Область техники, к которой относится изобретение
Изобретение относится к раку и другим заболеваниям и нарушениям, например, воспалительным заболеваниям и нарушениям, и их терапевтической модуляции. В частности, изобретение относится к соединениям на основе коротких пептидов, способным модулировать запрограммированную гибель клеток (PCD) и пролиферацию раковых клеток и провоспалительных/аутоиммунных клеток.
Уровень техники
Индукцию апоптоза давно считают способом направленного воздействия на раковые клетки, а также провоспалительные, аутоиммунные клетки и другие больные клетки. Существует ряд клеточных путей, участвующих в запуске гибели клеток, включая путь c-Jun N-концевой киназы JNK. Киназы JNK чувствительны к цитокинам и стрессовым стимулам, таким как ультрафиолетовое излучение, тепловой шок и осмотический шок. Кроме того, в ответ на цитокины и клеточный стресс происходит активация пути NF-κВ. Путь NF-κВ может ингибировать путь JNK посредством взаимного влияния, опосредованного Gadd45β и киназой JNK, активированной митогеном киназы 7 протеинкиназы (NFKK7/JNKK2). Активность MKK7 ингибируется Gadd45β, членом семейства индуцируемых факторов Gadd45 и непосредственной транскрипционной мишенью NF-κВ. Это означает, что Gadd45β опосредует супрессию посредством NF-κВ передачи сигнала JNK путем связывания с MKK7 и ингибирования его активности. См. статью Papa, et al. 2004, Nature Cell Biology 6(2):1462153.
Предложено использование ингибиторов NF-κВ при лечении рака и воспалительных заболеваний. Однако, поскольку NF-κВ обладает рядом активностей, включая участие в PCD, иммунитете, воспалении и развитии тканей, предпочтительно ингибировать специфические функции NF-κВ, а не саму NF-κВ.
Настоящее изобретение относится к ингибированию Gadd45β, которая, как известно, регулируется положительным образом при ряде форм рака и, кроме того, при хронических воспалительных и наследственных нарушениях.
Множественная миелома (ММ), известная также как миелома клеток плазмы или болезнь Калера, представляет собой рак клеток плазмы. Множественная миелома в настоящее время неизлечима, хотя можно вызвать временные ремиссии при использовании стероидов, химиотерапии, талидомида, ингибиторов протеасом (Pis), например, бортецомиба, мелфалана и трансплантатов стволовых клеток. Согласно данным Американского общества борьбы с раковыми заболеваниями в США живет приблизительно 45000 больных множественной миеломой при том, что в США каждый год диагностируют приблизительно 15000 новых случаев. Средний период выживания от времени постановки диагноза составляет приблизительно три года. Множественная миелома является второй наиболее распространенной формой рака крови после лимфомы не-Ходжкина и составляет приблизительно 1% от всех форм рака и приблизительно 2% от всех смертей от рака. Частота множественной миеломы, по-видимому, будет возрастать, и имеется ряд доказательств снижения возраста, в котором начинается развитие заболевания. Таким образом существует явная необходимость в усовершенствованных препаратах для лечения множественной миеломы.
Почти все первичные опухоли множественной миеломы и клеточные линии множественной миеломы проявляют конститутивную активность NF-κВ. Блокирование активности NF-κВ вызывает гибель клеток множественной миеломы. Главным препятствием в достижении долговременных результатов лечения рака при использовании стратегий с направленностью на NF-κВ является отсутствие специфичности и, следовательно, неудовлетворительная переносимость лечения. Это обусловлено плейотропными функциями NF-κВ и протеасом. Существует необходимость в принципиально новом подходе к лечению, который является более специфическим, безопасным и, вследствие этого, более эффективным.
Одной из ключевых функций NF-κВ при множественной миеломе является стимуляция выживаемости. Показано (см. статью De Smaele, et al. (2001) Nature 414:306-313), что NF-κВ обеспечивает цитопротекцию, подавляя каскад JNK МАРК посредством Gadd45β, члена семейства индуцируемых факторов Gadd45. NF-κВ регулирует положительным образом Gadd45β в ответ на различные стимулы и способствует выживаемости путем направленного воздействия на киназу JNK MKK7 (см. статьи Papa, et al. 2004 Nature Cell Biology 6:146-153, Papa, et al. 2007, J. Biol. Chem. 282: 19029-19041, Papa, et al. (2008) J. Cliri. Invest. 118:191-1923).
Ингибиторы протеасом (PIs) и прямые ингибиторы NF-κВ уничтожают клетки множественной миеломы путем активации пути JNK, но непригодны для терапевтического лечения множественной миеломы вследствие их неизбирательных эффектов на NF-κВ и/или неизбирательных эффектов на протеасому, которые препятствуют их использованию в полностью ингибирующих лечебных дозах.
Кроме множественной миеломы Gadd45β экспрессируется на высоких уровнях в других опухолях, включая диффузную круп но клеточную В-клеточную лимфому, лимфому Беркетта, промоноцитарный лейкоз и другие формы лейкоза, а также некоторые солидные опухоли, включая гепатоцеллюларную карциному, рак мочевого пузыря, рак головного мозга и центральной нервной системы, рак молочной железы, рак головы и шеи, рак легкого и рак простаты. Вследствие этого ингибирование Gadd45β в данных опухолях может индуцировать гибель раковых клеток и, таким образом, имеет благоприятные терапевтические эффекты. Известно также, что многие гематологические злокачественные заболевания (в том числе множественная миелома, лимфома из клеток мантии, лимфома MALT, диффузная крупноклеточная В-клеточная лимфома, лимфома Ходжкина, миелодиспластический синдром, Т-клеточный лейкоз взрослых (HTLV-1), хронический лимфоцитарный лейкоз, хронический миелоидный лейкоз, острый миелогенный лейкоз и острый лимфоцитарный лейкоз) и солидные опухоли (включая рак молочной железы, рак шейки матки, рак почки, рак легкого, рак толстой кишки, рак печени, рак пищевода, рак желудка, рак гортани, рак щитовидной железы, рак паращитовидной железы, рак мочевого пузыря, рака яичников, рак простаты, рак поджелудочной железы и многие другие формы рака) демонстрируют конститутивную активацию NF-κВ, давая направленные на выживаемость сигналы клеткам посредством PCD, которые могли бы в другом случае привести к повышенному уровню гибели опухолевых клеток (см. статью V.Baud and M.Karin 2009, Nat. Rev. Drug Disc. 8:33-40). Конститутивную активность NF-κВ обнаруживают также в меланоме, цилиндроме, карциноме сквамозных клеток (кожи, а также головы и шеи), карциноме полости рта, эндометриальной карциноме, ретинобластоме, астроцитоме и глиобластоме (см. статью V.Baud and M.Karin 2009, Nat. Rev. Drug Disc. 8: 33-40). Ингибирование Gadd45β в данных опухолях, характеризующихся патологически высокой конститутивной активностью NF-κВ, также может давать благоприятные терапевтические эффекты посредством индукции запрограммированной гибели клеток среди раковых клеток. Настоящее изобретение основано на реализации того факта, что направленность отдельных направленных на выживаемость функций NF-κВ, опосредованная Gadd45β, дает более безопасный, более эффективный способ лечения, чем прямая направленность NF-κВ на ряд заболеваний и нарушений, включая рак, а также другие болезни, характеризующиеся патологической выживаемостью клеток, или болезни, которые можно было бы лечить индукций повышенного уровня PCD (такие как аутоиммунные заболевания, хронические воспалительные заболевания, дегенеративные заболевания, а также ишемические и сосудистые заболевания).
Широкий круг заболеваний и нарушений зависит от активности NF-κВ. В самом деле в настоящее время считают, что патогенез практически каждого известного человеческого заболевания или нарушения зависит от воспаления и, следовательно, включает NF-κВ. Она действует как групповой переключатель воспалительного ответа, координации экспрессии последовательности из более 200 генов, кодирующих цитокины, рецепторы, факторы транскрипции, хемокины, провоспалительные ферменты и другие факторы, включая направленные на выживаемость факторы, которые инициируют и замедляют воспаление. Соединения, соответствующие изобретению, ингибируют отдельную направленную на выживаемость активность NF-κВ при воспалении. Вследствие этого заболевания и нарушения, поддающиеся лечению данными соединениями, включают, кроме распространенных хронических воспалительных заболеваний (таких как воспалительная болезнь кишки, ревматоидный артрит и псориаз), другие заболевания и нарушения, которые в существенной степени зависят от воспалительного компонента. Примеры данных заболеваний и нарушений, которые лечатся противовоспалительными препаратами или препаратами, ингибирующими NF-κВ, или таких, которые, как предполагают, подходят для лечения ингибиторами NF-κВ и которые можно было бы лечить соединением, соответствующим изобретению, включают:
1. Дыхательные пути: обструктивные заболевания дыхательных путей, включая: астму, в том числе бронхиальную, аллергическую, эндогенную, экзогенную, вызываемую физической нагрузкой, вызываемую лекарственными препаратами (включая вызываемую аспирином и NSAID (нестероидными противовоспалительными лекарственными препаратами)) и вызываемую пылью астму, как интермиттирующую, так и постоянную, и всех степеней тяжести, а также другие случаи гиперчувствительности дыхательных путей; хроническое обструктивное легочное заболевание (COPD); бронхит, в том числе инфекционный и эозинофильный бронхит; эмфизему; бронхоэктазию; муковисцидоз; саркоидоз; "легкие фермера" (экзогенный аллергический альвеолит) и подобные заболевания; пневмонит, связанный с гиперчувствительностью; фиброз легкого, в том числе криптогенный фиброзирующий альвеолит, идиопатическое интерстициальное воспаление легких, фиброз как осложнение противоопухолевой терапии и хроническую инфекцию, включая туберкулез и аспергиллез и другие грибковые инфекции; осложнения при трансплантации легкого; васкулитные и тромботические нарушения легочной сосудистой сети и легочную гипертензию; противокашлевую активность, в том числе лечение хронического кашля, ассоциированного с воспалительными и секреторными состояниями дыхательных путей, и ятрогенный кашель; острый и хронический ринит, включая медикаментозный ринит и вазомоторный ринит; круглогодичный и сезонный аллергический ринит, в том числе нейрогенный ринит (сенная лихорадка); назальный полипоз; острую вирусную инфекцию, включая насморк, и инфекцию респираторно-синцитиального вируса, гриппа, коронавируса (в том числе SARS (тяжелый острый респираторный сидром) или аденовируса; либо эозинофильный эзофагит;
2. Кости и суставы: поражения ревматического или подагрического характера, ассоциированные с остеоартритом/остеоартрозом или включающие их, как первичные, так и вторичные, например, врожденная дисплазия тазобедренного сустава; шейный и поясничный спондилит и боль в нижнем отделе спины и шее; остеопороз; ревматоидный артрит и болезнь Стилла; серонегативные спондилоартропатии, включая анкилозирующий спондилит, псориатический артрит, реактивный артрит и недифференцированную спондартропатию; септический артрит и другие связанные с инфекцией артропатии и нарушения костей, такие как туберкулез, включая болезнь Пота и синдром Понсе; острый и хронический кристаллический синовит, включая подагру, заболевание, связанное с отложением пирофосфата кальция, и обусловленное апатитом кальция воспаление сухожилия, бурсы и синовии; болезнь Бехчета; первичный и вторичный синдром Шегрена, системный склероз и ограниченная склеродермия; системная красная волчанка, заболевание комбинированной соединительной ткани и заболевание недифференцированной соединительной ткани; воспалительные миопатии, в том числе дерматомиозит и полимиозит; ревматическая полимиалгия; ювенильный артрит, в том числе идиопатический воспалительный артрит, распространяющийся на любые суставы, и ассоциированные синдромы, и ревматическая лихорадка и ее системные осложнения; васкулиты, включая артериит гигантских клеток, артериит Такаяши, синдром Чин-Страусса, узелковый полиартериит, микроскопический полиартериит и васкулиты, ассоциированные с вирусной инфекцией, реакции гиперчувствительности, криоглобулины и парапротеины, боль в нижней области спины; семейная средиземноморская лихорадка, синдром Макла-Уэллса, семейная ирландская лихорадка, болезнь Кикучи; вызываемые лекарственными препаратами арталгии, тендиниты и миопатии;
3. Боль и ремоделирование соединительных тканей при скелетно-мышечных нарушениях, обусловленных повреждением [например, спортивной травмой]
или заболеванием: артритом (например, ревматоидным артритом, остеоартритом, подагрой или кристаллической артропатией), другим заболеванием суставов (таким как дегенерация межпозвоночного диска или дегенерация височно-нижнечелюстного сустава), заболевание, связанное с ремоделированием костей (такое как остеопороз, болезнь Педжета или остеонекроз), полихондрит (воспаление нескольких хрящей), склеродермия, нарушение комбинированной соединительной ткани, спондилоартропатии или заболевание периодонта (такое как периодонтит;
4. Кожа: псориаз, атопический дерматит, контактный дерматит или другие экзематозные дерматозы и реакции гиперчувствительности замедленного типа; фито- и фотодерматит; себорейный дерматит, герпетиформный дерматит, плоский лишай, склеротический атофический лишай, гангренозная пиодермия, кожный саркоид, дискоидная красная волчанка, пемфигус, пемфигоид, буллезный эпидермолиз, крапивница, ангионевротический отек (болезнь Квинке), васкулит, токсическая эритема, кожные эозинофилии, очаговая алопеция, облысение по мужскому типу, синдром Свита, синдром Вебера-Христиана, мультиформная эритема; целлюлит, как инфекционный, так и неинфекционный; панникулит; кожные лимфомы, рак кожи, отличный от меланомы и другие диспластические повреждения; вызываемые лекарственными препаратами нарушения, включая постоянную лекарственную сыпь;
5. Глаза: блефарит, конъюнктивит, включая круглогодичный и весенний аллергический конъюнктивит, ирит; передний и задний увеит; хориоидит; аутоиммунные, дегенеративные или воспалительные нарушения, поражающие сетчатку; офтальмит, в том числе симпатический офтальмит; саркоидоз; инфекции, включая вирусную, грибковую и бактериальную;
6. Желудочно-кишечный тракт: глоссит, гингивит, периодонтит, эзофагит, включая рефлюкс; эозинофильный гастроэнтерит, мастоцитоз, болезнь Крона, колит, в том числе язвенный колит, проктит, анальный зуд; заболевания брюшной полости, синдром раздраженной кишки и пищевые аллергии, которые могут иметь эффекты, отдаленные от кишки (например, мигрень, ринит или экзему);
7. Брюшная полость: гепатит, включая аутоиммунный, алкогольный и вирусный; фиброз и цирроз печени; холецистит, панкреатит, как острый, так и хронический;
8. Мочеполовая система: нефрит, включая интерстициальный и гломерулонефрит; нефротический синдром; цистит, включая острый и хронический (интерстициальный) цистит и язву Ханнера; острый и хронический уретрит, простатит, эпидидимит, оофорит и сальпингит; вульвовагинит; болезнь Пейрони; эректильная дисфункция (как у мужчин, так и у женщин);
9. Отторжение аллотрансплантата: острое и хроническое после, например, трансплантации почки, сердца, печени, легкого, костного мозга, кожи или роговицы, или после переливания крови, либо хроническая болезнь трансплантат против хозяина;
10. ЦНС (центральная нервная система): болезнь Альцгеймера и другие нарушения, связанные с деменцией, включая БКЯ (болезнь Крейтцфельда-Якоба) и нвБКЯ (болезнь Крейтцфельда-Якоба, новый вариант); амилоидоз; рассеянный склероз и другие демиелинизирующие синдромы; церебральный атеросклероз и васкулит; височный артериит; злокачественная миастения; острая и хроническая боль (острая, временная или хроническая, центральной или периферической природы), включая боли в животе, головную боль, мигрень, невралгию тройничного нерва, атипическую лицевую боль, боль в суставах и костях, боль, вызываемую раком и инвазией опухоли, невропатические болевые синдромы, включая диабетическую, пост-герпетическую и связанную с ВИЧ невропатию; нейросаркоидоз; осложнения на центральную и периферическую нервную систему злокачественных, инфекционных или аутоиммунных процессов;
11. Другие аутоиммунные и аллергические нарушения, в том числе тиреоидит Хашимото, болезнь Грейвса, болезнь Аддисона, сахарный диабет, идиопатическая тромбоцитопеническая пурпура, эозинофильный фасцит, синдром гипер-IgE, антифосфолипидный синдром;
12. Другие заболевания с воспалительным или иммунологическим компонентом, включая синдром приобретенного иммунодефицита (СПИД), лепру, синдром Сезари и паранеопластические синдромы;
13. Сердечно-сосудистая система: атеросклероз, поражающий коронарное и периферическое кровообращение; перикардит; миокардит, воспалительная и аутоиммунная кардиомиопатия, включая саркоид миокарда; ишемические реперфузионные повреждения; эндокардит, вальвулит и аортит, включая инфекционный (например, сифилитический); васкулит; нарушения проксимальных и периферических вен, включая флебит и тромбоз, в том числе тромбоз глубоких вен и осложнения варикоза вен;
14. Желудочно-кишечный тракт: Глютеновая болезнь, проктит, эозинофильный гастроэнтерит, мастоцитоз, болезнь Крона, язвенный колит, микроскопический колит, индетерминантный колит, нарушение раздраженной кишки, синдром раздраженной кишки, невоспалительная диарея, пищевые аллергии, которые могут иметь эффекты, отдаленные от кишки, например, мигрень, ринит или экзему.
Настоящее изобретение относится к новым ингибиторам комплекса Gadd45β/MKK7 и/или передачи сигнала данного комплекса, которые можно использовать для подавления функции, направленной на выживаемость NF-κВ при онкологических, воспалительных, аутоиммунных и дегенеративных, ишемических и сосудистых нарушениях.
Раскрытие изобретения
Согласно первому аспекту изобретения предусматривают соединение формулы I:
I: X1-A-X2,
А представляет собой А””, или А”-[М-А'-]n М-А”';
А”” представляет собой А”, А”' или Z1-Y2-Y3-Z4, где Y2-Y3 представляет собой олигопептидную группу или олигопептоидную группу, имеющую остатки Y2-Y3, и Z, присоединено к N-концевому атому азота Y2-Y3, и Z4 присоединено к С-концевому атому углерода Y2-Y3;
А” представляет собой А' или Y1-Y2-Y3-Y4, где Y1-Y2-Y3 представляет собой олигопептоидную группу или олигопептоидную группу, содержащую остатки Y1-Y2-Y3, и Z4 присоединено к С-концевому атому углерода Y1-Y2-Y3;
А”' представляет собой А' или Z1-Y2-Y3-Y4, где Y2-Y3-Y4 представляет собой олигопептоидную группу или олигопептоидную группу, содержащую остатки Y2-Y3-Y4, и Z1 присоединено к N-концевому атому азота Y2-Y3-Y4;
каждое присутствующее А' независимо представляет собой олигопептидную группу или олигопептоидную группу, содержащую остатки Y1-Y2-Y3-Y4;
n представляет собой целое число от 0 до 18,
Y1 и Y4 независимо представляют собой остатки аминокислот или -13-остатки производных аминокислот, имеющих ароматические боковые цепи,
Y2 представляет собой остаток аминокислоты или остаток производного аминокислоты или отсутствует,
Y3 представляет собой остаток аминокислоты или остаток производного аминокислоты или отсутствует;
Z1 представляет собой группу формулы II:
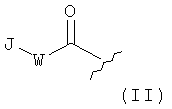
которая связана с N-концевым атомом азота Y2,
W отсутствует или представляет собой кислород или азот или алкиленовую группу из одного-трех атомов углерода, причем алкиленовая группа из одного-трех атомов углерода необязательно замещена по меньшей мере одним заместителем, выбранным из алкила с одним-четырьмя атомами углерода или 5-10-членной карбоциклической или гетероциклической ароматической группы;
J представляет собой 5-10-членную карбоциклическую или гетероциклическую ароматическую группу, причем ароматическая группа необязательно замещена по меньшей мере одним заместителем, выбранным из гидроксила, галогена, алкила из одного-четырех атомов углерода или алкокси из одного-четырех атомов углерода;
Z4 представляет собой группу формулы III:
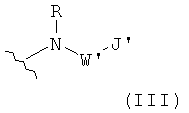
которая связана с С-концевым атомом углерода Y3,
R представляет собой водород или алкил из одного-четырех атомов
углерода;
W' отсутствует или представляет собой алкиленовую группу из
одного-трех атомов углерода, причем алкиленовая группа из одного-трех атомов углерода необязательно замещена по меньшей мере одним заместителем, выбранным из алкила из одного-четырех атомов углерода или 5-10-членной карбоциклической или гетероциклической ароматической группой;
J' представляет собой 3-10-членную алифатическую карбоциклическую группу или 5-10-членную карбоциклическую или гетероциклическую ароматическую группу, причем алифатическая или ароматическая группа необязательно замещена по меньшей мере одним заместителем, выбранным из гидроксила, галогена, алкила из одного-четырех атом углерода или алкокси из одного-четырех атомов углерода;
М представляет собой пептидную связь между предшествующей олигопептидной или олигопептидоидной группой (А', А” или А”') и следующей олигопептидной или олигопептидоидной группой (А', А" или А”') либо линкерную группу, присоединенную посредством амидной связи, сложно-эфирной связи, эфирной связи или тиоэфирной связи к концевой карбоксильной группе предшествующей олигопептидной или олигопептидоидной группы (А', А" или А"') и посредством амидной связи, сложно-эфирной связи, эфирной связи или тиоэфирной связи к концевой аминогруппе следующей олигопептидоидной группы (А', А” или А”');
Х1 отсутствует или представляет собой группу, введенную в аминоконец А с целью блокировать свободные аминогруппы;
X2 отсутствует или представляет собой группу, введенную в - карбоксильный конец А с целью блокировать свободные карбоксильные группы;
при условии, что Х1 отсутствует, если А содержит Z1 и X2 отсутствует, если А содержит Z4,
или их производные, причем указанные производные выбраны из группы, состоящей из:
a) олигомеров или мультимеров молекул соединения формулы I, причем указанные олигомеры и мультимеры содержат две или более молекул соединения формулы I, каждая их которых соединена с обычной группой основы посредством амидной связи между амино- или группой карбоновой кислоты, присутствующей в молекулах соединения формулы I и противоположной амино- или группой карбоновой кислоты на группе основы, причем указанная группа основы участвует по меньшей мере в 2 амидных связях,
b) производных, содержащих молекулу соединения формулы I или его олигомер либо мультимер, как определено выше в разделе а), конъюгированные посредством амидной связи, сложно-эфирной связи, эфирной связи или тиоэфирной связи с:
ПЭГ,
соединениями на основе ПЭГ,
проникающими в клетку пептидами,
флуоресцентными красителями,
биотином или другой группой метки,
жирными кислотами,
наночастицами дискретного размера
или
хелатирующими лигандами, комплексованными с ионами металлов или
ионами радиоактивных веществ.
c) производных, содержащих молекулу соединения формулы I или его олигомер либо мультимер, как определено выше в разделе а), которые модифицированы амидированием, гликозилированием, карбамилированием, ацилированием, сульфатированием, фосфорилированием, циклизацией, липидированием, пегилированием или связыванием с пептидным или пептидоидным партнером слияния с целью получения слитого пептида или слитого пептидоида и
d) солей и сольватов молекулы соединения формулы 1 или его производного, как определено выше в разделе а).
Согласно второму аспекту изобретения предусматривают фармацевтическую композицию, содержащую соединение, соответствующее первому аспекту изобретения, и фармацевтически приемлемый носитель.
Согласно третьему аспекту изобретения представлен способ лечения заболевания или нарушения, характеризующегося повышенной активностью и/или экспрессией NF-κВ и/или повышенной активностью и/или экспрессией Gadd45β, предусматривающий введение терапевтически эффективного количества соединения, соответствующего первому аспекту изобретения, или фармацевтической композиции, соответствующей второму аспекту изобретения, нуждающемуся в этом пациенту.
Согласно четвертому аспекту изобретения предусматривают соединение, соответствующее первому аспекту изобретения, или композиции, соответствующей второму аспекту изобретения, предназначенные для использования в качестве лекарственного средства.
Согласно пятому аспекту изобретения предусматривают применение соединения, соответствующего первому аспекту изобретения, или фармацевтической композиции, соответствующей второму аспекту изобретения, для изготовления лекарственного средства для лечения заболевания или нарушения, характеризующегося повышенной активностью и/или экспрессией NF-κВ и/или повышенной активностью и/или экспрессией Gadd45β.
Краткое описание чертежей
Фигура 1. Показано схематическое представление защитного взаимного влияния путей NF-κВ и JNK в контексте передачи сигнала ФНО-R1 (рецептора 1 фактора некроза опухоли). Можно видеть, что Gadd45β опосредует взаимное влияние пути выживаемости, индуцируемого NF-κВ, и пути гибели, индуцируемого MKK7 и JNK. Ингибирование данного взаимного влияния путем блокирования Gadd45β позволяет MKK7 активировать JNK, запуская, таким образом, путь гибели в опухолевых клетках.
Фигура 2. Представлена модель комплекса Gadd45β-MKK7. Модель строят, как описано в ссылке Papa S. et al. 2007, J Biol Chem 282: 19029-19041. Далее модель уточняют, используя кристаллографическую структуру MKK7 (pdb (база данных процесса): 2DYL) и моделируют структуру Gadd45β на кристаллографической структуре Gadd45β (pdb: 3FFM). Структура Gadd45β показана синим цветом (ленты), тогда как структура MKK7 показана желтым цветом (ленты с поверхностью Ван дер Ваальса также представлены). Ингибирующие кислые петли 60-71 и 104-118 Gadd45β (см. статью Papa S. et al 2007, J Biol Chem 282: 19029-19041) выделены.
Фигура 3. Представлен скрининг с помощью ELISA (иммуноферментный твердофазный анализ), используемый для выделения соединяющих D-тетрапептидов (DTPs) 1 и 2. Антагонисты взаимодействия Gadd45β/MKK7 отбирают путем скрининга упрощенной комбинаторной библиотеки пептидов общей формулы Fmoc-(βA1a)2-X1-X2-X3-X4-CONH2 (см. ссылку на статью Marasco et al. 2008, Curr. Protein Pept. Sci. 9:447-67), полученной с использованием одной из 12 аминокислот в каждом положении от Х1 до Х4. Данную библиотеку, содержащую в целом 124=20736 различных пептидов, многократно подвергают деконволюционному анализу в четырех стадиях конкурентных анализов ELISA, используя на каждой стадии покрытый человеческий MKK7 (42 нМ), растворимый меченный биотином человеческий (h) Gadd45β (21 нм) и каждую из 12 суббиблиотек в номинальной концентрации 42 нМ (не показано).
Фигура 4. Представлен скрининг библиотеки второго поколения, полученной из наиболее активного пептида первого поколения. Скрининг данной библиотеки дает два высокоактивных пептида (отмечены на Фиг.4 как 1 и 8).
Фигура 5. Представлена активность изомеров оптимизированных пептидов. Так оптимизированные пептиды освобождают от метки Fmoc-(βAla)2 и синтезируют в виде D-изомеров, получая DTP1 и DTP2, которые нарушают взаимодействие Gadd45β/MKK7 с IC50 (50% ингибирующая концентрация) 0,22 нм и 0,19 нМ, соответственно. Можно также видеть, что L-изомеры данных пептидов (т.е. LTP1 и LTP2) демонстрируют 1650, близкую к данным значениям для DTP в конкурентных анализах ELISA, тогда как пептиды отрицательного контроля, LNC и DNC, не проявляют определяемый ингибирующий эффект на формирование комплекса Gadd45β/MKK7. LNC - отрицательный контроль L-изомера; LTP1 - тетрапептид L-изомера 1; LTP2 - тетрапептид L-изомера 2; DNC - отрицательный контроль D-изомера.
Фигура 6. Показана стабильность Z-DTPs в биологических жидкостях. Результаты конкурентных анализов ELISA показывают, что Z-защищенные DTP (Z-DTP1, Z-DTP2) сохраняют полную ингибирующую активность после 48-часового инкубирования с человеческой сывороткой при 37°С (IC50=0,19 нМ, Z-DTP1; IC50=0,18 нМ, ZDTP2), тогда как Z-защищенные LTP (Z-LTP1, Z-LTP2) почти полностью инактивируются после данной обработки (1050>10 мкМ). Анализы проводят, как показано на Фиг.3С, используя покрытый MKK7, растворимый биотин-hGadd45β и указанные концентрации тетрапептидов. Z-LNC и Z-DNC - отрицательные контроли L- и D-изомеров, соответственно.
Фигура 7. Представлены анализы соиммунопреципитации (co-IP), показывающие эффективное и специфическое нарушение взаимодействия Gadd45β/MMK7D-тетрапептидами 1 и 2 (DTP1 и DTP2), но не D-тетрапептидами отрицательного контроля (NC) (NC1, NC2, NC3 и NC4). Co-IP проводят при использовании антитела к FLAG (MKK7), и результаты вестерн-блоттинга проявляют с помощью антител к НА (детекция HA-Gadd45β) (верх) или к MKK7 (низ), как указано.
Фигура 8. Представлены анализы киназы MKK7, показывающие эффективное и специфическое нарушение взаимодействия Gadd45β/MMK7 и восстановление каталитической активности MKK7 посредством D-тетрапептидов 1 и 2 (DTP1 и DTP2), но не D-тетрапептидами отрицательного контроля (NC) (NC1, NC2, NC3 и NC4). Активную MKK7 подвергают иммунопреципитации с использованием антитела к FLAG из клеток НЕК293Т, обработанных форбол 12-миристат 13-ацетатом (РМА)/иономицином (P/I) и инкубируют с D-тетрапептидами в присутствии (верхняя панель) или в отсутствие (нижняя панель) рекомбинантного человеческого (h) Gadd45β, как указано. Как показано, ни активные соединяющие D-тетрапептиды, ни контрольные NC тетрапептиды не ингибируют каталитическую активность MKK7 при инкубировании с киназой в отсутствие Gadd45β (нижняя панель).
Фигуры 9-11. Представлены анализы включения [3H]тимидина, показывающие, что Z-защищенные производные DTP2 (Z-DTP2) (Фиг.9), но не ацетиловые производные DTP2 (Ac-DTP2) (Фиг.10) или Z-защищенные производные L-изомеров DTP2 (Z-LTP2) (Фиг.11), обладают существенной активностью уничтожения опухолевых клеток в опухолевых клеточных линиях. Результаты выражают как доля выживаемости/пролиферации опухолевых клеток после обработки либо 10 мкМ Z-DTP2 (Фиг.9), Ac-DTP2 (Фиг.10) или Z-LTP2 (Фиг.11) (закрашенные колонки) или Z-DNC (Фиг.9), Ac-DNC (Фиг.10) и Z-LNC (Фиг.11) (незакрашенные колонки) относительно уровня выживаемости/пролиферации необработанных клеток. Точки времени указаны. Показаны 3 из 8 тестированных чувствительных клеточных линий множественной миеломы (т.е. U266, KMS-11, NCI-H929) (Фиг.9-11) и клеточные линии лимфомы Беркетта (BJAB) (Фиг.9) и промоноцитарного лейкоза (U937) (Фиг.9). Данные результаты показывают высокую цитотоксическую активность Z-DTP2 (Фиг.9) по сравнению с отсутствием активности yAc-DTP2 (Фиг.10) и низкой активностью Z-LTP2 (Фиг.11) (см. также Фиг.12, 13 и 14 и Таблицу IV; дополнительные линии множественной миеломы). Отсутствие активности уничтожения опухолевых клеток Ac-DTP2 (Фиг.10) в клеточных линиях множественной миеломы коррелируется низким уровнем проницаемости клеток у данного соединения, как установлено в анализах CaCO2 (результаты не показаны). Выживаемость клеточных линий множественной миеломы после обработки другими, менее эффективными производными DTP (разработанными также для повышения поглощения DTP клетками), включая несущие метильную (Me), ацетиловую (Ас), миристиловую (Муг), 3-метокси,4-гидрокси-бензоиловую, бензоиловую, SCI-бензилоксикарбонильную (6CI-Z) и/или флуоренилметилоксикарбониловую (Fmoc) группу, не показана. Хотя активность in vitro и клеточное поглощение Z-LTP (Фиг.11) сравнимы с данными показателями Z-DTP (см. Фиг.5; результаты также не показаны), Z-LTP2 демонстрируют низкую активность в клетках множественной миеломы вследствие низкой стабильности в биологических жидкостях (см. Фиг.6).
Фигуры 12-14. Представлена проапоптотическая активность Z-DTP избирательна в отношении опухолевых клеточных линий с конститутивной активностью NF-κВ. Анализы включения [3H]тимидина, проведенные, как описано на Фиг.9-11, показывающие выживаемость клеток на панели опухолевых клеточных линий после обработки 10 мкМ Z-DTP или Z-DNC в следующие периоды времени: 144 часа (Фиг.12); 24 часа, 72 часа или 144 часа (Фиг.13 и Фиг.14), как указано. На Фиг.12 показана высокая активность уничтожения опухолевых клеток Z-DTP2 в 8 из 9 клеточных линий множественной миеломы, 1 из 2 клеточных линий диффузной крупноклеточной В-клеточной лимфомы (DLBCL; LY3), 1 из 1 клеточной линии промоноцитарного лейкоза (U937) и в 1 из 6 клеточных линий лимфомы Беркетта (BJAB), которые тестируют (см. также Фиг.15). Интересно, что Z-DTP2 показывает цитотоксическую активность только в клеточной линии DLBCL (АВС)-подобного подтипа активированных В-клеток (т.е. LYg) и не показывает в подтипе, подобном В-клеткам зародышевого центра (GCB) (т.е. SUDHL6), который не характеризуется конститутивной активацией NF-κВ (см. статью Ngo VN, et al. Nature 441(7089): 106-10; см. также Фиг.22 и Фиг.23, уровни экспрессии Gadd45β). Они также проявляют активность в клеточных линиях множественной миеломы, фактически все из которых характеризуются конститутивной активацией NF-κВ. Активность уничтожения опухолевых клеток Z-DTP2 (Фиг.13) и Z-DTP1 (Фиг.14) в клеточных линиях множественной миеломы и DLBCL после обработки 10 мкМ Z-DTP1, Z-DTP2 и Z-DNC в указанные точки времени (т.е. 24, 72 или 144 часа, как показано). Результаты подтверждают анализами исключения трипанового синего (результаты не показаны) и анализами с использованием йодида пропидия (PI) (см. Фиг.16-20; результаты также не показаны).
Фигура 15. Представлены анализы включения [3H]тимидина, показывающие отсутствие цитотоксичности Z-DTP2 на панели из 22 устойчивых опухолевых клеточных линий после обработки Z-DTP2 в течение 144 часов, даже когда данное соединение используют в очень высоких концентрациях, т.е. 100 мкМ. ZDNC означает Z-защищенный D-отрицательный контроль. Кроме того, показаны чувствительные клеточные линии BJAB (лимфомы Беркетта), KMS-11 и KMS-12 (множественная миелома). Таким образом, существует сильная корреляция среди данных клеточных линий между чувствительностью к индуцированной Z-DTP2 гибели и уровнями эндогенной экспрессии Gadd45β (см. Фиг.22 и 23).
Фигуры 16-20. Показано, что индуцированная Z-DTP2 гибель в клеточных линиях множественной миеломы обусловлена апоптозом. Анализы на основе окрашивания ядра йодидом пропидиума (PI) показывают индукцию апоптоза (т.е. содержание ДНК в фазе суб-G1; см. FL2-A) в репрезентативных клеточных линиях множественной миеломы ARH-77 (Фиг.16), KMS-11 (Фиг.17), JJN-3 (Фиг 18), NCI-H929 (Фиг 19) и U266 (Фиг.20) после обработки 10 мкМ Z-DTP2 или ZDNC1, как показано для точек 72 или 144 час. Показано также содержание ДНК в необработанных клетках, культивируемых в тех же условиях. Доли апоптотических клеток представлены на гистограммах.
Фигура 21. Показано, что обработка Z-DTP2 вызывает сильную активацию JNK в клеточных линиях множественной миеломы. Клетки KMS11 и NCI-H929 обрабатывают 10 мкМ Z-DTP2 или Z-DNC, как показано, и мониторируют активацию JNK в указанных точках времени с помощью вестерн-блоттинга с использованием специфического антитела к фосфо (P)-JNK. Повышенный уровень фосфорилирования JNK (маркер активации JNK) наблюдают только после обработки Z-DTP2, но не после обработки Z-защищенным пептидом отрицательного контроля (Z-DNC). Стимуляцию α-ФНО (2000 ед/мл) используют в качестве положительного контроля при активации JNK. Важно, что аналогичные эффекты ZDTP2 наблюдают в отношении активации MKK7 (результаты не показаны). Более того, как видно по биологической активности Gadd45β (см. ссылки на статьи: De Smaele, et al. (2001) Nature 414:306313; Papa, S et al., (2004) Nat. Cell Biol. 6, 146-153; Papa, et al. 2007 J. Biol. Chem. 282:19029-19041; Papa, et al. (2008) J.CIin.Invest. 118:191-1923), эффекты Z-DTP в клеточных линиях множественной миеломы специфичны в отношении пути MKK7/JNK, поскольку при использовании данных соединений не наблюдают никаких эффектов на активацию путей IKK/NF-κВ, ERK и р38 в данных клеточных линиях (результаты не показаны).
Фигуры 22 и 23. Показана сильная корреляция в опухолевых клеточных линиях между чувствительностью клеток к индуцированной Z-DTP гибели и уровнями экспрессии Gadd45β. На верхней панели Фиг.22 показывают экспрессию Gadd45β на панели из 29 раковых клеточных линий (кОТ-ПЦР (количественная ПЦР с использованием обратной полимеразы); колонки красного цвета), тогда как нижняя панель Фиг.22 показывает долю гибели клеток в тех же клеточных линиях после обработки 10 мкМ Z-DTP2 в течение 144 час (включение [3H]тимидина; колонки черного цвета). На Фиг.23 показана схема корреляции экспрессии Gadd45β относительно процента выживаемости клеток после обработки Z-DTP2 в том же эксперименте, что показан на Фиг.23. Достоверность коэффициента корреляции между двумя параметрами очень высокая (р<0,01) (Коэффициент корреляции Пирсона, который количественно определяет связь двух переменных величин, рассчитанный с использованием пакета программ GraphPad). Данные результаты подтверждают высокую специфичность Z-DTP в отношении мишени в клетках. Значения Фиг.22 (верхняя панель) нормализованы относительно β-актина.
Фигуры 24 и 25. Показаны химические структуры подходящих соединений, раскрытых в данном патенте, и описание возможных фармакофоров и стратегий их оценки. На Фиг.24 показаны химические структуры исходного соединения Z-DTP2 (Z-D-Tyr-D-Glu-D-Arg-DPhe-NH2) [SEQ ID NO:1]
и производных Z-DTP2: mDTP1 (п-гидроксибензойная кислота-D-Glu-Arg-фенетиламина), mDTP2 (Ac-D-Tyr-D-Glu-D-Phe-NH2) и mDTP3 (Ac-D-Tyr-D-Arg-D-Phe-NH2). Данные модифицированные соединения Z-DTP2 (далее называемые mDTP) тестируют на активность как in vitro (ELISA), так и на клетках (анализы гибели). Представлены также молекулярные массы (MM), IC50 in vitro и на клетках, а также эффективность Z-DPT2 как лигандов данных репрезентативных модифицированных соединений (см. также Таблицу V). На Фиг.25 описаны основные стадии способа, разработанного для идентификации возможного фармакофора биоактивных соединений (см. статью Geeson МР. 2008 J Med Chem. 51:817834). Большинство из предложенных изменений уже используют: М-концевые группы (см. Таблицу III); замена Tyr циклооксиаланином, Phe циклооксиаланином, удаление внутреннего Glu и/или Arg, замена Glu на Asp, сложно-эфирные пролекарственные формы на Asp боковой цепи (см. Таблицу V); замена Tyr на Phe, замена Arg на His, Lys или Pro (см. Таблицу VI). Одновременно результаты показывают, что биоактивный фармакофор можно описать следующим образом: тирозин или подобный ароматический цикл с донорами/акцепторами Н-связи необходим в положении Y1; по меньшей мере одна a-аминокислота необходима в положении Y2 и/или Y3, предпочтительно в основной группой для улучшения клеточного всасывания. Пролин, аспарагины или лейцин в положении Y2 при наличии или в отсутствие аргинина в положении Y3 также позволяют сохранить биоактивность. Расстояние, превышающее приблизительно 7 Å, между двумя ароматическими циклами (т.е. расстояние, превышающее занимаемое одной а-аминокислотой) вызывает снижение биоактивности; ароматический цикл необходим в положении Y4 при наличии или в отсутствие групп доноров/акцепторов Н-связи для сохранения биоактивности (см. Таблицу VI).
Фигуры 26-30. Показана цитотоксическая активность Z-DTP в первичных клетках множественной миеломы, выделенных у 5 репрезентативных пациентов. Каждая панель представляет результаты, полученные на клетках, выделенных у разных пациентов, т.е. пациента 1 (Фиг.26), пациента 2 (Фиг.27), пациента 3 (Фиг.28), пациента 4 (Фиг.29), и пациента 5 (Фиг.30). Обработки Z-DTP2, Z-DTP1 и Z-DNC проводят в указанных концентрациях в течение 48 часов. Показаны также необработанные клетки каждого пациента (-). Анализы проводят с использованием исключения трипанового синего и подсчета клеток. Значения представляют собой долю живых клеток, наблюдаемых после обработки Z-DTP2, Z- DTP1 или Z-DNC относительно выживаемости необработанных контрольных клеток.
Фигуры 31 и 32. Показано отсутствие цитотоксической активности Z-DTP в первичных нетрансформированных клетках, выделенных у пациентов с отсутствием множественной миеломы, включая клетки стромы костного мозга (BMSCs) (Фиг.31), мононуклеарные клетки периферической крови (PBMNCs) (Фиг.31) и мезенхимные стволовые клетки (MSCs) (Фиг.32) или очищенные первичные В- и Т-лимфоциты мышей (Фиг.32). Обработку Z-DTP2, Z-DTP1 и Z-DNC проводят в указанных концентрациях, в течение: 48 час (BMSCs, PBMNCs) (Фиг.31), 72 час (мышиные В- и Т-клетки) (Фиг.32) либо 144 час MSCs (Фиг.32). Анализы проводятся использованием исключения трипанового синего и подсчета клеток (Фиг.31) или включения [3H]тимидина (Фиг.32).
Фигуры 33-35. Показана индукция гибели клеток в репрезентативных клеточных линиях множественной миеломы после опосредованного shPHK сайленсинга экспрессии Gadd45β. Чувствительные kz-dtp клеточные линии множественной миеломы ARH-77 (Фиг.33) и NCI-H929 (Фиг.34) устойчивую к Z-DTP клеточную линию множественной миеломы RPMI-8226 (Фиг.35), инфицируют лентивирусом, экспрессирующим либо shPHK, специфическими в отношении Gadd45β (т.е. sh-Gadd45β-1, sh-Gadd45β-2 или sh-Gadd45β-3), shPHK, специфическими в отношении MKK7 (т.е. sh-MKK7-1 или sh-MKK7-2), либо неспецифическими shPHK (т.е. sh-NS-1 или sh-NS-2) и мониторируют выживаемость инфицированных клеток в течение 8 дней при использовании проточной цитометрии, обнаруживают клетки, экспрессирующие на повышенном уровне белок зеленой флуоресценции (eGFP), которые являются инфицированными клетками, и подсчитывают клетки. Показывают процент выживаемости eGFP+ (т.е. инфицированных) клеток множественной миеломы в указанные точки времени относительно выживаемости клеток множественной миеломы eGFP+ в той же культуре в 0. Клетки инфицируют лентивирусом pLentiLox.3.7, экспрессирующим указанные sh-PHK, а также eGFP, используя стандартные способы (как описано в ссылке Yang H et al., Рrос NatI Acad Sci USA. 5 июля 2006; 103(27):10397-402). Через 5 дней клетки eGFP+ подвергают сортингу, используя клеточный сортер BD FACSAria™ II, затем оставляют в покое в течение 2 дней перед началом анализов выживаемости клеток. Данная точка времени (т.е. начало анализов выживаемости) отмечена на графиках как день 0. Результаты показывают, что ингибирование экспрессии Gadd45β вызывает быструю гибель клеток в клеточных линиях множественной миеломы, которые чувствительны к вызываемой Z-DTP токсичности (т.е. в клеточных линиях ARH-77, NCI-H929) (Фиг.33 и 34), но не в клеточной линии множественной миеломы RPM18226 (Фиг.35), которая устойчива к данной токсичности. Данные результаты далее показывают специфичность в отношении мишени Z-DTP в отношении комплекса Gadd45β/MKK7 в клетках множественной миеломы (см. также Фиг.9-15, 22 и 23; гибель и анализы кОТ-ПЦР). Они также демонстрируют важную роль, которую играет Gadd45β в выживаемости клеток множественной миеломы, таким образом далее подтверждая правильность выбора Gadd45β в качестве терапевтической мишени при множественной миеломе.
Фигуры 36 и 37. Представлены результаты анализов включения [3H]тимидина, показывающие, что опосредованный sh-PHK сайленсинг Gadd45β, но не MKK7 имеет сильную активность уничтожения раковых клеток в клеточных линиях множественной миеломы, которые чувствительны к вызываемой Z-DTP гибели (т.е. клеточные линии ARH-77 и NCI-H929; см. также Фиг.9-14, чувствительность к вызываемой Z-DTP гибели). Выживаемость устойчивой kz-dtp клеточной линии множественной миеломы RPM18226 используют вместо клеток, на которые не действует опосредованное sh-PHK ингибирование Gadd45β. На Фиг.36 показана выживаемость трех репрезентативных клеточных линий множественной миеломы, RPMI-8226, NCI-H929 и ARH-77, после сайленсинга Gadd45β или MKK7. На Фиг.37 показана выживаемость клеточной линии множественной миеломы ARH-77 после сайленсинга Gadd45β или MKK7 при использовании трех различных специфических в отношении Gadd45β sh-PHK (т.е. sh-Gadd45β-1, sh-Gadd45β-2 или sh-Gadd45β-3), двух различных специфических в отношении MKK7 sh-PHK (т.е. sh-MKK7-1 или sh-MKK7-2) и двух различных неспецифических sh-PHK (т.е. sh-NS-1 или sh-NS-2). Клеточные линии множественной миеломы инфицируют лентивирусом pLentiLox.3.7, экспрессирующим указанные sh-PHK, затем проводят сортинг клеток множественной миеломы eGFP+ (т.е. клеток, инфицированных лентивирусом), используя клеточный сортер BD FACSAria™ II, как показано на Фиг.33-35. Анализы включения [3H]тимидина проводят через 10 дней после сортинга клеток, что соответствует дню 8 на Фиг.33-35. Показан процент включения [3H]тимидина (т.е. импульсы в мин), мера пролиферации клеток, в день 8 (т.е. через 10 дней после сортинга клеток) относительно включения [3H]тимидина, происходящего в тех же клетках в день 0 (т.е. через 2 дня после сортинга клеток). Данные результаты, кроме того, устанавливают специфичность в отношении мишени Z-DTP к комплексу Gadd45β/MKK7 в клетках множественной миеломы (см. также Фиг.9-15, 22 и 23, вызываемая Z-DTP гибель и экспрессия Gadd45β; см. Фиг.33-35, сайленсинг генов Gadd45β и MKK7) и подтверждают важную роль, которую Gadd45β играет в выживаемости клеток множественной миеломы. Вместе взятые, они, кроме того, также подтверждают роль Gadd45β как терапевтической мишени при множественной миеломе.
Фигуры 38-43. Представлены результаты анализов окрашивания йодидом пропидиума (PI), показывающие, что опосредованный sh-PHK сайленсинг Gadd45β индуцирует апоптоз в чувствительных к Z-DTP клеточных линиях множественной миеломы, ARH-77 (Фиг.38-41) и NCI-H929 (Фиг.42), но не в устойчивой к Z-DTP клеточной линии множественной миеломы RPMI-8226 (Фиг.43) (см. также Фиг.33-37, опосредованный sh-PHK сайленсинг; Фиг.9-14, 22 и 23 чувствительность клеточной линии множественной миеломы к индуцируемой Z-DTP гибели и экспрессии Gadd45β). Никакую существенную индукцию апоптоза не наблюдают в тех же клеточных линиях множественной миеломы в отсутствие лентивирусной инфекции (неинфицированные) или после экспрессии sh-PHK, специфических в отношении MKK7 (т.е. sh-MKK7-1 и sh-MKK7-2) или неспецифических sh-PHK (т.е. sh-NS-1 и sh-NS-2). Клеточные линии множественной миеломы инфицируют экспрессирующими sh-PHK лентивирусами pLentiLox.3.7 и проводят сортинг клеток множественной миеломы eGFP+ (т.е. клеток, инфицированных лентивирусом) с использованием клеточного сортера BD FACSAria™ II, как показано на Фиг.33-35. Анализы ядерного окрашивания PI проводят через 10 дней после сортинга клеток, что соответствует дню 8 на Фиг.33-35. На гистограммах показана доля апоптотических клеток (т.е. клеток, демонстрирующих содержание ДН К в фазе суб-G1). Важно, что уровни апоптоза, индуцированного различными специфическими в отношении Gadd45β sh-PHK (т.е. sh Gadd45y&, sh-Gadd45β-2 и sh-Gadd45β) коррелирует с уровнями понижающей регуляции Gadd45β, индуцируемой каждой из данных специфических в отношении Gadd45β sh-PHK (результаты не показаны). Данные результаты, кроме того, устанавливают специфичность в отношении мишени Z-DTP в отношении комплекса Gadd45β/MKK7 в клетках множественной миеломы (см. также Фиг.9-15, анализы гибели с использованием Z-DTP; см. Фиг.22 и Фиг.23, статистически достоверная корреляция между экспрессией Gadd45β и чувствительностью раковых клеток к вызываемой Z-DTP гибели; Фиг.33-37, индукция гибели клеток множественной миеломы путем понижающей регуляции Gadd45β, но не MKK7), и подтверждение важной роли, которую Gadd45β играет в выживаемости клеток множественной миеломы. Вместе взятые, они, кроме того, также подтверждают роль Gadd45β как терапевтической мишени при множественной миеломе.
