СВЯЗАННЫЕ ИЗОБРЕТЕНИЯ
[001] Данное изобретение связано с заявкой РСТ PCT/US12/034575, поданной 20 апреля 2012 года, под названием «Prostate-Specific Membrane Antigen Binding Proteins And Related Compositions And Methods», которая заявляет приоритет Временной патентной заявки США 61/478449, поданной 22 апреля 2011 года, а также связано с Заявкой США №61/636557, все три из которых включены в данный документ посредством ссылки в полном объеме.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[002] Настоящее изобретение связано с моноспецифичными и полиспецифичными белковыми терапевтическими средствами, которые связываются или взаимодействуют с CD3. Оно включает антитела, их фрагменты, scFv, Fab, однодоменные антитела ди-scFv, и полипептиды, содержащие антитела или их фрагменты.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[003] Содержимое текстового файла (название: «2479_1 13PC03_SequenceListing_ascii.txt», размер: 459 722 байта; дата создания: 16 апреля 2013 года), поданного в электронном виде с данной заявкой, который включен в данное описание посредством ссылки в полном объеме.
УРОВЕНЬ ТЕХНИКИ
[004] Нацеливание комплекса TCR на Т-клетки человека с помощью моноклональных анти-СВ3 антител применялось или предлагалось для лечения аутоиммунного заболевания и связанных расстройств, например, в лечении отторжения аллотрансплантанта органа. Моноклональные антитела мыши, специфичные в отношении человеческого CD3, такие как ОКТ3 (Kung et al. (1979) Science 206: 347-9), представляли собой первое поколение таких терапевтических средств. Хотя ОКТ3 обладает выраженной иммуносупрессорной активностью, его клиническое применение было затруднено серьезными побочными эффектами, связанными с его иммуногенным и митогенным потенциалом (Chatenoud (2003) Nature Reviews 3:123-132). Оно индуцировало антиглобулиновый ответ, что способствовало его быстрому клиренсу и нейтрализации (Chatenoud et al. (1982) Eur. J. Immunol. 137:830-8). Кроме того, ОКТ3 индуцирует пролиферацию Т-клеток и выработку цитокинов in vitro, а также приводит к высвобождению цитокинов in vivo в больших масштабах (Hirsch et al. (1989) J. Immunol 142:737-43, 1989). Высвобождение цитокинов (также обозначаемое как «цитокиновый шторм»), в свою очередь, приводит к «гриппоподобному» синдрому, который характеризуется лихорадкой, ознобом, головной болью, тошнотой, рвотой, диареей, дыхательной недостаточностью, септическим менингитом и гипотензией (Chatenoud, 2003). Такие серьезные побочные эффекты ограничили более широкое применение ОКТЗ при трансплантации, а также расширение его применения на другие клинические области, например, аутоиммунные явления (Id.).
[005] С целью уменьшения побочных эффектов первого поколения моноклональных анти-CD3 антител, второе поколение генетически сконструированных моноклональных анти-CD3 антител было разработано не только путем пересадки определяющих комплементарность участков (CDR) мышиных моноклональных анти-CD3 антител в человеческие последовательности IgG, но и введения не-FcR-связывающих мутаций в Fc, с целью снижения частоты возникновения цитокинового шторма (Cole et al. (1999) Transplantation 68: 563; Cole et al. (1997) J. Immunol. 159: 3613). См. также Публикацию PCT WO 2010/042904, которая включена в данное описание посредством ссылки в полном объеме.
[006] В дополнение к моноспецифичным терапевтическим средствам, нацеленным на CD3, полиспецифичные полипептиды, которые избирательно связываются с Т-клетками и клетками опухоли, могли бы предложить механизм перенаправления цитотоксичности Т-клеток на клетки опухоли и лечение рака. Однако одной из проблем при конструировании биспецифичного или полиспецифичного антитела, рекрутирующего Т-клетки, состояла в сохранении специфичности при одновременном подавлении регулирования активации Т-клеток множественными регуляторными путями.
[007] Остается потребность в улучшенных моноспецифичных и полиспецифичных анти-CD3 молекулах. Хотя предыдущие улучшения были произведены в части Fc нацеленной на CD3 молекулы, с целью снижения вероятности цитокинового шторма, по-прежнему существует потребность в анти-CD3 терапевтических средствах с увеличенным периодом полувыведения по сравнению с предшествующими молекулами из уровня техники, которые могли бы эффективно производиться, демонстрировать улучшенное связывание с Т-клетками и/или улучшенное перенаправление цитотоксичности Т-клеток (в случае полиспецифичных молекул, разработанных для лечения рака).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[008] В одном из аспектов настоящее изобретение включает полипептид, связывающийся с CD3, который содержит домен связывания с CD3 со сниженной изоэлектрической точкой, по сравнению с доменом связывания с последовательностью аминокислот SEQ ID NO: 41. В некоторых вариантах реализации домен связывания с CD3 включает участок VH и VL, причем каждый участок VH и VL содержит каркасные участки. Для снижения изоэлектрической точки (пи) домена связывания и/или полипептида, две или более аминокислот могут быть модифицированы в каркасных участках посредством замены положительно заряженных аминокислот нейтральными аминокислотами и/или замены нейтральных аминокислот отрицательно заряженными аминокислотами. Например, R и Т могут быть заменены S, Q может быть заменен Е, Y может быть заменен F, и Т может быть заменен V. В одном варианте реализации, например, с целью снижения риска иммуногенности, аминокислоты, которые заменены в последовательности, преобладают в последовательности IgG зародышевой линии человека или содержатся в последовательности зародышевой линии человека в таком же или ближайшем положении, например, в соответствующей последовательности IgG зародышевой линии человека. В некоторых вариантах реализации последовательность IgG зародышевой линии человека включает SEQ ID NO: 43 или 44.
[009] В другом варианте реализации изобретения полипептид, связывающийся с CD3, включает модификации в предшарнирном участке, которые приводят к снижению изоэлектрической точки предшарнирного участка или полного полипептида. Предшарнирный участок находится в соединении между доменом связывания и шарнирным участком. Например, предшарнирный участок из 3-х аминокислот с последовательностью RRT может быть заменен последовательностью SSS, с целью снижения изоэлектрической точки. В некоторых вариантах реализации предшарнирный участок обладает сниженной изоэлектрической точкой по сравнению с полипептидом, связывающимся с CD3, с предшарнирным участком RRT.
[0010] Полипептиды, связывающиеся с CD3, могут быть сконструированы таким образом, чтобы снизить изоэлектрическую точку, например, посредством мутации каркасного участка(-ов) участков VH и/или VL и/или предшарнирного участка. Снижение изоэлектрической точки может быть на 0,5-2,5 или более единиц.
[0011] В одном варианте реализации изобретения, полипептид, связывающийся с CD3, представляет собой антитело, например, гуманизированное. В другом варианте реализации он представляет собой низкомолекулярный модульный иммунофармацевтический белок (SMIP). В некоторых вариантах реализации полипептид, связывающийся с CD3, содержит одноцепочечный вариабельный фрагмент (scFv).
[0012] В другом варианте реализации он представляет собой полиспецифичный или биспецифичный полипептид. Например, полипептид, связывающийся с CD3, может содержать домен связывания с CD3 и домен связывания с антигеном опухоли. В одном варианте реализации биспецифичная или полиспецифичная молекула перенаправляет цитотоксичность Т-клеток на клетку опухоли.
[0013] В другом варианте реализации изобретения полипептид CD3 существует, как гомодимер или гетеродимер.
ОПИСАНИЕ ФИГУР
[0014] Фиг. 1 представляет собой график, на котором проиллюстрирована теоретическая изоэлектрическая точка гуманизированных полипептидов Cris-7 SMIP, сконструированных ранее, по сравнению с теоретической изоэлектрической точкой около 200 других полипептидов SMIP.
[0015] Фиг. 2А и 2В иллюстрируют отсечение SMIP полипептидов, связывающихся с CD3, с более высокой изоэлектрической точкой, по сравнению с полипептидами CD3 по изобретению. Фиг. 2А иллюстрирует капиллярный электрофорез в натрия лаурилсульфате, с помощью которого количественно определяется содержание низкомолекулярного отсечения очищенных белков. Фиг. 2В иллюстрирует отсеченные молекулы CD3 SMIP. Последовательности, представленные на Фиг. 2В, соответствуют аминокислотам 252-260 SEQ ID NO: 4, 243-252 SEQ ID NO: 4 и 251-260 SEQ ID NO: 240.
[0016] Фиг. 3 иллюстрирует выравнивание последовательностей аминокислот VH Cris-7 мыши и производных гуманизированных последовательностей VH. CrisT VH мыши, H1, Н2, Н3, Н4, Н5, Н6 и консенсусная последовательность соответствуют SEQ ID NO: 45,22,24, 26,28, 30, 32 и 229, соответственно.
[0017] Фиг. 4 иллюстрирует выравнивание последовательностей аминокислот VL Cris-7 мыши и производных гуманизированных последовательностей VL. Cris7 VL мыши, LI, L2, L3, L4 и консенсусная последовательность соответствуют SEQ ID NO: 46, 34, 36, 38,40 и 230, соответственно.
[0018] Фиг. 5 иллюстрирует выравнивание последовательностей аминокислот Н3 и последовательностей зародышевой линии IGHV1-69*01 и IGHV3-30*01. Н3, IGHV1-69*01, IGHV3-30*01 и консенсусные последовательности соответствуют SEQ ID NO: 26,43,44 и 331, соответственно.
[0019] Фиг. 6 иллюстрирует выравнивание последовательностей аминокислот участка каппа J участка VL последовательности зародышевой линии IGKV1-5*01 и L1. Участок J каппа участка VL (IGKVI-5*01), цепи L1 и консенсусных последовательностей соответствует SEQ ID NO: 232, 34 и 233, соответственно. Перечисленные последовательности IGKJ1*01, IGKJ2*01, IGKJ3*01, IGKJ4*01 и IGKJ5*01 соответствуют SEQ ID NO: 234-238, соответственно.
[0020] Фиг. 7A и 7B иллюстрируют выравнивание последовательностей аминокислот различных SMIP молекул полипептида, связывающегося с CD3, со сниженным теоретическим значением изоэлектрической точки, по сравнению с DRA209. Последовательности, приведенные для DRA209, DRA219, DRA222, DRA221, DRA223, DRA224, DRA225 и консенсусная последовательность соответствуют аминокислотам 1-423 SEQ ID NO: 4, аминокислотам 1-423 SEQ ID NO: 6, аминокислотам 1-423 SEQ ID NO: 8, аминокислотам 1-423 SEQ ID NO: 10, аминокислотам 1-423 SEQ ID NO: 12, аминокислотам 1-426 SEQ ID NO: 14, аминокислотам 1-426 SEQ ID NO: 16 и SEQ ID NO: 239, соответственно.
[0021] Фиг. 8 иллюстрирует выравнивание последовательностей аминокислот вариантов изоэлектрической точки DRA233 и DRA234, по сравнению с DRA161. Последовательности, приведенные для DRA161, DRA233, DRA234, и консенсусная последовательность соответствуют SEQ ID NO: 240,18, 20 и 241, соответственно.
[0022] Фиг. 9 иллюстрирует, каким образом были определены эмпирические значения изоэлектрической точки.
[0023] Фиг. 10 представляет собой график, иллюстрирующий эмпирическое значение изоэлектрической точки DRA209 SMIP и scFv.
[0024] Фиг. 11 представляет собой график, иллюстрирующий эмпирическое значение изоэлектрической точки DRA161 SMIP по сравнению с вариантами изоэлектрической точки DRA233 и DRA234.
[0025] Фиг. 12 представляет собой график, иллюстрирующий такое же связывание вариантов изоэлектрической точки SMIP с Т-клетками, как и в случае DRA209 и DRA161, за исключением DRA227. DRA227 SMIP используется в качестве контроля и не содержит домена связывания с CD3 или Т-клетками. Буква «а» после каждого обозначения молекулы указывает, что конкретная молекула была получена в клетках яичника китайского хомяка (ЯКХ), а четырехзначное число представляет собой номер серии.
[0026] На Фиг. 13 представлены изображения невосстанавливающего НЛС-ПАГЕ, демонстрирующие, что связывающиеся с CD3 конструкты SMIP DRA233 и DRA234 менее склонны к фрагментации, чем SMIP DRA161 (который разделяет VH и VL с DRA209).
[0027] На Фиг. 14 представлены изображения восстанавливающего НЛС-ПАГЕ, демонстрирующие, что связывающиеся с CD3 конструкты SMIP DRA233 и DRA234 менее склонны к фрагментации, чем SMIP DRA161 (который разделяет VH и VL с DRA209).
[0028] Фиг. 15 представляет собой графики и таблицу, которые иллюстрируют, что варианты изоэлектрической точки DRA233 и DRA234 преимущественно экспрессируются интактными (например, отсутствует отсечение).
[0029] На Фиг. 16 представлены изображения, иллюстрирующие невосстановленный КЭ-НЛС DRA161 (очищенный).
[0030] Фиг. 17 иллюстрирует данные исследования ФК DRA234 и DRA233.
[0031] На Фиг. 18 проиллюстрированы данные анализа WinNonLin для DRA233 и DRA234, как описано в Примере 14.
[0032] Фиг. 19 представляет собой таблицу ФК оценок для DRA233 и DRA234.
[0033] Фиг. 20 представляет собой таблицу параметров ФК для DRA161, по сравнению с DRA233 (со временной точкой 504 часа и без нее) и DRA234.
[0034] Фиг. 21 иллюстрирует результаты зависимых от мишени анализов пролиферации Т-клеток с использованием различных гомодимерных полипептидных биспецифичных молекул, нацеленных на CD19. Фиг. 21А и 21В иллюстрируют результаты пролиферации Т-клеток CD4+ и пролиферации Т-клеток CD8+, соответственно, как описано в Примере 10.
[0035] Фиг. 22 иллюстрирует результаты зависимого от мишени анализа пролиферации Т-клеток с использованием различных гетеродимерных молекул биспецифичного полипептида, нацеленных на CD 19. Фиг. 22А и 22В иллюстрируют результаты пролиферации Т-клеток CD8+ и пролиферации Т-клеток CD4+, соответственно, как описано в Примере 10.
[0036] Фиг. 23 иллюстрирует результаты анализа перенаправленной цитотоксичности Т-клеток с использованием различных полипептидных гетеродимеров и гомодимеров, нацеленных на CD19. Фиг. 23А иллюстрирует результаты для гомодимерных полипептидов биспецифичного TSC129a, TSC233 и TSC234. Фиг. 23В иллюстрирует результаты для гетеродимерных биспецифичных полипептидов TSC127, TSC227 и TSC228.
[0037] Фиг. 24 иллюстрирует результаты анализа перенаправленной цитотоксичности Т-клеток с использованием TSC275, TSC277, TSC278 и TSC279, которые представляют собой различные гомодимеры биспецифичных полипептидов, нацеленные на RON.
[0038] Фиг. 25 иллюстрирует результаты связывания с Т-клетками гетеродимеров и гомодимеров биспецифичных полипептидов. Фиг. 25А и 25В иллюстрируют дозозависимое связывание биспецифичных молекул TSC228 (гетеродимер) и TSC249 (гомодимер) с клетками Jurkat.
[0039] Фиг. 26 иллюстрирует результаты зависимой от мишени пролиферации Т-клеток под действием гомодимеров полипептида, нацеленных на простатоспецифичный мембранный антиген (ПСА). Фиг. 26А иллюстрирует результаты для клеток С4-2 В, экспрессирующих целевой простатоспецифический мембранный антиген (ПСМА) при обработке TSC249. Фиг. 26В иллюстрирует результаты для клеток DU-145, которые не экспрессировали целевого антигена ПСМА при обработке TSC249. Фиг. 26С иллюстрирует результаты после обработки выделенных Т-клеток TSC249.
[0040] Фиг. 27 иллюстрирует результаты анализа перенаправленной цитотоксичности Т-клеток с использованием гомодимерных биспецифичных полипептидов TSC194 и TSC249, оба из которых нацелены на ПСМА.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0041] В изобретении предложены полипептиды, связывающиеся с CD3, с улучшенными характеристиками, по сравнению с полипептидами, связывающимися с CD3, из уровня техники. Молекулы по изобретению сконструированы таким образом, чтобы продемонстрировать сниженную изоэлектрическую точку, посредством модификации аминокислот в каркасных участках тяжелой и/или легкой цепи и/или предшарнирном участке, с целью снижения изоэлектрической точки молекулы. В одном варианте реализации модификации произведены на участке каппа J участка VL. В некоторых вариантах реализации модификации произведены посредством замены положительно заряженных аминокислот на аминокислоты с нейтральным зарядом и/или замены аминокислот с нейтральным зарядом на отрицательный заряд.
[0042] В некоторых вариантах реализации полипептид по изобретению, связывающийся с CD3, обладает теоретическим значением изоэлектрической точки на около 0,5-2,5 единиц меньше, чем анти-CD3 DRA209 SMIP (SEQ ID NO: 4). Например, DRA209 обладает теоретическим значением изоэлектрической точки 9, тогда как вариант изоэлектрической точки DRA219 (SEQ ID NO: 6) обладает теоретическим значением изоэлектрической точки 8,4, DRA221 (SEQ ID NO: 10) обладает теоретическим значением изоэлектрической точки 8,2, DRA222 (SEQ ID NO: 8) обладает теоретическим значением изоэлектрической точки 7,5, DRA223 (SEQ ID NO: 12) обладает теоретическим значением изоэлектрической точки 7,2 и DRA224 (SEQ ID NO: 14) обладает теоретическим значением изоэлектрической точки 6,8. В некоторых вариантах реализации полипептид, связывающийся с CD3, обладает эмпирической изоэлектрической точкой, по меньшей мере на 1 единицу меньшей, чем полипептид SEQ ID NO: 4.
[0043] Неожиданно, варианты изоэлектрической точки демонстрируют улучшенный период полувыведения и лучше экспрессируются в клетках СНО, чем DRA209, например, см. Пример 4 ниже, в котором описан способ измерения периода полувыведения. Кроме того, полиспецифичные конструкты, содержащие DRA222 scFv, продемонстрировали улучшенные свойства, по сравнению с подобными конструктами с другими доменами связывания анти-CD3, такими как домен связывания DRA209 (см. связанную Публикацию РСТ WO 2012/145714).
[0044] Заголовки разделов, используемые в настоящем документе, предназначены только для организационных целей и не должны интерпретироваться как ограничивающие описанный объект. Все документы или части документов, цитируемые в настоящем документе, в том числе, но не ограничиваясь ими, патенты, патентные заявки, статьи, книги и трактаты, таким образом, явно включены в данный документ посредством ссылки в полном объеме для любой цели. В случае если один или более включенных документов или частей документов определяют термин, который противоречит определению данного термина в заявке, преобладает определение, приведенное в данной заявке.
[0045] В настоящем документе любой интервал концентрации, процентный интервал, интервал соотношения или целочисленный интервал следует интерпретировать как включающий значение любого целого числа в пределах заявляемого интервала и, в соответствующих случаях, его частей (например, одна десятая и одна сотая часть целого числа), если не указано иное. В данном документе, «около» обозначает ±20% номинального интервала, значения или структуры, если не указано иное. Следует понимать, что указание на форму единственного числа в настоящем документе обозначает «один или более» перечисленных компонентов, если не указано иное. Использование альтернативы (например, «или») следует понимать, как подразумевающее любую одну, обе, или любую комбинацию указанных альтернатив. В данном документе, термины «включают» и «содержат» используются синонимическим образом. Кроме того, следует понимать, что полипептиды, содержащие различные комбинации компонентов (например, домены или участки) и заместителей, описанных в настоящем документе, раскрыты в настоящей заявке в такой же степени, как если бы каждый полипептид был указан индивидуально. Таким образом, выбор конкретных компонентов индивидуальных полипептидов находится в пределах контекста настоящего описания.
[0046] В данном документе термин «домен связывания» или «участок связывания» обозначает домен, участок, часть или сайт белка, полипептида, олигопептида или пептида, который обладает способностью специфично распознавать и связываться с молекулой мишени, такой как антиген, лиганд, рецептор, субстрат или ингибитор (например, CD3 или ассоциирующийся с опухолью антиген, такой как RON, CD19, CD37 или ПСМА). Примеры доменов связывания включают одноцепочечные вариабельные участки (например, доменные антитела, sFv, scFv и scFab) антитела, рецепторные эктодомены и лиганды (например, цитокины, хемокины). В некоторых вариантах реализации домен связывания включает или состоит из сайта связывания с антигеном (например, содержащего вариабельную последовательность тяжелой цепи и вариабельную последовательность легкой цепи или три определяющих комплементарность участка (CDR) легкой цепи и три CDR тяжелой цепи от антитела, размещенные в альтернативных каркасных участках (FR) (например, человеческие FR, необязательно содержащие одну или более замен аминокислот). Различные анализы известны для идентификации доменов связывания по настоящему изобретению, которые специфично связываются с конкретной мишенью, в том числе, вестерн-блоттинг, ТИФА, скрининг библиотеки показа фага и анализ взаимодействия BIACORE®. В данном документе, полипептид, связывающийся с CD3, может содержать «домен связывания с CD3» и, необязательно, «второй домен связывания». Домен связывания с CD3 может быть размещен на амино- или карбокси-конце. В некоторых вариантах реализации домен связывания с CD3 включает гуманизированный scFv, происходящий от моноклонального антитела мыши (например, Cris-7 или HuM291). Например, домен связывания с CD3 может состоять из участка VH и участка VL, происходящего от моноклонального антитела мыши, причем участки VH и VL соединены линкером, таким как линкер (Gly4Ser)3 (SEQ ID NO: 76). В других вариантах реализации домен связывания с CD3 состоит по существу из или состоит из гуманизированного scFv, происходящего от моноклонального антитела мыши. В некоторых вариантах реализации домен связывания с CD3 по изобретению содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи мышиного антитела Cris-7 или HuM291.
[0047] Домен связывания «специфично связывается» с мишенью, если он связывается с мишенью с аффинностью или Ка (т.е., константа ассоциации равновесия конкретного взаимодействия связывания с единицами 1/М), больше или равной 105 М-1, и, в то же время, в существенной мере не связывается с другими компонентами, присутствующими в испытуемом образце. Домены связывания могут быть классифицированы как домены связывания с «высокой аффинностью» и домены связывания с «низкой аффинностью». Домены связывания с «высокой аффинностью» обозначают связывающие домены с Ка по меньшей мере 107 М-1, по меньшей мере 108 М-1, по меньшей мере 109 М-1, по меньшей мере 1010 М-1, по меньшей мере 1011 М-1, по меньшей мере 1012 М-1 или по меньшей мере 1013 М-1. Домены связывания с «низкой аффинностью» обозначают домены связывания с Ка до 107 М-1, до 106 М-1, до 105 М-1. Альтернативно, аффинность может определяться по константе равновесной диссоциации (Kd) конкретного взаимодействия связывания с единицами М (например, от 10-5 М до 10-13 М). Значения аффинности полипептидов с доменом связывания и одноцепочечных полипептидов в соответствии с настоящим изобретением могут быть легко определены с использованием традиционных техник (см., например, Scatchard et al. (1949) Ann. N Y. Acad. Sci. 51:660; и патенты США №№5283173, 5468614 или эквивалентные).
[0048] «CD3» известен в уровне техники как полибелковый комплекс из 6 цепей (см., например, Abbas and Lichtman, 2003; Janeway et al., pp.172 and 178, 1999), которые являются субъединицами комплекса рецептора Т-клетки. У млекопитающих, субъединицы CD3 комплекса рецептора Т-клетки представляют собой цепь CD3γ, цепь CD3δ, две цепи CD3ε и гомодимер цепей CD3ζ. Цепи CD3γ, CD3δ и CD3ε представляют собой в высокой степени родственные белки иммуноглобулинового суперсемейства, содержащие единственный иммуноглобулиновый домен. Трансмембранные участки цепей CD3γ, CD3δ и CD3ε заряжены отрицательно, и это является характеристикой, позволяющей таким цепям ассоциироваться с положительно заряженными рецепторными цепями Т-клеток. Каждый из внутриклеточных хвостов цепей CD3γ, CD3δ и CD3ε содержит единственный консервативный мотив, известный как иммунорецепторный активационный мотив на основе тирозина или ИТАМ, тогда как каждая цепь CD3ζ содержит три. Считается, что ИТАМ важны для емкости проведения сигнала комплекса TCR. CD3, использующиеся в документе данного изобретения, могут происходить от различных видов, в том числе, человека, обезьяны, мыши, крысы или других млекопитающих.
[0049] В данном документе «консервативной заменой», признанной в уровне техники, является замена одной аминокислоты другой аминокислотой, которая обладает подобными свойствами. Примеры консервативных замен известны в данной области (см., например, WO 97/09433, стр. 10, опубликована 13 марта 1997 года; Lehninger, Biochemistry, Second Edition; Worth Publishers, Inc. NY: NY (1975), pp.71-77; Lewin, Genes IV, Oxford University Press, NY and Cell Press, Cambridge, MA (1990), p. 8). В некоторых вариантах реализации консервативная замена включает замену лейцина серином.
[0050] В данном документе термин «производное» обозначает модификацию одного или более остатков аминокислот в пептиде химическими или биологическими средствами, с применением фермента или без него, например, гликозилированием, алкилированием, ацилированием, образованием эфира или образованием амида.
[0051] В данном документе полипептид или последовательность аминокислот, «производная от» обозначенного полипептида или белка, обозначает происхождение полипептида. В одном варианте реализации полипептид или последовательность аминокислот, которая происходит от конкретной последовательности, содержат последовательность аминокислот, по существу идентичную такой последовательности или ее части, где часть состоит по меньшей мере из 10-20 аминокислот, по меньшей мере из 20-30 аминокислот или по меньшей мере из 30-50 аминокислот, или по меньшей мере из 50-150 аминокислот, или которая иным способом может быть идентифицирована специалистом, имеющим средний уровень квалификации в данной области, как происходящая от последовательности. В одном варианте реализации гуманизированный или химерный домен связывания происходит от VH и/или VL антитела, генерируемого другим животным. Например, гуманизированный домен связывания или химерный домен связывания может происходить от участков VH и VL мышиного антитела. Например, гуманизация мышиных антител может быть осуществлена посредством модификации каркасных участков способами, известными из уровня техники.
[0052] Полипептиды, происходящие от другого полипептида, могут содержать одну или более мутаций относительно исходного полипептида, например, один или более остатков аминокислот, которые заменены другим остатком аминокислоты или которые содержат одну или более вставок или делеций остатка аминокислоты. Полипептид может содержать последовательность аминокислот, которая не существует в природе. Такие вариации обязательно обладают менее чем 100% идентичности последовательности или сходства с исходным полипептидом. В одном варианте реализации вариант будет содержать последовательность аминокислот с идентичностью последовательности аминокислот или сходством с последовательностью аминокислот исходного полипептида от около 60% до менее чем 100%. В другом варианте реализации вариант будет содержать последовательность аминокислот с идентичностью последовательностей аминокислот или сходством с последовательностью аминокислот исходного полипептида от около 75% до менее чем 100%, от около 80% до менее чем 100%, от около 85% до менее чем 100%, от около 90% до менее чем 100%, от около 95% до менее чем 100%.
[0053] В данном документе, если не указано иное, положение остатка аминокислоты в вариабельном участке молекулы иммуноглобулина нумеруется согласно конвенции о нумерации Kabat (Kabat, Sequences of Proteins of Immunological Interest, 5th ed. Bethesda, MD: Public Health Service, National Institutes of Health (1991)), и положение остатка аминокислоты в константном участке молекулы иммуноглобулина нумеруется согласно номенклатуре ЕС (Ward et al, 1995 Therap. Immunol. 2:77-94).
[0054] В данном документе термин «димер» обозначает биологический объект, состоящий из двух субъединиц, связанных друг с другом посредством одного или более видов внутримолекулярных сил, в том числе, ковалентных связей (например, дисульфидные связи) и других взаимодействий (например, электростатические взаимодействия, солевые мостики, водородная связь и гидрофобные взаимодействия), и стабильный в соответствующих условиях (например, в физиологических условиях, в водном растворе, пригодном для экспрессии, очистки и/или хранения рекомбинантных белков, или в условиях неденатурирующего и/или невосстанавливающего электрофореза). «Гетеродимер» или «гетеродимерный белок» в настоящем документе обозначает димер, образованный двумя различными полипептидами. Гетеродимер не включает антитело, образованное четырьмя полипептидами (т.е., две легких цепи и две тяжелых цепи). В одном варианте реализации изобретения гетеродимер создан с использованием платформы аварийного перехватчика, которая включает домен гетеродимеризации. «Гомодимер» или «гомодимерный белок» в настоящем документе обозначает димер, образованный двумя идентичными полипептидами. В некоторых вариантах изобретения гомодимер создан с использованием платформы аварийного SMIP, PIMS или Scorpion.
[0055] В данном документе «шарнирный участок» или «шарнир» обозначает полипептид, происходящий от (а) междоменного участка трансмембранного белка (например, трансмембранный белок типа I); или (б) стебелькового участка С-лектина типа II. Например, шарнирный участок может происходить от междоменного участка члена суперсемейства иммуноглобулина; пригодные шарнирные участки в пределах данного конкретного класса включают (i) иммуноглобулиновые шарнирные участки (состоящие, например, из верхнего и/или ядерного шарнирного участка(-ов)) иммуноглобулина или их функциональные варианты, в том числе, дикий тип и модифицированные шарниры иммуноглобулина, и (ii) участки (или их функциональные варианты), которые соединяют иммуноглобулиновые V-подобные или иммуноглобулиновые С-подобные домены.
[0056] «Шарнирный участок иммуноглобулина дикого типа» обозначает природные верхние и средние последовательности аминокислот шарнира, вставленные между и соединенные с доменами СН1 и СН2 (для IgG, IgA и IgD) или вставленные между и соединенные с доменами СН1 и СН3 (для IgE и IgM), найденные в тяжелой цепи антитела. В некоторых вариантах реализации последовательность шарнирного участка иммуноглобулина дикого типа является человеческой и может включать шарнирный участок IgG человека.
[0057] «Модифицированный шарнирный участок иммуноглобулина дикого типа» или «модифицированный шарнирный участок иммуноглобулина» обозначает (а) шарнирный участок иммуноглобулина дикого типа, содержащий до 30% модификаций аминокислот (например, до 25%, 20%, 15%, 10%, или 5% замен или делеций аминокислот), или (б) часть шарнирного участка иммуноглобулина дикого типа, длина которого составляет от около 5 аминокислот (например, около 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот) до около 120 аминокислот (например, с длиной от около 10 до около 40 аминокислот или от около 15 до около 30 аминокислот, или от около 15 до около 20 аминокислот, или от около 20 до около 25 аминокислот), содержащий до около 30% модификаций аминокислот (например, до около 25%, 20%, 15%, 10%, 5%, 4%, 3%, 2%, или 1% замен или делеций аминокислот или их комбинаций), и содержит основной шарнирный участок IgG, как раскрыто в Публикациях РСТ WO 2011/090762 и WO 2011/090754.
[0058] В данном документе термин «гуманизированный» обозначает способ создания антитела или связывающихся с иммуноглобулином белков и полипептидов, происходящих из негуманоидных видов (например, мышь или крыса), менее иммуногенных для человека, при сохранении свойств связывания с антигеном оригинального антитела, с применением методов генной инженерии. В некоторых вариантах реализации домен(-ы) связывания антитела или связывающихся с иммуноглобулином белков и полипептидов (например, вариабельные участки легкой и тяжелой цепи, Fab, scFv) гуманизирован(-ы). Негуманоидные домены связывания могут быть гуманизированы с применением методов, известных как пересадка CDR (Jones et al., Nature 321:522 (1986)), и их варианты, в том числе, «реконструирование» (Verhoeyen, et al., 1988 Science 239:1534-1536; Riechmann, et al., 1988 Nature 332:323-337; Tempest, et al., BioTechnol 1991 9:266-271), «гиперхимеризация» (Queen, et al., 1989 Proc Natl Acad Sci USA 86:10029-10033; Co, et al., 1991 Proc Natl Acad Sci USA 88:2869-2873; Co, et al, 1992 J Immunol 148:1149-1154) и «облицовка» (Mark, et al., "Derivation of therapeutically active humanized and veneered anti-CD 18 antibodies. B: Metcalf BW, Dalton BJ, eds. Cellular adhesion: molecular definition to therapeutic potential. New York: Plenum Press, 1994: 291-312). Другие участки антитела или связывающихся с иммуноглобулином белков и полипептидов, такие как шарнирный участок и домены константного участка, также могут быть гуманизированы, если они происходят из негуманоидного источника.
[0059] «Константный участок иммуноглобулина» или «константный участок» представляет собой термин, определенный в настоящем документе таким образом, чтобы обозначать последовательность пептида или полипептида, которая соответствует или происходит от части или полноразмерного, одного или более доменов константного участка. В одном варианте реализации константный участок содержит все домены константного участка исходного антитела. В одном варианте реализации константный участок содержит домены СН2 и СН3 IgG, например, домены СН2 и СН3 lgG1. В некоторых вариантах реализации константный участок не содержит домена СН1. В некоторых вариантах реализации домены константного участка, составляющие константную подобласть, являются человеческими. В некоторых вариантах реализации домены константного участка слитого белка в соответствии с данным описанием не обладают или обладают минимальными эффекторными функциями антителозависимой клеточно-опосредованной цитотоксичности (АЗКЦ) и активации комплемента и комплементзависимой цитотоксичности (КЗЦ), при сохранении способности связываться с некоторыми рецепторами Fc (такими как FcRn, рецептор Fc новорожденных) и сохранении относительно длинного периода полувыведения in vivo.
[0060] В других вариациях, слитый белок в соответствии с описанием содержит константные домены, которые сохраняют такую эффекторную функцию или обе АЗКЦ и КЗЦ. В некоторых вариантах реализации связывающий домен в соответствии с настоящим описанием слит с человеческим константным участком IgG (например, lgG1), причем константный участок содержит одну или более из следующих мутировавших аминокислот: лейцин в положении 234 (L234), лейцин в положении 235 (L235), глицин в положении 237 (G237), глутамат в положении 318 (Е318), лизин в положении 320 (К320), лизин в положении 322 (К322), или любую их комбинацию (нумерация ЕС). Например, любая одна или более из указанных аминокислот может быть заменена аланином. В другом варианте реализации домен Fc IgG, такого как lgG1, содержит каждый из L234, L235, G237, Е318, К320 и К322 (нумерация ЕС), мутировавший, например, до аланина (т.е., L234A, L235A, G237A, Е318А, К320А и К322А, соответственно), и необязательно также мутацию N297A (т.е., гликозилирование домена СН2 по существу исключается).
[0061] В настоящем документе термин «низкомолекулярные модульные иммунофармацевтические белки» или «SMIP» используется для обозначения платформы белка как в общем раскрыто, например, в Публикации Патентных заявок США №№2003/0133939, 2003/0118592 и 2005/0136049, которые включены в данное описание посредством ссыпки в полном объеме. «Молекулы SMIP», описанные в Примерах и описании настоящего изобретения, следует понимать как связывающиеся белки, содержащие платформу SMIP, например, в порядке от амино- к карбокси-концу, первый домен связывания, шарнирный участок и константный участок иммуноглобулина. «CD3-специфичные молекулы» SMIP следует понимать как CD3-связывающие белки, содержащие платформу SMIP.
[0062] В настоящем документе термин «PIMS» используется для обозначения платформы белка как раскрыто в общем, например, в Публикации Патентной заявки США №2009/0148447, которая включена в данный документ посредством ссылки в полном объеме. «Молекулы PIMS», описанные в Примерах и настоящем описании следует понимать как связывающиеся белки, содержащие платформу PIMS, например, в порядке от амино- к карбокси-концу, константный участок иммуноглобулина, шарнирный участок и первый домен связывания. «CD3-специфичные молекулы» PIMS следует понимать как CD3-связывающие белки, содержащие платформу PIMS.
[0063] «Участок Fc» или «домен Fc» обозначает последовательность полипептида, соответствующую или происходящую от части исходного антитела, которая ответственна за связывание с рецепторами антитела на клетках и компоненте C1q комплемента. Fc поддерживает «кристаллический фрагмент», фрагмент антитела, который легко образует белковый кристалл. Различные фрагменты белка, которые были первоначально описаны с помощью протеолитического расщепления, могут определять общую полную структуру белка иммуноглобулина. Как первоначально определялось в литературе, фрагмент Fc состоит из связанных дисульфидной связью шарнирных участков тяжелой цепи, доменов СН2 и СН3. Однако позже термин стали применять к единой цепи, состоящей из СН3, СН2, и по меньшей мере части шарнира, достаточной для образования связанного дисульфидными связями димера со второй такой цепью. Обзор структуры и функций иммуноглобулина см. в Putnam, The Plasma Proteins, Vol. V (Academic Press, Inc., 1987), pp. 49-140; и Padlan, Mol. Immunol. 31:169-217, 1994. В данном документе термин Fc включает варианты природных последовательностей.
[0064] В данном документе «изоэлектрическая точка» или pI обозначает значение рН, при котором сетевой заряд равен нулю.
[0065] В данном документе термин «перехватчик» используется для обозначения моноспецифичной или полиспецифичной платформы белка, как в общем раскрыто в Публикациях РСТ WO 2011/090762 и WO 2011/090754. Молекулы перехватчика, описанные в настоящем документе, следует понимать как белки, связывающиеся с CD3, которые содержат две неидентичные полипептидной цепи, причем каждая полипептидная цепь содержит домен гетеродимеризации иммуноглобулина. Взаимодействующие на поверхности домены гетеродимеризации иммуноглобулина являются различными. В одном варианте реализации домен гетеродимеризации иммуноглобулина включает домен СН1 или его производное. В другом варианте реализации домен гетеродимеризации иммуноглобулина включает домен CL или его производное. В одном варианте реализации домен CL представляет собой изотип Сκ или Сλ или его производное.
[0066] В данном документе «стебельковый участок» С-лектина типа II обозначает часть внеклеточного домена С-лектина типа II, который размещен между лектин-подобным доменом С-типа (СТЛД; например, подобный СТЛД рецепторов природных клеток-киллеров) и трансмембранным доменом. Например, в молекуле CD94 человека (GenBank, номер доступа ААС50291.1, PRI 30 ноября 1995 года), внеклеточный домен соответствует остаткам аминокислот 34-179, тогда как СТЛД соответствует остаткам аминокислот 61-176. Соответственно, стебельковый участок молекулы CD94 человека включает остатки аминокислот 34-60, которые найдены между мембраной и СТЛД (см. Boyington et al., Immunity 10:75, 1999; для описаний других стебельковых участков см. также Beavil et al., Proc, Natl. Acad. Sci. USA 89:753, 1992; и Figdor et al, Nature Rev. Immunol. 2:77, 2002). Указанные С-лектины типа П также могут содержать стебельковый участок длиной от 6 до 10 соединяющих аминокислот и трансмембранный участок или СТЛД. В другом примере, белок NKG2A человека длиной 233 аминокислоты (GenBank номер доступа Р26715.1, PRI 15 июня 2010 года) содержит трансмембранный домен, варьирующий от аминокислот 71-93, и внеклеточный домен, варьирующий от аминокислот 94-233. СТЛД содержит аминокислоты 119-231, и стебельковый участок содержит аминокислоты 99-116, обрамленные соединяющими 5 и 2 аминокислотами. Другие С-лектины типа II, а также их внеклеточные лигандсвязывающие домены, междоменные или стебельковые участки и СТЛД известны из уровня техники (см., например, GenBank номер доступа NP_001993.2; ААН07037.1, PRI 15 июля 2006 года; NP_001773.1, PRI 20 июня 1010 года; AAL65234.1, PRI 17 января 2002 года; и САА04925.1, PRI 14 ноября 2006 года, для последовательностей CD23, CD69, CD72, NKG2A и NKG2D человека, а также из описания, соответственно).
[0067] В данном документе, «междоменный участок» трансмембранного белка (например, трансмембранный белок типа I) обозначает часть внеклеточного домена трансмембранного белка, которая расположена между двумя смежными доменами. Примеры междоменных участков включают участки, связывающие смежные домены Ig членов суперсемейства иммуноглобулина (например, шарнирный участок иммуноглобулина от IgG, IgA, IgD или IgE; участок, связывающий домены IgV и lgC2 CD2; или участок, связывающий домены IgV и IgC CD80 или CD86). Другим примером междоменного участка является участок, связывающий не-Ig домен и lgC2 CD22, связывающийся с сиаловой кислотой Ig-подобный лектин типа I.
[0068] Участок полипептида, «происходящий от» стебелькового участка С-лектина типа II, или «происходящий от» междоменного участка (например, шарнирный участок иммуноглобулина) трансмембранного белка, обозначает последовательность длиной от около 5 до около 150 аминокислот, все или по меньшей мере часть из которых включают (i) стебельковый участок дикого типа или последовательность междоменного участка; (ii) фрагмент стебелькового участка дикого типа или последовательности междоменного участка; (iii) полипептид, обладающий по меньшей мере 80%, 85%, 90% или 95% идентичностью последовательности аминокислот по отношению к (i), или (ii); или (iv) или (i), или (ii), в котором одна, две, три, четыре или пять аминокислот содержат делецию, вставку, замену, или любую их комбинацию, например, одна или более модификаций представляют собой замены или одна или более мутаций включают только одну делецию. В некоторых вариантах реализации производное стебелькового участка более устойчиво к протеолитическому расщеплению, по сравнению с последовательностью стебелькового участка дикого типа, такой как происходящие от около 8 до около 20, от около 10 до около 25 или от около 15 до около 25 аминокислот NKG2A, NKG2D, CD23, CD64, CD72 или CD94.
[0069] В данном документе термин «соединяющие аминокислоты» или «остатки соединяющих аминокислот» обозначает один или более (например, около 2-10) остатков аминокислот между двумя смежными участками или доменами полипептида, например, между шарниром и смежной константной подобластью иммуноглобулина или между шарниром и смежным доменом связывания, или между пептидным линкером, который соединяет два вариабельных домена иммуноглобулина и смежный вариабельный домен иммуноглобулина. Соединяющие аминокислоты могут быть результатом дизайна полипептидного конструкта (например, остатки аминокислот, образовавшиеся в результате использования сайта рестрикционного фермента в ходе конструирования молекулы нуклеиновой кислоты, кодирующей полипептид). Соединяющие аминокислоты между доменом связывания и шарнирным участком здесь и в дальнейшем обозначаются как предшарнирный участок, например, могут образовываться в результате добавления рестрикционного сайта в кодирующую нуклеотидную последовательность. В одном варианте реализации предшарнирный участок состоит из около1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или 15 аминокислот. Последовательность предшарнирного участка DRA209 и DRA161 не является частью последовательности мышиного Cris-7 или иммуноглобулина зародышевой линии человека. Предшарнирный участок DRA209 и DRA161 представляет собой последовательность аминокислот RRT, но, при этом, он представляет собой SSS для DRA219, DRA221, DRA222, DRA223, DRA224, DRA225, DRA228, DRA229, DRA233, DRA234, TSC249, TSC250, TSC251, TSC252, TSC295, TSC296, TSC301 и TSC302. Связывающие молекулы по изобретению могут содержать предшарнирный участок или не содержать его.
