновскими лучами в дозе 900 рентген и тимэктомии (Rees R. J.W., Path F.C., Recent bacteriologic, immunologic and pathologic studies on experimental human leprosy in the mous foot pad. Int. J. Leprosy. 1965. Vol. 33, N 3. P.646-652.). При этом авторы получали большой прирост биомассы в подушечках лап мышей, зараженных микобактериями лепры или микобактериями туберкулеза по методу Шепарда (Shepard C.C. The experimental disease that follows the injection of human leprosy bacilli into foot-pads of mice. J. Exper. Med. 1960. Vol.112. N 3. P.445-454.). К недостаткам приведенного способа следует отнести трудоемкость исполнения, вязанного со сложными процедурами, и отсутствие избирательного воздействия на определенное звено иммунитета.
Известен способ искусственного снижения моноцитопоэза и тем самым создания недостаточности СМФ путем многократного дренирования брюшной полости мышей (Кравцов В.Д., Зорина Т.Д., Аркадьева Г.Е., Фрейдлин И.С. Новая модель недостаточности мононуклеарных фагоцитов. Иммунология. 1985. N 5. С.48-50). В брюшную полость животных при помощи шприца вводят 5 мл раствора Хенкса и после непродолжительного массажа брюшной полости жидкость отсасывают пастеровской пипеткой с диаметром капилляра 1,5-2 мм. Недостатками способа являются трудоемкость операций и большая вероятность нарушения стерильности.
Известен способ удлинения периода персистенции микобактерий лепры в клетках макрофагоподобной линии P.388D1 и, следовательно, снижения функциональной способности клеток путем ежедневного добавления в среду для культивирования клеток тетрапептида тафцина (1 мкг/мл) (Вишневецкий Р.Ф. Длительное персистирование Mycobacterium leprae в клетках макрофагоподобной линии P. 3888D1. Автореф. дисс. канд. мед. наук. Санкт-Петербург, 1993 г.). Длительное персистирование микобактерий лепры в клетках макрофагоподобной линии, находящихся в питательной среде, обогащенной тафцином, приводит к уменьшению содержания лизосом в клетках, и, по мнению автора, тафцин снижает продукцию лизосом. Недостатком данного способа является низкая воспроизводимость результатов. Тафцин известен как стимулятор фагоцитирующих клеток и обладает протективным эффектом при ряде инфекционных заболеваний, а по данным Iver R.R., Prasad H.K., Bhutani L.K., Rao D.N. Effect of tuftsin stimulation on the microbial activity exerted by blood monocytes-macrophages of leprosy patients. Int. J. Immunopharmac. 1990. Vol. 12. P. 859-869 макрофаги, полученные от здоровых лиц и больных погранично-туберкулоидной и туберкулоидной лепрой (ВТ, ТТ), демонстрируют устойчивое усиление фагоцитоза при стимуляции тафцином и медленное снижение микробицидного ответа.
Этот способ принят в качестве прототипа.
Целью предлагаемого способа является повышение воспроизводимости моделей.
Поставленная цель достигается тем, что перитонеальные макрофаги стимулированных животных (мыши) за 2 ч до заражения однократно насыщают 0,6%-ным раствором перекиси водорода. При этом достигается достоверное снижение содержания активной пероксидазы перитонеальных макрофагов (ПМ) и их функциональной активности (показатели поглотительной способности ПМ). На фоне снижения функциональной активности достигается более длительное по сравнению с контролем персистирование в ПМ патогенных и условно патогенных микобактерий (M. tuberculosis, лабораторного вирулентного штамма, H37Rv, M.tuberculosis, полученных непосредственно от больных туберкулезом, пассивированных штаммов M. leprae, полученных из подушечек лап зараженных мышей, а также условно патогенного штамма M.lufu). При осуществлении предлагемого способа требуется лишь однократное насыщение ПМ 0,6%-ным раствором перекиси водорода, что выгодно отличает предлагаемый способ от прототипа. При этом обеспечивается лучшая воспроизводимость моделей. Изложенная сущность способа поясняется в описании двумя таблицами и электронограммами.
С целью изучения иммунитета при лепре возникает необходимость искусственного дефекта различных звеньев иммунной системы. Известно, что при лепре клетки системы мононуклеарных фагоцитов (СМФ) являются основным резервуаром для выживания и размножения Mycobacterium leprae. Ранее нами (Маслов А.К., Ющенко А.А. Оценка функционального состояния лепрозных макрофагов. Арх. патол. 1988. N 11. С.51-54) было установлено, что различия в степени активности и в локализации пероксидазы лепрозных макрофагов гранулем больных лепрой коррелируют с их функциональным состоянием и завершенностью фагоцитоза M.leprae. Электронно-цитохимический анализ цитоархитектоники и активности пероксидазы (ПО) лепрозных макрофагов гранулем больных лепрой, длительно наблюдавшихся до и в процессе лечения (до 16 лет), показал, что при низкой активности ПО фагоцитов регресс заболевания протекает медленнее. Больные чаще подвержены риску развития рецидива заболевания. При высокой активности ПО фагоцитов регресс заболевания протекает быстрее и более выражен. Риск развития рецидива заболевания минимален (Маслов А.К., Ющенко А.А. Прогнозирование эффективности терапии лепры на основе данных электронной микроскопии. В кн.: Тез. докл. XIV конфер. по электронной микроскопии. М., 1992. C.60. Маслов А. К., Ющенко А.А. АС N 1636717).
Настоящие результаты эксперимента по искусственному снижению активности ПО в клетках СМФ и установлению связи между степенью фагоцитарных реакций и активностью фермента были выполнены на перитонеальных макрофагах 300 беспородных мышей.
Известно, что основной функцией ПО является связывание перекиси водорода, препятствуя ее накоплению в клетке (Гайер Г. Электронная гистохимия. 1974. М.: Мир, с. 336). Учитывая этот факт, мы решили снизить активность ПО в клетках СМФ, насыщая последние раствором перекиси водорода.
Мышей стимулировали введением внутрибрюшинно 2%-ного раствора пептон-казеина (фирма Serva) в количестве 5 мл. Одним из этапов разработки способа явился подбор концентрации перкиси водорода, снижающий активность фермента, но не оказывающий отрицательного воздействия на ультраструктуру клеток. Испытывались 0,3%, 0,6% и 1% растворы перекиси водорода, свежеприготовленные из 30%, которые вводились ровно через 2 ч после стимуляции внутрибрюшинно в количестве 2 мл. Через 2 ч после введения H2O2 мышам также внутрибрюшинно вводили разведенную до конечной концентрации 106 в 1 мл взвесь культур патогенных и условнопатогенных микобактерий (M.tuberculosis, H37 Rv, M.tuberculosis, выделенных от больных туберкулезом, M.lufu), а также очищенную от тканей суспензию M.leprae, пассируемых интраплантарно на мышах. Животным контрольной серии перекись водорода не вводилась. Мышей забивали через 2 ч, 1, 2, 3, 4, 5 и 6 сут после заражения.
Перитонеальные макрофаги получали по общепринятой методике (Использование культуры мышиных перитонеальных макрофагов в качестве модели для изучения клеток мононуклеарной фагоцитирующей системы организма и их изменение под влиянием биологически активных веществ: Методические рекомендации. Л., 1975) промыванием брюшной полости мышей средой 199 с добавлением гепарина (0,2 ед. на 1 мл среды). Экссудат инкубировали в течение 1 ч при 37oC на чашках Петри. Неприкрепленные клетки смывали 0,85% NaCl.
Функциональную активность ПМ оценивали как с помощью электронной микроскопии (определение цитоархитектоники и подсчет процента пероксидазоактивных митохондрий), так и теста определения поглотительной способности (Шатров В.А., Кузнецова Л.В., Беляновская Т.И. Оценка функционального состояния моноцитов периферической крови у больных туберкулезом легких. Ж. микробиол. 1985. N 5. С. 76-78). Для определения поглотительной способности ПМ учитывали фагоцитарный показатель (ФП), представляющий собой процент фагоцитирующих ПМ к общему числу сосчитанных клеток, и фагоцитарное число (ФЧ) - среднее количество микобактерий, поглощенных одной клеткой. Для подсчета этих показателей в чашку Петри перед инкубацией помещали тонкую стеклянную пластинку, изготовленную из покровного стекла, размерами 1 х 2 см. После инкубации пластинку с прикрепленными клетками извлекали, промывали, высушивали и окрашивали по методу Циля-Нильсена.
Для электронно-микроскопического исследования ПМ прикрепившиеся клетки снимали кисточкой со дна чашки с 2 мл физиологического раствора, добавляли 2 мл фиксирующего раствора (4%-ный раствор глютарового альдегида) и дальнейшую обработку материала с заключением в эпоксидные смолы проводили как обычно. Кроме того, для контроля снижения активности ПО проводили электронно-цитохимическое изучение клеток СМФ. Пероксидазу выявляли по методу (Graham R.C., Karnovsky M.J. The early stages of absorption of injected horseradish peroxidase in the proximal tubule of the mouse kidney. J.Hist, Cytoch. 1966, N 14. P. 291-302). Учитывали процент митохондрий с активностью ПО в 20 срезах ПМ (А.К. Маслов, А.А. Ющенко. АС N 1636717). Вычисление значимости различий проводили по методу углового преобразования Фишера. Достоверность различий вычисляли по таблице: Определение значимости различий "p" по известному аргументу нормального распределения UP(двусторонний критерий) (Гублер Е.В. Вычислительные методы анализа и распознавания патологических процессов. Л., 1978. С. 269).
Предлагаемый способ был успешно апробирован в НИИ по изучению лепры на протяжении 1991-1993 гг. Ниже приводятся результаты апробации, иллюстрируемые двумя таблицами и 20-ю электронограммами, отражающими морфологию, взаимоотношение клеток и их функциональную активность. Учитывая приблизительное сходство результатов в отношении различных видов микобактерий, все показатели приведены по отношению к шт. M.tuberculosis.
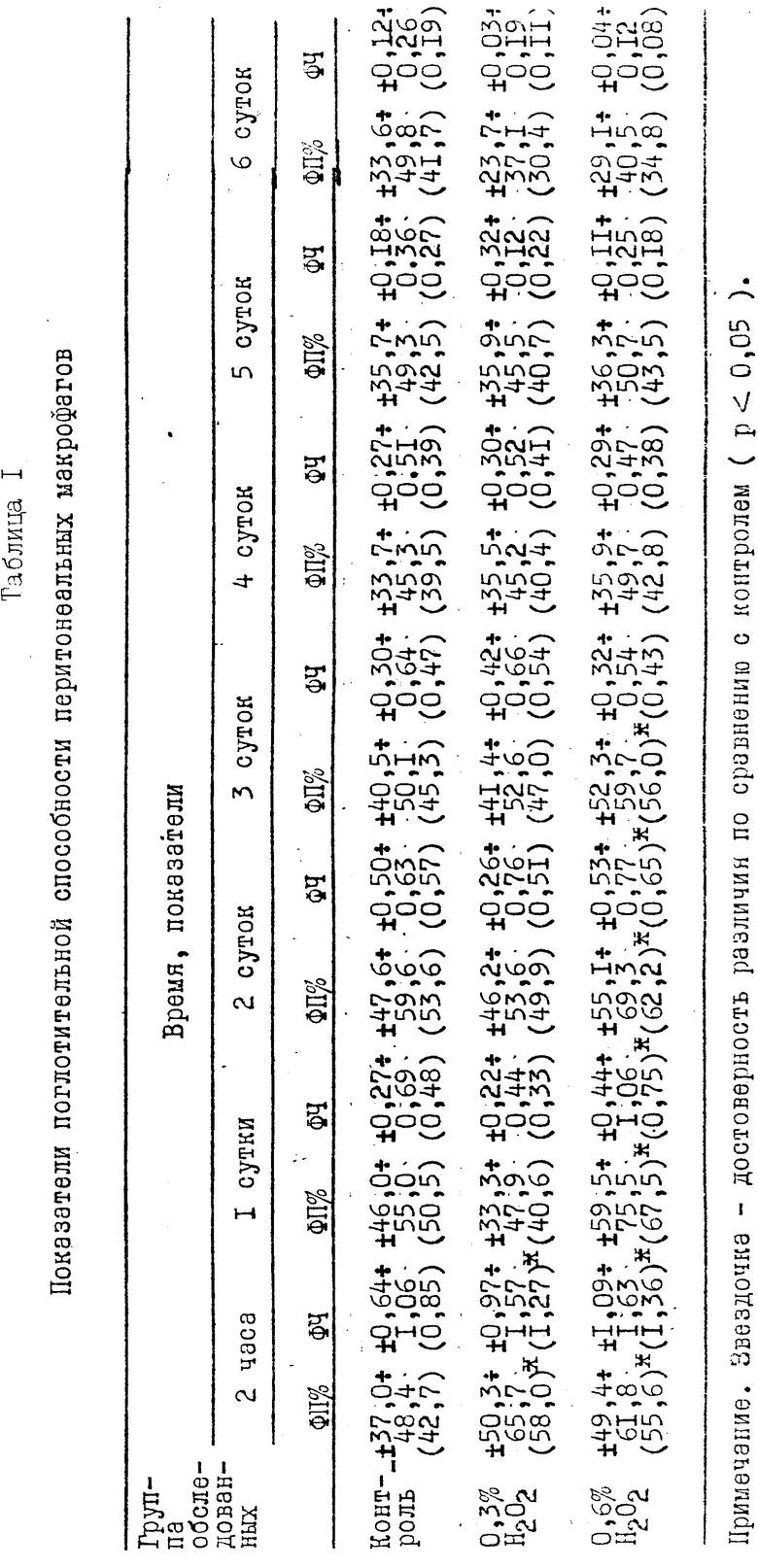
Результаты, отражающие показатели поглотительной способности ПМ по отношению к M.tuberculosis, представлены в табл. 1. Изучая поглотительную способность ПМ при воздействии концентрации H2O2 0,6%, мы выявили повышение показателей ФП и ФЧ по сравнению с контролем в течение первых суток после введения перекиси водорода. При концентрации H2O2 0,3%, напротив, в течение первых суток отмечалось снижение ФП и ФЧ по сравнению с контролем. В течение последующих трех суток показатели выравнивались и приближались к контрольным. В продолжение 4-6 сут со дня введения перекиси водорода показатели ФП и ФЧ в опыте снижались по сравнению с контролем. При концентрации H2O2 1% в течение первых суток после введения перекиси водорода отмечалось подавление поглотительной функции ПМ.
