СВЯЗАННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет предварительных заявок США №№ 61/179850, поданной 20 мая 2009, 61/287336, поданной 17 декабря 2009 и 61/314794, поданной 17 марта 2010, содержание которых приведено в настоящем описании в качестве ссылки в полном объеме.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к получению библиотек последовательностей ДНК, кодирующих гомологичные полипептиды, и к применению таких библиотек. В частности, изобретение относится к получению коллекций синтетических фрагментов антител, в которых один или несколько гипервариабельных участков (CDR) заменены коллекцией соответствующих CDR, полученных из природного источника. Изобретение также относится к получению коллекций фрагментов антител, содержащих CDR, полученных от иммунизированного животного, и к их применению в качестве усовершенствованного источника для получения фрагментов антител, обладающих высокой аффинностью. Изобретение также относится к диверсификации фрагмента полипептида путем вставки диверсифицированной последовательности синтетического или природного происхождения, не требующей модификации последовательности, кодирующей исходный полипептид.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Антитело состоит из четырех полипептидов: двух тяжелых цепей и двух легких цепей. Антиген-связывающий фрагмент антитела образован вариабельным доменом легкой цепи (VL) и вариабельным доменом тяжелой цепи (VH). На одном краю указанных доменов находятся шесть петель, также называемые гипервариабельные участки, которые образуют антиген-связывающий участок (CDR). Три CDR расположены на домене VH (H1, H2 и H3) и три других CDR расположены на домене VL (L1, L2 и L3). В процессе развития B-клетки уникальный участок, кодирующий иммуноглобулин, образуется в результате соматической рекомбинации, известной как V(D)J-рекомбинация. Вариабельные участки иммуноглобулина тяжелой и легкой цепи кодируются разными генными сегментами. Тяжелая цепь кодируется тремя сегментами, включающими вариабельный сегмент (V), сегмент разнообразия (D) и соединительный сегмент (J), тогда как вариабельный участок легкой цепи образуется в результате рекомбинации только двух сегментов V и J. В результате рекомбинации среди нескольких копий V-, D- и J-сегментов, присутствующих в геноме, образуется большое число паратопов антитела. V-сегмент кодирует CDR1 и CDR2, тогда как CDR3 образуется в результате событий рекомбинации. В ходе развития иммунного ответа вводится дополнительное разнообразие антиген-связывающих участков посредством процесса, называемого соматической гипермутацией (SHM). Этот процесс включает введение точечных мутаций в гены вариабельных участков тяжелых и легких цепей и, в особенности, в участки, кодирующие CDR. Такое дополнительное разнообразие обеспечивает возможность отбора и размножения B-клеток, экспрессирующих варианты антитела с повышенной аффинностью к распознаваемому ими антигену.
В последние годы было разработано несколько технологий с применением дисплеев, которые позволяют проводить скрининг больших коллекций белков или пептидов. Указанные дисплеи включают фаговый дисплей, бактериальный дисплей, дрожжевой дисплей и рибосомальный дисплей (Smith GP. Science. 1985 Jun 14;228(4705):1315-7; Hanes J and Plückthun A. Proc Natl Acad Sci USA. 1997 May 13;94(10):4937-42.; Daugherty PS et al., Protein Eng. 1998 Sep; 11(9):825-32.; Boder ET and Wittrup KD. Nat Biotechnol. 1997 Jun; 15(6):553-7). В частности, указанные способы широко используется для изучения антител и их фрагментов. Описан ряд способов получения библиотек полипептидов и скрининга таких библиотек с выявлением членов, обладающих желательными характеристиками связывания.
Первый способ включает улавливание реаранжированных иммуноглобулиновых генов из природных репертуаров методом амплификации с использованием в качестве источника генетического разнообразия тканей или клеток людей или других млекопитающих. Указанные коллекции реаранжированных тяжелых и легких цепей (VH и VL) объединяют с получением библиотек связывающих пар, которые можно отобразить на бактериофаге, или на другом средстве отображения, таком как бактерии, дрожжи или клетки млекопитающих. В данном случае улавливают большую фракцию иммуноглобулинового репертуара донора. Например, в таких репертуарах можно обнаружить все каркасные участки, кодируемые зародышевыми генами донора, а также разнообразие, образующееся в результате рекомбинации V(D)J и соматической гипермутации (Marks JD et al., J Mol Biol. 1991 Dec 5;222(3):581-97; McCaffety US Patent No. 5969108).
Недостатком природных репертуаров является то, что природные антитела могут содержать каркасные участки с низкой собственной стабильностью, которая ограничивает уровень их экспрессии, срок хранения и возможность использования в качестве реагентов или терапевтических молекул. Для преодоления указанных недостатков разработан ряд способов, включающих получение библиотек синтетических антител. В этих способах используют определенное или ограниченное число выбранных каркасных участков антител, кодируемых соответствующими зародышевыми генами. Выбор указанных каркасных участков обычно основан на их биохимической стабильности и/или частоте экспрессии в природных репертуарах антител. Затем, чтобы получить коллекцию связывающих белков, во все или в часть CDR вводят синтетическое разнообразие. Как правило, с помощью разных способов подвергают диверсификации весь участок CDR или часть участка CDR. В некоторых случаях диверсификацию осуществляют по выбранным положениям CDR (Knappik A et al., J Mol Biol. 2000 Feb 11;296(l):57-86). Остатки-мишени могут представлять собой остатки, которые с высокой частотой участвуют в контакте с антигеном, остатки, характеризующиеся максимальным разнообразием в природных репертуарах антител, или даже остатки, которые являются предпочтительной мишенью клеточного механизма генерирования соматических гипермутаций в процессе естественного созревания аффинности (Balint RF, Larrick JW. Gene. 1993 Dec 27;137(1):109-18).
Для диверсификации CDR антител используют несколько способов. ПЦР по методу перекрывающегося расширения с использованием вырожденных олигонуклеотидов широко используется для сборки каркасных участков и элементов CDR с целью воссоздания генов антител. В другом способе с помощью рекомбинантных методов в каркасные участки встраивают уникальные участки распознавания ферментами рестрикции на границе с каждым CDR, что позволяет вводить диверсифицированные CDR путем клонирования с использованием ферментов рестрикции. В любом случае, поскольку все члены библиотеки получают с использованием каркасных участков с выбранными и предпочтительными характеристиками, можно ожидать, что антитела из указанных репертуаров являются более стабильными и представляют собой улучшенный источник полезных реагентов. (Knappik, US 6696248; Sidhu SS, et al., Methods Enzymol. 2000;328:333-63; Lee CV et al., J Mol Biol. 2004 Jul 23;340(5): 1073-93).
Однако существенным недостатком указанных синтетических библиотек является неспособность к экспрессии значительной части членов библиотек, поскольку случайно диверсифицированные последовательности не обеспечивают соответствующую экспрессию и/или укладку белка. Эта проблема имеет особое значение для CDR3 тяжелой цепи. Действительно, такой CDR зачастую значительно влияет на энергию связывания с антигеном и сильно варьирует по длине и последовательности. В то время, как другие CDR (H1, H2, L1, L2 и L3) могут иметь только ограниченное число трехмерных конформаций, известных как традиционные складки, число конформаций, которые может принимать CDR3 тяжелой цепи, является слишком большим, чтобы предсказать его конформацию (Al-Lazikani B et al., J Mol Biol. 1997 Nov 7;273(4):927-48). Кроме того, применение длинных вырожденных олигонуклеотидов, используемых для перекрывания длинных CDRH3, часто приводит к делециям одиночных пар оснований. Указанные факторы значительно снижают функциональный размер синтетических репертуаров.
И природные и синтетические репертуары имеют преимущества и недостатки. С одной стороны, способы, основанные на улавливании естественно реаранжированных генов вариабельных участков антител, не являются оптимальными, поскольку они приводят к получению библиотеки с потенциально низким числом желательных каркасных участков. Преимуществом является то, что указанные реаранжированные гены вариабельных участков содержат CDR, совместимые с правильной укладкой домена, поскольку они экспрессируются в среде природного антитела. С другой стороны, преимущество стратегий, основанных на выборе каркасных участков и введении синтетического разнообразия, заключается в повышенной стабильности каркасных участков, а их недостатком является большое число последовательностей CDR, которые препятствуют укладке и/или экспрессии и могут дестабилизировать целый домен (фигура 1A). Следовательно, существует необходимость в новых способах, которые могли бы объединить преимущества использования выбранных каркасных участков с желательными характеристиками и CDR, совместимых с правильной укладкой, например, полученных из природных репертуаров.
Все описанные способы получения библиотек, основанные либо на улавливании последовательностей антител, полученных в результате естественной реаранжировки, либо на генерировании разнообразия синтетическими способами, имеют недостаток, заключающийся в появлении мутаций, приводящих к сдвигу рамки считывания и образованию нефункциональных последовательностей антител. Указанные мутации могут появляться на нескольких стадиях манипуляций с молекулами ДНК, кодирующими антитела, таких как амплификация ПЦР и сборка фрагмента ДНК, а также молекулярное клонирование. Встречаемость нефункциональных членов библиотек антител обычно изменяется от 15% до 45% в зависимости от используемых способов, основанных на улавливании или генерировании разнообразия антител (Persson MA et al., Proc Natl Acad Sci USA. 1991 Mar 15;88(6):2432-6; Schoonbroodt S, et al., Nucleic Acids Res. 2005 May 19;33(9):e81; Soderling E et al., Nat Biotechnol. 2000 Aug; 18(8):852-6; Rothe et al., J Mol Biol. 2008 Feb 29;376(4):1182-200). Встречаемость последовательностей, кодирующих не функциональные антитела, оказывает большое влияние на процесс идентификации антител. Во-первых, уменьшается функциональный размер библиотеки и, поскольку нефункциональные клоны часто характеризуются преимущественным ростом в процессе размножения библиотек, они размножаются быстрее и могут препятствовать процессу идентификации антител-кандидатов (De Bruin R et al., Nat Biotechnol 1999 April 17: 397-399). Указанные проблемы считаются серьезными ограничениями для полноценной эксплуатации потенциала библиотек антител. Получение высоко функциональных библиотек остается сложной задачей в данной области, которая заставляет предпринимать многочисленные исследования по улучшению процесса. Например, чтобы более эффективно отслеживать огромное разнообразие возможных сочетаний последовательностей, кодируемых синтетическими генами CDR, используют несколько стратегий диверсификации, целью которых является имитация применения аминокислот, присутствующих в природных последовательностях CDR (de Kruif J et al., J Mol Biol. 1995 Apr 21; 248(1):97-105; Sidhu SS et al., J Mol Biol. 2004 Apr 23;338(2):299-310). Другой подход, потенциальной издержкой которого является утрата разнообразия, включает зачистку исходной библиотеки с целью удаления нефункциональных клонов. Указанный способ используют для предварительного выбора синтетических репертуаров путем связывания библиотеки антител с общим лигандом. Эту стадию, позволяющую обогащать библиотеку членами, способными к экспрессии и правильной укладке, можно использовать для воссоздания более функциональной библиотеки (Winter and Tomlinson, US 6696245 B2). Независимо от способа качество любой библиотеки зависит от эффективности методов молекулярной биологии, используемых для ее получения, которые, как правило, позволяют получать библиотеки, содержащие от 15% до 45% нефункциональных членов. Следовательно, существует потребность в новых и высокоэффективных способах, способных минимизировать встречаемость нефункциональных генов, образующихся в результате сдвигов рамки считывания в процессе стадий молекулярного клонирования, и максимально повысить функциональность библиотек в результате улавливания участков CDR, способных к правильной укладке в каркасных участках антител, обладающих желательными свойствами. Кроме того, существует потребность в способах, которые позволяют улавливать последовательности CDR из иммунного репертуара животного в терапевтически полезной среде, такой как каркасные участки антител человека, с целью усовершенствования процесса генерирования антител с высокой аффинностью.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам получения библиотек последовательностей нуклеиновых кислот, которые объединяют преимущества выбора стабильных каркасных участков и вставки кодируемых природными генами гипервариабельных участков (CDR) или выбранных аминокислотных последовательностей, которые могут выполнять роль CDR в природной среде функционального полипептида, такого как антитело. Такой способ позволяет выделять длинные CDR, или аминокислотные последовательности, способные выполнять роль CDR, которые трудно получить с помощью синтетических кодирующих последовательностей. Путем объединения стабильных каркасных участков и характеризующихся правильной укладкой CDR, или аминокислотных последовательностей, способных выполнять роль CDR, настоящее изобретение максимально повышает долю функциональных антител в библиотеке и, как следствие, выход процесса селекции, а также качество выбранных клонов. Настоящее изобретение относится к способу улавливания CDR, экспрессирующихся естественным образом в разных видах, и их вставки в каркасные участки антител человека. Этот способ позволяет использовать репертуары CDRH3, которые сильно отличаются по длине и составу от человеческого репертуара. Изобретение позволяет получить фрагменты антител человека, отражающих структурные репертуары, полученные из других видов, и, следовательно, возможность анализировать разные структурные пространства. Способы по настоящему изобретению также позволяют вводить CDR синтетического происхождения, или аминокислотные последовательности, способные выполнять роль CDR, с более высокой частотой благоприятного результата, чем альтернативные способы, с меньшим числом ошибок, вызывающих сдвиги рамок считывания в кодирующей последовательности. Библиотеки, полученные с использованием способов настоящего изобретения, характеризуются высокой встречаемостью функциональных вариантов. Библиотеки вариантов, полученные с помощью указанного способа, используют для селекции и скрининга с применением любых описанных технологий дисплеев, селекции и скрининга.
Анализ иммунных репертуаров, полученных из разных видов, или из одного вида, на разных стадиях развития, показывает, что существуют убедительные различия в характеристиках состава и длины CDRH3. Например, средняя длина CDRH3 у взрослых людей больше, чем в эмбриональном периоде, или у новорожденных (Schroeder Jr, HW et al., 2001 Blood 98; 2745-2751). Интересно, что, несмотря на большое сходство зародышевых генов антител у людей и приматов, изменение длины CDRH3 в процессе развития может быть разным (Link JM et al., Molecular Immunol. 2005 42; 943-955). Сравнение последовательностей CDRH3, обнаруженных у мышей и людей, отчетливо демонстрирует, что у мышей средняя длина значительно меньше (Rock EP et al., J Exp Med 1994 179; 323- 328). В процессе раннего развития B-клеток в костном мозге средняя длина CDRH3 у мышей увеличивается, а у людей она имеет тенденцию уменьшаться, кроме того аминокислотные композиции репертуаров CDRH3 мыши и человека различаются (Zemlin M et al., 2003 J Mol Biol 334; 733-749; Ivanov I et al., 2005 J Immunol 174; 7773-7780). Приведенные примеры свидетельствуют о том, что разные виды экспрессируют разные диапазоны репертуаров CDRH3, несмотря на то, что в целом они специфичны к подобным классам антигенов, причем биологическое значение указанных фактов еще подлежит изучению. Показано, что форма участка связывания у антител, направленных против маленьких антигенов, таких как гаптены или пептиды, отличается от формы участка связывания антител, направленных против больших белков, причем форма участка связывания зависит от длины и состава CDR (Collis A et al., J Mol Biol 2003 325; 337-354). Приведенные факты позволяют предположить, что репертуары CDRH3, экспрессируемые разными видами, будут иметь разные способности к эффективному взаимодействию с разными классами мишеней.
В описанных в настоящем документе способах и библиотеках антител, используемых для получения терапевтических антител, применяются разные репертуары, экспрессируемые разными видами. Указанные репертуары, в которых выявлены разные трехмерные пространства, могут обеспечивать получение антител против широкого ряда классов и эпитопов мишеней. Подробно описаны способы получения библиотек из наивных или иммунизированных животных, которые обеспечивают улавливание соответствующих репертуаров и получение антител. Однако антитела, полученные из указанных библиотек, не являются человеческими и, следовательно, плохо подходят для лечения человека без дополнительной генно-инженерной обработки, такой как гуманизация. Таким образом, существует потребность в новых способах использования разнообразия, присутствующего в репертуаре, полученном из разных видов, и применения данного разнообразия в терапевтически полезной среде антитела человека.
Описанные в настоящем документе способы и библиотеки антител позволяют решить некоторые из указанных выше проблем и имеют преимущества по сравнению с существующим уровнем техники. Во-первых, описанные в настоящем документе способы объединяют выбор стабильного каркасного участка и вставку естественно кодируемых CDR, выбранных из природной среды функционального антитела. Во-вторых, указанные способы позволяют осуществлять высокоэффективную вставку синтетических или природных последовательностей CDR в каркасный участок антитела, которая в значительной степени минимизирует число сдвигов рамки считывания в библиотеке и, следовательно, повышает ее качество. И наконец, изобретение относится к новому способу применения естественного структурного разнообразия антител путем улавливания репертуаров CDRH3, естественно экспрессирующихся в разных видах, и их вставки в каркасные участки антител человека. Следовательно, указанные репертуары, обладающие структурным разнообразием, можно использовать для получения антител, применяющихся для лечения человека.
Описанные в настоящем документе способы позволяют получать антитела, которые содержат стабильный каркасный участок и CDR, обеспечивающие правильную укладку, или аминокислотные последовательности, способные выполнять роль CDR. Указанные способы позволяют улавливать природное разнообразие последовательностей в стабильных каркасных участках.
В описанных в настоящем документе способах зародышевые последовательности каркасных участков 1, 2 и 3 (FR1, FR2 и FR3) выбирают из зародышевых последовательностей желательного организма, например, из генома человека (см., например, фигуры 2 и 6). В одном из вариантов осуществления данного способа вариабельные домены выбранного антитела модифицируют путем введения "лишней" последовательности, которая служит участком интеграции диверсифицированных последовательностей. Разнообразие вводят в последовательность, находящуюся вне участка, кодирующего иммуноглобулин, путем введения участков, распознаваемых ферментами рестрикции, например, участков рестрикции типа IIs, в желательном положении, таком как гипервариабельный участок тяжелой цепи 3 (CDRH3), гипервариабельный участок легкой цепи 3 (CDR L3), гипервариабельный участок тяжелой цепи 1 (CDR H1), гипервариабельный участок легкой цепи 1 (CDR L1), гипервариабельный участок тяжелой цепи 2 (CDR H2) или гипервариабельный участок легкой цепи 2 (CDR L2). Хотя приведенные в настоящем документе примеры демонстрируют разнообразие по участку CDR3 (входящему в состав вариабельного домена тяжелой цепи и/или вариабельного домена легкой цепи), следует понимать, что разнообразие можно вводить по любому желательному положению, включающему, без ограничения, участок CDR1 (входящий в состав вариабельного домена тяжелой цепи и/или вариабельного домена легкой цепи) или участок CDR2 (входящий в состав вариабельного домена тяжелой цепи и/или вариабельного домена легкой цепи). Диверсифицированные последовательности ДНК получают вместе с фланкирующими последовательностями, которые включают участки рестрикции типа IIs. Описанные в настоящем документе способы обеспечивают получение совместимых липких концов с помощью ферментов рестрикции и сохранение рамки считывания, что позволяет лигировать диверсифицированные фрагменты ДНК в акцепторный каркасный участок.
Описанные в настоящем документе способы также можно использовать для получения кодируемых указанными последовательностями аминокислотных последовательностей, содержащих диверсифицированные участки. Например, в описанных в настоящем документе способах недиверсифицированные фрагменты кодируемых аминокислотных последовательностей выбирают из последовательностей желательного организма, например, из последовательностей человека. Фрагмент кодируемой аминокислотной последовательности модифицируют путем введения "лишней" последовательности, которая служит участком интеграции диверсифицированных последовательностей. Разнообразие вводят в последовательность в желательном положении (положениях) путем введения участков, распознаваемых ферментами рестрикции, например, участков рестрикции типа IIs, в соответствующее положение кодируемой аминокислотной последовательности. Диверсифицированные последовательности ДНК получают вместе с фланкирующими последовательностями, которые включают участки рестрикции типа IIs. Описанные в настоящем документе способы обеспечивают получение совместимых липких концов с помощью ферментов рестрикции и сохранение рамки считывания, что позволяет лигировать диверсифицированные фрагменты ДНК в акцепторный каркасный участок.
В описанных в настоящем документе способах "акцепторный каркасный участок" получают с использованием "лишнего" фрагмента ДНК, который содержит, или, предпочтительно, граничит с двумя участками ферментативной рестрикции типа IIs. (см., например, фигуру 6). Предпочтительно, два указанных участка ферментативной рестрикции типа IIs обеспечивают расщепление последовательностей по границе участка целевой диверсификации, такого как, например, участок CDRH3, участок CDR L3, участок CDR H1, участок CDR L1, участок CDR H2 или участок CDR L2. В настоящем описании термин "акцепторный каркасный участок" относится к последовательности нуклеиновых кислот, которая содержит последовательности, кодирующие участки FR1, FR2, FR3 и FR4, последовательности нуклеиновых кислот, кодирующие два CDR, или аминокислотные последовательности, которые могут выполнять роль этих CDR, и "лишний" фрагмент, который служит участком интеграции диверсифицированной последовательности нуклеиновых кислот. Например, в вариантах осуществления, в которых желательно проводить диверсификацию по участку CDR3 (входящему в состав вариабельного домена тяжелой цепи и/или вариабельного домена легкой цепи), акцепторный каркасный участок содержит последовательности нуклеиновых кислот, кодирующие участки FR1, FR2, FR3 и FR4, последовательности нуклеиновых кислот, кодирующие участки CDR1 и CDR2, и "лишний" фрагмент, который служит участком интеграции диверсифицированной последовательности нуклеиновых кислот. Например, в вариантах осуществления, в которых желательно проводить диверсификацию по участку CDR2 (входящему в состав вариабельного домена тяжелой цепи и/или вариабельного домена легкой цепи), акцепторный каркасный участок содержит последовательности нуклеиновых кислот, кодирующие участки FR1, FR2, FR3 и FR4, последовательности нуклеиновых кислот, кодирующие участки CDR1 и CDR3, и "лишний" фрагмент, который служит участком интеграции диверсифицированной последовательности нуклеиновых кислот. Например, в вариантах осуществления, в которых желательно проводить диверсификацию по участку CDR1 (входящему в состав вариабельного домена тяжелой цепи и/или вариабельного домена легкой цепи), акцепторный каркасный участок содержит последовательности нуклеиновых кислот, кодирующие участки FR1, FR2, FR3 и FR4, последовательности нуклеиновых кислот, кодирующие участки CDR2 и CDR3, и "лишний" фрагмент, который служит участком интеграции диверсифицированной последовательности нуклеиновых кислот.
Термины "лишний фрагмент", "лишний фрагмент ДНК" и "лишняя последовательность", а также их любые грамматические эквиваленты, используемые в настоящем описании как взаимозаменяемые, относятся к последовательности нуклеиновых кислот, которая содержит по меньшей мере два участка распознавания типа IIs и диверсифицированную последовательность. Акцепторный каркасный участок может представлять собой акцепторный каркасный участок вариабельного домена тяжелой цепи (VH) или акцепторный каркасный участок вариабельного домена легкой цепи (VL). Применение акцепторных каркасных участков и входящих в их состав лишних фрагментов позволяет проводить интеграцию последовательности CDR (природной или синтетической), или аминокислотной последовательности, которая может выполнять роль CDR, в акцепторный каркасный участок, в котором нуклеотиды или остатки донорного каркасного участка отсутствуют, или в котором для интеграции не требуется присутствие указанных нуклеотидов или остатков. Например, применение акцепторных каркасных участков и входящих в их состав лишних фрагментов позволяет проводить интеграцию последовательности CDR (природной или синтетической), выбранной из CDRH3, CDR L3, CDR H2, CDR L2, CDR H1 и CDR L1, или аминокислотной последовательности, которая может выполнять роль CDR, выбранного из CDRH3, CDR L3, CDR H2, CDR L2, CDR H1 и CDR L1, в акцепторный каркасный участок, в котором нуклеотиды или остатки донорного каркасного участка отсутствуют, или в котором для интеграции не требуется присутствие указанных нуклеотидов или остатков. Таким образом, после интеграции лишний фрагмент полностью удаляют, после чего кодирующий участок акцепторного белка и вставленные фрагменты белков (т.е. CDR) являются интактными.
В описанных в настоящем документе способах используют праймеры, которые конструируют так, чтобы обеспечить введение участков расщепления ферментами рестрикции типа IIs на границе с участком, по которому желательно введение разнообразия, таким как участок CDRH3, участок CDR L3, участок CDR H2, участок CDR L2, участок CDR H1 или участок CDR L1. Произвольные природные клоны CDR (см., например, фигуру 10), или синтетические последовательности CDR (см. пример 6), или аминокислотные последовательности, которые могут выполнять роль CDR, улавливаются описанными в настоящем документе акцепторными каркасными участками. Например, в вариантах осуществления, в которых желательно достичь разнообразия по участку CDR3 (входящему в состав вариабельного домена тяжелой цепи и/или вариабельного домена легкой цепи), произвольные природные клоны CDR3 (см., например, фигуру 10) или синтетические последовательности CDR3 (см. пример 6), или аминокислотные последовательности, которые могут выполнять роль CDR3, улавливаются описанными в настоящем документе акцепторными каркасными участками. Например, в вариантах осуществления, в которых желательно достичь разнообразия по участку CDR2 (входящему в состав вариабельного домена тяжелой цепи и/или вариабельного домена легкой цепи), произвольные природные клоны CDR2 (см., например, способы, показанные на фигуре 10) или синтетические последовательности CDR2 (см., например, способы, показанные в примере 6), или аминокислотные последовательности, которые могут выполнять роль CDR2, улавливаются описанными в настоящем документе акцепторными каркасными участками. Например, в вариантах осуществления, в которых желательно достичь разнообразия по участку CDR1 (входящему в состав вариабельного домена тяжелой цепи и/или вариабельного домена легкой цепи), произвольные природные клоны CDR1 (см., например, способы, показанные на фигуре 10) или синтетические последовательности CDR1 (см., например, способы, показанные в примере 6), или аминокислотные последовательности, которые могут выполнять роль CDR1, улавливаются описанными в настоящем документе акцепторными каркасными участками. В качестве примера, конструируют олигонуклеотидные праймеры, специфичные к участкам, фланкирующим последовательность ДНК, кодирующую CDRH3 иммуноглобулинов, т.е., специфичные к FR3 и FR4 вариабельного участка. Также можно сконструировать олигонуклеотидные праймеры, специфичные к участкам, фланкирующим последовательности ДНК, кодирующие другие участки, такие как, например, CDR L3, CDR H1, CDR L1, CDR H2 или CDR L2. Указанные олигонуклеотиды содержат на 5'-конце участок распознавания ферментом рестрикции типа IIs, а их 3'-конец совпадает с целевой последовательностью ДНК.
В некоторых вариантах осуществления праймер представляет собой нуклеиновую кислоту, выбранную из группы, включающей SEQ ID NO: 120-254.
В описанных в настоящем документе способах используют ферменты рестрикции типа IIs, такие как, например, FokI, чтобы вставить природные последовательности CDR, такие как, например, природные последовательности CDRH3, CDR L3, CDR H1, CDR L1, CDR H2 или CDR L2, в описанные в настоящем документе акцепторные каркасные участки. В описанных в настоящем документе способах используют ферменты рестрикции типа IIs, такие как, например, FokI, чтобы вставить синтетические последовательности CDR, такие как, например, синтетические последовательности CDRH3, CDR L3, CDR H1, CDR L1, CDR H2 или CDR L2, в описанные в настоящем документе акцепторные каркасные участки. В описанных в настоящем документе способах используют ферменты рестрикции типа IIs, такие как, например, FokI, чтобы вставить аминокислотные последовательности, которые могут выполнять роль желательного участка CDR, такие как, например, аминокислотная последовательность, которая может выполнять роль природного или синтетического участка CDRH3, CDR L3, CDR H1, CDR L1, CDR H2 или CDR L2, в описанные в настоящем документе акцепторные каркасные участки. Ферменты рестрикции типа IIs представляют собой ферменты, которые производят отщепление распознаваемой ими последовательности с одной стороны. Указанные ферменты являются промежуточными по размеру, как правило, содержат 400-650 аминокислот в длину и распознают непрерывные и асимметричные последовательности. Подходящие ферменты рестрикции типа IIs, также известные как рестрикционные эндонуклеазы типа IIs, а также распознаваемые ими последовательности описаны, например, в Szybalski et al., "Class-IIS Restriction Enzymes - a Review." Gene, vol. 100: 13-26 (1991), содержание которой включено в настоящее описание в качестве ссылки во всей полноте.
Первичные библиотеки включают акцепторный каркасный участок VH и стационарную последовательность VL (также называемую "холостая" последовательность VL) или акцепторный каркасный участок VL и стационарную последовательность VH (также называемую "холостая" последовательность VH). Таким образом, первичные библиотеки характеризуются разнообразием только в одной из тяжелой и легкой цепей. Вторичные библиотеки получают путем лигирования акцепторного каркасного участка VH с акцепторным каркасным участком VL (см. пример 7). Вторичные библиотеки характеризуются разнообразием как в тяжелой, так и в легкой цепях.
Изобретение относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен тяжелой цепи иммуноглобулина человека, содержащий несколько гипервариабельных участков 3 тяжелой цепи (CDRH3), выделенных из репертуара вариабельных доменов иммуноглобулинов отличных от человека видов. В некоторых вариантах осуществления способ включает следующие стадии: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены тяжелой цепи иммуноглобулина человека, причем каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот; (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих последовательности гипервариабельных участков 3 тяжелой цепи (CDRH3), выделенные из репертуара иммуноглобулинов отличных от человека видов, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот на каждом конце содержит участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDRH3, с помощью рестрикционного фермента типа IIs, который связывается с участком распознавания рестрикционным ферментом типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью рестрикционного фермента типа IIs, который связывается с участком распознавания рестрикционным ферментом типа IIs, указанным на стадии (a); и (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDRH3, или аминокислотных последовательностей, указанных на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR3 и FR4 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDRH3 или аминокислотную последовательность, которая может выполнять роль участка CDR3, с восстановлением последовательностей, кодирующих полноразмерный вариабельный домен иммуноглобулина, которые не содержат участки распознавания рестрикционными ферментами типа IIs, указанные на стадиях (a) и (b).
В некоторых вариантах осуществления описанную выше стадию (b) проводят путем амплификации последовательности CDRH3, полученной из отличных от человека видов, с использованием олигонуклеотидных праймеров, содержащих участок рестрикции типа IIs. В некоторых вариантах осуществления стадию (b) проводят путем амплификации последовательности CDRH3, полученной из отличных от человека видов, с использованием олигонуклеотидных праймеров, содержащих участок рестрикции типа IIs FokI. В некоторых вариантах осуществления отличные от человека виды включают отличных от человека приматов, грызунов, собак, кошек, овец, коз, крупный рогатый скот, лошадей или свиней.
Изобретение относится к способам получения библиотеки нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина, указанные способы включают (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулина, причем каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот; (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих участки гипервариабельного участка 3 (CDR3), или кодирующих аминокислотные последовательности, которые могут выполнять роль участка CDR3, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот на каждом конце содержит участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR3, или аминокислотные последовательности, которые могут выполнять роль участка CDR3, с помощью рестрикционного фермента типа IIs, который связывается с участком распознавания рестрикционным ферментом типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью рестрикционного фермента типа IIs, который связывается с участком распознавания рестрикционным ферментом типа IIs, указанным на стадии (a); и (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR3 или аминокислотные последовательности, которые могут выполнять роль участка CDR3, указанных на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR3 и FR4 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR3 или аминокислотную последовательность, которая может выполнять роль участка CDR3, с восстановлением последовательностей, кодирующих полноразмерный вариабельный домен иммуноглобулина, которые не содержат участки распознавания рестрикционными ферментами типа IIs, указанные на стадиях (a) и (b).