[0070] В данном документе определение «линкер между СН3 и СН1 или CL» обозначает один или более (например, около 2-12) остатков аминокислот между С-концом (например, СН3 дикого типа или мутантный СН3) СН3 и N-концом домена СН1 или домена CL (например, Ck).
[0071] В данном документе термин «пациент, нуждающийся в» обозначает больного, подверженного риску или страдающего от заболевания, расстройства или состояния, отвечающего на лечение или облегчение с помощью белка, связывающегося с CD3, или его полипептида или их составов, раскрытых в настоящем документе.
[0072] В данном документе термин «пептидный линкер» обозначает последовательность аминокислот, которая соединяет вариабельный участок тяжелой цепи с вариабельным участком легкой цепи и обеспечивает функцию распорки, совместимую со взаимодействием двух субдоменов связывания, таким образом, что полученный полипептид сохраняет специфичную аффинность связывания в отношении той же самой молекулы-мишени, что и антитело, которое содержит такие же вариабельные участки легкой и тяжелой цепи. В некоторых вариантах реализации длина линкера составляет от около 5 до около 35 аминокислот, например, от около 15 до около 25 аминокислот.
[0073] В данном документе термин «фармацевтически приемлемый» обозначает молекулярные объекты и составы, которые не вызывают аллергических или других серьезных неблагоприятных реакций у большинства субъектов, при введении их способами, хорошо известными из уровня техники. Молекулярные объекты и составы, одобренные регуляторным ведомством Федерального правительства или правительства штата, или перечисленные в Фармакопее США или другой общепринятой фармакопее для применения у животных, и, более конкретно, у человека, рассматриваются как «фармацевтически приемлемые».
[0074] В данном документе термин «промотор» обозначает участок ДНК, принимающий участие в связывании с РНК полимеразой для инициации транскрипции.
[0075] В данном документе термины «нуклеиновая кислота», «молекула нуклеиновой кислоты» или «полинуклеотид» обозначают дезоксирибонуклеотиды или рибонуклеотиды и их полимеры, в одно- или двухцепочечной форме. Если нет конкретных ограничений, термины охватывают нуклеиновые кислоты, содержащие аналоги природных нуклеотидов, которые обладают сходными свойствами связывания с референтной нуклеиновой кислотой и метаболизируются способом, до некоторой степени сходным с природными нуклеотидами. Если не указано иное, конкретная последовательность нуклеиновой кислоты также безоговорочно включает ее консервативно модифицированные варианты (например, замены с вырождением кодонов) и комплементарные последовательности, а также явно указанную последовательность. Конкретно, замены с вырождением кодонов могут быть осуществлены посредством генерации последовательностей, в которых третье положение одного или более выбранных (или всех) кодонов замещено остатками смешанных оснований и/или дезоксиинозина (Batzer et al. (1991) Nucleic Acid Res. 19:5081; Ohtsuka et al. (1985) J. Biol. Chem. 260:2605-2608; Cassol et al. (1992); Rossolini et al. (1994) Mol. Cell. Probes 8:91-98). Термин нуклеиновая кислота используется взаимозаменяемым образом с терминами, обозначающими ген, кДНК и мРНК, кодируемую геном. В данном документе термины «нуклеиновая кислота», «молекула нуклеиновой кислоты» или «полинуклеотид» предназначены включать молекулы ДНК (например, кДНК или геномную ДНК), молекулы РНК (например, мРНК), аналоги ДНК или РНК, сгенерированные с применением нуклеотидных аналогов, и их производные, фрагменты и гомологи.
[0076] Термин «экспрессия» обозначает биосинтез продукта, кодируемого нуклеиновой кислотой. Например, в случае сегмента нуклеиновой кислоты, кодирующего целевой полипептид, экспрессия включает транскрипцию сегмента нуклеиновой кислоты в мРНК и трансляцию мРНК в один или более полипептидов.
[0077] Термины «модуль экспрессии» и «кассета экспрессии» используются в настоящем документе взаимозаменяемым образом и обозначают сегмент нуклеиновой кислоты, кодирующий целевой полипептид и способный обеспечить экспрессию сегмента нуклеиновой кислоты в клетке-хозяине. Модуль экспрессии обычно содержит промотор транскрипции, открытую рамку считывания, кодирующую целевой полипептид, и терминатор транскрипции, все в действующей конфигурации. В дополнение к транскрипциональному промотору и терминатору, модуль экспрессии может дополнительно содержать другие сегменты нуклеиновых кислот, например, энхансер или сигнал полиаденилирования.
[0078] Термин «вектор экспрессии» в настоящем документе обозначает молекулу нуклеиновой кислоты, линейную или круговую, содержащую один или более модулей экспрессии. В дополнение к одному или более модулей экспрессии, вектор экспрессии также может содержать дополнительные сегменты нуклеиновых кислот, например, один или более источников репликации или один или более селекционных маркеров. Векторы экспрессии, в общем, происходят из ДНК плазмиды или вируса, или могут содержать элементы обоих.
[0079] В данном документе, Scorpion представляет собой термин, используемый для обозначения полиспецифичной платформы белка связывания. Полиспецифичные связывающиеся белки и полипептиды раскрыты, например, в Публикации заявки РСТ № WO 2007/146968, Публикации Патентной заявки США №2006/0051844, Публикации заявки РСТ WO 2010/040105, Публикации заявки РСТ WO 2010/003108 и патенте США 7166707. Полипептид Scorpion содержит два домена связывания (домены могут быть сконструированы таким образом чтобы специфично связываться с одной и той же или различными мишенями), два линкера, и константную субобласть иммуноглобулина. Линкеры для молекул Scorpion описаны в Публикации заявки РСТ WO 2010/003108. В некоторых вариантах реализации длина линкерной последовательности составляет около 2-45 аминокислот или 2-38 аминокислот или 5-45 аминокислот. В некоторых вариантах реализации линкер представляет собой шарнирный участок (например, от IgG) антитела или стебельковый участок С-лектина. Белки Scorpion представляют собой гомодимерные белки, содержащие два идентичных, связанных дисульфидной связью полипептида Scorpion.
[0080] В данном документе термин «идентичность последовательностей» обозначает взаимоотношение между двумя или более последовательностей полинуклеотидов или между двумя или более последовательностями полипептидов. Если положение в одной последовательности занято таким же основанием нуклеиновой кислоты или остатком аминокислоты в соответствующем положении последовательности сравнения, последовательности называют «идентичными» по этому положению. «Процент идентичности последовательностей» вычисляют посредством определения количества положений, в которых идентичное основание нуклеиновой кислоты или аминокислотный остаток находится в обеих последовательностях, чтобы получить количество «идентичных» положений. В дальнейшем количество «идентичных» положений делят на общее количество положений в окне сравнения и умножают на 100, с получением «процента идентичности последовательностей». Процент «идентичности последовательностей» определяют посредством сравнения двух оптимально выровненных последовательностей в окне сравнения. Длина окна сравнения для последовательностей нуклеиновых кислот может составлять, например, по меньшей мере 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 300, 400, 500, 600, 700, 800, 900 или 1000, или более нуклеиновых кислот. Окно сравнения для полипептидных последовательностей может составлять, например, по меньшей мере 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 300 или более аминокислот. Для того чтобы оптимально выровнять последовательности для сравнения, часть полинуклеотида или последовательности полипептида в окне сравнения может содержать дополнения или делеций, называемые промежутками, тогда как референтная последовательность остается постоянной. Оптимальным выравниванием является такое выравнивание, которое, даже с промежутками, дает наибольшее возможное количество «идентичных» положений между референтными и компараторными последовательностями. Процент «идентичности последовательностей» для двух последовательностей может быть определен с помощью версии программы «BLAST 2 Sequences)), которая была доступна в Национальном Центре биотехнологической информации по состоянию на 1 сентября 2004 года, причем указанная программа объединяет программы BLASTN (для сравнения нуклеотидных последовательностей) и BLASTP (для сравнения последовательностей полипептидов), и при этом указанные программы основаны на алгоритме Karlin и Altschul (Proc. Natl. Acad. Sci. США 90(12): 5873-5877, 1993). При использовании «BLAST 2 Sequences)) параметры, которые использовались по умолчанию, по состоянию на 1 сентября 2004 года, могут использоваться для размера слова (3), штрафа за открытие промежутка (11), штрафа за продление промежутка (1), сокращение промежутка (50), ожидаемого значения (10) и любого другого обязательного параметра, включая, без ограничений, опцию матрицы. Две нуклеотидных или аминокислотных последовательности считаются «обладающими в существенной мере подобной идентичностью» или «в существенной мере идентичными последовательностями», если идентичность двух последовательностей друг относительно друга составляет по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99%.
[0081] В данном документе «полипептид» или «полипептидная цепь» представляет собой единое, линейное и смежное расположение ковалентно соединенных аминокислот. Оно не включает две полипептидных цепи, которые соединены нелинейным образом, например, с помощью межцепочечной дисульфидной связи (например, половина молекулы иммуноглобулина, в которой легкая цепь соединена с тяжелой цепью посредством дисульфидной связи). Полипептиды могут содержать или образовывать одну или более внутрицепочечных дисульфидных связей. Относительно полипептидов, описанных в настоящем документе, ссылка на остатки аминокислот, соответствующие указанным SEQ ID NO, включает посттрансляционные модификации таких остатков.
[0082] «Белок» представляет собой макромолекулу, содержащую одну или более полипептидных цепей. Белок также может содержать непептидные компоненты, такие как углеводные группы. Углеводы и другие непептидные заместители могут быть добавлены к белку клеткой, в которой вырабатывается белок, и будут варьировать в зависимости от вида клеток. Белки определены в настоящем документе в терминах их структуры аминокислотного скелета; заместители, такие как углеводные группы, в общем не указаны, но тем не менее могут присутствовать.
[0083] Термины «амино-концевой» (N-концевой) и «карбокси-концевой» (С-концевой) используются в настоящем документе для обозначения положений в пределах полипептидов. Если контекст позволяет, указанные термины используются со ссылкой на конкретную последовательность или часть полипептида для обозначения близости или относительного положения. Например, определенная последовательность, расположенная в направлении карбокси-конца по отношению к референтной последовательности в пределах полипептида, размещена проксимально по отношению к карбокси-концу референтной последовательности, но не обязательно расположена на карбокси-конце всего полипептида.
[0084] «Рецептор Т-клеток (TCR)» представляет собой молекулу, найденную на поверхности Т-клеток, которые, наряду с CD3, в общем ответственны за распознавание антигенов, связанных с основным комплексом гистосовместимости (ОКГ) молекулы. Он состоит из связанного дисульфидными связями гетеродимера в высокой степени вариабельных α и β цепей в большинстве Т-клеток. В других Т-клетках экспрессируется альтернативный рецептор, образованный вариабельными γ и δ цепями. Каждая цепь TCR является членом суперсемейства иммуноглобулинов и содержит один N-концевой вариабельный домен иммуноглобулина, один константный домен иммуноглобулина, трансмембранный участок и короткий цитоплазматический хвост на С-конце (см. Abbas and Lichtman, Cellular and Molecular Immunology (5th Ed.), Editor: Saunders, Philadelphia, 2003; Janeway et al., Immunobiology: The Immune System in Health and Disease, 4th Ed., Current Biology Publications, pl48, 149, and 172, 1999). TCR в настоящем документе может происходить из различных видов животных, в том числе, человека, мыши, крысы или других млекопитающих.
[0085] Комплекс «TCR» в настоящем документе обозначает комплекс, образованный посредством соединения цепей CD3 с другими цепями TCR. Например, комплекс TCR может состоять из цепи CD3γ, цепи CD3δ, двух цепей CD3ε, гомодимера цепей CD3ζ, цепи TCRα и цепи TCRβ. Альтернативно, комплекс TCR может состоять из цепи CD3γ, цепи CD3δ, двух цепей CD3ε, гомодимера цепей CD3ζ, цепи TCRγ и цепи TCRδ.
[0086] «Компонент комплекса TCR» в настоящем документе обозначает цепь TCR (например, TCRα, TCRβ, TCRγ или TCRδ), цепь CD3 (например, CD3γ, CD3δ, CD3ε или CD3ζ, или комплекс, образованный двумя или более цепей TCR или цепей CD3 (например, комплекс TCRα и TCRβ, комплекс TCRγ и TCRδ, комплекс CD3ε и CD3δ, комплекс CD3γ и CD3ε или суб-TCR комплекс TCRα, TCRβ, CD3γ, CD3δ и две цепи CD3ε).
[0087] «Антителозависимая клеточно-опосредованная цитотоксичность» и «АЗКЦ» в настоящем документе обозначают опосредованный клетками процесс, в ходе которого неспецифичные цитотоксические клетки, которые экспрессируют FcγR (например, моноцитарные клетки, такие как природные клетки-киллеры (ПК) и макрофаги), распознают связанное антитело (или другой белок, способный к связыванию с FcγR) на клетке-мишени и впоследствии вызывают лизис клетки-мишени. В принципе, любая эффекторная клетка с активизацией FcγR может быть запущена в режим опосредования АЗКЦ. Основными клетками для опосредования АЗКЦ являются клетки ПК, которые экспрессируют только FcγRIII, тогда как моноциты, в зависимости от их состояния активации, локализации или дифференциации, могут экспрессировать FcγRI, FcγRII и FcγRIII. Обзор экспрессии FcγR на гемопоэтических клетках см., например, в Ravetch et al., 1991, Annu. Rev. Immunol, 9:457-92.
[0088] В данном документе термин «обладающий активностью АЗКЦ», со ссыпкой на полипептид или белок, означает, что полипептид или белок (например, содержащий шарнирный участок иммуноглобулина и константную подобласть иммуноглобулина, которая содержит домены СН2 и СН3, например, полученные из IgG (такого как lgG1)), способен опосредовать антителозависимую клеточно-опосредованную цитотоксичность (АЗКЦ) путем связывания с цитолитическим рецептором Fc (например, FcγRIII) на цитолитической иммунной эффекторной клетке, экспрессирующей рецептор Fc (например, клетке ПК).
[0089] «Комплементзависимая цитотоксичность» и «КЗЦ» в настоящем документе обозначают процесс, в ходе которого компоненты нормальной сыворотки («комплемент»), вместе с антителом или другим C1q-комплемент-связывающим белком, связанным с целевым антигеном, осуществляют лизис клетки-мишени, экспрессирующий целевой антиген. Комплемент состоит из группы белков сыворотки, которые действуют совместно и в точной последовательности для достижения эффекта.
[0090] Термины «классический путь комплемента» и «классическая система комплемента» в настоящем документе являются синонимическими и обозначают конкретный путь активации комплемента. Классический путь требует комплексов антиген-антитело для инициации и включает активацию, в точной последовательности, девяти основных белковых компонентов, обозначенных С1-С9. Продуктом нескольких стадий в процессе активации является фермент, который катализирует следующую стадию. Такой каскад обеспечивает амплификацию и активацию больших количеств комплемента под действием относительно небольшого начального сигнала.
[0091] В данном документе термин «обладающий активностью КЗЦ», со ссылкой на полипептид или белок, означает, что полипептид или белок (например, содержащий шарнирный участок иммуноглобулина и константную подобласть иммуноглобулина, которая содержит домены СН2 и СН3, например, полученные из IgG (такого как lgG1)) способен опосредовать комплементзависимую цитотоксичность (КЗЦ) путем связывания с белком комплемента C1q и активации классической системы комплемента.
[0092] В данном документе «перенаправленная цитотоксичность Т-клеток» и «ПЦТК» обозначает опосредованный Т-клетками процесс, в ходе которого цитотоксическая Т-клетка рекрутируется к клетке-мишени с помощью полиспецифичного белка, способного специфично связываться как с цитотоксической Т-клеткой, так и с клеткой-мишенью, посредством чего мишень-зависимая цитотоксическая реакция Т-клеток проявляется против клетки-мишени.
[0093] В данном документе термин «лечение», «терапия» или «облегчение» обозначает терапевтическое лечение или профилактическое/превентивное лечение. Лечение является терапевтическим, если по меньшей мере один симптом заболевания у индивидуума, получающего лечение, улучшается, или лечение может отодвигать ухудшение прогрессирующего заболевания у индивидуума или отодвигать начало дополнительных сопутствующих заболеваний.
[0094] В данном документе термин «трансформация», «трансфекция» и «трансдукция» обозначает перенос нуклеиновой кислоты (например, нуклеотидного полимера) в клетку. В данном документе термин «генетическая трансформация» обозначает перенос и инкорпорацию ДНК, особенно рекомбинантной ДНК, в клетку. Перенесенная нуклеиновая кислота может быть введена в клетку с помощью вектора экспрессии.
[0095] В данном документе термин «вариант» или «варианты» обозначает нуклеиновую кислоту или полипептид, отличающийся от референтной нуклеиновой кислоты или полипептида, но сохраняющий их существенные свойства. В общем, варианты в высокой степени подобны, и многие их участки идентичны референтной нуклеиновой кислоте или полипептиду. Например, вариант может демонстрировать идентичность последовательностей по меньшей мере около 70%, по меньшей мере около 80%, по меньшей мере около 90%, по меньшей мере около 95%, по меньшей мере около 96%, по меньшей мере около 97%, по меньшей мере около 98% или по меньшей мере около 99%, по сравнению с активной частью или полноразмерной нуклеиновой кислотой или полипептидом.
[0096] Термины «вариабельный участок легкой цепи» (также обозначается как «вариабельный домен легкой цепи» или «VL») и «вариабельный участок тяжелой цепи» (также обозначается как «вариабельный домен тяжелой цепи» или «VH») обозначают вариабельный участок связывания легкой и тяжелой цепи антитела, соответственно. Вариабельные участки связывания состоят из дискретных, четких подобластей, известных как «определяющие комплементарность участки (CDR)» и «каркасные участки» (FR). В одном варианте реализации FR гуманизированы. Термин «CL» обозначает «константный участок легкой цепи иммуноглобулина» или «константный участок легкой цепи», т.е., константный участок легкой цепи антитела. Термин «СН» обозначает «константный участок тяжелой цепи иммуноглобулина» или «константный участок тяжелой цепи», который дополнительно подразделяется, в зависимости от изотипа антитела на домены CH1, СН2 и СН3 (IgA, IgD, IgG), или CH1, СН2, СН3 и СН4 (IgE, IgM). «Fab» (антигенсвязывающий фрагмент) представляет собой часть антитела, которая связывается с антигенами и содержит вариабельный участок и домен СН1 тяжелой цепи, соединенный с легкой цепью посредством межцепочечной дисульфидной связи.
[0097] В настоящем документе раскрыты, среди прочего, полипептиды и белки, содержащие домены связывания, в частности, первый домен связывания, который специфично связывается с CD3. Полипептиды и белки, содержащие домены связывания, в соответствии с настоящим описанием, могут содержать одно или более из следующего: константную подобласть(-и) иммуноглобулина, линкерный пептид(-ы), шарнирный участок(-ки), домен(-ы) гетеродимеризации иммуноглобулина, домен(-ы) димеризации иммуноглобулина, одну или более соединяющих аминокислот, метку(-и), и т.п. Указанные компоненты раскрытых полипептидов и белков описаны более подробно в настоящем документе далее.
[0098] Дополнительно, полипептиды, связывающиеся с CD3, и белки, раскрытые в настоящем документе, могут существовать в форме антитела или слитого белка в любом из множества различных форматов (например, слитый белок может существовать в форме белка SMIP, белка PIMS, белка Scorpion или белка-перехватчика).
[0099] В некоторых вариантах реализации полипептид, связывающийся с CD3, содержит второй домен связывания. В некоторых вариантах реализации полипептид, связывающийся с CD3, содержит, от амино- к карбокси-концу, домен связывания с CD3, шарнирный участок, константный участок и второй домен связывания. В других вариантах полипептид, связывающийся с CD3, содержит, от амино- к карбокси-концу, второй домен связывания, шарнирный участок, константный участок и домен связывания с CD3.
[00100] Белок, связывающийся с CD3, в соответствии с настоящим изобретением в общем содержит (а) по меньшей мере одну полипептидную цепь, связывающуюся с CD3, содержащую домен связывания с CD3, как было указано ранее в настоящем документе. В некоторых вариантах, полипептид, связывающийся с CD3, дополнительно содержит (б) шарнирный участок, расположенный в направлении карбокси-конца по отношению к домену связывания с CD3, и (в) константный участок иммуноглобулина (например, полипептид SMIP). В дальнейших вариантах, связывающийся с CD3 полипептид дополнительно содержи (г) второй шарнирный участок, расположенный в направлении карбокси-конца по отношению к константной подобласти иммуноглобулина, и (д) второй домен связывания, расположенный в направлении карбокси-конца по отношению ко второму шарнирному участку (например, полипептид Scorpion). В некоторых вариантах реализации полипептид, связывающийся с CD3, содержит домен связывания с CD3, содержащий каркасные участки, предшарнирный участок и шарнирный участок.
[00101] В некоторых вариантах реализации полипептид, связывающийся с CD3, содержит три или более модифицированных аминокислот по сравнению с SEQ ID NO: 42, например, модификации, которые представляют собой замены положительно заряженных аминокислот нейтральными аминокислотами и/или замены нейтральных аминокислот отрицательно заряженными аминокислотами, например, в каркасных и/или предшарнирных участках. В некоторых вариантах реализации модифицированы по меньшей мере два, по меньшей мере три, по меньшей мере четыре, 3-5 или 3-10 аминокислот в каркасных участках VH и/или VL, посредством замены положительно заряженных аминокислот на нейтральные аминокислоты и/или замены нейтральных аминокислот отрицательно заряженными аминокислотами.
[00102] В одном из вариантов реализации изобретения полипептид, связывающийся с CD3, содержит модификации в предшарнирном участке, например, которые приводят к снижению изоэлектрической точки предшарнирного участка или всего полипептида. Предшарнирный участок расположен в соединении между доменом связывания и шарнирным участком. Например, три аминокислоты предшарнирного участка с последовательностью RRT могут быть заменены последовательностью SSS или SST, с целью снижения изоэлектрической точки. В некоторых вариантах реализации предшарнирный участок обладает сниженной изоэлектрической точкой по сравнению с полипептидом, связывающимся с CD3, причем предшарнирный участок представляет собой RRT.
[00103] В одном варианте реализации, с целью снижения риска иммуногенности, в последовательности заменяют аминокислоты, которые являются преобладающими в последовательности IgG зародышевой линии человека или содержатся в последовательности зародышевой линии человека в таком же или ближайшем положении, например, в соответствующей последовательности IgG зародышевой линии человека. В некоторых вариантах реализации последовательность IgG зародышевой линии человека содержит SEQ ID NO: 43 или 44.
[00104] В некоторых вариантах реализации по меньшей мере одна модификация аминокислоты введена в пределах участка J каппа (Jk) участка VL, по сравнению с мышиной последовательностью LQIT участка Cris-7 VL Jk (SEQ ID NO: 252). В некоторых вариантах реализации участок Jk содержит последовательность аминокислот VEIK (SEQ ID NO: 253), например, вместо LQIT (SEQ ID NO: 252).
[00105] В некоторых вариантах реализации полипептид, связывающийся с CD3, содержит шарнирный участок и константный участок, например, полипептид, связывающийся с CD3, может содержать (а) шарнирный участок, амино-концевой по отношению к домену связывания с CD3, и (б) константный участок иммуноглобулина, расположенный в амино-концевом направлении по отношению к шарнирному участку (например, полипептид PIMS). В некоторых вариантах реализации полипептид, связывающийся с CD3, содержит, от амино-конца к карбокси-концу, домен связывания с CD3, шарнирный участок и константный участок. В некоторых вариантах реализации полипептид, связывающийся с CD3, содержит, от амино-конца к карбокси-концу, константный участок, шарнирный участок и домен связывания с CD3.
[00106] Обычно, связывающиеся с CD3 полипептиды упомянутых выше форматов (SMIP, Scorpion или PIMS) способны к гомодимеризации, обычно посредством дисульфидной связи, через константный участок иммуноглобулина и/или шарнирный участок (например, через константный участок иммуноглобулина, содержащий домены СН2 и СН3 IgG и шарнирный участок IgG). Таким образом, в некоторых аспектах настоящего изобретения, два идентичных полипептида, связывающиеся с CD3, гомодимеризуются с образованием димерного белка, связывающегося с CD3.
[00107] В других вариантах реализации полипептид, связывающийся с CD3, дополнительно содержит домен гетеродимеризации, способный к гетеродимеризации с другим доменом гетеродимеризации во второй, неидентичной полипептидной цепи. В некоторых вариациях, вторая полипептидная цепь для гетеродимеризации содержит второй домен связывания. Соответственно, в некоторых аспектах настоящего изобретения, две неидентичных полипептидных цепи, одна из которых содержит домен связывания с CD3, и вторая необязательно содержит второй домен связывания, димеризуются с образованием гетеродимерного белка с 1-4 доменами связывания. В некоторых вариантах реализации полипептид связывающийся с CD3, содержит, от амино-конца к карбокси-концу, домен связывания с CD3, шарнирный участок, константный участок, домен гетеродимеризации и необязательно второй домен связывания.
[00108] В некоторых вариантах реализации второй домен связывания связывается или взаимодействует с молекулой-мишенью, и связывающийся с CD3 полипептид индуцирует цитотоксичность Т-клеток. Он может существовать в недимеризованной форме, в формате гетеродимера или гомодимера. В некоторых вариантах реализации второй домен связывания связывается или взаимодействует с ассоциированным с опухолью антигеном. В некоторых вариантах реализации полипептид, связывающийся с CD3, индуцирует лизис Т-клетками опухолевых клеток и/или индуцирует поликлональную активацию Т-клеток и экспансию вблизи опухоли.
[00109] Полипептиды, содержащие анти-CD3 домен связывания по изобретению, могут представлять собой или содержать антитела или производные антител, включая функциональные фрагменты антитела или производные фрагментов, которые сохраняют специфичность связывания. В некоторых вариантах реализации изобретение включает слитые белки и другие полипептиды, которые содержат вариабельные домены тяжелой и/или легкой цепи.
[00110] В некоторых вариантах реализации изобретения полипептид, связывающийся с CD3, содержит домен связывания с CD3, который содержит последовательность участка VH или последовательность участка VL, выбранную из группы, состоящей из SEQ ID NO: 28, 30, 32, 38 и 40. В некоторых вариантах реализации домен связывания с CD3 содержит участок VH, выбранный из группы, состоящей из SEQ ID NO: 22, 24, 26, 28, 30 и 32, и участок VL, выбранный из группы, состоящей из SEQ ID NO: 38 и 40. В некоторых вариантах реализации домен связывания с CD3 содержит участок VH, выбранный из группы, состоящей из SEQ ID NO: 28, 30 и 32, и участок VL, выбранный из группы, состоящей из SEQ ID NO: 34, 36, 38 и 40. В некоторых вариантах реализации участок VH содержит SEQ ID NO: 28 и участок VL содержит SEQ ID NO: 34; участок VH содержит SEQ ID NO: 28 и участок VL содержит SEQ ID NO: 38; участок VH содержит SEQ ID NO: 26 и участок VL содержит SEQ ID NO: 38; или участок VH содержит SEQ ID NO: 26 и участок VL содержит SEQ ID NO: 34. В некоторых вариантах реализации полипептид, связывающийся с CD3, содержит домен связывания с CD3, который содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, причем CDR3 тяжелой цепи содержит SEQ ID NO: 51. В некоторых вариантах реализации полипептид, связывающийся с CD3, содержит CDR1 тяжелой цепи содержащий SEQ ID NO: 49, CDR2 тяжелой цепи, содержащий SEQ ID NO: 50, и CDR3, тяжелой цепи содержащий SEQ ID NO: 51, а также CDR1 легкой цепи, содержащий SEQ ID NO: 52, CDR2 легкой цепи, содержащий SEQ ID NO: 53, и CDR3 легкой цепи, содержащий SEQ ID NO: 54.
[00111] В некоторых вариантах реализации полипептид, связывающийся с CD3, содержит последовательность аминокислот, выбранную из группы, состоящей из SEQ ID NO: 6, 8, 10, 12, 14, 16, 18 и 20.
[00112] Изобретение включает полиспецифичный связывающийся белок, содержащий димеризованные одноцепочечные полипептиды, причем каждый одноцепочечный полипептид содержит, от амино-конца к карбокси-концу, первый домен связывания, N-концевой линкер, константный участок иммуноглобулина, С-концевой линкер и домен связывания с CD3 по изобретению. Дополнительно, изобретение включает полиспецифичный связывающийся белок, содержащий димеризованные одноцепочечные полипептиды, каждый из которых содержит, от амино-конца к карбокси-концу, домен связывания с CD3 по изобретению, N-концевой линкер, константный участок иммуноглобулина, С-концевой линкер и первый домен связывания. В некоторых вариантах реализации N-концевой линкер может содержать или может по существу состоять из шарнирного участка иммуноглобулина.
[00113] В другом аспекте изобретения, полиспецифичный связывающийся белок содержит одноцепочечный полипептид, содержащий от амино-конца к карбокси-концу, первый домен связывания, N-концевой линкер, константный участок иммуноглобулина, С-концевой линкер и домен связывания с CD3. Дополнительно, изобретение включает полиспецифичный связывающийся белок, содержащий от амино-конца к карбокси-концу домен связывания с CD3, N-концевой линкер, константный участок иммуноглобулина, С-концевой линкер и первый домен связывания.
[00114] Изобретение включает полиспецифичный связывающийся белок, содержащий первый домен связывания, соединенный через линкерный домен со вторым доменом связывания (например, scFv, соединенный через линкер с другим scFv). Например, изобретение включает полиспецифичный связывающийся белок, содержащий первый домен связывания (в ориентации VH-линкер-VL или VL-линкер-VH), соединенный через пептидный линкерный домен с доменом связывания с CD3 (в ориентации VH-линкер-VL или VL-линкер-VH). В некоторых вариантах реализации биспецифичный белок в формате scFv-линкер-scFv может содержать вариабельные домены тяжелой и легкой цепи, происходящие от антитела к Т-клеточному антигену (например, CD3), включая, без ограничений, вариабельные домены, раскрытые в настоящем документе. Линкер, разделяющий домены scFv, может содержать ((Gly4)Ser)n, где n=1-5 (SEQ ID NO: 74-78). В одном варианте реализации линкер представляет собой ((Gly4)Ser)3 (SEQ ID NO: 76). Дополнительно, линкер может содержать около 8-12 аминокислот. В некоторых вариантах реализации белок по изобретению в данном формате («одноцепочечное биспецифичное антитело») не содержит участка Fc и в результате не обладает связанной с Fc эффекторной функцией.
[00115] Изобретение включает по существу любой вид полипептида, содержащего домен или домены связывания, как раскрыто в настоящем документе. Сюда включены антитела, их фрагменты, scFv, Fab, однодоменные ди-scFv, диатела, связывающиеся белки с двойным вариабельным доменом и полипептиды, содержащие фрагменты антител или антитела. Дополнительные виды полипептидов, которые включены в изобретение, раскрыты в настоящем документе.
[00116] В некоторых вариантах реализации домены VH и VL используются в качестве scFv и присоединены/слиты с другими полипептидами или последовательностями аминокислот таким способом, который позволяет scFv и VH/VL связываться с мишенью. В некоторых вариантах реализации домен VH и VL, как раскрыто в настоящем документе, используется в антителе или его фрагменте. Например домены VH и VL введены в природное положение доменов VH и VL в антителе. В некоторых вариантах реализации полипептид по изобретению, связывающийся с CD3, представляет собой антитело, например, с цепью VL и цепью VH, как раскрыто в настоящем документе.
[00117] В одном варианте реализации изобретения, полиспецифичный белок представляет собой димер scFv или диатело вместо цельного антитела. Диатела и димеры scFv могут быть сконструированы без участка Fc, с использованием только вариабельных доменов. Диатела представляет собой двухвалентные, биспецифичные антитела, в которых домены VH и VL экспрессируются на единой полипептидной цепи, но с использованием пептидного линкера, слишком короткого, чтобы позволить спаривание между двумя доменами на одной цепи, таким образом, вынуждая домены спариваться с комплементарными доменами другой цепи и образовывать два сайта связывания с антигеном (см., например, Holliger, P., et al. (1993) Proc. Natl. Acad. Sci. USA 90:6444-6448; Poljak, R.J., et al. (1994) Structure 2:1121-1123). Такие части связывания с антителом известны из уровня техники (Kontermann and Dubel eds., Antibody Engineering (2001) Springer-Verlag. New York. 790 pp.(ISBN 3-540-41354-5).
[00118] В одном варианте реализации изобретения, полиспецифичный белок представляет собой диатело, стабилизированное дисульфидными связями. Например, полиспецифичное антитело может содержать два различных полипептида, которые соэкспрессируются для генерации ковалентно связанного гетеродимерного комплекса с одним сайтом связывания для каждого из 2-х видов специфичности. В данном варианте реализации каждый Fv образован ассоциацией партнера VL на одной цепи с партнером VH на второй цепи в конфигурации VLA-VHB (первая цепь) и VLB-VHA (вторая цепь). Она может быть стабилизирована любым из двух альтернативных карбоксиконцевых доменов гетеродимеризации: спаривание VEPKSC (SEQ ID NO: 254) на одной цепи и FNRGEC (SEQ ID NO: 255) на другой, или спаривание противоположно заряженных биспиральных доменов. См., например, Moore et al., 2011, Blood. 1 17:4542-4551. В данном варианте реализации полиспецифичный связывающийся белок может содержать первую цепь, содержащую домен связывания с CD3 VH, соединенный с первым доменом связывания VL, и вторую цепь, содержащую домен связывания с CD3 VL, соединенный с первым доменом связывания VH, причем две цепи соединены посредством дисульфидной связи на С-концах. Стабилизированное дисульфидными связями диатело может быть сконструировано с использованием вариабельных тяжелых и легких цепей, происходящих от известных антител, включая, например, вариабельные тяжелые и легкие цепи, раскрытые в настоящем документе.
[00119] В другом варианте реализации полиспецифичный связывающийся белок представляет собой связывающийся белок с двумя вариабельными доменами, способный к связыванию с первым сайтом связывания (например, антиген опухоли) и комплексом TCR со специфичностью. В данном варианте реализации рекомбинантный белок содержит полипептидную цепь, которая содержит VD1-(X1)n-VD2-C-(X2)n, где VD1 - первый вариабельный домен, VD2 - второй вариабельный домен, С - константный домен, X1 - линкер (например, полипептидный линкер длиной около 10-20 аминокислот), Х2 представляет собой участок Fc, и n равно 0 или 1. См., например, патент США №8258268.
[00120] В одном варианте реализации изобретения полиспецифичный связывающийся белок содержит одну, две, три или более полипептидных цепей. Например, изобретение включает полиспецифичный белок с первой цепью, содержащей VH1-VL2, второй цепью, содержащей CH2-CH3-VL1-VH2 и третьей цепью, содержащей СН2-СН3. В данном варианте реализации VH1 и VL1 могут соответствовать вариабельным доменам первого домена связывания, и VH2 и VL2 могут соответствовать анти-CD3 вариабельным доменам. Альтернативно, VH1 и VL1 могут соответствовать анти-CD3 вариабельным доменам, и VH2 и VL2 могут соответствовать вариабельным доменам первого домена связывания.
[00121] В других вариантах реализации полиспецифичный связывающийся белок состоит из димеризованных одноцепочечных полипептидных цепей, содержащих различные одноцепочечные полипептиды (гетеродимер). В таком формате («полиспецифичный гетеродимер») каждая полипептидная цепь содержит домен связывания, N-концевой линкер, константный участок, С-концевой линкер и необязательно другой домен связывания, и дополнительно содержит домен гетеродимеризации. В некоторых вариациях вторая полипептидная цепь для гетеродимеризации содержит дополнительный домен связывания. Соответственно, в некоторых вариантах реализации настоящего изобретения гетеродимерный рекомбинантный белок может содержать два, три или четыре различных домена связывания. См. публикацию РСТ WO 2011/090762 относительно некоторых примеров и связанных способов.
[00122] Как указано выше, полипептид, связывающийся с CD3, в соответствии с настоящим описанием, содержит домен связывания, который специфично связывается с CD3. В некоторых вариациях, домен связывания с CD3 способен конкурировать за связывание с CD3 с одноцепочечным Fv (scFv), содержащим последовательность аминокислот, как показано в SEQ ID NO: 41. В конкретных вариантах реализации домен связывания с CD3 содержит (i) вариабельный участок тяжелой цепи (VH) иммуноглобулина, содержащий участки CDR HCDR1, HCDR2 и HCDR3, и (ii) вариабельный участок легкой цепи иммуноглобулина (VL), содержащий участки CDR LCDR1, LCDR2 и LCDR3. Пригодный домен связывания с CD3 включают такие, которые содержат участки VL и VH, происходящие от мышиного моноклонального антитела Cris-7 (Reinherz, Е.L. et al. (eds.), Leukocyte typing II., Springer Verlag, New York, (1986)). В некоторых из таких вариантов реализации LCDR3 содержит последовательность аминокислот, представленную в SEQ ID NO: 54, и/или HCDR3, содержит последовательность аминокислот, представленную в SEQ ID NO: 51; и LCDR1 и LCDR2 необязательно содержат последовательности аминокислот, представленные в SEQ ID N0:52 и SEQ ID NO: 53, соответственно, причем HCDR1 и HCDR2 необязательно содержат последовательности аминокислот, представленные в SEQ ID NO: 49 и SEQ ID NO: 50, соответственно. Например, в некоторых вариантах реализации LCDR1, LCDR2 и LCDR3 содержат последовательности аминокислот, соответственно представленные в SEQ ID NO: 52, 53 и 54; и/или HCDR1, HCDR2 и HCDR3 содержат последовательности аминокислот, представленные в SEQ ID NO: 49, 50 и 51, соответственно.
[00123] Примеры анти-CD3 антител, из которых домен связывания в соответствии с данным описанием может быть получен, включают моноклональное антитело Cris-7 (Reinherz, Е.L. et al. (eds.), Leukocyte typing II., Springer Verlag, New York, (1986)).

моноклональное антитело BC3 (Anasetti et al. (1990) J. Exp. Med. 172: 1691); моноклональное антитело ОКТ3 (Ortho multicenter Transplant Study Group (1985) N. Engl. J. Med. 313:337) и его производные, такие как ОКТЗ ala-ala (также называемый ОКТ3 AA-FL или ОКТ3 FL), гуманизированный вариант Fc с заменой аланина в положениях 234 и 235 (Herold et al. (2003) J. Clin. Invest. 11: 409); визилизумаб (Carpenter et al. (2002) Blood 99:2712), моноклональное антитело G19-4 (Ledbetter et al., 1986, J. Immunol. 136:3945) и моноклональное антитело 145-2C11 (Hirsch et al. (1988) J. Immunol. 140: 3766). Примером анти-TCR антитела является моноклональное антитело ВМА031 (Borst et al. (1990) Human Immunology 29: 175-188).
[00124] В некоторых вариантах реализации домен связывания представляет собой одноцепочечной фрагмент Fv (scFv), который включает участки VH и VL, специфичные в отношении целевой мишени. В некоторых вариантах реализации участки VH и VL являются человеческими.
[00125] В одном варианте реализации изобретения полипептиды, связывающиеся с CD3, обладают эмпирической и/или теоретической изоэлектрической точкой, которая по меньшей мере на 0,25, 0,5, 0,75, 1, 1,25, 1,5, 2, 2,5 или более единиц ниже изоэлектрической точки SMIP DRA209. В некоторых вариантах реализации полипептид по изобретению, связывающийся с CD3, обладает эмпирической и/или теоретической изоэлектрической точкой, которая от около 0,25 до около 2,5, от 0,5 до около 2,5, от около 0,5 до около 2,0, от около 0,5 до около 1,5, от около 0,5 до около 1,25, от около 0,5 до около 1,0, от около 0,5 до около 0,75, от около 0,75 до около 1,0, от около 0,75 до около 1,25, от около 0,75 до около 1,5, от около 0,75 до около 2,0, от около 1,0 до около 1,5, от около 1,0 до около 2,0, или от около 1,0 до около 2,5 единиц ниже изоэлектрической точки SMIP DRA209. В некоторых вариантах реализации полипептиды CD3 по изобретению обладают эмпирической или теоретической изоэлектрической точкой, которая составляет менее, чем около 8,9, 8,8, 8,7, 8,6, 8,5, 8,4, 8,3, 8,2, 8,1, 8, 7,5, 7,25, 7,0 или 6,75. В некоторых вариантах реализации полипептиды CD3 по изобретению обладают эмпирической или теоретической изоэлектрической точкой, которая составляет около 8,9, 8,8, 8,7, 8,6, 8,5, 8,4, 8,3, 8,2, 8,1, 8, 7,5, 7,25, 7,2 7,0, 6,8 или 6,75. В некоторых вариантах реализации полипептиды CD3 по изобретению обладают эмпирической или теоретической изоэлектрической точкой, которая составляет от около 7,9 до около 8,7, от 7,9 до около 8,6, от 8,0 до около 8,6, от 8,0 до около 8,5, от 8,0 до около 8,4, от 8,0 до около 8,3, от 8,0 до около 8,2, от 8,0 до около 8,1, от около 8,1 до около 8,6, от около 8,2 до около 8,6, от около 8,3 до около 8,6, от около 8,4 до около 8,6, от около 8,5 до около 8,6, от около 8,5 до около 8,3, от около 8,4 до около 8,2, или от около 8,3 до около 8,1.
[00126] В некоторых вариантах реализации домен связывания с CD3 содержит или представляет собой scFv со сниженной изоэлектрической точкой (эмпирической и/или теоретической) по меньшей мере на около 0,25, 0,5, 1, 1,25, 1,5, 2, 2,5 или более единиц, по сравнению с scFv SEQ ID NO: 41. В некоторых вариантах реализации домен связывания с CD3 содержит или представляет собой scFv со сниженной изоэлектрической точкой (эмпирической и/или теоретической), которая составляет от около 0,25 до около 2,5, от 0,5 до около 2,5, от около 0,5 до около 2,0, от около 0,5 до около 1,5, от около 0,5 до около 1,25, от около 0,5 до около 1,0, от около 0,5 до около 0,75, от около 0,75 до около 1,0, от около 0,75 до около 1,25, от около 0,75 до около 1,5, от около 0,75 до около 2,0, от около 1,0 до около 1,5, от около 1,0 до около 2,0, или от около 1,0 до около 2,5 единиц ниже изоэлектрической точки scFv SEQ ID NO: 41. Такие scFv включают scFv, представленные в SEQ ID NO: 6, 8, 10, 12, 14, 16, 18 и 20, или scFv, который по меньшей мере на около 90%, по меньшей мере около на 91%, по меньшей мере около на 92%, по меньшей мере около на 93%, по меньшей мере около на 94%, по меньшей мере около на 95%, по меньшей мере около на 96%, по меньшей мере около на 97%, по меньшей мере около на 98%, по меньшей мере около на 99%, по меньшей мере около на 99,5%, или 100% идентичен последовательности аминокислот scFv, представленных в SEQ ID NO: 6, 8,10, 12,14, 16,18 и 20.
