Изобретение относится к области иммунологии и медицины и касается искусственных пептидных мини-антигенов (ПМА), которые могут применяться для индукции контролируемого протективного гуморального IgG-опосредованного иммунного ответа против аллергена с целью замены патогенного IgE-опосредованного иммунного ответа.
ПМА является мини-аналогом гаптена на белке-носителе и состоит из пептидов, происходящих из разных белков.
ПМА является безопасной заменой экстрактов аллергенов для проведения специфической иммунотерапии (СИТ).
Областью медицинского применения являются все виды пыльцевой, пищевой, грибной и бытовой аллергии.
Биомишенью ПМА является адаптивный (антиген-специфический) иммунный ответ, включающий антигенпредставляющие клетки (макрофаги, дендритные и B-клетки), T- и B-клетки.
Ключевые слова: аллергия, пептиды, T- и B-эпитопы, наночастицы, структура вакцины, специфическая иммунотерапия, IgE, IgG.
Список сокращений
2. Введение
Для объяснения сути изобретения требуется схематичное понимание механизма формирования Антиген-специфического Иммунного Ответа (АСИО). Следует отметить, что АСИО всегда направлен на распознавание чужеродного белка. Именно белки несут в себе генетическую информацию в наиболее концентрированном виде. Фрагмент белка (пептид) длиной в 9 аминокислот (аа) может распознаваться иммунной системой как генетически чужеродный и против него может формироваться АСИО. Механизмов АСИО несколько. Для изложения сути данного изобретения мы остановимся только на одном механизме, называемом гуморальным АСИО (ГАСИО), поскольку именно формирование гуморального АСИО в виде молекул IgE является механизмом патогенеза аллергии I типа, для лечения которой может использоваться данное изобретение.
3. Биомишень I. Для формирования любого ГАСИО необходим белок-антиген (белок из аллергена или любой другой белок) и клетки иммунной системы, которые его распознают и реагируют формированием иммуноглобулинов. Такие иммуноглобулины, называемые также антителами, могут формировать стабильные комплексы с этим антигеном при встрече с ним в любых жидкостях и тканях организма. Существует несколько типов иммуноглобулинов: D, M, G1, G2, G3, G4, E и A. У каждого из типов иммуноглобулинов своя функция в иммунной системе. В идеале эти молекулы созданы природой для удаления из организма генетически чужеродного материала, попадающего в организм с патогенами: бактериями, вирусами, грибами, паразитами. Каждый из типов антител может оказывать как защитный (протективный) эффект, так и патогенный. Так, IgE-опосредованный ГАСИО на аллергены является патологической реакцией иммунитета, а такой же IgE ГАСИО на паразитов - протективной. Имеется ряд аутоиммунных заболеваний, где патогенный эффект оказывают IgG или IgA антитела.
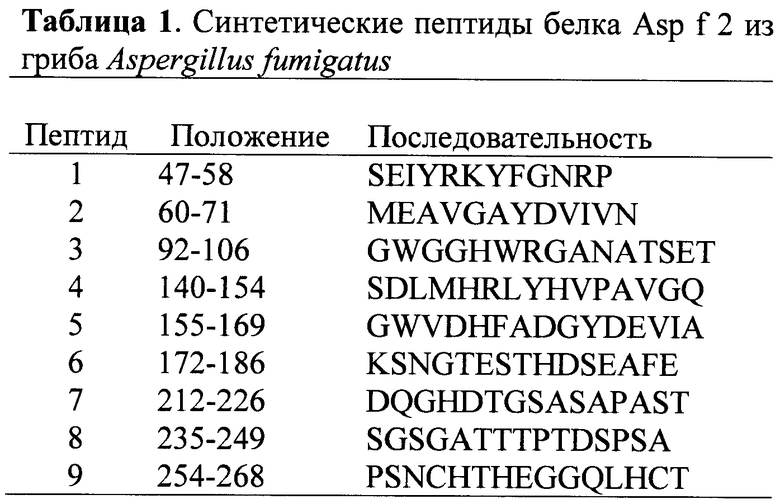
Для формирования ГАСИО необходимо, чтобы иммунная система опознала патоген, что на первом этапе происходит с помощью врожденной состемы иммунитета, узнающей не белки, а характерные структуры патогенов, например клеточную стенку бактерий, состоящую из полисахаридов. Это приводит к захвату патогена специализированными клетками-фагоцитами (фагоцитозу). После фагоцитоза патогены попадают в специализированные органелы фагоцитов (эндосомы и лизосомы), где патоген убивается и фракционируется на отдельные молекулы. Среди фагоцитов часть клеток имеет специализированную функцию для передачи информации клеткам адаптивного иммунитета с целью дальнейшего формирования АСИО. Дендритные клетки и макрофаги (в меньшей степени также и В-клетки) обладают такой функцией, за что получили название антигенпредставляющих клеток (АПК). Особенностью АПК является наличие эндосом, куда попадает патоген после фагоцитоза и куда клетки транспортируют специальные молекулы, называемые молекулами главного комплекса гистосовместимости II класса (ГКГСII). Эти молекулы играют доминирующую роль в формировании АСИО и являются первой биомишенью данного изобретения (Рис.1).
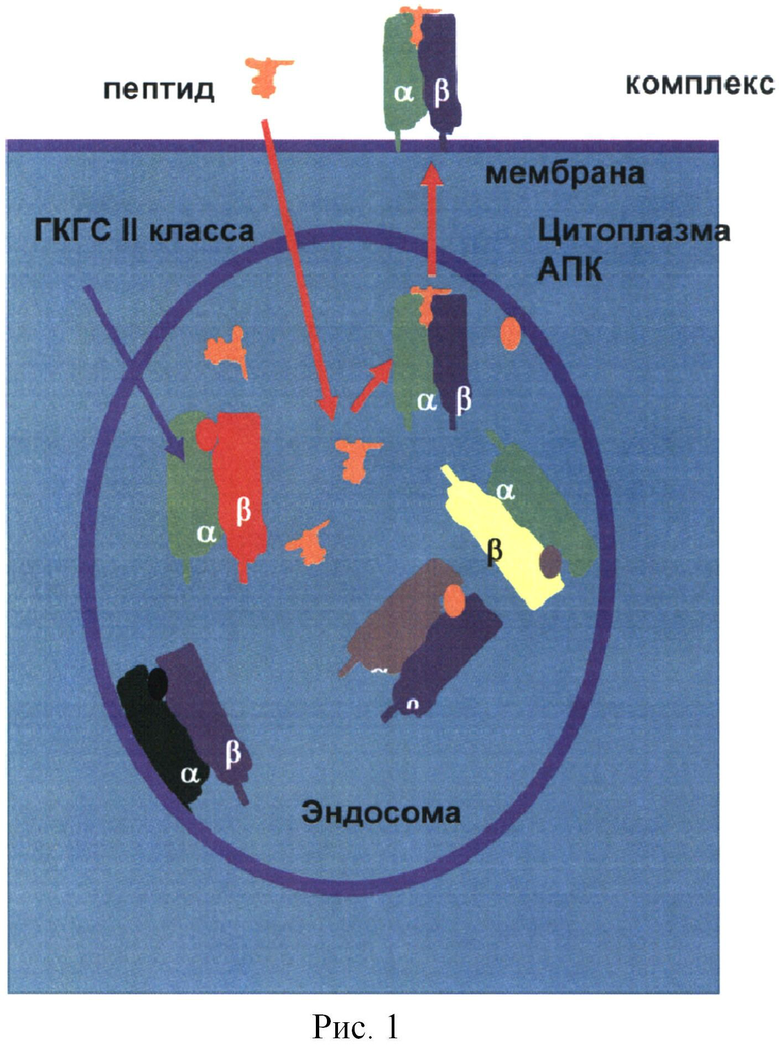
В процессе переваривания патогена в эндосомах его белки высвобождаются и фрагментируются на пептиды. Молекулы ГКГСII формируют комплекс с пептидами белков-антигенов за счет наличия специализированного участка-ловушки (кармана) на их поверхности, в который могут попасть пептиды только определенной структуры (Рис.2).
В организме человека молекул ГКГСII всего 3 (DR, DQ, DP), но имеется вариабельность этих молекул у разных людей (полиморфизм). Каждая молекула состоит из 2 цепей - α и β (Рис.1, 2), которые могут собираться из разных вариантов (аллелей). В результате вариабельности отдельный индивидуум может иметь до 12 различных комбинаций молекул ГКГСII. Общее количество вариантов в популяции людей ограничено. Все они классифицированы в системе HLA-DR, DQ, DP (от human leukocyte antigens DR, DQ, DP). Структура молекул HLA-DR охарактеризована для многих аллелей (см., например, базу данных http://www.syfpeithi.de). Для DQ и DP данных пока меньше.
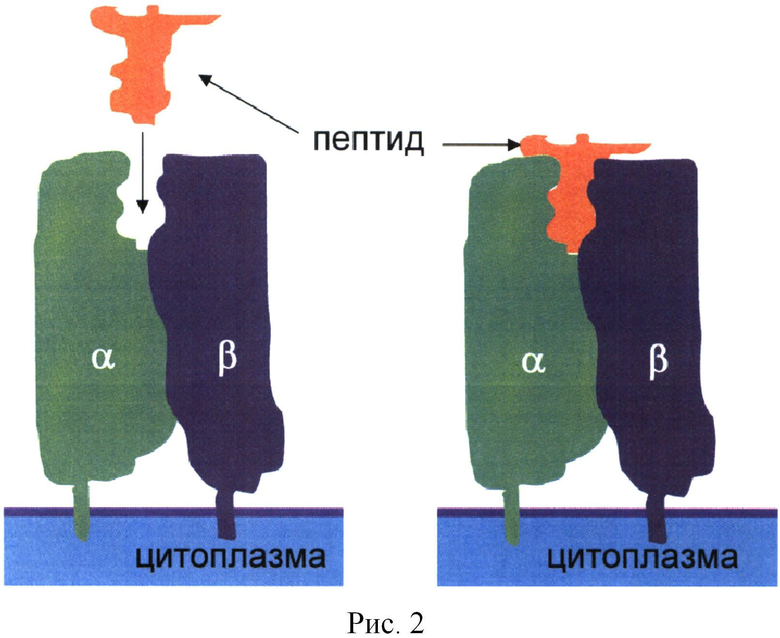
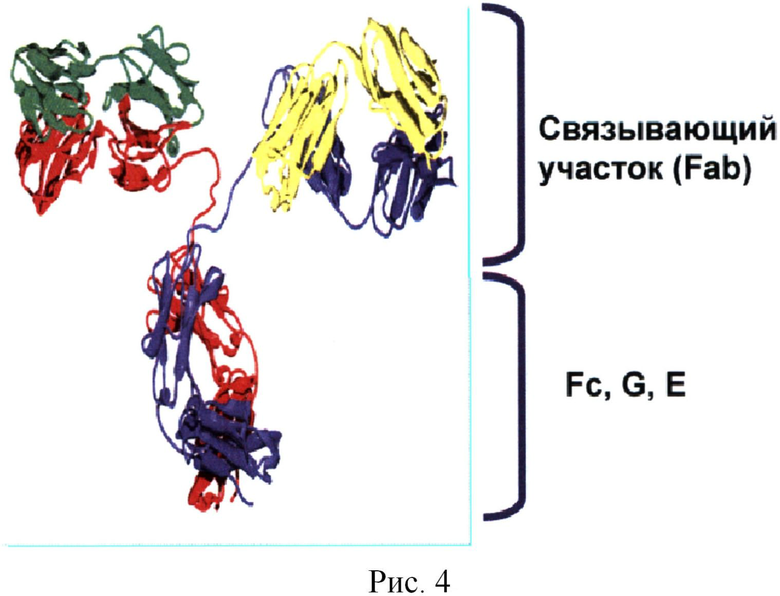
Комплекс чужеродного пептида, связанного в кармане ГКГСII, экспрессируется на поверхности АПК и является мощным стимулятором Т-клеток (Т-хелперов, CD4+ клеток) к активации, синтезу различных молекул, пролиферации (Рис.3) и активации других клеток АСИО, в случае ГАСИО - в стимулиции B-клеток к продукции антител (Рис.4). Такой пептид называется Т-эпитопом.
Зная структуру кармана определенной молекулы из любого белка, можно подобрать Т-эпитоп, который подходит по форме к карману ГКГСII. Зная, что тот или иной индивидуум имеет ту или иную молекулу ГКГСII, можно вводить в организм данный пептид и получить на него ответ, который выражается в пролиферации Т-клеток, распознающих именно тот пептид, что введен в организм.
После окончания иммунного ответа часть Т-клеток, специфичных к данному пептиду, сохраняется в виде Т-клеток памяти (ТКП). ТКП значительно быстрее активируются, чем наивные Т-клетки, впервые встретившие антиген. Когда человек часто болеет, например, гриппом определенного распространенного штамма, например, N1H1, то в его организме имеется достаточно много ТКП, которые обеспечивают реактивацию АСИО при новой инфекции - это и есть иммунитет, защищающий нас от знакомой инфекции. На принципе стимуляции ТКП (В-клетки памяти тоже имеются) основаны все современные вакцины.
Формулой изобретения, часть 1, является использование для индукции ГАСИО к аллергену пула предсуществующих ТКП, неспецифичных к аллергену. Специфичность ТКП может быть направлена на белки вируса гриппа, вируса Эпштейн-Барра, герпесов 1 и 2 типа и др, то есть на белки патогенов, вызывающих часто случающуюся или хроническую персистирующую инфекцию. Для индукции ГАСИО этим путем необходимо провести HLA-DR типирование конкретного больного (что делает множество фирм) и выбрать подходящий пептид для данной молекулы из базы данных.
При использовании полноразмерных белков рестрикция молекулами ГКГСII не является детерминирующей. Однако при работе с пептидными конструкциями этот вопрос является одним из наиболее важных и лимитирующих применение пептидных вакцин, поскольку в составе коротких фрагментов может не оказаться ни одного Т-клеточного эпитопа, способного связываться с конкретными молекулами ГКГС.
4. Биомишень II. Т-клетки, начинающие иммунный ответ, называются Т-клетками-хелперами. Их функцией является помощь другим клеткам, которые называются эффекторами. Эффекторами ГАСИО являются В-клетки. Функция B-клеток - продуцировать антитела высокой специфичности. В-клетки изначально распознают совсем другие пептиды, чем Т-клетки. На поверхности всех В-клеток имеются молекулы, являющиеся составной частью антитела, которая собственно и должна связываться с антигеном (IgD). Эта молекула называется В-клеточным рецептором (ВКР). В организме имеется большое разнообразие ВКР, за счет чего в норме иммунная система может распознавать все имеющиеся в природе патогены. В процессе формирования ГАСИО В-клетка проходит ряд преобразований, что в итоге приводит к двум принципиальным событиям: 1) аффинность взаимодействия ВКР с распознаваемым пептидом резко возрастает (константа диссоциации уменьшается с 10-6 до 10-9 и выше) и 2) В-клетка формирует растворимую (гуморальную) форму антитела, состоящую из высокоаффинного ВКР и константной части антитела (Fc, от fragment constant). Именно Fc определяет тип антитела: M, G1, G2, G3, G4, E или A (Рис.4).
Для формирования протективного противоаллергенного ГАСИО требуется, чтобы появились В-клетки памяти (ВКП), продуцирующие IgG антитела к белкам аллергена. В этом случае говорят, что антитела распознают В-эпитопы белков аллергена. Получить IgG антитела на В-эпитопы аллергена очень просто. Можно, например, проиммунизировать больного с аллергией экстрактом аллергена, что и делается при проведении лечения с помощью Специфической Иммунотерапии (СИТ). Однако при этом возникает много проблем, изложенных далее.
1. Опасность лечения методом СИТ. Можно спровоцировать обострение заболевания за счет активации тучных клеток, на которых уже имеются IgE антитела к данному аллергену. В ряде случаев может последовать анафилактический шок и опасность для жизни больного.
2. Экстенсивная иммунизация. В экстракте аллергенов много белков, иммунная система будет распознавать несколько белков, представленных в наибольшем количестве. В результате будет формироваться ГАСИО на пул белков. В случае успеха это защитит больного от дальнейшей аллергии на этот аллерген. При этом иммунная система подвергается значительной нагрузке. Любой иммунный ответ обходится организму очень дорого - при селекции Т- и В-клеток 90-99% пула селектируемых клеток погибает.
3. Неоправданная нагрузка. При формировании АСИО на патогены экстенсивный иммунный ответ необходим: иммунная система ищет ключевой АСИО, который блокирует размножение патогена в организме. В отличие от патогенов - большинство аллергенов не размножаются в организме! Это безвредные объекты: пыльца, пылевые клещи, кусочки эпидермиса животных и птиц, пищевые белки и др. Для их удаления из организма не нужно искать самый главный белок. Достаточно иметь антитела к белку, экспонированному на поверхности аллергенного комплекса. За этот белок антитело свяжется, а затем такой комплекс выведется из организма с помощью обычных механизмов, существующих для выведения комплексов антиген-антитело. При проведении СИТ, соответственно, лучше вызвать ответ на один-два таких поверхностных белка, чтобы решить проблему. С учетом того, что у аллергических больных IgE ответ часто наблюдается ко многим аллергенам, необходимо щадить иммунную систему. Им потребуется пройти несколько курсов десенсибилизации к разным аллергенам для того, чтобы выздороветь.
4. Нет стандартных экстрактов. Большой проблемой разработки препаратов для СИТ (а это экстракты аллергенов) является нестандартизованность таких препаратов. Каждая фирма получает свой экстракт. Даже при использовании одного протокола выделения аллергены различаются между собой из-за различий в исходном сырье.
5. Рекомбинантные аллергены. Альтернативой нестандартности сырья является получение рекомбинантных аллергенов. В мире ведется такая работа. Аллергенов много. В составе каждого из аллергенов также много белков. В настоящий момент проводят тестирование на «основные аллергены», то есть определяют, какие из белков конкретного аллергена чаще распознается IgE больных. Эти белки в дальнейшем планируется использовать для СИТ. Рекомбинантные белки дороги в производстве. Соответственно, дорогим будет и лечение.
6. Пептидные вакцины. В мире также разрабатываются пептидные вакцины для лечения аллергии. Это наиболее близкие к нашему изобретению препараты. Они состоят из пептидов основных белков аллергенов. Эффективность пептидных вакцин пока мала, что связано с рядом проблем, о которых будет идти речь далее.
Формулой изобретения, часть 2, является индукция ГАСИО на 5-8 любых гидрофильных пептидов длиной 10-15 аминокислот из поверхностно экспонированных на диком аллергене белков (можно из нескольких разных белков), поскольку иммунная система умеет формировать антитела к любым чужеродным пептидам. Не требуется определять природные В-эпитопы. Достаточно, чтобы формировались IgG к любым участкам белка-аллергена. В примере 1 показано, что при индукции IgG на произвольно выбранный гидрофильный пептид белка ДСГЗ полученные антитела связывают полноразмерный белок, что и требовалось доказать.
Требование экспонирования белков на поверхности аллергена дикого типа необходимо для доступа антител к этому белку. Требование гидрофильности к используемым пептидам является нежестким и связано с лучшей экспонированностью гидрофильных пептидов в водную среду при создании вакцины.
