Область техники, к которой относится настоящее раскрытие
Настоящее изобретение относится к области медицины и предусматривает композиции для генной терапии, предназначенные для применения в лечении метаболического нарушения, применения в лечении воспаления и/или фиброза печени, применения в лечении рака и/или применения в продлении жизни в здоровом состоянии у млекопитающих, в особенности, у людей.
Предшествующий уровень техники настоящего изобретения
Распространенность сахарного диабета растет с вызывающей тревогу скоростью и является основной проблемой здравоохранения в общемировом масштабе. Ожирение четко ассоциировано с резистентностью к инсулину и сахарным диабетом 2 типа (T2D) (Moller, D.Е., and Flier, J.S., 1991. N. Engl. J. Med. 325:938-948). Более того, ожирение повышает риск смерти (Peeters, A. et al., 2003. Ann. Intern. Med. 138:24-32), а также является существенным фактором риска для заболевания сердца, дисфункции иммунной системы, гипертензии, артрита, нейродегенеративных заболеваний и определенных типов рака (Roberst, D. L. et al., 2010. Annu. Rev. Med. 61:301-316; Spiegelman, В.M. et al., 1993. J. Biol. Chem. 268:6823-6826; Whitmer, R.A., 2007. Curr. Alzheimer Res. 4:117-122). Несмотря на клиническую значимость T2D и ожирения, не существует доступных эффективных методов лечения. Следовательно, существует насущная потребность в новых и безопасных подходах для предупреждения и борьбы с современной эпидемией T2D-ожирения. Недавно получило широкое признание утверждение, что ожирение является важным фактором риска для рака (Roberst, D.L. etal., 2010. Annu. Rev. Med. 61:301-316). С учетом современной эпидемии ожирения риски связанного с ожирением рака являются важной с клинической точки зрения проблемой, для решения которой срочно требуются новые и безопасные подходы. Увеличение массы и резистентность к инсулину также ассоциированы со старением. Следовательно, для продления жизни в здоровом состоянии необходимы новые и безопасные подходы к предупреждению и обращению развития ожирения и сахарного диабета. Фиброз печени представляет собой избыточное накопление белков внеклеточного матрикса, например, коллагена, преимущественно являющееся результатом хронического воспаления печени. Запущенный фиброз печени будет приводить к циррозу печени, портальной гипертензии и печеночной недостаточности. Таким образом, требуются новые и безопасные противофиброзные терапевтические средства.
Фактор роста фибробластов 21 (FGF21), фактор роста, преимущественно секретируемый печенью, а также жировой тканью и поджелудочной железой (Muise, Е.S. et al., 2008. Mot. Pharmacol. 74:403-412), как было показано, повышает рост бурой жировой ткани (ВАТ) и экспрессию термогенных генов в ВАТ и белой жировой ткани (WAT), стимулируя потребление энергии (Coskun, Т. et al., 2008. Endocrinology 149:6018-6027; Fisher, F.M. et al., 2012. Genes Dev. 26:271-281; Kharitonenkov, A. et al., 2005. J. Clin. Invest 115:1627-1635; Konishi, M. et al., 2000. J. Biol. Chem. 275:12119-12122; Tomlinson, E. et al., 2002. Endocrinology 143:1741-1747; Xu, J. et al., 2009. Diabetes 58:250-259).
Сверхэкспрессия FGF21 у трансгенных мышей защищала их от алиментарного ожирения (Kharitonenkov, A. et al., 2005. J. Clin. Invest 115:1627-1635). и введение FGF21 ob/ob мышам, db/db мышам или мышам, которых кормили рационом с высоким содержанием жиров (HFD), или ZDF крысам с ожирением стимулировало сильное снижение ожирения, значительно снижало уровень глюкозы и триглицеридов в крови, снижало уровни инсулина натощак и улучшало чувствительность к инсулину (Coskun, Т. et al., 2008. Endocrinology 149:6018-6027; Kharitonenkov, A. et al., 2005. J. Clin. Invest 115:1627-1635; Xu, J. et al., 2009. Diabetes 58:250-259; Adams, A.C. et al., 2012. PLoS. One. 7:e38438.; Berglund, E.D. et al., 2009. Endocrinology 150:4084-4093). Более того, введение FGF21 макакам-резусам с ожирением и диабетом резко снижало уровни глюкозы, фруктозамина, триглицеридов, инсулина и глюкагона в плазме крови натощак и индуцировало небольшую, но значимую потерю массы (Kharitonenkov, A. et al., 2007. Endocrinology 148:774-781).
Нативный белок FGF21 проявляет плохие фармакокинетические характеристики. Он характеризуется коротким периодом полужизни и является чувствительным к протеолитическому разрушению in vivo и агрегации in vitro (Huang, J. et al., 2013. J Pharmacol Exp Ther. 346(2):270-80; So. W.Y. and Leung. P.S. 2016. Med Res Rev. 36(4):672-704; Zhang, J. and Li, Y. 2015. Front Endocrinol (Lausanne). 6:168). Различные генно-инженерные подходы были разработаны для продления периода полужизни и улучшения стабильности и растворимости FGF21. В настоящее время два сконструированных миметика FGF21 (LY2405319 и PF-05231023) проходят исследования у людей. Тем не менее эти миметики FGF21 требуют нескольких введений, что накладывает значительную нагрузку на пациентов. Более того, сконструированные миметики/аналоги FGF21 могут проявлять более высокий риск иммуногенности, чем нативный FGF21, например, у пациентов, получавших лечение с использованием LY2405319, развивались реакции в месте инъекции, появлялись антитела к лекарственному средству и развивалась серьезная реакция гиперчувствительности (Gaich, G. et al., 2013. Cell Metab. 18(3):333-40).
Следовательно, все еще существует потребность в новых методах лечения сахарного диабета, и/или ожирения, и/или воспаления, и/или фиброза, и/или рака печени, и/или продления жизни в здоровом состоянии, которые не имеют всех недостатков существующих методов лечения.
Раскрытие настоящего изобретения
Авторы настоящего изобретения разработали улучшенные стратегии генной терапии, основанной на опосредуемом вектором на основе аденоассоциированного вируса (AAV) переносе гена FGF21 в печень, жировую ткань и/или скелетную мышцу, для противодействия метаболическим нарушениям, предпочтительно, сахарному диабету и/или ожирению. Генную терапию согласно настоящему изобретению также можно применять для противодействия воспалению и/или фиброзу печени. Кроме того, генную терапию согласно настоящему изобретению также можно применять для продления жизни в здоровом состоянии посредством противодействия метаболическим нарушениям, ассоциированным со старением, предпочтительно, сахарному диабету и/или ожирению. Кроме того, генную терапию согласно настоящему изобретению также можно применять для противодействия раку, предпочтительно, раку печени.
Создание одновекторных генных конструкций обеспечивает возможность продуцирования in vivo нативного FGF21, что должно привести в результате к уменьшенному риску иммуногенности или других проявлений токсичности.
Тем не менее, квалифицированному специалисту известно, что нативный FGF21 может быть чувствительным к протеолитическому разрушению in vivo и/или может иметь более быстрый клиренс in vivo. Все векторы, исследуемые в экспериментальной части, оказались способными обеспечивать возможность продолжительной секреции стабильного нативного FGF21 в кровоток. Эффективность сохраняется даже при однократном введении векторов, переносящих ген.
Таким образом, создание таких векторов на основе AAV для продуцирования in vivo нативного FGF21 не является стандартной практикой для специалиста в данной области техники, что демонстрируется в экспериментальной части.
Вирусная экспрессионная конструкция
В соответствии с первым аспектом предусмотрена вирусная экспрессионная конструкция, подходящая для экспрессии у млекопитающего и содержащая нуклеотидную последовательность, которая кодирует фактор роста фибробластов 21 (FGF21), подлежащий экспрессии в печени, жировой ткани и/или скелетной мышце.
Определения терминов «вирусная экспрессионная конструкция», «подходящий для экспрессии у млекопитающего», «печень», «жировая ткань» и «скелетная мышца» были представлены в части описания, озаглавленной «Общие определения».
Предпочтительная нуклеотидная последовательность, кодирующая FGF21, которая присутствует в вирусной экспрессионной конструкции согласно настоящему изобретению, является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 4, 5, 6, 7, 8, 9, 10 или 11. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения».
Более предпочтительная нуклеотидная последовательность, кодирующая человеческий FGF21, которая присутствует в вирусной экспрессионной конструкции согласно настоящему изобретению, является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 4, 5, 6 или 7. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения». SEQ ID NO: 4 представляет собой нуклеотидную последовательность, кодирующую человеческий FGF21. SEQ ID NO: 5 представляет собой оптимизированную по кодону нуклеотидную последовательность, кодирующую человеческий FGF21, вариант 1. SEQ ID NO: 6 представляет собой оптимизированную по кодону нуклеотидную последовательность, кодирующую человеческий FGF21, вариант 2. SEQ ID NO: 7 представляет собой оптимизированную по кодону нуклеотидную последовательность, кодирующую человеческий FGF21, вариант 3. Вариант 1, вариант 2 и вариант 3 кодируют одинаковый белок человеческого FGF21, и они были получены с помощью разных алгоритмов оптимизации по кодону. Еще одна предпочтительная нуклеотидная последовательность, кодирующая мышиный FGF21, которая присутствует в вирусной экспрессионной конструкции согласно настоящему изобретению, является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 8 или 9. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения». SEQ ID NO: 8 представляет собой нуклеотидную последовательность, кодирующую мышиный FGF21. SEQ ID NO: 9 представляет собой оптимизированную по кодону нуклеотидную последовательность, кодирующую мышиный FGF21. Еще одна предпочтительная нуклеотидная последовательность, кодирующая собачий FGF21, которая присутствует в вирусной экспрессионной конструкции согласно настоящему изобретению, является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 10 или 11. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения». SEQ ID NO: 10 представляет собой нуклеотидную последовательность, кодирующую собачий FGF21. SEQ ID NO: 11 представляет собой оптимизированную по кодону нуклеотидную последовательность, кодирующую собачий FGF21. Нуклеотидная последовательность, кодирующая FGF21, может быть получена из любого гена FGF21 или кодирующей последовательности FGF21, предпочтительно, от человека, мыши или собаки; или из мутированного гена FGF21 или кодирующей последовательности FGF21, или оптимизированного по кодону гена FGF21 или кодирующей последовательности FGF21, предпочтительно, от человека, мыши или собаки.
В контексте данного документа FGF21 проявляет по меньшей мере выявляемый уровень активности FGF21, как известно квалифицированному специалисту. Активность FGF21 заключается в повышении чувствительности к инсулину. Эту активность можно оценить с применением теста толерантности к инсулину, который описан в экспериментальной части, предпочтительно, в примере 8 или 9.
В соответствии с вариантом осуществления предусмотрена вирусная экспрессионная конструкция, описанная выше, в которой нуклеотидная последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, является выбранной из группы, состоящей из:
(a) нуклеотидной последовательности, кодирующей полипептид, содержащий аминокислотную последовательность, которая является по меньшей мере на 60% идентичной аминокислотной последовательности SEQ ID NO: 1, 2 или 3.
(b) нуклеотидной последовательности, которая является по меньшей мере на 60% идентичной нуклеотидной последовательности SEQ ID NO: 4, 5, 6, 7, 8, 9, 10 или 11.
(c) нуклеотидной последовательности, последовательность которой отличается от последовательности нуклеотидной последовательности из (b) вследствие вырожденности генетического кода.
Предпочтительная нуклеотидная последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, кодирует полипептид, содержащий аминокислотную последовательность, которая является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 1, 2 или 3. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения». SEQ ID NO: 1 представляет собой аминокислотную последовательность человеческого FGF21. SEQ ID NO: 2 представляет собой аминокислотную последовательность мышиного FGF21. SEQ ID NO: 3 представляет собой аминокислотную последовательность собачьего FGF21.
В соответствии с вариантом осуществления предусмотрена вирусная экспрессионная конструкция, которая описана выше, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и по меньшей мере один из элементов а), b), с), d) и е):
(a) специфический для печени промотор;
(b) специфический для жировой ткани промотор;
(c) комбинация убиквитарного промотора, и по меньшей мере одной нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, которая экспрессируется в печени, и по меньшей мере одной нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, которая экспрессируется в сердце, при этом указанная комбинация обеспечивает возможность специфической экспрессии в жировой ткани;
(d) активный в скелетных мышцах промотор и
(e) комбинация убиквитарного промотора и последовательности вектора на основе аденоассоциированного вируса (AAV), при этом указанная комбинация обеспечивает возможность специфической экспрессии в скелетной мышце.
Выражения «последовательность-мишень микроРНК, которая экспрессируется в печени», или «последовательность-мишень miRNA, которая экспрессируется в печени», или «сайт-связывания микроРНК, которая экспрессируется в печени», относятся к нуклеотидной последовательности, которая является комплементарной или частично комплементарной по меньшей мере части микроРНК, которая экспрессируется в печени. Аналогично, выражения «последовательность-мишень микроРНК, которая экспрессируется в сердце», или «последовательность-мишень miRNA, которая экспрессируется в сердце», или «сайт связывания микроРНК, которая экспрессируется в сердце», относятся к нуклеотидной последовательности, которая является комплементарной или частично комплементарной по меньшей мере части микроРНК, которая экспрессируется в сердце. Часть микроРНК, которая экспрессируется в печени, или часть микроРНК, которая экспрессируется в сердце, как определено в данном документе, означает нуклеотидную последовательность по меньшей мере из пяти или по меньшей мере из шести последовательных нуклеотидов в указанной микроРНК. Последовательность сайта связывания может характеризоваться идеальной комплементарностью по меньшей мере части экспрессируемой микроРНК, что означает, что последовательности могут идеально соответствовать, несоответствия могут не встречаться. В качестве альтернативы, последовательность сайта связывания может являться частично комплементарной по меньшей мере части экспрессируемой микроРНК, что означает, что может встречаться одно несоответствие/пять, шесть последовательных нуклеотидов. Частично комплементарные сайты связывания предпочтительно содержат участок, идеально или почти идеально комплементарный затравочному участку микроРНК, что означает, что между затравочным участком микроРНК и сайтом ее связывания могут не встречаться несоответствия (идеальная комплементарность) или может встречаться одно несоответствие/пять, шесть последовательных нуклеотидов (почти идеальная комплементарность). Затравочный участок микроРНК состоит из 5' участка микроРНК от приблизительно нуклеотида 2 до приблизительно нуклеотида 8 в микроРНК (т.е. 6 нуклеотидов). Часть, которая определена в данном документе, предпочтительно представляет собой затравочный участок указанной микроРНК. Разрушение информационной РНК (мРНК), содержащей последовательность-мишень для микроРНК, которая экспрессируется в печени, или микроРНК, которая экспрессируется в сердце, может происходить посредством пути РНК-интерференции или посредством прямого трансляционного контроля (ингибирования) мРНК. Настоящее изобретение не ограничивается путем, который в конечном счете используется miRNA в ингибировании экспрессии трансгена или кодируемого белка.
В контексте настоящего изобретения нуклеотидная последовательность, кодирующая последовательность-мишень микроРНК, которая экспрессируется в печени, может быть заменена на нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: 12 или 14-23. Более предпочтительная нуклеотидная последовательность является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 12 или 14-23. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения». В соответствии с одним вариантом осуществления нуклеотидная последовательность, кодирующая последовательность-мишень микроРНК, которая экспрессируется в печени, может быть заменена на нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: 12. Более предпочтительная нуклеотидная последовательность является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 12. В соответствии с дополнительным вариантом осуществления по меньшей мере одна копия нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, которая экспрессируется в печени, определенной в SEQ ID NO: 12 или 14-23, присутствует в вирусной экспрессионной конструкции согласно настоящему изобретению. В соответствии с дополнительным вариантом осуществления две, три, четыре, пять, шесть, семь или восемь копий нуклеотидной последовательности, кодирующей последовательность-мишень специфической для печени микроРНК, которая определена в SEQ ID NO: 12 или 14-23, присутствуют в вирусной экспрессионной конструкции согласно настоящему изобретению. В соответствии с предпочтительным вариантом осуществления одна, две, три, четыре, пять, шесть, семь или восемь копий нуклеотидной последовательности, кодирующей miRT122a (SEQ ID NO: 12), присутствуют в вирусной экспрессионной конструкции согласно настоящему изобретению.
Последовательность-мишень микроРНК, которая экспрессируется в печени, в данном случае проявляет по меньшей мере выявляемый уровень активности последовательности-мишени микроРНК, которая экспрессируется в печени, как известно квалифицированному специалисту. Активность последовательности-мишени микроРНК, которая экспрессируется в печени, заключается в связывании с ее когнатной микроРНК, которая экспрессируется в печени, и, когда она функционально связана с трансгеном, в опосредовании нарушения нацеливания экспрессии трансгена в печени. Эту активность можно оценить посредством измерения уровней экспрессии трансгена в печени с помощью qPCR, как описано в экспериментальной части.
В контексте настоящего изобретения нуклеотидная последовательность, кодирующая последовательность-мишень микроРНК, которая экспрессируется в сердце, может быть заменена на нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: 13 или 23-30. Предпочтительная нуклеотидная последовательность является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 13 или 23-30. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения». В соответствии с одним вариантом осуществления нуклеотидная последовательность, кодирующая последовательность-мишень микроРНК, которая экспрессируется в сердце, может быть заменена на нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: 13. Предпочтительная нуклеотидная последовательность является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 13. В соответствии с дополнительным вариантом осуществления по меньшей мере одна копия нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, которая экспрессируется в сердце, определенной в SEQ ID NO: 13 или 23-30, присутствует в вирусной экспрессионной конструкции согласно настоящему изобретению. В соответствии с дополнительным вариантом осуществления две, три, четыре, пять, шесть, семь или восемь копий нуклеотидной последовательности, кодирующей последовательность-мишень специфической для сердца микроРНК, которая определена в SEQ ID NO: 13 или 23-30, присутствуют в вирусной экспрессионной конструкции согласно настоящему изобретению. В соответствии с предпочтительным вариантом осуществления одна, две, три, четыре, пять, шесть, семь или восемь копий нуклеотидной последовательности, кодирующей miRT1 (SEQ ID NO: 13), присутствуют в вирусной экспрессионной конструкции согласно настоящему изобретению.
Активность последовательности-мишени микроРНК, которая экспрессируется в сердце, заключается в связывании с ее когнатной микроРНК, которая экспрессируется в сердце, и, когда она функционально связана с трансгеном, в опосредовании нарушения нацеливания экспрессии трансгена в сердце. Эту активность можно оценить посредством измерения уровней экспрессии трансгена в сердце с помощью qPCR, как описано в экспериментальной части.
В соответствии с вариантом осуществления предусмотрена вирусная экспрессионная конструкция, описанная выше, в которой нуклеотидная последовательность, кодирующая последовательность-мишень микроРНК, которая экспрессируется в печени, и нуклеотидная последовательность, кодирующая последовательность-мишень микроРНК, которая экспрессируется в сердце, является выбранной из группы, состоящей из последовательностей SEQ ID NO: 12-30 и/или их комбинаций.
В соответствии с одним вариантом осуществления по меньшей мере одна копия нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, которая экспрессируется в печени, определенной в SEQ ID NO: 12 или 14-23, и по меньшей мере одна копия нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, которая экспрессируется в печени, определенной в SEQ ID NO: 13 или 23-30, присутствуют в вирусной экспрессионной конструкции согласно настоящему изобретению. В соответствии с дополнительным вариантом осуществления две, три, четыре, пять, шесть, семь или восемь копий нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, которая экспрессируется в печени, определенной в SEQ ID NO: 12 или 14-23, и две, три, четыре, пять, шесть, семь или восемь копий нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, которая экспрессируется в сердце, определенной в SEQ ID NO: 13 или 23-30, присутствуют в вирусной экспрессионной конструкции согласно настоящему изобретению. В соответствии с дополнительным вариантом осуществления одна, две, три, четыре, пять, шесть, семь или восемь копий нуклеотидной последовательности, кодирующей miRT122a (SEQ ID NO: 12), и одна, две, три, четыре, пять, шесть, семь или восемь копий нуклеотидной последовательности, кодирующей miRT1 (SEQ ID NO: 13), объединены в вирусной экспрессионной конструкции согласно настоящему изобретению. В соответствии с дополнительным вариантом осуществления четыре копии нуклеотидной последовательности, кодирующей miRT122a (SEQ ID NO: 12), и четыре копии нуклеотидной последовательности, кодирующей miRT1 (SEQ ID NO: 13), объединены в вирусной экспрессионной конструкции согласно настоящему изобретению.
Определения терминов «промотор», «специфический для печени промотор», «специфический для жировой ткани промотор», «убиквитарный промотор», «активный в скелетных мышцах промотор» были представлены в части описания, озаглавленной «Общие определения».
Предпочтительный убиквитарный промотор представляет собой промотор CAG.
В контексте настоящего изобретения нуклеотидная последовательность промотора CAG может быть заменена на нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: 44. Предпочтительная нуклеотидная последовательность является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 44. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения».
Еще один предпочтительный убиквитарный промотор представляет собой промотор (CMV) цитомегаловируса.
В контексте настоящего изобретения нуклеотидная последовательность промотора CMV может быть заменена на нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: 45. Предпочтительная нуклеотидная последовательность является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 45. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения».
Предпочтительно, указанный промотор CMV применяют вместе с интронной последовательностью. В этом контексте интронная последовательность может быть заменена на нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: 43. Предпочтительная нуклеотидная последовательность является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 43. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения».
Предпочтительный специфический для печени промотор представляет собой промотор человеческого al-антитрипсина (hAAT).
В контексте настоящего изобретения нуклеотидная последовательность промотора hAAT может быть заменена на нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: 47. Предпочтительная нуклеотидная последовательность является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 47. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения».
Предпочтительно, указанный промотор hAAT применяют вместе с интронной последовательностью. Предпочтительная интронная последовательность представляет собой энхансер гепатоцитарного регуляторного участка (HCR) из аполипопротеина Е. Наиболее предпочтительная интронная последовательность представляет собой энхансер HCR из аполипопротеина Е, который определен в SEQ ID NO: 53. В этом контексте интронная последовательность может быть заменена на нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: 53. Предпочтительная нуклеотидная последовательность является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 53. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения». В соответствии с вариантом осуществления, указанный промотор hAAT применяют вместе с одной, двумя, тремя, четырьмя или пятью копиями интронной последовательности. В соответствии с предпочтительным вариантом осуществления указанный промотор hAAT применяют вместе с одной, двумя, тремя, четырьмя или пятью копиями энхансера HCR из аполипопротеина Е, который определен в SEQ ID NO: 53.
Другие специфические для печени промоторы представляют собой альбуминовый промотор, промотор основного белка мочи, промотор фосфоенолпируват-карбоксикиназы (PEPCK), промотор печеночного белка-активатора (liver-enriched protein activator), транстиретиновый промотор, промотор тироксин-связывающего глобулина, промотор аполипопротеина А1, промотор печеночной формы белка, связывающего жирные кислоты, и промотор фенилаланин-гидроксилазы.
Специфические для жировой ткани промоторы представляют собой промотор белка адипоцитов 2 (аР2, также известный как белок 4, связывающий жирные кислоты (FABP4)), промотор PPARy, адипонектиновый промотор, промотор фосфоенолпируват-карбоксикиназы (PEPCK), промотор, полученный из человеческого цитохрома р450 ароматазы (p450arom), промотор mini/aP2 (состоящий из специфического для жировой ткани энхансера аР2 и базового промотора аР2), промотор разобщающего белка 1 (UCP1), промотор mini/UCP1 (состоящий из специфического для жировой ткани энхансера UCP1 и базового промотора UCP1), адипсиновый промотор, лептиновый промотор или промотор Foxa-2. Предпочтительные специфические для жировой ткани промоторы представляют собой промотор mini/aP2 (SEQ ID NO: 54) и промотор mini/UCP1 (SEQ ID NO 55). В этом контексте последовательность специфического для жировой ткани промотора может быть заменена на нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: 53 или SEQ ID NO: 54. Предпочтительная нуклеотидная последовательность является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 53 или SEQ ID NO: 54. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения».
Предпочтительные активные в скелетных мышцах промоторы представляют собой промотор легкой цепи миозина, промотор тяжелой цепи миозина, десминовый промотор, промотор мышечной креатин-киназы (MCK), промотор гладкомышечного альфа-актина, промотор CK6, промотор миозинового шаперона Unc-45 В, базовый промотор MCK в комбинации с копиями энхансера MCK, промотор Enh358MCK (комбинация энхансера MCK с 358 п.о. проксимальным промотором гена МСК). Наиболее предпочтительный активный в скелетных мышцах промотор представляет собой промотор С5-12, который определен в SEQ ID NO: 56. В этом контексте последовательность активного в скелетных мышцах промотора может быть заменена на нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: 56. Предпочтительная нуклеотидная последовательность является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 56. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в части описания, озаглавленной «Общие определения».
Промотор, который применяют в контексте данного документа (особенно, когда промоторная последовательность определена как характеризующаяся минимальной процентной идентичностью с заданной SEQ ID NO), должен проявлять по меньшей мере активность промотора, которая известна квалифицированному специалисту. Пожалуйста, обратитесь к части описания, озаглавленной «Общие определения», для определения такой активности. Предпочтительно, промотор, определенный как характеризующийся минимальной процентной идентичностью с заданной SEQ ID NO, должен управлять транскрипцией нуклеотидной последовательности, он является функционально связанным с ней (т.е. нуклеотидной последовательностью, кодирующей FGF21), что оценивают в анализе, известном квалифицированному специалисту. В контексте настоящего изобретения указанный промотор является функционально связанным с нуклеотидной последовательностью FGF21, определенной выше. В соответствии с одним вариантом осуществления промотор является специфическим для клеток и/или специфическим для ткани, предпочтительно, специфическим для печени, жировой ткани и/или скелетной мышцы.
Таким образом, настоящим изобретением охватываются несколько вирусных экспрессионных конструкций:
вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и содержащая элемент а),
вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и содержащая элемент b),
вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и содержащая элемент с),
вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и содержащая элемент d),
вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и содержащая элемент е),
вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и содержащая элемент b) и нуклеотидную последовательность элемента с),
вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и содержащая элемент е) и нуклеотидную последовательность элемента с).
В соответствии с вариантом осуществления предусмотрена вирусная экспрессионная конструкция, которая описана в данном документе, в которой специфический для печени промотор представляет собой промотор человеческого α1-антитрипсина (hAAT), и/или специфический для жировой ткани промотор представляет собой промотор mini/ap2 и/или промотор mini/UCP1, и/или активный в скелетных мышцах промотор представляет собой промотор С5-12, и/или убиквитарный промотор представляет собой промотор цитомегаловируса (CMV) и/или промотор CAG.
В соответствии с вариантом осуществления предусмотрена вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и элемент а), при этом специфический для печени промотор представляет собой промотор hAAT (SEQ ID NO: 47).
В соответствии с предпочтительным вариантом осуществления предусмотрена вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и элемент а), при этом указанная конструкция представляет собой AAV8-hAAT-moFGF21. Эта конструкция содержит, например, вирусную экспрессионную конструкцию, которая изображена на фиг. 6А: ITR2-hAAT-moFGF21-polyA-ITR2; последовательность этой экспрессионной конструкции содержится в SEQ ID NO: 34.
Для этой конструкции в примере 3 неожиданно выявили высокую и стабильную специфическую для печени экспрессию после внутривенного введения. Было показано, что экспрессия является стабильной в течение периода до 1 года (пример 12). Всесторонние благоприятные терапевтические эффекты в отношении обращения развития и лечения ожирения и сахарного диабета продемонстрированы у ob/ob мышей (примеры 3 и 11), мышей, которых кормили рационом с высоким содержанием жиров (HFD), (примеры 4, 12-14) и старых мышей, которых кормили HFD (примеры 5, 12-14). Примеры 11 и 16 также выявили заметное улучшение при стеатозе печени, воспалении печени и фиброзе печени. В примере 15 показано улучшение при воспалении WAT, ассоциированном с ожирением. Пример 17 указывает на долговременную безопасность терапии. Пример 18 выявил благоприятный эффект в предупреждении возникновения опухолей печени. В примере 19 показан терапевтический потенциал в модели сахарного диабета I типа.
В соответствии с вариантом осуществления предусмотрена вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и элемент b), при этом специфический для жировой ткани промотор представляет собой промотор mini/aP2 (SEQ ID NO: 54) и/или промотор mini/UCP1 (SEQ ID NO 55).
В соответствии с вариантом осуществления предусмотрена вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и элемент с), при этом убиквитарный промотор представляет собой промотор CAG (SEQ ID NO: 44), и при этом по меньшей мере одна нуклеотидная последовательность, кодирующая последовательность-мишень микроРНК, которая экспрессируется в печени, является выбранной из группы, состоящей из SEQ ID NO: 12 или 14-23, и по меньшей мере одна нуклеотидная последовательность, кодирующая последовательность-мишень микроРНК, которая экспрессируется в сердце, является выбранной из группы, состоящей из SEQ ID NO: 13 или 23-30.
В соответствии с предпочтительным вариантом осуществления предусмотрена вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и элемент с), при этом указанная конструкция представляет собой AAV9-CAG-moFGF21-dmiRT или AAV8-CAG-moFGF21-dmiRT. Обозначения dmiRT и doublemiRT являются эквивалентными. Эти конструкции, например, содержат вирусную экспрессионную конструкцию, которая изображена на фиг. 1А: ITR2-CAG-moFGF21-4x miRT122a-4x miRT1-polyA-ITR2; последовательность этой экспрессионной конструкции содержится в SEQ ID NO: 32.
Для этих конструкций в примерах 1-2 неожиданно выявили высокую и стабильную специфическую для жировой ткани экспрессию после введения в eWAT. Всесторонние благоприятные терапевтические эффекты в отношении предупреждения, обращения развития и лечения ожирения и сахарного диабета показаны у нормальных мышей (пример 1) и ob/ob мышей (примеры 2 и 10). Пример 10 также выявил улучшение при стеатозе печени.
В соответствии с вариантом осуществления предусмотрена вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и элемент d), при этом активный в скелетных мышцах промотор представляет собой промотор С5-12 (SEQ ID NO: 56).
В соответствии с вариантом осуществления предусмотрена вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и элемент е), при этом убиквитарный промотор представляет собой промотор CMV (SEQ ID NO: 45), и серотип AAV представляет собой AAV1.
В соответствии с предпочтительным вариантом осуществления предусмотрена вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и элемент е), при этом указанная конструкция представляет собой AAV1-CMV-moFGF21. Эта конструкция содержит, например, вирусную экспрессионную конструкцию, которая изображена на фиг. 11А: ITR2-CMV-moFGF21-polyA-ITR2; последовательность этой экспрессионной конструкции содержится в SEQ ID NO: 36.
Для этой конструкции в примере 20 выявили высокую и стабильную специфическую для скелетной мышцы экспрессию после внутримышечного введения. Всесторонние благоприятные терапевтические эффекты в отношении предупреждения, обращения развития и лечения ожирения и сахарного диабета показаны у мышей, получавших HFD, и ob/ob мышей (примеры 6 и 21). Пример 20 выявил благоприятный эффект в продлении жизни в здоровом состоянии посредством предупреждения ожирения и сахарного диабета.
В соответствии с вариантом осуществления предусмотрена вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и элемент b) и нуклеотидную последовательность элемента с), при этом специфический для жировой ткани промотор представляет собой промотор mini/aP2 (SEQ ID NO: 54) и/или промотор mini/UCP1 (SEQ ID NO 55).
В соответствии с вариантом осуществления предусмотрена вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и элемент е) и нуклеотидную последовательность элемента с), при этом убиквитарный промотор представляет собой промотор CMV (SEQ ID NO: 45), и серотип AAV представляет собой AAV1.
Все конструкции согласно настоящему изобретению являются более привлекательными, чем конструкции из предшествующего уровня техники, такие как раскрытая в Zhang et al., EBioMedicine 15 (2017) 173-183, в особенности, конструкции, содержащие элемент а), который представляет собой специфический для печени промотор, предпочтительно, hAAT, и/или элемент с), который представляет собой комбинацию убиквитарного промотора и по меньшей мере одной нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, которая экспрессируется в печени, предпочтительно, miRT122a, и по меньшей мере одной нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, которая экспрессируется в сердце, предпочтительно, miRT1, при этом указанная комбинация обеспечивает возможность специфической экспрессии в жировой ткани, и/или элемент е), который представляет собой комбинацию убиквитарного промотора, предпочтительно, CMV, и последовательности вектора на основе аденоассоциированного вируса (AAV), предпочтительно, AAV1, при этом указанная комбинация обеспечивает возможность специфической экспрессии в скелетной мышце. Zhang и соавт. раскрыта кодирующая последовательность мышиного FGF21 дикого типа под контролем промотора фактора элонгации 1a (EF1a) (EF1a-mFGF21) (Zhang et al., EBioMedicine 15 (2017) 173-183). Эту конструкцию сравнивали с конструкциями согласно настоящему изобретению в примерах 23 и 24. Во всех экспериментах in vitro и in vivo все экспрессионные кассеты и векторы на основе AAV согласно настоящему изобретению опосредовали более высокий уровень экспрессии FGF21 в целевой ткани или типе клеток и более низкий уровень экспрессии FGF21 в нецелевых тканях, демонстрируя высокую эффективность экспрессионных кассет и векторов на основе AAV согласно настоящему изобретению, а также более высокую тканеспецифичность. Кроме того, конструкции CMV-moFGF21 и CAG-moFGF21-double miRT также опосредовали более высокий уровень продуцирования и секреции белка в культуральной среде в клетках HEK293 по сравнению с EF1a-mFGF21. Более того, hAAT-moFGF21 и AAV8-hAAT-moFGF21 также опосредовали более высокий уровень секреции FGF21 в кровоток, чем EF1a-mFGF21 и AAV8-EF1a-mFGF21.
В вирусной экспрессионной конструкции согласно настоящему изобретению могут присутствовать дополнительные последовательности, как подробно объяснено в части описания, озаглавленной «Общие определения». Предпочтительные дополнительные последовательности включают в себя инвертированные концевые повторы (ITR), сигнал полиаденилирования SV40 (SEQ ID NO: 50), сигнал полиаденилирования кроличьего β-глобина (SEQ ID NO: 51), последовательность энхансера CMV (SEQ ID NO: 46) и энхансер HCR из аполипопротеина Е (SEQ ID NO: 53). В контексте настоящего изобретения предусмотрено, что «ITR» охватывают один 5'ITR и один 3'ITR, каждый из которых получен из генома AAV. Предпочтительные ITR происходят из AAV2 и представлены SEQ ID NO: 48 (5'ITR) и SEQ ID NO: 49 (3'ITR). В контексте настоящего изобретения предусмотрено применение последовательности энхансера CMV (SEQ ID NO: 46) и последовательности промотора CMV (SEQ ID NO: 45) в виде двух отдельных последовательностей или в виде единой последовательности (SEQ ID NO: 52).
Каждая из этих дополнительных последовательностей может присутствовать в вирусной экспрессионной конструкции согласно настоящему изобретению (см., например, как изображено на фиг. 1, 2, 3, 4, 5, 6, 7, 8 и 9, а также как изображено на фиг. 11, 31 и 32).
В соответствии с вариантом осуществления вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и по меньшей мере один из элементов а), и/или b), и/или с), и/или d), и/или е), которые определены выше, дополнительно содержит:
- ITR, которые фланкируют экспрессионную кассету в указанной конструкции,
- сигналы полиаденилирования SV40 или кроличьего β-глобина, которые расположены в направлении 3' относительно нуклеотидной последовательности, кодирующей FGF21, и/или
- последовательность энхансера CMV или последовательность энхансера HCR, которая располагается в направлении 5' относительно нуклеотидной последовательности, кодирующей FGF21.
В соответствии с предпочтительным вариантом осуществления вирусная экспрессионная конструкция, содержащая нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и по меньшей мере один из элементов а), и/или b), и/или с), и/или d), и/или е), которые определены выше, дополнительно содержит ITR, которые фланкируют экспрессионную кассету в указанной конструкции, и, необязательно,
- сигналы полиаденилирования SV40 или кроличьего β-глобина, которые расположены в направлении 3' относительно нуклеотидной последовательности, кодирующей FGF21, и/или
- последовательность энхансера CMV или последовательность энхансера HCR, которая располагается в направлении 5' относительно нуклеотидной последовательности, кодирующей FGF21.
Эти последовательности применяли в экспериментальной части в некоторых из конструкций, идентифицированных в данном документе.
Таким образом, в соответствии с одним вариантом осуществления в случае каждой из этих предпочтительных вирусных экспрессионных конструкций, определенных выше, могла присутствовать дополнительная последовательность, выбранная из группы, состоящей из: ITR, сигнала полиаденилирования SV40, сигнала полиаденилирования кроличьего β-глобина, последовательности энхансера CMV, последовательности энхансера HCR из аполипопротеина Е.
В соответствии с предпочтительным вариантом осуществления вирусная экспрессионная конструкция содержит нуклеотидную последовательность, которая кодирует FGF21, подходящий для экспрессии у млекопитающего, и по меньшей мере один из элементов а), и/или b), и/или с), и/или d), и/или е), при этом присутствует дополнительная последовательность, которая является выбранной из группы, состоящей из: ITR, сигнала полиаденилирования SV40, сигнала полиаденилирования кроличьего β-глобина, последовательности энхансера CMV, последовательности энхансера HCR. Предпочтительные ITR представляют собой таковые из AAV2, которые представлены SEQ ID NO: 48 (5'ITR) и SEQ ID NO: 49 (3'ITR).
Предпочтительные вирусные экспрессионные конструкции содержат элементы а), и/или b), и/или с), и/или d), и/или е) и представляют собой конструкции, в которых экспрессионная кассета фланкирована 5'ITR и 3'ITR.
Другие предпочтительные вирусные экспрессионные конструкции содержат элементы а), и/или b), и/или с), и/или d), и/или е) и представляют собой конструкции, в которых экспрессионная кассета фланкирована 5'ITR и 3'ITR. Кроме того, присутствуют сигналы полиаденилирования SV40.
Другие предпочтительные вирусные экспрессионные конструкции содержат элементы а), и/или b), и/или с), и/или d), и/или е) и представляют собой конструкции, в которых экспрессионная кассета фланкирована 5'ITR и 3'ITR. Кроме того, присутствуют сигналы полиаденилирования кроличьего β-глобина.
Другие предпочтительные вирусные экспрессионные конструкции содержат элементы а), и/или b), и/или с), и/или d), и/или е) и представляют собой конструкции, в которых экспрессионная кассета фланкирована 5'ITR и 3'ITR. Кроме того, присутствует последовательность энхансера CMV.
Другие предпочтительные вирусные экспрессионные конструкции содержат элементы а), и/или b), и/или с), и/или d), и/или е) и представляют собой конструкции, в которых экспрессионная кассета фланкирована 5'ITR и 3'ITR. Кроме того, присутствует последовательность энхансера HCR из аполипопротеина Е.
Наиболее предпочтительные сконструированные вирусные экспрессионные конструкции включают в себя:
конструкцию В (представленную нуклеотидной последовательностью, содержащей SEQ ID NO: 32),
конструкцию D (представленную нуклеотидной последовательностью, содержащей SEQ ID NO: 34),
конструкцию F (представленную нуклеотидной последовательностью, содержащей SEQ ID NO: 36),
конструкцию G (представленную нуклеотидной последовательностью, содержащей SEQ ID NO: 37),
конструкцию Н (представленную нуклеотидной последовательностью, содержащей SEQ ID NO: 38),
конструкцию I (представленную нуклеотидной последовательностью, содержащей SEQ ID NO: 39),
конструкцию J (представленную нуклеотидной последовательностью, содержащей SEQ ID NO: 40),
конструкцию K (представленную нуклеотидной последовательностью, содержащей SEQ ID NO: 41).
конструкцию L (представленную нуклеотидной последовательностью, содержащей SEQ ID NO: 42).
Как будет понятно квалифицированному специалисту, каждая из этих вирусных экспрессионных конструкций уже содержит два ITR из AAV2 (т.е. SEQ ID NO: 48 (5'ITR) и SEQ ID NO: 49 (3'ITR)).
Конструкции В и G содержат сигнал полиаденилирования кроличьего β-глобина. Конструкция F содержит сигнал полиаденилирования SV40, последовательность энхансера CMV и нуклеотидную последовательность химерного интрона (состоящего из интронов из генов человеческого β-глобина и тяжелой цепи иммуноглобулина). Конструкции D, H-L содержат сигнал полиаденилирования SV40, последовательность энхансера HCR и нуклеотидную последовательность химерного интрона (состоящего из интронов из генов человеческого β-глобина и тяжелой цепи иммуноглобулина).
Как объясняется в общей части, озаглавленной «Общие определения», в настоящей заявке при каждом упоминании, относящемся к конкретной нуклеотидной последовательности SEQ ID NO (обозначенной SEQ ID NO: А, В или С), представляющей предпочтительные конструкции, сконструированные в данном документе, их можно заменить на:
i. нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: А, В или С;
ii. нуклеотидную последовательность, последовательность которой отличается от последовательности молекулы нуклеиновой кислоты из (i) вследствие вырожденности генетического кода.
Каждая нуклеотидная последовательность, описанная в данном документе посредством процентной идентичности (по меньшей мере 60%) с заданной нуклеотидной последовательностью, согласно дополнительному предпочтительному варианту осуществления, соответственно, является по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99% идентичной или более идентичной заданному нуклеотиду, соответственно. В соответствии с предпочтительным вариантом осуществления идентичность последовательности определяют посредством сравнения полной длины последовательностей, которые идентифицированы в данном документе. Если в данном документе не указано иное, идентичность заданной SEQ ID NO означает идентичность или сходство на основании полной длины указанной последовательности (т.е. по всей ее длине или целиком).
Конструкция, определенная посредством своей минимальной идентичности (т.е. по меньшей мере 60%) заданной SEQ ID NO, которая идентифицирована выше, включена в объем настоящего изобретения, когда эта конструкция, или вирусная экспрессионная конструкция, или вирусный вектор, содержащий эту конструкцию, или композиция, содержащая эту конструкцию или вектор, являются способными индуцировать экспрессию FGF21 в клетке, предпочтительно, в клетке печени, клетке жировой ткани или в клетке скелетной мышцы. Экспрессию FGF21 можно оценить с применением методик, известных квалифицированному специалисту. В соответствии с предпочтительным вариантом осуществления указанную экспрессию оценивают таким образом, как это выполнено в экспериментальной части.
В соответствии с предпочтительным вариантом осуществления вирусная экспрессионная конструкция является такой, чтобы конструкция была представлена нуклеотидной последовательностью, содержащей SEQ ID NO: 4, 5, 6, 7, 8, 9, 10 или 11, или последовательностью, по меньшей мере на 60% идентичной SEQ ID NO: 4, 5, 6, 7, 8, 9, 10 или 11, или последовательностью, по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 4, 5, 6, 7, 8, 9, 10 или 11.
Вирусный вектор
В соответствии с дополнительным аспектом предусмотрен вирусный вектор, содержащий вирусную экспрессионную конструкцию, которая определена выше, при этом указанный вирусный вектор представляет собой аденовирусный вектор, вектор на основе аденоассоциированного вируса, ретровирусный вектор или лентивирусный вектор, предпочтительно, вектор на основе аденоассоциированного вируса, выбранный из группы, состоящей из вектора на основе аденоассоциированного вируса 1 (AAV1), вектора на основе аденоассоциированного вируса 8 (AAV8) и вектора на основе аденоассоциированного вируса 9 (AAV9).
«Вирусный вектор» и «вектор на основе аденоассоциированного вируса (вектор на основе AAV)» дополнительно определены в части описания, озаглавленной «Общие определения».
В соответствии с вариантом осуществления применяют вектор на основе AAV, содержащий каждый из элементов, определенных выше в данном документе, и геном на основе рекомбинантного AAV (rAAV), содержащий ITR или его часть. Предпочтительные ITR представляют собой таковые из AAV2, которые представлены SEQ ID NO: 48 (5'ITR) и SEQ ID NO: 49 (3'ITR).
Предпочтительно, указанный вектор на основе AAV представляет собой вектор на основе AAV1, вектор на основе AAV8 или вектор на основе AAV9.
Вирусная экспрессионная конструкция и вирусный вектор согласно настоящему изобретению предпочтительно предназначены для применения в качестве лекарственного препарата. Лекарственный препарат предпочтительно предназначен для предупреждения, задержки, излечения, обращения развития и/или лечения метаболического нарушения, предпочтительно, сахарного диабета и/или ожирения. Сахарный диабет может представлять собой сахарный диабет 1 типа, сахарный диабет 2 типа или моногенный сахарный диабет. В соответствии с еще одним предпочтительным вариантом осуществления лекарственный препарат предназначен для предупреждения, задержки, излечения, обращения развития и/или лечения воспаления и/или фиброза печени. В соответствии с еще одним предпочтительным вариантом осуществления лекарственный препарат предназначен для продления жизни в здоровом состоянии, предпочтительно, посредством предупреждения, задержки, излечения, обращения развития и/или лечения метаболического нарушения, ассоциированного со старением, предпочтительно, сахарного диабета и/или ожирения. В соответствии с еще одним предпочтительным вариантом осуществления лекарственный препарат предназначен для предупреждения, задержки, излечения, обращения развития и/или лечения рака, предпочтительно, рака печени.
Получающий лечение субъект может представлять собой высшее млекопитающее, например, кошек, грызунов (предпочтительно, мышей, крыс, песчанок и морских свинок и, более предпочтительно, мышей и крыс), или собак, или людей.
Молекула нуклеиновой кислоты
В соответствии с дополнительным аспектом предусмотрена молекула нуклеиновой кислоты, подходящая для экспрессии у млекопитающего и представленная оптимизированной по кодону для экспрессии у млекопитающего нуклеотидной последовательностью, которая кодирует FGF21, подлежащий экспрессии в печени, жировой ткани и/или скелетной мышце.
Определение «оптимизация по кодону» было представлено в части описания, озаглавленной «Общие определения».
В соответствии с вариантом осуществления предусмотрена молекула нуклеиновой кислоты, которая описана выше, при этом нуклеотидная последовательность является по меньшей мере на 60% идентичной нуклеотидной последовательности SEQ ID NO: 4, 5, 6, 7, 8, 9, 10 или 11. Предпочтительная нуклеотидная последовательность является по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: 4, 5, 6, 7, 8, 9, 10 или 11.
Композиция
В соответствии с дополнительным аспектом предусмотрена композиция, содержащая вирусную экспрессионную конструкцию, которая определена выше, и/или вирусный вектор, который определен выше, и/или молекулу нуклеиновой кислоты, которая определена выше, вместе с одним или несколькими фармацевтически приемлемыми вспомогательными средствами или средами.
Эту композицию предпочтительно называют композицией для генной терапии. Предпочтительно, композиция представляет собой фармацевтическую композицию, при этом указанная фармацевтическая композиция содержит фармацевтически приемлемый носитель, вспомогательное вещество, разбавители, солюбилизаторы, наполнитель, консервант и/или вспомогательное средство.
Такой фармацевтически приемлемый носитель, наполнитель, консервант, солюбилизатор, разбавитель и/или вспомогательное средство можно найти, например, в Remington: The Science and Practice of Pharmacy, 20th Edition. Baltimore, MD: Lippincott Williams & Wilkins, 2000.
В соответствии с предпочтительным вариантом осуществления указанная композиция предназначена для применения в качестве лекарственного препарата, предпочтительно, для предупреждения, задержки, излечения, обращения развития и/или лечения метаболического нарушения, предпочтительно, сахарного диабета и/или ожирения. Сахарный диабет может представлять собой сахарный диабет 1 типа, сахарный диабет 2 типа или моногенный сахарный диабет. В соответствии с еще одним предпочтительным вариантом осуществления лекарственный препарат предназначен для предупреждения, задержки, излечения, обращения развития и/или лечения воспаления и/или фиброза печени. В соответствии с еще одним предпочтительным вариантом осуществления лекарственный препарат предназначен для продления жизни в здоровом состоянии, предпочтительно, посредством предупреждения, задержки, излечения, обращения развития и/или лечения метаболического нарушения, ассоциированного со старением, предпочтительно, сахарного диабета и/или ожирения. В соответствии с еще одним предпочтительным вариантом осуществления лекарственный препарат предназначен для предупреждения, задержки, излечения, обращения развития и/или лечения рака, предпочтительно, рака печени. Получающий лечение субъект может представлять собой высшее млекопитающее, например, кошек, грызуна (предпочтительно, мышей, крыс, песчанок и морских свинок и, более предпочтительно, мышей и крыс), или собак, или человека.
Предпочтительно, считают, что указанную вирусную экспрессионную конструкцию, вирусный вектор, и/или молекулу нуклеиновой кислоты, и/или композицию возможно применять для предупреждения, задержки, обращения развития, излечения и/или лечения метаболического нарушения, предпочтительно, сахарного диабета и/или ожирения, когда указанная вирусная экспрессионная конструкция, вирусный вектор, и/или молекула нуклеиновой кислоты, и/или композиция способны проявлять противодиабетический эффект и/или эффект, направленный против ожирения.
Предпочтительно, считают, что указанную вирусную экспрессионную конструкцию, вирусный вектор, и/или молекулу нуклеиновой кислоты, и/или композицию возможно применять для предупреждения, задержки, излечения, обращения развития и/или лечения воспаления и/или фиброза печени, когда указанная вирусная экспрессионная конструкция, вирусный вектор, и/или молекула нуклеиновой кислоты, и/или композиция способны проявлять противофиброзный эффект.
Предпочтительно, считают, что указанную вирусную экспрессионную конструкцию, вирусный вектор, и/или молекулу нуклеиновой кислоты, и/или композицию возможно применять для продления жизни в здоровом состоянии, предпочтительно, посредством предупреждения, задержки, излечения, обращения развития и/или лечения метаболического нарушения, ассоциированного со старением, предпочтительно, сахарного диабета и/или ожирения, когда указанная вирусная экспрессионная конструкция, вирусный вектор, и/или молекула нуклеиновой кислоты, и/или композиция способны проявлять противодиабетический эффект и/или эффект, направленный против ожирения, при старении.
Предпочтительно, считают, что указанную вирусную экспрессионную конструкцию, вирусный вектор, и/или молекулу нуклеиновой кислоты, и/или композицию можно применять для предупреждения, задержки, излечения, обращения развития и/или лечения рака, предпочтительно, рака печени, когда указанная вирусная экспрессионная конструкция, вирусный вектор, и/или молекула нуклеиновой кислоты, и/или композиция способны проявлять противораковый эффект.
Противодиабетический эффект может быть достигнут, когда повышается утилизация глюкозы в крови, и/или когда улучшается толерантность к глюкозе, и/или когда повышается чувствительность к инсулину. Его можно оценить с применением методик, известных квалифицированному специалисту, или таким образом, как выполнено в экспериментальной части, предпочтительно, как оценено в примере 8 или 9. В этом контексте «повышение» (соответственно, «улучшение») означает по меньшей мере выявляемое повышение (соответственно, выявляемое улучшение) с применением анализа, известного квалифицированному специалисту, или с применением анализов, которые выполнены в экспериментальной части, таких как измерение гликемии, инсулинемии и/или проведение теста толерантности к инсулину и/или теста толерантности к глюкозе. Повышение может представлять собой повышение по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90% или по меньшей мере на 100% при применении таких анализов, как измерение гликемии, инсулинемии и/или проведение теста толерантности к инсулину и/или теста толерантности к глюкозе.
Эффект, направленный против ожирения, может быть достигнут, когда снижается масса тела, увеличение массы тела и/или процентное содержание жира в организме. Эффект, направленный против ожирения, также может быть достигнут, когда снижается индекс массы тела (BMI), окружность талии, соотношение окружностей талии и бедер (WHR) и/или соотношение окружности талии к росту (WHtR). Его можно оценить с применением методик, известных квалифицированному специалисту, или таким образом, как выполнено в экспериментальной части. В этом контексте «снижение» (соответственно, «улучшение») означает по меньшей мере выявляемое снижение (соответственно, выявляемое улучшение) с применением анализа, известного квалифицированному специалисту, или с применением анализов, которые выполнены в экспериментальной части. Эффекты, направленные против ожирения, включают в себя как предупреждение ожирения, так и обращение развития ожирения, что оценивают посредством измерения массы тела индивида, BMI и/или массы тканей.
Противовоспалительный эффект в печени может быть достигнут посредством снижения инфильтрации макрофагов, сниженного уровня провоспалительных цитокинов. Его можно оценить с применением методик, известных квалифицированному специалисту, или таким образом, как выполнено в экспериментальной части. В этом контексте «снижение» (соответственно, «улучшение») означает по меньшей мере выявляемое снижение (соответственно, выявляемое улучшение) с применением анализа, известного квалифицированному специалисту, или с применением анализов, которые выполнены в экспериментальной части.
Противофиброзный эффект в печени может быть достигнут посредством снижения уровней накопленных белков внеклеточного матрикса, маркеров в крови (например, в том числе уровней N-концевого пропептида коллагена III типа, гиалуроновой кислоты, тканевого ингибитора металлопротеиназы 1 типа (TIMP-1), YKL-40, сывороточной глутамат-оксалоацетат-трансаминазы (SGOT), сывороточной глутамат-пируват-трансаминазы (SGPT) в плазме крови). Противофиброзный эффект может быть достигнут посредством улучшения оценки согласно системе оценки фиброза, такой как Metavir или Ishak. Его можно оценить с применением методик, известных квалифицированному специалисту, или таким образом, как выполнено в экспериментальной части. В этом контексте «снижение» (соответственно, «улучшение») означает по меньшей мере выявляемое снижение (соответственно, выявляемое улучшение) с применением анализа, известного квалифицированному специалисту, или с применением анализов, которые выполнены в экспериментальной части.
Эффект продления жизни в здоровом состоянии может быть достигнут, когда противодиабетический эффект и/или эффект, направленный против ожирения, которые определены выше в данном документе, применяют для предупреждения, задержки, излечения, обращения развития или лечения проявления или прогрессирования метаболического нарушения, ассоциированного со старением, предпочтительно, сахарного диабета и/или ожирения. Эффект продления жизни в здоровом состоянии также может быть достигнут посредством повышения продолжительности жизни в здоровом состоянии, при этом симптомы, ассоциированные с метаболическими нарушениями, предпочтительно, с сахарным диабетом и/или ожирением, отсутствуют или уменьшены. Эффект продления жизни в здоровом состоянии также может быть достигнут посредством улучшенной координации и равновесия (оцениваемой с помощью теста на вращающемся стержне Rota-Rod), памяти (оцениваемой с помощью теста распознавания объекта) и/или нервно-мышечной координации (оцениваемой с помощью теста на натянутом канате (Tightrope Test)), уменьшенного ухудшения митохондриальной и метаболической функций (оцениваемого с помощью измерения уровней экспрессии генов, вовлеченных в метаболизм и функционирование митохондрий, таких как PGC-1-альфа, АТФ-синтаза и ERR-альфа). Его можно оценить с применением методик, известных квалифицированному специалисту, или таким образом, как выполнено в экспериментальной части. В этом контексте «повышение» (соответственно, «улучшение») означает по меньшей мере выявляемое повышение (соответственно, выявляемое улучшение) с применением анализа, известного квалифицированному специалисту, или с применением анализов, которые выполнены в экспериментальной части.
Противораковый эффект может быть достигнут посредством снижения кумулятивной частоты возникновения рака за время жизни. Его можно оценить с применением методик, известных квалифицированному специалисту, или таким образом, как выполнено в экспериментальной части. В этом контексте «снижение» (соответственно, «улучшение») означает по меньшей мере выявляемое снижение (соответственно, выявляемое улучшение) с применением анализа, известного квалифицированному специалисту, или с применением анализов, которые выполнены в экспериментальной части.
Противодиабетический эффект и/или эффект, направленный против ожирения, также можно наблюдать, когда прогрессирование типичного симптома (например, инсулита, потери бета-клеток, увеличения массы тела) замедлилось согласно оценке врача. Уменьшение типичного симптома может означать замедление прогрессирования развития симптома или полное исчезновение симптомов. Симптомы и, следовательно, также уменьшение симптомов можно оценивать с применением ряда способов, по существу, таких же способов, которые применяют в диагностике сахарного диабета и/или ожирения, в том числе с применением клинического исследования и стандартных лабораторных тестов. Такие способы включают в себя как макроскопические, так и микроскопические методы, а также молекулярные методы, рентгенологические, биохимические, иммуногистохимические и другие методы.
Противовоспалительный эффект в печени также можно наблюдать, когда прогрессирование типичного симптома (например, усталость, гриппоподобные симптомы, темная моча, бледный стул, боль в животе, потеря аппетита, необъяснимая потеря массы, разлитие желчи) замедлилось согласно оценке врача. Уменьшение типичного симптома может означать замедление прогрессирования развития симптома или полное исчезновение симптомов. Симптомы и, следовательно, также уменьшение симптомов можно оценивать с применением ряда способов, по существу, таких же способов, которые применяют в диагностике фиброза печени, в том числе с применением клинического исследования и стандартных лабораторных тестов. Такие способы включают в себя как макроскопические, так и микроскопические методы, а также молекулярные методы, методы визуализации (эластография, рентгенография, MPT, КТ, ультразвуковое исследование, ангиография), биохимические, иммуногистохимические и другие методы.
Противофиброзный эффект в печени также можно наблюдать, когда прогрессирование типичного симптома (например, жесткость печени, разлитие желчи, потеря аппетита, трудность с ясностью мышления, накопление жидкости в ногах или желудке, тошнота, необъяснимая потеря массы, слабость) замедлилось согласно оценке врача. Уменьшение типичного симптома может означать замедление прогрессирования развития симптома или полное исчезновение симптомов. Симптомы и, следовательно, также уменьшение симптомов можно оценивать с применением ряда способов, по существу, таких же способов, которые применяют в диагностике фиброза печени, в том числе с применением клинического исследования и стандартных лабораторных тестов. Такие способы включают в себя как макроскопические, так и микроскопические методы, а также молекулярные методы, методы визуализации (эластография, рентгенография, МРТ, КТ, ультразвуковое исследование, ангиография), биохимические, иммуногистохимические и другие методы.
Эффект продления жизни в здоровом состоянии также можно наблюдать, когда прогрессирование типичного симптома метаболических нарушений, ассоциированных со старением (например, резистентность к инсулину, нарушение толерантности к глюкозе, увеличение массы тела), замедлилось согласно оценке врача. Уменьшение типичного симптома может означать замедление прогрессирования развития симптома или полное исчезновение симптомов. Симптомы и, следовательно, также уменьшение симптомов можно оценивать с применением ряда способов, по существу, таких же способов, которые применяют в диагностике сахарного диабета и/или ожирения, в том числе с применением клинического исследования и стандартных лабораторных тестов. Такие способы включают в себя как макроскопические, так и микроскопические методы, а также молекулярные методы, рентгенологические, биохимические, иммуногистохимические и другие методы.
Противораковый эффект также можно наблюдать, когда прогрессирование типичного симптома (например, размер опухоли, необъяснимая потеря массы, потеря аппетита, ощущение переполненности в желудке после небольшой порции пищи, тошнота или рвота, увеличенная печень, увеличенная селезенка, боль в животе или вблизи правой лопатки, потливость или накопление жидкости в животе, зуд, разлитие желчи) замедлилось согласно оценке врача. Уменьшение типичного симптома может означать замедление прогрессирования развития симптома или полное исчезновение симптомов. Симптомы и, следовательно, также уменьшение симптомов можно оценивать с применением ряда способов, по существу, таких же способов, которые применяют в диагностике рака, в том числе с применением клинического исследования и стандартных лабораторных тестов. Такие способы включают в себя как макроскопические, так и микроскопические методы, а также молекулярные методы, методы визуализации (рентгенография, MPT, КТ, ультразвуковое исследование, ангиография), биохимические, иммуногистохимические и другие методы.
Лекарственный препарат, который определен в данном документе (вирусная экспрессионная конструкция, вирусный вектор, молекула нуклеиновой кислоты, композиция), предпочтительно, способен облегчить один симптом или одну характеристику пациента или клетки, ткани или органа указанного пациента, если после по меньшей мере одной недели, одного месяца, шести месяцев, одного года или большего периода лечения с применением вирусной экспрессионной конструкции, и/или вирусного вектора, и/или молекулы нуклеиновой кислоты, и/или композиции согласно настоящему изобретению указанные симптом или характеристика уменьшились (например, уже не выявляются или замедлились), как определено выше.
Вирусная экспрессионная конструкция, и/или вирусный вектор, и/или молекула нуклеиновой кислоты, и/или композиция, которые определены в данном документе для применения в соответствии с настоящим изобретением, могут быть подходящими для введения в клетку, ткань и/или орган in vivo индивидов, пораженных метаболическим нарушением или имеющим риск развития метаболического нарушения, такого как сахарный диабет и/или ожирение, воспаление и/или фиброз печени, метаболическое нарушение, ассоциированное со старением, и/или рак, и их можно вводить in vivo, ex vivo или in vitro. Указанную вирусную экспрессионную конструкцию, и/или вирусный вектор, и/или молекулу нуклеиновой кислоты, и/или композицию можно вводить непосредственно или опосредованно в клетку, ткань и/или орган in vivo индивида, пораженного метаболическим нарушением или имеющего риск развития метаболического нарушения, такого как сахарный диабет и/или ожирение, воспаление и/или фиброз печени, метаболическое нарушение, ассоциированное со старением, и/или рак, и их можно вводить непосредственно или опосредованно in vivo, ex vivo или in vitro. Способ введения может быть внутривенным, подкожным, внутримышечным, интратекальным, внутрисуставным, интравентрикулярным, внутрибрюшинным, в жировую ткань, посредством ингаляции, пероральным, интраназальным, внутрипеченочным, во внутренние органы, внутриглазным, внутриушным, местным введением и/или посредством ретроградного внутрипротокового введения в поджелудочную железу. Предпочтительным способом введения является внутримышечный, внутривенный или введение в жировую ткань, как описано в разделе «Общие процедуры для примеров» в рамках настоящей заявки.
Вирусную экспрессионную конструкцию, и/или вирусный вектор, и/или молекулу нуклеиновой кислоты, и/или композицию согласно настоящему изобретению можно вводить непосредственно или опосредованно с применением подходящих средств, известных в уровне техники. Принимая во внимание прогресс, который уже достигнут на данный момент, предполагаются усовершенствования в средствах для обеспечения индивида или клетки, ткани, органа указанного индивида вирусной экспрессионной конструкцией, и/или вирусным вектором, и/или молекулой нуклеиновой кислоты, и/или композицией согласно настоящему изобретению. Такие будущие усовершенствования, разумеется, могут быть включены для достижения упомянутого эффекта настоящего изобретения. Вирусную экспрессионную конструкцию, и/или вирусный вектор, и/или молекулу нуклеиновой кислоты, и/или композицию можно доставлять без дополнительной обработки индивиду, в клетку, ткань или орган указанного индивида. В зависимости от заболевания или состояния клетка, ткань или орган указанного индивида могут быть такими, как определено выше в данном документе. При введении вирусной экспрессионной конструкции, и/или вирусного вектора, и/или молекулы нуклеиновой кислоты, и/или композиции согласно настоящему изобретению предпочтительно, чтобы такая вирусная экспрессионная конструкция, и/или вектор, и/или нуклеиновая кислота, и/или композиция были растворены в растворе, который является совместимым со способом доставки.
Как предусмотрено в данном документе, терапевтически эффективную дозу вирусной экспрессионной конструкции, вектора, молекулы нуклеиновой кислоты и/или композиции, которые упомянуты выше, предпочтительно вводят в однократной и индивидуальной дозе, таким образом избегая повторного периодического введения. Более предпочтительно, однократную дозу вводят в скелетную мышцу, в жировую ткань или внутривенно.
Дополнительное соединение может присутствовать в композиции согласно настоящему изобретению. Указанное соединение может помочь в доставке композиции. Ниже представлен перечень подходящих соединений: соединения, способные к образованию комплексов, наночастицы, мицеллы и/или липосомы, которые доставляют каждую составляющую, определенную в данном документе, заключенную в комплекс или захваченную в пузырьке или липосоме, через клеточную мембрану. Многие из этих соединений являются известными в уровне техники. Подходящие соединения содержат полиэтиленимин (PEI) или подобные катионные полимеры, в том числе полипропиленимин или сополимеры полиэтиленимина (РЕС) и производные, синтетические амфифильные вещества (SAINT-18), lipofectin™, DOTAP.
В зависимости от их идентификационных данных квалифицированному специалисту будет известно, какой тип состава является наиболее подходящим для композиции, которая определена в данном документе.
Способ/применение
В соответствии с дополнительным аспектом предусмотрена вирусная экспрессионная конструкция, которая определена выше, и/или вирусный вектор, который определен выше, и/или молекула нуклеиновой кислоты, которая определена выше, и/или композиция, которая определена выше, для применения в качестве лекарственного препарата.
В соответствии с вариантом осуществления предусмотрена указанная вирусная экспрессионная конструкция, и/или вирусный вектор, и/или молекула нуклеиновой кислоты, и/или композиция для применения в лечении и/или предупреждении метаболического нарушения, предпочтительно, сахарного диабета и/или ожирения. Также могут охватываться осложнения метаболического нарушения.
В соответствии с еще одним вариантом осуществления предусмотрена указанная вирусная экспрессионная конструкция, и/или вирусный вектор, и/или молекула нуклеиновой кислоты, и/или композиция для применения в лечении и/или предупреждении воспаления и/или фиброза печени. Также могут охватываться осложнения и/или фиброз печени.
В соответствии с еще одним вариантом осуществления предусмотрена указанная вирусная экспрессионная конструкция, и/или вирусный вектор, и/или молекула нуклеиновой кислоты, и/или композиция для применения в продлении жизни в здоровом состоянии, предпочтительно, посредством предупреждения, задержки, излечения, обращения развития и/или лечения метаболического нарушения, ассоциированного со старением, предпочтительно, сахарного диабета и/или ожирения.
В соответствии с еще одним вариантом осуществления предусмотрена указанная вирусная экспрессионная конструкция, и/или вирусный вектор, и/или молекула нуклеиновой кислоты, и/или композиция для применения в лечении и/или предупреждении рака, предпочтительно, рака печени. Также могут охватываться осложнения рака.
В соответствии с дополнительным аспектом предусмотрен способ предупреждения, задержки, обращения развития, излечения и/или лечения метаболического нарушения, предпочтительно, сахарного диабета и/или ожирения, а также их осложнений, предусматривающий применение вирусной экспрессионной конструкции, которая определена выше, и/или вирусного вектора, который определен выше, и/или молекулы нуклеиновой кислоты, которая определена выше, и/или композиции, которая определена выше.
Такой способ предпочтительно предназначен для облегчения одного или нескольких симптомов метаболического нарушения, такого как сахарный диабет и/или ожирение, у индивида, в клетке, ткани или органе указанного индивида, или чтобы облегчить одну или несколько характеристик или симптомов в клетке, ткани или органе указанного индивида, при этом способ предусматривает введение указанному индивиду вирусной экспрессионной конструкции, и/или вирусного вектора, и/или молекулы нуклеиновой кислоты, и/или композиции, которые определены в данном документе.
В соответствии с дополнительным аспектом предусмотрен способ предупреждения, задержки, обращения развития, излечения и/или лечения воспаления и/или фиброза печени, а также их осложнений, предусматривающий применение вирусной экспрессионной конструкции, которая определена выше, и/или вирусного вектора, который определен выше, и/или молекулы нуклеиновой кислоты, которая определена выше, и/или композиции, которая определена выше.
Такой способ, предпочтительно предназначен для облегчения одного или нескольких симптомов воспаления и/или фиброза печени у индивида, в клетке, ткани или органе указанного индивида, или чтобы облегчить одну или несколько характеристик или симптомов в клетке, ткани или органе указанного индивида, при этом способ предусматривает введение указанному индивиду вирусной экспрессионной конструкции, и/или вирусного вектора, и/или молекулы нуклеиновой кислоты, и/или композиции, которые определены в данном документе.
В соответствии с дополнительным аспектом предусмотрен способ продления жизни в здоровом состоянии, предпочтительно, посредством предупреждения, задержки, излечения, обращения развития и/или лечения метаболического нарушения, ассоциированного со старением, предпочтительно, сахарного диабета и/или ожирения, предусматривающий применение вирусной экспрессионной конструкции, которая определена выше, и/или вирусного вектора, который определен выше, и/или молекулы нуклеиновой кислоты, которая определена выше, и/или композиции, которая определена выше.
Такой способ, предпочтительно предназначен для облегчения одного или нескольких симптомов метаболического нарушения, ассоциированного со старением, такого как сахарный диабет и/или ожирение, у индивида, в клетке, ткани или органе указанного индивида, или чтобы облегчить одну или несколько характеристик или симптомов в клетке, ткани или органе указанного индивида, при этом способ предусматривает введение указанному индивиду вирусной экспрессионной конструкции, и/или вирусного вектора, и/или молекулы нуклеиновой кислоты, и/или композиции, которые определены в данном документе.
В соответствии с дополнительным аспектом предусмотрен способ предупреждения, задержки, обращения развития, излечения и/или лечения рака, предпочтительно, рака печени, а также их осложнений, предусматривающий применение вирусной экспрессионной конструкции, которая определена выше, и/или вирусного вектора, который определен выше, и/или молекулы нуклеиновой кислоты, которая определена выше, и/или композиции, которая определена выше.
Такой способ предпочтительно предназначен для облегчения одного или нескольких симптомов рака, такого как рак печени, у индивида, в клетке, ткани или органе указанного индивида, или чтобы облегчить одну или несколько характеристик или симптомов в клетке, ткани или органе указанного индивида, при этом способ предусматривает введение указанному индивиду вирусной экспрессионной конструкции, и/или вирусного вектора, и/или молекулы нуклеиновой кислоты, и/или композиции, которые определены в данном документе.
В контексте настоящего изобретения предусмотрено применение вирусной экспрессионной конструкции, и/или вирусного вектора, и/или молекулы нуклеиновой кислоты, и/или композиции, которые определены в данном документе, для производства лекарственного препарата для предупреждения, задержки, обращения развития, излечения и/или лечения метаболического нарушения, предпочтительно, сахарного диабета и/или ожирения.
В контексте настоящего изобретения предусмотрено применение вирусной экспрессионной конструкции, и/или вирусного вектора, и/или молекулы нуклеиновой кислоты, и/или композиции, которые определены в данном документе, для производства лекарственного препарата для предупреждения, задержки, излечения, обращения развития и/или лечения воспаления и/или фиброза печени.
В контексте настоящего изобретения предусмотрено применение вирусной экспрессионной конструкции, и/или вирусного вектора, и/или молекулы нуклеиновой кислоты, и/или композиции, которые определены в данном документе, для производства лекарственного препарата для продления жизни в здоровом состоянии, предпочтительно, посредством предупреждения, задержки, излечения, обращения развития и/или лечения метаболического нарушения, ассоциированного со старением, предпочтительно, сахарного диабета и/или ожирения.
В контексте настоящего изобретения предусмотрено применение вирусной экспрессионной конструкции, и/или вирусного вектора, и/или молекулы нуклеиновой кислоты, и/или композиции, которые определены в данном документе, для производства лекарственного препарата для предупреждения, задержки, обращения развития, излечения и/или лечения рака, предпочтительно, рака печени.
Метаболические нарушения включают в себя метаболический синдром, сахарный диабет, ожирение, связанные с ожирением сопутствующие заболевания, связанные с сахарным диабетом сопутствующие заболевания, гипергликемию, резистентность к инсулину, нарушение толерантности к глюкозе, стеатоз печени, алкогольные болезни печени (ALD), неалкогольную жировую болезнь печени (NAFLD), неалкогольный стеатогепатит (NASH), ишемическую болезнь сердца (CHD), гиперлипидемию, атеросклероз, патологии эндокринной системы, синдром остеосаркопенического ожирения (OSO), диабетическую нефропатию, хроническое заболевание почек (CKD), гипертрофию сердца, диабетическую ретинопатию, диабетическую нефропатию, диабетическую невропатию, артрит, сепсис, неоваскуляризацию в глазу, нейродегенерацию, деменцию, а также могут включать в себя депрессию, аденому, карциному.
Сахарный диабет включает в себя преддиабет, гипергликемию, сахарный диабет 1 типа, сахарный диабет 2 типа, сахарный диабет взрослого типа у молодых (MODY), моногенный сахарный диабет, неонатальный сахарный диабет, гестационный сахарный диабет, сахарный лабильный диабет, идиопатический сахарный диабет, индуцированный лекарственным средством или химическим соединением сахарный диабет, синдром мышечной скованности, липоатрофический сахарный диабет, латентный аутоиммунный сахарный диабет у взрослых (LADA).
Ожирение включает в себя избыточный вес, ожирение центральной/верхней части тела, периферическое ожирение/ожирение нижней части тела, морбидное ожирение, синдром остеосаркопенического ожирения (OSO), детское ожирение, менделевское (моногенное) синдромное ожирение, менделевское несиндромное ожирение, полигенное ожирение.
Выше в данном документе были определены метаболические нарушения, сахарный диабет, ожирение, а также тип субъекта, получающего лечение.
Воспаление и/или фиброз печени включают в себя аутоиммунный гепатит, вирусный гепатит, в том числе гепатит А, В, С, D и Е, алкогольный гепатит, неалкогольный стеатогепатит (NASH) и цирроз печени.
Рак включает в себя астроцитому, глиому, лейкоз, лимфому, меланому, миелому, нейробластому, саркому (в том числе хондросаркому, фибросаркому, рабдомиосаркому и остеосаркому), невриному, семиному и карциномы мочевого пузыря, молочной железы, шейки матки, ободочной кишки, эндометрия, пищевода, желчного пузыря, почки, печени, легкого, яичника, предстательной железы, поджелудочной железы, прямой кишки, кожи, желудка и щитовидной железы. Предпочтительный рак представляет собой рак печени, предпочтительно, печеночно-клеточную карциному. В соответствии с одним вариантом осуществления указанный способ или применение осуществляют in vitro, например, с применением клеточной культуры. Предпочтительно, указанный способ или применение осуществляют in vivo. Каждый признак этих способов/применений уже был определен в данном документе.
В способе согласно настоящему изобретению вирусная экспрессионная конструкция, и/или вектор, и/или молекула нуклеиновой кислоты, и/или композиция могут быть объединены с дополнительным соединением, которое, как известно, применяют для лечения метаболических нарушений, предпочтительно, сахарного диабета и/или ожирения, у индивида.
В еще одном способе согласно настоящему изобретению вирусная экспрессионная конструкция, и/или вектор, и/или молекула нуклеиновой кислоты, и/или композиция могут быть объединены с дополнительным соединением, которое, как известно, применяют для лечения воспаления и/или фиброза печени.
В еще одном способе согласно настоящему изобретению вирусная экспрессионная конструкция, и/или вектор, и/или молекула нуклеиновой кислоты, и/или композиция могут быть объединены с дополнительным соединением, которое, как известно, применяют для продления жизни в здоровом состоянии.
В еще одном способе согласно настоящему изобретению вирусная экспрессионная конструкция, и/или вектор, и/или молекула нуклеиновой кислоты, и/или композиция могут быть объединены с дополнительным соединением, которое, как известно, применяют для лечения рака, предпочтительно, рака печени.
В соответствии с предпочтительным вариантом осуществления лечение в применении или в способе согласно настоящему изобретению не должно повторяться. В качестве альтернативы, в применении или способе согласно настоящему изобретению указанное введение вирусной экспрессионной конструкции или указанной композиции можно повторять каждый год или каждые 2, 3, 4, 5, 6 лет.
Общие определения
Идентичность/сходство
В контексте настоящего изобретения фрагмент белка, или полипептид, или пептид, или производное пептида в качестве фактора роста фибробластов 21 (FGF21) представлены аминокислотной последовательностью.
В контексте настоящего изобретения молекула нуклеиновой кислоты в качестве молекулы нуклеиновой кислоты, кодирующей FGF21, представлена нуклеиновой кислотой или нуклеотидной последовательностью, которая кодирует фрагмент белка, или полипептид, или пептид, или производное пептида. Молекула нуклеиновой кислоты может содержать регуляторный участок.
Следует понимать, что каждая молекула нуклеиновой кислоты, или фрагмент белка, или полипептид, или пептид, или производное пептида, или конструкция, которые идентифицированы в данном документе с помощью заданного номерного идентификатора последовательности (SEQ ID NO), не ограничены этой конкретной последовательностью, которая раскрыта. Каждая кодирующая последовательность, которая идентифицирована в данном документе, кодирует заданный фрагмент белка, или полипептид, или пептид, или производное пептида, или конструкцию, или сама представляет собой фрагмент белка, или полипептид, или конструкцию, или пептид, или производное пептида. В настоящей заявке при каждом упоминании, относящемся к конкретной нуклеотидной последовательности SEQ ID NO (обозначенной SEQ ID NO: X в качестве примера), кодирующей заданный фрагмент белка, или полипептид, или пептид, или производное пептида, ее можно заменить на:
i. нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с SEQ ID NO: X;
ii. нуклеотидную последовательность, последовательность которой отличается от последовательности молекулы нуклеиновой кислоты из (i) вследствие вырожденности генетического кода; или
iii. нуклеотидную последовательность, которая кодирует аминокислотную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством аминокислот с аминокислотной последовательностью, кодируемой нуклеотидной последовательностью SEQ ID NO: X.
В настоящей заявке при каждом упоминании, относящемся к конкретной аминокислотной последовательности SEQ ID NO (обозначенной SEQ ID NO: Y в качестве примера), ее можно заменить на: полипептид содержащий, аминокислотную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательностью с аминокислотной последовательностью SEQ ID NO: Y.