[00127] В некоторых вариантах реализации степень идентичности SEQ ID NO: 41 домена связывания с CD3 по изобретению составляет от около 80% до около 99%, от около 82% до около 99%, от около 84% до около 99%, от около 86% до около 99%, от около 88% до около 99%, от около 90% до около 99%, от около 92% до около 99%, от около 94% до около 99%, от около 96% до около 99%, от около 97% до около 99%, от около 98% до около 99%, от около 80% до около 85%, от около 85% до около 90%, от около 85% до около 95%, от около 90% до около 97%, от около 90% до около 95%, от около 90% до около 93%, от около 90% до около 91%, от около 95% до около 96%, от около 96% до около 97%, от около 97% до около 98%, от около 96% до около 97% или от около 96% до около 98%.
[00128] В некоторых вариантах реализации домен связывания с CD3 по изобретению представляет собой вариант SEQ ID NO: 41, в котором одна или более аминокислот в последовательности аминокислот модифицированы таким образом, чтобы снизить изоэлектрическую точку домена связывания. Мутации включают замену аминокислоты(-от) другой аминокислотой(-ами), с целью снижения изоэлектрической точки домена связывания, вставку аминокислоты(-от), с целью снижения изоэлектрической точки молекулы связывания и/или делецию аминокислоты(-от), с целью снижения изоэлектрической точки домена связывания. В одном варианте реализации мутации представляют собой замены. В некоторых вариантах реализации мутация(-и) находятся в каркасных участках вариабельной цепи(-ей). В некоторых вариантах реализации вариант содержит мутацию 1,2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот.
[00129] В одном варианте реализации домен связывания с CD3 содержит или представляет собой scFv, содержащий тяжелую цепь или легкую цепь, выбранную из группы, состоящей из SEQ ID NO: 28, 30, 32, 38 и 40. В другом варианте реализации домен связывания с CD3, содержащий участок VH, выбранный из группы, состоящей из SEQ ID NO: 22, 24, 26, 28, 30 и 32, и участок VL, выбранный из группы, состоящей из SEQ ID NO: 38 и 40. В другом варианте реализации домен связывания с CD3 содержит участок VH, выбранный из группы, состоящей из SEQ ID NO: 28,30и32и участок VL, выбранный из группы, состоящей из SEQ ID NO: 34, 36, 38 и 40. В другом варианте реализации домен связывания с CD3 содержит SEQ ID NO: 28 и 34. В другом варианте реализации домен связывания с CD3 содержит SEQ ID NO: 28 и 38. В другом варианте реализации домен связывания с CD3 содержит SEQ ID NO: 26 и 38. В другом варианте реализации домен связывания с CD3 содержит SEQ ID NO: 26 и 34. В некоторых вариантах реализации домен связывания с CD3 содержит участок VL, выбранный из группы, состоящей из SEQ ID NO: 38 и 40 и участок VH, выбранный из группы, состоящей из SEQ ID NO: 22, 24, 26, 28, 30 и 32. В дополнительных вариантах реализации каждый CDR содержит не более одной, двух, или трех замен, вставок или делеций, по сравнению с участком моноклонального антитела или его фрагмента или производного, который специфично связывается с целевой мишенью (например, CD3).
[00130] В некоторых вариантах реализации домен связьюания с CD3 связывается или взаимодействует с субъединицей CD3ε комплекса рецептора Т-клеток на Т-клетках. В некоторых вариантах реализации домен связывания с CD3 конкурирует за связывание с CD3ε с моноклональным антителом Cris-7 или HuM291.
[00131] В некоторых вариантах реализации полипептид, связывающийся с CD3, индуцирует интернализацию комплекса рецептора Т-клеток.
[00132] В некоторых вариациях домен связывания представляет собой одноцепочечный Fv (scFv), содержащий участки VL и VH иммуноглобулина, соединенные пептидным линкером. Использование пептидных линкеров для соединения участков VL и VH известно из уровня техники, и существует большое количество публикаций в пределах данной конкретной области. Широко используемый пептидный линкер представляет собой 15mer, состоящий из трех повторов Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 74) последовательности аминокислот (Gly4Ser)3 (SEQ ID NO: 76). В некоторых вариантах реализации пептидный линкер содержит или состоит из любой из SEQ ID NO: 74-78. Применялись другие линкеры и технология показа фага, а также технология селективного инфекционного фага применялась с целью диверсификации и отбора пригодных линкерных последовательностей (Tang et al., J. Biol. Chem. 271, 15682-15686, 1996; Hennecke et al, Protein Eng. 11, 405-410, 1998). В некоторых вариантах реализации участки VH и VL соединяют пептидным линкером, содержащим последовательность аминокислот формулы (Gly4Ser)n, в которой n=1-5 (SEQ ID NO: 74-78, соответственно). Другие пригодные линкеры могут быть получены посредством оптимизации простого линкера (например, (Gly4Ser)n) с помощью случайного мутагенеза.
[00133] В конкретных вариантах реализации домен связывания содержит гуманизированные участки VL и/или VH иммуноглобулина. Методы гуманизации участков VL и VH иммуноглобулина известны из уровня техники и обсуждаются, например, в Публикации патентной заявки США №2006/0153837.
[00134] Ожидается, что «гуманизация» будет давать менее иммуногенное антитело с полным сохранением свойств связывания с антигеном оригинальной молекулы. Для того, чтобы сохранить все свойства связывания с антигеном оригинального антитела, структуру его сайта связывания с антигеном необходимо воспроизвести в «гуманизированной» версии. Это может быть достигнуто с помощью пересадки только нечеловеческих CDR на человеческие вариабельные домены каркаса и константные участки, с сохранением или без сохранения критических остатков каркаса (Jones et al., Nature 321:522 (1986); Verhoeyen et al., Science 239:1539 (1988)), или рекомбинацией полноразмерных нечеловеческих вариабельных доменов (для сохранения свойств связывания лиганда), но с «маскировкой» их сходной с человеческой поверхностью посредством рациональной замены контактирующих остатков (для снижения антигенности) (Padlan, Molec. Immunol. 28:489 (1991)).
[00135] По существу, гуманизация путем пересадки CDR включает рекомбинацию только CDR нечеловеческого антитела на человеческий каркас вариабельного участка и человеческий константный участок. Теоретически, это должно в существенной мере снизить или исключить иммуногенность (кроме случаев существования аллотипических или идиотипических отличий). Однако сообщалось о том, что также может возникнуть необходимость в сохранении некоторых остатков каркаса оригинального антитела (Reichmann et al., Nature, 332:323 (1988); Queen et al., Proc. Natl. Acad. Sci. USA, 86:10,029 (1989)).
[00136] Остатки каркаса, которые следует сохранить, поддаются идентификации посредством компьютерного моделирования. Альтернативно, критические остатки каркаса потенциально могут быть идентифицированы путем сравнения известных структур сайта связывания с антигеном (Padlan, Molec. Immunol., 31 (3): 169-217 (1994), включена в данное описание посредством ссылки).
[00137] Остатки, которые потенциально воздействуют на связывание с антигеном, делятся на несколько групп. Первая группа включает остатки, смежные с поверхностью антигенного сайта, которые могут таким образом осуществлять прямой контакт с антигенами. Такие остатки включают амино-концевые остатки и смежные с CDR. Вторая группа включает остатки, которые могут изменить структуру или относительное выравнивание CDR путем контакта с CDR или другой пептидной цепью антитела. Третья группа включает аминокислоты со скрытыми боковыми цепями, которые могут повлиять на структурную целостность вариабельных доменов. Остатки в указанных группах обычно находят в одних и тех же положениях (Padlan, 1994, выше), хотя их идентифицированные положения могут отличаться в зависимости от системы нумерации (см. Kabat et al., "Sequences of proteins of immunological interest, 5th ed., Pub. No. 91-3242, U.S. Dept. Health и Human Services, NIH, Bethesda, Md., 1991).
[00138] Хотя некоторые примеры, раскрытые в настоящем документе, включают гуманизацию scFv, SMIP, Scorpion и молекул перехватчика, но не антитела, знание о гуманизированных антителах в данной области применимо к полипептидам по изобретению.
[00139] В некоторых вариантах реализации шарнир представляет собой шарнирный участок иммуноглобулина человека дикого типа. В некоторых других вариантах реализации один или более остатков аминокислот могут быть добавлены к амино- или карбокси-концу шарнирного участка иммуноглобулина дикого типа как часть дизайна конструкта слитого белка. Например, дополнительные остатки соединяющих аминокислот на амино-конце шарнира могут представлять собой «RT», «RSS», «TG» или «Т», или на карбокси-конце шарнира могут представлять собой «SG», или делеция шарнира может сочетаться с добавлением, например, АР и «SG» добавляют на карбокси-конце.
[00140] В некоторых вариантах реализации шарнир представляет собой модифицированный шарнир иммуноглобулина, в которой один или более остатков цистеина в шарнирном участке иммуноглобулина дикого типа заменены одним или более других остатков аминокислот (например, серии или аланин).
[00141] Примеры модифицированных шарниров иммуноглобулина включают, без ограничений, шарнирный участок иммуноглобулина lgG1 человека, содержащий замену одного, двух или трех остатков цистеина, найденных в шарнире lgG1 дикого типа человека, на один, два или три различных остатка аминокислот (например, серина или аланина). Модифицированный шарнир иммуноглобулина дополнительно может содержать замену пролина на другую аминокислоту (например, серии или аланин). Например, раскрытый выше модифицированный шарнир lgG1 человека может дополнительно содержать замену пролина, размещенного в направлении карбокси-конца по отношению к трем остаткам цистеина шарнирного участка lgG1 дикого типа человека, на другой остаток аминокислоты (например, серии, аланин). В одном варианте реализации замены остатков пролина в ядерном шарнирном участке отсутствуют.
[00142] В некоторых вариантах реализации полипептид шарнира содержит или представляет собой последовательность, которая по меньшей мере на 80%, по меньшей мере на 81%, по меньшей мере на 82%, по меньшей мере на 83%, по меньшей мере на 84%, по меньшей мере на 85%, по меньшей мере на 86%, по меньшей мере на 87%, по меньшей мере на 88%, по меньшей мере на 89%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентична шарнирному участку иммуноглобулина дикого типа, такому как шарнир lgG1 человека дикого типа, шарнир lgG2 человека дикого типа или шарнир lgG4 человека дикого типа.
[00143] В дополнительных вариантах реализации шарнир, который присутствует в полипептиде, связывающемся с CD3, может представлять собой шарнир, который не основывается или не происходит от шарнира иммуноглобулина (т.е., шарнир иммуноглобулина дикого типа или модифицированный шарнир иммуноглобулина). Примеры таких шарниров включают пептиды длиной от около 5 до около 150 аминокислот, происходящие от междоменного участка трансмембранного белка или стебелькового участка С-лектина типа II, например, пептиды длиной от около 8 до 25 аминокислот и пептиды длиной от около 7 до 18 аминокислот.
[00144] В некоторых вариантах реализации междоменные шарниры или шарниры стебелькового участка содержат от 7 до 18 аминокислот и могут образовывать α-спиральную биспиральную структуру. В некоторых вариантах реализации междоменные шарниры или шарниры стебелькового участка содержат 0, 1, 2, 3 или 4 остатка цистеина. Примерами междоменных шарниров или шарниров стебелькового участка являются пептидные фрагменты междоменного или стебелькового участков, такие как фрагменты стебельковых участков CD69, CD72, CD94, NKG2A и NKG2D длиной от 10 до 150 аминокислот.
[00145] В некоторых вариантах реализации последовательности шарнира содержат около 5-150 аминокислот, 5-10 аминокислот, 10-20 аминокислот, 20-30 аминокислот, 30-40 аминокислот, 40-50 аминокислот, 50-60 аминокислот, 5-60 аминокислот, 5-40 аминокислот, 8-20 аминокислот или 10-15 аминокислот. Шарнир может быть в основном гибким, но может также обеспечивать более жесткие характеристики или может содержать в основном α-спиральную структуру с минимальной β-листовой структурой. Длина или последовательности шарниров могут влиять на аффинность связывания доменов связывания, с которыми шарниры соединены прямо или косвенно (через другой участок или домен, такой как домен гетеродимеризации), а также один или более видов активности участка Fc, с которым шарниры прямо или непрямо соединены.
[00146] В некоторых вариантах реализации последовательности шарниров стабильны в плазме и сыворотке, а также устойчивы к протеолитическому расщеплению. Первый остаток лизина в верхнем шарнирном участке lgG1 может быть модифицирован таким образом, чтобы минимизировать протеолитическое расщепление, например, лизин может быть заменен метионином, треонином, аланином или глицином, или удален.
[00147] В некоторых вариантах реализации изобретения, полипептид, связывающийся с CD3, способен к образованию гетеродимера со второй полипептидной цепью и включает шарнирный участок: (а) непосредственно амино-концевой по отношению к константному участку иммуноглобулина (например, амино-концевой по отношению к домену СН2, причем константный участок иммуноглобулина содержит домены СН2 и СН3, или амино-концевой по отношению к домену СН3, причем участки иммуноглобулина содержат домены СН3 и СН4), (б) вставленный между и соединенный с доменом связывания (например, scFv) и доменом гетеродимеризации иммуноглобулина, (в) вставленный между и соединенный с доменом гетеродимеризации иммуноглобулина и константным участком иммуноглобулина (например, если константный участок иммуноглобулина содержит домены СН2 и СН3 или домены СН3 и СН4), (г) вставленный между и соединенный с константным участком иммуноглобулина и доменом связывания, (д) на амино-конце полипептидной цепи, или (е) на карбокси-конце полипептидной цепи. Полипептидная цепь, содержащая шарнирный участок, как раскрыто в настоящем документе, может быть способна связываться с другой полипептидной цепью, чтобы образовать гомодимерный или гетеродимерный белок, раскрытый в настоящем документе, причем образованный димер будет содержать домен связывания, который сохраняет свою специфичность в отношении мишени и/или свою специфичную аффинность связывания с мишенью.
[00148] В некоторых вариантах реализации шарнир, присутствующий в полипептиде, который образует гетеродимер с другой цепью полипептида, может представлять собой шарнир иммуноглобулина, например, шарнирный участок иммуноглобулина дикого типа или модифицированный шарнирный участок такого иммуноглобулина. В некоторых вариантах реализации шарнир одной полипептидной цепи гетеродимерного белка идентичен соответствующему шарниру другой полипептидной цепи гетеродимера. В некоторых других вариантах реализации шарнир одной цепи отличается от шарнира другой цепи (например, длиной и/или последовательностью). Различные шарниры в различных цепях позволяют осуществить другой способ манипулирования аффинностью связывания доменов связывания, с которыми соединены шарниры, таким образом, что гетеродимер может связываться преимущественно с мишенью одного домена связывания, по сравнению с мишенью другого домена связывания. Например, в некоторых вариантах реализации гетеродимерный белок содержит CD3- или TCR-связывающийся домен в одной цепи и второй домен связывания (например, домен связывания с антигеном опухоли) в другой цепи. Присутствие двух различных шарниров в двух цепях может позволять гетеродимеру связываться вначале с антигеном опухоли, а затем с CD3 или другим компонентом TCR. Таким образом, гетеродимер может рекрутировать CD3+ Т-клетки к клеткам опухоли, которые экспрессируют антиген (например, ПСМА-экспрессирующие клетки опухоли), которые, в свою очередь, могут, повреждать или уничтожать клетки опухоли, экспрессирующие антиген.
[00149] Примеры шарнирных участков, пригодных для использования в соответствии с настоящим изобретением, приведены в Табл. 1 и 2 ниже.
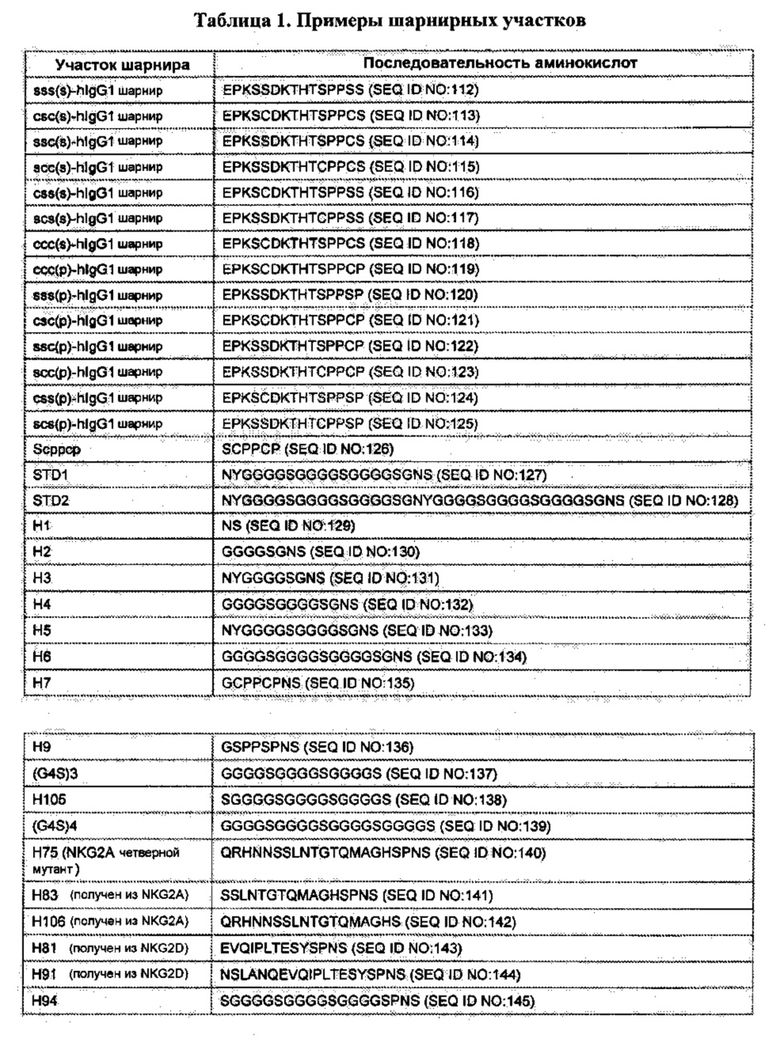
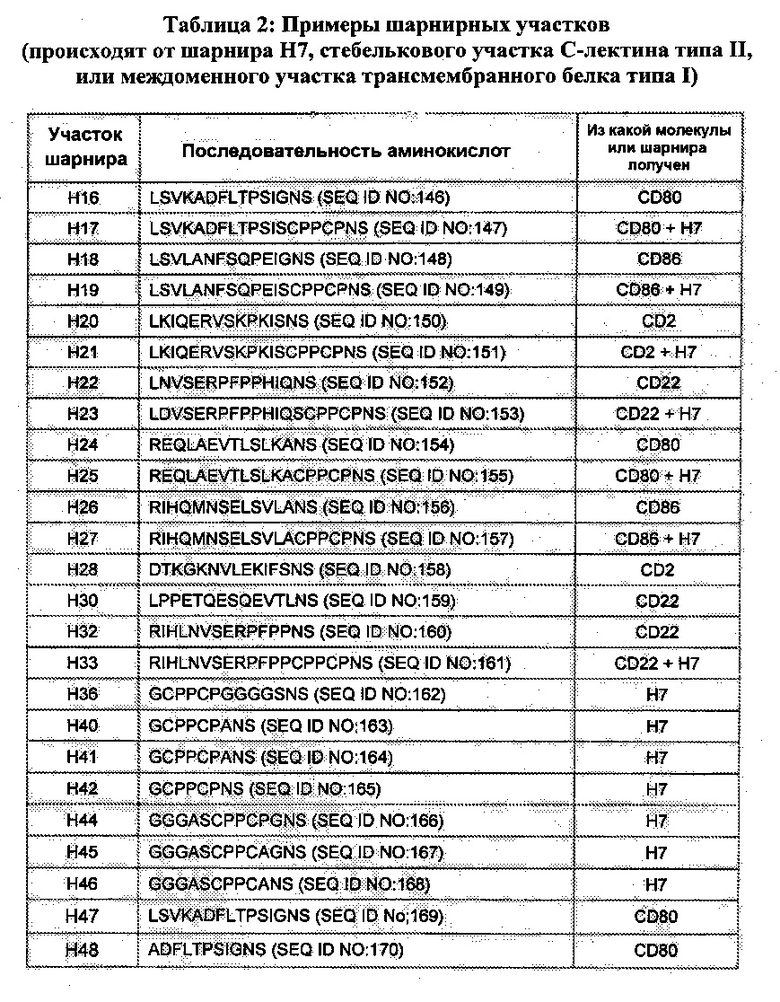
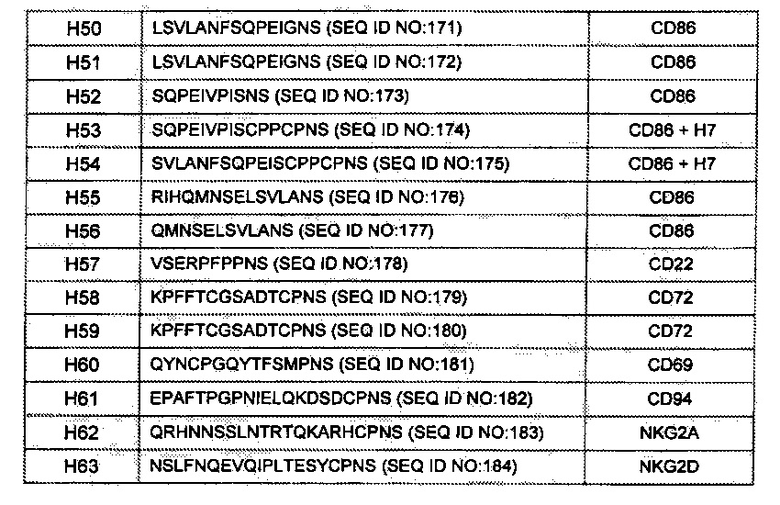
[00150] В некоторых вариантах реализации полипептид или белок, связывающийся с CD3, по изобретению, может содержать «домен димеризации иммуноглобулина» или «домен гетеродимеризации иммуноглобулина».
[00151] «Домен димеризации иммуноглобулина» или «домен гетеродимеризации иммуноглобулина» в настоящем документе обозначает домен полипептидной цепи иммуноглобулина, который взаимодействует или связывается со вторым доменом второй полипептидной цепи иммуноглобулина, причем взаимодействие первых и вторых доменов гетеродимеризации иммуноглобулина в существенной мере содействует или эффективно способствует гетеродимеризации первых и вторых полипептидных цепей (т.е., образованию димера между двумя различными полипептидными цепями, которые также обозначаются как «гетеродимер» или «гетеродимерный белок»). Взаимодействие между доменами гетеродимеризации иммуноглобулина «в существенной мере содействует или эффективно способствует» гетеродимеризации первых и вторых полипептидных цепей, если присутствует статистически значимое уменьшение димеризации между первыми и вторыми полипептидными цепями в отсутствие домена гетеродимеризации иммуноглобулина первой полипептидной цепи и/или домена гетеродимеризации иммуноглобулина второй полипептидной цепи. В некоторых вариантах реализации, если первые и вторые полипептидные цепи соэкспрессируются, по меньшей мере 60%, по меньшей мере от около 60% до около 70%, по меньшей мере от около 70% до около 80%, по меньшей мере от около 80% до около 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% первых и вторых полипептидных цепей образуют гетеродимеры друг с другом. Характерные домены гетеродимеризации иммуноглобулина включают домен СН1 иммуноглобулина, домен CL иммуноглобулина (например, изотипы СК или СА) или их производные, в том числе, домены СН1 и CL иммуноглобулина дикого типа и модифицированные (или мутантные) домены СН1 и CL иммуноглобулина, такие как раскрыты в настоящем документе.
[00152] Домены димеризации/гетеродимеризации могут использоваться в случае, если желательно сформировать гетеродимеры из двух неидентичных полипептидных цепей, где одна или обе полипептидных цепи содержат домен связывания. В некоторых вариантах реализации один член полипептидной цепи конкретных гетеродимеров, раскрытых в настоящем документе, не содержит домена связывания. Как указано выше, гетеродимерный белок в соответствии с настоящим документом содержит домен гетеродимеризации иммуноглобулина в каждой полипептидной цепи. Иммуноглобулиновые домены гетеродимеризации в полипептидных цепях гетеродимера отличаются друг от друга и, таким образом, могут быть дифференциально модифицированы с целью облегчения гетеродимеризации обоих цепей, а также минимизации гомодимеризации каждой из цепей. Как продемонстрировано в Публикации РСТ WO 2011/090762, домены гетеродимеризации иммуноглобулина, раскрытые в настоящем документе, позволяют эффективную гетеродимеризацию между различными полипептидами и облегчают очистку полученного гетеродимерного белка.
[00153] Как предусмотрено в настоящем документе, домены гетеродимеризации иммуноглобулина, пригодные для содействия гетеродимеризации двух различных одноцепочечных полипептидов (например, один короткий и один длинный) в соответствии с настоящим документом, содержат домены СН1 и CL иммуноглобулина, например, домены СН1 и CL человека. В некоторых вариантах реализации домен гетеродимеризации иммуноглобулина представляет собой домен СН1 дикого типа, например, домен CHI lgG1, lgG2, lgG3, lgG4, IgA1, lgA2, IgD, IgE или IgM дикого типа. В дополнительных вариантах реализации домен гетеродимеризации иммуноглобулина представляет собой домен CHI lgG1, lgG2, lgG3, lgG4, IgA1, lgA2, IgD, IgE или IgM дикого типа человека, как было представлено ранее в SEQ ID NO: 114, 186-192 и 194, соответственно, Публикации РСТ WO 2011/090762 (указанные последовательности включены в настоящий документ посредством ссылки). В некоторых вариантах реализации домен гетеродимеризации иммуноглобулина представляет собой домен lgG1 CH1 дикого типа человека, как представлено в SEQ ID NO: 114 WO 2011/090762 (указанная последовательность включена в настоящий документ посредством ссылки), которая является такой же, как и SEQ ID NO: 80 настоящей заявки.
[00154] В дополнительных вариантах реализации домен гетеродимеризации иммуноглобулина представляет собой модифицированный домен СН1 иммуноглобулина, такой как модифицированный домен CH1 lgG1, lgG2, lgG3, lgG4, IgA1, lgA2 IgD, IgE или IgM. В некоторых вариантах реализации домен гетеродимеризации иммуноглобулина представляет собой модифицированный домен CH1 lgG1, lgG2, lgG3, lgG4, IgA1, lgA2, IgD, IgE или IgM человека. В других вариантах реализации остаток цистеина в домене СН1 дикого типа (например, СН1 человека), принимающий участие в образовании дисульфидной связи с доменом CL иммуноглобулина дикого типа (например, CL человека) удален или заменен в модифицированном домене СН1 иммуноглобулина таким образом, что дисульфидная связь между модифицированным доменом СН1 и доменом CL дикого типа не образуется.
[00155] В некоторых вариантах реализации домен гетеродимеризации иммуноглобулина представляет собой домен CL дикого типа, например, домен Сκ дикого типа или домен Сλ, дикого типа. В конкретных вариантах реализации домен гетеродимеризации иммуноглобулина представляет собой домен Сκ дикого типа человека или домен Сλ человека, как было представлено ранее в SEQ ID NO: 112 и 113, соответственно, WO 2011/090762 (указанные последовательности включены в настоящий документ посредством ссылки), которые являются такими же, как SEQ ID NO: 81 и 82 настоящей заявки, соответственно. В дополнительных вариантах реализации домен гетеродимеризации иммуноглобулина представляет собой модифицированный домен CL иммуноглобулина, например, модифицированный домен Сκ или Сλ, такой как модифицированный домен Сκ или Сλ человека.
[00156] В некоторых вариантах реализации остаток цистеина домена CL (например, CL человека) дикого типа, принимающий участие в образовании дисульфидной связи с доменом СН1 (например, СН1 человека) иммуноглобулина дикого типа, удаляют или замещают в модифицированном домене CL иммуноглобулина. Такие модифицированные домены CL могут дополнительно содержать делецию аминокислоты на амино-конце. Пример домена Сκ представлен в SEQ ID NO: 141 WO 2011/090762 (указанная последовательность включена в настоящий документ посредством ссыпки и представляет собой SEQ ID NO: 83 настоящей заявки), в которой как первый остаток аргинина, так и последний остаток цистеина домена Сκ дикого типа человека удалены. В некоторых вариантах реализации только последний остаток цистеина дикого типа в домене Сκ человека удален в модифицированном домене Сκ, поскольку первый остаток аргинина, удаленный из домена Сκ дикого типа человека может быть поставлен линкером, который содержит аргинин на карбокси-конце и связывает амино-конец модифицированного домена Сκ с другим доменом (например, подобласть иммуноглобулина, например подобласть, содержащая домены СН2 и СН3 иммуноглобулина). Пример домена Сλ представлен в SEQ ID NO: 140 WO 2011/090762 (указанная последовательность включена в настоящий документ посредством ссылки и представляет собой SEQ ID NO: 84 настоящей заявки), в которой первый остаток аргинина в домене Сλ дикого типа человека удален, и цистеин, принимающий участие в образовании дисульфидной связи с цистеином в домене СН1, заменен серином.
[00157] В дополнительных вариантах реализации домен гетеродимеризации иммуноглобулина представляет собой модифицированный домен Сκ, который содержит одну или более замен аминокислоты, по сравнению с доменом Сκ дикого типа, в положениях, которые могут принимать участие в образовании сети межцепочечных водородных связей в интерфейсе Сκ-Сκ. Например, в некоторых вариантах реализации домен гетеродимеризации иммуноглобулина представляет собой модифицированный домен Сκ человека, содержащий замены одной или более аминокислот в положениях N29, N30, Q52, V55, Т56, S68 или Т70 различными аминокислотами. Нумерация аминокислот основана на их положении в модифицированной последовательности Сκ человека, как представлено в SEQ ID NO: 141 WO 2011/090762 (указанная последовательность включена в настоящий документ посредством ссылки и представляет собой SEQ ID NO: 83 настоящей заявки). В некоторых вариантах реализации домен гетеродимеризации иммуноглобулина представляет собой модифицированный домен Сκ человека, содержащий одну, две, три или четыре замены аминокислот в положениях N29, N30, V55 или Т70. Аминокислота, используемая для замены в вышеуказанных положениях, может представлять собой аланин или остаток аминокислоты с объемным фрагментом боковой цепи, таким как аргинин, триптофан, тирозин, глутамат, глутамин или лизин. Дополнительные остатки аминокислот, которые могут использоваться для замены остатков аминокислот в последовательности Сκ дикого типа человека в вышеуказанных положениях (например, N30), включают аспартат, метионин, серии и фенилаланин. Примеры модифицированных доменов Сκ человека представлены в SEQ ID NO: 142-178 WO 2011/090762 (указанные последовательности включены в настоящий документе посредством ссылки). Модифицированные домены Сκ человека представляют собой такие, которые облегчают гетеродимеризацию с доменом СН1, но минимизируют гомодимеризацию с другим доменом Сκ. Характерные модифицированные домены Сκ человека представлены в SEQ ID NO: 160 (N29W V55A Т70А), 161 (N29Y V55A Т70А), 202 (Т70Е N29A N30A V55A), 167 (N30R V55A Т70А), 168 (N30K V55A Т70А), 170 (N30E V55A Т70А), 172 (V55R N29A N30A), 175 (N29W N30Y V55A Т70Е), 176 (N29Y N30Y V55A Т70Е), 177 (N30E V55A Т70Е), 178 (N30Y V55A Т70Е), 838 (N30D V55A Т70Е), 839 (N30M V55A Т70Е), 840 (N30S V55A Т70Е), и 841 (N30F V55A Т70Е) WO 2011/090762 (указанные последовательности включены в настоящий документ посредством ссылки).
[00158] В некоторых вариантах реализации, в дополнение или в качестве альтернативы мутациям в доменах Сκ, раскрытых в настоящем документе, оба домена гетеродимеризации иммуноглобулина (например, домены СН1 и CL иммуноглобулина) полипептидного гетеродимера содержат мутации, таким образом, что результирующие домены гетеродимеризации иммуноглобулина образуют солевые мостики (т.е., ионные взаимодействия) между остатками аминокислот на модифицированных сайтах. Например, домены гетеродимеризации иммуноглобулина полипептидного гетеродимера могут представлять собой модифицированный домен СН1 в комбинации с модифицированным доменом Сκ. В мутантном домене СН1, валин в положении 68 (V68) домена СН1 дикого типа человека заменен остатком аминокислоты с отрицательным зарядом (например, аспартатом или глутаматом), тогда как лейцин в положении 29 (L29) мутантного домена Сκ человека, в котором первый остаток аргинина и последний остаток цистеина удалены, заменен остатком аминокислоты с положительным зарядом (например, лизин, аргинин или гистидин). Взаимодействие заряд-заряд между остатком аминокислоты с отрицательным зарядом полученного мутантного домена СН1 и остатком аминокислоты с положительным зарядом полученного мутантного домена Сκ приводит к образованию солевого мостика, который стабилизирует гетеродимерный интерфейс между мутантными доменами СН1 и Сκ. Альтернативно, V68 СН1 дикого типа может быть заменен на остаток аминокислоты с положительным зарядом, тогда как L29 мутантного домена Сκ человека, в котором первый остаток аргинина и последний остаток цистеина удалены, может есть быть заменен на остаток аминокислоты с отрицательным зарядом. Примеры мутантных последовательностей СН1, в которых V68 заменен на аминокислоту с отрицательным или положительным зарядом, представлены в SEQ ID NO: 844 и 845 WO 2011/090762 (указанные последовательности включены в настоящий документ посредством ссылки). Примеры мутантных последовательностей Сκ, в которых L29 заменен аминокислотой с отрицательным или положительным зарядом, представлены в SEQ ID NO: 842 и 843 WO 2011/090762 (указанные последовательности включены в настоящий документ посредством ссылки).
[00159] Остатки в положениях, кроме V68 домена СН1 человека и L29 домена Сκ человека, могут быть заменены аминокислотами с противоположным зарядом, чтобы обеспечить ионные взаимодействия между аминокислотами в дополнение или в качестве альтернативы мутациям в V68 домена СН1 и L29 домена Сκ. Такие положения могут быть идентифицированы любым пригодным способом, в том числе, случайным мутагенезом, анализом кристаллической структуры пары СН1-Сκ, с целью идентификации остатков аминокислот в интерфейсе СН1-Сκ и дальнейшей идентификации пригодных положений среди остатков аминокислот в интерфейсе СН1-Сκ с использованием набора критериев (например, склонность принимать участие в ионных взаимодействиях, близость к потенциальному остатку-партнеру, и т.п.).
[00160] В некоторых вариантах реализации полипептидные гетеродимеры в соответствии с настоящим документом содержат только одну пару доменов гетеродимеризации иммуноглобулина. Например, первая цепь полипептидного гетеродимера может содержать домен СН1 в качестве домена гетеродимеризации иммуноглобулина, тогда как вторая цепь может содержать домен CL (например, Сκ или Сλ) в качестве домена гетеродимеризации иммуноглобулина. Альтернативно, первая цепь может содержать домен CL (например, Сκ или Сλ) в качестве домена гетеродимеризации иммуноглобулина, тогда как вторая цепь может содержать домен СН1 в качестве домена гетеродимеризации иммуноглобулина. Как раскрыто в настоящем документе, домены гетеродимеризации иммуноглобулина первых и вторых цепей способны ассоциироваться с образованием гетеродимерного белка в соответствии с настоящим документом.
[00161] В некоторых других вариантах реализации гетеродимерные белки в соответствии с настоящим документом могут содержать две пары доменов гетеродимеризации иммуноглобулина. Например, первая цепь гетеродимера может содержать два домена СН1, тогда как вторая цепь может содержать два домена CL, которые связываются с двумя доменами СН1 в первой цепи. Альтернативно, первая цепь может содержать два домена CL, тогда как вторая цепь может содержать два домена СН1, которые связываются с двумя доменами CL в первой цепи. В некоторых вариантах реализации первая полипептидная цепь содержит домен СН1 и домен CL, тогда как вторая полипептидная цепь содержит домен CL и домен СН1, которые связываются с доменом СН1 и доменом CL первой полипептидной цепи, соответственно.
[00162] В тех вариантах реализации, в которых гетеродимерный белок содержит только одну пару гетеродимерных (например, один домен гетеродимеризации иммуноглобулина в каждой цепи), домен гетеродимеризации иммуноглобулина в каждой цепи может быть расположен в направлении амино-конца по отношению к константной подобласти иммуноглобулина указанной цепи. Альтернативно, домен гетеродимеризации иммуноглобулина в каждой цепи может быть расположен в направлении карбокси-конца по отношению к константной подобласти иммуноглобулина указанной цепи.
[00163] В тех вариантах реализации, в которых гетеродимерный белок содержит две пары гетеродимеризации (например, два домена гетеродимеризации иммуноглобулина в каждой цепи), оба домена гетеродимеризации иммуноглобулина в каждой цепи могут быть расположены в направлении амино-конца по отношению к константной подобласти иммуноглобулина в указанной цепи. Альтернативно, оба домена гетеродимеризации иммуноглобулина в каждой цепи могут быть расположены в направлении карбокси-конца по отношению к константной подобласти иммуноглобулина указанной цепи. В дополнительных вариантах реализации один домен гетеродимеризации иммуноглобулина в каждой цепи может быть размещен в направлении амино-конца по отношению к константной подобласти иммуноглобулина указанной цепи, тогда как другой домен гетеродимеризации иммуноглобулина в каждой цепи может быть расположен в направлении карбокси-конца по отношению к константной подобласти иммуноглобулина указанной цепи. Другими словами, в таких вариантах реализации константная подобласть иммуноглобулина вставлена между двумя доменами гетеродимеризации иммуноглобулина в каждой цепи.
[00164] Как раскрыто в настоящем документе, в некоторых вариантах реализации связывающиеся с CD3 полипептиды в соответствии с настоящим документом (например, низкомолекулярные модульные иммунофармацевтические белки (SMIP), биспецифичные гомодимеры (например, Scorpion), гетеродимерные моно- и полиспецифичные терапевтические средства (например, перехватчик) содержат константный участок иммуноглобулина в каждой полипептидной цепи. В настоящем документе термин «константный участок» используется взаимозаменяемым образом с термином «константная подобласть» и «константный участок» иммуноглобулина. Введение константного участка иммуноглобулина может замедлить клиренс гомодимерных и гетеродимерных белков, образованных двумя цепями полипептида, связывающегося с CD3, из кровотока после их введения субъекту.
[00165] Путем мутаций или других модификаций, константный участок иммуноглобулина в дальнейшем позволяет относительно легкую модуляцию эффекторных функций димерного полипептида (например, АЗКЦ, АЗКФ, КЗЦ, фиксация комплемента и связывание с рецепторами Fc), которые могут быть увеличены или уменьшены в зависимости от подлежащего лечению заболевания, как известно из уровня техники и раскрыто в настоящем документе.
[00166] В некоторых вариантах реализации константный участок иммуноглобулина одной или обеих полипептидных цепей полипептидных гомодимеров и гетеродимеров в соответствии с настоящим документом будет способен к опосредованию одной или более таких эффекторных функций.
[00167] В других вариантах реализации одна или более из указанных эффекторных функций снижена или отсутствует в константном участке иммуноглобулина или одной или обеих полипептидных цепях полипептидных гомодимеров и гетеродимеров в соответствии с настоящим документом, по сравнению с константным участком иммуноглобулина соответствующего дикого типа. Например, для моноспецифичных полипептидов, связывающихся с CD3, а также полипептидов, связывающихся с CD3, которые содержат второй домен связывания, например, сконструированных таким образом, чтобы вызывать перенаправленную токсичность Т-клеток (ПТТК), будет особенно предпочтительным содержать константный участок иммуноглобулина со сниженной или отсутствующей эффекторной функцией относительно константного участка иммуноглобулина соответствующего дикого типа. В одном варианте реализации константный участок модифицирован таким образом, чтобы он не фиксировал комплемент. Дополнительно, константный участок может быть модифицирован таким образом, чтобы он не связывался с одним или более рецепторов Fcγ, таких как CD16, CD32 и CD64.
[00168] Константная подобласть иммуноглобулина, которая присутствует в связывающихся с CD3 полипептидах согласно настоящему документу, может содержать или происходить от части или всех из: домена СН2, домена СН3, домена СН4 или любой их комбинации. Например, константная подобласть иммуноглобулина может содержать домен СН2, домен СН3, домены СН2 и СН3, домены СН3 и СН4, два домена СН3, домен СН4, два домена СН4, а также домен СН2 и часть домена СН3. В некоторых вариантах реализации константный участок содержит домены СН2 и СН3 lgG1, lgG2, lgG3, lgG4, IgA1, lgA2, IgD или любой их комбинации; домен СН3 иммуноглобулина lgG1, lgG2, lgG3, lgG4, IgA1, lgA2, IgD, IgE, IgM или любой их комбинации; или домены СН3 и СН4 иммуноглобулина IgE, IgM или их комбинации. В некоторых вариантах реализации константный участок по существу состоит из домена СН2 и домена СН3.
[00169] Домен СН2, который может быть образован константным участком иммуноглобулина связывающегося с CD3 полипептида в соответствии с настоящим документом, может представлять собой домен СН2 иммуноглобулина дикого типа или модифицированный домен СН2 иммуноглобулина, происходящий из некоторых классов или подклассов иммуноглобулина (например, lgG1, lgG2, lgG3, lgG4, IgA1, lgA2 или IgD) и от различных видов (включая, человека, мышь, крысу и других млекопитающих). В некоторых вариантах реализации константный участок модифицирован таким образом, чтобы уменьшить или исключить эффекторную функцию, уменьшить или исключить фиксацию комплемента и/или уменьшить или исключить связывание с рецепторами Fcγ. В некоторых вариантах реализации рецептор Fcγ выбран из группы, состоящей из CD16, CD32 и CD64.
[00170] В некоторых вариантах реализации домен СН2 представляет собой домен СН2 иммуноглобулина дикого типа человека, например, домен СН2 lgG1, lgG2, lgG3, lgG4, IgA1, lgA2 или IgD дикого типа человека, как было представлено ранее в SEQ ID NO: 115, 199-201 и 195-197 Публикации РСТ WO 2011/090762, соответственно (указанные последовательности включены в настоящий документ посредством ссылки). В некоторых вариантах реализации домен СН2 представляет собой домен СН2 lgG1 дикого типа человека, как было представлено ранее в SEQ ID NO: 115 WO 2011/090762 (указанная последовательность включена в настоящий документ посредством ссылки и представляет собой SEQ ID NO: 85 настоящей заявки). В некоторых вариантах реализации константный участок содержит или состоит из последовательности аминокислот SEQ ID NO: 55.
[00171] В некоторых вариантах реализации домен СН2 представляет собой модифицированный участок СН2 иммуноглобулина (например, модифицированный домен СН2 lgG1 человека), который содержит замену аминокислоты аспарагин в положении 297 (например, аспарагин на аланин). Такая замена аминокислоты уменьшает или исключает гликозилирование на данном сайте и исключает эффективное связывание Fc с FcγR и C1q. Последовательность модифицированного домена СН2 lgG1 человека, с заменой Asn на Ala в положении 297, представлена в SEQ ID NO: 324 WO 2011/090762 (указанная последовательность включена в настоящий документ посредством ссылки и представляет собой SEQ ID NO: 86 настоящей заявки).
[00172] В некоторых вариантах реализации домен СН2 представляет собой модифицированный участок СН2 иммуноглобулина (например, модифицированный домен СН2 lgG1), который содержит по меньшей мере одну замену или делецию в положениях 234-238. Например, участок СН2 иммуноглобулина может содержать замену в положении 234, 235, 236, 237 или 238, положениях 234 и 235, положениях 234 и 236, положениях 234 и 237, положениях 234 и 238, положениях 234-236, положениях 234, 235 и 237, положениях 234, 236 и 238, положениях 234, 235, 237 и 238, положениях 236-238, или любую другую комбинацию двух, трех, четырех или пяти аминокислот в положениях 234-238. Дополнительно или в качестве альтернативы, модифицированный участок СН2 может содержать одну или более (например, две, три, четыре или пять) делеций аминокислот в положениях 234-238, например, в одном из положений 236 или 237, тогда как другое положение содержит замену. Вышеуказанная мутация(-и) уменьшает или исключает активность антителозависимой клеточно-опосредованной цитотоксичности (АЗКЦ) или возможность связывания с рецептором Fc полипептидного гетеродимера, который содержит модифицированный домен СН2. В некоторых вариантах реализации остатки аминокислот в одном или более положений 234-238 заменены одним или более остатков аланина. В дополнительных вариантах реализации только один из остатков аминокислот в положениях 234-238 удален, тогда как один или более из остальных остатков аминокислот в положениях 234-238 могут быть заменены другой аминокислотой (например, аланином или серином).
[00173] В некоторых других вариантах реализации домен СН2 представляет собой модифицированный участок СН2 иммуноглобулина (например, модифицированный домен СН2 lgG1 человека), который содержит одну или более замен аминокислот в положениях 253, 310, 318, 320, 322 и 331. Например, участок СН2 иммуноглобулина может содержать замену в положении 253, 310, 318, 320, 322 или 331, положениях 318 и 320, положениях 318 и 322, положениях 318, 320 и 322, или любую другую комбинацию двух, трех, четырех, пяти или шести аминокислот в положениях 253, 310, 318, 320, 322 и 331. Вышеуказанная мутация(-и) уменьшает или исключает комплементзависимую цитотоксичность (КЗЦ) полипептидного гетеродимера, который содержит модифицированный домен СН2.
[00174] В некоторых других вариантах реализации, в дополнение к замене аминокислоты в положении 297, модифицированный участок СН2 (например, модифицированный домен СН2 lgG1 человека) может содержать одну или более (например, две, три, четыре или пять) дополнительных замен в положениях 234-238. Например, участок СН2 иммуноглобулина может содержать замену в положениях 234 и 297, положениях 234, 235 и 297, положениях 234, 236 и 297, положения 234-236 и 297, положениях 234, 235, 237 и 297, положениях 234, 236, 238 и 297, положениях 234, 235, 237, 238 и 297, положениях 236-238 и 297, или любую комбинацию двух, трех, четырех или пяти аминокислот в положениях 234-238 в дополнение к положению 297. Дополнительно или альтернативно, модифицированный участок СН2 может содержать одну или более (например, две, три, четыре или пять) делеций аминокислот в положениях 234-238, например, в положении 236 или положении 237. Дополнительная мутация(-и) уменьшает или исключает активность антителозависимой клеточно-опосредованной цитотоксичности (АЗКЦ) или возможность связывания с рецептором Fc полипептидного гетеродимера, который содержит модифицированный домен СН2. В некоторых вариантах реализации остатки аминокислот в одном или более положений 234-238 заменены одним или более остатков аланина. В дополнительных вариантах реализации только один из остатков аминокислот в положениях 234-238 удален, тогда как один или более из остальных остатков аминокислот в положениях 234-238 могут быть заменены другой аминокислотой (например, аланином или серином).
[00175] В некоторых вариантах реализации, в дополнение к одной или более (например, 2, 3, 4, или 5) заменам аминокислот в положениях 234-238, мутантный участок СН2 (например, модифицированный домен СН2 lgG1 человека) в слитом белке согласно настоящему документу может содержать одну или более (например, 2, 3, 4, 5 или 6) дополнительных замен аминокислот (например, замену аланином) в одном или более положениях, принимающих участие в фиксации комплемента (например, в положениях 1253, Н310, Е318, К320, К322 или Р331). Примеры мутантных участков СН2 иммуноглобулина включают участки СН2 lgG1, lgG2, lgG4 человека и lgG2a мыши с заменой аланина в положениях 234, 235, 237 (если они присутствуют), 318, 320 и 322. Примером мутантного участка СН2 иммуноглобулина является участок СН2 IGHG2c мыши с заменой аланина в положениях L234, L235, G237, Е318, К320 и К322.
[00176] В других вариантах реализации, в дополнение к замене аминокислоты в положении 297 и дополнительной делеции(-ям) или замене(-ам) в положениях 234-238, модифицированный участок СН2 (например, модифицированный домен СН2 lgG1 человека) может дополнительно содержать одну или более (например, две, три, четыре, пять или шесть) дополнительных замен в положениях 253, 310, 318, 320, 322 и 331. Например, участок СН2 иммуноглобулина может содержать (1) замену в положении 297, (2) одну или более замен или делеций или их комбинацию в положениях 234-238, и одну или более (например, 2, 3, 4, 5 или 6) замен аминокислот в положениях 1253, Н310, Е318, К320, К322 и Р331, например, одну, две, три замены в положениях Е318, К320 и К322. Аминокислоты в вышеуказанных положениях могут быть заменены аланином или серином.
[00177] В некоторых вариантах реализации полипептид участка СН2 иммуноглобулина содержит: (i) замену аминокислоты аспарагин в положении 297 и одну замену аминокислоты в положении 234, 235, 236 или 237; (ii) замену аминокислоты аспарагин в положении 297 и замену аминокислот в двух из положений 234-237; (iii) замену аминокислоты в положении аспарагина 297 и замену аминокислот в трех из положений 234-237; (iv) замену аминокислот в положении аспарагина 297, замену аминокислот в положениях 234, 235 и 237, а также делецию аминокислоты в положении 236; (v) замену аминокислот в трех из положений 234-237 и замену аминокислот в положениях 318, 320 и 322; или (vi) замену аминокислот в трех из положений 234-237, делецию аминокислоты в положении 236, а также замену аминокислот в положениях 318, 320 и 322.
[00178] Примеры модифицированных участков СН2 иммуноглобулина с заменой аминокислоты аспарагин в положении 297 включают: участок СН2 lgG1 человека с заменой аланина в положениях L234, L235, G237 и N297 и делецией в положении G236 (SEQ ID NO: 325 WO 2011/090762, указанная последовательность включена в настоящий документ посредством ссылки), участок СН2 lgG2 человека с заменой аланина в положениях V234, G236 и N297 (SEQ ID NO: 326 WO 2011/090762, указанная последовательность включена в настоящий документ посредством ссылки), участок СН2 lgG4 человека с заменой аланина в положениях F234, L235, G237 и N297 и делецией G236 (SEQ ID NO: 322 WO 2011/090762, указанная последовательность включена в настоящий документ посредством ссылки), участок СН2 lgG4 человека с заменой аланина в положениях F234 и N297 (SEQ ID NO: 343 WO 2011/090762, указанная последовательность включена в настоящий документ посредством ссылки), участок СН2 lgG4 человека с заменой аланина в положениях L235 и N297 (SEQ ID NO: 344 WO 2011/090762, указанная последовательность включена в настоящий документ посредством ссылки), участок СН2 lgG4 человека с заменой аланина в положениях G236 и N297 (SEQ ID NO: 345 WO 2011/090762, указанная последовательность включена в настоящий документ посредством ссылки), и участок СН2 lgG4 человека с заменой аланина в G237 и N297 (SEQ ID NO: 346 WO 2011/090762, указанная последовательность включена в настоящий документ посредством ссылки).
[00179] В некоторых вариантах реализации в дополнение к заменам аминокислот, раскрытым выше, модифицированный участок СН2 (например, модифицированный домен СН2 lgG1 человека) может содержать одну или более дополнительных замен аминокислот в одном или более положениях, кроме вышеуказанных положений. Такие замены аминокислот могут представлять собой консервативные или неконсервативные замены аминокислот. Например, в некоторых вариантах реализации Р233 может быть модифицирован до Е233 в модифицированном участке СН2 lgG2 (см., например, SEQ ID NO: 326 WO 2011/090762, указанная последовательность включена в настоящий документ посредством ссылки). Дополнительно или альтернативно, в определенных вариантах реализации модифицированный участок СН2 может содержать одну или более из вставок, делеций аминокислот или обеих. Вставка(-и), делеция(-и) или замена(-ы) могут быть произведены в любом месте участка СН2 иммуноглобулина, например, на N- или С-конце участка СН2 иммуноглобулина дикого типа, полученного в результате связи участка СН2 с другим участком (например, доменом связывания или доменом гетеродимеризации иммуноглобулина) посредством шарнира.
[00180] В некоторых вариантах реализации модифицированный участок СН2 в полипептиде в соответствии с настоящим документом содержит или представляет собой последовательность, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичную участку СН2 иммуноглобулина дикого типа, такому как участок СН2 lgG1, lgG2, или lgG4 дикого типа человека или lgG2a мыши дикого типа (например, IGHG2c).
[00181] Модифицированный участок СН2 иммуноглобулина в связывающемся с CD3 полипептиде в соответствии с настоящим документом может происходить от участка СН2 различных изотипов иммуноглобулина, таких как lgG1, lgG2, lgG3, lgG4, IgA1, lgA2 и IgD, из различных видов (включая человека, мышь, крысу и других млекопитающих). В определенных вариантах реализации модифицированный участок СН2 иммуноглобулина в слитом белке в соответствии с настоящим документом может происходить от участка СН2 lgG1, lgG2 или lgG4 человека или CD3 мыши (например, IGHG2c), последовательности которых представлены в SEQ ID NO: 115, 199, 201 и 320 WO 2011/090762 (указанные последовательности включены в настоящий документ посредством ссылки).
[00182] В некоторых вариантах реализации модифицированный домен СН2 представляет собой домен СН2 lgG1 человека с заменой аланина в положениях 235, 318, 320 и 322 (т.е., домен СН2 lgG1 человека с заменой L235A, Е318А, К320А и К322А) (SEQ ID NO: 595 WO 2011/090762, указанная последовательность включена в настоящий документ посредством ссылки) и необязательной мутацией N297 (например, до аланина). В некоторых других вариантах реализации модифицированный домен СН2 представляет собой домен СН2 lgG1 человека с заменой аланина в положениях 234, 235, 237, 318, 320 и 322 (т.е., домен СН2 lgG1 человека с заменой L234A, L235A, G237A, Е318А, К320А и К322А) (SEQ ID NO:596 WO 2011/090762, указанная последовательность включена в настоящий документ посредством ссылки) и необязательной мутацией N297 (например, до аланина).
[00183] В некоторых вариантах реализации модифицированный домен СН2 представляет собой модифицированный домен СН2 lgG1 человека с известными из уровня техники мутациями, которые повышают иммунологическую активность, такими как АЗКЦ, антителозависимый клеточный фагоцитоз (АЗКФ), КЗЦ, фиксация комплемента, связывание с рецептором Fc или любая их комбинация.
[00184] Домен СН3, который может образовывать константный участок иммуноглобулина связывающегося с CD3 полипептида в соответствии с настоящим документом, может представлять собой домен СН3 иммуноглобулина дикого типа или модифицированный домен СН3 иммуноглобулина из некоторых классов или подклассов иммуноглобулина (например, lgG1, lgG2, lgG3, lgG4, IgA1, IgA2, IgD, IgE, IgM) различных видов (включая человека, мышь, крысу и других млекопитающих). В некоторых вариантах реализации домен СН3 представляет собой домен СН3 иммуноглобулина человека дикого типа, например, домен СН3 lgG1, lgG2, lgG3, IgG4, IgA1, lgA2, IgD, IgE или IgM человека, как представлено в SEQ ID NO: 116, 208-210, 204-207 и 212 WO 2011/090762, соответственно (указанные последовательности включены в настоящий документ посредством ссылки). В некоторых вариантах реализации домен СН3 представляет собой домен lgG1 СН3 дикого типа человека, как ранее было представлено в SEQ ID NO: 116 WO 2011/090762 (указанная последовательность включена в настоящий документ посредством ссылки). В некоторых вариантах реализации домен СН3 представляет собой модифицированный домен СН3 иммуноглобулина человека, например, модифицированный домен СН3, базирующийся или происходящий от домена СН3 антител lgG1, lgG2, lgG3, lgG4, IgA1, lgA2, IgD, IgE или IgM дикого типа человека. Например, модифицированный домен СН3 может представлять собой домен СН3 lgG1 человека с одной или двумя мутациями в положениях Н433 и N434 (положения нумеруются согласно нумерации ЕС). Мутации в таких положениях могут принимать участие в фиксации комплемента. В некоторых других вариантах реализации модифицированный домен СН3 может представлять собой домен СН3 lgG1 человека, но с одной или двумя заменами аминокислот в положениях F405 или Y407. Аминокислоты в указанных положениях принимают участие во взаимодействии с другим доменом СН3. В некоторых вариантах реализации модифицированный домен СН3 может представлять собой модифицированный домен СН3 lgG1 человека, в котором последний участок лизина удален. Последовательность указанного модифицированного домена СН3 представлена в SEQ ID NO: 761 WO 2011/090762 (указанная последовательность включена в настоящий документ посредством ссылки).
[00185] В некоторых вариантах реализации связывающиеся с CD3 полипептиды образуют полипептидный гетеродимер, содержащий пару СН3, которая включает так называемые мутации «выступ-во-впадину» (см. Marvin and Zhu, Acta Pharmacologica Sinica 26:649-58, 2005; Ridgway et al., Protein Engineering 9:617-21, 1966). Более конкретно, мутации могут быть введены в каждый из двух доменов СН3 каждой полипептидной цепи, таким образом, что стерическая комплементарность, необходимая для ассоциации СН3/СН3, обязывает указанные два домена СН3 спариваться друг с другом. Например, домен СН3 в одном одноцепочечном полипептиде полипептидного гетеродимера может содержать мутацию T366W (мутация «выпуклости», при которой аминокислота небольшого размера заменяется большей), и домен СН3 в другом одноцепочечном полипептиде полипептидного гетеродимера может содержать мутацию Y407A (мутация «впадины», при которой аминокислота большего размера заменяется меньшей). Другие примеры мутаций выступ-по-впадину включают (1) мутацию T366Y в одном домене СН3 и Y407T в другом домене СН3, и (2) мутацию T366W в одном домене СН3 и мутацию T366S, L368A и Y407V в другом домене СН3.
[00186] Домен СН4, который может образовывать константный участок иммуноглобулина связывающихся с CD3 полипептидов согласно настоящему документу, может представлять собой домен СН4 иммуноглобулина дикого типа или модифицированный домен СН4 иммуноглобулина из молекул IgE или IgM. В некоторых вариантах реализации домен СН4 представляет собой домен СН4 иммуноглобулина дикого типа человека, например, домен СН4 молекул IgE и IgM дикого типа человека, как ранее было представлено в SEQ ID NO: 213 и 214 WO 2011/090762, соответственно (указанные последовательности включены в настоящий документ посредством ссылки). В некоторых вариантах реализации домен СН4 представляет собой модифицированный домен СН4 иммуноглобулина человека, например, модифицированный домен СН4, базирующийся или происходящий от домена СН4 молекул IgE или IgM человека, которые содержат мутации, повышающие или снижающие иммунологическую активность, которая доказанно ассоциируется с участком Fc IgE или IgM.
[00187] В некоторых вариантах реализации константный участок иммуноглобулина связывающихся с CD3 полипептидов согласно настоящему документу содержит комбинацию доменов СН2, СН3 или СН4 (т.е., более чем один домен константного участка, выбранный из СН2, СН3 и СН4). Например, константная подобласть иммуноглобулина может содержать домены СН2 и СН3 или домены СН3 и СН4. В некоторых других вариантах реализации константный участок иммуноглобулина может содержать два домена СН3 и не содержать доменов СН2 или СН4 (т.е., только два или более СН3). Множественные домены константного участка, которые образуют константную подобласть иммуноглобулина, могут базироваться или происходить от такой же молекулы иммуноглобулина или такого же класса или подкласса молекул иммуноглобулина. В определенных вариантах реализации константная подобласть иммуноглобулина представляет собой СН2СН3 IgG (например, СН2СН3 lgG1, СН2СН3 lgG2 и СН2СН3 lgG4), и может представлять собой СН2СН3 человека (например, lgG1, lgG2 и lgG4 человека). Например, в некоторых вариантах реализации константная подобласть иммуноглобулина содержит (1) домены СН2 и СН3 lgG1 дикого типа человека, (2) СН2 lgG1 человека с заменой N297A (т.е., CH2(N297A)) и СН3 lgG1 дикого типа человека, или (3) CH2(N297A) lgG1 человека и модифицированный СН3 lgG1 человека, в котором последний остаток лизина удален.
[00188] Альтернативно, множественные домены константного участка могут базироваться или происходить от различных молекул иммуноглобулина или различных классов или подклассов молекул иммуноглобулина. Например, в определенных вариантах реализации константная подобласть иммуноглобулина содержит как домен СН3 IgM человека, так и домен СН3 lgG1 человека. Множественные домены константного участка, которые образуют константную подобласть иммуноглобулина, могут быть непосредственно соединены вместе или могут быть соединены друг с другом с помощью одной или более (например, около 2-10) аминокислот.
[00189] Примеры константных подобластей иммуноглобулина представлены в SEQ ID NO: 305-309, 321, 323, 341, 342 и 762 WO 2011/090762 (указанные последовательности включены в настоящий документ посредством ссылки).
[00190] В определенных вариантах реализации константные участки иммуноглобулина обеих цепей гомодимера или гетеродимера связывающегося с CD3 полипептида идентичны друг другу. В других определенных вариантах реализации константная подобласть иммуноглобулина одной полипептидной цепи гетеродимерного белка отличается от константной подобласти иммуноглобулина другой полипептидной цепи гетеродимера. Например, одна константная подобласть иммуноглобулина гетеродимерного белка может содержать домен СН3 с мутацией «выступа», тогда как другая константная подобласть иммуноглобулина гетеродимерного белка может содержать домен СН3 с мутацией «впадины».
[00191] Некоторые варианты реализации изобретения включают использование полиспецифичных или биспецифичных молекул связывания, которые (i) нацелены на комплекс TCR (например, CD3) на Т-клетки человека и (ii) белок на поверхности клетки, например, с целью перенаправления цитотоксичности Т-клеток на клетку с белком на поверхности клетки, например, клетку опухоли, экспрессирующую антиген опухоли, с целью лечения рака.
[00192] В некоторых вариантах реализации белок, связывающийся с CD3, может содержать один или более дополнительных доменов связывания (например, второй домен связывания), которые связываются с другой мишенью, кроме CD3. Указанные другие молекулы-мишени могут содержать, например, ассоциированный с опухолью антиген. В одном варианте реализации изобретения, один или более дополнительных доменов связывания связываются или взаимодействуют с одним или более из следующих опухолевых антигенов: RON, c-Met, СЕАСАМ-6, ПСМА, ЕрСАМ, СЕА, РСТА-1, STEAP-1, STEAP-2, PSCA, PSA, PAP, ALCAM (CD 166), РЕСАМ-1, ЕрЬА2, CD151, CA-125/MUC16, MUC-1, MAGE-1, TROP2, IGF1R, TGFBR2, GHRHR, GHR, IL-6R, gpl30, TNFR2, OSMRβ, Patched-1, Frizzled, Robol, LTpR, CD19, CD25, CD26, CD27, CD30, CD33, CD44, CD44v6, CD63, CD80, CD81, CD86, CD100, CD151, CXCR4, CCR5, HER-2/ErbBl, HER-3/ErbB3, HER-4/ErbB4, EGFR/ErbB1, изоформа EGFRvlll, MUC2, MUC3, MUC4, MUC5AC, MUC5b, MUC7, βhCG, Lewis-Y, ганглиозид GD3, 9-О-ацетил-GD3, GM2, Globo H, фукозил GM1, поли SA, GD2, карбоангидраза IX (MN/CA IX), еж Соник (Shh), Wue-1, антиген клеток плазмы, (связанный с мембраной) IgE, хондроитина сульфат протеогликан меланомы (ХСПМ), CCR8, прекурсор опухолевого некротического фактора-альфа, мезотелин, антиген А33, Ly-6; десмоглеин 4, Е-кадгерин неоэпитоп, фетальный рецептор ацетилхолина, маркер СА19-9, рецептор мюллеровой ингибирующей субстанции (МИС) типа II, sTn (сталированный антиген Tn; TAG-72), ААФ (антиген активации фибробластов), эндосиалин, LG, SAS, ВСМА, TWEAKR/Fn14, FGFR4, VEGFR1, VEGFR2, SSX1 и SSX2.
[00193] В некоторых вариантах реализации домен связывания с CD3 связывается с доменом связывания с TCR с целью рекрутинга Т-клеток для нацеливания на клетки, экспрессирующие антиген опухоли. В некоторых вариантах реализации полипептид, связывающийся с CD3, может содержать домен связывания с CD3, который специфично связывается с комплексом TCR или его компонентом (например, TCRα, TCRβ, CD3γ, CD3δ и CD3ε), и другой домен связывания, который специфично связывается с ассоциированным с опухолью антигеном.
[00194] ПСМА представляет собой в высокой степени ограниченный, связанный с раком предстательной железы антиген клеточной мембраны. В клетках рака предстательной железы, экспрессия ПСМА в 1000 раз выше, чем на нормальном эпителии предстательной железы (Su et al., Cancer Res. 1995 44:1441-1443). Экспрессия ПСМА возрастает с прогрессированием рака предстательной железы, и обычно наиболее высока при метастатическом заболевании, в гормонрезистентых случаях, и при поражении более высокой степени (Israeli et al., Cancer Res. 1994, 54:1807-1811; Wright et al., Urologic Oncology: Seminars and Original Investigations 1995 1: 18-28; Wright et al., Urology 1996 48:326-332; Sweat et al., Urology 1998 52:637-640). Дополнительно, ПСМА обильно экспрессируется на вновь образованных сосудах во множестве других солидных опухолей, в том числе, мочевого пузыря, поджелудочной железы, меланоме, раке легкого и почки, но не на вновь образованных нормальных сосудах (Chang et al., Urology 2001 57:801-805; Divgi et al., Clin. Cancer Res. 1998 4:2729-3279). Домены связывания для нацеливания на ПСМА включают, без ограничений, раскрытые в Публикации РСТ WO 2012/145714, которая включена в настоящий документ посредством ссылки в полном объеме.
[00195] В некоторых вариантах реализации домен связывания с ПСМА содержит последовательность аминокислот, которая по меньшей мере на 90%, по меньшей мере на 95% или на 100% идентична последовательности аминокислот, выбранной из группы, состоящей из (i) аминокислот 1-107 и 124-243 SEQ ID NO: 212; (ii) аминокислот 1-107 и 124-243 SEQ ID NO: 226; или (iii) аминокислот 1-107 и 124-243 SEQ ID NO: 216. В некоторых вариантах реализации белок или молекула по изобретению содержит последовательность аминокислот, выбранную из группы, состоящей из SEQ ID NO: 212, 214, 216 и 226. Домены связывания с ПСМА, которые могут использоваться в соответствии с изобретением, также описаны в Публикации РСТ WO 2012/145714, включая домены связывания с ПСМА, содержащие последовательности аминокислот SEQ ID NO: 19, 21, 30, 31, 34 или 35 WO 2012/145714, или содержащие цепь VL, содержащую последовательность аминокислот, выбранную из группы, состоящей из SEQ ID NO: 5 и 23, и цепь VH, содержащую последовательность аминокислот, выбранную из группы, состоящей из SEQ ID NO: 2, 25 и 27 WO 2012/145714.
[00196] RON (рецептор Нантского происхождения, также известный как MST1R) представляет собой протеинтирозинкиназу рецепторного типа, которая имеет большое значение для эмбрионального развития, а также играет важную роль в воспалительных реакциях (Camp et al. Ann. Surg. Oncol. 12:273-281 (2005)). RON по большей части экспрессируется в происходящих из эпителия типах клеток, и высказано предположение о том, что RON, подобно целому ряду других тирозинкиназ рецепторного типа, может играть роль в прогрессировании эпителиальных злокачественных опухолей (Wang et al. Carcinogenesis 23:1291-1297 (2003)). Активация RON инициирует нижележащие пути проведение сигнала, которые вовлечены в туморигенную активность, такую как пролиферация клеток, ингибирование апоптоза и подвижность клеток. RON представляет терапевтическую мишень при раке эпителия, среди прочего, вследствие его свойств проведения сигнала и/или чрезмерной экспрессии RON при карциномах ободочной и прямой кишки, молочной железы, яичника и поджелудочной железы.
[00197] Домены связывания для нацеливания RON включают, без ограничений, раскрытые в разделе Примеров и раскрытые в Публикации РСТ WO 2011/090761, которая включена в настоящий документ посредством ссылки в полном объеме. В некоторых вариантах реализации анти-RON домен связывания содержит (а) домен VL, содержащий i. последовательность аминокислот CDR1 SEQ ID NO: 87, последовательность аминокислот CDR2 SEQ ID NO: 88, и последовательность аминокислот CDR3 SEQ ID NO: 89; или ii. последовательность аминокислот CDR1 SEQ ID NO: 90, последовательность аминокислот CDR2 SEQ ID NO: 91 и последовательность аминокислот CDR3 SEQ ID NO: 92; или (б) домен VH, содержащий i. последовательность аминокислот CDR1 SEQ ID NO: 93, последовательность аминокислот CDR2 SEQ ID NO: 94 и последовательность аминокислот CDR3 SEQ ID NO: 95; или ii. последовательность аминокислот CDR1 SEQ ID NO: 96, последовательность аминокислот CDR2 SEQ ID NO: 97, и последовательность аминокислот CDR3 SEQ ID NO: 98; или (с) VL (а) и VH (б). В одном варианте реализации домен VL содержит последовательность аминокислот согласно любой из SEQ ID NO: 99 или 100, и домен VH содержит последовательность аминокислот согласно любой из SEQ ID NO: 101, 102 и 103. В другом варианте реализации домены VL и VH гуманизированы. В определенных вариантах реализации гуманизированный VL содержит последовательность аминокислот любой из SEQ ID NO: 104, 105 и 106, и гуманизированный домен VH содержит последовательность аминокислот любой из SEQ ID NO: 107-111. В некоторых вариантах реализации анти-RON домен связывания содержит домен VL, содержащий последовательность аминокислот любой из SEQ ID NO: 99 и 100, и домен VH, содержащий последовательность аминокислот любой из SEQ IDNO: 101, 102 и 103. В некоторых вариантах реализации домен анти-RON связывания содержит домен VL, содержащий последовательность аминокислот любой из SEQ ID NO: 104, 105 и 106, и домен VH, содержащий последовательность аминокислот любой из SEQ ID NO: 107-111.
[00198] В некоторых вариантах реализации домен связывания с RON содержит последовательность аминокислот, которая по меньшей мере на 90%, 92%, 94%, 96%, 97%, 98%, 99% или 100% идентична SEQ ID NO: 187, и/или содержит последовательность аминокислот, которая по меньшей мере на 90%, 92%, 94%, 96%, 97%, 98%, 99% или 100% идентична SEQ ID NO: 188. В некоторых вариантах реализации домен связывания с RON содержит (i) SEQ ID NO: 187, в которой аминокислота 43 (аланин) заменена другой аминокислотой, например, лизином (А43К) или треонином (А43Т), (ii) SEQ ID NO: 188, в которой аминокислота 38 (глутамин) и/или 113 (глутамин) заменена другой аминокислотой, такой как глутаминовая кислота и/или аргинин, например, Q38R и/или Q113E, или (iii) любая их комбинация. В некоторых вариантах реализации биспецифичная молекула анти-RON/anra-CD3 содержит или состоит из последовательности аминокислот, которая по меньшей мере на 90%, 92%, 94%, 96%, 97%, 98%, 99% или 100% идентична любой из SEQ ID NO: 186, 190, 192 или 194. В некоторых вариантах реализации изобретения раскрыта нуклеиновая кислота, которая кодирует биспецифичную молекулу анти-RON/анти-CD3, содержащая нуклеотидную последовательность, которая по меньшей мере на 90%, 92%, 94%, 96%, 97%, 98%, 99% или 100% идентична любой из SEQ ID NO: 185,189,191 или 193.
[00199] CD19 найден на поверхности В-клеток, включая большинство злокачественных В-клеток. Нацеливание на CD19 может использоваться для подавления и лечения связанного с В-клетками заболевания, лейкозов и лимфом.
[00200] В некоторых вариантах реализации домен связывания с CD19 содержит последовательность аминокислот, которая по меньшей мере на 90%, 92%, 94%, 96%, 97%, 98%, 99% или 100% идентична аминокислотам 1-111 SEQ ID NO: 196 и/или содержит последовательность аминокислот, которая по меньшей мере на 90%, 92%, 94%, 96%, 97%, 98%, 99% или 100% идентична аминокислотам 128-251 SEQ ID NO: 196. В некоторых вариантах реализации биспецифичная молекула анти-CD19/анти-CD3 содержит или состоит из последовательности аминокислот, которая по меньшей мере на 90%, 92%, 94%, 96%, 97%, 98%, 99% или 100% идентична любой из SEQ ID NO: 196, 198, 200, 202, 204 или 206. В некоторых вариантах реализации изобретения раскрыта нуклеиновая кислота, которая кодирует молекулу биспецифичного анти-CD19/анти-CD3, причем нуклеиновая кислота содержит нуклеотидную последовательность, которая по меньшей мере на 90%, 92%, 94%, 96%, 97%, 98%, 99% или 100% идентична любой из SEQ ID NO: 195, 197, 199, 201, 203 или 206.
[00201] HER2 также известен как ErbB2 человека. Чрезмерная экспрессия HER2 (часто, но не однородно, вследствие амплификации гена) также наблюдается при раке молочной железы, раке яичника и других карциномах, включая карциномы желудка, слюнной железы, эндометрия, легкого, почки, ободочной кишки, щитовидной железы, поджелудочной железы и мочевого пузыря. Домены связывания для нацеливания на HER2 включают, без ограничений, раскрытые в Публикации РСТ WO 2009/055074, которая включена в настоящий документ посредством ссылки в полном объеме.
[00202] В некоторых вариантах реализации полипептид, связывающийся с CD3, содержит последовательность аминокислот, которая по меньшей мере на 90%, по меньшей мере на 95% или 100% идентична SEQ ID NO: 57, 59, 61, 63, 65, 67, 69, 71, 186,190, 192, 194, 196, 198, 200, 202, 204 или 206.
[00203] В некоторых вариантах реализации полипептид, связьшающийся с CD3, представляет собой часть гетеродимера. В некоторых вариантах реализации гетеродимер содержит пару одноцепочечных полипептидов, которая содержит последовательности аминокислот, которые по меньшей мере на 90%, по меньшей мере на 95% или 100% идентичны парам, выбранным из SEQ ID NO: 210 и 247, SEQ ID NO: 210 и 218, SEQ ID NO: 210 и 220, SEQ ID NO: 208 и 249, SEQ ID NO: 208 и 222, или SEQ ID NO: 208 и 224, SEQ ID NO: 212 и 218, SEQ ID NO: 216 и 222, SEQ ID NO: 228 и 226, или SEQ ID NO: 214 и 218.
[00204] Изобретение также включает нуклеиновые кислоты (например, ДНК или РНК), кодирующие полипептид, связывающийся с CD3, как раскрыто в настоящем документе, или одну или более полипептидных цепей димерного или гетеродимерного белка, связывающегося с CD3, как раскрыто в настоящем документе. Нуклеиновые кислоты по изобретению включают нуклеиновые кислоты, содержащие участок, который в существенной мере идентичен полинуклеотиду, как представлено в SEQ ID NO: 5, 7, 9, 11, 13, 15, 17, 19, 27, 29, 31, 37 и 39, или нуклеиновые кислоты, содержащие кодирующий участок, который кодирует такой же или подобный полипептид. В некоторых вариантах реализации нуклеиновая кислота в соответствии с настоящим изобретением по меньшей мере на 80%, обычно по меньшей мере около на 90%, и чаще по меньшей мере около на 95% или по меньшей мере около на 98% идентична кодирующему полипептид полинуклеотиду, как представлено в SEQ ID NO: 5, 7, 9, 11, 13, 15, 17, 19, 27, 29, 31, 37 и 39. Нуклеиновые кислоты по изобретению также включают дополнительные нуклеиновые кислоты. В некоторых случаях, последовательности будут полностью комплементарными (отсутствие несовпадений) при выравнивании. В других случаях может существовать около 20% несовпадений в последовательностях. В некоторых вариантах изобретения раскрываются нуклеиновые кислоты, кодирующие как первые, так и вторые полипептидные цепи гетеродимерного белка, связывающегося с CD3, по изобретению. Последовательности нуклеиновых кислот, раскрытые в настоящем документе, могут эксплуатироваться с использованием кодон-оптимизации, вырожденной последовательности, «молчащих» мутаций и других ДНК-техник для оптимизации экспрессии в конкретном хозяине, и настоящее изобретение включает такие модификации последовательностей.
[00205] Молекулы полинуклеотида, содержащие желательную последовательность полинуклеотида, разводят, помещая молекулу в вектор. Изобретение также включает векторы экспрессии, содержащие нуклеиновые кислоты по изобретению, и/или экспрессирующие определенные полипептиды по изобретению. Используются вирусные и невирусные векторы, в том числе, плазмиды. Выбор плазмиды будет зависеть от вида клетки, в которой разведение является желательным, и цели разведения. Некоторые векторы пригодны для амплификации и создания больших количеств желательной последовательности ДНК. Другие векторы пригодны для экспрессии в клетках культуры. Дополнительные векторы пригодны для переноса и экспрессии в клетках цельного организма животного или человека. Выбор пригодного вектора находится в пределах квалификации специалиста в данной области. Множество таких векторов коммерчески доступны. Часть или полноразмерный полинуклеотид вставляют в вектор, обычно с помощью присоединения ДНК-лигазой к расщепленному сайту рестрикционного фермента в векторе. Альтернативно, желательная нуклеотидная последовательность может быть вставлена рекомбинацией in vivo. Обычно это достигается путем присоединения участков гомологии к вектору по обе стороны желательной нуклеотидной последовательности. Участки гомологии добавляют посредством лигирования олигонуклеотидов или полимеразной цепной реакцией, с использованием праймеров, например, содержащих как участок гомологии, так и часть желательной нуклеотидной последовательности.
[00206] Для экспрессии может применяться кассета экспрессии или система экспрессии. Для экспрессии нуклеиновой кислоты, кодирующей полипептид, раскрытый в настоящем документе, кодирующую полипептид молекулу нуклеиновой кислоты, функционально связанную с регуляторными последовательностями, которые управляют транскрипциональной экспрессией в векторе экспрессии, вводят в клетку-хозяина. Изобретение включает клетки-хозяева, содержащие нуклеиновую кислоту и/или вектор экспрессии по изобретению, например, кодирующую полипептид по изобретению. В дополнение к последовательностям регуляции транскрипции, таким как промоторы и энхансеры, векторы экспрессии могут содержать последовательности регуляции трансляции и ген маркера, пригодного для селекции клеток, которые несут вектор экспрессии. Продукт гена, кодируемый полинуклеотидом по изобретению, экспрессируется в любой пригодной системе экспрессии, в том числе, например, системы бактерии, дрожжей, насекомого, амфибии и млекопитающего. В векторе экспрессии полинуклеотид, кодирующий полипептид, при необходимости, связан с регуляторной последовательностью для получения желательных свойств экспрессии. Это может включать промоторы, энхансеры, терминаторы, операторы, репрессоры и индукторы. Промоторы могут регулироваться (например, промотор индуцибельного стероидом вектора pIND (Invitrogen)) или быть конститутивными (например, промоторы из ЦМВ, SV40, фактор удлинения или последовательности LTR). Их связывают с желательной нуклеотидной последовательностью с применением методов, раскрытых выше для соединения с векторами. Могут применяться любые методы, известные из уровня техники. Соответственно, вектор экспрессии, в общем, обеспечивает участок инициации транскрипции и трансляции, который может быть индуцибельным или конститутивным, причем кодирующий участок функционально связан под транскрипциональным контролем участка инициации транскрипции и участка терминации транскрипции и трансляции.
[00207] Кассета экспрессии («модуль экспрессии») может быть введена во множество векторов, например, плазмиду, ВАС, YAC, бактериофаг, такой как лямбда, P1, М13, и т.п., вирусные векторы из вирусов растений или животных (например, базирующиеся на ретровирусах векторы, аденовирусные векторы), и т.п., причем векторы обычно характеризуются способностью обеспечить селекцию клеток, содержащих векторы экспрессии. Векторы могут быть предложены для внехромосомной поддержки, особенно в форме плазмид или вирусов, или для интеграции в хромосому хозяина. Если желательна внехромосомная поддержка, обеспечивается последовательность источника для репликации плазмиды, которая может давать низкое или высокое количество копий. Широкий спектр маркеров доступен для селекции, особенно таких, которые защищают от токсинов, более конкретно, от антибиотиков. Конкретный выбираемый маркер, выбирают в соответствии с природой хозяина, где, в некоторых случаях, комплементация может применяться для ауксотрофных хозяев. Для введения конструкта ДНК может применяться любой пригодный способ, в том числе, например, конъюгация, бактериальная трансформация, осаждение ДНК кальцием, электропорация, слияние, трансфекция, инфекция вирусными векторами, баллистическая трансфекция, и т.п.
[00208] Соответственно, белки для использования в пределах настоящего изобретения могут вырабатываться в генетически модифицированных клетках-хозяевах в соответствии с обычными методами. Пригодными клетками-хозяевами являются такие типы клеток, которые могут быть трансформированы или трансфецированы экзогенной ДНК и выращены в культуре, и включают бактерии, грибковые клетки и клетки высших эукариот в культуре (в том числе, клетки многоклеточных организмов в культуре), особенно клетки млекопитающих в культуре. Методы манипулирования клонированными молекулами ДНК и введения экзогенной ДНК в широкий спектр клеток-хозяев раскрыты Sambrook and Russell, Molecular Cloning: A Laboratory Manual (3rd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 2001), и Ausubel et al., Short Protocols in Molecular Biology (4th ed., John Wiley and Sons, 1999).
[00209] Например, для рекомбинантной экспрессии гомодимерного белка, связывающегося с CD3, который содержит две идентичные полипептидные цепи, как раскрыто в настоящем документе, вектор экспрессии, в общем, будет содержать сегмент нуклеиновой кислоты, кодирующий полипептид, который связывается с CD3, функционально связанный с промотором. Для рекомбинантной экспрессии гетеродимерного белка, связывающегося с CD3, который содержит различные первую и вторую полипептидные цепи, первая и вторая полипептидные цепи могут соэкспрессироваться из отдельных векторов в клетке-хозяине, с целью экспрессии полноразмерного гетеродимерного белка. Альтернативно, для экспрессии гетеродимерных белков, связывающихся с CD3, первая и вторая полипептидные цепи соэкспрессируются из отдельных модулей экспрессии в одном и том же векторе в клетке-хозяине, для экспрессии полноразмерного гетеродимерного белка. Вектор(-ы) экспрессии переносят в клетку-хозяина обычными методами, и далее трансфецированные клетки культивируют обычными методами для выработки кодируемого полипептида(-ов), с целью выработки соответствующего белка, связывающегося с CD3.
[00210] Чтобы направить рекомбинантный белок в секреторный путь клетки-хозяина, секреторную сигнальную последовательность (также известную как лидерная последовательность) вводят в вектор экспрессии. Секреторная сигнальная последовательность может быть последовательностью из природной формы рекомбинантного белка или может происходить из другого выделенного белка, или может быть синтезирована de novo. Секреторную сигнальную последовательность функционально связывают с кодирующей полипептид последовательностью ДНК, т.е., две последовательности соединяют в корректной рамке считывания и размещают таким образом, чтобы направить недавно синтезированный полипептид в секреторный путь клетки-хозяина. Секреторные сигнальные последовательности обычно размещают в направлении 5' по отношению к последовательности ДНК, кодирующей целевой полипептид, хотя некоторые сигнальные последовательности могут быть размещены где-нибудь в другом месте целевой последовательности ДНК (см., например, Welch с соавт., патент США №5037743; Holland с соавт., патент США №5143830). В конкретных вариациях секреторная сигнальная последовательность для использования в соответствии с настоящим изобретением содержит последовательность аминокислот MEAPAQLLFLLLLWLPDTTG (SEQ ID NO: 79).
[00211] Клетки млекопитающих в культуре являются пригодными хозяевами для выработки рекомбинантных белков с целью использования в настоящем изобретении. Способы введения экзогенной ДНК в клетки-хозяева млекопитающих включают опосредованную кальция фосфатом трансфекцию (Wigler et al., Cell 14:725, 1978; Corsaro and Pearson, Somatic Cell Genetics 7:603, 1981: Graham and Van der Eb, Virology 52:456, 1973), электропорацию (Neumann et al., EMBO J. 1:841-845, 1982), опосредованную ДЭАЭ-декстраном трансфекцию (Ausubel et al., выше) и опосредованную липосомами трансфекцию (Hawley-Nelson et al., Focus 15:73, 1993; Ciccarone et al., Focus 15:80, 1993). Выработка рекомбинантных полипептидов в культивируемых клетках млекопитающих раскрыта, например, Levinson с соавт., патент США №4713339; Hagen с соавт., патент США №4784950; Palmiter с соавт., патент США №4579821; и Ringold, патент США №4656134. Примеры пригодных клеток-хозяев млекопитающих включают почечные клетки африканской зеленой макаки (Vero; АКТК CRL 1587), почечные клетки эмбриона человека (293-НЕК; АКТК CRL 1573), почечные клетки детеныша хомяка (ВНК-21, ВНК-570; АКТК CRL 8544, АКТК CRL 10314), почечные клетки собаки (MDCK; АКТК CCL 34), клетки яичника китайского хомяка (СНО-К1; АКТК CCL61; СНО DG44; СНО DXB11 (Hyclone, Logan, UT); см. также, например, Chasin et al., Som. Cell. Molec. Genet. 12:555, 1986)), гипофизарные клетки крысы (GH1; АКТК CCL82), клетки HeLa S3 (АТСС CCL2.2), клетки гепатомы крысы (H-4-II-E; АКТК CRL 1548), трансформированные SV40 почечные клетки макаки (COS-1; АКТК CRL 1650) и мышиные эмбриональные клетки (NIH-3T3; АКТК CRL 1658). Дополнительные пригодные линии клеток известны из уровня техники и доступны в публичных депозитариях, таких как Американская коллекция типовых культур, Манассас, Вирджиния. Могут использоваться высокоактивные промоторы транскрипции, например, промоторы SV-40 или цитомегаловируса. См., например, патент США №4956288. Другие пригодные промоторы включают промоторы генов металлотионеина (патенты США №№4579821 и 4601978) и основной поздний промотор аденовируса.
[00212] Селекция с помощью лекарственных средств в общем используется для отбора в культуре клеток млекопитающих, в которые вставлена экзогенная ДНК. Такие клетки обычно называют «трансфектантами». Клетки, культивируемые в присутствии селективного агента, которые способны передавать целевой ген потомку, называют «стабильными трансфектантами». Примеры селекционных маркеров включают ген, кодирующий устойчивость к антибиотику неомицину, который позволяет осуществлять селекцию в присутствии лекарственного средства неомицинового типа, такого как G-418 или подобное; ген gpt для ксантин-гуанин фосфорибозилтрансферазы, который позволяет рост клетки-хозяина в присутствии микофенольной кислоты/ксантина; и маркеры, которые обеспечивают резистентность к зеоцину, блеомицину, бластоцидину и гигромицину (см., например, Gatignol et al., Mol. Gen. Genet. 207:342, 1987; Drocourt et al., Nucl. Acids Res. 18:4009, 1990). Дополнительно, системы селекции могут применяться с целью повышения уровня экспрессии целевого гена; такой процесс называют «амплификацией». Амплификацию осуществляют путем культивирования трансфектантов в присутствии низкого уровня селекционного агента, с последующим увеличением количества селекционного агента для селекции клеток, которые вырабатывают высокие уровни выработки введенных генов. Примерами поддающегося амплификации селекционного маркера являются дигидрофолатредуктаза, которая обеспечивает резистентность к метотрексату. Дополнительно могут применяться другие гены резистентности к лекарственным средствам (например, гигромицину, многим лекарственным средствам, пуромицин ацетилтрансферазе).
[00213] Другие высшие эукариотические клетки также могут использоваться в качестве хозяев, в том числе, клетки насекомых, клетки растений и клетки птиц. Использование Agrobacterium rhizogenes в качестве вектора для экспрессии генов в клетках растений рассмотрено Sinkar et al., J. Biosci. (Bangalore) 11:47-58, 1987. Трансформация клеток насекомого и выработка экзогенных полипептидов в них раскрыты Guarino с соавт., патент США №5162222 и публикация WIPO WO 94/06463.
[00214] Клетки насекомого могут быть инфицированы рекомбинантным бакуловирусом, обычно происходящим от вируса ядерного полиэдроза Autographa californica (AcNPV). См. King and Possee, The Baculovirus Expression System: A Laboratory Guide (Chapman and Hall, London); O'Reilly et al., Baculovirus Expression Vectors: A Laboratory Manual (Oxford University Press., New York 1994); и Baculovirus Expression Protocols. Methods in Molecular Biology (Richardson ed., Humana Press, Totowa, NJ, 1995). Дополнительно, рекомбинантный бакуловирус может быть получен с использованием системы на основе транспозона, раскрытой Luckow с соавт.(J. Virol. 67:4566-4579, 1993). Данная система, в которой используются векторы переноса, коммерчески доступна в форме набора (набор ВАС-ТО-ВАС; Life Technologies, Гейтерсберг, Мэриленд). Вектор переноса (например, PFASTBAC1; Life Technologies) содержит транспозон Tn7, с целью переноса ДНК, кодирующей целевой белок, в геном бакуловируса, поддерживаемый в Е. coli в форме крупной плазмиды, называемой «бакмидой». См. Hill-Perkins and Possee, J. Gen. Virol. 71: 971-976, 1990; Bonning et al., J. Gen. Virol. 75: 1551-1556, 1994; и Chazenbalk and Rapoport, J. Biol. Chem. 270: 1543-1549, 1995. Кроме того, векторы переноса могут включать слияние с сохранением рамки считывания с ДНК, кодирующей удлинение полипептида или метку аффинности, как было раскрыто выше. С применением методов, известных из уровня техники, вектор переноса, содержащий кодирующую белок последовательность ДНК, трансформируют в клетки хозяева Е. coli, и осуществляют скрининг клеток на предмет бакмид, которые содержат перемежающийся ген lacZ, указывающий на рекомбинантный бакуловирус. Бакмиду ДНК, содержащую рекомбинантный геном бакуловируса, выделяют с применением общих методов и используют для трансфекции клеток Spodoptera frugiperda, таких как клетки Sf9. Впоследствии вырабатывается рекомбинантный вирус, который экспрессирует целевой белок. Запасы рекомбинантного вируса получают способами, обычно применяемыми в данной области.
[00215] С целью выработки белка, рекомбинантный вирус используется для инфицирования клеток-хозяев, - обычно линии клеток, полученной от совки травяной, Spodoptera frugiperda (например, клетки Sf9 или Sf21) или Trichoplusia ni (например, клетки HIGH FIVE; Invitrogen, Карлсбад, Калифорния). См. в общем Glick and Pasternak, Molecular Biotechnology, Principles & Applications of Recombinant DNA (ASM Press, Washington, D.C., 1994). См. также патент США №5300435. Свободные от сыворотки среды используются для выращивания и содержания клеток. Пригодные препараты сред известны из уровня техники и могут быть приобретены у коммерческих поставщиков. Клетки выращивают от плотности инокуляции около 2-5×105 клеток до плотности 1-2×106 клеток, и в этой временной точке добавляют запасной препарат рекомбинантного вируса с кратностью инфицирования (КИ) 0,1-10, чаще около 3. Применяемые методики в общем описаны в доступных лабораторных руководствах (см., например, King и Possee, выше; O'Reilly с соавт., выше; Richardson, выше).
[00216] Дополнительно, грибковые клетки, в том числе клетки дрожжей, могут использоваться в пределах настоящего изобретения. Виды дрожжей, представляющие особый интерес в этом отношении, включают Saccharomyces cerevisiae, Pichia pastor is и Pichia methanolica. Способы трансформации клеток S. cerevisiae экзогенной ДНК и выработка ими рекомбинантных полипептидов раскрыты, например, в Kawasaki, патент США №4599311; Kawasaki с соавт., патент США №4931373; Brake, патент США №4870008; Welch с соавт., патент США №5037743; и Murray с соавт., патент США №4845075. Трансформированные клетки отбирают на основании фенотипа, определенного селекционным маркером, обычно резистентность к лекарственным средствам или способность расти в отсутствие конкретного питательного вещества (например, лейцина). Примером системы вектора для использования в Saccharomyces cerevisiae является система вектора РОТ1, раскрытая Kawasaki с соавт.(патент США №4931373), которая позволяет производить отбор трансформированных клеток по их росту в содержащих глюкозу средах. Пригодные промоторы и терминаторы для использования в дрожжах включают полученные из генов гликолитических ферментов (см., например, Kawasaki, патент США №4599311; Kingsman с соавт., патент США №4615974; и Bitter, патент США №4977092) и генов алкогольдегидрогеназы. См. также патенты США №№4990446; 5063154; 5139936; и 4661454. Системы трансформации для других дрожжей, в том числе, Hansenula polymorpha, Schizosaccharomyces pombe, Kluyveromyces lactis, Kluyveromyces fragilis, Ustilago maydis, Pichia pastoris, Pichia methanolica, Pichia guillermondii и Candida maltosa известны из уровня техники. См., например, Gleeson et al., J. Gen. Microbiol. 132:3459-3465, 1986; Cregg, патент США №4882279; и Raymond et al., Yeast 14:11-23, 1998. Клетки Aspergillus могут применяться в соответствии со способами, описанными McKnight с соавт., патент США №4935349. Способы трансформации Acremonium chrysogenum раскрыты Sumino с соавт., патент США №5162228. Способы преобразования Neurospora раскрыты Lambowitz, патент США №4486533. Выработка рекомбинантных белков в Pichia methanolica раскрыта в Патентах США №№5716808; 5736383; 5854039; и 5888768.
[00217] Дополнительно, прокариотные клетки-хозяева, в том числе, штаммы бактерий Escherichia coli, Bacillus и другие виды, являются пригодными клетками-хозяевами в пределах настоящего изобретения. Способы трансформации таких хозяев и экспрессии экзогенных последовательностей ДНК, клонированных в соответствии с настоящим документом, известны из уровня техники (см., например, Sambrook и Russell, выше). При экспрессии рекомбинантного белка в бактериях, таких как Е. coli, белок может сохраняться в цитоплазме, обычно в форме нерастворимых гранул, или может быть направлен в периплазматическое пространство последовательностью бактериальной секреции. В первом случае, клетки лизируют, гранулы выделяют и денатурируют с применением, например, изотиоцианат гуанидина или мочевины. В дальнейшем денатурированный белок может быть подвергнут рефолдингу и димеризован путем разбавления денатурирующего вещества, например, диализом против раствора мочевины и комбинации восстановленного и окисленного глутатиона, с последующим диализом против буферизованного солевого раствора. Альтернативно, белок может быть добыт из цитоплазмы в растворимой форме и выделен без применения денатурирующих веществ. Белок добывают из клетки в форме водного экстракта, например, в буферизованном фосфатом солевом растворе. Чтобы захватить целевой белок, экстракт наносят непосредственно на среду хроматографирования, например, иммобилизированное антитело или колонку гепарин-Сефарозы. Выделенные белки могут быть добыты из периплазматического пространства в растворимой и функциональной форме, путем разрушения клеток (например, с помощью обработки ультразвуком или осмотического шока), чтобы высвободить содержимое периплазматического пространства и добыть белок, таким образом, исключая потребность в денатурации и рефолдинге. Антитела, в том числе, одноцепочечные антитела, могут вырабатываться бактериальными клетками-хозяевами согласно известным способам. См., например, Bird et al., Science 242:423-428, 1988; Huston et al., Proc Natl. Acad Sci. USA 85:5879-5883, 1988; и Pantoliano et al., Biochem. 30:10117-10125,1991.
[00218] Трансформированные или трансфецированные клетки-хозяева культивируют в соответствии с обычными методиками в питательной среде, содержащей нутриенты и других компонентах, необходимых для роста выбранных клеток-хозяев. Множество пригодных сред, включая, определенные среды и сложные среды, известны из уровня техники и, в общем, содержат источник углерода, источник азота, эссенциальные аминокислоты, витамины и минералы. Дополнительно, среды могут содержать такие компоненты, как факторы роста или сыворотка, при необходимости. Питательную среду, в общем, выбирают для клеток, содержащих экзогенно введенную ДНК, например, путем селекции с применением лекарственного средства или дефицита эссенциального нутриента, которое дополняют селекционным маркером, расположенным в векторе экспрессии, или которым сотрансфецирована клетка-хозяин.
[00219] Белки, связывающиеся с CD3, могут быть очищены традиционными методами очистки белка, обычно комбинацией хроматографических методов. См., в общем, Affinity Chromatography: Principles & Methods (Pharmacia LKB Biotechnology, Uppsala, Sweden, 1988); Scopes, Protein Purification: Principles and Practice (Springer-Verlag, New York 1994). Белки, содержащие участок Fc иммуноглобулина, могут быть очищены аффинной хроматографией на иммобилизированном белке А или белке G. Дополнительные стадии очистки, например, гель-фильтрация, могут применяться для достижения желательного уровня чистоты или обеспечения обессоливания, обмена буфера, и т.п.
[00220] Изобретение также включает составы, содержащие полипептид, связывающийся с CD3, по изобретению и фармацевтически приемлемый носитель, разбавитель и/или вспомогательное вещество.
[00221] В одном варианте реализации изобретения моноспецифичный полипептид, связывающийся с CD3, вводят пациенту, страдающему от аутоиммунного заболевания, такого как ревматоидный артрит.В другом варианте реализации изобретения, моноспецифичный полипептид, связывающийся с CD3, по изобретению вводят субъекту, которому в скором времени предстоит пересадка органа.
[00222] В другом аспекте настоящего изобретения раскрыт способ лечения расстройства, характеризующегося чрезмерной экспрессией опухолевого антигена, например, ракового. В общем, такие способы включают введение субъекту, нуждающемуся в таком лечении, терапевтически эффективного количества белка, связывающегося с CD3, который содержит второй домен связывания, связывающийся с антигеном опухоли, как раскрыто в настоящем документе. В некоторых вариантах реализации белок, связывающийся с CD3, индуцирует перенаправленную цитотоксичность Т-клеток (ПЦТК) против экспрессирующих антиген клеток опухоли у субъекта.
[00223] В некоторых вариациях способа расстройство представляет собой рак. В других вариациях расстройство представляет собой аутоиммунное заболевание. В изобретении также раскрыты способы лечения рака или аутоиммунного расстройства, включающие введение терапевтически эффективного количества составов или связывающихся с CD3 полипептидов, раскрытых в настоящем документе, нуждающемуся полипептидов больному.
[00224] В каждом из вариантов способов лечения, раскрытых в настоящем документе, белок, связывающийся с CD3, вводят способом, согласующимся с обычными методиками, которые применяются для контроля заболевания или расстройства, для которого найдено лечение. В соответствии с настоящим документом, эффективное количество белка, связывающегося с CD3, вводят субъекту, нуждающемуся в таком лечении, в течение периода времени и в условиях, достаточных для предупреждения или лечения заболевания или расстройства.
[00225] Для введения белок, связывающийся с CD3, вводят в фармацевтический состав. Фармацевтический состав, содержащий белок, связывающийся с CD3, может быть получен в соответствии с известными способами приготовления фармацевтически приемлемых составов, в которых терапевтическую молекулу объединяют в смеси с фармацевтически приемлемым носителем. Состав считается «фармацевтически приемлемым носителем», если его введение может переноситься больным-реципиентом. Стерильный, буферизованный фосфатом солевой раствор является одним из примеров фармацевтически приемлемого носителя. Другие пригодные носители известны специалисту из уровня техники. (См., например, Gennaro (ed.), Remington's Pharmaceutical Sciences (Mack Publishing Company, 19th ed. 1995)). Дополнительно, препараты могут содержать одно или более вспомогательных веществ, консерванты, солюбилизаторы, буферные агенты, альбумин, с целью предупреждения потери белка на поверхностях флакона, и т.п.
[00226] Фармацевтический состав, содержащий белок, связывающийся с CD3, вводят субъекту в терапевтически эффективном количестве. Согласно способам по настоящему изобретению, белок, связывающийся с CD3, можно вводить субъектам с помощью различных способов введения, в том числе, например, внутримышечным, подкожным, внутривенным, внутрипредсердным, внутрисуставным, парентеральным, интраназальным, внутрилегочным, трансдермальным, внутриплевральным, интратекальным и пероральным способами введения. Для целей предотвращения и лечения, введение субъекту может осуществляться, например, в форме введения единичного болюса, посредством непрерывного введения (например, непрерывное трансдермальное введение) на протяжении длительного периода времени, или в схеме с повторным введением (например, каждый час, каждый день или каждую неделю).
[00227] «Терапевтически эффективное количество (или доза)» или «эффективное количество (или доза)» молекулы или состава представляет собой такое количество, которое производит статистически значимый эффект с точки зрения облегчения одного или более симптомов расстройства, например, статистически значимое замедление прогрессирования заболевания или статистически значимое улучшение функции органа. Точная доза будет определяться клиницистом согласно общепринятым стандартам, принимая во внимание природу и тяжесть состояния, которое подлежит лечению, особенности пациента, и т.п. Определение дозы находится в пределах компетенции среднего специалиста в данной области. При ссылке на комбинацию, терапевтически эффективная доза обозначает объединенные количества активных ингредиентов, которые приводят к терапевтическому эффекту, вводимые последовательно или одновременно (например, в одном и том же препарате или параллельно в разных препаратах).
[00228] Определение эффективных доз в данном контексте обычно базируется на экспериментальных исследованиях на животных, за которыми следуют клинические испытания, и осуществляется путем определения эффективных доз и схем введения, которые существенно уменьшают частоту или тяжесть указанного расстройства у модельных субъектов. Эффективные дозы составов по настоящему изобретению варьируют в зависимости от множества различных факторов, в том числе, способа введения, сайта-мишени, физиологического состояния больного, является ли больной человеком или животным, других вводимых терапевтических средств, профилактического или терапевтического характера лечения, а также специфической активности непосредственно состава и его способности вызывать желательную реакцию у индивидуума. Обычно, больной является человеком, но в случае некоторых заболеваний больной может быть негуманоидным млекопитающим. Обычно, схемы лечения корректируют таким образом, чтобы обеспечить оптимальную терапевтическую реакцию, например, для оптимизации безопасности и эффективности.
ПРИМЕРЫ
ПРИМЕР 1: ДИЗАЙН И КОНСТРУИРОВАНИЕ МОЛЕКУЛ АНТИ-CD3 SMIP СО СНИЖЕННЫМ ТЕОРЕТИЧЕСКИМ ЗНАЧЕНИЕМ ИЗОЭЛЕКТРИЧЕСКОЙ ТОЧКИ
[00229] L1, L2, H1, Н2 и Н3 (см. Фиг. 3 и 4) представляют собой тяжелые и легкие цепи гуманизированного Cris-7, которые были предварительно сконструированы и проанализированы на предмет связывания и активности. H3L1 используют для получения SMIP DRA209 (SEQ ID NO: 3 и 4; lgG4 AA АЗКЦ-КЗЦ нулевой Fc) и SMIP DRA161 (lgG4 AA N297A Fc). Хотя указанные конструкты демонстрируют улучшенные характеристики, по сравнению с ОКТЗ и другими анти-CD3 терапевтическими средствами из уровня техники, конструкты демонстрируют значительно более короткий период полувыведения, более высокое теоретическое значение изоэлектрической точки (см. Фиг. 1) и «отсечение» в домене связывания или предшарнирном участке, при экспрессии в клетках СНО (см. Фиг. 2А и 2В), по сравнению с другими молекулами SMIP.
[00230] Варианты изоэлектрической точки анти-CD3 были сконструированы таким образом, чтобы снизить изоэлектрическую точку домена связывания и/или предшарнирного участка. Для получения вариантов изоэлектрической точки анти-CD3, новые тяжелые и легкие цепи (L3, L4, Н4, Н5 и Н6) были синтезированы по заказу Blue Heron. См. Табл. 3 и Фиг. 3 и 4.

[00231] Н4 содержит 2 точечных мутации, по сравнению с Н3. Одна мутация представляет собой замену положительно заряженного остатка (К на Q), тогда как другая мутация вводит отрицательный заряд (замена Q на Е). Обе модификации были спроектированы таким образом, чтобы снизить изоэлектрическую точку белка (посредством устранения положительного заряда или введения отрицательного заряда). Мутации осуществлялись на последовательности каркаса, во избежание утраты аффинности, и введенные новые остатки были проверены на предмет присутствия в последовательности зародышевой линии в том же положении, чтобы избежать риска иммуногенности. С использованием последовательности зародышевой линии в качестве руководства, с целью снижения изоэлектрической точки уже гуманизированного домена связывания, могут быть осуществлены замены аминокислот, с целью добавления аминокислот, преобладающих в последовательности зародышевой линии. Указанные аминокислоты могут находиться в таком же положении в последовательности зародышевой линии, или ближайшем положении. См. Фиг. 5.
[00232] Н5 конструируют, повторно «гуманизируя» последовательность тяжелой цепи (по сравнению с DRA209 и DRA161), таким образом, чтобы получить более низкую изоэлектрическую точку, по сравнению с DRA209 и DRA161. Это осуществляют посредством проектирования тяжелой цепи в сравнении с каркасом VH1 последовательности тяжелой цепи зародышевой линии IGHV1-3*01.
[00233] Н6 конструируют, повторно «гуманизируя» последовательность тяжелой цепи (по сравнению с DRA209 и DRA161), таким образом, чтобы получить более низкую изоэлектрическую точку, по сравнению с DRA209 и DRA161. Иными словами, исходная последовательность, которую используют для конструирования Н6, представляет собой последовательность зародышевой линии вместо предварительно гуманизированной последовательности. Конкретно, это осуществляют посредством проектирования тяжелой цепи с использованием каркаса VH3 последовательности тяжелой цепи IGHV3-21*01.
[00234] L3 содержит 6 точечных мутаций, по сравнению с L1, где RW мутировал (т.е., содержит замену остатка с положительным зарядом, R на L) до LL (используют LL, так как LL чаще встречается в последовательности зародышевой линии, чем LW) и часть участка J каппа (Jk) (LQIT; SEQ ID NO: 252) заменена последовательностью Jk4 (VEIK; SEQ ID NO: 253). Jk4 представляет лучше соответствующий участок Jk, базирующийся на последовательности Cris-7 H CDR3. См. Фиг. 6. Снова, мутации производились только в каркасном участке, и введенные новые остатки были проверены на предмет присутствия в таком же положении в последовательности зародышевой линии, чтобы избежать риска иммуногенности.
[00235] L4 конструируют, повторно «гуманизируя» последовательность легкой цепи (по сравнению с DRA209 и DRA161) таким способом, чтобы получить более низкую изоэлектрическую точку, по сравнению с DRA209 и DRA161. L4 повторно гуманизируют, используя каркас Vk3 последовательности легкой цепи IGKV3-11*01 в качестве руководства.
[00236] Ожидается, что все новые легкие и тяжелые цепи будут обладать более низкой изоэлектрической точкой, на основании теоретических вычислений. Вектор NTI (Invitrogen) используют, чтобы облегчить соединение последовательностей (последовательности присоединены). Последовательности зародышевой линии получены с веб-сайта НЦБИ (http://vvww.ncbi.nlm.nih.gov/igblast/).
[00237] Варианты конструируют таким образом, чтобы они содержали замененные аминокислоты в предшарнирном участке молекул SMIP, который во всех остальных отношениях идентичен DRA209 и DRA161. Предшарнирный участок представляет собой короткую последовательность аминокислот, которая соединяет домен связывания с шарнирным участком, например, может быть результатом добавления рестрикционного сайта в кодирующую нуклеотидную последовательность.
[00238] Получен вариант реализации, содержащий замену последовательности RRT в предшарнирном участке DRA209 SSS с помощью ПЦР мутагенеза. Это было сделано намеренно, чтобы удалить два положительно заряженных остатка (аргинин). Нами экспериментально показано, что такая модификация транслируется в снижение изоэлектрической точки белка. Данный конструкт по-прежнему содержит комбинацию H3L1 и получил новое название, DRA219. За исключением удаления положительно заряженных остатков (два остатка аргинина), такая мутация SSS также вводит рестрикционный сайт Xhol, который позволяет легкую сборку более новых конструктов с вариантами изоэлектрической точки.
[00239] При использовании такого же хвоста Fc, как и в DRA209, следующие комбинации Н и L применялись для конструирования конструктов SMIP, с использованием стандартных методов молекулярной биологии: а) Н4 L1 (содержащий lgG4 AA АЗКЦ-КЗЦ нулевой Fc; также обозначенный в настоящем документе как DRA222; SEQ ID NO: 7 и 8), б) H4L3 (содержащий lgG4 AA АЗКЦ-КЗЦ нулевой Fc; также обозначенный в настоящем документе как DRA221; SEQ ID NO: 9 и 10), в) H3L3 (содержащий lgG4 AA АЗКЦ-КЗЦ нулевой Fc; также обозначенный в настоящем документе как DRA223; SEQ ID NO: 11 и 12), г) H5L4 (содержащий lgG4 AA АЗКЦ-КЗЦ нулевой Fc; также обозначенный в настоящем документе как DRA224; SEQ ID NO: 13 и 14), и д) H6L4 (содержащий lgG4 AA АЗКЦ-КЗЦ нулевой Fc; также обозначенный в настоящем документе как DRA225; SEQ ID NO: 15 и 16). См. Фиг. 7А и7В.
[00240] Все указанные конструкты были трансфецированы в клетки НЕК, с целью генерации белка в исследованиях связывания in vitro. Все конструкты экспрессировали желательный белок, кроме конструкта DRA225, который не проявлял видимой экспрессии. Все другие конструкты (i) связывались с CD3 в различной степени (DRA222 продемонстрировал несколько лучшее связывание) и (ii) обладали более низким значением изоэлектрической точки, чем родительская молекула, DRA209. Измеренные значения изоэлектрической точки приведены в Табл. 4.
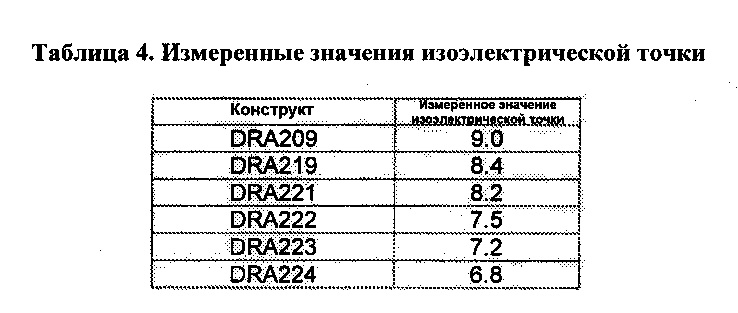
[00241] DRA219, DRA221, DRA222, DRA223 и DRA224 все ингибировали смешанные реакции лимфоцитов человека (СРЛ; данные не показаны), причем степень ингибирования коррелирует с их измеренной способностью связываться с CD3.
[00242] DRA219 (НЗ LI lgG4 AA АЗКЦ-КЗЦ нулевой Fc; SEQ ID NO: 5 и 6) и DRA222 (SEQ ID NO: 7 и 8) сохраняют связывание к CD3, а также обладают более низкой измеренной изоэлектрической точкой, причем их дополнительно клонируют в вектор рЕЕ12.4, который кодирует другой хвост Fc (lgG4 AA N297A), чтобы генерировать два дополнительных конструкта, - DRA233 (H3L1; SEQ ID NO: 17 и 18) и DRA234 (H4L1; SEQ ID NO: 19 и 20), соответственно. Дополнительно, DRA219 и DRA222 клонировали в вектор рЕЕ12.4, с целью экспрессии в СНО и назвали DRA228 и DRA229, соответственно. По существу, DRA219 равноценен DRA228, и DRA222 равноценен DRA229. Все конструкты (DRA228, DRA229, DRA233 и DRA234) впоследствии трансфецируют в клетки СНО, чтобы генерировать белки для исследования связывания, характеристики белка и изучения ФК. См. Фиг. 8.
[00243] Белки очищают из супернатанта клеток аффинной хроматографией на Белке А, с использованием системы ВЭЖХ АКТА Purifier. Предварительно заполненные колонки POROS A (Life Technologies) используют для препаратов in vitro, и предварительно заполненные колонки Белка A MabSuRe (GE Healthcare) используют для материала in vivo. Образцы загружают в присутствии ФБР и элюируют цитратным буфером. Элюенты нейтрализуют буфером ТРИС.
[00244] Капиллярную изоэлектрическую фокусировку выполняют на приборе Beckman РА 800 и используют для определения изоэлектрической точки (pl) очищенных белков. Анализ проводят в соответствии со стандартным протоколом Beckman clEF. Образцы готовят в 3 М растворе мочевины в геле clEF (Beckman), с добавлением амфолитов Pharmalyte 3-10 (Healthcare GE), катодных и анодных стабилизаторов и синтетических маркеров изоэлектрической точки пептида (Beckman). Образцы вводят в нейтральный капилляр (Beckman), а затем фокусируют путем погружения капиллярного входа в растворе анолита, выхода в раствор католита, с приложением электрического потенциала 25 кВ через капилляр в течение 15 минут. После фокусировки, образец мобилизируют химическими средствами, чтобы пропустить все разделенные компоненты сквозь детектор с фиксированной длиной волны, измеряющий оптическую плотность на длине волны 280 нм. Изоэлектрическую точку (pl) вычисляют для всех образцов на основе стандартной кривой, построенной с учетом результатов для синтетических маркеров изоэлектрической точки пептида. См. Фиг. 9-11. Конструкты содержали следующие измеренные значения изоэлектрической точки: DRA228=8,4; DRA229=7,5; DRA233=8,6; DRA234=8,0, тогда как DRA161=9,2 и DRA209=8,8. Таким образом, все варианты изоэлектрической точки, которые экспрессировались, обладали более низкой изоэлектрической точкой, чем соответствующие родительские конструкты DRA161 или DRA209.
ПРИМЕР 2: МОЛЕКУЛЫ AHTH-CD3 SMIP СВЯЗЫВАЮТСЯ С Т-КЛЕТКАМИ
[00245] Моноядерные клетки периферической крови человека (МЯПК) выделяют из свежей цельной крови с применением стандартного центрифугирования градиента плотности и трижды промывают ФБР, содержащим 0,2% альбумина телячьей сыворотки. Клетками покрывают 96-луночные планшеты с U-дном при плотности 200000 МЯПК/лунку и вводят метку с помощью 50 мкл антител в нМ концентрации, как проиллюстрировано на Фиг. 12, в течение 30 минут на льду. Планшеты трижды промывают и вводят метку в течение 30 минут с использованием 50 мкл коктейля антитела, содержащего оптимальные концентрации анти-CD5-АРС плюс козий F(ab')2 анти-человеческий IgG Fc-PE. Планшеты трижды промывают и фиксируют в 120 мкл 1% параформальдегида в ФБР, при 4С на протяжении ночи. Все циклы инкубации антитела и промывания выполняют на льду, с холодным ФБР, 0,2% АТС, 2 нМ ЭДТА. Образцы анализируют с помощью проточного питометра BD LSRII. Из каждой лунки отбирают 60 мкл. Собранные образцы анализируют с использованием программного обеспечения Flowjo путем открытия мембранного канала на Т-клетках (СТЭ против натрия хлорида/натрия цитрата, клетки CD5+). Средняя интенсивность флуоресценции (СИФ) на канале РЕ проиллюстрирована на графике. DRA227 SMIP используется только в качестве контроля и не содержит CD3 или домена связывания с Т-клетками, а также не был модифицирован для снижения изоэлектрической точки. См. Фиг. 12. Все проверенные варианты изоэлектрической точки связывались с CD3, и DRA228 и DRA229 сохраняли активность связывания, в высокой степени сходную с их родительскими конструктами.
ПРИМЕР 3: МОЛЕКУЛЫ AHTH-CD3 SMIP СО СНИЖЕННОЙ ИЗОЭЛЕКТРИЧЕСКОЙ ТОЧКОЙ ЛУЧШЕ ЭКСПРЕСИРУЮТСЯ В КЛЕТКАХ СНО
[00246] Эксклюзионная хроматография: эксклюзионную хроматографию используют для оценки содержания высокомолекулярного агрегата очищенных белков. Система ВЭЖХ представляет собой серию Agilent 1200, оборудованную четвертичным насосом, дегазатором растворителя, диодно-матричным детектором и аутосемплером с контролем температуры. Колонку Tosoh TSKgel G3000SWxl, уравновешенную в буфере анализа, содержащем 200 мМ калия фосфата и 250 мМ калия хлорида, рН 7,2, используют для хроматографического разделения. Скорость потока растворителя составляет 1 мл/мин, и масса инъекции растворителя составляет 50 мкг. Выход колонки контролируют с помощью диодно-матричного детектора Agilent на длине волны 280 нм.
[00247] НЛС-ПАГЕ: электрофорез на денатурирующем полиакриламидном геле натрия лаурилсульфата (НЛС-ПАГЕ) применяют для оценки целостности очищенных белков. Образцы готовят с помощью буфера для образцов Novex (Invitrogen) и нагревают до 85°С, выдерживая при этой температуре в течение 3 минут, чтобы гарантировать полную денатурацию до загрузки на 4-20% гель Трис-глицина Novex (Invitrogen). Образцы необязательно восстанавливают, добавляя восстанавливающий агент Novex перед стадией нагревания. Белки мобилизуют и разделяю, применяя 125 В на гели в течение ПО минут в камере для электрофореза, наполненной буфером Трис-глицин (Invitrogen, из 10Х запасного раствора). Полосы белка визуализируют окрашиванием SimplyBlue Safe Stain (Invitrogen). См. Фиг. 13-14. Проверенные варианты изоэлектрической точки продемонстрировали уменьшение отсечения по сравнению с родительской молекулой DRA161.
[00248] Капиллярный электрофорез в присутствии натрия лаурилсульфата (КЭ-НЛС): Капиллярный электрофорез в НЛС выполняют на приборе Beckman РА 800 и используют для количественного определения содержания низкомолекулярного отсечения очищенных белков. Образцы готовят в концентрации 1 мг/мл в 50% буфере для образцов НЛС (Beckman) и нагревают до 85°С, выдерживая при этой температуре в течение 3 минут, чтобы гарантировать полную денатурацию до проведения анализа. Образцы электрокинетически вводят (10 кВ в течение 20 секунд) в капилляр из кварцевого стекла без покрытия, наполненный буфером геля НЛС (Beckman), и отделяют, применяя напряжение 15 кВ через капилляр (обратная полярность) в течение 30 минут. Оптическую плотность на длине волны 220 нм и 280 нм измеряют в окне детектора. См. Фиг. 15-16. Это показывает, что качество белка вариантов изоэлектрической точки выше, чем родительской молекулы, например, вследствие более низкого очевидного протеолиза, иногда называемого в настоящем документе «отсечением».
ПРИМЕР 4: ВАРИАНТЫ ИЗОЭЛЕКТРИЧЕСКОЙ ТОЧКИ МОЛЕКУЛ АНТИ-CD3 SMIP ДЕМОНСТРИРУЮТ УВЕЛИЧЕННЫЙ ПЕРИОД ПОЛУВЫВЕДЕНИЯ
[00249] Самкам мышей BALB/c в точке времени 0 вводят внутривенной (в/в) инъекцией 200 мкл ФБР, содержащего 200 мкг DRA234 или DRA233. Трем мышам в каждой группе осуществляют инъекцию в каждой точке времени. У анестезированных мышей собирают кровь пункцией сердца в указанных точках времени после инъекции, и сыворотку собирают как раскрыто ниже. Концентрации DRA234 и DRA233 в сыворотке определяют с использованием твердофазного иммуноферментного анализа (ТИФА). Фармакокинетические параметры диспозиции для каждого белка оценивают некомпартментальным анализом с помощью программного обеспечения WinNonlin™ Professional (версия 5.3), применяя предварительно компилированную модель 201 для в/в введения болюса и редкого отбора проб.
[00250] В точке времени 0 мышам вводят в/в инъекцией 200 мкл ФБР, содержащего 200 пг (-10 мг/кг) DRA233 или DRA234. Трем мышам в каждой группе проводят инъекции в каждой временной точке. У анестезированных мышей отбирают кровь пункцией сердца, сыворотку собирают в t=15 минут, и спустя 2, 6, 24, 48, 72, 96, 168, 336 и 504 часа после инъекции. Концентрацию сыворотки для каждого белка определяют, используя специфический сандвичевый ТИФА. Результаты выражают, как среднюю концентрацию сыворотки (г/мл) ± стандартное отклонение во времени. Данные приведены в линейном (А) и логарифмическом (В) формате. См. Фиг. 17.
[00251] Анти-лекарственные антитела (АДА) обнаруживают с помощью сандвичевого ТИФА, с использованием планшетов, покрытых DRA161 (оригинальная форма DRA233 и DRA234), чтобы захватить АДА, и анти-мышиным IgG (H&L)-пероксидазой хрена (ПХ), чтобы обнаружить связанные АДА. Количество связанных комплексов измеряют, используя субстрат пероксидазы, со считыванием результатов с помощью устройства для считывания флуоресценции планшетов. Результаты выражают, как средние единицы флуоресценции (ЕФ) против разведения сыворотки. Уровни АДА явно наблюдались через 504 часа для DRA233, с возможными низкими уровнями, начиная с 336 часов; однако АДА, по-видимому, отсутствовали в любом из образцов DRA234 (Данные не показаны).
[00252] Среднюю концентрацию в сыворотке против профиля времени наносят на график для всех временных точек DRA233 ФК с данными и без данных для 504 часов. Концентрация сыворотки против профиля времени для DRA234 проиллюстрирована на Фиг. 18С.Результаты выражают, как наблюдаемый набор данных и предсказанные значения, вычисленные с помощью программного обеспечения WinNonLin™. Значение Rsq и скорректированные значения Rsq определяют пригодность аппроксимационной статистики для фазы терминальной элиминации, до и после коррекции для множества точек, используемых для оценки HL Lambda z. Приблизительный период полувыведения для DRA233 составляет 95 часов с использованием только временных точек, без обнаружимых уровней антилекарственных антител, и 84 часов для DRA234. См. Фиг. 18.
[00253] Оценка ФК по данным программы WinNonLin в табличном формате: фармакокинетический анализ проводят с помощью программного обеспечения WinNonlin™ (версия 5.3), применяя некомпартментальный анализ с использованием редкого отбора проб и предварительно компилированной модели 201 для в/в введения болюса с равномерным взвешиванием. Площадь под кривой (AUC) концентрация в сыворотке-время от временной точки 0 до последней измеримой концентрации (Tlast) оценивают с использованием линейного метода трапеций с линейно/логарифмическим способом вычисления интерполяции. Оценивают следующие фармакокинетические параметры диспозиции: максимальная наблюдаемая концентрация в сыворотке (Сшах), концентрация в точке последней измеримой концентрации (Clast), время достижения максимальной наблюдаемой концентрации в сыворотке (Ттах), очевидный период терминального элиминационного полувыведения (HL лямбда z), площадь под кривой времени введения дозы до последней измеримой концентрации (AUCall), AUC с момента введения дозы со временем, экстраполированным к бесконечности (AUCiNF_obs), клиренс из сыворотки (CL_obs), объем распределения, основанный на терминальной фазе (Vz_obs) и среднее время удерживания (СВУ). См. Фиг. 19 и 20. Данный Пример иллюстрирует, что проверенные варианты изоэлектрической точки демонстрируют более длительный период полувыведения из сыворотки in vivo и более медленный клиренс у мышей по сравнению с родительской молекулой, DRA161.
ПРИМЕР 5: БИСПЕЦИФИЧНЫЕ МОЛЕКУЛЫ ГОМОДИМЕРА АНТИ-CD3 И АНТИ-ПСМА
[00254] Биспецифичные гомодимеры, связывающиеся с CD3 и ПСМА конструируют, используя стандартные методы. Указанные конструкты, TSC249, TSC250, TSC251, TSC252, TSC295, TSC296, TSC301 и TSC302 раскрыты в Табл. 5. Вставка N-концевого домена связывания с scFv осуществляется посредством расщепления родительского шаблона и вставки scFv с помощью любого рестрикционного фермента Hindlll и Xhol или Agel и Xhol, причем желательные фрагменты идентифицируют и выделяют очисткой на агарозном геле, и лигирования. Вставку С-концевого домена связывания с scFv осуществляют посредством расщепления родительского шаблона и вставки scFv с помощью рестрикционных ферментов EcoRI и Notl, причем желательные фрагменты идентифицируют и выделяют очисткой на агарозном геле, и лигирования.


ПРИМЕР 6: БИСПЕЦЕФИЧНЫЕ ГОМОДИМЕРНЫЕ МОЛЕКУЛЫ - АНТИ-CDR3 И АНТИ-RON
[00255] Биспецифичные молекулы с гомодимерами, связывающимися с CD3 и RON, конструируют с применением стандартных методов. Указанные конструкты, TSC275, TSC277, TSC278 и TSC279 раскрыты в Табл. 6. TSC275 содержит цепи VL и VH hu4C04, SEQ ID NO: 187 и 188, соответственно, которые являются частью домена связывания с RON. TSC277, TSC278 и TSC279 являются такими же, как TSC275, за исключением мутаций/замен, приведенных в Табл. 6.
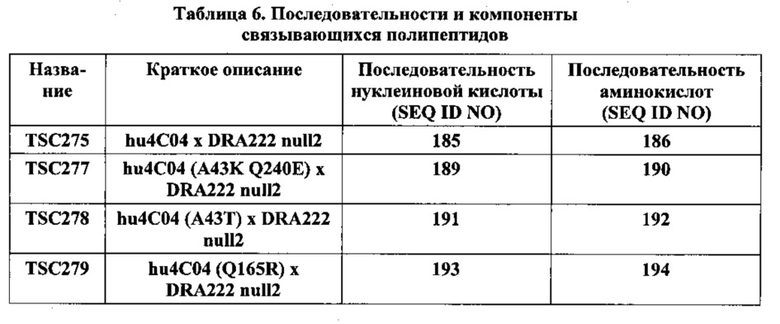
ПРИМЕР 7: БИСПЕЦИФИЧНЫЕ ГОМОДИМЕРНЫЕ МОЛЕКУЛЫ АНТИ-CD3 И АНТИ-CD 19
[00256] Биспецифичные гомодимерные молекулы, связывающиеся как с CD3, так и с CD 19, конструируют с применением стандартных методов, например, см. Публикацию РСТ WO2007/146968. Указанные конструкты, TSC233, TSC234, TSC235, TSC240, TSC241 и TSC242 раскрыты в Табл.7. TSC233 содержит цепи VL (аминокислоты 1-111 SEQ ID NO: 196) и VH (аминокислоты 128-251 SEQ ID NO: 196) HD37, которые являются частью домена связывания с CD 19.
