Изобретение относится к области биотехнологии, медицины, иммунологии; оно может быть использовано в фармацевтической промышленности и молекулярной фармакологии и представляет собой способ доставки и представления иммуногенов иммунной системе организма.
Достижения в области генетики, молекулярной биологии и биотехнологии определили современную стратегию разработки вакцин. На сегодняшний день одним из перспективных и наиболее разрабатываемым направлением является создание генно-инженерных вакцин при помощи наработки антигенов микробиологическим синтезом с последующей их очисткой до гомогенного состояния. Оно предпочтительнее, чем усовершенствование вакцин на основе живых аттенюированных или инактивированных возбудителей. Это обусловлено тем, что присутствие части белков инфекционного агента в вакцине нежелательно [1, 2]. Так, например, известно, что среди белков вируса HIV есть и В-клеточные эпитопы, стимулирующие продукцию усиливающих инфекционность антител, и Т-клеточные эпитопы, стимулирующие супрессирующие клетки, и области, перекрестно реагирующие с нормальными белками клеток, и иммуносупрессивные районы. Поэтому для создания эффективных и безопасных вакцин нового поколения перспективно создавать синтетические пептидные вакцины, которые, как ожидается, будут свободны от многих недостатков, присущих живым и инактивированным вакцинам [3, 4]. Наиболее перспективными могут быть вакцины, содержащие несколько Т и В-клеточных эпитопов инфекционного агента. К недостаткам генно-инженерных вакцин следует отнести сравнительно небольшие молекулярные массы синтезируемых полипептидов-иммуногенов, от тысячи до нескольких десятков тысяч дальтон. Поэтому при вакцинации используют различные адъюванты, что не всегда желательно и вызывает побочные эффекты. Наиболее сильным из адъювантов является адъювант Фрейнда, который для людей использовать запрещено.
Известны способы экспонирования эпитопов инфекционного агента на поверхности капсидных структур HBcAg, синтезированных в E.coli [5-9] и дрожжах S. cerevisiae [10] и получение на них иммунного ответа. HBcAg представляет собой нуклеокапсид вируса гепатита В и состоит из идентичных белковых субъединиц размером 21 кД, имеет диаметр около 25-27 нм [5, 6] или 30-34 нм [9] по данным разных авторов и является иммуногеном. Для реализации иммунного ответа на встроенные эпитопы необходимо выполнение трех основных условий [5, 6] : встроенная последовательность а) не нарушает капсидообразующих свойств химеры, б) экспонируется на ее наружной поверхности, в) сохраняет в составе химеры свою нативную конформацию. Наиболее перспективным по устойчивости к внедрению чужеродных последовательностей является район 144-й аминокислоты гена. Внедрение чужеродных последовательностей длиной 40-50 аминокислот в этом районе не приводит к нарушению процесса самосборки капсид, которые сохраняют морфологию немутантного HBcAg и экспонируют при этом на своей поверхности внедренные последовательности с сохранением их антигенной и иммуногенной активности [5, 6].
К недостаткам такой конструкции следует отнести ограниченность по размерам встраиваемых чужеродных эпитопов (порядка 40-50, до 100 аминокислот [5, 6]). К тому же, не все встроенные эпитопы проявляли свою антигенность в составе гибридных частиц [9, 11]. Например, было обнаружено, что эпитоп вируса иммунодефицита обезьян, встроенный в С-конец после 183-й аминокислоты, недоступен для антител и иммунологически не активен [11].
Технической задачей изобретения является создание высокоэффективной молекулярной конструкции вирусоподобного вида (вакцины) как средства доставки иммуногенов. Для вакцинации людей перспективным представляется создание такой конструкции вакцины, которая имела бы и антигенные свойства, и стимулятор иммунного ответа, и, в то же время имела молекулярную массу, сравнимую с молекулярными массами вирусов, т.е. являлась бы полным антигеном.
В качестве стимуляторов иммунного ответа применяются полисахариды, липидные (в том числе липосомы) и полинуклеотидные комплексы (двуспиральная РНК, фрагментированная ДНК) и др.
Поставленная задача решается путем создания вакцины нового поколения - молекулярной конструкции, которая представляет собой вирусоподобные частицы, в центре которых находится полинуклеотидный комплекс (рекомбинантная плазмидная ДНК или двуспиральная РНК), а на поверхности - гибридные белки, содержащие эпитопы инфекционного агента (фиг.1). Рекомбинантная плазмидная ДНК может содержать различные гены инфекционного агента, тем самым превращая конструкцию также в "ДНК-вакцину".
Сущность изобретения заключается в следующем.