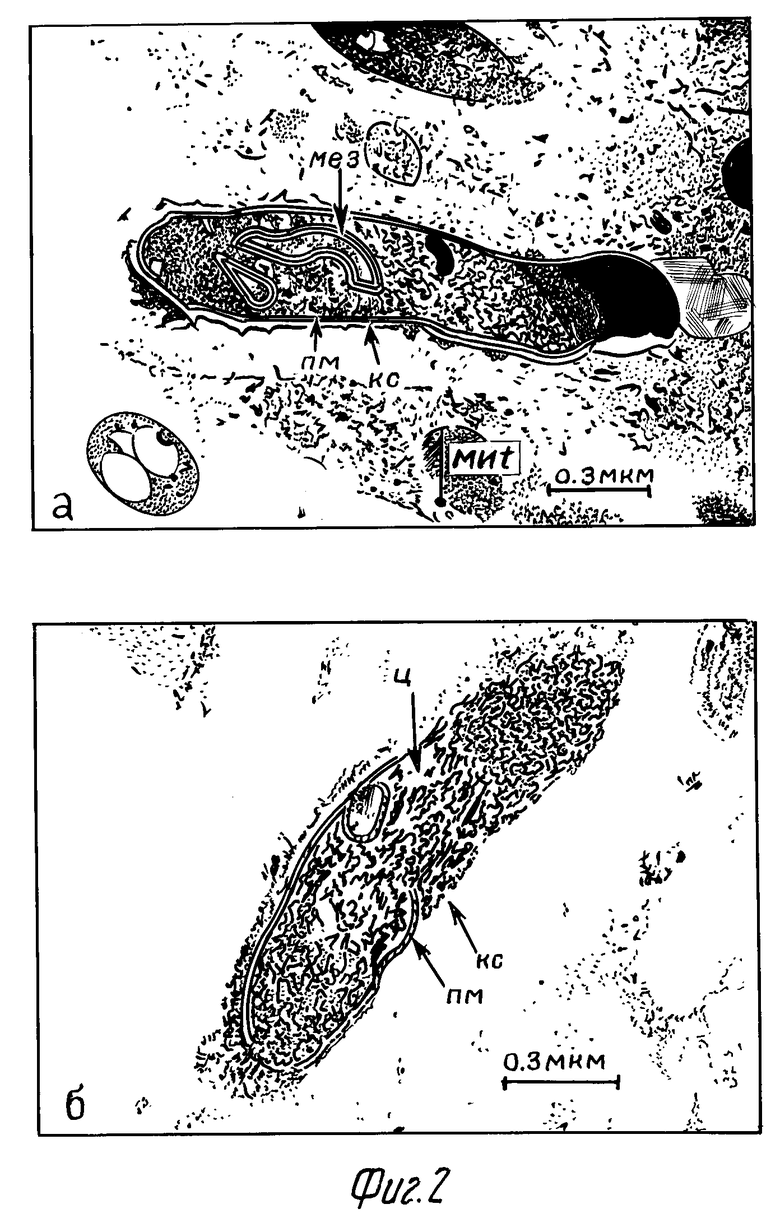
При электронно-микроскопическом изучении ПМ, предварительно обработанных 0,3% и 0,6% перекисью водорода и через 2 ч после заражения M.tuberculosis, было установлено, что большая часть микобактерий располагалась внутриклеточно. Через сутки после заражения ультраструктура микобактерий, расположенных в цитоплазме макрофагов опытной серии, в основном сохранялась: клеточная стенка микроорганизмов выражена, цитоплазма клеток без изменений, органоиды сохранены (фиг. 1а). В тоже время в контрольной серии ультраструктура части захваченных бактерий подвергалась некоторой деструкции: электронно-плотная часть клеточной стенки микроорганизмов размыта, слабо электронно-плотная. Цитоплазма разрежена. Наблюдалась деструкция внутрицитоплазматических органелл, конденсация нуклеотида. При этом ультраструктура плазматической мембраны практически не менялась (фиг.1б).
Через 2 сут в опытной серии структура большей части микобактериальных стенок практически не изменялась (фиг.2а). В контрольной группе структура преимущественного числа микобактерий изменялась значительно: у большинства клеток цитоплазма разрежена, иногда до полного просветления. Исчезают органоиды. Разрушению подвергалась плазматическая мембрана (фиг. 2б). Обнаружено много запустевших клеток.
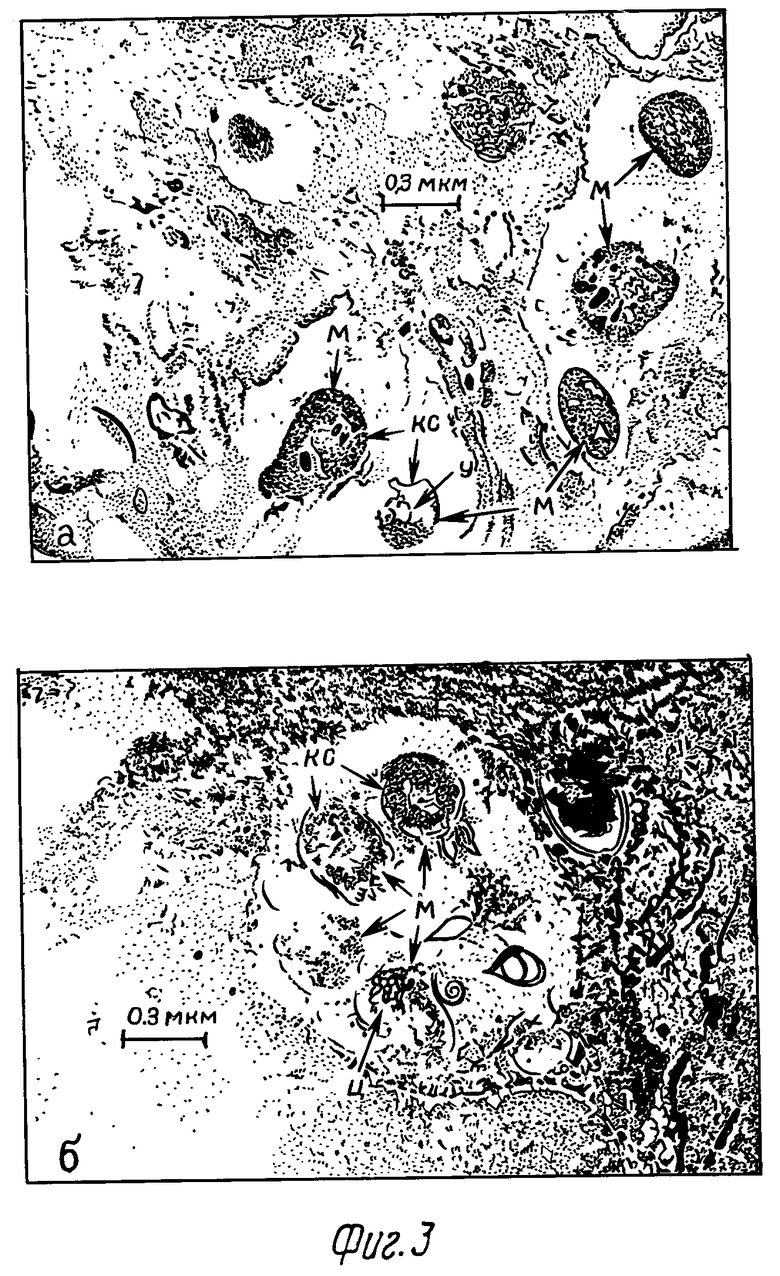
Через 3 сут ультраструктура микобактериальных клеток, расположенных в ПМ опытной серии, изменялась. Появились признаки деструкции клеточной стенки, разрежения цитоплазмы (фиг.3).
На этом этапе исследований нами обнаружены различия в ультраморфологии бактериальных клеток, находящихся в цитоплазме ПМ, подвергавшихся воздействию разных концентраций перекиси водорода. В случае первоначального воздействия на фагоциты 0,6% раствора H2O2 через 3 сут в их цитоплазме выявлялось большое количество микобактериальных клеток с сохраненной структурой органоидов (фиг. 3а). В случае воздействия 0,3% раствора перекиси водорода интактных микобактериальных клеток в этот период оказалось значительно меньше (фиг. 3б). Та же картина сохранялась на 4-е сутки персистенции микобактерий в ПМ мышей. В контрольной серии интактных клеток в этот период практически не обнаруживалось.