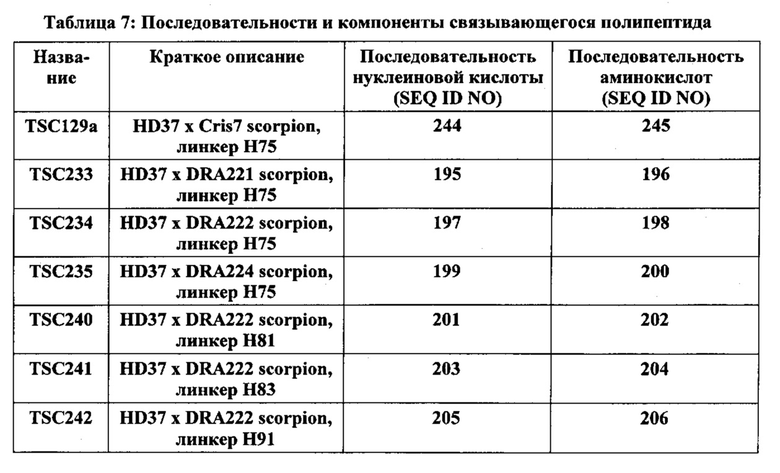
ПРИМЕР 8: БИСПЕЦИФИЧЫЕ ГЕТЕРОДИМЕРНЫЕ МОЛЕКУЛЫ АНТИ-CD3 И АНТИ-CD19
[00257] Различные биспецифичные гетеродимерные молекулы, содержащие домен связывания с CD19 и домен связывания с CD3, конструируют подобно тому, как раскрыто в Публикации РСТ WO 2011/090762, осуществляя сотрансфекцию нуклеиновых кислот, кодирующих последовательности аминокислот, как указано в Табл. 8. Гетеродимеры получают соэкспрессией двух различных полипептидных цепей, причем одна полипептидная цепь, содержит домен гетеродимеризации иммуноглобулина СН1, а другая полипептидная цепь, содержит домен гетеродимеризации иммуноглобулина CL.


[00258] TSC049 содержит, от амино-конца к карбокси-концу: HD37 (анти-CD19) scFv, шарнир SCC-P lgG1 человека, СН2 lgG1 человека, СН3 lgG1 человека и СН1 человека. TSC096 содержит, от амино-конца к карбокси-концу: HD37 (анти-CD19) scFv, шарнир lgG1 SCC-P, lgG1 СН2 человека, lgG1 СН3 и Cκ(YAE) человека. TSC125 содержит, от амино-конца к карбокси-концу: Cris7 (анти-CD3) scFv, шарнир lgG1 SCC-Р человека, lgG1 СН2 человека, lgG1 СН3 человека и СН1 человека. TSC218 содержит, от амино-конца к карбокси-концу: DRA221 (анти-CD3) scFv, шарнир lgG1 SCC-P человека, lgG1 СН2 человека, lgG1 СН3 человека и СН1 человека. TSC219 содержит, от амино-конца к карбокси-концу: DRA222 (анти-CD3) scFv, шарнир lgG1 SCC-P человека, lgG1 СН2 человека, lgG1 СН3 человека и СН1 человека. TSC220 содержит, от амино-конца к карбокси-концу: DRA224 (анти-CD3) scFv, шарнир lgG1 SCC-P человека, lgG1 СН2 человека, lgG1 СН3 человека и СН1 человека. TSC221 содержит, от амино-конца к карбокси-концу: DRA221 (анти-CD3) scFv, шарнир lgG1 SCC-P человека, lgG1 СН2 человека, lgG1 СН3 человека и Cκ(YAE) человека. TSC222 содержит, от амино-конца к карбокси-концу: DRA222 (анти-CD3) scFv, шарнир lgG1 SCC-P человека, lgG1 СН2 человека, lgG1 СН3 человека и Cκ(YAE) человека. TSC223 содержит, от амино-конца к карбокси-концу: DRA224 (анти-CD3) scFv, шарнир lgG1 SCC-P человека, человеческий lgG1 СН2, lgG1 СН3 человека и Cκ(YAE) человека.
ПРИМЕР 9: ГЕТЕРОДИМЕРНЫЕ БИСПЕЦИФИЧНЫЕ МОЛЕКУЛЫ АНТИ-CD3 И АНТИ-ПСМА
[00259] Различные гетеродимерные биспецифичные молекулы, содержащие домен связывания с ПСМА и домен связывания с CD3, конструируют подобно тому, как раскрыто в Публикации РСТ WO 2011/090762, осуществляя сотрансфекцию нуклеиновых кислот, кодирующих последовательности аминокислот, как указано в Табл. 9. Гетеродимеры получают соэкспрессией двух различных полипептидных цепей, причем одна полипептидная цепь содержит домен гетеродимеризации СН1 иммуноглобулина, и другая полипептидная цепь содержит домен гетеродимеризации иммуноглобулина CL. Некоторые домены связывания с ПСМА раскрыты в Публикации РСТ WO 2011/090761.

[00260] «Гуманизированный 107-1А4 VL-VH#2 scFv» и «гуманизированный 107-1А4 VL-VH#1 scFv» представляют собой гуманизированные scFv, базирующиеся на моноклональном анти-человеческом-ПСМА антителе 107-1А4, и также раскрыты в Публикации РСТ WO 2011/090761. «Гуманизированный 107-1А4 VL-VH#2 scFv» и «гуманизированный 107-1А4 VL-VH#1 scFv» соответствуют аминокислотам 1-243 SEQ ID NO: 216 и 226, соответственно.
[00261] TSC192 содержит, от амино-конца к карбокси-концу: гуманизированный 107-1А4 (анти-ПСМА) VL-VH#2 scFv, шарнир lgG1 SCC-P человека, lgG1 СН2 человека, lgG1 СН3 человека и Cκ(YAE) человека. TSC195 содержит, от амино-конца к карбокси-концу: гуманизированный 107-1А4 (анти-ПСМА) VL-VH#2 scFv, шарнир lgG1 SCC-P человека, lgG1 СН2 человека, lgG1 СН3 человека и СН1 человека. TSC193 содержит, от амино-конца к карбокси-концу: гуманизированный 107-1А4 (анти-ПСМА) VL-VH#1 scFv, шарнир lgG1 SCC-P человека, lgG1 СН2 человека, lgG1 СН3 человека и Cκ(YAE) человека.
[00262] TSC219 содержит, от амино-конца к карбокси-концу: DRA222 (анти-CD3) scFv, шарнир lgG1 SCC-P человека, lgG1 СН2 человека, lgG1 СН3 человека и СН1 человека. TSC222 содержит, от амино-конца к карбокси-концу: DRA222 (анти-CD3) scFv, шарнир lgG1 SCC-P человека, lgG1 СН2 человека, lgG1 СН3 человека и Cκ(YAE) человека. TSC254 содержит, от амино-конца к карбокси-концу: гуманизированный 107-1А4 (анти-ПСМА) VL-VH#1 scFv, шарнир lgG1 SCC-P человека, lgG1 СН2 человека, lgG1 СН3 человека, линкер и СН1 человека. TSC258 содержит, от амино-конца к карбокси-концу: DRA222 (анти-CD3) scFv, шарнир lgG1 SCC-P человека, lgG1 СН2 человека, lgG1 СН3 человека, линкер и Cκ(YAE) человека.
ПРИМЕР 10: ЗАВИСИМАЯ ОТ МИШЕНИ ПРОЛИФЕРАЦИЯ ПОЛИПЕПТИДОМ Т-КЛЕТКИ ПОД ДЕЙСТВИЕМ ГЕТЕРОДИМЕРОВ И ГОМОДИМЕРОВ. НАЦЕЛЕННЫХ НА CD19
[00263] С целью сравнения эффективности различных гетеродимеров биспецифичных молекул полипептида с точки зрения индуцирования зависимой от мишени активации и пролиферации Т-клеток, сравнивали три различных гомодимерных биспецифичных молекулы (TSC129a, TSC233 и TSC234 (SEQ ID NO: 245, 196 и 198, соответственно)) с общим доменом связывания анти-CD19 (HD37) и тремя различными доменами связывания анти-CD3 (Cris7 для TSC129a, домен связывания scFv от DRA221 (аминокислоты 1-244 SEQ ID NO: 10) для TSC233, домен связывания scFv от DRA222 (аминокислоты 1-244 SEQ ID NO: 8) для TSC234). Дополнительно сравнивали три различных гетеродимерных биспецифичных молекулы (TSC127, TSC227, TSC228) с тремя различными доменами связывания анти-CD3 (Cris7 для TSC127, домен связывания scFv от DRA221 (аминокислоты 1-244 SEQ ID NO: 10) для TSC227 и домен связывания scFv от DRA222 (аминокислота 1-244 SEQ ID NO: 8) для TSC228).
[00264] Клетки Дауди лимфомы Беркитта (CD 19+) получают из Американской коллекции типовых культур (АКТК, Манассас, Вирджиния) и культивируют в соответствии с прилагаемым протоколом. Моноядерные клетки периферической крови (МЯПК) выделяют из крови человека с использованием стандартных градиентов фиколла. Выделенные клетки промывают солевым буфером. Т-клетки дополнительно выделяют из МЯПК с помощью набора для выделения Т-клеток с ванночкой Miltenyi Biotec (Bergisch Gladbach, Германия) с использованием протокола производителя.
[00265] Пролиферацию оценивают посредством введения метки в популяции выделенных Т-клеток с помощью карбоксифлуоресцеин диацетат сукцинимидильного эфира (КФСЭ). Меченными КФСЭ Т-клетками покрывают 96-луночные планшеты с U-дном с плотностью Т-клеток 100000/лунку, при плотности опухолевых клеток 33 000/лунку, чтобы обеспечить соотношение Т-клеток к опухолевым клеткам приблизительно 3:1. Исследуемые молекулы в концентрациях, варьирующих от 1 нМ до 5 фМ, добавляют к смесям клетки в общем количестве 200 мкл/лунку в среде RPMI 1640, содержащей 10% человеческой или телячьей сыворотки, натрия пируват и заменимые аминокислоты. Планшеты инкубируют при 37°С, 5% СО2 в увлажненных инкубаторах. Через 3 дня клетки метят антителами для анализа методом проточной цитометрии. Клетки метят и промывают в первоначальных планшетах, чтобы минимизировать потери клеток в ходе переноса, и весь процесс введения метки осуществляют в солевом буфере, содержащем 0,2% альбумина телячьей сыворотки. Вначале клетки предварительно инкубируют с 100 мкг/мл IgG человека при комнатной температуре на протяжении 15 минут.Впоследствии, клетки инкубируют со смесью (общий объем 50 мкл) со следующими меченными красителем антителами: CD5-PE, CD4-APC, CD8-тихоокеанскйи голубой, CD25-PE-Cy7, а также 7-аминоактиномицин D (в дальнейшем 7AAD) в течение 40 минут.Планшеты дважды промывают, ресуспендируют в объемах от 80 до 120 мкл и немедленно анализируют на проточном цитометре BD LSRII, чтобы получить 80% содержимого каждой лунки. Файлы образцов анализируют с помощью программного обеспечения FlowJo для вычисления процента и количества клеток, для которых произошло по меньшей мере одно деление клетки, согласно их профилю КФСЭ, путем открытия мембранного канала последовательно на активированных, живых Т-клетках CD4+ или CD8+ (7AAD-, CD5+ CD25+ CD4+ или 7AAD-CD5+ CD25+ CD8+, соответственно). Средние значения и среднее отклонение вычисляют с помощью программного обеспечения Microsoft Excel. Графики строят с помощью Microsoft Excel или Graphpad Prism.
[00266] Анализ популяций живых CD4+ и CD8+ из клеток Дауди, обработанных цельными Т-клетками, выявил существенное увеличение как общего количества клеток, так и процента размножающихся клеток в присутствии клеток Дауди, показывающих целевой антиген CD19 (Фиг. 21 и 22).
[00267] В случае гомодимерных биспецифичных полипептидов (TSC129a, TSC233, TSC234), сравнивали пролиферацию между клетками CD4+ и CD8+ (Фиг. 21).
Пролиферация, индуцированная TSC129a и TSC234, была сравнимой для клеток CD4+ и CD8+ (Фиг. 21), свидетельствуя о том, что вариант изоэлектрической точки домена связывания анти-CD3 scFv от DRA222 обладает по существу равноценной активностью относительно родительского домена связывания с Cris7. Профиль пролиферации Т-клеток, индуцированной TSC233, слегка отличался для TSC129a и TSC234, со сниженной индукцией пролиферации CD4+ (Фиг. 21 А) и слегка увеличенной индукцией пролиферации CD8+ (Фиг. 21В). Это наводит на мысль о том, что вариант изоэлектрической точки домена связывания анти-CD3 scFv от DRA221 не полностью эквивалентен родительскому домену связывания с Cris7 с точки зрения активности.
[00268] В случае гетеродимерных биспецифичных полипептидов (TSC127, TSC227, TSC228), пролиферация была выше для Т-клеток CD8+, чем для Т-клеток CD4+ в присутствии клеток Дауди (Фиг. 22). Пролиферация, индуцированная TSC127 и TSC228, была сравнимой для клеток CD4+ и CD8+ (Фиг. 22), наводя на мысль о том, что вариант изоэлектрической точки домена связывания анти-CD3 scFv от DRA222 обладает активностью, по существу равноценной родительскому домену связывания с Cris7. Профиль пролиферации Т-клеток, индуцированной TSC227, слегка отличался от TSC127 и TSC228, с уменьшенной индукцией пролиферации CD4+ и сравнимой индукцией пролиферации CD8+. Снова, это наводит на мысль о том, что вариант изоэлектрической точки домена связывания анти-CD3 scFv от DRA221 не является полностью эквивалентным родительскому домену связывания с Cris7 с точки зрения активности.
ПРИМЕР 11: ПЕРЕНАПРАВЛЕНИЕ ЦИТОТОКСИЧНОСТИ Т-КЛЕТОК ПОЛИПЕПТИДНЫМИ ГЕТЕРОДИМЕРАМИ И ГОМОДИМЕРАМИ, НАЦЕЛЕННЫМИ НА CD19
[00269] С целью сравнения эффективности различных гетеродимеров биспецифичных молекул полипептида с точки зрения индуцирования зависимой от мишени цитотоксичности Т-клеток, сравнивали три различных гомодимерных биспецифичных молекулы (TSC129a, TSC233 и TSC234 (SEQ ID NO: 245, 196 и 198, соответственно)) с общим доменом связывания анти-CD19 (HD37) и тремя различными доменами связывания анти-CD3 (Cris7 для TSC129a, домен связывания scFv от DRA221 (аминокислоты 1-244 SEQ IDNO: 10) для TSC233, домен связывания scFv от DRA222 (аминокислоты 1-244 SEQ ID NO: 8) для TSC234)). Дополнительно, также сравнивались три различных гетеродимерных биспецифичных молекулы (TSC127, TSC227, TSC228) с тремя различными доменами связывания анти-CD3 (Cris7 для TSC127, домен связывания scFv от DRA221 (аминокислоты 1-244 SEQ IDNO: 10) для TSC227, и домен связывания scFv от DRA222 (аминокислоты 1-244 SEQ ID NO: 8) для TSC228).
[00270] Клетки Дауди лимфомы Беркитта (CD19+) получают из АКТК (Манассас, Вирджиния) и культивируют в соответствии с прилагаемым протоколом производителя. Моноядерные клетки периферической крови человека (МЯПК) выделяют из свежей цельной крови с применением стандартных градиентов фиколла и промывают в солевом буфере. Т-клетки дополнительно выделяют из МЯПК с помощью набора для выделения Т-клеток с ванночкой Miltenyi Biotec (Bergisch Gladbach, Германия) с использованием протокола производителя.
[00271] Цитотоксичность оценивают по высвобождению 51Cr Около 5×106 клеток Дауди обрабатывают 0,3 мКи 51Cr и инкубируют в течение 75 минут при 37°С. Через 75 минут клетки трижды промывают средой (RPMI + 10% сыворотки телячьего эмбриона) и ресуспендируют в 11,5 мл среды. 50 мкл полученной суспензии помещают в каждую лунку 96-луночных U-образных планшетов (около 20000 клеток/лунку). Биспецифичные молекулы в концентрациях от 500 пМ до 0,1 пМ добавляют к клеткам Дауди, доводя общий объем до 100 мкл/лунку. Клетки-мишени инкубируют при комнатной температуре на протяжении 15 минут. Затем добавляют 100 мкл выделенных Т-клеток (около 200000), чтобы получить соотношение Т-клеток к клеткам-мишеням 10:1. 50 мкл 0,8% NP-40 добавляют к контрольной лунке, содержащей клетки-мишени, оставляют на 15 минут, и далее добавляют по 100 мкл среды, чтобы обеспечить контроль общего лизиса.
[00272] Планшеты инкубируют на протяжении 4 часов, затем центрифугируют при 1500 об/мин в течение 3 минут, и 25 мкл супернатанта переносят из каждой лунки в соответствующую лунку 96-луночного планшета для образцов Luma. Планшетам для образцов дают высохнуть на воздухе под безопасной химической вытяжкой в течение 18 часов, после чего считывают радиоактивность на сцинтилляционном счетчике с использованием стандартного протокола.
[00273] Анализ данных цитотоксичности показал направляемую клетками Дауди цитотоксичность Т-клеток в присутствии Т-клеток и нацеленных биспецифичных молекул анти-CD19 (Фиг. 23).
[00274] В случае гомодимерных биспецифичных полипептидов (TSC129a, TSC233, TSC234), наблюдаемая цитотоксичность была сравнимой для TSC129a и TSC234 (Фиг. 23А), наводя на мысль о том, что вариант изоэлектрической точки DRA222 обладает равноценной активностью с родительским доменом связывания с Cris7. Незначительное уменьшение цитотоксичности наблюдалось в случае TSC233, наводя на мысль о том, что вариант изоэлектрической точки DRA221 обладает меньшей активностью, чем родительский домен связывания Cris7.
[00275] В случае гетеродимерных биспецифичных полипептидов (TSC127, TSC227, TSC228), наблюдаемая цитотоксичность была сравнимой для TSC127 и TSC228 (Фиг. 23В), наводя на мысль о том, что вариант изоэлектрической точки домена связывания DRA222 scFv обладает равноценной активностью с родительским доменом связывания с Cris7. Легкое уменьшение цитотоксичности наблюдалось для TSC227, наводя на мысль о том, что вариант изоэлектрической точки домена связывания DRA221 scFv обладает меньшей активностью, чем родительский домен связывания Cris7.
ПРИМЕР 12: ЦИТОТОКСИЧНОСТЬ Т-КЛЕТОК, ПЕРЕНАПРАВЛЕННАЯ ПОЛИПЕПТИДНЫМИ ГОМОДИМЕРАМИ, НАЦЕЛЕННЫМИ НА RON
[00276] Для сравнения эффективности различных гетеродимерных биспецифичных молекул полипептида с точки зрения индуцирования цитотоксичности Т-клеток, зависимой от мишени, сравнивают четыре различных гомодимерных биспецифичных молекулы (TSC275, TSC277, TSC278 и TSC279) с общим доменом связывания анти-CD3 (домен связывания scFv от DRA222 (аминокислоты 1-244 SEQ ID NO: 8)) и четырьмя вариантами одного и того же домена связывания анти-RON (hu4C04 scFv для TSC275, hu4C04 scFv (A43K/Q240E) для TSC277, hu4C04 scFv (A43T) для TSC278 и hu4C04 scFv (Q 165R) для TSC279).
[00277] Клетки рака молочной железы MDA-MB-453 (RON+) получают в АКТК (Манассас, Вирджиния) и культивируют в соответствии с протоколом, предложенным АКТК. Моноядерные клетки периферической крови человека (МЯПК) выделяют из свежей цельной крови человека с применением стандартных градиентов фиколла и промывают в солевом буфере. Т-клетки дополнительно выделяют из МЯПК с помощью набора для выделения Т-клеток с ванночкой Miltenyi Biotec (Bergisch Gladbach, Германия) с использованием протокола производителя.
[00278] Цитотоксичность оценивают по высвобождению 51Cr Около 5×106 клеток Дауди обрабатывают 0,3 мКи 51Cr и инкубируют в течение 75 минут при 37°С. Спустя 75 минут клетки трижды промывают средой (RPMI + 10% сыворотки телячьего эмбриона) и ресуспендируют в 11,5 мл среды. 50 мкл полученной суспензии помещают в каждую лунку 96-луночных U-образных планшетов (около 20000 клеток/лунку). Биспецифичные молекулы в концентрациях от 10 нМ до 0,005 фМ добавляют к клеткам Дауди, доводя общий объем до 100 мкл/лунку. Клетки-мишени инкубируют при комнатной температуре на протяжении 15 минут. Затем добавляют 100 мкл выделенных Т-клеток (около 100 000), чтобы получить соотношение Т-клеток к клеткам-мишеням 5:1. 50 мкл 0,8% NP-40 добавляют к контрольной лунке, содержащей клетки-мишени, оставляют на 15 минут, и далее добавляют по 100 мкл среды, чтобы обеспечить полный контроль лизиса.
[00279] Планшеты инкубируют на протяжении 24 часов, затем центрифугируют при 1500 об/мин в течение 3 минут, и 25 мкл супернатанта переносят из каждой лунки в соответствующую лунку 96-луночного планшета для образцов Luma. Планшетам для образцов дают высохнуть на воздухе под безопасной химической вытяжкой в течение 18 часов, после чего считывают радиоактивность на сцинтилляционном счетчике с использованием стандартного протокола.
[00280] Анализ данных цитотоксичности показал направляемую клетками MDA-МВ-453 цитотоксичность Т-клеток в присутствии Т-клеток и нацеленных биспецифичных молекул анти-RON (Фиг. 24).
[00281] В случае гомодимерных биспецифичных полипептидов (TSC275, TSC277, TSC278 и TSC279), наблюдаемая цитотоксичность в общем была сравнимой (Фиг. 24), наводя на мысль о том, что мутации в пределах домена связывания с ORN196 не воздействуют существенно на общую активность. Однако, несколько более низкий уровень максимального лизиса наблюдается для TSC278, по сравнению с тремя другими вариантами (Фиг. 24). Максимальный лизис наблюдается при концентрациях 10 пМ и 10 нМ.
ПРИМЕР 13: СВЯЗЫВАНИЕ ГЕТЕРОДИМЕРОВ И ГОМОДИМЕРОВ БИСПЕЦИФИЧНОГО ПОЛИПЕПТИДА С Т-КЛЕТКАМИ
[00282] Для сравнения эффективности молекул биспецифичного полипептида, демонстрирующих вариант изоэлектрической точки домена связывания с CD3 (scFv найден в DRA222 (аминокислоты 1-244 SEQ ID NO: 8)) при связывании с Т-клетками, характеристики связывания на клетке гетеродимера биспецифичного полипептида анти-CD19 х анти-CD3, TSC228 (как раскрыто в Примере 8, Табл. 8), сравнивают с характеристиками гомодимера биспецифичного полипептида анти-ПСМА х анти-CD3 TSC249 (как раскрыто в Примере 5, Табл. 5) в двух независимых экспериментах.
[00283] Клетки Т-клеточного лейкоза Jurkat (CD3+) получают из АКТК (Манассас, Вирджиния) и культивируют в соответствии с протоколом, предложенным АКТК. Связывание оценивают, инкубируя 5×105 клеток Jurkat в течение 30 минут при 4°С с серийными разведениями молекул биспецифичного TSC228 или TSC249, в концентрациях от 250 нМ до 0,125 нМ (TSC249) или от 125 нМ до 1 пМ (TSC228). Клетки трижды промывают, после чего инкубируют с козьим анти-человеческим IgG-ФИТЦ (разведение 1:200) еще в течение 30 минут при 4°С. Затем клетки снова трижды промывают, фиксируют в 1% растворе параформальдегида и считывают на инструменте FACS-Calibur.
[00284] Анализ методом прямого светорассеяния, подмножество с высоким боковым светорассеянием в FlowJo, версия 7.5, показал дозозависимое связывание (Tree Star, Inc, Ашленд, Орегон) биспецифичных молекул TSC228 и TSC249 с клетками Jurkat (Фиг. 25А и В, соответственно). Гетеродимер биспецифичного полипептида TSC228, который содержит только один домен связывания анти-CD3, связывался с более низкой очевидной аффинностью (ЕС50=24,5 нМ), чем гомодимер биспецифичного полипептида TSC249 (ЕС50=1,9 нМ), который содержит два домена связывания анти-CD3. Это наводит на мысль о том, что гетеродимер биспецифичного полипептида TSC249 связывается с некоторой степенью авидности.
ПРИМЕР 14: ЗАВИСИМАЯ ОТ МИШЕНИ ПРОЛИФЕРАЦИЯ Т-КЛЕТОК ПОД ДЕЙСТВИЕМ ГОМОДИМЕРОВ БИСПЕЦИФИЧНОГО ПОЛИПЕПТИДА, НАЦЕЛЕННЫХ НА ПСМА
[00285] Для определения эффективности молекул другого гетеродимера биспецифичного полипептида с точки зрения индуцирования зависимой от мишени активации и пролиферации Т-клеток, исследуют гомодимерную биспецифичную молекулу (TSC249, см. Пример 5, Табл. 5) с доменом связывания анти-ПСМА (hu107-1А4) и доменом связывания анти-CD3 (scFv от DRA222 (аминокислоты 1-244 SEQ ID NO: 8)) на линиях клеток рака предстательной железы, экспрессирующих ПСМА, линиях клеток рака предстательной железы, не экспрессирующих ПСМА, и Т-клетках в отсутствие клеток-мишеней.
[00286] Клетки карциномы предстательной железы С4-2 В (ПСМА+) получают из Онкологического центра М.Д, Андерсона (Хьюстон, Техас) и культивируют в соответствии с прилагаемым протоколом. Клетки карциномы предстательной железы DU-145 (ПСМА) получают из АКТК (Манассас, Вирджиния) и культивируют в соответствии с прилагаемым протоколом. Моноядерные клетки периферической крови (МЯПК) выделяют из крови человека с использованием стандартных градиентов фиколла. Выделенные клетки промывают солевым буфером. Т-клетки дополнительно выделяют из МЯПК с помощью набора для выделения Т-клеток с ванночкой Miltenyi Biotec (Bergisch Gladbach, Германия) с использованием протокола производителя.
[00287] Пролиферацию оценивают посредством введения метки в популяции выделенных Т-клеток с помощью карбоксифлуоресцеин диацетат сукцинимидильного эфира (КФСЭ). Одновременно проводят 3 независимых эксперимента. Меченные КФСЭ Т-клетки смешивают только с молекулой TSC249; меченные КФСЭ Т-клетки смешивают с клетками DU-145 и молекулой TSC249; и меченные КФСЭ Т-клетки смешивают с клетками С4-2 В и молекулой TSC249. Меченными КФСЭ Т-клетками покрывают 96-луночные планшеты с U-дном с плотностью Т-клеток 100 000/лунку, при плотности опухолевых клеток 33000/лунку, чтобы обеспечить соотношение Т-клеток к опухолевым клеткам около 3:1. Исследуемые молекулы в концентрациях, варьирующих от 20 нМ до 5 фМ, добавляют к смесям клетки в общем количестве 200 мкл/лунку в среде RPMI 1640, содержащей 10% человеческой или телячьей сыворотки, натрия пируват и заменимые аминокислоты. Планшеты инкубируют при 37°С, 5% СО2 в увлажненных инкубаторах. Спустя 3 дня клетки метят антителами для анализа методом проточной цитометрии. Клетки метят и промывают в первоначальных планшетах, чтобы минимизировать потери клеток в ходе переноса, и весь процесс введения метки осуществляют в солевом буфере, содержащем 0,2% альбумина телячьей сыворотки. Вначале клетки предварительно инкубируют с 100 мкг/мл IgG человека при в комнатной температуре на протяжении 15 минут. Впоследствии, клетки инкубируют со смесью (общий объем 50 мкл) со следующими антителами, меченными красителем: CD5-PE, CD4-APC, CD8-тихоокеанскйи голубой, CD25-PE-Cy7, а также 7-амино актиномицин D (в дальнейшем 7AAD) в течение 40 минут. Планшеты дважды промывают, ресуспендируют в объемах от 80 до 120 мкл и немедленно анализируют на проточной цитометре BD LSRII, чтобы получить 80% содержимого каждой лунки. Файлы образцов анализируют с помощью программного обеспечения FlowJo для вычисления процента и количества клеток, для которых произошло по меньшей мере одно деление клетки, согласно их профилю КФСЭ, путем открытия мембранного канала последовательно на активированных, живых Т-клетках CD4+ или CD8+ (7AAD-, CD5+ CD25+ CD4+ или 7AAD-CD5+ CD25+ CD8+, соответственно). Средние значения и среднее отклонение вычисляют с помощью программного обеспечения Microsoft Excel. Графики строят с помощью Microsoft Excel или Graphpad Prism.
[00288] Анализ популяций живых CD4+ и CD8+ из клеток С4-2В, обработанных цельными Т-клетками, выявляет существенное увеличение общего количества клеток в присутствии клеток С4-2В, показывающих целевой антиген при обработке TSC249 (Фиг. 26А). Такого увеличения не наблюдается в присутствии DU-145, которые не экспрессируют целевой антиген ПСМА, после обработки TSC249 (Фиг. 26В). В конечном итоге, указанное возрастание также не наблюдается, если выделенные Т-клетки обрабатывают TSC249 (Фиг. 26С). Это подтверждает зависимую от мишени природу пролиферации Т-клеток, индуцированную TSC249 в присутствии клеток опухоли, экспрессирующих целевой антиген.
ПРИМЕР 15: ТЕСТИРОВАНИЕ IN VIVO БИСПЕЦИФИЧНОГО НАЦЕЛИВАНИЯ НА CD3 И ПСМА
[00289] Для подтверждения эффективности биспецифичных молекул анти-CD3 и анти-ПСМА с точки зрения подавления роста опухоли in vivo, ПСМА-нацеленная молекула может быть оценена с применением любого одного или более из следующего.
[00290] Профилактическое лечение, или предотвращение приживления опухоли для подкожных опухолей: культивируемые, ПСМА-экспрессирующие линии опухолевых клеток (такие как LNCaP, LNCaP С4-2, LNCaP С4-2 В, VCaP, CWR22Rvl, LAPC4, MDA-PCa-2b, LuCaP 23,1, LuCaP 58, LuCaP 70, LuCaP 77) смешивают с лимфоцитами человека (моноядерными клетками периферической крови человека или очищенными Т-клетками) и вводят подкожной инъекцией иммунодефицитным мышам (например, SCID, NOD/SCID, и т.п.). Биспецифичную молекулу анти-ПСМА вводят внутривенной инъекцией в день инъекции и несколько последующих дней. Дозозависимое ингибирование разрастания опухоли, оцениваемое по объему опухоли, указывает на то, что соответствующая молекула обладает эффективностью против ПСМА-экспрессирующих опухолей in vivo.
[00291] Терапевтическое лечение или регресс ранее прижившихся подкожных опухолей: культивируемые, ПСМА-экспрессирующие линии опухолевых клеток (такие как LNCaP, LNCaP С4-2, LNCaP С4-2 В, VCaP, CWR22Rvl, LAPC4, MDA-PCa-2b, LuCaP 23ДА1, LuCaP 58, LuCaP 70 и LuCaP 77) вводят подкожной инъекцией иммунодефицитным мышам (например, SCID, NOD/SCID, и т.п.). Рост опухоли контролируют, и исследование начинают, когда опухоли демонстрируют признаки стабильного роста (обычно объем ~200 мм3). Лимфоциты человека (моноядерные клетки периферической крови человека или очищенные Т-клетки) вводят внутривенной инъекцией наряду с молекулой биспецифичного анти-ПСМА в день инъекции. Молекулу биспецифичного анти-ПСМА вводят инъекционно на протяжении нескольких последующих дней. Дозозависимое ингибирование роста опухоли, оцениваемое по объему опухоли, указывает на то, что соответствующая молекула обладает эффективностью против ПСМА-экспрессирующих опухолей in vivo.
[00292] Профилактическое лечение или предотвращение приживления опухоли для интратибиальных опухолей: культивируемые, ПСМА-экспрессирующие линии опухолевых клеток (такие как LNCaP С4-2, LNCaP С4-2 В, VCaP, CWR22Rv1, LAPC4, MDA-PCa-2b, LuCaP 23,1, LuCaP 58, LuCaP 70, LuCaP 77) смешивают с лимфоцитами человека (моноядерными клетками периферической крови человека или очищенными Т-клетками) и вводят интратибиально иммунодефицитным мышам (например, SCID, NOD/SCID, и т.п.). Биспецифичную молекулу анти-ПСМА вводят внутривенной инъекцией в день инъекции и несколько последующих дней. Дозозависимое ингибирование разрастания опухоли, оцениваемое по биомаркерам сыворотки, данным радиографии, флуоресцентной визуализации, потере массы и другим показательным параметрам объема опухоли, указывает на то, что соответствующая молекула обладает эффективностью против ПСМА-экспрессирующих опухолей in vivo.
[00293] Терапевтическое лечение или регресс ранее прижившихся интратибиальных опухолей: культивируемые, ПСМА-экспрессирующие линии опухолевых клеток (такие как LNCaP С4-2, LNCaP С4-2 В, VCaP, CWR22Rvl, LAPC4, MDA-PCa-2b, LuCaP 23, 1A1, LuCaP 58, LuCaP 70, LuCaP 77) вводят интратибиально иммунодефицитным мышам (например, SCID, NOD/SCID, и т.п.). Рост опухоли контролируют, и исследование начинают, когда опухоли демонстрируют признаки стабильного роста (обычно объем ~200 мм3). Лимфоциты человека (моноядерные клетки периферической крови человека или очищенные Т-клетки) вводят внутривенной инъекцией наряду с молекулой биспецифичного анти-ПСМА в день инъекции. Молекулу биспецифичного анти-ПСМА вводят инъекционно на протяжении нескольких последующих дней. Дозозависимое ингибирование роста опухоли, оцениваемое по биомаркерам сыворотки, данным радиографии, флуоресцентной визуализации, потере массы и другим показательным параметрам объема опухоли, указывает на то, что соответствующая молекула обладает эффективностью против ПСМА-экспрессирующих опухолей in vivo.
ПРИМЕР 16: ЦИТОТОКСИЧНОСТЬ Т-КЛЕТОК, ПЕРЕНАПРАВЛЕННАЯ ПОЛИПЕПТИДНЫМИ ГОМОДИМЕРАМИ, НАЦЕЛЕННЫМИ НА ПСМА
[00294] Для сравнения эффективности различных гетеродимерных биспецифичных молекул полипептида с точки зрения индуцирования зависимой от мишени цитотоксичности Т-клеток, сравнивают две различных гомодимерных биспецифичных молекулы (TSC194, TSC249) с общим доменом связывания анти-ПСМА (hul07-lA4) и двумя различными доменами связывания анти-CD3 (Cris7 scFV для TSC194, scFv от DRA222 для TSC249). TSC194 представляет собой гомодимерный биспецифичный ((huVL-VH#2 107-1А4 scFv-Fc-Cris7 scFv) (SEQ ID NO: 250 и 251).
[00295] Клетки C4-2B (ПСМА+) получают из Онкологического центра М.Д. Андерсона (Хьюстон, Техас) и культивируют в соответствии с опубликованным протоколом культивирования в среде RPMI-1640 (Life Technologies, Карлсбад, Калифорния), содержащей 10% сыворотки телячьего эмбриона. Моноядерные клетки периферической крови (МЯПК) выделяют из крови человека с использованием стандартных градиентов фиколла. Выделенные клетки промывают солевым буфером. Т-клетки дополнительно выделяют из МЯПК с помощью набора для выделения Т-клеток с ванночкой Miltenyi Biotec (Bergisch Gladbach, Германия) с использованием протокола производителя.
[00296] Цитотоксичность оценивают по высвобождению 51Cr. Около 5×106 клеток С4-2 обрабатывают 0,3 мКи 51Cr и инкубируют в течение 75 минут при 37°С. Спустя 75 минут клетки трижды промывают средой (RPMI + 10% сыворотки телячьего эмбриона) и ресуспендируют в 11,5 мл среды. 50 мкл полученной суспензии помещают в каждую лунку 96-луночных планшетов с U-образным дном (около 20000 клеток/лунку). Биспецифичные молекулы в концентрациях от 500 нМ до 0,2 пМ добавляют к клеткам С4-2, доводя общий объем до 100 мкл/лунку. Клетки-мишени инкубируют при комнатной температуре на протяжении 15 минут. Затем добавляют 100 мкл выделенных Т-клеток (около 200000), чтобы получить соотношение Т-клеток к клеткам-мишеням 10:1. 50 мкл 0,8% NP-40 добавляют к контрольной лунке, содержащей клетки-мишени, оставляют на 15 минут, и далее добавляют по 100 мкл среды, чтобы обеспечить полный контроль лизиса.
[00297] Планшеты инкубируют на протяжении 4 часов, затем центрифугируют при 1500 об/мин в течение 3 минут, и 25 мкл супернатанта переносят из каждой лунки в соответствующую лунку 96-луночного планшета для образцов Luma. Планшетам для образцов дают высохнуть на воздухе под безопасной химической вытяжкой в течение 18 часов, после чего считывают радиоактивность на сцинтилляционном счетчике с использованием стандартного протокола.
[00298] Анализ данных цитотоксичности показал направляемую клетками С4-2 цитотоксичность Т-клеток в присутствии Т-клеток и нацеленных биспецифичных молекул анти-ПСМА (Фиг. 27). В случае гомодимерных биспецифичных полипептидов (TSC194, TSC249), наблюдаемая цитотоксичность в общем была сравнимой для TSC194 и TSC249 (Табл. 10), наводя на мысль о том, что вариант изоэлектрической точки домена связывания анти-CD3 scFv DRA222 обладает равноценной активностью родительскому домену связывания Cris7.

