Область техники
Группа изобретений относится к биотехнологии и медицине, а именно к получению вирусоподобных частиц вируса гепатита С и их дальнейшему использованию, такому как профилактика вирусного гепатита С и лечение хронического гепатита С путем изготовления новой вакцины.
Предшествующий уровень техники
Вирус гепатита С (HCV) рассматривается как очень опасный патоген человека. В настоящее время это основная причина хронических заболеваний печени, часто приводящих к циррозу и раку печени. 170 млн. (3%) человек во всем мире хронически инфицированы гепатитом С и до 4 млн. новых случаев регистрируется каждый год (Liver Int. 2011 Jan; 31 Suppl 1: 68-77). В 75% инфицирование приводит к развитию хронического гепатита, который в 25% случаев развивается бессимптомно в течение нескольких десятилетий и приводит к циррозу и/или гепатоцеллюлярной карциноме. Передача инфекции HCV происходит в основном через кровь и другие жидкости организма человека. Антивирусная терапия в виде длительного курса сочетания рибавирина и интерферона не обеспечивает устойчивое выздоровление. Также эффективность лечения зависит от генотипа вируса. К сожалению, лечение не всегда хорошо переносится в силу токсичности препаратов, и не все пациенты могут завершить курс лечения (Antivir Ther. 2008; 13 Suppl 1: 3-8).
Эпидемиологическая ситуация в мире в отношении вирусного гепатита С (ВГС) ухудшается с каждым годом. Причина - отсутствие вакцин, с помощью которых можно было бы контролировать инфекцию путем массовой иммунизации населения, как это успешно происходит в отношении вирусного гепатита В.
Потенциальная возможность защиты от инфицирования вирусом гепатита С была показана в остром эксперименте на шимпанзе при иммунизации животных белками Е1 и Е2 перед заражением (Рrос. Nati. Acad. Sci. USA, 1994, Vol.91, pp.1294-1298). Защитная роль core-антигена, как компонента перспективной вакцины, опосредована его высокой межтиповой консервативностью, что проявляется в перекрестной иммуноспецифичности, составляющей 97% по данным распознавания core-антигена широкой панелью сывороток больных хроническим гепатитом С (Virology Journal, 2009, 6:84), а также его активностью в отношении индукции как гуморального, так и клеточного иммунитета (Journal of virology, Sept. 1995, p.5859-5863; Hepatology, Volume 24, Issue 1, 1996, p.14-20).
Биологическая сущность любой вакцины состоит в том, чтобы максимально близко воспроизвести модель природного процесса инфицирования человека, не допустив при этом развития инфекции. В результате формируется защитный иммунитет, но препарат должен быть безвредным и не нести потенциальную опасность в перспективе своего применения, даже если это весьма отдаленные побочные негативные осложнения.
Совершенно очевидно, что чем больше структурное сходство вакцины с природным вирусом, тем более она эффективна. Включение в состав препарата всех компонентов вируса, сохранив их вирус-нейтрализующие эпитопы, за исключением частей, ответственных за воспроизведение вируса (кодирующие последовательности ДНК или РНК) - задача конструирования эффективной безвредной вакцины. Благодаря успехам техники рекомбинантных ДНК и накопленному опыту, это стало реально возможным.
С этой точки зрения очевидно стремление многих исследователей в качестве основы вакцины против вируса гепатита С использовать его структурные антигены.
Гены вируса гепатита С клонировали и экспрессировали в прокариотических и эукариотических клетках. Есть сообщение об экспрессии всех трех структурных белков HCV - core, E1 и Е2 - в культуре клеток насекомых с образованием вирусоподобных частиц (J. Virol. 1998; 72 (5): 3827-3836).
Известна вакцинная композиция против вируса гепатита С, раскрытая в патенте RU 2351363 С2, опубликованном 10.04.2009, где в качестве клетки-хозяина используются клетки метилотрофных дрожжей Pichia pastoris. Система основана на применении промотора гена фермента алькогольоксидазы I (АОХ1). Промотор АОХ1 жестко регулируется и обеспечивает высокие уровни экспрессии фермента (до 30%), когда клетки выращивают в присутствии метанола, но не в присутствии глюкозы (Ellis S.B. etal. Molecular and Cellular Biology, 1985, 5: 1111; Couderc R. et al. Agric Biol Chem. 1980, 44: 2259). Дрожжи Pichia pastoris используются для экспрессии различных чужеродных белков, в том числе ВГС, а именно Е1 и Е2 белков (Antiviral Research (2010), Volume: 88, Issue: 1, p.80-85). Также существует патент на раздельную экспрессию белков E1, Е2 и core в клетках E.coli и Pichia pastoris, с последующим смешиванием очищенных продуктов для использования в качестве вакцины.
Известно использование структурных антигенов вируса гепатита С для создания ДНК-вакцин (Virology Journal, 2011, 8: 506; Biotechnol Appl Biochem, 2004, Apr, 39 (Pt 2), p.249-255).
Однако практическое применение известных технологий для создания вакцины против гепатита С сопряжено с рядом ограничений, связанных с их высокой стоимостью, а также проблемой безопасности применения у человека.
Наиболее близким аналогом заявленного изобретения является заявка WO 2011/042551 А1, 14.04.2011, раскрывающая способ получения вирусоподобных частиц вируса гепатита С в клетках дрожжей Schizosaccharomyses путем ко-экспрессии двух независимых последовательностей core и Е1-Е2, экспрессируемых раздельно с помощью разных плазмид совместно трансформированных в клетки дрожжей. Однако известный способ имеет ряд недостатков, связанных с тем, что продукты генов core и Е1-Е2, экспрессируемые раздельно, а не с общей рамки считывания, не дают возможности получить полноценные вирусоподобные частицы в клетках низших эукариот: полученные авторами частицы не проявляют Е1-Е2 антигенной активности. Кроме того, в связи с тем, что клетки дрожжей Schizosaccharomyses не являются высокопродуктивной системой экспрессии, их использование не может обеспечить достаточную продуктивность для массового производства вакцины.
Раскрытие изобретения
Задачей изобретения является создание экономичной и безопасной вакцины для профилактики и/или лечения заболеваний, вызываемых вирусом гепатита С, обеспечивающей выраженный специфический иммунный ответ на ее введение.
Технический результат, достигаемый при реализации заявленной группы изобретений, заключается в получении высокопродуктивной системы экспрессии Hansenula polymorpha вирусоподобных частиц, пригодных для использования в качестве вакцины для профилактики и/или лечения заболеваний, вызываемых вирусом гепатита С, и обладающих выраженными антигенными и иммуногенными свойствами, за счет содержания в них всех трех структурных антигенов (core, E1 и Е2) вируса гепатита С.
Указанный технический результат достигается за счет того, что способ получения вирусоподобных частиц, содержащих структурные антигены core, E1 и Е2 вируса гепатита С, включает трансформацию клеток дрожжей Hansenula polymorpha рекомбинантной плазмидой, содержащей единую открытую рамку считывания, несущую гены соге - Е1-Е2 вируса гепатита С, при прямой экспрессии которой образуются антигены core, E1 и Е2, последующее культивирование и выделение полученных частиц.
Указанный технический результат достигается также за счет того, что способ очистки полученных вирусоподобных частиц включает осветление гомогената разрушенных клеток, диафильтрацию, концентрирование, преципитацию при пониженном рН, ионообменную хроматографию, седиментационное центрифугирование в градиенте плотности, гидрофобную хроматографию и гель-хроматографию.
Указанный технический результат достигается также за счет того, что вирусоподобные частицы, содержащие антигены структурных белков core, E1 и Е2 вируса гепатита С, получены вышеуказанным способом.
Указанный технический результат достигается также за счет того, что вакцина для профилактики и/или лечения гепатита С, содержащая вирусоподобные частицы, получена вышеописанным способом. При этом вакцина может дополнительно содержать фармацевтически приемлемый адъювант.
В уровне техники отсутствуют сведения, указывающие на возможность получения вакцины с помощью синтеза всех структурных белков-антигенов вируса гепатита С в клетках дрожжей Hansenula polymorpha в виде вирусоподобных частиц путем прямой экспрессии генов всех структурных белков - E1, Е2 и core.
Настоящей группой изобретений показано, что совместный синтез белков-антигенов E1, Е2 и core в клонированных клетках дрожжей Hansenula polymorpha приводит к накоплению вирусоподобных частиц (ВПЧ), проявляющих антигенные и иммуногенные свойства всех исходных компонентов.
Использование предлагаемого способа позволяет получить рекомбинантную вакцину, содержащую синтезированные ВПЧ, полученные по технологии, в основе которой лежит высокоэффективный синтез антигенных продуктов в системе экспрессии Hansenula polymorpha. При этом высокоэффективная технология производства с ее низкой себестоимостью выводит способ в разряд перспективных разработок в области создания новой вакцины для борьбы с гепатитом С, вплоть до искоренения этой инфекции.
Полученные ВПЧ могут быть использованы как для профилактического, так и для терапевтического применения.
Для этого полученные ВПЧ используют в качестве активного начала для получения вакцин (профилактических и терапевтических вакцинных композиций). В дополнение к активному началу вакцинные композиции могут содержать фармацевтически приемлемый носитель и/или фармацевтически приемлемый адъювант. Типичные носители обычно представляют собой крупные медленно метаболизируемые макромолекулы, такие как белки, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, сополимеры аминокислот и неактивные вирусные частицы. Такие носители хорошо известны специалистам. Примеры фармацевтически приемлемых адъювантов для усиления эффективности композиции включают в себя следующие: гидроокись алюминия, алюминий в комбинации с 3-О-деацилированным монофосфориллипидом А, фосфат алюминия, N-ацетил-мурамил-L-треонил-D-изоглутамин, N-ацетил-нормурамил-L-аланил-D-изоглутамин, N-ацетилмурамил-L-аланил-D-изоглутамил-L-аланин-2-(1',2'-дипальмитоил-sn-глицеро-3-гидроксифосфорил-окси)-этиламин. Кроме того, можно применять такие адъюванты, как Stimulon (Cambridge Bioscience, Worcester, MA, USA), SAF-1 (Syntex) или бактериальные адъюванты на основе ДНК, такие как ISS (Dynavax) или CpG (Coley Pharmaceuticals), или адъюванты на основе блок-сополимеров, такие как Optivax (Vaxcel, Cythx), или адъюванты на основе инулина, такие как Algammulin и Gammalnulin (Anutech), неполный адъювант Фрейнда (IFА) или препараты Gerbu (Gerbu Biotechnik), но не ограничиваясь ими. Вакцинная композиция может также дополнительно содержать эксципиенты и разбавители, которые по своей сути являются нетоксичными и не терапевтическими, такие как вода, физиологический раствор, глицерин, этанол, увлажняющие и эмульгирующие агенты, вещества, забуферивающие рН, консерванты и тому подобное. В типичном случае вакцинную композицию готовят как инъецируемую в виде водного раствора или суспензии. Инъекция может быть подкожной, внутримышечной, внутривенной, внутрибрюшинной, внутрикожной. Другие типы введения включают в себя имплантацию, суппозитории, пероральный прием внутрь, введение в тонкий кишечник, ингаляцию, аэрозоли или назальные спреи или капли. Также могут быть приготовлены твердые формы, пригодные для растворения или суспендирования в жидких носителях перед инъекцией. Эти препараты можно также эмульгировать или инкапсулировать в липосомы для усиления адъювантного эффекта. Вакцинные композиции содержат эффективное количество активного вещества, а также любой из вышеупомянутых компонентов. Под эффективным количеством активного вещества понимается то, что введение этого количества индивиду эффективно для предотвращения или лечения заболевания или для индукции желаемого эффекта. Это количество варьирует в зависимости от здоровья и физического состояния индивида, подлежащего лечению, способности иммунной системы индивида давать эффективный иммунный ответ желаемой степени защиты, препарата вакцины, оценки лечащим врачом, штамма инфекционного патогена и других факторов. Предполагается, что это количество должно попасть в относительно широкий диапазон, который можно определить обычными испытаниями. Обычно это количество варьирует от 0,01 до 1000 мкг/дозу, более конкретно от 0,1 до 100 мкг/дозу. Режим дозирования может представлять собой схему разовых доз или схему множественных доз. Вакцину можно вводить в сочетании с другими терапевтическими агентами.
Еще в одном аспекте изобретение касается способа индуцирования HCV-специфичного иммунного ответа у млекопитающего, при котором указанному млекопитающему вводят эффективное количество ВПЧ, возможно в сочетании с фармацевтически приемлемым адъювантом и/или фармацевтически приемлемым носителем (в виде вакцинной композиции). Указанный способ, при котором указанному млекопитающему вводят эффективное количество ВПЧ, можно также применять для индукции HCV-специфичных антител у млекопитающего или для индукции специфичной Т-клеточной функции у млекопитающего. В указанных способах указанное введение может осуществляться для профилактических целей, то есть являться профилактическим введением, или для терапевтических целей, то есть являться терапевтическим введением.
Еще один аспект настоящего изобретения относится к способу иммунизации млекопитающего, при котором указанному млекопитающему вводят эффективное количество ВПЧ, возможно в сочетании с фармацевтически приемлемым адъювантом и/или фармацевтически приемлемым носителем.
Настоящее изобретение также относится к способу лечения млекопитающего, инфицированного HCV, при котором указанному млекопитающему вводят эффективное количество ВПЧ, возможно в сочетании с фармацевтически приемлемым адъювантом и/или фармацевтически приемлемым носителем.
Путь введения, доза и количество введений, как для профилактических, так и для терапевтических целей, могут быть оптимизированы специалистом в данной области известным образом.
К состояниям, для профилактики и/или лечения которых могут быть использованы полученные ВПЧ, возможно в сочетании с фармацевтически приемлемым адъювантом и/или фармацевтически приемлемым носителем, относятся HCV-инфекции (такие как гепатит С и хронический гепатит С) и ассоциированные с ними заболевания, в частности хронические заболевания печени, такие как цирроз, и опухоли печени, такие как гепатоцеллюлярная карцинома.
В наиболее предпочтительном варианте указанным млекопитающим является человек.
Краткое описание чертежей
Сущность заявленной группы изобретений поясняется чертежами, где:
на Фиг.1 изображена структура экспрессирующей плазмиды, в которой использованы следующие общепринятые биологические сокращения:
МОХ - метанол оксидаза (промотор),
LEU2 - дрожжевой ген, участвующий в синтезе лейцина,
ColE1 - участок инициации репликации плазмиды в бактериях,
Ampr - бактериальный ген устойчивости к ампицилину,
HARS - область автономной системы репликации Hansenula polymorpha;
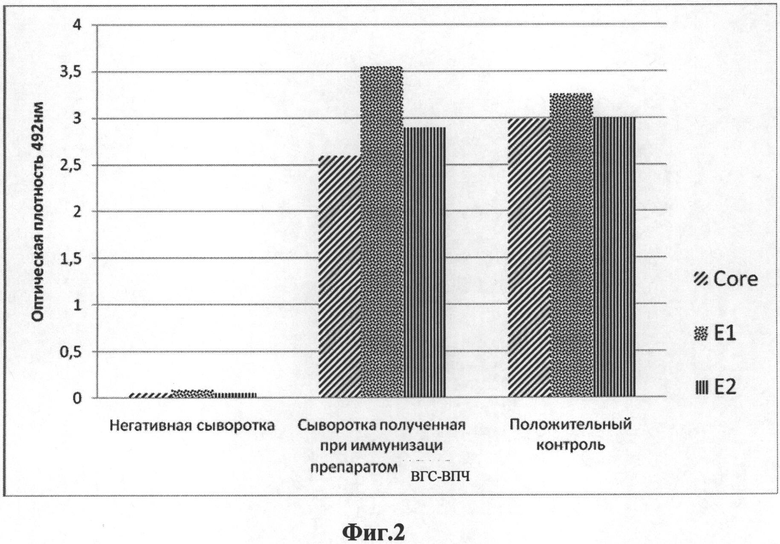
на Фиг.2 приведены диаграммы данных результатов иммуноферментного анализа, демонстрирующие детекцию специфических антител в сыворотке мышей, иммунизированных вакцинными препаратом ВГС-ВПЧ;
на Фиг.3 приведен электрофорез в полиакриламидном геле препаратов ВПЧ в денатурированных, восстановленных условиях:
A) Полоса, соответствующая Е2≥76 кДа
B) Полоса, соответствующая E1≈40 кДа
C) Полоса, соответствующая соrе≈28 кДа
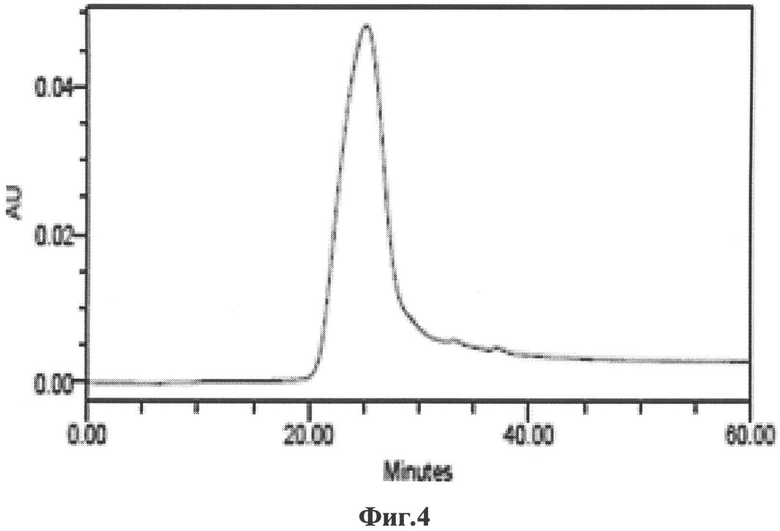
на Фиг.4 изображены результаты жидкостной хроматографии высокого давления очищенных образцов ВПЧ. По оси абсцисс отложено время прохода фракции через детектор оптической плотности. По оси ординат отложена оптическая плотность фракции;
на Фиг.5 изображена электронная микроскопия очищенных образцов ВПЧ.
Осуществление изобретения
Вирусоподобные частицы, содержащие структурные антигены core, E1 и Е2 вируса гепатита С, получают путем трансформации клеток дрожжей Hansenula polymorpha рекомбинантной плазмидой, содержащей единую открытую рамку считывания, несущую гены соrе - Е1-Е2 вируса гепатита С, при прямой экспрессии которой образуются антигены core, E1 и Е2; а также последующего культивирования клеток и выделения полученных частиц.
Особенности процесса ферментации и состава примененной питательной среды позволяют эффективно создавать оптимальные физиологические условия, необходимые для роста микроорганизмов и для предпочтительной экспрессии клонируемого гена, а также добиваться высокой степени агрегирования получаемого продукта в частицы.
Выделение и очистка целевых продуктов разработана целенаправленно для получения вакцинного препарата, соответствующего требованиям, предъявляемым к рекомбинантным вакцинам этого класса.
Очистка полученных ВПЧ, включает осветление гомогената разрушенных клеток, диафильтрацию, концентрирование, преципитацию при пониженном рН, ионообменную хроматографию, выполненную на DEAE Sepharose, седиментационное центрифугирование в градиенте плотности сахарозы, гидрофобную хроматографию и гель-хроматографию.
С помощью предлагаемого способа, включающего соблюдение специфических условий ферментации и очистки, рекомбинантные антигены core, E1 и Е2 получают при культивировании клеток дрожжей Hansenula polymorpha (Pichia angusta), при этом они отличаются высокой степенью агрегирования в частицы и большой однородностью, что придает им более высокие иммуногенные свойства.
Для получения вакцины (вакцинной композиции) для профилактики и/или лечения гепатита С, полученные ВПЧ могут быть сорбированы на адъюванте и/или смешаны с фармацевтически приемлемым носителем.
Более подробно в некоторых вариантах осуществления изобретение раскрыто в приведенных ниже примерах.
Пример 1. Получение стабильного клона штамма-продуцента
Конструирование плазмиды для экспрессии.
Структурные гены вируса гепатита С (core, E1 и Е2) были клонированы в экспрессионный вектор pSL. Вектор представляет собой плазмиду, содержащую промотор метанол оксидазы (МОХ) и автономно реплицирующуюся последовательность (ARS) Hansenula polymorpha, дрожжевой ген LEU2 (маркер селекции по лейцину), последовательность бактериального инициатора репликации и ген устойчивости к ампициллину.
