Настоящее изобретение относится к молекулам антител, ингибирующим тромбин.
Свертывание крови является ключевым процессом для предотвращения кровотечений из поврежденных кровеносных сосудов (гемостазе). Тем не менее, тромб, который блокирует течение крови через сосуд (при тромбозе) или отрывается, застревая в сосуде в другой части тела (при тромбоэмболии), может представлять серьезную угрозу для здоровья.
Для лечения патологической свертываемости крови доступен ряд способов антикоагулянтной терапии. Общим недостатком этих способов лечения является повышенный риск кровотечения (Mackman (2008) Nature 451(7181):914-918). Многие антикоагулянты характеризуются узким терапевтическим окном между дозой, предотвращающей тромбоз и дозой, вызывающей кровотечение. Это окно часто дополнительно ограничивается различиями в реакции у отдельных пациентов.
Настоящее изобретение относится к неожиданному наблюдению, заключающемуся в том, что молекулы антител, распознающие эпитоп экзосайта 1 тромбина, селективно ингибируют тромбин, не вызывая кровотечения. Указанные молекулы антител могут быть полезны при лечении и профилактике тромбоза, эмболии и других состояний, опосредованных тромбином.
Согласно одному из аспектов настоящего изобретения предложена молекула антитела, специфически связывающаяся с экзосайтом 1 тромбина.
Выделенные молекулы антител против экзосайта 1 могут ингибировать тромбин in vivo без стимуляции или существенной стимуляции кровотечения или кровоизлияния, т.е. указанные молекулы антител не ингибируют или практически не ингибируют нормальные физиологические реакции на повреждение сосудов (т.е. гемостаз). Например, молекулы антител могут не ингибировать или в минимальной степени ингибировать гемостаз (т.е. ингибировать в незначительной степени, которая не влияет на благополучие пациента или не требует дополнительного вмешательства). Указанные молекулы антител могут не увеличивать вероятность кровотечения или могут увеличивать вероятность кровотечения в минимальной степени.
Экзосайт 1 (также известный как "анион-связывающий экзосайт 1" и "экзосайт распознавания фибриногена") является хорошо описанным вторичным сайтом связывания молекулы тромбина (см., например, James A. Huntington, 2008, Structural Insights into the Life History of Thrombin, in Recent Advances in Thrombosis and Hemostasis 2008, editors; K. Tanaka and E.W. Davie, Springer Japan KK, Tokyo, pp. 80-106). Экзосайт 1 образуется в зрелом тромбине, но не в протромбине (см., например, Anderson et al. (2000) JBC 2775 16428-16434).
Экзосайт 1 участвует в распознавании субстратов тромбина, например, фибриногена, но удален от каталитического активного центра. С экзосайтом 1 связываются различные факторы связывания тромбина, в том числе антикоагулянтный додекапептид гируген (Naski et al. 1990 JBC 265 13484-13489), фактор V, фактор VIII, тромбомодулин (кофактор белка C и активации АТИФ), фибриноген, PAR1 и фибрин (кофактор активации фактора XIII).
Антитело против экзосайта 1 может связываться с экзосайтом 1 зрелого тромбина человека. Последовательность препротромбина человека приведена в SEQ ID NO: 1. Протромбин человека обладает последовательностью остатков 44-622 SEQ ID NO: 1. Зрелый тромбин человека обладает последовательностью остатков 314-363 (легкая цепь) и остатков 364-622 (тяжелая цепь).
В некоторых вариантах реализации настоящего изобретения антитело против экзосайта 1 также может связываться с экзосайтом 1 зрелого тромбина других видов животных. Последовательности тромбина других видов животных известны в данной области техники и доступны в базах данных общего пользования, например, Genbank. Соответствующие остатки последовательностей тромбина других видов животных легко идентифицировать с помощью инструментов для выравнивания последовательностей.
Схема нумерации остатков аминокислот в молекуле тромбина, представленная в настоящем документе, является общепринятой в данной области техники и основана на шаблоне химотрипсина (Bode W et al. EMBO J. 1989 Nov.; 8(11):3467-75). Тромбин содержит инсерционные петли по сравнению с химотрипсином, которые последовательно обозначаются строчными буквами.
Экзосайт 1 зрелого тромбина человека подчеркнут в SEQ ID NO: 1 и может включать следующие остатки аминокислот: M32, F34, R35, K36, S36a, P37, Q38, E39, L40, L65, R67, S72, R73, T74, R75, Y76, R77a, N78, E80, K81, 182, S83, M84, K109, K110, K149e, G150, Q151, S153 и V154. В некоторых вариантах реализации другие остатки аминокислот тромбина, расположенные вблизи (т.е. в пределах 0,5 нм или 1 нм) от любого из указанных остатков, также могут рассматриваться как часть экзосайта 1.
Антитело против экзосайта 1 может связываться с эпитопом, содержащим 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более 20 остатков аминокислот экзосайта 1. Предпочтительно, антитело против экзосайта 1 связывается с эпитопом, полностью состоящим из остатков аминокислот экзосайта 1.
Например, антитело против экзосайта 1 может связываться с эпитопом, содержащим 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или все 16 остатков аминокислот, выбранных из группы, состоящей из M32, F34, S36a, Р37, Q38, E39, L40, L65, R67, R73, T74, R75, Y76, R77a, 182 и Q151 тромбина человека или эквивалентных остатков аминокислот тромбина других видов животных. В еще одном варианте реализации эпитоп может содержать остатки аминокислот тромбина Q38, R73, Т74, Y76 и R77a и, необязательно, один или более дополнительных остатков аминокислот.
Молекулы антител против экзосайта 1, описанные в настоящем документе, специфичны по отношению к экзосайту 1 тромбина и связываются с указанным эпитопом с высоким сродством по сравнению с другими эпитопами, например, эпитопами белков млекопитающих, не являющихся зрелым тромбином. Например, молекула антитела против экзосайта 1 может проявлять сродство связывания по отношению к экзосайту 1 тромбина, по меньшей мере в 500 раз, по меньшей мере в 1000 раз или по меньшей мере в 2000 раз выше, чем по отношению к другим эпитопам.
Предпочтительно, молекула антитела, описанная в настоящем документе, специфичная по отношению к экзосайту 1, может связываться со зрелым тромбином, но не проявляет связывания или практически не проявляет связывания с протромбином.
Без привязки к конкретной теории, антитела против экзосайта 1 могут не иметь доступа к тромбину в ядре кровоостанавливающего тромба, и, следовательно, не иметь возможности влиять на гемостаз путем прерывания нормальной функции тромбина в местах повреждения сосудов. Однако поскольку антитела против экзосайта 1 связываются с тромбином на поверхности тромба и во внешней оболочке тромба, тромбоз предотвращается, т.е. предотвращается негемостатическое распространение тромба.
Молекула антитела против экзосайта 1 может обладать константой диссоциации по отношению к экзосайту 1 менее 50 нМ (наномоль), менее 40 nM, менее 30 нМ, менее 20 нМ, менее 10 нМ или менее 1 нМ. Например, молекула антитела может обладать сродством к экзосайту 1 от 0,1 до 50 нМ, например, от 0,5 до 10 нМ. Подходящая молекула антитела против экзосайта 1 может, например, обладать сродством к экзосайту 1 тромбина, равным приблизительно 1 нМ.
Кинетику связывания и сродство (выраженное как константа равновесной диссоциации, Kd) молекул антител против экзосайта 1 можно определить с помощью стандартных методик, например, поверхностного плазмонного резонанса, например, с помощью анализа BIAcore.
Молекула антитела против экзосайта 1, описанная в данном документе, может быть иммуноглобулином или его фрагментом, и может быть природной или частично или полностью синтетической, например, рекомбинантной молекулой.
Молекулы антител против экзосайта 1 могут включать любой полипептид или белок, содержащий антигенсвязывающий сайт антитела, в том числе Fab, Fab2, Fab3, диатела, триотела, тетратела, миниантитела и однодоменные антитела, в том числе нанотела, а также полные антитела любого изотипа или подкласса. Молекулы антител и способы их конструирования и применения описаны, например, в статье Holliger & Hudson, Nature Biotechnology 23(9):1126-1136 (2005).
В некоторых вариантах реализации настоящего изобретения молекула антитела против экзосайта 1 является полноразмерным антителом. Например, молекула антитела против экзосайта 1 может являться IgG, IgA, IgE или IgM или относиться к любому из подклассов изотипа, в частности, IgG1 и IgG4. Молекулы антител против экзосайта 1 могут являться моноклональными антителами. В других предпочтительных вариантах реализации молекула антитела против экзосайта 1 является фрагментом антитела.
Молекулы антител против экзосайта 1 могут представлять собой химерные, гуманизированные антитела или антитела человека.
Молекулы антител против экзосайта 1, описанные в настоящем документе, могут быть выделенными, т.е. не содержать примесей, например, антител, способных связывать другие полипептиды и/или компоненты сыворотки. Для некоторых целей предпочтительны моноклональные антитела, хотя поликлональные антитела также можно использовать.
Молекулы антител против экзосайта 1 можно получить с помощью способов, стандартно применяемых в данной области техники. Способы получения антител включают иммунизацию млекопитающего (например, мыши, крысы, кролика, лошади, козы, овцы или обезьяны) белком или его фрагментом. Антитела можно получить из иммунизированных животных с помощью любой из множества методик, известных в данной области техники, и подвергнуть скринингу, предпочтительно с помощью связывания антитела с представляющим интерес антигеном. Например, можно применить методики вестерн-блоттинга или иммунопреципитации (Armitage et al., 1992, Nature 357:80-82). Выделение антител и/или клеток, продуцирующих антитела, из организма животного может сопровождаться этапом умерщвления животного.
В качестве альтернативы или дополнения к иммунизации млекопитающего пептидом, антитело, специфичное по отношению к белку, можно получить из рекомбинантной библиотеки экспрессируемых вариабельных доменов иммуноглобулина, например, с помощью бактериофага лямбда или нитевидного бактериофага, отображающих функциональные связывающие домены иммуноглобулинов на своей поверхности; см., например, WO 92/01047. Библиотека может быть «наивной», т.е. сконструированной из последовательностей, полученных из организма, не иммунизированого белками (или фрагментами), или может быть сконструирована с использованием последовательностей, полученных из организма, подвергшегося воздействию антигена, представляющего интерес.
Другие молекулы антител против экзосайта 1 можно идентифицировать путем скрининга сыворотки крови пациента на антитела, связывающиеся с экзосайтом 1.
В некоторых вариантах реализации молекулы антител против тромбина можно получить любым удобным способом, например, способом, описанным выше, а затем скринингом на дифференциальное связывание со зрелым тромбином по сравнению с тромбином с мутацией экзосайта 1, гамма-тромбином (дефектным по экзосайту 1 вследствие автолиза по R75 и R77a) или протромбином. Подходящие способы скрининга известны в данной области техники.
Антитело, демонстрирующее повышенное связывание со зрелым тромбином, по сравнению с белками, не являющимися тромбином, тромбином с мутациями экзосайта 1, гамма-тромбином или протромбином, например, антитело, связывающееся со зрелым тромбином, но не связывающееся с тромбином с мутацией экзосайта I, гамма-тромбином или протромбином, можно идентифицировать как молекулу антитела против экзосайта 1.
После получения и/или выделения можно протестировать биологическую активность молекулы антитела против экзосайта 1. Например, можно определить способность молекулы указанного антитела ингибировать связывание и/или расщепление тромбином субстрата, кофактора или ингибитора тромбина и/или способность молекулы антитела ингибировать тромбоз, не вызывая кровотечения.
Активность подходящих молекул антител можно протестировать с использованием свертывания фибриногена или анализа тромбинового времени. Подходящие способы анализа известны в данной области техники.
Влияние молекулы антитела на свертываемость крови и кровотечение можно определить с помощью стандартных методик. Например, влияние молекулы антитела на тромбоз можно определить на животной модели, например, модели мыши с индукцией тромбов в кровеносных сосудах с помощью хлорида железа (III). Влияние на гемостаз также можно определить на животной модели, например, путем измерения кровотечения из хвостовой вены мыши.
Молекулы антител обычно включают антигенсвязывающий домен, содержащий вариабельный домен тяжелой цепи иммуноглобулина (VH) и вариабельный домен легкой цепи иммуноглобулина (VL), хотя возможны и антигенсвязывающие домены, содержащие только вариабельный домен тяжелой цепи (VH) (например, антитела верблюдовых или акулы).
Каждый из доменов VH и VL обычно содержит три гипервариабельных участка (CDR), ответственных за связывание антигена, перемежающихся с каркасными областями.
В некоторых вариантах реализации связывание с экзосайтом 1 может происходить полностью или в значительной степени через VHCDR3 молекулы антитела против экзосайта 1.
Например, молекула антитела против экзосайта 1 может содержать домен VH, содержащий HCDR3 с аминокислотной последовательностью SEQ ID NO: 5 или последовательностью SEQ ID NO: 5 с 1 или более, например, 2, 3, 4 или 5 или более заменами, делециями или инсерциями аминокислот. Указанные замены могут представлять собой консервативные замены аминокислот. В некоторых вариантах реализации HCDR3 может содержать аминокислотные остатки в положениях 4-9 SEQ ID NO: 5 (SEFEPF), или, более предпочтительно, аминокислотные остатки в положениях 2 и 4-10 SEQ ID NO: 5 (D и SEFEPFS) с заменами, делециями или инсерциями по одному или более другим положениям в SEQ ID NO: 5. HCDR3 может быть единственной или практически единственной областью молекулы антитела, взаимодействующей с эпитопом экзосайта 1 тромбина. Таким образом, HCDR3 может определять специфичность и/или сродство молекулы антитела по отношению к области экзосайта 1 тромбина.
Домен VH молекулы антитела против экзосайта 1 может дополнительно содержать HCDR2 с аминокислотной последовательностью SEQ ID NO: 4 или последовательностью SEQ ID NO: 4 с 1 или более, например, с 2, 3, 4 или 5 или более заменами, делециями или инсерциями аминокислот. В некоторых вариантах реализации HCDR3 может содержать аминокислотные остатки в положениях 3-7 SEQ ID NO: 4 (DPQDG), или аминокислотные остатки в положениях 2 и 4-7 SEQ ID NO: 4 (L и PQDG) SEQ ID NO: 4 с заменами, делециями или инсерциями по одному или более другим положениям в SEQ ID NO: 4.
Домен VH молекулы антитела против экзосайта 1 может дополнительно содержать HCDR1 с аминокислотной последовательностью SEQ ID NO: 3 или последовательностью SEQ ID NO: 3 с 1 или более, например, 2, 3, 4 или 5 или более заменами, делециями или инсерциями аминокислот. В некоторых вариантах реализации HCDR1 может содержать аминокислотный остаток T в положении 5 SEQ ID NO: 3 с заменами, делециями или инсерциями по одному или более других положений в SEQ ID NO: 3.
