Изобретение относится к целенаправленной блокировке генетической информации вирусной мРНК для репликации ретровирусов, например, HIV, в трансфецированных гематопоэтических клетках, более конкретно, к плазмидам для экспрессии анти-мРНК ретровируса HIV-1.
Изобретение относится к плазмидам, кодирующим последовательности анти-мРНК, которые в отличие от уровня техники обеспечивают полное торможение репликации вируса HIV-1 в измененных в отношении генов клетках человека. Описанные в рамках данной заявки плазмиды в особенности годятся для соматической генной терапии заболеваний СПИДом, так как они защищают гематопоэтические клетки от индуцируемых вирусом HIV-1 цитопатических эффектов. Таким образом можно предотвращать иммунную недостаточность у людей, зараженных вирусом HIV. Размножение вируса в зараженной клетке управляется генами вирусного генома. При этом этими генами транскрипцией и трансляцией образуются регуляторные генные продукты и структурные белки для построения инфекционных вирусных частиц, обеспечивающих размножение вируса. Играющим основную роль в этом процессе продуктом является мРНК, которая дает информации для образования регуляторных белков и структурных белков.
Высокоспецифичная блокировка мРНК, основанная исключительно на неповторимости молекулярной структуры вирусной мРНК, а именно специфичной для вируса последовательности нуклеооснований этой молекулы, может осуществляться с помощью комплементарной к рибонуклеиновой кислоте последовательности оснований. В результате специфичного в отношении последовательности спаривания нуклеооснований, как это аналогично описано для дезоксирибонуклеиновой кислоты, однонитевая мРНК комплексируется в двухнитевой РНК, так что генетическая информация последовательности нуклеооснований для осуществления биосинтеза белков становится недоступной. Тем самым вторая стадия экспрессии гена, то есть трансляция, задерживается. Комплементарная рибонуклеиновая кислота обозначается как анти-мРНК.
Механизм торможения экспрессии гена в результате взаимодействий анти-мРНК с целевой мРНК является комплексным и в основном еще не разъяснен. Одним объяснением является задержка сплайсинга и переработки РНК в клеточном ядре. Но детальные исследования в этом отношении еще не проводились. Есть публикация об обмене аденозина на инозин в двухнитевой РНК, результатом которого является задержка трансляции. Возможно, что затрудняемый перенос РНК через перинуклеарную мембрану в место биосинтеза белков и маскирование двухнитевой РНК-ой взаимодействия между рибосомами и РНК могут далее содействовать торможению экспрессии гена, вызываемому анти-мРНК.
Известна задержка процессов обмена веществ воздействием анти-мРНК в природных биологических системах, таких, как, например, бактерии и некоторые эукариотные многоклеточные организмы [см. К.М. Такаяма и М. Инуйе, Critical Reviews in Biochemistry and Molecular Biology, 25, 155 (1990)], а также в искусственно созданных переносом генов трансгенных организмах [см. Й. Моль и др. FEBS Letters 268, стр. 427 (1900)].
Есть и публикации, касающиеся создания в растениях и клетках животных устойчивых к воздействию вирусов организмов за счет включения гена, экспримирующего анти-мРНК (см. М. Куоццо и др. Biotechology 6, 549 (1988), а также Т. Рюден и Е. Гильбоа, J. Virol. 63, стр. 677, 1989 и ЕР 252440]. Но, как указывают сами авторы в этих публикациях, убедительное научное обоснование связи между экспрессией анти-мРНК и устойчивостью к воздействию вирусов до сих пор еще не удалось. Так, например, в частности, в случае микроиньицированных, экспримирующих анти-мРНК плазмид при временной экспрессии антивирусное действие является слабым или же приводит к неподдающимся обоснованной оценке результатам [(К. Риттнер и др., Nucleic Acids Resarch 19, стр. 1421 (1991)] . В литературе есть и сведения о экспримирующих анти-мРНК ретровирусных векторах для образования устойчивых к воздействию вирусов клеток (см. Р. И-Л. , То и др. в Gene Regulation: Biology of Antisense RNA and DNA. изд. Р. П. Эриксон и И.Г. Изант, Равен Пресс Лтд. Нью Йорк 1992]. Но эти ретровирусные векторы могут вызывать рост злокачественных опухолей, так как они содержат подобные онкогенам генные последовательности. Зависимость онкогенеза от ретровирусов широко известна.
Задачей изобретения является разработка плазмид для высокоэффективной экспрессии анти-мРНК ретровируса HIV-1.
Поставленная задача решается предлагаемой плазмидой pSXK1 для экспрессии анти-мРНК ретровируса HIV-1, включающей следующие элементы:
- генный фрагмент HIV-1 681 - 3825;
- промотор цитомегалии;
- ранний промотор SV40;
- сигнал полиаденилирования;
- неомициновый селекционный маркер;
- сайт рестрикции Hind III;
- ген устойчивости к ампициллину;
- возможный хозяин - E.coli.
