Настоящее изобретение относится к способу определения лигандов рецептора лептина путем использования переноса энергии между cлитыми протеинами, состоящими из рецепторов лептина и протеинов, являющихся донорами или акцепторами энергии.
Оно относится также к слитым протеинам, используемым для осуществления этого способа.
Лептин представляет собой протеин с молекулярной массой 16 кДа, который секретируется адипоцитами. Этот протеин связан с чувством насыщения и играет важную роль в контроле за прибавлением веса, потреблением энергии, в образовании костной ткани, ангиогенезе, а также в других физиологических процессах, таких как начало полового созревания и контроль за репродукцией или регуляцией иммунного ответа, регулируемого Т-лимфоцитами.
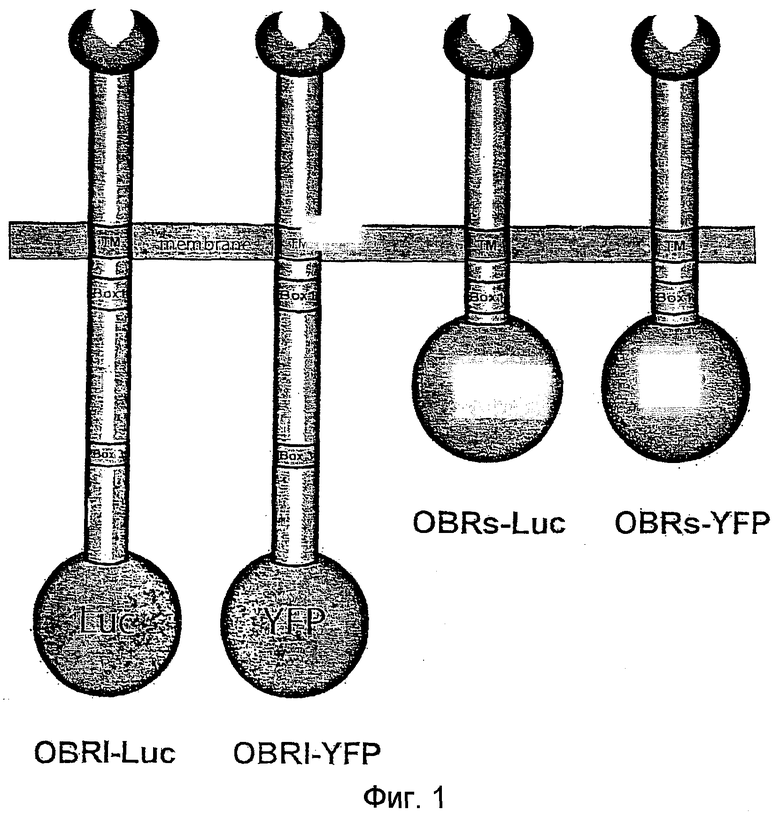
Рецептор лептина (OBR) относится к семейству цитокиновых рецепторов. Он состоит, как показано на фиг.1, из единственной полипептидной цепи, включающей трансмембранный домен (Тartaglia et al., J.Biol.Chem, 272, 6093-6096, 1995). В международной заявке на патент WO 97/19952 описан этот рецептор.
Описаны также шесть различных изоформ рецептора OBR, имеющих С-концевые участки различной длины. Все эти изоформы получены путем альтернативного сплайсинга из единственного гена. Существует также растворимая форма OBR, содержащая сайт связывания с лептином, соответствующий внеклеточному домену мембраносвязанных форм. Эта посттрансляционная растворимая форма, вырабатываемая в плазматической мембране в результате протеолиза из мембраносвязанных форм, находится в крови. В очень редких случаях обнаруживается другая растворимая форма OBR, являющаяся результатом мутации, вырабатывающей стоп кодон перед трансмембранным доменом.
Слитый протеин, состоящий из длинной формы рецептора лептина (OBRI), слитого с протеином EGFP (Enhanced Green Fluorescenсe Protein), используется согласно Lundin et al. (Biochemica et Biophysica Acta, 1499, 130-138, 2000) для изучения локализации рецептора.
Активация рецептора OBR протекает, по видимому, посредством тетрамерного комплекса, состоящего из двух janus киназ 2 (JAK2) и двух OBR. Предполагается, что активация рецептора, индуцированная лептином, приводит к изменению конформации рецептора OBR, в результате чего он сам активирует одну киназу JAK2, которая в свою очередь трансфосфорилирует другую киназу JAK2, затем рецептор OBR.
Активация рецептора OBR оказывается ответственной за все известные проявления лептина, такие как потеря веса и все явления, связанные с нарушением веса.
В последнее время были обнаружены ингибирующие свойства лептина в отношении синтеза костной ткани. Лептин действует как ингибитор активности остеобластов, т.е. популяции клеток, ответственных за формирование костной ткани.
Путем изменения содержания лептина можно было бы лечить болезни, связанные с уменьшением плотности костной ткани, такие как, например, остеопороз или, наоборот, болезни, связанные со значительной кальцификацией.
В 1999 г. авторами Хu et al (Proc. Natl Acad Sсi USA A 96, 151-156), был описан метод определения межбелкового взаимодействия в живых клетках. Этот метод стал объектом международной заявки на патент WO 99/66324.
Этот метод, называемый BRET (аббревиатура от Bioluminescent Resonance Energy Transfer или перенос энергии биолюминесцентного резонанса) основан на природном явлении, при котором морские организмы испускают флуоресцентное свечение. Ферментативная трансформация, под действием люциферазы Renilla (Luc), субстрата, который может проходить через мембрану, генерирует биолюминесцентную энергию, которая в свою очередь возбуждает акцептор энергии, такой как желтый флуоресцирующий белок (YFP). Этот метод соответствует LRET (аббревиатура Luminеscent Resonance Energу Transfer), описанный Wang et al (Mol Gen Genet 264: 578-587(2001)).
Эффективность переноса энергии зависит от физического расстояния и от взаимного расположения акцептора и донора энергии. Поэтому недостаточно совместной экспрессии люциферазы и YFP для индуцирования переноса энергии, т.к. расстояние между двумя участниками переноса должно быть менее 100 Ǻ. Для исследования взаимодействия между двумя потенциальными участниками этого взаимодействия первый протеин был слит с люциферазой, а второй протеин слит с YFP. Если между двумя протеинами происходит взаимодействие, то может наблюдаться передача энергии.
С тех пор метод, основанный на явлении BRET, применялся для исследования некоторых рецепторов, чья структура значительно отличалась от структуры рецептора лептина.
Так, ряд авторов описывает применение этого метода к рецепторам, относящимся к семейству рецепторов, связанных с протеинами G (GPCR), таких как 2 бета-адренергические рецепторы (Angers et al, 2000, Proc. Natl. Acad. Sci. USA, 10.1073), холецистокины типа А (ССК; Cheng et al, 2001. Biol. Chem. 276: 48040-48047), и рецепторов тиреотропин-рилизинг гормона (Kroeger et al, 2001, J.Biol. Chem. 276: 12736-12743).
Эти рецепторы крупного размера имеют структуру комплекса, содержащего 7 трансмембранных доменов.
Кроме того, авторы Boute et al (2001, Mol. Pharmacol.60: 640-645) описали процесс активации рецептора инсулина с использованием явления BRET.
Рецептор инсулина состоит из ковалентных димеров, в отличие от нековалентных комплексов, как в рецепторе лептина. Кроме того, он имеет довольно длинную внутриклеточную часть. В заключение авторы показали, что изменение BRET, индуцированное инсулином, может быть осуществлено только с растворимым рецептором.
В последние десятилетия ожирение стало главной проблемой в области здравоохранения в промышленных странах, в которых эта проблема в настоящее время затрагивает 20-30% населения. Эти цифры, по-видимому, будут угрожающим образом расти в последующие годы. Поскольку многочисленные причины ожирения, источниками которых являются в большей или меньшей степени, с одной стороны, факторы окружающей среды (состояние пищевых продуктов, доступность продуктов питания, энергетические затраты…) и, с другой стороны, многочисленные генетические факторы создают настоящую проблему для медицины.
С другой стороны, костные заболевания, такие как остеопороз, также затрагивают все более и более значительную часть населения.
Таким образом, открытие новых молекул, позволяющих лечить разные заболевания, в которых задействован рецептор лептина, такие как ожирение и остеопороз, было бы большим шагом вперед в области здравоохранения.
В то же время не существует способа специфического скрининга агонистов или антагонистов рецептора лептина, который был бы высокоэффективным.
Заявитель поставил задачу разработать тест для быстрого, специфического и эффективного скрининга агонистов или антагонистов рецептора лептина.
Заявитель неожиданно установил, что изменение BRET, индуцированное лептином, может быть осуществлено с применением одной из изоформ рецептора лептина, но не может быть осуществлено со всеми изоформами.
Заявитель также установил, что применение BRET к рецептору лептина оптимально при некоторых определенных рабочих условиях.
Таким образом, настоящее изобретение относится к способу определения лигандов рецептора лептина путем использования переноса энергии резонанса между первым слитым протеином, состоящим из рецептора лептина или основной части рецептора лептина, и из протеина, являющегося донором энергии, или из основной и активной части протеина, являющегося донором энергии, и вторым слитым протеином, состоящим из рецептора лептина или основной части рецептора лептина, и из протеина, являющегося акцептором энергии, или основной и активной части протеина, являющегося акцептором энергии.
Изобретение относится также к слитым протеинам, используемым для осуществления этого способа, а также к нуклеиновым кислотам, кодирующим эти протеины.
Изобретение относится также к способу лечения или профилактики заболеваний, в которых задействован лептин, состоящему во введении лиганда, выбранного согласно описанному выше способу, пациенту, который страдает указанным выше заболеванием.
Таким образом, первым объектом настоящего изобретения является слитый протеин, отличающийся тем, что он состоит из рецептора лептина или основной части рецептора лептина, и из протеина, являющегося акцептором или донором энергии, или основной и активной части протеина, являющегося акцептором или донором энергии.
