Область техники, к которой относится изобретение
Изобретение относится к фармацевтической композиции, содержащей связывающий гепарин белок, и ее использованию для лечения заболеваний или состояний, вызванных бактериальным эндотоксином.
Уровень техники
Частота встречаемости септического шока в результате системной реакции на тяжелую инфекцию возрастала в течение последних 50 лет и в настоящее время является наиболее широко распространенной причиной смерти в отделениях интенсивной терапии в США.
Причинами этого увеличения распространенности и высокой частоты встречаемости септического шока считаются возросшее применение инвазивных приспособлений, как, например, внутрисосудистых катетеров, возросшее применение цитотоксических и иммуносуппрессивных препаратов, увеличение продолжительности жизни больных, склонных к развитию сепсиса, и учащение инфекций, вызванных резистентными к антибиотикам организмами.
Патологическими состояниями, связанными с сепсисом, являются бактеремия (известная также как септицемия), характеризующаяся положительными результатами посева культур крови; сепсис, характеризующийся системной реакцией на инфекцию в форме учащения дыхания, тахикардии, гипертермии или гипотермии; синдром сепсиса, при котором имеются клинические проявления сепсиса и признаки нарушенной перфузии органов в виде патологического возрастания уровня лактата, олигурии или острого нарушения психического состояния; ранний септический шок, при котором имеются клинические проявления синдрома сепсиса, а также гипотония, продолжающаяся менее одного часа и реагирующая на обычные методы лечения; и стойкий септический шок, при котором имеются клинические проявления синдрома сепсиса и гипотония, продолжающаяся более одного часа, несмотря на обычные методы лечения. Известные медиаторы сепсиса взаимодействуют сложным образом для развития сепсиса или цитокинового каскада, начинающегося в участке инфекции или травмы (например, в брюшной полости). Фактором, инициирующим цитокиновый каскад (который является обычной причиной сепсиса при грамотрицательной бактеремии), является эндотоксин (кроме того, известный как липополисахарид, сокращенно LPS), выделяемый бактериями в участке инфекции или воспаления, где он вызывает освобождение фактора опухолевого некроза α (TNFα), интерлейкина-1, интерлейкина-6, интерлейкина-8 и фактора, активирующего тромбоциты (PAF) из макрофагов и других клеток. После освобождения TNFα, интерлейкина-1 и PAF арахидоновая кислота метаболизируется с образованием лейкотриенов, тромбоксана А2 и простагландинов. Интерлейкин-1 и интерлейкин-6 активируют Т клетки для выработки интерферона-γ, интерлейкина-2, интерлейкина-4 и фактора, стимулирующего колонии гранулоцитов-моноцитов. Нейтрофильные лейкоциты могут активироваться непосредственно большинством из этих медиаторов. Таким образом, может происходить вызванное нейтрофилами повреждение во время дегрануляции под влиянием освобождения свободных радикалов кислорода и лизосомальных ферментов во время агрегации в участке инфекции или воспаления.
Присутствие эндотоксина или других медиаторов сепсиса в очагах инфекции или воспаления, хотя и потенциально способно запустить цитокиновый каскад, само по себе не обязательно приведет к развитию сепсиса, поскольку цитокиновый каскад имеет определенные наследственные защитные механизмы против неконтролируемой активации цитокинового каскада. Так простагландин может тормозить освобождение цитокинов из макрофагов, а макрофаги могут быть способны подавлять Т клетки. Как представляется, клинические симптомы сепсиса появляются тогда, когда происходит освобождение слишком большого количества эндотоксинов или других медиаторов сепсиса или когда отсутствуют медиаторы, способные вмешаться в цитокиновый каскад, например, когда ослаблена иммунная система. Более подробное описание развития септических расстройств и участвующих в нем медиаторов дано в работе vide R.С. Bone, "The Pathogenesis of Sepsis", Ann. Int. Med., 1991, pp.457-469.
Продолжающаяся высокая смертность и частота осложнений, связанные с грамотрицательным сепсисом, побудили к интенсивному поиску лечебных средств, способных противодействовать потенциально летальным эффектам циркулирующего бактериального LPS. Во многих публикациях сообщают о значительном лечебном эффекте высоких доз внутривенно вводимого иммуноглобулина. Однако этот способ лечения требует IgG, полученного из плазмы доноров, подвергнутых скринингу для выявления естественно встречающихся высоких уровней антител к сердцевинному LPS или от очень больших пулов доноров (>1000). Моноклональные антитела против LPS, которые также были предложены для лечения бактеремии (например, WО 88/03211), оказались малоэффективными или неэффективными, вероятно, потому, что они не ингибируют цитокиновый каскад, вызванный LPS.
Недавно была определена ковалентная структура двух близко родственных белков, выделенных из нейтрофильных лейкоцитов периферической крови, полученных от человека и свиньи (см. H. Flodgaard et al., Eur. J. Biochem., 1991, vol. 197, pp.535-547; J. Pohl et al., FEBS Lett., 1990, vol. 272, p. 200 ff.). Оба белка имеют высокую аналогию с эластазой нейтрофилов, но благодаря селективным мутациям активного серина 195 и гистидина 57 (нумерация химотрипсина (В.S. Hartley, "Homologies in Serine Proteinases", Phil. Trans. Roy. Soc. Series, 1970, vol. 257, p.77 ff.)) у белков отсутствует протеазная активность. Благодаря их высокому сродству с гепарином белки были названы соответственно белком, связывающим человеческий гепарин (hHBP), и белком, связывающим свиной гепарин (рНВР); Schafer et al. (W.M. Schafer et al., Infect. Immun. , 1986, vol. 53, p. 651 ff.), назвали белок катионным антимикробным белком (САР37) благодаря его антимикробной активности. Было также показано, что белок хемотоксичен для моноцитов в диапазоне от 1,3•10-9 М до 10-8 М (Н. А. Pereira et al., J. Clin. Invest., 1990, vol.85, p.l468 ff.), что согласуется с результатами Flodgaard et al., op. cit.
Кроме того, было показано, что НВР является медиатором отслойки и сокращения эндотелиальных клеток и фибробластов при его добавлении к таким клеткам, выращенным в монослойной культуре. НВР также стимулирует выживание моноцитов и секрецию тромбоспондина (E. Ostergaard и Н. Flodgaard, J. Leukocyte Biol., 1992, vol.51, p.316 ff).
Из азурофильных гранул был также выделен белок с первыми 20-ю N-терминальными аминокислотными остатками, идентичными таковым hHBP и САР37, и названный азуроцидином (J.E. Gabay et al., Proc. Natl. Acad. Sci. USA, 1989, vol. 86, p.5610 ff.; C.G. Wilde et al., J. Biol. Chem. 1990, vol.265, p.2038 ff. ) и сообщалось о его антимикробных свойствах (D. Capanelli et al., J. Clin. Invest., 1990, vol.85, p.904 ff.).
Присутствие hHBP в нейтрофильных лейкоцитах и тот факт, что при фагоцитировании лейкоцитами Staph. aureus освобождается 89% САР37 (который идентичен hHBP) (H.A.Pereira et al., op. cit.), указывают на то, что функцией hHBP может быть его вовлечение в воспалительный процесс, поскольку белок, очевидно, освобождается из активированных нейтрофилов. H.A. Pereira et al., op. cit. предположили, что САР37 действует в участке воспаления, где он может специфично привлечь моноциты и, таким образом, быть одним из факторов, ответственных за приток моноцитов во второй волне воспаления. Ostergaard и Flodgaard, op. cit. предполагают, что, кроме важного значения, которое НВР имеет для мобилизации моноцитов, он может играть ключевую роль в механизме экстравазации нейтрофилов, а также моноцитов.
Структура НВР раскрыта в описании патента WO 89/08666 и в данных Н.Flodgaard et al. , op. cit. Иначе НВР был назван САР37 (описание патента WО 91/00907) и азуроцидином (см. C.G. Wilde et al., J. Biol. Chem., 1990, vol. 265, p.2038).
Краткое изложение сущности изобретения
Ранее не предполагалось, что НВР может применяться в лечении заболеваний или состояний, вызванных присутствием бактериального эндотоксина в участках инфекции или травмы.
Соответственно настоящее изобретение относится к фармацевтической композиции для профилактики или лечения заболеваний или состояний, связанных с индукцией цитокинового каскада липополисахаридом (LPS), причем композиция включает связывающий гепарин белок (НВР), который в гликозилированной форме имеет истинный молекулярный вес 28 кД (по данным метода SDS-PAGE в восстанавливающих условиях), т. е. белок, продуцируемый в азурофильных гранулах полиморфоядерных лейкоцитов, наряду с фармацевтически приемлемым носителем или растворителем.
Как было обнаружено, НВР связывается с LPS (см. работу Н.А. Pereira et al. , Proc. NatI. Acad. Sci. USA, 1993, vol.90, pp.4733-4737), где эту способность связывают с антибактериальным эффектом белка. Есть данные, что бактерицидное действие НВР уменьшается при рН 7 (т.е. физиологическом рН). Кроме того, септические состояния не вызываются грамотрицательными бактериями как таковыми, а эндотоксином, освобождаемым из мертвых бактериальных клеток, активирующим рецептор CD14 на моноцитах, и таким образом вызывающим цитокиновый каскад. Поэтому, казалось бы, антибактериальный эффект НВР не имеет значения для лечения септических состояний. В этом контексте следует отметить, что связывание с LPS как таковое, не является показателем какой-либо способности нейтрализовать эффекты эндотоксина, вызывающие цитокиновый каскад, что доказывается фактом, что антитела, выработанные против LPS, имеют незначительный или отсутствующий эффект в этом плане (см. Е.Th. Rietschel и M. Schlaak, Immun. Infect., 1993, vol.21, pp.25-36).
В то время как ранее опубликованные данные об усилиях, направленных на борьбу с септическими состояниями, были сконцентрированы на нейтрализации эндотоксина в циркуляции больных сепсисом (например, как предложено в описании патента WО 93/19087, в котором САР37 (т.е. НВР) предложен для лечения эндотоксемии путем связывания с LPS и обеспечения выведения LPS из циркуляции), большинство этих усилий были напрасными, потому что у больных с сепсисом, в основном, не бывает повышенных уровней циркулирующего эндотоксина. Однако НВР может быть эффективным на уровне локальных участков воспаления или инфекции после проявления феноменов сепсиса, то есть при процессах, развивающихся после эндотоксемии. В настоящее время предполагается, что НВР может оказывать свой благоприятный эффект не только посредством специфического связывания и высокого сродства с Липидом А. (токсической частью LPS) и, вследствие этого, подавления продолжающейся индукции цитокинового каскада, но также путем защиты моноцитов от апоптоза, возникающего в результате стрессового повреждения, вызванного освобождением свободных кислородных радикалов и цитокинов из фагоцитов (например, нейтрофилов, моноцитов, макрофагов) как части защитных механизмов организма против травм и инфекций.
Моноциты мигрируют к участкам повреждения или инфекции, где они фагоцитируют инфекционные микроорганизмы и вредные вещества, как, например, эндотоксин. НВР может также стимулировать фагоцитарное действие моноцитов. Способность НВР мобилизировать моноциты, как оказывается, зависит от его способности связываться с LPS, которое происходит пока еще неустановленным способом.
Кроме того, к удивлению было установлено, что НВР необратимо связывается с LPS при 37-45oС (т.е. при температуре тела от нормальной до повышенной). Это тоже может влиять на нейтрализацию LPS под действием НВР.
В другом аспекте изобретение относится к способу профилактики и лечения заболеваний или состояний, связанных с индукцией цитокинового каскада LPS, причем способ включает введение больному, которому показано такое лечение, эффективного количества связывающего гепарин белка (НВР), который в гликозилированной форме имеет истинный молекулярный вес 28 кД (по данным SDS-PAGE в условиях восстановления), т.е. белка, продуцируемого в азурофильных гранулах полиморфоядерных лейкоцитов.
В дальнейшем аспекте изобретение относится к использованию связывающего гепарин белка (НВР), который в гликозилированной форме имеет истинный молекулярный вес 28 кД, (по данным метода SDS-PAGE в условиях восстановления), т. е. белка, продуцируемого в азурофильных гранулах полиморфоядерных лейкоцитов, для производства лекарственного препарата для профилактики и лечения заболеваний или состояний, связанных с индукцией цитокинового каскада LPS.
Подробное изложение сущности изобретения
Пригодным может быть НВР, полученный от млекопитающих, в частности от человека или свиньи. В частности, НВР является человеческим НВР с аминокислотной последовательностью, показанной в Списке Последовательностей, представленной как "SEQ ID 1", или свиным НВР с аминокислотной последовательностью, показанной в Списке Последовательностей, представленной как "SEQ id 2", или функциональным аналогом, или его пептидным фрагментом, способным связываться с частью липида А LPS. Примеры таких функциональных аналогов включают производные нативного белка, полученные путем добавления одного или более аминокислотных остатков к какому-либо или обоим С- или N-концам нативного белка, замещения одного или более аминокислотных остатков на одном из или обоих концах нативного белка, делеции одного или более аминокислотных остатков на одном из или более концах нативного белка, или в одном или более участках внутри аминокислотной последовательности, или вставки одного или более аминокислотных остатков в одном или более участках в нативной аминокислотной последовательности. Термин выбран специально для включения пептидных фрагментов НВР, в частности фрагментов, обладающих одинаковой способностью с НВР уменьшать вызванный LPS цитокиновый каскад из мононуклеарных клеток человека. Примером такого фрагмента, способным связывать часть липида А LPS, является пептидный фрагмент, включающий аминокислотные остатки 20-53, в частности аминокислотные остатки 26-42 SEQ ID 1 или аминокислотные остатки 20-53 SEQ ID 2.
Последовательность ДНК, кодирующая НВР, может быть получена синтетическим путем в соответствии с установленными стандартными способами, например, фосфоамидитным методом, описанным S.L. Beaucage и M.H. Caruthers, Tetrahedron Letters, 1981, vol.22, pp.1859-1869, или методом, описанным Matthes et al. , EMBO Journal, 1984, vol.3, pp.801-805. В соответствии с фосфоамидитным методом олигонуклеотиды являются синтезированными, например, в автоматическом ДНК-синтезаторе, очищенными, ренатурированными, сшитыми и клонированными в соответствующих векторах.
Последовательность ДНК может быть также геномного происхождения или кДНК, например, полученная путем приготовления библиотеки генов или кДНК, и скрининга для выявления последовательностей ДНК, кодирующих весь или часть НВР, с помощью гибридизации с использованием синтетических олигонуклеотидных зондов в соответствии со стандартными методиками (см. Sambrook et al., Molecular cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor, 1989). Последовательность ДНК может быть также приготовлена посредством полимеразной цепной реакции с использованием специфичных праймеров, например, как описано в патенте США 4,685,202 или в работе R.K. Saiki et al., Science, 1988, vol. 239, pp. 487-491.
Затем последовательность ДНК встраивается в рекомбинантный вектор экспрессии, который может быть любым вектором, который может быть соответствующим образом подвержен процедурам рекомбинантной ДНК. Выбор вектора часто будет зависеть от клетки-хозяина, в которую он будет введен. Таким образом, вектором может быть автономно реплицирующий вектор, т.е. вектор, который существует как внехромосомная сущность, репликация которого независима от репликации хромосом, например плазмида. В альтернативном варианте вектором может быть вектор, который при введении в клетку-хозяин интегрируется в геном клетки-хозяина и реплицируется вместе с хромосомой (хромосомами), в который он был интегрирован.
В векторе последовательность ДНК, кодирующая НВР, должна быть функционально связана с подходящей последовательностью промотора. Промотором может быть любая последовательность ДНК, которая проявляет транскрипционную активность в выбранной клетке-хозяине, и может быть получен из генов, кодирующих белки, которые являются гомологичными или гетерологичными для клетки-хозяина. Примерами подходящих промоторов для направления транскрипции ДНК, кодирующей НВР в клетках млекопитающих, являются промо-тор SV40 (Subramani et al., Mol. Cell. Biol., 1981, vol.1, pp.854-864), промотор МТ-1 (ген металлотионеина) (Palmiter et al., Science, 1983, vol.22, pp.809-814) или главный поздний промотор аденовируса 2. Подходящим промотором для использования в клетках насекомых является полиэдриновый промотор (Vasuvedan et al., FEBBS Lett. , 1992, vol. 311, pp.7-11). Подходящие промоторы для использования в дрожжевых клетках-хозяевах включают промоторы из гликолитических генов дрожжей (Hitzeman et al., J. Biol. Chem., 1980, vol.255, pp.12073-12080); Alber и Kawasaki, J. Mol. Appl. Gen., 1982, vol.1, pp.419-434) или промоторы генов алкоголь дегидрогеназы (Young et al., в Genetic Engineering of Microorganisms for Chemicals (Hollaender et al., eds.). Plenum Press, New York, 1982), или ТРI1 (патент США 4,599,311), или ADH2-4c (Russel et al., Nature, 1983, vol.304, pp.652-654). Подходящими промоторами для использования в клетках-хозяевах нитчатых грибах являются, например, промотор ADH4 (McKnight et al., The EMBO J., 1985, vol.4, pp. 2093-2099) или промотор tpiA.
Последовательность ДНК, кодирующая НВР, может быть также функционально связана с подходящим терминатором, как например, терминатор гормона роста человека (Palmiter et al., op. cit.) или промоторами (для грибковых хозяев) ТРIl (Alber и Kawasaki, op. cit.) или ADH3 (McKnight et al., op. cit.). Кроме того, вектор может включать элементы, как например, сигналы полиаденилирования (например, из региона SV 40 или 5 Elb аденовируса), последовательности усилителя транскрипции (например, усилитель SV 40) и последовательности усилителя трансляции (например, последовательности, кодирующие РНК аденовируса VA).
Кроме того, вектор рекомбинантной экспрессии может включать последовательность ДНК, обеспечивающую возможность репликации вектора рассматриваемой клетки-хозяина. Примером такой последовательности (когда клеткой-хозяином является клетка млекопитающего) является ориджин репликации из SV 40. Вектор может также включать селектируемый маркер, например ген, продукт которого комплементирует дефект в клетке-хозяине, например ген, кодирующий дигидрофолат редуктазу (DHFR), или ген, который обеспечивает устойчивость к препарату, например неомицину или гидромицину.
Процедуры, используемые для сшивания последовательностей ДНК, кодирующих НВР, промотор и терминатор соответственно, и для вставки их в подходящие векторы, содержащие информацию, необходимую для репликации, хорошо известны специалистам в этой области (см., например, Sambrook et al., op. cit.).
Клеткой-хозяином, в которую вводится вектор экспрессии, может быть любая клетка, которая способна продуцировать НВР, и является предпочтительно эукариотической клеткой, как, например, клетки беспозвоночных (насекомых) или клетки позвоночных, например ооциты Xenopus laevis, или клетки млекопитающих, в частности клетки насекомых и млекопитающих. Примерами подходящих линий клеток млекопитающих являются линии клеток COS (например, АТСС CRL 1650), ВНК (например, АТСС CRL 1632, АТСС CCL 10) или СНО (например, АТСС CCL 61). Способы трансфекции клеток млекопитающих и экспрессии последовательностей ДНК, введенных в клетки, описаны, например, в работах Kaufman и Sharp, J. Mol. Biol., 1982, vol.159, pp.601-621; Sothern и Berg, J. Mol. Appl. Genet., 1982, vol. 1, pp. 327-341; Loyter et al., Proc. Natl. Acad. Sci. USA, 1982, vol. 79, pp. 422-426, Wigler et al., Cell, 1978, vol.14, p. 725; Corsaro и Pearson, Somatic Cell Genetics, 1981, vol.7, p.603; Graham и van der Eb, Virology, 1973, vol.52, p.456; и Neumann et al., EMBO J., 1982, vol.1, pp. 841-845.
В альтернативном варианте в качестве клеток-хозяев могут использоваться грибковые клетки (включая дрожжевые клетки). Примеры подходящих дрожжевых клеток включают клетки видов Saccharomyces или видов Schizoaccharomyces, в частности штаммы Saccharomyces cerevisiae. Примерами других грибковых клеток являются клетки нитчатых грибов, например виды Aspergillus или виды Neurospora, в частности штаммы Aspergillus oryzae или Aspergillus niger. Использование видов Aspergillus для экспрессии белков описано, например, в патенте ЕР 238 023.
Средой, используемой для культивирования клеток, может быть любая обычная среда, пригодная для выращивания клеток млекопитающих, как, например, содержащая сыворотку, или бессывороточная среда, содержащая соответствующие добавки, или подходящая среда для выращивания клеток насекомых, дрожжей или грибов. Подходящие среды можно приобрести у коммерческих поставщиков или их можно приготовить в соответствии с опубликованными прописями (например, в каталогах "American Type. Culture Collection").
НВР, продуцируемый клетками, затем может быть выделен из культуральной среды с помощью обычных процедур, включая отделение клеток-хозяев от среды путем центрифугирования или фильтрации, преципитации белковых компонентов супернатанта или фильтрата с помощью соли, например сульфата аммония, очистки с помощью различных хроматографических методов, например ионообменной хроматографии, аффинной хроматографии или им подобных.
К удивлению было обнаружено, что особенно хороший выход НВР может быть получен, когда НВР экспрессирован в виде про-НВР. Соответственно при специфической реализации настоящее изобретение относится к технологии производства НВР, в которой клетки-хозяева, содержащие последовательность ДНК, кодирующую зрелый НВР с предшествующим удлинением N-конца, культивируются в соответствующей культуральной среде в условиях, обеспечивающих возможность экспрессии НВР, и полученный в результате НВР выделяется из культуральной среды в виде НВР с удлиненным N-концом.
Удлинение N-конца может быть последовательностью, состоящей от около 5 до около 25 аминокислотных остатков, в частности от приблизительно 8 до приблизительно 15 аминокислотных остатков. Природу аминокислотных остатков в удлинении N-конца не считают принципиальной. Удлинение N-конца может соответственно быть пропептидом НВР с аминокислотной последовательностью Gly-Ser-Ser-Pro-Leu-Asp.
С целью облегчения продукции НВР, как правило, предпочтительно, чтобы последовательность ДНК, кодирующая N-кольцевое удлинение НВР, включала последовательность ДНК, кодирующую сайт расщепления протеазой, локализованный между последовательностью ДНК, кодирующей удлинение N-конца, и последовательностью ДНК, кодирующей зрелый НВР.
Примерами подходящих сайтов протеазного расщепления является сайт расщепления энтерокиназой с аминокислотной последовательностью (Asp)4-Lys или участок расщепления Фактором Ха с аминокислотной последовательностью Ile-Glu-Gly-Arg.
После выделения из культуральной среды удлиненный на N-конце НВР может предпочтительно быть расщеплен соответствующей протеазой для получения зрелого (и активного) НВР.
Примерами соответствующих ферментов являются энтерокиназа или Фактор Ха.
Благоприятными клетками-хозяевами, используемыми для производства НВР, являются клетки насекомых, например клетки Lepidoptera или Drosophila. В этом случае клетки могут соответственно трансфецироваться бакуловирусным вектором, например, как описано в патенте США 4,745,051 или патенте WO 92/01801.
Кроме того, было обнаружено, что с целью избежать связывания НВР с липополисахаридом, присутствующим в культуральной среде, предпочтительно вырабатывать НВР в присутствии гепарина или другого сульфированного полисахарида. Соответственно в специфическом воплощении настоящее изобретение относится к способу продукции НВР, в котором клетки-хозяева, содержащие последовательность ДНК, кодирующую НВР, культивируются в соответствующей культуральной среде, содержащей сульфированный полисахарид, в условиях, обеспечивающих возможность экспрессии НВР, и полученный в результате НВР выделяется из культуральной среды. НВР будет связываться с сульфированным полисахаридом, присутствующим в культуральной среде, и должен в последующем освобождаться из полисахарида в соответствующих условиях, например с помощью элюирования НВР солью, как, например, хлористым натрием.
Сульфированным полисахаридом предпочтительно является гепарин, хотя также могут использоваться другие сульфированные полисахариды, как, например, гепарин сульфат, декстран сульфат, дерматан сульфат, пентозан полисульфат или протамин сульфат.
Для облегчения отделения НВР, связанного с полисахаридом, из культуральной среды предпочтительно, чтобы сульфированный полисахарид был иммобилизован на инертном носителе. Инертным носителем может, например, быть агароза (например, Сефароза).
В этом воплощении также последовательности ДНК, кодирующей зрелый НВР, может предшествовать удлинение N-конца, как описано выше.
Еще в одном воплощении настоящее изобретение относится к способу продукции НВР, в котором клетки-хозяева, содержащие последовательность ДНК, кодирующую зрелый НВР, которой предшествует и которая слита с последовательностью ДНК, кодирующей другой белок, культивируются в соответствующей культуральной среде в условиях, обеспечивающих возможность экспрессии белка слияния НВР и другого белка, и полученный в результате белок слияния выделяется из культуральной среды. Другим белком может быть, например, легкая цепь Фактора VIII (последовательность ДНК, кодирующая легкую цепь Фактора VIII, может, например, быть получена, как описано в патенте ЕР 232 112).
В этом воплощении также может иметь преимущество введение последовательности ДНК, кодирующей сайт протеазного расщепления, как описано выше, расположенный между удлинением N-конца и зрелым НВР.
В фармацевтической композиции НВР может быть приготовлен с помощью любого из установленных способов приготовления фармацевтических композиций, например, как описано в руководстве Remington "Pharmaceutical Sciences", 1985. Обычно композиция может быть в форме, предназначенной для местного применения, или системных инъекций, или инфузии, и, как таковой, может быть приготовлена со стерильной водой, или изотоническим солевым раствором, или раствором глюкозы. Композиции могут стерилизоваться обычными методами стерилизации, которые хорошо известны в этой области. Полученные водные растворы могут упаковываться для использования или фильтроваться в асептических условиях и подвергнуты лиофильной сушке, причем в лиофилизированный препарат перед введением добавлялся стерильный водный раствор. Композиция может содержать фармацевтически приемлемые дополнительные вещества, требуемые для приближения физиологических условий, как, например, забуферивающие агенты, агенты, регулирующие концентрацию, и им подобные, например ацетат натрия, лактат натрия, хлористый натрий, хлористый калий, хлористый кальций и т.д. Концентрация НВР может колебаться в широких пределах, т.е. от менее чем около 0,5%, как от 1%, до величины 15-20% по весу. Единичная доза композиции обычно может содержать от приблизительно 10 мг до около 1 г НВР.
Фармацевтическая композиция настоящего изобретения, как считают, имеет преимущества при использовании в терапевтических целях, как, например, лечение сепсиса, септического шока, диссеминированного внутрисосудистого свертывания или менингококкового менингита. Как указано выше, настоящая композиция может быть особенно пригодна для местного применения в участках воспаления или инфекции (например, в брюшной полости), поскольку важно защитить моноциты, мигрирующие в такие участки, и гарантировать их способность включать эндотоксин и инфекционные бактерии. В настоящее время подходящей дозой считается ежедневная доза НВР 0,1-100 мг/кг веса тела в зависимости от тяжести состояния, подлежащего лечению, и состояния больного.
Композиция по данному изобретению может дополнительно содержать антимикробный агент для лечения этиологической бактериальной инфекции. Примерами подходящих антимикробных агентов являются бета-лактамные антибиотики (например, пенициллин), аминогликозиды, тетрациклины, бацитрацин, полимиксин, сульфониламиды, нитрофураны, налидиксиновая кислота и т.д. Для системного применения композиция может также содержать гепарин или другой сульфированный глюкозаминогликан в качестве антикоагулянтного агента, поскольку к удивлению было обнаружено, что гепарин не подавляет способность НВР связываться с LPS. Помимо действия в качестве антикоагулянта присутствие гепарина может также привести к удлинению периода полураспада НВР в плазме, так как он предотвращает связь НВР с глюкозаминогликанами на сосудистом эндотелии.
Краткое описание чертежей
Настоящее изобретение, кроме того, иллюстрируется в следующих примерах со ссылкой на прилагаемые чертежи, в которых
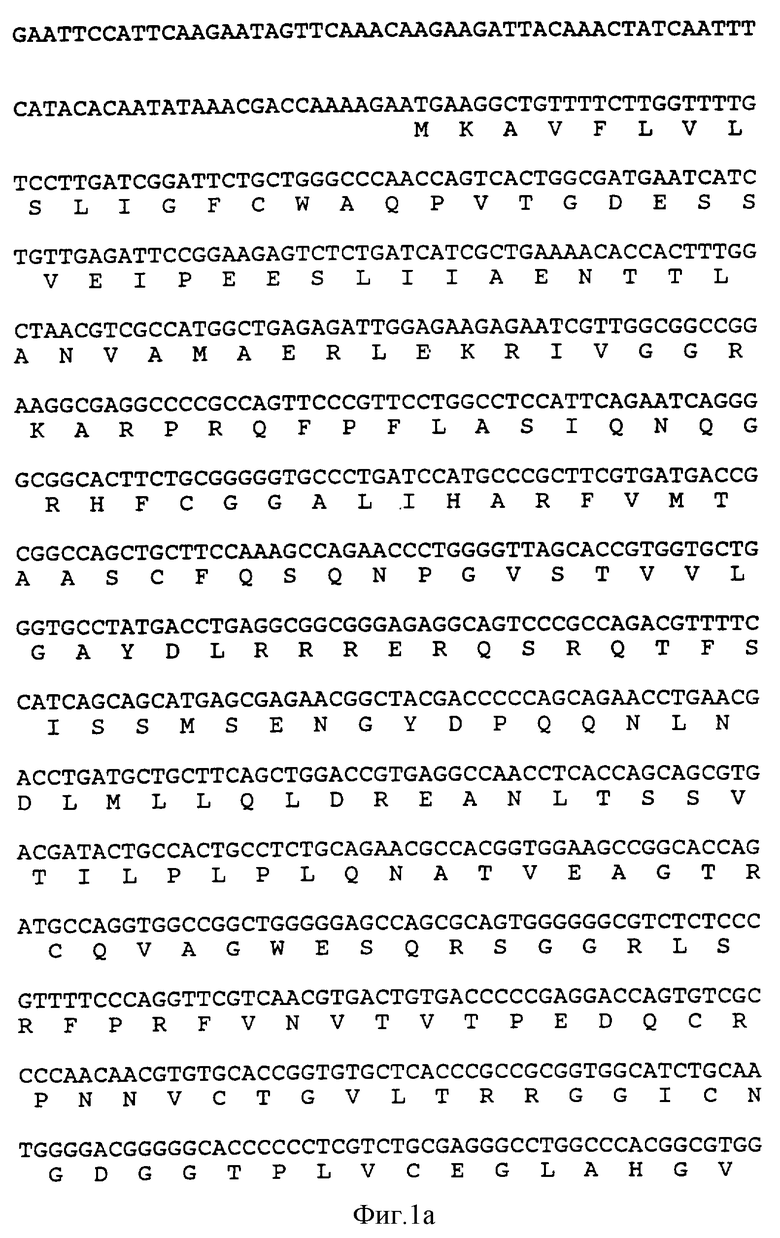
на фиг. 1 показана последовательность ДНК и полученная аминокислотная последовательность EcoRI-Xbal фрагмента 901 bp плазмиды pKFN-1783, кодирующей hHBP;
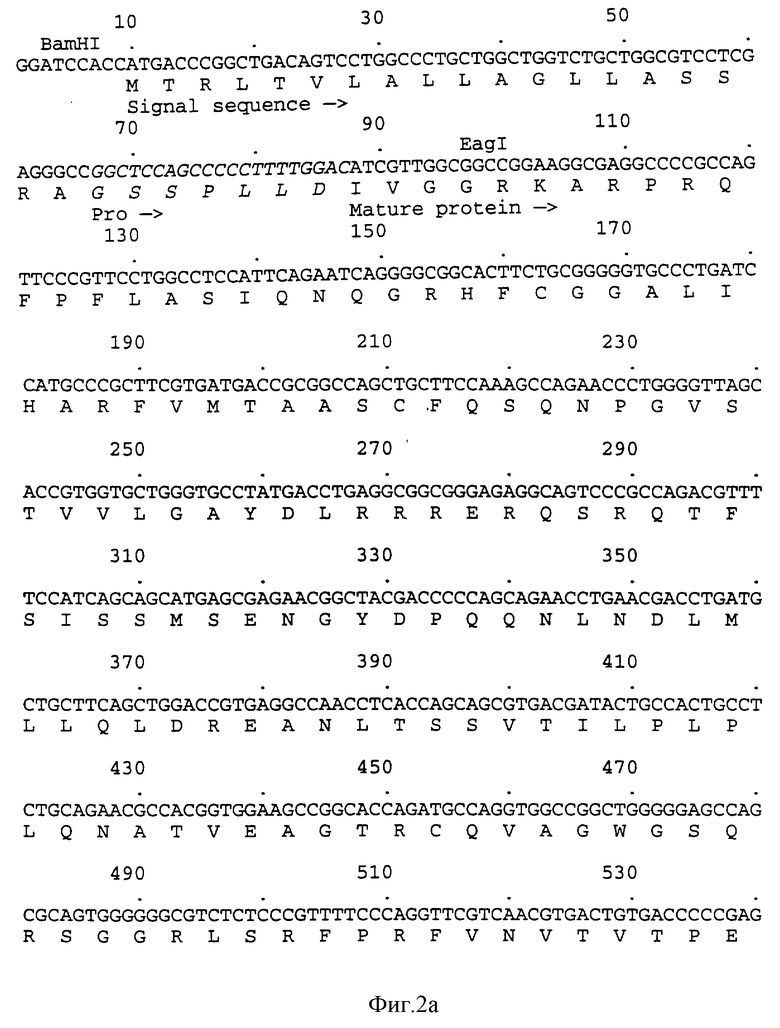
на фиг.2 показана последовательность ДНК и полученная аминокислотная последовательность BamHI-HindIII фрагмента 772 bp, вставленного в плазмиду pSX221, кодирующую hHBP;
