Изобретение относится к области биотехнологии, медицины, иммунологии; оно может быть использовано в фармацевтической промышленности и молекулярной фармакологии и представляет собой способ целенаправленной доставки генов в клетки-мишени.
Достижения в области генетики, молекулярной биологии и биотехнологии определяют разработку стратегии и тактики введения ДНК в клетки как для целей генной терапии, так и при создании ДНК-вакцин. ВОЗ в начале 90-х годов составила и утвердила рекомендации по проведению работ в этих направлениях. В постановлении восьмой сессии общего собрания Российской академии медицинских наук в плане развития приоритетных направлений фундаментальных медицинских исследований на первом месте стоит генодиагностика и генотерапия [1] . Большое внимание созданию и развитию ДНК-вакцин было отведено на международной конференции "Вакцины и иммунизация" 1998 г. [2]. Обсуждались как созданные препараты, так и механизмы их действия.
Из числа основных проблем, связанных с безопасностью этих методов, две являются основными: возможное мутагенное действие ДНК, встроенной в геном клеток человека, и иммунопатология, которая может возникать при длительной выработке антигена в организме [3]. Эти проблемы решаются как созданием специальных рекомбинантных векторов, содержащих гены, так и разработкой способов доставки генов в клетки. На сегодняшний день одним из перспективных и исследуемых направлением является разработка способов целенаправленной доставки генов в клетки-мишени организма. Суть заключается в том, чтобы доставить ген именно в те клетки, в которых он и должен функционировать.
Известны способы создания рекомбинантных векторов и введения их в организм [4] . Первоначально вводили "незащищенные" рекомбинантные ДНК, которые быстро инактивировались под действием нуклеаз крови и эффективность доставки ДНК в клетки была низкой [5-7].
В настоящее время разрабатываются способы защиты вводимых ДНК (липосомы и т.п.) и способы доставки ДНК к клеткам-мишеням и проникновения в них. Так в работе [8] (прототип) описывается использование комплекса белка с поликатионными нуклеиновыми кислотами. В качестве белка авторы использовали трансферритин и показали, что комплекс взаимодействует с рецепторами трансферритина, проникает в клетки посредством пиноцитоза и происходит экспрессия ДНК в клетках. Недостатком этого способа является отсутствие универсальности доставки ДНК (в другие клетки-мишени) и относительно низкая эффективность проникновения ДНК в клетки.
Технической задачей изобретения является создание высокоэффективного молекулярного вектора как средства целенаправленной доставки генов в клетки-мишени. Для генной терапии и ДНК-вакцинации людей перспективным представляется создание такой конструкции вектора, который защищал бы рекомбинантную плазмиду с встроенным геном от действия нуклеаз крови, и, в то же время, имел бы стимулятор для проникновения ДНК в клетки. В качестве стимуляторов проникновения ДНК в эукариотические клетки применяются: ДЕАЕ-декстран, кальций-фосфат и некоторые другие [9].
Поставленная задача решается путем создания молекулярного вектора - конструкции, которая представляет собой вирусоподобные частицы, в центре которых находится рекомбинантная плазмидная ДНК с доставляемым геном, а на поверхности антитела - к клеткам-мишеням. Антитела входят в состав конъюгата: спермидин-полиглюкин, который несет две функции: удерживает конъюгат на отрицательно заряженной ДНК за счет положительного заряда спермидина и в то же время является аналогом ДЕАЕ-декстрана, стимулятора проникновения молекул ДНК в эукариотические клетки.
Сущность изобретения заключается в следующем.
Молекулярный вектор для доставки генов в клетки-мишени представляет собой частицы, в центре которых находится рекомбинантная плазмидная ДНК, содержащая доставляемые гены, а на поверхности - антитела к клеткам-мишеням.
Для сборки молекулярного вектора создают рекомбинантную плазмидную ДНК, содержащую доставляемый ген, очищают ее известными способами [10]. Антитела к клеткам-мишеням получают, используя стандартные способы иммунизации антигенами (клетками-мишенями или их компонентами) с последующей очисткой антител из сыворотки крови [11].