Слитые протеины согласно изобретению состоят по существу из части, которая соответствует частичной или полной последовательности рецептора лептина, и из части, соответствующей протеину - донору или протеину-акцептору энергии. Они могут также включать другие последовательности аминокислот, происходящие из других протеинов, такие как сигнальная последовательность. Так, последовательность SEQ ID N 4 состоит из частичной последовательности SEQ ID N 2 и других последовательностей, в частности из сигнальной последовательности интерлейкина 3 мыши.
Преимущественно, протеином, являющимся донором энергии, является люцифераза Renilla. Однако этим протеином может быть любой другой протеин - донор энергии, у которого спектр излучения перекрывается в достаточной мере со спектром возбуждения акцептора для того, чтобы обеспечить эффективный перенос энергии между двумя партнерами. Таким образом, протеином - донором знергии может быть протеин GFP, если перенос энергии представляет собой FRET, или же акворин, если перенос энергии представляет собой CRET. Акворин может быть получен и использован по методу, описанному в заявке на патент ЕР 0 187 519 или в статье Inouye et al. (PNAS USA 82: 3154-3158 (1985)).
Флуоресцирующим протеином - акцептором энергии, является предпочтительно DsRed, GFP или его мутант, такой как YFP, EYFP, GFP дикого типа, GFPS65T или Тоpаz.
Он может быть представлен любым другим флуоресцирующим протеином - акцептором энергии, у которого спектр возбуждения перекрывался бы в достаточной мере со спектром излучения донора для эффективного переноса энергии между двумя партнерами.
Эти протеины известны специалисту, их последовательности он сможет найти в литературе, в частности, в обзоре Blinks et al. (Pharmacol. Rev. 28: 1-93 (1976)). В частности, протеин GFP описан Tsien (Annu. Rev. Biochem. 67: 509-544 (1998)), а его клонирование описано Prasher et al. (Genе 111: 229-233 (1992)). Клонирование DsRed описано Matz et al. (Nat. Biotechnol. 17: 969-973 (1999)). В отношении Rluc специалист найдет информацию в работе Blinks et al. (Pharmacol. Rev. 28: 1-93 (1976)) или в работе Lorenz et al. (PNAS 88: 4438-4442 (1991)).
Преимущественно, изоформа рецептора лептина, входящая полностью или частично в слитый протеин, представляет собой короткую изоформу или имеет короткий внутриклеточный домен.
Такая изоформа содержит преимущественно внутриклеточный домен Box 1, но не содержит внутриклеточный домен Box 3.
Предпочтительно, эта изоформа является изоформой OBRs, более предпочтительно, изоформой OBRs человека. Эта изоформа может иметь и другое происхождение.
Она может быть также любой другой изоформой, предпочтительно короткой, и более предпочтительно, содержащей по меньшей мере внеклеточный домен рецептора OBR, такой как растворимая форма рецептора OBR, содержащая сайт связывания с лептином, описанная авторами Lee et al. (Nature 379, 632-635 (1996)), Gavrilova et al. (JBC 272 : 30546-30551 (1997)), Maamr. еt al. (Endocrinology 142 : 4389-4393 (2001)) или Clement et al. (Nature 392 : 398-401 (1998)).
Согласно особенно предпочтительному варианту осуществления изоформа представляет собой изоформу OBRs человека с последовательностью SEQ ID N 2. Она может представлять собой вариант этого протеина, последовательность которого идентична последовательности SEQ ID N 2 по меньшей мере на 65%, предпочтительно по меньшей мере на 75%, еще более предпочтительно по меньшей мере на 85% или 95%.
Слитый протеин может содержать только часть изоформы OBRs человека. Преимущественно, он содержит часть, находящуюся между 46 и 866 аминокислотами в последовательности SEQ ID N 2.
Таким образом, часть, соответствующая рецептору лептина, может иметь последовательность SEQ ID N 4 или ее вариант, который идентичен по меньшей мере на 65%, предпочтительно по меньшей мере на 75% и еще более предпочтительно по меньшей мере на 85% или 95%.
Особенно предпочтительно слитый протеин-донор имеет последовательность SEQ ID No6 или ее вариант, который идентичен этой последовательности по меньшей мере на 65%.
Особенно предпочтительно слитый протеин-акцептор имеет последовательность SEQ ID N 8 или ее вариант, который идентичен этой последовательности по меньшей мере на 65%.
Другими объектами изобретения являются нуклеиновые кислоты, кодирующие эти протеины. Такими нуклеиновыми кислотами могут быть комплементарные или геномные ДНК, а также РНК. Эти нуклеиновые кислоты или полинуклеотиды могут находиться в форме одинарной цепи или в форме дуплекса.
Особенно предпочтительны комплементарные ДНК.
Предпочтительно изобретение относится к нуклеиновой кислоте, степень идентичности которой нуклеиновой кислоте с последовательностью SEQ ID N 5 или SEQ ID N 7 по нуклеотидам составляет по меньшей мере 65%, предпочтительно, по меньшей мере 75%, еще более предпочтительно по меньшей мере 85% или 95%.
Согласно другому аспекту изобретение относится к нуклеиновой кислоте, гибридизующейся в жестких условиях гибридизации с нуклеиновой кислотой, такой как описана выше, более конкретно, с нуклеиновой кислотой, имеющей нуклеотидные последовательности SEQ ID N 5 и SEQ ID N 7, или с нуклеиновой кислотой, имеющей комплементарную последовательность.
“Степень идентичности“ двух нуклеотидных последовательностей или аминокислот может быть определена согласно изобретению при сравнении двух оптимально выровненных последовательностей в окне сравнения.
Так, часть нуклеотидной или полипептидной последовательности в окне сравнения может содержать вставки или делеции (например “гэпы”) по сравнению со ссылочной последовательностью (которая не содержит эти вставки или эти делеции) для того, чтобы добиться оптимального выравнивания двух последовательностей.
Результат в процентах рассчитывают путем определения числа положений, в которых идентичное основание нуклеиновой кислоты или идентичный аминокислотный остаток обнаружен для двух сравниваемых последовательностей (нуклеиновой или пептидной), затем делят число положений, в которых имеется идентичность между двумя основаниями или аминокислотными остатками, на общее число положений в окне сравнения, затем умножают результат на 100, чтобы получить степень идентичности последовательности в процентах.
Оптимальное выравнивание сравниваемых последовательностей может быть осуществлено на компьютерных системах с помощью известных алгоритмов, содержащихся в пакете программ фирмы WISCONSIN GENETICS SOFTWARE PACKAGE, GENETICS CONPUTER GROUP (GCG), 575 Science Doctor, Madison, WISCONSIN.
Например, степень идентичности последовательности может быть рассчитана с помощью программного обеспечения BLAST (версии BLAST 1.4.9 март 1996, BLAST 2.0.4 февраль 1998 и BLAST 2.0.6 сентябрь 1998), с использованием исключительно параметров по умолчанию (S.F.Altschul et al, J.Mol.Biol. 1990, 215: 403-410, S.F.Altschul et al, Nucleic Acids Res. 1997, 25: 3389-3402). BLAST-поиск последовательностей, подобных или гомологичных последовательности, указанной в качестве ссылки, осуществляют с помощью алгоритма Altschul et al. Соответствующая ссылочная последовательность и заданные основания могут быть пептидными или нуклеиновыми последовательностями или представлять собой любую их комбинацию.
Под “жесткими условиями гибридизации“ понимают в данном описании следующие условия:
1. Конкурентное связывание с мембранами и предгибридизация:
- Смешивание 40 мкл ДНК спермы лосося (10 мг/мл) и 40 мкл ДНК плаценты человека (10 мг/мл),
- Денатурация в течение 5 минут при 96°С с последующим погружением смеси в лед,
- Удаление SSC 2X и вливание 4 мл формамидной смеси в пробирку для гибридизации, содержащую мембраны,
- Добавление смеси двух денатурированных ДНК,
- Инкубация при 42°С в течение 5-6 часов с перемешиванием в ротационной мешалке.
2. Конкурентное связывание меченого зонда:
- Добавление к меченому и очищенному зонду от 10 до 50 мкл ДНК Cot I в зависимости от количества повторов,
- Денатурация 7-10 мин при 95°С,
- Инкубация при 65°С в течение 2-5 часов.
3. Гибридизация:
- Удаление пред-гибридизованной смеси,
- Смешивание 40 мкл ДНК спермы лосося и 40 мкл ДНК плаценты человека и денатурация в течение 5 минут при 96°С с последующим погружением в лед,
- Добавление в пробирку для гибридизации 4 мл формамидной смеси, смеси двух ДНК и меченого зонда с денатурированной ДНК Cot I,
- Инкубация в течение 15-20 часов при 42оС с перемешиванием в ротационной мешалке.
4. Промывки:
- Промывка при комнатной темпeратуре в SSC 2X для отмывки,
- Промывка 2 раза по 5 минут при комнатной температуре при помощи SSC 2X и SDS 0,1% при 65°С,
- Промывка 2 раза по 15 минут при 65оС при помощи SSC 1X при 65оС и SDS 0,1% при 65°С.
Упаковывают мембраны в Saran и оставляют на хранение.
Условия гибридизации, описанные выше, пригодны для гибридизации в жестких условиях молекулы нуклеиновой кислоты с длиной цепи в интервале от 20 нуклеотидов до нескольких сотен нуклеотидов.
Разумеется, указанные выше условия могут быть соответственно скорректированы согласно знаниям специалиста в зависимости от длины нуклеиновой кислоты, которая подвергается гибридизации, или с учетом выбранного типа маркировки.
Соответствующие условия гибридизации могут быть выбраны, например, в соответствии со сведениями, содержащимися в работе HAMES et HIGGINS (1985, «Nucleic acid hydridization: a prасtical approach”, Hames and Higgins Ed., IRL Press, Oxford) или работой F.AUSUBEL et al. (1989, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y.).
