Изобретение относится к биотехнологии, в частности к генетической инженерии, и представляет собой сконструированную in vitro рекомбинантную плазмидную ДНК, содержащую кДНК процессированной формы лептина человека, trc-промотор Escherichia coli и синтетический участок - усилитель трансляции, обусловливающие биосинтез полипептида со свойствами лептина человека, а также штамм Escherichia coli - продуцент этого полипептида.
Лептин человека представляет собой полипептидный гормон с молекулярной массой 16000 Да [1]. Лептин синтезируется преимущественно жировыми клетками в виде предшественника, состоящего из 167 аминокислотных остатков (а.о.). При секреции происходит удаление лидерного пептида с образованием зрелой формы белка, состоящей из 146 а.о. Показано, что лептин не содержит гликозидных остатков [2].
Физиологическая функция лептина заключается в поддержании энергетического баланса у млекопитающих. Связывание лептина с рецептором в гипоталамусе уменьшает потребление пищи, стимулирует обмен веществ и увеличивает расходование энергии [3-5].
Рекомбинантный лептин находит широкое применение в лабораторных и клинических исследованиях. В работе [6] показано, что подкожное введение в течение 4 недель рекомбинантного лептина, содержащего на N-конце метиониновый остаток, в количестве 0,1 мг/кг веса в день приводит к потере веса у людей в среднем на 1,9 кг.
Известны способы получения лептина человека, основанные на экспрессии в дрожжах [7] и в бакуловирусной системе [8]. Недостатком обоих методов является недостаточно высокий выход целевого продукта, а также длительная процедура очистки белкового препарата.
Основным способом получения лептина человека является микробиологический синтез [9, 10] . Синтезируемый белок накапливается в нерастворимом виде в телах включения, что, с одной стороны, требует введения стадии ренатурации для получения биологически активной формы полипептида, а с другой стороны, упрощает процедуру очистки целевого белка от полипептидных компонентов клетки. Недостатками описанных в работах [9, 10] способов являются весьма умеренный для микробиологического синтеза выход целевого белка, а также большой расход индуктора, требуемый для достижения экспрессии.
Наиболее близким к заявляемому техническому решению (прототипом) является способ, описанный в работе [11]. Рекомбинантная плазмидная ДНК содержит кДНК зрелого лептина человека под контролем промотора фага Т7. Синтезируемый целевой белок содержит на N - конце 11 аминокислотных остатков гистидина. Синтез белка осуществляют при добавлении индуктора, изопропил-β-D-тиогалактопиранозида (ИПТГ), до конечной концентрации 1 мМ. Полученный белок очищают с помощью металлоаффинной хроматографии, после чего остатки гистидина удаляют тромбиновым гидролизом. В результате получают зрелый лептин человека с общим выходом 45 нг/л культуральной жидкости.
Недостатком способа-прототипа является недостаточно высокий уровень синтеза лептина, использование больших количеств ИПТГ для индукции, а также высокий базальный уровень синтеза белка, свойственный системам экспрессии на основе промотора фага Т7 [12].
Изобретение решает задачу получения полипептида со свойствами лептина человека путем индуцибельного синтеза, а также увеличение уровня его биосинтеза при одновременном снижении количества добавляемого индуктора.
Поставленная задача решается путем конструирования рекомбинантной плазмидной ДНК pTrcTE-Lep, кодирующей индуцибельный синтез полипептида со свойствами лептина человека, и штамма Escherichia coli W3110/pTrcTE-Lep, обеспечивающего синтез этого полипептида с уровнем экспрессии не ниже 43% суммарного клеточного белка. Индуцибельный высокий уровень синтеза целевого полипептида обеспечивается тем, что плазмида pTrcTE-Lep содержит trc-промотор Е. coli и синтетический усилитель трансляции гена 10 бактериофага Т7.