Вакцина (молекулярная конструкция вирусоподобного вида) представляет собой частицы (фиг. 1) 25-30 нм в диаметре, содержащие в центре полинуклеотидный комплекс (рекомбинантную плазмидную ДНК, содержащую гены инфекционного агента или двуспиральную РНК - стимулятор неспецифической резистентности организма), а на поверхности - гибридные белки, содержащие эпитопы (полипептиды) инфекционного агента и фермент (например глутатион-S-трансферазу или β-галактозидазу). Связь между полинуклеотидным комплексом и гибридными белками осуществляется посредством конъюгата: спермидин (для связи конъюгата с полинуклеотидным комплексом) - полиглюкин - субстрат для фермента (например, глутатион или галактопиранозид) для аффинной сорбции гибридных белков на конструкцию. При иммунизации созданной конструкцией титр антител выше и сохраняется более длительное время по сравнению с таковым при иммунизации исходным белком-иммуногеном.
Для сборки молекулярной конструкции получают рекомбинантную плазмидную ДНК, выделенную щелочным способом [12] , и содержащую гены инфекционного агента или двуспиральную РНК, выделенную из дрожжей, например из штамма Saccharomyces cerevisiae M437 Y116 [13], используемую как основной компонент лекарственного препарата для стимуляции неспецифической резистентности организма "Ридостин" [14, 15].
Полинуклеотидный комплекс покрывают слоем конъюгата (фиг.1):
- полиглюкин с молекулярной массой около 60000 Д активируют периодатом натрия, который затем удаляют гель-фильтрацией на колонке с сефадексом G-50 в калий-фосфатном буфере;
- к активированному полиглюкину добавляют спермидин и субстрат для фермента гибридного белка, например глутатион или аминофенилгалактопиранозид;
- после инкубации несвязавшиеся компоненты удаляют гель-фильтрацией на сефадексе G-50 в буфере: 0,1 М Трис-HCl, рН 8,3;
- полученный конъюгат в избытке добавляют к полинуклеотидному комплексу и после инкубации освобождаются от избытка конъюгата гель-фильтрацией на колонке с сефарозой CL-6B в физиологическом растворе.
Далее к полученному комплексу добавляют гибридные белки, содержащие эпитопы инфекционного агента и фермент, например глутатион-S-трансферазу или β-галактозидазу, и после инкубации полученную молекулярную конструкцию отделяют гель-фильтрацией на сефарозе CL-6B, используя для элюции физиологический раствор.
Гибридные белки получают по стандартной процедуре клонирования генов в одной рамке считывания с маркерными ферментами (глутатион-S-трансферазой, β-галактозидазой и т.д.) в разработанных или коммерческих векторах [16, 17]. Выделение и очистку белков проводят по разработанной методике или по рекомендациям фирм.
Процесс получения конструкции представлен на фиг.2. Полученную конструкцию используют для иммунизации. Титр нарабатываемых антител, силу и длительность иммунного ответа определяют по стандартным методикам.
Новыми, по сравнению с известными иммуногенными конструкциями, признаками являются: использование для иммунизации молекулярной конструкции вирусоподобного вида, содержащей в центре полинуклеотидный комплекс (рекомбинантную плазмидную ДНК, содержащую гены инфекционного агента или двуспиральную РНК - стимулятор неспецифической резистентности организма), а на поверхности - гибридные белки, содержащие эпитопы (полипептиды) инфекционного агента и глутатион-S-трансферазу (или β-галактозидазу). Связь между полинуклеотидным комплексом и гибридными белками осуществляется посредством полианиона - конъюгата: спермидин (для связи конъюгата с полинуклеотидным комплексом) - полиглюкин - глутатион (или галактопиранозид) (для аффинной сорбции гибридных белков на конструкцию). Именно эта совокупность признаков обеспечивает более высокий титр антител и более длительный иммунный ответ при иммунизации созданной молекулярной конструкцией по сравнению с аналогичными при иммунизации просто исходным белком-антигеном. Молекулярная конструкция позволяет экспонировать на поверхности частиц эпитопы любых инфекционных агентов продолжительностью от десятков до тысяч аминокислот (в аналогах - до 50-100 аминокислот) и создавать поливалентные вакцины.
Перечень графических материалов:
Фиг.1. Молекулярная конструкция вирусоподобного вида:
1 - плазмидная ДНК, 2 - конъюгат: спермидин-полиглюкин-глутатион, 3 - гибридный белок глутатион-S-трансфераза - TBI (GST-TBI).