В некоторых вариантах реализации молекула антитела может включать домен VH, содержащий HCDR1, HCDR2 и HCDR3, обладающие последовательностями SEQ ID NO: 3, 4 и 5, соответственно. Например, молекула антитела может содержать домен VH с последовательностью SEQ ID NO: 2 или последовательностью SEQ ID NO: 2 с 1 или более, например, 2, 3, 4 или 10 или более заменами, делециями или инсерциями аминокислот в SEQ ID NO: 2.
Молекула антитела против экзосайта 1 может дополнительно включать домен VL, например, домен VL, содержащий LCDR1, LCDR2 и LCDR3, обладающие последовательностями SEQ ID NO: 7, 8 и 9, соответственно, или последовательностями SEQ ID NO: 7, 8 и 9, соответственно, независимо содержащими одну или несколько, например, 2, 3, 4 или 5 или более замен, делеций или инсерций аминокислот. Указанные замены могут представлять собой консервативные замены аминокислот. Например, молекула антитела может содержать домен VL с последовательностью SEQ ID NO: 6 или последовательностью SEQ ID NO: 6 с 1 или более, например, 2, 3, 4 или 10 или более заменами, делециями или инсерциями аминокислот в SEQ ID NO: 6.
В некоторых вариантах реализации домен VL может содержать Tyr49.
Молекула антитела против экзосайта 1 может, например, содержать одну или несколько замен, делеций или инсерций аминокислот, улучшающих одно или более свойств антитела, например, сродство, функциональное время полужизни, скорость ассоциации и диссоциации.
Методики, необходимые для внедрения замен, делеций или инсерций в последовательности аминокислот CDR, VH- или VL-доменов и антител, в целом, доступны в данной области техники. Можно получить варианты последовательностей с заменами, делециями или инсерциями, которые могут или не могут предсказуемо оказывать минимальное или благоприятное влияние на активность, и протестировать их способность связывать экзосайт 1 тромбина и/или любое другое желательное свойство.
В некоторых вариантах реализации молекула антитела против экзосайта 1 может включать домен VH, содержащий HCDR1, HCDR2 и HCDR3, обладающие последовательностями SEQ ID NO: 3, 4 и 5, соответственно, и домен VL, содержащий LCDR1, LCDR2 и LCDR3, обладающие последовательностями SEQ ID NOS 7, 8 и 9, соответственно.
Например, VH- и VL-домены могут обладать аминокислотными последовательностями SEQ ID NO: 2 и SEQ ID NO: 6, соответственно; либо могут обладать аминокислотными последовательностями SEQ ID NO: 2 и SEQ ID NO: 6, независимо содержащими 1 или более, например, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более замен, делеций или инсерций аминокислот. Замены могут быть консервативными заменами.
В некоторых вариантах реализации антитело может содержать одну или несколько замен, делеций или вставок, удаляющих сайт гликозилирования. Например, сайт гликозилирования в домене VL SEQ ID NO 6 можно мутировать путем внедрения замены по N28 или S30.
Молекула антитела против экзосайта 1 может быть представлена в любом формате, как описано выше. В некоторых предпочтительных вариантах реализации молекула антитела против экзосайта 1 может представлять собой полноразмерное антитело, например, IgG, например, IgG1 или IgG4, IgA, IgE или IgM.
Молекула антитела против экзосайта 1 согласно настоящему изобретению может представлять собой молекулу, конкурирующую за связывание с экзосайтом 1 с молекулой антитела, описанной выше, например, молекулой антитела, которая
(i) связывает экзосайт 1 тромбина и
(ii) содержит VH-домен SEQ ID NO: 2 и/или VL-домен SEQ ID NO: 6; HCDR3 SEQ ID NO: 5; HCDR1, HCDR2, LCDR1, LCDR2 или LCDR3 SEQ ID NOS: 3, 4, 7, 8 или 9, соответственно; VH-домен, содержащий последовательности HCDR1, HCDR2 и HCDR3 SEQ ID NOS: 3, 4 и 5, соответственно; и/или VH-домен, содержащий последовательности HCDR1, HCDR2 и HCDR3 SEQ ID NOS: 3, 4 и 5, и VL-домен, содержащий последовательности LCDR1, LDR2 и LCDR3 SEQ ID NOS: 7, 8 и 9, соответственно.
Конкуренцию между молекулами антител можно легко проанализировать in vitro, например, с помощью твердофазного ИФА и/или путем мечения определенной репортерной молекулой молекул одного антитела, которые можно обнаружить в присутствии одной или более других немеченых молекул антител с целью обеспечить выявление молекул антител, связывающихся с тем же эпитопом или перекрывающимся эпитопом. Такие способы хорошо известны специалистам в данной области техники. Таким образом, в дополнительном аспекте настоящего изобретения предложена молекула антитела, содержащая антигенсвязывающий сайт антитела, конкурирующая с молекулой антитела, например, содержащей VH- и/или VL-домен, CDR, например, HCDR3, или набор CDR исходного антитела, описанного выше, за связывание с экзосайтом 1 тромбина. Подходящая молекула антитела может содержать антигенсвязывающий сайт антитела, конкурирующий с антигенсвязывающим сайтом антитела за связывание с экзосайтом 1, причем антигенсвязывающий сайт антитела состоит из VH-домена и VL-домена, и указанные VH- и VL-домены содержат последовательности HCDR1, HCDR2 и HCDR3 SEQ ID NO: 3, 4 и 5, и последовательности LCDR1, LDR2 и LCDR3 SEQ ID NOS: 7, 8 и 9, соответственно, например, VH- и VL-домены SEQ ID NOS: 2 и 6.
Молекула антитела против экзосайта 1, описанная в настоящем документе, может ингибировать связывание факторов, связывающихся с тромбином, в том числе факторов, связывающихся с экзосайтом 1. Например, молекула антитела может конкурентно или неконкурентно ингибировать связывание одного или более из fV, fVIII, тромбомодулина, фибриногена или фибрина, PAR1 и/или аналогов гиругена и гирудина с тромбином.
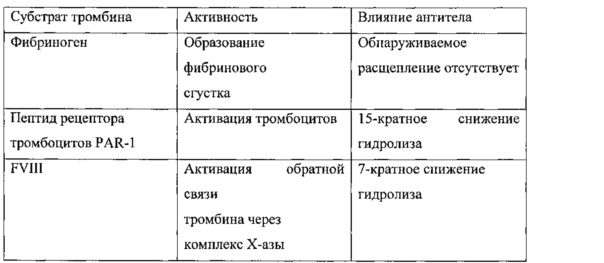
Молекула антитела против экзосайта 1, описанная в настоящем документе, может ингибировать одну или более активностей тромбина. Например, молекула антитела против экзосайта 1 может ингибировать гидролитическое расщепление одного или более субстратов тромбина, например, фибриногена, рецептора тромбоцитов PAR-1 и фактора свертывания FVIII. Например, связывание молекулы антитела с тромбином может привести к по меньшей мере 5-кратному, по меньшей мере 10-кратному или по меньшей мере 15-кратному снижению гидролиза фибриногена, PAR-1, фактора FVIII или других субстратов тромбина, например, фактора V, фактора XIII в присутствии фибрина и белка C и/или АТИФ в присутствии тромбомодулина. В некоторых вариантах реализации связывание тромбина молекулой антитела против экзосайта 1 может привести к отсутствию обнаруживаемого расщепления субстрата тромбина тромбином.
Методики измерения активности тромбина, например, путем измерения гидролиза субстратов тромбина in vitro, являются стандартными в данной области техники и описаны в настоящем документе.
Молекулы антител против экзосайта 1 можно дополнительно модифицировать путем химической модификации, например, ПЭГилирования или включения в липосомы, с целью улучшения их фармацевтических свойств, например, путем увеличения периода полужизни in vivo.
Влияние молекулы антитела против экзосайта 1 на свертываемость крови и кровотечение можно определить с помощью стандартных методик. Например, можно определить влияние антитела на модель тромбоза. Подходящие модели включают индукцию тромба в кровеносных сосудах с помощью хлорида железа (III) на модели мыши, с последующей оценкой кровотечения из хвостовой вены для проверки нормального гемостаза. Другие подходящие модели тромбоза хорошо известны в данной области техники (см., например, Westrick et al. ATVB (2007) 27:2079-2093)
Молекулы антител против экзосайта 1 могут быть включены в фармацевтические композиции с фармацевтически приемлемым вспомогательным веществом.
Фармацевтически приемлемое вспомогательное вещество может быть соединением или комбинацией соединений, входящих в фармацевтическую композицию, не вызывающим вторичных реакций и обеспечивающих, например, облегчение введения молекулы антитела против экзосайта 1, увеличение ее продолжительности жизни и/или ее эффективности в организме или увеличения ее растворимости в растворе. Эти фармацевтически приемлемые носители хорошо известны и должны быть приспособлены специалистом в данной области техники в зависимости от режима введения молекулы антитела против экзосайта 1.
В некоторых вариантах реализации молекулы антител против экзосайта 1 могут быть представлены в лиофилизированной форме для восстановления перед введением. Например, молекулы лиофилизированных антител можно восстановить в стерильной воде и смешать с физиологическим раствором перед введением индивиду.
Молекулы антител против экзосайта 1 обычно следует вводить в форме фармацевтической композиции, которая может содержать по меньшей мере один компонент в дополнение к указанной молекуле антитела. Таким образом, фармацевтические композиции могут содержать, в дополнение к молекуле антитела против экзосайта 1, фармацевтически приемлемый наполнитель, носитель, буфер, стабилизатор или другие материалы, хорошо известные специалистам в данной области техники. Такие материалы должны быть нетоксичными и не должны мешать эффективности молекулы антитела против экзосайта 1. Точная природа носителя или другого материала зависит от пути введения, которое может осуществляться путем болюса, вливания, инъекции или любым другим подходящим путем, как описано ниже.
Для парентерального, например, подкожного или внутривенного введения, например, путем инъекции, фармацевтическая композиция, содержащая молекулу антитела против экзосайта 1, может быть представлена в виде парентерально приемлемого апирогенного водного раствора с подходящим pH, изотоничностью и стабильностью. Специалисты в данной области техники легко могут изготовить подходящие растворы, используя, например, изотонические носители, например, раствор хлорида натрия для инъекций, раствор Рингера для инъекций, раствор Рингера с лактатом для инъекций. При необходимости можно использовать консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки, в том числе такие буферные вещества, как фосфат, цитрат и другие органические соли; антиоксиданты, например, аскорбиновую кислоту и метионин; консерванты (например, хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензэтония; фенол; бутиловый или бензиловый спирт; алкилпарабены, например метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и m-крезол); низкомолекулярные полипептиды; белки, например сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, например поливинилпирролидон; аминокислоты, например глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, например ЭДТА; сахара, например сахарозу, маннит, трегалозу или сорбит; солеобразующие противоионы, например, натрия; металлсодержащие комплексы (например, комплексы Zn-белок) и/или неионогенные ПАВ, например TWEEN™, плюроники (PLURONICS™) или полиэтиленгликоль (ПЭГ).
Фармацевтическую композицию, содержащую молекулу антитела против экзосайта 1, можно вводить отдельно или в сочетании с другими способами лечения, одновременно или последовательно, в зависимости от состояния, подлежащего лечению.
Молекулу антитела против экзосайта 1, описанная в настоящем документе, можно использовать в способе лечения человека или животного, в том числе профилактического или предохранительного лечения (например, лечения до наступления состояния у индивида с целью снижения риска указанного состояния у индивида; задержки его начала; или снижения его тяжести после начала). Способ лечения может включать введение молекулы антитела против экзосайта 1 индивиду, нуждающемуся в этом.
Введение, как правило, осуществляют в "терапевтически эффективном количестве", которое является достаточным для оказания благоприятного действия на пациента. Такое благоприятное действие может представлять собой, по меньшей мере, улучшение по меньшей мере одного симптома. Фактическое вводимое количество, скорость и период действия при введении зависят от природы и тяжести заболевания, подлежащего лечению, конкретного млекопитающего, подлежащего лечению, клинического состояния отдельного пациента, причины расстройства, области доставки композиции, способа введения, схемы введения и других факторов, известных врачам-практикам. За назначение лечения, например, решение по дозировке и т.д., отвечают врачи общей практики и другие врачи; оно может зависеть от тяжести симптомов и/или прогрессирования заболевания, подлежащего лечению. Подходящие дозы молекул антител хорошо известны в данной области техники (Ledermann J.A. et al. (1991) Int. J. Cancer 47:659-664; Bagshawe K.D. et al. (1991) Antibody, Immunoconjugates and Radiopharmaceuticals 4:915-922). Конкретные дозировки, которые можно использовать, могут быть указаны в настоящем документе или в Physician's Desk Reference (2003) в зависимости от вводимого типа лекарственного средства. Терапевтически эффективное количество или подходящую дозу молекулы антитела можно определить путем сравнения его активности in vitro и in vivo на животной модели. Известны методы экстраполяции эффективных дозировок у мышей и других экспериментальных животных на человека. Точная доза зависит от ряда факторов, в том числе предназначения антитела для профилактики или лечения, размера и местоположения зоны, подлежащей обработке, точной природы антитела (например, полноразмерное антитело, фрагмент) и природы обнаруживаемой метки или другой молекулы, присоединенной к антителу.
Типичная доза антитела находится в диапазоне от 100 мкг до 1 г при системном применении и от 1 мкг до 1 мг при местном применении. Можно вводить начальную повышенную нагрузочную дозу, а затем одну или большее количество пониженных доз. Обычно антитело представляет собой полноразмерное антитело, например, антитело изотипа IgG1 или IgG4. Дозу для однократного лечения взрослого пациента можно пропорционально корректировать для лечения детей и младенцев, а также корректировать с поправкой на другие форматы антител пропорционально их молекулярной массе. Лечение можно повторить с ежедневными, два раза в неделю, еженедельными или ежемесячными интервалами по усмотрению врача. График лечения индивида может зависеть от фармакодинамических и фармакодинамических свойств композиции антитела, пути введения и природы состояния, подлежащего лечению.
Лечение может быть периодическим, и период между введениями может составлять приблизительно две недели или более, например, приблизительно три недели или более, приблизительно четыре недели или более, приблизительно месяц или более, приблизительно пять недель или более, или приблизительно шесть недель или более. Например, лечение можно осуществлять раз в две-четыре недели или раз в четыре-восемь недель. Лечение можно проводить до и/или после операции, и/или можно вводить или наносить непосредственно на анатомическую область хирургического лечения или инвазивных процедур. Подходящие составы и пути введения описаны выше.
