Область изобретения
Изобретение относится, в общем, к областям действия лекарственных препаратов, клеточной регуляции и генной терапии. Более конкретно, настоящее изобретение касается нового открытия факта, что глюкокортикоиды усиливают активность репортерного гена при трансфекции катионным липидом или липосомами.
Описание прототипов
Как показано, за последние два года получили развитие и стали многообещающими клинические испытания генной терапии на людях для лечения заболеваний, вызванных генетическими дефектами или нарушением генной регуляции. Предметом генной терапии являлись некоторые заболевания, проявляющие значительную симптомологию в легких, в том числе муковисцидоз и рак легких. В этих исследованиях применяли рекомбинантные ретровирусные или аденовирусные векторы, а также катионные липиды для транспорта и доставки гена в клетку. Тем не менее клеточный транспорт и последующая экспрессия гена низки и, следовательно, терапевтические уровни генной экспрессии могут отсутствовать. Кроме того, иммунные реакции, развивающиеся против вирусных векторов, могут ограничивать их применение. Хотя катионные липиды являются менее эффективными при доставке, чем аденовирусные векторы, более новые химические конструкции обеспечили катионные липиды, которые являются значительно улучшенными по сравнению с исходными конструкциями. Некоторые испытания на животных и человеке показали, что при концентрациях катионных липидов, типичных для трансфекции, не развиваются отрицательные побочные эффекты или иммунные реакции.
Доставка средств генной терапии в легкие посредством аэрозоля позволяет доставлять гены непосредственно к ткани-мишени. Несколько групп продемонстрировали доставку аэрозольным способом и трансфекцию легких животных in vivo с использованием ДНК рецепторного гена, присоединенной к катионным липосомам. Выдающимися отличительными признаками, отмеченными в этих исследованиях, являются отсутствие токсичности и продолжительность экспрессии гена, равная примерно 1 месяцу. Экспрессия гена была по-прежнему низкой; даже умеренная трансфекция в легких мышей требует, по крайней мере, от 0,5 до 12 мг ДНК высокой чистоты. Как сообщалось, такие способы генной терапии невозможны для человека.
Другим подходом к повышению эффективности трансфекции является большее понимание поглощения плазмид и факторов, которые влияют на экспрессию трансфицированного гена в ткани-мишени. Что касается роли воспаления в трансфекции генов в клетках легких, было обнаружено, что подвергание клеток карциномы легких человека А549 воздействию иммунного стимулятора липополисахарида или цитокина IL-1β перед трансфекцией посредством pCMVβ-gal-DMRIE/DOPE снижало уровень белка β-gal ниже уровня, наблюдаемого в клетках, обработанных только средой.
Недостаток предыдущих исследований заключается в отсутствии эффективных средств доставки терапевтических уровней трансфицированных генов. Настоящее изобретение обеспечивает эту давнишнюю потребность в данной области знаний.
Краткое содержание изобретения
О механизме, при помощи которого клетка поглощает комплексы катионной липид-ДНК, или о том, какова судьба этих комплексов внутри клетки, известно немного. Еще меньше известно о факторах in situ, которые могут влиять на поглощение или последовательную доставку и экспрессию ДНК в ткани, в особенности у пациентов с хроническим воспалением легких или другим иммунным процессом. Настоящее изобретение демонстрирует два открытия, которые оказывают существенное влияние на трансфекцию в клеточной культуре и которые имеют параллели in vivo. Во-первых, настоящее изобретение показывает, что цитокин IL-1β и иммунный стимулятор липополисахарид (LPS) подавляют трансфекцию/экспрессию pCMVβ-gal, трансфецированного посредством катионного липида в А549 клеточные линии рака легких человека или первичные культуры клеток легких крысы. Во-вторых, противовоспалительные глюкокортикоиды локального действия, такие как беклометазондипропионат (ВЕС), обращают ингибиторное действие IL-1β и липополисахарида и даже усиливают экспрессию репортерных генов выше экспрессии, наблюдаемой в необработанных трансфицированных клетках, то есть не обработанных липополисахаридом или IL-1β и т.д. Этот эффект специфичен для глюкокортикоидов в противоположность другим типам стероидов, но не только для конкретного глюкокортикоида. Этот эффект может быть специфичен для глюкокортикоидов как противовоспалительных агентов, так как этот эффект не наблюдается, когда клетки предварительно обрабатывают другим иммуносупрессором циклоспорином А. Гликокортикоидопосредованное усиление трансгенной активности не зависит от использованных промотора, репортерного гена и катионного липида. Механизм, по которому глюкокортикоиды усиливали экспрессию репортерных генов, не включает в себя повышенное поглощение плазмиды липида, а скорее представляет собой внутриклеточный механизм, который не включает в себя нового белка. Кроме того, предварительная обработка первичных клеток легкого крысы синтетическими глюкокортикоидами локального действия в отсутствие липополисахарида или IL-1β перед трансфекцией повышает уровень β-gal протеина выше уровня в необработанных контролях. Настоящее изобретение описывает исследования механизма усиленной глюкокортикоидами трансфекции плазмидной ДНК. Таким образом, настоящее изобретение имеет прямое отношение к использованию генной терапии in vivo.
В одном варианте настоящего изобретения разработан способ повышения клеточной экспрессии гена в биологической ткани после доставки указанного гена в соответствующем векторе животному, включающий стадию введения указанному животному фармакологически эффективной дозы глюкокортикоида в количестве, достаточном для повышения клеточной экспрессии указанного гена.
В другом варианте настоящего изобретения разработан способ лечения патофизиологического состояния человека посредством повышения клеточной экспрессии гена в соответствующем векторе после доставки указанного гена в биологическую ткань нуждающегося в таком лечении человека, включающий стадию введения указанному человеку фармакологически эффективной дозы глюкокортикоида в количестве, достаточном для повышения клеточной экспрессии указанного гена.
Другие и дополнительные аспекты, отличительные признаки и преимущества настоящего изобретения будут ясны из следующего описания предпочтительных в настоящее время вариантов данного изобретения, приведенных для целей описания.
Краткое описание чертежей
Для того чтобы материал, в котором достигнуты указанные выше отличительные признаки, преимущества и цели данного изобретения, а также другие особенности, которые станут ясными, мог быть воспринят и понят в деталях, более подробные описания данного изобретения, кратко суммированные выше, можно подкрепить ссылками на определенные варианты изобретения, которые иллюстрируются прилагаемыми чертежами. Эти чертежи составляют часть описания. Однако необходимо отметить, что прилагаемые чертежи иллюстрируют предпочтительные варианты данного изобретения и, следовательно, их нельзя считать ограничительными для объема изобретения.
На фигуре 1 показано, что липополисахарид (LPS) и IL-1β подавляют активность β-галктозидазы (β-gаl) в трансфицированных комплексом pCMVβ-gal-катионный липид клетках А549. Клетки А549 в течение 4 часов обрабатывали средой 0,5 μг/мл липополисахарида или 100 Е/мл рекомбинантного человеческого IL-1-β. После обработки клетки трансфицировали 1 μг/мл pCMVβ-gal- 4 μг/мл DMRIE/DOPE. Активность β-gal определяли при помощи (CPRG) способа колориметирческого микротитрования, специфически измеряющего активность β-галктозидазы. Результаты представляют среднее значение и стандартное отклонение из 3 экспериментов. Относительное изменение (изменение в указанное число раз) = β-gal-активность в образцах липополисахарида (LPS) или IL-1/β-gal-активность в образцах, обработанных средой.