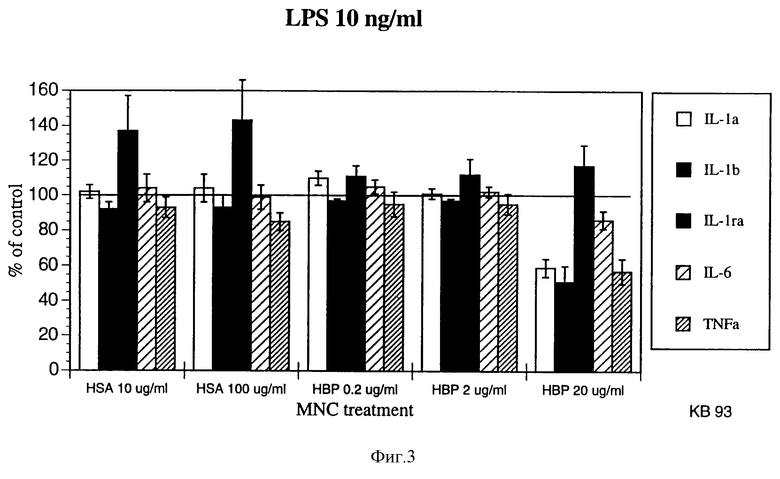
фиг. 3 является графиком, показывающим уровни цитокина в мононуклеарных клетках крови, обработанных HSA в концентрации 10 мкг/мл или меняющимися уровнями НВР в присутствии LPS (10 нг/мл);
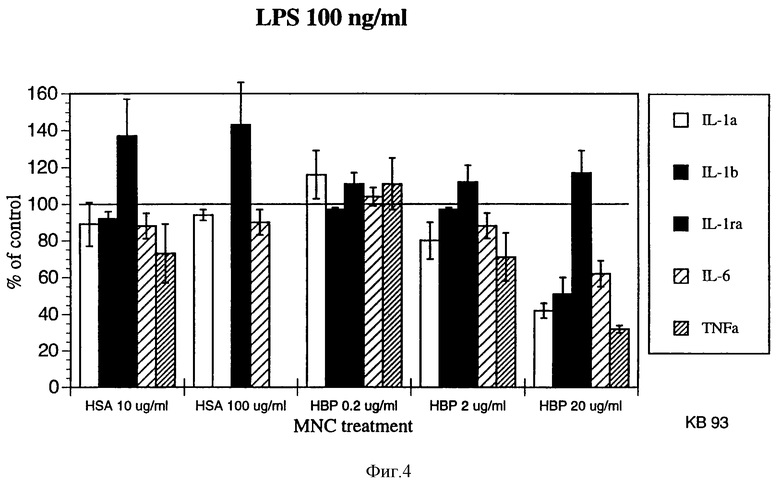
фиг. 4 является графиком, показывающим уровни цитокина в мононуклеарных клетках крови, обработанных HSA в концентрации 10 мкг/мл или меняющимися уровнями НВР в присутствии 100 нг/мл LPS.
Пример 1
Получение белка, связывающего человеческий гепарин, из дрожжей штамма KFN-1775.
Общие методы
Применялись стандартные ДНК-методы, описанные в руководстве (Sambrook, J., Fritch, E.F., и Maniatis, Т., (1989) Molecular Cloning: A Laboratory Manual, 2nd edn. , Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY). Синтетические олигонуклеотиды были приготовлены на автоматическом синтезаторе ДНК (380В, Applied Biosystems) с использованием имеющихся в продаже реактивов.
Определения последовательности ДНК выполнялись с помощью метода терминации дидезокси-цепи (Sanger, F., Micklen, S., и Coulson, A.R., Proc. NatI. Acad. Sci. USA, 1977, vol.74, pp.5463-5467).
Полимеразная цепная реакция (PCR) выполнялась на устройстве для термоциклической обработки ДНК (Perkin Elmer Cetus).
Анализ аминокислотной последовательности N-конца был проведен с помощью автоматической деградации по Эдману с использованием газофазового секвенатора "Applied Biosystems 477А". Проводилась обратнофазовая ВЭЖХ в реальном масштабе времени для выявления и количественного определения освобожденных РТН аминокислот из каждого цикла секвенатора.
1 мкл (106 pfu) библиотеки с ДНК костного мозга человека (Clontech, Palo Alto, , CA, USA) использовали в качестве матрицы в реакции PCR, содержащей 100 пикомолей каждого из праймеров NOR-3176.
(GTTGGAATTCAT (А/Т/С )CA(A/G)AA( Т/С )CA(A/G)GGN (А/С )G)
и
NOR-3174(CA(T/C)TG(A/G)TC(T/C)TCNGGNGT),
где N является смесью всех четырех нуклеотидов. Часть 5' первого праймера NOR-3176 содержит сайт EcoRI, а часть 3' соответствует аминокислотным остаткам 18-23, и обратный праймер NOR-2174 соответствует аминокислотным остаткам 149-154 в аминокислотной последовательности hHBP [Flodgaard, Н., Ostergaard, Е., Bayne, S., Svendsen, A., Thomson, J., Engels, M. и Wollmer, A., Eur. J. Biochem., 1991, vol.197, pp.535-547].
Реакция PCR выполнялась в объеме 100 мкл с использованием коммерческого набора (GeneAmp, Perkin Elmer Cetus) и следующего цикла: 94o в течение 1 мин, 50o в течение 2 мин и 72o в течение 3 мин. Через 35 циклов выполнялся последний цикл, при котором этап 72o поддерживался в течение 10 мин. Продукт PCR, фрагмент 420 bp, был выделен с помощью электрофореза на 1% агарозном геле.
Аналогичным образом в реакции PCR, как описано выше, использовались праймеры NOR-3173
(ACNCCNGA(A/G)GA(T/C)CA(A/G)TG) и NOR-3175
(GGTTTCTAGATTATCCCGG (A/G) TT (A/G) TTNA (A/G) NACNCC) с той же матрицей кДНК. Первый праймер NOR-3173 комплементарен NOR-2174, а обратный праймер NOR-3175 соответствует аминокислотным остаткам 215-221 в аминокислотной последовательности НВР со следующим за ней стоп-кодоном и участком Xbal. Реакция PCR выполнялась, как описано выше, и ее продукт, фрагмент 232 bp, был выделен с помощью электрофореза на агарозном геле.
Два описанных выше фрагмента PCR соединялись с помощью PCR удлинения с перекрытием [Horton, R.M., Hunt, H.D., Но, S.N., Pullen, J.K. и Pease, L.R., Gene, 1989, vol.77, pp.61-68] с использованием в качестве праймеров NOR-3176 и NOR-3175.
Полученный в результате фрагмент 635 bp был выделен и затем подвержен расщеплению EcoRI и Xbal с получением фрагмента 621 bp., который был лигирован с фрагментом 2,8 kb EcoRI-Xbal из плазмиды pTZ19R (Mead, D.A., Szczesna-Skorupa, E. и Kemper, В., Prot. Engin., 1986, vol.1, pp. 67-74).
Полученную в результате лигирования смесь использовали для трансформации компетентного штамма E. coli (r-, m+), селектируя на устойчивость к ампициллину. Определение последовательности ДНК показало, что плазмида pKFN-1726 из одной из полученных колоний кодировала ожидаемую аминокислотную последовательность hHBP18-21.
Плазмида pKFN-1726 подвергалась расщеплению PstI и Xbal, и полученный фрагмент 329 bp выделяли и использовали, как описано ниже.
N-терминальная часть НРБ выделялась в виде фрагмента 379 bp с помощью PCR с использованием той же матрицы кДНК, как описано выше, и праймеров MHJ-206
(GCTGAGAGATTGGAGAAGAGAATCGTTGGCGGCCGGAAGGCGAG)
и MHJ-200
(CGGCTTCCACCGTGGCGTTCTG)
Часть 3' MHJ-206 идентична N-терминальной кодирующей части гена НВР человека, а часть 5' MHJ-206 идентична С-терминальной части описанного ранее гибридного дрожжевого лидерного гена доминирующего гена гибридных дрожжей [Thim, L., Norris, K., Nielsen, Р. F., Bjorn, S.E., Christensеn и Petersеn, J., FEBS Lett., 1993, voi.318, pp.345-352].
Слияние в рамке гибридного лидерного гена и фрагмента 379 bp PCR, кодирующего N-терминальную часть НРБ, было получено с помощью PCR удлинения с перекрыванием [Horton, R.M., Hunt, H.D., Но, S.N., Pullen, J.K. и Pease, L.R., Gene, 1989, vol. 77, pp. 61-68]. Продукт подвергали перевариванию EcoRI и PstI и выделяли в виде фрагмента 572 bp.
Фрагмент 572 bp EcoRI-PstI и фрагмент 329 bp Pstl-Xbal, описанные выше, были лигированы С фрагментом 2,8 kb EcoRI-Xbal из плазмиды pTZ19R (Mead, D. A. , Szczesna-Skorupa, Е. и Kemper, В., Prot. Engin., 1986, vol.1, pp. 67-74). Лигирующая смесь использовалась для трансформации компетентного штамма E. coli (r-, m+), селектируя на устойчивость к ампициллину. Плазмида pKFN-1780 из одной из полученных в результате колоний была отобрана для дальнейшего использования.
Плазмида pKFN-1780 подвергалась перевариванию EcoRI и Xbal. Полученный в результате фрагмент 901 bр был лигирован с фрагментом 9,5 kb NcoI-Xbal из рМТ636 и фрагментом 1,4 kb NcoI-EcoRI из рМТ636. Плазмида рМТ636 описана в международной патентной заявке WO 89/01968.
РМТ636 является челночным вектором E.coli-S. cerevisae, содержащим ген TPI Schizosaccharomyces pombe (РОТ) (Russel, P.R., Gene, 1985, vol.40, pp. 125-130), промотор и терминатор триозофосфатизомеразы S.cerevisiae,TPIp и ТРIт (Alber, Т. и Kawasaki, G., J. Mol. Appl. Gen., 1982, vol.1, pp.419-434).
Лигирующую смесь использовали для трнсформации компетентного штамма E. coli (r-, m+), селектируя на устойчивость к ампициллину. Определение последовательности ДНК показало, что плазмиды из полученных в результате колоний содержали правильную последовательность ДНК для НРБ человека, правильно слитую с синтетическим дрожжевым сигнально-лидерным геном.
Одна плазмида pKFN-1783 была выбрана для дальнейшего использования.
Кассета экспрессии плазмиды pKFN-1783 содержит следующую последовательность: TPIp - сигнал-лидер-НВР-ТРIт.
Последовательность ДНК фрагмента 901 bp EcoRI-Xbal pKFN-1783 показана на фиг.1.
Трансформация дрожжей: штамм МТ663 В. cerevisiae (Е-2-7В XEll-36а/α, Δtpi/Δtpi, pep 4-3/pep 4-3) выращивался на среде YPGaL (1% Бакто-дрожжевой экстракт, 2% Бакто пептон, 2% галактоза, 1% лактат) до O.D., равной 0,6 при 600 нм.
100 мл культуры были собраны центрифугированием, промывали 10 мл воды, повторно центрифугировали и ресуспендировали в 10 мл раствора, содержащего 1,2 М сорбитол, 25 мМ Na2 ЭДТА, рН 8,0 и 6,7 мг/мл дитиотреитол. Суспензию инкубировали при 30oС в течение 15 минут, центрифугировали, и клетки ресуспендировали в 10 мл раствора, содержащего 1,2 М сорбитола, 10 мМ 1,2 мМ Na2 ЭДТА, 0,1 М цитрат натрия, рН 5,8 и 2 мг Novozym® 234. Суспензию инкубировали при 30oС в течение 30 минут, клетки собирали центрифугированием, отмывали в 10 мл 1,2 М сорбитола и 10 мл CAS (1,2 М сорбитол, 10 мМ СаСl2, 10 мМ Tris-HCl (Тris=Трис(гидроксиметил)аминометан) рН 7,5) и ресуспендировали в 2 мл CAS. Для трансформации 0,1 мл
ресуспендированных в CAS клеток смешивали приблизительно с 1 мкг плазмиды pKFN-1783 и оставляли при комнатной температуре на 15 минут. Добавляли 1 мл (20% полиэтиленгликоля 4000, 20 мМ CaCl2, 10 мМ Tris HC1, pH 7,5) и смесь оставляли еще на 30 минут при комнатной температуре. Смесь центрифугировали, а осадок ресуспендировали в 0,1 мл SOS (1,2 М сорбитол, 33% о/о YPD, 6,7 мМ CaCl2, 14 мкг/мл лейцин) и инкубировали при 30oС в течение 2 часов. Суспензию затем центрифугировали, и осадок ресуспендировали в 0,5 мл 1,2 М сорбитола. Затем 6 мл верхнего слоя агара (среда SC Sherman et al., (Methods in Yeast Genetics, Cold Spring Harbor Laboratory, 1982)), содержащая 1,2 М сорбитол плюс 2,5% агар) добавляли при 52oС и суспензию заливали сверху в чашки, содержащие ту же самую затвердеваемую через посредство агара, содержащую сорбитол среду.
Трансформированные колонии отбирали через трое суток при 30oС, повторно выделяли и использовали для приготовления жидких культур. Для дальнейшей характеристики был отобран один такой трансформант KFN-1775.
Ферментация: Штамм дрожжей KFN-1775 выращивался на среде YPD (1% экстракта дрожжей, 2% пептона (из Difco Laboratories) и 3% глюкозы). 1 литр культуры штамма встряхивали при 30oС до оптической плотности, равной 24, при 650 нм. После центрифугирования выделяли супернатант.
НВР очищали из сунернатанта, в основном, как описано в патенте WO 89/08666. Анализ аминокислотной последовательности остатков 1-20 показал идентичность N-терминальной последовательности SEQ ID 1.
Пример 2
Для экспрессии НВР (в качестве проформы) в клетках насекомых с использованием бакуловирусной системы были получены следующие праймеры PCR:
MHJ 2087: 5'- FFF AAG GAT CCA CCA TGA CCC GGC TGA CAG TCC TGG CCC TGC TGG CNG GTC TGC TGG CGT CCT CGA GGG CCG GCT CCA GCC CCC TTT TGG АСА TCG TTG GCG GCC GGA AGG C-3'
MHJ 2069: 5' -AAA AAA GCT TCC TAG GCT GGC CCC GGT CCC GGA TTG TTT AAA ACG CCA TC -3'
MHJ 2087 кодирует сайт BamHI, кодон инициации и препрочасть кДНК человека (Morgan, J.G. et al., J., Immun., 1991, vol.147, pp.3210-3214) с последующими первыми 20 нуклеотидами из начала зрелой части гена на pKFN1780.
MHJ 2089 комплементарен последним 8 кодонам из кодирующей части гена НВР на pKFN1780 плюс два дополнительных кодона в соответствии с указанной выше последовательностью кДНК. Его конец является HindIII-сайтом.
PCR выполнялась с использованием двух праймеров и pKFN1780 в качестве матрицы по следующей схеме:
3 цикла 95oС 60 сек, 50oС 120 сек, 72oС 120 сек
12 циклов 95oС 30 сек, 65oС 60 сек, 72oС 90 сек
Продукт PCR, фрагмент 760 bр, был выделен с помощью электрофореза на 1% агарозном геле, фракционирован с помощью BamHI и HindIII и встроен в рSХ221, разрезанную с использованием тех же двух ферментов (рSХ221 является производным pUC19 (Yannisch-Perron, С. et al., Gene, 1985, vol.33, pp.103-119)). Клонированная ДНК верифицировалась путем определения последовательности, и фрагмент BamHI-HindIII (показанный на фиг.2) отсекался и вставлялся в pBlueBacIII (Invitrogen Corporation) для экспрессии в клетках насекомых. Полученная в результате плазмида была названа pSX556.
Для получения рекомбинантного бакуловируса, экспрессирующего НВР, использовался набор МАХВАС, приобретенный в Invitrogen Corp. (San Diego, CA), и все манипуляции производились в соответствии с приложенным руководством "Baculovirus Expression System Manual" (версия 1.5.5). Вкратце 1 мкг линеаризированной AcMNPV ДНК и 3 мкг pSX556 были трансфецированы в клетки насекомых SF9 (2•106 клеток в 60 мм чашках). Полученный в результате культуральный супернатант собирали через 7 суток. Свежие монослои клеток SF9 в 100 мм чашках инфицировались супернатантом вируса при различных разведениях, а затем на них наслаивали 1,5% агарозу, содержащую полную среду TNM-FH со 150 мкг/мл Х-gal. Через 8 суток 6 предполагаемых рекомбинантных бляшек идентифицировали по их синему ободку и использовали для инфекции клеток SF9, находящихся в чашке с 6-ю отсеками. Через 5 дней соответствующую вирусную ДНК очищали и подвергали реакции PCR с прямым и обратным праймерами, находящимися по обе стороны от участка рекомбинации в вирусной ДНК. После оценки продуктов PCR на агарозном геле соответствующий наиболее чистый рекомбинантный вирус подвергался еще одному циклу очистки бляшки для гарантии того, что окончательный штамм рекомбинантного вируса будет свободен от вируса дикого типа. Выработка рекомбинантного НВР проводилась в клетках насекомых (SF9 и SF21), адаптированных к росту в бессывороточной среде SF900-II (Gibco BRL/Life-Technologies). Обычно перемешиваемые культуры объемом 5 л или ферментер объемом 10 л при плотности клеток в обоих типах, составляющей 1•106 /мл, инфицировались при множественности заражения, равной 1, и среду собирали через 3 дня после инфекции. Очистку НВР проводили, как описано в патенте WO 89/08666.
Пример 3
НВР без прорайона
Олигонуклеотидный линкер (см. ниже) был приготовлен, покрывая первые 99 bp последовательности, показанной на фиг. 2 (от BamHI до Еаgl), исключая часть, покрывающую прорегион (с 73 до 87, курсив), и он был подставлен вместо первоначального BamHI-EagI в pSX556, образуя pSX559. Ожидают, что эта конструкция при экспрессии в системе бакуловируса должна производить зрелый НВР.
Линкер состоит из четырех олигонуклеотидов, ренатурированных попарно, с получением следующих дуплексов:
MHJ2568/LWN5746:
5'-GATCCACCATGACCCGGCTGACAGTCCTGGCCC-3'
3'-GTGGTACTGGGCCGACTGTCAGGACCGGGACGACC-P-5'
LWN5745/MHJ2566:
5 5'-P-TGCTGGCTGGTCTGCTGGCGTCCTCGAGGGCCATCGTTGGC-3'
3'-GACCAGACGACCGCAGGAGCTCCCGGTAGCAACCGCCGG-5'
Пример 4
Экспрессия производного НВР с участком расщепления бычьей энтерокиназы, расположенным между пропептидом и зрелым НВР
Клетки насекомых, инфицированные бакуловирусным вектором, кодирующим НВР (сигнал-пропептид-зрелый НВР), не способны отщепить пропептид. Клетки насекомых, инфицированные вирусом, кодирующим НВР без пропептида (сигнальная последовательность, слитая непосредственно с последовательностью зрелого НВР), вырабатывают зрелую форму, но в очень маленьких количествах. Для увеличения выхода зрелого НВР был получен бакуловирусный вектор, кодирующий производное НВР с участком расщепления бычьей энтерокиназы (Asp4-Lys), вставленным между пропептидом и зрелым НВР, обеспечивающий возможность получения зрелого НВР путем процессинга in vitro полученной удлиненной формы НВР:
Фрагмент кДНК, кодирующий сигнал-Gly-Ser-Ser-Pro-Leu-Leu-Asp-Asp3-Lys-зрелый НВР, был генерирован с помощью PCR с полимеразой Pfu и прзймерами PBRa241.
(CCGGGATCCGATGACCCGGCTGACAGTCCTGGCCCTGCTGGCTGGTCTGCT GGCGTCCTCGAGGGCCGGCTCCAGCCCCCTTTTGGACGACGACGACAAGATCGTTG GCGGC) и PBRa 246
(CCGGGGATCCAACTAGGCTGGCCCCGGTCCCGG).
Продукт PCR подвергался перевариванию BamHI и клонировался в трансферный вектор pVL1393 (Invitrogen, San Diego, CA). Размножение рекомбинантного бакуловируса и получение белка производилось, как описано в примере 2 (однако в этом случае предполагаемые рекомбинантные бляшки идентифицировались с помощью их окклюзионно-отрицательного фенотипа).
Для получения формы НВР с удлиненным N-концом, 4•108 клеток SF9, растущих в бессывороточной среде SF900II (Gibco), осаждали ценгрифугированием и ресуспендировали в образце из штамма вируса, дающего MOI (множественность инфекции), равную 1. Клетки с вирусом переносились во флакон центрифуги "Belco" емкостью 0,5 л (#1965-00500), и свежая среда SF900II добавлялась до конечного объема 400 мл. Наконец в культуру добавлялись 1,5 г гепарина-Сефарозы (GL-6B, Pharmacia), который автоклавировался в 25 мл стерильного 0,9% раствора NaCl. Культура инкубировалась при 27oС в течение 3 суток.
Для выделения бусин гепарина-Сефарозы из культуры клеток насекомых, 400 мл ферментационной среды добавляли в кратном объеме в 8 пробирок емкостью 50 мл и центрифугировали в центрифуге TECHNOSPIN R (Sorvall Instruments) при 300 об/мин в течение 3 минут. Супернатанты с клетками отсасывались, и осажденные бусины гепарина-Сефарозы отделялись от остающихся загрязняющих клеток путем ресуспендирования в 30 мл стерильного раствора NaCl, добавляемого в каждую пробирку, с последующим ценгрифугированием при 300 об/мин.
Вся процедура повторялась дважды. Бусины окончательно промывали в 20 мл стерильного 0,5 М раствора NaCl, добавляемого в каждую трубку. Затем бусины собирали в одну трубку емкостью 50 мл в небольшом объеме 0,5 М стерильного раствора NaCl (20-30 мл) и переносили в стерильный стеклянный воронкообразный фильтр. Раствору давали возможность стечь с бусин и, наконец, rНВР элюировали 30 мл стерильного 3 М раствора NaCl. НВР очищали из 3 М NaCI элюата в соответствии со способом, описанным в патенте WO 89/08666.
Двести микрограмм очищенного НВР, содержащего пропоследовательность и участок расщепления энтерокиназы между пропоследовательностью и зрелым белком, были растворены в 50 мМ Na-ацетатном буфере, содержащем 25 мМ CaCl2, рН 5,1. Четыреста единиц энтерокиназы, приобретенной у компании "Biozyme Laboratories Limited", GB (партия 18•2 Код ЕКЗ), добавляли в объеме 1,2 микролитра, и смесь инкубировали в течение 1 часа при 37oС. НВР окончательно очищали из инкубационной смеси с помощью ВЭЖХ с высоким разрешением в соответствии со способом, описанным в патенте WO 89/08666.
Аликвотная проба очищенного НВР (1 нмоль) подвергалась определению последовательности N-конца, которое показало, что главная последовательность представляет правильный зрелый НВР, начинающийся с Ilе, свидетельствуя о том, что расщепление было успешным.
Пример 5
Экспрессия производных НВР с участком расщепления Фактора Xa, расположенным между пропептидом и зрелым НВР.
Альтернативной стратегией генерирования зрелого НВР было использование фактора Ха для расщепления in vitro полученного материала.
Участком распознавания/расщепления фактора Ха является Ile-Glu-Gly-Arg. Т.J. Gardella et al. (J. Biol. Chem., 1990, vol. 26, pp. 15854-15859) сообщили, что помещение трех маленьких аминокислот (Gly-Gly-Ser) непосредственно около участка распознавания фактора Ха со стороны N-конца вероятно свело к минимуму эффекты стерической помехи и, таким образом, может быть увеличена эффективность расщепления фактором Ха. Для генерации фрагментов кДНК, кодирующих сигнал-пропептид-Ile-Glu-Gly- Arg-зрелый НВР и сигнал-пропептид-Gly-Gly-Ser-Ile-Glu-Gly-Arg-зрелый НВР, были синтезированы два праймера, соответственно PBRa 249
(CCGGGGATCCGATGACCCGGCTGACAGTCCTGGCCCTGCTGG
CTGGTCTGCTGGCGTCCTCGAGGGCCGGCTCCAGCCCCCTTTTG
GACATCGAGGGTAGGATCGTTGGCGGC)
и PBRa 250
(CCGGGGATCCGATGACCCGGCTGACAGTCCTGGCCCTGCTGG
CTGGTCTGCTGGCGTCCTCGAGGGCCGGCTCCAGCCCCCTTTTG
GACGGTGGTTCCATCGAGGGTAGGATCGTTGGCGGC).
Совместно с праймером PBRa 226 (пример 4) эти два праймера были использованы при реакциях PCR с полимеразой Pfu. Эти два фрагмента были переварены BamHI и вставлены в pVL1393. Размножение рекомбинантных бакуловирусов и производство белков производились, как описано в примере 4.
Пример 6
Мононуклеарные клетки периферической крови (BMNC) получали от здоровых взрослых доноров крови (Банк крови, Rigshospitalet) путем центрифугирования надосадочного слоя лейкоцитов с цитратным антикоагулянтом на приборе Lymphoprep® (Nycomed, Осло, Норвегия). В некоторых экспериментах BMNC предварительно инкубировали с РТХ перед добавлением липополисахарида (LPS) (E. coli 055: В5; Difco Laboratories, Detroit MI) при конечной концентрации 1 мкг/мл, или РНА (Difco) при конечной концентрации 20 мкг/мл, или очищенного белкового производного туберкулина (PPD) (Государственный сывороточный институт, Копенгаген, Дания) при конечной концентрации 25 мкг/мл. Инкубацию проводили в среде RPMI-1640 (BRL-Gibco, Roskilde, Denmark), содержащей 3% нормальную человеческую сыворотку (NHS), инактивированную теплом при 56oС в течение 30 мин с добавлением 0,8 мМ глютамина, пенициллина/стрептомицина (BRL-Gibco, Дания) по 20 ЕД/мл каждого. После инкубации клетки собирали, и супернатанты были немедленно заморожены при -20oС; осажденные клетки быстро замораживались на жидком азоте и хранились при -80oС в течение более 1 недели перед тем, как была произведена экстракция РНК.
Содержание IL-1α, IL-1β, IL-2, IL-6, IFNτ, TNFα и TNFβ определяли с помощью двойной сэндвич-ELISA с использованием неспецифических поликлональных кроличьих антител к очищенным рекомбинантным цитокинам (М.В. Наnsen et аl., Immunol. Lett., 1991, vol.30, p.156). Чашки "Immuno-Maxisorp)" ("Nunc", Roskilde, Denmark) покрывались IgG, очищенным по сродству к белку А. Неприкрепленные участки блокировались 5% сывороточным альбумином человека в фосфатно-солевом буфере (PBS). Образцы растворялись в PBS с добавлением 2% нормальной кроличьей сыворотки ("Dako", Glostrup, Denmark), 10 мМ ЭДТА, 2000 KIE/мл апротинина и 5 мМ DL-дитиотреитола. Анализы калибровались международными стандартами соответствующих цитокинов (National Institute for Biological Standards and Controls, Potters Bar, Hertfordshire, UK). Биотинилированные поликлональные кроличьи антитела использовались в качестве выявляющих антител наряду со стрептавидин-пероксидазой (Kirkegaard and Perry La., Gaitherburg, MD). В качестве субстрата использовали 1,2-фенилендиамин-дигидрохлорид, а интенсивность окрашивания определяли с помощью фотометра при 492 нм. Вариационный коэффициент между определениями для диапазона концентрации между 8 нг/мл и 1 нг/мл был менее 15%. Предел чувствительности этих ELISA был 8-10 пг/мл.
BMNC оттаивали и обрабатывали сывороточным альбумином человека в количествах 1-мкг/мл и 100 мкг/мл, а также НВР в количествах 0,2 мкг/мл и 20 мкг/мл в присутствии соответственно 10 и 100 нг/мл LPS.
Результаты показаны на прилагаемых фигурах 3 и 4. На фигурах видно, что в присутствии НВР освобождение цитокинов из BMNC значительно снижено.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРИМЕНЕНИЕ ГЕПАРИНСВЯЗЫВАЮЩИХ АНТАГОНИСТОВ В ИНГИБИРОВАНИИ ВЫСВОБОЖДЕНИЯ БРАДИКИНИНА | 2000 |
|
RU2251433C2 |
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКОВ В ТРАНСФОРМИРОВАННЫХ ДРОЖЖЕВЫХ КЛЕТКАХ | 1999 |
|
RU2238323C2 |
| НОВЫЙ ЛИГАНД РЕЦЕПТОРА ЦИТОКИНА ZCYTOR17 | 2003 |
|
RU2490276C2 |
| ТРОМБОПОЭТИН | 1994 |
|
RU2245365C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ПРОТЕАЗНОЙ АКТИВНОСТЬЮ, И НУКЛЕИНОВЫЕ КИСЛОТЫ, КОДИРУЮЩИЕ УКАЗАННЫЕ ПОЛИПЕПТИДЫ | 2002 |
|
RU2318018C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГЕТЕРОЛОГИЧНОГО БЕЛКА | 1994 |
|
RU2143495C1 |
| ВАРИАНТЫ ГЛЮКОАМИЛАЗЫ | 2006 |
|
RU2439153C2 |
| ПОЛИПЕПТИД ТРОМБОПОЭТИН (ТРО), ДНК, КОДИРУЮЩАЯ ПОЛИПЕПТИД ТРО (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДА (ВАРИАНТЫ), ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СПОСОБ ЛЕЧЕНИЯ (ВАРИАНТЫ), АНТИТЕЛО К ПОЛИПЕПТИДУ ТРО | 1995 |
|
RU2194074C2 |
| ВАРИАНТЫ АЛЬФА-АМИЛАЗЫ | 2000 |
|
RU2231547C2 |
| БЕЛОК IFNAB-BPII, ОБЛАДАЮЩИЙ СПОСОБНОСТЬЮ СВЯЗЫВАТЬ ИНТЕРФЕРОН-α/β, ЕГО ПРЕДШЕСТВЕННИК И СЛИТЫЕ БЕЛКИ, СПОСОБ ПОЛУЧЕНИЯ IFNAB-BPII, МОЛЕКУЛЫ ДНК, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 1995 |
|
RU2232811C2 |
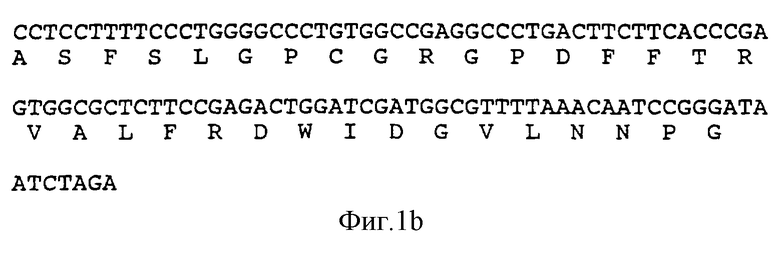
Изобретение относится к медицине и касается применения гепаринсвязывающего белка (НВР), который (а) лишен протеиназной активности, (б) присутствует в периферических нейтрофильных лейкоцитах и (в) является хемотоксическим агентом для моноцитов, для изготовления лекарственного средства для предотвращения или лечения сепсиса, грамотрицательного сепсиса, септического шока или диссеминированного внутрисосудистого свертывания. Изобретение позволяет использовать НВР после проявления феноменов сепсиса, т.е. при процессах, развивающихся после эндотоксемии. 12 з.п. ф-лы, 4 ил.
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| СПОСОБ ЛЕЧЕНИЯ СЕПТИЧЕСКИХ ЗАБОЛЕВАНИЙ У НОВОРОЖДЕННЫХ И ДЕТЕЙ ГРУДНОГО ВОЗРАСТА | 1989 |
|
RU2008009C1 |
| СПОСОБ ЛЕЧЕНИЯ СЕПСИСА У НОВОРОЖДЕННЫХ ДЕТЕЙ | 1991 |
|
RU2012341C1 |