Рекомбинантная плазмидная ДНК pTrcTE-Lep, кодирующая полипептид со структурой лептина человека, характеризуется следующими признаками:
имеет молекулярную массу 3,09 Md (4,682 т.п.о.);
кодирует аминокислотную последовательность зрелого лептина человека;
состоит из Cla I/Hind III - фрагмента ДНК плазмиды pTrcTEGF [13] длиной 4,232 т.п.о., содержащего trc-промотор Е. coli, синтетический усилитель трансляции TREN гена 10 бактериофага Т7, терминатор транскрипции фага лямбда term, ген bla β-лактамазы, ген lac Iq lac-репрессора и участок ori инициации репликации; а также из кДНК зрелого лептина человека, фланкированной сайтами рестрикции Cla I и Hind III;
содержит: trc-промотор Е. coli, синтетический усилитель трансляции TREN гена 10 бактериофага Т7, кДНК зрелого лептина человека, терминатор транскрипции фага лямбда term, ген bla β-лактамазы, определяющий устойчивость трансформированных плазмидой pTrcTE-Lep клеток к ампициллину, ген lac Iq lac-репрессора, участок ori инициации репликации; уникальные сайты узнавания рестрикционными эндонуклеазами, имеющими следующие координаты: Nco I - 265, Eco RI - 270, Kpn I - 286, Xba I - 340, Cla I - 377, Hind III - 828.
Особенностью предложенной плазмидной конструкции является то, что кДНК зрелого лептина человека находится под контролем trc-промотора Е. coli, a для усиления трансляции используется синтетический усилитель трансляции, что в совокупности обеспечивает индуцибельный синтез целевого белка с надежной регуляцией и высоким выходом, достигаемым при малых концентрациях индуктора.
Для получения штамма-продуцента полипептида со структурой лептина человека трансформируют компетентные клетки Escherichia coli W3110 рекомбинантной плазмидой pTrcTE-Lep.
Полученный штамм Escherichia coli W3110/pTrcTE-Lep характеризуется следующими признаками.
Морфологические признаки. Клетки мелкие палочковидной формы, грамотрицательные, неспороносные, 1х3-5 мкм, подвижные.
Культуральные признаки. При росте на плотной среде LA колонии круглые, гладкие, полупрозрачные, блестящие, серые, край ровный, диаметр колоний 1-3 мм; консистенция пастообразная. Рост в жидкой среде LB характеризуется ровным помутнением с образованием легкого осадка.
Физико-биохимические признаки. Клетки растут при температуре 4-42oС при оптимуме pH 6,8-7,2. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот. В качестве источника углерода используют аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к ампициллину (до 200 мкг/мл), обусловленную наличием в плазмиде гена бета-лактамазы.
Штамм Е. coli W3110/pTrcTE-Lep обеспечивает индуцибельный синтез полипептида со свойствами лептина человека в количестве не менее 43% от суммарного клеточного белка при концентрации индуктора 0,05 мМ. В отличие от прототипа в данном штамме достигается в 1,5 раза более высокий выход целевого белка при использовании лишь 1/20 от указанного в прототипе количества индуктора. Совокупность перечисленных свойств штамма Е. coli W3110/pTrcTE-Lep обусловливает большую технологичность процесса получения рекомбинантного полипептида.
Полученный штамм депонирован в коллекции культур микроорганизмов Института биоорганической химии им. М.М. Шемякина - Ю.А. Овчинникова Российской Академии Наук под номером 20.