Каждая нуклеотидная последовательность или аминокислотная последовательность, описанная в данном документе посредством ее процентной идентичности или сходства (по меньшей мере 60%) с заданной нуклеотидной последовательностью или аминокислотной последовательностью, соответственно, в соответствии с дополнительным предпочтительным вариантом осуществления характеризуется идентичностью или сходством, составляющими по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99% или более, с заданной нуклеотидной или аминокислотной последовательностью, соответственно. В соответствии с предпочтительным вариантом осуществления идентичность или сходство последовательности определяют посредством сравнения полной длины последовательностей, которые идентифицированы в данном документе. Если в данном документе не указано иное, идентичность или сходство с заданной SEQ ID NO означает идентичность или сходство на основании полной длины указанной последовательности {т.е. по всей ее длине или целиком).
Каждую некодирующую нуклеотидную последовательность (т.е. промотора или другого регуляторного участка) можно заменить на нуклеотидную последовательность, содержащую нуклеотидную последовательность, которая характеризуется по меньшей мере 60% идентичностью или сходством последовательности с конкретной нуклеотидной последовательностью SEQ ID NO (обозначенной SEQ ID NO: А в качестве примера). Предпочтительная нуклеотидная последовательность является по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 100% идентичной SEQ ID NO: А. Идентичность можно оценивать относительно целой SEQ ID NO или относительно ее части, как объясняется в данном документе. В соответствии с предпочтительным вариантом осуществления такая некодирующая нуклеотидная последовательность, такая как промотор, демонстрирует или проявляет по меньшей мере активность такой некодирующей нуклеотидной последовательности, такую как активность промотора, которая известна квалифицированному специалисту.
«Идентичность последовательности» в данном документе определена как взаимосвязь двух или более аминокислотных последовательностей (полипептида или белка) или двух или более последовательностей нуклеиновой кислоты (полинуклеотида), которую определяют посредством сравнения последовательностей. В соответствии с предпочтительным вариантом осуществления идентичность последовательности рассчитывают на основании полной длины двух заданных SEQ ID NO или их части. Их части предпочтительно означают по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% обеих SEQ ID NO. В уровне техники «идентичность» также означает степень связанности последовательностей между аминокислотными последовательностями или последовательностями нуклеиновой кислоты в зависимости от конкретного случая, которую определяют на основании совпадения между нитями таких последовательностей.
«Сходство» между двумя аминокислотными последовательностями определяют посредством сравнения аминокислотной последовательности и ее консервативных аминокислотных замен у одного полипептида с последовательностью второго полипептида. «Идентичность» и «сходство» можно легко рассчитать с помощью известных способов, в том числе, без ограничения, описанных в Computational Molecular Biology, Lesk, A. M., ed., Oxford University Press, New York, 1988; Biocomputing: Informatics and Genome Projects, Smith, D. W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A. M., and Griffin, H. G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heine, G., Academic Press, 1987; и Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991; и Carillo, H., and Lipman, D., SIAM J. Applied Math., 48:1073 (1988).
Предпочтительные способы определения идентичности предназначены для получения наибольшего совпадения между исследуемыми последовательностями. Способы определения идентичности и сходства систематизированы в общедоступных компьютерных программах. Предпочтительные способы с применением компьютерных программ для определения идентичности и сходства между двумя последовательностями включают в себя, например, программный пакет GCG (Devereux, J., et at, Nucleic Acids Research 12 (1): 387 (1984)), BestFit, BLASTP, BLASTN и FASTA (Altschul, S. F. etal, J. Mol. Biol. 215:403-410 (1990). Программа BLAST X является общедоступной от NCBI и других источников (BLAST Manual, Altschul, S., et al, NCBI NLM NIH Bethesda, MD 20894; Altschul, S., et al., J. Mol. Biol. 215:403-410 (1990). Хорошо известный алгоритм Смита-Ватермана также можно применять для определения идентичности.
Предпочтительные параметры для сравнения полипептидных последовательностей включают в себя следующие: Алгоритм: Needleman and Wunsch, J. Mol. Biol. 48:443-453 (1970); матрица сравнения: BLOSSUM62 из Hentikoff and Hentikoff, Proc. Natl. Acad. Sci. USA. 89:10915-10919 (1992); штраф за открытие гэпа: 12; и штраф за продолжение гэпа: 4. Программа, пригодная для использования с этими параметрами, общедоступна в виде программы «Ogap» от Genetics Computer Group, находящейся в Мадисоне, Висконсин. Вышеупомянутые параметры представляют собой параметры по умолчанию для сравнения аминокислот (вместе с отсутствием штрафа за закрытие гэпа).
Предпочтительные параметры для сравнения нуклеиновой кислоты включают в себя следующие: Алгоритм: Needleman and Wunsch, J. Mol. Biol. 48:443-453 (1970); матрица сравнения: совпадения = +10, несовпадение = 0; штраф за открытие гэпа: 50; штраф за продолжение гэпа: 3. Доступна в виде программы Gap от Genetics Computer Group, находящейся в Мадисоне, Висконсин. Выше приведены параметры по умолчанию для сравнений нуклеиновых кислот.
Необязательно, при определении степени сходства аминокислот квалифицированный специалист также может принимать во внимание так называемые «консервативные» аминокислотные замены, что будет ясно квалифицированному специалисту. Консервативные аминокислотные замены относятся к взаимозаменяемости остатков, имеющих подобные боковые цепи. Например, группа аминокислот, имеющих алифатические боковые цепи, представляет собой глицин, аланин, валин, лейцин и изолейцин; группа аминокислот, имеющих алифатические боковые цепи с гидроксильной группой, представляет собой серии и треонин; группа аминокислот, имеющих содержащие амидную группу боковые цепи, представляет собой аспарагин и глутамин; группа аминокислот, имеющих ароматические боковые цепи, представляет собой фенилаланин, тирозин и триптофан; группа аминокислот, имеющих основные боковые цепи, представляет собой лизин, аргинин и гистидин; и группа аминокислот, имеющих серосодержащие боковые цепи, представляет собой цистеин и метионин. Предпочтительные группы консервативных аминокислотных замен представляют собой: валин-лейцин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин и аспарагин-глутамин. Варианты замен в аминокислотной последовательности, раскрытой в данном документе, представляют собой те, при которых по меньшей мере один остаток в раскрытых последовательностях был удален, а на его место вставлен отличающийся остаток. Предпочтительно, аминокислотная замена является консервативной.
Предпочтительные консервативные замены для каждой из встречающихся в естественных условиях аминокислот являются следующими: Ala на Ser; Arg на Lys; Asn на Gln или His; Asp на Glu; Cys на Ser или Ala; Gln на Asn; Glu на Asp; Gly на Pro; His на Asn или Gln; Ile на Leu или Val; Leu на Ile или Val; Lys на Arg; Gln или Glu; Met на Leu или Ile; Phe на Met, Leu или Tyr; Ser на Thr; Thr на Ser; Trp на Tyr; Tyr на Trp или Phe; и Val на Ile или Leu.
Ген или кодирующая последовательность
Термины «ген», или «кодирующая последовательность», или «нуклеиновая кислота», или «нуклеотидная последовательность», или «нуклеиновый» относятся к участку ДНК или РНК (транскрибируемый участок), который «кодирует» конкретный белок, такой как FGF21. Кодирующая последовательность транскрибируется (ДНК) и транслируется (РНК) в полипептид, когда она помещена под управление соответствующего регуляторного участка, такого как промотор. Ген может содержать несколько функционально связанных фрагментов, таких как промотор, 5' лидерная последовательность, интрон, кодирующая последовательность и 3' нетранслируемая последовательность или 3' нетранслируемый участок (3' UTR), содержащий сайт полиаденилирования или сигнальную последовательность. Химерный или рекомбинантный ген (такой как ген FGF21) представляет собой ген, который в норме обнаруживается в естественных условиях, такой как ген, в котором, например, промотор не является ассоциированным в естественных условиях с частью или всем транскрибируемым участком ДНК. «Экспрессия гена» относится к процессу, при котором ген транскрибируется в РНК и/или транслируется в активный белок.
Промотор
В контексте данного документа термин «промотор» относится к фрагменту нуклеиновой кислоты, функцией которого является контроль транскрипции одного или нескольких генов (или кодирующей последовательности), расположенному выше по направлению транскрипции относительно сайта инициации транскрипции гена, и его идентифицируют на основании структуры по присутствию сайта связывания для ДНК-зависимой РНК-полимеразы, сайтов инициации транскрипции и любых других последовательностей ДНК, в том числе, без ограничения, сайтов связывания для транскрипционных факторов, сайтов связывания для белков-репрессоров и белков-активаторов и любых других последовательностей из нуклеотидов, которые, как известно специалисту в данной области техники, действуют непосредственно или опосредованно, регулируя величину транскрипции с промотора. «Конститутивный» промотор представляет собой промотор, который является активным в большинстве физиологических условий и условий развития. «Индуцируемый» промотор представляет собой промотор, который регулируется в зависимости от физиологических условий или условий развития. «Специфический для органа» или «специфический для ткани» промотор представляет собой промотор, который является активным в конкретном типе органа или ткани, соответственно. Специфические для органа и специфические для ткани промоторы регулируют экспрессию одного или нескольких генов (или кодирующей последовательности) преимущественно в одном органе или ткани, но также могут позволять выявляемый уровень («утечки») экспрессии в других органах или тканях. «Утечка» экспрессии в других органах или тканях означает, что экспрессия по меньшей мере однократно, по меньшей мере двукратно, по меньшей мере трехкратно, по меньшей мере четырехкратно или по меньшей мере пятикратно ниже, но при этом все еще является выявляемой, по сравнению со специфической для органа или специфической для ткани экспрессией, что оценивают с помощью стандартных анализов, известных квалифицированному специалисту (например, ПЦР, анализ методом вестерн-блоттинга, ELISA). Максимальное число органов или тканей, в которых может быть выявлена «утечка» экспрессии, составляет пять, шесть, семь или восемь. «Специфический для жировой ткани промотор» представляет собой промотор, который является способным к инициации транскрипции в жировой ткани, при этом все еще обеспечивая возможность какой-либо «утечки» экспрессии в других (максимум пяти, шести, семи или восьми) органах и частях тела. Транскрипцию в жировой ткани можно выявить в жировой ткани и жировых клетках, таких как белые адипоциты, бурые адипоциты, бежевые адипоциты, преадипоциты, стромальные сосудистые клетки. «Специфический для печени промотор» представляет собой промотор, который является способным к инициации транскрипции в печени, при этом все еще обеспечивая возможность какой-либо «утечки» экспрессии в других (максимум пяти, шести, семи или восьми) органах и частях тела. Транскрипцию в печени можно выявить в ткани печени и клетках печени, таких как гепатоциты, купферовские клетки и/или овальные клетки. Аналогично, «активный в скелетных мышцах промотор» представляет собой промотор, который является способным к инициации транскрипции в скелетной мышце, при этом все еще обеспечивая возможность какой-либо «утечки» экспрессии в других (максимум пяти, шести, семи или восьми) органах и частях тела. Транскрипцию в скелетной мышце можно выявить в клетках скелетной мышцы, таких как миоциты, миобласты, миосателлитоциты.
«Убиквитарный промотор» является активным практически во всех тканях, органах и клетках организма.
Подходящие промоторы для органоспецифической и/или тканеспецифической экспрессии нуклеотидной последовательности, кодирующей FGF21, включают в себя промотор человеческого α1-антитрипсина, промотор α1-антитрипсина в комбинации с энхансером гепатоцитарного регуляторного участка (HCR) из аполипопротеина Е, альбуминовый промотор, промотор основного белка мочи, промотор фосфоенолпируват-карбоксикиназы (РЕРСК), промотор печеночного белка-активатора (liver-enriched protein activator), транстиретиновый промотор, промотор тироксин-связывающего глобулина, промотор аполипопротеина А1, промотор печеночной формы белка, связывающего жирные кислоты, промотор фенилаланин-гидроксилазы, промотор белка адипоцитов 2 (аР2, также известный как белок 4, связывающий жирные кислоты, (FABP4)), промотор PPARy, адипонектиновый промотор, промотор фосфоенолпируват-карбоксикиназы (РЕРСК), промотор, полученный из человеческого цитохрома р450 ароматазы (p450arom), промотор mini/aP2 (состоящий из специфического для жировой ткани энхансера аР2 и базового промотора аР2), промотор разобщающего белка 1 (UCP1), промотор mini/UCP1 (состоящий из специфического для жировой ткани энхансера UCP1 и базового промотора UCP1), адипсиновый промотор, лептиновый промотор, промотор Foxa-2, промотор легкой цепи миозина, промотор тяжелой цепи миозина, десминовый промотор, промотор С5-12, промотор мышечной креатин-киназы (МСК), промотор гладкомышечного альфа-актина, промотор CK6, промотор миозинового шаперона Unc-45 В, базовый промотор MCK в комбинации с копиями энхансера МСК, промотор Enh358MCK (комбинация энхансера MCK с 358 п.о. проксимальным промотором гена MCK).
Функционально связанный
Термин «функционально связанный» определен в данном документе как конфигурация, при которой управляющая последовательность, такая как промоторная последовательность или регуляторная последовательность, размещена надлежащим образом в положении относительно нуклеотидной последовательности, представляющей интерес, предпочтительно, кодирующей FGF21, в результате чего промотор или управляющая или регуляторная последовательность управляет или воздействует на транскрипцию, и/или продукцию, или экспрессию представляющей интерес нуклеотидной последовательности, предпочтительно, кодирующей FGF21, в клетке и/или у субъекта. Например, промотор является функционально связанным с кодирующей последовательностью, если промотор способен инициировать или регулировать транскрипцию или экспрессию кодирующей последовательности, при этом следует понимать, что в данном случае кодирующая последовательность находится «под управлением» промотора. Когда одна или несколько нуклеотидных последовательностей и/или элементов, содержащихся в конструкции, определены в данном документе как «сконфигурированные таким образом, чтобы быть функционально связанными с необязательной нуклеотидной последовательностью, представляющей интерес», понятно, что указанные нуклеотидные последовательности и/или элементы сконфигурированы в указанной конструкции таким образом, что все эти нуклеотидные последовательности и/или элементы являются функционально связанными с указанной нуклеотидной последовательностью, представляющей интерес, когда указанная нуклеотидная последовательность, представляющая интерес, присутствует в указанной конструкции.
Вирусная экспрессионная конструкция
Экспрессионная конструкция несет геном, который способен стабилизировать и сохранять ее эписомально в клетке. В контексте настоящего изобретения клетка может означать включение клетки, применяемой для получения конструкции, или клетки, в которую будут вводить конструкцию. В качестве альтернативы, конструкция способна интегрироваться в геном клетки, например, посредством гомологичной рекомбинации или иным способом. Особенно предпочтительная экспрессионная конструкция представляет собой такую, в которой нуклеотидная последовательность, кодирующая FGF21, который определен в данном документе, является функционально связанной с промотором, который определен в данном документе, при этом указанный промотор способен управлять экспрессией указанной нуклеотидной последовательности (т.е. кодирующей последовательности) в клетке. Предпочтительно, указанный промотор управляет экспрессией указанной нуклеотидной последовательности по меньшей мере в одной клетке конкретного органа и/или конкретной ткани. Предпочтительно, указанный промотор управляет экспрессией указанной нуклеотидной последовательности по меньшей мере в одной клетке печени, жировой ткани и/или скелетной мышцы. Предпочтительно, указанный промотор управляет экспрессией по меньшей мере в 10%, 20%, 30%, 40%, 40%, 60%, 70%, 80%, 90% или 100% клеток печени, жировой ткани и/или скелетной мышцы. В контексте настоящего изобретения FGF21, подлежащий экспрессии в печени, жировой ткани или скелетной мышце, относится к предпочтительной или преимущественной (по меньшей мере на 10% выше, по меньшей мере на 20% выше, по меньшей мере на 30% выше, по меньшей мере на 40% выше, по меньшей мере на 50% выше, по меньшей мере на 60% выше, по меньшей мере на 70% выше, по меньшей мере на 80% выше, по меньшей мере на 90% выше, по меньшей мере на 100% выше, по меньшей мере на 150% выше, по меньшей мере на 200% выше или более высокому уровню) экспрессии FGF21 в печени, жировой ткани или скелетной мышце по сравнению с другими органами или тканями. В данной заявке в случаях, когда выражения «специфический для печени», или «специфический для жировой ткани», или «специфический для скелетной мышцы» упоминаются в контексте экспрессии, также предусмотрена специфическая для типа клеток экспрессия в типе (типах) клеток, составляющих печень, жировую ткань или скелетную мышцу, соответственно.
Вирусные экспрессионные конструкции согласно настоящему изобретению содержат нуклеотидную последовательность в форме, «подходящей для экспрессии у млекопитающего», что означает, что вирусные экспрессионные конструкции включают в себя одну или несколько регуляторных последовательностей, выбранных, исходя из клеток-хозяев млекопитающего, подлежащих применению для экспрессии, которые являются функционально связанными с нуклеотидной последовательностью, подлежащей экспрессии. Предпочтительно, указанные клетки-хозяева млекопитающего, подлежащие применению для экспрессии, представляют собой человеческие, мышиные или собачьи клетки.
Вирусные экспрессионные конструкции согласно настоящему изобретению содержат нуклеотидную последовательность, подлежащую экспрессии в печени, жировой ткани и/или скелетной мышце.
В контексте данного документа термин «жировая ткань» относится к ткани, состоящей из зрелых адипоцитов (т.е. жировых клеток) и комбинации мелких кровеносных сосудов, нервной ткани, лимфатических узлов и стромальной сосудистой фракции (SVF). SVF состоит из эндотелиальных клеток, фибробластов, клеток-предшественников адипоцитов (т.е. преадипоцитов) и иммунных клеток, таких как макрофаги и Т-клетки. У млекопитающих традиционно различают два отличающихся типа жировых тканей: белую жировую ткань (WAT) и бурую жировую ткань (ВАТ). У млекопитающих жировая ткань содержится в нескольких органах-депо. Жировые депо включают в себя, без ограничения, эпидидимальную WAT (eWAT), паховую WAT (iWAT), забрюшинную WAT (rWAT), мезентериальную WAT (mWAT), межлопаточную BAT (iBAT).
В контексте данного документа «скелетная мышца» относится к ткани, состоящей из мышечных волокон. Мышечное волокно, также известное как волокно мышцы, представляет собой одну многоядерную или синцитиальную клетку, которая является результатом слияния многих сотен миобластов, некоторые из которых остаются в зрелой мышце в виде недифференцированных клеток, известных как миосателлитоциты. Отдельные мышечные волокна окружены соединительной тканью, называемой эндомизием. Около 10-100 мышечных волокон образуют пучки, которые сами окружены еще одним слоем соединительной ткани, называемой перимизием. Наконец, скелетная мышца формируется группами пучков, которые также окружены еще одним слоем соединительной ткани, называемом эпимизием. Помимо мышечных волокон скелетная мышца также состоит из многочисленных кровеносных сосудов и нервов. Концы мышц сходятся в плотных соединительнотканных структурах, сухожилиях и апоневрозах, которые опосредуют прикрепление мышц к надкостнице костей или к соединительной ткани других мышц.
В контексте данного документа «печень» относится к ткани, состоящей из гепатоцитов. Гепатоциты составляют приблизительно 50-70% клеток в печени. Помимо гепатоцитов печень состоит из эндотелиальных клеток, перисинусоидальных клеток, овальных клеток, купферовских клеток и звездчатых клеток (клеток Ито). При активации купферовскими клетками звездчатые клетки превращаются в миофибробласты. Также в печени находятся центральные вены и портальные тракты (портальные триады), которые содержат предконцевые ветви печеночной артерии, воротная вена печени, желчные проточки и лимфатические сосуды.
Указано, что такая предпочтительная экспрессионная конструкция содержит экспрессионную кассету. Экспрессионная кассета в контексте данного документа содержит или состоит из нуклеотидной последовательности, кодирующей FGF21, которая функционально связана с промотором, при этом указанный промотор способен управлять экспрессией указанной нуклеотидной последовательности. В соответствии с вариантом осуществления экспрессионная кассета в контексте данного документа содержит или состоит из нуклеотидной последовательности, кодирующей FGF21, промотора и по меньшей мере одной нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, которая экспрессируется в печени, а также по меньшей мере одной нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, которая экспрессируется в сердце. В соответствии с одним вариантом осуществления описанные экспрессионные кассеты содержат нуклеотидные последовательности, кодирующие последовательности-мишени для микроРНК, которая экспрессируется в печени, и/или микроРНК, которая экспрессируется в сердце, с идеальной комплементарностью их когнатными микроРНК. В соответствии с еще одним вариантом осуществления описанные экспрессионные кассеты содержат одну или несколько нуклеотидных последовательностей, кодирующих сайты связывания микроРНК с неидеальной комплементарностью (одно несоответствие/пять последовательных нуклеотидов). В соответствии с еще одним вариантом осуществления экспрессионные кассеты могут содержать обе нуклеотидные последовательности, кодирующие идеальные и неидеальные сайты связывания микроРНК. Таким образом, экспрессионные кассеты можно адаптировать с получением в результате переменных уровней регуляции с применением нуклеотидных последовательностей, кодирующих один идеальный, несколько идеальных, один неидеальный, несколько неидеальных или комбинацию идеальных и неидеальных сайтов-мишеней для микроРНК. Кроме того, можно применять нуклеотидную последовательность, кодирующую сайты-мишени для разных микроРНК, тем самым допуская регуляцию гена с помощью нескольких микроРНК. Предпочтительным положением для нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, является 3'UTR. Тем не менее, также можно применять нуклеотидные последовательности (кодирующие последовательности-мишени), вставленные либо в кодирующую последовательность, либо в последовательности 5'UTR.
Выбор нуклеотидной последовательности, кодирующей последовательность-мишень микроРНК, определяется желаемым характером экспрессии. Присутствие эндогенной микроРНК в клетке будет ингибировать экспрессию гена или кодирующей последовательности с экспрессионной конструкции, которая содержит нуклеотидную последовательность, кодирующую последовательность-мишень для указанной микроРНК. Для ингибирования экспрессии гена или кодирующей последовательности, представляющей интерес, в заданном типе клеток выбирают нуклеотидную последовательность, кодирующую последовательность-мишень, которая распознается микроРНК, присутствующей в этом типе клеток.
Вирусная экспрессионная конструкция представляет собой экспрессионную конструкцию, которая предназначена для применения в генной терапии. Ее конструируют таким образом, чтобы она содержала часть вирусного генома, как определено ниже в данном документе.
Экспрессионные конструкции, раскрытые в данном документе, можно получить с применением методик на основе рекомбинации, при которых нуклеотидная последовательность, кодирующая указанный FGF21, экспрессируется в подходящей клетке, например, в культивируемых клетках или клетках многоклеточного организма, например, как описано в Ausubel et al., "Current Protocols in Molecular Biology", Greene Publishing and Wiley-Interscience, New York (1987); и в Sambrook and Russell (2001, выше); при этом оба из этих источников включены в данный документ посредством ссылки во всей своей полноте. См. также, Kunkel (1985) Proc. Natl. Acad. Sci. 82:488 (описывающая сайт-направленный мутагенез); и Roberts et al. (1987) Nature 328:731-734; или Wells, J.A., et al. (1985) Gene 34: 315 (описывающая кассетный мутагенез).
Как правило, последовательность нуклеиновой кислоты или нуклеотидную последовательность, кодирующую FGF21, применяют в экспрессионной конструкции или экспрессионном векторе. Фраза «экспрессионный вектор» или «вектор» обычно относится к нуклеотидной последовательности, которая способна к осуществлению экспрессии гена или кодирующей последовательности в хозяине, совместимом с такими последовательностями. Эти экспрессионные векторы, как правило, включают в себя по меньшей мере подходящие промоторные последовательности и, необязательно, сигналы терминации транскрипции. Дополнительный фактор, необходимый или полезный в осуществлении экспрессии, также можно применять, как описано в данном документе. Нуклеиновая кислота, или ДНК, или нуклеотидная последовательность, кодирующая FGF21, включена в ДНК-конструкцию, которую можно вводить в культуру клеток in vitro, и которая может экспрессироваться в культуре клеток in vitro. В частности, ДНК-конструкция является подходящей для репликации в хозяине-прокариоте, таком как бактерия, например, Е. coli, или ее можно вводить в культивируемые линии клеток млекопитающего, растения, насекомого, {например, Sf9), дрожжей, грибов или другие линии эукариотических клеток.
ДНК-конструкция, приготовленная для введения в конкретного хозяина, может включать в себя репликационную систему, распознаваемую хозяином, заданный сегмент ДНК, кодирующий желаемый полипептид, и регуляторные последовательности для инициации и терминации транскрипции и трансляции, функционально связанные с кодирующим полипептид сегментом. Термин «функционально связанный» уже был определен в данном документе. Например, промотор или энхансер является функционально связанным с кодирующей последовательностью, если он стимулирует транскрипцию последовательности. ДНК для сигнальной последовательности является функционально связанной с ДНК, кодирующей полипептид, если она экспрессируется в виде пребелка, который участвует в секреции полипептида. Обычно последовательность ДНК, которая является функционально связанной, является смежной, а в случае сигнальной последовательности и смежной, и находящейся в рамке считывания. Тем не менее, энхансеры не обязательно должны быть смежными с кодирующей последовательностью, транскрипцией которой они управляют. Связывание осуществляют посредством лигирования в подходящих рестрикционных сайтах или в адаптерах или линкерах, вставленных вместо них, или с помощью синтеза генов.
Выбор подходящей промоторной последовательности обычно зависит от клетки-хозяина, выбранной для экспрессии сегмента ДНК. Примеры подходящих промоторных последовательностей включают в себя прокариотические и эукариотические промоторы, хорошо известные в уровне техники (см., например, Sambrook and Russell, 2001, выше). Транскрипционная регуляторная последовательность, как правило, включает в себя гетерологичный энхансер или промотор, который распознается хозяином. Выбор соответствующего промотора зависит от хозяина, но такие промоторы, как промоторы trp, lac и фаговые промоторы, промоторы тРНК и промоторы гликолитических ферментов, являются известными и доступными (см., например, Sambrook and Russell, 2001, выше). Экспрессионный вектор включает в себя репликационную систему и транскрипционные и трансляционные регуляторные последовательности вместе с сайтом вставки для кодирующего полипептид сегмента. В большинстве случаев репликационная система является функциональной только в клетке, которую применяют для получения вектора (бактериальная клетка в виде Е. coli). Большинство плазмид и векторов не реплицируются в клетках, инфицированных вектором. Примеры подходящих для работы комбинаций клеточных линий и экспрессионных векторов описаны в Sambrook and Russell (2001, выше); и в Metzger et at. (1988) Nature 334: 31-36. Например, подходящие экспрессионные векторы могут экспрессироваться в дрожжах, например, S. cerevisiae, например, клетках насекомого, например, клетках Sf9, клетках млекопитающего, например, клетках СНО, и бактериальных клетках, например, Е. coli. Таким образом, клетка может представлять собой прокариотическую или эукариотическую клетку-хозяин. Клетка может представлять собой клетку, которая является подходящей для культивирования в жидких средах или на плотных средах.
В качестве альтернативы, клетка-хозяин представляет собой клетку, которая является частью многоклеточного организма, такого как трансгенное растение или животное.
Вирусный вектор
Вирусный вектор или вирусный вектор для генной терапии представляет собой вектор, который содержит вирусную экспрессионную конструкцию, которая определена выше.
Вирусный вектор или вирусный вектор для генной терапии представляет собой вектор, который является подходящим для генной терапии. Векторы, которые являются подходящими для генной терапии, описаны в Anderson 1998, Nature 392: 25-30; Walther and Stein, 2000, Drugs 60: 249-71; Kay et at., 2001, Nat. Med. 7: 33-40; Russell, 2000, J. Gen. Virol. 81: 2573-604; Amado and Chen, 1999, Science 285: 674-6; Federico, 1999, Curr. Opin. Biotechnol. 10: 448-53; Vigna and Naldini, 2000, J. Gene Med. 2: 308-16; Marin et al, 1997, Mol. Med. Today 3: 396-403; Peng and Russell, 1999, Curr. Opin. Biotechnol. 10: 454-7; Sommerfelt, 1999, J. Gen. Virol. 80: 3049-64; Reiser, 2000, Gene Ther. 7: 910-3; и в источниках, цитируемых в них.
Особенно подходящий вектор для генной терапии включает в себя аденовирусный вектор и вектор на основе аденоассоциированного вируса (AAV). Эти векторы инфицируют широкий спектр типов делящихся и неделящихся клеток, в том числе синовиальные клетки и клетки печени. Эписомальная природа аденовирусных векторов и векторов на основе AAV после попадания в клетку делает эти векторы подходящими для терапевтических применений (Russell, 2000, J. Gen. Virol. 81: 2573-2604; Goncalves, 2005, Virol J. 2(1):43), как указано выше. Векторы на основе AAV являются еще более предпочтительными, поскольку известно, что они приводят в результате к очень стабильной долговременной экспрессии трансгена (до 9 лет у собаки (Niemeyer et al, Blood. 2009 Jan 22; 113(4):797-806) и ~10 лет у человека (Buchlis, G. et al., Blood. 2012 Mar 29; 119(13):3038-41). Предпочтительные аденовирусные векторы модифицируют для снижения ответа хозяина, как рассмотрено в Russell (2000, выше). Способ генной терапии с применением векторов на основе AAV описан в Wang et al, 2005, J Gene Med. March 9 (Epub перед поступлением в печать), Mandel et al., 2004, Curr Opin Mol Ther. 6(5):482-90, и Martin et al., 2004, Eye 18(11):1049-55, Nathwani et al., N Engl J Med. 2011 Dec 22; 365(25):2357-65, Apparailly et al., Hum Gene Ther. 2005 Apr; 16(4):426-34.
Еще один подходящий вектор для генной терапии включает в себя ретровирусный вектор. Предпочтительный ретровирусный вектор для применения в настоящем изобретении представляет собой экспрессионную конструкцию на основе лентивируса. Лентивирусные векторы обладают способностью инфицировать и стабильно интегрироваться в геном делящихся и неделящихся клеток (Amado and Chen, 1999 Science 285: 674-6). Способы конструирования и применения экспрессионных конструкций на основе лентивирусов описаны в патентах США №№6165782, 6207455, 6218181, 6277633 и 6323031 и в Federico (1999, Curr Opin Biotechnol 10: 448-53) и Vigna et al. (2000, J Gene Med 2000; 2: 308-16).
Другие подходящие векторы для генной терапии включают в себя вектор на основе аденовируса, вектор на основе вируса герпеса, вектор на основе полиомавируса или вектор на основе вируса коровьей оспы.
Вектор для генной терапии содержит нуклеотидную последовательность, кодирующую FGF21, подлежащий экспрессии, при этом указанная нуклеотидная последовательность является функционально связанной с соответствующими регуляторными последовательностями. Такая регуляторная последовательность будет содержать по меньшей мере промоторную последовательность. Подходящие промоторы для экспрессии нуклеотидной последовательности, кодирующей FGF21, с вектора для генной терапии включают в себя, например, промотор CMV, промоторы длинных концевых повторов (LTR) вирусов, таких как промоторы из вируса лейкоза мышей Молони (MMLV), вируса саркомы Рауса или HTLV-1, ранний промотор вируса обезьян 40 (SV 40), промотор CAG, промотор α1-антитрипсина, промотор mini/aP2, промотор mini/UCP1, промотор С5-12 и промотор гена тимидинкиназы вируса простого герпеса.
Были описаны несколько индуцируемых промоторных систем, которые могут быть индуцированы посредством введения малых органических или неорганических веществ. Такие индуцируемые промоторы включают в себя промоторы, управляемые тяжелыми металлами, такие как металлотионеиновый промотор (Brinster et al. 1982 Nature 296: 39-42; Mayo et al. 1982 Cell 29: 99-108), управляемые RU-486 (антагонист прогестерона) (Wang et al. 1994 Proc. Natl. Acad. Sci. USA 91: 8180-8184), управляемые стероидами (Mader and White, 1993 Proc. Natl. Acad. Sci. USA 90: 5603-5607), управляемые тетрациклином (Gossen and Bujard 1992 Proc. Natl. Acad. Sci. USA 89: 5547-5551; патент США №5464758; Furth et al. 1994 Proc. Natl. Acad. Sci. USA 91: 9302-9306; Howe et al. 1995 J. Biol. Chem. 270: 14168-14174; Resnitzky et al. 1994 Mol. Cell. Biol. 14: 1669-1679; Shockett et al. 1995 Proc. Natl. Acad. Sci. USA 92: 6522-6526), и систему tTAER, которая основывается на мультихимерном трансактиваторе, состоящем из полипептида tetR в качестве активирующего домена VP16, и лиганд-связывающего домена эстрогенового рецептора (Yee et al., 2002, патентный документ US 6432705).
Вектор для генной терапии, необязательно, может содержать дополнительную нуклеотидную последовательность, кодирующую дополнительный полипептид.
Вектор для генной терапии предпочтительно составляют в композицию или фармацевтическую композицию, как определено в данном документе. В этом контексте композиция или фармацевтическая композиция могут содержать подходящий фармацевтический носитель, как определено выше в данном документе.
Вектор на основе аденоассоциированного вируса (вектор на основе AAV)
Предпочтительный вирусный вектор или предпочтительный вектор для генной терапии представляет собой вектор на основе AAV. Вектор на основе AAV в контексте данного документа предпочтительно содержит вектор на основе рекомбинантного AAV (вектор на основе rAAV). «Вектор на основе rAAV» в контексте данного документа относится к рекомбинантному вектору, содержащему часть генома AAV, заключенного в белковую оболочку из белка капсида, полученного из серотипа AAV, как объяснено в данном документе. Часть генома AAV может содержать инвертированные концевые повторы (ITR), полученные из такого серотипа аденоассоциированного вируса, как AAV1, AAV2, AAV3, AAV4, AAV5 и другие. Предпочтительные ITR представляют собой таковые из AAV2, которые представлены последовательностями, содержащими или состоящими из SEQ ID NO: 48 (5' ITR) и SEQ ID NO: 49 (3' ITR). Предпочтительно, настоящим изобретением также предусмотрено применение последовательности, по меньшей мере на 80% (или по меньшей мере на 81%, 82%, 83%, 84%, 85%, 86%, 87%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичной SEQ ID NO: 48, в качестве 5'ITR и последовательности, по меньшей мере на 80% идентичной SEQ ID NO: 49, в качестве 3'ITR.
Белковую оболочку, состоящую из белка капсида, можно получить из такого серотипа AAV, как AAV1, 2, 3, 4, 5 и другие. Предпочтительный капсид AAV представляет собой капсид AAV1, AAV3, AAV8, AAV9. Предпочтительный ITR происходит из AAV2. Белковую оболочку также можно назвать оболочкой из белков капсида. У вектора на основе rAAV может быть удален один или, предпочтительно, все гены AAV дикого типа, но он может все еще содержать функциональные последовательности нуклеиновой кислоты ITR. Функциональные последовательности ITR являются необходимыми для репликации, спасения и упаковки вирионов AAV. Последовательности ITR могут представлять собой последовательности дикого типа или могут являться по меньшей мере на 80%, 85%, 90%, 95%, 97%, 98%, 99% или 100% идентичными последовательностям дикого типа, или они могут быть изменены, например, посредством вставки, мутации, делеции или замены нуклеотидов, при условии, что они остаются функциональными. В этом контексте функциональность относится к способности управлять упаковкой генома в оболочку капсида, а затем обеспечивать возможность экспрессии в клетке-хозяине, подлежащей инфицированию, или в клетке-мишени. В контексте настоящего изобретения белковая оболочка капсида может происходить из другого серотипа, нежели геномный ITR вектора на основе rAAV.
Молекула нуклеиновой кислоты, представленная предпочтительной последовательностью нуклеиновой кислоты, предпочтительно вставлена между геномом rAAV или последовательностями ITR, как идентифицировано выше, например, экспрессионная конструкция, содержащая экспрессионный регуляторный элемент, функционально связанный с кодирующей последовательностью и 3' последовательностью терминации. Указанную молекулу нуклеиновой кислоты также можно назвать трансгеном.
«Функции помощника AAV» обычно относятся к соответствующим функциям AAV, требующимся для репликации и упаковки rAAV и обеспечиваемым вектору на основе rAAV in trans (внешним фактором). Функции помощника AAV дополняют функции AAV, которые отсутствуют у вектора на основе rAAV, но в них отсутствуют ITR AAV (которые обеспечиваются геномом вектора rAAV). Функции помощника AAV включают в себя две основных ORF AAV, а именно кодирующий участок rep и кодирующий участок cap или функциональные последовательности, по существу, идентичные им. Rep и Сар участки являются хорошо известными в уровне техники, см., например, Chiorini et at. (1999, J. of Virology, Vol 73(2): 1309-1319) или патентный документ US 5139941, которые включены в данный документ посредством ссылки. Функции помощника AAV могут обеспечиваться на конструкции-помощнике AAV. Введение конструкции-помощника в клетку-хозяин может происходить, например, посредством трансформации, трансфекции или трансдукции до или одновременно с введением генома rAAV, присутствующего в векторе на основе rAAV, который идентифицирован в данном документе. Таким образом, конструкции-помощники на основе AAV согласно настоящему изобретению могут быть выбраны таким образом, чтобы они продуцировали желаемую комбинацию серотипов для оболочки из белков капсида вектора на основе rAAV с одной стороны, и для репликации и упаковки генома rAAV, присутствующего в указанном векторе на основе rAAV, с другой стороны.
«Вирус-помощник AAV» обеспечивает дополнительные функции, требующиеся для репликации и упаковки AAV. Подходящие вирусы-помощники AAV включают в себя аденовирусы, вирусы простого герпеса (такие как HSV 1 и 2 типов) и вирусы коровьей оспы. Дополнительные функции, обеспечиваемые вирусом-помощником, также могут быть введены в клетку-хозяина посредством плазмид, как описано в патентном документе US 6531456, включенном в данный документ посредством ссылки.
«Трансген» определен в данном документе как ген, или кодирующая последовательность, или молекула нуклеиновой кислоты (т.е. молекула, кодирующая FGF21), которые были недавно введены в клетку, т.е. ген, который может присутствовать, но в норме может не экспрессироваться в клетке или экспрессироваться на недостаточном уровне. В этом контексте «недостаточный» означает, что несмотря на то, что указанный FGF21 экспрессируется в клетке, состояние и/или заболевание, которое определено в данном документе, все еще может проявляться. В этом случае настоящее изобретение обеспечивает возможность сверхэкспрессии FGF21. Трансген может содержать последовательности, которые являются нативными для клетки, последовательности, которые в естественных условиях не встречаются в клетке, и он может содержать комбинации и тех, и других. Трансген может содержать последовательности, кодирующие FGF21 и/или дополнительные белки, которые идентифицированы выше в данном документе, которые могут быть функционально связаны с соответствующими регуляторными последовательностями для экспрессии последовательностей, кодирующих FGF21, в клетке. Предпочтительно, трансген не интегрируется в геном клетки-хозяина.
«Трансдукция» относится к доставке FGF21 в реципиентную клетку-хозяин с помощью вирусного вектора. Например, трансдукция клетки-мишени вектором на основе rAAV согласно настоящему изобретению приводит к переносу генома rAAV, содержащегося в этом векторе, в подвергшуюся трансдукции клетку. «Клетка-хозяин» или «клетка-мишень» относится к клетке, в которую происходит доставка ДНК, как например, к мышечным клеткам субъекта. Векторы на основе AAV способны трансдуцировать как делящиеся, так и неделящиеся клетки.
Получение вектора на основе AAV
Получение рекомбинантного AAV (rAAV) для векторизации трансгенов было описано ранее. См. Ayuso Е, et al., Curr. Gene Ther. 2010; 10:423-436, Okada T, et al., Hum. Gene Ther. 2009; 20:1013-1021, Zhang H, et al., Hum. Gene Ther. 2009; 20:922-929, и Virag T, et al., Hum. Gene Ther. 2009; 20:807-817. Эти протоколы можно применять или адаптировать для получения AAV согласно настоящему изобретению. В соответствии с одним вариантом осуществления линию клеток-продуцентов временно трансфицируют полинуклеотидом согласно настоящему изобретению (содержащим экспрессионную кассету, фланкированную ITR) и конструкцией(конструкциями), которая(которые) кодирует(кодируют) белки rep и cap и обеспечивает(обеспечивают) функции помощника. В соответствии с еще одним вариантом осуществления клеточная линия стабильно обеспечивает функции помощника, и ее временно трансфицируют полинуклеотидом согласно настоящему изобретению (содержащим, экспрессионную кассету, фланкированную ITR) и конструкцией(конструкциями), которая(которые) кодирует(кодируют) белки rep и cap. В соответствии с еще одним вариантом осуществления клеточная линия стабильно обеспечивает белки rep и cap и функции помощника, и ее временно трансфицируют полинуклеотидом согласно настоящему изобретению. В соответствии с еще одним вариантом осуществления клеточная линия стабильно обеспечивает белки rep и cap, и ее временно трансфицируют полинуклеотидом согласно настоящему изобретению и полинуклеотидом, кодирующим функции помощника. В соответствии с еще одним вариантом осуществления клеточная линия стабильно обеспечивает полинуклеотид согласно настоящему изобретению, белки rep и cap и функции помощника. Способы получения и применения этих и других продуцирующих систем на основе AAV были описаны в уровне техники. См. Muzyczka N, и соавт., патентный документ US 5139941, Zhou X, и соавт., патентный документ US 5741683, Samulski R, и соавт., патентный документ US 6057152, Samulski R, и соавт., патентный документ US 6204059, Samulski R, и соавт., патентный документ US 6268213, Rabinowitz J, и соавт., патентный документ US 6491907, Zolotukhin S, и соавт., патентный документ US 6660514, Shenk Т, и соавт., патентный документ US 6951753, Snyder R, и соавт., патентный документ US 7094604, Rabinowitz J, и соавт., патентный документ US 7172893, Monahan Р, и соавт., патентный документ US 7201898, Samulski R, и соавт., патентный документ US 7229823, и Ferrari F, и соавт., патентный документ US 7439065.
Геном rAAV, присутствующий в векторе на основе rAAV, содержит по меньшей мере нуклеотидные последовательности участков инвертированных концевых повторов (ITR) из одного из серотипов AAV (предпочтительно, повторов из серотипа AAV2, который раскрыт выше в данном документе), или нуклеотидные последовательности, по существу, идентичные им, или нуклеотидные последовательности, по меньшей мере на 60% идентичные им, а также нуклеотидную последовательность, кодирующую FGF21 (под управлением подходящего регуляторного элемента), вставленную между двумя ITR. Геном вектора требует применения фланкирующих последовательностей 5' и 3' ITR для обеспечения возможности эффективной упаковки генома вектора в капсид rAAV.
Полный геном нескольких серотипов AAV и соответствующие ITR были секвенированы (Chiorini et al. 1999, J. of Virology Vol. 73, No. 2, p 1309-1319). Их можно либо клонировать, либо получить посредством химического синтеза, как известно в уровне техники, с применением, например, синтезатора олигонуклеотидов, который поставляется, например, Applied Biosystems Inc. (Фостерс, Калифорния, США), или с помощью стандартных методик молекулярной биологии. ITR можно клонировать из вирусного генома AAV или вырезать из вектора, содержащего ITR AAV. Нуклеотидные последовательности ITR можно либо лигировать на одном из концов в нуклеотидную последовательность, кодирующую один или несколько терапевтических белков, с применением стандартных методик молекулярной биологии, либо последовательность AAV между ITR можно заменить на желаемую нуклеотидную последовательность.
Предпочтительно, геном rAAV, который присутствует в векторе на основе rAAV, не содержит никаких нуклеотидных последовательностей, кодирующих вирусные белки, такие как гены rep (репликации) или cap (капсида) AAV. Этот геном rAAV может дополнительно содержать маркерный или репортерный ген, такой как ген, кодирующий, например, устойчивость к антибиотику, флуоресцентный белок {например, gfp), или ген, кодирующий выявляемый и/или селектируемый с помощью химических, ферментативных или иных методов продукт (например, lacZ, aph и т.д.), известный в уровне техники.
Геном rAAV, который присутствует в указанном векторе на основе rAAV, дополнительно содержит промоторную последовательность, функционально связанную с нуклеотидной последовательностью, кодирующей FGF21. Предпочтительные промоторные последовательности представляют собой промоторы, которые обеспечивают экспрессию в клетках скелетной мышцы и/или скелетной мышце, в клетках печени и/или печени и в жировых клетках и/или жировой ткани. Примеры таких промоторов включают в себя промотор CMV, CAG, mini/aP2, mini/UCP1, С5-12 и hAAT, как определено выше в данном документе.
Подходящая 3' нетранслируемая последовательность также может быть функционально связанной с нуклеотидной последовательностью, кодирующей FGF21. Подходящие 3' нетранслируемые участки могут представлять собой те, которые в естественных условиях асоциированы с нуклеотидной последовательностью, или они могут быть получены из отличающихся генов, таких как, например, сигнал полиаденилирования SV40 (SEQ ID NO: 50) и сигнал полиаденилирования кроличьего (3-глобина (SEQ ID NO: 51).
Необязательно, дополнительные нуклеотидные последовательности могут быть функционально связанными с нуклеотидной(нуклеотидными) последовательностью(последовательностями), кодирующей(кодирующими) FGF21, как например, нуклеотидные последовательности, кодирующие сигнальные последовательности, сигналы ядерной локализации, энхансеры экспрессии и т.п.
Оптимизация по кодону
«Оптимизация по кодону» в контексте данного документа относится к процессам, используемым для модификации существующей кодирующей последовательности или для конструирования кодирующей последовательности, например, для улучшения трансляции в клетке-хозяине или организме-хозяине экспрессии молекулы транскрипта РНК, транскрибируемой с кодирующей последовательностью, или для улучшения транскрипции кодирующей последовательности. Оптимизация по кодону включает в себя, без ограничения, процессы, включающие подбор кодонов для кодирующей последовательности для обеспечения соответствия предпочтению кодонов для организма-хозяина экспрессии. Например, для обеспечения соответствия предпочтению кодонов у хозяев экспрессии млекопитающих, предпочтительно, у хозяев экспрессии в виде мышей, собак или человека. Оптимизация по кодону также исключает элементы, которые потенциально оказывают отрицательное воздействие на стабильность и/или трансляцию РНК (например, последовательности терминации, ТАТА-боксы, сайты сплайсинга, сайты посадки рибосом, повторяющиеся и/или богатые GC последовательности и мотивы, обеспечивающие вторичные структуры РНК, или мотивы, обеспечивающие не стабильность).
В данном документе и в пунктах формулы изобретения глагол «содержать» и сочетания с ним используют в его неограничивающем смысле, что означает, что элементы после слова включены, но элементы, не упоминаемые специально, не исключаются. Кроме того, глагол «состоять из» можно заменить на «состоять, по существу, из», что означает, что вирусная экспрессионная конструкция, вирусный вектор, композиция, композиция для генной терапии, которые определены в данном документе, могут содержать дополнительный(дополнительные) компонент(компоненты) помимо специально идентифицированных компонентов, при этом указанный(указанные) дополнительный(дополнительные) компонент(компоненты) не изменяют уникальные характеристики настоящего изобретения.
Кроме того, упоминание элемента в форме единственного числа не исключает возможности того, что присутствует более чем один элемент, если контекстом явно не предписывается, чтобы присутствовал один и только один из элементов. Следовательно, единственное число обычно означает «по меньшей мере один».
Слово «примерно» или «приблизительно» при использовании совместно с численным значением (примерно 10, приблизительно 10) предпочтительно означает, что значение может представлять собой заданное значение 10 и может быть больше или меньше на 1% от значения.
Все патентные и литературные источники, упоминаемые в данном описании, тем самым включены посредством ссылки в своей полноте. Каждый вариант осуществления, который идентифицирован в данном документе, можно комбинировать, если не указано иное.
Настоящее изобретение дополнительно объясняется в следующих примерах. Эти примеры не ограничивают объем настоящего изобретения, а служат только для разъяснения настоящего изобретения.
Условные обозначения к фигурам
Фиг. 1. Предупреждение ожирения при введении в eWAT векторов AAV9-CAG-moFGF21-dmiRT С57В16 мышам. (А) Схематическое изображение векторов AAV-CAG-moFGF21-doublemiRT. Экспрессионная кассета содержала промотор С AG, оптимизированную по кодону кодирующую последовательность мышиного FGF21 и четыре тандемных повтора последовательности miRT122a и четыре тандемных повтора последовательности miRT1, клонированные в 3' нетранслируемый участок экспрессионной кассеты. ITR из AAV2 фланкировали экспрессионную кассету. Схематическое изображение представлено без соблюдения масштаба. CAG: промотор куриного β-актина/энхансер CMV; рА: полиА. (В) Уровни экспрессии FGF21 в тканях, участвующих в метаболизме. Уровни экспрессии оптимизированной по кодону кодирующей последовательности мышиного FGF21 измеряли с помощью RTqPCR в eWAT, iWAT, iBAT и печени С57В16 мышей и нормализовали к значениям Rplp0 (n=8-11 животных/группа). (С) Уровни циркулирующего в крови FGF21 (n=8-11 животных/группа). (D-E) Уровни экспрессии FGF21R1 (D) и β-клото (Е) в тканях, участвующих в метаболизме. Уровни экспрессии рецептора 1 FGF21 (FGF21R1) и β-клото измеряли с помощью RTqPCR в eWAT, iWAT, iBAT и печени С57В16 мышей и нормализовали к значениям Rplp0 (n=7 животных/группа). (F) Динамика изменения массы тела. Массу тела измеряли еженедельно (n=8-11 животных/группа). (G) Типичное изображение животных. (Н) Масса тканей. Масса eWAT, iWAT, rWAT, mWAT, iBAT и печени у получавших корм и HFD С57В16 мышей, которых обрабатывали векторами на основе AAV посредством введения в eWAT (n=8-11 животных/группа). Анализы осуществляли через 14 недель после введения в eWAT 1012 vg (вирусных геномов) векторов AAV9-CAG-moFGF21-doublemiRT или AAV9-CAG-null. Результаты представлены в виде среднего значения ±SEM. ND, не выявлялось. HFD, рацион с высоким содержанием жиров. AU, условные единицы. eWAT, эпидидимальная белая жировая ткань. iWAT, паховая белая жировая ткань. rWAT, забрюшинная белая жировая ткань. mWAT, мезентериальная белая жировая ткань. iBAT межлопаточная бурая жировая ткань. * р<0,05 в сравнении с AAV9-CAG-null с кормом, ** р<0,01 в сравнении с AAV9-CAG-null с кормом, *** р<0,001 в сравнении с AAV9-CAG-null с кормом, $ р<0,05 в сравнении с AAV9-CAG-null с HFD, $$ р<0,01 в сравнении с AAV9-CAG-null с HFD, $$$ р<0,001 в сравнении с AAV9-CAG-null с HFD.
Фиг. 2. Гистологический анализ жировой ткани и печени С57В16 мышей, обработанных посредством введения в eWAT векторов AAV9-CAG-moFGF21 doublemiRT. (А) Типичные изображения окрашенных гематоксилином и эозином срезов эпидидимальной белой жировой ткани (eWAT), паховой белой жировой ткани (iWAT) межлопаточной бурой жировой ткани (iBAT) и печени получавших корм и получавших HFD С57В16 мышей, которых обрабатывали посредством введения в eWAT векторов AAV9-CAG-moFGF21-doublemiRT или AAV9-CAG-null. Исходное увеличение ×100. (В) Средняя площадь белых адипоцитов в eWAT (n=4 животных/группа). (С) Частотное распределение площади белых адипоцитов в eWAT (n=4 животных/группа). Анализы осуществляли через 14 недель после введения в eWAT 1012 vg (вирусных геномов) векторов AAV9-CAG-moFGF21-doublemiRT или AAV9-CAG-null. Результаты представлены в виде среднего значения ±SEM. HFD, рацион с высоким содержанием жиров. ** р<0,01 в сравнении с AAV9-CAG-null с кормом, *** р<0,001 в сравнении с AAV9-CAG-null с кормом, $$ р<0,01 в сравнении с AAV9-CAG-null с HFD, $$$ р<0,001 в сравнении с AAV9-CAG-null с HFD.
Фиг. 3. Повышенное потребление энергии и чувствительность к инсулину у С57 В16 мышей, обработанных посредством введения в eWAT векторов AAV9-CAG-moFGF21-doublemiRT. (А-В) Уровни экспрессии UCP1 (А) и Dio2 (В). Уровни экспрессии UCP1 и Dio2 измеряли с помощью RTqPCR в iWAT и нормализовали к значениям Rplp0 (n=7 животных/группа). (С) Энергетический метаболизм. Потребление энергии (ЕЕ) измеряли с использованием калориметра непрямого действия с открытым контуром. Потребление кислорода и выработку диоксида углерода отслеживали одновременно. Данные получали через 9 недель после введения AAV во время дневной части (базальное состояние) и ночной части (фаза активности) цикла чередования света и темноты и корректировали на массу тела (n=8-11 животных/группа). (D) Содержание триглицеридов в печени (n=8-10 животных/группа). (E-F) Уровни триглицеридов (Е) и холестерина (F) в сыворотке крови (n=8-11 животных/группа). (G) Внутрибрюшинный тест толерантности к инсулину. Мышам давали внутрибрюшинную инъекцию 0,75 Ед. инсулина/кг массы тела и уровни глюкозы в крови измеряли в указанные моменты времени (n=6-11 животных/группа). Тесты осуществляли через 11 недель после введения AAV. (Н) Уровни циркулирующего в крови инсулина натощак. Если не указано иное, анализы осуществляли через 14 недель после введения в eWAT 1012 vg векторов AAV9-CAG-moFGF21-doublemiRT или AAV9-CAG-null. Результаты представлены в виде среднего значения ±SEM. HFD, рацион с высоким содержанием жиров. TG, триглицериды. Хол., холестерин. * р<0,05 в сравнении с AAV9-CAG-null с кормом, ** р<0,01 в сравнении с AAV9-CAG-null с кормом, *** р<0,001 в сравнении с AAV9-CAG-null с кормом, $ р<0,05 в сравнении с AAV9-CAG-null с HFD, $$ p<0,01 в сравнении с AAV9-CAG-null с HFD, $$$ р<0,001 в сравнении с AAV9-CAG-null с HFD.
Фиг. 4. Обращение развития ожирения при введении в eWAT векторов AAV8-CAG-moFGF21-dmiRT ob/ob мышам. (А) Уровни экспрессии FGF21 в тканях, участвующих в метаболизме. Уровень экспрессии оптимизированной по кодону кодирующей последовательности мышиного FGF21 измеряли с помощью RTqPCR в eWAT, iWAT, iBAT и печени ob/ob мышей и нормализовали к значениям Rplp0. (В) Уровни циркулирующего в крови FGF21. (C-D) Динамика изменения массы тела (С) и увеличения массы тела (D). Массу тела измеряли еженедельно. (Е) Масса тканей. Масса eWAT, iWAT, rWAT, mWAT, iBAT и печени ob/ob мышей, которых обрабатывали посредством введения в eWAT векторов на основе AAV. Анализы осуществляли через 16 недель после введения в eWAT 1010 vg, 5×1010 vg, 2×1011 vg или 1012 vg вектора AAV8-CAG-moFGF21-doublemiRT или 1012 vg вектора AAV8-CAG-null. Результаты представлены в виде среднего значения ±SEM. n=7-8 животных/группа. ND, не выявлялось. AU, условные единицы. eWAT, эпидидимальная белая жировая ткань. iWAT, паховая белая жировая ткань. rWAT, забрюшинная белая жировая ткань. mWAT, мезентериальная белая жировая ткань. iBAT межлопаточная бурая жировая ткань. * р<0,05 в сравнении с AAV8-CAG-null, ** р<0,01 в сравнении с AAV8-С AG-null, *** р<0,001 в сравнении с AAV8-CAG-null.
Фиг. 5. Улучшенная чувствительность к инсулину у ob/ob мышей, обработанных посредством введения в eWAT векторов AAV8-CAG-moFGF21-doublemiRT. (А) Внутрибрюшинный тест толерантности к инсулину. Ob/ob мышам давали внутрибрюшинную инъекцию 0,75 Ед. инсулина/кг массы тела и уровни глюкозы в крови измеряли в указанные моменты времени. Тесты осуществляли через 9 недель после введения AAV. (В) Уровни циркулирующего в крови инсулина натощак через 2 месяца после введения AAV. Результаты представлены в виде среднего значения ±SEM, n=7-8 животных/группа. * р<0,05 в сравнении с AAV8-CAG-null, ** р<0,01 в сравнении с AAV8-CAG-null, *** р<0,001 в сравнении с AAV8-CAG-null.
Фиг. 6. Обращение развития ожирения и ослабление метаболизма глюкозы при внутривенном введении векторов AAV8-hAAT-moFGF21 ob/ob мышам. (А) Схематическое изображение векторов AAV-hAAT-moFGF21. Экспрессионная кассета содержала промотор человеческого а 1-антитрипсина (hAAT) и оптимизированную по кодону кодирующую последовательность мышиного FGF21. ITR из AAV2 фланкировали экспрессионную кассету. Схематическое изображение представлено без соблюдения масштаба. рА: полиА. (В) Уровни экспрессии FGF21. Уровни экспрессии оптимизированной по кодону кодирующей последовательности мышиного FGF21 измеряли с помощью RTqPCR в печени ob/ob мышей и нормализовали к значениям Rplp0. (С) Уровни циркулирующего в крови FGF21. (D-E) Динамика изменения массы тела (С) и увеличения массы тела (D). Массу тела измеряли еженедельно. (F) Типичное изображение животных. (G) Масса тканей. Масса eWAT, iWAT, rWAT, mWAT, iBAT и печени ob/ob мышей, которых обрабатывали внутривенно векторами на основе AAV. (Н) Внутрибрюшинный тест толерантности к инсулину. Ob/ob мышам давали внутрибрюшинную инъекцию 0,75 Ед. инсулина/кг массы тела и уровни глюкозы в крови измеряли в указанные моменты времени. Тесты осуществляли через 9 недель после введения AAV. (I) Уровни циркулирующего в крови инсулина натощак через 3 месяца после введения AAV. Если не указано иное, анализы осуществляли через 20 недель после внутривенного введения 1011 vg или 5×1011 vg вектора AAV8-hAAT-moFGF21 или 5×1011 vg вектора AAV8-hAAT-null. Результаты представлены в виде среднего значения ±SEM. n=9-10 животных/группа. ND, не выявлялось. AU, условные единицы. eWAT, эпидидимальная белая жировая ткань. iWAT, паховая белая жировая ткань. rWAT, забрюшинная белая жировая ткань. mWAT, мезентериальная белая жировая ткань. iBAT межлопаточная бурая жировая ткань. * p<0,05 в сравнении с AAV8-hAAT-null, ** р<0,01 в сравнении с AAV8-hAAT-null, *** р<0,001 в сравнении с AAV8-hAAT-null.
Фиг. 7. Долговременное обращение развития ожирения при внутривенном введении векторов AAV-hAAT-moFGF21 C57bl6 мышам, которых кормили HFD. (А)
Уровни циркулирующего в крови FGF21. (В-С) Динамика изменения массы тела (С) и увеличения массы тела (D). Массу тела измеряли еженедельно. Анализы осуществляли через 52 недели после внутривенного введения 1010 vg или 5×1010 vg вектора AAV8-hAAT-moFGF21 или 5×1010 vg вектора AAV8-hAAT-null. Результаты представлены в виде среднего значения ±SEM, n=9-12 животных/группа. *** р<0,001 в сравнении с AAV8-hAAT-null с кормом, $$ р<0,01 в сравнении с AAV8-hAAT-null с HFD, $$$ p<0,001 в сравнении с AAV8-hAAT-null с HFD.
Фиг. 8. Долговременное повышенное потребление энергии и чувствительность к инсулину при внутривенном введении векторов AAV-hAAT-moFGF21 C57Bl6 мышам, которых кормили HFD. (А) Энергетический метаболизм. Потребление энергии (ЕЕ) измеряли с использованием калориметра непрямого действия с открытым контуром. Потребление кислорода и выработку диоксида углерода отслеживали одновременно. Данные получали через 4 недели после введения AAV во время дневной части (базальное состояние) и ночной части (фаза активности) цикла чередования света и темноты и корректировали на массу тела. (В) Внутрибрюшинный тест толерантности к инсулину. С57 В16 мышам давали внутрибрюшинную инъекцию 0,75 Ед. инсулина/кг массы тела и уровни глюкозы в крови измеряли в указанные моменты времени. Тесты осуществляли через 7 недель после введения AAV. (С) Уровни циркулирующего в крови инсулина натощак и после кормления. Результаты представлены в виде среднего значения ±SEM, n=9-12 животных/группа. HFD, рацион с высоким содержанием жиров. * р<0,05 в сравнении с AAV8-hAAT-null с кормом, ** р<0,01 в сравнении с AAV8-hAAT-null с кормом, *** р<0,001 в сравнении с AAV8-hAAT-null с кормом, $ р<0,05 в сравнении с AAV8-hAAT-null с HFD, $$ р<0,01 в сравнении с AAV8-hAAT-null с HFD, $$$ р<0,001 в сравнении с AAV8-hAAT-null с HFD.
Фиг. 9. Обращение развития ожирения при внутривенном введении векторов AAV-hAAT-moFGF21 старым мышам, которых кормили HFD. (А) Уровни циркулирующего в крови FGF21. (В-С) Динамика изменения массы тела (В) и увеличения массы тела (С). Массу тела измеряли еженедельно. Анализ осуществляли через 21 неделю после внутривенного введения 1010 vg, 2×1010 vg или 5×1010 vg вектора AAV8-hAAT-moFGF21 или 5×1010 vg вектора AAV8-hAAT-null. Результаты представлены в виде среднего значения ±SEM, n=7-8 животных/группа. HFD, рацион с высоким содержанием жиров. *** р<0,05 в сравнении с AAV8-hAAT-null с кормом, $ р<0,05 в сравнении с AAV8-hAAT-null с HFD, $$ р<0,01 в сравнении с AAV8-hAAT-null с HFD. $$$ р<0,001 в сравнении с AAV8-hAAT-null с HFD.
Фиг. 10. Повышенное потребление энергии и чувствительность к инсулину при внутривенном введении векторов AAV-hAAT-moFGF21 старым мышам, которых кормили HFD. (А) Энергетический метаболизм. Потребление энергии (ЕЕ) измеряли с использованием калориметра непрямого действия с открытым контуром. Потребление кислорода и выработку диоксида углерода отслеживали одновременно. Данные получали через 6 недель после введения AAV во время дневной части (базальное состояние) и ночной части (фаза активности) цикла чередования света и темноты и корректировали на массу тела. (В) Внутрибрюшинный тест толерантности к инсулину. Старым C57Bl6 мышам давали внутрибрюшинную инъекцию 0,75 Ед. инсулина/кг массы тела и уровни глюкозы в крови измеряли в указанные моменты времени. Тесты осуществляли через 9 недель после введения AAV. (С) Уровни циркулирующего в крови инсулина натощак и после кормления. Результаты представлены в виде среднего значения ±SEM, n=7-8 животных/группа. HFD, рацион с высоким содержанием жиров. ** р<0,01 в сравнении с AAV8-hAAT-null с кормом, *** р<0,001 в сравнении с AAV8-hAAT-null с кормом, $ p<0,05 в сравнении с AAVS-hAAT-null с HFD, $$ р<0,01 в сравнении с AAV8-hAAT-null с HFD, $$$ р<0,001 в сравнении с AAV8-hAAT-null с HFD.
Фиг. 11. Потеря массы тела при внутримышечном введении векторов AAV-CMV-moFGF21 C57Bl6 мышам. (А) Схематическое изображение векторов AAV-CMV-moFGF21. Экспрессионная кассета содержала промотор цитомегаловируса (CMV) и оптимизированную по кодону кодирующую последовательность мышиного FGF21. ITR из AAV2 фланкировали экспрессионную кассету. Схематическое изображение представлено без соблюдения масштаба. рА: полиА. (В) Уровни циркулирующего в крови FGF21. (C-D) Динамика изменения массы тела (С) и увеличения массы тела (D). Массу тела измеряли еженедельно. Результаты представлены в виде среднего значения ±SEM. n=6-7 животных/группа. * р<0,05 в сравнении с AAV1-CMV-null, ** р<0,01 в сравнении с AAV1-CMV-null. Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Фиг. 12. Повышенная продукция белка FGF21 при оптимизации по кодону нуклеотидных последовательностей, кодирующих человеческий FGF21. (А) Уровни белка hFGF21 в культуральной среде НЕК293 клеток, трансфицированных hFGF21 дикого типа или тремя различными вариантами оптимизированных по кодону последовательностей человеческого FGF21. Результаты представлены в виде среднего значения ±SEM. N=3 лунки/группа. ND, не выявлялось. * р<0,05 в сравнении с нетрансфицированными клетками.
Фиг. 13. Введение в eWAT векторов AAV8-CAG-moFGF21-dmiRT ob/ob мышам.
А, В Типичные изображения окрашивания гематоксилином-эозином срезов ткани (А) eWAT и (В) печени, полученных от ob/ob животных, которым вводили инъекцией в eWAT либо нулевой вектор на основе AAV8, либо кодирующий FGF21 вектор на основе AAV8 во всех исследуемых дозах. Масштабные метки: 100 мкм для eWAT и 200 мкм для печени.
С Гликемия в сытом состоянии.
D Инсулинемия в сытом состоянии через 3 месяца после введения AAV.
Пометки для FGF21 на фигуре относятся к moFGF21.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (А, В) n=6-9 животных/группа. В (С-Н) n=4-8 животных/группа. В (I) n=6-8 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой, которым вводили инъекцией нулевой вектор.
Фиг. 14. Воздействие переноса гена FGF21 в eWAT ob/ob мышей.
А Уровни адипонектина в сыворотке крови у ob/ob животных возрастом 25 недель, которым вводили инъекцией в eWAT в возрасте 11 недель либо векторы AAV8-CAG-null, либо векторы AAV8-CAG-moFGF21 - dmiRT в 4 различных дозах (1×1010, 5×1010, 2×1011, 1×1012 vg/мышь).
В Количественное определение с помощью qRT-PCR экспрессии маркера макрофагов F4/80 в тех же группах животных, что и в (А).
С Типичные изображения иммуноокрашивания срезов eW AT от ob/ob мышей, которые получали векторы AAV8-CAG-moFGF21-dmiRT, в отношении специфического для макрофагов маркера Мас2. n=4-8/группа. Масштабные метки: 200 мкм.
D Масса печени во всех группах обработки внутри eWAT.
Е, F Содержание триглицеридов и холестерина в печени в сытом состоянии в тех же когортах, что и в (А).
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (А, В, D) n=4-8 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой ob/ob мышей, которым вводили инъекцией нулевой вектор.
Фиг. 15. Уменьшенное ожирение и улучшенная чувствительность к инсулину у ob/ob мышей, обработанных векторами AAV8-hAAT-moFGF21.
А Типичные изображения окрашивания гематоксилином-эозином срезов ткани eWAT, полученных от ob/ob животных, которым вводили инъекцией либо нулевой вектор на основе AAV, либо кодирующий FGF21 вектор на основе AAV в количестве 1×1011 или 5×1011 vg/мышь.
В Уровни адипонектина в сыворотке крови во всех группах.
С Типичные изображения окрашивания гематоксилином-эозином срезов ткани печени, полученных от ob/ob животных, которым вводили инъекцией либо нулевой вектор на основе AAV, либо кодирующий FGF21 вектор на основе AAV в количестве 1×1011 или 5×1011 vg/мышь.
D Уровни глюкозы в крови в сытом состоянии.
Е Уровни инсулина в сыворотке крови в сытом состоянии через 5 месяцев после введения AAV.
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все данные представляют собой среднее значение ±SEM. В (А-С, Е, G-H) n=9-10 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой ob/ob мышей, которым вводили инъекцией нулевой вектор.
Фиг. 16. Воздействия переноса гена FGF21 в печень на ob/ob мышей.
А Иммуногистохимический анализ в отношении специфического для макрофагов маркера Мас2 в срезах eWAT от ob/ob мышей, которые получали векторы AAV8-hAAT-moFGF21. Масштабные метки: 500 мкм.
В, С Количественное определение с помощью qRT-PCR экспрессии маркеров воспаления F4/80 (В) и TNF-α (С) в тех же когортах мышей.
D, Е Масса (D) и типичные изображения печени (Е), полученные от животных, принадлежащих к тем же экспериментальным группам, как и в (А).
F, G Содержание триглицеридов и холестерина в печени в сытом состоянии в тех же когортах, что и в (А).
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (В, D-F, H-I) n=9-10 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой ob/ob мышей, которым вводили инъекцией нулевой вектор.
Фиг. 17. Обработка AAV8-hAAT-moFGF21 повышает экспрессию генов, вовлеченных в усвоение глюкозы и термогенез, в жировой ткани ob/ob мышей.
А, В Количественное определение с помощью qRT-PCR экспрессии PEPCK и G6Разы (глюкоза-6-фосфатаза) в печени у ob/ob мышей, которым вводили инъекцией в возрасте 2 месяца либо векторы AAV8-hAAT-null, либо векторы AAV8-hAAT-moFGF21.
C-F Количественное определение с помощью qRT-PCR экспрессии GLUT1 (С), GLUT4 (D), HKI (Е) и HKII (F) в eWAT, iWAT и iBAT у тех же животных, что и в (А).
G Относительная экспрессия UCP1 в iBAT в тех же когортах, что и в (А).
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (A-G) n=9-10 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой ob/ob мышей, которым вводили инъекцией нулевой вектор.
Фиг. 18. Опосредуемый AAV8 перенос гена FGF21 в печени противодействует индуцируемому HFD ожирению.
А Масса эпидидимального (eWAT), пахового (iWAT) и забрюшинного (rWAT) депо белой жировой ткани, печени и четырехглавой мышцы, полученных от мышей, обработанных векторами AAV8-hAAT-moFGF21 в молодом возрасте {верхняя секция) или во взрослом возрасте {нижняя секция).
В Уровни циркулирующего в крови FGF21 в различные моменты времени после введения вектора.
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (A-D) n=7-10 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой получавших корм животных, которым вводили инъекцией нулевой вектор. #Р<0,05, ##Р<0,01 и ###Р<0,001 в сравнении с группой получавших HFD животных, которым вводили инъекцией нулевой вектор. HFD, рацион с высоким содержанием жиров.
Фиг. 19. Перенос гена FGF21 в печень противодействует индуцированному HFD ожирению.
А, В Типичные изображения животных, принадлежащих ко всем экспериментальным группам в исследованиях, осуществляемых у молодых (А) или у взрослых (В) животных.
С Типичные изображения подушки эпидидимальной белой жировой ткани (eWAT), полученной при умерщвлении от животных, обработанных несколькими дозами AAV8-hAAT-moFGF21 в молодом возрасте (слева) или во взрослом возрасте (справа).
D Типичные изображения печени, полученной от животных, обработанных в молодом возрасте (слева) или во взрослом возрасте (справа).
Е Достигнутая с помощью AAV экспрессия FGF21 в печени животных, обработанных в молодом возрасте или во взрослом возрасте. qPCR осуществляли с использованием праймеров, которые специфически выявляли оптимизированную по кодону кодирующую последовательность мышиного FGF21 (coFGF21).
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (Е) n=7-10 животных/группа. HFD, рацион с высоким содержанием жиров. ND, не выявлялось.
Фиг. 20. Опосредованное AAV8-hAAT-moFGF21 повышенное потребление энергии и пониженное накопление жира в iBAT и iWAT.
А Оценка двигательной активности посредством теста открытого поля у животных, которые подверглись кормлению HFD с возраста ~2 месяцев, и которых обрабатывали либо нулевым вектором, либо вектором, кодирующим FGF21, 2 месяца спустя (в молодом возрасте).
В Окрашивание гематоксилином-эозином срезов ткани iBAT, полученных от животных, обработанный в молодом возрасте (слева) или во взрослом возрасте (справа).
С Анализ методом вестерн-блоттинга содержания UCP1 в iBAT от той же когорты животных, что и в (А). Показан типичный иммуноблот (слева). Гистограмма описывает денситометрический анализ двух отличающихся иммуноблотов (справа).
D Окрашивание гематоксилином-эозином срезов ткани iWAT, полученных от животных, обработанных в молодом возрасте (слева) или во взрослом возрасте (справа).
Е Количественное определение с помощью qRT-PCR экспрессии Phosphol в iWAT в группах животных, которых начинали кормить HFD, и которые получали векторы, кодирующие FGF21, в молодом возрасте или во взрослом возрасте.
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (А-С) n=7-10 животных/группа. В (Е) n=4 животных/группа. В (G) n=7-10 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой получавших корм животных, которым вводили инъекцией нулевой вектор. #Р<0,05 и ##Р<0,001 в сравнении с группой получавших HFD животных, которым вводили инъекцией нулевой вектор. HFD, рацион с высоким содержанием жиров.
Фиг. 21. Потребление энергии через 10 месяцев после переноса гена в печень.
А Потребление энергии измеряли через 10 месяцев после доставки вектора AAV8-hAAT-null или AAV8-hAAT-moFGF21 в когорте животных, которых начинали кормить HFD в возрасте 2 месяцев. Данные получали во время циклов чередования света и темноты.
В Анализ методом вестерн-блоттинга содержания UCP1 в iWAT от той же когорты животных. Показан типичный иммуноблот (слева). На графике показан денситометрический анализ двух отличающихся иммуноблотов (справа).
С Относительная экспрессия Serca2b и RyR2 в iWAT в группах животных, которых начинали кормить HFD, и которые получали векторы, кодирующие FGF21, в молодом возрасте или во взрослом возрасте.
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (А) n=7-10 животных/группа. В (В) n=4 животных/группа. В (С) n=7-10 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой получавших корм животных, которым вводили инъекцией нулевой вектор. ###Р<0,001 в сравнении с группой получавших HFD животных, которым вводили инъекцией нулевой вектор. HFD, рацион с высоким содержанием жиров.
Фиг. 22. Опосредованное AAV8-hAAT-moFGF21 обращение развития гиперплазии островков.
А Уровни глюкагона натощак в группе животных, которых начинали кормить HFD, и которые получали векторы, кодирующие FGF21, в молодом возрасте.
В Масса β-клеток в группе животных, которых начинали кормить HFD, и которые получали векторы, кодирующие FGF21, во взрослом возрасте.
С Типичные изображения результатов иммуноокрашивания в отношении инсулина в срезах поджелудочной железы от животных, которые получали 5×1010 vg/мышь AAV8-hAAT-moFGF21 во взрослом возрасте. Масштабные метки: 400 мкм. Масштабные метки на вкладках: 100 мкм.
D Типичные изображения результатов двойного иммуноокрашивания в отношении инсулина (темно-серый) и глюкагона (светло-серый) в срезах поджелудочной железы от животных, которые получали 5×1010 vg/мышь AAV8-hAAT-moFGF21 в молодом возрасте (верхняя секция) или во взрослом возрасте (нижняя секция).
Масштабные метки: 100 мкм.
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (А-С) n=7-10 животных/группа. В (D) n=4-5 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой получавших корм животных, которым вводили инъекцией нулевой вектор. #Р<0,05, ##Р<0,01 и ###Р<0,001 в сравнении с группой получавших HFD животных, которым вводили инъекцией нулевой вектор. HFD, рацион с высоким содержанием жиров.
Фиг. 23. Обработка AAV8-hAAT-moFGF21 улучшает толерантность к глюкозе.
А Толерантность к глюкозе исследовали в группе мышей, которых начинали кормить HFD, и которые получали векторы, кодирующие FGF21 в молодом возрасте, после внутрибрюшинной инъекции глюкозы (2 г/кг массы тела).
В Уровни инсулина в сыворотке крови во время теста толерантности к глюкозе, представленного в (А).
Информация о данных: все данные представляют собой среднее значение ±SEM. В (A-D) n=7-10 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой получавших корм животных, которым вводили инъекцией нулевой вектор. #Р<0,05, ##Р<0,01 и ###Р<0,001 в сравнении с группой получавших HFD животных, которым вводили инъекцией нулевой вектор. HFD, рацион с высоким содержанием жиров.
Фиг. 24. Обращение развития гипертрофии и воспаления WAT при обработке AAV8-hAAT-moFGF21.
А Типичные изображения результатов окрашивания гематоксилином-эозином eWAT от животных, которых кормили кормом или HFD и которым вводили либо векторы AAV8-hAAT-null, либо 5×1010 vg/мышь векторов AAV8-hAAT-moFGF21 в молодом возрасте (левые секции) или во взрослом возрасте (правые секции). В то время как мыши, которых кормили HFD и которым вводили инъекцией нулевой вектор имели более крупные адипоциты, животные, которых кормили HFD, и которые получали обработку FGF21, имели адипоциты уменьшенного размера. Масштабные метки: 100 мкм.
В Морфометрический анализ площади адипоцитов в WAT у животных, обработанных в молодом возрасте или во взрослом возрасте.
С, D Уровни циркулирующего в крови адипонектина (С) и лептина (D).
Е Иммуногистохимический анализ в отношении макрофаг-специфического маркера Мас2 в срезах eWAT для животных, которые получали 5×1010 vg/мышь AAV8-hAAT-moFGF21 во взрослом возрасте. На микрофотографиях иллюстрируется присутствие коронообразных структур {стрелки и вкладка) в eWAT у животных, которых кормили HFD и которым вводили инъекцией нулевой вектор, но такие структуры отсутствовали в eWAT у мышей, которых кормили HFD и обрабатывали FGF21. Масштабные метки: 200 мкм и 50 мкм (вкладка).
F-H Количественное определение с помощью qRT-PCR экспрессии маркеров воспаления F4/80 (F), IL1-β (G) и TNF-α (Н) в группе животных, которых начинали кормить HFD, и которые получали векторы, кодирующие FGF21, во взрослом возрасте.
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (В) n=4 животных/группа. В (F-H) n=7-10 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой получавших корм животных, которым вводили инъекцией нулевой вектор. #Р<0,05, ##Р<0,01 и ###Р<0,001 в сравнении с группой получавших HFD животных, которым вводили инъекцией нулевой вектор. HFD, рацион с высоким содержанием жиров.
Фиг. 25. Размер адипоцитов и воспаление у обработанных AAV8-hAAT-moFGF21 животных.
А Частотное распределение площади адипоцитов в группах животных, которых начинали кормить кормом или HFD и которые получали либо вектор AAV8-hAAT-null, либо 5×1010 vg/мышь AAV8-hAAT-moFGF21 векторов в молодом возрасте (верхний график) или во взрослом возрасте (нижний график).
В Иммуногистохимический анализ Мас2 в eWAT у животных, у которых исследование начинали в молодом возрасте. Коронообразные структуры образовывались инфильтрирующими макрофагами в eWAT у мышей, которых кормили HFD и которым вводили инъекцией нулевой вектор, указаны стрелками. Масштабные метки: 200 мкм и 50 мкм (вкладка).
С-Е Относительная экспрессия маркеров воспаления F4/80, CD68 и TNF-α, исследуемая с помощью qRT-PCR в той же когорте животных, что и в (В).
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (А) n=4 животных/группа. В (С-Е) n=7-10 животных/группа. ***Р<0,001 в сравнении с группой получавших корм животных, которым вводили инъекцией нулевой вектор. ###Р<0,001 в сравнении с группой получавших HFD животных, которым вводили инъекцией нулевой вектор. HFD, рацион с высоким содержанием жиров.
Фиг. 26. Обработка кодирующими FGF21 векторами обращает развитие стеатоза и воспаления печени.
А Типичные изображения результатов окрашивание гематоксилином-эозином срезов печени, полученных от животных, которых кормили кормом или HFD и которым вводили либо вектор AAV8-hAAT-null, либо 5×1010 vg/мышь векторов AAV8-hAAT-moFGF21. HFD явно индуцировал отложение капелек липидов в печени, и образование этого отложения обращалось под действием обработки AAVS-hAAT-moFGF21 как у животных в молодом возрасте, так и у взрослых животных. Масштабные метки: 100 мкм.
В, С Содержание триглицеридов и холестерина в печени в сытом состоянии в тех же когортах животных.
D Результаты иммуноокрашивания в отношении макрофаг-специфического маркера Мас-2 срезов печени от животных, которых кормили HFD, и которые получали либо вектор AAV8-hAAT-null, либо 5×1010 vg/мышь векторов AAVS-hAAT-moFGF21. Стрелками указано присутствие коронообразных структур. Масштабные метки: 200 мкм и 50 мкм (вкладка).
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (В-С) n=7-10 животных/группа. **Р<0,01 и ***Р<0,001 в сравнении с группой получавших корм животных, которым вводили инъекцией нулевой вектор. ##Р<0,01 в сравнении с группой получавших HFD животных, которым вводили инъекцией нулевой вектор. HFD, рацион с высоким содержанием жиров.
Фиг. 27. Опосредованное AAV8-hAAT-moFGF21 ослабление фиброза печени.
Анализ фиброза печени посредством трихром-окрашивания по Массону у животных, которых кормили HFD и которые получали 5×1010 vg/мышь либо вектора AAV8 hAAT-null, либо вектора AAV8-hAAT-moFGF21. Обработка AAV8-hAAT-moFGF21 (правые секции) заметно снижала выявление коллагеновых волокон, которые были легко выявлялись (показаны синим) у животных, обработанных нулевым вектором (левые секции). Масштабные метки: 50 мкм. Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Фиг. 28. Обработка AAV8-hAAT-moFGF21 обеспечивает улучшение при фиброзе печени.
А Анализ фиброза печени посредством окрашивания пикросириусом у животных, которых кормили HFD и которые получали 5×1010 vg/мышь либо вектора AAV8-hAAT-null, либо вектора AAV8-hAAT-moFGF21. Обработка AAV8-hAAT-moFGF21 (правые секции) заметно снижала выявление коллагеновых волокон, которые были легко выявлялись (показаны черным) у животных, обработанных нулевым вектором (левые секции). Масштабные метки: 50 мкм.
В, С Количественное определение с помощью qRT-PCR экспрессии коллагена 1 в печени в группе животных, которых начинали кормить HFD и которые получали векторы, кодирующие FGF21, в молодом возрасте (В) или во взрослом возрасте (С).
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ±SEM. В (В-С) n=7-10 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой получавших корм животных, которым вводили инъекцией нулевой вектор. #Р<0,05 и ###Р<0,001 в сравнении с группой получавших HFD животных, которым вводили инъекцией нулевой вектор. HFD, рацион с высоким содержанием жиров.
Фиг. 29. Наблюдали отсутствие аномалий костей у обработанных AAV8-hAAT-moFGF21 животных. Долговременные эффекты переноса гена FGF21 в отношении костей исследовали посредством сравнения мышей, которых кормили HFD, обработанных самой высокой дозой (5×1010 vg/мышь) векторов AAV8-hAAT-moFGF21 в молодом возрасте или во взрослом возрасте, с животными, которым вводили инъекцией нулевой вектор и которых кормили либо кормом, либо HFD.
А Общая назоанальная длина.
В Длина большеберцовой кости.
С-О Анализ методом компьютерной микротомографии (μСТ) эпифиза (C-J) и диафиза (K-О) большеберцовых костей, полученных в момент умерщвления, т.е. когда возраст животных составлял 18 месяцев, из мышей, которых кормили HFD и которым вводили либо нулевые векторы, либо векторы на основе AAV, кодирующие FGF21.
Р, Q Уровни циркулирующих в крови IGFBP1 (Р) и IGF1 (Q), измеренные с помощью ELISA.
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все данные представляют собой среднее значение ±SEM. В (A, P-Q) n=7-10 животных/группа. В (В-О) n=4 животных/группа. **Р<0,01 и ***Р<0,001 в сравнении с группой получавших корм животных, которым вводили инъекцией нулевой вектор. HFD, рацион с высоким содержанием жиров; BMD, минеральная плотность кости; ВМС, содержание минералов в кости; BV, объем кости; BV/TV, соотношение объем кости/объем ткани; BS/BV, соотношение поверхность кости/объем кости; Tb.N, число трабекул; Tb.Th, толщина трабекул; Tb.Sp, расстояние между трабекулами.
Фиг. 30. Анализ гликемических профилей у C57Bl6 мышей, обработанных векторами AAV8-hAAT-moFGF21. Уровни глюкозы в крови оценивали в сытом состоянии. AAV, IV введение 5×1010 vg или 2×1011 vg векторов AAV8-hAAT-moFGF21 (n=13 и 15, соответственно) или 2×1011 vg векторов AAV8-null (n=15). STZ, обработка стрептозоцином (5×50 мг/кг). Результаты представлены в виде среднего значения +SEM. * р<0,05; *** р<0,001 в сравнении с AAV8-hAAT-Null. Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Фиг. 31. Перенос гена FGF21 в скелетную мышцу здоровых животных.
А Уровни циркулирующего в крови FGF21, измеренные через 40 недель после инъекции ЗхЮ11 vg/мышь одного из векторов AAV1-CMV-Null или AAV1-CMV-moFGF21 в скелетную мышцу здоровых животных, которых кормили рационом в виде корма.
В Достигнутая с помощью AAV экспрессия FGF21 в мышцах и печени здоровых животных, которым вводили инъекцией внутримышечно векторы AAV1-CMV-Null или AAVl-CMV-moFGF21.
С Динамика изменений масса тела в 40-недельный период последующего наблюдения.
D Масса во влажном состоянии ткани различных мышц, жировых подушек и печени.
Е, F Содержание триглицеридов и холестерина в печени в сытом состоянии.
G Уровни инсулина в сыворотке крови в сытом состоянии.
Н Чувствительность к инсулину, оцениваемая посредством внутрибрюшинной инъекции инсулина (0,75 единиц/кг массы тела) и представлена в виде процента от исходного уровня глюкозы в крови.
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения ± SEM. В (А-Н) n=5-7 животных/группа. *Р<0,05, **Р<0,01 и ***Р<0,001 в сравнении с группой, которой вводили инъекцией нулевой вектор.
Фиг. 32. Опосредованный AAV1 перенос генов FGF21 в скелетную мышцу противодействует индуцируемому HFD ожирению и резистентности к инсулину.