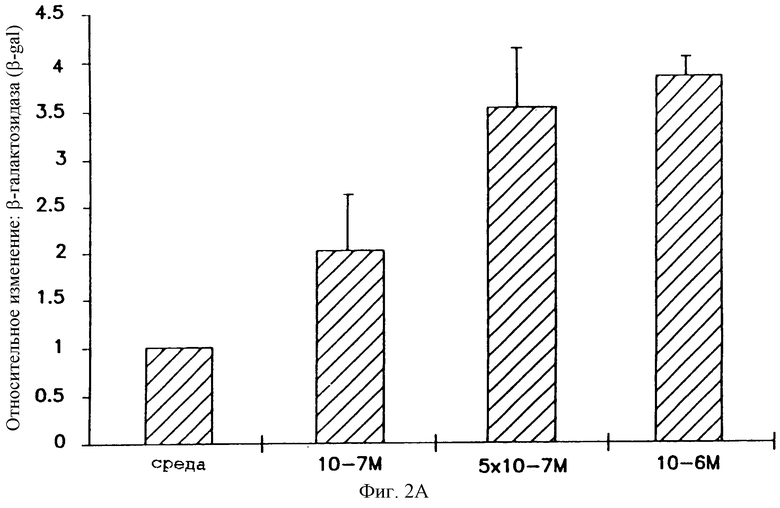
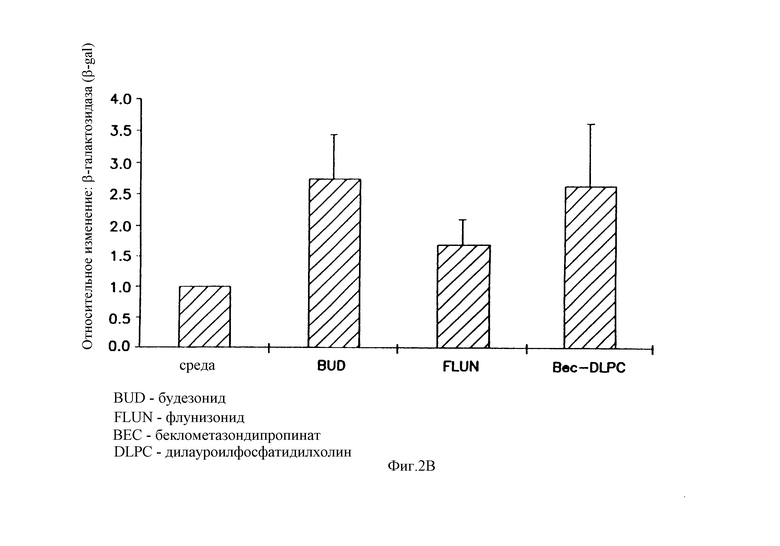
На фигуре 2 показано, что глюкокортикоиды усиливают β-gal-активность в транфицированных клетках А549. Клетки А549 оборабатывали в течение 4 часов глюкокортикоидами. На фигуре 2А показана реакция трансфицированных клеток на различные дозы беклометазона, диапазон варьирования дозы составляет от 10-7 до 10-6 М. На фигуре 2В показано, что некоторые другие глюкокортикоиды локального действия также вызывают усиление β-gal активности при 10-6 М: Budesonide (BUD); Flunisolide (FLUN); беклометазон-дипропионатдилауроилфосфатидилхолин, беклометазон-DLPC (beclomethasone-dipropionate-dilauroyl Phosphatidylcholine). После обработки клетки транфицировали 1 μг/мл pCMVβ-gal- 4 μг/мл DMRIE/DOPE. Активность β-gal определяли при помощи способа микротитрования. Результаты представляют средние и стандартные отклонения из 3 экспериментов. Относительное изменение = β-gal активность в клетках, обработанных глюкокортикоидом/β-gal активность в клетках, обработанных средой.
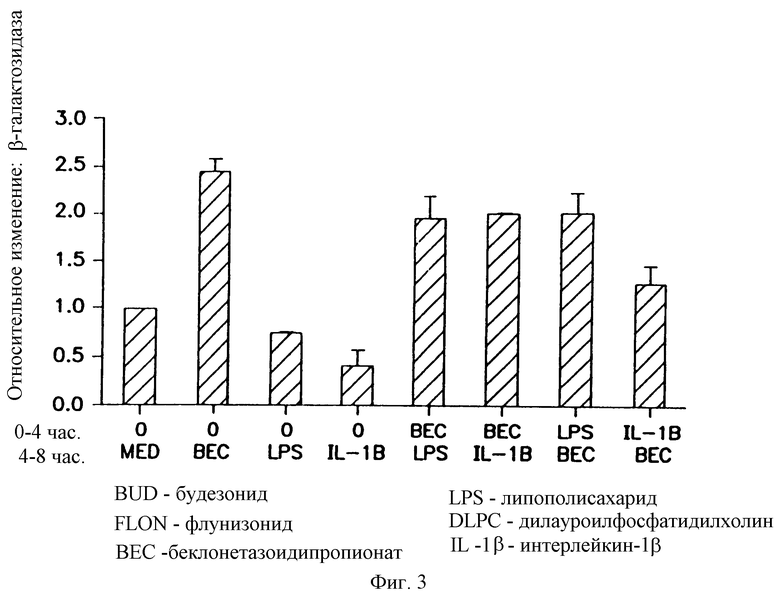
На фигуре 3 показано, что беклометазон обращает ингибиторное действие липополисахарида и IL-1β на β-gal активность в трансфицированных клетках А549. Клетки А549 обрабатывали в течение 4 часов только средой, беклометазоном (10-6 М), IL-1β (100 U/мл) или липополисахаридом (0,5 μг/мл). По прошествии первых 4 часов удаляли супернатант и клетки обрабатывали еще в течение 4 часов средой IL-1β, липополисахаридом или беклометазоном (то есть беклометазон, липополисахарид; беклометазон в течение первых 4 часов, липополисахарид в течение дополнительных 4 часов). После второй обработки клетки трансфицировали 1 μг/мл pCMVβ-gal- 4 μг/мл DMRIE/DOPE. Активность β-gal определяли при помощи способа микротитрования. Результаты представляют средние значения и стандартные отклонения (SD) из 2 экспериментов. Относительное изменение = β-gal активность в обработанных беклометазоном, липополисахаридом, IL-1 или их комбинацией клетках/β-gal активность в клетках, обработанных средой.
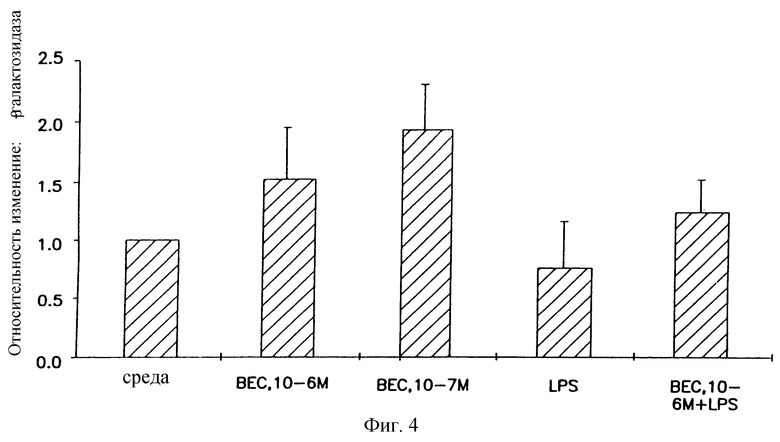
На фигуре 4 показано, что беклометазон усиливает β-gal активность в первичных клетках легких крысы. Первичные клетки легких крысы выделяли из ферментативно переваренных легких крысы и высевали в чашки для культивирования тканей при 1,0•105 клеток/на лунку. Клетки предварительно обрабатывали беклометазоном, липополисахаридом (0,5 μг/мл) или беклометазон + липополисахарид (4 часа беклометазон + 4 часа липополисахарид) в диапазоне концентраций 10-7-10-6 М в течение 4 часов, затем трансфицировали 3 μг ДНК и 12 μг DMRIE/DOPE. Через 48 часов определяли β-gal активность при помощи способа микротитрования CPRG. Результаты представляют средние значения и стандартные отклонения (SD) из 3 (*2) экспериментов. Относительное изменение = β-gal активность в обработанных беклометазоном, липополисахаридом, беклометазон + липополисахарид клетках/β-gal активность в клетках, обработанных средой.