Протеины согласно изобретению могут быть получены в соответствии с методами, известными специалисту. Однако преимущественно их получают путем экспрессии нуклеиновых кислот, описанных выше, кодирующих эти протеины, возможно встроенных в векторы экспрессии, в соответственно отобранных клетках с последующей, при необходимости, экстракцией и очисткой, которая может быть полной или частичной.
Изобретение относится также к рекомбинантному вектору, содержащему нуклеиновую кислоту согласно изобретению.
Преимущественно, этот рекомбинантный вектор содержит нуклеиновую кислоту, выбранную из следующих нуклеиновых кислот:
а) нуклеиновой кислоты, кодирующей протеин, идентичность которого, по аминокислотам, последовательности SEQ ID N 6 или SEQ ID N 8 или пептидному фрагменту или варианту этой последовательности составляет не менее 65%,
б) нуклеиновой кислоты, содержащей полинуклеотид с последовательностью SEQ ID N 5 или SEQ ID N 7, или фрагмент или вариант последней,
в) нуклеиновой кислоты, идентичность которой, по нуклеотидам, нуклеиновой кислоте с последовательностью SEQ ID N 5 или SEQ ID N 7 или фрагменту или варианту последней составляет не менее 65%,
г) нуклеиновой кислоты, гибридизующейся в условиях жесткой гибридизации, с нуклеиновой кислотой, имеющей последовательности SEQ ID N 5 или SEQ ID N 7 или фрагмент или вариант последней.
Под “вектором“ понимают в настоящем изобретении кольцевую или линейную молекулу ДНК или РНК, которая независимо может находиться как в форме одинарной цепи, так и в форме двойной цепи.
Согласно варианту осуществления, вектор экспрессии включает помимо нуклеиновой кислоты согласно изобретению, регуляторные последовательности, направляющие транскрипцию и/или трансляцию.
Согласно предпочтительному варианту, рекомбинантный вектор согласно изобретению включает, в частности, следующие элементы:
(1) элементы регуляции экспрессии встраиваемой нуклеиновой кислоты, такие как промоторы и энхансеры;
(2) кодирующую последовательность, входящую в нуклеиновую кислоту согласно изобретению, подлежащую встраиванию в указанный вектор, причем указанная кодирующая последовательность локализуется регуляторными сигналами, указанными в (1) и
(3) соответствующие последовательности инициации и остановки транскрипции.
Кроме того, рекомбинантные векторы согласно изобретению могут включать один или несколько сайтов инициации репликации в клетках-хозяевах, в которых их амплификация или экспрессия необходима, и маркеры или маркеры селекции.
Например, промоторы для эукариотических клеток включают промотор тимидинкиназы вируса HSV или же промотор L-металлотионина мыши.
Обычно для выбора соответствующего промотора специалист обращается к работе SAMBROOK et al. (1989, “Мolecular Cloning: A Laboratory Manual”, 2nd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.) или к методам, описанным FULLER et al. (1996, Immunology in Current Protocols in Molecular Biology, Ausubel et al).
Предпочтительными векторами согласно изобретению являются плазмиды, такие как например, векторы pCDNA3 (Invitrogen), pQE70, pQE60, pQE9 (Qiagen), psiX174. pBluescript SA, pNH8A, pNH16A, pNH18A, pNH46A, pWLNE0, pSV2CAT, pOG44, pXTI, pSG(Stratagene).
Пригодны также векторы типа baculovirus, такие как вектор pVL1392/1393 (Pharmingen), используемый для трансфекции клеток линии Sf9(ATCC N CRL 1711), полученных из Spodoptera frugiperda.
Могут быть пригодны также аденовирусные векторы, такие как аденовирус человека типа 2 или 5.
Рекомбинантным вектором согласно изобретению может быть также ретровирусный вектор или же адено-ассоциированный вектор (AAV). Такие адено-ассоциированные векторы описаны например, FLOTTE et al. (1992, Am.J.Resрir.Cell. Mol.Biol., 7: 349-356.
Настоящее изобретение относится также к клеткам, содержащим протеин, нуклеиновую кислоту или вектор, описанные выше, или фрагменты этих клеток, лизаты этих клеток или же мембраны этих клеток.
Такие клетки могут представлять собой клетки, выделенные из организма и выращенные в соответствующей культуральной среде. Однако такими клетками являются предпочтительно линии клеток. Особенно предпочтительны линии клеток HEK 293, COS (ATCC N CRL 1650), COS-M6 и HeLa(ATCC N CCL2) или же Cv I (ATCC N CСL 70), Sf-9 (ATCC N CRL 1711), CHO(ATCC N CCL-61) или 3T3 (ATCC N CRL-6361).
Мембраны этих клеток могут быть получены любым известным специалисту способом.
Предпочтительно, мембраны получают механическим измельчением клеток с последующим центрифугированием полученных эмульсий, как это показано в нижеследующих примерах.
Настоящее изобретение относится также к композициям, содержащим клетки, которые описаны выше, и сапонин.
Настоящее изобретение относится также к способу определения связывания соединений с рецептором лептина, включающему стадии, заключающиеся в том, что:
вводят в контакт указанное соединение со слитым протеином, являющимся донором энергии, таким как описано выше, и со слитым протеином, являющимся акцептором энергии, таким как описано выше, или с клетками, или с фрагментами, или с лизатами, или с клеточными мембранами, содержащими такой протеин, и с соответствующим ферментным субстратом, и
- измеряют перенос энергии.
Предпочтительно, способ осуществляют с использованием клеток, обработанных сапонином.
Слитые протеины, являющиеся донорами энергии, и слитые протеины, являющиеся акцепторами энергии, выбирают таким образом, чтобы энергия, выделяющаяся в результате активации донора, могла бы наиболее эффективно передаваться акцептору.
В преимущественном варианте осуществления указанного способа слитый протеин, который является донором энергии, представляет собой протеин, полученный в результате слияния рецептора лептина или основной части рецептора лептина, с люциферазой или основной частью люцеферазы, и в этом случае субстратом преимущественно является коэлентеразин.
В предпочтительном варианте осуществления способа слитый протеин, который является акцептором энергии, представляет собой протеин, полученный в результате слияния рецептора лептина или основной части рецептора лептина с YFP или основной частью YFP.
Согласно преимущественному варианту осуществления способа, перенос энергии, измеряемый в присутствии исследуемого соединения, сравнивают с переносом энергии, измеряемым в отсутствие исследуемого соединения.
Предпочтительно, способ осуществляют на мембранах клеток, которые описаны выше.
Предпочтительно, протеины, доноры и акцепторы энергии, согласно изобретению, выбирают таким образом, чтобы перенос энергии происходил посредством резонанса BRET или LRET. Однако такой перенос энергии может быть осуществлен и посредством резонанса FRET (Fluorescent Resonance Energy Transfer или перенос энергии флуоресцентного резонанса) или СRET (Chemioluminescent Resonance Energy Transfer или перенос энергии хемиолюминесцентого резонанса).
Независимо от типа переноса энергии протеиновые пары, состоящие из слитого протеина-донора/слитого протеина-акцептора, выбирают таким образом, чтобы обеспечить такой перенос.
CRET-перенос состоит в передаче энергии между акворином, который представляет собой люциферазу, и GFP.
FRET-перенос состоит в переносе энергии между двумя протеинами семейства GFP, имеющих разные спектры.
Для осуществления этих переносов специалист может воспользоваться работой Baubet et al. (PNAS USA 97: 7260-7265 (2000)) в случае переноса типа CRET, работой Matyus (J.Photochem. Photobiol. В 12: 323-337 (1992)) и работой Pollok et Heim (Trends Cell. Biol. В: 57-60 (1999)) в случае переноса типа FRET.
Другим объектом изобретения является способ скрининга или определения соединений, предназначенных для профилактики и/или лечения патологий, в которых задействован лептин, включающий стадии, заключающиеся в том, что:
- осуществляют контакт указанного соединения со слитым протеином-донором энергии, описанным выше, и со слитым протеином-акцептором энергии, описанным выше, или с клетками в присутствии сапонина или без него, или с фрагментами, или с лизатами, или с мембранами клеток, содержащими такие протеины, и, возможно, с соответствующим ферментным субстратом, и
- измеряют перенос энергии.
Этот способ может быть использован для скрининга агонистов или антагонистов рецептора лептина.
Способ согласно изобретению пригоден для 96-ти или 384-х луночных планшетов, которые обычно используются. Этот способ не требует применения радиоактивных молекул, он является чувствительным, воспроизводимым, быстрым и результат легко считывается. Так, способ дает удовлетворительное соотношение сигнал/фоновые шумы и низкую перекрестную реактивность с другими лигандами, не являющимися лептином. Это объясняется, по меньшей мере частично, тем, что активация рецептора OBR детектируется непосредственно на уровне рецептора, что позволяет исключить возможные источники перекрестной реактивности на других уровнях путей сигнализации, как можно наблюдать в случае анализа, основанного на репортерных генах или на росте клеток.
Кроме того, способ не ограничивается одним путем трансдукции, имеющим специфический сигнал, напротив, он способен определять все молекулы, взаимодействующие с рецепторами OBR.
Это способность особенно важна для осуществления крупномасштабного скрининга, поскольку все большее число лигандов мембранных рецепторов становится способными активировать некоторые определенные пути, и никакие другие.
Настоящее изобретение относится, кроме того, к применению соединений, отобранных согласно способу, состоящему в том, что
- вводят в контакт указанное соединение со слитым протеином-донором энергии, и со слитым протеином-акцептором энергии, описанными выше, или с клетками, или с фрагментами, или с лизатами, или с мембранами клеток, включающими такой протеин, и, возможно, с соответствующим ферментным субстратом, и
- измеряют перенос энергии,
для получения лекарственных средств, предназначенных для лечения или профилактики болезней, в которых задействован лептин или его рецептор.