Фигуры 44-47. Представлены результаты анализов ядерного окрашивания PI, показывающие, что опосредованный sh-РНК сайленсинг либо MKK7 либо Gadd45β не влияет на распределение клеточного цикла в клеточных линиях множественной миеломы. Показанные репрезентативные инфицированные лентивирусом клеточные линии множественной миеломы, т.е. ARH-77 (Фиг.44 и Фиг.45), NCI-H929 (Фиг.46) и RPMI-8226 (Фиг.47) берут из того же эксперимента, что представлен на Фиг.38-43. В отличие от результатов, показанных на Фиг.38-43 (где профили окрашивания PI представляют в логарифмическом масштабе, который выделяет апоптоз), окрашивание PI (т.е. FL2-A) на данной фигуре представляют в линейном масштабе, что выделяет распределение клеточного цикла. Доли клеток множественной миеломы, находящихся в разных фазах клеточного цикла (т.е. G1, S и Gg/M), представлены на гистограммах. Анализы клеточного цикла невозможно было провести со специфическими в отношении Gadd45β sh-PHK в клеточных линиях множественной миеломы ARH-77 (Фиг.44 и Фиг.45) и NCI-H929 (Фиг.46) вследствие индукции массивного апоптоза в данных клетках (см. Фиг.38-42).
Фигуры 48-50. Показано, что опосредованный sh-РНК сайленсинг MKK7 делает репрезентативную чувствительную к Z-DTP клеточную линию ARH-77 устойчивой к индуцированной Z-/mDTP гибели. Представлены анализы включения [3H]тимидина, показывающие IC50 D-изомера тетрапептида отрицательного контроля (Z-DNC) (Фиг.48-50), Z-DTP 1 (Фиг.48), Z-DTP2 (Фиг.49) или mDTP3 (Фиг.50) в клетках множественной миеломы ARH-77, экспрессирующих либо специфические в отношении MKK7 (sh-MKK7), либо неспецифические shRNAs (sh-NS). Обработку клеток ARH-77 Z-DNC, Z-DTP1, Z-DTP2 или mDTP3 проводят в течение 3 дней. Можно видеть, что клетки ARH-77, экспрессирующие sh-NS, высокочувствительны к вызываемой Z-/mDTP гибели, что показано по значениям 1050 1,4 мкМ (Z-DTP1; Фиг.48), 302 нМ (Z-DTP2; Фиг.49) и 303 нМ (mDTP3; Фиг.50), близким значениям, наблюдаемым в неинфицированных исходных клетках ARH-77 (см. Таблицу IV). Напротив, клетки ARH77, экспрессирующие sh-MKK7, становятся полностью устойчивыми к вызываемой Z-/mDTP гибели, что показано по значениям IC50>10 мкМ, близким значениям, наблюдаемым в клетках ARH-77, обработанных Z-DNC. 1050 рассчитывают, как описано в Примерах, используя повышающиеся концентрации Z-DNC (Фиг.48-50), Z-DTP1 (Фиг.48), Z-DTP2 (Фиг.49) и mDTP3 (Фиг.50), лежащие в интервале от 0,01 до 10 мкМ. На графиках показаны проценты импульсов в минуту (с.р.m.), результаты измерения пролиферации клеток, полученные для клеток, обработанных пептидом, относительно значений с.р.m., полученных для необработанных клеток. Аналогичные результаты получают для дополнительных чувствительных к Z-/mDTP клеточных линий множественной миеломы, включая клеточные линии U266, KMS-11 и KMS-12 (результаты не показаны). Данные результаты демонстрируют очень высокую специфичность в отношении мишени Z-/mDTP для комплекса Gadd45β/MKK7 в клетках множественной миеломы (см. также Фиг.22 и Фиг.23, корреляция между уровнем экспрессии Gadd45β и чувствительностью раковых клеток к вызываемой Z-DTP гибели).
Фигуры 51-54. Показано, что соединения, соответствующие изобретению, не связываются ни с Gadd45β, ни с MKK7 при выделении; напротив, для связывания им необходимо формирование комплекса Gadd45β/MKK7, как определяют в анализах системы biacore. На Фиг, 51 показано связывание Gadd45β с доменом киназы MKK7 (MKK7KD), иммобилизованном на чипе. Различные концентрации Gadd45β (лежащие в интервале от 20 до 200 нМ) наносят на чип, на котором предварительно иммобилизуют MKK7KD. Регистрируют дозозависимое связывание Gadd45β с MKK7KD и кривые диссоциации комплекса Gadd45β/MKK7KD и определяют значение константы равновесия диссоциации (KD) 4,0±0,7 нМ путем усреднения значений, определенных по кинетическим параметрам каждой отдельной кривой. Вкратце, термодинамический параметр константы равновесия диссоциации (KD) рассчитывают, учитывая фазы ассоциации (ka) и диссоциации (kd), соответствующие повышению или снижению сигнала SPR (выражаемого как единицы ответа, RU), соответственно. На Фиг.52 показано связывание MKK7KD с Gadd45β, когда Gadd45β иммобилизован на чипе. В данном случае значения KD определяют по нанесению MKK7KD в различных концентрациях (лежащих в интервале от 1 до 25 нМ) на чип с иммобилизованным Gadd45β. Кривые дозозависимости регистрируют для всех исследованных концентраций MKK7KD. На основании данных анализов получают значение KD 3,4±0,6 нМ, которое очень близко значению KD, полученному из Фиг.51. На Фиг.53 показано нанесение mDTP3 на чип, содержащий либо Gadd45β, либо MKK7KD. Для того, чтобы определить, связывается ли mDTP3 с Gadd45β и/или с MKK7KD, раствор, содержащий mDTP3 в концентрации, лежащей в интервале от 1 нМ до 10 мкМ, наносят на чип, дериватизированный либо одним, либо другим белком. Как можно видеть, никакое связывание mDTP3 либо с Gadd45β, либо с MKK7 не регистрируют даже при самой высокой из использованных концентраций mDTP3 (т.е. 10 мкМ). На Фиг.54 показано связывание mDTP3 с полученным комплексом Gadd45β/MKK7. Gadd45β в концентрации 100 нМ наносят на чип, дериватизированный MKK7KD (60 мкл; время контакта 3 мин). Белкам Gadd45β позволяют диссоциировать в течение приблизительно 10 мин и когда приблизительно 50% Gadd45β еще остается связанным с MKK7KD, наносят mDTP3 в концентрации либо 10 нМ, 100 нМ, либо 1 мкМ. Как можно видеть, при использовании в концентрации, равной или ниже 100 нМ, mDTP3 вызывает быструю диссоциацию комплекса Gadd45β/MKK7KD. Как также можно видеть, образование комплекса Gadd45β/MKK7KD быстро регистрируют после отмывания mDTP3. Однако, при более высоких концентрациях (например 1 мкМ), mDTP3 дает дозозависимое связывание и кривые диссоциации, указывающие на то, что это связывание либо с Gadd45β и/или с MKK7KD, либо с комплексом из двух белков. Данные результаты поддерживают ту точку зрения, что DTP не связываются с белками Gadd45β или MKK7 отдельно; напротив, они связываются с одним и/или другим белком либо с комплексом из двух белков только в случае, когда два белка находятся в контакте друг с другом.
Замечание относительно номенклатуры, используемой в данном контексте.
В разных разделах данного описания соединения обозначают символическим кодом, например, LTP, DTP, LNC, DTP1 и т.п. Коды, содержащие "NC", описывают соединения, которые являются отрицательными контролями, не входящими в объем изобретения. Коды, содержащие "ТР" (что означает сокращение от тетра или трипептидов/пептидоидов, хотя следует заметить, что основой некоторых соединений являются дипептидные/пептоидные мотивы), входят в объем изобретения. Префикс "L" или "D" означает остатки в L- или D-оптической конфигурации. Цифровой суффикс означает соединение со специальным номером, подробно описанное в другом месте. Префикс "Z", как в "Z-DTP" означает бензилоксикарбониловую N-концевую группу. Префикс "m", как в "mDTP", означает любую модификацию DTP, цель которой состоит в повышении уровня клеточного поглощения, клеточной активности и/или профиля РК, например, в удалении N- и/или С-конца (например, как в mDTP1), удалении группы Z и остатков Arg или Glu в Z-DTP2, как в mDTP2 и mDTP3, соответственно (дальнейшие примеры представляют на Фиг.24 и Фиг.25).
Осуществление изобретения
Стратегия, лежащая в основе настоящего изобретения, происходит из понимания того, что взаимное влияние NF-κB-JNK контролирует также выживаемость относительно запрограммированной гибели клеток, включая раковые клетки, которые погибли бы в ином случае. Важно, что Gadd45β регулируется положительным образом в раковых клетках в ответ на активацию NF-κВ и конститутивно экспрессируется на высоких уровнях в клетках множественной миеломы и других опухолей, включая диффузную крупноклеточную В-клеточную лимфому, лимфому Беркетта, промоноцитарный лейкоз и другие формы лейкоза, а также некоторые солидные опухоли, включая гепатоцеллюларную карциному, рак мочевого пузыря, рак головного мозга и центральной нервной системы, рак молочной железы, рака головы и шеи, рак легкого и рак простаты. Настоящее изобретение основано на способе стимуляции запрограммированной гибели клеток путем доставки соединений, направленных на Gadd45β/MKK7, которые препятствуют перекрестному влиянию NF-κВ-JNK, усиливая, таким образом, передачу цитотоксического сигнала JNK в клетках. Продукты и способы, соответствующие настоящему изобретению, могут особенно подходить для лечения нарушений, характеризующихся патологической повышающей регуляцией Gadd45β. Кроме того, они подходят в случаях заболеваний и нарушений, при которых Gadd45β необязательно может патологически регулироваться повышающим образом, но при которых NF-κВ патологически регулируется повышающим образом или активируется и при которых индуктор запрограммированной гибели клеток посредством передачи сигнала Gadd45β-MKK7 могут обеспечить лечение.
Примеры данных заболеваний, характеризующихся патологической повышающей регуляцией или активацией NF-κВ и в которых индуктор запрограммированной гибели клеток посредством передачи сигнала Gadd45β-MKK7 могут обеспечить лечение, включают; гематологические злокачественные заболевания (такие как множественная миелома, лимфома из клеток мантии, лимфома MALT, диффузная крупноклеточная В-клеточная лимфома, лимфома Ходжкина, миелодиспластический синдром, Т-клеточный лейкоз взрослых (HTLV-1), хронический лимфоцитарный лейкоз, хронический миелоидный лейкоз, острый миелогенный лейкоз и острый лимфоцитарный лейкоз), солидные опухоли (такие как рак молочной железы, рак шейки матки, рак почки, рак легкого, рак толстой кишки, рак печени, рак пищевода, рак желудка, рак гортани, рак щитовидной железы, рак паращитовидной железы, рак мочевого пузыря, рак яичников, рак простаты, рак поджелудочной железы и многие другие формы рака), другие формы рака (такие как меланома, цилиндрома, карцинома сквамозных клеток (кожи, а также головы и шеи), карцинома полости рта, эндометриальная карцинома, ретинобластома, астроцитома и глиобластома) и другие заболевания и нарушения, такие как аутоиммунные заболевания, хронические воспалительные заболевания, дегенеративные заболевания, а также ишемические и сосудистые заболевания.
Широкий круг заболеваний и нарушений зависит от активности NF-κВ. В самом деле в настоящее время считают, что патогенез практически каждого известного человеческого заболевания или нарушения зависит от воспаления и, следовательно, включает NF-κВ. Она действует как групповой переключатель воспалительного ответа, координации экспрессии последовательности из более 200 генов, кодирующих цитокины, рецепторы, факторы транскрипции, хемокины, провоспалительные ферменты и другие факторы, включая направленные на выживаемость факторы, которые инициируют и замедляют воспаление. Соединения, соответствующие изобретению, ингибируют отдельную направленную на выживаемость активность NF-κВ при воспалении. Вследствие этого заболевания и нарушения, поддающиеся лечению данными соединениями, включают, кроме распространенных хронических воспалительных заболеваний (таких как воспалительная болезнь кишки, ревматоидный артрит и псориаз), другие заболевания и нарушения, которые в существенной степени зависят от воспалительного компонента. Примеры данных заболеваний и нарушений, которые лечатся противовоспалительными препаратами или препаратами, ингибирующими NF-κВ, или таких, которые, как предполагают, подходят для лечения ингибиторами NF-κВ и которые можно было бы лечить соединением, соответствующим изобретению, включают:
1. Дыхательные пути: обструктивные заболевания дыхательных путей, включая: астму, в том числе бронхиальную, аллергическую, эндогенную, экзогенную, вызываемую физической нагрузкой, вызываемую лекарственными препаратами (включая вызываемую аспирином и NSAID) и вызываемую пылью астму, как интермиттирующую, так и постоянную, и всех степеней тяжести, а также другие случаи гиперчувствительности дыхательных путей; хроническое обструктивное легочное заболевание (COPD); бронхит, в том числе инфекционный и эозинофильный бронхит; эмфизему; бронхоэктазию; муковисцидоз; саркоидоз; "легкие фермера" (экзогенный аллергический альвеолит) и подобные заболевания; пневмонит, связанный с гиперчувствительностью; фиброз легкого, в том числе криптогенный фиброзирующий альвеолит, идиопатическое интерстициальное воспаление легких, фиброз как осложнение противоопухолевой терапии и хроническую инфекцию, включая туберкулез и аспергиллез и другие грибковые инфекции; осложнения при трансплантации легкого; васкулитные и тромботические нарушения легочной сосудистой сети и легочную гипертензию; противокашлевую активность, в том числе лечение хронического кашля, ассоциированного с воспалительными и секреторными состояниями дыхательных путей, и ятрогенный кашель; острый и хронический ринит, включая медикаментозный ринит и вазомоторный ринит; круглогодичный и сезонный аллергический ринит, в том числе нейрогенный ринит (сенная лихорадка); назальный полипоз; острую вирусную инфекцию, включая насморк, и инфекцию респиратрно-синцитиального вируса, гриппа, коронавируса (в том числе SARS (тяжелый острый респираторный сидром) или аденовируса; либо эозинофильный эзофагит;
2. Кости и суставы: поражения ревматического или подагрического характера, ассоциированные с остеоартритом/остеоартрозом или включающие их, как первичные, так и вторичные, например, врожденная дисплазия тазобедренного сустава; шейный и поясничный спондилит и боль в нижнем отделе спины и шее; остеопороз; ревматоидный артрит и болезнь Стилла; серонегативные спондилоартропатии, включая анкилозирующий спондилит, псориатический артрит, реактивный артрит и недифференцированную спондартропатию; септический артрит и другие связанные с инфекцией артропатии и нарушения костей, такие как туберкулез, включая болезнь Пота и синдром Понсе; острый и хронический кристаллический синовит, включая подагру, заболевание, связанное с отложением пирофосфата кальция, и обусловленное апатитом кальция воспаление сухожилия, бурсы и синовии; болезнь Бехчета; первичный и вторичный синдром Шегрена, системный склероз и ограниченная склеродермия; системная красная волчанка, заболевание комбинированной соединительной ткани и заболевание недифференцированной соединительной ткани; воспалительные миопатии, в том числе дерматомиозит и полимиозит; ревматическая полимиалгия; ювенильный артрит, в том числе идиопатический воспалительный артрит, распространяющийся на любые суставы, и ассоциированные синдромы, и ревматическая лихорадка и ее системные осложнения; васкулиты, включая артериит гигантских клеток, артериит Такаяши, синдром Чин-Страусса, узелковый полиартериит, микроскопический полиартериит и васкулиты, ассоциированные с вирусной инфекцией, реакции гиперчувствительности, криоглобулины и парапротеины, боль в нижней области спины; семейная средиземноморская лихорадка, синдром Макла-Уэллса, семейная ирландская лихорадка, болезнь Кикучи; вызываемые лекарственными препаратами арталгии, тендиниты и миопатии;
3. боль и ремоделирование соединительных тканей при скелетно-мышечных нарушениях, обусловленных повреждением [например, спортивной травмой]
или заболеванием: артритом (например, ревматоидным артритом, остеоартритом, подагрой или кристаллической артропатией), другим заболеванием суставов (таким как дегенерация межпозвоночного диска или дегенерация височно-нижнечелюстного сустава), заболевание, связанное с ремоделированием костей (такое как остеопороз, болезнь Педжета или остеонекроз), полихондрит (воспаление нескольких хрящей), склеродермия, нарушение комбинированной соединительной ткани, спондилоартропатии или заболевание периодонта (такое как периодонтит);
4. Кожа: псориаз, атопический дерматит, контактный дерматит или другие экзематозные дерматозы и реакции гиперчувствительности замедленного типа; фито- и фотодерматит; себорейный дерматит, герпетиформный дерматит, плоский лишай, склеротический атопический лишай, гангренозная пиодермия, кожный саркоид, дискоидная красная волчанка, пемфигус, пемфигоид, буллезный эпидермолиз, крапивница, ангионевротический отек (болезнь Квинке), васкулит, токсическая эритема, кожные эозинофилии, очаговая алопеция, облысение по мужскому типу, синдром Свита, синдром Вебера-Христиана, мультиформная эритема; целлюлит, как инфекционный, так и неинфекционный; панникулит; кожные лимфомы, рак кожи, отличный от меланомы и другие диспластические повреждения; вызываемые лекарственными препаратами нарушения, включая постоянную лекарственную сыпь;
5. Глаза: блефарит, конъюнктивит, включая круглогодичный и весенний аллергический конъюнктивит, ирит; передний и задний увеит; хориоидит; аутоиммунные, дегенеративные или воспалительные нарушения, поражающие сетчатку; офтальмит, в том числе симпатический офтальмит; саркоидоз; инфекции, включая вирусную, грибковую и бактериальную;
6. Желудочно-кишечный тракт: глоссит, гингивит, периодонтит, эзофагит, включая рефлюкс; эозинофильный гастроэнтерит, мастоцитоз, болезнь Крона, колит, в том числе язвенный колит, проктит, анальный зуд; заболевания брюшной полости, синдром раздраженной кишки и пищевые аллергии, которые могут иметь эффекты, отдаленные от кишки (например, мигрень, ринит или экзему);
7. Брюшная полость: гепатит, включая аутоиммунный, алкогольный и вирусный; фиброз и цирроз печени; холецистит, панкреатит, как острый, так и хронический;
8. Мочеполовая система: нефрит, включая интерстициальный и гломерулонефрит; нефротический синдром; цистит, включая острый и хронический (интерстициальный) цистит и язву Ханнера; острый и хронический уретрит, простатит, эпидидимит, оофорит и сальпингит; вульвовагинит; болезнь Пейрони; эректильная дисфункция (как у мужчин, так и у женщин);
9. Отторжение аллотрансплантата: острое и хроническое после, например, трансплантации почки, сердца, печени, легкого, костного мозга, кожи или роговицы, или после переливания крови, либо хроническая болезнь трансплантат против хозяина;
10. ЦНС (центральная нервная система): болезнь Альцгеймера и другие нарушения, связанные с деменцией, включая БКЯ и нвБКЯ; амилоидоз; рассеянный склероз и другие демиелинизирующие синдромы; церебральный атеросклероз и васкулит; височный артериит; злокачественная миастения; острая и хроническая боль (острая, временная или хроническая, центральной или периферической природы), включая боли в животе, головную боль, мигрень, невралгию тройничного нерва, атипическую лицевую боль, боль в суставах и костях, боль, вызываемую раком и инвазией опухоли, невропатические болевые синдромы, включая диабетическую, пост-герпетическую и связанную с ВИЧ невропатию; нейросаркоидоз; осложнения на центральную и периферическую нервную систему злокачественных, инфекционных или аутоиммунных процессов;
11. Другие аутоиммунные и аллергические нарушения, в том числе тиреоидит Хашимото, болезнь Грейвса, болезнь Аддисона, сахарный диабет, идиопатическая тромбоцитопеническая пурпура, эозинофильный фасцит, синдром гипер-IgE, антифосфолипидный синдром;
12. Другие заболевания с воспалительным или иммунологическим компонентом, включая синдром приобретенного иммунодефицита (СПИД), лепру, синдром Сезари и паранеопластические синдромы;
13. Сердечно-сосудистая система: атеросклероз, поражающий коронарное и периферическое кровообращение; перикардит; миокардит, воспалительная и аутоиммунная кардиомиопатия, включая саркоид миокарда; ишемические реперфузионные повреждения; эндокардит, вальвулит и аортит, включая инфекционный (например, сифилитический); васкулит; нарушения проксимальных и периферических вен, включая флебит и тромбоз, в том числе тромбоз глубоких вен и осложнения варикоза вен;
14. Желудочно-кишечный тракт: Глютеновая болезнь, проктит, эозинофильный гастроэнтерит, мастоцитоз, болезнь Крона, язвенный колит, микроскопический колит, индетерминантный колит, нарушение раздраженной кишки, синдром раздраженной кишки, невоспалительная диарея, пищевые аллергии, которые могут иметь эффекты, отдаленные от кишки, например, мигрень, ринит и экзема.
В результате авторы создают ряд синтетических молекул на основе D-энантиомеров тетрапептидов, трипептидов, дипептидов и подобных пептидомиметиков, включающих пептидоидные группы, которые нарушают взаимодействие Gadd45β/MKK7. Важно, что данные соединения проявляют активность ингибирования Gadd45β без подавления функции киназы MKK7. Это важно, поскольку подтверждает то, что соединения, соответствующие изобретению, могут индуцировать передачу цитотоксического сигнала JNK посредством ингибирования комплексов Gadd45β/MKK7.
Синтетические молекулы не связывают ни Gadd45β, ни MKK7 по отдельности, но они связываются с одним или другим белком, когда белки находятся в контакте друг с другом в связанном или несвязанном состоянии, вероятно, путем распознавания поверхности, которая становится доступной на Gadd45β, MKK7 и/или комплексе двух данных белков только когда Gadd45β и MKK7 приходят в контакт друг с другом, и, следовательно, вызывают конформационную модификацию в одном из двух белков или в комплексе в целом, что запускает диссоциацию комплекса. Данное свойство представляет особенный интерес, поскольку обеспечивает то, что соединения обладают очень высокой специфичностью в отношении мишени (т.е. комплекса Gadd45β/MKK7) и уменьшают вероятность того, что соединения, соответствующие изобретению, могут взаимодействовать и, таким образом, воздействовать на белки, которые имеют структуру, близкую структуре Gadd45β или MKK7. Данное свойство, которое определяет то, что терапевтическая мишень соединений, соответствующих изобретению, представляет собой границу раздела между двумя белками (т.е. Gadd45β и MKK7), обусловливает также, что соединения, соответствующие изобретению, не будут блокировать общие биологические активности Gadd45β или MKK7 in vivo, но скорее будут избирательно нарушать биологические функции, которыми обладают Gadd45β или MKK7, как часть комплекса Gadd45β/MKK7.
Примечательно, что соединения, соответствующие изобретению, как показано, индуцируют апоптоз в клеточных линиях множественной миеломы и первичных опухолевых клетках, а также других опухолевых В-клеточных линиях, включая клеточные линии диффузной круп но клеточной В-клеточной лимфомы и лимфомы Беркетта, а также другие формы рака, такие как промоноцитарный лейкоз, с IC50, лежащими в низком наномолярном интервале, но не обладают активностью в отношении опухолевых Т-клеточных линий или в отношении нормальных клеток, таких как нетрансформированные фибробласты, клетки стромы костного мозга (BMSCs), мононуклеарные клетки периферической крови (PBMNCs) и мезенхимные стволовые клетки (MSCs) или в очищенных первичных В- и Т-лимфоцитах мышей, даже при использовании в очень высоких концентрациях (т.е. 100 мкМ). Этот факт является доказательством того, что они обладают специфичностью в плане цитотоксической активности в отношении клеток с патологически конститутивно активной NF-κВ. Важно, что соединения, соответствующие изобретению, устойчивы к протеолизу, растворимы и стабильны в биологических жидкостях, сохраняя полную активность ингибирования после длительного инкубирования с человеческой сывороткой, и, вследствие этого, представляются подходящими кандидатами для системного применения.
Соединения, соответствующие изобретению, демонстрируют высокую специфичность направленности в отношении комплекса Gadd45β/MKK7 в клетках. Это показано на основании данных о том, что: 1) На большой панели опухолевых клеточных линий существует высоко достоверная статистическая корреляции между уровнями экспрессии Gadd45β и чувствительностью раковых клеток к вызываемой Z-/mDTP гибели; 2) опосредованная sh-PHK понижающая регуляция Gadd45β индуцирует апоптоз в чувствительных к Z-/mDTP, но не в устойчивых к Z/m DTP раковых клеточных линиях, и кинетика индукции апоптоза посредством специфических в отношении Gadd45β sh-PHK в данных клеточных линиях аналогична результатам, наблюдаемым с Z-/mDTP; 3) опосредованная sh-PHK понижающая регуляция MKK7 делает чувствительные к Z-/mDTP раковые клеточные линии полностью устойчивыми к индуцируемой Z-/mDTP гибели; 4) терапевтической мишенью, соответствующей изобретению, является граница раздела между двумя белками, Gadd45β и MKK7, что, кроме того, создает потенциальную возможность высокой специфичности мишени, ключевого преимущества решения авторов изобретения относительно имеющихся лекарственных препаратов. Данные результаты вместе с низкой токсичностью Z-/mDTP в отношении нормальных клеток и полученными данными о том, что выбивающее исключение Gadd45β хорошо переносится мышами, указывают на то, что направленность на отдельные способствующие выживаемости функции NF-κВ в выживаемости клеток с помощью опосредованного Z-/mDTP ингибирования Gadd45β/MKK7 могут привести к получению лекарственного препарата, который более специфичен, менее токсичен и, следовательно, более эффективен, чем лекарственные препараты, направленные на путь NF-κВ и/или протеасомы.
Более того, соединения, соответствующие изобретению, не обладают токсичностью в отношении нормальных клеток, и ингибирование Gadd45β, по-видимому, не имеет или имеет незначительные побочные эффекты, поскольку мыши с выбитым Gadd45β жизнеспособны и по всей видимости здоровы, показывая, что полная инактивация Gadd45β хорошо переносится in vivo. Кроме того, соединения, соответствующие изобретению, стабильны, растворимы, проникают в клетку и, вследствие этого, подходят для лечения множественной миеломы, диффузной крупноклеточной В-клеточной лимфомы и других форм рака, выживаемость которых зависит от NF-κВ, Их также используют для лечения хронических воспалительных и аутоиммунных заболеваний, особенно тех, которые опосредованы NF-κВ. Соединения, соответствующие изобретению, также имеют профили, которые привлекательны для терапевтического применения.
Изобретение также относится к разработке клинически пригодных анализов для предсказания ответа на лечение Z/mDTP у пациентов. Результаты, полученные на большой панели опухолевых клеточных линий, показывают, что чувствительность к вызываемой Z-/mDTP гибели коррелирует с высокой степенью достоверности в уровнями экспрессии Gadd45β (р<0,01), доказывая, таким образом, высокую специфичность цитотоксического действия Z-/mDTP в отношении Gadd45β. Более того, выбивание Gadd45β индуцирует апоптоз в клетках множественной миеломы, тогда как выбивание MKK7 делает данные клетки полностью устойчивыми к вызываемой Z-/mDTP-гибели. Вместе взятые, данные результаты показывают, что если бы лечение Z-/mDTP вошло в клиническую практику, было бы возможно предсказывать отвечающие популяции пациентов посредством простого и рентабельного анализа кОТ-ПЦР.
Согласно первому аспекту изобретения предусматривают соединение формулы I:
I: X1-A-Х2
где
А представляет собой А”” или А”-[М-А'-]n М-А”';
А”” представляет собой А”, А”'
или Z1-Y2-Y3-Z4, где Y2-Y3 представляет собой олигопептидную группу или олигопептоидную группу, имеющую остатки Y2-Y3, и Z1 присоединено к N-концевому атому азота Y2-Y3, и Z4 присоединено к С-концевому атому углерода Y2-Y3;
А” представляет собой А'
или Y1-Y2-Y3-Z4, где Y1-Y2-Y3 представляет собой олигопептоидную группу или олигопептоидную группу, содержащую остатки Y1-Y2-Y3 и Z4 присоединено к С-концевому атому углерода Y1-Y2-Y3;
А”' представляет собой А',
или Z1-Y2-Y3-Y4, где Y2-Y3-Y4 представляет собой олигопептоидную группу или олигопептоидную группу, содержащую остатки Y2-Y3-Y4, и Z1 присоединено к N-концевому атому азота Y2-Y3-Y4;
каждое присутствующее А' независимо представляет собой олигопептидную группу или олигопептоидную группу, содержащую остатки Y1-Y2-Y3-Y4;
n представляет собой целое число от 0 до 18,
Y1 и Y4 независимо представляют собой остатки аминокислот или остатки производных аминокислот, имеющих ароматические боковые цепи, согласно ряду вариантов осуществления каждая боковая цепь содержит алкиленовую группу из одного - трех атомов углерода, которые один или два раз замещены 5-10-членной карбоциклической или гетероциклической ароматической группой и необязательно далее замещена алкилом из 1-4 атомов углерода, причем указанная ароматическая группа необязательно замещена по меньшей мере одним заместителем, выбранным из гидроксила, галогена или С1-С4 алкила или С1-С4 алкокси.
Y2 отсутствует или представляет собой остаток аминокислоты или остаток производного аминокислоты, предпочтительно любой из 20 природных аминокислот в L- или D-конфигурации и/или предпочтительно остаток аминокислоты или остаток производного аминокислоты, имеющей боковую цепь, несущую предпочтительно отрицательный заряд в водном растворе при рН 7;
Y3 представляет собой остаток аминокислоты или остаток производного аминокислоты, предпочтительно любой из 20 природных аминокислот в L- или D-конфигурации и/или предпочтительно остаток аминокислоты или остаток производного аминокислоты, имеющей боковую цепь, несущую предпочтительно отрицательный заряд в водном растворе при рН 7;
Когда в ряде вариантов осуществления присутствуют оба, Y2 и Y3, предпочтительно, когда они такие, что способны к формированию солевого мостика между соответствующими положительным и отрицательными зарядами боковых цепей и/или такие, что расстояние между ароматическими центрами на Y1 и Y4 или на X1 и Х4 или на Х1 и Y4 или на Y1 и Х4 не превышает 10 или 20 Å и составляет не меньше, чем 3 Å. Предпочтительно, когда боковые цепи Y2 и Y3 состоят из не меньше, чем 30 атомов. Y2 и Y3 могут представлять собой природные аминокислоты или N-метил-аминокислоты в L- или D-конфигурации.
Z1 представляет собой группу формулы II:
которая связана с N-концевым атомом азота Y2,
W отсутствует или представляет собой кислород или азот или алкиленовую группу из одного-трех атомов углерода, причем алкиленовая группа из одного-трех атомов углерода необязательно замещена по меньшей мере одним заместителем, выбранным из алкила с одним-четырьмя атомами углерода или 5-10-членной карбоциклической или гетероциклической ароматической группы;
J представляет собой 5-10-членную карбоциклическую или гетероциклическую ароматическую группу, причем ароматическая группа замещена по меньшей мере одним заместителем, выбранным из гидроксила, галогена, алкила из одного-четырех атомов углерода или алкокси из одного-четырех атомов углерода;
Z4 представляет собой группу формулы III:
которая связана с С-концевым атомом углерода Y3,
R представляет собой водород или алкил из одного-четырех атомов углерода;
W отсутствует или представляет собой алкиленовую группу из одного-трех атомов углерода, причем алкиленовая группа из одного-трех атомов углерода необязательно замещена по меньшей мере одним заместителем, выбранным из алкила из одного-четырех атомов углерода или 5-10-членной карбоциклической или гетероциклической ароматической группой;
J' представляет собой 3-10-членнуюалифатическуюкарбоциклическую группу или 5-10-членную карбоциклическую или гетероциклическую ароматическую группу,
причем алифатическая или ароматическая группа необязательно замещена по меньшей мере одним заместителем, выбранным из гидроксила, галогена, алкила из одного-четырех атом углерода или алкокси из одного-четырех атомов углерода;
М представляет собой пептидную связь между предшествующей олигопептидной или олигопептидоидной группой (А', А” или А”')и последующей олигопептидной или олигопептидоидной группой (А', А” или А”') либо линкерную группу, присоединенную посредством амидной связи, сложно-эфирной связи, эфирной связи или тиоэфирной связи к концевой карбоксильной группе предшествующей олигопептидной или олигопептидоидной группы (А', А” или А”') и посредством амидной связи, сложно-эфирной связи, эфирной связи или тиоэфирной связи к концевой аминогруппе следующей олигопептидоидной группы (А', А” или А”');
Х1 отсутствует или представляет собой группу, введенную в -аминоконец А с целью блокировать свободные аминогруппы;
X2 отсутствует или представляет собой группу, введенную в -карбоксильный конец А с целью блокировать свободные карбоксильные группы;
Согласно ряду вариантов осуществления W отсутствует или представляет собой алкилен из 1-3 атомов углерода.
Предпочтительно, когда Х1 и X2 представляют собой группы из не более, чем 30 (или более предпочтительно 20 или 10) атомов при условии, что Х1 отсутствует, если А содержит Z1 и X2 отсутствует, если А содержит Z4 (т.е., если не имеется свободных амино или карбоксильных групп на концах молекулы, Х1 и X2 не требуются);
или их производные, причем указанные производные выбраны их группы, состоящей из:
a) олигомеров или мультимеров молекул соединения формулы I, причем указанные олигомеры и мультимеры содержат две или более молекул соединения формулы I, каждая их которых соединена с обычной группой основы посредством амидной связи между амино- или группой карбоновой кислоты, присутствующей в молекулах соединения формулы I и противоположной амино- или группой карбоновой кислоты на группе основы, причем указанная группа основы участвует по меньшей мере в 2 амидных связях,
b) производных, содержащих молекулу соединения формулы I или его олигомер либо мультимер, как определено выше в разделе а), конъюгированные посредством амидной связи, сложно-эфирной связи, эфирной связи или тиоэфирной связи с:
ПЭГ,
соединениями на основе ПЭГ,
проникающими в клетку пептидами,
флуоресцентными красителями,
биотином или другой группой метки,
жирными кислотами,
наночастицами дискретного размера
или
хелатирующими лигандами, комплексированными с ионами металлов или радиоактивных веществ.
c) производных, содержащих молекулу соединения формулы 1 или его олигомер либо мультимер, как определено выше в разделе а), которые модифицированы амидированием, гликозилированием, карбамилированием, ацилированием, сульфатированием, фосфорилированием, циклизацией, липидированием, пегилированием или связыванием с пептидным или пептидоидным партнером слияния с целью получения слитого пептида или слитого пептидоида.
и
d) солей и сольватов молекулы соединения формулы 1 или его производного, как определено выше в разделе а).
Согласно ряду вариантов осуществления:
Y1 представляет собой
D-триптофан,
L-триптофан,
D-тирозин,
L-тирозин,
D-3,3-дифенилаланин,
L-3,3-дифенилаланин,
D-Н-3-(4-пиридил)аланин,
L-Н-3-(4-пиридил)аланин,
D-Н-3-(3-пиридил)аланин,
L-Н-3-(3-пиридил)аланин,
D-Н-3-(2-пиридил)аланин,
L-Н-3-(2-пиридил)аланин,
D-2-амино-4-фенил-масляную кислоту,
L-2-амино-4-фенил-масляную кислоту,
D-H-4-гидрокси-фенилглицин,
L-H-4-гидрокси-фенилглицин,
D-3-(2-фурил)-аланин,
L-3-(2-фурил)-аланин,
L-гомофенилаланин,
D-гомофенилаланин,
D-3-(4-хинолил)-аланин,
L-3-(4-хинолил)-аланин;
D-нафтил-аланин;
L-нафтил-аланин;
п-гидрокси-бензойную кислоту;
п-гидрокси-фенил-уксусную кислоту;
3-(п-гидрокси-фенил)-пропионовую кислоту;
или N-метил-производные в L- или D-конфигурации любого из вышеприведенных соединений.
Альтернативно Y1 может представлять собой:
D-фенилаланин,
L-фенилаланин,
D-триптофан,
L-триптофан,
D-тирозин,
L-тирозин,
D-3,3-дифенилаланин,
L-3,3-дифенилаланин,
D-Н-3-(4-пиридил)аланин,
L-Н-3-(4-пиридил)аланин,
D-Н-3-(3-пиридил)аланин,
L-H-3-(3-пиридил)аланин,
D-Н-3-(2-пиридил)аланин,
L-Н-3-(2-пиридил)аланин,
D-2-амино-4-фенил-масляную кислоту,
L-2-амино-4-фенил-масляную кислоту,
D-фенилглицин,
L-фенилглицин,
D-H-4-гидрокси-фенилглицин,
L-H-4-гидрокси-фенилглицин,
D-3-(2-фурил)-аланин,
L-3-(2-фурил)-аланин,
L-циклогексилаланин,
D-циклогексилаланин,
L-гомофенилаланин,
D-гомофенилаланин,
D-3-(4-хинолил)-аланин,
L-3-(4-хинолил)-аланин;
D-нафтил-аланин или
L-нафтил-аланин.
Согласно ряду вариантов осуществления:
Y2 отсутствует или представляет собой
D-глутаминовая кислота,
L-глутаминовая кислота,
D-аспарагиновая кислота,
L-аспарагиновая кислота,
L-лейцин,
D-лейцин,
L-глутамин,
D-глутамин,
L-метионин,
D-метионин,
D-2-амино-гептандиоевая кислота,
L-2-амино-гептандиоевая кислота,
сложный метиловый или этиловый эфир любого из них,
L-гомосерин,
D-гомосерин;
или N-метил-производные любого из вышеуказанных соединений в L- или D-конфигурации.
Альтернативно Y2 может представлять собой:
D-глутаминовую кислоту,
L-глутаминовую кислоту,
D-аспарагиновую кислоту,
L-аспарагиновую кислоту,
D-2-амино-гептандиоевую кислоту,
L-2-амино-гептандиоевую кислоту,
сложный метиловый или этиловый эфир любого из них,
L-гомосерин,
или D-гомосерин;
Согласно ряду вариантов осуществления:
Y3 представляет собой
D-аргинин,
L-аргинин,
L-пролин,
D-пролин,
D-гистидин,
L-гистидин,
D-лизин,
D-α,β-диаминопропионовую кислоту (D-Dap),
L-α,β-диаминопропионовую кислоту (L-Dap),
L-α,β-диаминомасляную кислоту (L-Dab),
L-α,β-диаминомасляную кислоту (L-Dab),
L-орнитин,
D-орнитин,
L-лизин;
или N-метил-производные любого из вышеуказанных соединений в L- или D-конфигурации.
Альтернативно Y3 может представлять собой
D-аргинин,
L-аргинин,
D-гистидин,
L-гистидин,
D-лизин,
D-α,β-диаминопропионовую кислоту (D-Dap),
L-α,β-диаминопропионовую кислоту (L-Dap),
L-α,δ-диаминомасляную кислоту (L-Dab),
L-α,δ-диаминомасляную кислоту (L-Dab),
L-орнитин,
D-орнитин,
или L-лизин;
Согласно ряду вариантов осуществления:
Y4 представляет собой
D-фенилаланин,
L-фенилаланин,
D-триптофан,
L-триптофан,
D-тирозин,
L-тирозин,
D-3,3-дифенилаланин,
L-3,3-дифенилаланин,
D-Н-3-(4-пиридил)аланин,
L-Н-3-(4-пиридил)аланин,
D-Н-3-(3-пиридил)аланин,
L-Н-3-(3-пиридил)аланин,
D-H-3-(2-пиридил)аланин,
L-Н-3-(2-пиридил)аланин,
D-2-амино-4-фенил-масляную кислоту,
L-2-амино-4-фенил-масляную кислоту,
D-фенилглицин,
L-фенилглицин,
D-H-4-гидрокси-фенилглицин,
L-H-4-гидрокси-фенилглицин,
D-3-(2-фурил)аланин,
L-3-(2-фурил)аланин,
L-гомофенилаланин,
D-гомофенилаланин,
D-3-(4-хинолил)аланин,
L-3-(4-хинолил)-аланин;
D-нафтилаланин,
L-нафтилаланин,
Их N-метил-производные в L- или D-конфигурации,
Анилин,
Бензиламин,
или 2-фенил-этиламин.
Альтернативно Y4 может представлять собой
D-фенилаланин,
L-фенилаланин,
D-триптофан,
L-триптофан,
D-тирозин,
L-тирозин,
D-3,3-дифенилаланин,
L-3,3-дифенилаланин,
D-Н-3-(4-пиридил)аланин,
L-Н-3-(4-пиридил)аланин,
D-Н-3-(3-пиридил)аланин,
L-Н-3-(3-пиридил)аланин,
D-Н-3-(2-пиридил)аланин,
L-Н-3-(2-пиридил)аланин,
D-2-амино-4-фенилмасляную кислоту,
L-2-амино-4-фенилмасляную кислоту,
D-фенилглицин,
L-фенилглицин,
D-H-4-гидрокси-фенилглицин,
L-H-4-гидрокси-фенилглицин,
D-3-(2-фурил)аланин,
L-3-(2-фурил)аланин,
L-циклогексилаланин,
D-циклогексилаланин,
L-гомофенилаланин,
D-гомофенилаланин,
D-3-(4-хинолил)аланин,
L-3-(4-хинолил)аланин;
D-нафтилаланин
или L-нафтил-аланин.
Согласно ряду предпочтительных вариантов осуществления все Y1, Y2, Y3 и Y4 такие, как описано выше. Согласно ряду вариантов осуществления все Y1, Y2, Y3 и Y4 такие, как описано выше при условии, что Y2 представляет собой
D-глутаминовую кислоту,
L-глутаминовую кислоту,
D-аспарагиновую кислоту,
L-аспарагиновую кислоту,
D-2-амино-гептандиоевую кислоту,
L-2-амино-гептандиоевую кислоту,
сложный метиловый или этиловый эфир любого из них;
L-гомосерин,
L-лейцин,
D-лейцин,
L-глутамин,
D-Глутамин,
L-Метионин,
D-Метионин,
D-гомосерин,
Или N-метил-производные любого из вышеуказанных соединений в L- или
D-конфигурации
и Y3 представляет собой
D-аргинин,
L-аргинин,
D-гистидин,
L-гистидин,
D-лизин,
L-лизин,
L-пролин,
D-пролин,
D-α,β-диаминопропионовую кислоту (D-Dap),
L-α,β-диаминопропионовую кислоту (L-Dap),
L-α,δ-диаминомасляную кислоту (L-Dab),
L-α,δ-диаминомасляную кислоту (L-Dab),
D-орнитин,
L-орнитин
или N-метил-производные любого из вышеуказанных соединений в L- или D-конфигурации.
Согласно ряду вариантов осуществления оба, Y1 и Y2, такие, как описано выше, но одно или оба из Y2 и Y3 отсутствуют. Согласно ряду вариантов осуществления М представляет собой пептидную связь.
Согласно ряду вариантов осуществления Х1 представляет собой водород или Х1 представляет собой одну из следующих групп, введенных с аминоконец олигопептидной последовательности таким образом, чтобы сформировать амидную связь:
ацетил,
бензилоксикарбонил,
2-хлор-бензилоксикарбонил,
3-метокси,4-гидрокси-бензоил,
3-гидрокси,4-метокси-бензоил,
бензоил,
или флуоренилметоксикарбонил;
X2 представляет собой гидроксильную группу или одну из следующих групп, введенных в конец карбоновой кислоты олигопептидной последовательности таким образом, чтобы сформировать амидную связь:
амин,
D-фенилаланин,
L-фенилаланин,
D-триптофан,
L-триптофан,
D-тирозин,
L-тирозин,
D-3,3-дифенилаланин,
L-3,3-дифенилаланин,
D-Н-3-(4-пиридил)аланин,
L-Н-3-(4-пиридил)аланин,
D-Н-3-(3-пиридил)аланин,
L-Н-3-(3-пиридил)аланин,
D-Н-3-(2-пиридил)аланин,
L-Н-3-(2-пиридил)-аланин,
D-2-амино-4-фенилмасляную кислоту,
L-2-амино-4-фенилмасляную кислоту,
D-фенилглицин,
L-фенилглицин,
D-H-4-гидрокси-фенилглицин,
L-H-4-гидрокси-фенилглицин,
D-3-(2-фурил)аланин,
L-3-(2-фурил)аланин,
L-циклогексилаланин,
D-циклогексилаланин,
L-гомофенилаланин,
D-гомофенилаланин,
D-3-(4-хинолил)аланин,
L-3-(4-хинолил)аланин;
D-нафтилаланин,
L-нафтилаланин,
или N-метил-производные любого из вышеуказанных соединений в L- или D-конфигурации.
Согласно ряду вариантов осуществления:
Z1 представляет собой
4-гидрокси-бензоил,
(4-гидрокси-фенил)-ацетил,
3-(4-гидрокси-фенил)-пропионил,
бензоил,
бензилоксикарбонил,
2-фенил-ацетил,
3-фенилпропионил,
3,3-дифенилпропионил,
3-(1Н-индол-3-ил)пропионил,
(1Н-индол-3-ил)-ацетил,
фуран-2-ил-ацетил,
фуран-3-ил-ацетил,
3-пиридин-4-ил-пропионил,
3-пиридин-3-ил-пропионил,
3-пиридин-2-ил-пропионил,
3-пиримидин-4-ил-пропионил,
3-пиридазин-4-ил-пропионил,
3-[1,3,5]триазин-2-ил-пропионил,
2-пиридин-4-ил-ацетил,
2-пиридин-3-ил-ацетил,
2-пиридин-2-ил-ацетил,
2-пиримидин-4-ил-ацетил,
2-пиридазин-4-ил-ацетил,
2[1,3,5]триазин-2-ил-ацетил,
нафталин-1-ил-ацетил,
нафталин-2-ил-ацетил,
2-нафталин-1-ил-пропионил
или 2-нафталин-2-ил-пропионил,
Y2 представляет собой D-глутаминовую кислоту,
L-глутаминовую кислоту,
D-аспарагиновую кислоту,
L-аспарагиновую кислоту, L-лейцин,
D-лейцин,
L-Глутамин,
D-Глутамин,
L-Метионин,
D-Метионин,
D-2-амино-гептандиоевую кислоту,
L-2-амино-гептандиоевую кислоту,
сложный метиловый или этиловый эфир любого из них;
L-гомосерин,
D-гомосерин;
или N-метил-производные любого из вышеуказанных соединений в L- или D-конфигурации.
Y3 представляет собой
D-аргинин,
L-аргинин,
D-гистидин,
L-гистидин,
L-пролин,
D-пролин,
D-лизин,
L-лизин;
D-α,β-диаминопропионовую кислоту (D-Dap),
L-α,β-диаминопропионовую кислоту (L-Dap),
L-α,δ-диаминомасляную кислоту (L-Dab),
L-α,δ-диаминомасляную кислоту (L-Dab),
D-орнитин
L-орнитин
или N-метил-производные любого из вышеуказанных соединений в L- или D-конфигурации.
Z4 представляет собой фениламин,
бензиламин,
фенетиламин,
циклогексиламин,
2-циклогексил-этиламин,
3-циклогексил-пролиламин,
4-(2-амино-этил)фенол,
4-аминофенол,
4-аминометилфенол,
1 Н-индол-3-ил-амин,
2-(1Н-индол-3-ил)-этиламин,
С-(1Н-индол-3-ил)-метиламин,
2,2-дифенилэтиламин,
2,2-дипиридин-4-ил-этиламин,
2-пиридин-4-ил-этиламин,
2-пиридин-3-ил-этиламин,
2-пиридин-2-ил-этиламин,
2-пиримидин-4-ил-этиламин,
2-[1,3,5]триазин-2-ил-этиламин,
С-фуран-2-ил-метиламин,
С-фуран-3-ил-метиламин
или С-нафталин-2-ил-метиламин.
В соответствии с соглашением все пептиды и пептоиды и их участки описаны от N-конца к С-концу.
n может означать 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17 или 18. Согласно ряду предпочтительных вариантов осуществления n=0.
Согласно ряду предпочтительных вариантов осуществления А представляет собой А'. В данных вариантах осуществления соединение, вследствие этого, представляет собой тетрапептид, трипептид или дипептид (либо соответствующий пептоид) с необязательными блокирующими группами Х1 и X2 на одном или более концов.
Олигопептиды
Олигопептиды представляют собой короткие полимеры, образованные путем конденсации су-аминокислот (обозначаемых в данном контексте как просто "аминокислоты"). Связь между одним остатком аминокислоты и следующим известна как пептидная связь или амидная связь.
Аминокислоты
Как используют в данном контексте, термин "аминокислота" включает 20 стандартных аминокислот (Изолейцин, Аланин, Лейцин, Аспарагин, Лизин, Аспарагиновая кислота, Метионин, Цистеин, Фенилаланин, Глутаминовая кислота, Треонин, Глутамин, Триптофан, Глицин, Валин, Пролин, Серин, Тирозин, Аргинин и Гистидин) в своих обеих, D- и L-оптических конфигурациях. Он также включает синтетические ст-аминокислоты в обеих, D- и L-формах. Согласно ряду вариантов осуществления D-конфигурация предпочтительна.
Производные аминокислот
Как используют в данном контексте, данный термин включает N-замещенные глицины, которые отличаются от α-аминокислот тем, что их боковые цепи присоединены к атомам азота, расположенным вдоль скелета молекулы, а не к атомам α-углерода (как они присоединены в аминокислотах). В термин также входят сложные метиловые и этиловые эфиры α-аминокислот, β-аминокислоты и N-метилированные α-аминокислоты.
Олигопептоиды
Строго говоря, термин "олигопептид" относится только к олигомерам α-аминокислот. Аналогичный олигомер, включающий (во всех или некоторых положениях остатков) производное аминокислоты (например, N-замещенный глицин), известен как олигопептоид.
Производные
Предпочтительно, когда производные соединений, соответствующих первому аспекту изобретения, представляют собой функциональные производные. Термин "функциональное производное" используют в данном контексте для обозначения химического производного соединения формулы (I), имеющего такую же физиологическую функцию (как соответствующие немодифицированные соединения формулы (I) или, альтернативно, имеющие такую же функцию in vitro в функциональном анализе (например, в одном из анализов, описанных в одном из примеров, раскрытых в данном контексте).
Производные соединения, соответствующего изобретению, могут содержать структуру формулы (I), модифицированную хорошо известными способами, включая амидирование, гликозилирование, карбамилирование, ацилирование, например ацетилирование, сульфатирование, фосфорилирование, циклизацию и пегилирование. Структура формулы (I) может быть модифицирована по случайным положениям в молекуле или по заданным положениям в молекуле и может включать одну, две, три или более присоединенных химических групп. Производные включают соединения, в которых N-концевая группа NH2 замещена другой группой, например, метоксигруппой. Соединение, соответствующее изобретению, может представлять собой слитый белок, в котором структура формулы (I) слита с другим белком или полипептидом (партнером слияния) при использовании способов, известных у уровне техники. Любой подходящий пептид или белок можно использовать в качестве партнера слияния (например, сывороточный альбумин, углеродангидразу, глутатион-8-трансферазу или тиоредоксин и т.п.). Предпочтительные партнеры слияния не будут обладать вредной биологической активностью in vivo. Данные слитые белки можно получить путем присоединения карбокси-конца партнера слияния с аминоконцом структуры формулы (I) или наоборот. Необязательно можно использовать расщепляемый линкер для присоединения структуры формулы (I) к партнеру слияния. Полученный в результате расщепляемый слитый белок может быть расщеплен in vivo, так что происходит высвобождение активной формы соединения, соответствующего изобретению. Примеры данных расщепляемых линкеров включают, но без ограничения перечисленным, линкеры D-D-D-D-Y [SEQ ID NO.:227], G-P-R, A-G-G и H-P-F-H-L [SEQ ID NO.:228], которые могут расщепляться энтерокиназой, тромбином, убиквитин-расщепляющим ферментом и ренином, соответственно. См., например, Патент США No.6410707.
Соединение, соответствующее изобретению, может представлять собой физиологически функциональное производное структуры формулы (I). Термин физиологически функциональное производное" используют в данном контексте для определения химического производного соединения формулы (I), имеющего ту же самую физиологическую функцию, что соответствующее немодифицированное соединение формулы (I). Например, физиологически функциональное производное может быть превращаемым в организме в соединение формулы (I). Согласно настоящему изобретению примеры физиологически функциональных производных включают сложные эфиры, амиды и карбаматы, предпочтительно сложные эфиры и амиды.
Фармацевтически приемлемые сложные эфиры и амиды соединений, соответствующих изобретению, могут включать С1-20 алкил-, C2-20 алкенил-, C5-10 арил-, C5-10 или С1-20 алкил- или сложный эфир либо амид аминокислоты, присоединенный к подходящему центру, например, кислотной группе. Примерами подходящих групп являются гидрофобные заместителями из 4-26 атомы углерода, предпочтительно 5-19 атомами углерода. Подходящие липидные группы включают, но без ограничения перечисленным, следующие: лауроил (С12Н23), пальмитил (С15Н31), олеил (C15H29), стеарил (C17H35), холат и дезоксихолат.
Способы липидизации соединений, содержащих сульфгидрил, производными жирных кислот описаны в Патенте США No.5936092; Патенте США No.6093692 и Патенте США No.6225445. Производные жирных кислот соединения, соответствующего изобретению, содержащие соединение, соответствующее изобретению, присоединенное к жирной кислоте посредством дисульфидной связи, можно использовать для доставки соединения, соответствующего изобретению, в нервные клетки и ткани. Липидизация заметно повышает уровень всасывания соединений относительно уровня всасывания соответствующих нелипидизированных соединений, а также пролонгирует сохранение соединений в крови и тканях. Более того, дисульфидная связь в липидизированном производном относительно лабильна в клетках и, таким образом, способствует внутриклеточному высвобождению молекулы из групп жирных кислот. Подходящими липидсодержащими группами являются гидрофобные заместители с 4-26 атомами углерода, предпочтительно с 5-19 атомами углерода. Подходящие липидные группы включают, но без ограничения перечисленным, следующее: пальмитил (C15H31), олеоил (C15H29), стеарил (С17Н35). холат и дезоксихолат.
Способы циклизации включают циклизацию посредством формирования дисульфидного мостика и циклизацию типа "голова к хвосту" с использованием циклизационной смолы. Циклизованные петиды могут обладать повышенной стабильностью, включая повышенную устойчивость к ферментному разложению, в результате их конформационных ограничений. Циклизация может быть, в частности, целесообразной, когда нециклизованный пептид включает N-концевую группу цистеина. Подходящие циклизованные пептиды включают мономерные и димерные структуры, циклизованные по типу "голова к хвосту". Циклизованные пептиды могут включать один или более дополнительных остатков, особенно дополнительный цистеин, введенный с целью формирования дисульфидной связи, или боковую цепь, введенную с целью циклизации на основе смолы.
Соединение, соответствующее изобретению, может представлять собой пегилированную структуру формулы (I). Пегилированные соединения, соответствующие изобретению, могут давать дополнительные преимущества, такие как повышенная растворимость, стабильность и время циркуляции в кровотоке полипептида или пониженная иммуногенность (см. Патент США No.4179337).
Химические группы для дериватизации соединения, соответствующего изобретению, могут быть также выбраны из водорастворимых полимеров, таких как полиэтиленгликоль, сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлоза, декстран, поливиниловый спирт и т.п. Структура полимера, предназначенная для дериватизации соединения, соответствующего изобретению, может иметь любую молекулярную массу и может быть разветвленной или неразветвленной. Могут быть использованы полимеры другой молекулярной массы в зависимости от требуемого терапевтического профиля, например, требуемой продолжительности замедленного высвобождения, эффектов (при их наличии) на биологическую активность, простоты обращения, степени или отсутствия антигенности и других известных эффектов полиэтиленгликоля на терапевтический белок или аналог. Например, полиэтиленгликоль может иметь среднюю молекулярную массу приблизительно 200, 500, 1000, 1500, 2000, 2500, 3000, 3500, 4000, 4500, 5000, 5500, 6000, 6500, 7000, 7500, 8000, 8500, 9000, 9500, 10000, 10500, 11000,11500, 12000, 12500,13000, 13500, 14000, 14500, 15000, 15500, 16000, 16500, 17000, 17500, 18000, 18500, 19000, 19500, 20000, 25000, 30000, 35000, 40000, 50000, 55000, 60000, 65000, 70000, 75000, 80000, 85000, 90000, 95000 или 100000 кД.
Соли и сольваты соединений, соответствующих изобретению, которые подходят для использования в лекарственном средстве, представляют собой те, в которых противоион или ассоциированный растворитель фармацевтически приемлемы. Однако, соли и сольваты, имеющие фармацевтически неприемлемые противоионы или ассоциированные растворители, входят в объем настоящего изобретения, например, для применения в качестве промежуточных продуктов при получении соединений формулы (I) и их фармацевтически приемлемых солей или сольватов.
Подходящие соли, соответствующие изобретению, включают образованные органическими или неорганическими кислотами или основаниями. Фармацевтически приемлемые кислотно-аддитивные соли включают те, которые сформированы с использованием хлористоводородной, бромистоводородной, серной, азотной, лимонной, винной, уксусной, фосфорной, молочной, пировиноградной, уксусной, трифторуксусной, янтарной, перхлорной, фумаровой, малеиновой, гликолевой, молочной, салициловой, оксалоуксусной, метансульфоновой, этансульфоновой, п-толуолсульфоновой, муравьиной, бензойной, малоновой, нафталин-2-сульфоновой, бензолсульфоновой и изетионовой (2-гидроксиэтансульфоновой) кислот. Другие кислоты, такие как щавелевая, хотя сами не являются фармацевтически приемлемыми, могут быть использованы в качестве промежуточных продуктов при получении соединений, соответствующих изобретению, и их фармацевтически приемлемых солей. Фармацевтически приемлемые соли с основаниями включают соли аммония, соли щелочных металлов, например, соли калия и натрия, соли щелочноземельных металлов, например, соли кальция и магния, и соли с органическими основаниями, например, дициклогексиламином и N-метил-D-глюкомином.
Компетентные специалисты в области органической химии будут иметь в виду, что многие органические соединения могут образовывать комплексы с растворителями, в которых они реагируют или из которых они осаждаются или кристаллизуются. Данные комплексы известны как "сольваты". Например, комплекс в водой известен как "гидрат". Настоящее изобретение предусматривает сольваты соединений, соответствующих изобретению.
Согласно ряду предпочтительных вариантов осуществления соединение имеет полупериод существования в кровотоке человека по меньшей мере 0,1, 0,2, 0,5, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или, наиболее предпочтительно, по меньшей мере 12 часов.
Предпочтительно, когда соединение сохраняет по меньшей мере 20, 30, 40, 50, 60, 70, 80, 90 или, наиболее предпочтительно, 99% своей способности связываться с Gadd45β и/или MKK7 (и/или их ассоциации), как оценивают в анализе связывания in vitro, или по меньшей мере 20, 30, 40, 50, 60, 70, 80, 90 или, наиболее предпочтительно, 99% своей способности блокировать взаимодействие Gadd45β с MKK7, как оценивают в конкурентном анализе связывания in vitro после инкубирования в нормальной человеческой сыворотке в течение 24 часов при 37°С.
Альтернативно или дополнительно соединение обладает по меньшей мере одной из следующих активностей:
a) Способность ингибировать по меньшей мере 20, 30, 40, 50, 60, 70, 80, 90 или, наиболее предпочтительно, 99% взаимодействий MKK7 Gadd45β в условиях анализа, описанных в примерах.
b) Способность in vitro уничтожать по меньшей мере 20, 30, 40, 50, 60, 70, 80, 90 или, наиболее предпочтительно, 99% клеток в культуре клеточной линии человеческой миеломы, выбранной из группы, состоящей из U266, KMS-11, NCI-H929, ARH-77, JJN-3, KMS-12, KMS-18 и KMS-27, или культуры клеточной линии DLBCL LY-3, или культуры промоноцитарной клеточной линии U937, или культуры клеточной линии лимфомы Беркетта BJAB или культуры первичных опухолевых клеток (например, первичных опухолевых клеток множественной миеломы) в условиях, при которых по меньшей мере 90% Т-клеточной линии JURKAT не погибает.
Согласно ряду предпочтительных вариантов осуществления группа олигопептидного ядра соединения, идентифицированного как А в Формуле I, имеет последовательность аминокислот, выбранную из группы, состоящей из:
(L-Tyr)-(L-Asp)-(L-His)-(L-Phe), [SEQ ID NO.: 2]
(L-Tyr)-(L-Glu)-(L-Arg)-(L-Phe), [SEQ ID NO.: 3]
(L-Tyr)-(L-Glu)-(L-His)-(L-Phe), [SEQ ID NO.: 4]
(L-Trp)-(L-Asp)-(L-His)-(L-Phe), [SEQ ID NO.: 5]
(L-Trp)-(L-Glu)-(L-His)-(L-Phe), [SEQ ID NO.: 6]
(L-Tyr)-(L-Asp)-(L-Arg)-(L-Phe), [SEQ ID NO.: 7]
(L-Tyr)-(L-Asp)-(L-Lys)-(L-Phe), [SEQ ID NO.: 8]
(L-Tyr)-(L-Glu)-(L-Lys)-(L-Phe), [SEQ ID NO.: 9]
(L-Trp)-(L-Glu)-(L-Lys)-(L-Phe), [SEQ ID NO.: 10]
(L-Trp)-(L-Glu)-(L-Arg)-(L-Phe), [SEQ ID NO.: 11]
(L-Trp)-(L-Asp)-(L-Lys)-(L-Phe), [SEQ ID NO.: 12]
(L-Trp)-(L-Asp)-(L-Arg)-(L-Phe), [SEQ ID NO.: 13]
(L-Tyr)-(L-Asp)-(L-His)-(L-Trp), [SEQ ID NO.: 14]
(L-Tyr)-(L-Glu)-(L-His)-(L-Trp), [SEQ ID NO.: 15]
(L-Trp)-(L-Asp)-(L-His)-(L-Trp), [SEQ ID NO.: 16]
(L-Trp)-(L-Glu)-(L-His)-(L-Trp), [SEQ ID NO.: 17]
(L-Tyr)-(L-Asp)-(L-Arg)-(L-Trp), [SEQ ID NO.: 18]
(L-Tyr)-(L-Asp)-(L-Lys)-(L-Trp), [SEQ ID NO.: 19]
(L-Tyr)-(L-Glu)-(L-Lys)-(L-Trp), [SEQ ID NO.: 20]
(L-Tyr)-(L-Glu)-(L-Arg)-(L-Trp), [SEQ ID NO.: 21]
(L-Trp)-(L-Glu)-(L-Lys)-(L-Trp), [SEQ ID NO.: 22]
(L-Trp)-(L-Glu)-(L-Arg)-(L-Trp), [SEQ ID NO.: 23]
(L-Trp)-(L-Asp)-(L-Lys)-(L-Trp), [SEQ ID NO.: 24]
(L-Trp)-(L-Asp)-(L-Arg)-(L-Trp), [SEQ ID NO.: 25]
(L-Tyr)-(L-Asp)-(L-His)-(L-Tyr), [SEQ ID NO.: 26]
(D-Tyr)-(D-Glu)-(D-Arg)-(D-Phe), [SEQ ID NO.: 27]
(D-Tyr)-(D-Asp)-(D-His)-(D-Phe), [SEQ ID NO.: 28]
(D-Trp)-(D-Glu)-(D-Arg)-(D-Phe), [SEQ ID NO.: 29]
(D-Trp)-(D-Asp)-(D-His)-(D-Phe), [SEQ ID NO.: 30]
(D-Tyr)-(D-Asp)-(D-Arg)-(D-Phe), [SEQ ID NO.: 31]
(D-Tyr)-(D-Asp)-(D-His)-(D-Tyr), [SEQ ID NO.: 32]
(D-Tyr)-(D-Glu)-(D-Arg)-(D-Tyr), [SEQ ID NO.: 33]
(D-Trp)-(D-Asp)-(D-His)-(D-Typ), [SEQ ID NO.: 34]
(D-Trp)-(D-Glu)-(D-Arg)-(D-Typ), [SEQ ID NO.: 35]
(D-Tyr)-(D-Asp)-(D-Lys)-(D-Phe), [SEQ ID NO.: 36]
(D-Tyr)-(D-Glu)-(D-His)-(D-Phe), [SEQ ID NO.: 208]
(D-Tyr)-(D-Asp)-(D-Lys)-(D-Phe), [SEQ ID NO.: 209]
(D-Trp)-(D-Glu)-(D-His)-(D-Phe), [SEQ ID NO.: 210]
(D-Tyr)-(D-Glu)-(D-Lys)-(D-Phe), [SEQ ID NO.: 211]
(D-Trp)-(D-Glu)-(D-Lys)-(D-Phe), [SEQ ID NO.: 212]
(D-Trp)-(D-Asp)-(D-Lys)-(D-Phe), [SEQ ID NO.: 213]
(D-Tyr)-(D-Asp)-(D-His)-(D-Trp), [SEQ ID NO.: 214]
(D-Tyr)-(D-Glu)-(D-His)-(D-Trp), [SEQ ID NO.: 215]
(D-Trp)-(D-Asp)-(D-His)-(D-Trp), [SEQ ID NO.: 216]
(D-Trp)-(D-Glu)-(D-His)-(D-Trp), [SEQ ID NO.: 217]
(D-Tyr)-(D-Asp)-(D-Arg)-(D-Trp), [SEQ ID N0.: 218]
(D-Tyr)-(D-Asp)-(D-Lys)-(D-Trp), [SEQ ID NO.: 219]
(D-Tyr)-(D-Glu)-(D-Lys)-(D-Trp), [SEQ ID NO.: 220]
(D-Tyr)-(D-Glu)-(D-Arg)-(D-Trp), [SEQ ID NO.: 221]
(D-Trp)-(D-Glu)-(D-Lys)-(D-Trp), [SEQ ID NO.: 222]
(D-Trp)-(D-G1u)-(D-Arg)-(D-Trp), [SEQ ID NO.: 223]
(D-Trp)-(D-Asp)-(D-Lys)-(D-Trp), [SEQ ID NO.: 224]
(D-Trp)-(D-G1n)-(D-Arg)-(D-Trp), [SEQ ID NO.: 225]
(D-Trp)-(D-Asn)-(D-Lys)-(D-Trp), [SEQ ID NO.: 226]
(L-Tyr)-(L-Asp)-(L-Phe),
(D-Tyr)-(D-Asp)-(D-Phe),
(L-Tyr)-(L-Glu)-(L-Phe),
(L-Tyr)-(L-Arg)-(L-Phe),
(D-Tyr)-(D-Arg)-(D-Phe),
(D-Tyr)-(D-Glu)-(D-Phe),
(D-Tyr)-(D-Pro)-(D-Phe),
(D-Tyr)-(D-Leu)-(D-Phe),
(D-Tyr)-(D-Asp)-(D-Tyr),
(D-Tyr)-(D-Glu)-(D-Tyr),
(D-Tyr)-(D-Arg)-(D-Tyr),
(D-Tyr)-(D-Pro)-(D-Tyr),
(D-Tyr)-(D-Leu)-(D-Tyr),
(D-Phe)-(D-Pro)-(D-Phe),
(D-Phe)-(D-Leu)-(D-Phe),
(D-Phe)-(D-Arg)-(D-Tyr),
(D-Phe)-(D-,G1u)-(D-Tyr),
(D-Phe)-(D-Asp)-(D-Tyr),
(D-Phe)-(D-Pro)-(D-Tyr),
(D-Phe)-(D-Leu)-(D-Tyr),
(D-Tyr)-(D-Pro)-(D-Trp),
(D-Tyr)-(D-Leu)-(D-Trp),
(D-Tyr)-(D-Asp)-(D-Trp),
(D-Tyr)-(D-Glu)-(D-Trp),
(D-Tyr)-(D-Arg)-(D-Trp),
(D-Tyr)-(D-Pro)-(D-Trp),
(D-Tyr)-(D-Leu)-(D-Trp),
(D-Phe)-(D-Pro)-(D-Trp),
(D-Phe)-(D-Leu)-(D-Trp),
(D-Phe)-(D-Arg)-(D-Trp),
(D-Phe)-(D-Glu)-(D-Trp),
(D-Phe)-(D-Asp)-(D-Trp),
(D-Phe)-(D-Pro)-(D-Trp) и
(D-Phe)-(D-Leu)-(D-Trp).
В других вариантах осуществления структура А выбрана из группы, состоящей из:
п-гидроксибензойная кислота-(L-Glu)-(L-Arg)-анилина
п-гидроксибензойная кислота-(D-Glu)-(L-Arg)-анилина
п-гидроксибензойная кислота-(L-Glu)-(D-Arg)-анилина
п-гидроксибензойная кислота-(D-Glu)-(D-Arg)-анилина
п-гидроксибензойная кислота-(L-Glu)-(L-Arg)-бензиламина
п-гидроксибензойная кислота-(D-Glu)-(L-Arg)-бензиламина
п-гидроксибензойная кислота-(L-Glu)-(D-Arg)-бензиламина
п-гидроксибензойная кислота-(D-Glu)-(D-Arg)-бензиламина
п-гидроксибензойная кислота-(L-Glu)-(L-Arg)-2-фенил-этил-амина
п-гидроксибензойная кислота-(D-Glu)-(L-Arg)-2-фенил-этил-амина
п-гидроксибензойная кислота-(L-Glu)-(D-Arg)-2-фенил-этил-амина
п-гидроксибензойная кислота-(D-Glu)-(D-Arg)-2-фенил-этил-амина
2-(4-гидрокси-фенил)уксусная кислота-(L-Glu)-(L-Arg)-анилина
2-(4-гидрокси-фенил)уксусная кислота-(D-Glu)-(L-Arg)-анилина
2-(4-гидрокси-фенил)уксусная кислота-(L-Glu)-(D-Arg)-анилина
2-(4-гидрокси-фенил)уксусная кислота-(D-Glu)-(D-Arg)-анилина
2-(4-гидрокси-фенил)уксусная кислота-(L-Glu)-(L-Arg)-бензиламина
2-(4-гидрокси-фенил)уксусная кислота-(D-Glu)-(L-Arg)- бензиламина
2-(4-гидрокси-фенил)уксусная кислота-(L-Glu)-(D-Arg)-бензиламина
2-(4-гидрокси-фенил)уксусная кислота-(D-Glu)-(D-Arg)-бензиламина
2-(4-гидрокси-фенил)уксусная кислота-(L-Glu)-(L-Arg)-2-фенил-этил-амина
2-(4-гидрокси-фенил)уксусная кислота-(D-Glu)-(L-Arg)-2-фенил-этил-амина
2-(4-гидрокси-фемил)уксусная кислота-(L-Glu)-(D-Arg)-2-фемил-этил-амина
2-(4-гидрокси-фенил)уксусная кислота-(D-Glu)-(D-Arg)-2-фенил-этил-амина
3-(4-гидрокси-фенил)пропионовая кислота-(L-Glu)-(L-Arg)-анилина
3-(4-гидрокси-фенил)пропионовая кислота-(D-Glu)-(L-Arg)-анилина
3-(4-гидрокси-фенил)пропионовая кислота-(L-Glu)-(D-Arg)-анилина
3-(4-гидрокси-фенил)пропионовая кислота-(D-Glu)-(D-Arg)-анилина
3-(4-гидрокси-фенил)пропионовая кислота-(L-Glu)-(L-Arg)-бензиламина
3-(4-гидрокси-фенил)пропионовая кислота-(D-Glu)-(L-Arg)-бензиламина
3-(4-гидрокси-фенил)пропионовая кислота-(L-Glu)-(D-Arg)-бензиламина
3-(4-гидрокси-фенил)пропионовая кислота-(D-Glu)-(D-Arg)-бензиламина
3-(4-гидрокси-фенил)пропионовая кислота-(L-Glu)-(L-Arg)-2-фенил-этил-амина
3-(4-гидрокси-фенил)пропионовая кислота-(D-Glu)-(L-Arg)-2-фенил-этил-амина
3-(4-гидрокси-фенил)пропионовая кислота-(L-Glu)-(D-Arg)-2-фенил-этил-амина
3-(4-гидрокси-фенил)пропионовая кислота-(D-Glu)-(D-Arg)-2-фенил-этил-амина
п-гидроксибензойная кислота-(L-Arg)-анилина
п-гидроксибензойная кислота-(D-Arg)-анилина
п-гидроксибензойная кислота-(L-Glu)-анилина
п-гидроксибензойная кислота-(D-Glu)-анилина
п-гидроксибензойная кислота-(L-Arg)-бензиламина
п-гидроксибензойная кислота-(D-Arg)-бензиламина
п-гидроксибензойная кислота-(L-Glu)-бензиламина
п-гидроксибензойная кислота-(D-Glu)-бензиламина
п-гидроксибензойная кислота-(L-Arg)-2-фенил-этил-амина
п-гидроксибензойная кислота-(D-Arg)-2-фенил-этил-амина
п-гидроксибензойная кислота-(D-Glu)-2-фенил-этил-амина
п-гидроксибензойная кислота-(L-Glu)-2-фенил-этил-амина
2-(4-гидрокси-фенил)уксусная кислота-(L-Arg)-анилина
2-(4-гидрокси-фенил)уксусная кислота-(D-Arg)-анилина
2-(4-гидрокси-фенил)уксусная кислота-(L-Glu)-анилина
2-(4-гидрокси-фенил)уксусная кислота-(D-Glu)-анилина
2-(4-гидрокси-фенил)уксусная кислота-(D-Arg)-бензиламина
2-(4-гидрокси-фенил)уксусная кислота-(L-Arg)-бензиламина
2-(4-гидрокси-фенил)уксусная кислота-(D-Glu)-бензиламина
2-(4-гидрокси-фенил)уксусная кислота-(L-Glu)-бензиламина
2-(4-гидрокси-фенил)уксусная кислота-(L-Arg)-2-фенил-этил-амина
2-(4-гидрокси-фенил)уксусная кислота-(D-Arg)-2-фенил-этил-амина
2-(4-гидрокси-фенил)уксусная кислота-(L-Glu)-2-фенил-этил-амина
2-(4-гидрокси-фенил)уксусная кислота-(D-Glu)-2-фенил-этил-амина
3-(4-гидрокси-фенил)пропионовая кислота-(L-Arg)-анилина
3-(4-гидрокси-фенил)пропионовая кислота-(D-Arg)-анилина
3-(4-гидрокси-фенил)пропионовая кислота-(L-Glu)-анилина
3-(4-гидрокси-фенил)пропионовая кислота-(D-Glu)-анилина
3-(4-гидрокси-фенил)пропионовая кислота-(L-Arg)-бензиламина
3-(4-гидрокси-фенил)пропионовая кислота-(D-Arg)-бензиламина
3-(4-гидрокси-фенил)пропионовая кислота-(L-Glu)-бензиламина
3-(4-гидрокси-фенил)пропионовая кислота-(D-Glu)-бензиламина
3-(4-гидрокси-фенил)пропионовая кислота-(L-Arg)-2-фенил-этил-амина
3-(4-гидрокси-фенил)пропионовая кислота-(D-Arg)-2-фенил-этил-амина
3-(4-гидрокси-фенил)пропионовая кислота-(L-Glu)-2-фенил-этил-амина
3-(4-гидрокси-фенил)пропионовая кислота-(D-Glu)-2-фенил-этил-амина
Альтернативно группа, обозначенная как А' в формуле I может представлять собой олигопептид, имеющий последовательность аминокислот, выбранный из группы, которая представлена непосредственно выше.
Согласно ряду вариантов осуществления группа А' представляет собой пептидную или пептоидную группу, имеющую остатки Хаа1-Хаа2- Хаа3-Хаа4,
где:
Хаа1 представляет собой L-Tyr, D-Tyr, N- метил -L-Tyr, N-метил-О-Tyr, п-гидроксибензойную кислоту, 2-(4-гидрокси-фенил)уксусную кислоту, 3-(4-гидрокси-фенил)пропионовую кислоту или ацетил.
Хаа2 представляет собой L-Glu, D-Glu, L-Asp или D-Asp, N-метил-L-GlLi, N-метил-D-Glu, N-метил-L-Asp, N-метил-D-Asp, L-Pro, D-Pro, N-метил-L-Pro, N-метил-D-Pro, L-Leu, D-Leu, N-метил-L-Leu, N-метил-D-Leu или отсутствует.
Xaa3 представляет собой L-Arg, D-Arg, L-His или D-His, L-Lys, D-Lys, N-метил-L-Arg, N-метил-D-Arg, N-метил-L-l-lis, N-метил-D-His, N-метил-L-Lys, N-метил-D-Lys или отсутствует; и
Хаа4 представляет собой анилин, бензиламин, 2-фенил-этиламин, L-Phe или D-Phe, N-метил-L-Phe, N-метил-О-Phe, L-Trp, D-Trp, N-метил-L-Trp, N-метил-D-Trp.
Согласно ряду вариантов осуществления либо Хаа2, либо Хаа3 отсутствуют, но не оба, Хаа2 и Хаа3. Согласно другим вариантам осуществления оба, Хаа2 и Хаа3 отсутствуют.
М может представлять собой просто амидную связь между соседними пептидными или пептоидными группами. Альтернативно оно может представлять собой молекулярную группу, введенную в качестве спейсера и присоединенную к соседним пептидным или пептоидным группам амидными связями.
М может представлять собой дополнительную аминокислоту. Предпочтительно, когда оно представляет собой дополнительную аминокислоту с необъемной боковой цепью, например глицин, аланин или серин либо производные любой из них. Альтернативно М может представлять собой неаминокислотную группу, например, ε-аминокапроновую кислоту, Заминопропионовую кислоту, 4-аминомасляную кислоту. Другие группы могут быть представлены метиламином, этиламином, пропиламином, бутиламином, метиленом, диметиленом, триметиленом или тетраматиленом. Во всех случаях М должно быть таким, чтобы его присутствие физически не мешало связыванию группы А” и Gadd45β и/или MKK7. Степень возможного нарушения можно оценить с помощью анализа связывания in vitro, как раскрывают в данном контексте.
Олигомеры и мультимеры
Первый аспект изобретения охватывает олигомеры или мультимеры молекул соединения формулы I, причем указанные олигомеры и мультимеры содержат две или более молекул соединения формулы I, каждая из которых связана с обычной структурой основы через амидную связь, сформированную между амином или группой карбоновой кислоты, присутствующих в молекулах соединения формулы 1 и противоположной аминогруппой или группой карбоновой кислоты на структуре основы, причем указанная структура основы участвует по меньшей мере в 2 амидных связях.
Согласно ряду вариантов осуществления обычной основой может быть аминокислота лизин. Лизин представляет собой трифункциональную аминокислоту, имеющую кроме функциональных групп, которые определяют его как аминокислоту, аминогруппу на боковой цепи. Данная трифункциональная природа позволяет ему формировать три амидные связи с пептидами, пептоидами или подобными молекулами. Другие трифункциональные аминокислоты, которые можно использовать в качестве обычной основы включают D-α,β-диаминопропионовую кислоту (D-Dap), L-α,β-диаминопропионовую кислоту (L-Dap), D-α,δ-диаминомасляную кислоту (L-Dab), L-α,δ-диаминомасляную кислоту (L-Dab) и L-орнитин, D-орнитин. Другие трифункциональные нестандартные аминокислоты также могут быть использованы в соответствии с изобретением. Обычная основа может также содержать разветвленные пептиды, пептоиды или подобные молекулы, которые включают в свою последовательность трифункциональные аминокислоты и имеют по меньшей мере три функционально активные концевые группы, способные к формированию амидных связей.
Пептиды, проникающие в клетку.
Согласно ряду вариантов осуществления соединения формулы 1 конъюгируют с проникающим в клетку пептидом (СРР).
Данные пептиды можно присоединить к соединению формулы 1 либо посредством одной или более ковалентных связей, либо путем нековалентных связей.
СРР могут либо непосредственно проникать в плазмалемму, например, СРР может представлять собой Tat или производное, пептиды, выделенные из последовательности Antennapedia, либо полиаргининовую метку, пептид PTD-4, или функционально эквивалентный проникающий в клетку пептид (см. статью Но A, Schwarze SR, Mermelstein SJ, Waksman G, Dowdy SF 2001 Synthetic protein transduction domains: enhanced transduction potential in vitro and in vivo (Синтетические домены белка трансдукции повышают потенциал трансдукции in vitro и in vivo) Cancer Res 61:474-477).
Альтернативно СРР могут войти в клетку, опосредуя эндоцитоз или опосредуя формирование временных структур, стягивающих мембраны. В плане обсуждения пептидов, проникающих в клетку читателя отсылают к статье Wagstaff et al. (2006). Curr. Med. Chem. 13:171-1387 и приведенным в ней ссылкам.
Согласно ряду вариантов осуществления соединения, соответствующие изобретению, можно конъюгировать с наночастицами (например, наночастицами золота) с целью стимуляции клеточного поглощения
Флуоресцентные красители, группы меток и липидированные производные
Соединения формулы I можно конъюгировать с флуоресцентными красителями, чтобы мониторировать их проникновение в ткани- или клетки-мишени. Флуоресцентные красители можно получить с аминогруппами (т.е., сукцинимидами, изотиоцианатами, гидразинами), карбоксильными группами (т.е., карбодиимидами), тиоловыми группами (т.е., малеимидами и ацетилбромидами) и азидными группами, которые можно использовать для избирательной реакции с пептидами группами соединений формулы I. Примеры флуоресцентных красителей включают флуоресцеин и его производные, родамин и его производные.
Соединения формулы I можно конъюгировать с наночастицами дискретного размера, такими, как описано в статье Chithrani DB, Mol Membr Biol. 7 октября 2010 (в печати), имеющим дискретный размер до 100 нм, где пептиды или их производные могут быть присоединены дисульфидным мостиком, чтобы обеспечить специфическое высвобождение при восстановлении среды цитозоля. Кроме того пептид-наночастицы, конъюгированные посредством амидных, эфирных, сложно-эфирных, тиоэфирных связей, можно использовать с той же целью с получением данных соединений с низкой токсичностью. Наночастицы будут создавать благоприятные условия для поглощения клетками, а также обеспечивать средство визуализации и количественной оценки поглощения клетками при использовании способов флуоресценции (см. статью Schrand AM, Lin JB, Hens SC, Hussain SM., Nanoscale. 27 сентября 2010, в печати).
Группы меток можно присоединить аналогичными средствами и аналогичным образом обеспечить мониторирование успешного введения метки в ткани и клетки.
Производные жирных кислот соединения, соответствующего изобретению, содержащие соединение формулы I, связанное с жирной кислотой дисульфидной связью, можно использовать для доставки соединения, соответствующего изобретению, в клетки и ткани. Липидизация заметно повышает поглощение соединения относительно уровня поглощения соответствующего нелипидизированного соединения, а также пролонгирует удерживание соединений в крови и ткани. Более того, дисульфидная связь в липидизированном производном относительно лабильна в клетках и, таким образом, способствует внутриклеточному высвобождению молекулы из структуры жирной кислоты. Подходящие липидсодержащие структуры представляют собой гидрофобные заместители с 4-26 атомами углерода, предпочтительно 5-19 атомами углерода. Подходящие липидные группы включают, но без ограничения перечисленным, следующее: пальмитоил (C15H31), олеил (C15H29), стеарил (С17Н35), холат, линолат и дезоксихолат.
Ионные конъюгаты
Изобретение также охватывает соединения формулы I, функционально присоединенные к ионам металлов или радиоактивным ионам. Данное присоединение, как правило, осуществляют конъюгированием хелатирующего ион реактива (например, ЭДТА), который хелатируется с ионом. Таким образом радиоактивные ионы (например 99mTc, 111In, 64Cu, 67Cu, 89Sr, 90Y, 117mSn, 153Sm, 186Re, 188Re или 177Lu) можно доставить в клетки-мишени в качестве радиотерапии. Нерадиоактивные ионы металлов (например, ионы гадолиния) можно использовать как маркер, определяемый ЯМР-спектроскопией.
Соли и сольваты
Соли и сольваты соединений, соответствующих изобретению, которые подходят для использования в лекарственных средствах, представляют собой те, в которых противоион или ассоциированный растворитель фармацевтически приемлемы. Однако, соли и сольваты, имеющие фармацевтически неприемлемые противоионы или ассоциированные растворители, входят в объем настоящего изобретения, например, для использования в качестве промежуточных продуктов для получения соединений формулы (I) и их фармацевтически приемлемых солей или сольватов.
Подходящие соли согласно изобретению включают образованные с органическими или неорганическими кислотами или основаниями. Фармацевтически приемлемые кислотно-аддитивные соли включают те, которые сформированы с использованием хлористоводородной, бромистоводородной, серной, азотной, лимонной, винной, уксусной, фосфорной, молочной, пировиноградной, трифторуксусной, янтарной, перхлорной, фумаровой, малеиновой, гликолевой, салициловой, оксалоуксусной,метансульфоновой,этансульфоновой,п-толуолсульфоновой, муравьиной, бензойной, малоновой, нафталин-2-сульфоновой, бензолсульфоновой и изетионовой кислот. Другие кислоты, такие как щавелевая, хотя сами не являются фармацевтически приемлемыми, могут быть использованы в качестве промежуточных продуктов при получении соединений, соответствующих изобретению, и их фармацевтически приемлемых солей. Фармацевтически приемлемые соли с основаниями включают соли аммония, соли щелочных металлов, например, соли калия и натрия, соли щелочноземельных металлов, например, соли кальция и магния, и соли с органическими основаниями, например, дициклогексиламином и N-метил-D-глюкомином.
Компетентные специалисты в области органической химии будут иметь в виду, что многие органические соединения могут образовывать комплексы с растворителями, в которых они реагируют или из которых они осаждаются или кристаллизуются. Данные комплексы известны как "сольваты". Например, комплекс в водой известен как "гидрат". Настоящее изобретение предусматривает сольваты соединений, соответствующих изобретению.
Примеры предпочтительных молекул формулы 1 приводят ниже. Если не указана L-/D-конфигурация остатка аминокислоты, значит, включены обе конфигурации.
Ацетил-Tyr-Glu-Arg-Phe-NH2 [SEQ ID NO.:37]
пара-гидроксибензойная кислота-С1u-Arg-анилин
пара-гидроксибензойная кислота-Glu-Arg-бензиламин
пара-гидроксибензойная кислота-Glu-Arg-2-фенил-этиламин
2-(4-гидроксифенил)уксусная кислота-Glu-Arg-анилин
2-(4-гидроксифенил)уксусная кислота-Glu-Arg-бензиламин
2-(4-гидроксифенил)уксусная кислота-Glu-Arg-2-фенил-этиламин
3-(4-гидроксифенил)уксусная кислота-Glu-Arg-3-анилин
3-(4-гидроксифенил)уксусная кислота-Glu-Arg-бензиламин
3-(4-гидроксифенил)уксусная кислота-Glu-Arg-2-фенил-этил-амин
Ацетил-Tyr-Asp-His-Phe-NH2 [SEQ ID NO.:38]
пара-гидроксибензойная кислота-Asp-Hus-анилин
пара-гидроксибензойная кислота-Asp-His-бензиламин
пара-гидроксибензойная кислота-Asp-His-3-фенил-пропиламино
2-(4-гидроксифенил)уксусная кислота-Asp-His-анилин
2-(4-гидроксифенил)уксусная кислота-Asp-His-бензиламин
2-(4-гидроксифенил)уксусная кислота-Asp-His-2-фенил-этиламин
3-(4-гидроксифенил)пропионовая кислота-Asp-His-анилин
3-(4-гидроксифенил)пропионовая кислота-Asp-His-бензиламин
3-(4-гидроксифенил)пропионовая кислота-Asp-His-2-фенил-этил-амин
Ацетил-Tyr-Asp-Lys-Phe-NH2, [SEQ ID NO.:39]
Ацетил-Tyr-Glu-Lys-Phe-NH2, [SEQ ID N0.:40]
Ацетил-Tyr-Glu-His-Phe-NH2, [SEQ ID NO.:41]
Ацетил-Tyr-Asp-Arg-Phe-NH2, [SEQ ID NO.:42]
Ацетил-Trp-Glu-His-Phe-NH2, [SEQ ID NO.:43]
Ацетил-Trp-Glu-Lys-Phe-NH2, [SEQ ID NO.:44]
Ацетил-Trp-Asp-His-Phe-NH2, [SEQ ID NO.:45]
Ацетил-Trp-Asp-Lys-Phe-NH2, [SEQ ID NO.:46]
Ацетил-Tyr-Glu-Arg-Tyr-NH2, [SEQ ID NO.:47]
Ацетил-Tyr-Asp-Lys-Tyr-NH2, [SEQ ID NO.:48]
Ацетил-Tyr-Glu-Lys-Tyr-NH2, [SEQ ID NO.:49]
Ацетил-Tyr-Glu-His-Tyr-NH2, [SEQ ID NO.:50]
Ацетил-Tyr-Asp-Arg-Tyr-NH2, [SEQ ID NO.:51]
Ацетил-Trp-Glu-His-Tyr-NH2, [SEQ ID NO.:52]
Ацетил-Trp-Glu-Lys-Tyr-NH2, [SEQ ID NO.:53]
Ацетил-Trp-Asp-His-Tyr-NH2, [SEQ ID NO.:54]
Ацетил-Trp-Asp-Lys-Tyr-NH2, [SEQ ID NO.:55]
Внутренний пактам ацетил-Tyr-Glu-Lys-Phe-NH2, [SEQ ID NO.:56]
Ацетил-Tyr-Gln-Arg-Phe-NH2, [SEQ ID NO.:57]
Ацетил-Tyr-Met-Arg-Phe-NH2, [SEQ ID NO.:58]
Ацетил-Tyr-Leu-Arg-Phe-NH2, [SEQ ID NO.:59]
Ацетил-Tyr-Arg-Phe-NH2,
Ацетил-Tyr-Arg-Tyr-NH2,
Ацетил-Tyr-Glu-Phe-NH2,
Ацетил-Tyr-Glu-Tyr-NH2,
Ацетил-Tyr-Asp-Phe-NH2,
Ацетил-Tyr-Asp-Tyr-NH2,
Ацетил-Tyr-Pro-Phe-NH2,
Ацетил-Tyr-Lys-Phe-NH2,
Ацетил-Tyr-His-Phe-NH2,
H-Tyr-Arg-Phe-NH2,
H-Tyr-Arg-Tyr-NH2,
H-Tyr-Glu-Phe-NH2,
H-Tyr-Glu-Tyr-NH2,
H-Tyr-Asp-Phe-NH2,
H-Tyr-Asp-Tyr-NH2,
H-Tyr-Pro-Phe-NH2,
H-Tyr-Lys-Phe-NH2,
H-Tyr-His-Phe-NH2,
Бензилоксикарбонил-Tyr-Arg-Phe-NH2,
Бензилоксикарбонил-Tyr-Arg-Tyr-NH2,
Бензилоксикарбонил-Tyr-Glu-Phe-NH2,
Бензилоксикарбонил-Tyr-Glu-Tyr-NH2,
Бензилоксикарбонил-Tyr-Asp-Phe-NH2,
Бензилоксикарбонил-Tyr-Asp-Tyr-NH2,
Бензилоксикарбонил-Tyr-Pro-Phe-NH2,
Бензилоксикарбонил-Tyr-Lys-Phe-NH2,
Бензилоксикарбонил-Tyr-His-Phe-NH2,
Бензилоксикарбонил-Tyr-Glu-Arg-Phe-NH2, [SEQ ID NO.:60]
Бензилоксикарбонил-Tyr-Asp-His-Phe-NH2, [SEQ ID NO.:61]
Бензилоксикарбонил-Tyr-Asp(ОМе)-His-Phe-NH2, [SEQ ID NO.:62]
Бензилоксикарбонил-Tyr-Arg-Phe-NH2,
Бензилоксикарбонил-Tyr-Glu-Phe-NH2,
Бензилоксикарбонил-(N-метил)Tyr-(N-метил)Arg-(N-метил)Phe-NH2,
Бензилоксикарбонил-(N-метил)Tyr-Glu-(N-метил)Phe-NH2,
Бензилоксикарбонил-Tyr-(N-метил)Arg-(N-метил)Phe-NH2,
Бензилоксикарбонил-(N-метил)Tyr-(N-метил)Arg-Phe-NH2,
Бензилоксикарбонил-Tyr-Glu-(N-метил)Phe-NH2,
Бензилоксикарбонил-Tyr-(N-метил)Glu-Phe-NH2,
Бензилоксикарбонил-(N-метил)Tyr-Glu-Phe-NH2,
Ацетил-(N-метил)Tyr-(N-метил)Arg-(N-метил)Phe-NH2,
Ацетил-(N-метил)Tyr-Glu-(N-метил)Phe-NH2,
Ацетил-Tyr-(N-метил)Arg-(N-метил)Phe-NH2,
Ацетил-(N-метил)Tyr-(N-метил)Arg-Phe-NH2,
Ацетил-Tyr-Glu-(N-метил)Phe-NH2,
Ацетил-Tyr-(N-метил)Glu-Phe-NH2,
Ацетил-(N-метил)Tyr-Glu-Phe-NH2,
H-(N-метил)Tyr-(N-метил)Arg-(N-метил)Phe-NH2,
Н-(N-метил)Tyr-Glu-(N-метил)Phe-NH2,
Н-Tyr-(N-метил)Arg-(N-метил)Phe-NH2,
Н-(N-метил)Tyr-(N-метил)Arg-Phe-NH2,
Н-Tyr-Glu-(N-метил)Phe-NH2,
H-Tyr-(N-метил)Glu-Phe-NH2,
Н-(N-метил)Tyr-Glu-Phe-NH2,
Ацетил-Tyr-Glu-(β-гомо)Phe-NH2,
Ацетил-Tyr-(β-гомо)Glu-Phe-NH2,
Ацетил-(β-гомо)Tyr-Glu-Phe-NH2,
Ацетил-Tyr-Phe-NH2,
Ацетил-Phe-Tyr-NH2,
Бензилоксикарбонил-Tyr-Phe-NH2,
Бензилоксикарбонил-Phe-Tyr-NH2,
H-Tyr-Phe-NH2,
H-Phe-Tyr-NH2,
(3-Метокси,4-гидрокси-бензоил)-Tyr-Glu-Arg-Phe-NH2, [SEQ ID NO.:63]
(3-Метокси,4-гидрокси-бензоил)-Tyr-Asp-His-Phe-NH2, [SEQ ID NO.:64]
(3-Метокси,4-гидрокси-бензоил)-Tyr-Asp(ОМе)-His-Phe-NH2, [SEQ ID NO.:65]
(3-Метокси,4-гидрокси-бензоил)-Tyr-Arg-Phe-NH2,
(3-Метокси,4-гидрокси-бензоил)-Tyr-Glu-Phe-NH2,
Флуоренилметилоксикарбонил-Tyr-Glu-Arg-Phe-NH2, [SEQ ID NO.:66]
Флуоренилметилоксикарбонил-Tyr-Asp-His-Phe-NH2, [SEQ ID NO.:67]
Флуоренилметилоксикарбонил-Tyr-Asp(ОМе)-His-Phe-NH2, [SEQ ID NO.:68]
Флуоренилметилоксикарбонил-Tyr-Asp(ОМе)-His-Phe-NH2 [SEQ ID NO.:69]
Флуоренилметилоксикарбонил-Tyr-Arg-Phe-NH2,
Флуоренилметилоксикарбонил-Tyr-Glu-Phe-NH2,
Миристил-Tyr-Glu-Arg-Phe-NH2, [SEQ ID NO.:70]
Миристил-Tyr-Asp-His-Phe-NH2, [SEQ ID NO.:71]
Миристил-Tyr-Arg-Phe-NH2,
Миристил-Tyr-Glu-Phe-NH2,
Миристил-Tyr-Asp(OMe)-His-Phe-NH2, [SEQ ID NO.:72]
Ацетил-Tyr-Glu-Arg-Phe-Gly-Tyr-Glu-Arg-Phe-NH2, [SEQ ID NO.:73]
Ацетил-Tyr-Asp-His-Phe-Gly-Tyr-Asp-His-Phe-NH2, [SEQ ID NO.:74]
Ацетил-Tyr-Arg-Phe-Gly-Tyr-Arg-Phe-NH2, [SEQ ID NO.:75],
Ацетил-Tyr-Asp(OMe)-His-Phe-Gly-Tyr-Asp(OMe)-His-Phe-NH2, [SEQ ID NO.:76]
бензилоксикарбонил-Tyr-Glu-Arg-Phe-Gly-Tyr-Glu-Arg-Phe-NH2, [SEQ ID NO.:77]
бензилоксикарбонил-Tyr-Asp-His-Phe-Gly-Tyr-Asp-His-Phe-NH2, [SEQ ID NO.:78]
бензилоксикарбонил-Tyr-Arg-Phe-Gly-Tyr-Arg-Phe-NH2, [SEQ ID NO.:79]
бензилоксикарбонил-Tyr-Asp(OMe)-His-Phe-Gly-Tyr-Asp(OMe)-His-Phe-NH2, [SEQ ID NO.:80].
Другие примеры соединений, соответствующих изобретению, включают:
Соединение А1
Ac-Tyr-Phe-NH2
Соединение A3
Ac-Tyr-bAla-Phe-NH2
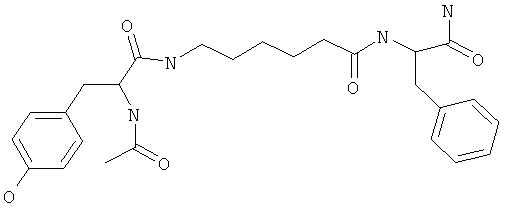
Соединение А6
Ас-Tyr-(6-амино-капроновая кислота)-Phe-NH2
Соединение А7
Ac-Tyr-Tyr-NH2
Соединение А8
Ac-Phe-Tyr-NH2
Соединение А9
Ac-Phe-Arg-Phe-NH2
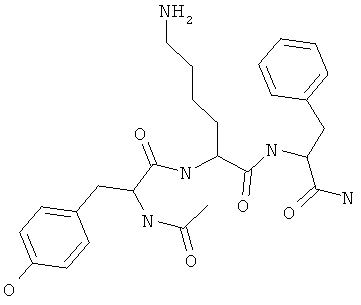
Соединение В2
Ac-Tyr-Lys-Phe-NH2
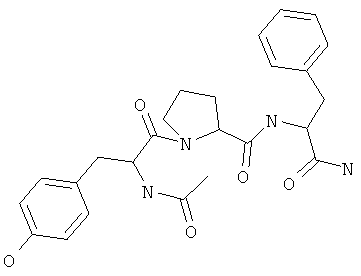
Соединение В13
Ас-Tyr-Pro-Phe-NH2
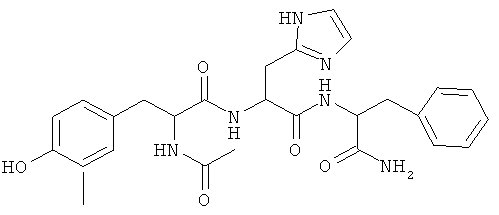
Соединение В16
Ac-Tyr-His-Phe-NH2
Соединение Н1
L-3,3-дифенил-аланин
Соединение Н2
L-Н-3(4-пиридил) аланин
Соединение Н3
L-H-4-гидрокси-фенилглицин
Соединение Н4
L-2-амино-4-фенилмасляная кислота
Соединение Н5
L-фенилглицина
Соединение Н6
L-H-4-гидрокси-фенилглицин
Соединение Н7
L-гомофенилаланин
Соединение Н8
L-3-(2-фурил)-аланин
Соединение Н9
L-3-(4-хинолил)-аланин
Соединение H10
L-нафтилаланин
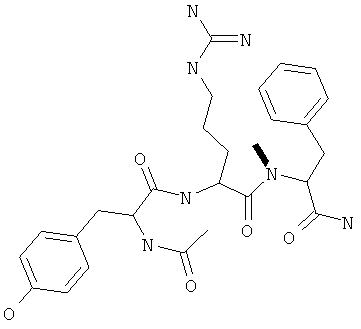
Соединение I1
Ac-Tyr-Arg-(N-Me)Phe-NH2
Соединение I2
Ac-Tyr-(N-Me)Arg-Phe-NH2,
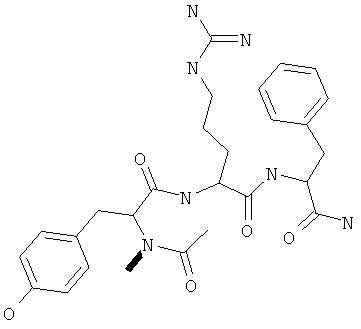
Соединение I3
Ac-(N-Me)Tyr-Arg-Phe-NH2,
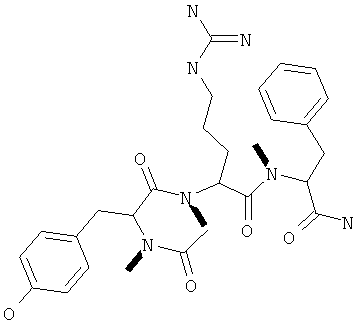
Соединение I4
Ac-(N-Me)Tyr-(N-Me)Arg-(N-Me)Phe-NH2
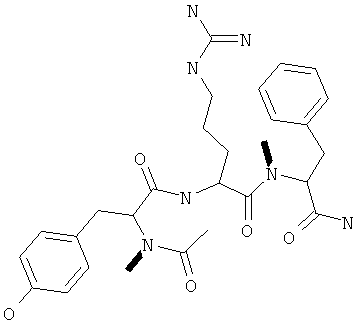
Соединение I5
Ac-(N-Me)Tyr-Arg-(N-Me)Phe-NH2
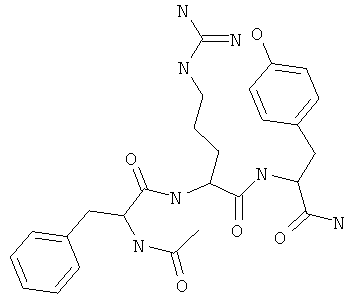
Соединение O1
Ac-Phe-Arg-Tyr-NH2
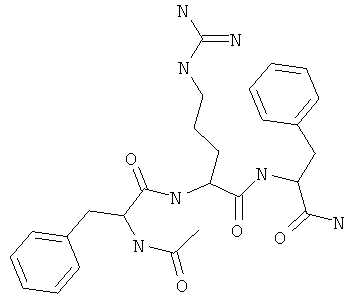
Соединение O2
Ac-Phe-Arg-Phe-NH2
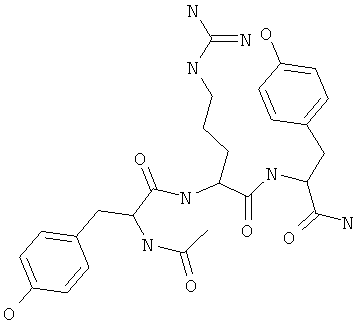
Соединение O3
Ac-Tyr-Arg-Tyr-NH2
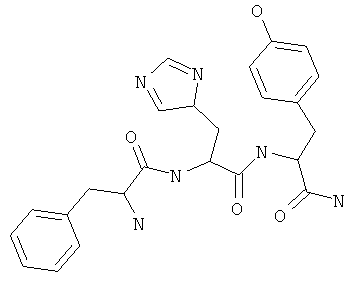
Соединение O5
H-Phe-His-Tyr-NH2
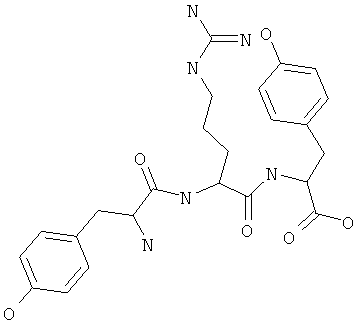
Соединение O7
H-Phe-His-Phe-NH2
Соединение O8
H-Phe-Arg-Tyr-NH2,
Соединение O9
H-Phe-Arg-Phe-NH2
Соединение O10
H-Tyr-Arg-Tyr-NH2
Соединение Р1
4-(гидроксил)-фенилуксусная кислота-Arg-3-фенил-этиламин
Соединение Р2
4-(гидроксил)-фенил-уксусная кислота-His-3-фенил-этиламин
Соединение Р3
4-(гидроксил)-фенил-уксусная кислота-Glu-3-фенил-этиламин
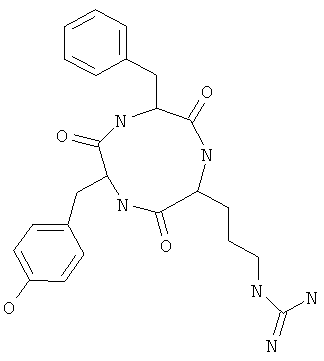
Соединение G1
Цикло(Tyr-Arg-Phe)
Соединение G2
Цикло(Рhе-Arg-Tyr)
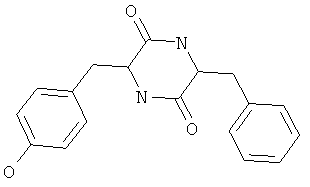
Соединение G3
Цикло(Tyr-Phe)
Соединение N1
Наночастицы золота-Tyr-Arg-Phe-NH2
Согласно ряду вариантов осуществления соединения, специально раскрытые в данном контексте, включая приведенные в примерах, представляют собой предпочтительные соединения или предпочтительные варианты осуществления группы А' формулы I. Настоящее изобретение предусматривает мультимерные варианты или специальные соединения, детально описанные в данном контексте. Например, настоящее изобретение предусматривает состоящие из 3 или 4 остатков пептидные или пептидоидные структуры специальных соединений, описанных в данном контексте в качестве соответствующих структуре А, А', А', А”' или А”” соединений формулы I.
Фармацевтические композиции
Согласно второму аспекту изобретения предусматривают фармацевтическую композицию, содержащую соединение, соответствующее первому аспекту изобретения, и фармацевтически приемлемый носитель.
Хотя возможно введение активного ингредиента в виде монопрепарата, предпочтительно, чтобы он находился в фармацевтическом препарате или композиции. Соответственно, изобретение предусматривает фармацевтический препарат, содержащий соединение формулы (I) или его производное либо его соль или сольват, как определено выше, и фармацевтически приемлемый носитель. Фармацевтические композиции, соответствующие изобретению, могут принимать форму фармацевтического препарата, как описано ниже.
Фармацевтические препараты, соответствующие изобретению, включают подходящие для перорального, парентерального (включая подкожное, внутрикожное, внутримышечное, внутривенное или внутрисуставное), ингаляционного (включая пылевидные препараты или аэрозоли из мелких частиц, которые можно получить с использованием различных типов дозированных аэрозолей под давлением, распылителей или инсуффляторов), ректального и местного (включая накожное, чрескожное, через слизистые, защечное, подъязычное и внутриглазное) введения, хотя наиболее подходящий путь может зависеть, например, от состояния или нарушения у пациента.
Препараты можно удобно представить в дозированной лекарственной форме и можно изготовить любым способом, хорошо известным в области фармации. Все способы в ключа ют стадию приведения активного ингредиента в контакт с носителем, который содержит один или более дополнительных ингредиентов. Как правило, препараты получают путем однородного и тщательного перемешивания активного ингредиента с жидкими носителями или тонко разделенными твердыми носителями либо обоими с последующим (при необходимости) формованием продукта с получением требуемого препарата.
Препараты, соответствующие настоящему изобретению, подходящие для перорального применения, могут быть представлены в виде отдельных элементов, таких как капсулы, крахмальные капсулы или таблетки, каждая из которых содержит заданное количество активного ингредиента, в виде порошка или гранул, в виде раствора или суспензии в водной жидкости или неводной жидкости либо в виде жидкой эмульсии типа масло в воде или жидкой эмульсии типа вода в масле. Активный ингредиент может также присутствовать в виде болюса, электуария или пасты. Различные фармацевтически приемлемые носители и их получение описаны в стандартных справочниках по препаратам, например, Remington's Pharmaceutical Sciences (Справочник по фармацевтическим наукам Ремингтона) by E.W.Martin. См. также статью Wang, Y.J. and Hanson, M.A., Journal of Parenteral Science and Technology, Technical Report No.10, Supp. 42:2S, 1988.
Таблетка может быть изготовлена путем прессования или формования, необязательно с одним или более вспомогательных ингредиентов. Прессованные таблетки можно изготовить путем прессования в подходящем устройстве активного ингредиента в сыпучем виде, таком как порошок или гранулы, необязательно смешанного со связующим компонентом, скользящим веществом, инертным разбавителем, смазывающим, поверхностно-активным или диспергирующим компонентом. Формованные таблетки можно изготовить путем формования в подходящем устройстве смеси порошкового соединения, увлажненного инертным жидким разбавителем. Таблетки необязательно могут быть покрытыми или иметь насечки и могут иметь состав, обеспечивающий замедленное или контролируемое высвобождение находящегося в них активного ингредиента. Настоящие соединения можно, например, вводить в форме, подходящей для немедленного высвобождения или замедленного высвобождения. Немедленное высвобождение или замедленное высвобождение может быть достигнуто при использовании подходящих фармацевтических композиций, содержащих настоящие соединения, или, особенно в случае замедленного высвобождения, при использовании таких устройств, как подкожные имплантаты или осмотические насосы. Настоящие соединения можно также вводить липосомально.
Предпочтительно, когда композиции, соответствующие изобретению, пригодны для подкожного введения, например, путем инъекции.
Примеры композиций, предназначенных для перорального введения, включают суспензии, которые могут содержать, например, микрокристаллическую целлюлозу для придания объема, альгиновую кислоту или альгинат натрия в качестве суспендирующего компонента, метилцеллюлозу в качестве компонента, повышающего вязкость, и подсластители или вкусовые добавки такие, которые известны в уровне техники, и таблетки с немедленным высвобождением, которые могут содержать, например, микрокристаллическую целлюлозу, дикальцийфосфат, крахмал, стеарат магния и/или лактозу и/или другие наполнители, связующие компоненты, сухие наполнители, разрыхлители, разбавители и смазывающие вещества такие, которые известны в уровень техники. Соединения формулы (I) или их вариант, производное, соль или сольват также можно доставлять через полость рта путем подъязычного и/или защечного введения. Формованные таблетки, прессованные таблетки или лиофилизированные таблетки представляют собой примеры форм, которые могут быть использованы. Примеры композиций включают такие, в которых настоящее соединение(я) находится в составе с быстрорастворимыми разбавителями, такими как маннит, лактоза, сахароза и/или циклодекстрины. В данные препараты могут быть также включены высокомолекулярные наполнители, такие как целлюлозы (avicel) или полиэтиленгликоли (ПЭГ). Данные препараты могут также включать наполнитель, способствующий адгезии к слизистым, такой как гидроксипропилцеллюлоза (НРС), гидроксипропилметилцеллюлоза (НРМС), натрий карбоксиметилцеллюлоза (SCMC), сополимер малеинового ангидрида (например, Gantrez) и ингредиенты для контроля высвобождения, такие как полиакриловый сополимер (например Carbopol 934). Для простоты изготовления и применения могут быть также добавлены смазывающие вещества, скользящие вещества, красители и стабилизаторы.
Препараты для парентерального введения включают водные и неводные стерильные инъекционные растворы, которые могут содержать антиоксиданты, буферы, бактериостатики и растворенные вещества, которые делают препарат изотоническим относительно крови предусматриваемого реципиента, а также водные и неводные стерильные суспензии, которые могут включать суспендированные компоненты и сгустители. Препараты могут быть представлены в однодозных и многодозных контейнерах, например, запаянных ампулах и флаконах, и могут храниться в сублимационно высушенном (лиофилизированном) состоянии, требующем только добавления стерильного жидкого носителя, например, солевого раствора или воды для инъекций, непосредственно перед применением. Приготовленные для немедленного применения инъекционные растворы и суспензии можно получить из стерильных порошков, гранул и таблеток ранее описанного типа. Примеры композиций для парентерального введения включают инъекционные растворы или суспензии, которые могут содержать, например, подходящие нетоксичные приемлемые для парентерального применения разбавители или растворители, такие как маннит, 1,3-бутандиол, вода, раствор Рингера, изотонический раствор хлорида натрия, или другие подходящие диспергирующие или смачивающие и суспендирующие компоненты, включая синтетические моно- или диглицериды и жирные кислоты, в том числе олеиновую кислоту, или Кремафор (Cremaphor). Водный носитель может представлять собой, например, изотонический буферный раствор с рН от приблизительно 3,0 до приблизительно 8,0, предпочтительно с рН от приблизительно 3,5 до приблизительно 7,4, например от 3,5 до 6,0, например, от 3,5 до приблизительно 5,0. Используемые буферы включают буферы на основе цитрата натрия-лимонной кислоты и фосфата натрия-фосфорной кислоты, а также ацетата натрия/уксусной кислоты. Предпочтительно, когда композиция не включает окислители и другие соединения, которые, как известно, вредны для соединения формулы I и родственных молекул. Наполнители, которые могут быть включены, представляют собой, например, другие белки, такие как человеческий сывороточный альбумин или препараты плазмы. При необходимости фармацевтическая композиция может также содержать минимальные количества нетоксичных вспомогательных субстанций, таких как смачивающие или эмульгирующие вещества, консерванты и забуферивающие рН компоненты и т.п., например, ацетат натрия или сорбитанмонолаурат.
Примеры композиций для назального аэрозольного или ингаляционного введения включают растворы в солевом растворе, которые могут содержать, например, бензиловый спирт или другие подходящие консерванты, вещества, способствующие всасыванию, предназначенные для повышения биодоступности, и/или солюбилизирующие либо диспергирующие компоненты, такие как известные в области техники. Удобно, когда в композициях для назального аэрозольного или ингаляционного введения соединение, соответствующее изобретению, доставляют в форме эерозольного спрея из упаковки или разбрызгивателя под давлением при использовании подходящего пропеллента, например, дихлородифторметана, трихлорфторметана, дихлортетрафторэтана, диоксида углерода или другого подходящего газа. В случае аэрозоля под давлением дозированную форму можно получить, снабдив его клапаном для доставки отмеренного количества. Можно изготовить капсулы и картриджи, например, из желатина, предназначенные для использования в ингаляторе или инсуффляторе, которые содержат порошковую смесь соединения и подходящей порошковой основой, например, лактозой или крахмалом. В одном специальном неограничивающем примере соединение, соответствующее изобретению, вводят в виде аэрозоля из дозирующего клапана через аэрозольный адаптер, известный также исполнительный механизм клапана. Необязательно включены также стабилизатор и/или пористые частицы для глубокой легочной доставки (например, см. Патент США No.6447743).
Препараты для ректального применения могут быть представлены в форме удерживающей клизмы или суппозитория с обычными носителями, таким как масло какао, синтетические сложные эфиры глицеридов или полиэтиленгликоль. Данные носители, как правило, твердые при обычных температурах, но разжижаются и/или растворяются в прямой кишке, высвобождая лекарственный препарат.
Препараты для местного применения в полости рта, например, защечно или подъязычно, включают лепешки, содержащие активный ингредиент в основе с вкусовой добавкой, такой как сахароза и акация или трагакант, и пастилки, содержащие активный ингредиент в основе, такой как желатин и глицерин или сахароза и акация. Примеры композиций для местного применения включают носитель для местного применения, такой как Plastibase (минеральное масло, желированное полиэтиленом).
Предпочтительными дозированными препаратами являются те, которые содержат эффективную дозу активного ингредиента, как указано выше в данном материале, или ее подходящую фракцию.
Следует иметь в виду, что, в дополнение к ингредиентам, конкретно упомянутым выше, препараты, соответствующие данному изобретению, могут включать другие компоненты, принятые в уровне техники, имеющие отношение к типу рассматриваемого препарата, например, те, которые предназначены для перорального введения, могут включать вкусовые добавки.
Соединения, соответствующие изобретению, удобно, кроме того, вводить в виде систем с замедленным высвобождением. Подходящие примеры систем с замедленным высвобождением, соответствующих изобретению, включают подходящие полимерные материалы, например, полупроницаемые полимерные матрицы в форме формованных изделий, например, пленки или микрокапсулы, подходящие гидрофобные материалы, например, в виде эмульсии в приемлемом масле, или ионообменные смолы и малорастворимые производные соединения, соответствующего изобретению, например, малорастворимую соль. Системы с замедленным высвобождением можно вводить перорально, ректально, парентерально, интравагинально, внутрибрюшинно, местно, например в виде порошка, мази, геля, капель или чрескожного пластыря, защечно или в виде спрея для рта или носа.
Препараты для введения могут быть соответствующим образом изготовлены так, чтобы они обеспечивали контролируемое высвобождение соединений, соответствующих изобретению. Например, фармацевтические композиции могут находиться в форме частиц, содержащих один или более биоразрушаемых полимеров, желирующих полисахариды и/или биоадгезивных полимеров, амфифильных полимеров, средств, способных модифицировать свойства границ раздела частиц соединения формулы (I). Данные композиции проявляют ряд свойств биосовместимости, которые дают возможность контролируемого высвобождения активной субстанции. См. Патент США No.5700486.
Соединение, соответствующее изобретению, можно доставить с помощью насоса (см. статьи Langer, supra; Sefton, CRC Crit. Ref Biomed. Eng. 14:201, 1987; Buchwald et al., Surgery 88:507, 1980; Saudek et al., N. Engl. J. Med. 321:574, 1989) или посредством непрерывных подкожных инфузий, например, с использованием мининасоса. Можно также использовать упаковку для внутривенного раствора. Другие системы с контролируемым высвобождением обсуждают в обзоре Langer (см. Science 249:1527-1533, 1990). В другом аспекте описания соединения, соответствующие изобретению, доставляются помощью имплантированного насоса, описанного, например, в Патенте США No.6436091, Патенте США No.5939380, Патенте США No.5993414.
Имплантируемые устройства для инфузий лекарственных препаратов используют для обеспечения пациентов постоянным и длительным дозированием или инфузией лекарства или любого другого лекарственного препарата. Как правило, данное устройство можно отнести как к категории активных, так и пассивных. Соединение, соответствующее настоящему изобретению, можно изготовить в виде депо-препарата. Данный депо-препарат длительного действия можно ввести путем имплантации, например, подкожно или внутримышечно, или путем внутримышечной инъекции. Так, например, соединения можно изготовить с подходящими полимерными или гидрофобными материалами, например, в виде эмульсии в приемлемом масле, или ионообменными смолами, либо в виде слаборастворимых производных, например, в виде слаборастворимой соли.
Терапевтически эффективное количество соединения, соответствующего изобретению, можно ввести как одноимпульсную дозу, как болюсную дозу или импульсные дозы, вводимые в течение времени. Так, при импульсных дозах проводят болюсное введение соединения, соответствующего изобретению, за которым следует период времени, когда никакое соединение, соответствующее изобретению, не вводят пациенту, с последующим вторым болюсным введением. В специальных неограничивающих примерах импульсные дозы соединения, соответствующего изобретению, вводят в течение дня, в течение недели или в течение месяца.
В одном варианте осуществления терапевтически эффективное количество соединения, соответствующего изобретению, вводят с терапевтически эффективным количеством другого препарата, например, другого противоопухолевого химиотерапевтического препарата (например, талидомида, дексаметазона, бортецомиба, леналидомида, мелфалана, цисплатина, доксорубицина, 5-ФУ (5-фторурацила) и т.п.) или препарата для лечения анемии (например, эритропоэтина) или препарата для предупреждения переломов костей (например, бисфосфоната, такого как памидронат или цолендроновая кислота).
Терапевтически эффективное количество соединения, соответствующего изобретению, будет зависеть от используемой молекулы, пациента, проходящего лечение, тяжести и типа поражения и способа и пути введения.
Согласно третьему аспекту изобретения предложен способ лечения
нарушения или заболевания, предусматривающий введение соединения, соответствующего первому и второму аспектам изобретения, или фармацевтической композиции, соответствующей второму аспекту изобретения, введение терапевтически эффективного количества соединения, соответствующего первому аспекту изобретения, или фармацевтической композиции, соответствующей второму аспекту изобретения, нуждающемуся в этом пациенту.
Нарушения и заболевания
Соединения, композиции и способы, соответствующие изобретению, подходят для лечения или предупреждения заболеваний и нарушений, которые характеризуются либо патологической повышенной экспрессией или активностью Gadd45β, либо характеризуются патологической активацией пути NF-κВ и подлежат лечению индукцией запрограммированной гибели клеток путем ингибирования активности Gadd45β.
Заболевания, подходящие для лечения или профилактики, включают рак. Предпочтительно, когда как представляет собой форму рака, экспрессирующую повышенные уровни Gadd45β относительно уровней, соответствующих нормальным здоровым клеткам или тканям. Формы рака, которые, как известно, экспрессируют высокие уровни Gadd45, и, таким образом, подходят для лечения соединениями, соответствующими изобретению, включают: множественную миелому, диффузную крупноклеточную В-клеточную лимфому, лимфому Беркетта, промоноцитарный лейкоз и другие формы лейкоза, а также солидные опухоли, такие как гепатоцеллюлярная карцинома, рак мочевого пузыря, рак головного мозга и центральной нервной системы, рак молочной железы, рак головы и шеи, рак легкого и рак простаты. Согласно ряду вариантов осуществления рак представляет собой форму рака, которая зависит от NF-κВ в плане выживаемости. Такие специфические формы рака, которые зависят от NF-κВ в плане выживаемости и, таким образом, подходят для лечения или предупреждения, включают: множественную миелому, лимфому из клеток мантии, лимфому MALT, лимфому Ходжкина, диффузную крупноклеточную В-клеточную лимфому, лимфому Беркетта, промоноцитарный лейкоз, миелодиспластитеческий синдром, Т-клеточный лейкоз взрослых (HTLV-1), хронический лимфоцитарный лейкоз, хронический миелогенный лейкоз, острый миелогенный лейкоз, острый лимфобластный лейкоз, рак, ассоциированный с колитом, рак толстой кишки, рак печение (например, гепатоцеллюлярная карцинома), рак шейки матки, рак почки, рак легкого, рак пищевода, рак желудка, рак глотки, рак простаты, рак поджелудочной железы, рак щитовидной железы, рак паращитовидной железы, рак мочевого пузыря, рак яичников, рак молочной железы, меланому, цилиндрому, карциному сквамозных клеток (кожи, а также головы и шеи), карциному полости рта, эндометриальную карциному, ретинобластому, астроцитому и глиобластому. Согласно ряду предпочтительных вариантов осуществления рак представляет собой множественную миелому. Согласно ряду вариантов осуществления клетки, взятые у пациента (например полученные из биоптата рака пациента или выделенные из крови или другой жидкости тела пациентов, в которую они могут выделяться опухолью) можно тестировать на активацию NF-κВ и/или повышенный уровень активности Gadd45^, чтобы определить подходит ли рак для лечения способами, соединениями и композициями, соответствующими изобретению.
Другие заболевания и нарушения, подходящие для лечения или предупреждения, включают аутоиммунное заболевание (например, рассеянный склероз, волчанка, диабет типа I), аллергическое заболевание (например, астму), хроническое воспалительное заболевание (например, воспалительную болезнь кишки, ревматоидный артрит, псориаз, язвенный колит), генетическое заболевание (например, недержание пигмента, ангидротическая эктодермальная дисплазия с иммунодефицитом и цилиндроматозом), ишемическое и сосудистое заболевание (например, атеросклероз, стенокардия, инсульт, инфаркт миокарда) и дегенеративное заболевание (например, болезнь Альцгеймера и Паркинсона), заболевания печени, такие как фиброз печени и цирроз печени.
Широкий круг заболеваний и нарушений зависит от активности NF-κВ. В самом деле в настоящее время считают, что патогенез практически каждого известного человеческого заболевания или нарушения зависит от воспаления и, следовательно, включает NF-κВ. Она действует как групповой переключатель воспалительного ответа, координации экспрессии последовательности из более 200 генов, кодирующих цитокины, рецепторы, факторы транскрипции, хемокины, провоспалительные ферменты и другие факторы, включая направленные на выживаемость факторы, которые инициируют и замедляют воспаление. Соединения, соответствующие изобретению, ингибируют отдельную направленную на выживаемость активность NF-κВ при воспалении. Вследствие этого заболевания и нарушения, поддающиеся лечению данными соединениями, включают, кроме распространенных хронических воспалительных заболеваний (таких как воспалительная болезнь кишки, ревматоидный артрит и псориаз), другие заболевания и нарушения, которые в существенной степени зависят от воспалительного компонента. Примеры данных заболеваний и нарушений, которые лечатся противовоспалительными препаратами или препаратами, ингибирующими NF-κВ, или таких, которые, как предполагают, подходят для лечения ингибиторами NF-κВ и которые можно было бы лечить соединением, соответствующим изобретению, включают:
1. Дыхательные пути: обструктивные заболевания дыхательных путей, включая: астму, в том числе бронхиальную, аллергическую, эндогенную, экзогенную, вызываемую физической нагрузкой, вызываемую лекарственными препаратами (включая вызываемую аспирином и NSAID (нестероидными противовоспалительными лекарственными препаратами)) и вызываемую пылью астму, как интермиттирующую, так и постоянную, и всех степеней тяжести, а также другие случаи гиперчувствительности дыхательных путей; хроническое обструктивное легочное заболевание (COPD); бронхит, в том числе инфекционный и эозинофильный бронхит; эмфизему; бронхоэктазию; муковисцидоз; саркоидоз; "легкие фермера" (экзогенный аллергический альвеолит) и подобные заболевания; пневмонит, связанный с гиперчувствительностью; фиброз легкого, в том числе криптогенный фиброзирующий альвеолит, идиопатическое интерстициальное воспаление легких, фиброз как осложнение противоопухолевой терапии и хроническую инфекцию, включая туберкулез и аспергиллез и другие грибковые инфекции; осложнения при трансплантации легкого; васкулитные и тромботические нарушения легочной сосудистой сети и легочную гипертензию; противокашлевую активность, в том числе лечение хронического кашля, ассоциированного с воспалительными и секреторными состояниями дыхательных путей, и ятрогенный кашель; острый и хронический ринит, включая медикаментозный ринит и вазомоторный ринит; круглогодичный и сезонный аллергический ринит, в том числе нейрогенный ринит (сенная лихорадка); назальный полипоз; острую вирусную инфекцию, включая насморк, и инфекцию респираторно-синцитиального вируса, гриппа, коронавируса (в том числе SARS (тяжелый острый респираторный сидром) или аденовируса; либо эозинофильный эзофагит;
2. Кости и суставы: поражения ревматического или подагрического характера, ассоциированные с остеоартритом/остеоартрозом или включающие их, как первичные, так и вторичные, например, врожденная дисплазия тазобедренного сустава; шейный и поясничный спондилит и боль в нижнем отделе спины и шее; остеопороз; ревматоидный артрит и болезнь Стилла; серонегативные спондилоартропатии, включая анкилозирующий спондилит, псориатический артрит, реактивный артрит и недифференцированную спондартропатию; септический артрит и другие связанные с инфекцией артропатии и нарушения костей, такие как туберкулез, включая болезнь Пота и синдром Понсе; острый и хронический кристаллический синовит, включая подагру, заболевание, связанное с отложением пирофосфата кальция, и обусловленное апатитом кальция воспаление сухожилия, бурсы и синовии; болезнь Бехчета; первичный и вторичный синдром Шегрена, системный склероз и ограниченная склеродермия; системная красная волчанка, заболевание комбинированной соединительной ткани и заболевание недифференцированной соединительной ткани; воспалительные миопатии, в том числе дерматомиозит и полимиозит; ревматическая полимиалгия; ювенильный артрит, в том числе идиопатический воспалительный артрит, распространяющийся на любые суставы, и ассоциированные синдромы, и ревматическая лихорадка и ее системные осложнения; васкулиты, включая артериит гигантских клеток, артериит Такаяши, синдром Чин-Страусса, узелковый полиартериит, микроскопический полиартериит и васкулиты, ассоциированные с вирусной инфекцией, реакции гиперчувствительности, криоглобулины и парапротеины, боль в нижней области спины; семейная средиземноморская лихорадка, синдром Макла-Уэллса, семейная ирландская лихорадка, болезнь Кикучи; вызываемые лекарственными препаратами арталгии, тендиниты и миопатии;
3. боль и ремоделирование соединительных тканей при скелетно-мышечных нарушениях, обусловленных повреждением [например, спортивной травмой] или заболеванием: артритом (например, ревматоидным артритом, остеоартритом, подагрой или кристаллической артропатией), другим заболеванием суставов (таким как дегенерация межпозвоночного диска или дегенерация височно-нижнечелюстного сустава), заболевание, связанное с ремоделированием костей (такое как остеопороз, болезнь Педжета или остеонекроз), полихондрит (воспаление нескольких хрящей), склеродермия, нарушение комбинированной соединительной ткани, спондилоартропатии или заболевание периодонта (такое как периодонтит;
4. Кожа: псориаз, атопический дерматит, контактный дерматит или другие экзематозные дерматозы и реакции гиперчувствительности замедленного типа; фито- и фотодерматит; себорейный дерматит, герпетиформный дерматит, плоский лишай, склеротический атофический лишай, гангренозная пиодермия, кожный саркоид, дискоидная красная волчанка, пемфигус, пемфигоид, буллезный эпидермолиз, крапивница, ангионевротический отек (болезнь Квинке), васкулит, токсическая эритема, кожные эозинофилии, очаговая алопеция, облысение по мужскому типу, синдром Свита, синдром Вебера-Христиана, мультиформная эритема; целлюлит, как инфекционный, так и неинфекционный; панникулит; кожные лимфомы, рак кожи, отличный от меланомы и другие диспластические повреждения; вызываемые лекарственными препаратами нарушения, включая постоянную лекарственную сыпь;
5. Глаза: блефарит, конъюнктивит, включая крулогодичный и весенний аллергический конъюнктивит, ирит; передний и задний увеит; хориоидит; аутоиммунные, дегенеративные или воспалительные нарушения, поражающие сетчатку; офтальмит, в том числе симпатический офтальмит; саркоидоз; инфекции, включая вирусную, грибковую и бактериальную;
6. Желудочно-кишечный тракт: глоссит, гингивит, периодонтит, эзофагит, включая рефлюкс; эозинофильный гастроэнтерит, мастоцитоз, болезнь Крона, колит, в том числе язвенный колит, проктит, анальный зуд; заболевания брюшной полости, синдром раздраженной кишки и пищевые аллергии, которые могут иметь эффекты, отдаленные от кишки (например, мигрень, ринит или экзему);
7. Брюшная полость: гепатит, включая аутоиммунный, алкогольный и вирусный; фиброз и цирроз печени; холецистит, панкреатит, как острый, так и хронический;
8. Мочеполовая система: нефрит, включая интерстициальный и гломерулонефрит; нефротический синдром; цистит, включая острый и хронический (интерстициальный) цистит и язву Ханнера; острый и хронический уретрит, простатит, эпидидимит, оофорит и сальпингит; вульвовагинит; болезнь Пейрони; эректильная дисфункция (как у мужчин, так и у женщин);
9. Отторжение аллотрансплантата: острое и хроническое после, например, трансплантации почки, сердца, печени, легкого, костного мозга, кожи или роговицы, или после переливания крови, либо хроническая болезнь трансплантат против хозяина;
10. ЦНС (центральная нервная система): болезнь Альцгеймера и другие нарушения, связанные с деменцией, включая БКЯ (болезнь Крейтцфельда-Якоба) и нвБКЯ (болезнь Крейтцфельда-Якоба, новый вариант); амилоидоз; рассеянный склероз и другие демиелинизирующие синдромы; церебральный атеросклероз и васкулит; височный артериит; злокачественная миастения; острая и хроническая боль (острая, временная или хроническая, центральной или периферической природы), включая боли в животе, головную боль, мигрень, невралгию тройничного нерва, атипическую лицевую боль, боль в суставах и костях, боль, вызываемую раком и инвазией опухоли, невропатические болевые синдромы, включая диабетическую, пост-герпетическую и связанную с ВИЧ невропатию; нейросаркоидоз; осложнения на центральную и периферическую нервную систему злокачественных, инфекционных или аутоиммунных процессов;
11. Другие аутоиммунные и аллергические нарушения, в том числе тиреоидит Хашимото, болезнь Грейвса, болезнь Аддисона, сахарный диабет, идиопатическая тромбоцитопеническая пурпура, эозинофильный фасцит, синдром гипер-IgE, антифосфолипидный синдром;
12. Другие заболевания с воспалительным или иммунологическим компонентом, включая синдром приобретенного иммунодефицита (СПИД), лепру, синдром Сезари и паранеопластические синдромы;
13. Сердечно-сосудистая система: атеросклероз, поражающий коронарное и периферическое кровообращение; перикардит; миокардит, воспалительная и аутоиммунная кардиомиопатия, включая саркоид миокарда; ишемические реперфузионные повреждения; эндокардит, вальвулит и аортит, включая инфекционный (например, сифилитический); васкулит; нарушения проксимальных и периферических вен, включая флебит и тромбоз, в том числе тромбоз глубоких вен и осложнения варикоза вен;
14. Желудочно-кишечный тракт: Глютеновая болезнь, проктит, эозинофильный гастроэнтерит, мастоцитоз, болезнь Крона, язвенный колит, микроскопический колит, индетерминантный колит, нарушение раздраженной кишки, синдром раздраженной кишки, невоспалительная диаррея, пищевые аллергии, которые могут иметь эффекты, отдаленные от кишки, например, мигрень, ринит и экзему.
Согласно четвертому аспекту изобретения предусматривают соединение, соответствующее первому аспекту изобретения, или композицию, соответствующую второму аспекту изобретения, предназначенные для применения в качестве лекарственного средства.
Согласно пятому аспекту изобретения предусматривают применение соединения, соответствующего первому аспекту изобретения, или фармацевтической композиции, соответствующей второму аспекту изобретения, для получения лекарственного средства для лечения заболевания или нарушения. Данное заболевание или нарушение и пациент определены в ряде предпочтительных вариантов осуществления, как описано выше, относительно третьего аспекта изобретения.
Предпочтительно, когда продукты, способы, соответствующие изобретению, предназначены для лечения заболеваний и нарушений у человека.
Терапевтические и диагностические (тераностические) аспекты изобретения
Изобретение охватывает в различных вариантах осуществления способы лечения, применение соединений или композиций, соответствующих изобретению, для получения лекарственного средства и соединения или композиции, соответствующие изобретению, предназначенные для терапевтического применения.
Согласно ряду вариантов осуществления изобретение может также охватывать:
a) Способы лечения или предупреждения заболевания или нарушения, как определено выше, где заболевание или нарушение представляет собой рак у отдельного пациента и где предварительно получен образец предполагаемого рака пациента (например, путем взятия биоптата или жидкости тела, такой как кровь или мокрота) и проведен анализ, чтобы показать повышенные уровни экспрессии и/или активности Gadd45β и/или повышенные уровни экспрессии и/или активности NF-κВ.
b) Соединения или композиции, соответствующие изобретению, предназначенные для применения в качестве лекарственного средства для лечения тканей пациента, которые, как выявлено ранее, демонстрируют повышенные уровни экспрессии и/или активности Gadd45β и/или повышенные уровни экспрессии и/или активности NF-κВ.
с) Применение соединений или композиций, соответствующих изобретению, для изготовления лекарственного средства для лечения заболевания или нарушения, которое либо характеризуется патологической повышенной экспрессией и/или активностью Gadd45β, либо характеризуется патологической активацией пути NF-κВ и поддается лечению посредством индукции запрограммированной гибели клеток путем ингибирования активности Gadd45β, причем заболевание или нарушение представляет собой рак, причем предварительно выявлено, что клетки указанной формы рака демонстрируют повышенные уровни экспрессии и/или активности Gadd45β
Термин "повышенные уровни" может означать повышенные по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 50%, по меньшей мере на 75%, по меньшей мере на 100%, по меньшей мере на 200%, по меньшей мере на 300%, по меньшей мере на 400%, по меньшей мере на 500%, по меньшей мере на 600%, по меньшей мере на 700%, по меньшей мере на 800%, по меньшей мере на 900% или по меньшей мере на 1000% по сравнению с уровнями в контрольной здоровой ткани той же природы и необязательно получены у того же пациента или у здорового пациента. Уровни экспрессии и активности можно определить любым способом, известным в уровне техники, включая ОТ-ПЦР (полимеразная цепная реакция с использованием обратной транскриптазы), саузерн-блоттинг, нозерн-блоттинг, вестерн-блоттинг, ELISA, радиоиммунный анализ, анализ киназ и другие анализы связывания, функциональные анализы и/или анализы экспрессии.
Данный тераностический аспект изобретения в основном иллюстрируют результатами, представленными на Фиг.22 и Фиг.23. Результаты, приведенные в данном контексте, демонстрируют, что на панели из 29 раковых клеточных линий разных по природе тканей чувствительность раковых клеток к вызываемой Z-DTP гибели коррелирует с очень высокой степенью статистической достоверности с уровнями эндогенной экспрессии Gadd45β, как показано в анализах кОТ-ПЦР. В действительности график корреляции уровня экспрессии Gadd45β относительно процента выживаемости/пролиферации клеток после обработки Z-DTP2 показывает, что достоверность коэффициента корреляции в области значений 2 параметров является очень высокой (р<0,01) (Корреляция Пирсона). Удивительно, что единственной клеточной линией множественной миеломы (в целом из 9 тестированных клеточных линий множественной миеломы), которая устойчива к вызываемой Z-/mDTP гибели, а также к гибели клеток, вызываемой опосредованным sh-PHK сайленсингом Gadd45β (см. Фиг.33-43), является клеточная линия RPMI-8226, которая экспрессирует самые низкие, почти неопределяемые, уровни Gadd45β (см. Фиг.22). Данные результаты показывают, что если бы препарат на основе DTP нашел клиническое применение, было бы возможным предсказать популяции отвечающих на лечение пациентов с помощью простого и рентабельного анализа кОТ-ПЦР. Например, первичную клетку, полученную от пациентов с множественной миеломой, можно проанализировать на уровни экспрессии Gadd45β, и пациентов с высокими уровнями данной экспрессии можно считать теми, кто получит максимальный положительный эффект от лечения соединениями, соответствующими изобретению. Следовательно, важным аспектом изобретения является тераностический аспект, т.е. применение клинически эффективного анализа для предсказания ответа на лечение DTP у пациентов.
Данный тераностический аспект изобретения поддерживается, кроме того, очень высокой специфичностью в отношении мишени соединений, соответствующих изобретению, в клетках в отношении комплекса Gadd45β/MKK7. Данный факт показывает, что чем выше уровни экспрессии мишени (т.е. Gadd45β) в клетках, тем выше будет вероятность того, что данные клетки будут зависеть от Gadd45β в плане выживаемости, следовательно, чем выше будет вероятность того, что данные клетки будут чувствительны к вызываемой Z-/mDTP гибели. Указанную высокую специфичность Z-/mDTP демонстрируют обнаружением того, что: 1) На большой панели опухолевых клеточных линий существует высоко достоверная статистическая корреляция между уровнями экспрессии Gadd45β и чувствительностью раковых клеток к вызываемой Z-/mDTP гибели (см. Фиг.22 и Фиг.23); 2) опосредованная sh-PHK понижающая регуляция Gadd45β быстро индуцирует апоптоз в чувствительных к Z-/mDTP, но не в устойчивых к Z-/mDTP раковых клеточных линиях (см. Фиг.33-43), и кинетика индукции апоптоза посредством специфических в отношении Gadd45β sh-PHK в данных клеточных линиях аналогична процессам, наблюдаемым с Z-/mDTP (см. Фиг.9, 13 и 14); 3) опосредованная sh-PHK понижающая регуляция MKK7 делает чувствительные к Z-/mDTP раковые клеточные линии полностью устойчивыми к индуцируемой Z-/mDTP гибели (см. Фиг.48-50), 4) терапевтической мишенью изобретения является граница раздела между белками Gadd45β и MKKI (см. Фиг.51-54), которая, кроме того, обеспечивает возможность высокой избирательности в отношении мишени, ключевого преимущества решения авторов изобретения относительно имеющихся лекарственных препаратов. Данные результаты вместе с низкой токсичностью Z-/mDTP в отношении нормальных клеток и полученными данными о том, что выбивающее исключение Gadd45β хорошо переносится мышами (см. ссылку Papa, et al. (2008) J. Clin. Invest. 118:191-1923), указывают на то, что направленность на отдельные способствующие выживаемости функции NF-κВ в выживаемости клеток с помощью опосредованного Z-/mDTP ингибирования Gadd45β/lvlKK7 могут привести к получению лекарственного препарата, который более специфичен, менее токсичен и, следовательно, более эффективен, чем лекарственные препараты, направленные на путь NF-κВ и/или протеасомы.
Примеры
Следующие неограничивающие примеры иллюстрируют изобретение.
Пример 1. Синтез Z-DTP2
В качестве примера описан синтез Z-DTP2. Z-DTP2 содержит тетрапептидное ядро из D-тирозина, D-глутамина, D-аргинина, D-фенилаланина с бензилоксикарбонилом (который представляет собой группу Z), связанным с N-концом посредством амидной связи, и аминогруппой, связанной с С-концом посредством амидной связи.
Материалы и способы
Z-DTP2 получают вручную в соответствии с твердофазным способом Fmoc/tBu (см. статьи Fields G.B. and Noble R.L 1990 Int J Pept Protein Res; 35: 161-214; Bodansky, M. and Bodansky A. 1995, The practice of peptide synthesis (Методы синтеза пептидов), 2 изд., Springer Verlag, Berlin) и начинают с 500 гмоль (1000 мг) амидной полистироловой смолы Rink (Fmoc-RINK-AMresin, GL Biochem, Shangai, China, номер по каталогу 49001), имеющей замещение 0,50 ммоль/г. Смолу помещают в полипропиленовую емкость объемом 30 мл, снабженную тефлоновой перегородкой массой 20 г, полипропиленовой верхней крышкой и нижней полипропиленовой крышкой с наконечником Люэра. Смоле дают набухнуть с использованием 10,0 мл смеси 50:50 дихлорметан(ДХМ):диметилформамид(ДМФ) (оба реагента формы LabScan, Stillorgan, Ireland; ДХМ N° по каталогу H6508L; ДМФ N° по каталогу Н33Н11Х) в течение 20 минут. Затем после удаления растворителя под вакуумом отщепляют группу Fmoc обработкой 5,0 мл смеси ДМФ-пиперидин 8:2 (Пиперидин фирмы Pip, N° по каталогу 80641; Sigma-Aldrich, Milan, Italy) в течение 20 минут при комнатной температуре (КТ). Реагент удаляют под вакуумом и смолу трижды промывают 5,0 мл ДМФ. Затем 2,5 ммоль, 0,97 г, Fmoc-D-Phe-OH (GL Biochem, Shangai. N° по каталогу 35702) растворяют в 5,0 мл ДМФ (конечная концентрация 0,5 М) и активируют 5,0 мл 0,5 М раствора бензотриазол-1-ил-окси-трис-пирролидинофосфонийгексафторфосфата (PyBOP, Novabiochem, N° по каталогу 01-62-0016) в ДХМ и 0,90 мл диизопропил-этиламина (5,0 ммоль; DIEA, Sigma-Aldrich, N° по каталогу D-3887). Раствор активированной аминокислоты выливают на смолу и выдерживают при энергичном перемешивании в течение 30 минут. Раствор осушают под вакуумом и трижды промывают смолу 5,0 мл ДМФ. Группу Fmoc на α-NH2 удаляют, как описано ранее при использовании раствора 8:2 ДМФ-Pip (5,0 мл) в течение 20 минут и тщательно промывают 5,0 мл ДМФ (3 раза). Раствор Fmoc-D-Arg(Pbf)-OH (2,5 ммоль, 1,6 г в 5,0 мл ДМФ; GL Biochem, Shangai, N° по каталогу 36404) активируют, как описано, с помощью 2,5 ммоль PyBOP и 5,0 ммоль чистого DIEA. Раствор переносят на смолу и оставляют при перемешивании на 30 минут. После отщепления групп Fmoc с использованием 5,0 мл раствора 8:2 ДМФ-Pip и промывания ДМФ (3 раза, 5,0 мл), раствор Fmoc-(D)-Glu(tBu)-OH 0,50 М в ДМФ (2,5 ммоль, 1,1 г в 5,0 мл ДМФ; GL Biochem, Shangai, N° по каталогу 36605), предварительно активированный PyBOP и DIEA, как описано выше, добавляют к смоле и дают реакции протекать в течение 30 минут при комнатной температуре. После отмывании аминокислоты группу Fmoc удаляют, как описано выше (обработкой в течение 20 минут, 8:2 ДМФ:Pip, 5,0 мл) и смолу трижды промывают 5,0 мл ДМФ. 2,5 ммоль Fmoc-(D)-Tyr(tBu)-OH (1,2 г, GL Biochem, Shangai, N° по каталогу 36906), растворенного в 5,0 мл ДМФ, предварительно активируют с помощью PyBOP и DIEA, как описано выше, переносят на смолу и оставляют при перемешивании на 45 минут. Раствор аминокислоты удаляют осушением под вакуумом, затем смолу 5 раз промывают 5,0 мл ДМФ. 5 ммоль Z-OSu (бензилоксикарбонил-N-гидрокси-сукцинимид, GL Biochem, Shangai, N° по каталогу 10502) растворяют в 10 мл ДМФ и добавляют к смоле. Добавляют 2,4 мл DIEA и выдерживают реакцию при перемешивании в течение ночи. После высушивания раствора смолу обильно промывают ДМФ, ДХМ, метиловым спиртом (МеОН, LabScan, N° по каталогу С2517) и этиловым эфиром (Et2O, LabScan, N° по каталогу А3509Е) и сушат под вакуумом и взвешивают. Масса составляет 1,1 г. Для отщепления пептида смолу обрабатывают 10,0 мл смеси, состоящей из смеси ТФА-H2O-TIS 90:5:5 (об/об/об) (ТФА, трифторуксусная кислота, Sigma-Aldrich, Italy N° по каталогу 91700; TIS, три-изопропилсилан, Sigma-Aldrich, N° по каталогу 23378-1), в течений 3 часов при КГ. Смолу удаляют фильтрацией, затем добавляют 20 мл холодного Et2O к раствору трифторуксусной кислоты, что приводит к образованию осадка белого цвета. После удаления растворителей центрифугированием осадок промывают 10,0 мл холодного Et2O, растворяют в 10,0 мл H2O/CH3CN 50:50 (об/об) и лиофилизируют. Пептид характеризуют ЖХ-МС (жидкостная хроматография-масс-спектрометрия) с использованием колонки С18 с узким каналом 50×2 мм ID ONYX (Phenomenex, Torrance, CA, USA), уравновешенной при 600 мкл/мин 5% CH3CN, 0,05% ТФА. Анализ проводят с использованием градиента CH3CN, 0,05% ТФА от 5% до 70% в течение 3 минут. Пептид очищают полупрепаративной ВЭЖХ с обращенной фазой при использовании колонки С18 10×1 см ONYX (Phenomenex, Torrance, CA, USA), уравновешенной при скорости 20 мл/мин с инъекцией 20 мг в каждом проведении. Для элюции пептида используют градиент от 5% до 65% в течение 8 минут. Очищенные фракции объединяют и характеризуют ЖХ-МС. Установленная ММ (молекулярная масса) пептида А составляет 746,8 аем (атомных единиц массы) (теоретически 746,83 аем) и продукт имеет чистоту больше 95% (ВЭЖХ). Выход приблизительно 60% достигают после очистки всего сырца.
Пример 2. Дозозависимое ингибирование взаимодействия Gadd45β и MKK7 при отборе соединений общей формулы (I)
Для оценки ингибирующих свойств пептидов проводят анализы на основе ELISA. В данных анализах слитым белком глутатион S-трансферазы (GST) и активированной митогеном киназы протеинкиназы 7 (MKK7) покрывают стенки 96-луночного планшета, тогда как биотинилированный hGadd45β используют в растворе. hGadd45β биотинилируют с использованием набора EZ Link NHS-LC-биотин (Pierce, Rockford, IL) в соответствии с работой Tornatore et al. (см. статью Tornatore L, et al. (2008). J Mol Bio1;378:97-111). Материалы и способы
Наконец, связь между Gadd45β и MKK7 исследуют в анализах ELISA, как также описано в статье Tornatore et al. (см. статью Tornatore L, et al. (2008). J Mol Bio1; 378:97-111). Слитой с GST киназой полной длины покрывают в течение 16 часов при 4°С в концентрации 42 нМ в буфере А (25 мМ Трис рН 7,5, 150 мМ NaCl, 1 мМ DTT и 1 мМ ЭДТА) лунки 96-луночного планшета для микротитрования. Некоторые лунки наполняют только буфером и используют как пустые. После инкубирования в течение 16 час при 4°С растворы удаляют и лунки наполняют 350 мкл 1% (масс/об) раствора NFDM (обезжиренного сухого молока) в PBS (забуференном фосфатом солевом растворе). Планшет инкубируют в течение 1 часа при 37°С в темноте. После промывания буфером T-PBS (PBS с добавлением 0,004% (об/об) детергента Tween) лунки наполняют 100 мкг биотинилированного Gadd45(в концентрациях, лежащих в интервале от 8,4 нМ до 168 нМ. В каждой точке результат получают в трех повторностях. После инкубирования в течение 1 часа в темноте при 37°С растворы удаляют и лунки снова промывают T-PBS. Затем 100 мкл разведения 1:10000 стрептавидина, конъюгированного с пероксидазой хрена, растворенного в буфере, добавляют в каждую лунку и инкубируют планшет в течение 1 часа при 37°С в темноте. После удаления раствора фермента и промывания добавляют 100 мкл хромогенного субстрата о-фенилендиамина (0,4 мг/мл в 50 мМ буфера на основе фосфата-цитрата натрия, содержащего 0,4 мг/мл мочевины в пероксиде водорода) и дают появиться окрашиванию, выдерживая в темноте в течение 5 минут. Реакцию останавливают добавлением 50 мкл 2,5 М H2SO4. Поглощение при длине волны 490 нм измеряют во всех лунках и усредняют значения после вычитания соответствующих значений в пустых лунках. Затем связанный белок определяют, как описано выше. Молярная концентрация биотинилированного Gadd45β, при которой определяют полумаксимальный сигнал ELISA, соответствует константе диссоциации (KD) (см. статью Friguet В, Chaffotte AF, Djavadi-Ohaniance L, Goldberg ME. J Immunol Methods. 1985 Mar 18;77(2):305-19). Конкурентные анализы связывания проводят с покрытием GST-MKK7 в концентрации 42 нМ, как описано, концентрацией биотинилированного Gadd45β 21 нМ (условия пренасыщения соотношение 1:0,5 моль/моль) и, в первом тесте, с использованием конкурентных реагентов в концентрации 21 н. Связывание биотинилированного Gadd45β с GST-MKK7 анализируют в присутствии возрастающих количеств конкурирующего пептида (концентрации лежат в интервале от 0,01 нМ до 100 нМ) и значения, полученные с конкурирующим реагентом, выражают как процент от связывания, определенного в отсутствие конкурирующего реагента. Данные по активности, выраженные как процент от способности ингибирования при концентрации 21 нМ в условиях анализа, представляют в следующей Таблице I для отобранной группы соединений, соответствующих изобретению. Согласно условиям, принятым в таблице, термины "L-Xaa" и "D-Xaa" относятся к L- и D-формам аминокислоты Хаа. Данные по IC50 отобранных соединений (т.е. доза соединения, необходимая для достижения 50% снижения уровня связывания Gadd45β с MKK7) приводят на Фиг.5.
Пример 3. Выделение образцов тетрапептидов
Материалы и способы
Скрининг с использованием ELISA используют для идентификации образцов D-тетрапептидов, из которых можно выделить предпочтительные соединения, соответствующие изобретению. В упрощенной комбинаторной библиотеке пептидов (см. статью Marasco et al. 2008, Curr. Protein Pept. Sci. 9:447-67) проводят скрининг антагонистов взаимодействия Gadd45β/MKK7. Данная библиотека содержит в целом 124=20736 различных тетрапептидов, образованных комбинациями следующих остатков аминокислот Gin (Q), Ser (S), Arg (R), Ala (A), Tyr (Y), Pro (P), Met (M), Cys (C), Phe (F), Leu (L), His (H), Asp (D) и многократно деконволюционируют в четырех стадиях конкурентных анализов ELISA при использовании на каждой стадии покрытого MKK7 (42 нМ), растворимого биотин-hGadd45β (21 нМ) и каждой из 12 суббиблиотек (42 нМ). Результаты каждого скрининга показаны в Таблице I выше (где используют стандартные однобуквенные коды остатков аминокислот и X2, X3 и Х4 представляют собой смеси вышеуказанных 12 остатков) (см. также Фиг.3). Полученный в результате наиболее активный пептид описанный в Таблице I (т.е. Fmoc-(βAla)2-YDHF-NH2, также обозначенный как Fmoc-LTPI) затем подвергают нескольким циклам оптимизации и удалению метки Fmoc-(βAla)2 с получением Ac-LTPI и AC-LTP2 (см. Таблицу II). Затем данные тетрапептиды повторно синтезируют с использованием D-изомеров тех же аминокислоты, получая в итоге образцы тетрапептидов 1 и 2 (DTP1 и DTP2), которые нарушают взаимодействие Gadd45β/MKK7 с IC50 0,22 нМ и 0,19 нМ, соответственно (см. фиг.5).
Последовательность DTP1: Ацетил-(D-Tyr)-(D-Asp)-(D-His)-(D-Phe)-NH2 [SEQ ID NO.:38]
Последовательность DTP2: Ацетил-(D-Tyr)-(D-Glu)-(D-Arg)-(D-Phe)-NH2 [SEQ ID NO.:37]
Кроме того, следующие последовательности выбирают в качестве отрицательных контролей (NC):
Последовательность NC1: Ацетил-(D-Tyr)-(D-Asp)-(D-His)-(D-Gln)-NH2 [SEQ ID NO.:81]
Последовательность NC2: Ацетил-(L-Tyr)-(L-Asp)-(L-His)-(L-Ala)-NH2 [SEQ ID NO.:82]
Последовательность NC3: Ацетил-(L-Tyr)-(L-Glu)-(L-Lys)-(L-Trp)-NH2, [SEQ ID NO.:83]
Последовательность NC4: Ацетил-(L-Tyr)-(L-Asp)-(L-Lys)-(L-Trp)-NH2 [SEQ ID NO.:84]
На Фиг.3-5 показывают конкурентные анализы связывания ELISA. Процент ингибирования ацетилированных пептидов и/или модифицированных пептидов (т.е. пептидов, конъюгированных либо с ацетилом, либо с другими группами) показывают, соответственно, в Таблицах II и III (в которых используют стандартные однобуквенные коды остатков аминокислот).
Пример 4. Анализы иммунопреципитации
Материалы и способы
Человеческую эмбриональную почку (HEK-293) культивируют в среде Игла, модифицированной Дульбекко (DMEM) с добавлением 10% сыворотки телячьих эмбрионов (FCS), 100 Ед./мл пенициллина, 100 мг/мл стрептомицина и 1% глутамина. Клетки HEK-293 (2,2×106) высевают на чашки для культивирования тканей площадью 10 см2 и на следующий день трансфицируют плазмидами pcDNA-FLAG-MKK7 и pcDNA-HA-Gadd45β с использованием стандартного способа преципитации Са3(PO4)3 (см. статью Papa, S et al., (2004) Nat. Cell Biol. 6, 146-153). Через сорок восемь часов после трансфекции клетки однократно промывают PBS, затем ресуспендируют и инкубируют в течение 30 мин при 4°С в лизирующем буфере (20 мМ HEPES, 350 мМ NaCl, 20% глицерин, 1 мМ MgCl2, 0,2 мМ EGTA, 1 мМ DTT (дитиотрейтол), 1 мМ Na3VO4 и 50 мМ NaF) с добавлением ингибиторов протеазы (1 мМ фенилметилсульфонилфторида, 10 мкМ химостатина, 2 мкг/мл апротинина и 2 мг/мл лейпептина) при редком осторожном встряхивании. Лизированные клетки собирают и центрифугируют при 45000×g в течение 40 мин. Полученные в результате осветленные лизаты клеток используют для дальнейшего анализа.
Образцы тетрапептидов DTP1 и DTP2, выделенные в Примере 3, вместе с тетрапептидами отрицательного контроля (NCI, NC2, NC3 и NC4) совместно инкубируют с Gadcl45β/MKK7, чтобы продемонстрировать, что активные D-тетрапептиды, но не тетрапептиды отрицательного контроля, нарушают взаимодействие Gadd45β/MKK7. Иммунопреципитацию проводят с использованием в основном тех же условий, что описаны в статье Papa, S et al., (2004) Nat. Cell Biol. 6, 146-153 и ссылках в ней, и антитела к FLAG, которое вызывает преципитацию меченного FLAG MKK7. Результаты вестерн-блоттинга проявляют с использованием антител к MKK7 или антител к НА (связывающихся к НА-Gadd45β), как указано на Фиг.7 (нижняя и верхняя панели, соответственно).
Результаты
Результаты представлены на Фиг.7. Из вестерн-блотов, представленных на Фиг.7, можно видеть, что существует сильное взаимодействие между Gadd45β и MKK7, когда проводят иммунопреципитацию с лизатами клеток HEK-293, временно экспрессирующих HA-Gadd45β и FLAG-MKK7, и антителами к FLAG (специфически связывающимися с меченным FLAG MKK7). Данный результат получают, когда проводят иммунопреципитацию либо в отсутствие тетрапептидов, либо в присутствии D-тетрапептидов отрицательного контроля (NC) NCI, NC2, NC2 или NC4. Однако, когда соиммунопреципитацию проводят в присутствии 1 или 5 нМ DTP1 или DTP2, преципитированный комплекс, не содержит или содержит очень мало Gadd45β, указывая на то, что взаимодействие MKK7 и Gadd45β нарушают активные соединения DTP, приводя таким образом к уменьшению количества Gadd45β в соиммунопрециптатах. Данные результаты подтверждают и расширяют результат, наблюдаемый в конкурентных анализах ELISA, показанный на Фиг.3-6.
Пример 5. Стабильность DTP в человеческой сыворотке
Материалы и способы
На Фиг.6, представляющей связывание Gadd45β/MKK7, показано, что конкурентные анализы ELISA проводят с целью определения стабильности конъюгированных с Z D-тетрапептидов в человеческой сыворотке. Для данной цели активности наиболее активных антагонистов Gadd45β/MKK7, выбранных при скрининге комбинаторных библиотек, описанном в Примере 3 (т.е. Z-LTP1, Z-LTP2, Z-DTP1 и Z-DTP2), а также одного L-тетрапептида отрицательного контроля (т.е. Z-LNC) и соответствующего D-энантиомера (т.е. Z-DNC) сравнивают до и после 48-часового предварительного инкубирования с человеческой сывороткой при 37°С в конкурентных анализах ELISA. Анализ ELISA проводят, как описано на Фиг.5. Вкратце, 100 мкл 42 нМ рекомбинантным GST-MKK7 в буфере для ELISA (25 мМ Трис рН 7,5, 150 мМ NaCl, 1 мМ DTT, 1 мМ ЭДТА) покрывают лунки 96-луночных планшетов путем инкубирования в течение ночи при 4°С. После блокирования с 2% NFDM в течение 1 час при 37°С планшеты промывают TPBS и затем в лунки добавляют 21 нМ рекомбинантного биотинилированного человеческого (h) Gadd45β вместе с повышающимися концентрациями тетрапептидов, которые либо подвергают, либо не подвергают предварительному инкубированию с человеческой сывороткой, как указано. В плане дальнейшего обсуждения условий, используемых для конкурентного анализа ELISA, читателя адресуют к статье Tornatore et al 2008 JMB, 378: 97-111 и приведенным в ней ссылкам.
Результаты
На Фиг.5 показывают, что активности DTP1 и DTP2 сравнимы с активностями соответствующих им L-энантиомеров (т.е. LTP1 и LTP2, соответственно) при ингибировании формирования комплекса Gadd45β/MMK7, как показывают при сравнении IC50 DTP и LTP в конкурентных анализах ELISA. На Фиг.6 показано, что не происходит никакой потери активности после 48-часового инкубирования Z-DTP1 или Z-DTP2 с человеческой сывороткой при 37°С. Фактически результаты показывают, что после данного предварительного инкубирования Z-DTP, но не Z-LTP полностью сохраняют свою способность нарушать взаимодействие Gadd45β-MICK7 в конкурентных анализах ELISA. При сравнении IC50 тетрапептидов после предварительного инкубирования с сывороткой и без предварительного инкубирования с сывороткой можно видеть, что Z-DTP1 и Z-DTP2 полностью стабильны после предварительного инкубирования с сывороткой (IC50=0,19 нМ для Z-DTP1 и IC50=0,18 нМ для Z-DTP2), тогда как Z-LTP1 и Z-LTP2 нестабильны, поскольку последние тетрапептиды показывают существенную потерю активности после предварительного инкубирования с сывороткой (см. их IC50>10 мкМ). При всех тестированных концентрациях ингибирующие активности D-тетрапептидов, которые предварительного инкубируют с сывороткой не отличаются в данных анализах от активностей D-тетрапептидов, которые не подвергают данному предварительному инкубированию (см. Фиг.5 и Фиг.6).
Сравнение дозозависимых типов, показанных на Фиг.5 и Фиг.6, указывает на то, что Z-DTP1 и Z-DTP2 стабильны в человеческой сыворотке при 37°С и, таким образом, подходят для системного применения, тогда как Z-LTP1 и Z-LTP2 не подходят. Можно также видеть, что у тетрапептидов отрицательного контроля (например, Z-DNC и Z-LNC) отсутствует какая-либо активность в вышеупомянутых конкурентных анализах ELISA независимо от того, были они предварительно инкубированы с человеческой сывороткой или нет (см. Фиг.5 и Фиг.6). Результаты, приведенные на Фиг.5 и Фиг.6, также показывают, что N-концевое введение бензилоксикарбониловой (Z) группы (вместо ацетильной группы) не подрывает способность как DTP1, так и DTP2 или как LTP1, так и LTP2 ингибировать формирование комплекса Gadd45β/MKK7, хотя введение группы Z заметно повышает поглощение DTP клетками (результаты не показаны), следовательно, заметно повышает клеточную активность DTP в анализах уничтожения опухолей (см. Фиг.9 и Фиг.10).
Пример 6. Определение IC50 Z-DTP на панели клеточных линий множественной миеломы
Материалы и способы
Для дальнейшего исследования эффектов обработки D-тетрапептидом на выживаемость/пролиферацию клеточных линий множественной миеломы клетки 8 клеточных линий множественной миеломы (из 9 тестированных клеточных линий множественной миеломы), которые чувствительны к вызываемой Z-DTP гибели (т.е.клетки U266, KMS-11, NC1-H929, ARH-77, JJN-3, KMS-12, KMS-18, KMS-27; см. также Фиг.12-14, 22 и 23) обрабатывают повышающимися концентрациями (лежащими в интервале от 0,01 до 10 мкМ) Z-DTP1 или Z-DTP2 в течение 24, 72 или 144 час, как показывают в Таблице IV. Культуры клеток множественной миеломы и обработки Z-DTP ведут, как описано в Примере 8 (см. ниже). Эффекты Z-DTP на выживаемость/пролиферацию клеток множественной миеломы оценивают с помощью анализов включения [3H]тимидина, проводимых таким же образом, как описано в Примере 8. Количество пролиферации клеток, измеренное для каждой из используемых концентраций Z-DTP и для необработанных культур (т.е. культур, инкубируемых в среде без добавок), выражают как количество импульсов в минуту (с.р.m.), что прямо коррелирует со степенью пролиферации клеток. Все эксперименты проводят в трех повторностях. Затем определяют средние концентрации Z-DTP1 и Z-DTP2, а также их производных (например, mDTP3), которые приводят в результате к 50% ингибированию клеточной пролиферации (т.е. IC50) относительно пролиферации клеток, зарегистрированной для необработанных культур. IC50 Z-DTP1 и Z-DTP2, рассчитанные для 8 тестированных чувствительных клеточных линий множественной миеломы в показанные точки времени (т.е. день 1, 3 и 6), приводят в Таблице IV. IC50 данных двух соединений, а также данные величины 31 дополнительного соединения (включая результаты для производных Z-DTP2, таких как mDTP3), рассчитанные в клетках множественной миеломы KMS-11 и/или KMS-12 в день 1, 3 и 6, приводят в Таблице V.
Как можно видеть из Таблицы IV, Z-DTP1 и Z-DTP2 заметно снижают поглощение [3H]-TdR во всех тестированных клеточных линиях множественной миеломы дозозависимым образом (за исключением клеточной линии RPMI-8226, которая демонстрирует очень низкие уровни Gadd45β, дальнейшее обсуждение ниже, см, Фиг.22 и 23) (см. также Фиг.12-14, 22 и 23). Аналогичные результаты получают в данных клеточных линиях множественной миеломы, когда 1050 Z-DTP1 и ZDTP2 рассчитывают, используя анализы исключения трипанового синего (измерение выживаемости клеток) (результаты не показаны).
Результаты
Как показано в Таблице IV, все тестированные клеточные линии множественной миеломы проявляют высокую чувствительность к вызываемому Z-DTP ингибированию выживаемости/пролиферации клеток (см. также Фиг.12-14, 22 и 23). Однако, как также можно видеть из Таблицы IV, данные клеточные линии проявляют вариабельную чувствительность KZ-DTP1 и Z-DTP2. Фактически некоторые клеточные линии уже высокочувствительны к вызываемому Z-DTP ингибированию выживаемости/пролиферации клеток после 24-часовой обработки данными соединениями (например, см. IC50=1,3 мкМ для клеток KMS-12 и IC50=2,88 мкМ для клеток KMS-11 при обработке Z-DTP2 в точке 24 час), и все из них высокочувствительны как к Z-DTP1, так и к Z-DTP2 после обработки в течение 144 час при IC50, лежащем в интервале от 10,1 нМ до 4,9 мкМ для Z-DTP1 и от 10 нМ до 4,5 мкМ для Z-DTP2 в данной точке времени (см. Таблицу IV). Известно, что производное Z-DTP2, mDTP3 (соединение 17), тестируют на клеточных линиях KMS-11 и KMS-12 и показывают повышенную клеточную активность в данных клеточных линиях по сравнению с Z-DTP1 и Z-DTP2 при IC50 16 нМ и 25 нМ, соответственно, в день 6 по сравнению с 1650 Z-DTP1 (т.е. 316 нМ и 10,1 нМ, соответственно) и Z-DTP2 (т.е. 66 нМ и 10 нМ, соответственно) (см. Таблицу V) (см. также Фиг.48-50). Следовательно, активные DTP, но не Z-DNC отрицательного контроля обладают сильной цитотоксической активностью в большинстве клеточных линий множественной миеломы. Более того, полученные авторами самые последние производные (например mDTP3) сохраняют высокую активность in vitro, но демонстрируют повышенную клеточную активность в клетках множественной миеломы при в существенной степени пониженной молекулярной массе (-500 относительно >700), следовательно, повышенной эффективности как лигандов (см. Таблицу V) (см. также Фиг.24 и Фиг.25).
Пример 7. Восстановление ингибируемой Gadd45β каталитической активности MKK7 посредством тетрапептидов
Материалы и способы
На фиг.8 показано проведение временной трансфекции pcDNA-FIAG- MKK7 клеток HEK-293 при использовании способа преципитации Са3(PO4)2 в основном, как описано в статье Papa, S et al., (2004) Nat. Cell Biol. 6, 146-153 и приведенных в ней ссылках. Через 36 час после трансфекции клетки обрабатывают 100 нг/мл форбол 12-миристат 13-ацетата (РМА) и 1 мкМ иономицина в течение 30 мин при 37°С. Клеточные экстракты получают, как описано в примере 4 и используют для иммунопреципитации с антителами к FLAG (связывающимися с FLAG-MKK7), как описано в статье Papa, S et al., (2004) Nat. Cell Biol. 6, 146-153 и приведенных в ней ссылках. Вкратце, 50 мкг клеточного лизата из РМА-иономицина (обработанные или необработанные P/D клетки HEK-293, временно экспрессирующие FLAG-MKK7 инкубируют с 10 мкл аффинного геля М2 к FLAG (SIGMA) в течение 4 час при 4°С при перемешивании. Затем иммунопреципитаты промывают 3 раза в лизирующем буфере и еще дважды в буфере для киназы (10 мМ HEPES, 5 мМ MgCl2, 1 мМ MnCl2, 12,5 мМ β-глицерофосфата, 2 мМ DTT, 4 мМ NaF и 0,1 мМ Na3VO4). Наконец, каталитическую активность MKK7 измеряют в анализе киназ путем инкубирования продуктов иммунопреципитации FLAG-MKK7 при 30°С for 20 мМ с 20 мкл буфера для киназы, содержащего 2 мкМ рекомбинантного GST-JNK1 и 5 мкКи [γ-32P]АТФ) (реакция киназы), как описано в статье Papa, S et al., (2004) Nat. Cell Biol. 6, 146153 и приведенных в ней ссылках.
В некоторых реакциях для тестирования способности антагонистов D-тетрапептида нарушать взаимодействие Gadd45β-MKK7 и, таким образом, высвобождать каталитическую активность MKK7 от вызываемого Gadd45β ингибирования, продукты иммунопреципитации FLAG-MKK7 1) сначала предварительно инкубируют в течение 10 мин при 30°С либо с 1 нМ, либо с 5 нМ DTP1, DTP2 или D-тетрапетидами отрицательного контроля (NC) NC1, NC2, NC3 и NC4, и 2) затем инкубируют в течение следующих 10 мин при 30°С с добавлением или без добавления 5 мкМ слитого с GST белка или рекомбинантного человеческого (h)Gadd45β (GSThGadd45β; очищенный из бактериальных лизатов, как описано в статье Papa, S., (2007) J. Biol. Chem. 282, 19029-19041), перед тем, как использовать их для вышеописанной реакции киназы, как показано на Фиг.8.
Во всех случаях реакции киназы останавливают добавлением буфера для образцов Лэммли. Затем белки разделяют с помощью 10% SDS-PAGE (электрофорез в полиактиламидном геле с использованием додецилсульфата натрия) и активность киназы MKK7 показывают с помощью авторадиографии. В плане дальнейшего обсуждения условий анализа киназы MKK7 читателя адресуют к статьям Papa, S et al., (2007) J. Biol. Chem. 282, 19029-19041 и Papa, S et al., (2004) Nat. Cell Biol. 6, 146-153 и приведенным в них ссылках.
Результаты
Результаты показаны на Фиг.8, где интенсивность полосы соответствует стелен и измеряемой активности киназы MKK, поскольку данная интенсивность пропорциональна количеству [γ-32Р]АТФ), включенной посредством MKK7 в ее субстрат, GST-JNK1. Как можно видеть из Фиг.5-7, инкубирование с DTP1 или DTP2 эффективно и специфически нарушает взаимодействие Gadd45β с MKK7 и, вследствие этого, как можно видеть на Фиг.8, полностью восстанавливает каталитическую активность MKK7, тогда как инкубирование с тетрапептидами отрицательного контроля (NC) NC1, NC2, NC3 или NC4 не дает данного эффекта (см. Фиг.8, верхние панели). На Фиг.8 можно также видеть, что ни контрольные тетрапептиды NC1, NC2, NC3 и NC4, ни активные тетрапептиды DTP1 и DTP2, не вызывают никакого ингибирования каталитической активности MKK7 при инкубировании с MKK7 в отсутствие рекомбинантного GST-hGadd45β (см. Фиг.8, нижние панели). Данные результаты согласуются с представленными на Фиг.5-7, где показано, что только DTP1 и DTP2, но не NC1, NC2, NC3 или NC4 способны нарушать взаимодействие MKK7-Gadd45β либо в ELISA, либо в анализах соиммунопреципитации.
Пример 8. Специфическое уничтожение опухолевых клеточных линий, характеризующихся конститутивной активностью NF-κВ и/или высокими уровнями экспрессии Gadd45β, тетрапептидами
Материалы и способы
В данном примере исследуют использование контрольных тетрапептидов (т.е. Z-DNC, Z-LNC и Ac-DNC) и биоактивных in vitro иллюстративных тетрапептидов (т.е. Z-DTP1, Z-DTP2, Z-LTP2 и AC-DTP2) для уничтожения большой панели человеческих и мышиных опухолевых клеточных линий тканей различной природы. Тестированные опухолевые клеточные линии включают: клеточные линии множественной миеломы U266, KMS-11, NC1-H929, ARH-77, JJN-3, KMS-12, KMS-18, KMS-27, RPMI8226; клеточные линии диффузной крупноклеточной В-клеточной лимфомы LY-3 и SUDHL6, клеточные линии лимфомы Беркетта BJAB, ST486, RAJI, RAMOS, Namalwa и HS-SULTAN, клеточную линию промоноцитарного лейкоза U937, клеточные линии Т-клеточного лейкоза и лимфомы JURKAT, HUT-78, MT-2, MT-4, MOLT4, MT2-HTLV-I и СЕМ, клеточные линии рака молочной железы MCF7, MD-MDA-231 и MD-MDA-486, клеточные линии пре-В-клеточной лимфомы NALM-6 (человеческая) и 70Z/3 (мышиная), клеточную линию хронического миелогенного лейкоза К652, клеточные линии В-клеточной лимфомы KARPAS (человеческая) и А20 (мышиная), клеточную линию человеческой эмбриональной почки HEK-293Т. Опухолевые клеточные линии культивируют, как описано ранее (см. статью Zazzeroni et al. 2003, Blood 102: 3270-3279), в RPMI-1640 (клеточные линии множественной миеломы, диффузной крупноклеточной В-клеточной лимфомы, лимфомы Беркетта, промоноцитарного лейкоза, Т-клеточного лейкоза и лимфомы, пре-В-клеточной лимфомы, хронического миелогенного лейкоза и В-клеточной лимфомы) или среде DMEM (клеточные линии рака молочной железы и эмбриональной почки) с добавлением 10% сыворотки телячьих эмбрионов (FBS), 1% глутамина, 100 Ед/мл пенициллина и 100 мкг/мл стрептомицина в увлаженной атмосфере 5% CO2 при 37°С.
Для анализов ингибирования пролиферации (см. Фиг.9-15) и анализов гибели клеток (см. Фиг.16-20) клетки высевают в лунки 96-луночных планшетов в концентрации 1,0×104 клеток/мл (анализы пролиферации) или в лунки 24-луночных планшетов в концентрации 4×105 клеток/мл (анализы апоптоза) и культивируют до 6 дней. В этот период времени клетки культивируют в среде без добавок (необработанные культуры) или в среде с добавлением либо контрольных (например, Z-DNC), либо активных (например, Z-DTP2) тетрапептидов (обработанные культуры)для достижения конечной концентрации тетрапептидов в культурах 10 мкМ или 100 мкМ, как указано. Для анализов ингибирования пролиферации, направленных на оценку эффектов тетрапептидов на выживаемость/пролиферацию опухолевых клеток, культуры оценивают ежедневно либо по исключению трипанового синего (дифференцирующему живые и мертвые клетки) и подсчету клеток (результаты не показаны), либо по данным анализа включения [3H]тимидина (см. Фиг.9-15), как указано. В данных последних анализах эффекты Z-DTP, Ac-dtp и Z-LTP на выживаемость/пролиферацию опухолевых клеточных линий исследуют измерением синтеза ДНК с использованием стандартных анализов поглощения меченного тритием тимидина ([3H]тимидина; [3H]-TdR). В показанных анализах клетки инкубируют в течение 24, 72, 120 или 144 час при 37°С в присутствии или в отсутствие контрольных или биоактивных тетрапептидов, как указано, затем дополнительно инкубируют в течение 18 час с [3H]-TdR (0,037 Мбк/лунку, что равно 0,5 мкКи/лунку). Затем клетки собирают на плоских стекловолоконных фильтрах, используя автоматизированное устройство для сбора клеток для 96-луночных планшетов, после чего добавляют сцинтилляционную жидкость и измеряют включение [3H]тимидина, измеряемое с помощью жидкой сцинтилляционной спектроскопии на β-счетчике. Результаты выражают как проценты импульсов в минуту (с.р.m.) (прямо коррелируют с уровнем пролиферации клеток), измеряемые в обработанных тетрапептидами культурах относительно с.р. т., измеряемых в соответствующих культурах, инкубируемых с среде без добавок (необработанные клетки). Все эксперименты проводят в трех повторностях. Как дополнительно обсуждают ниже, Z-DTP2, Z-LTP2 и Ac-DTP2 дают близкие результаты с Z-DTP1, Z-LTP1 и Ac-DTPI, соответственно в данных анализах выживаемости/пролиферации, хотя Z-DTP2 проявляет активность немного выше, чем Z-DTP1 (результаты не показаны; см. также Таблицу IV).
Апоптоз клеток в культуре измеряют в указанных точках времени с использованием окрашивания ядра йодидом пропидия (PI) и проточной цитометрии (FC), проводимых в основном так, как описано ранее (см. статью Riccardi and Nicoletti (2006) Nature Protocols 1, - 1458-1461), с целью идентифицировать клетки с содержанием ДНК в фазе суб-G1 (т.е. апоптотические клетки) (см. Фиг.16-20). Для данных анализов клетки (4×105 клеток/мл) культивируют в 24-луночных планшетах в течение 72 или 144 час, как указано на Фиг.16-20, затем дважды промывают в 1Х солевом растворе, забуференном фосфатом (PBS) и фиксируют 70% ледяным этанолом в течение 16 час. при -20°С, после чего их центрифугируют и затем ресуспендируют в 1×PBS, содержащем 100 мкг/мл РНКазы А. После данной стадии клетки инкубируют при комнатной температуре в течение 30 мин и центрифугируют, затем ресуспендируют в 50 мкг/мл PI и инкубируют в течение дополнительных 45 минут при 4°С в темноте. Наконец, проводят проточную цитометрию (FC) с использованием автоматизированной системы FACsCalibur и анализируют результаты с помощью программы FlowJo.
Для определения основы различной чувствительности опухолевых клеточных линий к вызываемой Z-DTP гибели измеряют уровни экспрессии Gadd45β на панели из 29 опухолевых клеточных линий или тканей различной природы при использовании количественной полимеразной цепной реакции с использованием обратной транскриптазы (кОТ-ПЦР) и соотносят данные уровни со степенью чувствительности данных клеточных линий к цитотоксической активности Z-DTPs. Для данных анализов, которые показаны на Фиг.22 и 23, клеточные линия рака молочной железы и HEK-293Т культивируют в колбах объемом 75 см2 (5×106 клеток/колбу) в полной среде DM ЕМ, тогда как все другие клеточные линии культивируют в лунках 6-луночных планшетов при густоте 5×105 клеток/лунку в полной среде RPMI-1640, как описано выше. Общую РНК экстрагируют тризолом и очищают, используя мининабор для PHK Pure Like RNA (Invitrogen). 1 мкг РНК добавляют в качестве матрицы в реакции обратной трансферазы (ОТ), проводимые при использовании набора для ПЦР PHK GeneAmp (Applied Biosystems). кОТ-ПЦР проводят с полученными в результате кДНК в трех повторностях при использовании смеси SYBR Green PCR Master Mix (Applied Biosystems), Gadd45β-специфических праймеров, перечисленных с Таблице VII и устройства для проведения ПЦР в режиме реального времени ABI 7900. Экспериментальные значения Ct нормализуют относительно β-актина и рассчитывают относительную экспрессию мРНК сравнительно эталонного образца (т.е. мРНК из клеток HEK-293Т). Чувствительность раковых клеток к уничтожению Z-DTP анализируют, как описано выше, проводя анализы включения [3H]тимидина после обработки клеток 10 мкМ Z-DTP2 в течение 144 час. На Фиг.22 и 23 показана также кривая корреляции экспрессии мРНК Gadd45β относительно процента выживаемости клеток после обработки Z-DTP2. Достоверность коэффициента корреляции в области значений 2 параметров рассчитывают по корреляции Пирсона, которая количественно выражает связь между двумя переменными, при использовании программы GraphPad.
Для того, чтобы определить, обусловлена ли вызываемая Z-DTP гибель раковых клеточных линий индукцией передачи цитотоксического сигнала JNK, мониторируют активацию JNK после обработки двух репрезентативных чувствительных клеточных линий множественной миеломы (т.е. клеточных линий KMS11 и NCI-H929) Z-DTP2 (см. Фиг.21). В данном случае используют анализы с помощью вестерн-блоттинга для оценки фосфорилирования JNK как показателя активации JNK. Клеточные линии множественной миеломы KMS11 и NCI-H929 культивируют в 6-луночных планшетах с густотой 5×105 клеток/лунку в полной среде RPMI-1640, как описано выше, и обрабатывают 10 мкМ Z-DTP2 или тетрапептидом отрицательного контроля Z-DNC в течение 3, 6, 12 или 24 час (см. Фиг.21). После обработки тетрапептидом получают клеточные лизаты в основном как описано в Примере 4, и вестерн-блоттинг проводят с использованием антитела к фосфо(P)-JNK. Способ, используемый в анализах с использованием вестерн-блоттинга, описан в ссылках De Smaele, et al. (2001) Nature 414:306-313; Papa, S et al., (2004) Nat. Cell Biol. 6, 146-153; Papa, et al. 2007 J. Biol. Chem. 282:19029-19041; Papa, et al. (2008) J. Clin. Invest. 118:1911923. Уровни β-актина определяют с использованием антитела к β-актину, и они служат контрольной нагрузкой (см. Фиг.21). Стимуляцию с помощью α-ФНО (2000 Ед/мл) клеток KMS11 и NCI-H929 проводят в течение 5, 10 или 30 мин и используют в качестве положительного контроля для активации JNK (см. Фиг.21). Данные анализы показывают, что активацию JNK вызывает только обработка Z-DTP2, но не обработка Z-DNC. Аналогичные эффекты Z-DTP2 показаны при активации MKK7 (результаты не приводят). Важно что, как видно по биологической активности Gadd45β (см. ссылки De Smaele, et al. (2001) Nature 414:306-313; Papa, S et al., (2004) Nat. Cell Biol. 6, 146-153; Papa, et al. 2007 J. Biol. Chem. 282:19029-19041; Papa, et al. (2008) J. Clin. Invest. 118:191-1923), эффекты Z-DTP2 в клетках множественной миеломы специфичны в отношении пути MKK7/JNK, поскольку данные соединения не проявляют никакого эффекта на активацию путей IKK/NF-κВ, ERK и р38 (результаты не приводят).
Результаты
На Фиг.12-14 показывают, что Z-защищенные производные DTP2 (Z-DTP2), но не ацетильные производные (Ac-DTP2) или L-изомеры Z-DTP2 (Z-LTP2) и не тетрапептиды отрицательного контроля Z-DNC, Ac-DNC и Z-LNC в существенной степени ингибируют пролиферацию трех репрезентативных клеточных линий множественной миеломы из 8 протестированных чувствительных клеточных линий множественной миеломы (т.е. U266, KMS-11 и NCI-H929), клеточной линии лимфомы Беркетта, BJAB и клеточной линии промоноцитарного лейкоза U937 (см, также Фиг.12-14, 22 и 23 и Таблицу IV; дополнительные линии множественной миеломы). Клетки культивируют с 10 мкМ либо Z-DTP2, либо Z-DNC (см. Фиг.9), Ac-DTP2 или Ac-DNC (см. Фиг.10) и Z-LTP2 или Z-LNC (см. Фиг.11), как указано. Измеряют включение [3H]tTdR (измеряя синтез ДНК) обработанных клеток и сравнивают с данными, полученными для клеток, культивируемых в среде без добавок. Результаты выражают как процент с.р.m., наблюдаемый в опухолевых клетках после обработки Z-DTP2, Ac-DTP2 или Z-LTP2 (закрашенные колонки) или Z-DNC, Ac-DNC или Z-LNC (незакрашенные колонки) относительно с.р.m., измеряемому в клетках, культивируемых в среде без добавок (необработанные клетки). Заметное ингибирование пролиферации клеток наблюдают в клеточных линиях множественной миеломы и других опухолевых клеточных линиях, обработанных Z-DTP2, но не в клеточных линиях, обработанных Z-DNC. На Фиг.10 показано, что отсутствие активности уничтожения опухолевых клеток у Ac-DTP2 в клеточных линиях множественной миеломы коррелирует с низкой клеточной проницаемостью данного соединения, как устанавливают в анализах с использованием CaCO2 (результаты не приводят). Выживаемость клеточных линий множественной миеломы после обработки другими, менее эффективными производными DTP (также созданными для улучшения клеточного поглощения DTP), включая несущие метил (Me), ацетил (Ас), миристил (Myr), 3-метокси,4-гидроксибензоил, бензоил, 6CI-бензилоксикарбонил (6CI-Z) и/или флуоренилметилоксикарбонил (Fmoc) не показана. Хотя активность и клеточное поглощение Z-LTP in vitro сравнимы сданными показателями Z-DTP (см. Фиг.5; результаты также не приводят), Z-LTP2 демонстрирует низкую активность в клетках множественной миеломы (см. Фиг.9) вследствие низкой стабильности в биологических жидкостях (см. Фиг.6). Аналогичное ингибирование пролиферации клеток наблюдают в опухолевых клеточных линиях, обработанных Z-DTP1, но не в обработанных Ac-DTP1 или Z-LTP1 (результаты не приводят), несмотря на это (как также видно с производными DTP2) данные два последних соединения демонстрируют сравнимую с Z-DTP1 активность in vitro (см. Фиг.5 и 6). В совокупности данные результаты показывают высокую цитотоксическую активность Z-DTP (см. Фиг.9 и результаты не приводят) по сравнению с отсутствием активности Ac-DTP (см. Фиг.10 и результаты не приводят) и низкой активностью Z-LTP (см. Фиг.11 и результаты не приводят).
На Фиг.12-14, показано, что исследуют эффекты обработки D-тетрапептидом на пролиферацию увеличенной панели клеточных линий множественной миеломы (т.е. U266, KMS-11, JJN-3, NCI-H929, ARH-77, KMS-27, KMS-18, KMS-12 и RPMI-8226). Другие тестированные клеточные линии включают клеточные линии диффузной крупноклеточной В-клеточной лимфомы LY-3 и SUDHL6, клеточные линии лимфомы Беркетта BJAB, ST486 и RAJI и клеточную линию промоноцитарного лейкоза U937. Клетки обрабатывают 10 мкМ либо Z-DTP2, Z-DTP1, либо Z-DNC, как показано, в указанные точки времени (т.е. 24, 72 или 144 час). Измеряют включение [3H]тимидина обработанных клеток, как на Фиг.9-11, и сравнивают с данными для клеток, культивируемых в среде без добавок. Результаты выражают как процент с.р.m., наблюдаемый с опухолевыми клетками, обработанными Z-DTP2 или Z-DTP1 (закрашенные колонки) или Z-DNC (незакрашенные колонки) относительно с.р.m., измеряемым в необработанных клетках. На Фиг.12 показывают, что Z-DTP2, но не Z-DNC в существенной степени ингибирует выживаемость/пролиферацию 8 из 9 протестированных клеточных линий множественной миеломы (т.е. U266, KMS-11, JJN-3 и NCI-H929, ARH-77, KMS-27, KMS-18, KMS-12), клеточной линии лимфомы Беркетта BJAB, клеточной линии диффузной крупноклеточной В-клеточной лимфомы (DLBCL) LY-3 и клеточной линии промоноцитарного лейкоза U937 (см. также Фиг.22 и 23). Отмечают, что Z-DTP2 показывает цитотоксическую активность только в клеточной линии DLBCL подтипа, подобного активированным В-клеткам (АВС) (т.е. LY3), которая зависит от NF-κВ в плане выживаемости и не показывает в подтипе, подобном В-клеткам зародышевого центра (GCB) (т.е. SUDHL6), который не характеризуется конститутивной активацией NF-κВ (см. статью Ngo VN, et al. Nature 441(7089):106-10; см. также Фиг.22 и 23). Она показывает также сильную цитотоксическую активность Z-DTP в подавляющем большинстве множества протестированных клеточных линий, все из которых зависят от NF-κВ в плане выживаемости. Как показано на Фигурах 13 и 14, ингибирующие эффекты Z-DPT1 и Z-DTP2 на пролиферацию опухолевых клеток повышаются с течением времени, максимальное ингибирование пролиферации наблюдают после обработки тетрапептидом в течение 144 час, хотя данные эффекты уже очевидны после обработки в течение 24 час. Данные результаты согласуются в результатами, полученными при подсчете клеток в анализах исключения трипанового синего (результаты не приводят) и в анализах окрашивания ядер PI на содержание ДНК (см. Фиг.16-20; результаты также не приводят). В совокупности данные и другие результаты показывают, что цитотоксическая активность Z-DTP избирательна в отношении опухолевых клеток, проявляющих конститутивную активность NF-κВ (см. также Фиг.22 и 23; результаты также не приводят).
Специфичность цитотоксической активности Z-DTP далее подтверждают анализами пролиферации с использованием [3H]тимидина, показанными на Фиг.15. На данной Фиг. показывают отсутствие вызываемой Z-DTP2 цитотоксичности на панели из 22 устойчивых опухолевых клеточных линий после обработки в течение 144 часов, даже когда данное соединение используют в очень высоких концентрациях, т.е 100 мкМ. Анализы пролиферации с использованием [3H]тимидина, показанные на Фиг.15, проводят, как описано на Фиг.12-14.
Как можно видеть на Фиг.15, Z-DTP2 не проявляет цитотоксичность в клеточных линиях Т-клеточного лейкоза и клеточных линиях лимфомы JURKAT, HUT-78, MT-2, MT-4, MOLT4, MT2-HTLV-I и СЕМ, клеточных линиях лимфомы Беркетта BJAB, ST486, RAH, RAMOS, Namalwa и HS-SULTAN, клеточных линиях рака молочной железы MCF7, MD-MDA-231 и MD-MDA-486, клеточных линиях пре-В-клеточной лимфомы NALM-6 и 70Z/3, клеточных линиях В-клеточной лимфомы KARPAS и А20, клеточной линии хронического миелогенного лейкоза К652, клеточной линии человеческой эмбриональной почки HEK-293Т и клеточной линии множественной миеломы RPMI-8226 (см. также Фиг.22 и 23). На Фиг.15 также показаны чувствительные клеточные линии BJAB (лимфомы Беркетта), KMS-11 и KMS-12 (множественной миеломы). Отмечают, что существует сильная корреляция в данных клеточных линиях между чувствительностью к вызываемой Z-DTP2 гибелью и уровнями эндогенной экспрессии Gadd45β (см. Фиг.22 и 23). Заметим, что клеточная линия RPM 1-8226, которая является единственной из протестированных клеточных линий множественной миеломы, устойчивой к гибели, вызываемой Z-/mDTP (см. Фиг.12 и 15), проявляет очень низкие уровни Gadd45β (см. Фиг.22 и 23), дополнительно подтверждая, что цитотоксическая активность DTP при раке зависит от уровней конститутивной экспрессии Gadd45β.
На Фиг.16-20 представлено, что помещенные панели показывают процент клеток, демонстрирующих окрашивание йодидом пропидия (PI), указывающее количество ДНК в фазе суб-G1 (т.е. клеток, которые либо погибли, либо погибают от апоптоза) после обработки в течение указанных периодов времени (т.е. 72 или 144 час) либо средой для культивирования без добавок (необработанные), либо средой для культивирования, доставляющей конечную концентрацию 10 мкМ либо Z-DTP2 либо Z-NC1. Проценты апоптотических клеток представлены на гистограммах. Показано пять репрезентативных чувствительных клеточных линий множественной миеломы, NCI-H929, KMS-11, ARH-77, JJN-3 и U266. как можно видеть, вызываемая Z-DTP2 гибель клеток множественной миеломы обусловлено запуском апоптоза, и часть апоптотических клеток, наблюдаемая после воздействия на клетки данного соединения, возрастает со временем обработки.
На Фиг.21 показывают, что обработка Z-DTP2 вызывает сильную активацию JNK в клеточных линиях множественной миеломы. Две репрезентативные чувствительные клеточные линии множественной миеломы, KMS11 и NCI-H929, обрабатывают 10 мкМ Z-DTP2 или Z-DNC, как показано. Активацию JNK мониторируют в указанные точки времени вестерн-блоттингом с использованием специфического антитела к фосфо(Р)-ЛМК. Можно видеть, что фосфорилирование ЛМК (маркер активации JN К) повышается только после обработки Z-DTP2, но не после обработки Z-защищенным пептидов отрицательного контроля (Z-DNC). Фактически Z-DTP2 вызывает даже более сильную активацию JNK, чем стимуляция с помощью α-ФНО (2000 Ед/мл), используемого в качестве положительного контроля. Обнаружены аналогичные эффекты Z-DTP2 на активацию MKK7, и анализы киназы используют для мониторирования активностей JNK и MKK7 (результаты не приводят). Отмечают, что, как видно по биологической активности Gadd45((см. ссылки De Smaele, et al. (2001) Nature 414:306-313; Papa, S et al. (2004) Nat. Cell Biol. 6, 146-153; Papa, et al. (2007) J. Biol. Chem. 282:1902919041; Papa, et al. (2008) J. Clin. Invest. 118:191-1923), эффекты Z-DTP2, а также Z-DTP1 и mDTP3 (результаты не приводят) в клетках множественной миеломы специфичны в отношении пути MKK7/JNK, поскольку с данными соединениями не наблюдают никакие эффекты на пути IKK/NFKB, ERK и р38 (результаты не приводят). Важно, что обработка Z-DTP не приводит к активации JNK в клеточной линии множественной миеломы RPMI-8226, которая устойчива к вызываемой Z-DTP гибели (см. Фиг.12 и 15). Данные и другие результаты (см. также Фиг.48-50) поддерживают точку зрения о том, что Z-DTP индуцируют апоптоз в опухолевых клеточных линиях путем активации передачи цитотоксического сигнала JNK. Важно, что результаты, представленные на Фиг.22 и 23 показывают, что чувствительность раковых клеточных линий к вызываемой Z-DTP гибели коррелирует с очень высокой степенью статистической достоверности в уровнями эндогенной экспрессии Gadd45β (p<0,01). Экспрессию мРНК Gadd45β оценивают на панели из 29 раковых клеточных линий при использовании анализов кОТ-ПЦР (см. Фиг.22, верхняя панель, красные колонки). Значения нормализуют относительно β-актина. Выживаемость/пролиферацию в тех же раковых клеточных линиях определяют, проводя включение [3H]тимидина после обработки 10 мкМ Z-DTP2 в течение 144 часов. Данные результаты показывают на нижней панели Фиг.22 (колонки черного цвета). Приведенные в данном контексте значения представляют собой процент с.р.m., измеренный в клетках, обработанных Z-DTP2, относительно с.р.m., измеренному в необработанных клетках. На Фиг.23 показана кривая корреляции экспрессии Gadd45β относительно процента выживаемости/пролиферации, наблюдаемого после обработки Z-DTP2, в том же самом эксперименте, показанном на Фиг.22. Как можно видеть, достоверность коэффициента корреляции в области значений 2 параметров является очень высокой (р<0,01) (Корреляция Пирсона, которая количественно оценивает связь между двумя переменными, рассчитанная с использованием программы GraphPad). Это является ключевым фактором в плане создания успешного терапевтического препарат для человека. Данные результаты демонстрируют высокую специфичность к мишени Z-DTP в клетках в отношении Gadd45β. Дополнительно поддерживает данное заключение то, что опосредованный shРНК сайленсинг Gadd45β индуцирует апоптоз в клетках множественной миеломы, тогда как опосредованный shРНК сайленсинг MKK7 делает данные клетки полностью устойчивыми к вызываемой Z-DTP гибели (см. Фиг.33-50). В совокупности данные результаты также показывают, что если бы лечение на основе DTP вошло в клиническую практику, было бы возможно предсказывать отвечающие популяции пациентов посредством простого и рентабельного анализа кОТ-ПЦР. Соответственно, из этого следует, что первичную клетку, полученную от пациентов с множественной миеломой, можно проанализировать на уровни экспрессии Gadd45β, и пациентов с высокими уровнями данной экспрессии можно считать теми, кто получит максимальный благоприятный эффект от лечения соединениями, соответствующими изобретению. Следовательно важным аспектом изобретения является тераностический аспект, т.е. применение клинически эффективного анализа для предсказания ответа на лечение DTP у пациентов.
Пример 9. IC50 vitro и в клетках панели производных Z-DTP
Разработан всесторонний план направленной оптимизации доставки безопасного и эффективного нового препарата для лечения рака и других заболеваний и нарушений при использовании современных достижений в качестве отправной точки. Z-DTP2 уже показывает высокую стабильность, высокую растворимость, активность в суб-нМ диапазоне in vitro и высокую активность в клетках множественной миеломы (первичных и клеточных линий) и других раковых клетках при высокой специфичности в отношении мишени и отсутствии токсичности в нормальных клетках (см. Фиг.5, 6, 12-15, 22, 23, 26-32; см. также Таблицу IV; результаты не приводят). Соединение также проявляет высокий исходный уровень DMPK и профили безопасности in vivo (однократная внутривенная болюсная доза у мышей) (см. Таблицы VIII и IX). Для получения производных DTP с усовершенствованными свойствами ADMET, но при сохранении высокой биоактивности используют рациональное молекулярное конструирование (см. статью Geeson МР. 2008 J Med Chem. 51:817-834). В данном подходе модифицируют размер (молекулярную массу), липофильность (LogP) и состояние ионизации (молекулярный заряд), т.е. ключевые связанные с объемом свойства молекул, которые влияют на свойства ADMET, при использовании созданной авторами модели фармакофора для сохранения структурных элементов, отвечающих за биоактивность in vitro (см. Фиг.24 и 25). Как описано в данном Примере, самые последние из полученных авторами производных (например, mDTP3 и mDTP4) сохраняют высокую активность in vitro, но показывают повышенную активность уничтожения в клетках множественной миеломы при в существенной степени пониженной молекулярной массе (-500 относительно >700), но при повышенной эффективности лиганда (см. Фиг.24 и 25). Используют также дополнительные средства повышения клеточной активности пептидов и значений РК, включая циклизацию, введение блокирующих групп для интернализации чувствительных амидов и/или замещение неамидными связями.
Материалы и способы
Сконструировано 33 соединения на основе образцов последовательностей тетрапептидов: Tyr-Glu-Arg-Phe и Tyr-Asp-His-Phe, полученных при скрининге библиотеки (см. Фиг.3-5). Все соединения за исключением соединений 2, 3, 4, 5, 6, 7, 10, 11, 12, 13, 14, 15 и 16 (см. Таблицу V) получают твердофазным способом в соответствии с классическими химическими способами с использованием Fmoc/tBu (как описано в ссылке Fields GB, Noble RL Solid phase peptide synthesis utilizing 9-fluorenylmethoxy-carbonil amino acids (Твердофазный синтез пептидов с использованием 9-флуоренилметоксикарбониламинокислот). - Int J Pept Protein Res 1990; 35:161-214). Только аминокислоты в D-конфигурации используют для сборки пептидов, показанных в Таблице V. N-концевое ацетилирование осуществляют обработкой 10% уксусным ангидридом в диметилформамиде (ДМФ), содержащем 5% DIEA (диизопропилэтиламин). При необходимости группу Z вводят обработкой на смоле Z-OSu (Бензилоксикарбонил-N-гидроксисукцинимидом) 0,5 М в ДМФ/5% DIEA. Соединения отщепляют от смолы с использованием ТФА (трифторуксусной кислоты) и обработки очистителями, затем очищают до однородности препаративной обращенно-фазовой (RP)-PLC (препаративной жидкостной хроматографией). Синтез соединений 2, 3, 4, 5, 6, 7, 10, 11, 12, 13, 14, 15 и 16 (см. таблицу V) осуществляют в других фирмах. Идентичность и чистоту соединение оценивают с использованием анализов ЖХ-МС и ЯМР-спектроскопии. Х1 представляет собой бензойную кислоту, X2 представляет собой бензиловую кислоту, Y1 представляет собой анилин, Y2 представляет собой бензиламин и Y3 представляет собой фенетиламин. Соединения растворяют в ДМСО (диметилсульфоксиде) при исходной концентрации 5 мМ и затем аликвоты серийно разводят в буфере для достижения концентраций, необходимых для проведения конкурентных анализов ELISA. Белки получают, как описано в статье Tornatore L, et al. (2008). J Mol Biol; 378:97-111.
Конкурентные анализы связывания ELISA проводят, как описано в ссылке Tornatore L, et al. (2008). J Mol Biol; 378:97-111 (см. только способы, описанные в Примерах 2 и 3), с использованием пептидов в возрастающих концентрациях, лежащих в интервале от 0,01 нМ до 10 нМ. Вкратце, GST-MKK7 иммобилизуют в концентрации 42 нМ на лунках 96-луночных планшетов для микротитрования. Конкурирующие соединения предварительно инкубируют в биотин-hGadd45β (21 нМ) и затем инкубируют с покрытием киназой. Для каждого соединения рассчитывают IC50 in vitro, как концентрацию, приводящую в результате к 50% снижению связывания Gadd45β с MKK7 относительно связывания, наблюдаемого в отсутствие конкурирующих соединений.
Исследуют эффекты каждого соединения на выживаемость/пролиферацию производных DTP производные на двух репрезентативных, чувствительных клеточных линиях множественной миеломы, KMS12 и KMS11. Анализы включения [3H]тимидина на клеточных линиях множественной миеломы KMS11 и KMS12 проводят, как описано в примерах 6 и 8 (см. Фиг.9-14 и Таблицу IV). Вкратце, клетки культивируют в 96-луночных планшетах и обрабатывают раздельно указанным соединением в лунках 96-луночных планшетов при использовании возрастающих концентраций соединения, лежащих в интервале от 0,1 нМ до 10 мкМ. Культивирование клеток и обработку соединениями проводят, как описано на Фиг.9-14 и в Таблице IV. Поглощение [3Н]тимидина, измеряющее выживаемость/пролиферацию клеток, определяют после обработки соединениями в течение 1, 3 или 6 дней, как указано. В данные точки времени IC50 каждого соединения рассчитывают, как описано в Примере 6 путем определения концентрации, дающей в результате 50% ингибирования выживаемости/пролиферации клеток относительно выживаемости/пролиферации, наблюдаемой у необработанных клеток.
Значения IC50 in vitro (ELISA) и в клетках (клетки KMS-11 и KMS-12) для 33 соединений, описанных в данном Примере, приводят в Таблице V.
Результаты
В таблице V показаны значения IC50 in vitro и в клетках панели тетра- и трипептидов, созданных на основе консенсусных последовательностей Tyr-Glu-Arg-Phe и TyrAsp-His-Phe, полученных при скрининге библиотеки и химическими способами оптимизации образцов.
Данные соединения подвергают скринингу in vitro с использованием конкурентного анализа ELISA, где замещение связывания биотин-Cadd45β с покрытием GST-MKK7 определяют тестированием активностей соединений в различных концентрациях. In vivo 1050 для группы выбранных соединений определяют с использованием анализов включения [3H]тимидина в клеточных линиях миеломы KMS-11 и KMS-12 с целью оценки активности соединений уничтожать раковые клетки. 1050 указанных соединений в клетках определяют после обработки в течение 1, 3 или 6 дней. Z означает бензилоксикарбониловую группу. Как можно видеть из Таблицы V, наиболее активными соединениями в клетках являются соединение 9, обозначенное как Z-DTP2 (IC50=10 нМ в клетках KMS-11; IS50=66 нМ в KMS-12) и соединение 17, обозначенное как mDTP3 (IC50=25 нМ в клетках KMS-11, IC50=16 нМ в KMS-12).
Все 33 соединения, описанные в данном Примере, подвергают скринингу in vitro в конкурентных анализах ELISA на их способность нарушать взаимодействие Gadd45β/MKK7 (см. таблицу V). Большинство из данных соединений за исключением соединений 18, 20, 21, 22, 32 и 33 также подвергают скринингу в клетках с использованием анализов включения [3Н]тимидина в клеточных линиях множественной миеломы KMS-11 и/или KMS-12, и их IC50 в данных клетках определяют в день 1, 3 и 6. Как можно видеть из таблицы V, соединения 1, 2, 3, 4, 5, 6, 7, 9, 15 и 17 тестируют на обеих клеточных линиях. Соединения 15 и 19 тестируют только на клетках KMS-12. Соединения 10, 11, 12, 13, 14, 23, 24, 25, 26, 27, 28, 29, 30 и 31 тестируют только на клеточной линии KMS-11. Соединения 18, 20, 21, 22, 32 и 33 не тестируют на клетках вследствие их относительно низкой активности in vitro.
В Таблице V показывают, что IC50 in vitro тестированных соединений лежат в интервале от 100 рМ (см. соединение 7, X2-Asp-His-Y3; соединение 15, Х2-Glu-Arg-Y3 и соединение 19, Z-Tyr-Arg-Phe) до >10 нМ (см. соединения 24, 27, 30, 31, 32 и 33). Как можно видеть, активности соединений, которые получены in vitro, часто отражают их активности в клетках, хотя некоторые из соединений, активных in vitro, обладают относительно низкой активностью в клетках, предположительно вследствие низкого уровня их клеточного поглощения, например, сравним соединение 15 (показывающее in vitro IC50=100 пМ и IC50=263 нМ в клетках KMS-11) с соединением 9 (Z-DTP2; показывающим in vitro IC50=190 пМ и IC50=10 нМ в клетках KMS11). Результаты, полученные на клетках, показывают также, что присутствие группы Z на N-конце и/или основные боковые цепи дают в результате повышенную активность в клетках, обусловленную повышенным уровнем клеточного поглощения. В качестве примеров важности основной боковой цепи сравним IC50 в клетках соединения 19 (Z-Tyr-Arg-Phe-NH2; IC50=81 нМ в клетках KMS-12 в день 3) с данной величиной для соединения 8 (Z-Tyr-Asp-Phe-NH2; IC50>10 мкМ в клетках KM S-11 в день 3) (т.е. замена Arg на Asp) или с данной величиной для соединения 16 (Z-Tyr-Glu-Phe-NH2; IC50=3,0 мкМ в клетках KMS11 в день 3) (т.е. замена Arg на Glu), кроме того, отметим сравнимые низкие IC50 in vitro данных трех соединений, все из которых лежат в интервале суб-нМ (см. Таблицу V). В качестве примеров важности группы Z сравним IC50 в клетках U266, KMS-11 и NCI-H929 для Z-DTP2(CM. Фиг.9) сданной величиной для Ac-DTP2 (см. Фиг.10); см. также близкие значения IC50 in vitro для данных двух соединений (см. Фиг.5 и 6; результаты не приводят). Данные также показывают, что соединения без ароматических циклов по обоим концам (например соединения 20 и 21) неактивны как in vitro, так и в клетках (см. таблицу V), указывая на то, что данные ароматические циклы абсолютно необходимы для проявления биоактивности. Интересно, что присутствие 2 тирозинов на N-конце также приводит в результате к потере активности (см. таблицу V).
Пример 10. IC50 in vitro панели дополнительных производных Z-DTP Материалы и способы
Панель из 18 дополнительных соединений разрабатывают на основе образца трипептидной последовательности Tyr-Arg-Phe (т.е. mDTP3) с целью исследования важности для биоактивности: 1) расстояния между двумя ароматическими циклами; 2) свойствами аминокислоты в центральном положении; 3) присутствия ацетильной группы на N-конце; 4) и присутствия заместителей ароматических циклов (см. Таблицу VI). Все соединения получают способом твердофазного синтеза в соответствии с классическими химическими способами с использованием Fmoc/tBu (как описано в ссылке Fields GB, Noble RL. Solid phase peptide synthesis utilizing 9-fluorenylmethoxy-carbonil amino acids (Твердофазный синтез пептидов с использованием 9-флуоренилметоксикарбониламинокислот). Int. J. Pept. Protein Res. 1990:35:161-214). N-концевое ацетилирование проводят путем обработки 10% уксусным ангидридом в диметилформамиде (ДМФ), содержащем 5% DIEA (диизопропилэтиламин). Соединения отщепляют от смолы с использованием ТФА (трифторуксусной кислоты) и обработки очистителями, затем очищают до однородности препаративной обращенно-фазовой(RP)-PLC. Идентичность и чистоту соединения оценивают с помощью анализов ЖХ-МС и ЯМР-спетроскопии. Соединения очищают с помощью ВЭЖХ с обращенной фазой, затем все растворяют в DMSO в исходной концентрации 5 мМ и хранят до использования. Затем аликвоты серийно разводят в буфере для достижения концентраций, требующихся для конкурентных анализов ELISA. Конкурентные анализы связывания ELISA проводят, как описано в ссылке Tornatore L, et al. (2008). J Mol Biol; 378:97-111 (см. также раздел Способы в Примерах 2 и 3), используя пептиды в возрастающих концентрациях, лежащих в интервале от 0,01 нМ до 10 нМ. Вкратце, GST-MKK7 иммобилизуют в концентрации 42 нМ на лунках 96-луночных планшетов для микротитрования. Конкурирующие соединения предварительно инкубируют в биотин-hCadd45β (21 нМ) и затем инкубируют с покрытием киназой. Для каждого соединения рассчитывают IC50 in vitro, как концентрацию, приводящую в результате к 50% снижению связывания Gadd45β с MKK7 относительно связывания, наблюдаемого в отсутствие конкурирующих соединений.
Результаты
В Таблице VI показывают значения IC50 панели из 18 трипептидов и дипептидов, созданных на основе mDTP3 (Ac-D-Tyr-D-Arg-D-Phe). Соединения создают для исследования влияния на биоактивность следующих параметров: 1) расстояния между двумя ароматическими циклами на N- и С-концах (см. соединения А1, А1бис, A3, А6, А7 и А8); 2) свойств аминокислоты в центральном положении (см. соединения В2, В13, В16, В16бис, O5 и O5 бис); 3) присутствие или отсутствие гидроксильной группы на остатках ароматического цикла в положениях 1 и 3 (см. соединения А9, O1, O3, O5, O5бис, O6, O7 и O8); присутствие ацетильной группы на N-конце (см. соединения А9 и O7; В16 и В16 бис; O1 и O8; O3 и O6; O5 и O5 бис).
18 дополнительных соединений тестируют на активность in vitro, используя конкурентные анализы ELISA и повышающиеся концентрации соединений, лежащие в интервале от 0,01 нМ до 100 нМ. Как можно видеть из таблицы VI, все протестированные дипептиды неактивны независимо от присутствия аминокислоты Phe или Tyr либо на N-конце, либо на С-конце (см. соединения А1, А1бис, А7 и А8). Введение спейсеров более длинных, чем α-аминокислота в центральном положении трипептидов также приводит к потере активности in vitro (см. соединения A3 и А6, несущие β-аланин и ε-капроновую кислоту в середине, соответственно). Это неверно для тетрапептидов, где положения Y2 и Y3 заняты Asp/Glu или His/Arg - сравним IC50 in vitro соединения 9 (т.е. ZDTP2) с данным значением соединения 16 (т.е. mDTP2) и значений соединения 1 (т.е. Z-DTP1) со значениями соединения 8 (т.е. Z-Tyr-Asp-Phe-NH2). Это обусловлено тем, что Z-DPT2 и Z-DTP1, которые содержат дополнительную аминокислоту между двумя активными ароматическими группами, сохраняют высокую активность in vitro (см. IC50 в Таблице V). Важно, как показано в Таблице VI, что удаление гидроксильной группы на N-концевом тирозине также приводит в результате к полной потере биоактивности in vitro (см. соединения А9, O1, O5, O5бис, O7 и O8) независимо от присутствия ацетильной группы. Существенно, что данное наблюдение указывает на важный вклад гидроксильной группы на взаимодействие активных соединений с белками-мишенями. В действительности данная группа, вероятно, участвует в формировании Н-связи или полярного взаимодействия. Напротив, присутствие гидроксильной группы на ароматическом цикле на С-конце не влияет на активность соединений (см. соединения А9, O1, O3, O5, O5бис, O6, O7 и O8). Аналогично замена аргинина другой основной аминокислотой, такой как гистидин или лизин, либо пролин не изменяет биоактивность in vitro (см. соединения В2, В13, В16, В16бис, 05 и 05бис), позволяя предположить минимальную роль боковой цепи данного остатка в способности соединений нарушать взаимодействие Gadd45β/MKK7.
Пример 11. Лентивирусные инфекции, устанавливающие важную роль Gadd45β в выживаемости клеток множественной миеломы
Материал и способы
Для определения роли Gadd45β и MKK7 в выживаемости клеточных линий множественной миеломы исследуют эффекты понижающей регуляции экспрессии Gadd45β или MKK7 в данных клетках (см. Фиг.33-47). В данном случае осуществляют инфекцию лентивирусами, экспрессирующими направленные на Gadd45β и MKK7 sh-PHK, которая приводит в результате к сайленсингу генов Gadd45β и MKK7, соответственно. Последовательности ДНК, кодирующие направленные маленькие шпилечные (sh)-PHK, перечислены в таблице VII. Направленные последовательности sh-PHK (т.е. sh-Gadd45β-1, sh-Gadd45β-2, sh-Gadd45β-3, sh-MKK7-1 и sh-MKK7-2) и неспецифические контрольные последовательности sh-NS-1 и sh-NS-2 интродуцируют между рестрикционными сайтами BamH1 и HpaI лентивирусного вектора, LentiLox3.7 (см. ссылку Yang et all 2006 PNAS 103, 10397-10402). Продукцию лентивирусного препарата с высоким титром в клетках HEK-293Т проводят с использованием в основном таких же условий, что описаны в ссылках Pham et all 2004 Cell 116, 529-542 и Yang et all 2006 PNAS 103, 10397-10402. Для интродукции последовательностей sh-PHK, направленных на Gadd45β и MKK7, и неспецифических контрольных последовательностей sh-PHK пять репрезентативных чувствительных к ZDTP клеточных линий множественной миеломы, ARH-77, NCI-H929, U266, KMS11 и KMS12 и устойчивую к Z-DTP клеточную линию множественной миеломы RPMI-8226 инфицируют лентивирусами LentiLox3.7, как представлено в опубликованных протоколах в основном, как описано в ссылке Yang et all 2006 PNAS 103, 10397-10402. Через 5 дней после инфекции клетки множественной миеломы eGFP подвергают сортингу с использованием клеточного сортера BD FACSAria™ II, затем оставляют в покое на 2 дня перед началом анализов выживаемости клеток и клеточной пролиферации. Выживаемость инфицированных клеток множественной миеломы мониторируют в течение 8 дней с помощью проточной цитометрии, измеряя экспрессию белка повышенной зеленой флуоресценции (eGFP) (метка инфицированных клеток), и подсчитывают клетки (см. Фиг.33-37). Уровень апоптоза (см. Фиг.38-43) и распределение клеточного цикла (см. Фиг.44-47) измеряют, проводя анализы окрашивания ядер PI, как описано в статье Riccardi С.and Nicoletti I 2006 Nature Protocols 1, 1458-1461 (см. также раздел Способы, приведенный в Примере 8).
Результаты
На Фиг.33-35 показано, что опосредованный sh-PHK сайленсинг экспрессии Gadd45β дает в результате быструю индукцию гибели клеток, приводящую к пониженному уровню пролиферации в репрезентативных чувствительных к Z-DTP клеточных линиях множественной миеломы ARH-77 (см. Фиг.33) и NCI-H929 (см. Фиг.34), но не в устойчивой к Z-DTP клеточной линии множественной миеломы RPMI-8226 (см. Фиг.35). В эксперименте, показанном на Фиг.33-35, клеточные линии множественной миеломы инфицируют лентивирусом, экспрессирующим либо специфические в отношении Gadd45β sh-PHK (sh-Gadd45β-1, sh-Gadd45β-2 или sh-Gadd45β-3), специфические в отношении MKK7 shPHK (sh-MKK7-1 или sh-MKK7-2), либо неспецифические sh-PHK (sh-NS-1 или sh-NS-2), и выживаемость инфицированных клеток мониторируют в течение 8 дней при использовании проточной цитометрии, показывающей клетки, экспрессирующие белок с повышенной зеленой флуоресценцией (eGFP), т.е. инфицированные клетки, и подсчитывают клетки. Показан процент выживаемости eGFP+ (т.е. инфицированных лентивирусом) клеток множественной миеломы в указанные точки времени, указанный относительно выживаемости eGFP+ клеток множественной миеломы в той же культуре в день 0. Клетки инфицируют лентивирусами pLentiLox.3.7, экспрессирующими указанные sh-PHK и eGFP, используя стандартные способы (как описано в статье Yang H et al., Proc Natl Acad Sci US A. 5 июля 2006; 103(27): 10397-402). Через 5 дней клетки eGFP+ подвергают сортингу с использованием клеточного сортера BD FACSAria™ II, затем оставляют в покое на 2 дня перед началом анализов выживаемости клеток. Данную точку времени (т.е. начало анализов выживаемости) отмечают на Фиг.33-35 как день 0. Результаты показывают, что ингибирование экспрессии Gadd45β, но не ингибирование экспрессии MKK7 быстро вызывает гибель клеток в клеточных линиях множественной миеломы которые чувствительны к вызываемой Z-DTP токсичности (т.е. в клеточных линиях ARH-77 и NCI-H929) (см. Фиг.33 и 34, соответственно), но не в клеточной линии множественной миеломы RPMI-8226 (см. Фиг.35), которая устойчива к данной токсичности. Данные результаты, кроме того, устанавливают специфичности направленности Z-DTP в отношении комплекса Gadd45β/MKK7 в клетках множественной миеломы (см. также Фиг.9-15, 22 и 23; гибель и анализы кОТ-ПЦР). В действительности в согласии сданным заключением также находится тот факт, что кинетика ингибирования пролиферации клеток множественной миеломы, наблюдаемая после сайленсинга Gadd45β, очень близка к наблюдаемой после обработки данных клеток Z-DTP (см. Фиг.9, 13 и 14). Результаты также демонстрируют важную роль, которую Gadd45β играет в выживаемости клеток множественной миеломы, таким образом дополнительно подтверждая правильность выбора Gadd45β в качестве терапевтической мишени при множественной миеломе.
На Фиг.36 и 37 показывают, что опосредованный sh-PHK сайленсинг Gadd45β, но не сайленсинг MKK7 обладает высокой активностью ингибирования выживаемости/пролиферации только в отношении клеточных линий множественной миеломы, которые чувствительны к вызываемой Z-DTP гибели (например, клеточные линии ARH-77 и NCI-H929, см. также Фиг.9-14, чувствительность к гибели, вызываемой Z-DTP). Прямо противоположно то, что на выживаемость устойчивой к Z-DTP клеточной линии множественной миеломы RPMI8226 совершенно не влияет опосредованное sh-PHK ингибирование Gadd45β. Пролиферацию/выживаемость клеток на Фиг.36 и 37 определяют при использовании анализов включения [3H]тимидина, проводимых, как описано в Примерах 6 и 8. На Фиг.36 показывают выживаемость трех репрезентативных клеточных линий множественной миеломы RPMI-8226, NCI-H929 и ARH-77, после сайленсинга Gadd45β или MKK7. На Фиг.37 показывают выживаемость/пролиферацию клеточной линии множественной миеломы ARH-77 после сайленсинга Gadd45β или MKK7 при использовании трех различных специфических в отношении Gadd45β shPHK (sh-Gadd45β-1, sh-Gadd45β-2 или sh-Gadd45β-3), двух различных специфических в отношении MKK7sh-PHK (sh-MKK7-1 или sh-MKK7-2) и двух различных неспецифических sh-PHK (sh-NS1 или sh-NS-2). Клеточные линии множественной миеломы инфицируют экспрессирующим указанную sh-PHK лентиврусом pLentiLox.3.7, затем клетки множественной миеломы eGFP+ (т.е. клетки, инфицированные лентивирусом) подвергают сортингу с использованием клеточного сортера BD FACSAria™ II, как на Фиг.33-35. Анализы включения [3H]тимидина, представленные на Фиг.36 и 37, проводят через 10 дней после сортинга, что соответствует дню 8 на Фиг.33-35. Показывают процент включения [^тимидина (т.е. с.р.m.), количественный показатель пролиферации клеток в день 8 (т.е. через 10 дней после сортинга клеток) относительно включения [3H]тимидина, происходящего в тех же клетках в день 0 (т.е. через 2 дня после сортинга). Данные результаты, кроме того, устанавливают специфичность к мишени Z-DTP в отношении комплекса Gadd45β/MKK7 в клетках множественной миеломы (см. также Фиг.9-15, 22, 23 и 33-35) и подтверждают важную роль, которую играет Gadd45β в выживаемости клеток множественной миеломы. В совокупности они дополнительно подтверждают правильность выбора Gadd45β в качестве терапевтической мишени при множественной миеломе.
На Фиг.38-43 показывают, что опосредованный sh-PHK сайленсинг Gadd45β эффективно индуцирует апоптоз в чувствительных к Z-DT клеточных линиях множественной миеломы, ARH-77 (см. Фиг.38-41) и NCI-H929 (см. Фиг.42), но не в устойчивой к Z-DTP клеточной линии множественной миеломы RPMI-8226 (см. Фиг.43) (см. также Фиг.33-37, опосредованный sh-PHK сайленсинг, Фиг.9-14, 22 и 23, чувствительность клеточной линии множественной миеломы к вызываемой Z-DTP гибели и уровни экспрессии Gadd45β). Индукцию апоптоза на Фиг.38-43 определяют при использовании анализов окрашивания ядер PI, проводимых, как описано в Примере 8. Данные результаты демонстрируют, что ингибирование выживаемости/пролиферации клеток множественной миеломы вызвано понижающей регуляцией экспрессии Gadd45β, наблюдаемой на Фиг.33-37, обусловлено индукцией запрограммированной гибели клеток, опосредованной путем апоптоза. Заметим, что никакой существенной индукции апоптоза в тех же клеточных линиях множественной миеломы не наблюдают в отсутствие лентивирусной инфекции (неинфицированные) или после экспрессии специфической в отношении MKK7 sh-PHK (sh-MKK7-1 и sh-MKK7-2) или неспецифической sh-PHK (sh-NS-1 и sh-NS-2) (см. Фиг.38-43). Клеточные линии множественной миеломы инфицируют экспрессирующими sh-PHK лентивирусами pLentiLox.3.7 и проводят сортинг клеток множественной миеломы eGFP+ (т.е. клеток, инфицированных лентивирусом) с использованием клеточного сортера BD FACSAria™ II, как на Фиг.33-35. Анализы ядерного окрашивания PI проводят через 10 дней после сортинга клеток, что соответствует дню 8 на Фиг.33-35. Проценты апоптотических клеток (т.е. клеток, демонстрирующих содержание ДНК в фазе суб-G1) показаны на гистограммах. Важно, что уровни апоптоза, индуцированные различными специфическими в отношении Gadd45β sh-PHK (т.е. sh-Gadd45β-1, sh-Gadd45β-2 и sh-Gadd45β-3) коррелируют с уровнями понижающей регуляции Gadd45β, индуцируемой каждой из данных специфических в отношении Gadd45β sh-PHK (см. Фиг.38-41; кроме того, результаты не приводят). Данные, представленные на Фиг.38-43, кроме того, устанавливают специфичность к мишени Z-DTPs в отношении комплекса Gadd45β/MKK7 в клетках множественной миеломы (см. также Фиг.9-15, анализы гибели с использованием Z-DTP; Фиг.22 и 23, статистически достоверная корреляция между уровнем экспрессии Gadd45β и чувствительностью раковых клеток к гибели, вызываемой Z-DTP, Фиг.33-37, индукция гибели клеточной линии множественной миеломы путем понижающей регуляции Gadd45β, но не MKK7) и подтверждают важную роль, которую Gadd45β играет в выживаемости клеток множественной миеломы. В совокупности они дополнительно подтверждают правильность выбора Gadd45β в качестве терапевтической мишени при множественной миеломе.
На Фиг.44-47 показывают, что опосредованный sh-RNA сайленсинг либо MKK7, либо Gadd45β не влияет на распределение клеточного цикла в клеточных линиях множественной миеломы. Показанные репрезентативные инфицированные лентивирусом клеточные линии множественной миеломы, т.е ARH-77 (см. Фиг.44 и 45), NCI-H929 (см. Фиг.46) и RPMI-8226 (см. Фиг.47), получены в том же самом эксперименте, что представлен на Фиг.38-43. Анализы клеточного цикла, представленные на Фиг.44-47, проводят при использовании анализов окрашивания ядер PI, которые осуществляют, как описано в Примере 8 (см. также Фиг.38-43). В отличие от результатов, показанных на Фиг.38-43 (на которой профили окрашивания PI представляют в логарифмическом масштабе, который выделяет апоптоз), окрашивание PI (т.е. FL2-A) на данных Фиг. представляют в линейном масштабе, который выделяет распространение клеточного цикла. Проценты клеток множественной миеломы в различных фазах клеточного цикла (т.е. G1, S и G2/M) показаны на гистограммах. Анализы клеточного цикла не могли быть проведены со специфическими в отношении Gadd45β sh-PHK в случае клеточных линий множественной миеломы ARH-77 (см. Фиг.44 и 45) и NCI-H929 (см. Фиг.46) вследствие индукции массивного апоптоза после экспрессии данных sh-PHK (см. Фиг.38-42). тем не менее, как можно видеть на Фиг.44 и 45, понижающая регуляция Gadd45β не влияет на распределение клеточного цикла в устойчивой к Z-DTP клеточной линии RPMI-8229.
Пример 12. Понижающая регуляция экспрессии MKK7 делает клеточные линии множественной миеломы с нормальной чувствительностью полностью устойчивыми к вызываемой Z-DTP гибели
Материалы и способы
Для оценки специфичности к мишени Z-/mDTP в отношении комплекса Gadd45β/MKK7 исследуют эффекты понижающей регуляции экспрессии MKK7 на чувствительность к вызываемой Z-/mDTP гибели чувствительных клеточных линий множественной миеломы (см. Фиг.48-50). В данном случае репрезентативную клеточную линию множественной миеломы ARH77 инфицируют лентивирусами, экспрессирующими специфическую в отношении MKK7 sh-PHK, что приводит в результате к сайленсингу гена MKK7, или контрольной неспецифической sh-РНК. Последовательности ДНК, кодирующие направленную маленькую шпилечную (sh)-PHK, перечислены в таблице VII. Направленные на MKK7 последовательности shRNA и неспецифические контрольные последовательности интродуцируют между сайтами рестрикции BamH1 and HpaI лентивирусного вектора LentiLox3.7, как описано в Примере 11 (см. ссылку Yang et al 2006 PNAS 103, 10397-10402). Продукцию лентивирусного препарата с высоким титром в клетках HEK-293Т осуществляют с использованием в основном тех же условий, что описаны в ссылке Yang et all 2006 PNAS 103, 10397-10402. Для интродукции направленных на MKK7 и неспецифических контрольных последовательностей sh-PHK репрезентативную чувствительную к Z-DTP клеточную линию множественной миеломы ARH-77 инфицируют лентивирусами LentiLox3.7, экспрессирующими либо специфическую в отношении MKK7 sh-PHK (sh-MKK7), либо неспецифическую sh-PHK (sh-NS), как представлено в опубликованных протоколах, в основном как описано в ссылке Yang et all 2006 PNAS 103, 10397-10402. Через 5 дней после инфекции клетки ARH-77 eGFP+ подвергают сортингу с использованием клеточного сортера BD FACSAria™ II. Затем через 10 дней после сортинга клеток инфицированные лентивирусом клетки множественной миеломы ARH-77 обрабатывают либо Z-DTP1, Z-DTP2, mDTP3, либо Z-NC в течение 72 час при 37°С или культивируют в тех же условиях в отсутствие обработки пептидом, как описано в Примере 8. Обработки Z-DTP1, Z-DTP2, mDTP3 и Z-NC осуществляют в следующих конечных концентрациях пептидов: 0,01 мкМ, 0,03 мкМ, 0,1 мкМ, 0,3 мкМ, 1 мкМ, 3 мкМ и 10 мкМ. После данных обработок выживаемость/пролиферацию ARH-77 определяют проведением анализов включения [3H]тимидина, как описано в Примерах 6 и 8. Результаты, полученные в данных экспериментах, выражают как проценты выживаемости/пролиферации (т.е. с.р.m.), наблюдаемой в инфицированных лентивирусом клетках множественной миеломой, обработанных Z-DTP1, Z-DTP2, mDTP3 или Z-NC, относительно выживаемости/пролиферации соответствующих инфицированных лентвирусом клеток в отсутствие обработки пептидом. Средние концентрации Z-DTP1, Z-DTP2, mDTPS и Z-DNC, приводящие в результате к 50% (IC50) ингибированию выживаемости/пролиферации клеток, определяют, проводя анализы инкорпорации [3H]тимидина и рассчитывают, как описано в Примере 6. Результаты, полученные в данных экспериментах, показывают на Фиг.48-50.
Результаты
На Фиг.48-50 показывают, что опосредованный sh-RNA сайленсинг MKK7 делает репрезентативную чувствительную к Z-AnDTP клеточную линию ARM-77 полностью устойчивой к вызываемой Z/mDTP гибели. Анализы включения [3H]тимидина, представленные на данных Фиг., показывают IC50 D-изомеров тетрапептидов отрицательного контроля (Z-DNC) (см. Фиг.48-50), Z-DTP1 (см. Фиг.48), Z-DTP2 (см. Фиг.49) и mDTP3 (см. Фиг.50) в клетках множественной миеломы ARH-77, экспрессирующих либо специфическую в отношении MKK7 (sh-MKK7), либо неспецифическую sh-PHK (sh-NS). Обработки клеток ARH-77 осуществляют с разными концентрациями данных пептидов и выживаемость/пролиферацию клеток анализируют с помощью анализов включения [3H]тимидина через 3 дня. Можно видеть, что экспрессирующие sh-NS клетки ARH-77 высокочувствительны к вызываемой Z-/mDTP гибели, как показано на основании значений IC50 1,4 мкМ (Z-DTP1; см. Фиг.48), 302 нМ (Z-DTP2; см. Фиг.49) и 303 нМ (mDTP3; см. Фиг.50), аналогичных тому, что наблюдают в неинфицированных исходных клетках ARH-77 (см. Таблицу IV). Однако, напротив, экспрессирующие sh-MKK7 клетки ARH-77 становятся устойчивыми к вызываемой Z-/mDTP гибели, как показано на основании значений IC50>10 мкМ Z-DTP1, Z-DTP2 и mDTP3, аналогично тому, что наблюдают в обработанных Z-DNC клетках ARH-77 (см. Фиг.48, 20В и 20С). IC50 рассчитывают, как описано в Примере 6, используя возрастающие концентрации Z-DNC (см. Фиг.48-50), ZDTP1 (см. Фиг.48), Z-DTP2 (см. Фиг.49) и mDTP3 (см. Фиг.50), лежащие в интервале от 0,01 до 10 мкМ. На графиках представлены проценты импульсов в минуту (с.р.m.), измерение выживаемости/пролиферации клеток, полученное для обработанных клеток относительно значений с.р.m., полученных с необработанными клетками. Аналогичные данные получают с дополнительными чувствительными к Z-/mDTP клеточными линиями множественной миеломы, включая клеточные линии U266, KMS-11 и KMS-12 (результаты не приводят). Данные результаты (т.е. потерю чувствительности к Z-/mDTP в чувствительной клетке множественной миеломы путем сайленсинга MKK7) в сочетании с результатами, показанными на Фиг.22 и 23 (т.е. сильная корреляция между экспрессией Gadd45β и чувствительностью раковых клеток к вызываемой Z-DTP гибели), заключительно демонстрирует очень высокую специфичность к мишени Z-/mDTPs в отношении комплекса Gadd45β/MKK7 в клетках множественной миеломы.
Пример 13. Z-DTPs сохраняет высокую и специфическую цитотоксическую активность в первичных клетках множественной миеломы, полученных от пациентов
Материалы и способы
Для подтверждения того, что Z-/mDTP сохраняют цитотоксическую активность в первичных клетках множественной миеломы, исследуют эффекты Z-DTP1 и Z-DTP2 на выживаемость клеток множественной миеломы, выделенных у пациентов с клиническим диагнозом множественная миелома. В данном случае клетки множественной миеломы очищают из аспиратов костного мозга (ВМ) пациентов с множественной миеломой путем отрицательного отбора с использованием конъюгированных с CD138 магнитных частиц, в основном, как описано в ссылке Hideshima Т. et all 2006, Blood 107: 4053-4062. Чистоту клеток множественной миеломы подтверждают проточной цитометрией с использованием как CD138, так и антител к CD45, также в основном в соответствии с процедурой, описанной в ссылке Hideshima Т. et all 2006, Blood 107: 4053-4062. Затем очищенные клетки ВМ CD138+ культивируют в концентрации 4×105 клеток/мл в лунках 96-луночных планшетов и обрабатывают либо 1 мкМ, либо 10 мкМ Z-DTP1, Z-DTP2 или Z-DNC в течение 48 час. Выживаемость клеток измеряют подсчетом клеток с использованием анализов исключения трипанового синего (см. Фиг.26-30).
Для определения in vitro терапевтического индекса Z-/mDTP проводят также анализы выживаемости и пролиферации с первичными нетрансформированными клетками как человеческой, так и мышиной природы после обработки либо 10 мкМ, либо 100 мкМ Z-DTP1, Z-DTP2 и Z-DNC. В данном случае клетки стромы костного мозга (BMSC) мононуклеарные клетки периферической крови (PBMNC) и мезенхимные стволовые клетки (MSC) очищают у здоровых людей после разделения по плотности в Ficoll-Hypaque в соответствии с протоколами, приведенными в ссылке Piva R. et all 2008 Blood 111: 2765-2775). Клетки BMSC, PBMNC и MSC затем обрабатывают в течение указанного времени и концентрациями пептидов, определенными на Фиг.31 и 32. Для дальнейшего установления специфичности цитотоксической активности Z-/mDTP в отношении раковых клеток используют также первичные В- и Т-лимфоциты, очищенные из селезенки и лимфатических узлов мышей, соответственно, в основном, как описано в ссылке Shirakawa et al 2010 Cell Mal immunology 1-12. Затем В- и Т-клетки активируют стимуляцией 1 нг/мл LPS в течение 16 час с последующей обработкой 100 мкМ Z-DTP1, Z-DTP2 или Z-DNC в течение 72 час, как показано на Фиг.32.
Результаты
На Фиг.26-30 показывают, что Z-DTP1 и Z-DTP2, но не Z-DNC, проявляют сильную цитотоксическую активность в первичных клетках множественной миеломы, выделенных у 5 репрезентативных пациентов. Каждая панель показывает результаты, полученные с клетками множественной миеломы, выделенными у разных пациентов, т.е. пациента 1 (см. Фиг.26), пациента 2 (см. Фиг.27), пациента 3 (см. Фиг.28), пациента 4 (см. Фиг.29) и пациента 5 (см. Фиг.30). Обработки Z-DTP2, Z-DTP 1 и Z-DNC проводят в указанных концентрациях (т.е. 1 мкМ или 10 мкМ) в течение 48 час. Анализы проводят при использовании исключения трипанового синего. Значения представляют собой процент живых клеток после обработки Z-DTP2, Z-DTP1 или Z-DNC относительно выживаемости необработанных контрольных клеток. Сильную цитотоксическую активность, сравнимую с активностью Z-DTP2 и Z-DTP1, наблюдают также в первичных клетках миеломы, полученных от пациентов, с mDTP3 в аналогичных условиях эксперимента (результаты не приводят). Данные результаты демонстрируют, что Z-/mDTP сохраняет активность в первичных клетках множественной миеломы и показывают, что лечение на основе Z-/mDTP можно использовать для лечения у пациентов множественной миеломы.
На Фиг.31 и 32 показывают, что Z-DTP1 и Z-DTP2 не проявляют токсичность в отношении нормальных первичных клеток либо мышиной, либо человеческой природы, даже при использовании в очень высоких концентрациях, т.е. 100 мкМ. Тестированные первичные клетки включают нормальные клетки стромы костного мозга (BMSC) (см. Фиг.31), мононуклеарные клетки периферической крови (PBMNC) (см. Фиг.31) и мезенхимные стволовые клетки (MSC) (см. Фиг.32), выделенные у пациентов, не страдающих множественной миеломой, и очищенные первичные В- и Т-лимфоциты, выделенные у мышей (см. Фиг.32). Обработку Z-DTP2, Z-DTP1 и Z-DNC проводят в указанных концентрациях в течение либо 48 час (BMSCs, PBMNCs) (см. Фиг.31), 72 час (мышиные В- и Т-клетки) (см. Фиг.32), либо 144 час MSC (см. Фиг.32). Анализы проводят при использовании исключения трипанового синего (см. Фиг.31) или включения [3H]тимидина (см. Фиг.32). Результаты, представленные на Фиг.26-32, показывают, что Z-DTP имеют высокие терапевтические индексы in vitro (т.е. отсутствие токсичности в нормальных клетках относительно высокой токсичности в раковых клетках). В действительности Z-DTP1 и Z-DTP2, но не Z-DNC, показывают сильную активность уничтожения опухолевых клеток в клетках множественной миеломы, полученных у пациентов (см. Фиг.26-30), но не проявляют токсичность в первичных нормальных клетках, полученных у здоровых людей или мышей (см. Фиг.31), даже при использовании очень высоких концентраций, таких как 100 мкМ (см. Фиг.32). Данные результаты демонстрируют, что Z-DTP не обладают неизбирательными цитотоксическими эффектами в клетках, скорее их цитотоксические эффекты специфичны в отношении раковых клеток и/или клеток, характеризующихся высокими уровнями экспрессии или активности Gadd45β и/или конститутивной высокой экспрессии или активности NF-κВ.
Высокая активность Z-/mDTP в клетках множественной миеломы и других раковых клетках в сочетании с отсутствием токсичности в первичных нормальных клетках, включая первичные человеческие BMSC, MSC, PBMNC и мышиные В- и Т-лимфоциты, даже при использовании в высоких концентрациях (т.е. 100 мкМ), демонстрирует, что соединения, соответствующие изобретению, обладают благоприятными терапевтическими индексами in vitro (см. Фиг.15, отсутствие токсичности в клеточных линиях, которое не зависит от NF-κВ в плане выживаемости, см. Фиг.22 и 23, корреляция между экспрессией Gadd45β и чувствительность раковых клеток к Z-DTP; см. Фиг.26-32, анализы гибели в первичных клетках), ключевым преимуществом данного изобретения относительно имеющихся лекарственных препаратов. У соединений, соответствующих изобретению, также отсутствует токсичность в опухолевых клеточных линиях, таких как Т-клеточный лейкоз, лимфома Беркетта и многие другие, которые не зависят от NF-κВ в плане выживаемости (даже при использовании в концентрации 100 мкМ; см. Фиг.15), показывая, что их активность имеет присущую им специфичность в отношении клеток с конститутивно активной NF-κВ. Более того, на большой панели опухолевых клеточных линий тканей различной природы существует высоко статистически достоверная корреляция между уровнями экспрессии Gadd45β и чувствительностью к вызываемой Z-/mDTP гибели (р<0,01; см. Фиг.22 и 23), устанавливая тем самым высокую специфичность цитотоксического действия Z-/mDTP в отношении Gadd45β. Важно, что опосредованная sh-РНК понижающая регуляция Gadd45β вызывает апоптоз в чувствительных к Z-/mDTP клеточных линиях множественной миеломы (например ARH-77 и NCIH929) с кинетикой, аналогичной параметрам, наблюдаемым с Z-/mDTP, но не в устойчивых к Z-/mDTP клеточных линиях множественной миеломы (например, RPMI-8226), и опосредованная sh-PHК понижающая регуляция MKK7 при водит в результате к утрате чувствительности к вызываемой Z-/mDTP гибели в чувствительных клеточных линиях множественной миеломы (например ARH-77) (см. Фиг.48-50). В совокупности данные результаты показывают, что цитотоксическая активность Z-/mDTP ограничена опухолевыми клетками, характеризующимися конститутивно активной NF-κВ и/или высокими уровнями экспрессии или активности Gadd45β, т.е. Z-/mDTPs проявляет активность на уровнях нМ в чувствительных клеточных линиях множественной миеломы, но не обладает токсичностью в устойчивых опухолевых линиях, которые не зависят от NF-κВ в плане выживаемости или которые проявляют низкие уровни экспрессии Gadd45β даже при использовании в концентрации 100 мкМ. Более того, в противоположность мышам с отсутствием компонентов ядра пути IKK/NF-κВ мыши Gadd45β-/- жизнеспособны и, вероятно, здоровы (см. статью Papa et all 2008 J Clin Invest 118, 1911-1923), показывая, что (в отличие от полной блокады протеасома/NF-κB) полная инактивация Gadd45β хорошо переносится in vivo (см. статью Papa et all 2008 J Clin Invest 118, 1911-1923). В совокупности данные результаты показывают, что лекарственный препарат на основе Z-/mDTP будет безопасным и специфичным (см. Фиг.15, 31 и 32 отсутствие токсичности в NF-κВ-независимых опухолевых клеточных линиях и нормальных первичных клетках; см. Фиг.22 и 23, корреляция между экспрессией Gadd45β и чувствительность раковых клеток к вызываемой Z-mDTP гибели; см Фиг.26-30, вызываемая Z-/mDTP гибель первичных клеток множественной миеломы).
Ингибиторы протеасом (PI), такие как бортецомиб и другие препараты для лечения множественной миеломы также уничтожают клетки множественной миеломы путем активации JNK (см. статью Chauhan et al 2008 Blood 111, 1654 1664), но не могут лечить вследствие низких терапевтических индексов (см. статьи Lauback et al 2009 Leukemia 23, 2222-2232; Ludwing et a12010 Oncologist 15, 6-25 и www.cancecare.on.ca/). Направленность на отдельные функции NF-κВ в процессе выживаемости множественной миеломы через Gadd45β сделает возможным разделить функции NF-κВ в направлениях иммунитета, воспаления и выживаемости, что дает, таким образом, более безопасный, более специфический препарат, который может переноситься в дозах, необходимых для лечения. Z/mDTP очерчивают совершенно новый класс лекарственных препаратов, направленных на новый путь во множественной миеломе и потенциально других формах рака, а также заболеваниях или нарушении, которые зависят от NF-κВ в плане выживаемости.
Пример 14. Характеристики связывания mDTP3 с белками Gadd45β и MKK7 в виде отдельных молекул и как части комплекса Gadd45β/MKK7
В качестве примера эксперименты по связыванию проводят с mDTP3 в отношении Gadd45β, домена киназы MKK7 (MKK7KD) и комплекса Gadd45β/MKK7 с использованием способа поверхностного плазменного резонанса.
Материалы и способы
Для того, чтобы определить, как DTP связывается с комплексом Gadd45β/MKK7, проводят эксперименты с помощью прибора Biacore3000 SPR (GE Healthcare, Milan, Italy) при использовании 4-канальных сенсорных чипов СМ5 (GE Healthcare, Milan, Italy). Человеческий Gadd45β полной длины получают и очищают, как описано в ссылке Tornatore L, et al. (2008). J Mol Biol; 378:97-111. Конститутивно активный домен киназы MKK7, охватывающий остатки 101-405 и несущий мутации S287D и Т291 D (MKK7KD), экспрессируют в Е. coli в виде слитого белка His6. Белок очищают до однородности в двух последовательных стадиях аффинной хроматографии (основа Ni-NTA) и гель-фильтрации (Superdex G75) и затем характеризуют с помощью SDS-PAGE, ЖХ-МС для подтверждения идентичности и чистоты, а также кругового дихроизма с целью оценки укладки.
MKK7KD иммобилизуют на сенсорном чипе Biacore классическими химическими способами присоединения EDC/NHS при рН 5 (pi белка ?9) при скорости потока 5 мкл/мин. Достигают уровня иммобилизации приблизительно 8000 Единиц ответа. Gadd45β, который по природе является кислым белком с pI приблизительно 4,5, иммобилизуют при рН 3,5 (уровни иммобилизации 6000 RU) на отдельном канале. Остаточные реакционные группы на обоих, Gadd45β и MKK7KD, каналах в итоге инактивируют обработкой этаноламином. На другом канале проводят такую же процедуру активации с помощью EDC/NHS и инактивацию этаноламином. Данный канал используют в качестве эталона и получаемый от него сигнал считают пустым и, соответственно, вычитают значения из значений для экспериментальных каналов, определяющих белки Gadd45(или МKK7KD, с целью удаления неспецифического связывания с поверхностью чипа. Для того, чтобы определить, эффективно ли иммобилизованы два белка, проводят повторные введения Gadd45β (20-200 нМ) и МKK7KD (1-25 нМ) с повышающимися концентрациями белков (время контакта 3 мин; 60 мкл). Регенерацию достигают при использовании ввода либо 1М NaCl (1 мМ, канал, дериватизированный MKKKD), либо 20 мМ NaOH (30 сек, канал, дериватизированный Gadd45β).
Наконец, возрастающие концентрации трипептида mDTP3 (Ac-D-Tyr-D-Arg-D-Phe-NH2) наносят на чип, причем концентрации лежат в интервале от 1 нМ до 10 мкМ. В отдельном эксперименте mDTP3 вводят во время фазы диссоциации либо Gadd45(из иммобилизованного MKK7KD, либо MKK7KD из иммобилизованного Gadd45β. Результаты данных анализов приводят на Фиг.51-54.
Результаты
Как можно видеть на Фиг.51, связывание Gadd45β с иммобилизованным MKK7KD очень эффективно. Дозозависимое связывание и кривые диссоциации наблюдают при всех используемых концентрациях. Константу диссоциации KD взаимодействия Gadd45β/MKK7KD оценивают путем усреднения значений, рассчитанных относительно каждой из различных кривых, и определяют, что она составляет 4,0±0,7 нМ (см. Фиг.51). Аналогично повторные вводы MKK7«о на канал Gadd45β дают дозозависимые кривые ассоциации и диссоциации (см. Фиг.52), на основании которых выводят KD 3,4±0,6 нМ.
Для того, чтобы определить, связывается ли mDTP3 с МKK7KD и/или с Gadd45β, образцы пептида (т.е. mDTP3) вводят в каналы, дериватизированные Gadd45β и MKK7KD. Неожиданно, как можно видеть из Фиг.53, результаты показывают, что данный пептид не связывается ни с Gadd45β, ни с MKK7KD в растворе. Удивительно, однако, что, когда mDTP3 вводят во время фазы диссоциации Gadd45β из MKK7KD (см. Фиг.54) или фазы диссоциации МKK7KD из Gadd45β (результаты не приводят), связывание наблюдают и можно зарегистрировать кривые дозозависимой ассоциации и диссоциации. На Фиг.54 показывают, что, когда mDTP3 вводят в низких концентрациях либо 10 нМ либо 100 нМ, данный пептид вызывает быструю диссоциацию комплекса Gadd45β/MKK7KD. Как можно также видеть, образовавшийся комплекс Gadd45β/MKK7KD быстро выделяют после отмывания пептидов. На Фиг.54 также показывают, что, когда mDTP3 вводят в более высоких концентрациях (например 1 мкМ), регистрируют дозозависимое связывание и диссоциацию, указывающие на то, что mDTP3 связывается либо с Gadd45β и/или с MKK7KD либо с комплексом двух данных белков. В совокупности данные результаты показывают, что mDTP3 не способен связываться либо с Gadd45β, либо с MKK7KD по отдельности даже при использовании в высоких концентрациях, скорее его связывание либо с Gadd45β, MKK7KD либо с поверхностью, образованной при взаимодействии двух белков, требует формирования комплекса Gadd45β/MKK7. Данные результаты важны, поскольку они показывают, что терапевтической мишенью является граница между двумя белками (т.е. Gadd45β и MKK7), которая обеспечивает возможность высокой избирательности в клетках, ключевого преимущества данного изобретения относительно имеющихся лекарственных препаратов.
Пример 15. Фармакокинетические профили (DMPK) in vivo Z-DTP2 и mDTP3
Для оценки пригодности Z-DTP2 и mDTP3 для терапевтического применения in vivo проводят фармакокинетические анализы на мышах.
Материалы и способы
Исследование фармакокинетики на мышах:
Краткое изложение протокола:
Z-DTP2 и Z-mDTP3 вводят мышам внутривенно. Пробы крови собирают в 7 точках времени после внутривенной (i.v.) инъекции соединений в течение 8 час и анализируют плазму с помощью ЖХ-МС/МС с целью определения концентрации соединений в каждой точке времени.
Ход эксперимента:
Трем самцам мышей CD1 массой 25-30 г каждый вводят дозу в зависимости от пути введения, точки времени, соединения. Тестируемое соединение вводят внутривенно (на типичном уровне дозы 10 мг соединения на кг массы тела). Животным создают свободный доступ к пище на протяжении исследования.
Животных подвергают анестезии, забирают пробы крови в гепаринизированные пробирки и животных умерщвляют в следующие точки времени:
Внутривенное введение дозы: 0,08, 0,25, 0,5, 1, 2, 4 час 8 после введения дозы.
Получение образцов:
Пробы крови центрифугируют с целью получения плазмы, которую переносят в отдельный контейнер, снабженный этикеткой. Аликвоты, полученные в отдельные точки времени у трех животных, анализируют раздельно. Белки осаждают добавлением трех объемов метанола и центрифугированием в течение 30 мин при 4°С. Аликвоты по 100 мкл полученных в результате супернатантов разводят 200 мкл воды качества для проведения ВЭЖХ в лунках 96-луночного планшета.
Количественный анализ:
Стандартные кривые получают в матрицах с плазмой без добавок и обрабатывают идентично образцам. Образцы плазмы количественно оценивают ЖХ-МС/МС и концентрацию каждого соединения в плазме выражают в м кг/мл.
Фармакокинетический анализ:
Фармакокинетические параметры рассчитывают с использованием анализа на некомпартментной модели, как описано на веб-сайте http://www.pharsight.com/main.php
Биоанализ:
Краткое изложение протокола:
Концентрацию тестируемого соединения в образцах плазмы измеряют с помощью ЖХ-МС/МС. Результаты количественно оценивают с использованием пятиточечной стандартной кривой в интервале 3-3000 нг/мл.
Ход эксперимента:
Белки осаждают из аликвот по 50 мкл отдельных образцов плазмы добавлением 150 мкл метанола. После осаждения белка образцы плазмы центрифугируют в течение 30 мин при 4°С. Аликвоты по 100 мкл полученного в результате супернатанта разводят 200 мкл воды качества для проведения ВЭЖХ в лунках 96-луночного планшет. Тестируемое соединение количественно оценивают с помощью ЖХ-МС/МС по пятиточечной стандартной кривой, полученной путем введения в плазму различных концентраций тестируемого соединения, растворенного в ДМСО при интервале конечных концентраций 3-3000 нг/мл (конечная концентрация ДМСО 1%), и обрабатывают идентично тест-образцам, как описано выше.
Результаты
Фармакокинетические исследования на самцах мышей CD1 показывают, что как Z-DTP2, так и mDTP3 имеют профили DMPK in vivo, подходящие для введения путем внутривенной (i.v.) инфузии (см. Таблицы VIII, IX [А] и IX [В]) в отсутствие острой токсичности у мышей. В Таблице VIII приводят значения наиболее важных in vivo фармакокинетических параметров, полученных с Z-DTP2 и mDTP3, включая полупериод существования в плазме (Т1/2), гомеостаз (Vss) и конечные (Vβ) объемы распределения, а также общий клиренс (tot CL), площадь под кривой зависимости концентрации в плазме от времени (AUC) и концентрация в точке времени 0 (С0). Значения рассчитывают на основании данных кривых зависимости концентрации в плазме относительно времени, полученных на основе некомпартментных и компартментных способов анализа (см. статьи Groulx A. 2006 ScianNew 9: 1-5 и DiStefano 3rd 1982 Am J Physiol Regul Integr Comp Physiol 243: 1-6) (результаты не приводят). Каждый из показанных параметров представляет собой среднее из экспериментальных значений, полученных в трех различных группах самцов мышей CD1 после однократной внутривенной (i.v.) инъекции соединения в дозе 10 мг/кг массы тела. Трем самцам мышей CD1 (массой тела 25-30 г) внутривенно вводят дозу либо Z-DTP2, либо mDTP3. Пробы крови собирают в 7 точках времени, как показано (т.е. через 0,08, 0,25, 0,5, 1, 2, 4 и 8 час после инъекции) и плазму анализируют жидкостной хроматографией с масс-спектрометрией (ЖХ-МС) с целью определения концентраций в крови двух соединений в каждой точке времени. Графики профиля концентрации в плазме относительно времени экстраполируют как для Z-DTP2, так и для mDTP3. Результаты показывают, что оба, Z-DTP2 и mDTP3, имеют мультифазный характер размещения после внутривенной инъекции (результаты не приводят). В действительности кривые зависимости концентрации от времени внутривенно введенных соединений демонстрируют явный биоэкспоненциальный профиль с крутой фазой начального распределения и длинным конечным Т1/2 (результаты не приводят).
Основные фармакокинетические параметры, экстраполированные из данных кривых зависимости концентрации в плазме от времени (т.е. СО, AUC до последней точки, Т1/2, Vβ, Vss и CL), важны для расчета уровней дозирования и схемы введения, необходимых для достижения требуемых системных равновесных концентраций лекарственного препарата (т.е. терапевтических системных концентраций). Как видно из таблицы VIII, Z-DTP2 и mDTP3 демонстрируют полупериоды существования in vivo приблизительно 2 час и приблизительно 1 час и 20 мин, соответственно.
Интересно, что оба, Z-DTP2 и mDTP3, показывают исходный распределительный полупериод существования приблизительно 5 мин, при котором можно было бы предположить быстрое тканевое/клеточное поглощение, но альтернативно можно было бы предположить связывание с белками плазмы. Наиболее важно, что оба соединения демонстрируют очень медленное выведение из тканей, которое отражается в конечном полупериоде существования приблизительно 8 час (см. Таблицу VIII и результаты не приводят) (см. веб-сайты http://www.pharsight.com/main.php и http://www.meds.com/leukemia/idamycin/adriamycin.html и статью Kupperman et al 2010 Cancer Res 70 1970-1980). Результаты также демонстрируют, что как Z-DTP2, так и mDTP3 соответствуют общей линейной фармакокинетической системе (см. статью Berezhkovskiy (2007) J Pharm Sci. 96, 1638-52), как показывает тот факт, что их значения общего объемного распределения превышают значения равновесного объемного распределения (т.е. Vβ>Vss).
Оба, конечное и равновесное распределения, а также конечные полупериоды существования двух соединений синергически участвуют в установлении количества лекарственного препарата, необходимого в организме для получения постоянной скорости вливания.
Важно, что Z-DTP2 и mDTP3 показывают значения общего клиренса, лежащие в интервале 66-90 мл/мин/кг и 22-27 мл/мин/кг, соответственно, позволяя предположить медленные скорости метаболического выведения и выведения с желчью обоих соединений (см. таблицу VIII и результаты не приводят).
В таблицах IX [А] и IX [В] показывают предсказанное дозирование для введения in vivo ZDTP2 и mDTP3, соответственно, необходимое для достижения системных терапевтических концентраций двух соединений. Значения представляют дозирование, выраженное в мг/час, необходимые для получения требуемых постоянных концентраций в плазме 1, 5 или 10 мкМ для обоих, Z-DTP (см. Таблицу IX [А]) или mDTP3 (см. Таблицу IX [В]). Существенно, что, несмотря на сравнимые полупериоды существования, а также сравнимый конечный полупериод существования с Z-DTP2, mDTP3 демонстрирует значение общего клиренса, которое в три раза ниже, чем данное значение у Z-DTP2 (см. таблицу VIII и результаты не приводят). Заметим, что даже небольшое различие в данном фармакокинетическом параметре может в существенной степени воздействовать на размер дозы и схему, необходимые для достижения требуемой постоянной концентрации соединения в плазме, как видно по разнице в дозах, предсказанных для Z-DTP2 и mDTP3 (см. Таблицы IX [А] и IX [В], соответственно). В действительности Таблицы IX [А] и IX [В] (анализы на модели) показывают, что для достижения постоянной концентрации в плазме 1, 5 или 10 мкМ доза, необходимая для mDTP3, существенно ниже, чем необходимая для Z-DTP2. Таким образом, основываясь на данных фармакокинетических результатах и значениях IC50, установленных для двух соединений в клеточных линиях множественной миеломы (см. Таблицу IV), для достижения постоянной концентрации в плазме до 10 мкМ будет необходимо вводить Z-DTP2 и mDTP3 путем постоянной внутривенной инфузии со скоростью 0,976 мг/час и 0,218 мг/час, соответственно (см. Таблицы IX [А] и IX [В]).
Отметим, что синтез Z-/mDTP не требует много времени и несложен, следовательно, препарат рентабелен даже при постоянном применении. Таким образом, даже при низком Т1/2, лечение Z-/mDTP путем инфузии будет приемлемо для находящихся в больничных условиях пациентов, уже проходящих химиотерапию. Соединения, соответствующие изобретению, также имеют высокую растворимость и обладают высокой специфичностью и благоприятными профилями безопасности, поэтому их можно доставлять в высоких дозах в небольших объемах для максимального повышения лечебных эффектов, как успешно используют в существующих пептидных препаратах.
Таблицы
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К BCMA И BCMA-CAR-T-КЛЕТКИ | 2020 |
|
RU2762942C1 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К BCMA И BCMA-CAR-NK-КЛЕТКИ | 2020 |
|
RU2795443C2 |
| ВИДЫ КОМБИНИРОВАННОЙ ТЕРАПИИ РАКА, НАЦЕЛИВАЮЩИЕСЯ НА CD38 И TGF-БЕТА | 2019 |
|
RU2808632C2 |
| КОНЪЮГАТЫ АНТИТЕЛ С ЛЕКАРСТВЕННЫМИ ВЕЩЕСТВАМИ И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ РАКОВЫХ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2799547C2 |
| АНТИТЕЛА К CD38 И СОСТАВЫ | 2020 |
|
RU2826957C2 |
| Способ и композиция для лечения рака, уничтожения метастатических раковых клеток и профилактики метастазов рака, используя антитела к конечным продуктам повышенного гликирования ( AGE) | 2017 |
|
RU2728964C2 |
| АГЕНТЫ ДЛЯ ЛЕЧЕНИЯ ЭКСПРЕССИРУЮЩИХ КЛАУДИН РАКОВЫХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2798990C2 |
| Т-КЛЕТКИ С ДВУМЯ ХИМЕРНЫМИ АНТИГЕННЫМИ РЕЦЕПТОРАМИ CAR | 2021 |
|
RU2839662C1 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ CD269 (BCMA) | 2015 |
|
RU2749041C2 |
| АНТИ-FcRH5 АНТИТЕЛА | 2014 |
|
RU2687132C2 |











































































































































































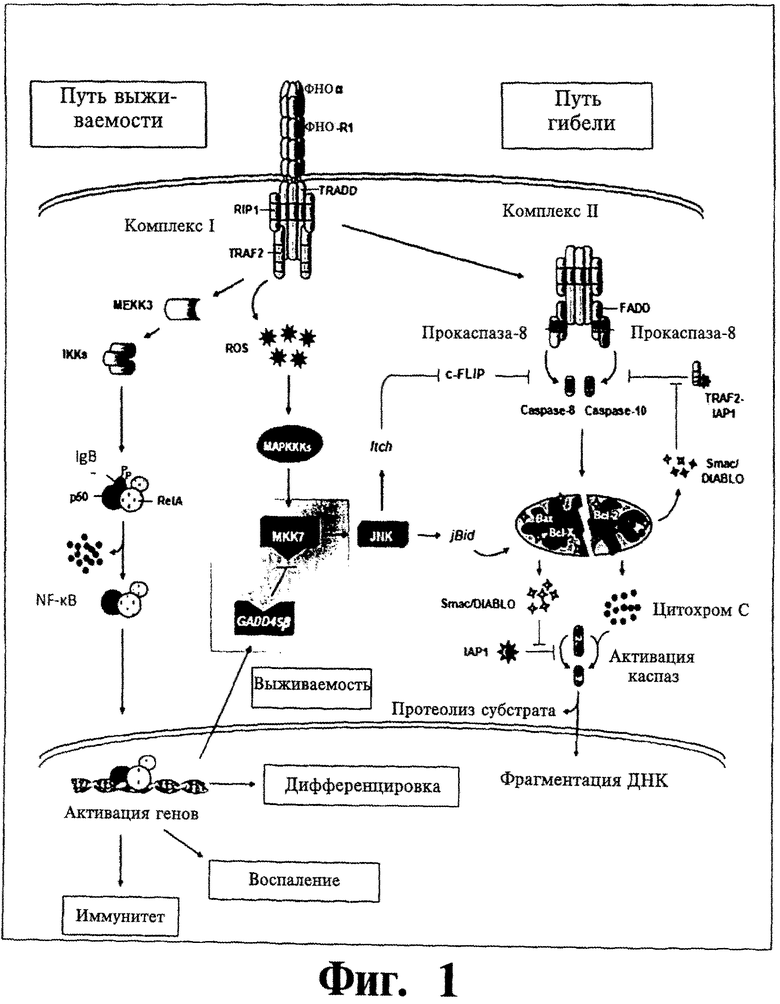
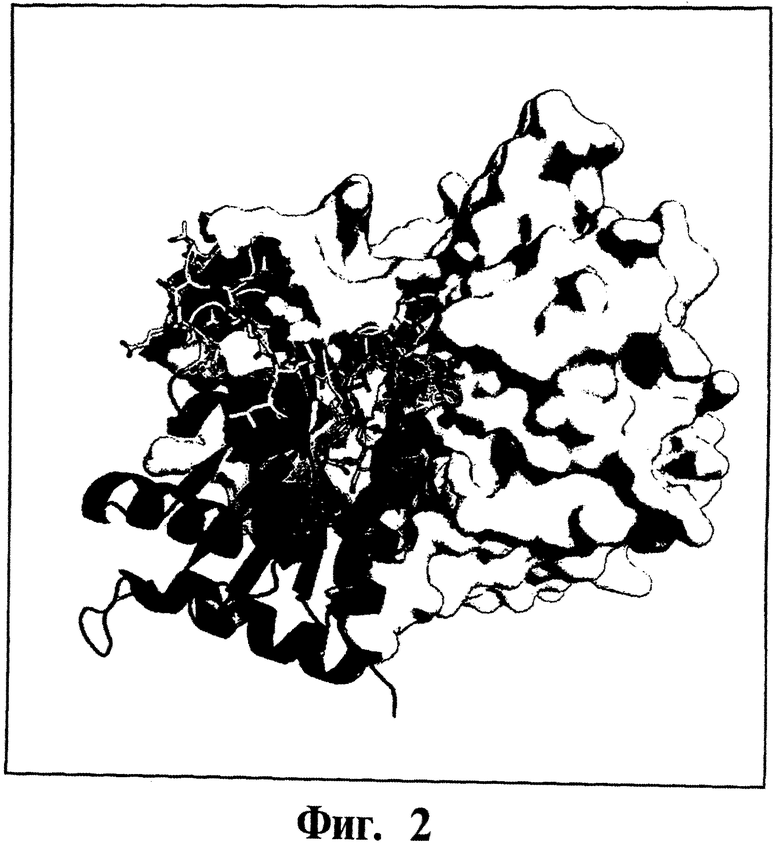
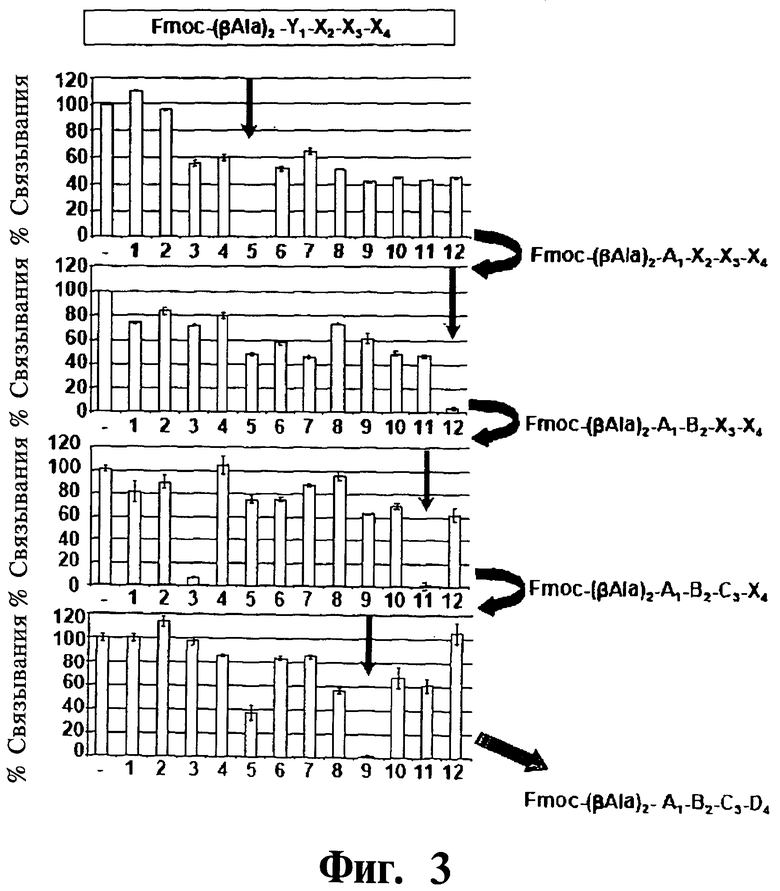
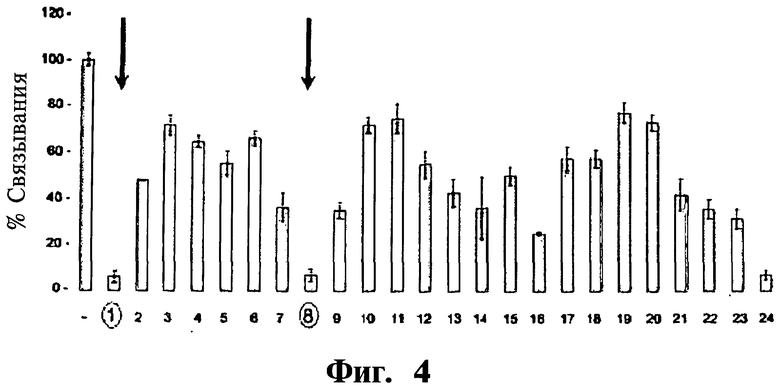
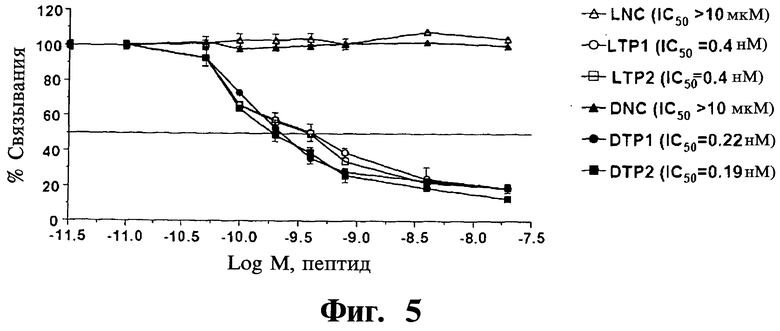
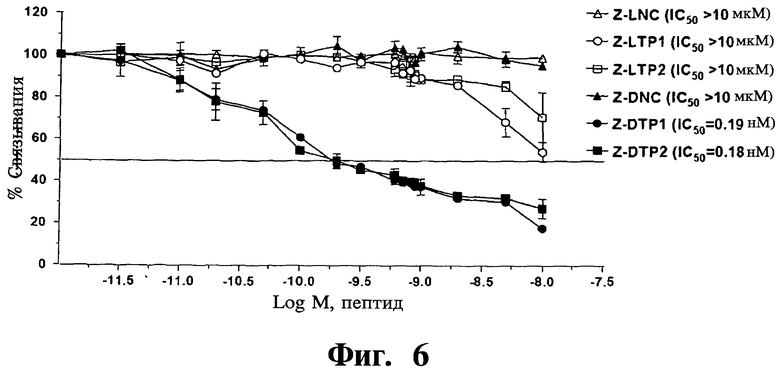
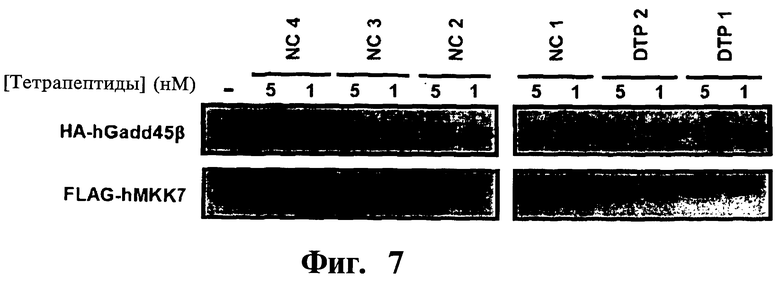
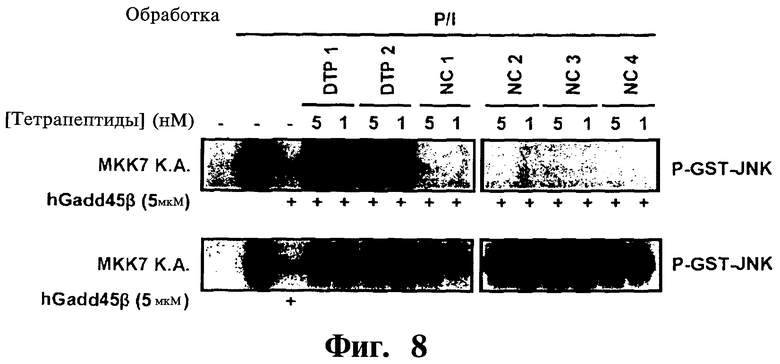
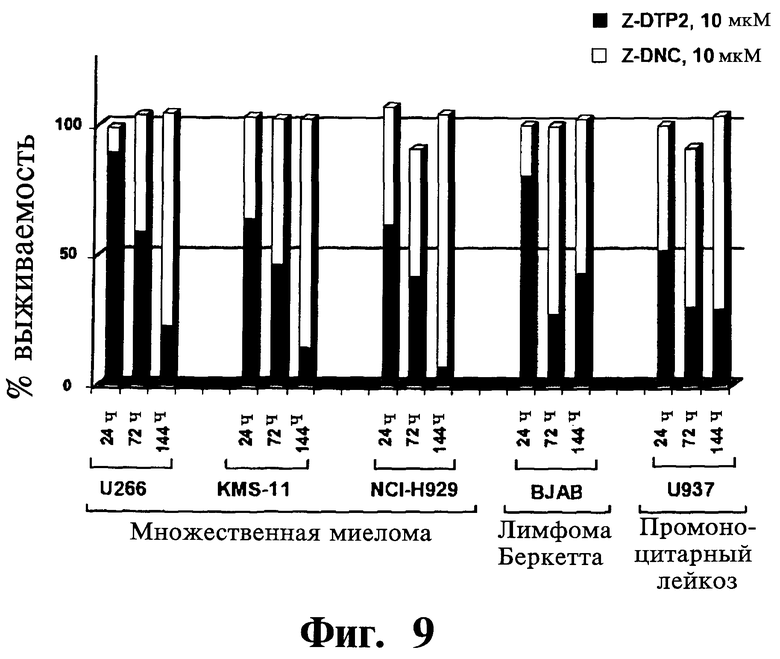

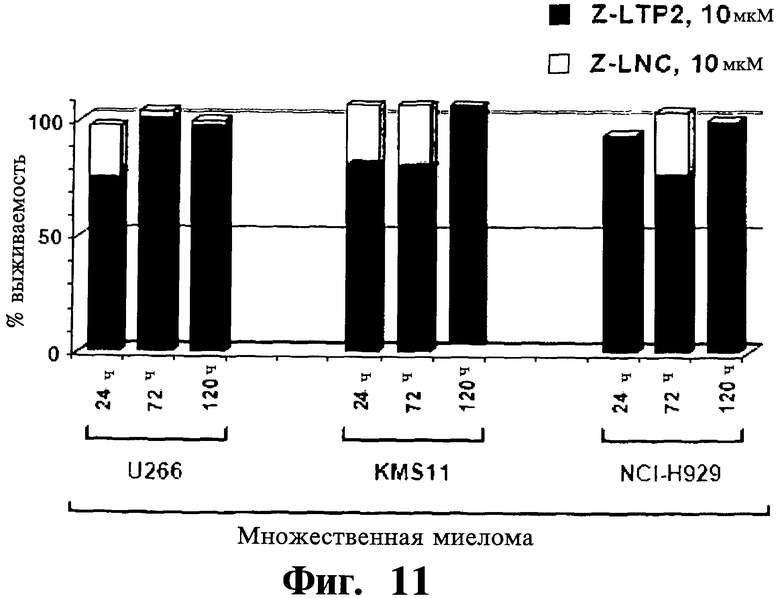
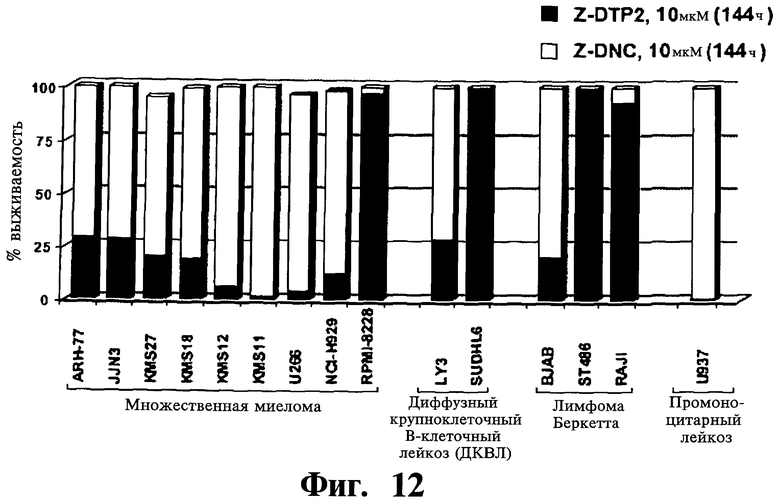
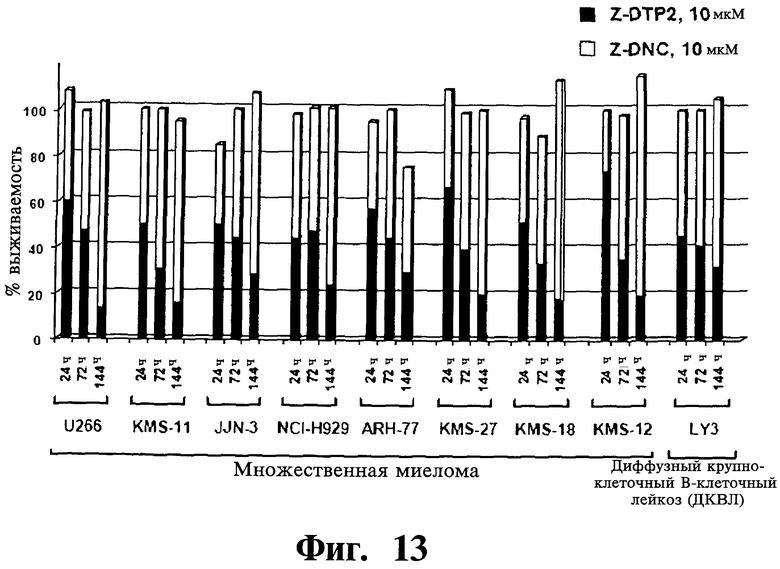
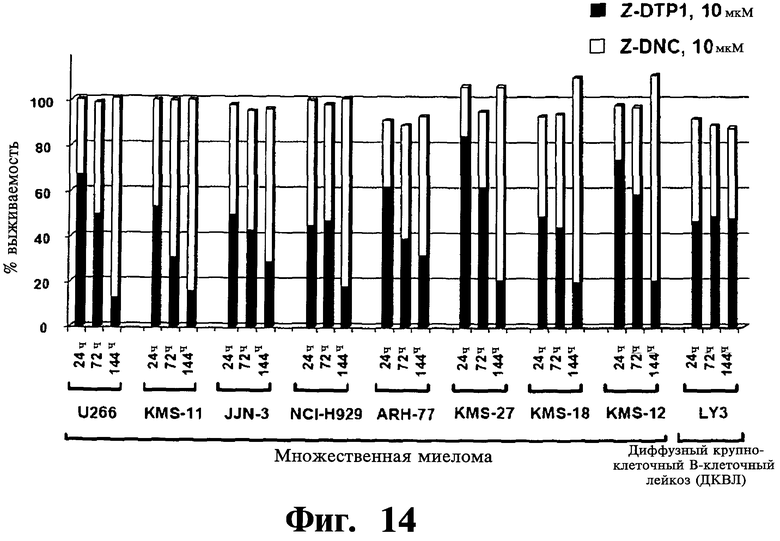
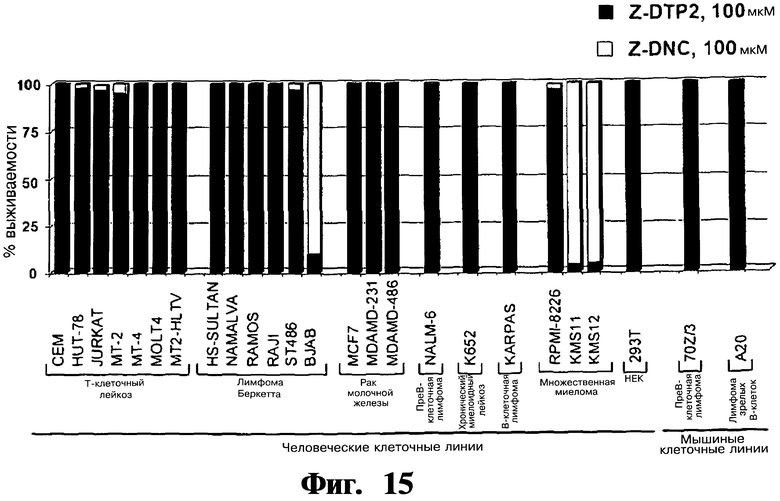

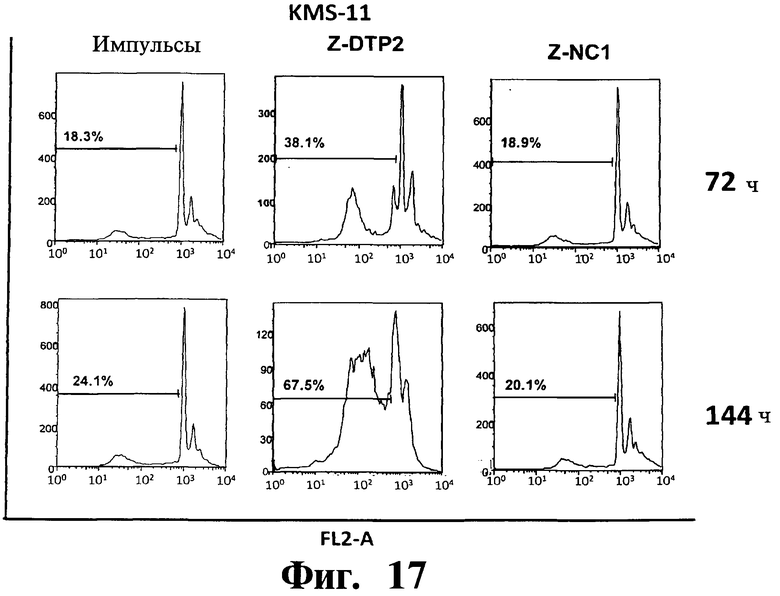
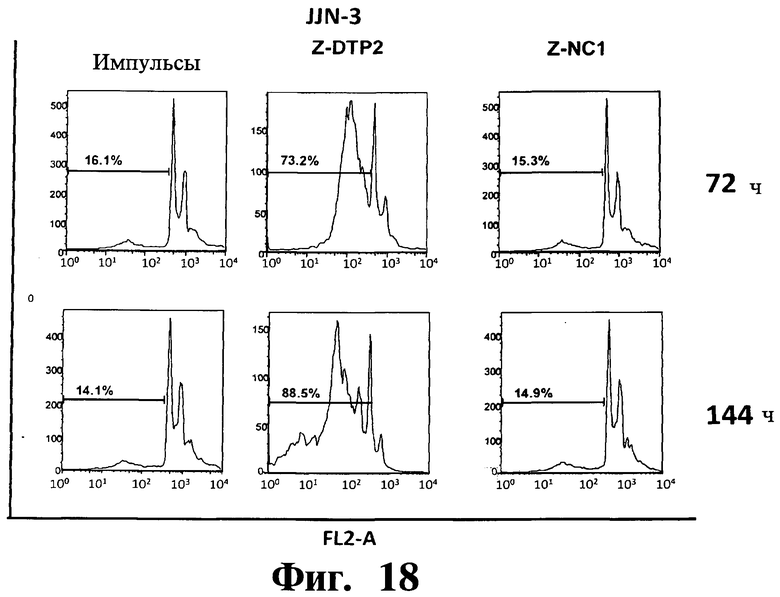
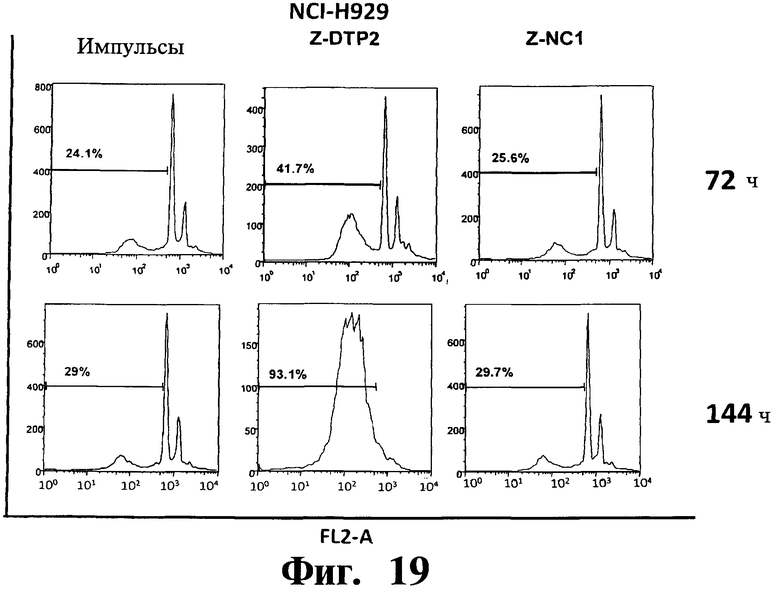
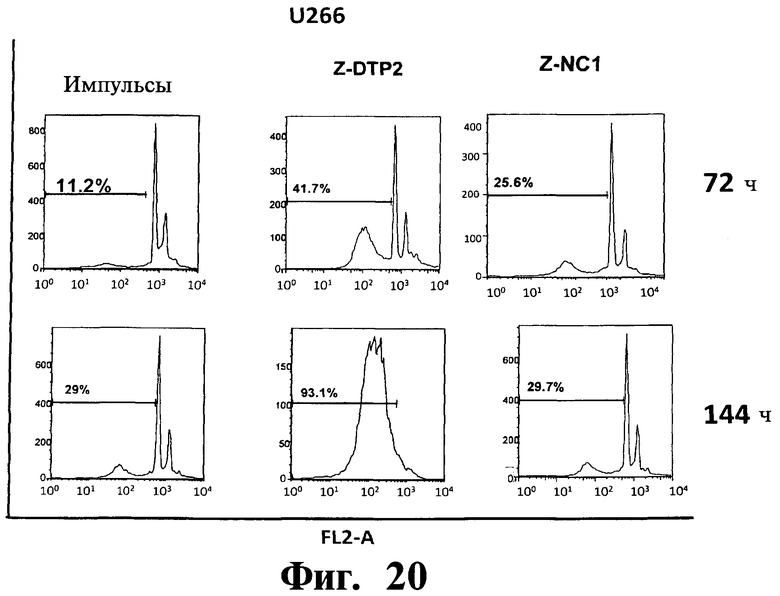
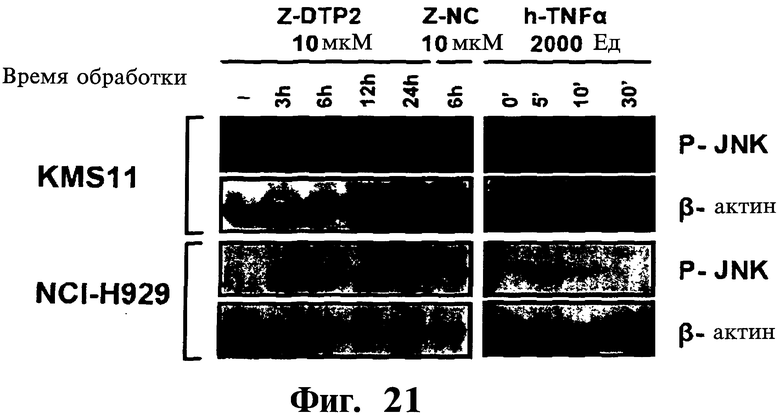
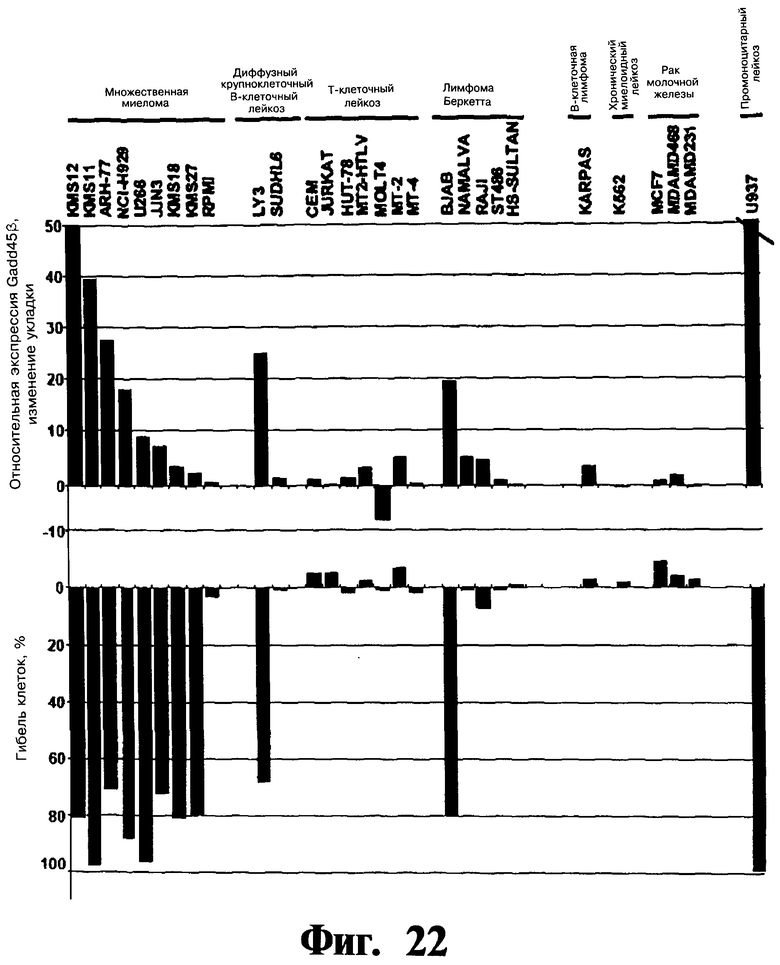
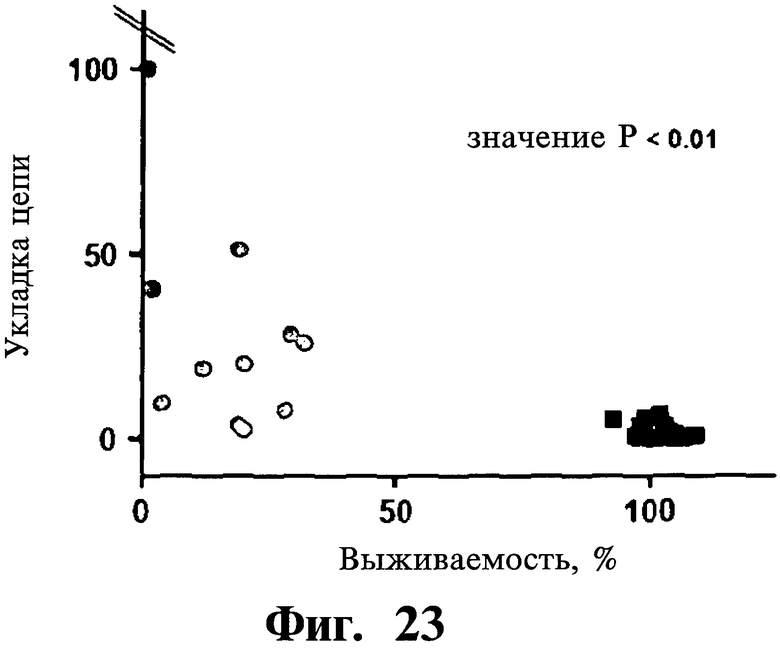
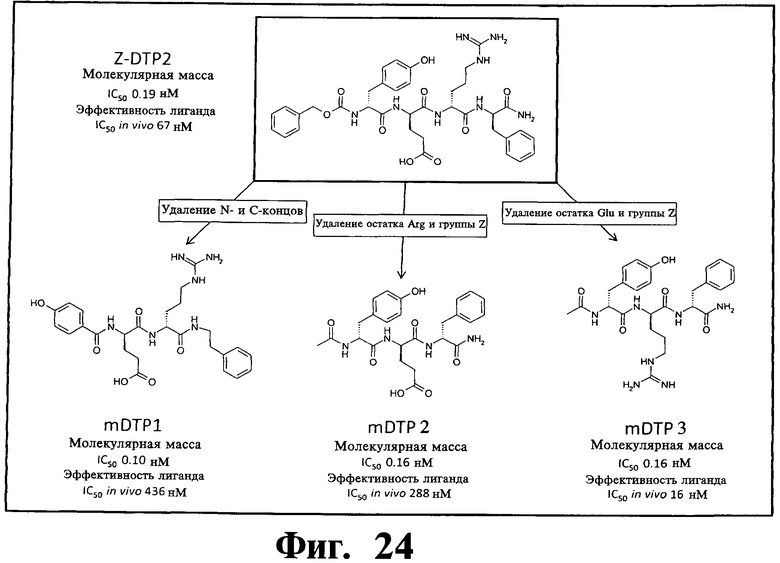
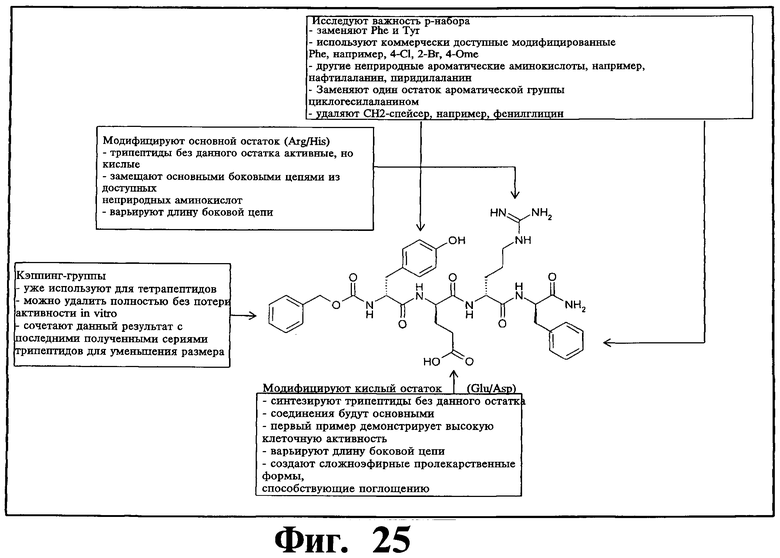
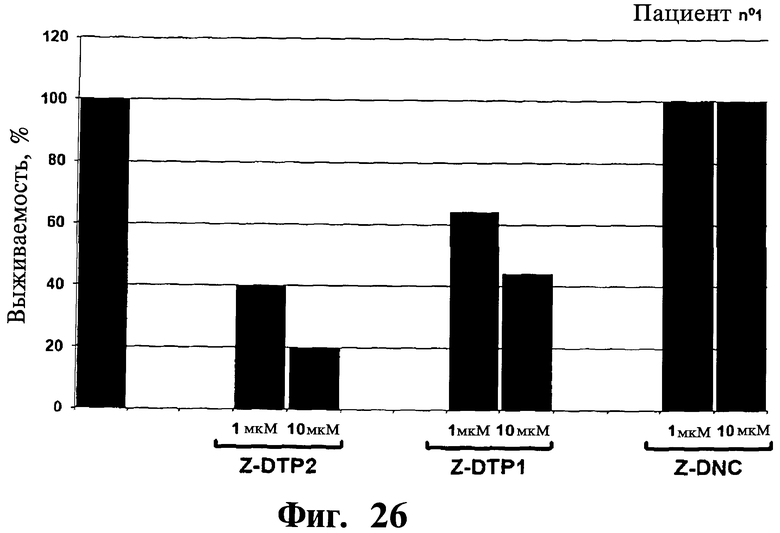
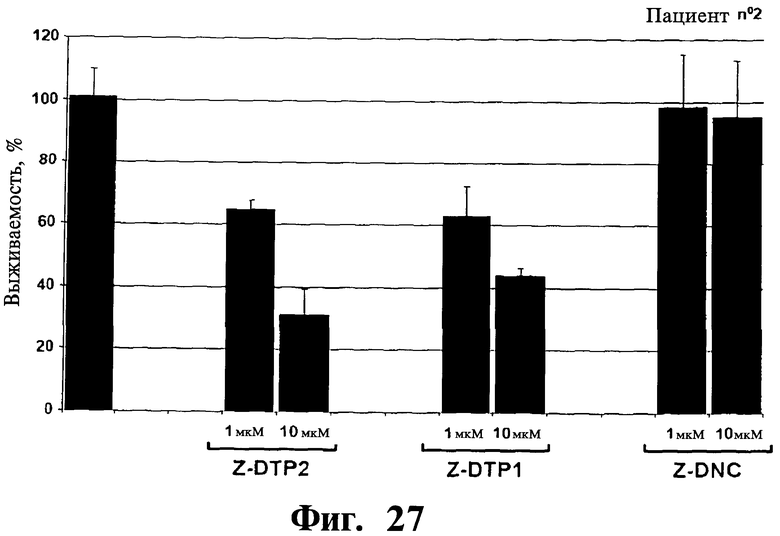
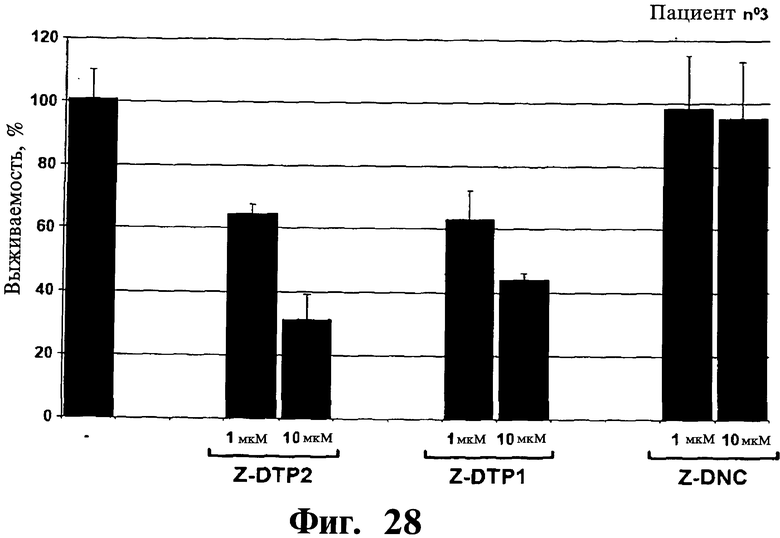
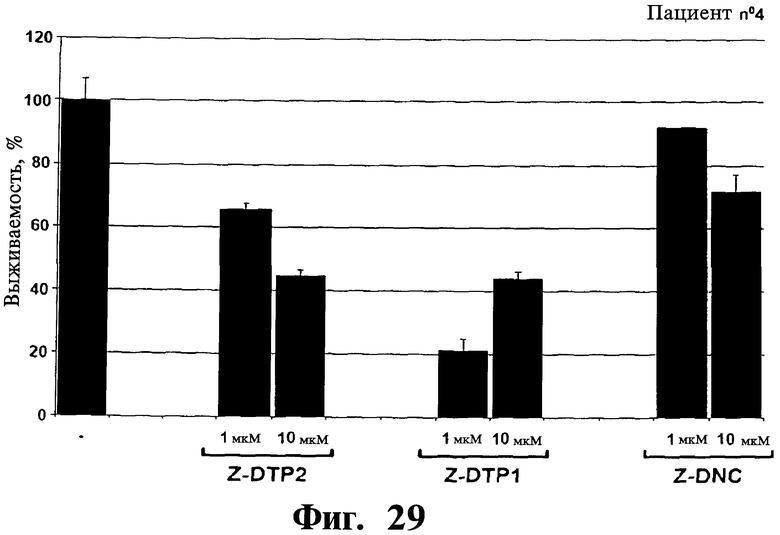
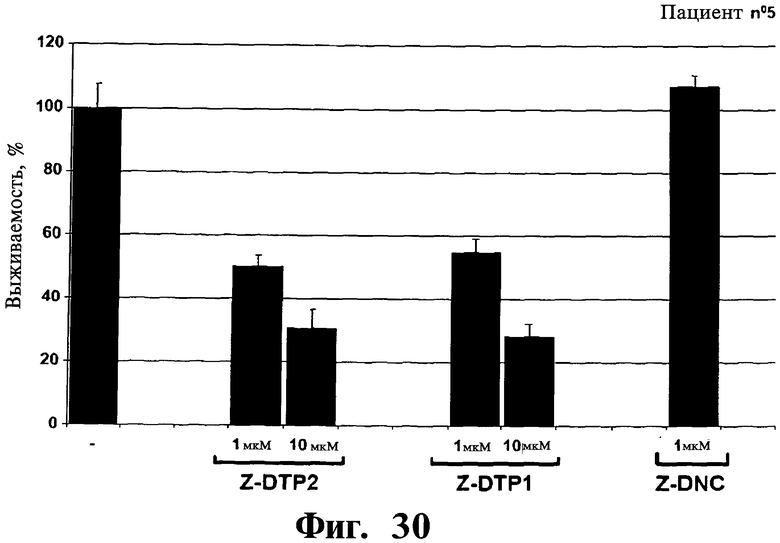
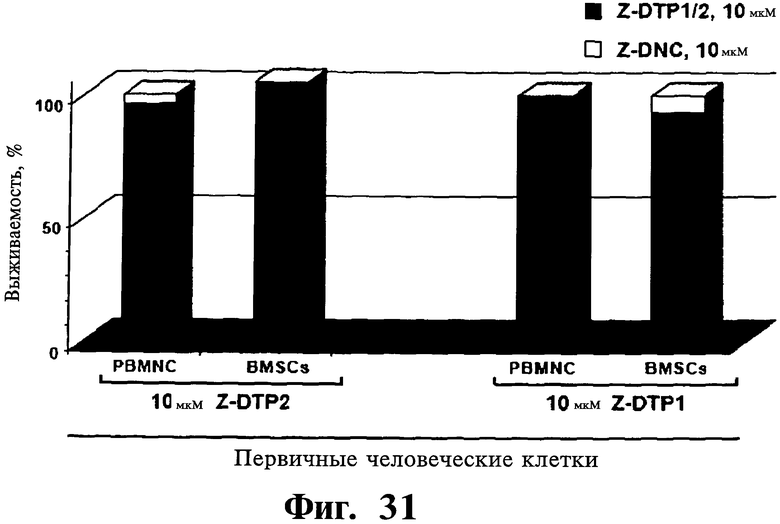
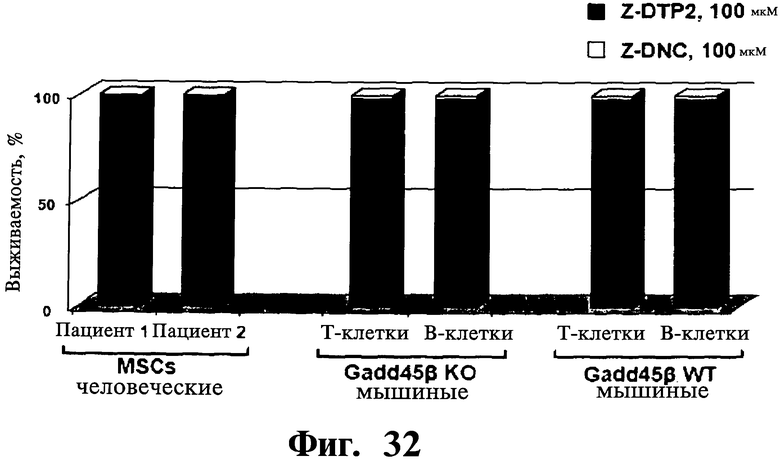

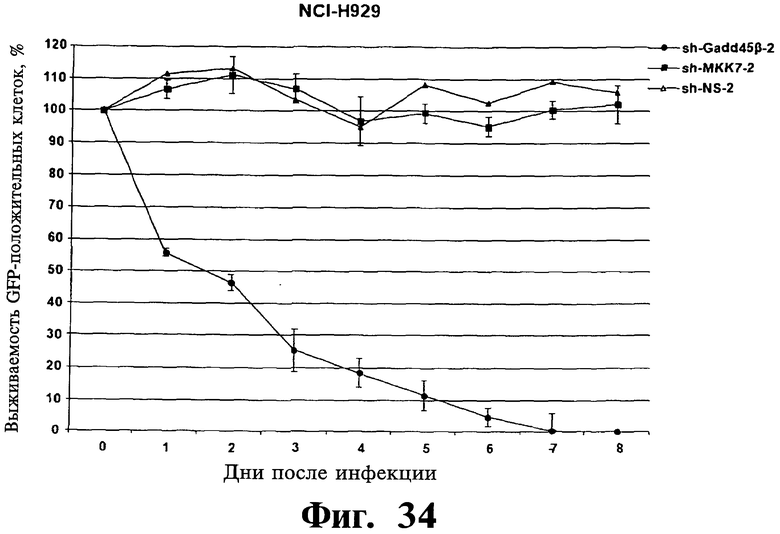
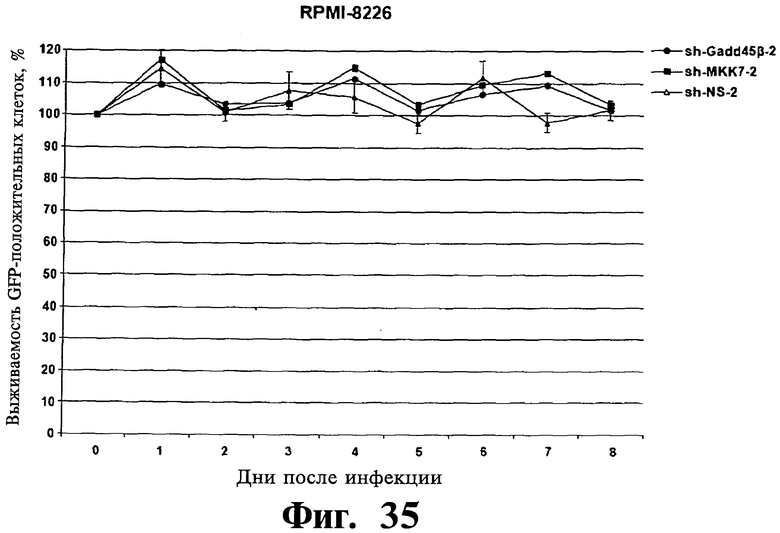
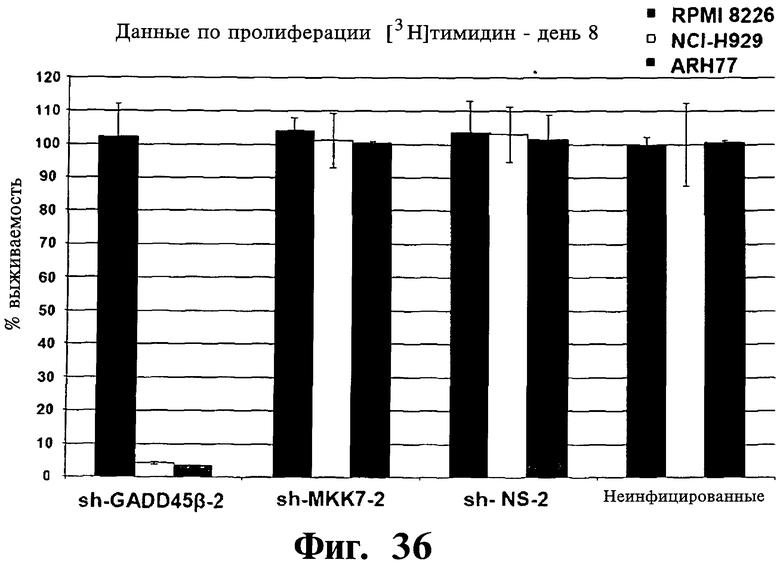
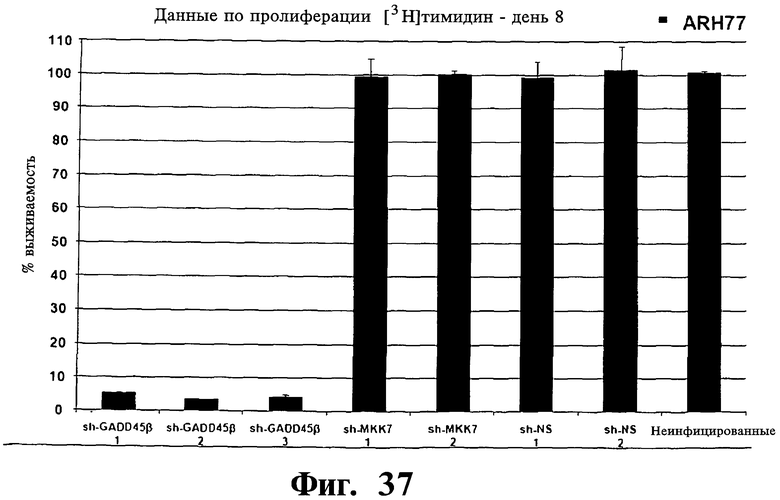

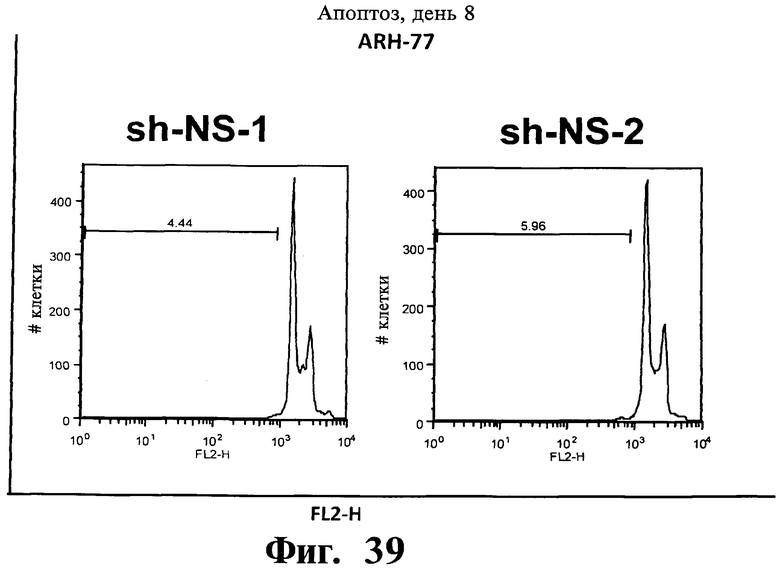
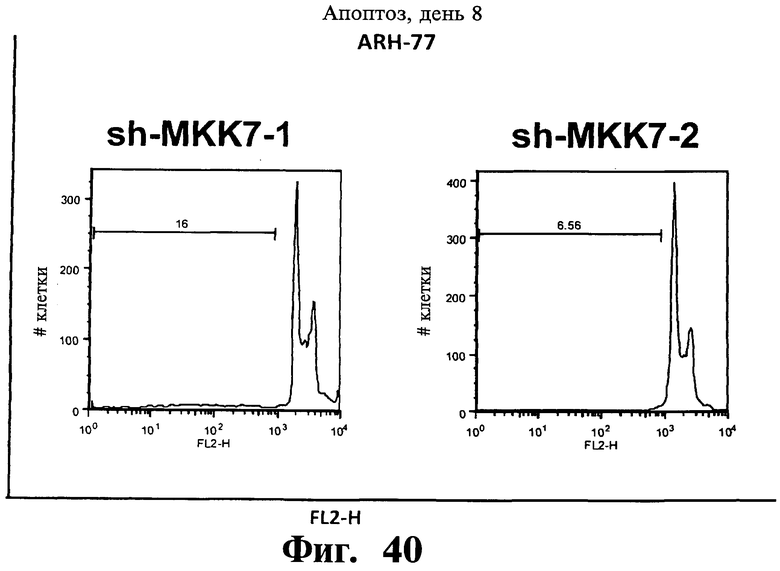
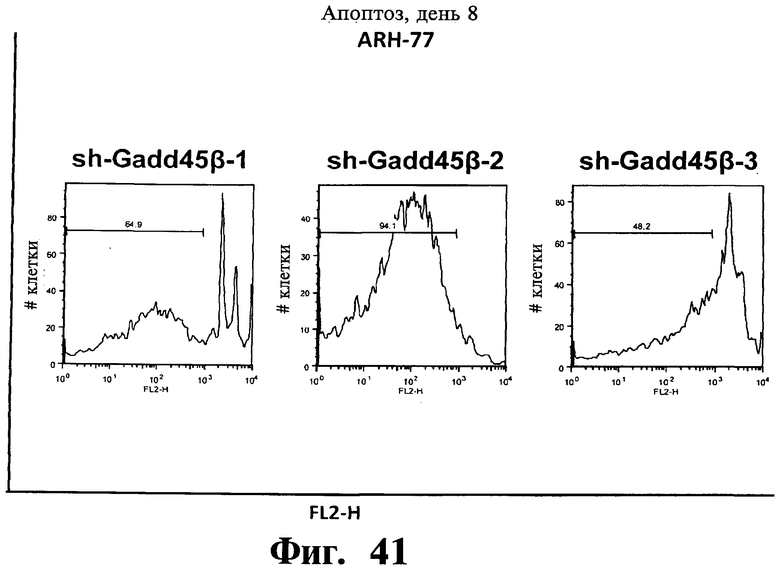
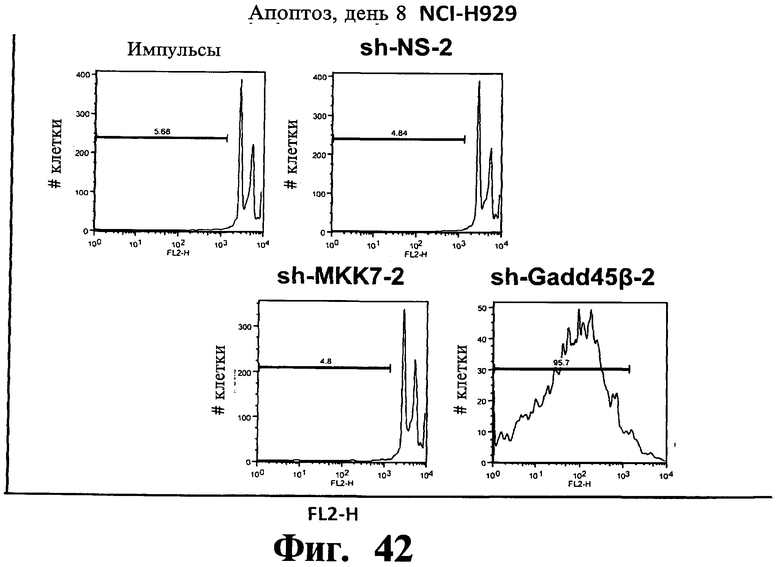
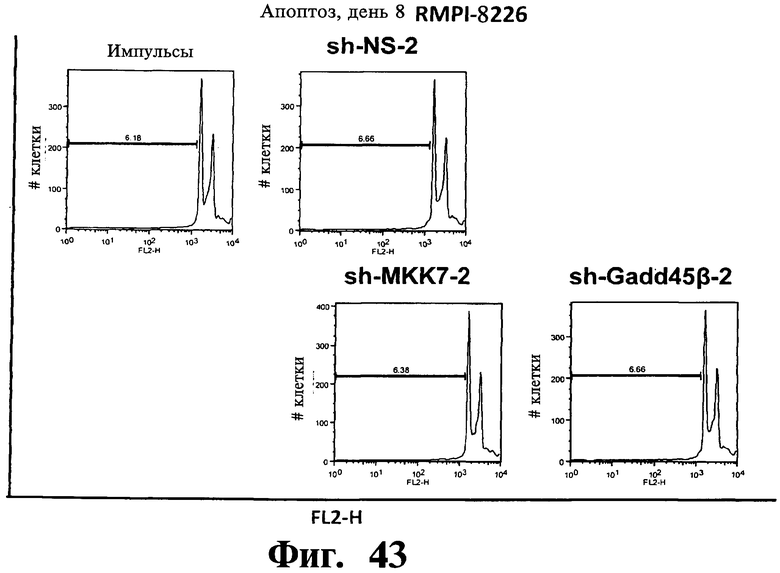
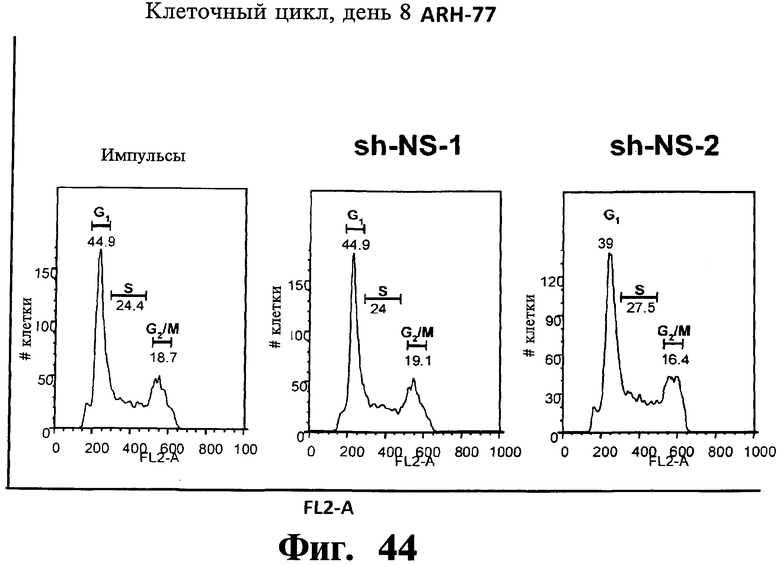
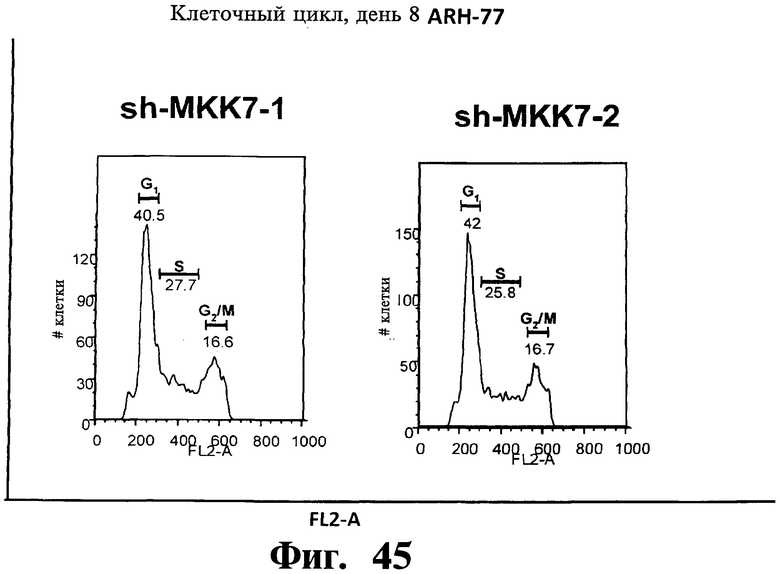
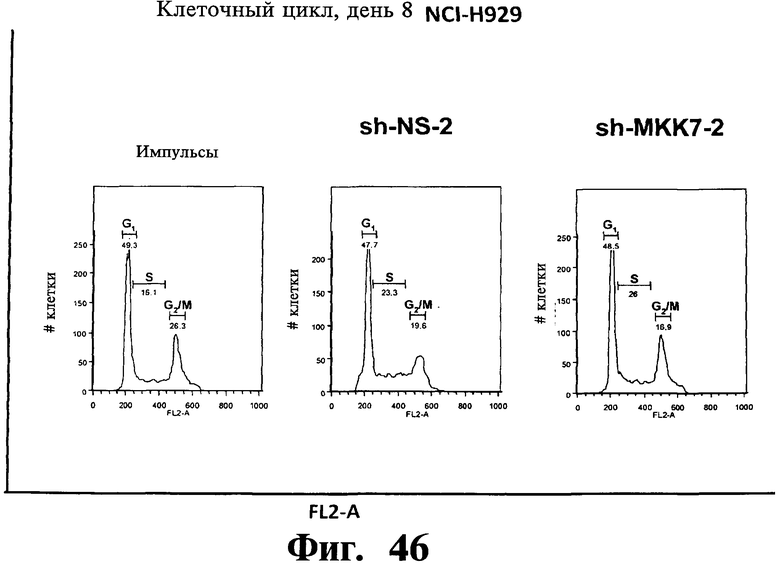
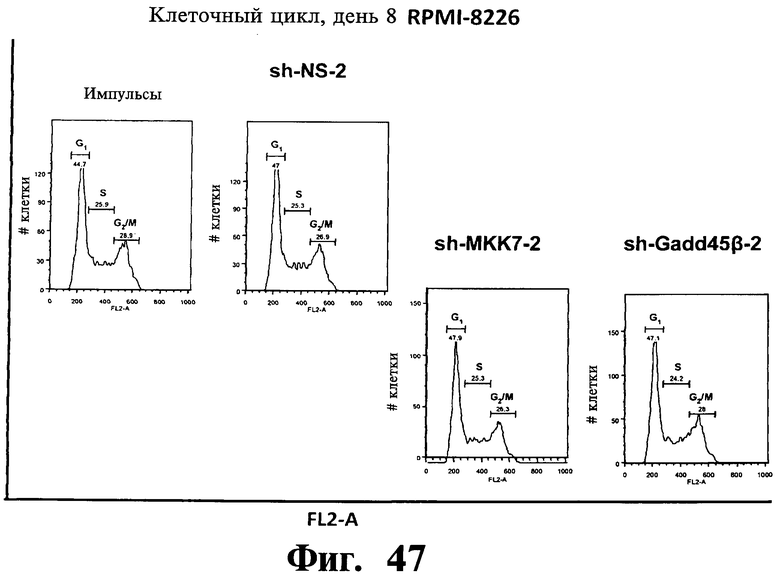
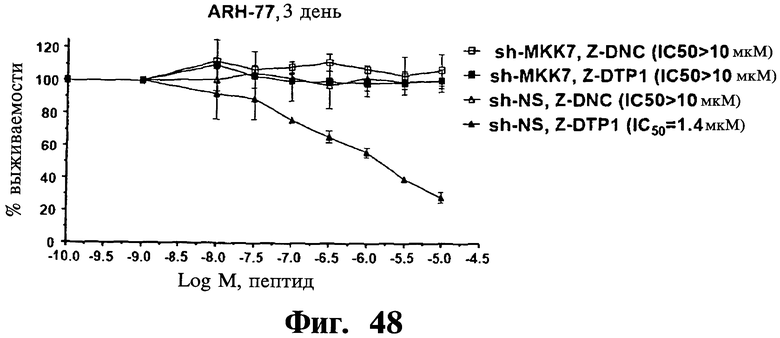
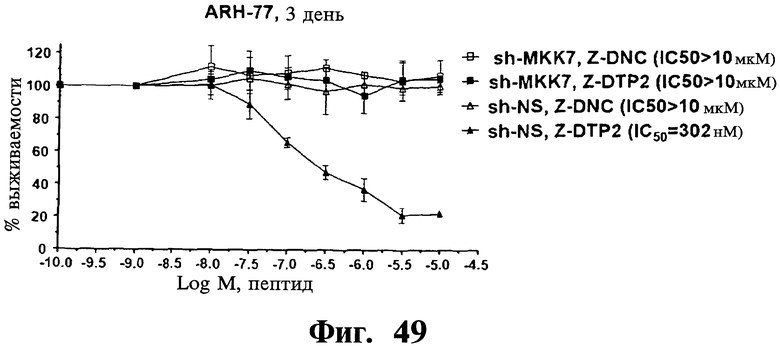
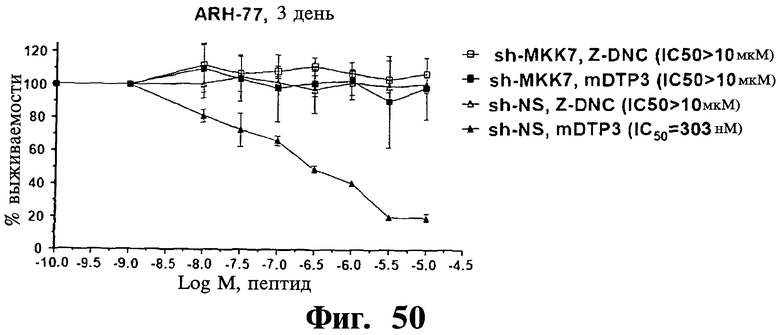
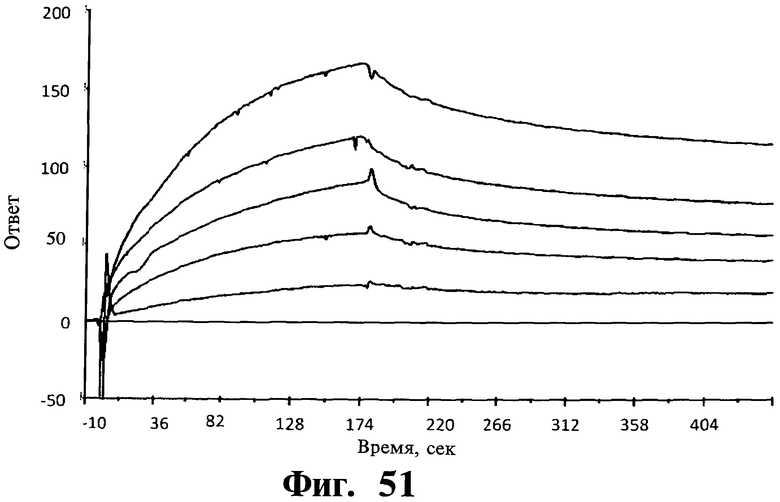
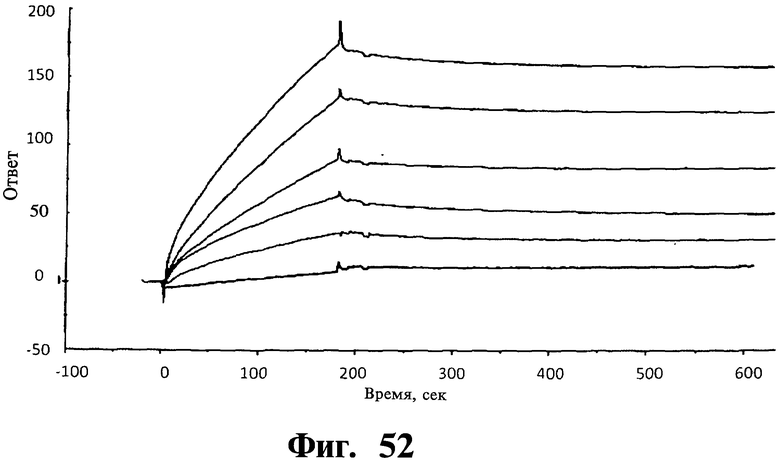
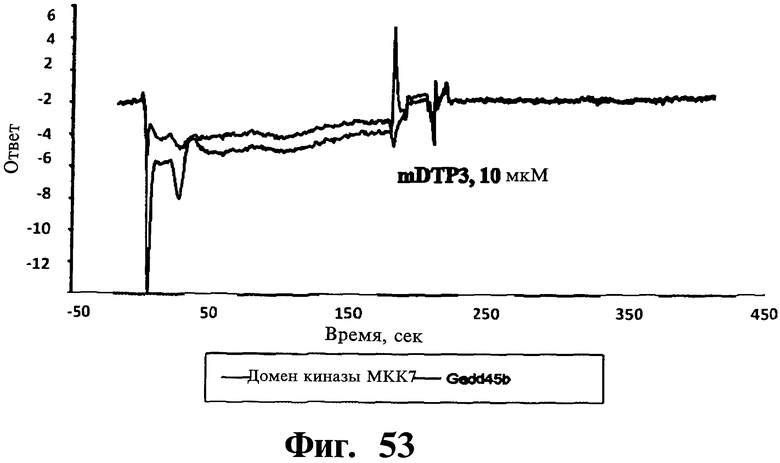
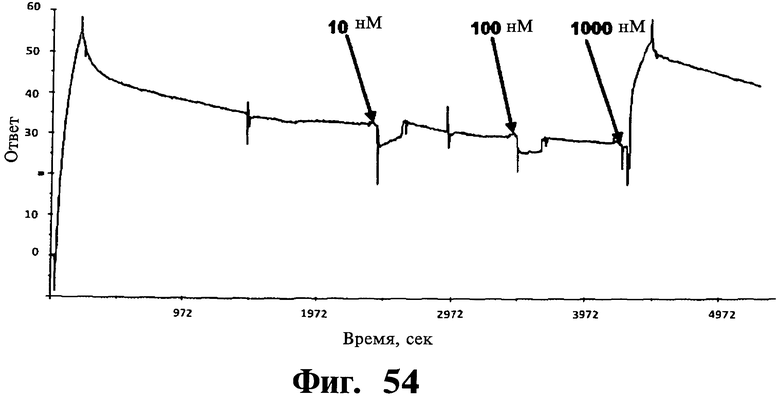
Изобретение относится к применению соединений на основе тетрапептидных и трипептидных групп и соответствующих пептоидных групп при лечении или профилактике ракового заболевания у человека, которое характеризуется повышенным уровнем экспрессии или активности Gadd45β по сравнению с обычными здоровыми клетками, в случае зависимости жизнеспособности и/или роста раковых клеток от NF-κB. 8 н. и 6 з.п. ф-лы, 54 ил., 9 табл., 15 пр.
1. Применение соединений, охватываемых общей структурной формулой I:
I: X1-A-X2,
где
А представляет собой олигопептидную группу или олигопептоидную группу, имеющую остатки Y1-Y2-Y3-Y4,
Y1 представляет собой
D-триптофан,
L-триптофан,
D-тирозин,
L-тирозин,
D-фенилаланин,
L-фенилаланин,
или N-метиловые производные в L- или D-конфигурации любого из вышеперечисленных;
Y2 отсутствует или представляет собой
D-глутаминовую кислоту,
L-глутаминовую кислоту,
D-аспарагиновую кислоту,
L-аспарагиновую кислоту,
L-лейцин,
D-лейцин,
L-глутамин,
D-глутамин,
L-метионин,
D-метионин,
D-2-аминогептандиоевую кислоту,
L-2-аминогептандиоевую кислоту,
сложный метиловый или этиловый эфир любого из вышеперечисленных,
L-гомосерин,
D-гомосерин,
или N-метиловые производные в L- или D-конфигурации любого из вышеперечисленных;
Y3 отсутствует или представляет собой
D-аргинин,
L-аргинин,
L-пролин,
D-пролин,
D-гистидин,
L-гистидин,
D-лизин,
D-α,β-диаминопропионовую кислоту (D-Dap),
L-α,β-диаминопропионовую кислоту (L-Dap),
L-α,δ-диаминомасляную кислоту (L-Dab),
L-α,δ-диаминомасляную кислоту (L-Dab),
L-орнитин,
D-орнитин,
L-лизин,
или N-метиловые производные в L- или D-конфигурации любого из вышеперечисленных;
при условии, что если Y2 отсутствует, то Y3 обязательно представляет собой аминокислотный остаток или остаток производного аминокислоты из указанных выше значений для Y3 и, наоборот, если Y3 отсутствует, то Y2 обязательно представляет собой аминокислотный остаток или остаток производного аминокислоты из указанных выше значений для Y2;
Y4 представляет собой
D-фенилаланин,
L-фенилаланин,
D-триптофан,
L-триптофан,
D-тирозин,
L-тирозин,
N-метиловые производные в L- или D-конфигурации вышеперечисленных,
Х1 отсутствует или представляет собой группу, введенную в амино- конец последовательности олигопептида или олигопептоида А для блокировки свободных аминогрупп, включающей
ацетил,
бензилоксикарбонил,
2-хлор-бензилоксикарбонил,
3-метокси-4-гидрокси-бензоил,
3-гидрокси-4-метокси-бензоил,
бензоил и
флуоренилметоксикарбонил;
Х2 отсутствует или представляет собой группу NHR, введенную в карбоксильный конец олигопептидной последовательности с образованием амидной связи, в которой R выбран из водорода, C1-C4-алкила, необязательно замещенного фенильной группой, и фенила;
или их производные, которые модифицированы липидированием или пегилированием, и фармацевтически приемлемые соли соединения формулы I или его вышеуказанных производных при лечении или профилактике ракового заболевания у человека, которое характеризуется повышенным уровнем экспрессии или активности Gadd45β по сравнению с обычными здоровыми клетками, в случае зависимости жизнеспособности и/или роста раковых клеток от NF-κB.
2. Применение по п. 1, в котором А представляет собой олигопептид, имеющий аминокислотную последовательность, выбранную из группы, включающей
(D-Tyr)-(D-Glu)-(D-Arg)-(D-Phe), [SEQ ID NO.: 27]
(D-Tyr)-(D-Asp)-(D-His)-(D-Phe), [SEQ ID NO.: 28]
(D-Tyr)-(D-Glu)-(D-Arg)-(D-Phe), [SEQ ID NO.: 29]
(D-Tyr)-(D-Asp)-(D-His)-(D-Phe), [SEQ ID NO.: 30]
(D-Tyr)-(D-Glu)-(D-Arg)-(D-Phe), [SEQ ID NO.: 31]
(D-Tyr)-(D-Asp)-(D-His)-(D-Tyr), [SEQ ID NO.: 32]
(D-Tyr)-(D-Glu)-(D-Arg)-(D-Tyr), [SEQ ID NO.: 33]
(D-Trp)-(D-Asp)-(D-His)-(D-Typ), [SEQ ID NO.: 34]
(D-Trp)-(D-Glu)-(D-Arg)-(D-Typ), [SEQ ID NO.: 35]
(D-Tyr)-(D-Asp)-(D-Lys)-(D-Phe), [SEQ ID NO.: 36]
(D-Tyr)-D-Glu)-(D-His)-(D-Phe), [SEQ ID NO.: 208]
(D-Tyr)-(D-Asp)-(D-Lys)-(D-Phe), [SEQ ID NO.: 209]
(D-Trp)-(D-Glu)-(D-His)-(D-Phe), [SEQ ID NO.: 210]
(D-Tyr)-(D-Glu)-(D-Lys)-(D-Phe), [SEQ ID NO.: 211]
(D-Trp)-(D-Glu)-(D-Lys)-(D-Phe), [SEQ ID NO.: 212]
(D-Trp)-(D-Asp)-(D-Lys)-(D-Phe), [SEQ ID NO.: 213]
(D-Tyr)-(D-Asp)-(D-His)-(D-Trp), [SEQ ID NO.: 214]
(D-Try)-(D-Glu)-(D-His)-(D-Trp), [SEQ ID NO.: 215]
(D-Trp)-(D-Asp)-(D-His)-(D-Trp), [SEQ ID NO.: 216]
(D-Trp)-(D-Glu)-(D-His)-(D-Trp), [SEQ ID NO.: 217]
(D-Tyr)-(D-Asp)-(D-Arg)-(D-Trp), [SEQ ID NO.: 218]
(D-Tyr)-(D-Asp)-(D-Lys)-(D-Trp), [SEQ ID NO.: 219]
(D-Tyr)-(D-Glu)-(D-Lys)-(D-Trp), [SEQ ID NO.: 220]
(D-Tyr)-(D-Glu)-(D-Arg)-(D-Trp), [SEQ ID NO.: 221]
(D-Trp)-(D-Glu)-(D-Lys)-(D-Trp), [SEQ ID NO.: 222]
(D-Trp)-(D-Glu)-(D-Arg)-(D-Trp), [SEQ ID NO.: 223]
(D-Trp)-(D-Asp)-(D-Lys)-(D-Trp), [SEQ ID NO.: 224]
(D-Trp)-(D-Gln)-(D-Arg)-(D-Trp), [SEQ ID NO.: 225]
(L-Tyr)-(L-Asp)-(L-Phe),
(D-Tyr)-(D-Arg)-(D-Phe),
(D-Tyr)-(D-Asp)-(D-Phe),
(L-Tyr)-(L-Glu)-(L-Phe),
(L-Tyr)-(L-Arg)-(L-Phe),
(D-Tyr)-(D-Arg)-(D-Phe),
(D-Tyr)-(D-Glu)-(D-Phe),
(D-Tyr)-(D-Pro)-(D-Phe),
(D-Tyr)-(D-Leu)-(D-Phe),
(D-Tyr)-(D-Asp)-(D-Tyr),
(D-Tyr)-(D-Glu)-(D-Tyr),
(D-Tyr)-(D-Pro)-(D-Tyr),
(D-Tyr)-(D-Leu)-(D-Tyr),
(D-Phe)-(D-Pro)-(D-Phe),
(D-Phe)-(D-Leu)-(D-Phe),
(D-Phe)-(D-Arg)-(D-Tyr),
(D-Phe)-(D-Glu)-(D-Tyr),
(D-Phe)-(D-Asp)-(D-Tyr),
(D-Phe)-(D-Pro)-(D-Tyr),
(D-Phe)-(D-Leu)-(D-Tyr),
(D-Tyr)-(D-Pro)-(D-Trp),
(D-Tyr)-(D-Leu)-(D-Trp),
(D-Tyr)-(D-Asp)-(D-Trp),
(D-Tyr)-(D-Glu)-(D-Trp),
(D-Tyr)-(D-Arg)-(D-Trp),
(D-Tyr)-(D-Pro)-(D-Trp),
(D-Tyr)-(D-Leu)-(D-Trp),
(D-Phe)-(D-Pro)-(D-Trp),
(D-Phe)-(D-Leu)-(D-Trp),
(D-Phe)-(D-Arg)-(D-Trp),
(D-Phe)-(D-Glu)-(D-Trp),
(D-Phe)-(D-Asp)-(D-Trp),
(D-Phe)-(D-Pro)-(D-Trp),
(D-Phe)-(D-Leu)-(D-Trp),
Ацетил-Tyr-Asp-Lys-Phe-NH2, [SEQ ID NO.: 39]
Ацетил-Tyr-Glu-Lys-Phe-NH2, [SEQ ID NO.: 40]
Ацетил-Tyr-Glu-His-Phe-NH2, [SEQ ID NO.: 41]
Ацетил-Tyr-Asp-Arg-Phe-NH2, [SEQ ID NO.: 42]
Ацетил-Trp-Glu-His-Phe-NH2, [SEQ ID NO.: 43]
Ацетил-Trp-Glu-Lys-Phe-NH2, [SEQ ID NO.: 44]
Ацетил-Trp-Asp-His-Phe-NH2, [SEQ ID NO.: 45]
Ацетил-Trp-Asp-Lys-Phe-NH2, [SEQ ID NO.: 46]
Ацетил-Tyr-Glu-Arg-Tyr-NH2, [SEQ ID NO.: 47]
Ацетил-Tyr-Asp-Lys-Tyr-NH2, [SEQ ID NO.: 48]
Ацетил-Tyr-Glu-Lys-Tyr-NH2, [SEQ ID NO.: 49]
Ацетил-Tyr-Glu-His-Tyr-NH2, [SEQ ID NO.: 50]
Ацетил-Tyr-Asp-Arg-Tyr-NH2, [SEQ ID NO.: 51]
Ацетил-Trp-Glu-His-Tyr-NH2, [SEQ ID NO.: 52]
Ацетил-Trp-Glu-Lys-Tyr-NH2, [SEQ ID NO.: 53]
Ацетил-Trp-Asp-His-Tyr-NH2, [SEQ ID NO.: 54]
Ацетил-Trp-Asp-Lys-Tyr-NH2, [SEQ ID NO.: 55]
Внутренний лактам ацетил-Tyr-Glu-Lys-Phe-NH2, [SEQ ID NO.: 56]
Ацетил-Tyr-Gln-Arg-Phe-NH2, [SEQ ID NO.: 57]
Ацетил-Tyr-Met-Arg-Phe-NH2, [SEQ ID NO.: 58]
Ацетил-Tyr-Leu-Arg-Phe-NH2, [SEQ ID NO.: 59]
Ацетил-Tyr-Arg-Phe-NH2,
Ацетил-D-Tyr-D-Arg-D-Phe-NH2,
Ацетил-Tyr-Glu-Phe-NH2,
Ацетил-Tyr-Glu-Tyr-NH2,
Ацетил-Tyr-Asp-Phe-NH2,
Ацетил-Tyr-Asp-Tyr-NH2,
H-Tyr-Arg-Phe-NH2,
H-Tyr-Glu-Phe-NH2,
H-Tyr-Glu-Tyr-NH2,
H-Tyr-Asp-Phe-NH2,
H-Tyr-Asp-Tyr-NH2,
H-Tyr-Pro-Phe-NH2,
H-Tyr-Lys-Phe-NH2,
Бензилоксикарбонил-Tyr-Arg-Phe-NH2,
Бензилоксикарбонил-Tyr-Arg-Tyr-NH2,
Бензилоксикарбонил-Tyr-Glu-Phe-NH2,
Бензилоксикарбонил-Tyr-Glu-Tyr-NH2,
Бензилоксикарбонил-Tyr-Asp-Phe-NH2,
Бензилоксикарбонил-Tyr-Asp-Tyr-NH2,
Бензилоксикарбонил-Tyr-Pro-Phe-NH2,
Бензилоксикарбонил-Tyr-Lys-Phe-NH2,
Бензилоксикарбонил-Tyr-His-Phe-NH2,
Бензилоксикарбонил-Tyr-Glu-Arg-Phe-NH2, [SEQ ID NO.: 60]
Бензилоксикарбонил-Tyr-Asp-His-Phe-NH2, [SEQ ID NO.: 61]
Бензилоксикарбонил-Tyr-Asp(ОМе)-His-Phe-NH2, [SEQ ID NO.: 62]
Бензилоксикарбонил-Tyr-Arg-Phe-NH2,
Бензилоксикарбонил-Tyr-Glu-Phe-NH2,
Бензилоксикарбонил-(N-метил)Tyr-(N-метил)Arg-(N-метил)Phe-NH2,
Бензилоксикарбонил-(N-метил)Tyr-Glu-(N-метил)Phe-NH2,
Бензилоксикарбонил-Tyr-(N-метил)Arg-(N-метил)Phe-NH2,
Бензилоксикарбонил-(N-метил)Tyr-(N-метил)Arg-Phe-NH2,
Бензилоксикарбонил-Tyr-Glu-(N-метил)Phe-NH2,
Бензилоксикарбонил-Tyr-(N-метил)Glu-Phe-NH2,
Бензилоксикарбонил-(N-метил)Tyr-Glu-Phe-NH2,
Ацетил-(N-метил)Tyr-Glu-(N-метил)Phe-NH2,
Ацетил-Tyr-(N-метил)Arg-(N-метил)Phe-NH2,
Ацетил-(N-метил)Tyr-(N-метил)Arg-Phe-NH2,
Ацетил-Tyr-Glu-(N-метил)Phe-NH2,
Ацетил-Tyr-(N-метил)Glu-Phe-NH2,
Ацетил-(N-метил)Tyr-Glu-Phe-NH2,
Н-(N-метил)Tyr-(N-метил)Arg-(N-метил)Phe-NH2,
Н-(N-метил)Tyr-Glu-(N-метил)Phe-NH2,
Н-Tyr-(N-метил)Arg-(N-метил)Phe-NH2,
Н-(N-метил)Tyr-(N-метил)Arg-Phe-NH2,
H-Tyr-Glu-(N-метил)Phe-NH2,
H-Tyr-(N-метил)Glu-Phe-NH2,
H-(N-метил)Tyr-Glu-Phe-NH2,
Ацетил-Tyr-Glu-(β-гомо)Phe-NH2,
Ацетил-Tyr-(β-гомо)Glu-Phe-NH2,
Ацетил-(β-гомо)Tyr-Glu-Phe-NH2,
(3-Метокси,4-гидрокси-бензоил)-Tyr-Glu-Arg-Phe-NH2, [SEQ ID NO.: 63]
(3-Метокси,4-гидрокси-бензоил)-Tyr-Asp-His-Phe-NH2, [SEQ ID NO.: 64]
(3-Метокси,4-гидрокси-бензоил)-Tyr-Asp(OMe)-His-Phe-NH2, [SEQ ID NO.: 65]
(3-Метокси,4-гидрокси-бензоил)-Tyr-Arg-Phe-NH2,
(3-Метокси,4-гидрокси-бензоил)-Tyr-Glu-Phe-NH2,
Флуоренилметилоксикарбонил-Tyr-Glu-Arg-Phe-NH2, [SEQ ID NO.: 66]
Флуоренилметилоксикарбонил-Tyr-Asp-His-Phe-NH2, [SEQ ID NO.: 67]
Флуоренилметилоксикарбонил-Tyr-Asp(OMe)-His-Phe-NH2, [SEQ ID NO.: 68]
Флуоренилметилоксикарбонил-Tyr-Asp(OMe)-His-Phe-NH2, [SEQ ID NO.: 69]
Флуоренилметилоксикарбонил-Tyr-Arg-Phe-NH2,
Флуоренилметилоксикарбонил-Tyr-Glu-Phe-NH2,
Миристил-Tyr-Glu-Arg-Phe-NH2, [SEQ ID NO.: 70]
Миристил-Tyr-Asp-His-Phe-NH2, [SEQ ID NO.: 71]
Миристил-Tyr-Arg-Phe-NH2,
Миристил-Tyr-Glu-Phe-NH2,
Миристил-Tyr-Asp(OMe)-His-Phe-NH2, [SEQ ID NO.: 72]
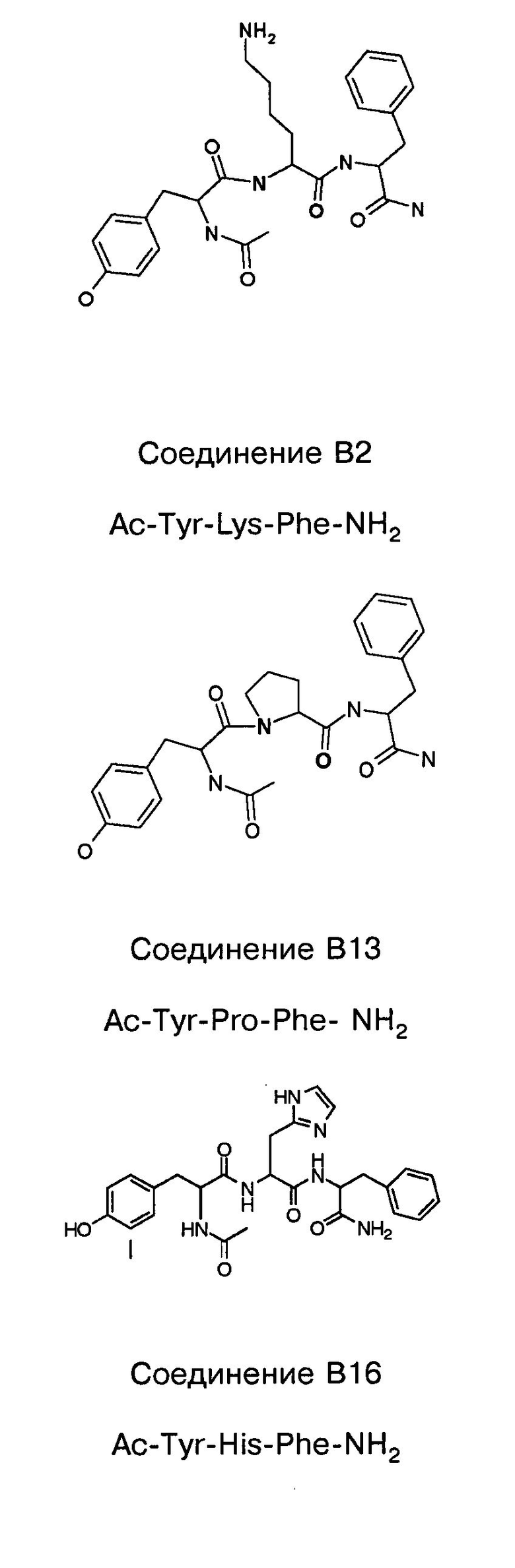
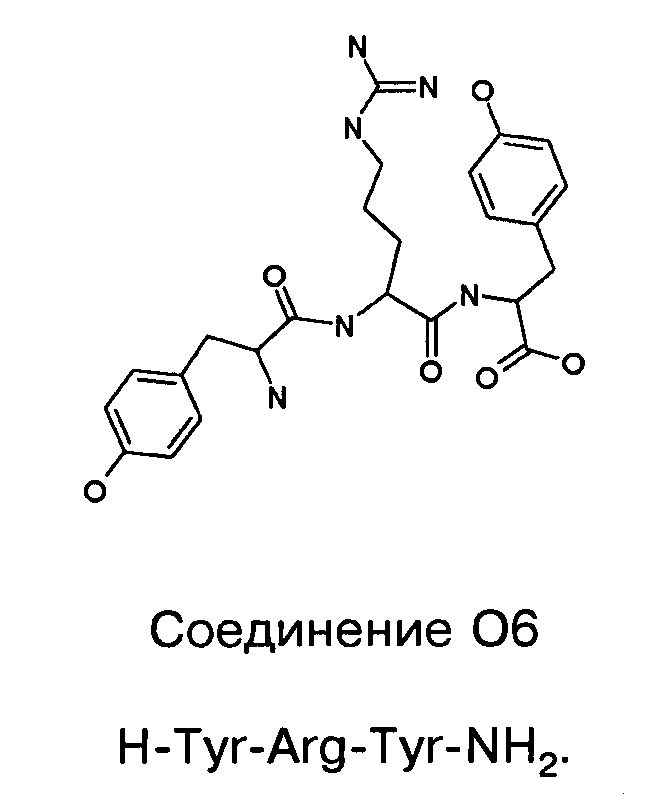
3. Применение по любому из пп. 1-2, в котором соединения характеризуются полупериодом существования в кровотоке человека по меньшей мере 15 минут.
4. Применение по любому из пп. 1-2, в котором соединения или сохраняют по меньшей мере 20% своей способности связываться с Gadd45β и/или MKK7 в соответствии с результатами анализов на связывание in vitro, или по меньшей мере 20% своей способности блокировать взаимодействие Gadd45β с MKK7 в соответствии с результатами анализов на связывание in vitro, или обладают способностью восстанавливать по меньшей мере 20% ингибированной Gadd45β каталитической активности MKK7 в соответствии с результатами анализов на условия, которые не ингибируют активность MKK7.
5. Применение по любому из пп. 1-2, в котором соединения обладают способностью уничтожать in vitro по меньшей мере 20% клеток в культуре клеточной линии человеческой миеломы, выбранной из группы, включающей клетки U266, KMS-11, NCl-H929, ARH-77 и JJN3, KMS-12, KMS-18 и KMS-27, в условиях, при которых 80% Т-клеточной линии JURKAT не погибает.
6. Применение по любому из пп. 1-2, в котором заболевание или нарушение выбрано из группы, включающей множественную миелому, лимфому Беркетта, промоноцитарный лейкоз или диффузную крупноклеточную В-клеточную лимфому.
7. Применение по п. 1, в котором А представляет собой олигопептид, имеющий аминокислотную последовательность (D-Tyr)-(D-Arg)-(D-Phe).
8. Трипептид, включающий аминокислотную последовательность (D-Tyr)-(D-Arg)-(D-Phe).
| WO 2006049597 A1, 11.05.2006 | |||
| WO 2009073725 A2, 11.06.2009 | |||
| WO 2004094471 A2, 04.11.2004 | |||
| US 2008146496 A1 19.06.2008 | |||
| US 20070032430 A1 08.02.2007 | |||
| WO 2004013306 A2, 12.02.2004 | |||
| US 2008095821 A1, 24.04.2008 | |||
| US 2005265970 A1,01.12.2005 | |||
| ПЕПТИД, СТИМУЛИРУЮЩИЙ ПРОТИВООПУХОЛЕВЫЙ ИММУННЫЙ ОТВЕТ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ, СПОСОБ ЛЕЧЕНИЯ МЛЕКОПИТАЮЩЕГО И СПОСОБ МОДУЛЯЦИИ ИММУННОГО ОТВЕТА | 2002 |
|
RU2305107C2 |

