Предпосылки создания изобретения
Эффективное встраивание (инсерция) экзогенной нуклеиновой кислоты в хромосомную и нехромосомную нуклеиновую кислоту клеток необходимо в области молекулярной биологии для идентификации областей хромосомы, участвующих в экспрессии или регуляции экспрессии пептидов и протеинов. Эту же технологию целесообразно использовать при создании новых терапевтических и фармакологических агентов.
Один из общепринятых методов основан на применении транспозонного мутагенеза in vivo с использованием Тn5 (Тn5-мутагенез) для встраивания представляющих интерес полинуклеотидов в клеточную ДНК и конструирования банков клеток, которые содержат встроенные полинуклеотиды в случайных или псевдослучайных положениях. Существующие методы Тn5-мутагенеза in vivo требуют наличия клеток-мишеней, кодирующих транспозазу либо нативно, либо с помощью встроенной экспрессионной конструкции. Таким образом, может оказаться необходимым конструировать пригодную систему экспрессии, соответствующую каждому типу клеток-мишеней. Для этого может требоваться много времени и наличие многочисленных сведений о характеристиках каждого типа клеток.
Во многих случаях ген, кодирующий транспозазу, кодируется активным транспозоном, который может обеспечивать продолжение транспозиции в клетке-мишени после завершения начальной требуемой стадии мутагенеза. Такая неплановая остаточная транспозиция является нежелательной вследствие того, что она осложняет анализ банков инсерционных мутантов.
Кроме того, для осуществления многих методов Тn5-мутагенеза in vivo требуется сложный биологический механизм для интродукции экзогенной ДНК в клетки-мишени, такой как трансдукция с использованием фага, например бактериофага лямбда, или конъюгация с использованием плазмиды. Целесообразно избегать применения таких сложных биологических систем.
Shoji-Tanaka А. с соавторами в B.B.R.C. 203: 1756-1764 (1994) описали применение очищенной ретровирусной интегразы для опосредованного переноса гена в клетки мышей.
Kuspa А. и W.F. Loomis в P.N.A.S. U.S.A., 89: 8803-8807 (1992) и в других публикациях описали специфичную интеграцию плазмиды, линеаризованной с помощью рестриктазы, в геномный сайт рестрикции клетки-мишени путем электропорации с использованием расщепленной ферментативным путем нуклеиновой кислоты вместе с отщепляющим ферментом.
Краткое изложение сущности изобретения
Настоящее изобретение в целом относится к способу эффективной инсерции способного к транспозиции (мобильного) полинуклеотида в случайные или псевдослучайные положения в хромосомной или внехромосомной нуклеиновой кислоте клетки-мишени, предусматривающему стадии объединения в клетке-мишени клеточной нуклеиновой кислоты с синаптическим комплексом, который включает (а) протеин Тn5-транспозазы, объединенный в комплекс с (б) полинуклеотидом, который включает пару нуклеотидных последовательностей, адаптированных к функциональному взаимодействию с Тn5-транспозазой, и мобильную нуклеотидную последовательность между ними, в условиях, которые опосредуют транспозиции в клеточной ДНК. Согласно этому способу синаптический комплекс формируют in vitro в условиях, которые являются неблагоприятными или препятствуют тому, чтобы синаптические комплексы, которые могут применяться для транспозиции, фактически подвергались продуктивной транспозиции. Согласно способу частота продуктивной транспозиции мобильной нуклеотидной последовательности в нуклеиновой кислоте-мишени может быть повышена либо с помощью гиперактивной транспозазы, либо мобильного полинуклеотида, который включает последовательности, особенно хорошо адаптированные для эффективной транспозиции в присутствии Тn5-транспозазы, либо с помощью обоих этих факторов.
Настоящее изобретение также в целом относится к способу формирования банка клеток, которые содержат инсерционные мутации, предусматривающему стадии объединения в многочисленных клетках-мишенях клеточной нуклеиновой кислоты с описанным выше синаптическим комплексом, и отбора клеток, которые содержат инсерционные мутации.
Другим объектом изобретения является банк клеток, которые содержат инсерционные мутации, созданный согласно указанному выше способу. Такие популяции клеток, которые содержат случайные и независимые мутантные инсерции в их геномах, могут быть подвергнуты скринингу с целью отбора тех клеток, которые включают инсерционные мутации, вызывающие фенотипическое или генотипическое изменение по сравнению с клетками, которые не подвергались инсерционному мутагенезу.
Преимуществом настоящего изобретения является то, что мобильные полинуклеотиды, применяемые для формирования синаптических комплексов, могут состоять из ДНК транспозона, помимо любых фланкирующих последовательностей. Это является важным потому, что уменьшает вероятность внутримолекулярной транспозиции и повышает вероятность транспозиции в геноме-мишени. Кроме того, отсутствие донорных каркасных (ДК) последовательностей полинуклеотида упрощает получение последовательностей транспозона, которые могут применяться согласно способу по изобретению.
Еще одним преимуществом настоящего изобретения является то, что синаптический комплекс может быть сформирован в условиях, которые являются неблагоприятными для возникновения случаев непродуктивной внутримолекулярной транспозиции. Это преимущество обусловливает то, что практически все синаптические комплексы могут подвергаться транспозиции при объединении с клеточной ДНК. Очень небольшое количество нуклеиновых кислот в синаптических комплексах, если таковые вообще имеются, являются неактивными.
Отличительной особенностью настоящего изобретения является то, что усиливающие транспозицию условия начинают проявляться только после того, как синаптический комплекс приходит в контакт с нуклеиновой кислотой-мишенью в клетке-мишени.
Другие объекты, преимущества и отличительные особенности настоящего изобретения станут очевидными после ознакомления с приведенным ниже подробным описанием изобретения.
Краткое описание некоторых аспектов чертежей
На фиг.1 показан перенос синаптического комплекса в клетку-мишень с последующим отбором инсерционных мутантов.
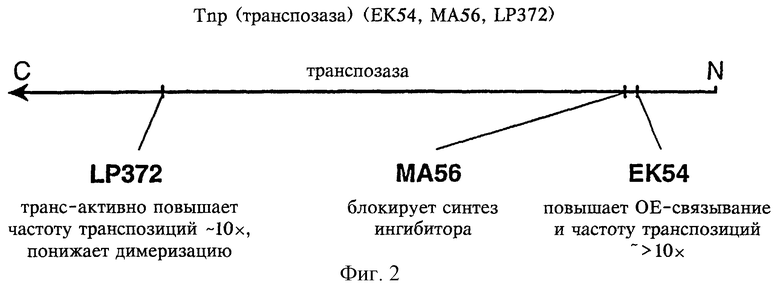
На фиг.2 представлен предпочтительный для применения согласно способу фермент транспозаза.
На фиг.3 представлены концевые последовательности транспозона, которые могут применяться в мобильной полинуклеотидной последовательности по настоящему изобретению.
На фиг.4 представлен транспозон или мобильная полинуклеотидная последовательность, имеющая мозаичные концы, и мобильная нуклеотидная последовательность между этими концами.
Подробное описание изобретения
В системе для транспозиции in vitro с использованием модифицированной Тn5-транспозазы, описанной в PCT/US7/15941 (WO 98/10077), которая полностью включена в настоящее описание в качестве ссылки, заявители продемонстрировали продуктивную транспозицию мобильного полинуклеотида in vitro с помощью модифицированной мутантной Тn5-транспозазы и модифицированных полинуклеотидных последовательностей Тn5, фланкирующих мобильную нуклеотидную последовательность.
Настоящее изобретение отличается от системы, известной из уровня техники, тем, что согласно настоящему изобретению синаптические комплексы формируют in vitro, при этом транспозиция in vitro практически отсутствует. При этом после того как синаптические комплексы интродуцируют в клетки-мишени, быстро происходит транспозиция in vivo. На фиг.1 представлено схематическое изображение одного из аспектов основанного на применении Тn5 способа эффективной интродукции по настоящему изобретению инсерционных мутаций в случайные и псевдослучайные положения в хромосомной или внехромосомной нуклеиновой кислоты клетки-мишени. Согласно этому способу клеточную нуклеиновую кислоту в клетке-мишени объединяют с синаптическим комплексом, который включает (а) протеин Тn5-транспозазы (показанный на фиг.1 в виде пары примыкающих друг к другу окружностей) и (б) полинуклеотид, который включает пару инвертированных нуклеотидных последовательностей (показанных в виде стрелок на фиг.1), адаптированных к функциональному взаимодействию с Тn5-транспозазой, и мобильную нуклеотидную последовательность между ними, в условиях, которые опосредуют транспозиции в клеточной ДНК. Структура синаптического комплекса является вспомогательным средством при транспозиции мобильного полинуклеотида в клеточной нуклеиновой кислоте. В настоящем описании представлен способ, с помощью которого можно отделить формирование синаптического комплекса от транспозиции посредством воздействия на способность синаптического комплекса участвовать в реакции переноса полинуклеотидной цепи. Согласно этому способу синаптический комплекс получают in vitro в условиях, которые являются неблагоприятными или препятствуют тому, чтобы синаптические комплексы подвергались продуктивной транспозиции. В изобретении описано использование методов Тn5-мутагенеза применительно, например, к бактериям, которые не являются близкородственными Е. coli и о структуре которых мало известно, и определены дополнительные требования к промоторам и другим регуляторным последовательностям. В таких клетках транспозаза может не экспрессироваться в необходимых количествах или может быть нестабильной. Для способа по настоящему изобретению не требуется получать эндогенную молекулу транспозазы. Кроме того, поскольку интродуцируемая транспозаза входит в состав комплекса с полинуклеотидом транспозона, она может действовать в присутствии любой ДНК-мишени и сама по себе является достаточно стабильной для того, чтобы облегчать транспозицию полинуклеотида, с которым она образует комплекс. Представленный в настоящем описании способ, с помощью которого можно препятствовать тому, чтобы в синаптических комплексах происходил перенос полинуклеотидной цепи, достигается путем снижения или полного устранения ионов магния (Mg++) из реакционной смеси.
Таким образом, согласно предпочтительному варианту осуществления способа пригодную транспозазу и приемлемый мобильный элемент ДНК объединяют в реакции in vitro, где реакционная смесь содержит такую концентрацию магния, которая недостаточна для того, чтобы поддерживать перенос полинуклеотидной цепи, или более предпочтительно смесь вообще не содержит ионы магния. Приемлемым реакционным буфером для образования синоптических комплексов может служить буфер, описанный на странице 19 включенной в настоящее описание в качестве ссылки международной заявки на патент, который модифицируют, удаляя ацетат магния. Из реакционного буфера также могут быть удалены БСА и спермидин. Из буфера может быть удалена тРНК, если в реакционной смеси присутствуют нуклеазы, в этом случае тРНК может быть добавлена. Также можно еще более упростить реакционный буфер. Типичная реакция описана ниже в примере.
Синаптические комплексы предназначены для того, чтобы опосредовать эффективную транспозицию и предпочтительно они могут храниться до применения в холодных условиях. Синаптические комплексы могут быть интродуцированы в пригодные клетки-мишени, обладающие способностью обеспечивать условия, благоприятные для транспозиции, после чего происходит эффективная транспозиция. Путем интродукции синаптических комплексов в многочисленные пригодные клетки-мишени и отбора тех клеток, которые включают инсерционные мутации, может быть создан банк клеток, которые содержат случайные или псевдослучайные инсерционные мутации. Для создания банка инсерционных мутантов мобильная нуклеотидная последовательность предпочтительно содержит соответствующий селектируемый маркер, такой как ген, обусловливающий устойчивость к антибиотику, благодаря чему клетки, лишенные инсерционной мутации в их клеточной ДНК, могут быть легко отличены от клеток, в которых может происходить продуктивная транспозиция. Созданные согласно способу банки могут быть подвергнуты скринингу в отношении генотипических или фенотипических изменений после транспозиции. На молекулярном генетическом уровне могут применяться стандартные аналитические методы, включая гибридизацию, картирование с помощью рестрикционных фрагментов, секвенирование нуклеотидных последовательностей и сочетание этих методов, или другие методы для идентификации генетических изменений. На фенотипическом уровне можно оценить представителей банка транспозиционных мутантов в отношении отдельных мутантов, имеющих измененные характеристики роста или другой фенотип.
Набор синаптических комплексов, включающий мобильные полинуклеотидные последовательности, которые имеют требуемые характеристики, такие как описанные ниже, может поступать в продажу с целью облегчения быстрого получения банков инсерционных мутаций, для которого встраиваемые последовательности создают с определенной, описанной ниже целью. Синаптические комплексы по настоящему изобретению практически не содержат полинуклеотидных молекул, которые уже подверглись продуктивной транспозиции. Кроме того, поскольку синаптические комплексы получают в реакции in vitro, комплексы могут рассматриваться как практически чистый препарат, не содержащий других протеинов, генетического материала и т.п.
Тn5-транспозаза в синаптических комплексах может представлять собой Тn5-транспозазу, которая эффективно образует синаптические комплексы in vitro (например, 25% или более ДНК транспозона превращается в синаптические комплексы с помощью транспозазы). Транспозаза может представлять собой гиперактивную Тn5-транспозазу, такую как транспозаза, описанная в международной заявке PCT/US97/15941. Предпочтительная мутантная Тn5-транспозаза представляет собой мутантную Тn5-транспозазу, модифицированную относительно Тn5-транспозазы дикого типа, причем мутантная транспозаза несет мутацию в положении 54 и мутацию в положении 372, мутантная транспозаза обладает большей авидностью по отношению к инвертированным повторяющимся последовательностям на концах Тn5 ДНК-донора и меньшей способностью образовывать мультимеры по сравнению с Тn5-транспозазой дикого типа. Мутация в положении 54, которая обусловливает более высокую авидность по отношению к инвертированным повторяющимся последовательностям на концах Тn5, представляет собой замену глутаминовой кислоты на лизин в транспозазе дикого типа. Мутация в положении 372, которая приводит к пониженной способности образовывать непродуктивные мультимеры, представляет собой замену лизина на пролин в транспозазе дикого типа.
Также предпочтительно, чтобы транспозаза не содержала так называемый протеин-ингибитор, т.е. протеин, кодируемый последовательностью, частично перекрывающейся последовательностью транспозазы, который может оказывать влияние на активность транспозазы. Согласно этому способу транспозазу применяют в очищенной или частично очищенной форме, и если фермент транспозазу получают из клеток (используя общепринятые методы), то должна существовать возможность отделения транспозазы от фермента-ингибитора до применения ее согласно способу по изобретению. Однако также возможно генетически устранять возможность присутствия какого-либо загрязняющего протеина-ингибитора простым удалением его стартового кодона из гена, который кодирует транспозазу.
Кодон AUG в гене Тn5-транспозазы дикого типа, который кодирует метионин в положении 56 на аминокислотной последовательности транспозазы, представляет собой первый кодон протеина-ингибитора. Однако уже было установлено, что замена метионина в положении 56 не оказывает заметного воздействия на активность транспозазы, но в то же самое время препятствует трансляции протеина-ингибитора, приводя тем самым к некоторому увеличению скорости транспозиции (Weigand T.W. и W.S.Reznikoff, "Characterization of Two Hypertransposing Tn5 Mutants", J.Bact. 174: 1229-1239), публикация включена в настоящее описание в качестве ссылки). В частности, авторы настоящего изобретения в предпочтительном варианте осуществления заменили метионин на аланин (и заменили кодирующий метионин кодон AUG на кодирующий аланин кодон GCG). Таким образом, предпочтительная транспозаза по настоящему изобретению включает в положении 56 аминокислотной последовательности аминокислоту, отличную от метионина, хотя эта замена главным образом может дать только техническое преимущество (поскольку она гарантирует отсутствие ингибитора в системе in vitro), и не является существенной для изобретения (поскольку для элиминации протеина-ингибитора из системы in vitro могут использоваться другие методы). На фиг.2 схематически изображен предпочтительный фермент транспозаза, имеющий мутации в положениях 54, 56 и 372 относительно Тn5-транспозазы дикого типа.
Мобильный полинуклеотид в синаптических комплексах характеризуется как нуклеотидная последовательность, фланкированная парой инвертированных полинуклеотидов, включающая нуклеотидную последовательность длиной 18 или 19 пар оснований, которая может функционировать в системе транспозиции Tn5 in vitro. Полинуклеотид (или его любая часть) может быть синтезирован с помощью методов, хорошо известных специалистам в данной области, или может быть получен с использованием методов генетической инженерии путем создания фрагментов нуклеиновых кислот. Как показано на фиг.3, известная нуклеиновая последовательность наружный конец (ОЕ) концевой последовательности транспозона Tn5 дикого типа (5'-CTGACTCTTATACACAAGT-3') может применяться в качестве фланкирующих полинуклеотидов, так же как и внутренний конец (IE) концевая последовательности транспозона Tn5 дикого типа (5'-CTGTCTCTTGATCAGATCT-3'). Хотя может применяться ОЕ-концевая последовательность дикого типа, заявители продемонстрировали, что частота транспозиций является по меньшей мере столь же высокой, а как правило, существенно более высокой по сравнению с той, которая может быть получена при использовании ОЕ дикого типа, если окончания в конструкции представляют собой мозаичные промежуточные последовательности между ОЕ и IE. Предпочтительные мозаичные концевые последовательности включают основания АТА в положениях 10, 11 и 12 соответственно, а также нуклеотиды, общие с нуклеотидами ОЕ и IE дикого типа (например, в положениях 1-3, 5-9, 13, 14, 16 и необязательно 19). Нуклеотиды в положениях 4, 15, 17 и 18 могут соответствовать нуклеотидам, обнаруженным в этих положениях либо в ОЕ дикого типа, либо в IE дикого типа. Следует отметить, что частота транспозиций может быть увеличена по сравнению с частотой при использовании ОЕ дикого типа, если нуклеотид в положении 4 представляет собой Т, в сочетании с наличием указанных выше нуклеотидов. Предпочтительные мозаичные последовательности включают CTGTCTCTTATACACATCT-3' и CTGTCTCTTATACAGATCT-3'. Также могут применяться комбинации неидентичных окончаний.
Мобильный полинуклеотид предпочтительно представляет собой линейный полинуклеотид, содержащий на концах инвертированные полинуклеотидные последовательности, необходимые для транспозиции Тn5. Также можно, но менее предпочтительно, применять мобильный полинуклеотид, который является линейным, но который содержит последовательности, отличные от последовательностей мобильной нуклеиновой кислоты, и инвертированные полинуклеотидные последовательности. Такие дополнительные последовательности могут быть локализованы на любой стороне мобильного полинуклеотида, хотя такое строение является менее предпочтительным, поскольку оно затрудняет получение ДНК транспозона. Определенная транспозиция может иметь место, когда мобильный полинуклеотид находится на кольцевой, суперспиральной молекуле ДНК, хотя частота транспозиций при этом является не столь высокой, как в случае, когда мобильная последовательность представляет собой линейную молекулу.
Мобильная нуклеотидная последовательность между требуемыми концевыми последовательностями может включать любую последовательность, которую необходимо встроить в геном-мишень. На фиг.4 представлен схематический транспозон, имеющий так называемые мозаичные концы и мобильную полинуклеотидную последовательность между ними. Специалист в данной области может легко сконструировать требуемую мобильную нуклеотидную последовательность, соответствующую конкретной цели.
Изобретение не ограничено какими-либо определенными нуклеиновыми кислотами, расположенными между инвертированными полинуклеотидными последовательностями. Наоборот, последовательность может представлять собой любую обнаруживаемую последовательность или последовательность, которая кодирует продукт, который может быть обнаружен с использованием методов, известных в данной области. В качестве примера, который не ограничивает объем изобретения, мобильная нуклеотидная последовательность может обеспечивать клетку-мишень селектируемым маркером, который может представлять собой пептид или протеин, кодируемый мобильной нуклеотидной последовательностью. Последовательность может кодировать протеин, который придает клетке устойчивость к антибиотику. В альтернативном варианте мобильный полинуклеотид может включать последовательность, присутствие которой может быть обнаружено в клетках-мишенях. Такая последовательность может включать последовательность, расщепляемую редкой рестриктазой, или любую последовательность, для которой существует зонд.
Мобильный полинуклеотид также может включать последовательности, которые регулируют экспрессию (транскрипцию или трансляцию) соседней последовательности. Регуляторные последовательности могут облегчать экспрессию протеина или пептида, кодируемого кодирующей последовательностью мобильного полинуклеотида. В другом варианте мобильный полинуклеотид может включать регуляторные последовательности помимо любой кодирующей последовательности или кодирующие последовательности помимо любой регуляторной последовательности. В предыдущем варианте случаи транспозиции против хода транскрипции геномных кодирующих последовательностей могут приводить к появлению регуляторных элементов, необходимых для модуляции транскрипции и/или трансляции эндогенной кодирующей последовательности. В последнем случае перенесенная последовательность может позволить выявить ранее не известные регуляторные последовательности в геноме путем интродукции маркерного гена, продукт которого образуется только тогда, когда ген переносят в положение, соседнее относительно регуляторной последовательности в клеточной нуклеиновой кислоте.
После образования из указанных выше компонентов in vitro синаптический комплекс может быть интродуцирован в клетки-мишени с помощью методов, известных специалистам в данной области. В другом варианте синаптические комплексы могут храниться до применения, например, при -40°С. Предпочтительным методом интродукции синаптического комплекса в клетку является электропорация, например, с помощью метода, описанного у Dauer W.J. и др., Nucleic Acids Research, 16: 6127 (1998), публикация полностью включена в настоящее описание в качестве ссылки. Транспозиция происходит после введения комплекса в нуклеиновую кислоту клетки без дополнительного вмешательства. Если синаптические комплексы присутствуют в объеме, превышающем несколько микролитров, то предпочтительно подвергать синаптические комплексы диализу в противотоке буфера с низким содержанием солей, например, такого как 6 мМ Трис, рН 7,5 с 7-10% глицерина (не ограничиваясь таким буфером), до применения реакции электропорации для уменьшения количества солей в реакционной смеси до столь низкого уровня, при котором они не могут образовывать электрическую дугу между электродами. Другие приемлемые методы интродукции синаптических комплексов в клетки-мишени известны и могут включать трансформацию, трансфекцию и методы с использованием липосом.
Система обеспечивает быстрый и эффективный способ введения мутаций в клетки. Поскольку для этого применяется мобильная полинуклеотидная последовательность, система не является специфичной для определенного хозяина и может работать в любой клетке-мишени. Изобретение, прежде всего, может найти применение для клеток, особенно бактериальных клеток, которые не кодируют транспозазу, поскольку требуемая молекула транспозазы вносится непосредственно как часть синаптического комплекса. Продемонстрировано, что способ может работать в несодержащих ядра системах-мишенях, таких как бактериальные клетки. В частности, синаптические комплексы интродуцировали в клетки Е. coli (штамм MG1655) и наблюдали продуктивную транспозицию. С научной точки зрения не известно никаких причин, препятствующих использованию способа применительно к ядерным клеткам, таким как архебактерии, клетки растений и животных, прежде всего, клетки нематод, амфибий и млекопитающих, включая (но не ограничиваясь ими) клетки грызунов и людей. При использовании способов интродукции синаптических комплексов в ядерные клетки может оказаться предпочтительным встраивать в синаптический комплекс сигнал ядерной локализации, предпочтительно в виде части генетически модифицированного протеина транспозазы.
Изобретение должно стать более понятным при рассмотрении следующих примеров, которые приведены только для иллюстрации и не ограничивают объем изобретения.
Примеры
В реакционной смеси объемом 1 мкл объединяли 0,05 мкг очищенной гиперактивной транспозазы (EK54/MA56/LP372), аминокислотная последовательность которой описана в международной заявке PCT/US97/15941, которая включена в настоящее описание в качестве ссылки, с 0,1 мкг мобильного полинуклеотида, включающего кассету экспрессии, которая кодирует протеин, придающий клетке-мишени устойчивость к канамицину.
Кассету экспрессии фланкировали мозаичными концами, также описанными в указанной международной заявке, включенной в настоящее описание в качестве ссылки, которые показаны на фиг.3. В разных реакциях полинуклеотид использовали в виде суперспиральной плазмиды, линеаризованной плазмиды или в виде полинуклеотидного фрагмента, включающего его концевые инвертированные последовательности, которые необходимы для транспозиции, опосредованной Тn5-транспозазой.
Смесь инкубировали в течение 1 ч в присутствии ионов магния (Mg++) или без них в указанном выше реакционном буфере. В отсутствии ионов магния синаптические комплексы образовывались, но транспозиция in vitro не происходила.
После инкубации реакционную смесь смешивали с 40 мкл штамма Е. coli MG1655 и подвергали электропорации согласно методу, описанному у Dauer. Клетки (1,4×109) высевали на планшеты со средой LB-Kan и подсчитывали количество устойчивых к канамицину колоний. Когда использовали 10 мкл реакционной смеси, то смесь подвергали диализу в противотоке пригодного буфера для предотвращения образования электрической дуги между электродами.

Из таблицы видно, что транспозиция мобильного полинуклеотида, придающего клетке устойчивость к канамицину, выше в варианте, в котором реакционную смесь инкубировали без ионов магния. Наиболее высокую частоту транспозиций наблюдали в варианте, в котором мобильный полинуклеотид не содержал последовательности, фланкирующие мобильную область ("высвобождаемый элемент"). Более чем 10-кратное уменьшение устойчивых к канамицину КОЕ наблюдали при добавлении в реакционную смесь ионов магния, что, вероятно, является результатом непродуктивной внутримолекулярной транспозиции, которая происходит в присутствии ионов магния.
При создании банка для достижении 99%-ной вероятности полного охвата (т.е. встраивания в каждую ОРС) необходимо примерно 20000 случаев транспозиции. При этих расчетах предполагалось, что транспозиция действительно является случайной в отношении выбранной ОPC-мишени и что каждая ОРС имеет одинаковый размер. Хотя Тn5-вставки были обнаружены во всех генах, для которых они предназначались, для Тn5 обнаружены определенные смещения в отношении последовательностей-мишеней, что может отражать случайный характер распределения генов. Также могут существовать неизвестные смещения, влияющие на транспозицию путем конденсации/организации тела нуклеоида. Кроме того, не все ОРС имеют одинаковый размер, их размер в Е.coli, например, находится в диапазоне от 100 кодонов до 2383 кодонов. С другой стороны, результаты описанной выше электропорации/транспозиции синаптического комплекса отражают его воздействия в результате встраивания только на несущественные (не жизненно важные) функции, поскольку при этом образуются только жизнеспособные колониеобразующие единицы. Это четко характеризует полноту транспозиции встроенного пула. Таким образом, для получения полностью репрезентативного банка необходимо учитывать от 50000 до 500000 жизнеспособных случаев транспозиции. Представленное в настоящем описании изобретение обеспечивает достаточную эффективность, позволяющую получить банк указанного размера.
Настоящее изобретение не ограничено приведенным выше описанием, оно включает все вариации и модификации, подпадающие под объем приведенной ниже формулы изобретения.



Изобретение относится к биотехнологии, в частности к способу получения инсерционных мутаций в случайных или псевдослучайных положениях в хромосомной или внехромосомной нуклеиновой кислоты клетки-мишени. Проводят стадию объединения в клетке-мишени клеточной нуклеиновой кислоты с синаптическим комплексом, который включает (а) протеин Tn5-транспозазы и (б) полинуклеотид. Последний включает пару нуклеотидных последовательностей, адаптированных к функциональному взаимодействию с Tn5-транспозазой, и мобильную нуклеотидную последовательность между ними в условиях, которые опосредуют транспозиции в клеточной ДНК. Согласно способу синаптический комплекс получают in vitro в условиях, которые являются неблагоприятными или препятствуют тому, чтобы синаптические комплексы подвергались продуктивной транспозиции. Способ позволяет повысить частоту продуктивной транспозиции. 13 з.п. ф-лы, 4 ил., 1 табл.

| GORYSHIN et al., “Tn5 in Vitro Transposition”, The Journal of Biological Chemistry 273:7367-7374 (1998).US 5925545 A, 20.07.1999.US 5948622 A, 07.09.1999.US 5965443 A, 12.10.1999.MIZUUCHI, KIYOSHI, “In Vitro Transposition of Bactetiophage Mu: A Biochemical Approach to a Novel Replication Reaction,” Cell, 35, P.785-794 (1983).MORITA et al., “Nested Deletions from a Fixed Site as an Aid to Nucleotide Seguencing: An in vitro System Using Tn3 Transposase”, DNA Research, 3, p.431-433 (1996).TOMACSANYI et al., “Intramoleclar Transposition by a Synthetic IS50 (Tn5) Derivative”, J | |||
| Bacteriology | |||
| Приспособление для воспроизведения изображения на светочувствительной фильме при посредстве промежуточного клише в способе фотоэлектрической передачи изображений на расстояние | 1920 |
|
SU172A1 |
| ROSS A | |||
| et al., “The Organization of the Outside End of Transposon Tn5”, Journal of Bacteriology, 178(6), p.1671-1679 (Mar | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |
| и др | |||
| Методы генетической инженерии | |||
| Молекулярное клонирование, М., Мир, 1984, стр | |||
| Счетный сектор | 1919 |
|
SU107A1 |