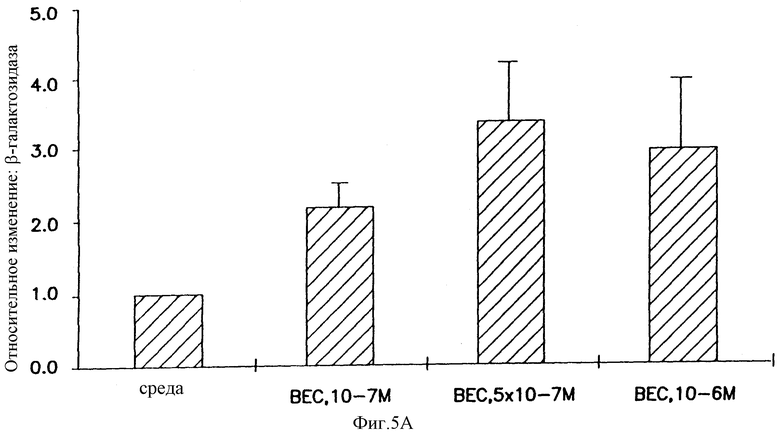
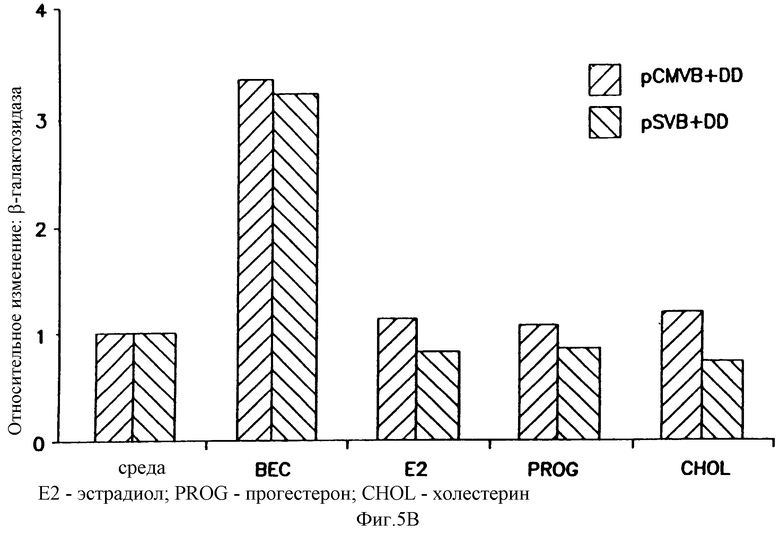
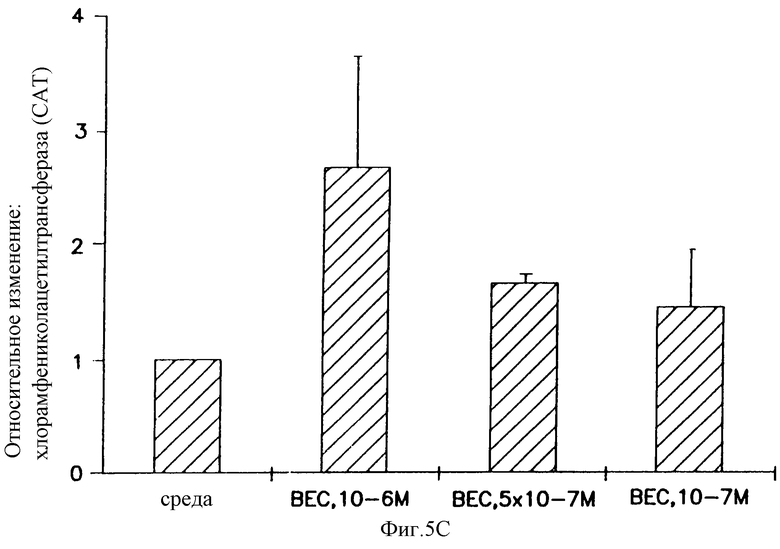
На фигуре 5 показано, что беклометазонопосредованное усиление не ограничено катионным липидом, векторным ором или векторными репортерными генами и специфично относительно глюкокортикоидов. Клетки А549 обрабатывали в течение 4 часов беклометазоном или другими стероидами (на фигуре 5В стероиды: эстроген, Е2; прогестерон, PROG; холестерол, CHOL). После обработки клетки трансфицировали: на фигуре 5А 1 μг/мл pCMVβ-gal и 3 μг/мл DOSPA/DOPE, на фигуре 5В 1 μг/мл pSVβ и 4 μг/мл DMRIE/DOPE или 1 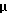 г/мл pCMVβ-gal и 4 μг/мл DMRIE и на фигуре 5С 1 μг/мл pCMVHICAT и 4 μг/мл DMRIE/DOPE. Активность β-gal определяли при помощи способа микротитрования. Активность хлорамфениколацетилтрансферазы (CAT) определяли способом тонкослойной хроматографии (TLC) хлорамфениколацетилтранферазы, где 14С-изотопные ацетилированные формы хлорамфеникола разделяли способом TLC, а 14С-включенис определяли при помощи бета-скеннера, детектируя 14С. Результаты на фигуре 5А и фигуре 5С представляют средние значения и стандартные отклонения (SD) из 3 экспериментов. Результаты на фигуре 5В представляют данные 3 экспериментов. Относительное изменение = активность β-gal или CAT в клетках, обработанных беклометазоном, или стероид/активность β-gаl, или хлорамфениколацетилтрансферазы в клетках, обработанных средой.
г/мл pCMVβ-gal и 4 μг/мл DMRIE и на фигуре 5С 1 μг/мл pCMVHICAT и 4 μг/мл DMRIE/DOPE. Активность β-gal определяли при помощи способа микротитрования. Активность хлорамфениколацетилтрансферазы (CAT) определяли способом тонкослойной хроматографии (TLC) хлорамфениколацетилтранферазы, где 14С-изотопные ацетилированные формы хлорамфеникола разделяли способом TLC, а 14С-включенис определяли при помощи бета-скеннера, детектируя 14С. Результаты на фигуре 5А и фигуре 5С представляют средние значения и стандартные отклонения (SD) из 3 экспериментов. Результаты на фигуре 5В представляют данные 3 экспериментов. Относительное изменение = активность β-gal или CAT в клетках, обработанных беклометазоном, или стероид/активность β-gаl, или хлорамфениколацетилтрансферазы в клетках, обработанных средой.
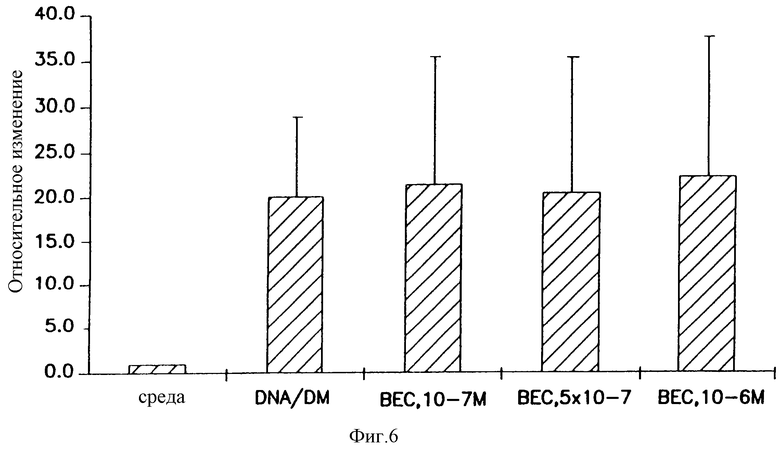
На фигуре 6 показано, что беклометазон не повышает поглощение плазмид в клетках А549. Клетки А549 обрабатывали в течение 4 часов средой или указанной дозой беклометазона, затем трансфицировали 3H-тимидинмеченым pCMVβ-gal. Через 2 часа после трансфекции клетки промывали, слегка обрабатывали трипсином и центрифугировали. Осадки после центрифугирования лизировали, повторно суспендировали и добавляли к ним жидкий коктейль для сцинтилляционного счета радиоактивности. Определяли поглощение 3H-тимидинмеченым pCMVβ-gal (имп/мин) на образец. Результаты представляют средине значения и стандартные отклонения (SD) из 4 экспериментов, и каждая экспериментальная точка получена в двух повторностях. Относительное изменение = имп/мин образцов, предварительно обработанных беклометазоном/имп/мин образцов, предварительно обработанных средой.
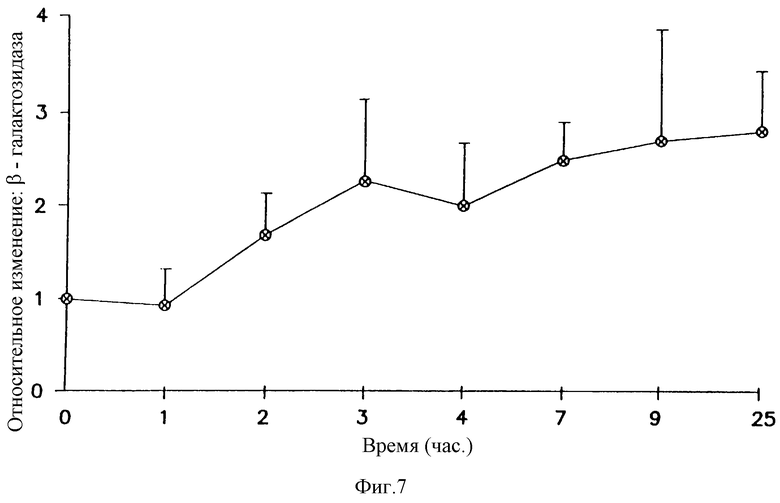
На фигуре 7 показана кинетика экспрессии репортерного гена, усиленной беклометазоном. Клетки А549 обрабатывали в течение различных периодов времени белометазоном 10-6 М или средой. Клетки трансфицировали 1 μг/мл pCMVβ-gal- 4 μг/мл DMRIE/DOPE. β-gal активность определяли при помощи способа микротитрования CPRG. Результаты представляют средние значения и стандартные отклонения (SD) из 2 экспериментов. Относительное изменение = β-gal активность в клетках, обработанных беклометазоном/β-gаl активность в клетках, обработанных средой.
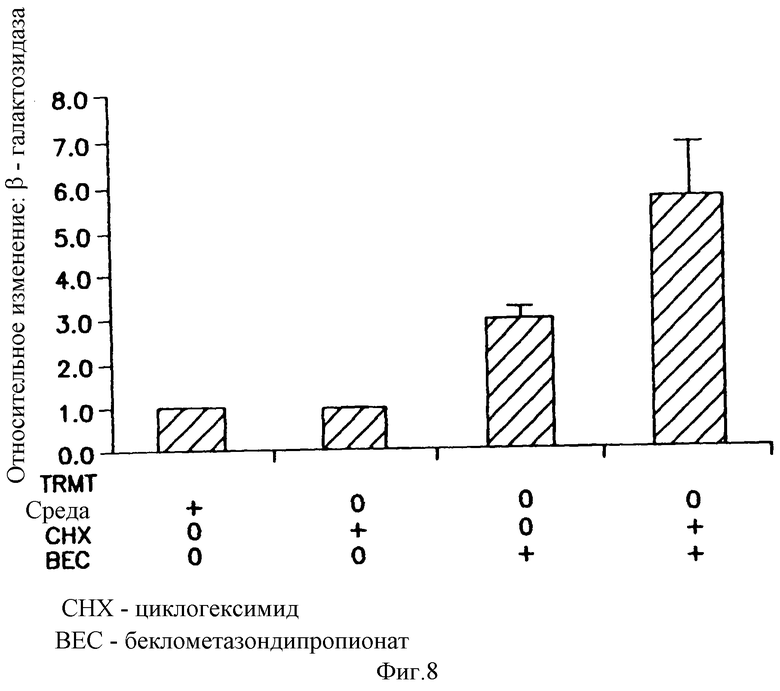
На фигуре 8 показано, что для беклометазонопосредованного усиления β-gal активности не требуется белковый синтез. Клетки обрабатывали в течение 30 минут либо средой, либо СНХ (10 μг/мл). Клетки, предварительно обработанные средой, затем обрабатывали в течение 4 часов либо средой, либо беклометазоном 10-6 М. Клетки, предварительно обработанные СНХ, затем обрабатывали в течение 4 часов либо СНХ, либо беклометазоном 10-6 М+СНХ. Клетки трансфицировали 1 μг/мл pCMVβ-gal- 4 μг/мл DMRIE/DOPE. β-gal активность определяли при помощи способа CPRG микротитрования. Результаты представляют средние значения и стандартные отклонения (SD) из 2 экспериментов. Относительное изменение ВЕС = β-gal активность в клетках, обработанных беклометазоном, /β-gal активность в клетках, обработанных средой. Относительное изменение беклометазон + СНХ = β-gal активность в клетках, обработанных беклометазоном + СНХ, /β-gal активность в клетках, обработанных СНХ.