В некоторых вариантах реализации молекулы антител против экзосайта 1, описанные в настоящем документе, можно вводить путем подкожных инъекций. Подкожные инъекции можно вводить с помощью автоинжектора, например, при долгосрочной профилактике/лечении.
В некоторых предпочтительных вариантах реализации терапевтическое действие молекулы антитела против экзосайта 1 может присутствовать на протяжении нескольких времен полужизни в зависимости от дозы. Например, терапевтический эффект однократного введения дозы молекулы антитела против экзосайта 1 может сохраняться в организме индивида в течение 1 месяца или более, 2 месяцев или более, 3 месяцев или более, 4 месяцев или более, 5 месяцев или и более, или 6 месяцев или более.
Молекулы антител против экзосайта 1, описанные в настоящем документе, ингибируют тромбин и могут применяться при лечении состояний, опосредованных тромбином.
Гемостаз является нормальной реакцией свертывания, т.е. профилактикой кровотечений или кровоизлияний, например, из поврежденного кровеносного сосуда. Гемостаз останавливает кровотечение и кровоизлияние из кровеносных сосудов в организме.
Молекулы антител против экзосайта 1 могут не оказывать влияния или практически не оказывать влияния на гемостаз, т.е. не вызывать кровотечения или кровоизлияния.
В аспектах настоящего изобретения предложены; молекула антитела против экзосайта 1, описанная в настоящем документе, для применения в рамках способа лечения человека или животного; молекула антитела против экзосайта 1, описанная в настоящем документе, для применения рамках способа лечения расстройства, опосредованного тромбином; применение молекулы антитела против экзосайта 1, описанной в настоящем документе, при изготовлении лекарственного средства для лечения состояния, опосредованного тромбином; а также способ лечения состояния, опосредованного тромбином, включающий введение молекулы антитела против экзосайта 1, описанной в настоящем документе, индивиду, нуждающемуся в этом.
Ингибирование тромбина антителами против экзосайта 1, описанными в настоящем документе, может обеспечивать благоприятный эффект при лечении состояний, опосредованных тромбином. Состояние, опосредованное тромбином, может включать расстройства, связанные с образованием или активностью тромбина.
Тромбин играет ключевую роль в гемостазе, свертывании и тромбозе. Состояния, опосредованные тромбином, включают тромботические состояния, например, тромбоз и эмболию.
Тромбоз представляет собой свертывание, происходящее в избытке, превышающем потребности гемостаза (т.е. избыточное свертывание), или не требующееся для гемостаза (т.е. внегемостатическое или негемостатическое свертывание).
Тромбоз представляет собой свертывание крови в просвете кровеносного сосуда. Он характеризуется образованием сгустка (тромба), превышающего потребности или не требующегося для гемостаза. Сгусток может препятствовать кровотоку через кровеносный сосуд, что ведет к медицинским осложнениям. Сгусток может оторваться от места своего образования, что приводит к эмболии в другом месте кровеносной системы. В артериальной системе тромбоз, как правило, является результатом разрыва атеросклеротической бляшки.
В некоторых вариантах реализации тромбоз может возникнуть после первоначальной физиологической гемостатической реакции, например, повреждения эндотелиальных клеток в кровеносном сосуде. В других вариантах реализации тромбоз может происходить в отсутствие какой-либо физиологической гемостатической реакции.
Тромбоз может происходить у людей с внутренней предрасположенностью к тромбозу (т.е. тромбофилии) или у "здоровых" людей без внутренней предрасположенности к тромбозам, например, в ответ на внешний раздражитель.
Тромбоз и эмболия могут произойти в любой вене, артерии или другом кровеносном сосуде в кровеносной системе и может включать капиллярный тромбоз.
Тромбоз и эмболия могут быть связаны с хирургической операцией (возникать во время операции или после нее) или внедрением в организм пациента посторонних предметов, например, коронарных стентов.
Например, антитела против экзосайта 1, описанные в настоящем документе, можно применять при хирургических и других процедурах, при которых кровь подвергается воздействию искусственных поверхностей, например, открытой операции на сердце и диализе.
Тромботические состояния могут включать тромбофилию, тромботический инсульт и окклюзию коронарной артерии.
Пациенты, подходящие для лечения, описанного в настоящем документе, включают пациентов с состояниями, при которых тромбоз является симптомом или побочным эффектом лечения, или сопровождающихся повышенным риском тромбоза, или пациентов, предрасположенных или подверженных повышенному риску тромбоза по сравнению с общей популяцией. Например, молекулу антитела против экзосайта 1, описанную в настоящем документе, также можно применять при лечении или профилактике венозного тромбоза у больных раком, а также при лечении или профилактике нозокомиального тромбоза, который вызывает 50% случаев венозной тромбоэмболии.
Молекулы антител против экзосайта 1, описанные в настоящем документе, могут оказывать терапевтическое или другое благоприятное действие при состояниях, опосредованных тромбином, например, тромботических состояниях, без существенного ингибирования или помех для гемостаза. Например, риск кровотечения у пациентов, получавших молекулы антител против экзосайта 1, может не возрастать или практически не возрастать по сравнению с индивидами, не получавшими лечения.
У индивидов, получавших обычные антикоагулянты, например, натуральный и синтетический гепарин, варфарин, прямые ингибиторы сериновых протеаз (например, аргатробан, дабигатран, апиксабан и ривароксабан), гирудин и его производные (например, лепирудин и бивалирудин) и антитромбоцитарные препараты (например, клопидогрель, тиклопидин и абциксимаб), наблюдались кровотечения. Риск кровотечения у пациентов, получавших молекулы антител против экзосайта 1, описанные в настоящем документе, может быть понижен по сравнению с индивидами, получавшими обычные антикоагулянты.
Состояния, опосредованные тромбином, включают нетромботические состояния, связанные с активностью тромбина, в том числе воспаление, инфекцию, опухолевый рост и метастазирование, отторжение органов и деменцию (сосудистую и несосудистую, например, болезнь Альцгеймера) (Licari et al. J Vet Emerg Crit Care (San Antonio). 2009 Feb; 19(1):11-22; Tsopanoglou et al. Eur Cytokine Netw. 2009 Dec 1; 20(4):171-9).
Молекулы антител против экзосайта 1, описанные в настоящем документе, также можно применять при тестировании in vitro, например, при анализе и оценке характеристик свертывания, например, в образце, полученном от пациента.
Молекулы антител против экзосайта 1 можно применять при измерении образования тромбина. Анализ образования тромбина технически проблематичен, поскольку превращение фибриногена в фибрин вызывает помутнение образца, исключающее использование простого хромогенного анализа по конечной точке.
Добавление молекулы антитела против экзосайта 1, описанной в настоящем документе, в образец крови предотвращает или ингибирует образование фибрина и, следовательно, помутнение, и обеспечивает измерение образования тромбина с помощью хромогенного субстрата, без необходимости дефибринизации.
Например, способ измерения образования тромбина может включать контакт образца крови с хромогенным субстратом тромбина в присутствии молекулы антитела против экзосайта 1, описанной в настоящем документе, и измерение хромогенного сигнала субстрата;
причем хромогенный сигнал указывает на образование тромбина в образце.
Хромогенный сигнал можно измерять непосредственно без дефибринизации образца.
Подходящие субстраты хорошо известны в данной области техники и включают S2238 (H-D-Phe-Pip-Arg-pNa), β-Ala-Gly-Arg-p-нитроанилида диацетат (Prasa, D. et al. (1997) Thromb. Haemost. 78, 1215; Sigma Aldrich Inc) и Tos-Gly-Pro-Arg-pNa.AcOH (Biophen CS-01(81); Aniara Inc OH USA).
Молекулы антител против экзосайта 1 также можно применять при ингибировании или предотвращении свертывания крови, как описано выше, в экстракорпоральном кровотоке, например, при гемодиализе и экстракорпоральной мембранной оксигенации.
Например, способ ингибирования или предотвращения свертывания крови in vitro или ex vivo может включать введение молекулы антитела против экзосайта 1, описанной в настоящем документе, в образец крови. Образец крови можно ввести в экстракорпоральный кровоток до, одновременно с или после введения антитела против экзосайта 1 и, необязательно, подвергать обработке, например, гемодиализу или оксигенации. В некоторых вариантах реализации обработанную кровь затем вводят индивиду. В других вариантах реализации предложены молекула антитела против экзосайта 1, описанная в настоящем документе, для применения в рамках способа ингибирования или предотвращения свертывания крови в образце крови ex vivo, и применение молекулы антитела против экзосайта 1, описанной в настоящем документе, при изготовлении лекарственного средства для применения в рамках способа ингибирования или предотвращения свертывания крови в образце крови ex vivo.
Другие аспекты изобретения относятся к получению молекул антител, которые связываются с эпитопом экзосайта 1 тромбина и могут применяться, например, при лечении патологического свертывания крови или тромбоза.
Способ получения антигенсвязывающего домена антитела к эпитопу экзосайта 1 тромбина, может включать;
обеспечение VH-домена, являющегося вариантом аминокислотной последовательности исходного VH-домена путем добавления, делеций, замены или инсерций одной или нескольких аминокислот в аминокислотной последовательности исходного VH-домена, включающего HCDR1, HCDR2 и HCDR3, причем HCDR1, HCDR2 и HCDR3 имеют аминокислотные последовательности SEQ ID NOS: 3, 4 и 5 соответственно и;
в некоторых случаях объединение полученного таким образом VH-домена с одним или несколькими VL-доменами с получением одной или более комбинации VH/VL; и
тестирование указанного VH-домена, являющегося вариантом аминокислотной последовательности исходного VH-домена или комбинации или комбинаций VH/VL, с целью выявления антигенсвязывающего домена антитела к эпитопу экзосайта 1 тромбина.
VH-домен, являющийся вариантом аминокислотной последовательности исходного VH-домена, может содержать последовательность HCDR3 SEQ ID NO: 5 или вариант с добавлением, делецией, заменой или инсерцией одной, двух, трех или более аминокислот.
VH-домен, являющийся вариантом аминокислотной последовательности исходного VH-домена, может содержать последовательности HCDR1 и HCDR2 SEQ ID NO: 3 или 4, соответственно, или варианты указанных последовательностей с добавлением, делецией, заменой или инсерцией одной, двух, трех или более аминокислот.
Способ получения молекулы антитела, специфически связывающейся с эпитопом экзосайта 1 тромбина, может включать;
обеспечение исходной нуклеиновой кислоты, кодирующей VH-домен, или исходного репертуара нуклеиновых кислот, каждая из которых кодирует VH-домен, причем VH-домен или VH-домены содержат HCDR1, HCDR2 и/или HCDR3, подлежащие замене, или не содержат области, кодирующей HCDR1, HCDR2 и/или HCDR3;
объединение указанной исходной нуклеиновой кислоты или исходного репертуара нуклеиновых кислот с донорной нуклеиновой кислотой или донорными нуклеиновыми кислотами, кодирующими или полученными путем мутации аминокислотной последовательности HCDR1, HCDR2 и/или HCDR3, обладающих аминокислотными последовательностями SEQ ID NOS: 3, 4 и 5, соответственно, так что указанная донорная нуклеиновая кислота или донорные нуклеиновые кислоты встраиваются в участки CDR1, CDR2 и/или CDR3 исходной нуклеиновой кислоты или исходного репертуара с получением продукта - репертуара нуклеиновых кислот, кодирующих VH домены;
экспрессию нуклеиновых кислот указанного полученного репертуара для продукции полученных VH-доменов;
необязательное объединение указанных полученных VH-доменов с одним или более VL-доменами;
выбор молекулы антитела, связывающегося с экзосайтом 1 тромбина, причем молекула антитела содержит полученный VH-домен и, необязательно, VL-домен; и
выделение указанной молекулы антитела или кодирующей ее нуклеиновой кислоты.
Подходящие методики созревания и оптимизации молекул антител хорошо известны в данной области техники.
Антигенсвязывающие домены антител и молекулы антител к эпитопу экзосайта 1 тромбина можно тестировать, как описано выше; Например, можно определить способность связываться с тромбином и/или ингибировать расщепление субстратов тромбина.
Влияние молекулы антитела на свертывание и кровотечение можно определить с помощью стандартных методик. Например, можно использовать модель тромбоза мыши за счет индукции тромба с помощью хлорида железа (III) в кровеносных сосудах, например, бедренной вене или сонной артерии, с последующей оценкой кровотечения из хвостовой вены для проверки нормального гемостаза.
Различные дополнительные аспекты и варианты реализации настоящего изобретения очевидны для специалистов в данной области техники в свете настоящего описания.
Все документы, упомянутые в настоящем описании, полностью включены в настоящую заявку посредством ссылок.
Если не указано иное, остатки антитела пронумерованы в настоящем документе в соответствии со схемой нумерации Кабата.
Фразу "и/или", используемую в настоящем документе, следует рассматривать как специфическое описание каждой из двух указанных особенностей или компонентов друг с другом или по отдельности. Например, "A и/или B" следует рассматривать как специфическое описание каждого из (i) A, (ii) B и (iii) A и B, как если бы каждый из них по отдельности описывался в настоящем документе.
Если иное не следует из контекста, описания и определения особенностей, изложенных выше, не ограничиваются каким-либо конкретным аспектом или вариантом реализации настоящего изобретения и в равной степени применимы ко всем описанным аспектам и вариантам реализации. Таким образом, функции, изложенные выше, описаны во всех комбинациях и сочетаниях.
Некоторые аспекты и варианты реализации настоящего изобретения будут проиллюстрированы посредством примеров со ссылкой на фигуры и таблицы, описанные ниже.
