Область техники
Данное изобретение относится к способу интеграции генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего, включающему введение по меньшей мере одного экспрессионного вектора, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, в суспензионную клетку млекопитающего; и к способу получения белка, включающему суспензионное культивирование суспензионной клетки млекопитающего, которая продуцирует белок, суспензионной клетки млекопитающего, которая экспрессирует белок; а также к экспрессионному вектору, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
Уровень техники
Получение экзогенных белков методами рекомбинантных ДНК используется в различных отраслях промышленности, таких как фармацевтическая промышленность и пищевая промышленность. В большинстве случаев рекомбинантные белки получают путем введения экспрессионного вектора, содержащего нуклеотидную последовательность, кодирующую интересующий белок, в хозяина, например, Escherichia coli, дрожжи, клетку насекомого, растительную клетку и животную клетку, отбора трансформанта, в котором экспрессионный вектор интегрирован в хромосому, и последующего культивирования трансформированной линии клеток в соответствующих условиях культивирования.
Однако, чтобы создать хозяина, способного эффективно продуцировать экзогенный белок, необходимо отбирать клетку-хозяина, обладающую хорошей продуктивностью для каждого представляющего интерес белка, так что существует потребность в дальнейших технических инновациях в области методов производства экзогенного белка для каждого хозяина.
В бактериальных системах, таких как Escherichia coli, и дрожжевых системах, отличающихся от животных клеток, во многих случаях трудно добиться посттрансляционных модификаций, таких как модификации сахарной цепи, и, таким образом, возникает проблема с получением белка, обладающего присущей ему активностью.
Поскольку в системе клеток насекомых продуцируемый белок подвергается посттрансляционным модификациям, таким как фосфорилирование и добавление сахарных цепей, эта система имеет то преимущество, что может экспрессироваться белок, обладающий присущей ему физиологической активностью. Однако, поскольку структура сахарной цепи секретируемого белка отличается от таковой в клетках млекопитающих, антигенность и тому подобное становится проблемой, когда белок применяют в фармацевтических целях.
Кроме того, поскольку в системе клеток насекомых используют рекомбинантный вирус при введении экзогенного гена, с точки зрения безопасности существует проблема, связанная с необходимостью инактивации и локализации вируса.
В системе животных клеток посттрансляционные модификации, такие как фосфорилирование, добавление сахарной цепи и фолдинг, можно проводить с белками высших животных, включая человека, в большей степени аналогично тому, как это происходит с белками, продуцируемыми в живом организме. Такие точные посттрансляционные модификации необходимы для воссоздания в рекомбинантном белке физиологической активности, изначально присущей белку, и систему производства белка, в которой в качестве хозяина используется клетка млекопитающего, как правило, применяют для получения фармацевтических продуктов и тому подобного, когда необходима такая физиологическая активность.
Однако система экспрессии белка, в которой в качестве хозяина используется клетка млекопитающего, как правило, обладает низкой продуктивностью и, кроме того, во многих случаях создает проблему стабильности введенных генов. Повышение продуктивности для белка при использовании культуры клеток млекопитающих в качестве хозяина не только очень важно при производстве медикаментов для лечения, диагностических средств и тому подобного, но и вносит значительный вклад в их исследование и разработку. Таким образом, необходимо в срочном порядке разработать систему экспрессии генов, которая легко позволяет получать линию клеток с высокой продуктивностью при использовании культуры клеток млекопитающих, в частности, клеток яичника китайского хомячка (клетки CHO), в качестве хозяина.
Транспозон представляет собой мобильный генетический элемент, который может перемещаться из одного локуса в другой локус на хромосоме. Транспозон является сильным инструментом для исследований в области молекулярной биологии и генетики и используется для таких целей, как мутагенез, улавливание гена и получение трансгенных особей, у насекомых или нематод (например, Drosophila melanogaster или Caenorhabditis elegans) и растений. Однако разработка такого метода отставала в случае позвоночных животных, включая клетки млекопитающих.
Тем не менее, в последние годы появились сообщения о транспозонах, которые обладают активностью также и в организме позвоночных животных, и некоторые из них, как было показано, обладают активностью в клетках млекопитающих, например, клетках мыши и человека. Типичные примеры включают транспозоны Tol1 (патентная ссылка 1) и Tol2 (непатентная ссылка 1), клонированные из оризии (рыбы семейства карпозубых), транспозон «спящая красавица», восстановленный из неавтономного транспозона, существующего в геноме лососевых рыб (непатентная ссылка 2), искусственный транспозон «лягушачий принц» (непатентная ссылка 3), полученный из лягушки, и транспозон piggyBac (непатентная ссылка 4), полученный из насекомого.
Эти ДНК транспозоны использовали для мутагенеза, улавливания генов, получения трансгенных особей, экспрессии устойчивых к лекарственным средствам белков и тому подобного, в качестве инструмента для введения генов с целью привнесения нового фенотипа в геном клетки млекопитающего (непатентные ссылки 5-12).
В случае насекомых был изучен способ, в котором экзогенный ген вводят в хромосому шелкопряда, используя транспозон piggyback, полученный из чешуекрылого насекомого, для экспрессии белка, кодируемого указанным экзогенным геном, и был описан способ получения белка с использованием вышеуказанного метода (патентная ссылка 2).
Однако, поскольку интересующий белок не экспрессируется на достаточном уровне и производится во всем теле шелкопряда, это вызывает экономические проблемы в связи с необходимостью усовершенствованных методов очистки для извлечения экспрессированного экзогенного белка в высокоочищенной форме из жидкостей организма, содержащих большое количество примесных белков.
Кроме того, известен пример того, как белок, связанный с устойчивостью к G418, экспрессируется в клетке млекопитающего с использованием полученного из оризии транспозона Tol2 (непатентные ссылки 12 и 13).
В случае получения белкового лекарственного средства для медицинского применения с использованием культивируемой клетки млекопитающего важно, чтобы в процессе его производства отсутствовал компонент животного происхождения, с целью предотвращения неожиданного загрязнения неизвестным вирусом или патогенным полипептидом. Клетку CHO наиболее часто используют в качестве животной клетки для получения белкового лекарственного средства, и в результате исследований последних лет также была создана суспензионная линия клеток CHO, которую можно культивировать в безопасной среде без применения сыворотки или компонента животного происхождения. Однако продуктивность клеточной линии, в которую ген был введен в бессывороточных или безбелковых условиях, ограничена наполовину по сравнению с клеточной линией, в которую ген был введен в условиях наличия сыворотки (непатентная литература 14). Показано, что трансдукция генов в бессывороточных или безбелковых условиях технически сложна.
Как правило, селектируемый маркер для скрининга клеток, экспрессирующих интересующий белок, находится в том же экспрессионном векторе. Это основано на предположении, что существует область, в которой ген, присутствующий в геноме, легко экспрессируется и область, в которой ген, присутствующий в геноме, экспрессируется плохо (это называют эффектами положения, непатентная литература 15), и что представляющий интерес белок также экспрессируется, когда экспрессируется селектируемый маркер.
С другой стороны, известно также, что, когда интересующий белок состоит из двух или более полипептидов, например, антитело и тому подобное, каждый полипептид экспрессируется при помощи различных векторов. В случае антитела показано, что продуктивность возрастает, когда экспрессия тяжелой цепи антитела выше, чем экспрессия легкой цепи (непатентная литература 16). Поскольку предсказано, что экспрессия тяжелой цепи и легкой цепи становится постоянной на одном и том же векторе. Стало возможным получение линии клеток, которые экспрессируют тяжелую цепь и легкую цепь в оптимальном соотношении, путем намеренной экспрессии тяжелой цепи и легкой цепи при помощи различных векторов для достижения высокой продуктивности. Однако, если белок экспрессируется с использованием двух или более различных векторов, также необходимы два или более селектируемых маркера.
В качестве способа преодоления этого, сообщалось о случае, когда ген dhfr, исходно соответствующий одной полипептидной цепи, был разделен на две полипептидных цепи, и одна из них была размещена на экспрессионном векторе тяжелой цепи, а другая была размещена на экспрессионном векторе легкой цепи (непатентная литература 17).
Однако клетка, описанная в непатентной литературе 17, является клеткой CHO, при этом клетка зависима от наличия белкового компонента, добавленного к среде, и, как описано выше, существует вероятность того, что эффективность введения гена является высокой в отличие от случая введения гена в бессывороточных или безбелковых условиях. По прогнозам, отбор высокопродуктивной клетки по-прежнему будет труден, если ген введен в бессывороточных или безбелковых условиях, имеющих высокую степень безопасности при отсутствии опасности вирусной инфекции и тому подобного.
Перечень ссылок
Патентная литература
Патентная литература 1: W02008/072540.
Патентная литература 2: Японская опубликованная нерассмотренная патентная заявка № 2001-532188.
Непатентная литература
Непатентная литература 1: Nature 383, 30 (1996).
Непатентная литература 2: Cell 91, 501-510 (1997).
Непатентная литература 3: Nucleic Acids Res, 31, 6873-6881 (2003).
Непатентная литература 4: Insect Mol. Biol. 5, 141-151 (1996).
Непатентная литература 5: Genetics. 166, 895-899 (2004).
Непатентная литература 6: PLoS Genet, 2, el69 (2006).
Непатентная литература 7: Proc. Natl. Acad. Sei. USA 95, 10769-10773 (1998).
Непатентная литература 8: Proc. Natl. Acad. Sei. USA 98:6759-6764 (2001).
Непатентная литература 9: Nature 436, 221-22 6 (2005).
Непатентная литература 10: Nucleic Acids Res., 31, 6873-6881 (2003).
Непатентная литература 11: Nucleic Acids Res., 35, e87 (2007).
Непатентная литература 12: Proc Natl. Acad. Sei. USA, 103, 15008-15013 (2006).
Непатентная литература 13: Plos Genetics, 2,1715-1724(2006).
Непатентная литература 14: Biotech. Bioeng. 96, 1118-1126.
Непатентная литература 15: Nature Biotech. 22, 1393-1398.
Непатентная литература 16: Biotech. Bioeng. 96, 337-348 (2007).
Непатентная литература 17: Biotech. Bioeng. 84, 439-444 (2003).
Раскрытие изобретения
Проблемы, решаемые при помощи изобретения
Для получения и анализа интересующего белка необходимо выбрать линию клеток, которые стабильно и на высоком уровне экспрессируют интересующий белок, используя культивируемые клетки животного происхождения. Однако подготовка и культивирование клеток, продуцирующих интересующий белок, требует значительных усилий и времени.
Кроме того, хотя известно, что интересующий белок экспрессируется в клетке млекопитающего с использованием транспозонной последовательности, способ получения клетки, которая способна экспрессировать интересующий белок на высоком уровне и, следовательно, может быть использована в качестве системы производства белка с использованием транспозонной последовательности; способ получения клетки млекопитающего, которая способна продуцировать интересующий белок на высоком уровне с использованием транспозонной последовательности; а также способ получения белка с использованием такой клетки остаются неизвестными.
Как описано выше, существует потребность в экспрессии интересующего белка в большом количестве путем создания системы производства белка, способной продуцировать интересующий белок на высоком уровне с использованием культивируемой клетки млекопитающего эффективно и за короткий период времени. Кроме того, было бы желательно создать продуцирующую клетку, для которой не требуются какие-либо компоненты животного происхождения в период от введения гена до создания продуцирующей клетки.
Таким образом, целями изобретения являются предложение клетки, способной экспрессировать интересующий белок на высоком уровне, которую можно эффективно создавать, и способ получения интересующего белка с помощью данной клетки.
Способы решения проблем
Для решения вышеупомянутых проблем авторы настоящего изобретения провели интенсивные исследования и в результате обнаружили, что интересующий белок можно эффективно получать путем введения по меньшей мере одного экспрессионного вектора, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, в суспензионную клетку млекопитающего; и интеграции генного фрагмента, вставленного между парой (двумя) транспозонных последовательностей, в хромосому клетки млекопитающего. Кроме того, было установлено, что интересующий белок можно эффективно получать, используя такую клетку, и, таким образом, было совершено изобретение.
В частности, изобретение относится к следующему:
1. Способ получения интересующего белка, включающий введение по меньшей мере одного экспрессионного вектора, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, в суспензионную клетку млекопитающего; интеграцию генного фрагмента, содержащего ДНК, кодирующую интересующий белок, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего для получения клетки млекопитающего, которая экспрессирует интересующий белок; и суспензионное культивирование клетки млекопитающего.
2. Способ получения интересующего белка, включающий следующие этапы (A)-(C):
(A) этап одновременного введения следующих экспрессионных векторов (a) и (b) в суспензионную клетку млекопитающего:
(a) по меньшей мере один экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) экспрессионный вектор, который содержит ДНК, кодирующую транспозазу, которая узнает транспозонные последовательности и обладает способностью перемещать генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому;
(B) этап получения суспензионной клетки млекопитающего, которая экспрессирует интересующий белок в результате временной экспрессии транспозазы с экспрессионного вектора, введенного в суспензионную клетку млекопитающего на этапе (A) с целью интеграции генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего, и
(C) этап суспензионного культивирования суспензионной клетки млекопитающего, которая экспрессирует интересующий белок, полученной на этапе (B), для получения интересующего белка.
3. Способ получения суспензионной клетки млекопитающего, которая экспрессирует интересующий белок, включающий введение по меньшей мере одного экспрессионного вектора, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, в суспензионную клетку млекопитающего и интеграцию генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего.
4. Способ, описанный в любом из вышеприведенных пунктов 1-3, в котором по меньшей мере один из экспрессионных векторов, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, представляет собой экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
5. Способ, описанный в любом из вышеприведенных пунктов 1-4, включающий введение экспрессионного вектора, который содержит генный фрагмент, содержащий селектируемый маркер, и содержит пару транспозонных последовательностей на обоих концах генного фрагмента, в клетку млекопитающего в дополнение к экспрессионному вектору, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
6. Способ, описанный в любом из вышеприведенных пунктов 1-5, в котором ДНК, кодирующая интересующий белок, представляет собой ДНК, кодирующую антитело.
7. Способ, описанный в вышеприведенном пункте 6, в котором ДНК, кодирующая антитело, представляет собой по меньшей мере одну из ДНК, кодирующей H-цепь антитела, и ДНК, кодирующей L-цепь антитела.
8. Способ, описанный в любом из вышеприведенных пунктов 4-7, в котором экспрессионный вектор, выбранный из следующих пунктов (a)-(d), вводят в суспензионную клетку млекопитающего:
(a) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(c) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и
(d) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь и L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
9. Способ, описанный в любом из вышеприведенных пунктов 1-8, в котором суспензионная клетка млекопитающего представляет собой клетку, способную выживать и пролиферировать в бессывороточной среде.
10. Способ, описанный в любом из вышеприведенных пунктов 1-9, в котором суспензионная клетка млекопитающего представляет собой по меньшей мере клетку, выбранную из суспензионной клетки CHO, при этом клетка CHO адаптирована к суспензионному культивированию, клетки PER.C6, клетки крысиной миеломы YB2/3HL.P2.G11.16Ag.20 (или также называемой YB2/0) и суспензионной клетки мышиной миеломы NS0, адаптированной к суспензионному культивированию.
11. Способ, описанный в вышеприведенном пункте 10, в котором клетка CHO представляет собой по меньшей мере клетку, выбранную из CHO-K1, CHO-K1SV, DUKXB11, CHO/DG44, Pro-3 и CHO-S.
12. Способ, описанный в любом из вышеприведенных пунктов 4-11, в котором ген селектируемого маркера представляет собой ген устойчивости к циклогексимиду.
13. Способ, описанный в вышеприведенном пункте 12, в котором ген устойчивости к циклогексимиду соответствует рибосомному белку.
14. Способ, описанный в любом из вышеприведенных пунктов 1-13, в котором пара транспозонных последовательностей представляет собой нуклеотидные последовательности, полученные из пары транспозонов ДНК-типа, которые функционируют в клетке млекопитающего.
15. Способ, описанный в вышеприведенном пункте 14, в котором нуклеотидные последовательности, полученные из пары транспозонов ДНК-типа, представляют собой нуклеотидные последовательности, полученные из пары транспозонов Tol1, или нуклеотидные последовательности, полученные из пары транспозонов Tol2.
16. Способ, описанный в вышеприведенном пункте 15, в котором нуклеотидные последовательности, полученные из пары транспозонов Tol2, представляют собой нуклеотидную последовательность, содержащую нуклеотидную последовательность, приведенную в SEQ ID NO: 2, и нуклеотидную последовательность, приведенную в SEQ ID NO: 3.
17. Способ, описанный в вышеприведенном пункте 15, в котором нуклеотидные последовательности, полученные из пары транспозонов Tol1, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 14, и нуклеотидную последовательность, приведенную в SEQ ID NO: 15.
18. Суспензионная клетка млекопитающего, которая имеет хромосому, в которую интегрирован генный фрагмент, вставленный между парой транспозонов, и которая продуцирует интересующий белок, получаемый путем одновременного введения по меньшей мере одного экспрессионного вектора (a), который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионного вектора (b), который содержит ДНК, кодирующую транспозазу (трансферазу), способную узнавать транспозонные последовательности и обладающую способностью переносить генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому.
19. Клетка млекопитающего, описанная в вышеприведенном пункте 18, в которой по меньшей мере один из экспрессионного вектора (a), который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, представляет собой экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
20. Клетка млекопитающего, описанная в вышеприведенном пункте 18 или 19, которая представляет собой клетку, полученную в результате дополнительного введения экспрессионного вектора (c), который содержит генный фрагмент, содержащий ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, в дополнение к экспрессионным векторам (a) и (b).
21. Клетка млекопитающего, описанная в любом из вышеприведенных пунктов 18-20, в которой ДНК, кодирующая интересующий белок, представляет собой ДНК, кодирующую антитело.
22. Клетка млекопитающего, описанная в вышеприведенном пункте 21, в которой ДНК, кодирующая антитело, представляет собой по меньшей мере одну из ДНК, кодирующей H-цепь антитела, и ДНК, кодирующей L-цепь антитела.
23. Клетка млекопитающего, описанная в любом из вышеприведенных пунктов 18-22, в которую введен экспрессионный вектор, выбранный из следующих (a)-(d):
(a) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(c) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и
(d) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь и L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
24. Клетка млекопитающего, описанная в любом из вышеприведенных пунктов 18-23, представляющая собой клетку, способную выживать и пролиферировать в бессывороточной среде.
25. Клетка млекопитающего, описанная в любом из вышеприведенных пунктов 18-24, представляющая собой любую суспензионную клетку млекопитающего, выбранную из суспензионной клетки CHO, при этом клетка CHO адаптирована к суспензионному культивированию, клетки PER.C6, клетки крысиной миеломы YB2/3HL.P2.G11.16Ag.20 (или также называемой YB2/0) и суспензионной клетки мышиной миеломы NS0, адаптированной к суспензионному культивированию.
26. Клетка млекопитающего, описанная в вышеприведенном пункте 25, при этом клетка CHO представляет собой любую клетку, выбранную из CHO-K1, CHO-K1SV, DUKXB11, CHO/DG44, Pro-3 и CHO-S.
27. Клетка млекопитающего, описанная в любом из вышеприведенных пунктов 19-26, в которой ген селектируемого маркера представляет собой ген устойчивости к циклогексимиду.
28. Клетка млекопитающего, описанная в вышеприведенном пункте 27, в которой ген устойчивости к циклогексимиду представляет собой ген, кодирующий мутант человеческого рибосомного белка L36a.
29. Клетка млекопитающего, описанная в любом из вышеприведенных пунктов 19-28, в которой пара транспозонных последовательностей представляет собой нуклеотидные последовательности, полученные из пары транспозонов ДНК-типа, которые функционируют в клетке млекопитающего.
30. Клетка млекопитающего, описанная в вышеприведенном пункте 29, в которой нуклеотидные последовательности, полученные из пары транспозонов ДНК-типа, представляют собой нуклеотидные последовательности, полученные из пары транспозонов Tol1, или нуклеотидные последовательности, полученные из пары транспозонов Tol2.
31. Клетка млекопитающего, описанная в вышеприведенном пункте 30, в которой нуклеотидные последовательности, полученные из пары транспозонов Tol2, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 2, и нуклеотидную последовательность, приведенную в SEQ ID NO: 3.
32. Клетка млекопитающего, описанная в вышеприведенном пункте 30, в которой нуклеотидные последовательности, полученные из пары транспозонов Tol1, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 14, и нуклеотидную последовательность, приведенную в SEQ ID NO: 15.
33. Экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
34. Экспрессионный вектор, описанный в вышеприведенном пункте 33, в котором пара транспозонных последовательностей представляют собой нуклеотидные последовательности, полученные из пары транспозонов Tol1, или нуклеотидные последовательности, полученные из пары транспозонов Tol2.
35. Экспрессионный вектор для белка, описанный в вышеприведенном пункте 34, в котором нуклеотидные последовательности, полученные из пары транспозонов Tol2, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 2, и нуклеотидную последовательность, приведенную в SEQ ID NO: 3.
36. Экспрессионный вектор, описанный в вышеприведенном пункте 34, в котором нуклеотидные последовательности, полученные из пары транспозонов Tol1, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 14, и нуклеотидную последовательность, приведенную в SEQ ID NO: 15.
Эффект изобретения
Согласно способу получения белка по изобретению, интересующий белок можно эффективно производить с использованием суспензионной клетки млекопитающего. Кроме того, клетку по настоящему изобретению можно использовать в качестве продуцирующей клетки для получения рекомбинантного белка или рекомбинантного полипептида с высокой эффективностью.
Краткое описание чертежей
На фигуре 1 представлено схематическое изображение транспозонного вектора для экспрессии антитела против М2 человеческого вируса гриппа. Tol2-L представляет транспозон Tol2 с левого края (SEQ ID NO: 2), Tol2-R представляет транспозон Tol2 с правого края (SEQ ID NO: 3), CMV представляет промотор CMV, поли-A представляет сайт полиаденилирования, Hc представляет кДНК H-цепи человеческого антитела, Lc представляет кДНК L-цепи человеческого антитела и CHX-r представляет ген устойчивости к циклогексимиду.
На фигуре 2 представлено схематическое изображение экспрессионного вектора для антитела против М2 человеческого вируса гриппа. CMV представляет промотор CMV, поли-A представляет сайт полиаденилирования, Hc представляет кДНК H-цепи человеческого антитела, Lc представляет кДНК L-цепи человеческого антитела и CHX-r представляет ген устойчивости к циклогексимиду.
На фигуре 3 представлено схематическое изображение экспрессионного вектора для Tol2 транспозазы. CAGGS представляет промотор CAGGS, поли-A представляет сайт полиаденилирования и TPase кДНК представляет кДНК Tol2 транспозазы.
На фигуре 4 приведен результат исследования уровня экспрессии антитела против М2 человеческого вируса гриппа в суспензионной клетке CHO-K1 и прикрепляющейся клетке CHO-K1 при использовании Tol2 транспозонного вектора для экспрессии антитела против М2 человеческого вируса гриппа. На фигуре 4A приведен результат для суспензионной клетки CHO-K1, а на фигуре 4B приведен результат для прикрепляющейся клетки CHO-K1. На обеих фигурах на оси ординат указано количество продуцируемого антитела (мкг/мл), а на оси абсцисс указано количество трансгенных клонов каждой клетки.
На фигуре 5 представлено схематическое изображение Tol1 транспозонного вектора для экспрессии антитела против М2 человеческого вируса гриппа. Tol1-L представляет транспозон Tol1 с левого края (SEQ ID NO: 14), Tol1-R представляет транспозон Tol1 с правого края (SEQ ID NO: 15), CMV представляет промотор CMV, поли-A представляет сайт полиаденилирования, Hc представляет кДНК H-цепи человеческого антитела, Lc представляет кДНК L-цепи человеческого антитела и CHX-r представляет ген устойчивости к циклогексимиду.
На фигуре 6 представлено схематическое изображение экспрессионного вектора для Tol1 транспозазы. CAGGS представляет промотор CAGGS, поли-A представляет сайт полиаденилирования и TPase кДНК представляет кДНК Tol1 транспозазы.
На фигуре 7 приведен результат исследования уровня экспрессии антитела против М2 человеческого вируса гриппа в суспензионной клетке CHO-K1 при использовании Tol1 транспозонного вектора для экспрессии антитела против М2 человеческого вируса гриппа. На оси ординат указано количество продуцируемого антитела (мкг/мл), а на оси абсцисс указано количество трансгенных клонов каждой клетки.
На фигуре 8 представлено схематическое изображение транспозонного вектора для экспрессии тяжелой цепи антитела против CD98 человека. Tol2-L представляет транспозон Tol2 с левого края (SEQ ID NO: 2), Tol2-R представляет транспозон Tol2 с правого края (SEQ ID NO: 3), Pmo представляет промотор вируса лейкемии мышей Молони, поли-A представляет сайт полиаденилирования и Hc представляет кДНК тяжелой цепи антитела против CD98 человека (SEQ ID NO: 18).
На фигуре 9 представлено схематическое изображение транспозонного вектора для экспрессии легкой цепи антитела против CD98 человека. Tol2-L представляет транспозон Tol2 с левого края (SEQ ID NO: 2), Tol2-R представляет транспозон Tol2 с правого края (SEQ ID NO: 3), CMV представляет промотор CMV, поли-A представляет сайт полиаденилирования и Lc представляет кДНК легкой цепи антитела против CD98 человека (SEQ ID NO: 21).
На фигуре 10 представлено схематическое изображение транспозонного вектора для экспрессии гена устойчивости к циклогексимиду. Tol2-L представляет транспозон Tol2 с левого края (SEQ ID NO: 2), Tol2-R представляет транспозон Tol2 с правого края (SEQ ID NO: 3), CMV представляет промотор CMV, поли-A представляет сайт полиаденилирования и CHX-r представляет ген устойчивости к циклогексимиду. (SEQ ID NO: 7).
На фигуре 11 представлено продуцируемое количество антитела против TNFα человека при введении в клетку CHO-K1 тандемного вектора TNFα-CHX или вектора TNFαH-CHX и вектора TNFαL. На оси ординат указана концентрация антитела (мкг/мл), продуцируемого в среду, контрольная схема обозначена как «Контроль», а экспериментальная схема обозначена как «Эксперимент».
На фигуре 12 представлено продуцируемое количество антитела против CD20 человека при введении в клетку CHO-K1 тандемного вектора CD20-CHX или вектора CD20H-CHX и вектора CD20L. На оси ординат указана концентрация антитела (мкг/мл), продуцируемого в среду, контрольная схема обозначена как «Контроль», а экспериментальная схема обозначена как «Эксперимент».
На фигуре 13 представлена структура экспрессионного вектора A для антитела. На фигуре 13 Tol2-L представляет фрагмент ДНК, содержащий последовательность Tol2-L (SEQ ID NO: 2), а Tol2-R представляет фрагмент ДНК, содержащий последовательность Tol2-R (SEQ ID NO: 3), CMV представляет промотор CMV, поли-A представляет сайт полиаденилирования, Hc представляет ген тяжелой цепи антитела к CD98, Lc представляет ген легкой цепи антитела против CD98 человека, SO представляет промотор SV40, SV представляет сайт полиаденилирования SV40 и Neo-r представляет ген устойчивости к неомицину.
Варианты осуществления изобретения
Данное изобретение относится к способу получения интересующего белка, включающему введение по меньшей мере одного экспрессионного вектора, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит транспозонные последовательности на обоих концах генного фрагмента, в суспензионную клетку млекопитающего; интеграцию генного фрагмента, вставленного между парой (двумя) транспозонных последовательностей, в хромосому клетки млекопитающего для получения суспензионной клетки млекопитающего, которая экспрессирует указанный интересующий белок; и суспензионное культивирование клетки млекопитающего.
Примеры способа получения интересующего белка в настоящем изобретении (далее в данном документе называемого способом по настоящему изобретению) включают способ получения интересующего белка, включающий следующие этапы (A)-(C):
(A) этап одновременного введения следующих экспрессионных векторов (a) и (b) в суспензионную клетку млекопитающего:
(a) по меньшей мере один экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) вектор, который содержит ДНК, кодирующую транспозазу, которая узнает транспозонные последовательности и обладает способностью перемещать генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому;
(B) этап временной экспрессии транспозазы с экспрессионного вектора (b), который введен в суспензионную клетку млекопитающего на этапе (A) для интеграции генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего с целью получения суспензионной клетки млекопитающего, которая экспрессирует интересующий белок, и
(C) этап суспензионного культивирования суспензионной клетки млекопитающего, которая экспрессирует интересующий белок, полученной на этапе (B), с целью получения интересующего белка.
Кроме того, настоящее изобретение относится к суспензионной клетке млекопитающего, в которую введен по меньшей мере один экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, для интеграции генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому, и которая продуцирует интересующий белок.
В настоящем изобретении интересующий белок представляет собой белок, состоящий из одного или более полипептидов, и в соответствии со способом по изобретению можно осуществлять любую экспрессию по меньшей мере одного интересующего белка и/или экспрессию по меньшей мере одного полипептида.
По меньшей мере один из экпрессионных векторов, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, означает один или два или более видов экспрессионного вектора. В частности, для экспрессии интересующего белка, состоящего из двух или более полипептидов, необходимо использовать два или более экспрессионных вектора, которые содержат генный фрагмент, содержащий ДНК, кодирующую соответствующие полипептиды, и также содержат пару транспозонных последовательностей на обоих концах генного фрагмента.
Более конкретно, например, если вышеуказанный интересующий белок, состоящий из двух или более полипептидов, представляет собой антитело, H-цепь и L-цепь антитела могут быть экспрессированы с помощью одного экспрессионного вектора или могут быть экспрессированы с помощью двух экспрессионных векторов, вектора, который экспрессирует H-цепь, и вектора, который экспрессирует L-цепь, соответственно.
Способом по настоящему изобретению можно получать интересующий белок с использованием суспензионной клетки млекопитающего, продуцирующей интересующий белок, в которой генный фрагмент, вставленный между парой транспозонных последовательностей, интегрирован в хромосому, путем введения экспрессионного вектора, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
Ген селектируемого маркера, используемый в качестве показателя встраивания гена, можно интегрировать в тот же вектор, что и экспрессионный вектор, который содержит ДНК, кодирующую интересующий белок, или можно интегрировать в другой вектор.
То есть, по меньшей мере один из экспрессионных векторов, которые содержат генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержат пару транспозонных последовательностей на обоих концах генного фрагмента, можно использовать в качестве экспрессионного вектора, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и ген селекционного маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
Кроме того, в дополнение к экспрессионному вектору, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, в клетку млекопитающего можно дополнительно вводить экспрессионный вектор, который содержит генный фрагмент, содержащий ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
В частности, примеры способа получения интересующего белка по настоящему изобретению включают способ, включающий следующие этапы (A)-(C):
(A) этап одновременного введения следующих экспрессионных векторов (a) и (b) в суспензионную клетку млекопитающего:
(a) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) вектор, который содержит ДНК, кодирующую транспозазу, которая узнает транспозонные последовательности и обладает способностью переносить генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому;
(B) этап временной экспрессии транспозазы с экспрессионного вектора (b), который введен в суспензионную клетку млекопитающего на этапе (A) для интеграции генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего, и получения суспензионной клетки млекопитающего, которая экспрессирует интересующий белок, и
(C) этап суспензионного культивирования суспензионной клетки млекопитающего, которая экспрессирует интересующий белок, полученной на этапе (B), для получения интересующего белка.
Кроме того, примеры способа получения интересующего белка по настоящему изобретению включают способ, включающий следующие этапы (A)-(C):
(A) этап одновременного введения следующих экспрессионных векторов (a), (b) и (с) в суспензионную клетку млекопитающего:
(a) по меньшей мере один экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) экспрессионный вектор, который содержит селектируемый маркер и пару транспозонных последовательностей на обоих концах селектируемого маркера;
(с) вектор, который содержит ДНК, кодирующую транспозазу, которая узнает транспозонные последовательности и обладает способностью переносить генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому;
(B) этап временной экспрессии транспозазы с экспрессионного вектора (c), который введен в суспензионную клетку млекопитающего на этапе (A) для интеграции генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего, и получения суспензионной клетки млекопитающего, которая экспрессирует интересующий белок, и
(C) этап суспензионного культивирования суспензионной клетки млекопитающего, которая экспрессирует интересующий белок, полученной на этапе (B), для получения интересующего белка.
Настоящее изобретение относится к суспензионной клетке млекопитающего, в которую по меньшей мере один экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит селектируемый маркер и пару транспозонных последовательностей на обоих концах селектируемого маркера, введены для интеграции генного фрагмента и селектируемого маркера, вставленных между парой транспозонных последовательностей, в хромосому, и которая продуцирует интересующий белок.
Кроме того, настоящее изобретение относится к суспензионной клетке млекопитающего, в которую введен экспрессионный вектор для белка, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и селектируемый маркер, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, для интеграции генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому, и которая продуцирует интересующий белок.
Кроме того, примеры суспензионной клетки млекопитающего, которая продуцирует интересующий белок по настоящему изобретению, включают суспензионную клетку млекопитающего, в которую одновременно введены экспрессионный вектор (a), который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и ген селектируемого маркера, и также содержит транспозонные последовательности на обоих концах генного фрагмента, и вектор (b), содержащий ДНК, кодирующую транспозазу (трансферазу), которая узнает транспозонные последовательности и обладает способностью переносить генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому для интеграции генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому, и которая продуцирует интересующий белок.
По настоящему изобретению, число экспрессионных векторов, которые содержат генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержат пару транспозонных последовательностей на обоих концах генного фрагмента, предназначенных для введения в суспензионную клетку млекопитающего, не имеет конкретных ограничений при условии, что клетка млекопитающего может экспрессировать и продуцировать интересующий белок, и примеры включают предпочтительно от 1 до 20 видов экспрессионных векторов, более предпочтительно можно упомянуть от 2 до 10 видов экспрессионных векторов и, например, от 3 до 8 видов экспрессионных векторов, от 4 до 7 видов экспрессионных векторов, от 1 до 6 видов экспрессионных векторов, от 1 до 5 видов экспрессионных векторов, от 1 до 4 видов экспрессионных векторов, и от 1 до 3 видов экспрессионных векторов являются предпочтительными.
Кроме того, примеры вариантов осуществления настоящего изобретения включают способ увеличения интеграции генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего путем одновременного введения в суспензионную клетку млекопитающего (a) по меньшей мере одного экспрессионного вектора, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и (b) вектора, который содержит ДНК, кодирующую транспозазу, способную узнавать транспозонные последовательности и обладающую способностью встраивать генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому, способ интеграции ДНК, кодирующей интересующий белок, в хромосому клетки млекопитающего с высокой частотой и суспензионную клетку млекопитающего, которая получена данными способами и может продуцировать интересующий белок.
Термин «транспозон» в настоящей спецификации относится к мобильному генетическому элементу и означает генетическую единицу, которая перемещается по хромосоме или с хромосомы на другую хромосому (транспозиция), сохраняя при этом определенную структуру.
Транспозон содержит повторяющиеся транспозонные последовательности (также называемые инвертированной повторяющейся последовательностью (IR последовательностью) или концевой инвертированной повторяющейся последовательностью (TIR последовательностью)), которые располагаются в том же направлении или в обратном направлении на обоих концах генетической единицы, и нуклеотидную последовательность, кодирующую транспозазу, которая узнает транспозонную последовательность для вставки гена, находящегося между транспозонными последовательностями.
Транспозаза, транслируемая с транспозона, может встраивать ДНК путем узнавания транспозонных последовательностей на обоих концах транспозона, вырезания фрагмента ДНК, вставленного между парой транспозонных последовательностей, и вставки фрагмента в сайт, в который он будет встроен.
Термин «транспозонная последовательность» в настоящей спецификации означает нуклеотидную последовательность транспозона, узнаваемую транспозазой, и имеет то же значение, что и IR последовательность или TIR последовательность. ДНК, содержащая нуклеотидную последовательность, может содержать несовершенный повторяющийся фрагмент при условии, что она может быть встроена (вставлена в другое место в геноме) в результате активности транспозазы, и существует транспозонная последовательность, специфичная для транспозазы.
В качестве транспозонной последовательности, используемой по изобретению, предпочтительной является нуклеотидная последовательность, полученная из транспозона ДНК-типа, и нуклеотидная последовательность, полученная из пары природных или искусственных транспозонов ДНК-типа, которая может узнаваться транспозазой и подвергаться перемещениям в клетках млекопитающих, является более предпочтительной.
Примеры нуклеотидной последовательности, полученной из транспозона ДНК-типа, включают нуклеотидные последовательности, полученные из транспозона Tol1 и транспозона Tol2 из рыбы оризии, транспозон «спящая красавица», восстановленный из неавтономного транспозона, существующего в геноме лососевых рыб, искусственный транспозон «лягушачий принц» из лягушки и транспозон PiggyBac из насекомого.
В частности, среди них нуклеотидные последовательности, полученные из транспозона Tol2 из рыбы оризии, содержащие нуклеотидную последовательность, приведенную в SEQ ID NO: 6, и транспозона Tol2 из рыбы оризии, содержащие нуклеотидную последовательность, приведенную в SEQ ID NO: 13, являются предпочтительными.
Примеры нуклеотидной последовательности, полученной из пары транспозонов Tol2, включают нуклеотидную последовательность в положениях 1-2229 и нуклеотидную последовательность в положениях 4148-4682 в нуклеотидной последовательности транспозона Tol2, приведенной в SEQ ID NO: 6 в списке последовательностей.
В качестве нуклеотидной последовательности, полученной из пары транспозонов Tol2, нуклеотидная последовательность в положениях 1-200 (SEQ ID NO: 2) (далее в данном документе называемая «последовательность Tol2-L») и нуклеотидная последовательность в положениях 2285-2788 (SEQ ID NO: 3) (далее в данном документе называемая «последовательность Tol2-R») в нуклеотидной последовательности транспозона Tol2, приведенной в SEQ ID NO: 1 в списке последовательностей, являются более предпочтительными.
В качестве транспозонной последовательности, полученной из пары транспозонов Tol1, пример включает нуклеотидную последовательность, содержащую нуклеотидную последовательность в положениях 1-157 и нуклеотидную последовательность в положениях 1748-1855 в нуклеотидной последовательности транспозона Tol1, приведенной в SEQ ID NO: 13 в списке последовательностей.
В качестве транспозонной последовательности, полученной из пары транспозонов Tol1, нуклеотидная последовательность в положениях 1-200 (SEQ ID NO: 14) (далее в данном документе называемая «последовательность Tol1-L») и нуклеотидная последовательность в положениях 1351-1855 (SEQ ID NO: 15) (далее в данном документе называемая «последовательность Tol1-R») в нуклеотидной последовательности транспозона Tol1, приведенной в SEQ ID NO: 13 в списке последовательностей, являются более предпочтительными.
Примеры транспозонной последовательности для использования по изобретению включают транспозонные последовательности, для которых реакции транспозиции контролируются путем использования частичной последовательности транспозонной последовательности, полученной из вышеуказанного транспозона, путем регулирования длины нуклеотидной последовательности и путем модифицирования нуклеотидной последовательности за счет добавления, делеции или замены.
В качестве способа получения интересующего белка по настоящему изобретению примеры также включают способ, в котором по меньшей мере один из интересующих белков продуцируется с использованием по меньшей мере двух транспозонных последовательностей и по меньшей мере двух транспозаз.
В частности, примеры включают способ получения белка, который включает этапы введения вектора, содержащего ДНК, кодирующую первый интересующий белок, вставленную в две транспозонные последовательности Tol1, вектора, содержащего ДНК, кодирующую второй интересующий белок, вставленную в две транспозонные последовательности Tol2, экспрессионного вектора для Tol1 транспозазы и экспрессионного вектора для Tol2 транспозазы одновременно или по порядку в хромосому клетки млекопитающего и, таким образом, получения клетки млекопитающего, которая продуцирует два интересующих белка.
Кроме того, первый интересующий белок и второй интересующий белок могут быть одинаковыми, и продуктивность для интересующего белка можно также повышать, увеличивая число копий гена, который вводят в клетку.
Что касается контроля реакции транспозиции транспозона, реакцию транспозиции можно ускорять или подавлять, ускоряя или подавляя узнавание транспозонной последовательности транспозазой, соответственно. Кроме того, что касается реакции транспозиции транспозона, реакцию транспозиции можно усиливать путем укорачивания длины нуклеотидной последовательности, вставленной между парой (двумя) транспозонных последовательностей, и реакцию транспозиции можно снижать путем увеличения этой длины. Вследствие этого, в случае экспрессии и получения интересующего белка, содержащего несколько белков, интересующие белки можно получать путем встраивания ДНК, кодирующей каждый белок, в разные экспрессионные векторы, интеграции ДНК в хромосому клетки-хозяина и получения суспензионной клетки млекопитающего, способной создавать интересующий белок, чтобы получать интересующий белок с использованием клетки.
Термин «транспозаза» в настоящей спецификации означает фермент, который узнает нуклеотидные последовательности, имеющие транспозонные последовательности, и перемещает генный фрагмент, находящийся между нуклеотидными последовательностями по хромосоме или с хромосомы на другую хромосому.
Примеры транспозазы включают ферменты из Tol1 и Tol2, полученных из рыбы оризии, транспозона «спящая красавица» (SB), восстановленного из неавтономного транспозона, существующего в геноме лососевых рыб, транспозона «спящая красавица 11» (SB11), искусственного транспозона «лягушачий принц» (FP), полученного из лягушки, и транспозона PiggyBac (PB), полученного из насекомого.
В качестве транспозазы можно использовать природный фермент, а также любую транспозазу, в которой часть ее аминокислот заменены, делетированы, вставлены и/или добавлены, при условии, что сохраняется такая же транспозиционная активность, что и у транспозазы. Контролируя ферментативную активность транспозазы, можно контролировать реакцию транспозиции ДНК, находящейся между транспозонными последовательностями.
Для анализа того, присутствует ли транспозиционная активность, подобная активности транспозазы, ее можно измерять при помощи 2-компонентной аналитической системы, раскрытой в японской опубликованной нерассмотренной патентной заявке № 235575/2003.
В частности, способен ли неавтономный элемент Tol2 переноситься и встраиваться в хромосому клетки млекопитающего за счет активности транспозазы, можно анализировать путем раздельного использования плазмиды, содержащей лишенный Tol2 транспозазы транспозон Tol2 (полученный из Tol2 неавтономный транспозон), и плазмиды, содержащей Tol2 транспозазу.
Термин «неавтономный транспозон» в настоящей спецификации означает транспозон, который утратил транспозазу, существующую внутри транспозона, и, вследствие этого, не в состоянии осуществлять собственную автономную транспозицию. Неавтономный транспозон может переносить ДНК, вставленную между транспозонными последовательностями неавтономного транспозона, в хромосому клетки-хозяина при одновременном наличии в клетке белка транспозазы, мРНК, кодирующей белок транспозазу, или ДНК, кодирующей белок транспозазу.
Ген транспозазы означает ген, кодирующий транспозазу. Для повышения экспрессионной эффективности в клетке млекопитающего, последовательность, которая регулирует пространство между консенсусной последовательностью Козака (Kozak М., Nucleic Acids Res., 12, 857-872 (1984)) или последовательностью связывания рибосомы, последовательностью Шайна-Дальгарно и инициирующим кодоном до соответствующего расстояния (например, от 6 до 18 оснований), можно присоединять к сайту, расположенному выше от кодона инициации трансляции ATG гена.
В соответствии со способом по изобретению, для того, чтобы интегрировать генный фрагмент, содержащий ДНК, кодирующую интересующий белок, в по меньшей мере одном экспрессионном векторе в хромосому клетки-хозяина, экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, вводят в клетку-хозяина и дают возможность транспозазе проявлять свою активность в отношении транспозонных последовательностей, содержащихся в экспрессионном векторе, который введен в клетку.
Для того, чтобы позволить транспозазе проявлять свою активность в отношении транспозонных последовательностей, содержащихся в экспрессионном векторе, который введен в клетку, транспозазу можно инъецировать в клетку, или экспрессионный вектор, содержащий ДНК, кодирующую по меньшей мере один интересующий белок, или ДНК, кодирующую интересующий белок, можно вводить в клетку-хозяина вместе с экспрессионным вектором, содержащим ДНК, кодирующую интересующий белок и ген селектируемого маркера. Кроме того, при введении РНК, кодирующей транспозазу, в клетку-хозяина транспозаза может экспрессироваться в клетке.
Не существует конкретных ограничений для экспрессионного вектора. Любой экспрессионный вектор может быть использован при произвольном выборе среди экспрессионных векторов, известных специалистам в данной области, в зависимости от клетки-хозяина, в которую вводят экспрессионный вектор, содержащий ген транспозазы; от применения и тому подобного.
В случае, когда интересующий белок, состоящий из двух или более полипептидов, или два или более интересующих белка получают способом по изобретению, продуцирующую белок клетку, в которой ДНК, кодирующая каждый белок, встроена в хромосому клетки-хозяина, можно получать путем вставки ДНК, кодирующей каждый белок, в один и тот же экспрессионный вектор, или вставки ДНК в соответствующие различные экспрессионные векторы и введения экспрессионных векторов в клетку-хозяина.
Транспозазу можно вводить в экспрессионный вектор для экспрессии совместно с интересующим белком или можно вводить в вектор, отличный от экспрессионного вектора. Можно позволять транспозазе действовать временно или можно ей позволять действовать постоянно, однако предпочтительно позволять транспозазе действовать временно, чтобы подготовить клетку к стабильной продукции.
В качестве способа создания условий для временной активности транспозазы примеры включают способ, включающий создание экспрессионного вектора, содержащего ДНК, кодирующую транспозазу, и экспрессионного вектора, содержащего ДНК, кодирующую интересующий белок, и последующее введение обеих экспрессионных плазмид одновременно в клетку-хозяина.
Термин «экспрессионный вектор» в настоящей спецификации означает экспрессионный вектор, используемый с целью введения в клетку млекопитающего и экспрессии интересующего белка. Экспрессионный вектор, используемый по изобретению, имеет структуру, в которой по меньшей мере пара транспозонных последовательностей присутствует на обоих концах экспрессионной кассеты.
Термин «экспрессионная кассета» в настоящей спецификации означает нуклеотидную последовательность, в которой имеется область, контролирующая экспрессию гена, необходимая для экспрессии интересующего белка, и последовательность, кодирующая интересующий белок. Примеры области, контролирующей экспрессию гена, включают энхансер, промотор и терминатор. Экспрессионная кассета может включать ген селектируемого маркера.
Можно использовать любой промотор, при условии, что он может функционировать в животной клетке. Примеры включают промотор IE (предраннего) гена цитомегаловируса (CMV), ранний промотор SV40, промотор ретровируса, металлотионеиновый промотор, промотор теплового шока, SRα промотор, вирус лейкемии мышей Молони, энхансер и тому подобное. Кроме того, энхансер IE гена человеческого CMV можно использовать вместе с промотором.
Термин «ген селектируемого маркера» означает дополнительный ген другого маркера, который можно использовать, чтобы отличить клетку, в которую введен плазмидный вектор, от клетки без вектора.
Примеры гена селектируемого маркера включают ген устойчивости к лекарственному средству (ген устойчивости к неомицину, ген дигидрофолатредуктазы (DHFR), ген устойчивости к пуромицину, ген устойчивости к бластицидину, ген устойчивости к зеоцину, ген устойчивости к гигромицину и ген устойчивости к циклогексимиду (японская опубликованная нерассмотренная патентная заявка № 262879/2002)), гены флуоресцентных и биолюминесцентных маркеров (например, зеленый флуоресцентный белок GFP) и тому подобное.
В данном изобретении предпочтительным селектируемым маркером является ген устойчивости к лекарственному средству, и особенно предпочтительным селектируемым маркером является ген устойчивости к циклогексимиду. Кроме того, свойство устойчивости к лекарственному средству и люминесцентное свойство белка селектируемого маркера можно также изменять, создавая вариант с модифицированными аминокислотами путем генетической модификации гена селектируемого маркера или контролируя транскрипцию или трансляцию гена селектируемого маркера (например, модификация промотора, модификация аминокислотного кодона и тому подобное). Кроме того, клетки с введенным геном селектируемого маркера, обладающие различной степенью устойчивости к лекарственному средству, можно также отбирать, регулируя концентрацию лекарственного средства.
Для контроля свойства устойчивости к лекарственному средству и люминесцентного свойства белка селектируемого маркера предпочтительно использовать ослабленный ген селектируемого маркера. Ослабленный ген селектируемого маркера представляет собой ген селектируемого маркера, который модифицирован таким образом, что активность белка, кодируемого геном селектируемого маркера, внутри клетки понижена.
Примеры гена селектируемого маркера, модифицированного таким образом, что активность в клетке становится пониженной, включают (A) ген селектируемого маркера, у которого аминокислотная последовательность белка, кодируемого геном селектируемого маркера, модифицирована так, что активность белка в клетке снижена, и (B) ген селектируемого маркера, у которого нуклеотидная последовательность, контролирующая экспрессию гена селектируемого маркера, модифицирована или нуклеотидная последовательность внутри ORF (открытой рамки считывания) модифицирована так, что экспрессия гена селектируемого маркера снижена.
Примеры гена селектируемого маркера, у которого аминокислотная последовательность белка, кодируемого геном селектируемого маркера, модифицирована таким образом, что активность белка в клетке снижена, включают ген устойчивости к неомицину, описанный Sauter et al. [Biotech. Bioeng., 89, 530-538 (2005)] или Chen et al. [.Journal of Immunological Methods, 295, 49-56 (2004)].
Примеры метода снижения уровня экспрессии белка в клетке путем модификации нуклеотидной последовательности, контролирующей экспрессию гена селектируемого маркера, включают метод модификации последовательности промотора, последовательности терминатора, последовательности энхансера, консенсусной последовательности Козака или последовательности Шайна-Дальгарно, которая контролирует экспрессию гена селектируемого маркера. Более конкретно, примеры включают метод, в котором промоторная последовательность, которая контролирует экспрессию гена селектируемого маркера, заменена более слабой промоторной последовательностью.
Примеры метода снижения уровня экспрессии белка в клетке путем модификации нуклеотидной последовательности в ORF гена селектируемого маркера включают метод, в котором кодон в ORF заменен синонимичным кодоном с более низкой частотой использования кодона в клетке.
Примеры ослабленного гена селектируемого маркера по изобретению включают селектируемый маркер, в котором вышеуказанный кодон в ORF гена заменен синонимичным кодоном с более низкой частотой использования кодона в клетке.
В клетках различных биологических видов синонимичный кодон с более низкой частотой использования среди всех синонимичных кодонов можно выбирать на основании известной литературы, баз данных и тому подобного.
В качестве такой замены синонимичным кодоном с более низкой частотой использования, в частности, в случае клетки CHO, примеры включают замену кодона лейцина на TTA, замену кодона аргинина на CGA или CGT, замену кодона аланина на GCG, замену кодона валина на GTA, замену кодона серина на TCG, замену кодона изолейцина на ATA, замену кодона треонина на ACG, замену кодона пролина на CCG, замену кодона глютаминовой кислоты на GAA, замену кодона тирозина на TAT, замену кодона лизина на AAA, замену кодона фенилаланина на TTT, замену кодона гистидина на CAT, замену кодона глютамина на CAA, замену кодона аспарагина на AAT, замену кодона аспарагиновой кислоты на GAT, замену кодона цистеина на TGT и замену кодона глицина на GGT.
В ослабленном гене селектируемого маркера число заменяемых кодонов по сравнению с геном селектируемого маркера до модификации не имеет конкретных ограничений при условии, что можно эффективно получать продуцирующую белок клетку, однако предпочтительно заменять кодоны, соответствующие 20 или более аминокислотным остаткам.
В ослабленном гене селектируемого маркера число модифицируемых оснований по сравнению с геном селектируемого маркера до модификации не имеет конкретных ограничений, однако предпочтительно модифицировать 10% или более нуклеотидной последовательности, кодирующей ген селектируемого маркера.
Кроме того, в случае ослабленного гена селектируемого маркера аминокислотные остатки, кодируемые заменяемыми кодонами, не имеют конкретных ограничений, но предпочтительные примеры включают лейцин, аланин, серин и валин.
В случае ослабленного гена селектируемого маркера, случаи замены кодонов, соответствующих лейцину, не имеют конкретных ограничений, однако предпочтительно заменять кодоны, соответствующие 70% или более остатков лейцина, среди кодонов, соответствующих всем остаткам лейцина, содержащимся в гене селектируемого маркера.
Кроме того, в случае ослабленного гена селектируемого маркера случаи замены кодонов, соответствующих аланину, не имеют конкретных ограничений, однако предпочтительно заменять кодоны, соответствующие 70% или более остатков аланина среди кодонов, соответствующих всем остаткам аланина, содержащимся в гене селектируемого маркера.
Конкретные примеры ослабленного гена селектируемого маркера, полученного при помощи такой модификации, в котором кодоны заменены синонимичными кодонами с меньшей частотой использования, включают ген устойчивости к неомицину, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 37, 38 или 39, ген устойчивости к пуромицину, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 41, 43 или 44, ген устойчивости к зеоцину, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 45 или 46, и ген устойчивости к гигромицину, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 47 или 48.
Кроме того, можно ослаблять ген селектируемого маркера также путем значительного увеличения концентрации лекарственного средства по сравнению с обычно используемой концентрацией, когда устойчивую к лекарственному средству клетку отбирают при создании продуцирующей антитело клетки, или проводя дополнительное введение до того, как ген устойчивости к лекарственному средству обеспечит преобразование и распад лекарственного средства.
Циклогексимид (далее в данном документе иногда называемый CHX) представляет собой ингибитор белкового синтеза, и примеры использования гена устойчивости к CHX в качестве гена селектируемого маркера включают известные случаи с дрожжами [Kondo K., J. Bacteriol., 177, 24, 7171-7177 (1995)] и животными клетками (JP-A-2002-262879).
В случае животных клеток было установлено, что трансформант, экспрессирующий белок, кодируемый нуклеотидной последовательностью, представленной SEQ ID NO: 7 в списке последовательностей, в котором пролин в положении 54 субъединицы L36a человеческого рибосомного белка, кодируемой нуклеотидной последовательностью, представленной SEQ ID NO: 5 в списке последовательностей, заменен глютамином, обеспечивает устойчивость к циклогексимиду. Кроме того, примеры маркера устойчивости к циклогексимиду включают мутантную субъединицу L44 человеческого рибосомного белка, когда пролин в положении 54 субъединицы L44 человеческого рибосомного белка заменен глютамином.
Метод введения вышеупомянутого экспрессионного вектора для белка, содержащего транспозонную последовательность, плазмидного вектора для экспрессии транспозазы или РНК не имеет конкретных ограничений. Примеры включают кальций-фосфатную трансфекцию, электропорацию, липосомный метод, метод генной пушки, липофекцию и тому подобное.
Примеры метода прямого введения транспозазы в форме белка включают метод микроинъекции или попадание в клетку путем эндоцитоза. Введение гена можно проводить методом, описанным в Shin Idenshi Kogaku Handbook (New Genetic Engineering Handbook) (Новый справочник по генетической инженерии), под редакцией Masami Muramatsu и Tadashi Yamamoto, опубликованном Yodo-sha, ISBN 9784897063737.
Клетка-хозяин может быть любой клеткой млекопитающего при условии, что она может быть субкультивирована и может стабильно экспрессировать интересующий белок. Примеры клетки-хозяина включают клетку PER.C6, клетку Namalwa лейкемии человека, клетку COS обезьяны, клетку YB2/3HL.P2.G11.16Ag.20 (также называемую YB2/0) крысиной миеломы, клетку NS0 мышиной миеломы, клетку SP2/0-Ag14 мышиной миеломы, клетку BHK сирийского хомячка, HBT5637 (японская нерассмотренная патентная заявка № 1998-000299), клетку CHO яичника китайского хомячка (Journal of Experimental Medicine, 108, 945 (1958); Proc. Natl. Acad. Sci. USA., 601275 (1968); Genetics, 55, 513 (1968); Chromosoma, 41, 129 (1973); Methods in Cell Science, 18, 115 (1996); Radiation Research, 148, 260 (1997); Proc. Natl. Acad. Sci. USA., 11, 4216 (1980); Proc. Natl. Acad. Sci., 60, 1275 (1968); Cell, 6, 121 (1975); Molecular Cell Genetics, Appendix I, II (pp. 883-900)), CHO/DG44, CHO-K1 (ATCC CCL-61), DUKXB11 (ATCC CCL-9096), Pro-5 (ATCC CCL-1781), CHO-S (Life Technologies, каталожный № 11619), Pro-3 и субклональную клеточную линию клетки CHO.
Кроме того, вышеуказанную клетку-хозяина можно также использовать в способе получения белка по изобретению, модифицируя клетку таким образом, чтобы она подходила для производства белка, за счет модификации хромосомной ДНК, введения экзогенного гена и тому подобного.
Кроме того, для контроля структуры сахарной цепи, связанной с продуцируемым интересующим белком, в качестве клетки-хозяина можно также использовать Lec13 с приобретенной устойчивостью к лектину [Somatic Cell and 20 Molecular Genetics, 12, 55 (1986)] и клетку CHO, из которой удален ген α1,6-фукозилтрансферазы (WO2005/35586, WO2002/31140), клетку, дефицитную по ГДФ-маннозо-4,6-дегидратазе (GMD), и клетку, дефицитную по белку Fx.
В настоящем изобретении интересующий белок также включает любой белок, состоящий из по меньшей мере одного полипептида, и сложный белок, состоящий из двух или более полипептидов белка. Кроме того, белок и полипептид являются синонимами в данном изобретении, однако белковая молекула с относительно низкой молекулярной массой или белок, являющийся составной частью сложного белка, иногда могут быть названы полипептидами.
Интересующий белок в данном изобретении может быть любым белком или полипептидом при условии, что он может быть экспрессирован способом по изобретению. В частности, примеры интересующего белка включают человеческий сывороточный белок, альбумин-связывающий белок, пептидный гормон, фактор роста, цитокин, фактор свертывания крови, фибринолитический белок, антитело, белок селектируемого маркера, мембранный белок, частичные фрагменты различных белков и тому подобное. В частности, примеры интересующего белка включают человеческий внутривенный иммуноглобулин (IVIG), эритропоэтин (EPO), альбумин, гормон роста (GH), фолликулостимулирующий гормон (FSH), фактор роста гепатоцитов (HGF), инсулин, инсулиноподобный фактор роста-I (IGF-I), интерферон (INF), Fas-лиганд, факторы свертывания крови (II, VII, VIII, IX, X), протромбин, фибриноген, протеин С, протеин S, антитромбин III (ATIII), тканевой активатор плазминогена (tPA), моноклональное антитело, поликлональное антитело и тому подобное.
Антитело представляет собой молекулу, состоящую из полипептида тяжелой цепи (H-цепи) антитела и двух полипептидов легкой цепи (L-цепи) антитела, и как подклассы известны IgA, IgD, IgE, IgG и IgM. Кроме того, IgG подразделяется на IgG1, IgG2, IgG3 и IgG4.
IgG антитело представляет собой гетеротетрамерную молекулу, состоящую из двух полипептидов H-цепи и двух полипептидов L-цепи. Каждая из H-цепи и L-цепи состоит из вариабельной области (V), имеющей отношение к связыванию антигена, и константной области (C), и каждая из них называется VH, CH, VL или CL, соответственно. Область CH дополнительно подразделяется на CH1, CH2 и CH3 области, и CH2 и CH3 области в сочетании называются Fc-областью или просто Fc.
Антитело включает моноклональное антитело, которое реагирует с единственным эпитопом, поликлональное антитело, которое реагирует с двумя или более эпитопами, и рекомбинантное антитело.
Моноклональное антитело представляет собой антитело, которое секретируется одной клональной антитело-продуцирующей клеткой и узнает только один эпитоп (также называемый антигенной детерминантой), и аминокислотная последовательность (первичная структура), составляющая моноклональное антитело, единообразна.
Поликлональное антитело представляет собой смесь моноклональных антител и может реагировать с двумя или более эпитопами.
Примеры рекомбинантного антитела включают химерное антитело, гуманизированное антитело, человеческое антитело, Fc-слитый белок, антитело с модифицированными аминокислотами в Fc и мультивалентное антитело, а также частичный фрагмент антитела. Антитело с модифицированными аминокислотами может иметь аминокислотную модификацию либо в вариабельной области, либо в константной области, и активность антитела контролируется.
Мультивалентное антитело включает мультивалентное антитело, которое реагирует с двумя или более различными эпитопами на одном антигене, мультивалентное антитело, которое реагирует с двумя или более различными антигенами, и тому подобное, но может включать любое мультивалентное антитело. Кроме того, мультивалентное антитело может быть любым мультивалентным антителом, имеющим любую структуру, при условии, что оно сохраняет способность связывать антиген (WO2001/77342, патент США № 7612181 и WO2009/131239).
В соответствии со способом получения по настоящему изобретению, любой из вышеуказанных интересующих белков и/или интересующих пептидов может быть экпрессирован и продуцирован.
Примеры клетки, в которую введена ДНК, кодирующая по меньшей мере один интересующий белок по настоящему изобретению, включают антитело-продуцирующую клетку, полученную в результате следующих этапов (A) и (B).
Этап (A) - этап одновременного введения обоих из сочетания экспрессионного вектора, выбранного из следующих (a)-(c), или экспрессионного вектора (d) и экспрессионного вектора (e) в суспензионную клетку млекопитающего:
(a) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ген селектируемого маркера и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(c) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и
(d) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь и L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(e) вектор, содержащий ДНК, кодирующую транспозазу, которая узнает транспозонные последовательности и обладает способностью переносить генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому; и
Этап (B) - этап отбора суспензионной клетки млекопитающего, экспрессирующей антитело, в которой гены вышеуказанных H-цепи, L-цепи и селектируемого маркера, вставленные между парой транспозонных последовательностей, интегрированы в хромосому вышеуказанной клетки млекопитающего в результате временной экспрессии транспозазы с экспрессионного вектора (e), который введен в суспензионную клетку млекопитающего на этапе (A).
Примеры способа получения антитела по настоящему изобретению включают способ получения интересующего белка, включающий следующие этапы (A)-(C).
Этап (A) - этап одновременного введения сочетания экспрессионного вектора, выбранного из следующих (a)-(c), или экспрессионного вектора (d) и экспрессионного вектора (e) в суспензионную клетку млекопитающего:
(a) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ген селектируемого маркера и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(c) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и
(d) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь и L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(e) вектор, содержащий ДНК, кодирующую транспозазу, которая узнает транспозонные последовательности и обладает способностью переносить генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому;
Этап (B) - этап получения суспензионной клетки млекопитающего, экспрессирующей антитело, в которой гены вышеуказанных H-цепи, L-цепи и селектируемого маркера, вставленные между парой транспозонных последовательностей, интегрированы в хромосому вышеуказанной клетки млекопитающего в результате временной экспрессии транспозазы с экспрессионного вектора (e), который введен в суспензионную клетку млекопитающего на этапе (A); и
Этап (C) - этап продуцирования антитела культивируемой в суспензии суспензионной клеткой млекопитающего, полученной на этапе (B), которая экспрессирует антитело.
Кроме того, настоящее изобретение включает способ получения линии клеток, обладающих высокой продуктивностью в отношении антитела, и способ скрининга линии клеток, включающий следующие этапы (A) и (B).
Этап (A) - этап одновременного введения сочетания экспрессионного вектора, выбранного из следующих (a)-(c), или экспрессионного вектора (d) и экспрессионного вектора (e) в суспензионную клетку млекопитающего:
(a) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(c) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и
(d) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь и L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(e) вектор, содержащий ДНК, кодирующую транспозазу, которая узнает транспозонные последовательности и обладает способностью переносить генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому; и
Этап (B) - этап отбора суспензионной клетки млекопитающего, экспрессирующей антитело на высоком уровне, в которой гены вышеуказанных H-цепи, L-цепи и селектируемого маркера, вставленные между парой транспозонных последовательностей, интегрированы в хромосому вышеуказанной клетки млекопитающего в результате временной экспрессии транспозазы с экспрессионного вектора (e), который введен в суспензионную клетку млекопитающего на этапе (A).
Кроме того, настоящее изобретение включает способ получения антитела, включающий следующие этапы (A), (B) и (C).
Этап (A) - этап одновременного введения сочетания экспрессионного вектора, выбранного из следующих (a)-(c), или экспрессионного вектора (d) и экспрессионного вектора (e) в суспензионную клетку млекопитающего:
(a) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(c) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и
(d) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую H-цепь и L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(e) вектор, содержащий ДНК, кодирующую транспозазу, которая узнает транспозонные последовательности и обладает способностью переносить генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому;
Этап (B) - этап получения суспензионной клетки млекопитающего, экспрессирующей антитело, в которой гены вышеуказанных H-цепи, L-цепи и селектируемого маркера, вставленные между парой транспозонных последовательностей, интегрированы в хромосому вышеуказанной клетки млекопитающего в результате временной экспрессии транспозазы с экспрессионного вектора (e), который введен в суспензионную клетку млекопитающего на этапе (A); и
Этап (C) - этап продуцирования антитела культивируемой в суспензии суспензионной клеткой млекопитающего, полученной на этапе (B), которая экспрессирует антитело.
Примеры клетки млекопитающего, в которую введена ДНК, кодирующая по меньшей мере один интересующий белок по настоящему изобретению, включают продуцирующую поликлональное антитело клетку, в которую введены несколько различных генов антител, продуцирующую сложную молекулу клетку и тому подобное.
Примеры продуцирующей поликлональное антитело клетки включают клетку, в которую введены по меньшей мере два или более различных генов моноклональных антител, клетку, в которую введены гены нескольких моноклональных антител против нескольких антигенов, клетку, которая иммунизирована антигеном и в которую введена генная библиотека антитела, отличного от человеческого, клетку, в которую введена генная библиотека антитела, полученного от пациента, и тому подобное.
Клетка, продуцирующая сложную молекулу, может быть любой клеткой при условии, что введены ДНК, кодирующие соответствующие белки, которые совместно экспрессируются в клетке с образованием сложной молекулы. Конкретные примеры включают клетку, в которую совместно трансфицированы FcγRIII (CD 16) и общая γ-цепь, клетку, в которой совместно экспрессируются неонатальный Fc-рецептор (FcRn) и β2-макроглобулин, клетку, в которую совместно трансфицированы CD98 и LAT1 (WO2007/114496), и тому подобные.
Антитело, которое получают способом получения антитела по настоящему изобретению, может быть любым антителом, и примеры включают антитело, которое узнает связанный с опухолью антиген, антитело, которое узнает связанный с аллергией или воспалением антиген, антитело, которое узнает связанный с сердечно-сосудистым заболеванием антиген, антитело, которое узнает антиген, связанный с аутоиммунными заболеваниями, антитело, которое узнает связанный с вирусной или бактериальной инфекцией антиген, и тому подобные.
Примеры связанного с опухолью антигена включают CD1a, CD2, CD3, CD4, CD5, CD6, CD7, CD9, CD10, CD13, CD19, CD20, CD21, CD22, CD25, CD28, CD30, CD32, CD33, CD38, CD40, лиганд CD40 (CD40L), CD44, CD45, CD46, CD47, CD52, CD54, CD55, CD55, CD59, CD63, CD64, CD66b, CD69, CD70, CD74, CD80, CD89, CD95, CD98, CD105, CD134, CD137, CD138, CD147, CD158, CD160, CD162, CD164, CD200, CD227, адреномедуллин, ангиопоэтин-родственный белок 4 (ARP4), аврору, B7-H1, B7-DC, интеглин, стромальный антиген 2 костного мозга (BST2), CA125, CA19.9, карбоангидразу 9 (CA9), кадгерин, cc-хемокиновый рецептор (CCR) 4, CCR7, карциноэмбриональный антиген (CEA), богатый цистеином рецептор 1 фактора роста фибробластов (CFR-1), c-Met, c-Myc, коллаген, СТА, фактор роста соединительной ткани (CTGF), CTLA-4, цитокератин-18, DF3, E-катерин, рецептор эпидермального фактора роста (EGFR), EGFRvIII, EGFR2 (HER2), EGFR3 (HER3), EGFR4 (HER4), эндоглин, эпителиальную молекулу клеточной адгезии (EpCAM), эндотелиальный рецептор протеина С (EPCR), эфрин, эфриновый рецептор (Eph), EphA2, эндотелиазу-2 (ET2), FAM3D, белок, активирующий фибробласты (FAP), гомолог 1 Fc-рецептора (FcRH1), ферритин, фактор роста фибробластов 8 (FGF-8), рецептор FGF8, основной FGF (bFGF), рецептор bFGF, рецептор FGF (FGFR) 3, FGFR4, FLT1, FLT3, фолатный рецептор, гомолог 10 Frizzled (FZD10), рецептор 4 frizzled (FZD-4), G250, рецептор G-CSF, ганглиозид (такой как GD2, GD3, GM2 и GM3), глобо H, gp75, gp88, GPR-9-6, гепараназу I, фактор роста гепатоцитов (HGF), рецептор HGF, HLA антиген (такой как HLA-DR), HM1.24, глобулу человеческого молочного жира (HMFG), hRS7, белок теплового шока 90 (hsp90), идиотипический эпитоп, инсулин-подобный фактор роста (IGF), рецептор IGF (IGFR), интерлейкин (такой как IL-6 и IL-15), рецептор интерлейкина (такой как IL-6R и IL-15R), интегрин, связанный с транслокацией иммунного рецептора белок 4 (IRTA-4), калликреин 1, KDR, KIR2DL1, KIR2DL2/3, KS1/4, lamp-1, lamp-2, ламинин-5, Lewis y, сиалил-Lewis x, рецептор лимфотоксина-бета (LTBR), LUNX, связанный с меланомой хондроитин сульфат протеогликан (MCSP), мезотелин, MICA, рецептор II типа ингибирующего вещества Мюллера (MISIIR), муцин, молекулу адгезии нервных клеток (NCAM), Necl-5, Notch 1, остеопонтин, фактор роста тромбоцитов (PDGF), рецептор PDGF, тромбоцитарный фактор 4 (PF-4), фосфатидилсерин, простатический специфический антиген (PSA), антиген стволовых клеток простаты (PSCA), простатический специфический мембранный антиген (PSMA), белок/пептид, родственный паратиреоидному гормону (PTHrP), активатор рецептора лиганда NF-каппаВ (RANKL), рецептор для опосредованной гиалуроновой кислотой подвижности (RHAMM), ROBO1, SART3, семафорин 4B (SEMA4B), ингибитор секреторной лейкоцитарной протеазы (SLPI), SM5-1, сфингозин-1-фосфат, связанный с опухолью гликопротеин 72 (TAG-72), рецептор трансферрина (TfR), TGF бета, Thy-1, Tie-1, рецептор Tie2, T-клеточный иммуноглобулиновый домен и муциновый домен 1 (TIM-1), тканевой фактор человека (hTF), антиген Tn, фактор некроза опухоли (TNF), антиген Thomsen-Friedenreich (антиген TF), рецептор TNF, родственный фактору некроза опухоли апоптоз-индуцирующий лиганд (TRAIL), рецептор TRAIL (такой как DR4 и DR5), переносчик аминокислот 2 системы ASC (ASCT2), trkC, TROP-2, рецептор Fn14 TWEAK, коллагеназу IV типа, рецептор урокиназы, сосудистый эндотелиальный фактор роста (VEGF), рецептор VEGF (такой как VEGFR1, VEGFR2 и VEGFR3), виментин, VLA-4 и тому подобные, а также антитела против вышеуказанных антигенов.
Кроме того, примеры антитела, которое узнает связанный с опухолью антиген, включают [AntiCancer Res., 13, 331 (1993)], анти-GD3 антитело [Cancer Immunol. Immunother., 36, 260 (1993)], анти-GM2 антитело [Cancer Res., 54, 1511 (1994)], анти-CD52 антитело [Proc. Natl. Acad. Sci. USA, 89, 4285 (1992)], анти-MAGE антитело [British J. Cancer, 83, 493 (2000)], анти-HM1.24 антитело [Molecular Immunol., 36, 387 (1999)], антитело против белка, родственного паратиреоидному гормону (PTHrP) [Cancer, 88, 2909 (2000)], анти-bFGF антитело, анти-FGF-8 антитело [Proc. Natl. Acad. Sci. USA, 86, 9911 (1989)], анти-bFGFR антитело, анти-FGFR1 антитело (WO2005/037235), анти-FGF-8R антитело [J. Biol. Chem., 265, 16455 (1990)], анти-IGF антитело [J. Neurosci. Res., 40, 647 (1995)], анти-IGF-IR антитело [J. Neurosci. Res, 40, 647 (1995)], анти-PSMA антитело [J. Urology, 160, 2396 (1998)], анти-VEGF антитело [Cancer Res, 57, 4593 (1997), авастин (R)], анти-VEGFR антитело [Oncogene, 19, 2138 (2000), WO96/30046], анти-CD20 антитело [Curr. Opin. Oncol., 10, 548 (1998), патент США 5736137, ритуксан (R), окрелизумаб, офатумумаб], анти-EGFR антитело (эрбитукс (R), вектибикс (R)), анти-HER2 антитело [Proc. Natl. Acad. Sci. USA, 89, 4285 (1992), патент США 5725856, герцептин (R), пертузумаб), анти-HER3 антитело (US2008/0124345), c-Met антитело (патент США 6468529), анти-CD10 антитело, анти-EGFR антитело (WO96/402010), анти-Apo-2R антитело (WO98/51793), anti-ASCT2 антитело (WO2010/008075), анти-CEA антитело [Cancer Res., 55 (23 suppl): 5935s-5945s, (1995)], анти-CD38 антитело, анти-CD33 антитело, анти-CD22 антитело, анти-CD20 антитело с модифицированными аминокислотами [Immunology, 115, 4393, 2010), анти-EpCAM антитело, анти-А33 антитело, антитело против фолатного рецептора (MRAb-003) и тому подобные.
Примеры антитела, которое узнает связанный с аллергией или воспалением антиген, включают антитело против интерлейкина 6 [Immunol. Rev., 127, 5 (1992)], антитело против рецептора интерлейкина 6 [Molecular Immunol., 31, 371 (1994)], антитело против интерлейкина 5 [Immunol. Rev., 127, 5 (1992)], антитело против рецептора интерлейкина 5, антитело против интерлейкина 4 [Cytokine, 3, 562 (1991)], антитело против рецептора интерлейкина 4 [J. Immunol. Meth, 217, 41 (1998)], антитело против фактора некроза опухоли [Hybridoma, 13, 183 (1994)], антитело против рецептора фактора некроза опухоли [Molecular Pharmacol., 58, 237 (2000)], анти-CCR4 антитело [Nature, 400, 776 (1999)], антитело против хемокина [Peri et al., J. Immuno. Meth., 174, 249-257 (1994)], антитело против рецептора хемокина [J. Exp. Med., 186, 1373 (1997)] и тому подобные. Примеры антитела, которое узнает связанный с сердечно-сосудистым заболеванием антиген, включают анти-GpIIb/IIIa антитело [J. Immunol., 152, 2968 (1994)], антитело против фактора роста тромбоцитов [Science, 253, 1129 (1991)], антитело против рецептора фактора роста тромбоцитов [J. Biol. Chem., 272, 17400 (1997)], антитело против фактора свертывания крови [Circulation, 101, 1158 (2000)], анти-IgE антитело, анти-αVβ3 антитело, анти-α4β7 антитело, и тому подобные.
Примеры антитела, которое узнает связанные с вирусной или бактериальной инфекцией антигены, включают анти-gp120 антитело [Structure, 8, 385 (2000)], анти-CD4 антитело [J. Rheumatology, 25, 2065 (1998)], анти-CCR5 антитело, антитело против веротоксина [J. Clin. Microbiol, 37, 396 (1999)], анти-M2 антитело (JP2003-235575), и тому подобные.
Эффекторную активность моноклонального антитела, полученного способом по настоящему изобретению, можно контролировать различными методами. Примеры известных методов включают метод контроля количества фукозы (далее в данном документе также называемой «коровая фукоза»), которая связана с N-ацетилглюкозамином (GlcNAc) посредством α-1,6 связи в восстанавливающем конце N-связанной сахарной цепи сложного типа, связанной с аспарагином (Asn) в положении 297 Fc-области антитела (WO2005/035586, WO2002/31140 и WO00/61739), метод контроля эффекторной активности путем модифицирования аминокислотного остатка(остатков) в Fc-области антитела, и тому подобные. Эффекторную активность моноклонального антитела, полученного способом по настоящему изобретению, можно контролировать, используя любой из данных методов.
«Эффекторная активность» означает зависимую от антитела активность, которая индуцируется с помощью Fc-области антитела. В качестве эффекторной активности известна антитело-зависимая клеточная цитотоксичность (ADCC активность), комплемент-зависимая цитотоксичность (CDC активность), антитело-зависимый фагоцитоз (ADP активность) фагоцитирующими клетками, такими как макрофаги или дендритные клетки, и тому подобные.
Кроме того, контролируя содержание коровой фукозы N-связанной сахарной цепи сложного типа в Fc-области моноклонального антитела, которое получают способом по настоящему изобретению, эффекторную активность антитела можно увеличивать или уменьшать.
В качестве метода снижения содержания фукозы, которая связана с N-связанной сахарной цепью сложного типа, связанной с Fc-областью антитела, можно получать антитело, с которым фукоза не связана, путем экспрессии антитела с использованием клетки CHO, дефицитной по гену, кодирующему α1,6-фукозилтрансферазу. Антитело, с которым фукоза не связана, обладает высокой ADCC активностью.
С другой стороны, в качестве метода увеличения содержания фукозы, которая связана с N-связанной сахарной цепью сложного типа, связанной с Fc-областью антитела, можно получать антитело, с которым фукоза связана, путем экспрессии антитела с использованием клетки-хозяина, в которую введен ген, кодирующий α1,6-фукозилтрансферазу. Антитело, с которым фукоза связана, обладает более низкой ADCC активностью, чем антитело, с которым фукоза не связана.
Кроме того, модифицируя аминокислотный остаток(остатки) в Fc-области антитела, ADCC активность или CDC активность можно увеличивать или уменьшать. Например, CDC активность антитела можно увеличивать, используя аминокислотную последовательность Fc-области антитела, описанную в US2007/0148165.
Кроме того, ADCC активность или CDC активность антитела можно увеличивать или уменьшать, осуществляя модификацию аминокислот, описанную в патентах США №№ 6737056 или 7297775 или 7317091.
Термин «суспензионная клетка млекопитающего» в настоящем изобретении означает клетку, которая не прикрепляется к опоре для клеточных культур, имеющей покрытие для облегчения прикрепления культивируемых клеток, такой как микрогранулы, контейнер для культивирования тканевой культуры (также называемый контейнером для культивирования тканей или контейнером для прикрепляющихся культур и тому подобное), и способна выживать и расти, будучи суспендировнной в культуральном растворе.
Поскольку клетка не прикрепляется к опоре для клеточных культур, клетка может выживать и расти в состоянии одиночной клетки в культуральном растворе или выживать и расти в состоянии массы клеток, образованной в результате слипания двух или более клеток.
Кроме того, в качестве суспензионной клетки млекопитающего, используемой по настоящему изобретению, предпочтительной является клетка, способная выживать и расти в бессывороточной среде, не содержащей эмбриональной телячьей сыворотки (далее в данном документе обозначаемой FCS), и тому подобного, при этом будучи суспендированной в культуральном растворе без прикрепления к опоре для клеточных культур, и клетка млекопитающего, способная выживать и расти, будучи суспендированной в безбелковой среде, не содержащей белка, является более предпочтительной.
Культуральный контейнер для культивирования тканей может быть любым, например, колбой, чашкой Петри и тому подобным, при условии, что на него нанесено покрытие для прикрепляющихся культур. В частности, например, является ли клетка суспензионной клеткой млекопитающего, можно подтвердить, используя коммерчески доступную колбу для культивирования тканей (производитель Greiner), колбу для прикрепляющихся культур (производитель Sumitomo Bakelite) и тому подобное.
В качестве суспензионной клетки млекопитающего, используемой по настоящему изобретению, может служить либо клетка, полученная в результате дополнительной адаптации клетки, исходно имеющей суспензионные свойства, к суспензионному культивированию, либо суспензионная клетка млекопитающего, полученная в результате адаптации прикрепляющейся клетки млекопитающего к условиям суспензионного культивирования.
Примеры клетки, исходно имеющей суспензионные свойства, включают клетку PER.C6, клетку YB2/3HL.P2.G11.16Ag.20 (или также называемую YB2/0) крысиной миеломы, клетку CHO-S (производитель Invitrogen) и тому подобные.
Вышеуказанную «суспензионную клетку млекопитающего, полученную путем адаптации прикрепляющейся клетки млекопитающего к условиям суспензионного культивирования», можно получать методом, описанным в Mol. Biotechnol., 2000, 15(3), 249-57, или методом, описанным далее, и можно получать путем создания клетки, которая демонстрирует свойство пролиферации и свойство выживания, аналогичные тем, которые существовали до адаптации к суспензионному культивированию, или превосходящие те, которые существовали до адаптации к суспензионному культивированию (J. Biotechnol., 2007, 130(3), 282-90).
Термин «аналогичные тем, которые существовали до адаптации к суспензионному культивированию» означает, что коэффициент выживаемости, скорость пролиферации (время удвоения) и тому подобное у клетки, адаптированной к суспензионному культивированию, являются практически такими же, что и у клетки до адаптации к суспензионному культивированию.
В настоящем изобретении примеры метода адаптации прикрепляющейся клетки млекопитающего к условиям суспензионного культивирования включают следующий метод. Содержание сыворотки в содержащей сыворотку среде снижают до 1/10 и повторяют субкультивирование при относительно высокой концентрации клеток. Если клетка млекопитающего становится способной выживать и пролиферировать, содержание сыворотки вновь снижают и субкультивирование повторяют. Таким методом можно получать суспензионную клетку млекопитающего, которая способна выживать и пролиферировать в бессывороточных условиях.
Кроме того, суспензионную клетку млекопитающего можно также получать методом, включающим культивирование с добавлением соответствующего неионного сурфактанта, такого как Pluronic-F68 или тому подобные, в культуральный раствор.
Примеры прикрепляющейся клетки млекопитающего, которая приобретает суспензионные свойства в результате адаптации к условиям суспензионного культивирования, включают клетку NS0 мышиной миеломы, клетку CHO и тому подобные.
В настоящем изобретении в качестве свойства, имеющегося у суспензионной клетки млекопитающего, суспензионное культивирование проводят в условиях плотности клеток 2×105 клеток/мл, а затем концентрация клеток после культивирования в течение 3 или 4 дней становится предпочтительно 5×105 клеток/мл или более, более предпочтительно 8×105 клеток/мл или более, особенно предпочтительно 1×106 клеток/мл или более, наиболее предпочтительно 1,5×106 клеток/мл или более.
Кроме того, время удвоения суспензионной клетки млекопитающего по настоящему изобретению составляет предпочтительно 48 часов или менее, более предпочтительно 24 часа или менее, особенно предпочтительно 18 часов или менее, наиболее предпочтительно 11 часов или менее.
Примеры среды для суспензионного культивирования включают коммерчески доступную среду, такую как среда CD-CHO (Invitrogen), среда EX-CELL 325-PF (SAFC Biosciences), среда SFM4CHO (HyClone) и тому подобные. Кроме того, ее также можно получать путем смешивания сахаридов, аминокислот и тому подобного, которые необходимы для культивирования клеток млекопитающих.
Суспензионную клетку млекопитающего можно культивировать, используя культуральный контейнер, который можно использовать для суспензионного культивирования в условиях культивирования, позволяющих культивировать в суспензии. Примеры культурального контейнера включают 96-луночный планшет для суспензионной клеточной культуры (производитель Corning), T-колбу (производитель Becton Dickinson), коническую колбу (производитель Corning) и тому подобное.
Что касается условий культивирования, например, можно статически культивировать в атмосфере 5% CO2 при температуре культивирования 37°C. Можно также использовать оборудование для встряхивания культуры, такое как культивационное оборудование, предназначенное исключительно для суспензионных культур, например, волновой биореактор (производитель GE Healthcare Bioscience).
Что касается условий для суспензионного культивирования суспензионной клетки млекопитающего с использованием оборудования волнового биореактора, клетку можно культивировать методом, описанным на домашней странице компании GE Healthcare Bioscience .
В дополнение к культивированию при встряхивании, можно также использовать культивирование с помощью оборудования для орбитального встряхивания, такого как биореактор. Культивирование с использованием биореактора можно проводить по методу, описанному в Cytotechnology, (2006) 52: 199-207, и тому подобным.
В настоящем изобретении при использовании линии клеток, отличных от суспензионных клеток млекопитающего, можно использовать любую линию клеток при условии, что она является линией клеток млекопитающего, адаптированной к суспензионному культивированию с помощью вышеупомянутого метода, и представляет собой линию клеток, которую можно использовать в способе получения белка по настоящему изобретению.
Очистку интересующего белка, полученного при помощи суспензионной клетки млекопитающего, проводят путем отделения интересующего белка от примесей, отличных от интересующего белка, в культуральном растворе или клеточном гомогенате, содержащем интересующий белок. Примеры методов разделения включают центрифугирование, диализ, осаждение сульфатом аммония, колоночную хроматографию, фильтрование и тому подобное. Разделение можно проводить на основании разницы в физико-химических свойствах интересующего белка и примесей, либо разницы в их авидности по отношению к самому колоночному носителю.
В качестве метода очистки интересующего белка проводят очистку методом, описанным в Protein Experimentation Note (первый том) - Extraction, Separation and Expression of Recombinant Protein (перевод учебника, написанного на японском языке) (под редакцией Masato Okada и Kaori Miyazaki, опубликовано Yodo-sha, ISBN 9784897069180), и тому подобными.
Полное содержание всех литературных источников, таких как научные документы, патенты, патентные заявки, цитируемых в данном документе, включено в данный документ посредством ссылки в той же степени, что и иллюстративно описанных источников, соответственно.
Настоящее изобретение описано выше путем демонстрации предпочтительных вариантов его осуществления ради простоты понимания. Далее в данном документе настоящее изобретение описано более конкретно на основании примеров, однако вышеупомянутые объяснения и следующие далее примеры приведены исключительно с целью иллюстрации и не предназначены для целей ограничения изобретения. Соответственно, объем изобретения не ограничен вариантами осуществления и примерами, которые конкретно описаны в настоящей спецификации, но ограничен только формулой изобретения.
Различные экспериментальные методы, относящиеся к рекомбинации, которые описаны далее, такие как клонирование и тому подобное, осуществляли в соответствии с методами генетической инженерии, описанными в Molecular Cloning, 2е издание, под редакцией J. Sambrook, E. F. Frisch и T. Maniatis, Current Protocols in Molecular Biology, под редакцией Frederick M. Ausubel et al., опубликованных Current Protocols, и тому подобного.
ПРИМЕРЫ
Пример 1
Получение транспозонного вектора для экспрессии антитела против М2 вируса гриппа человека
Плазмиду, содержащую генную экспрессионную кассету для клеток млекопитающего, включающую произвольный ген человеческого антитела и ген маркера устойчивости к лекарственному средству, вставленные между парой транспозонных последовательностей Tol2, использовали в качестве плазмидного вектора для экспрессии белка.
Каждая ДНК используемых генов была химически и искусственно синтезирована на основании известной нуклеотидной последовательности или получена путем создания праймеров для обеих ее концевых последовательностей с последующим проведением ПЦР при использовании соответствующего источника ДНК в качестве матрицы. Для последующего проведения манипуляций с генами сайт рестрикции для фермента рестрикции был добавлен к концу праймера.
В нуклеотидной последовательности (SEQ ID NO: 1) неавтономного транспозона Tol2, раскрытой в японской опубликованной нерассмотренной патентной заявке № 235575/2003, нуклеотидную последовательность в положениях 1-200 (последовательность Tol2-L) (SEQ ID NO: 2) и нуклеотидную последовательность в положениях 2285-2788 (последовательность Tol2-R) (SEQ ID NO: 3) использовали в качестве транспозонных последовательностей.
Каждый фрагмент синтетической ДНК, содержащий пару транспозонных последовательностей (производитель TAKARA BIO INC.), получали следующим методом. Получали фрагмент ДНК, содержащий нуклеотидную последовательность, в которой последовательность узнавания для фермента рестрикции NruI была присоединена как к 5'-концу, так и к 3'-концу последовательности Tol2-R. Затем получали фрагмент ДНК, содержащий нуклеотидную последовательность, в которой последовательность узнавания для фермента рестрикции FseI была присоединена к 5'-концу последовательности Tol2-L, а для фермента рестрикции AscI была присоединена к ее 3'-концу.
Затем полученные таким образом фрагменты ДНК, содержащие последовательность Tol2-R и последовательность Tol2-L, встраивали в экспрессионный вектор N5LG1_M2_Z3 (WO2006/061723), содержащий нуклеотидную последовательность, кодирующую аминокислотную последовательность антитела Z3G1 против М2 вируса гриппа человека.
Вектор N5LG1_M2_Z3 (WO2006/061723), в который нуклеотидная последовательность (SEQ ID NO: 9), кодирующая H-цепь (SEQ ID NO: 10), и нуклеотидная последовательность (SEQ ID NO: 11), кодирующая L-цепь (SEQ ID NO: 12) антитела Z3G1 против М2 вируса гриппа человека (депозитный № в ATCC PTA-5968: депонировано 13 марта 2004 г., Американская коллекция типовых культур, Manassas, VA, USA), были встроены под контролем энхансера/промотора CMV, использовали в качестве экспрессионной кассеты гена антитела.
Фрагмент ДНК, содержащий последовательность Tol2-R, вставляли в сайт фермента рестрикции NruI, расположенный на 5'-конце генного фрагмента, содержащего экспрессионную кассету гена антитела и ген селектируемого маркера в векторе N5LG1_M2_Z3. Затем фрагмент ДНК, содержащий последовательность Tol2-L, вставляли в сайты фермента рестрикции FseI и AscI, расположенные на 3'-конце.
Кроме того, был сконструирован транспозонный вектор для экспрессии антитела против М2 вируса гриппа человека (фигура 1) путем встраивания экспрессионной кассеты гена устойчивости к циклогексимиду, в которой нуклеотидная последовательность (SEQ ID NO: 5), кодирующая ген устойчивости к циклогексимиду (ген, в котором пролин в положении 54 человеческого рибосомного белка L36a заменен глютамином), введена под контролем энхансера/промотора CMV в сайт узнавания FseI вектора N5LG1_M2_Z3, связанного с транспозонной последовательностью Tol2.
С другой стороны, вектор, не содержащий транспозонных последовательностей, был назван экспрессионным вектором антитела против М2 вируса гриппа человека и использован в качестве контрольного вектора (фигура 2).
Пример 2
Получение экспрессионного вектора транспозазы
Транспозазу экспрессировали, используя экспрессионный вектор, независимый от экспрессионного вектора для интересующего антитела. То есть ген, кодирующий Tol2 транспозазу (SEQ ID NO: 4) из рыбы оризии, был вставлен ниже CAGGS промотора в векторе pCAGGS (Gene, 108, 193-200, 1991) для получения экспрессионного вектора Tol2 транспозазы (далее в данном документе называемого вектором Tol2) (фигура 3).
Пример 3
Получение трансформанта с использованием клетки животного-млекопитающего
(1) Получение суспензионной клетки CHO
Прикрепляющиеся клетки CHO, которые культивировали, используя среду α-MEM (Invitrogen), содержащую 10% сыворотки (FCS), отделяли от подложки обработкой трипсином, а затем извлекали в свежую среду α-MEM, содержащую 10% сыворотки, с последующим встряхиванием культуры при 37°C в инкубаторе с 5% CO2. Через несколько дней подтверждали рост этих клеток, а затем проводили культивирование при встряхивании путем инокуляции их в среду α-MEM, содержащую 5% FCS, в концентрации 2×105 клеток/мл с последующим встряхиванием культуры.
Спустя еще несколько дней после этого, проводили инокуляцию аналогичным образом, используя среду α-MEM, содержащую 5% FCS. В конечном итоге получали клетки, адаптированные к суспензионному культивированию, путем повторного субкультивирования и встряхивания культуры, используя бессывороточную среду α-MEM и подтверждая, что клетки обладают такой же способностью к росту, что и в случае их культивирования в присутствии сыворотки.
(2) Получение клетки CHO, продуцирующей антитело
В качестве экспрессионного вектора использовали транспозонный вектор для экспрессии антитела против М2 вируса гриппа человека, полученный в примере 1 и примере 2 (далее в данном документе называемый транспозонным вектором), и вектор Tol2 pCAGGS-T2TP (фигура 3, Kawakami K. & Noda Т., Genetics, 166, 895-899 (2004)). Кроме того, экспрессионный вектор антитела против М2 вируса гриппа человека, лишенный транспозонных последовательностей, использовали в качестве контроля.
Вводя вышеуказанные экспрессионные векторы в адаптированную к суспензионному культивированию клетку CHO-K1 (Американская коллекция типовых культур, каталожный № CCL-61) или клетку HEK293 (Invitrogen, клетка FreeStyle 293F), сравнивали частоты получения устойчивых к циклогексимиду клонов.
Клетки каждого вида (4×106 клеток) суспендировали в 400 мкл PBS, и транспозонный вектор для экспрессии антитела против М2 вируса гриппа человека (10 мкг) и вектор Tol2 (25 мкг) совместно трансфицировали напрямую в форме кольцевой ДНК методом электропорации. В связи с этим, для того, чтобы экспрессировать Tol2 транспозазу временно, вектор Tol2 был напрямую введен в форме кольцевой ДНК с целью предотвращения интеграции в хромосому хозяина.
Кроме того, в качестве контроля, экспрессионный вектор антитела против М2 вируса гриппа человека (10 мкг) был линеаризован при помощи фермента рестрикции, а затем введен в клетки каждого вида стандартным методом введения генов при помощи электропорации.
Электропорацию проводили, используя кювету с шириной зазора 4 мм (производитель Bio-Rad), при помощи электропоратора (Gene Pulser Xcell System (производитель Bio-Rad)) в условиях напряжения 300 В, электростатической мощности 500 мкФ и комнатной температуры.
После введения генов электропорацией клетки каждого вида высевали в три 96-луночных планшета и культивировали в CO2-инкубаторе в течение 3 дней, используя среду EX-CELL 325-PF от производителя SAFC Biosciences для клеток CHO и среду FreeStyle-293 (производитель Invitrogen) для клеток HEK293.
Затем, начиная со дня смены среды на 4-й день после введения генов, 3 мкг/мл циклогексимида добавляли к среде так, что клетки культивировались в присутствии циклогексимида, после чего культивировали в течение 3 недель, еженедельно производя смену среды.
После культивирования в течение 3 недель подсчитывали число лунок, в которых были обнаружены устойчивые к циклогексимиду колонии. Результаты приведены в таблице 1 и таблице 2.
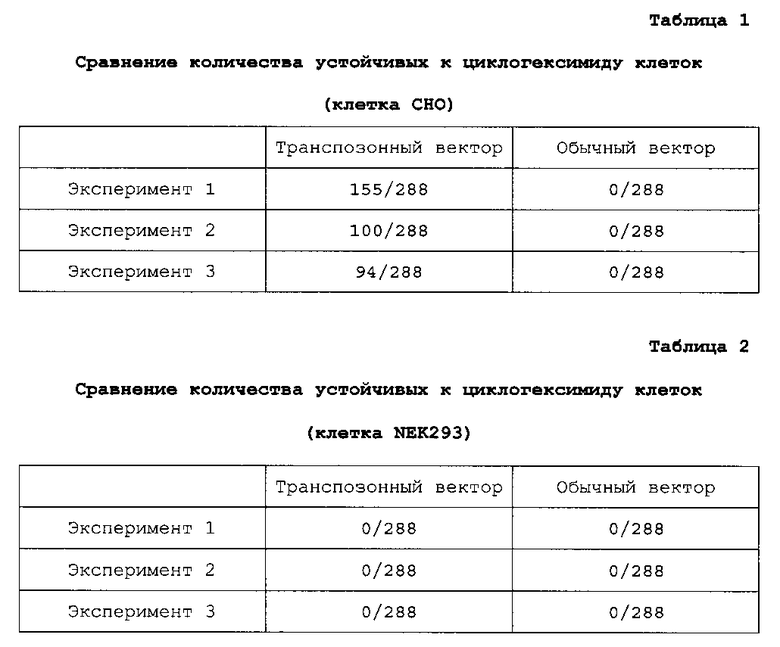 Как видно из таблицы 1, каждый из экспрессионного транспозонного вектора антитела против М2 вируса гриппа человека или экспрессионного вектора антитела против М2 вируса гриппа человека вводили в суспензионную клетку CHO-К1. В результате, устойчивые к циклогексимиду трансформанты не были получены из клеток, в которые был введен экспрессионный вектор антитела против М2 вируса гриппа человека, как и в других клеточных линиях, однако устойчивые к циклогексимиду трансформанты были получены с высокой частотой из клеток, в которые был введен транспозонный вектор для экспрессии антитела против М2 вируса гриппа человека.
Как видно из таблицы 1, каждый из экспрессионного транспозонного вектора антитела против М2 вируса гриппа человека или экспрессионного вектора антитела против М2 вируса гриппа человека вводили в суспензионную клетку CHO-К1. В результате, устойчивые к циклогексимиду трансформанты не были получены из клеток, в которые был введен экспрессионный вектор антитела против М2 вируса гриппа человека, как и в других клеточных линиях, однако устойчивые к циклогексимиду трансформанты были получены с высокой частотой из клеток, в которые был введен транспозонный вектор для экспрессии антитела против М2 вируса гриппа человека.
С другой стороны, как видно из таблицы 2, устойчивые к циклогексимиду трансформанты не были получены, когда либо транспозонный вектор для экспрессии антитела против М2 вируса гриппа человека, либо экспрессионный вектор антитела против М2 вируса гриппа человека были введены в клетки HEK293.
На основании данных результатов было установлено, что ген, кодирующий интересующий белок, и ген устойчивости к циклогексимиду, которые были вставлены между парой транспозонных последовательностей, эффективно встроились в хромосому клетки-хозяина в суспензионной клетке млекопитающего.
(3) Изучение продукции антитела в суспензионной клетке CHO и прикрепляющейся клетке CHO
Чтобы изучить эффективность продукции антитела в суспензионной клетке CHO или прикрепляющейся клетке CHO, изучали количество антител, продуцируемых каждой клеточной линией. В качестве суспензионной клетки CHO использовали суспензионную клетку CHO-K1, адаптированную к суспензионному культивированию. Кроме того, в качестве прикрепляющейся клетки CHO использовали прикрепляющуюся клетку CHO-K1 до адаптации к суспензионному культивированию.
Экспрессионный транспозонный вектор антитела против М2 вируса гриппа человека (10 мкг) и вектор Tol2 (25 мкг) вводили электропорацией в суспензионную клетку CHO-K1 и прикрепляющуюся клетку CHO-K1, соответственно. Затем суспензионную клетку CHO-K1 и прикрепляющуюся клетку CHO-K1 инокулировали в три 96-луночных планшета для каждого вида клеток.
Среду для суспензионных клеток (EX-CELL 325-PF, производитель SAFC Biosciences) использовали для суспензионной клетки CHO-K1 и среду α-MEM, содержащую 10% сыворотки, использовали для прикрепляющейся клетки CHO-K1. Клетку каждого вида культивировали в CO2-инкубаторе в течение 3 дней. Начиная со дня смены среды на 4-й день после электропорации, 3 мкг/мл циклогексимида добавляли к среде так, что клетки культивировались в присутствии циклогексимида, и клетки дополнительно культивировали в течение 3 недель. В этом случае смену среды производили каждую неделю.
В случае суспензионной клетки CHO-K1 1×106 клеток высевали в 6-луночный планшет с последующим культивированием при встряхивании в CO2-инкубаторе в течение 3 дней, и количество белка антитела измеряли методом ВЭЖХ, используя культуральный супернатант.
В случае прикрепляющейся клетки CHO-K1 смену среды производили, когда клетки достигали конфлюэнтности в 6-луночном планшете (2×106 клеток), и после статического культивирования в течение 3 дней количество белка антитела измеряли методом ВЭЖХ, используя культуральный супернатант.
Концентрацию антитела в культуральном супернатанте измеряли по методу, описанному в Yeast Res., 7 (2007), 1307-1316. Результаты приведены на фигуре 4.
Как показано на фигуре 4A, большое количество клеток, демонстрирующих заметно высокий уровень экспрессии антитела, было получено при использовании клетки CHO-K1, адаптированной для суспензионного культивирования. С другой стороны, как показано на фигуре 4B, только клетки, демонстрирующие уровень экспрессии антитела на пределе обнаружения методом ВЭЖХ (5 мкг/мл) или менее, были получены при использовании прикрепляющейся клетки CHO-K1.
На основании данных результатов установлено, что если нужно экспрессировать интересующий белок с использованием транспозонного вектора, интересующий белок может быть экспрессирован на высоком уровне, если использовать суспензионную клетку млекопитающего.
Кроме того, исходя из результатов примеров 1-3, установлено, что способ по изобретению можно использовать в качестве нового способа получения интересующего белка путем эффективного создания продуцирующей клетки, которая способна на высоком уровне экспрессировать экзогенный ген, с использованием суспензионной клетки млекопитающего, адаптированной к суспензионному культивированию.
Пример 4
Получение экспрессирующей антитело клетки с использованием Tol1 транспозона и получение антитела
(1) Получение Tol1 транспозонного вектора для экспрессии антитела против М2 вируса гриппа человека
Таким же образом, как и в примере 1, плазмиду, которая содержала генную экспрессионную кассету для клеток млекопитающего, содержащую произвольный ген человеческого антитела и ген маркера устойчивости к лекарственному средству, вставленные между парой Tol1 транспозонных последовательностей, использовали в качестве экспрессионного плазмидного вектора для белка.
Каждую ДНК используемых генов химически синтезировали искусственным образом на основании известной информации о последовательности, либо получали путем создания праймеров для обеих ее концевых последовательностей с проведением ПЦР при использовании соответствующего источника ДНК в качестве матрицы. С целью последующего проведения манипуляций с генами, сайт расщепления для ферментов рестрикции был добавлен к концу праймера.
В нуклеотидной последовательности неавтономного транспозона Tol1, представленной SEQ ID NO: 13 в списке последовательностей (WO2008/072540), нуклеотидную последовательность в положениях 1-200 (последовательность Tol1-L) (SEQ ID NO: 14) и нуклеотидную последовательность в положениях 1351-1855 (последовательность Tol1-R) (SEQ ID NO: 15) использовали в качестве транспозонных последовательностей.
Каждый из фрагментов синтетической ДНК, содержащих пару транспозонных последовательностей, получали следующим методом. Получали фрагмент ДНК, содержащий нуклеотидную последовательность, в которой последовательность узнавания для фермента рестрикции NruI была присоединена как к 5'-концу, так и к 3'-концу последовательности Tol1-R. Затем получали фрагмент ДНК, содержащий нуклеотидную последовательность, в которой последовательность узнавания для фермента рестрикции FseI была присоединена к 5'-концу последовательности Tol1-L, а для фермента рестрикции AscI была присоединена к ее 3'-концу.
Затем полученные таким образом фрагменты ДНК, содержащие последовательность Tol1-R и последовательность Tol1-L, встраивали в экспрессионный вектор N5LG1_M2_Z3. Фрагмент ДНК, содержащий последовательность Tol1-R, встраивали в сайт фермента рестрикции NruI, находящийся на 5'-конце генного фрагмента, содержащего экспрессионную кассету гена антитела и ген селектируемого маркера в векторе N5LG1_M2_Z3, а фрагмент ДНК, содержащий последовательность Tol1-L, встраивали в сайты ферментов рестрикции FseI и AscI, находящиеся на 3'-конце.
Кроме того, был сконструирован Tol1 транспозонный вектор для экспрессии антитела против М2 вируса гриппа человека (фигура 5) путем встраивания экспрессионной кассеты гена устойчивости к циклогексимиду, в которой ген устойчивости к циклогексимиду (ген, в котором пролин в положении 54 человеческого рибосомного белка L36a был заменен глютамином) введен под контролем энхансера/промотора CMV в сайт узнавания FseI вектора N5LG1_M2_Z3, связанного с Tol1 транспозонной последовательностью.
(2) Получение экспрессионного вектора Tol1 транспозазы
Транспозазу экспрессировали, используя экспрессионный вектор, независимый от экспрессионного вектора для интересующего антитела. То есть, экспрессионную кассету гена Tol1 транспозазы, в которой фрагмент ДНК, кодирующий Tol1 транспозазу из рыбы оризии (SEQ ID NO: 17), состоящий из нуклеотидной последовательности, представленной SEQ ID NO: 16, был введен под контролем энхансера/промотора CMV, встраивали в pBluescriptII SK (+) (производитель Stratagene), который затем использовали в качестве экспрессионного вектора Tol1 транспозазы pTol1ase (фигура 6).
(3) Получение клетки CHO, продуцирующей антитело
Используя экспрессионные векторы, полученные в вышеуказанных пунктах (1)-(3), эффективность введения экспрессионного вектора при помощи транспозона Tol1 изучали таким же образом, что и в примере 3. Результат приведен в таблице 3.
Как видно из таблицы 3, когда Tol1 транспозонный вектор для экспрессии антитела против М2 вируса гриппа человека был введен в суспензионную клетку CHO-K1, устойчивые к циклогексимиду трансформанты были получены с высокой частотой, как и в случае примера 3, в котором был введен Tol2 транспозонный вектор для экспрессии антитела против М2 вируса гриппа человека.
На основании данных результатов было установлено, что ген антитела и ген устойчивости к циклогексимиду, вставленные между парой транспозонных последовательностей, эффективно трансдуцируются в хромосому клетки-хозяина, а именно, суспензионной клетки млекопитающего, также и в случае использования Tol1 транспозона.
(4) Изучение продукции антитела суспензионной клеткой CHO
Эффективность продукции антитела суспензионной клеткой CHO изучали, используя транспозон Tol1 таким же образом, что и в примере 3(3).
Концентрацию антитела в культуральном супернатанте измеряли методом, описанным в FEMS Yeast Res., 7 (2007), 1307-1316. Результаты приведены на фигуре 7.
Как видно из фигуры 7, большое число клеток, демонстрирующих заметно высокий уровень экспрессии антитела, было также получено в случае использования транспозона Tol1. На основании данного результата было установлено, что аналогично случаю использования нуклеотидной последовательности, полученной из транспозона Tol2, суспензионную клетку млекопитающего, способную на высоком уровне экспрессировать интересующий белок, можно также получать, если в качестве транспозонной последовательности использовать нуклеотидную последовательность, полученную из транспозона Tol1.
Пример 5
Получение антитела против CD98 человека
(1) Получение экспрессионного транспозонного вектора тяжелой цепи антитела против CD98 человека и экспрессионного транспозонного вектора легкой цепи антитела против CD98 человека
Для получения антитела против CD98 человека, имеющего H-цепь и L-цепь с вариабельной областью, представленные аминокислотными последовательностями SEQ ID NO: 20 и 23, соответственно, аминокислотные последовательности H-цепи и L-цепи были получены путем соединения аминокислотной последовательности константной области антитела IgG1 человека с каждой вариабельной областью антитела.
С использованием последовательностей, встроенных в вектор (N5KG1-Val C2IgG1NS/I117L), раскрытых в японском патенте № 4324637 в качестве генных последовательностей (SEQ ID NO: 18 и 21, соответственно) вариабельной области тяжелой цепи и вариабельной области легкой цепи антитела против CD98 человека, к которым присоединена сигнальная последовательность, и с использованием транспозонной последовательности и промотора, аналогичных тем, что были использованы в примере 1, были, соответственно, сконструированы экспрессионный транспозонный вектор тяжелой цепи антитела против CD98 человека (далее в данном документе называемый вектором CD98H) и экспрессионный транспозонный вектор легкой цепи антитела против CD98 человека (далее в данном документе называемый вектором CD98L), (фигуры 8 и 9).
Используемый фрагмент ДНК был химически синтезирован искусственным образом на основании общеизвестной последовательности или получен путем создания праймеров для обеих ее концевых последовательностей и проведения ПЦР с использованием соответствующего источника ДНК в качестве матрицы. Сайт расщепления для фермента рестрикции был присоединен к концу каждого праймера для последующих рекомбинационных процедур с генами.
(2) Получение экспрессионного транспозонного вектора гена устойчивости к циклогексимиду
Экспрессионный транспозонный вектор гена устойчивости к циклогексимиду (далее в данном документе называемый вектором CHX) был сконструирован путем присоединения последовательности, кодирующей ген устойчивости к циклогексимиду (SEQ ID NO: 7), под контролем энхансера/промотора CMV, описанного в примере 1, и встраивания пары транспозонных последовательностей (Tol2-L, Tol2-R) на обоих концах экспрессионной кассеты гена устойчивости к циклогексимиду (фигура 10).
Используемый фрагмент ДНК был искусственным образом химически синтезирован на основании общеизвестной последовательности, либо получен путем создания праймеров для обеих ее концевых последовательностей с последующим проведением ПЦР при использовании соответствующего источника ДНК в качестве матрицы. Сайт расщепления для фермента рестрикции был присоединен к концу каждого праймера для последующих рекомбинационных процедур с генами.
(3) Получение клетки CHO, продуцирующей антитело против CD98 человека
Вектор CD98H (фигура 8), вектор CD98L (фигура 9) и вектор CHX (фигура 10), полученные в вышеуказанных пунктах (1) и (2), и вектор Tol2 (фигура 3), полученный в примере 2, вводили в клетку CHO-K1, адаптированную к суспензионному культивированию, и сравнивали количество появившихся клеток, способных на высоком уровне экспрессировать антитело.
В экспериментальной схеме 4×106 клеток клетки CHO-K1 суспендировали в 400 мкл PBS, и вектор CD98H (10 мкг), вектор CD98L (10 мкг), вектор CHX (10 мкг) и вектор Tol2 (10 мкг) напрямую совместно трансфицировали в форме кольцевой ДНК методом электропорации. Для того чтобы экспрессия Tol2 транспозазы была временной и чтобы предотвратить интеграцию в хромосому хозяина, вектор Tol2 вводили напрямую в форме кольцевой ДНК. Электропорацию проводили при помощи электропоратора (Gene Pulser Xcell System (производитель Bio-Rad)) в условиях напряжения 300 В, электростатической мощности 500 мкФ и комнатной температуры, используя кювету с шириной зазора 4 мм (производитель Bio-Rad).
Кроме того, в контрольной схеме каждый из вектора CD98H (10 мкг), вектора CD98L (10 мкг) и вектора CHX (10 мкг) линеаризовали, используя фермент рестрикции PciI (Takara Bio Inc.), а затем проводили электропорацию таким же образом, как описано выше.
После введения генов электропорацией клетки в каждой кювете суспендировали в среде CD OptiCHO с добавлением 0,5% гидролизата соевых бобов (далее в данном документе называемой средой 0,5CD), инокулировали в один 96-луночный планшет и культивировали в течение 4 дней в CO2-инкубаторе. Затем, начиная со смены среды через 5 дней после введения генов, проводили культивирование в присутствии циклогексимида, используя среду 0,5CD с добавлением 3 мкг/мл циклогексимида (C4859, Sigma-Aldrich), и продолжали культивирование в течение 4 недель, меняя среду с интервалами в одну неделю.
Через 4 недели культивирования экспрессию антитела определяли сэндвич-методом (LENCETM, Perkin-Elmer Corp), используя FRET (резонансный перенос энергии флуоресценции). Что касается клеток, экспрессирующих антитело на высоком уровне, то клоны, экспрессирующие антитело с концентрацией в культуральном супернатанте, составляющей 5,0 мкг/мл или более, считали экспрессирующими антитело клетками, результаты приведены в таблице 4.
Как видно из таблицы 4, большое количество клеток, экспрессирующих антитело против CD98 человека, было обнаружено в экспериментальной схеме, в которой вектор Tol2 был ко-трансфицирован в суспензионную клетку CHO-K1 совместно с экспрессионным транспозонным вектором тяжелой цепи антитела против CD98 человека, экспрессионным транспозонным вектором легкой цепи антитела против CD98 человека и вектором гена устойчивости к циклогексимиду, однако клетки, экспрессирующие антитело против CD98 человека, не были обнаружены в контрольной схеме, в которой вектор Tol2 не был совместно трансфицирован, несмотря на то, что векторы были преобразованы в линейные цепи.
Пример 6
Получение антитела против CD98 человека
(1) Получение экспрессионного транспозонного вектора, содержащего генный фрагмент тяжелой цепи антитела против CD98 человека, генный фрагмент легкой цепи антитела против CD98 человека и ген устойчивости к циклогексимиду
Экспрессионный транспозонный вектор, содержащий генный фрагмент тяжелой цепи антитела против CD98 человека, генный фрагмент легкой цепи антитела против CD98 человека и ген устойчивости к циклогексимиду (далее в данном документе называемый тандемным вектором CD98-CHX), был сконструирован с использованием синтетической ДНК и метода ПЦР таким же образом, как описано выше, путем соединения экспрессионного транспозонного вектора тяжелой цепи антитела против CD98 человека, полученного в примере 5(1), с экспрессионной кассетой гена легкой цепи антитела против CD98 человека, полученной в примере 5(1), и кассетой гена устойчивости к циклогексимиду, полученной в примере 5(2).
(2) Получение экспрессионного транспозонного вектора, содержащего генный фрагмент тяжелой цепи антитела против CD98 человека и ген устойчивости к циклогексимиду
Экспрессионный транспозонный вектор, содержащий генный фрагмент тяжелой цепи антитела против CD98 человека и ген устойчивости к циклогексимиду (далее в данном документе называемый экспрессионным транспозонным вектром CD98H-CHX), был сконструирован с использованием синтетической ДНК и метода ПЦР таким же образом, как описано выше, путем соединения экспрессионного транспозонного вектора тяжелой цепи антитела против CD98 человека, полученного в примере 5(1), с кассетой гена устойчивости к циклогексимиду, полученной в примере 5(2).
(3) Получение клетки CHO, продуцирующей антитело против CD98 человека
Используя экспрессионные транспозонные векторы, полученные в описанном выше примере 5(1) и (2) и в описанном выше примере 6(1) и (2), частоту появления клеток, способных на высоком уровне экспрессировать антитело против CD98 человека, сравнивали в случае переноса генов H-цепи и L-цепи антитела против CD98 человека с использованием одного и того же экспрессионного вектора (контрольная схема), в случае переноса генов H-цепи или L-цепи антитела против CD98 человека или гена устойчивости к циклогексимиду, соответственно, с использованием различных экспрессионных векторов (экспериментальная схема 1) и в случае переноса генов H-цепи или L-цепи с использованием различных экспрессионных векторов (экспериментальная схема 2).
В экспериментальной схеме 1 4×106 клеток клетки CHO-K1 суспендировали в 400 мкл PBS, и вектор CD98H (10 мкг), вектор CD98L (10 мкг), вектор CHX (10 мкг) и вектор Tol2 (10 мкг) напрямую совместно трансфицировали в виде кольцевой ДНК методом электропорации.
В экспериментальной схеме 2 4×106 клеток клетки CHO-K1 суспендировали в 400 мкл PBS, и вектор CD98H-CHX (10 мкг), вектор CD98L (10 мкг) и вектор Tol2 (10 мкг) напрямую совместно трансфицировали в виде кольцевой ДНК методом электропорации.
В контрольной схеме 4×106 клеток клетки CHO-K1 суспендировали в 400 мкл PBS, и тандемный вектор CD98-CHX (10 мкг) и вектор Tol2 (20 мкг) напрямую совместно трансфицировали в виде кольцевой ДНК методом электропорации. Кроме того, во всех экспериментах, для того, чтобы экспрессия Tol2 транспозазы была временной и чтобы предотвратить интеграцию в хромосому хозяина, вектор Tol2 был введен напрямую в форме кольцевой ДНК.
В следующем методе частоту появления продуцирующих антитело клеток подтверждали таким же образом, как и в примере 5(3). Что касается клеток, продуцирующих антитело, то клоны, у которых концентрация антитела в культуральном супернатанте составляла 3,0 мкг/мл или более, считали экспрессирующими антитело клетками. Результаты приведены в таблице 5.
В экспериментальной схеме 1, в которой вводили вектор CD98H, вектор CD98L и вектор CHX, и экспериментальной схеме 2, в которой вводили вектор CD98H и вектор CD98L, частота появления клеток, способных на высоком уровне экспрессировать антитело против CD98 человека, заметно увеличилась.
Приведенные выше результаты свидетельствуют о том, что клетки, обладающие высокой продуктивностью в отношении антитела, можно легко получать и производить, когда различные экспрессионные векторы, в которых ген тяжелой цепи антитела и ген легкой цепи антитела, соответственно, вставлены между транспозонными последовательностями, совместно трансфицированы в суспензионную клетку CHO, по сравнению со случаем, когда экспрессионный вектор, полученный путем интеграции гена тяжелой цепи антитела и гена легкой цепи антитела в один и тот же экспрессионный вектор, вводят в суспензионную клетку CHO. Кроме того, результаты экспериментальной схемы 1 и экспериментальной схемы 2 показали, что даже когда векторов для введения два или более, достаточно по меньшей мере одного гена устойчивости к лекарственному средству (гена селектируемого маркера). Кроме того, было обнаружено, что ген устойчивости к лекарственному средству может присутствовать на экспрессионном векторе, в который встроен ген тяжелой цепи антитела, либо на другом независимом векторе.
Приведенные выше результаты демонстрируют, что транспозонный вектор эффективен в качестве средства для эффективного введения генов, расположенных на двух или более векторах, в суспензию клеток млекопитающего, что обычно достигается с трудом. Кроме того, показано, что с целью достижения высокой продуктивности в отношении белка, содержащего более одного полипептида, или более чем одного белка, эффективно вводить полипептиды и белки, используя различные транспозонные векторы.
(4) Культивирование клетки CHO, продуцирующей антитело против CD98 человека
Лучшие три линии клеток с высокой продуктивностью для антитела были выбраны из клеток, в которые был введен тандемный вектор CD98-CHX, полученный в вышеупомянутом примере 6(3), и клеток, в которые были введены вектор CD98H-CHX и вектор CD98L, и было проведено сравнение их уровней экспрессии антитела. Подробности экспериментов приведены ниже.
Клетку CHO-K1, полученную в примере 6(3), выбранную на основании устойчивости к циклогексимиду и также экспрессирующую анти-CD98 антитело, культивировали для наращивания, используя 96-луночный планшет, 24-луночный планшет и 6-луночный планшет (Corning Glassworks) именно в таком порядке. После наращивания измеряли концентрацию антитела в супернатанте каждой культуры и выбирали наилучшие три клеточных линии клеток CHO с высоким уровнем экспрессии анти-CD98 антитела. Затем каждую из выбранных таким образом трех клеточных линий суспендировали в 3 мл среды 0,5% CD (Invitrogen), а именно среды 0,5CD, до плотности 2×105 клеток/мл и культивировали на шейкере в течение 5 дней при 37°C в атмосфере 5% CO2, используя 6-луночный планшет. Количество антитела в среде через 5 дней культивирования определяли методом ВЭЖХ (Waters Associates, Inc.). Результаты приведены в таблице 6.
Как видно из таблицы 6, клетка CHO-K1, в которую были совместно трансфицированы вектор CD98H и вектор CD98L, демонстрирует высокий уровень продукции антитела по сравнению с клеткой CHO-K1, в которую был введен тандемный вектор CD98-CHX.
Приведенные выше результаты свидетельствуют не только о том, что можно с легкостью получать и производить клеточную линию, продуцирующую антитело на высоком уровне, но также о том, что полученная таким образом клетка имеет высокую продуктивность для антитела, когда различные экспрессионные векторы, в которых, соответственно, ген тяжелой цепи антитела и ген легкой цепи антитела вставлены между парой транспозонных последовательностей, совместно трансфицированы в суспензионную клетку CHO.
Пример 7
Получение антитела против фактора некроза опухоли-альфа (TNFα) человека
(1) Получение экспрессионного транспозонного вектора, содержащего генный фрагмент тяжелой цепи антитела к TNFα, генный фрагмент легкой цепи антитела к TNFα и ген устойчивости к циклогексимиду
Для получения антитела против TNFα человека с аминокислотной последовательностью SEQ ID NO: 26 и SEQ ID NO: 29 был сконструирован экспрессионный транспозонный вектор с генным фрагментом тяжелой цепи антитела против TNFα человека, генным фрагментом легкой цепи антитела против TNFα человека и геном устойчивости к циклогексимиду (далее в данном документе называемый тандемным вектором TNFα-CHX) путем замены генных фрагментов VH и VL экспрессионного транспозонного вектора, содержащего генный фрагмент тяжелой цепи и генный фрагмент легкой цепи антитела против CD98 человека и ген устойчивости к циклогексимиду, полученного в примере 6(1) (тандемный вектор CD98-CHX) на VH и VL из антитела против TNFα человека, соответственно.
Последовательности гена тяжелой цепи и гена легкой цепи антитела против TNFα человека получали с использованием синтетической ДНК, получая аминокислотные последовательности (SEQ ID NO: 26 и 29), в которых сигнальная последовательность присоединена к аминокислотным последовательностям (SEQ ID NO: 25 и 28) субъединицы вариабельной области тяжелой цепи или субъединицы вариабельной области легкой цепи адалимумаба (рекомбинантного), изображенных на фигуре 1 и фигуре 2, соответственно, в инспекционном отчете о подкожной инъекции 40 мг HUMIRA(R) (Агентство фармацевтических препаратов и медицинских устройств, 14 февраля 2008 г.), и определяя нуклеотидные последовательности таким образом, что аминокислотные последовательности не менялись (SEQ ID NO: 24 и 27). Для последующих манипуляций с генами на конце искусственных последовательностей добавляли сайт расщепления для ферментов рестрикции.
(2) Получение экспрессионного транспозонного вектора, содержащего фрагмент тяжелой цепи антитела против TNFα человека и ген устойчивости к циклогексимиду
Экспрессионный транспозонный вектор, содержащий фрагмент тяжелой цепи антитела против TNFα человека и ген устойчивости к циклогексимиду (далее в данном документе называемый вектором TNFαH-CHX), был сконструирован путем изменения области генного фрагмента VH экспрессионного транспозонного вектора, содержащего генный фрагмент тяжелой цепи антитела против CD98 человека и ген устойчивости к циклогексимиду (вектор CD98H-CHX), полученного в примере 6(2), на генный фрагмент VH антитела к TNFα. В качестве гена тяжелой цепи антитела против TNFα человека использовали последовательность той же последовательности, которая приведена в данном пункте (1).
(3) Получение экспрессионного транспозонного вектора гена легкой цепи антитела против TNFα человека
Экспрессионный транспозонный вектор гена легкой цепи антитела против TNFα человека (далее в данном документе называемый вектором CD98L) был сконструирован путем изменения области гена легкой цепи экспрессионного транспозонного вектора гена легкой цепи антитела против CD98, полученного в примере Example 6(1), на легкую цепь антитела против TNFα человека. В качестве гена VL антитела против TNFα человека использовали ту же последовательность, что и последовательность, приведенная в данном пункте (1).
(4) Получение клетки CHO, продуцирующей антитело против TNFα человека
Для получения клетки CHO-K1, продуцирующей антитело против TNFα человека, тандемный вектор TNFα-CHX (20 мкг), полученный в вышеуказанном пункте (1), и экспрессионный вектор Tol2 транспозазы (вектор Tol2) (10 мкг), полученный в примере 2, вводили в клетку CHO-K1, адаптированную к суспензионному культивированию, полученную в примере 3 (контрольная схема).
Таким же образом, вектор TNFαH-CHX (10 мкг), вектор TNFαL (10 мкг) и вектор Tol2 (10 мкг), полученные в вышеуказанных пунктах (2) и (3), были напрямую совместно трансфицированы в форме кольцевой ДНК (экспериментальная схема). Частоту появления клеток, способных на высоком уровне экспрессировать антитело, сравнивали, проводя введение генов, культивирование клеток и тому подобное таким же образом, как в примере 6, за исключением того, что культивирование клеток с введенными генами проводили на пяти 96-луночных планшетах. Что касается клеток с высокой продуктивностью для антитела, то клоны, у которых концентрация антитела в культуральном супернатанте составляла 3,0 мкг/мл или более, считали экспрессирующими антитело клетками. Результаты приведены в таблице 7.
Как видно из таблицы 7, как и в случае клетки, продуцирующей антитело против CD98 человека, полученной в примере 6, для клетки CHO-K1, в которую вектор TNFαH-CHX и вектор TNFαL были совместно трансфицированы, наблюдали примерно в 4 раза более высокую частоту появления клеток, в которых антитело против TNFα человека экспрессировалось на высоком уровне, по сравнению с клеткой CHO-K1, в которую был введен тандемный вектор TNFα-CHX.
Данные результаты свидетельствуют о том, что в любом случае с антителом, линию клеток с высокой продуктивностью для антитела можно легко получать и производить путем совестного трансфицирования встроенных в различные экспрессионные векторы гена тяжелой цепи антитела и гена легкой цепи антитела, которые соответственно вставлены между парой транспозонных последовательностей, в суспензионную клетку CHO.
(5) Культивирование клетки CHO, продуцирующей антитело против TNFα человека
Клетки, выбранные на основании устойчивости к циклогексимиду из клеток с введенным тандемным вектором TNFα-CHX, полученные в вышеуказанном пункте (4), и клетки, в которые были совместно трансфицированы вектор TNFαH-CHX и вектор TNFαL, и также экспрессирующие антитело против TNFα человека, отбирали и культивировали для наращивания, используя 96-луночный планшет, 24-луночный планшет и 6-луночный планшет, именно в таком порядке. Что касается 4 клеточных линий из клеток, в которые был введен тандемный вектор TNFα-CHX, успешно размноженных при культивировании, и 52 клеточных линий, в которые были совместно трансфицированы вектор TNFαH-CHX и вектор TNFαL, эти клетки культивировали таким же образом, как в примере 6(4), за исключением того, что период культивирования составлял 7 дней, и измеряли уровни экспрессии антител. Результаты приведены на фигуре 11.
В результате, клетка CHO-K1, в которую были совместно трансфицированы вектор TNFαH-CHX и вектор TNFαL, продемонстрировала примерно в 2,4 раза более высокую продуктивность для антитела, чем продуктивность клетки CHO-K1, в которую был введен тандемный вектор TNFα-CHX.
Этот результат показывает, что, как и в случае примера 6(4), не только можно получать и производить клетку с высокой продуктивностью для антитела, но также полученная таким образом клетка имеет высокую продуктивность для антитела, когда различные экспрессионные векторы, в которых каждый из гена тяжелой цепи антитела и гена легкой цепи антитела, соответственно, вставлен между парой транспозонных последовательностей, совместно трансфицированы в суспензионную клетку CHO.
Пример 8
Получение антитела против CD20 человека
(1) Получение экспрессионного транспозонного вектора, содержащего генный фрагмент тяжелой цепи антитела против CD20 человека, генный фрагмент легкой цепи антитела против CD20 человека и ген устойчивости к циклогексимиду
Для получения антитела против CD20 человека, содержащего VH и VL, представленные аминокислотными последовательностями SEQ ID NO: 32 и 35, соответственно, экспрессионный транспозонный вектор, содержащий генный фрагмент тяжелой цепи антитела против CD20 человека, генный фрагмент легкой цепи антитела против CD20 человека и ген устойчивости к циклогексимиду (далее в данном документе называемый тандемным вектором CD98-CHX), был сконструирован путем замены областей генов VH и VL антитела в тандемном векторе CD98-CHX, полученном в примере 6(1) на VH и VL из антитела против CD20 человека, соответственно.
Последовательности генов области VH и области VL антитела против CD20 человека получали с использованием синтетической ДНК, получая нуклеотидную последовательность, описанную в регистрационном № GenBank AR000013, и аминокислотные последовательности (SEQ ID NO: 31 и 34, соответственно), в которых сигнальная последовательность была присоединена к аминокислотным последовательностям (SEQ ID NO: 32 и 35, соответственно) VH и VL ритуксимаба, описанным в сопроводительном листе к инспекционному отчету об инъекции 10 мг/мл Rituxan(R) (сообщение Национального института наук о здоровье, № 3395, 28 августа 2003 г.), и определяя нуклеотидные последовательности таким образом, что аминокислотные последовательности не менялись (SEQ ID NO: 30 и 33). Для последующих манипуляций с генами на конце искусственных последовательностей добавляли сайт расщепления для ферментов рестрикции.
(2) Получение экспрессионного транспозонного вектора, содержащего генный фрагмент тяжелой цепи антитела против CD20 человека и ген устойчивости к циклогексимиду
Экспрессионный транспозонный вектор, содержащий генный фрагмент тяжелой цепи антитела против CD20 человека и ген устойчивости к циклогексимиду (далее в данном документе называемый вектором CD20H-CHX), был сконструирован путем изменения области гена VH антитела в векторе CD98H-CHX, полученном в примере 6(2), на VH из антитела против CD20 человека. В качестве гена тяжелой цепи антитела против CD20 человека использовали ту же последовательность, что и последовательность, приведенная в вышеуказанном пункте (1).
(3) Получение экспрессионного транспозонного вектора гена легкой цепи антитела против CD20 человека
Экспрессионный транспозонный вектор гена легкой цепи антитела против CD20 человека (далее в данном документе называемый вектором CD20L) был сконструирован путем изменения областей гена VL антитела против CD98 человека, полученного в примере 6(1), на VL из антитела против CD20 человека. В качестве генов тяжелой и легкой цепи антитела против CD20 человека использовали те же последовательности, что и последовательность, приведенная в вышеуказанном пункте (1).
(4) Получение клетки CHO, продуцирующей антитело против CD20 человека
Для получения клетки CHO-K1, продуцирующей антитело против CD20 человека, тандемный вектор CD20-CHX, полученный в вышеуказанном пункте (1), и экспрессионный вектор Tol2 транспозазы (вектор Tol2), полученный в примере 2, были введены в клетку CHO-K1, адаптированную к суспензионному культивированию, полученную в примере 3(1) (контрольная схема).
Аналогичным образом, вектор CD20H-CHX (10 мкг) и вектор CD20L (10 мкг), полученные в вышеуказанных пунктах (2) и (3), были совместно трансфицированы в клетку CHO-K1, вместе с вектором Tol2 (10 мкг) (экспериментальная схема). Частоту появления клеток, способных на высоком уровне экспрессировать антитело, сравнивали, проводя введение генов, культивирование клеток и тому подобное таким же образом, что и в примере 6, за исключением того, что культивирование клеток с введенными генами проводили на пяти 96-луночных планшетах. Кроме того, лунки с концентрацией антитела 3,0 мкг/мл или более считали лунками с экспрессированным антителом. Результаты приведены в таблице 8.
В результате, клетка CHO-K1, в которую были совместно трансфицированы вектор CD20H-CHX и вектор CD20L, демонстрировала примерно в 3 раза более высокую частоту появления клеток, экспрессирующих на высоком уровне антитело против CD20 человека, по сравнению с клеткой CHO-K1, в которую был введен тандемный вектор CD20-CHX.
Этот результат аналогичен результату в случаях с антителом против CD98 человека и антителом против TNFα человека, описанных в примере 6(3) или примере 7(3), и свидетельствует о том, что линию клеток, продуцирующих антитело на высоком уровне, можно легко получать и производить в случае каждого из антител, когда различные экспрессионные векторы, в которые каждый из гена тяжелой цепи антитела и гена легкой цепи антитела, соответственно, вставлен между транспозонными последовательностями, совместно трансфицированы в суспензионную клетку CHO.
(5) культивирование клетки CHO, продуцирующей антитело против CD20 человека
Клетки, выбранные на основе устойчивости к циклогексимиду из клеток с введенным тандемным вектором CD20-CHX, полученные в вышеуказанном пункте (3), и клетки, в которые были совместно трансфицированы вектор CD20H-CHX и вектор CD20L, и также экспрессирующие антитело против CD20 человека, отбирали и культивировали для наращивания, используя 96-луночный планшет, 24-луночный планшет и 6-луночный планшет, именно в таком порядке. Что касается 4 клеточных линий из клеток контрольной схемы, успешно размноженных при культивировании, и 50 клеточных линий из клеток экспериментальной схемы, эти клетки культивировали таким же образом, как в примере 6(4), за исключением того, что период культивирования составлял 7 дней, и измеряли их уровни экспрессии антител. Результаты приведены на фигуре 12.
Как видно из фигуры 12, установлено, что клетка CHO-K1, в которую были совместно трансфицированы вектор CD20H-CHX и вектор CD20L, демонстрировала примерно в 1,6 раза более высокую продуктивность для антитела, чем клетка CHO-K1, в которую был введен тандемный вектор CD20-CHX.
Этот результат аналогичен результату в случаях с антителом против CD98 человека и антителом против TNFα человека, описанных в примере 6(4) или примере 7(5), и свидетельствует не только о том, что клеточную линию, имеющую высокую продуктивность для антитела, можно легко получать и производить, если различные экспрессионные векторы, в которых ген тяжелой цепи антитела и ген легкой цепи антитела, соответственно, вставлены между транспозонными последовательностями, совместно трансфицированы в суспензионную клетку CHO, но и о том, что полученная таким образом клетка имеет высокую продуктивность для антитела.
Пример 9
Получение транспозонного вектора, экспрессирующего ген устойчивости к неомицину и антитело против CD98 человека
(1) Получение транспозонного вектора, экспрессирующего ген устойчивости к неомицину дикого типа и антитело против CD98 человека
Плазмиду, которая содержала генную экспрессионную кассету для использования в клетке млекопитающего, содержащую произвольный ген антитела человека и ген маркера устойчивости к лекарственному средству, вставленные между парой полученных из Tol2 нуклеотидных последовательностей, использовали в качестве плазмидного вектора для экспрессии белка.
ДНК используемого гена получали, проводя химический синтез искусственным образом на основании общеизвестной нуклеотидной последовательности, либо путем создания праймеров для обеих ее концевых последовательностей с последующим проведением ПЦР при использовании соответствующего источника ДНК. Для последующих манипуляций с генами на конце праймера добавляли сайт расщепления для ферментов рестрикции.
В нуклеотидной последовательности неавтономного транспозона Tol2 (SEQ ID NO: 1), раскрытой в JP-A-2003-235575, нуклеотидную последовательность в положениях 1-200 (последовательность Tol2-L) (SEQ ID NO: 2) и нуклеотидную последовательность в положениях 2285-2788 (последовательность Tol2-R) (SEQ ID NO: 3) использовали в качестве транспозонных последовательностей.
Синтезировали фрагмент ДНК, содержащий каждую из последовательности Tol2-R и последовательности Tol2-L.
Получали фрагмент ДНК, содержащий нуклеотидную последовательность (SEQ ID NO: 18), кодирующую H-цепь антитела, под контролем промотора CMV, амплифицированный на основе вектора N5KG1-Val C2IgG1NS/I117L антитела против CD98 человека (японский патент № 4324637), в качестве кассеты гена тяжелой цепи антитела, и фрагмент ДНК, содержащий нуклеотидную последовательность (SEQ ID NO: 21), кодирующую легкую цепь антитела, под контролем промотора SV40, амплифицированный на основе вектора N5KG1-Val C2IgG1NS/I117L антитела против CD98 человека, в качестве кассеты гена легкой цепи антитела.
В качестве кассеты гена устойчивости к неомицину получали фрагмент ДНК, содержащий ДНК, содержащую нуклеотидную последовательность, кодирующую ген устойчивости к неомицину, под контролем промотора SV40 (ДНК, кодирующую неомицинфосфотрансферазу, состоящую из нуклеотидной последовательности, представленной SEQ ID NO: 36, и с регистрационным № GenBank U47120.2).
Экспрессионный вектор A анти-CD98 антитела получали, соединяя вышеуказанную экспрессионную кассету гена тяжелой цепи антитела, экспрессионную кассету гена легкой цепи антитела и экспрессионную кассету гена устойчивости к неомицину, и дополнительно соединяя оба его конца с фрагментом ДНК, содержащим последовательность Tol2-R, и фрагментом ДНК, содержащим последовательность Tol2-L (фигура 13).
(2) Получение экспрессионного транспозонного вектора антитела против CD98 человека, содержащего модифицированный ген устойчивости к неомицину 1
Получали экспрессионный транспозонный вектор В антитела против CD98 человека, в котором ген устойчивости к неомицину экспрессионного транспозонного вектора А антитела против CD98 человека, полученного в пункте (1), содержащего ген устойчивости к неомицину дикого типа, был заменен на модифицированный ген устойчивости к неомицину 1, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 37.
Модифицированный ген устойчивости к неомицину 1 кодирует аминокислотную последовательность, идентичную последовательности гена устойчивости к неомицину дикого типа, и был изменен для получения нуклеотидной последовательности, в которой модифицированы 167 оснований, соответствующих 22% всех оснований. В частности, среди в общей сложности 32 остатков лейцина, кодоны, соответствующие 25 остаткам лейцина, были изменены на ТТА.
(3) Получение экспрессионного транспозонного вектора антитела против CD98 человека, содержащего модифицированный ген устойчивости к неомицину 2
Получали экспрессионный транспозонный вектор С антитела против CD98 человека, в котором ген устойчивости к неомицину экспрессионного транспозонного вектора А антитела против CD98 человека, полученного в пункте (1), содержащего ген устойчивости к неомицину дикого типа, был заменен на модифицированный ген устойчивости к неомицину 2, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 38.
Модифицированный ген устойчивости к неомицину 2 кодировал аминокислотную последовательность, идентичную последовательности гена устойчивости к неомицину дикого типа, и имел нуклеотидную последовательность, в которой 180 оснований, соответствующих 23% всех оснований, были модифицированы. В частности, среди в общей сложности 32 остатков лейцина, кодоны, соответствующие 28 остаткам лейцина, были изменены на ТТА.
(4) Получение экспрессионного транспозонного вектора антитела против CD98 человека с модифицированным геном устойчивости к неомицину 3
Получали экспрессионный транспозонный вектор D антитела против CD98 человека, в котором ген устойчивости к неомицину экспрессионного транспозонного вектора А антитела против CD98 человека, полученного в пункте (1), содержащего ген устойчивости к неомицину дикого типа, был заменен на модифицированный ген устойчивости к неомицину 3, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 39.
Модифицированный ген устойчивости к неомицину 3 кодировал аминокислотную последовательность, идентичную последовательности гена устойчивости к неомицину дикого типа, и имел нуклеотидную последовательность, в которой 203 основания, соответствующих 26% всех оснований, были модифицированы. В частности, среди в общей сложности 32 остатков лейцина, кодоны, соответствующие 30 остаткам лейцина, были измерены на ТТА.
Пример 10
Получение антитела с помощью продуцирующей антитело клетки CHO, экспрессирующей модифицированный ген устойчивости к неомицину
Получали продуцирующие антитело клетки A-D путем введения каждого из экспрессионных транспозонных векторов A-D антитела против CD98 человека, полученных в примере 9(1)-(4), в суспензионную клетку CHO-K1 вместе с вектором pCAGGS-T2TP, экспрессирующим Tol2 транспозазу с аминокислотной последовательностью, представленной SEQ ID NO: 40 [Kwakami K. & Noda T., Genetics, 166, 895-899 (2004)].
Введение векторов в суспензионную клетку CHO проводили путем суспендирования клетки CHO (4×106 клеток) в 400 мкл буфера PBS и совместного трансфицирования экспрессионного транспозонного вектора антитела против CD98 человека (10 мкг) и экспрессионного вектора Tol2 транспозазы pCAGGS-T2TP (20 мкг) напрямую в форме кольцевой ДНК методом электропорации.
В данном случае экспрессионный вектор Tol2 транспозазы был также введен в форме кольцевой ДНК для того, чтобы Tol2 транспозаза экспрессировалась временно.
Кроме того, в качестве контроля, в котором не использовали Tol2 транспозазу, экспрессионный транспозонный вектор D антитела против CD98 человека (10 мкг) из примера 19(4) был линеаризован при помощи фермента рестрикции PciI (TARA BIO INC.), а затем введен в суспензионную клетку CHO-K1 методом электропорации.
Электропорацию проводили при помощи электропоратора [Gene Pulser Xcell System (производитель Bio-Rad)] в условиях напряжения 300 В, электростатической мощности 500 мкФ и комнатной температуры, используя кювету с шириной зазора 4 мм (производитель Bio-Rad).
После введения генов электропорацией клетки в каждой кювете инокулировали в один 96-луночный планшет и культивировали в течение 3 дней в CO2-инкубаторе, используя среду CD OptiCHO (Invitrogen) с добавлением 5% гидролизата соевых бобов.
Затем, начиная со смены среды через 4 дня после введения генов, проводили культивирование в присутствии G418 (Geneticin(R), Invitrogen), добавляя G418 до получения конечной концентрации 500 мкг/мл, и культивирование продолжали в течение 3 недель, меняя среду с интервалами в одну неделю.
После культивирования экспрессию антитела определяли сэндвич-методом при помощи анализа LANCE(R) (Perkin-Elmer Corp) с применением FRET (флуоресцентного резонансного переноса энергии). Результаты приведены в таблице 9.
Как видно из таблицы 9, уровни экспрессии антитела против CD98 человека клеток В-D, экспрессирующих модифицированные гены устойчивости к неомицину, были выше, чем уровни клетки A, экспрессирующей ген устойчивости к неомицину дикого типа.
В частности, в случае клетки D, продуцирующей антитело против CD98 человека, которая экспрессирует модифицированный ген устойчивости к неомицину 3, была получена клеточная линия, демонстрирующая в 10 раз более высокий уровень экспрессии, чем таковой у клетки A, продуцирующей антитело против CD98 человека, которая экспрессирует ген устойчивости к неомицину дикого типа.
Кроме того, даже при использовании модифицированного гена устойчивости к неомицину 3, не удалось получить клетку, экспрессирующую антитело против CD98 человека, из контрольной клетки, в которую экспрессионный вектор Tol2 транспозазы не был совместно трансфицирован, несмотря на приведение вектора в линейную форму.
Пример 11
Получение транспозонного вектора, экспрессирующего ген устойчивости к пуромицину и антитело против CD98 человека
(1) Получение экспрессионного транспозонного вектора антитела против CD98 человека, содержащего модифицированный ген устойчивости к пуромицину 1
Получали экспрессионный транспозонный вектор E антитела против CD98 человека, в котором ген устойчивости к неомицину экспрессионного транспозонного вектора A антитела против CD98 человека, полученного в примере 9(1), который содержал ген устойчивости к неомицину дикого типа, был заменен на модифицированный ген устойчивости к пуромицину 1 с нуклеотидной последовательностью, представленной SEQ ID NO: 41.
Модифицированный ген устойчивости к пуромицину 1 кодировал аминокислотную последовательность, идентичную последовательности гена устойчивости к пуромицину дикого типа с нуклеотидной последовательностью, представленной SEQ ID NO: 42 (ген пуромицин-N-ацетилтрансферазы с нуклеотидной последовательностью, описанной в регистрационном № GenBank U07648.1), и имел нуклеотидную последовательность, в которой 17 оснований, соответствующих 3% всех оснований, модифицированы. В частности, среди в общей сложности 28 остатков аланина, содержащихся в гене устойчивости к пуромицину, кодоны, соответствующие 17 остаткам аланина, были изменены на GCG в результате модификации, вместе с кодонами, которые уже были кодонами GCG в диком типе, кодоны, соответствующие всем остаткам аланина, были измерены на GCG.
(2) Получение экспрессионного транспозонного вектора антитела против CD98 человека, содержащего модифицированный ген устойчивости к пуромицину 2
Получали экспрессионный транспозонный вектор F антитела против CD98 человека, в котором ген устойчивости к неомицину экспрессионного транспозонного вектора A антитела против CD98 человека, полученного в примере 9(1), содержащего ген устойчивости к неомицину дикого типа, был заменен на модифицировнный ген устойчивости к пуромицину 2 с нуклеотидной последовательностью, представленной SEQ ID NO: 43. Модифицированный ген устойчивости к пуромицину 2 кодирует аминокислотную последовательность, идентичную последовательности гена устойчивости к пуромицину дикого типа, и имеет нуклеотидную последовательность, в которой 79 оснований, соответствующих 14% общего количества оснований, модифицированы. В частности, в дополнение к модификации кодонов, соответствующих остаткам аланина модифицированного гена устойчивости к пуромицину 1, кодоны, соответствующие остаткам лейцина, были изменены на ТТА и кодоны, соответствующие остаткам валина, были изменены на GTA, и кодоны для серина были изменены на TCG.
Пример 12
Получение антитела с помощью продуцирующей антитело клетки СНО, экспрессирующей модифицированный ген устойчивости к пуромицину 1
Получали продуцирующие антитело клетки Ε и F путем введения экспрессионного транспозонного вектора Ε антитела против CD98 человека из примера 11(1), содержащего модифицированный ген устойчивости к пуромицину 1, экспрессионного транспозонного вектора F антитела против CD98 человека из примера 11(2), содержащего модифицированный ген устойчивости к пуромицину 2, и экспрессионного вектора Tol2 транспозазы pCAGGS-T2TP в суспензионную клетку CHO-К1.
Введение векторов в суспензионную клетку проводили путем суспендирования суспензионной клетки СНО (4×106 клеток) в 400 мклбуфера PBS и совместного трансфицирования экспрессионного транспозонного вектора антитела против CD98 человека, содержащего модифицированный ген устойчивости к пуромицину, в форме кольцевой ДНК (10 мкг) и pCAGGS-T2TP (20 мкг) напрямую методом электропорации.
В этом случае, экспрессионный вектор Tol2 транспозазы pCAGGS-T2TP также вводили напрямую в форме кольцевой ДНК для временной экспрессии Tol2 транспозазы.
Электропорацию проводили при помощи электропоратора [Gene Pulser Xcell System (производитель Bio-Rad)] в условиях напряжения 300 В, электростатической мощности 500 мкФ и комнатной температуры, используя кювету с шириной зазора 4 мм (производитель Bio-Rad).
После введения генов электропорацией клетки в каждой кювете инокулировали в один 96-луночный планшет и культивировали в течение 3 дней в CO2-инкубаторе, используя среду CD OptiCHO (Invitrogen) с добавлением 5% гидролизата соевых бобов.
Затем, начиная со смены среды через 2 дня после введения генов, проводили культивирование в течение 4 недель, добавляя пуромицин (P9620, Sigma-Aldrich) до конечной концентрации 5 мкг/мл и меняя среду на содержащую пуромицин среду с интервалами в одну неделю.
После культивирования уровень экспрессии антитела определяли сэндвич-методом при помощи анализа LANCE(R) (Perkin-Elmer Corp) с применением FRET (флуоресцентного резонансного переноса энергии). Результаты приведены в таблице 10.
Как видно из таблицы 10, продуцирующая антитело клетка F, экспрессирующая модифицированный ген устойчивости к пуромицину 2, демонстрировала в два или более раза более высокую продуктивность для антитела, чем продуцирующая антитело клетка E, экспрессирующая модифицированный ген устойчивости к пуромицину 1.
Пример 13
Получение антитела с помощью продуцирующей антитело клетки CHO, экспрессирующей модифицированный ген устойчивости к пуромицину 2
Продуцирующую антитело клетку F, полученную в примере 12, экспрессирующую модифицированный ген устойчивости к пуромицину 2, культивировали в конической колбе для получения антитела против CD98 человека.
В частности, продуцирующую антитело клетку F культивировали для наращивания, используя 96-луночный планшет, 24-луночный планшет и 6-луночный планшет именно в таком порядке. Выбрали две линии клеток продуцирующей антитело клетки F, в которых число клеток было значительно повышено (клеточная линия 1 и клеточная линия 2), их, соответственно, суспендировали в 35 мл среды CD OptiCHO (Invitrogen) с добавлением 5% гидролизата соевых бобов так, чтобы получить плотность клеток 2×105 клеток/мл, и культивировали в течение 1 недели на шейкере, используя коническую колбу емкостью 125 мл (с изогнутой крышкой, Corning Glassworks), при 37°C в атмосфере 5% CO2, получая таким образом антитело против CD98 человека.
Количество антитела в среде после культивирования определяли методом ВЭЖХ (Waters Associates, Inc.). Результаты приведены в таблице 11.
Приведенные выше результаты свидетельствуют о том, что в суспензионной клетке CHO ген антитела, вставленный между парой транспозонных последовательностей, и модифицированный ген устойчивости к лекарственному средству успешно встроены в хромосому хозяина и также эффективны для отбора экспрессирующей на высоком уровне клетки. Кроме того, установлено, что полученную таким образом клетку можно культивировать для наращивания и производство интересующего белка в условиях суспензионного культивирования возможно.
При помощи способа получения белка по настоящему изобретению интересующий белок можно эффективно производить, используя суспензионную клетку млекопитающего. Клетку по настоящему изобретению можно использовать в качестве продуцирующей белок клетки для получения рекомбинантного белка.
Список последовательностей
SEQ ID NO: 1 - Описание искусственной последовательности; нуклеотидная последовательность неавтономного транспозона Tol2.
SEQ ID NO: 2 - Описание искусственной последовательности; последовательность Tol2-L.
SEQ ID NO: 3 - Описание искусственной последовательности; последовательность Tol2-R.
SEQ ID NO: 7 - Описание искусственной последовательности; нуклеотидная последовательность гена устойчивости к циклогексимиду.
SEQ ID NO: 8 - Описание искусственной последовательности; аминокислотная последовательность белка, кодируемого геном устойчивости к циклогексимиду.
SEQ ID NO: 9 - Описание искусственной последовательности; нуклеотидная последовательность, кодирующая H-цепь антитела M2Z3.
SEQ ID NO: 10 - Описание искусственной последовательности; аминокислотная последовательность H-цепи антитела M2Z3.
SEQ ID NO: 11 - Описание искусственной последовательности; нуклеотидная последовательность, кодирующая L-цепь антитела M2Z3.
SEQ ID NO: 12 - Описание искусственной последовательности; аминокислотная последовательность L-цепи антитела M2Z3.
SEQ ID NO: 13 - Описание искусственной последовательности; нуклеотидная последовательность неавтономного Tol1.
SEQ ID NO: 14 - Описание искусственной последовательности; последовательность Tol1-L.
SEQ ID NO: 15 - Описание искусственной последовательности; последовательность Tol1-R.
SEQ ID NO: 18 - Описание искусственной последовательности; нуклеотидная последовательность, кодирующая вариабельную область тяжелой цепи анти-CD98 антитела.
SEQ ID NO: 19 - Описание искусственной последовательности; аминокислотная последовательность вариабельной области тяжелой цепи анти-CD98 антитела.
SEQ ID NO: 20 - Описание искусственной последовательности; аминокислотная последовательность вариабельной области тяжелой цепи анти-CD98 антитела.
SEQ ID NO: 21 - Описание искусственной последовательности; нуклеотидная последовательность, кодирующая вариабельную область легкой цепи анти-CD98 антитела.
SEQ ID NO: 22 - Описание искусственной последовательности; аминокислотная последовательность вариабельной области легкой цепи анти-CD98 антитела.
SEQ ID NO: 23 - Описание искусственной последовательности; аминокислотная последовательность вариабельной области легкой цепи анти-CD98 антитела.
SEQ ID NO: 24 - Описание искусственной последовательности; нуклеотидная последовательность, кодирующая вариабельную область тяжелой цепи антитела против TNFα человека.
SEQ ID NO: 25 - Описание искусственной последовательности; аминокислотная последовательность вариабельной области тяжелой цепи антитела против TNFα человека.
SEQ ID NO: 26 - Описание искусственной последовательности; аминокислотная последовательность вариабельной области тяжелой цепи антитела против TNFα человека.
SEQ ID NO: 27 - Описание искусственной последовательности; нуклеотидная последовательность, кодирующая вариабельную область легкой цепи антитела против TNFα человека.
SEQ ID NO: 28 - Описание искусственной последовательности; аминокислотная последовательность вариабельной области легкой цепи антитела против TNFα человека.
SEQ ID NO: 29 - Описание искусственной последовательности; аминокислотная последовательность вариабельной области легкой цепи антитела против TNFα человека.
SEQ ID NO: 30 - Описание искусственной последовательности; нуклеотидная последовательность, кодирующая вариабельную область тяжелой цепи антитела против CD20 человека.
SEQ ID NO: 31 - Описание искусственной последовательности; аминокислотная последовательность вариабельной области тяжелой цепи антитела против CD20 человека.
SEQ ID NO: 32 - Описание искусственной последовательности; аминокислотная последовательность вариабельной области тяжелой цепи антитела против CD20 человека.
SEQ ID NO: 33 - Описание искусственной последовательности; нуклеотидная последовательность, кодирующая вариабельную область легкой цепи антитела против CD20 человека.
SEQ ID NO: 34 - Описание искусственной последовательности; аминокислотная последовательность вариабельной области легкой цепи антитела против CD20 человека.
SEQ ID NO: 35 - Описание искусственной последовательности; аминокислотная последовательность вариабельной области легкой цепи антитела против CD20 человека.
SEQ ID NO: 36 - Описание искусственной последовательности; нуклеотидная последовательность гена устойчивости к неомицину дикого типа.
SEQ ID NO: 37 - Описание искусственной последовательности; нуклеотидная последовательность модифицированного гена устойчивости к неомицину.
SEQ ID NO: 38 - Описание искусственной последовательности; нуклеотидная последовательность модифицированного гена устойчивости к неомицину.
SEQ ID NO: 39 - Описание искусственной последовательности; нуклеотидная последовательность модифицированного гена устойчивости к неомицину.
SEQ ID NO: 41 - Описание искусственной последовательности; нуклеотидная последовательность модифицированного гена устойчивости к пуромицину.
SEQ ID NO: 42 - Описание искусственной последовательности; нуклеотидная последовательность гена устойчивости к пуромицину дикого типа.
SEQ ID NO: 43 - Описание искусственной последовательности; нуклеотидная последовательность модифицированного гена устойчивости к пуромицину.
SEQ ID NO: 44 - Описание искусственной последовательности; нуклеотидная последовательность модифицированного гена устойчивости к пуромицину.
SEQ ID NO: 45 - Описание искусственной последовательности; нуклеотидная последовательность модифицированного гена устойчивости к зеоцину.
SEQ ID NO: 46 - Описание искусственной последовательности; нуклеотидная последовательность модифицированного гена устойчивости к зеоцину.
SEQ ID NO: 47 - Описание искусственной последовательности; нуклеотидная последовательность модифицированного гена устойчивости к гигромицину.
SEQ ID NO: 48 - Описание искусственной последовательности; нуклеотидная последовательность модифицированного гена устойчивости к гигромицину.
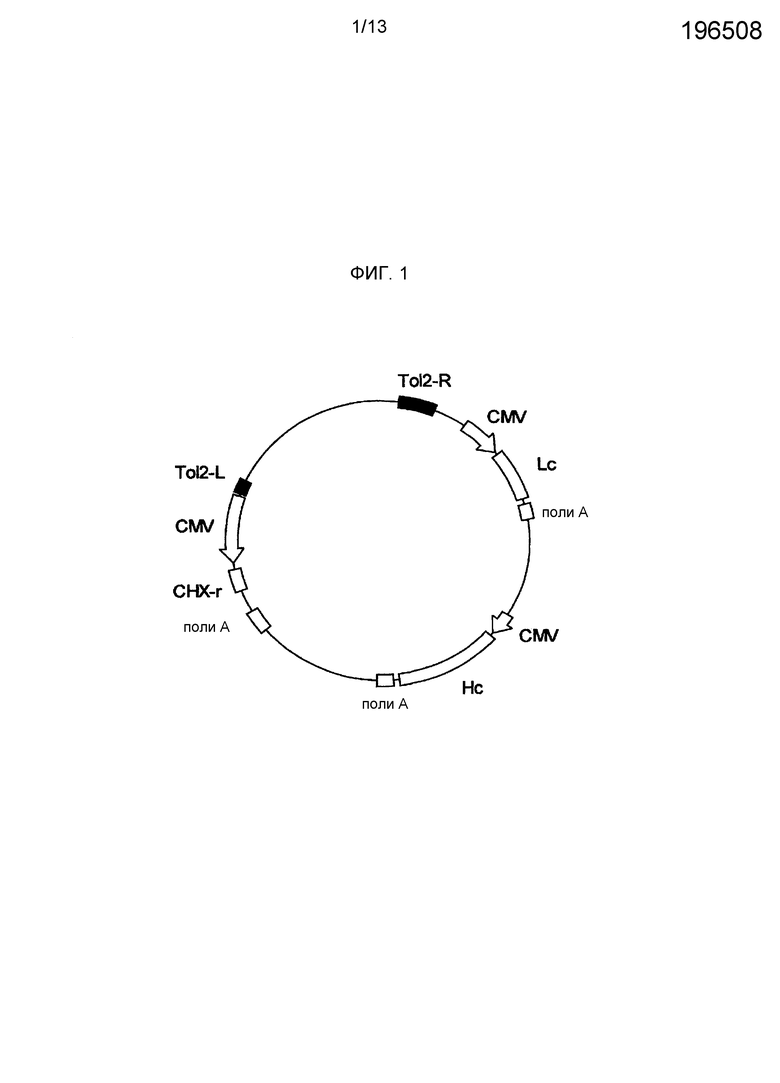
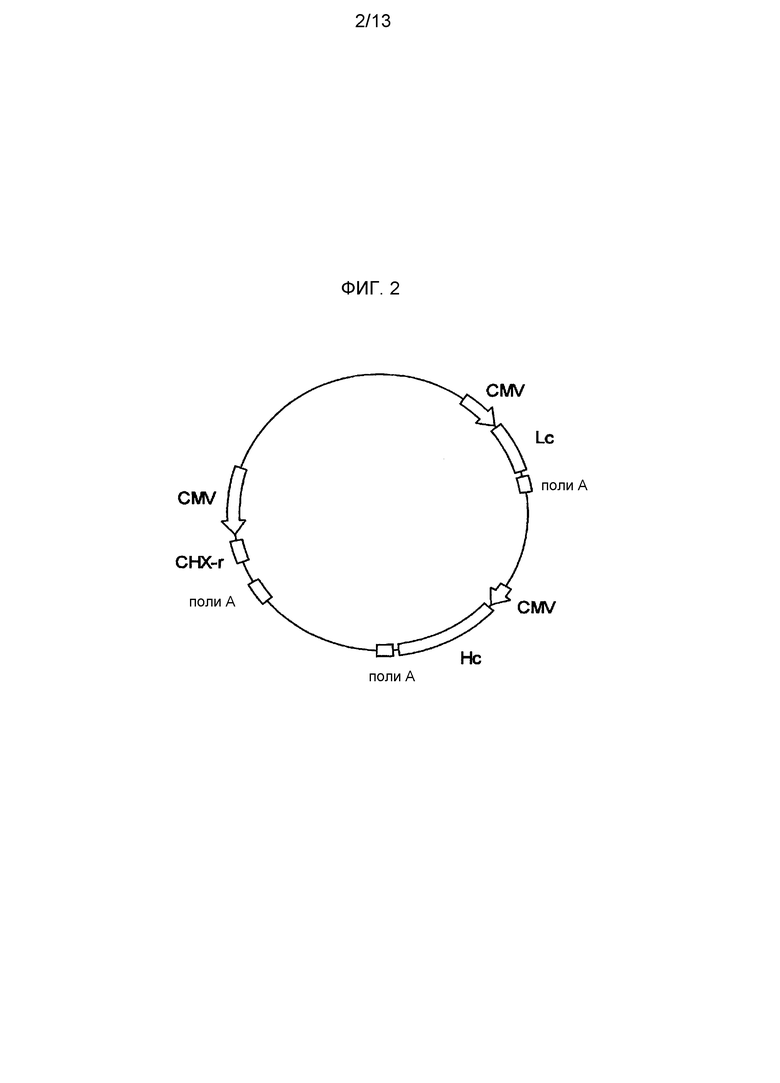
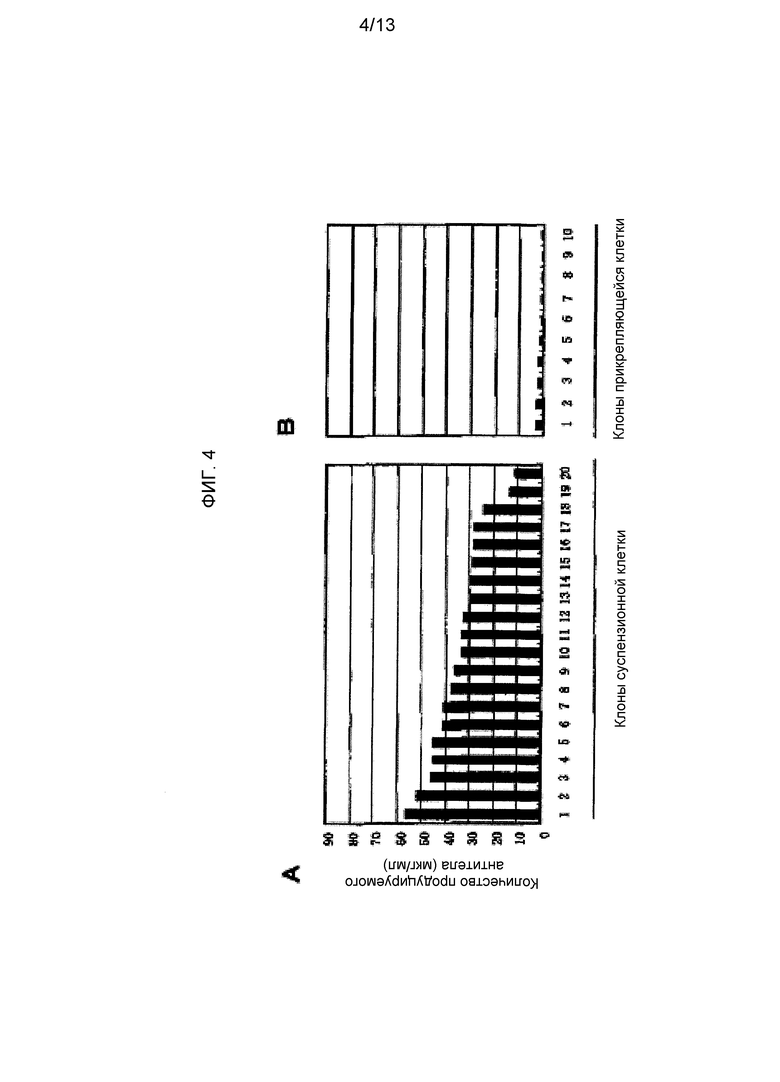
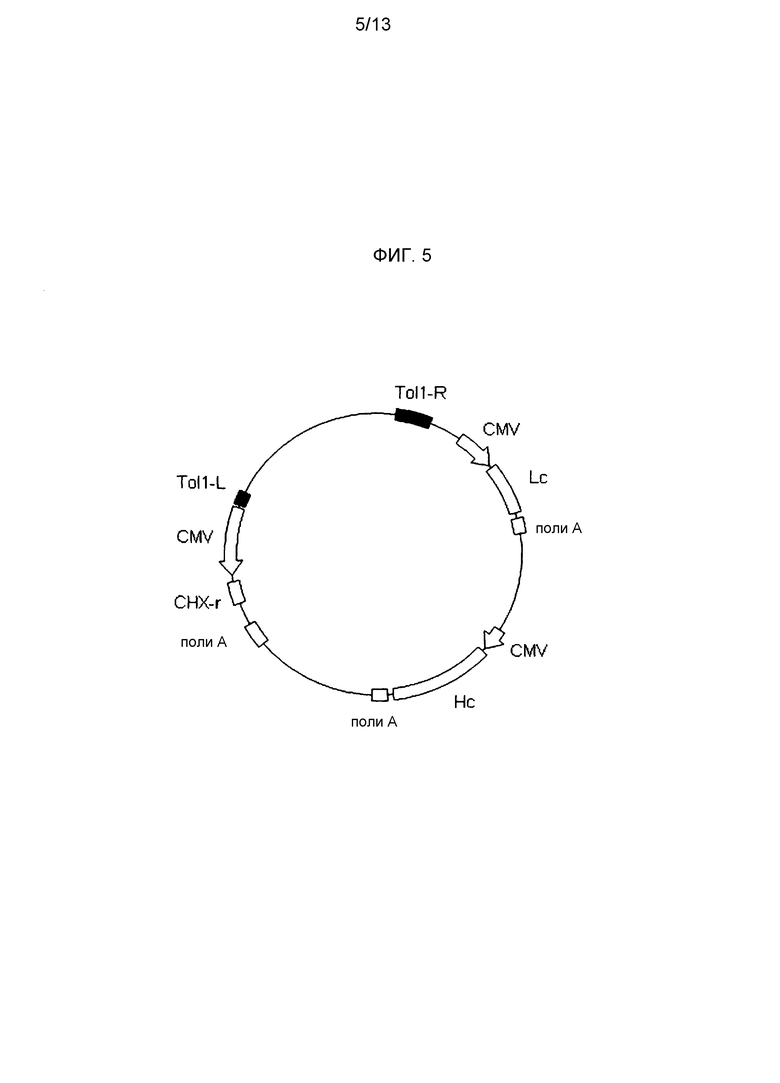
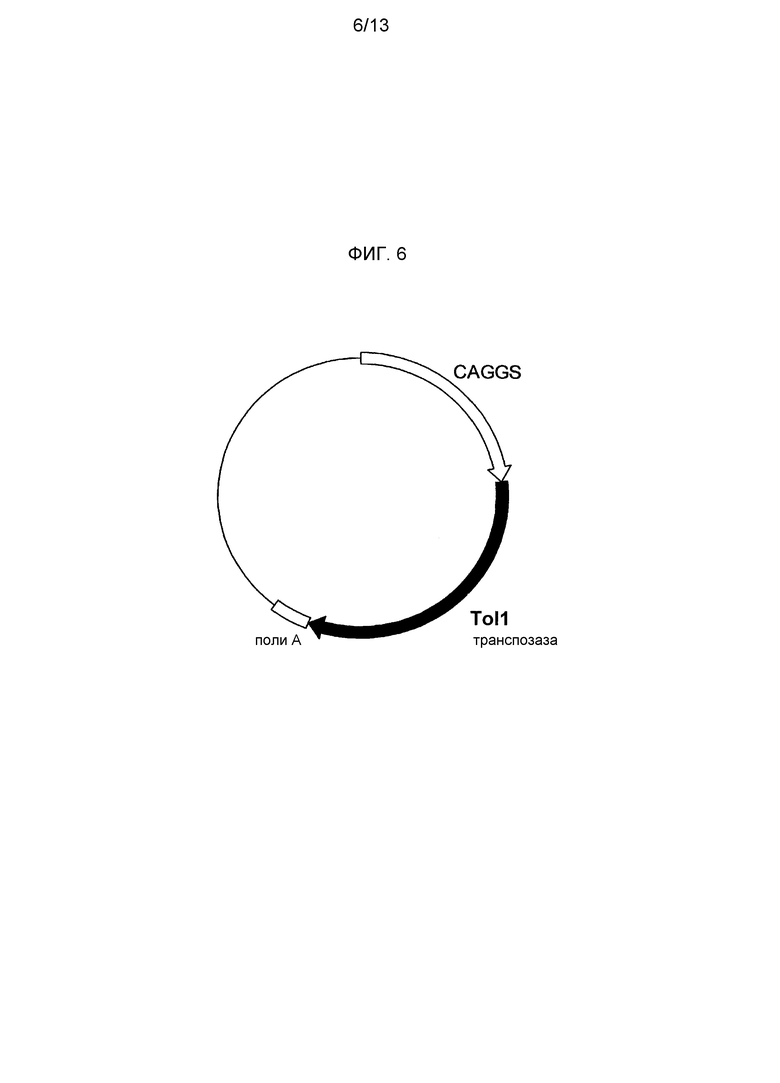
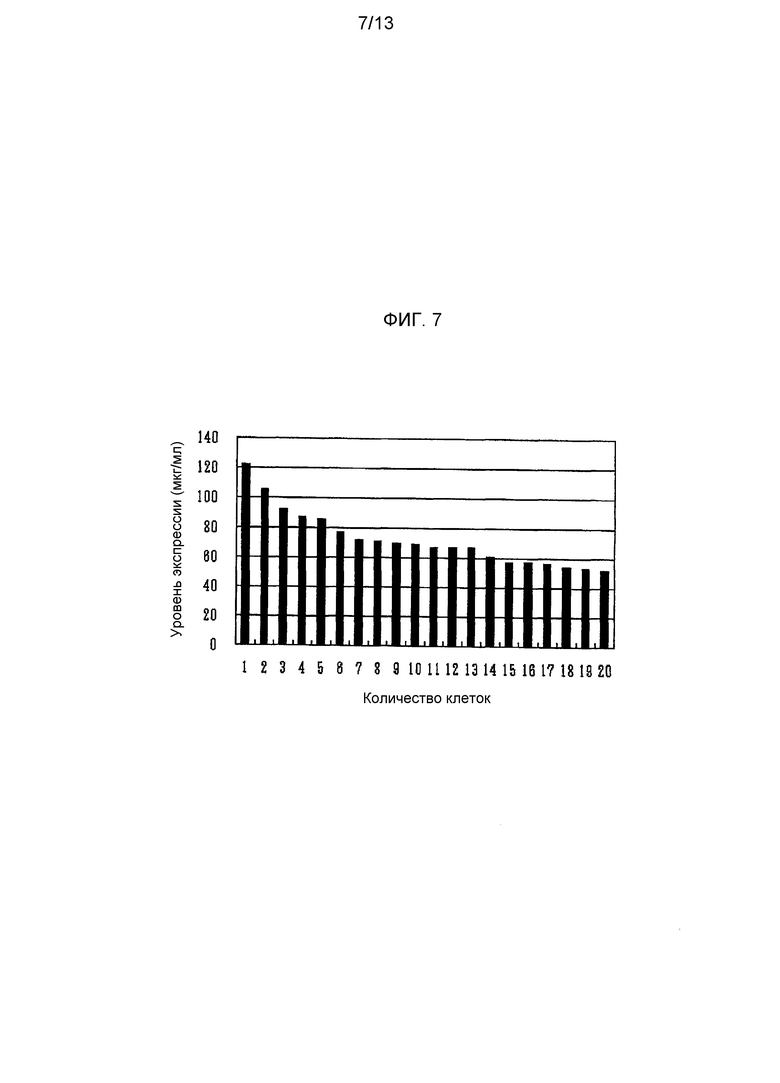
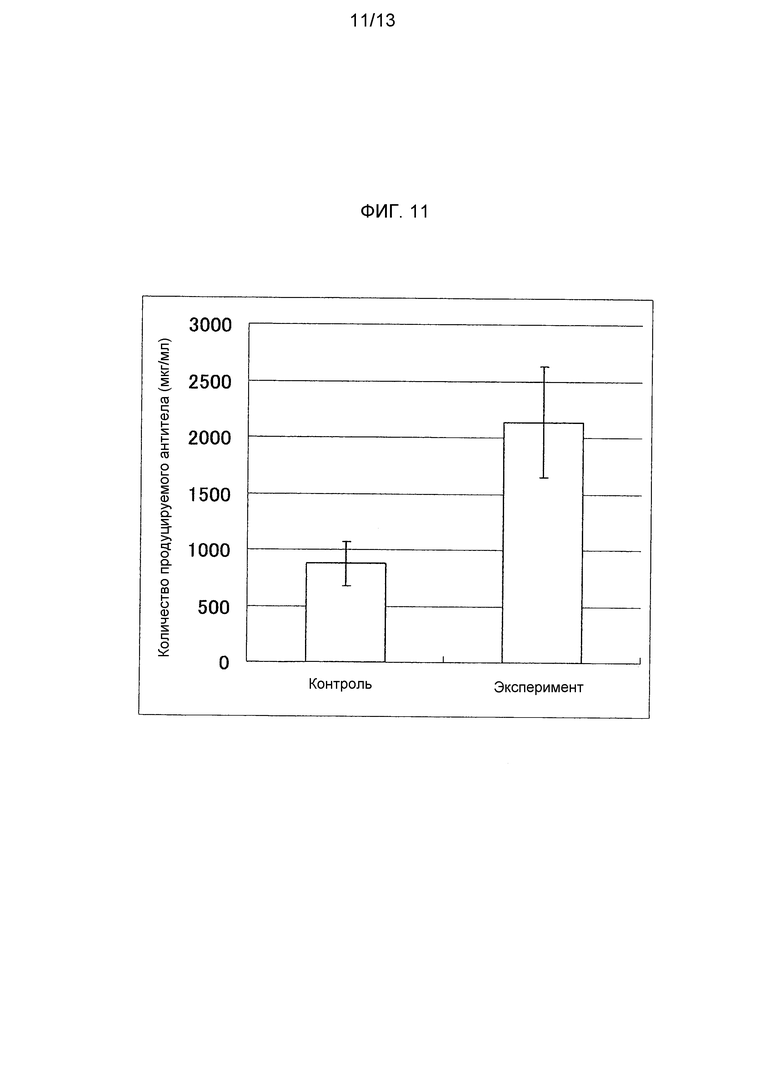
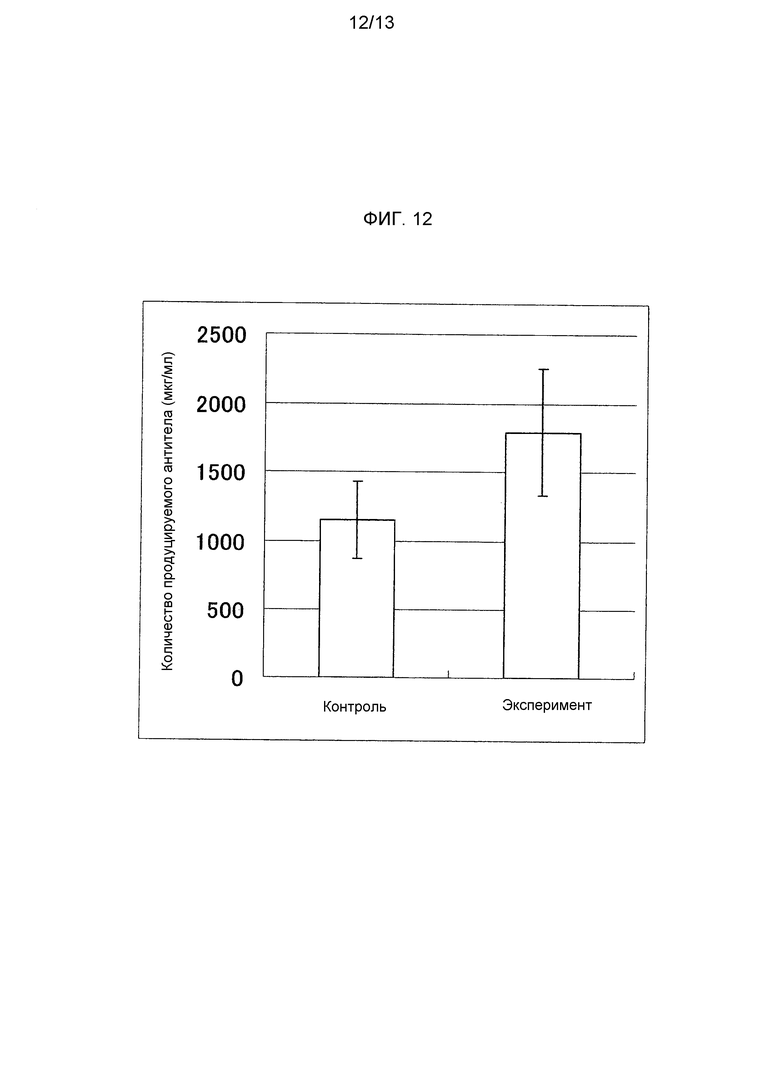
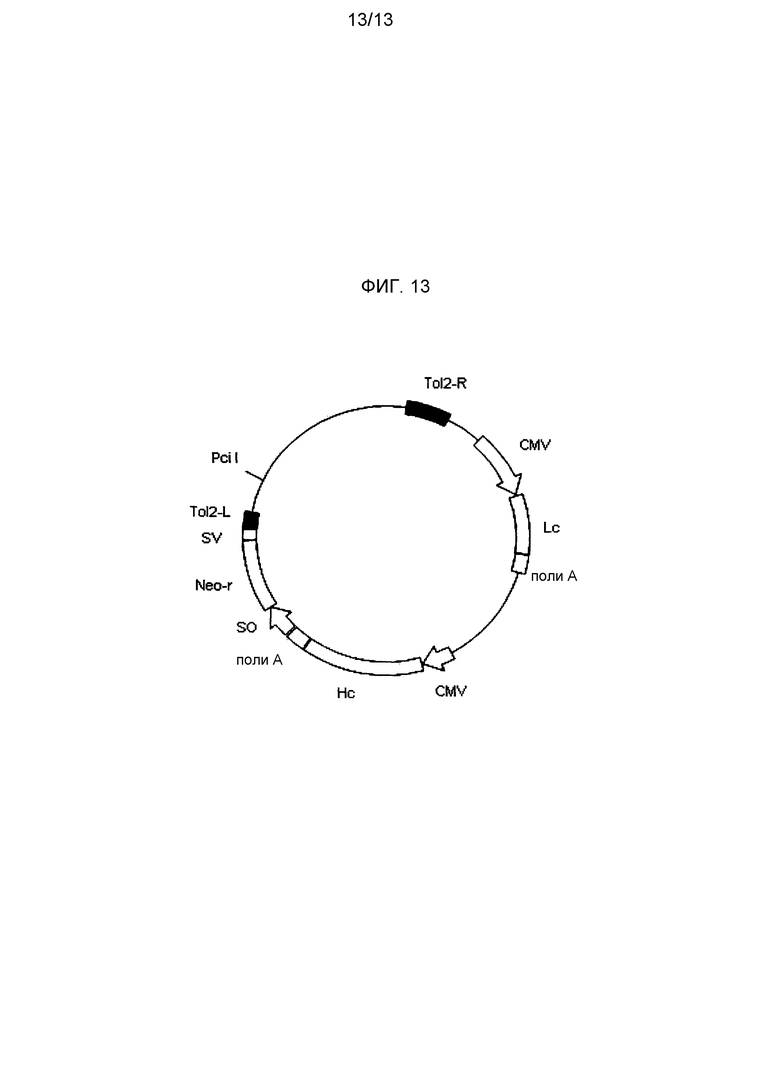

























































Изобретение относится к биохимии. Описан способ получения интересующего белка, включающий введение по меньшей мере одного экспрессионного вектора, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также пару транспозонных последовательностей Tol1 или Tol2 на обоих концах генного фрагмента, в суспензионную клетку млекопитающего СНО и суспензионное культивирование клетки млекопитающего, где суспензионная клетка млекопитающего представляет собой суспензионную клетку СНО, которая способна выживать и пролиферировать в бессывороточной среде. Также представлены соответствующая суспензионная клетка и вектор. Изобретение позволяет за короткий период времени эффективно и на высоком уровне продуцировать интересующий белок. 5 н. и 26 з.п. ф-лы, 13 ил., 11 табл., 13 пр.
1. Способ получения интересующего белка, включающий введение по меньшей мере одного экспрессионного вектора, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, в суспензионную клетку млекопитающего; интеграцию генного фрагмента, содержащего ДНК, кодирующую интересующий белок, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего для получения клетки млекопитающего, которая экспрессирует интересующий белок; и суспензионное культивирование клетки млекопитающего,
где суспензионная клетка млекопитающего представляет собой суспензионную клетку СНО, которая способна выживать и пролиферировать в бессывороточной среде, и где пара транспозонных последовательностей представляет собой нуклеотидные последовательности, полученные из пары транспозонов Tol1, или нуклеотидные последовательности, полученные из пары транспозонов Tol2.
2. Способ получения интересующего белка, включающий следующие этапы (А)-(С):
(А) этап одновременного введения следующих экспрессионных векторов (а) и (b) в суспензионную клетку млекопитающего:
(а) по меньшей мере один экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) экспрессионный вектор, который содержит ДНК, кодирующую транспозазу, которая узнает транспозонные последовательности и обладает способностью перемещать генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому;
(B) этап получения суспензионной клетки млекопитающего, которая экспрессирует интересующий белок в результате временной экспрессии транспозазы с экспрессионного вектора, введенного в суспензионную клетку млекопитающего на этапе (А), с целью интеграции генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего, и
(C) этап суспензионного культивирования суспензионной клетки млекопитающего, которая экспрессирует интересующий белок, полученной на этапе (В), для получения интересующего белка, где суспензионная клетка млекопитающего представляет собой суспензионную клетку СНО, которая способна выживать и пролиферировать в бессывороточной среде, и где пара транспозонных последовательностей представляет собой нуклеотидные последовательности, полученные из пары транспозонов Tol1, или нуклеотидные последовательности, полученные из пары транспозонов Tol2.
3. Способ получения суспензионной клетки млекопитающего, которая экспрессирует интересующий белок, включающий введение по меньшей мере одного экспрессионного вектора, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, в суспензионную клетку млекопитающего и интеграцию генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего,
где суспензионная клетка млекопитающего представляет собой суспензионную клетку СНО, которая способна выживать и пролиферировать в бессывороточной среде, и где пара транспозонных последовательностей представляет собой нуклеотидные последовательности, полученные из пары транспозонов Tol1, или нуклеотидные последовательности, полученные из пары транспозонов Tol2.
4. Способ по любому одному из пп. 1-3, в котором по меньшей мере один из экспрессионных векторов, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, представляет собой экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
5. Способ по любому одному из пп. 1-3, включающий введение экспрессионного вектора, который содержит генный фрагмент, содержащий селектируемый маркер, и содержит пару транспозонных последовательностей на обоих концах генного фрагмента, в клетку млекопитающего в дополнение к экспрессионному вектору, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
6. Способ по любому одному из пп. 1-3, в котором ДНК, кодирующая интересующий белок, представляет собой ДНК, кодирующую антитело.
7. Способ по п. 6, в котором ДНК, кодирующая антитело, представляет собой по меньшей мере одну из ДНК, кодирующей Н-цепь антитела, и ДНК, кодирующей L-цепь антитела.
8. Способ по любому одному из пп. 1-3, в котором экспрессионный вектор, выбранный из следующих пунктов (а)-(d), вводят в суспензионную клетку млекопитающего:
(a) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую Н-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую Н-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(c) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую Н-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и
(d) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую Н-цепь и L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
9. Способ по любому одному из пп. 1-3, в котором клетка СНО представляет собой по меньшей мере клетку, выбранную из CHO-K1, CHO-K1SV, DUKXB11, CHO/DG44, Pro-3 и CHO-S.
10. Способ по п. 4, в котором ген селектируемого маркера представляет собой ген устойчивости к циклогексимиду.
11. Способ по п. 10, в котором ген устойчивости к циклогексимиду представляет собой ген, кодирующий мутант человеческого рибосомного белка L36a.
12. Способ по п. 5, в котором ген селектируемого маркера представляет собой ген устойчивости к циклогексимиду.
13. Способ по п. 12, в котором ген устойчивости к циклогексимиду представляет собой ген, кодирующий мутант рибосомного белка L36a человека.
14. Способ по п. 8, в котором ген селектируемого маркера представляет собой ген устойчивости к циклогексимиду.
15. Способ по п. 14, в котором ген устойчивости к циклогексимиду представляет собой ген, кодирующий мутант рибосомного белка L36a человека.
16. Способ по любому одному из пп. 1-3, в котором нуклеотидные последовательности, полученные из пары транспозонов Tol2, представляют собой нуклеотидную последовательность, содержащую нуклеотидную последовательность, приведенную в SEQ ID NO: 2, и нуклеотидную последовательность, приведенную в SEQ ID NO: 3.
17. Способ по любому одному из пп. 1-3, в котором нуклеотидные последовательности, полученные из пары транспозонов Tol1, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 14, и нуклеотидную последовательность, приведенную в SEQ ID NO: 15.
18. Суспензионная клетка млекопитающего, продуцирующая интересующий белок, которая имеет хромосому, в которую интегрирован генный фрагмент, вставленный между парой транспозонов, и которая продуцирует интересующий белок, получаемый путем одновременного введения по меньшей мере одного экспрессионного вектора (а), который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионного вектора (b), который содержит ДНК, кодирующую транспозазу (трансферазу), способную узнавать транспозонные последовательности и обладающую способностью переносить генный фрагмент, вставленный между парой транспозонных последовательностей, в хромосому, где суспензионная клетка млекопитающего представляет собой суспензионную клетку СНО, которая способна выживать и пролиферировать в бессывороточной среде, и где пара транспозонных последовательностей представляет собой нуклеотидные последовательности, полученные из пары транспозонов Tol1, или нуклеотидные последовательности, полученные из пары транспозонов Tol2.
19. Клетка млекопитающего по п. 18, в которой по меньшей мере один из экспрессионного вектора (а), который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, представляет собой экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
20. Клетка млекопитающего по п. 18, которая представляет собой клетку, полученную в результате дополнительного введения экспрессионного вектора (с), который содержит генный фрагмент, содержащий ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, в дополнение к экспрессионным векторам (а) и (b).
21. Клетка млекопитающего по любому одному из пп. 18-20, в которой ДНК, кодирующая интересующий белок, представляет собой ДНК, кодирующую антитело.
22. Клетка млекопитающего по п. 21, в которой ДНК, кодирующая антитело, представляет собой по меньшей мере одну из ДНК, кодирующей Н-цепь антитела, и ДНК, кодирующей L-цепь антитела.
23. Клетка млекопитающего по любому одному из пп. 18-20, в которую введен экспрессионный вектор, выбранный из следующих пунктов (а)-(d):
(a) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую Н-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(b) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую Н-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента;
(c) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую Н-цепь антитела, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, и
(d) экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую Н-цепь и L-цепь антитела, и ген селектируемого маркера, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента.
24. Клетка млекопитающего по любому одному из пп. 18-20, при этом клетка СНО представляет собой любую клетку, выбранную из СНО-K1, CHO-K1SV, DUKXB11, CHO/DG44, Pro-3 и CHO-S.
25. Клетка млекопитающего по п. 19 или 20, в которой ген селектируемого маркера представляет собой ген устойчивости к циклогексимиду.
26. Клетка млекопитающего по п. 25, в которой ген устойчивости к циклогексимиду представляет собой ген, кодирующий мутант человеческого рибосомного белка L36a.
27. Клетка млекопитающего по п. 18, в которой нуклеотидные последовательности, полученные из пары транспозонов Tol2, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 2, и нуклеотидную последовательность, приведенную в SEQ ID NO: 3.
28. Клетка млекопитающего по п. 18, в которой нуклеотидные последовательности, полученные из пары транспозонов Tol1, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 14, и нуклеотидную последовательность, приведенную в SEQ ID NO: 15.
29. Экспрессионный вектор, который содержит генный фрагмент, содержащий ДНК, кодирующую интересующий белок, и также содержит пару транспозонных последовательностей на обоих концах генного фрагмента, где пара транспозонных последовательностей представляет собой нуклеотидные последовательности, полученные из пары транспозонов Tol1, или нуклеотидные последовательности, полученные из пары транспозонов Tol2.
30. Экспрессионный вектор для белка по п. 29, в котором нуклеотидные последовательности, полученные из пары транспозонов Tol2, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 2, и нуклеотидную последовательность, приведенную в SEQ ID NO: 3.
31. Экспрессионный вектор по п. 29, в котором нуклеотидные последовательности, полученные из пары транспозонов Tol1, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 14, и нуклеотидную последовательность, приведенную в SEQ ID NO: 15.
| SUMIYAMA K, A simple and hihly efficient transgenesis method in mice with the Tol2 transposon system and cytoplasmic microinjection, Genomics, 22.02.2010, 95(2010) 306-311 | |||
| WO2003102191 A, 11.12.2003 | |||
| PARK CW, DNA methylation of Sleeping Beauty with transposition into the mouse genome, Genes Cells 2005, 10(8), реф | |||
| ZHONG B, Comparison of |