На 5-е сутки отмечались признаки деструкции большей части микобактерий в опытных сериях. Обнаружено много запустевших клеток (фиг. 4). Однако следует отметить, что большое количество относительно интактных клеток микобактерий наблюдалось в опыте с предварительным введением в брюшную полость мышей 0,6%-ного раствора H2O2. При воздействии этих 2-х концентраций перекиси водорода цитоархитектоника перитонеальных макрофагов мышей существенно не изменялась.
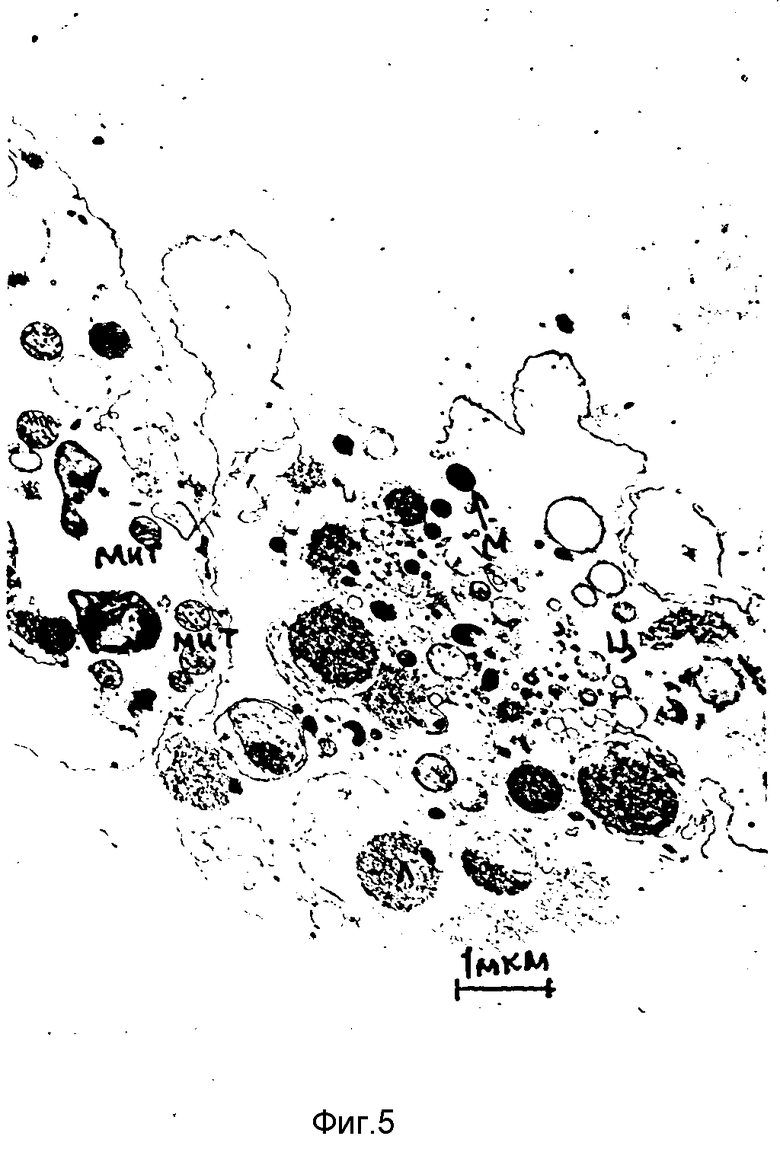
При введении в брюшную полость мышей 1%-ного раствора перекиси водорода в экссудате через 2 ч обнаруживалось большое количество ПМ, ультратонкое строение которых изменялось значительно. Цитоплазма ПМ просветлена. Наблюдалось разрушение органелл и мембранных структур ПМ. В цитоплазме немногочисленных ПМ размещались лишь единичные микобактериальные клетки (фиг. 5). Эти данные совпали с показателями ФП и ФЧ фагоцитов при воздействии H2O2 в данной концентрации.
Таким образом, предварительно вводимая в брюшную полость стимулированных животных перекись водорода в концентрации 0,6% обеспечивает начальное повышение поглотительной способности фагоцитов по сравнению с контролем и при этом не влияет на ультраструктуру фагоцитов. При воздействии перекиси водорода в этой концентрации наблюдалось персистирование микобактерий в стимулированных ПМ в течение 5-6 сут после заражения. В то же время в контроле за этот период времени микобактериальные клетки лизируются полностью.
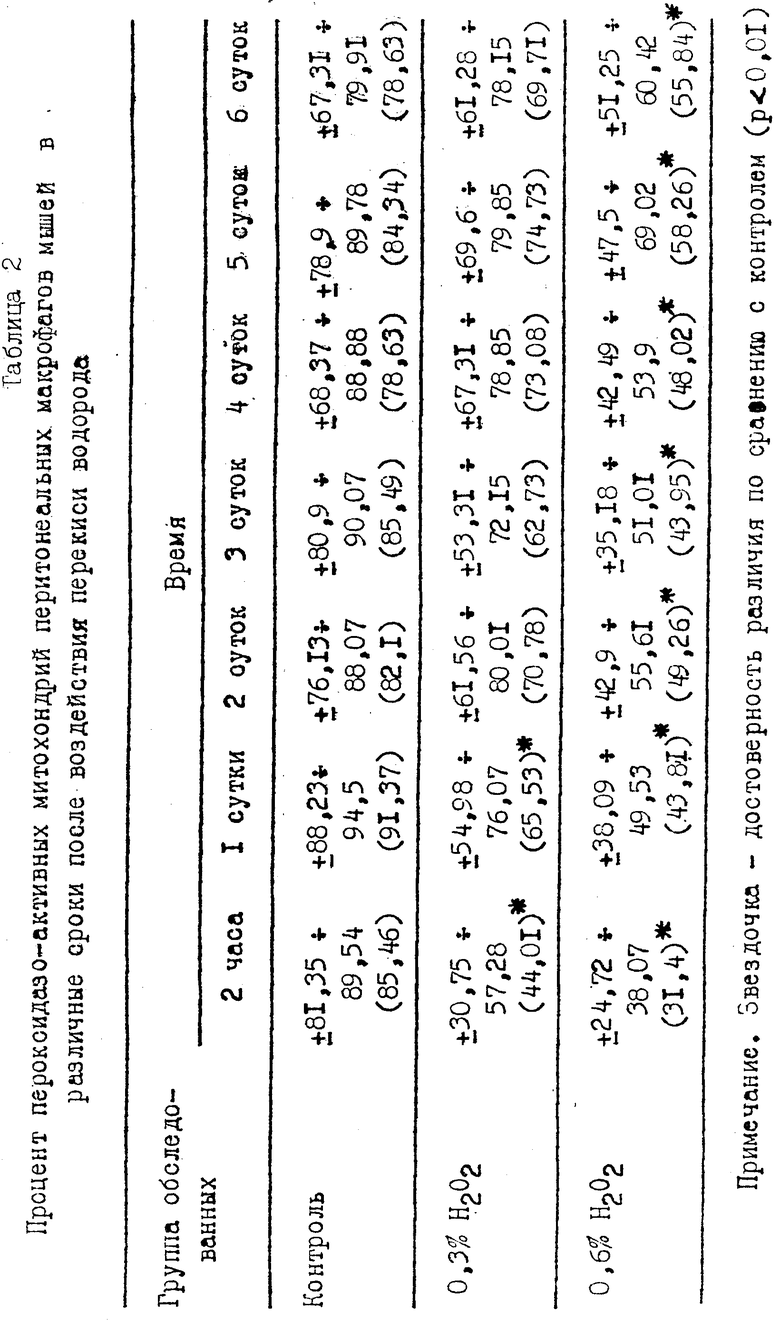
Результаты проведенного исследования активности пероксидазы перитонеальных макрофагов мышей в различные периоды после воздействия перекиси водорода показали, что в опытных сериях по прошествии 2 ч с момента воздействия H2O2 количество пероксидазоактивных митохондрий достоверно снижается, причем наиболее заметно после воздействия H2O2 в концентрации 0,6% (см. табл. 2).