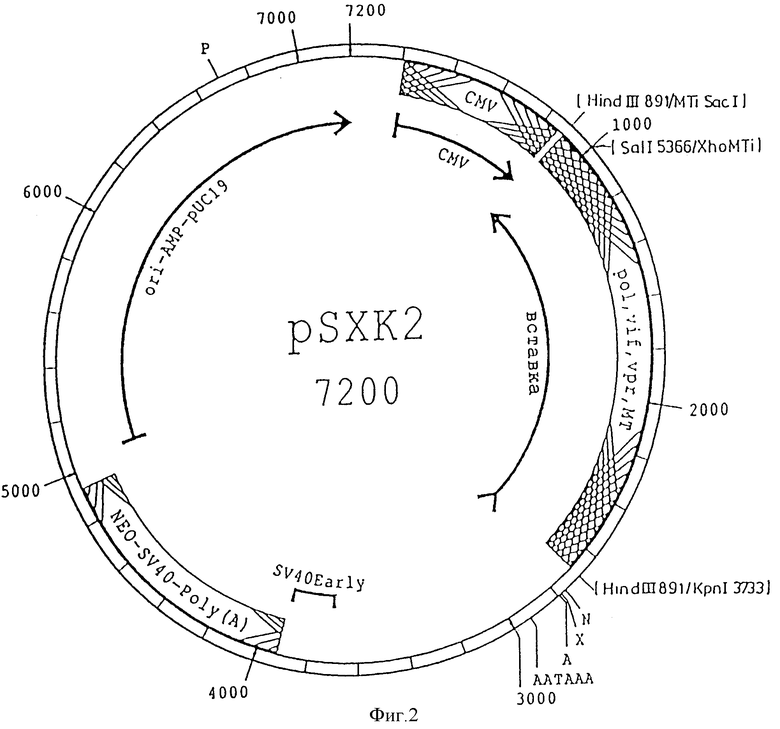
Поставленная задача также решается предлагаемой плазмидой pSXK2 для экспрессии анти-мРНК ретровируса HIV-1, включающей следующие элементы:
- генный фрагмент HIV-1 4158 - 5786;
- гибридный промотор, состоящий из нуклеотидов 209 - 865 промотора цитомегалии и нуклеотидов 150 - 369 металлотионеинового промотора;
- сайт рестрикции Hind III;
- ген устойчивости к ампициллину;
- возможный хозяин - E.coli.
В релевантных для репликации вируса HIV-1 клетках-хозяевах человека плазмиды согласно изобретению показывают антивирусное действие, которое обеспечивает полную защиту от репликации ретровируса HIV-1 в человеческих клетках в течение более 60 дней.
Изобретение далее иллюстрируется следующими примерами.
Пример 1
Плазмида pSXK1
Плазмида pSXK1 для экспрессии анти-мРНК ретровируса HIV-1 сконструирована с тем, чтобы показать экспрессию анти-мРНК под контролем сильного конститутивно экспримирующего промотора, в данном случае промотора вируса цитомегалии. В рестрикционный сайт Hind III у нуклеотида 891 (вектор pRc/CMV, N V750-20 в каталоге фирмы Инвайтроджин, Сан Диего, CA 92121, США) вставлен в ориентации 3' ---> 5' генный фрагмент HIV-1, включающий нуклеотиды 681 - 3825. Сигнал полиаденилирования является производным гена гормона роста крупного рогатого скота. Впрочем плазмида тоже включает неомициновый селекционный маркер и фрагмент ДНК pBR322 для репликации и селекции в клетках E.coli.
Рестрикционная карта плазмиды pSXK1 представлена на фиг. 1.
Пример 2
Плазмида pSXK 2
Плазмида pSXK 2 для экспрессии анти-мРНК ретровируса HIV отличается от плазмиды pSXK 1 тем, что имеет полученный в результате сочетания промоторные последовательности, включающие нуклеотиды 209 - 865 промотора CMV (см. пример 1) и нуклеотиды 150 - 369 металлотионеинового промотора (UWGCG GenEMBL, дело: MUSMETI. RO). Таким образом, он также включает гибридный промотор. Кроме того, включающая нуклеотиды 4158 - 5786 провирусная последовательность ДНК размещена в ориентации 3' ---> 5' в качестве матрицы для экспрессии анти-мРНК под контролем гибридного промотора CMV и металлотионеина. Рестрикционная карта плазмиды pSXK2 представлена на фиг. 2.
Пример 3
Трансфекция и клонирование лимфоцитов человека предлагаемыми плазмидами для экспрессии анти-мРНК
В качестве примеров трансфецированных гематопоэтических клеточных линий человека, которые проявляют устойчивость к воздействию HIV-1 эндогенно экспримируемой анти-мРНК-ой, выбирают растущие без сцепления с субстратом, подобную моноцитам клеточную линию U 937 и Т- лимфоцитную клеточную линию HUT 78. Обе клеточные линии получены от американской коллекции АТСС (12301 Парклаун Драйв, г. Роквилл, MD 20852 - 1776, США).
Клеточные линии культивируют в рекомендуемой АТСС среде. Трансфекцию плазмидами осуществляют нижеуказанными методами, из которых электропорация дает наилучшие результаты.
1) Метод с применением фосфата кальция (аналогично Виглеру и др., 1977, CELL 11, стр. 223).
2) Метод с применением содержащего диэтиламиноэтиловые группы декстрана.
За день до трансфекции клетки доводят до плотности 3-4•105 /мл питательной среды. За 2-4 часа до трансфекции клетки центрифугируют и подают в свежую среду. 0,5 - 20 мкг ДНК смешивают с 1070 мкл буфера PBS и к полученной смеси добавляют 120 мкл раствора, содержащего 10 мг/мл раствора содержащего диэтиламиноэтиловые группы декстрана и полученный таким образом раствор используют для трансфекции. Перед трансфекцией 40 мл клеток центрифугируют, подают в 10 мл буфера PBS и еще раз центрифугируют. Полученный при этом центрифугат суспендируют в 1,2 мл раствора буфера PBS и вышеуказанного декстрана и инкубируют в течение 30 минут, после чего к клеткам добавляют 0,8 мл холодного 20%-ного раствора диметилсульфоксида в буфере TBS. По истечении трех минут добавляют 10 мл TBS и центрифугируют. Клетки промывают средой RPMI, после чего подают в 40 мл той же среды.