А, В Динамика изменений массы тела (А) и увеличения массы тела (В) у животных, обработанных AAV1-CMV-moFGF21. С57В16 мышей кормили HFD в течение ~12 недель, а затем вводили 3×1011 vg/мышь векторов AAV1-CMV-moFGF21. Контрольные мыши с ожирением и контрольные мыши, которых кормили кормом, получали 3×1011 vg AAV1-CMV-null.
С Уровни циркулирующего в крови FGF21 в различные моменты времени после введения вектора.
D, Е Уровни глюкозы в крови натощак (D) и уровни инсулина в сыворотке крови в сытом состоянии (Е) в тех же группах животных, что и в (А, В).
F Чувствительность к инсулину определяли во всех экспериментальных группах после внутрибрюшинной инъекции инсулина (0,75 единицы/кг массы тела). Результаты рассчитывали в виде процента от исходных уровней глюкозы в крови.
Пометки для FGF21 на фигуре относятся к moFGF21 в соответствии с условными обозначениями к фигурам.
Информация о данных: все значения представлены в виде среднего значения±SEM. В (A-F) мыши, которых кормили HFD, n=10 животных/группа; мыши, которых кормили кормом, n=5 животных/группа. ***Р<0,001 в сравнении с группой животных, которых кормили HFD и которым вводили инъекцией нулевой вектор.
Фиг. 33. Повышенные уровни циркулирующего в крови FGF21 in vivo при оптимизации по кодону нуклеотидных последовательностей, кодирующих человеческий FGF21. Уровни циркулирующего в крови hFGF21 у С57В16 мышей, которым вводили с помощью гидродинамических методов плазмиды, кодирующие hFGF21 дикого типа или три различных варианта оптимизированных по кодону последовательностей человеческого FGF21. Результаты представлены в виде среднего значения±SEM. n=9-10 мышей/группа. ND, не выявлялось. Отрицательный контроль, необработанные мыши. * р<0,05 в сравнении с необработанными мышами.
Фиг. 34. Повышенные уровни экспрессии FGF21 in vitro под действием экспрессионных кассет hAAT-moFGF21, С AG-moFGF21 - doublemiRT и CMV-moFGF21. (А) Уровни экспрессии FGF21 в клетках НЕК293, трансфицированных плазмидами, которые кодируют кодирующую последовательность мышиного FGF21 WT под управлением промотора EF1a (EF1a-mFGF21) или оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора CMV (CMV-moFGF21) или промотора СAG в сочетании с четырьмя тандемными повторами последовательности miRT122a и четырьмя тандемными повторами последовательности miRT1 (CAG-moFGF21 -doublemiRT). (В и С) Внутриклеточное содержание белка FGF21 (В) и уровни белка FGF21 в культуральной среде (С) в тех же клетках, что и в (A). (D) Уровни экспрессии FGF21 в клетках С2С12, трансфицированных плазмидами, которые кодируют кодирующую последовательность мышиного FGF21 WT под управлением промотора EF1a (EF1a-mFGF21) или оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора CMV (CMV-moFGF21). (Е) Уровни экспрессии FGF21 в клетках HepG2, трансфицированных плазмидами, которые кодируют кодирующую последовательность мышиного FGF21 WT под управлением промотора EF1a (EF1a-mFGF21) или оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора hAAT (hAAT-moFGF21). qPCR осуществляли с использованием праймеров, которые выявляли как кодирующие последовательности FGF21 wt, так и оптимизированные по кодону кодирующие последовательности FGF21. Результаты представлены в виде среднего значения±SEM. N=3 лунки/группа. ND, не выявлялось. * р<0,05 в сравнении с контролем. ### р<0,001 в сравнении с EF1a-mFGF21.
Фиг. 35. Повышенная экспрессия FGF21 в печени in vivo и уровни циркулирующего в крови FGF21 под действием экспрессионных кассет hAAT-moFGF21 и CMV-moFGF21. (А) Уровни экспрессии FGF21 в печени С57В16 мышей, которым вводили с помощью гидродинамическим методов плазмиды, которые кодируют кодирующую последовательность мышиного FGF21 WT под управлением промотора фактора элонгации 1a (EF1a) (EF1a-mFGF21) или оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора CMV (CMV-moFGF21) или промотора hAAT (hAAT-moFGF21). qPCR осуществляли с использованием праймеров, которые выявляли как кодирующие последовательности FGF21 wt, так и оптимизированные по кодону кодирующие последовательности FGF21. (В) Уровни циркулирующего в крови FGF21 в тех же когортах, что и в (А). Результаты представлены в виде среднего значения±SEM. n=5 мышей/группа. ** р<0,01 в сравнении с EF1a-mFGF21
Фиг. 36. Повышенная экспрессия FGF21 в печени in vivo и уровни циркулирующего в крови FGF21 под действием AAV8-hAAT-moFGF21. (А) Уровни экспрессии FGF21 в печени С57 В16 мышей, которым внутривенно вводили 1×1010 vg, 2×1010 vg или 5×1010 vg векторов на основе AAV8, которые кодируют кодирующую последовательность мышиного FGF21 WT под управлением промотора фактора элонгации 1a (EF1a) (AAV8-EF1a-mFGF21) или оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора hAAT (AAV8-I1AAT-moFGF21). qPCR осуществляли с использованием праймеров, которые выявляли как кодирующие последовательности FGF21 wt, так и оптимизированные по кодону кодирующие последовательности FGF21. (В) Уровни циркулирующего в крови FGF21 в тех же когортах, что и в (А). Анализы осуществляли через две недели после введения AAV. Результаты представлены в виде среднего значения±SEM. n=4-5 мышей/группа. Контроль, необработанные мыши. ** р<0,01 и *** р<0,001 в сравнении с контролем. ## р<0,01 и ### р<0,001 в сравнении с AAV8-EF1a-mFGF21
Фиг. 37. Повышенная экспрессия FGF21 в жировой ткани in vivo под действием AAV8-CAG-moFGF21-dmiRT. (А-В) Уровени экспрессии FGF21 в eWAT (А) или печени (В) С57 В16 мышей, которым вводили в eWAT 2×1010 vg, 5×1010 vg или 1×1011 vg либо векторов на основе AAV8, которые кодируют кодирующую последовательность мышиного FGF21 WT под управлением промотора фактора элонгации 1a (EF1a) (AAV8-EF1a-mFGF21), либо векторов на основе AAV8, которые кодируют оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора С AG в сочетании с четырьмя тандемными повторами последовательности miRT122a и четырьмя тандемными повторами последовательности miRT1 (AAV8-CAG-moFGF21-doublemiRT). qPCR осуществляли с использованием праймеров, которые выявляли как кодирующие последовательности FGF21 wt, так и оптимизированные по кодону кодирующие последовательности FGF21. Анализы осуществляли через две недели после введения AAV. Результаты представлены в виде среднего значения±SEM. n=4-5 мышей/группа. Контроль, необработанные мыши. eWAT, эпидидимальная белая жировая ткань. * р<0,05, ** р<0,01 и
Фиг. 38. Повышенная экспрессия FGF21 в скелетной мышце in vivo под действием AAV1-CMV-moFGF21. (А-В) Уровни экспрессии FGF21 в четырехглавой мышце (А) или печени (В) С57В16 мышей, которым вводили внутримышечно 5×1010 vg, 1×1011 vg или 3×1011 vg либо векторов на основе AAV8, которые кодируют кодирующую последовательность мышиного FGF21 WT под управлением промотора фактора элонгации 1a (EF1a) (AAV8-EF1a-mFGF21), либо векторов на основе AAV1, которые кодируют оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора CMV (AAV1-CMV-FGF21). qPCR осуществляли с использованием праймеров, которые выявляли как кодирующие последовательности FGF21 wt, так и оптимизированные по кодону кодирующие последовательности FGF21. Анализы осуществляли через две недели после введения AAV. Результаты представлены в виде среднего значения±SEM. n=4-5 мышей/группа. Контроль, необработанные мыши. * р<0,05, ** р<0,01 и *** р<0,001 в сравнении с контролем. # р<0,05, ## р<0,01 и ### р<0,001 в сравнении с AAV8-EF1a-mFGF21.
ПРИМЕРЫ
Общие процедуры для примеров
Характеристики субъектов
Использовали самцов C57B1/6J мышей и B6.V-Lepob/OlaHsd (ob/ob) мышей. Мышей кормили ad libitum стандартным рационом (2018S Tek1ad Global Diets®, Harlan Labs., Inc., Мадисон, Висконсин, США) или рационом с высоким содержанием жиров (TD.88137 Har1an Tek1ad, Мадисон, Висконсин, США) и содержали в условиях цикла чередования 12 часов света и темноты (освещение включали в 8:00 до полудня) и при стабильной температуре (22°С±2). Для получения образцов ткани мышей подвергали анестезии посредством ингаляционной анестезии изофлураном (IsoFlo®, Abbott Laboratories, Эббот-Парк, Иллинойс, США) и декапитировали. Представляющие интерес ткани вырезали и хранили при -80°С или с формалином до анализа. Все экспериментальные процедуры были одобрены Комитетом по этике для экспериментов на животных и человеке (Ethics Committee for Animal and Human Experimentation) Автономного университета Барселоны (Universitat Autonoma de Barcelona).
Векторы па основерекомбинантного AAV
Векторы на основе одноцепочечного AAV серотипа 1, 8 или 9 получали посредством тройной трансфекции клеток НЕК293 в соответствии со стандартными способами (Ayuso, Е. et al., 2010. Curr Gene Ther. 10(6):423-36). Клетки культивировали в 10 роллерных бутылях (850 см2, плоские; Corning™, Sigma-Aldrich Co., Сент-Луис, Миссури, США) в DMEM с 10% FBS до 80% конфлюентности и совместно трансфицировали с помощью способа с использованием фосфата кальция плазмидой, несущей экспрессионную кассету, фланкированную ITR AAV2, плазмидой-помощником, несущей ген rep AAV2 и ген cap AAV серотипов 1, 8 или 9, и плазмидой, несущей функции помощника аденовируса. Используемые трансгены представляли собой: оптимизированные по кодону или wt кодирующие последовательности мышиного, собачьего или человеческого FGF21, управляемые 1) энхансером ранних генов цитомегаловируса (СМУ)/промотором куриного бета-актина (CAG) с добавлением четырех тандемных повторов последовательности miRT122a (5'CAAACACCATTGTCACACTCCA3') (SEQ ID NO:12) и четырех тандемных повторов последовательности miRT1 (5'ТТАСАТАСТТСТТТАСАТТССА3') (SEQ ID NO: 13), клонированных в 3' нетранслируемый участок экспрессионной кассеты; 2) промотором CMV или 3) промотором человеческого α1-атитрипсина (hAAT). Некодирующие плазмиды, несущие проморы CAG, hAAT или CMV использовали для получения нулевых векторов. AAV очищали с использованием оптимизированного способа, основанного на стадии осаждения полиэтиленгликолем и двумя последовательными процедурами выделения в градиентах хлорида цезия (CsCl). Этот основанный на использовании CsCl протокол второго поколения резко снижал содержание пустых капсидов AAV и примесей ДНК и белков (Ayuso, Е. et al., 2010. Curr Gene Ther. 10(6):423-36). Очищенные векторы на основе AAV подвергали диализу против PBS, фильтровали и хранили при -80°С. Титры вирусных геномов определяли с помощью количественной ПЦР, следуя протоколу, описанному для стандартного образца материала AAV2 с использованием линеаризованной плазмидной ДНК в качестве стандарта для построения кривой (Lock М, et al., Hum. Gene Ther. 2010; 21:1273-1285). Векторы конструировали в соответствии с методиками молекулярной биологии, хорошо известными в уровне техники.
Введение в eWAT in vivo векторов на основе AAV
Мышей подвергали анестезии с помощью внутрибрюшинной инъекции кетамина (100 мг/кг) и ксилазина (10 мг/кг). Лапаротомию осуществляли для того, чтобы получить доступ к эпидидимальной белой жировой ткани. Векторы на основе AAV ресуспендировали в PBS с 0,001% Pluronic® F68 (Gibco) и вводили инъекцией непосредственно в эпидидимальную жировую подушку. Каждая эпидидимальная жировая подушка получала дважды инъекцию 50 мкл раствора AAV (одна инъекция вблизи яичка, а другая в середину жировой подушки). Брюшную полость промывали стерильным солевым раствором и закрывали с доступом через два слоя.
Системное введение векторов на основе AAV
Подходящее количество раствора AAV разводили в 200 мкл PBS с 0,001% Pluronic® и вводили инъекцией вручную в латеральную хвостовую вену без приложения давления в момент доставки. Перед инъекцией животных помещали под 250 Вт лампу с излучением тепла в инфракрасном диапазоне (Philips NV, Амстердам, Нидерланды) на несколько минут для того, чтобы расслабить кровеносные сосуды, облегчить визуальное наблюдение и упростить доступ к хвостовой вене. Пластиковое приспособление для фиксирования лап (Harvard Apparatus, Холлистон, Массачусетс, США) использовали для фиксирования животного для инъекции. Анестезию не использовали, поскольку использовали соответствующее приспособление для фиксирования лап. Для инъекции животным использовали иглу 30 калибра.
Внутримышечное введение векторов на основе AAV
Мышей подвергали анестезии с помощью внутрибрюшинной инъекции кетамина (100 мг/кг) и ксилазина (10 мг/кг). Задние конечности брили и векторы вводили посредством внутримышечной инъекции в общем объеме 180 мкл, разделенном на шесть мест инъекции, распределенных в четырехглавой мышце, икроножной мышце и передней краниальной болынеберцовой мышце каждой задней конечности.
Иммуногистохимический и морфометрический анализ
Ткани фиксировали в течение 24 часов в формалине (Panreac Quimica), заливали в парафин и делали срезы. Образцы тканей окрашивали гематоксилином-эозином. Площадь адипоцитов определяли на 12 изображениях окрашенной гематоксилином/эозином WAT на животное, полученных при 10-кратном увеличении с использованием микроскопа Nikon Eclipse Е800 (Nikon, Токио, Япония), соединенного с видеокамерой с монитором при использовании программного обеспечения для анализа изображений (analySIS 3.0; Soft Imaging System, Сентер Вэлли, Пенсильвания. США, EEUU), и площадь каждого адипоцита количественно определяли в мкм2. Рассчитывали среднюю площадь адипоцитов для каждой экспериментальной группы, и распределение адипоцитов по категориям согласно размеру представлено в виде гистограммы. Использовали четыре животных на группу и анализировали по меньшей мере 250 адипоцитов на животное.
Иммуногистохимический анализ
Ткани фиксировали в течение 12-24 часов в 10% формалине, заливали в парафин и делали срезы. Срезы инкубировали в течение ночи при 4°С с крысиным антителом к Мас2 (1:50; CL8942AP; Cedarlane), антителом морской свинки к инсулину (1:100; 1-8510; Sigma-Aldrich) или кроличьим антителом к глюкагону (1:100; 219-01; Signet Labs). В качестве вторичных антител использовали биотинилированное кроличье антитело к крысиному антителу (1:300; Е0467; Dako), козье антитело к кроличьему IgG (конъюгированное с флуорофором Alexa Fluor 568) (1:200; A11011; ThermoFisher), козье антитело к IgG морской свинки (конъюгированное с флуорофором Alexa Fluor 488) (1:300; А11073; ThermoFisher) или кроличье антитело к антителу морской свинки, конъюгированное с пероксидазой (1:300; Р0141; Dako). Набор для выявления пероксидазы ABC peroxidase kit (Pierce) использовали для выявления при иммунологическом анализе и срезы подвергали контрастному окрашиванию гематоксилином по Майеру. Краситель Хехста (В2261; Sigma-Aldrich) использовали для контрастного окрашивания ядер в образцах при флуоресцентном окрашивании. Окрашивание пикросириусом красным и трихром-окрашивание по Массону использовали для оценки фиброза. Выраженную в процентах площадь β-клеток в поджелудочной железе анализировали на двух окрашенных в отношении инсулина срезах, расположенных на расстоянии 200 мкм, посредством деления площади всех инсулин + клеток на одном срезе на общую площадь ткани поджелудочной железы на этом срезе. Массу β-клеток рассчитывали посредством умножения массы поджелудочной железы на выраженную в процентах площадь β-клеток, как описано ранее (Jimenez et al, 2011).
Анализ РНК
Общую РНК получали из жировых депо или печени с применением реактива для лизиса QIAzol Lysis Reagent (Qiagen NV, Венло, Нидерланды) или реактива для выделения Tripure iso1ation reagent (Roche Diagnostics Corp., Индианаполис, Индиана, США), соответственно, и набора для выделения РНК RNeasy Lipid Tissue Minikit (Qiagen NV, Венло, Нидерланды). Для того чтобы удалить оставшиеся вирусные геномы, общую РНК обрабатывали ДНКазой I (Qiagen NV, Венло, Нидерланды). Для RT-PCR (ПНР с обратной транскрипцией) 1 мкг образцов РНК подвергали обратной транскрипции с использованием набора для синтеза кДНК Transcriptor First Strand cDNA Synthesis Kit (04379012001, Roche, Калифорния, США). Количественную ПЦР в реальном времени осуществляли в SmartCyclerll® (Cepheid, Саннивейл, США) с использованием супермикса EXPRESS SYBRGreen qPCR supermix (Invitrogen™, Life Technologies Corp., Карлсбад, Калифорния, США). Данные нормализовали к значениям RplpO и анализировали, как описано ранее (Pfaffl, М., Nucleic Acids Res. 2001; 29(9):е45).
Анализы гормонов и метаболитов
Уровни глюкозы в крови измеряли с использованием анализатора Glucometer Elite™ (Bayer, Леверкузен, Германия). Уровни циркулирующего в крови FGF21 определяли с помощью количественного иммуноферментного сендвич-анализа с набором Mouse/Rat FGF-21 ELISA kit (MF2100, R&Dsystems, Абингдон, Великобритания). Концентрации инсулина в сыворотке крови определяли с помощью сендвич-анализа крысиного инсулина методом ELISA (90010, Crystal Chem INC. Даунерс-Гроув, Иллинойс, 60515, США). Для экстракции липидов из ткани замороженные образцы массой примерно 100 мг взвешивали и гомогенизировали в 15 мл смеси хлороформ:метанол (2:1). Липидную и водную фазы затем разделяли посредством добавления 3 мл 0,05% H2SO4 и сохранения их в течение ночи при 4°С. После того как фазы были разделены, верхнюю водную фазу удаляли с использованием пипетки Пастера и 1 мл нижней липидной фазы улавливали в стеклянной пробирке. 1 мл хлороформа и Triton Х-100 в виде 1% раствора добавляли в стеклянную пробирку и ее инкубировали при 90°С на бане для выпаривания хлороформа. Все оставшиеся водные частицы удаляли из липидной фазы при использовании смеси хлороформа и Triton Х-100. После выпаривания хлороформом промывали стенки пробирки для концентрирования образца и его опять подогревали до 90°С для выпаривания хлороформа. После того как осадок становился полностью сухим и концентрированным, его ресуспендировали посредством добавления 500 мкл H20 miliQ при 37°С. Наконец, определяли количество триглицеридов с использованием коммерческого продукта GPO-РАР (Roche Diagnostics, Базель, Швейцария). Сывороточные триглицериды и холестрин количественно определяли спектрофотометрически с использованием набора для ферментного анализа (Horiba-ABX, Монпелье, Франция). Все биохимические параметры определяли с использованием анализатора Pentra 400 Analyzer (Horiba-АВХ).
Гликемию определяли с использованием Glucometer EliteTM (Bayer). Уровни глюкагона измеряли с использованием радиоиммунологического анализа глюкагона (#GL-32К, EMD Millipore). Адипонектин, лептин, IGFBP1 и IGF1 определяли с использованием набора для определения мышиного адипонектина методом ELISA (80569, Crystal Chem), набора для определения мышиного лептина методом ELISA (90030, Crystal Chem), набора для определения IGFBP1 (мышиного) методом ELISA (КА3054, Abnova) и набора m/r IGF-I-ELISA kit (Е25, Mediagnost), соответственно.
Тесты толерантности к инсулину
Для тестов толерантности к инсулину инсулин (0,75 МЕ/кг массы тела; Humulin Regu1ar; Eli Lilly, Индианаполис, Индиана, США) вводили инъекцией внутрибрюшинно бодрствующим сытым мышам. Концентрацию глюкозы определяли в образцах крови, полученных из хвостовой вены в указанные моменты времени после инъекции инсулина.
Тест толерантности к глюкозе
Бодрствующих мышей ограничивали в кормлении в течение ночи (16 часов) и вводили им в виде внутрибрюшинной инъекции глюкозу (2 г/кг массы тела). Гликемию измеряли в образцах крови из хвостовой вены в указанные моменты времени. Венозную кровь собирали из хвостовой вены в пробирки (Microvette® СВ 300, SARSTEDT) в те же моменты времени и немедленно центрифугировали для отделения сыворотки, которую использовали для измерения уровней инсулина.
Оксиметрия
Калориметр непрямого действия с открытым контуром (Oxylet, Pan1ab, Корнелья, Испания) использовали для мониторинга потребления кислорода, выработки диоксида углерода в восьми камерах для измерения метаболизма одновременно. Мышей размещали по одной и позволяли акклиматизироваться к камерам для измерения метаболизма в течение 24 часов и данные собирали каждые 15 мин. в течение 3 мин. в каждой клетке в течение еще 24 часов. Данные получали во время цикла чередования света и темноты и корректировали на массу тела. Для расчета потребления энергии использовали программное обеспечение для определения показателей метаболизма, которое обеспечено производителем.
Трансфекция НЕК293, С2С12 и HepG2 клеток
Клетки культивировали в 24-луночном планшете и трансфицировали 0,8 мкг ДНК на лунку с использованием липофектамина 2000, следуя инструкциям производителя (Thermo Fisher Scientific).
Анализ костей
Объем и архитектуру костей оценивали с помощью μСТ (микроКТ). Большеберцовые кости мышей фиксировали в стабилизированном буфером при нейтральном рН формалине (10%) и сканировали с использованием компьютерного томографа eXplore Locus СТ scanner (General Electric) при разрешении 27 микрон. Трабекулы анализировали в 1 мм3 проксимального эпифиза большеберцовой кости и в 1,8 мм3 кортикального слоя диафиза большеберцовой кости у 4 мышей/группа. Параметры костей рассчитывали с использованием инструмента для анализа Micro View 3D Image Viewer & Analysis Tool. Длину большеберцовой кости измеряли от межмыщелкового возвышения до медиальной лодыжки.
Анализ методом вестерн-блоттинга
iWAT и iBAT гомогенизировали в реактиве для лизиса QIAzol Lysis Reagent (Qiagen) и белковую фракцию выделяли из органической фазы, следуя инструкциям производителя. Белки разделяли с помощью электрофореза в полиакриламидном геле с 12% додецилсульфатом натрия и анализировали посредством иммуноблоттинга с использованием поликлонального кроличьего антитела к UCP1 (ab10983; Abcam) и поликлонального кроличьего антитела к α-тубулину (ab4074; Abcam). Выявление осуществляли с использованием реактива для выявления ECL Plus detection reagent (Amersham Biosciences).
Тест открытого поля
Тест открытого поля осуществляли между 9:00 до полудня и 1:00 после полудня, как сообщалось ранее (Haurigot et al, 2013). Вкратце, животных помещали в центр ярко освещенной камеры (41×41×30 см), которую пересекали 2 группы фотодетекторов (LE 8811; Panlab), которые выявляют горизонтальные и вертикальные движения. Двигательные и исследовательские активности оценивали в течение первых 6 минут. Общее покрытое расстояние оценивали с использованием видеосистемы для отслеживания движений (SMART Junior; Pan1ab).
Статистический анализ
Все значения представлены в виде среднего значения±SEM. Различия между группами сравнивали с помощью t-критерия Стьюдента. Различия считали значимыми при р<0,05.
Примеры
Пример 1. Предупреждение ожирения и сахарного диабета под действием введения в eWAT векторов AAV-CAG-moFGF21-dmiRT С57В16 мышам
Авторы настоящего изобретения оценивали терапевтический потенциал опосредованного AAV генно-инженерного воздействия на жировую ткань с использованием FGF21 для предупреждения ожирения и сахарного диабета у самцов С57В16 мышей возрастом 8 недель. Введение в eWAT (eWAT: эпидидимальная белая жировая ткань) 1012 вирусных геномов (vg) векторов на основе AAV9, кодирующих оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением убиквитарного промотора CAG, который включал в себя сайты-мишени miR122 и miR1 (AAV9-CAG-moFGF21 -doublemiRT) (фиг. 1А), опосредовало специфическую для жировой ткани сверхэкспрессию FGF21 (фиг. 1В), а также высокую секрецию белка в кровоток (фиг. 1С). Обработанные AAV9-CAG-moFGF21 -doublemiRT мыши также демонстрировали сверхэкспрессию рецептора 1 FGF21 (FGF21R1) в eWAT (FIG ID) и β-клото (корецептора FGF21) в жировой ткани и печени (фиг. 1Е) по сравнению с векторами AAV9-CAG-null (векторы, которые сохраняют равную инфекционную способность, но не кодируют какой-либо трансген). Конструкция CAG-moFGF21-doublemiRT содержится в SEQ ID NO: 32, и конструкция CAG-null содержится в SEQ ID NO:31.
После опосредованного AAV переноса гена FGF21 в eWAT мыши, которых кормили рационом в виде корма, демонстрировали потерю массы тела (фиг. 1F и 1G). При воздействии рационом с высоким содержанием жиров (HFD) животные, сверхэкспрессирующие FGF21 в жировой ткани, оставались худыми в течение периода проведения эксперимента, в то время как у обработанных AAV9-CAG-null мышей прогрессивно развивалось ожирение (фиг. 1F и 1G). С учетом их более низкой массы тела обработанные AAV9-CAG-moFGF21-doublemiRT мыши, которых кормили как кормом, так и HFD, демонстрировали уменьшенную массу жировых депо и печени (фиг. 1Н).
Гистологический анализ белой жировой ткани посредством окрашивания гематоксилином-эозином выявил уменьшенный размер белых апоцитов в eWAT и iWAT (iWAT: паховая белая жировая ткань) и многочисленные мультилокулярные адипоциты в iWAT, что говорит о том, что произошло «побурение» этого депо (фиг. 2А). Морфометрический анализ дополнительно подтвердил уменьшенную среднюю площадь белых адипоцитов у обработанных AAV9-CAG-moFGF21-doublemiRT мышей (фиг. 2 В). Частотное распределение площади белых адипоцитов также отличалось у групп. У обработанных AAV9-CAG-moFGF21-doublemiRT мышей, которых кормили как кормом, так и HFD, присутствовало повышенное количество мелких адипоцитов и меньше крупных адипоцитов (фиг. 2С). Примечательно, что частотное распределение площади белых адипоцитов обработанных AAV9-CAG-moFGF21-doublemiRT мышей, которых кормили HFD, было практически идентичным таковому у обработанных AAV9-CAG-null животных, которых кормили кормом (фиг. 2С). Таким образом, индуцируемая HFD гипертрофия адипоцитов, наблюдаемая у обработанных AAV9-CAG-null мышей, блокировалась у мышей, сверхэкспрессирующих FGF21. Сверхэкспрессия UCP1 и Dio2 в iWAT (фиг. 3А и 3В) дополнительно подтверждала «побурение» iWAT у обработанных AAV9-CAG-moFGF21-doublemiRT мышей, которых кормили кормом и HFD.
Гистологический анализ iBAT (iBAT: межлопаточная бурая жировая ткань) показал низкое накопление липидов в этом депо у обработанных AAV9-CAG-moFGF21-doublemiRT мышей, которых кормили кормом и HFD, по сравнению с AAV9-CAG-null мышами (фиг. 2А). В соответствии с этим результатом и «побурением» iWAT потребление энергии (фиг. 3С) у обработанных AAV9-CAG-moFGF21-doublemiRT мышей, которых кормили HFD, во время циклов чередования света и темноты было выше, чем потребление энергии у AAV9-CAG-null мышей, которых кормили HFD. В совокупности эти данные говорят о том, что обработанные AAV9-CAG-moFGF21-doublemiRT мыши характеризуются повышенной термогенной активностью.
Гистологические срезы печени показали пониженное накопление липидов в гепатоцитах у мышей, сверхэкспрессирующих FGF21, по сравнению с обработанными AAV9-CAG-null мышами под действием как корма, так и HFD (фиг. 2А). Соответственно, у обработанных AAV9-CAG-moFGF21-doublemiRT мышей, которых кормили HFD, нормализовалось содержание триглицеридов (TG) в их печени (фиг. 3D). Параллельно уровни циркулирующих в крови TG, общего холестерина, HDL-холестерина и LDL-холестерина нормализовались у мышей, которых кормили HFD, и у которых сверхэкспрессировался FGF21 (фиг. 3Е и 3F).
Мыши, которых кормили HFD, и у которых сверхэкспрессировался FGF21, были более чувствительны к инсулину, чем обработанные AAV9-null мыши, которых кормили HFD (фиг. 3G), и обработанные AAV9-CAG-moFGF21-doublemiRT мыши, которых кормили как кормом, так и HFD, демонстрировали пониженные уровни циркулирующего в крови инсулина по сравнению с их обработанными AAV9-CAG-null аналогами (фиг. 3Н).
Пример 2. Обращение развития ожирения и улучшение метаболизма глюкозы при введении в eWAT векторов AAV-CAG-moFGF21-dmiRT ob/ob мышам
Авторы настоящего изобретения оценивали противодиабетический и направленный против развития ожирения терапевтический потенциал опосредованного AAV генно-инженерного воздействия на жировую ткань с использованием FGF21 у самцов ob/ob мышей возрастом 11 недель, которые имеют дефектную передачу сигнала с участием лептина и представляют собой широко применяемую генетическую модель ожирения и сахарного диабета. С этой целью осуществляли исследование зависимости доза-эффект. Ob/ob мышам вводили локально в eWAT четыре разные дозы (1010 vg, 5×1010 vg, 2×1011 vg или 1012 vg) векторов AAV8-CAG-moFGF21-doublemiRT (фиг. 1А). В качестве контроля ob/ob животным вводили в eWAT 1012 vg векторов AAV8-CAG-null.
Введение в eWAT векторов AAV8-CAG-moFGF21-doublemiRT опосредовало специфическую сверхэкспрессию FGF21 в белой жировой ткани, а также высокую секрецию белка в кровоток дозозависимым образом (фиг. 4А и 4В). В частности, доза 1012vg векторов AAV8-CAG-moFGF21-doublemiRT опосредовала очень сильную сверхэкспрессию FGF21 в eWAT и iWAT (фиг. 4А) и достигала наиболее высоких уровней циркулирующего в крови FGF21 (фиг. 4В). Напротив, самая низкая вводимая доза, 1010vg векторов AAV8-CAG-moFGF21-doublemiRT, давала только очень умеренную сверхэкспрессию FGF21 в eWAT (фиг. 4А), и животные, обработанные этой дозой, не демонстрировали различий в уровнях FGF21 в сыворотке крови по сравнению с обработанными AAV8-CAG-null животными (фиг. 4 В), возможно, вследствие того, что FGF21 действовал паракринно-аутокринным образом. Соответственно, в то время как у обработанных AAV8-CAG-null животных прогрессивно увеличивалась их масса тела, животные, обработанные векторами AAV8-CAG-moFGF21-doublemiRT, демонстрировали уменьшенное увеличение массы тела, пропорциональное вводимой дозе векторов (фиг. 4С и 4D). Примечательно, что животные, обработанные 1012vg векторов AAV8-CAG-moFGF21-doublemiRT теряли примерно 15% массы в течение первых двух недель после введения AAV, а после этого увеличивали свою массу тела до тех пор, пока они не достигали исходной массы тела (фиг. 4С и 4D). Таким образом, животные, которым вводили в eWAT 1012 vg векторов AAV8-CAG-moFGF21-doublemiRT, демонстрировали 40% различие в общей массе тела по сравнению с обработанными AAV8-CAG-null животными в конце эксперимента (фиг. 4D). Соответственно, животные, обработанные 1012 vg векторов AAV8-CAG-moFGF21-doublemiRT, демонстрировали заметно уменьшенное ожирение и 60% уменьшение массы печени (фиг. 4Е). Масса iBAT увеличивалась в этой когорте мышей (фиг. 4Е), возможно, вследствие повышенной термогенной активности.
У животных, обработанных 5×1010 vg, 2×1011 vg или 1012 vg векторы AAV8-CAG-moFGF21-doublemiRT, присутствовала улучшенная чувствительность к инсулину по сравнению с обработанными AAV8-CAG-null мышами (фиг. 5А). Животные, обработанные 2×1011 vg или 1012 vg векторов AAV8-CAG-moFGF21-doublemiRT, также демонстрировали более низкие уровни циркулирующего в крови инсулина, чем ob/ob мыши, обработанные векторами AAV8-CAG-null (фиг. 5В).
Пример 3. Обращение развития ожирения и улучшение метаболизма глюкозы при внутривенном введении векторов AAV-hAAT-moFGF21 ob/ob мышам
Авторы настоящего изобретения также оценивали противодиабетические и направленные против развития ожирения эффекты, опосредованные повышенными уровнями циркулирующего в крови FGF21, в результате опосредованного AAV генно-инженерного воздействия на печень у самцов ob/ob мышей возрастом 8 недель. Ob/ob мышам вводили внутривенно (IV) 1011 vg или 5×1011 vg векторов на основе AAV8, кодирующих оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением специфического для печени промотора человеческого α1-антитрипсина (hAAT) (AAV8-hAAT-moFGF21) (фиг. 6А). В качестве контроля ob/ob животным вводили IV 5×1011 vg векторов AAV8-hAAT-null. Конструкция hAAT-moFGF21 содержится в SEQ ID NO: 34, и конструкция hAAT-null содержится в SEQ ID NO:33.
Внутривенное введение векторов AAV8-hAAT-moFGF21 опосредовало специфическую сверхэкспрессию FGF21 в печени, а также высокую секрецию белка в кровоток дозозависимым образом (фиг. 6 В и 6С). В частности, доза 5×1011 vg векторов AAV8-hAAT-moFGF21 опосредовала очень сильную сверхэкспрессию FGF21 в печени (фиг. 6В) и достигала наиболее высоких уровней циркулирующего в крови FGF21 (фиг. 6С). Масса тела животных, обработанных этой дозой векторов AAV8-hAAT-moFGF21, снижалась примерно на 7% в течение первых двух недель после введения AAV и после этого немного возрастала, в то время как обработанные AAV8-hAAT-null мыши прогрессивно прибавляли в массе (фиг. 6D, 6Е и 6F). Мыши, которым вводили 1011 vg векторов AAV8-hAAT-moFGF21, набирали заметно меньшую массу, чем обработанные AAV8-hAAT-null животные (фиг. 6D, 6Е и 6F). В частности, животные AAV8-hAAT-null демонстрировали 50% увеличение их массы тела в конце эксперимента по сравнению с 10% увеличением массы животных, обработанных 10nvg векторов AAV8-hAAT-moFGF21 (фиг. 6Е). В соответствии с их более низкой массой тела животные, сверхэкспрессирующие FGF21 в печени, демонстрировали значительно уменьшенное ожирение, в особенности, у тех животных, которые были обработаны наиболее высокой дозой векторов, и примерно 60% уменьшение массы печени (фиг. 6G). Подобным образом, масса iBAT увеличивалась в обеих группах обработанных AAV8-hAAT-moFGF21 мышей (фиг. 6G), возможно, вследствие более высокой термогенной активности у этих животных по сравнению с мышами, которым вводили векторы AAV8-hAAT-null.
Животные, обработанные векторами AAV8-hAAT-moFGF21, демонстрировали улучшенную чувствительность к инсулину и пониженные уровни циркулирующего в крови инсулина по сравнению с обработанными AAV8-hAAT-null мышами (фиг. 6Н и 61)
Пример 4. Долговременное обращение развития ожирения и сахарного диабета при внутривенном введении векторов AAV-hAAT-moFGF21 мышам, которых кормили HFD
Авторы настоящего изобретения также оценивали противодиабетические и направленные против развития ожирения эффекты, опосредованные повышенными уровнями циркулирующего в крови FGF21 в результате опосредованного AAV генно-инженерного воздействия на печень у С57В16 мышей с ожирением. Самцов С57В16 мышей возрастом девять недель (молодой возраст) кормили HFD в течение 9 недель, а затем вводили IV 1010vg или 5×1010vg векторов AAV8-hAAT-moFGF21 (фиг. 6А). После введения AAV обработанных AAV8-hAAT-moFGF21 мышей поддерживали на HFD в течение 52 недель. В качестве контролей 5×1010vg AAV8-hAAT-null вводили IV С57В16 мышам, которых кормили кормом и HFD. Эти две последние когорты мышей поддерживали после этого либо на рационе в виде корма, либо на HFD.
Внутривенное введение векторов AAV8-hAAT-moFGF21 у мышей, которых кормили HFD, опосредовало высокую секрецию FGF21 в кровоток дозозависимым образом (фиг. 7А).
Отсутствие различий в массе тела наблюдали между обработанными AAV8-null мышами, которых кормили HFD, и животными, которых кормили HFD и которым вводили 1010 vg векторов AAV8-hAAT-moFGF21 (фиг. 7 В и 1С). Тем не менее, животные, которых кормили HFD, обработанные 5×1010vg векторов AAV8-hAAT-moFGF21, вначале теряли 20% массы тела после введения AAV, а затем прогрессивно набирали массу аналогично обработанным AAV8-hAAT-null мышам, которых кормили кормом (фиг. 7В и 7С). Примечательно, что с 9 недели после введения AAV и позже не наблюдали статистически значимых различий в общей массе тела и увеличении массы тела между животными, которых кормили HFD и которым вводили 5×1010vg векторов AAV8-hAAT-moFGF21, и обработанными AAV8-hAAT-null мышами, которых кормили кормом (фиг. 7 В и 7С).
Потребление энергии у мышей, которых кормили HFD, обработанных 5×1010vg векторов AAV8-hAAT-moFGF21, во время циклов чередования света и темноты было выше, чем у получавших AAV8-hAAT-null мышей, которых кормили кормом и HFD (фиг. 8А). Отсутствие различий в потреблении энергии наблюдали между обработанными AAV8-hAAT-null животными, которых кормили кормом и HFD, и мышами, которым вводили 1010vg векторов AAV8-hAAT-moFGF21 (фиг. 8А). В совокупности эти данные говорят о том, что мыши, обработанные 5×1010vg AAV8-hAAT-moFGF21, характеризуются повышенной термогенной активностью.
У животных, обработанных 1010vg векторов AAV8-hAAT-moFGF21, присутствовала улучшенная чувствительность к инсулину по сравнению с мышами, которых кормили HFD и которым вводили векторы AAV8-hAAT-null, и их чувствительность к инсулину была подобной таковой у мышей, обработанных векторами AAV8-hAAT-null, которых кормили кормом (фиг. 8 В). Примечательно, что у животных, которым вводили 5×1010 vg векторов AAV8-hAAT-moFGF21, присутствовала улучшенная чувствительность к инсулину по сравнению мышами, с которых кормили кормом и которым вводили векторы AAV8-hAAT-null, и они демонстрировали нормализованные уровни циркулирующего в крови инсулина (фиг. 8 В и 8С).
Пример 5. Обращение развития ожирения и сахарного диабета при внутривенном введении векторов AAV-hAAT-moFGF21 старым мышам, которых кормили HFD
Авторы настоящего изобретения также оценивали противодиабетические и направленные против развития ожирения эффекты FGF21 у старых (взрослых) С57В16 мышей с ожирением. Самцов С57В16 мышей возрастом семь с половиной месяцев кормили HFD в течение 8 недель, а затем вводили IV 1010 vg, 2×1010 vg или 5×1010 vg векторов AAV8-hAAT-moFGF21 (фиг. 6А). После введения AAV обработанных AAV8-hAAT-moFGF21 мышей поддерживали на HFD в течение 22 недель. В качестве контролей 5×1010 vg AAV8-hAAT-null вводили IV старым С57В16 мышам, которых кормили кормом и HFD. Эти две последние когорты мышей поддерживали после этого либо на рационе в виде корма, либо на HFD.
Внутривенное введение векторов AAV8-hAAT-moFGF21 старым мышам, которых кормили HFD, опосредовало высокую секрецию FGF21 в кровоток дозозависимым образом (фиг. 9А).
Отсутствие различий в массе тела наблюдали между обработанными AAV8-null мышами, которых кормили HFD, и животными, которых кормили HFD и которым вводили 1010 vg векторов AAV8-hAAT-moFGF21 (фиг. 9 В и 9С). Тем не менее, животные, которых кормили HFD, обработанные либо 2×1010vg, либо 5×1010vg векторов AAV8-hAAT-moFGF21, вначале теряли соответственно 15 и 20% массы тела после введения AAV (фиг. 9 В и 9С). После этого у животных, обработанных 2×1010vg векторов AAV8-hAAT-moFGF21, прогрессивно увеличивалась масса аналогично обработанным AAV8-hAAT-null мышам, которых кормили кормом, в то время как наблюдали отсутствие значимых изменений в массе тела у животных, обработанных 5×1010vg векторов AAV8-hAAT-moFGF21 (фиг. 9 В и 9С). Примечательно, что с 3 недели после введения AAV и позже не наблюдали статистически значимых различий в общей массе тела и увеличении массы тела между животными, которых кормили HFD и которым вводили 5×1010 vg векторов AAV8-hAAT-moFGF21, и обработанными AAV8-hAAT-null мышами, которых кормили кормом (фиг. 9 В и 9С).
Потребление энергии у мышей, которых кормили HFD, обработанных 5×1010vg векторов AAV8-hAAT-moFGF21, во время циклов чередования света и темноты было выше, чем у получавших AAV8-hAAT-null мышей, которых кормили кормом и HFD (фиг. 10А). Животные, обработанные 2×1010vg векторов AAV8-hAAT-moFGF21, демонстрировали повышенное потребление энергии во время дневной части цикла и тенденцию к повышению потребления энергии во время ночной части цикла чередования света и темноты (фиг. 10А). Животные, обработанные 1010vg векторов AAVS-hAAT-moFGF21, демонстрировали повышенное потребление энергии во время ночной части цикла чередования света и темноты (фиг. 10А). Наблюдали отсутствие различий между обработанными AAV8-hAAT-null животными, которых кормили кормом и HFD, и мышами, которым вводили 1010vg векторов AAV8-hAAT-moFGF21 (фиг. 10А). В совокупности эти данные говорят о том, что старые мыши, обработанные AAV8-hAAT-moFGF21, характеризуются повышенной термогенной активностью.
У животных, обработанных 1010vg или 2×1010 vg векторов AAV8-hAAT-moFGF21, присутствовала улучшенная чувствительность к инсулину по сравнению с мышами, которых кормили HFD и которым вводили векторы AAV8-hAAT-null, и их чувствительность к инсулину была подобной таковой у мышей, обработанных векторами AAV8-hAAT-null, которых кормили кормом (фиг. 10 В). Примечательно, что у животных, которым вводили 5×1010 vg векторов AAV8-hAAT-moFGF21, присутствовала улучшенная чувствительность к инсулину по сравнению с мышами, которых кормили кормом и которым вводили векторы AAV8-hAAT-null (фиг. 10А). Животные, обработанные 1010vg, 2×1010vg или 5×1010vg векторов AAV8-hAAT-moFGF21, демонстрировали более низкие уровни циркулирующего в крови инсулина натощак и в сытом состоянии, чем обработанные AAV8-hAAT-null мыши, которых кормили HFD (фиг. 10С). Примечательно, что наблюдали отсутствие различий в уровнях циркулирующего в крови инсулина в сытом состоянии между старыми животными, которым вводили IV 2×1010vg или 5×1010vg векторов AAV8-hAAT-moFGF21, и обработанными AAV8-hAAT-null мышами, которых кормили кормом (фиг. 10С).
Пример 6. Оценка потери массы при внутримышечном введении векторов AAV-CMV-moFGF21 С57В16 мышам
Авторы настоящего изобретения также оценивали терапевтический потенциал повышения уровней циркулирующего в крови FGF21 под действием опосредованного AAV генно-инженерного воздействия на скелетные мышцв у С57В16 мышей. Для того чтобы целенаправленно воздействовать на скелетную мышцу выбрали промотор CMV и серотип AAV1. Несмотря на то что промотор CMV представляет собой убиквитарный промотор, его одновременное применение вместе с капсидами AAV1 обеспечивает возможность очень эффективного целенаправленного воздействия на скелетную мышцу без трансдукции печени, как опубликовано ранее (Mas et al., Diabetes 2006; Callejas et al., Diabetes 2013).
Дозу 3×1011 vg векторов на основе AAV1, кодирующих оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением убиквитарного промотора CMV (AAV1-CMV-moFGF21) (фиг. 11А), вводили посредством внутримышечной инъекции в четырехглавую мышцу, икроножную мышцу и переднюю краниальную болыпеберцовую мышцу каждой задней конечности (5×1010 vg/мышца) самцов C57BL6 мышей возрастом 6-12 недель. В качестве контроля животным С57В16 соответствующей возрастной группы вводили внутримышечно в те же мышцы 3×1011 vg векторов AAV1-CMV-null (5×1010 vg/мышца). Конструкция CMV-moFGF21 содержится в SEQ ID NO: 36, и конструкция CMV-null содержится в SEQ ID NO:35.
Внутримышечное введение векторов AAV1-CMV-moFGF21 опосредовало высокую секрецию FGF21 в кровоток (фиг. 11В). Животные, обработанные векторами AAV1-CMV-moFGF21, демонстрировали уменьшенную массу тела и увеличение массы тела по сравнению с обработанными AAV1-CMV-null мышами (фиг. 11С и 11D).
Пример 7. Повышенная продукция белка под действием оптимизированных по кодону нуклеотидных последовательностей человеческого FGF21
Чтобы оценить, была ли способна оптимизация по кодону опосредовать повышенную продукцию белка FGF21, клетки НЕК293 трансфицировали плазмидами, кодирующими три разные оптимизированные по кодону нуклеотидные последовательности человеческого FGF21 (SEQ ID NO: 40-42). В качестве контроля использовали нетрансфицированные клетки и клетки, трансдуцированные кодирующей последовательностью hFGF21 дикого типа. Экспрессия трех оптимизированных по кодону последовательностей человеческого FGF21 и последовательности человеческого FGF21 WT находилась под управлением промотора hAAT (SEQ ID NO:47). Клетки, трансдуцированные любым из варианта 1 или 3 оптимизированного по кодону человеческого FGF21, были способны секретировать более высокие уровни человеческого FGF21 в культуральную среду по сравнению с диким типом или вариантом 2 оптимизированного по кодону FGF21 (фиг. 12), таким образом демонстрируя повышенную продукцию белка FGF21 при оптимизации по кодону согласно вариантам 1 и 3.
Пример 8. Обращение развития ожирения и сахарного диабета у мышей при введении векторов на основе AAV, кодирующих человеческий FGF21 (эксперимент in vivo, доказывающий активность FGF21)
Мышей, которых кормили HFD, обрабатывали векторами на основе AAV, кодирующими человеческий FGF21. В качестве контролей такую же дозу векторов AAV-null вводили мышам, которых кормили кормом и HFD.
Для оценки способности человеческого FGF21 индуцировать «побурение» WAT и термогенную активность ВАТ, повышать потребление энергии и улучшать метаболизм глюкозы и энергетический метаболизм осуществляют следующие тесты:
- измерение массы тела и потребления корма и жидкости еженедельно;
- измерение температуры тела;
- измерение потребления энергии и дыхательного коэффициента с помощью непрямой калориметрии;
- измерение гликемии;
- оценка утилизации глюкозы во всем организме с помощью внутрибрюшинного теста толерантности к глюкозе;
- оценка чувствительности к инсулину с помощью внутрибрюшинного теста толерантности к инсулину;
- анализы в образцах ткани и сыворотки, включающие в себя
- оценку уровня сверхэкспрессии человеческого FGF21 в подвергнутой целенаправленному воздействию ткани и в кровоток,
- морфологический и гистологический анализ,
- определение уровней циркулирующих в крови гормонов и цитокинов,
- определение метаболических параметров сыворотки крови, таких как свободные жирные кислоты, глицерин, триглицериды, холестерин и кетоновые тела,
- оценку способности к «побурению» посредством оценки присутствия бежевых адипоцитов в паховой жировой подушке с помощью иммуногистохимического анализа и экспрессии генов маркеров классических белых, бурых и бежевых адипоцитов.
Пример 9. In vitro анализ для оценки активности FGF21
Ожидается, что FGF21 повышает усвоение глюкозы и экспрессию GLUT1 в клетках 3T3-L1 (Kharitonenkov, A. et al., 2005. J. Clin. Invest 115:1627-1635).
Пример 10. Обращение развития ожирения и улучшение метаболизма глюкозы при введении в eWAT векторов AAV8-CAG-moFGF21-dmiRT ob/ob мышам: дальнейшие наблюдения
Авторы настоящего изобретения дополнительно оценивали противодиабетический и направленный против развития ожирения терапевтический потенциал опосредованного AAV генно-инженерного воздействия на жировую ткань с использованием FGF21 у ob/ob мышей (см. пример 2).
Ob/ob мыши, которые получали инъекции в eWAT векторов AAV8-CAG-moFGF21-dmiRT, демонстрировали снижение размера белых адипоцитов в эпидидимальной подушке (фиг. 13А). Уровни циркулирующего в крови адипонектина также повышались с дозой (фиг. 14А). Воспаление eWAT, оцениваемое посредством окрашивания Мас2, также снижалось в зависимости от дозы вектора, как и экспрессия маркера макрофагов F4/80 (фиг. 14 В и С).
В печени ob/ob мышей, которым вводили инъекцией нулевые векторы или самую низкую дозу AAV8-CAG-moFGF21-dmirT, проявлялось накопление капелек липидов в гепатоцитах (фиг. 13 В). Введение кодирующих FGF21 векторов в дозах 5×1010vg/мышь или выше полностью предупреждало развитие стеатоза печени (фиг. 13 В), что коррелировало с массой органа (фиг. 14D) и общим содержанием триглицеридов и холестерина в нем (фиг. 14Е и F). Дополнительное доказательство того, что доза 5×1010 vg/мышь представляла собой пороговый уровень для терапевтической эффективности, получено в результате анализа гликемии и инсулинемии. В то время как доза 1×1010 vg/мышь не модифицировала уровни глюкозы в крови в сытом состоянии и только частично снижала уровни инсулина, дозы 5×1010vg/мышь и выше полностью нормализовали гликемию и инсулинемию (фиг. 13С и D). В целом, этот пример подтверждает терапевтический потенциал сверхэкспрессии FGF21 в жировой ткани.
Пример 11. Обращение развития ожирения и улучшение метаболизма глюкозы при внутривенном введении векторов AAV8-hAAT-moFGF21 ob/ob мышам: дальнейшие наблюдения
Авторы настоящего изобретения дополнительно оценивали противодиабетический и направленный против развития ожирения терапевтический потенциал внутривенного введения векторов AAV8-hAAT-moFGF21 у ob/ob мышей (см. пример 3).
В соответствии с их более низкой массой тела ob/ob животные, сверхэкспрессирующие FGF21 в печени, демонстрировали значительно уменьшенный размер белых адипоцитов, в особенности, те животные, которых обрабатывали 5×1011vg (фиг. 15А). Он изменялся параллельно с дозозависимым повышением уровней циркулирующего в крови адипонектина (фиг. 15В) и уменьшенным воспалением WAT, о чем свидетельствовало уменьшенное окрашивание в отношении Мас2 и экспрессия F4/80 и TNF-α в eWAT (фиг. 16А-С). Примечательно, что ob/ob мыши, обработанные 5×1011vg, демонстрировали значительное снижение содержания «коронообразных» структур в eWAT (фиг. 16А).
В то время как ob/ob мыши возрастом 7 месяцев демонстрировали заметный стеатоз печени, в печени обработанных FGF21 ob/ob мышей не проявлялось накопление липидов в гепатоцитах (фиг. 15С). Это согласовалось с 60% уменьшением массы этого органа (фиг. 16D и Е), а также с заметным снижением общего содержания триглицеридов и холестерина в печени (фиг. 16F и G) у ob/ob мышей, получавших терапевтические векторы. Ob/ob животные, обработанные обеими дозами AAV8-hAAT-moFGF21, также демонстрировали пониженную гликемию в сытом состоянии, а их инсулинемия в сытом состоянии снижалась на ~70% (фиг. 15D и Е).
Авторы настоящего изобретения оценивали, являлось ли снижение уровней циркулирующей в крови глюкозы, наблюдаемое у ob/ob мышей после обработки AAV8-hAAT-moFGF21, результатом подавления глюконеогенеза в печени, посредством измерения экспрессии фосфоенолпируват-карбоксикиназы (РЕРСК) и глюкоза-6-фосфатазы (G6Pase) с помощью qPCR. Наблюдали отсутствие изменений в экспрессии этих ферментов в печени обработанных AAV8-hAAT-moFGF21 ob/ob мышей, за исключением животных, обработанных 1×1011vg AAV8-hAAT-moFGF21, которые демонстрировали повышенную экспрессию РЕРСК (фиг. 17А и В). Эти результаты говорят о том, что опосредованная AAV долговременная экспрессия FGF21 в печени и последующее повышение уровня циркулирующего в крови FGF21 не снижали уровень глюкозы за счет ингибирования продукции глюкозы в печени.
Эффекты FGF21 в отношении снижения уровня глюкозы также объясняли повышенным усвоением глюкозы адипоцитами и усиленным потреблением энергии (Xu J. et al., 2009. AJP Endocrinol. Metab. 297:E1105-E1114; Ding X. et al., 2012. CellMetab. 16:387-393; Camacho R.C. et al., 2013. Eur. J. Pharmacol. 715:41-45; Emanuelli B. et al., 2014. Clin. Invest. 124:515-527; Kharitonenkov A. et al., 2005. Endocrinology 148: 774-781; Hondares E. et al., 2010. Cell Metab. 11:206-212; Samms R.J. et al., 2015. Cell Rep.11:991-999). Таким образом, авторы настоящего изобретения оценивали с помощью qPCR экспрессию ключевых компонентов аппарата усвоения глюкозы, таких как транспортеры глюкозы Glutl и Glut4, фосфорилирующие глюкозу ферменты гексокиназы I и II (HKI и HKI) и UCP1 в случае iBAT, в различных подушках жировой ткани (iWAT, eWAT и iBAT). У обработанных AAV8-FGF21 ob/ob мышей экспрессия Glutl повышалась в iWAT и iBAT (фиг. 17С), а экспрессия Glut4 повышалась в eWAT, iWAT и iBAT (фиг. 17D). Экспрессия HKI и HKII активировалась только в iBAT (фиг. 17Е и F). Более того, экспрессия UCP1 повышалась в iBAT ob/ob мышей, обработанных высокой дозой векторов AAV8-hAAT-moFGF21 (фиг. 17G). В совокупности эти результаты говорят о том, что долговременное ослабление гликемии, наблюдаемое у ob/ob мышей после обработки векторами AAV8-hAAT-moFGF21, возможно, является результатом повышенного усвоения глюкозы белыми и бурыми адипоцитами и усиленного термогенеза в iBAT.
Пример 12. Долговременное обращение развития ожирения и сахарного диабета при внутривенном введении векторов AAV8-hAAT-moFGF21 мышам, которых кормили HFD, и старым мышам, которых кормили HFD: уменьшенная масса ткани и стабильная экспрессия вплоть до 1 года
Типичные изображения животных, принадлежащих ко всем экспериментальным группам в исследованиях, осуществляемых на животных в молодом возрасте или во взрослом возрасте (см. примеры 4 и 5), представлены на фиг. 19А-В.
Обращение развития ожирения под действием обработки AAV8-hAAT-moFGF21 происходило параллельно с дозозависимым снижением массы основных депо белой жировой ткани (WAT), таких как эпидидимальная (eWAT), паховая (iWAT) и забрюшинная (rWAT) жировые подушки, в обеих группах животных, обработанных в молодом возрасте или во взрослом возрасте (фиг. 18А и фиг. 19С). Индуцированное HFD увеличение массы печени полностью нормализовалось под действием переноса гена FGF21 при самых высоких используемых дозах вектора, в то время как масса четырехглавой мышцы не изменялась под действием рациона или доставки AAV (фиг. 18А и фиг. 19D).
Обработанные AAV8-hAAT-moFGF21 мыши обоих возрастов демонстрировали специфическую сверхэкспрессию оптимизированного по кодону FGF21 в печени (фиг. 19Е), которая приводила в результате к дозозависимой секреции FGF21 в кровоток в обеих группах мышей, при этом уровни оставались стабильными в течение периода до 1 года после однократного введения вектора (фиг. 18В).
Пример 13. Долговременное обращение развития ожирения и сахарного диабета при внутривенном введении векторов AAV8-hAAT-moFGF21 мышам, которых кормили HFD, и старым мышам, которых кормили HFD: повышенная двигательная активность и изучение термогенного механизма
Повышенное потребление энергии (см. примеры 4 и 5) также наблюдали у животных, которых подвергали обработке в молодом возрасте, спустя 10 месяцев после доставки AAV8-hAAT-moFGF21 (фиг. 21 А).
Это наблюдение согласовалось с опосредованными AAV8-hAAT-moFGF21 эффектами в отношении двигательной активности. В отличие от пониженной активности, наблюдаемой в тесте открытого поля у животных, которых кормили HFD, и которые получали векторы AAV8-null, мыши, обработанные 5×1010vg AAV8-hAAT-moFGF21 в молодом возрасте, демонстрировали такую же степень спонтанной двигательной активности, что и животные, которых кормили кормом и которым вводили инъекцией нулевой вектор. Как показано на фиг. 20А, спустя 1 год после доставки AAV8-hAAT-moFGF21, обработанные животные преодолевали большее расстояние, меньше времени лежали и проводили больше времени, выполняя медленные и быстрые движения, чем необработанные контроли, которых кормили HFD.
С учетом того, что эти изменения в потреблении энергии могут отражать изменения в термогенезе, авторы настоящего изобретения оценивали степень активации бурой жировой ткани (ВАТ). Как мыши, обработанные в молодом возрасте, так и мыши, обработанные во взрослом возрасте 5×1010vg AAV8-hAAT-moFGF21, демонстрировали пониженное отложение липидов в iBAT (фиг. 20 В). Содержание белка UCP1 в ВАТ повышалось дозозависимо у мышей, обработанных векторами AAV8-hAAT-moFGF21 в молодом возрасте (фиг. 20С), что соответствовало повышению несократительного термогенеза, индуцируемому переносом гена FGF21 в печень.
«Побурение» подкожной WAT, характеризующееся появлением бежевых адипоцитов, также ассоциировано с повышениями потребления энергии (Harms & Seale, 2013). Для того чтобы оценить, ответственно ли «побурение» за усиление потребления энергии, наблюдаемое после обработки AAV8-hAAT-moFGF21, осуществляли гистологическую оценку iWAT. В соответствии с уменьшенной массой этой подушки (фиг. 18А) адипоциты обработанных AAV8-hAAT-moFGF21 животных, которых кормили HFD, были меньше, чем адипоциты животных, которых кормили HFD и которым вводили инъекцией нулевой вектор (фиг. 20D). Тем не менее, обработка векторами AAV8-hAAT-moFGF21 не приводила в результате к повышенному выявлению мультилокулярных бежевых адипоцитов в iWAT при любой из исследуемых доз у животных, обработанных либо в молодом возрасте, либо во взрослом возрасте (фиг. 20D). Соответственно, не существовало статистический значимых различий в уровнях белка UCP1 в iWAT между группами животных, которых кормили HFD (фиг. 21 В).
Запускаемый креатином субстратный цикл и опосредуемое Са2+-АТФазой 2b (Serca2b) сарко/эндоплазматического ретикулума циклическое изменение уровней кальция могут повышать термогенез в iWAT независимо от UCP1 (Kazak L. et al., 2015. Cell 163:643 655; Ikeda K. et al., 2017. Nat. Med. 23:1454-1465). Более высокие уровни экспрессии фосфатазы orphan 1 (Phospho1), фермента, вовлеченного в запускаемый креатином субстратный цикл, наблюдали в iWAT мышей, которых кормили HFD, обработанных 5×1010vg AAV8-hAAT-moFGF21, по сравнению с соответствующими по возрасту контрольными группами животных, которых кормили кормом и HFD (фиг. 20Е), что говорит о том, что активность запускаемого креатином субстратного цикла, возможно, повышалась в результате переноса гена FGF21. Что касается термогенного механизма, зависимого от циклических изменений уровней кальция, отсутствие различий в уровнях экспрессии Serca2b выявили в iWAT животных, обработанных векторами AAV8-hAAT-moFGF21, по сравнению с обработанными нулевым вектором животными, которых которых кормили кормом или HFD (фиг. 21С). С другой стороны, экспрессия рианодинового рецептора 2 (RyR2), другого фермента, вовлеченного в тот же цикл, в iWAT повышалась при кормлении HFD как у мышей, обработанных нулевым вектором, так и у обработанных AAV8-hAAT-moFGF21 мышей (фиг. 21С). В совокупности эти результаты говорят о том, что термогенный механизм, зависимый от циклического изменения уровней кальция, не вовлечен в улучшение энергетического гомеостаза всего организма, наблюдаемое после обработки AAV-FGF21.
Пример 14. Долговременное обращение развития ожирения и сахарного диабета при внутривенном введении векторов AAV8-hAAT-moFGF21 мышам, которых кормили HFD, и старым мышам, которых кормили HFD: уровни глюкагона, гиперплазия островков и толерантность к глюкозе
Кроме этого, животные, которых кормили HFD, обработанные в молодом возрасте векторами AAV8-hAAT-moFGF21, демонстрировали пониженные уровни циркулирующего в крови глюкагона по сравнению с обработанными нулевым вектором мышами, которых кормили HFD (фиг. 22А).
В то время как у обработанных вектором AAV8-null мышей развивалась гиперплазия островков как следствие кормления HFD, масса β-клеток у животных, обработанных векторами AAV8-hAAT-FGF21 (в дозах 2×1010 или 5×1010 vg/мышь), была подобной таковой у контрольных мышей, которых кормили рационом в виде корма (фиг. 22В и С). Двойное иммуноокрашивание в отношении инсулина и глюкагона срезов поджелудочной железы от обработанных AAV8-hAAT-moFGF21 мышей, которых кормили HFD, показывало нормальное распределение α- и β-клеток в островках этих животных с локализацией экспрессирующих глюкагон клеток на периферии островка, а экспрессирующих инсулин клеток - в сердцевине (фиг. 22D).
Для оценки толерантности к глюкозе у обработанных FGF21 мышей осуществляли внутрибрюшинный тест толерантности к глюкозе (GTT) (2 г глюкозы/кг массы тела) через 10 недель после введения AAV. Животные, которых кормили HFD и которым вводили инъекцией либо нулевые векторы, либо кодирующие FGF21 векторы в дозе 1×1010 vg/мышь, были нетолерантны к глюкозе и демонстрировали значительно повышенные уровни циркулирующего в крови инсулина во время GTT (фиг. 23А и В). В отличие от этого, животные, обработанные 5×1010 vg/мышь AAV8-hAAT-moFGF21, демонстрировали улучшенную толерантность к глюкозе по сравнению с контрольными мышами, которых кормили кормом (фиг. 23А). Уровни инсулина были неотличимы в этих двух экспериментальных группах (фиг. 23В). Эти результаты дополнительно подтверждали улучшенную чувствительность к инсулину у мышей, которых кормили HFD, обработанных 5 ×1010 vg/мышь AAV8-hAAT-moFGF21.
Пример 15. Обращение развития ассоциированной с HFD гипертрофии и воспаления WAT при внутривенном введении векторов AAV8-hAAT-moFGF21
Кормление HFD индуцирует увеличение размера адипоцитов WAT (Sattar N. & Gill J.M.R., 2014. ВМС Med. 12: 123). Введение кодирующих FGF21 векторов, противодействовало этому увеличению (фиг. 24А). Морфометрический анализ WAT выявил, что площадь белых адипоцитов у животных, обработанных в молодом возрасте 1×1010 или 5×1010 vg вектора, и у мышей, обработанных во взрослом возрасте 2×1010 или 5×1010 vg вектора, была подобной таковой у животных, которых кормили рационом в виде корма (фиг. 24В). В обеих группах обработанных FGF21 животных присутствовало перераспределение размера адипоцитов с большей долей более мелких адипоцитов (фиг. 25А). В соответствии со снижением ожирения и обращением развития гипертрофии WAT уровни адипонектина и лептина также нормализовались у животных, обработанных самыми высокими дозами векторов AAV8-hAAT-moFGF21, вне зависимости от возраста, в котором происходило начало обработки (фиг. 24С и D).
Ожирение также вызывает воспаление WAT (Hajer G.R. et al., 2008. Eur. Heart J. 29: 2959-2571). Следовательно, авторы настоящего изобретения анализировали воспаление в этой ткани посредством иммуноокрашивания в отношении специфического для макрофагов маркера Мас2 и по экспрессии провоспалительных молекул. В то время как мыши, которых кормили HFD, демонстрировали повышенное присутствие макрофагов, выявляемое в виде «коронообразных» структур, в eWAT, животные, обработанные в молодом возрасте или во взрослом возрасте 5×1010 vg AAV8-hAAT-moFGF21, не имели признаков инфильтрации макрофагов (фиг. 24Е и фиг. 25В). Это происходило параллельно с нормализацией экспрессии маркеров макрофагов F480 и CD68 и провоспалительных цитокинов TNFα и IL-1β (фиг. 24F-H и фиг. 25С-Е), указывая на то, что экспрессия FGF21 противодействовала воспалению WAT, ассоциированному с ожирением.
Пример 16. Обращение развития стеатоза, воспаления и фиброза печени при внутривенном введении векторов AAV8-hAAT-moFGF21
Гистологический анализ печени показал, что все обработанные нулевым вектором животные, которых кормили HFD, имели заметный стеатоз печени на момент умерщвления (фиг. 26A-D). В отличие от этого мыши, которых кормили HFD, и которые получали 5×1010 vg AAV8-hAAT-moFGF21 в молодом возрасте или во взрослом возрасте, подтверждали обращение развития этого патологического отложения липидов (фиг. 26А). Эти гистологические данные согласовались с заметным снижение общего содержания триглицеридов и холестерина в печени у обработанных 5×1010 vg AAV8-hAAT-moFGF21 животных (фиг. 26В и С). Кроме того, животные, которых кормили HFD и обрабатывали 5×1010 vg векторов AAV8-hAAT-moFGF21 в молодом возрасте или во взрослом возрасте, не демонстрировали признаков воспаления печени, о чем свидетельствует отсутствие окрашивания в отношении Мас2, которое выявляло присутствие макрофагов в печени обработанных нулевым вектором мышей, которых кормили HFD (фиг. 26D). Наконец, перенос гена FGF21 в печень обращал развитие фиброза печени. В то время как коллагеновые волокна легко выявлялись после окрашивания пикросириусом красным или трихром-окрашивания по Массону срезов печени от животных, которых кормили HFD и которым вводили инъекцией контрольные нулевые векторы, они не выявлялись в печени обработанных AAV8-hAAT-moFGF21 мышей (фиг. 28А и фиг. 27). Эти мыши также демонстрировали заметно сниженную экспрессию коллагена 1 в печени (фиг. 28В и С). В совокупности эти данные указывают на то, что обработка AAV8-hAAT-moFGF21 защищала от развития индуцированного HFD неалкогольного стеатогепатита (NASH).
Пример 17. Долговременная безопасность направленной на печень обработки AAV-FGF21
Фармакологическое лечение с использованием FGF21 или трансгенная сверхэкспрессия ассоциировали с нарушением гомеостаза кости вследствие повышенной резорбции кости, которая может вызвать рарефикацию кости (Wei W. et al., 2012. Ргос.Natl. Acad. Sci. 109: 3143-3148; WangX. et al., 2015. Cell Metab. 22: 811-824; Charoenphandhu N. et al., 2017. J. Bone Miner. Metab. 35: 142 149; Talukdar S. et al., 2016. Cell Metab. 23: 427 440; Kim A.M. et al., 2017. Diabetes, Obes. Metab). С учетом терапевтического потенциала AAV8-hAAT-moFGF21 для лечения ожирения и сахарного диабета авторы настоящего изобретения оценивали долговременные эффекты переноса гена в отношении костей животных, обработанных самой высокой дозой вектора. На момент умерщвления (в возрасте ~16,5 месяцев) назо-анальная длина и длина болынеберцовой кости были нормальными у животных, которым вводили векторы AAV8-hAAT-moFGF21 в возрасте 9 или 29 недель (фиг. 29А и В). Затем авторы настоящего изобретения изучали структуру кости с помощью компьютерной микротомографии (μСТ). Анализ проксимального эпифиза болынеберцовой кости выявил отсутствие значимых различий в трабекулярном и кортикальном веществе кости мышей, которых кормили HFD и которым вводили 5×1010 vg AAV8-hAAT-moFGF21, по сравнению с мышами в соответствующих возрастных группах, обработанных нулевыми векторами. В частности, отсутствие различий документально подтверждалось минеральной плотностью кости (BMD) (фиг. 29С), содержанием минералов в кости (ВМС) (фиг. 29D), объемом кости (BV) (фиг. 29Е), соотношением объем кости/объем ткани (BV/TV) (фиг. 29F), соотношением поверхность кости/объем кости (BS/BV) (фиг. 29G), числом трабекул (Tb.N) (FIG 29Н), толщиной трабекул (Tb.Th) (фиг. 291) или расстоянием между трабекулами (Tb.Sp) (фиг. 29J). Подобным образом, анализ компактного вещества кости в диафизе болынеберцовой кости показал отсутствие различий в ВМС, BMD, BV, BV/TV или BS/BV между группами животных, которых кормили HFD и которым вводили инъекцией нулевой вектор, или группами обработанных FGF21 животных (фиг. 29K-O).
Сообщалось, что патологические эффекты FGF21 опосредованы, по меньшей мере частично, повышенной продукцией связывающего белка 1 инсулиноподобного фактора роста (IGFPB1) печенью (Wang X. et al., 2015. Cell Metab. 22: 811-824). В соответствии с отсутствием изменений кости обработка высокой дозой AAV8-hAAT-moFGF21 не вела к повышению уровней циркулирующего в крови белка IGFBP1 у животных, обработанных 12 (в молодом возрасте) или 6 (во взрослом возрасте) месяцами ранее, по сравнению с мышами, которых кормили HFD и которым вводили инъекцией нулевой вектор (фиг. 29Р). Уровни циркулирующего в крови IGF1 также являлись нормальными во всех экспериментальных группах (фиг. 29Q). В совокупности эти результаты подтверждают безопасность для костной ткани опосредованного AAV переноса гена FGF21 в печень.
Пример 18. Предупреждение индуцированных HFD опухолей печени при внутривенном введении векторов AAV8-hAAT-moFGF21
Долговременное (>60 недель) кормление HFD ассоциировалось с повышенной частотой возникновения новообразований печени у C57BL/6J мышей (Hill-Baskin А.Е. et al., 2009. Hum. Mol. Genet. 18: 2975-2988; Nakagawa H., 2015. World J. Hepatol. 7: 2110). В исследовании авторов настоящего изобретения на животных, кормление которых HFD начинали в молодом возрасте и продолжали его в течение 60 недель, авторы настоящего изобретения обнаружили опухоли в печени у 66,7% (6/9) животных, которым вводили инъекцией нулевые векторы. Животные, обработанные векторами AAV8-hAAT-moFGF21, были защищены от индуцированного HFD развития новообразований печени: 0% (0/8) животных, обработанных 5×1010 vg кодирующих FGF21 векторов, демонстрировали опухоли, а частота возникновения составляла 40% (4/10) в когорте, обработанной самой низкой дозой (1×1010 vg). Ни у одной (0/11) из мышей, которых кормили кормом, не развились опухоли в тот же период времени (таблица 1).
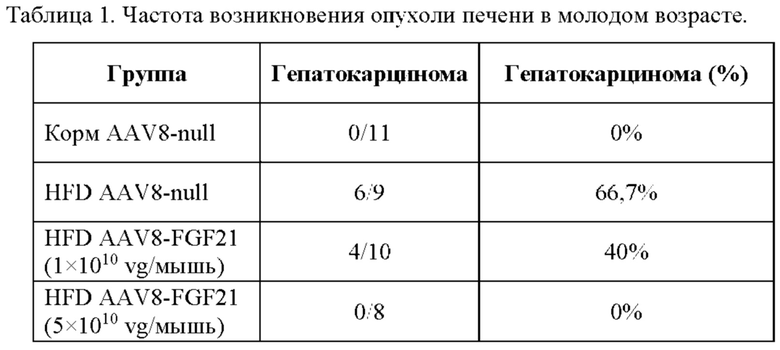
Пример 19. Ослабление индуцированной STZ гипергликемии под действием специфической для печени сверхэкспрессии FGF21, опосредованной AAV8
Материалы и способы
Животные
Авторы настоящего изобретения использовали самцов С57b16 мышей возрастом 9 недель. Мыши имели свободный доступ к стандартному рациону, и их содержали при цикле чередования 12 часов света и темноты (освещение включали в 08:00). Для индукции сахарного диабета мыши получали пять внутрибрюшинных инъекций, в последовательные дни, стрептозоцина (50 мг/кг), растворенного в 0,1 моль/л нитратном буфере (рН 4,5). Уровни глюкозы в крови оценивали с использованием анализатора (Glucometer Elite; Bayer, Леверкузен, Германия). Процедуры по уходу за животными и экспериментальные процедуры были одобрены Комитетом по этике в экспериментах на животных и человеке (Ethics Committee in Animal and Human Experimentation) Автономного университета Барселоны (Universitat  de Barcelona).
de Barcelona).
Введение векторов на основе AAV in vivo
Для системного введения векторы на основе AAV разводили в 200 мкл 0,001% F68 Pluronic® (Gibco) в PBS и вводили инъекцией через хвостовую вену.
Результаты
Для того чтобы исследовать защитный потенциал полученного с помощью AAV FGF21 против сахарного диабета 1 типа, 5×1010 vg или 2×1011 vg векторов на основе AAV8, кодирующих оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора hAAT (AAV8-hAAT-moFGF21), вводили IV самцам С57В16 мышей возрастом 9 недель. Контрольные мыши получали 2×1011 vg векторов AAV8-hAAT-null. Через две недели после введения AAV всех животных обрабатывали стрептозоцином (STZ) (5 доз по 50 мг/кг; 1 доза в сутки) для запуска диабетического процесса.
Анализ уровней глюкозы в крови выявил, что животные, обработанные векторами на основе AAV8, кодирующими moFGF21, демонстрировали более низкие уровни циркулирующей в крови глюкозы, чем С57В16 мыши, обработанные векторами AAV8-hAAT-null (фиг. 30).
Пример 20. Продление жизни в здоровом состоянии при внутримышечном введении векторов AAV-CMV-moFGF21 С57В16 мышам вследствие предупреждения увеличения массы и резистентности к инсулину, ассоциированных со старением
Скелетная мышца (Skm) является легкодоступной тканью, и ее применили для продуцирования секретируемых терапевтических белков (Haurigot V. et al., 2010. J. Clin. Invest. 123: 3254-3271; Callejas D. et al., 2013. Diabetes 62: 1718-1729; Jaen M.L. et al., 2017. Mol. Ther. Methods Clin. Dev. 6: 1-7). Чтобы изучить, может ли Skm являться приемлемым источником циркулирующего в крови FGF21, использовали векторы на основе AAV серотипа 1, которые демонстрируют высокий тропизм в отношении Skm (Chao L. et al., 2000. J. Clin. Invest. 106: 1221 1228; Wu Z. et al., 2006. J. Virol. 80: 9093 9103; Lisowski L. et al., 2015. Curr. Opin. Pharmacol. 24:59-67) и несут оптимизированный мышиный FGF21 под контролем промотора CMV (AAV1-CMV-moFGF21). Векторы вводили инъекцией в дозе 5×1010 vg/мышца в четырехглавую мышцу, икроножную мышцу и переднюю краниальную болынеберцовую мышцу обеих лап (общая доза 3×1011 vg/мышь) С57В16 мышей возрастом 8 недель. Контрольным животным вводили инъекцией векторы AAV1-CMV-Null в такой же дозе. Использование здоровых мышей, которых кормили стандартным рационом, дополнительно позволило авторам настоящего изобретения оценить долговременную безопасность генной терапии с использованием FGF21.
Животным возрастом одиннадцать месяцев, которым вводили инъекцией векторы, кодирующие FGF21, в возрасте 8 недель, демонстрировали заметное повышение циркулирующего в крови FGF21 (фиг. 31А), которое происходило параллельно с высокими уровнями экспрессии происходящего из вектора FGF21 в 3 получавших инъекцию мышцах (фиг. 31В). В соответствии с предшествующими сообщениями эта комбинация серотипа вектора, промотора и пути введения не приводила к экспрессии трансгена в печени (фиг. 31В).
В конце ~10-месячного периода последующего наблюдения мыши, которым вводили инъекцией внутримышечно AAV1-CMV-moFGF21, сохраняли массу тела, которую они имели на момент начала исследования, и были на ~38% более стройными, чем контроли, чья масса неуклонно увеличивалась по мере того, как животные старели (фиг. 31С). В то время как перенос гена FGF21 оказывал лишь незначительное воздействие на массу мышц, масса белых и бурых депо, а также печени печень существенно уменьшалась (фиг. 31D). Действительно, масса анализируемых подушек WAT уменьшалась на >50% (фиг. 31D). Более того, мыши, обработанные AAV1-CMV-moFGF21, демонстрировали заметное снижение общего содержания триглицеридов в печени (фиг. 31Е). Наблюдали отсутствие изменений уровней холестерина в печени (фиг. 31F). В противоположность животным, которым вводили инъекцией нулевой вектор, животные, обработанные AAV1-CMV-moFGF21, демонстрировали нормогликемию {данные не показаны) и пониженную инсулинемию, когда были в возрасте примерно 1 года (фиг. 31G). Соответственно, обработанные FGF21 мыши демонстрировали заметно улучшенную чувствительность к инсулину в конце исследования (фиг. 31H). В целом, это исследование демонстрирует, что введение векторов на основе AAV, которое приводит к значимым с терапевтической точки зрения уровням циркулирующего в крови FGF21, является безопасным в течение длительного периода времени в здоровом состоянии, и его можно применять для обращения развития увеличения массы тела и резистентности к инсулину, ассоциированных со старением.
Пример 21. Обращение развития ожирения и сахарного диабета при внутримышечном введении векторов AAV1-CMV-moFGF21 С57В16 мышам, которых кормили HFD
Авторы настоящего изобретения затем оценивали, способно ли im (внутримышечное) введение векторов AAV1-CMV-moFGF21 также обращать развитие ожирения и резистентности к инсулину. С этой целью С57В16 мышей возрастом два месяца кормили либо кормом, либо HFD в течение 12 недель. В течение этих первых 3 месяцев последующего наблюдения масса животных, которых кормили кормом, увеличилась на 20%, а у животных, которых кормили HFD, развилось ожирение (95% увеличение массы тела) (фиг. 32А и В). Затем векторы вводили инъекцией в дозе 5×1010 vg/мышца в четырехглавую мышцу, икроножную мышцу и переднюю краниальную болынеберцовую мышцу обеих лап (общая доза 3×1011 vg/мышь) С57В16 мышей с ожирением. В качестве контролей другая когорта мышей с ожирением и когорта мышей, которых кормили кормом, получали 3×1011 vg некодирующих нулевых векторов (AAV1-CMV-null). После доставки AAV мышей содержали с кормлением кормом или HFD. Животные, обработанные AAV1-CMV-moFGF21, испытывали прогрессирующую потерю массы тела (фиг. 32А и В). Обращение развития ожирения под действием обработки AAV1-CMV-FGF21 происходило параллельно с повышениями уровней циркулирующего в крови FGF21 (фиг. 32С).
Обработанные нулевым вектором мыши, которых кормили HFD, демонстрировали нормальную гликемию в сытом состоянии (фиг. 32D), но имели гиперинсулинемию (фиг. 32Е), что говорит о том, что у этих мышей развилась резистентность к инсулину. В отличие от этого, мыши, которых кормили HFD, обработанные AAV1-CMV-moFGF21, к концу исследования имели нормогликемию и нормоинсулинемию (фиг. 32D и Е). Более того, животные, которым вводили AAV1-CMV-moFGF21, демонстрировали большую чувствительность к инсулину, чем у контролей, которых кормили HFD (фиг. 32F).
Пример 22. Повышенные уровни циркулирующего в крови FGF21 под действием оптимизированных по кодону нуклеотидных последовательностей человеческого FGF21
Чтобы оценить, была ли способна оптимизация по кодону опосредовать повышенные уровни циркулирующего в крови FGF21 самцам С57В16 мышей возрастом 8 недель вводили инъекцией с помощью гидродинамических методов плазмиды, кодирующие три разные оптимизированные по кодону нуклеотидные последовательности человеческого FGF21 (SEQ ID NO: 40-42) под управлением промотора hAAT. В качестве контроля использовали необработанных мышей и мышей, которым вводили инъекцией с помощью гидродинамических методов плазмиду, которая кодирует кодирующую последовательность hFGF21 дикого типа под управлением промотора hAAT.
Материалы и способы
Доставка мышам плазмид in vivo с помощью гидродинамической инъекции в хвостовую вену
Плазмидную ДНК разводили в солевом растворе в объеме (мл), равном ~10% средней массы тела животного (граммы), и вручную вводили инъекцией в латеральную хвостовую вену менее чем за 5 секунд. Перед инъекцией животных помещали под 250 Вт лампу с излучением тепла в инфракрасном диапазоне (Philips NV) на несколько минут для того, чтобы расслабить кровеносные сосуды и облегчить визуальное наблюдение и упростить доступ к хвостовой вене. Пластиковое приспособление для фиксирования лап (Harvard Apparatus) использовали для фиксирования животного для инъекции. Анестезию не использовали, поскольку она не нужна при использовании соответствующего приспособления для фиксирования лап. Авторы настоящего изобретения использовали для инъекции животным иглы для подкожных инъекций калибром 26G и длиной 3/8 дюйма (BD), самый большой допустимый калибр иглы, который плотно входил в вену, к которой осуществляли доступ.
Результаты
Мыши, обработанные любым из варианта 2 или 3 оптимизированного по кодону человеческого FGF21, были способны секретировать более высокие уровни человеческого FGF21 в кровоток по сравнению с диким типом или вариантом 1 оптимизированного по кодону FGF21 (фиг. 33), таким образом демонстрируя повышенную продукцию белка FGF21 при оптимизации по кодону согласно вариантам 2 и 3.
Пример 23. Повышенные уровни экспрессии и продукции белка FGF21 in vitro и in vivo под действием экспрессионных кассет hAAT-moFGF21, CAG-moFGF21-doublemiRT и CMV-moFGF21
Материалы и способы
Доставка плазмид in vivo мышам с помощью гидродинамической инъекции в хвостовую вену
Плазмидную ДНК разводили в солевом растворе в объеме (мл), равном ~10% средней массы тела животного (граммы), и вручную вводили инъекцией в латеральную хвостовую вену менее чем за 5 секунд. Перед инъекцией животных помещали под 250 Вт лампу с излучением тепла в инфракрасном диапазоне (Philips NV) на несколько минут для того, чтобы расслабить кровеносные сосуды, облегчить визуальное наблюдение и упростить доступ к хвостовой вене. Пластиковое приспособление для фиксирования лап (Harvard Apparatus) использовали для фиксирования животного для инъекции. Анестезию не использовали, поскольку она не нужна при использовании соответствующего приспособления для фиксирования лап.Авторы настоящего изобретения использовали для инъекции животным иглы для подкожных инъекций калибром 26G и длиной 3/8 дюйма (BD), самый большой допустимый калибр иглы, который плотно входил в вену, к которой осуществляли доступ.
Результаты
In vitro
Клетки HEK293 трансфицировали плазмидами, которые кодировали кодирующую последовательность мышиного FGF21 WT под управлением промотора фактора элонгации la (EF1a) (EF1a-mFGF21) (Zhang et al., EBioMedicine 15 (2017) 173-183) (SEQ ID NO: 57) или оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора CMV (CMV-moFGF21) или промотора CAG в сочетании с четырьмя тандемными повторами последовательности miRT122a и четырьмя тандемными повторами последовательности miRT1 (CAG-moFGF21-doublemiRT). В качестве контроля использовали нетрансфицированные клетки. Клетки HEK293, трансдуцированные CAG-moFGF21-doublemiRT, экспрессировали более высокие уровни FGF21 по сравнению с клетками, трансдуцированными EF1a-mFGF21, или нетрансдуцированными клетками (фиг. 34А). Более того, клетки HEK293, трансдуцированные CAG-moFGF21-doublemiRT, также демонстрировали более высокое содержание внутриклеточного белка FGF21 и более высокие уровни белка FGF21 в культуральной среде (фиш. 34В и С). Несмотря на то что клетки HEK293, трансдуцированные EF1a-mFGF21 или CMV-moFGF21, экспрессировали сходные уровни FGF21 (фиг. 34А), клетки HEK293, трансдуцированные CMV-moFGF21, демонстрировали более высокое содержание внутриклеточного белка FGF21 и более высокие уровни белка FGF21 в культуральной среде (фиг. 34В и С).
Клетки С2С12 трансфицировали плазмидами, которые кодировали кодирующую последовательность мышиного FGF21 WT под управлением промотора EF1a (EF1a-mFGF21) (Zhang et al., EBioMedicine 15 (2017) 173-183) или оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора CMV (CMV-moFGF21). В качестве контроля использовали нетрансфицированные клетки. Клетки С2С12, трансдуцированные CMV-moFGF21, экспрессировали более высокие уровни FGF21 по сравнению с клетками, которые трансдуцировали EF1a-mFGF21, или нетрансдуцированными клетками (фиг. 34D).
Клетки HepG2 трансфицировали плазмидами, которые кодировали кодирующую последовательность мышиного FGF21 WT под управлением промотора EF1a (EF1a-mFGF21) (Zhang et al., EBioMedicine 15 (2017) 173-183) или оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора hAAT (hAAT-moFGF21). В качестве контроля использовали нетрансфицированные клетки. Клетки HepG2, трансдуцированные hAAT-moFGF21, экспрессировали более высокие уровни FGF21 по сравнению с клетками, которые трансдуцировали EF1a-mFGF21, или нетрансдуцированными клетками (фиг. 34Е).
In vivo
Самцам C57B16 мышей возрастом 8 недель вводили с помощью гидродинамических методов 5 мкг плазмид, которые кодируют кодирующую последовательность мышиного FGF21 WT под управлением промотора фактора элонгации la (EF1a) (EF1a-mFGF21) (Zhang et al., EBioMedicine 15 (2017) 173-183) или оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора CMV (CMV-moFGF21) или промотора hAAT (hAAT-moFGF21). Анализ уровней экспрессии FGF21 в печени через 24 часа после введения плазмид выявил, что животные, обработанные hAAT-moFGF21 или CMV-moFGF21, экспрессировали намного более высокие уровни FGF21, чем животные, получающие EF1a-mFGF21 (фиг. 35А). Кроме того, животные, обработанные hAAT-moFGF21 или CMV-moFGF21, демонстрировали более высокие уровни циркулирующего в крови FGF21, чем животные, получавшие EF1a-mFGF21 (фиг. 35В).
Пример 24. Повышенная экспрессия FGF21 в тканях-мишенях in vivo и уровни циркулирующего в крови FGF21 под действием AAV8-hAAT-moFGF21, AAV8-CAG-moFGF21-doublemiRT и AAV1-CMC-moFGF21 по сравнению с AAV8-EF1a-mFGF21
Экспрессия в печени
Самцам С57В16 мышей вводили внутривенно 1×1010 vg, 2×1010 vg или 5×1010 vg векторов на основе AAV8, которые кодируют кодирующую последовательность мышиного FGF21 WT под управлением промотора фактора элонгации la (EF1a) (AAV8-EF1a-mFGF21) или оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением специфического для печени промотора hAAT (AAV8-I1AAT-moFGF21). Через две недели после введения AAV животные, обработанные AAV8-I1AAT-moFGF21, демонстрировали как высокие уровни экспрессии FGF21 в печени, так и более высокие уровни циркулирующего в крови FGF21, чем животные, обработанные AAV8-EF1a-mFGF21, независимо от дозы вектора (фиг. 36А и В).
Экспрессия в жировой ткани
Самцам С57В16 мышей вводили в eWAT 2×1010 vg, 5×1010 vg или 1×1011 vg либо векторов на основе AAV8, которые кодируют кодирующую последовательность мышиного FGF21 WT под управлением промотора фактора элонгации la (EF1a) (AAV8-EF1a-mFGF21), либо векторов на основе AAV8, которые кодируют оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора CAG в сочетании с четырьмя тандемными повторами последовательности miRT122a и четырьмя тандемными повторами последовательности miRT1 (AAV8-CAG-moFGF21-doublemiRT). Через две недели после введения AAV животные, обработанные AAV8-CAG-moFGF21-doublemiRT, демонстрировали более высокие уровни экспрессии FGF21 в WAT, чем животные, которым вводили AAV8-EF1a-mFGF21 (фиг. 37А). Более того, животные, обработанные AAV8-CAG-moFGF21-doublemiRT, показывали намного более низкую экспрессию FGF21 в печени, чем животные, которым вводили AAV8-EF1a-mFGF21 (фиг. 37В), демонстрируя, что введение в eWAT векторов AAV8-CAG-moFGF21-doublemiRT эффективно препятствовало экспрессии трансгена в нецелевых тканях.
Экспрессия в скелетных мышцах
Самцам С57В16 мышей вводили внутримышечно 5×1010 vg, 1×1011 vg или 3×1011 vg либо векторов на основе AAV8, которые кодируют кодирующую последовательность мышиного FGF21 WT под управлением промотора фактора элонгации la (EF1a) (AAV8-EF1a-mFGF21), либо векторов на основе AAV1, которые кодируют оптимизированную по кодону кодирующую последовательность мышиного FGF21 под управлением промотора CMV (AAV1-CMV-FGF21). Через две недели после введения AAV животные, обработанные AAV1-CMV-FGF21, демонстрировали намного более высокие уровни экспрессии FGF21 в скелетной мышце, чем животные, которым вводили AAV8-EF1a-mFGF21 (фиг. 38А). Более того, животные, обработанные AAV8-EF1a-mFGF21, демонстрировали высокую экспрессию FGF21 в печени, в то время как внутримышечное введение векторов AAV1-CMV-FGF21 эффективно препятствовало экспрессии трансгена в печени (фиг. 38В).
ПОСЛЕДОВАТЕЛЬНОСТИ
SEQ ID NO: Тип последовательности
1 Аминокислотная последовательность FGF21 homo sapiens
2 Аминокислотная последовательность FGF21 mus musculus
3 Аминокислотная последовательность FGF21 canis lupus familiaris
4 Нуклеотидная последовательность FGF21 homo sapiens
5 Оптимизированная по кодону нуклеотидная последовательность FGF21 homo sapiens - вариант 1
6 Оптимизированная по кодону нуклеотидная последовательность FGF21 homo sapiens - вариант 2
7 Оптимизированная по кодону нуклеотидная последовательность FGF21 homo sapiens - вариант 3
8 Нуклеотидная последовательность FGF21 mus musculus
9 Оптимизированная по кодону нуклеотидная последовательность FGF21 mus musculus
10 Нуклеотидная последовательность FGF21 canis lupus familiaris
11 Оптимизированная по кодону нуклеотидная последовательность FGF21 canis lupus familiaris
12 Нуклеотидная последовательность, кодирующая miRT 122а
13 Нуклеотидная последовательность, кодирующая miRT 1
14 Нуклеотидная последовательность, кодирующая miRT 152
15 Нуклеотидная последовательность, кодирующая miRT 199а-5р
16 Нуклеотидная последовательность, кодирующая miRT 199а-3р
17 Нуклеотидная последовательность, кодирующая miRT215
18 Нуклеотидная последовательность, кодирующая miRT 192
19 Нуклеотидная последовательность, кодирующая miRT 148а
20 Нуклеотидная последовательность, кодирующая miRT 194
21 Нуклеотидная последовательность, кодирующая miRT 124
22 Нуклеотидная последовательность, кодирующая miRT 216
23 Нуклеотидная последовательность, кодирующая miRT 125
24 Нуклеотидная последовательность, кодирующая miRT 133а
25 Нуклеотидная последовательность, кодирующая miRT206
26 Нуклеотидная последовательность, кодирующая miRT 130
27 Нуклеотидная последовательность, кодирующая miRT99
28 Нуклеотидная последовательность, кодирующая miRT208-5p
29 Нуклеотидная последовательность, кодирующая miRT208a-3p
30 Нуклеотидная последовательность, кодирующая miRT499-5p
31 Конструкция А
32 Конструкция В
33 Конструкция С
34 Конструкция D
35 Конструкция Е
36 Конструкция F
37 Конструкция G
38 Конструкция Н
39 Конструкция I
40 Конструкция J
41 Конструкция K
42 Конструкция L
43 Нуклеотидная последовательность химерного нитрона, состоящего из интронов из генов человеческого β-глобина и тяжелой цепи иммуноглобулина
44 Нуклеотидная последовательность промотора CAG
45 Нуклеотидная последовательность промотора CMV
46 Нуклеотидная последовательность энхансера CMV
47 Нуклеотидная последовательность промотора hAAT
48 Усеченный 5'ITR AAV2
49 Усеченный 3'ITR AAV2
50 Сигнал полиаденилирования SV40
51 Сигнал полиаденилирования кроличьего β-глобина
52 Последовательность промотора CMV и энхансера CMV
53 Энхансер гепатоцитарного регуляторного участка (HCR) из аполипопротеина Е
54 Промотор mini/aP2
55 Промотор mini/UCP1
56 Промотор С5-12
57 pAAV-EF1a-mmFGF21-рА
Аминокислотная последовательность FGF21 homo sapiens (SEQ ID NO: 1)