Плазмидную ДНК с доставляемым геном покрывают слоем конъюгата (фиг.1):
- полиглюкин с молекулярной массой около 60000 Д активируют периодатом натрия, который затем удаляют гель-фильтрацией на колонке с сефадексом G-50 в калий-фосфатном буфере;
- к активированному полиглюкину добавляют спермидин и антитела в отношении на 10 молекул полиглюкина 100 молекул спермидина и 1 - антител;
- после инкубации несвязавшиеся компоненты удаляют гель-фильтрацией на сефадексе G-50 в буфере: 0,1 М Tpис-НCl, pH 8,3;
- полученный конъюгат в избытке добавляют к плазмидной ДНК и после инкубации освобождаются от избытка конъюгата гель-фильтрацией на колонке с сефарозой CL-6В в физиологическом растворе.
Процесс получения молекулярного вектора для доставки генов в клетки-мишени представлен на фиг.2. Стабильность плазмиды в клетках и экспрессию гена определяют по известным методикам (картирование плазмидной ДНК рестриктазами, методами ИФА, ПЦР и др.).
Новыми, по сравнению с известными конструкциями, признаками являются: использование для доставки генов молекулярного вектора, содержащего в центре рекомбинантную плазмидную ДНК с доставляемым геном, а на поверхности антитела - к клеткам-мишеням. Антитела удерживаются на поверхности за счет вхождения их в состав конъюгата: спермидин-полиглюкин, который несет две функции: удерживает конъюгат на отрицательно заряженной ДНК за счет положительного заряда спермидина и в то же время является аналогом ДЕАЕ-декстрана, стимулятора проникновения молекул ДНК в эукариотические клетки.
Именно эта совокупность признаков обеспечивает целенаправленную доставку ионов в клетки-мишени и более высокую степень трансформации клеток рекомбинантной ДНК. Согласно предложенной схеме была создана конструкция молекулярного вектора, которая представляет собой вирусоподобные частицы, в центре которых находится рекомбинантная плазмидная ДНК с доставляемым геном, а на поверхности - антитела к клеткам-мишеням и проверена ее биологическая активность. Показано, что вводимый ген доставляется и накапливается в тех клетках организма, антитела к которым находятся на поверхности конструкции. Таким образом можно сделать вывод, что предлагаемое техническое решение позволяет целенаправленно доставлять гены к клеткам-мишеням, защищать их от действия нуклеаз крови и, кроме того, комплекс полиглюкин-спермидин, являясь аналогом ДЕАЕ-декстрана, используемого in vitro для стимуляции трансформации эукариотических клеток, аналогично способствует проникновению доставляемых генов в клетки.
Перечень графических материалов:
фиг.1. Молекулярный вектор для доставки генов в клетки-мишени:
1 - плазмидная ДНК, содержащая доставляемый ген, 2 - конъюгат: спермидин-полиглюкин - антитела к клеткам-мишеням.
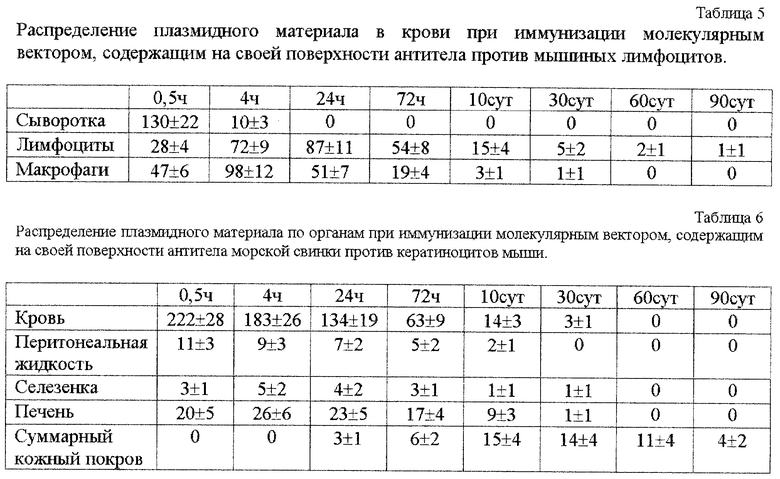
фиг.2. Гель-фильтрация на колонке с сефарозой CL-6B:
1 - плазмидная ДНК, 2 - то же, после инкубации с конъюгатом: спермндин-полиглюкин - антитела.