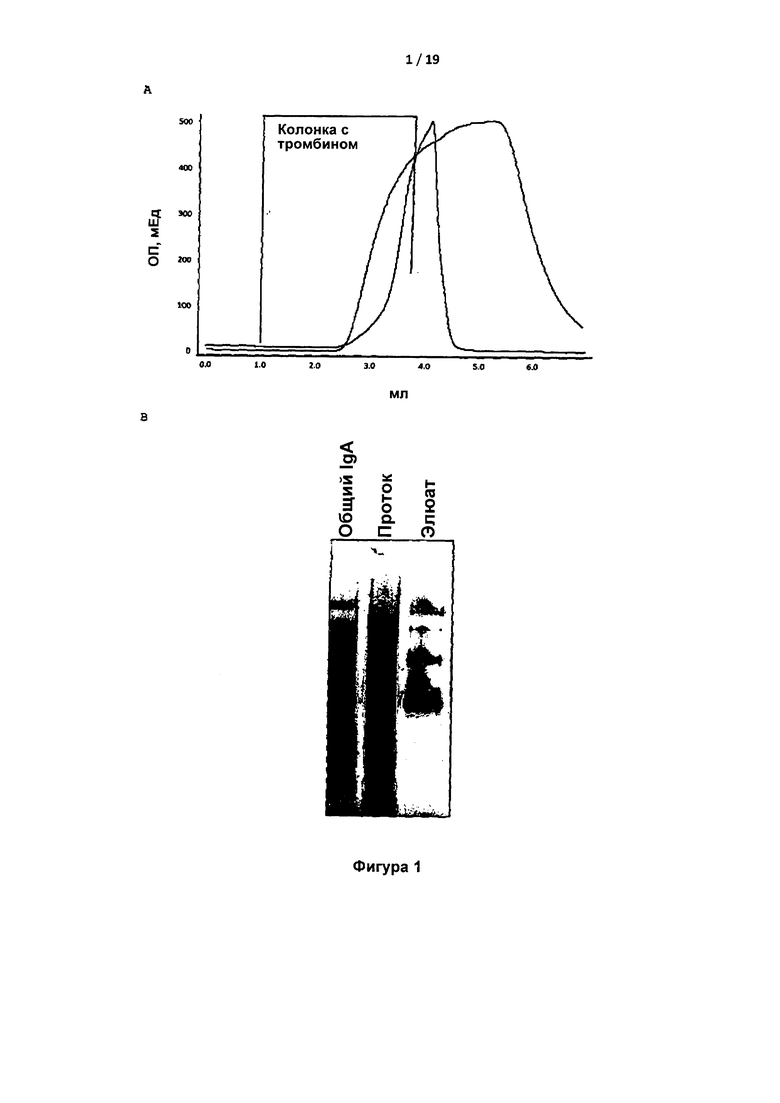
На Фигуре 1 показано связывание и элюирование IgA на колонке с тромбином человека-сефарозой. На Фигуре 1A показан профиль элюирования IgA (узкий пик) с колонки с тромбином человека-сефарозой градиентом pH (от нейтрального к низкому, показанным восходящей наклонной линией). На Фигуре 1B показан нативный голубой гель с показанной полной загрузкой IgA, элюат с колонки с тромбином человека и элюат после элюирования при низком pH.
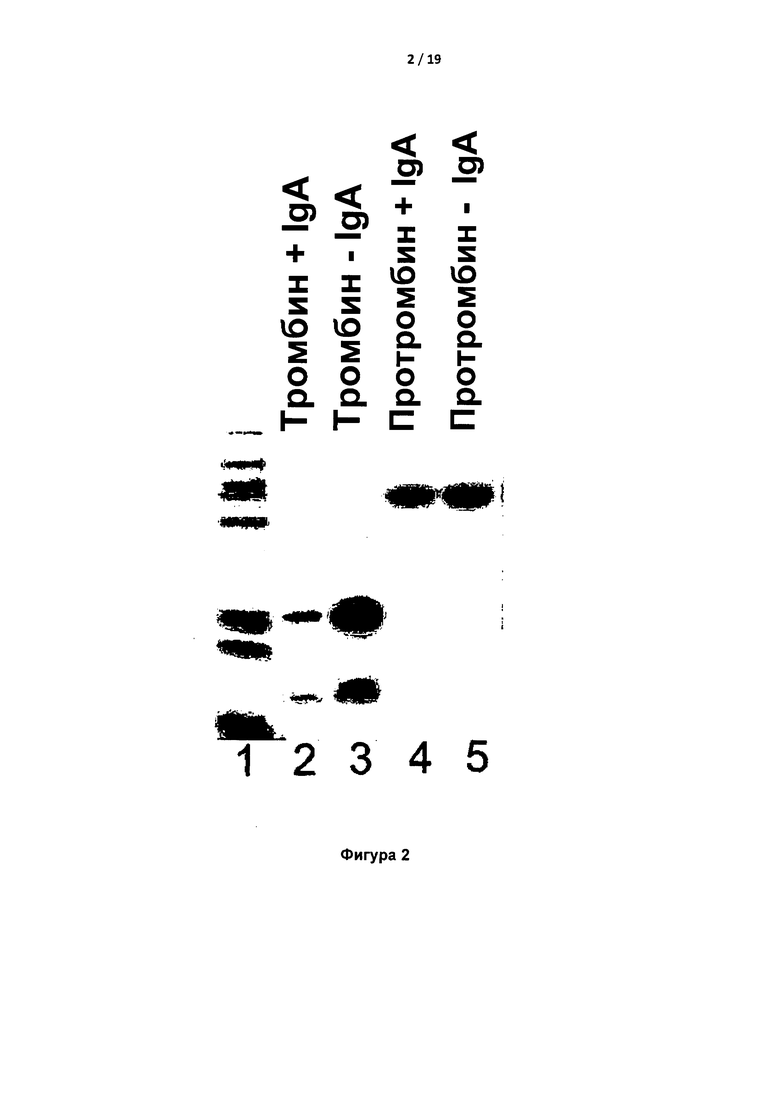
На Фигуре 2 показан невосстанавливающий гель для электрофореза в ДСН-ПААГ, демонстрирующий, что IgA связывало тромбин, но не протромбин. В этом понижающемся анализе для связывания IgA в присутствии тромбина или протромбина использовали лектин-агарозу. Супернатант подвергали электрофорезу в геле с ДСН. Дорожка 1 представляет собой стандарты размера; дорожка 2 показывает истощение тромбина в супернатанте; дорожка 3 показывает, что истощение зависит от присутствия IgA; дорожки 3 и 4 показывают, что протромбин не исчерпывается и поэтому не связывается с IgA.
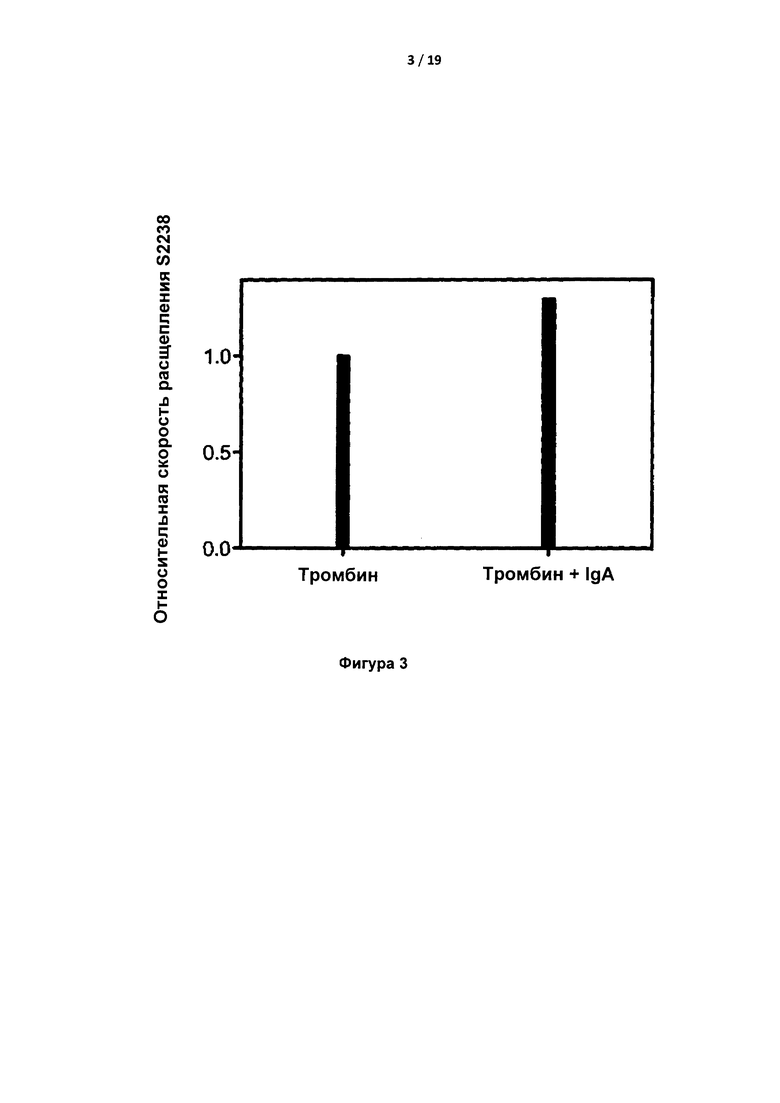
На Фигуре 3 показана относительная скорость расщепления S2238 тромбином в присутствии или в отсутствие IgA (т.е. одиночный наклон Abs405 со временем для гидролиза S2238). Это означает, что IgA не связывается в активном центре тромбина.
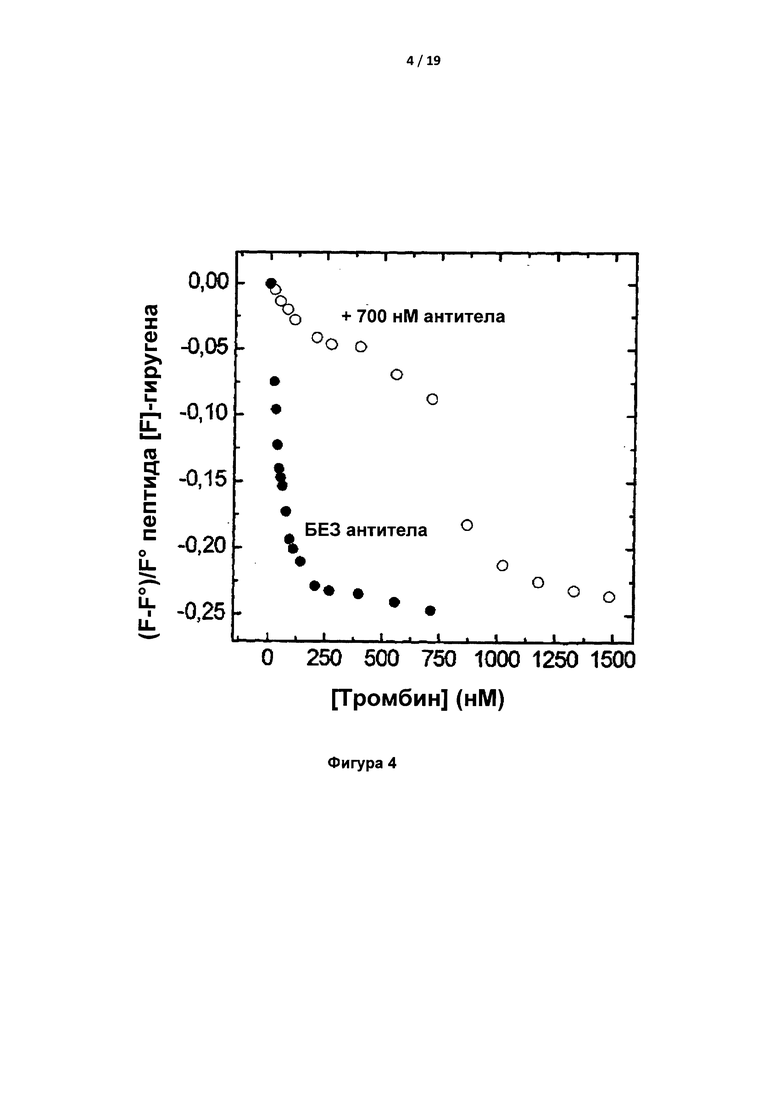
На Фигуре 4 показаны результаты исследований связывания, которые показывают, что IgA конкурирует с флуоресцентно-меченым додекапептидом гиругеном за связывание с тромбином.
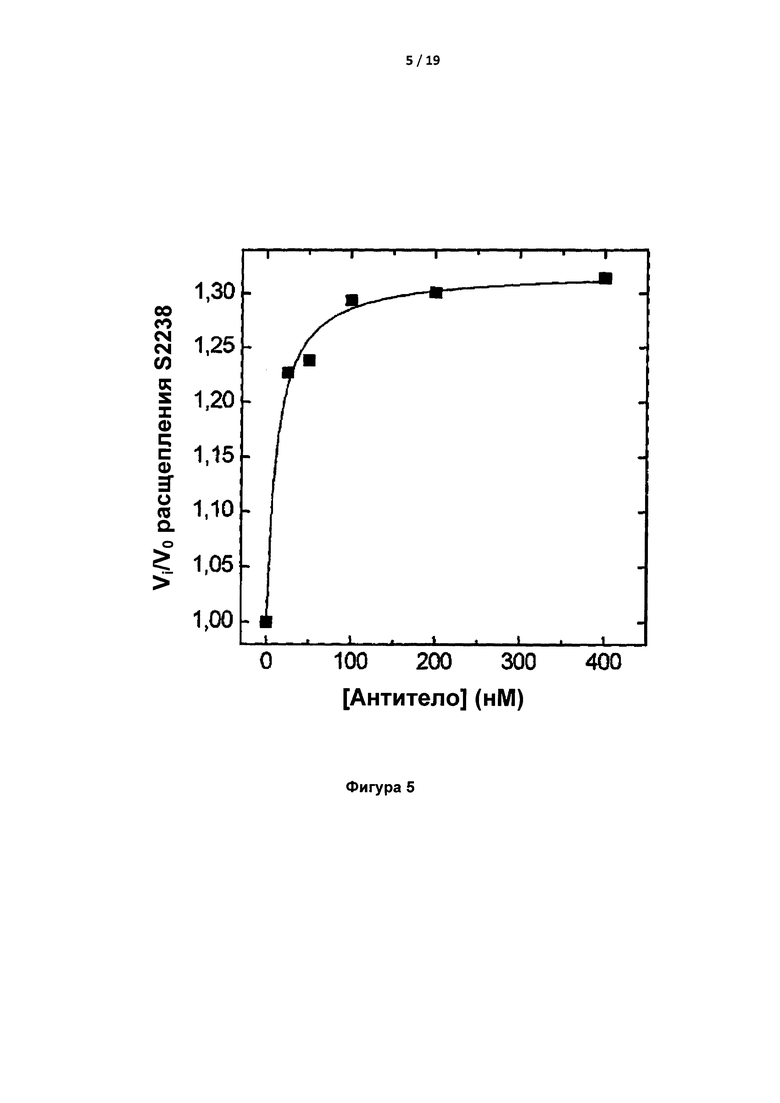
На Фигуре 5 показано влияние IgA на расщепление S2238 тромбином. Этот анализ позволяет оценить Kd для взаимодействия IgA-тромбин, равную 12 нМ.