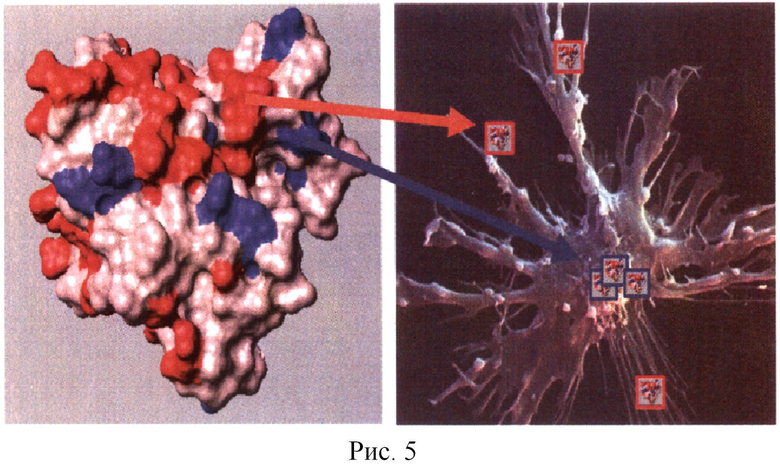
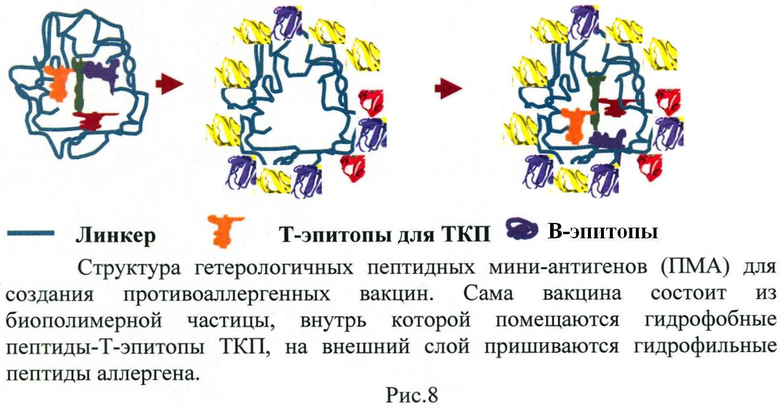
5. Структура вакцины. Для индукции эффективного ГАСИО необходимо выбранные пептиды упаковать специальным образом, а именно физически связать T- и B-эпитопы между собой. Это требование связано с тем, что механизм активации ГАСИО связан с одновременным присутствием Т- и В-эпитопов одного белка в одной АПК (Рис.5). Каждый природный белок имеет много потенциальных Т- и В-эпитопов. На рис.5 приведена кристаллографическая структура белка, на котором разными оттенками серого отмечены участки в структуре молекулы белка, соответствующие В- и Т-эпитопам белка.
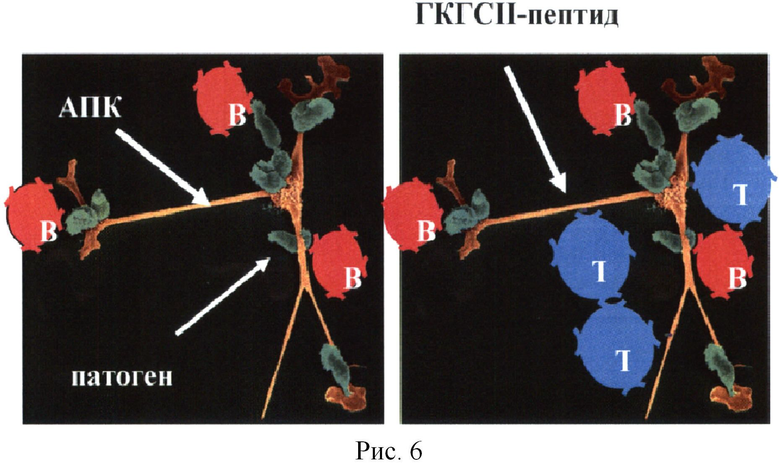
При формировании АСИО принципиальным является участие АПК на этапе возникновения центра, в котором проходит селекция Т- и В-клеток (герминальные центры в лимфатических узлах и селезенке). Такие АПК собирают антиген на периферии за счет липких рецепторов (маннозные рецепторы, мусорные рецепторы и др). Патогены налипают на поверхность АПК. Часть из них фагоцитируется и попадает внутрь. Такой «ежик», набрав антиген, продвигается в ближайший лимфатический узел, где и оседает, формируя вокруг себя «шубу» из В-клеток, отловленных из потока лимфы за счет разпознавая пока с малой аффинностью В-эпитопов белков прилипших патогенов (Рис.6, слева).
Одновременно АПК процессирует белки патогена внутри в эндосомах и формирует комплексы ГКГСII-пептиды, которые затем траспортирует на свою поверхность. За эти комплексы, как за крючки, тормозятся рядом Т-клетки данной специфичности. Формируется терминальный центр, где потом будет происходить формирование АСИО.
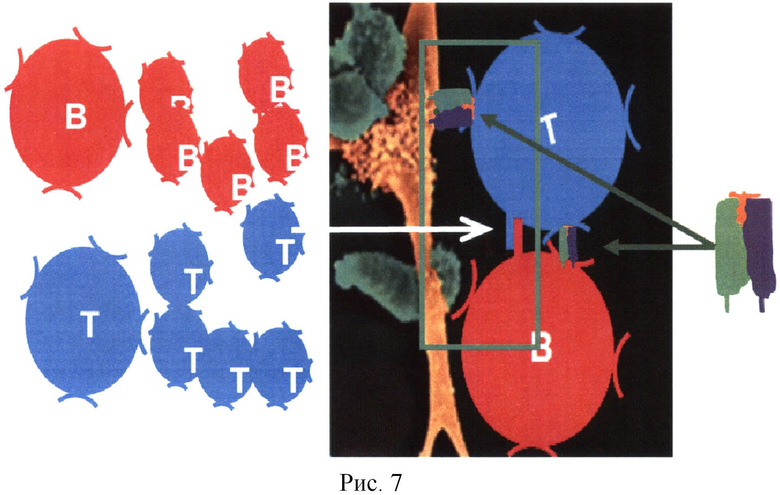
Для завершения процесса активации и формирования клеток-эффекторов гуморального ответа (созревание ГАСИО) требуется взаимодействие между T- и B-клетками, специфичными к одному и тому же белку (когнатное взаимодействие) (Рис.7).
Для этого природа создала специальный механизм: все активированные B-клетки становятся АПК, то есть начинают процессировать антиген и представлять пептиды белков антигена в комплексе с ГКГСII. T- и B-клетки размножаются (пролиферируют) и, в процессе размножения, на каждом цикле деления отбираются только те, что смогли вступить в контакт друг с другом и с материнской АПК (Рис.7). При этом между T- и B-клетками появляется второй контакт через специальные молекулы CD40-CD40L, без сигнала от которого В-клетка не может начать синтезировать IgG (не переключает тип антитела, что связано с генными перестроениями). Похоже, что именно этот контакт нарушен при аллергической реакции из-за недостаточного градиента концентрации антигена. Кроме двух контактов (ГКГСII-пептид-антигена-ТКР и CD40-CD40L) для оптимального ГАСИО требуется еще и сигнал через гуморальные факторы (цитокины), которые продуцируются АПК и T-клетками локально и связываются В-клетками также локально. Формируется трехсигнальная система запуска ГАСИО. Все это происходит в непосредственной близости трех клеток: АГГК-Т-клетка-В-клетка. При их разделении в пространстве координированность нарушается и ГАСИО не формируется. Именно требование сближенности в пространстве этих клеток требует, чтобы Т- и В-эпитопы антигена были связаны друг с другом физически. Особый способ связи (линкер) является третьей сутью изобретения, о чем пойдет речь ниже.