Наконец, изобретение относится к способу лечения или профилактики болезней, связанных с лептином или его рецептором, включающему стадии:
- отбора соединения методом, состоящим в:
+ контактировании указанного соединения со слитым протеином- донором и со слитым протеином - акцептором энергии, или с клетками, или с фрагментами, или с лизатами, или с мембранами клеток, содержащими такой протеин, и с соответствующим ферментным субстратом, и
+ в измерении переноса энергии, и
- введения указанного соединения пациенту, страдающему указанным заболеванием.
Такими болезнями могут быть болезни, связанные с уменьшением плотности костной ткани, как например, остеопороз или, наоборот, болезни, связанные со значительной кальцификацией.
Такими болезнями могут быть также болезни, оказывающие влияние на вес, такие как ожирение, диабет или анорексия.
Соединения согласно изобретению могут входить в состав фармацевтических композиций для введения их топическим, пероральным, парентеральным, носовым, внутривенным, внутримышечным, подкожным, интраокулярным и т.д. путем. Предпочтительно, фармацевтические композиции для инъецируемых составов содержат фармацевтически приемлемые носители. Подразумеваются под ними, в частности, стерильные, изотонические солевые растворы (однонатриевый фосфат, двунатриевый фосфат, хлорид натрия, калия, кальция или магния и т.д. или смеси таких солей), или сухие композиции, в частности, лиофилизованные, которые, после добавления, в зависимости от необходимости, стерилизованной воды или физиологической сыворотки, создают инъецируемые растворы.
Наконец, способ согласно изобретению позволяет также осуществить скрининг сыворотки у тучных пациентов для определения наличия или отсутствия нефункционального лептина или же скрининг молекул, воздействующих на димеризацию OBR.
На фиг.1 схематически представлены слитые протеины. Box 1 обозначает сайт связывания JAK2; Box 3 обозначает сайт связывания протеинов STAT; TM обозначает транс-мембранный домен.
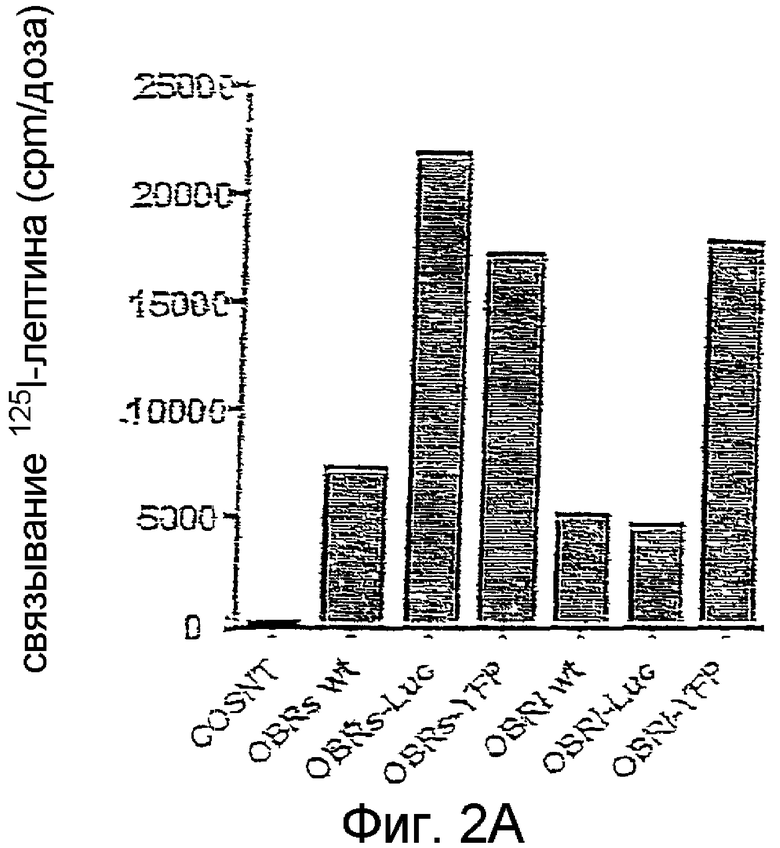
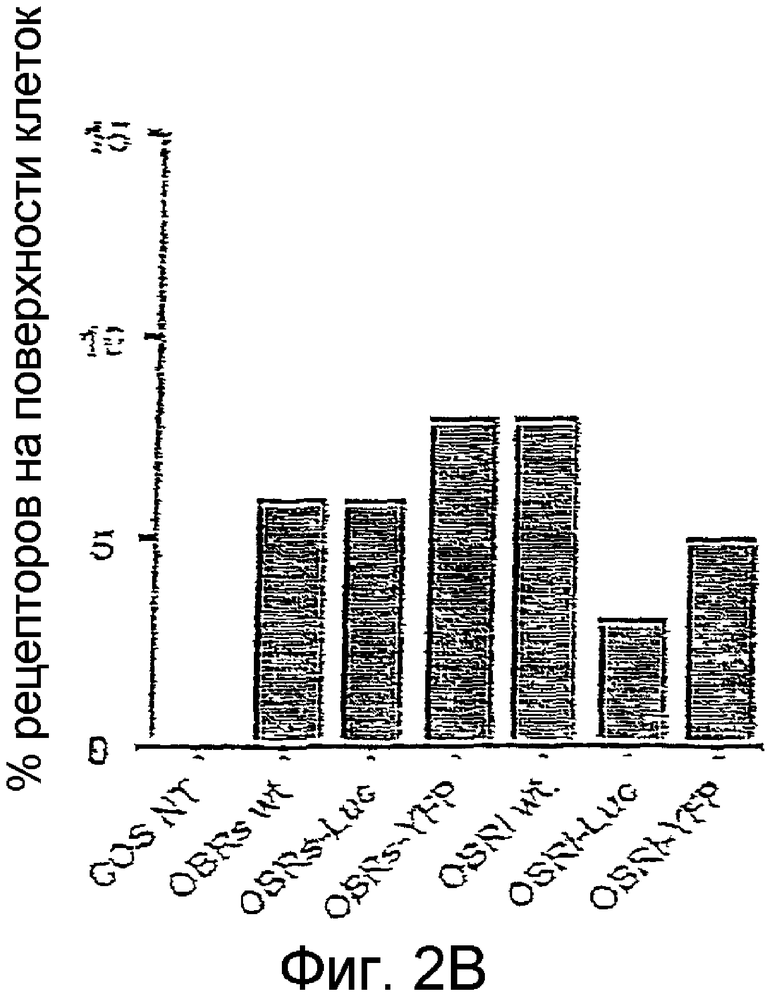
На фиг. 2А и 2B иллюстрирована экспрессия конструкций OBR в клетках COS, экспериментально осуществленная с помощью радиометки с использованием 125I-лептина в качестве радио-лиганда. На фиг.2А и 2B указаны соответственно общее содержание в клетках OBR и количество сайтов связывания с поверхностью клеток.
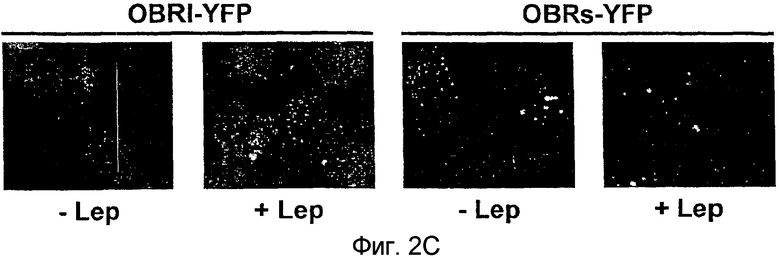
На фиг.2С показана локализация в клетках экспрессии конструкций OBRI-YFP и OBRs-YFP в присутствии и в отсутствие лептина.
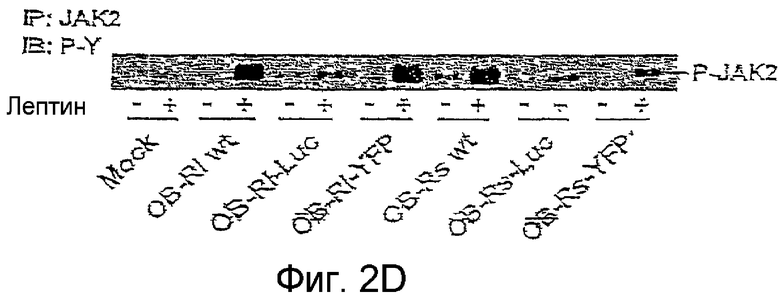
На фиг. 2D иллюстрируется активация JAK2 различными конструкциями OBR.
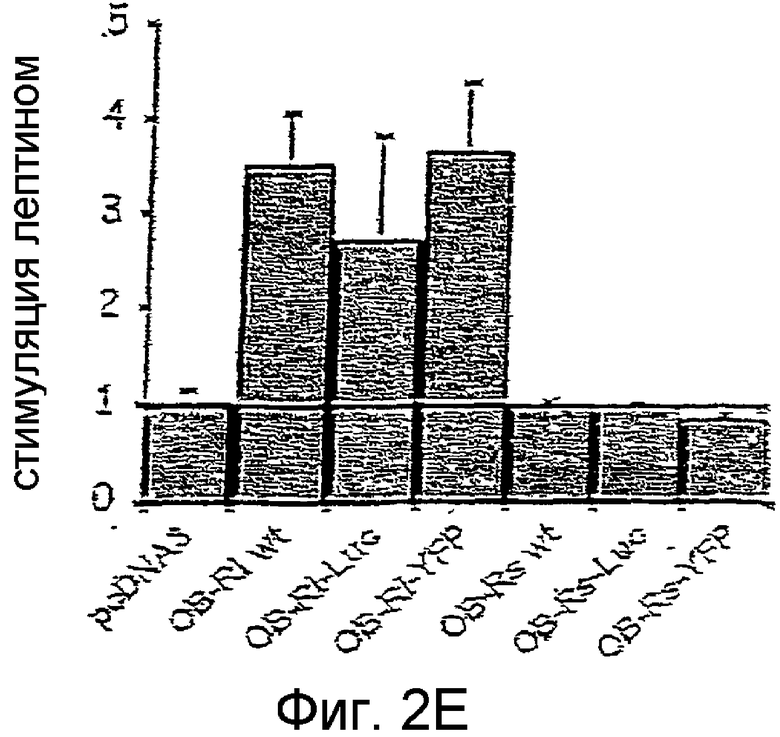
На фиг.2Е иллюстрируется эффект стимуляции лептином клеток, коэкспрессирующих репортерный ген для STAT3 и различныe конструкции OBR.
