Ссылка на родственные заявки
Согласно настоящей заявке испрашивается преимущество в соответствии с 35 U.S.C. § 119(e) согласно предварительным заявкам на выдачу патента США №№61/955362, поданной 19 марта 2014 г., 61/955358, поданной 19 марта 2014 г., и 62/043042, поданной 28 августа 2014 г., содержания которых полностью включены в настоящий документ посредством ссылки.
Область техники, к которой относится настоящее изобретение
Описанная в настоящем документе технология относится к получению плюрипотентных клеток.
Предшествующий уровень техники настоящего изобретения
Современные способы получения плюрипотентных клеток основаны преимущественно на тканях с ограниченной доступностью (например, эмбриональная ткань или пуповинная кровь) или добавлении перепрограммирующих факторов (Hanna, J. et al. Cell 2008 133, 250-264; Hockemeyer, D. et al. Cell stem cell 2008 3, 346-353; Kim, D. et al. Cell stem cell 2009 4, 472-476; Kim, J.B. Nature 2009 461, 649-643; Okabe, M. et al. Blood 2009 114, 1764-1767), которое предусматривает введение экзогенных нуклеиновых кислот. Способы легкого получения стволовых клеток, особенно аутологичных стволовых клеток, без осложнений, обусловленных добавлением экзогенных перепрограммирующих факторов, ускорят исследование в области клеточной дифференцировки и разработке видов терапии на основе стволовых клеток. Наряду с выдвинутым предположением о том, что повреждение клеток в результате воздействия на них таких раздражителей, как ожоги, химическое повреждение, травма и облучение, может изменять нормальные соматические клетки так, что они становятся злокачественными клетками, не существует прямого доказательства того, что здоровые взрослые соматические клетки можно превратить в другие состояния без специфической манипуляции с помощью перепрограммирующих факторов.
Ранее исследователи сообщили об обнаружении "взрослых стволовых клеток" во взрослых тканях (Reynolds, В.А. & Weiss, S. Science 1992 255, 1707-1710; Megeney, L.A. et al., Genes & development 1996 10, 1173-1183; Caplan, A.I. Journal of orthopaedic research 1991 9, 641-650; Lavker, R.M. & Sun, Т.T. The Journal of investigative dermatology 1983 81, 121s-127s). Такие сообщения остаются противоречивыми. Например, исследователи, ищущие клетки, экспрессирующие маркер стволовых клеток Oct4, не смогли обнаружить экспрессирующие Oct4 клетки во взрослом костном мозге при нормальном гомеостазе (Lengner, С.J. et al. Cell Cycle 2008 7, 725-728; Berg, J.S. & Goodell, M.A. Cell stem cell 2007 1, 359-360), тогда как другие исследователи сообщают о способности выделить экспрессирующие Oct4 клетки из различных взрослых тканей (Jiang, Y. et al. Nature 2010 418, 41-49; D'Ippolito, G. et al. Journal of cell science 2004 117, 2971-2981; Johnson, J. et al. Cell 2005 122, 303-315; Kucia, M. et al. Leukemia 2006 20, 857-869; Kuroda, Y. et al. PNAS 2011 107, 8639-8643; Obokata, H. et al. Tissue engineering. 2011 Part A 17, 607-615; Rahnemai-Azar, A. et al. Cytotherapy 2011 13, 179-192; Huang, Y. et al. Transplantation 2010 89, 677-685; Zuba-Surma, E.K. et al. Journal of cellular and molecular medicine 2011 15, 1319-1328; Paczkowska, E. et al. Annals of transplantation 2011 16, 59-71). Была выдвинута гипотеза о том, что указанные клетки представляют либо популяцию взрослых стволовых клеток, либо являются исключительно артефактом используемых техник. В любом случае они остаются редкими и не представляют собой адекватный источник плюрипотентных клеток для исследовательских и терапевтических целей.
Краткое раскрытие настоящего изобретения
В настоящем документе описаны улучшенные способы создания плюрипотентных клеток, например, клеток STAP, которые обеспечивают увеличенную эффективность, выход и/или качество по сравнению со способами, раскрытыми в международной патентной публикации WO 2013/163296 и статье Obokata et al. Nature 2014 505: 641-647; каждая из которых включена в настоящий документ посредством ссылки. Кроме того, в настоящем документе описаны способы и применения, относящиеся к клеткам, полученным с помощью настоящих способов.
Краткое описание графических материалов
На фиг. 1A-1D показано получение экспрессирующих Oct4 клеток из CD45-положительных соматических клеток. На фиг. 1А показана экспрессия Oct4-GFP подвергнутых воздействию стресса клеток. Подвергнутые воздействию стресса клетки экспрессируют Oct4-GFP, тогда как необработанные контроли нет. Увеличение экспрессирующей Oct4 колонии показано вверху справа в подвергнутой воздействию стресса группе. Масштабная полоска равна 100 мкм. На фиг. 1В показана анализ популяции подвергнутых воздействию стресса клеток и не подвергнутого воздействию стресса контроля. Экспрессирующая GFP клеточная популяция наблюдается только в подвергнутой воздействию стресса группе в день 5. На фиг. 1С показано анализ размера клеток CD45-положительных клеток до и после воздействия стресса в день 7. На фиг. 1D показано хронологическое изменение CD45-положительных клеток после воздействия стресса.
На фиг. 2А-2В показано определение характеристик клеток животного каллюса (АСС). На фиг. 2А показано хронологическое изменение экспрессии плюрипотентных маркерных генов. Содержания иРНК нормировали с помощью GAPDH. (n=3, среднее + S.D. (среднеквадратичное отклонение)). На фиг. 2В показано анализ метилирования промоторных генов Oct4 и Nanog.
На фиг. 3A-3D показаны клеточные модификации после воздействия стресса. На фиг. 3А показана относительная экспрессия генов защиты от стресса в течение фазы образования АСС. Образцы собирали в день 3 и день 7 и сравнивали с CD45-положительными клетками. (n=3, среднее + S.D.) На фиг. 3В показано измерение общего клеточного АТР. (n=3, среднее + S.D.) На фиг. 3С показано измерение ROS. Планки погрешностей показывают SD. На фиг. 3D показана относительная экспрессия генов факторов репликации мтДНК. (n=3, среднее + S.D.)
На фиг. 4А-4В показано получение химерной мыши из АСС. На фиг. 4А показана схема получения химерной мыши. На панели (i) продемонстрировано, что АС диссоциировали до отдельных клеток с помощью трипсина или (панель ii) АС разрезали на небольшие фрагменты, затем вводили с помощью инъекции в бластоцисты. На фиг. 4 В показан анализ вклада в организм химеры. Ткани от 9 детенышей анализировали с помощью FACS.
На фиг. 5А-5С показаны эксперименты с условиями получения АСС. На фиг. 5А продемонстрировано, что CD45-положительные клетки подвергали воздействию различных стрессов и экспрессию Oct4-GFP анализировали с помощью FACS. Процентное отношение экспрессирующих Oct4-GFP клеток к выжившим клеткам после воздействия стресса. (n=3, среднее + S.D.) На фиг. 5В показано определение условия рН. CD45-положительные клетки подвергали воздействию растворов с различным значением рН. Через 3 дня после воздействия стресса экспрессию Oct4-GFP анализировали с помощью FACS. На фиг. 5С показано определение условия культивирования. Подвергнутые воздействию стресса клетки культивировали в различных средах. Количество экспрессирующих GFP АС подсчитывали на 14 день. (n=3, среднее + S.D.)
На фиг. 6А-6В показано получение АСС из CD45-положительных клеток, полученных от мышей ICR. На фиг. 6А показано хронологическое изменение CD45-положительных клеток после воздействия стресса. Экспрессию Е-кадгерина и SSEA-1 анализировали с помощью FACS. На фиг. 6В продемонстрировано, что экспрессию генов Oct4 дважды положительных в отношении Е-кадгерина/SSEA1 клеток подтверждали с помощью RT-PCR. (n=3, среднее + S.D.)
На фиг. 7А-7В показано получение АСС из различных тканей, полученных от мышей GOF. На фиг. 7А показано отношение экспрессирующих Oct4-GFP клеток после воздействия стресса. Соматические клетки выделяли из различных тканей и подвергали воздействию различных видов стресса. Экспрессию Oct4-GFP анализировали с помощью FACS. На фиг. 7В показана экспрессия эмбриональных генов АСС, происходящих из различных тканей. Экспрессию генов нормировали с помощью GAPDH. (n=3, среднее + S.D.)
На фиг. 8 показана относительная экспрессия генов защиты от стресса в течение первых 7 дней. После воздействия стресса клетки собирали в 1, 3 и 7 день и экспрессию генов сравнивали с нативными CD45-положительными клетками. На графиках синим цветом показана экспрессия генов белков теплового шока. На графиках зеленым цветом показана экспрессия генов репарации ДНК. На графиках красным цветом показана экспрессия окислительно-восстановительных генов. На оси Y показаны относительная кратность изменения экспрессии.
На фиг. 9 показана дифференцировка АСС. На графике показан анализ вклада в формирование химеры. Признаки химеры, созданные с помощью АСС, происходящих из различных соматических клеток, анализировали с помощью FACS. На графике показано среднее 5 признаков химеры при Е13.5-15.5.
На фиг. 10 продемонстрировано, что воздействие стресса вызывало перепрограммирование соматических клеток путем мезенхимально-эпителиальной передачи (МЕТ). Экспрессия связанных с МЕТ генов показана в нативных клетках и в клетках через 3 и 7 дней после начала воздействия стресса. На оси Y показана экспрессия в %, нормированная к уровню в образце с уровнем экспрессии для указанного гена.
На фиг. 11 показан анализ FACS клеточных популяций до и после стресса. Экспрессия GFP являлась очевидной, указывая на образование плюрипотентных клеток, в клеточных популяциях из каждого исследуемого типа ткани.
На фиг. 12А-12Е продемонстрировано, что обработка с помощью низких значений рН индуцировала изменение судьбы в коммитированных соматических клетках. На фиг. 12А показана схема экспериментального протокола. На фиг. 12В показан анализ с помощью проточной цитометрии (верхний ряд: oct3/4::GFP+/CD45-; нижний ряд: необработанные CD45+ клетки). На оси Y представлено количество Oct3/4:GFP клеток и на оси X представлено количество CD45+ клеток. На обеих осях обозначены основные деления 0, 100, 1000 и 10000. На фиг. 12С показан график жизнеспособных oct3/4::GFP+ и oct3/4::GFP - клеток с течением времени в культуре. На фиг. 12D показан график клеточного размера Oct3/4::GFP+ клеток (левый пик) и CD45+ клеток (правый пик). На фиг. 12Е показаны результаты анализа геномных перестроек tcrβ в выделенных oct3/4::GFP+ сферах с помощью геномной ПЦР.
На фиг. 13А-13В продемонстрировано, что индуцированные низким значением рН Oct3/4+ клетки характеризуются плюрипотентностью. На фиг. 13А показан график анализа экспрессии генов с помощью кПЦР в индуцированных низким значением рН oct3/4::GFP+ клетках на 7 день по сравнению с CD45+ клетками (серии представляют слева направо экспрессию oct3/4, nanog, sox2, ecat1, esg1, dax1 и klf4). Образцы собирали в 3 день и 7 день и сравнивали с CD45-положительными клетками. (n=3, среднее + S.D.) На фиг. 13В показаны результаты бисульфитного секвенирования промоторных областей oct3/4 и nanog. CD45+ клетки, с дополнительной культурой или без нее, проявляли сильно метилированные паттерны на обоих промоторах.
На фиг. 14А-14В продемонстрировано, что клетки STAP можно получить из других тканевых источников. На фиг. 14А показан график скорости продукции oct3/4::GFP+ клеток на 7 день культивирования для ряда тканей (серии представляют слева направо CD45+ клетки, костный мозг, головной мозг, легкое, мышца, адипоциты, фибробласты, печень и хондроциты). На фиг. 14В показан график анализа экспрессии генов в кластерах клеток oct3/4::GFP+ (серии представляют слева направо экспрессию Oct3/4, Nanog, Sox2, Klf4 и Rex1).
На фиг. 15А-15В показано определение характеристик клеток STAP в качестве плюрипотентных клеток. На фиг. 15А показан график экспрессии генов маркеров ES клеток в клетках STAP (серии представляют слева направо ES, EpiSC, STAP и CD45). На фиг. 15В показан график % Х-хромосомной инактивации в клетках STAP.
На фиг. 16А показан график экспрессии Oct4-GFP, проанализированной с помощью FACS в CD45-положительных клетках, подвергнутых воздействию различных видов стресса. Процентное отношение экспрессирующих клеток Oct4-GFP к выжившим клеткам после воздействия стресса. (n=3, среднее + S.D.) На фиг. 16В показан график определения условия рН. CD45-положительные клетки подвергали воздействию растворов с различными значениями рН. На 3 день после воздействия стресса экспрессию Oct4-GFP анализировали с помощью FACS. (n=3, среднее + S.D.) На фиг. 16С показан график определения условия культивирования. Подвергнутые воздействию стресса клетки культивировали в различных средах. Количество измененной под воздействием стресса массы экспрессирующих GFP клеток подсчитывали на 14 день. (n=3, среднее + S.D.)
На фиг. 17А-17В показано получение SAC из CD45-положительных клеток, полученных от мышей ICR. На фиг. 17А показано хронологическое изменение CD45-положительных клеток после воздействия стресса. Экспрессию Е-кадгерина и SSEA-1 анализировали с помощью FACS. На фиг. 17В показан график экспрессии гена Oct4 дважды положительных в отношении Е-кадгерина/SSEA1 клеток, подтвержденной с помощью ОТ-ПЦР. (n=3, среднее + S.D.)
На фиг. 18А-18В показано получение SAC из различных тканей, полученных от мышей GOF. На фиг. 18А показан график отношения экспрессирующих Oct4-GFP клеток после воздействия стресса. Соматические клетки выделяли из различных тканей и подвергали воздействию различных видов стресса. Экспрессию Oct4-GFP анализировали с помощью FACS. Серии представляют слева направо ВМ, головной мозг, легкое, мышцу, жировую ткань, фибробласт и печень. На фиг. 18В показан график экспрессии эмбриональных генов SAC, полученных из различных тканей. Показатели экспрессию генов нормировали с помощью GAPDH. (n=3, среднее + S.D.) Серии представляют слева направо Oct4, Nanog, Sox2, Klf4 и Ecat1.
На фиг. 19 показан график относительной экспрессии генов защиты от стресса в течение первых 7 дней. После воздействия стресса клетки собирали в 1, 3 и 7 день и экспрессию генов сравнивали с нативными CD45-положительными клетками. На оси Y показана относительная кратность изменения экспрессии.
На фиг. 20 показаны анализы реаранжировки цепи TCRβ SAC и химерных мышей, полученных из SAC из CD45+ клеток. Химерные мыши 2N №1, №2, №3, №5, №6, №7, №8 и №9 экспрессировали реаранжированную ДНК.
На фиг. 21 показан анализ генотипирования химерных мышей 4N. Генотипирование проводили, чтобы доказать, что химерные мыши 4N, полученные с помощью SAC, происходящих из бластоцист 129/Sv×B6GFP F1 и 4N, полученных из ICR, экспрессировали специфический для SAC (129/Sv×B6GFP) ген.
На фиг. 22 продемонстрировано, что клетки STAP вносят вклад как в эмбриональную, так и плацентарную ткань in vivo. На графике показано соотношение признаков, в которые введенные инъекцией клетки вносили вклад только в отношении эмбриональной части, а также в отношении тканей плаценты и желточного мешка.
На фиг. 23А-23С продемонстрировано, что обработка FGF4 индуцировала некоторый характер трофобластной линии в клетках STAP. На фиг. 23А показана схема обработки FGF4 для индукции TS-подобных (F4I) клеток из клеток STAP. На фиг. 23В показан график анализа кПЦР экспрессии маркеров. На фиг. 23С показан график количественного определения плацентарного вклада с помощью анализа FACS. В отличие от клеток F4I, ES клетки не вносили вклад в плацентарные ткани на обнаруживаемом уровне.
На фиг. 24A-24D продемонстрировано, что подобные клеткам ES стволовые клетки можно получить из клеток STAP. На фиг. 24А показана схема индукции линий стволовых клеток из клеток STAP. На фиг. 24В показан график, демонстрирующий активный рост клеток STAP-S в поддерживающей культуре в течение 120 дней. Сходные результаты получали с 16 независимыми линиями. Напротив, исходные клетки STAP быстро уменьшались в количестве. На фиг. 24С показан график анализа кПЦР экспрессии маркерных генов. Клетки ES и STAP-S экспрессировали связанные с плюрипотентностью гены, которые не экспрессировались в CD45+ клетках. На фиг. 24D показано схематическое представление исследования метилирования ДНК с помощью бисульфатного секвенирования.
На фиг. 25А-25В продемонстрировано, что стволовые клетки STAP являются плюрипотентными и совместимыми с передачей зародышевой линии и тетраплоидной комплементацией. На фиг. 25А показан график вклада клеток STAPS в различные ткани у химерных мышей в анализах инъекции в бластоцисты (2N). На фиг. 25В показан график вклада в плацентарные ткани. В отличие от исходных клеток STAP и клеток TS, клетки STAPS не сохраняли способность вносить плацентарные вклады. Исследовали три независимых линии, и все показали значительные вклады в эмбриональные части.
На фиг. 26 продемонстрирована плюрипотентность, индуцированная отмывкой с помощью кислоты.
На фиг. 27 показано сверху: механическая гипералгезия, на которую указывает снижение порога одергивания лапы после инъекции капсаицина, снижается у крыс, получивших лечение с помощью интратекального SSP-SAP. На последующих графиках показан ответ через 10 мин после инъекции капсаицина, когда наблюдается наибольшая разница. Внизу: через пять недель после имплантации стволовых клеток спинного мозга сохранялась индуцированная капсаицином гипералгезия.
На фиг. 28 показаны тактильные (вверху) и термические (внизу) ответы после инъекций капсаицина в лапу крыс, которым вначале ввели инъекцию и/т (интратекально) SSP-SAP, которая сильно снижает состояние гипералгезии (ср. с фиг. 27), и затем обрабатывали с помощью стволовых клеток, люмбальной и/т инъекции. "Ранее не подверженный экспериментам ответ" показывает гипералгезический ответ на капсаицин до любых манипуляций. "BL1" представляет собой фоновый ответ до инъекции капсаицина у крыс, которым за 2 недели ранее ввели или SSP-SAP, или неактивную холостую пробу-SAP. "BL2" представляет собой фоновый ответ, без инъекции капсаицина, через 1-2 дня после доставки стволовых клеток. Следует обратить внимание на способность имплантата стволовых клеток возвращать гипералгезический ответ получивших лечение SSP-SAP крыс к значению не подвергнутых экспериментам крыс и получивших холостую пробу-SAP контролей.
На фиг. 29 продемонстрировано, что эффективность специфического антагониста NK1-R увеличивается у крыс, если чувствительность к капсаицину сохранялась с помощью имплантатов стволовых клеток. IC50 L-733.060 составляет ~ 0,3 мМ (30 мкл и/т инъекции) для обеих типов гипералгезия у не подвергнутых экспериментам крыс (О,  ; левая панель; и у тех крыс, которые получили холостую пробу-SAP с последующими стволовыми клетками, не показано), тогда как у сохранивших стволовые клетки крыс (правая панель) IC50 составляет ~30 мкМ для тактильной гипералгезия (
; левая панель; и у тех крыс, которые получили холостую пробу-SAP с последующими стволовыми клетками, не показано), тогда как у сохранивших стволовые клетки крыс (правая панель) IC50 составляет ~30 мкМ для тактильной гипералгезия ( ) и ~5 мкМ для термической гипералгезии (
) и ~5 мкМ для термической гипералгезии ( ).
).
Подробное раскрытие настоящего изобретения
Аспекты описанной в настоящем документе технологии относятся к получению или созданию плюрипотентных клеток из клеток. Аспекты описанной в настоящем документе технологии основаны на открытии авторами настоящего изобретения того, что стресс может индуцировать продукцию плюрипотентных стволовых клеток из клеток без необходимости вводить экзогенный ген, транскрипт, белок, ядерный компонент или цитоплазму в клетку или без необходимости слияния на клеток. Согласно некоторым вариантам осуществления стресс индуцирует снижение количества цитоплазмы и/или митохондрий в клетке; запуская процесс дедифференцировки и давая в результате плюрипотентные клетки. Согласно некоторым вариантам осуществления стресс вызывает разрушение клеточной мембраны, например, по меньшей мере у 10% клеток, подвергнутых воздействию стресса. Указанные плюрипотентные клетки характеризуются одним или несколькими из следующего: способность дифференцироваться в каждый из трех зародышевых листков (in vitro и/или in vivo), создание тератома-подобных клеточных масс in vivo и способность создавать жизнеспособных эмбрионов и/или химерных мышей.
В настоящем документе описаны эксперименты, демонстрирующие, что воздействие на клетки определенных относящихся к окружающей среде видов стресса, включая в себя без ограничения стрессовые воздействия, которые снижают количество цитоплазмы и/или митохондрий в клетке, может снижать митохондриальную активность, деметилировать области генома, ассоциированные с дедифференцировкой, вызывать проявление клетками маркеров известных путей дедифференцировки. Соответственно, согласно некоторым вариантам осуществления в настоящем документе предусмотрены способы создания плюрипотентных клеток из клеток, причем способы предусматривают удаление по меньшей мере приблизительно 40% цитоплазмы и/или митохондрий из клетки и выбор плюрипотентности или клеток, проявляющих маркеры плюрипотентности, причем клетка не присутствует в ткани. Кроме того, в настоящем документе описаны другие стрессовые воздействия, которые могут создавать плюрипотентные клетки из клеток.
Для удобства определенные термины, используемые в настоящем документе в описании настоящего изобретения, примерах и прилагаемой формуле изобретения, собраны в настоящем документе. Если не указано иное или не вытекает из контекста, следующие термины и фразы включают в себя значения, предусмотренные ниже. Если явно не указано иное или не очевидно из контекста, термины и фразы ниже не исключают значения, которое термин или фраза приобрели в настоящей области техники, к которой они принадлежат. Определения предусмотрены для содействия в описании конкретных вариантов осуществления, и не предусмотрено, что они ограничивают заявленное изобретение, поскольку объем настоящего изобретения ограничен только формулой изобретения. Если не установлено иное, все технические и научные термины, используемые в настоящем документе, имеют такое же значение, как то, которое подразумевает под ними специалист в настоящей области техники, к которой принадлежит настоящее изобретение.
Используемый в настоящем документе термин "содержащий" или "содержит" используют со ссылкой на композиции, способы и их соответствующий(е) компонент(ы), которые являются необходимыми для способа или композиции, но он открыт для включения не указанных элементов, будь то необходимых или необязательных.
Используемый в настоящем документе термин "состоящий по существу из" относится к тем элементам, которые необходимы для данного варианта осуществления. Термин допускает присутствие элементов, которые фактически не оказывают влияния на основную(ые) и новую(ые) или функциональную(ые) характеристику(и) настоящего варианта осуществления.
Термин "состоящий из" относится к композициям, способам и их соответствующим компонентам, описанным в настоящем документе, которые исключают любой элемент, не перечисленный в настоящем описании варианта осуществления.
Используемый в настоящем описании изобретения и прилагаемой формулы изобретения формы единственного числа включают в себя ссылки на формы множественного числа, если только контекст ясно не диктует иное. Таким образом, например, ссылки на "способ" включают в себя один или несколько способов и/или стадий типа, описанных в настоящем документе и/или которые станут очевидными специалистам в настоящей области техники при чтении настоящего раскрытие и т.д. Аналогично, предусмотрено, что слово "или" включает в себя "и", если в контексте ясно не указано иное. Несмотря на то, что способы и материалы, сходные или эквивалентные описанным в настоящем документе, можно использовать в осуществлении на практике или испытании настоящего раскрытия, подходящие способы и материалы описаны ниже. Термин "например" используется в настоящем документе для указание на неограничивающий пример.
Определения общих терминов в клеточной биологии и молекулярной биологии можно найти в "The Merck Manual of Diagnosis and Therapy", 19-е издание, опубликованный Merck Research Laboratories, 2006 (ISBN 0-911910-19-0); Robert S. Porter et al. (eds.), и The Encyclopedia of Molecular Biology, опубликованный Blackwell Science Ltd., 1994 (ISBN 0-632-02182-9). Определения общих терминов в молекулярной биологии также можно найти в Benjamin Lewin, Genes X, опубликованный Jones & Bartlett Publishing, 2009 (ISBN-10: 0763766321); Kendrew et al. (eds.), Molecular Biology and Biotechnology: a Comprehensive Desk Reference, опубликованный VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8) и Current Protocols in Protein Sciences 2009, Wiley Intersciences, Coligan et al., eds.
Если не указано иное, настоящее изобретение проводили с использованием стандартных процедур, описанных, например, в Sambrook et al., Molecular Cloning: A Laboratory Manual (3 ed.), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., USA (2001); Davis et al., Basic Methods in Molecular Biology, Elsevier Science Publishing, Inc., New York, USA (1995); Current Protocols in Cell Biology (CPCB) (Juan S. Bonifacino et. al. ed., John Wiley and Sons, Inc.), и Culture of Animal Cells: A Manual of Basic Technique by R. Ian Freshney, Publisher: Wiley-Liss; 5th edition (2005), Animal Cell Culture Methods (Methods in Cell Biology, Vol. 57, Jennie P. Mather and David Barnes editors, Academic Press, 1st edition, 1998), которые полностью включены посредством ссылки в настоящий документ.
Термины "уменьшать", "снижать", "сниженный" и "снижение" используют в настоящем документе, как правило, для обозначения уменьшения на статистически значимое количество относительно эталонного значения. Тем не менее, во избежание сомнения, "снижать", "снижение" или "уменьшать", как правило, означают уменьшение по меньшей мере на 10% по сравнению с отсутствием данной обработки и могут включать в себя, например, уменьшение по меньшей мере приблизительно на 20%, по меньшей мере приблизительно на 25%, по меньшей мере приблизительно на 30%, по меньшей мере приблизительно на 35%, по меньшей мере приблизительно на 40%, по меньшей мере приблизительно на 45%, по меньшей мере приблизительно на 50%, по меньшей мере приблизительно на 55%, по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 65%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 75%, по меньшей мере приблизительно на 80%), по меньшей мере приблизительно на 85%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 98%, по меньшей мере приблизительно на 99%, вплоть до и включая в себя, например, полное отсутствие данной величины или параметр по сравнению с отсутствием данной обработки или любое уменьшение от 10 до 99% по сравнению с отсутствием данной обработки.
Термины "увеличенный", "увеличивать" или "усиливать" используют в настоящем документе, как правило, для обозначения увеличения на статистически значимое количество; во избежание сомнения, термины "увеличенный", "увеличивать" или "усиливать" означают увеличение по меньшей мере на 10% по сравнению с эталонным уровнем, например, увеличение по меньшей мере приблизительно на 20%, или по меньшей мере приблизительно на 30%, или по меньшей мере приблизительно на 40%, или по меньшей мере приблизительно на 50%, или по меньшей мере приблизительно на 60%, или по меньшей мере приблизительно на 70%, или по меньшей мере приблизительно на 80%) или по меньшей мере приблизительно на 90% или вплоть до и включая в себя 100% увеличение или любое увеличение от 10 до 100% по сравнению с эталонным уровнем или по меньшей мере приблизительно в 2 раза, или по меньшей мере приблизительно в 3 раза, или по меньшей мере приблизительно в 4 раза, или по меньшей мере приблизительно в 5 раз, или по меньшей мере приблизительно в 10 раз или любое увеличение от 2-кратного до 10-кратного или больше по сравнению с эталонным уровнем.
Используемые в настоящем документе термины "лечить", "лечение", "осуществление лечения" или "облегчение" при использовании в отношении заболевания, нарушения или медицинского состояния означают терапевтические виды лечения состояния, при которых у объекта происходит обратное развитие, улучшение, облегчение, ингибирование, замедление или прекращение прогрессирования или тяжести симптома или состояния. Термин "обеспечение лечения" включает в себя снижение или облегчение по меньшей мере одного неблагоприятного эффекта или симптома состояния. Лечение является, как правило, "эффективным", если один или несколько симптомов или клинических маркеров снижаются. Альтернативно, лечение является "эффективным", если прогрессирование состояния снижается или прекращается. Иными словами, "лечение" включает в себя не только улучшение симптомов или маркеров, но также прекращение или по меньшей мере замедление прогрессирования или ухудшения симптомов, что можно ожидать при отсутствии лечения. Благоприятные или требуемые клинические результаты включают в себя без ограничения облегчение одного или нескольких симптомов, уменьшение степени недостаточности, стабилизированное (т.е. не ухудшающееся) состояние здоровья, задержку или замедление прогрессирования заболевания и уменьшение интенсивности или временное облегчение симптомов. Лечение также может включать в себя выживаемость субъекта, превышающую предполагаемую статистически смертность.
Используемый в настоящем документе термин "введение" относится к помещению плюрипотентной клетки, полученной согласно описанным в настоящем документе способам, и/или по меньшей мере частично дифференцированного потомства такой плюрипотентной клетки в организм субъекта с помощью способа или пути, который приводит по меньшей мере к частичной локализации клеток в требуемом месте. Фармацевтическая композиция, содержащая плюрипотентную клетку, полученную согласно описанным в настоящем документе способам, и/или по меньшей мере частично дифференцированного потомства такой плюрипотентной клетки можно вводить с помощью любого подходящего пути, который приводит к эффективному лечению субъекта.
Используемый в настоящем документе термин "субъект" означает человека или животного. Как правило, животное представляет собой такое позвоночное, как примат, грызун, домашнее животное или промысловое животное. Приматы, например, включают в себя следующих: шимпанзе, яванские макаки, паукообразные обезьяны и макаки, например, макаки-резусы. Грызуны включают в себя мышей, крыс, лесных сурков, хорьков, кроликов и хомяков. Домашние и промысловые животные включают в себя коров, лошадей, свиней, оленей, бизонов, буйволов, кошачьих, например, домашнюю кошку, псовых, например, собаку, лису, волка, виды птиц, например, курицу, эму, страуса, и рыбу, например, форель, сома и лосося. Пациент или субъект включает в себя любой подкласс перечисленного выше, например, всех из вышеперечисленных. Согласно определенным вариантам осуществления субъект представляет собой млекопитающее, например, примата, например, человека.
Субъект предпочтительно представляет собой млекопитающее. Млекопитающее может представлять собой человека, не являющегося человеком примата, мышь, крысу, собаку, кошку, лошадь или корову без ограничения указанными примерами. Млекопитающие, отличные от людей, можно успешно использовать в качестве субъектов, которые представляют животные модели заболевания, ассоциированного с отсутствием, дисфункцией и/или недостаточностью данной клетки или ткани или отсутствием, дисфункцией или недостаточностью компартмента стволовой клетки. Кроме того, описанные в настоящем документе способы можно использовать для лечения одомашненных животных и/или питомцев. Субъект может являться самцом или самкой. Субъект может представлять собой субъекта, у которого ранее диагностировали или идентифицировали как страдающего или характеризующегося отсутствием, дисфункцией и/или недостаточностью клеточного типа, ткани или компартмента стволовой клетки или одним или несколькими заболеваниями или состояниями, ассоциированными с таким состоянием, и необязательно, но мог не быть к настоящему времени подвергнут лечению в отношении такого состояния. Субъект также может представлять собой субъекта, у которого диагностировали или идентифицировали как страдающего от состояния, включая в себя дефицит, дисфункцию или недостаточность клеточного типа или ткани или компартмента стволовой клетки, но который демонстрирует улучшения в отношении известных факторов риска в результате получения одного или нескольких типов лечения в отношении такого состояния. Альтернативно, субъект также может представлять собой субъекта, которого ранее не диагностировали как характеризующегося таким состоянием. Например, субъект может представлять собой субъекта, который проявляет один или несколько факторов риска в отношении такого состояния, или субъекта, который не проявляет факторы риска в отношении таких состояний.
Используемый в настоящем документе термин "выбирать" при использовании со ссылкой на клетку или популяцию клеток, относится к отбору, отделению, выделению и/или селективному размножению одной или нескольких клеток, характеризующихся требуемой характеристикой. Используемый в настоящем документе термин "выбирать" необязательно подразумевает, что клетки без требуемой характеристики невозможно размножить в предусмотренных условиях.
Используемый в настоящем документе термин "поддерживать" относится к поддержанию жизнеспособности клетки или популяции клеток. Поддерживаемая популяция будет содержать ряд метаболически активных клеток. Ряд указанных клеток может являться приблизительно стабильными в течение периода, составляющего по меньшей мере один день или могут расти.
Используемый в настоящем документе термин "обнаруживаемый уровень" относится к уровню вещества или активности в образце, который обеспечивает возможность отличить количество вещества или активности от референсного уровня, например, содержания вещества или уровня активности в клетке, которую не подвергали воздействию стресса. Согласно некоторым вариантам осуществления обнаруживаемый уровень может представлять собой уровень по меньшей мере на 10% больше, чем референсный уровень, например, на 10% больше, на 20% больше, на 50% больше, на 100% больше, на 200% больше или на 300% или больше.
Термин "статистически значимое" или "значимо" относится к статистическому уровню значимости и, как правило, означает разницу в два среднеквадратичных отклонения (2SD) выше или ниже референсного значения, например, концентрации или численности маркера, например, маркера стволовых клеток или маркера дифференцировки. Термин относится к статистическому доказательству того, что имеет место разница. Ее определяют как вероятность принятия решения отклонить нулевую гипотезу, когда нулевая гипотеза действительно верна. Решение зачастую принимают с использованием р-значения.
За исключением рабочих примеров или там, где указано иное, все числа, выражающие количества ингредиентов или реакционные условия, используемые в настоящем документе, следует понимать как модифицированные во всех случаях с помощью термина "приблизительно." Термин "приблизительно" при использовании в связи с процентными отношениями может означать ±1%.
Другие термины определены в настоящем документе в описании различных аспектов описанной в настоящем документе технологии.
Аспекты описанной в настоящем документе технологии относятся к способам создания плюрипотентной клетки из клетки, а также применениям и способам применения указанных плюрипотентных клеток. В отличие от существующих способов создания плюрипотентных клеток (т.е. индуцированных плюрипотентных стволовых клеток или клеток iPS), которые основаны на увеличении экспрессии перепрограммирующих факторов, например, путем введения нуклеиновокислотных конструктов, кодирующих один или несколько перепрограммирующих факторов (например, Oct4), описанные в настоящем документе способы подвергают клетки стрессу, но не требуют введения чужеродных перепрограммирующих агентов.
Согласно некоторым вариантам осуществления стресс сокращает объем клеточной цитоплазмы и/или количество клеточных митохондрий. Сокращение объема клеточной цитоплазмы или количества клеточных митохондрий индуцирует ответ на стресс, во время которого клетка приобретает по меньшей мере плюрипотентные способности. Согласно одному аспекту в настоящем документе описан способ создания плюрипотентной клетки, предусматривающий удаление по меньшей мере приблизительно 40% цитоплазмы из клетки и выбор клеток, проявляющих плюрипотентность, причем клетка не присутствует в ткани. Согласно одному аспекту описанное в настоящем документе настоящее изобретение относится к способу создания плюрипотентной клетки, предусматривающему удаление по меньшей мере приблизительно 40% митохондрий из клетки и выбор клеток, проявляющих плюрипотентность, причем клетка не присутствует в ткани.
Клетки, используемые в способах, анализах и композициях, описанных в настоящем документе, могут относиться к любому типу клеток, например, взрослая клетка, эмбриональная клетка, дифференцированная клетка, стволовая клетка, клетка-предшественник и/или соматическая клетка. Клетку можно описать с помощью комбинаций описанных выше терминов, например, клетка может представлять собой эмбриональную стволовую клетку или дифференцированную соматическую клетку. Клетка, используемую в способах, анализах и композициях, описанных в настоящем документе, можно получить от субъекта. Согласно некоторым вариантам осуществления клетка представляет собой клетку млекопитающего. Согласно некоторым вариантам осуществления клетка представляет собой клетку человек. Согласно некоторым вариантам осуществления клетка представляет собой взрослую клетку. Согласно некоторым вариантам осуществления клетка представляет собой неонатальную клетку. Согласно некоторым вариантам осуществления клетка представляет собой фетальную клетку. Согласно некоторым вариантам осуществления клетка представляет собой амниотическую клетку. Согласно некоторым вариантам осуществления клетка представляет собой клетку пуповинной крови.
"Взрослая" относится к тканям и клеткам, полученным от или находящимся внутри животного-субъекта в любое время после рождения. "Эмбриональная" относится к тканям и клеткам, полученным от или находящимся внутри животного-субъекта в любое время до рождения.
Используемый в настоящем документе термин "соматическая клетка" относится к любой клетке, отличной от половой клетки, клетки, присутствующей в или полученной из предимплантационного эмбриона, или клетки, полученной путем пролиферации такой клетки in vitro. Иначе говоря, соматическая клетка относится к любым клеткам, образующим тело организма, в отличие от зародышевых клеток. У млекопитающих зародышевые клетки (также известные как "гаметы") представляют собой сперматозоиды и яйцеклетки, которые сливаются во время оплодотворения с образованием клети, которая называется зигота, из которой развивается весь эмбрион млекопитающего. Каждый другой тип клеток в теле млекопитающего, за исключением сперматозоидов и яйцеклеток, клеток, из которых они образованы (гаметоцитов) и недифференцированных стволовых клеток, представляет собой соматическую клетку: внутренние органы, кожа, кости, кровь и соединительная ткань образованы из соматических клеток. Согласно некоторым вариантам осуществления соматическая клетка представляет собой "неэмбриональную соматическую клетку", причем под этим термином подразумевают соматическую клетку, которая не присутствует или не получена из эмбриона и не является результатом пролиферации такой клетки in vitro. Согласно некоторым вариантам осуществления соматическая клетка представляет собой "взрослую соматическую клетку", причем под этим термином подразумевают клетку, которая присутствует или получена из организма, отличного от эмбриона или плода или является результатом пролиферации такой клетки in vitro. Следует отметить, что взрослые и неонатальные или эмбриональные клетки можно различить по структурным отличиям, например, эпигенетической организации, такой как паттерны метилирования. Согласно некоторым вариантам осуществления соматическая клетка представляет собой соматическую клетку млекопитающего. Согласно некоторым вариантам осуществления соматическая клетка представляет собой соматическую клетку человека. Согласно некоторым вариантам осуществления соматическая клетка представляет собой взрослую соматическую клетку. Согласно некоторым вариантам осуществления соматическая клетка представляет собой неонатальную соматическую клетку.
Используемый в настоящем документе термин "дифференцированная клетка" относится к клетке, которая является более специализированной в своем направлении развития или функции, чем в предыдущий момент своего развития, и включает в себя как клетки, которые являются терминально дифференцированными, так и клетки, которые, хотя и не являются терминально дифференцированными, являются более специализированными, чем в предыдущий момент своего развития. Развитие клетки из некоммитированной клетки (например, стволовой клетки) в клетку с увеличенной степенью направленности на конкретный тип дифференцированных клеток и, в итоге, в терминально дифференцированную клетку известно как прогрессирующая дифференцировка или прогрессирующее коммитирование. В контексте клеточного онтогенеза прилагательное "дифференцированный" или "дифференцирующий" представляет собой относительный термин. "Дифференцированная клетка" представляет собой клетку, которая прошла дальше по пути развития, чем клетка с которой ее сравнивают. Таки образом, стволовые клетки могут дифференцироваться до линиеспецифических клеток-предшественников (таких как мезодермальная стволовая клетка), которые в свою очередь могут дифференцироваться в другие типы клеток-предшественников дальше по пути развития (такие как предшественник кардиомиоцита), и затем до окончательно дифференцированной клетки, которая играет характерную роль в определенном типе ткани и может сохранять или не сохранять способность пролиферировать в дальнейшем.
Используемый в настоящем документе термин "стволовая клетка" относится к клетке в недифференцированном или частично дифференцированном состоянии, которая характеризуется свойством самообновления и характеризуется дифференцировочным потенциалом естественным образом дифференцироваться в более дифференцированный тип клеток, без специального подразумеваемого значения в отношении дифференцировочного потенциала (т.е. тотипотентные, плюрипотентные, мультипотентные и т.д.). Под самообновлением подразумевают, что стволовая клетка способна к пролиферации и порождению большего количества таких стволовых клеток, при этом сохраняя свой дифференцировочный потенциал. Соответственно, термин "стволовая клетка" относится к любому подклассу клеток, которые характеризуются дифференцировочным потенциалом, при определенных обстоятельствах, дифференцироваться до более специализированного или дифференцированного фенотипа, и которые сохраняют способность, при определенных обстоятельствах, пролиферировать без существенной дифференцировки. Термин "соматическая стволовая клетка" используют в настоящем документе для обозначения любой стволовой клетки, происходящей из неэмбриональной ткани, включая в себя фетальную, ювенильную и взрослую ткань. Природные соматические стволовые клетки были выделены из большого разнообразия взрослых тканей, включая в себя кровь, костный мозг, головной мозг, ольфакторный эпителий, кожу, поджелудочную железу, скелетную мышцу и сердечную мышцу. Иллюстративные встречающиеся в природе соматические стволовые клетки включают в себя без ограничения мезенхимальные стволовые клетки и гемопоэтические стволовые клетки. Согласно некоторым вариантам осуществления стволовые клетки или клетки-предшественники могут представлять собой эмбриональные стволовые клетки. Используемый в настоящем документе термин "эмбриональные стволовые клетки" относится к стволовым клеткам, происходящим из ткани, образованной после оплодотворения, но до окончания гестации, включая в себя преэмбриональную ткань (такую как, например, бластоцисту), эмбриональную ткань или фетальную ткань, взятую в любое время в течение гестации, как правило, ног не обязательно, приблизительно до 10-12 недель гестации. Чаще эмбриональные стволовые клетки представляют собой тотипотентные клетки, происходящие из раннего эмбриона или бластоцисты. Эмбриональные стволовые клетки можно получить напрямую из подходящей ткани, включая в себя без ограничения ткань человека, или из установленных эмбриональных клеточных линий. Согласно одному варианту осуществления эмбриональные стволовые клетки получают, как описано Thomson et al. (патенты США №№5843780 и 6200806; Science 282:1145, 1998; Curr. Top. Dev. Biol. 38:133 ff, 1998; Proc. Natl. Acad. Sci. U.S.A. 92: 7844, 1995, которые полностью включены в настоящий документ посредством ссылки).
Иллюстративные стволовые клетки включают в себя эмбриональная стволовые клетки, взрослые стволовые клетки, плюрипотентные стволовые клетки, нейрональные стволовые клетки, стволовые клетки печени, стволовые клетки мышцы, стволовые клетки предшественника мышцы, эндотелиальные клетки-предшественники, стволовые клетки костного мозга, хондрогенные стволовые клетки, лимфоидные стволовые клетки, мезенхимальные стволовые клетки, гемопоэтические стволовые клетки, стволовые клетки центральной нервной системы, стволовые клетки периферической нервной системы и подобное. Описания стволовых клеток, включая в себя способы их выделения и культивирования, можно найти, среди прочего, в Embryonic Stem Cells, Methods and Protocols, Turksen, ed., Humana Press, 2002; Weisman et al., Annu. Rev. Cell. Dev. Biol. 17: 387 403; Pittinger et al., Science, 284: 143 47, 1999; Animal Cell Culture, Masters, ed., Oxford University Press, 2000; Jackson et al., PNAS 96(25): 14482 86, 1999; Zuk et al., Tissue Engineering, 7: 211 228, 2001 ("Zuk et al."); Atala et al., особенно Главы 33 41; и патенты США №№5559022, 5672346 и 5827735. Описания стромальных клеток, включая в себя способы их выделения, можно найти, среди прочего, в Prockop, Science, 276: 71 74, 1997; Theise et al., Hepatology, 31:235 40, 2000; Current Protocols in Cell Biology, Bonifacino et al., eds., John Wiley & Sons, 2000 (включая в себя обновления в марте 2002 г.); и патент США №4963489.
Используемый в настоящем документе термин "клетки-предшественники" относится к клеткам в недифференцированном или частично дифференцированном состоянии и которые характеризуются дифференцировочным потенциалом дифференцироваться по меньшей мере до одного более дифференцированного фенотипа, без специального подразумеваемого значения в отношении дифференцировочного потенциала (т.е. тотипотентные, плюрипотентные, мультипотентные и т.д.) и которые не характеризуются свойством самообновления. Соответственно, термин "клетка-предшественник" относится к любому подклассу клеток, который характеризуется дифференцировочным потенциалом, при конкретных условиях, дифференцироваться до более специализированного или дифференцированного фенотипа. Согласно некоторым вариантам осуществления стволовые клетки или клетки-предшественники представляют собой плюрипотентные стволовые клетки. Согласно некоторым вариантам осуществления стволовые клетки или клетки-предшественники представляют собой тотипотентные стволовые клетки.
Термин "тотипотентная" относится к стволовой клетке, которая может давать начало любому типу ткани или клеток в организме. "Плюрипотентные" стволовые клетки могут давать начало любому типу клеток в организме, за исключением половых клеток. Стволовые клетки, которые могут давать начало меньшему или ограниченному количеству различных типов клеток, как правило, называют "мультипотентные." Таким образом, тотипотентные клетки дифференцируются в плюрипотентные клетки, которые могут давать начало большинству, но не всем тканям, необходимым для развития плода. Плюрипотентные клетки подвергаются дополнительной дифференцировке в мультипотентные клетки, которые отвечают за то, чтобы давать начало клеткам, которые характеризуются конкретной функцией. Например, мультипотентные гемопоэтические стволовые клетки дают начало эритроцитам, лейкоцитам и тромбоцитам в крови.
Используемый в настоящем документе термин "плюрипотентная" относится к клетке со способностью, при различных условиях, дифференцироваться до типов клеток, характерных для всех трех зародышевых листков (т.е. эндодерма (например, ткань кишечника), мезодерма (например, кровь, мышцы и сосуды) и эктодерма (например, кожа и нервы)). Плюрипотентные клетки главным образом характеризуются своей способностью дифференцироваться до всех трех зародышевых листков, с использованием, например, анализа образования тератомы у бестимусных мышей с мутацией nude («голых» мышей). Плюрипотентность также доказывают с помощью экспрессии маркеры эмбриональных стволовых (ES) клеток, хотя предпочтительное исследование на плюрипотентность представляет собой демонстрацию способности дифференцироваться в клетки каждого из трех зародышевых листков.
Описанные в примерах в настоящем документе клетки "АСС" и "STAP" представляют собой неограничивающие примеры плюрипотентных клеток. "STAP стволовые клетки" представляют собой неограничивающие примеры плюрипотентных стволовых клеток. Термин "плюрипотентная клетка" и термин "плюрипотентная стволовая клетка" можно использовать в настоящем документе взаимозаменяемо, поскольку обе клетки можно использовать подходящим образом для цели настоящего изобретения.
Используемый в настоящем документе термин "плюрипотентность" или "плюрипотентное состояние" относится к клетке со способностью дифференцироваться в три эмбриональных зародышевых листка: эндодерму (ткань кишечника), мезодерму (включая в себя кровь, мышцу и сосуды) и эктодерму (такую как кожу и нервы).
Термин "мультипотентная" при использовании со ссылкой на "мультипотентную клетку" относится к клетке, которая способна дифференцироваться в некоторые, но не все клетки, происходящие из трех зародышевых листков. Таким образом, мультипотентная клетка представляет собой частично дифференцированную клетку. Мультипотентные клетки хорошо известны в настоящей области техники, и неограничивающие примеры мультипотентных клеток могут включать в себя взрослые стволовые клетки, такие как, например, гемопоэтические стволовые клетки и нейрональные стволовые клетки. Мультипотентная означает, что стволовая клетка может образовывать многие типы клеток в данной линии дифференцировки, но не клетки других линий дифференцировки. Например, мультипотентная стволовая клетка крови может образовывать многие различные типы клеток крови (эритроциты, лейкоциты, тромбоциты и т.д.), но она не может образовать нейроны. Термин "мультипотентность" относится к клетке со степенью многосторонности развития, которая меньше, чем у тотипотентных и плюрипотентных клеток.
Термин "тотипотентность" относится к клетке со степенью дифференцировки, описывающей способность создавать все клетки во взрослом организме, а также экстраэмбриональные ткани, включая в себя плаценту. Оплодотворенная яйцеклетка (зигота) является тотипотентной, равно как и образованные в ходе раннего дробления (бластомеры)
Клетка, используемая в описанных в настоящем документе способах, может представлять собой клетку, которая не присутствует в ткани. Используемый в настоящем документе термин "ткань" относится к организованному биоматериалу (например, группе, слою или агрегации) сходным образом специализированных клеток, объединенных в выполнении по меньшей мере одной конкретной функции. Если клетки удаляют из организованной суперструктуры или иным образом отделяют от организованной суперструктуры, которая существует in vivo, они больше не присутствуют в ткани. Например, если образец крови разделяют на две или больше неидентичных фракции или селезенку разрезают на мелкие фрагменты и механически диссоциируют с помощью пипеток Пастера, клетки больше не присутствуют в ткани. Согласно некоторым вариантам осуществления клетки, которые не присутствуют в ткани, представляют собой выделенные клетки. Термин "выделенные", используемый в настоящем документе со ссылкой на клетки, относится к клетке, которая механически или физически отделена от другой группы клеток, с которыми она ассоциирована в норме in vivo. Способы отделения одной или нескольких клеток от другой группы клеток хорошо известны в настоящей области техники. См., например, Culture of Animal Cells: a manual of basic techniques (3rd edition), 1994, R.I. Freshney (ed.), Wiley-Liss, Inc.; Cells: a laboratory manual (vol. 1), 1998, D.L. Spector, R.D. Goldman, L.A. Leinwand (eds.), Cold Spring Harbor Laboratory Press; Animal Cells: culture and media, 1994, D.C. Darling, S.J. Morgan, John Wiley and Sons, Ltd. Необязательно выделенную клетку культивируют in vitro, например, в присутствии других клеток.
Согласно некоторым вариантам осуществления клетка, не присутствуя в ткани, присутствует в популяции клеток. Согласно некоторым вариантам осуществления популяция клеток представляет собой популяцию клеток. Используемый в настоящем документе термин "популяция клеток" относится к группе, составляющей по меньшей мере 2 клетки, например, 2 клетки, 3 клетки, 4 клетки, 10 клеток, 100 клеток, 1000 клеток, 10000 клеток, 100000 клеток или любое значение в пределах этих диапазонов или больше клеток. Необязательно популяция клеток может представлять собой клетки, которые характеризуются общим происхождением, например, они могут происходить из одной и той же родительской клетки, они могут являться клональными, они могут быть выделены или происходить из клеток, выделенных из одной и той же ткани, или они могут быть выделены или происходить из клеток, выделенных из одного и того же образца ткани. Популяция клеток может содержать 1 или больше типов клеток, например, 1 тип клеток, 2 типа клеток, 3 типа клеток, 4 типа клеток или больше типов клеток. Популяция клеток может являться гетерогенной или гомогенной. Популяция клеток может являться по существу гомогенной, если она содержит по меньшей мере 90% одного и того же типа клеток, например, 90%, 92%, 95%, 98%, 99% или больше клеток в популяции принадлежат одному типу клеток. Популяция клетки может являться гетерогенной, если меньше чем 90% клеток, присутствующих в популяции, принадлежат к одному типу клеток.
Согласно некоторым вариантам осуществления описанные в настоящем документе способы могут относиться к тому, чтобы заставить неплюрипотентную клетку (например, дифференцированную клетку) приобрести плюрипотентный фенотип. Согласно некоторым вариантам осуществления создание плюрипотентной клетки может предусматривать создание клетки в более плюрипотентным фенотипом, т.е. создание условий, при которых клетка приобретает фенотип, который характеризуется более широким дифференцировочным потенциалом. В качестве неограничивающего примера подобные очень малым эмбриональным клеткам (VSEL) клетки могут являться унипотентными вместо плюрипотентных и/или могут являться ограниченными в своей способности дифференцироваться в определенные типы дифференцированных клеток (возможно, вследствие эпигенетического состояния VSEL, больше напоминающего дифференцированные клетки, чем эмбриональные стволовые клетки). Согласно способам, описанным в настоящем документе, унипотентную клетку и/или клетку с ограниченной способностью к дифференцировке можно заставить приобрести более плюрипотентный фенотип. Более плюрипотентный фенотип может представлять собой фенотип, который способен дифференцироваться в большее количество типов дифференцированных клеток, например, из двух унипотентных клеток, та, которая может дифференцироваться в большее количество типов дифференцированных клеток указанной линии дифференцировки, является более плюрипотентной, и/или плюрипотентная клетка является более плюрипотентной, чем унипотентная клетка.
Способы создания плюрипотентной клетки (или более плюрипотентной клетки), описанные в настоящем документе, могут предусматривать, например, удаление части цитоплазмы из клетки и/или удаление митохондрий из клетки. Согласно некоторым вариантам осуществления удаление части цитоплазмы или митохондрий из клетки удаляет частичный эпигенетический контроль клетки. Согласно некоторым вариантам осуществления удаляют по меньшей мере приблизительно 40% цитоплазмы, например, удаляют по меньшей мере приблизительно 40%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 90% или больше цитоплазмы клетки. Согласно некоторым вариантам осуществления удаляют от 60% до 80% цитоплазмы клетки. Согласно некоторым вариантам осуществления удаляют по меньшей мере приблизительно 40% митохондрий, например, удаляют по меньшей мере приблизительно 40%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 90% или больше митохондрий клетки. Согласно некоторым вариантам осуществления удаляют от 50% до 90% митохондрий клетки.
Способ воздействия на клетку стресса и/или удаления части цитоплазмы или митохондрий из клетки может предусматривать любой относящийся к окружающей среде стимул, который будет вызывать образование пор и/или разрывов в мембране клетки ниже порога летальности. Стресс может включать в себя нефизиологический стрессовое воздействие на ткань или клеточную культуру. Неограничивающие примеры подходящих стимулов окружающей среды включают в себя следующее: травма, механические стимулы, химическое воздействие, ультразвуковая стимуляция, кислородная недостаточность, недостаточность питательный веществ, облучение, воздействие экстремальных температур, диссоциация, суспендирование, физический стресс, гиперосмос, гипоосмос, повреждение мембран, токсин, экстремальная концентрация ионов, активный кислород, УФ-воздействие, яркий свет видимого спектра, недостаточность необходимого питания или нефизиологически кислотная среда. Согласно некоторым вариантам осуществления один стимул окружающей среды можно применить к клетке. Согласно некоторым вариантам осуществления многочисленные стимулы окружающей среды можно применить к клетке, например, можно применить 2 стимула, 3 стимула, 4 стимула или больше стимулов. Многочисленные стимулы окружающей среды можно применить одновременно или отдельно.
Согласно некоторым вариантам осуществления стресс может представлять собой стресс, который будет вызывать разрушение мембраны по меньшей мере у 10% клеток, подвергнутых воздействию стресса. Используемый в настоящем документе термин "разрушение мембраны" относится к дезинтеграции, разрыву или разрушению мембраны так, чтобы формировались поры или разрывы, достаточные для высвобождения обнаруживаемого количества органелл и/или клеточного материала, включая в себя без ограничения митохондрии и ДНК, во внеклеточное окружение. Способы обнаружения высвобождения клеточного материала, например, митохондрий, известны в настоящей области техники и описаны в настоящем документе. Высвобожденный клеточный материал может являться свободным или инкапсулированным или окруженным мембранами.
Стресс может вызывать разрушение мембраны по меньшей мере у 10% клеток, подвергнутых воздействию стресса, например, у 10% или больше, у 20% или больше, у 30% или больше, у 40% или больше, у 50% или больше, у 60% или больше, у 70% или больше, у 80% или больше или у 90% или больше. Согласно некоторым вариантам осуществления клетки, подвергнутые воздействию стресса, могут представлять собой клетки одного и того же типа и характеристик, как клетки, которые необходимо сделать более плюрипотентными согласно описанию, представленному в настоящем документе, например, стресс, подходящий для одного типа клеток, может являться не подходящим для другого типа клеток.
Продолжительность времени, в течение которого клетки подвергают воздействию стресса, может варьировать в зависимости от используемого стимула. Например, при использовании условий с низким содержанием питательных веществ для воздействия на клетки согласно способам, описанным в настоящем документе, клетки можно культивировать при условиях с низким содержанием питательных веществ в течение 1 недели или больше, например, 1 недели, 2 недель или 3 недель или дольше. Согласно некоторым вариантам осуществления клетки культивируют при условиях с низким содержанием питательных веществ в течение приблизительно 3 недель. Согласно другому неограничивающему примеру клетки, подвергнутые воздействию условий с низким значением рН или гипоксических условий согласно описанным в настоящем документе способам, можно подвергать воздействию в течение нескольких минут или дольше, например, включая в себя в течение нескольких часов, например, в течение по меньшей мере 2 минут, в течение по меньшей мере 5 минут, в течение по меньшей мере 20 минут, в течение по меньшей мере 1 часа, в течение по меньшей мере 2 часов, в течение по меньшей мере 6 часов или дольше.
Механические стимулы, которые индуцируют создание плюрипотентных клеток, могут включать в себя любую форму контакта вещества или поверхности с клеточной мембраной, который будет механически разрушать целостность мембраны. Механический стимул может включать в себя воздействие на клетку напряжения сдвига и/или высокого давления. Иллюстративная форма механического стимула представляет собой суспендирование. Суспендирование представляет собой процесс растирания и/или истирания поверхности частицы посредством трения. Неограничивающий пример процесса суспендирования клетки предусматривает то, что клетку заставляют пройти через прибор, причем прибор содержит отверстие меньше, чем размер клетки. Например, клетку можно заставить, с помощью вакуумметрического давления и/или потока жидкости, пройти через пипетку, в которой по меньшей мере часть внутреннего пространства пипетки характеризуется диаметром, меньшим чем диаметр клетки. Согласно некоторым вариантам осуществления клетку пропускают через по меньшей мере один прибор с меньшим отверстием, чем размер клетки. Согласно некоторым вариантам осуществления клетку пропускают через несколько приборов, характеризующихся постепенно уменьшающимися отверстиями. Согласно некоторым вариантам осуществления клетки можно суспендировать в течение 5 или больше минут, например, 5 минут, 10 минут, 20 минут, 30 минут или 60 минут. Согласно некоторым вариантам осуществления клетки можно суспендировать путем пропускания их через пипетку Пастера с внутренним диаметром, составляющим 50 мкм. Согласно некоторым вариантам осуществления клетки можно суспендировать путем пропускания их через пипетку Пастера с внутренним диаметром, составляющим 50 мкм в течение 20 минут.
Другие способы приложения стресса, необходимого, чтобы индуцировать клетки создать плюрипотентные клетки, включают в себя, например, воздействие определенных химических средств или физико-химических условий (например, высокого или низкого значения рН, осмотического шока, температурных экстремумов, кислородной недостаточности и т.д.). Обработки указанного типа и других типов, которые индуцируют создание плюрипотентных клеток, обсуждают дополнительно ниже. Химическое воздействие может включать в себя, например, любую комбинацию рН, осмотического давления и/или порообразующих соединений, которые разрушают или нарушают целостность клеточной мембраны. В качестве неограничивающего примера, клетки можно подвергнуть действию нефизиологически кислотной среды или низкого значения рН, стрептолизина О или дистиллированной воды (т.е. осмотическому шоку).
Низкое значение рН может включать в себя значение рН, ниже чем 6,8, например, 6,7, 6,5, 6,3, 6,0, 5,8, 5,4, 5,0, 4,5, 4,0 или ниже. Согласно некоторым вариантам осуществления низкое значение рН составляет от приблизительно 3,0 до приблизительно 6,0. Согласно некоторым вариантам осуществления низкое значение рН составляет от приблизительно 4,5 до приблизительно 6,0. Согласно некоторым вариантам осуществления низкое значение рН составляет от 5,4 до 5,8. Согласно некоторым вариантам осуществления низкое значение рН составляет от 5,4 до 5,6. Согласно некоторым вариантам осуществления низкое значение рН составляет приблизительно 5,6. Согласно некоторым вариантам осуществления низкое значение рН составляет приблизительно 5,7. Согласно некоторым вариантам осуществления низкое значение рН составляет приблизительно 5,5. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию условий с низким значением рН в течение вплоть до нескольких дней, например, в течение 6 дней или меньше, в течение 4 дней или меньше, в течение 3 дней или меньше, в течение 2 дней или меньше, в течение 1 дня или меньше, в течение 12 часов или меньше, в течение 6 часов или меньше, в течение 3 часов или меньше, в течение 2 часов или меньше, в течение 1 часа или меньше, в течение 30 минут или меньше, в течение 20 минут или меньше или меньше чем 10 минут. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию рН от 5,4 до 5,6 в течение 3 дней или меньше. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию рН от приблизительно 5,6 до 6,8 в течение 3 дней или меньше. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию рН от приблизительно 5,6 до 6,8 в течение 1 часа или меньше. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию рН от приблизительно 5,6 до 6,8 в течение приблизительно 30 минут. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию рН от приблизительно 5,6 до 6,8 в течение приблизительно 20 минут. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию рН от приблизительно 5,6 до 5,8 в течение 3 дней или меньше. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию рН от приблизительно 5,6 до 5,8 в течение 1 часа или меньше. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию рН от приблизительно 5,6 до 5,8 в течение приблизительно 30 минут. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию рН от приблизительно 5,6 до 5,8 в течение приблизительно 20 минут.
Согласно некоторым вариантам осуществления клетки можно подвергнуть действию АТР, чтобы индуцировать создание плюрипотентных клеток. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию АТР в концентрациях, составляющих от приблизительно 20 мкМ до приблизительно 200 мМ. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию АТР в концентрациях, составляющих от приблизительно 200 мкМ до приблизительно 20 мМ. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию АТР в концентрациях, составляющих приблизительно 2,4 мМ. Согласно некоторым вариантам осуществления клетка можно подвергнуть действию АТР, разбавленного в HBSS. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию АТР в течение 1 минуты или дольше, например, по меньшей мере 1 минуту, по меньшей мере 2 минуты, по меньшей мере 5 минут, по меньшей мере 15 минут, по меньшей мере 30 минут, по меньшей мере 45 минут, по меньшей мере 1 час или дольше. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию АТР в течение от приблизительно 5 минут до приблизительно 30 минут. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию АТР в течение приблизительно 15 минут. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию приблизительно 2,4 мМ АТР в течение приблизительно 15 минут.
Согласно некоторым вариантам осуществления клетки можно подвергнуть действию CaCl2, чтобы индуцировать создание плюрипотентных клеток. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию CaCl2 в концентрациях, составляющих от приблизительно 20 мкМ до приблизительно 200 мМ. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию CaCl2 в концентрациях, составляющих от приблизительно 200 мкМ до приблизительно 20 мМ. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию CaCl2 в концентрациях, составляющих приблизительно 2 мМ. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию CaCl2, разбавленного в HBSS. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию CaCl2 в течение 1 дня или дольше, например, по меньшей мере 1 день, по меньшей мере 2 дня, по меньшей мере 1 неделю, по меньшей мере 2 недели, по меньшей мере 3 недели или дольше. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию CaCl2 в течение от приблизительно 1 недели до 3 недель. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию CaCl2 в течение приблизительно 2 недель. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию приблизительно 2 мМ CaCl2 в течение приблизительно 2 недели. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию приблизительно 2 мМ CaCl2 в течение приблизительно 1 недели.
Примеры порообразующих соединений включают в себя стрептолизин О (SLO), сапонин, дигитонин, филипин, Ае I, цитолизин морской анемоны, аэролизин, аматоксин, амебапор, гомолог амебапора из Entamoeba dispar, бревинин-1Е, бревинин-2Е, барбатолизин, цитолизин Entervcoccus faecalis, дельта-гемолизин, дифтерийный токсин, цитолизин El Tor Vibrio cholerae, эквинатоксин, энтеротоксин Aeromonas hydrophila, эскулентин, гранулизин, гемолизин Vibrio parahaemolyticus, интермедилизин Streptococcus intermedins, лентивирусный литический пептид, лейкотоксин Actinobacillus actinomycetemcomitans, магаинин, мелиттин, мембраноассоциированный лимфотоксин, Мет-энкефалин, неокиоторфин, фрагмент 1 неокиоторфина, фрагмент 2 неокиоторфина, фрагмент 3 неокиоторфина, фрагмент 4 неокиоторфина, NK-лизин, парадаксин, альфа-цитолизин Staphylococcus aureus, альфа-цитолизин Clostridium septicum, токсин Bacillus thuringiensis, колицин, комплемент, дефензин, гистолизин, листериолизин, магаинин, мелиттин, пневмолизин, дрожжевой киллерный токсин, валиномицин, краун-эфиры Петерсона, перфорин, перфринголизин О, тета-токсин Clostridium perfringens, фаллолизин, фаллотоксин и другие молекулы, такие как те, что описаны в Regen et al. Biochem Biophys Res Commun 1989 159: 566-571; которая полностью включена в настоящий документ посредством ссылки. Способы очистки или синтезирования порообразующих соединений хорошо известны специалисту в настоящей области техники. Кроме того, порообразующие соединения являются коммерчески доступными, например, стрептолизин О (№ по кат. S5265; Sigma-Aldrich; Сент-Луис, Миссури). В качестве неограничивающего примера, клетки можно подвергнуть действию SLO в течение приблизительно 5 минут или больше, например, по меньшей мере 5 минут, по меньшей мере 10 минут, по меньшей мере 20 минут, по меньшей мере 30 минут, по меньшей мере 45 минут, по меньшей мере 1 час, по меньшей мере 2 часа, по меньшей мере 3 часа или дольше. Согласно некоторым вариантам осуществления клетки подвергают воздействию SLO в течение от приблизительно 30 минут до 2 часов. Согласно некоторым вариантам осуществления клетки подвергают воздействию SLO в течение приблизительно 50 минут. В качестве неограничивающего примера, клетки можно подвергнуть действию SLO в концентрациях, составляющих от приблизительно 10 нг/мл до 1 мг/мл. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию SLO в концентрациях, составляющих от приблизительно 1 мкг/мл до 100 мкг/мл. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию SLO в концентрации, составляющей приблизительно 10 мкг/мл. Согласно некоторым вариантам осуществления клетки можно подвергнуть действию SLO в концентрации, составляющей приблизительно 10 мкг/мл в течение приблизительно 50 минут.
Условия кислородной недостаточности, которые индуцируют создание плюрипотентных клеток, могут включать в себя культивирование клеток при условиях сниженного содержания кислорода, например, культивирование клеток при 10% кислорода или меньше. Согласно некоторым вариантам осуществления клетки культивируют при 5% кислорода или меньше. Продолжительность культивирования при условиях сниженного содержания кислорода может составлять 1 час или дольше, например, 1 час, 12 часов, 1 день, 2 дня, 1 неделю, 2 недели, 3 недели, 1 месяц, 2 месяца или дольше. Согласно некоторым вариантам осуществления клетки можно культивировать при условиях сниженного содержания кислорода в течение от 1 недели до 1 месяца. Согласно некоторым вариантам осуществления клетки можно культивировать при условиях сниженного содержания кислорода в течение приблизительно 3 недель.
Условия недостаточности питательный веществ, которые индуцируют создание плюрипотентных клеток, могут включать в себя отсутствие любого фактора или питательного вещества, которое является благоприятным для роста клеток. Согласно некоторым вариантам осуществления условия недостаточности питательный веществ предусматривают культивирование клеток в базовой среде культивирования, например, F12 или DMEM без дополнительных добавок, таких как FBS или факторы роста. Продолжительность культивирования в условиях недостаточности питательный веществ может составлять 1 час или дольше, например, 1 час, 12 часов, 1 день, 2 дня, 1 неделю, 2 недели, 3 недели, 1 месяц, 2 месяца или дольше. Согласно некоторым вариантам осуществления клетки можно культивировать в условиях недостаточности питательный веществ в течение от 1 недели до 1 месяца. Согласно некоторым вариантам осуществления клетки можно культивировать в условиях недостаточности питательный веществ в течение приблизительно 2 недель. Согласно некоторым вариантам осуществления клетки можно культивировать в условиях недостаточности питательный веществ в течение приблизительно 3 недель. Согласно некоторым вариантам осуществления условия недостаточности питательный веществ могут включать в себя условия без факторов роста или условия с меньше чем 50% от стандартной концентрации одного или нескольких факторов роста для данного типа клеток.
Воздействие экстремальных температур, которое индуцирует создание плюрипотентных клеток, может включать в себя воздействие либо низких температур, либо высоких температур. Для клетки млекопитающего экстремальная низкая температура может представлять собой температуру ниже 35°С, например, 34°С, 33°С, 32°С, 31°С или ниже. Согласно некоторым вариантам осуществления экстремальная низкая температура может представлять собой температуру ниже замораживания. Замораживание клеток может вызвать появление перфораций в мембране кристаллами льда и обеспечит путь для сокращения цитоплазмы. Для клетки млекопитающего экстремальная высокая температура может представлять собой температуру выше 42°С, например, 43°С, 44°С, 45°С, 46°С или выше. Согласно некоторым вариантам осуществления экстремальная высокая температура может представлять собой температуру, составляющую приблизительно 85°С или выше. Продолжительность культивирования при экстремальных температурах может составлять 20 минут или дольше, например, 20 минут, 30 минут, 1 час, 12 часов, 1 день, 2 дня, 1 неделю, 2 недели, 3 недели, 1 месяц, 2 месяца или дольше. Ясно, что чем выше температура, тем короче воздействие, которое будет, как правило, являться переносимым для обеспечения возможности создания плюрипотентных клеток.
Дополнительные примеры стрессовых воздействий, которые можно использовать в описанных в настоящем документе способах, включают в себя без ограничения ультразвуковую стимуляцию и обработку облучением.
Согласно некоторым вариантам осуществления после того, как клетки подвергли воздействию стресса, клетки можно культивировать до отбора согласно описанным ниже в настоящем документе способам. Клетки можно культивировать в течение по меньшей мере 1 часа перед отбором, например, стрессовый стимул удаляют и клетки культивируют в течение по меньшей мере 1 часа, по меньшей мере 2 часов, по меньшей мере 6 часов, по меньшей мере 12 часов, по меньшей мере 1 дня, по меньшей мере 2 дней, по меньшей мере 7 дней или дольше перед отбором согласно приведенному в настоящем документе описанию. В качестве неограничивающего примера, клетки можно подвергнуть действию SLO в течение приблизительно 50 минут и затем культивировать в среде культивирования без SLO в течение приблизительно 7 дней перед отбором. Согласно некоторым вариантам осуществления среда культивирования, используемая для культивирования клеток перед отбором, не содержит факторы дифференцировки или не стимулируют дифференцировку. Согласно некоторым вариантам осуществления среда культивирования представляет собой среду, подходящую для культивирования стволовых клеток и/или плюрипотентных клеток. Примеры таких сред описаны ниже в настоящем документе.
Согласно некоторым вариантам осуществления количество цитоплазмы в клетке сокращается. Сокращение цитоплазмы в клетке можно определить с помощью мониторинга размера клетки. Способы определения размера клетки хорошо известны специалисту в настоящей области техники и включают в себя, в качестве неограничивающего примера, цитофлуориметрический анализ. Кратко, отдельные клетки окрашивают с помощью пропидиумйодида, фильтруют и измеряют, например, на анализаторе DAKO GALAXY™ (DAKO) с использованием программного обеспечения FLOMAX™. Цитофлуориметрический анализ затем можно провести для установления размера клеток. Микросферы заданных размеров ресуспендируют в изотоническом фосфатно-солевом буфере (рН 7,2) и используют в качестве стандарта, с которым сравнивают размер клеток, содержащихся в сферах с использованием цитофлуориметрического анализа. Как клетки, так и сферы анализируют с использованием одной и той же установки приборов (прямое светорассеяние, представляющее размер клеток и сфер, и боковое светорассеяние, представляющее гранулометрический состав клеток). Размер клеток можно рассчитать на кривой с использованием размера сфер на оси X и значений прямого светорассеяния на оси Y.
Согласно некоторым вариантам осуществления количество митохондрий клетке сокращается. Способы определения количества митохондрий в клетке хорошо известны специалисту в настоящей области техники и включают в себя окрашивание с помощью специфического в отношении митохондрий красителя и подсчет количества видимых митохондрий на клетку под микроскопом. Специфические в отношении митохондрий красители являются коммерчески доступными, например, MITOTRACKER™ (№ по кат. М7512 Invitrogen; Гранд Айленд, Нью-Йорк). Согласно некоторым вариантам осуществления количество митохондрий или интенсивность сигнала от специфически в отношении митохондрий красителей можно уменьшить по меньшей мере на 40% после обработки с помощью способов, описанных выше в настоящем документе. Согласно некоторым вариантам осуществления выбирают клетки, в которых количество митохондрий или интенсивность сигнала от специфически в отношении митохондрий красителей уменьшаются по меньшей мере на 40% после обработки с помощью способов, описанных выше в настоящем документе.
Количество митохондрий и/или разрушение мембраны также можно обнаружить путем измерения окислительно-восстановительной активности во внеклеточном окружении. Как только митохондрии высвобождаются во внеклеточное окружение с помощью стресса, описанного в настоящем документе, содержание ROS во внеклеточном окружении может увеличивать и его можно использовать для измерения эффективности данного стресса.
Согласно некоторым вариантам осуществления любого из аспектов, описанных в настоящем документе, клетку можно подвергнуть воздействию стресса в присутствии LIF (ингибирующего лейкоз фактора).
Согласно некоторым аспектам после удаления части цитоплазмы и/или митохондрий клетки, способ дополнительно предусматривает выбор проявляющих плюрипотентность клеток. Плюрипотентные клетки можно выбрать путем выбора клеток, которые проявляют маркеры, фенотипы или функции плюрипотентных клеток. Выбор клеток может предусматривать выделение и размножение клеток, проявляющих требуемые характеристики, или культивирование популяции клеток с неизвестными характеристиками при таких условиях, что клетки с требуемой(ыми) характеристикой(ами) будут выживать и/или размножаться с большей скоростью, чем те клетки, которые не характеризуются требуемой(ыми) характеристикой(ами). Неограничивающие примеры маркеров и характеристик плюрипотентных клеток описаны в настоящем документе ниже. Согласно некоторым вариантам осуществления выбор клеток в отношении плюрипотентности предусматривает, по меньшей мере частично, выбор клеток, которые экспрессируют Oct4. Согласно некоторым вариантам осуществления выбор клеток в отношении плюрипотентности предусматривает, по меньшей мере частично, выбор клеток, которые экспрессируют Nanog. Согласно некоторым вариантам осуществления выбор клеток в отношении плюрипотентности предусматривает, по меньшей мере частично, выбор клеток, которые экспрессируют Oct4, Nanog, Е-кадгерин и/или SSEA. Согласно некоторым вариантам осуществления плюрипотентные клетки можно выбрать путем выбора клеток, экспрессирующих SSEA-1 и Е-кадгерин с использованием антител, специфических в отношении указанных маркеров, и FACS. Согласно некоторым вариантам осуществления клетки можно выбирать на основании размера с использованием FACS или других приборов для сортировки клеток, известных в настоящей области техники и/или описанных в настоящем документе. Клетки также можно выбирать по их неспособности прикрепляться к культуральным чашкам.
Клетки также можно выбирать на основании меньшего размера после воздействия на них стресса. Иными словами, повергнутые воздействию стресса клетки, которые прогрессируют до плюрипотентности, характеризуются меньшим размером, чем их неплюрипотентные соматические предшественники. Согласно некоторым вариантам осуществления выбирают клетки с диаметром меньше чем 8 мкм, например, клетки с диаметром 8 мкм или менее, 7 мкм или менее, 6 мкм или менее, 5 мкм или менее или меньше. Клетки можно выбирать на основании размера после их культивирования в течение короткого периода (например, несколько минут - несколько дней) или после того, как их оставили в состоянии покоя после воздействия стресса. Согласно некоторым вариантам осуществления клетки можно выбирать на основании размера непосредственно после воздействия стресса. Клетки можно выбирать на основании размера с помощью любого способа, известного в настоящей области техники, например, применение фильтра или с помощью FACS.
Согласно некоторым вариантам осуществления описанных в настоящем документе способов плюрипотентную клетку, созданную согласно способам, описанными в настоящем документе, можно культивировать для обеспечения возможности размножения указанной плюрипотентной клетки (т.е. размножения стволовой клетки). Согласно некоторым вариантам осуществления плюрипотентную, созданную согласно описанным в настоящем документе способам, можно сдержать in vitro. Согласно одному аспекту описанная в настоящем документе технология относится к композиции, содержащей плюрипотентную клетку и/илие по меньшей мере ее частично дифференцированное потомство. Согласно некоторым вариантам осуществления плюрипотентную клетку и/или по меньшей мере ее частично дифференцированное потомство можно содержать in vitro, например, в виде клеточной линии. Клеточные линии можно использовать для скрининга и/или исследования кандидатных средств, например, терапевтических средств для данного заболевания и/или средств, которые модулируют стволовые клетки, как описано ниже в настоящем документе. Согласно некоторым вариантам осуществления плюрипотентную клетку и/или по меньшей мере ее частично дифференцированное потомство можно получить из клетки, полученной от субъекта с заболеванием, например, заболеванием, ассоциированным с недостаточностью встречающейся в природе типа клеток или ткани или встречающейся в природе плюрипотентной и/или мультипотентной клетки (как описано в настоящем документе ниже), и/или заболеванием, в которое вовлечены клетки, которые содержат генетические мутации, например, злокачественная опухоль. Описанные в настоящем документе композиции можно использовать, например, в моделировании заболевания, разработке лекарственных средств, диагностике и индивидуализированной терапии.
Условия, подходящие для размножения и/или содержания стволовых и/или плюрипотентных клеток, известны в настоящей области техники. Размножение стволовых клеток обеспечивает увеличение количества клеток без существенной индукции или обеспечения возможности дифференцировки В качестве неограничивающего примера, условия, подходящие для размножения плюрипотентных клеток, включают в себя помещение клеток с плотностью 1×106 клеток/см2 в F12/DMEM (1:1, объем/объем), дополненную с помощью 2% В27, 20 нг/мл основного фактора роста фибробластов и 10 нг/мл эпидермального фактора роста. Приблизительно 50% среды можно заменять каждые 2-3 дня в течение продолжительности культивирования. Согласно некоторым вариантам осуществления условия, подходящие для размножения стволовых и/или плюрипотентных клеток, предусматривают культивирование клеток в B27-LIF (т.е. бессывороточная среда, содержащая LIF (1×103 Ед/мл, Chemicon; № по кат.: ESG1107 EMD Millipore, Биллерика, Массачусетс) и добавку В27 (№ по кат.: 0080085-SA; Invitrogen; Гранд Айленд, Нью-Йорк), как описано в Hitoshi, S et al. Genes & development 2004 18, 1806-1811; которая полностью включена посредством ссылки в настоящий документ. Другие среды, подходящие для культивирования клеток, описанных в настоящем документе, описаны в примерах в настоящем документе, например, среда культивирования для формирования ES, 2i, 3i и АСТН, условие культивирования ES, ES-LIF, условие культивирования эмбриональных нейрональных стволовых клеток и условие культивирования EpiSC. Согласно некоторым вариантам осуществления условия размножения или содержания плюрипотентных клеток могут включать в себя культивирование клеток в присутствии LIF (ингибирующего лейкоз фактора).
Во время размножение плюрипотентная клетка, созданная согласно описанным в настоящем документе способам, будет продолжать экспрессировать одинаковый(е) маркер(ы) плюрипотентной стволовой клетки. Неограничивающие примеры маркеров плюрипотентных стволовых клеток включают в себя SSEA-1, SSEA-2, SSEA-3, SSEA-4 (которые обобщенно в настоящем документе называются SSEA), АР, антиген Е-кадгерина, Oct4, Nanog, Ecat1, Rex1, Zfp296, GDF3, Dppa3, Dppa4, Dppa5, Sox2, Esrrb, Dnmt3b, Dnmt31, Utf1, Tcl1, Bat1, Fgf4, Neo, Cripto, Cdx2 и Slc2a3. Способы определения, экспрессирует ли клетка маркер плюрипотентных стволовых клеток, хорошо известны специалисту в настоящей области техники и включают в себя, например, ОТ-ПЦР, применение конструктов репортерных генов (например, экспрессия конструкта Oct4-GFP, описанного в настоящем документе, объединенная с FACS или флуоресцентной микроскопией), и FACS или флуоресцентную микроскопию с использованием антител, специфических в отношении представляющих интерес маркеров клеточной поверхности.
Маркеры плюрипотентных клеток также включают в себя удлиненные теломеры по сравнению с клетками. Длину теломеры можно определить, например, путем выделения геномной ДНК, обработки гДНК с помощью рестрикционных ферментов, таких как Hinf1 и Rsa1, и обнаружения теломер с помощью реагента для анализа длины теломер. Такие реагенты известны в настоящей области техники и являются коммерчески доступными, например, набор TELOMERE LENGTH ASSAY TELOTAGGG™ (№ по кат.. 12209136001 Roche; Индианаполис, Индиана).
Согласно некоторым вариантам осуществления клетку, обработанную согласно описанным в настоящем документе способам, можно изменить так, чтобы она больше напоминала эпигенетическое состояние эмбриональной стволовой клетки по сравнению с этой же клеткой до обработки согласно раскрытым способам. Эпигенетическое состояние клетки относится к химической маркировке генома в противоположность изменениям в нуклеотидной последовательности генома. Виды эпигенетической маркировки могут включать в себя метилирование ДНК (импринты), а также метилирование и ацетилирование белков, ассоциированных с ДНК, таких как гистонов. Термин 'метилирование ДНК' относится к добавлению метильной (СН3) группы к конкретному основанию в ДНК. У млекопитающих метилирование происходит почти исключительно в 5'-положении на цитозине, когда за ним следует гуанин (CpG). Согласно некоторым вариантам осуществления эпигенетическое состояние может содержать эпигенетические паттерны метилирования, например, паттерны метилирования ДНК. Анализы для определения присутствия и расположения эпигенетических маркировок известны в настоящей области техники и могут включать в себя бисульфитное секвенирование, например, как описано в примере 2 в настоящем документе. Кратко, ДНК обрабатывают с помощью набора для модификации ДНК CpGenome™ DNA Modification Kit (Chemicon, Темекула, Калифорния) и представляющие интерес области (например, гены Nanog и Oct4) амплифицируют и секвенируют.
Некоторые аспекты описанной в настоящем документе технологии относятся к анализам с использованием плюрипотентной стволовой клетки, полученной с помощью описанных в настоящем документе способов. Например, плюрипотентную стволовую клетку, полученную с помощью описанных в настоящем документе способов, можно использовать для скрининга и/или идентификации средств, которые модулируют жизнеспособность, дифференцировку или размножение плюрипотентных стволовых клеток. Такие анализы могут предусматривать приведение в контакт плюрипотентной клетки, полученной согласно способам, описанным в настоящем документе, с кандидатным средством и определение того, отличается ли жизнеспособность, дифференцировка и/или размножение плюрипотентной клетки, контактирующей с кандидатным средством, от жизнеспособности, дифференцировки и/или размножения плюрипотентной клетки, не контактирующей с кандидатным средством. Согласно некоторым вариантам осуществления средство может увеличивать жизнеспособность, дифференцировку и/или размножение плюрипотентной стволовой клетки. Согласно некоторым вариантам осуществления средство может уменьшать жизнеспособность, дифференцировку и/или размножение плюрипотентной стволовой клетки. Согласно некоторым вариантам осуществления плюрипотентная стволовая клетка может контактировать с многочисленными кандидатными средствами, например, для определения синергических или антагонистических эффектов или для скрининга кандидатных средств в пулах.
Кандидатное средство идентифицируют как средство, которое модулирует жизнеспособность полученной плюрипотентной клетки, если число плюрипотентных клеток, которые являются жизнеспособными, т.е. живыми, является выше или ниже в присутствии кандидатного средства по сравнению с его отсутствием. Способы определения жизнеспособности клетки хорошо известны в настоящей области техники и включают в себя, в качестве неограничивающего примера, определение количества жизнеспособных клеток по меньшей мере в двух временных точках, путем обнаружения силы сигнала от маркера живой клетки, или количества или пропорции клеток, окрашенных с помощью маркера живых клеток. Маркеры живых клеток являются коммерчески доступными, например, PRESTO BLUE™ (№ по кат. А-13261; Life Technologies; Гранд Айленд, Нью-Йорк). Кандидатное средство идентифицируют как средство, которое модулирует размножение полученной плюрипотентной клетки, если скорость размножения плюрипотентной клетки изменяется, т.е. число полученных клеток-потомства в данный момент времени является выше или ниже в присутствии кандидатного средства. Способы определения скорости размножения клетки известны в настоящей области техники и включают в себя, в качестве неограничивающего примера, определение увеличения количества живых клеток с течением времени.
Кандидатное средство идентифицируют как средство, которое модулирует дифференцировку плюрипотентной клетки, если скорость или характер дифференцировки плюрипотентной клетки является выше или ниже в присутствии кандидатного средства. Способы определения скорости или характера дифференцировки клетки известны в настоящей области техники и включают в себя, в качестве неограничивающего примера, обнаружение маркеров или морфологии конкретной линии дифференцировки и сравнение числа клеток и/или скорости появления клеток с такими маркерами или морфологией в популяции, контактирующей с кандидатным средством, с популяцией, не контактирующей с кандидатным средством. Маркеры и морфологические характеристики различных линий дифференцировки клеток и типов зрелых клеток известны в настоящей области техники. В качестве неограничивающего примера, мезодермальные клетки отличают от плюрипотентных клеток по экспрессии актина, миозина и десмина. Хондроциты можно отличить от типов их клеток-предшественников по окрашиванию с помощью сафранина-O и/или красителей FASTGREEN™ (Fisher; Питтсбург, Филадельфия, F99). Остеоциты можно отличить от типов их клеток-предшественников по окрашиванию с помощью ализаринового красного красителя (Sigma; Сент-Луис, Миссури: № по кат. А5533).
Согласно некоторым вариантам осуществления кандидатное средство может представлять собой потенциальный ингибитор опухолевых стволовых клеток, например, описанные в настоящем документе способы можно использовать для создания плюрипотентных клеток из зрелых опухолевых клеток и использовать для скрининга в отношении средств, которые ингибируют образование и/или жизнеспособность опухолевых клеток. Описанные в настоящем документе способы также можно использовать для скрининга средств, которые уничтожают зрелые опухолевые клетки, но которые не стимулируют развитие и/или выживаемость опухолевых стволовых клеток.
Согласно некоторым вариантам осуществления плюрипотентные клетки приводят в контакт с одним или несколькими кандидатными средствами и культивируют при условиях, которые стимулируют дифференцировку до конкретной клеточной линии дифференцировки или типа зрелых клеток. Условия, подходящие для дифференцировки, известны в настоящей области техники. В качестве неограничивающего примера, условия, подходящие для дифференцировки до мезодермальной линии дифференцировки, включают в себя DMEM, дополненную 20% фетальной телячьей сывороткой (FCS), с заменой среды каждые 3 дня. В качестве дополнительного неограничивающего примера, условия, подходящие для дифференцировки до нейрональной линии дифференцировки, включают в себя помещение клеток на покрытые орнитином слайд-камеры в F12/DMEM (1:1, объем/объем), дополненной 2% В27, 10% FCS, 10 нг/мл bFGF и 20 нг/мл LEGF. Среду можно заменять каждые 3 дня.
Используемый в настоящем документе термин "кандидатное средство" относится к любому соединению, которое в норме не присутствует или не присутствует в содержаниях, подлежащих введению в клетку, ткань или субъект. Кандидатное средство можно выбрать из группы, содержащей следующее: химические средства; низкомолекулярные органические или неорганические молекулы; последовательности нуклеиновой кислоты; аналоги нуклеиновой кислоты; белки; пептиды; аптамеры; пептидомиметик, пептидное производное, пептидные аналоги, антитела; интратела; биологические макромолекулы, экстракты, полученные из биологических материалов, таких как бактерии, растения, грибы или клетки или ткани животных; встречающиеся в природе или синтетические композиции или их функциональные фрагменты. Согласно некоторым вариантам осуществления кандидатное средство представляет собой любое химическое соединение или фрагмент, включая в себя без ограничение синтетические и встречающиеся в природе небелковые соединения. Согласно определенным вариантам осуществления кандидатное средство представляет собой низкомолекулярную молекулу, содержащую химический фрагмент. Например, химические фрагменты включают в себя незамещенные или замещенные алкильные, ароматические или гетероциклические фрагменты, включая в себя макролиды, лептомицины и родственные природные продукты или их аналоги. Кандидатные средства, как известно, могут характеризоваться требуемой активностью и/или свойством или их можно выбрать из библиотеки разнообразных соединений.
Кандидатные средства можно подвергнуть скринингу в отношении их способности модулировать жизнеспособность, размножение и/или дифференцировку плюрипотентной клетки. Согласно одному варианту осуществления кандидатные средства подвергают скринингу с использованием анализов на жизнеспособность, дифференцировку и/или размножение, описанных выше и в примерах в настоящем документе.
Как правило, соединения можно исследовать в любой концентрации, которая может модулировать клеточную функцию, экспрессию генов или активность белка относительно контроля с течением соответствующего периода времени. Согласно некоторым вариантам осуществления соединения исследуют в концентрациях в диапазоне от приблизительно 0,1 нМ до приблизительно 1000 мМ. Согласно одному варианту осуществления соединение исследуют в диапазоне от приблизительно 0,1 мкМ до приблизительно 20 мкМ, от приблизительно 0,1 мкМ до приблизительно 10 мкМ или от приблизительно 0,1 мкМ до приблизительно 5 мкМ.
В зависимости от конкретного варианта осуществления подлежащего осуществлению на практике, кандидатные или исследуемые средства можно предоставить в свободном виде в растворе или их можно прикрепить к носителю или твердой подложке, например, сферам. Ряд подходящих твердых подложек можно использовать для иммобилизации исследуемых средств. Примеры подходящих твердых подложек включают в себя агарозу, целлюлозу, декстран (коммерчески доступный, например, в виде сефадекса, сефарозы), карбоксиметилцеллюлозу, полистирол, полиэтиленгликоль (ПЭГ), фильтровальную бумагу, нитроцеллюлозу, ионообменные смолы, полимерные пленки, сополимер полиаминметилвинилового эфира и малеиновой кислоты, стеклянные бусины, аминокислотный сополимер, сополимер этилена и малеиновой кислоты, нейлон, шелк и т.д. Кроме того, для описанных в настоящем документе способов исследуемые средства можно подвергнуть скринингу индивидуально или в группах или пулах. Групповой скрининг является особенно применимым, если предполагают, что коэффициенты успеха для эффективных исследуемых средств будут низкими, так что не предполагают больше одного положительного результата для данной группы.
Способы разработки библиотек низкомолекулярных молекул, полимерных и основанных на геноме библиотек описаны, например, в Ding, et al. J Am. Chem. Soc. 124: 1594-1596 (2002) и Lynn, et al., J. Am. Chem. Soc. 123: 8155-8156 (2001). Коммерчески доступные библиотеки соединений можно получить, например, от ArQule (Уоберн, Массачусетс), Invitrogen (Карлсбад, Калифорния), Ryan Scientific (Маунт Плезант, Южная Каролина) и Enzo Life Sciences (Фармингдейл, Нью-Йорк). Указанные библиотеки можно подвергнуть скринингу в отношении способности представителей модулировать жизнеспособность, размножение и/или дифференцировку плюрипотентных стволовых клеток. Кандидатные средства могут представлять собой встречающиеся в природе белки или их фрагменты. Такие кандидатные средства можно получить из природного источника, например, клеточного или тканевого лизата. Библиотеки полипептидных средств также можно получить, например, из библиотеки кДНК, коммерчески доступной или созданной рутинными способами. Кандидатные средства также могут представлять собой пептиды, например, пептиды, составляющие от приблизительно 5 до приблизительно 30 аминокислот, при этом от приблизительно 5 до приблизительно 20 аминокислот являются предпочтительными и от приблизительно 7 до приблизительно 15 являются особенно предпочтительными. Пептиды могут представлять собой продукт ферментативной обработки встречающихся в природе белков, статистических пептидов или "смещенных" статистических пептидов. Согласно некоторым способам кандидатные средства представляют собой полипептиды или белки. Пептидные библиотеки, например, комбинаторные библиотеки пептидов или других соединений могут являться полностью рандомизированными, без эталонов или постоянных признаков последовательностей в любом положении. Альтернативно, библиотека может быть смещенной, т.е., некоторые положения в пределах последовательности либо сохраняются постоянными, либо их выбирают из ограниченного числа возможностей. Например, в некоторых случаях нуклеотиды или аминокислотные остатки рандомизируют в пределах заданного класса, например, гидрофобных аминокислот, гидрофильных остатков, стерически смещенных (либо небольших, либо больших) остатков, в направлении образования цистеинов, для перекрестных сшивок, пролинов для доменов SH-3, серинов, треонинов, тирозинов или гистидинов для сайтов фосфорилирования или до пуринов.
Кандидатные средства также могут представлять собой нуклеиновые кислоты. Нуклеиновокислотные кандидатные средства могут представлять собой встречающиеся в природе нуклеиновые кислоты, статистические нуклеиновые кислоты или "смещенные" статистические нуклеиновые кислоты. Например, продукты ферментативной обработки прокариотических или эукариотических геномов можно использовать сходным с представленным выше описанием для белков.
Согласно некоторым вариантам осуществления кандидатное средство, которое подвергают скринингу и идентифицируют в отношении модуляции жизнеспособности, размножения и/или дифференцировки плюрипотентной клетки согласно описанным в настоящем документе способам, может увеличивать жизнеспособность, размножение и/или дифференцировку плюрипотентной клетки по меньшей мере на 5%, предпочтительно по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, в 1 раз, в 1,1 раза, в 1,5 раза, в 2 раза, в 3 раза, в 4 раза, в 5 раз, в 10 раз, в 50 раз, в 100 раз или больше относительно необработанного контроля. Согласно некоторым вариантам осуществления кандидатное средство, которая подвергают скринингу и идентифицируют в отношении модуляции жизнеспособности, размножения и/или дифференцировки плюрипотентной клетки согласно описанным в настоящем документе способам, может уменьшать жизнеспособность, размножение и/или дифференцировку плюрипотентной клетки по меньшей мере на 5%, предпочтительно по меньшей мере на 10%, 20%, 30%, 40%, 50%, 50%, 70%, 80%, 90%, 95%, 97%, 98%, 99% или больше, вплоть до и включая в себя полное сокращение (т.е. нулевую жизнеспособность, рост, размножение или дифференцировку) относительно необработанного контроля.
Согласно некоторым вариантам осуществления кандидатное средство функционирует напрямую в форме, в которой его вводят. Альтернативно, кандидатное средство можно модифицировать или использовать внутриклеточно для получения формы, которая модулирует требуемую активность, например, введение последовательности нуклеиновой кислоты в клетку и его транскрипция, приводящая к получению ингибитора или активатора экспрессии генов или активности белка внутри клетки.
Предусмотрено, что описанные в настоящем документе способы и композиции можно использовать, например, в разработке противораковых вакцин. Создание по меньшей мере частично дифференцированного потомства плюрипотентных опухолевых клеток, полученных согласно представленному в настоящем документе описанию (например, путем обработки зрелой опухолевой клетки согласно описанным в настоящем документе способам), может обеспечить разнообразный и изменяющийся антигенный профиль, который может обеспечить возможность разработки более действенных противораковых вакцин на основе АПК (антигенпредставляющих клеток).
Согласно некоторым вариантам осуществления описанные в настоящем документе способы относятся к увеличении эффективности трансформации клетки. Подвергая клетки воздействию стресса, например, индуцируя плюрипотентность согласно представленному в настоящем документе описанию, можно сделать клетки более восприимчивыми к способам генетической модификации, включая в себя без ограничения трансгенную вставку, вирусные векторы и/или эндонуклеазы «цинковые пальцы». Предусмотрено, что описанные в настоящем документе способы могут обеспечить возможность модифицировать клетки до генетически восприимчивого состояния, так чтобы голую ДНК можно было использовать для трансформации полученных плюрипотентных клеток.
Некоторые аспекты описанной в настоящем документе технологии относятся к способам клеточной терапии, предусматривающим введение плюрипотентной клетки, полученной с помощью описанных в настоящем документе способов, или по меньшей мере частично дифференцированного потомства такой клетки субъекту, нуждающемуся в клеточной терапии. Согласно некоторым вариантам осуществления предусмотрено терапевтически эффективное количество плюрипотентных клеток или по меньшей мере частично дифференцированное потомство плюрипотентной клетки. Согласно некоторым вариантам осуществления плюрипотентные клетки и/или их потомство являются аутологичными. Согласно некоторым вариантам осуществления плюрипотентные клетки и/или их потомство являются аллогенными. Согласно некоторым вариантам осуществления плюрипотентные клетки и/или их потомство являются аутологичными. Согласно некоторым вариантам осуществления плюрипотентные клетки и/или их потомство являются HLA-совпадающими аллогенными. Согласно некоторым вариантам осуществления плюрипотентные клетки и/или их потомство являются сингенными. Согласно некоторым вариантам осуществления плюрипотентные клетки и/или их потомство являются ксеногенными. Согласно некоторым вариантам осуществления клеточная терапия может представлять собой аутологичную терапию, например, клетку от субъекта можно использовать для создания плюрипотентной клетки согласно описанным в настоящем документе способам, и плюрипотентную клетку и/или по меньшей мере частично дифференцированное потомство такой плюрипотентной клетки можно вводить субъекту. Используемый в настоящем документе термин "нуждающийся в клеточной терапии субъект" относится к субъекту, которого диагностировали как характеризующегося или подверженного риску наличия или развития заболевания, ассоциированного с недостаточностью встречающегося в природе типа клеток или ткани или встречающейся в природе плюрипотентной и/или мультипотентной клетки (например, стволовой клетки).
Согласно некоторым вариантам осуществления описанные в настоящем документе способы можно использовать для лечения генетических нарушений, например, Тея-Сакса или гемофилии, например, путем введения аллогенных плюрипотентных клеток и/или их потомства, полученного согласно представленному в настоящем документе описанию.
Согласно одному аспекту в настоящем документе представлен способ получения клетки или ткани, которые совместимы с клеточной терапией, подлежащей введению субъекту, предусматривающий: создание плюрипотентной клетки (или более плюрипотентной клетки) из клетки согласно описанным в настоящем документе способам, причем клетка представляет собой аутологичную клетку совпадающую по HLA аллогенную клетку. Согласно некоторым вариантам осуществления плюрипотентную клетку (или более плюрипотентную клетку) можно дифференцировать по заданной линии дифференцировки клеток перед введением клетки или ткани субъекту.
Плюрипотентные клетки, например, плюрипотентные стволовые клетки, созданные согласно описанным в настоящем документе способам можно использовать в противораковой терапии. Например, высокодозная химиотерапия вместе с трансплантацией гемопоэтических стволовых клеток для регенерации гемопоэтической системы костного мозга могут извлечь полезный эффект из использования плюрипотентных клеток, созданных согласно представленному в настоящем документе описанию.
Неограничивающие примеры заболеваний, ассоциированных с недостаточностью встречающегося в природе типа клеток или тканей или встречающейся в природе плюрипотентной и/или мультипотентной клетки, включают в себя следующее: апластическая анемия, анемия Фанкони и пароксизмальная ночная гемоглобинурия (PNH). Другие заболевания включают в себя, например, следующее: острые лейкозы, включая в себя острый лимфобластный лейкоз (ALL), острый миелогенный лейкоз (AML), острый бифенотипический лейкоз и острый недифференцированный лейкоз; хронические лейкозы, включая в себя хронический миелогенный лейкоз (CML), хронический лимфоцитарный лейкоз (CLL), ювенильный хронический миелогенный лейкоз (JCML) и ювенильный миеломоноцитарный лейкоз (JMML); миелопролиферативные нарушения, включая в себя острый миелофиброз, ангиогенную миелоидную метаплазию (миелофиброз), истинную полицитемию и эссенциальную тромбоцитемию; лизосомные болезни накопления, включая в себя мукополисахаридозы (MPS), синдром Гурлер (MPS-IH), синдром Шейе (MPS-IS), синдром Гунтера (MPS-II), синдром Санфилиппо (MPS-III), синдром Моркио (MPS-IV), синдром Марото-Лами (MPS-VI), синдром Слая, недостаточность бета-глюкуронидазы (MPS-VII), адренолейкодистрофию, муколипидоз II (заболевание I-клеток), болезнь Краббе, болезнь Гоше, болезнь Ниманна-Пика, болезнь Вольмана и метохроматическую лейкодистрофию; гистиоцитозы, включая в себя семейный эритрофагоцитарный лимфогистиоцитоз, гистиоцитоз-Х и гемофагоцитоз; фагоцитарные нарушения, включая в себя синдром Чедиака-Хигаши, хроническое грануломатозное заболевание, недостаточность актина нейтрофилов и ретикулярную дисгенезию; наследственные тромоцитарные аномалии, включая в себя амегакариоцитоз/врожденную тромбоцитопению; нарушения плазматических клеток, включая в себя множественную миелому, плазмоклеточный лейкоз и макроглобулинемию Вальденстрема. Другие злокачественные опухоли, подлежащие лечению с помощью видов терапии стволовыми клетками, включают в себя без ограничения злокачественную опухоль молочной железы, саркому Юинга, нейробластому и почечно-клеточную карциному, среди прочего. Кроме того, подлежащими лечению с помощью терапии стволовыми клетками являются следующие нарушения: нарушения в легких, включая в себя COPD и бронхиальную астму; врожденные иммунные нарушения, включая в себя атаксию-телеангиэктазию, синдром Костаманна, недостаточность адгезии лейкоцитов, синдром ДиДжорджи, синдром «голых» лимфоцитов, синдром Оменна, тяжелый комбинированный иммунодефицит (SCID), SCID с недостаточностью аденогзиндеаминазы, SCID с отсутствием Т- и В-клеток, отсутствие Т-клеток, SCID с нормальными В-клетками, вариабельный неклассифицируемый иммунодефицит и связанное с Х-хромосомой лимфопролиферативное нарушение; другие наследственные нарушения, включая в себя синдром Леш-Нихана, хрящево-волосяную гипоплазию, тромбастению Гланцманна и остеопетроз; неврологические состояния, включая в себя острый и хронический инсульт, травматическое повреждение головного мозга, церебральный паралич, рассеянный склероз, амиотрофический латеральный склероз и эпилепсию; кардиологические состояния, включая в себя атеросклероз, врожденную сердечную недостаточность и инфаркт миокрада; метаболические нарушения, включая в себя сахарный диабет; и нарушения зрения, включая в себя макулярную дегенерацию и атрофию зрительного нерва. Такие заболевания или нарушения можно лечить или путем введения плюрипотентных клеток самих по себе, обеспечивая in vivo дифференцировку до требуемого типа клеток с введением или без введения средств для стимуляции требуемой дифференцировки, и/или путем введения плюрипотентных клеток, дифференцированных до или по меньшей мере частично дифференцированных в направлении требуемого типа клеток in vitro. Способы диагностики таких состояний хорошо известны клиническим врачам, квалифицированным в настоящей области техники. Согласно некоторым вариантам осуществления субъект может представлять собой субъекта, получившего лечение с помощью лучевой терапии или других видов терапии, которые подвергали абляции популяцию клеток или стволовых клеток, например, субъект может представлять собой субъект со злокачественной опухолью, чей костный мозг был подвергнут абляции с помощью лучевой терапии.
Согласно некоторым вариантам осуществления плюрипотентные клетки вводят субъекту. Согласно некоторым вариантам осуществления по меньшей мере частично дифференцированную клетку вводят субъекту. Согласно некоторым вариантам осуществления способ клеточной терапии может дополнительно предусматривать дифференцировку плюрипотентной клетки по заданной линии дифференцировки клеток перед введением клетки. Способы дифференцировки плюрипотентной клетки по заданным линиям дифференцировки клеток известны в настоящей области техники и примеры описаны в настоящем документе.
Согласно некоторым вариантам осуществления композицию, содержащую плюрипотентную клетку, полученную согласно описанным в настоящем документе способам, или по меньшей мере частично дифференцированную клетку, которая представляет собой потомство плюрипотентной клетки, вводят субъекту.
Согласно некоторым вариантам осуществления композиция, содержащая плюрипотентную клетку, полученную согласно описанным в настоящем документе способам, или по меньшей мере частично дифференцированную клетку, которая представляет собой потомство плюрипотентной клетки, может необязательно дополнительно содержать G-CSF, GM-CSF и/или M-CSF, и/или ее можно вводить субъекту, которому ввели или будут вводить G-CSF, GM-CSF и/или M-CSF в отдельной композиции. Введение G-CSF, GM-CSF и/или M-CSF, например, может индуцировать состояние воспаления, благоприятное для регенерации органов и удаления тканевого детрита, запавшей или наслоившейся ткани.
Согласно некоторым вариантам осуществления введение плюрипотентных клеток и/или их по меньшей мере частично дифференцированного потомства может происходить в пределах относительно короткого периода времени после получения плюрипотентной клетки в культуре согласно описанным в настоящем документе способам (например, через 1, 2, 5, 10, 24 или 48 часов после получения). Согласно некоторым вариантам осуществления введение по меньшей мере частично дифференцированного потомства может происходить в пределах относительно короткого периода времени после дифференцировки плюрипотентной клетки в культуре согласно описанным в настоящем документе способам (например, через 1, 2, 5, 10, 24 или 48 часов после получения). Согласно некоторым вариантам осуществления плюрипотентные клетки и/или их по меньшей мере частично дифференцированное потомство можно криоконсервировать перед введением.
Согласно некоторым аспектам описанная в настоящем документе технология относится к композиции, содержащей плюрипотентную клетку, созданную согласно описанным в настоящем документе способам, и/или по меньшей мере частично дифференцированное потомство плюрипотентной клетки. Согласно некоторым вариантам осуществления фармацевтическая композиция содержит плюрипотентную клетку, созданную согласно описанным в настоящем документе способам, и/или по меньшей мере частично дифференцированное потомство плюрипотентной клетки и необязательно фармацевтически приемлемый носитель. Композиции могут дополнительно содержать по меньшей мере одно фармацевтически приемлемое вспомогательное вещество.
Фармацевтическая композиция может включать в себя подходящие вспомогательные вещества или стабилизаторы и может представлять собой, например, растворы, суспензии, гели или эмульсии. Как правило, композиция будет содержать от приблизительно 0,01 до 99 процентов, предпочтительно от приблизительно 5 до 95 процентов клеток, вместе с носителем. Клетки, будучи комбинированными с фармацевтически или физиологически приемлемыми носителями, вспомогательными веществами или стабилизатором, можно вводить парентерально, подкожно, с помощью имплантата или инъекции. Для большинства терапевтических целей клетки можно вводить посредством инъекции в виде раствора или суспензии в жидкой форме. Термин "фармацевтически приемлемый носитель" относится к носителю для введения плюрипотентной клетки, созданной согласно описанным в настоящем документе способам, и/или по меньшей мере частично дифференцированного потомства плюрипотентной клетки. Такие носители включают в себя без ограничения физиологический раствор, забуференный физиологический раствор, декстрозу, воду, глицерин и их комбинации. Каждый носитель должен являться "приемлемым" в смысле совместимости с другими ингредиентами состава, например, носитель не уменьшает воздействие средства на субъекта. Иными словами, носитель является фармацевтически инертным и совместимым с живыми клетками.
Подходящие составы также включают в себя водные и неводные стерильные инъекционные растворы, которые могут содержать антиоксиданты, буферы, бактериостатические средства, бактерицидные антибиотики и растворенные вещества, которые делают состав изотоническим по отношению к жидкостям организма предусмотренного реципиента. Водные и неводные стерильные суспензии могут включать в себя суспендирующие средства и загустители. Составы могут быть представлены в контейнерах для однократного или многократного приема.
Примеры парентеральных лекарственных форм включают в себя без ограничения готовые для инъекции растворы, готовые для инъекции суспензии и эмульсии. Парентеральные лекарственные формы можно получить, например, с использованием саморассасывающихся каркасных материалов для удержания плюрипотентных клеток, созданных согласно описанным в настоящем документе способам, и/или по меньшей мере частично дифференцированного потомства плюрипотентной клетки.
Термин 'эпигенетическая модификация' относится к химической маркировке генома. Эпигенетические метки могут включать в себя метилирование ДНК (импринты), а также метилирование и ацетилирование белков, ассоциированных с ДНК, таких как гистоны. Экспрессия гена, специфического для родителя, от которого он происходит (или из материнской, или из отцовской хромосомы) часто наблюдается у млекопитающих и обусловлена эпигенетическими модификациями. В родительских зародышевых линиях эпигенетическая модификация может приводить к стабильному сайленсингу или активации генов.
Используемый в настоящем документе термин "вводить" или "трансплантировать" относится к помещению клеток в организм субъекта с помощью способа или пути, которые приводят по меньшей мере к частичной локализации клеток в требуемом месте так, чтобы производился требуемый эффект.
Описанные в настоящем документе плюрипотентные стволовые клетки и/или их по меньшей мере частично дифференцированное потомство можно вводить любым способом, который считается приемлемым лечащим врачом и может включать в себя локальное введение, например, с помощью инъекции суспензии клеток или, например, с помощью имплантации препарата клеток, размещенного или выращенного на или в пределах имплантируемого каркаса или подложки. Имплантируемые каркасы могут включать в себя любое число разлагаемых или рассасывающихся полимеров, или, например, шелковый каркас, среди прочего. Подходящие пути введения фармацевтической композиции, содержащей плюрипотентные стволовые клетки, описанные в настоящем документе, и/или их по меньшей мере частично дифференцированное потомство, включают в себя без ограничения локальное введение, например, интраперитонеальное, парентеральное, внутриполостное или подкожное введение. Используемые в настоящем документе фразы "парентеральное введение" и "введенный парентерально" относятся к способам введения, отличным от энтерального и местного введения, как правило, с помощью инъекции, и включают в себя без ограничения интраперитонеальную, интрадермальную, подкожную инъекцию и инфузию. Введение может предусматривать применение игл, катетеров и шприцов, подходящих для инъекции, или хирургической имплантации. Применение комбинации средств доставки и мест доставки предусмотрено для достижения требуемого клинического эффекта.
Термин 'эпигенетическая модификация' относится к химической маркировке генома. Эпигенетические метки могут включать в себя метилирование ДНК (импринты), а также метилирование и ацетилирование белков, ассоциированных с ДНК, таких как гистоны. Экспрессия гена, специфического для родителя, от которого он происходит (или из материнской, или из отцовской хромосомы) часто наблюдается у млекопитающих и обусловлена эпигенетическими модификациями. В родительских зародышевых линиях эпигенетическая модификация может приводить к стабильному сайленсингу или активации генов.
Согласно одному варианту осуществления субъекту вводят терапевтически эффективное количество плюрипотентных стволовых клеток, описанных в настоящем документе, и/или их по меньшей мере частично дифференцированное потомство. "Терапевтически эффективный количество" представляет собой количество описанных в настоящем документе плюрипотентных стволовых клеток и/или их по меньшей мере частично дифференцированного потомства, достаточное для получения измеряемого улучшения симптома или маркера состояния, подлежащего лечению. Фактические уровни дозировки клеток в терапевтической композиции могут варьировать так, чтобы вводить количество клеток, которое является эффективным для достижения требуемого терапевтического ответа для конкретного субъекта. Выбранный уровень дозировки будет зависеть от разнообразных факторов, включая в себя без ограничения активность терапевтической композиции, состав, путь введения, комбинацию с другими лекарственными средствами или видами лечения, тяжесть состояния, подлежащего лечению, физическое состояние субъекта, предыдущую медицинскую историю субъекта, подлежащего лечению, и опыт и решение лечащего врача или специалиста-практика, вводящего терапию. Как правило, доза и схема введения должны являться достаточными для того, чтобы привести к замедлению и предпочтительно ингибированию прогрессирования состояния, а также предпочтительно вызывая уменьшение одного или нескольких симптомов или маркеров состояния. Определение и регулирование терапевтически эффективной дозы, а также оценка того, когда и как производить такие регулирования, известны специалистам в настоящей области медицины.
Дозировку описанных в настоящем документе плюрипотентных стволовых клеток и/или их по меньшей мере частично дифференцированного потомства, введенных согласно описанным в настоящем документе способам, может определить лечащий врач и отрегулировать при необходимости так, чтобы она соответствовала наблюдаемым эффектам лечения. В отношении длительности и частоты лечения, как правило, квалифицированные клинические врачи осуществляют мониторинг субъектов для определения того, когда лечение обеспечивает терапевтический эффект, и для определения, следует ли вводить следующую дозу клеток, увеличить или уменьшить дозировку, прервать лечение, возобновить лечение или произвести другое изменение в режиме лечения. Если предполагают, что введенные клетки внедрятся и приживутся в среде на длительное время, могут потребоваться повторные дозировки. Тем не менее, введение можно повторить при необходимости и в случае, если это переносится субъектом. Дозировка не должна быть настолько большой, чтобы вызвать значительные неблагоприятные побочные эффекты. Кроме того, в случае любого осложнения лечащий врач отдельно сможет отрегулировать дозировку. Как правило, тем не менее, дозировка может находиться в диапазоне от 100 до 1×109 плюрипотентных стволовых клеток согласно представленному в настоящем документе описанию и/или их по меньшей мере частично дифференцированного потомства для взрослого человека, например, 100-10000 клеток, 1000-100000 клеток, 10000-1000000 клеток или 1000000-1×109 клеток. Эффективные дозы можно экстраполировать из кривых зависимости ответа от дозы, полученных, например, из исследуемых биоанализов или систем на животных моделях.
Терапевтические композиции, содержащая описанные в настоящем документе плюрипотентные стволовые клетки и/или их по меньшей мере частично дифференцированное потомство, полученное согласно представленному в настоящем документе описанию, необязательно исследуют в одной или нескольких соответствующих in vitro и/или in vivo животных моделях заболевания, таких как SCID мышиная модель, для подтверждения эффективности, оценки in vivo роста трансплантированных клеток и установки дозировок, согласно способам, хорошо известным в настоящей области техники. В частности, дозировки вначале можно определить по активности, стабильности или другим подходящим параметрам лечения по сравнению с отсутствием лечения (например, сравнение получивших лечение с не получившими лечение животными моделями), в релевантном анализа. В определении эффективного количества описанных в настоящем документе плюрипотентных стволовых клеток и/или их по меньшей мере частично дифференцированного потомства, лечащий врач оценивает среди прочих критериев рост и объем трансплантированных клеток и прогрессирование состояния, подлежащего лечению. Дозировка может варьировать в зависимости от используемой формы дозировки и используемого пути введения.
В отношении описанных в настоящем документе терапевтических способов, не предусмотрено, что введение описанных в настоящем документе плюрипотентных стволовых клеток и/или их по меньшей мере частично дифференцированного потомства ограничено конкретным способом введения, дозировкой или частотой введения доз. Предусмотрены все способы введения, включая в себя внутримышечный, внутривенный, интраперитонеальный, интравезикулярный, интраартикулярный пути введения, введение в место поражения, подкожный или любой другой путь, достаточный для обеспечения дозы, адекватной для лечения состояния, подлежащего лечению.
Согласно некоторым вариантам осуществления описанные в настоящем документе способы можно использовать для создания плюрипотентных клеток in vivo, например, клетку, присутствующую в организме субъекта, можно подвергнуть действию стресса согласно представленному в настоящем документе описанию так, чтобы она приобрела плюрипотентный фенотип. Способы приложения стрессовых воздействий, описанных в настоящем документе, к клеткам in vivo являются очевидными, например, слабокислые растворы можно вводить к ткань посредством инъекции и/или прямого нанесения, температуры можно изменять с помощью зондов, которые могут нагревать или охлаждать окружающую ткань или посредством применения неинвазивных способов, например, облучение сфокусированным пучком. In vivo модуляцию плюрипотентности можно использовать, например, для увеличения регенерации ткани или заживления ран. Неограничивающие примеры могут включать в себя инъекцию слабой кислоты в артритный коленный сустав, чтобы индуцировать клетки коленного сустава (например, синовиальные или хрящевые клетки) приобретать плюрипотентный фенотип и создавать новые ткани. Дополнительный неограничивающий пример может включать в себя лечение субъекта с инсультом или повреждением центральной нервной системы (например, повреждением спинного мозга). После снятия воспаления, клетки, смежные с пораженной областью можно обработать стрессовым воздействием согласно представленному в настоящем документе описанию, создавая плюрипотентные клетки, которые могут заново населять поврежденную ткань и/или регенерировать или восстанавливать поврежденную ткань.
В дополнительном неограничивающем примере изменения в эпигенетическом статусе (например, путем обработки деметилазой) могут вызвать превращение не секретирующих инсулин клеток (например, секретирующих альфа-глюкагон клеток поджелудочной железы) в секретирующие инсулин клетки (например, бета-клетки). Соответственно, обработка не секретирующей инсулин клетки (например, секретирующей альфа-глюкагон клетки поджелудочной железы) в соответствии с описанными в настоящем документе способами может дать в результате клетку, становящуюся секретирующей инсулин клеткой, например, бета-подобной клеткой, либо in vivo, либо in vitro.
Кроме того, предусмотрено, что описанные в настоящем документе плюрипотентные клетки могут сливаться с другими клетками (т.е. "клетками-реципиентами"), например, клетками, не обработанными согласно описанным в настоящем документе способам, неплюрипотентными клетками, зрелыми клетками, злокачественными клетками и/или поврежденными клетками. Слияние клеток может привести к увеличенному уровню экспрессии фермента клеточной репарации и/или активности клетки-реципиента по сравнению с состоянием перед слиянием. Это может увеличивать здоровье и/или функцию клетки-реципиента, например, путем увеличения репарации клеточного повреждения, мутаций и/или модификации эпигенетического статуса клетки-реципиента.
Согласно некоторым вариантам осуществления путем увеличения плюрипотентности клеток in vivo можно модулировать эпигенетические маркеры (например, статус метилирования, деметилирования и/или гидроксиметилирования ДНК) указанных клеток. Обнаружили, что модуляция эпигенетических маркеров вовлечена, например, в развитие злокачественной опухоли, артрита, аутоиммунного заболевания, старения и т.д., и предусмотрено лечение таких эпигенетически-связанных состояний в соответствии с описанными в настоящем документе способами.
Согласно некоторым вариантам осуществления одновременно можно обработать множество тканей in vivo, например, слабокислое условие можно индуцировать в многочисленных органах, например, последовательно или синхронно (например, в головном мозге, сердце, печени, легком и/или щитовидной железе) для лечения распространенного повреждения или старения.
Кроме того, предусмотрено, что in vivo обработку клеток согласно представленному в настоящем документе описанию можно комбинировать с введением плюрипотентных клеток и/или по меньшей мере их частично дифференцированного потомства, которое получили согласно представленному в настоящем документе описанию.
В настоящем документе предусмотрено, что описанные в настоящем документе способы можно использовать для лечения, например, плода или эмбриона в матке.
Эффективность лечения можно оценить, например, путем измерения маркера, индикатора, симптома или частоты возникновения состояния, подлежащего лечению согласно представленному в настоящем документе описанию, или любого другого измеряемого параметра, соответствующего, например, количеству потомства плюрипотентной клетки. В пределах компетенции специалиста в настоящей области техники находится осуществления мониторинга эффективности лечения или профилактики измерения любого из таких параметров или любой комбинации параметров.
Эффективное лечение является очевидным, если существует статистически значимое улучшение одного или нескольких маркеров, индикаторов или симптомов состояния, подлежащего лечению, или имеет место невозможность ухудшения или развития симптомов там, где это предполагали в ином случае. В качестве примера, благоприятное изменение по меньшей мере приблизительно на 10% в измеряемом параметре состояния и предпочтительно по меньшей мере приблизительно на 20%, приблизительно на 30%, приблизительно на 40%, приблизительно на 50% или больше может являться показателем эффективного лечения. Эффективность для плюрипотентных клеток, созданных согласно описанным в настоящем документе способам, и/или по меньшей мере частично дифференцированного потомства плюрипотентной клетки также можно оценить с использованием экспериментальной животной модели, известной в настоящей области техники для состояния, описанного в настоящем документе. При использовании экспериментальной животной модели, эффективность лечения является очевидной, если наблюдается статистически значимое изменение маркера, например, количество гемопоэтических клеток, присутствующих в организме мыши после абляции костного мозга и лечения с помощью плюрипотентных клеток согласно представленному в настоящем документе описанию.
Согласно одному аспекту в настоящем документе описан способ получения плюрипотентной клетки, способной дифференцироваться в плацентарную клетку, причем способ предусматривает культивирование плюрипотентной клетки, полученной согласно описанным в настоящем документе способам, в присутствии FGF4. Согласно некоторым вариантам осуществления плюрипотентная клетка способна дифференцироваться в эмбриональную стволовую клетку. Согласно некоторым вариантам осуществления концентрация FGF4 составляет от приблизительно 1 нМ до приблизительно 1 мкМ. Согласно некоторым вариантам осуществления концентрация FGF4 составляет от 1 нМ до 1 мкМ. Согласно некоторым вариантам осуществления концентрация FGF4 составляет от приблизительно 5 нМ до приблизительно 500 нМ. Согласно некоторым вариантам осуществления концентрация FGF4 составляет от приблизительно 10 нМ до приблизительно 100 нМ.
Согласно некоторым аспектам описанная в настоящем документе технология относится к системе создания плюрипотентной клетки из клетки, содержащая удаление части цитоплазмы и/или митохондрий из клетки.
Система для создания плюрипотентной клетки из клетки согласно описанным в настоящем документе способам может содержать контейнер, в котором клетки подвергают воздействию стресса. Контейнер может являться подходящим для культивирования соматических и/или плюрипотентных клеток, как, например, в случае, когда клетки культивируют в течение нескольких дней или дольше при условиях низкого содержания кислорода для снижения количества цитоплазмы и/или митохондрий согласно описанным в настоящем документе способам. Альтернативно, контейнер может являться подходящим для воздействия на клетки стресса, но не для культивирования клеток, как, например, в случае, когда клетки суспендируют в приборе, характеризующимся узкой щелью, в течение ограниченного периода, например, меньше чем 1 час. Контейнер может представлять собой, например, сосуд, пробирку, микроструйный прибор, пипетки, биореактор или чашку для культивирования клеток. Контейнер можно содержать в окружающей среде, которая обеспечивает условия, подходящие для культивирования соматических и/или плюрипотентных клеток (например, содержать внутри инкубатора) или в окружающей среде, которая обеспечивает условия, которые будут вызывать связанное с окружающей средой стрессовое воздействие на клетку (например, содержать внутри инкубатора, обеспечивающего окружающую среду с низким содержанием кислорода). Контейнер может характеризоваться конфигурацией, обеспечивающей воздействие 1 или несколько воздействий связанных с окружающей средой стрессовых воздействий, описанных выше в настоящем документе, например, 1 стрессового воздействия, 2 стрессовых воздействий, 3 стрессовых воздействий или больше. Контейнеры, подходящие для манипуляции и/или культивирования соматических и/или плюрипотентных клеток, хорошо известны специалисту в настоящей области техники и являются коммерчески доступными (например, № по кат. CLS430597 Sigma-Aldrich; Сент-Луис, Миссури). Согласно некоторым вариантам осуществления контейнер представляет собой микроструйный прибор. Согласно некоторым вариантам осуществления контейнер представляет собой чашку для культивирования клеток, колбу или планшет.
Согласно некоторым вариантам осуществления система дополнительно может содержать средства для выбора плюрипотентных клеток, например, система может содержать систему FACS, которая может выбирать клетки, экспрессирующие маркер плюрипотентности (например, Oct4-GFP) или выбирать по размеру, как описано выше в настоящем документе. Способы и приборы для выбора клеток хорошо известны специалисту в настоящей области техники и являются коммерчески доступными, например, прибор BD FACSARIA SORP™, объединенный с программных обеспечением BD LSRII™ и BD FACSDIVA™ (№ по кат. 643629), произведенные BD Biosciences; Франклин Лейке, Нью-Джерси.
Согласно некоторым вариантам осуществления в систему поставляют клетки, которые не присутствуют в ткани. Согласно некоторым вариантам осуществления в систему предоставляют ткани, и система дополнительно содержит средство выделения одного или нескольких типов клеток. В качестве неограничивающего примера, система может содержать тканевой гомогенизатор. Тканевые гомогенизаторы и способы их использования известны в настоящей области техники и являются коммерчески доступными (например, FASTH21™, № по кат. 21-82041 Omni International; Кеннесо, Джорджия). Альтернативно, система может содержать центрифугу для обработки образцов крови или жидкости.
Согласно некоторым вариантам осуществления система может являться автоматизированной. Способы автоматизированного выделения клеток, культивирования клеток и приборы для выбора клеток известны в настоящей области техники и являются коммерчески доступными. Например, тканевой гомогенизатор FASTH21™ (№ по кат. 21-82041 Omni International; Кеннесо, Джорджия) и BD FACSARIA SORP™.
Согласно некоторым вариантам осуществления система может являться стерильной, например, с ней можно работать в стерильной окружающей среде или с системой можно работать как с закрытой, стерильной системой.
Согласно одному аспекту в настоящем документе описан способ увеличения способности плюрипотентной клетки к самообновлению, причем способ предусматривает культивирование клетки в присутствии адренокортикотропного гормона (АСТН), среды 2i или 3i. Используемый в настоящем документе термин "способность к самообновлению" относится к продолжительности времени, в течение которого клетку можно культивировать и пересевать in vitro, например, количество пассажей, которым можно подвергнуть клетку и ее потомство и они продолжат производить жизнеспособные клетки. Клетка, у которой вызвали появление увеличенной способности к самообновлению согласно описанному в настоящем документе способу, может представлять собой, например, тотипотентную клетку и/или клетку, созданную путем воздействия на нее стресса согласно приведенному в настоящем документе описанию.
Согласно некоторым вариантам осуществления культивирование в присутствии АСТН может предусматривать культивирование клетки в клеточной среде, содержащей от приблизительно 0,1 мкМ до приблизительно 1,000 мкМ, например, от приблизительно 0,1 мкМ до приблизительно 100 мкМ, от приблизительно 0,1 мкМ до приблизительно 10 мкМ, или приблизительно 10 мкМ. Согласно некоторым вариантам осуществления культивирование клетки в присутствии АСТН может предусматривать культивирование клетки в среде LIF, содержащей АСТН. LIF, АСТН, 2i и 3i являются коммерчески доступными и хорошо известными в настоящей области техники, например, АСТН можно приобрести в Sigma-Aldrich (№ по кат. А0673; Сент-Луис, Миссури) и среды LIF можно приобрести в Millipore (например, № по кат. ESG1107; Биллерика, Массачусетс), и 3i можно приобрести в Stem Cells Inc. (например, в виде "iSTEM Stem Cell Culture Medium, № по кат. SCS-SF-ES-01; Ньюарк, Калифорния).
Согласно некоторым вариантам осуществления стадия культивирования может продолжаться в течение по меньшей мере 3 дней, например, по меньшей мере 3 дня, по меньшей мере 4 дня, по меньшей мере 5 дней, по меньшей мере 6 дней, по меньшей мере 7 дней или дольше. После стадии культивирования клетки можно содержать при условиях, подходящих для содержания плюрипотентных клеток согласно представленному в настоящем документе описанию.
Согласно некоторым вариантам осуществления после стадии культивирования клетка может экспрессировать обнаруживаемое и/или увеличенное содержание маркера стволовой клетки. Маркеры стволовых клеток и способы их обнаружения описаны в настоящем документе. Согласно некоторым вариантам осуществления маркер стволовой клетки можно выбрать из группы, состоящей из Oct3/4; Nanog; Rex1; Klf4; Sox2; Klf2; Esrr-бета; Tbx3 и Klf5.
Согласно одному аспекту в настоящем документе предусмотрены способы создания плюрипотентных клеток или клеток STAP, которые представляют собой улучшение по сравнение с предшествующими способами, например, обеспечивают увеличенную эффективность, качество и/или выход.
Согласно одному варианту осуществления в настоящем документе предусмотрен способ создания плюрипотентных клеток или клеток STAP, например, из клеточной суспензии и/или условия культивирования ткани.
В качестве первой стадии, исходную клетку (например, начальный материал) в суспензии можно осадить и/или удалить из раствора. В качестве только одного примера, клетку можно осадить путем центрифугирования в центрифужной пробирке при от приблизительно 800 об/мин до приблизительно 1600 об/мин в течение от приблизительно 1 минуты до приблизительно 20 минут. В качестве только одного примера, клетку можно осадить путем центрифугирования в центрифужной пробирке при приблизительно 1200 об/мин в течение 5 минут. Согласно некоторым вариантам осуществления клетку в суспензии можно привести в контакт с таким пищеварительным ферментом, как трипсин, перед ее осаждением и/или удалением из раствора. В качестве только одного примера, трипсин-EDTA, в концентрации от приблизительно 0,01% до приблизительно 0,5% (Gibco: 25300-054) можно добавить к чашке для культивирования ткани, содержащей клетки, в течение от приблизительно 1 до приблизительно 20 минут, для высвобождения прикрепленных клеток, которые необходимо добавить к центрифужной пробирке. В качестве только одного примера, трипсин-EDTA, 0,05% (Gibco: 25300-054) можно добавить к чашке для культивирования ткани, содержащей клетки, в течение приблизительно 3-5 минут, для высвобождения прикрепленных клеток, подлежащих добавлению к центрифужной пробирке. Согласно вариантам осуществления предусматривающим центрифугирование, супернатант можно аспирировать вплоть до клеточного осадка после центрифугирования.
Согласно некоторым вариантам осуществления первую стадию проводят на популяции, составляющей по меньшей мере 1 миллион жизнеспособных клеток. Согласно некоторым вариантам осуществления первую стадию проводят на популяции, составляющей по меньшей мере 5 миллионов жизнеспособных клеток. Согласно некоторым вариантам осуществления первую стадию проводят на популяции, составляющей по меньшей мере 10 миллионов жизнеспособных клеток.
В качестве второй стадии, клетки можно ресуспендировать в физиологическом солевом растворе, например, HBSS (сбалансированный соляной раствор Хэнка) (например, HBSS, не содержащий Са+ Mg+: Gibco 14170-112). Согласно некоторым вариантам осуществления клетку можно ресуспендировать в концентрации, составляющей от приблизительно 1×103 клеток/мл до приблизительно 1×109 клеток/мл.
Согласно некоторым вариантам осуществления клетку можно ресуспендировать в концентрации, составляющей от приблизительно 1×105 клеток/мл до приблизительно 1×107 клеток/мл. Согласно некоторым вариантам осуществления клетку можно ресуспендировать в концентрации, составляющей приблизительно 1×106 клеток/мл. Согласно некоторым вариантам осуществления клетки можно ресуспендировать в 50 мл пробирке. Согласно некоторым вариантам осуществления клетки можно ресуспендировать в 2-3 мл HBSS в 50 мл пробирке.
В качестве третьей стадии, клетку, в суспензии/растворе, можно суспендировать, например, пропустить через щель, отверстие и/или просвет, достаточно маленькие для создания, например, напряжений сдвига. Согласно иллюстративным вариантам осуществления описанным ниже в настоящем документе, щель, отверстие и/или просвет содержится в стеклянной пипетке, характеризующейся отверстием такого размера, как описано ниже в настоящем документе. Суспендирование можно осуществить с помощью ряда альтернативных средств. Неограничивающие примеры могут включать в себя щели, просветы или каналы в микроструйном приборе, приборе для манипуляций с клетками, содержащем насос и систему трубок, пропускающем клеточную суспензию через сетку или фильтр, заставляя клеточную суспензию протекать через барьеры или частицы, и подобное. Специалист в настоящей области техники может эмпирически определить соответствующие значения давления, скорости потока, напряжения сдвига и т.д. для различных систем суспендирования, основанных на настоящем раскрытии. Дополнительное обсуждение жидкостных стрессовых воздействий и расчеты, относящиеся к таким стрессовым воздействиям, можно найти, например, в Fournier "Basic Transport Phenomena in Biomedical Engineering" Taylor & Francis, 1999; которая полностью включена в настоящий документ посредством ссылки.
Согласно некоторым вариантам осуществления суспендирование может продолжаться в течение от приблизительно 10 минут до приблизительно 2 часов, например, от приблизительно 20 минут до приблизительно 1 часа или приблизительно 30 минут. Согласно некоторым вариантам осуществления суспендирование может продолжаться в течение по меньшей мере 10 минут, например, 10 минут или больше, 20 минут или больше, 30 минут или больше, 40 минут или больше, 50 минут или больше или 60 минут или больше. Согласно некоторым вариантам осуществления суспендирование может продолжаться до тех пор, пока суспензия не будет легко суспендироваться через отверстие или просвет. Согласно некоторым вариантам осуществления суспендирование в последней щели или просвете можно продолжать до тех пор, пока суспензия не буде легко проходить через щель или просвет. Согласно некоторым вариантам осуществления суспендирование в каждой щели или просвете можно продолжать до тех пор, пока суспензия не будет легко проходить через данную щель или просвет.
Согласно некоторым вариантам осуществления суспендирование может предусматривать суспендирование через серию отверстий или просветов, например, серию постепенно уменьшающихся отверстий или просветов. Согласно некоторым вариантам осуществления серия отверстий или просветов содержит по меньшей мере 2 отверстия или просвета, например, 2, 3, 4, 5, 10, 20, 50 или больше отверстий или просветов. Согласно некоторым вариантам осуществления на одно или несколько отверстий или просветов можно предварительно нанести, например, HBSS или воду.
В качестве исключительно иллюстративного варианта осуществления клетки можно суспендировать через множественные, например, три отверстия или просвета. Согласно некоторым вариантам осуществления первое отверстие или просвет может характеризоваться внутренним диаметром, составляющим от приблизительно 0,5 мм до приблизительно 2,0 мм. Согласно некоторым вариантам осуществления первое отверстие или просвет может характеризоваться внутренним диаметром, составляющим от приблизительно 0,7 мм до приблизительно 1,5 mm. Согласно некоторым вариантам осуществления первое отверстие или просвет может характеризоваться внутренним диаметром, составляющим приблизительно 1,1 мм. Согласно некоторым вариантам осуществления суспендирование через первую щель или просвет можно проводить в течение от приблизительно 1 минуты до приблизительно 10 минут. Согласно некоторым вариантам осуществления суспендирование через первую щель или просвет можно проводить в течение приблизительно 5 минут. В качестве исключительно иллюстративного варианта осуществления первое отверстие или просвет находится в стандартной 9'' стеклянной пипетке (например, например, 9'' одноразовые пипетки Пастера Fisher (Fisher brand 9'' Disposable Pasteur Pipettes): 13-678-20D), и клеточную суспензию можно пипетировать в пипетку и из пипетки в течение 5 минут с помощью достаточного значения силы для диссоциации клеточных агрегатов и всего сопутствующего дебриса.
Согласно некоторым вариантам осуществления последние две щели или просвета в серии могут характеризоваться внутренними диаметрами, составляющими от приблизительно 90 до приблизительно 200 мкм и от приблизительно 25 мкм до приблизительно 90 мкм. Согласно некоторым вариантам осуществления последние две щели или просвета в серии могут характеризоваться внутренними диаметрами, составляющими от приблизительно 100 до приблизительно 150 мкм и от приблизительно 50 мкм до приблизительно 70 мкм. Согласно некоторым вариантам осуществления суспендирование может предусматривать от приблизительно 5 до приблизительно 20 минут суспендирования через от второй до последней щели или просвета и от приблизительно 5 до приблизительно 20 минут суспендирования в последней щели или просвете. Согласно некоторым вариантам осуществления суспендирование может предусматривать приблизительно 10 минут суспендирования через от второй до последней щели или просвета и приблизительно 15 минут суспендирования в последней щели или просвете.
В качестве исключительно иллюстративного варианта осуществления последние два отверстия или просвета могут быть образованы пипетками, модифицированными следующим образом: сделать две оплавленных пипетки с очень маленькими выходными отверстиями следующим образом: нагреть стандартную 9'' (22,86 см) стеклянную пипетку над горелкой Бунзена, и затем тянуть и растягивать дальний (расплавленный) конец пипетки, пока просвет не схлопнется, и конец не оторвется, оставляя закрытый, заостренный стеклянный кончик. Подождать, пока пипетка охладится, и затем отламывать закрытый дальний конец, пока нельзя будет идентифицировать очень маленький просвет. Повторить этот процесс со второй пипеткой, но отломить кончик немного более близко, получая немного больший просвет на дальнем конце. Больший просвет должен составлять приблизительно 100-150 микрон в диаметре, в то время как другая пипетка должна иметь меньший просвет приблизительно 50-70 микрон. Клеточную суспензию можно суспендировать через пипетку с большим просветом в течение 10 минут. После этого можно осуществить суспендирование через пипетку, характеризующуюся меньшим просветом (50-70 мкм), в течение дополнительных 15 минут. Продолжать суспендировать суспензию, пока она не станет легко проходить вверх и вниз по оплавленной пипетке с меньшим внутренним диаметром. Каждую пипетку можно предварительно покрыть средой. Кроме того, во время суспендирования следует избегать забора воздуха и создания пузырей или пены в суспензии клеток.
Согласно некоторым вариантам осуществления суспендирование можно проводить со скоростью, составляющей от приблизительно 1 до приблизительно 200 циклов в минуту, например, всю суспензию пропускают через щель, просвет или отверстие 1-100 раз в минуту. Согласно некоторым вариантам осуществления суспендирование можно проводить со скоростью, составляющей от приблизительно 10 до приблизительно 60 циклов в минуту. Согласно некоторым вариантам осуществления суспендирование можно проводить со скоростью, составляющей приблизительно 40 циклов в минуту. Согласно некоторым вариантам осуществления в которых для суспендирования используют пипетку, суспензию могут выпускать из и впускать обратно в пипетку приблизительно 20 раз в минуту.
На следующей стадии, суспендированные клетки можно выделить из суспензии. Согласно некоторым вариантам осуществления приблизительно 0-50 объемов HBSS можно добавить к суспендированной суспензии и суспензию центрифугируют при от приблизительно 800-1600 об/мин в течение от приблизительно 1 минуты до приблизительно 30 минут и затем супернатант отбирают с помощью аспирации. Согласно некоторым вариантам осуществления приблизительно 9 объемов HBSS можно добавить к суспендированной суспензии и суспензию центрифугируют при приблизительно 1200 об/мин в течение приблизительно 5 минут и затем супернатант отбирают с помощью аспирации.
На следующей стадии, клетки можно ресуспендировать в HBSS, при этом полученная суспензия характеризуется рН от приблизительно 5,0 до приблизительно 6,0. Согласно некоторым вариантам осуществления полученная суспензия может характеризоваться рН от приблизительно 5,4 до приблизительно 5,8. Согласно некоторым вариантам осуществления полученная суспензия может характеризоваться рН от приблизительно 5,6 до приблизительно 5,7. Согласно некоторым вариантам осуществления полученная суспензия может характеризоваться рН, составляющим приблизительно 5,6. Согласно некоторым вариантам осуществления раствор HBSS перед смешиванием с клетками может характеризоваться рН от приблизительно 5,0 до приблизительно 5,7. Согласно некоторым вариантам осуществления раствор HBSS перед смешиванием с клетками может характеризоваться рН от приблизительно 5,3 до приблизительно 5,6. Согласно некоторым вариантам осуществления раствор HBSS перед смешиванием с клетками может характеризоваться рН, составляющим приблизительно 5,4. Согласно некоторым вариантам осуществления клетки можно ресуспендировать при концентрации, составляющей от приблизительно 2×104 клеток/мл до приблизительно 2×108 клеток/мл. Согласно некоторым вариантам осуществления клетки можно ресуспендировать при концентрации, составляющей приблизительно 2×106.
В качестве исключительно иллюстративного примера, стадию ресуспендирования согласно предыдущему абзацу можно проводить следующим образом: когда подкисляете раствор, мягко суспендировать его с использованием 5 мл пипетки в течение 10 секунд сразу после добавления кислоты к раствору Хенкса. HBSS обладает очень слабой буферной емкостью, так что любой раствор, перенесенный из супернатанта предшествующей суспензии будет существенно влиять на рН HBSS. Инструкции ниже показывают, как получить HBSS с оптимальным рН, составляющим 5,6-5,7 для получения клеток STAP в соответствии с этим экспериментальным вариантом осуществления. Сначала следует титровать рН предварительно охлажденного HBSS (при 4°С) с помощью 12 н. HCl до рН 5,6. Это осуществляют посредством медленного добавления 11,6 мкл 12 н. HCl к 50 мл HBSS. После подтверждения указанного значения рН, следует стерилизовать раствор фильтрацией через шприц с фильтром 0,2 микрон или через фильтр на горле бутыли, в новый стерильный контейнер для хранения. Необходимо подтвердить конечный рН 5,6-5,7 в ходе предварительного тестового эксперимента с соответствующим количеством клеток. Поскольку рН HBSS является настолько важным, рН раствора следует проверять, повторно титровать и повторно стерилизовать перед каждым применением.
На следующей стадии, клетки в суспензии HBSS можно инкубировать приблизительно при их in vivo температуре. Например, клетки млекопитающего можно инкубировать приблизительно при 37°С. Согласно некоторым вариантам осуществления инкубацию можно проводить в течение от приблизительно 5 минут до приблизительно 3 часов. Согласно некоторым вариантам осуществления инкубацию можно проводить в течение от приблизительно 10 минут до приблизительно 1 часа. Согласно некоторым вариантам осуществления инкубацию можно проводить в течение от приблизительно 15 минут до приблизительно 40 минут. Согласно некоторым вариантам осуществления инкубацию можно проводить в течение приблизительно 25 минут.
На следующей стадии, клетки выделяют из кислого раствора HBSS. В качестве только одного примера, клетки можно осадить с помощью центрифугирования в центрифужной пробирке при от приблизительно 800 об/мин до приблизительно 1600 об/мин в течение от приблизительно 1 минуты до приблизительно 20 минут. В качестве только одного примера, клетки можно осадить с помощью центрифугирования в центрифужной пробирке при приблизительно 1200 об/мин в течение 5 минут. Согласно некоторым вариантам осуществления супернатант затем можно отобрать с помощью аспирации.
На следующей стадии, клетки можно ресуспендировать в средах, подходящих для содержания и/или выбора плюрипотентной клетки. Согласно некоторым вариантам осуществления среда представляет собой среду для образования клеточных сфер. Используемый в настоящем документе термин "среда для образования клеточных сфер" относится к DMEM/F12 с 1% антибиотиком и 2% В27 Gibco 12587-010. Согласно некоторым вариантам осуществления среда может дополнительно содержать факторы роста, например, b-FGF (20 нг/мл), EGF (20 нг/мл), гепарин (0,2%, Stem Cell Technologies 07980). Указанные факторы приводят в соответствие к используемому типу клеток. Например, согласно некоторым вариантам осуществления LIF (1000 ед.) можно добавить, если клетки являются мышиными). Согласно некоторым вариантам осуществления такие добавки, как bFGF, EGF и гепарин могут не являться обязательными. Согласно некоторым вариантам осуществления клетки можно ресуспендировать в средах при концентрации, составляющей 105 клеток/см3.
На следующей стадии, клетки можно культивировать и/или содержать, например, культивировать при 37°C с 5% СО2. Согласно некоторым вариантам осуществления клетки можно перемешивать во время культивирования/содержания для предотвращения прилипания к контейнеру для культивирования клеток. Согласно некоторым вариантам осуществления клетки можно аккуратно пипетировать с использованием, например, 5 мл пипетки, дважды в день в течение 2 минут, в течение первой недели, чтобы препятствовать их прикреплению ко дну чашек. Согласно некоторым вариантам осуществления это может способствовать хорошему формированию сфер. Согласно некоторым вариантам осуществления среду для образования клеточных сфер, необязательно содержащую добавки, можно добавлять через день. Например, добавить 1 мл/день в 10 см чашку для культивирования или 0,5 мл/день в 6 см чашку.
Согласно второму варианту осуществления в настоящем документе предусмотрен способ создания плюрипотентных клеток или клеток STAP из, например, мягкой ткани, которая может содержать эритроциты (RBC). Такие ткани могут включать в себя без ограничения печень, селезенку и легкое.
На первой стадии, мягкую ткань, (например, отрезанную, отмытую, стерильную кань органа) механически разрезают на тонкие срезы, измельчают, скоблят и/или вымачивают. Согласно некоторым вариантам осуществления указанную стадию можно проводить в присутствии пищеварительных ферментов и/или ферментов, которые расщепляют ЕСМ. Согласно некоторым вариантам осуществления фермент может представлять собой коллагеназу. В настоящем документе предусмотрено, что различные типы коллагеназы или ферментов лучше подходят для обработки тканей различных органов, на основе компонентов ЕСМ указанной ткани и соединительных тканей. Специалист в настоящей области техники сможет легко определить соответствующие ферменты для каждого типа ткани. Согласно некоторым вариантам осуществления ткань представляет собой селезенку, и нет необходимости в использовании фермента. В качестве исключительно иллюстративного примера, ткань можно измельчать и соскабливать в течение от приблизительно 1 минуты до приблизительно 30 минут с использованием скальпелей и/или ножниц для увеличения площади поверхности, которую подвергают воздействию коллагеназы, пока ткань не станет гелеобразной по консистенции. В качестве исключительно иллюстративного примера, ткань можно измельчать и соскабливать в течение от приблизительно 10 минут с использованием скальпелей и/или ножниц. Согласно некоторым вариантам осуществления можно добавить дополнительный фермент и инкубировать ткань с ферментом, необязательно при перемешивании. В качестве только одного примера, ткань можно хранить в инкубаторе/встряхивателе в течение 30 минут при 37°С при 90 об/мин. Согласно некоторым вариантам осуществления ткань можно разбавить в HBSS после воздействия фермента и/или механического разрушения.
На следующей стадии, клетку, в суспензии, можно суспендировать, например, пропускать через отверстие или просвет, достаточно маленькие, чтобы создавать, например, напряжения сдвига. Согласно некоторым вариантам осуществления суспендирование может продолжаться в течение от приблизительно 10 минут до приблизительно 2 часов, например, от приблизительно 20 минут до приблизительно 1 часа или приблизительно 30 минут. Согласно некоторым вариантам осуществления суспендирование может продолжаться в течение по меньшей мере 10 минут, например, 10 минут или больше, 20 минут или больше, 30 минут или больше, 40 минут или больше, 50 минут или больше или 60 минут или больше. Согласно некоторым вариантам осуществления суспендирование можно продолжать, пока суспензия не будет легко суспендироваться через отверстие или просвет. Согласно некоторым вариантам осуществления суспендирование в последней щели или просвете можно продолжать, пока суспензия не будет легко проходить через щель или просвет. Согласно некоторым вариантам осуществления суспендирование в каждой щели или просвете можно продолжать, пока суспензия не будет легко проходить через такую щель или просвет.
Согласно некоторым вариантам осуществления суспендирование может предусматривать суспендирование через серию отверстий или просветов, например, серия постепенно уменьшающихся отверстий или просветов. Согласно некоторым вариантам осуществления серия отверстий или просветов содержит по меньшей мере 2 отверстия или просвета, например, 2, 3, 4, 5, 10, 20, 50 или больше отверстий или просветов. Согласно некоторым вариантам осуществления одно или несколько отверстий или просветов можно предварительно покрыть, например, с помощью HBSS или водой.
В качестве исключительно иллюстративного варианта осуществления клетки можно суспендировать через три отверстия или просвета. Согласно некоторым вариантам осуществления первое отверстие или просвет может характеризоваться внутренним диаметром, составляющим от приблизительно 0,5 мм до приблизительно 2,0 мм. Согласно некоторым вариантам осуществления первое отверстие или просвет может характеризоваться внутренним диаметром, составляющим от приблизительно 0,7 мм до приблизительно 1,5 мм. Согласно некоторым вариантам осуществления первое отверстие или просвет может характеризоваться внутренним диаметром, составляющим приблизительно 1,1 мм. Согласно некоторым вариантам осуществления суспендирование через первую щель или просвет можно проводить в течение от приблизительно 1 минуты до приблизительно 10 минут. Согласно некоторым вариантам осуществления суспендирование через первую щель или просвет можно проводить в течение приблизительно 5 минут. В качестве исключительно иллюстративного варианта осуществления первое отверстие или просвет находится в стандартной 9'' стеклянной пипетке (например, 9'' одноразовые пипетки Пастера Fisher (Fisher brand 9'' Disposable Pasteur Pipettes): 13-678-20D), и клеточную суспензию можно пипетировать в пипетку и из пипетки в течение 5 минут с помощью достаточного значения силы для диссоциации клеточных агрегатов и всего сопутствующего дебриса.
Согласно некоторым вариантам осуществления последние две щели или просвета в серии могут характеризоваться внутренними диаметрами, составляющими от приблизительно 90 до приблизительно 200 мкм и от приблизительно 25 мкм до приблизительно 90 мкм. Согласно некоторым вариантам осуществления последние две щели или просвета в серии могут характеризоваться внутренними диаметрами, составляющими от приблизительно 100 до приблизительно 150 мкм и от приблизительно 50 мкм до приблизительно 70 мкм. Согласно некоторым вариантам осуществления суспендирование может предусматривать приблизительно 5 до приблизительно 20 минут суспендирования через от второй до последней щели или просвета и приблизительно 5 до приблизительно 20 минут суспендирования в последней щели или просвете. Согласно некоторым вариантам осуществления суспендирование может предусматривать приблизительно 10 минут суспендирования через от второй до последней щели или просвета и приблизительно 15 минут суспендирования в последней щели или просвете.
В качестве исключительно иллюстративного варианта осуществления последние два отверстия или просвета могут быть образованы пипетками, модифицированными следующим образом: сделать две оплавленных пипетки с очень маленькими выходными отверстиями следующим образом: нагреть стандартную 9'' (22,86 см) стеклянную пипетку над горелкой Бунзена, и затем тянуть и растягивать дальний (расплавленный) конец пипетки, пока просвет не схлопнется, и конец не оторвется, оставляя закрытый, заостренный стеклянный кончик. Подождать, пока пипетка охладится, и затем отламывать закрытый дальний конец, пока нельзя будет идентифицировать очень маленький просвет. Повторить этот процесс со второй пипеткой, но отломить кончик немного более близко, получая немного больший просвет на дальнем конце. Больший просвет должен составлять приблизительно 100-150 микрон в диаметре, в то время как другая пипетка должна иметь меньший просвет приблизительно 50-70 микрон. Клеточную суспензию можно суспендировать через пипетку с большим просветом в течение 10 минут. После этого можно осуществить суспендирование через пипетку, характеризующуюся меньшим просветом (50-70 мкм), в течение дополнительных 15 минут. Продолжать пипетиовать суспензию, пока она не станет легко проходить вверх и вниз по оплавленной пипетке с меньшим внутренним диаметром. Каждую пипетку можно предварительно покрыть средой. Кроме того, во время суспендирования следует избегать забора воздуха и создания пузырей или пены в суспензии клеток.
Согласно некоторым вариантам осуществления суспендирование можно проводить со скоростью, составляющей от приблизительно 1 до приблизительно 200 циклов в минуту, например, всю суспензию пропускают через щель, просвет или отверстие 1-100 раз в минуту. Согласно некоторым вариантам осуществления суспендирование можно проводить со скоростью, составляющей от приблизительно 10 до приблизительно 60 циклов в минуту. Согласно некоторым вариантам осуществления суспендирование можно проводить со скоростью, составляющей приблизительно 40 циклов в минуту. Согласно некоторым вариантам осуществления в которых для суспендирования используют пипетку, суспензию можно выпускать из пипетки и обратно в пипетку приблизительно 20 раз в минуту.
На следующей стадии, не являющиеся эритроцитами суспендированные клетки можно выделить из красной крови. Согласно некоторым вариантам осуществления приблизительно 0-50 объемов HBSS можно добавить к суспендированной суспензии и затем добавить 0,1-20 объемов раствора для выделения эритроцитов. Специалисту в настоящей области техники знакомы растворы для выделения эритроцитов, например, лимфолит или бусины со специфическими в отношении эритроцитов антителами.
В качестве исключительно иллюстративного варианта осуществления после завершения суспендирования дополнительный HBSS можно добавить к клеткам, затем 1 объема лимфолита можно добавить на дно пробирки для создания хорошего бислоя. Согласно некоторым вариантам осуществления следует избегать смешивания двух растворов. Эту смесь можно центрифугировать при 1000 g в течение 10 мин. Вращать пробирку на 180° и заново центрифугировать при 1000 g в течение дополнительных 10 мин. Это будет вызывать образование эритроцитами осадка на дне пробирки. С использованием стандартной 9'' стеклянной пипетки аспирировать клеточные суспензии, слой между HBSS и лимфолитом удаляют и помещают в новую 50 мл пробирку. HSBB можно добавить к суспензии до общего объема, составляющего 20 мл HBSS, и затем суспензию перемешивают с помощью 5 мл пипетки в течение 1 минуты.
На следующей стадии, клетки выделяют из раствора HBSS. В качестве только одного примера, клетки можно осадить с помощью центрифугирования в центрифужной пробирке при от приблизительно 800 об/мин до приблизительно 1600 об/мин в течение от приблизительно 1 минуты до приблизительно 20 минут. В качестве только одного примера, клетки можно осадить с помощью центрифугирования в центрифужной пробирке при приблизительно 1200 об/мин в течение 5 минут. Согласно некоторым вариантам осуществления на следующей стадии клетки можно ресуспендировать в HBSS, при этом полученная суспензия характеризуется рН, составляющим от приблизительно 5,0 до приблизительно 6,0. Согласно некоторым вариантам осуществления полученная суспензия может характеризоваться значением рН, составляющим от приблизительно 5,4 до приблизительно 5,8. Согласно некоторым вариантам осуществления полученная суспензия может характеризоваться значением рН, составляющим от приблизительно 5,6 до приблизительно 5,7. Согласно некоторым вариантам осуществления полученная суспензия может характеризоваться значением рН, составляющим приблизительно 5,6. Согласно некоторым вариантам осуществления раствор HBSS перед смешиванием с клетками может характеризоваться значением рН, составляющим от приблизительно 5,0 до приблизительно 5,7. Согласно некоторым вариантам осуществления раствор HBSS перед смешиванием с клетками может характеризоваться рН, составляющим от приблизительно 5,3 до приблизительно 5,6. Согласно некоторым вариантам осуществления раствор HBSS перед смешиванием с клетками может характеризоваться рН, составляющим приблизительно 5,4. Согласно некоторым вариантам осуществления клетки можно ресуспендировать при концентрации, составляющей от приблизительно 2×104 клеток/мл до приблизительно 2×108 клеток/мл. Согласно некоторым вариантам осуществления клетки можно ресуспендировать при концентрации, составляющей приблизительно 2×106.
В качестве исключительно иллюстративного примера, стадию ресуспендирования согласно предыдущему абзацу можно проводить следующим образом: когда подкисляете раствор, мягко суспендировать его с использованием 5 мл пипетки в течение 10 секунд сразу после добавления кислоты к раствору Хенкса. HBSS обладает очень слабой буферной емкостью, так что любой раствор, перенесенный из супернатанта предшествующей суспензии будет существенно влиять на рН HBSS. Инструкции ниже показывают, как получить HBSS с оптимальным рН, составляющим 5,6-5,7 для получения клеток STAP в соответствии с этим экспериментальным вариантом осуществления. Сначала следует титровать рН предварительно охлажденного HBSS (при 4°С) с помощью 12 н. HCl до рН 5,6. Это осуществляют посредством медленного добавления 11,6 мкл 12 н. HCl к 50 мл HBSS. После подтверждения указанного значения рН, следует стерилизовать раствор фильтрацией через шприц с фильтром 0,2 микрон или через фильтр на горле бутыли, в новый стерильный контейнер для хранения. Необходимо подтвердить конечный рН 5,6-5,7 в ходе предварительного тестового эксперимента с соответствующим количеством клеток. Поскольку рН HBSS является настолько важным, рН раствора следует проверять, повторно титровать и повторно стерилизовать перед каждым применением.
На следующей стадии, клетки в суспензии HBSS можно инкубировать приблизительно при их in vivo температуре. Например, клетки млекопитающего можно инкубировать при приблизительно 37°С. Согласно некоторым вариантам осуществления инкубацию можно осуществлять в течение от приблизительно 5 минут до приблизительно 3 часов. Согласно некоторым вариантам осуществления инкубацию можно осуществлять в течение от приблизительно 10 минут до приблизительно 1 часа. Согласно некоторым вариантам осуществления инкубацию можно осуществлять в течение от приблизительно 15 минут до приблизительно 40 минут. Согласно некоторым вариантам осуществления инкубацию можно осуществлять в течение приблизительно 25 минут.
На следующей стадии, клетки выделяют из кислого раствора HBSS. В качестве только одного примера, клетки можно осадить с помощью центрифугирования в центрифужной пробирки при от приблизительно 800 об/мин до приблизительно 1600 об/мин в течение от приблизительно 1 минуты до приблизительно 20 минут. В качестве только одного примера, клетки можно осадить с помощью центрифугирования в центрифужной пробирке при приблизительно 1200 об/мин в течение 5 минут. Согласно некоторым вариантам осуществления супернатант затем можно отобрать с помощью аспирации.
На следующей стадии, клетки можно ресуспендировать в средах, подходящих для содержания и/или выбора плюрипотентной клетки. Согласно некоторым вариантам осуществления среда представляет собой среду для образования клеточных сфер. Используемый в настоящем документе термин "среда для образования клеточных сфер" относится к DMEM/F12 с 1% антибиотиком и 2% В27 Gibco 12587-010. Согласно некоторым вариантам осуществления среда может дополнительно содержать факторы роста, например, b-FGF (20 нг/мл), EGF (20 нг/мл), гепарин (0,2%, Stem Cell Technologies 07980). Указанные факторы приводят в соответствие к используемому типу клеток. Например, согласно некоторым вариантам осуществления LIF (1000 ед.) можно добавить, если клетки являются мышиными). Согласно некоторым вариантам осуществления такие добавки, как bFGF, EGF и гепарин могут не являться необходимыми. Согласно некоторым вариантам осуществления клетки можно ресуспендировать в средах при концентрации, составляющей 105 клеток/см3.
На следующей стадии, клетки можно культивировать и/или содержать, например, культивировать при 37°C с 5% СО2. Согласно некоторым вариантам осуществления клетки можно перемешивать во время культивирования/содержания для предотвращения прилипания к контейнеру для культивирования клеток. Согласно некоторым вариантам осуществления клетки можно аккуратно пипетировать с использованием, например, 5 мл пипетки, дважды в день в течение 2 минут, в течение первой недели, чтобы препятствовать их прикреплению к дну чашек. Согласно некоторым вариантам осуществления это может стимулировать образование хороших сфер. Согласно некоторым вариантам осуществления среду для образования клеточных сфер, необязательно содержащую добавки, можно добавлять через день. Например, добавить 1 мл/день в 10 см чашку для культивирования или 0,5 мл/день в 6 см чашку.
Согласно одному аспекту в настоящем документе предусмотрен способ лечения неврологического повреждения у позвоночного, причем способ предусматривает введение позвоночному плюрипотентных (включая в себя "более плюрипотентные" клетки согласно представленному в настоящем документе описанию) клеток или клеток STAP согласно представленному в настоящем документе описанию позвоночному, нуждающемуся в лечении неврологического повреждения. Согласно некоторым вариантам осуществления вводимые клетки представляют собой клетки, созданные с помощью улучшенных способов, описанных в настоящем документе, например, способов согласно двум аспектам, описанным непосредственно выше, и/или примеру 5. Согласно некоторым вариантам осуществления клетки можно вводить в каркасе, гидрогеле или составе отсроченного высвобождения. Согласно некоторым вариантам осуществления клетки могут являться аутологичными по отношению к позвоночному. Согласно некоторым вариантам осуществления клетки создают из нервной ткани. Согласно некоторым вариантам осуществления позвоночное нуждается в лечении от воздействия нейротоксина, острого неврологического повреждения, хронического неврологического повреждения и/или дегенеративного неврологического заболевания. Согласно некоторым вариантам осуществления неврологическое повреждение может включать в себя повреждение спинного мозга, нервов и/или головного мозга. Согласно некоторым вариантам осуществления позвоночное может представлять собой грызуна, например, мышь или крысу. Согласно некоторым вариантам осуществления позвоночное может представлять собой представителя семейства псовых, представителя семейства кошачьих, собаку, кошку, одомашненное животное, лошадь или примата, например, человека. Согласно некоторым вариантам осуществления способ может предусматривать повторные введения, например, 2 или больше, 3 или больше, 4 или больше или больше введений. Согласно некоторым вариантам осуществления клетки можно вводить в место повреждения, например, хирургически имплантировать и/или вводить с помощью инъекции.
Согласно одному аспекту в настоящем документе предусмотрен набор, содержащий пипетку, характеризующуюся отверстием, составляющим от приблизительно 90 до приблизительно 200 мкм в диаметре, и/или пипетку, характеризующуюся отверстием, составляющим от приблизительно 25 мкм до приблизительно 90 мкм в диаметре. Согласно некоторым вариантам осуществления первая пипетка содержит отверстие, составляющее от приблизительно 100 до приблизительно 150 мкм в диаметре, и вторая пипетка содержит отверстие, составляющее от приблизительно 50 мкм до приблизительно 70 мкм в диаметре.
Согласно некоторым вариантам осуществления набор может дополнительно содержать дополнительную пипетку, характеризующуюся отверстием, составляющим от приблизительно 0,5 мм до приблизительно 2,0 мм в диаметре. Согласно некоторым вариантам осуществления пипетка может характеризоваться отверстием, составляющим от приблизительно 0,7 мм до приблизительно 1,5 мм в диаметре. Согласно некоторым вариантам осуществления пипетка может характеризоваться отверстием, составляющим приблизительно 1,1 мм в диаметре.
Согласно некоторым вариантам осуществления наборы альтернативно могут быть предоставлены вместе с приборами, характеризующимися щелями и/или просветам с диаметрами, описанными выше для пипеток, например, микроструйными приборами, характеризующимися каналами со щелями или просветами с описанными внутренними диаметрами.
Согласно некоторым вариантам осуществления набор может дополнительно содержать HBSS. Согласно некоторым вариантам осуществления HBSS может характеризоваться рН, составляющим от приблизительно 5,0 до приблизительно 5,7. Согласно некоторым вариантам осуществления HBSS может характеризоваться рН, составляющим от приблизительно 5,3 до приблизительно 5,6. Согласно некоторым вариантам осуществления HBSS может характеризоваться рН, составляющим приблизительно 5,4. Согласно некоторым вариантам осуществления набор может дополнительно содержат кислоту для титрования рН HBSS. Согласно некоторым вариантам осуществления кислота представляет собой HCl. Согласно некоторым вариантам осуществления набор может дополнительно содержать среда для образования клеточных сфер, и необязательно факторы роста.
Набор представляет собой любое готовое изделие (например, упаковку или контейнер), содержащее по меньшей мере одну мультиэлектродную матрицу согласно различным вариантам осуществления в настоящем документе, причем готовое изделие рекламируют, распространяют или продают как установку для проведения способов или анализов, описанных в настоящем документе. Описанные в настоящем документе наборы включают в себя реагенты и/или компоненты, которые обеспечивают возможность создания, культивирования и/или выбора плюрипотентных клеток. Описанные в настоящем документе наборы необязательно могут содержать дополнительные компоненты, применимые для проведения способов и анализов, описанных в настоящем документе. Такие реагенты могут включать в себя, например, среды для культивирования клеток, факторы роста, факторы дифференцировки, буферные растворы, метки, визуализирующие реагенты и подобное. Такие ингредиенты известны специалисту в настоящей области техники и могут варьировать в зависимости от конкретных клеток и способов или анализа, подлежащего проведению. Кроме того, набор может содержать листок-вкладыш с инструкцией и/или может обеспечивать информацию относительно соответствия полученных результатов.
Не предусмотрено, что описание вариантов осуществления настоящего раскрытия является исчерпывающим или ограничивает раскрытие конкретной раскрытой формой. Наряду с тем, что конкретные варианты осуществления и примеры настоящего раскрытия описаны в настоящем документе для иллюстративных целей, различные эквивалентные модификации возможны в пределах объема настоящего раскрытия, которые смогут установить специалисты в настоящей области техники. Например, наряду с тем, что стадии или функции способов представлены в определенном порядке, согласно альтернативным вариантам осуществления можно осуществлять функции в отличающемся порядке или функции можно осуществлять по существу одновременно. Идеи раскрытия, предусмотренные в настоящем документе, можно применить к другим процедурам или способам в соответствующих случаях. Различные описанные в настоящем документе варианты осуществления можно комбинировать для обеспечения дополнительных вариантов осуществления. Аспекты настоящего раскрытия можно модифицировать при необходимости для использования композиций, функций и понятий представленных выше ссылок и применения для обеспечения дополнительных вариантов осуществления настоящего раскрытия. Указанные и другие изменения можно осуществить в отношении раскрытия в свете подробного описания.
Конкретные элементы любого из вышеописанных вариантов осуществления можно комбинировать или замещать элементами в других вариантах осуществления. Кроме того, наряду с тем, что преимущества, связанные с определенными вариантами осуществления настоящего раскрытия, были описаны в контексте указанных вариантов осуществления другие варианты осуществления также могут проявлять такие преимущества, и нет необходимости, чтобы все варианты осуществления проявляли такие преимущества для попадания в объем настоящего раскрытия.
Все патенты и другие указанные публикации полностью включены в настоящий документ посредством ссылки с целью описания и раскрытия, например, методик, описанных в этих публикациях, которые можно использовать применительно к настоящему изобретению. Указанные публикации представлены исключительно для их раскрытия до даты подачи настоящей заявки. Ничто в этом отношении не должно быть истолковано как признание, что авторы изобретения не наделены правом предшествовать такому раскрытию на основании предшествующего изобретения или ни по какой другой причине. Все заявления относительно даты или представления относительно содержания указанных документов основаны на информации, доступной заявителям, и не составляют признания правильности дат или содержания указанных документов.
Настоящее изобретение дополнительно проиллюстрировано следующими примерами, которые не следует рассматривать как ограничивающие.
Некоторые варианты осуществления описанной в настоящем документе технологии можно определить согласно любому из следующих пронумерованных абзацев:
1. Способ создания плюрипотентной клетки, предусматривающий воздействие на клетку стресса.
2. Способ согласно абзацу 1, при котором плюрипотентную клетку создают без введения экзогенного гена, транскрипта, белка, ядерного компонента или цитоплазмы или без слияния клеток.
3. Способ по любому из абзацев 1-2, дополнительно предусматривающий выбор клетки, проявляющей плюрипотентность.
4. Способ по любому из абзацев 1-3, при котором клетка не присутствует как часть ткани.
5. Способ по любому из абзацев 1-4, при котором клетка представляет собой соматическую клетку, стволовую клетку, клетку-предшественник или эмбриональную клетку.
6. Способ по любому из абзацев 1-5, при котором клетка представляет собой выделенную клетку.
7. Способ по любому из абзацев 1-6, при котором клетка присутствует в гетерогенной популяции клеток.
8. Способ по любому из абзацев 1-7, при котором клетка присутствует в гомогенной популяции клеток.
9. Способ по любому из абзацев 1-8, при котором выбор проявляющей плюрипотентность клетки предусматривает выбор клетки, экспрессирующей маркер стволовой клетки.
10. Способ по абзацу 9, при котором маркер стволовой клетки выбирают из группы, состоящей из следующего:
Oct4; Nanog; Е-кадгерин и SSEA4.
11. Способ по любому из абзацев 1-10, при котором выбор проявляющей плюрипотентность клетки предусматривает выбор клетки, которая не является адгерентной.
12. Способ по любому из абзацев 1-11, при котором стресс включает в себя нефизиологический стресс на ткань или клеточную культуру.
13. Способ по любому из абзацев 1-12, при котором стресс включает в себя воздействие на клетку по меньшей мере одного стимула окружающей среды, выбранного из следующего: травма, механические стимулы, химическое воздействие, ультразвуковая стимуляция, кислородная недостаточность, облучение, воздействие экстремальных температур, диссоциация, суспендирование, физический стресс, гиперосмос, гипоосмос, повреждение мембраны, токсин, экстремальная концентрация ионов, активный кислород, воздействие УФ, яркий свет видимого спектра, недостаточность необходимых питательных веществ или нефизиологически кислотная среда.
14. Способ по любому из абзацев 1-13, при котором стресс предусматривает воздействие на клетку рН, составляющего от приблизительно 3,0 до приблизительно 6,8.
15. Способ по любому из абзацев 1-14, при котором стресс предусматривает воздействие на клетку рН, составляющего от приблизительно 4,5 до приблизительно 6,0.
16. Способ по абзацу 15, при котором стресс предусматривает воздействие на клетку рН, составляющего от приблизительно 5,4 до приблизительно 5,8.
17. Способ по любому из абзацев 12-16, при котором клетку подвергают воздействию в течение 2-3 дней.
18. Способ по любому из абзацев 12-17, при котором клетку подвергают воздействию в течение 1 дня или меньше.
19. Способ по любому из абзацев 12-18, при котором клетку подвергают воздействию в течение 1 часа или меньше.
20. Способ по любому из абзацев 12-19, при котором клетку подвергают воздействию в течение приблизительно 30 минут.
21. Способ по абзацу 13, при котором воздействие экстремальных температур предусматривает воздействие на клетку температур ниже 35°С или выше 42°С.
22. Способ по абзацу 21, при котором воздействие экстремальных температур предусматривает воздействие на клетку температур замораживания или ниже замораживания или воздействие на клетку температур по меньшей мере приблизительно 85°С.
23. Способ по абзацу 13, при котором механический стимул предусматривает воздействие на клетку напряжения сдвига и/или высокого давления.
24. Способ по абзацу 23, при котором механический стимул предусматривает пропускание клетки через по меньшей мере один прибор со щелью, меньше чем размер клетки.
25. Способ по абзацу 23, при котором механический стимул предусматривает пропускание клетки через несколько приборов, характеризующихся постепенно уменьшающимися щелями.
26. Способ по любому из абзацев 1-25, дополнительно предусматривающий культивирование плюрипотентной клетки для обеспечения размножения плюрипотентной клетки.
27. Способ по любому из абзацев 1-26, при котором плюрипотентная клетка экспрессирует маркер стволовой клетки.
28. Способ по абзацу 27, при котором маркер стволовой клетки выбирают из группы, состоящей из следующего:
Oct4; Nanog; Е-кадгерин и SSEA4.
29. Способ по любому из абзацев 1-28, при котором клетка представляет собой клетку млекопитающего.
30. Способ по любому из абзацев 1-29, при котором клетка представляет собой клетку человека.
31. Способ по любому из абзацев 1-30, при котором клетка представляет собой взрослую клетку, неонатальную клетку, фетальную клетку, амниотическую клетку или клетку пуповинной крови.
32. Способ по любому из абзацев 1-31, дополнительно предусматривающий содержание плюрипотентной клетки in vitro.
33. Способ по любому из абзацев 1-32, при котором эпигенетическое состояние клетки изменяют, чтобы оно приобретало большее сходство с эпигенетическим состоянием эмбриональной стволовой клетки.
34. Способ по абзацу 33, при котором эпигенетическое состояние включает в себя паттерны метилирования.
35. Способ по любому из абзацев 1-34, при котором стресс включает в себя удаление по меньшей мере приблизительно 40% цитоплазмы из клетки.
36. Способ по абзацу 35, при котором по меньшей мере приблизительно 50% цитоплазмы удаляют из клетки.
37. Способ по абзацу 36, при котором по меньшей мере приблизительно 60% цитоплазмы удаляют из клетки.
38. Способ по абзацу 37, при котором 60-80% цитоплазмы удаляют из клетки.
39. Способ по абзацу 37, при котором по меньшей мере приблизительно 80% цитоплазмы удаляют из клетки.
40. Способ по абзацу 39, при котором по меньшей мере приблизительно 90% цитоплазмы удаляют из клетки.
41. Способ по любому из абзацев 1-40, при котором стресс включает в себя удаление по меньшей мере приблизительно 40% митохондрий из клетки.
42. Способ по абзацу 41, при котором удаление части цитоплазмы удаляет по меньшей мере приблизительно 50% митохондрий из цитоплазмы.
43. Способ по абзацу 42, при котором удаление цитоплазмы или митохондрий удаляет приблизительно 50%-90% митохондрий из цитоплазмы.
44. Способ по абзацу 42, при котором удаление цитоплазмы или митохондрий удаляет больше чем 90%) митохондрий из цитоплазмы.
45. Способ по любому из абзацев 1-44, при котором стресс является достаточным, чтобы разрушить клеточную мембрану по меньшей мере 10% клеток, подвергнутых воздействию стресса.
46. Анализ, предусматривающий:
приведение в контакт плюрипотентной клетки, полученной с помощью способа по любому из абзацев 1-45, с кандидатным средством.
47. Анализ по абзацу 46 для применения в идентификации средств, которые влияют на одно или несколько из жизнеспособности, дифференцировки, пролиферации плюрипотентной клетки.
48. Применение плюрипотентной клетки, полученной с помощью способа по любому из абзацев 1-45, в способе клеточной терапии субъекта.
49. Способ получения клетки или ткани, которая является совместимой с клеточной терапией, подлежащей введению субъекту, предусматривающий:
создание плюрипотентной клетки из клетки по любому из абзацев 1-45;
причем клетка представляет собой аутологичную клетку совпадающую по HLA аллогенную клетку.
50. Способ по абзацу 49, дополнительно предусматривающий дифференцировку плюрипотентной клетки по заданной линии дифференцировки перед введением клетки или ткани субъекту.
51. Композиция, содержащая плюрипотентную клетку, в которой плюрипотентная клетка создана из клетки с помощью способов по любому из абзацев 1-45.
52. Способ получения плюрипотентной стволовой клетки, причем способ предусматривает культивирование клетки в присутствии адренокортикотропного гормона (АСТН), среды 2i или 3i.
53. Способ по абзацу 52, при котором клетку культивируют в среде LIF, содержащей АСТН.
54. Способ по абзацу 52 или 53, при котором АСТН присутствует в концентрации, составляющей от приблизительно 0,1 мкМ до приблизительно 100 мкМ.
55. Способ по любому из абзацев 52-54, при котором клетка представляет собой клетку, созданную с помощью способа по любому из абзацев 1-45.
56. Способ по любому из абзацев 52-55, при котором клетка представляет собой тотипотентную клетку.
57. Способ по любому из абзацев 52-56, при котором клетку культивируют в присутствии АСТН, среды 2i или 3i в течение по меньшей мере 3 дней.
58. Способ по любому из абзацев 52-57, при котором клетку культивируют в присутствии АСТН, среды 2i или 3i в течение по меньшей мере 5 дней.
59. Способ по любому из абзацев 52-58, при котором клетку культивируют в присутствии АСТН, среды 2i или 3i в течение по меньшей мере 7 дней.
60. Способ по любому из абзацев 52-59, при котором после стадии культивирования клетка экспрессирует обнаруживаемое содержание маркера стволовой клетки, выбранного из группы, состоящий из следующего:
Oct3/4; Nanog; Rex1; Klf4; Sox2; Klf2; Esrr-бета; Tbx3 и Klf5.
61. Способ увеличения способности плюрипотентной клетки к самообновлению, причем способ предусматривает культивирование клетки в присутствии адренокортикотропного гормона (АСТН), среды 2i или 3i.
62. Способ по абзацу 61, при котором клетку культивируют в среде LIF, содержащей АСТН.
63. Способ по любому из абзацев 61-62, при котором АСТН присутствует в концентрации, составляющей от приблизительно 0,1 мкМ до приблизительно 100 мкМ.
64. Способ по любому из абзацев 61-63, при котором клетка представляет собой клетку, созданную с помощью способа по любому из абзацев 1-45.
65. Способ по любому из абзацев 61-64, при котором клетка представляет собой тотипотентную клетку.
66. Способ по любому из абзацев 61-65, при котором клетку культивируют в присутствии АСТН, среды 2i или 3i в течение по меньшей мере 3 дней.
67. Способ по любому из абзацев 61-66, при котором клетку культивируют в присутствии АСТН, среды 2i или 3i в течение по меньшей мере 5 дней.
68. Способ по любому из абзацев 61-67, при котором клетку культивируют в присутствии АСТН, среды 2i или 3i в течение по меньшей мере 7 дней.
69. Способ по любому из абзацев 61-68, при котором после стадии культивирования клетка экспрессирует обнаруживаемое содержание маркера стволовой клетки, выбранного из группы, состоящей из следующего:
Oct3/4; Nanog; Rex1; Klf4; Sox2; Klf2; Esrr-бета; Tbx3 и Klf5.
70. Способ аутологичной клеточной терапии у субъекта, нуждающегося в клеточной терапии, предусматривающий:
a. создание плюрипотентной клетки из клетки по любому из абзацев 1-45, причем клетку получают от субъекта, и
b. введение композиции, содержащей плюрипотентную клетку или ее дифференцированное потомство, субъекту.
71. Способ по абзацу 70, дополнительно содержащая дифференцировку плюрипотентная клетка по заданной линии дифференцировки перед введением композиции субъекту.
72. Способ получения плюрипотентной клетки, способной к дифференцировке ва плацентарную клетку, причем способ предусматривает культивирование плюрипотентной клетки, созданные с помощью способа по любому из абзацев 1-45 в присутствии FGF4.
73. Способ по абзацу 72, при котором концентрация FGF4 составляет 1 нМ - 1 мкМ.
74. Способ по абзацу 72 или 73, при котором плюрипотентная клетка способна к дифференцировке в эмбриональную стволовую клетку.
Некоторые варианты осуществления описанной в настоящем документе технологии можно определить согласно любому из следующих пронумерованных абзацев:
1. Способ создания плюрипотентной клетки, предусматривающий следующее:
a. Выделение исходной клетки из раствора;
b. Ресуспендирование клетки, полученной на стадии, в сбалансированном солевом растворе Хенкса (HBSS);
c. Суспендирование клетка суспензия полученной на стадии b;
d. Добавление от приблизительно 2 до приблизительно 20 объемов HBSS к клеточной суспензии;
e. Выделение клетки из суспензии, полученной на стадии d;
f. Ресуспендирование клетки, полученной на стадии е, в HBSS, характеризующимся рН, составляющим от приблизительно 5,0 до приблизительно 6,0;
g. Инкубация клеток приблизительно при их естественной in vivo температуре;
h. Выделение клетки из суспензии, полученной на стадии g; и
i. Ресуспендирование клеточного осадка, полученного на стадии h, в среде.
2. Способ по абзацу 1, при котором выделение предусматривает центрифугирование.
3. Способ по любому из абзацев 1-2, дополнительно предусматривающий приведение исходной клетки в контакт с трипсином в течение от приблизительно 1 минуты до приблизительно 10 минут перед стадией а.
4. Способ по абзацу 3, дополнительно предусматривающий приведение исходной клетки в контакт с трипсином в течение от приблизительно 3 минут до приблизительно 5 минут перед стадией а.
5. Способ по любому из абзацев 3-4, при котором трипсин дезактивируют путем приведения клеточного осадка в контакт с минимальной эссенциальной средой Игла в модификации Дюльбекко (DMEM)/F-12, содержащей 10% термически инактивированной фетальной бычьей сыворотки (FBS).
6. Способ по любому из абзацев, 1-5, при котором стадия суспендирования с предусматривает суспендирование клеток через серию щелей или просветов постепенно уменьшающихся диаметров.
7. Способ по абзацу 6, при котором серия содержит по меньшей мере 3 щели или просвета.
8. Способ по любому из абзацев 6-7, при котором по меньшей мере первую щель или просвет предварительно покрывают HBSS или водой.
9. Способ по любому из абзацев 6-8, при котором первая щель или просвет характеризуются внутренним диаметром, составляющим от приблизительно 0,5 мм до приблизительно 2,0 мм.
10. Способ по абзацу 9, при котором первая щель или просвет характеризуются внутренним диаметром, составляющим от приблизительно 0,7 мм до приблизительно 1,5 мм.
11. Способ по абзацу 10, при котором первая щель или просвет характеризуются внутренним диаметром, составляющим приблизительно 1,1 мм.
12. Способ по любому из абзацев 6-11, при котором суспендирование через первую щель или просвет проводят в течение от приблизительно 1 минуты до приблизительно 10 минут.
13. Способ по абзацу 12, при котором суспендирование через первую щель или просвет проводят в течение приблизительно 5 минут.
14. Способ по любому из абзацев 6-13, при котором последние две щели или просвета в серии характеризуются внутренними диаметрами, составляющими от приблизительно 90 до приблизительно 200 мкм и от приблизительно 25 мкм до приблизительно 90 мкм.
15. Способ по абзацу 14, при котором последние две щели или просвета в серии характеризуются внутренними диаметрами, составляющими от приблизительно 100 до приблизительно 150 мкм и от приблизительно 50 мкм до приблизительно 70 мкм.
16. Способ по любому из абзацев 1-15, при котором суспендирование предусматривает от приблизительно 5 до приблизительно 20 минут суспендирования через от второй до последней щели или просвета и от приблизительно 5 до приблизительно 20 минут суспендирования в последней щели или просвете.
17. Способ по абзацу 16, при котором суспендирование предусматривает от приблизительно 10 минут суспендирования через от второй до последней щели или просвета и от приблизительно 15 минут суспендирования в последней щели или просвете.
18. Способ по любому из абзацев 6-17, при котором суспендирование в последней щели или просвете продолжают, пока суспензия не будет легко проходить через щель или просвет.
19. Способ по любому из абзацев 6-18, при котором суспендирование в каждой щели или просвете продолжают, пока суспензия не будет легко проходить через такую щель или просвет.
20. Способ по любому из абзацев 1-19, при котором общее время суспендирования составляет приблизительно 30 минут.
21. Способ по любому из абзацев 1-20; при котором приблизительно 5 до приблизительно 15 объемов HBSS добавляют на стадии d.
22. Способ по любому из абзацев 1-21; при котором приблизительно 10 объемов HBSS добавляют на стадии d.
23. Способ по любому из абзацев 1-22, при котором рН HBSS на стадии f составляет от приблизительно 5,1 до приблизительно 5,7.
24. Способ по любому из абзацев 1-23, при котором рН HBSS на стадии f составляет приблизительно 5,4.
25. Способ по любому из абзацев 1-24, при котором рН клеточной суспензии в HBSS, полученной на стадии f, составляет от приблизительно 5,0 до приблизительно 6,0.
26. Способ по любому из абзацев 1-25, при котором рН клеточной суспензии в HBSS, полученной на стадии f, составляет от приблизительно 5,6 до приблизительно 5,7.
27. Способ по любому из абзацев 1-26, при котором стадия f предусматривает ресуспендирование клеток до концентрации, составляющей от приблизительно 0,5 миллиона клеток/мл до приблизительно 4 миллионов клеток/мл.
28. Способ по любому из абзацев 1-27, при котором стадия f предусматривает ресуспендирование клеток до концентрации, составляющей приблизительно 2 миллиона клеток/мл.
29. Способ по любому из абзацев 1-28, при котором стадия g предусматривает инкубацию клеток при приблизительно 37°С в течение приблизительно 25 минут.
30. Способ по любому из абзацев 1-29, при котором объединенные стадии g и h продолжаются от приблизительно 10 минут до приблизительно 1 часа.
31. Способ по любому из абзацев 1-30, при котором объединенные стадии g и h продолжаются в течение приблизительно 30 минут.
32. Способ по любому из абзацев 1-31, при котором среда стадии i представляет собой среду для образования клеточных сфер, содержащую DMEM/F12, приблизительно 1% антибиотика, приблизительно 2% В27 и необязательно один или несколько факторов роста.
33. Способ по абзацу 32, при котором факторы роста содержат bFGF, EGF и гепарин.
34. Способ создания плюрипотентной клетки, при котором исходная клетка присутствует в ткани, содержащей эритроциты, причем способ предусматривает следующее:
a. Механическое разрезание ткани на тонкие срезы в присутствии одного или нескольких расщепляющих ЕСМ ферментов;
b. Инкубация образца, полученного на стадии а, при приблизительно естественной для ткани in vivo температуры ткани при перемешивании ткани;
c. Разбавление клеточной суспензии, полученной на стадии b, в HBSS;
d. Выделение клетки из суспензии, полученной на стадии с;
e. Ресуспендирование а клетка полученной на стадии d in HBSS;
f. Суспендирование клеточной суспензии, полученной на стадии е;
g. Добавление от приблизительно 0,1 до приблизительно 10 объемов раствора для выделения эритроцитов к клеточной суспензии, полученной на стадии f, для создания бислоя;
h. Отделение слоя HBSS, полученного на стадии g, от слоя раствора для выделения эритроцитов;
i. Выделение клетки из суспензия HBSS, полученной на стадии h;
j. Ресуспендирование клетки, полученной на стадии i в HBSS с рН, составляющим от приблизительно 5,0 до приблизительно 6,0;
k. Инкубация клеток при приблизительно естественной для ткани in vivo температуре;
l. Выделение клетки из суспензии, полученной на стадии k; и
m. Ресуспендирование клетки, полученной на стадии l, в среде.
35. Способ по абзацу 34, при котором ткань, содержащую эритроциты, выбирают из группы, состоящей из следующего:
легкое; селезенка и печень.
36. Способ по любому из абзацев 34-35, при котором ткань представляет собой легкое, и расщепляющий ЕСМ фермент представляет собой коллагеназу Р.
37. Способ по любому из абзацев 34-36, при котором разрезание на тонкие срезы на стадии а продолжают в течение приблизительно 10 минут.
38. Способ по любому из абзацев 34-37, при котором суспендирование на стадии f предусматривает суспендирование клеток через серию щелей или просветов постепенно уменьшающихся диаметров.
39. Способ по абзацу 38, при котором серия содержит по меньшей мере 3 щели или просвета.
40. Способ по любому из абзацев 38-39, при котором по меньшей мере первую щель или просвет предварительно покрывают HBSS или водой.
41. Способ по любому из абзацев 38-40, при котором первая щель или просвет характеризуются внутренним диаметром, составляющим от приблизительно 0,5 мм до приблизительно 2,0 мм.
42. Способ по абзацу 41, при котором первая щель или просвет характеризуются внутренним диаметром, составляющим от приблизительно 0,7 мм до приблизительно 1,5 мм.
43. Способ по абзацу 42, при котором первая щель или просвет характеризуются внутренним диаметром, составляющим приблизительно 1,1 мм.
44. Способ по любому из абзацев 38-43, при котором суспендирование через первую щель или просвет проводят в течение от приблизительно 1 минуты до приблизительно 10 минут.
45. Способ по абзацу 44, при котором суспендирование через первую щель или просвет проводят в течение приблизительно 5 минут.
46. Способ по любому из абзацев 38-45, при котором последние две щели или просвета в серии характеризуются внутренними диаметрами, составляющими от приблизительно 90 до приблизительно 200 мкм и от приблизительно 25 мкм до приблизительно 90 мкм.
47. Способ по абзацу 46, при котором последние две щели или просвета в серии характеризуются внутренними диаметрами, составляющими от приблизительно 100 до приблизительно 150 мкм и от приблизительно 50 мкм до приблизительно 70 мкм.
48. Способ по любому из абзацев 38-47, при котором суспендирование предусматривает от приблизительно 5 до приблизительно 20 минут суспендирования через от второй до последней щели или просвета и от приблизительно 5 до приблизительно 20 минут суспендирования в последней щели или просвете.
49. Способ по абзацу 48, при котором суспендирование предусматривает приблизительно 10 минут суспендирования через от второй до последней щели или просвета и приблизительно 15 минут суспендирования в последней щели или просвете.
50. Способ по любому из абзацев 34-49, при котором суспендирование в последней щели или просвете продолжают, пока суспензия не будет легко проходить через щель или просвет.
51. Способ по любому из абзацев 34-50, при котором суспендирование в каждой щели или просвете продолжают, пока суспензия не будет легко проходить через такую щель или просвет.
52. Способ по любому из абзацев 34-51, при котором общее время суспендирования составляет приблизительно 30 минут.
53. Способ по любому из абзацев 34-52, при котором рН HBSS на стадии j составляет от приблизительно 5,1 до приблизительно 5,7.
54. Способ по абзацу 53, при котором рН HBSS на стадия j составляет приблизительно 5,4.
55. Способ по любому из абзацев 34-54, при котором рН клеточной суспензии в HBSS, полученной на стадии j, составляет от приблизительно 5,0 до приблизительно 6,0.
56. Способ по абзацу 55, при котором рН клеточной суспензии в HBSS, полученной на стадии j, составляет от приблизительно 5,6 до приблизительно 5,7.
57. Способ по любому из абзацев 34-56, при котором стадия к предусматривает инкубацию клеток приблизительно при 37°С в течение приблизительно 25 минут.
58. Способ по любому из абзацев 34-57, при котором объединенные стадии к и 1 продолжаются от приблизительно 10 минут до приблизительно 1 часа.
59. Способ по любому из абзацев 34-58, при котором среда на стадии m представляет собой среду для образования клеточных сфер, содержащую DMEM/F12, 1% антибиотика, 1% В27 и необязательно один или несколько факторов роста.
60. Способ по абзацу 59, при котором факторы роста содержат bFGF, EGF и гепарин.
61. Способ по любому из абзацев 1-60, при котором исходная клетка представляет собой мышиную клетку, и среда для образования клеточных сфер содержит LIF.
62. Способ по любому из абзацев 1-61, дополнительно предусматривающий стадию культивирования полученных клеток в течение по меньшей мере одной недели, причем культивирование предусматривает:
a. Добавление среды для образования клеточных сфер, необязательно содержащей факторы роста;
b. Перемешивание клеток для препятствия прикреплению к дну чашки.
63. Способ по абзацу 62, при котором среду для образования клеточных сфер добавляют каждые 1-4 дня.
64. Способ по абзацу 63, при котором среду для образования клеточных сфер добавляют каждые 2 дня.
65. Способ по любому из абзацев 62-64, при котором перемешивание предусматривает пипетирование клеток с помощью пипетки.
66. Способ по абзацу 65, при котором пипетка содержит отверстие, составляющее приблизительно 1,1 мм в диаметре.
67. Способ по любому из абзацев 65-66, при котором пипетирование проводят по меньшей мере один раз в день.
68. Способ по абзацу 67, при котором пипетирование проводят по меньшей мере два раза в день.
69. Способ по любому из абзацев 1-68, дополнительно предусматривающий выбор клетки с плюрипотентностью, при котором выбор предусматривает способ, выбранный из группы, состоящей из следующего:
выбор клеток с низким прилипанием; выбор клеток, которые представляют собой компонент сферы; и выбор клеток с маленьким относительным размером.
70. Способ по любому из абзацев 1-69, дополнительно предусматривающий конечную стадию выбора клетки, проявляющей плюрипотентность.
71. Способ по любому из абзацев 1-70, при котором исходная клетка не присутствует как часть ткани.
72. Способ по любому из абзацев 1-71, при котором исходная клетка представляет собой соматическую клетку, стволовую клетку, клетку-предшественник или эмбриональную клетку.
73. Способ по любому из абзацев 1-72, при котором исходная клетка представляет собой выделенную клетку.
74. Способ по любому из абзацев 1-73, при котором исходная клетка присутствует в гетерогенной популяции клеток.
75. Способ по любому из абзацев 1-74, при котором исходная клетка присутствует в гомогенной популяции клеток.
76. Способ по любому из абзацев 1-75, при котором выбор клетки, проявляющей плюрипотентность, предусматривает выбор клетки, экспрессирующей маркер стволовой клетки.
77. Способ по любому из абзаца 76, при котором маркер стволовой клетки выбирают из группы, состоящей из следующего:
Oct4; Nanog; Е-кадгерин и SSEA4.
78. Способ по любому из абзацев 1-77, при котором выбор клетки, проявляющей плюрипотентность, предусматривает выбор клетки, которая не является адгерентной.
79. Способ по любому из абзацев 1-78, дополнительно предусматривающий культивирование плюрипотентной клетки для обеспечения размножения плюрипотентной клетки.
80. Способ по любому из абзацев 1-79, при котором плюрипотентная клетка экспрессирует маркер стволовой клетки.
81. Способ по абзацу 80, при котором маркер стволовой клетки выбирают из группы, состоящей из следующего:
Oct4; Nanog; Е-кадгерин и SSEA4.
82. Способ по любому из абзацев 1-81, при котором исходная клетка представляет собой клетку млекопитающего.
83. Способ по любому из абзацев 1-82, при котором исходная клетка представляет собой клетку человек.
84. Способ по любому из абзацев 1-83, при котором исходная клетка представляет собой взрослую клетку, неонатальную клетку, фетальную клетку, амниотическую клетку или клетку пуповинная кровь.
85. Способ по любому из абзацев 1-84, дополнительно предусматривающий содержание плюрипотентная клетка in vitro.
86. Анализ, предусматривающий:
приведение в контакт плюрипотентной клетки, полученной с помощью способа по любому из абзацев 1-85, с кандидатным средством.
87. Анализ по абзацу 86 для применения в идентификации средств, которые влияют на одно или несколько из жизнеспособности, дифференцировки, пролиферации плюрипотентной клетки.
88. Применение плюрипотентной клетки, полученной с помощью способа по любому из абзацев 1-85, в способе клеточной терапии для субъекта.
89. Способ получения клетки или ткани, которые являются совместимыми с клеточной терапией, подлежащей введению субъекту, предусматривающий следующее:
создание плюрипотентной клетки из клетки по любому из абзацев 1-85;
причем клетка представляет собой аутологичную клетку совпадающую по HLA аллогенную клетку.
90. Способ по абзацу 89, дополнительно предусматривающий дифференцировку плюрипотентной клетки по заданной линии дифференцировки перед введением клетки или ткани субъекту.
91. Композиция, содержащая плюрипотентную клетку, в которой плюрипотентная клетка создана из клетки с помощью способов по любому из абзацев 1-85.
92. Способ аутологичной клеточной терапии у субъекта, нуждающегося в клеточной терапии, предусматривающий следующее:
a. создание плюрипотентной клетки из клетки по любому из абзацев 1-85, причем клетку получают от субъекта, и
b. введение композиции, содержащей плюрипотентную клетку или ее дифференцированное потомство, субъекту.
93. Способ по абзацу 92, дополнительно предусматривающий дифференцировку плюрипотентной клетки по заданной линии дифференцировки перед введением композиции субъекту.
94. Набор, содержащий две пипетки, причем первая пипетка характеризуется отверстием, составляющим от приблизительно 90 до приблизительно 200 мкм в диаметре, и вторая пипетка характеризуется отверстием, составляющим от приблизительно 25 мкм до приблизительно 90 мкм в диаметре.
95. Набор по абзацу 94, при котором первая пипетка характеризуется отверстием, составляющим от приблизительно 100 до приблизительно 150 мкм в диаметре, и вторая пипетка характеризуется отверстием, составляющим от приблизительно 50 мкм до приблизительно 70 мкм в диаметре.
96. Набор по любому из абзацев 94-95, дополнительно содержащий HBSS.
97. Набор по абзацу 94-96, при котором HBSS характеризуется рН, составляющим приблизительно 5,4.
98. Набор по любому из абзацев 96-97, дополнительно содержащий кислоты для титрования рН HBSS.
99. Набор по абзацу 98, при котором кислота представляет собой HCl.
100. Набор по любому из абзацев 94-99, дополнительно содержащий среду для образования клеточных сфер и необязательно факторы роста.
Некоторые варианты осуществления описанной в настоящем документе технологии можно определить согласно любому из следующих пронумерованных абзацев:
1. Способ создания плюрипотентной клетки, предусматривающий следующее:
a. Выделение исходной клетки из раствора;
b. Ресуспендирование клетки, полученной на стадии а, в растворе сбалансированного солевого раствора Хенкса (HBSS) и АТР, характеризующемся рН, составляющим от приблизительно 4,0 до приблизительно 6,5;
c. Суспендирование клеточной суспензии, полученной на стадии b;
d. Ресуспендирование клетки, полученной на стадии с, в HBSS;
e. Выделение клетки из суспензии, полученной на стадии d; и
f. Ресуспендирование клеточного осадка, полученного на стадии g, в среде.
2. Способ создания плюрипотентной клетки, предусматривающий следующее:
a. Выделение исходной клетки из раствора;
b. Ресуспендирование клетки, полученной на стадии а, в растворе сбалансированного солевого раствора Хенкса (HBSS) и АТР, характеризующемся рН, составляющим от приблизительно 4,0 до приблизительно 6,5;
c. Выделение клетки из суспензии, полученной на стадии b;
d. Ресуспендирование клетки, полученной на стадии с, в растворе HBSS и АТР, характеризующемся рН, составляющим от приблизительно 4,0 до приблизительно 6,5, до клеточной суспензии;
e. Суспендирование клеточной суспензии, полученной на стадии d;
f. Ресуспендирование клетки, полученной на стадии е, в HBSS;
g. Выделение клетки из суспензии, полученной на стадии f; и
h. Ресуспендирование клеточного осадка, полученного на стадии g, в среде.
3. Способ по любому из пп. 1-2, при котором АТР присутствует в растворе HBSS и АТР в концентрации, составляющие от приблизительно 1,5 до приблизительно 5 мг/см3.
4. Способ по п. 3, при котором АТР присутствует в растворе HBSS и АТР в концентрации, составляющей от приблизительно 2,7 мМ до приблизительно 9 мМ.
5. Способ по любому из пп. 1-4, при котором раствор HBSS и АТР, характеризующийся рН, составляющим от приблизительно 4,0 до приблизительно 6,5, получают путем титрования раствора HBSS с раствором АТР до достижения требуемого рН.
6. Способ по п. 5, при котором раствор АТР характеризуется концентрацией, составляющей от приблизительно 50 мМ до приблизительно 500 мМ.
7. Способ по п. 6, при котором раствор АТР характеризуется концентрацией, составляющей приблизительно 200 мМ.
8. Способ по любому из пп. 1-7, при котором раствор HBSS и АТР характеризуется рН, составляющим от приблизительно 5,0 до приблизительно 5,7.
9. Способ по любому из пп. 1-8, при котором раствор HBSS и АТР характеризуется рН, составляющим приблизительно 5,0.
10. Способ по любому из пп. 1-9, при котором HBSS согласно стадии f по п. 62 и стадии d по п. 61 не содержит АТР.
11. Способ по любому из пп. 1-10, при котором выделение предусматривает центрифугирование.
12. Способ по любому из пп. 1-11, дополнительно предусматривающий приведение исходной клетки в контакт с трипсином в течение от приблизительно 1 минуты до приблизительно 10 минут перед стадией а.
13. Способ по п. 12, дополнительно предусматривающий приведение исходной клетки в контакт с трипсином в течение от приблизительно 3 минут до приблизительно 5 минут перед стадией а.
14. Способ по любому из пп. 12-13, при котором трипсин дезактивируют путем приведения клеточного осадка в контакт с минимальной эссенциальной средой Игла в модификации Дюльбекко (DMEM)/F-12, содержащей 10% термически инактивированной фетальной бычьей сыворотки (FBS).
15. Способ по любому из пп. 1-14, при котором суспендирование предусматривает суспендирование клеток через серию щелей или просветов постепенно уменьшающихся диаметров.
16. Способ по п. 15, при котором серия содержит по меньшей мере 3 щели или просвета.
17. Способ по любому из пп. 15-16, при котором по меньшей мере первую щель или просвет предварительно покрывают HBSS или водой.
18. Способ по любому из пп. 15-17, при котором первая щель или просвет характеризуются внутренним диаметром, составляющим от приблизительно 0,5 мм до приблизительно 2,0 мм.
19. Способ по п. 18, при котором первая щель или просвет характеризуются внутренним диаметром, составляющим от приблизительно 0,7 мм до приблизительно 1,5 мм.
20. Способ по п. 19, при котором первая щель или просвет характеризуются внутренним диаметром, составляющим приблизительно 1,1 мм.
21. Способ по любому из пп. 15-20, при котором суспендирование через первую щель или просвет проводят в течение от приблизительно 1 минуты до приблизительно 10 минут.
22. Способ по п. 21, при котором суспендирование через первую щель или просвет проводят в течение приблизительно 5 минут.
23. Способ по любому из пп. 15-22, при котором последние две щели или просвета в серии характеризуются внутренними диаметрами, составляющими от приблизительно 90 до приблизительно 200 мкм и от приблизительно 25 мкм до приблизительно 90 мкм.
24. Способ по п. 23, при котором последние две щели или просвета в серии характеризуются внутренними диаметрами, составляющими от приблизительно 100 до приблизительно 150 мкм и от приблизительно 50 мкм до приблизительно 70 мкм.
25. Способ по любому из пп. 1-24, при котором суспендирование предусматривает от приблизительно 5 до приблизительно 20 минут суспендирования через от второй до последней щели или просвета и от приблизительно 5 до приблизительно 20 минут суспендирования в последней щели или просвете.
26. Способ по п. 25, при котором суспендирование предусматривает приблизительно 10 минут суспендирования через от второй до последней щели или просвета и приблизительно 15 минут суспендирования в последней щели или просвете.
27. Способ по любому из пп. 15-26, при котором суспендирование в последней щели или просвете продолжают, пока суспензия не будет легко проходить через щель или просвет.
28. Способ по любому из пп. 15-27, при котором суспендирование в каждой щели или просвете продолжают, пока суспензия не будет легко проходить через такую щель или просвет.
29. Способ по любому из пп. 1-28, при котором общее время суспендирования составляет приблизительно 30 минут.
30. Способ по любому из пп. 1-29; при котором приблизительно 5 до приблизительно 15 объемов HBSS добавляют после суспендирования.
31. Способ по любому из пп. 1-30; при котором приблизительно 10 объемов HBSS добавляют после суспендирования.
32. Способ по любому из пп. 1-31, при котором рН HBSS, добавленного после суспендирования, составляет от приблизительно 5,1 до приблизительно 5,7.
33. Способ по любому из пп. 1-32, при котором рН HBSS, добавленного после суспендирования, составляет приблизительно 5,4.
34. Способ по любому из пп. 1-33, при котором рН клеточной суспензии в HBSS, полученной на стадии f по п. 62 или стадии d по п. 61, составляет от приблизительно 5,0 до приблизительно 6,0.
35. Способ по любому из пп. 1-33, при котором рН клеточной суспензии в HBSS, полученной на стадии f по п. 62 или стадии d по п. 61, составляет от приблизительно 5,6 до приблизительно 5,7.
36. Способ по любому из пп. 1-35, при котором после добавления HBSS после суспендирования клетки присутствуют в концентрации, составляющей от приблизительно 0,5 миллиона клеток/мл до приблизительно 4 миллионов клеток/мл.
37. Способ по любому из пп. 1-36, при котором после добавления HBSS после суспендирования клетки присутствуют в концентрации, составляющей приблизительно 2 миллиона клеток/мл.
38. Способ по любому из пп. 1-37, при котором среда представляет собой среду для образования клеточных сфер, содержащую DMEM/F12, приблизительно 1% антибиотика, приблизительно 2% В27 и необязательно один или несколько факторов роста.
39. Способ по п. 38, при котором факторы роста содержат bFGF, EGF и гепарин.
40. Способ создания плюрипотентной клетки, при котором исходная клетка присутствует в ткани, содержащей эритроциты, причем способ предусматривает следующее:
a. Механическое разрезание ткани на тонкие срезы в присутствии одного или нескольких расщепляющих ЕСМ ферментов;
b. Инкубация образца, полученного на стадии а, при приблизительно естественной для ткани in vivo температуре при перемешивании ткани;
c. Разбавление клеточной суспензии, полученной на стадии b, в растворе сбалансированного солевого раствора Хенкса (HBSS) и АТР, характеризующемся рН, составляющим от приблизительно 4,0 до приблизительно 6,5;
d. Суспендирование клеточной суспензии, полученной на стадии с;
e. Ресуспендирование клетки, полученной на стадии d, в HBSS;
f. Выделение клетки из суспензии, полученной на стадии е; и
g. Ресуспендирование клеточного осадка, полученного на стадии g, в среде.
41. Способ создания плюрипотентной клетки, при котором исходная клетка присутствует в ткани, содержащей эритроциты, причем способ предусматривает следующее:
a. Механическое разрезание ткани на тонкие срезы в присутствии одного или нескольких расщепляющих ЕСМ ферментов;
b. Инкубация образца, полученного на стадии а, при приблизительно естественной для ткани in vivo температуре при перемешивании ткани;
c. Разбавление клеточной суспензии, полученной на стадии b, в растворе сбалансированного солевого раствора Хенкса (HBSS) и АТР, характеризующемся рН, составляющем от приблизительно 4,0 до приблизительно 6,5;
d. Выделение клетки из суспензии, полученной на стадии b;
e. Ресуспендирование клетки, полученной на стадии с, в растворе HBSS и АТР, характеризующемся рН, составляющим от приблизительно 4,0 до приблизительно 6,5, до клеточной суспензии;
f. Суспендирование клеточной суспензии, полученной на стадии d;
g. Ресуспендирование клетки, полученной на стадии е, в HBSS;
h. Выделение клетки из суспензии, полученной на стадии f; и
i. Ресуспендирование клеточного осадка, полученного на стадии g, в среде.
42. Способ по любому из пп. 40-41, при котором ткань, содержащую эритроциты, выбирают из группы, состоящей из следующего:
легкое; селезенка и печень.
43. Способ по любому из пп. 40-42, при котором ткань представляет собой легкое, и расщепляющий ЕСМ фермент представляет собой коллагеназу Р.
44. Способ по любому из пп. 40-43, при котором разрезание на тонкие срезы на стадии а продолжают в течение приблизительно 10 минут.
45. Способ по любому из пп. 40-44, при котором АТР присутствует в растворе HBSS и АТР в концентрации, составляющей от приблизительно 1,5 до приблизительно 5 мг/см3.
46. Способ по п. 45, при котором АТР присутствует в растворе HBSS и АТР в концентрации, составляющей от приблизительно 2,7 мМ до приблизительно 9 мМ.
47. Способ по любому из пп. 40-46, при котором раствор HBSS и АТР, характеризующийся рН, составляющим от приблизительно 4,0 до приблизительно 6,5, получают путем титрования раствора HBSS с раствором АТР до достижения требуемого рН.
48. Способ по п. 47, при котором раствор АТР характеризуется концентрацией, составляющей от приблизительно 50 мМ до приблизительно 500 мМ.
49. Способ по п. 48, при котором раствор АТР характеризуется концентрацией, составляющей приблизительно 200 мМ.
50. Способ по любому из пп. 40-49, при котором раствор HBSS и АТР характеризуется рН, составляющим от приблизительно 5,0 до приблизительно 5,7.
51. Способ по любому из пп. 40-50, при котором раствор HBSS и АТР характеризуется рН, составляющим приблизительно 5,0.
52. Способ по любому из пп. 40-50, при котором HBSS согласно стадии g по п. 101 и стадии е по п. 100 не содержит АТР.
53. Способ по любому из пп. 40-50, при котором выделение предусматривает центрифугирование.
54. Способ по любому из пп. 40-53, дополнительно предусматривающий приведение исходной клетки в контакт с трипсином в течение от приблизительно 1 минуты до приблизительно 10 минут перед стадией с.
55. Способ по п. 54, дополнительно предусматривающий приведение исходной клетки в контакт с трипсином в течение от приблизительно 3 минут до приблизительно 5 минут перед стадией с.
56. Способ по любому из пп. 54-55, при котором трипсин дезактивируют путем приведения клеточного осадка в контакт с минимальной эссенциальной средой Игла в модификации Дюльбекко (DMEM)/F-12, содержащей 10% термически инактивированной фетальной бычьей сыворотки (FBS).
57. Способ по любому из пп. 40-57, при котором суспендирование предусматривает суспендирование клеток через серию щелей или просветов постепенно уменьшающихся диаметров.
58. Способ по п. 57, при котором серия содержит по меньшей мере 3 щели или просвета.
59. Способ по любому из пп. 40-58, при котором по меньшей мере первую щель или просвет предварительно покрывают HBSS или водой.
60. Способ по любому из пп. 40-59, при котором первая щель или просвет характеризуются внутренним диаметром, составляющим от приблизительно 0,5 мм до приблизительно 2,0 мм.
61. Способ по п. 60, при котором первая щель или просвет характеризуются внутренним диаметром, составляющим от приблизительно 0,7 мм до приблизительно 1,5 мм.
62. Способ по п. 61, при котором первая щель или просвет характеризуются внутренним диаметром, составляющим приблизительно 1,1 мм.
63. Способ по любому из пп. 40-62, при котором суспендирование через первую щель или просвет проводят в течение от приблизительно 1 минуты до приблизительно 10 минут.
64. Способ по п. 40-63, при котором суспендирование через первую щель или просвет проводят в течение приблизительно 5 минут.
65. Способ по любому из пп. 40-64, при котором последние две щели или просвета в серии характеризуются внутренними диаметрами, составляющими от приблизительно 90 до приблизительно 200 мкм и от приблизительно 25 мкм до приблизительно 90 мкм.
66. Способ по п. 65, при котором последние две щели или просвета в серии характеризуются внутренними диаметрами, составляющими от приблизительно 100 до приблизительно 150 мкм и от приблизительно 50 мкм до приблизительно 70 мкм.
67. Способ по любому из пп. 40-66, при котором суспендирование предусматривает от приблизительно 5 до приблизительно 20 минут суспендирования через от второй до последней щели или просвета и от приблизительно 5 до приблизительно 20 минут суспендирования в последней щели или просвете.
68. Способ по п. 67, при котором суспендирование предусматривает приблизительно 10 минут суспендирования через от второй до последней щели или просвета и приблизительно 15 минут суспендирования в последней щели или просвете.
69. Способ по любому из пп. 40-68, при котором суспендирование в последней щели или просвете продолжают, пока суспензия не будет легко проходить через щель или просвет.
70. Способ по любому из пп. 40-69, при котором суспендирование в каждой щели или просвете продолжают, пока суспензия не будет легко проходить через такую щель или просвет.
71. Способ по любому из пп. 40-70, при котором общее время суспендирования составляет приблизительно 30 минут.
72. Способ по любому из пп. 40-71, при котором приблизительно 5 до приблизительно 15 объемов HBSS добавляют после суспендирования.
73. Способ по любому из пп. 40-72; при котором приблизительно 10 объемов HBSS добавляют после суспендирования.
74. Способ по любому из пп. 40-73, при котором рН HBSS, добавленного после суспендирования, составляет от приблизительно 5,1 до приблизительно 5,7.
75. Способ по любому из пп. 40-74, при котором рН HBSS, добавленного после суспендирования, составляет приблизительно 5,4.
76. Способ по любому из пп. 40-75, при котором рН клеточной суспензии в HBSS, полученной на стадии g по п. 101 или стадии е по п. 100, составляет от приблизительно 5,0 до приблизительно 6,0.
77. Способ по любому из пп. 40-76, при котором рН клеточной суспензии в HBSS, полученной на стадии g по п. 101 или стадии е по п. 100, составляет от приблизительно 5,6 до приблизительно 5,7.
78. Способ по любому из пп. 40-77, при котором после добавления HBSS после суспендирования клетки присутствуют в концентрации, составляющей от приблизительно 0,5 миллиона клеток/мл до приблизительно 4 миллиона клеток/мл.
79. Способ по любому из пп. 40-78, при котором после добавления HBSS после суспендирования клетки присутствуют в концентрации, составляющей приблизительно 2 миллиона клеток/мл.
80. Способ по любому из пп. 40-79, при котором среда представляет собой среду для образования клеточных сфер, содержащую DMEM/F12, приблизительно 1% антибиотика, приблизительно 2% В27 и необязательно один или несколько факторов роста.
81. Способ по п. 80, при котором факторы роста содержат bFGF, EGF и гепарин.
82. Способ по любому из пп. 1-81, при котором исходная клетка представляет собой мышиную клетку, и среда для образования клеточных сфер содержит LIF.
83. Способ по любому из пп. 1-81, дополнительно предусматривающий стадию культивирования полученных клеток в течение по меньшей мере одной недели, причем культивирование предусматривает следующее:
j. Добавление среды для образования клеточных сфер, необязательно содержащей факторы роста;
k. Перемешивание клеток для препятствия прикрепления ко дну чашки.
84. Способ по п. 83, при котором среду для образования клеточных сфер добавляют каждые 1-4 дня.
85. Способ по п. 83, при котором среду для образования клеточных сфер добавляют каждые 2 дня.
86. Способ по любому из пп. 82-85, при котором перемешивание предусматривает пипетирование клеток с помощью пипетки.
87. Способ по п. 86, при котором пипетка содержит отверстие, составляющее приблизительно 1,1 мм в диаметре.
88. Способ по любому из пп. 86-87, при котором пипетирование проводят по меньшей мере один раз в день.
89. Способ по п. 88, при котором пипетирование проводят по меньшей мере два раза в день.
90. Способ по любому из пп. 1-89, дополнительно предусматривающий выбор клетки с плюрипотентностью, причем выбор предусматривает способ, выбранный из группы, состоящей из следующего:
выбор клеток с низким прилипанием; выбор клеток, которые представляют собой компонент сферы; и выбор клеток с маленьким относительным размером.
91. Способ по любому из пп. 1-90, дополнительно предусматривающий конечную стадию выбора клетки, проявляющей плюрипотентность.
92. Способ по любому из пп. 1-91, при котором исходная клетка не присутствует как часть ткани.
93. Способ по любому из пп. 1-92, при котором исходная клетка представляет собой соматическую клетку, стволовую клетку, клетку-предшественник или эмбриональную клетку.
94. Способ по любому из пп. 1-93, при котором исходная клетка представляет собой выделенную клетку.
95. Способ по любому из пп. 1-94, при котором исходная клетка присутствует в гетерогенной популяции клеток.
96. Способ по любому из пп. 1-95, при котором исходная клетка присутствует в гомогенной популяции клеток.
97. Способ по любому из пп. 1-96, при котором выбор клетки, проявляющей плюрипотентность, предусматривает выбор клетки, экспрессирующей маркер стволовой клетки.
98. Способ по п. 97, при котором маркер стволовой клетки выбирают из группы, состоящей из следующего:
Oct4; Nanog; Е-кадгерин и SSEA4.
99. Способ по любому из пп. 1-98, при котором выбор клетки, проявляющей плюрипотентность, предусматривает выбор клетки, которая не является адгерентной.
100. Способ по любому из пп. 1-99, дополнительно предусматривающий культивирование плюрипотентной клетки для обеспечения размножения плюрипотентной клетки.
101. Способ по любому из пп. 1-100, при котором плюрипотентная клетка экспрессирует маркер стволовой клетки.
102. Способ по п. 101, при котором маркер стволовой клетки выбирают из группы, состоящей из следующего:
Oct4; Nanog; Е-кадгерин и SSEA4.
103. Способ по любому из пп. 1-102, при котором исходная клетка представляет собой клетку млекопитающего.
104. Способ по любому из пп. 1-103, при котором исходная клетка представляет собой клетку человека.
105. Способ по любому из пп. 1-104, при котором исходная клетка представляет собой взрослую клетку, неонатальную клетку, фетальную клетку, амниотическую клетку или клетку пуповинной крови.
106. Способ по любому из пп. 1-105, дополнительно предусматривающий содержание плюрипотентной клетки in vitro.
107. Анализ, предусматривающий:
приведение в контакт плюрипотентной клетки, полученной с помощью способа по любому из пп. 1-106, с кандидатным средством.
108. Анализ по п. 107 для применения в идентификации средств, которые влияют на одно или несколько из жизнеспособности, дифференцировки, пролиферации плюрипотентной клетки.
109. Применение плюрипотентной клетки, полученной с помощью способа по любому из пп. 1-106, в способе клеточной терапии для субъекта.
110. Способ получения клетки или ткани, которые являются совместимыми с клеточной терапией, подлежащей введению субъекту, предусматривающий:
создание плюрипотентной клетки из клетки по любому из пп. 1-106;
причем клетка представляет собой аутологичную клетку совпадающую по HLA аллогенную клетку.
111. Способ по п. 110, дополнительно предусматривающий дифференцировку плюрипотентной клетки по заданной линии дифференцировки перед введением клетки или ткани субъекту.
112. Композиция, содержащая плюрипотентную клетку, причем плюрипотентная клетка создана из клетки с помощью способов по любому из пп. 1-106.
113. Способ аутологичной клеточной терапии у субъекта, нуждающегося в клеточной терапии, предусматривающий следующее:
c. создание плюрипотентной клетки из клетки по любому из пп. 1-106, причем клетку получают от субъекта, и
d. введение композиции, содержащей плюрипотентную клетку или ее дифференцированное потомство, субъекту.
114. Способ по п. 113, дополнительно предусматривающий дифференцировку плюрипотентной клетки по заданной линии дифференцировки перед введением композиции субъекту.
Некоторые варианты осуществления описанной в настоящем документе технологии можно определить согласно любому из следующих пронумерованных абзацев:
1. Способ лечения неврологического повреждения, причем способ предусматривает введение плюрипотентной клетки или клетки STAP субъекту, нуждающемуся в лечении неврологического повреждения.
2. Способ по п. 1, при котором плюрипотентную клетку или клетку STAP создают с помощью описанных в настоящем документе способов, например, согласно абзацам, начиная со второго абзаца на странице 62 и по третий абзац на странице 56, или согласно приведенным выше пронумерованным абзацам, примеру 5 или примеру 7.
3. Способ по любому из пп. 1-2, при котором клетка является аутологичной субъекту.
4. Способ по любому из пп. 1-3, при котором неврологическое повреждение выбирают из группы, состоящей из следующего:
острое неврологическое повреждение; хроническое неврологическое повреждение; дегенеративное неврологическое заболевание; повреждение нерва или повреждение спинного мозга.
ПРИМЕРЫ
ПРИМЕР 1
Все организмы обладают первобытным инстинктом выживания. Если растения подвергают значительным внешним стрессовым воздействиям, они активируют механизм выживания, который вызывает дедифференцировку клеток и обеспечивает регенерацию поврежденной области или всего организма. Наряду с тем, что такие механизмы, вероятно, присущи низшим организмам для выживания при экстремальных изменениях окружающей среды, они документально подтверждены у млекопитающих.
Авторы настоящего изобретения выдвинули гипотезу о том, что физический стресс может вызывать переход зрелых клеток млекопитающего обратно в состояние стволовой клетки, аналогично тому, что наблюдается у растений и низших организмов. Для исследования указанной гипотезы изучали зрелые клетки, полученные из взрослых соматических тканей. Для исследования первого фокуса, на котором физические стрессовые воздействия могут являться наиболее эффективными в изменении зрелых клеток для их обратного развития до менее зрелого состояния, изучали CD45-положительные лимфоциты, собранные от мышей Oct4-GFP. Клетки от указанных мышей предоставляют информацию в отношении обратного развития до фенотипа стволовой клетки при активации специфического в отношении стволовой клетки промотора Oct4. Зрелые, полностью дифференцированные клетки подвергали воздействию нескольких значительных внешних стимулов.
Например, CD45-положительные лимфоциты подвергали воздействию раствора с низким значением рН для обеспечения сильного химического стресса. В пределах 3 дней воздействия наблюдали появление экспрессирующих GFP клеток и в пределах 5 дней наблюдали появление сферических колоний, состоящих из экспрессирующих GFP клеток. Клетки, созданные таким образом, в указанном примере называют измененные стрессом стволовые клетки (SASC, или SAC). SAC также можно назвать обновленными стволовыми клетками (RSC), или клетки животного каллюса (АСС). SAC экспрессировали несколько маркеров, в норме ассоциированные с эмбриональными стволовыми клетками. SAC проявляли дифференцировочный потенциал, эквивалентный клеткам ES клетки, вносили вклад в создание химерных мышей и являлись способными к созданию целого плода при инъекции в бластоцисты 4N. Клетки, созданные таким образом, исходно показали низкую митохондриальную активность и другие состояния, в норме ассоциированные с индукцией клетки на основе механизмов борьбы с повреждением Затем они проявляли деметилирование промоторов генов Oct4 и Nanog. Перепрограммирование измененных под действием стресса клеток, вероятно, было индуцировано посредством мезенхимальные-эпителиального перехода. Данные согласуются с описаниями клеток, содержащихся в растительном каллусе, в ответ на повреждение (внешние стимулы). Растительный каллус образован из индуцированного стрессом превращения клеток в плюрипотентные растительные стволовые клетки, способные к образованию клональных организмов. Такая сферическая колония, созданная из зрелых полностью дифференцированных соматических клеток млекопитающего в ответ на значительные внешние стимулы, в настоящем документе называется животный каллюс, а измененные под действием стресса клетки, содержащиеся в такой колонии или каллюсе, "клетки животного каллюса" (АСС), или SAC.
Таким образом, значительные физические и химические стрессовые воздействия, вызывали перепрограммирование нормальных зрелых взрослых клеток в плюрипотентные стволовые клетки, способные к эмбриогенезу. Не связываясь с какой-либо теорией, механизм перепрограммирования, вероятно, включает в себя индукцию клеточной выживаемости и процесса репарации, в норме наблюдаемые в ответ на повреждение. В настоящем документе продемонстрировано, что клетки млекопитающего обладают механизмом выживания, очень сходным с таковым у растений, с обратным развитием до перепрограммированного состояния в ответ на значительные стрессогенные внешние стимулы.
Как сообщалось, различные типы клеток подвергли перепрограммированию до состояния плюрипотентной стволовой клетки посредством индукции или усиленной экспрессии конкретных генов1-5. Также полагают, что повреждение клеток в результате воздействия таких раздражающих факторов, как ожоги, химическое повреждение, травма и облучение, может изменять нормальные клетки так, что они становятся злокачественными клетками.
Введение
Вероятно, все организмы обладают общими инстинктом выживания при повреждении, связанном со стрессогенными стимулами, путем собственной адаптации к окружающей среде и регенерации своего организма. У растений онтогенез наблюдается не только в зиготах, но также в полностью дифференцированных клетках и незрелой пыльце. У позвоночных тритоны являются способными к регенерации нескольких анатомических структур и органов, включая в себя их конечности1. Особо следует отметить, что существенную регенеративную способность продемонстрировали как растения, так и тритоны, индуцированную внешними стимулами, которые вызвали клеточную дедифференцировку ранее полностью дифференцированных соматических клеток. Наряду с тем, что прошли миллиарды лет, начиная с ранних форм жизни, и различные организмы развились по своему уникальному пути, это инстинкт выживания может наследоваться от общего предка до организмов современбной эпохи. Несмотря на то, что считается, что окончательно дифференцированные клетки млекопитающего в норме являются неспособными к изменению процесса дифференцировки на противоположный, млекопитающие могут сохранять ранее недооцененную программу избегать гибели в ответ на сильнодействующие изменения окружающей среды.
Растительный каллюс представляет собой массу пролиферирующих клеток, образованные в ответ на такие внешние стимулы, как ранение, которые можно стимулировать в культуре с помощью растительных гормонов2. Каллюс содержит перепрограммированные соматические клетки, которые называются каллюсные клетки, каждая из которых способна клонально регенерировать до целого организма. Каллюсные клетки не являются присущими растениям, но они создаются из соматических клеток в ответ на внешние стимулы. Несмотря на то, что недавние исследования продемонстрировали, что соматические клетки млекопитающего можно перепрограммировать за счет экзогенных процессов, таких как генная индукция3-7, не сообщалось о перепрограммировании соматических клеток млекопитающего в ответ на внешние физические и/или химические стимулы, таким способом, который параллелен тому, что имеет место у растений. Интересно, что, как полагают, экстремальные внешние стимулы, такие как воздействие раздражающие средства, включая в себя ожоги, химическое повреждение, травму и облучение, могут изменять нормальные соматические клетки так, что они превращаются в злокачественные клетки. Такие наблюдения, как предполагают, указывают на то, что внешние стимулы будут приводить к изменению клеток млекопитающего.
В настоящем исследовании авторы выдвинули гипотезу о том, что клетки млекопитающего сохраняют механизм выживания в ответ на воздействие значительного внешнего стресса, таким же образом, как это наблюдается у растений. В настоящем документе представлено доказательство того, что приложение значительного физических и химических стимулов может вызывать перепрограммирование зрелых, полностью дифференцированных соматических клеток млекопитающего, происходящих из различных тканей, и что такие измененные под действием стресса клетки способны образовывать животный каллюс, содержащий "клетки животного каллюса", которые могут регенерировать клональный организм.
Результаты
Значительные физические и химические стимулы, приложенные к зрелым соматическим клеткам. Поскольку эмбриональный фактор транскрипции Oct4, как полагают, является ключевым в регуляции плюрипотентного статуса клеток, исходная стратегия состояла в идентификации таких внешних стимулов, которые наиболее эффективно изменяют зрелые клетки так, чтобы они становились перепрограммированными для экспрессии Oct4. CD45-положительные клетки гемопоэтической линии дифференцировки вначале исследовали, чтобы избежать загрязнения недифференцированными клетками. CD45-положительные клетки, собранные из селезенок, полученных от мышей Oct4-GFP (GOF)8, подвергали воздействию различных значительных физических и химических стимулов. Воздействия включали в себя: обработку осмотическим давлением, обработку с помощью значительной механической суспендирования, воздействие низкого значения рН, применение повреждения мембраны клетки с использованием стрептолизина О (SLO), воздействие недостаточного содержания питательных веществ и воздействие гипоксии и высокой концентрации Са2+. Далее экспрессирующие GFP клетки идентифицировали, сортировали и собирали с использованием FACS. Экспрессию гена Oct4 подтверждали с помощью ОТ-ПЦР. Воздействие каждого из приложенных стимулов приводило к перепрограммированию зрелых клеток с экспрессией GFP до некоторой степени (фигура 5А). Воздействие на зрелые клетки химического стресса с помощью низкого значения рН и физического стресса за счет значительной механической суспендирования, как оказалось, являются наиболее эффективными обработками в изменении зрелых клеток для экспрессии Oct4. Для определения оптимального значения рН для индукции превращения в экспрессирующие Oct4 клетки, CD45-положительные клетки подвергали воздействию растворов с изменяющейся кислотностью от рН 4,0 до рН 6,8. Через 3 дня после воздействия кислого раствора экспрессию GFP клеток анализировали с использованием FACS. Кислый раствор со значением рН 5,4-5,6, наиболее эффективно изменял клетки с экспрессией GFP (фигура 5В). Следовательно, на воздействии с помощью низкого значения рН сфокусировались в качестве стрессового воздействия выбора для остального исследования.
Затем определяли оптимальные условия культивирования для содержания измененных под действием стресса экспрессирующих Oct4 клеток. Исследовали несколько ранее описанных сред культивирования, включая в себя: среда культивирования для формирования ES, 3i9 и АСТН10, условие культивирования ES, ES-LIF11, условие культивирования эмбриональных нейрональных стволовых клеток, B27-LIF12 и условие культивирования EpiSC13. Клетки помещали в каждую сред и подсчитывали экспрессирующие GFP колонии (фигура 5С). Среда B27-LIF оказалась наиболее эффективной в создании экспрессирующих GFP сферических колоний. Следовательно, среду B27-LIF использовали для культивирования обработанных клеток.
Подвергнутые воздействию стресса CD45-положительные клетки культивировали в среде B27-LIF и в пределах 5 дней наблюдали появление экспрессирующих GFP сферических колоний, тогда как в необработанном контроле не наблюдали экспрессирующих GFP колоний (фигура 1А). Сферические колонии росли до приблизительно 70 мкм в диаметре в течение первых 7 дней и сферические колонии могли сохраняться в течение следующих 7 дней при указанном условии культивирования. Конфигурация колоний характеризовалась слегка неправильной формы, проявляя более сходную форму с каллюсом, наблюдаемым в ботанике, чем со сферами. Клеточную колонию, созданную под воздействием стресса, в связи с этим, в настоящем документе называли животный каллюс (АС). Культивированные клетки диссоциировали и затем проводили анализ популяции с использованием FACS. Анализ выявил, что приложение определенных значительных стимулов приводило к созданию измененных под действием стресса клеток, которые теперь называют клетки животного каллюса (АСС), которые не существовали ранее в популяции CD45-положительных клеток (фигура 1В). Фенотипическое изменение CD45-положительных клеток в результате воздействия стресса наблюдали на уровне отдельной клетки. Тогда как CD45-положительные клетки не экспрессировали GFP, АСС экспрессировали GFP, ассоциированный со сниженной экспрессией CD45 (данные не показаны). Исследование отдельных клеток выявило, что размер клетки обработанных клеток оказался меньше, чем у необработанных клеток. В связи с этим, размер клеток в популяции АСС анализировали с помощью FACS. Размер клеток АСС являлся достаточно маленьким, при этом 80% клеток составляли меньше чем 8 мкм в диаметре (фигура 1С).
Для исследования хронологического фенотипического изменения, ассоциированного с уменьшением CD45 и экспрессией Oct4, подвергнутые воздействию стресса CD45-положительные клетки анализировали в 1 день, 3 день и 7 день. В 1 день большинство клеток все еще экспрессировали CD45, но не Oct4. На 3 день экспрессия маркера стала выявлять CD45-отрицательные клетки или CD45-отрицательные/Осt4-положительные (dim) клетки. На 7 день экспрессия CD45 исчезла, и появились экспрессирующие Oct4 клетки (фигура 1D). Следует отметить, что во время первых 7 дней культивирования количество PI-положительных клеток (мертвых клеток) постепенно увеличивалось (данные не показаны), что указывало на то, что воздействие стресса и условия культивирования постепенно изменяло характер клеток и выбирало успешно изменившиеся клетки, которые экспрессировали Oct4.
Определение характеристик АСС. Для подтверждения перепрограммирования соматических клеток в результате воздействия внешних стимулов исследовали экспрессию генов-маркеров раннего эмбриогенеза АСС. В качестве положительного контроля раннего эмбриогенеза в следующих экспериментах использовали клетки ES. Характеристики экспрессии маркера и метилирования ДНК определяли следующим образом: иммунофлуоресцентное окрашивание на 7 день показало, что сферические колонии, содержащие АСС, однородно экспрессировали маркеры плюрипотентных клеток, антиген Е-кадгерина, Nanog, SSEA-1, РСАМ-1 и АР, и являлись положительными в отношении Oct4-GFP (данные не показаны). Анализ экспрессии генов показал, что клетки АСС и ES, но не первичные CD45-положительные клетки, экспрессировали сопоставимые содержания генов Oct4, Nanog, Sox2, Ecat1, Esg1, Dax1, Fgf5, Klf4 и Rex1 (фигура 2A). Экспрессия специфических в отношении ES генов в АСС достигала пика на 7 день (фигура 2А). Бисульфитное секвенирование проводили для определения статуса метилирования промоторов генов Oct4 и Nanog в АСС. Нативные лимфоциты и культивированные контрольные образцы лимфоцитов проявляли выраженное метилирование на обоих промоторах, тогда как АСС показали широко распространенное деметилирование указанных областей, что аналогично тому, что наблюдается в клетках ES (фигура 2В). Таким образом, продемонстрировали, что соматические клетки млекопитающего подвергли перепрограммированию под действием внешнего стресса.
Для подтверждения того, что экспрессия гена Oct4 является результатом воздействия стресса на зрелые клетки не только у мышей GOF, но и у мышей дикого типа, CD45-положительные лимфоциты собирали из селезенок, полученных от мышей ICR. Лимфоциты затем подвергали воздействию стресса и хронологически анализировали до 7 дня с использованием FACS. SSEA-1-положительную/Е-кадгерин-положительную клеточную популяцию наблюдали в подвергнутой воздействию стресса группе, тогда как экспрессию SSEA-1/Е-кадгерина не наблюдали в не подвергнутой воздействию стресса контрольной группе (фигура 6А). Указанные дважды положительные клетки экспрессировали ген Oct4, что подтверждали с помощью ОТ-ПЦР (фигура 6В). Указанные результаты продемонстрировали, что в результате воздействия стресса АСС, Осt4-положительные и экспрессирующая плюрипотентный маркер клетки создавали из CD45-положительных клеток независимо от линии мыши.
Указанные результаты означают, что зрелые полностью дифференцированные взрослые соматические клетки изменяют свое развитие на противоположное до "стволовости" в результате воздействия стресса.
Для оценки стволовости АСС исследовали их потенциал к самообновлению и их дифференцировочный потенциал. Для исследования их потенциала к самообновлению колонии АСС, полученные из ранее созревших CD45-положительных лимфоцитов, диссоциировали до отдельных клеток и помещали на 96-луночные планшеты по одной клетке на лунку в попытке создать происходящие клонально популяции. Через десять дней после помещения на планшеты наблюдали появление сферических колоний в 4 из 96 лунок. Время деления АСС варьировало от лунки к лунке. Некоторые делились за 12-16 ч, а другие делились за 30-34 ч. АСС пересевали по меньшей мере 5 раза, при этом наблюдали непрерывную экспрессию Oct4. Следовательно, АСС продемонстрировали потенциал к дифференцировке в клетки из всех трех зародышевых листков in vitro.
АС, полученные из зрелых лимфоцитов GOF, снова диссоциировали до отдельных клеток, сортировали так, чтобы они содержали только популяцию клеток, которая экспрессировала GFP, и затем культивировали в среде дифференцировки. Через 14-21 дней после помещения на планшеты клетки экспрессировали эктодермальный маркер, βIII-тубулин и GFAP, мезодермальный маркер, α-актин гладких мышц, и эндодермальный маркер, α-фетопротеин и цитокератин 7 (данные не показаны). Таким образом, АСС дифференцировались в клетки, представляющие все три зародышевых листка in vitro.
Вызванное стрессом изменение зрелых соматических клеток, происходящих из различных взрослых тканей. Для исследования того, можно ли АСС создать не только из зрелых лимфоцитов, но также из других типов соматических клеток, головной мозг, кожу, мышцу, жировую ткань, костный мозг, легкое и печень собирали от мышей Oct4-GFP (GOF)8. Клетки выделяли из образцов ткани, диссоциировали до отдельных клеток и обрабатывали с помощью различных физических и/или химических стрессовых условий. Эффективность способа изменения клеток отличалась в зависимости как от источника клеток, так и стрессового(ых) условия(й), которому(ым) подвергали клетки (фигура 7А). Способность стресса изменять зрелые клетки с экспрессией Oct4, различалась в зависимости от происхождения клеток, но стресс был способен изменить клетки с экспрессией Oct4 до некоторой степени в зрелых клетках, происходящих их всех трех зародышевых листков (фигура 7А). Колонии АСС, происходящие из любой зрелой ткани, экспрессировали плюрипотентные маркеры, Е-кадгерин, Nanog, РСАМ-1 и АР (данные не показаны), и специфические для ES маркерные гены (фигура 7В). Значительные физические и химические стрессовые воздействия изменяли зрелые соматические клетки с обратным развитием до состояния стволовой клетки независимо от источника тканей и происхождения зародышевых листков.
Клеточная модификация в начальной фазе создания АСС. Указанные результаты продемонстрировали, что сильные физические и химические стимулы приводят к перепрограммированию соматических клеток. Получили наблюдения, что подвергнутые воздействию стресса лимфоциты образуют АС в пределах 5 дней. Выдвинули гипотезу о том, что сильнодействующее изменение молекулярных событий происходило в результате стрессового воздействия. В связи с этим, исследования сосредоточили на начальной фазе перепрограммирования, которая происходила в течение первых 7 дней после воздействия стимулов.
Поскольку АСС выживали после значительного стрессового воздействия, предположили, что механизмы выживания, в норме включающиеся для репарации клеточного повреждения, были индуцированы во время создания АСС. Вначале экспрессию ряда кандидатных генов, вовлеченных в клеточная ответ на стресс и репарацию ДНК14, сравнивали в нативных CD45-положительных клетках и подвергнутых воздействию стресса CD45-положительных клетках в 1 день, 3 день и 7 день. Экспрессию генов клеточного ответа наблюдали уже в 1 день, и указанные гены были подвергнуты положительной регуляции в течение 7 дней при анализе смесей создающих АСС клеток и других клеток (фигура 8). Поскольку положительная регуляция генов клеточного ответа коррелировала с созданием АСС, АСС на 3 день и 7 день сортировали и анализировали экспрессию генов. За исключением Hif3a, все кандидатные гены подвергались положительной регуляции до различных степеней во время создания АСС (фигура 3А). Обнаружили, что 4 гена теплового шока и один ген репарации ДНК были подвергнуты положительной регуляции во время создания АСС. Кроме того, 7 из подвергнутых положительной регуляции генов, как известно, напрямую вовлечены в регуляцию окислительно-восстановительного состояния клетки. Эти результаты указывают на то, что потенциал к саморепарации или самозащите был индуцирован во время создания АСС.
Поскольку АСС проявляли положительную регуляцию ассоциированных с окислительно-восстановительным статусом клетки генов, далее исследовали митохондриальную функцию АСС. Митохондрии представляют собой органеллы, отвечающие за продукцию подавляющего большинства АТР посредством окислительно-восстановительной реакции с использованием кислорода внутри эукариотических клеток. Экспрессия GFP сферических колоний АСС постепенно уменьшалась от расположенных на периферии клеток через 7 дней, если колонии культивировали без пересева. АСС на 10 день содержали экспрессирующие GFP центральные клетки и не-GFP дифференцированные периферические клетки (данные не показаны). Морфологию митохондрий оценивали в АСС и дифференцированных клетках с помощью окрашивания специфическим для митохондрий красителя, MitoTracker Red. Митохондрии АСС наблюдали в виде околоядерных кластеров, которые выглядели пятнистыми и глобулярными, тогда как дифференцированные клетка содержали множество митохондрий, которые являлись нитевидными и были широко распределены в цитоплазме. Продукция АТР в АСС была меньше, чем таковая в нативных CD45-положительных клетках (фигура 3В). Кроме того, продукция активных формы кислорода (ROS) АСС была меньше, чем в нативных CD45-положительных клетках (фигура 3С). В итоге, оценивали ключевые факторы, вовлеченные в репликацию мтДНК, которые представляли собой митохондриальный фактор транскрипции A (Tfam), специфическую для митохондрий ДНК-пролимеразу гамма (Polg) и ее вспомогательную структурную единицу (Polg2). Экспрессия генов Tfam, Polg и Polg2 в АСС была ниже, чем таковая в дифференцированных клетках (фигура 3D). Следовательно, АСС содержали небольшие количества митохондрий, и митохондриальная активность АСС была ниже, чем в дифференцированных клетках. Указанные результаты указывают на то, что АСС приобретали метаболическую систему, отличную от дифференцированных клеток, для выживания в ответ на тяжелое стрессовое воздействие.
Дифференцировочный потенциал АСС. В итоге, оценивали, обладают ли АСС дифференцировочным потенциалом, сходным с таковым у клеток растительного каллюса. В качестве начального исследования в отношении способности к развитию исследовали АСС, имплантированные подкожно мышам с иммунодефицитом (SCID). Через шесть недель после трансплантации АСС создавали ткани, представляющие все три зародышевых листка (данные не показаны).
АСС дифференцировались в клетки, представляющие все три зародышевых листка in vivo и in vitro. Следовательно, оценивали потенциал АСС вклада в создание химеры. АСС для применения в исследованиях по созданию химеры получали с использованием CD45-положительных клеток, происходящих из F1 GFP (C57BL/6GFP×DBA/2 или 129/SvGFP×C57BL/6GFP) или GOF. Поскольку анализ экспрессии генов выявил, что на 7 день АСС экспрессировали самый высокий уровень генов-плюрипотентных маркеров, то АСС на 7 день использовали для исследования по созданию химерной мыши. Вначале использовали общепринятые способы создания химеры. АС диссоциировали до отдельных клеток путем обработки трипсином. АСС затем вводили с помощью инъекции в бластоцисты (фигура 4А). С использованием указанного подхода вклад диссоциированных АСС в создание химеры был достаточно низким (таблица 1). Следовательно, АСС без предварительной обработки трипсином, которая часто вызывает клеточное повреждение15, вводили с помощью инъекции в бластоцисты. АС разрезали на маленькие кластеры с использованием микроскальпеля под микроскопом. Маленькие кластеры АС затем вводили с помощью инъекции в бластоцисты (фигура 4А). С использованием указанного подхода вклад АСС в образование химеры сильно увеличивался (данные не показаны). Химерные мыши, созданные с помощью АСС, вырастали здоровыми (данные не показаны), и наблюдали передачу зародышевой линии. Коэффициент вклада в создание химеры каждой ткани анализировали с помощью FACS. Результаты показали, что АСС, происходящие из лимфоцитов, вносили свой вклад во все ткани (фигура 4В).
Как продемонстрировано выше, АСС можно создать из различных клеток, происходящих из всех трех зародышевых листков (фигура 7А-7В). Для исследования того, характеризуются ли АСС, происходящие из различных тканей, различными тенденциями в дифференцировке, АСС создавали из различных тканей, происходящих от мышей F1GFP, и вводили с помощью инъекции в бластоцисты ICR. Затем с использованием FACS анализировали соотношение вклада каждой ткани в созданных химерных мышей. Обнаружили, что АСС, происходящие из любой ткани, вносили свой вклад в создание химерной мыши (фигура 9). Кроме того, соотношение вклада в отношении кожи, головного мозга, мышцы, жировой ткани, печени и легкого анализировали у химерных мышей, созданных с использованием АСС, происходящих из различных тканей. АСС, происходящие из любой ткани, вносили свой вклад в создание всех трех зародышевых листков, и в дифференцировке не наблюдали никакой тенденции (фигура 9).
Создание мышей с помощью тетраплоидной комплементации, которая предусматривает инъекцию плюрипотентных клеток в 4N бластоцисты-хозяева, представляет наиболее строгое исследование дифференцировочного потенциала, поскольку полученные эмбрионы происходят только из введенных с помощью инъекции донорных клеток16. АСС создавали из лимфоцитов, происходящих от мышей DBA×B6GFP F1 или 129/SvGFP×B6GFP F1. АСС приводили к созданию (mid) 'полностью АСС эмбрионов' на поздних сроках беременности после инъекции в бластоцисты 4N (данные не показаны). Анализ генотипирования продемонстрировал, что 'полностью АСС эмбрионы' содержали специфические гены линии, которую использовали для создания АСС. Таким образом, АСС обладали потенциалом создавать клональный организм аналогично клеткам растительного каллюса.
Обсуждение
Соматические клетки млекопитающего проявляют способность к образованию животного каллюса (АС) в результате воздействия значительных внешних стимулов, очень сходным с растениями образом. Клетки, содержащиеся в этих каллюсах (клетки животного каллюса, АСС) обладают способность создавать химерных мышей и создавать новые эмбрионы, полностью состоящие только из клеток, созданных из АСС. Описанные в настоящем документе результаты продемонстрировали, что соматические клетки млекопитающего восстанавливают способность дифференцироваться в любой из трех зародышевых листков под действием внешних стимулов. Это указывает на то, что соматические клетки обладают большей пластичностью, чем считали ранее. Кроме того, настоящее исследование продемонстрировало потенциал к перепрограммированию соматической клетки без введения генов или введения чужеродных белков, и обеспечивает новое понимание потенциала взрослых стволовых клеток; представляя значительный этап в прояснении биологии стволовых клеток.
Материалы и методы
Сбор тканей и культивирование клеток. Для выделения зрелых лимфоцитов, селезенки, полученные от мышей GOF или мышей ICR в возрасте 1 недели, нарезали ножницами и подвергали механической диссоциации с помощью пастеровской пипетки. Диссоциированные селезенки пропускали через клеточное сито (BD Biosciences, Сан-Хосе). Собранные клетки ресуспендировали в среде DMEM и добавляли такой же объем лимфолита (CEDARLANE®, Онтарио, Канада), затем центрифугировали при 1000g в течение 15 мин. Слой лимфоцитов отбирали и окрашивали с помощью антитела к CD45 (ab25603, abeam, Кембридж, Массачусетс). CD45-положительные клетки сортировали с помощью FACS Aria (BD Biosciences). Затем CD45-положительные клетки подвергали воздействию стресса (раствор с рН 5,5 в течение 15 мин) и помещали в среду В27, дополненную 1000 ед LIF (Sigma) и 10 нг/мл FGF 2 (Sigma).
Воздействие внешних стимулов - воздействие стресса. Для получения механического стресса на зрелые клетки пипетку Пастера нагревали и затем растягивали для создания просветов, составляющих приблизительно 50 мкм в диаметрах, и затем обламывали. Зрелые соматические клетки затем суспендировали через указанные пипетки в течение 20 мин и культивировали в течение 7 дней. Для обеспечения гипоксического стимула в отношении зрелых клеток клетки культивировали в инкубаторе с 5% кислорода в течение 3 недель. Стимул недостаточного содержания питательных веществ обеспечивали в отношении зрелых клеток путем культивирования клеток в базовой среде культивирования в течение 3 недель. Для воздействия на зрелые клетки физиологического стресса их обрабатывали с помощью раствора с низким значением рН (рН 5,5) и культивировали в течение 7 дней. Кроме того, клетки подвергали более серьезному повреждению. Для создания пор в мембранах зрелых клеток клетки обрабатывали с помощью SLO (стрептолизин О).
Обработанные SLO клетки инкубировали в HBSS, содержащем 10 мкг/мл SLO при 37°С в течение 50 мин и затем культивировали в среде культивирования без SLO в течение 7 дней. Подвергнутые воздействию недостаточного содержания питательных веществ клетки культивировали в базовой среде в течение 2-3 недель. Подвергнутые воздействию "АТР" стресса клетки инкубировали в HBSS, содержащем 2,4 мМ АТР при 37°С в течение 15 мин и затем культивировали в среде культивирования в течение 7 дней. Подвергнутые воздействию "Са" стресса клетки культивировали в среде культивирования, содержащей 2 мМ CaCl2 в течение 2 недель.
Бисульфитное секвенирование. Клетки, происходящих от мышей GOF, диссоциировали до отдельных клеток. GFP-положительные клетки собирали с использованием FACS Aria. Геномную ДНК экстрагировали из АСС и исследовали. Обработку ДНК бисульфитом выполняли с использованием набора для модификации ДНК CpGenome DNA Modification Kit (Chemicon, Темекула, Калифорния, http://www.chemicon.com) согласно инструкциям производителя. Полученную модифицированную ДНК амплифицировали с помощью гнездовой полимеразной цепной реакции с использованием двух прямых (F) праймеров и одного обратного (R) праймера: Oct4 (F1, GTTGTTTTGTTTTGGTTTTGGATAT (SEQ ID NO: 1; F2, ATGGGTTGAAATATTGGGTTTATTTA (SEQ ID NO: 2); R,CCACCCTCTAACCTTAACCTCTAAC (SEQ ID NO: 3)). и Nanog (F1, GAGGATGTTTTTTAAGTTTTTTTT (SEQ ID NO:4); F2, AATGTTTATGGTGGATTTTGTAGGT (SEQ ID NO: 5); R, CCCACACTCATATCAATATAATAAC (SEQ ID NO: 6)). ПЦР проводили с использованием варианта Taq для горячего старта TaKaRa Ex Taq Hot Start Version (RR030A). Секвенирование ДНК проводили с использованием праймера М13 при содействии GRAS (The Genome Resource and Analysis Unit).
Иммуногистохимия. Культивированные клетки фиксировали с помощью 4% параформальдегида и пермеабилизировали с помощью 0,1% Triton X-100/PBS перед блокированием с помощью 1% раствора BSA (Life Technology, Токио, Япония). Вторичные антитела представляли собой антитела козы против антител мыши или антител кролика с присоединенным Alexa-488 или -594 (Invitrogen). Ядра клеток визуанализировали с помощью DAPI (Sigma). Предметные стекла фиксировали с помощью средства, препятствующего выгоранию флуоресценции SlowFade Gold (Invitrogen).
Активированная флуоресценцией сортировка клеток и проточная цитометрия. Клетки получали согласно стандартным протоколам и суспендировали в 0,1% BSA/PBS на льду перед анализом с помощью FACS. PI (BD Biosciences) использовали для исключения мертвых клеток. Клетки сортировали на BD FACSAria SORP и анализировали на BD LSRII с помощью программного обеспечения BD FACSDiva (BD Biosciences).
Получение РНК и анализ ОТ-ПЦР. РНК выделяли с помощью набора RNeasy Micro kit (QIAGEN). Обратную транскрипцию проводили с помощью набора для синтеза первой цепи SupeSACript III First Strand Synthesis kit (Invitrogen). SYBR Green Mix I (Roche Diagnostics) использовали для амплификации и образцы анализировали на приборе Lightcycler-II (Roche Diagnostics).
Исследования на животных. Для исследований онкогенности клетки, суспендированные в 100 мл PBS, вводили с помощью инъекции подкожно в паховые области совпадающих по возрасту мышей с иммунодефицитом SCID. Мышей умерщвляли и подвергали вскрытию через 6 недель.
Анализ АТР и ROS. Межклеточное содержание АТР измеряли с помощью набора для биолюминесцентного анализа АТР - АТР Bioluminescence Assay Kit HS II (Roche) согласно протоколу поставщика. Интенсивность люминесценции измеряли с использованием микропланшетного люминометра Gelomax 96 Microplate Luminometer (Promega, Мадисон, Висконсин) и показатели люминесценции нормализовали по количеству клеток. Для измерения содержаний ROS клетки инкубировали в среде, содержащей 2 мкМ дигидроэтидия (Molecular Probes) при 37°С в темноте в течение 15 минут. Клетки затем отмывали с помощью PBS и суспендировали в PBS, содержащем 0,5% BSA. Интенсивность флуоресценции 30000 клеток регистрировали с помощью BD Biosciences LSR II (BD Bioscience, Спаркс, Мэриленд).
Создание и анализы химерных мышей. Получение диплоидных и тетраплоидных химер. Диплоидных эмбрионов получали от самок линии ICR, спаренных с самцами линии ICR, и тетраплоидных эмбрионов получали от самок линии BDF1, спаренных с самцами линии BDF1. Тетраплоидные эмбрионов получали с помощью электрослияния 2-клеточных эмбрионов17. В настоящем исследовании поскольку обработка трипсином вызывала низкий химеризм, сферические колонии АСС эмбрионов с использованием микроскальпеля под микроскопом, затем небольшие кластеры АСС инъецировали в бластоцисты на 4,5 сутки посредством большой пипетки. На следующий день химерные бластоцисты переносили псевдобеременным 2,5 суток самкам.
Ссылки
1. Brockes, J.P. & Kumar, A. Plasticity and reprogramming of differentiated cells in amphibian regeneration. Nature reviews. Molecular cell biology 3, 566-574, doi:10.1038/nrm881 (2002).
2. Sinnott, J.J. & Burklund, C.W. The treatment of carotid insufficiency. The Nebraska state medical journal 45, 357-359 (1960).
3. Hanna, J. et al. Direct reprogramming of terminally differentiated mature В lymphocytes to pluripotency. Cell 133, 250-264, doi:10.1016/j.cell.2008.03.028 (2008).
4. Hockemeyer, D. et al. A drug-inducible system for direct reprogramming of human somatic cells to pluripotency. Cell stem cell 3, 346-353, doi:10.1016/j.stem.2008.08.014 (2008).
5. Kim, D. et al. Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins. Cell stem cell 4, 472-476, doi:10.1016/j.stem.2009.05.005 (2009).
6. Kim, J.B. et al. Direct reprogramming of human neural stem cells by OCT4. Nature 461, 649-643, doi:10.1038/nature08436 (2009).
7. Okabe, M. et al. Definitive proof for direct reprogramming of hematopoietic cells to pluripotency. Blood 114, 1764-1767, doi:10.1182/blood-2009-02-203695 (2009).
8. Ohbo, K. et al. Identification and characterization of stem cells in prepubertal spermatogenesis in mice small star, filled. Developmental biology 258, 209-225 (2003).
9. Ying, Q.L. et al. The ground state of embryonic stem cell self-renewal. Nature 453, 519-523, doi:10.1038/nature06968 (2008).
10. Ogawa, K., Matsui, H., Ohtsuka, S. & Niwa, H. A novel mechanism for regulating clonal propagation of mouse ES cells. Genes to cells: devoted to molecular & cellular mechanisms 9, 471-477, doi:10.1111/j.1356-9597.2004.00736.x (2004).
11. Gough, N.M. et al. LIF: a molecule with divergent actions on myeloid leukaemic cells and embryonic stem cells. Reproduction, fertility, and development 1, 281-288 (1989).
12. Hitoshi, S. et al. Primitive neural stem cells from the mammalian epiblast differentiate to definitive neural stem cells under the control of Notch signaling. Genes & development 18, 1806-1811, doi:10.1101/gad. 1208404 (2004).
13. Tesar, P.J. et al. New cell lines from mouse epiblast share defining features with human embryonic stem cells. Nature 448, 196-199, doi:10.1038/nature05972 (2007).
14. Saretzki, G., Armstrong, L., Leake, A., Lako, M. & von Zglinicki, T. Stress defense in murine embryonic stem cells is superior to that of various differentiated murine cells. Stem Cells 22, 962-971, doi:10.1634/stemcells.22-6-962 (2004).
15. Mitalipova, M.M. et al. Preserving the genetic integrity of human embryonic stem cells. Nature biotechnology 23, 19-20, doi:10.1038/nbt0105-19 (2005).
16. Nagy, A., Rossant, J., Nagy, R., Abramow-Newerly, W. & Roder, J.C. Derivation of completely cell culture-derived mice from early-passage embryonic stem cells. Proceedings of the National Academy of Sciences of the United States of America 90, 8424-8428 (1993).
17. Nagy, A. et al. Embryonic stem cells alone are able to support fetal development in the mouse. Development 110, 815-821 (1990).

* Все плоды собирали на 13,5 dpc - 15,5 dpc, и долю вклада SAC в каждый орган оценивали посредством FACS
** Вклад SAC в каждую химеру оценивали как высокий (>50% окраса шерстяного покрова с экспрессией GFP)
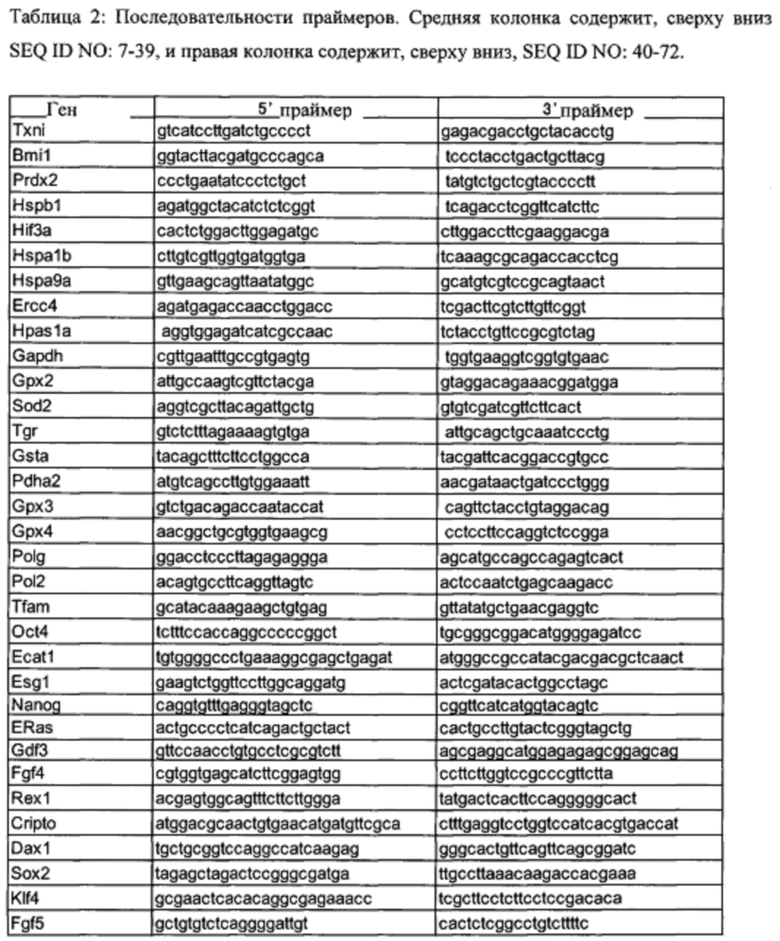
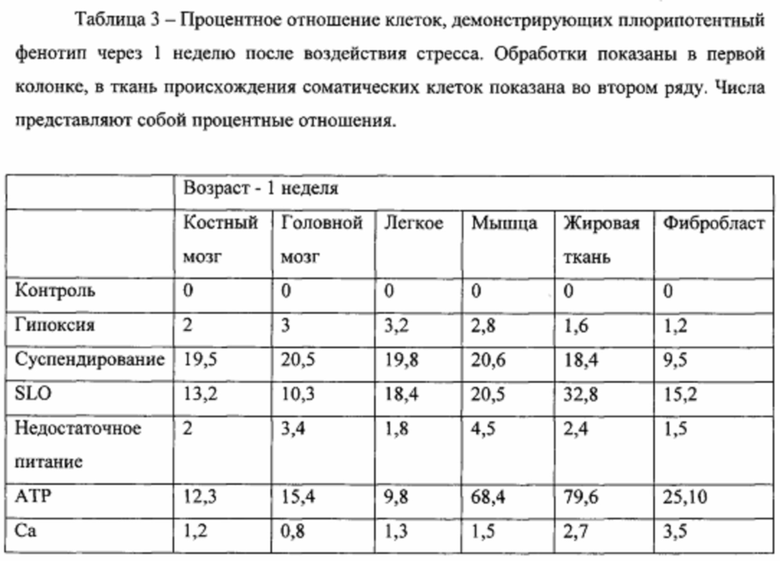
ПРИМЕР 2: Вызванное стимулом изменение судьбы соматических клеток до плюрипотентности
В настоящем документе описан феномен инициированной в ядре, «вызванной стимулом приобретенной плюрипотентности» (STAP), когда сильные внешние стимулы в достаточной мере перепрограммируют соматические клетки млекопитающих в плюрипотентные клетки, без использования переноса ядра или введения факторов транскрипции. В присутствии LIF, временный стресс при низком рН вызывает дедифференцировку CD45+ гемопоэтических клеток в клетки, которые экспрессируют маркеры плюрипотентных клеток, такие как Oct3/4, и обладают компетентностью для дифференцировки в клетки трех зародышевых листков. В этих клетках STAP, подобно клеткам ES, наблюдают значительное деметилирование в промоторных областях oct3/4 и nanog. Происходящие из гемопоэтических клеток клетки STAP несут перестройки гена Т-клеточного рецептора, что указывает на то, что эти коммитированные соматические клетки дают начало клеткам STAP посредством линейной конверсии. Инъекция в бластоцисты показывает, что клетки STAP эффективно вносят вклад в образовании химеры, даже в анализе тетраплоидной комплементации, и в образование потомства посредством передачи зародышевой линии. Таким образом, эпигенетическое состояние предопределения судьбы клеток можно радикальным образом инициализировать зависимым от контекста образом посредством сильных внешних стимулов.
При рассмотрении каналов эпигенетического ландшафта Уоддингтона, судьба соматических клеток постепенно определяется, по мере того, как клеточная дифференцировка катится вниз с холма. Как правило, считают, что обратное движение от статуса дифференцированных клеток требует искусственных, физических или генетических, манипуляций с функциями их ядер, таких как перенос ядра1 и введение множества факторов транскрипции2. Остается без ответа вопрос, могут ли соматические клетки подвергаться инициации их ядерной программы просто в ответ на внешние стимулы без этих непосредственных манипуляций с ядром. Известно, что такие ситуации возникают в растениях; резкие изменения условий культивирования могут изменять судьбу зрелых соматических клеток, например, от диссоциированных клеток моркови до незрелых клеток бластемы, из которых структура целого растения, включая стебли и корни, развивается в присутствии ауксинов. Трудным вопросом является то, могут ли соматические клетки животных обладать сходным потенциалом, который возникает по меньшей мере в специальных условиях. На протяжении последнего десятилетия, присутствие плюрипотентных клеток (или близкородственных типов клеток) во взрослых тканях являлось предметом споров, для которого противоречивые заключения опубликованы различными группами. Однако, ни одна из них не показала, что такие плюрипотентные клетки могут возникать из дифференцированных соматических клеток.
Гемопоэтические клетки, положительные по CD45 (общему лейкоцитарному антигену), представляют собой типичные линейно-коммитированные соматические клетки, которые часто используют в качестве исходных типов клеток для исследований перепрограммирования, таких как получение производных клеток iPS. Они никогда не экспрессируют связанные с плюрипотентностью маркеры, такие как Oct3/4, если не являются перепрограммированными. В частности, большинство CD45+ клеток из тканей селезенки считают составляющими популяции не относящихся к стволовым лейкоцитов (созревающих клеток или предшественников), и перевод в клетки iPS лимфоцитов, несущих геномные перестройки гена β-цепи (tcrβ) Т-клеточного рецептора, рассматривают как подлинный критерий перепрограммирования коммитированных соматических клеток. Авторы изобретения, таким образом, заинтересовались вопросом, можно ли перевести CD45+ клетки селезенки к приобретению плюрипотентности посредством резких изменений внешнего окружения, таких как изменения, вызванные простыми химическими возмущениями.
Результаты
Индуцированное воздействием низкого рН изменение судьбы коммитированных соматических клеток. CD45+ клетки, собранные из взрослых селезенок от мышей oct3/4::gfp В615, подвергали воздействию различных типов сильных временных стимулов, включая физические и химические стимулы, и исследовали по активации промотора осt3/4 после культивирования в суспензии с использованием содержащей LIF среды В27 в течение нескольких дней. Среди различных возмущений, сфокусировались на возмущениях посредством низкого рН. Как показано ниже, этот тип возмущений оказался наиболее эффективным для индукции oct3/4.
Без подвергания воздействию стимулов, ни одна из клеток, отсортированных по CD45, не экспрессировала oct3/4::GFP, вне зависимости от периода культивирования в содержащей LIF среде, позволяющей выживание отсортированных клеток. В отличие от этого, 30-минутная обработка CD45+ клеток селезенки средой с низким рН (рН4,5-6,0; фигура 12А) вызывала появление значительного количества oct3/4::GFP+ клеток на сутки 7 (d7) культивирования (фигура 12В; наиболее эффективный диапазон составлял рН5,4-5,8; фигура 16В). Эти клетки сохраняли экспрессию oct3/4::GFP без пассирования по меньшей мере в течение дополнительных 7 дней (всего 14 дней). На d7 в этой неадгезионной культуре, индуцированные низким рН oct3/4::GFP+ клетки формировали сферические (или обладающие немного неправильной формой) кластеры (данные не представлены; включающие от нескольких до нескольких дюжин клеток), которые больше не экспрессировали CD45 (фигура 12С). Интересно, что размер клеток для индуцированных низким рН oct3/4::GFP+ клеток являлся значительно меньшим, чем размер необработанных CD45+ клеток (см. иммунное окрашивание oct3/4::GFP и CD45 в отдельных клетках; фигура 12С); размер 80% упомянутых первыми клеток составлял менее 8 мкм в диаметре, в то время как размер контрольных CD45+ клеток лежал в диапазоне от 8 до 10 мкм (фигура 12D (левый пик показывает Oct3/4::GFP+ клетки, и правый пик показывает CD45+ клетки), как определено по анализу прямого светорассеяния в FACS). Эти наблюдения позволяют предполагать значительные различия между oct3/4::GFP+ и CD45+ популяциями, помимо различий в экспрессии двух маркеров.
Анализ зависимости от времени (фигура 12С) показал динамическое изменение популяций клеток в течение d1-d3. Большинство из выживших клеток на d1 (количество выживших клеток соответствовало ~85% от популяции на d0) все еще являлись CD45+ и oct3/4::GFP-. На d2 и d3, значительная популяция (21% и 34%, соответственно) общего количества выживших клеток становилась oct3/4::GFP+ и обладала слабым свечением для CD45 (фигура 12С; ~50-60% высеянных клеток были потеряны к тому времени). На d7, значительное количество oct3/4::GFP+/CD45- клеток (54% от общего количества выживших клеток) составляли популяцию, отдельную от популяции oct3/4::GFP-/CD45- (фигура 12В, сверху; общее количество клеток на d7 являлось сходным с количеством на d3). Не наблюдали очевидного образования oct3/4::GFP+/CD45- популяций в культуре необработанных CD45+ клеток (фигура 12В, снизу). Таким образом, размер oct3/4::GFP+/CD45- популяции в подвергнутой воздействию низкого рН группе являлся довольно значительным и соответствовал приблизительно половине общего количества выживших клеток на d7. Фактически, когда сигналы oct3/4::GFP впервые появлялись на d2, количество GFP+ клеток соответствовало ~8% от первоначально высеянных CD45+ клеток. Таким образом, кажется маловероятным, чтобы очень незначительная популяция (например, загрязняющая CD45- клетки) быстро выросла до формирования такой значительной oct3/4::GFP+ популяции в течение первых двух дней после воздействия низкого рН.
При анализе видеонаблюдения (данные не представлены), подвергнутые воздействию низкого рН CD45+ клетки, но не клетки без обработки, проявляли тенденцию формировать небольшие кластеры, которые постепенно начинали подавать сигналы GFP через первые несколько дней. Затем, эти небольшие oct3/4::GFP+ кластеры часто сливались и формировали более крупные сферы к d5, что указывает на то, что кластеры являются мультиклональными. Интересно, что эти GFP+ кластеры (но не GFP- клетки) являлись достаточно подвижными, и часто происходило выпячивание клеток (данные не представлены).
Чтобы протестировать, вносят ли вклад линейно-коммитированные CD45+ клетки селезенки, в частности, популяции Т-клеток, в oct3/4::GFP+ клетки, геномные перестройки tcrβ исследовали в выделенных oct3/4::GFP+ сферах посредством геномной ПЦР, и обнаружили, что каждая сфера содержала клетки с перестройками гена tcrβ (данные не представлены). Чтобы исключить возможность детекции перестройки в загрязняющих oct3/4::GFP-/CD45+ клетках, oct3/4::GFP+/CD45- клетки подвергали сортировке посредством FACS на d7 и подвергали анализу перестройки гена tcrβ. В этом случае, также явно наблюдали перестройки гена tcrβ (фигура 12Е). Эти данные демонстрируют, что эти популяции коммитированных соматических клеток в клетках селезенки (по меньшей мере, Т-клеток) вносили вклад в oct3/4::GFP+ клетки посредством изменения их судьбы от CD45+ до oct3/4::GFP+.
Индуцированные воздействием низкого рН Oct3/4+ клетки обладают плюрипотентностью. Затем исследовали, представляет ли экспрессия oct3/4::GFP+ в индуцированных стимулом клетках плюрипотентное состояние этих клеток или просто специфическое изменение характера экспрессии гена (в этом случае, oct3/4 и cd45) без приобретения плюрипотентности. Иммунологическое окрашивание показало, что на d7 oct3/4::GFP+ сферы экспрессировали связанные с плюрипотентностью маркеры, такие как Oct3/4, SSEA-1, Nanog, Е-кадгерин и АР (данные не представлены). Анализ экспрессии генов посредством qPCR показал, что эти индуцированные воздействием низкого рН oct3/4::GFP+ клетки на d7, в отличие от CD45+ клеток, экспрессировали уровни генов oct3/4, nanog, sox2, ecat1, esg1, dax1 и klf4, сравнимые с уровнями в ES клетках (фигура 13А (серии представляют собой, слева направо, экспрессию oct3/4, nanog, sox2, ecat1, esg1, dax1 и klf4); эти маркеры являлись положительными уже на d3), что указывает на то, что индуцированные воздействием низкого рН oct3/4::GFP+ клетки экспрессируют набор истинных маркерных генов, характерных для плюрипотентности, которые никогда не экспрессируются в клетках CD45+.
Затем исследовали, сопровождалось ли это сильное изменение характера экспрессии генов изменениями в эпигенетической модификации связанных с плюрипотентностью генов. С этой целью, проводили бисульфитное секвенирование для проверки статуса метилирования промоторных областей oct3/4 и nanog. Для CD45+ клеток, после дополнительного культивирования или без него, показан сильно метилированный характер обоих промоторов. В отличие от этого, для индуцированных воздействием низкого рН oct3/4::GFP+ клеток показано обширное деметилирование в этих областях, подобно ES клеткам (фигура 13В), демонстрирующее, что клетки подверглись значительному перепрограммированию эпигенетического статуса этих ключевых для плюрипотентности генов.
Затем проверяли, обладают ли индуцированные воздействием низкого рН клетки компетентностью для образования производных трех зародышевых листков, что является общепринятым критерием для плюрипотентного характера. Как анализы дифференцировки in vitro (данные не представлены), так и тест образования тератомы (данные не представлены), показали, что эти клетки могут давать начало эктодермальным (например, β-тубулин III+), мезодермальным (например, актин гладких мышц+) и эндодермальным (например, альфа-фетопротеин+) клеткам.
Совместно, эти данные демонстрируют, что состояние дифференцировки линии коммитированных соматических клеток можно переводить в состояние плюрипотентности посредством сильных стимулов, поданных извне. Далее в настоящем документе, изменение судьбы от соматических клеток до плюрипотентных клеток посредством сильных внешних стимулов, таких как низкий рН, обозначают как «вызванную стимулом приобретенную плюрипотентность» (STAP), и полученные клетки - как клетки STAP.
Клетки STAP из других тканей-источников. Другим важным вопросом относительно клеток STAP является то, ограничен ли феномен вызванного воздействием низкого рН перехода CD45+ лейкоцитами. Для изучения этого вопроса, сходные эксперименты по переходу проводили с соматическими клетки, собранными из тканей головного мозга, кожи, мышц, жировой ткани, тканей костного мозга, легкого и печени от мышей oct3/4::gfp.
Клетки из образцов тканей подвергали диссоциации до отдельных клеток, подвергали временному воздействию низкого рН и культивировали в содержащей LIF среде. Хотя эффективность перехода менялась среди тканей в зависимости от их происхождения, oct3/4::GFP+ клетки воспроизводимо наблюдали в культуре на d7 (фигура 14А (серии представляют собой, слева направо, CD45+ клетки, клетки костного мозга, головного мозга, легкого, мышц, жировой ткани, фибробластов, печени и хондроциты). Примечательно, что клетки STAP эффективно развивались из мезенхимальных клеток жировой ткани (данные не представлены), где CD45+ клетки являлись редкими, а также из первичной культуры клеток хондроцитов, что указывает на то, что не относящиеся к CD45+ популяции клеток могут давать начало клеткам STAP. Эти кластеры oct3/4::GFP+ клеток также экспрессировали связанные с плюрипотентностью маркеры (фигура 14В (серии представляют собой, слева направо, экспрессию Oct3/4, Nanog, Sox2, Klf4 и Rex1) и фигура 18В, данные не представлены) и специфические для клеток ES маркерные гены (фигура 14В и фигура 18В).
Характеристики клеток STAP как плюрипотентных клеток. Таким образом, клетки STAP экспрессируют специфические для клеток ES гены и имеют сходный характер метилирования в генах oct3/4 и nanog. Кроме того, клетки STAP можно получать в культуральной среде для клеток ES мыши, такой как содержащие LIF среды, но не в среде для EpiSC мыши (данные не представлены).
Однако, несмотря на то, что клетки STAP демонстрируют значительное сходство с клетками ES мыши, обнаружено также несколько отличных признаков. Например, для клеток STAP показана ограниченная способность к самообновлению. В отличие от клеток ES мыши (данные не представлены), когда сферы клеток STAP подвергали ферментативной диссоциации до отдельных клеток для клонального культивирования в каждой лунке 96-луночного планшета, колонии (АР+ или oct3/4::GFP+) не формировались после дополнительного 10-суточного культивирования в содержащей LIF среде (на основе G-MEM или В27), в адгезионных или в неадгезионных условиях (данные не представлены). В то время как нечасто наблюдали формирование сферических колоний (как правило, в 2-4 лунках из 96 лунок), эти колонии все были АР- и oct3/4::GFP-. Даже когда сферы клеток STAP подвергали частичной диссоциации и культивировали в условиях высокой плотности клеток (данные не представлены; предположительно, с лучшей поддержкой для самообновления), количество клеток начинало уменьшаться после двух пассажей, и oct3/4::GFP+ клетки невозможно было поддерживать дольше пяти пассажей. Эти характеристики роста и поддержания позволяют предполагать, что клетки STAP представляют популяцию плюрипотентных клеток, свойства которых частично отличаются от свойств клеток ES и iPS мыши.
Мыши EpiSC представляют собой другую категорию плюрипотентных стволовых клеток, которые, как считают, немного более продвинуты по стадиям дифференцировки. Клетки STAP, по-видимому, отличаются по поведению от клеток EpiSC в нескольких аспектах. В адгезионной культуре, подобно клеткам ES мыши, oct3/4::GFP+ клетки формировали полусферические колонии посредством наслоения, в отличие от монослойных плоских колоний, наблюдаемых для EpiSC мыши. Клетки STAP невозможно было также поддерживать в среде EpiSC, что позволяет предполагать, что они являются отличными от EpiSC (данные не представлены). Кроме того, обработка ингибитором ROCK, улучшающим пассирование отдельных клеток для EpiSC (ссылка; Ohgushi), не стимулировала формирования колоний из диссоциированных клеток STAP (данные не представлены).
Иммунное окрашивание показало, что клетки STAP являлись отрицательными по маркерам EpiSC клаудину 7 и ZO-1 и положительными по Klf2/4 (данные не представлены). Распределение по группам клеток ES, клеток STAP и EpiSC может оказаться не таким простым, поскольку экспрессия маркера клеток ES Esrrβ являлась низкой как в клетках STAP, так и в EpiSC, в то время как экспрессия elf5 являлась особенно низкой в клетках STAP (фигура 15А (серии представляют собой, слева направо, ES, EpiSC, STAP и CD45)). При кластерном анализе полногеномного транскриптома, клетки STAP являлись наиболее близкими к клеткам ES и обладали значительным сходством с бластоцистами по экспрессии РНК, в то время как они являлись наиболее удаленными от родительских CD45+ клеток (данные не представлены). Ситуация с инактивацией Х-хромосомы в клетках STAP вызывала интерес; ~40% женских клеток STAP (d7) обладали инактивированной хромосомой, в то время как инактивация Х-хромосомы была отменена у остальных (~60%) (фигура 15В).
Эти данные указывают на возможность того, что состояние дифференцировки клеток STAP может представлять собой новое метастабильное плюрипотентное состояние, близкородственное, но отличающееся от состояния клеток ES.
Формирование химер и передача зародышевой линии у мышей. Наконец, способность клеток STAP к формированию химер оценивали посредством анализа инъекции в бластоцисты. В отличие от клеток ES, когда клетки STAP (на фоне В6) подвергали диссоциации до отдельных клеток и инъецировали в бластоцисты ICR, не рождалось химерных мышей, обладающих темным окрасом шерстного покрова (таблица 4). Поскольку отдельные клетки STAP было очень сложно поддерживать in vitro, сделали вывод, что диссоциация клеток каким-то образом изменяет их активность. Таким образом, кластеры клеток STAP вручную разрезали на небольшие фрагменты с использованием микроскальпеля под микроскопом, и инъецировали целиком в бластоцисты (данные не представлены). После этой манипуляции, химерные мыши рождались со значительной частотой, и все развивались нормально (данные не представлены). Затем проверяли распределение по тканям инъецированных клеток STAP, полученных из CD45+ клеток мышей, конститутивно экспрессирующих GFP (F1 of C57BL/6GFP, скрещенных с DBA/2 или 129/Sv). От высокого до умеренного вклада экспрессирующих GFP клеток наблюдали в химерных эмбрионах после инъекции кластеров клеток STAP (данные не представлены).
Долю вклада GFP+ клеток в каждой ткани анализировали в этих химерных эмбрионах посредством FACS. Происходящие из CD45+ клеток клетки STAP вносили вклад во все исследованные ткани (данные не представлены). Более того, потомство, происходящее от клеток STAP, рождалось у химерных мышей (таблица 5). Этот потенциал клеток STAP является важным и демонстрирует истинную природу этих плюрипотентных клеток, поскольку передачу зародышевой линии рассматривают как строгий критерий плюрипотентности, так же как генетической и эпигенетической нормальности22. Затем проводили анализ тетраплоидной (4N) комплементации посредством инъекции клеток в 4N бластоцисты, который считают наиболее строгим тестом для способности к развитию инъецированных клеток, поскольку полученные эмбрионы развиваются только из этих донорных клеток23 (данные не представлены). При инъекции в 4N бластоцисты, происходящие из CD45+ клеток клетки STAP (из мышей DBA×B6 GFP или 129/Sv×B6 GFP F1) образовывали «полностью GFP+ эмбрионы» на E10,5 (данные не представлены), что показывает, что одних клеток STAP было достаточно для конструирования структуры целого эмбриона.
Совместно, эти данные явно показывают, что клетки STAP обладают способностью к развитию для дифференцировки во все линии соматических клеток и зародышевые линии в контексте эмбрионального окружения.
Обсуждение
Данные, описанные в настоящем документе, выявили неожиданно гибкую пластичность, которой соматические клетки обладают в латентном состоянии. Эта динамическая пластичность, даже переходящая в плюрипотентные клетки, возникает, когда клетки подвергают временному воздействию сильных стимулов, которое они в норме не испытывают в условиях их жизнедеятельности.
На переход от CD45+ клеток к клеткам STAP значительно не влияло по меньшей мере воздействие ингибиторов HDAC (например, трихостатина А) или 5-аза-цитидина.
В настоящем документе показано, что подвергание воздействию низкого рН значительно уменьшало количество клеток в культуре. Однако, фактически, уменьшение количества выживших клеток в течение первых 24 часов являлось ограниченным, что позволяет предполагать, что это воздействие маловероятно оказывало сильный летальный эффект на большинство клеток. Вместо этого, отсроченная потеря клеток происходила постепенно в течение d2-d5. В соответствии с этим, данные показывают, что ряд генов, вовлеченных в клеточный ответ на стресс и репарацию ДНК 21, был сильно индуцирован в испытавших воздействие низкого рН oct3/4::GFP+ клетках на d3, но не в контрольных клетках, культивированных в той же самой среде, что позволяет предполагать, что клетки отвечали на стимул как на опасный для жизни, или сублетальный, стресс. Интересно, что уровни экспрессии их генов становились даже выше на d7; таким образом, вызывает интерес исследование в будущем не только ролей индуцированных стрессом генов для выживания клеток, возможно, но также их возможного вовлечения в процесс перепрограммирования.
Другим открытым вопросом является то, можно ли перепрограммирование клеток инициировать специфически посредством воздействия низкого рН, или также посредством некоторых других типов сублетального стресса, таких как физическое повреждение, перфорация плазматической мембраны, шок из-за осмотического давления, недостаточность факторов роста, гипоксия и воздействие среды с высоким содержанием Са2+. Примечательно, что по меньшей мере некоторые из них, в частности, физическое повреждение посредством интенсивного суспендирования и перфорация мембраны посредством стрептолизина О, индуцировали образование oct3/4::GFP+ клеток из CD45+ клеток (фигура 18А). Эти данные указывают на возможность того, что конкретные общие регуляторные модули, стоящие ниже этих отдаленных сублетальных стрессов, действуют как ключевые для высвобождения соматических клеток из строго фиксированного эпигенетического состояния дифференцировки, приводя к глобальному изменению эпигенетической регуляции. Принимая во внимание, что некоторые oct3/4::GFP+ клетки появляются на d2, такой механизм перепрограммирования может начинать функционирование в течение первых двух дней.
Ссылки
1. Wakayama, Т., Perry, А.С., Zuccotti, М., Johnson, K.R. & Yanagimachi, R. Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature 394, 369-374, doi:10.1038/28615 (1998).
2. Takahashi, K. & Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663-676, doi:10.1016/j.cell.2006.07.024 (2006).
3. Jiang, Y. et al. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 418, 41-49, doi:10.1038/nature00870 (2002).
4. D'Ippolito, G. et al. Marrow-isolated adult multilineage inducible (MIAMI) cells, a unique population of postnatal young and old human cells with extensive expansion and differentiation potential. Journal of cell science 117, 2971-2981, doi:10.1242/jcs.01103 (2004).
5. Johnson, J. et al. Oocyte generation in adult mammalian ovaries by putative germ cells in bone marrow and peripheral blood. Cell 122, 303-315, doi:10.1016/j.cell.2005.06.031 (2005).
6. Kucia, M. et al. A population of very small embryonic-like (VSEL) CXCR4(+)SSEA-1(+)Oct-4+ stem cells identified in adult bone marrow. Leukemia : official journal of the Leukemia Society of America, Leukemia Research Fund, U.K 20, 857-869, doi:10.1038/sj.leu.2404171 (2006).
7. Kuroda, Y. et al. Unique multipotent cells in adult human mesenchymal cell populations. Proceedings of the National Academy of Sciences of the United States of America 107, 8639-8643, doi:10.1073/pnas.0911647107 (2010).
8. Obokata, H. et al. The potential of stem cells in adult tissues representative of the three germ layers. Tissue engineering. Part A 17, 607-615, doi:10.1089/ten.TEA.2010.0385 (2011).
9. Rahnemai-Azar, A. et al. Human marrow-isolated adult multilineage-inducible (MIAMI) cells protect against peripheral vascular ischemia in a mouse model. Cytotherapy 13, 179-192, doi:10.3109/14653249.2010.515579 (2011).
10. Huang, Y. et al. Bone marrow transplantation temporarily improves pancreatic function in streptozotocin-induced diabetes: potential involvement of very small embryonic-like cells. Transplantation 89, 677-685, doi:10.1097/TP.0b013e3181c9dc7d (2010).
11. Zuba-Surma, E.K. et al. Transplantation of expanded bone marrow-derived very small embryonic-like stem cells (VSEL-SCs) improves left ventricular function and remodelling after myocardial infarction. Journal of cellular and molecular medicine 15, 1319-1328, doi:10.1111/j.1582-4934.2010.01126.x (2011).
12. Paczkowska, E. et al. Aldehyde dehydrogenase (ALDH) - a promising new candidate for use in preclinical and clinical selection of pluripotent very small embryonic-like stem cells (VSEL SCs) of high long-term repopulating hematopoietic potential. Annals of transplantation: quarterly of the Polish Transplantation Society 16, 59-71 (2011).
13. Lengner, C.J., Welstead, G.G. & Jaenisch, R. The pluripotency regulator Oct4: a role in somatic stem cells? Cell Cycle 7, 725-728 (2008).
14. Berg, J.S. & Goodell, M.A. An argument against a role for Oct4 in somatic stem cells. Cell stem cell 1, 359-360, doi:10.1016/j.stem.2007.09.007 (2007).
15. Ohbo, K. et al. Identification and characterization of stem cells in prepubertal spermatogenesis in mice small star, filled. Developmental biology 258, 209-225 (2003).
16. Ying, Q.L. et al. The ground state of embryonic stem cell self-renewal. Nature 453, 519-523, doi:10.1038/nature06968 (2008).
17. Ogawa, K., Matsui, H., Ohtsuka, S. & Niwa, H.A novel mechanism for regulating clonal propagation of mouse ES cells. Genes to cells : devoted to molecular & cellular mechanisms 9, 471-477, doi:10.1111/j.1356-9597.2004.00736.x (2004).
18. Gough, N.M. et al. LIF: a molecule with divergent actions on myeloid leukaemic cells and embryonic stem cells. Reproduction, fertility, and development 1, 281-288 (1989).
19. Hitoshi, S. et al. Primitive neural stem cells from the mammalian epiblast differentiate to definitive neural stem cells under the control of Notch signaling. Genes & development 18, 1806-1811, doi:10.1101/gad.1208404 (2004).
20. Tesar, P.J. et al. New cell lines from mouse epiblast share defining features with human embryonic stem cells. Nature 448, 196-199, doi:10.1038/nature05972 (2007).
21. Saretzki, G., Armstrong, L., Leake, A., Lako, M. & von Zglinicki, T. Stress defense in murine embryonic stem cells is superior to that of various differentiated murine cells. Stem Cells 22, 962-971, doi:10.1634/stemcells.22-6-962 (2004).
22. Surani, M.A. & Barton, S.C. Development of gynogenetic eggs in the mouse: implications for parthenogenetic embryos. Science 222, 1034-1036 (1983).
23. Nagy, A., Rossant, J., Nagy, R., Abramow-Newerly, W. & Roder, J.C. Derivation of completely cell culture-derived mice from early-passage embryonic stem cells. Proceedings of the National Academy of Sciences of the United States of America 90, 8424-8428 (1993).
24. A.P. Dyban, V. S. B. Cytogenetics of Mammalian Embryonic Development. Oxford Univ. Press, New York (1987).
25. Gropp, A., Winking, H., Herbst, E.W. & Claussen, C.P. Murine trisomy: developmental profiles of the embryo, and isolation of trisomic cellular systems. The Journal of experimental zoology 228, 253-269, doi:10.1002/jez.1402280210 (1983).
Материалы и методы
Сбор тканей и культивирование клеток. Для выделения зрелых лимфоцитов, селезенки, полученные от мышей GOF или мышей ICR в возрасте 1 недели, нарезали ножницами и подвергали механической диссоциации с помощью пастеровской пипетки. Диссоциированные селезенки пропускали через клеточное сито (BD Biosciences, Сан-Хосе). Собранные клетки ресуспендировали в среде DMEM и добавляли такой же объем лимфолита (CEDARLANE®, Онтарио, Канада), затем центрифугировали при 1000 g в течение 15 мин. Слой лимфоцитов отбирали и окрашивали с помощью антитела против CD45 (ab25603, abeam, Кембридж, Массачусетс). Положительные по CD45 клетки сортировали посредством FACS Aria (BD Biosciences). Затем положительные по CD45 клетки обрабатывали с помощью стрессового воздействия (раствор с рН5,5 в течение 15 мин) и высевали в среду В27, дополненную 1000 ед. LIF (Sigma).
Воздействие внешних стимулов - стрессовое воздействие. Чтобы подвергнуть зрелые клетки механическому стрессу, пастеровские пипетки нагревали и затем вытягивали, чтобы получить просветы приблизительно 50 микрон в диаметре, и затем отламывали. Затем зрелые соматические клетки ресуспендировали через эти пипетки в течение 20 мин и культивировали в течение 7 дней. Для воздействия гипоксических стимулов на зрелые клетки, клетки культивировали в инкубаторе с 5% кислородом в течение 3 недель. Зрелые клетки подвергали воздействию стимула недостатка питательных веществ, посредством культивирования клеток в базовой культуральной среде в течение 3 недель. Высокую концентрацию Са в культуре обеспечивали для зрелых клеток посредством культивирования клеток в среде, содержащей 2 мМ CaCl2, в течение 7 дней. Чтобы подвергать зрелые клетки физиологическому стрессу, их обрабатывали раствором с низким рН (рН 5,5), и культивировали в течение 7 дней. Клетки подвергали также более серьезному повреждению. Для получения пор в мембранах зрелых клеток, клетки обрабатывали с помощью 230 нг/мл SLO (стрептолизин О) (S5265, Sigma) в течение 2 час, затем культивировали в течение 7 дней.
Бисульфитное секвенирование. Клетки, полученные от мышей GOF, подвергали диссоциации до отдельных клеток. Положительные по GFP клетки собирали с использованием FACS Aria™. Геномную ДНК выделяли из SAC и исследовали. Обработку ДНК бисульфитом проводили с использованием набора для модификации ДНК CpGenome™ DNA Modification Kit (Chemicon, Темекула, Калифорния, http://www.chemicon.com), следуя инструкциям производителя.
Полученную модифицированную ДНК амплифицировали посредством гнездовой полимеразной цепной реакции ПЦР с использованием двух прямых (F) праймеров и одного обратного (R) праймера: Oct4 (F1, GTTGTTTTGTTTTGGTTTTGGATAT; F2, ATGGGTTGAAATATTGGGTTTATTTA; R, ССАСССТСТААССТТААССТСТААС). И Nanog (F1, GAGGATGTTTTTTAAGTTTTTTTT; F2, AATGTTTATGGTGGATTTTGTAGGT; R, CCCACACTCATATCAATATAATAAC). ПЦР проводили с использованием варианта Taq для горячего старта TaKaRa Ex Taq Hot Start Version (RR030A). Секвенирование ДНК проводили с использованием праймера М13 при содействии GRAS (The Genome Resource and Analysis Unit).
Иммуногистохимия. Культивированные клетки фиксировали с помощью 4% параформальдегида и пермеабилизовали с помощью 0,1% Triton X-100/PBS перед блокированием с помощью 1% раствора BSA (Life Technology, Токио, Япония). Вторичные антитела представляли собой антитела козы против антител мыши или антител кролика с присоединенным Alexa-488 или -594 (Invitrogen). Ядра клеток визуанализировали с помощью DAPI (Sigma). Стекла фиксировали с помощью средства, препятствующего выгоранию флуоресценции, SlowFade Gold (Invitrogen).
Активированная флуоресценцией сортировка клеток и проточная цитометрия. Клетки получали в соответствии со стандартными способами и суспендировали в 0,1% BSA/PBS на льду перед FACS. PI™ (BD Biosciences) использовали для исключения мертвых клеток. Для отрицательного контроля, первичное антитело заменяли на IgG для отрицательного контроля такого же изотипа для обеспечения специфичности. Клетки сортировали на BD FACSAria SORP™ и анализировали на BD LSRII™ с помощью программного обеспечения BD FACSDiva™ (BD Biosciences).
Получение РНК и анализ ОТ-ПЦР. РНК выделяли с помощью набора для выделения РНК в микромасштабе RNeasy™ Micro kit (QIAGEN). Обратную транскрипцию проводили с помощью набора для синтеза первой цепи SupeSACript III First Strand Synthesis kit (Invitrogen). Смесь SYBR Green™ Mix I (Roche Diagnostics) использовали для амплификации, и образцы анализировали в устройстве Lightcycler-II™ (Roche Diagnostics).
Исследования на животных. Для исследований онкогенности клетки, суспендированые в 100 мл PBS, инъецировали подкожно в паховые области совпадающих по возрасту иммунодефицитных мышей SCID. Мышей умерщвляли и подвергали вскрытию через 6 недель.
Анализ АТР и ROS. Межклеточный уровень АТР измеряли посредством набора для биолюминесцентного анализа АТР - АТР Bioluminescence Assay Kit HS II™ (Roche), в соответствии с протоколом поставщика. Интенсивность люминесценции измеряли с использованием микропланшетного люминометра Gelomax™ 96 Microplate Luminometer (Promega, Мадисон, Висконсин), и показатели люминесценции нормализовали по количеству клеток. Для измерения уровней ROS, клетки инкубировали в среде, содержащей 2 мкМ дигидроэтидий (Molecular Probes) при 37°С в темноте в течение 15 минут. Затем клетки промывали PBS и суспендировали в PBS, содержащем 0,5% BSA. Интенсивность флуоресценции 30000 клеток регистрировали с помощью BD Biosciences LSR II (BD Bioscience, Spark, MD).
Получение и анализы химерных мышей
Получение диплоидных и тетраплоидных химер. Диплоидные эмбрионы получали от самок линии ICR, спаренных с самцами ICR, и тетраплоидные эмбрионы получали от самок линии BDF1, спаренных с самцами BDF1. Тетраплоидные эмбрионы получали посредством электрослияния 2-клеточных эмбрионов. В этом исследовании, поскольку обработка трипсином приводила к низкому уровню химеризма, сферические колонии SAC эмбрионов с использованием микроскальпеля под микроскопом, затем небольшие кластеры SAC инъецировали в бластоцисты на сутки 4,5 посредством большой пипетки. На следующие сутки, химерные бластоцисты переносили псевдобеременным самкам на сутки 2,5.
Анализ дифференцировки in vitro.
Анализ дифференцировки линий клеток мезодермы. Измененные под воздействием стресса клеточные массы собирали через 7 дней и подвергали диссоциации до отдельных клеток, затем собирали только положительные по Oct4-GFP клетки посредством сортировщика клеток. Собранные клетки культивировали в DMEM, дополненной 20% FCS. Среду меняли каждые 3 дня. Через 7-14 дней, мышечные клетки окрашивали с помощью антитела против α-актина гладких мышц (N1584, DAKO). В отрицательном контроле, первичное антитело заменяли на IgG для отрицательного контроля такого же изотипа для обеспечения специфичности.
Анализ дифференцировки линий нейрональных клеток. Измененные под воздействием стресса клеточные массы собирали через 7 дней и подвергали диссоциации до отдельных клеток, затем собирали только положительные по Oct4-GFP клетки посредством сортировщика клеток. Собранные клетки высевали в покрытые орнитином предметные стекла с лунками (Nalge Nunc International) в F12/DMEM (1:1, об./об.), дополненных 2% В27 (Invitrogen), 10% FCS, 10 нг/мл bFGF (R&D Systems) и 20 нг/мл EGF (R&D Systems). Среду меняли каждые 3 дня. Через 10-14 дней, клетки фиксировали 4% параформальдегидом в течение 30 минут при 4°С, промывали PBS, содержащим 0,2% Triton Х-100 в течение 15 минут при комнатной температуре, инкубировали с PBS, содержащим 2% FCS в течение 20 минут для блокирования неспецифических реакций, и инкубировали с моноклональным антителом против тубулина βIII мыши (G7121, Promega) и моноклональным антителом против GFAP мыши (АВ5804, CHEMICON). В отрицательном контроле, первичное антитело заменяли на IgG для отрицательного контроля такого же изотипа для обеспечения специфичности.
Анализ дифференцировки клеток печени. Измененные под воздействием стресса клеточные массы собирали через 7 дней и подвергали диссоциации до отдельных клеток, затем собирали только положительные по Oct4-GFP клетки посредством сортировщика клеток. Собранные клетки высевали на 2-камерные предметные стекла (Nalge Nunc International) в культуральной среде для гепатоцитов, состоящей из 500 мл базовой среды для гепатоцитов (Lonza, Вупперталь, Германия), 0,5 мл аскорбиновой кислоты, 10 мл BSA-FAF (не содержащей жирных кислот), 0,5 мл гидрокортизона, 0,5 мл трансферрина, 0,5 мл инсулина, 0,5 мл EGF и 0,5 мл гентамицина-амфотерицина (GA-1000; все из Lonza), дополненных 10% FCS, 1% пенициллином/стрептомицином (Sigma). Дифференцированные клетки детектировали посредством иммуногистохимии с использованием следующих антител; моноклонального антитела против α-фетопротеина мыши (МАВ1368, R&D System) и моноклонального антитела против цитокератина 7 мыши (ab668, abeam). В отрицательном контроле, первичное антитело заменяли на IgG для отрицательного контроля такого же изотипа для обеспечения специфичности.
Анализ дифференцировки in vivo: Измененные под воздействием стресса клеточные массы собирали через 7 дней и подвергали диссоциации до отдельных клеток, затем собирали только положительные по Oct4-GFP посредством сортировщика клеток. Собранные клетки ресуспендировали в 50 мкл DMEM с 10% FBS. Этот раствор высевали на лист 3×3×1 мм, состоящий из нетканой сетки из волокон полигликолевой кислоты, 200 микрон в диаметре, и имплантировали подкожно в заднюю часть бока мышей NOD/SCID в возрасте 4 недели. Через четыре недели имплантаты собирали и анализировали с использованием иммуногистохитмических способов. Имплантаты фиксировали с помощью 10% формальдегида, погружали в парафин и общепринятым образом нарезали толщиной 4 мкМ. Срезы окрашивали с использованием гематоксилина и эозина. Эндодермальные ткани идентифицировали с помощью маркера эндодермы моноклонального антитела против α-фетопротеина мыши (МАВ1368, R&D System). Эктодермальные ткани идентифицировали с помощью моноклонального антитела против тубулина βIII мыши (G7121, Promega). Мезодермальные ткани идентифицировали с помощью антитела против α-актина гладких мышц (N1584, DAKO). В отрицательном контроле, первичное антитело заменяли на IgG для отрицательного контроля такого же изотипа для обеспечения специфичности.
Анализ перестройки TCRβ -цепи. гДНК выделяли из SAC и кончиков хвостов химерных мышей, полученных с использованием SAC, полученных из положительных по CD45 клеток. ПЦР проводили с 50 нг гДНК с использованием следующих праймеров. (5'-GCACCTGTGGGGAAGAAACT-3' и 5'-TGAGAGCTGTCTCCTACTATCGATT-3') Амплифицированную ДНК анализировали электрофорезом в 1,5% агарозном геле.
Генотипирование химерных мышей. гДНК выделяли из кончиков хвостов 4N химерных мышей. Генотипирование проводили с использованием следующих праймеров. (GFP: F-AGAACTGGGACCACTCCAGTG и R-TTCACCCTCTCCACTGACAGATCT. IL-2: F-CTAGGCCACAGAATTGAAAGATCT и R-GTAGGTGGAAATTCTAGCATCATCC)
Затем определяли оптимальные условия культивирования для поддержания измененных под воздействием стресса экспрессирующих Oct4 клеток. Исследовали несколько описанных ранее культуральных сред, включая: культуральную среду для получения ES, 3i16 и АСТН17, условия культивирования ES, ES-LIF 18, условия культивирования эксперссирующих Oct4 примитивных нейрональных стволовых клеток, B27-LIF 19, и условия культивирования EpiSC20. Клетки высевали в каждую среду и подсчитывали экспрессирующие GFP колонии (фиг. S1C). Среда B27-LIF, по-видимому, являлась наиболее эффективной для получения экспрессирующих GFP сферических колоний. Таким образом, авторы настоящего изобретения использовали среду B27-LIF для культивирования обработанных клеток.
Чтобы исследовать, проявляют ли SAC, полученные из клеток, полученных из различных тканей, тенденцию к различной дифференцировке, SAC получали из различных тканей, полученных от мышей F1 GFP, инъецировали их в бластоцисты ICR. Затем, с использованием FACS анализировали соотношение распределения в каждой ткани полученных химерных мышей. Обнаружено, что SAC, полученные из любой ткани, вносили вклад в получение химерных мышей (данные не представлены). Кроме того, долю вклада в кожу, головной мозг, мышцы, жировую ткань, печень и легкие анализировали у химерных мышей, полученных с использованием SAC, полученных из различных тканей. SAC, полученные из любой ткани, вносили вклад в образование тканей, репрезентативных для всех трех зародышевых листков, и не наблюдали какой-либо тенденции к различиям (данные не представлены).
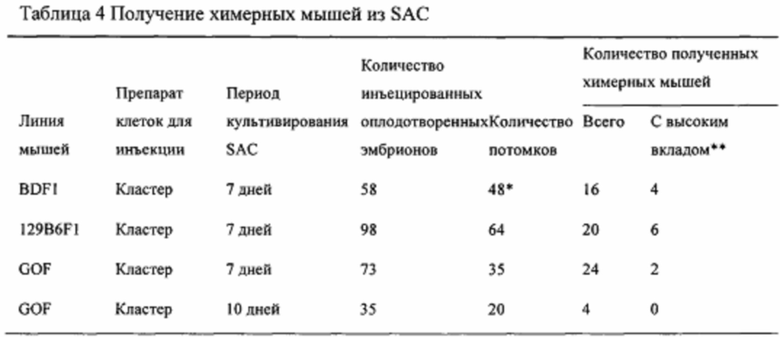
* Все плоды собирали на 13,5 dpc - 15,5 dpc, и долю вклада SAC в каждый орган оценивали посредством FACS
** Вклад SAC в каждую химеру оценивали как высокий (>50% окраса шерстяного покрова с экспрессией GFP)
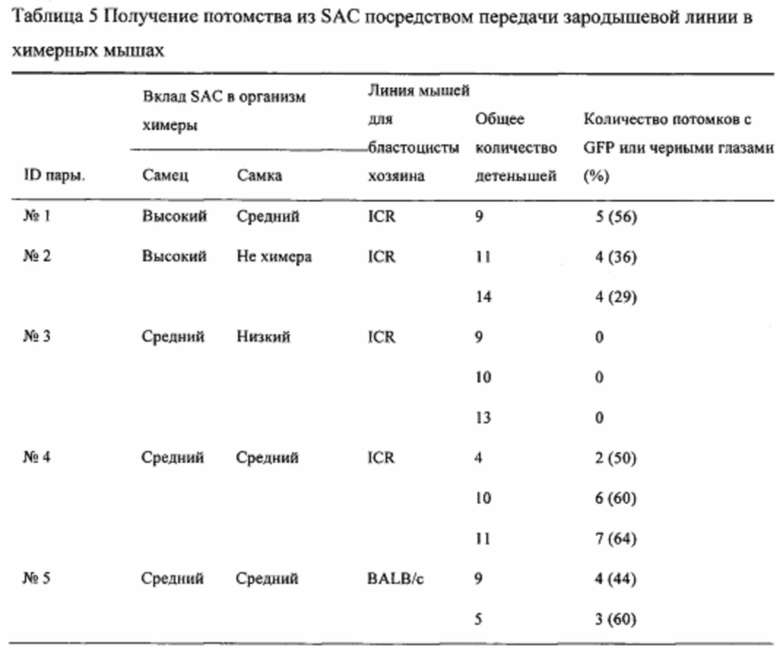
ПРИМЕР 3
Без желания быть связанными с теорией, предусматривают, что способы, описанные в настоящем документе, активируют процесс, родственный апоптозу, или контролируемой гибели клеток. Умеренное повреждение клеток может индуцировать активацию генов репарации. Серьезное повреждение клеток может активировать ранее не определенный механизм выживания. Предусматривают, что когда клетки подвергают значительному стрессу, такому как стрессы, описанные в настоящем документе, клеточные компоненты (например, митохондрии, пузырьки, ядра, рибосомы, эндоплазматический ретикулум, экзосомы, эндосомы, клеточные мембраны, митохондрии, лизосомы, АТР, белки, ферменты, углеводы, липиды и т.д.) высвобождаются из поврежденных клеток в «хранилище». Данные, описанные в настоящем документе, указывают на то, что это «хранилище» может являться способным к восстановлению клеток и/или стимуляции выживания клеток. Кроме того, предусматривают, без желания быть связанными с теорией, что митохондрии (и другие органеллы) являются способными направлять восстановление клеток. Из-за их небольшого размера, простоты, способности направлять дифференцировку и подобной прокариотической природы, митохондрии могут переживать стресс, показанный как летальный для родительской клетки. Митохондрии могут высвобождаться из клетки свободными, инкапсулированными в мембрану и/или связанными с другими клеточными компонентами.
Альтернативно, без желания быть связанными с теорией, ядра могут оставаться интактными, инкапсулированными в клеточную мембрану, которая может содержать некоторые митохондрии. Эти поврежденные клетки с очень небольшим количеством цитоплазмы и очень немногими органеллами, утратившие эпигенетический контроль ядра, могут затем взаимодействовать и, возможно, сливаться с органеллами, которые были вытеснены. Это приводит к клеткам с субклеточными компонентами, необходимыми для роста и репликации, но клетки теряют эпигенетический контроль, и таким образом, индуцируется более примитивное (например, более плюрипотентное) состояние.
ПРИМЕР 4: Потенциал развития для эмбриональных и плацентарных линий в перепрограммированных клетках с приобретенной плюрипотентностью
Как правило, судьбы постнатальных соматических клеток фиксированы и не меняются, если их не подвергают переносу ядра1,2 или генетическим манипуляциям с ключевыми факторами транскрипции3. Как показано в настоящем документе, авторы настоящего изобретения открыли неожиданный феномен перепрограммирования соматических клеток в плюрипотентные клетки посредством сублетальных стимулов, названный вызванной стимулом приобретенной плюрипотентностью (STAP)4. В настоящем документе описана также демонстрация того, что пеепрограммированные клетки STAP обладают уникальной способностью к дифференцировке, отличающейся от способности клеток ES. Клетки STAP могут вносить вклад не только в эмбриональные ткани, но также в плацентарную систему, как видно в анализе инъекции в бластоцисты. Их эффективность для вклада в плаценту дополнительно усиливали посредством культивирования с FGF4. И наоборот, при культивировании в течение дополнительных пассажей в среде для поддержания клеток ES, клетки STAP, для которых исходно показана ограниченная способность к самообновлению, образовывали активно пролиферирующие линии клеток, обладающие характеристиками, подобными клеткам ES, но не подобными трофобластам. Эти измененные клетки STAP (стволовые клетки STAP) приводили к рождению мышей в анализе тетраплоидной комплементации, но больше не вносили вклад в плацентарные ткани. Таким образом, клетки STAP, в отличие от клеток iPS, могут представлять собой новое метастабильное состояние плюрипотентности,6 которое отличается от состояния клеток ES. Технология стволовых клеток STAP может открывать универсальный, мощный ресурс для регенеративной медицины нового поколения.
В настоящем документе описан вызывающий интерес феномен изменения судьбы клеток: соматические клетки приобретают плюрипотентность после воздействия сублетальных стимулов, таких как воздействие низкого рН4. Когда CD45+ клетки селезенки (включая коммитированные Т клетки) подвергают воздействию рН5,7 в течение 30 мин и затем культивируют в присутствии LIF, значительная доля выживших клеток начинают экспрессировать маркер плюрипотентных клеток Oct3/4 на сутки 2 (d2). На d7, формируются кластеры плюрипотентных клеток с профилем маркеров истинной плюрипотентности и компетентностью к дифференцировке в клетки трех зародышевых листков (например, как показано по формированию тератом). Эти клетки STAP также могут вносить эффективный вклад в ткани химерных мышей и подвергаться передачи зародышевой линии в анализе инъекции в бластоцисты. В то время как эти характеристики напоминают характеристики клеток ES, клетки STAP, по-видимому, отличаются от клеток ES, по меньшей мере, по их ограниченной способностью к самообновлению(как правило, максимум 3-5 пассажей) и по их устойчивости к диссоциированной культуре4.
В настоящем примере, авторы изобретения далее исследовали уникальную природу клеток STAP, фокусируясь на их потенциалу к дифференцировке в две главные категории клеток в бластоцисте7-9: клетки типа внутренней клеточной массы (или подобные клеткам ES) и клетки трофобластной/плацентарной линии, после анализа инъекции в бластоцисты выявили неожиданные данные. Как правило, потомство инъецированных клеток ES обнаруживают в эмбриональной части химеры, но редко в плацентарной части7 (данные не представлены). Неожиданно, инъецированные клетки STAP вносили вклад не только в эмбрион, но также в плаценту и внеэмбриональные мембраны (фигура 22). Этот вклад в две линии наблюдали приблизительно у 60% химерных эмбрионов.
Эти данные содействовали исследованию способности к дифференцировке в трофобласты клеток STAP. Известно, что трофобластные линии клеток (трофобластные стволовые клетки; клетки TS)8,9 можно получать в длительной адгезионной культуре бластоцист в присутствии FGF4. Когда кластеры клеток STAP культивировали в таких же условиях (фигура 23А; один кластер на лунку в 96-луночном планшете), сфероидные кластеры клеток STAP постепенно исчезали, и клетки плоского вида, отличные от клеток STAP, росли и формировали колонии на d7-d10 (данные не представлены). В отличие от клеток STAP, обладающих высоким уровнем экспрессии oct3/4::GFP, эти плоские клетки (адгерентные ко дну планшета) проявляли умеренные сигналы GFP на сутки 7 культивирования с FGF4 (данные не представлены). Иммунное окрашивание показало, что индуцированные FGF4 (F4I) клетки сильно экспрессировали трофобластные маркеры10-12 интегрин альфа 7 и эомезодермин (данные не представлены) в дополнение к умеренным уровням oct3/4::GFP. Экспрессия Nanog являлась поддающейся детекции, но достаточно низкой (данные не представлены). В соответствии с этим, анализ qPCR показал, что клетки F4I экспрессировали значительные уровни маркерных генов трофобластных линий (например, cdx2), в то время как их экспрессия oct3/4 и nanog являлась более низкой, чем наблюдали в родительских клетках STAP (фигура 23В). Эти клетки F4I можно было эффективно размножать посредством пассирования с расщеплением трипсином каждые третьи сутки, и они оставались стабильными в течение более 30 пассажей в присутствии of FGF4 (в его отсутствие, они останавливали пролиферацию). В то время как их получение и размножение являлось возможным проводить как на клетках MEF, так и на покрытой желатином подложке, клетки, культивированные на фидерных клетках MEF, имели тенденцию проявлять более явный внешний вид эпителиальных клеток (данные не представлены).
В анализе инъекции в бластоцисты, вклад в плаценту клеток F4I наблюдали часто (50-60%) (данные не представлены). В химерных плацентах, клетки F4I, как правило, вносили вклад в ~10% от общего количества плацентарных клеток (фигура 23С, дорожки 1-3; следует отметить, что контрольные клетки ES не вносили значительного вклада в плаценту, дорожки 4-6). Эти данные позволяют предполагать, что клетки STAP обладают компетентностью для образования подобных TS клеток в ходе обработки FGF4, по меньшей мере, в свете экспрессии маркеров трофобластов и вклада в плаценту. Поскольку этот тип развития в подобные TS клетки не является общим с клетками ES (если не подвергать их генетической манипуляции)11, такая компетентность может представлять собой другой признак клеток STAP, отличающийся от клеток ES.
С другой стороны, клетки F4I, полученные из клеток STAP, могут обладать также характеристиками, отличающимися от характеристик полученных из бластоцист клеток TS. Во-первых, в отличие от общепринятых клеток TS13, клетки F4I экспрессировали умеренный уровень oct3/4 (данные не представлены). Более того, в отличие от клеток TS, инъецированные в бластоцисты клетки F4I также вносили вклад в эмбриональные части (в всех случаях это затрагивало химерные плаценты), хотя степень вклада, как правило, являлась низкой (данные не представлены).
Совместно, эти данные показывают, что популяция клеток STAP качественно отличается от клеток ES по отношению к их компетентности для дифференцировки в ткани плаценты.
Принимая это во внимание, исследовали дифференцировку в эмбриональные линии, другой тип клеток, присутствующий в бластоцистах. В отличие от клеток ES, клетки STAP обладают ограниченной способностью к самообновлению, и их невозможно размножать из отдельных клеток. Клетки STAP было невозможно поддерживать в течение более 5 пассажей (даже при частично диссоциированной культуре кластеров) в общепринятых содержащих LIF средах (включая среду B27+LIF, использованную для получения клеток STAP). Однако, содержащая АСТН среда с LIF15 (среда АСТН, далее в настоящем документе) оказывала относительно хорошие поддерживающие эффекты на скорость роста колоний клеток STAP (данные не представлены). При культивировании в этой среде на фидерных клетках MEF или на желатине в среде АСТН (фигура 24А), некоторая часть кластеров клеток STAP (как правило, обнаруженная в 20-50% лунок в культуре отдельных кластеров с использованием 96-луночных планшетов) продолжала расти (данные не представлены). Эти растущие колонии являлись сходными с колониями клеток ES мыши и экспрессировали высокий уровень oct3/4::GFP. В отличие от родительских клеток STAP, клетки в этих размножившихся колониях, после культивирования в этой среде в течение семи дней, становились устойчивыми к диссоциации, и их можно было пассировать в форме отдельных клеток (данные не представлены). В отличие от клеток STAP, эти измененные клетки можно было размножать экспоненциально, вплоть до по меньшей мере 120 дней культивирования (фигура 24В), подобно клеткам ES. Эта улучшенная способность к размножению не сопровождалась хромосомными аномалиями, как показано по многоцветному анализу FISH16 (данные не представлены). После размножения в течение семи дней, клетки росли, и их можно было поддерживать в любой из тестированных сред для клеток ES, в то время как первоначальное 7-суточное размножение наиболее эффективно выполняли с использованием среды АСТН (например, колонии формировались медленно и менее часто в среде 3i17; данные не представлены).
Далее в настоящем документе, пролиферативные клетки, полученные из клеток STAP, обозначены как стволовые клетки STAP. В отличие от клеток STAP, стволовые клетки STAP не образовывали подобные TS клетки в культуре с FGF4 (данные не представлены). В ходе иммунного окрашивания, обнаружено, что инактивацию Х-хромосомы18, которая обнаружена в значительной доле женских клеток STAP (ссылка), больше не наблюдали в стволовых клетках STAP (данные не представлены). Стволовые клетки STAP экспрессировали различные РНК-маркеры (фигура 24С) и белковые (данные не представлены) маркеры для клеток ES. Уровни метилирования ДНК в локусах oct3/4 и nanog, которые становятся деметилированными при переходе от CD45+ клеток к клеткам STAP, оставались низкими (фигура 24D). В дифференцирующей культуре19-21, стволовые клетки STAP образовывали эктодермальные, мезодермальные и эндодермальные производные (данные не представлены). Эти данные показывают, что стволовые клетки STAP обладают признаками, неотличимыми от признаков клеток ES.
В соответствии с этим, стволовые клетки STAP, даже после множества пассажей, могут формировать тератомы (данные не представлены) и, посредством инъекции в бластоцисты, эффективно вносят вклад в ткани химерных мышей (данные не представлены). Заметную эффективность стволовых клеток STAP в их распределение в эмбрионах было явно продемонстрировано тем фактом, что в анализе тетраплоидной комплементации5, эти клетки могли приводить к рождению мышей, способных вырастать до взрослого состояния и даже к производить потомство (данные не представлены). Принимая во внимание, что для восьми независимых линий стволовых клеток STAP воспроизводимо показана эта способность (следует отметить, что эта полная комплементация часто затруднена даже с использованием общепринятых линий клеток ES), авторы настоящего изобретения заключили, что клетки STAP, происходящие из взрослых соматических клеток, могут представлять собой привлекательный источник для получения линий плюрипотентных стволовых клеток, эквивалентных самим бластоцистам (или возможно, превосходящих их) в этом аспекте.
Важно, что в отличие от клеток STAP и F4I, стволовые клетки STAP, по-видимому, теряют свою способность вносить вклад в плацентарные ткани (данные не представлены), в то время как они дают начало различным тканям в химерах (фигуры 25А-25В). Таким образом, различие между клетками STAP и стволовыми клетками STAP не ограничено только активностью самообновления, но также затрагивает потерю компетентности для дифференцировки в плацентарные линии.
Эти данные указывают на уникальное плюрипотентное состояние клеток STAP. В то время как неспособность клонировать клетки STAP из отдельных клеток (как описано выше) затрудняет анализ коммитирования на уровне отдельных клеток, стоит отметить, что способом STAP можно переводить соматические клетки в плюрипотентную популяцию клеток с компетентностью как для эмбриональных, так и для плацентарных линий. Глубокое понимание состояния дифференцировки клеток STAP является важной темой для будущих исследований. В частности, будет интересно исследовать, представляют ли клетки STAP более незрелое состояние, чем клетки ES, как можно предполагать по их компетентности для плацентарных линий, которая напоминает эмбриональные клетки на стадии морулы. В недавнем исследовании опубликовано, что общепринятая культура клеток ES содержит также очень незначительную популяцию Oct3/4- клеток с отличающимися характеристиками, напоминающими признаки эмбрионов на очень ранних стадиях22. Клетки STAP могут обладать сходным метастабильным состоянием, обеспечивающим способность к двойной компетенции, однако, в отличие от клеток ES, оно обнаружено в большей части популяции клеток.
В настоящем документе показано, что клетки STAP обладают способностью к трансформации в подобные ES линии плюрипотентных стволовых клеток. Стоит отметить, что клетки STAP (из самок мышей) являются в некоторой степени мозаичными по инактивации Х-хромосомы; инактивация исчезает в ~40% клеток STAP4, в то время как остальные ее сохраняют. В клетках ES, напротив, обе Х-хромосомы являются воспроизводимо активированными. Интересно, что после получения, для «стволовых» клеток STAP не показано инактивации Х-хромосомы, подобно клеткам ES, что позволяет предполагать, что эпигенетический контроль в родительских клетках STAP является сходным, но не идентичным с эпигенетическим контролем в клетках ES мыши, также в этом смысле.
Настоящие результаты демонстрируют неожиданную «способность к спонтанному переходу» коммитированных соматических клеток для перепрограммирования их собственной судьбы в наивные клетки при воздействии сублетальных стимулов. Это вызывает множество любопытных и сложных биологических вопросов, включая описанные выше. Наряду с этим, этот впервые открытый феномен STAP может в корне изменить способы медицины с использованием стволовых клеток. Предусматривают, что может быть предоставлена возможность получения различных типов тканей посредством управляемой дифференцировки из клеток STAP или стволовых клеток STAP, полученных из соматических клеток без переноса генов (который может увеличивать риск злокачественной трансформации). Более того, в отличие от перехода в клетки iPS, переход в STAP происходи со значительно высокой частотой и продолжается посредством конкретных эндогенных программ, запускаемых сильными стимулами, такими как воздействие низкого рН. Поскольку стволовые клетки STAP, подобно клеткам ES, легко поддаются размножению и клонированию, они могут быть более пригодными, чем клетки STAP, для крупномасштабного получения тканей, которые можно использовать в медицине, со строгим контролем качества. В предварительном исследовании авторов настоящего изобретения, они преуспели в демонстрации эффективной дифференцировки стволовых клеток STAP в ретинальные клетки-предшественники23, кортикальные клетки-предшественники24 и сократительные кардиомиоциты25 (данные не представлены).
Методы
Культивирование клеток. Клетки STAP получали из CD45+ клеток посредством временного воздействия раствора с низким рН, с последующим культивированием в среде В27 + LIF (Obokata et al, 2013; подано совместно). Для получения линии клеток F4I, кластеры клеток STAP переносили в содержащую FGF4 среду TS на фидерные клетки MEF в 96-луночных планшетах. Клетки подвергали первому пассированию в течение d7-d10 с использованием общепринятого способа с трипсином. Для получения линии стволовых клеток STAP (STAPS), сферы STAP переносили в содержащую АСТН среду на фидерные клетки MEF или в покрытую желатином чашку. Через четыре - семь дней, клетки подвергали первому пассированию с использованием общепринятого способа с трипсином, и суспендированные клетки высевали в среду для поддержания ES, содержащую 5%FCS в 1%KSR.
Получение и анализы химерных мышей. Для инъекции стволовых клеток STAP, клеток F4I и клеток ES, использовали общепринятый способ инъекции в бластоцисты. Для инъекции клеток STAP, кластеры клеток STAP инъецировали целиком, поскольку обработка трипсином приводила к низкому уровню химеризма. Сферические колонии STAP эмбрионов с использованием микроскальпеля под микроскопом, затем небольшие кластеры колонии STAP инъецировали в бластоцисты на сутки 4,5 посредством большой пипетки. На следующие сутки, химерные бластоцисты переносили псевдобеременным самкам на сутки 2,5. Тетраплоидные эмбрионы получали посредством электрослияния 2-клеточных эмбрионов.
Анализ дифференцировки in vitro и in vivo: Формирование тератомы исследовали посредством инъекции 1×105 клеток из клеток STAPS подкожно в заднюю часть бока мышей NOD/SCID в возрасте 4 недели. Нейрональную дифференцировку in vitro индуцировали посредством способов SDIA и SFEBq24,26. Эндомезодермальную дифференцировку in vitro25 индуцировали посредством культивирования агрегатов клеток STAPS с факторами роста (активином) или 10% FCS.
Анализ кариотипа. Субконфлюэнтные клетки STAPS подвергали аресту в метафазе посредством колцемида и подвергали анализу многоцветной FISH (M-FISH). Специфические для хромосом мыши окрашивающие зонды метили комбинированным образом с использованием семи различных флуорохромов и гибридизовали, как описано ранее (Jentsch et al., 2003).
Культивирование клеток. Клетки STAP получали из CD45+ клеток, с последующим культивированием в среде В27 + LIF в течение 7 дней, как описано (Obokata et al, 2013; подано совместно). Для получения линии клеток F4I, кластеры клеток STAP переносили в содержащую FGF4 среду TS на фидерные клетки MEF в 96-луночные планшеты. Клетки подвергали первому пассированию в течение d7-d10 с использованием общепринятого способа с трипсином. Последующее пассирование проводили на каждые третьи сутки.
Для получения линии стволовых клеток STAP (STAPS), сферы STAP переносили в содержащую АСТН среду на фидерные клетки MEF. Через четыре - семь дней, клетки подвергали первому пассированию с использованием общепринятого способа с трипсином, и суспендированные клетки высевали в среду для поддержания ES, содержащую 5% FCS и 1% KSR. Последующее пассирование проводили на каждые вторые сутки.
Получение и анализы химерных мышей. Для получения диплоидных и тетраплоидных химер, диплоидные эмбрионы получали от самок линии ICR, спаренных с самцами ICR, и тетраплоидные эмбрионы получали от самок линии BDF1, спаренных с самцами BDF1. Тетраплоидные эмбрионы получали посредством электрослияния 2-клеточных эмбрионов. Для инъекции стволовых клеток STAP, клеток F4I и клеток ES, использовали общепринятый способ инъекции в бластоцисты. Для инъекции клеток STAP, кластеры клеток STAP инъецировали целиком, поскольку обработка трипсином приводила к низкому уровню химеризма. Сферические колонии STAP эмбрионов с использованием микроскальпеля под микроскопом, затем небольшие кластеры колонии STAP инъецировали в бластоцисты на сутки 4,5 посредством большой пипетки. На следующие сутки, химерные бластоцисты переносили псевдобеременным самкам на сутки 2,5.
Анализ дифференцировки in vitro и in vivo: 1×105 клеток из клеток STAP-S инъецировали подкожно в заднюю часть бока мышей 4 NOD/SCID в возрасте 4 недели. Через шесть недель, имплпантаты собирали и подвергали гистологическому анализу. Имплантаты фиксировали с помощью 10% формальдегида, погружали в парафин и общепринятым образом нарезали толщиной 4 мкМ. Срезы окрашивали с использованием гематоксилина и эозина.
Нейрональную дифференцировку in vitro индуцировали посредством способов SDIA и SFEBq. Эндомезодермальную дифференцировку in vitro индуцировали посредством культивирования агрегатов клеток STAPS с факторами роста (активином) или 10% FCS.
Иммунное окрашивание. Клетки фиксировали с помощью 4% PFA в течение 15 мин и затем пермеабилизовали с помощью 0,5% Triton Х-100, и затем инкубировали с первичными антителами: против H3K27me3 (Millipore; 1:300), против Oct3/4 (Santa Cruz Biotechnology; 1:300), против Nanog (eBioscience; 1:300), против KLF2/4 (R&D System; 1:300) и против Esrrβ(R&D System; 1:300). После инкубации в течение ночи, связавшиеся антитела визуанализировали с помощью вторичного антитела, конъюгированного с Alexa546 (Molecular Probes). Ядра окрашивали с помощью DAPI (Molecular Probes).
Получение РНК и анализ ОТ-ПЦР. РНК выделяли с помощью набора для выделения РНК в минимасштабе RNeasy™ Mini kit (QIAGEN). Обратную транскрипцию проводили с помощью набора для синтеза первой цепи SupeSACript III First Strand Synthesis kit (Invitrogen). Смесь Power SYBR™ Green Mix (Roche Diagnostics) использовали для амплификации ПЦР, и образцы анализировали в устройстве Lightcycler-II™ (Roche Diagnostics).
Анализ кариотипа. Анализ кариотипа проводили посредством анализа многоцветной FISH (M-FISH). Субконфлюэнтные клетки STAPS подвергали аресту в метафазе посредством колцемида (конечная концентрация 0,270 мкг/мл) в культуральной среде в течение 2,5 час при 37°С в 5% CO2. Клетки промывали с помощью PBS, обрабатывали трипсином/этилендиаминтетрауксусной кислотой (ЭДТА), ресуспендировали в среде для клеток и центрифугировали в течение 5 мин при 1200 об/мин. К осадку клеток в 3 мл PBS добавляли 7 мл предварительно нагретого гипотонического раствора 0,0375 М KCl. Клетки инкубировали в течение 20 мин при 37°С. Клетки центрифугировали в течение 5 мин при 1200 об/мин, и осадок ресуспендировали в 3-5 мл раствора 0,0375 М KCl. Клетки фиксировали с помощью метанола/уксусной кислоты (3:1; об./об.) посредством осторожного суспендирования. Фиксацию проводили четыре раза перед распределением клеток на предметных стеклах. Для способа FISH, специфические для хромосом мыши окрашивающие зонды метили комбинированным образом с использованием семи различных флуорохромов и гибридизовали, как описано ранее (Jentsch et al., 2003). Для каждой линии клеток, получали 9-15 метафазных пластинок с использованием эпифлуоресцентного микроскопа Leica DM RXA RF8 (Leica Mikrosysteme GmbH, Бенсхайм, Германия), оборудованного камерой Sensys CCD (Photometries, Тусон, Аризона). Камерой и микроскопом управляли посредством программного обеспечения Leica Q-FISH (Leica Microsystems hanging solutions, Кембридж, Великобритания). Метафазные пластинки обрабатывали на основе программного обеспечения Leica МСК и представляли в форме многоцветных кариограмм.
Бисульфитное секвенирование.
Геномную ДНК выделяли из клеток STAPS. Обработку ДНК бисульфитом проводили с использованием набора для модификации ДНК CpGenome DNA Modification Kit (Chemicon, Темекула, Калифорния, http://www.chemicon.com), следуя инструкциям производителя.
Полученную модифицированную ДНК амплифицировали посредством гнездовой полимеразной цепной реакции ПЦР с использованием двух прямых (F) праймеров и одного обратного (R) праймера: oct3/4 (F1, GTTGTTTTGTTTTGGTTTTGGATAT (SEQ ID NO: 73); F2,ATGGGTTGAAATATTGGGTTTATTTA (SEQ ID NO: 74); R,CCACCCTCTAACCTTAACCTCTAAC (SEQ ID NO: 75)). И nanog (F1,GAGGATGTTTTTTAAGTTTTTTTT (SEQ ID NO: 76); F2,AATGTTTATGGTGGATTTTGTAGGT (SEQ ID NO: 77); R,CCCACACTCATATCAATATAATAAC (SEQ ID NO: 78)). ПЦР проводили с использованием варианта Taq для горячего старта TaKaRa Ex Taq Hot Start Version (RR030A). Секвенирование ДНК проводили с использованием праймера М13 в Genome Resource and Analysis Unit, RIKEN CDB.
Ссылки
1. Gurdon, JB. The developmental capacity of nuclei taken from intestinal epithelium cells of feeding tadpoles. J Embryol Exp Morphol. 10, 622-640 (1962)
2. Wakayama, Т., Perry, A.C., Zuccotti, M., Johnson, K.R. & Yanagimachi, R. Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature 394, 369-374 (1998).
3. Takahashi, K. & Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663-676 (2006).
4. Obokata et al, Stimulus-Triggered Fate Conversion of Somatic Cells into Pluripotency. Подано совместно с этим документом.
5. Nagy, A., Rossant, J., Nagy, R., Abramow-Newerly, W. & Roder, J.C. Derivation of completely cell culture-derived mice from early-passage embryonic stem cells. Proc Nation AcadSci USA 90, 8424-8428 (1993).
6. Ohgushi, M. and Sasai, Y. Lonely death dance of human pluripotent stem cells: ROCKing between metastable cell states. Trends in Cell Biology 21, 274-282 (2011).
7. Niwa, H. How is pluripotency determined and maintained? Development 134, 635-646 (2007)
8. Quinn J, Kunath T, Rossant J. Mouse trophoblast stem cells. Methods Mol Med. 121, 125-148 (2006).
9. Rossant J. Stem cells and lineage development in the mammalian blastocyst. Reprod Fertil Dev. 19, 111-8 (2007)
10. Tanaka TS, Kunath T, Kimber WL, Jaradat SA, Stagg СA, Usuda M, Yokota T, Niwa H, Rossant J, Ко MS. Gene expression profiling of embryo-derived stem cells reveals candidate genes associated with pluripotency and lineage specificity. Genome Res. 12, 1921-1928(2002)
11. Klaffky E, Williams R, Yao CC, Ziober B, Kramer R, Sutherland A. Trophoblast-specific expression and function of the integrin alpha 7 subunit in the peri-implantation mouse embryo. Dev Biol. 239, 161-175 (2001).
12. Russ AP, Wattler S, Colledge WH, Aparicio SA, Carlton MB, Pearce JJ, Barton SC, Surani MA, Ryan K, Nehls MC, Wilson V, Evans MJ. Eomesodermin is required for mouse trophoblast development and mesoderm formation. Nature 404, 95-99 (2000).
13. Niwa H, Toyooka Y, Shimosato D, Strumpf D, Takahashi K, Yagi R, Rossant J. Interaction between Oct3/4 and Cdx2 determines trophectoderm differentiation. Cell 123, 917-929(2005)
14. Toyooka Y, Shimosato D, Murakami K, Takahashi K, Niwa H. Identification and characterization of subpopulations in undifferentiated ES cell culture. Development 135, 909-918(2008).
15. Ogawa, K., Matsui, H., Ohtsuka, S. & Niwa, H. A novel mechanism for regulating clonal propagation of mouse ES cells. Genes to cells 9, 471-477 (2004).
16. Jentsch I, Geigl J, Klein CA, Speicher MR. Seven-fluorochrome mouse M-FISH for high-resolution analysis of interchromosomal rearrangements. Cytogenet Genome Res. 103, 84-88 (2003),
17. Ying, Q. L. et al. The ground state of embryonic stem cell self-renewal. Nature 453, 519-523 (2008).
18. Murakami K, Araki K, Ohtsuka S, Wakayama T, Niwa H. Choice of random rather than imprinted X inactivation in female embryonic stem cell-derived extra-embryonic cells. Development 138, 197-202 (2011).
19. Watanabe, K., Kamiya, D., Nishiyama, A., Katayama, Т., Nozaki, S., Kawasaki, H., Mizuseki, K., Watanabe, Y., and Sasai, Y. (2005) Directed differentiation of telencephalic precursors from embryonic stem cells. Nature Neurosci. 8, 288-296
20. Kawasaki, H., Mizuseki, K., Nishikawa, S., Kaneko, S., Kuwana, Y., Nakanishi, S., Nishikawa, S.-I. and Sasai, Y. (2000) Induction of midbrain dopaminergic neurons from ES cells by Stromal Cell-Derived Inducing Activity. Neuron 28, 31-40.
21. Gouon-Evans V, Boussemart L, Gadue P, Nierhoff D, Koehler CI, Kubo A, Shafritz DA, Keller G. (2006) BMP-4 is required for hepatic specification of mouse embryonic stem cell-derived definitive endoderm. Nat Biotechnol. 24, 1402-1411.
22. Macfarlan TS, Gifford WD, Driscoll S, Lettieri K, Rowe HM, Bonanomi D, Firth A, Singer O, Trono D, Pfaff SL. Embryonic stem cell potency fluctuates with endogenous retrovirus activity. Nature 487, 57-63 (2012).
23. Eiraku, M., Takata, N., Ishibashi, H., Kawada, M., Sakakura, E., Okuda, S., Sekiguchi, K., Adachi, T. and Sasai, Y. (2011) Self-organizing optic-cup morphogenesis in three-dimensional culture. Nature 472, 51-56.
24. Eiraku, M, Watanabe, K., Matsuo-Takasaki, M., Kawada, M., Yonemura, S., Matsumura, M., Wataya, Т., Nishiyama, A., Muguruma, K. and Sasai, Y. (2008) Self-Organized Formation of Polarized Cortical Tissues from ES cells and its Active Manipulation by Extrinsic Signals. Cell Stem Cell 3, 519-532
25. Boheler KR, Czyz J, Tweedie D, Yang HT, Anisimov SV, Wobus AM. Differentiation of pluripotent embryonic stem cells into cardiomyocytes. Circ Res. 91, 189-201 (2002).
26. Kawasaki, H., Mizuseki, K., Nishikawa, S., Kaneko, S., Kuwana, Y., Nakanishi, S., Nishikawa, S.-I. and Sasai, Y. Induction of midbrain dopaminergic neurons from ES cells by Stromal Cell-Derived Inducing Activity. Neuron 28, 31-40 (2000)

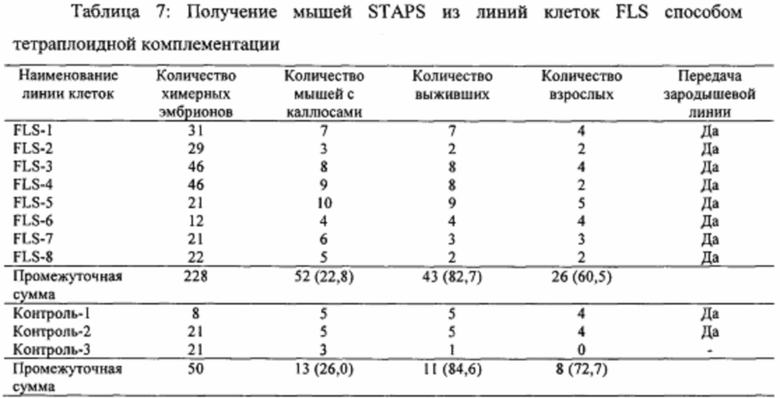
*: Потомство смешивали и подсаживали одной и той же матери из-за недостаточного количества кормящих матерей
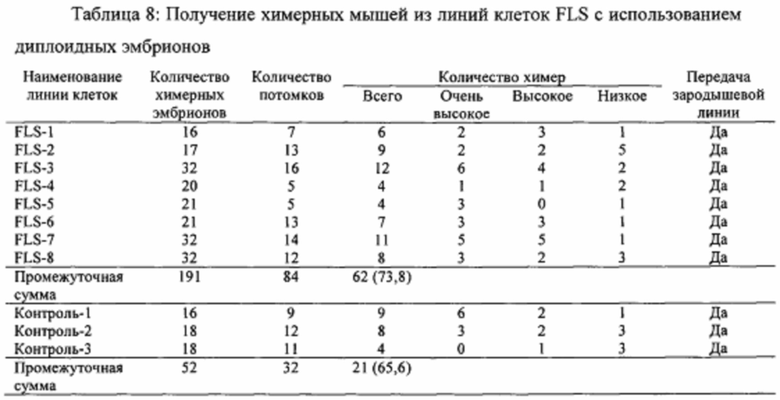
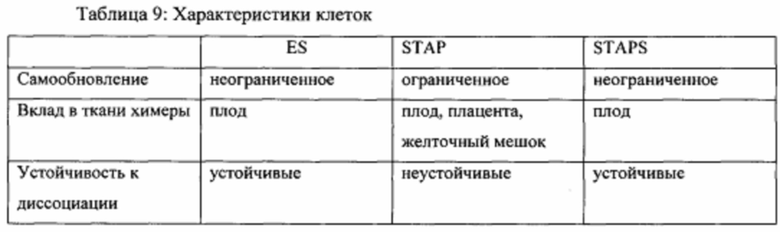
ПРИМЕР 5: Способ получения клеток STAP из зрелых соматических клеток.
В настоящем документе описан улучшенный способ получения клеток STAP, в зависимости от типа изучаемых клеток. Способ ниже представляет собой улучшение по сравнению со способами, описанными в статье авторов настоящего изобретения от 31 января 2014 г., опубликованной в Nature (Obokata et. Al., Stimulus triggered fate conversion of somatic cells into pluripotency. Nature 505. 641-647, 2014) и обеспечивает, например, увеличенную эффективность и выход. Способ является необычайно простым, но может немного меняться, если начинать с ткани, а не с суспензии клеток. Он также может меняться в зависимости от типа клеток или тканей, которые используют в качестве исходного материала.
В некоторых вариантах осуществления не следует пропускать никаких стадий. В некоторых вариантах осуществления следует суспендировать суспензию клеток в течение минимум 30 минут, пока суспензию невозможно будет легко суспендировать вверх и вниз через пипетки с уменьшающимся центральным отверстием до самых маленьких отверстий. Сначала описан способ, начиная с суспензии клеток, и затем описаны дополнительные стадии, необходимые при начале с использованием мягких тканей.
Получение клеток STAP, начиная с суспензии зрелых соматических клеток:
А1. Живые соматические клетки следует суспендировать в пробирке для центрифугирования и затем центрифугировать при 1200 об/мин в течение 5 минут. Примечание: трипсин-ЭДТА, 0,05% (Gibco: 25300-054), можно добавлять в чашку для культивирования клеток, содержащую клетки, на 3-5 минут, для высвобождения адгерентных клеток для добавления в пробирку для центрифугирования.
А2. Отобрать супернатант вплоть до осадка клеток.
A3. Ресуспендировать полученный осадок до концентрации 1×106 клеток/мл в сбалансированном солевом растворе Хенкса (HBSS без Ca+ Mg+: Gibco 14170-112) в 50 мл пробирке. Например, осадок можно ресуспендировать в 2-3 мл HBSS в 50 мл пробирке.
А4. Предварительно покрыть стандартную 9'' (22,86 см) стеклянную пипетку средой (так, чтобы клетки не прилипали к пипетке - иллюстративная пипетка представляет собой Fisher brand 9''Disposable Pasteur Pipettes: 13-678-20D). Суспендировать суспензию клеток в пипетку и из пипетки в течение 5 минут для диссоциации агрегатов клеток и всего сопутствующего дебриса. Это можно осуществлять со значительным усилием.
А5. В качестве конечной стадии процесса суспендирования, изготовить две оплавленные пипетки с очень маленькими отверстиями следующим образом:
Нагреть стандартную 9'' (22,86 см) стеклянную пипетку над горелкой Бунзена, и затем тянуть и растягивать дальний (расплавленный) конец пипетки, пока просвет не схлопнется и конец не оторвется, оставляя закрытый, заостренный стеклянный кончик. Подождать, пока пипетка охладится, и затем отламывать закрытый дальний конец, пока нельзя будет идентифицировать очень маленький просвет. Повторить этот процесс со второй пипеткой, но отломить кончик немного более близко, получая немного больший просвет на дальнем конце. Больший просвет должен составлять приблизительно 100-150 микрон в диаметре, в то время как другая пипетка должна иметь меньший просвет приблизительно 50-70 микрон.
Теперь суспендировать суспензию клеток через пипетку с большим просветом в течение 10 минут. За этим следует суспендирование через пипетку, имеющую меньший просвет (50-70 микрон) в течение дополнительных 15 минут. Продолжать пипетиовать суспензию, пока она не станет легко проходить вверх и вниз по оплавленной пипетке с меньшим отверстием. Предварительно покрыть каждую пипетку средой. Также, во время суспендирования следует избегать забора воздуха и создания пузырей или пены в суспензии клеток.
А6. Добавить HSBB к суспензии до общего объема 20 мл, центрифугировать при 1200 об/мин в течение 5 минут и затем отобрать супернатант.
А7. Ресуспендировать клетки в HBSS при рН 5,4, до концентрации клеток 2×106 клеток/мл, затем поместить в инкубатор при 37'С на 25 минут. рН HBSS будет увеличиваться при добавлении суспензии клеток, так что можно использовать раствор HBSS с более низким, чем желательный конечный рН 5,6.
При подкислении раствора, мягко суспендировать его с использованием 5 мл пипетки в течение 10 секунд сразу после добавления кислоты в раствор Хенкса. HBSS обладает очень слабой буферной емкостью, так что любой раствор, перенесенный из супернатанта предшествующей суспензии будет существенно влиять на рН HBSS. Инструкции ниже показывают, как получить HBSS с оптимальным рН, составляющим 5,6-5,7 для получения клеток STAP в соответствии с этим экспериментальным вариантом осуществления.
Сначала следует титровать рН предварительно охлажденного HBSS (при 4°С) с помощью 12 н. HCl до рН 5,6. Это осуществляют посредством медленного добавления 11,6 мкл 12 н. HCl к 50 мл HBSS. После подтверждения этого рН, следует стерилизовать раствор фильтрацией через шприц с фильтром 0,2 микрон или через фильтр на горле бутыли, в новый стерильный контейнер для хранения. Необходимо подтвердить конечный рН 5,6-5,7 в ходе предварительного тестового эксперимента с соответствующим количеством клеток. Поскольку рН HBSS является настолько важным, рН раствора следует проверять, повторно титровать и повторно стерилизовать перед каждым применением.
А8. Через 25 минут в кислом растворе, центрифугировать суспензию при 1200 об/мин в течение 5 минут.
А9. Отобрать супернатант и ресуспендировать полученный осадок в 5 мл того, что в настоящем документе обозначено «средой для образования клеточных сфер» (DMEM/F12 с 1% антибиотиком и 2% В27 Gibco 12587-010), и поместить в концентрации 105 клеток/см3, в культуральную чашку для неадгерентной культуры в присутствии следующих добавок: b-FGF (20 нг/мл), EGF (20 нг/мл), гепарин (0,2%, Stem Cell Technologies 07980). LIF (1000 ед.) следует добавлять, если клетки являются мышиными). В некоторых вариантах осуществления такие добавки, как bFGF, EGF и гепарин, могут не являться обязательными.
После помещения клеток в культуральные чашки, их можно осторожно пипетировать с использованием 5 мл пипетки, дважды в сутки в течение 2 минут, в течение первой недели, чтобы предотвратить их прикрепление ко дну чашек. Согласно некоторым вариантам осуществления это может способствовать хорошему формированию сфер. Следует добавлять среду для образования клеточных сфер, содержащую описанные добавки, на каждые вторые сутки. (Добавлять 1 мл/сутки на 10 см культуральную чашку или 0,5 мл/сутки на 6 см чашку)
В. Получение клеток STAP, начиная с мягких тканей, которые содержат большое количество эритроцитов.
B1. Поместить вырезанную, промытую, стерильную ткань органа в 60 мм чашку Петри, содержащую 50 мкл коллагеназы. (Может не являться необходимым подвергать селезенку воздействию каких-либо расщепляющих ферментов.) В настоящем документе предусматривают, что различные типы коллагеназы или ферментов являются лучшими для расщепления тканей различных органов.
B2. Нарезать и скоблить ткань в течение 10 минут с использованием скальпелей и ножниц для увеличения площади поверхности, подвергаемой воздействию коллагеназы, пока ткань не приобретет видимую гелеобразную консистенцию.
B3. Добавить дополнительные 450 мкл коллагеназы на чашку и поместить в инкубатор/встряхиватель на 30 минут при 37'C при 90 об/мин.
B4. Добавить 1,5 мл HBSS на чашку (получая общий объем 2,0 мл) и затем отобрать все содержимое через 5 мл пипетку и поместить в 50 мл пробирку.
B5. Теперь переходить к суспендированию, как описано ранее выше (стадии А4-5), начиная со зрелых соматических клеток.
B6. После завершения суспендирования (на стадии А5 при использовании культуральной чашки со зрелыми соматическими клетками), добавить 3 мл HBSS, получая объем 5 мл, в 15 мл пробирку и затем медленно добавляли 5 мл лимфолита на дно пробирки для получения хорошего бислоя. В некоторых вариантах осуществления смешивания двух растворов следует избегать.
B7. Центрифугировать эту пробирку при 1000 g в течение 10 мин. Повернуть пробирку на 180° и повторно центрифугировать при 1000 g в течение дополнительных 10 мин. Это заставит эритроциты формировать осадок на дне пробирки.
B8. С использованием стандартной 9'' (22,86 см) стеклянной пипетки отобрать слой суспензии клеток между HBSS и лимфолитом, и поместить в новую 50 мл пробирку.
89. Добавить HSBB в суспензию до общего объема 20 мл HBSS и затем тщательно перемешать суспензию посредством пипетирования через 5 мл пипетку в течение 1 минуты.
В10. Центрифугировать раствор при 1200 об/мин в течение 5 минут и отобрать супернатант.
В11. Следующие стадии см. в А7-9, как описано в этом примере.
ПРИМЕР 6: Восстановление посредством взрослых стволовых клеток STAP нормальных гипералгезических ответов, уменьшенных посредством химического подавления экспрессирующих NK-1 нейронов в спинном мозге крысы.
Повреждение спинного мозга появляется комплексом часто хронических неврологических сенсорных аномалий, включая онемение, парестезию и боль. Понимание механизмов, лежащих в основе любого из них, и разработка эффективных лекарственных средств осложняется обширными патологическими изменениями, возникающими в результате этих травматических повреждений. В настоящем документе описано очень специфическое цитологическое повреждение спинного мозга для получения ограниченной, но четко выраженной сенсорной недостаточности, которую затем обращают посредством имплантации подвергнутых воздействию стресса взрослых стволовых клеток (измененных посредством вызванной стимулом приобретенной плюрипотентности, STAP).
Высоко специфический цитотоксин SSP-SAP (20 мкл, 1 мкМ) инъецировали в интратекальное (i.t.) пространство спинного мозга самца крысы, чтобы подавить большинство экспрессирующих рецептор нейрокинина-1 (NK1R) нейронов. Через 2-3 недели сильные в норме гипералгезические ответы на инъекцию капсаицина (10 мкл, 0,1%) в подошву задней лапы, состоящие из механической гипералгезии на стимуляцию нитями фон Фрея и термической гипералгезии, проявляющейся как укорочение времени запаздывания отдергивания от источника теплового излучения, почти отсутствовали. Последующая i.t. инъекция стволовых клеток STAP, либо в форме суспензии индивидуальных клеток, либо в форме сферических агрегатов клеток, приводила к медленному восстановлению, в течение следующих 1-2 недель, индуцированной капсаицином механической и термической гипералгезии. Восстановленный ответ обладал такой же амплитудой и зависимостью от времени, как естественный ответ, до подавления, и являлся полностью ингибированным посредством i.t. инъекции антагониста NK-1R L-733,060 (при 300 мкМ). Иммуноцитохимия поясничного отдела спинного мозга от крыс с восстановленными функциями гипералгезии выявила окрашивание NK-1R на протяжении заднего рога. Таким образом, по-видимому, стволовые клетки STAP могут восстанавливать нормальную функцию после потери специфических нейронов спинного мозга и представляют собой модель для способов терапии повреждения спинного мозга.
Взрослых самцов крыс S-D сначала содержали в течение 4-5 дней для их ознакомления с участком для тестирования, для уменьшения индуцированной стрессом аналгезии и для получения данных по поведению наивных мышей и исходных данных по поведению. Способность к ответу на тактильное воздействие определяли посредством тестирования подошвенной поверхности задней лапы с помощью 15 г нити фон Фрея (VFH), 10 раз каждые 3 с. Исходная чувствительность, без обработок и без капсаицина равнялась ~1-2 отдергиваниям лапы на 10 тестов. Показателем термической чувствительности являлось времени запаздывания отдергивания лапы от источника теплового излучения (способ Харгрейвса: пороговое время устанавливали на 18 с.). Исходное запаздывание ~16 с. Определили, что до каких-либо интратекальных инъекций, ответы наивных крыс на инъекцию капсаицина в заднюю лапу составляли: тактильные: 6 отдергиваний/10 тестов VFH, и термические: запаздывание 6 с.
Интратекальные инъекции выполняли способом введения в область крестца, с использованием иглы 30 g, и вводили либо SSP-SAP (конъюгат модифицированного вещества Р - сапорина), выключающий большинство экспрессирующих NK1-R нейронов спинного мозга (Mantyh et al.,), либо его неактивный аналог, Blank-SAP (бессмысленный пептид, конъюгированный с сапорином).
Через несколько недель, когда острая гиперчувствительность, вызванная капсаицином, прекращалась у мышей после лечения SSP-SAP (но не у мышей после лечения Blank-SAP), стволовые клетки с вызванной стимулом приобретенной плюрипотентностью (STAP) (см. фигуру 26) инъецировали в ту же область крестцового отдела спинного мозга позвоночника, куда инъецировали конъюгаты SAP.
Ответы на тактильную и термическую стимуляцию после инъекции капсаицина отслеживали в течение следующих 5 недель, и в этот момент времени крыс подвергали анестезии с использованием пентобарбитала (75 мг/кг i.p.) и перфузии сердца холодным солевым раствором с 4% параформальдегидом. Получали срезы спинного мозга толщиной 50 мкм и окрашивали с использованием первичных антител против NK1-R и против нейронов: [антитело кролика против NK-1R (№ партии 011М4819, Sigma-Aldrich, Сент-Луис, Миссури) 1:5000 (2,3 мкг/мл) и антитело мыши против NeuN (№ партии LV1825845, Millipore, Биллерика, Массачусетс) 1:500 (2 мкг/мл), растворенные в PBS с 1% NDS и 0,3% Triton Х-100], затем интенсивно промывали и инкубировали в соответствующих 2-х антителах [антитело осла против антител кролика с Alexa Fluor 555 ((№ партии 819572) и антитело осла против антител мыши с Alexa Fluor 488 ((№ партии 1113537) (Invitrogen, Гранд Айленд, Нью-Йорк, США), оба 1:1000 (2 мкг/мл) и растворены в 1% NDS и 0,3% Triton Х-100 в PBS] перед рассмотрением с помощью флуоресцентного микроскопа.
Общее количество иммуноположительных по NK-1 и Neu-N клеточных тел на срез ткани подсчитывали для поверхностных (I и II) и глубоких (III-V) слоев для каждой из экспериментальных групп (n=3/группу). Результаты выражены как средний процент выживших нейронов в поверхностных и глубоких слоях у крыс, которым вводили SSP-SAP (или носитель), с обработкой или без обработки стволовыми клетками.
Механическая гипералгезия, показанная по снижению порога отдергивания лапы после инъекции капсаицина, уменьшалась у крыс после обработки интратекальным SSP-SAP (фигура 27). Через пять недель после имплантации стволовых клеток спинного мозга индуцированная капсаицином гипералгезия восстанавливалась. Имплантат стволовых клеток возвращал гипералгезический ответ обработанных SSP-SAP крыс к ответу наивных крыс и обработанного Blank-SAP контроля (фигура 28). Активность специфического антагониста NK1-R увеличивалась у крыс, у которых чувствительность к капсаицину была восстановлена посредством имплантатов стволовых клеток (фигура 29).
SSP-SAP является высоко эффективным для подавления экспрессирующих NK1R нейронов в спинном мозге и таким образом, для фактического прекращения ранних гипералгезических ответов на капсаицин, инъецированный в заднюю лапу. Введение стволовых клеток STAP восстанавливает «нормальные» гипералгезические тактильные и термические ответы на капсаицин у обработанных SSP-SAP крыс. У крыс, не испытывающих изменений в гипералгезической чувствительности из-за инъекции Blank-SAP, доставка стволовых клеток STAP не оказывала эффекта на ответы на капсаицин. Нормализацию гипералгезических ответов посредством стволовых клеток STAP осуществляли посредством возвращения NK1R-IR в спинной мозг. Активность антагониста NK1R для ингибирования вызванной капсаицином гипералгезии усиливалась в 10-60 у крыс, подвергнутых восстановлению с использованием стволовых клеток STAP, по сравнению с его активностью у наивных крыс или у крыс после введения Blank-SAP. Без желания быть связанными с теорией, предусматривают, что это может происходить из-за изменения аффинности антагониста для NK1R, индуцированного посредством стволовых клеток STAP, или из-за различий в связывании NK1R в общей схеме гипералгезических ответов у крыс после восстановления.
ПРИМЕР 7
В настоящем документе описан протокол с улучшенными результатами для получения плюрипотентных стволовых клеток STAP из зрелых соматических клеток, вне зависимости от источника клеток. Протокол переработан, чтобы отражать улучшенные способы. В этом протоколе используют комбинацию индивидуальных стрессов и способов, которые являются более эффективными для достижения желательного конечного результата; то есть, получения плюрипотентных клеток STAP.
Без желания быть связанными с теорией, в настоящем документе предусматривают, что в некоторых способах, описанных в настоящем документе, АТР используют в качестве источника энергии для улучшения жизнеспособности клеток и образованных сфер. Добавление АТР приводило к лучшему формированию сфер и являлось ассоциированным с заметным уменьшением рН раствора, воздействию которого подвергали зрелые клетки. Дальнейшее исследование полезности раствора с низким рН, содержащего АТР, для получения клеток STAP показывает, что в то время как рН сам по себе приводил к образованию клеток STAP, применение раствора с низким рН, содержащего АТР, существенно увеличивало эффективность этого перехода. Когда этот раствор использовали в комбинации с механическим суспендированием зрелых клеток, результаты являлись даже более значительными. Следовательно, в настоящем документе описан способ, включающий в себя эти данные для увеличения эффективности получения клеток STAP.
Описанный способ является эффективным для получения клеток STAP, вне зависимости от исследуемого типа клеток. В некоторых вариантах осуществления суспендирование суспензии зрелых клеток в растворе с низким рН, улучшенным с использованием АТР, продолжают в течение минимум 30 минут, например, пока суспензию невозможно будет легко суспендировать вверх и вниз через пипетки с уменьшающимся центральным отверстием до самых маленьких отверстий. Сначала описан способ для использования, начиная с суспензии клеток, и затем описаны дополнительные стадии, необходимые при начале с использованием мягких тканей.
А. Получение клеток STAP, начиная с суспензии зрелых соматических клеток:
А1. Приготовить раствор HBSS с низким рН, содержащий ACT, следующим образом и затем оставить для использования на стадии А4. Приготовить исходный раствор АТР, 200 мМ, для добавления к HBSS посредством добавления 110,22 мг порошка АТР (гидрат динатриевой соли аденозин-5'-трифосфата Sigma А2383) на каждый 1 мл воды (вода MilliQ). рН этого раствора составляет приблизительно 3,0.
Поместить 5 мл HBSS (с фенолом красным) [Life Technologies, 14170-161] в 15 мл пробирку. Поместить чистый датчик рН в HBSS. Титровать исходный раствор ACT в HBSS, по каплям, до получения желательного рН, например, 5,0. Регулярно перемешивать раствор, чтобы убедиться, что измерение является точным.
В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет от приблизительно 0,5 мг/см3 до приблизительно 100 мг/см3. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет от приблизительно 0,5 мг/см3 до приблизительно 20 мг/см3. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет от приблизительно 0,5 мг/см3 до приблизительно 10 мг/см3. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет от приблизительно 1,0 мг/см3 до приблизительно 7 мг/см3. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет от приблизительно 1,5 мг/см3 до приблизительно 5 мг/см3. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет от приблизительно 1 мМ до приблизительно 150 мМ. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет от приблизительно 1 мМ до приблизительно 50 мМ. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет от приблизительно 1 мМ до приблизительно 15 мМ. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет от приблизительно 2,0 мМ до приблизительно 10 мМ. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет от приблизительно 2,7 мМ до приблизительно 9 мМ.
В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет по меньшей мере приблизительно 0,5 мг/см3. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет по меньшей мере приблизительно 1,0 мг/см3. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет по меньшей мере приблизительно 1,5 мг/см3. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет по меньшей мере приблизительно 1 мМ. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет по меньшей мере приблизительно 2,0 мМ. В некоторых вариантах осуществления концентрация АТР в полученном растворе HBSS и АТР составляет по меньшей мере приблизительно 2,7 мМ.
В некоторых вариантах осуществления более высокой концентрации АТР можно достигать посредством забуферивания раствора до желательного рН и/или близкого к нему. В некоторых вариантах осуществления желательный рН составляет по меньшей мере 5,0. В некоторых вариантах осуществления желательный рН составляет по меньшей мере приблизительно 5,0. В некоторых вариантах осуществления желательный рН составляет приблизительно 5,0. В некоторых вариантах осуществления желательный рН составляет от приблизительно 4,0 до приблизительно 6,5. В некоторых вариантах осуществления желательный рН составляет от приблизительно 5,0 до приблизительно 5,7.
А2. Добавить живые соматические клетки, подлежащие обработке, в форме суспензии клеток, в пробирку для центрифугирования, и затем центрифугировать при 1200 об/мин в течение 5 минут. В некоторых вариантах осуществления трипсин-ЭДТА, 0,05% (Gibco: 25300-054), можно добавлять в культуральную чашку, содержащую клетки, на 3-5 минут, для высвобождения адгерентных клеток для добавления в пробирку для центрифугирования.
A3. Отобрать супернатант вплоть до осадка клеток.
А4. Ресуспендировать полученный осадок до концентрации 1×106 клеток/мл в сбалансированном солевом растворе Хенкса с низким рН, с АТР, (полученном выше на стадии 1А) в 50 мл пробирке. В некоторых вариантах осуществления можно использовать объем 2-3 мл суспензии клеток в 50 мл.
А5. Предварительно покрыть стандартную 9'' (22,86 см) стеклянную пипетку средой (так, чтобы клетки не прилипали к пипетке - например, Fisher brand 9'' Disposable Pasteur Pipettes: 13-678-20D). Суспендировать суспензию клеток в пипетку и из пипетки в течение 5 минут для диссоциации агрегатов клеток и всего сопутствующего дебриса. Это можно осуществлять со значительным усилием.
А6. В качестве конечной стадии процесса суспендирования, изготовить две оплавленные пипетки с очень маленькими отверстиями следующим образом: нагреть стандартную 9'' (22,86 см) стеклянную пипетку над горелкой Бунзена, и затем тянуть и растягивать дальний (расплавленный) конец пипетки, пока просвет не схлопнется и конец не оторвется, оставляя закрытый, заостренный стеклянный кончик. Подождать, пока пипетка охладится, и затем отламывать закрытый дальний конец, пока нельзя будет идентифицировать очень маленький просвет. Повторить этот процесс со второй пипеткой, но отломить кончик немного более близко, получая немного больший просвет на дальнем конце. Больший просвет должен составлять приблизительно 100-150 микрон в диаметре, в то время как другая пипетка должна иметь меньший просвет приблизительно 50-70 микрон. Теперь суспендировать суспензию клеток через пипетку с большим просветом в течение 10 минут. За этим следует суспендирование через пипетку, имеющую меньший просвет (50-70 микрон) в течение дополнительных 15 минут. Продолжать пипетиовать суспензию, пока она не станет легко проходить вверх и вниз по оплавленной пипетке с меньшим отверстием. Снова, следует помнить о предварительном покрытии каждой пипетки средой. Также, во время суспендирования следует избегать забора воздуха и создания пузырей или пены в суспензии клеток.
А7. Добавить обычный HBSS (не содержащий АТР) к суспензии до общего объема 20 мл, центрифугировать при 1200 об/мин в течение 5 минут и затем отобрать супернатант.
А8. Ресуспендировать полученный осадок в 5 мл того, что в настоящем документе обозначено «средой для образования клеточных сфер» (DMEM/F12 с 1% антибиотиком и 2% В27 Gibco 12587-010) и поместить в концентрации 105 клеток/мл, в культуральную чашку для неадгерентной культуры в присутствии следующих добавок: b-FGF (20 нг/мл), EGF (20 нг/мл), гепарин (0,2%, Stem Cell Technologies 07980). LIF (1000 ед.) следует добавлять, если клетки являются мышиными). В некоторых вариантах осуществления такие добавки, как bFGF, EGF и гепарин, могут не являться обязательными. После помещения клеток в культуральные чашки, их следует осторожно пипетировать с использованием 5 мл пипетки, дважды в сутки в течение 2 минут, в течение первой недели, чтобы предотвратить их прикрепление ко дну чашек. Это является важным для получения хорошего формирования сфер. Добавлять среду для образования клеточных сфер, содержащую описанные добавки, на каждые вторые сутки. (Добавлять 1 мл/сутки на 10 см культуральную чашку, или 0,5 мл/сутки на 6 см чашку.)
В. Получение клеток STAP, начиная с мягких тканей, которые содержат много эритроцитов.
В1. Поместить вырезанную, промытую, стерильную ткань органа в 60 мм чашку Петри, содержащую 50-500 мкл коллагеназы, в зависимости от размера ткани. Добавить достаточный объем коллагеназы, чтобы смочить всю ткань. Различные типы коллагеназ или других ферментов являются лучшими для расщепления тканей различных органов. (Может не являться необходимым подвергать селезенку воздействию каких-либо расщепляющих ферментов.)
B2. Нарезать и скоблить ткань в течение 10 минут с использованием скальпелей и ножниц для увеличения площади поверхности, подвергаемой воздействию коллагеназы, пока ткань не приобретет видимую гелеобразную консистенцию.
В настоящем документе конкретно предусмотрено, что в этом варианте осуществления способа, или в любом варианте осуществления способа, описанного в настоящем документе, соскабливание ткани можно осуществлять с помощью плоско обработанного лезвия и/или поверхности, например, скальпеля номер 11, в отличие от скальпеля с закругленным лезвием. Альтернативно, в любом варианте осуществления описанном в настоящем документе, применением звуковых волн высокой частоты можно заменять соскабливание, и/или их можно комбинировать (либо одновременно, либо последовательно) для разрушения ткани. Звуковые волны высокой частоты могут, например, разрушать мембраны, пробивать отверстия в ткани и/или мембранах, и/или вызывать протекание мембраны. Применение звуковых волн высокой частоты также легко поддается масштабированию. Специалистам в данной области известны способы воздействия звуковых волн высокой частоты на ткани, например, доступны коммерческие генераторы ультразвука (например, The Qsonica Q55 Sonicator, кат.No. UX-04712-52, доступный из Cole-Palmer; Верной Хилз, Иллинойс).
B3. Добавить дополнительную коллагеназу в чашку для получения общего объема = 0,5 мл, и поместить в инкубатор/встряхиватель на 30 минут при 37°С при 90 об/мин.
B4. Добавить 1,5 мл раствора HBSS/АТР с низким рН на чашку (получая общий объем 2,0 мл) и затем отобрать все содержимое через 5 мл пипетку и поместить в 50 мл пробирку.
B5. Теперь переходить к суспендированию, как описано ранее выше (стадии А4-5), начиная со зрелых соматических клеток.
B6. После завершения суспендирования (на стадии А5 при использовании культуральной чашки со зрелыми соматическими клетками), добавить 3 мл HBSS, получая объем 5 мл, в 15 мл пробирку и затем медленно добавлять 5 мл лимфолита на дно пробирки для получения хорошего бислоя. Раствор следует добавлять, как описано, для получения бислоя, и избегать смешивания двух растворов.
B7. Центрифугировать эту пробирку при 1000 g в течение 10 мин. Повернуть пробирку на 180° и повторно центрифугировать при 1000 g в течение дополнительных 10 мин. Это заставит эритроциты формировать осадок на дне пробирки.
B8. С использованием стандартной 9'' (22,86 см) стеклянной пипетки отобрать слой суспензии клеток между HBSS и лимфолитом, и поместить в новую 50 мл пробирку.
B9. Добавить HBSS в суспензию до общего объема 20 мл HBSS и затем тщательно перемешать суспензию посредством пипетирования через 5 мл пипетку в течение 1 минуты.
В10. Центрифугировать раствор при 1200 об/мин в течение 5 минут и отобрать супернатант.
В11. Следующие стадии см. в. А6-8.
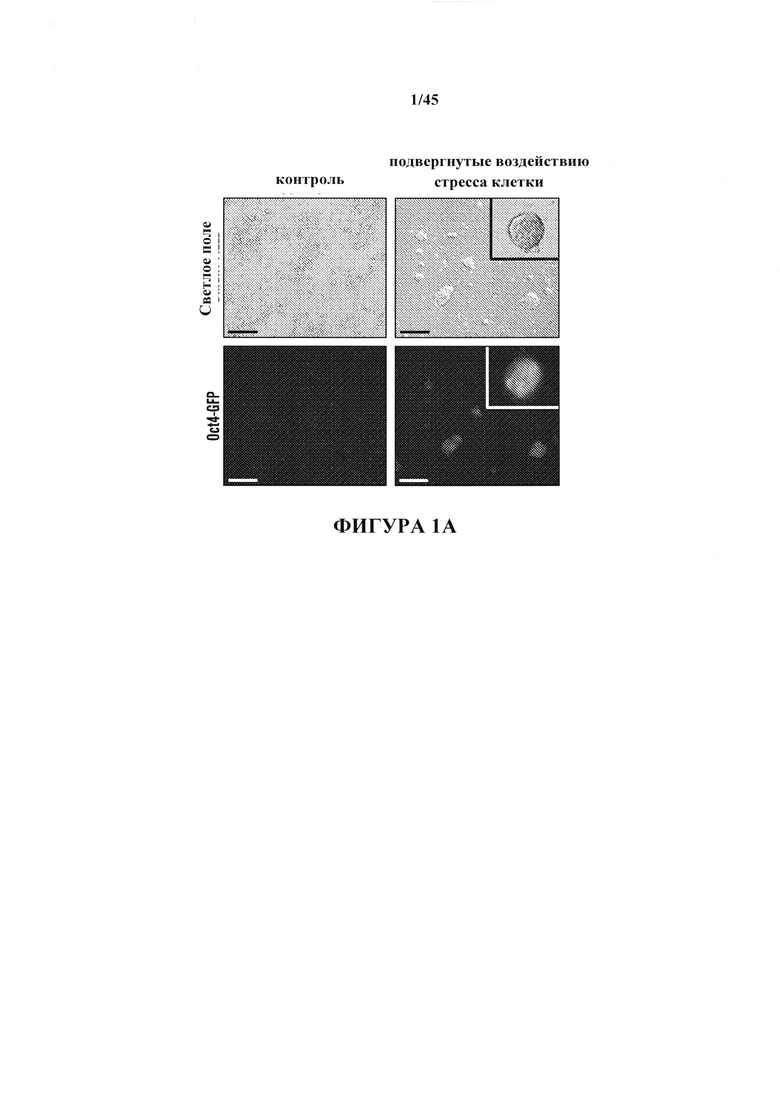
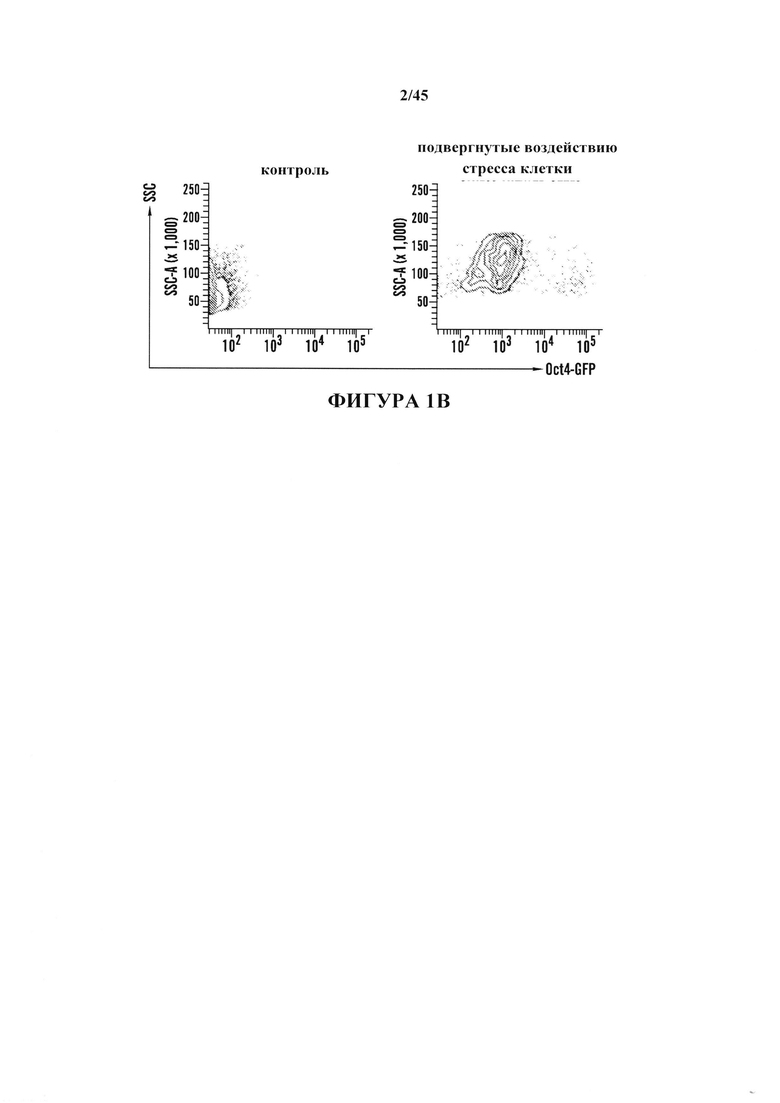
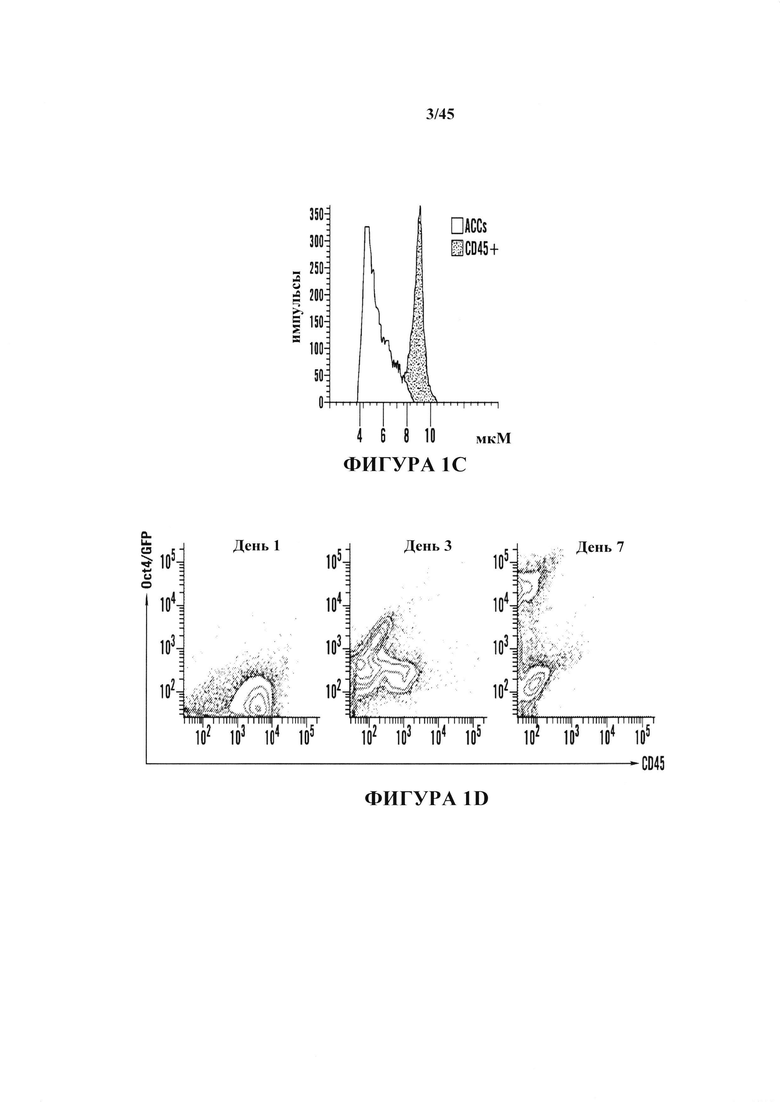
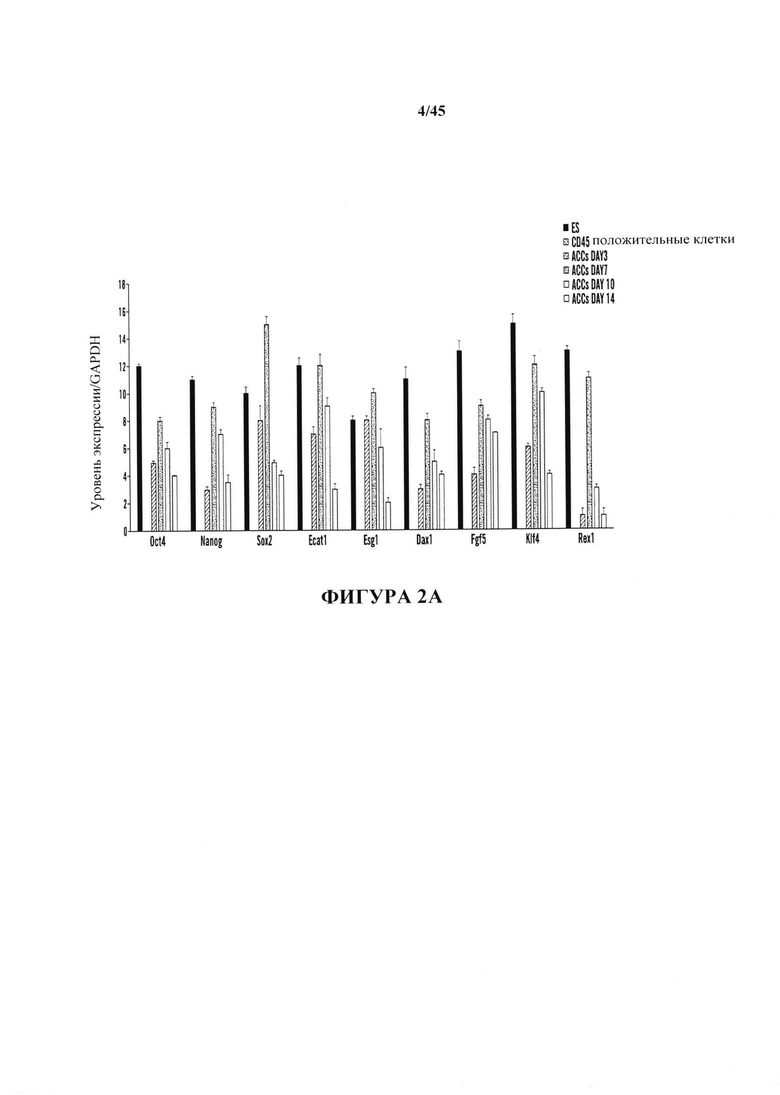
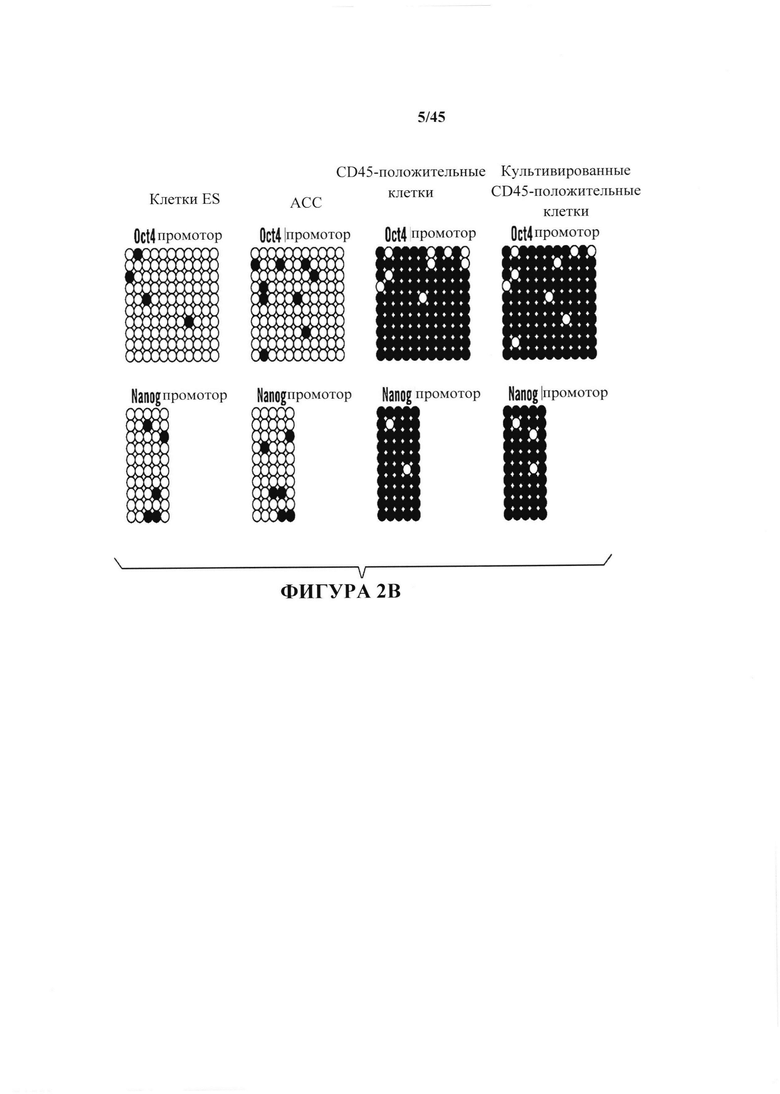

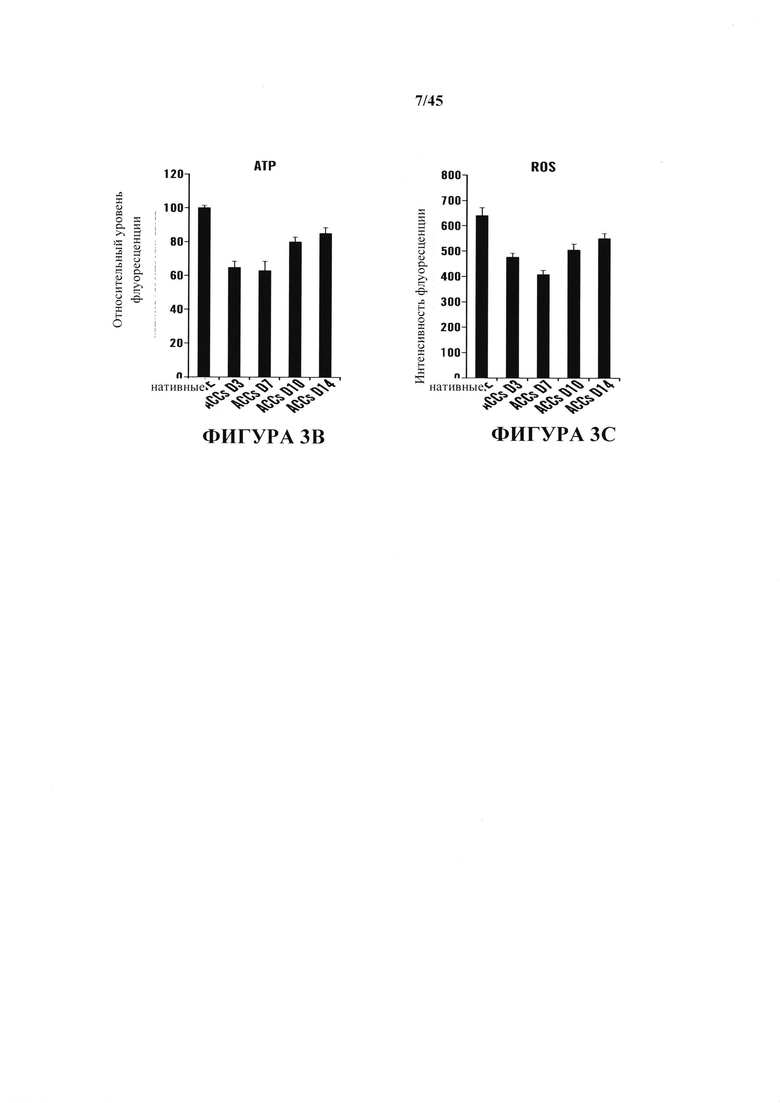
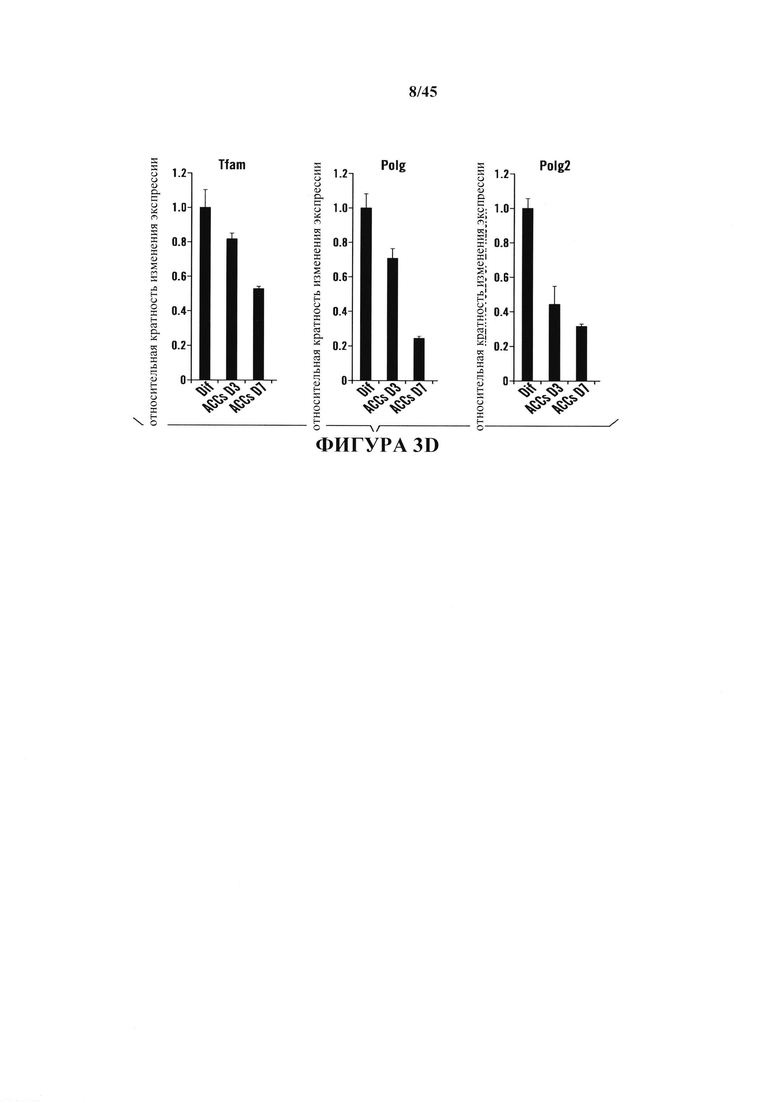
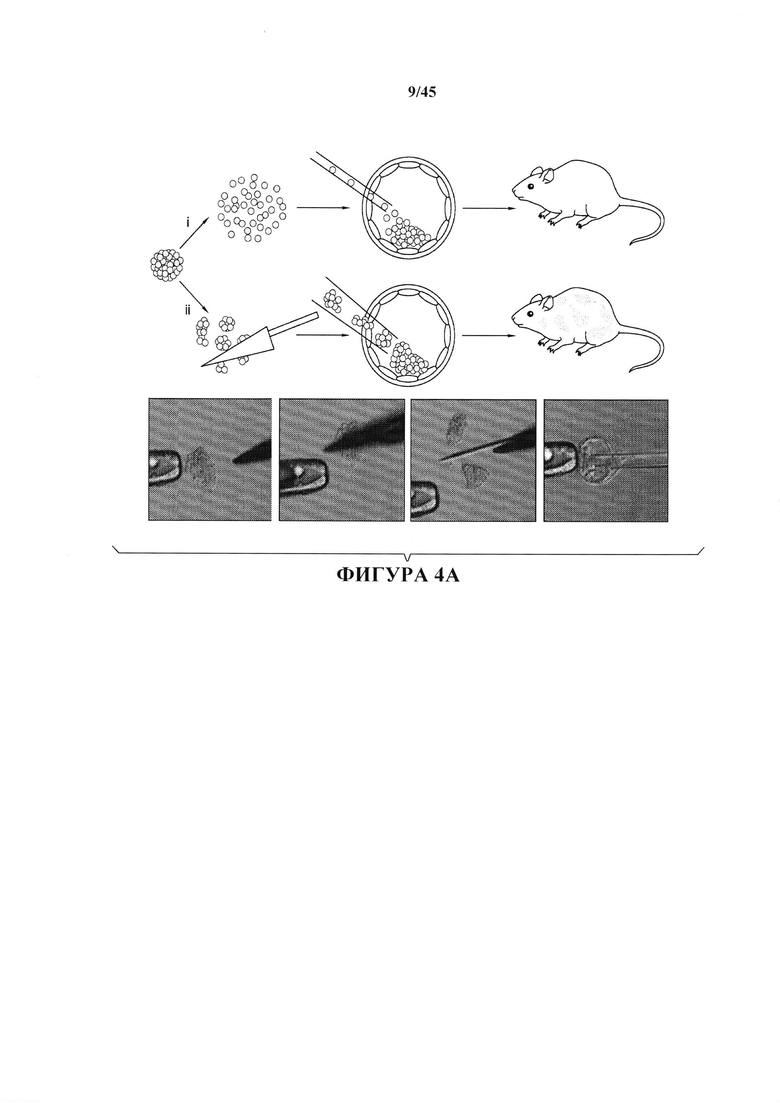

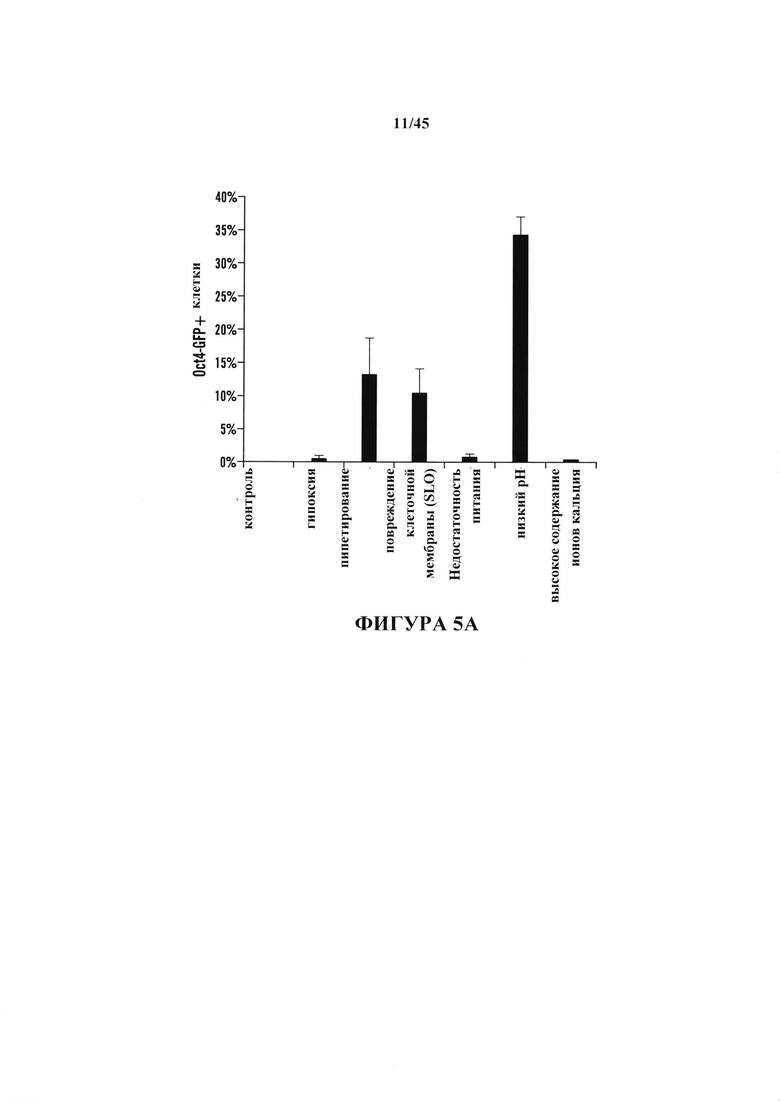
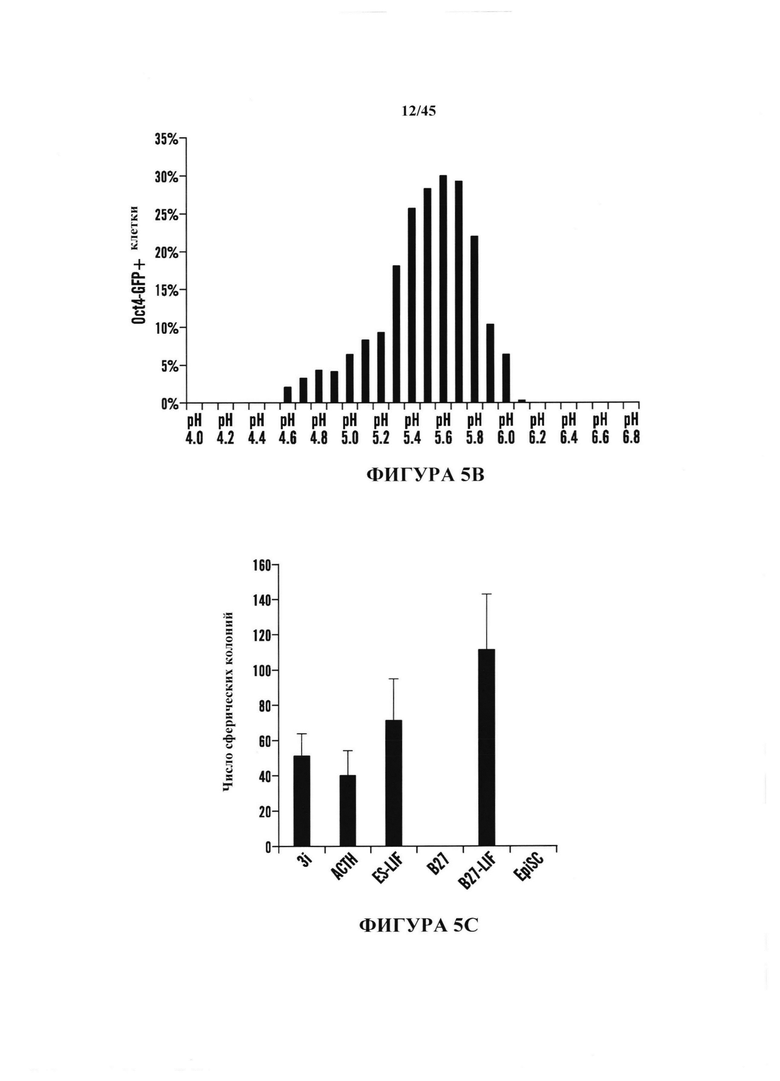
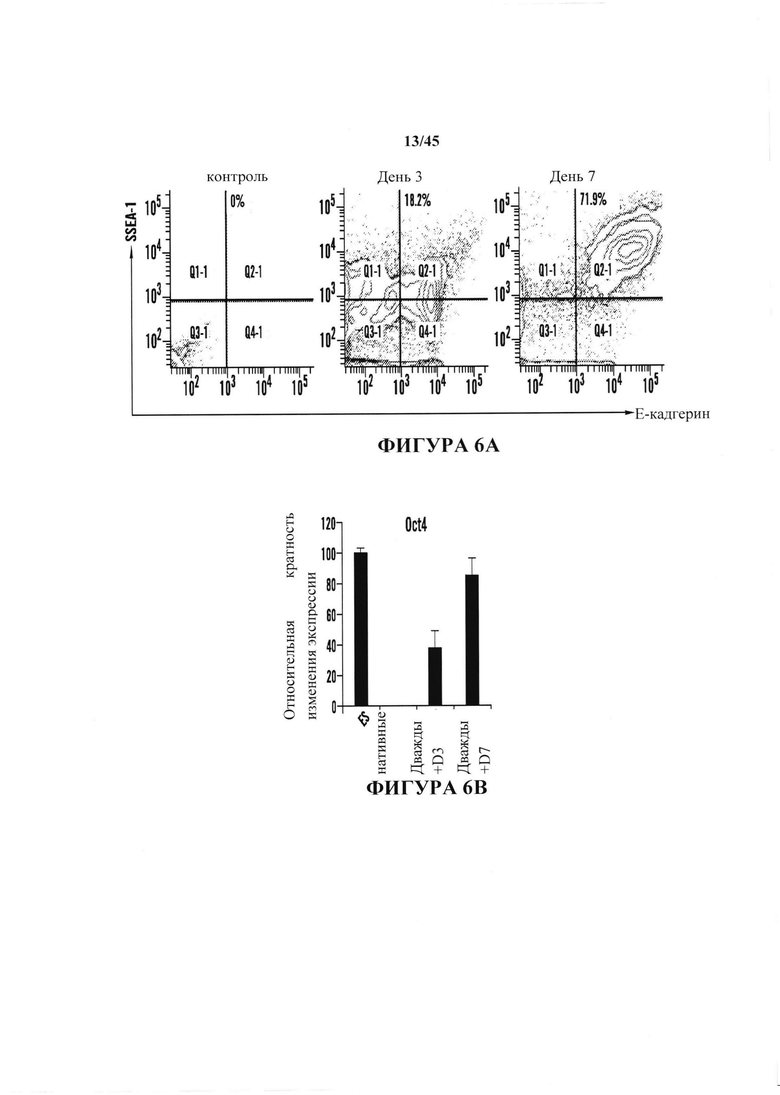
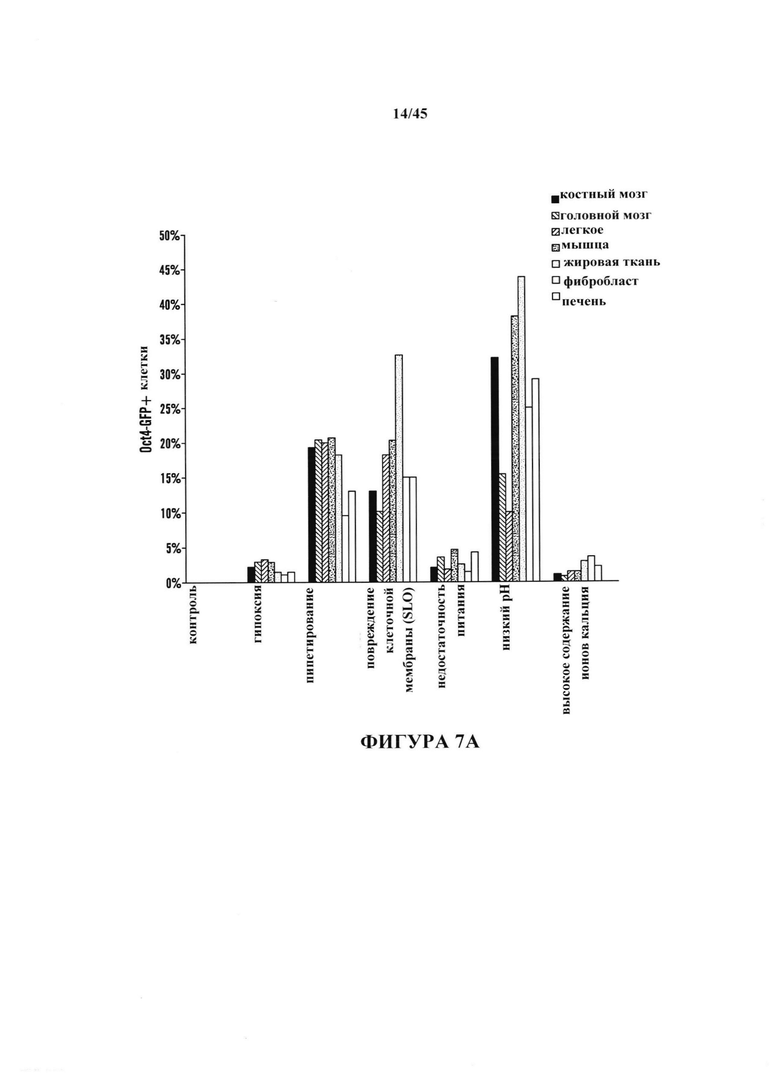
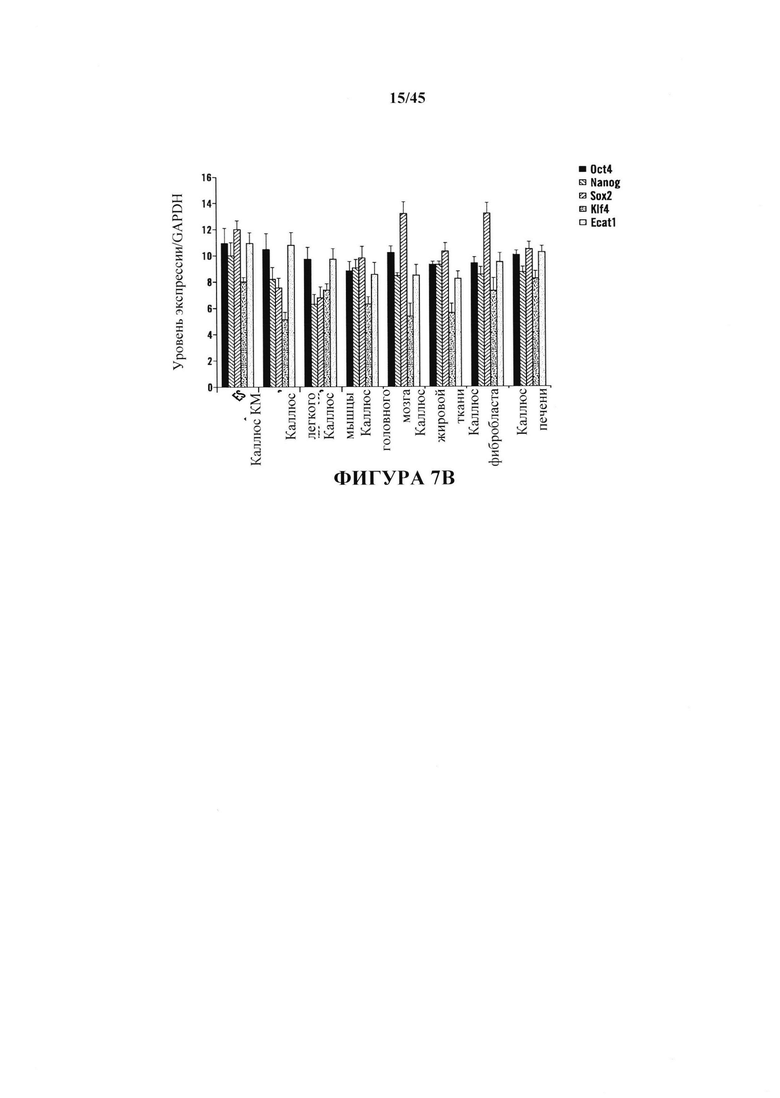
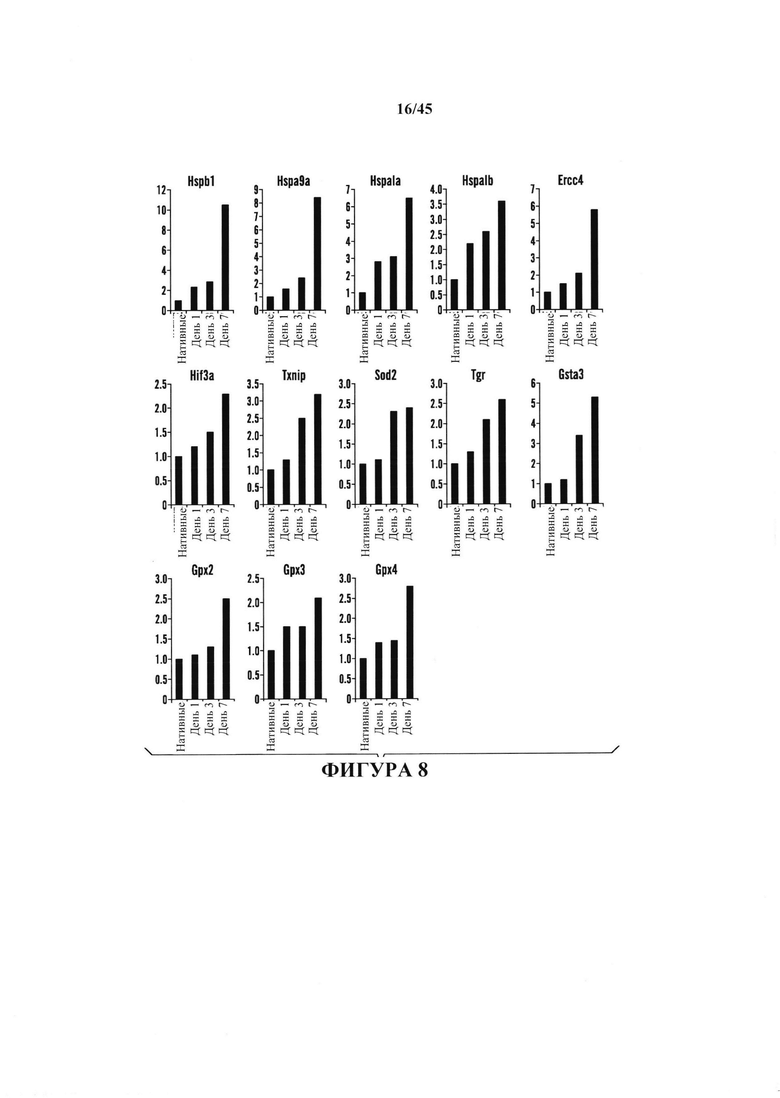
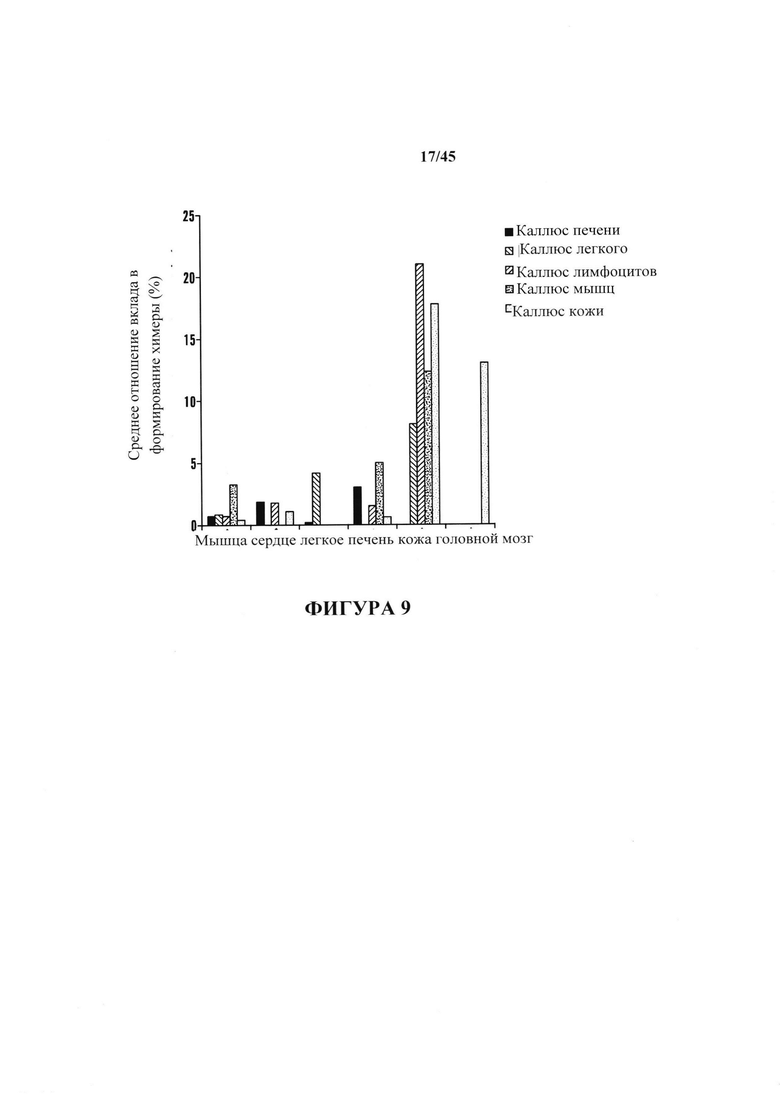
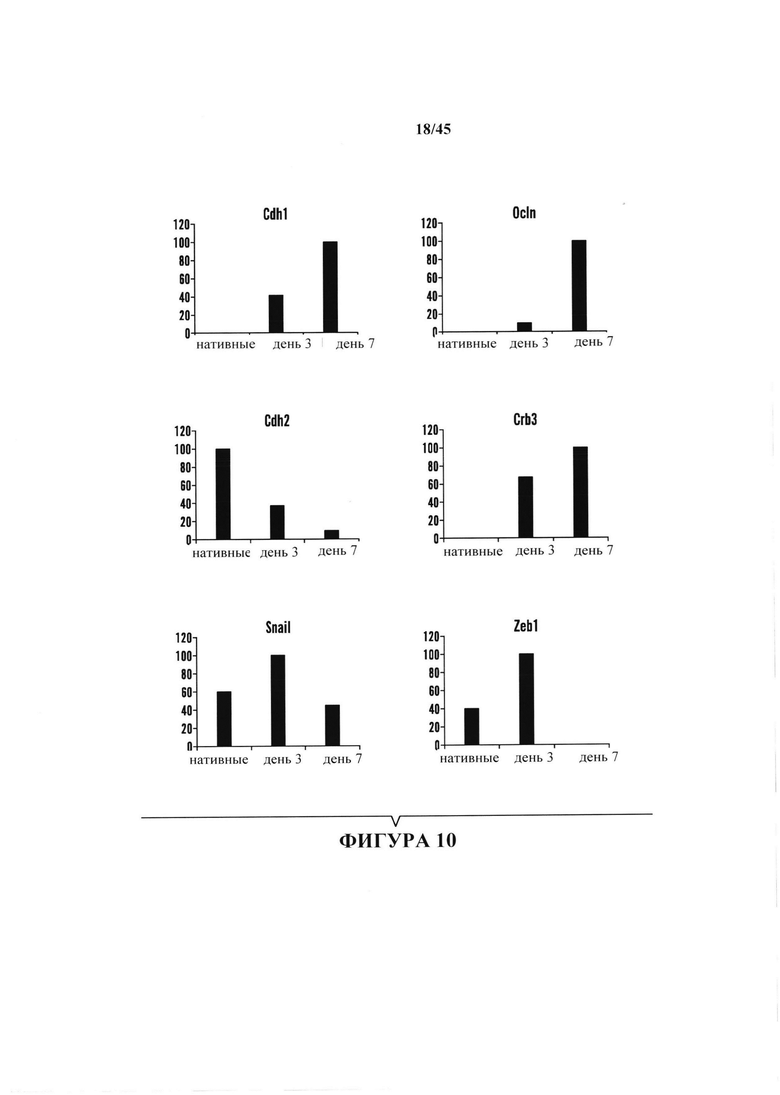
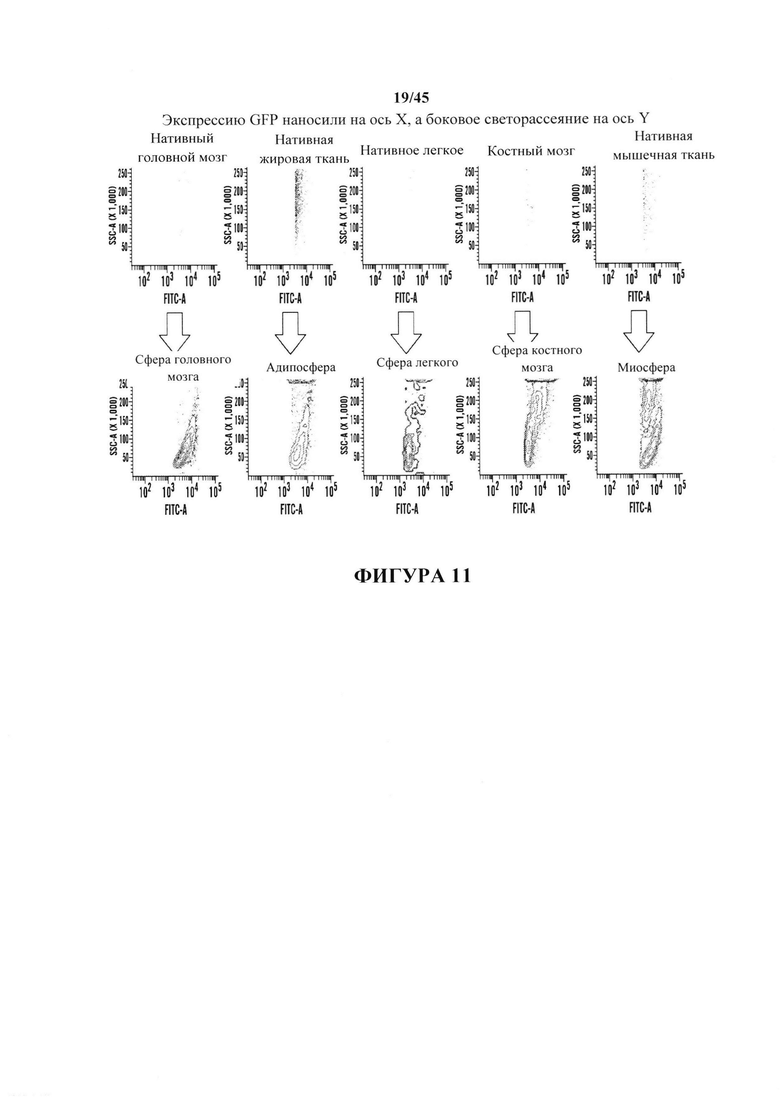
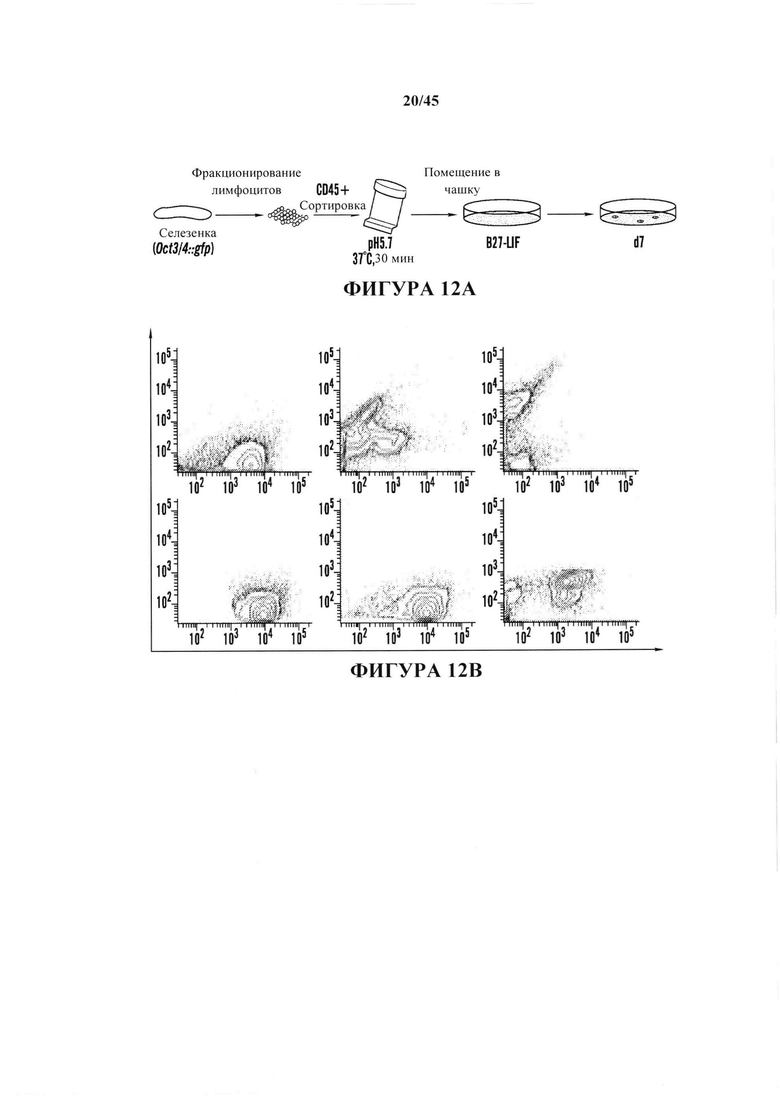
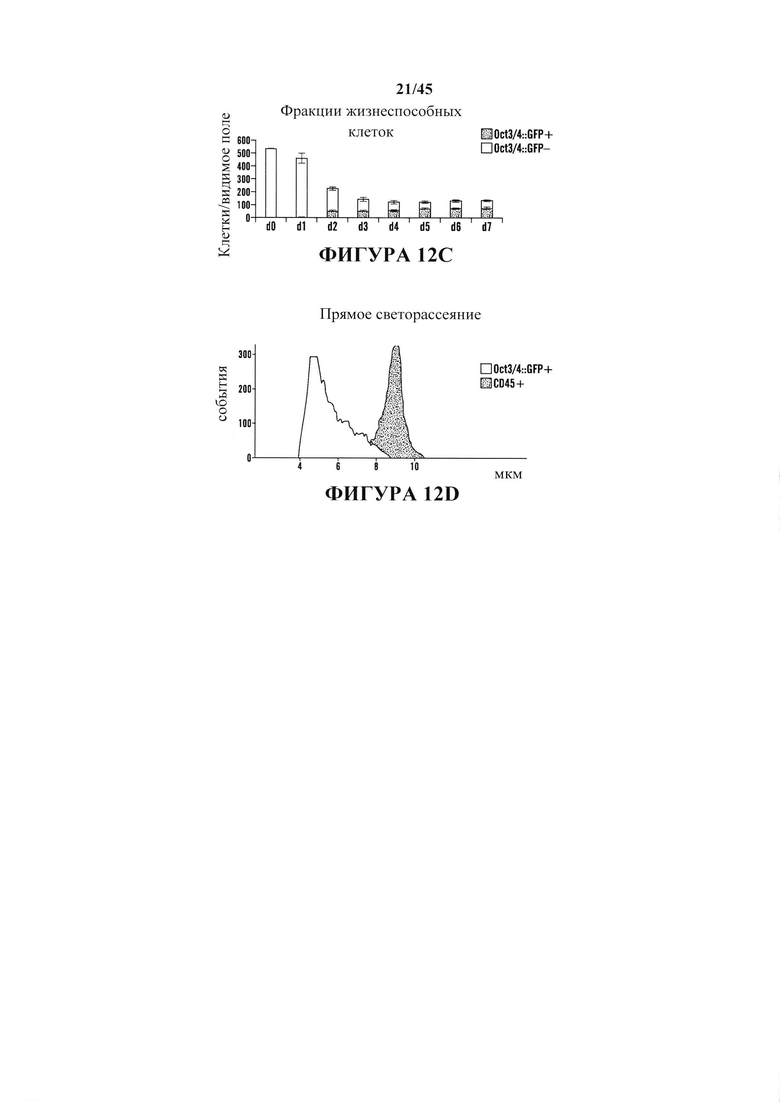
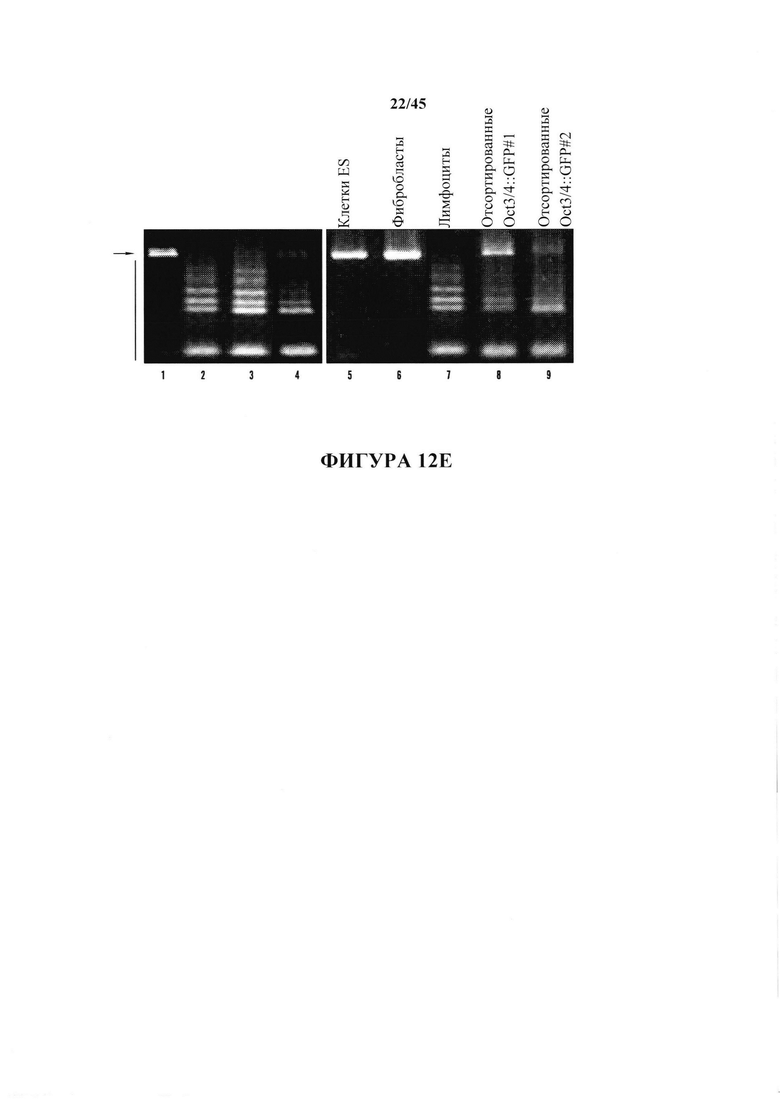

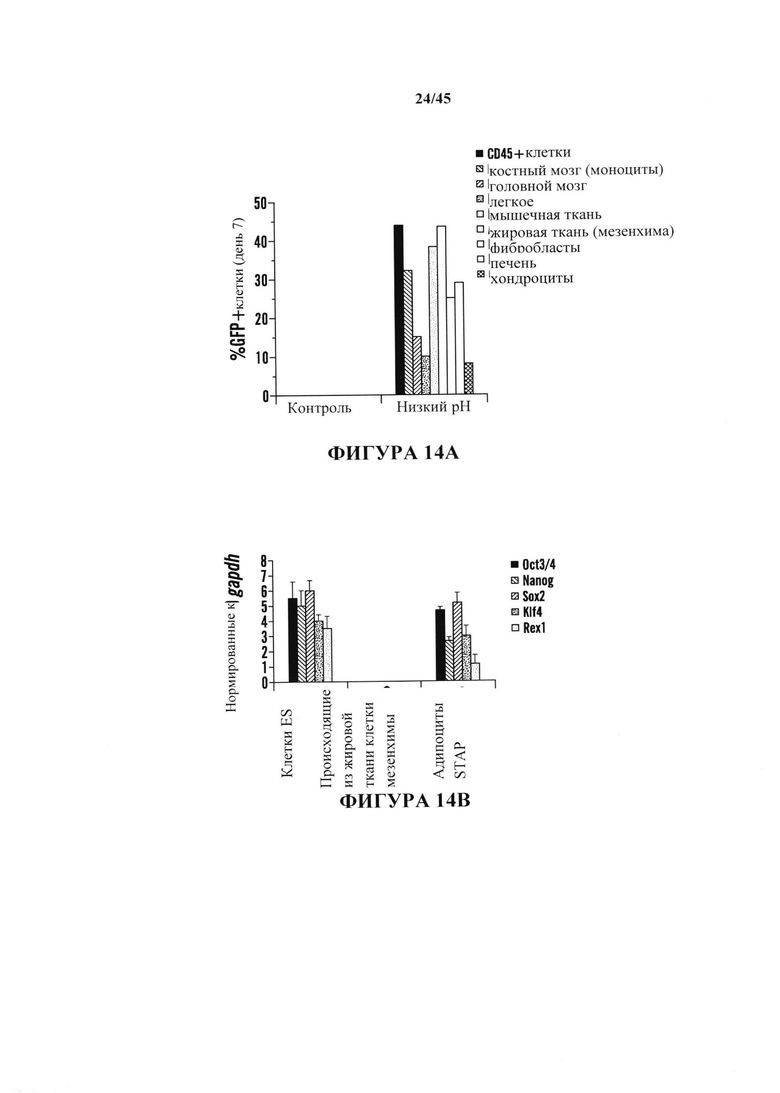
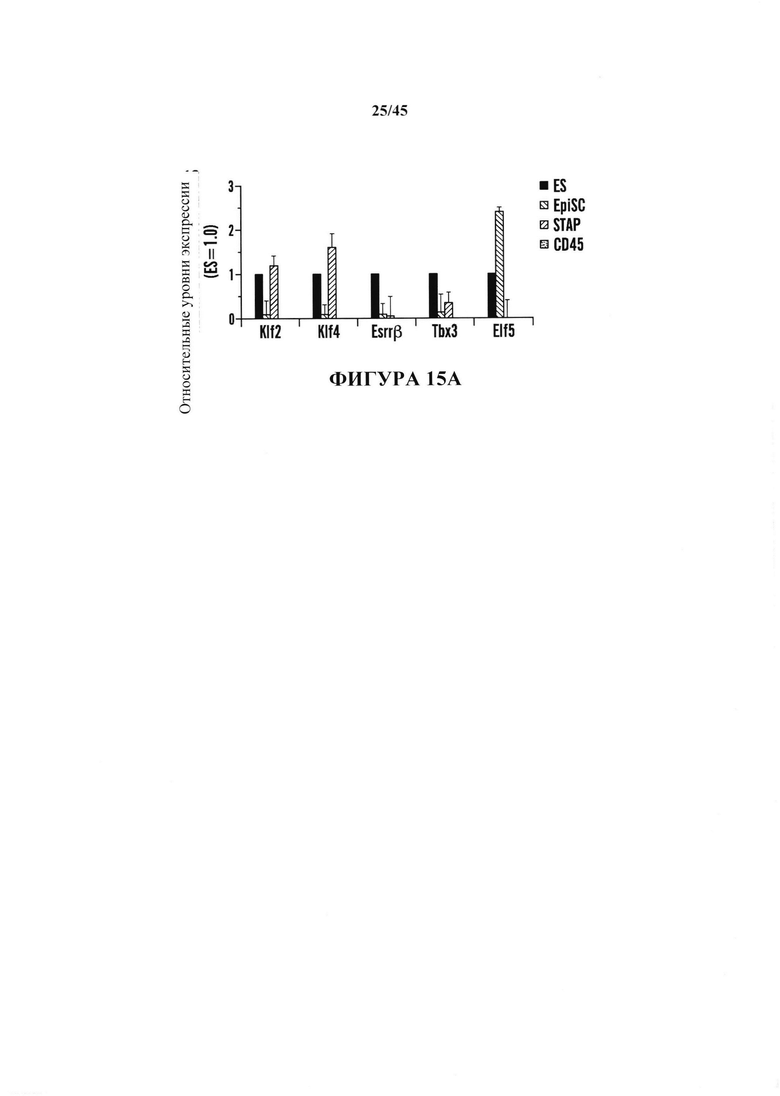
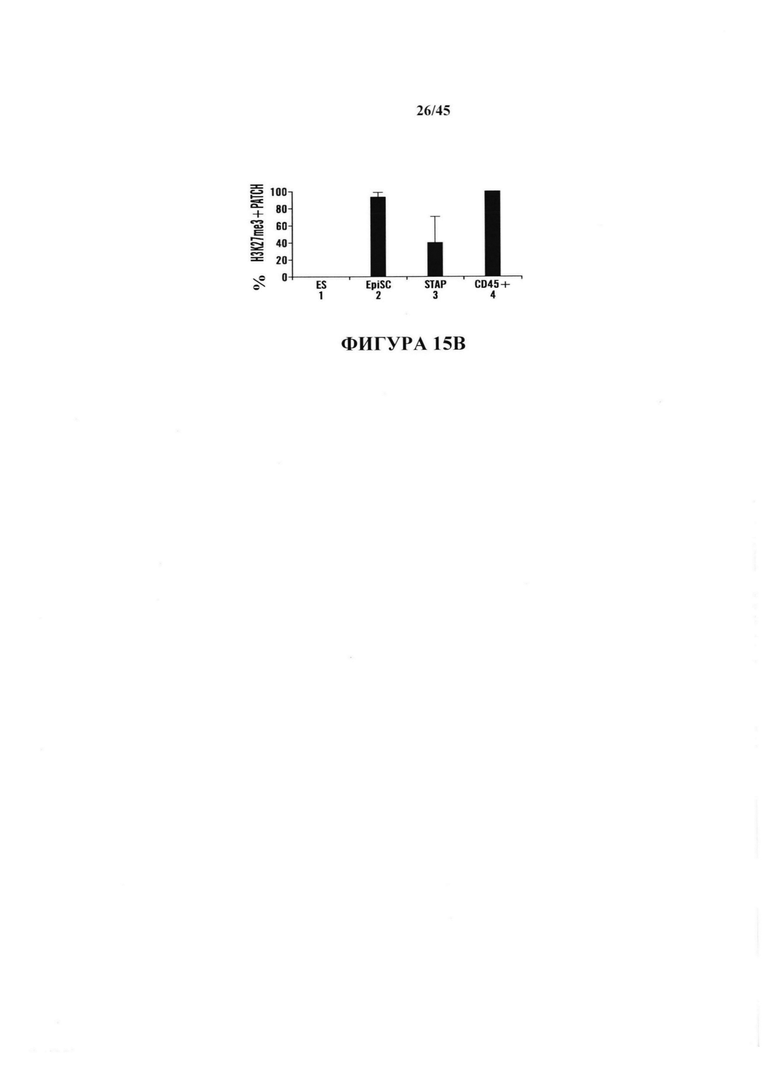
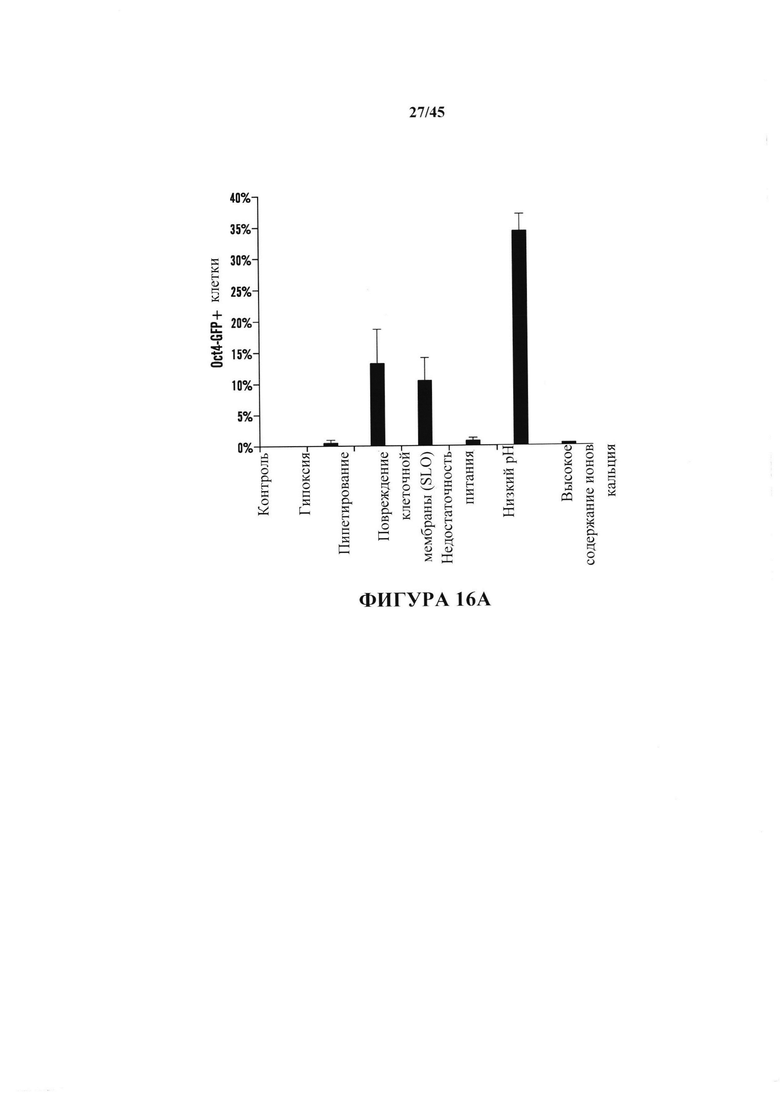
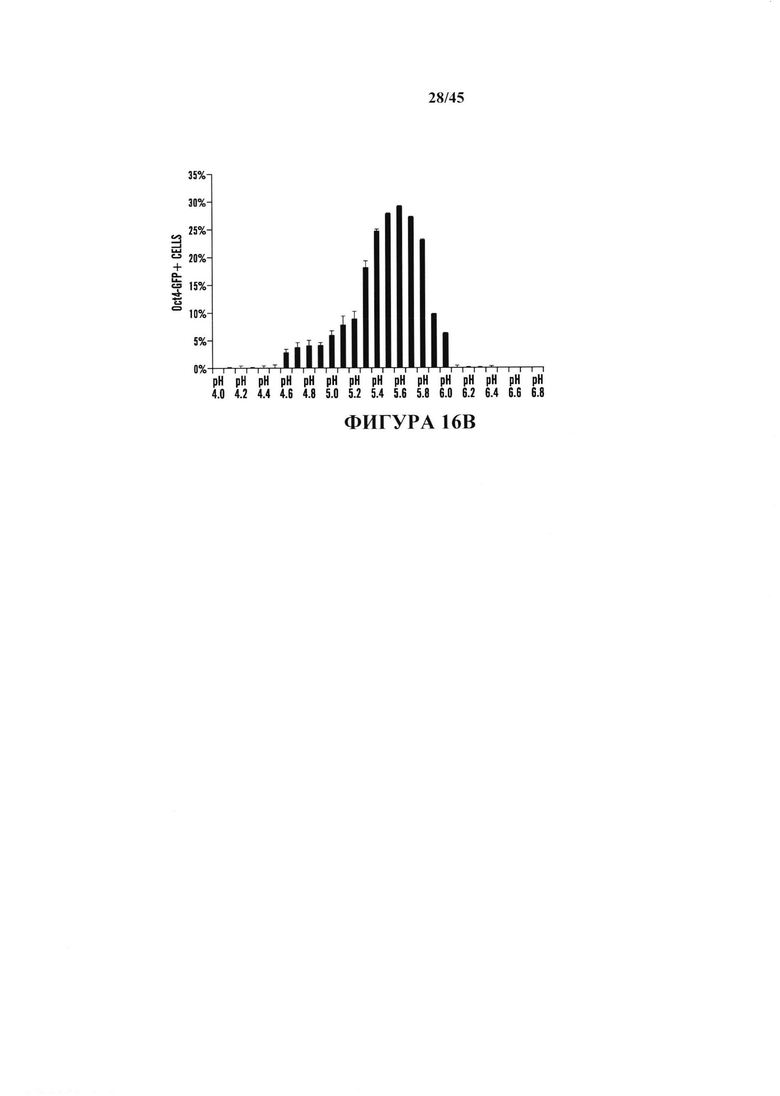
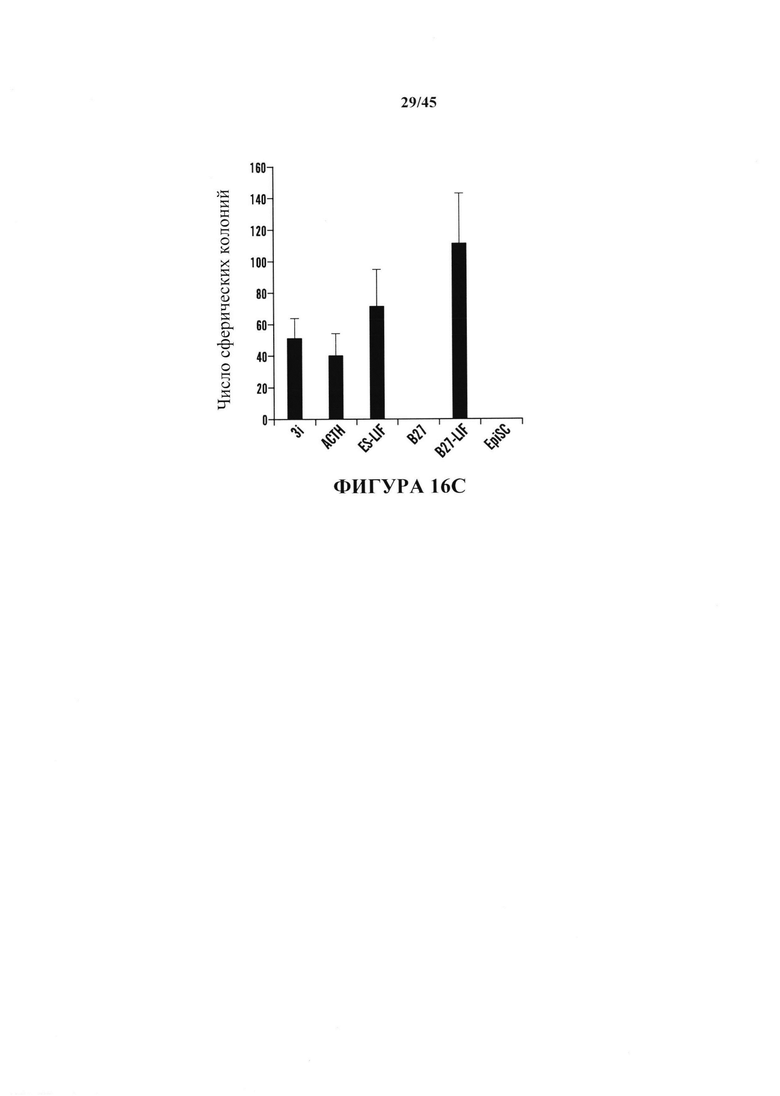

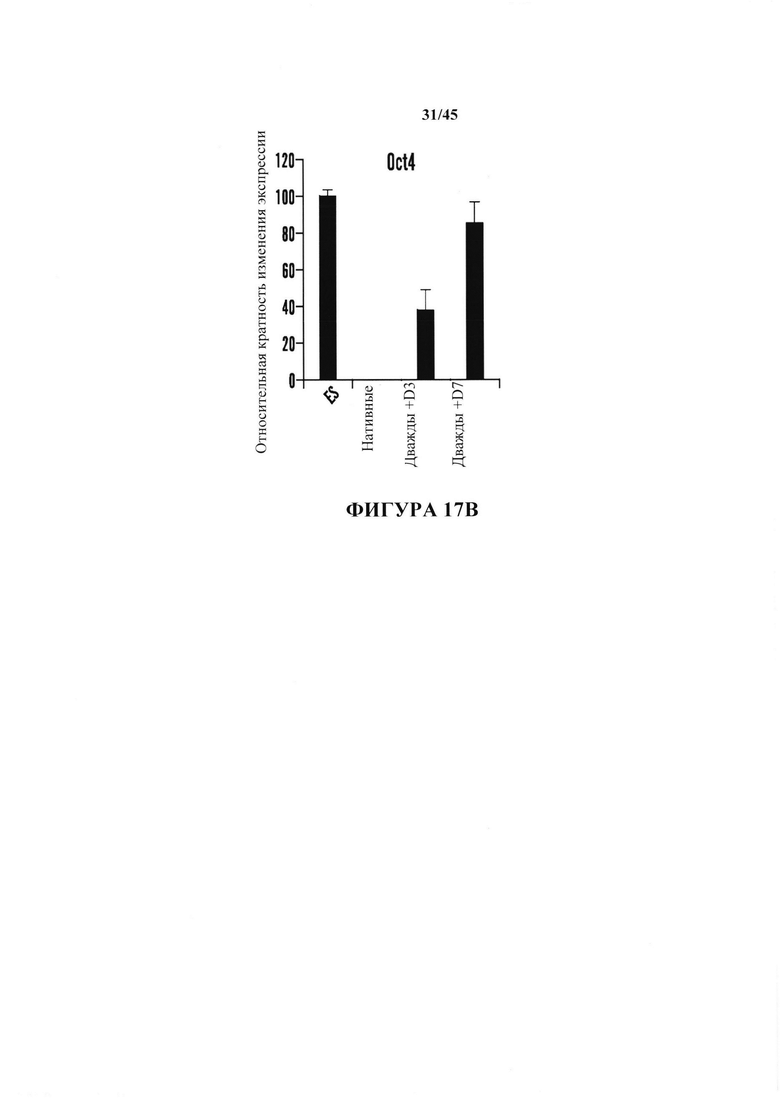
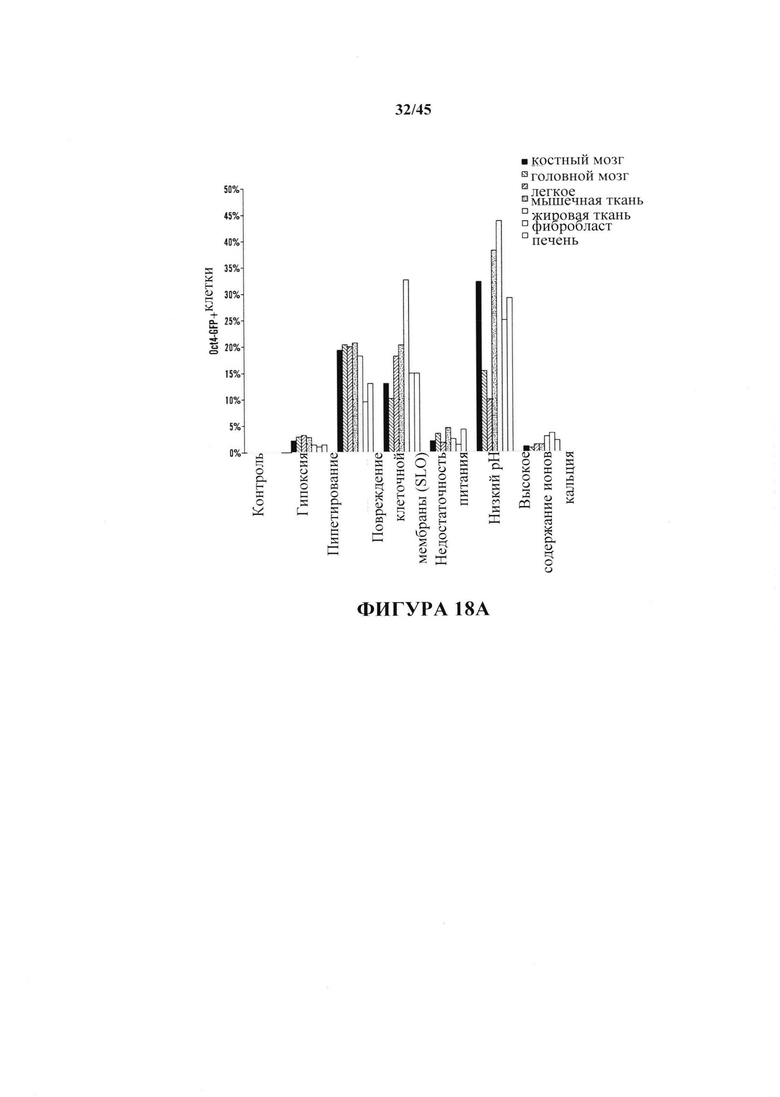
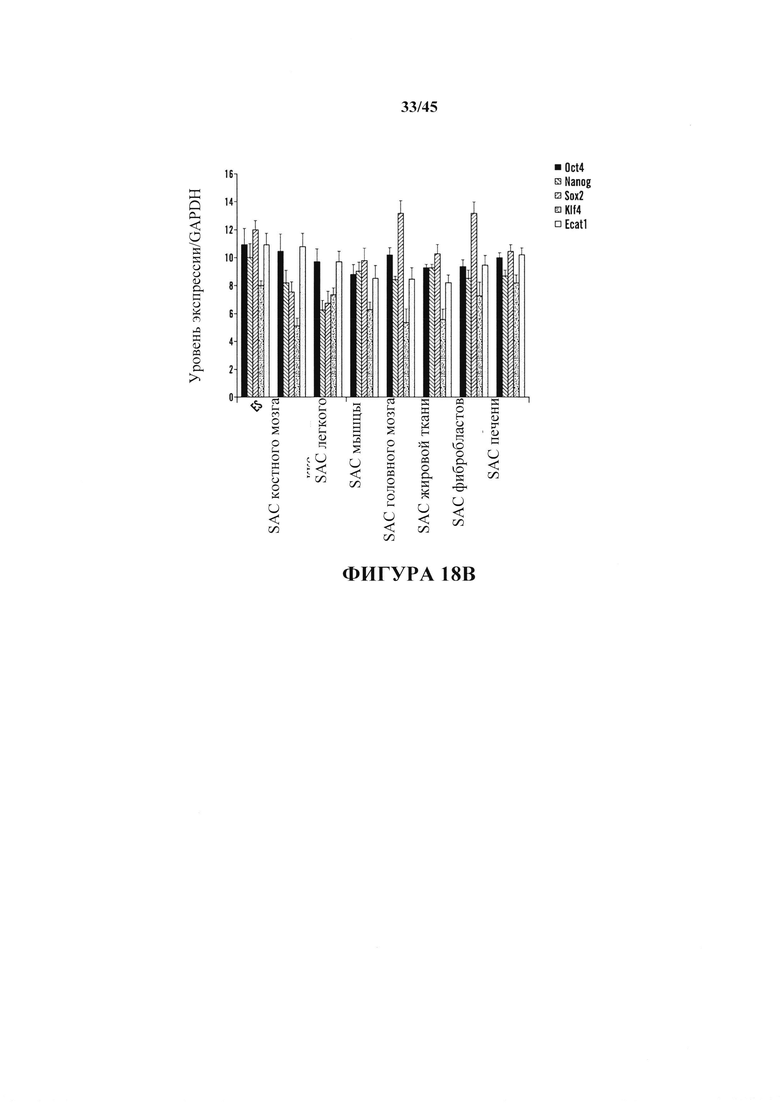
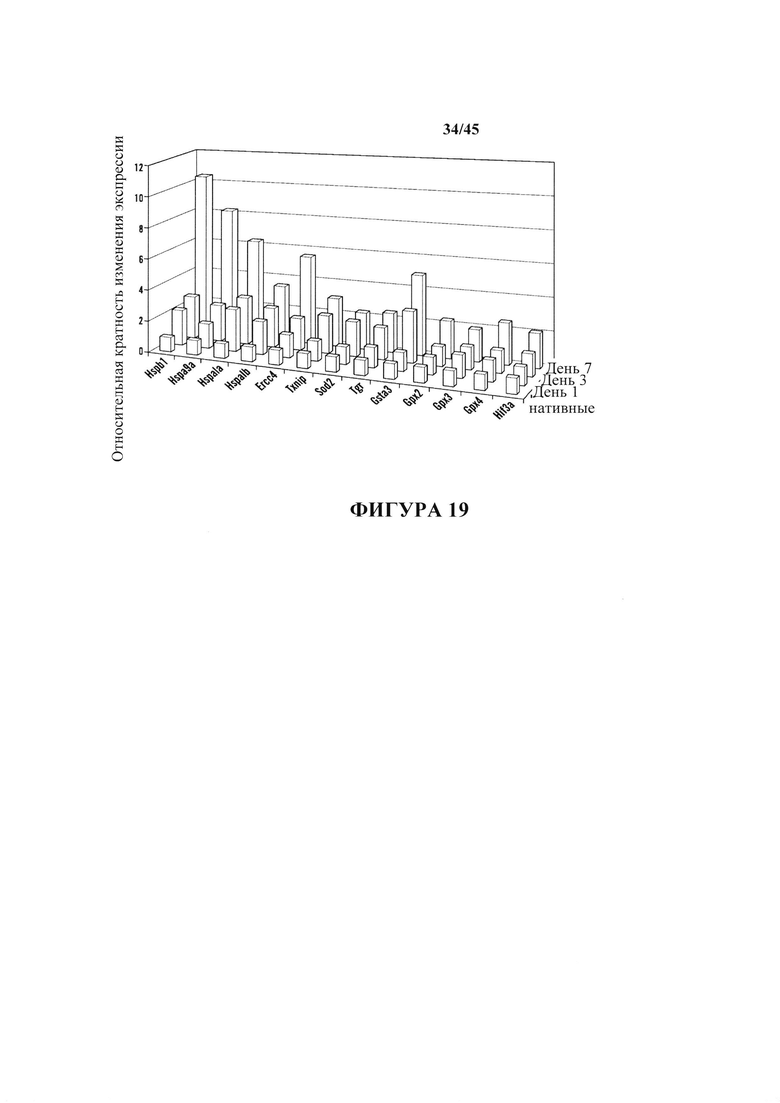
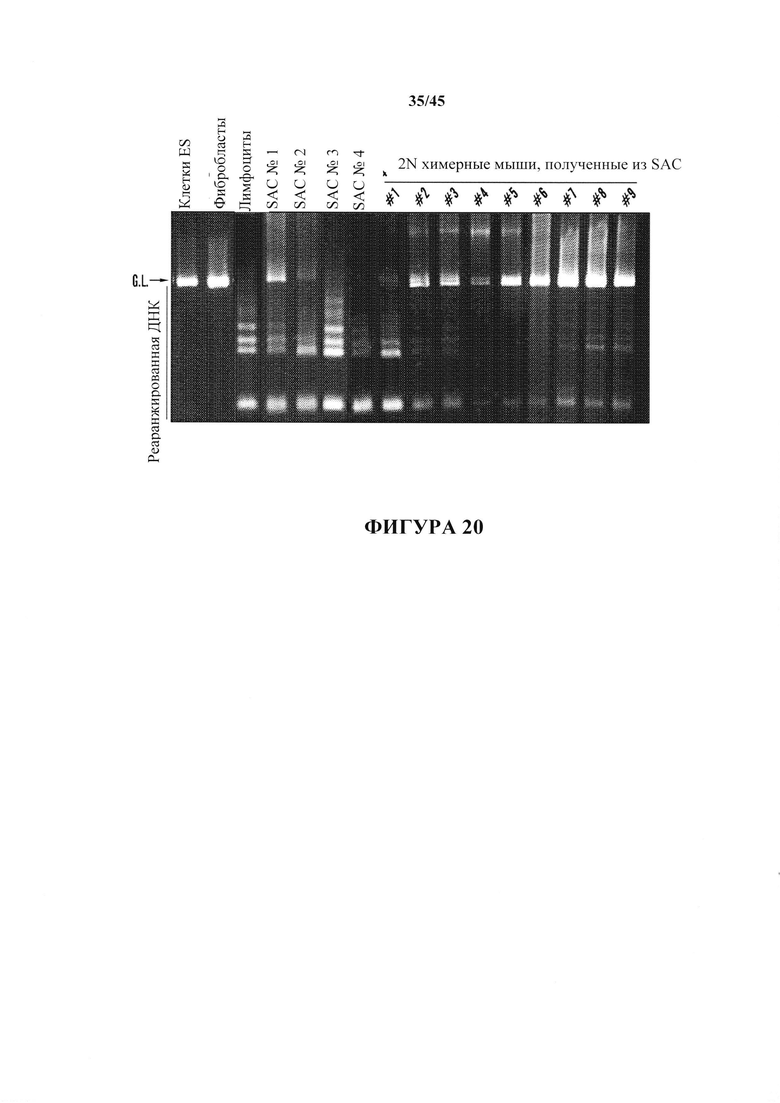


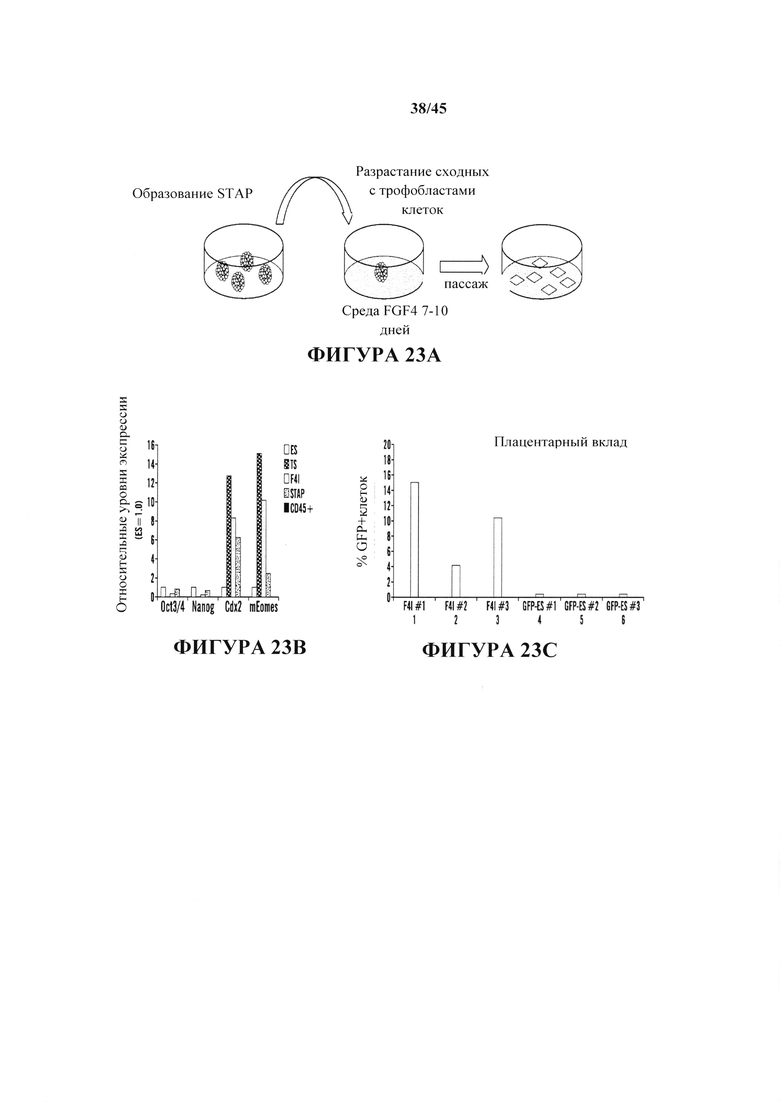
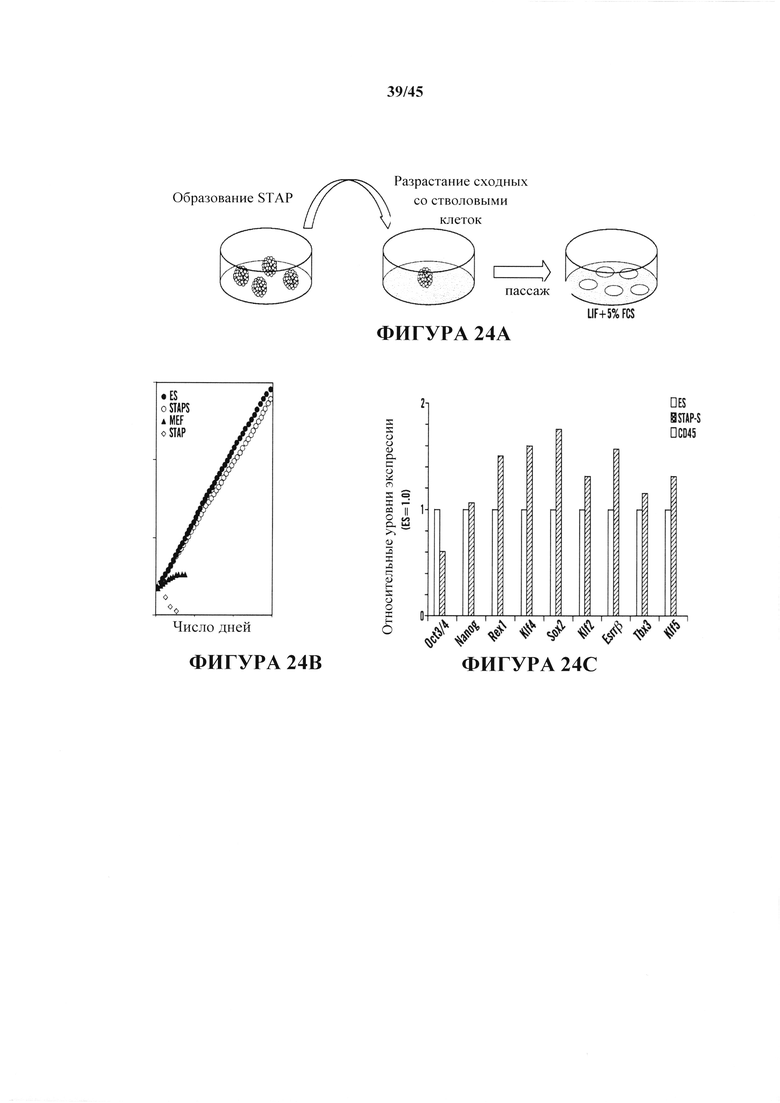

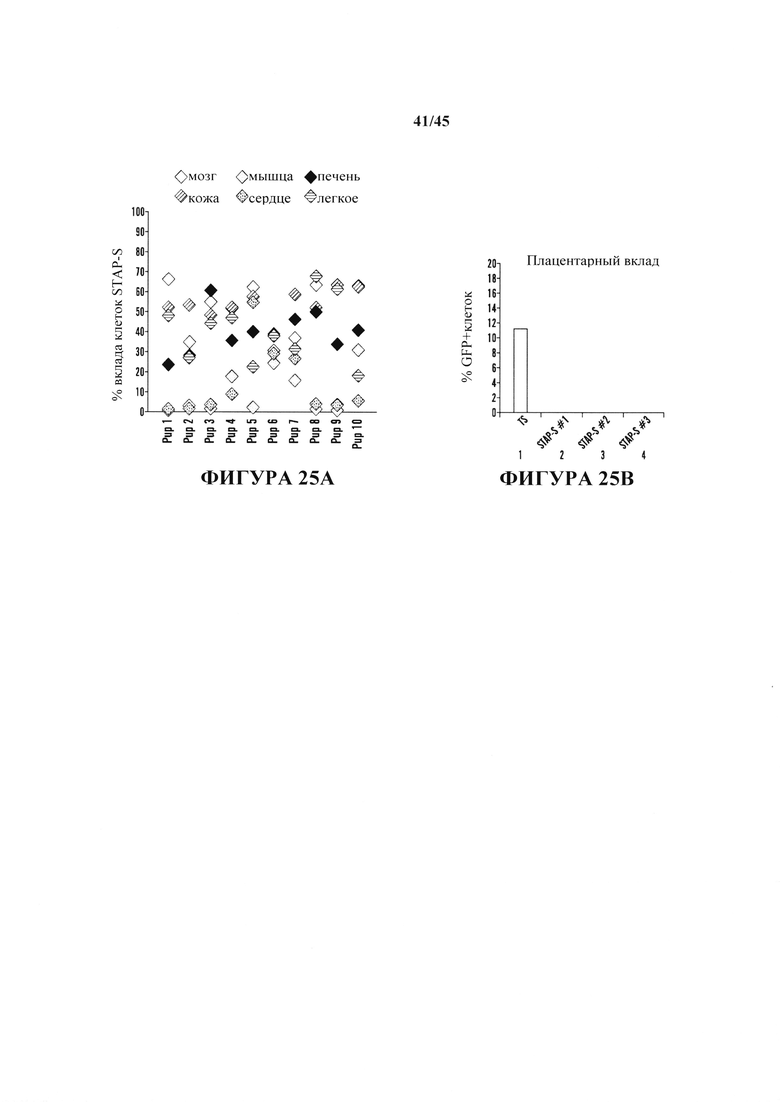

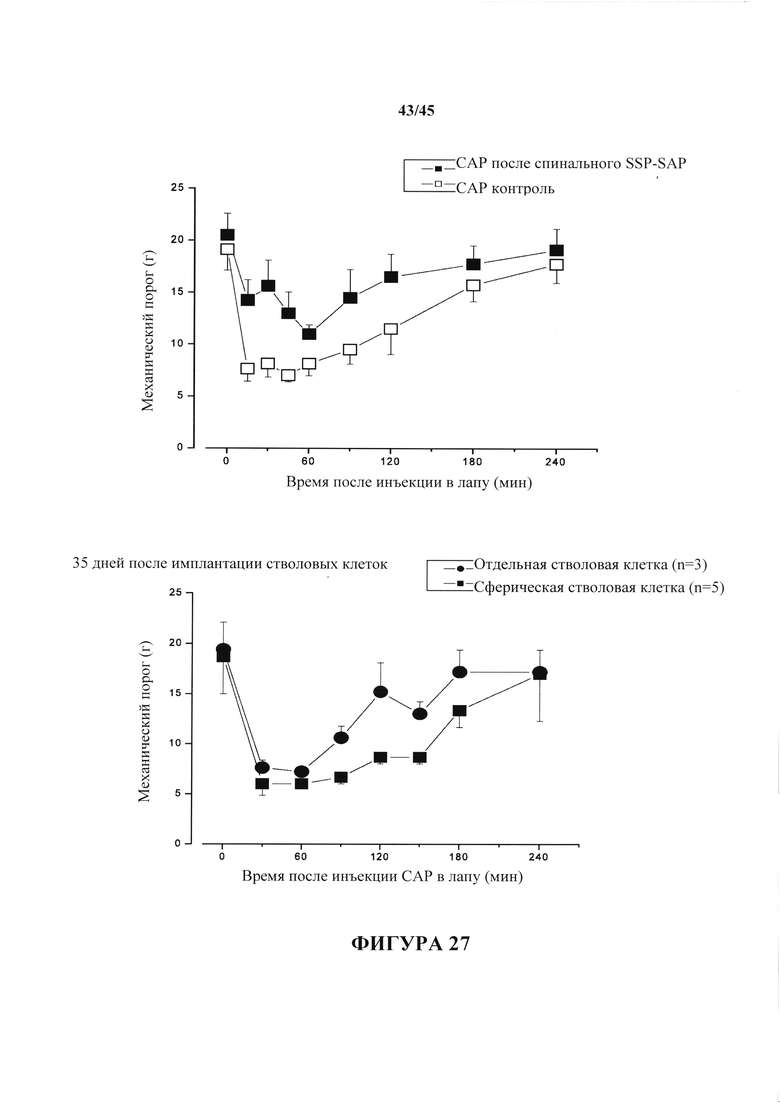
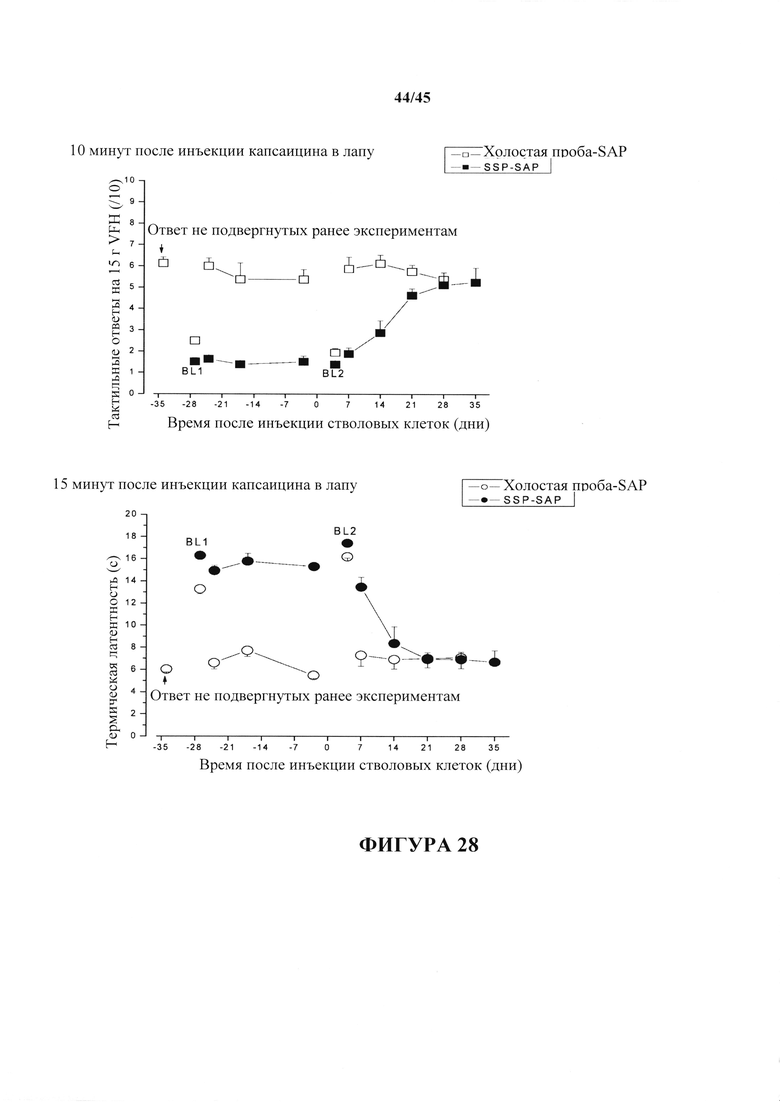
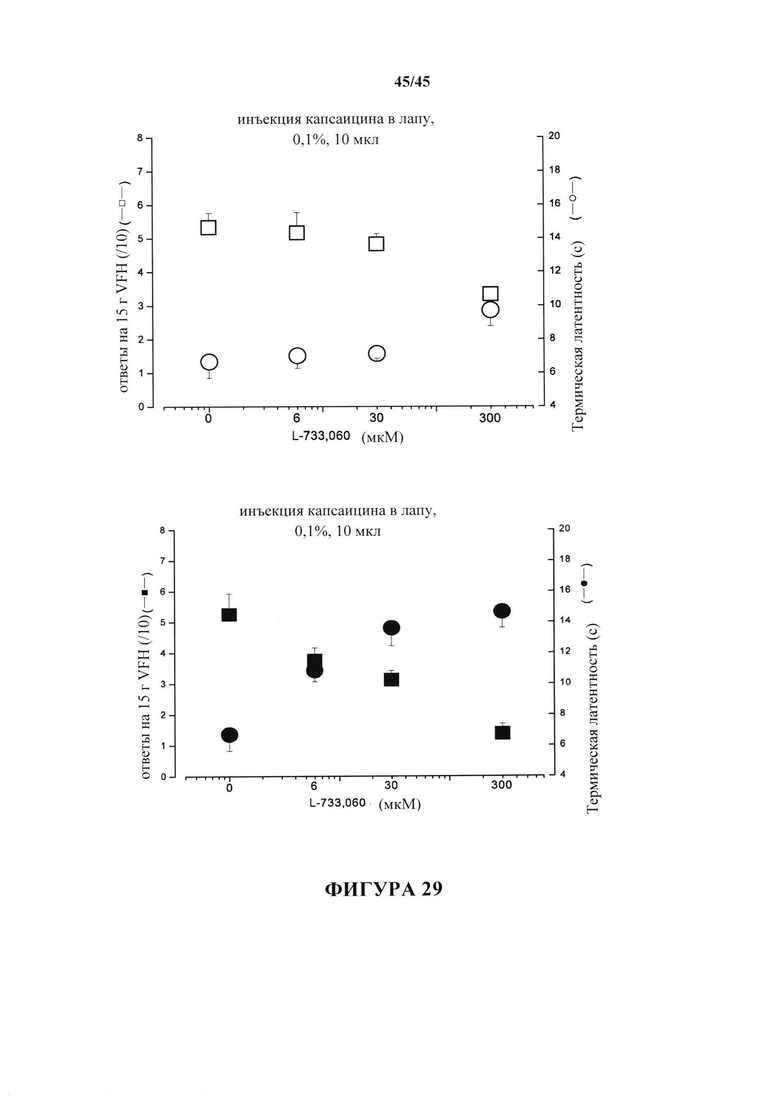
Изобретение относится к области биотехнологии, а именно к увеличению количества плюрипотентных клеток в популяции неэмбриональных дифференцированных соматических клеток и восстановлению нервной функции у субъекта с неврологическим повреждением. Способ включает воздействие на клеточную популяцию в виде механического разрушения пор клеточной мембраны с последующим воздействием рН от 5,4 до 5,6, в количестве и в течение времени, эффективных для увеличения количества клеток в клеточной суспензии, которые экспрессируют маркеры плюрипотентности, с последующим отбором клеток, экспрессирующих один или несколько маркеров плюрипотентности. Затем осуществляют введение полученных клеток в спинной мозг субъекта. 2 н. и 5 з.п. ф-лы, 9 табл., 29 ил., 7 пр.
1. Способ увеличения количества плюрипотентных клеток в популяции неэмбриональных дифференцированных соматических клеток, включающий воздействие на популяцию клеток эффективного количества механического и химического стресса для повышения уровня индуцированных стрессом генов,
где стресс представляет собой механическое разрушение пор клеточной мембраны с последующим воздействием рН от 5.4 до 5.6,
в количестве и в течение времени, эффективных для увеличения количества клеток в клеточной суспензии, которые экспрессируют маркеры плюрипотентности,
с последующим отбором клеток, экспрессирующих один или несколько маркеров плюрипотентности.
2. Способ по п. 1, при котором плюрипотентная клетка создана без введения экзогенного гена, транскрипта, белка, ядерного компонента или цитоплазмы или без слияния клеток.
3. Способ по п. 1, при котором клеточная популяция подвергается воздействию механического стресса, разрушающего поры, что приводит к потере по меньшей мере около 40% цитоплазмы или митохондрий из клетки.
4. Способ по любому из пп. 1-3, в котором механическое разрушение достигается посредством суспендирования клеток.
5. Способ по п. 1, при котором маркер плюрипотентности клетки выбирают из группы, состоящей из Oct4; Nanog; Е-кадгерин и SSEA.
6. Способ восстановления нервной функции у субъекта с неврологическим повреждением посредством использования плюрипотентных клеток, включающий:
получение плюрипотентных клеток в популяции неэмбриональных дифференцированных соматических клеток путем воздействия на клеточную популяцию эффективного количества механического и химического стресса для повышения уровня индуцированных стрессом генов, где стресс представляет собой механическое разрушение пор клеточной мембраны с последующим воздействием рН от 5,4 до 5,6, в количестве и в течение времени, эффективных для увеличения количества клеток в клеточной суспензии, которые экспрессируют маркеры плюрипотентности, с последующим отбором клеток, экспрессирующих один или несколько маркеров плюрипотентности;
введение полученных плюрипотентных клеток в спинной мозг субъекта.
7. Способ по п. 6, в котором неврологическое повреждение выбрано из группы, состоящей из:
острое неврологическое повреждение; хроническое неврологическое повреждение; дегенеративное неврологическое заболевание; травма нерва или повреждение спинного мозга.
| WO 2013163296 A1, 31.10.2013 | |||
| US 20060084167 A1, 20.04.2006 | |||
| WO 2013163296 A1, 31.10.2013 | |||
| НОВОСАДОВА Е.В | |||
| и др | |||
| "Индуцированные плюрипотентные стволовые клетки: от получения до применения в биохимических исследованиях", Успехи биологической химии | |||
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |