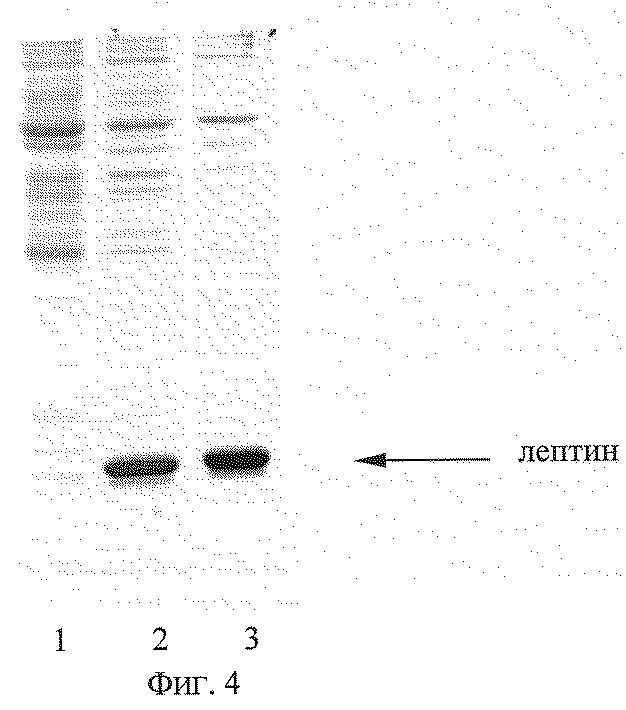
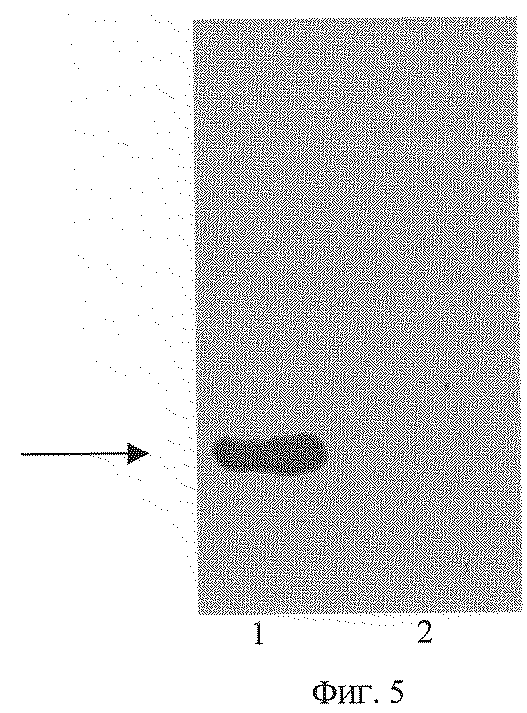
На фиг. 1 представлена физическая карта рекомбинантной плазмиды рТrсТЕ-Lep; на фиг.2(а) - нуклеотидная последовательность кДНК лептина человека с прилегающими регуляторными элементами: trc - промотор (1-30 п.о.), усилитель трансляции TREN (93-186 п.о.), ген лептина (190-630 п.о.); инициирующий и терминирующий кодоны подчеркнуты, в рамки взяты сайты рестриктаз: Eco RI, Kpn I, Cla I и Hind III; на фиг.2(б) - нуклеотидная последовательность олигонуклеотидных праймеров, использованных для амплификации кДНК лептина человека; на фиг.3 - аминокислотная последовательность полипептида лептина, кодируемого рекомбинантной плазмидой рТrсТЕ-Lep; на фиг.4 - электрофореграмма лизатов клеток штамма-реципиента Е. coli W3110 (дорожка 1), штамма-продуцента Е. coli W3110/pTrcTE-Lep при концентрации ИПТГ в среде 0,05 мМ (дорожка 2) и 1 мМ (дорожка 3) в 13%-ном полиакриламидном геле (стрелкой указан полипептид лептин); на фиг.5 - иммуноблот лизатов клеток штамма-продуцента Е. coli W3110/pTrcTE-Lep (дорожка 1) и штамма-реципиента Е. coli W3110 (дорожка 2) (стрелкой указан комплекс лептина с моноклональными антителами).
Изобретение иллюстрируется следующими примерами.
Пример 1. Конструирование промежуточной рекомбинантной плазмидной ДНК pSK-Lep.
Выделяют суммарную клеточную РНК из жировой ткани человека. Для этого 0,8 г жировой ткани размельчают в жидком азоте, после чего гомогенизируют в 10 мл реагента TRIZOL (Gibco BRL, США) с использованием гомогенизатора TISSUMIZER (Tekmar, США). К полученному гомогенату добавляют 2 мл хлороформа, перемешивают до образования суспензии и откручивают при 12000 g 15 мин при 4oС. Отбирают верхнюю водную фазу, содержащуюся в ней РНК осаждают добавлением 5 мл изопропилового спирта, выдерживают 10 мин при комнатной температуре, затем центрифугируют при 12000 g 10 мин при 4oС. Полученный осадок промывают 75%-ным этиловым спиртом, высушивают на воздухе и растворяют в 500 мкл воды. Концентрацию РНК определяют спектрофотометрически при длине волны 260 нм.
Суммарную клеточную РНК используют для получения кДНК лептина человека методом обратной транскрипции - полимеразной цепной реакции. Обратную транскрипцию проводят следующим образом: 5 мкг РНК в 10 мкл водного раствора инкубируют 5 мин при 75oC, после чего добавляют 50 пмоль праймера I (фиг.2 (б)), 4 мкл 5-кратного буфера (250 мМ трис-HCl, pH 8,3, 375 мМ КСl, 15 мМ MgCl2, 10 мМ дитиотреитол), 4 мкл смеси, содержащей 2,5 мМ каждого из дезоксинуклеозидтрифосфатов, 1 мкл (40 ед.) рекомбинантного ингибитора рибонуклеаз (Gibco BRL, США), 1,5 мкл (200 ед/мкл) M-MLV обратной транскриптазы (Gibco BRL, США) и воду до конечного объема 20 мкл. Реакционную смесь инкубируют 1 ч при 42oC, реакцию останавливают прогревом 5 мин при 75oС.
Синтез второй цепи и амплификацию кДНК лептина человека осуществляют следующим образом: к 5 мкл реакционной смеси, полученной после проведения обратной транскрипции, прибавляют по 100 пмоль праймеров I и II, 8 мкл смеси, содержащей 2,5 мМ каждого из дезоксинуклеозидтрифосфатов, 10 мкл 10-кратного буфера (100 мМ трис-HCl, pH 8,8, 500 мМ КСl, 15 мМ MgCl2), 2 ед. Taq ДНК-полимеразы (Fermentas, Литва) и воду до 100 мкл. Полимеразную цепную реакцию (ПЦР) проводят в следующем режиме: денатурация - 1 мин, 94oC; отжиг - 1 мин, 62oС; достройка - 1 мин, 72oС; количество циклов - 35. Праймер I представляет собой 36-звенный олигонуклеотид, в состав которого входят сайт узнавания рестриктазы Сlа I, стартовый ATG-кодон и 22 нуклеотида 5'-области кДНК зрелого лептина человека. Праймер II представляет собой 33-звенный олигонуклеотид, содержащий сайт узнавания рестриктазы Hind III, а также последовательность, комплементарную стоп-кодону гена лептина человека и прилегающим 18 нуклеотидам кодирующей области.
5 мкг плазмидной ДНК pBluescript (SK)+ ("Stratagene", США) обрабатывают 15 ед. рестриктазы Sma I (Fermentas, Литва) 1,5 ч при 30oС в 30 мкл буферного раствора, содержащего 33 мМ трис-ацетата (pH 7,9), 10 мМ ацетата магния, 66 мМ ацетата калия, и из полученного гидролизата выделяют линеаризованную плазмидную ДНК переносом на DEAE - мембрану NA-45 (Schleicher & Schuell, Германия) из 0,8%-ного агарозного геля.
0,5 мкг полученного в результате ПЦР фрагмента длиной 0,467 т.п.о. и 0,1 мкг векторной части плазмиды pBluescript (SK)+ длиной 2,961 т.п.о. сшивают при помощи лигазной реакции в течение 14 ч при 16oС в 15 мкл раствора, содержащего 40 мМ триса-НСl (pH 7,8), 10 мМ MgCl2, 10 мМ дитиотреитола, 0,5 мМ аденозинтрифосфата и 3 ед. Вейса Т4 ДНК-лигазы (Fermentas, Литва). 5 мкл реакционной смеси используют для трансформации 200 мкл компетентных клеток Escherichia coli XL-1 Blue ("Stratagene", США). 1/10 часть от общего количества клеток, использованных для трансформации, высевают на LB-агар, содержащий 75 мкг/мл ампициллина. В процессе рассева клеток на поверхность агара добавляют 100 мкл 0,1 М раствора ИПТГ и 20 мкл 4%-ного раствора 5-бромо-4-хлоро-3-индоксил-β-D-галактозида.
Конструирование промежуточной рекомбинантной плазмидной ДНК pSK-Lep позволяет использовать принцип цветной селекции для поиска клонов, содержащих встраиваемый фрагмент. Из выросших белых клонов выделяют плазмидную ДНК и анализируют рестрикционным анализом. Отбирают плазмидные ДНК, содержащие нужный набор рестрикционных фрагментов. Определяют нуклеотидную последовательность отобранных ДНК и окончательно отбирают плазмидные ДНК, в которых нуклеотидная последовательность кДНК лептина полностью соответствует данным, приведенным на фиг.2.
Пример 2. Конструирование рекомбинантной плазмидной ДНК pTrcTE-Lep.
10 мкг плазмидной ДНК pSK-Lep обрабатывают 20 ед. рестриктазы Сla I и 30 ед. Hind III (Fermentas, Литва) в течение 2 ч при 37oC в 50 мкл буферного раствора, содержащего 33 мМ трис-ацетата (pH 7,9), 10 мМ ацетата магния, 66 мМ ацетата калия, и из полученного гидролизата выделяют переносом из 2%-ного агарозного геля на DEAE-мембрану NA-45 фрагменты кДНК лептина человека длиной 0,351 т.п.о. (Cla I - Hind III фрагмент) и 0,099 т.п.о. (Hind III - Hind III фрагмент).
5 мкг плазмидной ДНК pTrcTEGF(b) размером 4,804 т.п.о., содержащей trc-промотор и усилитель трансляции TREN бактериофага Т7, обрабатывают 10 ед. рестриктазы Cla I и 15 ед. рестриктазы Hind III в течение 2 ч при 37oC в 30 мкл буферного раствора, содержащего 33 мМ трис-ацетата (pH 7,9), 10 мМ ацетата магния, 66 мМ ацетата калия, и из полученного гидролизата выделяют переносом из 0,8%-ного агарозного геля на DEAE-мембрану NA-45 векторную часть плазмидной ДНК длиной 4,232 т.п.о.
0,02 мкг Cla I - Hind III фрагмента кДНК лептина человека длиной 0,351 т.п.о. и 0,05 мкг векторной части плазмидной ДНК pTrcTEGF(b) длиной 4,232 т. п. о. соединяют при помощи лигазной реакции в течение 3 ч при 10oC в 15 мкл раствора, содержащего 40 мМ трис-НС1 (pH 7,8), 10 мМ MgCl2, 10 мМ дитиотреитола, 0,5 мМ аденозинтрифосфата и 2 ед. Вейса Т4 ДНК-лигазы. 5 мкл реакционной смеси используют для трансформации 200 мкл компетентных клеток XL-1 Blue. 1/10 клеток, использованных для трансформации, высевают на LB-агар, содержащий 75 мкг/мл ампициллина. Из выросших клонов выделяют плазмидную ДНК, содержащую фрагмент кДНК лептина человека.
5 мкг полученной плазмидной ДНК, содержащей фрагмент кДНК лептина человека длиной 0,351 т.п.о., обрабатывают 15 ед. рестриктазы Hind III в течение 1,5 ч при 37oC в 20 мкл буферного раствора, содержащего 33 мМ трис-ацетата (pH 7,9), 10 мМ ацетата магния, 66 мМ ацетата калия, и из гидролизата выделяют линеаризованную ДНК длиной 4,583 т.п.о. переносом из 0,8%-ного агарозного геля на DEAE-мембрану NA-45. 0,2 мкг полученной таким образом векторной ДНК длиной 4,583 т.п.о. соединяют с 0,02 мкг Hind III - Hind III фрагмента кДНК лептина длиной 0,099 т.п.о. посредством лигазной реакции в течение 3 ч при 10oC в 10 мкл раствора, содержащего 40 мМ трис-НСl (pH 7,8), 10 мМ MgCl2, 10 мМ дитиотреитола, 0,5 мМ аденозинтрифосфата и 2 ед. Вейса Т4 ДНК-лигазы. 2 мкл реакционной смеси используют для трансформации компетентных клеток XL-1 Blue. 1/10 клеток, использованных для трансформации, высевают на LB-агар, содержащий 75 мкг/мл ампициллина. Из выросших клонов выделяют плазмидную ДНК pTrcTE-Lep, содержащую Hind III - Hind III фрагмент кДНК лептина в требуемой ориентации.
Окончательно структуру рекомбинантной ДНК pTrcTE-Lep подтверждают определением нуклеотидной последовательности в области встроенной кДНК лептина человека.
Пример 3. Получение штамма-продуцента полипептида со свойствами лептина человека.
Рекомбинантной плазмидной ДНК pTrcTE-Lep трансформируют компетентные клетки Escherichia coli W3110 (American Type Culture Collection, 27325) и получают штамм-продуцент полипептида со свойствами лептина человека.
Пример 4. Определение продуктивности штамма-продуцента полипептида со свойствами лептина человека.
Ночную культуру клеток Е. coli W3110/pTrcTE-Lep в объеме 200 мкл переносят в 10 мл жидкой среды LB, содержащей 75 мкг/мл ампициллина, и выращивают до оптической плотности 0,7 (длина волны 550 нм) при 37oС при 175 об/мин. Добавляют ИПТГ до конечной концентрации 0,05 и 1 мМ, после чего клетки выращивают в течение 3 часов. Отбирают пробу 0,5 мл и центрифугируют 5 мин при скорости 6000 об/мин. Осажденные клетки суспендируют в 100 мкл буфера, содержащего 125 мМ трис-HCl, pH 6,8, 20% глицерин, 3% додецилсульфат натрия, 3% меркаптоэтанол, 0,005% бромфеноловый синий, инкубируют 10 мин в кипящей водяной бане, образцы объемом 5 мкл анализируют электрофорезом в 13% полиакриламидном геле с додецилсульфатом натрия. Гель окрашивают Кумасси R-250 (фиг. 4), сканируют и рассчитывают процентное содержание рекомбинантного белка в лизатах с использованием программы Scion Image (Scion Corp., США). По данным сканирования полипептид лептин составляет 43-46% от общего клеточного белка при концентрации ИПТГ 0,05 мМ и 50-54% при концентрации ИПТГ 1 мМ.
Пример 5. Иммуноблот лизатов штамма-продуцента Е. coli W3110/рТrсТЕ-Lep с моноклональными антителами к лептину человека.
Получают клетки штамма-реципиента Е. coli W3110 и штамма-продуцента Е. coli W3110/pTrcTE-Lep после индукции 0,05 мМ ИПТГ в течение 3 ч при 37oС при 175 об/мин. Для каждого штамма отбирают пробу 0,5 мл и центрифугируют 5 мин при скорости 6000 об/мин. Осажденные клетки суспендируют в 100 мкл буфера, содержащего 125 мМ трис-HCl, pH 6,8, 20% глицерин, 3% додецилсульфат натрия, 3% меркаптоэтанол, 0,005% бромфеноловый синий, инкубируют 10 мин в кипящей водяной бане, образцы объемом 5 мкл анализируют электрофорезом в 13% полиакриламидном геле с додецилсульфатом натрия, после чего осуществляют перенос белков из геля на нитроцеллюлозную мембрану. Проводят связывание белков на мембране с моноклональными антителами к лептину человека МАВ398 (R&D Systems, США) в течение 2 ч при концентрации антител 1,25 мкг/мл в 20 мМ трис-HCl, pH 7,6, 140 мМ NaCl. Мембрану отмывают от несвязавшихся антител, после чего мембрану инкубируют в присутствии конъюгата пероксидазы хрена и козлиных антител к антителам мыши (Sigma, США) в разведении 1:4000 в течение 2 ч в 20 мМ трис-HCl, pH 7,6, 140 мМ NaCl. Детекцию (фиг. 5) проводят с использованием набора для усиленной хемилюминесцентной детекции ECL (Amersham Life Science, Великобритания).
Пример 6. Выделение и характеризация рекомбинантного полипептида со свойствами лептина человека.
Проводят индукцию биосинтеза лептина в клетках штамма-продуцента Е. coli W3110/pTrcTE-Lep ИПТГ (конечная концентрация 0,05 мМ) в течение 3 ч. Клетки центрифугируют, после чего 1 г влажной биомассы обрабатывают лизоцимом и ультразвуком в 10 мл буферного раствора, содержащего 0,05 М трис-HCl, pH 8,0, 0,1 М NaCl и 0,001 М ЭДТА. Лизат центрифугируют в течение 10 мин при 10000 об/мин. Осадок, содержащий тельца включения, отмывают буферным раствором, содержащим 1% тритона Х-100, 0,05 М трис-HCl, pH 8,0, 0,025 М NaCl, 0,001 М ЭДТА, и тем же буферным раствором без тритона Х-100.
Для выделения лептина очищенные тельца включения растворяют в 10 мл 6 М раствора гуанидинхлорида, содержащего дитиотреитол, выдерживают в растворе окисленной и восстановленной формы глутатиона и выделяют хроматографическими методами: ион-обменной хроматографией и гель-фильтрацией. Лептин человека получают в 0,1 М уксусной кислоте, либо в 0,01 М трехзамещенном цитрате натрия, либо в 0,05 М бикарбонате аммония.
Описываемый способ выделения позволяет получить 34 мг лептина из 1 г влажной биомассы, что соответствует 34 мг белка из 100 мг лиофилизированной биомассы.
N-концевую аминокислотную последовательность определяют на секвенаторе 477А и ФТГ анализаторе 120А фирмы "Applied Biosystems", США. Препарат рекомбинантного лептина человека имеет следующую структуру N-конца молекулы: Met Val Pro Ile Gln Lys Val Gln Asp Asp..., т.е. соответствует структуре N-конца природного лептина человека с дополнительным метиониновым остатком, не влияющим на его биологические свойства [6].
Анализ вторичной структуры полипептида лептина осуществляют методом кругового дихроизма на приборе J500C фирмы JASCO (Япония). По данным анализа, доля аминокислотных остатков, входящих в состав α-спиральных участков составляет 64±2%, что в пределах погрешности опыта совпадает с данными рентгено-структурного анализа растворимой мутантной формы лептина, имеющей аминокислотную замену триптофана на аспарагиновую кислоту в 100-м положении [14] .
Таким образом, заявляемое техническое решение позволяет получить полипептид со структурой и свойствами, идентичными структуре и свойствам природного лептина человека; биосинтез полипептида индуцируется добавлением ИПТГ до конечной концентрации всего 0,05 мМ, и при этом уровень его синтеза составляет не менее 43% от суммарного клеточного белка за счет того, что ген лептина находится под контролем trc-промотора Е. coli, а трансляция белка усиливается за счет синтетического усилителя трансляции. Все это позволяет значительно повысить технологичность и экономичность процесса получения рекомбинантного лептина за счет значительного снижения расхода индуктора при одновременном увеличении выхода целевого продукта в 1,5 раза.
Источники информации
1. Zhang Y., Proenca R., Maffei M., Barone M., Leopold L., Friedman J. //Nature, 1994, v.372, p.425-432.
2. Cohen S. , Hallas J., Friedman J., Chait В., Benneth L., Chang D., Hecht R., Collins F.//Nature, 1996, v.382. N 6592, p.589.
3. Considine R., Caro J.//Hormone Research, 1996, v.46, p.249-256.
4. Spiegelman В., Flier J.//Cell, 1996, v.87, p.377-389.
5. Friedman J., Halaas J.//Nature, 1998, v.395, p.763-770.
6. Heymsfield S., Greenberg A., Fujioka К., Dixon R., Kushner R., Hunt Т., Lubina J., Patane J., Self В., Hunt P., Me Camish M.//JAMA, 1999, v.282, N 16, p.1568-1575.
7. Halaas J., Gajiwala K., Maffei M., Cohen S., Chait В., Rabinowitz D., Lallone R., Burley S., Friedman J.//Science, 1995, v.269, p.543-546.
8. Churgay L., Kovacevic S., Tinsley F., Kussow C., Millican R., Miller J., Hale J.//Gene, 1997, v.190, N 1, p.131-137.
9. Fawzi A. , Zhang H., van Heek M., Graziano M.//Horm. Metab. Res., 1996, v.28, p.694-697.
10. Altmann S. , Timans J., Rock F., Basan J., Kastelein R. //Protein Expression and Purification, 1995, v.6, p.722-726.
11. Патент США N 6048837, 2000.
12. Grossman Т. , Kawasaki E., Punreddy S., Osbume M. //Gene, 1998, v. 209, p.95-103.
13. Патент Российской Федерации 2113483.
14. Zhang F., Basinski M.B., Beals J.M., Briggs S.L., Churgay L.M., Clawson D.K., DiMarchi R.D., Furman T.C., Hale J.E., Hsiung H.M., Schoner B.E., Smith D.P., Zhang X.Y., Wery J.-P., Schevitz R.W. //Nature, v. 387, 1997, p. 206-209.


Изобретение относится к биотехнологии, в частности к генетической инженерии, и может быть использовано для получения полипептида со свойствами лептина человека. Рекомбинантная плазмидная ДНК содержит к ДНК процессированной формы лептина человека, trc-промотор Escherichia coli и синтетический участок - усилитель трансляции гена 10 бактериофага Т7, обусловливающие биосинтез полипептида со свойствами лептина человека. Esсherichia coli W3110/рTrcTE-Lep, трансформированный полученной плазмидой, обеспечивает синтез этого полипептида с уровнем экспрессии не ниже 43% суммарного клеточного белка при концентрации индуктора, изопропил-β-D-тиогалактопиранозида, 0,05 мМ. Изобретение позволяет увеличить уровень биосинтеза полипептида при одновременном снижении количества добавляемого индуктора. 2 с.п. ф-лы, 5 ил.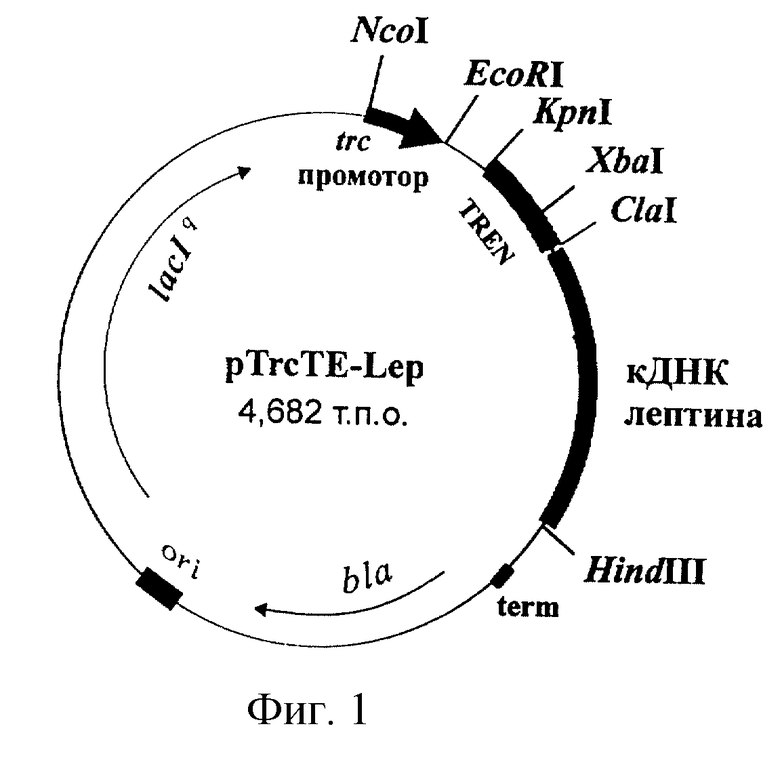
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PGGF 8, КОДИРУЮЩАЯ ПОЛИПЕПТИД СО СВОЙСТВАМИ ГРАНУЛОЦИТАРНОГО КОЛОНИЕСТИМУЛИРУЮЩЕГО ФАКТОРА ЧЕЛОВЕКА И ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ПОЛИПЕПТИДА СО СВОЙСТВАМИ ГРАНУЛОЦИТАРНОГО КОЛОНИЕСТИМУЛИРУЮЩЕГО ФАКТОРА ЧЕЛОВЕКА | 1996 |
|
RU2113483C1 |
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| US 6001816, 14.12.1999 | |||
| US 6087129, 11.07.2000. | |||