В последующие сутки количество пероксидазоактивных митохондрий возрастало как в контроле, так и в двух сериях опытов, причем наиболее быстро при воздействии H2O2 в концентрации 0,3%. В дальнейшем (до 6 сут) процент митохондрий, в которых выявлялась активность пероксидазы, в опытных сериях возрастал и к 6 суткам в макрофагах, подвергавшихся воздействию H2O2 в концентрации 0,3%, практически сравнивался с контрольным. В то же время в макрофагах, подвергавшихся воздействию H2O2 в концентрации 0,6%, процент пероксидазоактивных митохондрий оставался на уровне ниже контрольного.
Следующим этапом исследования явилось изучение цитоархитектоники перитонеальных макрофагов животных в опытной и контрольных сериях в различные сроки.
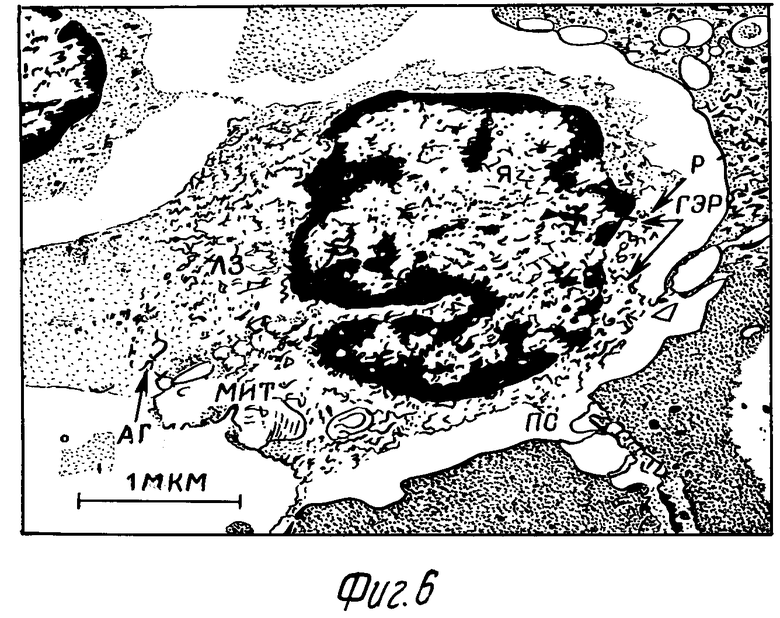
Морфологические исследования показали, что стимулированные макрофаги от интактных мышей, как обработанных 0,3% и 0,6% H2O2, так и интактные имели сферическую форму, почковидное или округлое ядро. У большинства обнаруживались многочисленные отростки (псевдоподии). Поверхностная мембрана трехслойна. Цитоплазма гранулярна. В цитоплазме имелись умеренно развитые мембранные структуры, состоящие из канальцев и вакуолей (гранулярный эндоплазматический ретикулум - ГЭР) с прикрепленными рибосомами, и пластинчатый аппарат Гольджи. Митохондрии вытянутой или овальной формы с матриксом небольшой электронной плотности и умеренным числом крист. В клетках имелись умеренно-электронно-оптически плотные гранулы - лизосомы. Хорошо выражены рибосомы и полисомы (фиг. 6).
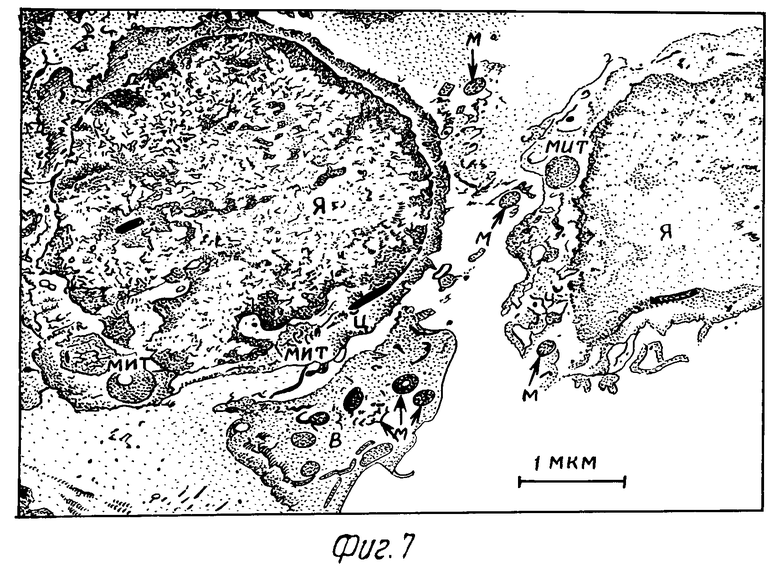
Через 2 ч после заражения культурами и суспензией микобактерий макрофаги несколько увеличивались в размерах. Ядро приобретает неправильную форму. Гранулярная эндоплазматическая сеть более выражена, чем в интактных клетках. В цитоплазме обнаруживались многочисленные вакуоли, содержащие фагоцитированные микобактерии (фиг. 7).
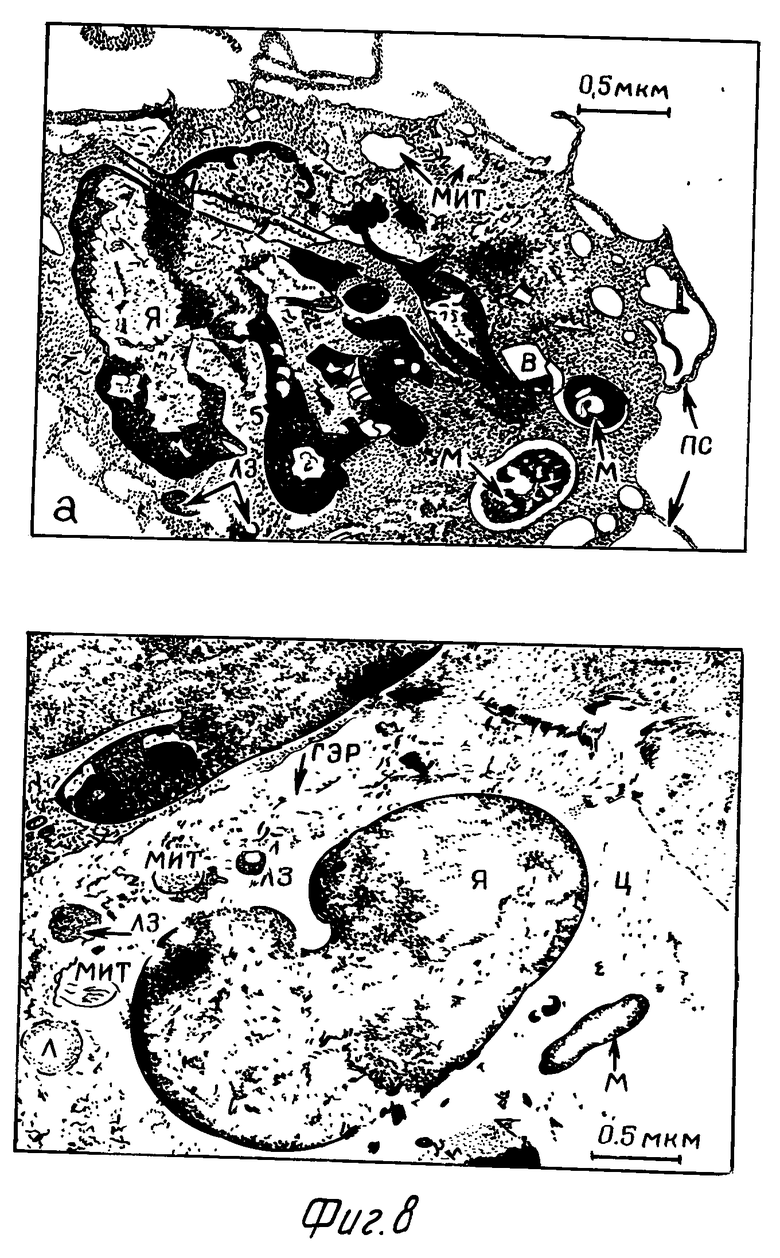
Через 1 сут в макрофагах опытных серий наблюдались выраженные длинные псевдоподии. Ядро неправильной формы. В цитоплазме кроме вакуолей значительной величины, содержащих интактные микобактериальные клетки, отмечались многочисленные мелкие округлые или овальные вакуоли, не содержащие какого-либо материала. Митохондрии мелкие, матрикс умеренно-электронно-оптически плотный. ГЭР слабо выражен, обнаруживался не во всех клетках. Лизосомы единичные, электронно-оптически плотные (фиг.8а). По сравнению с опытом в макрофагах контрольной серии отмечалось большее количество митохондрий и повышенное количество лизосом (фиг.8б).