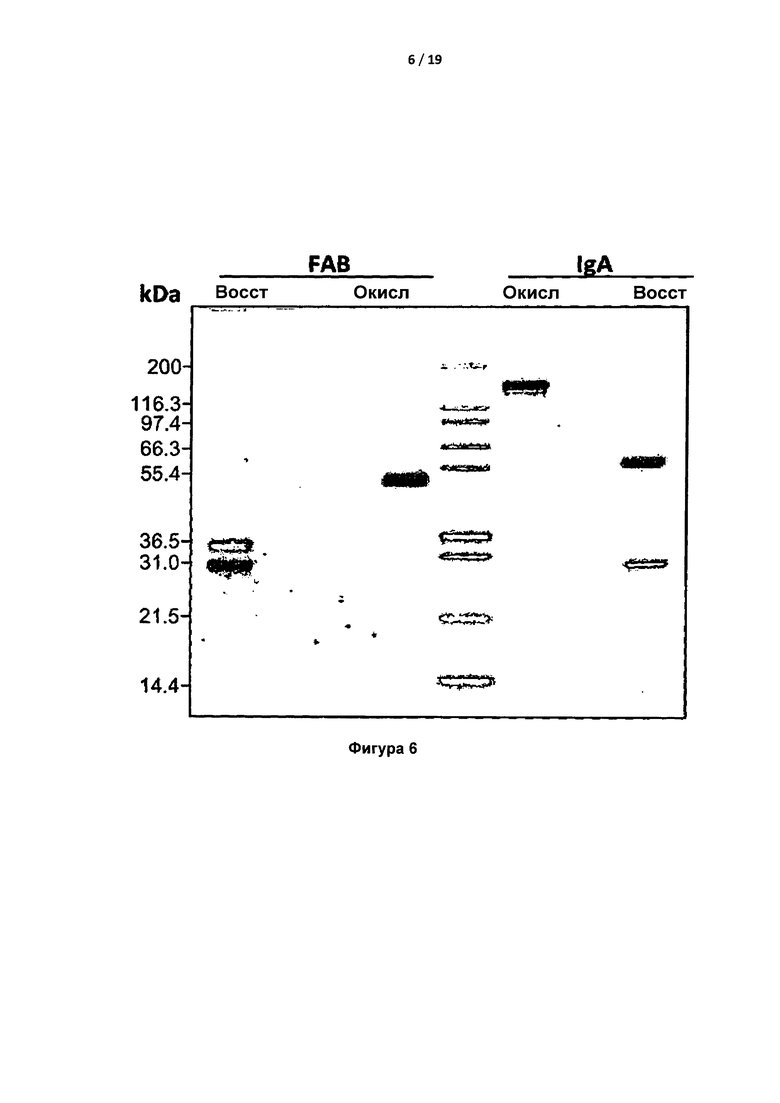
На Фигуре 6 показан электрофорез полноразмерных IgA и Fab-фрагментов в ДСН-ПААГ при восстанавливающих и невосстанавливающих (ox) условиях. Показано, что молекулярная масса невосстановленного IgA составляет 100-200 кДа, а молекулярная масса невосстановленного Fab - приблизительно 50 кДа.
На Фигуре 7 показана кристаллическая структура комплекса тромбин-Fab, показывающая взаимодействие между экзосайтом 1 тромбина и HCDR3 фрагмента Fab.
На Фигуре 8 показаны подробности кристаллической структуры, показывающие взаимодействие между конкретными остатками экзосайта 1 тромбина и HCDR3 фрагмента Fab.
На Фигуре 9 показаны флуоресцентные микрофотографии FeCl3-индуцированных тромбов в бедренной вене мышей C57BL/6 после введения фибриногена, меченного FITC, отобранных через 2-30 минут. Вводили 100 мкл PBS (контрольного носителя).
На Фигуре 10 показаны флуоресцентные микрофотографии FeCl3-индуцированных тромбов в бедренной вене мышей C57BL/6 после введения фибриногена, меченного FITC, и 40 нМ (конечная концентрация в крови мыши эквивалентна дозе приблизительно 0,6 мг/кг) IgA против экзосайта 1 (100 мкл в PBS).
На Фигуре 11 показаны флуоресцентные микрофотографии FeCl3-индуцированных тромбов в бедренной вене мышей C57BL/6 после введения фибриногена, меченного FITC, и 80 нМ (конечная концентрация в крови мыши эквивалентна дозе приблизительно 1,1 мг/кг) IgA против экзосайта 1 (100 мкл в PBS), и неповрежденной области для сравнения.
На Фигуре 12 показаны флуоресцентные микрофотографии FeCl3-индуцированных тромбов в бедренной вене мышей C57BL/6 после введения фибриногена, меченного FITC, и 200 нМ (конечная концентрация в крови мыши эквивалентна дозе приблизительно 3 мг/кг) IgA против экзосайта 1 (100 мкл в PBS), и неповрежденной области для сравнения.
На Фигуре 13 показаны флуоресцентные микрофотографии FeCl3-индуцированных тромбов в бедренной вене мышей C57BL/6 после введения фибриногена, меченного FITC, и 400 нМ (конечная концентрация в крови мыши эквивалентна дозе приблизительно 6 мг/кг) IgA против экзосайта 1 (100 мкл в PBS).
На Фигуре 14 показаны флуоресцентные микрофотографии FeCl3-индуцированных тромбов в бедренной вене мышей C57BL/6 после обработки фибриногеном, меченным FITC, и 4 мкМ (конечная концентрация в крови мыши эквивалентна дозе приблизительно 60 мг/кг) IgA против экзосайта 1 (100 мкл в PBS).
На Фигуре 15 показан количественный анализ ответа на дозу IgA против экзосайта 1 на основе флуоресцентных фотографий, показанных на фигурах 9-13.
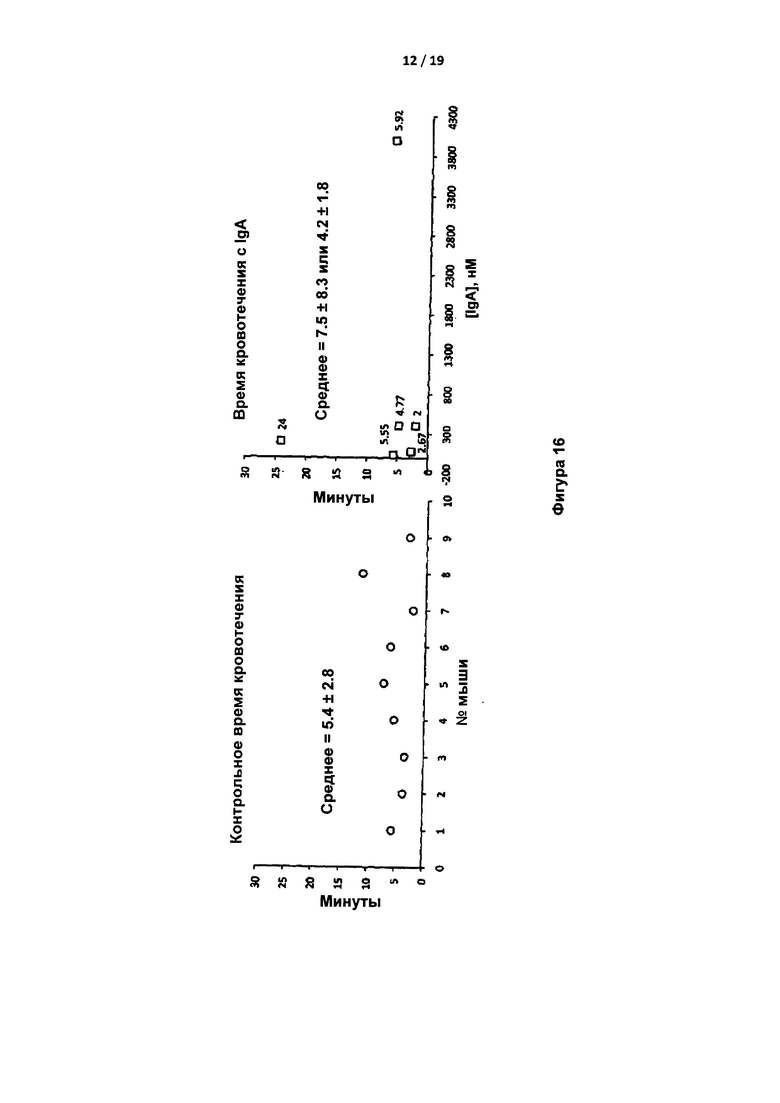
На Фигуре 16 показано время кровотечения из хвостовой вены у контрольных мышей C57BL/6 и у мышей, обработанных увеличивающимися количествами IgA против экзосайта 1. Во втором среднем исключены резко отклоняющиеся значения.
На Фигуре 17 показаны результаты анализов с укорачиванием хвоста самцов мышей дикого типа C57BL/6 (n=5) после инъекции в хвостовую вену IgA или PBS. Через 15 минут после инъекции хвосты обрезали по диаметру 3 мм и отслеживали потерю крови в течение 10 мин.
На Фигуре 18A-18D показаны результаты моделирования закупорки сонной артерии с помощью FeCl3 у 9-недельных самцов мышей C57BL/6 дикого типа, ранее получивших инъекцию 400 нМ IgA против тромбина (конечная концентрация в крови эквивалентна дозе приблизительно 6 мг/кг) или PBS за 15 мин до повреждения с помощью 5% FeCl3 в течение 2 мин. На Фигуре 18A показаны результаты типичных мышей, получивших инъекции PBS (закупорка в течение 20 минут), а на фигурах 18B, 18C и 18D показаны примеры результатов для мышей, получавших 400 нМ IgA против тромбина (закупорка отсутствовала).
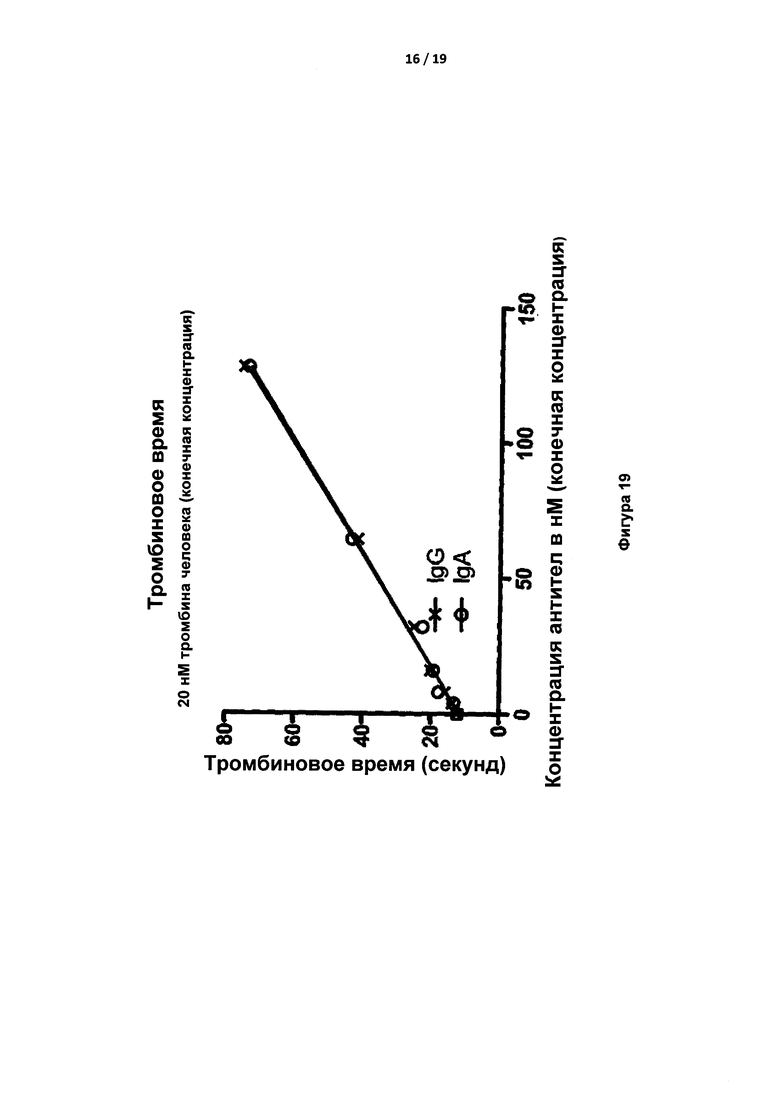
На Фигуре 19 показано тромбиновое время (т.е. свертывание объединенной плазмы) с увеличением концентрации IgG и IgA согласно изобретению при добавлении 20 нм тромбина человека.
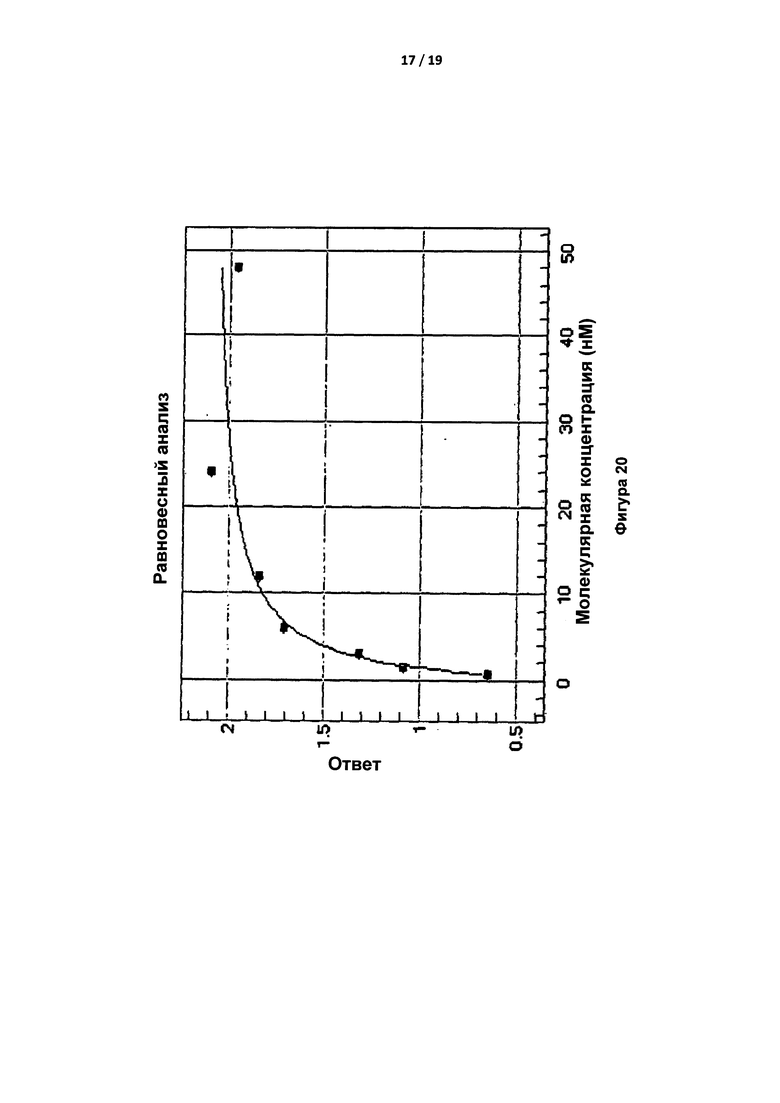
На Фигуре 20 показано связывание синтетического IgG с иммобилизованным тромбином (на приборе ForteBio Octet Red).