Каждая из T- и B-клеток, вступившая в такой контакт с АПК, перестает быть наивной и начинает активироваться. В результате активации появляются зрелые Т-клетки хелперы и высокоаффинные В-клетки, секретирующие антитела определенного класса.
Именно этот этап определяет, будет ли В-клетка продуцировать IgG или IgE. Это зависит от нескольких параметров, часть из которых пока плохо определена. Но точно известно, что для получения высокоаффинных антител G класса требуется:
1. Достаточно высокая пороговая концентрация антигена (очень малые концентрации иммунная система считает «шумом» и не реагирует на них);
2. Градиент антигена во времени (можно ввести сразу много антигена, который быстро выведется, в этом случае не будет ГАСИО);
3. Имеется некая оптимальная скорость поступления антигена, которая отобрана в процессе эволюции и связана со скоростью размножения патогенов и клеток иммунной системы;
4. Наличие сигнала через систему TLR врожденных рецепторов (Toll-like receptors).
Из трех необходимых условий аллергены удовлетворяют только одному: имеется градиент антигена во времени (сезон пыления деревьев и трав; постоянный контакт с домашними животными и пр). Пороговая концентрация белков аллергена ниже оптимума, так как аллергены не реплицируются в организме. Соответственно, скорость поступления аллергена ниже необходимой. Кроме того, аллергены не экспрессируют необходимых лигандов для системы TLR. А без этих сигналов меняется функционирование АПК. У людей, не склонных к аллергии, недостаток активации АПК приводит к игнорированию аллергена (толерантности). У 30% популяции людей имеется генетическая предрасположенность к распознаванию аллергенов, что выражается в аллергической реакции.
Нами проведены многочисленные эксперименты по анализу распознавания связанных между собой Т- и В-эпитопов. Оказалось, что при предъявлении иммунной системе «голого» Т-эпитопа к нему будут также формироваться антитела (пример 2). Этого, однако, хотелось бы избежать. Наличие в организме высокоаффинных антител к любому пептиду удалит этот пептид в любом окружении из организма в том случае, если такой пептид будет доступен антителам. Поскольку при проведении гипосенсибилизации против многих аллергенов хочется использовать ограниченный спектр Т-эпитопов, к которым имеется пул ТКП, мы постарались «спрятать» Т-эпитопы. Соответственно, мы разработали систему упаковки Т-эпитопов в гидрофобную частицу. Структура такой частицы является, по-видимому, уже запатентованным объектом. Так, аналогичную упаковку предлагает американская компания Selecta Biosciences. Область исследований Selecta Biosciences несколько иная, чем аллергия, но принцип упаковки пептидов один и тот же, а именно: пептиды, являющиеся Т-эпитопами, пакуются внутрь наночастицы, а гидрофильные пептиды, являющиеся В-эпитопами, располагаются на внешней поверхности частицы и доступны для иммунной системы и антител. Линкером может являться любой биоразлагаемый материал: полипептидный линкер, полисахариды, липиды. Структура комплекса показана на рис.8. Следует отметить, что пример 2 также демонстрирует отсутствие ответа при отсутствии в составе ПМА Т-эпитопов.
Один из наиболее важных параметров для новых аллерговакцин является отсутствие связывания IgE антител самой вакциной. Это позволит использовать высокую дозу для иммунизации больных, что сократит процесс лечения до 2-3 инъекций. Поскольку ПМА состоит из линейных пептидов аллергена, a IgE распознают преимущественно конформационные эпитопы (собирающиеся только в свернутом белке), то скорее всего такая конструкция не будет связывать IgE. Нами протестировано 7 сывороток больных с аллергией на гриб Aspergillus fumigatus (больные аллергическим бронхолегочным аспергиллезом, АБЛА) и показано, что сыворотки распознают основные белки, но не смесь линейных пептидов этих белков (Пример 3).
6. Ближайшие аналоги
Ближайшими аналогами предлагаемых ПМА являются: 1) пептидные вакцины для специфической иммунотерапии (ПИТ) и 2) пептидные конструкции с использованием «универсальных» Т-эпитопов.
1. Пептидные вакцины для специфической иммунотерапии
СИТ известна более 100 лет [1] и на настоящий момент является единственным способом патогенетического лечения аллергии. С учетом того, что число людей, страдающих аллергическими заболеваниями, постоянно растет, а для достижения состояния ремиссии требуется длительная СИТ, стоимость лечения выходит на первый план. Кроме того, из-за отсутствия стандартных экстрактов для СИТ не существует единых препаратов для лечения той или иной формы аллергии. Использование стандартизованных препаратов рекомбинантных аллергенов или пептидов из них дает надежду на решение данной проблемы [2-3]. При сравнении стоимости и безопасности пептидных и рекомбинантных препаратов для иммунотерапии преимущество имеет несомненно ПИТ. Существуют исследовательские препараты, направленные на разработку ПИТ, ориентированной на Т- и В-эпитопы белков аллергенов. Из них в клинических исследованиях изучены только пептиды, идентифицированные как Т-эпитопы аллергенов. К сожалению ПИТ, основанная на использовании Т-эпитопов, показала себя эффективной только при лечении аллергии на укусы пчел [4]. В лабораториях мира продолжаются исследования ПИТ, основанной на разных подходах [5]. При этом время от времени исследователи регистрируют формирование IgE у мышей на пептиды, что требуется учитывать при разработке вакцин [6]. Вакцин для ПИТ, основанных на гетерологичных пептидах, мы не встречали.
2. Пептидные конструкции с использованием «универсальных» Т-эпитопов
Поиск в базе данных PubMed по ключевым словам «universal T-cell epitopes allergy» не дал результатов.
Поиск в базе данных PubMed по ключевым словам «universal T-cell epitopes vaccines» дал несколько интересных результатов.
В статье Alexander J et al [7] авторы излагают часть нашей идеологии для создания вакцины против различных типов гриппа. В работе описан поиск универсальных Т-эпитопов вируса гриппа, которые распознаются (связываются) наиболее распространенными гаплотипами молекул ГКГСII HLA-DR. При создании противоаллергенных вакцин такие эпитопы могут быть частью противоаллергенной вакцины. Именно предсуществующие к ним T-клетки памяти будут помогать формировать ответ на В-эпитопы аллергенов.
В работе Gao J et al [8] описывается конструирование миниантигена, состоящего из T- и B-эпитопов вируса гепатита С. При этом все эпитопы происходят из одного антигенного комплекса, хотя и из разных белков. Подход близкий по сути, но сделан на основе получения слитого белка генноинженерными методами, что опять же даст более дорогой продукт, чем синтетические пептиды. Близкая идея описана в работах Calvo-Calle JM et al [9-10], где авторы получили пептидный «ежик» из одного T- и одного B-эпитопа Plasmodium falciparum для создания вакцины против малярии. Продукт получают уже синтетическим образом за счет синтеза разветвленных пептидов. Отсюда один шаг до наночастиц с нашитыми пептидами. Однако пока этот шаг не сделан.
Еще ближе к нашему изобретению лежит работа Batzloff MR et al [11], в которой «универсальный» Т-эпитоп GKLIPNASLIENCTKAEL из вируса кори собаки (!) сшивали с В-эпитопом белка M стрептококка группы A, получали липопептиды для интраназальной иммунизации. Один Т-эпитоп, один В-эпитоп, но уже из различных белков, то есть гетерологичные пептиды.
Нет работ, где бы авторы использовали пул предсуществующих Т-клеток памяти для поддержания de novo формирующегося ответа на безвредные антигены, такие как аллергены.
Суммируя приведенные сведения, суть данного изобретения может быть описана следующим образом:
Пептидный конъюгат любых гидрофобных пептидов, которые связываются с карманами наиболее распространенных молекул ГКГС II класса и к которым в популяции людей имеются Т-клетки памяти, с любыми гидрофильными пептидами аллергенов, представляет собой пептидный мини-антиген, который можно использовать для терапии аллергии. Пептиды должны быть упакованы специальным образом с тем, чтобы Т-эпитопы были свернуты внутри молекулы, а В-эпитопы экспонированы наружу с тем, чтобы Т-эпитопы не распознавались как В-эпитопы.
Такая упаковка возможна, но является предметом другого патента.
7. Экспериментальные доказательства положений изобретения
Пример 1. Иммунизация кролика произвольно выбранными пептидами белка адгезии десмоглеина 3 (ДСГЗ) приводит к появлению антител на этот пептид. Такие антитела распознают полноразмерный белок ДСГЗ в нативной конформации. При иммунизации комплексом, содержащим пептид вируса гриппа N1H1, аффинность антител резко возрастает.
ДСГЗ является белком адгезии кератиноцитов кожи. ДСГЗ является аутоантигеном при пузырном дерматозе аутоиммунного происхождения вульгарной пузырчатке (ВП). Этот белок экспрессируют клетки линии кератиноцитов человека НаСаТ.
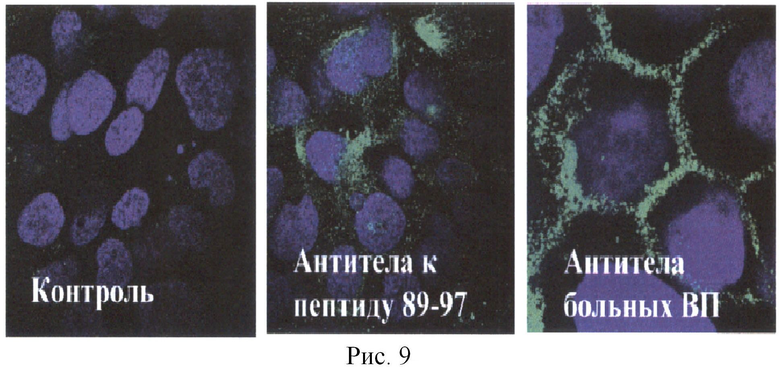
Кролика №1 иммунизировали пептидом 89-97 из белка человека десмоглеина 3. В-эпитопы этого белка не известны не только для кролика, но и для человека. В результате многократных иммунизаций получили антитела, которые распознавали клетки НаСаТ (Рис.9). Аффинность полученных антител не была высокой. Высокоаффинные антитела больных ВП связывали белок по всей поверхности клетки (правый рисунок), а сыворотки иммунного кролика связывались в основном с белком в цитоплазме клеток, и лишь некоторые - с десмосомами, где это белок концентрируется (центральная фотография). Сыворотка интактного кролика не связывалась с данными клетками (левый снимок). Поскольку произвольно выбранный пептид может не содержать Т-эпитопа или содержать не полный Т-эпитоп, то мы провели некоторую работу по подбору кролика, который отвечал на выбранный ранее нами пептид из белка вируса гриппа штамма N1H1 (RMQFSSLTVNVRGSGMR).
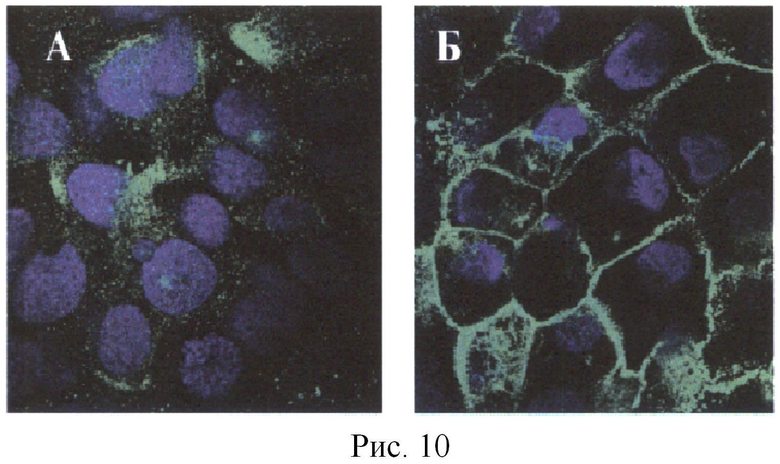
Данный пептид был ранее идентифицирован как Т-эпитоп в системе HLA-DR4 (Tan et al. Conservation and diversity of influenza A H1N1 HLA-restricted T cell epitope candidates for epitope-based vaccines. PLoS One. 2010 Jan 18; 5(1):e8754). Был получен конъюгат пептида ДСГЗ 89-97 с пептидом RMQFSSLTVNVRGSGMR. После иммунизации кролика конъюгатом, содержащим гетерологичные пептиды (Т-эпитоп - из белка вируса гриппа, В-эпитоп - из белка ДСГЗ), получили высокоаффинные антитела, связывающиеся с ДСГЗ внутри десмосом, что характерно для высокоаффинных антител таких, как антитела больных ВП (Рис.9, 10). Соответственно, нами показано, что введение пептида, распознаваемого иммунной системой как Т-эпитоп, в состав конъюгата с любым пептидом любого белка приводит к формированию антител на эти пептиды.