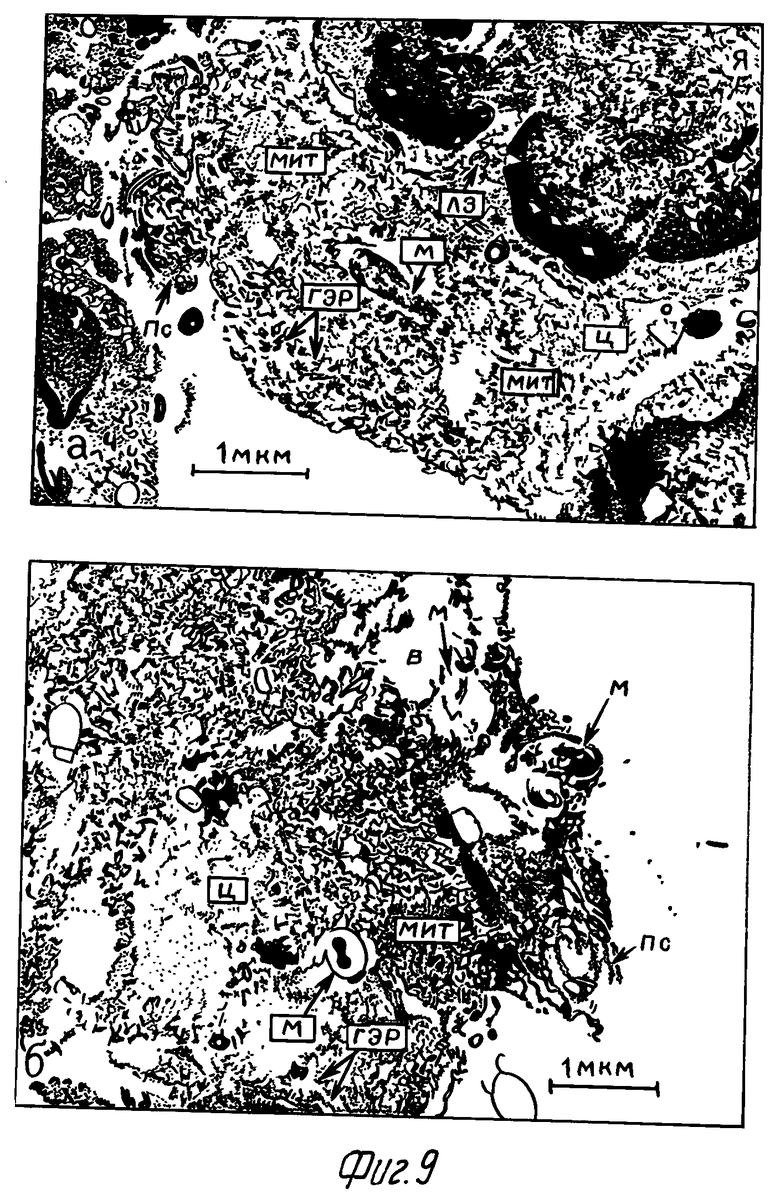
Через 2 сут после заражения макрофаги опытной серии большей частью имели округлую форму. У ряда клеток отмечались короткие толстые псевдоподии. Ядро правильной формы. Хроматин выражен, распределен равномерно в плоскости ядра. В цитоплазме отмечалось значительное количество митохондрий с выраженными кристами. Гранулярный эндоплазматический ретикулум выражен. Микобактерии большей частью расположены в вакуолях, некоторые непосредственно в цитоплазме фагоцита и не окружены мембраной фагосомы (фиг. 9а). В цитоплазме содержалось также небольшое количество гранул, по морфологии сходных с лизосомами. Слияние последних с фагосомами отмечалось в небольшом проценте случаев.
В контроле через 2 сут после заражения значительная часть макрофагов имела множество средней величины тонких псевдоподий. Ядро неправильной формы, нередко образовывало значительные выпячивания. Цитоплазма вакуолизирована. Наблюдался контакт лизосом с вакуолями, содержащими микобактериальные клетки. В цитоплазме отмечено также небольшое количество митохондрий с матриксом умеренной электронно-оптической плотности, почти сравнивающимся с электронно-оптической плотностью цитоплазмы (фиг. 9б).