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются одинаковыми ферментами рестрикции типа IIs. В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанными на стадии (a) и на стадии (b), распознаются разными ферментами рестрикции типа IIs. Например, участки распознавания ферментами рестрикции типа IIs включают участки распознавания FokI, участки распознавания BsaI и/или участки распознавания BsmBI.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка получают из последовательности гена человека. Например, последовательность человека представляет собой последовательность гена вариабельного домена тяжелой цепи человека, или последовательность, полученную из гена вариабельного домена тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена вариабельного домена тяжелой цепи человека выбрана из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи каппа человека, или последовательность, полученную из гена вариабельного домена легкой цепи каппа человека. Например, последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи лямбда человека, или последовательность, полученную из гена вариабельного домена легкой цепи лямбда человека. Например, последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR3 и включает природные последовательности, или последовательности, полученные от иммунизированных животных.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR3, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих и других естественно диверсифицированных полипептидных коллекций, или указанная совокупность получена из таких последовательностей.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR3 и включает последовательности природных иммуноглобулинов у людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанная совокупность получена из таких последовательностей.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR3, и включает синтетические последовательности.
В другом варианте осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR3, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка включает смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления предлагаемые способы включают дополнительную стадию (e) трансформации клетки-хозяина вектором экспрессии, полученным на стадии (d), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности последовательностей акцепторного каркасного участка. Например, клетка-хозяин может представлять собой E. coli. В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный вектор. Например, фагмидный вектор может представлять собой pNDS1.
Изобретение также относится к способам получения библиотеки нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина, которые включают (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулинов, причем каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), где участки FR1 и FR2 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот, участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены гипервариабельным участком 3 (CDR3); (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих участки гипервариабельного участка 1 (CDR1), или кодирующих аминокислотные последовательности, которые могут выполнять роль участка CDR1, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR1 или аминокислотные последовательности, способные выполнять роль участка CDR1, с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); и (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR1, или аминокислотные последовательности, которые могут выполнять роль участка CDR1, указанные на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR1 и FR2 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR1 или аминокислотные последовательности, которые могут выполнять роль участка CDR1, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b).
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются одинаковыми ферментами рестрикции типа IIs. В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются разными ферментами рестрикции типа IIs. Например, участки распознавания ферментами рестрикции типа IIs включают участки распознавания FokI, участки распознавания BsaI, и/или участки распознавания BsmBI.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка получают из последовательности гена человека. Например, последовательность человека представляет собой последовательность гена человека вариабельного домена тяжелой цепи или последовательность, полученную из последовательности гена вариабельного домена тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена вариабельного домена тяжелой цепи человека выбрана из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи каппа человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи каппа человека. Например, последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи лямбда человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи лямбда человека. Например, последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR1 и включает природные последовательности или последовательности, полученные из иммунизированных животных.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR1, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих и других естественно диверсифицированных полипептидных коллекций, или указанная совокупность получена из таких последовательностей.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR1 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанная совокупность получена из таких последовательностей.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR1, и включает синтетические последовательности.
В другом варианте осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR1, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка включает смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления предлагаемые способы включают дополнительную стадию (e) клонирования библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулинов, указанных на стадии (d), в векторе экспрессии и (f) трансформации клетки-хозяина вектором экспрессии, полученным на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности вариабельных доменов иммуноглобулинов, кодируемых библиотекой. Например, клетка-хозяин может представлять собой E. coli. В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный вектор. Например, фагмидный вектор может представлять собой pNDS1.
Изобретение также относится к способам получения библиотеки нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина, которые включают (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулинов, причем каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот, а участки FR3 и FR4 разделены гипервариабельным участком 3 (CDR3); (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих участки гипервариабельного участка 2 (CDR2) или аминокислотные последовательности, которые могут выполнять роль участка CDR2, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR2 или аминокислотные последовательности, способные выполнять роль участка CDR2 с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); и (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR2 или аминокислотные последовательности, которые могут выполнять роль участка CDR2, указанные на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR2 и FR3 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR2 или аминокислотные последовательности, которые могут выполнять роль участка CDR2, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b).
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются одинаковыми ферментами рестрикции типа IIs. В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются разными ферментами рестрикции типа IIs. Например, участки распознавания ферментами рестрикции типа IIs включают участки распознавания FokI, участки распознавания BsaI и/или участки распознавания BsmBI.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка получают из последовательности гена человека. Например, последовательность человека представляет собой последовательность гена вариабельного домена тяжелой цепи или последовательность, полученную из последовательности гена вариабельного домена тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена вариабельного домена тяжелой цепи человека выбрана из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи каппа человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи каппа человека. Например, последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи лямбда человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи лямбда человека. Например, последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR2 и включает природные последовательности или последовательности, полученные из иммунизированных животных.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR2, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих и других, естественно диверсифицированных полипептидных коллекций, или указанная совокупность получена из таких последовательностей.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR2 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанная совокупность получена из таких последовательностей.
В другом варианте осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR2, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка включает смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления предлагаемые способы включают дополнительную стадию (e) клонирования в векторе экспрессии библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулинов, указанных на стадии (d), и (f) трансформации клетки-хозяина вектором экспрессии, полученным на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности вариабельных доменов иммуноглобулинов, кодируемых библиотекой. Например, клетка-хозяин может представлять собой E. coli. В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный вектор. Например, фагмидный вектор может представлять собой pNDS1.
Изобретение также относится к способам получения мишень-специфичного антитела, вариабельного участка антитела или его фрагмента, которые включают (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулинов, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот; (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих участки гипервариабельного участка 3 (CDR3) или аминокислотные последовательности, которые могут выполнять роль участка CDR3, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR3 или аминокислотные последовательности, способные выполнять роль участка CDR3, с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); (d) клонирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR3 или аминокислотные последовательности, которые могут выполнять роль участка CDR3, в векторе экспрессии и лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR3 или аминокислотные последовательности, которые могут выполнять роль участка CDR,3 указанных на стадии (c), в акцепторный каркасный участок, так, чтобы участки FR3 и FR4 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR3 или аминокислотные последовательности, которые могут выполнять роль участка CDR3, с восстановлением полноразмерной последовательности гена вариабельного домена иммуноглобулина; (e) трансформацию клетки-хозяина вектором экспрессии, указанным на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности последовательностей акцепторного каркасного участка; (f) приведение в контакт клетки-хозяина с антигеном-мишенью; и (g) определение, какая из экспрессированных последовательностей акцепторных каркасных участков связывается с антигеном-мишенью.
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются одинаковыми ферментами рестрикции типа IIs. В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются разными ферментами рестрикции типа IIs. Например, участки распознавания ферментами рестрикции типа IIs включают участки распознавания FokI, участки распознавания BsaI, и/или участки распознавания BsmBI.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка получают из последовательности гена человека. Например, последовательность человека представляет собой последовательность гена вариабельного домена тяжелой цепи человека или последовательность, полученную из последовательности гена вариабельного домена тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена вариабельного домена тяжелой цепи человека выбрана из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи каппа человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи каппа человека. Например, последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи лямбда человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи лямбда человека. Например, последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR3 и включает природные последовательности или последовательности, полученные из иммунизированных животных.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR3, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих и других естественно диверсифицированных полипептидных коллекций, или указанная совокупность получена из таких последовательностей.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR3 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанная совокупность получена из таких последовательностей.
В другом варианте осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR3, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка включает смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный вектор. Например, фагмидный вектор представляет собой pNDS1. В некоторых вариантах осуществления клетка-хозяин представляет собой E. coli.
В некоторых вариантах осуществления способ включает дополнительную стадию (i) секвенирования последовательности, кодирующей вариабельный домен иммуноглобулина, который связывается с антигеном-мишенью.
Изобретение также относится к способам получения специфичного к мишени антитела, вариабельного участка антитела или его фрагмента, которые включают (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулинов, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), где участки FR1 и FR2 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот, участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены гипервариабельным участком 3 (CDR3); (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих участки гипервариабельного участка 1 (CDR1) или аминокислотные последовательности, которые могут выполнять роль участка CDR1, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR1 или аминокислотные последовательности, способные выполнять роль участка CDR1, с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); (d) клонирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR1 или аминокислотные последовательности, которые могут выполнять роль участка CDR1, в векторе экспрессии и лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR1 или аминокислотные последовательности, которые могут выполнять роль участка CDR1, указанные на стадии (c), в акцепторный каркасный участок так, чтобы участки FR1 и FR2 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR1, или аминокислотные последовательности, которые могут выполнять роль участка CDR1, с восстановлением полноразмерной последовательности, кодирующей ген вариабельного домена иммуноглобулина; (e) трансформацию клетки-хозяина вектором экспрессии, указанным на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности последовательностей акцепторного каркасного участка; (f) приведение в контакт клетки-хозяина с антигеном-мишенью; и (g) определение, какая из экспрессированных последовательностей акцепторных каркасных участков связывается с антигеном-мишенью.
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются одинаковыми ферментами рестрикции типа IIs. В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются разными ферментами рестрикции типа IIs. Например, участки распознавания ферментами рестрикции типа IIs включают участки распознавания FokI, участки распознавания BsaI, и/или участки распознавания BsmBI.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка получают из последовательности гена человека. Например, последовательность человека представляет собой последовательность гена вариабельного домена тяжелой цепи человека или последовательность, полученную из последовательности гена вариабельного домена тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена вариабельного домена тяжелой цепи человека выбрана из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи каппа человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи каппа человека. Например, последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи лямбда человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи лямбда человека. Например, последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR1 и включает природные последовательности или последовательности, полученные из иммунизированных животных.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR1, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих и других естественно диверсифицированных полипептидных коллекций, или указанная совокупность получена из таких последовательностей.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR1 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанная совокупность получена из таких последовательностей.
В другом варианте осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR1, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка включает смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный вектор. Например, фагмидный вектор представляет собой pNDS1. В некоторых вариантах осуществления клетка-хозяин представляет собой E. coli.
В некоторых вариантах осуществления способ включает дополнительную стадию (i) секвенирования последовательности, кодирующей вариабельный домен иммуноглобулина, который связывается с антигеном-мишенью.
Изобретение относится к способам получения мишень-специфичного антитела, вариабельного участка антитела или его фрагмента, которые включают (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулинов, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот, а участки FR3 и FR4 разделены гипервариабельным участком 3 (CDR3); (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих участки гипервариабельного участка 2 (CDR2), или кодирующих аминокислотные последовательности, которые могут выполнять роль участка CDR2, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR2 или аминокислотные последовательности, способные выполнять роль участка CDR2, с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR2 или аминокислотные последовательности, которые могут выполнять роль участка CDR2, указанные на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR2 и FR3 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR2 или аминокислотные последовательности, которые могут выполнять роль участка CDR2, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b); (e) клонирование библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулина, указанные на стадии (d), в векторе экспрессии; (f) трансформацию клетки-хозяина вектором экспрессии, указанным на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности вариабельных доменов иммуноглобулина, кодируемых библиотекой; (g) приведение в контакт совокупности вариабельных доменов иммуноглобулина, указанной на стадии (f), с антигеном-мишенью; и (h) определение, какая из экспрессированных последовательностей, кодирующих вариабельный домен иммуноглобулина, связывается с антигеном-мишенью.
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются одинаковыми ферментами рестрикции типа IIs. В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются разными ферментами рестрикции типа IIs. Например, участки распознавания ферментами рестрикции типа IIs включают участки распознавания FokI, участки распознавания BsaI и/или участки распознавания BsmBI.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка получают из последовательности гена человека. Например, последовательность человека представляет собой последовательность гена вариабельного домена тяжелой цепи человека или последовательность, полученную из последовательности гена вариабельного домена тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена вариабельного домена тяжелой цепи человека выбрана из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи каппа человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи каппа человека. Например, последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи лямбда человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи лямбда человека. Например, последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR2 и включает природные последовательности или последовательности, полученные из иммунизированных животных.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR2, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих и других естественно диверсифицированных полипептидных коллекций, или указанная совокупность получена из таких последовательностей.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR2 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанная совокупность получена из таких последовательностей.
В одном из вариантов осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR2, и включает синтетические последовательности.
В другом варианте осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR2, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка содержит смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления клетка-хозяин представляет собой E. coli. В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный вектор. Например, фагмидный вектор представляет собой pNDS1.
В некоторых вариантах осуществления способ включает дополнительную стадию (i) секвенирования последовательности, кодирующей вариабельный домен иммуноглобулина, который связывается с антигеном-мишенью.
Изобретение также относится к способам получения библиотеки нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина. Указанные способы включают следующие стадии: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка Ig, в которые вводят источник разнообразия в одном гипервариабельном участке (CDR), выбранном из группы, включающей гипервариабельный участок 1 (CDR1), гипервариабельный участок 2 (CDR2) и гипервариабельный участок 3 (CDR3), где последовательность акцепторного каркасного участка Ig содержит "лишнюю" последовательность нуклеиновых кислот, содержащую по меньшей мере два участка распознавания ферментами рестрикции типа IIs, и где источник разнообразия представляет собой CDR, выбранный из природных последовательностей CDR, которые содержат участки распознавания ферментами рестрикции типа IIs вне участка CDR, (b) введение источника разнообразия в каждый акцепторный каркасный участок Ig путем расщепления как источника разнообразия, так и акцепторного каркасного участка Ig ферментом рестрикции типа IIs; и (c) лигирование расщепленного источника разнообразия в акцепторный каркасный участок Ig с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b).
Последовательность природного участка CDR практически не отличается от последовательности дикого типа, т.е., находится в природном состоянии. Указанные природные последовательности участка CDR фланкируются аминокислотными последовательностями, полученными с помощью генно-инженерных методов (или путем иных искусственных манипуляций), так, что в результате они содержат два участка распознавания ферментами рестрикции типа IIs, причем с каждой стороны природной последовательности участка CDR находится один участок распознавания ферментами рестрикции типа типа IIs. Участки распознавания ферментами рестрикции типа IIs находятся вне участка, кодирующего CDR. Последовательности участков CDR не изменяются в пределах участка, кодирующего CDR - распознавание ферментами рестрикции и сплайсинг осуществляются по участкам, которые примыкают к границам участка, кодирующего CDR, но не находятся внутри участка, кодирующего CDR.
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, находящиеся в "лишних" последовательностях нуклеиновых кислот, и фланкирующие природные последовательности CDR, распознаются одинаковыми ферментами рестрикции типа IIs. В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, находящиеся в "лишних" последовательностях нуклеиновых кислот, и фланкирующие природные последовательности CDR, распознаются разными ферментами рестрикции типа IIs. Например, участки распознавания ферментами рестрикции типа IIs включают участки распознавания FokI, участки распознавания BsaI и/или участки распознавания BsmBI.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка Ig получают из последовательности гена человека. Например, последовательность человека представляет собой последовательность гена вариабельного домена тяжелой цепи человека или последовательность, полученную из последовательности гена вариабельного домена тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена вариабельного домена тяжелой цепи человека выбрана из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи каппа человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи каппа человека. Например, последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи лямбда человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи лямбда человека. Например, последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В некоторых вариантах осуществления комплект природных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR3, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих и других естественно диверсифицированных полипептидных коллекций, или указанный комплект получен из таких последовательностей.
В некоторых вариантах осуществления комплект природных нуклеиновых кислот кодирует участки CDR3 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанный комплект получен из таких последовательностей.
В некоторых вариантах осуществления комплект природных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR1, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих и других естественно диверсифицированных полипептидных коллекций, или указанный комплект получен из таких последовательностей.
В некоторых вариантах осуществления комплект природных нуклеиновых кислот кодирует участки CDR1 и включает последовательности иммуноглобулина, последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанный комплект получен из таких последовательностей.
В некоторых вариантах осуществления комплект природных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR2, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих и других естественно диверсифицированных полипептидных коллекций, или указанный комплект получен из таких последовательностей.
В некоторых вариантах осуществления комплект природных нуклеиновых кислот кодирует участки CDR2 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанный комплект получен из таких последовательностей.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка Ig включает смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи (VL).
В некоторых вариантах осуществления предлагаемые способы включают дополнительные стадии (e) клонирования библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулина, указанные на стадии (d), в векторе экспрессии и (f) трансформации клетки-хозяина вектором экспрессии, указанным на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности кодируемых библиотекой вариабельных доменов иммуноглобулина. Например, клетка-хозяин представляет собой E. coli. В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный вектор. Например, фагмидный вектор представляет собой pNDS1.
Изобретение также относится к способам получения библиотеки нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина. Указанные способы включают следующие стадии: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка Ig, в которые вводят источник разнообразия в одном гипервариабельном участке (CDR), выбранном из группы, включающей гипервариабельный участок 1 (CDR1), гипервариабельный участок 2 (CDR2) и гипервариабельный участок 3 (CDR3), где последовательность акцепторного каркасного участка Ig содержит "лишнюю" последовательность нуклеиновых кислот, содержащую по меньшей мере два участка распознавания ферментами рестрикции типа IIs, и где источник разнообразия представляет собой CDR, выбранный из синтетических последовательностей CDR, которые содержат участки распознавания ферментами рестрикции типа IIs вне участка CDR, (b) введение источника разнообразия в каждый акцепторный каркасный участок Ig путем расщепления как источника разнообразия, так и акцепторного каркасного участка Ig ферментом рестрикции типа IIs; и (c) лигирование расщепленного источника разнообразия в акцепторный каркасный участок Ig с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b).
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs в "лишних" последовательностях нуклеиновых кислот и синтетических последовательностях CDR распознаются одинаковыми ферментами рестрикции типа IIs. В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs в "лишних" последовательностях нуклеиновых кислот и синтетических последовательностях CDR распознаются разными ферментами рестрикции типа IIs. Например, участки распознавания ферментами рестрикции типа IIs включают участки распознавания FokI, участки распознавания BsaI и/или участки распознавания BsmBI.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка Ig получают из последовательности человека. Например, последовательность человека представляет собой последовательность гена вариабельного домена тяжелой цепи человека или последовательность, полученную из последовательности гена вариабельного домена тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена вариабельного домена тяжелой цепи человека выбрана из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи каппа человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи каппа человека. Например, последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи лямбда человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи лямбда человека. Например, последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR3, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR1, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR2, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка Ig включает смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления предлагаемые способы включают дополнительные стадии (e) клонирования библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулина, указанные на стадии (d), в векторе экспрессии и (f) трансформации клетки-хозяина вектором экспрессии, указанным на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности вариабельных доменов иммуноглобулина, кодируемых библиотекой. Например, клетка-хозяин представляет собой E. coli. В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный вектор. Например, фагмидный вектор представляет собой pNDS1.
Изобретение также относится к способам получения полипептида иммуноглобулина. Указанные способы включают следующие стадии: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка Ig, в которые вводят источник разнообразия в одном гипервариабельном участке (CDR), выбранном из группы, включающей гипервариабельный участок 1 (CDR1), гипервариабельный участок 2 (CDR2) и гипервариабельный участок 3 (CDR3), где последовательность акцепторного каркасного участка Ig содержит "лишнюю" последовательность нуклеиновых кислот, содержащую по меньшей мере два участка распознавания ферментами рестрикции типа IIs, и где источник разнообразия представляет собой CDR, выбранный из природных последовательностей CDR, которые содержат участки распознавания ферментами рестрикции типа IIs вне участка CDR, (b) введение источника разнообразия в каждый акцепторный каркасный участок Ig путем расщепления как источника разнообразия, так и акцепторного каркасного участка Ig ферментом рестрикции типа IIs; (c) лигирование расщепленного источника разнообразия в акцепторный каркасный участок Ig с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, (d) клонирование полноразмерной последовательности, кодирующей ген вариабельного домена иммуноглобулина, указанной на стадии (c), в векторе экспрессии; и (e) трансформация клетки-хозяина вектором экспрессии, указанным на стадии (d), и культивирование клетки-хозяина в условиях, достаточных для экспрессии полноразмерных последовательностей, кодирующих ген иммуноглобулина, которые не содержат участки распознавания ферментами рестрикции типа IIs.
В указанных вариантах осуществления последовательности природного участка CDR практически не отличаются от последовательностей дикого типа, т.е., находятся в природном состоянии. Указанные природные последовательности участка CDR фланкируются аминокислотными последовательностями, полученными с помощью генно-инженерных методов (или путем иных искусственных манипуляций), так, что в результате они содержат два участка распознавания ферментами рестрикции типа IIs, причем с каждой стороны природной последовательности участка CDR находится один участок распознавания ферментами рестрикции типа IIs.
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, находящиеся в "лишних" последовательностях нуклеиновых кислот, и фланкирующие природные последовательности CDR, распознаются одинаковыми ферментами рестрикции типа IIs. В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, находящиеся в "лишних" последовательностях нуклеиновых кислот, и фланкирующие природные последовательности CDR, распознаются разными ферментами рестрикции типа IIs. Например, участки распознавания ферментами рестрикции типа IIs включают участки распознавания FokI, участки распознавания BsaI и/или участки распознавания BsmBI.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка получают из последовательности гена человека. Например, последовательность человека представляет собой последовательность гена вариабельного домена тяжелой цепи человека или последовательность, полученную из последовательности гена вариабельного домена тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена вариабельного домена тяжелой цепи человека выбрана из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи каппа человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи каппа человека. Например, последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи лямбда человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи лямбда человека. Например, последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В некоторых вариантах осуществления комплект природных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR3, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих и других естественно диверсифицированных полипептидных коллекций, или указанный комплект получен из таких последовательностей.
В некоторых вариантах осуществления комплект природных нуклеиновых кислот кодирует участки CDR3 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена.
В некоторых вариантах осуществления комплект природных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR1, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих и других естественно диверсифицированных полипептидных коллекций, или указанная совокупность получена из таких последовательностей.
В некоторых вариантах осуществления комплект природных нуклеиновых кислот кодирует участки CDR1 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанный комплект получен из таких последовательностей.
В некоторых вариантах осуществления комплект природных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR2, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих и других естественно диверсифицированных полипептидных коллекций, или указанный комплект получен из таких последовательностей.
В некоторых вариантах осуществления комплект природных нуклеиновых кислот кодирует участки CDR2 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанный комплект получен из таких последовательностей.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка содержит смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный вектор. В некоторых вариантах осуществления клетка-хозяин представляет собой E. coli.
В некоторых вариантах осуществления способ также включает стадии приведения в контакт клетки-хозяина с антигеном-мишенью, определение, какая из клеток-хозяев, экспрессирующих полноразмерную последовательность, кодирующую ген вариабельного домена Ig, связывается с антигеном-мишенью, и посредством этого идентификации целевых специфических антител, вариабельных участков антител или их фрагментов. В некоторых вариантах осуществления способ включает дополнительную стадию (i) секвенирования последовательности, кодирующей вариабельный домен иммуноглобулина, который связывается с антигеном-мишенью.
Изобретение также относится к способам получения полипептида иммуноглобулина. Указанные способы включают следующие стадии: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка Ig, в которые вводят источник разнообразия в одном гипервариабельном участке (CDR), выбранном из группы, включающей гипервариабельный участок 1 (CDR1), гипервариабельный участок 2 (CDR2) и гипервариабельный участок 3 (CDR3), где последовательность акцепторного каркасного участка Ig содержит "лишнюю" последовательность нуклеиновых кислот, содержащую по меньшей мере два участка распознавания ферментами рестрикции типа IIs, и где источник разнообразия представляет собой CDR, выбранный из синтетических последовательностей CDR, которые содержат участки распознавания ферментами рестрикции типа IIs вне участка CDR, (b) введение источника разнообразия в каждый акцепторный каркасный участок Ig путем расщепления как источника разнообразия, так и акцепторного каркасного участка Ig ферментом рестрикции типа IIs; (c) лигирование расщепленного источника разнообразия в акцепторный каркасный участок Ig с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, (d) клонирование легированного акцепторного каркасного участка Ig, указанного на стадии (c), в векторе экспрессии; и (e) трансформация клетки-хозяина вектором экспрессии, указанным на стадии (d), и культивирование клетки-хозяина в условиях, достаточных для экспрессии полноразмерных последовательностей, кодирующих ген иммуноглобулина, которые не содержат участки распознавания ферментами рестрикции типа IIs.
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs в "лишних" последовательностях нуклеиновых кислот и синтетических последовательностях CDR распознаются одинаковыми ферментами рестрикции типа IIs. В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs в "лишних" последовательностях нуклеиновых кислот и синтетических последовательностях CDR распознаются разными ферментами рестрикции типа IIs. Например, участки распознавания ферментами рестрикции типа IIs включают участки распознавания FokI, участки распознавания BsaI и/или участки распознавания BsmBI.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка Ig получают из последовательности гена человека. Например, последовательность человека представляет собой последовательность гена вариабельного домена тяжелой цепи человека или последовательность, полученную из последовательности гена вариабельного домена тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена вариабельного домена тяжелой цепи человека выбрана из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи каппа человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи каппа человека. Например, последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20. В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи лямбда человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи лямбда человека. Например, последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR3, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR1, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR2, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка Ig содержит смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный вектор. В некоторых вариантах осуществления клетка-хозяин представляет собой E. coli.
В некоторых вариантах осуществления способ также включает стадии приведения в контакт клетки-хозяина с антигеном-мишенью, и определение какая из клеток-хозяев, экспрессирующих полноразмерную последовательность, кодирующую ген вариабельного домена Ig, связывается с антигеном-мишенью, и посредством этого идентификации целевых специфических антител, вариабельных участков антител или их фрагментов. В некоторых вариантах осуществления способ включает дополнительную стадию (i) секвенирования последовательности, кодирующей вариабельный домен иммуноглобулина, который связывается с антигеном-мишенью.
Изобретение относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 3 (CDR3), выделенных по отдельности из репертуара вариабельных доменов иммуноглобулинов млекопитающих. Изобретение также относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 2 (CDR2), выделенных по отдельности из репертуара вариабельных доменов иммуноглобулинов млекопитающих. Изобретение также относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 1 (CDR1), выделенных по отдельности из репертуара вариабельных доменов иммуноглобулинов млекопитающих.
Изобретение относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 3 (CDR3), выделенных по отдельности из репертуара вариабельных доменов иммуноглобулинов отличных от млекопитающих видов. Изобретение также относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 2 (CDR2), выделенных по отдельности из репертуара вариабельных доменов иммуноглобулинов отличных от млекопитающих видов. Изобретение также относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 1 (CDR1), выделенных по отдельности из репертуара вариабельных доменов иммуноглобулинов отличных от млекопитающих видов.
В некоторых вариантах осуществления к отличным от человека видам относятся отличные от человека приматы, грызуны, собаки, кошки, овцы, козы, крупный рогатый скот, лошади, члены семейства Camelidae, ламы, верблюды, одногорбые верблюды и свиньи.
Изобретение относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 3 (CDR3), выделенных по отдельности из репертуара вариабельных доменов иммуноглобулинов человека. Изобретение относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 2 (CDR2), выделенных по отдельности из репертуара вариабельных доменов иммуноглобулинов человека. Изобретение относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 1 (CDR1), выделенных по отдельности из репертуара вариабельных доменов иммуноглобулинов человека.
Изобретение относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 3 (CDR3), выделенных по отдельности из репертуара вариабельных доменов иммуноглобулинов отличных от человека видов.
В некоторых вариантах осуществления указанные способы включают следующие стадии: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулина человека, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот; (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих последовательности гипервариабельного участка 3 (CDR3), выделенных из репертуара иммуноглобулинов млекопитающих, где каждая из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR3, с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); и (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR3 или аминокислотные последовательности, указанные на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR3 и FR4 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR3 или аминокислотную последовательность, которая может выполнять роль участка CDR3, и была восстановлена полноразмерная последовательность, кодирующая вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b). Указанные стадии также можно проводить с использованием совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих последовательности гипервариабельного участка 2 (CDR2), выделенных из репертуара иммуноглобулинов млекопитающих. Указанные стадии также можно проводить с использованием совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих последовательности гипервариабельного участка 1 (CDR1), выделенных из репертуара иммуноглобулинов млекопитающих.
В некоторых вариантах осуществления стадию (b) проводят путем амплификации последовательности CDR3 млекопитающих с использованием олигонуклеотидных праймеров, содержащих участок рестрикции типа IIs. В некоторых вариантах осуществления олигонуклеотидный праймер конструируют так, чтобы повысить совместимость последовательности CDR3 млекопитающего и акцепторного каркасного участка, кодирующего вариабельный домен иммуноглобулина человека. В некоторых вариантах осуществления олигонуклеотидный праймер конструируют так, чтобы модифицировать последовательность, граничащую с последовательностью CDR3 млекопитающего, с целью обеспечения эффективного лигирования посредством совместимых липких концов в последовательность акцепторного каркасного участка, кодирующую вариабельный домен иммуноглобулина человека. В некоторых вариантах осуществления последовательности ДНК млекопитающих, фланкирующие участки CDR3, могут содержать липкие концы, совместимые с липкими концами расщепленных акцепторных каркасных участков до расщепления ферментами рестрикции типа IIs. В таких случаях олигонуклеотиды, используемые для амплификации, конструируют с целью модификации целевой последовательности млекопитающих так, чтобы после расщепления ферментом рестрикции типа IIs получить совместимые липкие концы и обеспечить эффективное лигирование. Указанные стадии также можно проводить путем амплификации последовательности CDR2 млекопитающих с использованием олигонуклеотидных праймеров, содержащих участок рестрикции типа IIs. Указанные стадии также можно проводить путем амплификации последовательности CDR1 млекопитающих с использованием олигонуклеотидных праймеров, содержащих участок рестрикции типа IIs.
В некоторых вариантах осуществления стадию (b) проводят путем амплификации последовательности CDR3 отличных от человека видов с использованием олигонуклеотидных праймеров, содержащих участок рестрикции FokI IIs. Указанные стадии также можно проводить путем амплификации последовательности CDR2 млекопитающих с использованием олигонуклеотидных праймеров, содержащих участок рестрикции FokI IIs. Указанные стадии также можно проводить путем амплификации последовательности CDR1 млекопитающих с использованием олигонуклеотидных праймеров, содержащих участок рестрикции FokI IIs.
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются разными ферментами рестрикции типа IIs. В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs включают участки распознавания BsmBI, участки распознавания BsaI, участки распознавания FokI, или их сочетания.
В некоторых вариантах осуществления диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR3, кодируют последовательности CDR3 тяжелой цепи (CDRH3). В некоторых вариантах осуществления диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR3, кодируют последовательности CDR3 легкой цепи (CDR L3). В некоторых вариантах осуществления диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR2, кодируют последовательности CDR2 тяжелой цепи (CDR H2). В некоторых вариантах осуществления диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR2, кодируют последовательности CDR2 легкой цепи (CDR L2). В некоторых вариантах осуществления диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR1, кодируют последовательности CDR1 тяжелой цепи (CDR H1). В некоторых вариантах осуществления диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR1, кодируют последовательности CDR1 легкой цепи (CDR L1).
