Область техники
В настоящем изобретении раскрыт способ продукции парвовируса с использованием существенно бессывороточной среды, которая позволяет увеличить продукцию парвовируса по сравнению со стандартной средой.
Уровень техники
Парвовирус H-1 (H-1PV) принадлежит к роду Protoparvovirus, входящему в подсемейство Parvovirinae семейства Parvoviridae (Cotmore et al., 2014). Вирус состоит из безоболочечного икосаэдрического капсида диаметром 25 нм и содержит одноцепочечный геном ДНК длиной около 5 тн, кодирующий неструктурные белки, а именно NS1 (83 кДа) и NS2 (25 кДа), и капсидные белки VP1 (81 кДа) и VP2 (65 кДа). Другой капсидный белок, VP3 (63 кДа), образуется путем посттрансляционного расщепления белка VP2 (Faisst et al., 1995; Halder et al., 2012; Hanson and Rhode, 1991; Toolan et al., 1960). Протопарвовирусы реплицируются в S-фазе клеточного цикла и проходят литический цикл после инфицирования пермиссивных клеток (Burnett et al., 2006).
Хотя естественным хозяином H-1PV являются крысы, в последнее время этот вирус вызывает большой интерес, поскольку он реплицируется преимущественно в трансформированных клетках, включая ряд опухолевых клеток человека. Таким образом вирус обладает онколитическими и онкосупрессивными свойствами, которые были продемонстрированы на различных клеточных культурах и животных моделях (Nuesch et al., 2012; Rommelaere et al., 2010). На моделях ксенотрансплантатов было показано, что H-1PV подавляет рост ряда опухолей человека, включая опухоль шейки матки, (Faisst et al., 1998; Li et al., 2013) опухоль поджелудочной железы (Angelova et al., 2009b; Grekova et al., 2011), рак, глиому и лимфому молочной железы (Dupressoir et al., 1989) (Geletneky et al., 2010; Kiprianova et al., 2011) (Angelova et al., 2009a). Кроме того, было показано, что H-1PV успешно уничтожает раковые стволовые клетки (EP 2 404 609 A1). На основе этих результатов доклинических исследований в 2011 году было начато первое клиническое исследование (фаза I/IIa) H-1PV на пациентах с рецидивирующей мультиформной глиобластомой (Geletneky et al., 2012).
Для испытания и последующего использования терапевтического потенциала H-1PV необходимо оптимизировать продукцию очищенных вирусных (векторных) материалов в среде, подходящей для доклинических, клинических применений или фундаментальных исследований.
Между тем составы сред для культивирования клеток широко описаны в литературе, и некоторые среды коммерчески доступны. Согласно обычной практике, продукция H-1PV осуществляется в различных клеточных культурах, таких как клетки NB-324K человека, с использованием стандартных сред для культивирования клеток.
Помимо основных питательных веществ, для клеточной культуры человека in vitro необходим комплексный ряд факторов роста (Werner et al., 1993). Добавление сыворотки животных в стандартные среды для культивирования клеток имеет важное значение для роста клеток, метаболизма и стимуляции пролиферации. Наиболее широко используются телячьи сыворотки взрослых или новорожденных животных или сыворотки эмбрионального происхождения в диапазоне 10-20% об./об. Эмбриональная телячья сыворотка (ЭТС) представляет собой смесь большинства факторов, необходимых для прикрепления, роста и пролиферации клеток, и, таким образом, используется в качестве почти универсальной ростовой добавки, эффективной для большинства типов клеток человека и животных (WO 2011/157447 A2; Allaume et al. al., 2012). Однако использование сыворотки животных в клеточной культуре также имеет ряд недостатков. Это недостатки с точки зрения биологии клетки, поскольку сыворотка в целом является плохо определяемой смесью компонентов в культуральных средах. Кроме того, в биотехнологическом производстве лекарственных препаратов для человека не допускается использовать компоненты животного происхождения в протоколах культивирования. Например, существует риск того, что культуральная среда или продукты, полученные из нее в результате очистки, могут быть иммуногенными, в частности если добавки получены от животного, отличного от источника культивируемых клеток, и вызывать иммунологический ответ.
Ряд бессывороточных сред коммерчески доступен, например, среды, разработанные для поддержки культуры эндотелиальных клеток (WO 2012/033328 A2). В бессывороточных средах содержание белка ниже, чем в средах с добавлением сыворотки, что позволяет упростить процесс очистки, увеличить выход целевого продукта, обеспечить улучшение контроля условий культивирования клеток, более высокую однородность характеристик от партии к партии и оптимизацию составов для определенных типов клеток. Кроме того, характеристики и композиции сред для культивирования клеток варьируют в зависимости от конкретных требований к клеткам. Важными параметрами являются осмолярность, pH и состав питательных веществ.
Чтобы преодолеть ограничения использования животных белков в бессывороточных средах, было предпринято несколько попыток создать среды для культивирования клеток животных, которые вовсе не содержат животных белков. Например, некоторые культуральные среды содержат внесенный в базальную среду экстракт дрожжевых клеток (патентная заявка Великобритании № GB 901673; Keay, L., Biotechnol. Bioengin. 17:745-764 (1975)) в качестве источников азота и других необходимых питательных веществ.
Однако ни один из этих подходов не обеспечил культуральную среду, которая была бы оптимальной для культивирования некоммерческих клеток NB-324K для крупномасштабной продукции H1-PV для доклинических или клинических применений. Поскольку клетки NB-324K трудно выращивать в целом, невозможно просто адаптировать среду, известную из опыта культивирования других коммерческих организмов.
Таким образом, сохраняется потребность в существенно бессывороточной культуральной среде, которая способствует росту клеток NB-324K и повышает уровень продуцирования H-1PV. Такие культуральные среды должны обеспечивать более простые и экономически эффективные продукцию и очистку парвовируса, продуцируемого культивируемыми клетками NB-324K, а также дать более надежные результаты.