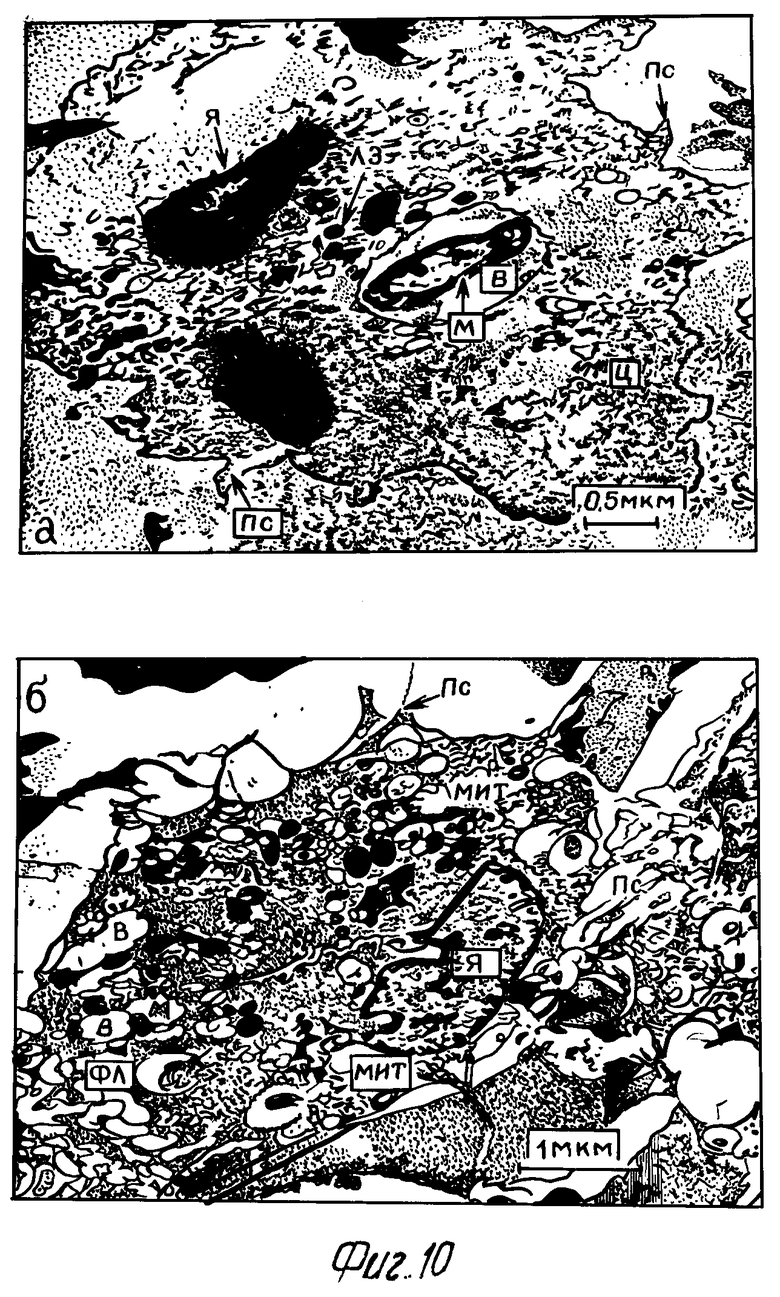
На 3-и сутки после введения микобактерий преимущественная часть макрофагов опытной серии претерпевала ультраструктурные изменения по сравнению с предыдущим сроком исследования. Псевдоподии макрофагов толстые и короткие. Цитоплазма разрежена. В ней определялись большие вакуоли, содержащие значительный процент интактных микобактерий (особенно в случае предварительного воздействия на фагоциты 0,6% H2O2) (фиг.10а). В цитоплазме фагоцитов увеличивалось количество мелких лизосом, однако в ряде случаев полноценного слияния их с вакуолями не наблюдается. Митохондрии малочисленны, матрикс не просветлен. Кристы митохондрий немногочисленны, большей частью разрушены.
В то же время в макрофагах контрольной серии псевдоподии длинные, тонкие, многочисленные. В цитоплазме наблюдалось большое количество лизосом, а также значительное число вакуолей, содержащих остатки фагоцитированных микобактериальных клеток. Отмечен полноценный контакт лизосом с фагосомами (фиг. 10б). Митохондрии многочисленные, полиморфные. Кристы большей частью сохранены.
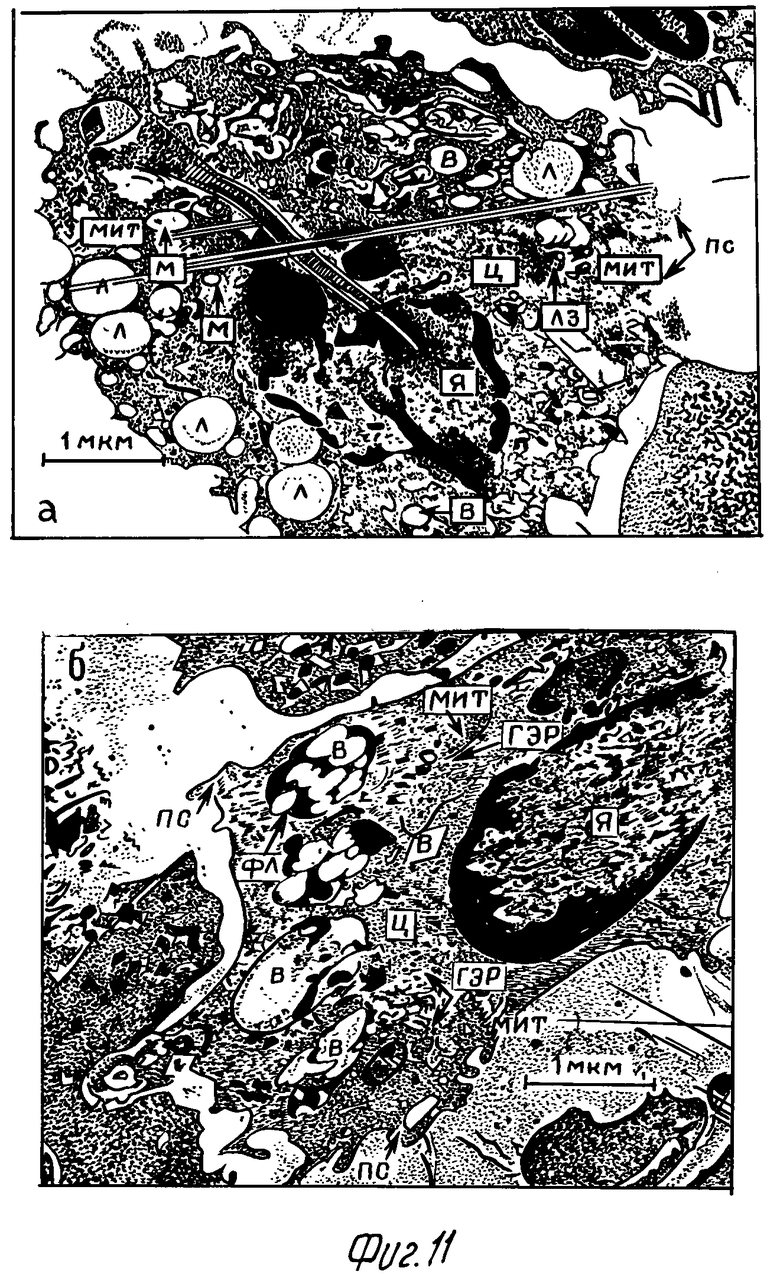
На 4-5-е сутки фагоцитарная активность макрофагов опытной серии отчетливо возрастала: увеличивается количество и длина псевдоподий. В цитоплазме отмечались вакуоли, содержащие как относительно интактные клетки микобактерий, так и их остатки. Характерной особенностью макрофагов в этот период являлось наличие в цитоплазме больших вакуолей, содержащих материал небольшой электронно-оптической плотности, по-видимому, соответствующий липидам (фиг. 11а).
В макрофагах контрольной серии в этот период отмечались большие вакуоли, либо не содержащие материала, либо содержащие остатки дезинтегрированных микобактериальных клеток (фиг. 11б). Псевдоподии макрофагов немногочисленные, короткие. Митохондрий немного, кристы сохранены. Ядро макрофагов округлой или овальной формы с преимущественно периферийным расположением хроматина.
На 6-е сутки в макрофагах опытной серии (при воздействии H2O2 в концентрации 0,6%) можно было обнаружить интактные микобактерии, расположенные в фаголизосомах (фиг. 12а). В цитоплазме фагоцитов отмечались многочисленные лизосомы, мелкие немногочисленные митохондрии преимущественно с разрушенными кристами и не ярко выраженный гранулярный эндоплазматический ретикулум.
Ультраструктура макрофагов контрольной серии в этот период по сравнению с предыдущим сроком наблюдения практически не изменялась. Ядро макрофагов округлой или овальной формы с небольшими выпячиваниями и периферийным расположением хроматина. В цитоплазме - большие вакуоли, содержащие остатки переваренных микобактериальных клеток, немногочисленные митохондрии и не резко выраженный гранулярный эндоплазматический ретикулум (фиг. 12б).
Таким образом, достоверное снижение уровня пероксидазы в перитонеальных макрофагах мышей обуславливает ряд ультраструктурных показателей, характеризующих изменение фагоцитарной активности клеток. В этом случае нарушается процесс внутриклеточного лизиса микроорганизмов. Процессы поглощения микобактериальных клеток в начальном периоде, напротив, активизируются. Наивысшие показатели поглотительной способности ПМ в опыте (как видно из табл. 1) приходились на первые 3 сут. В это же время (как видно из табл.2) отмечены низкие уровни митохондриальной пероксидазы фагоцитов.
Предполагаемым способом достигается повышение воспроизводимости моделей, причем достигнуто целенаправленное воздействие на клетки СМФ. При осуществлении способа не требуется ежедневных процедур, связанных с трудоемкостью и естественно большей вероятностью нарушения стерильности. Требуется лишь однократная процедура блокады миелопероксидазы перитонеальных макрофагов стимулированных мышей предложенной концентрацией (0,6%) перекиси водорода в количестве 2 мл, введенной внутрибрюшинно. Предлагаемый способ может быть легко воспроизведен в условиях экспериментальных лабораторий. Способ может быть рекомендован для оптимизации существующих моделей лепры (например, модель интраплантарного заражения мышей по методу Шепарда).
Размеры и условные обозначения, использованные на фигурах:
1 мкм - микрометр = 1000 нм = 10000  = 10-4 см = 10-6 м;
= 10-4 см = 10-6 м;
Н - нуклеоид микобактерий;
ПМ - плазматическая мембрана;
КС - клеточная стенка;
МЕЗ - мезосомы;
Р - рибосомы;
М - клетки микобактерий;
Ц - цитоплазма;
МИТ - митохондрии фагоцитов;
Я - ядро;
ПС - псевдоподии;
ЛЗ - лизосомы;
В - вакуоли;
Л - липиды;
ГЭР - гранулярный эндоплазматический ретикулум;
ФЛ - фаголизосомы;
АГ - аппарат Гольджи.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ МОДЕЛИРОВАНИЯ ЛЕПРОЗНОЙ ИНФЕКЦИИ | 1995 |
|
RU2102482C1 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ МЕДЛЕННОРАСТУЩИХ МИКОБАКТЕРИЙ | 1995 |
|
RU2106879C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ГРУППЫ РИСКА РЕЦИДИВА ЛЕПРЫ | 1993 |
|
RU2082979C1 |
| СПОСОБ МОДЕЛИРОВАНИЯ ЛЕПРЫ | 1993 |
|
RU2098866C1 |
| СПОСОБ ЛЕЧЕНИЯ ЭКСПЕРИМЕНТАЛЬНОЙ ЛЕПРОЗНОЙ ИНФЕКЦИИ | 2011 |
|
RU2467742C1 |
| СПОСОБ ДИАГНОСТИКИ ЛЕПРЫ | 1996 |
|
RU2124730C1 |
| СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ ТРОФИЧЕСКИХ ЯЗВ И ОЖОГОВ У БОЛЬНЫХ ЛЕПРОЙ | 1995 |
|
RU2095065C1 |
| СПОСОБ ЛЕЧЕНИЯ ЭКСПЕРИМЕНТАЛЬНОЙ ЛЕПРОЗНОЙ ИНФЕКЦИИ | 2006 |
|
RU2322991C1 |
| Способ прогнозирования рецидива лепрозного процесса | 1988 |
|
SU1636717A1 |
| СПОСОБ ИММУНОПРОФИЛАКТИКИ ЭКСПЕРИМЕНТАЛЬНОЙ ЛЕПРОЗНОЙ ИНФЕКЦИИ | 1996 |
|
RU2135196C1 |


Способ может быть использован в области медицины, а именно в лепрологии и фтизиатрии, и может применяться для оптимизации существующих моделей лепры и туберкулеза. Позволяет повысить воспроизводимость модели. Перитонеальные макрофаги стимулированных животных (мыши) за 2 ч до заражения взвесью микобактерий лепры или туберкулеза однократно насыщают введением в брюшную полость 0,6%-ного раствора перекиси водорода в количестве 2 мл на мышь. 2 табл., 12 ил.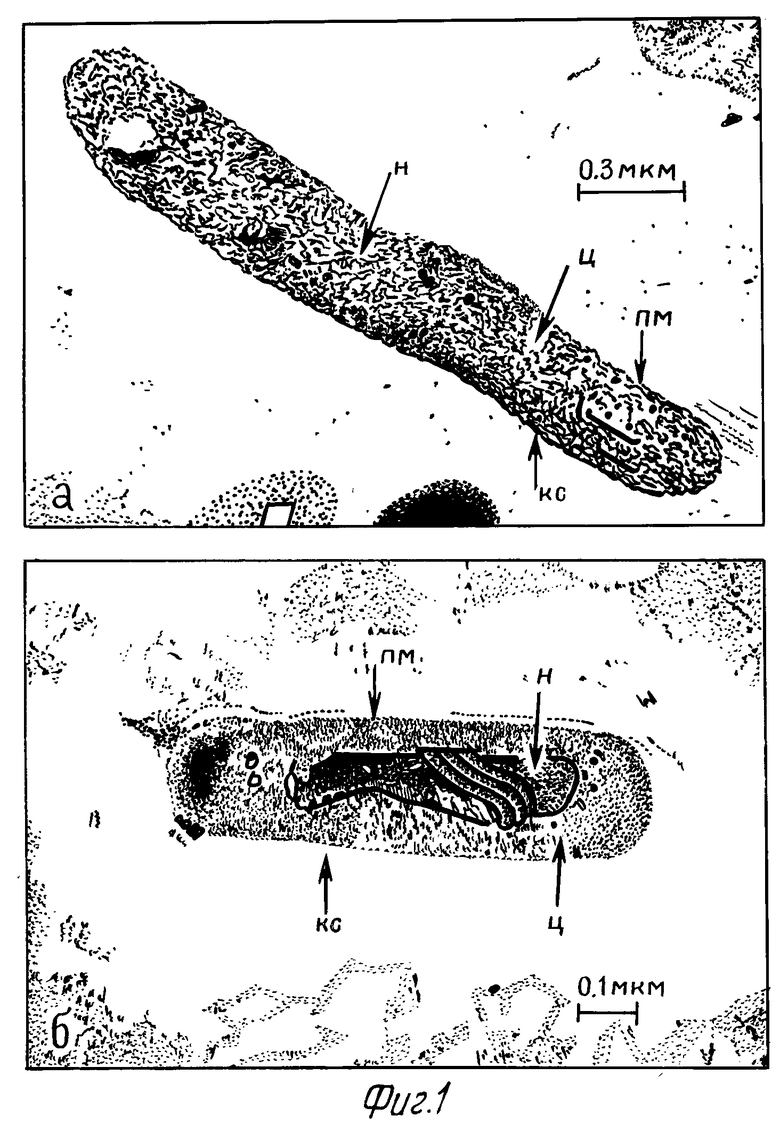
Способ моделирования дефекта макрофагов путем заражения мышей взвесью микобактерий лепры, отличающийся тем, что за 2 ч до внутрибрюшинного заражения мышей взвесью микобактерий лепры или туберкулеза в брюшную полость вводят однократно 0,6%-ный раствор перекиси водорода в количестве 2 мл на мышь.
| Вишневецкий Р.Ф | |||
| Уровень с пузырьком | 1922 |
|
SU388A1 |
| Автореф.дис | |||
| на соиск | |||
| степ.канд.мед.наук | |||
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |
