Область техники
Настоящее изобретение относится к специфическим связывающим молекулам, которые связываются с CD3, в частности, к антителам и их фрагментам с улучшенными свойствами.
Уровень техники
CD3 (кластер дифференцировки 3) является корецептором Т-клеток, который помогает активировать как цитотоксические Т-клетки (CD8+Т-клетки), так и хелперные Т-клетки (CD4+Т-клетки). CD3 ассоциирует с Т-клеточным рецептором (TCR) и ζ-цепью (зета-цепь, CD247) для генерации активирующего сигнала в Т-лимфоцитах. ζ-цепь TCR и молекулы CD3 вместе составляют комплекс TCR.
Антитела, которые связываются с CD3, известны и применяются в качестве иммуносупрессивных лекарственных средств. Примеры включают муромонаб-СD3 (Janssen-Cilag), отеликсизумаб (также известный как TRX4), теплизумаб (также известный как PRV-031) и визилизумаб.
Антитела к CD3 также применяют в качестве рекрутирующих Т-клетки агентов из класса терапевтических средств на основе белков, широко известных как биспецифические активаторы Т-клеток (Baeuerle et al, Cancer Res. 2009 Jun 15;69(12):4941-4). В таких терапевтических средствах домен распознавания клетки-мишени скомбинирован с направленным против CD3 (анти-СD3) доменом. Одновременный рекрутинг клеток-мишеней (например, раковой клетки) и CD3+ цитотоксических Т-клеток ведет к активации путей сигнализации через CD3 независимо от специфичности Т-клеточного рецептора, и в конечном итоге приводит к гибели клеток-мишеней. Биспецифическое антитело блинатумомаб (Amgen) является примером зарегистрированного для продажи терапевтического активатора Т-клеток для лечения острого лимфобластного лейкоза (ОЛЛ). Ряд других биспецифических активаторов Т-клеток исследуют в клинических испытаниях для лечения различных видов рака и инфекционных заболеваний (например, см. таблицы в источниках: Yuraszeck et al., Clin Pharmacol Ther. 2017 May;101(5):634-645, and Husain et al., BioDrugs. 2018 Oct;32(5):441-464). Большинство биспецифических активаторов Т-клеток распознают антигены клеточной поверхности на клетке-мишени. Кроме того, известны также биспецифические активаторы Т-клеток, которые распознают короткие пептиды, происходящие из внутриклеточных антигенов и представленные на поверхности клетки в комплексе с ГКГС (пГКГС, рМНС). (Liddy et al., Nat Med. 2012 Jun;18(6):980-7).
Антитело UCHT1 представляет собой клинически значимое антитело против CD3, известное в данной области техники (Shalaby et al., J Exp Med. 1992 Jan l;175(l):217-25; US5821337). ScFv-фрагмент гуманизированного антитела UCHT1 сливали с растворимым Т-клеточным рецептором для конструирования биспецифического активатора Т-клеток, который связывается с рМНС на клетке-мишени (например, см. WO2011001152).
Описание изобретения
Авторы настоящего изобретения неожиданно обнаружили, что введение определенных мутаций в аминокислотную последовательность UCHT1 приводит к образованию биспецифических активаторов Т-клеток с неожиданными свойствами, которые особенно благоприятны для клинического применения.
Согласно первому аспекту предлагается специфическая связывающая молекула, которая связывается с CD3 и содержит полипептид, имеющий иммуноглобулиновые домены VL и VH, при этом домен VL содержит определяющие комплементарность области (CDR) VLCDR1, VLCDR2 и VLCDR3, а домен VH включает определяющие комплементарность области (CDR) VHCDR1, VHCDR2, VHCDR3, каждая из которых имеет соответствующую аминокислотную последовательность, представленную ниже, где:
VLCDR1 представляет собой QDIRNY
VLCDR2 представляет собой YTS
VLCDR3 представляет собой QQGNTLPWT
VHCDR1 представляет собой GYSFTGYA
VHCDR2 представляет собой TNPYKGVS
VHCDR3 представляет собой ARSGYYGDSDWYFDV
или аминокислотную последовательность, по меньшей мере на 70% идентичную указанной последовательности.
CDR определены в соответствии с международной информационной системой ImMunoGeneTics (IMGT®) (LeFranc et al., Nucleic Acids Res. 2009 Jan;37(Database issue):D1006-12)
В частности, специфические связывающие молекулы согласно настоящему изобретению содержат аланин на С-конце VHCDR1 в положении 38 в соответствии с нумерацией IMGT® (в настоящем документе обозначен как положение 165, приведен в качестве примера и выделен жирным шрифтом в последовательности тяжелой цепи v1 на фиг. 1). Было обнаружено, что молекулы, имеющие эту мутацию, обладают улучшенными свойствами, включая улучшенное окно специфичности, по сравнению с молекулами без этой мутации.
CDR могут быть представлены в каркасной последовательности вариабельного домена антитела. Каркасная последовательность может быть каркасной последовательностью мыши или человека, гуманизированной каркасной последовательностью или любой другой подходящей каркасной последовательностью. Предпочтительно каркас представляет собой человеческую или гуманизированную каркасную последовательность. Человеческие или гуманизированные каркасные последовательности по существу имеют аминокислотную последовательность иммуноглобулина человека. В некоторых случаях каркасные последовательности мыши, человека и гуманизированные каркасные последовательности могут быть смешаны в любой комбинации.
Предпочтительно VL иммуноглобулина содержит общую последовательность VLFW1-VLCDR1-VLFW2-VLCDR2-VLFW3-VLCDR3-VLFW4, где VLFW1, VLFW2, VLFW3 и VLFW4 представляют собой каркасные последовательности VL (VLFW) 1-4, соответственно, VH иммуноглобулина содержит последовательность VHFW1-VHCDR1-VHFW2-VHCDR2-VHFW3-VHCDR3-VHFW4, где VHFW1, VHFW2, VHFW3 и VHFW4 представляют собой каркасные последовательности VH (VHFW) 14, соответственно; при этом необязательно последовательности VLFW и VHFW представляют собой каркасные последовательности мыши, человека или гуманизированные каркасные последовательности.
Предпочтительно VL иммуноглобулина содержит следующую общую последовательность: VLFW1-VLCDR1-VLFW2-VLCDR2-VLFW3-VLCDR3-VLFW4, где VLFW1, VLFW2, VLFW3 и VLFW4 представляют собой каркасные (FW) последовательности 1-4, соответственно, каждая из VLFW1, VLFW2, VLFW3 и VLFW4 имеет соответствующую аминокислотную последовательность, представленную ниже, где:
VLFW1 представляет собой AIQMTQSPSSLSASVGDRVTITCRAS или DIQMTQSPSSLSASVGDRVTITCRAS
VLFW2 представляет собой LNWYQQKPGKAPKLLIY
VLFW3 представляет собой RLESGVPSRFSGSGSGTDYTLTIS SLQPEDFATYYC
VLFW4 представляет собой FGQGTKVEIK
или аминокислотную последовательность, по меньшей мере на 70% идентичную указанной последовательности.
Предпочтительно VH иммуноглобулина содержит следующую общую последовательность: VHFW1-VHCDR1-VHFW2-VHCDR2-VHFW3-VHCDR3-VHFW4, где VHFW1, VHFW2, VHFW3 и VHFW4 представляют собой каркасные (FW) последовательности 1-4, соответственно, каждая из последовательностей VHFW1, VHFW2, VHFW3 и VHFW4 имеет соответствующую аминокислотную последовательность, представленную ниже, где:
VHFW1 - EVQLVESGGGLVQPGGSLRLSCAAS
VHFW2 - MNWVRQAPGKGLEWVAL
VHFW3 - TYNQKFKDRFTISVDKSKNTAYLQMNSLRАЕDTАVYYC или TYNQKFKDRFTFSVDKSKNTAYLQMNSLRAEDTAVYYC
VHFW4 - WGQGTLVTVSS
или аминокислотную последовательность, по меньшей мере на 70% идентичную указанной последовательности.
В частности, специфические связывающие молекулы согласно настоящему изобретению содержат фенилаланин в VHFW3 в положении 78 в соответствии с нумерацией IMGT® (в настоящем документе обозначен как положение 202, приведен в качестве примера и выделен жирным шрифтом в последовательности тяжелой цепи v2 на фиг. 1). Было обнаружено, что молекулы, имеющие эту мутацию в дополнение к аланину в положении 38, обладают повышенной эффективностью активации Т-клеток по сравнению с молекулами без этих мутаций.
Предпочтительно VL иммуноглобулина содержит следующую общую последовательность:
AIQMTQSPSSLSASVGDRVTITCRASQDIRNYLNWYQQKPGKАРKLLIYYTSRLESGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQGNTLPWTFGQGTKVEIK; или
DIQMTQSPSSLSASVGDRVTITCRASQDIRNYLNWYQQKPGKAPKLLIYYTSRLESGVPSRFSGSGSGTD YTLTISSLQPEDFATYYCQQGNTLPWTFGQGTKVEIK
или аминокислотную последовательность, по меньшей мере на 70% идентичную указанной последовательности.
Предпочтительно VH иммуноглобулина содержит следующую общую последовательность:
ЕVQLVESGGGLVQPGGSLRLSСAASGYSFTGYAMNWVRQAPGKGLEWVALINPYKGVSTYNQKFKDRFTISVDKSKNTAYLQMNSLRAEDTAVYYCARSGYYGDSDWYFDVWGQGTLVTVSS; или
ЕVQLVESGGGLVQPGGSLRLSСAASGYSFTGYAMNWVRQAPGKGLEWVALINPYKGVSTYNQKFKDRFTFSVDKSKNTAYLQMNSLRAEDTAVYYCARSGYYGDSDWYFDVWGQGTLVTVSS
или аминокислотную последовательность, по меньшей мере на 70% идентичную указанной последовательности.
Предпочтительно специфическая связывающая молекула находится в форме scFv-фрагмента.
Предпочтительно иммуноглобулиновые домены VL и VH соединены посредством линкера. Линкер может иметь любую аминокислотную последовательность, предпочтительно длиной 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 аминокислот. Предпочтительный линкер содержит последовательность с формулой (GGGGS)n, необязательно в дополнение к другим аминокислотам. Соответственно, предложена одноцепочечная специфическая связывающая молекула, имеющая последовательность:
AIQMTQSPSSLSASVGDRVTITCRASQDIRNYLNWYQQKPGKAPKLLIYYTSRLESGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQGNTLPWTFGQGTKVEIKGGGGSGGGGSGGGGSGGGGSGGGSEVQLVESGGGLVQPGGSLRLSCAASGYSFTGYAMNWVRQAPGKGLEWVALINPYKGVSTYNQKFKDRFTISVDKSKNTAYLQMNSLRAEDTAVYYCARSGYYGDSDWYFDVWGQGTLVTVSS; или
AIQMTQSPSSLSASVGDRVTITCRASQDIRNYLNWYQQKPGKAPKLLIYYTSRLESGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQGNTLPWTFGQGTKVEIKGGGGSGGGGSGGGGSGGGGSGGGSEVQLVESGGGLVQPGGSLRLSCAASGYSFTGYAMNWVRQAPGKGLEWVALINPYKGVSTYNQKFKDRFTFSVDKSKNTAYLQMNSLRAEDTAVYYCARSGYYGDSDWYFDVWGQGTLVTVSS
или аминокислотную последовательность, по меньшей мере на 70% идентичную указанной последовательности.
Специфическая связывающая молекула также может быть частью слитого белка, содержащего дополнительные домены. Слитые белки могут быть сконструированы посредством N-концевого или С-концевого слияния с доменами иммуноглобулина VL или VH. Дополнительные домены могут быть слиты при помощи линкеров. Линкерные последовательности обычно являются гибкими, поскольку состоят в основном из таких аминокислот, как глицин, аланин и серии, которые не имеют объемных боковых цепей, которые могут ограничивать гибкость. В альтернативном варианте желательными могут быть линкеры с большей жесткостью. Пригодные для применения или оптимальные длины линкерных последовательностей могут быть легко определены. Часто длина линкерной последовательности составляет менее 12, например, менее 10, или от 2 до 10 аминокислот. Линкер может представлять собой любую аминокислотную последовательность, предпочтительно длиной 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 аминокислот. Примеры подходящих линкеров включают, не ограничиваясь перечисленными: GGGSGGGG, GGGGS, GGGSG, GGSGG, GSGGG, GSGGGP, GGEPS, GGEGGGP и GGEGGGSEGGGS (как описано в WO2010/133828). Предпочтительный линкер содержит последовательность с формулой (GGGGS)n, необязательно в дополнение к другим аминокислотам.
Предпочтительно специфическая связывающая молекула демонстрирует одно или более улучшенных терапевтических свойств по сравнению с немутированной молекулой при использовании в качестве части биспецифической молекулы, предпочтительно биспецифического активатора Т-клеток согласно описанию в настоящем документе. Предпочтительно улучшенные терапевтические свойства выбирают из улучшенного терапевтического окна и/или увеличения максимальной активации Т-клеток при заданной концентрации. Улучшенное терапевтическое окно может позволять введение более высоких доз при минимизации токсичности в результате нецелевой активации. Увеличение максимальной активации Т-клеток при заданной концентрации биспецифической молекулы может обеспечивать более эффективный киллинг клеток-мишеней при заданной дозе лекарственного средства. Способы определения активации Т-клеток известны в данной области техники и включают высвобождение иммуноактивирующих цитокинов и опосредованной Т-клетками смерти клеток. Терапевтическое окно для биспецифического активатора Т-клеток может быть определено путем измерения активации Т-клеток в присутствии антиген-положительных клеток и антиген-отрицательных клеток и вычисления разницы между результатами указанных двух измерений. Подробнее предпочтительные методы описаны в примере 2.
Предпочтительно антитело против CD3 согласно настоящему изобретению связывает субъединицу s CD3.
Специфические связывающие молекулы согласно настоящему изобретению могут применяться в способе лечения или диагностики для организма человека или животного, таком как способ лечения состояния у пациента (предпочтительно человека), который включает введение указанному пациенту эффективного количества специфической связывающей молекулы согласно настоящему изобретению. Согласно настоящему изобретению также предложена специфическая связывающая молекула по настоящему изобретению для применения в медицине, а также применение специфической связывающей молекулы по настоящему изобретению при получении медикамента для диагностики или лечения опухоли.
Эти и другие аспекты настоящего изобретения подробнее описаны ниже.
В настоящем документе «лечение» включает любой режим, который может принести пользу человеку или животному, не являющемуся человеком, предпочтительно млекопитающему. Лечение может относиться к существующему состоянию или может быть профилактическим (профилактическое лечение).
Термин «антитело» в настоящем документе относится к молекулам иммуноглобулина и активным в иммунологическом отношении частям молекул иммуноглобулина, то есть молекулам, содержащим антигенсвязывающий сайт, который специфически связывает антиген, либо природным, либо частично или полностью синтезированным. Указанный термин также охватывает любой полипептид или белок, имеющий связывающий домен, который представляет собой связывающий домен антитела или гомологичен ему. Они могут быть получены из природных источников или могут быть частично или полностью синтезированы. Примерами антител являются изотипы иммуноглобулинов (например, IgG, IgE, IgM, IgD и IgA) и их изотипические подклассы; фрагменты, которые содержат антигенсвязывающий домен, такой как Fab, scFv, Fv, dAb, Fd; и диатела. Антитела могут быть поликлональными или моноклональными. Моноклональное антитело может быть обозначено в настоящем документе как «mAb».
Могут быть взяты моноклональные и другие антитела и использованы методы технологии рекомбинантной ДНК для получения других антител или химерных молекул, которые сохраняют специфичность исходного антитела. Такие методы могут включать введение ДНК, кодирующей вариабельную область иммуноглобулина или определяющие комплементарность области (CDR) антитела, в константные области или константные области плюс каркасные области другого иммуноглобулина. См., например, ЕР-А-184187, GB 2188638А или ЕР-А-239400. Гибридому или другую клетку, продуцирующую антитело, можно подвергнуть генетической мутации или другим изменениям, которые могут изменять или могут не изменять специфичность связывания продуцированных антител.
Поскольку антитела могут быть модифицированы несколькими способами, термин «антитело» следует понимать как охватывающий любую специфическую связывающую молекулу или вещество, имеющие связывающий домен с требуемой специфичностью. Таким образом, этот термин охватывает фрагменты антител, производные антител, функциональные эквиваленты и гомологи антител, гуманизированные антитела, в том числе любой полипептид, содержащий связывающий домен иммуноглобулина, как природный, так и полностью или частично синтетический. Следовательно, включены химерные молекулы, содержащие связывающий домен иммуноглобулина или его эквивалент, слитый с другим полипептидом. Клонирование и экспрессия химерных антител описаны в ЕР-А-0120694 и ЕР-А-0125023. Гуманизированное антитело может представлять собой модифицированное антитело, имеющее вариабельные области не являющегося человеческим антитела, например, антитела мыши, и константную область человеческого антитела. Способы получения гуманизированных антител описаны, например, в патенте США№5225539.
Было показано, что фрагменты целого антитела могут выполнять функцию связывания антигенов. Примерами связывающих фрагментов являются (i) Fab-фрагмент, состоящий из доменов VL, VH, CL и CHI; (ii) Fd-фрагмент, состоящий из доменов VH и СН1; (iii) Fv-фрагмент, состоящий из доменов VL и VH одного антитела; (iv) dAb-фрагмент (Ward, E.S. et al., Nature 341:544-546 (1989)), который состоит из домена VH; (v) выделенные области CDR; (vi) Е(аb')2-фрагменты, бивалентный фрагмент, содержащий два соединенных Fab-фрагмента; (vii) одноцепочечные молекулы Fv (scFv), где домены VH и VL соединены пептидным линкером, который позволяет двум доменам связываться с образованием антигенсвязывающего сайта (Bird et al., Science 242:423-426 (1988); Huston et al., PNAS USA 85:5879-5883 (1988)); (viii) биспецифические одноцепочечные димеры Fv (PCT/US92/09965) и (ix) «диатела», поливалентные или мультиспецифические фрагменты, сконструированные путем слияния генов (WO94/13804; P. Hollinger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993)).
Диатела представляют собой мультимеры полипептидов, причем каждый полипептид содержит первый домен, содержащий область связывания легкой цепи иммуноглобулина, и второй домен, содержащий область связывания тяжелой цепи иммуноглобулина, при этом указанные два домена соединены (например, с помощью пептидного линкера), но не способны связываться друг с другом с образованием антигенсвязывающего сайта: антигенсвязывающие сайты образуются в результате связывания первого домена одного полипептида в мультимере со вторым доменом другого полипептида в мультимере (WO94/13804).
Если необходимо использовать биспецифические антитела, они могут представлять собой обычные биспецифические антитела, которые могут быть получены различными способами (Hollinger & Winter, Current Opinion Biotechnol. 4:446-449 (1993)), например, получены химическим путем или из гибридных гибридом, или могут представлять собой любой из фрагментов биспецифических антител, упомянутых выше. Может быть предпочтительным применение димеров scFv или диател, а не целых антител. Диатела и scFv могут быть сконструированы без участка Fc с применением только вариабельных доменов, что потенциально уменьшает последствия антиидиотипической реакции. Другие формы биспецифических антител включают одноцепочечные «Янусины», описанные в источнике: Traunecker et al., EMBO Journal 10:3655-3659 (1991).
Помимо целых биспецифических антител могут также подходить для применения биспецифические диатела, поскольку их можно легко сконструировать и экспрессировать в Е. coli. Диатела (и многие другие полипептиды, такие как фрагменты антител) с подходящей специфичностью связывания можно легко отобрать из библиотек с применением фагового дисплея (WO94/13804). Если одно плечо диатела нужно сохранить неизменным, например, со специфичностью к антигену X, может быть создана библиотека с варьирующим другим плечом и выбрано антитело с подходящей специфичностью.
«Антигенсвязывающий домен» является частью антитела, которая содержит область, которая специфически связывается с частью антигена или со всем антигеном, или комплементарную части антигена или всему антигену. В случае значительного размера антигена антитело может связываться только с определенной частью антигена, которую называют эпитопом. Антигенсвязывающий домен может быть образован одним или несколькими вариабельными доменами антитела. Антигенсвязывающий домен может содержать вариабельную область легкой цепи антитела (VL) и вариабельную область тяжелой цепи антитела (VH).
Термин «специфический» обычно используется для обозначения ситуации, в которой один член специфически связывающейся пары не будет демонстрировать какого-либо значительного связывания с другими молекулами помимо его партнера (партнеров) для специфического связывания, и, например, будет иметь перекрестную реактивность в отношении любой другой молекулы менее чем приблизительно 30%, предпочтительно 20%, 10% или 1%. Указанный термин также относится к тем случаям, когда, например, антигенсвязывающий домен является специфическим для конкретного эпитопа, который имеется у ряда антигенов, и в таком случае специфическая связывающая молекула, несущая антигенсвязывающий домен, будет способна связываться с различными антигенами, несущими такой эпитоп.
Термин «выделенный» относится к состоянию, в котором предпочтительно будут находиться специфические связывающие молекулы или нуклеиновая кислота, кодирующая такие связывающие молекулы, в соответствии с настоящим изобретением. Молекулы и нуклеиновая кислота, как правило, не содержат или по существу не содержат материала, с которым они ассоциированы в природе, такого как другие полипептиды или нуклеиновые кислоты, с которыми они обнаруживаются в их естественной среде, или среде, в которой их получают (например, культуре клеток), когда такой состав получают с помощью технологии рекомбинантной ДНК, используемой in vitro или in vivo. Специфические связывающие молекулы и нуклеиновая кислота могут быть введены в составы с растворителями или адъювантами и все же могут быть выделенными для практических целей - например, молекулы обычно смешивают с желатином или другими носителями, если они применяются для покрытия микротитрационных планшетов для использования в иммуноанализах, или смешивают с фармацевтически приемлемыми носителями или разбавителями при использовании в диагностике или терапии. Специфические связывающие молекулы могут быть гликозилированы, как естественным путем, так и с помощью систем гетерологичных эукариотических клеток, или они могут быть негликозилированными (например, если они продуцируются путем экспрессии в прокариотической клетке).
Под термином «по существу как представленные» подразумевается, что области CDR согласно настоящему изобретению будут либо идентичны, либо высокогомологичны областям на фиг. 1а и 1cb. Под «высокогомологичными» областями подразумевается, что в CDR могут быть произведены от 1 до 5, от 1 до 4, от 1 до 3 замен, 2 замены или 1 замена.
В объем изобретения также входят полипептиды, имеющие аминокислотную последовательность, представленную на фиг.1 и 2, и последовательности, имеющие существенную идентичность с ними, например, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 99% идентичности с ними.
Вариабельные домены могут происходить из любой зародышевой линии или реаранжированного вариабельного домена человека или могут быть синтетическим вариабельным доменом на основе консенсусных последовательностей известных вариабельных доменов человека. Последовательности согласно настоящему изобретению могут быть введены в репертуар вариабельных доменов, в которых отсутствуют области CDR3, с использованием технологии рекомбинантной ДНК, таких как перестановка или комбинаторные методы, изложенные у Stemmer (Nature 370:389-391 (1994)), где автор описывает технологию, основанную на гене бета-лактамазы, но также отмечает, что этот подход может быть использован для получения антител.
Дополнительной альтернативой является получение новых областей VH или VL, несущих последовательности согласно настоящему изобретению, с использованием случайного мутагенеза, например, генов SC104 VH или VL для получения мутаций в пределах всего вариабельного домена. Такой метод описан Gram с соавторами (Gram et al. Proc. Natl. Acad. Sci. USA 89: 3576-3580(1992)), где автор использовал допускающую ошибки ПЦР.
Другой способ, который может быть использован, представляет собой направленный мутагенез в областях CDR генов VH или VL. Такие методы описаны в источниках: Barbas et al. (Proc. Natl. Acad. Sci. USA 91:3809-3813 (1994)) и Schier et al. (J. Mol. Biol. 263:551-567 (1996)).
Значительная часть вариабельного домена иммуноглобулина, обычно, содержит по меньшей мере три области CDR вместе с промежуточными каркасными областями. Эта часть также может включать по меньшей мере около 50% из первой или четвертой каркасных областей или из них обеих, причем в эти 50% входят С-концевые 50% первой каркасной области и N-концевые 50% четвертой каркасной области. Дополнительные остатки на N-конце или С-конце указанной значительной части вариабельного домена могут представлять собой остатки, обычно не ассоциированные с встречающимися в природе областями вариабельного домена. Например, конструирование специфических связывающих молекул по настоящему изобретению, полученных методами рекомбинантной ДНК, может приводить к введению N- или С-концевых остатков, кодируемых линкерами, введенными для облегчения клонирования или других этапов модификации, включая введение линкеров для присоединения вариабельных доменов по настоящему изобретению к дополнительным белковым последовательностям, включая тяжелые цепи иммуноглобулина, других вариабельных доменов (например, при продуцировании диател) или белковых меток, как более подробно обсуждается ниже.
Предпочтительно специфическая связывающая молекула содержит пару связывающих доменов на основе аминокислотных последовательностей для областей VL и VH, по существу представленных на фиг. 1. Одиночные связывающие домены на основе любой из этих последовательностей составляют дополнительные аспекты настоящего изобретения. В случае связывающих доменов на основе аминокислотной последовательности для области VH, по существу представленной на фиг. 1, такие связывающие домены могут быть использованы в качестве нацеливающих агентов, поскольку известно, что домены VH иммуноглобулина способны связывать антигены-мишени специфическим образом.
Специфические связывающие молекулы согласно настоящему изобретению могут дополнительно содержать константные области антитела или их части. Например, специфические связывающие молекулы на основе области VL, показанной на фиг. 1, могут быть присоединены на С-конце к константным доменам легкой цепи антитела, включающим цепи Сκ или Сλ человека. Аналогичным образом специфические связывающие молекулы на основе области VH, показанной на фиг. 1, могут быть присоединены на С-конце к полной тяжелой цепи иммуноглобулина или ее части, происходящей из антитела любого изотипа, например, IgG, IgA, IgE и IgM, и из антитела любого подкласса, в частности, IgG1 и IgG4.
Специфические связывающие молекулы согласно настоящему изобретению могут быть дополнительно помечены функциональной меткой. Такие функциональные метки включают токсины, такие как рицин, и ферменты, такие как бактериальная карбоксипептидаза или нитроредуктаза, которые способны превращать пролекарства в активные лекарственные средства. Кроме того, специфические связывающие молекулы могут быть присоединены или иным образом ассоциированы с химиотерапевтическими или цитотоксическими агентами, такими как калихеамицин, или радиоактивными метками, такими как 90Y или 131I.
Кроме того, специфические связывающие молекулы согласно настоящему изобретению могут быть ассоциированы с дополнительным терапевтическим агентом или нацеливающим фрагментом. Терапевтические агенты, которые могут быть ассоциированы со специфическими связывающими молекулами, включают иммуномодуляторы и эффекторы, радиоактивные соединения, ферменты (например, перфорин) или химиотерапевтические агенты (например, цисплатин). Чтобы гарантировать, что токсические эффекты проявляются в желаемом месте, токсин может находиться внутри липосомы, соединенной со специфическими связывающими молекулами, что обеспечивает медленное высвобождение соединения. Это предотвратит повреждающие эффекты при транспорте в организме и обеспечит максимальный эффект токсина после связывания специфических связывающих молекул с релевантными антиген-презентирующими клетками.
Примеры подходящих терапевтических агентов включают, не ограничиваясь перечисленными:
 Низкомолекулярные цитотоксические агенты, т.е. соединения, обладающие способностью убивать клетки млекопитающих, имеющие молекулярную массу менее 700 дальтон. Такие соединения могут также содержать токсичные металлы, способные оказывать цитотоксическое действие. Кроме того, следует понимать, что эти низкомолекулярные цитотоксические агенты также включают пролекарства, то есть соединения, которые распадаются или превращаются в физиологических условиях с высвобождением цитотоксических агентов. Примеры таких агентов включают: цисплатин, производные майтансина, рахелмицин (патент СС-1065), калихеамицин, доцетаксел, этопозид, гемцитабин, ифосфамид, иринотекан, мелфалан, митоксантрон, порфимер натрия фотофрин II, темозоломид, топотекан, триметрексата глюкуронат, ауристатин Е, винкристин и доксорубицин;
Низкомолекулярные цитотоксические агенты, т.е. соединения, обладающие способностью убивать клетки млекопитающих, имеющие молекулярную массу менее 700 дальтон. Такие соединения могут также содержать токсичные металлы, способные оказывать цитотоксическое действие. Кроме того, следует понимать, что эти низкомолекулярные цитотоксические агенты также включают пролекарства, то есть соединения, которые распадаются или превращаются в физиологических условиях с высвобождением цитотоксических агентов. Примеры таких агентов включают: цисплатин, производные майтансина, рахелмицин (патент СС-1065), калихеамицин, доцетаксел, этопозид, гемцитабин, ифосфамид, иринотекан, мелфалан, митоксантрон, порфимер натрия фотофрин II, темозоломид, топотекан, триметрексата глюкуронат, ауристатин Е, винкристин и доксорубицин;
 Пептидные цитотоксины, то есть белки или их фрагменты, обладающие способностью убивать клетки млекопитающих. Например, рицин, дифтерийный токсин, псевдомонадный бактериальный экзотоксин А, ДНКаза и РНКаза;
Пептидные цитотоксины, то есть белки или их фрагменты, обладающие способностью убивать клетки млекопитающих. Например, рицин, дифтерийный токсин, псевдомонадный бактериальный экзотоксин А, ДНКаза и РНКаза;
 Радионуклиды, т.е. нестабильные изотопы элементов, которые распадаются при одновременном излучении одной или нескольких α- или β-частиц или γ-лучей. Например, йод-131, рений-186, индий-111, иттрий-90, висмут-210 и висмут-213, актиний-225 и астатин-213; хелатирующие агенты могут быть использованы для облегчения ассоциации этих радионуклидов с высокоаффинными TCR или их мультимерами;
Радионуклиды, т.е. нестабильные изотопы элементов, которые распадаются при одновременном излучении одной или нескольких α- или β-частиц или γ-лучей. Например, йод-131, рений-186, индий-111, иттрий-90, висмут-210 и висмут-213, актиний-225 и астатин-213; хелатирующие агенты могут быть использованы для облегчения ассоциации этих радионуклидов с высокоаффинными TCR или их мультимерами;
 Иммуностимуляторы, т.е. иммунные эффекторные молекулы, которые стимулируют иммунный ответ.Например, цитокины, такие как ИЛ-2 и ИФН-γ;
Иммуностимуляторы, т.е. иммунные эффекторные молекулы, которые стимулируют иммунный ответ.Например, цитокины, такие как ИЛ-2 и ИФН-γ;
 Суперантигены и их мутантные формы;
Суперантигены и их мутантные формы;
 Слитые TCR-HLA, например, слияние с комплексом пептид-HLA, где пептид происходит из обычного патогена человека, такого как вирус Эпштейна-Барр (ВЭБ);
Слитые TCR-HLA, например, слияние с комплексом пептид-HLA, где пептид происходит из обычного патогена человека, такого как вирус Эпштейна-Барр (ВЭБ);
 Хемокины, такие как ИЛ-8, тромбоцитарный фактор 4, стимулирующий рост меланомы белок и т.д.;
Хемокины, такие как ИЛ-8, тромбоцитарный фактор 4, стимулирующий рост меланомы белок и т.д.;
 Антитела или их фрагменты, включая антитела против Т-клеточных или NK-клеточных детерминант (например, анти-СD3, анти-СD28 или анти-СD16);
Антитела или их фрагменты, включая антитела против Т-клеточных или NK-клеточных детерминант (например, анти-СD3, анти-СD28 или анти-СD16);
 Альтернативные белковые скаффолды с антитело-подобными характеристиками связывания;
Альтернативные белковые скаффолды с антитело-подобными характеристиками связывания;
 Активаторы комплемента;
Активаторы комплемента;
 Ксеногенные белковые домены, аллогенные белковые домены, вирусные/бактериальные белковые домены, вирусные/бактериальные пептиды.
Ксеногенные белковые домены, аллогенные белковые домены, вирусные/бактериальные белковые домены, вирусные/бактериальные пептиды.
Нацеливающие фрагменты, которые могут быть связаны со специфическими связывающими молекулами, включают:
 TCR (включая TCR альфа/бета и гамма/дельта);
TCR (включая TCR альфа/бета и гамма/дельта);
 Антитела или их фрагменты, которые распознают и связывают антигены, представленные на клетках-мишенях, включая антигены клеточной поверхности и пептиды, происходящие из внутриклеточных антигенов, которые представлены на клеточной поверхности в комплексе с MHC/HLA;
Антитела или их фрагменты, которые распознают и связывают антигены, представленные на клетках-мишенях, включая антигены клеточной поверхности и пептиды, происходящие из внутриклеточных антигенов, которые представлены на клеточной поверхности в комплексе с MHC/HLA;
 Альтернативные белковые скаффолды с антитело-подобными характеристиками связывания.
Альтернативные белковые скаффолды с антитело-подобными характеристиками связывания.
Далее, специфические связывающие молекулы по настоящему изобретению можно вводить отдельно или в комбинации с другими видами лечения, одновременно или последовательно, в зависимости от состояния, подлежащего лечению. Таким образом, согласно настоящему изобретению дополнительно предложены продукты, содержащие специфическую связывающую молекулу по настоящему изобретению и активный агент в качестве комбинированного состава для одновременного, раздельного или последовательного применения при лечении опухоли. Активные агенты могут включать химиотерапевтические или цитотоксические агенты, включая 5-фторурацил, цисплатин, митомицин С, оксалиплатин и тамоксифен, которые могут работать синергетически со связывающими молекулами согласно настоящему изобретению. Другие активные агенты могут включать подходящие дозы обезболивающих средств, таких как нестероидные противовоспалительные средства (например, аспирин, парацетамол, ибупрофен или кетопрофен) или опиаты, такие как морфин, или противорвотные средства.
Специфические связывающие молекулы согласно настоящему изобретению обычно вводят в форме фармацевтической композиции, которая может содержать по меньшей мере один ингредиент в дополнение к специфической связывающей молекуле. Фармацевтическая композиция может содержать, помимо активного ингредиента, фармацевтически приемлемое вспомогательное вещество, разбавитель, носитель, буфер, стабилизатор или другие материалы, хорошо известные специалистам в данной области техники. Такие материалы должны быть нетоксичными и не должны влиять на эффективность активного ингредиента. Точная природа носителя или другого материала зависит от пути введения, который может быть пероральным или инъекционным, например, внутривенным.
Предполагается, что инъекции будут основным путем для терапевтического введения указанных композиций, хотя также может быть использована доставка через катетер или другую хирургическую трубку. Некоторые подходящие пути введения включают внутривенное, подкожное и внутримышечное введение. Жидкие составы могут быть использованы после восстановления из порошковых составов.
Для внутривенной инъекции или инъекции в очаг поражения активный ингредиент находится в форме парентерально приемлемого водного раствора, который не содержит пирогенов и имеет подходящий рН, изотоничность и стабильность. Специалисты в данной области техники могут приготовить подходящие растворы с использованием, например, изотонических основ, таких как хлорид натрия для инъекций, раствор Рингера для инъекций, раствор Рингера с лактатом для инъекций. При необходимости могут быть включены консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки.
Фармацевтические композиции для перорального введения могут быть представлены в форме таблеток, капсул, порошков или жидкостей. Таблетка может содержать твердый носитель, такой как желатин или адъювант.Жидкие фармацевтические композиции обычно содержат жидкий носитель, такой как вода, продукты переработки нефти, животные или растительные масла, минеральное масло или синтетическое масло. Могут быть включены физиологический солевой раствор, раствор декстрозы или другого сахарида, или гликоли, такие как этиленгликоль, пропиленгликоль или полиэтиленгликоль. Если состав представляет собой жидкость, он может представлять собой, например, физиологический солевой раствор, содержащий буфер без фосфата с рН 6,8 7,6, или лиофилизированный порошок.
Указанную композицию также можно вводить в микросферах, липосомах, других системах доставки микрочастиц или составах с замедленным высвобождением, помещенных в определенные ткани, включая кровь. Подходящие примеры носителей с замедленным высвобождением включают полупроницаемые полимерные матрицы в форме общих изделий, например, суппозиториев или микрокапсул. Имплантируемые или микрокапсульные матрицы с замедленным высвобождением включают полилактиды (патент США №3773919; ЕР-А-0058481), сополимеры L-глутаминовой кислоты и гамма-этил-L-глутамата (Sidman et al., Biopolymers 22(1): 547-556, 1985), поли(2-гидроксиэтил-метакрилат) или этиленвинилацетат (Langer et al, J. Biomed. Mater. Res. 15: 167-277, 1981, and Langer, Chem. Tech. 12:98-105, 1982). Липосомы, содержащие полипептиды, получают хорошо известными способами: DE 3218121 A; Epstein et al., PNAS USA, 82: 3688-3692, 1985; Hwang et al, PNAS USA, 77: 4030-4034, 1980; EP-A-0052522; E-A-0036676; EP-A-0088046; EP-A-0143949; EP-A-0142541; JP-A-83-11808; патенты США №4485045 и №4544545. Обычно липосомы относятся к небольшим (около 200-800 ангстрем) липосомам однослойного типа, в которых содержание липидов выше около 30 мол. % холестерина, выбранную пропорцию корректируют для оптимальной скорости высвобождения полипептида.
Указанная композиция может быть введена локально в требуемое место или доставлена таким образом, чтобы она была нацелена на релевантные клетки.
Указанные композиции предпочтительно вводят индивидууму в «терапевтически эффективном количестве», то есть достаточном для того, чтобы у индивидуума наблюдался благоприятный эффект. Фактическое вводимое количество, а также скорость и временные характеристики введения зависят от природы и тяжести подлежащего лечению состояния. Назначение лечения, например, принятие решений о дозировке и т.п.входит в компетенцию врачей общей практики и других медицинских специалистов, и обычно учитываются заболевание, подлежащее лечению, состояние индивидуального пациента, место доставки, способ введения и другие факторы, известные лечащим врачам.
Оптимальная доза может быть определена лечащими врачами на основании ряда параметров, включая, например, возраст, пол, массу тела, тяжесть состояния, подлежащего лечению, вводимый активный ингредиент и путь введения. В целом, желательна концентрация полипептидов и антител в сыворотке, которая обеспечивает насыщение рецепторов. Обычно достаточно концентрации, превышающей приблизительно 0,1 нМ. Например, доза 100 мг/м2 антитела обеспечивает концентрацию в сыворотке приблизительно 20 нМ в течение приблизительно восьми дней.
Согласно приблизительной рекомендации дозы антител можно вводить еженедельно в количестве 10-300 мг/м2. Эквивалентные дозы фрагментов антител следует использовать с меньшими интервалами времени для поддержания в сыворотке уровня, превышающего концентрацию, допускающую насыщение сиалилтетраозил-карбогидратцерамида.
Доза композиции зависит от свойств связывающей молекулы, например, ее связывающей активности и периода полужизни в плазме in vivo, концентрации полипептида в составе, пути введения, места и скорости дозирования, клинической переносимости у пациента, патологического состояния, которым страдает пациент, и т.п., как хорошо известно практикующим врачам. Например, предпочтительными являются дозы 300 мкг антитела на пациента на одно введение, хотя дозировки могут варьировать от примерно 10 мкг до 6 мг на дозу. При проведении ряда последовательных инокуляций используют разные дозировки; лечащий врач может провести первичную инокуляцию, а затем проводить вторичную стимуляцию относительно более низкими дозами антитела.
Связывающие молекулы согласно настоящему изобретению могут быть получены полностью или частично путем химического синтеза. Связывающие молекулы могут быть легко получены в соответствии с хорошо известными, стандартными методами жидкофазного или предпочтительно твердофазного синтеза пептидов, общие описания которых широко доступны (см., например, J.M. Stewart and J.D. Young, Solid Phase Peptide Synthesis, 2nd edition, Pierce Chemical Company, Rockford, Illinois (1984), M. Bodanzsky and A. Bodanzsky, The Practice of Peptide Synthesis, Springer Verlag, New York (1984); и Applied Biosystems 430A Users Manual, ABI Inc., Foster City, California), или могут быть получены в растворе с помощью жидкофазного метода или с помощью любой комбинации твердофазного, жидкофазного методов и химии растворов, например, сначала путем завершения синтеза соответствующей части пептида, а затем, если это желательно и целесообразно, после удаления любых присутствующих защитных групп, путем введения остатка X путем реакции соответствующей угольной или сульфоновой кислоты, или их реакционноспособного производного.
Другим удобным способом получения связывающей молекулы в соответствии с настоящим изобретением является экспрессия кодирующей ее нуклеиновой кислоты за счет использования указанной нуклеиновой кислоты в экспрессионной системе.
Согласно настоящему изобретению дополнительно предложена выделенная нуклеиновая кислота, кодирующая специфическую связывающую молекулу согласно настоящему изобретению. Нуклеиновая кислота включает ДНК и РНК. В предпочтительном аспекте согласно настоящему изобретению предложена нуклеиновая кислота, которая кодирует специфическую связывающую молекулу согласно настоящему изобретению, как определено выше. Специалист в данной области техники сможет определить замены, делеции и/или добавления в таких нуклеиновых кислотах, которые все еще будут обеспечивать получение специфической связывающей молекулы согласно настоящему изобретению.
Согласно настоящему изобретению также предложены конструкции в форме плазмид, векторов, транскрипционных или экспрессионных кассет, которые содержат по меньшей мере одну нуклеиновую кислоту согласно описанию выше. Согласно настоящему изобретению также предложена рекомбинантная клетка-хозяин, которая содержит одну или более конструкций согласно описанию выше. Как уже упоминалось, нуклеиновая кислота, кодирующая специфическую связывающую молекулу согласно настоящему изобретению, образует аспект настоящего изобретения, как и способ получения специфической связывающей молекулы, включающий экспрессию кодирующей ее нуклеиновой кислоты. Экспрессия может быть удобным образом достигнута путем культивирования в подходящих условиях рекомбинантных клеток-хозяев, содержащих нуклеиновую кислоту. После продуцирования путем экспрессии специфическая связывающая молекула может быть выделена и/или очищена с использованием любой подходящей методики, а затем использована надлежащим образом.
Хорошо известны системы для клонирования и экспрессии полипептида в различных клетках-хозяевах. К подходящим клеткам-хозяевам относятся бактерии, клетки млекопитающих, дрожжевые и бакуловирусные системы. Линии клеток млекопитающих, доступные в данной области техники для экспрессии гетерологичного полипептида, включают клетки яичника китайского хомячка, клетки HeLa, клетки почек новорожденного хомяка, клетки меланомы мыши линии NS0 и многие другие. Обычным предпочтительным бактериальным хозяином является Е. coli. Экспрессия антител и фрагментов антител в прокариотических клетках, таких как клетки Е. coli, хорошо известна в данной области техники. Обзорную информацию можно найти, например, в источнике: Pluckthun, Bio/Technology 9:545-551 (1991). Экспрессия в эукариотических клетках в культуре также доступна специалистам в данной области в качестве варианта продуцирования специфической связывающей молекулы, см., например, недавнюю обзорную статью Reff, Curr. Opinion Biotech. 4:573-576 (1993); Trill et al., Curr. Opinion Biotech. 6:553-560 (1995).
Подходящие векторы могут быть выбраны или сконструированы, содержат соответствующие регуляторные последовательности, включая промоторные последовательности, терминирующие последовательности, последовательности полиаденилирования, энхансерные последовательности, маркерные гены и другие последовательности, по необходимости. Векторы могут представлять собой плазмиды, вирусные векторы, например, фаг или фагмиду, по необходимости. Для получения дополнительной информации см., например, Sambrook et al, Molecular Cloning: A Laboratory Manual: 2nd Edition, Cold Spring Harbor Laboratory Press (1989). Многие известные методики и протоколы для манипуляций с нуклеиновой кислотой, например, при получении нуклеиновокислотных конструкций, мутагенезе, секвенировании, введении ДНК в клетки и экспрессии генов, а также анализе белков, подробно описаны в источнике: Ausubel et al. eds., Short Protocols in Molecular Biology, 2nd Edition, John Wiley & Sons (1992).
Таким образом, в дополнительном аспекте настоящего изобретения предложена клетка-хозяин, содержащая нуклеиновую кислоту согласно описанию в настоящем документе. В другом дополнительном аспекте предложен способ, включающий введение такой нуклеиновой кислоты в клетку-хозяина. Для введения можно использовать любую доступную методику. Подходящие для эукариотических клеток методики могут включать кальций-фосфатную трансфекцию, трансфекцию с ДЭАЭ-декстраном, электропорацию, липофекцию и трансдукцию с использованием ретровируса или другого вируса, например, вируса осповакцины или, для клеток насекомых, бакуловируса. Подходящие для бактериальных клеток методики могут включать трансформацию с использованием хлорида кальция, электропорацию и трансдукцию с использованием бактериофага. После введения может быть вызвана или разрешена экспрессия нуклеиновой кислоты, например, путем культивирования клеток-хозяев в условиях, подходящих для экспрессии гена.
Нуклеиновая кислота согласно настоящему изобретению может быть интегрирована в геном (например, в хромосому) клетки-хозяина. Интеграции может способствовать включение последовательностей, стимулирующих рекомбинацию с геномом, в соответствии со стандартными методиками.
В рамках настоящего изобретения также предложен способ, который включает применение конструкции согласно описанию выше в системе для экспрессии специфической связывающей молекулы или полипептида согласно описанию выше.
В дополнительном аспекте предложена бифункциональная связывающая молекула, содержащая:
i) Т-клеточный рецептор (TCR) или антитело; и
ii) Специфическую связывающую молекулу, которая связывается с CD3 в соответствии с первым аспектом.
Следует понимать, что все признаки, описанные выше в отношении первого аспекта, в равной степени применимы к этому дополнительному аспекту.
Нацеливающий фрагмент может представлять собой Т-клеточный рецептор (TCR), антитело или фрагмент антитела.
Структура анти-CD3 и нацеливающего фрагмента может соответствовать любому известному формату (например, как описано в источнике: Brinkman et al., MAbs. 2017 Feb-Mar; 9(2): 182-212, фиг. 2).
Т-клеточный рецептор (TCR) может представлять собой гетеродимерную пару полипептидов TCR альфа/бета или гамма/дельта. Т-клеточный рецептор (TCR) может представлять собой одноцепочечный пептид TCR.
Специфическая связывающая молекула, которая связывается с CD3, может быть слита с С- или N-концом альфа- или бета-цепи Т-клеточного рецептора (TCR).
Предпочтительно эффектор CD3 слит с N-концом бета-цепи TCR. Бифункциональная связывающая молекула может быть в форме диатела, в котором область TCR-Va присоединена к анти-CD3-VL, a TCR-Vb присоединена к анти-CD3-VH, и наоборот.
В некоторых случаях специфическая связывающая молекула, которая связывается с CD3, может быть слита с С- или N-концом нацеливающего фрагмента. В других случаях специфическая связывающая молекула, которая связывается с CD3, слита с С- или N-концом нацеливающего фрагмента с помощью линкера.
Специфическая связывающая молекула, которая связывается с CD3, может быть слита с Т-клеточным рецептором (TCR) или TCR-подобным антителом с помощью линкера, который может быть полипептидным. Полипептидные линкерные последовательности обычно являются гибкими, поскольку они состоят в основном из аминокислот, таких как глицин, аланин и серии, которые не имеют объемных боковых цепей, способных ограничить гибкость. Пригодные для применения или оптимальные длины линкерных последовательностей могут быть легко определены. Часто линкерная последовательность имеет длину менее 12, например, менее 10, или от 5-10 аминокислот в длину. Длина линкера может составлять 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 аминокислот. Предпочтительный линкер содержит последовательность формулы (GGGGS)n, необязательно в дополнение к другим аминокислотам.
TCR может содержать не являющуюся нативной дисульфидную связь между константной областью альфа-цепи и константной областью бета-цепи.
TCR может связываться с ГКГС в комплексе с пептидным антигеном. Предпочтительно пептидный антиген представляет собой любой ассоциированный с заболеванием антиген. Предпочтительно пептидный антиген представляет собой любой опухолеассоциированный антиген. Предпочтительно пептидный антиген представляет собой пептид, происходящий из GP100, NYESO, MAGEA4 или PRAME согласно описанию в WO2011001152, WO2017109496, WO2017175006 и WO2018234319.
Подходящий TCR может иметь аминокислотную последовательность, определенную в WO2011001152, WO2017109496, WO2017175006 и WO2018234319.
TCR может иметь следующую аминокислотную последовательность:
DGGITQSPKYLFRKEGQNVTLSCEQNLNHDAMYWYRQDPGQGLRLIYYSQIMG DEQKGDIAEGYSVSREKKESFPLTVTSAQKNPTAFYLCASSWWTGGSAPIRFGPGTRLT VTEDLKNVFPPEVAVFEPSEAEISHTQKATLVCLATGFYPDHVELSWWVNGKEVHSGV CTDPQPLKEQPALNDSRYALSSRLRVSATFWQDPRNHFRCQVQFYGLSENDEWTQDRA KPVTQIVSAEAWGRAD
или аминокислотную последовательность, по меньшей мере на 70% идентичную указанной последовательности.
Бифункциональная связывающая молекула может иметь следующую аминокислотную последовательность:
AIQMTQSPSSLSASVGDRVTITCRASQDIRNYLNWYQQKPGKАРKLLIYYTSRLESGVPSRFSGSGSGTD LTISSLQPEDFATYYCQQGNTLPWTFGQGTKVEIKGGGGSGGGGSGGGGSGGGGSGGGSEVQLVESGGGLVQPGGSLRLSCAASGYSFTGYAMNWVRQAPGKGLEWVALINPYKGVSTYNQKFKDRFTISVDKSKNTAYLQMNSLRAEDTAVYYCARSGYYGDSDWYFDVWGQGTLVTVSSGGGGSDGGITQSPKYLFRKEGQNVTLSCEQNLNHDAMYWYRQDPGQGLRLIYYSQIMGDEQKGDIAEGYSVSREKKESFPLTVTSAQKNPTAFYLCASSWWTGGSAPIRFGPGTRLTVTEDLKNVFPPEVAVFEPSEAEISHTQKATLVCLATGFYPDHVELSWWVNGKEVHSGVCTDPQPLKEQPALNDSRYALSSRLRVSATFWQDP RNHFRCQVQFYGLSENDEWTQDRAKPVTQIVSAEAWGRAD
или аминокислотную последовательность, по меньшей мере на 70% идентичную указанной последовательности.
Или бифункциональная связывающая молекула может иметь следующую аминокислотную последовательность:
AIQMTQSPSSLSASVGDRVTITCRASQDIRNYLNWYQQKPGKАРKLLIYYTSRLESGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQGNTLPWTFGQGTKVEIKGGGGSGGGGSGGGGSGGGGSGGGSEVQLVESGGGLVQPGGSLRLSCAASGYSFTGYAMNWVRQAPGKGLEWVALINPYKGVSTYNQKFKDRFTFSVDKSKNTAYLQMNSLRAEDTAVYYCARSGYYGDSDWYFDVWGQGTLVTVSSGGGGSDGGITQSPKYLFRKEGQNVTLSCEQNLNHDAMYWYRQDPGQGLRLIYYSQIMGDEQKGDIAEGYSVSREKKESFPLTVTSAQKNPTAFYLCASSWWTGGSAPIRFGPGTRLTVTEDLKNVFPPEVAVFEPSEAEISHTQKATLVCLATGFYPDHVELSWWVNGKEVHSGVCTDPQPLKEQPALNDSRYALSSRLRVSATFWQDPRNHFRCQVQFYGLSENDEWTQDRAKPVTQIVSAEAWGRADили аминокислотную последовательность, по меньшей мере на 70% идентичную указанной последовательности.
В данном документе также описаны варианты специфических связывающих молекул или бифункциональных связывающих молекул с молчащим фенотипом. В настоящем документе термин «варианты с молчащим фенотипом» следует понимать как относящийся к специфической связывающей молекуле или вариабельному домену бифункциональной связывающей молекулы, который включает одно или более дополнительных аминокислотных изменений, включая замены, инсерции и делеции, при этом молекула имеет фенотип, аналогичный фенотипу соответствующей молекулы или полипептида без указанного изменения (изменений).
Варианты с молчащим фенотипом могут содержать одну или более консервативных замен и/или одну или более приемлемых замен. Под приемлемыми заменами подразумеваются замены, которые не подпадают под определение консервативных согласно описанию ниже, но, тем не менее, фенотипически не проявляются. Специалисту в данной области техники известно, что различные аминокислоты обладают аналогичными свойствами и, соответственно, являются «консервативными». Одна или более таких аминокислот белка, полипептида или пептида часто могут быть заменены одной или несколькими другими такими аминокислотами без устранения желаемой активности этого белка, полипептида или пептида.
Таким образом, часто аминокислоты глицин, аланин, валин, лейцин и изолейцин могут быть взаимно заменены (аминокислоты, имеющие алифатические боковые цепи). Для этих возможных замен предпочтительно использование глицина и аланина для взаимных замен (поскольку они имеют относительно короткие боковые цепи), а также валина, лейцина и изолейцина (поскольку они имеют более крупные алифатические боковые цепи, которые являются гидрофобными). Другие аминокислоты, которые часто могут быть взаимно заменены, включают: фенилаланин, тирозин и триптофан (аминокислоты, имеющие ароматические боковые цепи); лизин, аргинин и гистидин (аминокислоты, имеющие основные боковые цепи); аспартат и глутамат (аминокислоты, имеющие кислые боковые цепи); аспарагин и глутамин (аминокислоты, имеющие амидные боковые цепи); и цистеин и метионин (аминокислоты, имеющие серосодержащие боковые цепи). Подразумевается, что замены аминокислот в пределах объема настоящего изобретения могут быть осуществлены с использованием встречающихся или не встречающихся в природе аминокислот. Например, настоящим изобретением предусмотрено, что метальная группа на аланине может быть заменена этильной группой и/или что в остов пептида могут быть внесены незначительные изменения. Независимо от того, используются ли природные или синтетические аминокислоты, предпочтительно, чтобы присутствовали только L-аминокислоты.
Замены такого характера часто называют «консервативными» или «полуконсервативными» заменами аминокислот. Таким образом, настоящее изобретение охватывает применение специфической связывающей молекулы или бифункциональной связывающей молекулы, содержащей любую из аминокислотных последовательностей, описанных выше, но с одной или более консервативными заменами и/или одной или более приемлемыми заменами в последовательности, так что аминокислотная последовательность указанной специфической связывающей молекулы или бифункциональной связывающей молекулы имеет по меньшей мере 80% идентичности или по меньшей мере 90% идентичности, например, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичности со специфической связывающей молекулой или бифункциональной связывающей молекулой, описанной выше.
«Идентичность», как известно в данной области техники, представляет собой взаимосвязь между двумя или более полипептидными последовательностями или двумя или более полинуклеотидными последовательностями, определенную путем сравнения указанных последовательностей. В данной области техники идентичность также означает степень родства последовательностей, полипептидных или полинуклеотидных, в зависимости от обстоятельств, определяемую на основании совпадения строк таких последовательностей. Хотя существует ряд способов измерения идентичности двух полипептидных или двух полинуклеотидных последовательностей, способы, обычно используемые для определения идентичности, кодифицированы в виде компьютерных программ. Предпочтительные компьютерные программы для определения идентичности двух последовательностей включают, не ограничиваясь перечисленным, программный пакет GCG (Devereux, et al., Nucleic Acids Research, 12, 387 (1984)), BLASTP, BLASTN и FASTA (Atschul et al, J. Molec. Biol. 215, 403 (1990)).
Для сравнения аминокислотных последовательностей можно использовать программу, такую как программа CLUSTAL. Эта программа сравнивает аминокислотные последовательности и находит оптимальное выравнивание, вставляя пробелы в любую из последовательностей по мере необходимости. Можно рассчитать идентичность или сходство аминокислот (идентичность плюс сохранение типа аминокислоты) для оптимального выравнивания. Такая программа, как BLASTx, выравнивает самый длинный отрезок сходных последовательностей и присваивает значение соответствию. Таким образом, можно получить сравнение с несколькими найденными областями сходства, каждая из которых имеет отличающуюся оценку. Оба типа анализа идентичности предусмотрены настоящим изобретением.
Процент идентичности двух аминокислотных последовательностей или двух последовательностей нуклеиновых кислот определяют путем выравнивания указанных последовательностей с целью оптимального сравнения (например, в первую последовательность могут быть введены пропуски для наилучшего выравнивания с последовательностью) и сравнения остатков аминокислот или нуклеотидов в соответствующих положениях. «Наилучшее выравнивание» представляет собой выравнивание двух последовательностей, которое приводит к самому высокому проценту идентичности. Процент идентичности определяют по количеству идентичных остатков аминокислот или нуклеотидов в сравниваемых последовательностях (т.е. % идентичности = количество идентичных положений/общее количество положений х 100).
Определение процента идентичности двух последовательностей может быть выполнено с использованием математического алгоритма, известного специалистам в данной области техники. Примером математического алгоритма для сравнения двух последовательностей является алгоритм Карлина-Альтшула (Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA 87:2264-2268), модифицированный согласно Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-5877. Программы BLASTn и BLASTp (Altschul, et al. (1990) J. Mol. Biol. 215:403-410) включают такой алгоритм. Определение процента идентичности двух нуклеотидных последовательностей может быть выполнено с помощью программы BLASTn. Определение процента идентичности двух последовательностей белка может быть выполнено с помощью программы BLASTp.Чтобы получить выравнивания с пропусками в целях сравнения, можно использовать Gapped BLAST согласно описанию в Altschul et al. (1997) Nucleic Acids Res. 25:3389-3402. В качестве альтернативы можно использовать PSI-Blast для выполнения итерационного поиска, который обнаруживает отдаленные взаимосвязи между молекулами (там же). При использовании программ BLAST, Gapped BLAST и PSI-BLAST можно использовать установленные по умолчанию параметры соответствующих программ (например, BLASTn и BLASTp). Подробнее см. на http://www.ncbi.nlm.nih.gov. Общие параметры по умолчанию могут включать, например, размер слова = 3, ожидаемый порог = 10. Могут быть выбраны параметры для автоматического подстраивания под короткие входные последовательности. Другим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм Майерса и Миллера, CABIOS (1989). Программа ALIGN (версии 2.0), которая является частью пакета программного обеспечения для выравнивания последовательностей CGC, включает такой алгоритм. Другие алгоритмы для анализа последовательностей, известные в данной области техники, включают ADVANCE и ADAM, которые описаны в источнике: Torellis and Robotti (1994) Comput. Appl. Biosci., 10:3-5; и FASTA, описанный в источнике: Pearson and Lipman (1988) Proc. Natl. Acad. Sci. 85:2444-8. В FASTA ktup - это опция управления, которая задает чувствительность и скорость поиска. Для оценки процента идентичности согласно настоящему изобретению в качестве метода сравнения используют BLASTp с параметрами по умолчанию. Кроме того, когда указанный процент идентичности дает нецелое значение для аминокислот (т.е. последовательность из 25 аминокислот, идентичная на 90% другой последовательности, дает значение «22,5», которое округляют в меньшую сторону до ближайшего целого числа, соответственно, до «22»). Соответственно, в представленном примере % идентичности последовательности, имеющей 22 совпадающих аминокислоты из 25 аминокислот, находится в пределах 90% идентичности последовательности.
Как будет очевидно специалистам в данной области техники, возможно осуществить усечение или удлинение последовательностей на С-конце и/или N-конце специфической связывающей молекулы на 1, 2, 3, 4, 5 или более остатков, по существу не влияющее на ее функциональные характеристики. Последовательности на С-конце и/или N-конце специфической связывающей молекулы могут быть усечены или удлинены на 1, 2, 3, 4 или 5 остатков. Все такие варианты охвачены настоящим изобретением.
Мутации, включая консервативные и приемлемые замены, инсерции и делеции, могут быть введены в последовательности, полученные с применением любого подходящего способа, включая, но не ограничиваясь перечисленными, способы на основе полимеразной цепной реакции (ПЦР), клонирования с использованием эндонуклеаз рестрикции или процедур безлигазного клонирования (LIC). Эти способы подробно описаны во многих стандартных пособиях по молекулярной биологии. Для получения дополнительной информации о полимеразной цепной реакции (ПЦР) и клонировании с использованием рестрикционных эндонуклеаз см. руководство: Sambrook & Russell, (2001) Molecular Cloning - A Laboratory Manual (3rd Ed.) CSHL Press. Дополнительную информацию о процедурах безлигазного клонирования (LIC) можно найти в Rashtchian, (1995) Curr Opin Biotechnol 6(1): 30-6. Последовательности, предложенные согласно настоящему изобретению, могут быть получены с помощью твердофазного синтеза или любого другого подходящего способа, известного в данной области техники.
Нацеливающий фрагмент может представлять собой антитело или его фрагмент. Предпочтительно, чтобы антитела, включая их фрагменты, производные и варианты, связывались с антигенами, представленными на пораженных заболеванием или раковых клетках.
Специфические связывающие молекулы или бифункциональные связывающие молекулы согласно настоящему изобретению могут быть использованы в способах диагностики и лечения рака или инфекционных заболеваний у людей или животных. Примеры рака включают, не ограничиваясь перечисленными, жидкие опухоли, такие как лейкозы, лимфомы и миелома, и солидные опухоли, включая опухоли мочевого пузыря, молочной железы, шейки матки, ободочной и прямой кишки, пищевода, эндометрия и желудка, глиобластому, опухоли печени, меланому, опухоли легких, яичника, поджелудочной железы, предстательной железы, саркому, опухоли щитовидной железы. Примеры инфекционных заболеваний включают, не ограничиваясь перечисленными, ВИЧ, гепатит В, туберкулез, гепатит С.
При использовании в диагностике специфические связывающие молекулы или бифункциональные связывающие молекулы согласно настоящему изобретению могут быть помечены детектируемой меткой, например, радиоактивной меткой, такой как 131I или 99Iс, которые могут быть присоединены к специфическим связывающим молекулам согласно настоящему изобретению с использованием общепринятых химических способов, известных в области визуализации антител. Метки также включают ферменты, например, пероксидаза хрена. Метки также включают химические фрагменты, такие как биотин, которые могут быть детектированы за счет связывания со специфическим когнатным детектируемым фрагментом, например, меченым авидином.
Специфические связывающие молекулы или бифункциональная связывающая молекула согласно настоящему изобретению, в частности, специфические связывающие молекулы в растворимом формате, могут поддаваться высокоэффективному очищению. Выход может быть определен как на основании количества материала, сохраненного в процессе очищения (т.е. количества материала с корректной укладкой, полученного в конце процесса очищения, относительно количества солюбилизированного материала, полученного до рефолдинга), так и на основе количества материала с корректной укладкой, полученного в конце процесса очищения, относительно исходного объема культуры. Высокий выход означает более 1%, или более предпочтительно более 5%, или более высокий выход. Высокий выход означает более 1 мг/мл, или более предпочтительно более 3 мг/мл, или более 5 мг/мл, или более высокий выход.
Методы определения аффинности связывания (обратно пропорциональной константе равновесия KD) и периода полужизни (выраженного как  ) известны специалистам в данной области техники. Предпочтительно аффинность связывания и период полужизни определяют при помощи поверхностного плазмонного резонанса (ППР) или биослойной интерферометрии (ВLI), например, при помощи инструмента BIAcore или инструмента Octet, соответственно. Следует принимать во внимание, что удвоение аффинности приводит к уменьшению KD в два раза. рассчитывают как 1n2, разделенное на скорость диссоциации (koff). Таким образом, удвоение приводит к уменьшению koff в два раза. Значения KD и koff. Чтобы учесть различия между независимыми измерениями, в частности, для взаимодействий со временем диссоциации более 20 часов, аффинность связывания и/или период полужизни заданной молекулы могут быть измерены несколько раз, например, 3 или более раз, с использованием одного и того же протокола анализа и усреднением полученных результатов. Для сравнения данных связывания для двух образцов (т.е. двух разных молекул и/или двух составов с одной и той же молекулой) предпочтительно проводить измерения при одних и тех же условиях анализа (например, температуре), например, таких, как описанные в WO2018234319.
) известны специалистам в данной области техники. Предпочтительно аффинность связывания и период полужизни определяют при помощи поверхностного плазмонного резонанса (ППР) или биослойной интерферометрии (ВLI), например, при помощи инструмента BIAcore или инструмента Octet, соответственно. Следует принимать во внимание, что удвоение аффинности приводит к уменьшению KD в два раза. рассчитывают как 1n2, разделенное на скорость диссоциации (koff). Таким образом, удвоение приводит к уменьшению koff в два раза. Значения KD и koff. Чтобы учесть различия между независимыми измерениями, в частности, для взаимодействий со временем диссоциации более 20 часов, аффинность связывания и/или период полужизни заданной молекулы могут быть измерены несколько раз, например, 3 или более раз, с использованием одного и того же протокола анализа и усреднением полученных результатов. Для сравнения данных связывания для двух образцов (т.е. двух разных молекул и/или двух составов с одной и той же молекулой) предпочтительно проводить измерения при одних и тех же условиях анализа (например, температуре), например, таких, как описанные в WO2018234319.
TCR, описанные в настоящем документе, могут быть гетеродимерами ар. Альфа-бета гетеродимерные TCR обычно содержат последовательность константного домена альфа-цепи TRAC и/или последовательность константного домена бета-цепи TRBC1 или TRBC2. Константные домены могут быть полноразмерными, что означает присутствие внеклеточных, трансмембранных и цитоплазматических доменов, или они могут быть представлены в растворимом формате (т.е. не иметь транс мембранных или цитоплазматических доменов). Один или оба константных домена могут содержать мутации, замены или делеции относительно природных последовательностей TRAC и/или TRBC1/2. Термины TRAC и TRBC1/2 также охватывают природные полиморфные варианты, например, замену N на К в положении 4 TRAC (Bragado et al. International immunology. 1994 Feb;6(2):223-30).
Для растворимых TCR последовательности константных доменов альфа- и бета-цепей могут быть модифицированы путем усечения или замены с удалением природной дисульфидной связи между Cys4 экзона 2 TRAC и Cys2 экзона 2 TRBC1 или TRBC2. Последовательность (последовательности) константных доменов альфа- и/или бета-цепей может (могут) содержать введенную дисульфидную связь между остатками соответствующих константных доменов, как описано, например, в WO 03/020763. Константные домены альфа и бета могут быть модифицированы заменой Thr в положении 48 TRAC и Ser в положении 57 TRBC1 или TRBC2 на цистеины, причем указанные цистеины образуют дисульфидную связь между константными доменами альфа и бета TCR. TRBC1 или TRBC2 могут дополнительно содержать замену цистеина на аланин в положении 75 константного домена и замену аспарагина на аспарагиновую кислоту в положении 89 константного домена. Один или оба внеклеточных константных домена, присутствующих в гетеродимере ар, может (могут) быть усечен(ы) на С-конце или С-концах, например, до 15, или до 10, или до 8 или менее аминокислот. Один или оба внеклеточных константных домена, присутствующих в гетеродимере ар, может (могут) быть укорочен(ы) на С-конце или С-концах, например, до 15, или до 10, или до 8 аминокислот. С-конец внеклеточного константного домена альфа-цепи может быть усечен на 8 аминокислот. Растворимые TCR предпочтительно связаны с терапевтическими агентами и/или детектируемыми метками.
Константные домены ар гетеродимерного TCR могут быть полноразмерными и иметь как трансмембранные, так и цитоплазматические домены. Такие TCR могут содержать дисульфидную связь, соответствующую той, которая встречается в природе, между соответствующими константными доменами альфа и бета. Дополнительно или в качестве альтернативы, между внеклеточными константными доменами может присутствовать не встречающаяся в природе дисульфидная связь. Указанные не встречающиеся в природе дисульфидные связи более подробно описаны в WO03020763 и WO06000830. Не встречающаяся в природе дисульфидная связь может находиться между положением Thr 48 TRAC и положением Ser 57 TRBC1 или TRBC2. Один или оба константных домена могут содержать одну или более мутаций в виде замен или делеций относительно природных последовательностей TRAC и/или TRBC1/2. TCR с полноразмерными константными доменами предпочтительны для использования в адоптивной терапии.
TCR, описанные в настоящем документе, могут быть в одноцепочечном формате. Одноцепочечные форматы включают, не ограничиваясь перечисленными, полипептиды типа αβ TCR Vα-L-Vβ, Vβ-L-Vα, Vα-Cα-L-Vβ, Vα-L-Vβ-Cβ или Vα-Cα-L-Vβ-Cβ, где Vα и Vβ представляют собой вариабельные области TCR α и β, соответственно, Сα и Сβ представляют собой константные области TCR α и β, соответственно, a L представляет собой линкерную последовательность (Weidanz et al, (1998) J Immunol Methods. Dec l;221(l-2):59-76; Epel et al., (2002), Cancer Immunol Immunother. Nov;51(10):565-73; WO 2004/033685; W09918129). Там, где они присутствуют, один или оба константных домена могут быть полноразмерными, или они могут быть укорочены и/или содержать мутации согласно описанию выше. Предпочтительно одноцепочечные TCR являются растворимыми. Одноцепочечные TCR могут иметь введенную дисульфидную связь между остатками соответствующих константных доменов согласно описанию в WO 2004/033685. Одноцепочечные TCR более подробно описаны в WO2004/033685; W098/39482; WO01/62908; Weidanz et al. (1998) J Immunol Methods 221(1-2): 59-76; Hoo et al. (1992) Proc Natl Acad Sci U S A 89(10): 4759-4763; Schodin (1996) Mol Immunol 33(9): 819-829).
Специфические связывающие молекулы или бифункциональные связывающие молекулы могут быть связаны (ковалентно или иным образом) с фрагментом, модифицирующим фармакокинетику (ФК). Примеры модифицирующих ФК фрагментов включают, не ограничиваясь перечисленными, полиэтиленгликоль (ПЭГ) (Dozier et al., (2015) Int J Mol Sci. Oct 28;16(10):25831-64 and Jevsevar et al, (2010) Biotechnol J. Jan;5(l):113-28), модификацию остатками пролина, аланина и серина (PAS) (Schlapschy et al, (2013) Protein Eng Des Sel. Aug;26(8):489-501), альбумин и альбумин-связывающие домены (Dennis et al, (2002) J Biol Chem. Sep 20;277(38):35035-43) и/или неструктурированные полипептиды (Schellenberger et al, (2009) Nat Biotechnol. Dec;27(12): 1186-90). Дополнительные модифицирующие ФК фрагменты включают Fc-фрагменты антител.
В настоящем документе термин «антитело» охватывает такие фрагменты и варианты. Фрагменты и варианты/аналоги антител, которые пригодны для применения в композициях и методах, описанных в настоящем документе, включают минитела, Fab-фрагменты, Р(аb')2-фрагменты, dsFv- и scFv-фрагменты, диатела, нанотела (Nanobodies™) (эти конструкции, продаваемые Ablynx (Бельгия), содержат синтетический одиночный вариабельный домен тяжелой цепи иммуноглобулина, выделенный из антитела верблюдовых (например, верблюда или ламы)) и «доменные антитела» (Domantis (Бельгия), содержащие подвергнутый аффинному созреванию одиночный вариабельный домен тяжелой цепи иммуноглобулина или вариабельный легкий домен иммуноглобулина) или альтернативные белковые каркасы, которые проявляют характеристики связывания, подобные антителу, такие как аффитела (от Affibody (Швеция), содержащие скаффолд сконструированного белка А) или антикалины (от Pieris (Германия), содержащие сконструированные антикалины), и другие.
«Антитело» также включает TCR-подобные антитела (Chang et al, Expert Opin Biol Ther. 2016 Aug;16(8):979-87 and Dahan et al., Expert Rev Mol Med. 2012 Feb 24;14:e6).
Связь нацеливающего фрагмента и специфической связывающей молекулы по первому аспекту может быть осуществлена посредством ковалентного или нековалентного присоединения. Ковалентное присоединение может быть прямым или непрямым, через последовательность линкера. Линкерные последовательности обычно являются гибкими, поскольку они состоят в основном из таких аминокислот, как глицин, аланин и серии, которые не имеют объемных боковых цепей, способных ограничить гибкость. В альтернативных вариантах могут быть предпочтительными линкеры с большей жесткостью. Пригодные для применения или оптимальные длины линкерных последовательностей могут быть легко определены. Часто линкерная последовательность имеет длину менее 12, например, менее 10, или от 2-10 аминокислот, предпочтительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 аминокислот.Примеры подходящих линкеров, которые могут быть использованы в TCR согласно настоящему изобретению, включают, не ограничиваясь перечисленными: GGGSGGGG, GGGGS, GGGSG, GGSGG, GSGGG, GSGGGP, GGEPS, GGEGGGP и GGEGGGSEGGGS (как описано в WO2010/133828).
Как хорошо известно в данной области, специфические связывающие молекулы или бифункциональные связывающие молекулы могут подвергаться посттрансляционным модификациям. Одной из таких модификаций является гликозилирование, которое включает ковалентное присоединение олигосахаридных фрагментов к определенным аминокислотам в аминокислотной цепи. Например, остатки аспарагина или остатки серина/треонина являются хорошо известными сайтами прикрепления олигосахаридов. Статус гликозилирования конкретного белка зависит от ряда факторов, включая последовательность белка, конформацию белка и доступность определенных ферментов. Кроме того, статус гликозилирования (т.е. тип олигосахарида, ковалентная связь и общее количество присоединений) может влиять на функцию белка. Поэтому при получении рекомбинантных белков часто желательно контролировать гликозилирование. Контролируемое гликозилирование применялось для улучшения терапевтических средств на основе антител. (Jefferis et al., (2009) Nat Rev Drug Discov Mar;8(3):226-34.). Для растворимых TCR гликозилирование можно контролировать, например, за счет применения конкретных клеточных линий, например (включая, но не ограничиваясь перечисленными), клеточных линий млекопитающих, таких как клетки яичника китайского хомячка (СНО) или клетки эмбриональной почки человека (НЕK), или путем химической модификации. Такие модификации могут быть предпочтительными, поскольку гликозилирование может улучшать фармакокинетику, снижать иммуногенность и более точно имитировать природный белок человека (Sinclair and Elliott, (2005) Pharm Sci. Aug; 94(8): 1626-35).
Для введения пациентам специфические связывающие молекулы, бифункциональные связывающие молекулы, нуклеиновые кислоты, векторы экспрессии или клетки согласно настоящему изобретению могут быть предоставлены в составе стерильной фармацевтической композиции вместе с одним или несколькими фармацевтически приемлемыми носителями или вспомогательными веществами. Эта фармацевтическая композиция может быть представлена в любой подходящей форме (в зависимости от желаемого способа введения пациенту). Она может быть предоставлена в единичной лекарственной форме, как правило, предоставляется в герметичном контейнере и может быть предоставлена как часть набора. Такой набор обычно (хотя и необязательно) включает инструкции по применению. Он может включать в себя множество указанных единичных лекарственных форм.
Указанная фармацевтическая композиция может быть адаптирована для введения любым подходящим путем, таким как парентеральный (включая подкожный, внутримышечный, интратекальный или внутривенный), энтеральный (включая пероральный или ректальный), ингаляционный или интраназальный. Такие композиции могут быть получены любым способом, известным в области фармацевтики, например, путем смешивания активного ингредиента с носителем (носителями) или вспомогательным веществом (веществами) в стерильных условиях.
Дозировки веществ согласно настоящему изобретению могут варьировать в широких пределах в зависимости от заболевания или расстройства, подлежащего лечению, возраста и состояния индивидуума, подлежащего лечению, и т.д. Подходящий диапазон доз для молекулы согласно настоящему изобретению может находиться в диапазоне от 25 нг/кг до 50 мкг/кг или от 1 мкг до 1 г. В конечном счете подходящие дозы для применения определяет лечащий врач.
Специфические связывающие молекулы, бифункциональные связывающие молекулы, фармацевтические композиции, векторы, нуклеиновые кислоты и клетки согласно настоящему изобретению могут быть предоставлены по существу в чистой форме, например, чистой по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100%.
Способ лечения может дополнительно включать введение по отдельности, в комбинации или последовательно дополнительного противоопухолевого агента. Примеры
таких агентов известны в данной области техники и могут включать иммуностимулирующие агенты и/или модулирующие Т-клетки агенты.
Нуклеиновые кислоты, экспрессионные векторы, клетки-хозяева и способы получения, описанные выше применительно к первому аспекту, также предусмотрены и другими аспектами, описанными в настоящем документе.
Предпочтительными признаками каждого аспекта изобретения являются те же признаки, что и для всех других аспектов, с соответствующими изменениями. Упомянутые в настоящем документе документы предшествующего уровня техники включены в него посредством ссылки в максимальной возможной степени, разрешенной законом.
Описание чертежей
На фиг. 1 представлены аминокислотные последовательности VH и VL улучшенных вариантов UCHT1. CDR подчеркнуты. Мутации выделены жирным шрифтом.
На фиг. 2 показан пример последовательности аминокислот слитого белка TCR-анти-CD3, включающего улучшенные варианты scFv, направленного против CD3.
На фиг. 3 показано, что слияние TCR-CD3, включающее улучшенный вариант 1 scFv, направленного против CD3, имеет лучшее терапевтическое окно по сравнению с немутированным анти-СD3 (А), а слияние TCR-CD3, включающее улучшенный вариант 2 scFv, направленного против CD3, имеет более высокое значение Emax относительно немутированного анти-СD3 (В).
На фиг. 4 продемонстрированы улучшенные свойства Т-клеток в отношении киллинга клеток, опосредованные слиянием TCR-CD3, включающим вариант 1 и вариант 2 UCHT1.
Изобретение дополнительно описано в следующих неограничивающих примерах. Примеры
Следующие примеры описывают бифункциональные связывающие молекулы согласно настоящему изобретению, которые могут называться TCR-анти-СD3-биспецифическими белками.
1) Получение биспецифических слитых белков TCR-анти-CD3 с улучшенным анти-СD3 свойствами Слитые белки, содержащие TCR и направленный против CD3 scFv, известны в данной области техники (например, см. WO2011001152, WO2017109496, WO2017175006 и WO2018234319). Эти молекулы содержат scFv-фрагмент гуманизированного UCHT1.
В этом примере были получены варианты слитого белка TCR- анти -CD3, описанные в WO2018234319, которые включают либо анти-СD3 вариант 1 (Т165А), либо анти-СD3 вариант 2 (T165A+I202F). Мутации вводили с использованием стандартных способов мутагенеза и клонирования (таких, как описанные в руководстве: Sambrook, Joseph. (2001). Molecular cloning: a laboratory manual. Cold Spring Harbor, N.Y.: Cold Spring Harbor Laboratory Press). Аминокислотные последовательности VL и VH для T165A и T165A+I202F представлены на фиг. 1. Аминокислотные последовательности TCR-анти-CD3, включающие последовательности на фиг. 1, представлены на фиг.2.
Слитые белки TCR-анти-CD3, содержащие Т165А, T165A+I202F или немутированный (WT, ДТ) scFv-фрагмент UCHT1, экспрессировали в Е. coli в виде телец включения, а затем проводили рефолдинг и очищали с использованием способов, описанных в WO2018234319, см. пример 2.
2) Увеличенное терапевтическое окно и максимальная активация Т-клеток, опосредованные TCR-анти-CD3 с вариантами Т165А и T165A+I202F
А) Высвобождение ИФН-γ
Слитые белки TCR-анти-CD3, описанные выше, оценивали на способность опосредовать активацию CD3+ Т-клеток в присутствии антиген-положительных и антиген-отрицательных клеток. Высвобождение интерферона-γ (ИФН-γ) использовали в качестве сигнала об активации Т-клеток.
Анализы проводили с применением набора для анализа на ИФН-γ человека ELISpot (BD Biosciences) в соответствии с инструкциями производителя. Вкратце, клетки меланомы Ме1624 использовали в качестве антиген-положительных клеток-мишеней. Клетки В-клеточной лимфомы Granta-519 использовали в качестве антиген-отрицательных клеток-мишеней. Получали клетки-мишени с плотностью 1×106/мл в среде для анализа (RPMI1640, содержащей 10% инактивированной нагреванием ФБС и 1% пенициллин-стрептомицин-L-глутамина) и высевали по 50000 клеток на лунку в объеме 50 мкл. Мононуклеарные клетки периферической крови (МКПК), выделенные из свежей донорской крови, использовали в качестве эффекторных CD3+ клеток и высевали в количестве 35000 клеток на лунку в объеме 50 мкл. Белки TCR-анти-CD3 титровали до конечных концентраций от 10 нМ до 0,0001 нМ и добавляли в лунку в объеме 50 мкл.
Планшеты подготавливали и проявляли в соответствии с инструкциями производителя. Объем в лунках, содержащих клетки-мишени, эффекторные клетки и слитые белки, доводили до конечного объема 200 мкл средой для анализа. Все реакции проводили в трех повторностях. Также были приготовлены контрольные лунки либо без слитого белка, либо без эффекторных клеток или клеток-мишеней. Планшеты инкубировали в течение ночи (37°С/5% СО2). На следующий день планшеты трижды промывали промывочным буфером (пакет с 1хФСБ, содержащим 0,05% Tween-20, разведенный в деионизированной воде). Затем в каждую лунку добавляли первичное антитело для детекции в объеме 50 мкл. Планшеты инкубировали при комнатной температуре в течение 2 часов перед повторным трехкратным промыванием. Вторичную детекцию проводили путем добавления 50 мкл разбавленного конъюгата стрептавидин-HRP в каждую лунку и инкубирования при комнатной температуре в течение 1 часа, после чего повторяли этап промывания. Не более чем за 15 минут до использования добавляли одну каплю (20 мкл) хромогена АЕС на каждый 1 мл субстрата АЕС, перемешивали и добавляли по 50 мкл в каждую лунку. Регулярно отслеживали проявление пятен и промывали планшеты водопроводной водой для прекращения реакции проявления. Затем планшеты оставляли для просушивания при комнатной температуре по меньшей мере на 2 часа перед подсчетом пятен с использованием анализатора CTL с программным обеспечением Immunospot (Cellular Technology Limited). Данные были подготовлены и проанализированы с использованием программного обеспечения Prism.
Терапевтическое окно рассчитывали путем определения относительной эффективности активации Т-клеток, опосредованной вариантом TCR-anti-CD3 и вариантом дикого типа, направленной против антиген-положительных клеток, и сравнения значений Ес50 после подбор аппроксимирующей кривой в Prism. Для антиген-отрицательных клеток устойчивые значения Ес50 не могли быть получены, поэтому определяли «минимальную концентрацию для перекрестной реактивности» путем установки порогового количества пятен (например, 25 пятен на лунку) и определения, путем интерполяции, концентрации, которая первой обеспечит превышение этого количества.
Результаты Т165А
Для надежного определения терапевтического окна усредняли данные по 4 планшетам Ag+ и 4 планшетам Ag-, в каждом из которых рядом были расположены немутированный вариант и варианте Т165А. На фиг. 3А представлены кривые, полученные для одного планшета. Данные для каждого из индивидуальных планшетов приведены в таблицах ниже.


Среднее значение Относительная активность в отношении антиген-положительных клеток (т.е. Ес50 WT, разделенная на Ес50 Т165А) составило 0,41. Среднее значение относительной перекрестной реактивности против антиген-отрицательных клеток (т.е. концентрация WT, дающая 25 пятен, разделенная на концентрацию Т165А, дающую 25 пятен) составило 0,19. Эти данные демонстрируют, что терапевтическое окно Т165А приблизительно в 2 раза больше окна для WT.
T165A+I202F
На фиг. 3В показано, что вариант T165A+I202F приводит к более высокой максимальной активации Т-клеток (Emax) по сравнению с WT. В этом случае терапевтическое окно аналогично таковому для WT. Эти данные демонстрируют, что T165A+I202F более эффективен для активации Т-клеток.
В) Опосредованный Т-клетками киллинг
Способность слитых белков TCR-анти-CD3 опосредовать перенаправленный киллинг Т-клетками антиген-положительных и антиген-отрицательных опухолевых клеток исследовали с использованием платформы IncuCyte (Essen Bioscience). Этот анализ позволяет в режиме реального времени детектировать с помощью микроскопии высвобождение каспазы 3/7, маркера апоптоза.
Способ
Анализы проводили с использованием 96-луночного набора для анализа апоптоза с каспазой 3/7 CellPlayer (Essen Bioscience, кат.№4440) в соответствии с протоколом производителя. Вкратце, клетки-мишени Ме1624 (антиген-положительные) или MDA MB 231 (антиген-отрицательные) высевали в количестве 10000 клеток на лунку и инкубировали в течение ночи, чтобы позволить клеткам прикрепиться. Слитые белки TCR-анти-CD3 добавляли в концентрациях от 0,05 нМ до 0,0125 нМ (для антиген-положительных клеток) и от 20 нМ до 0,125 нМ (для антиген-отрицательных клеток). Эффекторные клетки CD3+(МКПК) использовали при соотношении эффекторных клеток и клеток-мишеней 10:1 (100000 клеток на лунку). Реагент для анализа NucView готовили в концентрации 30 мкМ и добавляли в объеме 25 мкл в каждую лунку, и доводили конечный объем до 150 мкл (с получением конечной концентрации 5 мкМ). Планшет помещали в инструмент IncuCyte и захватывали изображения через равные промежутки времени в течение 3 5 дней. Количество апоптотических клеток на каждом изображении определяли и записывали как количество объектов на мм2. Анализы проводили в трех повторностях. Графики были подготовлены с использованием программного обеспечения PRISM.
Результаты
Итоговые кривые киллинга для WT, Т165А и T165A+I202F против антиген-положительных и антиген-отрицательных клеток представлены на фиг. 4. Отметим, что для ясности представлены только кривые для указанных концентраций.
Т165А
Т165А демонстрирует небольшое снижение эффективности киллинга Т-клетками антиген-положительных клеток по сравнению с WT в концентрации 0,005 нМ. Киллинг антиген-отрицательных клеток не наблюдается до 20 нМ, тогда как для WT он наблюдается при концентрации 1 нМ. Эти данные подтверждают вышеприведенные выводы о том, что Т165А характеризуется улучшенным терапевтическим окном.
T165A+I202F
T165A+I202F демонстрирует увеличение опосредованного Т-клетками киллинга в заданной концентрации (например, 0,005 нМ) антиген-положительных клеток. Киллинг антиген-отрицательных клеток сравнимо с WT. Эти данные подтверждают вышеуказанные результаты и демонстрируют, что T165A+I202F более эффективен для активации Т-клеток.
Данные, представленные в указанном примере, демонстрируют, что слитые белки TCR-анти-CD3, содержащие варианты Т165А или T165A+I202F UCHT1, обладают улучшенными терапевтическими свойствами.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> IMMUNOCORE LIMITED/ ИММУНОКОР ЛИМИТЕД
<120> CD3-SPECIFIC BINDING MOLECULES / CD3-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ
<130> P121631WO
<150> GB1901305.1
<151> 2019-01-30
<160> 36
<170> PatentIn, версия 3.5
<210> 1
<211> 6
<212> PRT/Белок
<213> Homo sapiens
<400> 1
Gln Asp Ile Arg Asn Tyr
1 5
<210> 2
<211> 9
<212> PRT/Белок
<213> Homo sapiens
<400> 2
Gln Gln Gly Asn Thr Leu Pro Trp Thr
1 5
<210> 3
<211> 8
<212> PRT/Белок
<213> Homo sapiens
<400> 3
Gly Tyr Ser Phe Thr Gly Tyr Ala
1 5
<210> 4
<211> 8
<212> PRT/Белок
<213> Homo sapiens
<400> 4
Ile Asn Pro Tyr Lys Gly Val Ser
1 5
<210> 5
<211> 15
<212> PRT/Белок
<213> Homo sapiens
<400> 5
Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe Asp Val
1 5 10 15
<210> 6
<211> 26
<212> PRT/Белок
<213> Homo sapiens
<400> 6
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser
20 25
<210> 7
<211> 26
<212> PRT/Белок
<213> Homo sapiens
<400> 7
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser
20 25
<210> 8
<211> 17
<212> PRT/Белок
<213> Homo sapiens
<400> 8
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
1 5 10 15
Tyr
<210> 9
<211> 36
<212> PRT/Белок
<213> Homo sapiens
<400> 9
Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly
1 5 10 15
Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala
20 25 30
Thr Tyr Tyr Cys
35
<210> 10
<211> 10
<212> PRT/Белок
<213> Homo sapiens
<400> 10
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 11
<211> 25
<212> PRT/Белок
<213> Homo sapiens
<400> 11
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 12
<211> 17
<212> PRT/Белок
<213> Homo sapiens
<400> 12
Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala
1 5 10 15
Leu
<210> 13
<211> 38
<212> PRT/Белок
<213> Homo sapiens
<400> 13
Thr Tyr Asn Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys
1 5 10 15
Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 14
<211> 38
<212> PRT/Белок
<213> Homo sapiens
<400> 14
Thr Tyr Asn Gln Lys Phe Lys Asp Arg Phe Thr Phe Ser Val Asp Lys
1 5 10 15
Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 15
<211> 11
<212> PRT/Белок
<213> Homo sapiens
<400> 15
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 16
<211> 107
<212> PRT/Белок
<213> Homo sapiens
<400> 16
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 17
<211> 107
<212> PRT/Белок
<213> Homo sapiens
<400> 17
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 18
<211> 122
<212> PRT/Белок
<213> Homo sapiens
<400> 18
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 19
<211> 122
<212> PRT/Белок
<213> Homo sapiens
<400> 19
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Phe Thr Phe Ser Val Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 20
<211> 253
<212> PRT/Белок
<213> Homo sapiens
<400> 20
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
245 250
<210> 21
<211> 253
<212> PRT/Белок
<213> Homo sapiens
<400> 21
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Phe Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
245 250
<210> 22
<211> 8
<212> PRT/Белок
<213> искусственная последовательность
<220>
<223> линкерная последовательность
<400> 22
Gly Gly Gly Ser Gly Gly Gly Gly
1 5
<210> 23
<211> 5
<212> PRT/Белок
<213> искусственная последовательность
<220>
<223> линкерная последовательность
<400> 23
Gly Gly Gly Gly Ser
1 5
<210> 24
<211> 5
<212> PRT/Белок
<213> искусственная последовательность
<220>
<223> линкерная последовательность
<400> 24
Gly Gly Gly Ser Gly
1 5
<210> 25
<211> 5
<212> PRT/Белок
<213> искусственная последовательность
<220>
<223> линкерная последовательность
<400> 25
Gly Gly Ser Gly Gly
1 5
<210> 26
<211> 5
<212> PRT/Белок
<213> искусственная последовательность
<220>
<223> линкерная последовательность
<400> 26
Gly Ser Gly Gly Gly
1 5
<210> 27
<211> 6
<212> PRT/Белок
<213> искусственная последовательность
<220>
<223> линкерная последовательность
<400> 27
Gly Ser Gly Gly Gly Pro
1 5
<210> 28
<211> 5
<212> PRT/Белок
<213> искусственная последовательность
<220>
<223> линкерная последовательность
<400> 28
Gly Gly Glu Pro Ser
1 5
<210> 29
<211> 7
<212> PRT/Белок
<213> искусственная последовательность
<220>
<223> линкерная последовательность
<400> 29
Gly Gly Glu Gly Gly Gly Pro
1 5
<210> 30
<211> 12
<212> PRT/Белок
<213> искусственная последовательность
<220>
<223> линкерная последовательность
<400> 30
Gly Gly Glu Gly Gly Gly Ser Glu Gly Gly Gly Ser
1 5 10
<210> 31
<211> 244
<212> PRT/Белок
<213> Homo sapiens
<400> 31
Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys Glu Gly
1 5 10 15
Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp Ala Met
20 25 30
Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile Tyr Tyr
35 40 45
Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu Gly Tyr
50 55 60
Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val Thr Ser
65 70 75 80
Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser Trp Trp
85 90 95
Thr Gly Gly Ser Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg Leu Thr
100 105 110
Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe
115 120 125
Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val
130 135 140
Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp
145 150 155 160
Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Pro
165 170 175
Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu Ser Ser
180 185 190
Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro Arg Asn His Phe
195 200 205
Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr
210 215 220
Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp
225 230 235 240
Gly Arg Ala Asp
<210> 32
<211> 502
<212> PRT/Белок
<213> Homo sapiens
<400> 32
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
245 250 255
Gly Ser Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys
260 265 270
Glu Gly Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp
275 280 285
Ala Met Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile
290 295 300
Tyr Tyr Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu
305 310 315 320
Gly Tyr Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val
325 330 335
Thr Ser Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser
340 345 350
Trp Trp Thr Gly Gly Ser Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg
355 360 365
Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala
370 375 380
Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr
385 390 395 400
Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser
405 410 415
Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro
420 425 430
Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu
435 440 445
Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro Arg Asn
450 455 460
His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu
465 470 475 480
Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu
485 490 495
Ala Trp Gly Arg Ala Asp
500
<210> 33
<211> 502
<212> PRT/Белок
<213> Homo sapiens
<400> 33
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Phe Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
245 250 255
Gly Ser Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys
260 265 270
Glu Gly Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp
275 280 285
Ala Met Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile
290 295 300
Tyr Tyr Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu
305 310 315 320
Gly Tyr Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val
325 330 335
Thr Ser Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser
340 345 350
Trp Trp Thr Gly Gly Ser Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg
355 360 365
Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala
370 375 380
Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr
385 390 395 400
Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser
405 410 415
Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro
420 425 430
Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu
435 440 445
Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro Arg Asn
450 455 460
His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu
465 470 475 480
Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu
485 490 495
Ala Trp Gly Arg Ala Asp
500
<210> 34
<211> 202
<212> PRT/Белок
<213> Homo sapiens
<400> 34
Met Gly Asp Ala Lys Thr Thr Gln Pro Asn Ser Met Glu Ser Asn Glu
1 5 10 15
Glu Glu Pro Val His Leu Pro Cys Asn His Ser Thr Ile Ser Gly Thr
20 25 30
Asp Tyr Ile His Trp Tyr Arg Gln Leu Pro Ser Gln Gly Pro Glu Tyr
35 40 45
Val Ile His Gly Leu Thr Ser Asn Val Asn Asn Arg Met Ala Ser Leu
50 55 60
Ala Ile Ala Glu Asp Arg Lys Ser Ser Thr Leu Ile Leu His Arg Ala
65 70 75 80
Thr Leu Arg Asp Ala Ala Val Tyr Tyr Cys Ile Leu Ile Leu Gly His
85 90 95
Ser Arg Leu Gly Asn Tyr Ile Ala Thr Phe Gly Lys Gly Thr Lys Leu
100 105 110
Ser Val Ile Pro Asn Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu
115 120 125
Arg Asp Ser Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe
130 135 140
Asp Ser Gln Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile
145 150 155 160
Thr Asp Lys Cys Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn
165 170 175
Ser Ala Val Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala
180 185 190
Phe Asn Asn Ser Ile Ile Pro Glu Asp Thr
195 200
<210> 35
<211> 502
<212> PRT/Белок
<213> Homo sapiens
<400> 35
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Ile Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
245 250 255
Gly Ser Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys
260 265 270
Glu Gly Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp
275 280 285
Ala Met Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile
290 295 300
Tyr Tyr Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu
305 310 315 320
Gly Tyr Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val
325 330 335
Thr Ser Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser
340 345 350
Trp Trp Thr Gly Gly Ser Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg
355 360 365
Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala
370 375 380
Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr
385 390 395 400
Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser
405 410 415
Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro
420 425 430
Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu
435 440 445
Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro Arg Asn
450 455 460
His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu
465 470 475 480
Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu
485 490 495
Ala Trp Gly Arg Ala Asp
500
<210> 36
<211> 502
<212> PRT/Белок
<213> Homo sapiens
<400> 36
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe
145 150 155 160
Thr Gly Tyr Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ala Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn
180 185 190
Gln Lys Phe Lys Asp Arg Phe Thr Phe Ser Val Asp Lys Ser Lys Asn
195 200 205
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe
225 230 235 240
Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
245 250 255
Gly Ser Asp Gly Gly Ile Thr Gln Ser Pro Lys Tyr Leu Phe Arg Lys
260 265 270
Glu Gly Gln Asn Val Thr Leu Ser Cys Glu Gln Asn Leu Asn His Asp
275 280 285
Ala Met Tyr Trp Tyr Arg Gln Asp Pro Gly Gln Gly Leu Arg Leu Ile
290 295 300
Tyr Tyr Ser Gln Ile Met Gly Asp Glu Gln Lys Gly Asp Ile Ala Glu
305 310 315 320
Gly Tyr Ser Val Ser Arg Glu Lys Lys Glu Ser Phe Pro Leu Thr Val
325 330 335
Thr Ser Ala Gln Lys Asn Pro Thr Ala Phe Tyr Leu Cys Ala Ser Ser
340 345 350
Trp Trp Thr Gly Gly Ser Ala Pro Ile Arg Phe Gly Pro Gly Thr Arg
355 360 365
Leu Thr Val Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala
370 375 380
Val Phe Glu Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr
385 390 395 400
Leu Val Cys Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser
405 410 415
Trp Trp Val Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro
420 425 430
Gln Pro Leu Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Ala Leu
435 440 445
Ser Ser Arg Leu Arg Val Ser Ala Thr Phe Trp Gln Asp Pro Arg Asn
450 455 460
His Phe Arg Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu
465 470 475 480
Trp Thr Gln Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu
485 490 495
Ala Trp Gly Arg Ala Asp
500
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| УЛУЧШЕННЫЙ СИНТЕТИЧЕСКИЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР И АНТИГЕННЫЙ РЕЦЕПТОР | 2021 |
|
RU2820497C1 |
| РЕКРУТИРУЮЩИЕ Т-КЛЕТКИ ПОЛИПЕПТИДЫ, СПОСОБНЫЕ СВЯЗЫВАТЬ CD123 И TCR АЛЬФА/БЕТА | 2017 |
|
RU2775063C2 |
| СПОСОБЫ КУЛЬТИВИРОВАНИЯ КЛЕТОК И НАБОРЫ И УСТРОЙСТВО ДЛЯ НИХ | 2016 |
|
RU2761557C2 |
| МОЛЕКУЛЫ IMMTAC, СВЯЗЫВАЮЩИЕ РЕСТРИКТИРОВАННЫЙ ПО HLA-A*02 ПЕПТИД, С УВЕЛИЧЕННЫМ ПЕРИОДОМ ПОЛУВЫВЕДЕНИЯ | 2020 |
|
RU2833530C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2017 |
|
RU2775623C2 |
| НОВЫЕ БИСПЕЦИФИЧНЫЕ ПОЛИПЕПТИДНЫЕ КОМПЛЕКСЫ ПРОТИВ CD3/CD19 | 2018 |
|
RU2788127C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ КОМПЛЕКСА ОПУХОЛЕВЫЙ АНТИГЕН NY-ESO-1/HLA-A*02 | 2016 |
|
RU2775394C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) С АНТИГЕНСВЯЗЫВАЮЩИМИ ДОМЕНАМИ К КОНСТАНТНОЙ ОБЛАСТИ β Т-КЛЕТОЧНОГО РЕЦЕПТОРА | 2015 |
|
RU2830064C2 |
| СПЕЦИФИЧНЫЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2825837C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) С АНТИГЕНСВЯЗЫВАЮЩИМИ ДОМЕНАМИ К КОНСТАНТНОЙ ОБЛАСТИ β Т-КЛЕТОЧНОГО РЕЦЕПТОРА | 2015 |
|
RU2744046C2 |


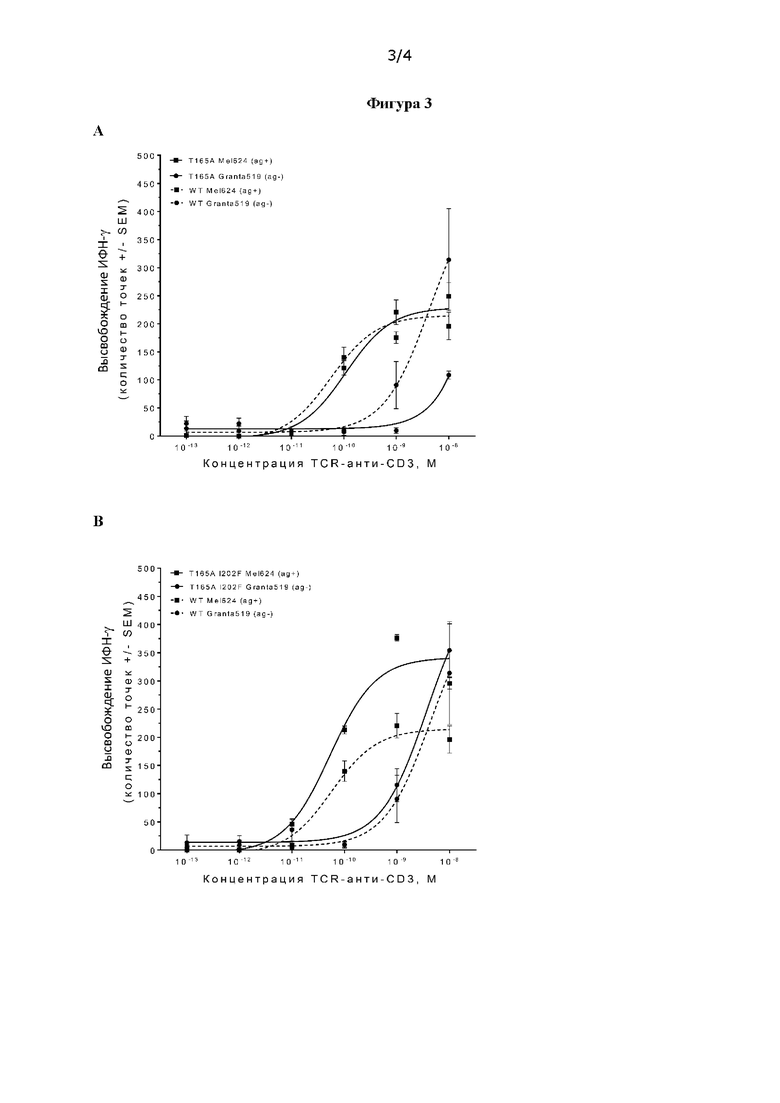

Группа изобретений относится к биотехнологии. Раскрыты: специфическая связывающая молекула, которая связывается с CD3, бифункциональная связывающая молекула, способная связываться с CD3 и клеткой-мишенью, и способы их получения, фармацевтические композиции для лечения рака и инфекционного заболевания, нуклеиновые кислоты, кодирующие специфическую и бифункциональную связывающие молекулы, экспрессионный вектор, клетки-хозяина для получения специфической и бифункциональной связывающих молекул. Также раскрыты способы лечения рака и инфекционного заболевания. Изобретение направлено на получение связывающих молекул, которые связываются с CD3, и их использование для лечения рака и инфекционного заболевания у субъекта. 13 н. и 14 з.п. ф-лы, 4 ил., 2 табл.
1. Специфическая связывающая молекула, которая связывается с CD3 и содержит полипептид, имеющий иммуноглобулиновый домен VL и иммуноглобулиновый домен VH, где домен VL содержит определяющие комплементарность области (CDR) VLCDR1, VLCDR2 и VLCDR3, а домен VH включает определяющие комплементарность области (CDR) VHCDR1, VHCDR2, VHCDR3, каждая из которых имеет соответствующую аминокислотную последовательность, представленную ниже, где:
VLCDR1 представляет собой QDIRNY (SEQ ID NO: 1)
VLCDR2 представляет собой YTS
VLCDR3 представляет собой QQGNTLPWT (SEQ ID NO: 2)
VHCDR1 представляет собой GYSFTGYA (SEQ ID NO: 3)
VHCDR2 представляет собой INPYKGVS (SEQ ID NO: 4)
VHCDR3 представляет собой ARSGYYGDSDWYFDV (SEQ ID NO: 5).
2. Специфическая связывающая молекула по п. 1, отличающаяся тем, что иммуноглобулиновый домен VL содержит общую последовательность VLFW1-VLCDR1-VLFW2-VLCDR2-VLFW3-VLCDR3-VLFW4, где VLFW1, VLFW2, VLFW3 и VLFW4 представляют собой каркасные последовательности VL (VLFW) 1–4, соответственно, иммуноглобулиновый домен VH содержит общую последовательность VHFW1-VHCDR1-VHFW2-VHCDR2-VHFW3-VHCDR3-VHFW4, где VHFW1, VHFW2, VHFW3 и VHFW4 представляют собой каркасные последовательности VH (VHFW) 1–4, соответственно.
3. Специфическая связывающая молекула по п. 2, отличающаяся тем, что последовательности VLFW и VHFW являются мышиными, человеческими или гуманизированными каркасными последовательностями.
4. Специфическая связывающая молекула по любому из предыдущих пунктов, отличающаяся тем, что иммуноглобулиновый домен VL содержит общую последовательность VLFW1-VLCDR1-VLFW2-VLCDR2-VLFW3-VLCDR3-VLFW4, где VLFW1, VLFW2, VLFW3 и VLFW4 представляют собой каркасные (FW) последовательности 1–4, соответственно, при этом каждая из последовательностей VLFW1, VLFW2, VLFW3 и VLFW4 имеет соответствующую аминокислотную последовательность, представленную ниже, где:
VLFW1 представляет собой AIQMTQSPSSLSASVGDRVTITCRAS (SEQ ID NO: 6) или
DIQMTQSPSSLSASVGDRVTITCRAS (SEQ ID NO: 7)
VLFW2 представляет собой LNWYQQKPGKAPKLLIY (SEQ ID NO: 8)
VLFW3 представляет собой RLESGVPSRFSGSGSGTDYTLTISSLQPEDFATYYC (SEQ ID NO: 9)
VLFW4 представляет собой FGQGTKVEIK (SEQ ID NO: 10).
5. Специфическая связывающая молекула по любому из предыдущих пунктов, отличающаяся тем, что иммуноглобулиновый домен VH содержит общую последовательность VHFW1-VHCDR1-VHFW2-VHCDR2-VHFW3-VHCDR3-VHFW4, где VHFW1, VHFW2, VHFW3 и VHFW4 представляют собой каркасные (FW) последовательности 1–4, соответственно, при этом каждая из последовательностей VHFW1, VHFW2, VHFW3 и VHFW4 имеет соответствующую аминокислотную последовательность, представленную ниже, где:
VHFW1 представляет собой EVQLVESGGGLVQPGGSLRLSCAAS (SEQ ID NO: 11)
VHFW2 представляет собой MNWVRQAPGKGLEWVAL (SEQ ID NO: 12)
VHFW3 представляет собой TYNQKFKDRFTISVDKSKNTAYLQMNSLRAEDTAVYYC (SEQ ID NO: 13) или TYNQKFKDRFTFSVDKSKNTAYLQMNSLRAEDTAVYYC (SEQ ID NO: 14)
VHFW4 представляет собой WGQGTLVTVSS (SEQ ID NO: 15).
6. Специфическая связывающая молекула по любому из предыдущих пунктов, отличающаяся тем, что иммуноглобулиновый домен VL содержит последовательность:
AIQMTQSPSSLSASVGDRVTITCRASQDIRNYLNWYQQKPGKAPKLLIYYTSRLESGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQGNTLPWTFGQGTKVEIK (SEQ ID NO: 16); или
DIQMTQSPSSLSASVGDRVTITCRASQDIRNYLNWYQQKPGKAPKLLIYYTSRLESGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQGNTLPWTFGQGTKVEIK (SEQ ID NO: 17).
7. Специфическая связывающая молекула по любому из предыдущих пунктов, отличающаяся тем, что иммуноглобулиновый домен VH содержит последовательность:
EVQLVESGGGLVQPGGSLRLSCAASGYSFTGYAMNWVRQAPGKGLEWVALINPYKGVSTYNQKFKDRFTISVDKSKNTAYLQMNSLRAEDTAVYYCARSGYYGDSDWYFDVWGQGTLVTVSS (SEQ ID NO: 18); или
EVQLVESGGGLVQPGGSLRLSCAASGYSFTGYAMNWVRQAPGKGLEWVALINPYKGVSTYNQKFKDRFTFSVDKSKNTAYLQMNSLRAEDTAVYYCARSGYYGDSDWYFDVWGQGTLVTVSS (SEQ ID NO: 19).
8. Специфическая связывающая молекула по любому из предыдущих пунктов, отличающаяся тем, что иммуноглобулиновый домен VL содержит последовательность:
AIQMTQSPSSLSASVGDRVTITCRASQDIRNYLNWYQQKPGKAPKLLIYYTSRLESGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQGNTLPWTFGQGTKVEIK (SEQ ID NO: 16); и
иммуноглобулиновый VH содержит последовательность:
EVQLVESGGGLVQPGGSLRLSCAASGYSFTGYAMNWVRQAPGKGLEWVALINPYKGVSTYNQKFKDRFTFSVDKSKNTAYLQMNSLRAEDTAVYYCARSGYYGDSDWYFDVWGQGTLVTVSS (SEQ ID NO: 19).
9. Специфическая связывающая молекула по любому из предшествующих пунктов, отличающаяся тем, что указанная молекула находится в форме scFv-фрагмента.
10. Специфическая связывающая молекула по любому из предшествующих пунктов, отличающаяся тем, что иммуноглобулиновые домены VL и VH соединены посредством линкера.
11. Специфическая связывающая молекула по любому из предшествующих пунктов, отличающаяся тем, что указанная молекула представляет собой одноцепочечную специфическую связывающую молекулу, имеющую последовательность:
AIQMTQSPSSLSASVGDRVTITCRASQDIRNYLNWYQQKPGKAPKLLIYYTSRLESGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQGNTLPWTFGQGTKVEIKGGGGSGGGGSGGGGSGGGGSGGGSEVQLVESGGGLVQPGGSLRLSCAASGYSFTGYAMNWVRQAPGKGLEWVALINPYKGVSTYNQKFKDRFTFSVDKSKNTAYLQMNSLRAEDTAVYYCARSGYYGDSDWYFDVWGQGTLVTVSS (SEQ ID NO: 21).
12. Бифункциональная связывающая молекула, способная связываться с CD3 и клеткой-мишенью, содержащая:
i) нацеливающий фрагмент, где указанный нацеливающий фрагмент представляет собой Т-клеточный рецептор (TCR), антитело или фрагмент антитела; и
ii) специфическую связывающую молекулу, которая связывается с CD3, по любому из пп. 1–11.
13. Бифункциональная связывающая молекула по п. 12, отличающаяся тем, что Т-клеточный рецептор (TCR) представляет собой гетеродимерную пару полипептидов TCR альфа/бета или одноцепочечный полипептид TCR альфа/бета.
14. Бифункциональная связывающая молекула по п. 12 или 13, отличающаяся тем, что гетеродимерная пара полипептидов TCR альфа/бета содержит не являющуюся нативной дисульфидную связь между константной областью альфа-цепи и константной областью бета-цепи.
15. Бифункциональная связывающая молекула по любому из пп. 12–14, отличающаяся тем, что специфическая связывающая молекула, которая связывается с CD3, по любому из пп. 1–11, слита с C- или N-концом нацеливающего фрагмента, необязательно посредством линкера.
16. Фармацевтическая композиция для лечения рака, содержащая специфическую связывающую молекулу, которая связывается с CD3, по любому из пп. 1–11, или бифункциональную связывающую молекулу по любому из пп. 12–15, где указанная специфическая связывающая молекула или бифункциональная связывающая молекула присутствует в эффективном количестве.
17. Фармацевтическая композиция для лечения инфекционного заболевания, содержащая специфическую связывающую молекулу, которая связывается с CD3, по любому из пп. 1–11, или бифункциональную связывающую молекулу по любому из пп. 12–15, где указанная специфическая связывающая молекула или бифункциональная связывающая молекула присутствует в эффективном количестве.
18. Нуклеиновая кислота, кодирующая специфическую связывающую молекулу, которая связывается с CD3, по любому из пп. 1–11.
19. Нуклеиновая кислота, кодирующая бифункциональную связывающую молекулу по любому из пп. 12–15.
20. Экспрессионный вектор, содержащий нуклеиновую кислоту по п. 18 или 19.
21. Клетка-хозяин для получения специфической связывающей молекулы, которая связывается с CD3, содержащая нуклеиновую кислоту по п. 18 или вектор по п. 20, отличающаяся тем, что нуклеиновая кислота, кодирующая специфическую связывающую молекулу, которая связывается с CD3, по любому из пп. 1–11 присутствует в виде одной открытой рамки считывания или двух отдельных открытых рамок считывания, кодирующих альфа-цепь и бета-цепь, соответственно.
22. Клетка-хозяин для получения бифункциональной связывающей молекулы, которая связывается с CD3, содержащая нуклеиновую кислоту по п. 19 или вектор по п. 20, где нуклеиновая кислота, кодирующая бифункциональную связывающую молекулу, которая связывается с CD3 в соответствии с любым из пп. 12–15, присутствует в виде одной открытой рамки или двух отдельных открытых рамок считывания, кодирующих альфа-цепь и бета-цепь, соответственно.
23. Способ получения специфической связывающей молекулы, которая связывается с CD3, по любому из пп. 1–11, включающий поддержание клетки-хозяина по п. 21 в оптимальных условиях для экспрессии нуклеиновой кислоты и выделение специфической связывающей молекулы, которая связывается с CD3.
24. Способ получения бифункциональной связывающей молекулы по любому из пп. 12–15, включающий поддержание клетки-хозяина по п. 22 в оптимальных условиях для экспрессии нуклеиновой кислоты и выделение бифункциональной связывающей молекулы.
25. Специфическая связывающая молекула, которая связывается с CD3, по любому из пп. 1–11, или бифункциональная связывающая молекула по любому из пп. 12–15 для применения в качестве медикамента для лечения рака или инфекционного заболевания.
26. Способ лечения рака, включающий введение специфической связывающей молекулы, которая связывается с CD3, по любому из пп. 1–11 или бифункциональной связывающей молекулы по любому из пп. 12–15 нуждающемуся в этом пациенту.
27. Способ лечения инфекционного заболевания, включающий введение специфической связывающей молекулы, которая связывается с CD3, по любому из пп. 1–11 или бифункциональной связывающей молекулы по любому из пп. 12–15 нуждающемуся в этом пациенту.
| WO 2018234319 A1, 27.12.2018 | |||
| БИСПЕЦИФИЧЕСКИЕ АСИММЕТРИЧНЫЕ ГЕТЕРОДИМЕРЫ, СОДЕРЖАЩИЕ АНТИ-CD3 КОНСТРУКЦИИ | 2013 |
|
RU2650868C2 |
| QI PAN, et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Веникодробильный станок | 1921 |
|
SU53A1 |
| BADRI H | |||
| et al., Optimization of radiation dosing schedules for proneural glioblastoma, J Math Bio, 2016, vol | |||
| Термосно-паровая кухня | 1921 |
|
SU72A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |

