Настоящее изобретение относится к scFv синтетическому аналогу (R6313/G2) вариабельных доменов моноклонального антитела 6313/G2 к рецептору ангиотензина II типа 1.
Ангиотензин II играет центральную роль в электролитном гомеостазе млекопитающих и регулировании кровяного давления (Peach Physiol. Rev 57 313-370 (1977); Vinson et al. "The Adrenal Cortex", Prentice Hall, Englefield Heights (1992)). Были обнаружены два основных типа рецепторов ангиотензина II, обозначенные как тип 1 и тип 2 (АТ1 и АТ2), однако большинство хорошо известных проявлений активности ангиотензина II происходит по механизму подтипа АТ1 (Herblin et al. Am. J. Hypertens. 4 299S-302S (1991); Ouali et al. J. Steroid. Biochem. Mol. Biol. 43 271-280 (1992)).
Для изучения действия рецептора (Vinson et al. Mol. Med. Today 1 35-38 (1995)) применяли моноклональное антитело 6313/G2 к рецептору подтипа АТ1 (Barker et al. J. Mol. Endocrinol. 11 241-245 (1993)). Было предложено применять моноклональное антитело в качестве терапевтического агента для контроля сужения кровеносных сосудов, например, при лечении гипертензии или сокращения других клеток гладких мышц (например, матки).
Указанное антитело применяли в качестве специфического визуализирующего агента в различных тканях, например, при раке гортани (Marsigliante et al. Cancer Letters 110 19-27 (1996)), почек (Harrison-Bernard et al. Am. J. Physiol. 42 F170-F177 (1997); Cheng et al. J. Physiol. 43 F10-F17 (1998)) и головного мозга (Yang et al. J. Neuroscience 17 1660-1669 (1997)). Было показано, что данное антитело блокирует вызванную ангиотензином-II интернализацию рецептора АТ1 и активацию РКС (протеинкиназы С), но с другой стороны, стимулирует чувствительность к кальцию (Kapas et al. Biochem. Biophys. Res. Comm. 204 1292-1298 (1994); Vinson et al. J. Endocrinol. 141 R5-R9 (1994)). Сообщалось о присутствии рецепторов АТ1 и АТ2 в опухолях молочной железы, связываемом с местной выработкой ангиотензина (Inwang et al. Brit. J. Cancer 75 1279-1283 (1997); Tahmasebi et al. Eur. J. Cancer 34 1777-1782 (1998)).
Моноклональное антитело 6313/G2 секретируется гибридомной клеточной линией, депонированной 22 июля 1993 г. согласно Будапештскому договору в Европейскую коллекцию клеточных культур животных, Porton Down, Великобритания, с присвоением номера доступа 93072117. Депонирование было произведено доктором Gavin P Vinson и доктором Stewart Barker, отделение биохимии, колледжа Queen Mary & Westfield College, Mile End Road, Лондон Е1 4NS. Депонент уполномочил заявителя передать на рассмотрение материал, депонируемый в заявке, и предоставил его безусловное и безотзывное согласие на общедоступность в соответствии с Правилом 28(1)(d) Европейской Патентной Конвенции.
Данная гибридомная клеточная линия продуцирует антитело, которое специфично связывает аминокислотные остатки 8-17 рецептора АТ1 гладких мышц кровеносных сосудов крысы, данная последовательность была также обнаружена в рецепторе АТ1 клеток человека и быка. Эпитопная последовательность представляет собой следующую:
EDGIKRIQDD или альтернативно представлена в виде:
NH2-Glu-Asp-Gly-Ile-Lys-Arg-Ile-Gln-Asp-Asp-COOH
Были получены моноклональные антитела к пептидной последовательности, содержащей N-концевую последовательность рецептора ангиотензина II типа 1 (Barker et al. Journal of Molecular Endocrinology 11 241-245 (1993); WO 95/09186). Сообщалось, что такие моноклональные антитела могут найти дополнительное терапевтическое применение при некоторых медицинских состояниях, где возможность такого терапевтического применения ранее не предполагалась или не была показана (WO 2004/018519). Предполагается, что такие терапевтические эффекты состоят в способности моноклональных антител блокировать вредное действие ангиотензина II при имеющих отношение к ангиотензину II медицинских состояниях, в то же время сохраняя полезные эффекты данной молекулы.
К настоящему времени было показано, что scFv синтетические аналоги моноклональных антител, специфичные к рецептору АТ1, обладают ценными и неожиданными свойствами, позволяющими предложить применение таких аналогов для терапии или лечения заболеваний. Авторы настоящего изобретения получили как аналоги scFv мышей, так и гуманизированные варианты аналогов scFv мышей.
Согласно первому аспекту настоящего изобретения, предложена специфически связывающаяся молекула, которая специфически связывается с пептидом, имеющим аминокислотную последовательность EDGIKRIQDD, и содержит полипептид, включающий VL домен иммуноглобулина, связанный с VH доменом иммуноглобулина, причем домен VL содержит участки, определяющие комплементарность (CDRs) VLCDR1, VLCDR2 и VLCDR3, и домен VH содержит участки, определяющие комплементарность (CDRs) VHCDR1, VHCDR2, VHCDR3, каждый из которых соответственно имеет следующую аминокислотную последовательность, где
VHCDR1 представляет собой GYSFTGYNMN
VHCDR2 представляет собой NIDPYYGGTTYNQKFKG
VHCDR3 представляет собой EVDY
VLCDR1 представляет собой RASKSVSTSGYSYMH
VLCDR2 представляет собой LVSNLES
VLCDR3 представляет собой QHIRELTRSEG
или аминокислотную последовательность, по меньшей мере на 70% идентичную указанным последовательностям.
Согласно одному из вариантов реализации настоящего изобретения, предложена специфически связывающаяся молекула, которая специфически связывается с пептидом, имеющим аминокислотную последовательность EDGIKRIQDD, и содержит полипептид, включающий домен иммуноглобулина VL, связанный с VH доменом иммуноглобулина, причем домен VL содержит участки, определяющие комплементарность (CDRs) VLCDR1, VLCDR2 и VLCDR3, и домен VH содержит участки, определяющие комплементарность (CDRs) VHCDR1, VHCDR2, VHCDR3, каждый из которых соответственно имеет следующую аминокислотную последовательность, где
VHCDR1 представляет собой GYSFTGYNMN или GYSFTGYNMS
VHCDR2 представляет собой NIDPYYGGTTYNQKFKG
VHCDR3 представляет собой EVDY
VLCDR1 представляет собой RASKSVSTSGYSYMH
VLCDR2 представляет собой LVSNLES
VLCDR3 представляет собой QHIRELTRSEG
или аминокислотную последовательность, по меньшей мере на 80% идентичную указанным последовательностям.
В другом варианте реализации настоящего изобретения CDRs специфически связывающиеся молекулы имеют следующие аминокислотные последовательности:
VHCDR1 представляет собой GYSFTGYNMN или GYSFTGYNMS
VHCDR2 представляет собой NIDPYYGGTTYNQKFKG
VHCDR3 представляет собой EVDY
VLCDR1 представляет собой RASKSVSTSGYSYMH
VLCDR2 представляет собой LVSNLES или LVSDLED
VLCDR3 представляет собой QHIRELTRSEG.
CDRs обозначены согласно определению комбинаций консервативных последовательностей (Kabat et al. in "Sequences of Proteins of Immunological Interest", Nat'l. Inst. Health, Bethesda, MD (1987)), и структурному определению (Chothia and Lesk J. Mol Biol. 196: 901-17 (1987)). Данные определения также впоследствии были описаны в источнике Carter et al, Proc Nat'l Acad Sci USA. 89: 4285-9 (1992).
Аминокислоты могут быть названы с применением трехбуквенной или однобуквенной кодировки, а также в следующем виде: глицин (G или Gly), аланин (А или Ala), валин (V или Val), лейцин (L или Leu), изолейцин (I или Не), пролин (Р или Pro), фенилаланин (F или Phe), тирозин (Y или Tyr), триптофан (W или Trp), лизин (К или Lys), аргинин (R или Arg), гистидин (Н или His), аспарагиновая кислота (D или Asp), глутаминовая кислота (Е или Glu), аспарагин (N или Asn), глутамин (Q или Gln), цистеин (С или Cys), метионин (М или Met), серин (S или Ser) и треонин (Т или Thr). Если остатком является аспарагиновая кислота или аспарагин, могут применяться символы Asx или В. Если остатком является глутаминовая кислота или глутамин, могут применяться символы Glx или Z. Ссылки на аспарагиновую кислоту включают аспартат, а ссылки на глутаминовую кислоту включают глутамат, если из контекста не следует обратное.
Настоящее изобретение также включает варианты вышеуказанных пептидных последовательностей. Примером варианта настоящего изобретения является гибридный белок, содержащий вышеуказанный пептид, в котором одна или более аминокислот заменена на одну или более из других аминокислот. Специалист в данной области знает, что различные аминокислоты имеют похожие свойства. Одна или более из таких аминокислот в составе вещества часто может быть заменена на одну или более из других таких аминокислот без потери необходимой активности такого вещества.
Так, аминокислоты глицин, аланин, валин, лейцин и изолейцин часто могут быть заменены на другие (аминокислоты, содержащие алифатические боковые цепи). Из указанных допустимых замещений предпочтительно, чтобы взаимозаменялись аминокислоты глицин и аланин (поскольку они содержат относительно короткие боковые цепи) и чтобы взаимозаменялись аминокислоты валин, лейцин и изолейцин (поскольку они содержат алифатические боковые цепи большего размера, которые являются гидрофобными). Другие аминокислоты, которые могут взаимозаменяться, включают: фенилаланин, тирозин и триптофан (аминокислоты, содержащие ароматические боковые цепи); лизин, аргинин и гистидин (аминокислоты, содержащие боковые цепи основного характера); аспартат и глутамат (аминокислоты, содержащие боковые цепи кислотного характера); аспарагин и глутамин (аминокислоты, содержащие амидные боковые цепи), и цистеин и метионин (аминокислоты, содержащие в боковых цепях серу).
Замены такого рода часто называют "консервативными" или "полуконсервативными" аминокислотными заменами.
В аминокислотной последовательности вышеуказанного гибридного белка также могут производиться делеции и вставки аминокислот. Так, например, аминокислоты, не оказывающие существенного влияния на активность полипептида или по меньшей мере те аминокислоты, удаление которых не устраняет указанную активность, могут быть удалены. Такие делеции могут быть ценными, поскольку они влекут уменьшение общей длины и молекулярной массы полипептида без ущерба для сохранения активности. Это может помочь уменьшить количество полипептида, требуемого для достижения конкретной цели, например могут быть уменьшены уровни дозировки.
В аминокислотной последовательности вышеуказанного гибридного белка могут производиться вставки. Это может быть. сделано для изменения свойств вещества согласно настоящему изобретению (например, для облегчения идентификации, очистки или экспрессии, как объяснено выше по отношению к гибридным белкам).
Изменения аминокислотной последовательности, представленные выше, могут быть произведены с применением любой подходящей технологии, например с применением направленного мутагенеза или твердофазного синтеза.
Считается предпочтительным, что замены или вставки аминокислот в пределах объема притязаний настоящего изобретения могут производиться с применением встречающихся в природе или не встречающихся в природе аминокислот. Независимо от того, применяли ли природные или синтетические аминокислоты, предпочтительно, чтобы присутствовали только L-аминокислоты.
"Идентичность", как известно специалистам в данной области техники, является соотношением между двумя или более полипептидными последовательностями или двумя или более полинуклеотидными последовательностями, определяемым путем сравнения последовательностей. В данной области техники идентичность также означает степень сходства последовательностей между двумя полипептидными или полинуклеотидными последовательностями, которую в зависимости от обстоятельств, можно определить сопоставлением цепочек таких последовательностей. Несмотря на то, что существует ряд способов для оценки идентичности двух полипептидных или двух нуклеотидных последовательностей, наиболее часто применяют способы, которые позволяют определить идентичность при помощи закодированных компьютерных программ. Предпочтительные компьютерные программы для определения идентичности двух последовательностей включают пакет программ GCG (Devereux, et al., Nucleic acids Research, 12, 387 (1984), BLASTP, BLASTN и FASTA (Atschul et al., J. Molec. Biol. 215, 403 (1990), но не ограничиваются им.
Предпочтительно аминокислотная последовательность CDRs согласно данному изобретению имеет идентичность по меньшей мере 70%, которая определена с примением параметров, принимающих значение по умолчанию для компьютерной программы BLAST (Atschul et al,, J. Mol. Biol. 215, 403-410 (1990), предлагаемой HGMP (Проект картирования генома человека), на аминокислотном уровне, к вышеописанной аминокислотной последовательности CDRs VH и VL.
Более предпочтительно последовательность CDR может иметь по меньшей мере 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% по меньшей мере 99% идентичности на аминокислотном уровне по отношению к последовательностям, показанным выше.
В одном из вариантов реализации настоящего изобретения специфически связывающаяся молекула может иметь следующее размещение CDRs
(R6313клон12D и гуманизированные варианты HuCY и вар3)
VHCDR1:GYSFTGYNMN
VHCDR2: NIDPYYGGTTYNQKFKG
VHCDR3: EVDY
VLCDR1: RASKSVSTSGYSYMH
VLCDR2: LVSNLES
VLCDR3: QHIRELTRSEG
или
(Р6313клон 11В)
VHCDR1: GYSFTGYNMS
VHCDR2: NIDPYYGGTTYNQKFKG
VHCDR3: EVDY
VLCDR1: RASKSVSTSGYSYMH
VLCDR2: LVSNLES
VLCDR3: QHIRELTRSEG
или
(гуманизированный вариант вар4)
VHCDR1: GYSFTGYNMN
VHCDR2: NIDPYYGGTTYNQKFKG
VHCDR3: EVDY
VLCDR1: RASKSVSTSGYSYMH
VLCDR2: LVSDLED
VLCDR3: QHIRELTRSEG.