Таким образом, техническая проблема, лежащая в основе настоящего изобретения, заключается в оптимизации крупномасштабной продукции парвовируса в существенно бессывороточной среде.
Решение данной технической проблемы достигается посредством раскрытия вариантов осуществления, отраженных в формуле изобретения.
Сущность изобретения
Исследования онколитического протопарвовируса достигли стадии перехода в клиническую практику, при этом первое клиническое исследование (фаза I/IIa) H-1PV проводилось на пациентах с рецидивирующей операбельной злокачественной глиомой (Geletneky et al., 2012). Ожидается, что за этим исследованием последуют дальнейшие клинические исследования, направленные на оценку эффективности и расширение применения данного подхода при других видах рака, таких как рак поджелудочной железы (клиническое исследование: "ParvOryx02"; https://clinicaltrials.gov/ct2/show/NCT02653313) или нейробластома (Lacroix et al., 2010; Li et al., 2013). Эти разработки полагаются на доступность робастных методик крупномасштабной продукции протопарвовируса.
В ходе экспериментов, приведших к настоящему изобретению, были введены различные инновации для оптимизации продукции H1-PV с устранением нежелательных контаминантов в средах для культивирования клеток посредством ЭТС. Авторы изобретения сфокусировались на разработке существенно бессывороточной среды, используемой в качестве основы для продукции H-1PV для доклинических и клинических применений в рамках противораковой виротерапии.
Способ по настоящему изобретению предполагает применение существенно бессывороточной среды для культивирования клеток, содержащей такие ингредиенты, как глюкоза, глутамин, глутамат, лактат, аммоний, белки (например, факторы роста, инсулин и т. д.), в котором указанная среда способна поддерживать культивирование эпителиальных клеток in vitro, предпочтительно клеток NB-324K, для продукции H-1PV.
Краткое описание чертежей
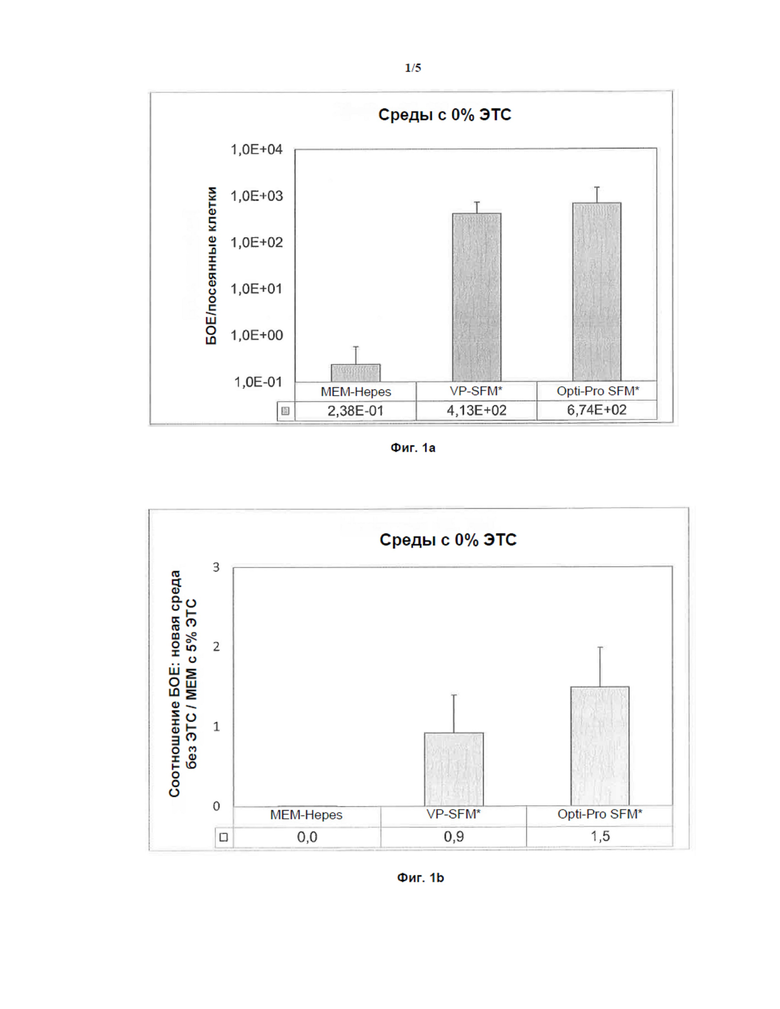
Фигуры 1a и 1b: Различные среды для более высокой наработки H1-PV без ЭТС
На Фиг. 1а показана гистограмма, демонстрирующая влияние сред MEM-Hepes, VP-SFM и Opti-Pro SFM на продукцию H-1PV в клетках NB-324K без ЭТС. Титры БОЕ определяли с помощью анализа бляшкообразования и выражали относительно количества посеянных клеток.
На Фиг. 1b представлена гистограмма, демонстрирующая влияние сред MEM-Hepes, VP-SFM и Opti-Pro SFM на продукцию H-1PV в клетках NB-324K без ЭТС. Соотношение БОЕ для сред VP-SFM и Opti-Pro SFM без ЭТС выражали относительно среды MEM-Hepes с 5% ЭТС.
Фигуры 2a и 2b: Различные среды для более высокой наработки H1-PV с 5% ЭТС
На Фиг. 2а показана гистограмма, демонстрирующая влияние сред MEM-Hepes, VP-SFM и Opti-Pro SFM на продукцию H-1PV в клетках NB-324K с 5% ЭТС. Титры БОЕ определяли с помощью анализа бляшкообразования и выражали относительно количества посеянных клеток.
На Фиг. 2b показана гистограмма, демонстрирующая влияние сред MEM-Hepes, VP-SFM и Opti-Pro SFM на продукцию H-1PV в клетках NB-324K с 5% ЭТС. Соотношение БОЕ для сред VP-SFM и Opti-Pro SFM с 5% ЭТС выражали относительно среды MEM-Hepes с 5% ЭТС.