Нуклеотидная последовательность FGF21 homo sapiens (SEQ ID NO: 4)

Оптимизированная по кодону нуклеотидная последовательность FGF21 homo sapiens - вариант 1 (SEQ ID NO: 5)

Оптимизированная по кодону нуклеотидная последовательность FGF21 homo sapiens - вариант 2 (SEQ ID NO: 6)

Оптимизированная по кодону нуклеотидная последовательность FGF21 homo sapiens - вариант 3 (SEQ ID NO: 7)

Нуклеотидная последовательность, кодирующая miRT122a (последовательность-мишень микроРНК 122а) (SEQ ID NO: 12)
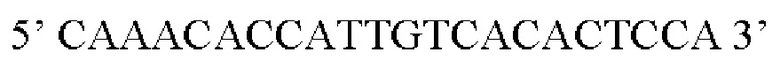
Нуклеотидная последовательность, кодирующая miRT1 (последовательность-мишень микроРНК 1) (SEQ ID NO: 13)
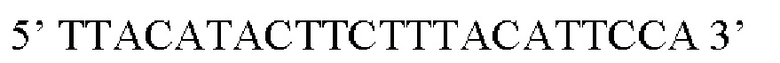
Нуклеотидная последовательность, кодирующая miRT152 (последовательность-мишень микроРНК 152) (SEQ ID NO: 14)
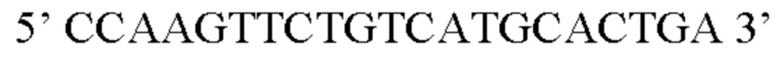
Нуклеотидная последовательность, кодирующая miRT199a-5р (последовательность-мишень микроРНК 199а) (SEQ ID NO: 15)
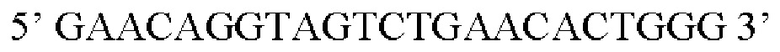
Нуклеотидная последовательность, кодирующая miRT199a-3р (последовательность-мишень микроРНК 199а) (SEQ ID NO: 16)
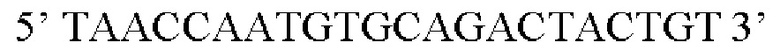
Нуклеотидная последовательность, кодирующая miRT215 (последовательность-мишень микроРНК 215) (SEQ ID NO: 17)

Нуклеотидная последовательность, кодирующая miRT192 (последовательность-мишень микроРНК 192) (SEQ ID NO: 18)

Нуклеотидная последовательность, кодирующая miRT 148а (последовательность-мишень микроРНК 148а) (SEQ ID NO: 19)
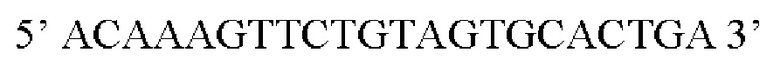
Нуклеотидная последовательность, кодирующая miRT 194 (последовательность-мишень микроРНК 194) (SEQ ID NO: 20)
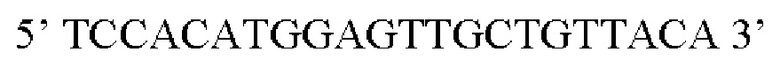
Нуклеотидная последовательность, кодирующая miRT124 (последовательность-мишень микроРНК 124) (SEQ ID NO: 21)
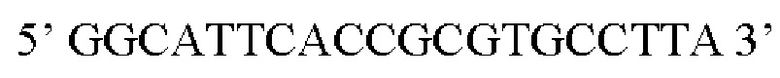
Нуклеотидная последовательность, кодирующая miRT216 (последовательность-мишень микроРНК 216) (SEQ ID NO: 22)
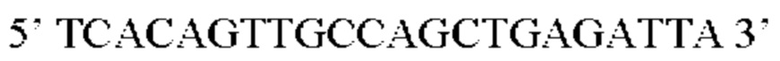
Нуклеотидная последовательность, кодирующая miRT125 (последовательность-мишень микроРНК 125) (SEQ ID NO: 23)
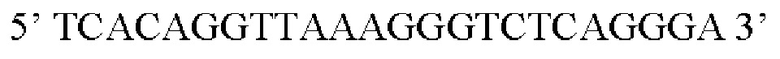
Нуклеотидная последовательность, кодирующая miRT 133 а (последовательность-мишень микроРНК 133а) (SEQ ID NO: 24)
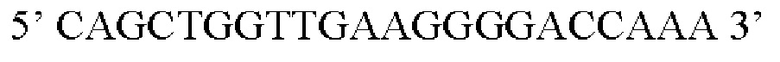
Нуклеотидная последовательность, кодирующая miRT206 (последовательность-мишень микроРНК 206) (SEQ ID NO: 25)
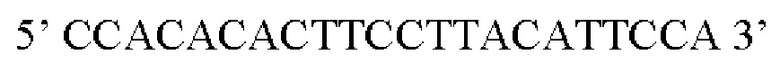
Нуклеотидная последовательность, кодирующая miRT130 (последовательность-мишень микроРНК 130) (SEQ ID NO: 26)
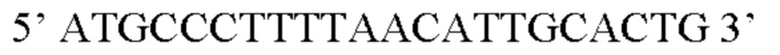
Нуклеотидная последовательность, кодирующая miRT99 (последовательность-мишень микроРНК 99) (SEQ ID NO: 27)
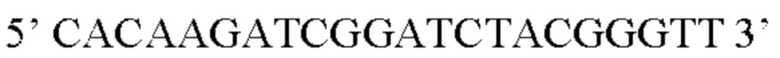
Нуклеотидная последовательность, кодирующая miRT208-5p (последовательность-мишень микроРНК 208а) (SEQ ID NO: 28)
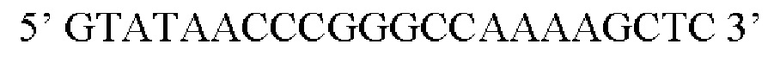
Нуклеотидная последовательность, кодирующая miRT208a-3p (последовательность-мишень микроРНК 208а) (SEQ ID NO: 29)
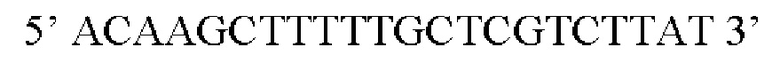
Нуклеотидная последовательность, кодирующая miRT499-5p (последовательность-мишень специфической для сердца микроРНК 499) (SEQ ID NO: 30)
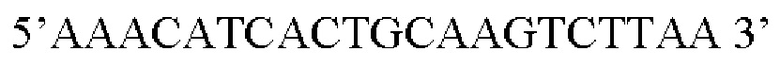
Нуклеотидная последовательность промотора CAG (SEQ ID NO: 44)

Нуклеотидная последовательность промотора CMV (SEQ ID NO: 45)

Нуклеотидная последовательность энхансера CMV (SEQ ID NO: 46)

Нуклеотидная последовательность промотора hAAT (SEQ ID NO: 47)

Усеченный 5'ITR AAV2 (SEQ ID NO: 48)
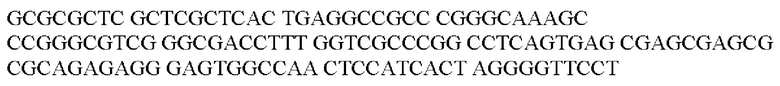
Усеченный 3'ITR AAV2 (SEQ ID NO: 49)
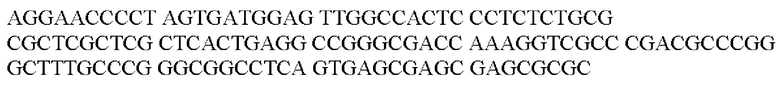
Сигнал полиаденилирования SV40 (SEQ ID NO: 50)
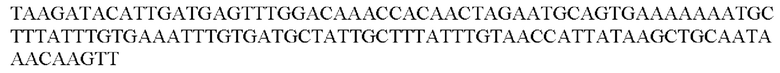
Сигнал полиаденилирования кроличьего β-глобина (SEQ ID NO: 51)

Последовательность промотора CMV и энхансера CMV (SEQ ID NO: 52)
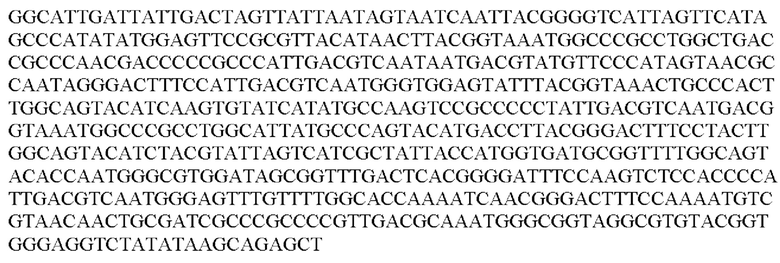
Энхансер гепатоцитарного регуляторного участка (HCR) из аполипопротеина Е (SEQ ID NO: 53)
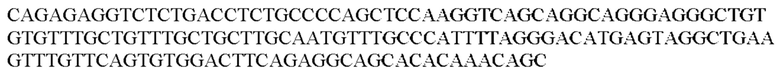
Промотор miniaP2 (SEQ ID NO: 54)

Промотор miniUCP1 (SEQ ID NO: 55)

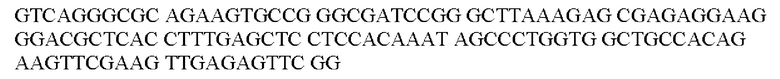
Промотор С5-12 (SEQ ID NO: 56)

pAAV-EF1a-mmFGF21-pA (SEQ ID NO: 57)