В одном из вариантов реализации изобретения специфически связывающаяся молекула может содержать вариабельную тяжелую цепь (VH) и вариабельную легкую цепь (VL), соединяемые пептидным линкером. Линкер может содержать от 1 до 20 аминокислот, таких как, например, 1, 2, 3 или 4 аминокислоты, 5, 10 или 15 аминокислот, или другое промежуточное количество в диапазоне от 1 до 20, в зависимости от необходимости. Пептидный линкер может быть образован остатками любых обычно применяемых аминокислот, таких как глицин и/или серин. Одним из примеров подходящего линкера является Gly4Ser. Можно применять мультимеры таких линкеров, такие как, например, димер, тример, тетрамер или пентамер, например (Gly4Ser)2, (Gly4Ser)3, (Gly4Ser)4 или (Gly4Ser)5. Однако в других вариантах реализации изобретения пептидный линкер может присутствовать, и домен VL может быть связан с доменом VH пептидной связью.
Специфически связывающаяся молекула может представлять собой одноцепочечный вариабельный аналог (scFv). Специфически связывающаяся молекула или scFv может быть связана с другими специфически связывающимися молекулами (например, с другими scFv, фрагментами антител, Fab, химерными антителами IgG (например, с человеческими каркасными участками)) или связана с другими scFv согласно настоящему изобретению таким образом, чтобы образовался мультимер, представляющий собой мультиспецифично связывающий белок, например, димер, тример или тетрамер Специфичные к двум молекулам scFv иногда называют диателами, специфичные к трем молекулам называют триателами и специфичные к четырем молекулам тетрателами, в которых каждый scFv в димере, тримере или тетрамере обладает различной специфичностью. Диатела, триатела и тетратела также могут являться моноспецифичными, если каждый scFv в димере, тримере или тетрамере обладает одинаковой специфичностью.
В одном из вариантов реализации настоящего изобретения специфически связывающаяся молекула может представлять собой scFv аналог моноклонального антитела, идентифицируемый как R6313/G2. В настоящей заявке данный scFv также обозначают как R6313клон120 или клон 12D с последовательностью, показанной на Фиг.14. В другом варианте реализации настоящего изобретения специфически связывающаяся молекула может представлять собой scFv аналог моноклонального антитела, идентифицируемый как К6313клон11В в настоящей заявке, обозначенный также клон как 11 В с последовательностью, показанной также на Фиг.14.
scFv может быть получен в соответствии с любой пригодной технологией с применением стандартных химических или молекулярно-биологических способов. В одном варианте реализации изобретения аналоги моноклонального антитела могут быть приготовлены в виде scFv из демонстрационной библиотеки фаговых антител человека, ранее не подвергавшихся экспериментам (McCafferty et al., Nature 348, 552-554 (1990); также описанной в WO 92/01047).
Аналог моноклонального антитела может быть гуманизирован за счет модификации аминокислотной последовательности scFv. Способы уменьшения иммуногенности специфично связывающих молекул согласно изобретению могут включать имплантацию CDR (участков, определяющих комплементарность) на подходящий участок трехмерной подложки каркаса антитела или реконструкцию вариабельных остатков на поверхности, например посредством сайт-специфичного мутагенеза или других технологий, общепринятых в молекулярной биологии (Roguska et al. Protein Eng. 9 895-904 (1996)).
Другие приемлемые способы могут включать идентификацию потенциальных эпитопов Т-клеток внутри молекулы с последующим их удалением, например, посредством сайт-специфичного мутагенеза (де иммунизация). Гуманизация специфично связывающейся молекулы может стать необходимой, если молекулу предполагается применять в качестве терапевтического агента. При необходимости может проводиться гуманизация CDR участков или окружающих их каркасных последовательностей.
Авторы настоящей заявки продуцировали, гуманизированные варианты scFv R6313/G2, как описано в примерах.
В одном из вариантов реализации изобретения специфически связывающаяся молекула может представлять собой одно или более гуманизированных scFv, идентифицируемых как HuCY, вар3 и/или вар4. Последовательности этих гуманизированных scFv показаны на Фиг.14.
Согласно другому аспекту настоящего изобретения предложена фармацевтическая композиция, содержащая вышеописанную специфически связывающуюся молекулу.
Композиция, применяемая в соответствии с данным аспектом изобретения, может быть приготовлена любым подходящим способом. Как правило, лекарственное средство будет поставляться как составная часть стерильной фармацевтической композиции, обычно содержащей фармацевтически приемлемый носитель. Композиции согласно изобретению можно применять в комбинации с фармацевтически приемлемым носителем или носителями, или адъювантом, и/или разбавителем. Такие носители могут включать, не ограничиваясь перечисленными: физиологический раствор, буферный физиологический раствор, декстрозу, липосомы, воду, глицерин, полиэтиленгликоль, этанол и их комбинации. Данная фармацевтическая композиция может находиться в любой подходящей форме (в зависимости от требуемого способа введения пациенту).
Указанная композиция может быть представлена в единичной лекарственной форме, как правило, в герметичном контейнере, а также может быть представлена в виде составной части набора. Такой набор будет обычно включать (хотя и не обязательно) инструкции по применению. Он может содержать множество указанных единичных лекарственных форм.
Фармацевтическая композиция может быть приспособлена для введения любым приемлемым способом, например, пероральным (включая буккальный или подъязычный), ректальным, назальным, местным (включая буккальный, подъязычный или чрескожный), вагинальным или парентеральным способом (включая подкожный, внутримышечный, внутривенный или внутрикожный). Такие композиции могут быть приготовлены любым способом, известным специалистам в фармации, например посредством смешивания активного ингредиента с носителем(носителями) или наполнителем(наполнителями) в стерильных условиях.
Фармацевтические композиции, предназначенные для перорального введения, могут быть представлены в виде дискретных единиц, таких как капсулы или таблетки; в виде порошков или гранул; в виде растворов, сиропов или суспензий (в водных или неводных жидкостях; или в виде съедобных пен или взбитых масс, или в виде эмульсий).
Подходящие наполнители для таблеток или твердых желатиновых капсул включают лактозу, кукурузный крахмал или их производные, стеариновую кислоту или ее соли.
Подходящие наполнители для применения в виде мягких желатиновых капсул включают, например, растительные масла, воски, жиры, полутвердые или жидкие полиолы и так далее.
Для приготовления растворов и сиропов, могут применяться наполнители, которые могут включать, например, воду, полиолы и сахара. Для приготовления суспензий (например, в растительных маслах) можно применять масла, получая суспензии типа масло-в-воде или вода-в-масле.
Фармацевтические композиции, приспособленные для чрескожного введения, могут быть представлены в виде дискретных пластырей, предназначенных для сохранения тесного контакта с эпидермисом реципиента в течение продолжительного периода времени. Например, активный ингредиент может доставляться из пластыря путем лекарственного электрофореза, как в общем виде описано в Pharmaceutical Research, 3(6):318 (1986).
Фармацевтические композиции, предназначенные для местного применения, можно получать в виде мазей, кремов, суспензий, лосьонов, порошков, растворов, паст, гелей, спреев, аэрозолей или масел. При инфекциях глаза и других внешних тканей, например, рта и кожи, данные композиции предпочтительно наносят местно в виде мази или крема. При приготовлении мази можно применять активный ингредиент совместно с парафиновой или смешиваемой с водой основой для мази. Альтернативно активный ингредиент может быть введен в состав крема с основой типа масло-в-воде или вода-в-масле. Фармацевтические композиции, приспособленные для местного введения в глаз, включают глазные капли, в которых активный ингредиент растворен или суспендирован в подходящем носителе, в особенности в водном растворителе. Фармацевтические композиции, приспособленные для местного введения в рот включают лепешки, пастилки и растворы для полоскания полости рта.
Фармацевтические композиции, приспособленные для ректального введения могут быть представлены в виде суппозиториев или клизм.
Фармацевтические композиции, приспособленные для назального введения, в которых носитель представляет собой твердое вещество, включают крупный порошок с диаметром частиц в диапазоне от 20 до 500 микрон, который вводится при вдыхании через нос, то посредством быстрой ингаляции через носовой проход из контейнера, удерживаемого вблизи носа. Подходящие композиции, в которых носитель является жидким, для введения в виде назального спрея или капель в нос, включают водные или масляные растворы активного ингредиента.
Фармацевтические композиции, приспособленные для введения при ингаляции, включают мелко размолотые пудры или аэрозоли, которые могут быть получены с применением различных типов ингаляторов отмеренных доз, работающих под давлением, распылителей, или инсуффляторов.
Фармацевтические композиции, приспособленные для вагинального введения, могут быть представлены в составе пессариев, тампонов, кремов, гелей, паст, пен или спреев.
Фармацевтические композиции, приспособленные для парентерального введения, включают водные и неводные стерильные растворы для инъекций, которые могут содержать антиоксиданты, буферы, бактериостатики и растворенные вещества, делающие композицию по существу изотоничной с кровью целевого реципиента, и водные и неводные стерильные суспензии; которые могут содержать суспендирующие агенты и загустители. Наполнители, которые можно применять в растворах для инъекций включают, например, воду, спирты, полиолы, глицерин и растительные масла. Композиции могут быть предоставлены в контейнерах для единичной дозы или контейнерах, содержащих многократные дозы, например в герметичных ампулах и флаконах, которые могут храниться в лиофилизованном состоянии, при применении которых требуется только добавление непосредственно перед использованием стерильного жидкого носителя, например воды для инъекций. Инъекционные растворы и суспензии для приготовления могут быть приготовлены из стерильных порошков, гранул и таблеток.
Фармацевтические композиции могут содержать консерванты, солюбилизаторы, стабилизаторы, смачивающие агенты, эмульгаторы, подсластители, красители, ароматизаторы, соли (вещества согласно настоящему изобретению могут сами непосредственно быть представлены в форме фармацевтически приемлемой соли), буферы, покрытия или антиоксиданты. Фармацевтические композиции могут также содержать терапевтически активные агенты дополнительно к веществу согласно настоящему изобретению.
В некоторых вариантах реализации композиция концентрата активного лекарственного средства может содержать агент для поддержания фармацевтически приемлемой тоничности, буферный агент и фармацевтически приемлемое поверхностно-активное вещество.
Альтернативно композиция может содержать активный ингредиент плюс фосфат натрия, одноосновный фосфат натрия, двухосновный фосфат натрия, хлорид натрия, полисорбат 80 или полисорбат 20 (поверхностно-активное вещество для минимизации риска агрегации, вызванной встряхиванием) и воду (Фармакопея США/Европейская Фармакопея), необязательно при значении рН, доведенном до от примерно 6,0 до 7,0, например около 6,5.
Концентрат активного лекарственного средства может быть лиофилизован или нелиофилизован.
Другие композиции могут содержать в качестве буферного агента ацетат натрия тригидрат, хлорид натрия для модифицирования тоничности, уксусную кислоту для доведения значения рН и воду для инъекций.
Концентрат активного лекарственного средства перед введением может быть разбавлен 0,9% хлористым натрием.
Дозы вещества согласно настоящему изобретению могут варьироваться в широких пределах в зависимости от заболевания или расстройства, подлежащего лечению, возраста и состояния субъекта, подлежащего лечению и других факторов, в итоге адекватные дозы, которые необходимо применить, определяет лечащий врач.
Введение дозы можно повторять столько раз, сколько это уместно. При развитии побочных эффектов дозировка и/или частота введения доз может быть уменьшена в соответствии с обычной клинической практикой.
При введении млекопитающим, и в частности людям, ожидается, что ежедневная доза активного агента будет составлять от 1 мкг/кг до 10 мг/кг массы тела, как правило, от примерно 10 мкг/кг до 1 мг/кг массы тела. Врач в любом случае определит фактическую дозу, наиболее подходящую для данного субъекта, которая будет зависеть от ряда факторов, включая возраст, массу тела, пол и чувствительность субъекта. Вышеприведенные дозы приведены в качестве примера для среднего случая. Конечно, могут возникать обстоятельства, когда будут предпочтительными более высокие или более низкие дозы, и как таковые они будут находиться в пределах объема настоящего изобретения.
Вышеописанные специфически связывающиеся молекулы могут найти применение в медицине, например, для лечения рака. Не желая углубляться в теорию, авторы считают, что виды рака, связанные с ангиотензиновым рецептором типа 1 (AT1R) могут оказаться особенно чувствительными к терапии с применением специфически связывающихся молекул согласно изобретению. Такие виды рака многочисленны и включают рак молочной железы, рак предстательной железы, рак яичников, рак матки, рак прямой и толстой кишки, рак поджелудочной железы, рак гипофиза, хориокарциному, болезнь Ходжкина, рак кожи, рак почек, опухоли надпочечников, рак печени, рак легких, лейкоз и клетки нейробластомы.
Таким образом, данный аспект изобретения также включает способ лечения рака у субъекта, включающий введение указанному субъекту специфически связывающейся молекулы, описанной выше. Поэтому данное изобретение также включает применение вышеописанной специфически связывающейся молекулы для получения лекарственного средства для лечения рака. Способ лечения может относиться к субъекту, такому как человек или животное, и изобретение в равной мере распространяется на медицину для людей и/или ветеринарную медицину.
Согласно другому аспекту настоящего изобретения предложен комбинированный препарат вышеописанной специфически связывающейся молекулы и ангиотензина II для раздельного, одновременного или последовательного введения для лечения рака у субъекта. Рак может быть таким, как описано выше.
Согласно настоящему изобретению также предложена композиция, содержащая специфически связывающуюся молекулу согласно изобретению, как определено выше, и ангиотензин II. Такие композиции могут быть получены в виде фармацевтической композиции, содержащей фармацевтически приемлемый адъювант и/или разбавитель.
Альтернативно, согласно настоящему изобретению также предложен набор составных частей, содержащих специфически связывающуюся молекулу согласно изобретению и ангиотензин II, причем каждая из составных частей включена для введения в состав фармацевтического препарата, включая, но не ограничиваясь перечисленными: таблетки для перорального введения, ингаляторы для введения в легкие и растворы для инъекций для внутривенного введения.
Варианты реализации настоящего изобретения, относящиеся к применению ангиотензина II, могут включать любую, удобную в практическом смысле форму ангиотензина, приемлемую для применения для лечения людей или ветеринарной медицине. Приемлемо, если ангиотензин II представлен в форме лиофилизованного продукта, в котором остаток образован из раствора, содержащего Ангиотензин II, трегалозу, альбумин сыворотки человека и уксусную кислоту. Одним из поставщиков ангиотензина II является NIBSC, South Mimms, Великобритания, который поставляет ангиотензин II в виде ампул, содержащих лиофилизованный остаток, приготовленный из 0,5 мл раствора, содержащего 2,5 мкг ангиотензина II (Ileu5), 3 мг трегалозы, 1 мг альбумина сыворотки человека, уксусной кислоты концентрации 2×10-3 моль/л.
Синтетические аналоги вариабельных участков моноклонального антитела 6313/G2, такие как scFv, называемые R6313/G2, обладают уникальными и ценными свойствами, в особенности по сравнению с исходным гибридомным антителом. Например:
1. R6313/G2 существенно ингибировал рост раковых клеток, содержащихся в системе полых волокон у интактных бестимусных мышей (nu/nu mice). Это неожиданно, потому что при концентрациях 13 нмоль/кг, вводимых дважды в сутки, не было обнаружено влияния на массу тела, концентрацию альдостерона в кровотоке или активность животного, то есть другие функции, связанные с ангиотензином, оказались назатронутыми. (Примечание: исходное гибридомное антитело увеличивало секрецию альдостерона in vitro).
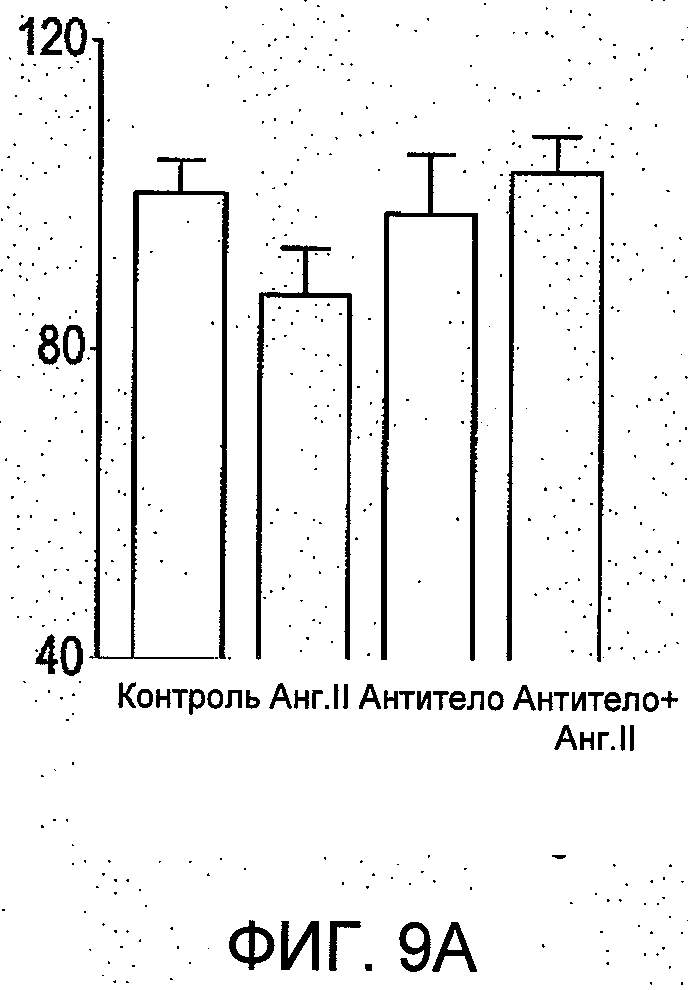
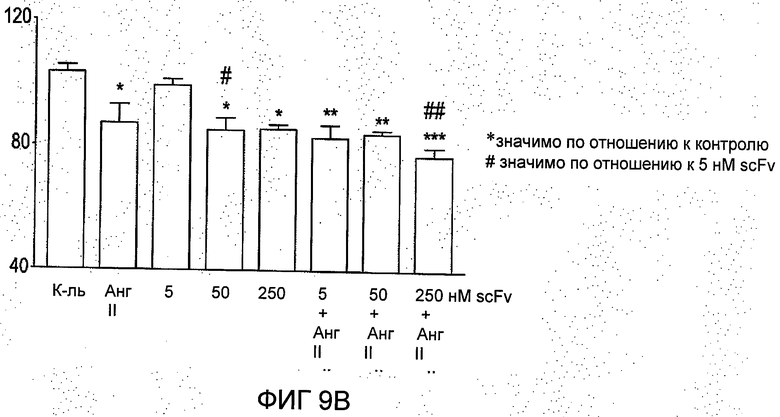
2. R6313/G2 в отдельности ингибирует инвазию клетки T-47D через воспроизведенные белки матрикса (ЕСМ) базальной мембраны, извлеченные из опухоли мышей Engelbroth-Holm-Swam (EHS) в концентрациях 50 и 250 нмоль/л (см. Фиг.9(b)). Очищенное моноклональное антитело (при 100 нмоль/л) не проявило значимого ингибирующего эффекта. Более того, в присутствии ангиотензина II в концентрации 100 нмоль/л (на Фиг.9(b)) эффект R6313/G2 становится значительно более выраженным при наибольшей концентрации R6313/G2 (250 нмоль/л) - до 25% ингибирования инвазии клеток в течение периода исследования более 24 часов.
3. В присутствии до 100 нмоль/л ангиотензина II R6313/G2 значительно ингибирует пролиферации клеток рака молочной железы. Это неожиданно, потому что требуется присутствие ангиотензина II, и R6313/G2 в отсутствие ангиотензина II неэффективен.
4. У интактных крыс in vivo, R6313/G2 существенно ингибирует повышение кровяного давления, вызванное стрессом. Это. неожиданно, поскольку R6313/G2 не уменьшает и даже может увеличить кровяное давление крови в состоянии покоя.
5. В присутствии, но не в отсутствие ангиотензина II до 100 нмоль/л R6313/G2 усиливает в клетках молочной железы апоптоз, приводимый в действие каспазой. Отсутствие ангиотензина блокирует данный эффект. По этой причине присутствие ангиотензина II неожиданно необходимо для проявления активности R6313/G2.
6. Неожиданно обнаружили, что R6313/G2 оказался намного более эффективным in vivo, чем in vitro. Так, при дозе 25 нмоль/кг in vivo показан противораковый эффект в клетках, эквивалентный эффекту, достижимому in vitro в концентрации 3,3 микромоль/л.
7. У интактных бестимусных мышей (nu/nu mice), содержащих ксенотрансплантат клеток раковых опухолей R6313/G2, существенно ингибировал рост клеток рака в дозе 13 нмоль/кг, также без проявления других эффектов.
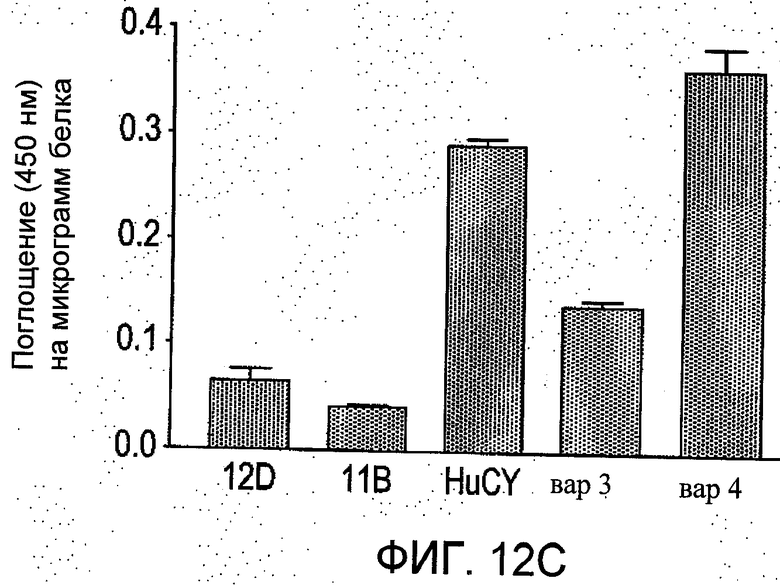
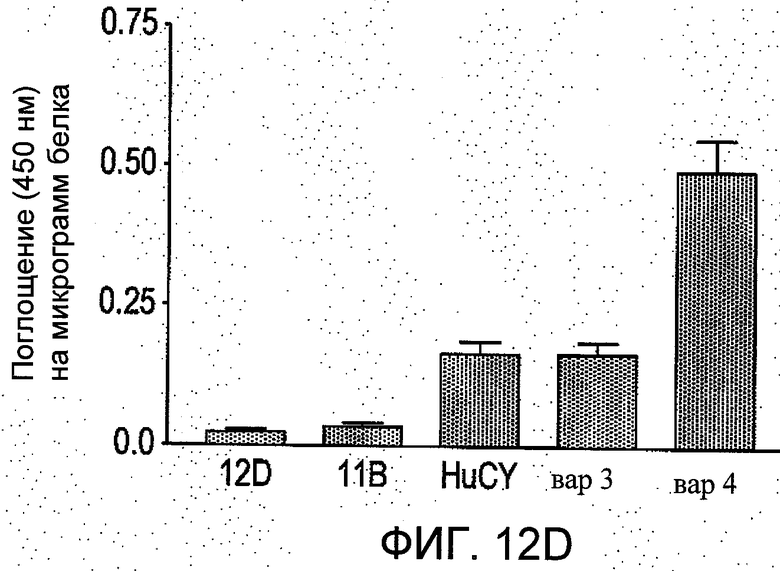
8. Как можно увидеть из Фиг.12, на повторно обработанной поверхности (гуманизированного) scFv наблюдалось повышенное связывание пептидного антигена по сравнению с scFv мыши. Следовательно, данное обстоятельство предоставляет преимущество как с точки зрения потенциальной стоимости лечения, так и с учетом выгоды от уменьшения потенциала активации иммунного ответа у человека. Более того, гуманизированный вариант вар4 показал гораздо более существенное увеличение связывания по сравнению с материнским scFv мыши. Данное обстоятельство было неожиданным, поскольку один из CDR (VLCDR2) указанного варианта был модифицирован относительно материнского scFv мышей.
Предпочтительные признаки второго и последующих аспектов настоящего изобретения представляют собой те же, что и для первого аспекта mutatis mutandis (с соответствующими изменениями).
Согласно одному из вариантов реализации настоящего изобретения предложена специфически связывающаяся молекула, содержащая полипептид с последовательностью, показанной на Фиг.14.
Настоящее изобретение будет дополнительно описано посредством следующих примеров, которые приводятся только в иллюстративных целях. В примерах делаются ссылки на ряд графических материалов, в которых:
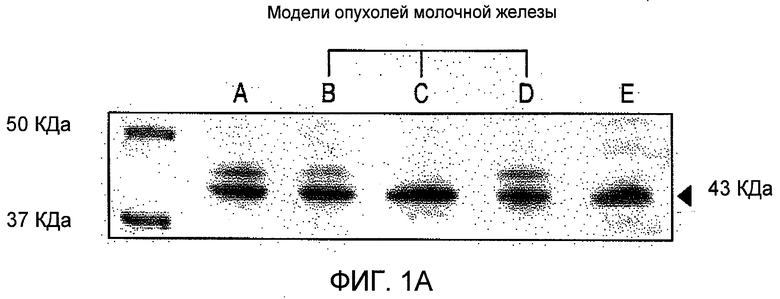
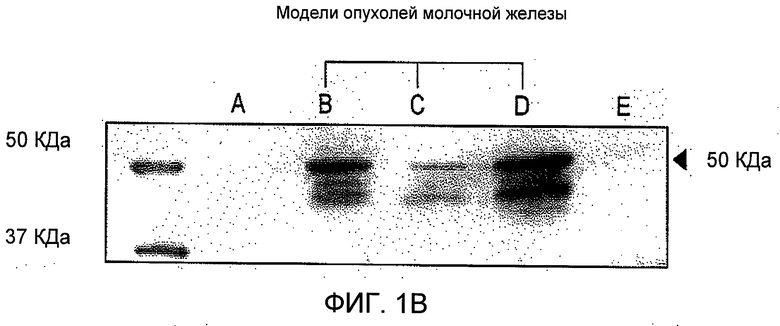
На Фиг.1 представлен иммуноблоттинг подтипов рецептора ангиотензина II. Содержание рецептора АТ1 (А) и рецептора АТ2 (В) анализировали в лизатах трех линий клеток рака молочной железы (Дорожка В, T47D; Дорожка С, MDA-MB-231; Дорожка D, MCF-7) и клетках гладк мышц (Дорожка А). Только лизаты трех линий рака молочной железы (Дорожки B-D) содержали рецептор АТ2, хотя клетки MDA-MB-231 содержали относительно меньшее количество: в RASMC (клетки гладких мышц аорты крысы) (Дорожка А) не было определимых количеств рецептора АТ2.