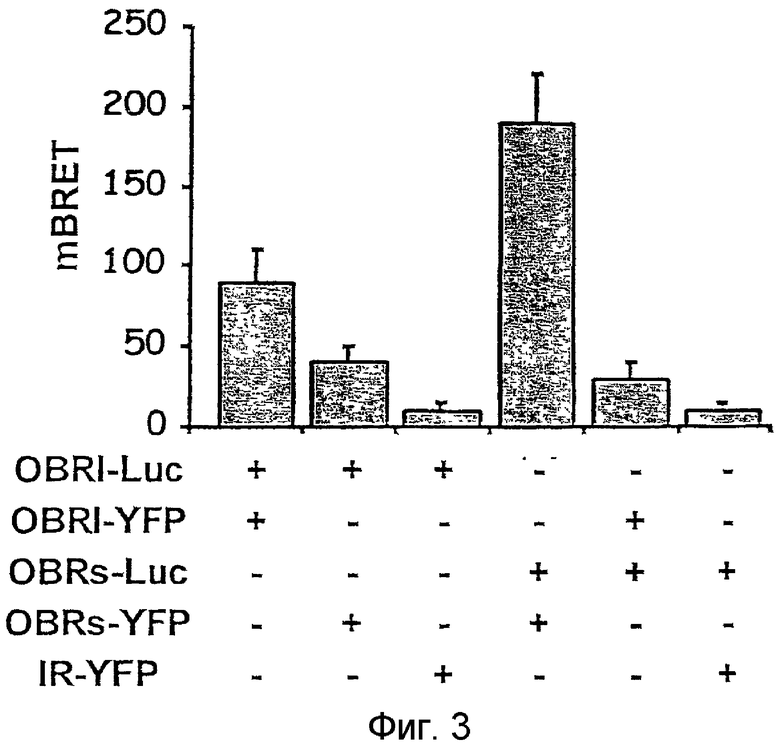
На фиг. 3 показана конститутивная димеризация OBR. Клетки HEK 293, экспрессирующие указанные различные конструкции OBR, инкубируют в присутствии коэлентеразина. Перенос энергии измерен с помощью люминометра.
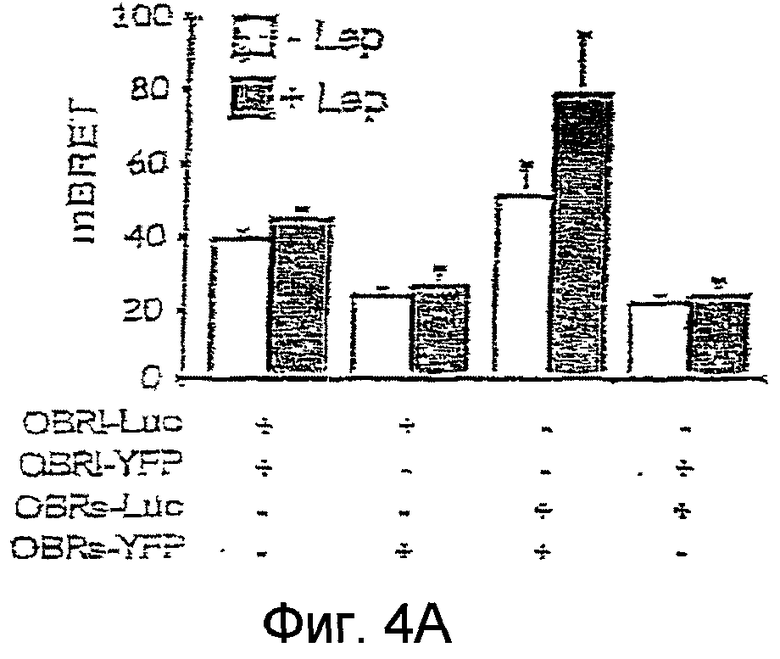
На фиг. 4А и 4B иллюстрируется эффект связывания лептина на конститутивный BRET рецепторов ORB.
На фиг.4А: клетки HeLа, экспрессирующие различные указанные конструкции OBR, инкубируют в присутствии лептина перед инициированием реакции люциферазы. Перенос энергии измерен с помощью люминометра.
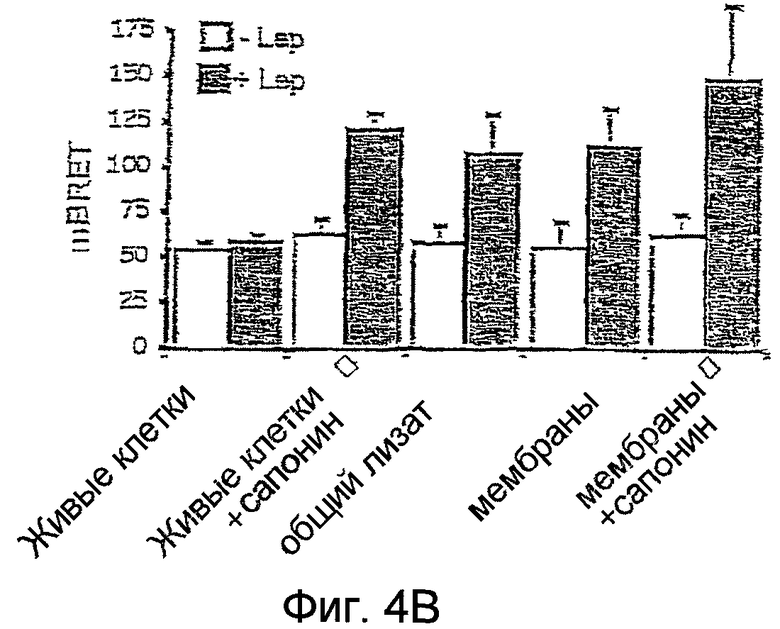
На фиг.4B: эффект лептина сравнивают в цельных клетках, коэкспрессирующих OBRs-Luc OBR-YFP, в присутствии или в отсутствие сапонина, в общих лизатах и в мембранных препаратах.
На фиг. 5А-5Е показаны оптимизация и характеристика изменения BRET, индуцированного лептином, с использованием рецепторов OBRs. Мембраны, полученные из клеток HeLa или COS, коэкспрессирующих OBRs-Luc и OBRs-YFP, прединкубированы с лептином или без него перед инициированием реакции с люциферазой.
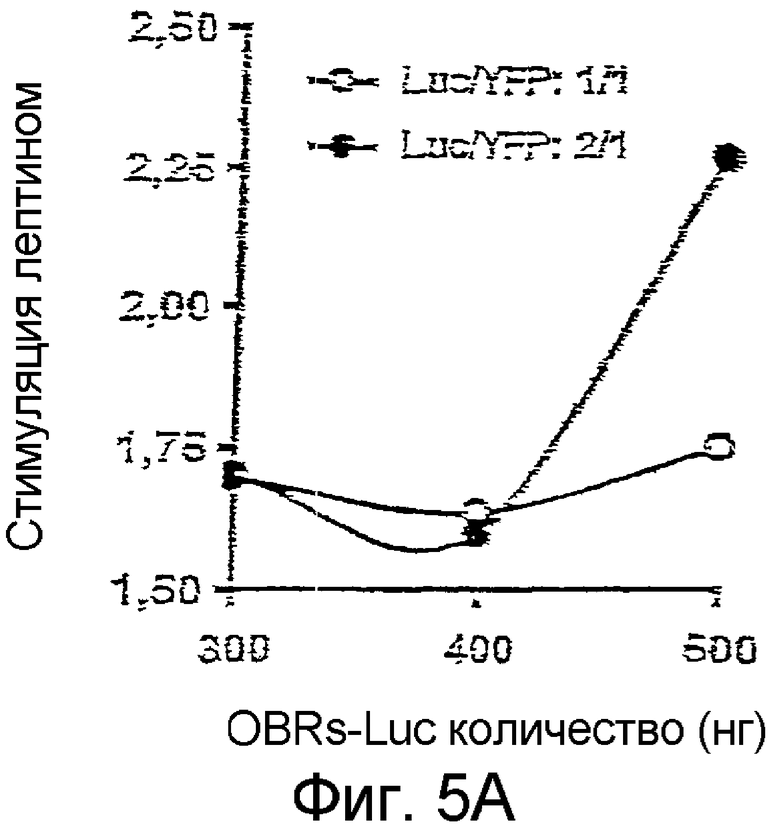
На фиг.5А: Оптимизация относительных и абсолютных уровней экспресии OBRs-Luc и OBRs-YFP.