| название | год | авторы | номер документа |
|---|---|---|---|
| БЕЛКИ, СВЯЗЫВАЮЩИЕ СПЕЦИФИЧЕСКИЙ МЕМБРАННЫЙ АНТИГЕН ПРОСТАТЫ, И СООТВЕТСТВУЮЩИЕ КОМПОЗИЦИИ И СПОСОБЫ | 2012 |
|
RU2632647C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ, СВЯЗЫВАЮЩИЕСЯ С 5T4 И 4-1BB, И ОТНОСЯЩИЕСЯ К НИМ КОМПОЗИЦИИ И СПОСОБЫ | 2018 |
|
RU2804458C2 |
| СПОСОБ ОТБОРА И ПОЛУЧЕНИЯ ВЫСОКОСЕЛЕКТИВНЫХ И МУЛЬТИСПЕЦИФИЧНЫХ НАЦЕЛИВАЮЩИХ ГРУПП С ЗАДАННЫМИ СВОЙСТВАМИ, ВКЛЮЧАЮЩИХ ПО МЕНЬШЕЙ МЕРЕ ДВЕ РАЗЛИЧНЫЕ СВЯЗЫВАЮЩИЕ ГРУППИРОВКИ, И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2639287C2 |
| БИСПЕЦИФИЧЕСКИЕ АСИММЕТРИЧНЫЕ ГЕТЕРОДИМЕРЫ, СОДЕРЖАЩИЕ АНТИ-CD3 КОНСТРУКЦИИ | 2013 |
|
RU2650868C2 |
| ТРИСПЕЦИФИЧНЫЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, КОТОРЫЕ СПЕЦИФИЧЕСКИ СВЯЗЫВАЮТ АНТИГЕНЫ МНОЖЕСТВА ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2718692C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2810924C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА-ИНГИБИТОРЫ КОНТРОЛЬНЫХ ТОЧЕК | 2017 |
|
RU2767357C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2012 |
|
RU2650775C2 |
| СОДЕРЖАЩИЕ СУПЕРСПИРАЛЬ И/ИЛИ ПРИВЯЗКУ БЕЛКОВЫЕ КОМПЛЕКСЫ И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2573915C2 |
| Биспецифическое антитело и его применение | 2019 |
|
RU2785292C2 |




































































































































































































































































































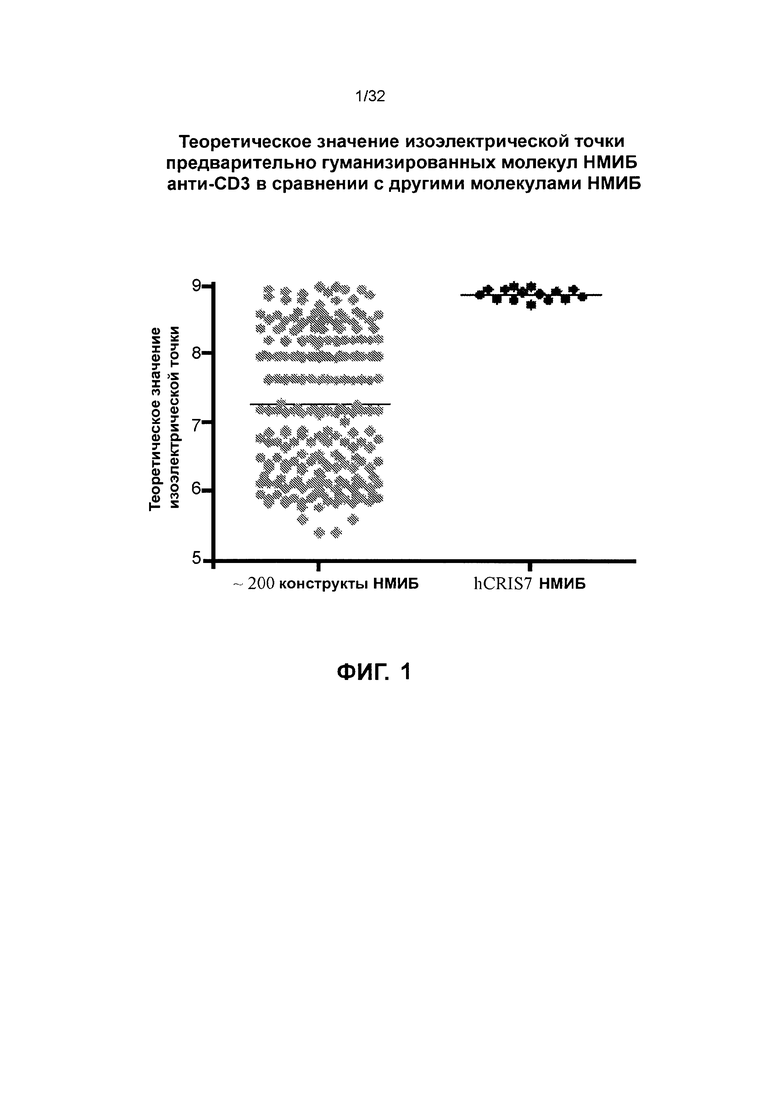
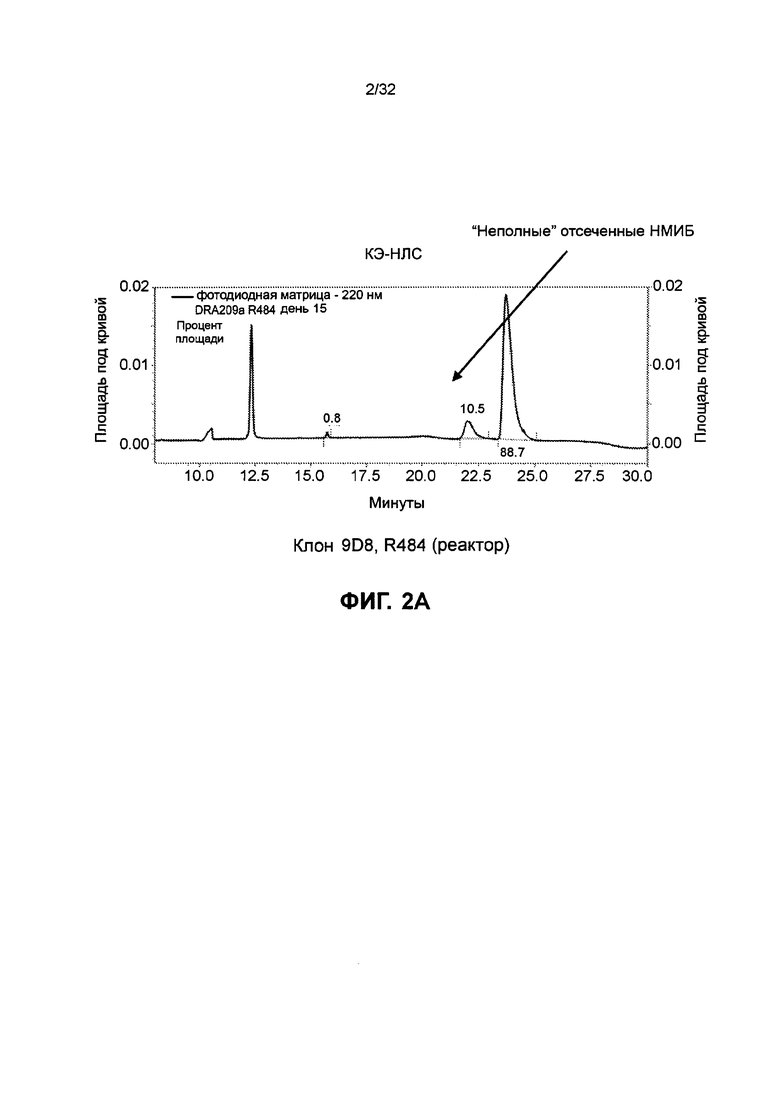
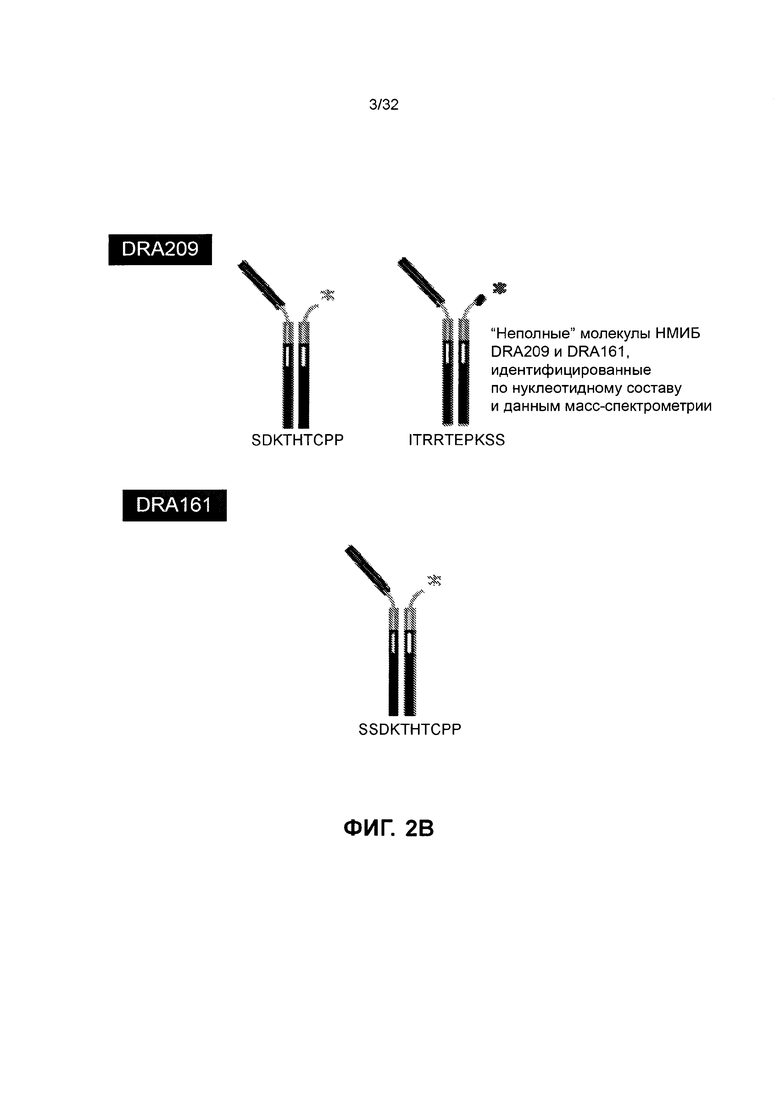



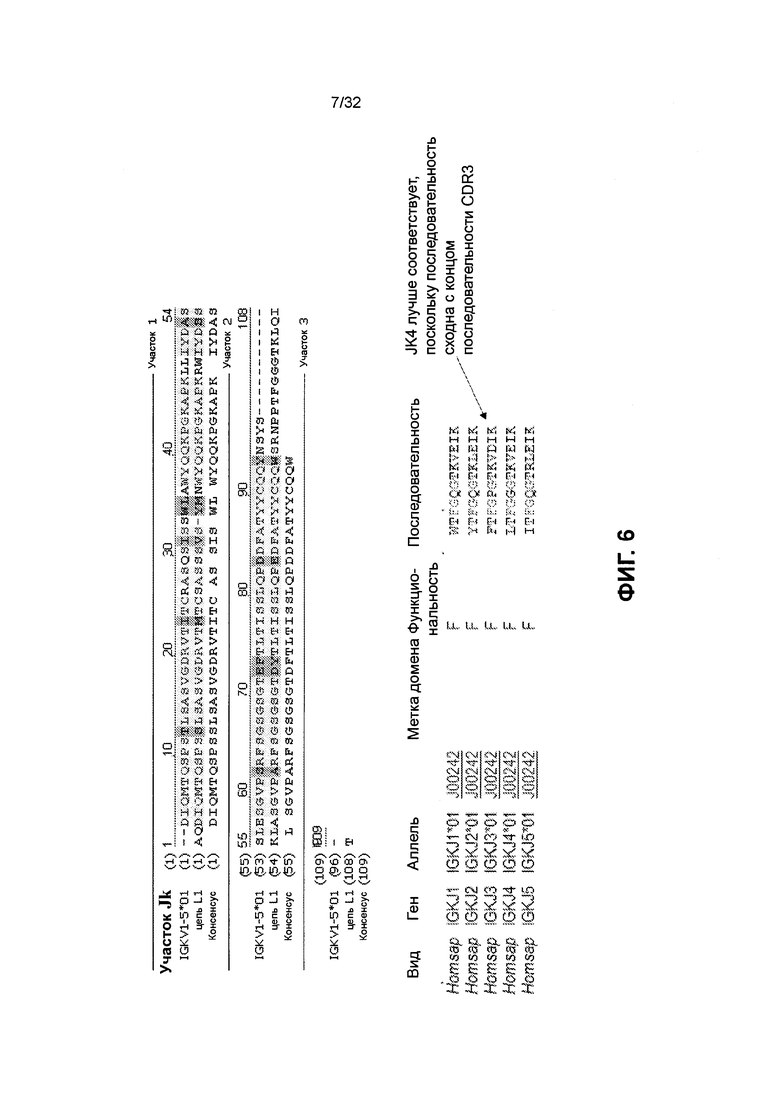
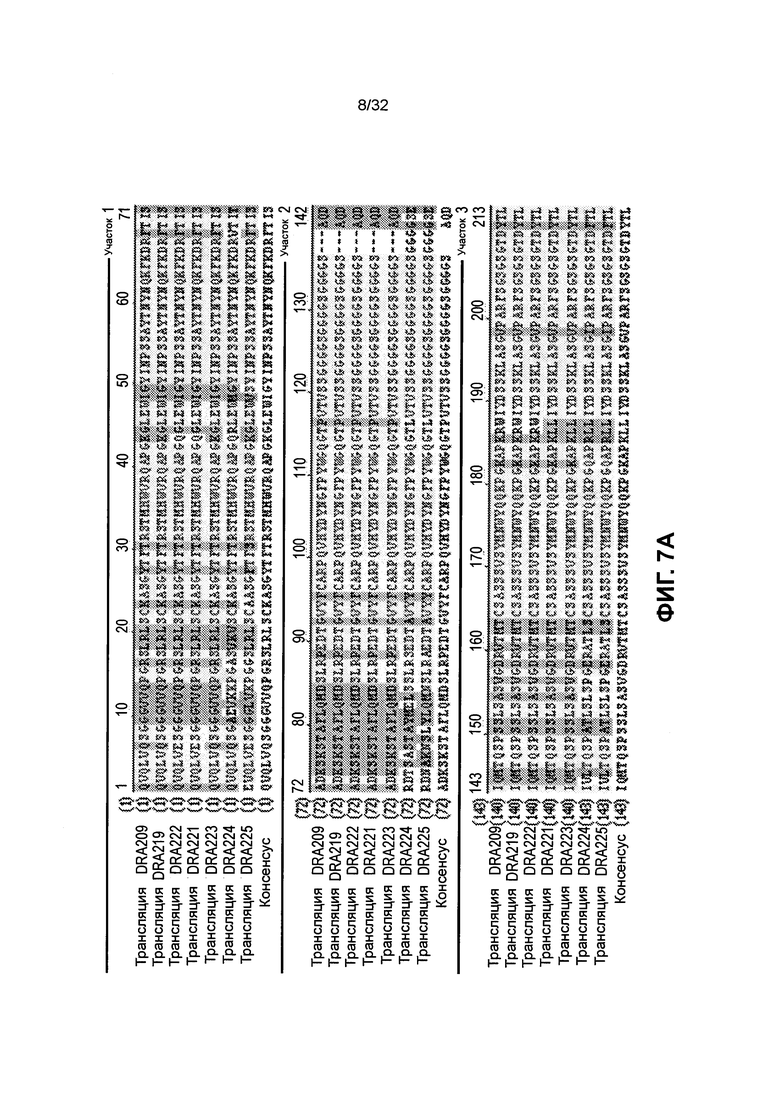
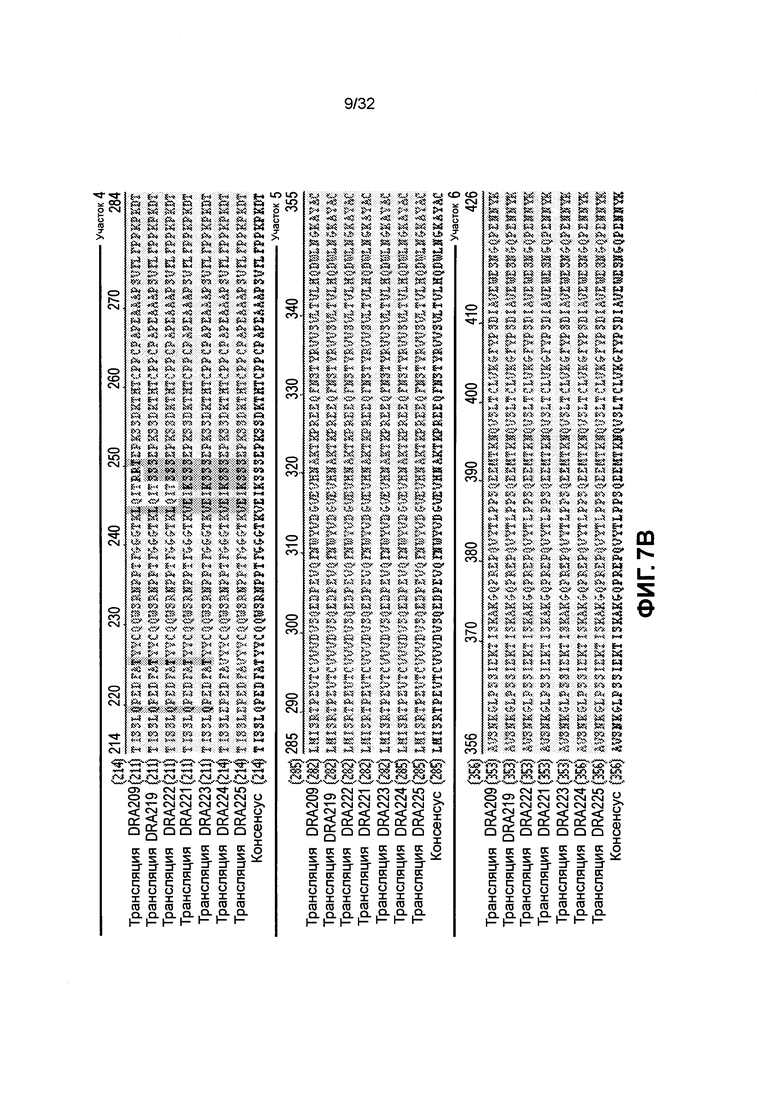

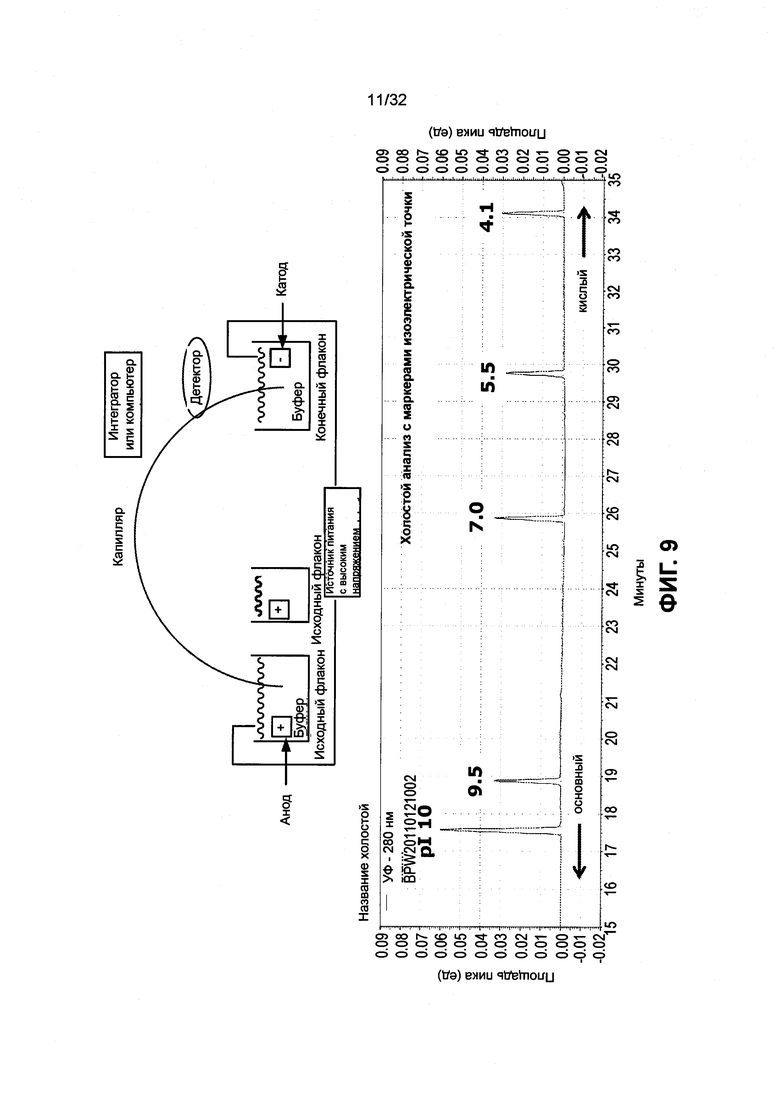
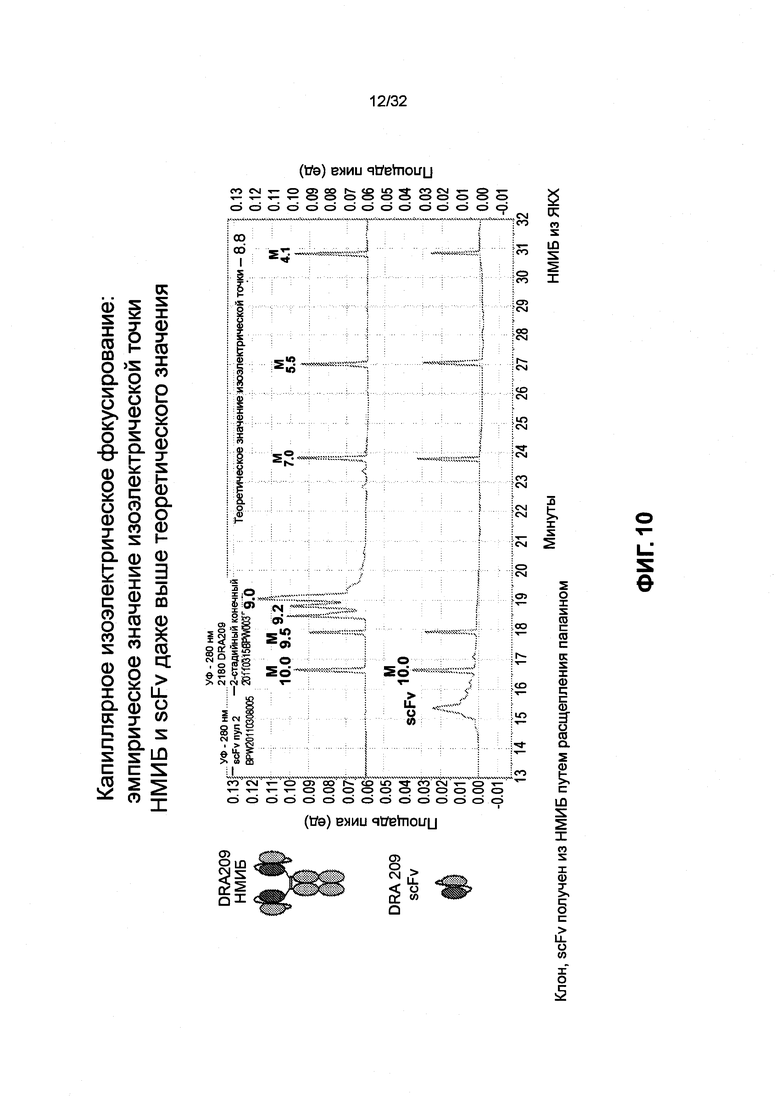
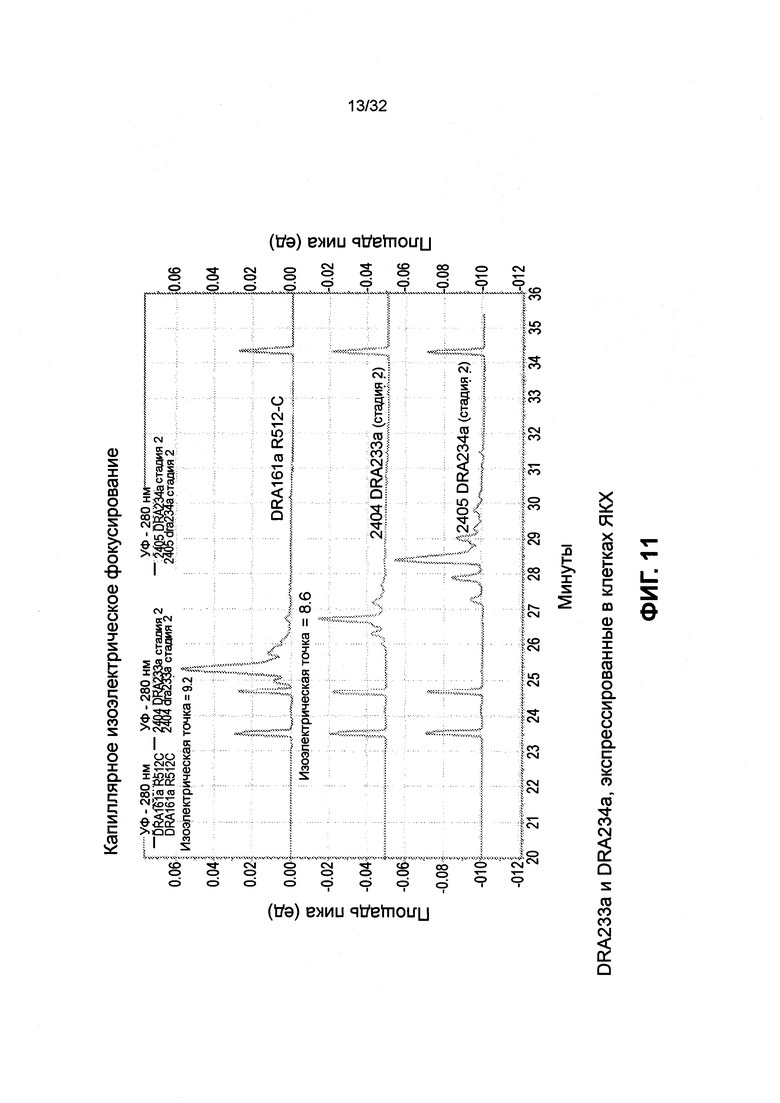
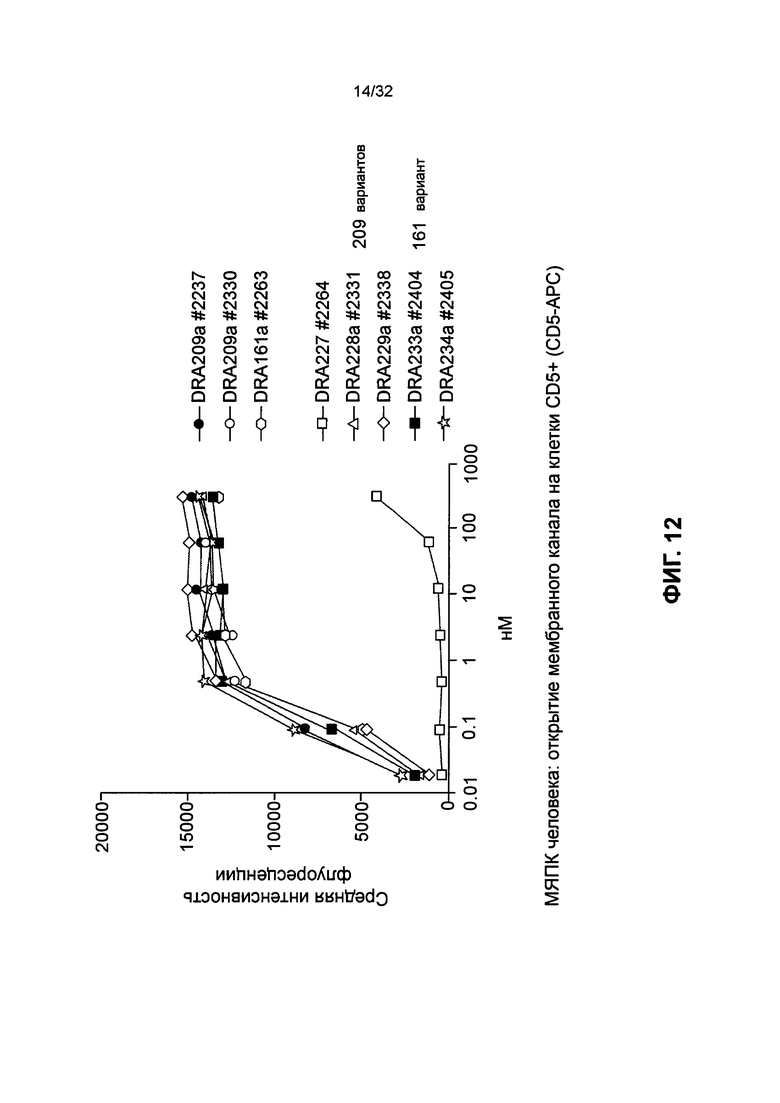
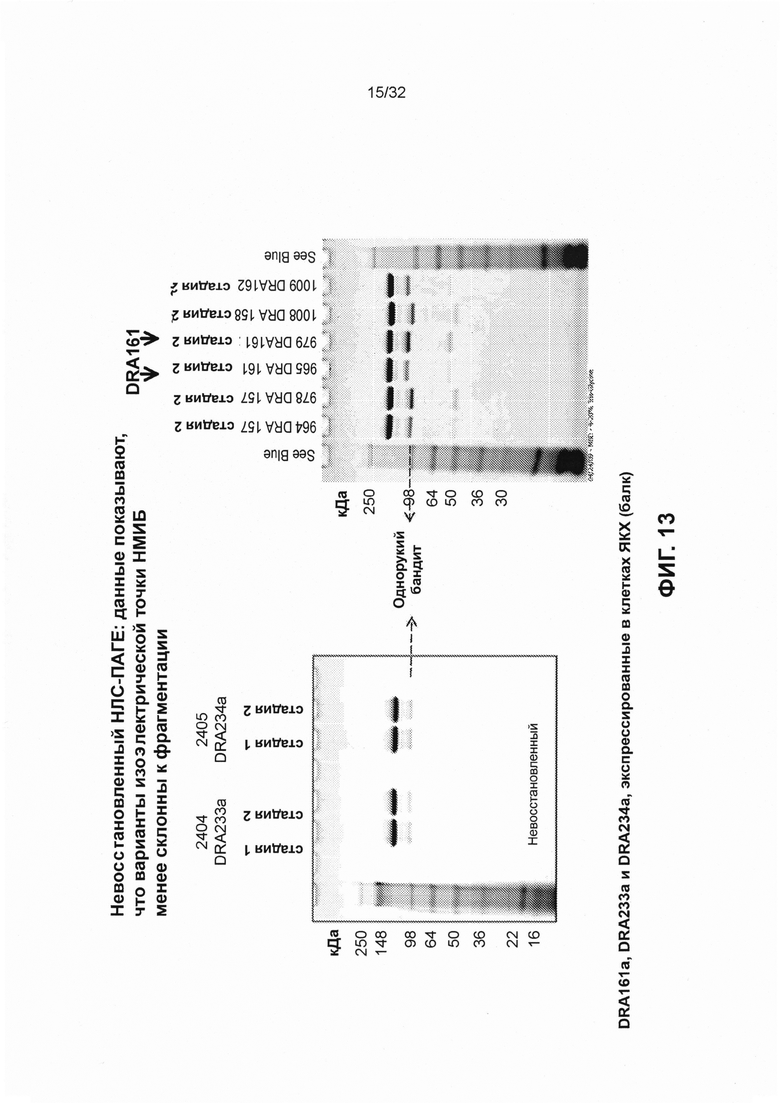
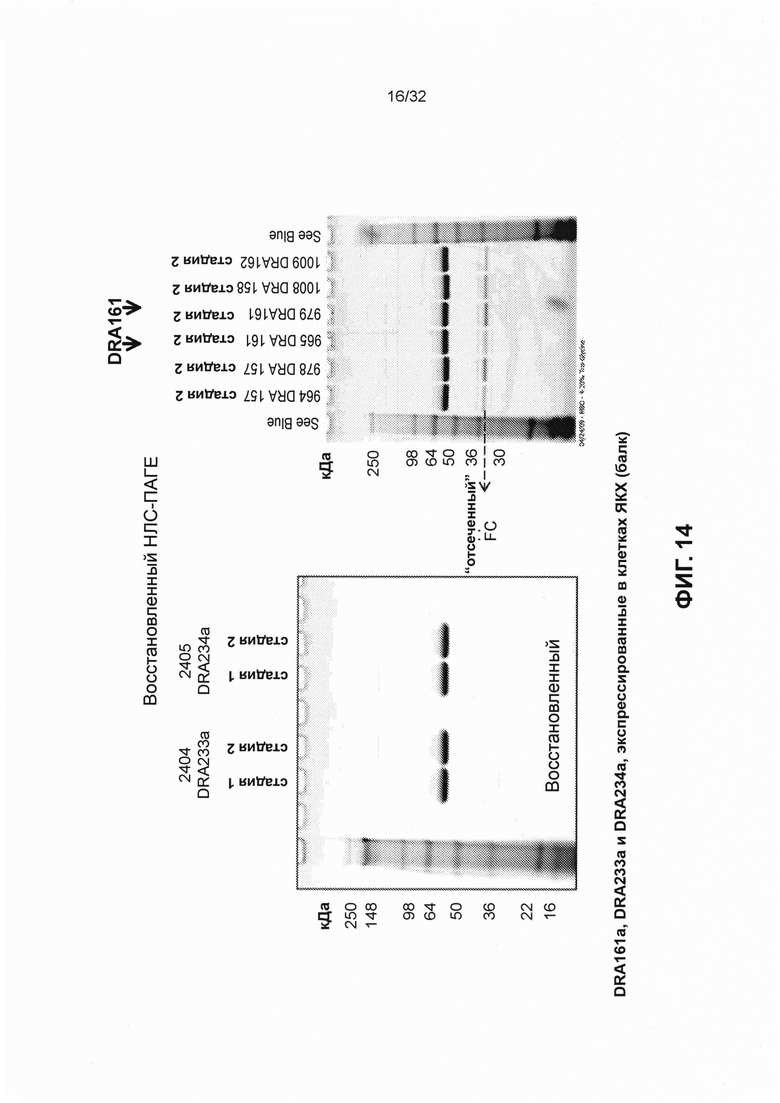
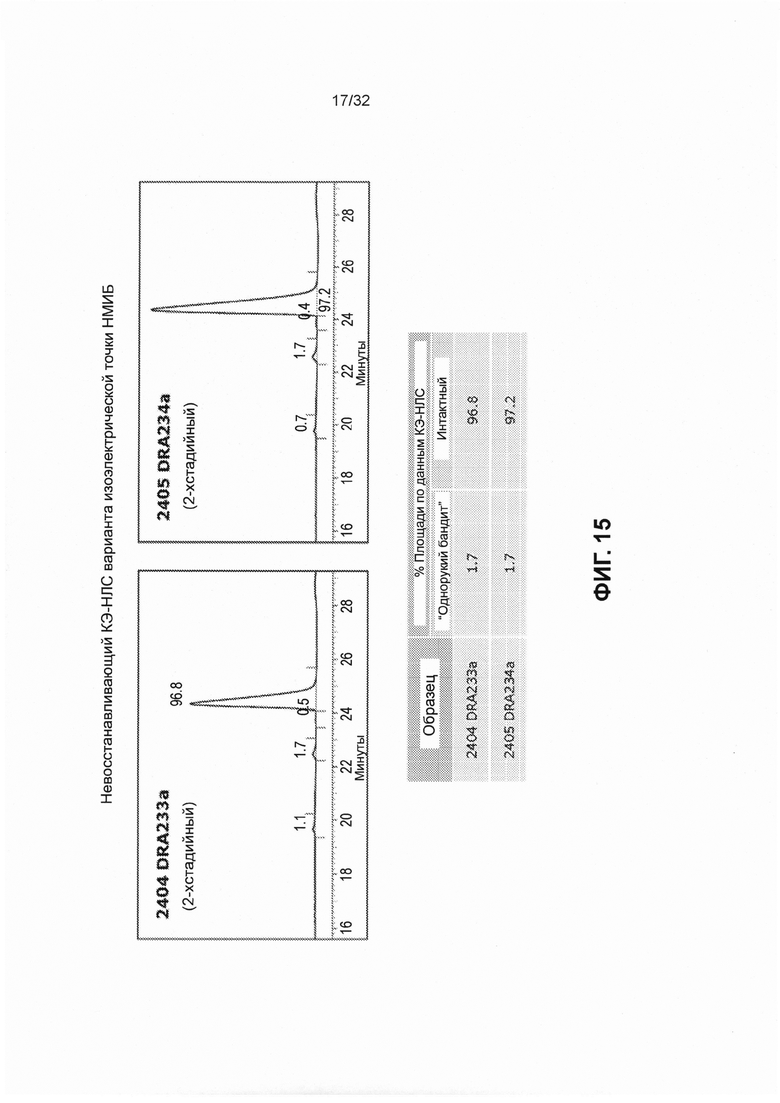
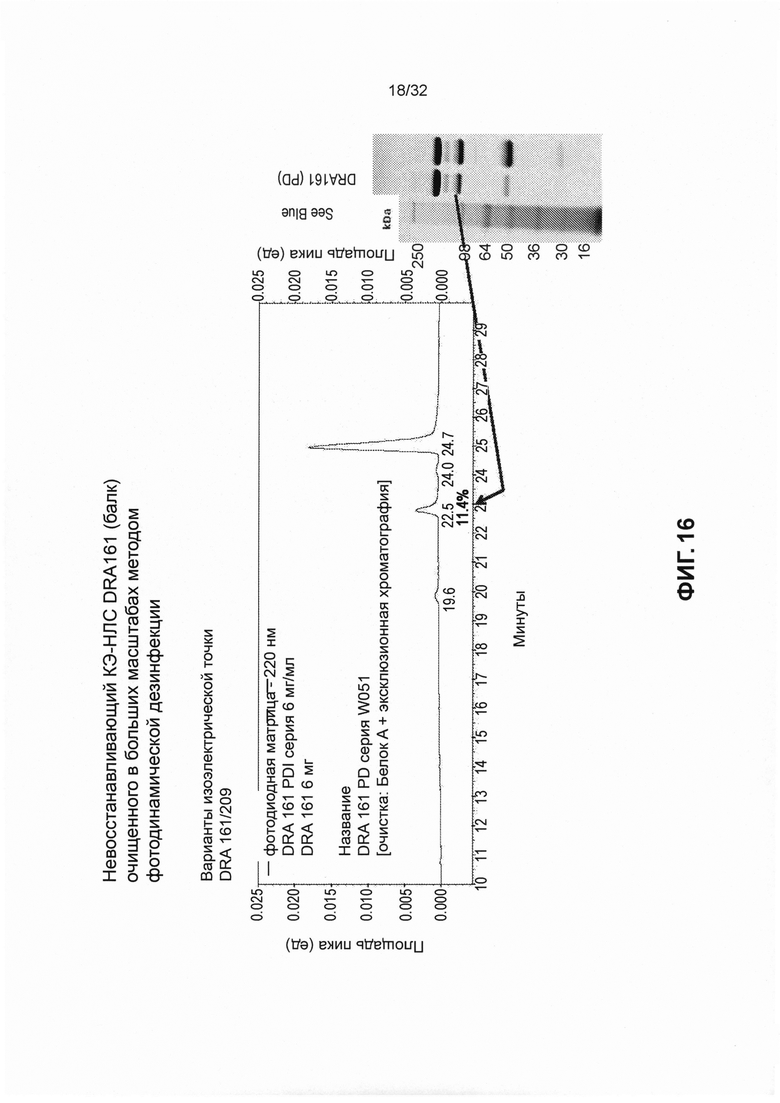
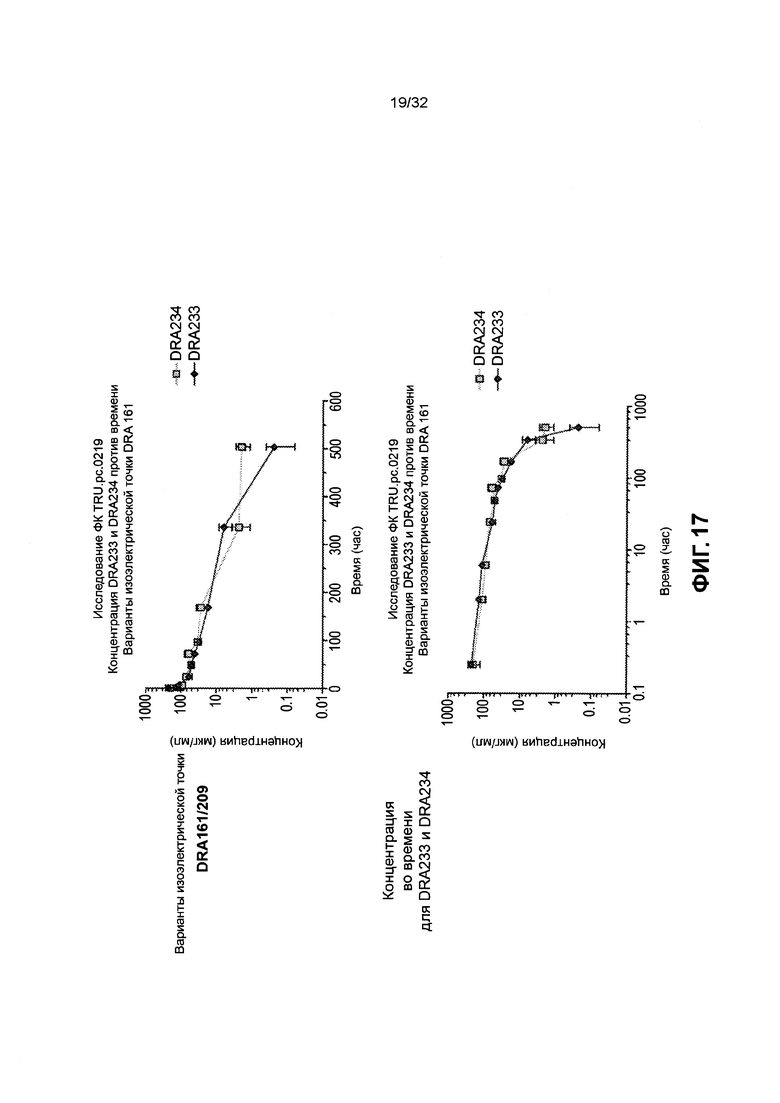
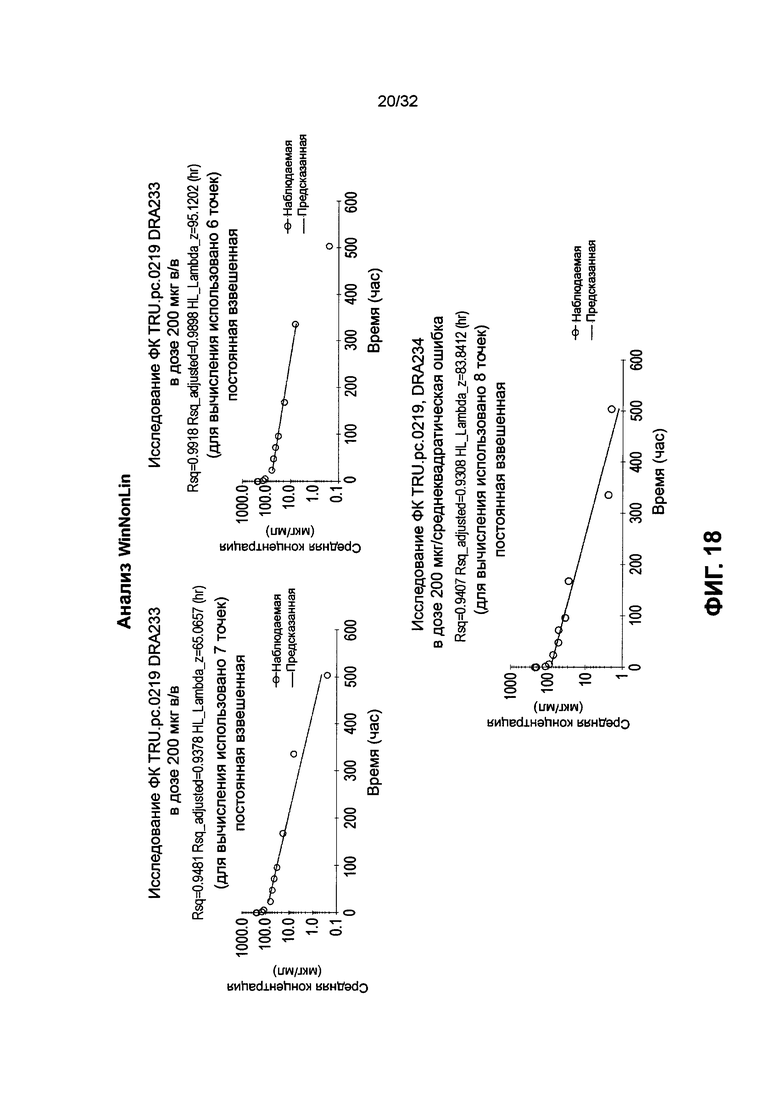
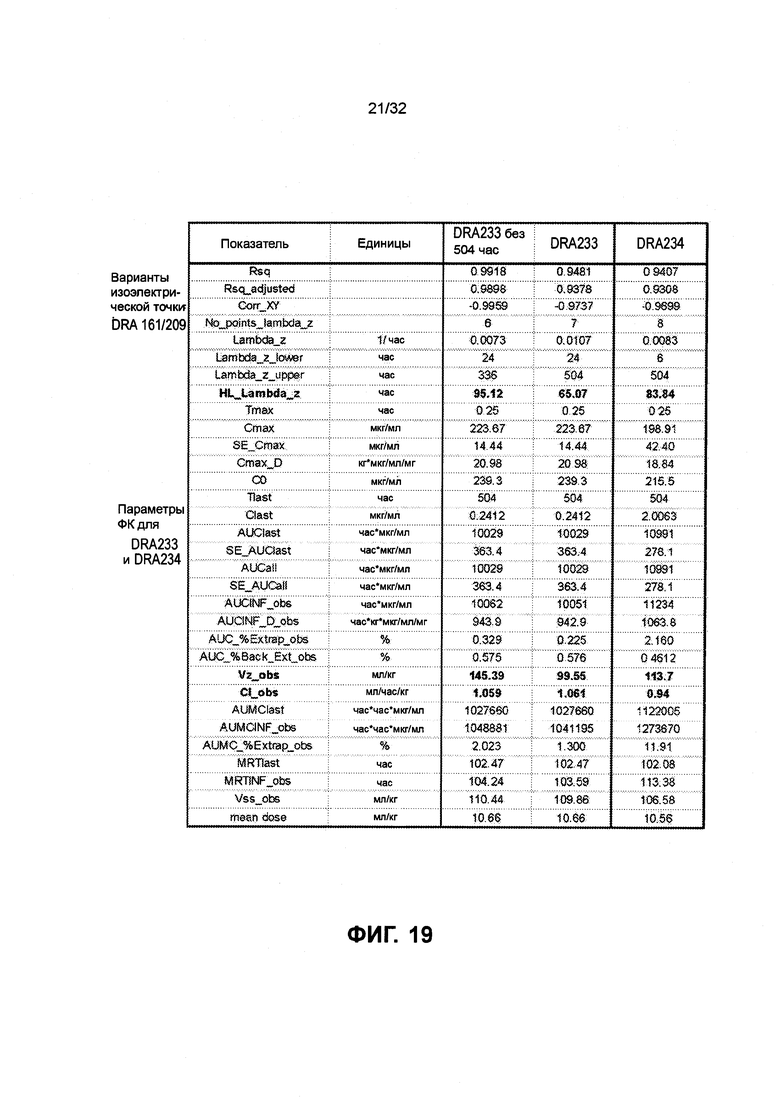

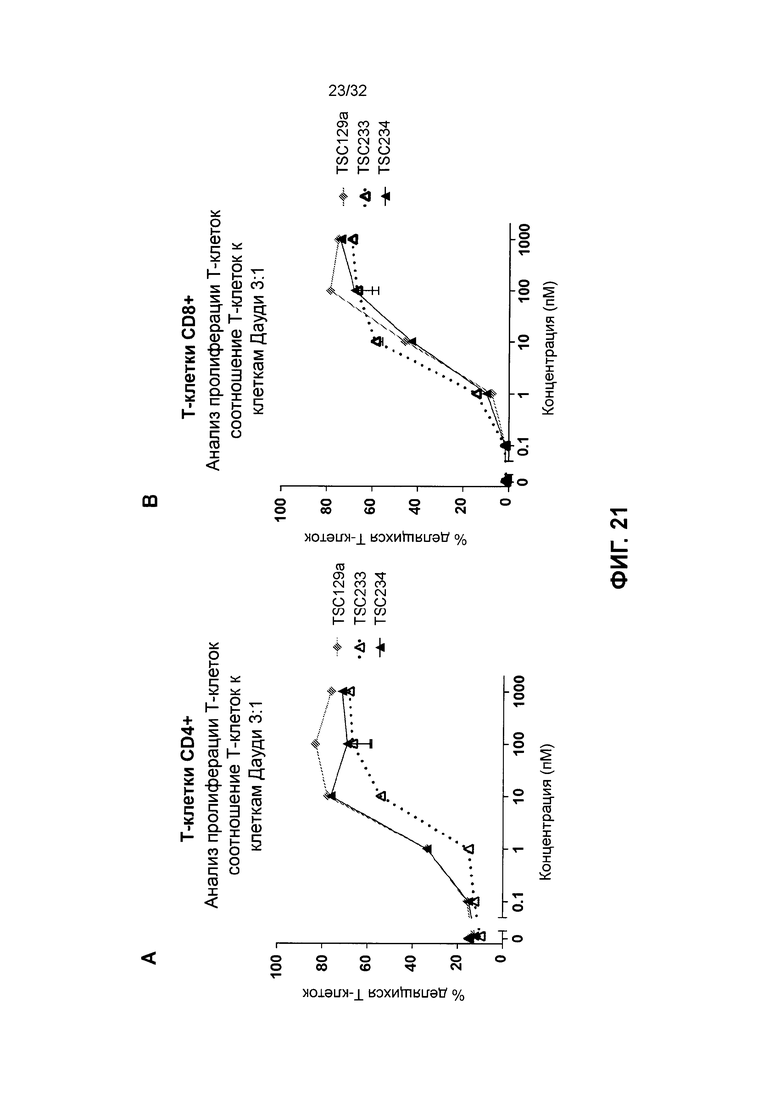

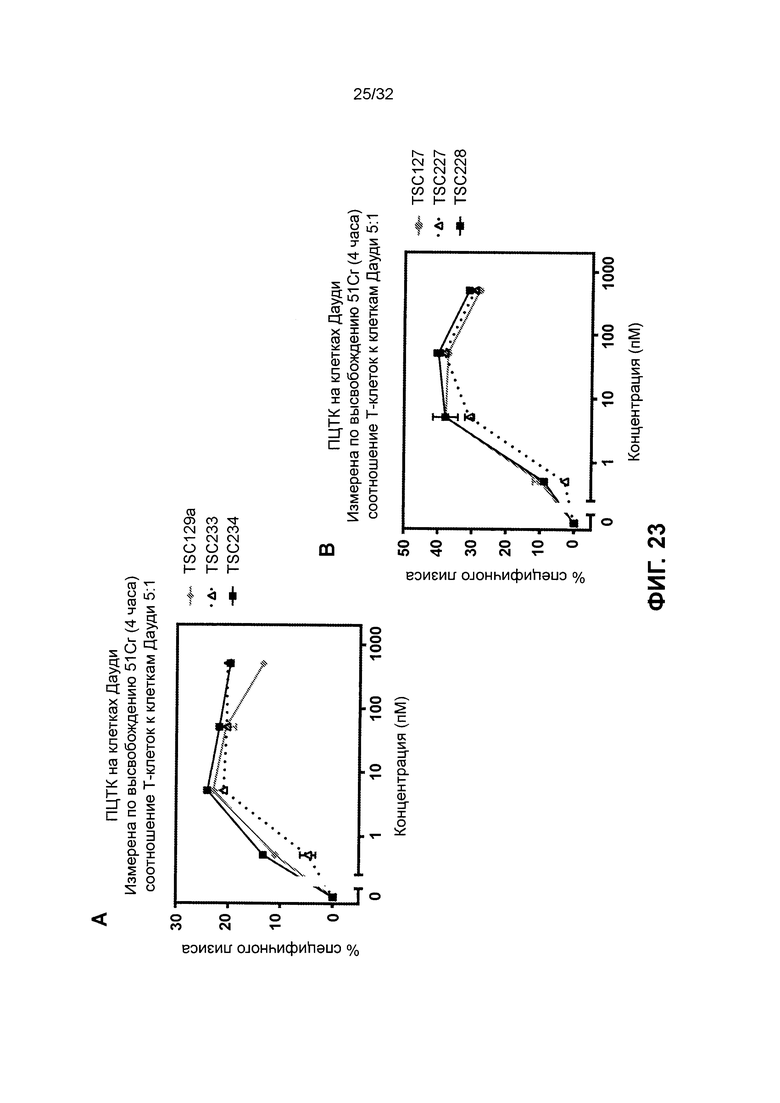
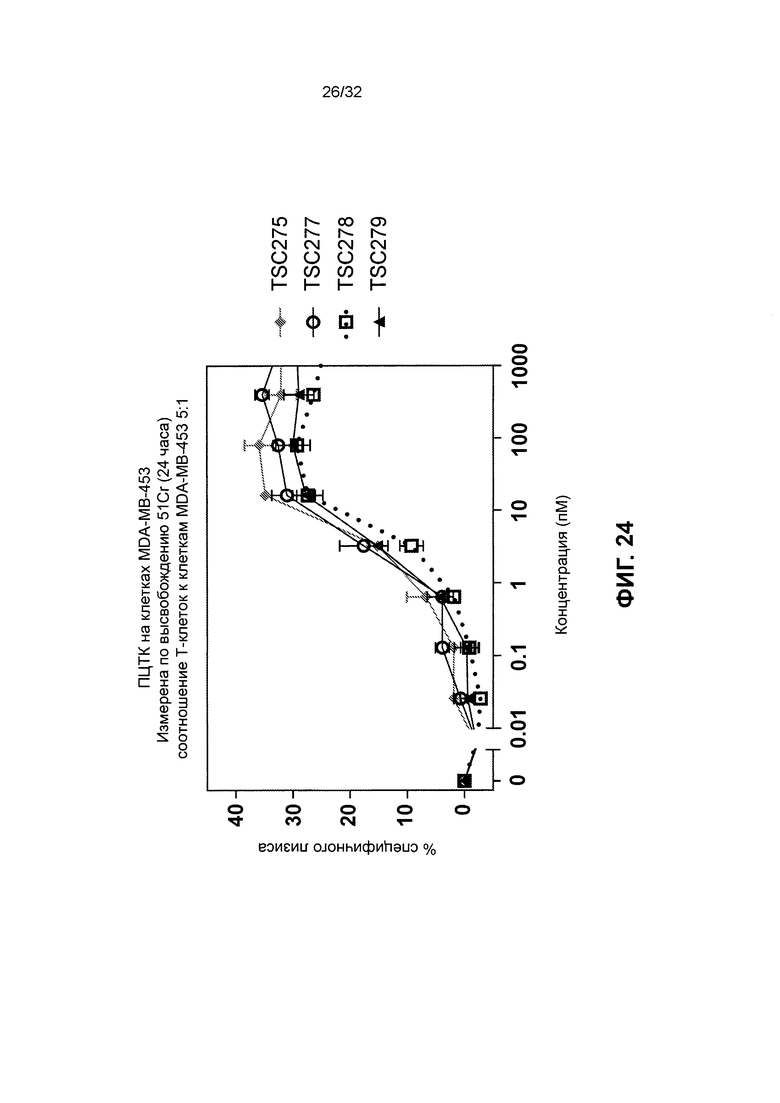
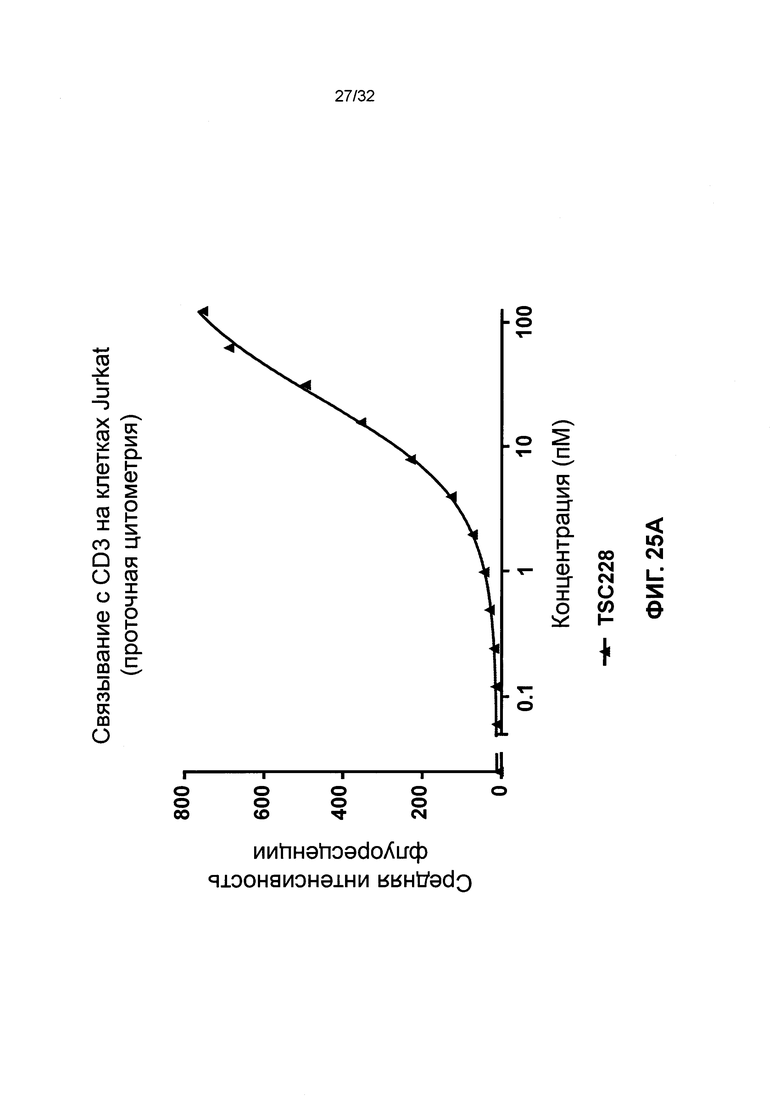
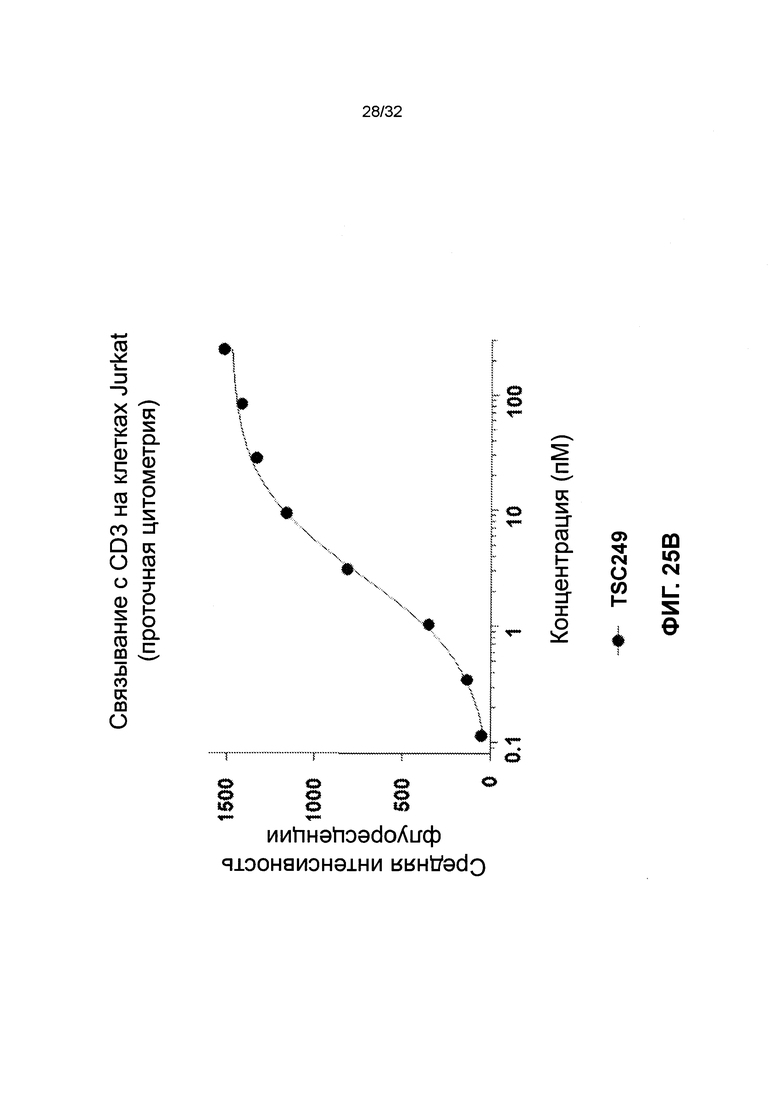
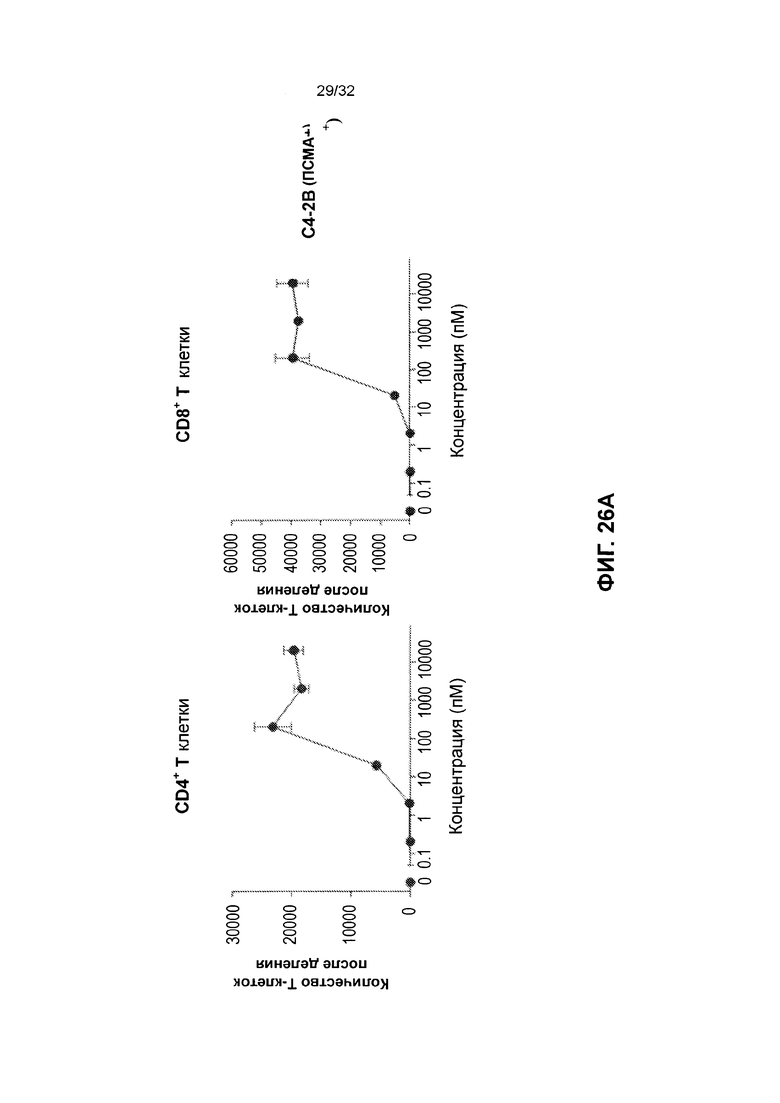

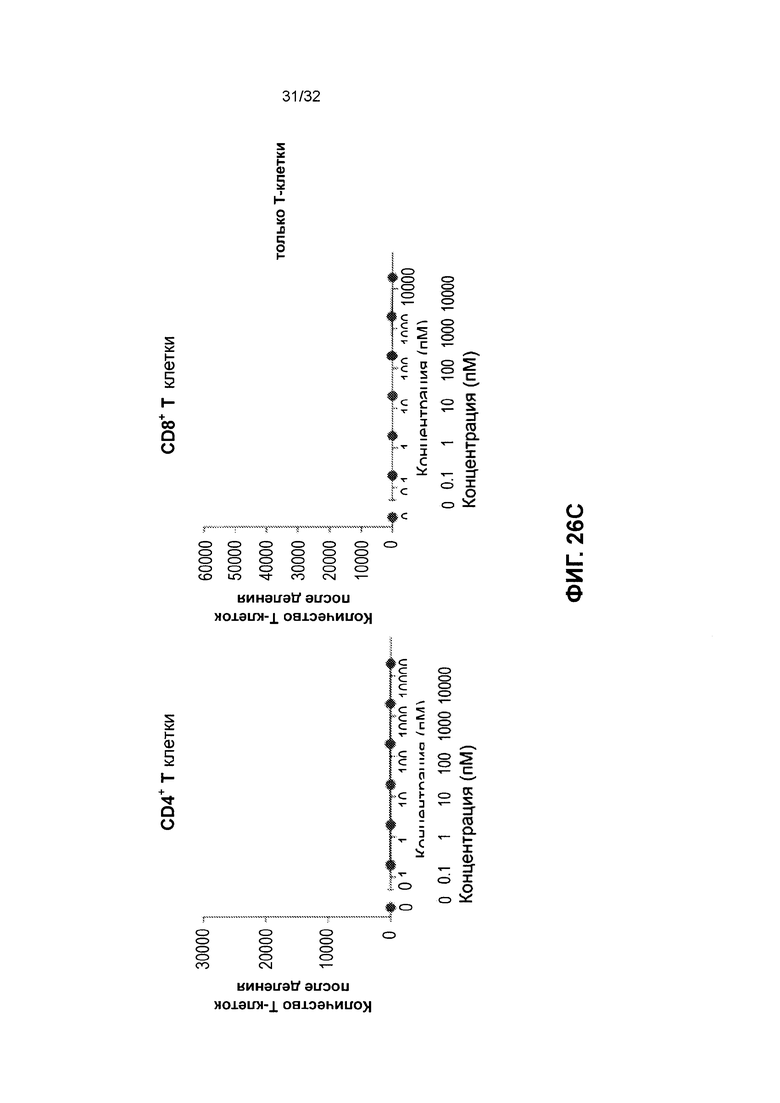
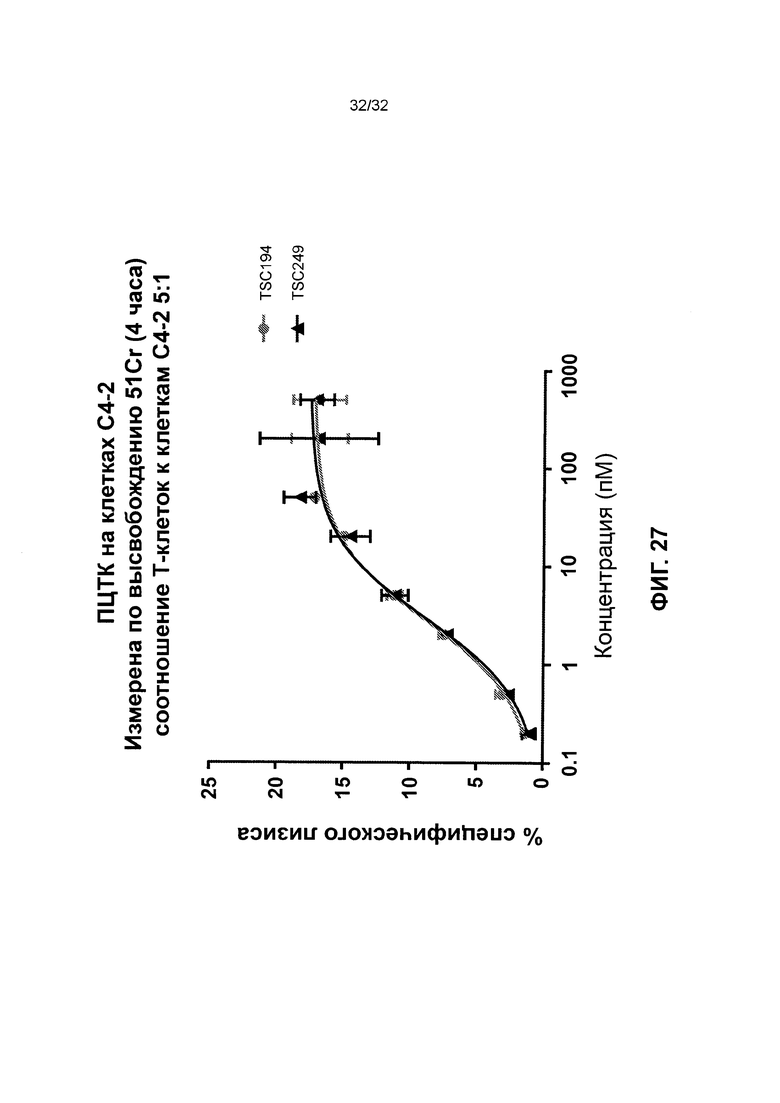
Изобретение относится к области биохимии, в частности к полипептиду, специфически связывающемуся с CD3, а также нуклеиновой кислоте, его кодирующей. Причем указанный полипептид содержит домен связывания с CD3 со сниженной изоэлектрической точкой. Также раскрыты вектор экспрессии и клетка-хозяин, содержащие вышеуказанную нуклеиновую кислоту. Изобретение также относится к фармацевтической композиции для лечения рака или аутоиммунного заболевания, а также к способу лечения опухоли, экспрессирующей простатоспецифический мембранный антиген с использованием вышеуказанного полипептида. Изобретение позволяет эффективно связывать как комплекс рецептора Т-клеток на Т-клетках, так и антиген опухоли, чтобы индуцировать зависимую от мишени цитотоксичность, активацию и пролиферацию Т-клеток. 13 н. и 47 з.п. ф-лы, 27 ил., 10 табл., 16 пр.
1. Полипептид, специфически связывающийся с CD3, причем указанный полипептид содержит домен связывания с CD3, причем указанный домен связывания с CD3 имеет сниженную изоэлектрическую точку по сравнению с изоэлектрической точкой домена связывания с последовательностью аминокислот SEQ ID NO: 41;
причем домен связывания с CD3 содержит гуманизированный вариабельный участок тяжелой цепи (VH) и гуманизированный вариабельный участок легкой цепи (VL), и при этом указанные гуманизированный участок VH и гуманизированный участок VL содержат каркасные участки; и
при этом участок VH содержит SEQ ID NO: 28 и участок VL содержит SEQ ID NO: 34 или последовательность аминокислот, которая идентична SEQ ID NO: 34 на от примерно 90% до менее 100%.
2. Полипептид по п. 1, отличающийся тем, что две или более аминокислоты в каркасных участках VL модифицированы по сравнению с SEQ ID NO: 34 посредством замены положительно заряженных аминокислот нейтральными аминокислотами и/или замены нейтральных аминокислот отрицательно заряженными аминокислотами.
3. Полипептид по п. 1, отличающийся тем, что по меньшей мере один из каркасных участков содержит последовательность аминокислот из последовательности IgG зародышевой линии человека.
4. Полипептид по п. 3, отличающийся тем, что последовательность IgG зародышевой линии человека содержит SEQ ID NO: 43.
5. Полипептид по п. 3, отличающийся тем, что последовательность IgG зародышевой линии человека содержит SEQ ID NO: 44.
6. Полипептид, специфически связывающийся с CD3, причем указанный полипептид содержит домен связывания с CD3 и предшарнирный участок, причем указанный домен связывания с CD3 имеет сниженную изоэлектрическую точку по сравнению с изоэлектрической точкой домена связывания с последовательностью аминокислот SEQ ID NO: 41;
причем домен связывания с CD3 содержит гуманизированный вариабельный участок тяжелой цепи (VH) и гуманизированный вариабельный участок легкой цепи (VL), и при этом указанные гуманизированный участок VH и гуманизированный участок VL содержат каркасные участки; и
при этом участок VH содержит SEQ ID NO: 28 и участок VL содержит SEQ ID NO: 34 или последовательность аминокислот, которая идентична SEQ ID NO: 34 на от примерно 90% до менее 100%.
7. Полипептид по п. 6, отличающийся тем, что предшарнирный участок имеет последовательность SSS и указанный полипептид имеет сниженную изоэлектрическую точку по сравнению с изоэлектрической точкой полипептида, идентичного указанному, за исключением того, что он имеет предшарнирный участок, имеющий последовательность RRT.
8. Полипептид, специфически связывающийся с CD3, причем указанный полипептид содержит домен связывания с CD3 с каркасными участками, предшарнирный участок и шарнирный участок,
причем домен связывания с CD3 специфически связывается с CD3;
при этом домен связывания с CD3 имеет сниженную изоэлектрическую точку по сравнению с изоэлектрической точкой домена связывания с последовательностью аминокислот SEQ ID NO: 41;
при этом домен связывания с CD3 содержит гуманизированный участок VH и гуманизированный участок VL;
при этом участок VH содержит SEQ ID NO: 28 и участок VL содержит SEQ ID NO: 34 или последовательность аминокислот, которая идентична SEQ ID NO: 34 на от примерно 90% до менее 100%.
9. Полипептид по п. 8, отличающийся тем, что по меньшей мере две или более аминокислоты в каркасных участках VL модифицированы по сравнению с SEQ ID NO: 34 путем замены положительно заряженных аминокислот нейтральными аминокислотами и/или замены нейтральных аминокислот отрицательно заряженными аминокислотами.
10. Полипептид по п. 2 или 8, отличающийся тем, что по меньшей мере четыре или более аминокислоты в каркасных участках VL модифицированы по сравнению с SEQ ID NO: 34 путем замены положительно заряженных аминокислот нейтральными аминокислотами и/или замены нейтральных аминокислот отрицательно заряженными аминокислотами.
11. Полипептид по п. 2 или 8, отличающийся тем, что от 3 до 5 аминокислот в каркасных участках VL модифицированы по сравнению с SEQ ID NO: 34 путем замены положительно заряженных аминокислот нейтральными аминокислотами и/или замены нейтральных аминокислот отрицательно заряженными аминокислотами.
12. Полипептид по п. 2 или 8, отличающийся тем, что от 3 до 10 аминокислот в каркасных участках VL модифицированы по сравнению с SEQ ID NO: 34 путем замены положительно заряженных аминокислот нейтральными аминокислотами и/или замены нейтральных аминокислот отрицательно заряженными аминокислотами.
13. Полипептид по п. 8, отличающийся тем, что его предшарнирный участок содержит последовательность аминокислот SSS или SST.
14. Полипептид, специфически связывающийся с CD3, причем указанный полипептид содержит домен связывания с CD3, шарнирный участок и константный участок, причем указанный домен связывания с CD3 имеет сниженную изоэлектрическую точку по сравнению с изоэлектрической точкой домена связывания с последовательностью аминокислот SEQ ID NO: 41;
причем домен связывания с CD3 содержит гуманизированный вариабельный участок тяжелой цепи (VH) и гуманизированный вариабельный участок легкой цепи (VL), и при этом указанные гуманизированный участок VH и гуманизированный участок VL содержат каркасные участки; и
при этом участок VH содержит SEQ ID NO: 28 и участок VL содержит SEQ ID NO: 34 или последовательность аминокислот, которая идентична SEQ ID NO: 34 на от примерно 90% до менее 100%.
15. Полипептид по любому из пп. 1-14, причем указанный полипептид имеет эмпирическую изоэлектрическую точку, которая на 0,5-2,5 единиц pi ниже, чем эмпирическая изоэлектрическая точка полипептида SEQ ID NO: 4.
16. Полипептид по п. 1, отличающийся тем, что указанный полипептид представляет собой гуманизированное антитело, одноцепочечный Fv (scFv) или SMIP.
17. Полипептид по п. 14, отличающийся тем, что указанный полипептид содержит от аминоконца к карбоксиконцу домен связывания с CD3, шарнирный участок и константный участок.
18. Полипептид по п. 14, отличающийся тем, что указанный полипептид содержит от аминоконца к карбоксиконцу константный участок, шарнирный участок и домен связывания с CD3.
19. Полипептид, специфически связывающийся с CD3, причем указанный полипептид содержит домен связывания с CD3, шарнирный участок и константный участок и второй домен связывания, причем указанный домен связывания с CD3 имеет сниженную изоэлектрическую точку по сравнению с изоэлектрической точкой домена связывания с последовательностью аминокислот SEQ ID NO: 41;
причем домен связывания с CD3 содержит гуманизированный вариабельный участок тяжелой цепи (VH) и гуманизированный вариабельный участок легкой цепи (VL), и при этом указанные гуманизированный участок VH и гуманизированный участок VL содержат каркасные участки; и
при этом участок VH содержит SEQ ID NO: 28 и участок VL содержит SEQ ID NO: 34 или последовательность аминокислот, которая идентична SEQ ID NO: 34 на от примерно 90% до менее 100%.
20. Полипептид по п. 19, отличающийся тем, что указанный связывающий полипептид содержит от аминоконца к карбоксиконцу домен связывания с CD3, шарнирный участок, константный участок и второй домен связывания.
21. Полипептид по п. 19, отличающийся тем, что указанный полипептид содержит от аминоконца к карбоксиконцу второй домен связывания, шарнирный участок, константный участок и домен связывания с CD3.
22. Полипептид, специфически связывающийся с CD3, причем указанный полипептид содержит домен связывания с CD3, шарнирный участок, константный участок и домен гетеродимеризации, причем указанный домен связывания с CD3 имеет сниженную изоэлектрическую точку по сравнению с изоэлектрической точкой домена связывания с последовательностью аминокислот SEQ ID NO: 41;
причем домен связывания с CD3 содержит гуманизированный вариабельный участок тяжелой цепи (VH) и гуманизированный вариабельный участок легкой цепи (VL), и
при этом указанные гуманизированный участок VH и гуманизированный участок VL содержат каркасные участки; и
при этом участок VH содержит SEQ ID NO: 28 и участок VL содержит SEQ ID NO: 34 или последовательность аминокислот, которая идентична SEQ ID NO: 34 на от примерно 90% до менее 100%.
23. Полипептид по п. 22, отличающийся тем, что указанный полипептид содержит от аминоконца к карбоксиконцу домен связывания с CD3, шарнирный участок, константный участок и домен гетеродимеризации.
24. Полипептид, специфически связывающийся с CD3, причем указанный полипептид содержит домен связывания с CD3, шарнирный участок, константный участок, домен гетеродимеризации и второй домен связывания, причем указанный домен связывания с CD3 имеет сниженную изоэлектрическую точку по сравнению с изоэлектрической точкой домена связывания с последовательностью аминокислот SEQ ID NO: 41;
причем домен связывания с CD3 содержит гуманизированный вариабельный участок тяжелой цепи (VH) и гуманизированный вариабельный участок легкой цепи (VL), и при этом указанные гуманизированный участок VH и гуманизированный участок VL содержат каркасные участки; и
при этом участок VH содержит SEQ ID NO: 28 и участок VL содержит SEQ ID NO: 34 или последовательность аминокислот, которая идентична SEQ ID NO: 34 на от примерно 90% до менее 100%.
25. Полипептид по п. 24, отличающийся тем, что второй домен связывания связывается или взаимодействует с молекулой-мишенью и указанный полипептид индуцирует Т-клеточную цитотоксичность.
26. Полипептид по п. 25, отличающийся тем, что второй домен связывания связывается или взаимодействует с ассоциированным с опухолью антигеном.
27. Полипептид по п. 26, отличающийся тем, что указанный полипептид индуцирует лизис клеток опухоли Т-клетками.
28. Полипептид по п. 26, отличающийся тем, что указанный полипептид индуцирует поликлональную активацию Т-клеток и экспансию вблизи опухоли.
29. Полипептид по любому из пп. 26-28, отличающийся тем, что ассоциированный с опухолью антиген выбран из группы, состоящей из RON, c-Met, СЕАСАМ-6, ПСМА, ЕрСАМ, СЕА, РСТА-1, STEAP-1, STEAP-2, PSCA, PSA, PAP, ALCAM (CD166), РЕСАМ-1, EphA2, CD151, CA-125/MUC16, MUC-1, MAGE-1, TROP2, IGF1R, TGFBR2, GHRHR, GHR, IL-6R, gp130, TNFR2, OSMRβ, Patched-1, Frizzled, Robol, LTβR, CD25, CD26, CD27, CD30, CD33, CD44, CD44v6, CD63, CD80, CD81, CD86, CD100, CD151, CXCR4, CCR5, HER-2/ErbB1, HER-3/ErbB3, HER-4/ErbB4, EGFR/ErbB1, изоформы EGFRvIII, MUC2, MUC3, MUC4, MUC5AC, MUC5b, MUC7, βhCG, Lewis-Y, ганглиозида GD3, 9-О-ацетил-CD3, GM2, Globo H, фукозила GM1, поли SA, GD2, карбоангидразы IX (MN/CA IX), ежа Соник (SHH), Wue-1, антигена клеток плазмы, (связанного с мембраной) IgE, хондроитина сульфат протеогликана меланомы (ХСПМ), CCR8, прекурсора опухолевого некротического фактора-альфа, мезотелина, антигена А33, Ly-6; десмоглеина 4, Е-кадгерин неоэпитопа, фетального рецептора ацетилхолина, маркера СА19-9, рецептора мюллеровой ингибирующей субстанции (МИС) типа II, sTn (сиалированного антигена Tn; TAG-72), ААФ (антигена активации фибробластов), эндосиалина, LG, SAS, ВСМА, TWEAKR/Fn14, FGFR4, VEGFR1, VEGFR2, SSX1 и SSX2.
30. Полипептид по любому из пп. 1-29, отличающийся тем, что домен связывания с CD3 связывается или взаимодействует с субъединицей CD3ɛ комплекса рецептора Т-клеток на Т-клетках.
31. Полипептид по любому из пп. 1-30, отличающийся тем, что указанный полипептид индуцирует интернализацию комплекса рецептора Т-клеток.
32. Полипептид по любому из пп. 1-31, отличающийся тем, что домен связывания с CD3 представляет собой гуманизированный домен связывания с CD3, происходящий от мышиного моноклонального антитела Cris-7 или HuM291.
33. Полипептид по п. 32, отличающийся тем, что домен связывания с CD3 содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи мышиного антитела Cris-7 или HuM291.
34. Полипептид по п. 1, отличающийся тем, что домен связывания с CD3 содержит участок VL, выбранный из группы, состоящей из SEQ ID NO: 36 и SEQ ID NO: 38.
35. Полипептид по п. 1, отличающийся тем, что участок VL содержит SEQ ID NO: 38.
36. Полипептид по п. 1, отличающийся тем, что указанный полипептид содержит последовательность аминокислот SEQ ID NO: 8.
37. Полипептид по п. 1, отличающийся тем, что домен связывания с CD3 содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, при том что CDR3 тяжелой цепи содержит SEQ ID NO: 51.
38. Полипептид по п. 37, отличающийся тем, что CDR2 тяжелой цепи содержит SEQ ID NO: 50 и CDR3 тяжелой цепи содержит SEQ ID NO: 51, и при этом CDR1 легкой цепи содержит SEQ ID NO: 52, CDR2 легкой цепи содержит SEQ ID NO: 53 и CDR3 легкой цепи содержит SEQ ID NO: 54.
39. Полипептид по п. 14, отличающийся тем, что константный участок содержит домены СН2 и СН3 IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgD или любую их комбинацию; домен СН3 иммуноглобулина IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgD, IgE, IgM или любую их комбинацию или домены СН3 и СН4 иммуноглобулина IgE, IgM или их комбинацию.
40. Полипептид по п. 14, отличающийся тем, что константный участок содержит СН2 и СН3.
41. Полипептид по п. 14, отличающийся тем, что константный участок по существу состоит из домена СН2 и домена СН3.
42. Полипептид по любому из пп. 39-41, отличающийся тем, что константный участок модифицирован таким образом, чтобы уменьшить или исключить эффекторную функцию.
43. Полипептид по любому из пп. 39-41, отличающийся тем, что константный участок модифицирован таким образом, чтобы он не фиксировал комплемент.
44. Полипептид по любому из пп. 39-41, отличающийся тем, что константный участок модифицирован таким образом, чтобы он не связывался с рецепторами Fcγ.
45. Полипептид по п. 44, отличающийся тем, что рецепторы Fcγ выбраны из группы, состоящей из CD16, CD32 и CD64.
46. Полипептид, специфически связывающийся с CD3, причем указанный полипептид содержит домен связывания с CD3, шарнирный участок, константный участок, домен димеризации иммуноглобулина и второй домен связывания, причем указанный домен связывания с CD3 имеет сниженную изоэлектрическую точку по сравнению с изоэлектрической точкой домена связывания с последовательностью аминокислот SEQ IDNO: 41;
причем домен связывания с CD3 содержит гуманизированный вариабельный участок тяжелой цепи (VH) и гуманизированный вариабельный участок легкой цепи (VL), и при этом указанные гуманизированный участок VH и гуманизированный участок VL содержат каркасные участки; и
при этом участок VH содержит SEQ ID NO: 28 и участок VL содержит SEQ ID NO: 34 или последовательность аминокислот, которая идентична SEQ ID NO: 34 на от примерно 90% до менее 100%.
47. Полипептид по п. 46, отличающийся тем, что указанный полипептид образует гомодимер или гетеродимер.
48. Полипептид по п. 2, отличающийся тем, что по меньшей мере одна модификация аминокислоты находится в пределах участка J каппа (Jk) участка VL.
49. Полипептид по п. 48, отличающийся тем, что участок Jk содержит последовательность аминокислот VEIK (SEQ ID NO: 253).
50. Полипептид по п. 24, отличающийся тем, что второй домен связывания содержит последовательность аминокислот, которая по меньшей мере на 90%, по меньшей мере на 95% или 100% идентична последовательности аминокислот, выбранной из группы, состоящей из аминокислот 1-107 и 124-243 SEQ ID NO: 212; аминокислот 1-107 и 124-243 SEQ ID NO: 226; аминокислот 1-107 и 124-243 SEQ ID NO: 216 и 1-111 и 128-251 SEQ ID NO: 196.
51. Полипептид по п. 1, отличающийся тем, что указанный связывающий полипептид содержит последовательность аминокислот, по меньшей мере на 90%, по меньшей мере на 95% или 100% идентичную SEQ ID NO: 57, 59, 61, 63, 65, 67, 69, 71, 186, 190, 192, 194, 196, 198, 200, 202, 204 или 206.
52. Полипептид по п. 1, отличающийся тем, что указанный полипептид представляет собой часть гетеродимера.
53. Полипептид по п. 52, отличающийся тем, что гетеродимер содержит пару одноцепочечных полипептидов, при том что одноцепочечная пара содержит последовательности аминокислот, которые по меньшей мере на 90%, по меньшей мере на 95% или 100% идентичны паре SEQ ID NO: 212 и 218.
54. Полипептид по п. 1, отличающийся тем, что степень идентичности домена связывания с CD3 SEQ ID NO: 41 составляет от около 90% до около 99%.
55. Нуклеиновая кислота, кодирующая любой из полипептидов по любому из пп. 1-54.
56. Нуклеиновая кислота по п. 55, причем указанная нуклеиновая кислота содержит SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 19, SEQ ID NO: 27 или SEQ ID NO: 37.
57. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 55 или 56.
58. Рекомбинантная клетка-хозяин, содержащая вектор экспрессии по п. 57, причем указанная клетка-хозяин экспрессирует любой из полипептидов по пп. 1-54.
59. Фармацевтическая композиция для лечения рака или аутоиммунного заболевания, содержащая эффективное количество полипептида по любому из пп. 1-54 и фармацевтически приемлемый носитель, разбавитель или вспомогательное вещество.
60. Способ лечения опухоли, экспрессирующей простатоспецифический мембранный антиген (ПСМА), включающий введение терапевтически эффективного количества биспецифического конструкта, содержащего ПСМА-связывающий полипептид и полипептид по любому из пп. 1-54, пациенту, нуждающемуся в этом.
| WO2010042904 A2, 15.04.2010 | |||
| WO2011090762 A1, 28.07.2011 | |||
| FLAVIA SERPIERI ET AL | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Железобетонный фасонный камень для кладки стен | 1920 |
|
SU45A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| FENG ZHOU ET AL | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Способ очищения сернокислого глинозема от железа | 1920 |
|
SU47A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| ЛЕКАРСТВЕННОЕ СРЕДСТВО ДЛЯ ПРЕДОТВРАЩЕНИЯ ОТТОРЖЕНИЯ ТРАНСПЛАНТАТА, МОНОКЛОНАЛЬНОЕ АНТИТЕЛО К CD3-АНТИГЕНУ Т-ЛИМФОЦИТОВ ЧЕЛОВЕКА, ГИБРИДОМА И СПОСОБ ЛЕЧЕНИЯ БОЛЬНЫХ, ИМЕЮЩИХ РЕАКЦИЮ ОСТРОГО ОТТОРЖЕНИЯ ТРАНСПЛАНТАТА ПОСЛЕ ПЕРЕСАДКИ ПОЧКИ | 2000 |
|
RU2179862C1 |

