Область техники
Настоящее изобретение относится к простому, эффективному и быстрому способу ферментативной модификации и синтеза нуклеиновой кислоты in vitro с использованием непрерывного и краткого микроволнового излучения с частотой от 2300 до 2500 мГц, выходной мощностью от 600 до 900 ватт и временем облучения от 5 до 120 секунд; а также к устройству, необходимому для проведения данного способа.
Предпосылки создания изобретения
Ферментативные реакции нуклеиновых кислот являются основой молекулярной биологии. Наиболее распространенными ферментативными реакциями в молекулярной биологии являются сайт-специфичное расщепление, лигирование, дефосфорилирование, фосфорилирование (кинирование) и реакции синтеза ДНК и РНК in vitro. Данные реакции проводят с использованием разных ферментов, таких как рестрикционная эндокуклеаза (рестриктаза), лигаза, фосфатаза, киназа, ДНК-полимераза, обратная транскриптаза и РНК-полимераза. Продукты упомянутых реакций широко применяются в молекулярной биологии.
Открытие нескольких рестрикционных эндокуклеаз (REN) дало начало современной биотехнологии. Эти ферменты катализируют сайт-специфическое гидролитическое расщепление двухцепочечной ДНК, индуцируя одиночные разрывы (ники) в противолежащих цепях.
Лигаза представляет собой другой важный фермент, который может модифицировать нуклеиновые кислоты путем образования фосфодиэфирной связи между двумя совместимыми концами двухцепочечной ДНК и цепей, образовавшихся в результате одноцепочечных разрывов гибридов ДНК-ДНК или ДНК-РНК [Engler, M.J. and Richardson, C.C. (1982) in The Enzymes (Boyer, P.D, ed.) Vol.5, p.3, Academic Press, San Diego]. Продукты реакций, катализируемых ДНК-лигазой, используются для проведения клонирования и манипулирования с генами.
Фосфатаза катализирует удаление концевых фосфатных групп нуклеиновых кислот [Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Molecular cloning, A Laboratory Manual, second edition. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York], она нужна для снижения фона вектора при клонировании и для введения концевой метки [γ32p] АТФ.
Полинуклеотидкиназа модифицирует нуклеиновые кислоты путем переноса или обмена фосфатной группы, находящейся в гамма-положении АТФ, в 5'-гидроксильный конец полинуклеотидов (двух- и одноцепочечных ДНК и РНК) и нуклеозид 3'-монофосфатов [Sambrook, J., Fritsch, E. F. and Maniatis, (1989) Molecular cloning, A Laboratory Manual, second edition, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York].
Синтез ДНК и РНК на матрице, катализируемый соответствующей полимеразой, обычно используют при проведении исследований, связанных с рекомбинантными ДНК. Синтез ДНК на матрице ДНК используется для получения радиоактивномеченных зондов для гибридизации посредством ник-трансляции [Rigby et al. (1977), J. Biol. Chem. 113:237], а также для реакций дополнения концов при лигировании и введении меток во фрагменты ДНК, синтеза второй цепи кДНК, полимеразной цепной реакции (ПЦР) и секвенирования ДНК.
Синтез ДНК на матрице РНК, катализируемый обратной транскриптазой, используется для синтеза кДНК и амплификации генов методом ПЦР-ОТ. Синтез РНК на матрице ДНК (транскрипция) используется для идентификации промотора, специфичного к РНК-полимеразе, для сопряженных реакций транскрипции и трансляции, получения зондов РНК с высоким содержанием метки и др.
Обычно вышеуказанные реакции проводят на водяной бане, в которой поддерживается оптимальная температура, необходимая для проявления максимальной активности фермента. Время инкубации варьирует от нескольких минут до нескольких часов, в зависимости от конкретного фермента и субстрата, а также от их концентраций. Для достижения определенной цели большинство данных реакций нужно проводить последовательно, поскольку продукт одной реакции является субстратом для другой реакции. Хотя проведение большинства указанных реакций занимает от нескольких минут до нескольких часов, для завершения конкретной совокупности реакций требуется очень много времени. Как правило, для гарантии того, что реакция завершена, на ее проведение предпочтительно нужно затратить большее время.
Основным недостатком традиционных методов является большая продолжительность реакции, и молекулярные биологи вынуждены тратить много своего ценного времени на проведение длительных экспериментов с использованием упомянутых ферментов. Чтобы ферментативная реакция протекала быстрее, нужно активировать или фермент, или субстрат, или и тот и другой. Скорость органической реакции, как правило, увеличивается при повышении температуры, но поскольку большинство ферментов инактивируется при нагревании, температуру не всегда можно увеличить. Альтернативным способом ускорения химической или биологической реакции является применение микроволнового излучения в качестве источника энергии. В действительности микроволновое излучение давно применяется для увеличения скорости органических реакций.
В 1986 г. Gedbye et. al и Giguere et. al продемонстрировали, что многие органические реакции протекают очень быстро под действием микроволнового излучения [Gedye, R.N. et. al., Tetrahedron Lett. (1986), 26, 279; Giguere, R. J et. al, Tetrahedron Lett. (1986), 27, 4945-4948]. Позднее универсальность химии "Активации органических реакций под действием микроволнового излучения" (MORE - Microwave-induced Organic Reactions Enhancement) была продемонстрирована многими авторами на ряде органических реакций, таких как каталитическое гидрирование, реакция Дильса-Альдера, свободнорадикальная реакция, ацилирование, деацилирование и др. [Bose, A.K, et al., Res. Chem. Intermed. (1994) 20, 1-11; Bose, A.K., et al., J. Org. Chem. (1991) 56, 6968-6970; Nahar, P. Tetrahedron Lett. (1997), 38, 7253-7254]. Практически все эти реакции проводили в бытовой микроволновой печи при частоте 2450 мГц и максимальной выходной мощности 700 ватт. Данные печи обычно имеют 10 разных уровней, соответствующих разной выходной мощности, десятый уровень микроволновой печи соответствует максимальной или самой высокой выходной мощности, а первый уровень - самой низкой. Хотя основным преимуществом реакций, проводимых с использованием микроволнового излучения, является значительное уменьшение времени их протекания, были также опубликованы данные по селективности или специфичности таких реакций [Nahar, P. Tetrahedron Lett. (1997), 38, 7253-7254].
Энергия микроволнового излучения также успешно использовалась для анализа белков [Akins, Jr. et al.; U.S. patent №№5403747 and 5478748] и иммуногистохимии [Boon, M.E. et al.; Am. J. Clin. Pathol. (1989), Boon, M.E. et al.; In Microwave Cookbook of Pathology: The art of Microscopic Visualisation]. Кроме того, изучался гидролиз аденозинтрифосфата под действием микроволнового излучения [Jahngen, E.G.E. et al. J. Org. Chem. (1990) 55, 3406-3409].
Некоторые исследователи изучали влияние микроволнового излучения на биополимеры, особенно, на ДНК и ферменты. Однако данные исследования имеют противоречивые результаты. В некоторых случаях микроволновое излучение использовалось для инактивации ферментов мозга мелких лабораторных животных [Galli Claudio and Racagni Giogio, Methods in enzymology, (1982) 86, 636-642]. Однако Byus, C.V. et al. обнаружили увеличение активности орнитиндекарбоксилазы в культивируемых клетках при использовании энергии микроволнового излучения [Byus et al., Cancer Res. (1988) 48, 4222-4226]. Кроме того, существуют публикации, в которых описано увеличение, а также подавление клеточной ферментативной активности [Dutta, S.K. and Verma, M. Current Scince (1989) 58, 58-63; Dutta, S.K. and Verma, M. Current Scince (1988) 57, 779-786]. Maleev, V. Ya. et al. не смогли обнаружить увеличения поглощения микроволнового излучения молекулами ДНК (Maleev, V. Ya et al. Biopolymers (1987) 26, 1965-1970). Однако, в соответствии с Narasimhan et al., в молекулах ДНК под действием микроволнового излучения может происходить одноцепочечный разрыв, двухцепочечный разрыв и локализованное разделение цепей [Narasimhan, V. and Huh, W.K.; Biochem. International (1991) 25,363-370].
Влияние микроволнового излучения на активацию ферментативных реакций синтеза и модификации нуклеиновых кислот ранее не было описано, хотя существуют такие данные для реакций расщепления нуклеиновых кислот под действием ферментов рестрикции (Jhingan, патент США №5350686). Автор осуществлял расщепление ДНК ферментом рестрикции с использованием энергии микроволнового излучения на самом низком уровне мощности, который составляет 10% от выходной мощности 700 ватт стандартной микроволновой печи. В процессе реакции автор также использовал ряд пауз, в течение которых реакционная смесь не подвергалась воздействию микроволнового излучения. В данном методе общее время реакции расщепления для большинства ферментов находилось в интервале от 6 до 15 минут. Однако в некоторых контрольных образцах плазмидной ДНК при инкубации при 37°С наблюдалось почти полное расщепление продуктов. Из этого эксперимента не ясно, насколько сильно микроволновое излучение влияет на вышеуказанные ферментативные реакции, поскольку большую часть времени фермент не получал соответствующего микроволнового излучения. По-видимому, в данных случаях время имеет более важное значение, чем микроволновое излучение.
Описанный метод (патент США №5350686) ферментативной модификации макромолекул с использованием микроволнового излучения имеет несколько недостатков, например (1) неэффективный выигрыш во времени; можно получить сравнимые результаты для некоторых реакций расщепления под действием ферментов рестрикции, проводимых вне микроволновой печи в течение такого же времени при 37°C; (2) при проведении реакции приходится делать несколько пауз, в течение которых реакционная смесь не подвергается воздействию микроволнового излучения, (3) не существует единообразных условий для ферментативных реакций; в большинстве случаев для реакции расщепления под действием ферментов рестрикции автор использовал разное время воздействия микроволнового излучения; и (4) эффективную автоматизацию процесса нельзя осуществить из-за (a) более длительного времени реакции и (b) отсутствия единообразия условий реакции.
Авторы настоящего изобретения преодолели все вышеуказанные недостатки. Для любой реакции нужны оптимальные условия. Оптимальное количество тепловой энергии ускоряет ферментативную реакцию, в мягких условиях (количество тепловой энергии ниже оптимального) реакция протекает медленнее, тогда как в жестких условиях (количество тепловой энергии выше оптимального) происходит дезактивация фермента. Вышесказанное справедливо и для реакций, протекающих с использованием энергии микроволнового излучения. Если не поддерживать оптимальные условия, ферментативные реакции могут завершиться неудачно или может получиться неполный продукт. В настоящем изобретении найдены подходящие условия, большинство из которых находится в противоречии с опубликованным методом или не известно в данной области.
Кроме того, протекание ферментативной реакции с использованием микроволнового излучения в большой степени зависит, помимо фермента и субстрата, от состава реакционной смеси. Буфер, его pH, концентрации солей и другие ингредиенты играют важную роль в ферментативных реакциях, особенно, если они проводятся с использованием микроволнового излучения. Авторы данного изобретения обнаружили, что в буфере для разбавления, содержащем 50% (об./об.) глицерина, фермент рестрикции инактивируется при воздействии непрерывного микроволнового излучения высокой мощности даже в течение короткого периода времени (15 секунд и больше), тогда как в обычных условиях при 37°C он остается активным. Буфер для разбавления поставщик рекомендует использовать для разбавления раствора с высокой концентрацией фермента, если это необходимо.
В противоположность данным, опубликованным Jhingan (патент №5350686), авторы данного изобретения обнаружили, что ферментативную модификацию биологической макромолекулы можно проводить в режиме самой высокой мощности микроволновой печи, при частоте 2450 мГц и мощности 700 ватт. Более того, также было обнаружено, что нет необходимости прерывать микроволновое облучение ферментативных реакций, что опять же находится в противоречии с приведенной выше публикацией.
Цели данного изобретения
Основной целью настоящего изобретения является предоставление быстрого и эффективного способа проведения ферментативных реакций с получением специфических продуктов данных ферментативных реакций, которые нужны для анализа генов и манипуляций с ними в молекулярной биологии, генной инженерии и других областях.
Другой целью настоящего изобретения является предоставление простого, воспроизводимого способа, который не требует дополнительных исследований или дорогого оборудования.
Очередной целью настоящего изобретения является предоставление способа, который можно легко автоматизировать для проведения большинства ферментативных реакций, использующихся в биотехнологии или молекулярной биологии, в одном устройстве.
Следующей целью настоящего изобретения является разработка способа, в котором используется облучение, не оказывающее неблагоприятного влияния на фермент.
Краткое описание изобретения
Настоящее изобретение относится к простому, эффективному и быстрому способу ферментативной модификации и ферментативного синтеза нуклеиновой кислоты in vitro с использованием непрерывного и краткого микроволнового излучения с частотой от 2300 до 2500 мГц, выходной мощностью от 600 до 900 ватт и временем облучения от 5 до 120 секунд; а также к устройству, необходимому для проведения данного способа.
Подробное описание настоящего изобретения
Соответственно, настоящее изобретение относится к простому, эффективному и быстрому способу ферментативной модификации и ферментативного синтеза нуклеиновой кислоты in vitro с использованием непрерывного и краткого микроволнового излучения.
В одном воплощении настоящего изобретения предоставляется реакционная смесь, содержащая (i) фермент, выбранный из группы, состоящей из рестрикционной эндонуклеазы, лигазы, фосфатазы, киназы, обратной транскриптазы, ДНК-полимеразы и РНК-полимеразы, (ii) биомолекулу, выбранную из ДНК, РНК или олигонуклеотидов, в качестве субстрата для выбранного фермента, (iii) буфер, подходящий для конкретного фермента, и (iv) другие ингредиенты, в маленькой пробирке Эппендорфа или на маленьком кусочке парафильма.
В другом воплощении настоящего изобретения указанные пробирку Эппендорфа или кусочек парафина с реакционной смесью помещают в микроволновую печь.
В следующем воплощении настоящего изобретения указанные пробирку Эппендорфа или кусочек парафильма непрерывно подвергают воздействию микроволнового излучения при частоте от 2300 до 2500 мГц и выходной мощности от 600 до 900 ватт.
В следующем воплощении настоящего изобретения реакционную смесь облучают в течение периода времени от 5 до 120 секунд.
В следующем воплощении настоящего изобретения реакцию останавливают добавлением тетраацетата этилендиамина (EDTA) и/или нагреванием на водяной бане при 75°С в течение 3-15 минут с последующим добавлением геля, несущего краситель.
В следующем воплощении настоящего изобретения продукт(ы) реакции анализируют методом электрофореза на агарозном геле, авторадиографии, подсчета радиоактивной метки и другими традиционными методами.
В следующем воплощении настоящего изобретения нуклеиновую кислоту выбирают из ДНК, РНК и олигонуклеотида.
В следующем воплощении настоящего изобретения другие ингредиенты выбирают из группы, состоящей из MgCl2, MgSO4, NaCl, KCl, CH3COOK, тетраацетата этилендиамина, дитиотреитола, спермидина, БСА, тритона Х-100, нуклеотидов, матрицы ДНК, матрицы РНК и олигонуклеотидных праймеров.
В следующем воплощении настоящего изобретения рестрикционную эндонуклеазу выбирают из Hind III, BamHI, EcoRI, Hae III, Bgl II, Pst I, Bst E II и Bst NI.
В следующем воплощении настоящего изобретения в качестве лигазы используют ДНК-лигазу T4.
В следующем воплощении настоящего изобретения в качестве киназы используют полинуклеотидкиназу T4 (TPNK).
В следующем воплощении настоящего изобретения в качестве фосфатазы используют щелочную фосфатазу из кишечника теленка (CIP).
В следующем воплощении настоящего изобретения полимеразу выбирают из группы, состоящей из ДНК-полимеразы I E.coli, фрагмента Кленова ДНК-полимеразы I E.coli, РНК-полимеразы T7 и РНК-полимеразы SP6.
В следующем воплощении настоящего изобретения в качестве обратной транскриптазы используют обратную транскриптазу вируса птичьего миелобластоза (AMV-RT).
В следующем воплощении настоящего изобретения используют ДНК, выбранную из группы, состоящей из геномной ДНК, ДНК лямбда-фага, плазмидной ДНК, ДНК бакуловирусов, ДНК растений и ДНК млекопитающих.
В следующем воплощении настоящего изобретения для двойного расщепления нуклеиновой кислоты используют более одной рестрикционной эндонуклеазы.
В следующем воплощении настоящего изобретения для двойного расщепления используют рестрикционные эндонуклеазы EcoRI и Hind III.
В следующем воплощении настоящего изобретения микроволновое облучение можно проводить в любом устройстве или в любой камере, где может быть сгенерировано микроволновое излучение.
В следующем воплощении настоящего изобретения упомянутый способ можно частично или полностью автоматизировать с помощью специально сконструированного устройства.
В следующем воплощении настоящего изобретения для проведения ферментативной реакции можно использовать указанное выше устройство.
В следующем воплощении настоящего изобретения предоставляется реакционная камера, состоящая из магнетрона, вытяжного вентилятора, волоконно-оптического термометра, охлаждающей системы и вращающейся платформы, на которой должна размещаться реакционная смесь.
В следующем воплощении настоящего изобретения предоставляются компьютерные средства на основе микропроцессора, обеспечивающие заранее запрограммированную команду для проведения различных реакций посредством подходящего аппаратного оборудования и программного обеспечения.
В следующем воплощении настоящего изобретения магнетрон является источником микроволнового излучения и работает при частоте приблизительно 2450 мГц и выходной мощности в интервале от 100 ватт до 800 ватт.
В следующем воплощении настоящее изобретение относится к быстрому и эффективному способу сайт-специфичного расщепления, модификации и синтеза биологических молекул. Более конкретно, описывается получение сконструированных биомолекул с помощью ферментативных реакций и с использованием краткого и непрерывного микроволнового излучения. Способ данного изобретения может применяться в биотехнологии, в частности в молекулярной биологии, в промышленных процессах, протекающих с использованием ферментов, и для очистки сточных вод, кроме того, данный способ можно автоматизировать для применения в родственной области.
В следующем воплощении настоящее изобретение в особенности относится к быстрому и эффективному способу сайт-специфичного расщепления, модификации и синтеза биологических молекул in vitro с помощью ферментативных реакций, активированных микроволновым излучением. Данное изобретение в особенности относится к быстрому и эффективному способу проведения сайт-специфичного расщепления, модификации и синтеза биологических молекул in vitro с помощью ферментативных реакций, в которых используется краткое и непрерывное микроволновое излучение без пауз. Предпочтительно частота микроволнового излучения составляет от 2000 до 2800 мГц при выходной мощности от 500 до 900 ватт. Продолжительность микроволнового излучения составляет от 5 до 120 секунд. Реакции расщепления, модификации и синтеза нуклеиновых кислот предпочтительно катализируются рестрикционной эндонуклеазой, лигазой, фосфатазой, киназой, обратной транскриптазой, ДНК-полимеразой и РНК-полимеразой. Специфические продукты данных ферментативных реакций нужны для анализа генов и манипуляций с ними в молекулярной биологии и генной инженерии.
В следующем воплощении настоящее изобретение относится к быстрому методу получения продуктов ферментативных реакцию. Данное изобретение в особенности относится к быстрому и эффективному способу проведения ферментативных реакций сайт-специфичного расщепления, модификации и синтеза нуклеиновых кислот in vitro. Реакции расщепления, модификации и синтеза нуклеиновых кислот предпочтительно катализируются рестрикционной эндонуклеазой, лигазой, фосфатазой, киназой, ДНК-полимеразой, обратной транскриптазой и РНК-полимеразой. Это очень быстрый способ проведения реакций сайт-специфичного расщепления, лигирования, дефосфорилирования, фосфорилирования (кинирования) и синтеза ДНК и РНК in vitro. Специфические продукты данных ферментативных реакций нужны для анализа генов и манипуляций с ними в молекулярной биологии и генной инженерии.
В следующем воплощении настоящего изобретения с точки зрения достижения цели настоящего изобретения и преодоления недостатков известных способов поведения ферментативных реакций предоставляется быстрый и эффективный способ проведения ферментативных реакций сайт-специфичного расщепления, модификации и синтеза нуклеиновых кислот in vitro с использованием микроволнового излучения. В настоящем изобретении используются ферментативные реакции, применяющиеся в молекулярной биологии, а именно реакции сайт-специфичного расщепления, лигирования, дефосфорилирования, фосфорилирования (кинирования), синтеза ДНК in vitro и синтеза РНК in vitro.
В следующем воплощении настоящее изобретение включает в себя (a) воздействие на реакционную смесь, содержащую фермент, субстрат, подходящий буфер и другие ингредиенты (как описано в примерах), микроволнового излучения в микроволновой печи при самом высоком значении выходной мощности микроволновой печи (700 ватт) в течение короткого периода времени и без пауз.
В следующем воплощении настоящего изобретения предоставляется способ, обеспечивающий сайт-специфическое расщепление, модификацию и синтез нуклеиновых кислот in vitro в течение интервала времени от 5 до 120 секунд. Ферментативные реакции, проводимые по способу данного изобретения, демонстрируют воспроизводимые результаты, что делает этот способ эксплуатационно гибким. Раскрывается новое открытие, заключающееся в том, что воздействие микроволнового излучения на реакционную смесь фермент-субстрат приводит к тому, что расщепление, модификация и синтез макромолекул, в особенности нуклеиновых кислот, завершается менее чем через 140 секунд.
В следующем воплощении настоящего изобретения предоставляется очень быстрый способ проведения известных на настоящий момент ферментативных реакций сайт-специфического расщепления, модификации и синтеза нуклеиновых кислот in vitro. Синтез in vitro в данном изобретении включает в себя синтез ДНК и РНК на матрице ДНК и синтез ДНК на матрице РНК (обратная транскрипция). Данный способ можно автоматизировать.
В следующем воплощении новизна настоящего изобретения заключается в том, что большинство реакций завершается в течение 5-150 секунд, и, следовательно, отчетливо демонстрируется влияние микроволнового излучения на время протекания реакции.
В следующем воплощении другой аспект новизны настоящего изобретения заключается в обнаружении, что ферменты рестрикции в разбавляющем буфере, содержащем 50% (об./об.) глицерина, утрачивают свои ферментативные свойства при кратковременном воздействии энергии микроволнового излучения с высокой выходной мощностью.
В следующем воплощении другой аспект новизны настоящего изобретения заключается в том, что ферменты рестрикции в буфере для расщепления остаются активными после воздействия энергии микроволнового излучения с высокой выходной мощностью (700 ватт) в течение короткого промежутка времени (по меньшей мере 140 секунд) и без пауз.
В следующем воплощении другой аспект новизны настоящего изобретения заключается в том, что условия реакции являются более единообразными в отличие от опубликованного способа, где в большинстве случаев расщепление ДНК под действием различных ферментов рестрикции проводят в разных условиях.
В следующем воплощении другой аспект новизны настоящего изобретения заключается в том, что все или почти все ферментативные реакции, обычно использующиеся в молекулярной биологии, можно проводить по способу данного изобретения.
В следующем воплощении другой аспект новизны настоящего изобретения заключается в том, что способ данного изобретения можно легко автоматизировать для проведения большинства ферментативных реакций, обычно использующихся в молекулярной биологии, в одном устройстве.
В следующем воплощении настоящего изобретения предоставляется быстрый и эффективный способ сайт-специфичного расщепления, модификации и синтеза нуклеиновых кислот in vitro с помощью ферментативных реакций, индуцируемых микроволновым излучением, а именно сайт-специфичного расщепления, лигирования, дефосфорилирования, фосфорилирования (кинирования), синтеза ДНК in vitro и синтеза РНК in vitro. Способ включает в себя воздействие на реакционную смесь, содержащую фермент, микроволнового излучения в микроволновой печи при высоком уровне мощности, в течение короткого периода времени и без пауз. Ферментативные реакции представляют собой деликатный метод, и неподходящие условия могут препятствовать получению нужных результатов.
В следующем воплощении настоящего изобретения авторы обнаружили, что ферментативная активность сохраняется после воздействия микроволнового излучения, соответствующего самому высокому уровню мощности микроволновой печи (700 ватт), в течение короткого периода времени от 5 до 120 секунд. Это открытие находится в противоречии с опубликованными данными (патент США №5350686), свидетельствующими о том, что дезактивация ферментов происходит после воздействия микроволнового излучения, соответствующего гораздо более низкому уровню мощности. Это может быть следствием того, что на предыдущем уровне технике использовалось гораздо более длительное облучение, от 15 до 31 минут, с разным числом пауз. Авторы данного изобретения проводят ферментативные реакции с использованием микроволнового излучения на самом высоком уровне мощности, в течение короткого периода времени и без пауз.
Подробное описание фигур
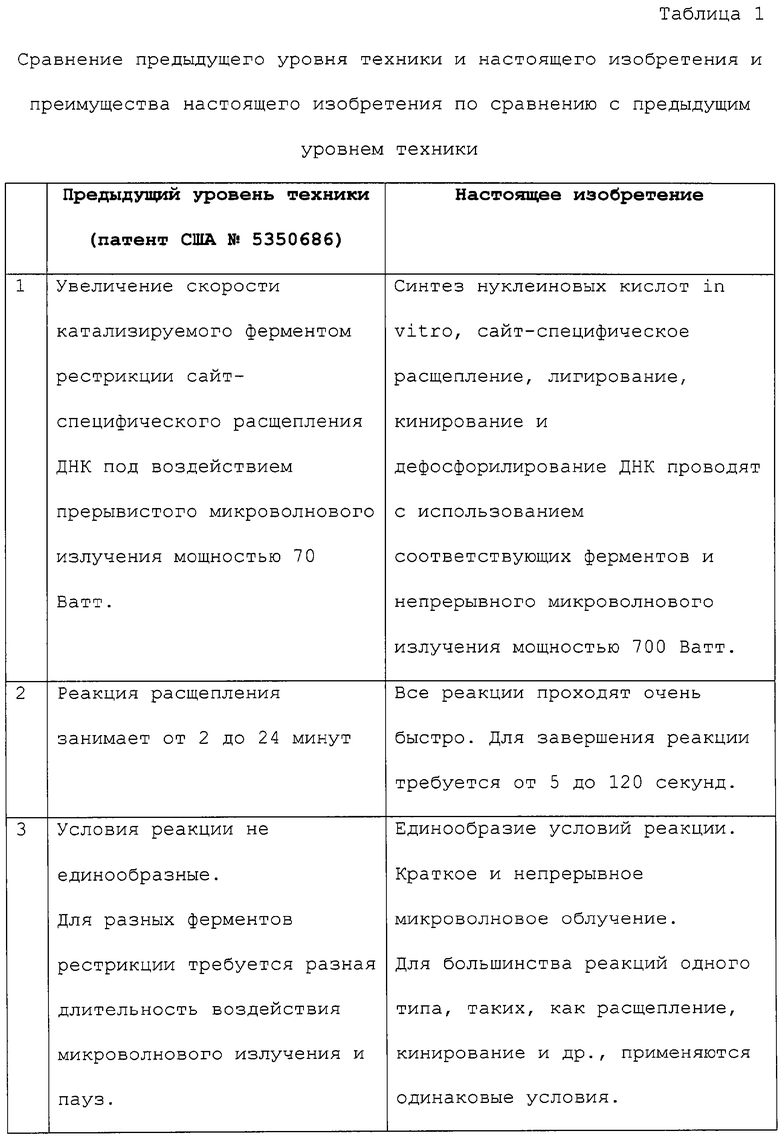
Фиг.1.
Стабильность Hind III при воздействии микроволнового излучения мощностью 700 ватт.
Hind III в соответствующем буфере для расщепления подвергают воздействию микроволнового излучения в течение разных периодов времени: 0 (полоса 1), 30 (полоса 2), 50 (полоса 3), 70 (полоса 4), 90 (полоса 5), 110 (полоса 6) и 130 (полоса 7) секунд. Hind III, облученную в течение разных периодов времени, анализируют по расщеплению лямбда-ДНК, подвергая воздействию микроволнового излучения мощностью 700 ватт в течение 50 секунд. Для получения каждой полосы в 1,0% агарозном геле наносят 1,0 мкг расщепленной ДНК (пример 1).
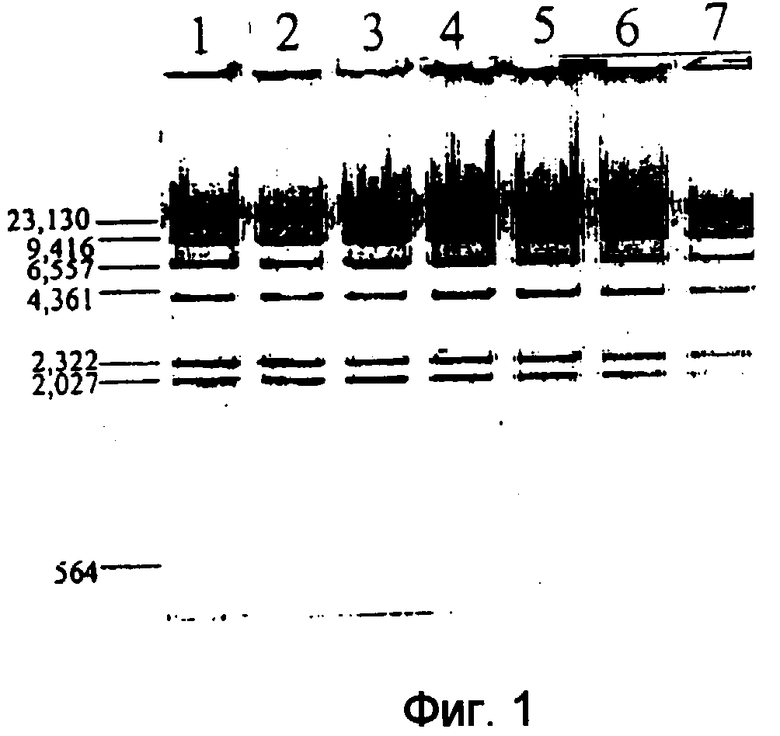
Фиг.2
Расщепление ДНК рестрикционной эндонуклеазой с использовнием микроволнового излучения мощностью 700 ватт.
Расщепление лямбда-ДНК под действием Hind III (полоса 2), BamHI (полоса 3), EcoRI (полоса 4), двойное расщепление лямбда-ДНК под действием EcoRI и Hind III (полоса 5) и расщепление клона кДНК лектина гороха по сайту Pst I вектора PUC19 под действием Pst I (полоса 6) проводят, используя 2,0 мкг ДНК, 1,0 мкл раствора каждого фермента в соответствующем буфере для расщепления и общего объема смеси 20,0 мкл. Двойное расщепление с использованием Hind III и EcoRI проводят в буфере для расщепления для EcoRI. В качестве контроля (полоса 1) используют 2,0 мкг лямбда-ДНК в 20,0 мкл буфера для расщепления для Hind III без фермента. Для получения каждой полосы в 1,0% агарозном геле наносят 1,0 мкг ДНК (пример 2).