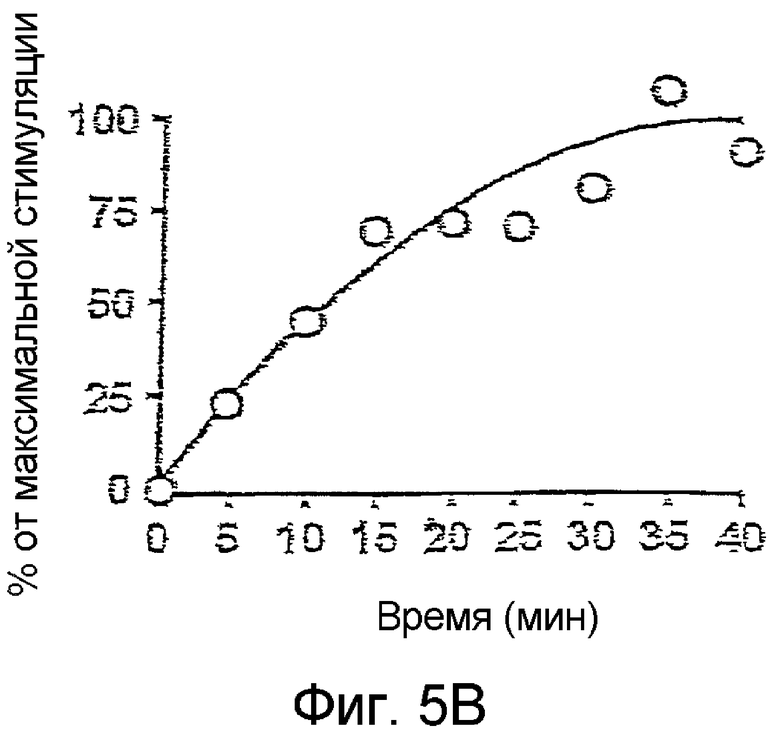
На фиг.5B: Изменение сигнала BRET, индуцированное лептином, в зависимости от времени.
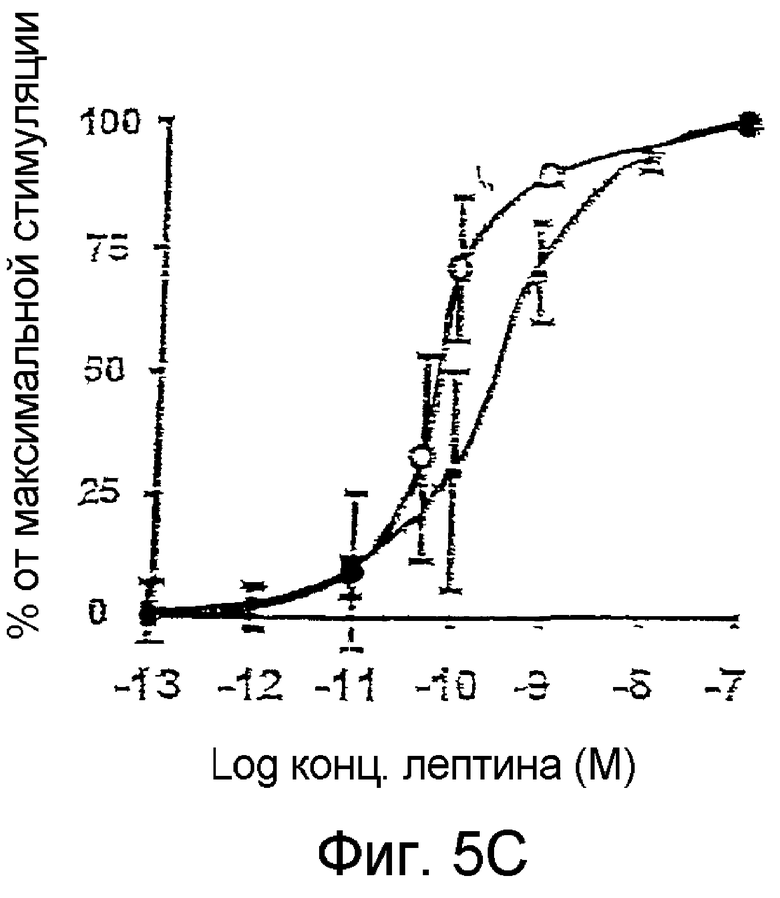
На фиг.5С: Кривые доза-ответ и BRET/концентрация лептина на мембране и в интактных клетках в присутствии сапонина (0,05%).
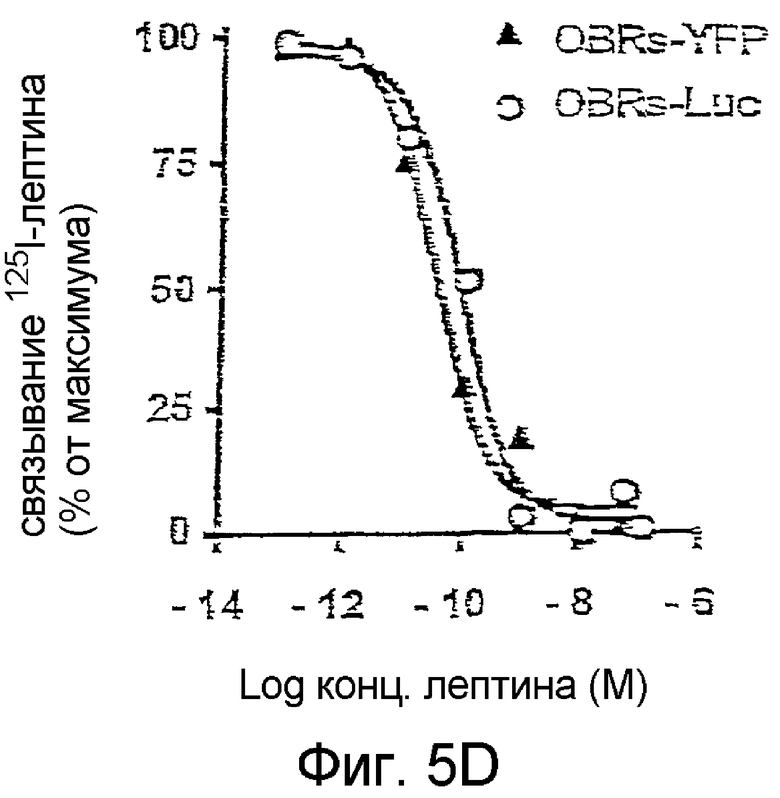
На фиг.5D: Конкурентное связывание 125I-лептина при увеличении возрастающих концентраций лептина.
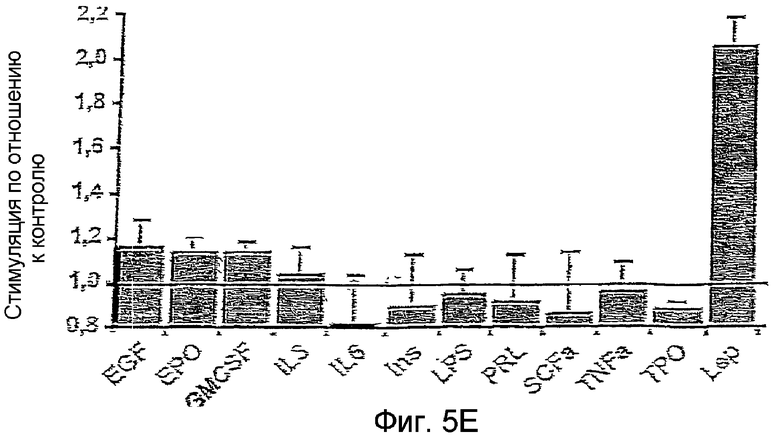
На фиг.5Е: Специфичность изменений BRET, индуцированных лептином. Мембраны прединкубированы в присутствии насыщенных концентраций эритропоэтина (EPO, 10U/мл), тромбопоэтина (ТРО, 10 нМ), колониестимулирующего фактора гранулоцитов/макрофагов (GM-CSF, 250 нг/мл), интерлейкина 3 (IL3, 280 нг/мл), интерлейкина 6 (IL6, 100 нг/мл), пролактина (PRL, 200 нг/мл), фактора α стволовых клеток (SCFα, 250 нг/мл), эпидермального фактора роста (EGF, 100 нг/мл), инсулина (Ins, 100 нМ), липополисахарида (LPS, 100 нг/мл) и фактора α некроза опухоли (TNFα, 50 нг/мл).
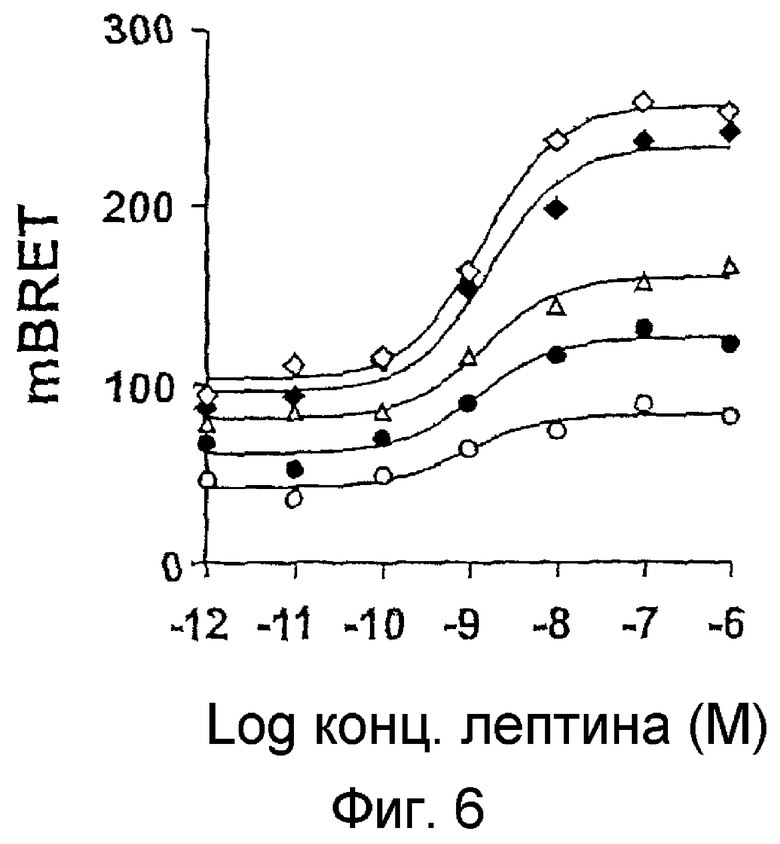
На фиг.6: Котрансфекция клеток COS постоянным количеством OBRs-Luc (50 нг) и возрастающим количеством OBRs-YFP: ο,200 нг; ●,400 нг; Δ,800; ♦1600; ◊,3200. Измерения BRET сделаны на клетках в присутствии сапонина (0,015%), инкубированных или не инкубированных в присутствии возрастающих доз лептина, и выражены в мBRET.
Настоящее изобретение иллюстрируется нижеследующими примерами, не ограничивающими его.
Материалы и методы, используемые в примерах
Конструкции плазмид, трансфекции и культивирование клеток
Слитые протеины OBR-YFP и OBR-Luc сконструированы путем сшивания YFP и Luc на С-конце рецепторов классическими методами молекулярной биологии. Кодируемые участки YFP, полученные из вектора pGFPtpz-N1 CytogemR - Topaze (Packard, Meriden, CT) были введены в сайт EcoRV вектора pcDNA3/CMV (Invitrogen, Groningen, Netherlands), который содержит модифицированный полилинкерный сайт. Кодируемый участок Renilla люциферазы получен из pRL-CМV (Promega, Madison, WI) и встроен в сайт EcoRV модифицированного вектора pcDNA3. Кодируемые участки OBRI и OBRs (от Dr.Gainsford, Royal Melbourne Hospital, Victoria, Australia) встроены в два вектора, описанные выше, соответственно в сайты клонирования EcoR1/BamH1 и Nhel. Стоп-кодоны делетировались путем направленного мутагенеза, и устанавливалась фаза слитого протеина.
Линии клеток HEK 293, COS-M6 и HeLa были культивированы в среде DMEM, дополненной следующими компонентами: 10% (об/об) FBS, 4,5 г/л глюкозы, 100 U/мл пенициллина, 0,1 мг/мл стрептомицина, 1 мМ глутамина (Life Technologies, Gaithersburg, MD).
Транзиторные транфекции осуществляли с использованием реактива трансфекции FuGene 6 (Roche, Bale, Швеция).
Флуоресцентная микроскопия
После двух дней трансфекции конструкциями YFP клетки были инкубированы в присутствии 100 нМ лептина в течение 60 мин и 0,01 мМ бис-бензамидина в течение 15 мин, затем промывали при помощи PBS и фиксировали в течение 20 мин. при комнатной температуре в 4%-м холодном растворе параформальдегида в PBS. Фракции исследовались путем флуоресцентной микроскопии с использованием фильтров FITC и DAPI.
Приготовление мембран и солюбилизация.
Клетки помещали на лед, дважды промывали в PBS при температуре льда и механически открепляли в буфере 1 (5 мМ Tris, 2 мМ EDTA, рН 7.4, 5 мг/л ингибитора трипсина сои, 5 мг/л лейпептина и 10 мг/л бензамидина) при температуре льда.
Суспензии клеток гомогенизировали три раза в течение 5 сек на гомогенизаторе Polytron (Janke and Кunkel Ultra-Turrax T25). Лизат центрифугировали при 450 х g в течение 5 мин при 4оС и надосадочную жидкость центрифугировали при 48000 х g в течение 30 мин. при 4°С. Конечный осадок промывали два раза буфером 1 и повторно суспендировали в растворе (75 мМ Tris (pH 7,4), 12.5 мМ MgCl2, 5 мМ EDTA с ингибиторами протеаз, описанными выше) и сразу же использовали в опытах на связывание радиоактивных лиганд или в опытах BRET.
Иммунопреципитация JAK2.
Клетки HeLa были котрансфицированы плазмидами, экспрессирующими меченый JAK2 посредством HA2 (от Dr.Wojchowsky, Pensylvania State University, Pensylvania, USA), и плазмидами, содержащими различные конструкции OBR.
Полученные клетки были лизированы в лизирующем буфере (10 мМ Tris, 150 мМ NaCl, 5 мМ EDTA, 5% глицерина, 0,02% NaN3, NP40 0,1%, ортованадат 1 мМ, 5 мг/л ингибитора трипсина сои и 10 мг/л бензамидина), затем их центрифугировали в течение 15 мин при 13000 об/мин. Растворимую фракцию иммуноприципитировали в течение 2 час поликлональным антителом к JAK2 (HR-758) (1 мкг/мл) (Santa-Cruz Biotechnology, Santa Cruz, CA).
Иммуноабсорбция ДСН-ЭФ на ПААГ
Иммунопреципитаты JAK2 денатурировали в растворе (62,5 мМ Tris/HCl (pH 6,8), 5% SDS, 10% глицерина и 0,05% голубого бромфенола) при 100оС в течение 10 мин. Протеины разделяли при помощи ДСН-ЭФ на ПААГ в 7%-ном полиакриламиде, и переносили на нитроцеллюлозу. Иммунодетекцию осуществляли антифосфотирозиновым антителом 4G10 (2 мкг/мл) (Upstate Biotechnology, Lake Placid, NY). Иммунореактивность была обнаружена при использовании соответствующего вторичного антитела, связанного с пероксидазой хрена и с хемилюминесцентным реагентом ECL (Amersham, Aylesbury, UK).
Тест связывания 125 I-лептина.
Трансфицированные клетки обеднялись сывороткой в среде DMEM (1% BSA) в течение 24 час перед опытами по связыванию. Для измерения связывания лептина с поверхностью клеток, клетки были дважды промыты при помощи PBS при температуре льда и инкубированы в буфере связывания (DMEM, 25 мМ Hepes, рН 7.4, 1% BSA), содержащем 100000 cpm/лунку 125I-лептина (PerkinElmer life sciences, Paris, France) в отсутствие или в присутствии 200 нМ нерадиоактивного лептина (рекомбинантный лептин человека (PeproTech Inc, USA)) в течение 4 час при 4°С. Клетки дважды промывали при помощи PBS при температуре льда, лизировали в 1N NaOH и подсчитывали радиоактивность на счетчике с гамма-излучением. Для измерения общего количества лептина, связанного в экстракте, клетки суспензировали в 1,5 мл буфера связывания, содержащего 0,15% дигитонина, в течение 2 час при 4°С. Экстракты центрифугировали 30 мин в центрифуге Eppendorf c максимальной скоростью при 4°С. Надосадочную жидкость (0,2 мл) инкубировали 125I-лептином с концентрацией 100000 срm в общем объеме 0,25 мл в отсутствие или в присутствии 200 нМ лептина при перемешивании в течение ночи при 4єС.
Добавляли 0,5 мл γ-глобулина (1,25 мг/мл) и 0,5 мл полиэтиленгликоля 6000 (25% вес/об.) для осаждения комплексов рецептор-лиганд, образующихся при центрифугировании (17000 х g в течение 3 мин). Осадок промывали 1 мл полиэтиленгликоля 6000 (12% вес/об.) и подсчитывали.
Активация репортерного гена.
Клетки HeLa ко-трансфицировали при помощи 2,6 мкг плазмиды, несущей репортерный ген STAT3 (от Dr. Levy, New York University, New York, USA), 200 пг pcDNA3, содержащего кодируемый участок Renilla люциферазы (используемой в качестве внутреннего контроля) и при помощи 1,4 мкг различных конструкций OBR или только носителем. Спустя 48 час после трансфекции, клетки обеднялись средой DMEM (1% BSA) в течение ночи, затем стимулировались при помощи 1 нМ лептина в течение 6-8 часов. Затем клетки были промыты и лизированы в пассивном лизирующем буфере (Promega Corporation, Madison, WI) в течение 15 минут при комнатной температуре. Общие лизаты центрифугировали в течение 2 минут при 15000 об/мин и надосадочные жидкости были использованы для количественного анализа Люциферазы ((Promega Corporation, Madison, WI) с использованием люминометра Berthold (Lumat LB 9507).
Результаты выражены в виде отношения активности люциферазы светлячка к активности люциферазы Renilla.
Измерение BRET
Спустя 48 час после трансфекции клеток HeLa, клетки COS и HEK 293, экспрессирующие OBR, открепляли и промывали при помощи PBS. Клетки в количестве 1-2х105 распределяли в оптических 96-ти луночных планшетах (Packard Instrument Company, Meriden, CT) в отсутствие или в присутствии лиганд при 25оС. Мембраны, полученные из клеток, эксперессирующих OBR, были использованы в тесте для измерения BRET. Субстрат коэлентеразин h (Molecular Probes, Eugene, OR) вводили до конечной концентрации 5 мкМ и осуществляли считывание на флюро/люминометре FusionTM (Packard Instrument Company, Meriden, CT), который обеспечивает последовательную интеграцию люминесцентных сигналов, улавливаемых с помощью двух фильтров (фильтр Luc: 485 +/- 10 нМ; фильтр YFP: 530 +/-12,5 нМ). Отношение BRET описывается как разность между излучением при 530 нм/485 нм котранфицированных слитых протеинов Luc и YFP и излучением при 530 нм/485 нм только одного слитого протеина Luc. Результаты выражаются в единицах миллиBRET (мBU), 1 единица мBRET соответствует значению отношения BRET, умноженному на 1000. Следующие лиганды были использованы для определения специфичности количественного анализа: рекомбинантный эритропоэтин человека (EPO), инсулин (Ins), липополисахарид (LPS, Sigma Aldrich, St.Lоuis, USA), рекомбинантный тромбопоэтин человека (TPO), GM-CSF, интерлейкин 3 (IL3), интерлейкин 6 (IL6), пролактин (PRL), SCF, EGF и TNFα.
Пример 1. Функциональная экcпрессия слитых протеинов OBR
Длинная форма (OBRI) и короткая форма (OBRs) рецепторов OBR были слиты своими С-концами с YFP или Luc (Фиг.1). Экспрессия этих слитых протеинов подтверждается на клетках COS, трансфицированных в экпериментах на связывание с 125I-лептином (Фиг.2А). Аналогичные результаты были получены на трансфицированных клетках HeLa.
Экспрессия на поверхности клеток слитых протеинов и рецепторов дикого типа, экспрессированных в клетках COS, варьируется в интервале от 5 до 10%, что согласуется с уже описанными значениями. Аналогичные данные получены с клетками HEK 293, экспрессирующими эндогенные OBR (14+/-3%).
Локализация слитых протеинов OBR в клетках HeLa изучалась по методу флуоресцентной микроскопии на протеинах, слитых с YFP. Флуоресценция, вызванная OBR1-YFP, точечно распределяется в клетках, в то время как флуоресценция, вызванная OBRs-YFP, локализуется в ламеллах. Стимуляция, вызванная лептином, локализует OBR1-YFP в больших внутриклеточных ламеллах, соответствующих, по-видимому, эндосомальному компартменту. Локализация OBRs-YFP не вносит каких-либо существенных изменений. Результаты, полученные флуоресцентной микроскопией, подтверждают локализацию OBR главным образом во внутриклеточном компартменте и согласуются с описанной локализацией слитого протеина OBR1-GFP в клетках COS.
Функциональная экспрессия слитых протеинов оценивается по величине активации пути JAK-STAT. Киназы JAK2 ассоциируются с внутриклеточными доменами OBRs и OBR1. Связывание лигандов индуцирует трансфосфорилирование JAK2 и фосфорилирование OBR1, но не OBRs. Фосфорилированый OBR1 cоздает сайт заякоревания для протеинов STAT, которые активируются путем фосфорилирования тирозина после связывания с рецептором. Активированные протеины STAT затем димеризуются и переходят в ядро, где они стимулируют транскрипцию генов посредством элементов, реагирующих на STAT, как описано Tartaglia (1997, J.Biol.Chem. 272, 6093-6096).
Как показано на фиг. 2С, все конструкции OBRs индуцируют фосфорилирование JAK2, что указывает на активацию JAK2. Активация репортерного гена STAT3 увеличивается 2-4 раза при помощи OBR1-wt и слитых протеинов OBR1, в то время как короткие изоформы не влияют на активность репортерного гена. Эти результаты показывают, что в клетках HeLa экспрессируются функциональные слитые протеины OBR.
Пример 2. Конститутивная димеризация OBR в живых клетках.
Димеризация OBR-Luc и OBR-YFP исследовалась на живых клетках. Наиболее заметный перенос энергии отмечается между OBRs-Luc и OBRs-YFP, а также между OBR1-Luc и OBR1-YFP, экспрессированных в эквимолярных количествах, что указывает на то, что конститутивные гомодимеры характерны для двух рецепторов (Фиг.3А,B). Существование гетеродимеров OBRs/OBR1 в живых клетках подтверждается тем, что обнаружен BRET-перенос между OBRs-Luc и OBR1-YFP, а также между OBR1-Luc и OBRs-YFP. О специфичности этих взаимодействий говорит отсутствие значимого переноса энергии между OBRs-Luc и OBR1-Luc и протеином слияния YFP и рецептора инсулина, описанным выше (BOUTE et al., 2001, цитированная выше).
Эти результаты свидетельствуют, что короткие и длинные изоформы включаются в гетеро- и гомокомплексы живых клеток.
Пример 3. Влияние связывания лептина на конститутивный BRET рецепторов OBR.
Для того чтобы оценить влияние агонистов на конститутивный BRET, клетки прединкубировали в присутствии лептина, затем инициировали реакцию люциферазы со своим субстратом.
Не было обнаружено никакого изменения конститутивного BRET в случае гомодимеров OBR1 и двух комбинаций гетеродимеров OBRs/OBR1, тогда как BRET возрастал в случае гомодимеров OBRs (Фиг.4 А).
Изменения BRET в случае гомодимеров OBRs, индуцированные лептином, исследовались на различных клеточных препаратах.
Механическое разрушение клеток в гипотоническом буфере заметно способствует увеличению BRET, вызванному лептином, тогда как начальный BRET остается без изменения.
Подобные результаты были получены с мембранной фракцией после выделения цитозола. Поскольку все пары OBRs-Luc/OBRs-YFP задействованы в формировании начального сигнала BRET, то только рецепторы, находящиеся на поверхности клеток (5-10%), могут стимулироваться лептином, который неспособен проходить через мембрану интактных клеток. Измельчение мембран клеток увеличивает фракцию OBR, проницаемую для лептина, и тем самым способствует увеличению BRET, индуцированному лептином.
Аналогичные результаты были получены на клетках, обработанных сапонином. Этот компонент проделывает отверстия в мембранах и обеспечивает проницаемость протеинов, как, например, лептина, во внутриклеточные компартменты, где находится большинство рецепторов OBRs.
В аналогичных экспериментах, осуществленных с использованием препаратов, полученных из клеток, эксперессирующих гомодимеры OBR1 или гетеродимеры OBRs/OBR1, не наблюдалось никакого изменения BRET, индуцированного лептином.
Затем были определены количества и соотношения OBRs-Luc и OBRs-YFP, позволяющие оптимизировать BRET, индуцированный лептином (Фиг.5А). Наилучшие результаты были получены в случае, когда использовались 500 нг ДНК, кодирующей OBRs-Luc и 250 нг ДНК, кодирующей OBRs-YFP.
При таких оптимальных условиях насыщенная концентрация лептина увеличивает в 2-2,5 раза начальный сигнал BRET в клетках, инкубированных в присутствии сапонина, или в мембранах, полученных из клеток, экспрессирующих гомодимеры OBRs. Это увеличение является функцией времени. Максимальные значения достигаются спустя 20 мин после инкубации с 1 нМ лептина при комнатной температуре (Фиг.5B). При высоких концентрациях лептина максимальные значения достигаются спустя 5 минут после инкубации при комнатной температуре.
Эффект лептина является дозозависимым при значении ЕС50 около 100 пМ (Фиг.5С), что согласуется со значениями Ki, полученными со слитыми протеинами OBRs-Luc (116 пМ) и OBRs-YFP (35 пМ) (Фиг.5D). Специфичность теста подтверждается отсутствием BRET, инуцированным лигандом при насыщенной концентрации нескольких цитокинов и других лигандов мембранных рецепторов, таких как эритропоэтин, тромбопоэтин, GM-CSF, IL3, IL6, PRL, SCFα, EGF, инсулин, LPS и TNFα.
Распределение рецепторов в виде димеров происходит согласно статистическому закону распределения и при численном соотношении рецепторов 1/1 можно получить следующее распределение рецепторов, если все они находятся в димерной форме: 1/4 Luc/Luc, 1/4 YFP/YFP и 1/2 рецепторов, способных запустить сигнал BRET (1/4 Luc/YFP, 1/4 YFP/Luc). Однако при измерении BRET совокупность молекул, слитых с Luc, дает сигнал люминесценции и, следовательно, при соотношении 1/1 наблюдается, что только половина всей популяции рецепторов способна запустить сигнал BRET. Поэтому для усиления сигнала BRET был проведен эксперимент с насыщением молекул Luc молекулами YFP так, чтобы совокупность молекул Luc находились в форме димеров с молекулами YFP (способными запустить сигнал BRET). Результаты, представленные на фиг.6, показывают, что начальный сигнал BRET усиливается при насыщении, и что стимуляция лептином пропорциональна начальному сигналу и усиливает в 2-2,5 раз начальный сигнал BRET. При насыщении получают наилучшее разрешение начального и индуцированного сигнала BRET, что позволяет осуществить более легкий отбор искомых молекул.
Изобретение относится к области биотехнологии и может быть использовано для скрининга соединений со свойствами агонистов или антагонистов рецептора лептина. Предложен способ определения лигандов рецептора лептина, предусматривающий контакт соединения-кадидата с клетками, экспрессирующими после их ко-трансфекции двумя экспрессирующими векторами, соответственно, два слитых протеина, один из которых состоит из короткой изоформы рецептора лептина (OBRs) и белка-донора энергии (предпочтительно люциферазы), а второй - из OBRs и белка-акцептора энергии (предпочтительно GFP или его мутантной формы); измерение переноса энергии (BRET) между слитыми протеинами; сравнение его величины с соответствующим показателем, измеренным в той же системе, но без тестируемого соединения, и оценку результата, где увеличение переноса энергии считается свидетельством связывания соединения-кандидата с рецептором лептина. Перспективы применения изобретения связаны с разработкой препаратов, предназначенных для профилактики или лечения болезней, в которых задействован лептин или его рецептор. 3 з.п. ф-лы, 15 ил.
1. Способ определения связывания соединений с рецептором лептина, включающий стадии, заключающиеся в том, что
a) получают клетки, совместно трансфицированные двумя векторами, один из которых обеспечивает экспрессию первого слитого протеина, отличающегося тем, что он состоит из короткой изоформы рецептора лептина (OBRs) и люциферазы, а второй вектор обеспечивает экспрессию второго слитого протеина, отличающегося тем, что он состоит из OBRs и флуоресцентного белка, представленного GFP или одной из его мутантных форм, и тем, что он характеризуется спектром возбуждения, перекрывающим спектр излучения люциферазы, являющейся частью первого слитого протеина;
b) осуществляют контакт указанного соединения с клетками, полученными на стадии а), и измеряют перенос энергии;
c) сравнивают перенос энергии, измеряемый в присутствии исследуемого соединения, с переносом энергии, измеряемым в отсутствие исследуемого соединения, и
d) анализируют результаты, полученные на стадии с), где увеличение переноса энергии коррелирует со связыванием соединения с рецептором лептина.
2. Способ по п.1, отличающийся тем, что его осуществляют с клетками, обработанными сапонином.
3. Способ по п.1, отличающийся тем, что субстратом является коэлентеразин.
4. Способ по п.1, отличающийся тем, что рецептор лептина представляет собой изоформу OBRs человека с последовательностью SEQ ID NO:2.
| LUNDIN A | |||
| et al | |||
| Biochim | |||
| et Biophys | |||
| Acta | |||
| Приспособление для регулирования подачи горючей жидкости | 1924 |
|
SU1499A1 |
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| BJORBAEK С | |||
| et al | |||
| J | |||
| Biol | |||
| Chem | |||
| Паровоз с приспособлением для автоматического регулирования подвода и распределения топлива в его топке | 1919 |
|
SU272A1 |