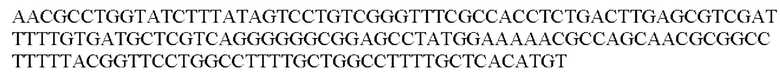
Промотор фактора элонгации 1-альфа: с 150 по 1327 (1178 п. о.) FGF21 Mus museums: с 1359 по 1991 (633 п.о.)
SEQ ID NO: 57 также содержит усеченные 5' и 3' ITR AAV2 и полиА SV40 (уже включенные в перечень последовательностей, SEQ ID NO: 48, 49 и 50)
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Университат Аутонома де Барселона
<120> ВИРУСНАЯ ЭКСПРЕССИОННАЯ КОНСТРУКЦИЯ, СОДЕРЖАЩАЯ КОДИРУЮЩУЮ
ПОСЛЕДОВАТЕЛЬНОСТЬ ФАКТОРА РОСТА ФИБРОБЛАСТОВ 21 (FGF21)
<130> P6066689PCT
<150> EP17172818.1
<151> 24-05-2017
<150> ES201700615
<151> 24-05-2017
<160> 57
<170> PatentIn version 3.5
<210> 1
<211> 209
<212> Белок
<213> Homo sapiens
<400> 1
Met Asp Ser Asp Glu Thr Gly Phe Glu His Ser Gly Leu Trp Val Ser
1 5 10 15
Val Leu Ala Gly Leu Leu Leu Gly Ala Cys Gln Ala His Pro Ile Pro
20 25 30
Asp Ser Ser Pro Leu Leu Gln Phe Gly Gly Gln Val Arg Gln Arg Tyr
35 40 45
Leu Tyr Thr Asp Asp Ala Gln Gln Thr Glu Ala His Leu Glu Ile Arg
50 55 60
Glu Asp Gly Thr Val Gly Gly Ala Ala Asp Gln Ser Pro Glu Ser Leu
65 70 75 80
Leu Gln Leu Lys Ala Leu Lys Pro Gly Val Ile Gln Ile Leu Gly Val
85 90 95
Lys Thr Ser Arg Phe Leu Cys Gln Arg Pro Asp Gly Ala Leu Tyr Gly
100 105 110
Ser Leu His Phe Asp Pro Glu Ala Cys Ser Phe Arg Glu Leu Leu Leu
115 120 125
Glu Asp Gly Tyr Asn Val Tyr Gln Ser Glu Ala His Gly Leu Pro Leu
130 135 140
His Leu Pro Gly Asn Lys Ser Pro His Arg Asp Pro Ala Pro Arg Gly
145 150 155 160
Pro Ala Arg Phe Leu Pro Leu Pro Gly Leu Pro Pro Ala Leu Pro Glu
165 170 175
Pro Pro Gly Ile Leu Ala Pro Gln Pro Pro Asp Val Gly Ser Ser Asp
180 185 190
Pro Leu Ser Met Val Gly Pro Ser Gln Gly Arg Ser Pro Ser Tyr Ala
195 200 205
Ser
<210> 2
<211> 210
<212> Белок
<213> Mus musculus
<400> 2
Met Glu Trp Met Arg Ser Arg Val Gly Thr Leu Gly Leu Trp Val Arg
1 5 10 15
Leu Leu Leu Ala Val Phe Leu Leu Gly Val Tyr Gln Ala Tyr Pro Ile
20 25 30
Pro Asp Ser Ser Pro Leu Leu Gln Phe Gly Gly Gln Val Arg Gln Arg
35 40 45
Tyr Leu Tyr Thr Asp Asp Asp Gln Asp Thr Glu Ala His Leu Glu Ile
50 55 60
Arg Glu Asp Gly Thr Val Val Gly Ala Ala His Arg Ser Pro Glu Ser
65 70 75 80
Leu Leu Glu Leu Lys Ala Leu Lys Pro Gly Val Ile Gln Ile Leu Gly
85 90 95
Val Lys Ala Ser Arg Phe Leu Cys Gln Gln Pro Asp Gly Ala Leu Tyr
100 105 110
Gly Ser Pro His Phe Asp Pro Glu Ala Cys Ser Phe Arg Glu Leu Leu
115 120 125
Leu Glu Asp Gly Tyr Asn Val Tyr Gln Ser Glu Ala His Gly Leu Pro
130 135 140
Leu Arg Leu Pro Gln Lys Asp Ser Pro Asn Gln Asp Ala Thr Ser Trp
145 150 155 160
Gly Pro Val Arg Phe Leu Pro Met Pro Gly Leu Leu His Glu Pro Gln
165 170 175
Asp Gln Ala Gly Phe Leu Pro Pro Glu Pro Pro Asp Val Gly Ser Ser
180 185 190
Asp Pro Leu Ser Met Val Glu Pro Leu Gln Gly Arg Ser Pro Ser Tyr
195 200 205
Ala Ser
210
<210> 3
<211> 209
<212> Белок
<213> Canis lupus familiaris
<400> 3
Met Gly Trp Ala Glu Ala Gly Phe Glu His Leu Gly Leu Trp Val Pro
1 5 10 15
Val Leu Ala Val Leu Leu Leu Glu Ala Cys Arg Ala His Pro Ile Pro
20 25 30
Asp Ser Ser Pro Leu Leu Gln Phe Gly Gly Gln Val Arg Gln Arg Tyr
35 40 45
Leu Tyr Thr Asp Asp Ala Gln Glu Thr Glu Ala His Leu Glu Ile Arg
50 55 60
Ala Asp Gly Thr Val Val Gly Ala Ala Arg Gln Ser Pro Glu Ser Leu
65 70 75 80
Leu Glu Leu Lys Ala Leu Lys Pro Gly Val Ile Gln Ile Leu Gly Val
85 90 95
Lys Thr Ser Arg Phe Leu Cys Gln Gly Pro Asp Gly Thr Leu Tyr Gly
100 105 110
Ser Leu His Phe Asp Pro Val Ala Cys Ser Phe Arg Glu Leu Leu Leu
115 120 125
Glu Asp Gly Tyr Asn Ile Tyr His Ser Glu Thr Leu Gly Leu Pro Leu
130 135 140
Arg Leu Arg Pro His Asn Ser Ala Tyr Arg Asp Leu Ala Pro Arg Gly
145 150 155 160
Pro Ala Arg Phe Leu Pro Leu Pro Gly Leu Leu Pro Ala Pro Pro Glu
165 170 175
Pro Pro Gly Ile Leu Ala Pro Glu Pro Pro Asp Val Gly Ser Ser Asp
180 185 190
Pro Leu Ser Met Val Gly Pro Ser Gln Gly Arg Ser Pro Ser Tyr Ala
195 200 205
Ser
<210> 4
<211> 630
<212> ДНК
<213> Homo sapiens
<400> 4
atggactcgg acgagaccgg gttcgagcac tcaggactgt gggtttctgt gctggctggt 60
cttctgctgg gagcctgcca ggcacacccc atccctgact ccagtcctct cctgcaattc 120
gggggccaag tccggcagcg gtacctctac acagatgatg cccagcagac agaagcccac 180
ctggagatca gggaggatgg gacggtgggg ggcgctgctg accagagccc cgaaagtctc 240
ctgcagctga aagccttgaa gccgggagtt attcaaatct tgggagtcaa gacatccagg 300
ttcctgtgcc agcggccaga tggggccctg tatggatcgc tccactttga ccctgaggcc 360
tgcagcttcc gggagctgct tcttgaggac ggatacaatg tttaccagtc cgaagcccac 420
ggcctcccgc tgcacctgcc agggaacaag tccccacacc gggaccctgc accccgagga 480
ccagctcgct tcctgccact accaggcctg ccccccgcac tcccggagcc acccggaatc 540
ctggcccccc agccccccga tgtgggctcc tcggaccctc tgagcatggt gggaccttcc 600
cagggccgaa gccccagcta cgcttcctga 630
<210> 5
<211> 630
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированный по кодону Homo sapiens FGF21 - вариант 1
<400> 5
atggattctg atgagacagg cttcgagcac agcggcctgt gggtttcagt tctggctgga 60
ctgctgctgg gagcctgtca ggcacaccct attccagata gcagccctct gctgcagttc 120
ggcggacaag tgcggcagag atacctgtac accgacgacg cccagcagac agaagcccac 180
ctggaaatca gagaggatgg cacagttggc ggagccgccg atcagtctcc tgaatctctg 240
ctccagctga aggccctgaa gcctggcgtg atccagatcc tgggcgtgaa aaccagccgg 300
ttcctgtgcc aaagacctga cggcgccctg tatggcagcc tgcactttga tcctgaggcc 360
tgcagcttca gagagctgct gcttgaggac ggctacaacg tgtaccagtc tgaggcccat 420
ggcctgcctc tgcatctgcc tggaaacaag agccctcaca gagatcccgc tcctagaggc 480
cctgccagat ttctgcctct tcctggattg cctcctgctc tgccagagcc tcctggaatt 540
ctggctcctc agcctcctga tgtgggcagc tctgatcctc tgagcatggt cggacctagc 600
cagggcagat ctcctagcta cgcctcttga 630
<210> 6
<211> 630
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированный по кодону Homo sapiens FGF21 - вариант 2
<400> 6
atggacagcg atgaaaccgg gttcgagcac agcggtctgt gggtgtccgt gctggccgga 60
ctgctcctgg gagcctgtca ggcgcacccc atccctgact cctcgccgct gctgcaattc 120
ggcggacaag tccgccagag atacctgtac accgacgacg cccagcagac cgaagcccac 180
ctggaaattc gggaggacgg gactgtggga ggcgctgcag atcagtcacc cgagtccctc 240
ctccaactga aggccttgaa gcccggcgtg attcagatcc tgggcgtgaa aacttcccgc 300
ttcctttgcc aacggccgga tggagctctg tacggatccc tgcacttcga ccccgaagcc 360
tgctcattcc gcgagctgct ccttgaggac ggctataacg tgtaccagtc tgaggcccat 420
ggactccccc tgcatctgcc cggcaacaag tcccctcacc gggatcctgc cccaagaggc 480
ccagctcggt ttctgcctct gccgggactg cctccagcgt tgcccgaacc ccctggtatc 540
ctggccccgc aaccacctga cgtcggttcg tcggacccgc tgagcatggt cggtccgagc 600
cagggaaggt ccccgtccta cgcatcctga 630
<210> 7
<211> 630
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированный по кодону Homo sapiens FGF21 - вариант 3
<400> 7
atggattccg acgaaactgg atttgaacat tcagggctgt gggtctctgt gctggctgga 60
ctgctgctgg gggcttgtca ggctcacccc atccctgaca gctcccctct gctgcagttc 120
ggaggacagg tgcggcagag atacctgtat accgacgatg cccagcagac agaggcacac 180
ctggagatca gggaggacgg aaccgtggga ggagcagccg atcagtctcc cgagagcctg 240
ctgcagctga aggccctgaa gcctggcgtg atccagatcc tgggcgtgaa gacatctcgg 300
tttctgtgcc agcggcccga cggcgccctg tacggctccc tgcacttcga tcccgaggcc 360
tgttctttta gggagctgct gctggaggac ggctacaacg tgtatcagag cgaggcacac 420
ggcctgccac tgcacctgcc tggcaataag tcccctcacc gcgatccagc acccaggggc 480
ccagcacgct tcctgcctct gccaggcctg ccccctgccc tgccagagcc acccggcatc 540
ctggcccccc agcctccaga tgtgggctcc agcgatcctc tgtcaatggt ggggccaagt 600
caggggcgga gtccttcata cgcatcataa 630
<210> 8
<211> 633
<212> ДНК
<213> Mus musculus
<400> 8
atggaatgga tgagatctag agttgggacc ctgggactgt gggtccgact gctgctggct 60
gtcttcctgc tgggggtcta ccaagcatac cccatccctg actccagccc cctcctccag 120
tttgggggtc aagtccggca gaggtacctc tacacagatg acgaccaaga cactgaagcc 180
cacctggaga tcagggagga tggaacagtg gtaggcgcag cacaccgcag tccagaaagt 240
ctcctggagc tcaaagcctt gaagccaggg gtcattcaaa tcctgggtgt caaagcctct 300
aggtttcttt gccaacagcc agatggagct ctctatggat cgcctcactt tgatcctgag 360
gcctgcagct tcagagaact gctgctggag gacggttaca atgtgtacca gtctgaagcc 420
catggcctgc ccctgcgtct gcctcagaag gactccccaa accaggatgc aacatcctgg 480
ggacctgtgc gcttcctgcc catgccaggc ctgctccacg agccccaaga ccaagcagga 540
ttcctgcccc cagagccccc agatgtgggc tcctctgacc ccctgagcat ggtagagcct 600
ttacagggcc gaagccccag ctatgcgtcc tga 633
<210> 9
<211> 633
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированный по кодону Mus musculus FGF21
<400> 9
atggaatgga tgagaagcag agtgggcacc ctgggcctgt gggtgcgact gctgctggct 60
gtgtttctgc tgggcgtgta ccaggcctac cccatccctg actctagccc cctgctgcag 120
tttggcggac aagtgcggca gagatacctg tacaccgacg acgaccagga caccgaggcc 180
cacctggaaa tccgcgagga tggcacagtc gtgggcgctg ctcacagaag ccctgagagc 240
ctgctggaac tgaaggccct gaagcccggc gtgatccaga tcctgggcgt gaaggccagc 300
agattcctgt gccagcagcc tgacggcgcc ctgtacggct ctcctcactt cgatcctgag 360
gcctgcagct tcagagagct gctgctggag gacggctaca acgtgtacca gtctgaggcc 420
cacggcctgc ccctgagact gcctcagaag gacagcccta accaggacgc cacaagctgg 480
ggacctgtgc ggttcctgcc tatgcctgga ctgctgcacg agccccagga tcaggctggc 540
tttctgcctc ctgagcctcc agacgtgggc agcagcgacc ctctgagcat ggtggaacct 600
ctgcagggca gaagccccag ctacgcctct tga 633
<210> 10
<211> 629
<212> ДНК
<213> Canis lupus familiaris
<400> 10
atgggctggg ccgaggccgg gttcgagcac ctgggactgt gggtccctgt gctggctgtg 60
cttttgctgg aagcctgccg ggcacatccg atccctgact ccagccccct cctacaattt 120
ggaggtcaag ttcgacagcg gtacctctac accgacgatg cccaggagac agaggcccac 180
ctagagatca gggccgatgg cacagggtgg gggctgcccg ccagagccct gaaagtctcc 240
tggagctgaa agccctaaag ccaggggtca ttcaaatctt gggagtcaaa acatccaggt 300
tcctgtgcca gggcccagat gggacactat atggctcgct ccatttcgac cctgtggcct 360
gcagtttccg agaactgctt cttgaggatg ggtacaacat ctaccactcc gagacccttg 420
gtctcccgct tcgcctgcgc ccccacaact ccgcataccg ggacttggca ccccgcgggc 480
ctgcccgctt cctgccactg ccaggcctgc ttccagcacc cccagagcct ccagggatcc 540
tggccccgga gcctcctgac gtgggctcct cggaccctct gagcatggtg gggccttcac 600
agggccggag tcccagctat gcttcctaa 629
<210> 11
<211> 630
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированный по кодону Canis lupus familiaris FGF21
<400> 11
atgggatggg ctgaggctgg attcgaacac ctgggactct gggtgcccgt cctggccgtg 60
ctgctcctgg aggcttgcag ggctcatccc atccctgaca gctccccact cctgcagttt 120
ggaggacagg tgaggcagcg gtacctgtat accgacgatg cccaggagac agaagctcac 180
ctggaaattc gggctgatgg aacagtggtc ggagctgccc gacagtcccc agagtctctc 240
ctggaactga aggccctcaa acccggagtg atccagattc tgggcgtcaa gacttctaga 300
ttcctgtgcc agggaccaga cggcaccctg tacggcagcc tgcatttcga tcctgtggcc 360
tgttcctttc gagagctcct gctcgaagac ggctacaaca tctatcactc tgagaccctg 420
ggactcccac tgcgactcag acctcataat agtgcctatc gagatctggc tcccaggggc 480
ccagctaggt ttctgccact ccccggactg ctccctgctc cacctgagcc acccggcatt 540
ctggctccag aacctccaga cgtgggctct agtgatccac tgagtatggt cggcccctca 600
caggggaggt cacctagcta cgccagctga 630
<210> 12
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT122a
<400> 12
caaacaccat tgtcacactc ca 22
<210> 13
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT1
<400> 13
ttacatactt ctttacattc ca 22
<210> 14
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT152
<400> 14
ccaagttctg tcatgcactg a 21
<210> 15
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT199a-5p
<400> 15
gaacaggtag tctgaacact ggg 23
<210> 16
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT199a-3p
<400> 16
taaccaatgt gcagactact gt 22
<210> 17
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT215
<400> 17
gtctgtcaat tcataggtca t 21
<210> 18
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT192
<400> 18
ggctgtcaat tcataggtca g 21
<210> 19
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT148a
<400> 19
acaaagttct gtagtgcact ga 22
<210> 20
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT194
<400> 20
tccacatgga gttgctgtta ca 22
<210> 21
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT124
<400> 21
ggcattcacc gcgtgcctta 20
<210> 22
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT216
<400> 22
tcacagttgc cagctgagat ta 22
<210> 23
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT125
<400> 23
tcacaggtta aagggtctca ggga 24
<210> 24
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT133a
<400> 24
cagctggttg aaggggacca aa 22
<210> 25
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT206
<400> 25
ccacacactt ccttacattc ca 22
<210> 26
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT130
<400> 26
atgccctttt aacattgcac tg 22
<210> 27
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT99
<400> 27
cacaagatcg gatctacggg tt 22
<210> 28
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT208a-5p
<400> 28
gtataacccg ggccaaaagc tc 22
<210> 29
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT208a-3p
<400> 29
acaagctttt tgctcgtctt at 22
<210> 30
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая miRT499-5p
<400> 30
aaacatcact gcaagtctta a 21
<210> 31
<211> 6343
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pAAV-CAG-null
<400> 31
aattcgagct cggtacccgg gaatcaattc actcctcagg tgcaggctgc ctatcagaag 60
gtggtggctg gtgtggccaa tgccctggct cacaaatacc actgagatct ttttccctct 120
gccaaaaatt atggggacat catgaagccc cttgagcatc tgacttctgg ctaataaagg 180
aaatttattt tcattgcaat agtgtgttgg aattttttgt gtctctcact cggaaggaca 240
tatgggaggg caaatcattt aaaacatcag aatgagtatt tggtttagag tttggcaaca 300
tatgcccata tgctggctgc catgaacaaa ggttggctat aaagaggtca tcagtatatg 360
aaacagcccc ctgctgtcca ttccttattc catagaaaag ccttgacttg aggttagatt 420
ttttttatat tttgttttgt gttatttttt tctttaacat ccctaaaatt ttccttacat 480
gttttactag ccagattttt cctcctctcc tgactactcc cagtcatagc tgtccctctt 540
ctcttatgga gatccctcga cctgcagccc aagctgtaga taagtagcat ggcgggttaa 600
tcattaacta caaggaaccc ctagtgatgg agttggccac tccctctctg cgcgctcgct 660
cgctcactga ggccgggcga ccaaaggtcg cccgacgccc gggctttgcc cgggcggcct 720
cagtgagcga gcgagcgcgc agctgcatta atgaatcggc caacgcgcgg ggagaggcgg 780
tttgcgtatt gggcgctctt ccgcttcctc gctcactgac tcgctgcgct cggtcgttcg 840
gctgcggcga gcggtatcag ctcactcaaa ggcggtaata cggttatcca cagaatcagg 900
ggataacgca ggaaagaaca tgtgagcaaa aggccagcaa aaggccagga accgtaaaaa 960
ggccgcgttg ctggcgtttt tccataggct ccgcccccct gacgagcatc acaaaaatcg 1020
acgctcaagt cagaggtggc gaaacccgac aggactataa agataccagg cgtttccccc 1080
tggaagctcc ctcgtgcgct ctcctgttcc gaccctgccg cttaccggat acctgtccgc 1140
ctttctccct tcgggaagcg tggcgctttc tcatagctca cgctgtaggt atctcagttc 1200
ggtgtaggtc gttcgctcca agctgggctg tgtgcacgaa ccccccgttc agcccgaccg 1260
ctgcgcctta tccggtaact atcgtcttga gtccaacccg gtaagacacg acttatcgcc 1320
actggcagca gccactggta acaggattag cagagcgagg tatgtaggcg gtgctacaga 1380
gttcttgaag tggtggccta actacggcta cactagaaga acagtatttg gtatctgcgc 1440
tctgctgaag ccagttacct tcggaaaaag agttggtagc tcttgatccg gcaaacaaac 1500
caccgctggt agcggtggtt tttttgtttg caagcagcag attacgcgca gaaaaaaagg 1560
atctcaagaa gatcctttga tcttttctac ggggtctgac gctcagtgga acgaaaactc 1620
acgttaaggg attttggtca tgagattatc aaaaaggatc ttcacctaga tccttttaaa 1680
ttaaaaatga agttttaaat caatctaaag tatatatgag taaacttggt ctgacagtta 1740
ccaatgctta atcagtgagg cacctatctc agcgatctgt ctatttcgtt catccatagt 1800
tgcctgactc cccgtcgtgt agataactac gatacgggag ggcttaccat ctggccccag 1860
tgctgcaatg ataccgcgag acccacgctc accggctcca gatttatcag caataaacca 1920
gccagccgga agggccgagc gcagaagtgg tcctgcaact ttatccgcct ccatccagtc 1980
tattaattgt tgccgggaag ctagagtaag tagttcgcca gttaatagtt tgcgcaacgt 2040
tgttgccatt gctacaggca tcgtggtgtc acgctcgtcg tttggtatgg cttcattcag 2100
ctccggttcc caacgatcaa ggcgagttac atgatccccc atgttgtgca aaaaagcggt 2160
tagctccttc ggtcctccga tcgttgtcag aagtaagttg gccgcagtgt tatcactcat 2220
ggttatggca gcactgcata attctcttac tgtcatgcca tccgtaagat gcttttctgt 2280
gactggtgag tactcaacca agtcattctg agaatagtgt atgcggcgac cgagttgctc 2340
ttgcccggcg tcaatacggg ataataccgc gccacatagc agaactttaa aagtgctcat 2400
cattggaaaa cgttcttcgg ggcgaaaact ctcaaggatc ttaccgctgt tgagatccag 2460
ttcgatgtaa cccactcgtg cacccaactg atcttcagca tcttttactt tcaccagcgt 2520
ttctgggtga gcaaaaacag gaaggcaaaa tgccgcaaaa aagggaataa gggcgacacg 2580
gaaatgttga atactcatac tcttcctttt tcaatattat tgaagcattt atcagggtta 2640
ttgtctcatg agcggataca tatttgaatg tatttagaaa aataaacaaa taggggttcc 2700
gcgcacattt ccccgaaaag tgccacctga cgtctaagaa accattatta tcatgacatt 2760
aacctataaa aataggcgta tcacgaggcc ctttcgtctc gcgcgtttcg gtgatgacgg 2820
tgaaaacctc tgacacatgc agctcccgga gacggtcaca gcttgtctgt aagcggatgc 2880
cgggagcaga caagcccgtc agggcgcgtc agcgggtgtt ggcgggtgtc ggggctggct 2940
taactatgcg gcatcagagc agattgtact gagagtgcac catatgcggt gtgaaatacc 3000
gcacagatgc gtaaggagaa aataccgcat caggcgattc caacatccaa taaatcatac 3060
aggcaaggca aagaattagc aaaattaagc aataaagcct cagagcataa agctaaatcg 3120
gttgtaccaa aaacattatg accctgtaat acttttgcgg gagaagcctt tatttcaacg 3180
caaggataaa aatttttaga accctcatat attttaaatg caatgcctga gtaatgtgta 3240
ggtaaagatt caaacgggtg agaaaggccg gagacagtca aatcaccatc aatatgatat 3300
tcaaccgttc tagctgataa attcatgccg gagagggtag ctatttttga gaggtctcta 3360
caaaggctat caggtcattg cctgagagtc tggagcaaac aagagaatcg atgaacggta 3420
atcgtaaaac tagcatgtca atcatatgta ccccggttga taatcagaaa agccccaaaa 3480
acaggaagat tgtataagca aatatttaaa ttgtaagcgt taatattttg ttaaaattcg 3540
cgttaaattt ttgttaaatc agctcatttt ttaaccaata ggccgaaatc ggcaaaatcc 3600
cttataaatc aaaagaatag accgagatag ggttgagtgt tgttccagtt tggaacaaga 3660
gtccactatt aaagaacgtg gactccaacg tcaaagggcg aaaaaccgtc tatcagggcg 3720
atggcccact acgtgaacca tcaccctaat caagtttttt ggggtcgagg tgccgtaaag 3780
cactaaatcg gaaccctaaa gggagccccc gatttagagc ttgacgggga aagccggcga 3840
acgtggcgag aaaggaaggg aagaaagcga aaggagcggg cgctagggcg ctggcaagtg 3900
tagcggtcac gctgcgcgta accaccacac ccgccgcgct taatgcgccg ctacagggcg 3960
cgtactatgg ttgctttgac gagcacgtat aacgtgcttt cctcgttaga atcagagcgg 4020
gagctaaaca ggaggccgat taaagggatt ttagacagga acggtacgcc agaatcctga 4080
gaagtgtttt tataatcagt gaggccaccg agtaaaagag tctgtccatc acgcaaatta 4140
accgttgtcg caatacttct ttgattagta ataacatcac ttgcctgagt agaagaactc 4200
aaactatcgg ccttgctggt aatatccaga acaatattac cgccagccat tgcaacggaa 4260
tcgccattcg ccattcaggc tgcgcaactg ttgggaaggg cgatcggtgc gggcctcttc 4320
gctattacgc cagctgcgcg ctcgctcgct cactgaggcc gcccgggcaa agcccgggcg 4380
tcgggcgacc tttggtcgcc cggcctcagt gagcgagcga gcgcgcagag agggagtggc 4440
caactccatc actaggggtt ccttgtagtt aatgattaac ccgccatgct acttatctac 4500
tcgacattga ttattgacta gttattaata gtaatcaatt acggggtcat tagttcatag 4560
cccatatatg gagttccgcg ttacataact tacggtaaat ggcccgcctg gctgaccgcc 4620
caacgacccc cgcccattga cgtcaataat gacgtatgtt cccatagtaa cgccaatagg 4680
gactttccat tgacgtcaat gggtggagta tttacggtaa actgcccact tggcagtaca 4740
tcaagtgtat catatgccaa gtacgccccc tattgacgtc aatgacggta aatggcccgc 4800
ctggcattat gcccagtaca tgaccttatg ggactttcct acttggcagt acatctacgt 4860
attagtcatc gctattacca tggtcgaggt gagccccacg ttctgcttca ctctccccat 4920
ctcccccccc tccccacccc caattttgta tttatttatt ttttaattat tttgtgcagc 4980
gatgggggcg gggggggggg gggggcgcgc gccaggcggg gcggggcggg gcgaggggcg 5040
gggcggggcg aggcggagag gtgcggcggc agccaatcag agcggcgcgc tccgaaagtt 5100
tccttttatg gcgaggcggc ggcggcggcg gccctataaa aagcgaagcg cgcggcgggc 5160
gggagtcgct gcgttgcctt cgccccgtgc cccgctccgc cgccgcctcg cgccgcccgc 5220
cccggctctg actgaccgcg ttactcccac aggtgagcgg gcgggacggc ccttctcctc 5280
cgggctgtaa ttagcgcttg gtttaatgac ggcttgtttc ttttctgtgg ctgcgtgaaa 5340
gccttgaggg gctccgggag ggccctttgt gcggggggag cggctcgggg ggtgcgtgcg 5400
tgtgtgtgtg cgtggggagc gccgcgtgcg gctccgcgct gcccggcggc tgtgagcgct 5460
gcgggcgcgg cgcggggctt tgtgcgctcc gcagtgtgcg cgaggggagc gcggccgggg 5520
gcggtgcccc gcggtgcggg gggggctgcg aggggaacaa aggctgcgtg cggggtgtgt 5580
gcgtgggggg gtgagcaggg ggtgtgggcg cgtcggtcgg gctgcaaccc cccctgcacc 5640
cccctccccg agttgctgag cacggcccgg cttcgggtgc ggggctccgt acggggcgtg 5700
gcgcggggct cgccgtgccg ggcggggggt ggcggcaggt gggggtgccg ggcggggcgg 5760
ggccgcctcg ggccggggag ggctcggggg aggggcgcgg cggcccccgg agcgccggcg 5820
gctgtcgagg cgcggcgagc cgcagccatt gccttttatg gtaatcgtgc gagagggcgc 5880
agggacttcc tttgtcccaa atctgtgcgg agccgaaatc tgggaggcgc cgccgcaccc 5940
cctctagcgg gcgcggggcg aagcggtgcg gcgccggcag gaaggaaatg ggcggggagg 6000
gccttcgtgc gtcgccgcgc cgccgtcccc ttctccctct ccagcctcgg ggctgtccgc 6060
ggggggacgg ctgccttcgg gggggacggg gcagggcggg gttcggcttc tggcgtgtga 6120
ccggcggctc tagagcctct gctaaccatg ttcatgcctt cttctttttc ctacagctcc 6180
tgggcaacgt gctggttatt gtgctgtctc atcattttgg caaagaattg attaattcga 6240
gcgaacgcgt cgagtcgctc ggtacgattt aaattgaatt ggcctcgagc gcaagcttga 6300
gctagctcga tatcggccta ggctggatcc gcgcggccgc aag 6343
<210> 32
<211> 7319
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pAAV-CAG-moFGF21-dmiRT
<400> 32
agtgagcgag cgagcgcgca gctgcattaa tgaatcggcc aacgcgcggg gagaggcggt 60
ttgcgtattg ggcgctcttc cgcttcctcg ctcactgact cgctgcgctc ggtcgttcgg 120
ctgcggcgag cggtatcagc tcactcaaag gcggtaatac ggttatccac agaatcaggg 180
gataacgcag gaaagaacat gtgagcaaaa ggccagcaaa aggccaggaa ccgtaaaaag 240
gccgcgttgc tggcgttttt ccataggctc cgcccccctg acgagcatca caaaaatcga 300
cgctcaagtc agaggtggcg aaacccgaca ggactataaa gataccaggc gtttccccct 360
ggaagctccc tcgtgcgctc tcctgttccg accctgccgc ttaccggata cctgtccgcc 420
tttctccctt cgggaagcgt ggcgctttct catagctcac gctgtaggta tctcagttcg 480
gtgtaggtcg ttcgctccaa gctgggctgt gtgcacgaac cccccgttca gcccgaccgc 540
tgcgccttat ccggtaacta tcgtcttgag tccaacccgg taagacacga cttatcgcca 600
ctggcagcag ccactggtaa caggattagc agagcgaggt atgtaggcgg tgctacagag 660
ttcttgaagt ggtggcctaa ctacggctac actagaagaa cagtatttgg tatctgcgct 720
ctgctgaagc cagttacctt cggaaaaaga gttggtagct cttgatccgg caaacaaacc 780
accgctggta gcggtggttt ttttgtttgc aagcagcaga ttacgcgcag aaaaaaagga 840
tctcaagaag atcctttgat cttttctacg gggtctgacg ctcagtggaa cgaaaactca 900
cgttaaggga ttttggtcat gagattatca aaaaggatct tcacctagat ccttttaaat 960
taaaaatgaa gttttaaatc aatctaaagt atatatgagt aaacttggtc tgacagttac 1020
caatgcttaa tcagtgaggc acctatctca gcgatctgtc tatttcgttc atccatagtt 1080
gcctgactcc ccgtcgtgta gataactacg atacgggagg gcttaccatc tggccccagt 1140
gctgcaatga taccgcgaga cccacgctca ccggctccag atttatcagc aataaaccag 1200
ccagccggaa gggccgagcg cagaagtggt cctgcaactt tatccgcctc catccagtct 1260
attaattgtt gccgggaagc tagagtaagt agttcgccag ttaatagttt gcgcaacgtt 1320
gttgccattg ctacaggcat cgtggtgtca cgctcgtcgt ttggtatggc ttcattcagc 1380
tccggttccc aacgatcaag gcgagttaca tgatccccca tgttgtgcaa aaaagcggtt 1440
agctccttcg gtcctccgat cgttgtcaga agtaagttgg ccgcagtgtt atcactcatg 1500
gttatggcag cactgcataa ttctcttact gtcatgccat ccgtaagatg cttttctgtg 1560
actggtgagt actcaaccaa gtcattctga gaatagtgta tgcggcgacc gagttgctct 1620
tgcccggcgt caatacggga taataccgcg ccacatagca gaactttaaa agtgctcatc 1680
attggaaaac gttcttcggg gcgaaaactc tcaaggatct taccgctgtt gagatccagt 1740
tcgatgtaac ccactcgtgc acccaactga tcttcagcat cttttacttt caccagcgtt 1800
tctgggtgag caaaaacagg aaggcaaaat gccgcaaaaa agggaataag ggcgacacgg 1860
aaatgttgaa tactcatact cttccttttt caatattatt gaagcattta tcagggttat 1920
tgtctcatga gcggatacat atttgaatgt atttagaaaa ataaacaaat aggggttccg 1980
cgcacatttc cccgaaaagt gccacctgac gtctaagaaa ccattattat catgacatta 2040
acctataaaa ataggcgtat cacgaggccc tttcgtctcg cgcgtttcgg tgatgacggt 2100
gaaaacctct gacacatgca gctcccggag acggtcacag cttgtctgta agcggatgcc 2160
gggagcagac aagcccgtca gggcgcgtca gcgggtgttg gcgggtgtcg gggctggctt 2220
aactatgcgg catcagagca gattgtactg agagtgcacc atatgcggtg tgaaataccg 2280
cacagatgcg taaggagaaa ataccgcatc aggcgattcc aacatccaat aaatcataca 2340
ggcaaggcaa agaattagca aaattaagca ataaagcctc agagcataaa gctaaatcgg 2400
ttgtaccaaa aacattatga ccctgtaata cttttgcggg agaagccttt atttcaacgc 2460
aaggataaaa atttttagaa ccctcatata ttttaaatgc aatgcctgag taatgtgtag 2520
gtaaagattc aaacgggtga gaaaggccgg agacagtcaa atcaccatca atatgatatt 2580
caaccgttct agctgataaa ttcatgccgg agagggtagc tatttttgag aggtctctac 2640
aaaggctatc aggtcattgc ctgagagtct ggagcaaaca agagaatcga tgaacggtaa 2700
tcgtaaaact agcatgtcaa tcatatgtac cccggttgat aatcagaaaa gccccaaaaa 2760
caggaagatt gtataagcaa atatttaaat tgtaagcgtt aatattttgt taaaattcgc 2820
gttaaatttt tgttaaatca gctcattttt taaccaatag gccgaaatcg gcaaaatccc 2880
ttataaatca aaagaataga ccgagatagg gttgagtgtt gttccagttt ggaacaagag 2940
tccactatta aagaacgtgg actccaacgt caaagggcga aaaaccgtct atcagggcga 3000
tggcccacta cgtgaaccat caccctaatc aagttttttg gggtcgaggt gccgtaaagc 3060
actaaatcgg aaccctaaag ggagcccccg atttagagct tgacggggaa agccggcgaa 3120
cgtggcgaga aaggaaggga agaaagcgaa aggagcgggc gctagggcgc tggcaagtgt 3180
agcggtcacg ctgcgcgtaa ccaccacacc cgccgcgctt aatgcgccgc tacagggcgc 3240
gtactatggt tgctttgacg agcacgtata acgtgctttc ctcgttagaa tcagagcggg 3300
agctaaacag gaggccgatt aaagggattt tagacaggaa cggtacgcca gaatcctgag 3360
aagtgttttt ataatcagtg aggccaccga gtaaaagagt ctgtccatca cgcaaattaa 3420
ccgttgtcgc aatacttctt tgattagtaa taacatcact tgcctgagta gaagaactca 3480
aactatcggc cttgctggta atatccagaa caatattacc gccagccatt gcaacggaat 3540
cgccattcgc cattcaggct gcgcaactgt tgggaagggc gatcggtgcg ggcctcttcc 3600
actgaggccc agctgcgcgc tcgctcgctc actgaggccg cccgggcaaa gcccgggcgt 3660
cgggcgacct ttggtcgccc ggcctcagtg agcgagcgag cgcgcagaga gggagtggcc 3720
aactccatca ctaggggttc cttgtagtta atgattaacc cgccatgcta cttatctact 3780
cgacattgat tattgactag ttattaatag taatcaatta cggggtcatt agttcatagc 3840
ccatatatgg agttccgcgt tacataactt acggtaaatg gcccgcctgg ctgaccgccc 3900
aacgaccccc gcccattgac gtcaataatg acgtatgttc ccatagtaac gccaataggg 3960
actttccatt gacgtcaatg ggtggagtat ttacggtaaa ctgcccactt ggcagtacat 4020
caagtgtatc atatgccaag tacgccccct attgacgtca atgacggtaa atggcccgcc 4080
tggcattatg cccagtacat gaccttatgg gactttccta cttggcagta catctacgta 4140
ttagtcatcg ctattaccat ggtcgaggtg agccccacgt tctgcttcac tctccccatc 4200
tcccccccct ccccaccccc aattttgtat ttatttattt tttaattatt ttgtgcagcg 4260
atgggggcgg gggggggggg ggggcgcgcg ccaggcgggg cggggcgggg cgaggggcgg 4320
ggcggggcga ggcggagagg tgcggcggca gccaatcaga gcggcgcgct ccgaaagttt 4380
ccttttatgg cgaggcggcg gcggcggcgg ccctataaaa agcgaagcgc gcggcgggcg 4440
ggagtcgctg cgttgccttc gccccgtgcc ccgctccgcg ccgcctcgcg ccgcccgccc 4500
cggctctgac tgaccgcgtt actcccacag gtgagcgggc gggacggccc ttctcctccg 4560
ggctgtaatt agcgcttggt ttaatgacgg cttgtttctt ttctgtggct gcgtgaaagc 4620
cttgaggggc tccgggaggg ccctttgtgc ggggggagcg gctcgggggg tgcgtgcgtg 4680
tgtgtgtgcg tggggagcgc cgcgtgcggc tccgcgctgc ccggcggctg tgagcgctgc 4740
gggcgcggcg cggggctttg tgcgctccgc agtgtgcgcg aggggagcgc ggccgggggc 4800
ggtgccccgc ggtgcggggg gctgcgaggg gaacaaaggc tgcgtgcggg gtgtgtgcgt 4860
gggggggtga gcagggggtg tgggcgcgtc ggtcgggctg caaccccccc tgcacccccc 4920
tccccgagtt gctgagcacg gcccggcttc gggtgcgggg ctccgtacgg ggcgtggcgc 4980
ggggctcgcc gtgccgggcg gggggtggcg gcaggtgggg gtgccgggcg gggcggggcc 5040
gcctcgggcc ggggagggct cgggggaggg gcgcggcggc ccccggagcg ccggcggctg 5100
tcgaggcgcg gcgagccgca gccattgcct tttatggtaa tcgtgcgaga gggcgcaggg 5160
acttcctttg tcccaaatct gtgcggagcc gaaatctggg aggcgccgcc gcaccccctc 5220
tagcgggcgc ggggcgaagc ggtgcggcgc cggcaggaag gaaatgggcg gggagggcct 5280
tcgtgcgtcg ccgcgccgcc gtccccttct ccctctccag cctcggggct gtccgcgggg 5340
ggacggctgc cttcgggggg gacggggcag ggcggggttc ggcttctggc gtgtgaccgg 5400
cggctctaga gcctctgcta accatgttca tgccttcttc tttttcctac agctcctggg 5460
caacgtgctg gttattgtgc tgtctcatca ttttggcaaa gaattgatta attcgagcga 5520
acgcgtcgag tcgctcggta cgatttaaat tgaattggcc tcgagcgcaa gcttgagcta 5580
gcgccaccat ggaatggatg agaagcagag tgggcaccct gggcctgtgg gtgcgactgc 5640
tgctggctgt gtttctgctg ggcgtgtacc aggcctaccc catccctgac tctagccccc 5700
tgctgcagtt tggcggacaa gtgcggcaga gatacctgta caccgacgac gaccaggaca 5760
ccgaggccca cctggaaatc cgcgaggatg gcacagtcgt gggcgctgct cacagaagcc 5820
ctgagagcct gctggaactg aaggccctga agcccggcgt gatccagatc ctgggcgtga 5880
aggccagcag attcctgtgc cagcagcctg acggcgccct gtacggctct cctcacttcg 5940
atcctgaggc ctgcagcttc agagagctgc tgctggagga cggctacaac gtgtaccagt 6000
ctgaggccca cggcctgccc ctgagactgc ctcagaagga cagccctaac caggacgcca 6060
caagctgggg acctgtgcgg ttcctgccta tgcctggact gctgcacgag ccccaggatc 6120
aggctggctt tctgcctcct gagcctccag acgtgggcag cagcgaccct ctgagcatgg 6180
tggaacctct gcagggcaga agccccagct acgcctcttg agaatgcggg cccggtaccc 6240
ccgacgcggc cgctaattct agatcgcgaa caaacaccat tgtcacactc cagtatacac 6300
aaacaccatt gtcacactcc agatatcaca aacaccattg tcacactcca aggcgaacaa 6360
acaccattgt cacactccaa ggctattcta gatcgcgaat tacatacttc tttacattcc 6420
agtatacatt acatacttct ttacattcca gatatcatta catacttctt tacattccaa 6480
ggcgaattac atacttcttt acattccaag gctacctgag gcccgggggt acctcttaat 6540
taactggcct catgggcctt ccgctcactg cccgctttcc agtcgggaaa cctgtcgtgc 6600
cagtcaggtg caggctgcct atcagaaggt ggtggctggt gtggccaatg ccctggctca 6660
caaataccac tgagatcttt ttccctctgc caaaaattat ggggacatca tgaagcccct 6720
tgagcatctg acttctggct aataaaggaa atttattttc attgcaatag tgtgttggaa 6780
ttttttgtgt ctctcactcg gaaggacata tgggagggca aatcatttaa aacatcagaa 6840
tgagtatttg gtttagagtt tggcaacata tgcccatatg ctggctgcca tgaacaaagg 6900
ttggctataa agaggtcatc agtatatgaa acagccccct gctgtccatt ccttattcca 6960
tagaaaagcc ttgacttgag gttagatttt ttttatattt tgttttgtgt tatttttttc 7020
tttaacatcc ctaaaatttt ccttacatgt tttactagcc agatttttcc tcctctcctg 7080
actactccca gtcatagctg tccctcttct cttatggaga tccctcgacc tgcagcccaa 7140
gctgtagata agtagcatgg cgggttaatc attaactaca aggaacccct agtgatggag 7200
ttggccactc cctctctgcg cgctcgctcg ctcactgagg ccgggcgacc aaaggtcgcc 7260
cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc gagcgcgcag ctggcgtaa 7319
<210> 33
<211> 5678
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pGG2-hAAT-null
<400> 33
cagcagctgc gcgctcgctc gctcactgag gccgcccggg caaagcccgg gcgtcgggcg 60
acctttggtc gcccggcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc 120
atcactaggg gttccttgta gttaatgatt aacccgccat gctacttatc tacgtagcca 180
tgctctagac atggctcgac agatctgata tcatcgatga attcgagctc ggtacccggc 240
cgcagattta ggtgacacta tagaatatgc atcactagta agcttgcgaa ttccagtcta 300
cagagaggtc tctgacctct gccccagctc caaggtcagc aggcagggag ggctgtgtgt 360
ttgctgtttg ctgcttgcaa tgtttgccca ttttagggac atgagtaggc tgaagtttgt 420
tcagtgtgga cttcagaggc agcacacaaa cagcaagctt gcgaattcca gtctacagag 480
aggtctctga cctctgcccc agctccaagg tcagcaggca gggagggctg tgtgtttgct 540
gtttgctgct tgcaatgttt gcccatttta gggacatgag taggctgaag tttgttcagt 600
gtggacttca gaggcagcac acaaacagca agcttgcgaa ttccagtcta cagagaggtc 660
tctgacctct gccccagctc caaggtcagc aggcagggag ggctgtgtgt ttgctgtttg 720
ctgcttgcaa tgtttgccca ttttagggac atgagtaggc tgaagtttgt tcagtgtgga 780
cttcagaggc agcacacaaa cagcaagctt tgctctagac tggaattcgt cgacgagctc 840
cctatagtga gtcgtattag aggccgactg acccggtacc cggggatctt gctaccagtg 900
gaacagccac taaggattct gcagtgagag cagagggcca gctaagtggt actctcccag 960
agactgtctg actcacgcca ccccctccac cttggacaca ggacgctgtg gtttctgagc 1020
caggtacaat gactcctttc ggtaagtgca gtggaagctg tacactgccc aggcaaagcg 1080
tccgggcagc gtaggcgggc gactcagatc ccagccagtg gacttagccc ctgtttgctc 1140
ctccgataac tggggtgacc ttggttaata ttcaccagca gcctcccccg ttgcccctct 1200
ggatccactg cttaaatacg gacgaggaca gggccctgtc tcctcagctt caggcaccac 1260
cactgacctg ggacagtgaa tgtccccctg atctgcggcc gtgactctct taaggtagcc 1320
ttgcagaagt tggtcgtgag gcactgggca ggtaagtatc aaggttacaa gacaggttta 1380
aggagaccaa tagaaactgg gcttgtcgag acagagaaga ctcttgcgtt tctgataggc 1440
acctattggt cttactgaca tccactttgc ctttctctcc acaggtgtcc actcccagtt 1500
caattacagc tcttaaggct agagtactta atacgactca ctataggcta gcctcgacct 1560
cgagacgcgt gatatcggat cccggccggc ggccgcttcc ctttagtgag ggttaatgct 1620
tcgagcagac atgataagat acattgatga gtttggacaa accacaacta gaatgcagtg 1680
aaaaaaatgc tttatttgtg aaatttgtga tgctattgct ttatttgtaa ccattataag 1740
ctgcaataaa caagttaaca acaacaattg cattcatttt atgtttcagg ttcaggggga 1800
gatgtgggag gttttttaaa gcaagtaaaa cctctacaaa tgtggtaaaa tccgataagg 1860
gactagagca tggctacgta gataagtagc atggcgggtt aatcattaac tacaaggaac 1920
ccctagtgat ggagttggcc actccctctc tgcgcgctcg ctcgctcact gaggccgggc 1980
gaccaaaggt cgcccgacgc ccgggctttg cccgggcggc ctcagtgagc gagcgagcgc 2040
gccagctggc gtaatagcga agaggcccgc accgatcgcc cttcccaaca gttgcgcagc 2100
ctgaatggcg aatggaattc cagacgattg agcgtcaaaa tgtaggtatt tccatgagcg 2160
tttttccgtt gcaatggctg gcggtaatat tgttctggat attaccagca aggccgatag 2220
tttgagttct tctactcagg caagtgatgt tattactaat caaagaagta ttgcgacaac 2280
ggttaatttg cgtgatggac agactctttt actcggtggc ctcactgatt ataaaaacac 2340
ttctcaggat tctggcgtac cgttcctgtc taaaatccct ttaatcggcc tcctgtttag 2400
ctcccgctct gattctaacg aggaaagcac gttatacgtg ctcgtcaaag caaccatagt 2460
acgcgccctg tagcggcgca ttaagcgcgg cgggtgtggt ggttacgcgc agcgtgaccg 2520
ctacacttgc cagcgcccta gcgcccgctc ctttcgcttt cttcccttcc tttctcgcca 2580
cgttcgccgg ctttccccgt caagctctaa atcgggggct ccctttaggg ttccgattta 2640
gtgctttacg gcacctcgac cccaaaaaac ttgattaggg tgatggttca cgtagtgggc 2700
catcgccctg atagacggtt tttcgccctt tgacgttgga gtccacgttc tttaatagtg 2760
gactcttgtt ccaaactgga acaacactca accctatctc ggtctattct tttgatttat 2820
aagggatttt gccgatttcg gcctattggt taaaaaatga gctgatttaa caaaaattta 2880
acgcgaattt taacaaaata ttaacgtcta caatttaaat atttgcttat acaatcttcc 2940
tgtttttggg gcttttctga ttatcaaccg gggtacatat gattgacatg ctagttttac 3000
gattaccgtt catcgattct cttgtttgct ccagactctc aggcaatgac ctgatagcct 3060
ttgtagagac ctctcaaaaa tagctaccct ctccggcatg aatttatcag ctagaacggt 3120
tgaatatcat attgatggtg atttgactgt ctccggcctt tctcacccgt ttgaatcttt 3180
acctacacat tactcaggca ttgcatttaa aatatatgag ggttctaaaa atttttatcc 3240
ttgcgttgaa ataaaggctt ctcccgcaaa agtattacag ggtcataatg tttttggtac 3300
aaccgattta gctttatgct ctgaggcttt attgcttaat tttgctaatt ctttgccttg 3360
cctgtatgat ttattggatg ttggaatcgc ctgatgcggt attttctcct tacgcatctg 3420
tgcggtattt cacaccgcat atggtgcact ctcagtacaa tctgctctga tgccgcatag 3480
ttaagccagc cccgacaccc gccaacaccc gctgacgcgc cctgacgggc ttgtctgctc 3540
ccggcatccg cttacagaca agctgtgacc gtctccggga gctgcatgtg tcagaggttt 3600
tcaccgtcat caccgaaacg cgcgagacga aagggcctcg tgatacgcct atttttatag 3660
gttaatgtca tgataataat ggtttcttag acgtcaggtg gcacttttcg gggaaatgtg 3720
cgcggaaccc ctatttgttt atttttctaa atacattcaa atatgtatcc gctcatgaga 3780
caataaccct gataaatgct tcaataatat tgaaaaagga agagtatgag tattcaacat 3840
ttccgtgtcg cccttattcc cttttttgcg gcattttgcc ttcctgtttt tgctcaccca 3900
gaaacgctgg tgaaagtaaa agatgctgaa gatcagttgg gtgcacgagt gggttacatc 3960
gaactggatc tcaacagcgg taagatcctt gagagttttc gccccgaaga acgttttcca 4020
atgatgagca cttttaaagt tctgctatgt ggcgcggtat tatcccgtat tgacgccggg 4080
caagagcaac tcggtcgccg catacactat tctcagaatg acttggttga gtactcacca 4140
gtcacagaaa agcatcttac ggatggcatg acagtaagag aattatgcag tgctgccata 4200
accatgagtg ataacactgc ggccaactta cttctgacaa cgatcggagg accgaaggag 4260
ctaaccgctt ttttgcacaa catgggggat catgtaactc gccttgatcg ttgggaaccg 4320
gagctgaatg aagccatacc aaacgacgag cgtgacacca cgatgcctgt agcaatggca 4380
acaacgttgc gcaaactatt aactggcgaa ctacttactc tagcttcccg gcaacaatta 4440
atagactgga tggaggcgga taaagttgca ggaccacttc tgcgctcggc ccttccggct 4500
ggctggttta ttgctgataa atctggagcc ggtgagcgtg ggtctcgcgg tatcattgca 4560
gcactggggc cagatggtaa gccctcccgt atcgtagtta tctacacgac ggggagtcag 4620
gcaactatgg atgaacgaaa tagacagatc gctgagatag gtgcctcact gattaagcat 4680
tggtaactgt cagaccaagt ttactcatat atactttaga ttgatttaaa acttcatttt 4740
taatttaaaa ggatctaggt gaagatcctt tttgataatc tcatgaccaa aatcccttaa 4800
cgtgagtttt cgttccactg agcgtcagac cccgtagaaa agatcaaagg atcttcttga 4860
gatccttttt ttctgcgcgt aatctgctgc ttgcaaacaa aaaaaccacc gctaccagcg 4920
gtggtttgtt tgccggatca agagctacca actctttttc cgaaggtaac tggcttcagc 4980
agagcgcaga taccaaatac tgtccttcta gtgtagccgt agttaggcca ccacttcaag 5040
aactctgtag caccgcctac atacctcgct ctgctaatcc tgttaccagt ggctgctgcc 5100
agtggcgata agtcgtgtct taccgggttg gactcaagac gatagttacc ggataaggcg 5160
cagcggtcgg gctgaacggg gggttcgtgc acacagccca gcttggagcg aacgacctac 5220
accgaactga gatacctaca gcgtgagcta tgagaaagcg ccacgcttcc cgaagggaga 5280
aaggcggaca ggtatccggt aagcggcagg gtcggaacag gagagcgcac gagggagctt 5340
ccagggggaa acgcctggta tctttatagt cctgtcgggt ttcgccacct ctgacttgag 5400
cgtcgatttt tgtgatgctc gtcagggggg cggagcctat ggaaaaacgc cagcaacgcg 5460
gcctttttac ggttcctggc cttttgctgg ccttttgctc acatgttctt tcctgcgtta 5520
tcccctgatt ctgtggataa ccgtattacc gcctttgagt gagctgatac cgctcgccgc 5580
agccgaacga ccgagcgcag cgagtcagtg agcgaggaag cggaagagcg cccaatacgc 5640
aaaccgcctc tccccgcgcg ttggccgatt cattaatg 5678
<210> 34
<211> 6363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pGG2-hAAT-moFGF21
<400> 34
cgcgtgatat cggatcccgg ccggcggccg cttcccttta gtgagggtta atgcttcgag 60
cagacatgat aagatacatt gatgagtttg gacaaaccac aactagaatg cagtgaaaaa 120
aatgctttat ttgtgaaatt tgtgatgcta ttgctttatt tgtaaccatt ataagctgca 180
ataaacaagt taacaacaac aattgcattc attttatgtt tcaggttcag ggggagatgt 240
gggaggtttt ttaaagcaag taaaacctct acaaatgtgg taaaatccga taagggacta 300
gagcatggct acgtagataa gtagcatggc gggttaatca ttaactacaa ggaaccccta 360
gtgatggagt tggccactcc ctctctgcgc gctcgctcgc tcactgaggc cgggcgacca 420
aaggtcgccc gacgcccggg ctttgcccgg gcggcctcag tgagcgagcg agcgcgccag 480
ctggcgtaat agcgaagagg cccgcaccga tcgcccttcc caacagttgc gcagcctgaa 540
tggcgaatgg aattccagac gattgagcgt caaaatgtag gtatttccat gagcgttttt 600
ccgttgcaat ggctggcggt aatattgttc tggatattac cagcaaggcc gatagtttga 660
gttcttctac tcaggcaagt gatgttatta ctaatcaaag aagtattgcg acaacggtta 720
atttgcgtga tggacagact cttttactcg gtggcctcac tgattataaa aacacttctc 780
aggattctgg cgtaccgttc ctgtctaaaa tccctttaat cggcctcctg tttagctccc 840
gctctgattc taacgaggaa agcacgttat acgtgctcgt caaagcaacc atagtacgcg 900
ccctgtagcg gcgcattaag cgcggcgggt gtggtggtta cgcgcagcgt gaccgctaca 960
cttgccagcg ccctagcgcc cgctcctttc gctttcttcc cttcctttct cgccacgttc 1020
gccggctttc cccgtcaagc tctaaatcgg gggctccctt tagggttccg atttagtgct 1080
ttacggcacc tcgaccccaa aaaacttgat tagggtgatg gttcacgtag tgggccatcg 1140
ccctgataga cggtttttcg ccctttgacg ttggagtcca cgttctttaa tagtggactc 1200
ttgttccaaa ctggaacaac actcaaccct atctcggtct attcttttga tttataaggg 1260
attttgccga tttcggccta ttggttaaaa aatgagctga tttaacaaaa atttaacgcg 1320
aattttaaca aaatattaac gtctacaatt taaatatttg cttatacaat cttcctgttt 1380
ttggggcttt tctgattatc aaccggggta catatgattg acatgctagt tttacgatta 1440
ccgttcatcg attctcttgt ttgctccaga ctctcaggca atgacctgat agcctttgta 1500
gagacctctc aaaaatagct accctctccg gcatgaattt atcagctaga acggttgaat 1560
atcatattga tggtgatttg actgtctccg gcctttctca cccgtttgaa tctttaccta 1620
cacattactc aggcattgca tttaaaatat atgagggttc taaaaatttt tatccttgcg 1680
ttgaaataaa ggcttctccc gcaaaagtat tacagggtca taatgttttt ggtacaaccg 1740
atttagcttt atgctctgag gctttattgc ttaattttgc taattctttg ccttgcctgt 1800
atgatttatt ggatgttgga atcgcctgat gcggtatttt ctccttacgc atctgtgcgg 1860
tatttcacac cgcatatggt gcactctcag tacaatctgc tctgatgccg catagttaag 1920
ccagccccga cacccgccaa cacccgctga cgcgccctga cgggcttgtc tgctcccggc 1980
atccgcttac agacaagctg tgaccgtctc cgggagctgc atgtgtcaga ggttttcacc 2040
gtcatcaccg aaacgcgcga gacgaaaggg cctcgtgata cgcctatttt tataggttaa 2100
tgtcatgata ataatggttt cttagacgtc aggtggcact tttcggggaa atgtgcgcgg 2160
aacccctatt tgtttatttt tctaaataca ttcaaatatg tatccgctca tgagacaata 2220
accctgataa atgcttcaat aatattgaaa aaggaagagt atgagtattc aacatttccg 2280
tgtcgccctt attccctttt ttgcggcatt ttgccttcct gtttttgctc acccagaaac 2340
gctggtgaaa gtaaaagatg ctgaagatca gttgggtgca cgagtgggtt acatcgaact 2400
ggatctcaac agcggtaaga tccttgagag ttttcgcccc gaagaacgtt ttccaatgat 2460
gagcactttt aaagttctgc tatgtggcgc ggtattatcc cgtattgacg ccgggcaaga 2520
gcaactcggt cgccgcatac actattctca gaatgacttg gttgagtact caccagtcac 2580
agaaaagcat cttacggatg gcatgacagt aagagaatta tgcagtgctg ccataaccat 2640
gagtgataac actgcggcca acttacttct gacaacgatc ggaggaccga aggagctaac 2700
cgcttttttg cacaacatgg gggatcatgt aactcgcctt gatcgttggg aaccggagct 2760
gaatgaagcc ataccaaacg acgagcgtga caccacgatg cctgtagcaa tggcaacaac 2820
gttgcgcaaa ctattaactg gcgaactact tactctagct tcccggcaac aattaataga 2880
ctggatggag gcggataaag ttgcaggacc acttctgcgc tcggcccttc cggctggctg 2940
gtttattgct gataaatctg gagccggtga gcgtgggtct cgcggtatca ttgcagcact 3000
ggggccagat ggtaagccct cccgtatcgt agttatctac acgacgggga gtcaggcaac 3060
tatggatgaa cgaaatagac agatcgctga gataggtgcc tcactgatta agcattggta 3120
actgtcagac caagtttact catatatact ttagattgat ttaaaacttc atttttaatt 3180
taaaaggatc taggtgaaga tcctttttga taatctcatg accaaaatcc cttaacgtga 3240
gttttcgttc cactgagcgt cagaccccgt agaaaagatc aaaggatctt cttgagatcc 3300
tttttttctg cgcgtaatct gctgcttgca aacaaaaaaa ccaccgctac cagcggtggt 3360
ttgtttgccg gatcaagagc taccaactct ttttccgaag gtaactggct tcagcagagc 3420
gcagatacca aatactgtcc ttctagtgta gccgtagtta ggccaccact tcaagaactc 3480
tgtagcaccg cctacatacc tcgctctgct aatcctgtta ccagtggctg ctgccagtgg 3540
cgataagtcg tgtcttaccg ggttggactc aagacgatag ttaccggata aggcgcagcg 3600
gtcgggctga acggggggtt cgtgcacaca gcccagcttg gagcgaacga cctacaccga 3660
actgagatac ctacagcgtg agctatgaga aagcgccacg cttcccgaag ggagaaaggc 3720
ggacaggtat ccggtaagcg gcagggtcgg aacaggagag cgcacgaggg agcttccagg 3780
gggaaacgcc tggtatcttt atagtcctgt cgggtttcgc cacctctgac ttgagcgtcg 3840
atttttgtga tgctcgtcag gggggcggag cctatggaaa aacgccagca acgcggcctt 3900
tttacggttc ctggcctttt gctggccttt tgctcacatg ttctttcctg cgttatcccc 3960
tgattctgtg gataaccgta ttaccgcctt tgagtgagct gataccgctc gccgcagccg 4020
aacgaccgag cgcagcgagt cagtgagcga ggaagcggaa gagcgcccaa tacgcaaacc 4080
gcctctcccc gcgcgttggc cgattcatta atgcagcagc tgcgcgctcg ctcgctcact 4140
gaggccgccc gggcaaagcc cgggcgtcgg gcgacctttg gtcgcccggc ctcagtgagc 4200
gagcgagcgc gcagagaggg agtggccaac tccatcacta ggggttcctt gtagttaatg 4260
attaacccgc catgctactt atctacgtag ccatgctcta gacatggctc gacagatctg 4320
atatcatcga tgaattcgag ctcggtaccc ggccgcagat ttaggtgaca ctatagaata 4380
tgcatcacta gtaagcttgc gaattccagt ctacagagag gtctctgacc tctgccccag 4440
ctccaaggtc agcaggcagg gagggctgtg tgtttgctgt ttgctgcttg caatgtttgc 4500
ccattttagg gacatgagta ggctgaagtt tgttcagtgt ggacttcaga ggcagcacac 4560
aaacagcaag cttgcgaatt ccagtctaca gagaggtctc tgacctctgc cccagctcca 4620
aggtcagcag gcagggaggg ctgtgtgttt gctgtttgct gcttgcaatg tttgcccatt 4680
ttagggacat gagtaggctg aagtttgttc agtgtggact tcagaggcag cacacaaaca 4740
gcaagcttgc gaattccagt ctacagagag gtctctgacc tctgccccag ctccaaggtc 4800
agcaggcagg gagggctgtg tgtttgctgt ttgctgcttg caatgtttgc ccattttagg 4860
gacatgagta ggctgaagtt tgttcagtgt ggacttcaga ggcagcacac aaacagcaag 4920
ctttgctcta gactggaatt cgtcgacgag ctccctatag tgagtcgtat tagaggccga 4980
ctgacccggt acccggggat cttgctacca gtggaacagc cactaaggat tctgcagtga 5040
gagcagaggg ccagctaagt ggtactctcc cagagactgt ctgactcacg ccaccccctc 5100
caccttggac acaggacgct gtggtttctg agccaggtac aatgactcct ttcggtaagt 5160
gcagtggaag ctgtacactg cccaggcaaa gcgtccgggc agcgtaggcg ggcgactcag 5220
atcccagcca gtggacttag cccctgtttg ctcctccgat aactggggtg accttggtta 5280
atattcacca gcagcctccc ccgttgcccc tctggatcca ctgcttaaat acggacgagg 5340
acagggccct gtctcctcag cttcaggcac caccactgac ctgggacagt gaatgtcccc 5400
ctgatctgcg gccgtgactc tcttaaggta gccttgcaga agttggtcgt gaggcactgg 5460
gcaggtaagt atcaaggtta caagacaggt ttaaggagac caatagaaac tgggcttgtc 5520
gagacagaga agactcttgc gtttctgata ggcacctatt ggtcttactg acatccactt 5580
tgcctttctc tccacaggtg tccactccca gttcaattac agctcttaag gctagagtac 5640
ttaatacgac tcactatagg ctagcctcga cctcgagcgc aagcttgagc tagcgccacc 5700
atggaatgga tgagaagcag agtgggcacc ctgggcctgt gggtgcgact gctgctggct 5760
gtgtttctgc tgggcgtgta ccaggcctac cccatccctg actctagccc cctgctgcag 5820
tttggcggac aagtgcggca gagatacctg tacaccgacg acgaccagga caccgaggcc 5880
cacctggaaa tccgcgagga tggcacagtc gtgggcgctg ctcacagaag ccctgagagc 5940
ctgctggaac tgaaggccct gaagcccggc gtgatccaga tcctgggcgt gaaggccagc 6000
agattcctgt gccagcagcc tgacggcgcc ctgtacggct ctcctcactt cgatcctgag 6060
gcctgcagct tcagagagct gctgctggag gacggctaca acgtgtacca gtctgaggcc 6120
cacggcctgc ccctgagact gcctcagaag gacagcccta accaggacgc cacaagctgg 6180
ggacctgtgc ggttcctgcc tatgcctgga ctgctgcacg agccccagga tcaggctggc 6240
tttctgcctc ctgagcctcc agacgtgggc agcagcgacc ctctgagcat ggtggaacct 6300
ctgcagggca gaagccccag ctacgcctct tgagaatgcg ggcccggtac ccccgacgcg 6360
gcc 6363
<210> 35
<211> 4403
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pAAV-CMV-null
<400> 35
cgacggtacc agcgctgtcg aggccgcttc gagcagacat gataagatac attgatgagt 60
ttggacaaac cacaactaga atgcagtgaa aaaaatgctt tatttgtgaa atttgtgatg 120
ctattgcttt atttgtaacc attataagct gcaataaaca agttaacaac aacaattgca 180
ttcattttat gtttcaggtt cagggggaga tgtgggaggt tttttaaagc aagtaaaacc 240
tctacaaatg tggtaaaatc gattaggatc ttcctagagc atggctacct agacatggct 300
cgacagatca gcgctcatgc tctggaagat ctcgatttaa atgcggccgc aggaacccct 360
agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg ccgggcgacc 420
aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc gagcgcgcag 480
ctgcctgcag gggcgcctga tgcggtattt tctccttacg catctgtgcg gtatttcaca 540
ccgcatacgt caaagcaacc atagtacgcg ccctgtagcg gcgcattaag cgcggcgggt 600
gtggtggtta cgcgcagcgt gaccgctaca cttgccagcg ccctagcgcc cgctcctttc 660
gctttcttcc cttcctttct cgccacgttc gccggctttc cccgtcaagc tctaaatcgg 720
gggctccctt tagggttccg atttagtgct ttacggcacc tcgaccccaa aaaacttgat 780
ttgggtgatg gttcacgtag tgggccatcg ccctgataga cggtttttcg ccctttgacg 840
ttggagtcca cgttctttaa tagtggactc ttgttccaaa ctggaacaac actcaaccct 900
atctcgggct attcttttga tttataaggg attttgccga tttcggccta ttggttaaaa 960
aatgagctga tttaacaaaa atttaacgcg aattttaaca aaatattaac gtttacaatt 1020
ttatggtgca ctctcagtac aatctgctct gatgccgcat agttaagcca gccccgacac 1080
ccgccaacac ccgctgacgc gccctgacgg gcttgtctgc tcccggcatc cgcttacaga 1140
caagctgtga ccgtctccgg gagctgcatg tgtcagaggt tttcaccgtc atcaccgaaa 1200
cgcgcgagac gaaagggcct cgtgatacgc ctatttttat aggttaatgt catgataata 1260
atggtttctt agacgtcagg tggcactttt cggggaaatg tgcgcggaac ccctatttgt 1320
ttatttttct aaatacattc aaatatgtat ccgctcatga gacaataacc ctgataaatg 1380
cttcaataat attgaaaaag gaagagtatg agtattcaac atttccgtgt cgcccttatt 1440
cccttttttg cggcattttg ccttcctgtt tttgctcacc cagaaacgct ggtgaaagta 1500
aaagatgctg aagatcagtt gggtgcacga gtgggttaca tcgaactgga tctcaacagc 1560
ggtaagatcc ttgagagttt tcgccccgaa gaacgttttc caatgatgag cacttttaaa 1620
gttctgctat gtggcgcggt attatcccgt attgacgccg ggcaagagca actcggtcgc 1680
cgcatacact attctcagaa tgacttggtt gagtactcac cagtcacaga aaagcatctt 1740
acggatggca tgacagtaag agaattatgc agtgctgcca taaccatgag tgataacact 1800
gcggccaact tacttctgac aacgatcgga ggaccgaagg agctaaccgc ttttttgcac 1860
aacatggggg atcatgtaac tcgccttgat cgttgggaac cggagctgaa tgaagccata 1920
ccaaacgacg agcgtgacac cacgatgcct gtagcaatgg caacaacgtt gcgcaaacta 1980
ttaactggcg aactacttac tctagcttcc cggcaacaat taatagactg gatggaggcg 2040
gataaagttg caggaccact tctgcgctcg gcccttccgg ctggctggtt tattgctgat 2100
aaatctggag ccggtgagcg tgggtctcgc ggtatcattg cagcactggg gccagatggt 2160
aagccctccc gtatcgtagt tatctacacg acggggagtc aggcaactat ggatgaacga 2220
aatagacaga tcgctgagat aggtgcctca ctgattaagc attggtaact gtcagaccaa 2280
gtttactcat atatacttta gattgattta aaacttcatt tttaatttaa aaggatctag 2340
gtgaagatcc tttttgataa tctcatgacc aaaatccctt aacgtgagtt ttcgttccac 2400
tgagcgtcag accccgtaga aaagatcaaa ggatcttctt gagatccttt ttttctgcgc 2460
gtaatctgct gcttgcaaac aaaaaaacca ccgctaccag cggtggtttg tttgccggat 2520
caagagctac caactctttt tccgaaggta actggcttca gcagagcgca gataccaaat 2580
actgtccttc tagtgtagcc gtagttaggc caccacttca agaactctgt agcaccgcct 2640
acatacctcg ctctgctaat cctgttacca gtggctgctg ccagtggcga taagtcgtgt 2700
cttaccgggt tggactcaag acgatagtta ccggataagg cgcagcggtc gggctgaacg 2760
gggggttcgt gcacacagcc cagcttggag cgaacgacct acaccgaact gagataccta 2820
cagcgtgagc tatgagaaag cgccacgctt cccgaaggga gaaaggcgga caggtatccg 2880
gtaagcggca gggtcggaac aggagagcgc acgagggagc ttccaggggg aaacgcctgg 2940
tatctttata gtcctgtcgg gtttcgccac ctctgacttg agcgtcgatt tttgtgatgc 3000
tcgtcagggg ggcggagcct atggaaaaac gccagcaacg cggccttttt acggttcctg 3060
gccttttgct ggccttttgc tcacatgtcc tgcaggcagc tgcgcgctcg ctcgctcact 3120
gaggccgccc gggcaaagcc cgggcgtcgg gcgacctttg gtcgcccggc ctcagtgagc 3180
gagcgagcgc gcagagaggg agtggccaac tccatcacta ggggttcctg cggccgcgat 3240
atctgtagtt aatgattaac ccgccatgct acttatctac agatctcaat attggccatt 3300
agccatatta ttcattggtt atatagcata aatcaatatt ggctattggc cattgcatac 3360
gttgtatcta tatcataata tgtacattta tattggctca tgtccaatat gaccgccatg 3420
ttggcattga ttattgacta gttattaata gtaatcaatt acggggtcat tagttcatag 3480
cccatatatg gagttccgcg ttacataact tacggtaaat ggcccgcctg gctgaccgcc 3540
caacgacccc cgcccattga cgtcaataat gacgtatgtt cccatagtaa cgccaatagg 3600
gactttccat tgacgtcaat gggtggagta tttacggtaa actgcccact tggcagtaca 3660
tcaagtgtat catatgccaa gtccgccccc tattgacgtc aatgacggta aatggcccgc 3720
ctggcattat gcccagtaca tgaccttacg ggactttcct acttggcagt acatctacgt 3780
attagtcatc gctattacca tggtgatgcg gttttggcag tacaccaatg ggcgtggata 3840
gcggtttgac tcacggggat ttccaagtct ccaccccatt gacgtcaatg ggagtttgtt 3900
ttggcaccaa aatcaacggg actttccaaa atgtcgtaac aactgcgatc gcccgccccg 3960
ttgacgcaaa tgggcggtag gcgtgtacgg tgggaggtct atataagcag agctcgttta 4020
gtgaaccgtc agatcactag gctagctatt gcggtagttt atcacagtta aattgctaac 4080
gcagtcagtg cttctgacac aacagtctcg aacttaagct gcagtgactc tcttaaggta 4140
gccttgcaga agttggtcgt gaggcactgg gcaggtaagt atcaaggtta caagacaggt 4200
ttaaggagac caatagaaac tgggcttgtc gagacagaga agactcttgc gtttctgata 4260
ggcacctatt ggtcttactg acatccactt tgcctttctc tccacaggtg tccactccca 4320
gttcaattac agctcttaag gctagagtac ttaatacgac tcactataga atacgactca 4380
ctatagggag acgctagcgt cga 4403
<210> 36
<211> 5073
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pAAV-CMV-moFGF21
<400> 36
ggggctagcg ccaccatgga atggatgaga agcagagtgg gcaccctggg cctgtgggtg 60
cgactgctgc tggctgtgtt tctgctgggc gtgtaccagg cctaccccat ccctgactct 120
agccccctgc tgcagtttgg cggacaagtg cggcagagat acctgtacac cgacgacgac 180
caggacaccg aggcccacct ggaaatccgc gaggatggca cagtcgtggg cgctgctcac 240
agaagccctg agagcctgct ggaactgaag gccctgaagc ccggcgtgat ccagatcctg 300
ggcgtgaagg ccagcagatt cctgtgccag cagcctgacg gcgccctgta cggctctcct 360
cacttcgatc ctgaggcctg cagcttcaga gagctgctgc tggaggacgg ctacaacgtg 420
taccagtctg aggcccacgg cctgcccctg agactgcctc agaaggacag ccctaaccag 480
gacgccacaa gctggggacc tgtgcggttc ctgcctatgc ctggactgct gcacgagccc 540
caggatcagg ctggctttct gcctcctgag cctccagacg tgggcagcag cgaccctctg 600
agcatggtgg aacctctgca gggcagaagc cccagctacg cctcttgaga atgcgggccc 660
ggtaccccct cgacggtacc agcgctgtcg aggccgcttc gagcagacat gataagatac 720
attgatgagt ttggacaaac cacaactaga atgcagtgaa aaaaatgctt tatttgtgaa 780
atttgtgatg ctattgcttt atttgtaacc attataagct gcaataaaca agttaacaac 840
aacaattgca ttcattttat gtttcaggtt cagggggaga tgtgggaggt tttttaaagc 900
aagtaaaacc tctacaaatg tggtaaaatc gattaggatc ttcctagagc atggctacct 960
agacatggct cgacagatca gcgctcatgc tctggaagat ctcgatttaa atgcggccgc 1020
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 1080
ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc 1140
gagcgcgcag ctgcctgcag gggcgcctga tgcggtattt tctccttacg catctgtgcg 1200
gtatttcaca ccgcatacgt caaagcaacc atagtacgcg ccctgtagcg gcgcattaag 1260
cgcggcgggt gtggtggtta cgcgcagcgt gaccgctaca cttgccagcg ccctagcgcc 1320
cgctcctttc gctttcttcc cttcctttct cgccacgttc gccggctttc cccgtcaagc 1380
tctaaatcgg gggctccctt tagggttccg atttagtgct ttacggcacc tcgaccccaa 1440
aaaacttgat ttgggtgatg gttcacgtag tgggccatcg ccctgataga cggtttttcg 1500
ccctttgacg ttggagtcca cgttctttaa tagtggactc ttgttccaaa ctggaacaac 1560
actcaaccct atctcgggct attcttttga tttataaggg attttgccga tttcggccta 1620
ttggttaaaa aatgagctga tttaacaaaa atttaacgcg aattttaaca aaatattaac 1680
gtttacaatt ttatggtgca ctctcagtac aatctgctct gatgccgcat agttaagcca 1740
gccccgacac ccgccaacac ccgctgacgc gccctgacgg gcttgtctgc tcccggcatc 1800
cgcttacaga caagctgtga ccgtctccgg gagctgcatg tgtcagaggt tttcaccgtc 1860
atcaccgaaa cgcgcgagac gaaagggcct cgtgatacgc ctatttttat aggttaatgt 1920
catgataata atggtttctt agacgtcagg tggcactttt cggggaaatg tgcgcggaac 1980
ccctatttgt ttatttttct aaatacattc aaatatgtat ccgctcatga gacaataacc 2040
ctgataaatg cttcaataat attgaaaaag gaagagtatg agtattcaac atttccgtgt 2100
cgcccttatt cccttttttg cggcattttg ccttcctgtt tttgctcacc cagaaacgct 2160
ggtgaaagta aaagatgctg aagatcagtt gggtgcacga gtgggttaca tcgaactgga 2220
tctcaacagc ggtaagatcc ttgagagttt tcgccccgaa gaacgttttc caatgatgag 2280
cacttttaaa gttctgctat gtggcgcggt attatcccgt attgacgccg ggcaagagca 2340
actcggtcgc cgcatacact attctcagaa tgacttggtt gagtactcac cagtcacaga 2400
aaagcatctt acggatggca tgacagtaag agaattatgc agtgctgcca taaccatgag 2460
tgataacact gcggccaact tacttctgac aacgatcgga ggaccgaagg agctaaccgc 2520
ttttttgcac aacatggggg atcatgtaac tcgccttgat cgttgggaac cggagctgaa 2580
tgaagccata ccaaacgacg agcgtgacac cacgatgcct gtagcaatgg caacaacgtt 2640
gcgcaaacta ttaactggcg aactacttac tctagcttcc cggcaacaat taatagactg 2700
gatggaggcg gataaagttg caggaccact tctgcgctcg gcccttccgg ctggctggtt 2760
tattgctgat aaatctggag ccggtgagcg tgggtctcgc ggtatcattg cagcactggg 2820
gccagatggt aagccctccc gtatcgtagt tatctacacg acggggagtc aggcaactat 2880
ggatgaacga aatagacaga tcgctgagat aggtgcctca ctgattaagc attggtaact 2940
gtcagaccaa gtttactcat atatacttta gattgattta aaacttcatt tttaatttaa 3000
aaggatctag gtgaagatcc tttttgataa tctcatgacc aaaatccctt aacgtgagtt 3060
ttcgttccac tgagcgtcag accccgtaga aaagatcaaa ggatcttctt gagatccttt 3120
ttttctgcgc gtaatctgct gcttgcaaac aaaaaaacca ccgctaccag cggtggtttg 3180
tttgccggat caagagctac caactctttt tccgaaggta actggcttca gcagagcgca 3240
gataccaaat actgtccttc tagtgtagcc gtagttaggc caccacttca agaactctgt 3300
agcaccgcct acatacctcg ctctgctaat cctgttacca gtggctgctg ccagtggcga 3360
taagtcgtgt cttaccgggt tggactcaag acgatagtta ccggataagg cgcagcggtc 3420
gggctgaacg gggggttcgt gcacacagcc cagcttggag cgaacgacct acaccgaact 3480
gagataccta cagcgtgagc tatgagaaag cgccacgctt cccgaaggga gaaaggcgga 3540
caggtatccg gtaagcggca gggtcggaac aggagagcgc acgagggagc ttccaggggg 3600
aaacgcctgg tatctttata gtcctgtcgg gtttcgccac ctctgacttg agcgtcgatt 3660
tttgtgatgc tcgtcagggg ggcggagcct atggaaaaac gccagcaacg cggccttttt 3720
acggttcctg gccttttgct ggccttttgc tcacatgtcc tgcaggcagc tgcgcgctcg 3780
ctcgctcact gaggccgccc gggcaaagcc cgggcgtcgg gcgacctttg gtcgcccggc 3840
ctcagtgagc gagcgagcgc gcagagaggg agtggccaac tccatcacta ggggttcctg 3900
cggccgcgat atctgtagtt aatgattaac ccgccatgct acttatctac agatctcaat 3960
attggccatt agccatatta ttcattggtt atatagcata aatcaatatt ggctattggc 4020
cattgcatac gttgtatcta tatcataata tgtacattta tattggctca tgtccaatat 4080
gaccgccatg ttggcattga ttattgacta gttattaata gtaatcaatt acggggtcat 4140
tagttcatag cccatatatg gagttccgcg ttacataact tacggtaaat ggcccgcctg 4200
gctgaccgcc caacgacccc cgcccattga cgtcaataat gacgtatgtt cccatagtaa 4260
cgccaatagg gactttccat tgacgtcaat gggtggagta tttacggtaa actgcccact 4320
tggcagtaca tcaagtgtat catatgccaa gtccgccccc tattgacgtc aatgacggta 4380
aatggcccgc ctggcattat gcccagtaca tgaccttacg ggactttcct acttggcagt 4440
acatctacgt attagtcatc gctattacca tggtgatgcg gttttggcag tacaccaatg 4500
ggcgtggata gcggtttgac tcacggggat ttccaagtct ccaccccatt gacgtcaatg 4560
ggagtttgtt ttggcaccaa aatcaacggg actttccaaa atgtcgtaac aactgcgatc 4620
gcccgccccg ttgacgcaaa tgggcggtag gcgtgtacgg tgggaggtct atataagcag 4680
agctcgttta gtgaaccgtc agatcactag gctagctatt gcggtagttt atcacagtta 4740
aattgctaac gcagtcagtg cttctgacac aacagtctcg aacttaagct gcagtgactc 4800
tcttaaggta gccttgcaga agttggtcgt gaggcactgg gcaggtaagt atcaaggtta 4860
caagacaggt ttaaggagac caatagaaac tgggcttgtc gagacagaga agactcttgc 4920
gtttctgata ggcacctatt ggtcttactg acatccactt tgcctttctc tccacaggtg 4980
tccactccca gttcaattac agctcttaag gctagagtac ttaatacgac tcactataga 5040
atacgactca ctatagggag acgctagcgt cga 5073
<210> 37
<211> 7292
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pAAV-CAG-cloFGF21-dmiRT
<400> 37
ggccgctaat tctagatcgc gaacaaacac cattgtcaca ctccagtata cacaaacacc 60
attgtcacac tccagatatc acaaacacca ttgtcacact ccaaggcgaa caaacaccat 120
tgtcacactc caaggctatt ctagatcgcg aattacatac ttctttacat tccagtatac 180
attacatact tctttacatt ccagatatca ttacatactt ctttacattc caaggcgaat 240
tacatacttc tttacattcc aaggctacct gaggcccggg ggtacctctt aattaactgg 300
cctcatgggc cttccgctca ctgcccgctt tccagtcggg aaacctgtcg tgccagtcag 360
gtgcaggctg cctatcagaa ggtggtggct ggtgtggcca atgccctggc tcacaaatac 420
cactgagatc tttttccctc tgccaaaaat tatggggaca tcatgaagcc ccttgagcat 480
ctgacttctg gctaataaag gaaatttatt ttcattgcaa tagtgtgttg gaattttttg 540
tgtctctcac tcggaaggac atatgggagg gcaaatcatt taaaacatca gaatgagtat 600
ttggtttaga gtttggcaac atatgcccat atgctggctg ccatgaacaa aggttggcta 660
taaagaggtc atcagtatat gaaacagccc cctgctgtcc attccttatt ccatagaaaa 720
gccttgactt gaggttagat tttttttata ttttgttttg tgttattttt ttctttaaca 780
tccctaaaat tttccttaca tgttttacta gccagatttt tcctcctctc ctgactactc 840
ccagtcatag ctgtccctct tctcttatgg agatccctcg acctgcagcc caagctgtag 900
ataagtagca tggcgggtta atcattaact acaaggaacc cctagtgatg gagttggcca 960
ctccctctct gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc 1020
cgggctttgc ccgggcggcc tcagtgagcg agcgagcgcg cagctggcgt aaagtgagcg 1080
agcgagcgcg cagctgcatt aatgaatcgg ccaacgcgcg gggagaggcg gtttgcgtat 1140
tgggcgctct tccgcttcct cgctcactga ctcgctgcgc tcggtcgttc ggctgcggcg 1200
agcggtatca gctcactcaa aggcggtaat acggttatcc acagaatcag gggataacgc 1260
aggaaagaac atgtgagcaa aaggccagca aaaggccagg aaccgtaaaa aggccgcgtt 1320
gctggcgttt ttccataggc tccgcccccc tgacgagcat cacaaaaatc gacgctcaag 1380
tcagaggtgg cgaaacccga caggactata aagataccag gcgtttcccc ctggaagctc 1440
cctcgtgcgc tctcctgttc cgaccctgcc gcttaccgga tacctgtccg cctttctccc 1500
ttcgggaagc gtggcgcttt ctcatagctc acgctgtagg tatctcagtt cggtgtaggt 1560
cgttcgctcc aagctgggct gtgtgcacga accccccgtt cagcccgacc gctgcgcctt 1620
atccggtaac tatcgtcttg agtccaaccc ggtaagacac gacttatcgc cactggcagc 1680
agccactggt aacaggatta gcagagcgag gtatgtaggc ggtgctacag agttcttgaa 1740
gtggtggcct aactacggct acactagaag aacagtattt ggtatctgcg ctctgctgaa 1800
gccagttacc ttcggaaaaa gagttggtag ctcttgatcc ggcaaacaaa ccaccgctgg 1860
tagcggtggt ttttttgttt gcaagcagca gattacgcgc agaaaaaaag gatctcaaga 1920
agatcctttg atcttttcta cggggtctga cgctcagtgg aacgaaaact cacgttaagg 1980
gattttggtc atgagattat caaaaaggat cttcacctag atccttttaa attaaaaatg 2040
aagttttaaa tcaatctaaa gtatatatga gtaaacttgg tctgacagtt accaatgctt 2100
aatcagtgag gcacctatct cagcgatctg tctatttcgt tcatccatag ttgcctgact 2160
ccccgtcgtg tagataacta cgatacggga gggcttacca tctggcccca gtgctgcaat 2220
gataccgcga gacccacgct caccggctcc agatttatca gcaataaacc agccagccgg 2280
aagggccgag cgcagaagtg gtcctgcaac tttatccgcc tccatccagt ctattaattg 2340
ttgccgggaa gctagagtaa gtagttcgcc agttaatagt ttgcgcaacg ttgttgccat 2400
tgctacaggc atcgtggtgt cacgctcgtc gtttggtatg gcttcattca gctccggttc 2460
ccaacgatca aggcgagtta catgatcccc catgttgtgc aaaaaagcgg ttagctcctt 2520
cggtcctccg atcgttgtca gaagtaagtt ggccgcagtg ttatcactca tggttatggc 2580
agcactgcat aattctctta ctgtcatgcc atccgtaaga tgcttttctg tgactggtga 2640
gtactcaacc aagtcattct gagaatagtg tatgcggcga ccgagttgct cttgcccggc 2700
gtcaatacgg gataataccg cgccacatag cagaacttta aaagtgctca tcattggaaa 2760
acgttcttcg gggcgaaaac tctcaaggat cttaccgctg ttgagatcca gttcgatgta 2820
acccactcgt gcacccaact gatcttcagc atcttttact ttcaccagcg tttctgggtg 2880
agcaaaaaca ggaaggcaaa atgccgcaaa aaagggaata agggcgacac ggaaatgttg 2940
aatactcata ctcttccttt ttcaatatta ttgaagcatt tatcagggtt attgtctcat 3000
gagcggatac atatttgaat gtatttagaa aaataaacaa ataggggttc cgcgcacatt 3060
tccccgaaaa gtgccacctg acgtctaaga aaccattatt atcatgacat taacctataa 3120
aaataggcgt atcacgaggc cctttcgtct cgcgcgtttc ggtgatgacg gtgaaaacct 3180
ctgacacatg cagctcccgg agacggtcac agcttgtctg taagcggatg ccgggagcag 3240
acaagcccgt cagggcgcgt cagcgggtgt tggcgggtgt cggggctggc ttaactatgc 3300
ggcatcagag cagattgtac tgagagtgca ccatatgcgg tgtgaaatac cgcacagatg 3360
cgtaaggaga aaataccgca tcaggcgatt ccaacatcca ataaatcata caggcaaggc 3420
aaagaattag caaaattaag caataaagcc tcagagcata aagctaaatc ggttgtacca 3480
aaaacattat gaccctgtaa tacttttgcg ggagaagcct ttatttcaac gcaaggataa 3540
aaatttttag aaccctcata tattttaaat gcaatgcctg agtaatgtgt aggtaaagat 3600
tcaaacgggt gagaaaggcc ggagacagtc aaatcaccat caatatgata ttcaaccgtt 3660
ctagctgata aattcatgcc ggagagggta gctatttttg agaggtctct acaaaggcta 3720
tcaggtcatt gcctgagagt ctggagcaaa caagagaatc gatgaacggt aatcgtaaaa 3780
ctagcatgtc aatcatatgt accccggttg ataatcagaa aagccccaaa aacaggaaga 3840
ttgtataagc aaatatttaa attgtaagcg ttaatatttt gttaaaattc gcgttaaatt 3900
tttgttaaat cagctcattt tttaaccaat aggccgaaat cggcaaaatc ccttataaat 3960
caaaagaata gaccgagata gggttgagtg ttgttccagt ttggaacaag agtccactat 4020
taaagaacgt ggactccaac gtcaaagggc gaaaaaccgt ctatcagggc gatggcccac 4080
tacgtgaacc atcaccctaa tcaagttttt tggggtcgag gtgccgtaaa gcactaaatc 4140
ggaaccctaa agggagcccc cgatttagag cttgacgggg aaagccggcg aacgtggcga 4200
gaaaggaagg gaagaaagcg aaaggagcgg gcgctagggc gctggcaagt gtagcggtca 4260
cgctgcgcgt aaccaccaca cccgccgcgc ttaatgcgcc gctacagggc gcgtactatg 4320
gttgctttga cgagcacgta taacgtgctt tcctcgttag aatcagagcg ggagctaaac 4380
aggaggccga ttaaagggat tttagacagg aacggtacgc cagaatcctg agaagtgttt 4440
ttataatcag tgaggccacc gagtaaaaga gtctgtccat cacgcaaatt aaccgttgtc 4500
gcaatacttc tttgattagt aataacatca cttgcctgag tagaagaact caaactatcg 4560
gccttgctgg taatatccag aacaatatta ccgccagcca ttgcaacgga atcgccattc 4620
gccattcagg ctgcgcaact gttgggaagg gcgatcggtg cgggcctctt ccactgaggc 4680
ccagctgcgc gctcgctcgc tcactgaggc cgcccgggca aagcccgggc gtcgggcgac 4740
ctttggtcgc ccggcctcag tgagcgagcg agcgcgcaga gagggagtgg ccaactccat 4800
cactaggggt tccttgtagt taatgattaa cccgccatgc tacttatcta ctcgacattg 4860
attattgact agttattaat agtaatcaat tacggggtca ttagttcata gcccatatat 4920
ggagttccgc gttacataac ttacggtaaa tggcccgcct ggctgaccgc ccaacgaccc 4980
ccgcccattg acgtcaataa tgacgtatgt tcccatagta acgccaatag ggactttcca 5040
ttgacgtcaa tgggtggagt atttacggta aactgcccac ttggcagtac atcaagtgta 5100
tcatatgcca agtacgcccc ctattgacgt caatgacggt aaatggcccg cctggcatta 5160
tgcccagtac atgaccttat gggactttcc tacttggcag tacatctacg tattagtcat 5220
cgctattacc atggtcgagg tgagccccac gttctgcttc actctcccca tctccccccc 5280
ctccccaccc ccaattttgt atttatttat tttttaatta ttttgtgcag cgatgggggc 5340
gggggggggg ggggggcgcg cgccaggcgg ggcggggcgg ggcgaggggc ggggcggggc 5400
gaggcggaga ggtgcggcgg cagccaatca gagcggcgcg ctccgaaagt ttccttttat 5460
ggcgaggcgg cggcggcggc ggccctataa aaagcgaagc gcgcggcggg cgggagtcgc 5520
tgcgttgcct tcgccccgtg ccccgctccg cgccgcctcg cgccgcccgc cccggctctg 5580
actgaccgcg ttactcccac aggtgagcgg gcgggacggc ccttctcctc cgggctgtaa 5640
ttagcgcttg gtttaatgac ggcttgtttc ttttctgtgg ctgcgtgaaa gccttgaggg 5700
gctccgggag ggccctttgt gcggggggag cggctcgggg ggtgcgtgcg tgtgtgtgtg 5760
cgtggggagc gccgcgtgcg gctccgcgct gcccggcggc tgtgagcgct gcgggcgcgg 5820
cgcggggctt tgtgcgctcc gcagtgtgcg cgaggggagc gcggccgggg gcggtgcccc 5880
gcggtgcggg gggctgcgag gggaacaaag gctgcgtgcg gggtgtgtgc gtgggggggt 5940
gagcaggggg tgtgggcgcg tcggtcgggc tgcaaccccc cctgcacccc cctccccgag 6000
ttgctgagca cggcccggct tcgggtgcgg ggctccgtac ggggcgtggc gcggggctcg 6060
ccgtgccggg cggggggtgg cggcaggtgg gggtgccggg cggggcgggg ccgcctcggg 6120
ccggggaggg ctcgggggag gggcgcggcg gcccccggag cgccggcggc tgtcgaggcg 6180
cggcgagccg cagccattgc cttttatggt aatcgtgcga gagggcgcag ggacttcctt 6240
tgtcccaaat ctgtgcggag ccgaaatctg ggaggcgccg ccgcaccccc tctagcgggc 6300
gcggggcgaa gcggtgcggc gccggcagga aggaaatggg cggggagggc cttcgtgcgt 6360
cgccgcgccg ccgtcccctt ctccctctcc agcctcgggg ctgtccgcgg ggggacggct 6420
gccttcgggg gggacggggc agggcggggt tcggcttctg gcgtgtgacc ggcggctcta 6480
gagcctctgc taaccatgtt catgccttct tctttttcct acagctcctg ggcaacgtgc 6540
tggttattgt gctgtctcat cattttggca aagaattgat taattcgagc gaacgcgtcg 6600
agtcgctcgg tacgatttaa attgaattgg cctcgagcgc aagcttgagc tagcgccacc 6660
atgggatggg ctgaggctgg attcgaacac ctgggactct gggtgcccgt cctggccgtg 6720
ctgctcctgg aggcttgcag ggctcatccc atccctgaca gctccccact cctgcagttt 6780
ggaggacagg tgaggcagcg gtacctgtat accgacgatg cccaggagac agaagctcac 6840
ctggaaattc gggctgatgg aacagtggtc ggagctgccc gacagtcccc agagtctctc 6900
ctggaactga aggccctcaa acccggagtg atccagattc tgggcgtcaa gacttctaga 6960
ttcctgtgcc agggaccaga cggcaccctg tacggcagcc tgcatttcga tcctgtggcc 7020
tgttcctttc gagagctcct gctcgaagac ggctacaaca tctatcactc tgagaccctg 7080
ggactcccac tgcgactcag acctcataat agtgcctatc gagatctggc tcccaggggc 7140
ccagctaggt ttctgccact ccccggactg ctccctgctc cacctgagcc acccggcatt 7200
ctggctccag aacctccaga cgtgggctct agtgatccac tgagtatggt cggcccctca 7260
caggggaggt cacctagcta cgccagctga gc 7292
<210> 38
<211> 6278
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pGG2-hAAT-cloFGF21
<400> 38
ggccgcttcc ctttagtgag ggttaatgct tcgagcagac atgataagat acattgatga 60
gtttggacaa accacaacta gaatgcagtg aaaaaaatgc tttatttgtg aaatttgtga 120
tgctattgct ttatttgtaa ccattataag ctgcaataaa caagttaaca acaacaattg 180
cattcatttt atgtttcagg ttcaggggga gatgtgggag gttttttaaa gcaagtaaaa 240
cctctacaaa tgtggtaaaa tccgataagg gactagagca tggctacgta gataagtagc 300
atggcgggtt aatcattaac tacaaggaac ccctagtgat ggagttggcc actccctctc 360
tgcgcgctcg ctcgctcact gaggccgggc gaccaaaggt cgcccgacgc ccgggctttg 420
cccgggcggc ctcagtgagc gagcgagcgc gccagctggc gtaatagcga agaggcccgc 480
accgatcgcc cttcccaaca gttgcgcagc ctgaatggcg aatggaattc cagacgattg 540
agcgtcaaaa tgtaggtatt tccatgagcg tttttccgtt gcaatggctg gcggtaatat 600
tgttctggat attaccagca aggccgatag tttgagttct tctactcagg caagtgatgt 660
tattactaat caaagaagta ttgcgacaac ggttaatttg cgtgatggac agactctttt 720
actcggtggc ctcactgatt ataaaaacac ttctcaggat tctggcgtac cgttcctgtc 780
taaaatccct ttaatcggcc tcctgtttag ctcccgctct gattctaacg aggaaagcac 840
gttatacgtg ctcgtcaaag caaccatagt acgcgccctg tagcggcgca ttaagcgcgg 900
cgggtgtggt ggttacgcgc agcgtgaccg ctacacttgc cagcgcccta gcgcccgctc 960
ctttcgcttt cttcccttcc tttctcgcca cgttcgccgg ctttccccgt caagctctaa 1020
atcgggggct ccctttaggg ttccgattta gtgctttacg gcacctcgac cccaaaaaac 1080
ttgattaggg tgatggttca cgtagtgggc catcgccctg atagacggtt tttcgccctt 1140
tgacgttgga gtccacgttc tttaatagtg gactcttgtt ccaaactgga acaacactca 1200
accctatctc ggtctattct tttgatttat aagggatttt gccgatttcg gcctattggt 1260
taaaaaatga gctgatttaa caaaaattta acgcgaattt taacaaaata ttaacgtcta 1320
caatttaaat atttgcttat acaatcttcc tgtttttggg gcttttctga ttatcaaccg 1380
gggtacatat gattgacatg ctagttttac gattaccgtt catcgattct cttgtttgct 1440
ccagactctc aggcaatgac ctgatagcct ttgtagagac ctctcaaaaa tagctaccct 1500
ctccggcatg aatttatcag ctagaacggt tgaatatcat attgatggtg atttgactgt 1560
ctccggcctt tctcacccgt ttgaatcttt acctacacat tactcaggca ttgcatttaa 1620
aatatatgag ggttctaaaa atttttatcc ttgcgttgaa ataaaggctt ctcccgcaaa 1680
agtattacag ggtcataatg tttttggtac aaccgattta gctttatgct ctgaggcttt 1740
attgcttaat tttgctaatt ctttgccttg cctgtatgat ttattggatg ttggaatcgc 1800
ctgatgcggt attttctcct tacgcatctg tgcggtattt cacaccgcat atggtgcact 1860
ctcagtacaa tctgctctga tgccgcatag ttaagccagc cccgacaccc gccaacaccc 1920
gctgacgcgc cctgacgggc ttgtctgctc ccggcatccg cttacagaca agctgtgacc 1980
gtctccggga gctgcatgtg tcagaggttt tcaccgtcat caccgaaacg cgcgagacga 2040
aagggcctcg tgatacgcct atttttatag gttaatgtca tgataataat ggtttcttag 2100
acgtcaggtg gcacttttcg gggaaatgtg cgcggaaccc ctatttgttt atttttctaa 2160
atacattcaa atatgtatcc gctcatgaga caataaccct gataaatgct tcaataatat 2220
tgaaaaagga agagtatgag tattcaacat ttccgtgtcg cccttattcc cttttttgcg 2280
gcattttgcc ttcctgtttt tgctcaccca gaaacgctgg tgaaagtaaa agatgctgaa 2340
gatcagttgg gtgcacgagt gggttacatc gaactggatc tcaacagcgg taagatcctt 2400
gagagttttc gccccgaaga acgttttcca atgatgagca cttttaaagt tctgctatgt 2460
ggcgcggtat tatcccgtat tgacgccggg caagagcaac tcggtcgccg catacactat 2520
tctcagaatg acttggttga gtactcacca gtcacagaaa agcatcttac ggatggcatg 2580
acagtaagag aattatgcag tgctgccata accatgagtg ataacactgc ggccaactta 2640
cttctgacaa cgatcggagg accgaaggag ctaaccgctt ttttgcacaa catgggggat 2700
catgtaactc gccttgatcg ttgggaaccg gagctgaatg aagccatacc aaacgacgag 2760
cgtgacacca cgatgcctgt agcaatggca acaacgttgc gcaaactatt aactggcgaa 2820
ctacttactc tagcttcccg gcaacaatta atagactgga tggaggcgga taaagttgca 2880
ggaccacttc tgcgctcggc ccttccggct ggctggttta ttgctgataa atctggagcc 2940
ggtgagcgtg ggtctcgcgg tatcattgca gcactggggc cagatggtaa gccctcccgt 3000
atcgtagtta tctacacgac ggggagtcag gcaactatgg atgaacgaaa tagacagatc 3060
gctgagatag gtgcctcact gattaagcat tggtaactgt cagaccaagt ttactcatat 3120
atactttaga ttgatttaaa acttcatttt taatttaaaa ggatctaggt gaagatcctt 3180
tttgataatc tcatgaccaa aatcccttaa cgtgagtttt cgttccactg agcgtcagac 3240
cccgtagaaa agatcaaagg atcttcttga gatccttttt ttctgcgcgt aatctgctgc 3300
ttgcaaacaa aaaaaccacc gctaccagcg gtggtttgtt tgccggatca agagctacca 3360
actctttttc cgaaggtaac tggcttcagc agagcgcaga taccaaatac tgtccttcta 3420
gtgtagccgt agttaggcca ccacttcaag aactctgtag caccgcctac atacctcgct 3480
ctgctaatcc tgttaccagt ggctgctgcc agtggcgata agtcgtgtct taccgggttg 3540
gactcaagac gatagttacc ggataaggcg cagcggtcgg gctgaacggg gggttcgtgc 3600
acacagccca gcttggagcg aacgacctac accgaactga gatacctaca gcgtgagcta 3660
tgagaaagcg ccacgcttcc cgaagggaga aaggcggaca ggtatccggt aagcggcagg 3720
gtcggaacag gagagcgcac gagggagctt ccagggggaa acgcctggta tctttatagt 3780
cctgtcgggt ttcgccacct ctgacttgag cgtcgatttt tgtgatgctc gtcagggggg 3840
cggagcctat ggaaaaacgc cagcaacgcg gcctttttac ggttcctggc cttttgctgg 3900
ccttttgctc acatgttctt tcctgcgtta tcccctgatt ctgtggataa ccgtattacc 3960
gcctttgagt gagctgatac cgctcgccgc agccgaacga ccgagcgcag cgagtcagtg 4020
agcgaggaag cggaagagcg cccaatacgc aaaccgcctc tccccgcgcg ttggccgatt 4080
cattaatgca gcagctgcgc gctcgctcgc tcactgaggc cgcccgggca aagcccgggc 4140
gtcgggcgac ctttggtcgc ccggcctcag tgagcgagcg agcgcgcaga gagggagtgg 4200
ccaactccat cactaggggt tccttgtagt taatgattaa cccgccatgc tacttatcta 4260
cgtagccatg ctctagacat ggctcgacag atctgatatc atcgatgaat tcgagctcgg 4320
tacccggccg cagatttagg tgacactata gaatatgcat cactagtaag cttgcgaatt 4380
ccagtctaca gagaggtctc tgacctctgc cccagctcca aggtcagcag gcagggaggg 4440
ctgtgtgttt gctgtttgct gcttgcaatg tttgcccatt ttagggacat gagtaggctg 4500
aagtttgttc agtgtggact tcagaggcag cacacaaaca gcaagcttgc gaattccagt 4560
ctacagagag gtctctgacc tctgccccag ctccaaggtc agcaggcagg gagggctgtg 4620
tgtttgctgt ttgctgcttg caatgtttgc ccattttagg gacatgagta ggctgaagtt 4680
tgttcagtgt ggacttcaga ggcagcacac aaacagcaag cttgcgaatt ccagtctaca 4740
gagaggtctc tgacctctgc cccagctcca aggtcagcag gcagggaggg ctgtgtgttt 4800
gctgtttgct gcttgcaatg tttgcccatt ttagggacat gagtaggctg aagtttgttc 4860
agtgtggact tcagaggcag cacacaaaca gcaagctttg ctctagactg gaattcgtcg 4920
acgagctccc tatagtgagt cgtattagag gccgactgac ccggtacccg gggatcttgc 4980
taccagtgga acagccacta aggattctgc agtgagagca gagggccagc taagtggtac 5040
tctcccagag actgtctgac tcacgccacc ccctccacct tggacacagg acgctgtggt 5100
ttctgagcca ggtacaatga ctcctttcgg taagtgcagt ggaagctgta cactgcccag 5160
gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga cttagcccct 5220
gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc ctcccccgtt 5280
gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc ctcagcttca 5340
ggcaccacca ctgacctggg acagtgaatg tccccctgat ctgcggccgt gactctctta 5400
aggtagcctt gcagaagttg gtcgtgaggc actgggcagg taagtatcaa ggttacaaga 5460
caggtttaag gagaccaata gaaactgggc ttgtcgagac agagaagact cttgcgtttc 5520
tgataggcac ctattggtct tactgacatc cactttgcct ttctctccac aggtgtccac 5580
tcccagttca attacagctc ttaaggctag agtacttaat acgactcact ataggctagc 5640
gccaccatgg gatgggctga ggctggattc gaacacctgg gactctgggt gcccgtcctg 5700
gccgtgctgc tcctggaggc ttgcagggct catcccatcc ctgacagctc cccactcctg 5760
cagtttggag gacaggtgag gcagcggtac ctgtataccg acgatgccca ggagacagaa 5820
gctcacctgg aaattcgggc tgatggaaca gtggtcggag ctgcccgaca gtccccagag 5880
tctctcctgg aactgaaggc cctcaaaccc ggagtgatcc agattctggg cgtcaagact 5940
tctagattcc tgtgccaggg accagacggc accctgtacg gcagcctgca tttcgatcct 6000
gtggcctgtt cctttcgaga gctcctgctc gaagacggct acaacatcta tcactctgag 6060
accctgggac tcccactgcg actcagacct cataatagtg cctatcgaga tctggctccc 6120
aggggcccag ctaggtttct gccactcccc ggactgctcc ctgctccacc tgagccaccc 6180
ggcattctgg ctccagaacc tccagacgtg ggctctagtg atccactgag tatggtcggc 6240
ccctcacagg ggaggtcacc tagctacgcc agctgagc 6278
<210> 39
<211> 6278
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pGG2-hAAT-hsFGF21
<400> 39
gatctgatat catcgatgaa ttcgagctcg gtacccggcc gcagatttag gtgacactat 60
agaatatgca tcactagtaa gcttgcgaat tccagtctac agagaggtct ctgacctctg 120
ccccagctcc aaggtcagca ggcagggagg gctgtgtgtt tgctgtttgc tgcttgcaat 180
gtttgcccat tttagggaca tgagtaggct gaagtttgtt cagtgtggac ttcagaggca 240
gcacacaaac agcaagcttg cgaattccag tctacagaga ggtctctgac ctctgcccca 300
gctccaaggt cagcaggcag ggagggctgt gtgtttgctg tttgctgctt gcaatgtttg 360
cccattttag ggacatgagt aggctgaagt ttgttcagtg tggacttcag aggcagcaca 420
caaacagcaa gcttgcgaat tccagtctac agagaggtct ctgacctctg ccccagctcc 480
aaggtcagca ggcagggagg gctgtgtgtt tgctgtttgc tgcttgcaat gtttgcccat 540
tttagggaca tgagtaggct gaagtttgtt cagtgtggac ttcagaggca gcacacaaac 600
agcaagcttt gctctagact ggaattcgtc gacgagctcc ctatagtgag tcgtattaga 660
ggccgactga cccggtaccc ggggatcttg ctaccagtgg aacagccact aaggattctg 720
cagtgagagc agagggccag ctaagtggta ctctcccaga gactgtctga ctcacgccac 780
cccctccacc ttggacacag gacgctgtgg tttctgagcc aggtacaatg actcctttcg 840
gtaagtgcag tggaagctgt acactgccca ggcaaagcgt ccgggcagcg taggcgggcg 900
actcagatcc cagccagtgg acttagcccc tgtttgctcc tccgataact ggggtgacct 960
tggttaatat tcaccagcag cctcccccgt tgcccctctg gatccactgc ttaaatacgg 1020
acgaggacag ggccctgtct cctcagcttc aggcaccacc actgacctgg gacagtgaat 1080
gtccccctga tctgcggccg tgactctctt aaggtagcct tgcagaagtt ggtcgtgagg 1140
cactgggcag gtaagtatca aggttacaag acaggtttaa ggagaccaat agaaactggg 1200
cttgtcgaga cagagaagac tcttgcgttt ctgataggca cctattggtc ttactgacat 1260
ccactttgcc tttctctcca caggtgtcca ctcccagttc aattacagct cttaaggcta 1320
gagtacttaa tacgactcac tataggctag cgccaccatg gactcggacg agaccgggtt 1380
cgagcactca ggactgtggg tttctgtgct ggctggtctt ctgctgggag cctgccaggc 1440
acaccccatc cctgactcca gtcctctcct gcaattcggg ggccaagtcc ggcagcggta 1500
cctctacaca gatgatgccc agcagacaga agcccacctg gagatcaggg aggatgggac 1560
ggtggggggc gctgctgacc agagccccga aagtctcctg cagctgaaag ccttgaagcc 1620
gggagttatt caaatcttgg gagtcaagac atccaggttc ctgtgccagc ggccagatgg 1680
ggccctgtat ggatcgctcc actttgaccc tgaggcctgc agcttccggg agctgcttct 1740
tgaggacgga tacaatgttt accagtccga agcccacggc ctcccgctgc acctgccagg 1800
gaacaagtcc ccacaccggg accctgcacc ccgaggacca gctcgcttcc tgccactacc 1860
aggcctgccc cccgcactcc cggagccacc cggaatcctg gccccccagc cccccgatgt 1920
gggctcctcg gaccctctga gcatggtggg accttcccag ggccgaagcc ccagctacgc 1980
ttcctgagcg gccgcttccc tttagtgagg gttaatgctt cgagcagaca tgataagata 2040
cattgatgag tttggacaaa ccacaactag aatgcagtga aaaaaatgct ttatttgtga 2100
aatttgtgat gctattgctt tatttgtaac cattataagc tgcaataaac aagttaacaa 2160
caacaattgc attcatttta tgtttcaggt tcagggggag atgtgggagg ttttttaaag 2220
caagtaaaac ctctacaaat gtggtaaaat ccgataaggg actagagcat ggctacgtag 2280
ataagtagca tggcgggtta atcattaact acaaggaacc cctagtgatg gagttggcca 2340
ctccctctct gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc 2400
cgggctttgc ccgggcggcc tcagtgagcg agcgagcgcg ccagctggcg taatagcgaa 2460
gaggcccgca ccgatcgccc ttcccaacag ttgcgcagcc tgaatggcga atggaattcc 2520
agacgattga gcgtcaaaat gtaggtattt ccatgagcgt ttttccgttg caatggctgg 2580
cggtaatatt gttctggata ttaccagcaa ggccgatagt ttgagttctt ctactcaggc 2640
aagtgatgtt attactaatc aaagaagtat tgcgacaacg gttaatttgc gtgatggaca 2700
gactctttta ctcggtggcc tcactgatta taaaaacact tctcaggatt ctggcgtacc 2760
gttcctgtct aaaatccctt taatcggcct cctgtttagc tcccgctctg attctaacga 2820
ggaaagcacg ttatacgtgc tcgtcaaagc aaccatagta cgcgccctgt agcggcgcat 2880
taagcgcggc gggtgtggtg gttacgcgca gcgtgaccgc tacacttgcc agcgccctag 2940
cgcccgctcc tttcgctttc ttcccttcct ttctcgccac gttcgccggc tttccccgtc 3000
aagctctaaa tcgggggctc cctttagggt tccgatttag tgctttacgg cacctcgacc 3060
ccaaaaaact tgattagggt gatggttcac gtagtgggcc atcgccctga tagacggttt 3120
ttcgcccttt gacgttggag tccacgttct ttaatagtgg actcttgttc caaactggaa 3180
caacactcaa ccctatctcg gtctattctt ttgatttata agggattttg ccgatttcgg 3240
cctattggtt aaaaaatgag ctgatttaac aaaaatttaa cgcgaatttt aacaaaatat 3300
taacgtctac aatttaaata tttgcttata caatcttcct gtttttgggg cttttctgat 3360
tatcaaccgg ggtacatatg attgacatgc tagttttacg attaccgttc atcgattctc 3420
ttgtttgctc cagactctca ggcaatgacc tgatagcctt tgtagagacc tctcaaaaat 3480
agctaccctc tccggcatga atttatcagc tagaacggtt gaatatcata ttgatggtga 3540
tttgactgtc tccggccttt ctcacccgtt tgaatcttta cctacacatt actcaggcat 3600
tgcatttaaa atatatgagg gttctaaaaa tttttatcct tgcgttgaaa taaaggcttc 3660
tcccgcaaaa gtattacagg gtcataatgt ttttggtaca accgatttag ctttatgctc 3720
tgaggcttta ttgcttaatt ttgctaattc tttgccttgc ctgtatgatt tattggatgt 3780
tggaatcgcc tgatgcggta ttttctcctt acgcatctgt gcggtatttc acaccgcata 3840
tggtgcactc tcagtacaat ctgctctgat gccgcatagt taagccagcc ccgacacccg 3900
ccaacacccg ctgacgcgcc ctgacgggct tgtctgctcc cggcatccgc ttacagacaa 3960
gctgtgaccg tctccgggag ctgcatgtgt cagaggtttt caccgtcatc accgaaacgc 4020
gcgagacgaa agggcctcgt gatacgccta tttttatagg ttaatgtcat gataataatg 4080
gtttcttaga cgtcaggtgg cacttttcgg ggaaatgtgc gcggaacccc tatttgttta 4140
tttttctaaa tacattcaaa tatgtatccg ctcatgagac aataaccctg ataaatgctt 4200
caataatatt gaaaaaggaa gagtatgagt attcaacatt tccgtgtcgc ccttattccc 4260
ttttttgcgg cattttgcct tcctgttttt gctcacccag aaacgctggt gaaagtaaaa 4320
gatgctgaag atcagttggg tgcacgagtg ggttacatcg aactggatct caacagcggt 4380
aagatccttg agagttttcg ccccgaagaa cgttttccaa tgatgagcac ttttaaagtt 4440
ctgctatgtg gcgcggtatt atcccgtatt gacgccgggc aagagcaact cggtcgccgc 4500
atacactatt ctcagaatga cttggttgag tactcaccag tcacagaaaa gcatcttacg 4560
gatggcatga cagtaagaga attatgcagt gctgccataa ccatgagtga taacactgcg 4620
gccaacttac ttctgacaac gatcggagga ccgaaggagc taaccgcttt tttgcacaac 4680
atgggggatc atgtaactcg ccttgatcgt tgggaaccgg agctgaatga agccatacca 4740
aacgacgagc gtgacaccac gatgcctgta gcaatggcaa caacgttgcg caaactatta 4800
actggcgaac tacttactct agcttcccgg caacaattaa tagactggat ggaggcggat 4860
aaagttgcag gaccacttct gcgctcggcc cttccggctg gctggtttat tgctgataaa 4920
tctggagccg gtgagcgtgg gtctcgcggt atcattgcag cactggggcc agatggtaag 4980
ccctcccgta tcgtagttat ctacacgacg gggagtcagg caactatgga tgaacgaaat 5040
agacagatcg ctgagatagg tgcctcactg attaagcatt ggtaactgtc agaccaagtt 5100
tactcatata tactttagat tgatttaaaa cttcattttt aatttaaaag gatctaggtg 5160
aagatccttt ttgataatct catgaccaaa atcccttaac gtgagttttc gttccactga 5220
gcgtcagacc ccgtagaaaa gatcaaagga tcttcttgag atcctttttt tctgcgcgta 5280
atctgctgct tgcaaacaaa aaaaccaccg ctaccagcgg tggtttgttt gccggatcaa 5340
gagctaccaa ctctttttcc gaaggtaact ggcttcagca gagcgcagat accaaatact 5400
gtccttctag tgtagccgta gttaggccac cacttcaaga actctgtagc accgcctaca 5460
tacctcgctc tgctaatcct gttaccagtg gctgctgcca gtggcgataa gtcgtgtctt 5520
accgggttgg actcaagacg atagttaccg gataaggcgc agcggtcggg ctgaacgggg 5580
ggttcgtgca cacagcccag cttggagcga acgacctaca ccgaactgag atacctacag 5640
cgtgagctat gagaaagcgc cacgcttccc gaagggagaa aggcggacag gtatccggta 5700
agcggcaggg tcggaacagg agagcgcacg agggagcttc cagggggaaa cgcctggtat 5760
ctttatagtc ctgtcgggtt tcgccacctc tgacttgagc gtcgattttt gtgatgctcg 5820
tcaggggggc ggagcctatg gaaaaacgcc agcaacgcgg cctttttacg gttcctggcc 5880
ttttgctggc cttttgctca catgttcttt cctgcgttat cccctgattc tgtggataac 5940
cgtattaccg cctttgagtg agctgatacc gctcgccgca gccgaacgac cgagcgcagc 6000
gagtcagtga gcgaggaagc ggaagagcgc ccaatacgca aaccgcctct ccccgcgcgt 6060
tggccgattc attaatgcag cagctgcgcg ctcgctcgct cactgaggcc gcccgggcaa 6120
agcccgggcg tcgggcgacc tttggtcgcc cggcctcagt gagcgagcga gcgcgcagag 6180
agggagtggc caactccatc actaggggtt ccttgtagtt aatgattaac ccgccatgct 6240
acttatctac gtagccatgc tctagacatg gctcgaca 6278
<210> 40
<211> 6278
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pGG2-hAAT-hsoFGF21 вариант 1
<400> 40
gatctgatat catcgatgaa ttcgagctcg gtacccggcc gcagatttag gtgacactat 60
agaatatgca tcactagtaa gcttgcgaat tccagtctac agagaggtct ctgacctctg 120
ccccagctcc aaggtcagca ggcagggagg gctgtgtgtt tgctgtttgc tgcttgcaat 180
gtttgcccat tttagggaca tgagtaggct gaagtttgtt cagtgtggac ttcagaggca 240
gcacacaaac agcaagcttg cgaattccag tctacagaga ggtctctgac ctctgcccca 300
gctccaaggt cagcaggcag ggagggctgt gtgtttgctg tttgctgctt gcaatgtttg 360
cccattttag ggacatgagt aggctgaagt ttgttcagtg tggacttcag aggcagcaca 420
caaacagcaa gcttgcgaat tccagtctac agagaggtct ctgacctctg ccccagctcc 480
aaggtcagca ggcagggagg gctgtgtgtt tgctgtttgc tgcttgcaat gtttgcccat 540
tttagggaca tgagtaggct gaagtttgtt cagtgtggac ttcagaggca gcacacaaac 600
agcaagcttt gctctagact ggaattcgtc gacgagctcc ctatagtgag tcgtattaga 660
ggccgactga cccggtaccc ggggatcttg ctaccagtgg aacagccact aaggattctg 720
cagtgagagc agagggccag ctaagtggta ctctcccaga gactgtctga ctcacgccac 780
cccctccacc ttggacacag gacgctgtgg tttctgagcc aggtacaatg actcctttcg 840
gtaagtgcag tggaagctgt acactgccca ggcaaagcgt ccgggcagcg taggcgggcg 900
actcagatcc cagccagtgg acttagcccc tgtttgctcc tccgataact ggggtgacct 960
tggttaatat tcaccagcag cctcccccgt tgcccctctg gatccactgc ttaaatacgg 1020
acgaggacag ggccctgtct cctcagcttc aggcaccacc actgacctgg gacagtgaat 1080
gtccccctga tctgcggccg tgactctctt aaggtagcct tgcagaagtt ggtcgtgagg 1140
cactgggcag gtaagtatca aggttacaag acaggtttaa ggagaccaat agaaactggg 1200
cttgtcgaga cagagaagac tcttgcgttt ctgataggca cctattggtc ttactgacat 1260
ccactttgcc tttctctcca caggtgtcca ctcccagttc aattacagct cttaaggcta 1320
gagtacttaa tacgactcac tataggctag cgccaccatg gattctgatg agacaggctt 1380
cgagcacagc ggcctgtggg tttcagttct ggctggactg ctgctgggag cctgtcaggc 1440
acaccctatt ccagatagca gccctctgct gcagttcggc ggacaagtgc ggcagagata 1500
cctgtacacc gacgacgccc agcagacaga agcccacctg gaaatcagag aggatggcac 1560
agttggcgga gccgccgatc agtctcctga atctctgctc cagctgaagg ccctgaagcc 1620
tggcgtgatc cagatcctgg gcgtgaaaac cagccggttc ctgtgccaaa gacctgacgg 1680
cgccctgtat ggcagcctgc actttgatcc tgaggcctgc agcttcagag agctgctgct 1740
tgaggacggc tacaacgtgt accagtctga ggcccatggc ctgcctctgc atctgcctgg 1800
aaacaagagc cctcacagag atcccgctcc tagaggccct gccagatttc tgcctcttcc 1860
tggattgcct cctgctctgc cagagcctcc tggaattctg gctcctcagc ctcctgatgt 1920
gggcagctct gatcctctga gcatggtcgg acctagccag ggcagatctc ctagctacgc 1980
ctcttgagcg gccgcttccc tttagtgagg gttaatgctt cgagcagaca tgataagata 2040
cattgatgag tttggacaaa ccacaactag aatgcagtga aaaaaatgct ttatttgtga 2100
aatttgtgat gctattgctt tatttgtaac cattataagc tgcaataaac aagttaacaa 2160
caacaattgc attcatttta tgtttcaggt tcagggggag atgtgggagg ttttttaaag 2220
caagtaaaac ctctacaaat gtggtaaaat ccgataaggg actagagcat ggctacgtag 2280
ataagtagca tggcgggtta atcattaact acaaggaacc cctagtgatg gagttggcca 2340
ctccctctct gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc 2400
cgggctttgc ccgggcggcc tcagtgagcg agcgagcgcg ccagctggcg taatagcgaa 2460
gaggcccgca ccgatcgccc ttcccaacag ttgcgcagcc tgaatggcga atggaattcc 2520
agacgattga gcgtcaaaat gtaggtattt ccatgagcgt ttttccgttg caatggctgg 2580
cggtaatatt gttctggata ttaccagcaa ggccgatagt ttgagttctt ctactcaggc 2640
aagtgatgtt attactaatc aaagaagtat tgcgacaacg gttaatttgc gtgatggaca 2700
gactctttta ctcggtggcc tcactgatta taaaaacact tctcaggatt ctggcgtacc 2760
gttcctgtct aaaatccctt taatcggcct cctgtttagc tcccgctctg attctaacga 2820
ggaaagcacg ttatacgtgc tcgtcaaagc aaccatagta cgcgccctgt agcggcgcat 2880
taagcgcggc gggtgtggtg gttacgcgca gcgtgaccgc tacacttgcc agcgccctag 2940
cgcccgctcc tttcgctttc ttcccttcct ttctcgccac gttcgccggc tttccccgtc 3000
aagctctaaa tcgggggctc cctttagggt tccgatttag tgctttacgg cacctcgacc 3060
ccaaaaaact tgattagggt gatggttcac gtagtgggcc atcgccctga tagacggttt 3120
ttcgcccttt gacgttggag tccacgttct ttaatagtgg actcttgttc caaactggaa 3180
caacactcaa ccctatctcg gtctattctt ttgatttata agggattttg ccgatttcgg 3240
cctattggtt aaaaaatgag ctgatttaac aaaaatttaa cgcgaatttt aacaaaatat 3300
taacgtctac aatttaaata tttgcttata caatcttcct gtttttgggg cttttctgat 3360
tatcaaccgg ggtacatatg attgacatgc tagttttacg attaccgttc atcgattctc 3420
ttgtttgctc cagactctca ggcaatgacc tgatagcctt tgtagagacc tctcaaaaat 3480
agctaccctc tccggcatga atttatcagc tagaacggtt gaatatcata ttgatggtga 3540
tttgactgtc tccggccttt ctcacccgtt tgaatcttta cctacacatt actcaggcat 3600
tgcatttaaa atatatgagg gttctaaaaa tttttatcct tgcgttgaaa taaaggcttc 3660
tcccgcaaaa gtattacagg gtcataatgt ttttggtaca accgatttag ctttatgctc 3720
tgaggcttta ttgcttaatt ttgctaattc tttgccttgc ctgtatgatt tattggatgt 3780
tggaatcgcc tgatgcggta ttttctcctt acgcatctgt gcggtatttc acaccgcata 3840
tggtgcactc tcagtacaat ctgctctgat gccgcatagt taagccagcc ccgacacccg 3900
ccaacacccg ctgacgcgcc ctgacgggct tgtctgctcc cggcatccgc ttacagacaa 3960
gctgtgaccg tctccgggag ctgcatgtgt cagaggtttt caccgtcatc accgaaacgc 4020
gcgagacgaa agggcctcgt gatacgccta tttttatagg ttaatgtcat gataataatg 4080
gtttcttaga cgtcaggtgg cacttttcgg ggaaatgtgc gcggaacccc tatttgttta 4140
tttttctaaa tacattcaaa tatgtatccg ctcatgagac aataaccctg ataaatgctt 4200
caataatatt gaaaaaggaa gagtatgagt attcaacatt tccgtgtcgc ccttattccc 4260
ttttttgcgg cattttgcct tcctgttttt gctcacccag aaacgctggt gaaagtaaaa 4320
gatgctgaag atcagttggg tgcacgagtg ggttacatcg aactggatct caacagcggt 4380
aagatccttg agagttttcg ccccgaagaa cgttttccaa tgatgagcac ttttaaagtt 4440
ctgctatgtg gcgcggtatt atcccgtatt gacgccgggc aagagcaact cggtcgccgc 4500
atacactatt ctcagaatga cttggttgag tactcaccag tcacagaaaa gcatcttacg 4560
gatggcatga cagtaagaga attatgcagt gctgccataa ccatgagtga taacactgcg 4620
gccaacttac ttctgacaac gatcggagga ccgaaggagc taaccgcttt tttgcacaac 4680
atgggggatc atgtaactcg ccttgatcgt tgggaaccgg agctgaatga agccatacca 4740
aacgacgagc gtgacaccac gatgcctgta gcaatggcaa caacgttgcg caaactatta 4800
actggcgaac tacttactct agcttcccgg caacaattaa tagactggat ggaggcggat 4860
aaagttgcag gaccacttct gcgctcggcc cttccggctg gctggtttat tgctgataaa 4920
tctggagccg gtgagcgtgg gtctcgcggt atcattgcag cactggggcc agatggtaag 4980
ccctcccgta tcgtagttat ctacacgacg gggagtcagg caactatgga tgaacgaaat 5040
agacagatcg ctgagatagg tgcctcactg attaagcatt ggtaactgtc agaccaagtt 5100
tactcatata tactttagat tgatttaaaa cttcattttt aatttaaaag gatctaggtg 5160
aagatccttt ttgataatct catgaccaaa atcccttaac gtgagttttc gttccactga 5220
gcgtcagacc ccgtagaaaa gatcaaagga tcttcttgag atcctttttt tctgcgcgta 5280
atctgctgct tgcaaacaaa aaaaccaccg ctaccagcgg tggtttgttt gccggatcaa 5340
gagctaccaa ctctttttcc gaaggtaact ggcttcagca gagcgcagat accaaatact 5400
gtccttctag tgtagccgta gttaggccac cacttcaaga actctgtagc accgcctaca 5460
tacctcgctc tgctaatcct gttaccagtg gctgctgcca gtggcgataa gtcgtgtctt 5520
accgggttgg actcaagacg atagttaccg gataaggcgc agcggtcggg ctgaacgggg 5580
ggttcgtgca cacagcccag cttggagcga acgacctaca ccgaactgag atacctacag 5640
cgtgagctat gagaaagcgc cacgcttccc gaagggagaa aggcggacag gtatccggta 5700
agcggcaggg tcggaacagg agagcgcacg agggagcttc cagggggaaa cgcctggtat 5760
ctttatagtc ctgtcgggtt tcgccacctc tgacttgagc gtcgattttt gtgatgctcg 5820
tcaggggggc ggagcctatg gaaaaacgcc agcaacgcgg cctttttacg gttcctggcc 5880
ttttgctggc cttttgctca catgttcttt cctgcgttat cccctgattc tgtggataac 5940
cgtattaccg cctttgagtg agctgatacc gctcgccgca gccgaacgac cgagcgcagc 6000
gagtcagtga gcgaggaagc ggaagagcgc ccaatacgca aaccgcctct ccccgcgcgt 6060
tggccgattc attaatgcag cagctgcgcg ctcgctcgct cactgaggcc gcccgggcaa 6120
agcccgggcg tcgggcgacc tttggtcgcc cggcctcagt gagcgagcga gcgcgcagag 6180
agggagtggc caactccatc actaggggtt ccttgtagtt aatgattaac ccgccatgct 6240
acttatctac gtagccatgc tctagacatg gctcgaca 6278
<210> 41
<211> 6278
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pGG2-hAAT-hsoFGF21 вариант 2
<400> 41
cagcagctgc gcgctcgctc gctcactgag gccgcccggg caaagcccgg gcgtcgggcg 60
acctttggtc gcccggcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc 120
atcactaggg gttccttgta gttaatgatt aacccgccat gctacttatc tacgtagcca 180
tgctctagac atggctcgac agatctgata tcatcgatga attcgagctc ggtacccggc 240
cgcagattta ggtgacacta tagaatatgc atcactagta agcttgcgaa ttccagtcta 300
cagagaggtc tctgacctct gccccagctc caaggtcagc aggcagggag ggctgtgtgt 360
ttgctgtttg ctgcttgcaa tgtttgccca ttttagggac atgagtaggc tgaagtttgt 420
tcagtgtgga cttcagaggc agcacacaaa cagcaagctt gcgaattcca gtctacagag 480
aggtctctga cctctgcccc agctccaagg tcagcaggca gggagggctg tgtgtttgct 540
gtttgctgct tgcaatgttt gcccatttta gggacatgag taggctgaag tttgttcagt 600
gtggacttca gaggcagcac acaaacagca agcttgcgaa ttccagtcta cagagaggtc 660
tctgacctct gccccagctc caaggtcagc aggcagggag ggctgtgtgt ttgctgtttg 720
ctgcttgcaa tgtttgccca ttttagggac atgagtaggc tgaagtttgt tcagtgtgga 780
cttcagaggc agcacacaaa cagcaagctt tgctctagac tggaattcgt cgacgagctc 840
cctatagtga gtcgtattag aggccgactg acccggtacc cggggatctt gctaccagtg 900
gaacagccac taaggattct gcagtgagag cagagggcca gctaagtggt actctcccag 960
agactgtctg actcacgcca ccccctccac cttggacaca ggacgctgtg gtttctgagc 1020
caggtacaat gactcctttc ggtaagtgca gtggaagctg tacactgccc aggcaaagcg 1080
tccgggcagc gtaggcgggc gactcagatc ccagccagtg gacttagccc ctgtttgctc 1140
ctccgataac tggggtgacc ttggttaata ttcaccagca gcctcccccg ttgcccctct 1200
ggatccactg cttaaatacg gacgaggaca gggccctgtc tcctcagctt caggcaccac 1260
cactgacctg ggacagtgaa tgtccccctg atctgcggcc gtgactctct taaggtagcc 1320
ttgcagaagt tggtcgtgag gcactgggca ggtaagtatc aaggttacaa gacaggttta 1380
aggagaccaa tagaaactgg gcttgtcgag acagagaaga ctcttgcgtt tctgataggc 1440
acctattggt cttactgaca tccactttgc ctttctctcc acaggtgtcc actcccagtt 1500
caattacagc tcttaaggct agagtactta atacgactca ctataggcta gcgccaccat 1560
ggacagcgat gaaaccgggt tcgagcacag cggtctgtgg gtgtccgtgc tggccggact 1620
gctcctggga gcctgtcagg cgcaccccat ccctgactcc tcgccgctgc tgcaattcgg 1680
cggacaagtc cgccagagat acctgtacac cgacgacgcc cagcagaccg aagcccacct 1740
ggaaattcgg gaggacggga ctgtgggagg cgctgcagat cagtcacccg agtccctcct 1800
ccaactgaag gccttgaagc ccggcgtgat tcagatcctg ggcgtgaaaa cttcccgctt 1860
cctttgccaa cggccggatg gagctctgta cggatccctg cacttcgacc ccgaagcctg 1920
ctcattccgc gagctgctcc ttgaggacgg ctataacgtg taccagtctg aggcccatgg 1980
actccccctg catctgcccg gcaacaagtc ccctcaccgg gatcctgccc caagaggccc 2040
agctcggttt ctgcctctgc cgggactgcc tccagcgttg cccgaacccc ctggtatcct 2100
ggccccgcaa ccacctgacg tcggttcgtc ggacccgctg agcatggtcg gtccgagcca 2160
gggaaggtcc ccgtcctacg catcctgagc ggccgcttcc ctttagtgag ggttaatgct 2220
tcgagcagac atgataagat acattgatga gtttggacaa accacaacta gaatgcagtg 2280
aaaaaaatgc tttatttgtg aaatttgtga tgctattgct ttatttgtaa ccattataag 2340
ctgcaataaa caagttaaca acaacaattg cattcatttt atgtttcagg ttcaggggga 2400
gatgtgggag gttttttaaa gcaagtaaaa cctctacaaa tgtggtaaaa tccgataagg 2460
gactagagca tggctacgta gataagtagc atggcgggtt aatcattaac tacaaggaac 2520
ccctagtgat ggagttggcc actccctctc tgcgcgctcg ctcgctcact gaggccgggc 2580
gaccaaaggt cgcccgacgc ccgggctttg cccgggcggc ctcagtgagc gagcgagcgc 2640
gccagctggc gtaatagcga agaggcccgc accgatcgcc cttcccaaca gttgcgcagc 2700
ctgaatggcg aatggaattc cagacgattg agcgtcaaaa tgtaggtatt tccatgagcg 2760
tttttccgtt gcaatggctg gcggtaatat tgttctggat attaccagca aggccgatag 2820
tttgagttct tctactcagg caagtgatgt tattactaat caaagaagta ttgcgacaac 2880
ggttaatttg cgtgatggac agactctttt actcggtggc ctcactgatt ataaaaacac 2940
ttctcaggat tctggcgtac cgttcctgtc taaaatccct ttaatcggcc tcctgtttag 3000
ctcccgctct gattctaacg aggaaagcac gttatacgtg ctcgtcaaag caaccatagt 3060
acgcgccctg tagcggcgca ttaagcgcgg cgggtgtggt ggttacgcgc agcgtgaccg 3120
ctacacttgc cagcgcccta gcgcccgctc ctttcgcttt cttcccttcc tttctcgcca 3180
cgttcgccgg ctttccccgt caagctctaa atcgggggct ccctttaggg ttccgattta 3240
gtgctttacg gcacctcgac cccaaaaaac ttgattaggg tgatggttca cgtagtgggc 3300
catcgccctg atagacggtt tttcgccctt tgacgttgga gtccacgttc tttaatagtg 3360
gactcttgtt ccaaactgga acaacactca accctatctc ggtctattct tttgatttat 3420
aagggatttt gccgatttcg gcctattggt taaaaaatga gctgatttaa caaaaattta 3480
acgcgaattt taacaaaata ttaacgtcta caatttaaat atttgcttat acaatcttcc 3540
tgtttttggg gcttttctga ttatcaaccg gggtacatat gattgacatg ctagttttac 3600
gattaccgtt catcgattct cttgtttgct ccagactctc aggcaatgac ctgatagcct 3660
ttgtagagac ctctcaaaaa tagctaccct ctccggcatg aatttatcag ctagaacggt 3720
tgaatatcat attgatggtg atttgactgt ctccggcctt tctcacccgt ttgaatcttt 3780
acctacacat tactcaggca ttgcatttaa aatatatgag ggttctaaaa atttttatcc 3840
ttgcgttgaa ataaaggctt ctcccgcaaa agtattacag ggtcataatg tttttggtac 3900
aaccgattta gctttatgct ctgaggcttt attgcttaat tttgctaatt ctttgccttg 3960
cctgtatgat ttattggatg ttggaatcgc ctgatgcggt attttctcct tacgcatctg 4020
tgcggtattt cacaccgcat atggtgcact ctcagtacaa tctgctctga tgccgcatag 4080
ttaagccagc cccgacaccc gccaacaccc gctgacgcgc cctgacgggc ttgtctgctc 4140
ccggcatccg cttacagaca agctgtgacc gtctccggga gctgcatgtg tcagaggttt 4200
tcaccgtcat caccgaaacg cgcgagacga aagggcctcg tgatacgcct atttttatag 4260
gttaatgtca tgataataat ggtttcttag acgtcaggtg gcacttttcg gggaaatgtg 4320
cgcggaaccc ctatttgttt atttttctaa atacattcaa atatgtatcc gctcatgaga 4380
caataaccct gataaatgct tcaataatat tgaaaaagga agagtatgag tattcaacat 4440
ttccgtgtcg cccttattcc cttttttgcg gcattttgcc ttcctgtttt tgctcaccca 4500
gaaacgctgg tgaaagtaaa agatgctgaa gatcagttgg gtgcacgagt gggttacatc 4560
gaactggatc tcaacagcgg taagatcctt gagagttttc gccccgaaga acgttttcca 4620
atgatgagca cttttaaagt tctgctatgt ggcgcggtat tatcccgtat tgacgccggg 4680
caagagcaac tcggtcgccg catacactat tctcagaatg acttggttga gtactcacca 4740
gtcacagaaa agcatcttac ggatggcatg acagtaagag aattatgcag tgctgccata 4800
accatgagtg ataacactgc ggccaactta cttctgacaa cgatcggagg accgaaggag 4860
ctaaccgctt ttttgcacaa catgggggat catgtaactc gccttgatcg ttgggaaccg 4920
gagctgaatg aagccatacc aaacgacgag cgtgacacca cgatgcctgt agcaatggca 4980
acaacgttgc gcaaactatt aactggcgaa ctacttactc tagcttcccg gcaacaatta 5040
atagactgga tggaggcgga taaagttgca ggaccacttc tgcgctcggc ccttccggct 5100
ggctggttta ttgctgataa atctggagcc ggtgagcgtg ggtctcgcgg tatcattgca 5160
gcactggggc cagatggtaa gccctcccgt atcgtagtta tctacacgac ggggagtcag 5220
gcaactatgg atgaacgaaa tagacagatc gctgagatag gtgcctcact gattaagcat 5280
tggtaactgt cagaccaagt ttactcatat atactttaga ttgatttaaa acttcatttt 5340
taatttaaaa ggatctaggt gaagatcctt tttgataatc tcatgaccaa aatcccttaa 5400
cgtgagtttt cgttccactg agcgtcagac cccgtagaaa agatcaaagg atcttcttga 5460
gatccttttt ttctgcgcgt aatctgctgc ttgcaaacaa aaaaaccacc gctaccagcg 5520
gtggtttgtt tgccggatca agagctacca actctttttc cgaaggtaac tggcttcagc 5580
agagcgcaga taccaaatac tgtccttcta gtgtagccgt agttaggcca ccacttcaag 5640
aactctgtag caccgcctac atacctcgct ctgctaatcc tgttaccagt ggctgctgcc 5700
agtggcgata agtcgtgtct taccgggttg gactcaagac gatagttacc ggataaggcg 5760
cagcggtcgg gctgaacggg gggttcgtgc acacagccca gcttggagcg aacgacctac 5820
accgaactga gatacctaca gcgtgagcta tgagaaagcg ccacgcttcc cgaagggaga 5880
aaggcggaca ggtatccggt aagcggcagg gtcggaacag gagagcgcac gagggagctt 5940
ccagggggaa acgcctggta tctttatagt cctgtcgggt ttcgccacct ctgacttgag 6000
cgtcgatttt tgtgatgctc gtcagggggg cggagcctat ggaaaaacgc cagcaacgcg 6060
gcctttttac ggttcctggc cttttgctgg ccttttgctc acatgttctt tcctgcgtta 6120
tcccctgatt ctgtggataa ccgtattacc gcctttgagt gagctgatac cgctcgccgc 6180
agccgaacga ccgagcgcag cgagtcagtg agcgaggaag cggaagagcg cccaatacgc 6240
aaaccgcctc tccccgcgcg ttggccgatt cattaatg 6278
<210> 42
<211> 6278
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pGG2-hAAT-hsoFGF21 вариант 3
<400> 42
cagcagctgc gcgctcgctc gctcactgag gccgcccggg caaagcccgg gcgtcgggcg 60
acctttggtc gcccggcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc 120
atcactaggg gttccttgta gttaatgatt aacccgccat gctacttatc tacgtagcca 180
tgctctagac atggctcgac agatctgata tcatcgatga attcgagctc ggtacccggc 240
cgcagattta ggtgacacta tagaatatgc atcactagta agcttgcgaa ttccagtcta 300
cagagaggtc tctgacctct gccccagctc caaggtcagc aggcagggag ggctgtgtgt 360
ttgctgtttg ctgcttgcaa tgtttgccca ttttagggac atgagtaggc tgaagtttgt 420
tcagtgtgga cttcagaggc agcacacaaa cagcaagctt gcgaattcca gtctacagag 480
aggtctctga cctctgcccc agctccaagg tcagcaggca gggagggctg tgtgtttgct 540
gtttgctgct tgcaatgttt gcccatttta gggacatgag taggctgaag tttgttcagt 600
gtggacttca gaggcagcac acaaacagca agcttgcgaa ttccagtcta cagagaggtc 660
tctgacctct gccccagctc caaggtcagc aggcagggag ggctgtgtgt ttgctgtttg 720
ctgcttgcaa tgtttgccca ttttagggac atgagtaggc tgaagtttgt tcagtgtgga 780
cttcagaggc agcacacaaa cagcaagctt tgctctagac tggaattcgt cgacgagctc 840
cctatagtga gtcgtattag aggccgactg acccggtacc cggggatctt gctaccagtg 900
gaacagccac taaggattct gcagtgagag cagagggcca gctaagtggt actctcccag 960
agactgtctg actcacgcca ccccctccac cttggacaca ggacgctgtg gtttctgagc 1020
caggtacaat gactcctttc ggtaagtgca gtggaagctg tacactgccc aggcaaagcg 1080
tccgggcagc gtaggcgggc gactcagatc ccagccagtg gacttagccc ctgtttgctc 1140
ctccgataac tggggtgacc ttggttaata ttcaccagca gcctcccccg ttgcccctct 1200
ggatccactg cttaaatacg gacgaggaca gggccctgtc tcctcagctt caggcaccac 1260
cactgacctg ggacagtgaa tgtccccctg atctgcggcc gtgactctct taaggtagcc 1320
ttgcagaagt tggtcgtgag gcactgggca ggtaagtatc aaggttacaa gacaggttta 1380
aggagaccaa tagaaactgg gcttgtcgag acagagaaga ctcttgcgtt tctgataggc 1440
acctattggt cttactgaca tccactttgc ctttctctcc acaggtgtcc actcccagtt 1500
caattacagc tcttaaggct agagtactta atacgactca ctataggcta gcgccaccat 1560
ggattccgac gaaactggat ttgaacattc agggctgtgg gtctctgtgc tggctggact 1620
gctgctgggg gcttgtcagg ctcaccccat ccctgacagc tcccctctgc tgcagttcgg 1680
aggacaggtg cggcagagat acctgtatac cgacgatgcc cagcagacag aggcacacct 1740
ggagatcagg gaggacggaa ccgtgggagg agcagccgat cagtctcccg agagcctgct 1800
gcagctgaag gccctgaagc ctggcgtgat ccagatcctg ggcgtgaaga catctcggtt 1860
tctgtgccag cggcccgacg gcgccctgta cggctccctg cacttcgatc ccgaggcctg 1920
ttcttttagg gagctgctgc tggaggacgg ctacaacgtg tatcagagcg aggcacacgg 1980
cctgccactg cacctgcctg gcaataagtc ccctcaccgc gatccagcac ccaggggccc 2040
agcacgcttc ctgcctctgc caggcctgcc ccctgccctg ccagagccac ccggcatcct 2100
ggccccccag cctccagatg tgggctccag cgatcctctg tcaatggtgg ggccaagtca 2160
ggggcggagt ccttcatacg catcataagc ggccgcttcc ctttagtgag ggttaatgct 2220
tcgagcagac atgataagat acattgatga gtttggacaa accacaacta gaatgcagtg 2280
aaaaaaatgc tttatttgtg aaatttgtga tgctattgct ttatttgtaa ccattataag 2340
ctgcaataaa caagttaaca acaacaattg cattcatttt atgtttcagg ttcaggggga 2400
gatgtgggag gttttttaaa gcaagtaaaa cctctacaaa tgtggtaaaa tccgataagg 2460
gactagagca tggctacgta gataagtagc atggcgggtt aatcattaac tacaaggaac 2520
ccctagtgat ggagttggcc actccctctc tgcgcgctcg ctcgctcact gaggccgggc 2580
gaccaaaggt cgcccgacgc ccgggctttg cccgggcggc ctcagtgagc gagcgagcgc 2640
gccagctggc gtaatagcga agaggcccgc accgatcgcc cttcccaaca gttgcgcagc 2700
ctgaatggcg aatggaattc cagacgattg agcgtcaaaa tgtaggtatt tccatgagcg 2760
tttttccgtt gcaatggctg gcggtaatat tgttctggat attaccagca aggccgatag 2820
tttgagttct tctactcagg caagtgatgt tattactaat caaagaagta ttgcgacaac 2880
ggttaatttg cgtgatggac agactctttt actcggtggc ctcactgatt ataaaaacac 2940
ttctcaggat tctggcgtac cgttcctgtc taaaatccct ttaatcggcc tcctgtttag 3000
ctcccgctct gattctaacg aggaaagcac gttatacgtg ctcgtcaaag caaccatagt 3060
acgcgccctg tagcggcgca ttaagcgcgg cgggtgtggt ggttacgcgc agcgtgaccg 3120
ctacacttgc cagcgcccta gcgcccgctc ctttcgcttt cttcccttcc tttctcgcca 3180
cgttcgccgg ctttccccgt caagctctaa atcgggggct ccctttaggg ttccgattta 3240
gtgctttacg gcacctcgac cccaaaaaac ttgattaggg tgatggttca cgtagtgggc 3300
catcgccctg atagacggtt tttcgccctt tgacgttgga gtccacgttc tttaatagtg 3360
gactcttgtt ccaaactgga acaacactca accctatctc ggtctattct tttgatttat 3420
aagggatttt gccgatttcg gcctattggt taaaaaatga gctgatttaa caaaaattta 3480
acgcgaattt taacaaaata ttaacgtcta caatttaaat atttgcttat acaatcttcc 3540
tgtttttggg gcttttctga ttatcaaccg gggtacatat gattgacatg ctagttttac 3600
gattaccgtt catcgattct cttgtttgct ccagactctc aggcaatgac ctgatagcct 3660
ttgtagagac ctctcaaaaa tagctaccct ctccggcatg aatttatcag ctagaacggt 3720
tgaatatcat attgatggtg atttgactgt ctccggcctt tctcacccgt ttgaatcttt 3780
acctacacat tactcaggca ttgcatttaa aatatatgag ggttctaaaa atttttatcc 3840
ttgcgttgaa ataaaggctt ctcccgcaaa agtattacag ggtcataatg tttttggtac 3900
aaccgattta gctttatgct ctgaggcttt attgcttaat tttgctaatt ctttgccttg 3960
cctgtatgat ttattggatg ttggaatcgc ctgatgcggt attttctcct tacgcatctg 4020
tgcggtattt cacaccgcat atggtgcact ctcagtacaa tctgctctga tgccgcatag 4080
ttaagccagc cccgacaccc gccaacaccc gctgacgcgc cctgacgggc ttgtctgctc 4140
ccggcatccg cttacagaca agctgtgacc gtctccggga gctgcatgtg tcagaggttt 4200
tcaccgtcat caccgaaacg cgcgagacga aagggcctcg tgatacgcct atttttatag 4260
gttaatgtca tgataataat ggtttcttag acgtcaggtg gcacttttcg gggaaatgtg 4320
cgcggaaccc ctatttgttt atttttctaa atacattcaa atatgtatcc gctcatgaga 4380
caataaccct gataaatgct tcaataatat tgaaaaagga agagtatgag tattcaacat 4440
ttccgtgtcg cccttattcc cttttttgcg gcattttgcc ttcctgtttt tgctcaccca 4500
gaaacgctgg tgaaagtaaa agatgctgaa gatcagttgg gtgcacgagt gggttacatc 4560
gaactggatc tcaacagcgg taagatcctt gagagttttc gccccgaaga acgttttcca 4620
atgatgagca cttttaaagt tctgctatgt ggcgcggtat tatcccgtat tgacgccggg 4680
caagagcaac tcggtcgccg catacactat tctcagaatg acttggttga gtactcacca 4740
gtcacagaaa agcatcttac ggatggcatg acagtaagag aattatgcag tgctgccata 4800
accatgagtg ataacactgc ggccaactta cttctgacaa cgatcggagg accgaaggag 4860
ctaaccgctt ttttgcacaa catgggggat catgtaactc gccttgatcg ttgggaaccg 4920
gagctgaatg aagccatacc aaacgacgag cgtgacacca cgatgcctgt agcaatggca 4980
acaacgttgc gcaaactatt aactggcgaa ctacttactc tagcttcccg gcaacaatta 5040
atagactgga tggaggcgga taaagttgca ggaccacttc tgcgctcggc ccttccggct 5100
ggctggttta ttgctgataa atctggagcc ggtgagcgtg ggtctcgcgg tatcattgca 5160
gcactggggc cagatggtaa gccctcccgt atcgtagtta tctacacgac ggggagtcag 5220
gcaactatgg atgaacgaaa tagacagatc gctgagatag gtgcctcact gattaagcat 5280
tggtaactgt cagaccaagt ttactcatat atactttaga ttgatttaaa acttcatttt 5340
taatttaaaa ggatctaggt gaagatcctt tttgataatc tcatgaccaa aatcccttaa 5400
cgtgagtttt cgttccactg agcgtcagac cccgtagaaa agatcaaagg atcttcttga 5460
gatccttttt ttctgcgcgt aatctgctgc ttgcaaacaa aaaaaccacc gctaccagcg 5520
gtggtttgtt tgccggatca agagctacca actctttttc cgaaggtaac tggcttcagc 5580
agagcgcaga taccaaatac tgtccttcta gtgtagccgt agttaggcca ccacttcaag 5640
aactctgtag caccgcctac atacctcgct ctgctaatcc tgttaccagt ggctgctgcc 5700
agtggcgata agtcgtgtct taccgggttg gactcaagac gatagttacc ggataaggcg 5760
cagcggtcgg gctgaacggg gggttcgtgc acacagccca gcttggagcg aacgacctac 5820
accgaactga gatacctaca gcgtgagcta tgagaaagcg ccacgcttcc cgaagggaga 5880
aaggcggaca ggtatccggt aagcggcagg gtcggaacag gagagcgcac gagggagctt 5940
ccagggggaa acgcctggta tctttatagt cctgtcgggt ttcgccacct ctgacttgag 6000
cgtcgatttt tgtgatgctc gtcagggggg cggagcctat ggaaaaacgc cagcaacgcg 6060
gcctttttac ggttcctggc cttttgctgg ccttttgctc acatgttctt tcctgcgtta 6120
tcccctgatt ctgtggataa ccgtattacc gcctttgagt gagctgatac cgctcgccgc 6180
agccgaacga ccgagcgcag cgagtcagtg agcgaggaag cggaagagcg cccaatacgc 6240
aaaccgcctc tccccgcgcg ttggccgatt cattaatg 6278
<210> 43
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Химерный интрон, состоящий из интронов из генов человеческого бета-глобина и тяжелой цепи иммуноглобулина
<400> 43
gtaagtatca aggttacaag acaggtttaa ggagaccaat agaaactggg cttgtcgaga 60
cagagaagac tcttgcgttt ctgataggca cctattggtc ttactgacat ccactttgcc 120
tttctctcca cag 133
<210> 44
<211> 1671
<212> ДНК
<213> Искусственная последовательность
<220>
<223> промотор CAG
<400> 44
gacattgatt attgactagt tattaatagt aatcaattac ggggtcatta gttcatagcc 60
catatatgga gttccgcgtt acataactta cggtaaatgg cccgcctggc tgaccgccca 120
acgacccccg cccattgacg tcaataatga cgtatgttcc catagtaacg ccaataggga 180
ctttccattg acgtcaatgg gtggagtatt tacggtaaac tgcccacttg gcagtacatc 240
aagtgtatca tatgccaagt acgcccccta ttgacgtcaa tgacggtaaa tggcccgcct 300
ggcattatgc ccagtacatg accttatggg actttcctac ttggcagtac atctacgtat 360
tagtcatcgc tattaccatg gtcgaggtga gccccacgtt ctgcttcact ctccccatct 420
cccccccctc cccaccccca attttgtatt tatttatttt ttaattattt tgtgcagcga 480
tgggggcggg gggggggggg gggcgcgcgc caggcggggc ggggcggggc gaggggcggg 540
gcggggcgag gcggagaggt gcggcggcag ccaatcagag cggcgcgctc cgaaagtttc 600
cttttatggc gaggcggcgg cggcggcggc cctataaaaa gcgaagcgcg cggcgggcgg 660
gagtcgctgc gttgccttcg ccccgtgccc cgctccgcgc cgcctcgcgc cgcccgcccc 720
ggctctgact gaccgcgtta ctcccacagg tgagcgggcg ggacggccct tctcctccgg 780
gctgtaatta gcgcttggtt taatgacggc ttgtttcttt tctgtggctg cgtgaaagcc 840
ttgaggggct ccgggagggc cctttgtgcg gggggagcgg ctcggggggt gcgtgcgtgt 900
gtgtgtgcgt ggggagcgcc gcgtgcggct ccgcgctgcc cggcggctgt gagcgctgcg 960
ggcgcggcgc ggggctttgt gcgctccgca gtgtgcgcga ggggagcgcg gccgggggcg 1020
gtgccccgcg gtgcgggggg ctgcgagggg aacaaaggct gcgtgcgggg tgtgtgcgtg 1080
ggggggtgag cagggggtgt gggcgcgtcg gtcgggctgc aaccccccct gcacccccct 1140
ccccgagttg ctgagcacgg cccggcttcg ggtgcggggc tccgtacggg gcgtggcgcg 1200
gggctcgccg tgccgggcgg ggggtggcgg caggtggggg tgccgggcgg ggcggggccg 1260
cctcgggccg gggagggctc gggggagggg cgcggcggcc cccggagcgc cggcggctgt 1320
cgaggcgcgg cgagccgcag ccattgcctt ttatggtaat cgtgcgagag ggcgcaggga 1380
cttcctttgt cccaaatctg tgcggagccg aaatctggga ggcgccgccg caccccctct 1440
agcgggcgcg gggcgaagcg gtgcggcgcc ggcaggaagg aaatgggcgg ggagggcctt 1500
cgtgcgtcgc cgcgccgccg tccccttctc cctctccagc ctcggggctg tccgcggggg 1560
gacggctgcc ttcggggggg acggggcagg gcggggttcg gcttctggcg tgtgaccggc 1620
ggctctagag cctctgctaa ccatgttcat gccttcttct ttttcctaca g 1671
<210> 45
<211> 212
<212> ДНК
<213> Искусственная последовательность
<220>
<223> промотор CMV
<400> 45
gtgatgcggt tttggcagta caccaatggg cgtggatagc ggtttgactc acggggattt 60
ccaagtctcc accccattga cgtcaatggg agtttgtttt ggcaccaaaa tcaacgggac 120
tttccaaaat gtcgtaacaa ctgcgatcgc ccgccccgtt gacgcaaatg ggcggtaggc 180
gtgtacggtg ggaggtctat ataagcagag ct 212
<210> 46
<211> 380
<212> ДНК
<213> Искусственная последовательность
<220>
<223> энхансер CMV
<400> 46
ggcattgatt attgactagt tattaatagt aatcaattac ggggtcatta gttcatagcc 60
catatatgga gttccgcgtt acataactta cggtaaatgg cccgcctggc tgaccgccca 120
acgacccccg cccattgacg tcaataatga cgtatgttcc catagtaacg ccaataggga 180
ctttccattg acgtcaatgg gtggagtatt tacggtaaac tgcccacttg gcagtacatc 240
aagtgtatca tatgccaagt ccgcccccta ttgacgtcaa tgacggtaaa tggcccgcct 300
ggcattatgc ccagtacatg accttacggg actttcctac ttggcagtac atctacgtat 360
tagtcatcgc tattaccatg 380
<210> 47
<211> 397
<212> ДНК
<213> Искусственная последовательность
<220>
<223> промотор hAAT
<400> 47
gatcttgcta ccagtggaac agccactaag gattctgcag tgagagcaga gggccagcta 60
agtggtactc tcccagagac tgtctgactc acgccacccc ctccaccttg gacacaggac 120
gctgtggttt ctgagccagg tacaatgact cctttcggta agtgcagtgg aagctgtaca 180
ctgcccaggc aaagcgtccg ggcagcgtag gcgggcgact cagatcccag ccagtggact 240
tagcccctgt ttgctcctcc gataactggg gtgaccttgg ttaatattca ccagcagcct 300
cccccgttgc ccctctggat ccactgctta aatacggacg aggacagggc cctgtctcct 360
cagcttcagg caccaccact gacctgggac agtgaat 397
<210> 48
<211> 128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Усеченный AAV2 5'ITR
<400> 48
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgacctttgg 60
tcgcccggcc tcagtgagcg agcgagcgcg cagagaggga gtggccaact ccatcactag 120
gggttcct 128
<210> 49
<211> 128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Усеченный AAV2 3'ITR
<400> 49
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc 120
gagcgcgc 128
<210> 50
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Сигнал полиаденилирования SV40
<400> 50
taagatacat tgatgagttt ggacaaacca caactagaat gcagtgaaaa aaatgcttta 60
tttgtgaaat ttgtgatgct attgctttat ttgtaaccat tataagctgc aataaacaag 120
tt 122
<210> 51
<211> 449
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Сигнал полиаденилирования бета-глобина кролика
<400> 51
gatctttttc cctctgccaa aaattatggg gacatcatga agccccttga gcatctgact 60
tctggctaat aaaggaaatt tattttcatt gcaatagtgt gttggaattt tttgtgtctc 120
tcactcggaa ggacatatgg gagggcaaat catttaaaac atcagaatga gtatttggtt 180
tagagtttgg caacatatgc ccatatgctg gctgccatga acaaaggttg gctataaaga 240
ggtcatcagt atatgaaaca gccccctgct gtccattcct tattccatag aaaagccttg 300
acttgaggtt agattttttt tatattttgt tttgtgttat ttttttcttt aacatcccta 360
aaattttcct tacatgtttt actagccaga tttttcctcc tctcctgact actcccagtc 420
atagctgtcc ctcttctctt atggagatc 449
<210> 52
<211> 592
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Промотор CMV и энхансер CMV
<400> 52
ggcattgatt attgactagt tattaatagt aatcaattac ggggtcatta gttcatagcc 60
catatatgga gttccgcgtt acataactta cggtaaatgg cccgcctggc tgaccgccca 120
acgacccccg cccattgacg tcaataatga cgtatgttcc catagtaacg ccaataggga 180
ctttccattg acgtcaatgg gtggagtatt tacggtaaac tgcccacttg gcagtacatc 240
aagtgtatca tatgccaagt ccgcccccta ttgacgtcaa tgacggtaaa tggcccgcct 300
ggcattatgc ccagtacatg accttacggg actttcctac ttggcagtac atctacgtat 360
tagtcatcgc tattaccatg gtgatgcggt tttggcagta caccaatggg cgtggatagc 420
ggtttgactc acggggattt ccaagtctcc accccattga cgtcaatggg agtttgtttt 480
ggcaccaaaa tcaacgggac tttccaaaat gtcgtaacaa ctgcgatcgc ccgccccgtt 540
gacgcaaatg ggcggtaggc gtgtacggtg ggaggtctat ataagcagag ct 592
<210> 53
<211> 154
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Энхансер области контроля гепатоцитов (HCR) из аполипопротеина E
<400> 53
cagagaggtc tctgacctct gccccagctc caaggtcagc aggcagggag ggctgtgtgt 60
ttgctgtttg ctgcttgcaa tgtttgccca ttttagggac atgagtaggc tgaagtttgt 120
tcagtgtgga cttcagaggc agcacacaaa cagc 154
<210> 54
<211> 651
<212> ДНК
<213> Искусственная последовательность
<220>
<223> промотор mini/aP2
<400> 54
gattaacccg ccatgctact tatctactcg acattgatta ttgactaggg gaattccagc 60
aggaatcagg tagctggaga atcgcacaga gccatgcgat tcttggcaag ccatgcgaca 120
aaggcagaaa tgcacatttc acccagagag aagggattga tgtcagcagg aagtcaccac 180
ccagagagca aatggagttc ccagatgcct gacatttgcc ttcttactgg atcagagttc 240
actagtggaa gtgtcacagc ccaaacactc ccccaaagct cagcccttcc ttgccttgta 300
acaatcaagc cgctcctgga tgaactgctc cgccctctgt ctctttggca gggttggagc 360
ccactgtggc ctgagcgact tctatggctc ccttttctgt gattttcatg gtttctgagc 420
tcttttcccc cgctttatga ttttctcttt ttgtctctct cttgctaaac ctccttcgta 480
tatatgccct ctcaggtttc atttctgaat catctactgt gaactattcc cattgtttgc 540
cagaagcccc ctggttcttc cttctagaca ccaggcaagg ggcaggaggt aagaggcagg 600
agtccataaa acagccctga gagcctgctg ggtcagtgcc tgctgtcaga a 651
<210> 55
<211> 722
<212> ДНК
<213> Искусственная последовательность
<220>
<223> промотор mini/UCP1
<400> 55
gacgtcacag tgggtcagtc acccttgatc acactgcacc agtcttcacc tttccacgct 60
tcctgccaga gcatgaatca ggctctctgg ggataccggc ctcaccccta ctgaggcaaa 120
ctttctccca cttctcagag gctctgaggg cagcaaggtc agccctttct ttggaatcta 180
gaaccactcc ctgtcttgag ctgacatcac agggcaggca gatgcagcag ggaagggcct 240
gggactggga cgttcatcct acaagaaagc tgtggaactt ttcagcaaca tctcagaaat 300
cagatcgcac ttattcaaag gagccaggcc ctgctctgcg ccctggtgga ggctcctcat 360
gtgaagagtg acaaaaggca ccatgttgtg gatacggggc gaagcccctc cggtgtgtcc 420
tccaggcatc atcaggaact agtgccaaag cagaggtgct ggccagggct ttgggagtga 480
cgcgcgtctg ggaggcttgt gcgcccaggg cacgcccctg ccgattccca ctagcaggtc 540
ttgggggacc tgggccggct ctgcccctcc tccagcaatc gggctataaa gctcttccaa 600
gtcagggcgc agaagtgccg ggcgatccgg gcttaaagag cgagaggaag ggacgctcac 660
ctttgagctc ctccacaaat agccctggtg gctgccacag aagttcgaag ttgagagttc 720
gg 722
<210> 56
<211> 326
<212> ДНК
<213> Искусственная последовательность
<220>
<223> промотор C5-12
<400> 56
cggccgtccg ccttcggcac catcctcacg acacccaaat atggcgacgg gtgaggaatg 60
gtggggagtt atttttagag cggtgaggaa ggtgggcagg cagcaggtgt tggcgctcta 120
aaaataactc ccgggagtta tttttagagc ggaggaatgg tggacaccca aatatggcga 180
cggttcctca cccgtcgcca tatttgggtg tccgccctcg gccggggccg cattcctggg 240
ggccgggcgg tgctcccgcc cgcctcgata aaaggctccg gggccggcgg cggcccacga 300
gctacccgga ggagcgggag gcgcca 326
<210> 57
<211> 4930
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pAAV-EF1a-mmFGF21-pA
<400> 57
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgcggccgcg gctccggtgc ccgtcagtgg gcagagcgca 180
catcgcccac agtccccgag aagttggggg gaggggtcgg caattgaacc ggtgcctaga 240
gaaggtggcg cggggtaaac tgggaaagtg atgtcgtgta ctggctccgc ctttttcccg 300
agggtggggg agaaccgtat ataagtgcag tagtcgccgt gaacgttctt tttcgcaacg 360
ggtttgccgc cagaacacag gtaagtgccg tgtgtggttc ccgcgggcct ggcctcttta 420
cgggttatgg cccttgcgtg ccttgaatta cttccactgg ctgcagtacg tgattcttga 480
tcccgagctt cgggttggaa gtgggtggga gagttcgagg ccttgcgctt aaggagcccc 540
ttcgcctcgt gcttgagttg aggcctggcc tgggcgctgg ggccgccgcg tgcgaatctg 600
gtggcacctt cgcgcctgtc tcgctgcttt cgataagtct ctagccattt aaaatttttg 660
atgacctgct gcgacgcttt ttttctggca agatagtctt gtaaatgcgg gccaagatct 720
gcacactggt atttcggttt ttggggccgc gggcggcgac ggggcccgtg cgtcccagcg 780
cacatgttcg gcgaggcggg gcctgcgagc gcggccaccg agaatcggac gggggtagtc 840
tcaagctggc cggcctgctc tggtgcctgg cctcgcgccg ccgtgtatcg ccccgccctg 900
ggcggcaagg ctggcccggt cggcaccagt tgcgtgagcg gaaagatggc cgcttcccgg 960
ccctgctgca gggagctcaa aatggaggac gcggcgctcg ggagagcggg cgggtgagtc 1020
acccacacaa aggaaaaggg cctttccgtc ctcagccgtc gcttcatgtg actccacgga 1080
gtaccgggcg ccgtccaggc acctcgatta gttctcgagc ttttggagta cgtcgtcttt 1140
aggttggggg gaggggtttt atgcgatgga gtttccccac actgagtggg tggagactga 1200
agttaggcca gcttggcact tgatgtaatt ctccttggaa tttgcccttt ttgagtttgg 1260
atcttggttc attctcaagc ctcagacagt ggttcaaagt ttttttcttc catttcaggt 1320
gtcgtgagga atttcgactg ctagcacgcg tgatatcaat ggaatggatg agatctagag 1380
ttgggaccct gggactgtgg gtccgactgc tgctggctgt cttcctgctg ggggtctacc 1440
aagcataccc catccctgac tccagccccc tcctccagtt tgggggtcaa gtccggcaga 1500
ggtacctcta cacagatgac gaccaagaca ctgaagccca cctggagatc agggaggatg 1560
gaacagtggt aggcgcagca caccgcagtc cagaaagtct cctggagctc aaagccttga 1620
agccaggggt cattcaaatc ctgggtgtca aagcctctag gtttctttgc caacagccag 1680
atggagctct ctatggatcg cctcactttg atcctgaggc ctgcagcttc agagaactgc 1740
tgctggagga cggttacaat gtgtaccagt ctgaagccca tggcctgccc ctgcgtctgc 1800
ctcagaagga ctccccaaac caggatgcaa catcctgggg acctgtgcgc ttcctgccca 1860
tgccaggcct gctccacgag ccccaagacc aagcaggatt cctgccccca gagcccccag 1920
atgtgggctc ctctgacccc ctgagcatgg tagagccttt acagggccga agccccagct 1980
atgcgtcctg agatatcaaa gaattctaag cttgtcgacg aatgcaattg ttgttaatta 2040
attgttaact tgtttattgc agcttataat ggttacaaat aaagcaatag catcacaaat 2100
ttcacaaata aagcattttt ttcactgcat tctagttgtg gtttgtccaa actcatcaat 2160
gtatcttagt cgagttaatt aacggcggcc gcaggaaccc ctagtgatgg agttggccac 2220
tccctctctg cgcgctcgct cgctcactga ggccgggcga ccaaaggtcg cccgacgccc 2280
gggctttgcc cgggcggcct cagtgagcga gcgagcgcgc agctgcctgc aggggcgcct 2340
gatgcggtat tttctcctta cgcatctgtg cggtatttca caccgcatac gtcaaagcaa 2400
ccatagtacg cgccctgtag cggcgcatta agcgcggcgg gtgtggtggt tacgcgcagc 2460
gtgaccgcta cacttgccag cgccctagcg cccgctcctt tcgctttctt cccttccttt 2520
ctcgccacgt tcgccggctt tccccgtcaa gctctaaatc gggggctccc tttagggttc 2580
cgatttagtg ctttacggca cctcgacccc aaaaaacttg atttgggtga tggttcacgt 2640
agtgggccat cgccctgata gacggttttt cgccctttga cgttggagtc cacgttcttt 2700
aatagtggac tcttgttcca aactggaaca acactcaacc ctatctcggg ctattctttt 2760
gatttataag ggattttgcc gatttcggcc tattggttaa aaaatgagct gatttaacaa 2820
aaatttaacg cgaattttaa caaaatatta acgtttacaa ttttatggtg cactctcagt 2880
acaatctgct ctgatgccgc atagttaagc cagccccgac acccgccaac acccgctgac 2940
gcgccctgac gggcttgtct gctcccggca tccgcttaca gacaagctgt gaccgtctcc 3000
gggagctgca tgtgtcagag gttttcaccg tcatcaccga aacgcgcgag acgaaagggc 3060
ctcgtgatac gcctattttt ataggttaat gtcatgataa taatggtttc ttagacgtca 3120
ggtggcactt ttcggggaaa tgtgcgcgga acccctattt gtttattttt ctaaatacat 3180
tcaaatatgt atccgctcat gagacaataa ccctgataaa tgcttcaata atattgaaaa 3240
aggaagagta tgagtattca acatttccgt gtcgccctta ttcccttttt tgcggcattt 3300
tgccttcctg tttttgctca cccagaaacg ctggtgaaag taaaagatgc tgaagatcag 3360
ttgggtgcac gagtgggtta catcgaactg gatctcaaca gcggtaagat ccttgagagt 3420
tttcgccccg aagaacgttt tccaatgatg agcactttta aagttctgct atgtggcgcg 3480
gtattatccc gtattgacgc cgggcaagag caactcggtc gccgcataca ctattctcag 3540
aatgacttgg ttgagtactc accagtcaca gaaaagcatc ttacggatgg catgacagta 3600
agagaattat gcagtgctgc cataaccatg agtgataaca ctgcggccaa cttacttctg 3660
acaacgatcg gaggaccgaa ggagctaacc gcttttttgc acaacatggg ggatcatgta 3720
actcgccttg atcgttggga accggagctg aatgaagcca taccaaacga cgagcgtgac 3780
accacgatgc ctgtagcaat ggcaacaacg ttgcgcaaac tattaactgg cgaactactt 3840
actctagctt cccggcaaca attaatagac tggatggagg cggataaagt tgcaggacca 3900
cttctgcgct cggcccttcc ggctggctgg tttattgctg ataaatctgg agccggtgag 3960
cgtgggtctc gcggtatcat tgcagcactg gggccagatg gtaagccctc ccgtatcgta 4020
gttatctaca cgacggggag tcaggcaact atggatgaac gaaatagaca gatcgctgag 4080
ataggtgcct cactgattaa gcattggtaa ctgtcagacc aagtttactc atatatactt 4140
tagattgatt taaaacttca tttttaattt aaaaggatct aggtgaagat cctttttgat 4200
aatctcatga ccaaaatccc ttaacgtgag ttttcgttcc actgagcgtc agaccccgta 4260
gaaaagatca aaggatcttc ttgagatcct ttttttctgc gcgtaatctg ctgcttgcaa 4320
acaaaaaaac caccgctacc agcggtggtt tgtttgccgg atcaagagct accaactctt 4380
tttccgaagg taactggctt cagcagagcg cagataccaa atactgtcct tctagtgtag 4440
ccgtagttag gccaccactt caagaactct gtagcaccgc ctacatacct cgctctgcta 4500
atcctgttac cagtggctgc tgccagtggc gataagtcgt gtcttaccgg gttggactca 4560
agacgatagt taccggataa ggcgcagcgg tcgggctgaa cggggggttc gtgcacacag 4620
cccagcttgg agcgaacgac ctacaccgaa ctgagatacc tacagcgtga gctatgagaa 4680
agcgccacgc ttcccgaagg gagaaaggcg gacaggtatc cggtaagcgg cagggtcgga 4740
acaggagagc gcacgaggga gcttccaggg ggaaacgcct ggtatcttta tagtcctgtc 4800
gggtttcgcc acctctgact tgagcgtcga tttttgtgat gctcgtcagg ggggcggagc 4860
ctatggaaaa acgccagcaa cgcggccttt ttacggttcc tggccttttg ctggcctttt 4920
gctcacatgt 4930
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВЕКТОРЫ И ПОСЛЕДОВАТЕЛЬНОСТИ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ | 2011 |
|
RU2588667C2 |
| АДЕНОАССОЦИИРОВАННЫЕ ВИРУСНЫЕ ВЕКТОРЫ ДЛЯ ЛЕЧЕНИЯ МУКОПОЛИСАХАРИДОЗОВ ТИПА IV A | 2019 |
|
RU2794960C2 |
| АДЕНОАССОЦИИРОВАННЫЕ ВИРУСНЫЕ ВЕКТОРЫ ДЛЯ ЛЕЧЕНИЯ ЛИЗОСОМНЫХ БОЛЕЗНЕЙ НАКОПЛЕНИЯ | 2015 |
|
RU2718248C2 |
| ВЕКТОРЫ НА ОСНОВЕ АДЕНОАССОЦИИРОВАННЫХ ВИРУСОВ ДЛЯ ЛЕЧЕНИЯ МУКОПОЛИСАХАРИДОЗОВ | 2016 |
|
RU2744593C2 |
| ЛЕЧЕНИЕ ЗАБОЛЕВАНИЙ ПОСРЕДСТВОМ ЭКСПРЕССИИ ФЕРМЕНТА, ОБЛАДАЮЩЕГО ДЕЗОКСИРИБОНУКЛЕАЗНОЙ (ДНКазной) АКТИВНОСТЬЮ, В ПЕЧЕНИ | 2019 |
|
RU2773691C2 |
| ГЕННАЯ ТЕРАПИЯ ОФТАЛЬМОЛОГИЧЕСКИХ НАРУШЕНИЙ | 2016 |
|
RU2762747C2 |
| ПОЛУЧЕНИЕ УЛУЧШЕННОЙ ЧЕЛОВЕЧЕСКОЙ PAH ДЛЯ ЛЕЧЕНИЯ ТЯЖЕЛОЙ PKU С ПОМОЩЬЮ ГЕННОЙ ЗАМЕСТИТЕЛЬНОЙ ТЕРАПИИ, НАПРАВЛЕННОЙ НА ПЕЧЕНЬ | 2019 |
|
RU2839587C2 |
| ВАРИАНТ СРЕДСТВА ДЛЯ RNAi | 2018 |
|
RU2789647C2 |
| Кодон-оптимизированная последовательность нуклеотидов, кодирующая hAIPL1, и её содержащий экспрессионный вектор | 2021 |
|
RU2785621C1 |
| ЛЕЧЕНИЕ КОМПЛЕМЕНТ-ОПОСРЕДУЕМЫХ РАССТРОЙСТВ | 2017 |
|
RU2768982C2 |
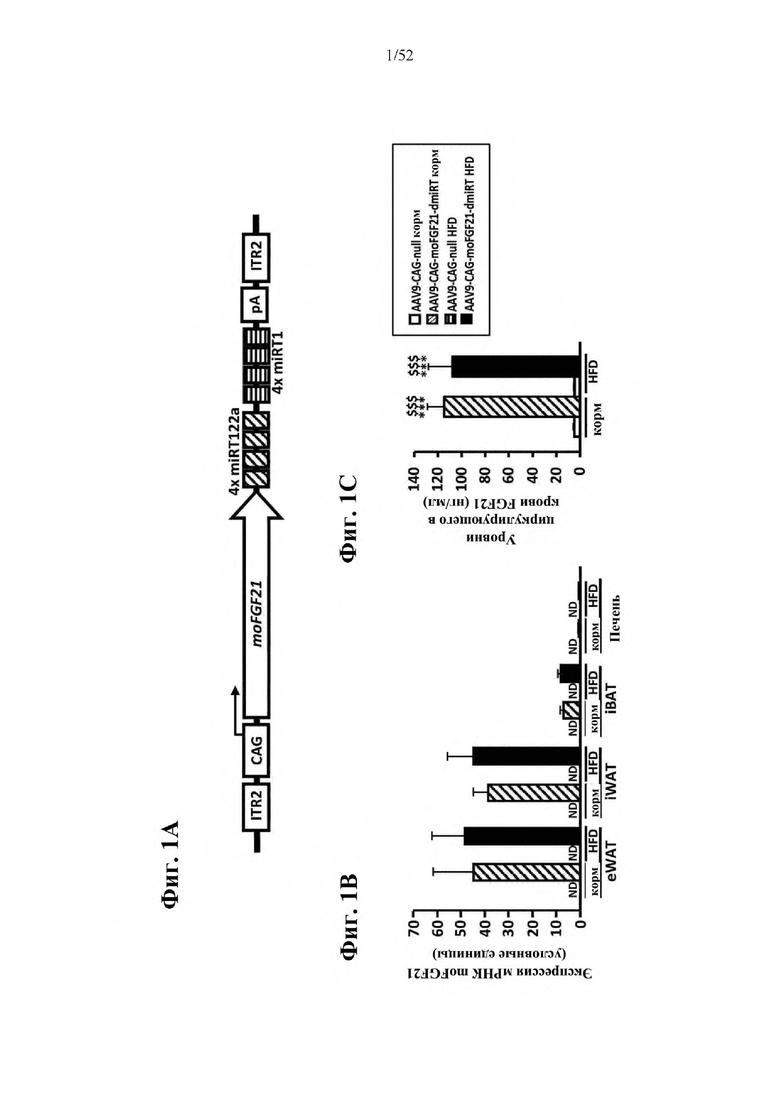
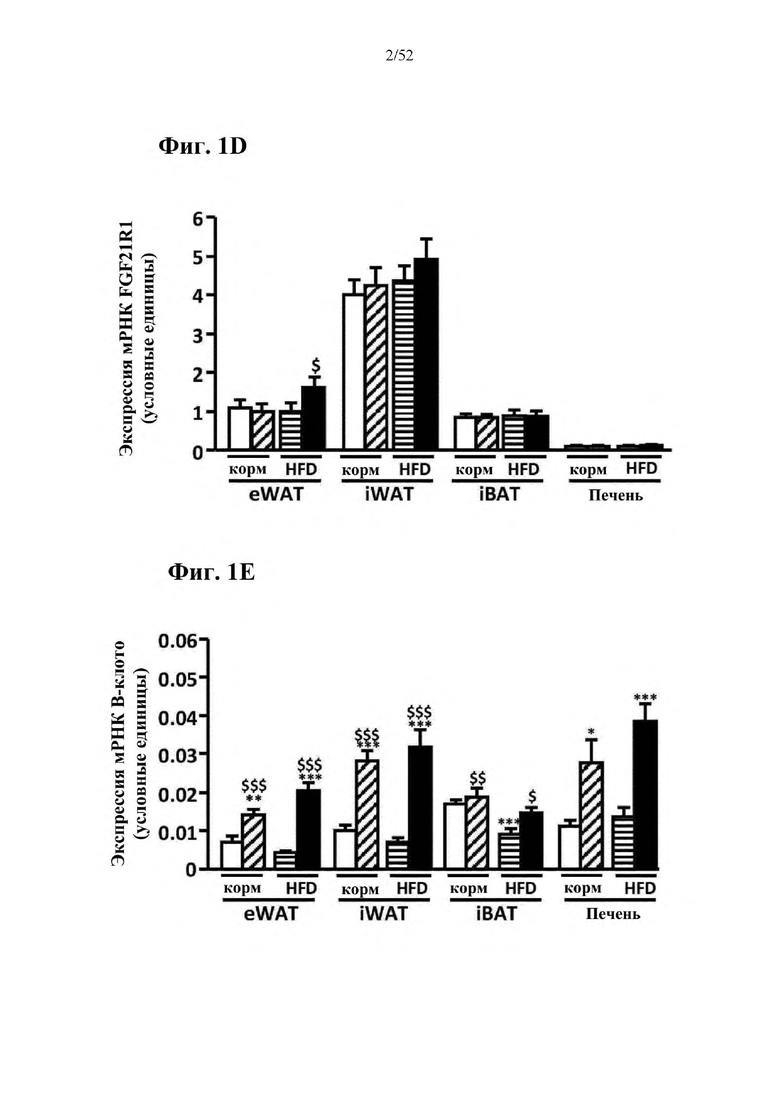
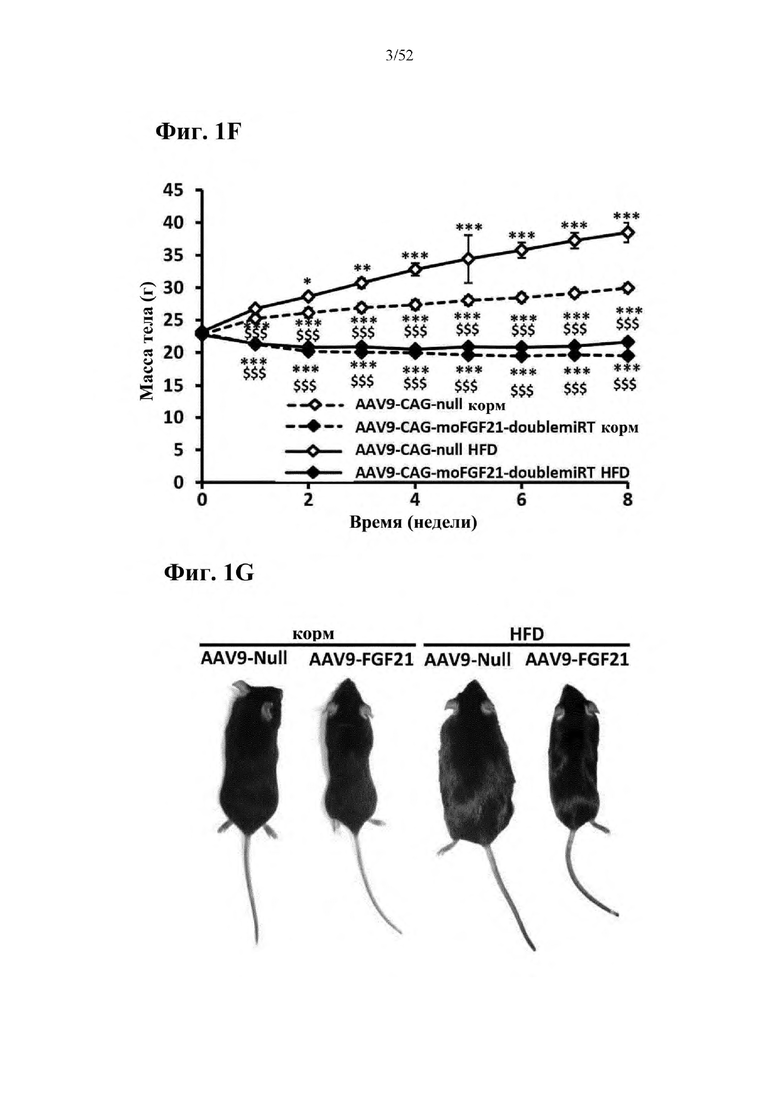
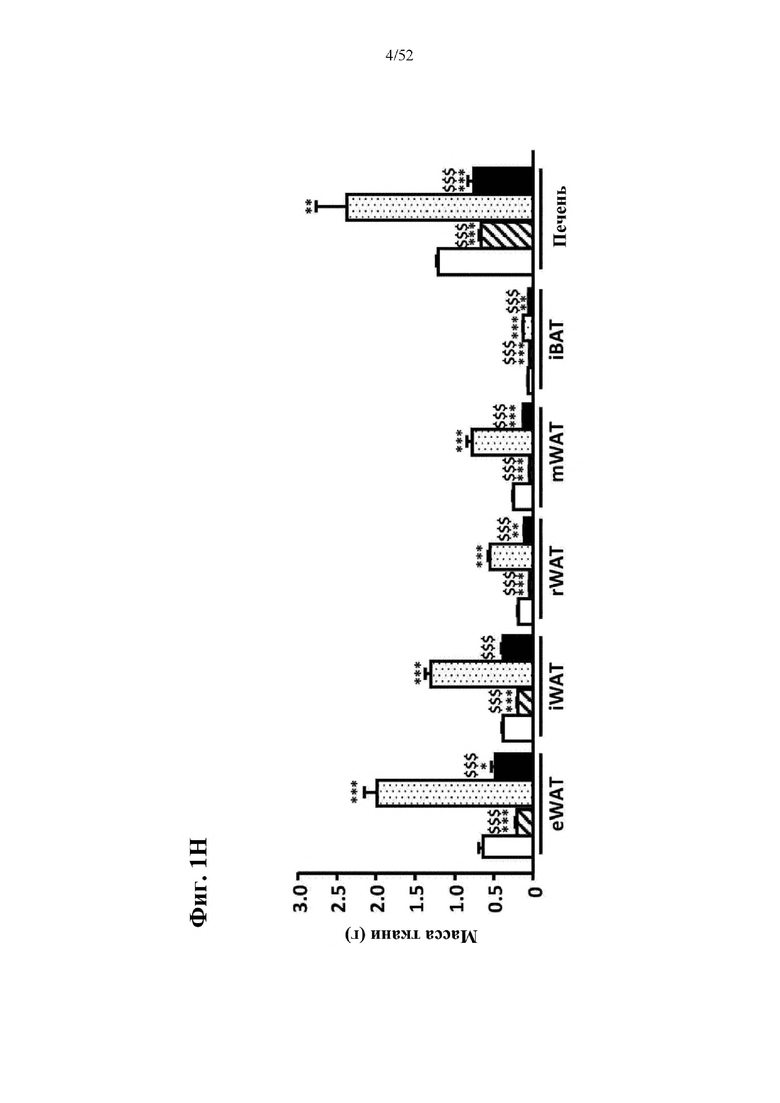