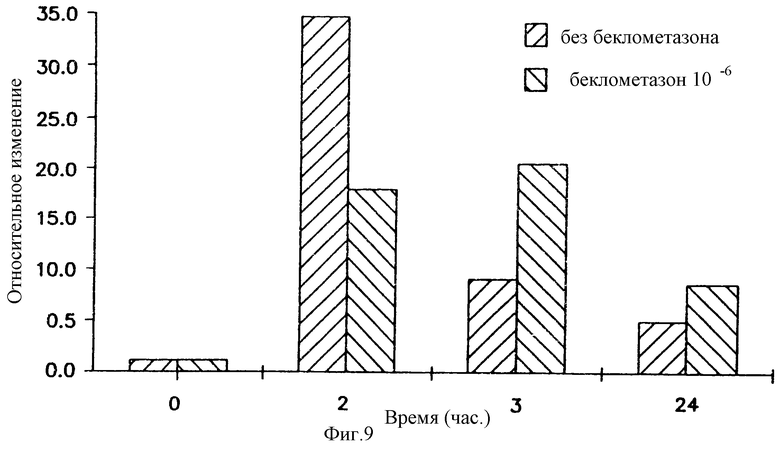
На фигуре 9 показано, что беклометазон повышает уровни мРНК устойчивого состояния в клетках А549. Клетки А549 обрабатывали в течение 4 часов беклометазоном 10-6 М или средой.
Клетки трансфицировали 1 μг/мл pCMV β-gal плюс 4 μг/мл DMRIE/DOPE. В указанные моменты времени (часы) после трансфекции собирали общую РНК, превращали в кДНК и специфические мессенджеры β-gal амплифицировали при помощи RS-PCR (РНК-специфической полимеразной цепной реакцией), как описано ниже. Пробы RS-PCR разделяли на 1%-ном геле ТАЕ-агарозы [ТАЕ=буфер 1X Трис-ацетат (40 мМ)-ЭДТА (2 мМ)] и переносили на найлоновый фильтр. Фильтр гибридизовали с 12Р-меченым зондом β-gal. Фильтр анализировали бета-скеннером и считали (на радиоактивность) полосы подходящего размера. Относительное изменение = имп/мин β-gal в клетках, обработанных беклометазоном, /имп/мин β-gal в клетках, обработанных средой.
Подробное описание изобретения
В настоящем изобретении могут быть использованы следующие сокращения: липополисахарид: липополисахарид Salmonella typhimurium; IL-1β: интерлейкин-1-β; GC: глюкокортикоиды; Е2: эстрадиол; PROG: прогестерон; CHOL: холестерин; BUD: будезонид; ВЕС: беклометазондипропионат; FLUN: флунизонид; DLPC: дилауроилфосфатидилхолин; β-gal: бета-галактозидаза Escherichia coli; CAT: хлорамфениколацетилтрансфераза; DMRIE/DOPE: бромид N-(2-гидроксиэтил)-N,N-диметил-2,3-бис-(тетрадецитокси)-1-пропанаминия/диолеилфосфатидилэтаноламин; DOSPA/DOPE: трифторацетат 2,3-диолеилокси-N-[2(сперминкарбоксамидо)этил]-N, N-диметил-1-пропанаминия/диолеилфосфатидилэтаноламин; DMEM: модифицированная по способу Дульбекко минимальная поддерживающая среда; FBS: фетальная телячья сыворотка; СНХ: циклогексимид; RS-PCR: РНК-специфическая полимеразная цепная реакция; CPRG: хлорфеноловый красный β-D-галактопиранозид; сАМР: циклический аденозинмонофосфат; CREB: белок, связывающий отвечающий на сАМР элемент.
Настоящее изобретение относится к способу повышения клеточной экспрессии гена в клетке после доставки указанного гена в животное, предусматривающему стадию введения данному животному фармакологически эффективной дозы глюкокортикоида в количестве, достаточном для усиления клеточной экспрессии указанного гена.
Настоящее изобретение относится также к способу лечения патофизиологического состояния человека путем усиления клеточной экспрессии гена после доставки указанного гена в биологическую ткань пациента, нуждающегося в таком лечении, отличающемуся тем, что он включает в себя стадию введения данному пациенту фармакологически эффективной дозы глюкокортикоида в количестве, достаточном для усиления клеточной экспрессии указанного гена.
Обычно в способе настоящего изобретения можно использовать любой глюкокортикоид, но, по-видимому, не любое противовоспалительное средство, так как в клетках, обработанных циклоспорином-А, эффект не был обнаружен. Типичные примеры применимых глюкокортикоидов включают гидрокортизон, преднизон, преднизолон, триамцинолон, бетаметазон, будезонид, флунизолид и дексаметазон. Глюкокортикоиды могут быть либо синтетическими, либо несинтетическими. Обычно глюкокортикоиды вводят в дозе примерно от 0,6 мг/кг до 50 мг/кг в зависимости от того, какой глюкокортикоид должен быть доставлен, поскольку существует различие в активности, и в зависимости от того, должна ли доставляться физиологическая или фармакологическая доза. В способах настоящего изобретения глюкокортикоид может находиться в виде растворимого в липиде вещества, вещества, растворимого в этаноле или воде, любая их этих форм может быть включена в липосому.
Обычно глюкокортикоид можно вводить таким способом, который оптимизирует способность глюкокортикоида усиливать активность доставленного гена. Например, можно вводить глюкокортикоид совместно с указанным геном, до или после доставки указанного гена. Способ введения может быть любым, какой требуется, включая аэрозольный, внутривенный, внутрибрюшинный и др.
Способы настоящего изобретения эффективны, независимо от биологической ткани, в которую доставляют ген. Например, активность гена может быть повышена, если его доставляют в такие ткани, как печень, лейкоциты, легкие, желудочно-кишечный тракт, почка, скелетные мышцы, гладкие мышцы, нервная ткань, клетки кожи, раковые клетки, глаз, костный мозг и опухоли.
Доставку гена можно осуществлять любым способом. Например, путем инъекции, пероральным способом, через кожу или при помощи аэрозоля. В другом варианте глюкокортикоид может быть включен в липосому, нейтральную или несущую активные вещества, как известно в этой области. Альтернативно ген может быть растворен в таком растворителе, как этанол.
Способы настоящего изобретения эффективны для любых животных, как для человека, так и для животных. И хотя способы настоящего изобретения применимы в первую очередь к людям, специалистам будут очевидны различные ветеринарные применения.
В способах настоящего изобретения трансфекцию гена можно производить различными известными способами. Типичные примеры способов трансфекции гена в биологическую ткань включают вирусную трансфекцию, трансфекцию катионным липидом и направленную генную терапию, использующую рецептор и катионный амин, такой как поли-L-лизин. На самом деле, активность любого гена можно усилить, применяя данную методологию к рекомбинантному гену, природному гену, кДНК или олигомеру. Глюкокортикоид усиливает активность вектора при промоторе или на стадии клеточной регуляции до трансляции.
Следующие далее примеры приведены с целью иллюстрации различных вариантов данного изобретения, но не для ограничения настоящего изобретения.
Пример 1
Клетки и клеточная культура
А549 получали из АТСС. А549 поддерживали в смеси: модифицированная по способу Дульбекко минимальная поддерживающая среда (DMEM, GIBCO-Life Technologies) плюс 10%-ная фетальная телячья сыворотка (FBS, Hyclone), содержащая низший липополисахарид, 200 мМ L-глутамин и 50 μг/мл гентамицина. Для большинства трансфекцией клетки высевали в 12-луночные пластины (Corning) при 3•104 клеток/мл. Такая концентрация клеток дает монослой, конфлюентный примерно на 30% через 24 часа после переноса клеток.
Первичные клетки легкого крысы готовили следующим образом. Легкие брали из взрослых самок Sprague-Daweley крыс и мелко размельчали. Размельченные легкие суспендировали в растворе 0,5Х трипсина в сбалансированном солевом растворе Хэнка в течение 1 часа, а затем в растворе, содержащем 100 Е/мл коллагеназы (С. perfringens, IV, Sigma Chemical), 100 Е/мл ДНКзы I (Sigma) в модифицированной по способу Дульбекко минимальной поддерживающей среде (DMEM) с 10%-ной фетальной телячьей сывороткой (FBS). Ткань инкубировали на качалке при 37oС в течение 1,5 часов. Диссоциированные клетки фильтровали через несколько слоев стерильной марли и высевали в DMEM + 10% FBS в 6-луночные планшеты (Corning) по 1,0•106 клеток/на лунку. Клетки инкубировали до трансфекции во влажном инкубаторе при 37oС, 5% CO2 в течение 13-24 часов.
Пример 2
Химикаты и реагенты
Циклогексимид, липополисахарид (липополисахарид S. Туphimurium). Будезонид (BUD) и флунизолид также приобретали у Sigma Chemical Co., St. Louis, МО. Все глюкокортикоиды растворяли в абсолютном этаноле. Беклометазондипропионат получали в качестве подарка из Orion Pharmaceuticals, Kuopio, Finland. Беклометазон в нейтральной липосоме, дилаурилфосфатидилхолин (DLPC, Avanti Polar Lipids, Birmingham, AL) готовили путем растворения 0,5 мг беклометазона и 25 мг DLPC в трет-бутаноле при 37oС. Затем образцы мгновенно замораживали в этаноле с сухим льдом и лиофилизировали. Липосомы восстанавливали в стерильной воде, не содержащей эндотоксина. Человеческий рекомбинантный IL-1β покупали из Genzyme Corp (Cambridge, MA).
Пример 3
кДНК
pCMVβ-gal и pCMVHICAT получали из Genzyme Corp., a pSVβ из Clontech (CA). Плазмидную ДНК экстрагировали и очищали, используя систему колоночной очистки Qiagen (Qiagen, Chatsworth, CA). Большинство липополисахарида удаляли из плазмидных препаратов, используя Е-ТОХ колонку (Sterogene, СА). Плазмиды оценивали на эндотоксин, используя набор LAL от Biowhitaker/Microbiological Associates (Bethesda, MD). Концентрацию ДНК определяли по поглощению при 260 нм (А260) и сравнением одинаковых концентраций ДНК, выделенных из плазмид, с CsCl2-oчищeннoй ДНК. Одна единица оптической плотности (OD) при 260 нм соответствует 50 μг/мл ДНК.
Пример 4
Липиды
Трифторацетат 2,3-диолеилокси-N- [2(сперминкарбоксамидо)этил]-N,N-диметил-1-пропанаминия (DOSPA)/диолеилфосфатидилэтаноламин (DOPE) (DOSPA/DOPE, липофектамин) получали из GIBCO/BRL. Бромид N-(2-гидроксиэтил)-N,N-диметил-2,3-бис(тетрадецитокси)-1-пропанаминия (DMRIE)/DOPE получали из Genzyme Corp. (Framingham, MA).
Пример 5
Получение препарата ДНК-катионная липосома
Соотношение ДНК-липид оптимизировали следующим образом. Плазмидную кДНК (2,5 μг) объединяли в воде с различными концентрациями катионного липида с целью получения набора соотношений ДНК:липид. Смеси ДНК:липид инкубировали в течение 15 минут при комнатной температуре и комплексы разделяли посредством электрофореза на 1%-ном геле агарозы в буфере IX Трис-ацетат (40 мМ) - ЭДТА (2 мМ) (ТАЕ). Оптимальными считали соотношения концентраций ДНК:липид, когда вся ДНК была связана липидом и, следовательно, удерживалась в начале геля. Соотношения подтверждали трансфекцией клеток А549, как описано ниже. Оптимальные соотношения ДНК: липид подтверждали для каждой комбинации липида и плазмиды и для каждой партии ДНК и липида. Для DOSPA/DOPE оптимальным соотношением по весу было соотношение 1 μг ДНК: 3 μг липида, а для DMRIE/DOPE 1 μг ДНК: 4 μг липида.
Пример 6
Трансфекция
Все трансфекции проводили в OPTIMEM (GIBCO-Life Technologies). Один μг pCMVβ-gal объединяли с 4 μг DMRIE/DOPE или 3 μг DOSPA/DOPE в 1 мл OPTIMEM и инкубировали в течение 15 минут при комнатной температуре. Клеточные монослои дважды промывали в бессывороточной среде, затем покрывали 1 мл трансфекционной смеси и инкубировали в течение 2,5 часов при 37oС во влажном инкубаторе при 5% CO2. ДНК-липосомный верхний слой заменяли DMEM с 10% FBS и инкубировали клетки в течение 48 часов. Клеточные лизаты собирали и анализировали на общий белок в соответствии с инструкциями изготовителей (БСА-тест, Pierce Chemical, Rockfied, IL) и определяли активность β-галактозидазы при помощи колориметрического способа с хлорфеноловым красным β-D-галактопиранозидом (CPRG), как описано (Boerhinger-Mannheim, Germany). Где указано, клеточные монослои промывали PBS, затем фиксировали смесью 2%-ного формальдегида - 0,2% глутаральдегида и окрашивали посредством X-gal (40 μг/мл, 5'-3' Inc. Boulder, CO) в 20 мМ феррицианиде калия, 20 мМ ферроцианиде калия и 2 мМ хлориде магния.
Пример 7
Исследование поглощения комплексов ДНК-липид
3Н-тимидинмеченую плазмиду pCMV β-gal получали, добавляя 1 мКи 3Н-TdR (3Н-TdR, 74 ГВк/ммоль, Amersham) к 25 мл выращиваемой в течение ночи культуры pCMVβ-gal-трасформированного штамма DH5α Escherichia coli. Меченую плазмиду выделяли, используя колонки типа Qiagen (размером 100 μг, Chatworth, CA) по инструкции изготовителя. Насыщаемое связывание меченой ДНК определяли следующим образом. Серийные разведения от 1 μг до 0,125 μг 3H-TdR-мeчeнoй ДНК добавляли к клеткам А549 и инкубировали 2, 6 или 24 часа. Монослои промывали, ополаскивали IX трипсином, слегка трипсинизировали и центрифугировали, получая осадок. Осадки лизировали в 100 μл буфера для лизиса (0,1 М Трис и 0,5% Тритон Х-100), добавляли жидкий сцинтилляционный коктейль (BCS, Amersham) и определяли количественно метку 3Н-плазмиды жидкостной сцинтилляцией. Для экспериментов по поглощению клетки А549 предварительно обрабатывали либо глюкокортикоидами, либо средой в течение 4 часов. Клетки обрабатывали насыщаемой концентрацией смеси 3Н-меченая плазмида плюс липид в течение 2,5 часов. Поглощение определяли, как описано выше.
Пример 8
РНК-специфическая полимеразная цепная реакция (RS-PCR)
RS-PCR проводили, как описано и хорошо известно в данной области. Вкратце, клетки предварительно обрабатывали беклометазоном при концентрации 10-6 М и проводили трансфекцию, как описано выше. После завершения инкубации клетки лизировали, используя подкисленный гуанидинийизотиоцианат (Fluka Chemical), содержащий 0,75 М цитрат натрия и 1% саркозил. РНК собирали, используя систему RNAeasy sistem (Qiegen, Chatsworth, CA). Общее количество РНК определяли по поглощению при 260 нм (А260) и 0,5 μг РНК подвергали электрофорезу на геле для определения целостности РНК. Используя 0,3 μг общей РНК, получали кДНК с применением обратной транскриптазы MMLV (GIBCO, Life Technologies), замещая праймером T30D20-gal (GAACATCGATGACAAGCTTAGGTATCGATACACCTCGCGGAAACCGAСАТ) олиго dТ. Этот праймер содержит 20 пар оснований, комплементарных 3' концу мРНК и 30 посторонних оснований. Кроме того, каждую реакцию снабжали 2,5 μКи 32Р α-dCTR в качестве изотопного индикатора. Реакцию инкубировали при 37oС в течение 1 часа. После завершения инкубации собирали гибриды ДНК-РНК, используя набор для Qiaquick spin PCR очистки (Qiagen, Chatsworth, CA) для удаления праймера и невключившегося изотопа. Один μл кДНК наносили в виде пятна на нитроцеллюлозные фильтры в 2-х повторностях и считали на жидкостном сцинтилляционном счетчике (LSC).
Для PCR использовали 25000 имп/мин для каждого образца. В остальном PCR осуществляли так, как описано. PCR-коктейль содержал 500 мМ КС1, 50 мМ Трис-HCl, рН 9,0, 50 мМ NaCl, 10 мМ MgCl2, 200 μМ dNTP и 1 единицу ДНК-полимеразы Tag (Promega, Madison WI).
Праймеры:
5β-gal (GAGAATCCGACGGGTTGTTACT) и
Т30-gal (GAACATCGATGAACAAGCTTAGGTATCGATA), который представляет 30 концевых нуклеотидов T30D20-gal олигомера, использовали при концентрации 1,25 μМ. Циклическое PCR оптимизировали для этой комбинации праймеров и матриц (33 цикла). По завершении PCR продукты разделяли в 1%-ном геле ТАЕ-агарозы и затем промокали (блоттировали) на найлоновый фильтр. Фильтры гибридизовали в течение ночи с 32Р-меченым pCMV β-gal-зондом для удаления негибридизированной метки. Блоты оценивали на бета-скеннере Betagen и определяли имп/мин для полос правильного размера.
Пример 9
Влияние бактериального липополисахарида и интерлейкина-1β на эффективность трансфекции
Липополисахарид в бактериальной клетке является эффективным иммуномодулятором, способным инициировать каскад событий, в том числе образование цитокина и рекрутмент клеток. В предыдущих исследованиях было показано, что клетки А549 реагируют на оба вещества, липополисахарид и IL-1β, позитивной регуляцией мРНК IL-1β (липополисахарид), а также индукцией мРНК IL-8 (IL-1β).
В настоящем изобретении клетки А549 обрабатывали в течение 4 часов либо 100 Е/мл IL-1β, либо 0,5 μг/мл липополисахарида, затем трансфицировали pCMVβ-gal-DMRIE/DOPE. Как можно видеть из Фигуры 1, как IL-1β, так и липополисахарид существенно снижали β-gal-активность относительно клеток, обработанных средой.
Пример 10
Активность β-галактозидазы, повышенная глюкокортикоидами локального действия, в А549 клетках, трансфицированных комплексом pCMV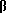 -gal-катионный липид
-gal-катионный липид
Клетки А549 предварительно перед трансфекцией обрабатывали в течение 4 часов синтетическим беклометазоном локального действия в диапазоне 10-7-10-6 М или средой. При дозе беклометазона в диапазоне 5•10-7-10-6 М наблюдали устойчивое усиление в 2-4 раза, и только слабое усиление наблюдали при дозе беклометазона 10-7 М (фигура 2А). Кроме того, другие стероиды локального действия, такие как будезонид, FLUN и нейтральная липосомная форма беклометазона, беклометазон-DLPC, также усиливали β-gal-активность в клетках А549 (фигура 2В). Беклометазон-DLPC при концентрации 10-6 М также повышал β-gal-активность в такой же степени, что и будезонид или беклометазон в этаноле (фигура 2А), что указывает на то, что липосомная часть не препятствует этому эффекту. FLUN устойчиво демонстрировал меньшее усиливающее действие, что согласуется с активностью этих глюкокортикоидов.
Пример 11
Обработка глюкокортикоидами обращает подавление экспрессии гена IL-1β и липопоплисахаридом
По-видимому, беклометазон обращает нежелательные эффекты двух указанных иммуномодуляторов. Для более подробной демонстрации этого явления клетки А549 предварительно обрабатывали в течение 4 часов 10-6 М беклометазоном с последующей обработкой липополисахаридом или IL-1β в течение 4 часов перед трансфекцией pCMV β-gal-DMRIE/DOPE. Как было показано раньше, IL-1 β и липополисахарид подавляли степень β-gal-экспрессии и усиленной беклометазоном β-gal-активности в 2,5 раза. В клетках перед обработкой беклометазоном, а затем обработанных IL-1β или липополисахаридом (беклометазон, липополисахарид; беклометазон, IL-1β) β-gal-экспрессия превышала экспрессию, наблюдаемую в необработанных клетках, в 2 раза и приближалась к уровню клеток, предварительно обработанных "только беклометазоном"; это дает основание предполагать, что беклометазон блокировал ингибиторные эффекты IL-1β или липополисахарида.
Так как многие из пациентов, подвергавшихся генной терапии, могут иметь хронические грамотрицательные бактериальные инфекции, ингибиторные эффекты липополисахарида и IL-1β были бы, наиболее вероятно, уместны перед доставкой гена или беклометазона. Для определения, может ли усиливающее действие беклометазона на β-gal-экспрессию преодолеть подавление β-gal-активности в клетках, предварительно обработанных липополисахаридом или IL-1β, клетки А549 обрабатывали в течение 4 часов липополисахаридом или IL-1β, затем обрабатывали в течение 4 часов беклометазоном перед трансфекцией pCMVβ-gal-DMRIE/DOPE (IL-1β, ВЕС; липополисахарид, ВЕС).
На фигуре 3 показано, что даже при обработке клеток липополисахаридом или IL-1β, так что процесс ингибирования уже устанавливается (см. обработку только липополисахаридом, IL-1β), обработка беклометазондипропионатом не только восстанавливала β-gal-активность, но все еще увеличивала β-gal-
активность больше, чем в обработанных средой трансфицированных клетках. Уровень β-gal-активности в предварительно обработанных липополисахаридом или IL-1β, а затем обработанных беклометазоном клетках не вполне достигал уровня усиленной β-gal-активности, обнаруженного в клетках, предобработанных только беклометазоном, что дает основание предполагать, что остается некоторая часть ингибиторной активности, индуцируемой липополисахаридом или IL-1β. Таким образом, настоящее изобретение демонстрирует, что даже во время существования инфекций обработка беклометазоном улучшает трансфекцию гена комплексом ДНК-катионный липид.
Пример 12
Характеристика глюкокортикоидопосредованного повышения активности репортерного гена
Для определения, является ли усиление глюкокортикоидами β-gal-активности более обобщенным явлением, выделяли первичные клетки легких крыс и обрабатывали их либо средой, либо глюкокортикоидами в течение 4 часов перед трансфекцией pCMVβ-gal-DMRIE/DOPE. Как можно видеть из фигуры 4, комбинированные результаты из нескольких различных препаратов клеток крысы дают 2-кратное увеличение β-gal-активности. По-видимому, первичные клетки легких крыс являются более чувствительными к беклометазону, чем клетки А549 с оптимальным усилением β-gal-активности при 10-7 М. Аналогично наблюдениям на клетках А549, липополисахарид проявлял β-gal-активность. Также обработка беклометазоном с последующий стимуляцией липополисахаридом восстанавливала β-gаl-активность до активности, соответствующей клеткам, предварительно обработанным средой; усилением выше контроля было незначительным. Таким образом, глюкокортикоиды также снимали ингибиторные действия липополисахарида на β-gal-активность в гетерогенной популяции первичных клеток легких крыс, что дает основание предполагать, что глюкокортикоиды могут также повышать трансфекцию аналогичных типов клеток in vivo.
Действие глюкокортикоидов не было специфическим по отношению к катионному липиду DMRIE/DOPE, так как экспрессия β-gal также усиливалась в клетках, предварительно обработанных беклометазоном, затем трансфицированных pCMVβ-gal- и DOSPA/DOPE, другим катионным липидом (фигура 5А). Эффект неспецифичен по отношению к промотору CMV в векторе, так как глюкокортикоидопосредованное усилие наблюдали и тогда, когда клетки предобрабатывали беклометазондипропионатом и трансфицировали pSVβ-gal-DMRIE/DOPE, вектором, который содержит промотор SV40 (фигура 5В). И, наконец, клюкокортикоидопосредованное усиление экспрессии гена не ограничено репортерным геном β-gal, так как аналогичные результаты наблюдали, когда рецепторным геном было хлорамфениколацетил-трансфераза (CAT) (фигура 5С).
Чтобы исключить возможность того, что другие стероиды также могут усиливать β-gаl-активность, клетки А549 обрабатывали различными дозами эстрогена (эстрадиола), прогестерона, холестерина или беклометазона в течение 4 часов перед трансфекцией pCMVβ-gal или pSVβ с использованием катионного липида DMRIE/DOPE. Усиление β-gal-активности наблюдали только в клетках, обработанных беклометазоном, что свидетельствует о том, что в А549 эффект усиления является глюкокортикоидспецифичным (фигура 5В). Зависимость от дозы беклометазонопосредованного усиления при использовании другого липида, промотора или репортерного гена была аналогична реакции, наблюдаемой при использовании в клетках А549 pCMVβ-gal плюс DMRIE/DOPE, причем увеличение активности репортерного гена было максимальным при 5•10-7 М-10-6 M.
Эти объединенные результаты исследований настоящего изобретения показывают, что этот эффект, относящийся к действию глюкокортикоидов на клетки, является общим явлением, а не специфическим для конкретных глюкокортикоида, репортерного гена, промотора, липида или клеточной линии. Действие зависело от дозы в первичных клетках легких, демонстрируя большую чувствительность к беклометазону. Так как эффект не был специфическим относительно отдельных глюкокортикоидов, для остальных описанных здесь исследований использовали беклометазон, растворенный в этаноле.
Пример 13
Механизм опосредованного беклометазондипропионатом усиления экспрессии репортерного гена: поглощение вектора
Одной из возможностей усиления экспрессии репортерного гена было увеличение поглощения вектора. К этой возможности обратились двумя способами: определением количества β-gаl-окрашенных клеток относительно активности и поглощением комплекса "радиоактивно-меченая плазмида - катионный липид".
Для окрашенных клеток дупликатные клеточные культуры клеток А549 обрабатывали в течение 4 часов беклометазондипропионатом или только средой, затем трансфицировали pCMVβ-gal с использованием DMRIE/DOPE. Через 48 часов один набор культур лизировали и определяли β-да1-активность, тогда как другой набор культур фиксировали нейтральным забуференным формалином и окрашивали β-gal (для каждого варианта по две лунки). Окрашенные клетки подсчитывали на площади 1 см2. Как можно видеть из таблицы, хотя в β-gаl-тесте β-gаl-активность возрастала, количество окрашенных на β-gal клеток было одинаково в образцах клеток, предобработанных средой, и образцах клеток, предобработанных беклометазондипропионатом, что дает основание предполагать, что повышенная β-gal-экспрессия не была обусловлена большим числом клеток, поглотивших плазмиды.
Так как возможно, что технология окрашивания не была недостаточно чувствительна для детектирования увеличенного поглощения плазмид, этот вопрос решали сравнением поглощения радиоактивно-меченой плазмиды в клетках, обработанных беклометазоном или средой. Для гарантии того, чтобы плазмида по существу не изменялась при введении метки, ее метили метаболическим способом путем добавления 3Н-тимидина к бактериальной культуре во время получения плазмиды (vide supra). После определения насыщаемого связывания и кинетики поглощения для меченого материала клетки А549 обрабатывали в течение 4 часов несколькими концентрациями беклометазондипропионата и затем трансфицировали 3Н-меченым ДНК-DMRIE/DOPE. Через 2 часа клетки обрабатывали трипсином, лизировали и определяли поглощение радиоактивно-меченых плазмид при помощи LSC. Результаты, представленные на фигуре 6, подтверждают то, что обнаружено при окрашивании; по-видимому, беклометазон не усиливает поглощение и, следовательно, для повышения β-да1-активности беклометазон должен действовать по другому механизму.
Пример 14
Кинетика GC-опосредованного усиления экспрессии трансфицированной β-gаl
Для определения временного хода опосредованного глюкокортикоидом усиления β-gal-активности клетки А549 предварительно обрабатывали в течение различного времени средой или 10-6 M беклометазоном для определения кинетики данного эффекта. Как можно видеть на фигуре 7, для увеличения β-gal-трансфекции требовалось действие беклометазона в течение минимум 3-4 часов. Повышенная β-gal-активность была максимальной через 8 часов после обработки и сохранялась максимальной в течение 24 часов наблюдения. Необходимость 3-4-часовой предварительной обработки плюс эксперименты по поглощению плазмиды предполагают, что скорее стадия внутриклеточного синтеза, чем событие на клеточной поверхности, участвует в опосредовании усиленного беклометазоном β-gal-ответа.
Пример 15
Внутриклеточное действие опосредованного глюкокортикоидами повышения векторной активности
Для демонстрации того, требует ли беклометазонопосредованное усиление β-gal-активности синтеза белков, использовали ингибитор белкового синтеза циклогексимид (СНХ). Культуры клеток, обозначенных А549, предобрабатывали средой или СНХ для ингибирования белкового синтеза. Через 30 минут обработанные средой клеточные культуры обрабатывали либо средой, либо беклометазоном. Культуры, уже обработанные СНХ, дополнительно обрабатывали в течение 4 часов клеток либо средой + СНХ, либо беклометазоном + СНХ. Все клетки трансфицировали pCMVβ-gal-DMRIE/DOPE и через 48 часов определяли β-gal-активность.
Как можно видеть на фигуре 8, СНХ умеренно подавлял β-gal-активность в контрольных клетках наиболее вероятно путем ингибирования синтеза β-gal-белка (сравните pCMVβ-gal плюс и минус СНХ). В противоположность этому, в клетках, обработанных беклометазоном + СНХ, все еще было видно опосредованное беклометазоном повышение β-gal-активности. В действительности, опосредованное беклометазоном повышение β-gal-активности было даже больше в присутствии СНХ. Эти результаты показывают, что синтез белков не был необходимым для повышения β-gal-активности беклометазоном.
Пример 16
Действие ВЕС на уровни мРНК pCMV β-gal в клетках А549
Для установления кинетики экспрессии гена β-gal клеток А549 трансфицировали pCMVβ-gal-DMRIE/DOPE. В различные моменты времени после завершения трансфекции (2,5 часа) собирали общую РНК (Т=2, 3, 7, 11 и 24 часа). мРНК являлась детектируемой при 2 часах и оставалась на высоком уровне в течение всего периода исследования (данные не показаны). Сразу же после установления кинетики экспрессии гена β-gal клетки А549 преедварительно обрабатывали средой или 10-6 М беклометазоном в течение 4 часов с последующей трансфекцией в течение 2,5 часов pCMV β-gal. Общую РНК собирали, когда Т=0, 2, 3 или 24 час, и после этого определяли уровни мРНК способом RS-PCR. Хотя уровни мРНК были высокими в необработанных клетках, а также в клетках, обработанных беклометазоном, при Т= 2 часам (38-и 18-кратная индукция соответственно) в клетках, обработанных беклометазоном, при Т=3 и Т=24 часам обнаруживалось явное увеличение уровней мРНК устойчивого состояния (фигура 9). Эти результаты показывают, что глюкокортикоиды либо повышали транскрипцию, либо стабилизировали мРНК в цитоплазме.
Настоящее изобретение демонстрирует, что глюкокортикоиды повышают экспрессию репортерных генов, трансфицированных в клетки посредством катионных липидов. Действие глюкокортикоидов не зависит от конкретных катионного липида, векторного промотора, векторного репортерного гена или типа клеток. Напротив, действие глюкокортикоидов является общим явлением для глюкокортикоидов, поскольку другие стероиды, такие как эстроген, прогестерон или холестерин, не повышали экспрессию репортерного гена с любым из двух промоторов, запускающих ген β-gаl. Глюкокортикоиды, по-видимому, действовали, не повышая поглощение плазмид клетками, а увеличивая каким-то образом экспрессию репортерного гена. Для обнаружения повышения экспрессии гена требовалось минимально 3-4 часа воздействия глюкокортикоидами. Для усиления активности репортерного гена беклометазоном не требовался синтез новых белков, и СНХ не повышал экспрессию гена в отсутствие беклометазондипропионата, давая основание полагать, что, наблюдаемая в присутствии СНХ, супериндукция может быть глюкокортикоидспецифической.
Действие глюкокортикоидов наблюдали в А549, клеточной линии рака легких человека, а также в первичных клеточных изолятах легких крысы. Глюкокортикоидопосредованного увеличения β-gаl-активности не наблюдали в клетках COS-1 почек обезьян, трансфицированных pCMVβ-gal-DMRIE/DOPE. Возможно, что клетки COS-1 испытывают недостаток рецепторов для глюкокортикоидов или недостаток какого-либо другого глюкокортикоидчувствительного внутриклеточного фактора, участвующего в усиленной реакции. Клеточная линия, которая не реагирует на действие глюкокортикоидов, может быть полезной в выяснении точной природы этого действия.
Наиболее вероятно, большее усиление β-gal-активности беклометазоном в присутствии липополисахарида (фигура 4) должно было бы наблюдаться, если использовали концентрацию беклометазона 10-7 М вместо концентрации 10-6 М, которая была менее эффективна, чем 10-7 М, в первичных культурах крыс. Эту модификацию проверяли вместе с концентрацией промежуточного продукта 5•10-7 М.
Липополисахарид присутствовал в использованных препаратах плазмид и фактически в каждом другом плазмидном препарате, получаемом в большинстве лабораторий. Удаление липополисахарида из плазмиды является препятствием, которое следует преодолеть, при клиническом применении генной терапии.
В любом случае, находится ли липополисахарид в плазмиде или предсуществует в воспаленных легких, способность беклометазона обращать (снимать) ингибиторные действия липополисахарида, или IL-1β, или, возможно, других цитокинов, или иммунных процессов должна иметь большое значение для генной терапии в клинике.
Механизм глюкокортикоидопосредованного усиления экспрессии репортерного гена только начал проясняться. Показано, что глюкокортикоиды обладают многочисленными регуляторными свойствами, которые зависят от концентрации глюкокортикоидов, типа клеток и гена-мишени или внутриклеточного сайта. Глюкокортикоиды могут оказывать сильное влияние на иммунную реакцию; гены, для которых известно, что они отрицательно регулируются глюкокортикоидами, включают ген IL-1β, TNF-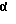 , фактор межклеточной адгезии молекулы -1 (ICAM-1), а также структурные гены, которые создают внеклеточный матрикс, такой как коллаген и стромелизин. Показано, что глюкокортикоиды регулируют гены по нескольким различным механизмам, таким как снижение транскрипции генов, как видно для гена IL-1β, но глюкокортикоиды также действуют на экспрессию IL-1β на посттранскрипционной стадии.
, фактор межклеточной адгезии молекулы -1 (ICAM-1), а также структурные гены, которые создают внеклеточный матрикс, такой как коллаген и стромелизин. Показано, что глюкокортикоиды регулируют гены по нескольким различным механизмам, таким как снижение транскрипции генов, как видно для гена IL-1β, но глюкокортикоиды также действуют на экспрессию IL-1β на посттранскрипционной стадии.
Глюкокортикоиды также позитивно регулируют экспрессию генов. Глюкокортикоидный рецептор после взаимодействия с глюкокортикоидами присоединяется непосредственно к регуляторным элементам, то есть специфической ДНК-последовательности, известной как отвечающий на глюкокортикоиды элемент (GRE). В промоторе, отвечающего на глюкокортикоид гена, связывание комплекса GC-глюкокортикоидный рецептор позитивно регулирует транскрипцию данного гена. Альтернативно было показано, что глюкокортикоидный рецептор связывается с другими факторами транскрипции, такими как c-fos или c-jun, которые в отсутствие глюкокортикоидного рецептора активируют транскрипцию гена. При ассоциации глюкокортикоидного рецептора с c-fos или c-jun, c-fos и c-jun не доступны для того, чтобы активировать чувствительные гены.
В настоящем изобретении ни один из использованных промоторов не имеет более общих GRE, как определено поиском в генном банке последовательностей. Период полураспада матричной РНК мог быть удлинен посредством глюкокортикоидопосредованного ингибирования мРНК-специфической эндонуклеазы, но так как в отсутствие беклометазона СНХ не оказывает супериндуцирующего влияния на экспрессию генов, ингибирование эндонуклеазы также должно иметь некоторую специфичность в отношении глюкокортикоидов. Альтернативно кофактор, который усиливает транскрипцию или трансляцию, кофактор, для которого не является необходимым синтез белка de novo, может позитивно модулироваться глюкокортикоидами, повышая таким образом образование белка репортерного гена.
Недавнее исследование Liu и др. показали усиление дексаметазоном (DEX) активности репортерного гена, когда он был присоединен к сегменту промотора соматостатина. Было показано, что этот механизм не содержит классического GRE в промоторе, но вместо этого глюкокортикоидный рецептор обнаружил кооперативную активность с белком, связывающим сАМР-регуляторный элемент (CREB), который, как известно, присоединяется к другому транскрипционному элементу, сАМР-регуляторному элементу (CRE). И сАМР, и протеинкиназа А участвовали в Dex-опосредованном усилении активности репортерного гена. Сходная ситуация была показана для гена РЕРСК; глюкокортикоиды позитивно регулировали и сАМР усиливал транскрипцию. В настоящем изобретении продемонстрированное действие глюкокортикоидов не было специфическим относительно единственного промоторного района. То есть глюкокортикоиды усиливали β-gаl-активность из векторов, которые имели либо промотор SV40, либо промотор CMV. Маловероятно, что оба эти промотора имеют одинаковые CRE последовательности в своих промоторах, но эта возможность неизвестна.
Другое исследование показало позитивные эффекты клеточной пролиферации на экспрессию трансфицированного гена. Подобно тому, что здесь наблюдалось, не отмечали возрастания поглощения репортерного гена, но в клетках, стимулированных к пролиферации повреждением клеток, наблюдали 10-кратное увеличение активности люциферазы. Маловероятно, что глюкокортикоиды могли индуцировать клеточную пролиферацию в исследованиях настоящей заявки, так как плотность клеток А549 или клеток легких была такова, что клетки находились в фазе логарифмического роста в течение всего периода инкубации 48 часов.
Глюкокортикоиды использовали в клинике в течение многих лет и их считали безопасными и эффективными. Пациенты с хроническим воспалением легких являются подходящими кандидатами для локального лечения глюкокортикоидами, доставляемыми в виде аэрозоля. Кроме того, показано, что лечение глюкокортикоидами улучшает общую функцию легких у пациентов с муковисцидозом, в особенности, если лечение дается на ранней стадии заболевания.
Безопасность глюкокортикоидов и стоимость генной терапии, которая включает дорогие препараты ДНК и катионного липида, и учет потенциальных побочных эффектов повторного приема больших количеств ДНК-липида делают настоящее изобретение все более привлекательным как в смысле генной терапии, так и с точки зрения молекулярной биологии глюкокортикоидов.
Любые патенты и публикации, упомянутые в данном описании, являются показателями уровня специалистов, к которым данное изобретение имеет отношение. Эти патенты и публикации включены здесь в виде ссылок в той же степени, как если бы каждая отдельная публикация специально и индивидуально была указана, как включенная в виде ссылки.
Специалисту в данной области должно быть понятно, что настоящее изобретение хорошо приспособлено для выполнения целей и получения упомянутых результатов и преимуществ, а также других присущих ему преимуществ. Приведенные примеры вместе с описанными здесь способами, процедурами, лечением, молекулами и специфическими соединениями представляют собой предпочтительные в настоящее время варианты, являются примерами и не предназначены для ограничения объема изобретения. Для специалиста в данной области ясны изменения и другие применения, которые соответствуют сущности изобретения, определенной формулой изобретения.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
(2) ИНФОРМАЦИЯ ДЛЯ SEQ ID NO:1:
(i) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 50
(B) ТИП: нуклеиновая кислота
(C) ТИП НИТИ: однонитевая
(D) Топология: линейная
(ii) ТИП МОЛЕКУЛЫ:
(A) Описание: другая нуклеиновая кислота
(iii) ГИПОТЕТИЧЕСКАЯ: нет
(iv) АНТИСМЫСЛОВАЯ: нет
(vi) ИСТОЧНИК ПРОИСХОЖДЕНИЯ:
(B) ШТАММ: N/A
(C) ИНДИВИДУАЛЬНЫЙ ИЗОЛЯТ: N/A
(D) СТАДИЯ РАЗВИТИЯ: N/A
(F) ТИП ТКАНИ: N/A
(G) КЛЕТОЧНЫЙ ТИП: N/A
(Н) КЛЕТОЧНАЯ ЛИНИЯ: N/A
(xi) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: SEQ ID NO:1: GAACATCGAT GACAAGCTTA GGTATCGATA CACCTCGCGG AAACCGACAT 50
(3) ИНФОРМАЦИЯ ДЛЯ SEQ ID NO:2:
(i) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 22
(B) ТИП: нуклеиновая кислота
(C) ТИП НИТИ: однонитевая
(D) Топология: линейная
(ii) ТИП МОЛЕКУЛЫ:
(A) Описание: другая нуклеиновая кислота
(iii) ГИПОТЕТИЧЕСКАЯ: нет
(iv) АНТИСМЫСЛОВАЯ: нет
(vi) ИСТОЧНИК ПРОИСХОЖДЕНИЯ:
(B) ШТАММ: N/A
(C) ИНДИВИДУАЛЬНЫЙ ИЗОЛЯТ: N/A
(D) СТАДИЯ РАЗВИТИЯ: N/A
(F) ТИП ТКАНИ: N/A
(G) КЛЕТОЧНЫЙ ТИП: N/A
(Н) КЛЕТОЧНАЯ ЛИНИЯ: N/A
(xi) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: SEQ ID NO:2: GAGAATCCGA CGGGTTGTTA CT 22
(4) ИНФОРМАЦИЯ ДЛЯ SEQ ID NО:3:
(i) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 31
(B) ТИП: нуклеиновая кислота
(C) ТИП НИТИ: однонитевая
(D) Топология: линейная
(ii) ТИП МОЛЕКУЛЫ:
(А) Описание: другая нуклеиновая кислота
(iii) ГИПОТЕТИЧЕСКАЯ: нет
(iv) АНТИСМЫСЛОВАЯ: нет
(vi) ИСТОЧНИК ПРОИСХОЖДЕНИЯ:
(B) ШТАММ:
(C) ИНДИВИДУАЛЬНЫЙ ИЗОЛЯТ:
(D) СТАДИЯ РАЗВИТИЯ:
(F) ТИП ТКАНИ:
(G) КЛЕТОЧНЫЙ ТИП:
(Н) КЛЕТОЧНАЯ ЛИНИЯ:
(xi) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: SEQ ID NO:3: GAACATCGAT GAACAAGCTT AGGTATCGAT A 31

Изобретение относится к области медицины и биотехнологии и касается усиления экспрессии генов глюкокортикоидами. Способ включает усиление экспрессии гена в клетке посредством введения в клетку экспрессирующего ген вектора и введение в указанную клетку фармакологически эффективной дозы глюкокортикоида в количестве, достаточном для усиления клеточной экспрессии указанного гена, где указанный ген не зависит от промотора, не имеющего глюкокортикоидреактивного элемента. Преимущество изобретения заключается в повышении эффективности доставки терапевтических уровней трансфицированных генов. 12 з.п.ф-лы, 1 табл., 9 ил.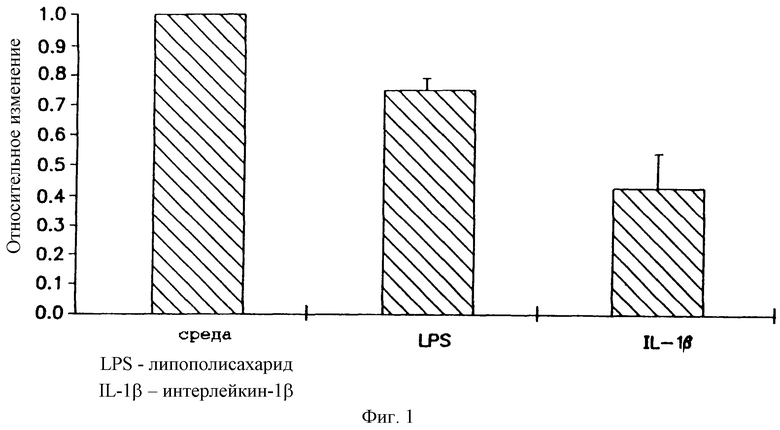
| MALONE et al | |||
| Dexamethasone enhancement of Gene Expression after hepatic DNA infection, J | |||
| of Biol | |||
| Chemistry, 1994, v.269, №47, рр.29903-29907 | |||
| SLATER et al | |||
| Regulation of transcription by steroid hormones | |||
| - Annals New York Academy of Sciences, 1994, v.733, рр.103-112 | |||
| JDO et al | |||
| Gene therapy for hepatoma cells using a retrovirus vector carrying Herpes Simplex Virus thymidine Kinase Gene under control of Human alpha - fetoprotein promotor | |||
| - Cancer Research, 15.07.1995, v.55, рр.3105-3109. |