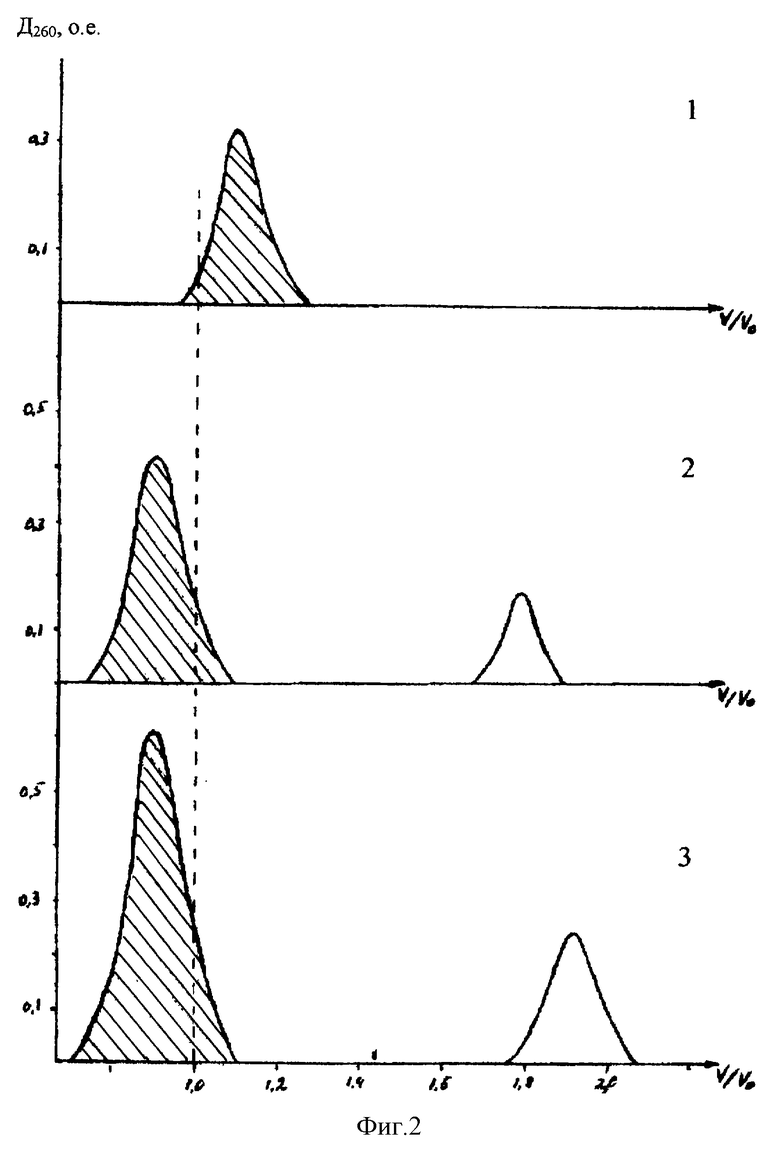
Фиг.2. Гель-фильтрация на колонке с сефарозой CL-6B:
1 - плазмидная ДНК,
2 - то же, после инкубации с конъюгатом: спермидин-полиглюкин-глутатион,
3 - после инкубации с гибридным белком глутатион-S-трансфераза - TBI.
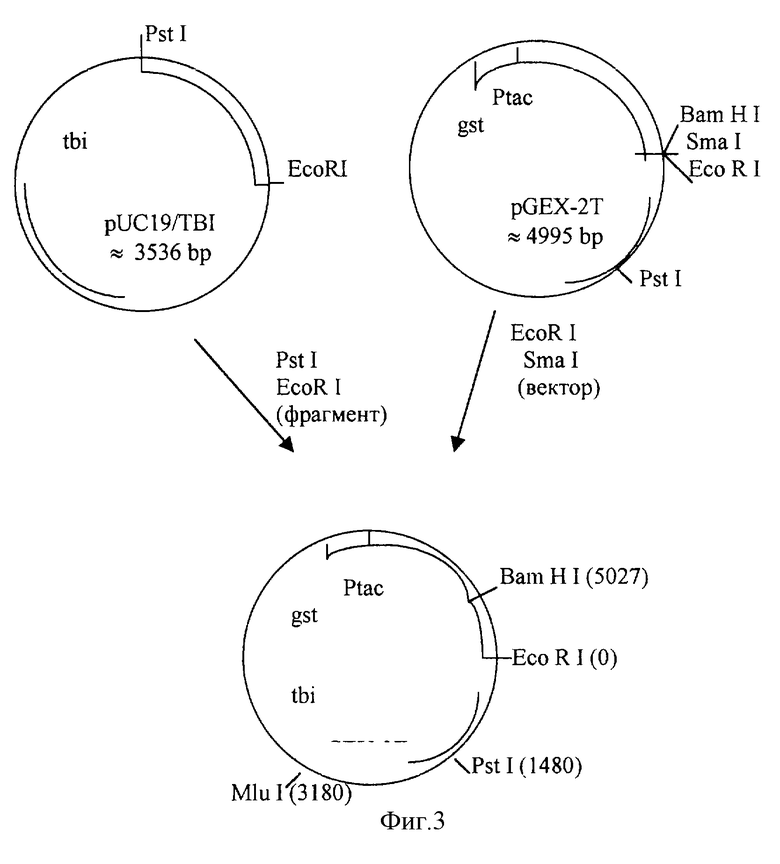
Фиг. 3. Схема конструирования рекомбинантной плазмиды: bla - ген β-лактамазы, кодирующий устойчивость к ампициллину (Арr); gst - ген, кодирующий глутатион-S-трансферазу; tbi - ген, кодирующий белок TBI.
Фиг.4. Электронограмма негативно окрашенного препарата микрочастиц вакцины. Увеличение 70000.
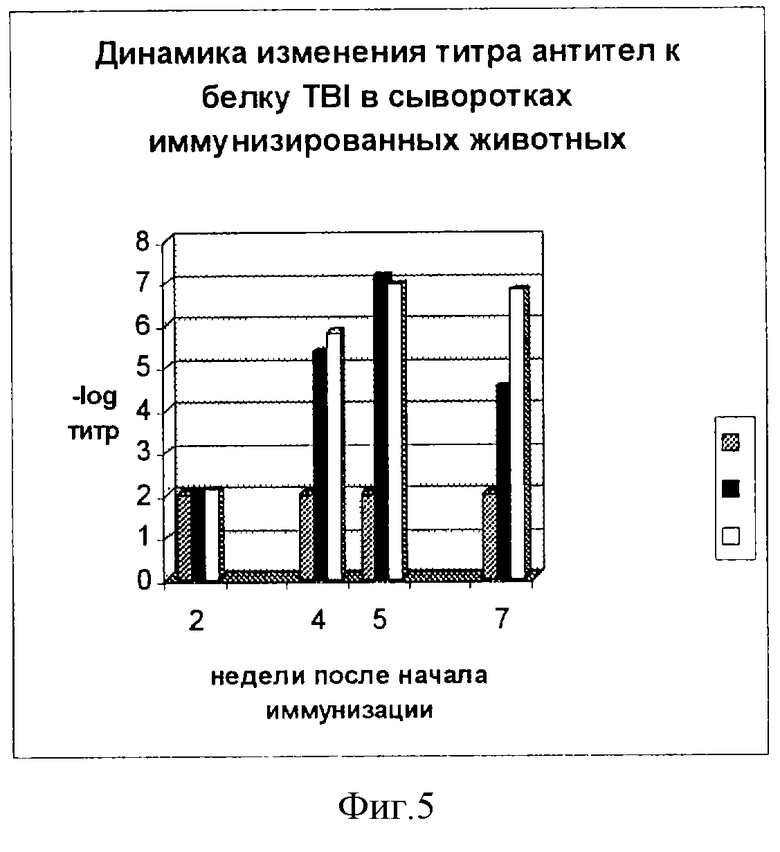
Фиг. 5. Динамика изменения титра антител к белку TBI в сыворотках животных, иммунизированных белком TBI-GST (черные столбцы); вирусоподобными частицами с белком TBI-GST (белые столбцы). Контрольные животные (серые столбцы). В каждом опыте использовали сыворотку от пяти животных.
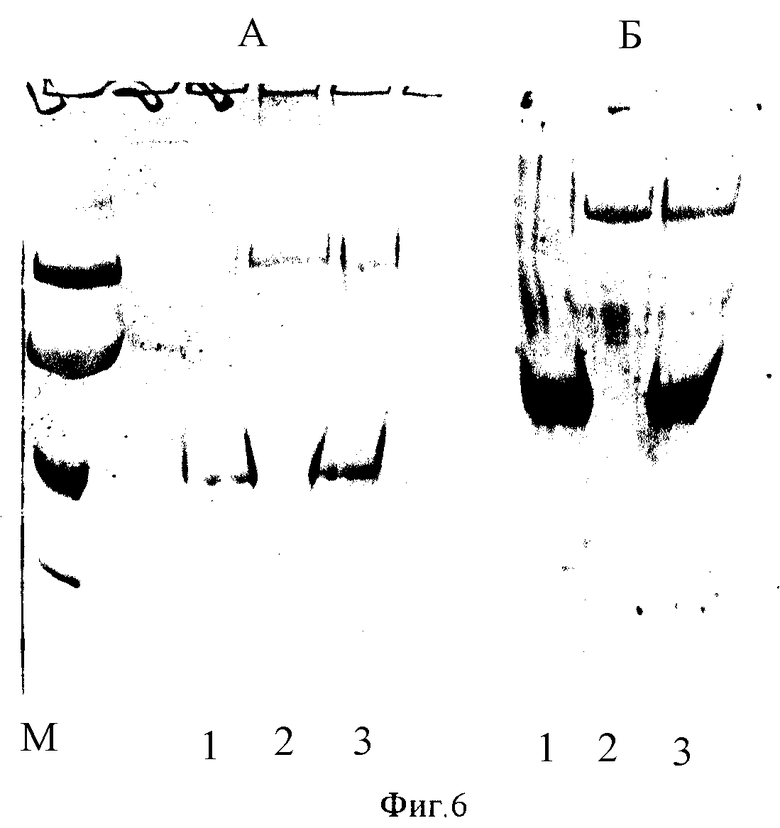
Фиг. 6. Электрофореграмма образцов вакцины в 10%-ном полиакриламидном геле: а - окраска кумасси R-250: 1 - конструкция с белком GST - TBI, 2 - белок gp41-β-гaлaктoзидaзa, 3 - конструкция с белками GST - TBI и gp41-β-галактозидаза; б - иммуноблоттинг с антителами положительной по HIV-I сывороткой человека. М - маркеры мол.масс.
Изобретение иллюстрируется следующими примерами конкретного выполнения.
Пример 1. Создание плазмид, несущих гены гибридных белков.
Целью создания плазмиды pGEX-2T-TBI было получение гибридного белка глутатион-S-трансфераза-ТВI (GST-TBI), способного одной своей половиной молекулы аффинно соединяться с конъюгатом глутатион-полисахарид, а второй половиной (TBI) - экспонировать эпитопы HIV-I на поверхности конструкции [18] . При создании плазмиды pGEX-2T-TBI исходную плазмиду pUC19/TBI [18] гидролизуют рестриктазами Pst I и EcoR I (фиг.3), выделяют фрагмент длиной 445 п. о., который затем лигируют с коммерческим вектором фирмы "Pharmacia" pGEX-2Т [16] , обработанным рестриктазами EcoR I и Sma I. Правильность встройки определяют секвенированием ДНК в районе соединения генов, кодирующих составные гибридного белка. Рамка считывания гена при этом не нарушается. В созданной плазмиде сайты узнавания для рестриктаз Pst I и Sma I отсутствуют.
Полученной плазмидой трансформируют компетентные клетки реципиентного штамма E. coli SG 200-50 и суспензию высевают на агаризованную среду, содержащую ампициллин (100 мкг/мл). Для наработки плазмиды культуру клеток выращивают в L-бульоне до оптической плотности Д550≅1,5-2,0 о.е., добавляют хлорамфеникол и инкубируют в течение 3-5 ч при 32oС. Плазмидную ДНК выделяют щелочным методом с последующей очисткой хроматографией на гидроксилапатите.
Плазмида pGP41-16 [19] , содержащая ген, кодирующий гибридный белок pGP41-16 - β-галактозидаза создана по традиционной технологии и любезно предоставлена авторами.
Пример 2. Выделение и очистка гибридных белков.
Для наработки биомассы рекомбинантных клеток выросшие на селективной (Ар) среде клоны вносят в 100 мл L-бульона; подращивают до оптической плотности Д550≅1,0 о.е., добавляют ампициллин до конечной концентрации 500 мкг/мл и инкубируют в течение 2 ч для лизиса безплазмидных клеток. После этого засевают 1,6 л L-бульона (8 колб по 200 мл). Индукцию биосинтеза целевого белка проводят добавлением в среду изопропилтиогалактопиранозида до конечной концентрации 0,1 мМ. Гибридный белок GST-TBI накапливается в клетках в нерастворимой форме, в тельцах включения.
Очистку белка GST-TBI проводят по методу, разработанному для искусственного белка TBI [20]. Для этого суспензию клеток обрабатывют ультразвуком и отмывают тельца включения. Гибридный белок растворяют в 6 М растворе мочевины в течение 2 ч, осадок удаляют центрифугированием, и затем проводят ренатурацию белка, последовательно инкубируя раствор при концентрациях мочевины 3 М и 1,5 М, после этого проводят диализ против буфера, не содержащего денатурирующего агента. При таком режиме ренатурации выпадения целевого белка в осадок не наблюдается. Для доочистки используют аффинную хроматографию на глутатион-сефарозе.
Очистку гибридного белка gp 41(607-620)-β-галактозидаза проводят согласно рекомендациям авторов [19].
Пример 3. Создание вирусоподобной конструкции (вакцины).
А) В вирусах молекулы ДНК и РНК не связаны с белками ковалентными связями. Подобную структуру создавали и мы. В центре конструкции вакцины находится интактный полинуклеотидный комплекс, а на поверхности - гибридный белок, содержащий антигены HIV-I. Для связи используют конъюгат полиглюкина со спермидином и глутатионом. Положительно заряженный спермидин обеспечивает связывание конъюгата с полинуклеотидным комплексом, глутатион - аффинно сорбирует гибридный белок TBI-GST. Схематично вид конструкции представлен на фиг.1.
В качестве "центрального ядра" конструкции используют плазмиду pGEX-2T-TBI (мол. массой около 3250000 Д), содержащую ген, кодирующий эпитопы HIV-I.
Для получения полисахаридной вставки 50 мг (0,8 мкМ) полиглюкина (60000 Д) обрабатывают 0,5 М раствором периодата натрия в течение 20-30 мин и затем проводят гель-фильтрацию на колонке с сефадексом G-50 в 20 мМ фосфатном буфере (рН 7,6). После этого в раствор активированного полиглюкина вносят 5 мг глутатиона (16 мкМ) и 2,5 мг спермидина (16 мкМ). Смесь инкубируют в течение ночи, затем добавляют боргидрид натрия, перемешивают еще 2 ч и непрореагировавшие компоненты удаляют гель-фильтрацией на сефадексе G-50, используя 100 мМ Трис-НСl (рН 8,3). Полученный таким образом препарат содежит 3,3 мг белка. Так как исходные компоненты (глутатион и спермидин) вносятся в эквимолярном соотношении, то можно предположить, что и полученный конъюгат содержит их одинаковые количества, т.е. на 0,8 мкМ полиглюкина по 7,2 мкМ глутатиона и спермидина (соотношение около 1:9:9).
Для сборки конструкции к раствору, содержащему 400 мкг нуклеотидного материала (около 0,12 нМ) в физиологическом растворе добавляют полисахаридную вставку (0,1 мг по белку), смесь инкубируют 2 ч при 4oС и образовавшийся комплекс отделяют гель-фильтрацией на сефарозе CL-6B. После этого добавляют гибридный белок (1,5 мг) в физ. растворе и также инкубируют в течение 2 ч. Полученную конструкцию отделяют от несвязавшихся компонентов гель-фильтрацией на сефарозе CL-6B (фиг.2).
Исследование физических свойств молекулярной конструкции.
При исследовании комплекса [нуклеотидный материал - полисахаридная вставка] было обнаружено, что на 0,12 нМ плазмидной ДНК содержалось 0,025 мг белка, т.е. на 1 молекулу - по 540 молекул глутатиона и спермидина или соответственно - по 60 молекул полисахаридной вставки.
Конечная конструкция содержала 0,96 мг белка (около 20 нМ гибридного белка при его мол. массе около 45000 Д). Таким образом конечная конструкция имела соотношение: на 1 молекулу нуклеотидного материала - 60 молекул полисахаридной вставки - 170 молекул гибридного белка. Ее мол. масса около: 3250000+60•60000+170•45000= 14500000 Д и при гель-хроматографии на колонке с сефарозой CL-6B она элюируется в свободном объеме (фиг.2).
Из теоретических расчетов следует, что шаровидная молекула нуклеиновой кислоты с молекулярной массой около 3250 кДа может сорбировать около 55 молекул полиглюкина. Комплекс [нуклеотидный материал - полисахаридная вставка] при молекулярной массе около 3250 кДа+55•60 кДа=6850 кДа может сорбировать примерно 120 шарообразных молекул гибридного белка (мол. массой 45 кДа). Так как в эксперименте это соотношение составило 1:60:170, то, вероятно, можно объяснить некоторой некомпактностью полисахаридного конъюгата и дипольной пространственной структурой гибридного белка. При дипольной пространственной структуре на поверхности конструкции находится только белок TBI (антиген).
Для проверки полноты упаковки нуклеотидного материала в собранной конструкции проводили ее обработку смесью ДНКазы и РНКазы. Опыт показал, что если для исходной плазмидной ДНК полная деградация наблюдалась уже через 30 мин инкубации, то в собранной конструкции нуклеотидный материал сохранялся интактным в течение суток.
По данным электронной микроскопии препарат молекулярной конструкции содержал шарообразные частицы, имеющие в основном диаметр от 10 до 25 им (фиг. 4). Встречались также более крупные частицы, диаметр которых достигал 50-100 нм. Возможно, это обусловлено тем, что полисахаридная вставка содержит на поверхности не одну молекулу спермидина, а девять, и происходит объединение ряда частиц. Тем не менее, размер полученной конструкции сравним с размером полиовируса и вируса полиомы, и приближается к размеру ВИЧ.
Б) Схожие результаты были получены при создании молекулярной конструкции, содержащей в качестве центрального ядра двуспиральную РНК из дрожжей штамма Saccharomyces cerevisiae M437 Y116.
В) При создании конструкции вакцины, содержащей два гибридных белка: GST-TBI и gp 41(607-620)-β-галактозидаза, на стадии получения полисахаридной вставки к активированному полиглюкину добавляют спермидин, а вместо глутатиона - аминофенилгалактопиранозид. И затем при сборке в раствор с плазмидной ДНК вносят на девять частей конъюгата [полиглюкин-спермидин-глутатион] одну часть конъюгата [полиглюкин-спермидин-галактопиранозид]. Затем в соответствующих отношениях вносят и гибридные белки. Конструкция проявляет суммарные антигенные свойства. Следует отметить, что искусственный белок TBI не имеет в своем составе последовательности gp 41(607-620) и сочетание обоих белков в конструкции вакцины должно увеличивать ее иммуногенные свойства.
Пример 4. Биологические испытания молекулярной конструкции.
Для иммунизации были использованы мыши линии BALB/c весом 18-20 грамм. Животные содержались на стандартном рационе. Все манипуляции с животными в эксперименте проводили с применением седативных средств в соответствии с ветеринарным законодательством.
Иммунизация. С целью изучения иммунного ответа на искусственные вирусоподобные частицы, экспонирующие белок TBI, была проведена иммунизация мышей линии BALB/c. Животным вводили внутримышечно трехкратно с интервалом через две недели по 45 мкг (по белку) собранной конструкции. В качестве положительного контроля использовали очищенный белок TBI-GST (80 мкг на мышь). Через каждые две недели проводили взятие крови и приготовление сыворотки для определения титра антител к белку TBI.
Для определения титра антител к эпитопам вируса ВИЧ-1 (белку TBI) использовали иммуноферментный анализ (ИФА). Антиген в конечной концентрации 5 мкг/мл в объеме 100 мкл сорбировали в течение ночи в полистироловых планшетах. Для титрования использовали последовательные трехкратные разведения сывороток, начиная с 1:15. Связавшиеся специфические антитела выявляли с помощью конъюгата антител кролика против IgG мыши с пероксидазой хрена ("Sigma").
Накопление антител в сыворотке мышей, иммунизированных чистым белком TBI и частицами, представлено на фиг.5. Как видно из графика, вирусоподобные частицы индуцируют более высокий титр антител по сравнению с титром, индуцируемым чистым белком TBI-GST, несмотря на то что количество вводимого животным белка TBI-GST в составе частиц было в два раза меньше. Две группы мышей отличаются также и по динамике изменения титров сыворотки. Группы мышей, иммунизированных TBI-GST и вирусоподобными частицами, отличаются и по динамике изменения титров сыворотки. Через неделю после последней иммунизации титр сыворотки животных, иммунизированных TBI-GST, достигал значения 1,8•107, но через три недели резко снижается до 1•105 (фиг.5). У мышей, иммунизированных вирусоподобными частицами, через неделю после последней иммунизации обратный титр достигает 0,9•107 и практически не изменяется через три недели. Таким образом, вирусоподобные частицы вызывают пролонгацию иммунного ответа, выступая в качестве своеобразного депо антигенов инфекционного агента. По-видимому, белок TBI в составе вирусоподобных частиц в организме животного более стабилен и меньше подвержен действию протеаз.
При постановке иммуноферментного анализа осуществляли перенос белков из геля на нитроцеллюлозный фильтр. Иммуноблот вели с сывороткой крови человека, содержащей антитела к белкам ВИЧ-1. Иммунные комплексы выявляли с помощью конъюгата белка А с пероксидазой. Как видно из фиг.6, первая конструкция вакцины содержит белок GST-TBI, вторая - оба белка. Обе конструкции проявляют антигенные свойства.
Таким образом, использование предлагаемой молекулярной конструкции по сравнению с аналогами позволяет:
- экспонировать на поверхности молекулярной конструкции любые эпитопы инфекционных агентов продолжительностью от десятков до тысяч аминокислот;
- использовать гены инфекционных агентов в составе рекомбинантных плазмидных ДНК ("центральное ядро" частиц) в качестве ДНК-вакцины;
- использовать двуспиральную РНК в молекулярной конструкции ("центральное ядро" частиц) в качестве неспецифического стимулятора иммунного ответа;
- используемый для получения конъюгата полиглюкин также является стимулятором иммунного ответа;
- сравнительно большие размеры частиц молекулярной конструкции (около 25 нм) позволяют ее использовать для иммунизации без адъювантов как полноценные антигены.
Литература
1. Bolognesi D.P. //Mol.Biol.Med. 1990. V.7. Р.1-15.
2. Ada G.L. //Nature. 1989. V.339. Р.331-332.
3. Steward M.W., Howard C.R. // Med. Lab. Sci. 1985. V.42. P.376-387.
4. Rowlands D.J. // Biochem.Soc.Trans. 1989. V.17. P.945-947.
5. Борисова Г.П., Берзинь И.Г., Осе В.П. и др. Кор-антиген вируса гепатита в качестве носителя чужеродных антигенных детерминант. // "Новые направления биотехнологии". Тез. докладов. - Пущино. - 1990. - С. 61.
6. Борисова Г.П., Берзинь И.Г., Цибиногин В.В. и др. // Докл. АН СССР. - 1990. - Т. 312. - С. 751-754.
7. Филатов Ф.П. Конструирование герпесвирусных векторов, экспрессирующих рекомбинантные гены на основе HBcAg. //"Новые направления биотехнологии". Тез. докладов. - Пущино. - 1990. - С. 70.
8. Цибиногин В.В., Лосева В.Я., Борисова Г.П. Иммунологические свойства HBcAg, содержащего чужеродные антигенные детерминанты. //"Новые направления биотехнологии". Тез. докладов. - Пущино. - 1990. - С. 71.
9. Патент РФ 2121504, кл. С 12 N 15/51, опубл. 1998.
10. Татьков С. И., Кувшинов В.Н., Комарских М.Е. и др. Конструирование вакцины нового поколения против вируса гепатита В. //"Новые направления биотехнологии". Тез. докладов. - Пущино. - 1990. - С. 68.
11. Yon J., Corcoran Т., Kent К. et al. //J.Gen.Virol. - 1992. - V.73. - P. 2569-2575.
12. Маниатис Т., Фрич Э., Сэмбрук Дж. // Молекулярное клонирование. - М. : Мир, 1984. - 479 с.
13. Дужак А.Б., Лобова Н.Н., Подгорный В.Ф. и др. II Антибиотики и мед. биотехнология. - 1985. - 1. - С. 19-21.
14. Патент РФ 2083221, кл. А 61 К 38/20, опубл. 1997.
15. ВФС 42-2457-94. Ридостин для инъекций.
16. Pharmacia Biotech "BioDirectory". - 1996.
17. Каталог фирмы "Promega". Promega protocols and applications guide. - 1997.
18. Eroshkin A.M., Karginova E.A., Gileva I.P. et al.//Protein Enginiring. - 1995. - V.8. - N.2. - P. 167-173.
19. Белявская В.А., Закабунин А.И., Долгова И.Н. и др.//Молекул. генет. микробиол. и вирусол. - 1994. - 6. - С.12-16.
20. Лебедев Л.Р., Ерошкин A.M., Гилева И.П. //Биотехнология. - 1997. - 7-8. - С. 38-42.

Изобретение относится к области биотехнологии, в частности к медицине, иммунологии. Вакцина представляет собой частицы 25-30 нм в диаметре, содержащие в центре полинуклеотидный комплекс (рекомбинантную плазмидную ДНК, содержащую гены инфекционного агента или двуспиральную РНК-стимулятор неспецифической резистентности организма), а на поверхности - гибридные белки, содержащие эпитопы (полипептиды) инфекционного агента и фермент (например, глутатион-S-трансферазу или β-галактозидазу). Связь между полинуклеотидным комплексом и гибридными белками осуществляется посредством коньюгата: спермидин (для связи коньюгата с полинуклеотидным комплексом) - полиглюкин-субстрат для фермента (например, глутатион или галактопиранозид) для аффинной сорбции гибридных белков на конструкцию. Вакцина обеспечивает более высокий титр антител и более длительный иммунный ответ при иммунизации. 6 ил.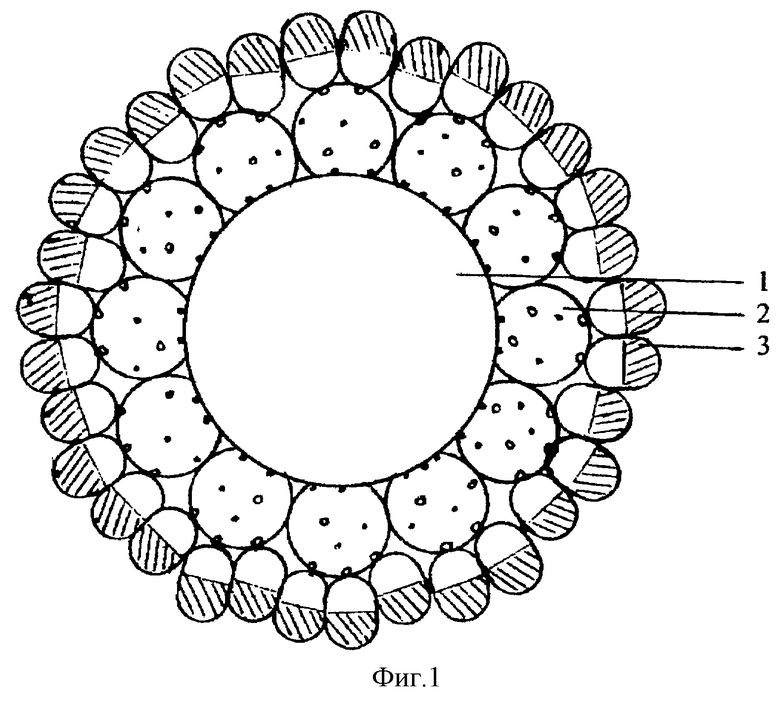
Вакцина против вирусных инфекций, отличающаяся тем, что представляет собой молекулярную конструкцию, в центре которой находится полинуклеотидный комплекс, представляющий собой рекомбинантулую плазмидную ДНК, содержащую гены инфекционного агента, или двуспиральную РНК, выделенную из дрожжей, при этом на поверхности полинуклеотидного комплекса экспонированы гибридные белки, включающие эпитопы инфекционного агента и фермент глутатион-S-трансферазу или β-галактозидазу, причем полинуклеотидный комплекс связан с гибридными белками посредством коньюгата, содержащего спермидин-полиглюкин-глутатион или спермидин-полиглюкин-галактопиранозид.
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| БОРИСОВА Г.П | |||
| и др | |||
| Кор-антиген вируса гепатита в качестве носителя чужеродных антигенных детерминант | |||
| Новые направления в биотехнологии./ Тезисы докладов | |||
| - Пущино, 1990, с | |||
| Устройство для сортировки каменного угля | 1921 |
|
SU61A1 |