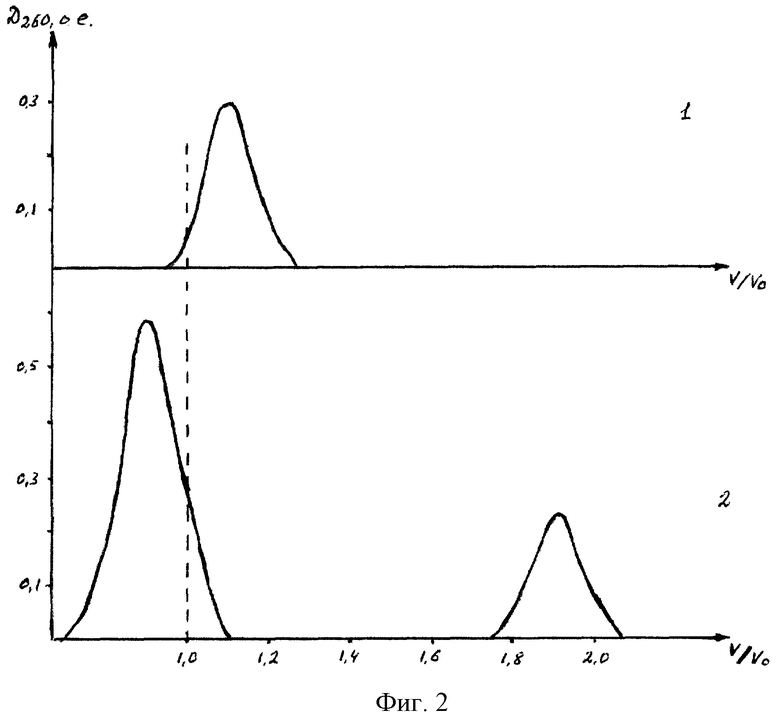
фиг. 3. Рестрикциоиный анализ (Нinf I) плазмидных ДНК. Дорожки электрофореграммы агарозного геля: 1 - исходная плазмидная ДНК, 2 - плазмидная ДНК после пассирования в лимфоцитах, 3 - то же, в кератиноцитах. Н - ДНК плазмиды pBR322+Hinf I.
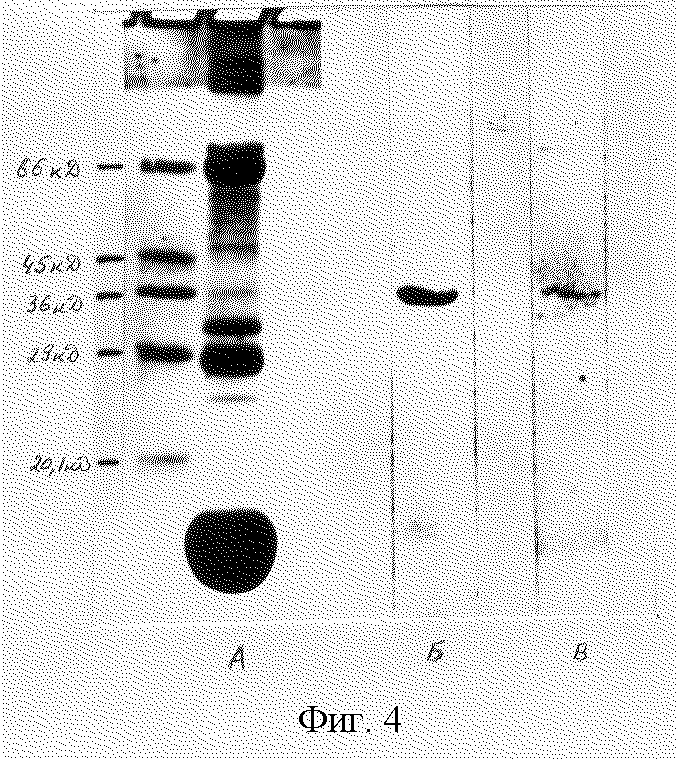
фиг. 4. Анализ белков лимфоцитов, полученных на 10-ые сутки после иммунизации мышей молекулярным вектором. А - электрофореграмма в 15%-ном поликриламидном геле, Б - иммуноблотинг с использованием поликлональных анти-α-ФНО-антител, В - то же, с использованием поликлональных анти-γ-ИФН-антител.
Изобретение иллюстрируется следующими примерами конкретного выполнения.
Пример 1. Плазмиды для доставки генов.
Используют плазмиду, содержащую ген, кодирующий биосинтез гибридного белка ннтерферон-гамма - фактор некроза опухолей-альфа [12].
Пример 2. Получение и очистка антител к клеткам-мишеням.
Антитела получают стандартным способом [11]. Для этого выделяют мышиные лимфоциты, очищают их центрифугованием в градиенте гипак-фиколл. Антигены, по 105 клеток в 0,1 мл физ. раствора, вводят трехкратно внутрибрюшинно с интервалом в две недели. Забор крови проводят через 10 дней после последней иммунизации. Для очистки антител из сывороток применяют фракционирование сульфатом аммония с последующей хроматографией на колонке с ДЕАЕ-целлюлозой. В качестве антител к лимфоцитам человека используют аптечный препарат антилимфоцитарного иммуноглобулина (лошадиный против лимфоцитов человека), дополнительно очищенный на колонке с ДЕАЕ-целлюлозой.
Пример 3. Создание молекулярного вектора для доставки генов в клетки-мишени.
В центре вектора находится интактная плазмидная ДНК, содержащая доставляемый ген, а на поверхности - антитела к клеткам-мишеням или белки, имеющие сродство к рецепторам клеток-мишеней. Для связи используют конъюгат полиглюкина со спермидином и антителом или белком. Положительно заряженный спермидин обеспечивает связывание конъюгата с плазмидной ДНК. Схематично вид конструкции представлен на фиг.1.
Для получения полисахаридной вставки 50 мг (0,8 мкМ) полиглюкина (60000 Д) обрабатывают 0,5 М раствором периодата натрия в течение 20-30 мин и затем проводят гель-фильтрацию на колонке с сефадексом G-50 в 20 мМ фосфатном буфере (рН 7,6). После этого в раствор активированного полиглюкина вносят 1,3 мг спермидина (8 мкМ) и 22 мг (0,17 мкМ) антител или белков. Смесь инкубируют в течение ночи, затем добавляют боргидрид натрия, перемешивают еще 2 ч и непрореагировавшие компоненты удаляют гель-фильтрацией на сефадексе G-50, используя 100 мМ Трис-НС1 (рН 8,3). Полученный таким образом препарат содержит 23,2 мг белка. Так как исходные компоненты (спермидин и антитела) вносятся в определенном соотношении, то можно предположить, что и полученный конъюгат содержит их в том же соотношении, т.е. около 1:10:0,1.
Для сборки конструкции к раствору, содержащему 400 мкг нуклеотидного материала (около 0,12 нМ) в физиологическом растворе добавляют конъюгат (200 мкг по белку), смесь инкубируют 2 ч при 4oC и полученную конструкцию отделяют от несвязавшихся компонентов гель-фильтрацией на сефарозе CL-6B (фиг. 2).
Исследование свойств молекулярной конструкции.
Конечные конструкции содержали примерно по 80 мкг белка (около 0,6 нМ антител при их мол. массе около 130000 Д на 0,12 нМ плазмиды). Таким образом они имели примерное соотношение: на 1 молекулу плазмиды - 50 молекул полисахаридной вставки - 5 молекул антител. Их мол. масса около 6700000 Д и при гель-хроматографии на колонке с сефарозой CL-6B они элюировались в свободном объеме (фиг.2).
Из теоретических расчетов на поверхности суперскрученной молекулы плазмиды мол. массой около 3000000-4000000 Д шарообразной формы может поместиться около 50-60 молекул полиглюкина (мол. массой 60000 Д). Так как в эксперименте это соотношение составляло 1:50, то можно предположить, что материал ДНК полностью "упакован".
Для проверки полноты упаковки нуклеотидного материала в собранной конструкции проводили ее обработку смесью ДНКазы и РНКазы (10 мкг/мл). Опыт показал, что если для исходной плазмидной ДНК полная деградация наблюдалась уже через 30 мин инкубации, то в собранной конструкции нуклеотидный материал сохранялся интактным в течение суток. При хранении конструкции в стерильных условиях при температуре 4oC ее свойства не изменялись по крайней мере в течение 1 мес.
Пример 4. Биологические испытания молекулярной конструкции.
А) Испытания in vitro. Первоначально исследования по доставке плазмидной ДНК, ее стабильности, способности проникать в клетки были проведены в опытах in vitro. При этом использовались клетки: лимфоциты, макрофаги, гепатоциты, кардиомиоциты, фибробласты, клетки мозга, легких. Использовались клетки двух видов животных: мышиные и 15-суточных эмбрионов кур.
К 0,3 мл клеточных суспензий с концентрацией 106 кл/мл вносили по 5 мкг препаратов чистой плазмиды и по 5 мкг "упакованной" в конструкции с двумя видами антител на поверхности. Первые - кроличьи антитела против мышиных лимфоцитов, вторые - антилимфоцитарный иммуноглобулин (лошадиный против лимфоцитов человека). Через различные периоды времени инкубации клетки собирали центрифугированием и анализировали на наличие в них плазмидного материала как описано выше. Клетки E.coli выросли на чашках с ампициллином только после контакта с нуклеотидным материалом, полученным из лимфоцитов, макрофагов и культуральной среды при использовании молекулярного вектора. Данные представлены в таблицах 1, 2. При использовании в опыте чистой плазмиды для введения гена роста клеток E.coli на селективной среде не наблюдали, выросло только несколько колоний из культуральной среды после 30-ти минутной инкубации.
Как видно из таблиц 1 и 2, если за первые 3 ч накопление плазмиды в лимфоцитах и макрофагах идет примерно с одинаковой скоростью, то в последующем в лимфоцитах плазмидного материала оказывается больше. К 72 ч инкубации в макрофагах плазмидного материала остается около 35% от максимального значения, тогда как в лимфоцитах его уровень сохраняется. Из таблиц также видно, что динамики накопления плазмидного материала в клетках при использовании молекулярных векторов с антителами против лимфоцитов, полученных из разных источников, кореллируют. Схожие данные были получены при использовании культур клеток эмбрионов кур для инкубации с плазмидным материалом.
Аналогичные эксперименты были проведены с молекулярным вектором, содержащим на поверхности антитела морской свинки к кератиноцитам мыши. При использовании чистой плазмиды для введения гена роста клеток E.coli на селективной среде также как в предыдущих опытах не наблюдали. Но наблюдался рост трансформированных клеток E.coli после инкубации с нуклеотидным материалом, полученным из фибробластов. Данные представлены в таблице 3.
Б) Испытания in vivo. Животным вводили внутрибрюшинно по 10 мкг, как препарата чистой плазмиды, так и по 10 мкг в конструкциях. Через определенные промежутки времени животных забивали цервикальной дислокацией, извлекали органы, отмывали от остатков крови, клетки тканей гомогенизировали, клетки макрофагов и лимфоцитов разделяли методами, описанными в [11]. Из культур клеток извлекали нуклеотидный материал, как описано выше, и анализировали его на наличие плазмиды. При иммунизации мышей препаратами чистой плазмиды роста клеток E.coli на селективной среде в последующем не наблюдали. Распределение плазмидного материала при иммунизации препаратами молекулярного вектора с антителами против лимфоцитов на своей поверхности представлено в таблице 4.
В клетках сердца, мышц, мозга, кожного покрова плазмидного материала не обнаружили. Наличие же плазмид в органах, приведенных в таблице 4 вероятно можно объяснить наличием лимфоцитов и макрофагов. Было исследовано распределение плазмидного материала среди компонентов крови. Результаты представлены в таблице 5.
Из таблицы 5 видно, что плазмидный материал накапливается и депонируется в основном в клетках лимфоцитов. Обнаружение его в макрофагах объясняется фагоцитозом, а снижение его уровня в макрофагах соответствующим киллингом попавшего в клетки материала. Снижение уровня плазмидного материала в фракции лимфоцитов крови к 60-ым и 90-ым суткам можно объяснить депонированием лимфоцитов в селезенке и лимфатических узлах, а также элиминацией части лимфоцитов из организма при их обновлении. Данные таблицы 4 показывают, что накопление плазмидного материала происходит в селезенке, месте депонирования лимфоцитов.
Динамика распределения плазмидного материала при использовании молекулярного вектора с антителами к кератиноцитам на своей поверхности в крови мышей сходна с отображенной в таблице 3. Распределение плазмидного материала в этом случае по органам представлено в таблице 6.
Из таблицы 6 видно, что плазмидный материал в процессе эксперимента накапливается в кожном покрове и к 60-ым, 90-ым суткам его уровень стекается.
Пример 5. Определение стабильности плазмиды с доставляемым геном после трансформации эукариотических клеток
Для проявления экспрессии доставляемого гена в эукариотические клетки существенную роль играет стабильность самой плазмиды, несущей ген. Известно, что чужеродный генетический материал в клетках может элиминироваться (удаляться) или подвергаться деградации, в том числе - с сохранением "маркеров", но с потерей доставляемого гена (делецией). Так как выделенные из эукариотических клеток плазмидный материал позволял расти трансформированным им клеткам Е. coli на селективной среде, то можно сделать вывод о наличии плазмиды и сохранении маркера. Для проверки сегрегационной стабильности плазмид проводили их физическое картирование (рестрикционный анализ). На фиг. 3 представлены элeктpoфopeграммы гидролиза образцов плазмиды, выделенных из клеток E.coli, трансформированных нуклеотидным материалом, полученным из лимфоцитов и фибробластов в процессе испытаний in vivo. Прямой рестрикционный анализ образцов плазмиды, выделенных из эукариотических клеток был затруднен, возможно из-за частичного метилирования ДНК. Как видно из фиг. 3 образцы плазмидной ДНК гидролизуются мелкощепящей эндонуклеазой рестрикции Нinf I на одинаковое количество фрагментов, размеры которых совпадают с аналогичными для фрагментов исходной плазмидной ДНК. Таким образом, можно сделать вывод, что плазмидная ДНК, используемая для экспериментов, в эукариотических клетках достаточно стабильна
Пример 6. Определение экспрессии доставленных генов в клетках
Так как в эксперименте использовался ген, встроенный под контроль прокариотических промоторов, то его экспрессии мы особо не ожидали. Тем не менее, при постановке иммуноблота с антителами к ФНО-α и γ-интерферону обнаружили, что в лимфоцитах, полученных из мышей на 10-ый день после введения молекулярной конструкции на иммуноблоте наблюдается полоса с мол. массой около 32000 Д, взаимодействующая с антителами как против ФНО, так и против интерферона (фиг. 4). При исследовании сыворотки соответствующие полосы визуально не обнаруживались. Таким образом, использование предлагаемого молекулярного вектора для доставки генов в клетки-мишени позволяет:
- целенаправленно доставлять гены в клетки-мишени, сводя до минимума их попадание в другие виды клеток;
- использовать гены инфекционных агентов в составе рекомбинантных плазмидных ДНК ("центральное ядро" частиц) в качестве ДНК-вакцины;
- использовать положительно заряженный комплекс полиглюкин-спермидин (аналог ДЕАЕ-декстрана) в качестве стимулятора проникновения ДНК в клетки.
Литература
1. Постановление восьмой (LXXI) сессии Общего собрания Российской академии медицинских наук "Новые технологии в медицине XXI века". //Вестник РАМН. - 1999. - 10. - С. 32-34.
2. Бектимиров Т.A., Баринский И.Ф. "Международная конференция "Вакцины и иммунизация" //Вопросы вирусологии. - 1999. - 2. - С. 95-96.
3. Воробьев А.А., Медуницин Н.В. Новые принципы и методы создания иммунобиологических препаратов. //Вестник РАМН. - 1999. - 10. - С. 16-17.
4. Ann. N.Y. Acad. Sci. V. 772: Vaccines. A New Era in Vaccinology./ Eds M.A. Lin, M.R., Hilleman, R. Kurth. New York, 1995.
5. Wolff J. A. , Malone R.W., Williams P. et al. //Science.-1990. - V. 247. - P. 1465-1468.
6. Tang D. C. , Devit M., Jonston S.A. //Nature - 1992. - V. 356. - P. 152-154.
7. Ulmer J.B., Donnelly J.J., Parker S.F. et al. //Science. - 1993. - V. 259. - P. 1745-1748.
8. Патент США 5792645A. Beug, Yartmut; Birnstiel, Max L. et al. "Комплексы белков с поликатионными нуклеиновыми кислотами и способы их использования". Опубликован 06.06.1994 г.
9. Щелкунов С.Н. Клонирование генов. //Новосибирск. - "Наука". - 1986. - 228 С.
10. Маниатис Т., Фрич Э., Сэмбрук Дж. //Молекулярное клонирование. - М.: Мир., 1984. - 479 с.
11. Иммунологические методы. Под ред. Г. Фримеля. - М. "Медицина". - 1987.
12. Коробко В.Г., Давыдов И.В., Шингарова Л.Н. и др. Гены гибридных лимфокинов человека. 1. Конструирование рекомбинантных плазмид, кодирующих гибриды иммунного интерферона и факторов некроза опухолей человека. //Биоорган. химия. - 1991. - Т. 17. - 2. - С. 189-196.
Изобретение относится к области биотехнологии, медицины, иммунологии и может быть использовано в фармацевтической промышленности и молекулярной фармакологии. Молекулярная конструкция представляет собой частицы, в центре которых находится рекомбинантная плазмидная ДНК с доставляемым геном, а на поверхности - антитела к клеткам-мишеням. Антитела входят в состав конъюгата - спермидин-полиглюкин, способного удерживать конъюгат на отрицательно заряженной ДНК за счет положительного заряда спермидина и стимулировать проникновение молекул ДНК в эукариотические клетки. Изобретение позволяет разрабатывать средства для доставки генов в клетки-мишени. 4 ил., 6 табл.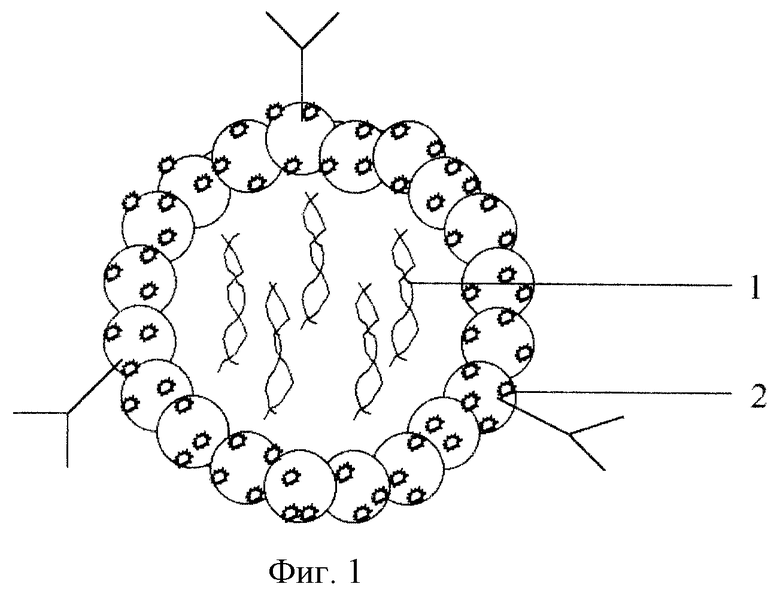
Молекулярный вектор для доставки гена в клетки-мишени, представляющий собой молекулярную конструкцию, содержащую в центре плазмиду, включающую доставляемый в клетки-мишени ген, покрытую конъюгатом: спермидин-полиглюкин - антитела к клеткам-мишеням.
| US 5792645, 11.08.1998 | |||
| Огнетушитель | 0 |
|
SU91A1 |
| WAGNER et al., Proc | |||
| Natl | |||
| Acad | |||
| Sci | |||
| USA, 1991, v | |||
| Шланговое соединение | 0 |
|
SU88A1 |
| ЩЕЛКУНОВ С.Н | |||
| Клонирование генов | |||
| - Новосибирск: Наука, 1986, с | |||
| Приспособление для нагрузки тендеров дровами | 1920 |
|
SU228A1 |