3) Сочетание (метод по Санари-Гольдину)
Логарифмически растущие клетки (1•107 клеток/мл) центрифугируют и промывают средой RPMI, после чего добавляют 2 мл суспензии протопластов E.coli (2•103 клеток/мл в 10% сахарозы и 10 мМ хлористого магния). Протопласты со служащей для трансфекции плазмидой получают по методу Санари-Гольдину и др. После 8-минутной инкубации центрифугируют и тщательно добавляют в течение двух минут 45%-ный раствор полиэтиленгликоля с молекулярным весом 400 в среде RPMI. После одноминутной инкубации добавляют в течение 7 минут 10 мл среды RPMI, которая не содержит телячьей эмбриональной сыворотки. Клетки центрифугируют, промывают средой RPMI и подают в 10 мл той же среды.
4) Электропорация
Для осуществления электропорации применяют аппарат Gene Pulser Apparatus and Capacitance Extender фирмы Био Рад. При этом логарифмически росшие клетки доводят до плотности 1•107 клеток/мл, промывают буфером PBS и повторно суспендируют в среде RPMI без телячьей эмбриональной сыворотки, содержащей 10 мМ декстрозы и 0,1 мМ дитиотреитола. 0,4 мл суспензии клеток подают в предназначенную для электропорации кювету и добавляют 10 -20 мкг ДНК. При этом ДНК растворяют в соотношении 1 1мкг/мкл буфера ТЕ. Электропорацию осуществляют в следующих условиях: емкость: 260 - 960 мкF; напряжение: 150 - 300 Bт; расстояние между электродами: 0,2/0,4 см; сопротивление 100 - ∞ Oм. До и после электропорации клетки инкубируют в течение 10 минут при комнатной температуре, после чего подают в 10 мл среды RPMI, содержащей 10% телячьей эмбриональной сыворотки.
Подбор продуктов трансфекции
Подбор продуктов трансфекции осуществляют в каждом случае с учетом устойчивости к воздействию антибиотика G418.
1) Селекция в вязких средах.
Трансфецированные клетки сначала инкубируют в содержащей антибиотик G 418 среде RPMI до тех пор, пока рост больше не может обнаруживаться. Затем клетки высеивают в вязкую среду. Для упрочнения среды служит либо агар, либо метилцеллюлоза. Предварительно селекционированные клетки высеивают в пластинку размером 10 см и добавляют 10 мл среды RPMI, содержащей 1 мг/мл антибиотика G 418, и 2 мл 2%-ного раствора разжиженного агара. После остывания агара осуществляют инкубацию при температуре 37oC в термостате. В дополнительном опыте агар заменяют на метилцеллюлозу.
2) Селекция с последующим клонированием в камере типа Transwell cell culture chamber TM фирмы Костар.
48 часов после трансфекции клетки высеивают в снабженную 96 микроячейками пластинку в количестве 1•105 на 0.2 мл ячейки. В случае клеток U937 антибиотик G418 добавляют в концентрации 0,7 мг/мл, а в случае клеток HUT 78 - в концентрации 1,0 мг/мл. 1/2 объема ячейки заполняют свежей средой после расхода среды. В случае клеток U 937 добавляют только 0,5 мг/мл антибиотика G418. Инкубацию проводят до появления в отдельных ячейках устойчивых клеток. Эти смешанные клоны выращивают до большего объема и затем клонируют. Для этого применяют специальную камеру роста фирмы Костар (тип: Transwell TM, No. каталога: 3425, адрес фирмы: 205 Бродвей, г. Кембридж, МА 02139, США).
Клетки небольшой плотностью (50 - 200 клеток на 6 мл) высеивают в мягком arape с антибиотиком G 418 (см. селекцию в вязкой среде) и 6 мл среды подают в центральную камеру, которая мембраной отделена от очень плотно растущей жидкой культуры клеток U 937 и HUT 78, соответственно (200 000 клеток/мл). После расхода жидкую среду частично заменяют, чтобы обеспечить дальнейший рост клеток. Как только в мягком агаре появятся устойчивые к антибиотику G 418 колоний, их выделяют при помощи пипетки и выращивают.
Примерно по 10 независимым трансфецированным клеточным линиям U 937 и HUT 78 клонируют вышеуказанным образом плазмидами pSXK1 и pSXK2, соответственно, и дальше обрабатывают с тем, чтобы доказать экспрессию анти-мРНК и устойчивость к ретровирусу HIV-1.
Пример 4
Доказательство интеграции интактных экспрессионных векторных конструктов и доказательство экспрессии анти-мРНК.
После обработки трансфецированных клеточных линий U 937 и HUT 78 известными приемами осуществляют градиентное центрифугирование с применением хлорида цезия, после чего ДНК и РНК выделяют из этих клеточных линий известными приемами [см. Л.Г. Дейвис и др., Basic Methods in Molecular Biology, изд. Эльсвир, Нью Йорк (1986)]. Доказательство интеграции интактных плазмид для экспресии анти-мРНК осуществляют при помощи сцепной реакции с применением полимеразы и олигонуклеотидных затравок, которые амплифицируют перекрывающиеся ДНК-фрагменты последовательностей промотора, вставки и окончания. Затем амплифицированную ДНК идентифицируют путем анализа согласно Соутерну с применением генных зондов вставки ДНК. Результаты анализа представлены на фиг. 3 - 5. Для предлагаемых плазмид применяют фланкирующие затравки 5'-CTT TCC AAA ATG TCG TAA CAA CTCC-З' из промоторной последовательности и 5'-ATT TAG GTG ACA CTA TAG AAT-З' из последовательности окончания. Во вставке (последовательности HIV) выбирают затравки, которые в зависимости от своих пар дают реакционные продукты длиной примерно 500 - 2100 пар оснований.
Доказательство экспрессии анти-мРНК. осуществляют путем анализа согласно Нортерну РНК клонированных трансфецированных клеточных линий.
Пояснения к упомянутым в данном примере фиг. 3 - 5.
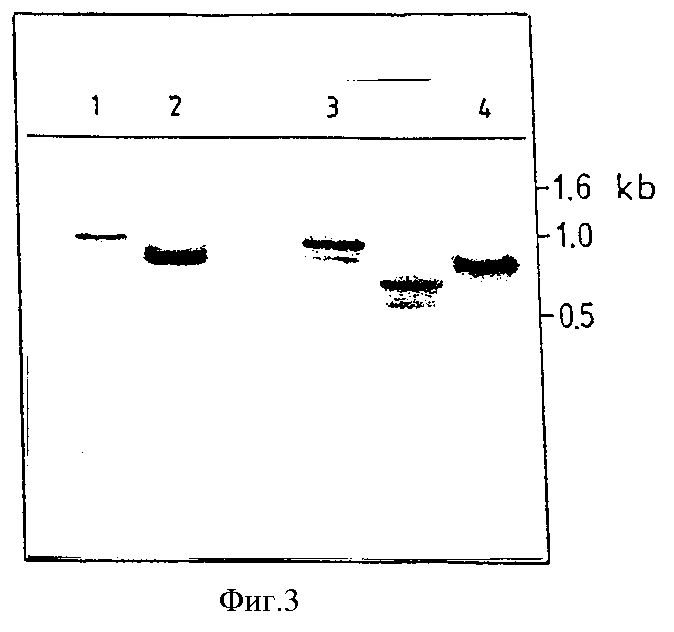
Фиг. 3
Анализ согласно Соутерну хромозомной ДНК из продукта трансфекции клонированной клеточной линии U 937 после вышеуказанной амплификации. Продукт содержит плазмиду pSXK 1, включенную в хромозомную ДНК. Величина фрагментов амплификации и результаты гибридизации генным зондом HIV, который не содержит последовательность затравки продукта амплификации, указывают на неприкосновенность последовательности промотора и вставки. Цифрой 1 обозначен контрольный опыт, 5'- выступ, фрагмент длиной 887 пар оснований (приведенное сокращение "kb" обозначает "килопар оснований"), цифрой 2 - контрольный опыт, 3'- выступ, фрагмент длиной 775 пар оснований, цифрой 3 - хромозомная ДНК, 5'-выступ, фрагмент длиной 887 пар оснований, цифрой 4 - хромозомная ДНК, 3'-выступ, фрагмент длиной 775 пар оснований.
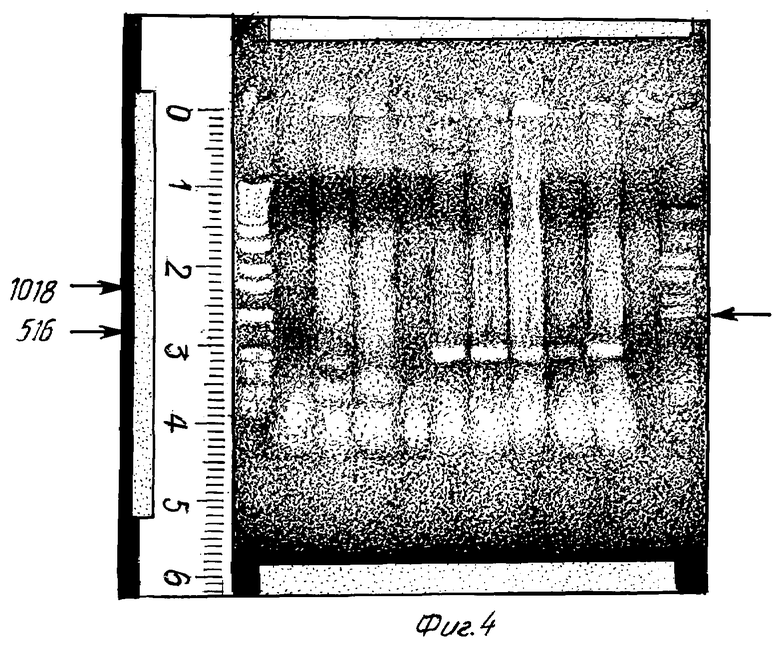
Фиг. 4
Доказательство генного фрагмента HIV-1 в хромозомной ДНК из продукта трансфекции клонированной клеточной линии HUT 78 после вышеуказанной амплификации. Продукт амплификации разделяют на агаровом геле и после окрашивания бромидом этидиа снимают под УФ-светом. Продукт реакции имеет длину 561 пару оснований и соответствует включающему нуклеотиды 699 - 1305 генному фрагменту HIV-1, который также является компонентом плазмиды pSXK 1.
Этот фрагмент указан стрелкой. Буквой М обозначена ДНК со стандартной длиной, отмечены размеры 516 пар оснований и 1018 пар оснований. Приведенные на фиг. 4 цифры имеют следующие значения.
1 = отрицательный контроль, на реакцию амплификации не подают ДНК
2 = хромосомная ДНК из клеток U 937 (дикий тип)
3 = хромосомная ДНК: из клеток HUT 78 (дикий тип)
4 = отрицательный контроль, на реакцию амплификации не подают ДНК
5 = положительный контроль
6 = хромосомная ДНК из продуктов трансформации HUT 78 (SSHUT 6/1 107 - 9/1)
7 = хромосомная ДНК из продуктов трансформации HUT 78 (SSHUT 6/1 107 - 9/1)
8 = хромосомная ДНК из продуктов трансформации HUT 78 (SSHUT 6/1 107 - 9/1)
9 = хромосомная ДНК из продуктов трансформации HUT 78 (SSHUT 6/1 107 - 9/1).
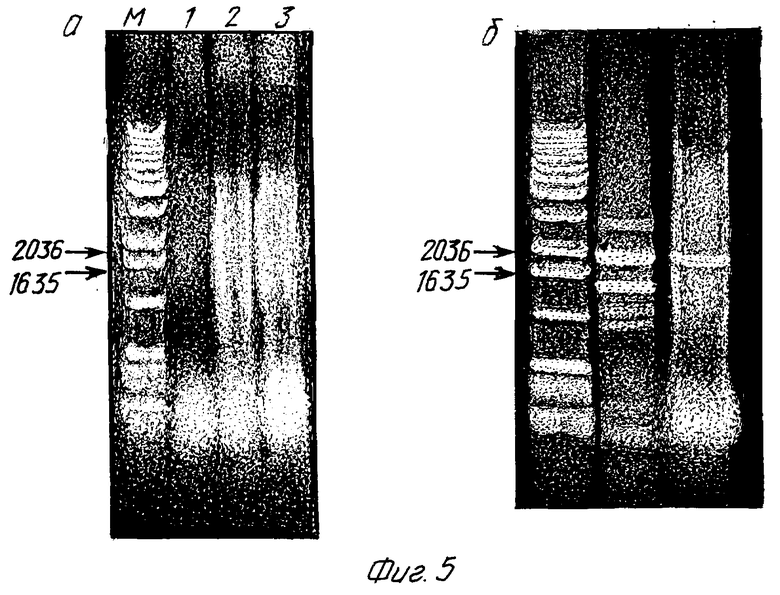
Фиг. 5 (a, b)
Доказательство включающего нуклеотиды 4158 - 5786 генного фрагмента HIV-1 в хромосомной ДНК из моноцитных клеток U 937 и Т- лимфоцитных клеток HUT 78 после вышеуказанной амплификации. Клетки предварительно трансферируют плазмидой pSXK2. Продукты реакции разделяют на агаровом геле и после окрашивания бромидом этидия снимают под УФ-светом. Продукт включает общий генный фрагмент HIV-1 в плазмиде pSXK2 (нуклеотиды 4158 - 5786) и имеет эту длину 1811 пар оснований. Этот продукт указан стрелкой. Буквой М обозначена ДНК со стандартной длиной, отмечены стандарты 1635 пар оснований и 2036 пар оснований. Приведенные цифры имеют следующие значения:
1 = отрицательный контроль, на реакцию амплификации не подают ДНК
2 = хромосомная ДНК из клеток U 937 (дикий тип)
3 = хромосомная ДНК из клеток HUT 78 (дикий тип)
4 = хромосомная ДНК из продуктов трансформации HUT 78 (SSHUT 2/1 107-12/1)
5 = хромосомная ДНК из продуктов трансформации U 937 (SSUK 3/1 107 -12/1).
Пример 5
Доказательство устойчивости к воздействию вирусов трансфецированных клонированных гематопоэтических клеточных линий
Клонированные, экспримирующие анти-мРНК клеточные линии HUT 78 и U 937 по примерам 3 и 4, зараженные HIV-1 [см. Л. Ратнер и др., Nature 313, стр. 277 (1985)] или фрагментом D 194, выделенным из HIV-2 [см. Х Кюнель и др., Nucleic Acids Research (1990) 18 стр. 6142], сравнивают по степени устойчивости к вирусам с взятыми в качестве контроля соответствующими исходными клеточными линиями HUT 78 и U 937, зараженными теми же вирусами.
1. Инфекция клеток HUT 78 и U 937 ретровирусом HIV-1 (контроль).
Осуществление опыта
По 10 мл суспензии клеток смешивают с имеющей различную степень разведения надосадочной жидкостью, содержащей HUT 78/HIV-1 (примерно 104 стандартных единиц/мл в культурах PBL, активность обратной транскриптазы замороженного основного раствора: 650 000 отсчетов в минуту/мл) в течение 60 минут. Затем клетки удаляют центрифугированием и три раза промывают с тем, чтобы удалить несвязанные вирусные частицы и вирусные белки. В заключение клетки повторно суспендируют в 10 мл питательной среде и культивируют в термостате. Каждые 3 - 4 дни 200 мкл свободной от клеток надосадочной жидкости проверяют на наличие антигенов HIV известным приемом. Для снабжения клеток отбирают по 5 мл суспензии, которые заменяют на 5 мл свежей питательной среды.
2. Инфекция экспримирующих анти-мРНК клеток HUT 78 и U 937 ретровирусом HIV-1
Осуществление опыта
По 10 мл суспензии клеток (примерно 4•105 клеток/мл) смешивают с разбавленной надосадочной жидкостью HUT 78/HIV-1 (примерно 104 стандартных единиц/мл в культурах PBL, активность обратной транскриптазы замороженного основного раствора: 650 000 отсчетов в минуту/мл) в течение 60 минут. При этом в случае клеток HUT 78 конечное разбавление основного раствора вируса составляет 1 : 1000, а в случае клеток U 937 - 1:50. Клетки удаляют путем центрифугирования и тщательно четыре раза промывают с тем, чтобы удалить несвязанные вирусные частицы и вирусные белки. В заключение клетки повторно суспендируют в 10 мл питательной среды (среде RPMI 1640, содержащей 16% телячьей эмбриональной сыворотки) и культивируют в термостате в течение 60 дней. Каждые 3 - 4 дни 160 мкл свободной от клеток надосадочной жидкости исследуют на наличие антигенов HIV известным приемом. Для снабжения клеток каждые 3 - 4 дни отбирают по 5 мл суспензии, которые заменяют на 5 мл свежей питательной среды.
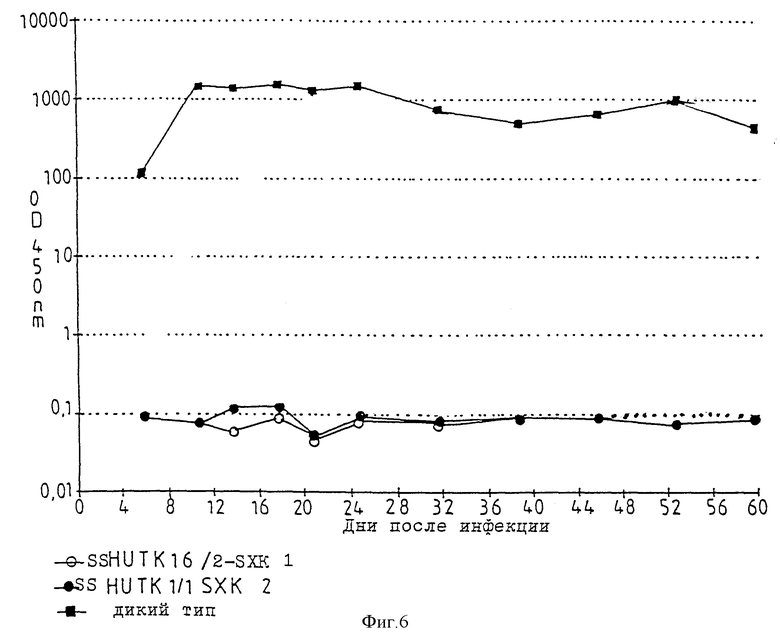
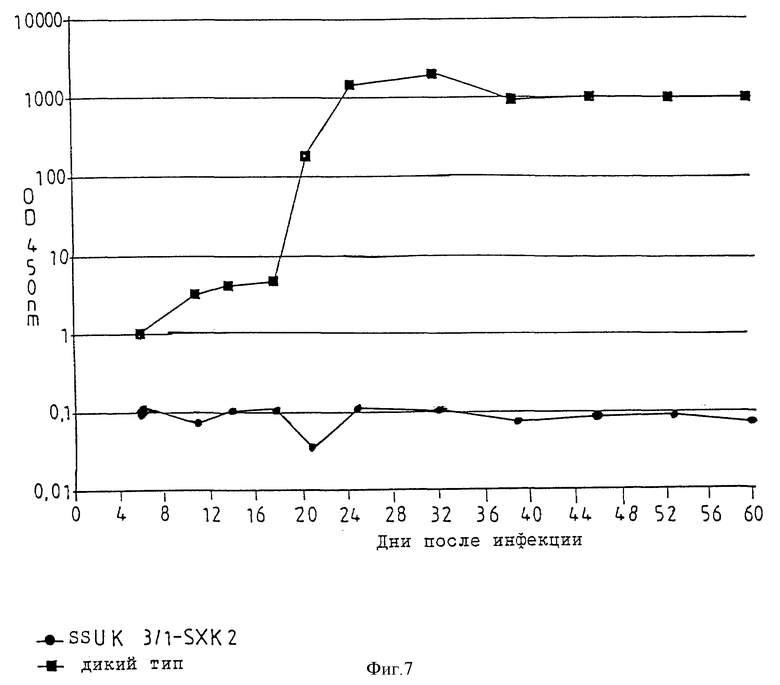
В опытах применяют два продукта трансфекции HUT 78 (SSHUTK16/2-SXK1, SSHUTKI/1-SXK2; см. фиг. 6) и один продукт трансфекции клеток U 937 (SSUK3/1-SXK2; см. фиг. 7). Как при определении активности обратной транскриптазы, так и при определении антигенов HIV в этих экспримирующих анти-мРНК клеточных линиях с плазмидами pSXKI, pSXK2 после инфекции за период инкубации до 60 дней не обнаруживают признаков размножения ретровируса HIV. В противоположность опытам по изобретению в контрольной клеточной линии определяется явное размножение ретровируса HIV. Результаты опытов представлены на фиг. 6 и 7, по которым даются следующие пояснения.
Фиг. 6
Представлен процесс образования антигена HIV-1 за 60 дней после инфекции (в день 0) клеточных линий HUT 78 фрагментом LAV/BRU, выделенным из ретровируса HIV-1. На ординату нанесена оптическая плотность при 450 нм (OD 450 nm), полученную в результате имунноферментного твердофазного анализа надосадочной жидкости клеточной культуры. При оптической плотности больше 2,0 надосадочную жидкость разбавляют и указанное значение оптической плотности рассчитывают по разбавленному раствору. Верхнее значение на ординате представляет собой предел обнаружения антигена HIV.
Как видно, в экспримирующих анти-мРНК клеточных линиях SSHUT K16/2-SXKI и SSHUT К1/1-SXK2 репликация ретровируса HIV полностью задерживается в течение 60 дней.
Фиг. 7
Представлен ход процесса образования ретровируса HIV-1 за 60 дней после инфекции (в день 0) клеточных линий U 937 фрагментом LAV/BRU, выделенным из ретровируса HIV-1. На ординату нанесена оптическая плотность при 450 нм (OD 450 nm), полученная в результате имунноферментного твердофазного анализа надосадочной жидкости клеточной культуры. При оптической плотности более 2,0 надосадочную жидкость разбавляют и указанное значение оптической плотности рассчитывают по разбавлению. Указанное верхнее значение представляет собой предел определения антигена HIV-1. В экспримирующей анти-мРНК клеточной линии SSUK3/1-SXK2 репликация ретровируса HIV полностью задерживается в течение 60 дней,
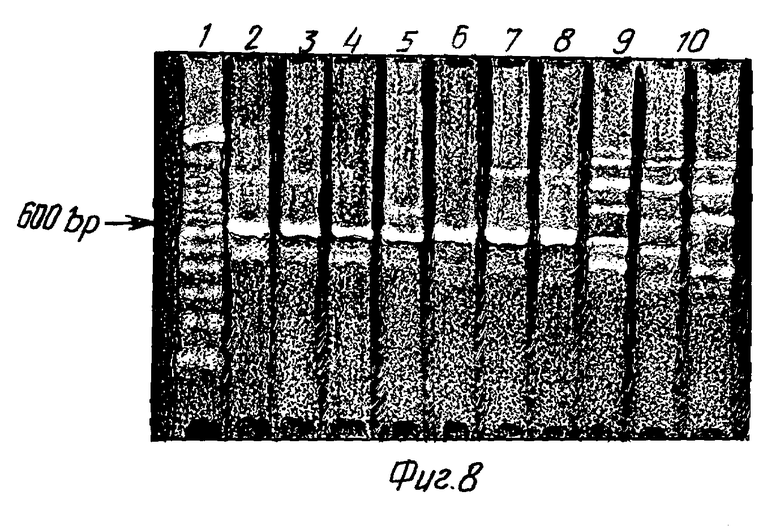
Факт инфекции HIV-1 в клеточных линиях SSUK3/1-SXK2, SSHUT K16/2- SXK1 и SSHUT К1/1-SXK2, которые после инкубации в течение 60 дней не проявляют ни активности обратной транскриптазы, ни наличие антигена HIV-1, доказывают, например, при помощи вышеупомянутой реакции. При этом в инкубированных в течение 60 дней клетках SSHUT K16/2-SXK1 и SSHUT K1/1-SXK2 можно доказывать TAT-последовательности и содержащиеся в оболочке последовательности HIV-1 (см. фиг. 8). В этой реакции применяют затравки 5'-ATG GAG CCA GTA GAT CCT-3' и 5'-TCT ACC ATG TCAT-3' для получения фрагмента длиной 690 пар оснований. Плазмиды pSXK1 и pSXK2 не содержат этих последовательностей и поэтому они не могут дать неправильно-положительный продукт. На фиг. 8 окрашенный бромидом этидия агаровый гель после электрофореза показывает продукты, полученные в результате вышеупомянутой реакции из хромосомной ДНК нижеследующих клеточных линий. Для сравнения с результатом данной реакции в скобках приведен результат определения антигена HIV соответствующей пробы клеточной культуры.
След М: фрагмент со стандартным молекулярным весом, размер 600 пар оснований отмечен.
След 1: SSHUT K16/2-SXK1 с 702 парами оснований положительно (опыт по определению антигена: отрицательный).
След 2: тот же продукт, что и по следу 1 (параллельный опыт).
След 3: тот же продукт, что и по следу 1 (параллельный опыт).
След 4: SSHUT K1/1-SXK2 с 702 парами оснований, положительно (опыт по определению антигена: отрицательный).
След 5: тот же продукт, что и по следу 4 (параллельный опыт).
След 6: тот же продукт, что и по следу 4 (параллельный опыт).
След 7: HUT 78, положительный контроль.
Следы 8-10: культуры SSUK3/1-SXK2: продукт с 703 парами оснований не может доказываться (опыт по определению антигена: отрицательный).
Пример 6
Контрольные опыты
Трансфекция клеточных линий U 937 и HUT 78 плазмидами по примеру 1, 2 без вставок провирусной ДНК из ретровируса HIV-1 в анти-м-ориентации не приводит к тому, что репликация вируса в этих клеточных линиях зараживается. Контрольные опыты с применением плазмид по примерам 1, 2, которые имеют вставку провирусной ДНК из ретровируса HIV-1 в м-ориентации, не проводились, так как соответствующие m-транскрипты могут связывать регуляторные белки репликации HIV, так, как, например, TAT, REV, и таким образом компетитивно задерживают репликацию вируса [см. Г. Й. Грэхем и др. PNAS 87, стр. 5817 (1990)]. Следовательно, контрольные опыты такого рода не имеют смысла.
Резюме результатов проведенных опытов
Плазмиды pSXK 1, pSXK 2 обеспечивают эндогенное образование анти-мРНК в отношении ретровируса HIV-1 (см. пример 4) в человеческих клеточных линиях крови после трансфекции и клонирования трансфецированных клеточных линий по примеру 3. Кроме того, согласно примеру 5 доказана их антивирусная активность.
Неожиданным является тот факт, что в трансфецированных клеточных линиях можно доказывать ярко выраженную задержку репликации ретровируса HIV-1. И в случае применения специально адаптированного к трансфецированным клеточным линиям вируса HIV-1 можно было равным образом доказывать устойчивость к вирусу экспримирующих анти-мРНК клеток (см. фиг. 6, 7 ).
Эти результаты свидетельствуют о том, что подходящие промоторы для экспрессии анти-мРНК и, конечно, вид последовательностей анти-мРНК обеспечивают придание генетически измененным клеткам устойчивости к заражению ретровирусом. В примере 4 показано, что в результате неизвестного до сих пор сплайсинга РНК-транскриптов (+) нити провирусной ДНК HIV в ориентации 3' ---> 5' образуются несколько молекул мРНК, которые обеспечивают антивирусную активность.
Экспериментальные работы, описанные в примерах 1 - 6, доказывают, что плазмиды, включающие кодирующие анти-мРНК фрагменты ДНК из ретровируса, обеспечивают создание антивирусных свойств в клетках, которые защищают от заражения ретровирусом.
На фиг. 6 - 8 видно, что 60 дней после заражения ретровирусом HIV клеточные линии SSHUT 16/2-SXK 1 и SSHUT K1/1-SXK2 проявляют положительную реакцию при определении провирусной ДНК HIV, тогда как определение гена HIV является отрицательным. По этому результату можно заключить, что после заражения HIV 1 на первой стадии цикла репликации HIV еще образуется провирусная ДНК в трансфецированных клеточных линиях HUT 78, однако транскрипция и трансляция провирусной ДНК не допускается анти-мРНК, так что белок HIV для репликации вируса не образуется в доказываемых количествах.
60 дней после инфекции HIV 1 в ДНК клеточной линии SSUK3/1-SXK2 нельзя было доказывать во всех опытах образование белков HIV и провирусной ДНК HIV. По этому результату можно заключить, что образуемая в клеточной линии SSUK3/1-SXK2 анти-мРНК не только задерживает первую стадию цикла репликации HIV, но и не допускает проведения второй стадии этого цикла (транскрипция и трансляция провирусной ДНК).
Образование провирусной ДНК из геномной (+) нитевой РНК в результате процесса обратной транскрипции очевидно не допускается анти-мРНК. В отношении этого действия анти-мРНК есть уже предположения в литературе (см.Р. И. Л. То и П. Е. Нейман в Gene Regulation: Biology of Antisense RNA and DNA, изд. P. П. Эриксон энд Й.Г. Изэнт, Рэвент Пресс Лтд, Нью Йорк, 1992), но оно до сих пор не могло показываться из-за отсутствия соответствующих высокоэффективных плазмид.
Изобретение относится к целенаправленной блокировке генетической информации вирусной мРНК для репликации ретровирусов, например HIV, в трансфецированных гематопоэтических клетках, более конкретно, к плазмидам для экспрессии анти-мРНК ретровируса HIV-1. Плазмиды, кодирующие последовательности анти-мРНК, обеспечивают полное торможение репликации вируса HIV-1 в измененных в отношении генов клетках человека. Плазмиды пригодны для соматической генной терапии СПИДа, так как они защищают гематопоэтические клетки от индуцируемых вирусом HIV-1 цитопатических эффектов. 2 с.п.ф-лы, 8 ил.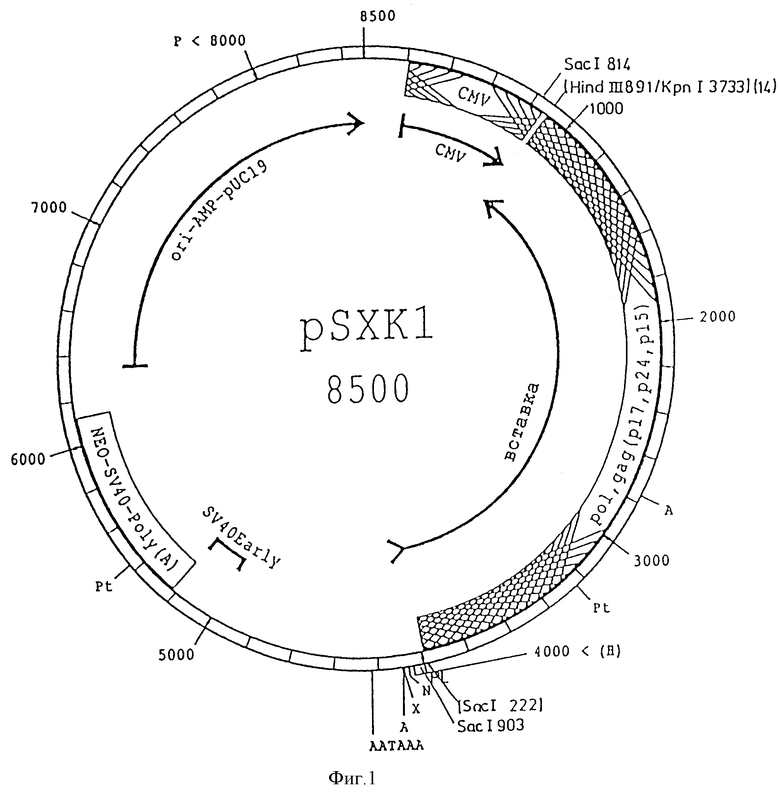
| PCT 9215680, 17.09.92 | |||
| PCT 8810311, 29.12.88 | |||
| ПОЛУАВТОМАТИЧЕСКАЯ ЛИНИЯ ДЛЯ ОТДЕЛКИ i" МЕБЕЛЬНЫХ ЩИТОВI | 0 |
|
SU331939A1 |
| EP 0240332, 07.10.87. | |||