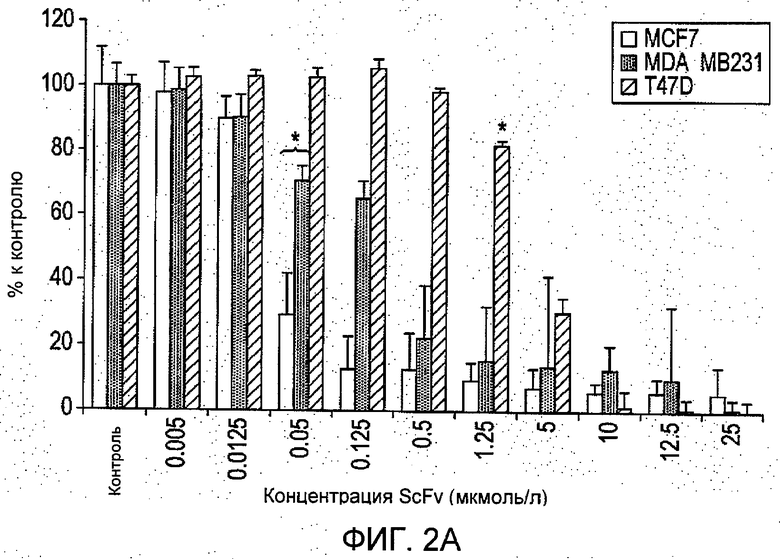
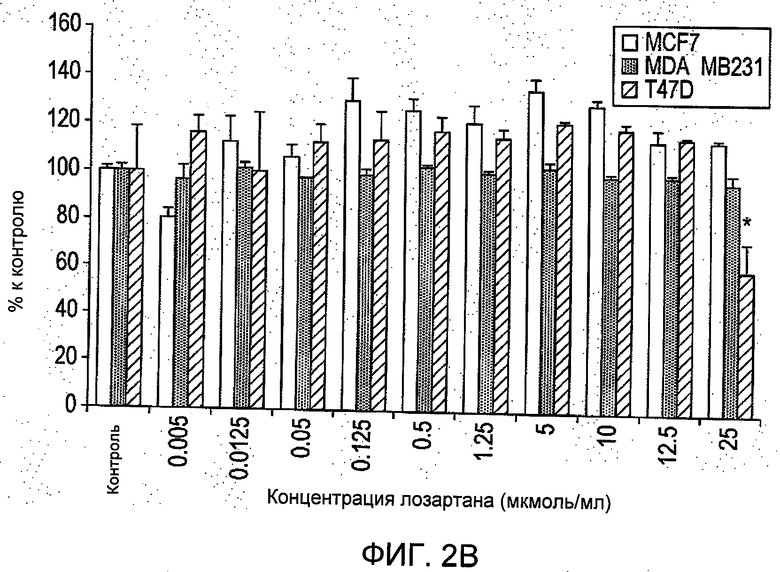
На Фиг.2 представлены (А) зависимое от дозы ингибирование выживания клеток при помощи R6313/G2 в присутствии 100 нмоль/л ангиотензина II, определенное посредством исследования с помощью ХТТ (набор для изучения пролиферации клеток) через 48 часов. Пороговые значения (*) для трех линий клеток (Р<0,05 или лучше) были 0,05 мкМ для MCF-7 и MDA-MB-231 клеток, и 1,25 мкМ для клеток T47D; значения IC50 составили: 2,8 мкмоль/л, 1,53 нмоль/л и 30 нмоль/л для T47D, MDA-MB-231 и MCF-7 клеток, соответственно. (В) Лозартан в отдельности демонстрировал ингибирование роста клеток T47D при 25 мкмоль/л на 43%, влияние на другие виды клеток в примененных дозах отсутствовало. Результаты являются средними для восьми образцов ±S.D. (квадратичное отклонение).
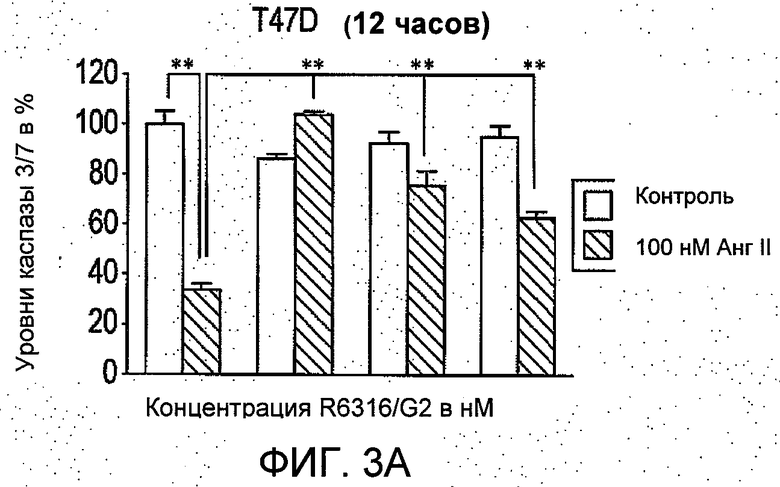
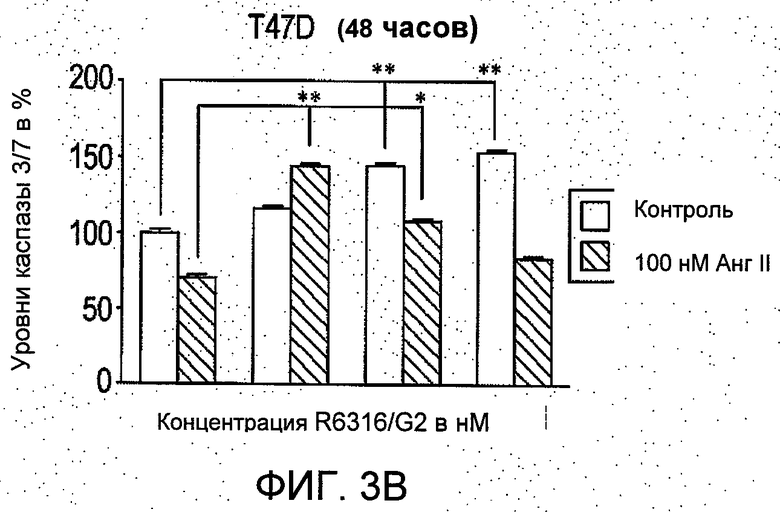
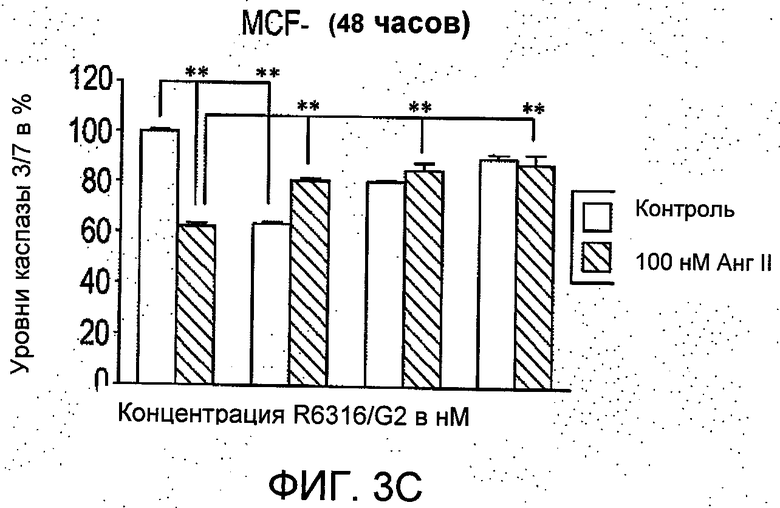
На Фиг.3 представлено (А и В) влияние R6313/G2 на активность каспазы -3/7 в клетках T47D в присутствии и при отсутствии 100 нмоль/л ангиотензина II через 12 часов и 48 часов. Через 12 часов активность каспазы 3/7, ингибируемая ангиотензином II, сравнивали с контрольными значениями для необработанных клеток, и это ингибирование было блокировано R6313/G2. Данное блокирующее действие R6313/G2 было менее выражено, если применяли более высокие дозы, но через 48 часов R6313/G2 сам по себе обусловил зависимое от дозы увеличение активности каспазы -3/7. (С) в клетках MCF7, R6313/G2 в отдельности уменьшал активность каспазы-3/7, но в присутствии ангиотензина II он зависимо от дозы увеличивал активность каспазы 3/7. Данные являются средними из восьми образцов ±S.D. *Р<0,05, **Р<0,01.
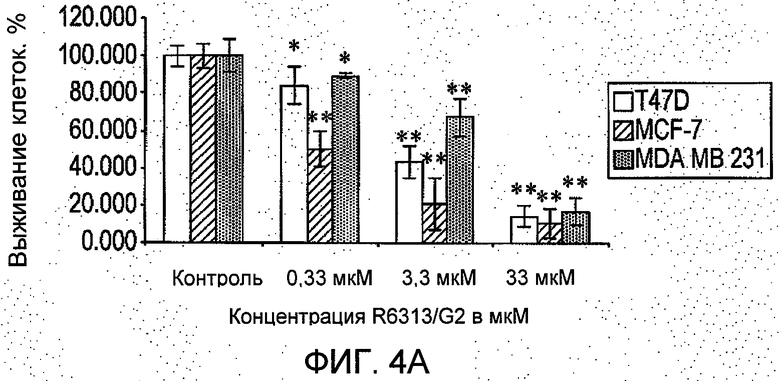
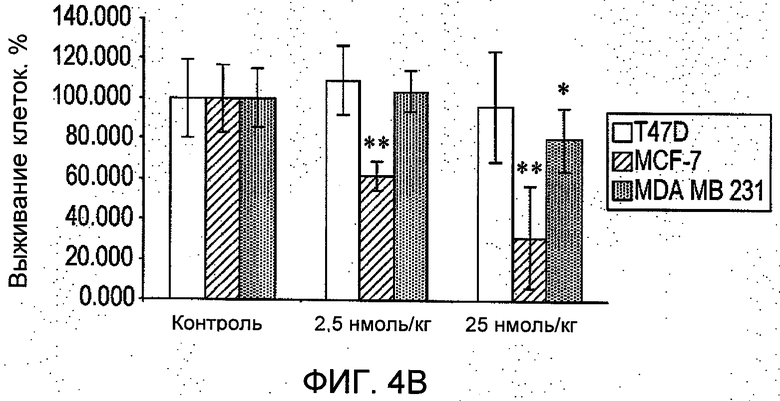
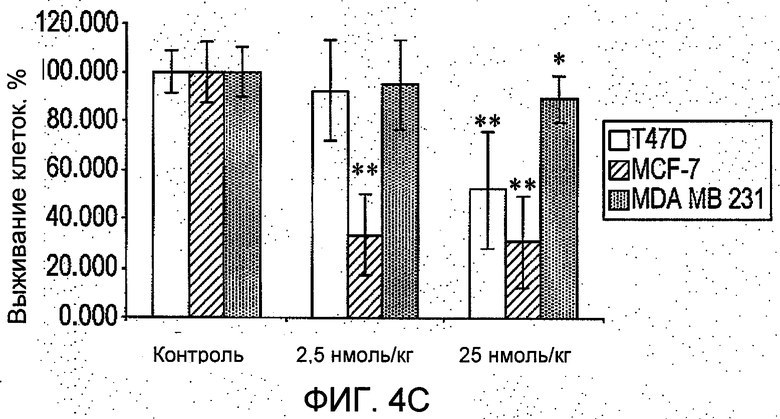
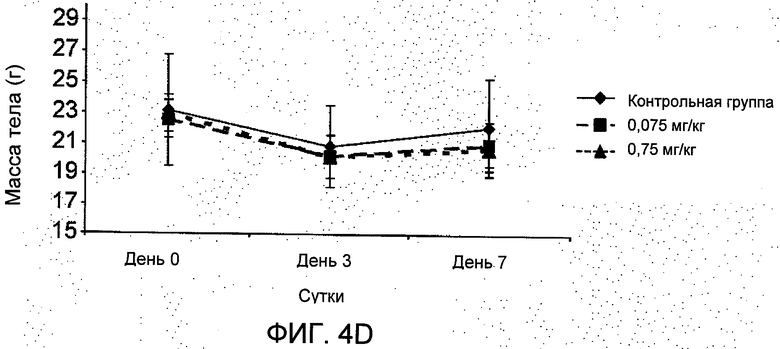
На Фиг.4 представлено: (А) в исследовании in vitro в системе полых волокон R6313/G2 в зависимости от дозы ингибировал выживание клеток рака молочной железы во всех трех линиях (T47D, MCF-7 и MDA-MB-231) в присутствии ангиотензина II через 48 часов. (В) в исследовании in vivo в системе полых волокон в месте подкожного введения R6313/G2 в зависимости от дозы ингибировал выживание клеток MCF-7, хотя клетки MDA MB 231 ингибировались только в самой высокой дозе. (С) в исследовании in vivo в системе полых волокон, в месте введения внутрибрюшинно R6313/G2, ингибировал выживание клеток MCF7 у животных, получавших 0,07 и 0,7 мг/кг (2,5 и 25 нмоль/кг) дважды в сутки, и клеток T47D и MBA MB 231 при 0,7 мг/кг (25 нмоль/кг) дважды в сутки. Значения выражены в виде средних значений ±S.D. *Р<0,05, **Р<0,001. (D) Показано, что за период лечения массы тела животных не изменились.