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка включает по меньшей мере часть последовательности гена вариабельного домена тяжелой цепи человека, выбранную из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51, или получена из такой последовательности. В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка включает по меньшей мере часть последовательности гена вариабельного домена легкой цепи каппа человека, или получена из такой последовательности. Например, последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20. В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка включает по меньшей мере часть последовательности гена вариабельного домена легкой цепи лямбда человека, или получена из такой последовательности. Например, последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка содержит смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления описанные в настоящем документе способы также включают следующие стадии: (e) клонирование библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулина, указанные на стадии (d), в векторе экспрессии и (f) трансформация клетки-хозяина вектором экспрессии, указанным на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности вариабельных доменов иммуноглобулина, кодируемых библиотекой. В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный или фаговый вектор. В некоторых вариантах осуществления клетка-хозяин представляет собой E. coli.
Изобретение относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 3 (CDR3), выделенных по отдельности из вариабельных доменов иммуноглобулинов иммунизированного отличного от человека млекопитающего или отличных от человека видов. Изобретение также относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 2 (CDR2), выделенных по отдельности из вариабельных доменов иммуноглобулинов иммунизированного отличного от человека млекопитающего. Изобретение также относится к способам получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий одну из совокупности последовательностей гипервариабельного участка 1 (CDR1), выделенных по отдельности из вариабельных доменов иммуноглобулинов иммунизированного отличного от человека млекопитающего.
В некоторых вариантах осуществления к отличным от человека видам относятся отличные от человека приматы, грызуны, собаки, кошки, овцы, козы, крупный рогатый скот, лошади, члены семейства Camelidae, ламы, верблюды, одногорбые верблюды и свиньи.
В некоторых вариантах осуществления способы включают следующие стадии: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулина человека, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот; (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих последовательности гипервариабельного участка 3 (CDR3), выделенные из иммунизированного отличного от человека млекопитающего, где каждая из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR3 с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); и (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR3 или аминокислотные последовательности, указанные на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR3 и FR4 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR3 или аминокислотную последовательность, которая может выполнять роль участка CDR3, и чтобы была восстановлена полноразмерная последовательность, кодирующая вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b). Указанные стадии также можно проводить с использованием совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих последовательности гипервариабельного участка 2 (CDR2), выделенных из иммунизированного отличного от человека млекопитающего. Указанные стадии также можно проводить с использованием совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих последовательности гипервариабельного участка 1 (CDR1), выделенных из иммунизированного отличного от человека млекопитающего.
В некоторых вариантах осуществления стадию (b) проводят путем амплификации последовательности CDR3 иммунизированного отличного от человека млекопитающего с использованием олигонуклеотидных праймеров, содержащих участок рестрикции типа IIs. В некоторых вариантах осуществления олигонуклеотидный праймер конструируют так, чтобы повысить совместимость последовательности CDR3 млекопитающего и акцепторного каркасного участка, кодирующего вариабельный домен иммуноглобулина человека. В некоторых вариантах осуществления олигонуклеотидный праймер конструируют так, чтобы модифицировать последовательность, граничащую с последовательностью CDR3 млекопитающих с целью обеспечения эффективного лигирования посредством совместимых липких концов в последовательность акцепторного каркасного участка, кодирующую вариабельный домен иммуноглобулина человека. В некоторых вариантах осуществления последовательности ДНК млекопитающих, фланкирующие участки CDR3, могут содержать липкие концы, совместимые с липкими концами расщепленных акцепторных каркасных участков до расщепления ферментами рестрикции типа IIs. В таких случаях олигонуклеотиды, используемые для амплификации, конструируют с целью модификации целевой последовательности млекопитающих так, чтобы после расщепления ферментом рестрикции типа IIs получить совместимые липкие концы и обеспечить эффективное лигирование. Указанные стадии также можно проводить путем амплификации последовательности CDR2 иммунизированных отличных от человека млекопитающих с использованием олигонуклеотидных праймеров, содержащих участок рестрикции типа IIs. Указанные стадии также можно проводить путем амплификации последовательности CDR1 иммунизированных отличных от человека млекопитающих с использованием олигонуклеотидных праймеров, содержащих участок рестрикции типа IIs.
В некоторых вариантах осуществления стадию (b) проводят путем амплификации последовательности CDRH3 отличного от человека млекопитающего с использованием олигонуклеотидных праймеров, содержащих участок рестрикции FokI IIs. Указанные стадии также можно проводить путем амплификации последовательности CDR2 отличного от человека млекопитающего с использованием олигонуклеотидных праймеров, содержащих участок рестрикции FokI IIs. Указанные стадии также можно проводить путем амплификации последовательности CDR1 отличного от человека млекопитающего с использованием олигонуклеотидных праймеров, содержащих участок рестрикции FokI IIs.
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются разными ферментами рестрикции типа IIs. В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs включают участки распознавания BsmBI, участки распознавания BsaI, участки распознавания FokI, или их сочетания.
В некоторых вариантах осуществления диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR3, кодируют последовательности CDR3 тяжелой цепи (CDRH3). В некоторых вариантах осуществления диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR3, кодируют последовательности CDR3 легкой цепи (CDR L3). В некоторых вариантах осуществления диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR2, кодируют последовательности CDR2 тяжелой цепи (CDR H2). В некоторых вариантах осуществления диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR2, кодируют последовательности CDR2 легкой цепи (CDR L2). В некоторых вариантах осуществления диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR1, кодируют последовательности CDR1 тяжелой цепи (CDR H1). В некоторых вариантах осуществления диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR1, кодируют последовательности CDR1 легкой цепи (CDR Ll).
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка включает по меньшей мере часть последовательности гена вариабельного домена тяжелой цепи человека, выбранной из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51, или она может быть получена из такой последовательности.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка включает по меньшей мере часть последовательности гена вариабельного домена легкой цепи каппа человека, или она может быть получена из такой последовательности. Например, последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20. В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка включает по меньшей мере часть последовательности гена вариабельного домена легкой цепи лямбда человека, или она может быть получена из такой последовательности. Например, последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка содержит смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления способы также включают следующие стадии: (e) клонирование библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулина, указанные на стадии (d), в векторе экспрессии и (f) трансформация клетки-хозяина вектором экспрессии, указанным на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности вариабельных доменов иммуноглобулина, кодируемых библиотекой. В некоторых вариантах осуществления клетка-хозяин представляет собой E. coli. В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный или фаговый вектор.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На фигуре 1A приведено схематическое изображение домена белка, в состав которого входят каркасный участок и петлеобразные участки, содержащие остатки, участвующие в осуществлении контакта с другим белком, или с другой молекулой. Изображено несколько случаев: стабильный домен белка и правильно уложенные петлеобразные участки; правильно уложенные петлеобразные участки, которые вставлены в домен с ограниченной собственной стабильностью; домен белка, обладающий собственной стабильностью, на которую влияют петлеобразные участки.
На фигуре 1B приведено схематическое изображение разных типов библиотек репертуаров белков, полученных с использованием разных стратегий диверсификации.
На фигуре 2 приведено схематическое изображение акцепторного каркасного участка вариабельного домена антитела. Указаны каркасные участки, CDR и участок рестрикции типа IIS-RM.
На фигуре 3 приведено схематическое изображение способа улавливания последовательностей CDRH3 из природных репертуаров.
На фигуре 4 приведено схематическое изображение преимущества применения праймеров, содержащих участки рестрикции типа IIS-RM, для амплификации и вставки природных участков CDR в акцепторные каркасные участки.
На фигуре 5 приведено иллюстративное изображение последовательности зародышевого гена вариабельного домена тяжелой и легкой цепи, выбранного для получения акцепторных каркасных участков.
На фигуре 6 приведено схематическое изображение способа амплификации, используемого для получения акцепторных каркасных участков путем добавления зародышевых последовательностей "лишнего" фрагмента и участка FR4.
На фигуре 7, вверху, приведено иллюстративное изображение элементов последовательностей "лишних" фрагментов акцепторного каркасного участка VH. Последовательности ДНК, распознаваемые и расщепляемые ферментом рестрикции BsmBI заключены в красные и черные рамки соответственно и показаны в нижней части фигуры. Рамка считывания, соответствующая последовательности вариабельного домена антитела, подчеркнута.
На фигуре 8 приведено иллюстративное изображение последовательностей 20 акцепторных каркасных участков.
На фигуре 9 приведено схематическое изображение вектора pNDS1 одного или в сочетании с контрольным вариабельным участком тяжелой цепи или контрольным вариабельным участком легкой цепи.
На фигуре 10 изображена таблица, в которой приведены последовательности CDRH3, полученные из кДНК человека в качестве источника и вставленные в акцепторные каркасные участки человека.
На фигуре 11 изображена таблица, в которой приведены конструкции синтетических последовательностей CDR для VH, VK и Vλ. Положения пронумерованы по системе Кабат. Указано теоретическое разнообразие CDR, полученное с использованием определенной стратегии диверсификации кодонов (NNS, DVK, NVT, DVT). Стратегии, используемые для синтеза CDR VH заключены в рамки.
На фигуре 12 приведено схематическое изображение вставки синтетического CDR в акцепторный каркасный участок и элементы последовательности.
На фигуре 13 приведено схематическое изображение первичных библиотек и рекомбинация цепей, проводимая для получения вторичных библиотек.
На фигуре 14 приведено схематическое изображение получения библиотек акцепторного VH, объединенных с синтетическими библиотеками VL, и улавливание репертуаров CDRH3 человеческого или нечеловеческого происхождения.
На фигуре 15 приведено схематическое изображение получения библиотек MnA, MiB и MiC с использованием в качестве источника разнообразия репертуара CDRH3, полученного из наивных мышей или мышей, иммунизированных hIFNγ или hCCL5/RANTES. Размер библиотек указан в верхних панелях. На нижних панелях показано распределение длины CDRH3, присутствующих в указанных библиотеках.
На фигуре 16 изображен ряд графиков, иллюстрирующих титрование емкости фага в процессе селекции против hIFNγ с использованием вторичных библиотек AD1 и AE1.
На фигуре 17 изображен ряд графиков, иллюстрирующих титрование выхода фага в процессе селекции против моноклонального антитела 5E3 с использованием вторичных библиотек AD1 и AE1.
На фигуре 18 изображен ряд графиков, иллюстрирующих встречаемость длин CDRH3, присутствующих в библиотеках AE1 и AD1, после трех циклов селекции против моноклонального антитела 5E3. Указано распределение каждой длины CDRH3 в разных семействах VH. Однако если длина CDRH3 составляет более 16 аминокислот, последовательности размером 70 п.о., предоставляемые платформой Illumina Sequencing, не охватывают каркасную последовательность в степени, достаточной для однозначной идентификации семейства VH1 и, следовательно, семейство VH указывается как неопределенное.
На фигуре 19 изображен ряд графиков, иллюстрирующих кривую доза-ответ, полученную методом ELISA с использованием 6 очищенных препаратов scFv против антитела 5E3 мыши или нерелевантного антитела 1A6 мыши. Семь клонов кодируют разные scFv. Клон A6, кодирующий scFv, специфичный к hIFNγ, используют в качестве отрицательного контроля.
На фигуре 20 изображен график, иллюстрирующий кривую доза-ответ, полученную методом ELISA с использованием очищенных препаратов scFv против hIFNγ по сравнению с используемым в качестве положительного контроля scFv, специфичным к hIFNγ (A6).
На фигуре 21 изображен график, иллюстрирующий ингибиторный эффект очищенных препаратов scFv в анализе, проводимом с использованием репортерного гена люциферазы, регулируемого hIFNγ. Нейтрализующую активность двух scFv-кандидатов (AD1R4P1A9 и AE14R3P2E4) сравнивают с активностью scFv (G9), используемого в качестве положительного контроля, и scFv (D11), используемого в качестве отрицательного контроля.
На фигуре 22 изображен график, иллюстрирующий ингибиторный эффект очищенных препаратов scFv в анализе индукции MHCII в ответ на hIFNγ. Нейтрализующую активность двух scFv-кандидатов (AD1R4P1A9 и AE14R3P2E4) сравнивают с активностью scFv (D11), используемого в качестве отрицательного контроля.
На фигуре 23 изображен ряд графиков, иллюстрирующих ингибиторный эффект двух кандидатов AD1R4P1A9 и AE14R3P2E4, преобразованных в IgG, в анализе, проводимом с использованием репортерного гена люциферазы, регулируемого hIFNγ. Нейтрализующую активность двух IgG сравнивают с активностью нерелевантного IgG в отношении RANTES человека (N1-0701).
На фигуре 24 изображен ряд графиков, иллюстрирующих кривую доза-ответ, полученную методом ELISA с использованием IgG G11 и DA4 против 5E3 мыши, химерного 5E3 крысы и соответствующих изотипов антител мыши и крысы.
На фигуре 25 изображен ряд графиков, иллюстрирующих детекцию 5E3 мыши методом ELISA в сыворотке мыши с разными разведениями, с использованием антиидиотипических IgG G11 и DA4 в качестве улавливающих антител.
На фигуре 26 изображен график, иллюстрирующий отношение фагового выхода/входа (phage output/input) в процессе селекции против hIFNγ с использованием библиотек MnA и MiB.
На фигуре 27 изображен график, иллюстрирующий наилучшие соотношения, полученные при скрининге scFv методом ELISA с использованием клонов, полученных из библиотек MnA, MiB и MiC после каждого цикла селекции против hIFNγ. Пороговым значением считают половину сигнала, полученного с использованием A6 в качестве контрольного scFv.
На фигуре 28 приведен график, изображающий кривую распределения scFv, дающих разные уровни сигнала в экспериментах по связыванию hIFNγ, которые были получены с использованием клонов из библиотек MnA и MiB.
На фигуре 29 приведен график, изображающий кривую доза-ответ, полученную методом ELISA с использованием очищенных препаратов scFv из библиотек MnA и MiB против hIFNγ по сравнению с используемым в качестве положительного контроля scFv, специфичным к hIFNγ (A6).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Преимущество библиотек синтетических белков и, в особенности, библиотек синтетических антител заключается в том, что в процессе получения библиотеки можно выбрать строительные блоки, составляющие указанные синтетические белки, и придать белкам желательные характеристики. Однако такие библиотеки имеют существенный недостаток, заключающийся в том, что рандомизация фрагментов указанных синтетических белков с получением коллекции вариантов часто приводит к образованию нефункциональных белков и, следовательно, может сильно уменьшать размер функциональной библиотеки и ухудшать ее рабочие характеристики. Другим недостатком синтетического разнообразия является то, что из-за практических ограничений трудно получить библиотеку с размером, охватывающим теоретическое разнообразие рандомизированных аминокислотных участков. Даже с использованием систем дисплеев, таких как рибосомальный дисплей, можно получить разнообразие 1013-1014 и выбрать образцы, которые могут максимально охватывать полную рандомизацию участков из 9 аминокислот. А поскольку средний размер природного CDRH3 (также называемого в настоящем документе CDR3 тяжелой цепи или CDR3 VH) больше 9 и может превышать 20 аминокислот в длину, синтетическое разнообразие не является подходящим способом для получения таких CDR.
Сочетание способов, обычно используемых для манипуляций с ДНК и получения библиотеки вариантов белка, привносит ошибки в последовательность ДНК. Указанные ошибки могут вызывать изменение рамки считывания ДНК и утрату способности ДНК кодировать функциональный полипептид. Как правило, библиотеки антител, полученные путем сборки фрагментов ДНК методом ПЦР и/или рестрикционного клонирования, содержат от 15% до 45% последовательностей, которые не находятся в правильной рамке считывания, обеспечивающей трансляцию белка. Указанные нефункциональные члены библиотеки могут снижать эффективность селекции антител и препятствовать процессу идентификации и, следовательно, они считаются недостатком в данной области. Описанные способы, основанные на стратегии альтернативного клонирования, позволяют вводить разнообразие в библиотеку антител с большей устойчивостью к ошибкам. Как правило, встречаемость последовательности в рамке считывания составляет примерно 90%. Другим преимуществом изобретения является объединение выбранных акцепторных каркасных участков вариабельного домена антитела с петлями CDR, которые обеспечивают высокую вероятность правильной укладки. Это позволяет улавливать длинные CDR, плохо охватываемые способами, основанными на синтетической рандомизации. Кроме того, в описанных способах клонирование диверсифицированных последовательностей проводят без модификации участка, кодирующего акцепторный вариабельный домен антитела. Другое преимущество данного способа заключается в том, что один и тот же набор акцепторных каркасных участков антитела может улавливать несколько источников разнообразия. Указанные источники включают, без ограничения: природные CDR антител человека или антител других млекопитающих, CDR антител кур, CDR антитело-подобных молекул, таких как VHH верблюдовых, IgNAR акул, вариабельные петли T-клеточных рецепторов. Кроме того, природные CDR можно получить из наивных или иммунизированных животных. В последнем случае CDR извлекают из последовательностей, участвующих в распознавании антигена, используемого для иммунизации.
Характерной особенностью описанных в настоящем документе способов является эффективное улавливание последовательностей, кодирующих CDR3 тяжелой цепи отличных от человека видов, и вставка указанных CDR3 в каркасные участки иммуноглобулина человека. Следовательно, с использованием указанных способов можно получить разные связывающие участки антитела, образованные уловленным репертуаром CDRH3 из других видов, и обеспечить выбор другого трехмерного пространства. Указанные способы позволяют получать антитела человека с новыми характеристиками и диапазоном классов и эпитопов мишеней, отличающимся от соответствующего диапазона, доступного для репертуара CDRH3 человека. Кроме того, указанные новые антитела содержат каркасный участок человека наряду с участками CDR1 и CDR2 и, следовательно, могут использоваться для лечения людей.
В указанном способе выбранные домены белка, например вариабельные домены антитела, модифицируют путем введения "лишней" последовательности, которая служит участком интеграции диверсифицированных последовательностей. После интеграции "лишний" фрагмент полностью удаляют, в результате чего остается интактный кодирующий участок акцепторного белка и вставленные фрагменты белков (т.е., CDR). Данное событие интеграции опосредуется ферментом рестрикции типа IIs, который распознает конкретный участок последовательности ДНК, но и расщепляет ДНК на некотором расстоянии от этого участка. Описанный способ имеет два основных преимущества: (1) он позволяет расщеплять акцепторный каркасный участок, не оказывая влияния на последовательность, кодирующую этот участок (нет необходимости конструировать молчащие участки рестрикции); и (2) он позволяет расщеплять и клонировать естественно диверсифицированные последовательности, которые по определению не содержат совместимые участки рестрикции.
Как указано выше, предпринятые ранее попытки получить библиотеки и/или дисплеи последовательностей антител отличаются от описанных в настоящем документе способов. Например, некоторые способы требуют прививания каждого CDR, как описано, например, в патенте США № 6300064, в котором участки ферментативной рестрикции вводят на границе каждого CDR, не только на участке CDRH3. В других способах последовательности CDR из природных источников амплифицируют и подвергают реаранжировке, как описано, например, в патенте США № 6989250. В некоторых способах, таких как описанные в публикации патентной заявки США № 20060134098, последовательности, полученные из мыши (или другого млекопитающего), добавляют к каркасному участку человека, так, чтобы полученное антитело содержало участки CDR1 и CDR2 мыши и участок CDR3 человека. С помощью других способов, например, описанных в публикации патентной заявки США № 20030232333, можно получить антитела, которые содержат синтетические участки CDR1 и/или CDR1/CDR2 наряду с природным участком CDR3. Однако эти способы не позволяют получать библиотеки, которые содержат стабильные каркасные участки и участки CDR с правильной укладкой.
Предложенные в настоящем документе способы позволяют конструировать акцепторные каркасные участки антитела для клонирования разнообразия. Разработана стратегия введения разнообразия в CDR3 выбранных доменов антитела человека, которая позволяет избежать модификации последовательности исходного каркасного участка. Стратегия основана на введении участков рестрикции типа IIs вне участка, кодирующего иммуноглобулин. Этот класс ферментов рестрикции распознает асимметричную и непрерывную последовательность из 4-7 пар оснований и расщепляет ДНК на определенном расстоянии от нее, составляющем до 20 оснований, независимо от последовательности ДНК, присутствующей в участке расщепления. Чтобы использовать преимущество данной системы для клонирования диверсифицированных последовательностей в выбранных каркасных участках, конструируют акцепторные каркасные участки, содержащие "лишний" фрагмент ДНК вместо CDR3, который включает два участка рестрикции типа IIs. Подобным образом, получают диверсифицированные последовательности ДНК, фланкирующие последовательности которых содержат участи типа IIs. При условии, что липкие концы, образующиеся в результате действия ферментов рестрикции, являются совместимыми и сохраняется рамка считывания, фрагменты ДНК можно лигировать в акцепторный каркасный участок и воссоздать кодируемые CDR3 в новой среде акцепторного каркасного участка антитела (фигура 2).
Предложенные в настоящем документе способы позволяют улавливать естественное разнообразие CDR. В стратегии, разработанной для улавливания естественно диверсифицированных фрагментов белков в качестве источника разнообразия, также используется преимущество ферментов рестрикции типа IIs. В качестве примера, конструируют олигонуклеотидные праймеры, специфичные к участкам, фланкирующим последовательность ДНК, которая кодирует CDRH3 иммуноглобулинов, т.е., специфичные к FR3 и FR4 вариабельного участка. Указанные олигонуклеотиды содержат на 5'-конце участок распознавания ферментом рестрикции типа IIs, а их 3'-фрагмент совпадает с целевой последовательностью ДНК. Фермент рестрикции, расщепляющий указанный участок, предпочтительно, представляет собой фермент, который расщепляет ДНК на большом расстоянии от участка распознавания, такой как FokI. Такой фермент является ключевым элементом способа и обеспечивает эффективную амплификацию природной последовательности ДНК, поскольку он позволяет вырезать последовательность, кодирующую CDRH3, путем расщепления ДНК на границе между CDR и каркасным участком, и при этом сохранить хорошее совпадение между 3'-концом праймера и ДНК, фланкирующей CDRH3 (фигура 3). Такое точное вырезание последовательности, кодирующей CDR, очень трудно осуществить с помощью ферментов типа II, которые расщепляют ДНК по участку распознавания, поскольку соответствующий участок рестрикции отсутствует в природной последовательности ДНК, а введение таких участков трудно осуществить в процессе амплификации вследствие плохого отжига праймера. Таким образом, указанный способ позволяет осуществлять амплификацию диверсифицированных белковых последовательностей и их вставку в любой акцепторный каркасный участок антитела независимо от происхождения амплифицируемого разнообразия (фигура 4).
Описанные в настоящем документе способы позволяют получить библиотеку нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина, и включают следующие стадии: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулинов, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот; (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих участки гипервариабельного участка 3 (CDR3), или кодирующих аминокислотные последовательности, которые могут выполнять роль участка CDR3, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR3, или аминокислотные последовательности, способные выполнять роль участка CDR3, с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); и (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR3, или аминокислотные последовательности, которые могут выполнять роль участка CDR3, указанные на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR3 и FR4 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR3, или аминокислотную последовательность, которая может выполнять роль участка CDR3, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b).
Настоящее изобретение относится к способам получения библиотеки нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина, который включает следующие стадии: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулинов, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот, участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены гипервариабельным участком 3 (CDR3); (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих участки гипервариабельного участка 1 (CDR1), или аминокислотные последовательности, которые могут выполнять роль участка CDR1, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR1, или аминокислотные последовательности, способные выполнять роль участка CDR1, с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); и (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR1, или аминокислотные последовательности, которые могут выполнять роль участка CDR1, указанные на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR1 и FR2 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR1, или аминокислотную последовательность, которая может выполнять роль участка CDR1, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b).
Описанные в настоящем документе способы позволяют получить библиотеку нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина, и включают: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулинов, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот, а участки FR3 и FR4 разделены гипервариабельным участком 3 (CDR3); (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих участки гипервариабельного участка 2 (CDR2), или аминокислотные последовательности, которые могут выполнять роль участка CDR2, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR2 или аминокислотные последовательности, способные выполнять роль участка CDR2, с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); и (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR2 или аминокислотные последовательности, которые могут выполнять роль участка CDR2, указанных на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR2 и FR3 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR2 или аминокислотные последовательности, которые могут выполнять роль участка CDR2, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b).
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b) описанных выше способов, распознаются разными ферментами рестрикции типа IIs. Например, в некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs представляют собой участки распознавания BsmBI, участки распознавания BsaI, участки распознавания FokI, или их сочетания.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка получают из последовательности гена человека. Например, в некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена тяжелой цепи человека или последовательность, полученную из последовательности гена вариабельного домена тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена вариабельного домена тяжелой цепи человека выбрана из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51.
В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи каппа человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи каппа человека. Например, в некоторых вариантах осуществления последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20.
В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи лямбда человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи лямбда человека. Например, в некоторых вариантах осуществления последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR3, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих, и других естественно диверсифицированных полипептидных коллекций, или она может быть получена из таких последовательностей.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR3 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанная совокупность получена из таких последовательностей.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR3, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR1, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих, и других естественно диверсифицированных полипептидных коллекций, или она может быть получена из таких последовательностей.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR1 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанная совокупность получена из таких последовательностей.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR1, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR2, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих, и других естественно диверсифицированных полипептидных коллекций, или она может быть получена из таких последовательностей.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR2 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанная совокупность получена из таких последовательностей.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR2, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка содержит смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления описанные в настоящем документе способы дополнительно включают следующие стадии: (e) клонирование библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулина, указанные на стадии (d), в векторе экспрессии и (f) трансформация клетки-хозяина вектором экспрессии, указанном на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности кодируемых библиотекой вариабельных доменов иммуноглобулина.
В некоторых вариантах осуществления клетка-хозяин представляет собой E. coli. В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный вектор.
Описанные в настоящем документе способы позволяют генерировать или иным образом получать специфичные к мишени антитела, вариабельные участки антител или их фрагменты и включают следующие стадии: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулинов, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот; (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих участки гипервариабельного участка 3 (CDR3) или аминокислотные последовательности, которые могут выполнять роль участка CDR3, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR3 или аминокислотные последовательности, способные выполнять роль участка CDR3, с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR3 или аминокислотные последовательности, которые могут выполнять роль участка CDR3, указанные на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR3 и FR4 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR3 или аминокислотные последовательности, которые могут выполнять роль участка CDR3, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b); (e) клонирование библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулина, указанные на стадии (d), в векторе экспрессии; (f) трансформация клетки-хозяина вектором экспрессии, указанным на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности кодируемых библиотекой вариабельных доменов иммуноглобулина; (g) приведение в контакт совокупности доменов иммуноглобулина, указанных на стадии (f), с антигеном-мишенью; и (h) определение, какой из экспрессированных вариабельных доменов иммуноглобулина, кодируемых последовательностью, связывается с антигеном-мишенью.
Описанные в настоящем документе способы позволяют генерировать или иным образом получать специфичные к мишени антитела, вариабельные участки антител или их фрагменты и включают следующие стадии: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулинов, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот, участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены гипервариабельным участком 3 (CDR3); (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих участки гипервариабельного участка 1 (CDR1) или аминокислотные последовательности, которые могут выполнять роль участка CDR1, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR1 или аминокислотные последовательности, способные выполнять роль участка CDR1, с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR1 или аминокислотные последовательности, которые могут выполнять роль участка CDR1, указанные на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR1 и FR2 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR1 или аминокислотные последовательности, которые могут выполнять роль участка CDR1, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b); (e) клонирование библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулина, указанные на стадии (d), в векторе экспрессии; (f) трансформация клетки-хозяина вектором экспрессии, указанным на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности кодируемых библиотекой вариабельных доменов иммуноглобулина; (g) приведение в контакт совокупности доменов иммуноглобулина, указанных на стадии (f), с антигеном-мишенью; и (g) определение, какой из экспрессированных вариабельных доменов иммуноглобулина, кодируемых последовательностью, связывается с антигеном-мишенью.
Описанные в настоящем документе способы позволяют генерировать или иным образом получать специфичные к мишени антитела, вариабельные участки антител или их фрагменты и включают следующие стадии: (a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулинов, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены "лишней" последовательностью нуклеиновых кислот, содержащей по меньшей мере два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот, а участки FR3 и FR4 разделены гипервариабельным участком 3 (CDR3); (b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих участки гипервариабельного участка 2 (CDR2) или аминокислотные последовательности, которые могут выполнять роль участка CDR2, где каждая последовательность из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs; (c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR2 или аминокислотные последовательности, способные выполнять роль участка CDR2, с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (a), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs, который связывается с участком распознавания ферментом рестрикции типа IIs, указанным на стадии (a); (d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR2 или аминокислотные последовательности, которые могут выполнять роль участка CDR2, указанные на стадии (c), в расщепленный акцепторный каркасный участок, указанный на стадии (c), так, чтобы участки FR2 и FR3 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR2 или аминокислотные последовательности, которые могут выполнять роль участка CDR2, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs, указанные на стадиях (a) и (b); (e) клонирование библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулина, указанные на стадии (d), в векторе экспрессии; (f) трансформация клетки-хозяина вектором экспрессии, указанным на стадии (e), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности кодируемых библиотекой вариабельных доменов иммуноглобулина; (g) приведение в контакт совокупности вариабельных доменов иммуноглобулина, указанных на стадии (f), с антигеном-мишенью; и (h) определение, какой из экспрессированных вариабельных доменов иммуноглобулина, кодируемых последовательностью, связывается с антигеном-мишенью.
В некоторых вариантах осуществления описанные в настоящем документе способы дополнительно включают стадию (i) секвенирования последовательности, кодирующей вариабельный домен иммуноглобулина, который связывается с антигеном-мишенью.
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs, указанные на стадии (a) и на стадии (b), распознаются разными ферментами рестрикции типа IIs.
В некоторых вариантах осуществления участки распознавания ферментами рестрикции типа IIs представляют собой участки распознавания BsmBI, участки распознавания BsaI, участки распознавания FokI, или их сочетания.
В некоторых вариантах осуществления последовательность нуклеиновых кислот акцепторного каркасного участка получают из последовательности гена человека. Например, в некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена тяжелой цепи человека или последовательность, полученную из последовательности гена вариабельного домена тяжелой цепи человека. Например, в некоторых вариантах осуществления последовательность гена вариабельного домена тяжелой цепи человека выбрана из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51.
В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи каппа человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи каппа человека. Например, в некоторых вариантах осуществления последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20.
В некоторых вариантах осуществления последовательность человека представляет собой последовательность гена вариабельного домена легкой цепи лямбда человека или последовательность, полученную из последовательности гена вариабельного домена легкой цепи лямбда человека. Например, в некоторых вариантах осуществления последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR3, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих, и других естественно диверсифицированных полипептидных коллекций, или она может быть получена из таких последовательностей.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR3 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанная совокупность получена из таких последовательностей.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR3, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR1, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих, и других естественно диверсифицированных полипептидных коллекций, или она может быть получена из таких последовательностей.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR1 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанная совокупность получена из таких последовательностей.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует аминокислотные последовательности, которые могут выполнять роль участка CDR1, и включает синтетические последовательности.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот включает последовательности, выбранные из природных последовательностей CDR2, природных последовательностей Ig человека, природных последовательностей Ig млекопитающих, природных последовательностей петлеобразного участка T-клеточного рецептора млекопитающих, и других естественно диверсифицированных полипептидных коллекций, или она может быть получена из таких последовательностей.
В некоторых вариантах осуществления совокупность диверсифицированных нуклеиновых кислот кодирует участки CDR2 и включает последовательности природных иммуноглобулинов людей, подвергшихся воздействию конкретного иммуногена, или последовательности, полученные у животных, которые были идентифицированы как животные, подвергшиеся воздействию конкретного антигена, или указанная совокупность получена из таких последовательностей.
В некоторых вариантах осуществления совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка содержит смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
В некоторых вариантах осуществления вектор экспрессии представляет собой фагмидный вектор. В некоторых вариантах осуществления клетка-хозяин представляет собой E. coli.
Если не указано иначе, научные и технические термины, используемые в применении к настоящему изобретению, имеют традиционные значения, хорошо известные рядовым специалистам в данной области. Кроме того, если контекст не указывает иначе, термины, используемые в единственном числе, включают множественное число, а термины, используемые во множественном числе, включают единственное число. Как правило, в настоящем изобретении используют хорошо известные и традиционно используемые в данной области номенклатуру и методы культивирования клеток и тканей, молекулярной биологии, белковой и олиго- или полинуклеотидной химии и гибридизации. Для получения рекомбинантных ДНК, олигонуклеотидного синтеза, культивирования и трансформации тканей используют стандартные методы (такие как электропорация, липофекция). Ферментные реакции и очистку проводят в соответствии с инструкциями производителя, или с помощью традиционно используемых в данной области или описанных в настоящем документе способов. Описанные выше методы и процедуры обычно проводят традиционными способами, хорошо известными в данной области и описанными в разных общих и более специальных справочных руководствах, которые цитируются и обсуждаются на протяжении настоящего описания. См., например, Sambrook et al. Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y. (1989)). В настоящем изобретении используют хорошо известные и традиционно используемые в данной области номенклатуру, лабораторные процедуры и методы аналитической химии, синтетической органической химии и медицинской и фармацевтической химии. Используют также стандартные методы химического синтеза, химического анализа, получения, составления и доставки фармацевтических средств, и лечения пациентов.
Если не указано иначе, используемые в настоящем описании термины имеют следующие значения:
В настоящем описании термин "антитело" относится к молекулам иммуноглобулинов и иммунологически активным фрагментам молекул иммуноглобулинов (Ig), т.е., к молекулам, которые содержат антиген-связывающий участок, способный к специфическому связыванию (иммунному взаимодействию) с антигеном. Фраза "специфическое связывание", или "иммунное взаимодействие", или "иммунноспецифическое связывание" означает, что антитело взаимодействует с одним или несколькими антигенными детерминантами целевого антигена и не взаимодействует с другими полипептидами, или взаимодействует с ними с гораздо меньшей аффинностью (Kd 10-6). Антитела включают, без ограничения, поликлональные антитела, моноклональные антитела, химерные антитела, dAb (доменные антитела), одноцепочечные антитела, фрагменты Fab, Fab' и F(ab')2, scFvs и экспрессионные библиотеки Fab.
Известно, что основной структурный элемент антитела включает тетрамер. Каждый тетрамер состоит из двух идентичных пар полипептидных цепей, причем каждая пара содержит одну "легкую" (примерно 25 кДа) и одну "тяжелую" цепь (примерно 50-70 кДа). Амино-концевой фрагмент каждой цепи содержит вариабельный участок размером примерно от 100 до 110 или более аминокислот, который в первую очередь отвечает за распознавание антигена. Карбокси-концевой участок каждой цепи включает константный участок, который в основном отвечает за эффекторную функцию. Как правило, молекулы антител человека подразделяют на классы IgG, IgM, IgA, IgE и IgD, которые отличаются друг от друга природой тяжелой цепи, входящей в состав молекулы. Некоторые классы также подразделяют на подклассы, такие как IgG1, IgG2 и другие. Кроме того, легкая цепь человека может представлять собой цепь каппа или цепь лямбда.
Термин "моноклональное антитело" (MAb), или "композиция моноклонального антитела", в настоящем описании относится к популяции молекул антител, которая включает только одну разновидность молекулы антитела, содержащую единственный продукт гена легкой цепи и единственный продукт гена тяжелой цепи. В частности, в популяции моноклональных антител все молекулы имеют одинаковые гипервариабельные участки (CDR). MAb содержит антиген-связывающий участок, способный к иммунному взаимодействию с конкретным эпитопом антигена и характеризующийся уникальной аффинностью связывания к нему.
Термин "антиген-связывающий участок" или "связывающий фрагмент" относится к фрагменту молекулы иммуноглобулина, который участвует в связывании антигена. В образовании антиген-связывающего участка участвуют аминокислотные остатки N-концевых вариабельных ("V") участков тяжелой ("H") и легкой ("L") цепей. Три высоко дивергентные последовательности, входящие в состав V-участков тяжелой и легкой цепей, называемые "гипервариабельные участки", располагаются между более консервативными фланкирующими последовательностями, известными как "каркасные участки" или "FR". Таким образом, термин "FR" относится к аминокислотным последовательностям, которые в природе находятся между гипервариабельными участками иммуноглобулинов и вплотную примыкают к ним. В молекуле антитела три гипервариабельных участка легкой цепи и три гипервариабельных участка тяжелой цепи, расположенные определенным образом относительно друг друга в трехмерном пространстве, образуют антиген-связывающую поверхность. Антиген-связывающая поверхность комплементарна трехмерной поверхности связываемого антигена и три гипервариабельных участка каждой из тяжелых и легких цепей называют "участки, определяющие комплементарность", или "CDR". Отнесение аминокислот к конкретным доменам осуществляют в соответствии с принципами, описанными в Kabat Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md. (1987 and 1991)), или Chothia & Lesk J. Mol. Biol. 196:901-917 (1987), Chothia et al. Nature 342:878-883 (1989).
В настоящем описании термин "эпитоп" включает любую белковую детерминанту, способную специфически связываться с иммуноглобулином, scFv или T-клеточным рецептором. Термин "эпитоп" включает любую белковую детерминанту, способнцую специфически связываться с иммуноглобулином или T-клеточным рецептором. Эпитопные детерминанты обычно состоят из совокупности химически активных поверхностных групп молекул, таких как аминокислоты или боковые цепи сахаров и, как правило, имеют специфические характеристики трехмерной структуры и распределения зарядов. Например, можно получить антитела против N-концевых или C-концевых участков полипептида. Считается, что антитело специфически связывается с антигеном, если константа диссоциации ≤1 мкМ; например, ≤100 нМ, предпочтительно, ≤10 нМ, более предпочтительно, ≤1 нМ.
В настоящем описании термины "иммунологическое связывание" и "характеристики иммунологического связывания" относятся к нековалентным взаимодействиям, которые наблюдаются между антигеном и специфичной к нему молекулой иммуноглобулина. Силу или аффинность взаимодействий, опосредующих иммунологическое связывание, можно выразить в виде константы диссоциации (Kd) взаимодействия, причем более низкое значение Kd соответствует более высокой аффинности. Характеристики иммунологического связывания выбранных полипептидов можно количественно определить с помощью хорошо известных в данной области способов. Один из таких способов включает измерение скоростей образования и диссоциации комплекса антиген-связывающий участок/антиген, где указанные скорости зависят от концентраций партнеров, участвующих в образовании комплекса, аффинность взаимодействия и геометрических параметров, которые равным образом влияют на скорость в обоих направлениях. Таким образом, и "константу скорости прямой реакции" (Kon) и "константу скорости обратной реакции" (Koff) можно определить путем расчета концентраций и фактических скоростей ассоциации и диссоциации. (см. Nature 361:186-87 (1993)). Отношение Koff/Kon, позволяющее аннулировать все параметры, не связанные с аффинностью, равно константе диссоциации Kd. (общую информацию можно найти в Davies et al. (1990) Annual Rev Biochem 59:439-473). Считается, что антитело по настоящему изобретению специфически связывается с мишенью, если равновесная константа связывания (Kd), измеренная с помощью таких анализов, как анализ радиолигандного связывания, или подобные анализы, известные специалистам в данной области, составляет ≤1 мкМ, например, ≤100 нМ, предпочтительно, ≤10 нМ, более предпочтительно, ≤1 нМ.
Термин "выделенный полинуклеотид" в данном описании относится к полинуклеотиду, имеющему геномное, кДНК, синтетическое или смешанное происхождение, который, вследствие его происхождения, (1) не связан с полноразмерным полинуклеотидом, или с частью полинуклеотида, в котором "выделенный полинуклеотид" присутствует в природе, (2) функционально связан с полинуклеотидом, с которым он не связан в природе, или (3) не встречается в природе как часть более крупной последовательности. В соответствии с настоящим изобретением полинуклеотиды включают описанные в настоящем документе молекулы нуклеиновых кислот, кодирующие молекулы тяжелой цепи иммуноглобулина, и молекулы нуклеиновых кислот, кодирующие молекулы легкой цепи иммуноглобулина.
Термин "выделенный белок" в настоящем описании относится к белку, кодируемому кДНК, рекомбинантной РНК, или полинуклеотидом синтетического происхождения, или полинуклеотидом смешанного происхождения, который, вследствие его происхождения или источника дериватизации, (1) не связан с белками, обнаруженными в природе, (2) не содержит других белков из того же источника, например, не содержит морских белков, (3) экспрессируется в клетках других видов или (4) не встречается в природе.
Термин "полипептид" используют в настоящем описании как общий термин, обозначающий нативный белок, фрагменты или аналоги полипептидной последовательности. Следовательно, фрагменты и аналоги нативного белка являются разновидностями класса полипептидов. В соответствии с настоящим изобретением полипептиды включают описанные в настоящем документе молекулы тяжелой цепи иммуноглобулина и молекулы легкой цепи иммуноглобулина, молекулы антител, образованные путем сочетания молекул тяжелой цепи иммуноглобулина с молекулами легкой цепи иммуноглобулина, такими как молекулы легкой цепи каппа иммуноглобулина, и наоборот, а также их фрагменты и аналоги.
Термин "природный" в применении к объекту в данном описании означает, что объект можно обнаружить в природе. Например, полипептидная или полинуклеотидная последовательность, которая присутствует в организме (включающем вирусы), которую можно выделить из природного источника и которая не была специально модифицирована человеком в лаборатории или иным образом, является природной.
Термин "функционально связанный" в настоящем описании относится к такому расположению указанных компонентов, которое обеспечивает их взаимосвязь и предполагаемое функционирование. Контролирующую последовательность, "функционально связанную" с кодирующей последовательностью, лигируют так, чтобы обеспечить экспрессию кодирующей последовательности в условиях, соответствующих контролирующей последовательности.
Термин "контролирующая последовательность" в данном описании относится к полинуклеотидной последовательности, необходимой для обеспечения экспрессии и процессинга кодирующей последовательности, с которой она лигирована. Природа такой контролирующей последовательности варьирует в зависимости от организма хозяина, у прокариотов такая контролирующая последовательность обычно включает промотор, участок связывания рибосомы и терминатор транскрипции, у эукариотов такая контролирующая последовательность обычно включает промотор и терминатор транскрипции. Подразумевается, что термин "контролирующая последовательность" включает, как минимум, все компоненты, необходимые для обеспечения экспрессии и процессинга, и, кроме того, он может включать другие компоненты, присутствие которых может оказывать благоприятный эффект, такие как лидерные последовательности и последовательности партнеров по слиянию. Термин "полинуклеотид" в настоящем описании используется для обозначения полимера из нуклеотидов, длина которого составляет по меньшей мере 10 оснований, включающих рибонуклеотиды или дезоксирибонуклеотиды, или модифицированные формы любых типов нуклеотидов. Данный термин включает одноцепочечные или двухцепочечные формы ДНК.
В настоящем описании двадцать незаменимых аминокислот и их аббревиатуры соответствуют традиционному применению. См. Immunology - A Synthesis (2nd Edition, E. S. Golub and D.R. Gren, Eds., Sinauer Associates, Sunderland Mass. (1991)). Полипептиды настоящего изобретения также могут содержать стереоизомеры (например, D-аминокислоты) двадцати традиционных аминокислот, неприродные аминокислоты, такие как α-,α-дизамещенные аминокислоты, N-алкил-аминокислоты, молочную кислоту и другие нетрадиционные аминокислоты. Примеры нетрадиционных аминокислот включают: 4-гидроксипролин, γ-карбоксиглутамат, ε-N,N,N-триметиллизин, ε-N-ацетиллизин, O-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, σ-N-метиларгинин и другие подобные аминокислоты и иминокислоты (например, 4-гидроксипролин). В соответствии со стандартным применением и соглашением при изображении полипептида слева указывают амино-конец, а справа - карбокси-конец.
В применении к полипептидам термин "практически идентичный" означает, что две пептидные последовательности при оптимальном выравнивании, например, с помощью программ GAP или BESTFIT, в которых используют вес гэпа по умолчанию, имеют по меньшей мере 80 процентную идентичность последовательностей, предпочтительно, по меньшей мере 90 процентную идентичность последовательностей, более предпочтительно, по меньшей мере 95 процентную идентичность последовательностей, и наиболее предпочтительно, по меньшей мере 99 процентную идентичность последовательностей.
Предпочтительно, положения, в которых остатки не являются идентичными, отличаются консервативными аминокислотными заменами.
Термин "консервативные замены аминокислот" относится к взаимозаменяемости остатков, содержащих подобные боковые цепи. Например, группа аминокислот, содержащих алифатические боковые цепи включает глицин, аланин, валин, лейцин и изолейцин; группа аминокислот, содержащих алифатические гидроксильные боковые цепи включает серин и треонин; группа аминокислот, содержащих амидные боковые цепи, включает аспарагин и глутамин; группа аминокислот, содержащих ароматические боковые цепи, включает фенилаланин, тирозин и триптофан; группа аминокислот, содержащих основные боковые цепи, включает лизин, аргинин и гистидин; и группа аминокислот, имеющих серосодержащие боковые цепи, включает цистеин и метионин. Предпочтительные консервативные замены аминокислот осуществляются в пределах следующих групп: валин-лейцин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин, глутаминовая кислота-аспарагиновая кислота и аспарагин-глутамин.
Как описано в данном документе, предполагается, что настоящее изобретение охватывает минорные вариации аминокислотных последовательностей антител или молекул иммуноглобулина, при условии, что вариации аминокислотной последовательности поддерживают по меньшей мере 75%, более предпочтительно, по меньшей мере 80%, 90%, 95% и, наиболее предпочтительно, 99%. В частности, используют консервативные замены аминокислот. Консервативные замены представляют собой замены в пределах семейства аминокислот, содержащих подобные боковые цепи. Генетически кодируемые аминокислоты обычно подразделяют на следующие семейства: (1) кислые аминокислоты, такие как аспартат, глутамат; (2) основные аминокислоты, такие как лизин, аргинин, гистидин; (3) неполярные аминокислоты, такие как аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан, и (4) незаряженные полярные аминокислоты, такие как глицин, аспарагин, глутамин, цистеин, серин, треонин, тирозин. Гидрофильные аминокислоты включают аргинин, аспарагин, аспартат, глутамин, глутамат, гистидин, лизин, серин и треонин. Гиброфобные аминокислоты включают аланин, цистеин, изолейцин, лейцин, метионин, фенилаланин, пролин, триптофан, тирозин и валин. Другие семейства аминокислот включают (i) серин и треонин, которые представляют собой алифатическое гидроксильное семейство; (ii) аспарагин и глутамин, которые представляют собой амид-содержащее семейство; (iii) аланин, валин, лейцин и изолейцин, которые представляют собой алифатическое семейство; и (iv) фенилаланин, триптофан и тирозин, которые представляют собой ароматическое семейство. Например, можно ожидать, что отдельная замена лейцина на изолейцин или валин, аспартата на глутамат, треонина на серин, или подобная замена аминокислоты на структурно родственную аминокислоту не будет оказывать большое влияние на связывание или свойства полученной молекулы, особенно, если замена не затрагивает аминокислоту, находящуюся в рамке считывания. Приводит ли замена аминокислоты к получению функционального пептида, можно легко определить с помощью анализа удельной активности полипептидного производного. Анализы подробно описаны в настоящем документе. Специалисты в данной области могут легко получить фрагменты или аналоги антител или молекул иммуноглобулинов. Предпочтительные амино- и карбокси-концы фрагментов или аналогов находятся вблизи границ функциональных доменов. Структурные и функциональные домены можно идентифицировать путем сравнения информации по последовательности нуклеиновых кислот и/или аминокислотной последовательности с общедоступной или частной базой данных. Предпочтительно, для идентификации мотивов последовательности или предсказания конформационных доменов белка, которые встречаются в других белках с известной структурой и/или функцией, используют компьютерные способы сравнения. Способы идентификации последовательности белка, имеющего известную трехмерную структуру, известны. Bowie et al. Science 253:164 (1991). Таким образом, вышеприведенные примеры демонстрируют, что специалисты в данной области могут определить мотивы и структурные конформации последовательности, которые можно использовать для идентификации структурных и функциональных доменов в соответствии с настоящим изобретением.
Предпочтительными являются аминокислотные замены, которые: (1) уменьшают чувствительность к протеолизу, (2) уменьшают чувствительность к окислению, (3) изменяют аффинность связывания при образовании белковых комплексов, (4) изменяют аффинность связывания и (4) придают таким аналогам другие физико-химические или функциональные свойства или модифицируют другие физико-химические или функциональные свойства таких аналогов. Аналоги могут включать разные последовательности-мутеины, отличные от природной пептидной последовательности. Например, в природную последовательность (предпочтительно, в участок полипептида, находящийся вне домена (доменов), участвующего в образовании межмолекулярных контактов) можно ввести одну или несколько аминокислотных замен (предпочтительно, консервативных аминокислотных замен). Консервативная аминокислотная замена не вызывает существенного изменения структурных характеристик исходной последовательности (например, замена аминокислоты не нарушает спираль, образованную исходной последовательностью, или не искажает другие типы вторичной структуры, характерные для исходной последовательности). Примеры известных в данной области вторичных и третичных структур полипептидов описаны в Proteins, Structures and Molecular Principles (Creighton, Ed., W. H. Freeman and Company, New York (1984)); Introduction to Protein Structure (C. Branden and J. Tooze, eds., Garland Publishing, New York, N.Y. (1991)); и Thornton et at. Nature 354:105 (1991).
В настоящем описании термины "метка" или "меченный" относятся к включению в состав полипептида детектируемого маркера, например, путем введения меченной радиоактивным изотопом аминокислоты или присоединения к полипептиду биотинилированных фрагментов, которые можно детектировать с помощью меченного авидина (такого как стрептавидин, содержащий флуоресцентный маркер или маркер, обладающий ферментативной активностью, которую можно детектировать оптическими или калориметрическими методами). В некоторых случаях метка или маркер также могут быть терапевтическими. Для мечения полипептидов и гликопротеинов можно использовать разные методы, известные в данной области. Примеры меток, используемых для мечения полипептидов включают, без ограничения: радиоизотопы или радионуклиды (такие как 3H, 14C, 15N, 35S, 90Y, 99Tc, 111In, 125I, 131I), флуоресцентные метки (такие как FITC, родамин, лантанидные люминофоры), ферментные метки (такие как пероксидаза хрена, β-галактозидаза, люцифераза, щелочная фосфатаза), хемилюминесцентные, биотинилированные группы, заранее идентифицированные полипептидные эпитопы, распознаваемые вторичным репортером (таким как последовательность, образующая пару лейциновой молнии, участки связывания вторичных антител, метал-связывающие домены, эпитопные маркеры). В некоторых вариантах осуществления метки присоединяют через спейсерные группы разной длины, чтобы уменьшить возможное стерическое затруднение. Термин "фармацевтический агент или фармацевтическое средство" в данном описании относится к химическим соединениям или композициям, способным вызывать желательный терапевтический эффект при соответствующем введении пациенту.
Другие химические термины используются в настоящем описании в соответствии с традиционной практикой, принятой в данной области, как описано, например, в The McGraw-Hill Dictionary of Chemical Terms (Parker, S., Ed., McGraw-Hill, San Francisco (1985)).
В настоящем описании термин "практически чистый" означает, что частицы объекта преобладают среди присутствующих частиц (т.е., в молярном отношении объект присутствует в более обильном количестве, чем любые другие отдельные частицы, входящие в состав композиции), предпочтительно, практически чистая фракция представляет собой композицию, где частицы объекта составляют по меньшей мере примерно 50 процентов (в молярном отношении) от всех присутствующих макромолекулярных частиц.
Как правило, практически чистая композиция составляет более чем примерно 80 процентов от всех макромолекулярных частиц, присутствующих в композиции, более предпочтительно, более чем примерно 85%, 90%, 95% и 99%. Наиболее предпочтительно, частицы объекта очищены практически до однородного состояния (частицы примесей в композиции невозможно детектировать традиционными способами), при котором композиция содержит практически один вид макромолекулярных частиц.
Термин пациент включает человека и животных.
Антитела очищают хорошо известными методами, такими как аффинная хроматография с использованием белка A или белка G, которая позволяет получить, в основном, фракцию IgG иммунной сыворотки. Затем, или альтернативно, специфический антиген, который является мишенью целевого иммуноглобулина, или его эпитоп, можно иммобилизовать на колонке с целью очистки иммунноспецифического антитела методом иммуноаффинной хроматографии. Очистка иммуноглобулинов описана, например, в D. Wilkinson (The Scientist, published by The Scientist, Inc., Philadelphia PA, Vol. 14, No. 8 (April 17, 2000), pp. 25-28).
Настоящее изобретение описывается нижеследующими примерами, которые не ограничивают объем изобретения, указанный в формуле изобретения.
ПРИМЕРЫ
ПРИМЕР 1: Клонирование зародышевых генов вариабельных доменов иммуноглобулинов
Для конструирования библиотек выбирают семь зародышевых генов вариабельных доменов тяжелой цепи человека (VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23, VH5-51), пять зародышевых генов вариабельных доменов легкой цепи каппа человека (VK1-33, VK1-39, VK3-11, VK3-15, VK3-20) и два зародышевых гена вариабельных доменов легкой цепи лямбда человека (VL1-44, VL1-51) (Lefranc, M.-P. et al., 1999 Nucleic Acids Research, 27, 209-212). Выбор указанных генов обусловлен тем, что они часто используются у людей для экспрессии репертуаров антител и кодируют каркасные участки, которые обладают подходящими стабильностью и профилем экспрессии в виде отдельных доменов и в составе пары VH/VL (Ewert S et al., J Mol Biol. 2003 Jan 17;325(3):531-53). Указанные гены амплифицируют методом гнездовой ПЦР с использованием двух наборов специфических праймеров и геномной ДНК человека. Данный способ необходим, поскольку 5'-последовательности зародышевых генов одного семейства являются идентичными или очень похожими. Для каждого гена конструируют первую пару праймеров, называемых геномные локаторы, которые являются специфичными к 5'- и 3'-нетранслируемым участкам, фланкирующим зародышевый ген. Затем конструируют вторую пару праймеров, специфичных к началу каркасного участка 1 (FR1) и к концу FR2. 14 независимых продуктов ПЦР клонируют в векторе pGEMT-easy (Promega, Madison WI) и секвенированием подтверждают их природу и целостность. Аминокислотные последовательности продуктов выбранных зародышевых генов показаны на фигуре 5.
Ниже приведены используемые праймеры и сочетания праймеров.
Геномные локаторы
5 K1-33 TGTTTCTAATCGCAGGTGCCAGATG (SEQ ID NO:120)
3 K1-33 ATTTATGTTATGACTTGTTACACTG (SEQ ID NO:121)
5 K1-39 TATTTGTTTTTATGTTTCCAATCTC (SEQ ID NO:122)
3 K1-39 CCTTGGAGGTTTATGTTATGACTTG (SEQ ID NO:123)
5 K3-11 TTATTTCCAATTTCAGATACCACCG (SEQ ID NO:124)
3 K3-11 TTGTTGGGGTTTTTGTTTCATGTGG (SEQ ID NO:125)
5 K3-15 TATTTCCAATTTCAGATACCACTGG (SEQ ID NO:126)
3 K3-15 ATGTTGAATCACTGTGGGAGGCCAG (SEQ ID NO:127)
5 K3-20 TTATTTCCAATCTCAGATACCACCG (SEQ ID NO:128)
3 K3-20 TTTTGTTTCAAGCTGAATCACTGTG (SEQ ID NO:129)
5 L1-44 ATGTCTGTGTCTCTCTCACTTCCAG (SEQ ID NO:130)
3 L1-44 TTCCCCATTGGCCTGGAGCACTGTG (SEQ ID NO:131)
5 L1-51 GTGTCTGTGTCTCTCCTGCTTCCAG (SEQ ID NO:132)
3 L1-51 CTTGTCTCAGTTCCCCATTGGGCTG (SEQ ID NO:133)
5 H1-2 ATCTCATCCACTTCTGTGTTCTCTC (SEQ ID NO:134)
3 H1-2 TTGGGTTTCTGACACCCTCAGGATG (SEQ ID NO:135)
5 H1-18 CAGGCCAGTCATGTGAGACTTCACC (SEQ ID NO:136)
3 H1-18 CTGCCTCCTCCCTGGGGTTTCTGAA (SEQ ID NO:137)
5 H1-69 CCCCTGTGTCCTCTCCACAGGTGTC (SEQ ID NO:138)
3 H1-69 CCGGCACAGCTGCCTTCTCCCTCAG (SEQ ID NO:139)
5 DP-47 GAGGTGCAGCTGTTGGAG (SEQ ID NO:140)
5 H3-23 TCTGACCAGGGTTTCTTTTTGTTTGC (SEQ ID NO:141)
3 H3-23 TTGTGTCTGGGCTCACAATGACTTC (SEQ ID NO:142)
5 H3-30 TGGCATTTTCTGATAACGGTGTCC (SEQ ID NO:143)
3 H3-30 CTGCAGGGAGGTTTGTGTCTGGGCG (SEQ ID NO:144)
5 H3-48 ATATGTGTGGCAGTTTCTGACCTTG (SEQ ID NO:145)
3 H3-48 GGTTTGTGTCTGGTGTCACACTGAC (SEQ ID NO:146)
5 H5-a GAGTCTGTGCCGGAAGTGCAGCTGG (SEQ ID NO:147)
Праймеры, специфичные к кодирующей последовательности
5 VH1 TATCAGGTGCAGCTGGTGCAG (SEQ ID NO:148)
5 VH3 TATCAGGTGCAGCTGGTGGAG (SEQ ID NO:149)
5 VH5 TATGAGGTGCAGCTGGTGCAG (SEQ ID NO: 150)
3 VH1/3 ATATCTCTCGCACAGTAATACAC (SEQ ID NO:151)
3 VH3 ATATCTCTCGCACAGTAATATAC (SEQ ID NO:152)
3 VH5 ATATGTCTCGCACAGTAATACAT (SEQ ID NO:153)
5 VK1 TATGACATCCAGATGACCCAGTCTCCATCCTC (SEQ ID NO:154)
3 DPK9 ATAGGAGGGGTACTGTAACT (SEQ ID NO:155)
3 DPK1 ATAGGAGGGAGATTATCATA (SEQ ID NO:156)
5 DPK22_L6 TATGAAATTGTGTTGACGCAGTCT (SEQ ID NO:157)
3 DPK22 ATAGGAGGTGAGCTACCATACTG (SEQ ID NO:158)
5 DPK21 TATGAAATAGTGATGACGCAGTCT (SEQ ID NO:159)
3 DPK21 ATAGGAGGCCAGTTATTATACTG (SEQ ID NO:160)
3 L6 CAGCGTAGCAACTGGCCTCCTAT (SEQ ID NO:161)
5 DPL2 TACAGTCTGTGCTGACTCAG (SEQ ID NO:162)
3 DPL2 ATAGGACCATTCAGGCTGTCATC (SEQ ID NO:163)
5 DPL5 TATCAGTCTGTGTTGACGCAG (SEQ ID NO:164)
3 DPL5 ATAGGAGCACTCAGGCTGCTAT (SEQ ID NO:165)
Сочетания праймеров, используемые для амплификации выбранных зародышевых генов.
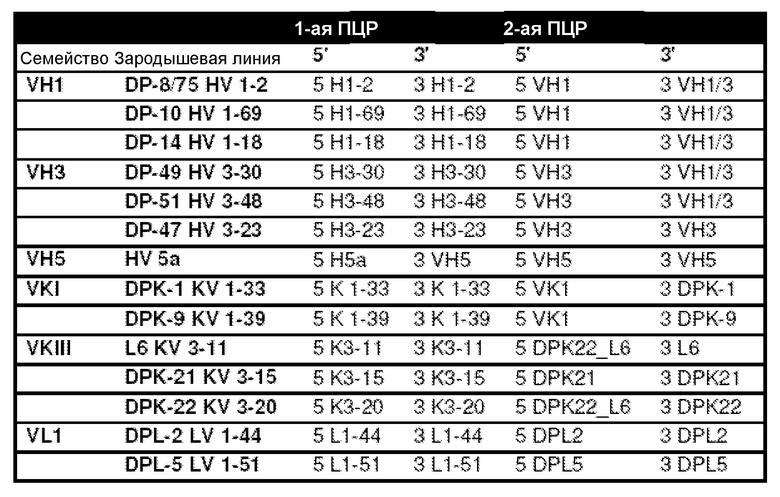
ПРИМЕР 2: Получение акцепторных каркасных участков
Последовательности выбранных зародышевых генов анализируют на присутствие участков рестрикции типа IIs. Участок BsmBI отсутствует в выбранных зародышевых генах вариабельного домена антитела. Два участка BsmBI обнаружены в скелете pNDS1, фагмидного вектора, используемого для клонирования акцепторного каркасного участка. Эти два участка удаляют путем направленного мутагенеза, чтобы можно было ввести единственные участки BsmBI в "лишнюю" последовательность ДНК акцепторных каркасных участков. Каждый зародышевый ген амплифицируют с помощью нескольких гнездовых ПЦР, чтобы добавить к 3'-концу последовательности FR3 "лишнюю" последовательность ДНК и затем последовательность, кодирующую FR4, специфичный для каждого соответствующего вариабельного сегмента (VH, Vκ, Vλ). Аминокислотная последовательность FR4 VH соответствует участку FR4, кодируемому зародышевыми J-генами JH1, JH3, JH4 и JH5. Аминокислотная последовательность FR4 VK соответствует участку FR4, кодируемому зародышевыми J-генами JK1. Аминокислотная последовательность FR4 Vλ соответствует участку FR4, кодируемому зародышевыми J-генами JL2 и JL3. Два варианта последовательности FR4 Vκ получают в результате одной аминокислотной замены в положении 106 (аргинин или глицин). Для получения акцепторного каркасного участка на основе зародышевого гена VH3-23 также конструируют два варианта, различающиеся одной аминокислотой (лизин или аргинин) в положении 94, последнем положении FR3. На последней стадии амплификации участки SfiI/NcoI и XhoI вводят в 5'- и 3'-конец VH, соответственно.
Подобным образом, участки SalI и NotI вводят в 5'- и 3'-конец VL, соответственно (фигура 6). "Лишний" фрагмент конструируют так, чтобы трансляционная рамка считывания была сдвинута и не обеспечивала бы экспрессию какого-либо функционального белка на последовательности, кодирующей акцепторный каркасный участок (фигура 7). Используемые в данном процессе праймеры приведены ниже.
VH
5 VH1 CAGCCGGCCATGGCCCAGGTGCAGCTGGTGCAG (SEQ ID NO:166)
5 VH3-30 CAGCCGGCCATGGCCCAGGTGCAGCTGGTGGAG (SEQ ID NO:167)
5 VH3-23 CAGCCGGCCATGGCCGAGGTGCAGCTGTTGGAG (SEQ ID NO:168)
5 VH3-48 CAGCCGGCCATGGCCGAGGTGCAGCTGGTGGAGTCTGGGGGAG (SEQ ID NO:169)
5 VH5-51 CAGCCGGCCATGGCCGAGGTGCAGCTGGTGCAG (SEQ ID NO:170)
3 VH1/3 CTTACCGTTATTCGTCTCATCTCGCACAGTAATACAC (SEQ ID NO:171)
3 VH3-23 CTTACCGTTATTCGTCTCATTTCGCACAGTAATATAC (SEQ ID NO:172)
3 VH3-48 CTCGCACAGTAATACACAGCCGTGTCCTCGGCTCTCAGGCTG (SEQ ID NO:173)
3 VH5-51 CTTACCGTTATTCGTCTCATCTCGCACAGTAATACAT (SEQ ID NO:174)
3 VHext1 CAATACGCGTTTAAACCTGGTAAACCGCCTTACCGTTATTCGTCTCA (SEQ ID NO:175)
3 VHext2 GTTCCCTGGCCCCAAGAGACGCGCCTTCCCAATACGCGTTTAAACCTG (SEQ ID NO:176)
3 VHext3 CCTCCACCGCTCGAGACTGTGACCAGGGTTCCCTGGCCCCAAGAG (SEQ ID NO:177)
VK
5 VK1 CGGGTCGACGGACATCCAGATGACCCAGTC (SEQ ID NO:178)
5 VK3-11 CGGGTCGACGGAAATTGTGTTGACACAGTCTCCAGC (SEQ ID NO:179)
5 VK3-15 CGGGTCGACGGAAATAGTGATGACGCAGTCTCCAGC (SEQ ID NO:180)
5 VK3-20 CGGGTCGACGGAAATTGTGTTGACGCAGTCTCCAGG (SEQ ID NO:181)
3 VK1-33 CCTTACCGTTATTCGTCTCGCTGCTGACAGTAATATGTTGCAATA (SEQ ID NO:182)
3 VK1-39 CCTTACCGTTATTCGTCTCGCTGCTGACAGTAGTAAGTTGCAAAA (SEQ ID NO:183)
3 VK3 CCTTACCGTTATTCGTCTCGCTGCTGACAGTAATAAACTGCAAAATC (SEQ ID NO:184)
3 VKext1 CCAATACGCGTTTAAACCTGGTAAACCGCCTTACCGTTATTCGTCTC (SEQ ID NO:185)
3 VKext2 GGTCCCTTGGCCGAATGAGACGCGCCTTCCCAATACGCGTTTAAAC (SEQ ID NO:186)
3 Vkext3R GTGCGGCCGCCCGTTTGATTTCCACCTTGGTCCCTTGGCCGAATG (SEQ ID NO:187)
3 VKext3G GTGCGGCCGCCCCTTTGATTTCCACCTTGGTCCCTTGGCCGAATG (SEQ ID NO:188)
Vλ
5 VL1-44 CGGGTCGACGCAGTCTGTGCTGACTCAGCCAC (SEQ ID NO:189)
5 VL1-51 CGGGTCGACGCAGTCTGTGTTGACGCAGCCGC (SEQ ID NO:190)
3 VL1-44 CCTTACCGTTATTCGTCTCCTGCTGCACAGTAATAATC (SEQ ID NO:191)
3 VL1-51 CCTTACCGTTATTCGTCTCCTGTTCCGCAGTAATAATC (SEQ ID NO:192)
3 Vlext2 CCCTCCGCCGAACACAGAGACGCGCCTTCCCAATACGCGTTTAAAC (SEQ ID NO:193)
3 Vlext3 GTGCGGCCGCCCCTAGGACGGTCAGCTTGGTCCCTCCGCCGAACACAGA (SEQ ID NO:194)
Последовательности 20 конечных собранных акцепторных каркасных участков приведены на фигуре 8.
ПРИМЕР 3: Получение фагмидных акцепторных векторов, содержащих неизменный вариабельный домен
Фагмидный вектор pNDS1, используемый для экспрессии scFv, вначале модифицируют, удаляя два участка BsmBI. Домен VH3-23, содержащий заданную последовательность CDR3, клонируют в модифицированном pNDS1, используя участки рестрикции SfiI и XhoI, с получением фагмидного вектора pNDS_VHdummy. Этот домен содержит участок BsmBI в участке FR4, который корректируют путем молчащий сайт-направленного мутагенеза. Затем домен VK1-39, содержащий заданную последовательность CDR3, клонируют в модифицированном pNDS1, используя участки рестрикции SalI и NotI, с получением фагмидного вектора pNDS_VKdummy (фигура 9). 8 акцепторных каркасных участков VH клонируют в векторе pNDS_VKdummy, используя участки рестрикции SalI и NotI. 12 акцепторных каркасных участков VL клонируют в векторе pNDS_VHdummy, используя участки рестрикции SfiI и XhoI. Полученные 20 фагмидных векторов pNDS, описанных ниже, на данной стадии можно использовать для клонирования диверсифицированных CDR3 с использованием участков BsmBI, присутствующих в "лишних" фрагментах ДНК.
Акцепторы VH: pNDS_VH1-2_VKd; pNDS_VH1-18_VKd; pNDS_VH1-69_VKd; pNDS_VH3-23R_VKd; pNDS_VH3-23K_VKd; pNDS_VH3-30_VKd; pNDS_VH5-51_VKd; pNDS_VH3-48_VKd.
Акцепторы VL: pNDS_VHd_VK1-33G; pNDS_VHd_VK1-33R; pNDS_VHd_VK1-39G; pNDS_VHd_VK1-39R; pNDS_VHd_VK3-11G; pNDS_VHd_VK3-11R; pNDS_VHd_VK3-15G; pNDS_VHd_VK3-15R; pNDS_VHd_VK3-20G; pNDS_VHd_VK3-20R; pNDS_VHd_VL1-44; pNDS_VHd_VK1-51.
ПРИМЕР 4: Улавливание природного разнообразия CDRH3 из человеческих репертуаров
В качестве матрицы для амплификации последовательностей CDRH3 используют несколько источников кДНК человека. Указанные источники включают фетальную селезенку человека, а также пулы выделенных клеток периферической крови нормальных взрослых мужского и женского пола. Для извлечения последовательностей CDRH3, полученных с использованием реаранжированной кДНК VH, кодируемой конкретным зародышевым геном, или последовательностей CDRH3, полученных с использованием любой кДНК VH, используют несколько способов амплификации.
Вначале смеси праймеров, совпадающих с 5'-кодирующими участками основных семейств VH человека, используют в сочетании со смесями праймеров, совпадающих со всеми участками JH человека. Это обеспечивает ПЦР-амплификацию большинства генов вариабельных доменов тяжелой цепи иммуноглобулина. Предполагаемые продукты амплификации размером примерно 400 пар оснований (п.о.) выделяют методом электрофореза на агарозном геле и очищают. Полученную ДНК используют в качестве матрицы на второй стадии ПЦР, проводимой с использованием праймеров, содержащих 13 п.о. и 14 п.о., совпадающих с концевым участком FR3 и началом FR4, соответственно. В большинстве случаев последний остаток FR3 представляет собой либо аргинин, либо лизин. Поскольку совпадение последней п.о. играет ключевую роль в удлинении праймера под действием полимеразы, используют два разных 5'-праймера: 5 VHR_FOK (SEQ ID NO: 205, показанная ниже) и 5 VHK_FOK (SEQ ID NO: 206, показанная ниже). Важно, что указанные праймеры также содержат участок рестрикции FokI, используемый для вырезания последовательности CDRH3 (фигура 4). Праймеры, используемые на второй стадии ПЦР, биотинилируют по 5'-концу, чтобы облегчить последующие стадии очистки (см. пример 5). Описанный двухстадийный способ позволяет проводить эффективную амплификацию последовательностей CDRH3, несмотря на ограниченное число пар оснований. Чтобы определить оптимальные условия, амплификацию проводят при разных температурах отжига (от 30°C до 70°C) с использованием нескольких термостабильных ДНК-полимераз. Обнаружено, что для данного набора праймеров оптимальными условиями являются температура отжига 55-58°C и применение полимеразы GoTaq (Promega). Продукт второй амплификации после разделения на 2% агарозном геле дает смазанную полосу в нижней части геля, сответствующую смеси CDRH3 разной длины. Чтобы обогатить смесь длинными CDRH3, используют либо препарат полноразмерной ДНК, экстрагированный из геля, либо участок, соответствующий более крупным фрагментам ДНК.
Альтернативно первую стадию амплификации проводят с использованием 5'-праймера 5 VH3-23H2 (SEQ ID NO: 201, показанная ниже), специфичного к последовательности, кодирующей CDRH2 зародышевого VH3-23. Поскольку разные зародышевые гены различаются по данному CDR, можно, предпочтительно, амплифицировать кДНК VH, кодируемые выбранным зародышевым геном. Последующие стадии очистки и амплификации идентичны описанным. С помощью данного способа можно выделить CDR из конкретной среды каркасного участка и снова ввести его в такой же, подобный или другой каркасный участок.
Ниже приведен список праймеров, используемых для амплификации природных репертуаров CDRH3 человека.
1-ая стадия ПЦР
5 VH1/5 CCGCACAGCCGGCCATGGCCCAGGTGCAGCTGGTGCAGTCTGG (SEQ ID NO:195)
5 VH3 CCGCACAGCCGGCCATGGCCGAGGTGCAGCTGGTGGAGTCTGG (SEQ ID NO:196)
5 VH2 CCGCACAGCCGGCCATGGCCCAGRTCACCTTGCTCGAGTCTGG (SEQ ID NO:197)
5 VH4 CCGCACAGCCGGCCATGGCCCAGGTGCAGCTGCAGGAGTCGGG (SEQ ID NO:198)
5 VH4DP64 CCGCACAGCCGGCCATGGCCCAGCTGCAGCTGCAGGAGTCCGG (SEQ ID NO:199)
5 VH4DP63 CCGCACAGCCGGCCATGGCCCAGGTGCAGCTACAGCAGTGGGG (SEQ ID NO:200)
5 VH3-23H2 TGGAGTGGGTCTCAGCTATTAGTGGTAGTGGT (SEQ ID NO:201)
3 HJ1/2 CGATGGGCCCTTGGTGGAGGCTGAGGAGACRGTGACCAGGGTGCC (SEQ ID NO:202)
3 HJ3/6 CGATGGGCCCTTGGTGGAGGCTGAAGAGACGGTGACCRTKGTCCC (SEQ ID NO:203)
3 HJ4/5 CGATGGGCCCTTGGTGGAGGCTGAGGAGACGGTGACCAGGGTTCC (SEQ ID NO:204)
2-ая стадия ПЦР
5 VHR_FOK GAGCCGAGGACACGGCCGGATGTTACTGTGCGAGA (SEQ ID NO:205)
5 VHK_FOK GAGCCGAGGACACGGCCGGATGTTACTGTGCGAAA (SEQ ID NO:206)
3 JH1_FOK GAGGAGACGGTGACGGATGTGCCCTGGCCCCA (SEQ ID NO:207)
3 JH2_FOK GAGGAGACGGTGACGGATGTGCCACGGCCCCA (SEQ ID NO:208)
3 JH3456_FOK GAGGAGACGGTGACGGATGTYCCTTGGCCCCA (SEQ ID NO:209)
ПРИМЕР 5: Получение первичных библиотек путем клонирования природных CDRH3 человека в акцепторных каркасных участках
Амплифицированные CDRH3 расщепляют с помощью FokI, после чего полученные в результате расщепления крайние участки и нерасщепленную ДНК удаляют, используя магнитные шарики, покрытые стрептавидином. Одновременно акцепторные векторы pNDS VH расщепляют с помощью BsmBI. Поскольку липкие концы, образовавшиеся в результате указанных расщеплений, являются совместимыми, коллекцию природных CDRH3 можно лигировать в акцепторный каркасный участок VH с восстановлением соответствующей рамки считывания. Лигированную ДНК очищают, концентрируют и используют для трансформации компетентных клеток E. coli XL1 Blue, после чего произвольные клоны анализируют секвенированием, чтобы подтвердить воссоздание последовательности CDRH3 и правильность соединения CDR и каркасного участка (фигура 10). Результаты показывают, что все клоны содержат последовательности CDRH3, рамка считывания восстанавливается и, следовательно, кодирует вариабельный домен тяжелой цепи иммуноглобулина. Кроме того, все CDR имеют разные последовательности, свидетельствуя о том, что с помощью данного способа можно улавливать большое разнообразие природных последовательностей. Длина CDRH3 также варьирует, причем обнаружены относительно длинные CDR, содержащие от 10 до 15 остатков, что подчеркивает преимущество данного способа, позволяющего отбирать длинные последовательности CDR, которые трудно улавливать с использованием синтетического разнообразия.
С помощью описанного способа, природные последовательности CDRH3, полученные либо из пула выделенных клеток периферической крови человека, либо из фетальной селезенки человека, клонируют в каждом из акцепторных каркасных участков pNDS VH и используют для трансформации электрокомпетентных клеток E. coli TG1, которые затем помещают в планшеты 2×TYAG Bioassay (среда 2×TY, содержащая 100 мкг/мл ампициллина и 2% глюкозы). После инкубации в течение ночи при 30°C в планшеты добавляют 10 мл жидкой среды 2×TYAG, клетки соскребают с поверхности и переносят в полипропиленовую пробирку объемом 50 мл. К суспензии клеток добавляют 2×TYAG, содержащую 50% глицерина, до получения конечной концентрации глицерина 17%. Аликвоты библиотек хранят при -80°C. С помощью данного способа получают 14 первичных библиотек, представляющих всего 8,1×109 трансформантов. Чтобы определить качество и степень разнообразия библиотек, секвенируют 180 произвольно выбранных клонов. Все клоны кодируют разные последовательности VH и >89% из них находятся в рамке считывания. Указанные первичные библиотеки содержат разнообразие только в CDRH3, поскольку они объединены с холостым доменом VL.
ПРИМЕР 6: Получение первичных библиотек путем клонирования синтетических CDR3 в акцепторных каркасных участках
Хотя способ имеет особое значение для получения природного разнообразия, его также можно использовать для интеграции синтетического разнообразия в акцепторные каркасные участки. Синтетические последовательности CDR3 конструируют как для VH, так и для VL. При конструировании учитывают встречаемость CDR определенной длины и стратегию диверсификации (кодоны NNS, DVK, NVT или DVT), которая обеспечивает полный охват теоретического разнообразия при соответствующем числе трансформантов в библиотеке (~5×109 трансформантов) (фигура 11). При конструировании CDR3 цепей Vκ и Vλ остатки, необходимые для поддержания канонической структуры CDR, оставляют неизменными. Для тяжелой цепи получают только CDR3, содержащие до 10 диверсифицированных положений, поскольку число клонов, необходимое для охвата разнообразия, обусловленного более длинными CDR, выходит за пределы практических ограничений эффективности трансформации.
Вырожденные олигонуклеотиды разной длины синтезируют, используя рандомизированные кодоны NNS, NVT, DVK или DVT. Для каждого CDRH3 синтезируют два олигонуклеотида, кодирующих либо метионин, либо фенилаланин в положении 100z (фигура 11). Каждый олигонуклеотид удлиняют и амплифицируют, используя два внешних биотинилированных праймера, с получением двухцепочечных фрагментов ДНК, кодирующих конструированные CDR. Указанные внешние праймеры содержат участки рестрикции BsmBI, позволяющие впоследствии вырезать последовательности CDR и вставить их в акцепторные каркасные участки (фигура 12). Собранные фрагменты ДНК обрабатывают, не используя очистку в геле, и расщепляют BsmBI. Полученные в результате расщепления крайние участки и нерасщепленную ДНК удаляют, используя магнитные шарики, покрытые стрептавидином. Расщепленные фрагменты ДНК концентрируют путем осаждения этанолом и лигируют в соответствующие акцепторные векторы pNDS VH, VK или Vλ. Продукты лигирования очищают, концентрируют и используют для трансформации электрокомпетентных клеток E. coli TG1, которые затем помещают в планшеты 2×TYAG Bioassay (среда 2×TY, содержащая 100 мкг/мл ампициллина и 2% глюкозы). После инкубации в течение ночи при 30°C в планшеты добавляют 10 мл жидкой среды 2×TYAG, клетки соскребают с поверхности и переносят в полипропиленовую пробирку объемом 50 мл. К суспензии клеток добавляют 2×TYAG, содержащую 50% глицерина, до получения конечной концентрации глицерина 17%. Аликвоты библиотек хранят при -80°C. Получают 24 первичные библиотеки тяжелых цепей, представляющие всего 1,6×1010 трансформантов. Подобным образом, получают 13 первичных библиотек легкой цепи, представляющие всего 6,9×109 трансформантов. Указанные первичные библиотеки содержат разнообразие только в CDRH3, поскольку они объединены с холостым доменом VL. Чтобы определить качество и степень разнообразия библиотек, секвенируют 330 произвольно выбранных клонов. Все клоны кодируют разные вариабельные домены и >90% из них находятся в рамке считывания. Такая низкая встречаемость последовательностей, содержащих сдвиги рамки считывания, резко контрастирует с результатами, обычно получаемыми при конструировании библиотек синтетических фрагментов антител с использованием методов ПЦР с перекрыванием, которые зачастую приводят к введению вставок и значительному уменьшению числа функциональных клонов (15-45%).
Разнообразие в указанных первичных библиотеках ограничено CDRH3 или CDRL3, поскольку они объединены с холостыми цепями VL или VH, соответственно.
Ниже перечислены праймеры, используемые для сборки синтетических CDR.
5 H3_R_biot ATGATGCTGCTGGCACGTCTCCGAGA (SEQ ID NO:210)
3 H3_M_biot CCACGTCATCCGATCCGTCTCCCCCAATAATCCAT (SEQ ID NO:211)
3 H3_F_biot CCACGTCATCCGATCCGTCTCCCCCAATAATCAAA (SEQ ID NO:212)
H3_4nnsF GCTGGCACGTCTCCGAGANNSNNSNNSNNSTTTGATTATTGGGGGAGACG (SEQ ID NO:213)
H3_4nnsM GCTGGCACGTCTCCGAGANNSNNSNNSNNSATGGATTATTGGGGGAGACG (SEQ ID NO:214)
H3_5nnsF GCTGGCACGTCTCCGAGANNSNNSNNSNNSNNSTTTGATTATTGGGGGAGACG (SEQ ID NO:215)
H3_5nnsM GCTGGCACGTCTCCGAGANNSNNSNNSNNSNNSATGGATTATTGGGGGAGACG (SEQ ID NO:216)
H3_6nnsF GCTGGCACGTCTCCGAGANNSNNSNNSNNSNNSNNSTTTGATTATTGGGGGAGACG (SEQ ID NO:217)
H3_6nnsM GCTGGCACGTCTCCGAGANNSNNSNNSNNSNNSNNSATGGATTATTGGGGGAGACG (SEQ ID NO:218)
H3_6dvkF GCTGGCACGTCTCCGAGADVKDVKDVKDVKDVKDVKTTTGATTATTGGGGGAGACG (SEQ ID NO:219)
H3_6dvkM GCTGGCACGTCTCCGAGADVKDVKDVKDVKDVKDVKATGGATTATTGGGGGAGACG (SEQ ID NO:220)
H3_7dvkF GCTGGCACGTCTCCGAGADVKDVKDVKDVKDVKDVKDVKTTTGATTATTGGGGGAGACG (SEQ ID NO:221)
H3_7dvkM GCTGGCACGTCTCCGAGADVKDVKDVKDVKDVKDVKDVKATGGATTATTGGGGGAGACG (SEQ ID NO:222)
H3_7nvtF GCTGGCACGTCTCCGAGANVTNVTNVTNVTNVTNVTNVTTTTGATTATTGGGGGAGACG (SEQ ID NO:223)
H3_7nvtM GCTGGCACGTCTCCGAGANVTNVTNVTNVTNVTNVTNVTATGGATTATTGGGGGAGACG (SEQ ID NO:224)
H3_8nvtF GCTGGCACGTCTCCGAGANVTNVTNVTNVTNVTNVTNVTNVTTTTGATTATTGGGGGAGACG (SEQ ID NO:225)
H3_8nvtM GCTGGCACGTCTCCGAGANVTNVTNVTNVTNVTNVTNVTNVTATGGATTATTGGGGGAGACG (SEQ ID NO:226)
H3_9nvtF GCTGGCACGTCTCCGAGANVTNVTNVTNVTNVTNVTNVTNVTNVTTTTGATTATTGGGGGAGAC
G (SEQ ID NO:227)
H3_9nvtM GCTGGCACGTCTCCGAGANVTNVTNVTNVTNVTNVTNVTNVTNVTATGGATTATTGGGGGAGACG (SEQ ID NO:228)
H3_9dvtF GCTGGCACGTCTCCGAGADVTDVTDVTDVTDVTDVTDVTDVTDVTTTTGATTATTGGGGGAGACG (SEQ ID NO:229)
H3_9dvtM GCTGGCACGTCTCCGAGADVTDVTDVTDVTDVTDVTDVTDVTDVTATGGATTATTGGGGGAGACG (SEQ ID NO:230)
H3_10dvtF GCTGGCACGTCTCCGAGADVTDVTDVTDVTDVTDVTDVTDVTDVTDVTTTTGATTATTGGGGGAGACG (SEQ ID NO:231)
H3_10dvtM GCTGGCACGTCTCCGAGADVTDVTDVTDVTDVTDVTDVTDVTDVTDVTATGGATTATTGGGGGAGACG (SEQ ID NO:232)
5 KL3_biot CCGGTGTAGCGAAGGCGTCTCAGCAG (SEQ ID NO:233)
3 KL3_ biot TAGGGTCGCCTTGATCGTCTCCCGAAGGTCGG (SEQ ID NO:234)
K_4nns GAAGGCGTCTCAGCAGNNSNNSNNSNNSCCGACCTTCGGGAGACG (SEQ ID NO:235)
K_5nns GAAGGCGTCTCAGCAGNNSNNSNNSNNSCCGNNSACCTTCGGGAGACG (SEQ ID NO:236)
K_6nns GAAGGCGTCTCAGCAGNNSNNSNNSNNSNNSCCGNNSACCTTCGGGAGACG (SEQ ID NO:237)
5 L44W_biot CGGTCAGTCGCAATACGTCTCCAGCATGGGAT (SEQ ID NO:238)
5 L44Y_biot CGGTCAGTCGCAATACGTCTCCAGCATATGAT (SEQ ID NO:239)
3 L_biot CAGGACCAGTCTCGTGAGGATCGTCTCAACAC (SEQ ID NO:240)
L44W_4nns CGTCTCCAGCATGGGATNNSNNSNNSNNSGTGTTGAGACGATCCTC (SEQ ID NO:241)
L44Y_4nns CGTCTCCAGCATATGATNNSNNSNNSNNSGTGTTGAGACGATCCTC (SEQ ID NO:242)
L44W_5nns CGTCTCCAGCATGGGATNNSNNSNNSNNSNNSGTGTTGAGACGATCCTC (SEQ ID NO:243)
L44Y_5nns CGTCTCCAGCATATGATNNSNNSNNSNNSNNSGTGTTGAGACGATCCTC (SEQ ID NO:244)
L44W_6nns CGTCTCCAGCATGGGATNNSNNSNNSNNSNNSNNSGTGTTGAGACGATCCTC (SEQ ID NO:245)
L44Y_6nns CGTCTCCAGCATATGATNNSNNSNNSNNSNNSNNSGTGTTGAGACGATCCTC (SEQ ID NO:246)
5 L51W_biot CGGTCAGTCGCAATACGTCTCGAACATGGGAT (SEQ ID NO:247)
5 L51Y_biot CGGTCAGTCGCAATACGTCTCGAACATATGAT (SEQ ID NO:248)
L51W_4nns CGTCTCGAACATGGGATNNSNNSNNSNNSGTGTTGAGACGATCCTC (SEQ ID NO:249)
L51Y_4nns CGTCTCGAACATATGATNNSNNSNNSNNSGTGTTGAGACGATCCTC (SEQ ID NO:250)
L51W_5nns CGTCTCGAACATGGGATNNSNNSNNSNNSNNSGTGTTGAGACGATCCTC (SEQ ID NO:251)
L51Y_5nns CGTCTCGAACATATGATNNSNNSNNSNNSNNSGTGTTGAGACGATCCTC (SEQ ID NO:252)
L51W_6nns CGTCTCGAACATGGGATNNSNNSNNSNNSNNSNNSGTGTTGAGACGATCCTC (SEQ ID NO:253)
L51Y_6nns CGTCTCGAACATATGATNNSNNSNNSNNSNNSNNSGTGTTGAGACGATCCTC (SEQ ID NO:254)
ПРИМЕР 7: Получение вторичных библиотек
Чтобы получить библиотеки scFv, несущие разнообразие как в тяжелых, так и в легких цепях, первичные библиотеки синтетических легких цепей объединяют либо с первичными библиотеками синтетических тяжелых цепей, либо с первичными библиотеками природных тяжелых цепей (фигура 13). Фагмидную ДНК, полученную из каждой первичной библиотеки, расщепляют ферментами рестрикции XhoI/NotI. Фрагменты ДНК, соответствующие линкеру и легким цепям, из первичных синтетических библиотек вставляют путем лигирования в расщепленные векторы первичных библиотек природных или синтетических тяжелых цепей. Альтернативно последовательность линкер-VL также можно амплифицировать с использованием специфических праймеров до расщепления ферментами XhoI/NotI и лигирования. Продукты лигирования после очистки методами фенол/хлороформной экстракции и преципитации используют для трансформации электрокомпетентных клеток E. coli TG1, которые затем помещают в планшеты 2×TYAG Bioassay (среда 2×TY, содержащая 100 мкг/мл ампициллина и 2% глюкозы). После инкубации в течение ночи при 30°C в планшеты добавляют 10 мл жидкой среды 2×TYAG, клетки соскребают с поверхности и переносят в полипропиленовую пробирку объемом 50 мл. К суспензии клеток добавляют 2×TYAG, содержащую 50% глицерина, до получения конечной концентрации глицерина 17%. Аликвоты библиотек хранят при -80°C. Чтобы ограничить число библиотек, подлежащих рекомбинации, их объединяют по подклассам цепей (т.е., VH1, VH3, VH5, VK1, VK3, Vλ1) с получением 9 сочетаний библиотек (т.е., VH1×VK1, VH1×VK3, VH1×Vλ1, VH3×VK1, VH3×VK3, VH3×Vλ1, VH5×VK1, VH5×VK3, VH5×Vλ1). Общий размер вторичных синтетических библиотек (несущих синтетическое разнообразие как в VH, так и в VL) составляет 7,3×109 трансформантов. Общий размер вторичных природных библиотек (несущих природное разнообразие в VH и синтетическое разнообразие в VL) составляет 1,5×1010 трансформантов.
ПРИМЕР 8: Получение библиотек антител человека, несущих репертуар CDRH3 из отличных от человека видов
Чтобы использовать альтернативные источники разнообразия, которые позволяют объяснить различие в трехмерных пространствах связывающих участках антител, библиотеку создают путем улавливания CDRH3 мыши и введения их в коллекцию каркасных участков антител человека. Для осуществления данного подхода конструируют акцепторную библиотеку, содержащую коллекцию генов VL с синтетическим разнообразием CDRL3, и объединяют ее с коллекцией акцепторных последовательностей, содержащих "лишнюю" последовательность ДНК, которая хорошо подходит для рестрикционного клонирования типа IIS, описанного в примере 2. Данная библиотека является исходным материалом для быстрого получения вторичных библиотек с использованием нескольких источников природных (человеческих и нечеловеческих) или синтетических CDRH3. В данном примере природное разнообразие CDRH3 улавливают из наивных мышей Balb/c и из мышей, иммунизированных hIFNγ или hCCL5 (hRANTES).
Первая стадия включает получение акцепторных библиотек путем клонирования коллекции VL, содержащих синтетическое разнообразие CDRL3, в векторах, несущих акцепторные каркасные участки VH (фигура 14). Последовательности VL получают из семи первичных синтетических библиотек, описанных в примере 6, методом ПЦР-амплификации с использованием праймеров 5'biot-VHdummy и 3'biot-fdtseq. Полученные VL, содержащие фрагменты размером примерно 400 п.о., расщепляют с использованием XhoI/NotI и очищают на центрифужных колонках, удаляя праймеры и ферменты. Подобным образом, акцепторные векторы pNDS VH, содержащие CDRH3, "лишнюю" последовательность и холостую легкую цепь, расщепляют с помощью XhoI/NotI и SwaI (SwaI разрезает последовательность внутри холостой VL) и очищают на колонках Chroma Spin TE с отсечением по 1000 п.о., избавляясь от холостого фрагмента VL. Затем расщепленные фрагменты VL лигируют в акцепторные векторы VH (фигура 14). Чтобы ограничить число библиотек, подлежащих рекомбинации, их объединяют по подклассам цепей (т.е., VH1, VH3, VH5, Vκ1, Vκ3, Vλ1) с получением 9 сочетаний библиотек (т.е., VH1×Vκ1, VH1×Vκ3, VH1×Vλ1, VH3×Vκ1, VH3×Vκ3, VH3×Vλ1, VH5×Vκ1, VH5×Vκ3, VH5×Vλ1). Продукты лигирования используют для трансформации электрокомпетентных клеток E. coli TG1, которые затем помещают в планшеты 2×TYAG Bioassay (среда 2×TY, содержащая 100 мкг/мл ампициллина и 2% глюкозы). После инкубации в течение ночи при 30°C в планшеты добавляют 6 мл жидкой среды 2×TYAG, клетки соскребают с поверхности и переносят в полипропиленовую пробирку объемом 50 мл. К суспензии клеток добавляют 50% глицерин до получения конечной концентрации глицерина 17%. Аликвоты библиотек хранят при -80°C. Общий размер полученной акцепторной библиотеки, несущей синтетическое разнообразие по CDRL3, составляет 1,9×109 трансформантов.
Следующая стадия включает выделение последовательности CDRH3 из отличного от человека источника. Клетки выделяют из селезенки пяти наивных или иммунизированных мышей Balb/c и получают общую РНК. кДНК получают методом ОТ-ПЦР, используя экстрагированную РНК в качестве матрицы. Полученную кДНК используют в качестве матрицы для выделения и амплификации VH мыши методом ПЦР. Ряд ПЦР проводят с использованием 15 разных 5'-праймеров (один для каждой подгруппы VH мыши), специфичных к началу участка FR1, и комплект 3'-праймеров (четыре праймера, охватывающих участок JH). Продукты первых ПЦР объединяют и очищают на 2% агарозном геле. Очищенную ДНК используют в качестве матрицы при проведении второй ПЦР с целью выделения участка CDRH3 мыши.
5'- и 3'-Праймеры, используемые во второй ПЦР, направлены на участки FR3 и FR4 VH мыши, соответственно. С помощью этих праймеров добавляют участок рестрикции FokI, чтобы обеспечить точное вырезание CDRH3 и клонирование в акцепторных векторах человека. Однако выравнивание последовательностей VH мыши показывает, что последовательность на 5'-конце CDR-H3 мыши, расположенная в участке расщепления FokI, почти всегда отличается от последовательности человека на одно основание, тогда как 3'-концы у этих двух видов одинаковые. Последовательности, расщепляемые FokI, заключены в рамки в приведенной ниже таблице:
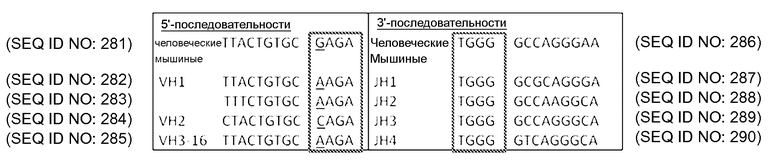
Следовательно, основание нужно корректировать в процессе второй стадии амплификации, чтобы получить липкие концы, совместимые с липкими концами, полученными в результате расщепления акцепторных каркасных участков. Наблюдающаяся эффективная амплификация позволяет предположить, что указанное превращение легко осуществляется. На 3'-конце последовательности мыши и человека, подлежащие разрезанию ферментами рестрикции типа IIS, являются идентичными, что позволяет избежать проблем, связанных с коррекцией.
Праймеры, используемые для второй амплификации, биотинилируют по 5'-концам, чтобы облегчит последующие стадии очистки. Акцепторные векторы расщепляют BsmBI и очищают на колонках Chroma Spin TE с отсечением по 1000 п.о. После расщепления и очистки девять разных сочетаний библиотек объединяют в эквимолярном соотношении для лигирования уловленных CDRH3 мыши.
Лигированную ДНК после очистки методом фенол/хлороформной экстракции и концентрирования методом преципитации используют для трансформации электрокомпетентных клеток E. coli TG1, которые затем помещают в планшеты 2×TYAG Bioassay (среда 2×TY, содержащая 100 мкг/мл ампициллина и 2% глюкозы). После инкубации в течение ночи при 30°C в планшеты добавляют 10 мл жидкой среды 2×TYAG, клетки соскребают с поверхности и переносят в полипропиленовую пробирку объемом 50 мл. К суспензии клеток добавляют 50% глицерин до получения конечной концентрации глицерина 17%. Аликвоты библиотек хранят при -80°C. Получают три библиотеки с похожими размерами: MnA, 2,5×108 трансформантов (несущих ограниченное природное разнообразие каркасных участков человека, разнообразие CDRH3 наивных мышей и синтетическое разнообразие CDRL3); MiB, 7,3×107 трансформантов (несущих ограниченное природное разнообразие каркасных участков человека, разнообразие CDRH3 мышей, иммунизированных против hIFNγ и синтетическое разнообразие CDRL3), и MiC, 1,8×108 трансформантов (несущих ограниченное природное разнообразие каркасных участков человека, разнообразие CDRH3 мышей, иммунизированных против hCCL5 и синтетическое разнообразие CDRL3). Произвольные клоны анализируют путем секвенирования, чтобы подтвердить восстановление последовательности CDRH3 и правильность соединения CDR с каркасными участками. Результаты показывают, что все клоны содержат последовательности CDRH3 и происходит восстановление рамки считывания, кодирующей вариабельный домен тяжелой цепи иммуноглобулина. Все CDR являются разными и имеют сходство с типичными последовательностями CDRH3 мыши, свидетельствуя о том, что описанный способ позволяет улавливать большое разнообразие природных последовательностей CDRH3 мыши. Кроме того, анализ профилей длин CDRH3 показывает, что в наивной библиотеке улавливается гауссово распределение, которое соответствует ожидаемому распределению длин в нормальном репертуаре мыши. И наоборот, в двух иммунных библиотеках профили отличаются, позволяя предположить, что улавливается другой репертуар CDRH3 (фигура 15).
Праймеры, используемые для амплификации CDRH3 мыши
1-ая ПЦР
5'-праймеры:
m5 VH1 ATGCGGCCCAGCCGGCCATGGCCSAGGTYCAGCTBCAGCAGTC (SEQ ID NO:256)
m5 VH2 ATGCGGCCCAGCCGGCCATGGCCCAGGTTCACCTGCAGCARTC (SEQ ID NO:257)
m5 VH3 ATGCGGCCCAGCCGGCCATGGCCCAGGTRCAGCTGAAGGAGTC (SEQ ID NO:258)
m5 VH4 ATGCGGCCCAGCCGGCCATGGCCCAGGTCCAACTVCAGCARCC (SEQ ID NO:259)
m5 VH5 ATGCGGCCCAGCCGGCCATGGCCCAGATCCAGTTGGTVCAGTC (SEQ ID NO:260)
m5 VH6 ATGCGGCCCAGCCGGCCATGGCCCAGGTGCAGCTGAAGSASTC (SEQ ID NO:261)
m5 VH7 ATGCGGCCCAGCCGGCCATGGCCGAGGTGCAGSKGGTGGAGTC (SEQ ID NO:262)
m5 VH8 ATGCGGCCCAGCCGGCCATGGCCGAAGTGAARSTTGAGGAGTC (SEQ ID NO:263)
m5 VH9 ATGCGGCCCAGCCGGCCATGGCCGAKGTSVAGCTTCAGGAGTC (SEQ ID NO:264)
m5 VH10 ATGCGGCCCAGCCGGCCATGGCCGAGGTGAASSTGGTGGAATC (SEQ ID NO:265)
m5 VH11 ATGCGGCCCAGCCGGCCATGGCCGAGGTGAAGCTGRTGGARTC (SEQ ID NO:266)
m5 VH12 ATGCGGCCCAGCCGGCCATGGCCGARGTGAAGCTGRTGGAGTC (SEQ ID NO:267)
m5 VH13 ATGCGGCCCAGCCGGCCATGGCCGAAGTGCAGCTGTTGGAGAC (SEQ ID NO:268)
m5 VH14 ATGCGGCCCAGCCGGCCATGGCCGARGTGAAGCTTCTCSAGTC (SEQ ID NO:269)
m5 VH15 ATGCGGCCCAGCCGGCCATGGCCCARGTTACTCTGAAAGAGT (SEQ ID NO:270)
3'-праймеры:
m3 HJ1 CCTGAACCGCCGCCTCCGCTCGAGACGGTGACCGTGGTCCC (SEQ ID NO:271)
m3 HJ2 CCTGAACCGCCGCCTCCGCTCGAGACTGTGAGAGTGGTGCC (SEQ ID NO:272)
m3 HJ3 CCTGAACCGCCGCCTCCGCTCGAGACAGTGACCAGAGTCCC (SEQ ID NO:273)
m3 HJ4 CCTGAACCGCCGCCTCCGCTCGAGACGGTGACTGAGGTTCC (SEQ ID NO:274)
2-ая ПЦР
5'-праймеры:
5 VHR_FOK_biot GAGCCGAGGACACGGCCGGATGTTACTGTGCGAGA (SEQ ID NO:275)
3'-праймеры:
3'mJH1_Fok_biot GGGGCGCAGGGACATCCGTCACCGTCTCCTC (SEQ ID NO:276)
3'mJH2_Fok_biot GAGGAGACTGTGAGGGATGTGCCTTGGCCCCA (SEQ ID NO:277)
3'JH1_Fok GAGGAGACGGTGACGGATGTGCCCTGGCCCCA (SEQ ID NO:278)
3'mJH4_Fok_biot GAGGAGACGGTGACGGATGTTCCTTGACCCCA (SEQ ID NO:279)
ПРИМЕР 9: Фаговое спасение библиотек
Каждую первичную и вторичную библиотеку спасают независимо в соответствии со стандартными способом фаговых дисплеев, кратко описанным ниже. Объем клеток из замороженных аликвот библиотек, достаточный для охватывания по меньшей мере 10-кратного теоретического разнообразия библиотеки, добавляют к 500 мл 2×TYAG и выращивают при 37°C при встряхивании (240 об/мин) до достижения значения OD600 от 0,3 до 0,5. Затем культуру суперинфицируют хелперным фагом MK13K07 и инкубируют в течение одного часа при 37°C (150 об/мин). Затем среду заменяют центрифугированием клеток при 2000 об/мин в течение 10 мин, чтобы удалить среду, с последующим ресуспендированием осадка в 500 мл 2×TY-AK (100 мкг/мл ампициллина; 50 мкг/мл канамицина). Затем культуру выращивают в течение ночи при 30°C (240 об/мин). Культуру центрифугируют при 4000 об/мин в течение 20 минут, чтобы осадить клетки. Супернатант собирают, добавляют 30% (об/об) PEG 8000 (20%)/2,5M NaCl и смесь инкубируют 1 час на льду, чтобы осадить фаговые частицы. Фаговые частицы собирают центрифугированием при 10000 об/мин в течение 30 мин и ресуспендируют в 10 мл буфера TE (10 мМ трис-HCl pH 8,0; 1 мМ EDTA). Ресуспендированный раствор центрифугируют при 10000 об/мин, чтобы удалить клеточные остатки, и снова повторяют процедуру осаждения. После конечного ресуспендирования фаг титруют путем инфицирования E. coli и измерения поглощения при 280 нм. Также определяют уровень проявления scFv на поверхности фага методом вестерн-блоттинга с использованием моноклонального антитела против c-myc. Очищенный фаг из разных библиотек хранят замороженным при -80°C после добавления глицерина до конечной концентрации 15% (масс/об).
Чтобы использовать в процессе селекции удобное число библиотек, очищенный фаг объединяют с 4 рабочими библиотеками: AA1 - фаг из всех первичных синтетических библиотек VH; AB1 - фаг из всех первичных синтетических библиотек VL; AC1 - фаг из всех первичных природных библиотек VH; AD1 - фаг из всех вторичных природных библиотек; AE1 - фаг из всех вторичных синтетических библиотек; MnA - библиотеки с разнообразием, полученным из наивных мышей; MiB - библиотеки с разнообразием, полученным из мышей, иммунизированных hIFNγ; MiC - библиотеки с разнообразием, полученным из мышей, иммунизированных hCCL5/RANTES.
ПРИМЕР 10. Высокопроизводительное секвенирование библиотек антител
Качество и разнообразие библиотеки можно оценить путем секвенирования ДНК произвольных членов библиотеки. В большинстве случаев секвенируют несколько сотен клонов, которые представляют собой лишь небольшую часть библиотеки (меньше 1 на 10000000 членов библиотеки). Чтобы определить эффективность описанных в настоящем документе способов, используют технологию секвенирования следующего поколения, позволяющую анализировать более представительное число членов библиотеки. ДНК, выделенную из библиотеки AE1, используют в качестве матрицы для высокопроизводительного секвенирования с использованием геномного анализатора illumina. Данная система секвенирования ДНК следующего поколения позволяет прочитать биллионы оснований за несколько дней. Считывания при секвенировании являются относительно короткими (примерно 70 оснований), но хорошо совмещаются с дизайном библиотек настоящего изобретения. Поскольку разнообразие ограничено участками CDR3, для идентификации семейства VH достаточно считывание 70 оснований, позволяющее охватить CDRH3 и часть каркасного участка 3. Данная технология применяется для секвенирования нескольких миллионов участков CDRH3 из библиотеки AE1. 5078705 последовательностей получают для общего числа оснований 365666760. Анализ результатов показывает, что 5007022 последовательностей (98,6% от общего числа) являются уникальными. Всего 4680882 последовательностей можно однозначно приписать к семейству VH (VH1, VH3 и VH5) и определить представительство семейств VH в библиотеке AE1 (41% VH1; 30% VH3; 29% VH5). Важным фактом является то, что доля вставок в рамку считывания варьирует от 88 до 91%. Эти данные подтверждают в более статистической манере результаты секвенирования 24 первичных синтетических библиотек VH, описанных в примере 6. Полученные результаты секвенирования вместе демонстрируют, что процесс рестрикционного клонирования типа IIs, используемый в данном способе, является очень надежным и приводит к эффективной и продуктивной вставке в конструкции 24 независимых библиотек, проводимой с целью генерирования разнообразия VH библиотеки AE1.
Секвенирование миллионов членов библиотеки является беспрецендентной стадией контроля качества библиотек антител. Результаты демонстрируют, что данный способ позволяет воспроизводимо и надежно получать высококачественные библиотеки с высокой степенью разнообразия.
ПРИМЕР 11: Селекция фаговых дисплеев с использованием вторичных библиотек
Жидкофазная селекция против интерферона гамма человека (hIFNγ):
Аликвоты фаговых библиотек AD1 и AE1 (1011-1012 БОЕ) блокируют PBS, содержащим 3% (масс/об) обезжиренного молока, в течение одного часа при комнатной температуре на вращающемся смесителе. Затем блокированный фаг отсеивают на магнитных гранулах, покрытых стрептавидином (Dynal M-280), в течение одного часа при комнатной температуре на вращающемся смесителе. Отсеянный фаг инкубируют с биотинилированным in vivo hIFNγ (100 нМ) в течение двух часов при комнатной температуре на вращающемся смесителе. Гранулы улавливают с помощью магнитной стойки, затем промывают четыре раза PBS/0,1% Tween 20 и 3 раза PBS. Затем гранулы непосредственно добавляют к 10 мл суспензии экспоненциально растущих клеток TG1 и инкубируют в течение одного часа при 37°C, медленно встряхивая (100 об/мин). Чтобы оттитровать продукт селекции, используют серийные разведения аликвоты инфицированных TG1. Остальные инфицированные TG1 центрифугируют при 3000 об/мин в течение 15 минут, снова суспендируют в 0,5 мл 2×TYAG (среда 2×TY, содержащая 100 мкг/мл ампициллина и 2% глюкозы) и вносят в планшеты 2×TYAG agar Bioassay. После инкубации в течение ночи при 30°C в планшеты добавляют 10 мл 2×TYAG, затем клетки соскребают с поверхности и переносят в полипропиленовые пробирки объемом 50 мл. К суспензии клеток добавляют среду 2×TYAG, содержащую 50% глицерина, до получения конечной концентрации глицерина 17%. Аликвоты продукта селекции держат при -80°C. Фаговые продукты титруют после каждого цикла и регистрируют прогрессирующее увеличение выхода, свидетельствующее о том, что происходит обогащение клонами, специфичными к мишени (фигура 16).
Селекция методом пэннинга против моноклонального антитела 5E3 крысы:
Иммунопробирки покрывают раствором 5E3 в PBS с концентрацией 10 мкг/мл в течение ночи при 4°C, а иммунопробирки для отсеивания фагов покрывают в тех же условиях нерелевантным антителом крысы. После промывания иммунопробирки блокируют PBS, содержащим 3% (масс/об) обезжиренного молока, в течение одного часа при комнатной температуре. Аликвоты фаговых библиотек AD1 и AE1 (1011-1012 БОЕ) блокируют PBS, содержащим 3% (масс/об) обезжиренного молока в течение одного часа при комнатной температуре на вращающемся смесителе. Затем блокированный фаг отсеивают в иммунопробирках, покрытых нерелевантным антителом крысы в течение одного часа при комнатной температуре на вращающемся смесителе. Отсеянный фаг переносят в иммунопробирки, покрытые 5E3 и инкубируют в течение двух часов при комнатной температуре на вращающемся смесителе. Пробирки промывают пять раз PBS/0,1% Tween 20 и 3 раза PBS. Фаг элюируют 100 мМ TEA в течение 10 минут и нейтрализуют 1M трис-HCl pH 7,5. Фаг добавляют к 10 мл суспензии экспоненциально растущих клеток TG1 и инкубируют в течение одного часа при 37°C, медленно встряхивая (100 об/мин). Чтобы оттитровать продукт селекции, используют серийные разведения аликвоты инфицированных TG1. Остальные инфицированные TG1 центрифугируют при 3000 об/мин в течение 15 минут, снова суспендируют в 0,5 мл 2×TYAG (среда 2×TY, содержащая 100 мкг/мл ампициллина и 2% глюкозы) и вносят в планшеты 2×TYAG agar Bioassay. После инкубации в течение ночи при 30°C в планшеты добавляют 10 мл 2×TYAG, затем клетки соскребают с поверхности и переносят в полипропиленовые пробирки объемом 50 мл. К суспензии клеток добавляют среду 2×TYAG, содержащую 50% глицерина, до получения конечной концентрации глицерина 17%. Аликвоты продукта селекции держат при -80°C. Циклы селекции проводят с чередованием 5E3 крысы и химерной версии 5E3, в которой вариабельный участок соединен с константными доменами мыши. Указанные чередующиеся циклы проводят с целью повышения доли клонов, специфичных к вариабельному участку 5E3, и получения антиидиотипических антител. Фаговые продукты титруют после каждого цикла и регистрируют прогрессирующее увеличение выхода, свидетельствующее о том, что происходит обогащение клонами, специфичными к мишени (фигура 17).
Фаговая помощь: 100 мкл клеточной суспензии, полученной в предыдущих циклах селекции, добавляют к 20 мл 2xTYAG и выращивают при 37°C при встряхивании (240 об/мин) до достижения значения OD600 от 0,3 до 0,5. Затем культуру суперинфицируют хелперным фагом MK13K07 3,3×1010 и инкубируют в течение одного часа при 37°C (150 об/мин). Затем среду заменяют путем центрифугирования клеток при 2000 об/мин в течение 10 мин, чтобы удалить среду, с последующим ресуспендированием осадка в 20 мл 2×TY-AK (100 мкг/мл ампициллина; 50 мкг/мл канамицина). Затем культуру выращивают в течение ночи при 30°C (240 об/мин)
Помощь моноклональных фагов для ELISA: Отдельные клоны помещают в титрационный микропланшет, содержащий 150 мкл среды 2×TYAG (2% глюкозы) на лунку и выращивают при 37°C (100-120 об/мин) в течение 5-6 ч. Хелперный фаг M13KO7 добавляют в каждую лунку, чтобы достичь множественность заражения (MOI) 10 (т.е., 10 фагов на каждую лунку в культуре) и инкубируют при 37°C (100 об/мин) в течение 1 ч. После выращивания планшеты центрифугируют при 3200 об/мин в течение 10 мин. Супернатант осторожно удаляют, клетки ресуспендируют в 150 мкл среды 2×TYAK и выращивают в течение ночи при 30°C (120 об/мин). Перед проведением ELISA фаг блокируют путем добавления 150 мкл PBS с удвоенной концентрацией, содержащего 5% обезжиренного порошкообразного молока, с последующей инкубацией в течение одного часа при комнатной температуре. Затем планшеты центрифугируют 10 минут при 3000 об/мин и фаг-содержащий супернатант используют для проведения ELISA.
Фаговый метод ELISA: Планшеты ELISA (Maxisorb, NUNC) покрывают в течение ночи раствором hIFNγ в PBS, 2 мкг/мл, или раствором 5E3 крысы в PBs, 2 мкг/мл. Контрольные планшеты покрывают 2 мкг/мл БСА или нерелевантным моноклональным антителом крысы. Затем планшеты блокируют 3% обезжиренным молоком/PBS при комнатной температуре в течение 1 ч. Планшеты промывают 3 раза PBS, содержащим 0,05% Tween 20, затем переносят супернатанты заранее блокированного фага и инкубируют в течение одного часа при комнатной температуре. Затем планшеты промывают 3 раза PBS, содержащим 0,05% Tween 20. В каждую лунку добавляют 50 мкл 3% обезжиренного молока/PBS, содержащего (HRP)-конъюгированное антитела против M13 (Amersham, разведение 1:10000). После инкубации при комнатной температуре в течение 1 ч планшеты промывают 5 раз PBS, содержащим 0,05% Tween 20. Затем результаты анализа ELISA проявляют путем добавления 50 мкл TMB (Sigma) и 50 мкл 2N H2SO4, чтобы остановить реакцию. Интенсивность абсорбции считывают при 450 нм. Можно идентифицировать клоны, специфичные к hIFNγ, причем коэффициент эффективности поиска варьирует от 10% до 30% после третьего цикла селекции. Также можно идентифицировать клоны, специфичные к вариабельному участку 5E3, где коэффициент эффективности поиска варьирует от 7 до 48% после третьего цикла селекции.
Секвенирование фагового клона: Отдельные клоны выращивают в 5 мл среды 2×TYAG (2% глюкозы) на лунку при 37°C (120 об/мин) в течение ночи. На следующий день фагмидную ДНК очищают и используют для секвенирования ДНК с использованием праймера, специфичного к pNDS1: mycseq, 5'-CTCTTCTGAGATGAGTTTTTG. (SEQ ID NO:255).
Крупномасштабная очистка scFv: Исходную культуру в 1 мл 2xTYAG инокулируют отдельной колонией, свежеполученной из полосы планшета, содержащего 2×TYAG-агар, и инкубируют при встряхивании (240 об/мин) при 37°C в течение 5 часов. 0,9 мл данной культуры используют для инокуляции 400 мл культуры в такой же среде и выращивают в течение ночи при 30°C при энергичном встряхивании (300 об/мин).
На следующий день культуру индуцируют путем добавления 400 мкл 1 M IPTG и продолжают инкубировать еще 3 часа. Клетки собирают центрифугированием при 5000 об/мин в течение 10 минут при 4°C. Осажденные клетки ресуспендируют в 10 мл охлажденного на льду буфера TES, дополненного ингибиторами протеаз, как описано выше. Осмотический шок достигают путем добавления 15 мл буфера TES, разбавленного 1:5, и инкубируют в течение 1 часа на льду. Клетки центрифугируют при 10000 об/мин в течение 20 минут при 4°C, чтобы осадить клеточные остатки. Супернатант осторожно переносят в новую пробирку. К супернатанту добавляют имидазол до конечной концентрации 10 мМ. В каждую пробирку добавляют 1 мл смолы Ni-NTA (Qiagen), уравновешенной в PBS, и инкубируют на вращающемся смесителе при 4°C (20 об/мин) в течение 1 часа. Пробирки центрифугируют при 2000 об/мин в течение 5 минут, после чего супернатант осторожно удаляют. Осажденную смолу ресуспендируют в 10 мл холодного (4°C) буфера для промывания 1 (50 мМ NaH2PO4, 300 мМ NaCl, 10 мМ имидазол, pH до 8,0). Суспензию наносят на полупрепаративную колонку (Biorad). Колонку промывают гравитационным течением, используя 8 мл холодного буфера для промывания 2 (50 мМ NaH2PO4, 300 мМ NaCl, 20 мМ имидазол, pH до 8,0). scFv элюируют с колонки 2 мл буфера для элюирования (50 мМ NaH2PO4, 300 мМ NaCl, 250 мМ имидазол, pH до 8,0). Фракции анализируют по поглощению при 280 нм, белок-содержащие фракции объединяют и затем меняют буфер на деминерализующей колонке PD10 (Amersham), уравновешенной PBS. scFv в PBS анализируют методом SDS-PAGE, а их количество определяют по поглощению при 280 нм. Аликвоты очищенных scFv хранят при -20°C и при 4°C.
ПРИМЕР 12. Анализ профилей CDR3, полученных в результате селекции с использованием высокопроизводительного секвенирования
С помощью технологии секвенирования следующего поколения, описанной в примере 10, анализируют распределение длин CDRH3 в каждом семействе VH библиотек AE1 и AD1, а также в продукте, полученном после третьего цикла селекции. Профили библиотек AE1 и AD1 несомненно являются разными (фигура 18). Распределение длин CDRH3 в библиотеке AE1 соответствует предполагаемому дизайну библиотеки, причем значения длин варьируют в диапазоне 9-15 аминокислот. И, наоборот, в библиотеке AD1 обнаружены гораздо более длинные CDRH3, содержащие до 22 аминокислот, причем профиль длин соответствует распределению, наблюдающемуся в природных репертуарах человека. Полученные результаты подтверждают, что природный репертуар CDRH3 человека улавливается в процессе конструирования библиотеки AD1. Подобный анализ, проведенный после трех циклов селекции против 5E3, демонстрирует, что получены совершенно разные профили длин CDRH3. В частности, после селекции, проведенной на библиотеке AD1, наблюдается сильное увеличение доли CDRH3, длина которых составляет 8 и 21 аминокислот. Ряд полученных данных демонстрирует, что разные профили CDRH3 из двух библиотек обогащаются после селекции против одной мишени. Кроме того, данный анализ показывает, что длинные CDRH3, которые очень трудно охватить с использованием синтетического разнообразия, с помощью настоящего изобретения можно уловить в выбранные каркасные участки человека с последующей селекцией.
ПРИМЕР 13. Анализ связывания идентифицированных scFv
Анализируют связывание очищенных препаратов scFv клонов, имеющих разные последовательности и положительно идентифицированных против вариабельного участка 5E3, с химерным 5E3 методом ELISA в дозо-зависимой манере. Указанные препараты также анализируют против нерелевантного антитела мыши (1A6). Планшеты для ELISA (Maxisorb, NUNC) покрывают в течение ночи раствором 5E3 мыши в PBS с концентрацией 2 мкг/мл. Контрольные планшеты покрывают моноклональным антителом 1A6, 2 мкг/мл. Затем планшеты блокируют 3% обезжиренным молоком/PBS при комнатной температуре в течение 1 ч. Планшеты промывают 3 раза PBS, содержащим 0,05% Tween 20, добавляют очищенные scFv в разных концентрациях и инкубируют в течение одного часа при комнатной температуре. Затем планшеты промывают 3 раза PBS, содержащим 0,05% Tween 20. В каждую лунку добавляют 50 мкл 3% обезжиренного молока/PBS, содержащего (HRP)-конъюгированное антитело против myc. После инкубации при комнатной температуре в течение 1 ч планшеты промывают 5 раз PBS, содержащим 0,05% Tween 20. Затем результаты ELISA проявляют путем добавления 50 мкл флуоресцентного субстрата Amplex Red и сигнал прочитывают на флуоресцентном спектрофотометре. Результаты показывают, что большая часть клонов обладает высокой специфичностью к 5E3, поскольку они не распознают 1A6, и что они направлены против вариабельных участков 5E3 (фигура 19).
Подобным образом, очищенные препараты scFv клонов, имеющих разные последовательности и идентифицированных методом фагового ELISA как способные связываться с hIFNγ, анализируют на связывание с hIFNγ в дозо-зависимой манере. Планшеты для ELISA (Maxisorb, NUNC) покрывают в течение ночи раствором hIFNγ в PBS с концентрацией 2 мкг/мл, а контрольные планшеты покрывают раствором БСА в PBS, 2 мкг/мл. Затем планшеты блокируют 3% обезжиренным молоком/PBS при комнатной температуре в течение 1 ч. Планшеты промывают 3 раза PBS, содержащим 0,05% Tween 20, добавляют очищенные scFv в разных концентрациях и инкубируют в течение одного часа при комнатной температуре. Затем планшеты промывают 3 раза PBS, содержащим 0,05% Tween 20. В каждую лунку добавляют 50 мкл 3% обезжиренного молока/PBS, содержащего (HRP)-конъюгированное антитело против myc. После инкубации при комнатной температуре в течение 1 ч планшеты промывают 5 раз PBS, содержащим 0,05% Tween 20. Затем результаты ELISA проявляют путем добавления 50 мкл субстрата TMB и 50 мкл 2N H2SO4, чтобы остановить реакцию. Сигнал прочитывают на спектрофотометре при 450 нм. Результаты показывают, что выбранные клоны связываются с hIFNγ в дозо-зависимой манере и дают хороший сигнал по сравнению с положительным контролем scFv A6, который обладает высокой аффинностью к hIFNγ (фигура 20).
ПРИМЕР 14: Ингибирование под действием ScFv интерферон гамма-индуцированной экспрессии репортерного гена
Ряд выбранных scFv, специфичных к hIFNγ, получают и очищают с помощью описанных выше способов и затем тестируют на способность блокировать биологическую активность hIFNγ. Репортерный ген (люциферазы светлячков), контролируемый hIFNγ-индуцируемым промотором GBP1, трансфицируют в клеточную линию меланомы человека, Me67.8. scFv в разных концентрациях инкубируют с hIFNγ, 2 нг/мл, и затем добавляют в клеточную культуру. После инкубации в течение 6 часов проводят анализ люциферазного репортера и измеряют интенсивность люминесценции. Активность сравнивают с активностью scFv, выделенных из другой библиотеки антител человека scFv, полученной путем традиционного улавливания репертуаров VH/VL от доноров-людей (клон G9). Результаты показывают, что scFv, выделенные как из синтетических, так и из природных библиотек человеческого разнообразия (AE1 и AD1), способны нейтрализовать биологическую активность hIFNγ в дозо-зависимой манере (фигура 21). Нейтрализующая способность этих scFv превосходит уровень scFv клона G9.
ПРИМЕР 15: Ингибирование под действием scFv интерферон гамма-индуцированной экспрессии MHC класса II
Анализ методом проточной цитометрии используют для идентификации полностью антител человека IgG или их фрагментов, способных блокировать экспрессию hIFNγ-индуцированных молекул MHC класса II. После внесения клеток Me67.8 в планшеты к культурам добавляют 5 нг/мл рекомбинантного hIFNγ человека в присутствии разных концентраций кандидатных полностью человеческих моноклональных антител против hIFNγ. После культивирования в течение 48 ч клетки окрашивают флуоресцентно меченным антителом против MHC класса II человека (HLA-DR) и анализируют с помощью FACSCalibur®. Для каждого кандидатного антитела измеряют IC50 (условия, при которых происходит 50% ингибирование hIFNγ-индуцированной экспрессии MHC класса II, т.е., концентрация, вызывающая 50% ингибирование).
Очищенные полностью человеческие scFv получают по описанному выше способу. Влияние выбранных scFv на hIFNγ-индуцированную экспрессию MHC класса II на клетках меланомы анализируют с помощью описанного выше метода проточной цитометрии. Указанные scFv ингибируют hIFNγ-индуцированную экспрессию MHC II на клетках меланомы (фигура 22).
ПРИМЕР 16: Реформатирование scFv в формат IgG
Последовательности VH и VL выбранных scFv амплифицируют с использованием специфических олигонуклеотидов, позволяющих ввести лидерную последовательность и участок рестрикции HindIII в 5'-концевую область. Участок ApaI вводят в 3'-концевую область тяжелой цепи, а участки AvrII и BsiWI вводят в 3'-концевую область последовательности легкой цепи лямбда и каппа, соответственно. Амплифицированные последовательности VH расщепляют HindIII/ApaI и клонируют в векторе экспрессии pCon_gamma1 (LONZA, Basel, Switzerland). Амплифицированные последовательности VL лямбда расщепляют HindIII/AvrII и клонируют в векторе экспрессии pCon_lambda2, а амплифицированные последовательности VL каппа расщепляют HindIII/BsiWI и клонируют в векторе экспрессии pCon_kappa (LONZA, Basel, Switzerland). Перед трансфекцией в клетки млекопитающих структуру конструкций подтверждают секвенированием.
Последовательности кДНК VH и VL в подходящих векторах экспрессии трансфицируют в клетки млекопитающих, используя реагент для трансфекции Fugene 6 (Roche, Basel, Switzerland). Коротко говоря, клетки Peak культивируют в 6-луночных планшетах в концентрации 6×105 клеток на лунку в 2 мл культуральной среды, содержащей фетальную бычью сыворотку. Векторы экспрессии, кодирующие кандидатные последовательности VH и VL совместно трансфицируют в клетки, используя реагент для трансфекции Fugene 6, в соответствии с инструкциями производителя. На следующий день после трансфекции культуральную среду отсасывают, к клеткам добавляют 3 мл свежей бессывороточной среды и культивируют три дня при 37°C. После культивирования в течение трех дней супернатант собирают для получения IgG, очищенных на быстропроточных колонках, содержащих белок G-сефароза 4B (Sigma, St. Louis, MO), в соответствии с инструкциями производителя. Коротко говоря, супернатанты из трансфицированных клеток инкубируют в течение ночи при 4°C с буфером для связывания IgG ImmunoPure (G) (Pierce, Rockford IL). Затем образцы пропускают через быстропроточные колонки, содержащие белок G-сефароза 4B и затем IgG очищают, используя буфер для элюирования. Затем элюированную фракцию IgG диализуют против PBS и определяют количество IgG по абсорбции при 280 нм. Чистоту и целостность IgG подтверждают методом SDS-PAGE.
ПРИМЕР 17: Ингибирование под действием IgG биологической активности интерферона гамма
Два scFv, AE1-4-R3-P2E4 (2E4) и A2-AD1-R4P1A9 (1A9), которые обладают ингибиторной активностью в отношении hIFNγ, подтвержденной в функциональных анализах, реформатируют в IgG, как описано в примере 16, и тестируют с помощью описанного в примере 14 анализа, использующего репортерный ген, индуцируемый интерфероном гамма. Результаты, приведенные на фигуре 23, показывают, что в формате IgG и 1A9, и 2E4 способны нейтрализовать активность hIFNγ с IC50 42 нМ и 10 нМ, соответственно, тогда как используемый в качестве отрицательного контроля IgG (NI-0701) не оказывает эффекта в данном анализе. Таким образом, два указанных кандидата, выделенные из библиотек как с синтетическим, так и с природным разнообразием, можно реформатировать в полноразмерные IgG, обладающие нейтрализующей активностью в отношении выбранной мишени.
ПРИМЕР 18: Разработка фармакокинетического анализа для детекции 5E3 в сыворотке мыши
Два scFv-кандидата, AD15E3R3P1_A4 и AD25E3R3P1_G11, которые специфически связываются с моноклональным антителом 5E3 мыши (фигура 19), реформатируют в полноразмерные IgG человека, как описано в примере 16. Специфичность соответствующих IgG DA4 и G11 подтверждают методом ELISA против 5E3 мыши и химерной версии данного моноклонального антитела, в котором вариабельные участки мыши соединены с константными участками IgG крысы. Результаты, приведенные на фигуре 24, демонстрируют, что IgG DA4 и G11 являются специфичными к вариабельному участку 5E3, поскольку они связываются как с мышиным, так и с химерным 5E3 крысы, и не связываются с мышиными и крысиными изотипическими контролями. Указанные два моноклональные антитела используют для разработки анализа для фармакокинетических исследований, позволяющего количественно определять 5E3 в сыворотке мыши. Несколько разведений сыворотки мыши смешивают с раствором антитела 5E3 мыши, 5 мкг/мл, и проводят серийные разведения так, чтобы концентрация сыворотки поддерживалась постоянной на протяжении серий разведения. Планшеты Maxisorb (Nunc, Denmark) покрывают в течение ночи 1 мкг/мл IgG DA4 или IgG G11. После блокирования PBS/1% БСА в лунки добавляют серийные разведения добавленных препаратов сыворотки. После инкбации и промывания регистрируют сигнал с использованием моноклонального антитела против легкой цепи каппа мыши, конъюгированного с пероксидазой хрена (HRP), и флуоресцентного субстрата (Amplex red; Invitrogen). Результаты показывают, что оба антитела можно использовать для специфической детекции моноклонального антитела 5E3 мыши в сыворотке мыши (фигура 25). Предел детекции 5E3 мыши в сыворотке составляет примерно 200 нг/мл и концентрация сыворотки не оказывает заметного влияния на результаты анализа, свидетельствуя о том, что IgG DA4 и IgG G11 являются высоко специфичными к 5E3 мыши и не связываются с другими иммуноглобулинами мыши. Описанные эксперименты демонстрируют, что высокоспецифичные антиидиотипические антитела нельзя выделить из природных или синтетических библиотек AE1 и AD1.
ПРИМЕР 19: Селекция фагов с использованием библиотек, содержащих разнообразие CDRH3, уловленное из наивных и иммунизированных мышей
Библиотеки MnA, MiB и MiC, описанные в примерах 8 и 9, используют параллельно для селекции фагов против hIFNγ по способу, описанному в примере 11. В процессе селекции наблюдаются похожие обогащения фагов (фигура 26).
Экспрессия scFv в формате титрационного микропланшета: Отдельные клоны помещают в титрационный микропланшет, содержащий 150 мкл среды 2xTYAG (2% глюкозы) на лунку и выращивают при 37°C (100-120 об/мин) в течение 5-6 ч. Планшеты центрифугируют при 280 об/мин, среду отбрасывают, а клеточный осадок ресуспендируют в 100 мкл среды 2xTYA, содержащей 1 мМ IPTG. Планшеты инкубируют в течение ночи при 30°C при встряхивании (100 об/мин). После выращивания планшеты центрифугируют при 3200 об/мин в течение 10 мин и супернатант осторожно переносят в планшет, содержащий 2×PBS, дополненный 5% порошкообразного обезжиренного молока для блокирования.
Анализ scFv методом ELISA: Планшеты ELISA (Maxisorb, NUNC) покрывают в течение ночи раствором hIFNγ в PBS, 2 мкг/мл. Контрольные планшеты покрывают раствором рекомбинантного БСА, 2 мкг/мл (Sigma). Затем планшеты блокируют 3% обезжиренным молоком/PBS при комнатной температуре в течение 1 ч. Планшеты промывают 3 раза PBS, содержащим 0,05% Tween 20, затем переносят супернатанты заранее блокированного scFv и инкубируют в течение одного часа при комнатной температуре. Затем планшеты промывают 3 раза PBS, содержащим 0,05% Tween 20. В каждую лунку добавляют 50 мкл 3% обезжиренного молока/PBS, содержащего (HRP)-конъюгированное антитело против cMyc (разведение 1:5000). После инкубации при комнатной температуре в течение 1 ч планшеты промывают 5 раз PBS, содержащим 0,05% Tween 20. Затем результаты анализа ELISA проявляют путем добавления 50 мкл Amplex Red (Invitrigen). Интенсивность флуоресценции измеряют при 590 нм с возбуждением при 530 нм. Частоту событий, дающих сигнал, составляющий половину от интенсивности контрольного клона A6, оценивают после каждого цикла селекции для трех библиотек (фигура 27). Коэффициент эффективности поиска, полученный для библиотеки MiB, значительно превышает аналогичный коэффициент для двух других библиотек и средний уровень сигнала превышает уровень, наблюдающийся у клонов, полученных из библиотеки MiB, свидетельствуя о том, что происходит обогащение scFv, обладающими повышенной аффинностью (фигура 28). Чтобы подтвердить данное наблюдение, положительные клоны секвенируют, экспрессируют в более крупном масштабе и очищают для применения в экспериментах по связыванию, описанных в примере 13. scFv, полученные из библиотеки MiB, имеют более высокую кажущуюся аффинность к hIFNγ, чем scFv, выделенные из наивной библиотеки MnA (фигура 29). Результаты показывают, что репертуар CDRH3 из мышей, иммунизированных белком, который может улавливаться средой каркасных участков антител человека эффективным способом с получением повышенной встречаемости фрагментов антител человека, обладающих высокой аффинностью. Таким образом, библиотеки, полученные с помощью способа настоящего изобретения, представляют собой мощное средство получения антител с терапевтическим потенциалом.
Другие варианты осуществления
Хотя изобретение описывается совместно с подробным описанием, приведенное выше описание предназначено для иллюстрации, но не для ограничения объема изобретения, который определяется объемом прилагающейся формулы изобретения. Другие аспекты, преимущества и модификации входят в объем нижеследующей формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДЫ И ИХ ПРИМЕНЕНИЯ | 2006 |
|
RU2470941C2 |
| СВЯЗЫВАЮЩИЕ ЭЛЕМЕНТЫ С ИЗМЕНЕННЫМИ ДИВЕРСИФИЦИРОВАННЫМИ ДОМЕНАМИ ОСТОВА | 2017 |
|
RU2778694C2 |
| АНТИТЕЛО ИЛИ ЕГО ФРАГМЕНТ, ИМЕЮЩИЕ НЕЙТРАЛИЗУЮЩУЮ АКТИВНОСТЬ В ОТНОШЕНИИ ВИЧ | 2005 |
|
RU2380378C2 |
| АНТИТЕЛА ПРОТИВ МИЕЛИН-АССОЦИИРОВАННОГО ГЛИКОПРОТЕИНА (MAG) | 2003 |
|
RU2303461C9 |
| ПОЛИПЕПТИДЫ, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2013 |
|
RU2673153C2 |
| АНТИТЕЛО ПРОТИВ С5 И СПОСОБ ПРЕДУПРЕЖДЕНИЯ И ЛЕЧЕНИЯ ОБУСЛОВЛЕННЫХ КОМПЛЕМЕНТОМ ЗАБОЛЕВАНИЙ | 2014 |
|
RU2663349C2 |
| АНТИТЕЛА ПРОТИВ СИГЛЕКА-15 ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С ПОТЕРЕЙ КОСТНОЙ МАССЫ | 2010 |
|
RU2596392C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ АНТИГЕН - ЛИГАНД CD30 ЧЕЛОВЕКА | 2013 |
|
RU2650800C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО К IL-5Rα | 2017 |
|
RU2698048C2 |
| СПОСОБЫ И КОМПОЗИЦИЯ ДЛЯ СЕКРЕЦИИ ГЕТЕРОЛОГИЧНЫХ ПОЛИПЕПТИДОВ | 2010 |
|
RU2585488C2 |




































































































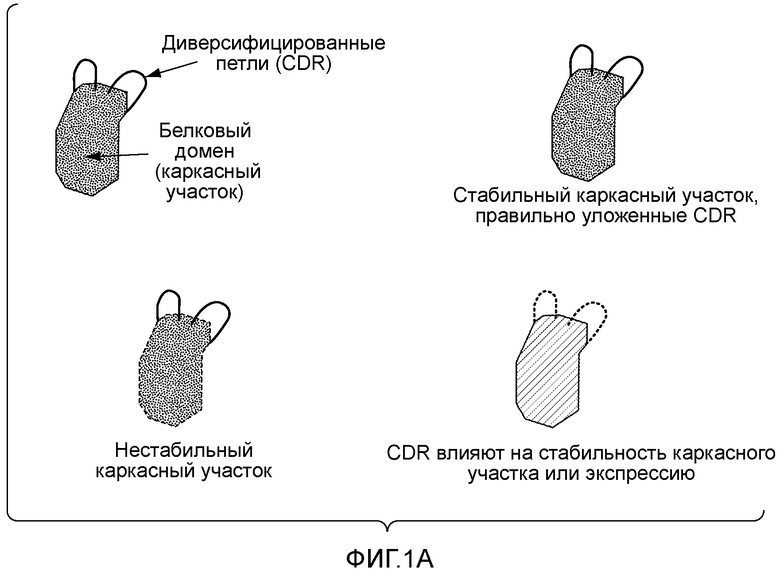
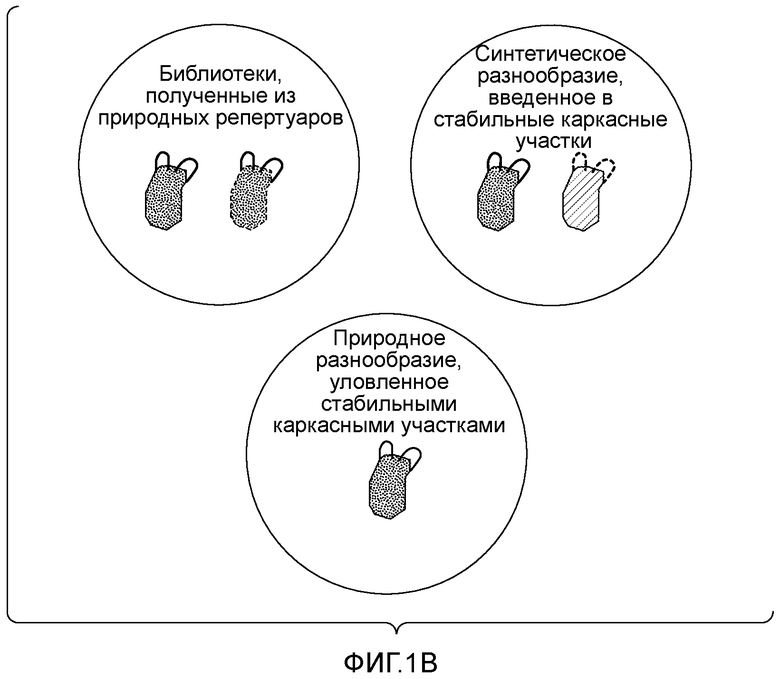

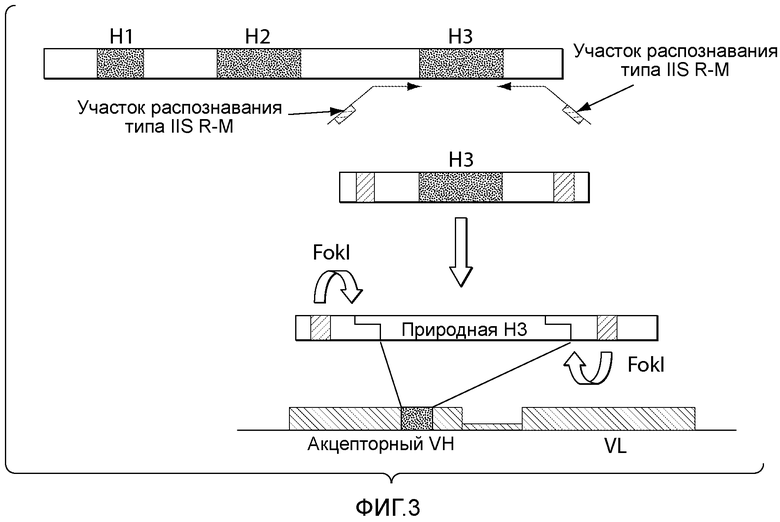
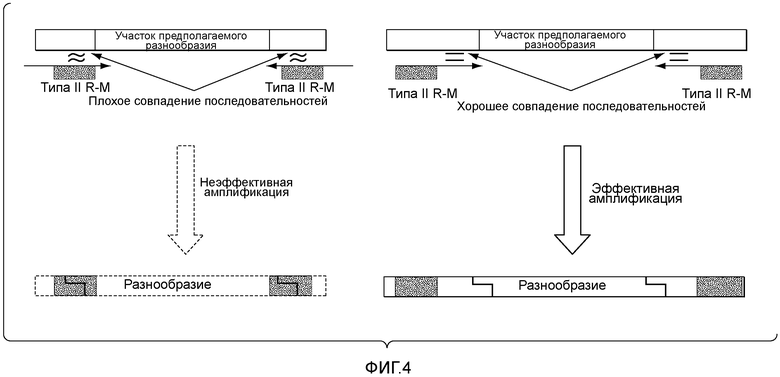

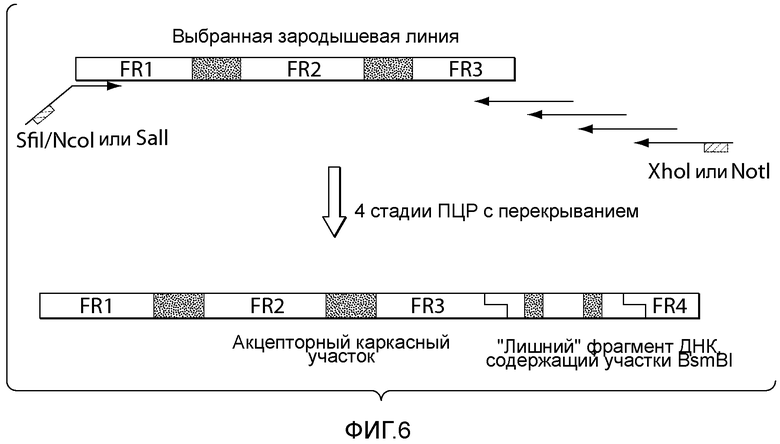
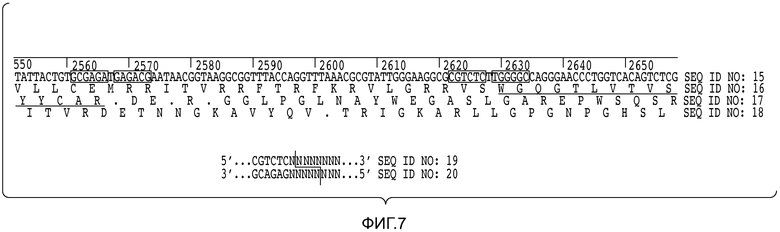




















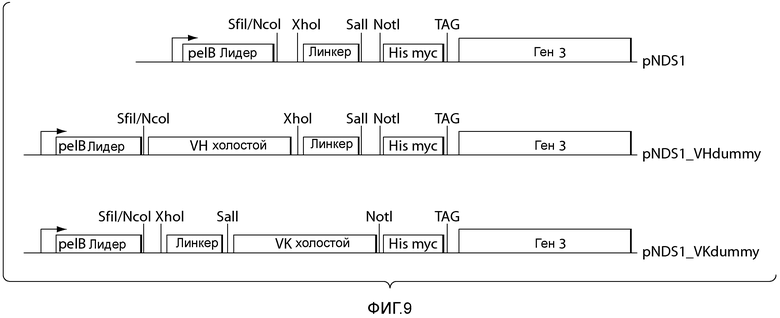
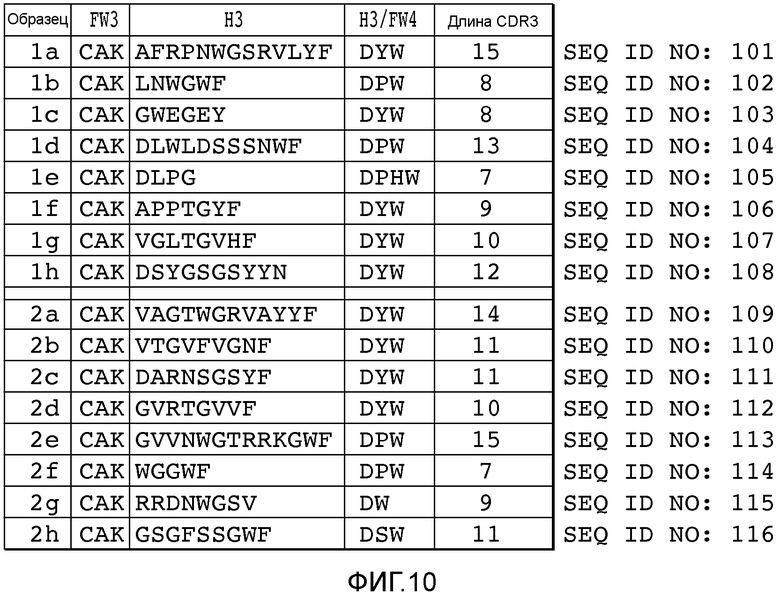
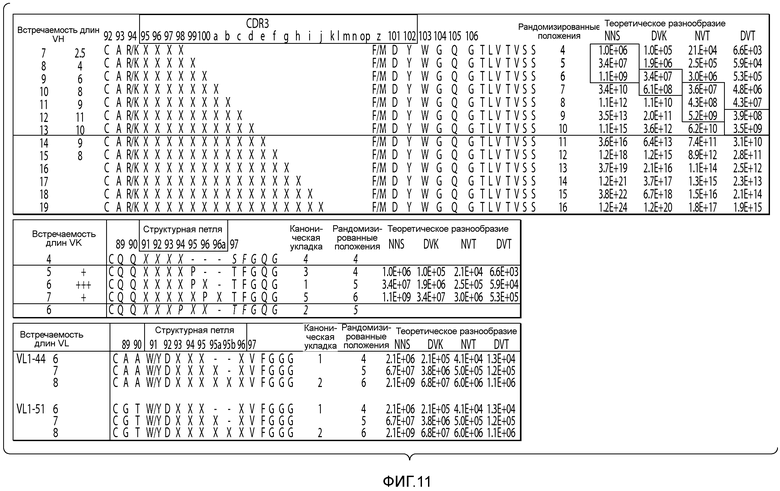
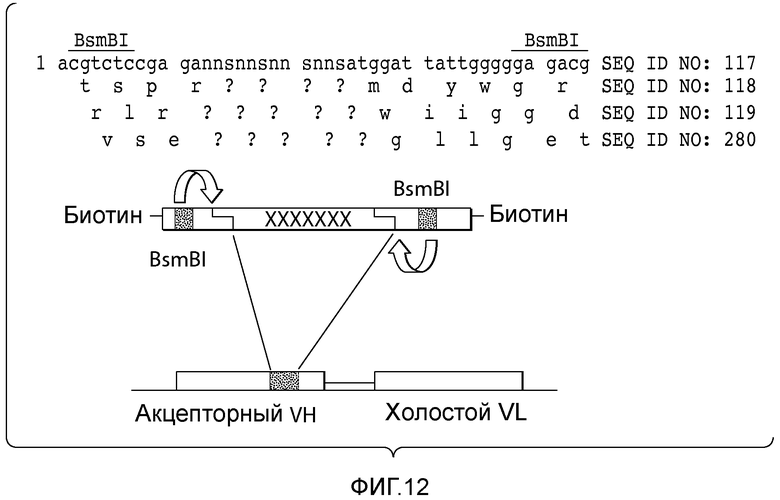
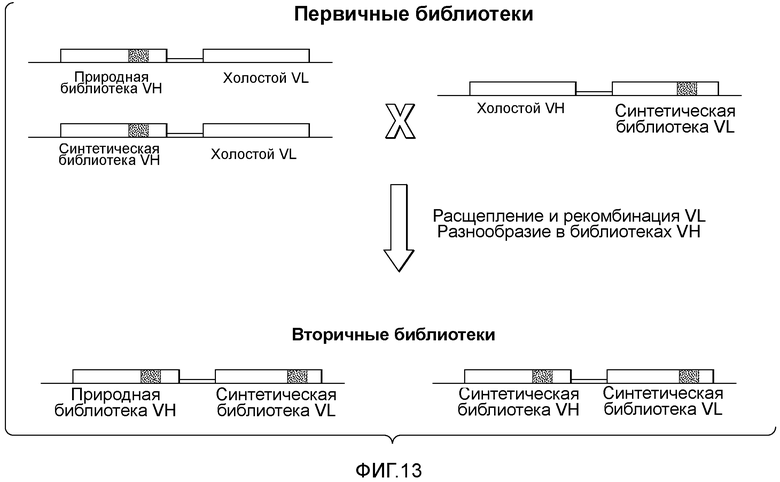
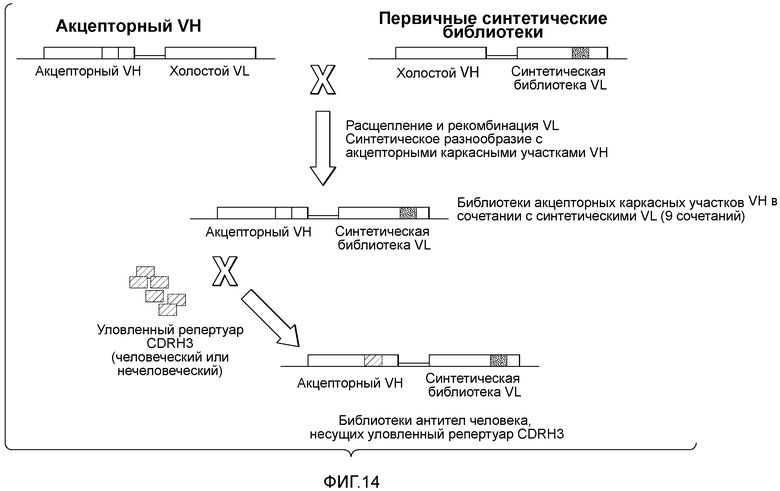
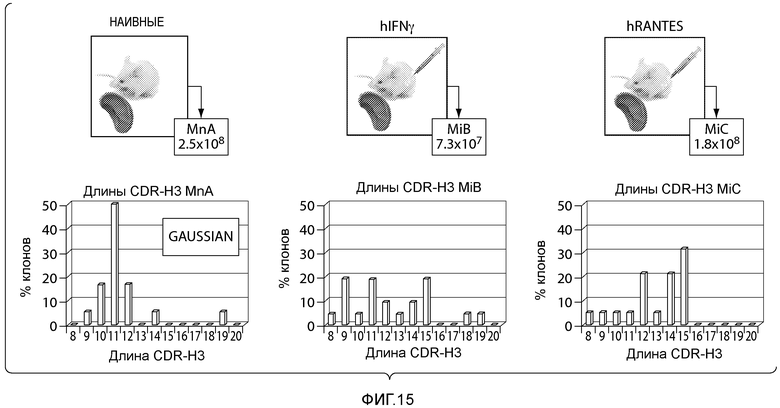
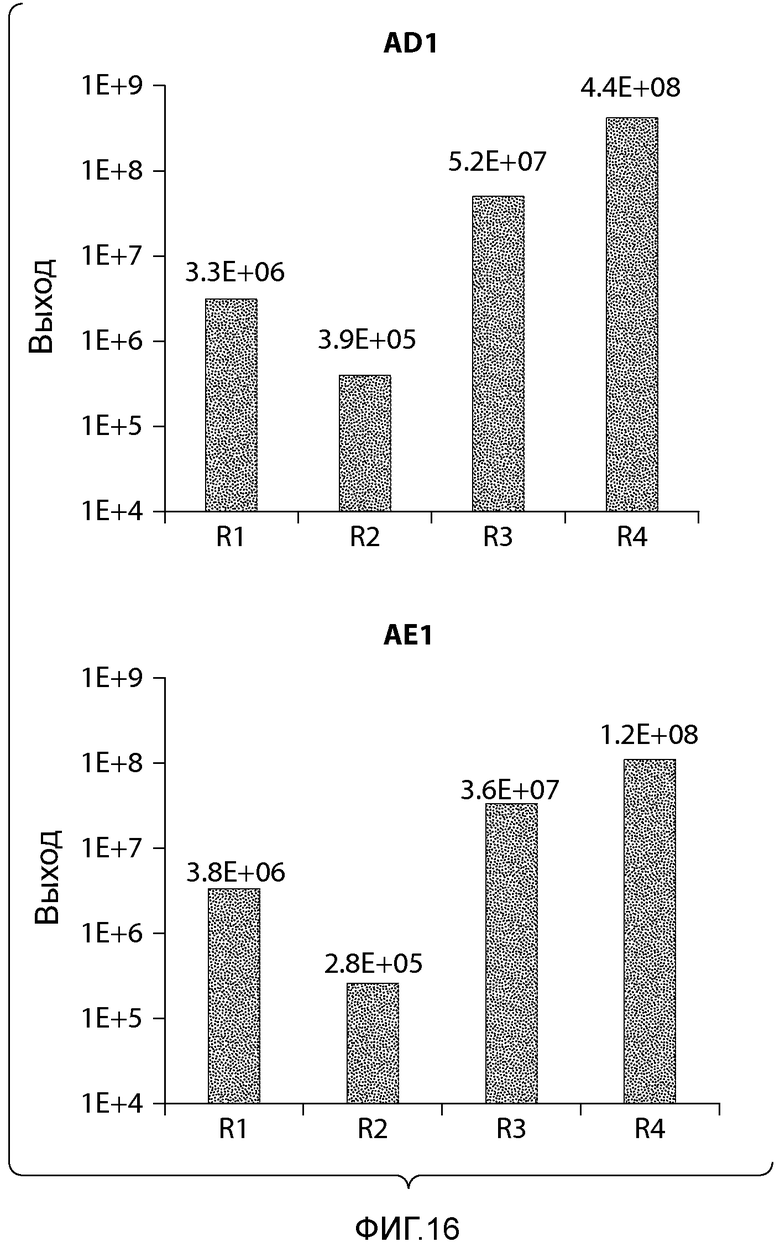
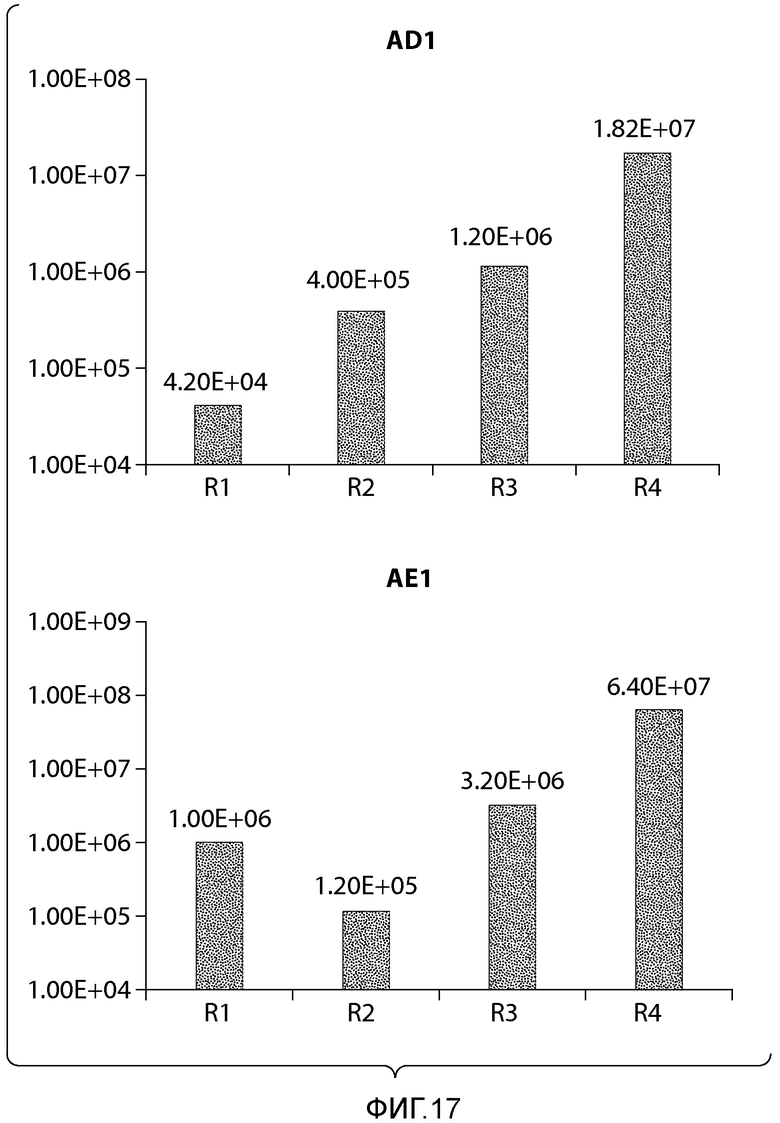
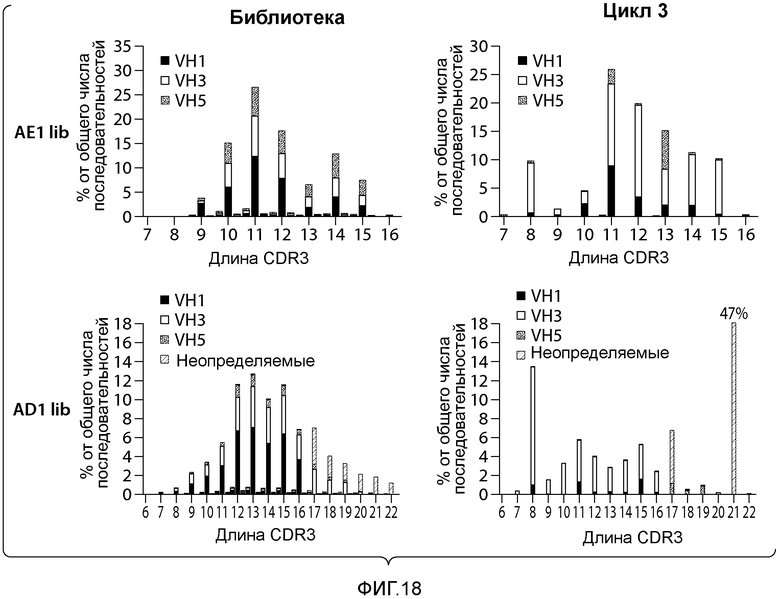
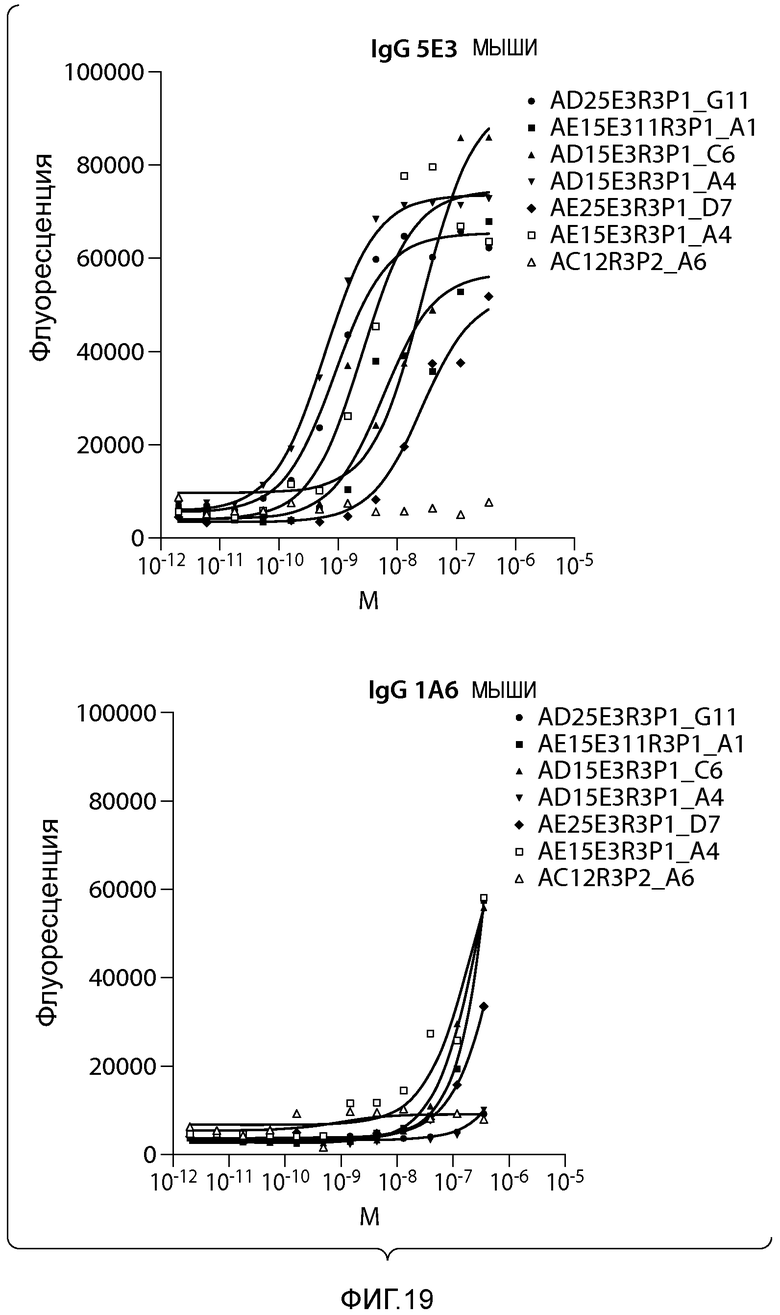
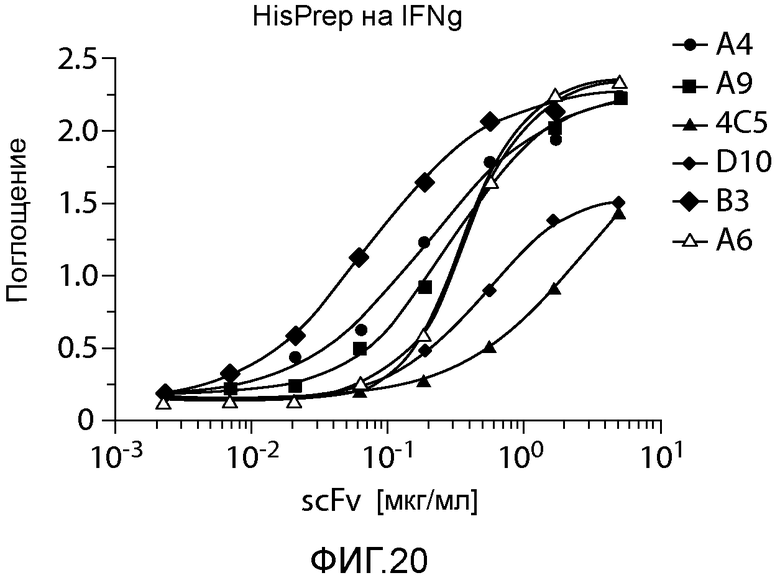
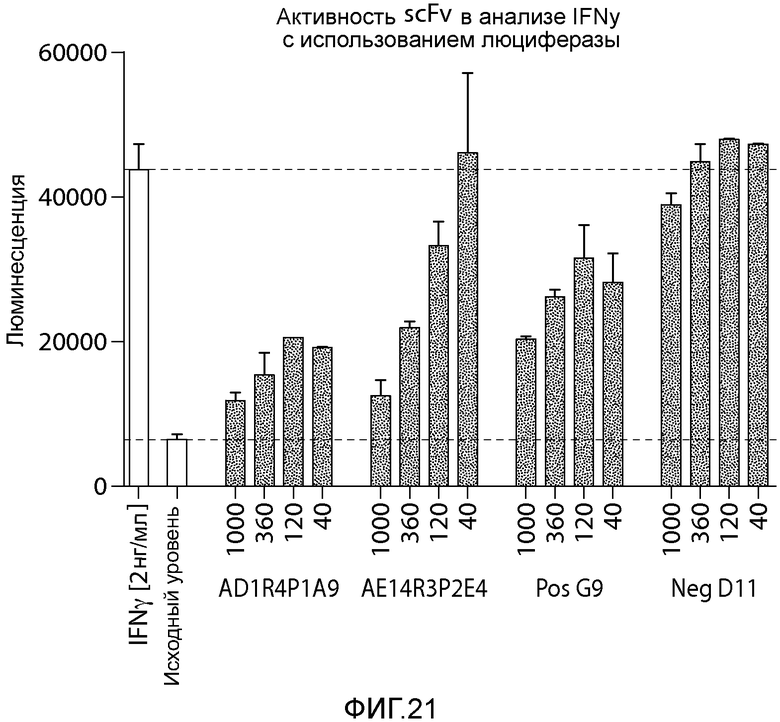
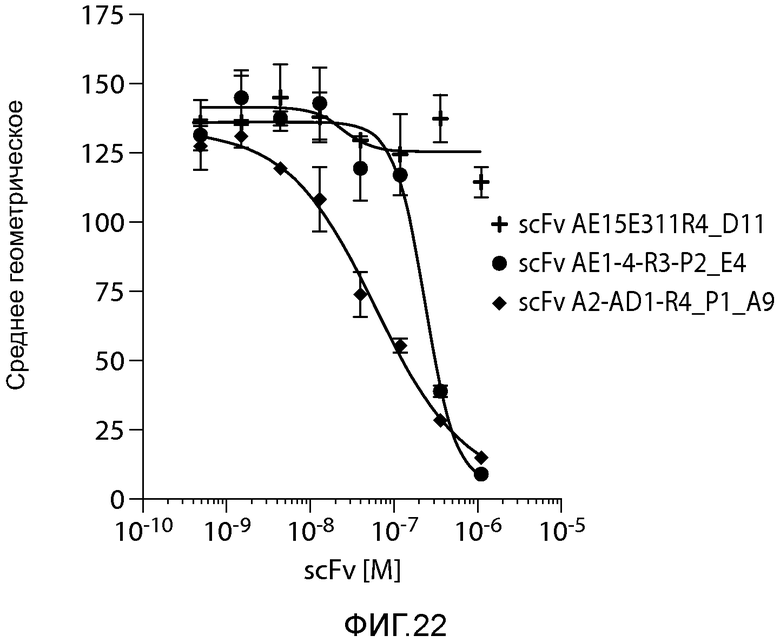
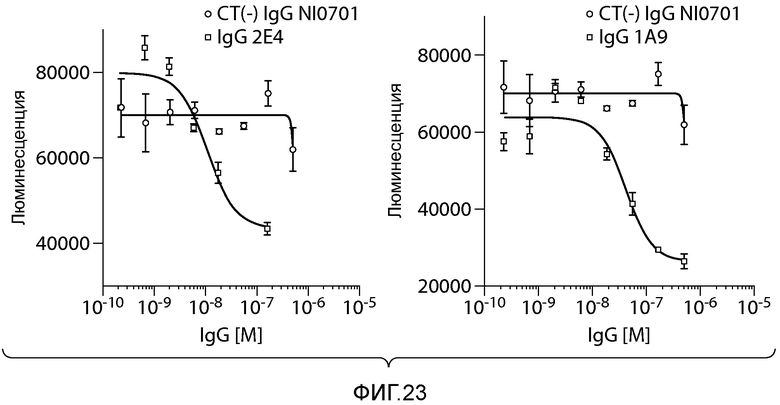
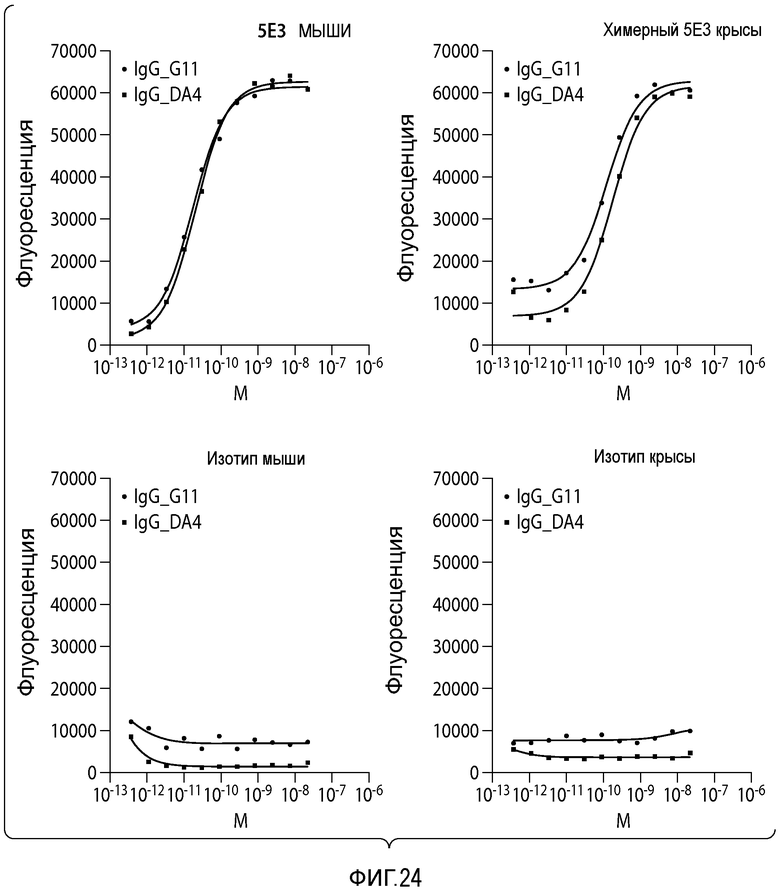

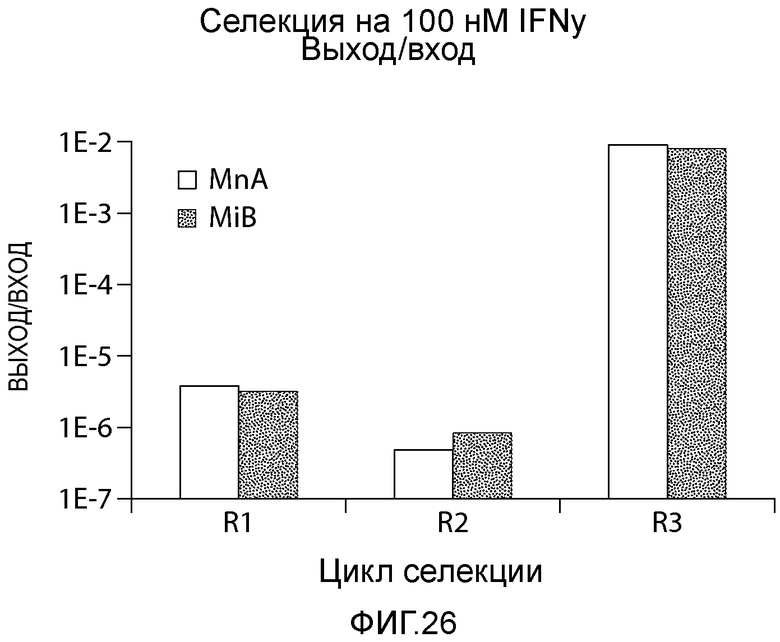
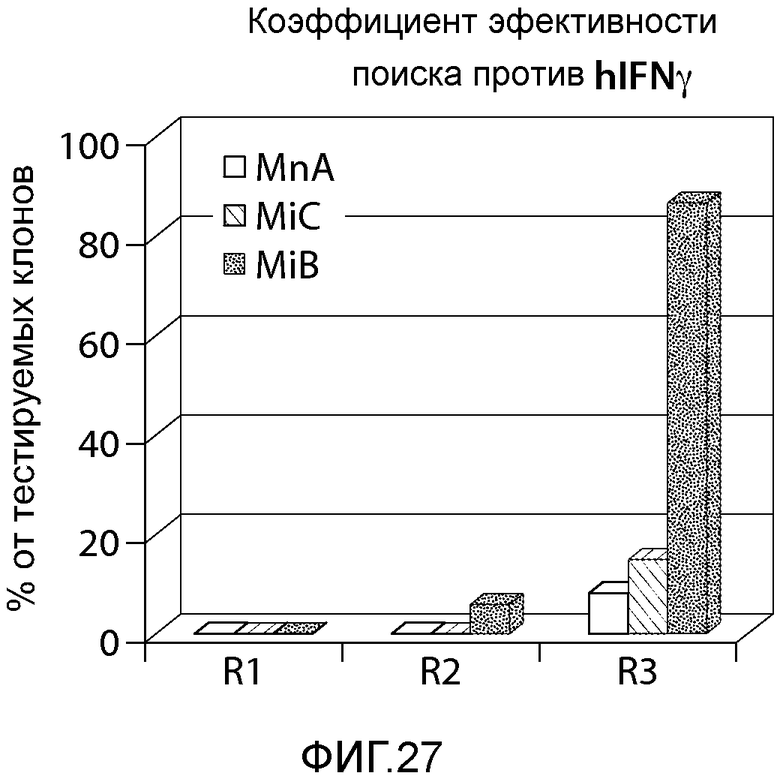
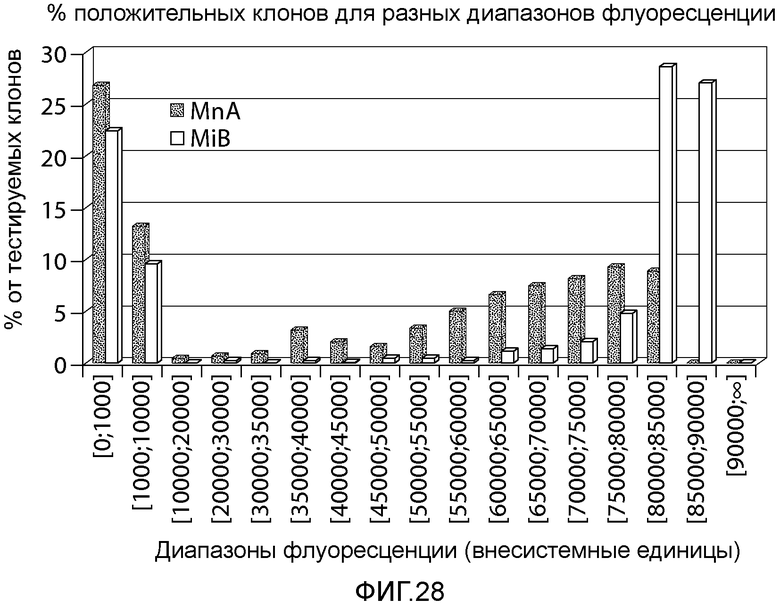
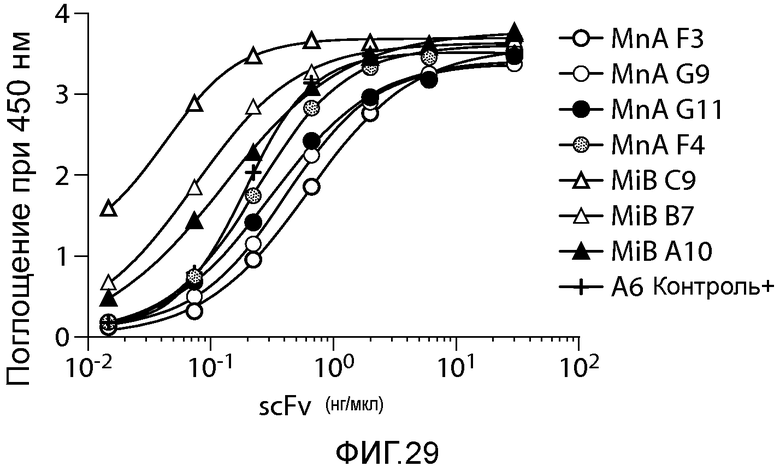
Изобретение относится к области биотехнологии, генной инженерии и иммунологии. Предложены способы получения библиотек последовательностей ДНК, кодирующих гомологичные полипептиды, и к применению таких библиотек для идентификации естественно диверсифицированных полипептидных вариантов. Изобретение также относится к способам, подходящим для получения коллекций синтетических фрагментов антител, в которых один или несколько гипервариабельных участков (CDR) заменены коллекцией соответствующих CDR, полученных из природного источника. Изобретение также относится к способам, подходящим для диверсификации фрагмента полипептида путем вставки диверсифицированной последовательности синтетического или природного происхождения, не требующей модификации последовательности, кодирующей исходный полипептид. Предложенная группа изобретений может быть использована в медицине. 4 н. и 32 з.п. ф-лы, 30 ил., 19 пр.
1. Способ получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий последовательности гипервариабельного участка 3 (CDR3), где каждая последовательность CDR3 в коллекции нуклеиновых кислот выделена по отдельности из репертуара вариабельных доменов иммуноглобулинов млекопитающих, где способ включает:
(a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулина человека, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены "лишней" последовательностью нуклеиновой кислоты, где «лишняя» последовательность нуклеиновой кислоты содержит два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот, где два участка распознавания ферментами рестрикции типа IIs представляют собой участки распознавания ферментами рестрикции типа IIa BsmBI;
(b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих последовательности гипервариабельного участка 3 (CDR3), выделенных из репертуара иммуноглобулинов млекопитающих, где каждая из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs, где каждый участок распознавания ферментами рестрикции типа IIs на каждом конце представляет собой участок распознавания ферментами рестрикции типа IIa FokI;
(c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR3, с помощью фермента рестрикции типа IIs FokI, который связывается с участком распознавания ферментом рестрикции типа IIs FokI, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (а), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs BsmBI, который связывается с участком распознавания ферментом рестрикции типа IIs BsmBI, указанным на стадии (а); и
(d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR3 или аминокислотные последовательности, указанные на стадии (с), в расщепленный акцепторный каркасный участок, указанный на стадии (с) так, чтобы участки FR3 и FR4 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR3 или аминокислотные последовательности, которые могут выполнять роль участка CDR3, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участка распознавания ферментами рестрикции типа IIs BsmBI, как указано на стадии (а), или участка распознавания ферментами рестрикции типа IIs FokI, как указано на стадии(b).
2. Способ получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий последовательность гипервариабельного участка 3 (CDR3), где каждая последовательность CDR3 в коллекции нуклеиновых кислот выделена по отдельности из репертуара вариабельных доменов иммуноглобулинов млекопитающих, отличных от человека, где способ включает:
(a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулина человека, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены "лишней" последовательностью нуклеиновых кислот, где «лишняя» последовательность нуклеиновой кислоты содержит два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот, где два участка распознавания ферментами рестрикции типа IIs представляют собой участки распознавания ферментами рестрикции типа IIa BsmBI;
(b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих последовательности гипервариабельного участка 3 (CDR3), выделенных из репертуара иммуноглобулинов млекопитающих, где каждая из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs, где каждый участок распознавания ферментами рестрикции типа IIs на каждом конце представляет собой участок распознавания ферментами рестрикции типа IIa FokI;
(c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR3, с помощью фермента рестрикции типа IIs FokI, который связывается с участком распознавания ферментом рестрикции типа IIs FokI, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (а), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs BsmBI, который связывается с участком распознавания ферментом рестрикции типа IIs BsmBI, указанным на стадии (а); и
(d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR3 или аминокислотные последовательности, как указано на стадии (с), в расщепленный акцепторный каркасный участок, указанный на стадии (с) так, чтобы участки FR3 и FR4 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR3 или аминокислотные последовательности, которые могут выполнять роль участка CDR3, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs BsmBI, как указано на стадиях (а) и (b), как указано на стадии (а), или участка распознавания ферментами рестрикции типа IIs FokI, как указано на стадии (b).
3. Способ получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий последовательность гипервариабельного участка 3 (CDR3), где каждая последовательность CDR3 в коллекции нуклеиновых кислот выделена по отдельности из репертуара вариабельных доменов иммуноглобулинов человека, где способ включает:
(а) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулина человека, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены "лишней" последовательностью нуклеиновых кислот, где «лишняя» последовательность нуклеиновой кислоты содержит два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот, где два участка распознавания ферментами рестрикции типа IIs представляют собой участки распознавания ферментами рестрикции типа IIa BsmBI;
(b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих последовательности гипервариабельного участка 3 (CDR3), выделенных из репертуара иммуноглобулинов млекопитающих, где каждая из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs, где каждый участок распознавания ферментами рестрикции типа IIs на каждом конце представляет собой участок распознавания ферментами рестрикции типа IIa FokI;
(c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR3, с помощью фермента рестрикции типа IIs FokI, который связывается с участком распознавания ферментом рестрикции типа IIs FokI, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (а), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs BsmBI, который связывается с участком распознавания ферментом рестрикции типа IIs BsmBI, указанным на стадии (а); и
(d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR3 или аминокислотные последовательности, указанные на стадии (с), в расщепленный акцепторный каркасный участок, указанный на стадии (с) так, чтобы участки FR3 и FR4 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR3 или аминокислотные последовательности, которые могут выполнять роль участка CDR3, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs BsmBI, как указано на стадии (а), или участка распознавания ферментами рестрикции типа IIs FokI, как указано на стадии(b).
4. Способ по любому из пп.1-3, где стадию (b) проводят путем амплификации последовательности CDR3 млекопитающих с использованием олигонуклеотидных праймеров, содержащих участок рестрикции типа IIs FokI.
5. Способ по п.4, где олигонуклеотидный праймер конструируют так, чтобы повысить совместимость последовательности CDR3 млекопитающего и акцепторного каркасного участка, кодирующего вариабельный домен иммуноглобулина человека.
6. Способ по п.5, где олигонуклеотидный праймер конструируют так, чтобы модифицировать последовательность нуклеиновых кислот на границе последовательности CDR3 млекопитающих с получением совместимой липкой нуклеотидной последовательности в акцепторном каркасном участке, кодирующей вариабельный домен иммуноглобулина человека.
7. Способ по п.4, где стадию (b) проводят путем амплификации последовательностей CDR3 отличных от человека видов с использованием олигонуклеотидных праймеров, содержащих участок рестрикции Fokl IIs.
8. Способ по п.2, где отличный от человека вид представляет собой грызунов.
9. Способ по любому из пп.1-3, где диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR3, кодируют последовательности CDR3 тяжелой цепи (CDRH3).
10. Способ по любому из пп.1-3, где диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR3, кодируют последовательности CDR3 легкой цепи (CDRL3).
11. Способ по любому из пп.1-3, где последовательность нуклеиновых кислот акцепторного каркасного участка содержит последовательность гена вариабельного домена тяжелой цепи человека, выбранного из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51.
12. Способ по любому из пп.1-3, где последовательность нуклеиновых кислот акцепторного каркасного участка содержит последовательность гена вариабельного домена легкой цепи каппа человека.
13. Способ по п.12, где последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20.
14. Способ по любому из пп.1-3, где последовательность нуклеиновых кислот акцепторного каркасного участка содержит последовательность гена вариабельного домена легкой цепи лямбда человека.
15. Способ по п.14, где последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
16. Способ по любому из пп.1-3, где совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка включает смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
17. Способ по любому из пп.1-3, дополнительно включающий следующие стадии:
(е) стадию клонирования библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулина, указанные на стадии (d), в векторе экспрессии и
(f) стадию трансформации клетки-хозяина вектором экспрессии, указанным на стадии (е), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности кодируемых библиотекой вариабельных доменов иммуноглобулина.
18. Способ по п.17, где клетка-хозяин представляет собой Е. coli.
19. Способ по п.17, где вектор экспрессии представляет собой фагмидный или фаговый вектор.
20. Способ получения коллекции нуклеиновых кислот, где каждая нуклеиновая кислота кодирует вариабельный домен иммуноглобулина человека, содержащий последовательность гипервариабельного участка 3 (CDR3), где каждая последовательность CDR3 в коллекции нуклеиновых кислот выделена по отдельности из вариабельных доменов иммуноглобулинов иммунизированного отличного от человека млекопитающего, где способ включает:
(a) получение совокупности последовательностей нуклеиновых кислот акцепторного каркасного участка, кодирующих разные вариабельные домены иммуноглобулина человека, где каждая последовательность нуклеиновых кислот акцепторного каркасного участка содержит первый каркасный участок (FR1), второй каркасный участок (FR2), третий каркасный участок (FR3) и четвертый каркасный участок (FR4), причем участки FR1 и FR2 разделены гипервариабельным участком 1 (CDR1), участки FR2 и FR3 разделены гипервариабельным участком 2 (CDR2), а участки FR3 и FR4 разделены "лишней" последовательностью нуклеиновых кислот, где «лишняя» последовательность нуклеиновой кислоты содержит два участка распознавания ферментами рестрикции типа IIs, разделенные произвольной последовательностью нуклеиновых кислот, где два участка распознавания ферментами рестрикции типа IIs представляют собой участки распознавания ферментами рестрикции типа IIa BsmBI;
(b) получение совокупности диверсифицированных последовательностей нуклеиновых кислот, кодирующих последовательности гипервариабельного участка 3 (CDR3), выделенные из иммунизированного отличного от человека млекопитающего, где каждая из совокупности диверсифицированных последовательностей нуклеиновых кислот содержит на каждом конце участок распознавания ферментом рестрикции типа IIs, где каждый участок распознавания ферментами рестрикции типа IIS на каждом конце представляет собой участок распознавания ферментами рестрикции типа IIa FokI;
(c) расщепление каждой последовательности из совокупности последовательностей нуклеиновых кислот, кодирующих участки CDR3, с помощью фермента рестрикции типа IIs FokI, который связывается с участком распознавания ферментом рестрикции типа IIs FokI, указанным на стадии (b), и отщепление "лишней" последовательности нуклеиновых кислот, указанной на стадии (а), от акцепторного каркасного участка с помощью фермента рестрикции типа IIs BsmBI, который связывается с участком распознавания ферментом рестрикции типа IIs BsmBI, указанным на стадии (а); и
(d) лигирование расщепленных последовательностей нуклеиновых кислот, кодирующих участки CDR3 или аминокислотные последовательности, указанные на стадии (с), в расщепленный акцепторный каркасный участок, указанный на стадии (с), так, чтобы участки FR3 и FR4 были разделены последовательностями нуклеиновых кислот, кодирующими участок CDR3 или аминокислотные последовательности, которые могут выполнять роль участка CDR3, с восстановлением полноразмерной последовательности, кодирующей вариабельный домен иммуноглобулина, которая не содержит участки распознавания ферментами рестрикции типа IIs BsmBI, как указано на стадии (а), или участка распознавания ферментами рестрикции типа IIs FokI, как указано на стадии (b).
21. Способ по п.20, где стадию (b) проводят путем амплификации последовательности CDR3 иммунизированного отличного от человека млекопитающего с использованием олигонуклеотидных праймеров, содержащих участок рестрикции типа IIs FokI.
22. Способ по п.21, где олигонуклеотидный праймер конструируют так, чтобы повысить совместимость последовательности CDR3 млекопитающего с акцепторным каркасным участком, кодирующим вариабельный домен иммуноглобулина человека.
23. Способ по п.22, где олигонуклеотидный праймер конструируют так, чтобы модифицировать последовательность нуклеиновых кислот на границе последовательности CDR3 млекопитающих с получением липкой нуклеотидной последовательности, совместимой с акцепторным каркасным участком, кодирующим вариабельный домен иммуноглобулина человека.
24. Способ по п.21, где стадию (b) проводят путем амплификации последовательности CDRH3 отличного от человека млекопитающего с использованием олигонуклеотидных праймеров, содержащих участок рестрикции FokI IIs.
25. Способ по п.20, где отличные от человека млекопитающие представляет собой грызунов.
26. Способ по п.20, где диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR3, кодируют последовательности CDR3 тяжелой цепи (CDRH3).
27. Способ по п.20, где диверсифицированные последовательности нуклеиновых кислот, кодирующие последовательности CDR3, кодируют последовательности CDR3 легкой цепи (CDR L3).
28. Способ по п.20, где последовательность нуклеиновых кислот акцепторного каркасного участка содержит последовательность гена вариабельного домена тяжелой цепи человека, выбранную из VH1-2, VH1-69, VH1-18, VH3-30, VH3-48, VH3-23 и VH5-51.
29. Способ по п.20, где последовательность нуклеиновых кислот акцепторного каркасного участка содержит последовательность гена вариабельного домена легкой цепи каппа человека.
30. Способ по п.29, где последовательность гена вариабельного домена легкой цепи каппа человека может быть выбрана из VK1-33, VK1-39, VK3-11, VK3-15 и VK3-20.
31. Способ по п.20, где последовательность нуклеиновых кислот акцепторного каркасного участка содержит последовательность гена вариабельного домена легкой цепи лямбда человека.
32. Способ по п.31, где последовательность гена вариабельного домена легкой цепи лямбда человека может быть выбрана из VL1-44 и VL1-51.
33. Способ по п.20, где совокупность последовательностей нуклеиновых кислот акцепторного каркасного участка содержит смесь по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена тяжелой цепи (VH) и по меньшей мере одной последовательности нуклеиновых кислот акцепторного каркасного участка вариабельного домена легкой цепи.
34. Способ по п.20, дополнительно включающий следующие стадии:
(е) стадию клонирования библиотеки нуклеиновых кислот, кодирующих вариабельные домены иммуноглобулина, как указано на стадии (d), в векторе экспрессии и
(f) стадию трансформации клетки-хозяина вектором экспрессии, указанным на стадии (е), и культивирование клетки-хозяина в условиях, достаточных для экспрессии совокупности кодируемых библиотекой вариабельных доменов иммуноглобулина.
35. Способ по п.34, где клетка-хозяин представляет собой Е. coli.
36. Способ по п.34, где вектор экспрессии представляет собой фагмидный или фаговый вектор.
| REITER Y | |||
| et al., "An antibody single-domain phage display library of a native heavy chain variable region: isolation of functional single-domain VH molecules with a unique interface", J Mol Biol | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |
| HOOGENBOOM H.R | |||
| et al., "Antibody phage display technology and its applications", Immunotechnology | |||
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| АНТИТЕЛА К Aβ-ПЕПТИДУ И ИХ ПРИМЕНЕНИЕ | 2003 |
|
RU2341533C2 |