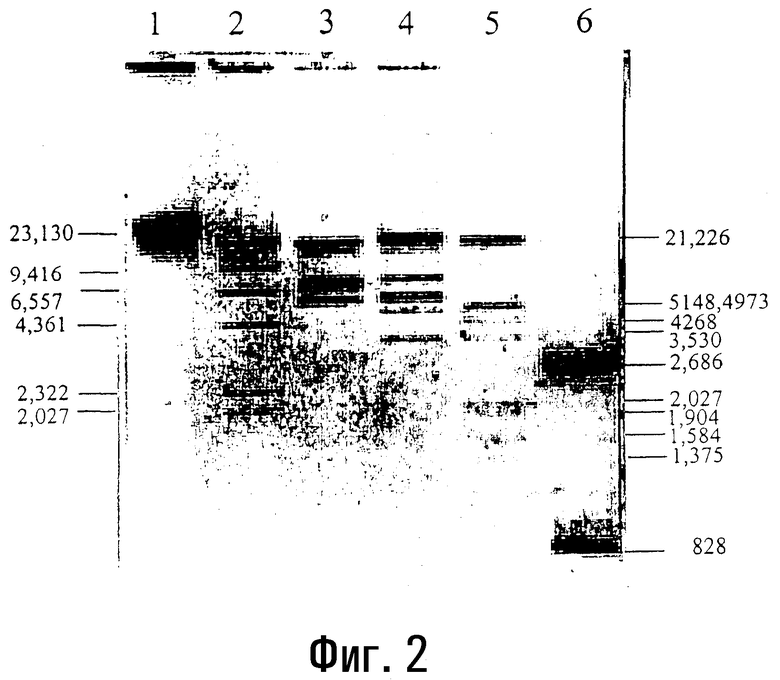
Фиг.3
Лигирование лямбда-фрагментов Hind III, катализируемое ДНК-лигазой T4, и фрагментация лигированной ДНК под действием Hind III в микроволновой печи.
На полосу 1 наносят неразрезанную лямбда-ДНК (1,0 мкг), обработанную микроволновым излучением в течение 50 секунд в буфере для расщепления для Hind III. На линию 2 наносят разрезанную Hind III лямбда-ДНК (1,0 мкг), полученную в результате воздействия на реакционную смесь микроволнового излучения в течение 50 секунд. На полосу 3 наносят продукты реакции лигирования разрезанной Hind III лямбда-ДНК, проводимой при 37°C в течение 45 минут. На полосу 5 наносят продукты реакции лигирования разрезанной Hind III лямбда-ДНК, проводимой с использованием микроволнового излучения в течение 20 секунд.
Лигированные ДНК полос 3 и 5 повторно разрезают Hind III с использованием микроволнового излучения в течение 50 секунд и наносят соответственно на полосы 4 и 6 1,0% агарозного геля. Каждая из этих полос содержит 1,5 мкг ДНК (пример 3).
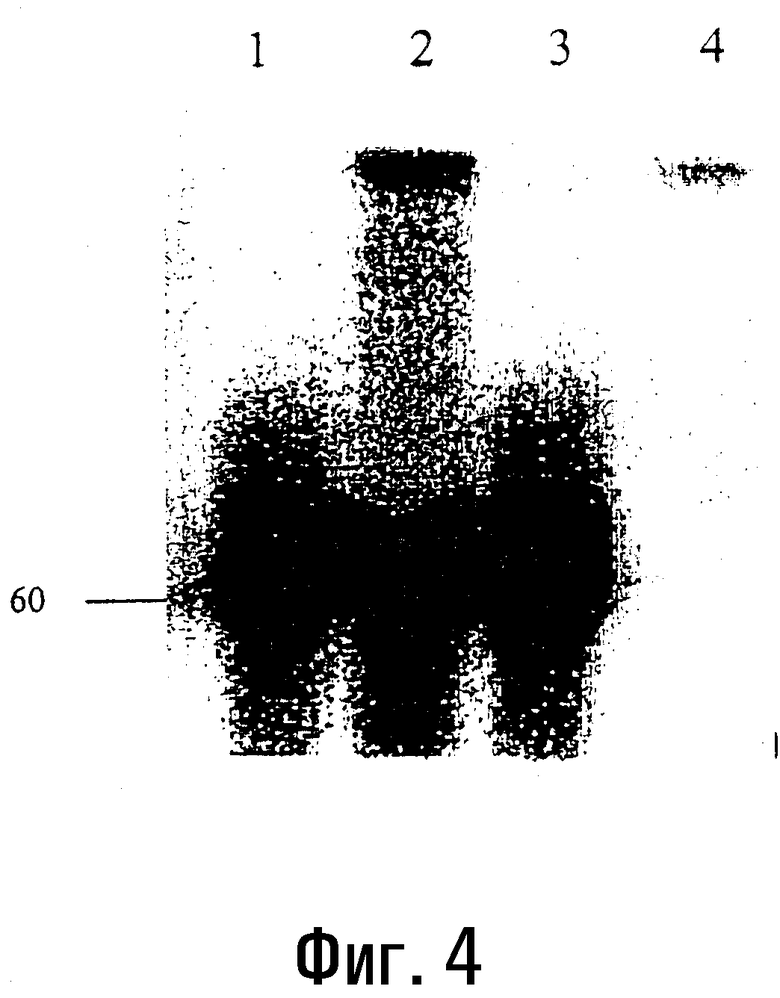
Фиг.4
Кинирование олигонуклеотида размером 60 оснований с помощью полинуклеотидкиназы T4.
Кинирование олигонуклеотида размером 60 оснований (5'-CCCCGCCCCGCG-3')5 с помощью полинуклеотидкиназы T4 и с использованием микроволнового излучения в течение 20 (полоса 1), 30 (полоса 2), 50 (полоса 3) и 0 (полоса 4) секунд (пример 5b).
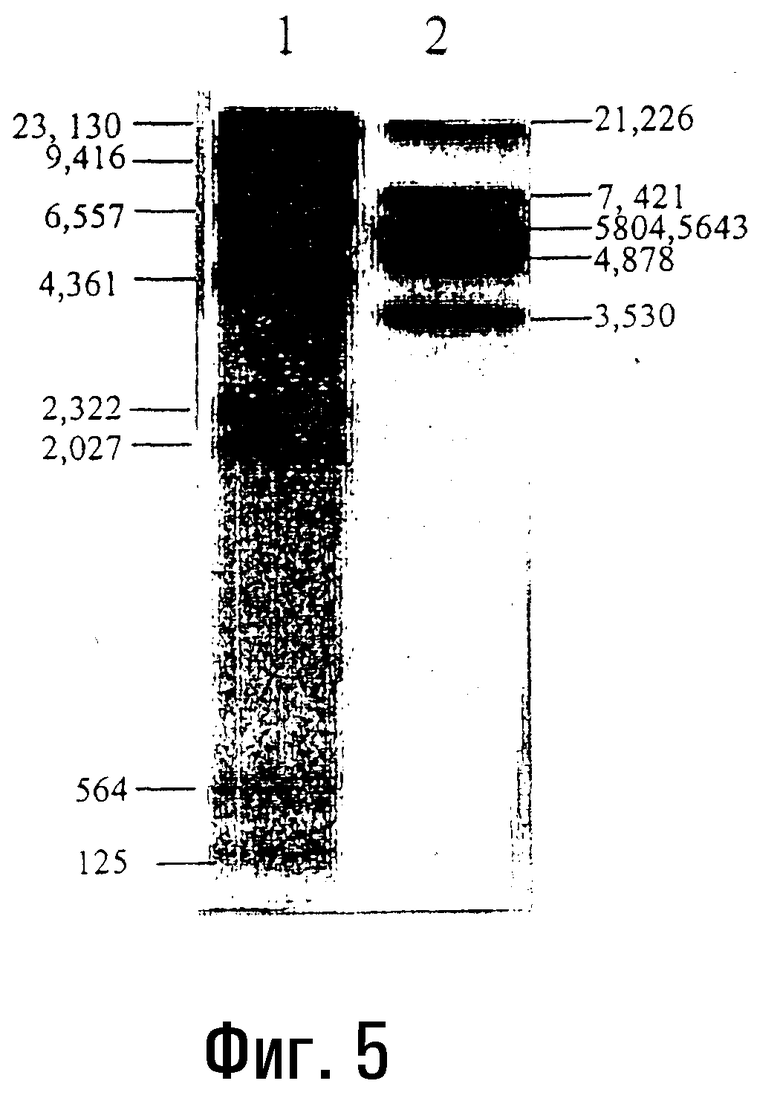
Фиг.5. Полоса 1.
Синтез ДНК с использованием фрагмента Кленова ДНК-полимеразы E.coli и микроволнового излучения.
Авторадиограмма продуктов, полученных в результате расщепления лямбда-ДНК с помощью Hind III с последующей реакцией дополнения концов с помощью ДНК-полимеразы I E.coli, на 1% агарозном геле. Обе реакции проводят с использованием микроволнового излучения мощностью 700 ватт в течение 50 и 20 секунд соответственно (пример 7).
Фиг.5. Полоса 2.
Реакции расщепления, дефосфорилирования и кинирования с использованием микроволнового излучения.
Катализируемое TPNK кинирование CIP-дефосфорилированных фрагментов EcoRI лямбда-ДНК.
Авторадиограмма продуктов, полученных в результате расщепления лямбда-ДНК с помощью EcoRI с последующим дефосфорилированием 5'-концов фрагментов EcoRI с помощью CIP и кинированием дефосфорилированных 5'-концов фрагментов EcoRI с помощью TPNK (пример 5a).
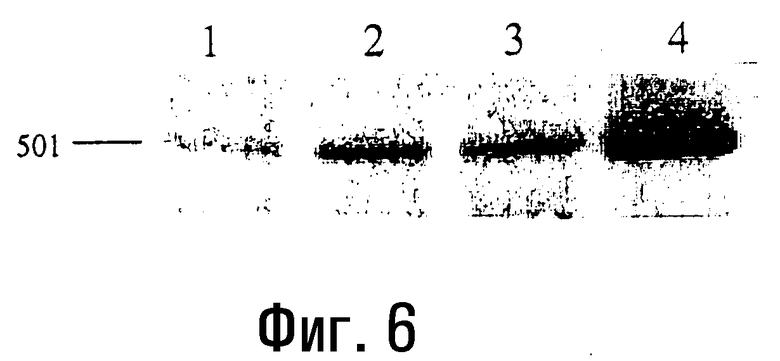
Фиг.6
Катализируемый AMV-RT синтез кДНК на матрице мРНК Е-селектина с использованием микроволнового излучения.
Продукты реакции, катализируемой AMV-RT и проводимой с использованием микроволнового излучения в течение 10 (полоса 1), 30 (полоса 3) и 50 (полоса 4) секунд, а также в течение 45 минут при 48°C (полоса 2), амплифицируют методом ПЦР, используя ДНК-полимеразу Tfl I, и получают специфическую кДНК размером 501 п.о. Продукты ПЦР-ОТ анализируют на 1,4% агарозном геле (пример 8).
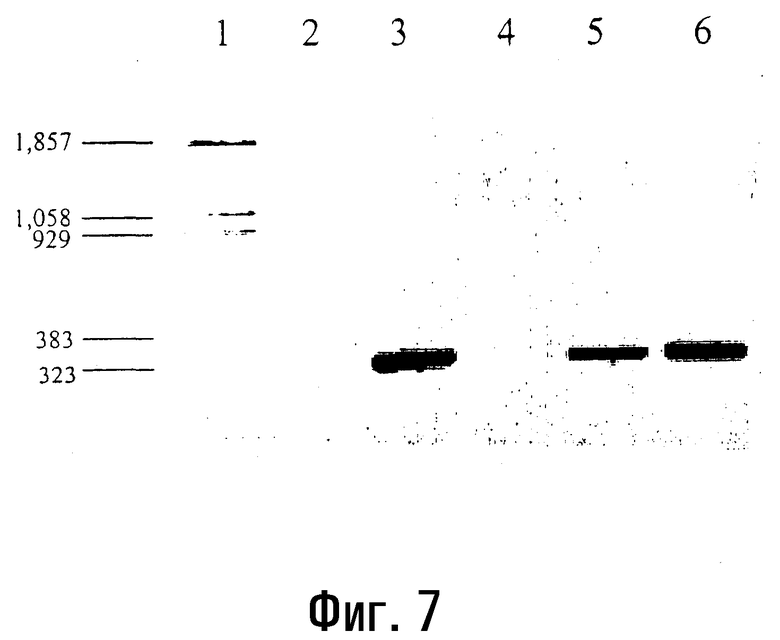
Фиг.7
Катализируемый AMV-RT синтез кДНК на матрице синтетической мРНК с использованием микроволнового излучения.
Продукты реакции, катализируемой AMV-RT и проводимой с использованием микроволнового излучения в течение 30 (полоса 5) и 50 (полоса 6) секунд, а также в течение 45 минут при 48°C (полоса 3), амплифицируют методом ПЦР, используя ДНК-полимеразу Tfl I, и получают специфическую кДНК размером 323 п.о. Продукты ПЦР-ОТ анализируют на 1,4% агарозном геле. В контрольных реакциях в образцы, соответствующие полосе 2 (45 мин при 48°C) и полосе 4 (воздействие микроволнового излучения в течение 50 сек), матрицу РНК (РНК 1151 nt, синтезированная in vitro, от Promega,) не добавляют. На полосу 1 наносят маркерную ДНК pBR 322 Bst NI (пример 9).




В следующем воплощении настоящее изобретение включает в себя быстрый и эффективный способ сайт-специфичного расщепления, модификации и синтеза нуклеиновых кислот in vitro с использованием микроволнового излучения, индуцирующего ферментативные реакции, а именно расщепление рестрикционной эндонуклеазой, лигирование, дефосфорилирование, фосфорилирование (кинирование), синтез ДНК in vitro, обратную транскрипцию и синтез РНК in vitro. Способ включает в себя воздействие микроволнового излучения на реакционную смесь, содержащую фермент и субстрат, в микроволновой печи в режиме максимальной мощности в течение короткого промежутка времени и без пауз.
В следующем воплощении настоящего изобретения способ обеспечивает высокоселективное специфическое расщепление, модификацию и синтез нуклеиновых кислот в течение периода времени от 5 до 120 секунд. Чтобы найти оптимальное время реакции, все реакции проводят с использованием микроволнового излучения разной длительности. Продукты реакций, в которых использовались соответствующие ферменты и микроволновое излучение, идентичны продуктам, полученным с помощью традиционных способов, проводимых при оптимальной температуре (37°С или ниже) в течение периода времени от 30 минут до нескольких часов. Контрольные реакции, проводимые вне микроволновой печи в течение такого же периода времени (как и в случае реакций, в которых используется микроволновое излучение), не дают желаемых продуктов реакции. Отсутствие неспецифических полос ДНК в присутствии или в отсутствие фермента показывает, что микроволновое излучение не индуцирует неспецифическую нуклеазную активность и не вызывает фрагментации ДНК в условиях реакции данного изобретения.
В следующем воплощении настоящего изобретения исследуют стабильность Hind III при воздействии микроволнового излучения самого высокого уровня мощности (700 ватт) в течение до 130 секунд. Было обнаружено, что Hind III в соответствующем буфере для разбавления, содержащем 50% глицерина (об./об.), теряет активность после облучения мощностью 700 ватт в течение времени свыше 15 секунд.
В следующем воплощении настоящего изобретения, наоборот, было обнаружено, что тот же фермент (Hind III) в соответствующем буфере для расщепления (общее содержание глицерина в реакционной смеси составляет 6,25% об./об.) является стабильным при воздействии микроволнового излучения мощностью 700 ватт в течение до 130 секунд (пример 1, фиг.1).
В следующем воплощении настоящего изобретения было обнаружено, что расщепление с помощью рестрикционной эндонуклеазы можно проводить по способу данного изобретения, т.е. воздействуя на реакционную смесь микроволновым излучением с самым высоким уровнем мощности в течение короткого времени и без пауз. Это находится в противоречии с опубликованным способом, где было обнаружено, что при использовании излучения с высоким уровнем мощности фермент теряет активность.
В следующем воплощении настоящего изобретения предоставляется способ, который обеспечивает высокоселективное расщепление нуклеиновых кислот с высокой специфичностью (расщепленная ДНК способна опять лигироваться) в течение гораздо более короткого периода времени. Эндонуклеазная активность ферментов рестрикции, таких как BamHI, EcoRI, Hind III, Hae III, Bgl II, Pst I и Bst E II, увеличивается, если реакционную смесь, содержащую субстрат ДНК, фермент и совместимый буфер, подвергают воздействию микроволнового излучения с самым высоким уровнем мощности. В типичном эксперименте 1 мкг лямбда-ДНК полностью расщепляется под действием 5-20 единиц (как определено New England Biolabs) каждого фермента рестрикции (объем реакционной смеси 20 мкл) при воздействии микроволнового излучения в течение 30-70 секунд (пример 2, фиг.2). Было обнаружено, что двойное расщепление ДНК-субстрата двумя ферментами, например, EcoRI и Hind III при воздействии микроволнового излучения завершается в течение такого же периода времени. Другое преимущество, касающееся расщепления рестрикционными эндонуклеазами, заключается в том, что данный способ является совершенно единообразным в широком диапазоне рестрикционных эндонуклеаз и ДНК-субстратов. ДНК-субстрат включает в себя лямбда-фаги, бакуловирусы, плазмиды и ДНК растений и млекопитающих. Сайт-специфическое расщепление ДНК-субстрата под действием ферментов рестрикции по способу данного изобретения подтверждают реакциями лигирования, проводимыми традиционными способами и по способу данного изобретения.
В следующем воплощении настоящего изобретения фрагменты ДНК, полученные в результате расщепления со сдвигом под действием Hind III, и фрагменты ДНК с тупыми концами, полученные в результате расщепления под действием Hae III, полностью лигируются ДНК-лигазой T4 в течение 10-15 секунд при воздействии микроволнового излучения. Контрольный эксперимент проводят при 16°C в течение 8 часов традиционным способом.
В следующем воплощении настоящего изобретения фрагменты ДНК, полученные в результате расщепления рестрикционной эндонуклеазой (проводимого традиционными способами и по способу данного изобретения), используют в качестве субстрата для реакций лигирования, и если лигированные ДНК снова подвергнуть расщеплению рестрикционной эндонуклеазой с использованием такого же фермента, то образуются такие же специфические фрагменты (пример 3, фиг.3).
В следующем воплощении настоящего изобретения модифицированные нуклеиновые кислоты, а именно дефосфорилированные, фосфорилированные и лигированные ДНК, являются важными инструментами молекулярной биологии. При воздействии микроволнового излучения на реакционную смесь все указанные продукты получают в достаточных количествах в течение 10-60 секунд, то есть происходит значительное уменьшение времени реакции по сравнению с традиционным способом. Так, дефосфорилирование концевых фосфатных групп фрагментов ДНК, полученных в результате действия рестрикционной эндонуклеазы, проводят с использованием щелочной фосфатазы кишечника теленка (CIP) в микроволновой печи в течение разных периодов времени, варьирующихся от 20 до 50 секунд. Получение продуктов реакции (дефосфорилированной ДНК), проводимой с использованием микроволнового излучения, подтверждается, если они не способны лигироваться ДНК-лигазой T4, но могут подвергаться кинированию под действием полинуклеотидкиназы Т4 (пример 4). Дефосфорилированные фрагменты ДНК, полученные в результате расщепления рестрикционной эндонуклеазой, фосфорилируют с помощью полинуклеотидкиназы T4 (TPNK) и с использованием [γ32P]АТФ в микроволновой печи в течение периода времени от 10 до 50 секунд. Фосфорилирование ДНК оценивают по количеству радиоактивности в веществе, осаждаемом трихлоруксусной кислотой, а также с помощью авторадиографии. Степень фосфорилирования, полученная в результате реакции, проводимой с использованием микроволнового излучения в течение 30 секунд, равна степени фосфорилирования, получаемой в результате традиционной реакции, проводимой при 37°C в течение 20 минут (пример 5).
В следующем воплощении настоящего изобретения в качестве субстрата для реакции кинирования и затем для реакции дефосфорилирования в микроволновой печи также используются синтетические олигонуклеотиды. Обнаружено, что обе реакции проходят с такой же эффективностью, как и в случае использования в качестве субстрата нуклеиновых кислот (пример 5b, фиг.4).
В следующем воплощении настоящего изобретения ферментативный синтез in vitro, проводимый по способу данного изобретения, включает в себя направленный синтез ДНК и РНК на матрице ДНК и направленный синтез ДНК на матрице РНК (обратная транскрипция).
В следующем воплощении настоящего изобретения ДНК-зависимый синтез ДНК, катализируемый ДНК-полимеразой I, проводят при помощи микроволнового излучения и с использованием в качестве матрицы никированной (содержащей разрывы) ДНК (ник-трансляция) и фрагментов, полученных в результате смещенного разреза рестрикционной эндонуклеазой и имеющих утопленные 3'-концы. Ник-трансляцию проводят в микроволновой печи в течение разных периодов времени. Синтез ДНК отслеживают по включению [α32P]dCTP, которое определяют по количеству радиоактивности в осадке, выпадающем при обработке трихлоруксусной кислотой. Было обнаружено, что после реакции, проводимой с использованием микроволнового излучения в течение 45 секунд, в веществе, осаждаемом ТХУ, наблюдается такое же количество радиоактивности на микрограмм никированной ДНК, как и после традиционной реакции, проводимой при 16°C в течение 60 минут. Однако если реакцию с использованием микроволнового излучения проводят в течение 30 секунд, то результат получается хуже, а если ее проводят в течение 60 секунд, то результат лучше, чем в традиционной реакции (пример 6).
В следующем воплощении настоящего изобретения точность воспроизведения структуры ДНК, синтезированной с помощью реакции ник-трансляции по способу данного изобретения, подтверждают путем гибридизации с матрицей ДНК (не никированной) с использованием саузерн-блоттинга. ДНК с затупленными концами получают через 20 секунд, если реакционную смесь, содержащую фрагмент Кленова ДНК-полимеразы I E.coli и матрицы ДНК с утопленными 3'-концами, подвергают воздействию микроволнового излучения. Было обнаружено, что после микроволнового облучения в течение 20 секунд наблюдается такая же степень завершения синтеза ДНК, как и в контрольном эксперименте, проводимом при 37°C в течение 30 минут. Воздействие микроволнового излучения в течение более короткого периода времени приводит к худшему результату, а в течение более длительного периода времени - к такому же или немного лучшему результату. Результаты синтеза ДНК, проведенного с использованием микроволнового излучения, подтверждают путем (i) включения [α32P]dCTP, (ii) авторадиографии и (iii) лигирования с векторной ДНК, имеющей тупые концы (пример 7; фиг.5, линия 1).
В следующем воплощении настоящего изобретения также обнаружено, что каталитическая активность РНК-полимеразы и обратной транскриптазы увеличивается под воздействием микроволнового излучения. Так, синтез ДНК (кДНК) на матрице РНК, катализируемый обратной транскриптазой вируса птичьего миелобластоза (AMV-RT), проводят с использованием двух матриц РНК и специфичных к матрице праймеров, при воздействии микроволнового излучения в течение разных периодов времени. Было обнаружено, что оптимальное время воздействия, при котором получают такой же результат, как в контрольной реакции, проводимой традиционным способом при 48°C в течение 45 минут, составляет 50 секунд. Заметная степень завершения реакции наблюдается уже после 10 секунд микроволнового облучения. Синтез кДНК отслеживают после амплификации с помощью полимеразы Tfl методом полимеразной цепной реакции с использованием обратной транскриптазы (ПЦР-ОТ) (пример 8, фиг.6 и пример 9, фиг.7).
В следующем воплощении настоящего изобретения для синтеза РНК на матрице ДНК (транскрипция) нужна специфическая последовательность ДНК (промотор), которая обеспечивает связывание РНК-полимеразы с матрицей ДНК и посредством этого инициирует транскрипцию. Синтез РНК in vitro, катализируемый РНК-полимеразой T7, проводят, используя матрицы pSPT 18 и pSPT 19-нео-ДНК, соответственно (Boehringer Mannheim Germany, номер по каталогу 999644). Продукты, полученные в результате микроволнового облучения реакционных смесей в течение разных периодов времени, варьирующих от 5 до 20 секунд, анализируют по включению [α32P]UTP и синтезу транскрипта определенного размера. Было обнаружено, что после реакции, проводимой с использованием микроволнового излучения в течение 5 секунд, в веществе, осаждаемом трихлоруксусной кислотой, наблюдается почти такое же количество радиоактивности, как и после традиционной реакции, проводимой при 37°C в течение 20 минут.
В следующем воплощении настоящего изобретения все реакции проводят в бытовой микроволновой печи (BPL, India), работающей при частоте приблизительно 2450 мГц, с максимальной выходной мощностью приблизительно 700 Вт. Все реакции проводят в микроволновой печи, установленной на максимальный уровень мощности (уровень 10), это означает, что коэффициент использования магнетрона при выходной мощности 700 ватт составляет 100%.
В следующем воплощении настоящего изобретения для проведения реакции последовательно смешивают буфер, субстрат(ы) и фермент в определенном объеме, как указано в стандартной инструкции. В следующем воплощении настоящего изобретения все ферментативные реакции проводят при таких значениях рН и ионной силы, которые используются в традиционных реакциях. Реакционную смесь вносят в пробирку Эппендорфа, которую помещают в центр вращающегося диска микроволновой печи. Реакционную смесь подвергают воздействию микроволнового излучения в течение периода, предпочтительно составляющего 5-120 секунд. В процессе микроволнового облучения пробирку Эппендорфа держат открытой. Реакции останавливают сразу после окончания микроволнового облучения добавлением тетраацетата этилендиамина (EDTA, pH 8,0) и/или нагреванием при 75°C в течение 3-15 минут. Продукты реакции анализируют традиционными методами, такими как электрофорез в агарозном геле, авторадиография, счет радиоактивности в веществе, осаждаемом трихлоруксусной кислотой (ТХУ), лигирование, кинирование, гибридизация и трансформация. Результаты всех реакций, проводимых по способам данного изобретения, сравнивают с результатами традиционных реакций, проводимых путем инкубации реакционной смеси на водяной бане при температуре, оптимальной для фермента.
Настоящее изобретение предлагает способ сайт-специфичного расщепления, модификации и синтеза нуклеиновых кислот, который обладает явными преимуществами по сравнению с существующими способами, известными до настоящего момента.
Нижеследующие примеры, описывающие данное изобретение в деталях, приведены только для иллюстрации и, следовательно, не должны рассматриваться как ограничивающие объем данного изобретения. Предполагается, что данное изобретение, описанное в соответствии с нижеприведенной формулой изобретения, может подвергаться различным модификациям в пределах его объема притязаний.
Пример 1
Стабильность Hind III в соответствующем буфере для расщепления после воздействия микроволнового излучения мощностью 700 ватт
Аликвоты раствора Hind III объемом 16 мкл помещают в пробирки Эппендорфа после разбавления 5 мкл (100 единиц) исходного раствора фермента смесью, содержащей 16 мкл 10×буфера для расщепления для Hind III (100 мМ Tris-HCl, pH 7,9, 500 мМ NaCl, 100 мМ MgCl2, 10 мМ DTT) и 107 мкл воды, с получением конечного объема 144 мкл. Каждую аликвоту подвергают воздействию микроволнового излучения мощностью 700 ватт (уровень мощности 10) в течение разных периодов времени: 0, 30, 50, 70, 90, 110 и 130 секунд. Каждый облученный фермент анализируют по расщеплению лямбда-ДНК традиционным способом и с использованием микроволнового излучения.
Расщепление с помощью облученного фермента проводят после добавления в каждую пробирку 4 мкл (2,0 мкг) лямбда-ДНК. Для проведения традиционного способа половину содержимого пробирки вносят в пробирку Эппендорфа и инкубируют при 37°C в течение 4 часов.
Для проведения реакции с использованием микроволнового излучения оставшуюся половину наносят на кусочек парафильма, помещают в центр микроволновой печи и подвергают микроволновому облучению мощностью 700 ватт в течение 50 секунд.
Все продукты реакции анализируют на 1,0% агарозном геле, где в каждую лунку вносят 1,0 мкг расщепленной ДНК (фиг.1).
Пример 2
Расщепление ДНК под действием рестрикционной эндонуклеазы с использованием микроволнового излучения мощностью 700 ватт
Все реакции расщепления проводят одновременно путем помещения реакционных смесей на парафильме в микроволновую печь на 50 секунд. Реакции расщепления лямбда-ДНК под действием Hind III, BamHI, EcoRI, двойного расщепления лямбда-ДНК под действием EcoRI и Hind III и расщепления клона кДНК лектина гороха по сайту Pst I вектора PUC19 под действием Pst I проводят, используя 2,0 мкг ДНК, 1,0 мкл раствора каждого фермента в соответствующем буфере для расщепления, так, что объем реакционной смеси составляет 20,0 мкл. Реакцию двойного расщепления под действием Hind III и EcoRI проводят в буфере для расщепления, подходящем для EcoRI. В контрольной реакции реакционная смесь содержит 2,0 мкг лямбда-ДНК в 20,0 мкл буфера для расщепления, подходящего для Hind III, и не содержит фермент. Каждый мкл раствора фермента содержит либо 20,0 (Hind III и EcoRI), либо 10,0 (BamHI и Pst I) единиц. На каждую полосу 1,0% агарозного геля наносят 1,0 мкг ДНК (фиг.2).
Пример 3
Расщепление лямбда-ДНК с помощью Hind III с последующим лигированием под действием ДНК-лигазы T4 с использованием микроволнового излучения
Фрагменты, полученные в результате расщепления с помощью Hind III, для реакций лигирования получают путем расщепления 8,0 мкг ДНК в 160,0 мкл реакционной смеси, содержащей 5,0 мкл (100 единиц) Hind III и 16,0 мкл 10×буфера для расщепления, подходящего для Hind III, в пробирке Эппендорфа объемом 1,5 мл в течение 50 секунд в микроволновой печи.
Реакционную смесь для лигирования объемом 20,0 мкл, содержащую 6,0 мкг фрагментов, полученных в результате расщепления с помощью Hind III, 2,0 мкл 10×буфера для лигирования (50 мМ Tris-HCl (pH 7,5), 10 мМ MgCl2, 10 мМ DDT, 1 мМ АТФ и 25 мкг/мл БСА) и 20 единиц способной к связыванию ДНК-лигазы T4 (New England Biolab. Inc. USA), помещают в две пробирки Эппендорфа объемом 0,5 мл, по 10,0 мкл в каждую. Одну из пробирок, содержащих реакционную смесь, подвергают воздействию микроволнового излучения в течение 20 секунд, а другую пробирку держат при 37°C в течение 45 минут. Реакции останавливают нагреванием при 65°C в течение 5 минут. Половину ДНК (1,5 мкг) из каждой пробирки Эппендорфа используют для повторного расщепления с помощью Hind III с использованием микроволнового излучения, как описано выше. Другую половину используют для того, чтобы подтвердить образование лигированных фрагментов с помощью электрофореза на агарозном геле.
В контрольном эксперименте неразрезанную лямбда-ДНК (1,0 мкг) подвергают воздействию микроволнового излучения мощностью 700 ватт в течение 50 секунд в буфере для расщепления, подходящем для Hind III, чтобы продемонстрировать неспецифическое расщепление, если оно присутствует (фиг.3).
Пример 4
Реакция дефосфорилирования с помощью щелочной фосфатазы
Дефосфорилирование 5'-фосфатных групп лямбда-фрагментов, полученных в результате расщепления с помощью EcoRI, проводят, используя 5 единиц щелочной фосфатазы кишечника телят (Boehringer Mannheim) в 20 мкл общего объема реакционной смеси, содержащей 1,0 мкг ДНК, 10 мМ Tris-HCl (pH 7,9), 10 мМ MgCl2, 50 мМ NaCl и 1 мМ DTT, в микроволновой печи. После воздействия микроволнового излучения в течение 30 секунд реакцию останавливают добавлением 2 мкл 0,5 мМ EDTA (pH 8,0) с последующей инкубацией при 75°C в течение 15 минут. ДНК освобождают от белков путем экстракции смесью фенол-хлороформ и последующего осаждения этанолом. Было обнаружено, что полученные таким способом фрагменты ДНК, обработанные фосфатазой, не способны лигироваться под действием ДНК-лигазы T4, но эффективно кинируются с помощью полинуклеотидкиназы, что подтверждает эффективность реакции дефосфорилирования, проведенной в течение 30 секунд.
Пример 5
Реакция кинирования с помощью полинуклеотидкиназы T4
a) Фрагментацию лямбда-ДНК под действием EcoRI проводят в течение 50 секунд с использованием микроволнового излучения, как описано в примере 2.
Реакцию дефосфорилирования с помощью CIP поводят в общем объеме 20,0 мкл, в пробирке Эппендорфа объемом 1,5 мл, в течение 30 секунд в микроволновой печи, используя 1,0 мкг ДНК и 1,0 единицу CIP, как описано выше.
Фосфорилирование 5'-гидроксильных групп дефосфорилированных фрагментов 0,5 мкг лямбда-ДНК, расщепленных EcoRI, проводят путем нанесения 20 мкл реакционной смеси, содержащей полинуклеотидкиназу T4 (Bangalore Genei), 70 мМ Tris-HCl (pH 7,6), 10 мМ MgCl2, 5 мМ DTT и 20 мкКи [γ32P]АТФ (уд. активность >4000 Ки/ммоль), на парафильм и воздействия на него микроволновым излучением в течение 20 секунд. Реакцию останавливают добавлением EDTA (pH 8,0) до получения конечной концентрации 5 мМ с последующим нагреванием при 65°C в течение 10 минут. Невключенную радиоактивность удаляют гель-фильтрацией на вращающейся колонке Sephadex G-50. Фосфорилирование фрагментов ДНК подтверждают авторадиографией, нанося объем реакционной смеси, равный 0,1 мкл.
b) Фосфорилирование 5'-гидроксильных групп 0,1 мкг олигонуклеотида размером 60 оснований (5'-CCCCGCCCCGCG-3')5 с помощью полинуклеотидкиназы T4 проводят с использованием микроволнового излучения в течение разных периодов времени (0, 50, 30 и 20 секунд), как описано выше (фиг.4).
Пример 6
Синтез ДНК с помощью полимеразы I E.coli
Реакции синтеза ДНК, катализируемые ДНК-полимеразой I E.coli, проводят в 50 мкл реакционной смеси, содержащей 1 мкг никированной ДНК лямбда-фага или матрицы ДНК вируса Autographa Californica nucleopolyhedrosis (AcNPV) (обработанной 5 фемтограммами ДНК-азы I), 5 единиц ДНК-полимеразы I E.coli (NEB), 50 мМ Tris-HCl (pH 7,5), 10 мМ MgSO4, 1 мМ дитиотриитол (DTT), 50 мкг/мл бычьего сывороточного альбумина (БСА, пентаксная фракция V), немеченные dATP, dTTP, dGTP (1 нМ каждого), 100 мкКи [α32P]dCTP (уд. активность >4000 Ки/ммоль) в микроволновой печи в течение разных периодов времени. После воздействия микроволнового излучения реакции останавливают добавлением 4 мкл 0,25 M EDTA (pH 8,0). Синтез ДНК на каждой матрице ДНК анализируют путем осаждения трихлоруксусной кислотой (ТХУ) после удаления невключенных радиоактивно меченных нуклеотидов через маленькую колонку Sephadex G-50 при центрифугировании. Количество ДНК, синтезированное в реакции с использованием микроволнового излучения в течение 45 секунд, равно количеству ДНК, синтезированному в традиционной реакции, проводимой при 16°C в течение 60 минут. Меченные ДНК, синтезированные по способу данного изобретения, эффективно гибридизуются с фрагментами, полученными в результате расщепления матриц ДНК рестрикционной эндонуклеазой.
Пример 7
Синтез ДНК с помощью фрагмента Кленова полимеразы I E.coli
Реакции синтеза ДНК, катализируемые фрагментом Кленова ДНК-полимеразы I E.coli, проводят в 20 мкл реакционной смеси, содержащей 1 мкг фрагментов ДНК лямбда-фага, полученных в результате расщепления Hind III (с использованием микроволнового излучения, как описано в примере 1), 1 мкл (8 единиц) раствора фрагмента Кленова ДНК-полимеразы I E.coli, 50 мМ Tris-HCl, pH 7,2, 10 мМ MgSO4, 1 мМ DTT, 50 мкг/мл БСА, 4 пмоль [α32Р]dCPT (уд. активность >4000 мкКи/ммоль) и 2 нмоль каждого из dTTP, dATP, dGTP, в микроволновой печи. Реакционную смесь наносят на парафильм и подвергают воздействию микроволнового излучения в течение 20 секунд, после чего реакцию останавливают добавлением 4 мкл 0,25 M EDTA (pH 8,0). Если используются радиоактивно меченные нуклеотиды, невключенные нуклеотиды удаляют с помощью вращающейся колонки Sephadex G-50. Дополнение утопленных 3'-концов матрицы ДНК путем синтеза ДНК анализируют по количеству радиоактивности в веществе, осаждаемом ТХУ, а специфичность синтеза ДНК определяют с помощью авторадиографии и лигирования с вектором ДНК, имеющим тупые концы. Количество ДНК, синтезированное по способу данного изобретения с использованием микроволнового излучения, равно количеству ДНК, синтезированному традиционным способом при 37°C в течение 30 минут (фиг.5, полоса 1).
Пример 8
Синтез кДНК на матрице мРНК Е-селектина, катализируемый AMV-RT-азой, с использованием микроволнового излучения
Три разные серии экспериментов проводят для синтеза кДНК, катализируемого обратной транскриптазой вируса птичьего миелобластоза (AMV-RT-аза), с использованием в качестве матрицы мРНК Е-селектина [Tucker et al. (1991) J. Biol. Chem., 266, 2466-2473], специфических прямых праймеров, путем микроволнового облучения реакционной смеси в течение 10, 30 и 50 секунд, соответственно. кДНК определенного размера идентифицируют с помощью электрофореза на агарозном геле после амплификации продукта реакции первой цепи кДНК с последующим добавлением специфичных к матрице обратного и прямого праймеров и ДНК-полимеразы Tfl. Прямой и обратный праймеры, используемые для матриц мРНК Е-селектина, имеют следующие последовательности: 5'-GAGTGGGCATGTGGAATGATG-3' и 3'-GGTCTCGGAAGTCACATGGA-5', соответственно. Синтез и ПЦР-амплификацию второй цепи кДНК после инактивации обратной транскриптазы (94°C в течение 2 мин) проводят, используя 40 циклов (94°C в течение 30 секунд/денатурация, 60°C в течение 1 мин/отжиг, 68°C в течение 2 мин/наращивание цепи). Реакционная смесь для ПЦР-ОТ, объем которой составляет 50 мкл, содержит 50 мМ Tris-HCl (pH 8,3), 50 мМ KCl, 10 мМ MgCl2, 0,5 мМ спермидина, 10 мМ DTT, 0,2 мМ dNTP (каждого), 1 мкМ прямого праймера, 1 мкМ обратного праймера, 1 мМ MgSO4, 5 единиц AMV-RT-азы и 5 единиц ДНК-полимеразы Tfl. Реакции инициируют добавлением РНК. Все компоненты реакционной смеси для ПЦР-ОТ, кроме мРНК Е-селектина и ее праймеров, получают от Promega.
Образование амплифицированных продуктов определенного размера, то есть 501 п.о., при использовании матрицы мРНК Е-селектина в реакции, проводимой как традиционным способом, так и по способу данного изобретения, подтверждают специфическим синтезом первой цепи кДНК с использованием мРНК. Количество амплифицированных кДНК, синтезированных в течение 50 секунд, было больше, чем в случае реакции, проводимой традиционным способом (фиг.6).
Пример 9
Синтез кДНК на синтетической мРНК в качестве матрицы, катализируемый AMV-RT-азой, с использованием микроволнового излучения
Три разные серии экспериментов проводят для синтеза кДНК, катализируемого обратной транскриптазой вируса птичьего миелобластоза (AMV-RT-аза), с использованием в качестве матрицы синтетической мРНК (РНК 1151 nt, синтезированная in vitro, от Promega,), специфических прямых праймеров, путем микроволнового облучения реакционной смеси в течение 10, 30 и 50 секунд, соответственно. кДНК определенного размера идентифицируют с помощью электрофореза на агарозном геле после амплификации продукта реакции первой цепи кДНК с последующим добавлением специфичных к матрице обратного и прямого праймеров и ДНК-полимеразы Tfl. Прямой и обратный праймеры, используемые для матриц синтетических мРНК, имеют следующие последовательности: 5'-GCCATTCTCACCGGATTCAGTCGTC-3' и 3'-GACTGAACTGCCGCCGA-5', соответственно. Образование амплифицированных продуктов определенного размера, то есть 323 п.о., при использовании матрицы синтетической мРНК в реакции, проводимой как традиционным способом, так и по способу данного изобретения, подтверждают специфическим синтезом первой цепи кДНК с использованием мРНК. Количество амплифицированных кДНК, синтезированных в течение 50 секунд, было больше, чем в случае реакции, проводимой традиционным способом.
Синтез и ПЦР-амплификацию второй цепи кДНК после инактивации обратной транскриптазы AMV (94°C в течение 2 мин) проводят, используя 40 циклов (94°C в течение 30 секунд/денатурация, 60°C в течение 1 мин/отжиг, 68°C в течение 2 мин/наращивание цепи). Реакционная смесь для ПЦР-ОТ, объем которой составляет 50 мкл, содержит 50 мМ Tris-HCl (pH 8,3), 50 мМ KCl, 10 мМ MgCl2, 0,5 мМ спермидина, 10 мМ DTT, 0,2 мМ dNTP (каждого), 1 мкМ прямого праймера, 1 мкМ обратного праймера, 1 мМ MgSO4, 5 единиц AMV-RT-азы и 5 единиц ДНК-полимеразы Tfl. Реакцию инициируют добавлением РНК. Все компоненты реакционной смеси для ПЦР-ОТ получают от Promega (фиг.7).
Пример 10
Синтез РНК с помощью РНК-полимеразы T7
Синтез РНК, катализируемый РНК-полимеразой SP6 и T7 поводят, используя pSPT18-нео и pSPT19-нео ДНК, полученные в результате расщепления EcoRI (Boehringer Mannheim, Germany), соответственно, в объеме 20 мкл. Каждая реакционная смесь содержит 0,5 мкг матрицы ДНК, 0,5 мМ каждого из ATP, GTP, CTP, UTP, 40 мМ Tris-HCl (pH 7,9), 6 мМ MgCl2, 2 мМ спермидина, 10 мМ DTT. Для проведения синтеза с использованием радиоактивной метки 0,5 мМ UTP заменяют на 50 мкКи [α32Р]UTP (уд. активность >400 Ки/ммоль). Реакционную смесь подвергают воздействию микроволнового излучения в течение 20 секунд, после чего реакцию останавливают добавлением 2 мкл 0,25 M EDTA (pH 8,0). Синтез РНК анализируют по включению [α32Р]UTP в вещество, осаждаемое ТХУ. По осаждению ТХУ обнаружено, что по способу данного изобретения синтезируется больше РНК, чем по традиционному способу, проводимому при 37°C в течение 45 минут. Размер РНК, синтезированной с использованием немеченных NTP, определяемый на формальдегид-агарозном геле, идентичен размеру РНК, полученной по традиционному способу.
Пример 11
Расщепление ДНК под действием рестрикционной эндонуклеазы с использованием микроволнового излучения мощностью 900 ватт
Реакцию расщепления проводят в пробирке Эппендорфа с использованием 1 мкг лямбда-ДНК, 5 Ед Hind III в соответствующем буфере для расщепления так, что объем реакционной смеси составлял 20,0 мкл. Пробирку Эппендорфа подвергают воздействию микроволнового излучения мощностью 900 ватт в течение 30 секунд (проба 1). В другом эксперименте длительность воздействия микроволнового излучения составила 40 секунд (проба 2). Контрольный эксперимент проводился при 37°С в течение 2 часов (проба 3). Через указанное время реакцию останавливали добавлением ЭДТА с последующей тепловой инактивацией. Продукты реакции контролировались при помощи электрофореза на фоне лямбда-ДНК, не подвергавшейся расщеплению (проба 4).
В результате контроля электрофореграмм установили, что все образцы, обозначенные как пробы 1-3, продемонстрировали сходные элекрофореграммы. Факт расщепления ДНК под действием микроволнового излучения мощностью 900 ватт доказан за счет наличия существенных отличий в электрофореграммах по отношению к нерасщепленной ДНК.
Обобщение результатов, представленных в примерах
1. Стабильность Hind III.
Hind III в буфере расщепления был подвергнут воздействию микроволнового излучения в течение 0-130 секунд. Стабильность фермента после воздействия на него излучения была проверена путем осуществления им реакции расщепления. Реакции расщепления были выполнены в обычном варианте и в реакциях с микроволновым воздействием. Пробы, отмеченные как "0 секунд" воздействия, не подвергались микроволновому облучению и использовались как контроль. Фиг.1 показывает, что все продукты расщепления имеют схожую картину, указывая на то, что фермент является устойчивым при короткой продолжительности воздействия, как в Примере 1.
2. Расщепление под действием рестрикционной эндонуклеазы. Реакции расщепления лямбда-ДНК под действием Hind III, BamHI, EcoRI, двойного расщепления лямбда-ДНК под действием EcoRI и Hind III и расщепления клона кДНК лектина гороха по сайту Pst I вектора PUC19 под действием Pst I проведены с использованием режима 50 секунд и мощности 700 ватт. Результаты, представленные на Фиг.2, показывают полное расщепление.
3. Расщепление лямбда-ДНК с помощью Hind III с последующим лигированием.
Фиг.3 показывает результаты, полученные для неразрезанной ДНК (полоса 1), разрезанной Hind III ДНК (полоса 2) при 50 секундном облучении, полосы 3 и 5 соответственно показывают результаты для ДНК, лигированной обычным способом, и при 20 секундном облучении, полосы 4 и 6 показывают результаты для лигированных ДНК, которые повторно подверглись разрезанию с помощью Hind III с использованием микроволнового излучения в течение 50 секунд. Последовательное расщепление и лигирование при микроволновом облучении подтверждает эффективность представленного способа, наряду с доказательствами эффективности, полученными традиционными методами.
4. Реакция дефосфорилирования.
Эта реакция была выполнена в режиме 30-секундного микроволнового облучения. Фрагменты ДНК, обработанные фосфатазой, не способны лигироваться под действием ДНК-лигазы Т4, но эффективно кинируются с помощью полинуклеотидкиназы, что подтверждает эффективность реакции дефосфорилирования, проведенной в течение 30 секунд.
5. Реакция кинирования.
Фиг.4 показывает эффективность реакции кинирования при воздействии излучения в течение 23-50 секунд, в то время как при значении "0 секунд" показано отсутствие кинирования.
6. Синтез ДНК с помощью полимеразы I Е. coli.
Количество ДНК, синтезированное в реакции с использованием микроволнового излучения в течение 20 секунд, равно количеству ДНК, синтезированному в традиционной реакции, проводимой при 37°С в течение 30 минут.
7. Синтез кДНК, катализируемый AMV-RT-азой.
Количество амплифицированных кДНК, синтезированных в течение 50 секунд, было больше, чем в случае реакции, проводимой традиционным способом (фиг.6).
Следовательно, настоящее изобретение предлагает простой, эффективный и ускоренный метод модификации и синтеза нуклеиновой кислоты.
Применение в промышленности
Данное изобретение можно использовать для быстрого и эффективного способа получения продуктов ферментативных реакций с использованием краткого и непрерывного микроволнового излучения. Способ данного изобретения можно использовать в биотехнологии, особенно в молекулярной биологии, для промышленной обработки и очистки сточных вод. Способ данного изобретения также можно автоматизировать для применения в родственной области.
Способ настоящего изобретения с использованием микроволнового излучения в особенности можно использовать для быстрого сайт-специфического расщепления ДНК, модификации ДНК и олигонуклеотидов, а также для синтеза нуклеиновых кислот in vitro. Данные реакции включают в себя расщепление ДНК с помощью рестрикционной эндонуклеазы, лигирование, дефосфорилирование, кинирование и синтез нуклеиновых кислот in vitro. Продукты данных реакций играют ключевую роль в молекулярной биологии и используются для геномного картирования, амплификации ДНК, секвенирования ДНК, экспрессии генов, очистки продукта, выделения продукта, манипуляции с генами в генной инженерии и др. Настоящее изобретение можно автоматизировать для применения в молекулярной биологии.
Преимущества
1. Значительное уменьшение времени реакции сайт-специфичного расщепления, модификации ДНК и олигонуклеотидов, а также синтеза нуклеиновых кислот является основным преимуществом настоящего изобретения. Время реакции уменьшается до 5-70 секунд для расщепления под действием рестрикционной эндонуклеазы, лигирования, дефосфорилирования, фосфорилирования (кинирования), обратной транскрипции, синтеза ДНК in vitro и синтеза РНК in vitro.
2. Почти все ферментативные реакции, используемые в молекулярной биологии, а также в другой области, могут быть проведены по способу данного изобретения в необычайно короткий период времени, что позволяет сэкономить много ценного времени и делает способ дешевым и технологичным.
3. Условия реакций очень простые и не требуют дорогого оборудования.
4. Способ данного изобретения можно автоматизировать для применения в молекулярной биологии.
5. Способ данного изобретения можно использовать в биотехнологической промышленности, в том числе в ферментативных реакциях.


Изобретение относится к области биотехнологии и может быть использовано для проведения ферментативных реакций, субстратом в которых служат нуклеиновые кислоты. Ферментативную обработку нуклеиновой кислоты проводят одним или двумя ферментами в присутствии микроволнового излучения, а также буфера и других ингредиентов, необходимых для проведения ферментативной реакции. Применение изобретения позволяет быстро, просто и эффективно провести ферментативную модификацию и синтез нуклеиновой кислоты in vitro. 11 з.п. ф-лы, 7 ил., 1 табл.
(a) получение реакционной смеси, содержащей один или два фермента, нуклеиновую кислоту, являющуюся субстратом для этого фермента(ов), буфер, подходящий для указанного фермента или указанных двух ферментов, и другие ингредиенты, необходимые для проведения ферментативной реакции;
(b) обработку реакционной смеси непрерывным микроволновым излучением с частотой 2300-2500 мГц и выходной мощностью от 600 до 900 Вт в устройстве или камере, в которых может быть генерировано указанное микроволновое излучение, в течение 5-120 с;
(c) остановку реакции и
(d) анализ продуктов реакции,
где указанный фермент выбирают из группы, состоящей из рестрикционной эндонуклеазы, лигазы, фосфатазы, киназы, обратной транскриптазы, ДНК-полимеразы и РНК-полимеразы, а указанную нуклеиновую кислоту выбирают из группы, состоящей из ДНК, РНК и олигонуклеотида.
| US 5350686, 27.09.1994 | |||
| СПОСОБ ПОЛУЧЕНИЯ АМИНОКИСЛОТНЫХ ИЛИ НУКЛЕИНОВОКИСЛОТНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ НА СУБСТРАТЕ, СПОСОБ СКРИНИНГА БОЛЬШОГО КОЛИЧЕСТВА АМИНОКИСЛОТНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, СУБСТРАТ ДЛЯ СКРИНИНГА | 1990 |
|
RU2107072C1 |
| MARIE-CLAIRE PARKER, THIERRY BESSON, SYLVAIN LAMARE AND MARIE-DOMINIQUE LEGOY, Microwave radiation can increase the rate of enzyme-catalysed reactions in organic media, Tetrahedron Letters, Volume 37, Issue 46, 11 November 1996, p | |||
| Способ обстрагивания брусьев по фигурному профилю | 1926 |
|
SU8383A1 |