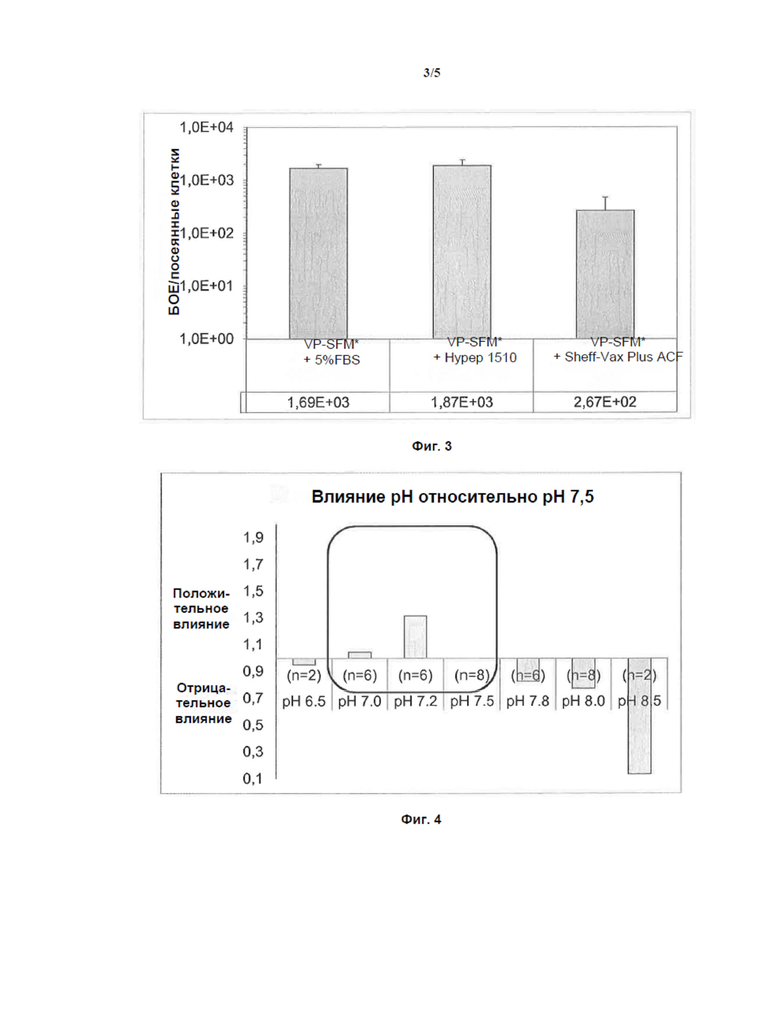
Фигура 3: Среда VP-SFM с и без ЭТС, дополненная Hypep 1510 или Sheff-Vax Plus ACF
На Фиг. 3 показана гистограмма, демонстрирующая влияние среды VP-SFM с добавлением 5% ЭТС, среды VP-SFM с добавлением Hypep 1510 и среды VP-SFM с добавлением Sheff-Vax Plus ACF на продукцию H-1PV в клетках NB-324K. Титры БОЕ определяли с помощью анализа бляшкообразования и выражали относительно количества посеянных клеток.
Фигура 4: влияние pH на наработку H-1PV
БОЕ определяли с помощью анализа бляшкообразования.
Фигура 5: Морфология клеток NB-234K в разных средах
На Фиг. 5 показана морфология клеток NB-324K в средах MEM-Hepes, VP-SFM и Opti-Pro SFM, определенная с помощью световой микроскопии.
Фигура 6: Иммунофлюоресцентное окрашивание
На Фиг. 6 показано, что главная линия клеток (главный банк клеток, ГБК) представляет собой линию эпителиальных клеток, подобную сравниваемой линии клеток эмбриональной почки человека 293T, и отличается от линии клеток фибробластов (HEF).
Подробное описание изобретения
Термин "клеточная культура" означает поддержание клеток в искусственной среде in vitro. Среда по настоящему изобретению может быть использована для культивирования адгезивных клеток млекопитающего, то есть клеток, которые прилипают к стенкам сосуда для культивирования, предпочтительно эпителиальных клеток, таких как клетки NB-324K.
Термин "культивирование" означает поддержание клеток in vitro в условиях, благоприятствующих росту, дифференцировке или продолжительной жизнеспособности (в активном или неактивном состоянии) клеток.
Фраза "среда для культивирования клеток" относится к питательному раствору для культивирования клеток.
Термин "EGF" (англ. Epidermal Growth Factor - эпидермальный фактор роста) относится к эпидермальному фактору роста, который стимулирует пролиферацию и дифференцировку путем связывания с его рецептором EGFR (англ. epidermal growth factor receptor - рецептор эпидермального фактора роста). Кроме того, EGF регулирует миграцию и клеточный рост эпителиальных клеток кишечника. EGF предпочтительно представляет собой EGF человека (hEGF).
Термин "ингредиент" относится к любому соединению химического или биологического происхождения, которое можно использовать в среде для культивирования клеток для поддержания или стимулирования роста, пролиферации клеток. Термины "компонент", "питательное вещество" и "ингредиент" могут использоваться взаимозаменяемо, и, подразумевается, что все они относятся к вышеуказанным соединениям. Типичные ингредиенты, которые используются в средах для культивирования клеток, включают глюкозу, глутамин, глутамат, лактат, аммоний, факторы роста, инсулин и белки.
"Бессывороточная" среда представляет собой среду, которая не содержит сыворотки. Бессывороточная среда отличается от среды с низким содержанием сыворотки и существенно бессывороточной среды, обе из которых содержат сыворотку.
Настоящее изобретение относится к способу продукции парвовируса H-1 путем выращивания H-1-инфицированной линии клеток-продуцентов NB-324K, ("главный банк клеток" или "главная линия клеток") в бессывороточной культуральной среде, при этом указанная среда содержит нижеуказанные ингредиенты. Как показано в разделе с примерами, настоящее изобретение позволяет получить в четыре раза более высокий титр парвовируса, чем обычно достигается с помощью стандартной ЭТС-содержащей среды.
Способ по настоящему изобретению состоит в том, что используют бессывороточную среду для культивирования клеток, содержащую такие ингредиенты, как глюкоза, глутамин, глутамат, лактат, аммоний, белки (например, факторы роста, инсулин и т. д.), в котором указанная среда способна поддерживать культивирование эпителиальных клеток in vitro, предпочтительно клеток NB-324K, для продукции H-1PV.
Предпочтительным фактором роста, используемым в средах по настоящему изобретению, является эпителиальный фактор роста (EGF), предпочтительно EGF человека (hEGF), в то время как предпочтительным инсулином является инсулин человеческий рекомбинантный. Белковые ингредиенты среды по настоящему изобретению не имеют человеческого или животного происхождения, и концентрация белка является сверхнизкой, как правило, менее 10 мкг/мкл.
Среду по настоящему изобретению можно использовать для выращивания эпителиальных клеток человека, предпочтительно клеток NB-324K, для получения высокой плотности и/или для увеличения продукции парвовируса. В одном предпочтительном варианте осуществления настоящего изобретения используют существенно бессывороточную среду для культивирования клеток, содержащую примерно 16-22 мМ глюкозы, 3-5 мМ глутамина, 0,1-0,6 мМ глутамата, 0,5-1,0 мМ лактата, менее 0,3 мМ аммония и 3-10 мкг/мкл белков, при этом среда способна поддерживать культивирование клеток NB-324K для продукции H-1PV.
Предпочтительно культуральная среда содержит 17-20 мМ глюкозы, примерно 4 мМ глутамина, примерно 0,15-0,5 мМ глутамата, примерно 0,7 мМ лактата, менее 0,2 мМ аммония и 4-8 мкг/мкл белков (например, добавки, эпителиальный фактор роста и инсулин).
В конкретном предпочтительном варианте осуществления культуральная среда содержит 19,14 мМ глюкозы, 4,25 мМ глутамина, 0,174 мМ глутамата, 0,669 мМ лактата, менее 0,05 мМ аммония и около 5 мкг/мкл белков. Эти белки включают EGF, инсулин и белковые добавки. Эта среда называется "VP-SFM".
В другом предпочтительном варианте осуществления культуральная среда содержит 17,49 мМ глюкозы, 4,25 мМ глутамина, 0,486 мМ глутамата, 0,641 мМ лактата, 0,18 мМ аммония, 7,5 нг/мкл rEGF (эпидермальный фактор роста рекомбинантный) и около 7,5 мкг/мкл белков. Эти белки включают EGF и белковые добавки. Эта среда называется "Opti-Pro SFM".
Добавки, которые могут быть добавлены к среде по настоящему изобретению, включают инсулин, соевые пептиды (например, Hypep 1510), Sheff-Vax Plus ACF, инсулин-трансферрин-селен A, инсулин-трансферрин-селен X и химически определяемый липидный концентрат. Инсулин является коммерчески доступным, например, от Life Technologies, Inc. (Роквилл, Мэриленд). Соевые пептиды являются коммерчески доступными, например, от Quest International (Норвич, Нью-Йорк).
В предпочтительном варианте осуществления среду по настоящему изобретению комбинируют с добавкой, предпочтительно с соевыми белками. Одним подходящим соевым белком является ультрафильтрованный ферментативный гидролизат сои, например, Hypep 1510® (Kerry Ingredients & Favors, Норвич, Нью-Йорк, США). Другой подходящей добавкой является Sheff-Vax Plus ACF® (Kerry, Inc., Килдэр, Ирландия), который представляет собой гидролизат на основе растительных компонентов, содержащий факторы роста и следовые количества элементов.
В еще одном предпочтительном варианте осуществления рН среды по настоящему изобретению необходимо довести до примерно до 7,0-7,5, предпочтительно примерно до 7,2.
Среда по настоящему изобретению может быть использована для облегчения культивирования эпителиальных клеток. В частности, эти среды могут быть использованы для культивирования главной линии клеток на основе клеток NB-324K, которая, как было установлено авторами изобретения, является линией эпителиальных клеток. Хотя среда по настоящему изобретению в частности пригодна для культивирования эпителиальных клеток, предполагается, что она может использоваться в любом стандартном протоколе культивирования клеток, где клетки могут выращиваться по той же методике, что и эпителиальные клетки in vitro.
Кроме того, поскольку среда по настоящему изобретению не содержит сыворотки и имеет низкую концентрацию белка, указанная среда может быть использована для быстрой продукции парвовируса, например, H-1PV. В настоящей заявке продемонстрировано, что продукция H1-PV в среде VP-SFM, дополненной глутамином и соевыми пептидами, показывает тот же выход, что и ЭТС-содержащая среда, и является подходящей альтернативой для бессывороточной продукции. Другой подходящей средой является среда Opti-Pro SFM, дополненная глутамином.
Следующие примеры предназначены для иллюстрации, но не ограничения объема настоящего изобретения. Несмотря на то, что такие примеры описывают общепринятые способы, специалисты в данной области техники могут альтернативно использовать и другие известные им способы.
Пример 1
Материал и методы
(A) Среда для культивирования клеток и продукции H-1PV
Среда MEM была дополнена 2 мМ L-глутамином. Каждая из сред VP-SFM и OptiPro-SFM была дополнена 4 мМ L-глутамином.
(=VP-SFM *)
(=Opti-Pro SFM*)
(B) Культивирование клеток и продукция H-1PV
Клетки почки новорожденного человека NB-324K (главный банк клеток), трансформированные обезьяньим вирусом SV40 (Tattersall and Bratton, 1983), культивировали при 37°C в различной среде (см. выше).
После переноса клеток ГБК в каждую среду клетки адаптировали к клеткам по крайней мере в течение 2 недель. Чтобы осуществить продукцию, клетки NB-324K высевали при плотности 3,6×104 кл/см2 и немедленно инфицировали H-1PV при множественности инфекции (МИ) 0,01 бляшкообразующих единиц (БОЕ) на клетку. Инфицированные клетки инкубировали в течение 4 дней при 37°C в атмосфере с 5% CO2 до тех пор, пока цитопатический эффект (ЦПЭ), определяемый как процент открепленных клеток, наблюдаемых под микроскопом, не достигал по крайней мере 30%. Плотность и жизнеспособность клеток определяли путем окрашивания живых клеток 0,4% трипановым синим (Invitrogen TM, Германия). Клетки подсчитывали с помощью счетчика клеток Countess® (Life Technologies, Германия), морфологию наблюдали под микроскопом.
Для сбора среду аспирировали, инфицированные клетки обрабатывали PBS/1 мМ ЭДТА. Полученные из среды супернатант и открепленные клетки центрифугировали в течение 5 минут при 5000×g. Осадок промывали PBS, ресуспендировали в буфере Virus Tris/ЭДТА, pH 8,7 (VTE), содержащем 0,05 М Трис-HCl, 0,5 мМ ЭДТА, и подвергали трем циклам замораживания/оттаивания. После центрифугирования в течение 5 минут при 5000xg клеточный дебрис отбрасывали. Затем клеточный лизат обрабатывали ультразвуком при 48 Вт в течение 1 мин в ультразвуковом гомогенизаторе Sonorex Super 10 P (Bandelin, Германия) и обрабатывали ДНКазой (50 ед/мл, Sigma, Германия) в течение 30 мин при 37°C.
(C) Анализ бляшкообразования (БОЕ)
Анализы бляшкообразования проводили, по существу, как описано у Tattersall and Bratton, 1983. Клетки NB-324K выращивали в однослойных культурах в среде MEM, содержащей 5% ЭТС, 100 мкг/мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ L-глутамина. Их инфицировали при 60% конфлуентности серийными разведениями H-1PV и инкубировали в течение 1 часа при 37°C. Затем инокулят заменяли верхним слоем бактоагара (1,7% в MEM, содержащей 5% ЭТС). На четвертый день после инфицирования живые клетки окрашивали в течение 18-24 часов путем добавления 0,02% раствора для окрашивания толуиленового красного (Sigma, Германия), содержащего бактоагар (Becton Dickinson, Германия). Чашки инкубировали при 37°°С в атмосфере с 5% СО2. Количество бляшкообразующих единиц оценивали через 5 дней после инфицирования на негатоскопе, концентрацию выражали в БОЕ/мл.
(D) Иммунофлюоресцентный анализ
Иммунофлюоресцентный анализ основан на использовании флюоресцентно-маркированных антител, которые связываются с соответствующим специфическим антигеном, при котором могут быть определены некоторые вне- и внутриклеточные структуры. Этот принцип используется для окрашивания кератина при помощи антитела к CH/HK-Keratin для обнаружения [антигена] в эпителиальных клетках. Таким образом клетки культивировали на предметных стеклах во флаконах для культивирования клеток 145 см² (Greiner Bio-One). Клетки почек новорожденных человека NB-324K (главный банк клеток) высевали при плотности 3E6 клеток/флакон. Для окрашивания на кератин HEF (human embryonic fibroblasts - эмбриональные фибробласты человека) и HEK293T (эпителиальные клетки почки человека) при плотности клеток 1,2 Е6 кл/флакон или 5E6 кл/флакон высевали в качестве контролей. После 24-часовой инкубации клетки на предметных стеклах промывали в PBS, фиксировали в течение 10 минут в метаноле и затем в течение 5 минут в ацетоне. Обе жидкости предварительно охлаждали до -20°С. Для детекции кератина предметные стекла предварительно блокировали PBS с 0,05% БСА и затем инкубировали с антителом к CH/HK-Keratin в течение 1 часа. Затем предметные стекла промывали в PBS с 0,1% Triton, перед тем как второе антитело Cy3gp (Dianova) инкубировали в течение 1 часа. Несвязанные вторые антитела удаляли тройной промывкой в PBS с 0,1% Triton. Предметные стекла погружали в этанол и затем сушили. Предметные стекла заливали заключающей средой для предметных стекол (CELL LAB), включающей 1 мкг/мл ДАФИ, окрашивающий ядро клеток. Анализ проводили с помощью флуоресцентного микроскопа BZ-9000 (Keyence).
(E) Микроскопия клеток
С помощью светового микроскопа (Axiovert 25) наблюдали морфологию, плотность и форму клеток.
Пример 2
Сравнение уровня продуцирования H-1PV в средах MEM-Hepes, VP-SFM и Opti-Pro SFM без ЭТС
Авторы изобретения сначала определяли уровень продуцирования H-1PV в различных средах, включая MEM-Hepes, VP-SFM с добавлением глутамина (VP-SFM *) и Opti-Pro SFM с добавлением глутамина (Opti-Pro SFM *), при этом каждая среда не содержала ЭТС (эмбриональной телячьей сыворотки). Как упомянуто в приведенной выше таблице VP-SFM * и Opti-Pro SFM * содержат ЭТС в качестве белкового ингредиента. Клетки собирали, как описано выше, и инфекционные частицы анализировали с помощью анализа бляшкообразования.
Как показано на Фиг. 1а, H-1PV-инфицированные клетки NB-324K, культивированные в средах VP-SFM * и Opti-Pro SFM * с 0% ЭТС, продуцировали более высокие титры H-1PV по сравнению со стандартной средой MEM-Hepes с 0% ЭТС.
Эти результаты демонстрируют, что среды VP-SFM* и Opti-Pro SFM* без ЭТС облегчают быструю продукцию H-1PV по сравнению со стандартной средой MEM-Hepes без ЭТС (см. Фиг. 1a). VP-SFM* с 0% ЭТС (0,9 ± 0,5) и Opti-Pro SFM* (1,5 ± 0,5) с 0% ЭТС показали такой же или даже больший уровень продуцирования по сравнению с MEM-Hepes с 5% ЭТС (сравнение Фиг. 2b с Фиг. 1b). Кроме того, стандартная среда MEM-Hepes с 0% ЭТС показала существенное снижение уровня продуцирования или почти отсутствие продуцирования (Фиг. 1а).
VP-SFM* и Opti-Pro SFM* отличаются от стандартной среды MEM-Hepes присутствием в них EGF. Это показывает, что эпидермальный фактор роста EGF может стимулировать пролиферацию клеток (Fallon et al., 1984), что может облегчить продуцирование H1-PV.
Пример 3
Сравнение уровня продуцирования H-1PV в средах MEM-Hepes, VP-SFM и Opti-Pro SFM с 5% ЭТС
Чтобы определить уровень продуцирования H-1PV в средах MEM-Hepes, VP-SFM* и Opti-Pro SFM*, где каждая среда содержит 5% ЭТС, клетки NB-324K собирали, как описано выше, и инфекционные частицы анализировали с помощью анализа бляшкообразования.
Как показано на Фиг. 2b, среда VP-SFM* (+5% ЭТС) была примерно на 4,0x ± 2,3 лучше по уровню продуцирования H-1PV, чем среда сравнения MEM-Hepes (+5% ЭТС). Среда Opti-Pro SFM* (+5% ЭТС) показала более низкое значение БОЕ, чем среда сравнения MEM-Hepes (+5% ЭТС) (около 10% от MEM-Hepes).
Эти результаты показывают, что среда Opti-Pro SFM* имеет более высокий уровень продуцирования без добавления ЭТС по сравнению с добавлением 5% ЭТС (Фиг. 1a и Фиг. 2a). Напротив, стандартная среда MEM-Hepes показала гораздо более высокий уровень продуцирования с 5% ЭТС по сравнению со средой без ЭТС (Фиг. 1a и Фиг. 2a).
Пример 4
Определение уровня продуцирования H-1PV в среде VP-SFM с Hypep 1510 или Sheff-Vax Plus ACF, дополненной и не дополненной ЭТС
Hypep 1510® и Sheff-Vax Plus ACF® использовали в качестве добавок в VP-SFM вместо ЭТС. Клетки культивировали с 5% ЭТС в VP-SFM и в бессывороточной VP-SFM, дополненной Hypep 1510® или Sheff-Vax Plus ACF®. Клетки собирали, как описано выше, и инфекционные частицы анализировали с помощью анализа бляшкообразования.
Как показано на Фиг. 3, VP-SFM, дополненная Hypep 1510 (1,87 * 103 БОЕ/кл), показала увеличение уровня продуцирования по сравнению с VP-SFM с 5% ЭТС (1,69 * 103 БОЕ/кл).
Эти результаты демонстрируют, что продукция H1-PV в VP-SFM, дополненной Hypep 1510, показала тот же выход, что и в среде VP-SFM, дополненной 5% ЭТС. Таким образом, среду VP-SFM по настоящему изобретению, при добавлении в нее Hypep 1510, можно использовать для культивирования H-1PV-инфицированных клеток NB-324K в бессывороточной среде с результатами, сопоставимыми с теми, полученными со средой, дополненной 5% ЭТС.
Пример 5
Влияние pH во время инфицирования на уровень продуцирования H-1PV
Необходимо определить идеальное значение pH для продукции H-1 PV. В связи с этим рН 6,5, 7,0, 7,2, 7,5, 7,8, 8,0 и 8,5 регулировали в среде MEM либо с помощью HCl, либо с NaOH. Для обеспечения стабильного pH во время инфицирования вирусом H-1PV использовали флаконы для культивирования клеток с закрытыми крышками, чтобы избежать газообмена в течение 4 часов. Поскольку для культивирования клеток необходима атмосфера с CO2, после 4 часов инкубации использовали крышки с фильтром в течение 4 дней инкубации. Клетки собирали, как описано выше, и инфекционные частицы анализировали с помощью анализа бляшкообразования.
Как показано на Фиг. 4, значение pH должно быть доведено до диапазона от 7,0 до 7,5, предпочтительно до примерно 7,2, для достижения [оптимального] выхода инфекционных частиц.
Пример 6
Морфология NB—324K в средах MEM-Hepes, VP-SFM и Opti-Pro SFM
Как показано на Фиг. 5 на изображениях со светового микроскопа, морфология характеризуется той же многоугольной формой со стандартными габаритами и размерами во всех трех средах.
Как показано на Фиг. 6 с помощью иммунофлюоресценции, главная линия клеток (ГБК) представляет собой линию эпителиальных клеток, подобную линии клеток эмбриональной почки человека 293T, и отличается от линии клеток эмбриональных фибробластов человека (HEF).
Источники
1. Angelova, A.L., Aprahamian, M., Balboni, G., Delecluse, H.J., Feederle, R., Kiprianova, I., Grekova, S.P., Galabov, A.S., Witzens-Harig, M., Ho, A.D., Rommelaere, J. and Raykov, Z., 2009a. Oncolytic rat parvovirus H-1PV, a candidate for the treatment of human lymphoma: In vitro and in vivo studies. Molecular therapy; The journal of the American Society of Gene Therapy 17, 1164-72.
2. Angelova, A.L., Aprahamian, M., Grekova, S.P., Hajri, A., Leuchs, B., Giese, N.A., Dinsart, C., Herrmann, A., Balboni, G., Rommelaere, J. and Raykov, Z., 2009b. Improvement of Gemcitabine-Based Therapy of Pancreatic Carcinoma by Means of Oncolytic Parvovirus H-1PV. Clinical Cancer Research 15, 511-519.
3. Burnett, E., Cotmore, S.F. and Tattersall, P., 2006. Segregation of a single outboard left-end origin is essential for the viability of parvovirus minute virus of mice. J Virol 80, 10879-83.
4. Cotmore, S.F., Agbandje-McKenna, M., Chiorini, J.A., Mukha, D.V., Pintel, D.J., Qiu, J., Soderlund-Venermo, M., Tattersall, P., Tijssen, P., Gatherer, D. and Davison, A.J., 2014. The family Parvoviridae. Archives of virology 159, 1239-47.
5. Dupressoir, T., Vanacker, J.M., Cornelis, J.J., Duponchel, N. and Rommelaere, J., 1989. Inhibition by parvovirus H-1 of the formation of tumors in nude mice and colonies in vitro by transformed human mammary epithelial cells. Cancer research 49, 3203-8.
6. Faisst, S., Faisst, S.R., Dupressoir, T., Plaza, S., Pujol, A., Jauniaux, J.C., Rhode, S.L. and Rommelaere, J., 1995. Isolation of a Fully Infectious Variant of Parvovirus H-1 Supplanting the Standard Strain in Human-Cells. Journal of Virology 69, 4538-4543.
7. Faisst, S., Guittard, D., Benner, A., Cesbron, J.Y., Schlehofer, J.R., Rommelaere, J. and Dupressoir, T., 1998. Dose-dependent regression of HeLa cell-derived tumours in SCID mice after parvovirus H-1 infection. International journal of cancer. Journal international du cancer 75, 584-9.
8. Fallon, A. E., Rozin, P., and Pliner, P., 1984. The child's conception of food: The development of food rejections with special reference to disgust and contamination sensitivity. Child Devel. 55: 566-575.
9. Geletneky, K., Huesing, J., Rommelaere, J., Schlehofer, J.R., Leuchs, B., Dahm, M., Krebs, O., von Knebel Doeberitz, M., Huber, B. and Hajda, J., 2012. Phase I/IIa study of intratumoral/intracerebral or intravenous/intracerebral administration of Parvovirus H-1 (ParvOryx) in patients with progressive primary or recurrent glioblastoma multiforme: ParvOryx01 protocol. BMC cancer 12, 99.
10. Geletneky, K., Kiprianova, I., Ayache, A., Koch, R., Herrero, Y.C.M., Deleu, L., Sommer, C., Thomas, N., Rommelaere, J. and Schlehofer, J.R., 2010. Regression of advanced rat and human gliomas by local or systemic treatment with oncolytic parvovirus H-1 in rat models. Neuro-oncology 12, 804-14.
11. Grekova, S.P., Aprahamian, M., Daeffler, L., Leuchs, B., Angelova, A., Giese, T., Galabov, A., Heller, A., Giese, N.A., Rommelaere, J. and Raykov, Z., 2011. Interferon gamma improves the vaccination potential of oncolytic parvovirus H-1PV for the treatment of peritoneal carcinomatosis in pancreatic cancer. Cancer biology & therapy 12, 888-95.
12. Griffith, O.M., 2006. Practical Techniques for centrifugal separations. FiberLite, Piramon Technologies, Inc.
1. Halder, S., Nam, H.J., Govindasamy, L., Vogel, M., Dinsart, C., Salome, N., McKenna, R. and Agbandje-McKenna, M., 2012. Production, purification, crystallization and structure determination of H-1 Parvovirus. Acta crystallographica. Section F, Structural biology and crystallization communications 68, 1571-6.
14. Hanson, N.D. and Rhode, S.L., 3rd, 1991. Parvovirus NS1 stimulates P4 expression by interaction with the terminal repeats and through DNA amplification. J Virol 65, 4325-33.
15. Keay, L., 1975. Biotechnol. Bioengign. 17:745-764.
16. Kiprianova, I., Thomas, N., Ayache, A., Fischer, M., Leuchs, B., Klein, M., Rommelaere, J. and Schlehofer, J.R., 2011. Regression of Glioma in Rat Models by Intranasal Application of Parvovirus H-1. Clinical Cancer Research 17, 5333-5342.
17. Kongsvik, J.R. and Toolan, H.W., 1972. Effect of proteolytic enzymes on the hemagglutinating property of the parvoviruses, H-1, H-3, and RV. Proceedings of the Society for Experimental Biology and Medicine. Society for Experimental Biology and Medicine 140, 140-4.
18. Lacroix, J., Leuchs, B., Li, J., Hristov, G., Deubzer, H.E., Kulozik, A.E., Rommelaere, J., Schlehofer, J.R. and Witt, O., 2010. Parvovirus H1 selectively induces cytotoxic effects on human neuroblastoma cells. International journal of cancer. Journal international du cancer 75, 584-9.
19. Li, J., Bonifati, S., Hristov, G., Marttila, T., Valmary-Degano, S., Stanzel, S., Schnolzer, M., Mougin, C., Aprahamian, M., Grekova, S.P., Raykov, Z., Rommelaere, J. and Marchini, A., 2013. Synergistic combination of valproic acid and oncolytic parvovirus H-1PV as a potential therapy against cervical and pancreatic carcinomas. EMBO molecular medicine 5, 1537-55.
20. Nuesch, J.P., Lacroix, J., Marchini, A. and Rommelaere, J., 2012. Molecular pathways: rodent parvoviruses--mechanisms of oncolysis and prospects for clinical cancer treatment. Clin Cancer Res 18, 3516-23.
21. Paradiso, P.R., 1981. Infectious process of the parvovirus H-1: correlation of protein content, particle density, and viral infectivity. J Virol 39, 800-7.
22. Rommelaere, J., Geletneky, K., Angelova, A.L., Daeffler, L., Dinsart, C., Kiprianova, I., Schlehofer, J.R. and Raykov, Z., 2010. Oncolytic parvoviruses as cancer therapeutics. Cytokine & growth factor reviews 21, 185-95.
23. Toolan, H.W., Dalldore, G., Barclay, M., Chandra, S. and Moore, A.E., 1960. An Unidentified, Filtrable Agent Isolated from Transplanted Human Tumors. Proceedings of the National Academy of Sciences of the United States of America 46, 1256-8.
24. Werner, R. G. et al, Mammalian Cell Cultures Part I: Characterization, morphology and metabolism, in: Arzneim.-Forsch./Drug Res. 43:1134-1139 (1993)).
25. Wrzesinski, C., L. Tesfay, N. Salome, J. C. Jauniaux, J. Rommelaere, J. Cornelis, and C. Dinsart. 2003. Chimeric and pseudotyped parvoviruses minimize the contamination of recombinant stocks with replication-competent viruses and identify a DNA sequence that restricts parvovirus H-1 in mouse cells. J. Virol. 77:3851-3858.)
26. Zolotukhin, S., Byrne, B.J., Mason, E., Zolotukhin, I., Potter, M., Chesnut, K., Summerford, C., Samulski, R.J. and Muzyczka, N., 1999. Recombinant adeno-associated virus purification using novel methods improves infectious titer and yield. Gene Ther 6, 973-85.
27. UK Patent Application No. GB 901676
28. Allaume, X., El-Andaloussi, N., Leuchs, B., Bonifati, S., Akulkrani, A., Marttila T., Kaufmann, J., Nettelbeck, D., Kleinschmidt, J., Rommelaere, J., Marchini, A., 2012. Retargeting of Rat Parvovirus H-1PV to Cancer Cells through Genetic Eneginering of the Viral Capsid, J. of Virology, Vol. 86, No. 7, 3452-3465.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ РАКА ПАРВОВИРУСОМ В КОМБИНАЦИИ С БЕВАЦИЗУМАБОМ | 2016 |
|
RU2679442C1 |
| СПОСОБ РЕПЛИКАЦИИ ВИРУСА В ПТИЧЬИХ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТКАХ | 2006 |
|
RU2457253C2 |
| ПРЕПАРАТ ПАРВОВИРУСА ДЛЯ ЛЕЧЕНИЯ ОПУХОЛЕЙ | 2016 |
|
RU2698717C2 |
| Бессывороточная питательная среда для культивирования клеток MDCK или Vero или вакцинных штаммов вирусов кори или гриппа | 2018 |
|
RU2703826C1 |
| БИОТРАНСПЛАНТАТ, СПОСОБ ЕГО ПОЛУЧЕНИЯ И СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ ПАРОДОНТА | 2009 |
|
RU2418571C1 |
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКА | 2010 |
|
RU2563514C2 |
| АНТИТЕЛО ПРОТИВ Siglec-15 | 2010 |
|
RU2539790C2 |
| Способ получения органоидов из опухолевой ткани органов головы и шеи | 2021 |
|
RU2787378C1 |
| Катионный разветвленный пептид для ингибирования респираторных вирусов | 2022 |
|
RU2810986C2 |
| Способ изготовления биотрансплантата, биотрансплантат для устранения рецессий десны и восстановления утраченного десной объема, способ устранения рецессий десны и восстановления утраченного десной объема | 2018 |
|
RU2700510C1 |



Изобретение относится к биотехнологии и представляет собой способ оптимизации крупномасштабной продукции парвовируса в существенно бессывороточной среде. Описан способ оптимизации продукции парвовируса, включающий существенно бессывороточную среду, которая позволяет увеличить продукцию парвовируса по сравнению со стандартной средой, предпочтительно для продукции H-1PV. 4 з.п. ф-лы, 6 ил., 1 табл., 6 пр.
1. Способ выращивания линии клеток-продуцентов NB-324K, инфицированных парвовирусом H-1, включающий культивирование инфицированных клеток NB-324K в бессывороточной среде, содержащей 16-22 мМ глюкозы, 3-5 мМ глутамина, 0,1-0,6 мМ глутамата, 0,5-1,0 мМ лактата, менее 0,3 мМ аммония и 3-10 мкг/мкл белков, включающих эпителиальный фактор роста и соевые пептиды, и при этом указанная среда имеет рН 7,0-7,5.
2. Способ по п. 1, в котором указанная среда содержит 19,14 мМ глюкозы, 4,25 мМ глутамина, 0,174 мМ глутамата, 0,669 мМ лактата, менее 0,05 мМ аммония и около 5 мкг/мкл белков, содержащих инсулин, эпителиальный фактор роста и соевые пептиды.
3. Способ по п. 1, в котором указанная среда содержит 17,49 мМ глюкозы, 4,25 мМ глутамина, 0,486 мМ глутамата, 0,641 лактата, 0,18 мМ аммония и 7,5 мкг/мкл эпителиального фактора роста и соевых пептидов.
4. Способ по п. 1, в котором указанный соевый пептид представляет собой Hypep 1510 ®.
5. Способ согласно любому из пп. 1-4, в котором рН составляет примерно 7,2.
| Шаманская Т | |||
| В., Осипова Е | |||
| Ю., Пурбуева Б | |||
| Б., Устюгов А | |||
| Ю., Астрелина Т | |||
| А., Яковлева М | |||
| В., Румянцев С | |||
| А | |||
| Культивирование мезенхимальных стволовых клеток ex vivo в различных питательных средах (обзор литературы и собственный опыт), Онкогематология, 2010, стр | |||
| Разборное приспособление для накатки на рельсы сошедших с них колес подвижного состава | 1920 |
|
SU65A1 |
| Mary P.Rosser et al., Transient transfection of CHO-K1-S using serum-free | |||