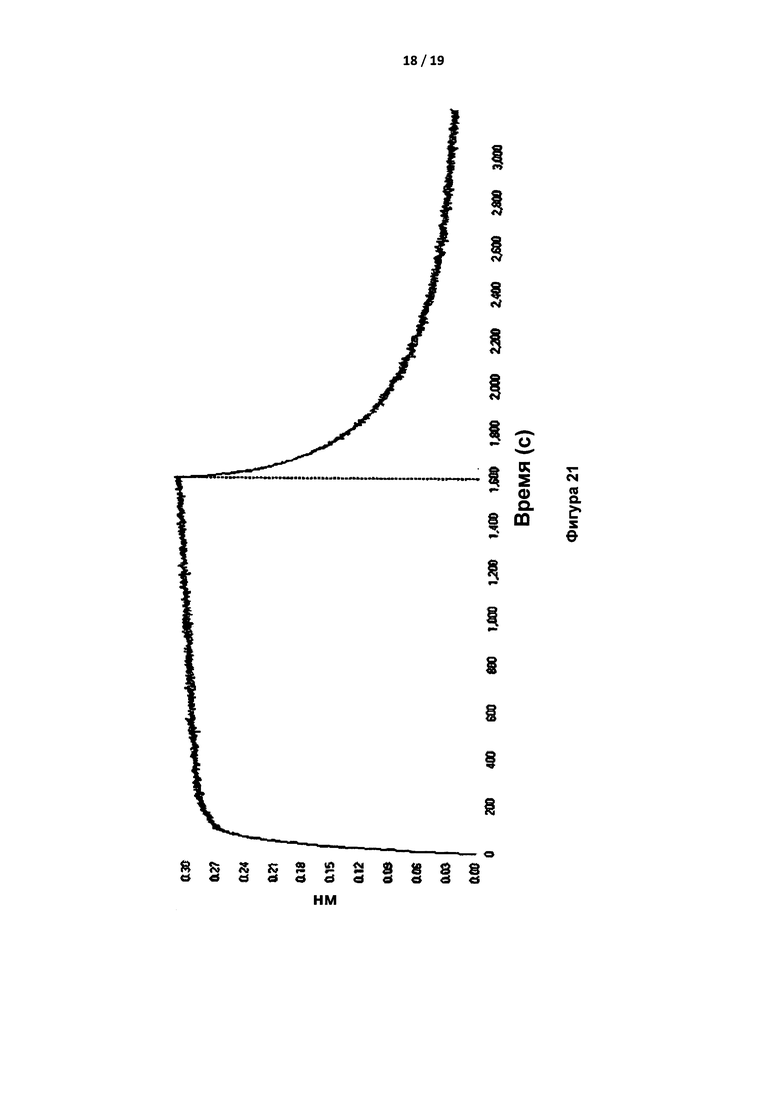
На Фигуре 21 показана типичная кривая Octet для связывания 24 нМ S195A тромбина с иммобилизованным IgG, на которой показана фаза ассоциации с последующей фазой диссоциации. Черная линия представляет собой аппроксимацию.
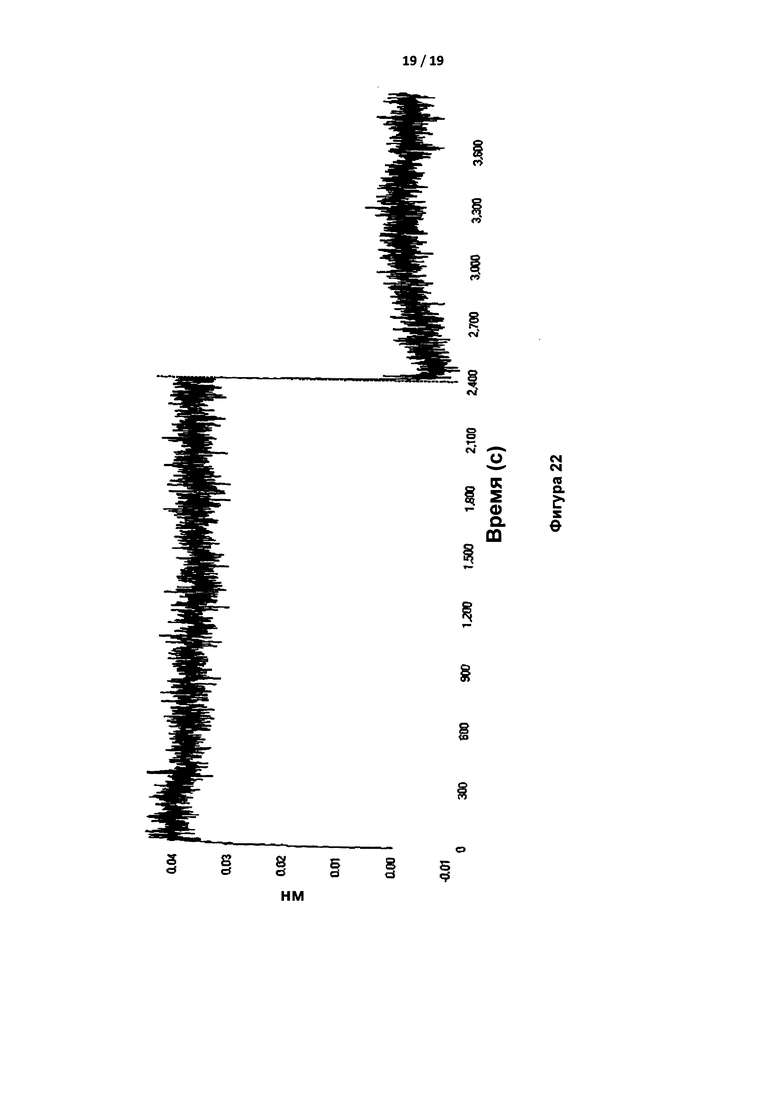
На Фигуре 22 показана кривая Octet 500 нМ протромбина с иммобилизованным IgG в наконечнике. Использовали концентрации, аналогичные концентрациям в эксперименте с тромбином на фиг. 21. Признаки связывания отсутствовали даже при указанной высокой концентрации.
Эксперименты
1. Выделение и оценка характеристик антитела
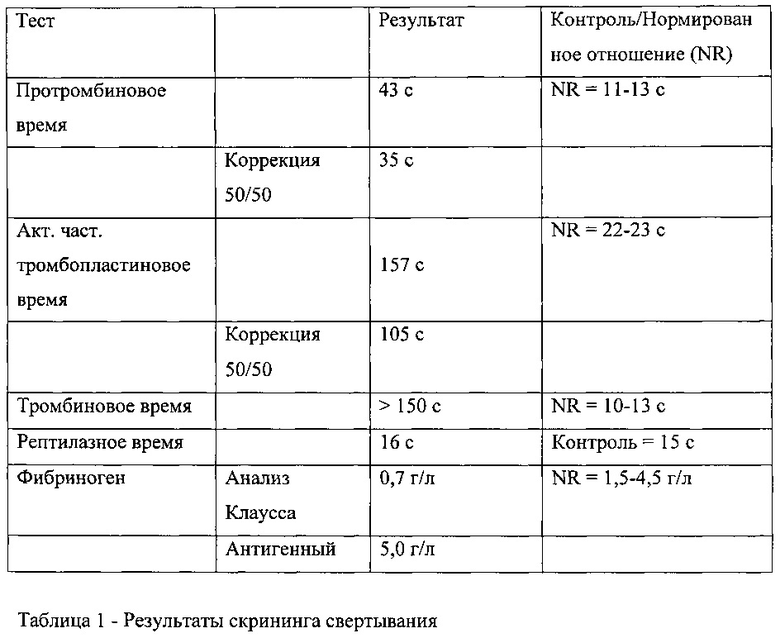
Скрининг свертывания проводили в образце плазмы крови пациента. Анализы свертывания проводили на пациенте, страдавшем субдуральной гематомой после травмы головы. Гематома спонтанно рассосалась без вмешательства. Кровотечения в анамнезе отсутствовали, в течение 4 лет после обращения пациента дальнейшие эпизоды кровотечений также отсутствовали. Результаты показаны в Таблице 1.
Протромбиновое время (ПВ), активированное частичное тромбопластиновое время (АЧТВ) и тромбиновое время (ТВ) были увеличены у пациента по сравнению с контрольной группой, но рептилазное время было нормальным.
Тромбиновое время не корректировалось с помощью гепариназы, что указывало на отсутствие реакции при лечении или загрязнении гепарином. Согласно твердофазному ИФА и рептилазному анализу, уровни фибриногена у пациента находились в пределах нормы. Анализ Клаусса выдавал заниженный уровень фибриногена в связи с присутствием ингибитора тромбина. Обнаружено, что времена свертывания ПВ и АЧТВ оставались увеличенными после смешанного теста с использованием смеси с объединенной плазмой здоровых индивидов в соотношении 50:50. Это указывало на присутствие ингибитора в образце пациента.
Обнаружено, что плазма крови пациента содержала высокий титр IgA. Обнаружено, что указанная молекула IgA связывалась с тромбином человека на колонке (Фигура 1). IgA-связывающая лектин-агароза снижала уровень тромбина в присутствии, но не в отсутствие IgA. Уровень протромбина не снижался за счет лектин-агарозы в присутствии IgA, что указывало на специфическое связывание IgA с тромбином, но не с протромбином (Фигура 2).
Затем исследовали сайт связывания IgA на молекуле тромбина.
В присутствии IgA скорость расщепления S2238 тромбином была несколько выше, что указывало на отсутствие блокирования активного центра тромбина IgA (Фигура 3).
Связывание флуоресцентно меченного гиругена с тромбином ингибировалось в присутствии 700 нМ IgA, что указывало на перекрывание эпитопа для антитела с сайтом связывания гиругена на тромбине, а именно экзосайтом 1 тромбина (рис. 4).
Протестировано влияние IgA на гидролиз некоторых прокоагулянтных субстратов тромбина. Результаты показаны в Таблице 2. Указанные результаты продемонстрировали, что молекула IgA, выделенная из образца пациента, ингибировала множественные варианты прокоагулянтной активности тромбина.
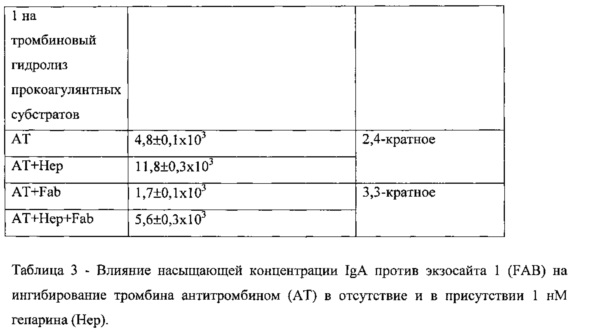
Ингибирование тромбина антитромбином (AT) в присутствии IgA менялось лишь в минимальной степени как в отсутствие, так и в присутствии гепарина (Таблица 3).
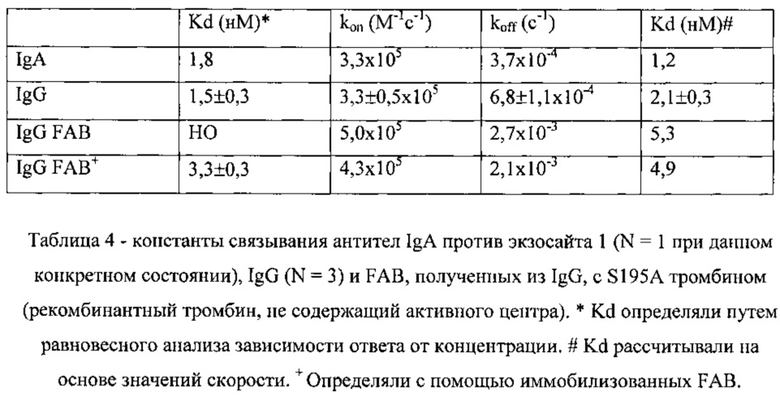
Константу диссоциации (Kd) IgA для тромбина был первоначально определяли на основании скорости гидролиза S2238; она составила приблизительно 12 нМ (Фигура 5). Kd связывания IgA с S195A тромбином (инактивированным путем мутации каталитического серина) составила 2 нМ при определении с помощью прибора ForteBio Octet Red (Таблица 4).
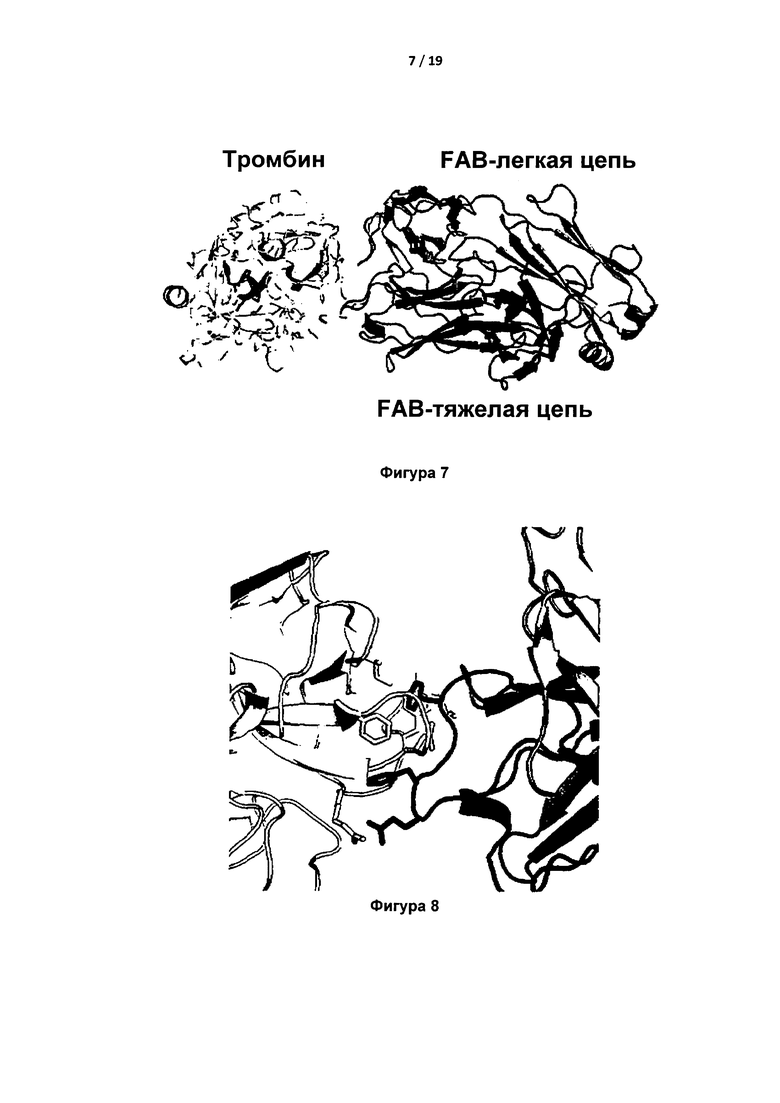
Очищенный IgA расщепляли папаином (Фигура 6), фрагмент Fab выделяли и объединяли с PPACK-тромбином человека (PPACK представляет собой ковалентный ингибитор активного центра). Комплекс PPACK-тромбин человека-FAB кристаллизовали и использовали для структурного анализа. Статистические параметры полученной структуры были следующими: разрешение 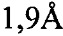 ; R-фактор = 19,43%; Rсвоб = 23,42%; один комплекс в асимметричной единице; карта Рамачандрана: благоприятные = 97,0%, выбросы = 0%. Кристаллическая структура показала тесную ассоциацию между HCDR3 Fab IgA и экзосайтом 1 тромбина (Фигура 7).
; R-фактор = 19,43%; Rсвоб = 23,42%; один комплекс в асимметричной единице; карта Рамачандрана: благоприятные = 97,0%, выбросы = 0%. Кристаллическая структура показала тесную ассоциацию между HCDR3 Fab IgA и экзосайтом 1 тромбина (Фигура 7).
В частности, остатки M32, F34, Q38, Е39, L40, L65, R67, R73, T74, R75, Y76, R77a и I82 экзосайта 1 непосредственно взаимодействовали с петлей HCDR3 Fab IgA (Фигура 8).
Анализ PISA взаимодействия антитело-тромбин показал, что общая закрытая площадь поверхности в комплексе составила 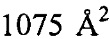 . Контактные остатки в тяжелой цепи IgA представляли собой (нумерация по Кабату): 30, 51, 52а, 53-55, 96, 98, 99, 100, 100а, 100b, 100c, 100d). Все они находились в CDR: CDRH1-GYTLTEAAIH; CDRH2-GLDPQDGETVYAQOFKG; CDRH3-GDFSEFEPFSMDYFHF (подчеркнутые остатки являлись контактными). Установлено, что CDRH3 был наиболее важным и обеспечивал 85% закрытой площади поверхности антитела. Легкая цепь осуществляла один минимальный контакт посредством Tyr49, непосредственно перед CDRL2 (с Ser36a тромбина). Вклад некоторых отдельных остатков в закрытую поверхность составлял:
. Контактные остатки в тяжелой цепи IgA представляли собой (нумерация по Кабату): 30, 51, 52а, 53-55, 96, 98, 99, 100, 100а, 100b, 100c, 100d). Все они находились в CDR: CDRH1-GYTLTEAAIH; CDRH2-GLDPQDGETVYAQOFKG; CDRH3-GDFSEFEPFSMDYFHF (подчеркнутые остатки являлись контактными). Установлено, что CDRH3 был наиболее важным и обеспечивал 85% закрытой площади поверхности антитела. Легкая цепь осуществляла один минимальный контакт посредством Tyr49, непосредственно перед CDRL2 (с Ser36a тромбина). Вклад некоторых отдельных остатков в закрытую поверхность составлял: 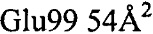 ,
, 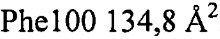 ,
, 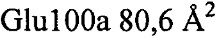 ,
, 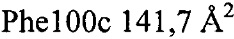 .
.
Установлено, что контактные остатки в тромбине представляли собой (нумерация аналогично химотрипсину): 32, 34, 36а-40, 65, 67, 73-76, 77а, 82 и 151. Отдельные остатки, вносившие наиболее важный вклад в закрытую поверхность, представляли собой: 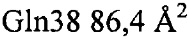 ,
, 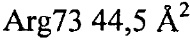 ,
, 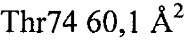 ,
, 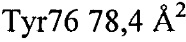 ,
, 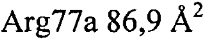 .
.
Пациент не демонстрировал повышенного или аномального кровотечения или геморрагии, несмотря на то, что уровень указанного IgA в кровотоке составлял 3 г/л, что демонстрировало ингибирование тромбина данным антителом без влияния на нормальный гемостаз.
2. Влияние IgA на животные модели тромбоза
Мышей C57BL/6 анестезировали. Катетер (для инъекции соединения) вставляли в сонную артерию. Через сонную артерию вводили фибриноген, меченный FITC (2 мг/мл). Через сонную артерию также вводили PBS (контроль) или IgA. Обнажали бедренную вену и наносили 10% FeCl3 (на фрагменте насыщенной промокательной бумаге 3 мм длиной) в течение 3 мин, чтобы вызвать свертывание.
Флуоресцентные микрофотографии получали по всей длине области повреждения через 0, 5, 10 и 20 мин после повреждения FeCl3 с использованием методик флуоресцентной микроскопии.
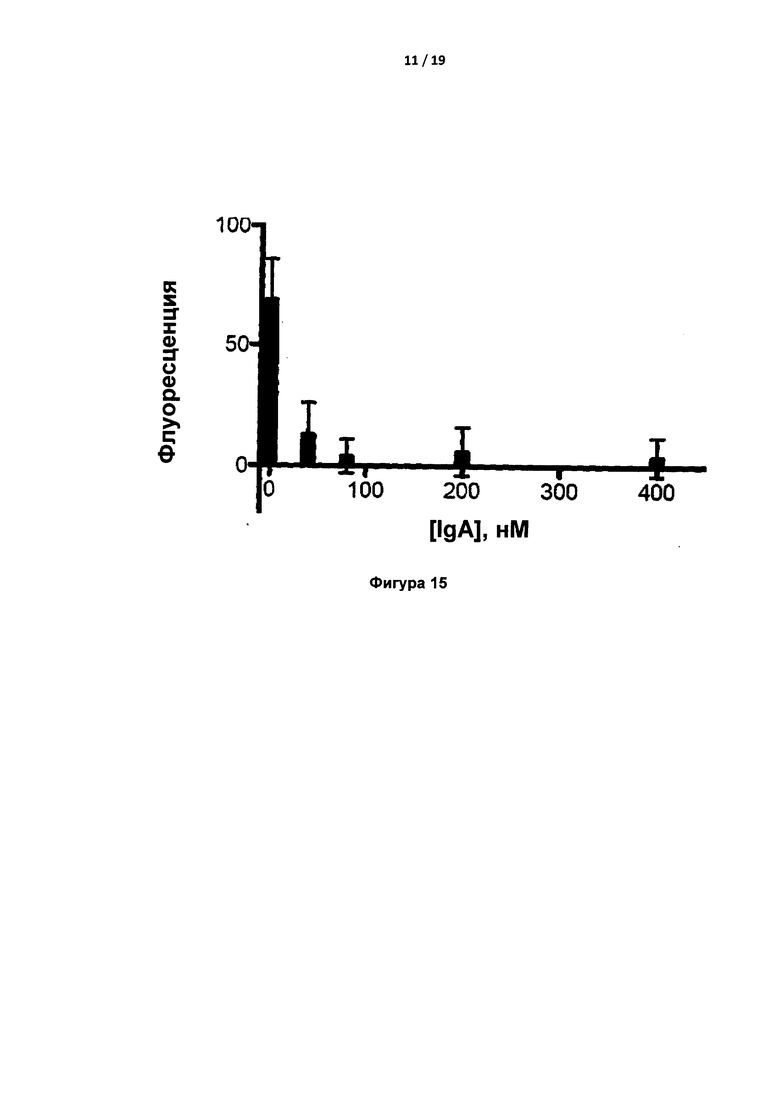
Сгустки (отложения фибрина) в бедренной вене были ясно видны в виде светлых областей (Фигура 9). Обнаружено, что наименьшая доза антитела вызывала значительное ингибирование свертывания, однако по мере увеличения дозы свертывание прекращалось (фигуры 10-15).
Кроме того, у мышей измеряли время кровотечения. Время кровотечения оценивали как время до прекращения кровотока после обрезания хвоста. Установлено, что, несмотря на присутствие одного аномального образца, время кровотечения не зависело от лечения с применением IgA против экзосайта 1 (фигура 16).
Указанные результаты показывают, что IgA-антитело против экзосайта 1 являлось мощным ингибитором тромбоза, но не оказывало влияния на время кровотечения.
3. Анализ с укорачиванием хвоста
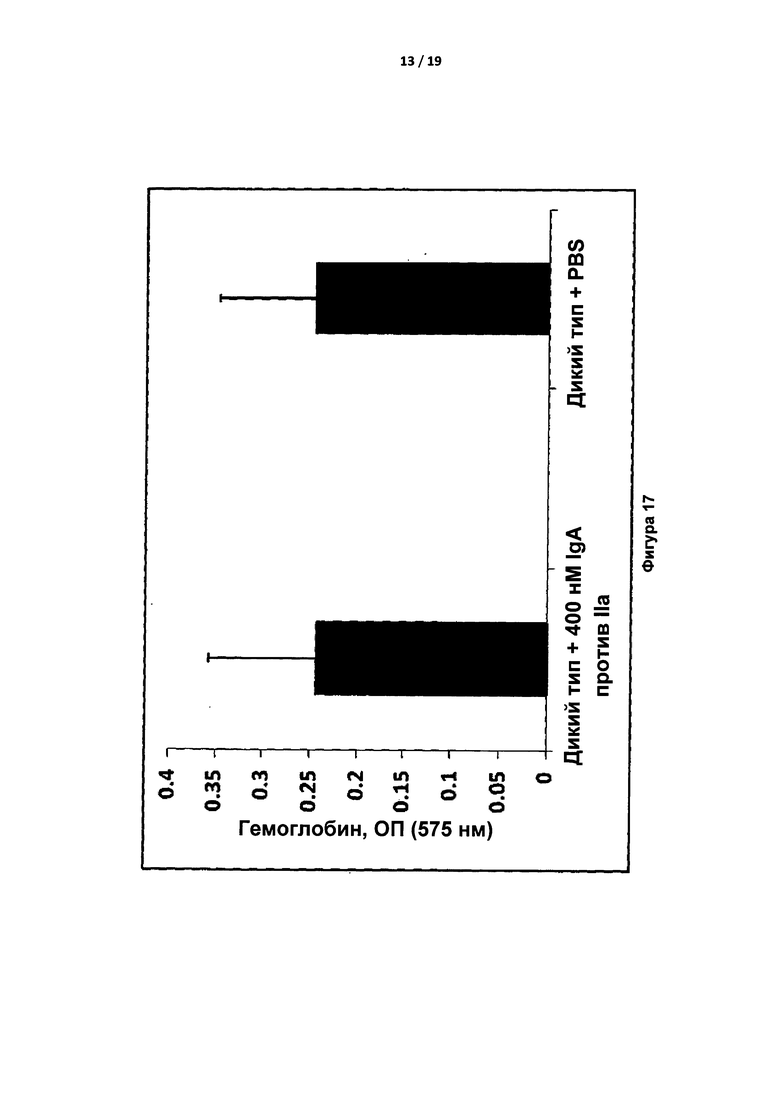
Анализ с укорачиванием хвоста проводили на самцах мышей C57BL/6 дикого типа, получивших инъекцию 400 нМ IgA (конечная концентрация в крови была эквивалентна дозе приблизительно 6 мг/кг) или PBS. Потерю крови отслеживали в течение 10 минут после обрезания хвоста по диаметру 3 мм через 15 минут после инъекции. Установлено, что полная потеря крови не зависела от лечения с применением IgA против экзосайта 1 (фигура 17).
4. Травматическая закупорка сонной артерии с применением FeCl3
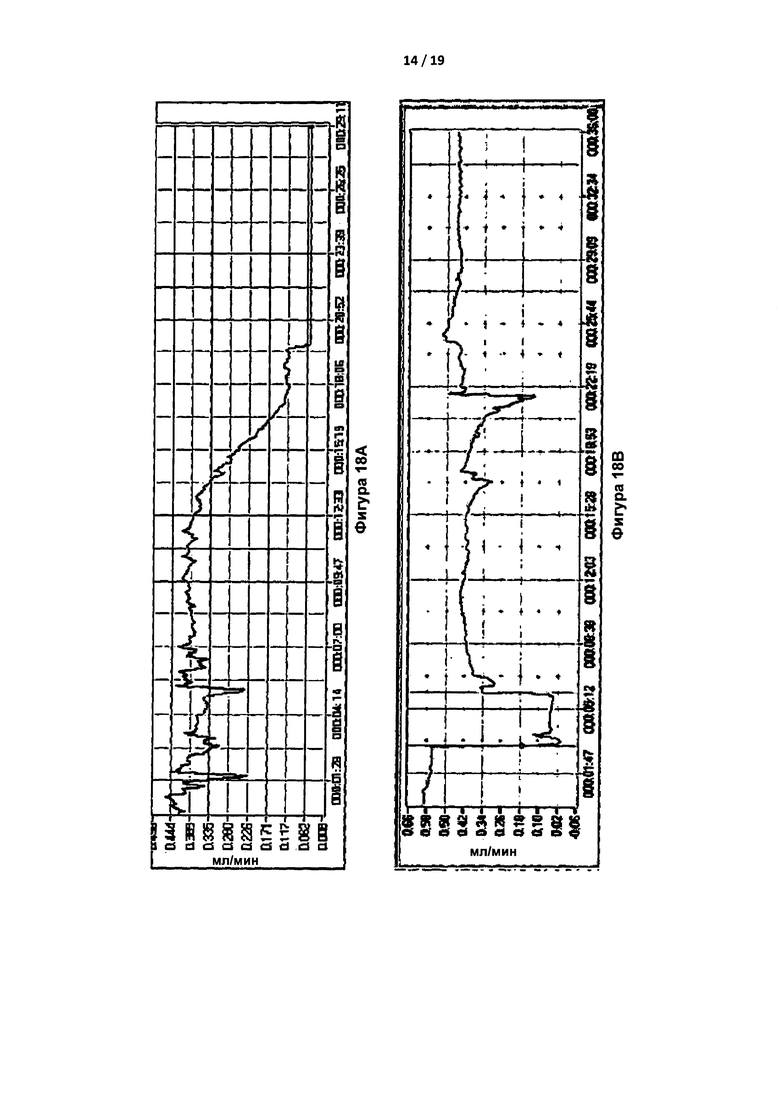
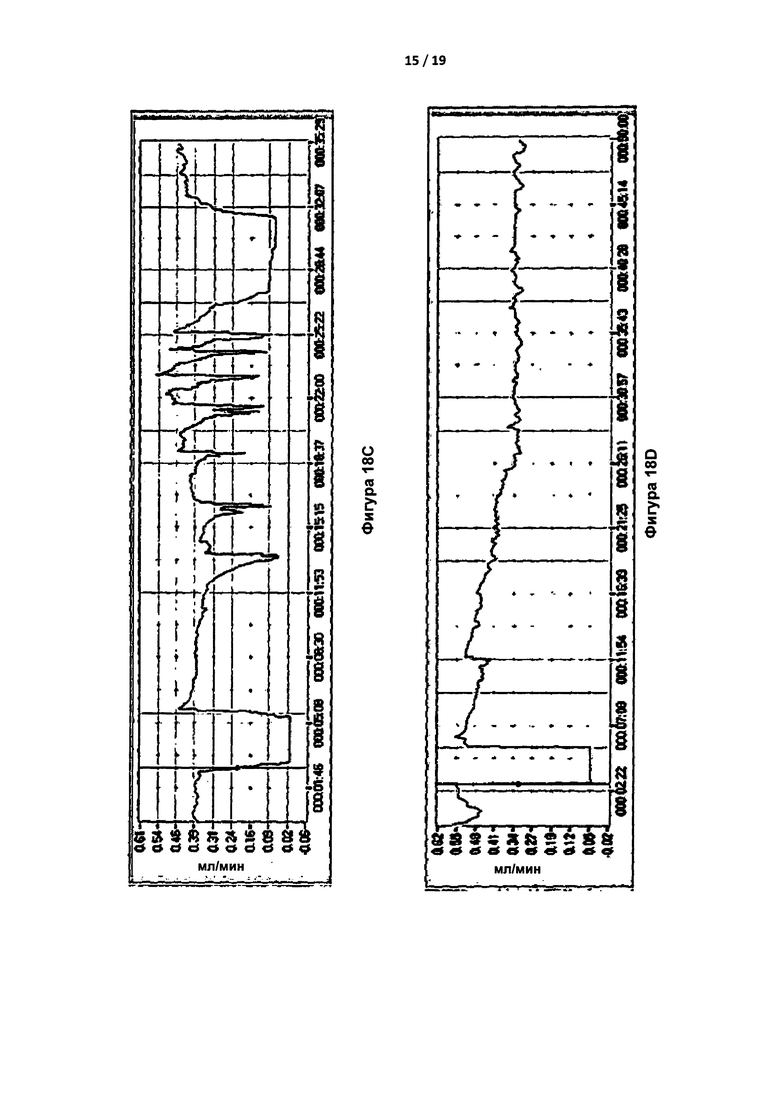
Исследования с травматической закупоркой сонной артерии с применением FeCl3 проводили на 9-недельных самцах мышей C57BL/6 дикого типа. Мышам вводили путем инъекции 400 нм IgA против IIa (конечная концентрация в крови была эквивалентна дозе приблизительно 6 мг/кг) или PBS за 15 мин до нанесения повреждения за счет 5% FeCl3 в течение 2 мин. Затем отслеживали кровоток с помощью допплеровской методики и измеряли время до закупорки. "Сгусток" определяли как стабильный окклюзионный тромб, при котором кровоток снижался до значений, как правило, менее 0,1 мл/мин, и оставался сниженным. У контрольных мышей формирование стабильного сгустка наблюдали приблизительно через 20 минут после повреждения (Фигура 18а). Вместе с тем, у большинства мышей, получивших 400 нМ IgA против IIa, устойчивые сгустки не образовывались; на их кривых сгустки быстро рассасывались, повторно рассасывались или не образовывались. Три типичные кривые показаны на Фигурах 18B-18D.
5. IgG против экзосайта 1
Молекулу IgA, обнаруженную у пациента, описанного выше, преобразовали в формат IgG с использованием стандартных методик.
Время свертывания объединенной плазмы человека с добавками возрастающих количеств исходного IgA и нового IgG тестировали при добавлении до 20 нМ тромбина человека (Фигура 19). И исходное IgA, и синтетическое IgG повышали время до образования сгустка аналогичным зависимым от концентрации образом, что подразумевает одинаковое сродство по отношению к тромбину.
Указанные результаты подтвердили путем измерения связывания синтетического IgG с иммобилизованным S195A тромбином с применением прибора ForteBio™ Octet Red. Тромбин присоединяли к зонду и отслеживали связывание антител (при различных концентрациях). Анализировали скорости ассоциации и диссоциации. Оба антитела обладали аналогичными скоростями ассоциации, составлявшими приблизительно 3×105 M-1с-1, скоростями диссоциации, составлявшими приблизительно 5×10-4 с-1, и константами диссоциации (Kd), составлявшими приблизительно 2 нМ. Kd, составлявшие приблизительно 2 нМ, также получили для IgA и IgG при равновесном анализе (Таблица 4). Типичная равновесная кривая показана на Фигуре 20. Таким образом, свойства IgA воспроизводились в каркасе IgG.
Связывание протромбина с антителом IgG тестировали с применением системы Octet путем иммобилизации IgG. Тромбин связывался с иммобилизованным IgG с скоростью и сродством, сопоставимыми с аналогичными параметрами, полученными при применении иммобилизованного тромбина (Таблица 4); протромбин не связывался с IgG. Фигура 21 представляет собой кривую связывания и диссоциации 24 нМ тромбина и иммобилизованного IgG. Фигура 22 представляет собой тот же самый эксперимент при применении 500 нМ протромбина, и демонстрирует отсутствие признаков связывания.
Последовательности
Аминокислотная последовательность препротромбина человека (SEQ ID NO: 1; GeneID: 2147; NP_000497.1 GI: 4503635; остатки экзосайта 1 подчеркнуты)

Аминокислотная последовательность VH-домена IgA и IgG против экзосайта 1 с нумерацией по Кабату (CDR подчеркнуты): (SEQ ID NO: 2).
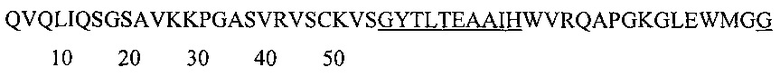
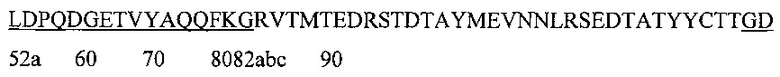

Аминокислотная последовательность HCDR1 IgA и IgG против экзосайта 1 (SEQ ID NO: 3).
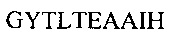
Аминокислотная последовательность HCDR2 IgA и IgG против экзосайта 1 (SEQ ID NO: 4).
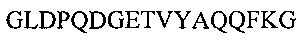
Аминокислотная последовательность HCDR3 IgA и IgG против экзосайта 1 (SEQ ID NO: 5).

Аминокислотная последовательность VL-домена IgA и IgG против экзосайта 1 с нумерацией по Кабату: (SEQ ID NO: 6).
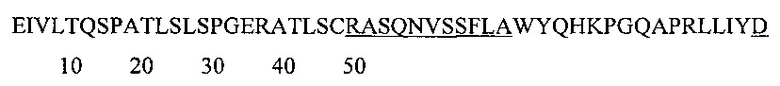
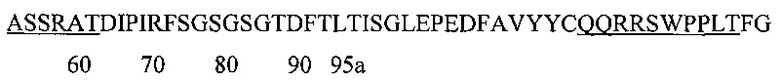

Аминокислотная последовательность LCDR1 IgA и IgG против экзосайта 1 (SEQ ID NO: 7).
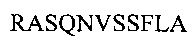
Аминокислотная последовательность LCDR2 IgA и IgG против экзосайта 1 (SEQ ID NO: 8).
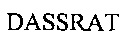
Аминокислотная последовательность LCDR3 IgA и IgG против экзосайта 1 (SEQ ID NO: 9).

| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К ФАКТОРУ XI И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2739946C2 |
| НЕЙТРАЛИЗУЮЩИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, НАПРАВЛЕННЫЕ ПРОТИВ ВИРУСА ГРИППА, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2721706C2 |
| ИНГИБИРУЮЩИЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ ФАКТОРА XII/XIIA И ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2660370C2 |
| МОЛЕКУЛЫ АНТИТЕЛ ПРОТИВ APRIL И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2793755C2 |
| Антитело против FXI/FXIA, его антигенсвязывающий фрагмент и фармацевтическое применение | 2021 |
|
RU2838408C1 |
| МОЛЕКУЛЫ АНТИТЕЛ ЧЕЛОВЕКА К IL-13 | 2004 |
|
RU2387667C2 |
| АНТИТЕЛА ПРОТИВ ИНГИБИТОРА ПУТИ ТКАНЕВОГО ФАКТОРА И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2705540C2 |
| ЧЛЕНЫ СВЯЗЫВАНИЯ ПРОТИВ IL-1R1 | 2009 |
|
RU2555532C2 |
| СОЕДИНЕНИЯ | 2007 |
|
RU2466139C2 |
| ИЗОЛИРОВАННОЕ АНТИТЕЛО ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, СПЕЦИФИЧЕСКОЕ(ИЙ) ДЛЯ ЧЕЛОВЕЧЕСКОГО ИММУНОГЛОБУЛИНА Е (ВАРИАНТЫ), КОДИРУЮЩАЯ ЕГО МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, КЛЕТКА-ХОЗЯИН, СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛА ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ПРИМЕНЕНИЕ ЕЕ ДЛЯ ЛЕЧЕНИЯ РАССТРОЙСТВ, АССОЦИИРОВАННЫХ С IgE | 2008 |
|
RU2466140C2 |



Настоящее изобретение относится к иммунологии. Предложена выделенная молекула антитела, специфически связывающаяся с областью экзосайта 1 тромбина, а также антигенсвязывающий фрагмент указанного антитела. Рассмотрено применение молекулы антитела при изготовлении медикаментов. Описана фармацевтическая композиция, а также способ лечения состояния, опосредованного тромбином. Данное изобретение может найти дальнейшее применение в терапии заболеваний, связанных с тромбином. 5 н. и 20 з.п. ф-лы, 22 ил., 4 табл.
1. Выделенная молекула антитела, специфически связывающаяся с областью экзосайта 1 тромбина, при этом указанная молекула антитела содержит вариабельный домен тяжелой цепи (VH), включающий гипервариабельные участки HCDR1, HCDR2, HCDR3, которые имеют последовательности SEQ ID NO: 3, 4 и 5, соответственно, и вариабельный домен легкой цепи (VL), включающий гипервариабельные участки LCDR2, LCDR3, которые имеют последовательности SEQ ID NO: 8 и 9, соответственно, и LCDR1, который имеет последовательность SEQ ID NO: 7 или LCDR1, который имеет последовательность SEQ ID NO: 7, в котором присутствует мутация, устраняющая сайт гликозилирования, посредством замены остатка аминокислоты в положении, соответствующем остатку S30 в последовательности SEQ ID NO: 6.
2. Молекула антитела по п. 1, которая ингибирует активность тромбина.
3. Молекула антитела по п. 2, которая вызывает минимальное ингибирование гемостаза и/или вызывает минимальное кровотечение.
4. Молекула антитела по п. 2 или 3, которая не ингибирует гемостаз и/или не вызывает кровотечение.
5. Молекула антитела по любому из пп. 1-3, отличающаяся тем, что указанная молекула антитела содержит VH-домен, который имеет аминокислотную последовательность SEQ ID NO: 2.
6. Молекула антитела по любому из пп. 1-3, отличающаяся тем, что указанная молекула антитела содержит VL-домен, который имеет аминокислотную последовательность SEQ ID NO: 6 или VL-домен, который имеет аминокислотную последовательность SEQ ID NO: 6, в которой присутствует мутация, устраняющая сайт гликозилирования, посредством замены остатка аминокислоты в положении остатка S30.
7. Молекула антитела по любому из пп. 1-3, которая представляет собой полноразмерное антитело.
8. Молекула антитела по п. 7, которая представляет собой IgA или IgG.
9. Молекула антитела по любому из пп. 1-3, которая представляет собой моноклональное антитело.
10. Антигенсвязывающий фрагмент молекулы антитела по любому из пп. 1-3, который специфически связывается с областью экзосайта 1 тромбина.
11. Антигенсвязывающий фрагмент по п. 10, отличающийся тем, что указанный фрагмент представляет собой Fab.
12. Фармацевтическая композиция для лечения опосредованного тромбином заболевания, причем указанная композиция содержит эффективное количество молекулы антитела по любому из пп. 1-3 и фармацевтически приемлемое вспомогательное вещество.
13. Молекула антитела по любому из пп. 1-3 для применения в способе лечения человека или животного.
14. Молекула антитела по любому из пп. 1-3 для применения в способе лечения состояния, опосредованного тромбином.
15. Молекула антитела по п. 14, отличающаяся тем, что указанное состояние, опосредованное тромбином, представляет собой тромботическое состояние.
16. Молекула антитела по п. 15, отличающаяся тем, что указанное тромботическое состояние представляет собой тромбоз или эмболию.
17. Молекула антитела по п. 14, отличающаяся тем, что указанное состояние, опосредованное тромбином, представляет собой воспаление, инфекцию, опухолевый рост, метастазирование опухоли или деменцию.
18. Применение молекулы антитела по любому из пп. 1-3 при изготовлении медикамента для лечения состояния, опосредованного тромбином.
19. Применение по п. 18, отличающееся тем, что указанное состояние, опосредованное тромбином, представляет собой тромботическое состояние.
20. Применение по п. 19, отличающееся тем, что указанное тромботическое состояние представляет собой тромбоз или эмболию.
21. Применение по п. 18, отличающееся тем, что указанное состояние, опосредованное тромбином, представляет собой воспаление, инфекцию, опухолевый рост, метастазирование опухоли или деменцию.
22. Способ лечения состояния, опосредованного тромбином, включающий введение эффективного количества молекулы антитела по любому из пп. 1-3 индивиду, нуждающемуся в этом.
23. Способ по п. 22, отличающийся тем, что указанное состояние, опосредованное тромбином, представляет собой тромботическое состояние.
24. Способ по п. 23, отличающийся тем, что указанное тромботическое состояние представляет собой тромбоз или эмболию.
25. Способ по п. 22, отличающийся тем, что указанное состояние, опосредованное тромбином, представляет собой воспаление, инфекцию, опухолевый рост, метастазирование опухоли или деменцию.
| WU QINGYU et al | |||
| "Аctivation-induced exposure of the thrombin anion-binding exosite | |||
| Interactions of recombinant mutant prothrombins with thrombomodulin and a thrombin exosite-specific antibody" | |||
| Journal of Biological Chemistry, 1994, 269(5): 3725-3730 | |||
| ARNAUD EMMANUEL et al | |||
| "An autoantibody directed against human thrombin anion-binding exosite in a patient with arterial thrombosis: effects on platelets, endothelial cells, and protein C activation" | |||
| Blood, 1994, 84(6): 1843-1850 | |||
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Экономайзер | 0 |
|
SU94A1 |
| US 5688507 A, 18.11.1997 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| МОИСЕЕНКО В.М | |||
| "Моноклональные антитела в лечении злокачественных опухолей" | |||
| Практическая онкология, 2003, т | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |