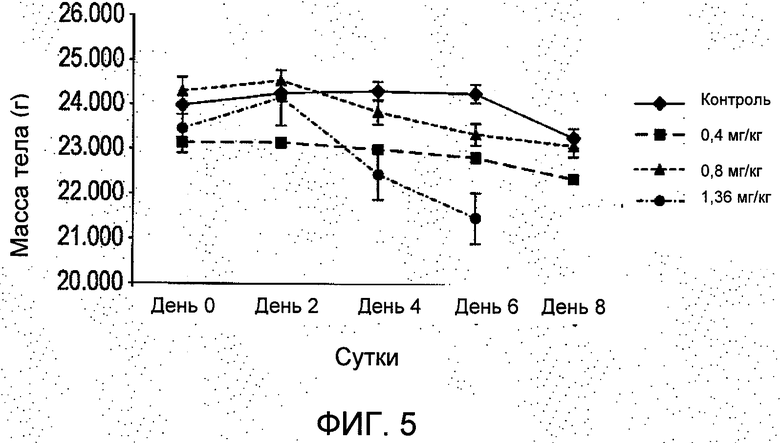
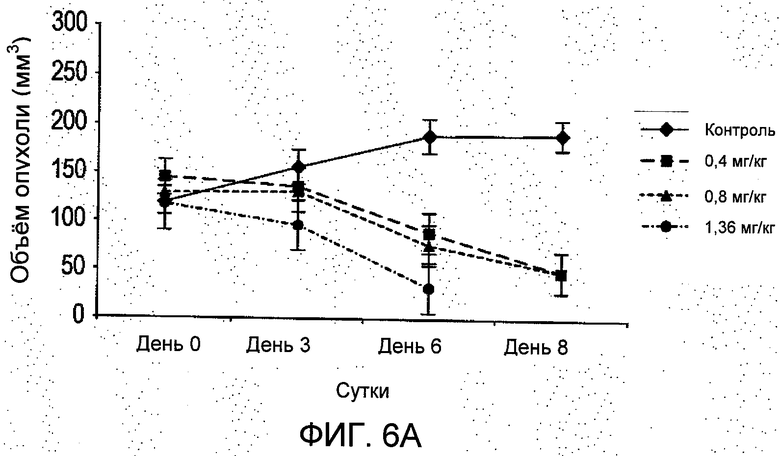
На Фиг.5 показано, что лечение R6313/G2 животных с имплантированными ксенотрансплантатами клеток MCF7 in vivo в дозах 0,4 мг/кг (13 нмоль/кг; -■-) и 0,8 мг/кг (27 нмоль/кг; -▲-); дважды за сутки не привело к уменьшению массы тела, и что за время лечения не было гибели животных, а в группе, получавшей 0,8 мг/кг group, четыре животных погибли на восьмые сутки. Масса тела существенно уменьшилась у животных, получавших 1,36 мг/кг (45,3 нмоль/кг; -•- *Р<0,05), и все они погибли на восьмые сутки 8. Данные являются средними ±S.E.M. (стандартная ошибка среднего), n=8, если не указано иное (значения n в скобках). Контроль (-♦-).
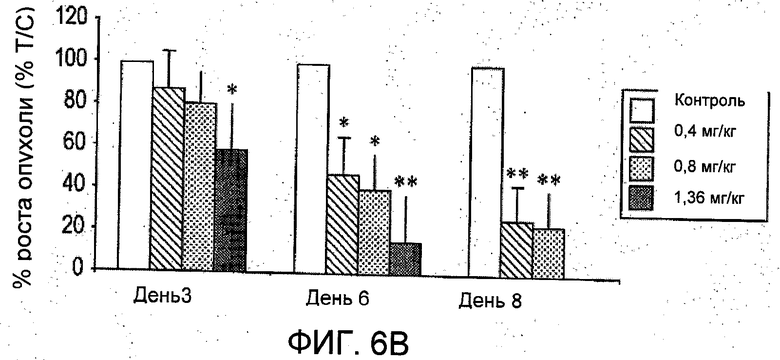
На Фиг.6 представлено влияние R6313/G2 на ксенотрансплантаты клеток MCF-7 in vivo. (А) объемы опухоли MCF-7, среднее значение ±S.E.M. (В) Те же данные, что и для (А), обработанные данные выражены в процентах от средних значений в контроле. *Р<0,05, **Р<0,001.
На Фиг.7 представлен образец ксенотрансплантата клеток MCF7 в контроле (вверху) и после обработки 0,4 мг/кг R6313/G2 в течение 7 суток, дважды в сутки, как на Фиг.6 (внизу).
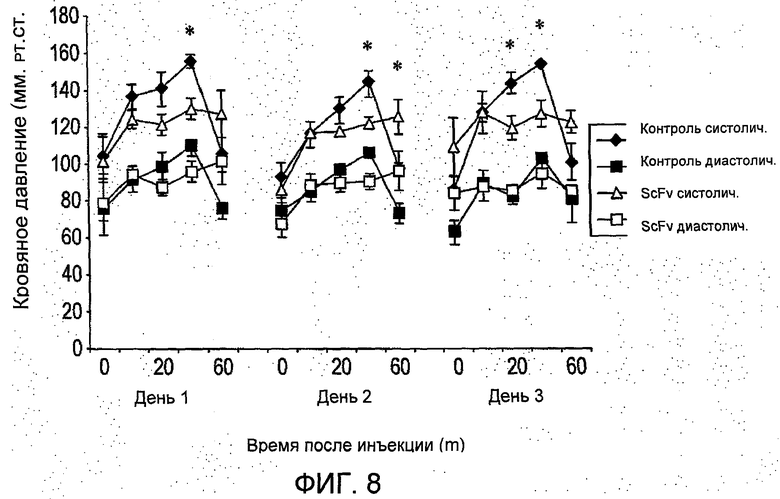
На Фиг.8 представлено значение кровяного давления у крыс, обработанных R6313/G2 (scFv, 0,4 мг/кг в сутки в течение трех суток) по сравнению с контролем (Con), который получал только фосфатно-солевой буферный раствор (PBS). *Р<0,05, сравнение диастолических давлений в Con и ScFv.
На Фиг.9 представлено сравнение очищенного моноклонального антитела из супенатанта гибридомы (Mab; Фиг.9а) и scFv, R6313/G2 клон12D (scFv; Фиг.9b), в исследованиях клеточной инвазии с применением клеток рака молочной железы T-47D. В данной модели клетки инвазии разрушают через общепринятую модель экстрацеллюларного матрикса. Фиг.9b показывает, что scFv сам по себе существенно ингибирует инвазию клеток T-47D через базальную мембрану из белков матрикса (ЕСМ), произведенную, извлеченную из опухоли мышей Engelbreth-Holm-Swam (EHS) в концентрациях 50 и 250 нмоль/л (b). Очищенное моноклональное антитело (при 100 нМ) не оказало существенного ингибирующего эффекта. Более того, в присутствии ангиотензина II (Ang II) при 100 нмоль/л (в b), влияние scFv становится значительно более выраженным при наивысшей концентрации scFv (250 нМ).
На Фиг.10 представлено выстраивание полных последовательностей клонов 12D и 11В. Эффективная аминокислотная последовательность вариантов scFv начинается у аминокислоты 9 и заканчивается на 9 остатке от конца последовательности, представленной на Фиг.10. Дополнительные аминокислоты у каждого конца включают часть pCANTAB 5E вектора сигнальной последовательности (N-конец к последовательности scFv) и пептидную последовательность, включающую часть пептида, экспрессирующего Е-маркерную последовательность (С-конец в scFv последовательности), экспрессирующийся через pCANTAB 5E и применяющуюся при афинной очистке scFv. В частности, эффективная аминокислотная последовательность начинается после МА сигнального сайта расщепления пептидазами, который составляет часть периплазматической направленной сигнальной последовательности и заканчивается перед тройной аланиновой мостиковой последовательностью и GAPVPY Е-маркерной последовательностью.
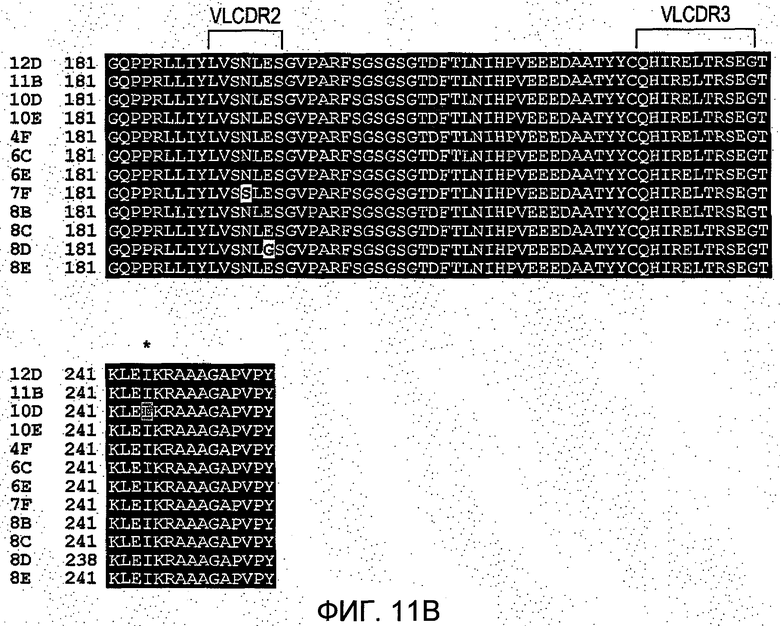
На Фиг.11 представлено выстраивание полных последовательностей клонов 12D, 11В, 10D, 10Е, 4F, 6С, 6Е, 7F, 8В, 8С, 8D и 8Е.
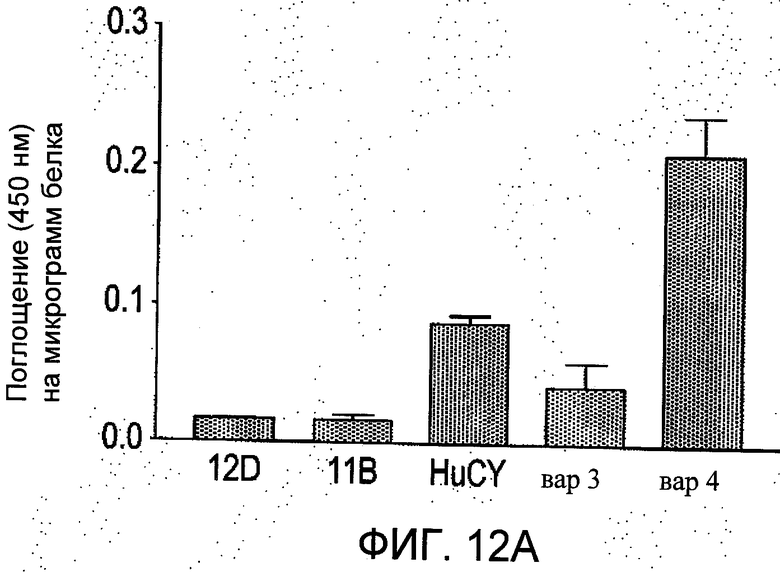
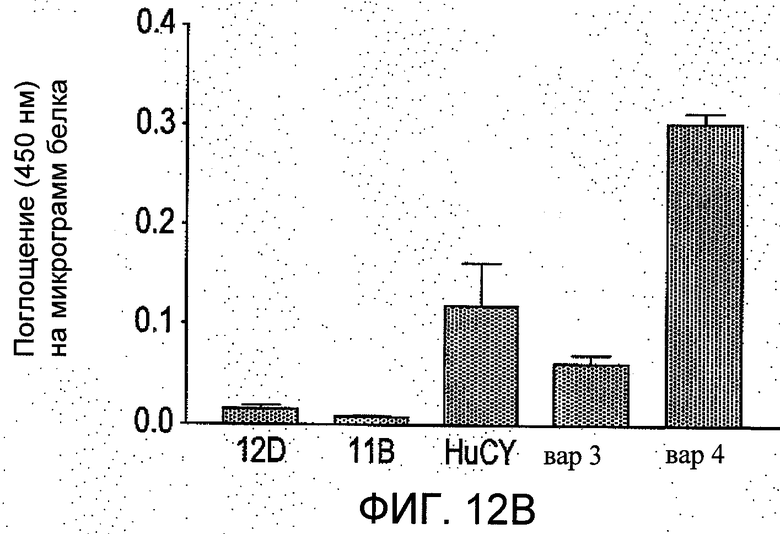
На Фиг.12 группы а-b отображают четыре независимых сравнения, проводимые методом ELISA (иммуноферментный твердофазный анализ) в трех повторностях. Показано количество связей пяти различных вариантов scFv с пептидным антигеном (из N-концевого региона рецеатора АТ1). Данные представлены как средние значения ±S.E.M. значений поглощения при 450 нм на 1 мкг белка в ячейке после вычитания фона.
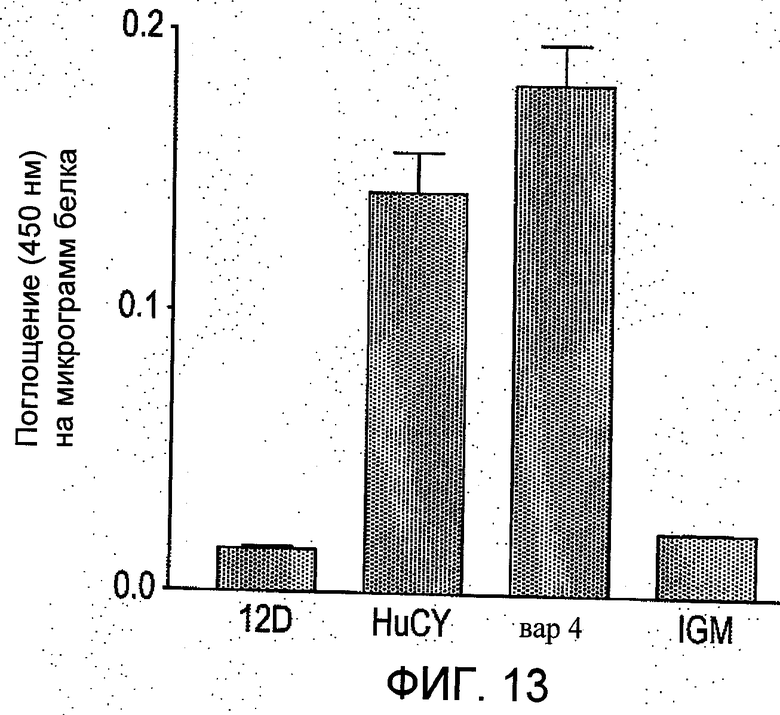
На Фиг.13 представлено косвенное сравнение мышиного scFv (12D) и варианта scFv, полученного генно-инженерными способами против очищенного IgM (иммуноглобулина М) из исходной гибридомы. Анализ ELISA проводили, применяя конъюгат вторичного антитела с антигистаминовым маркером пероксидазы (1:1000) для scFv и конъюгат вторичного антитела с анти-IgM пероксидазой (1:2500) для IgM полученного из гибридомы,.
На Фиг.14 представлено выстраивание последовательностей клонов 12D, 11В и гуманизированных вариантов HuCY, вариант 3 и вариант 4.
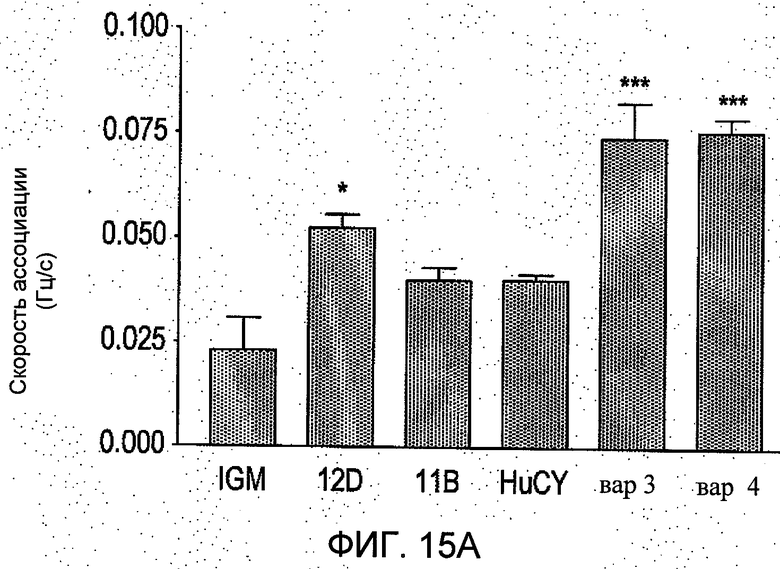
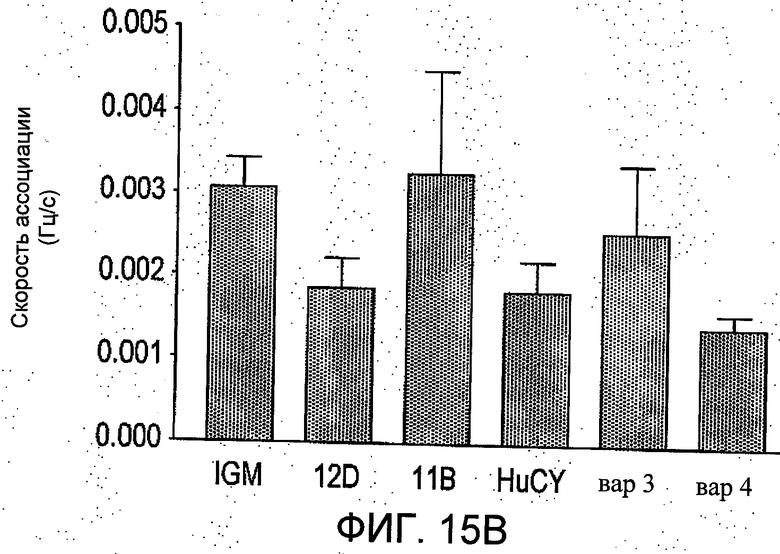
На Фиг.15 группа а) показывает сравнение изменения резонансной частоты наблюдаемой скорости ассоциации в Гц/секунда, между IgM и вариантами scFv. Эти данные были получены с применением биосенсора Attana 100 QCM Biosensor и выражены в виде средних значений ±S.E.M. от резонансной частоты, измеренной на покрытой стрептовидином поверхности чипа QCM (микровесы на кварцевых кристаллах) с применением в качестве связывающей мишени биотинилированного пептидного антигена. ScFvs 12D, вар3 и вар4 показали существенно более значительные изменения резонансной частоты за период инъекции 100 секунд. Группа b) показывает сравнение наблюдаемых скоростей диссоциации различных белков антител, определенных аналогично. В данных экспериментальных условиях не было обнаружено значимых различий в этих конечных скоростях диссоциации.
Пример 1. Получение scFv
Гибридома моноклонального антитела мышей 6313/G2 была выращена ранее описанным способом (Barker et al. J. Mol. Endocrinol. 11 241-245 (1993)). Для получения тяжелых и легких цепей посредством полимеразной цепной реакции использовали пул комплементарной ДНК, полученный из матричной РНК. Для создания библиотеки scFv вставок применяли кодировку фрагмента линкера (Gly4Ser)3 и для направленного клонирования данных вставок в фагмидный вектор рСАМТАВ 5 Е (Amersham Pharmacia, High Wycombe, ВЕЛИКОБРИТАНИЯ) создали индикаторную фаговую библиотеку. Е-маркер для экспрессируемой последовательности GAPVPYPDPLEPR встраивали в данный вектор и применяли в последующих стадиях пэннинга и очистки. Фагмидную библиотеку затем применяли для трансформирования TG1 E. coli и спасение фагмида проводили, применяя фаг-хелпер М13К07 с последующими циклами пэннинга. Положительно экспрессирующие клоны были идентифицированы ELISA, применяя планшеты с 96 ячейками, покрытыми оригинальным антигенным пептидом (EDGIKRIQDD) и анти-Е-маркерным антителом (Amersham Pharmacia), определяемым с применением вторичного антитела, связанного с пероксидазой хрена (HRP). Один конкретный клон подвергали дальнейшей экспрессии, очистке и функциональной оценке, если он показывал наиболее сильный сигнал антигена в ELISA.
6313/G2 scFv (R6313/G2, клон 12D) очищали на колонках HiTrap E-tag (Amersham Pharmacia) с последующей очисткой на колонке Protein L (BD Clontech, Cowley, Oxford, Великобритания), связывающей иммуноглобулины, включая scFv. В экспериментах in vitro и in vivo было необходимо получить среднюю степень очистки с последующим диализом в течение ночи против фосфатно-солевого буфера (PBS) и концентрированием с применением концентраторов, пропускающих вещества с молекулярной массой до 30 кДа (Millipore, Watford, Великобритания). Конечный раствор антитела в соответствии с установившейся практикой воссоздавали в PBS в концентрации 10 мг/мл.
Моноклональное антитело для сравнения в исследованиях инвазии очищали на колонке с иммобилизованным маннаном, связывающей белки (Perbio Science Великобритания, Ltd) с последующим концентрированием на фильтрах, пропускающих вещества с молекулярной массой до 100 кДа.
Предварительные исследования показали, что наивысшая активность относится к клонам Р6313клон120 (также описанный как R6313/G2) и Р6313клон11 В, в которых CDR представляют собой следующие:
R6313клон12D
VHCDR1: GYSFTGYNMN
VHCDR2: NIDPYYGGTTYNQKFKG
VHCDR3: EVDY
VLCDR1: RASKSVSTSGYSYMH
VLCDR2: LVSNLES
VLCDR3: QHIRELTRSEG
или
Р6313клон11В
VHCDR1: GYSFTGYNMS
VHCDR2: NIDPYYGGTTYNQKFKG
VHCDR3: EVDY
VLCDR1: RASKSVSTSGYSYMH
VLCDR2: LVSNLES
VLCDR3: QHIRELTRSEG
Согласно исследованиям методом ELISA на планшетах при пэннинге библиотеки комплементарной ДНК, полученной из РНК из клеточной популяции первоначальной гибридомы, имелись существенно более сильные агенты, связующие антиген. Единственным изменением в CDR является VHCDRH1. Однако также имеется и другое различие между двумя последовательностями в N-конце, KLQQ и QLQE, соответственно. Дополнительные десять клонов показывают другие изменения в CDR и других местах структуры (см. Фиг.11), однако в исследовании ELISA они связывались с антигенами на планшете ELISA менее сильно.
Полные последовательности клонов 12D и 11В выстроены, как показано на Фиг.10. Первые 6 аминокислот и последние 8 аминокислот являются ведущими последовательностями pCANTAB 5E вектора, причем последние 5 составляют часть Е-маркера в экспрессированном белковом продукте, применяемом в данных экспериментах.
Эффективные аминокислотные последовательности клонов 12D и 11В представлены на Фиг.14.
Пример 2. Исследования активности
Активность scFv R6313/G2 изучали следующими методами:
Методы обработки клеточных культур:
Клетки рака молочной железы MCF-7, T47D и MDA-MB-231 были получены из коллекции The American Tissue Culture Collection (LGC Promochem, Teddington, Великобритания). Клетки гладких мышц аорты крысы (RASMC) были получены из первичной культуры (Barker et al., 1996). Клетки MCF-7 выдерживали в минимально поддерживающей среде (MEM), клетки T47D и MDA-MB-231 в питательной среде RPMI 1640, и RASMC выдерживали в среде Игла, модифицированной Дульбекко (DMEM). Все среды были дополнены 2 мМ L-глутамина, 10% сывороткой плода теленка (FBS), 50 усл.ед/мл пенициллина и 0,05 мг/мл стрептомицина. Клетки выдерживали при 37°С в увлажненной атмосфере (95% кислорода, 5% СO2).
Исследование жизнеспособности клеток.
Конфлюэнтные монослои клеток были удалены из сосудов для клеточных культур с применением системы трипсин/ЭДТА (этилендиаминотетрауксусная кислота). Клетки (15×103 на ячейку) были посеяны на планшеты для тканей с 96 ячейками, содержащими соответствующую для каждой линии клеток среду. Через 24 часа клетки обрабатывали ангиотензином II (100 нМ) и R6313/G2 в диапазоне концентраций от 0,005 до 25 мкМ, или лозартаном в аналогичном диапазоне концентраций, и их дополнительно инкубировали в течение 48 часов. Жизнеспособность клеток определяли по способности метаболически активных клеток восстанавливать внутреннюю соль 2,3-бис-[2-метокси-4-нитро-5-сульфофенил]-2Н-тетразолий-5-карбоксианилида (ХТТ) в окрашенный формазановый продукт. Поглощение измеряли, применяя считыватель микропланшетов Multiskan Ascent (Thermo Labsystem, Хельсинки, Финляндия) при длине волны 450 нм и опорной длине волны 630 нм. Каждое измерение проводили в трехкратной повторности. 1С50 были рассчитаны с применением формулы нелинейной регрессии при помощи программного обеспечения GraphPad Prism v4.0 (GraphPad Software Inc. San Diego CA, США).
Экстракция белков и вестерн-блоттинг.
Клетки выращивали в присутствии и при отсутствии ангиотензина II (100 нмоль/л) в течение 24 часов, затем три раза промывали стерильным PBS (рН 7,4), инкубировали в течение 5 минут в лизисном буфере (PBS рН 7,4, 1% NP-40/тритон Х-100, 0,1% SDS (додецилсульфат натрия) и 0,5% дезоксихолата натрия с ингибиторами протеаз лейпептином 10 мкг/мл, апротинином 30 мкг/мл и 0,1 мМоль/л фенилметилсульфонилфторида) и собирали. Лизаты клеток гомогенизировали с применением ультразвукового гомогенизатора (циклами 2×5 с; Bandelin Sonoplus, SLS, Hessle ВЕЛИКОБРИТАНИЯ). После гомогенизации образцы центрифугировали при 20000 g в течение 10 минут при 4°С. Супернатанты отбирали и хранили при -80°С. Концентрации белков оценивали с применением устройства для исследования белков Bio-Rad (Bio-Rad Laboratories, Hemel Hempstead Великобритания). Для вестерн-блоттинга образцы, содержащие суммарно 50 мкг лизата клеток, помещали в 10% SDS-полиакриламидный гель и подвергали электрофорезу. Белки переносили на мембраны Hybond-C (Amersham Biosciences Ltd, Chalfont St Giles Великобритания) в буфере для переноса (39 мМол/л глицина, 48 мМоль/л основания триса (трис-гидроксиметиламинометана), 20% метанола и 0,037% SDS), применяя аппарат для переноса transBlot (Bio-Rad Laboratories, Hemel Hempstead Великобритания) при 120 мА в течение 1,5 часа при 4°С. Мембраны промывали и затем инкубировали в блокирующем буфере (1Х физиологический раствор, забуференный трисом, TBS), 0,1% твина 20 и 5% сухого молока) в течение 1 часа при комнатной температуре и после этого три раза промывали по 10 минут промывочным буфером (1Х TBS и 0,1% твина 20). Мембраны инкубировали с поликлональным антителом к рецепторам АТ1 кролика или антителом к рецептору АТ2 при разбавлении 1:500 в блокирующем буфере. Мембраны оставляли на ночь инкубироваться при 4°С, и затем промывали, как было описано выше и инкубировали с вторичным антителом антиIgG (иммуногглобулина G) кролика (Amersham Biosciences) (1:2000) в течение 1 часа при комнатной температуре. Дополнительно проводили промывки и проводили определение иммунности, инкубируя мембраны в течение 1 минуты в электрохемолюминисцентном (ECL) реагенте для детектирования вестерн-блоттинга (Amersham Biosciences) и экспонировали на Biomax пленке для определения хемолюминисценции (Kodak, Rochester NY, США).
Активность апоптоз-каспаза-3/7.
Активацию каспаз во время апоптоза определяли путем анализа каспазы 3/7 Apo-ONE (Promega Corp, Southampton Великобритания), согласно инструкциям производителя. Вкратце, клетки выращивали до 90% конфлюэнтности и три раза промывали стерильным PBS. Клетки собирали, применяя смесь трипсина и ЭДТА и подсчитывали. Клетки (104 на ячейку) высевали на планшет с 96 ячейками и инкубировали с R6313/G2 в концентрациях от 0,1 до 3 мкМ в присутствии или при отсутствии 100 нмоль/л ангиотензина II, общий объем 150 мкл, в течение 24 и 48 часов. После инкубации в каждую ячейку был добавлен субстрат Z-DEVD-R110 каспазы-3/7 (100 мкл). В «холостых» ячейках содержался только реагент, а в контрольных ячейках отсутствовало антитело и/или ангиотензин II. Флуоресценцию измеряли каждые 2 часа в течение периода 8 часов на спектрофлуориметре Fluostar Optima (BMG Laboratories, Offenburg Германия) с длиной волны возбуждения 485 нм и при длине волны эмиссии 535 нм.
Исследование в полых волокнах.
Методики, связанные с полыми волокнами были разработаны на основе способа Hollingshead. (Hollingshead et al. Life Sc/57 131-41(1995)).
Приготовление полых волокон.
Полые волокна из поливинилидендифторида (PVDF) (отсечка при 500 кДа, внутренний диаметр 1 мм; Spectrum Europe B.V., Breda, Нидерланды) промывали 70% этанолом с применением тупоносой иглы 21 калибра (0,724 мм) и шприца вместимостью 10 мл. Затем иглы оставляли в 70% этаноле на 72 часа, повторно промывали 70% этанолом, затем дистиллированной водой и затем автоклавировали при 131°С. Окончательно, перед загрузкой суспензий клеток, волокна промывали культуральной средой RPMI 1640. Клетки (MCF-7, T47D и MDA-MB-231) вводили в волокна при плотности клеток от 2,5 до 3,0×106 клетка/мл. Затем волокна запаивали теплом с интервалами 2 см и помещали на чашки Петри, содержащие 3 мл подходящей для данных клеток среды.
Для проверки эффективности способа до применения in vivo волокна с клетками инкубировали in vitro в течение 48 часов в присутствии или при отсутствии антитела в диапазоне концентраций от 0,33 мкМ до 33 мкмоль/л и ангиотензина II (100 нмоль/л).
Исследование с полыми волокнами in vivo.
Перед имплантацией бестимусным мышам чистопородной линии balb/c nu/nu mice, самкам возраста 5-6 недель сегменты полых волокон, содержащие клетки, оставляли инкубировать при 37°С в культуральной среде в течение ночи. Волокна имплантировали в животных под анестезией (2% изофлуоран). Каждому животному подкожно (s.c.) и внутрибрюшинно (i.p.) водили по три двухсантиметровых волокна, каждое из которых содержало одну из линий клеток MCF-7, T47D или MDA-MB-231. Для создания внутрибрюшинных имплантатов через кожу и мускулатуру абдоминальной стенки брюшины делали небольшой надрез. Волокна помещали в брюшную полость и оба надреза были закрыты металлическими скобками для сшивания ран (Harvard Instruments, Edenbridge Великобритания). Для подкожных имплантатов на спине делали маленький надрез. Волокна имплантировали слева от средней линии спины в направлении черепа. Маленький надрез закрывали металлическими скобками для сшивания ран.
Обработка антителами.
Мыши (n=5/группа) с имплантатами из полых волокон получали R6313/G2 (0,07 мг/кг (2,5 нмоль/кг) и 0,7 мг/кг (25 нмоль/кг) в 0,1 мл PBS, подкожно) два раза в сутки в течение шести суток. Контрольные животные (n=5) получали только носитель. Через 24 часа после заключительной инъекции животных умерщвляли смещением шейных позвонков и волокна извлекали и на 30 минут переносили в предварительно нагретую среду RPMI 1640, содержащую 20% FBS.
Оценка роста опухолевых клеток внутри полых волокон.
Жизнеспособность клеток оценивали, применяя модифицированный анализ с применением МТТ (бромида 3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолия). Волокна инкубировали в RPMI 1640 с 20% FBS, содержащим 1 мг/мл МТТ и инкубировали при 37°С в атмосфере 95% O2, 5% CO2 в течение 4 часов. Реагент отсасывали и добавляли 2 мл стерильно отфильтрованного 2,5% сульфата протамина (0,9 г NaCl, 2,5 г сульфата протамина в 100 мл воды). Образцы выдерживали при 4°С в темноте по меньшей мере в течение 24 часов для фиксации формазанового продукта. Добавляли свежий 2,5% сульфат протамина и волокна выдерживали при 4°С в течение 2-4 часов. Каждое волокно переносили в свою ячейку планшета с 24 ячейками, разрезали пополам и оставляли на ночь сушиться, предохраняя от света. В каждую ячейку добавляли диметилсульфоксид (ДМСО) (300 мкл) и экстрагировали формазановый продукт. Аликвоту (190 мкл) из каждой ячейки переносили на планшет с 96 ячейками и считывали поглощение при 540 нм на фотометрическом считывающем устройстве для микропланшетов Multiskan Ascent (Thermo labsystem). Обработанные значения рассчитывали в процентах к контролю.
Анализ ксенотрансплантата in vivo.
Мышам вводили подкожно в правый бок 150 мкл стерильного PBS, содержащего 7,5×106 клеток MCF-7. Опухолевые клетки выращивали в течение 4 недель без гормональной поддержки, после чего мыши еженедельно получали подкожные инъекции 17р-эстрадиолвалерата (0,1 мг/кг по массе тела) в кунжутном масле (Kasukabe et al., Breast Cancer Res. 7 (6) R1097-110 (2005)) дополнительно в течение 8 недель. Ежедневно обследовали состояние здоровья животных и два раза в неделю измеряли массу тела. Размеры опухолей измеряли три раза в неделю при помощи штангенциркулей, а объемы рассчитывали по формуле (L×W2)/2, где L и W представляют собой наибольшие и наименьший диаметры, соответственно. Как только объемы опухолей достигали 150-200 мм3, мышей разделяли случайным образом на опытные и контрольные группы по 8-10 животных на группу. Мышам дважды в сутки в течение семи суток вводили подкожные инъекции R6313/G2 в стерильном PBS (0,1 мл) в дозах 0,4 мг/кг (13 нмоль/кг), 0,8 мг/кг (27 нмоль/кг) и 1,36 мг/кг (45,3 нмоль/кг) по массе тела. Мыши контроля получали стерильный PBS. По завершении исследования животных умерщвляли смещением шейных позвонков. Относительные массы тела (%) рассчитывали как (Wt/Wi)×100, где Wt представляет собой массу тела в любое заданное время, a Wi представляет собой массу тела в начале лечения. Объем опухоли нетто рассчитывали как Vt-Vi, где Vt представляет собой объем опухоли в любое заданное время, a Vi представляет собой объем опухоли в начале лечения, и выражали в процентах OTVi.
Кровяное давление у крыс.
Для данной части исследований крысы были выбраны ввиду относительно большего удобства манипулирования при анализе кровяного давления. Первоначально, за 4-5 суток до начала эксперимента крыс самок Уистар приучали к манипуляциям и оборудованию для измерения кровяного давления. Кровяное давление у находящихся в сознании животных определяли с применением системы хвостовой манжеты Kent Scientific Corporation (Torrington CT, USA) Coda 6+, в которой во время измерения кровяного давления животных содержат в утепленных ограничителях. Сначала животных стабилизировали и измеряли фоновое кровяное давление перед лечением. Затем крыс на короткое время извлекали из ограничителей для подкожной инъекции R6313/G2, 0,4 мг/кг в стерильном PBS (0,1 мл). Контрольные животные получали только PBS. Значения давления крови определяли с интервалами за одночасовой период до их возвращения в клетки. Процедуры повторяли ежедневно в течение трех суток.
Статистический анализ.
Все данные были представлены в виде среднего значения±SE. Статистический анализ выполняли, применяя специализированную программу ANOVA (дисперсионный анализ). В случае значимого результата в дисперсионном анализе по кривым «доза-эффект» применяли f-критерий Стьюдента. Значение Р менее 0,05 считали статистически значимым.
Флуориметрический анализ инвазии.
Применяли прцедуру QCM™ Cell invasion assay (Chemicon Cat No.ECM555). В ней применяется флуориметрическое определение клеток, проникших через воспроизведенную базальную белковую мембрану матрикса (ЕСМ), полученную из опухоли мышей Engelbreth-Holm-Swam (EHS) (Repesh LA (1989) Invasion Metastasis 9: 192-208).
Вставки, покрытые ЕСМ, помещали в планшеты с 96 ячейками и на внутреннюю поверхность вставок, для гидратирования слоя ЕСМ добавляли 100 мкл предварительно нагретой среды, не содержащей сыворотку, и выдерживали в течение 1-2 ч при комнатной температуре. Среду удаляли и в ячейки планшета с 96 ячейками, в которые были помещены вставки, добавляли 150 мкл среды, не содержащей сыворотку. Затем вносили 100 мкл среды, содержащей 105 клеток на одну вставку. Планшет накрывали и инкубировали в течение 24 ч при 37°С в увлажненном инкубаторе 5% CO2/95% воздух. После удаления клеток с внутренней поверхности вставки и промывки PBS вставку помещали в планшет с 96 ячейками, содержащий раствор для отслоения клеток, и инкубировали при 37°С в течение 30 мин. Таким образом, клетки, проникшие через ЕСМ на низ вставки, были от нее оторваны с последующим лизисом и измерением флуоресценции согласно инструкциям изготовителя с применением фильтра 480/520, помещенного во флуориметр Fluostar Optima f (BMG Labtech).
Пример 3: Создание гуманизированных вариантов
Основой гуманизированных вариантов являлась вышеописанная последовательность 12D из мышей. Замены аминокислот были выбраны посредством комбинации ранее опубликованных подходов с некоторой степенью разумных расширений. Прежде всего последовательность scFv мышей применяли для обнаружения наиболее гомологичных вариабельных тяжелых и вариабельных легких цепей по базе данных NCBI (Национального центра биотехнологической информации), применяя поиск по BLAST (семейство компьютерных программ, служащих для поиска гомологов белков или нуклеиновых кислот). Во-вторых, применяли подход, описанный Padlan (Molecular Immunology 28 (4/5) 489-498 (1991)) для поиска аминокислотных остатков, которые, вероятно, должны быть экспонированы (то есть гидрофильные) или скрыты (гидрофобные) в целой молекуле scFv. Также со ссылкой на статью Padlan были предложены возможные варианты, согласно которым аминокислоты, завершающие линии зародышевых клеток человека, можно было бы применять для адекватной замены мышиных остатков, и таким образом обеспечить замену поверхности scFv, в которой, в общем плане, замена этих экспонированных мышиных остатков привела бы, в целом, к получению менее иммуногенного белка scFv.
На любых из двух сторон одного из CDR (начиная с данного места CY) были включены две дополнительные замены, которые, как считалось, являются чувствительными к изменениям, потому что остатки аминокислот, завершающих линии зародышевых клеток человека в этих двух позициях полностью сохранялись на концах четырех субгрупп вариабельных участков, в соответствии с данными Padlan (см. выше).
Все три гуманизированных варианта, идентифицированных посредством данного подхода, HuCY, вар3 и вар4, связываются с пептидным антигеном (из N-концевого региона рецептора АТ1) более прочно (ELISA), чем оба мышиных scFv, 12D и 11В.
Последовательность scFv белка была модифицирована только в каркасных участках, содержащихся внутри вариабельных тяжелых и вариабельных легких цепей, чтобы продуцировать гуманизированные варианты HuCY и вар3. Однако, гуманизированный вариант вар4 также имеет два дополнительных изменения в легкой цепи CDR2.
CDR гуманизированных вариантов представляют собой следующие:
HuCY и вар3
VHCDR1: GYSFTGYNMN
VHCDR2: NIDPYYGGTTYNQKFKG
VHCDR3: EVDY
VLCDR1: RASKSVSTSGYSYMH
VLCDR2: LVSNLES
VLCDR3: QHIRELTRSEG
вар4 VHCDR1: GYSFTGYNMN
VHCDR2: NIDPYYGGTTYNQKFKG
VHCDR3: EVDY
VLCDR1: RASKSVSTSGYSYMH
VLCDR2: LVSDLED
VLCDR3: QHIRELTRSEG
Последовательности клонов 12D, 11В и гуманизированных вариантов HuCY, вариант 3 и вариант 4 показаны на фиг.14.
Пример 4. Исследования активности гуманизированных вариантов
Активность гуманизированных вариантов изучали следующими методами.
Получение и очистка ScFv для сравнительных исследований связывания. Все последовательности генов синтезированы Blue Heron Biotechnology (Bothell, WA, USA) и интегрированы в вектор бактериальной белковой экспрессии, в направлении от кодирующей последовательности His-маркера под контролем промотора Т7/ас. Данный вектор включал периплазматическую позиционирующую лидерную последовательность, отщепляемую сигнальной пептидазой после достижения периплазматического пространства бактерии-хозяина. Специалист в данной области поймет, что для продукции scFv могут применяться также другие пригодные векторы.
Данные конструкции были трансформированы в Rosetta 2 (DE3) компетентные клетки (Merck-Novagen), согласно протоколу изготовителя. Штаммы выращивали согласно общепринятым способам в LB (лизогенном бульоне) или LB (лизогенном агаре) при 37°С, с 30 мг/литр канамицина и 34 мг/литр хлорамфеникола. Экспрессию белков проводили с применением 1 л бактериальных культур, выращенных в течение 24 часов в двухлитровых диафрагменных сосудах при 37°С. Продукцию белков инициировали добавлением IPTG (изопропил-β-D-галактопиранозид) в конечной концентрации 0,4 мМ, в течение 5 часов при 25°С. Осажденные бактериальные клетки собирали на центрифуге Beckman Coulter Avanti J-30I при 5000 g (rcf) в течение 20 минут при 4°С. Осадок клеток ресуспендировали в 10 мл (на 1 литр культуры) в нижеследующем буферном растворе, 0,4 М трис-HCL рН 8, 1 мМ ЭДТА, до очистки полученные осадки клеток затем сохраняли при -20°С.
Для приготовления периплазматической фракции 1L осадки оттаивали и добавляли следующие буферные растворы: 10 мл 1М сахарозы и 30 мл 1/5 буферного раствора TES (40 мМ трис-HCL, рН 8, 0,1 мМ ЭДТА, 0,1 М сахарозы и 5 мМ MgSO4). Затем данную клеточную суспензию встряхивали на льду в течение 40 минут и после этого центрифугировали при 17418 g (rcf) в течение 20 минут при 4°С для отделения растворимого периплазматического супернатанта. Этот осмотический шоковый протокол представляет собой модифицированную версию способа, описанного в руководстве, взятом из Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, New York (1989).
Периплазматические фракции затем фильтровали через фильтры с диаметров пор 0,45 мкм и наносили на предварительно очищенные 1 мл His-связывающие колонки, нагруженные NiSO4 (Merck-Novagen). Очистку проводили согласно инструкциям руководства для поли(этилентерефталатных) систем, Очистку проводили согласно, применяя буферные растворы, поставляемые в наборе His-Bind Buffer kit (Merck-Novagen). Буферную смесь в полученном элюате меняли пропусканием через колонку PD-10 (Pharmacia), предварительно уравновешенную фосфатно-солевым буферным раствором рН 7,4 (PBS; Sigma Р4417). Затем полученную фракцию scFv концентрировали на вращающемся концентраторе (Amicon) с отсечкой при молекулярной массе 10 кДа. Концентрацию scFv фракции в PBS устанавливали выполнением исследований в 10-15% SDS-полиакриламидных гелях, выполняя анализы белков по протоколу Bio-Rad и регистрируя УФ (ультрафиолетовое) поглощение при 280 нм, конвертируя последнее в концентрацию путем деления на коэффициент экстинкции 1,7 (молярный коэффициент экстинкции, деленный на молекулярную массу белка - приблизительно 25,7 кДа).
IgM для исследований сравнительного связывания очищали, как было предварительно описано для исследований инвазии в клетки. Коэффициент экстинкции для IgM составляет 1,18 (Johnstone A, Thorpe R. IмMunochemistry in practice. 2nd ed. Oxford: Blackwell Scientific Publications (1987)).
Анализ связывания антигена посредством ELISA.
Иммуносорбционный ферментный анализ (ELISA) проводили на планшетах Maxisorp с 96 ячейками, покрытыми пептидным антигеном EDGIKRIQDDC-биотин (2-8 мкг/мл в карбонатном буфере рН 9,6), оставляя на ночь при 4 или 37°С. Покрытые ячейки блокировали с применением 1% щелочерастворимого казеина (Blocking buffer) в течение 1 ч при комнатной температуре, затем три раза промывали PBS, содержащем 0,1% об./об. твина 20. Добавляли образцы ScFv, разбавленные между 1:1 до 1:100 в PBS-T и перед промывкой, описанной ранее, инкубировали в течение 1 ч при комнатной температуре. Затем в течение 1 ч при комнатной температуре добавляли вторичное HRP-конъюгированное антитело с анти-His-маркером (разбавленное 1:1000 в блокирующем буфере). Затем ячейки три раза промывали PBS-T, и после этого - дважды (без твина 20). Добавляли 100 мкл раствора субстрата в ТМВ (Тетраметилбензидин) и оставляли для проявления цвета на 30 мин, по истечении этого времени для остановки реакции добавляли 100 мкл 2М серной кислоты. На спектрофотометре для считывания планшетов считывали поглощение при 450 нм. Концентрации белков определяли с помощью биофотометра Eppendorf, считывая показания при 280 нм и коэффициент экстинкции для scFv 1,7 как описано выше. Результаты представлены на Фиг.12.
Как можно видеть из Фиг.12, в обновленных поверхностях scFv (HuCY, вар3 и вар4), имеет место повышенное связывание с пептидным антигеном по сравнению с мышиным scFv (12D и 11В). Неожиданно было обнаружено, что изменения, сделанные в каркасах мышиных scFv, приводящие к образованию HuCY, по данным ELISA, связываются с антигенным пептидом EDGIKRIQDDC-биотин в 5 раз более активно по сравнению с тем же количеством мышиного scFv той же степени очистки. Более того, гуманизированный вариант вар4 показал 5-10-кратное увеличение связывания с антигеном по сравнению с материнским мышиным scFv. Данное обстоятельство является неожиданным, поскольку один из CDR (VLCDR2) вар4 был модифицирован относительно материнского мышиного scFv.
На Фиг.13 показано косвенное сравнение мышиного scFv (12D) и сконструированного варианта scFv (HuCY и вар4) против очищенного IgM из исходной гибридомы. Как можно видеть из Фиг.13, вариантные scFv обладают более высокой активностью по сравнению с мышиным scFv.
Сравнение характеристик связывания при помощи микровесов на кварцевых кристаллах Quartz Crystal Microbalance
Получали сравнительные данные характеристик связывания вариантов scFv и IgM по отношению к иммобилизованному пептидному антигену с применением микровесов на кварцевых кристаллов Attana 100. Исследование включало применение покрытого стрептавидином (0,1 мг/мл) чипа с биотином (золоченый кварцевый кристалл) в биосенсоре Attana 100 (Микровесы на кварцевых кристаллах (QCM)) (Attana АВ, Стокгольм, Швеция). Пептидный антиген, соответствующий данной последовательности, применяемый для инициирования исходной мышиной гибридомы, биотинилированной у С-конца, в концентрации 4 мкг/мл вводили в контакт с чипом для создания мишени связывания для фрагментов scFv и впоследствии IgM вводили в контакт с поверхностью чипа QCM. Для отслеживания изменений резонансной частоты чипа пептидный антиген, соответствующий данной последовательности, в ответ на связывание тестируемого антитела с антигеном применяли программное обеспечение Atteste (Attana, Швеция). Значение отклонения, выраженное в Гц, за период введения образца 100 с в Гц/секунда для каждого антитела определяли в концентрации 9 мкг/мл. Также измеряли в Гц/секунда скорость, при которой антитело освобождается с чипа QCM из прямолинейной части записи результатов биосенсора, по мере того как антитело постепенно удалялось из чипа, и по мере того как уменьшалась резонансная частота чипа. Буфером при исследовании служил буфер PBS, содержащий 0,005% твина 20 (PBST), и его также применяли для разбавления антитела: в каждом случае конечная концентрация белка составляла 9 мкг/мл. Attana 100 устанавливали с инъекционной петлей объемом 50 мкл и проводили циклы исследований с постоянной скоростью насоса 20 мкл/мин. Образцы и контрольные буферные растворы PBST вводили за 100 секунд, что создавало объем образца в поперечнике чипа 33 мкл. Между каждым отдельным экспериментом для регенерирования стрептавидин-антигенной поверхности применяли 100 мМ (объем 6,6 мкл) фосфорную кислоту.
Результаты показаны на Фиг.15. За время инъекции ScFv 12D, вар3 и вар4, демонстрировали значительно большее возрастание резонансной частоты за время инъекции по сравнению с IgM, в то время в скорости удаления антитела с чипа QCM при данных экспериментальных условиях не было замечено значимой разницы.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ АНТИ-SIGLEC-9 АНТИТЕЛА | 2020 |
|
RU2833330C1 |
| АНТИТЕЛО К АННЕКСИНУ А1 | 2011 |
|
RU2596403C2 |
| МОЛЕКУЛЫ АНТИТЕЛ К CD73 И ПУТИ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2791192C2 |
| АНТИ-CD40-АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2012 |
|
RU2609647C2 |
| МОЛЕКУЛЫ АНТИТЕЛ ПРОТИВ ВИРУСА BK | 2021 |
|
RU2835801C1 |
| CD3-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2826453C2 |
| СВЯЗЫВАЮЩИЕ ILT7 МОЛЕКУЛЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2756109C2 |
| 5-БРОМ-2,6-ДИ-(1Н-ПИРАЗОЛ-1-ИЛ)ПИРИМИДИН-4-АМИН ДЛЯ ПРИМЕНЕНИЯ В ЛЕЧЕНИИ РАКА | 2016 |
|
RU2745560C2 |
| СЛИТЫЕ БЕЛКИ С АЛЬБУМИН-СВЯЗЫВАЮЩИМИ ДОМЕНАМИ | 2018 |
|
RU2786444C2 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕСЯ С PSL PSEUDOMONAS, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2708977C2 |


























Настоящее изобретение относится к биотехнологии и представляет собой гуманизированные синтетические аналоги (R6313/G2) вариабельных доменов моноклонального scFv антитела 6313/G2 к рецептору ангиотензина II типа 1, т.е. специфически связывающуюся молекулу, которая специфически связывается с пептидом, имеющим аминокислотную последовательность EDGIKRIQDD. Предложены также способ лечения рака, применение специфически связывающейся молекулы согласно изобретению при приготовлении средства для лечения рака, комбинированный препарат и фармацевтические композиции, содержащие специфически связывающуюся молекулу согласно изобретению и ангиотензин II. Настоящее изобретение позволяет блокировать вредное действие ангиотензина II и применять указанные аналоги для лечения рака у человека. 6 н. и 3 з.п. ф-лы, 15 ил., 4 пр.
1. Специфически связывающаяся молекула, которая
специфически связывается с пептидом, имеющим аминокислотную последовательность EDGIKRIQDD, и
состоит из полипептида, который представляет собой VL домен иммуноглобулина, связанный с VH доменом иммуноглобулина, причем
домен VL содержит участки, определяющие комплементарность (CDRs) VLCDR1, VLCDR2 и VLCDR3, и
домен VH содержит участки, определяющие комплементарность (CDRs) VHCDR1, VHCDR2, VHCDR3, в которой указанные CDRs имеют следующие аминокислотные последовательности:
VHCDR1 представляет собой GYSFTGYNMN
VHCDR2 представляет собой NIDPYYGGTTYNQKFKG
VHCDR3 представляет собой EVDY
VLCDR1 представляет собой RASKSVSTSGYSYMH
VLCDR2 представляет собой LVSNLES
VLCDR3 представляет собой QHIRELTRSEG;
или
VHCDR1 представляет собой GYSFTGYNMS
VHCDR2 представляет собой NIDPYYGGTTYNQKFKG
VHCDR3 представляет собой EVDY
VLCDR1 представляет собой RASKSVSTSGYSYMH
VLCDR2 представляет собой LVSNLES
VLCDR3 представляет собой QHIRELTRSEG;
или
VHCDR1 представляет собой GYSFTGYNMN
VHCDR2 представляет собой NIDPYYGGTTYNQKFKG
VHCDR3 представляет собой EVDY
VLCDR1 представляет собой RASKSVSTSGYSYMH
VLCDR2 представляет собой LVSDLED
VLCDR3 представляет собой QHIRELTRSEG.
2. Специфически связывающаяся молекула по п.1, состоящая из полипептида с аминокислотной последовательностью, которая выбрана из аминокислотных последовательностей, приведенных на Фиг.14.
3. Специфически связывающаяся молекула по п.1 или 2 для применения в медицине.
4. Специфически связывающаяся молекула по п.1 или 2 для применения при лечении рака.
5. Способ лечения рака у субъекта, включающий введение субъекту специфически связывающейся молекулы по п.1 или 2.
6. Применение специфически связывающейся молекулы по п.1 или 2 при приготовлении лекарственного средства для лечения рака.
7. Комбинированный препарат, содержащий специфически связывающуюся молекулу по п.1 или 2 и ангиотензин II для раздельного, одновременного или последовательного введения при лечении рака у субъекта.
8. Фармацевтическая композиция для блокирования вредного действия ангиотензина II, содержащая эффективное количество специфически связывающейся молекулы по п.1 или 2 и фармацевтически приемлемый носитель, адъювант и/или разбавитель.
9. Фармацевтическая композиция для блокирования вредного действия ангиотензина II, содержащая эффективное количество специфически связывающейся молекулы по п.1 или 2, ангиотензин II и фармацевтически приемлемый носитель, адъювант и/или разбавитель.
| WO 2004018519 A2, 04.03.2004 | |||
| BARKER S | |||
| et al., A monoclonal antibody to a conserved sequence in the extracellular domain recognizes the angiotensin II AT1 receptor in mammalian target tissues, Journal of Molecular Endocrinology, 1993, 11, pp.241-245 | |||
| US 6641811 B1, 04.11.2003 | |||
| ПРИМЕНЕНИЕ АНТАГОНИСТА РЕЦЕПТОРА AT-1 ИЛИ МОДУЛЯТОРА РЕЦЕПТОРА АТ-2 ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНЕЙ, СВЯЗАННЫХ С УВЕЛИЧЕНИЕМ СОДЕРЖАНИЯ РЕЦЕПТОРОВ АТ-1 ИЛИ АТ-2 | 1999 |
|
RU2271809C2 |