Для этого:
Плазмиду, содержащую полный геном HCV генотипа 1а (любезно предоставлена NL Biotechnologies LLC, USA), использовали для амплификации последовательности структурных генов вируса. Лидирующий полинуклеотидный праймер содержал последовательно сайт рестрикционной эндонукреазы BamHI, кодон инициации трансляции и последовательность нуклеотидов 5' конца core-белка вируса гепатита С. Обратный праймер содержал последовательно сайт рестрикционной эндонуклеазы EcoRI, комплиментарную стоп-кодону последовательность и последовательность нуклеотидов 3' конца белка Е2 вируса гепатита С. Ставилась полимеразная цепная реакция для амплификации и продукты амплифицикации подвергались электрофорезу в агарозном геле. Гели окрашивали 0,3 мкг/мл этидиум бромида и анализировали в ультрафиолетовом свете. Продукты амплификации очищали QIAquick PCR Purification Kit (QIAGEN Inc., Valencia, CA), подвергали рестрикции ферментами BamHI и EcoRI, а затем «сшивались» с линейным фрагментом вектора pSL BamHI-EcoRI.
Для получения рекомбинантного штамма дрожжей - продуцента HCV-LP проводили генетическую трансформацию реципиентного штамма Hansenula polymorpha DL-1 (GellissenG (ed) (2002) Hansenula polymorpha - biology and applications), несущего мутацию LEU, плазмидой: pSL-HCVLP-303 (фиг.1). Трансформацию клеток проводили в соответствии с методикой, описанной в Dohman, An efficient transformation procedure enabling long-term storage of competent cells of various yeast genera. Yeast. 1991 Oct; 7 (7): 691-692.
Для отбора клона, в котором произошла успешная интеграция плазмиды в геном, осуществляли генетическую стабилизацию культуры трансформированных клеток. Для этого проводили четыре пассажа колоний-трансформантов на минимальной агаризованой среде YNBD (Yeast Nitrogen Base (Difco, 239210) 0,7%, Д-Глюкоза 1,5%, агар-агар 1,5%). Колонии высевали методом штриха и инкубировали в течение 40-60 часов при температуре 25-30°С. Правильно сформировавшиеся изолированные колонии отбирали на следующий пассаж. После проведения четырех пассажей, колонии пересевали на жидкую богатую среду Y1P1D2 (дрожжевой экстракт 1%, бакто-пептон 1%, Д-глюкоза 2%) и инкубировали при температуре 25-30°С в течение 40-60 часов при принудительной аэрации (в орбитальном шейкере со скоростью вращения 150-220 об/мин) в колбах Эрленмейера, наполненных средой на 1/12. По окончании инкубации приготавливали серию разведений взвеси клеток дрожжей, высевали на чашки с агаризованой средой Y1P1D2 (с добавлением 1,5% агар-агара) и инкубировали в термостате в течение 60 часов при температуре 20-30°С. Отбирали 50 единичных колоний на агаризованую среду YNBD и 50 единичных колоний на агаризованую среду YNBD с добавлением лейцина (40 мг/л). Чашки инкубировали в термостате в течение 40-60 часов при температуре 30°С, после чего производили подсчет колоний и определяли эффективность интеграции фрагмента в хромосому по формуле
Ki=I_YNBD/I_(YNBD+L)×100Ki=I_YNBD/I_(YNBD+L)×100, где
Ki - коэффициент интеграции
IYNBD - Количество колоний, выросших на агаризованой среде YNBD
IYNBD+L - Количество колоний, выросших на агаризованой среде YNBD с добавлением лейцина.
В случае если коэффициент интеграции составлял менее 98%, процедуру генетической стабилизации повторяли еще раз.
В случае если коэффициент интеграции составлял 98% и более, производили закладку культуры на долгосрочное хранение.
Для долгосрочного хранения клеток одну из колоний, выросших на чашке с агаризованой средой YNBD, высевали в жидкую среду YNBD и выращивали при принудительной аэрации в колбах, наполненных средой на 1/12, в течение 12-24 часов при температуре 25-30°С. К полученной взвеси клеток добавляли глицерин до 20% (объем/объем), после чего аликвоты (0.5 мл) замораживали при температуре -80°С (банк культуры).
Пример 2. Получение биомассы клеток штамма-продуцента
Первый пассаж культуры проводили, высевая в жидкую среду YNBD 2,5% (объем/объем) размороженной суспензии клеток из банка культуры, с последующей инкубацией в течение 36 часов при температуре 20-30°С и принудительной аэрации.
Полученную суспензию клеток использовали в качестве посевного материала для второго пассажа, высевая в свежую среду YP2D (1% дрожжевого экстракта, 2% пептона, 1% Д-глюкоза) 20% суспензии (объем/объем) с последующей инкубацией в течение 12-36 часов при температуре 20-30°С и принудительной аэрации.
Для проведения третьего пассажа в свежую среду Y4P2G1 (4% дрожжевого экстракта, 2% пептона, 1% глицерина) высевали 12,5% (объем/объем) полученной на втором пассаже культуры и инкубировали при принудительной аэрации в течение 12-24 часов при температуре 20-30°С.
400 мл полученной культуры инокулировали в ферментер BioFlo 110 (New Brunswick Scientific), содержащий 3600 мл среды Y4P2M0,5 (4% дрожжевого экстракта, 2% бакто-пептона, 0,5% метанола) и проводили ферментацию при температуре 20-30°С, добавляя по 20 г метанола каждый раз, когда концентрация растворенного кислорода в среде поднималась от базового уровня (5-15%) и достигала не менее 40% от максимума (100%), что свидетельствовало о недостатке углеродного питания. Рост культуры контролировали, отбирая пробы клеточной суспензии и измеряя оптическую плотность при длине волны 600 нм и влажный вес биомассы (в г/л культуры). Влажный вес определяли, взвешивая осадок клеток, остающийся после центрифугирования взвеси клеток в течение 10 мин на микроцентрифуге Beckman при 10000 g. Окончание ферментации определяли по отсутствию роста культуры в течение 12-24 часов.
Биомассу собирали при помощи центрифугирования на центрифуге Avanti-J25 (Beckman) на роторе JLA 9. 1000 (7500 об/мин, 15 минут). Биомассу ресуспендировали в дистиллированной воде и центрифугировали в тех же условиях. Отмытую биомассу расфасовывали в контейнеры и хранили при температуре -80°С.
В результате получали с каждого литра среды 100-300 г биомассы клеток штаммов-продуцентов.
Пример 3. Выделение и очистка рекомбинантных антигенов в виде вирусоподобных частиц (ВПЧ) из клеток штаммов-продуцентов
Биомассу клеток штамма-продуцента ВГС-ВПЧ, полученную в результате ферментации, разрушали экструзионным способом. Для этого биомассу размораживали в течение 12 часов при температуре 4°С, после чего ресуспендировали в буфере DBN (3М мочевина, 0,05 MNa2HPO4, 0,01 М ЭДТА, 0,5% Tween-20, рН 8.0) из расчета 400 мл буфера на каждые 100 г биомассы (1/4 масса/объем). Суспензию охлаждали до 6°С и разрушали с помощью гомогенизатора Gaulin APV 2000, контролируя температуру и не допуская ее повышения выше 35°С. Контроль эффективности разрушения осуществляли путем центрифугирования проб суспензии по 1 мл при 9000 g в течение 15 минут и измерения оптической плотности супернатанта при длине волны 280 нм (OD280) в стандартных кюветах 1 см. Разрушение проводили до достижения OD280 образца, разведенного в 100 раз, значения не менее 0,68 единиц.
Гомогенизат осветляли путем центрифугирования при 9000 g в течение 15 минут. Супернатант фильтровали через мембрану с размером пор 0,45 мкм (Millipore, диаметр фильтра 44,5 мм).
Отфильтрованный гомогенизат концентрировали при помощи системы SartoriusSlice 200 с установленной мембраной Millipore 300 кДа до 1/3 начального объема, после чего диализовали против 10 объемов фосфатного буфера (10 mMNa2HPO4, рН 9.0), доводя конечную OD280 диализата до значения не менее 0,7 единиц при разведении в 100 раз.
Стандартизованный таким образом диализат очищали от посторонних белков путем преципитации с понижением рН. Для этого к раствору белка добавляли соляную кислоту, периодически отбирая пробы по 1 мл, центрифугируя их при 22000 rpm на центрифуге Microfuge 8 (Beckman) в течение 10 минут, с последующим измерением ОD280 супернатанта, разведенного в 100 раз. Процесс продолжали до достижения OD280 0,20-0,25 единиц. По достижении требуемых параметров, основной материал центрифугировали на ультрацентрифуге OptimaL-90K (Beckman) с ротором TYPETI-45, 22000 об/мин в течение 20-40 минут. Супернатант отбирали и доводили рН при помощи гидроксида натрия до значения 9,2±0,2.
Полученный раствор белка фильтровали через мембрану Millipore с диаметром пор 0,45 нм, после чего наносили на хроматографическую колонку ХК-16 (General Electric), заполненную сорбентом DEAE fast-flow (General Electric), предварительно сбалансированную фосфатным буфером. Прохождение белка через колонку контролировали путем измерения OD280 на выходе из колонки. Полученный раствор белка подвергали дальнейшей очистке.
Для проведения седиментационного центрифугирования в сахарозном градиенте плотности, раствор белка наслаивали на 4 ступени плотности - 30%, 40%, 50% и 60%. Поликарбонатные пробирки Beckman 38Х-102 мм заполняли ступенями по 16 мл 60%, 50%, 40%, 30% сахарозного градиента и сверху наслаивали 16 мл раствора белка, что в сумме составляло 80% максимального объема пробирки. Заполненные пробирки центрифугировали на ультрацентрифуге Beckman OptimaL-90K с ротором TYPETi-55 при 30000-42000 об/мин в течение 3 часов. Затем из пробирок отбирали фракции по 4 мл и определяли наличие антигена в них при помощи ИФА. Фракции высокой концентрации антигена объединяли и подвергали полученный раствор дальнейшей очистке. Полученный раствор белка фильтровали через мембрану Millipore с диаметром пор 0,45 нм, после чего наносили на хроматографическую колонку ХК-50 (GeneralElectric), заполненную сорбентом SephadexG-25 (TOSOH), предварительно сбалансированную фосфатным буфером. Прохождение белка через колонку контролировали путем измерения OD280 на выходе из колонки. Полученный раствор белка подвергали дальнейшей очистке.
Для проведения очистки методом гидрофобной хроматографии к раствору белка добавляли сухой сульфат аммония до конечной концентрации 0,5М, после чего раствор инкубировали 30 минут на ледяной бане, а затем пропускали через фильтр с диаметром пор 0,45 мкм (Millipore). Профильтрованный раствор белка наносили на колонку ХК-26 (General Electric), наполненную носителем Phenyl 650М (General Electric), уравновешенным фосфатным буфером с добавлением 0,5 М сульфата аммония. После нанесения белка, колонку промывали фосфатным буфером с добавлением 0,5 М сульфата аммония, контролируя OD280 на выходе колонки. После того как OD280 снижалась до уровня фона, проводили десорбцию материала при помощи фильтрованной воды, собирая элюат до достижения фоновых значений OD280.
Частично очищенные препараты антигенов, полученные после гидрофобной хроматографии, подвергались окончательной очистке методом гель-хроматографии. Для этого раствор белка наносили на колонку ХК-26 (General Electric), наполненную на высоту 30 см носителем HW-65 (General Electric) и уравновешенную фосфатным буфером 10 мМ. Отбор фракций (объемом по 5 мл) на выходе колонки начинали при повышении OD280 и завершали при достижении OD280 фоновых значений. Каждую фракцию оценивали на наличие антигена (методом ИФА) и на чистоту (методом денатурирующего электрофореза в полиакриламидном геле и Высокоэффективной Жидкостной Хроматографии (ВЭЖХ)). Фракции, имеющие наибольшую чистоту и активность антигена, объединяли для дальнейшей концентрации и хранения.
Белок концентрировали на системе Amicon 8400 (Millipore) до конечной концентрации белка 1-4 мг/мл, фильтровали через стерильную мембрану с размером пор 0,22 мкм (AcrodiscTuffryn, PallLifeSciences), добавляли стерильный глицерин до конечной концентрации 4% и разливали на аликвоты, которые хранили при -80°С.
Пример 4. Определение иммуногенности белков на мышах.
Для проведения исследований на животных, антиген в стерильных условиях сорбировали на гидроокиси алюминия. Для этого 100 мкл 3% суспензии Аl(ОН)3 суспендировали в 2 мл вакцинного буфера (0,9% NaCl, 6,1 mMNa2HPO4, 3,9 mMNaH2PO4) и добавляли к суспензии очищенный белок до конечной концентрации 20 мкг/мл. Пробирки инкубировали на ротаторе со скоростью 5-10 об/мин при комнатной температуре в течение 12 часов, после чего использовали для приготовления вакцины в рабочих дозах 10 мкг белка на животное. В качестве разбавителя использовали вакцинный буфер с добавлением 16,5 мл 3% суспензии гидроксида алюминия на литр.
Вакцинную композицию использовали для иммунизации 6-и недельных мышей породы BALB-C путем внутрибрюшинной инъекции 0,5 мл вакцины. Через 15 дней вводили таким же способом вторую такую же дозу вакцины. Еще через 15 дней производили забой животных и забор плазмы крови. Антитела в сыворотках крови иммунизированных мышей определяли с помощью ИФА.
Для этого на твердую фазу (планшет ThermoScientific #475094) сорбировали по 1 мкг рекомбинантных антигенов на лунку: сердцевинный белок (core) производства AustralBiologicals #НСА-070-4 для детекции анти-core антител, либо белок E1 (US Biologicals #H1920-17С) для детекции антител к Е1, либо Е2 (US Biologicals #H1920-19Q) для детекции антител к Е2. Сыворотки мышей добавляли в разведении 1:500 и инкубировали 2 часа при температуре 37°С. Позитивными контролями были моноклональные антитела Pierce #МА1-080 (1:5000), Austral Biologicals #HCM-081-S (1:1000), US Biologicals #H1920-19E (1:2000) соответственно. Мышиные антитела выявляли анти-видовым конъюгатом KPL #074-1806 в разведении 1:5000 с инкубацией 1,5 часа при 37°С с последующим окрашиванием ортофенилендиамином с перекисью водорода. На диаграмме представлены данные определения специфических антител к core, E1, Е2 у мышей, иммунизированных вакцинным препаратом ВГС-ВПЧ (Фиг.2).
Пример 5. Определение чистоты антигена в окончательной фракции продукта.
Анализ полученных продуктов осуществляли методами, установленными Всемирной Организацией Здравоохранения (ВОЗ) для минимального доступного уровня чистоты для рекомбинантных вакцин
(www.who.int/biologicals/HEP_B_Recomm_after_ECBS_endorsment_final.pdf).
Образцы очищенных ВПЧ исследовали:
ДСН-ПААГ-электрофорез в восстанавливающих условиях с окрашиванием нитратом серебра, который показал наличие полос, соответствующих мономерам E1, Е2 и core (Фиг.3). Расположение полос для каждого мономера подтверждено методом иммуноблотинга с соответствующими контролями и коммерческими моноклональными антигенами;
Жидкостная хроматография высокого давления (ВДЖХ) (Фиг.4), результаты которой свидетельствуют о надмолекулярной структуре продукта и высокой степени чистоты в нативных неденатурирующих условиях.
Полученные данные свидетельствуют о чистоте ≥95% полученных продуктов.
Электронная микроскопия показала четкие формирования рекомбинантных вирусоподобных частиц. (Фиг.5)
Представленные примеры демонстрируют, что совместный синтез белков-антигенов E1, Е2 и core в клонированных клетках дрожжей Hansenula polymorpha приводит к накоплению вирусоподобных частиц, проявляющих антигенные и иммуногенные свойства всех исходных компонентов.
Приведенные примеры также убедительно демонстрируют возможность индуцирования специфических антител у белых мышей при введении им полученных вирусоподобных частиц, что подтверждает иммуногенность полученных вирусоподобных частиц и возможность их использования в качестве компонентов вакцины для человека. Таким образом, заявленная полиэпитопная (несет эпитопы всех структурных антигенов вируса) вакцина и способ ее получения, а также применения для профилактики гепатита С являются новыми на основании существующего уровня техники.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИЭПИТОПНАЯ ВАКЦИНА 4-ГО ПОКОЛЕНИЯ ПРОТИВ ГЕПАТИТА В И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2011 |
|
RU2469741C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА Е У ЖИВОТНЫХ | 2012 |
|
RU2501568C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ПОВЕРХНОСТНОГО АНТИГЕНА ВИРУСА ГЕПАТИТА В СЕРОТИПА "ayw" | 2015 |
|
RU2586511C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ МУТАНТНОГО ПОВЕРХНОСТНОГО АНТИГЕНА ВИРУСА ГЕПАТИТА В (ВАРИАНТЫ) | 2015 |
|
RU2586513C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО КАПСИДНОГО БЕЛКА ВИРУСА ГЕПАТИТА Е И РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА Е | 2012 |
|
RU2501809C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ HANSENULA POLYMORPHA - ПРОДУЦЕНТ КАПСИДНОГО БЕЛКА ВИРУСА ГЕПАТИТА Е | 2012 |
|
RU2493249C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА В (ВАРИАНТЫ) | 2015 |
|
RU2603729C2 |
| Способ получения рекомбинантной вакцины для профилактики папилломавирусной инфекции человека, рекомбинантная вакцина | 2018 |
|
RU2681174C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ПАПИЛЛОМАВИРУСНОЙ ИНФЕКЦИИ ЧЕЛОВЕКА И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2014 |
|
RU2546243C1 |
| ВЫДЕЛЕННЫЙ ОБОЛОЧЕЧНЫЙ БЕЛОК HCV, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЛЕКАРСТВО, ВАКЦИНА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ (ВАРИАНТЫ) ЕГО СОДЕРЖАЩИЕ | 2002 |
|
RU2274643C2 |


Изобретение относится к биотехнологии и представляет собой способ получения вирусоподобных частиц, содержащих структурные антигены core, E1 и Е2 вируса гепатита С и способ очистки вирусоподобных частиц. Способ получения вирусоподобных частиц включает трансформацию клеток дрожжей Hansenula polymorpha рекомбинантной плазмидой, содержащей единую открытую рамку считывания, несущую гены соrе-Е1-Е2 вируса гепатита С, при прямой экспрессии которой образуются антигены core, E1 и Е2. Осуществляют последующее культивирование и выделение полученных частиц. Способ очистки вирусоподобных частиц включает осветление гомогената разрушенных клеток, диафильтрацию, концентрирование, преципитацию при пониженном рН, ионообменную хроматографию, седиментационное центрифугирование в градиенте плотности, гидрофобную хроматографию и гель-хроматографию. Предложенное изобретение позволяет получать вирусоподобные частицы, пригодные для использования в качестве вакцины для профилактики и/или лечения заболеваний, вызываемых вирусом гепатита С, с высоким выходом. 2 н.п. ф-лы, 5 ил., 5 пр.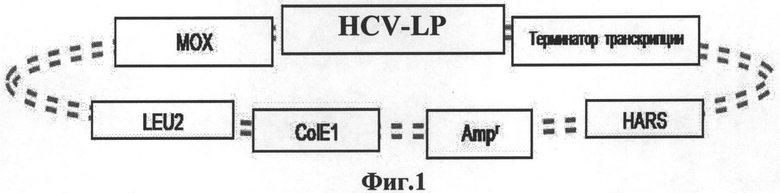
1. Способ получения вирусоподобных частиц, содержащих структурные антигены core, E1 и Е2 вируса гепатита С, включающий трансформацию клеток дрожжей Hansenula polymorpha рекомбинантной плазмидой, содержащей единую открытую рамку считывания, несущую гены соrе-Е1-Е2 вируса гепатита С, при прямой экспрессии которой образуются антигены core, E1 и Е2, последующее культивирование и выделение полученных частиц.
2. Способ очистки вирусоподобных частиц, полученных способом по п.1, включающий осветление гомогената разрушенных клеток, диафильтрацию, концентрирование, преципитацию при пониженном рН, ионообменную хроматографию, седиментационное центрифугирование в градиенте плотности, гидрофобную хроматографию и гель-хроматографию.
| ВАКЦИННАЯ КОМПОЗИЦИЯ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2005 |
|
RU2351363C2 |
| WO 2011042551 A1, 14.04.2011 | |||
| ОЛИГОМЕРНАЯ ЧАСТИЦА, ИНДУЦИРУЮЩАЯ ИММУНИТЕТ ПРОТИВ ВИРУСА ГЕПАТИТА С, СПОСОБ ПОЛУЧЕНИЯ ОЛИГОМЕРНОЙ ЧАСТИЦЫ, КОМПОЗИЦИЯ, СПЕЦИФИЧЕСКОЕ АНТИТЕЛО, НАБОР (ВАРИАНТЫ), ИММУНОЛОГИЧЕСКИЙ АНАЛИЗ И ВАКЦИНА ПРОТИВ ВИРУСА ГЕПАТИТА С | 1999 |
|
RU2247729C2 |
| EP 0001481983 A1, 01.12.2004 | |||
| BAUMERT T.F | |||
| ET AL., Hepatitis C virus structural proteins assemble into viruslike particles in insect cells, J Virol., 1998, v.72, no.5, p | |||
| Держатель для пачек трамвайных и т.п. билетов | 1925 |
|
SU3827A1 |