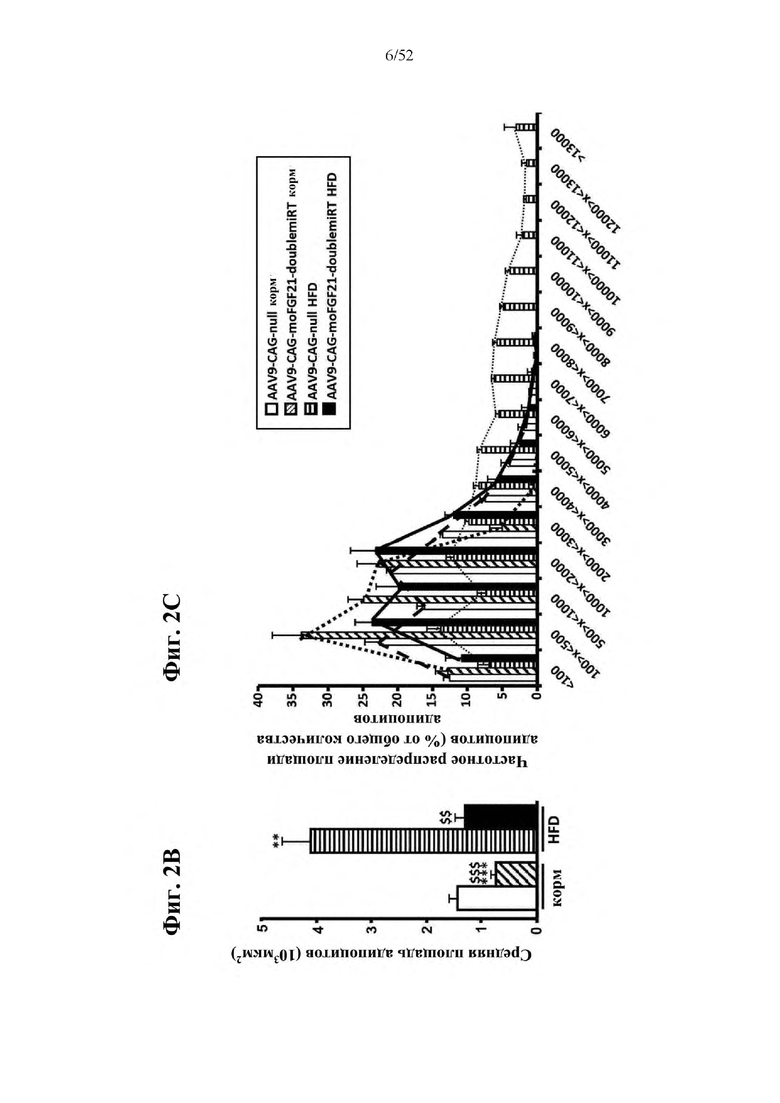
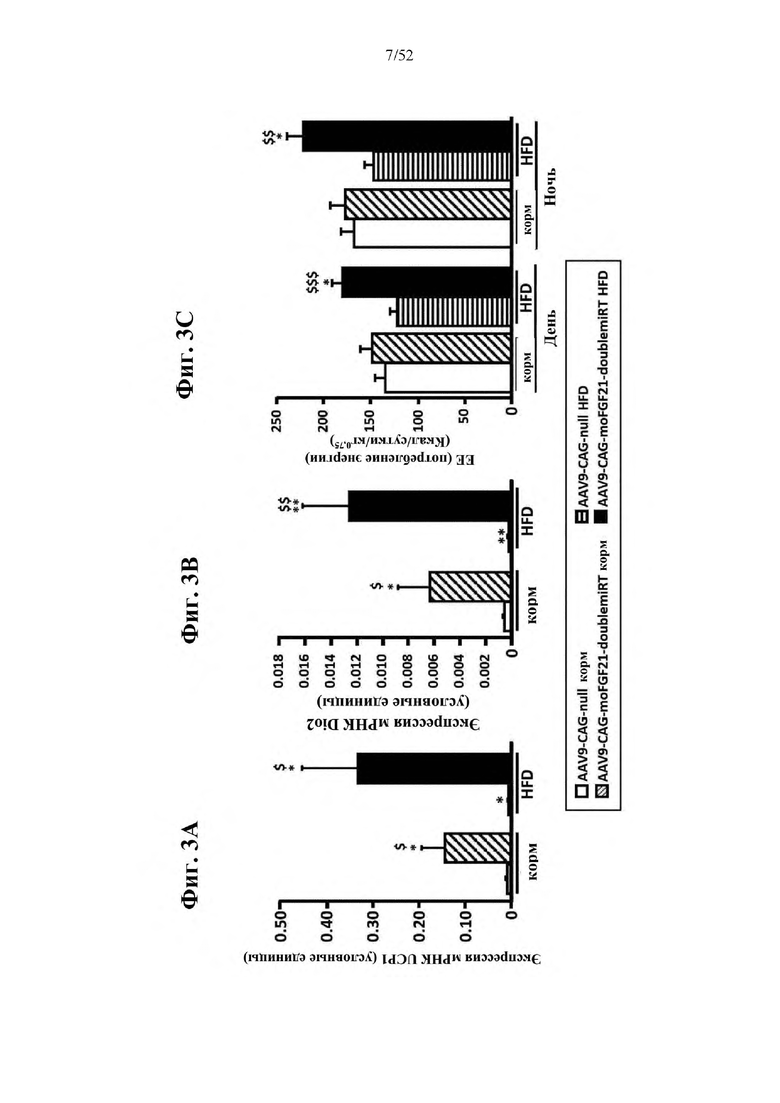
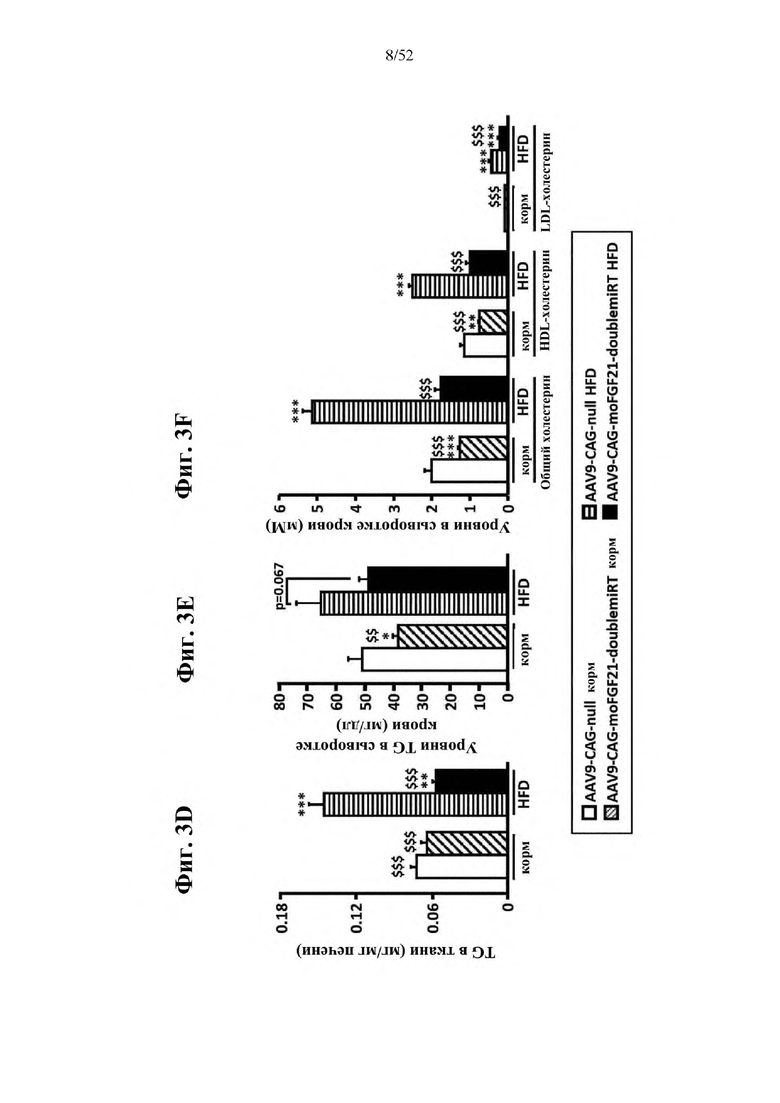
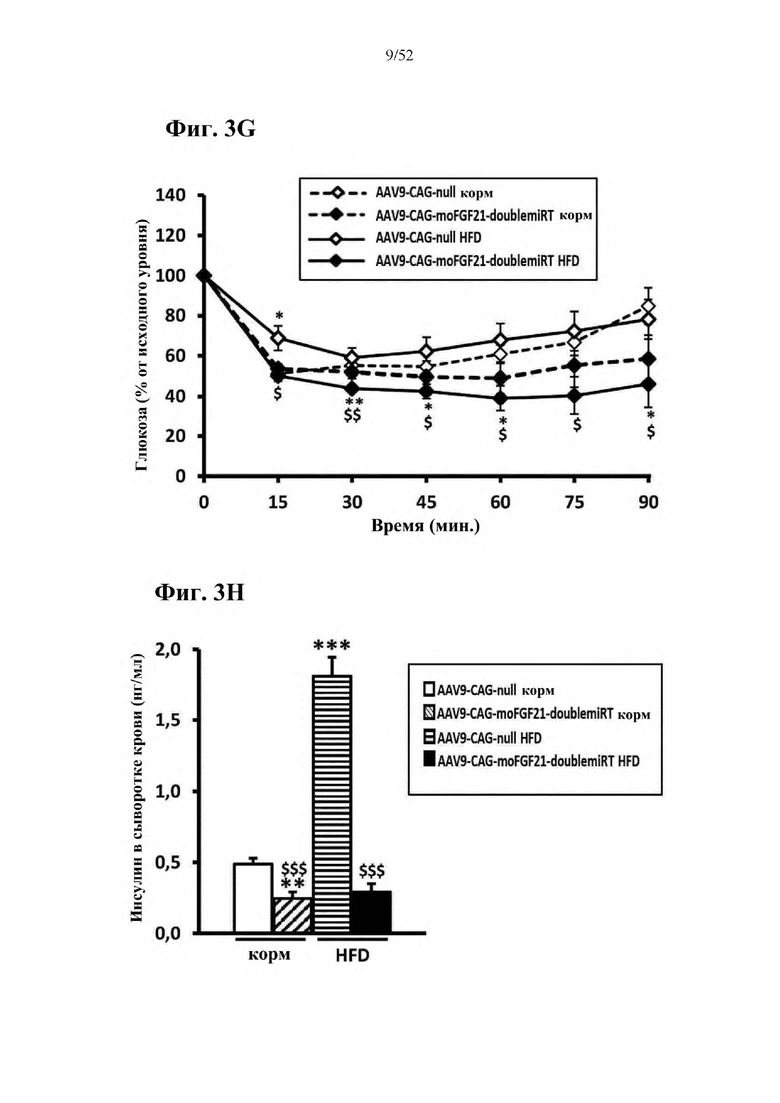
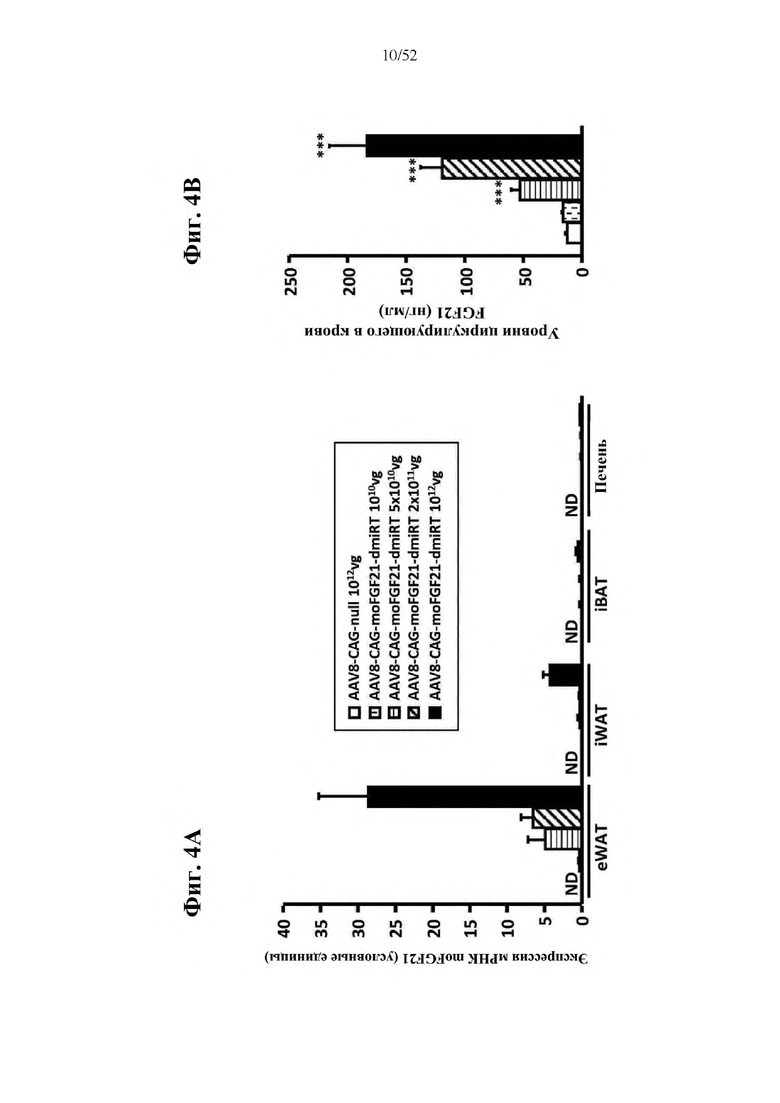
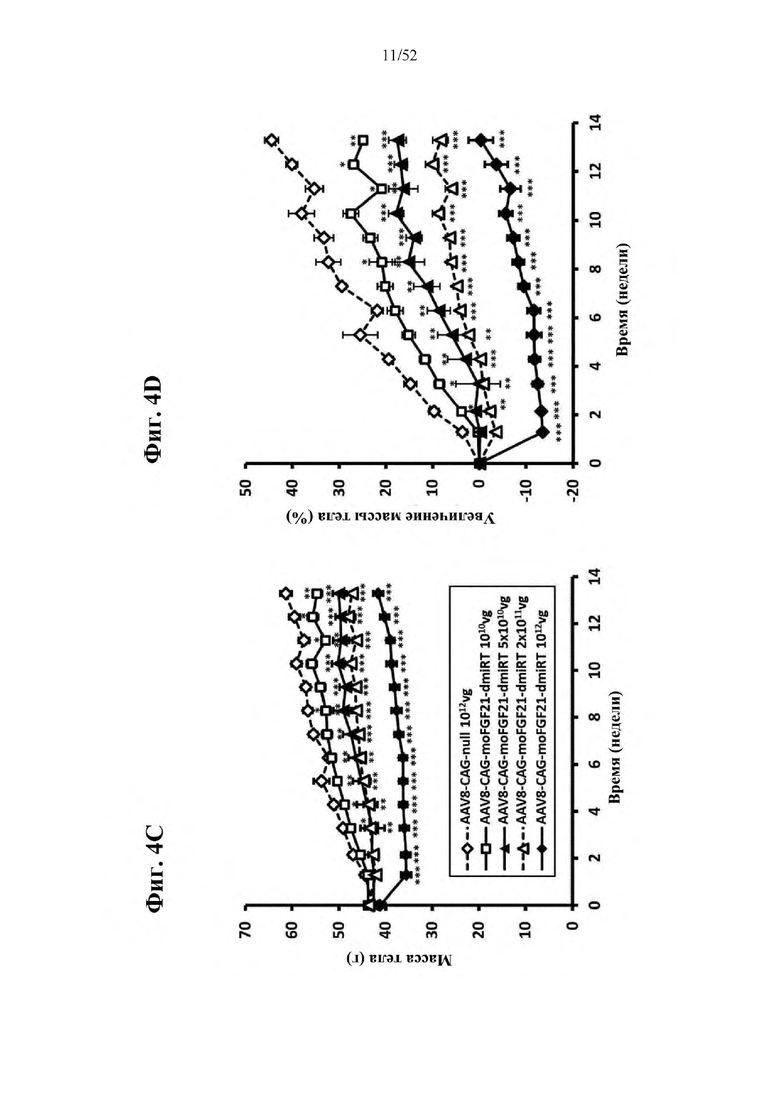
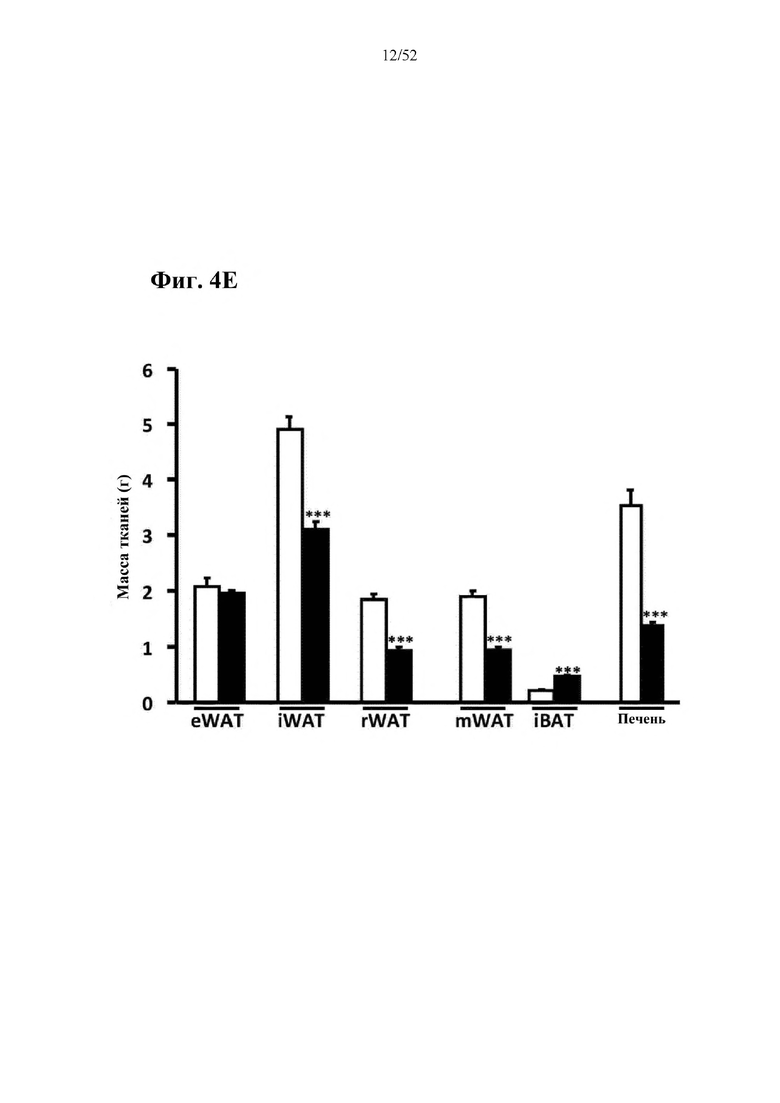
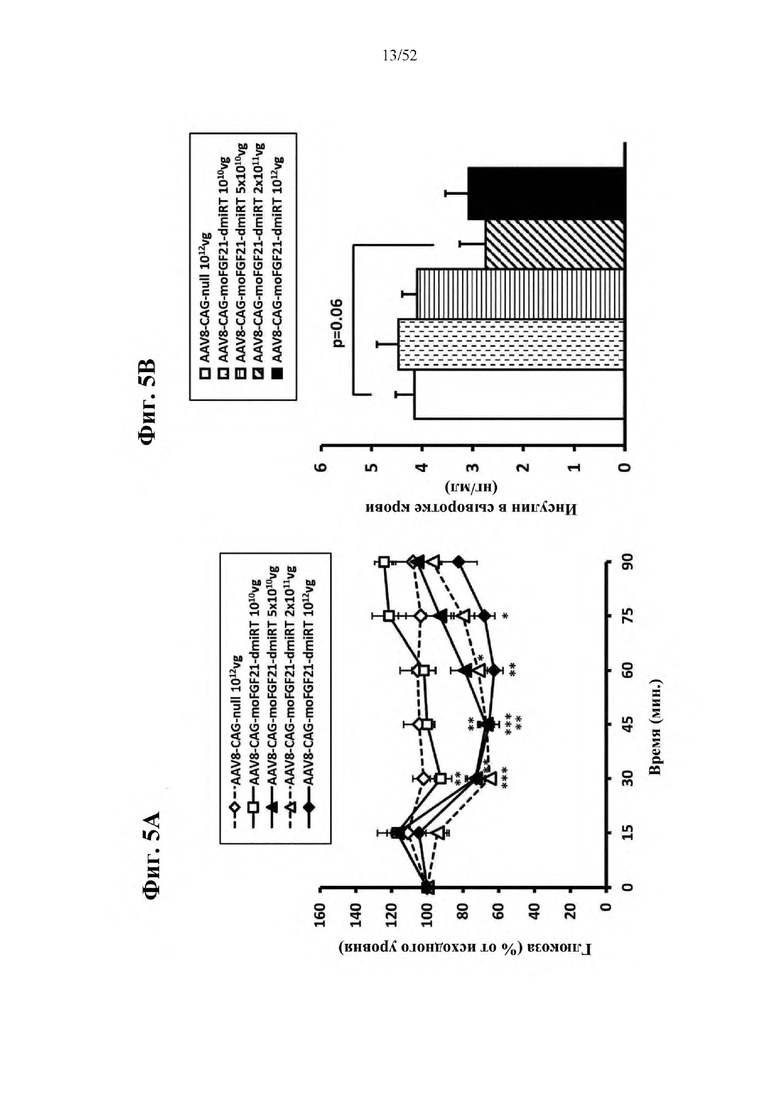
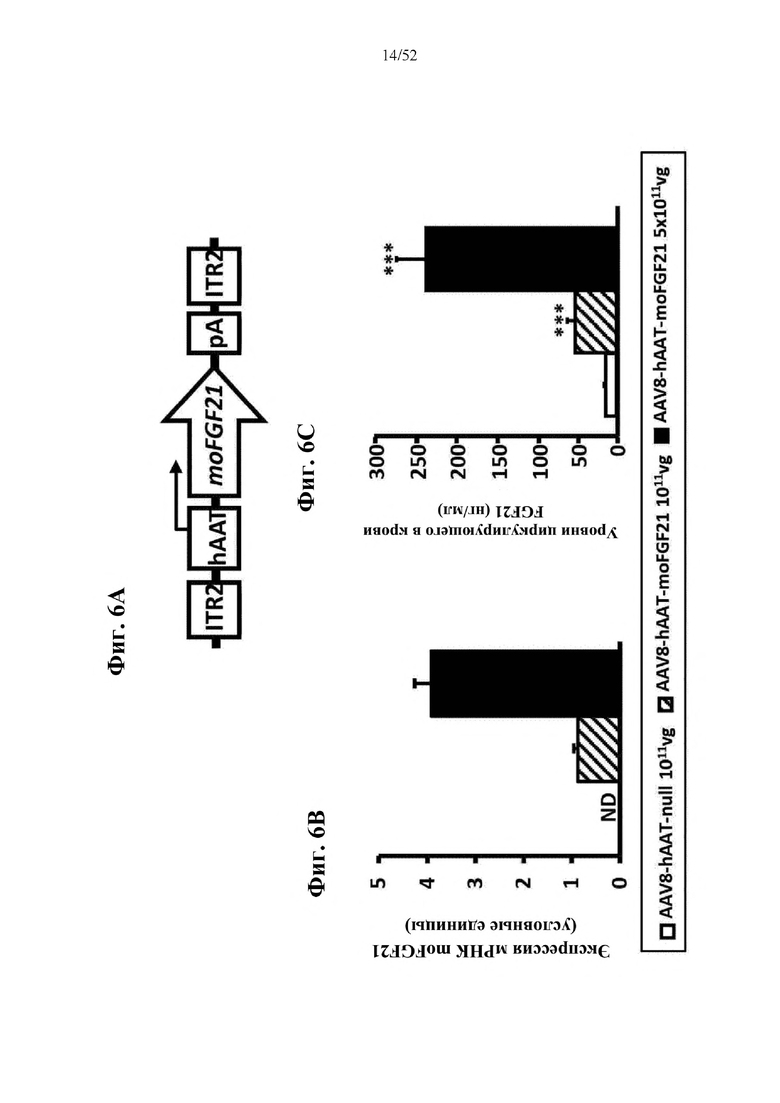
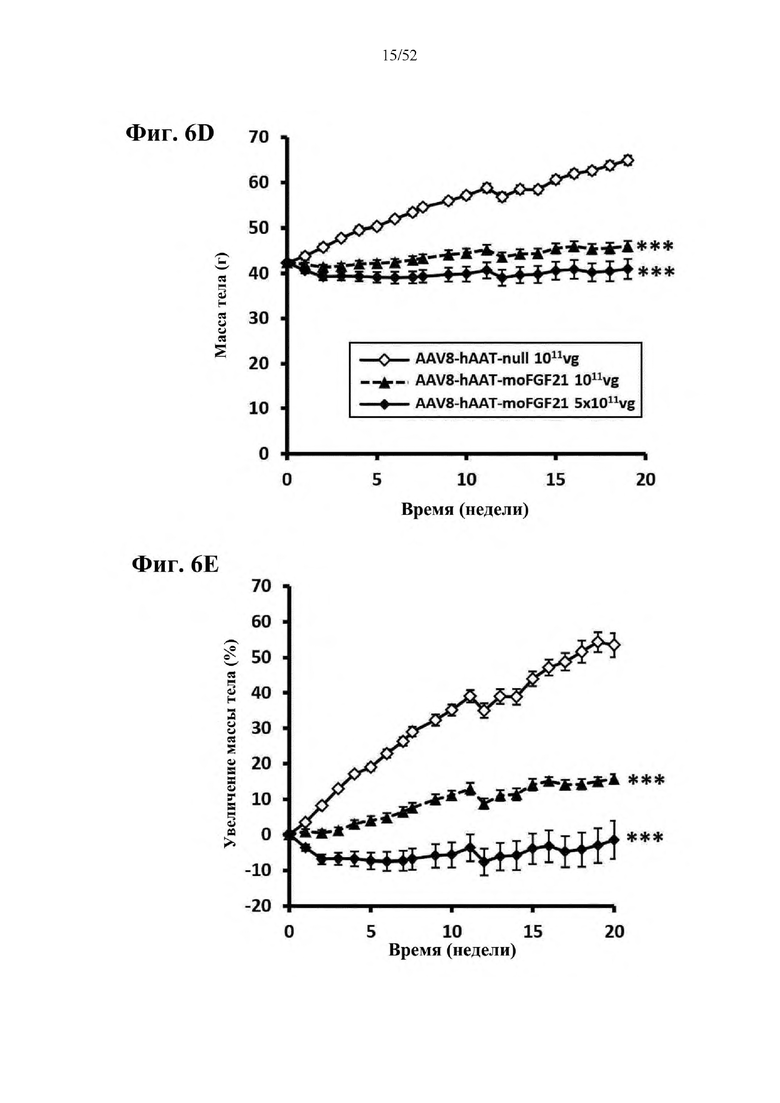
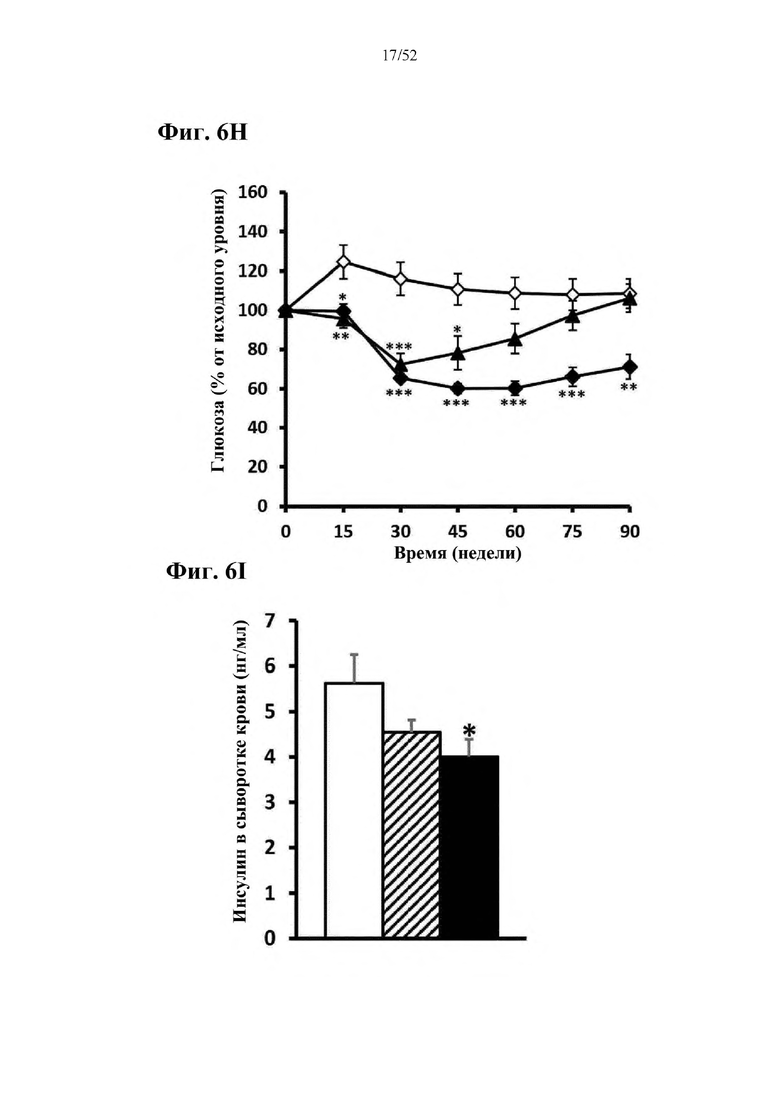
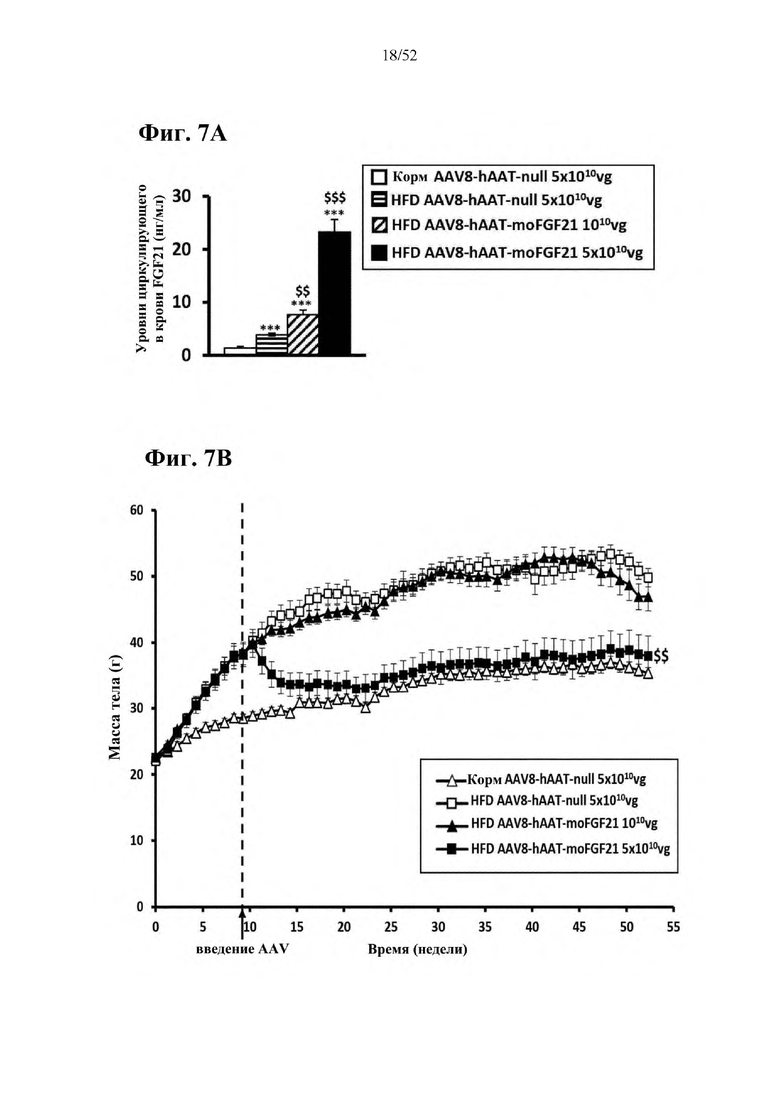
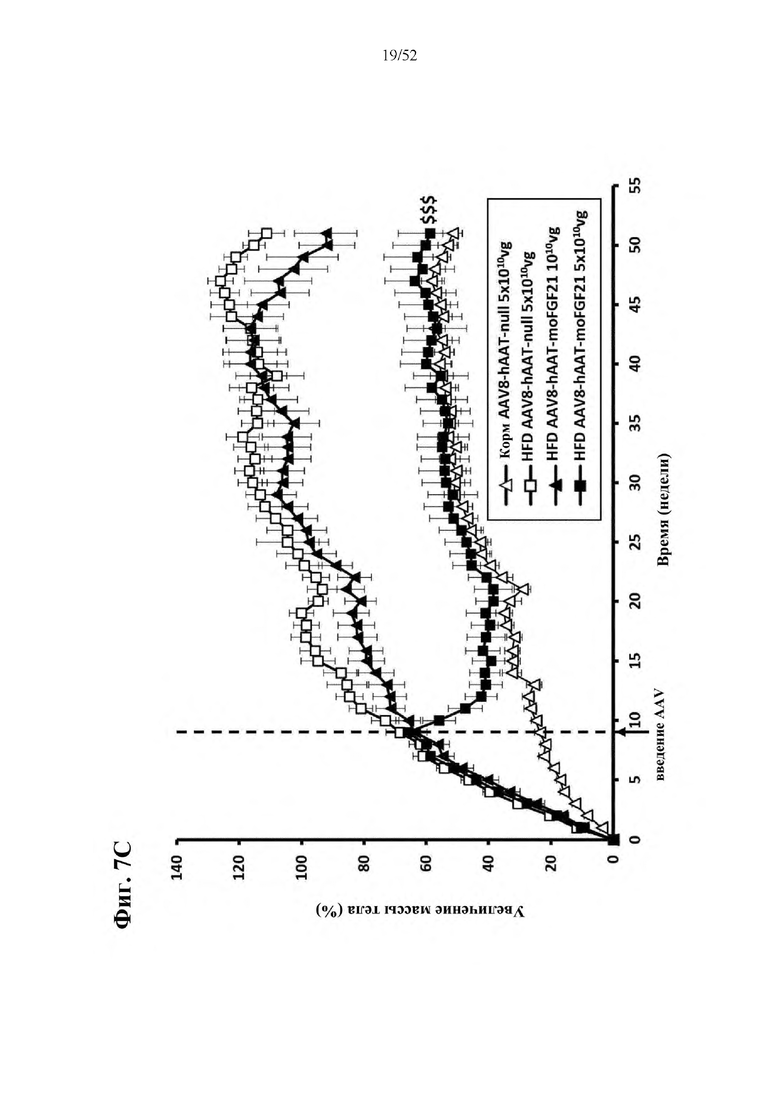
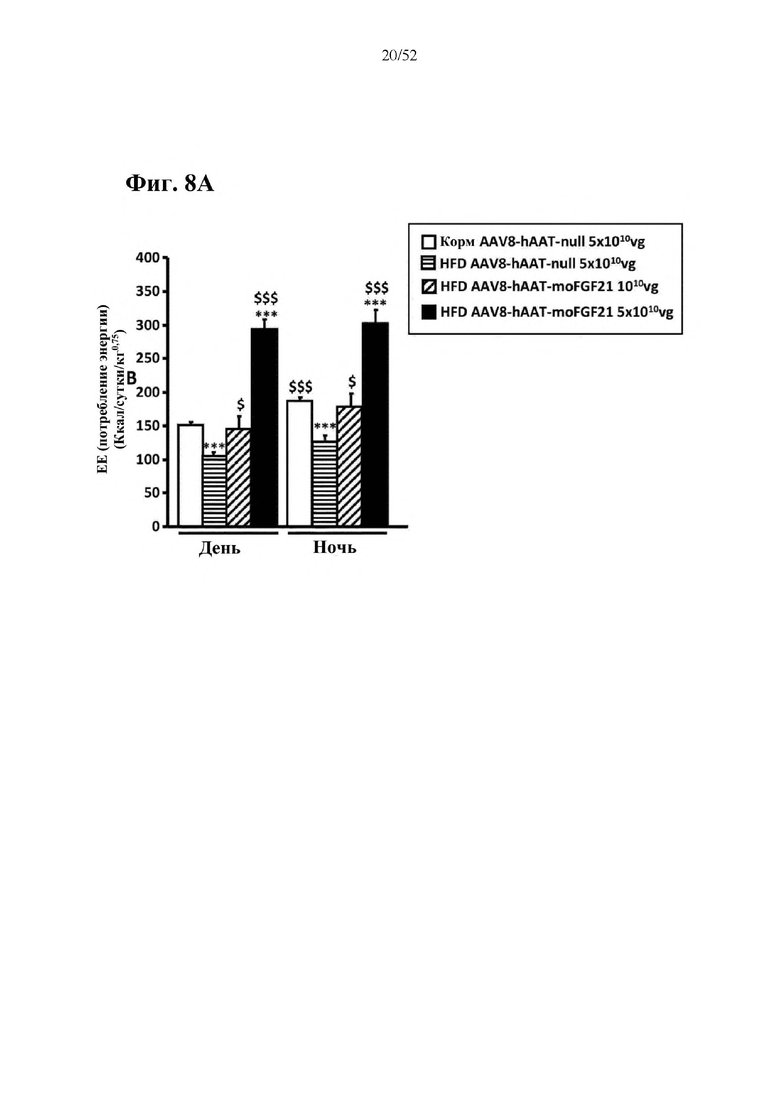
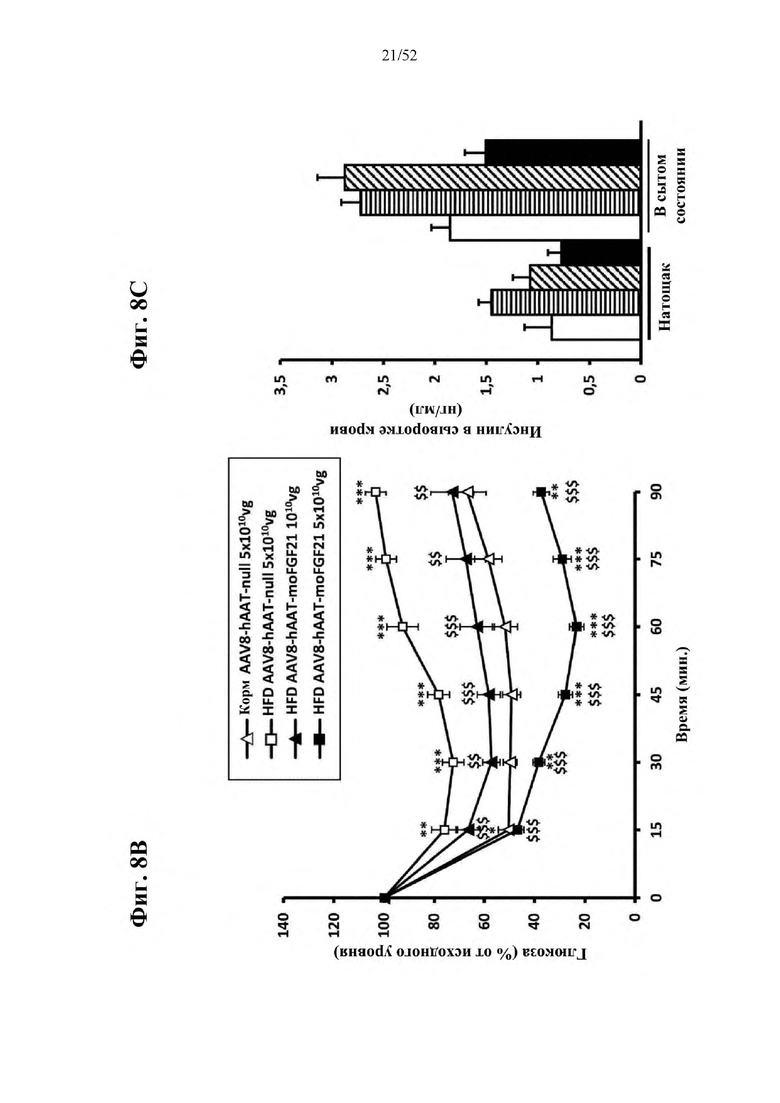
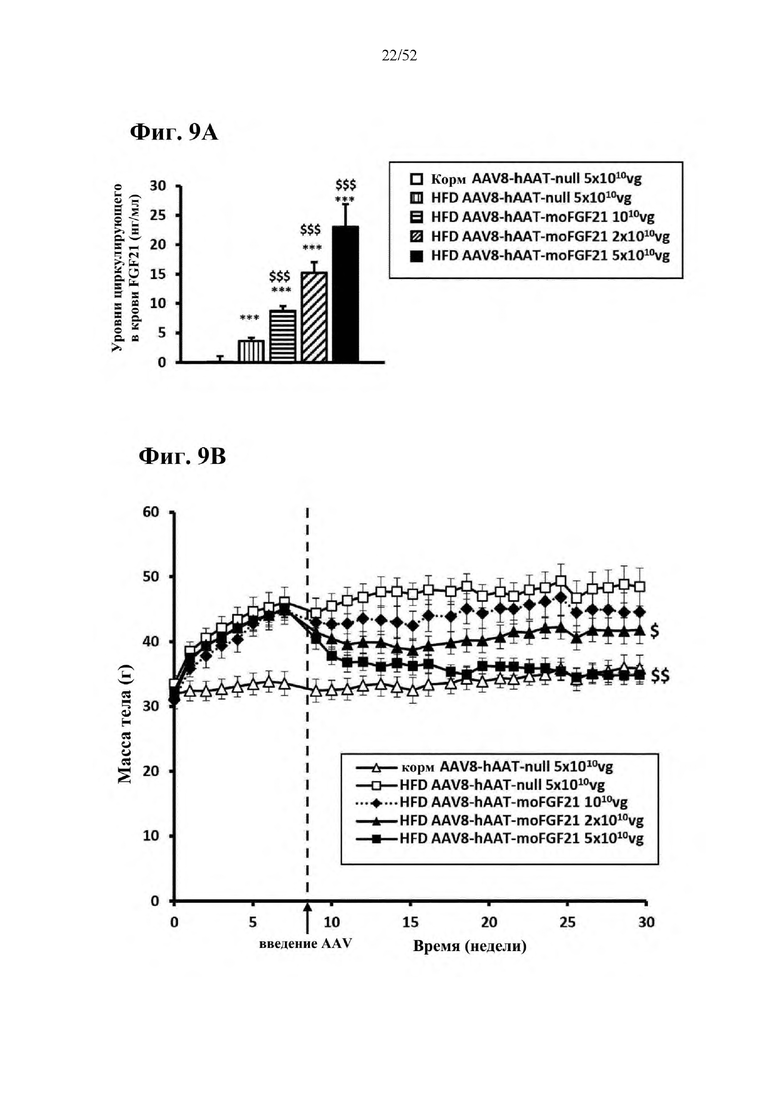
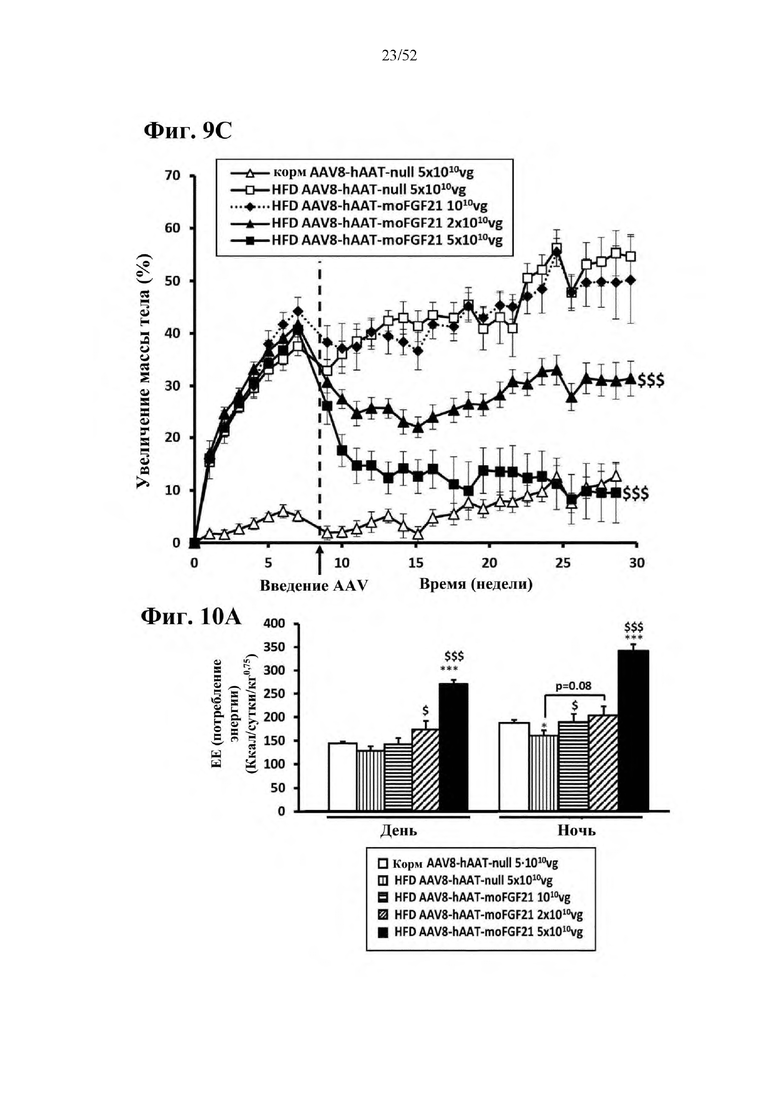
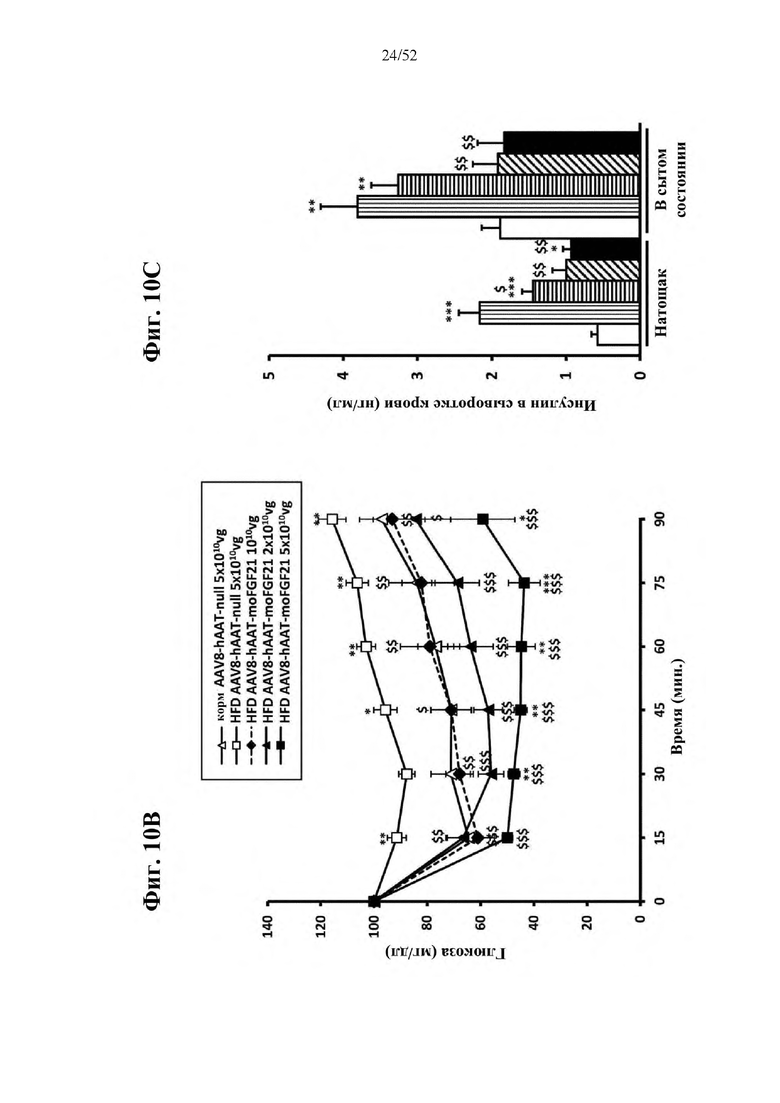
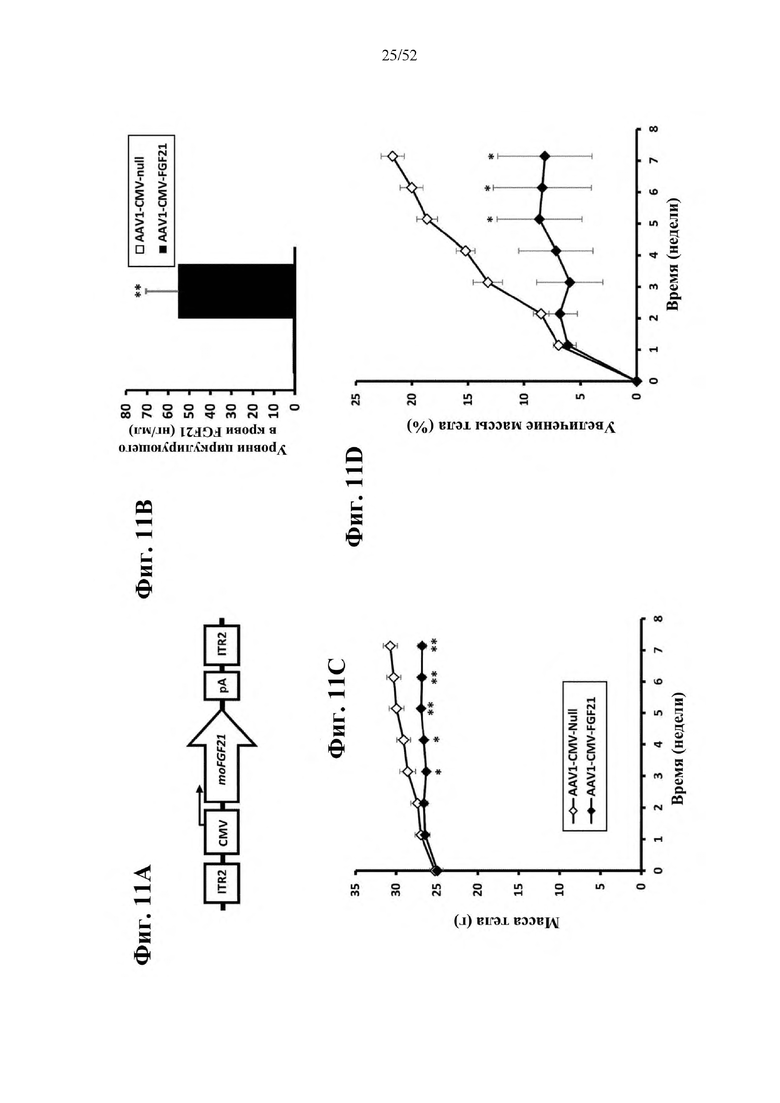
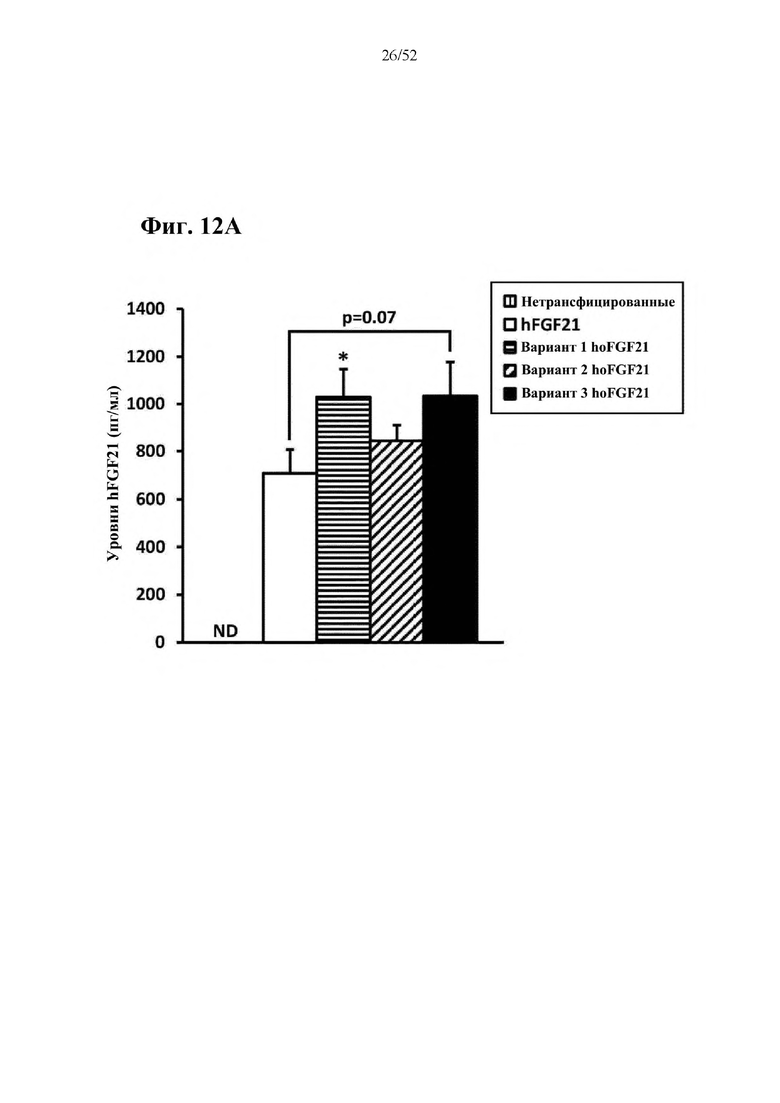
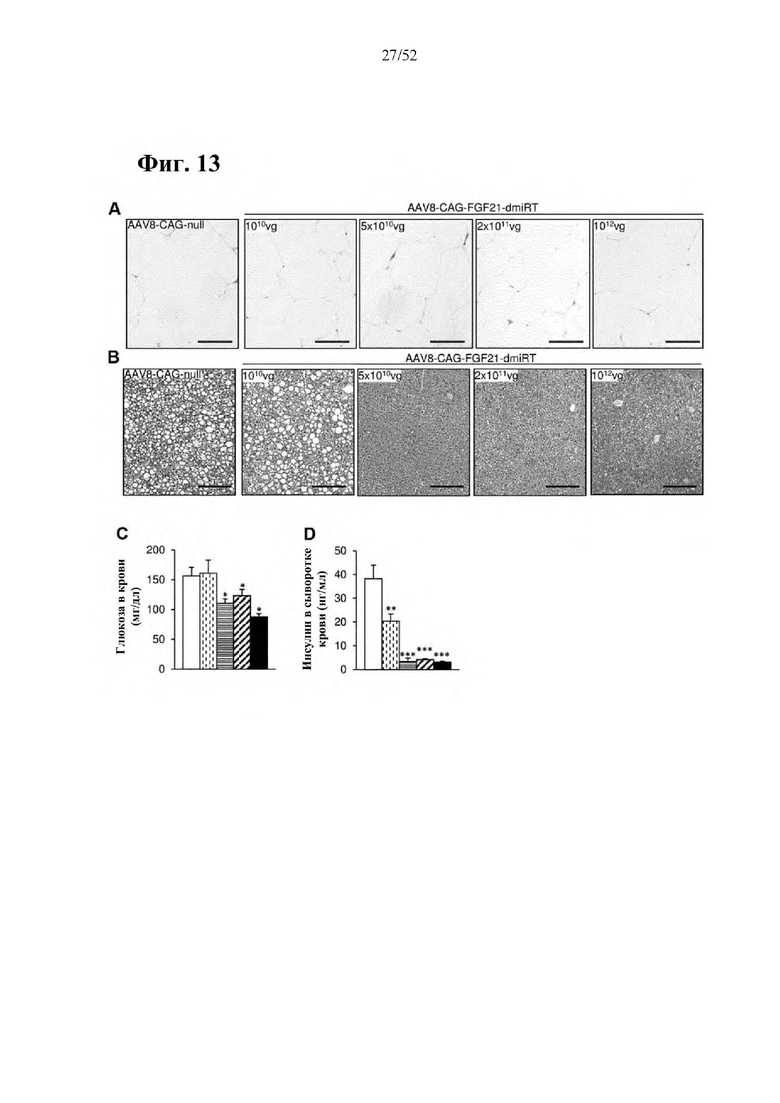
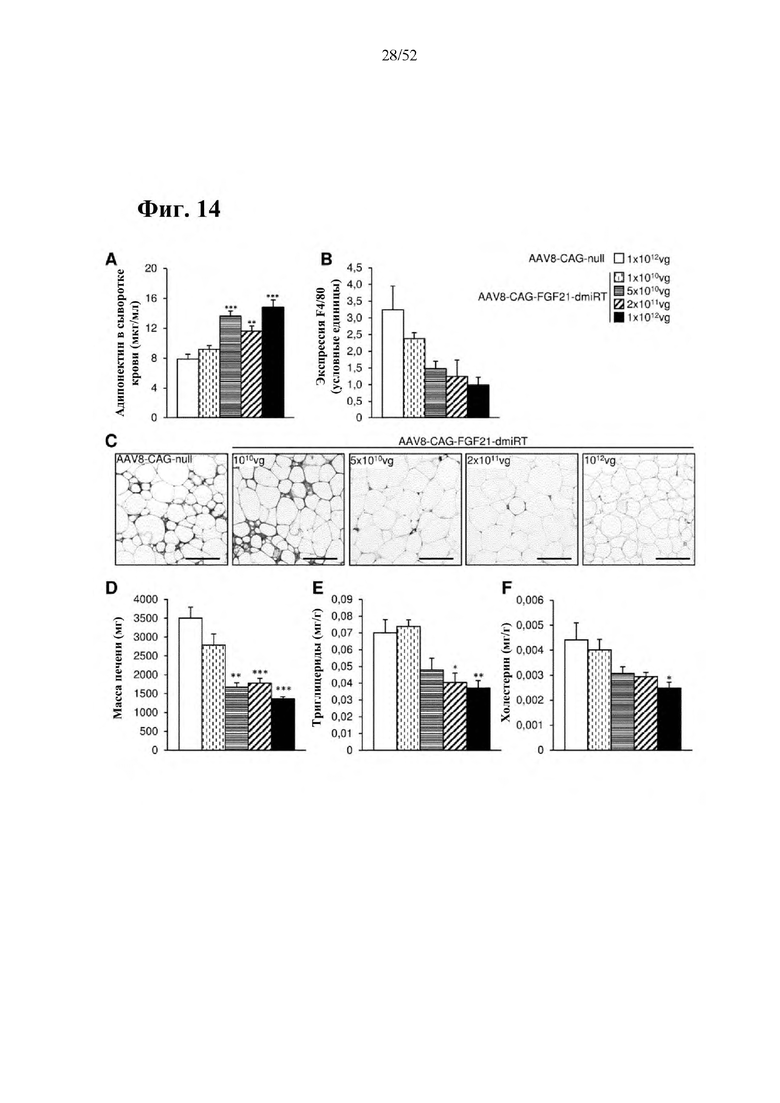
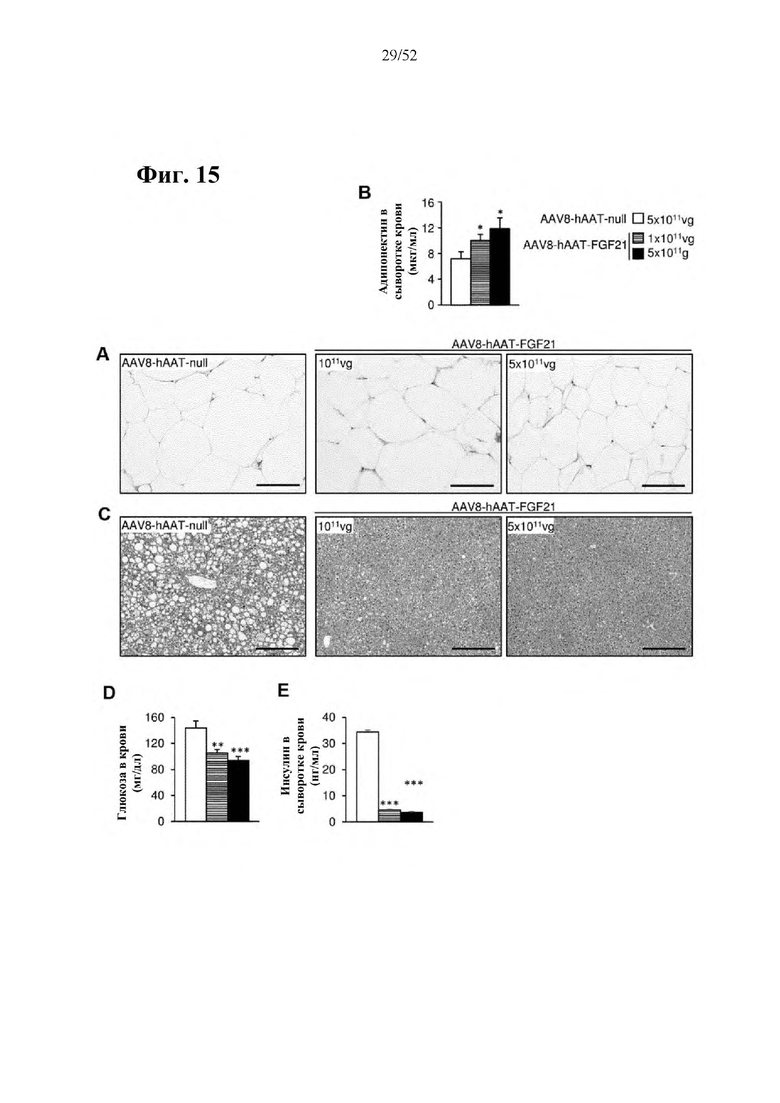
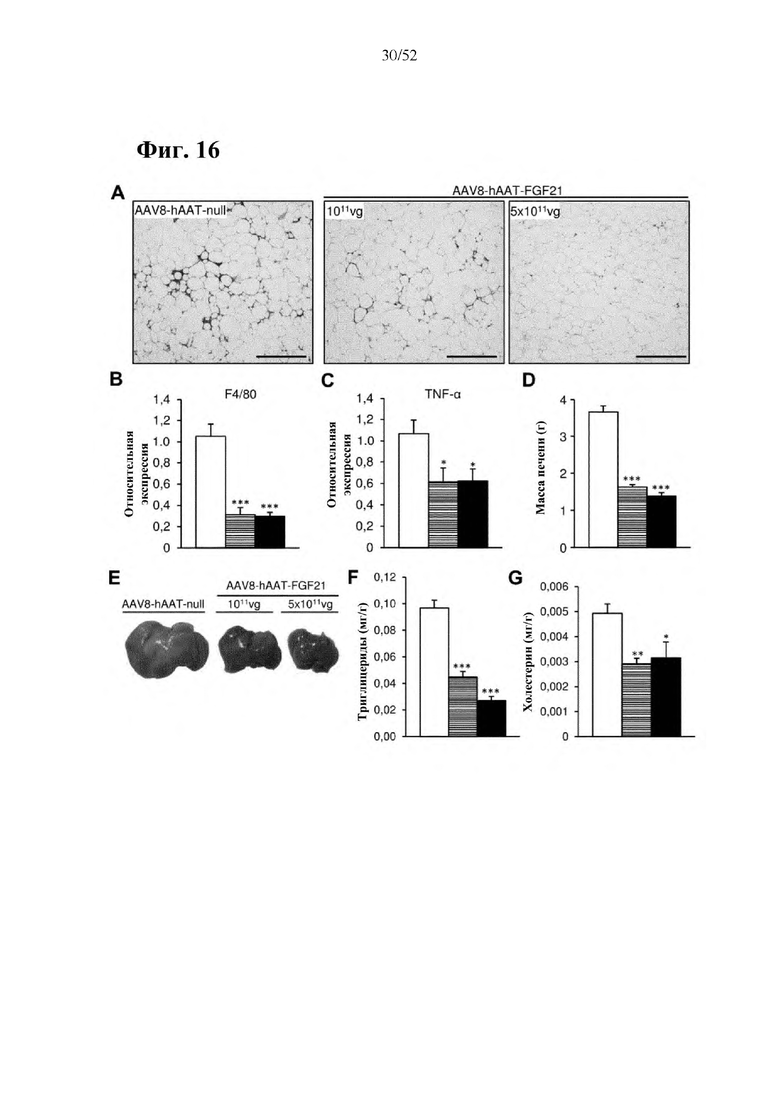
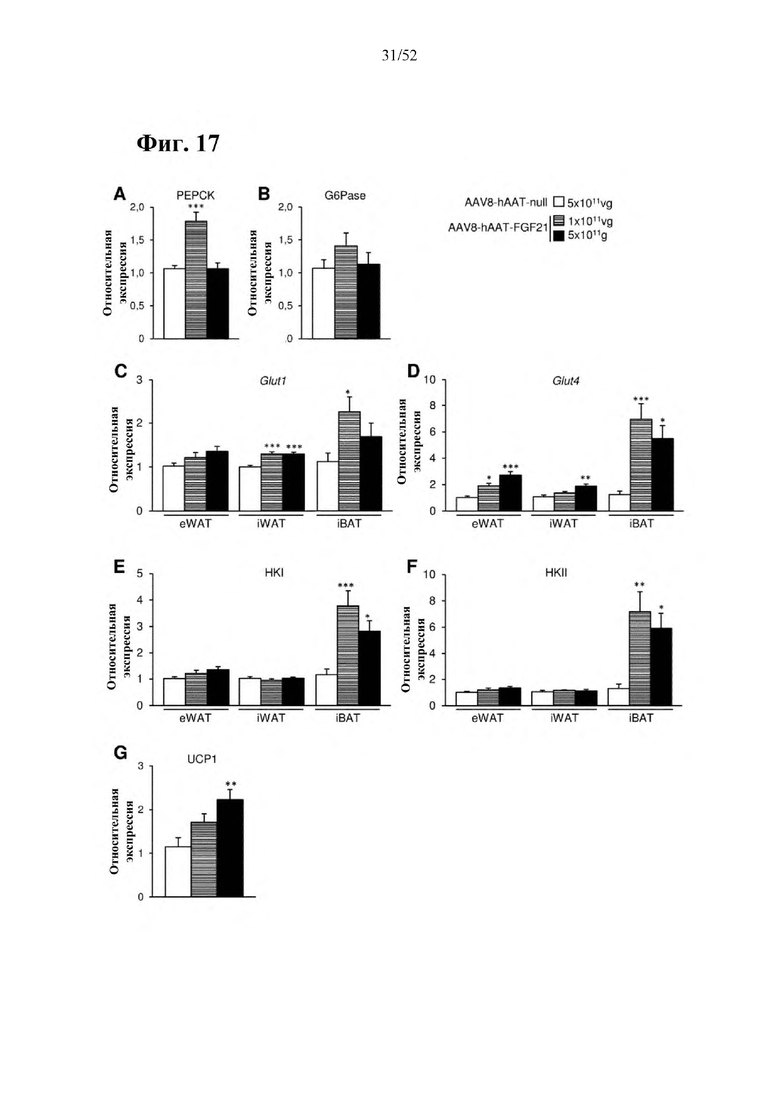
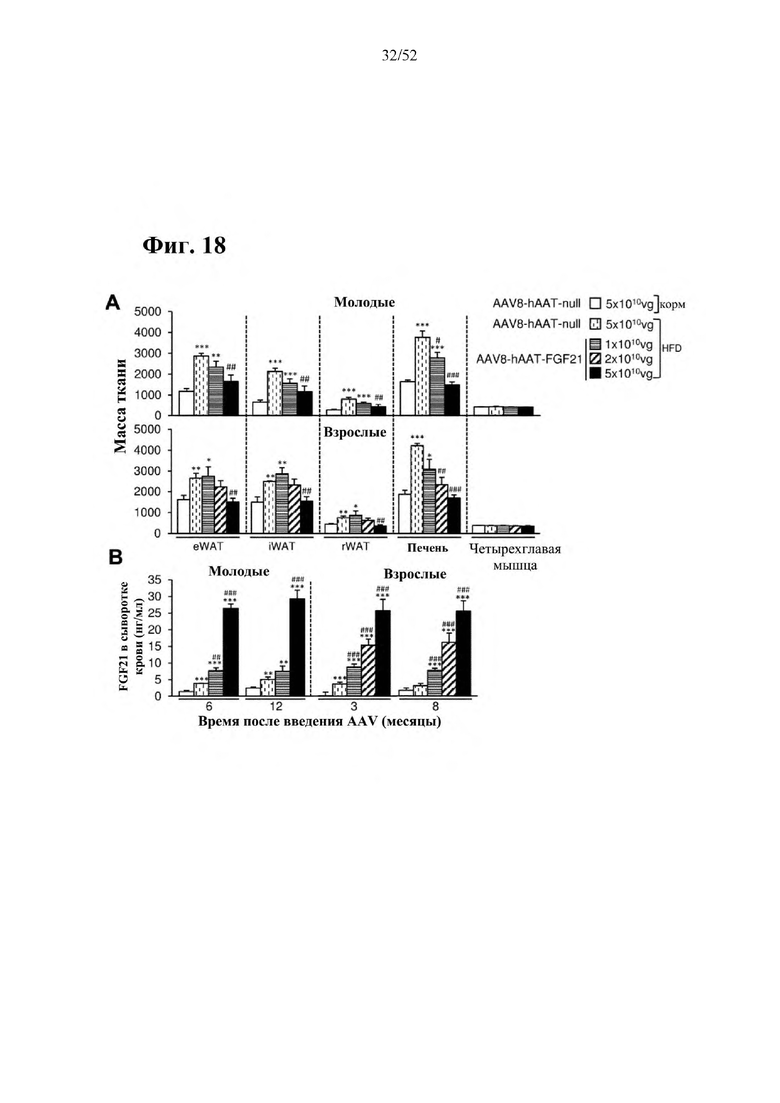
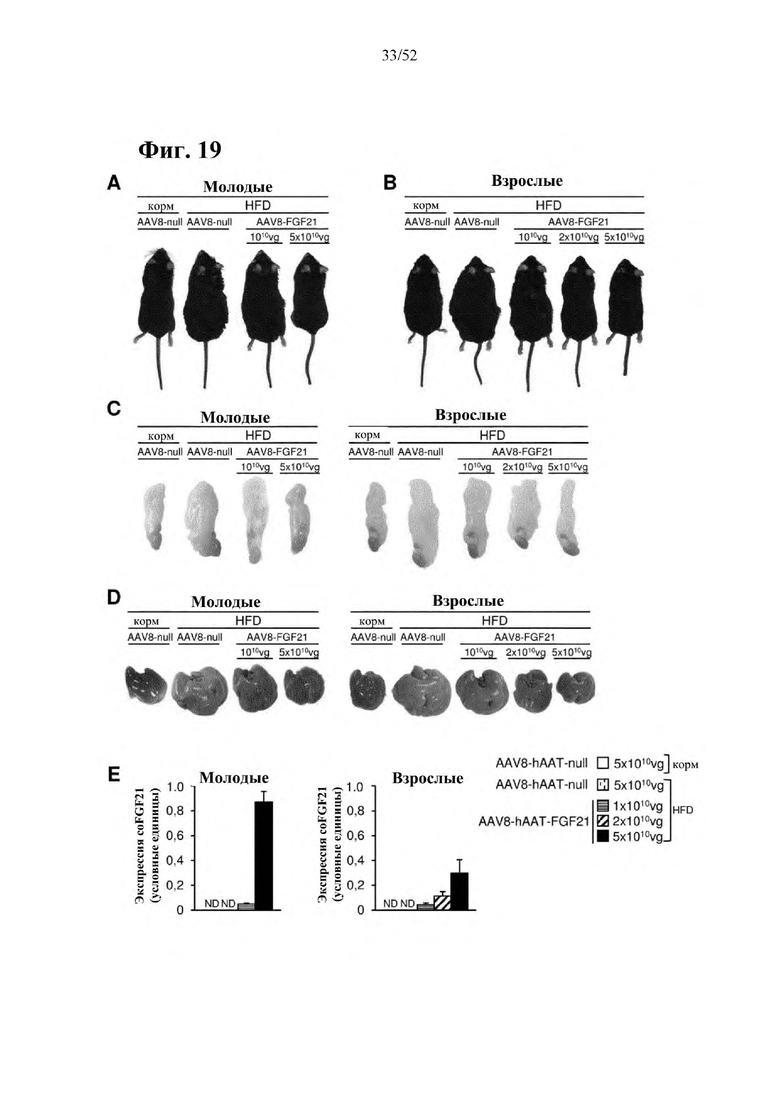
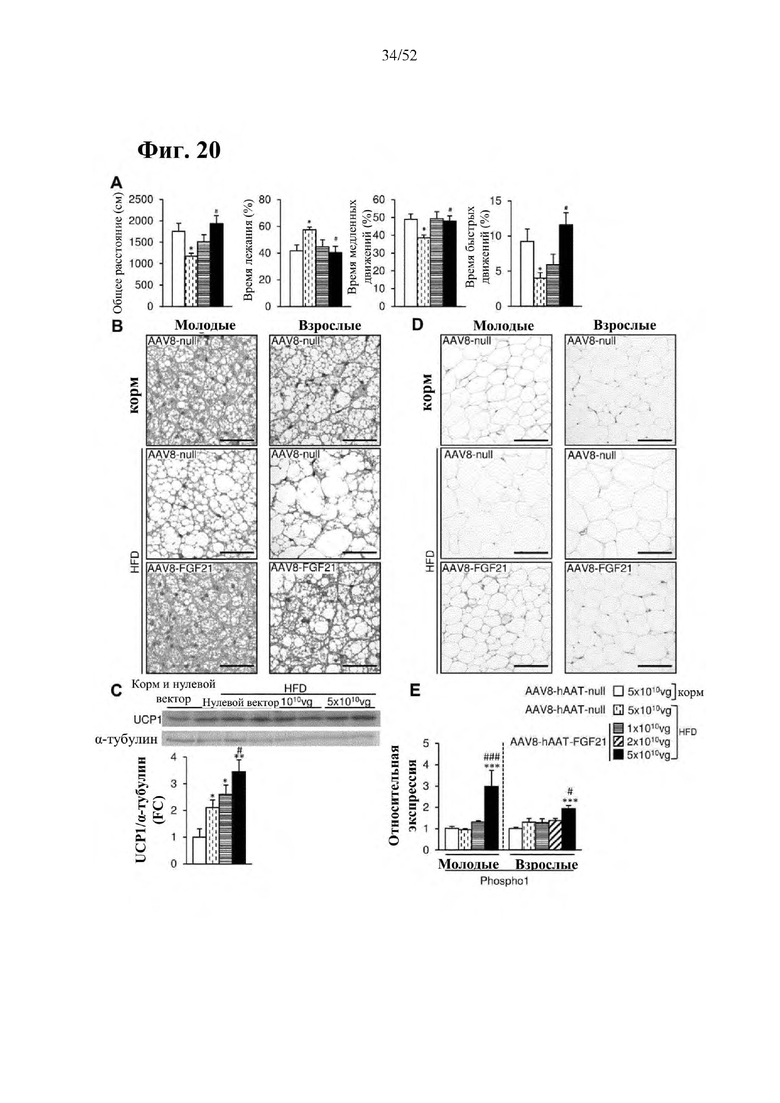
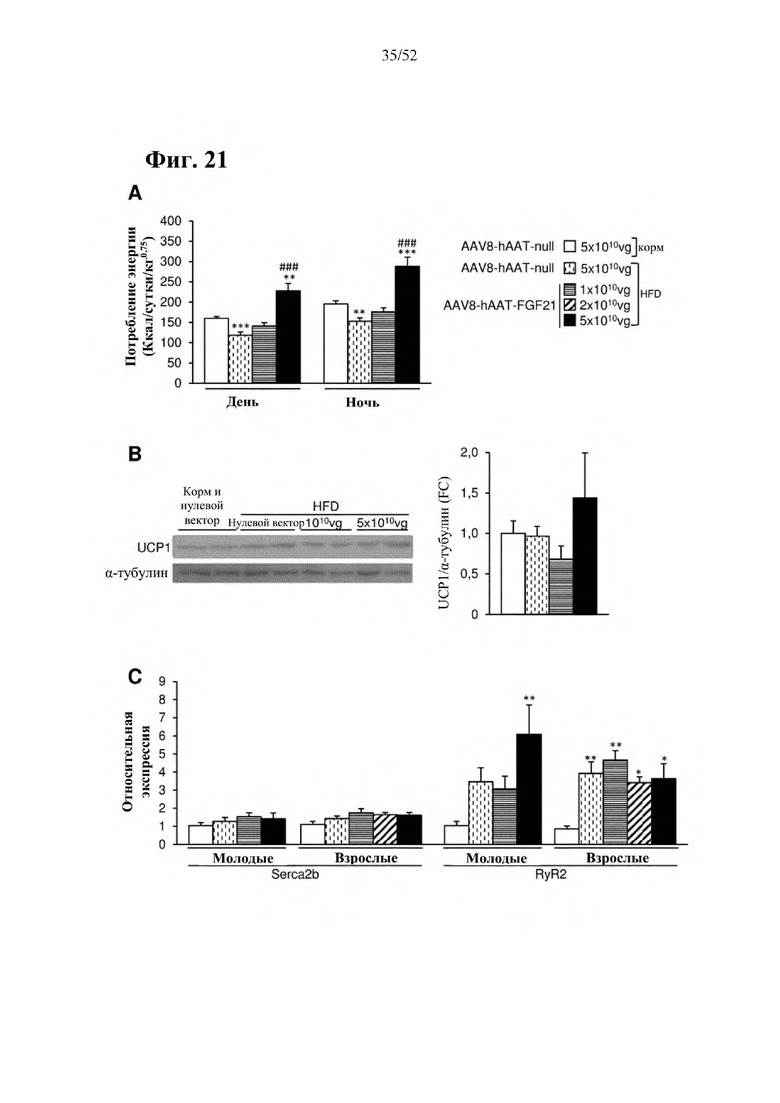
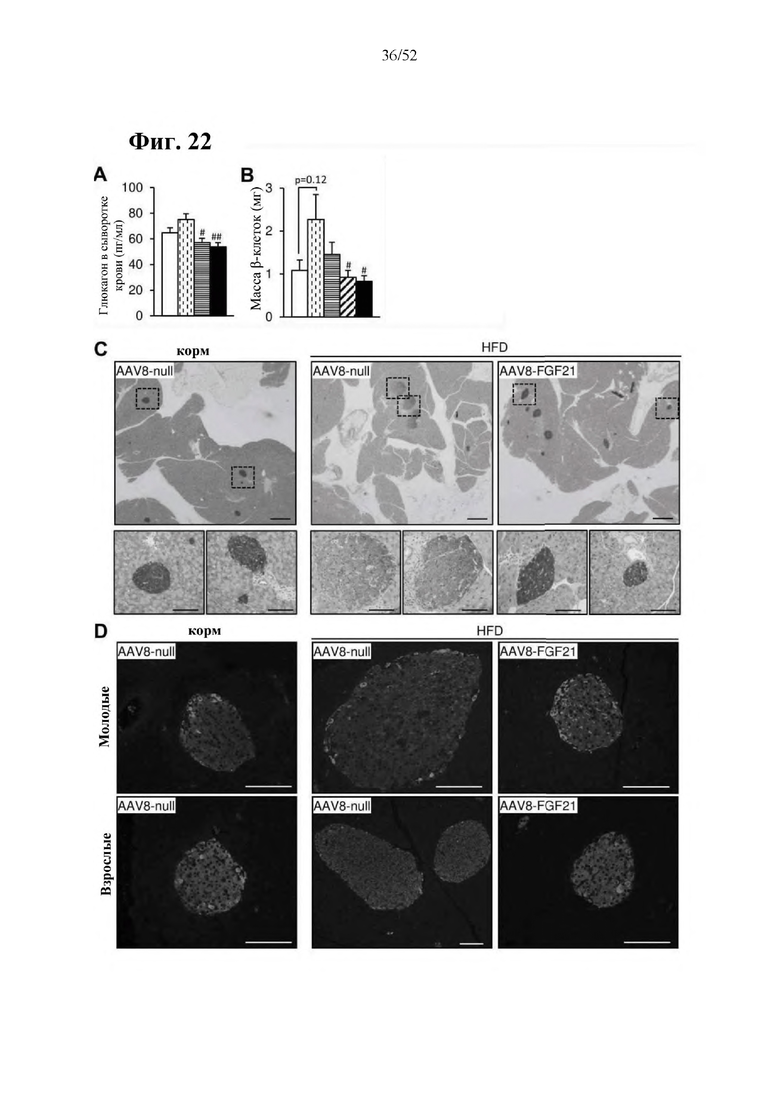
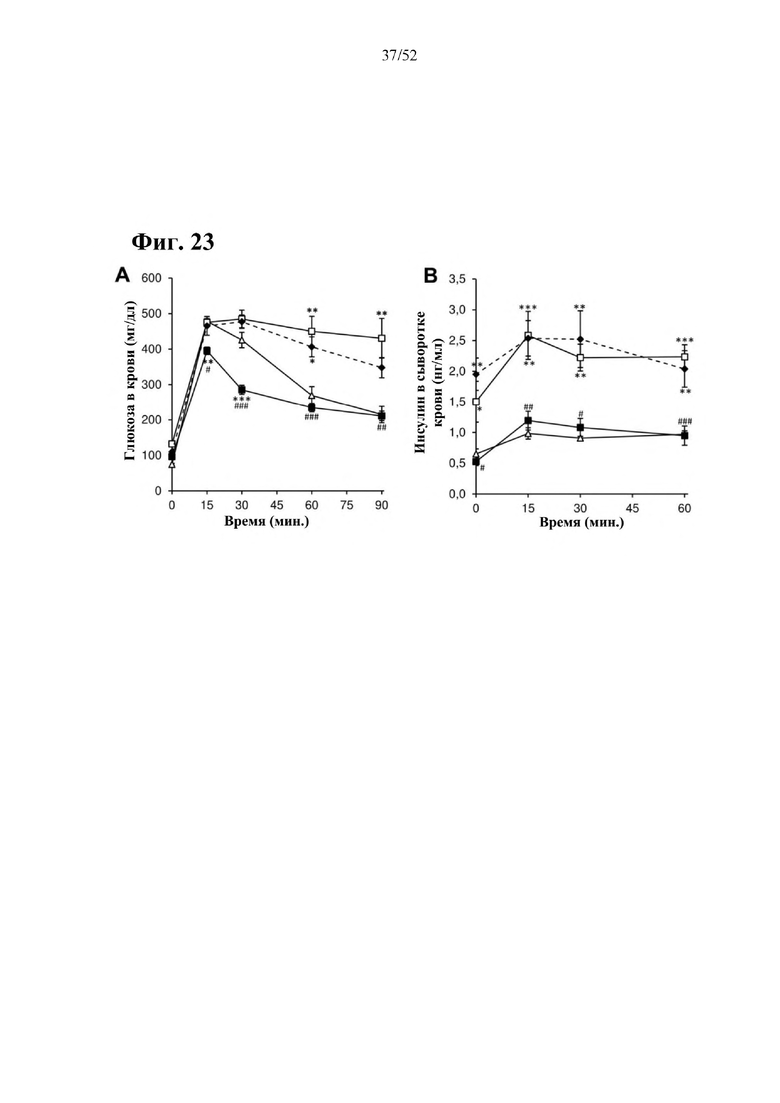
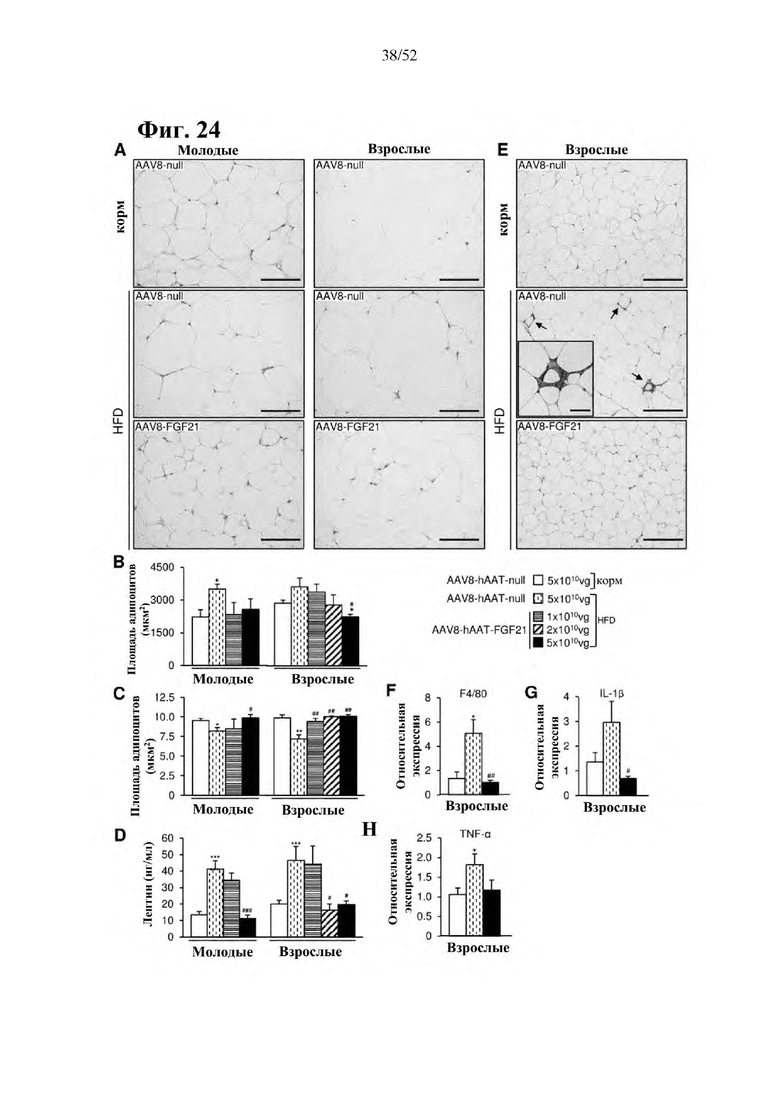
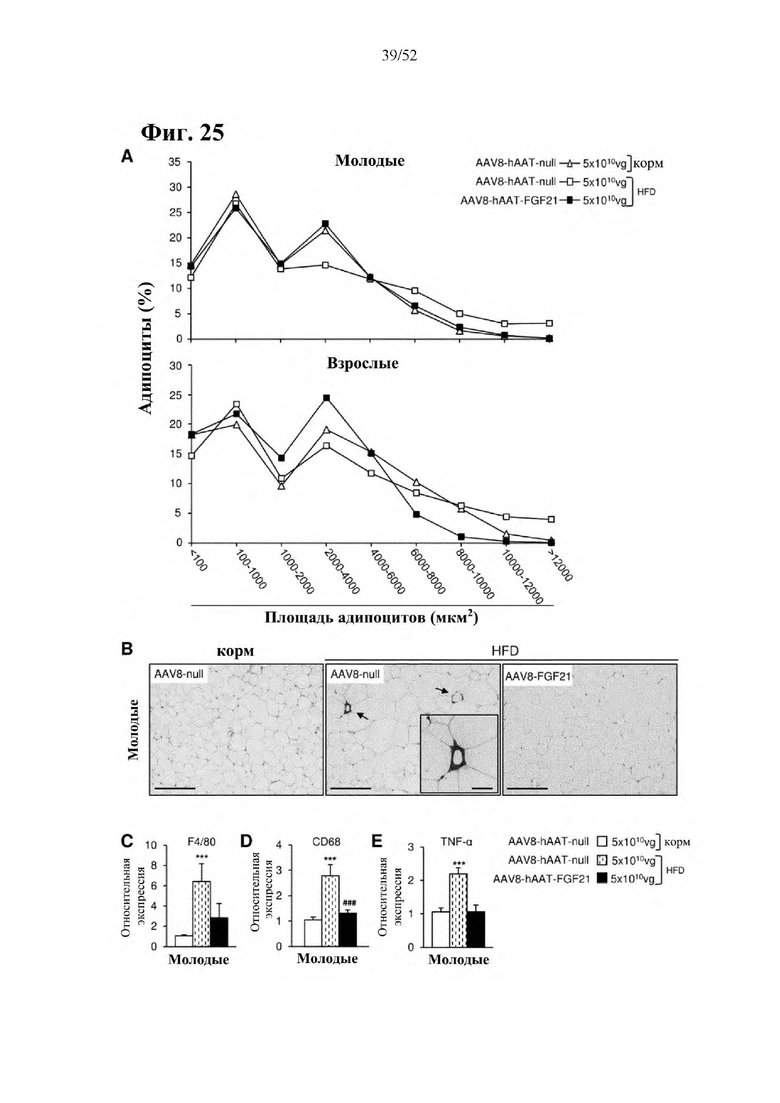
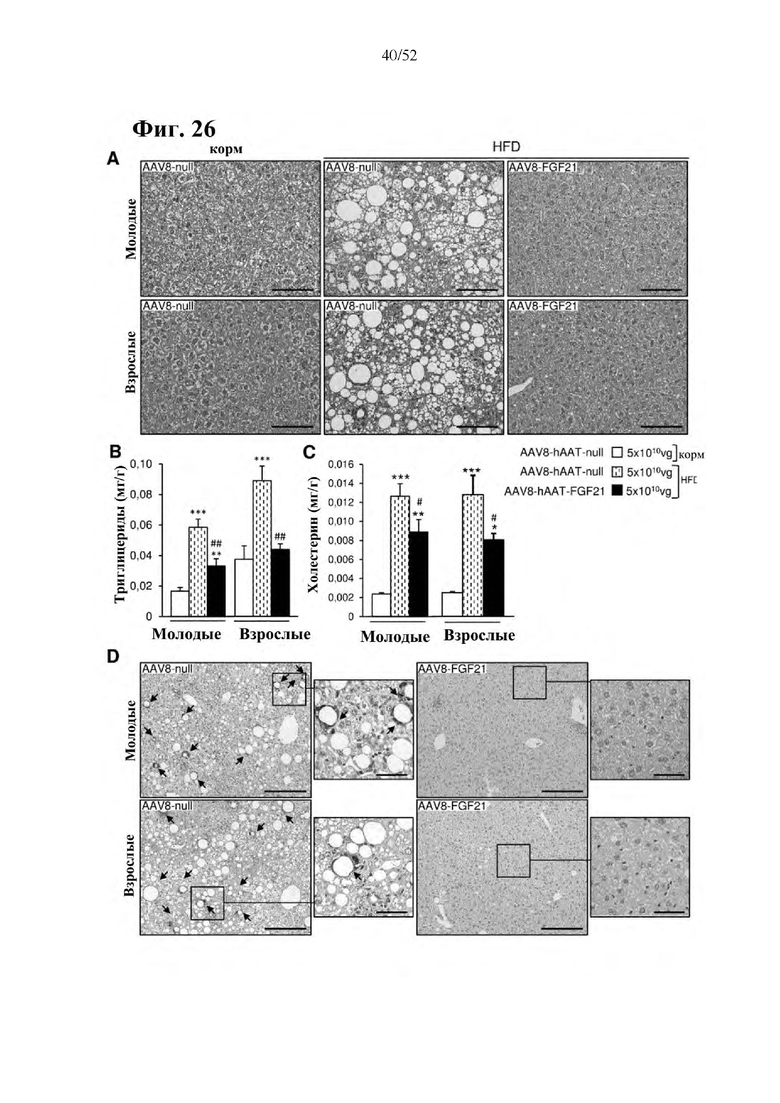
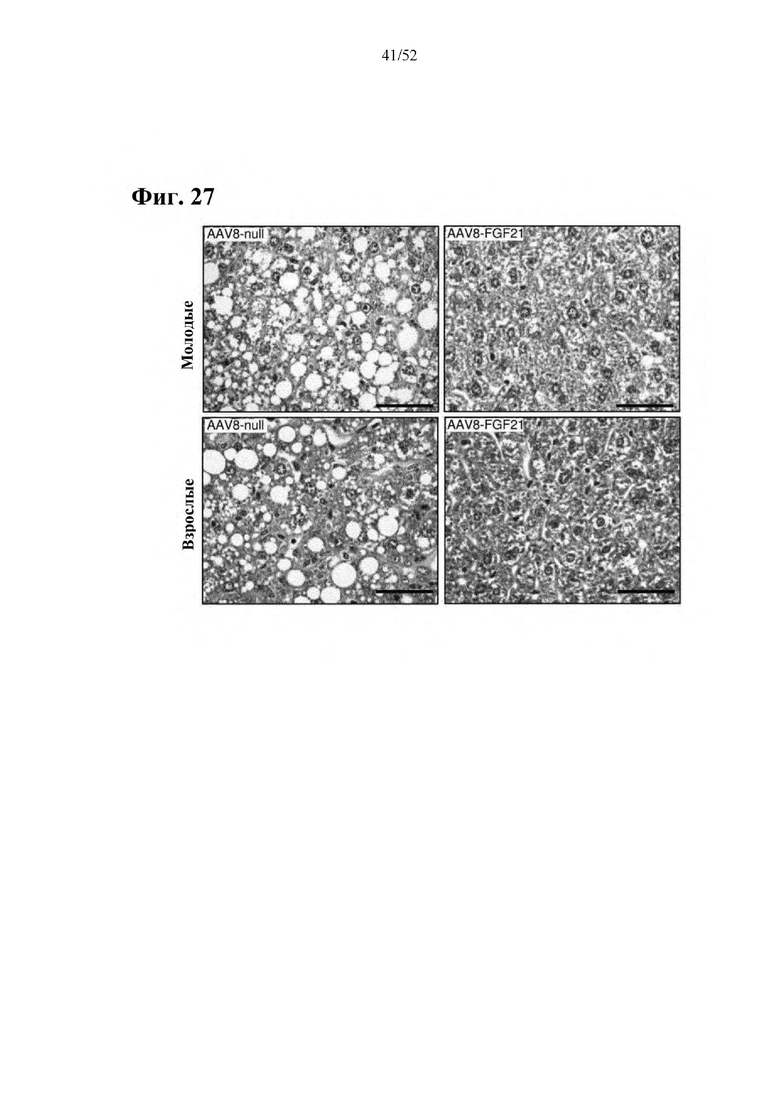
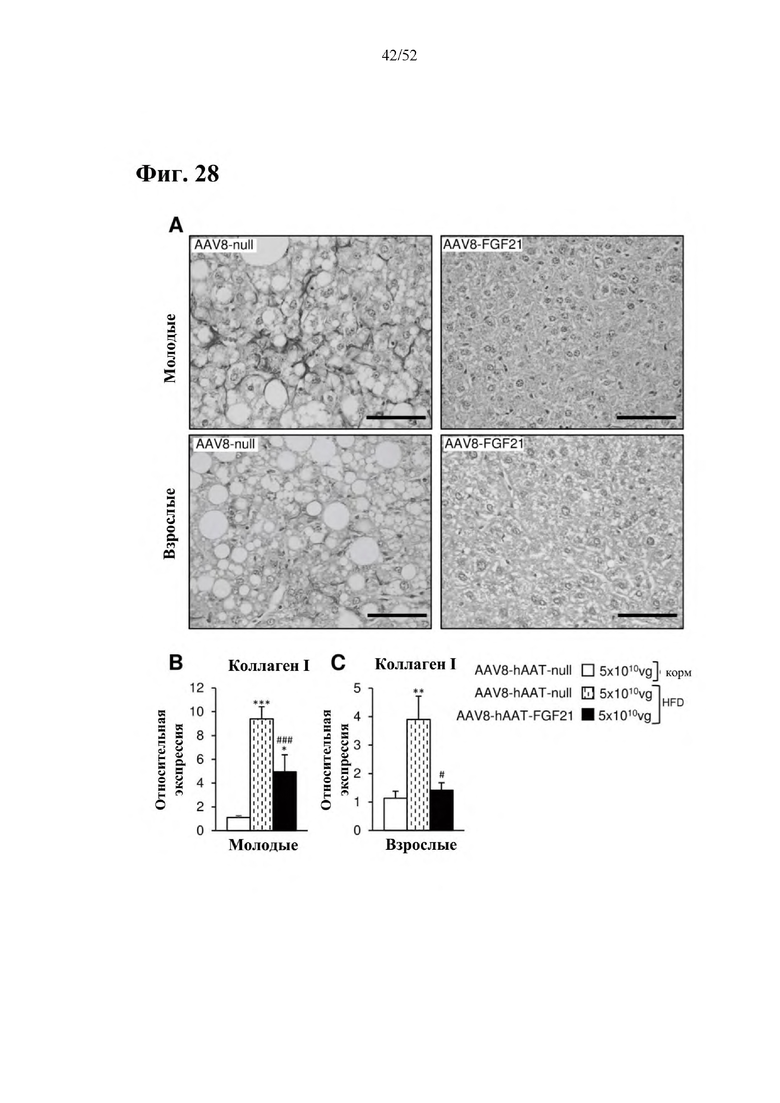
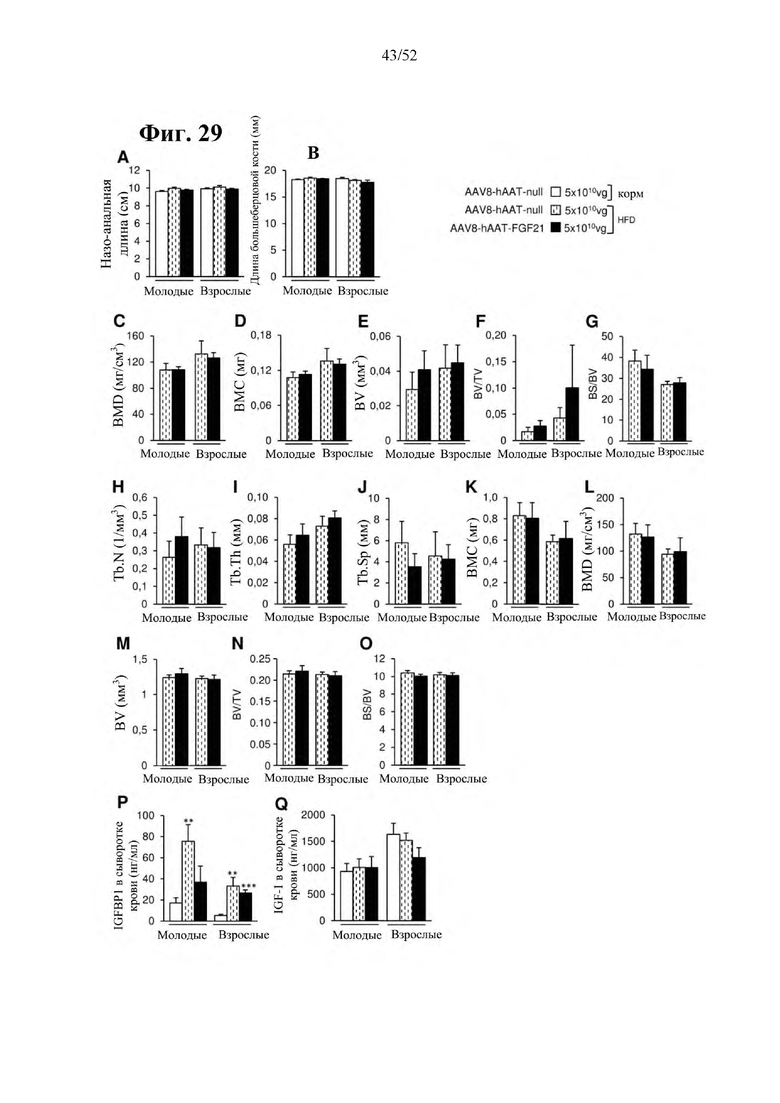
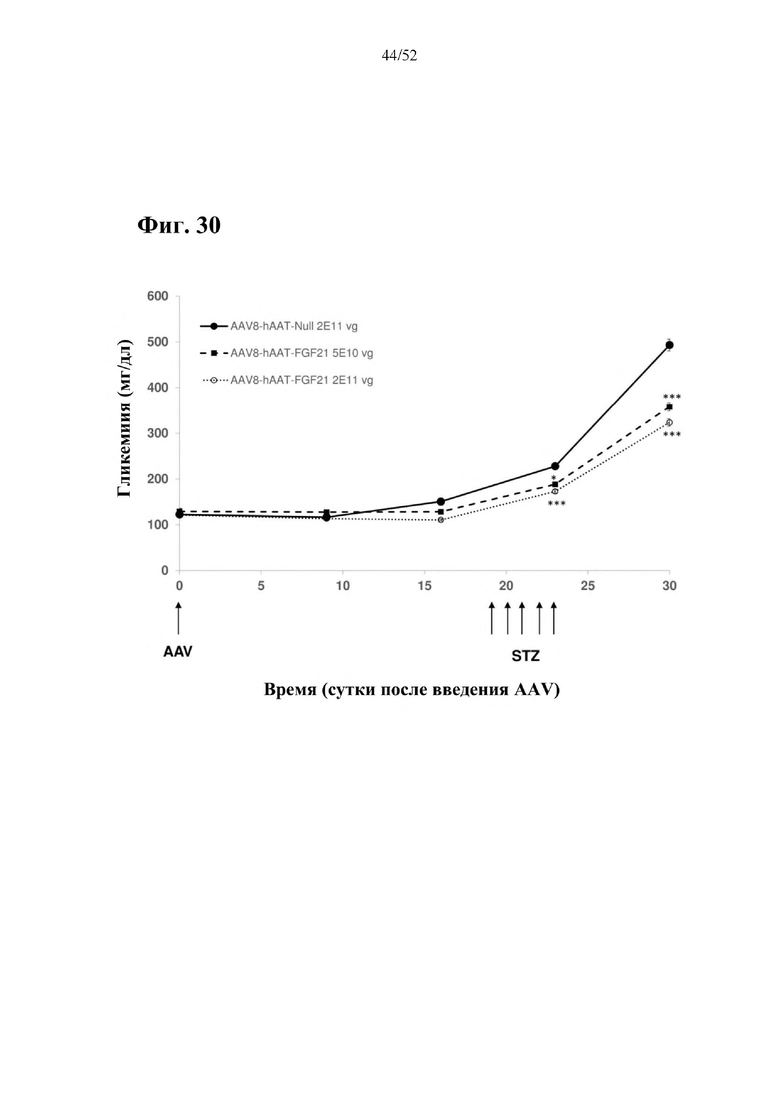
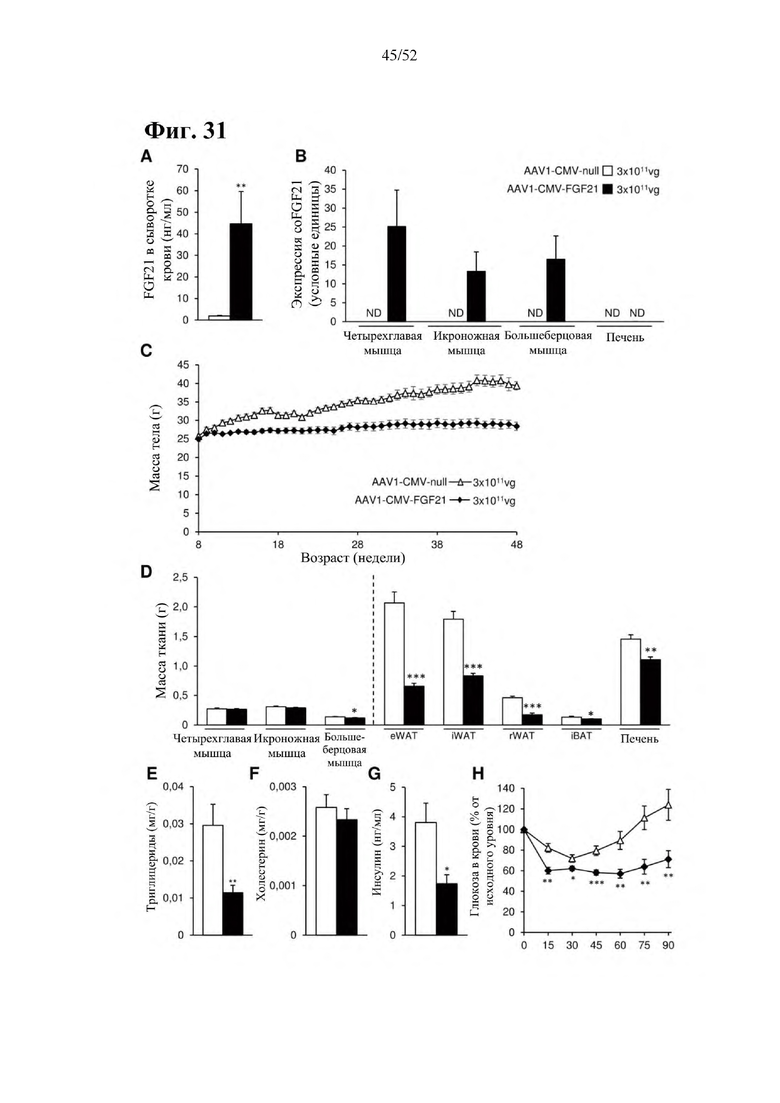
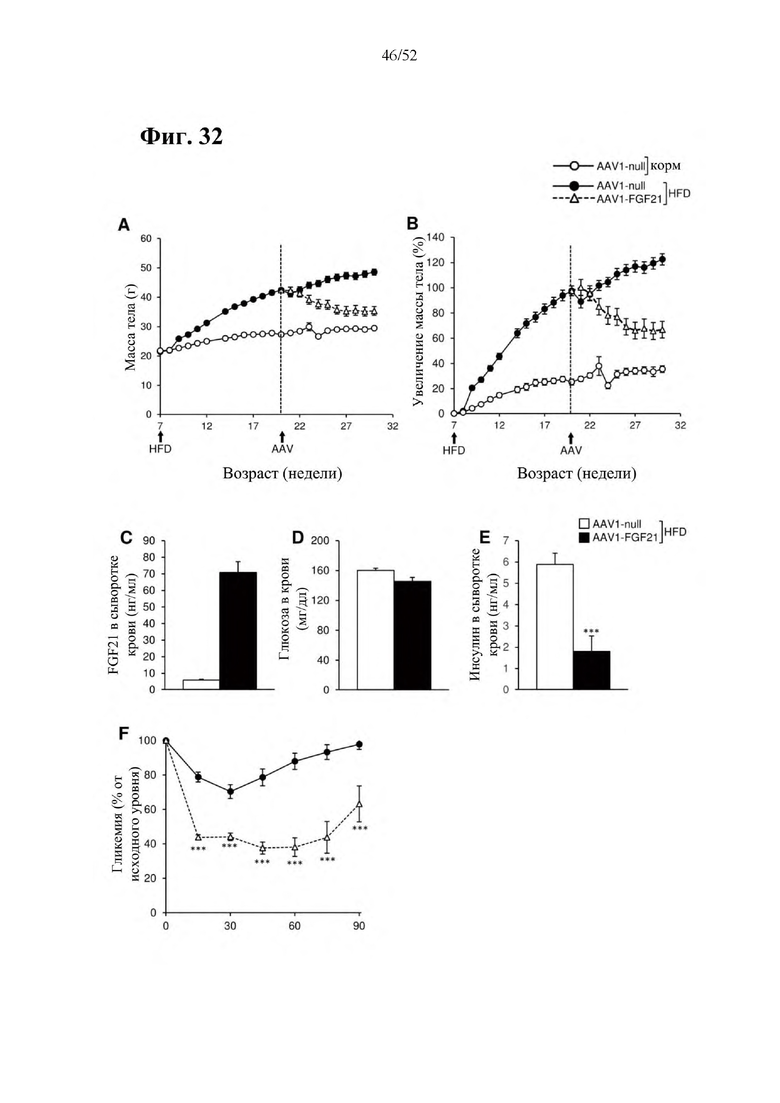
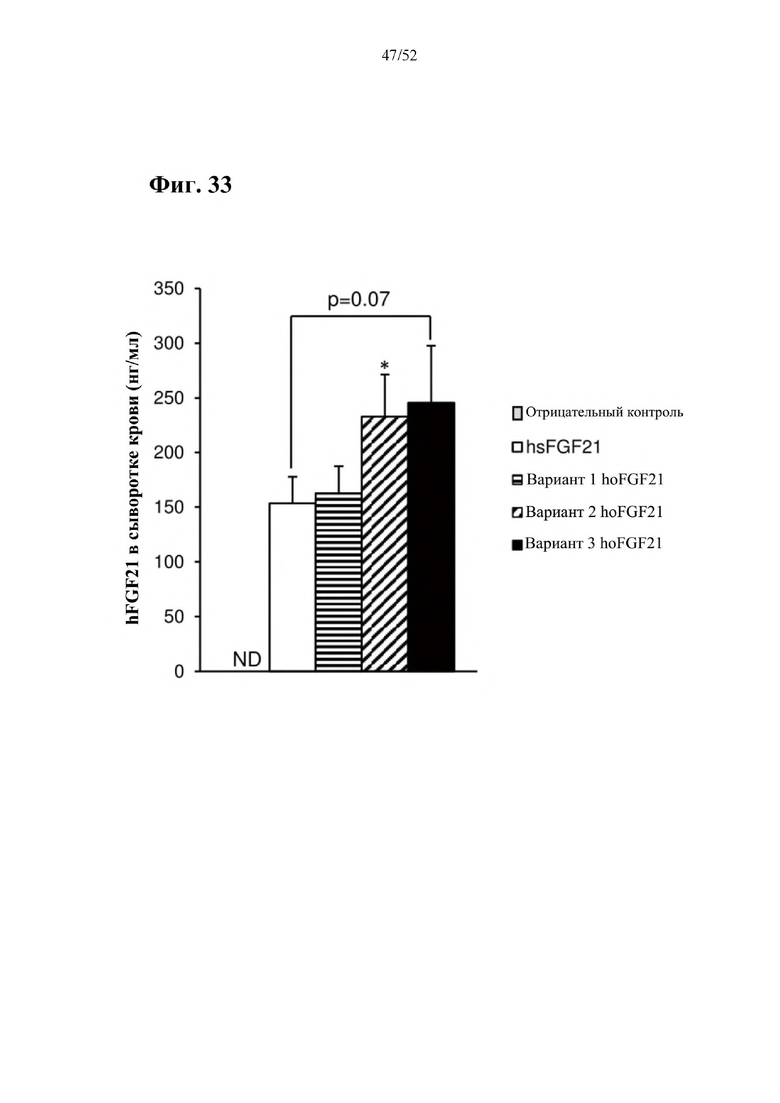
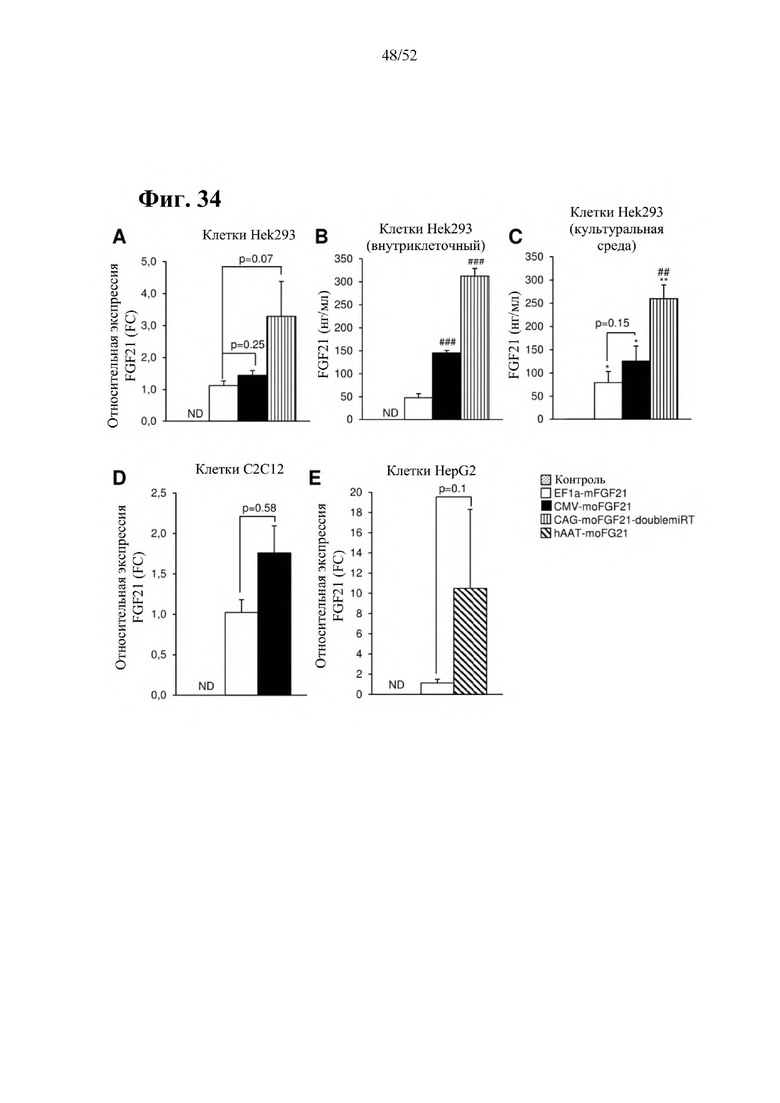
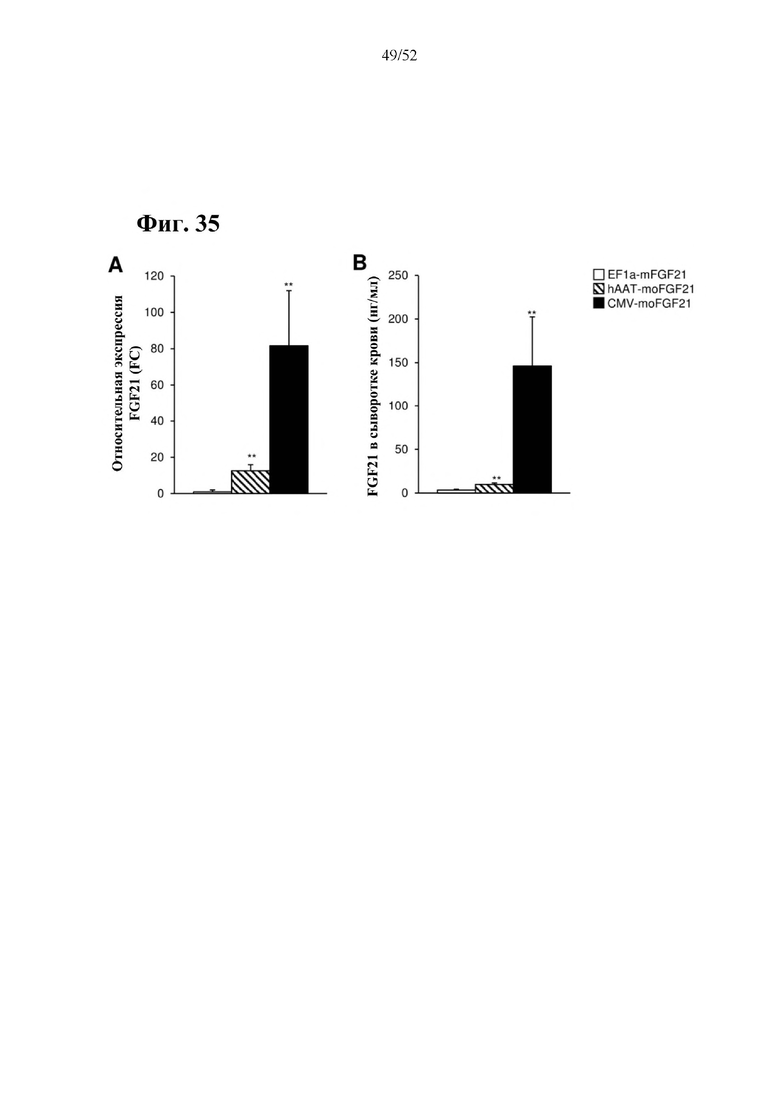
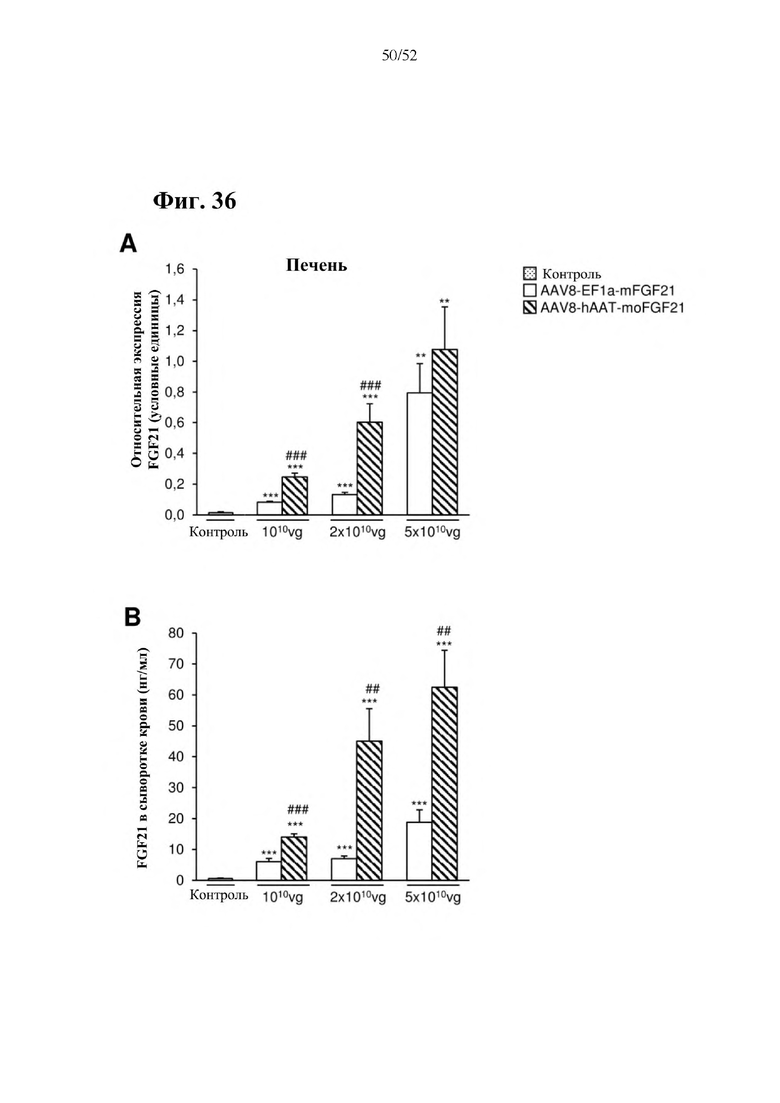
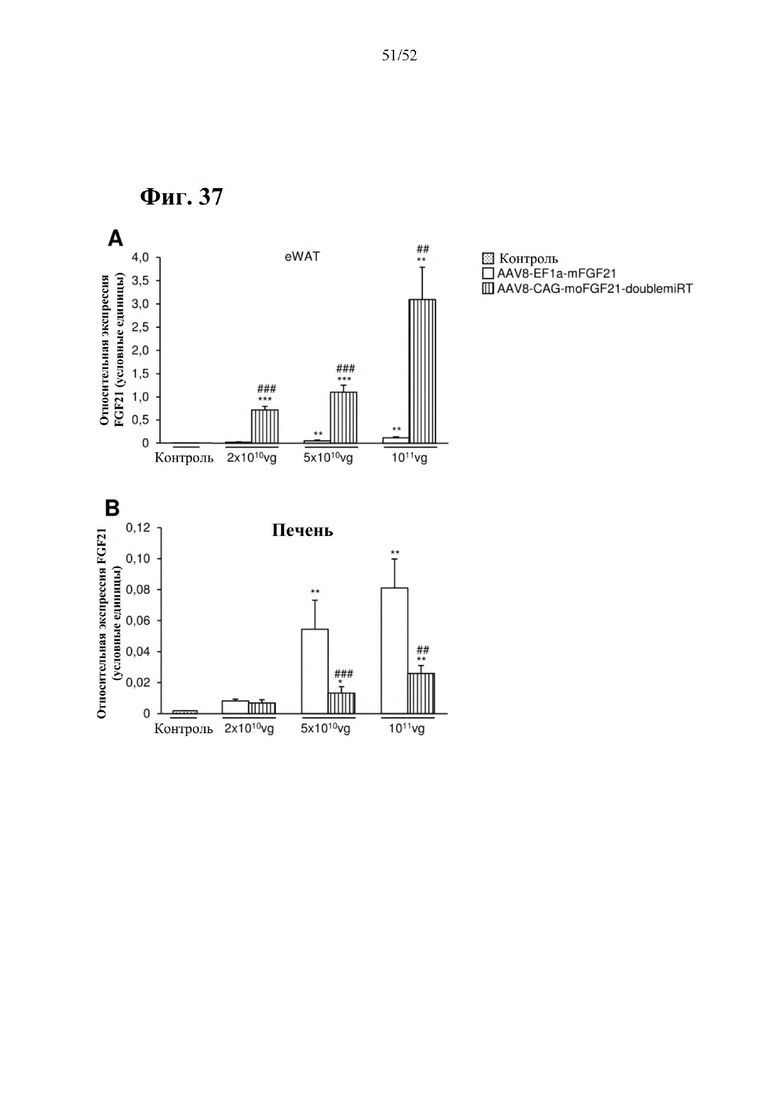
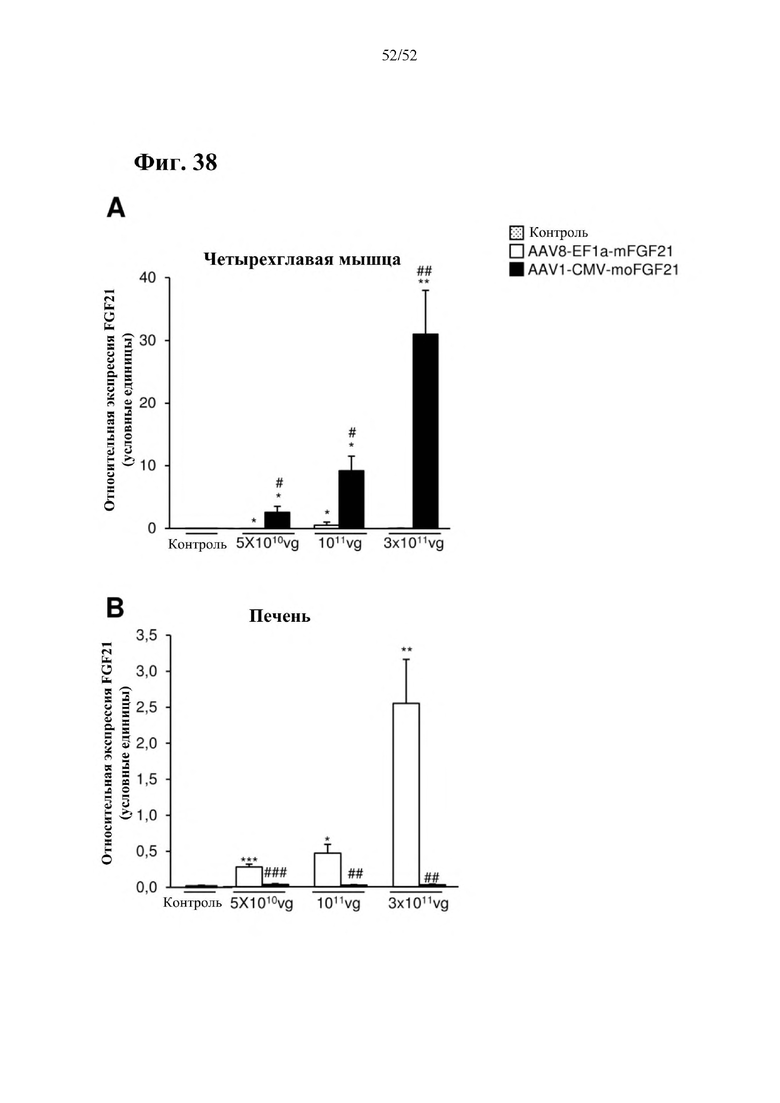
Группа изобретений относится к области медицины и предусматривает композиции для генной терапии, предназначенные для применения в лечении метаболического нарушения. Вектор на основе аденоассоциированного вируса 1 (AAV1) содержит вирусную экспрессионную конструкцию, содержащую убиквитарный промотор, функционально связанный с нуклеотидной последовательностью, которая кодирует фактор роста фибробластов 21 (FGF21), экспрессируемый в скелетной мышце. Молекула нуклеиновой кислоты представлена оптимизированной по кодону для экспрессии у млекопитающего нуклеотидной последовательностью, которая кодирует FGF21. Метаболическое нарушение представляет собой сахарный диабет, ожирение, неалкогольный стеатогепатит (NASH), воспаление или фиброз печени, метаболическое нарушение, ассоциированное со старением, рак. Группа изобретений обеспечивает возможность продолжительной секреции стабильного нативного FGF21 в кровоток. Эффективность сохраняется даже при однократном введении векторов, переносящих ген. 8 н. и 8 з.п. ф-лы, 77 ил., 1 табл., 24 пр.
1. Вектор на основе аденоассоциированного вируса 1 (AAV1), содержащий вирусную экспрессионную конструкцию, содержащую убиквитарный промотор, функционально связанный с нуклеотидной последовательностью, которая кодирует фактор роста фибробластов 21 (FGF21), экспрессируемый в скелетной мышце, при этом указанная нуклеотидная последовательность выбрана из группы, состоящей из:
(а) нуклеотидной последовательности, которая является по меньшей мере на 95% идентичной нуклеотидной последовательности SEQ ID NO: 5, 6 или 7; или
(b) нуклеотидной последовательности, последовательность которой отличается от последовательности нуклеотидной последовательности из (a) вследствие вырожденности генетического кода.
2. Вектор на основе AAV1 по п. 1, где убиквитарный промотор представляет собой промотор цитомегаловируса (CMV) или промотор CAG.
3. Молекула нуклеиновой кислоты, представленная оптимизированной по кодону для экспрессии у млекопитающего нуклеотидной последовательностью, которая кодирует FGF21, при этом нуклеотидная последовательность является по меньшей мере на 95% идентичной нуклеотидной последовательности SEQ ID NO: 5, 6 или 7.
4. Применение композиции в качестве лекарственного средства для генной терапии, причем указанная композиция содержит вектор на основе AAV1, который определен в п. 1, или молекулу нуклеиновой кислоты, которая определена в п. 3, вместе с одним или более фармацевтически приемлемыми вспомогательными веществами или средами.
5. Применение вектора на основе AAV1, который определен в п. 1, или молекулы нуклеиновой кислоты, которая определена в п. 3, в качестве лекарственного средства для генной терапии.
6. Применение вектора на основе AAV1, который определен в п. 1, или молекулы нуклеиновой кислоты, которая определена в п. 3, в лечении или предупреждении метаболического нарушения, причем указанное метаболическое нарушение представляет собой сахарный диабет, ожирение или неалкогольный стеатогепатит (NASH).
7. Применение вектора на основе AAV1, который определен в п. 1, или молекулы нуклеиновой кислоты, которая определена в п. 3, в лечении или предупреждении воспаления или фиброза печени.
8. Применение вектора на основе AAV1, который определен в п. 1, или молекулы нуклеиновой кислоты, которая определена в п. 3, в лечении или предупреждении метаболического нарушения, ассоциированного со старением.
9. Применение вектора на основе AAV1, который определен в п. 1, или молекулы нуклеиновой кислоты, которая определена в п. 3, в лечении или предупреждении рака, предпочтительно, рака печени.
10. Применение по п. 4 или 5, где лекарственное средство представлено в виде состава для внутримышечного введения.
11. Вектор на основе AAV1 по любому из пп. 1, 2, где вирусная экспрессионная конструкция содержит инвертированные концевые повторы (ITR), полученные из серотипа аденоассоциированного вируса AAV2.
12. Применение по любому из пп. 4-9, где вирусная экспрессионная конструкция содержит инвертированные концевые повторы (ITR), полученные из серотипа аденоассоциированного вируса AAV2.
13. Применение по п. 4, где лекарственное средство предназначено для лечения или предупреждения метаболического нарушения, при этом метаболическое нарушение представляет собой диабет, ожирение или неалкогольный стеатогепатит (NASH).
14. Применение по п. 4, где лекарственное средство предназначено для лечения или предупреждения воспаления или фиброза печени.
15. Применение по п. 4, где лекарственное средство предназначено для лечения или предупреждения метаболического нарушения, ассоциированного со старением.
16. Применение по п. 4, где лекарственное средство предназначено для лечения или предупреждения рака, предпочтительно рака печени.
| WO 2013033452 A2, 07.03.2013 | |||
| WO 2009149171 A2, 10.12.2009 | |||
| WO 2011127337 A2, 13.10.2011 | |||
| WO 2012066075 A1, 24.05.2012 | |||
| ЛЕЧЕНИЕ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ФАКТОРОМ РОСТА ФИБРОБЛАСТОВ 21 (FGF21), ПУТЕМ ИНГИБИРОВАНИЯ ПРИРОДНОГО АНТИСМЫСЛОВОГО ТРАНСКРИПТА К FGF21 | 2011 |
|
RU2610661C2 |