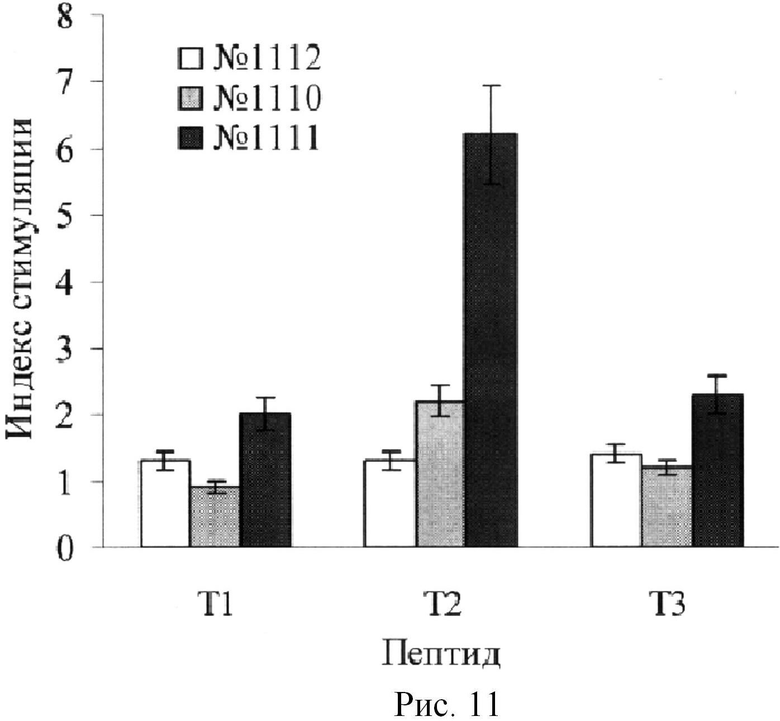
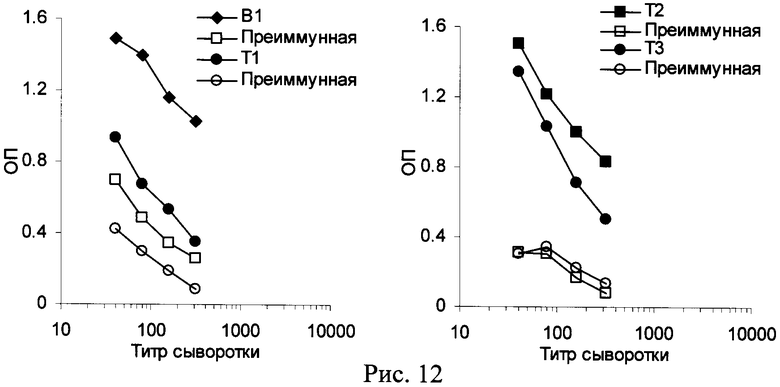
Пример 2. Поскольку природный Т-эпитоп обычно спрятан внутри молекулы белка из-за его гидрофобности, то к нему не образуются антитела. Мишенью антител являются гидрофильные экспонированные на поверхности пептиды. Однако при использовании мини-антигенов все пептиды, включая сам Т-эпитоп, оказываются доступны для антител. С большой вероятностью в этом случае антитела будут образовываться и к Т-эпитопу. Для проверки данного предположения был получен конъюгат, содержащий 4 различных пептида, один из которых был ранее идентифицирован как В-эпитоп (B1), а 3 - как потенциальные Т-эпитопы (T1, Т2, Т3). До иммунизации проверяли активность Т-эпитопов по индукции пролиферации лимфоцитов периферической крови кроликов №№1110-1112. Кролик №1111 ответил только на пептид Т2, остальные кролики вообще не ответили на пептиды (Рис.11). Поскольку мы отбирали Т-эпитопы по литературным данным для человека, то для кроликов мы посчитали удачей, что хотя бы один из 3-х кроликов ответил на 1 из 3-х эпитопов. Эти данные демонстрируют, что для подбора вакцины необходимо типирование HLA больного.
Кроликов №1111 и №1110 иммунизировали конъюгатом В1-Т1-Т2-Т3 в гидроокиси алюминия 3 раза. Анализ продукции IgG антител проводили иммуно-ферментным методом, где в качестве подложки использовали индивидуальные пептиды. Показали, что кролик №1110 не ответил формированием IgG ни на один пептид (нет Т-эпитопов, подходящих под ГКГСII кролика). Кролик №1111 ответил формированием IgG на пептиды В1, Т2 и Т3 (Рис.12). Антитела на Т1 не формировались. Полученные данные показали два факта: 1) при отсутствии подходящего Т-эпитопа в составе конъюгата иммунный ответ (ГАСИО) формироваться не будет; 2) «голые» Т-эпитопы могут становиться В-эпитопами, к которым формируются антитела.
Аналогичные данные были нами получены и на мышах. Ни один из выбранных Т-эпитопов не распознавался мышами линии BALB/c - ни одна мышь не формировала антител на конъюгаты. Мы заменили мышей на беспородных, у которых более рандомизированный ГКГСII. У 70% беспородных мышей формировался ГАСИО на все или почти все предъявленные пептиды (данные не приводятся).
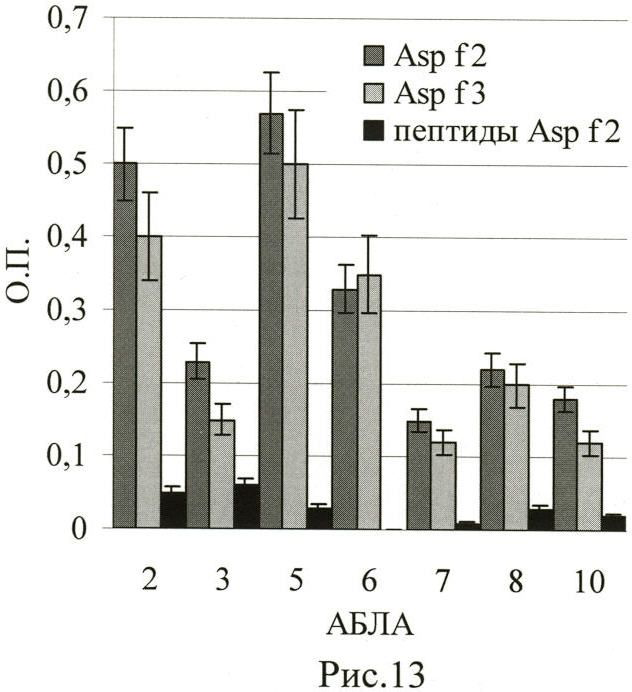
Пример 3. Линейные пептиды не содержат конформационных эпитопов, в то время как IgE чаще распознают именно конформационные эпитопы. Для проверки связывания пептидами IgE мы использовали пул синтетических пептидов Asp f2 и Asp f3, приведенных в таблицах 1 и 2. Сыворотки 7 из 10 больных АБЛА распознавали рекомбинантные белки Asp f2 и Asp f3, являющиеся основными аллергенами гриба Aspergillus fumigatus (Рис.13, серые столбики). При этом связывания с пулом линейных пептидов Asp f2 выявлено не было (Рис.13, черные столбики). Аналогичные данные были получены и для пула пептидов Asp f3 (данные не приводятся).


| название | год | авторы | номер документа |
|---|---|---|---|
| Способ капсулирования аллергенов в частицы хитозана для снижения распознавания аллергенов IgE антителами при проведении аллерген-специфической подкожной и трансэпидермальной иммунотерапии | 2020 |
|
RU2760672C1 |
| ГИБРИДНЫЕ БЕЛКИ-НОСИТЕЛИ ПЕПТИДОВ В КАЧЕСТВЕ ВАКЦИН ОТ АЛЛЕРГИИ | 2012 |
|
RU2630652C2 |
| Рекомбинантный полипептид на основе аллергена пыльцы березы и аллергена яблока в качестве вакцины от аллергии | 2020 |
|
RU2761431C1 |
| ВАКЦИНА, ОБЛАДАЮЩАЯ СПОСОБНОСТЬЮ ВЫЗЫВАТЬ ИММУННЫЙ ОТВЕТ ПРОТИВ СОБСТВЕННОГО JGE ОРГАНИЗМА, СПОСОБ ПОЛУЧЕНИЯ ВАКЦИНЫ И СПОСОБ ИММУНИЗАЦИИ МЛЕКОПИТАЮЩЕГО ПРОТИВ ОПОСРЕДУЕМЫХ JGE АЛЛЕРГИЧЕСКИХ РЕАКЦИЙ | 1992 |
|
RU2120805C1 |
| ИММУНОРЕГУЛЯТОРНАЯ ВАКЦИНА | 2013 |
|
RU2650709C2 |
| МОЛОЧНО-КИСЛЫЕ БАКТЕРИИ В КАЧЕСТВЕ АГЕНТОВ ДЛЯ ЛЕЧЕНИЯ И ПРЕДОТВРАЩЕНИЯ АЛЛЕРГИИ | 2002 |
|
RU2312892C2 |
| КЛЕЩЕВОЙ АЛЛЕРГЕН ДОМАШНЕЙ ПЫЛИ | 2007 |
|
RU2570563C2 |
| ПЕПТИДНЫЕ ИММУНОГЕНЫ И СОДЕРЖАЩИЕ ИХ СОСТАВЫ, НАЦЕЛЕННЫЕ НА МЕМБРАНОСВЯЗАННЫЕ IgE ДЛЯ ЛЕЧЕНИЯ ОПОСРЕДОВАННЫХ IgE АЛЛЕРГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2769983C2 |
| Иммуногенная композиция, включающая ее вакцина, набор для приготовления вышеуказанной композиции и способ лечения заболеваний, связанных с патологией секреции гастрина | 2014 |
|
RU2664197C2 |
| НОВЫЕ МУТАНТНЫЕ АЛЛЕРГЕНЫ | 2001 |
|
RU2285042C2 |
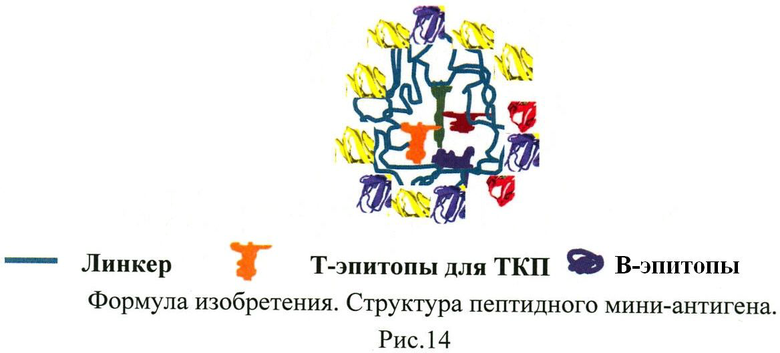
Изобретение раскрывает искусственные пептидные мини-антигены (ПМА), которые могут применяться для индукции контролируемого протективного гуморального IgG-опосредованного иммунного ответа против аллергена. Мини-антиген включает: 1) гидрофобные пептиды из различных белков и любых патогенов, которые связываются с карманами наиболее распространенных молекул ГКГС (главный комплекс гистосовместимости) II класса и к которым в популяции людей имеются Т-клетки памяти (Т-эпитопы Т-клеток памяти (ТКП)), 2) и гидрофильные пептиды из различных белков, находящихся на поверхности аллергена дикого типа, которые конъюгированы на носитель оболочки полимерной частицы и доступны для антител. (Структура мини-антигена приведена на рис.14 в тексте описания.) Особенностью мини-антигенов является возможность замены В-эпитопов на пептиды другого аллергена при сохранении ядра Т-эпитопов ТКП, что позволяет использовать пептидные мини-антигены для создания противоаллергенных вакцин против различных типов аллергии. Наличие Т-клеток памяти поможет значительно быстрее запустить иммунный ответ. Введение в конструкцию пептидов только основных белков аллергена позволит осуществить противоаллергенную защиту при минимальной нагрузке на иммунную систему. Отсутствие связывания IgE пептидами сделает вакцинирование безопасным и быстрым. 14 ил., 2 табл., 3 пр.
Гетерологичный пептидный мини-антиген в составе полимерной частицы для создания противоаллергенной вакцины, включающий:
- гидрофобные пептиды, которые связываются с карманами наиболее распространенных молекул ГКГС II класса (HLA-DR) и к которым в популяции людей имеются Т-клетки памяти, пептиды происходят из любых белков и любых патогенов, пептиды упакованы в ядро полимерного носителя и недоступны для антител (Рис.14);
- гидрофильные пептиды из различных белков одного аллергенного комплекса, происходящие из белков, находящихся на поверхности дикого аллергена, пептиды конъюгированы на носитель оболочки полимерной частицы и доступны для антител.
| ЕР 1547581 А1, 29.06.2005 | |||
| ВЕЗИКУЛЫ С РЕГУЛИРУЕМЫМ ВЫСВОБОЖДЕНИЕМ АКТИВНЫХ ИНГРЕДИЕНТОВ | 1994 |
|
RU2160093C2 |
| КОМПОЗИЦИИ ПОЛИМЕРНЫХ МИЦЕЛЛ | 2001 |
|
RU2308943C2 |
| ЛИПОСОМЫ, ОСУЩЕСТВЛЯЮЩИЕ ТИМУС-ЗАВИСИМУЮ ПОМОЩЬ "СЛАБЫМ" АНТИГЕНАМ, ИСПОЛЬЗУЕМЫМ ДЛЯ ПРИГОТОВЛЕНИЯ ВАКЦИНЫ | 1993 |
|
RU2107493C1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |

