Область изобретения
Настоящее изобретение относится к новым полипептидам, к способам их получения и к новым группам полипептидов-вариантов, основанных на общем каркасе. Эти группы можно, например, использовать для получения новых связывающих белков и полипептидов.
Предшествующий уровень техники
Были описаны разные способы конструирования новых связывающих белков (Nygren P.A. и  М. (1997) Curr Opin Struct Biol 7: 463-469). Одна стратегия заключалась в комбинировании формирования библиотеки и скрининга с отбором по необходимым свойствам.
М. (1997) Curr Opin Struct Biol 7: 463-469). Одна стратегия заключалась в комбинировании формирования библиотеки и скрининга с отбором по необходимым свойствам.
Полипептиды-варианты первого поколения Z, основанные на общем каркасе первого поколения, группы таких молекул и способы с их участием были описаны в WO 95/19374. Дополнительно, полипептиды-варианты Z, основанные на каркасе второго поколения, группы таких молекул и способы с их участием были описаны в WO 2009/080811. Сведения из этих двух описаний изобретений включены в данное описание изобретения посредством ссылки.
Для некоторых применений желательными являются полипептиды-варианты Z или их группы, имеющие улучшенные свойства, такие как более высокая щелочная стабильность, низкая антигенность, структурная стабильность, податливость химическому синтезу и гидрофильность. В WO 2009/080811 описаны Z-варианты, имеющие общий каркас с улучшенными свойствами, но не каждое необходимое свойство может быть получено у Z-вариантных полипептидов, как описано там.
Одним из ключевых факторов успеха полипептидных лекарственных средств является их стабильность. Полипептиды, демонстрирующие высокую структурную стабильность, будут скорее всего функционально противостоять химическим модификациям, изменениям физических условий и протеолизу как во время получения, так и в человеческом организме. Кроме того, стабильность будет влиять на активный срок годности полипептидных лекарственных средств, а также на продолжительность действия полипептидного лекарственного средства в человеческом организме.
Следовательно, остается потребность в улучшении стабильности Z-вариантных полипептидов.
Описание изобретения
Задачей настоящего изобретения является предложение полипептида с новым каркасом, полипептида, у которого уменьшены вышеуказанные и другие недостатки имеющихся в настоящее время Z-вариантных полипептидов.
Другой задачей настоящего изобретения является предложение способа получения полипептида на основе нового каркаса.
Также задачей настоящего изобретения является предложение группы таких улучшенных полипептидных вариантов, все из которых основаны на новом каркасе.
Другой задачей настоящего изобретения является предложение группы полинуклеотидов.
Еще одной задачей настоящего изобретения является предложение комбинации группы полипептидов и группы полинуклеотидов.
Еще одной задачей настоящего изобретения является предложение способа выбора необходимого полипептида, обладающего аффинностью к заданной мишени, из группы полипептидов.
Другая задача заключается в предложении способа выделения полинуклеотида, кодирующего необходимый полипептид, обладающий аффинностью к заданной мишени.
Другая задача заключается в предложении способа идентификации необходимого полипептида, обладающего аффинностью к заданной мишени.
Другая задача заключается в предложении способа выбора и идентификации необходимого полипептида, обладающего аффинностью к заданной мишени.
Связанная с этим задача заключается в предложении способа получения необходимого полипептида, обладающего аффинностью к заданной мишени.
Эти и другие цели могут быть достигнуты при помощи разных аспектов, раскрытых в настоящей заявке.
В первом аспекте раскрытия настоящего изобретения предложен полипептид, содержащий аминокислотную последовательность, выбранную из
1) EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q,
где каждый из Х2, Х3, Х4, Х6, Х7, Х10, Х11, Х17, X18, Х20, Х21, Х25 и Х28 независимо соответствует аминокислотному остатку; и
где, независимо друг от друга,
X16 выбран из N и Т;
Х26 выбран из K и S;
Х29Х30РХ32 выбран из DDPS и RQPE;
Х35 выбран из А и S;
Х36 выбран из Ε и N;
Х39 выбран из А, С и S;
Х45 выбран из Ε, N и S;
Х46 выбран из D, Ε и S, при условии, что Х46 не является D, когда Х45 представляет собой Ν;
Х47 выбран из А и S; и
2) аминокислотной последовательности, которая обладает по меньшей мере 91% идентичностью с последовательностью, определенной в (1), при условии, что Х46 не является D, когда Х45 представляет собой N.
В полипептидной последовательности (1), указанной выше, каждая аминокислота X, определенная как "независимо соответствующая любой аминокислоте", индивидуально соответствует аминокислотному остатку, который выбран из всех возможных аминокислот. Для ясности, это относится к положениям аминокислот, соответствующим положениям Х2, Х3, X4, Χ6, Х7, Х10, Χ11, Χ17, Х18, Х20, Х21, Х25 и Х28 в последовательности (1), указанной выше. Это означает, что каждый такой X может быть любым аминокислотным остатком, независимо от идентификации любого другого остатка, обозначенного как X в этой последовательности. В аминокислотной последовательности эти аминокислоты X могут быть выбраны из всех 20 природных аминокислотных остатков так, чтобы любой из этих 20 природных аминокислотных остатков мог присутствовать в соответствующем положении X в каждом отдельно взятом варианте. Выбор аминокислотного остатка в каждом положении может быть в той или иной степени случайным. Можно также ограничить группу, из которой выбирают разные варьирующиеся аминокислотные остатки, 19, 18, 17, 16 или менее 20 природными аминокислотными остатками. Вариабельность в разных положениях можно регулировать индивидуально, от одного, что означает отсутствие случайности, вплоть до всех 20 аминокислот. Случайное введение меньшего подмножества аминокислот может быть получено путем тщательного выбора вводимых дезоксирибонуклеотидных оснований, например, могут быть введены кодоны Т(А/С)С с получением случайного введения либо серина, либо тирозина в заданное положение полипептидной цепи. Аналогично, могут быть введены кодоны (T/C/A/G)CC с получением случайного введения фенилаланина, лейцина, аланина и валина в заданное положение полипептидной цепи. Специалистам известно много альтернативных комбинаций дезоксирибонуклеотидных оснований, которые можно использовать для получения разных комбинаций аминокислот в заданном положении полипептидной цепи. Совокупность аминокислот, которые могут появиться в заданном положении полипептидной цепи, также можно определить путем введения тринуклеотидов во время олигонуклеотидного синтеза вместо одного дезоксирибонуклеотидного основания за один раз. Определенная совокупность аминокислот также может быть получена с использованием сплит-пул синтеза, обеспечивающего возможность включения определенных кодонов в нужные положения при синтезе. Еще одна альтернатива получения случайных двунитевых линкеров заключается во включении случайных совокупностей тринуклеотидных структурных блоков с использованием лигирования и рестрикции сформированной затем двунитевой ДНК.
В одном воплощении настоящего изобретения предложен полипептид, обладающий аффинностью к заданной мишени. В одном таком воплощении аминокислотные остатки, которые обеспечивают специфичность связывания с мишенью, представляют собой остатки, находящиеся в положениях, соответствующих 2, 3, 4, 6, 7, 10, 11, 17, 18, 20, 21, 25 и 28 положениям в указанной выше последовательности (1). Аналогично, в таком полипептиде аминокислотные остатки, которые не обеспечивают специфичность связывания с мишенью, называются "аминокислотами каркаса" или просто "каркасом". Соответственно, в одном воплощении аминокислотные остатки каркаса, как определено в данном описании изобретения, являются аминокислотными остатками, которые находятся в положениях, соответствующих положениям 1, 5, 8, 9, 12-15, 19, 22-24, 27, 31, 33-34, 37-38, 40-44 и 48 в указанной выше последовательности (1). Специалисту понятно, что полезные свойства, обеспечиваемые каркасными аминокислотами полипептидов, как определено в данном описании изобретения, не зависят от специфичности связывания с мишенью указанного полипептида.
Специалисту понятно, что функция любого полипептида, такого как полипептид по настоящему изобретению, зависит от третичной структуры полипептида. Поэтому можно внести незначительные изменения в последовательность аминокислот полипептида без влияния на его функцию. Таким образом, описание изобретения включает модифицированные варианты указанного полипептида, которые не изменяют функциональные свойства полипептида, такие как его улучшенная стабильность и/или его аффинность связывания с заданной мишенью.
Таким образом, настоящее описание изобретения также включает полипептид, содержащий аминокислотную последовательность, обладающую 91%-ной или более идентичностью с последовательностью, определенной в (1). В некоторых воплощениях полипептид может содержать последовательность, которая по меньшей мере на 93%, например, по меньшей мере на 95%, например по меньшей мере на 97% идентична последовательности, определенной в (1).
В некоторых воплощениях такие различия между определениями последовательностей (1) и (2) могут быть обнаружены в любом положении последовательности полипептида, как описано в данном описании изобретения. В других воплощениях такие изменения могут быть обнаружены только в аминокислотных остатках каркаса. В других воплощениях указанные изменения могут быть обнаружены только в аминокислотных остатках, которые обеспечивают специфичность связывания с мишенью. Например, вполне возможно, что аминокислотный остаток, принадлежащий к определенной функциональной группе аминокислотных остатков (например, гидрофобный, гидрофильный, полярный и т.д.), может быть заменен другим аминокислотным остатком из той же функциональной группы.
Термин "% идентичности", как он использован в описании изобретения, может, например, быть рассчитан следующим образом. Искомую последовательность выравнивают с целевой последовательностью с использованием алгоритма CLUSTAL W (Thompson et al., Nucleic Acids Research, 22: 4673-4680 (1994)). Для сравнения использовали окно, соответствующее одной из выравниваемых последовательностей, например самой короткой. В некоторых случаях это окно может быть определено целевой последовательностью. В других случаях это окно может быть определено искомой последовательность. Сравнивают аминокислотные остатки в каждом положении и процент положений в искомой последовательности, которые имеют идентичные соответствия в целевой последовательности, регистрируют как % идентичности.
При использовании в качестве каркасов для связывающихся полипептидов, последовательности, описанные в данном описании изобретения, обеспечивали преимущества по сравнению с известными, аналогичными каркасами и были сконструированы для демонстрации высокой структурной стабильности и, следовательно, улучшенного срока годности при хранении. Эти преимущества также относятся к третьему аспекту настоящего раскрытия (смотрите ниже), который относится к группам полипептидных вариантов этого первого аспекта.
В одном воплощении настоящего изобретения Χ16 представляет собой Т.
В одном воплощении Х26 представляет собой K.
В одном воплощении Х29Х30РХ32 представляет собой DDPS.
В одном воплощении Х29Х30РХ32 представляет собой RQPE.
В одном воплощении Х35 представляет собой S.
В одном воплощении Х36 представляет собой Е.
В одном воплощении Х39 представляет собой S.
В одном воплощении Х45 выбран из Ε и S.
В одном воплощении Х45 представляет собой Е.
В одном воплощении Х45 представляет собой S.
В одном воплощении Х46 выбран из Ε и S.
В одном воплощении Х46 представляет собой Е.
В одном воплощении Х46 представляет собой S.
В одном воплощении Х46 представляет собой D.
В одном воплощении Х46 не является D или Е, когда Х45 представляет собой N.
В одном воплощении Х45Х46 выбран из ЕЕ, ES, SE и SS, например из ES и SE.
В одном воплощении Х45Х46 представляет собой ES.
В одном воплощении Х45Х46 представляет собой SE.
В одном воплощении Х45Х46 представляет собой SD.
В одном воплощении Х47 представляет собой S.
Термин "аффинность связывания с заданной мишенью", используемый в данном описании, относится к свойству полипептида, которое может быть проанализировано, например, путем применения технологии поверхностного плазмонного резонанса (SPR). Например, указанную аффинность связывания можно проанализировать в эксперименте, в котором заданную мишень или ее фрагмент иммобилизируют на сенсорный чип прибора и образец, содержащий тестируемый полипептид, пропускают над чипом. В качестве альтернативы, тестируемый полипептид иммобилизируют на сенсорный чип прибора и образец, содержащий заданную мишень или его фрагмент, пропускают над чипом. Специалист может затем интерпретировать результаты, полученные с помощью таких экспериментов, чтобы установить по меньшей мере качественную оценку аффинности связывания полипептида с заданной мишенью. Если необходимо количественное измерение, например, для определения значения KD взаимодействия, также можно использовать методы поверхностного плазмонного резонанса. Значения связывания могут, например, быть определены с помощью прибора Biacore (GE Healthcare) или ProteOn XPR 36 (Bio-Rad). Заданную мишень соответствующим образом иммобилизируют на сенсорный чип прибора и образцы полипептида, аффинность которого необходимо определить, готовят посредством серийного разведения и вводят в случайном порядке. Значения KD затем могут быть рассчитаны из этих результатов с использованием, например, 1:1 модели связывания Ленгмюра программного обеспечения BIAevaluation 4.1 или другого подходящего программного обеспечения, предоставляемого изготовителем прибора.
Термин "аффинность связывания с заданной мишенью" при использовании в данном описании изобретения также может относиться к свойству полипептида, которое может быть проанализировано с помощью ELISA. Например, аффинность связывания может быть проанализирована в эксперименте, в котором образцы полипептида захватывают покрытыми антителами планшетами ELISA и добавляют биотинилированную определенную мишень или ее фрагменты, с последующим добавлением конъюгата стрептавидина с HRP (пероксидазой хрена). Добавляют субстрат ТМВ (тетраметилбензидин) и измеряют поглощение при 450 нм, используя ридер для многолуночных планшетов, такой как Victor3 (Perkin Elmer). Специалист затем может интерпретировать полученные с помощью таких экспериментов результаты для установления по меньшей мере качественной меры аффинности связывания комплекса с заданной мишенью. Если необходимо количественное измерение, например для определения значения ЕС50 (половины максимальной эффективной концентрации) взаимодействия, также можно использовать ELISA. Ответ полипептида в зависимости от серийных разведений определенной мишени или ее фрагмента измеряют с использованием ELISA, как описано выше. Специалист может затем интерпретировать результаты, полученные с помощью таких экспериментов, и значения ЕС50 могут быть рассчитаны из результатов с использованием, например, GraphPad Prism 5 и нелинейной регрессии.
Как описано выше, считается, что Ζ полипептиды-варианты образуют или формируют часть трехспирального узла домена белка, мотив, обладающий аффинностью к определенной мишени, главным образом, формирующий часть двух альфа-спиралей с соединительной петлей в пределах указанного трехспирального узлового белкового домена.
Разные модификации и/или дополнения к полипептиду, как он определен выше, могут быть выполнены для того, чтобы приспособить полипептид к предполагаемому конкретному применению, не отступая от объема настоящего изобретения.
Такие модификации и дополнения описаны более подробно ниже и могут содержать дополнительные аминокислоты, включенные в ту же самую полипептидную цепь, или маркеры и/или терапевтические агенты, которые химически конъюгированы или иным образом присоединены к полипептиду.
Следовательно, в одном воплощении предложен полипептид, как он описан выше, содержащий дополнительные аминокислотные остатки. В некоторых воплощениях дополнительные аминокислотные остатки могут быть локализованы на С-конце полипептида. В некоторых воплощениях дополнительные аминокислотные остатки могут быть локализованы на N-конце полипептида.
В одном воплощении указанные дополнительные аминокислотные остатки на С-конце содержат АР.
В одном воплощении, указанные дополнительные аминокислотные остатки на N-конце содержат AEAKYAK.
В еще одном воплощении предложен полипептид, как он описан выше, который состоит из последовательности (1) или 2), имеющий от 0 до 7 дополнительных аминокислотных остатков на N-конце и от 0 до 3 дополнительных аминокислотных остатков на С-конце.
Дополнительные аминокислотные остатки могут играть роль в связывании полипептида, но в равной степени могут служить другим целям, связанным, например, с одним или более из получения, очистки, стабилизации, соединения или обнаружения полипептида. В некоторых воплощениях указанные дополнительные аминокислотные остатки образуют один или более полипептидных доменов.
Такие дополнительные аминокислотные остатки могут содержать один или более аминокислотных остатков, добавленных с целью химического связывания. Примером этого является добавление остатка цистеина в самом первом или самом последнем положении полипептидной цепи, т.е. на N- или С-конце. Остаток цистеина, который используется для химического связывания, может также быть введен посредством замены другой аминокислоты на поверхности белкового домена, предпочтительно на части поверхности, которая не вовлечена в связывание мишени. Такие дополнительные аминокислотные остатки могут также содержать "тег" для очистки или обнаружения полипептида, такие как гексагистидиновый (His6) тег или "myc" тег или "FLAG" тег для взаимодействия с антителами, специфичными к этому тегу. Специалисту известны и другие альтернативы.
"Дополнительные аминокислотные остатки", обсуждаемые выше, также могут составлять один или более полипептидный(х) домен(ов) с любой необходимой функцией, например с функцией другого связывания, или с функцией пролонгирования время полувыведения, или с ферментативной функцией, или с функцией хелатирования ионов металла, или с флуоресцентной функцией, или с любой их комбинацией.
В одном примере воплощения предложено соединение, обладающее аффинностью к определенной мишени, где указанное соединение содержит:
A. по меньшей мере один полипептид, как описано выше;
B. по меньшей мере один альбумин-связывающий домен стрептококкового белка G или его производное; и
C. возможно по меньшей мере одну связывающую группировку для связывания по меньшей мере одного указанного альбумин-связывающего домена или его производного с С- или N-концом по меньшей мере одного указанного полипептида.
Неограничивающие примеры производных альбумин-связывающего домена стрептококкового белка G раскрыты в WO 2009/016043 и WO 2012/004384.
Кроме того, в другом воплощении предложен полипептид, как описано выше, который содержит аминокислотную последовательность, выбранную из:
YAK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q АР; и
FNK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q АР.
где каждый Xy определен, как указано выше (и у обозначает аминокислотное положение остатка X в полипептидной последовательности, определенной в (1) выше.
В некоторых воплощениях предложен полипептид, который содержит аминокислотную последовательность, выбранную из
ADNNFNK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q APK;
ADNKFNK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q APK;
VDNKFNK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q APK;
VDAKYAK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q APK; и
AEAKYAK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q APK;
где Xy определен, как описано выше (и y обозначает аминокислотное положение остатка X в полипептидной последовательности, определяемой в (1) выше).
Варианты полипептидов, описанные в данном описании изобретения, можно получить, взяв Ζ вариант полипептида, например на основе известного каркаса и имеющего аффинность к заданной мишени и осуществив сайт-направленный мутагенез в выбранных положениях с получением полипептида, имеющего каркас по настоящему изобретению, сохраняющего целевую аффинность. Полипептид по настоящему изобретению может, в качестве альтернативы, быть изготовлен путем химического синтеза всей молекулы или с помощью других методов молекулярной биологии, известных специалисту в данной области техники, для присоединения связывающего мотива Ζ-вариантного полипептида на каркас, описанный в данном описании изобретения.
В качестве общей иллюстрации, исходные Ζ варианты полипептидов, содержащие следующую общую каркасную последовательность и обладающие специфичностью связывания, определяемой аминокислотной последовательности внутри связывающего мотива [ВМ]:
AEAKYAK-[BM]-DDPSQSSELL SEAKKLNDSQ APK
можно модифицировать, чтобы получить полипептид, как описано в данном описании изобретения.
В различных конкретных воплощениях этого аспекта описания изобретения представлены следующие полипептиды:
AEAKYAK-[BM]-RQPEQSSELL SEAKKLNDSQ APK
AEAKYAK-[BM]-DDPSQSSELL SEAKKLSESQ APK
AEAKYAK-[BM]-DDPSQSSELL SEAKKLESSQ APK
AEAKYAK-[BM]-DDPSQSSELL SEAKKLSDSQ APK
AEAKYAK-[BM]-DDPSQSSELL SEAKKLNESQ APK
AEAKYAK-[BM]-RQPEQSSELL SEAKKLSESQ APK
AEAKYAK-[BM]-RQPEQSSELL SEAKKLESSQ APK
AEAKYAK-[BM]-RQPEQSSELL SEAKKLSDSQ APK
AEAKYAK-[BM]-RQPEQSSELL SEAKKLNSSQ APK
Полипептиды, раскрытые в данном описании изобретения, имеют много применений, например применения терапевтического, диагностического или прогностического значения для заболевания. Неограничивающий перечень заболеваний, где указанные полипептиды могут найти терапевтическое, диагностическое или прогностическое применение, включают рак, воспалительные заболевания, аутоиммунное заболевание, инфекционные заболевания, неврологические заболевания, нейродегенеративные заболевания, глазные заболевания, заболевания почек, легочные заболевания, заболевания желудочно-кишечного тракта, сердечно-сосудистые заболевания, гематологические заболевания, дерматологические заболевания, аллергии и другие.
Таким образом, в одном воплощении предложен полипептид с аффинностью к заданной мишени. В более конкретных воплощениях указанная мишень выбрана из группы, состоящей из HER2, TNFα, EGFR, IGF1R, IgG, PDGFRβ, HER3, С5, FcRn, CAIX, амилоида β, CD4, IL8, IL6 и инсулина. В других воплощениях указанный полипептид может быть полезен для биотехнологического, промышленного и фармацевтического применения, например для применения в качестве аффинного лиганда в технологии разделения, в применении для очистки или в качестве агента для обнаружения. В более конкретном воплощении заданная мишень может быть альбумин-связывающим доменом ("ABD" или "модуль GA") из стрептококкового белка G или его производного.
Специалисту понятно, что список заданных мишеней следует рассматривать, как неограничивающий и, что полипептиды, определенные в данном описании изобретения, как обладающие аффинностью к другим заданным мишеням, подпадают в объем настоящего изобретения.
Неограничивающими примерами известных Z-вариантных полипептидов, основанными на известном каркасе и имеющими аффинность к разным мишеням, являются Z-варианты с аффинностью к рецептору EGF (раскрыт в WO 2007/065635), к рецептору HER2 (раскрыт в WO 2009/080810), к рецептору HER3 (раскрыт в WO 2010/056124), к рецептору IGF1 (раскрыт в WO 2009/019117), к β-рецептору PDGF (раскрыт в WO 2009/077175), к альбумин-связывающему домену (ABD) (раскрыт в WO 2014/064237), к неонатальному Fc рецептору (FcRn) (раскрыт в РСТ/ЕР 2014/055299) и к карбоангидразе IX (раскрыт в WO 2014/096163). Для ясности, следует иметь в виду, что в настоящем изобретении связывающий мотив [ВМ] Z-варианта соответствует первым 28 аминокислотным остаткам этих связывающих мотивов, описанных в документах, перечисленных выше, где определения связывающих мотивов представляют собой 29 аминокислотных остатков и соответствуют аминокислотным остаткам в положениях, соответствующих положениям 1-29 указанной выше последовательности (1).
В одном воплощении предложен полипептид с аффинностью к заданной мишени, который дополнительно содержит маркер, например маркер, выбранный из группы, состоящей из флуоресцентных красителей и металлов, хромофорных красителей, хемилюминесцентных соединений и биолюминесцентных белков, ферментов, радионуклидов и частиц. Такие маркеры могут быть использованы, например, для обнаружения полипептида.
В некоторых воплощениях полипептид присутствует как группировка в слитом полипептиде или конъюгате, также содержащем вторую группировку, обладающую нужной биологической активностью. Неограничивающие примеры такой нужной биологической активности включают терапевтическую активность, активность связывания и ферментативную активность.
В некоторых воплощениях указанная группировка также содержит маркер. Маркер в некоторых случаях может быть соединен только с полипептидом с аффинностью к заданной мишени, а в некоторых случаях и с полипептидом с аффинностью к заданной мишени, и со второй группировкой конъюгата или слитого полипептида. Кроме того, возможно, что метка может быть присоединена только ко второй группировке, а не к полипептиду с аффинностью к заданной мишени. Таким образом, в еще одном воплощении предложен полипептид с аффинностью к заданной мишени, содержащий вторую группировку, где указанный маркер соединен только со второй группировкой.
Раскрытые в данном описании изобретения полипептиды или слитые полипептиды можно использовать как реагенты для обнаружения, реагенты захвата, реагенты разделения, в качестве диагностических агентов для диагностики in vivo или in vitro, или в качестве терапевтических агентов. Методы с использованием полипептидов или слитых полипептиды по настоящему изобретению in vitro могут быть выполнены в разных форматах, таких как микротитрационные планшеты, белковые матрицы, на поверхностях биосенсоров, на срезах тканей и так далее.
Следует также понимать, что полипептид или слитые полипептиды по настоящему изобретению могут быть полезны в качестве терапевтического, диагностического или прогностического агента сами по себе или в качестве средства для нацеливания других терапевтических, диагностических или прогностических агентов, например с прямыми или опосредованными эффектами на указанную мишень. Прямой терапевтической эффект может быть, например, осуществлен путем ингибирования передачи сигнала указанной мишенью. Указанная мишень также может служить ценным маркером для прогноза течения некоторых заболеваний (например, заболеваний, перечисленных выше).
Следовательно, в одном воплощении предложен полипептид или слитый полипептид, как описано в данном описании изобретения, для применения в терапии или для применения в качестве диагностического агента. В другом воплощении указанный полипептид или слитый полипептид дополнительно содержит терапевтический агент. Неограничивающими примерами таких терапевтических агентов являются терапевтический агент, усиливающий эффект указанного полипептида или слитого полипептида, терапевтический агент, действующий синергично с указанным полипептидом или слитым полипептидом и терапевтический агент, оказывающий влияние на другой аспект заболевания, подлежащего лечению. Также предусмотрены фармацевтические композиции, содержащие полипептиды, как они раскрыты в данном описании изобретения, отдельно или вместе с дополнительными терапевтическими агентами.
Во втором аспекте настоящего описания изобретения предложен полинуклеотид, кодирующий полипептид или слитый полипептид, как описано в данном описании изобретения. Также включены в это описание изобретения способ получения полипептида или слитого полипептида, как описано выше, включающий экспрессию такого полинуклеотида; экспрессирующий вектор, содержащий полинуклеотид; и клетка-хозяин, содержащая указанный экспрессирующий вектор.
В третьем аспекте настоящего изобретения предложена группа полипептидных вариантов на основе общего каркаса, где каждый полипептид в группе содержит аминокислотную последовательность, выбранную из:
1) EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q,
где каждый из Х2, Х3, Х4, Х6, Х7, Х10, Х11, Х17, Χ18, Х20, Х21, Х25 и Х28 независимо соответствует любому аминокислотному остатку; и
где, независимо друг от друга,
Χ16 выбран из N и Т;
Х26 выбран из K и S;
Х29Х30РХ32 выбран из DDPS и RQPE;
Х35 выбран из А и S;
Х36 выбран из Ε и Ν;
Х39 выбран из А, С и S;
Х45 выбран из Ε, N и S;
Х46 выбран из D, Ε и S, при условии, что Х46 не является D, когда Х45 представляет собой Ν;
Х47 выбран из А и S; и
2) аминокислотная последовательность, которая обладает по меньшей мере 91% идентичностью с последовательностью, определенной в (1), при условии, что Х46 не является D, когда Х45 представляет собой N.
В вышеприведенной последовательности (1) каждый из Х2, Х3, Х4, Х6, Х7, Х10, Х11, Χ17, Χ18, Х20, Х21, Х25 и Х28 по отдельности соответствует аминокислотному остатку, который изменяется в указанной группе. Следовательно, каждый такой аминокислотный остаток может быть любым аминокислотным остатком независимо от идентификации любого другого остатка, обозначаемого как Xy в последовательности, как описано выше в связи с первым (полипептидным) аспектом описания изобретения. Неограничивающие варианты конкретных аминокислотных остатков Xy в группе полипептидов и любых дополнительных аминокислотных остатков в любом конце последовательности (1) или 2) являются такими же, как те, что приведены выше в качестве воплощений первого аспекта настоящего изобретения.
Как обсуждалось выше, полипептиды, содержащие незначительные изменения по сравнению с приведенными выше аминокислотными последовательностями без значительного влияния на их третичную структуру и функцию также входят в объем настоящего описания изобретения. Таким образом, настоящее описание изобретения охватывает группу вариантов полипептидов на основе общего каркаса, где каждый полипептид в группе содержит аминокислотную последовательность с 91%-ной или более высокой идентичностью последовательности, определенной в (1). В некоторых воплощениях каждый полипептид может содержать последовательность, которая по меньшей мере на 93%, например по меньшей мере на 95%, например по меньшей мере на 97% идентична последовательности, определенной в (1).
Группа, определенная в данном описании изобретения, состоит из большого количества уникальных и разных вариантов определенных полипептидных молекул. В данном контексте большое количество может означать, например, что эта группа содержит по меньшей мере 1×104 уникальных полипептидных молекул или по меньшей мере 1×106, по меньшей мере 1×108, по меньшей мере 1×1010, по меньшей мере 1×1012 или по меньшей мере 1×1014 уникальных полипептидных молекул. Специалисту понятно, что необходимо использовать группу, которая является достаточно большой, чтобы обеспечить необходимый размер группы. "Группа", описанная в данном описании изобретения, также может обозначать "библиотеку".
Для специалиста очевидно, что группа, описанная в данном описании изобретения, может быть полезной в качестве библиотеки для выбора новых связывающих молекул на основе полипептида, определенного в (1). Хорошо известно в данной области техники, что связывающие молекулы могут быть выделены из группы (или библиотеки) рандомизированных полипептидов. Эта технология была описана в общих чертах в РСТ публикации WO 95/19374, в Nord et al (1997) Nature Biotechnology 15: 772-777 и в WO 2009/080811, и успешно применялась для того, чтобы выбрать связывающие молекулы на основе общего Ζ домененного каркаса, к различным целевым молекулам посредством случайных изменений тринадцати разных положений связывания мишени и последующего выбора интересующих связывающих элементов с помощью фагового дисплея или другой системы выбора, основанной на взаимосвязи генотипа и фенотипа. Группа, как описано в данном описании изобретения, является группой полипептидов-вариантов, которые демонстрируют улучшенные свойства, в частности с точки зрения стабильности, по сравнению с группами из предшествующего уровня техники. Примеры Z-вариантов, выделенных из группы (или библиотеки) рандомизированных полипептидов, включают Z-варианты с аффинностью к рецептору EGF (раскрыты в WO 2007/065635), к рецептору HER2 (раскрыты в WO 2009/080810), к рецептору HER3 (раскрыты в WO 2010/056124), к рецептору IGF1 (раскрыты в WO 2009/019117), к β-рецептору PDGF (раскрыты в WO 2009/077175), к ABD (раскрыты в WO 2014/064237), к неонатальному Fc-рецептору (FcRn) (раскрыты в РСТ/ЕР 2014/055299) и к карбоангидразе IX (раскрыты в WO 2014/096163).
В четвертом аспекте настоящего описания изобретения предлагается группа полинуклеотидов. Каждый полинуклеотид в этой группе кодирует представителя группы полипептидов, как определено выше в связи с третьим аспектом.
В пятом аспекте настоящего изобретения предлагается комбинация полипептидной группы в соответствии с третьим аспектом и полинуклеотидной группы в соответствии с четвертым аспектом, в которой каждый представитель полипептидной группы физически и пространственно связан с соответствующим полинуклеотидом, кодирующим этого представителя посредством взаимосвязи генотипа и фенотипа. Это физическое или пространственное объединение будет более или менее жестким в зависимости от используемой системы.
Средства взаимосвязи генотипа и фенотипа могут включать систему фагового дисплея. Системы фаговых дисплеев хорошо известны специалисту и описаны, например, в Smith G.P. (1985) Science 228: 1315-1317 и Barbas C.F. et al. (1991) Proc Natl Acad Sci U S A 88: 7978-7982.
Кроме того, средства взаимосвязи генотипа и фенотипа могут включать систему дисплея на клеточной поверхности. Система дисплея на клеточной поверхности может включать прокариотические клетки, такие как грамположительные клетки или эукариотические клетки, такие как клетки дрожжей. Системы дисплея на клеточной поверхности хорошо известны специалисту. Прокариотические системы, например, описаны в Francisco J.A. et al. (1993) Proc Natl Acad Sci USA 90: 10444-10448 и Lee S.Y. et al. (2003) Trends Biotechnol 21:45-52. Эукариотические системы, например, описаны в Boder E.T. et al. (1997) Nat Biotechnol 15:553-557 и Gai S.A. et al. (2007) Curr Opin Struct Biol 17:467-473. В одном воплощении указанная взаимосвязь генотипа и фенотипа может включать систему фагового дисплея.
Кроме того, средства взаимосвязи генотипа и фенотипа могут включать систему бесклеточного дисплея. Система бесклеточного дисплея может включать систему рибосомного дисплея, или дисплейную систему с компартментализацией in vitro, или систему для cis дисплея, или систему дисплея на микрогранулах. Рибосомные дисплейные системы хорошо известны специалисту и описаны, например, в Mattheakis L.C. et al. (1994) Proc Natl Acad Sci USA 91: 9022-9026 и Zahnd С. et al. (2007) Nat Methods 4: 269-279. Системы компартментализации in vitro хорошо известны специалисту и описаны, например, в Sepp A. et al. (2002) FEBS Lett 532: 455-458. Системы cis дисплея хорошо известны специалисту и описаны, например, в Odegrip R. et al. (2004) Proc Natl Acad Sci USA 101: 2806-2810. Системы дисплея на микрогранулах хорошо известны специалисту и описаны, например, в Nord О. et al. (2003) J Biotechnol 106: 1-13.
Кроме того, средства взаимосвязи генотипа и фенотипа могут включать недисплейную систему, такую как структурный анализ комплементации фрагментов белка (РСА). РСА системы хорошо известны специалисту, и описаны, например, в Koch H. et al. (2006) J Mol Biol 357: 427-441.
В шестом аспекте настоящего изобретения предложен способ выбора искомого полипептида, обладающего аффинностью к заданной мишени из группы полипептидов, включающий стадии:
(а) обеспечения группы полипептидов в соответствии с третьим аспектом;
(б) приведение группы полипептидов в контакт с заданной мишенью в условиях, которые делают возможным специфическое взаимодействие между мишенью и по меньшей мере одним искомым полипептидом, обладающим аффинностью к мишени; и
(в) выбор, на основании указанного специфического взаимодействия, по меньшей мере одного искомого полипептида из остальной группы полипептидов.
Ниже этот способ называется "способ выбора" в соответствии с описанием изобретения.
Стадия (а) может включать подготовительные стадии обеспечения группы полинуклеотидов и экспрессирования указанной группы полинуклеотидов с получением указанной группы полипептидов. Средства получения группы полипептидов варьируются в зависимости от используемой дисплейной системы и примеры таких средств можно найти в приведенных выше ссылках о генотипе-фенотипе. Каждый представитель указанной группы полипептидов, используемой в способе выбора, может быть физически связан с полинуклеотидом, кодирующим этот представитель, при помощи средств взаимосвязи генотип-фенотип. Средством взаимосвязи генотипа и фенотипа может быть одно из обсуждаемых выше.
Стадия (б) включает стадии приведения группы полипептидов в контакт с заданной мишенью в условиях, которые делают возможным специфическое взаимодействие между мишенью и по меньшей мере одним искомым полипептидом, обладающим аффинностью к мишени. Диапазон применимых условий определяется устойчивостью мишени, устойчивостью дисплейной системы и желательными свойствами взаимодействия с мишенью. Например, может быть нужен конкретный метод разделения взаимодействия, например подкисление до заданного рН. Специалисту известно, какие эксперименты необходимы для определения подходящих условий.
Стадия (в) включает выделение по меньшей мере одного полипептида. Средства выделения искомого полипептида из остальной группы, основанные на специфическом взаимодействии между заданной мишенью и по меньшей мере одним искомым полипептидом, обладающим аффинностью к мишени, варьируются в зависимости от используемой дисплейной системы и их можно найти в вышеприведенных ссылках о генотипе-фенотипе. Например, дисплейные системы выбора in vitro являются бесклеточными, в отличие от таких систем, как фаговый дисплей и анализ компартментализации белковых фрагментов.
В седьмом аспекте настоящего изобретения предложен способ выделения полинуклеотида, кодирующего искомый полипептид, обладающий аффинностью к заданной мишени, включающий следующие стадии:
- выбор указанного искомого полипептида и полинуклеотида, кодирующего его, из группы полипептидов, используя способ выбора в соответствии с шестым аспектом; и
- выделение таким образом выделенного полинуклеотида, кодирующего искомый полипептид.
Ниже этот способ называется "способ выделения" в соответствии с описанием изобретения.
Отделение полинуклеотида от полипептида может быть осуществлено по-разному, в зависимости от используемой для выбора дисплейной системы. Например, в бесклеточных дисплейных системах, таких как cis-дисплей и рибосомный дисплей, полинуклеотид или соответствующую мРНК извлекают посредством эффективного элюирования из полипептида с использованием средств, описанных в вышеприведенных ссылках о генотипе-фенотипе.
Выделение полинуклеотида может быть осуществлено разными способами в зависимости от дисплейной системы, используемой для выбора. В большинстве из описанных выше систем выбора, например в анализе комплементации фрагментов белка, полинуклеотид может быть непосредственно выделен с помощью специфической ПЦР-амплификации с использованием соответствующих олигонуклеотидов. Кроме того, как в случае рибосомного дисплея, полинуклеотид может быть выделен из соответствующей мРНК с использованием обратной транскрипции. Различные средства выделения полинуклеотида можно найти в вышеприведенных ссылках о генотипе-фенотипе.
В восьмом аспекте настоящего изобретения предложен способ идентификации искомого полипептида, обладающего аффинностью к заданной мишени, включающей стадии:
- выделение полинуклеотида, кодирующего указанный искомый полипептид, с использованием способа в соответствии с седьмым аспектом; и
- секвенирование полинуклеотида для определения путем выведения (дедукции) аминокислотной последовательности указанного искомого полипептида.
Секвенирование полинуклеотида может быть сделано в соответствии со стандартными процедурами, хорошо известными специалистам в данной области.
В девятом аспекте настоящего изобретения предложен способ выбора и идентификации искомого полипептида, обладающего аффинностью к заданной мишени, из группы полипептидов, включающий следующие стадии:
(а) синтез каждого представителя группы полипептидов согласно третьему аспекту на отдельном носителе или грануле;
(б) селекция или обогащение носителей или гранул на основании взаимодействия полипептида с заданной мишенью; и
(в) идентификация полипептида посредством методологии определения характеристик белка.
На стадии (в), например, можно использовать масс-спектрометрический анализ.
Ниже этот способ называется "способ выбора и идентификации" в соответствии с описанием изобретения.
В десятом аспекте настоящего изобретения предложен способ получения искомого полипептида, обладающего аффинностью к заданной мишени, включающий следующие стадии:
- выбор и идентификация искомого полипептида с использованием способа выбора в соответствии с шестым аспектом или способа выбора и идентификации в соответствии с девятым аспектом; и
- получение указанного искомого полипептида.
Ниже этот способ называется "способ получения" в соответствии с описанием изобретения.
Что касается способа получения, то получение может быть выполнено с использованием рекомбинантной экспрессии полинуклеотида, кодирующего искомый полипептид. Получение может быть выполнено с использованием химического синтеза искомого полипептида de novo.
В одиннадцатом аспекте настоящего изобретения предложен способ получения искомого полипептида, обладающего аффинностью к заданной мишени, включающий следующие стадии:
(а1) выделение полинуклеотида, кодирующего указанный искомый полипептид с использованием способа выделения в соответствии с седьмым аспектом, или
(а2) обратная трансляция полипептида, идентифицированного с использованием способа выбора и идентификации в соответствии с девятым аспектом; и
(б) экспрессирование таким образом выделенного полинуклеотида с получением указанного искомого полипептида,
где стадию (б) выполняют после либо стадии (а1), либо стадии (а2).
Полипептиды, группы и способы по изобретению позволяют получить агенты с аффинностью к заданной мишени путем получения полипептида, который характеризуется специфическим связыванием с заданной мишенью.
Также возможно предложить полипептиды, связывающиеся с заданной мишенью, которые проявляют незначительное неспецифическое связывания или не проявляют его совсем.
Также возможно предложить полипептиды, связывающиеся с заданной мишенью, которые можно легко использовать в качестве группировки в слитом полипептиде.
Кроме того, возможно предложить полипептиды, связывающиеся с заданной мишенью, чтобы решить одну или более известных проблем, возникающих с существующими антительными реагентами.
Кроме того, возможно предложить полипептиды, связывающиеся с заданной мишенью, которые можно использовать в терапевтических и/или диагностических применениях.
Также можно предложить полипептиды, связывающиеся с заданной мишенью, которые легко получают посредством химического синтеза пептидов.
Кроме того, изобретение позволяет идентифицировать полипептиды, связывающиеся с заданной мишенью, которые проявляют улучшенную стабильность по сравнению с известным агентами, связывающимися с той же мишенью.
Также можно предложить полипептиды, связывающиеся с заданной мишенью, которые демонстрируют низкую антигенность при использовании in vivo у млекопитающих и/или которые проявляют улучшенное биораспределение при введении млекопитающему.
Рассмотренные выше модификации полипептидов, составляющих группу по настоящему изобретению, применимы также к полипептидам, полученным любым из вышеуказанных способов.
Полипептиды по настоящему изобретению могут быть получены любым известным способом, включая химический синтез или экспрессирование в разных прокариотических или эукариотических хозяевах, включающих бактериальные клетки, дрожжевые клетки, клетки растений, клетки насекомых, целые растения и трансгенные животные.
В то время как полипептиды, группы полипептидов и способы идентификации, выбора, выделения и получения, раскрытые в данном описании изобретения, были описаны со ссылкой на различные типичные аспекты и воплощения, специалистам в данной области понятно, что могут быть внесены различные изменения и могут быть использованы эквиваленты вместо их элементов без отступления от объема изобретения. Кроме того, многие модификации могут быть сделаны для адаптации конкретной ситуации или молекулы к идеям изобретения без отступления от его основного объема. Поэтому предполагается, что описание изобретения не ограничено каким-либо конкретным рассматриваемым воплощением, а включает все воплощения, подпадающие в объем прилагаемой формулы изобретения.
Краткое описание графических материалов
На Фиг. 1 представлен перечень аминокислотных последовательностей примеров полипептида, как описано в описании изобретения. Последовательности С5-связывающих Z-вариантных полипептидов, которые, как показано в Примерах 2-3, имеют улучшенную стабильность, представлены на Фиг. 1, как SEQ ID NO: 12, 17, 18 и 22, и их последовательности, соответствующие самой короткой последовательности, определенной в данном описании изобретения, представлены как SEQ ID NO: 19-21. Аминокислотные последовательности С5-связывающих полипептидов, слитых со альбумин-связывающими доменами, представлены на Фиг. 1 с идентификаторами последовательности SEQ ID NO: 4-11, 13-16 и 23-25. Последовательности Z-вариантных полипептидов с аффинностью к HER2, PDGF-Rβ, FcRn и CAIX, которые, как показано в Примере 12, имеют улучшенную стабильность, представлены как SEQ ID NO: 28-29, SEQ ID NO: 31-32, SEQ ID NO: 34-35 и SEQ ID NO: 37-42, соответственно, вместе с соответствующими контрольными полипептидами SEQ ID NO: 27, 30, 33 и 36. Последовательности указанных Z-вариантных полипептидов с аффинностью к HER2, ΡDGF-Rβ, FcRn и CAIX, соответствующих самой короткой последовательности, определенной в данном описании изобретения, представлены как SEQ ID NO: 43-54. Кроме того, аминокислотные последовательности контрольного С5-связывающего полипептида, контрольного С5-связывающего полипептида, слитого с альбумином, альбумин-связывающим доменом и человеческим С5, представлены как SEQ ID NO: 26, 1, 2 и 3, соответственно.
На Фиг. 2 представлено изображение геля SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), где первая дорожка содержит маркер размера SeeBlue 2Р и полосы представляют собой С5-связывающий полипептид PSI0242 (SEQ ID NO: 1) (0) до анализа стабильности; и (2н) после 2 недельного анализа стабильности.
На Фиг. 3 представлена хроматограмма обращенно-фазовой ВЭЖХ PSI0242 (SEQ ID NO: 1) до анализа стабильности (сплошная линия) и после 2 недельного анализа стабильности (пунктирная линия).
На Фиг. 4 представлено изображение SDS-PAGE геля, где первая дорожка содержит маркер размера SeeBlue 2Р и полосы, представляют собой (0) исходные образцы; и (2н) образцы после 2 недельного анализа стабильности. A: SEQ ID ΝΟ: 1; В: SEQ ID NO: 13; С: SEQ ID ΝO: 14; D: SEQ ID NO: 16.
На Фиг. 5 представлена хроматограмма обращенно-фазовой ВЭЖХ модифицированного С5 ингибитора (SEQ ID NO: 5) до анализа стабильности (сплошная линия) и после 2 недельного анализа стабильности (пунктирная линия).
На Фиг. 6 представлена хроматограмма обращенно-фазовой ВЭЖХ модифицированного С5 ингибитора (SEQ ID NO: 16) до анализа стабильности (сплошная линия) и после 2 недельного анализа стабильности (пунктирная линия).
На Фиг. 7 представлены CD (круговой дихроизм)-спектры, собранные для A: Z17351 (SEQ ID NO: 37); В: Z17352 (SEQ ID NO: 38); С: Z17355 (SEQ ID NO: 39); D: Z17357 (SEQ ID NO: 40); Ε: Ζ17359 (SEQ ID NO: 41); F: Z17360 (SEQ ID NO: 42); и G: Z09782 (SEQ ID NO: 36).
На Фиг. 8 представлены изображения SDS-PAGE гелей, показывающие исходные и являющиеся предметом изобретения полипептиды до (0) и после 2 недельного (2н) определения стабильности. А: Полипептиды, нацеленные на HER2: дорожка 1: Mw (Молекулярная масса), дорожка 2: Z02891 (0), дорожка 3: Z02891 (2н), дорожка 4: Mw, дорожка 5: Z17341 (0), дорожка 6: Z17341 (2н), дорожка 7: Z17342 (0), дорожка 8: Z17342 (2н); В: Полипептиды, нацеленные на PDGF-Rp: дорожка 1: Mw, дорожка 2: Z15805 (0), дорожка 3: Z15805 (2н), дорожка 4: Mw, дорожка 5: Z17343 (0), дорожка 6: Z17343 (2w), дорожка 7: Z17344 (0), дорожка 8: Z17344 (2н); С: Полипептиды, нацеленные на FcRn: дорожка 1: Z10103 (0), дорожка 2:Z10103 (2н), дорожка 3: Mw, дорожка 4: Z17347 (0), дорожка 5: Z17347 (2н), дорожка 6: Z17348 (0), дорожка 7: Z17348 (2н); и D: Полипептиды, нацеленные на CAIX: дорожка 1: Mw, дорожка 2: Z09782 (0), дорожка 3.Z09782 (2w), дорожка 4: Mw, дорожка 5: Z17351 (0), дорожка 6: Z17351 (2н), дорожка 7: Z17352 (0), дорожка 8: Z17352 (2н); дорожка 9: Z17355 (0), дорожка 10: Z17355 (2н), дорожка 11: Z17357 (0), дорожка 12: Z17357 (2н), дорожка 13: Z17359 (0), дорожка 14: Z17359 (2н), дорожка 15: Z17360 (0), дорожка 16: Z17360 (2w). Маркер размера молекулы (Mw) представлял собой Novex® Sharp предварительно окрашенный белковый стандарт (216, 160, 110, 80, 60, 50, 40, 30, 20, 15, 10, 3,5 кДа). (Диагональные полосы, видимые на Фиг. 8С, представляют собой артефакт, являющийся отпечатком второго геля, окрашиваемого в том же контейнере).
На Фиг. 9 показаны сенсограммы связывания Z-вариантов, содержащих аминокислотные замены ND на SE в положении 52-53 (черный) и исходные Z-варианты (серый) с аффинностью к той же мишени после 2 недельного анализа стабильности. А: Связывание Z017341 (SEQ ID NO: 28) и Z02891 (SEQ ID NO: 27) с HER2; В: Связывание Z017343 (SEQ ID NO: 31) и Z15805 (SEQ ID NO: 30) с PDGF-Rβ; С: Связывание Z017347 (SEQ ID NO: 34) and Z10130 (SEQ ID NO: 33) с FcRn и D: Связывание Z017351 (SEQ ID NO: 37) и Z09782 (SEQ ID NO: 36) с CAIX. Введенные концентрации каждого Z-варианта были такими же, как описано в Примере 13.
Примеры
В следующих Примерах раскрыты новые полипептиды-варианты Z, демонстрирующие улучшенную стабильность. Здесь свойства Z-вариантных полипептидов, основанных на предыдущих поколениях каркасов, сравнивали с Z-вариантными полипептидами, основанными на каркасе, раскрытом в данном описании изобретения.
Сравнительный пример 1
Анализ стабильности известного С5-связывающего Z-варианта
С5-связывающий Z-вариант, обозначенный как PSI0242 (SEQ ID NO: 1), был приготовлен в 25 мМ NaP/125 мМ NaCl рН 7,0 и подвергнут ускоренному изучению стабильности в течение 2 недель при 37°С. Стабильность определяли по появлению новых вариантов после тестирования стабильности с помощью SDS-PAGE и обращенно-фазовой ВЭЖХ (RPC). В обоих анализах исходный образец и образец, подвергнутый исследованию стабильности, исследовали параллельно. Для SDS-PAGE в каждую лунку загружали 7,5 мкг белка. RPC проводили на Agilent 1100 ВЭЖХ, используя подвижную фазу А, состоящую из 0,1% трифторуксусной кислоты (TFA) в воде, и подвижную фазу В, состоящую из смеси 0,1% TFA/45% МеОН/45% изопропиламина (IPA)/10% воды.
Результаты показывают, что в процессе инкубации образовались новые формы белка, которые были визуализированы в виде полос в SDS-PAGE (Фиг. 2) и как новые пики на хроматограммах обращенно-фазовой ВЭЖХ (RPC) (Фиг. 3). На Фиг. 3 основной пик после инкубации в течение 2 недель соответствует 57% исходного белкового образца.
Положения 1-60 в SEQ ID NO: 1 соответствуют полипептиду Z06175a, ранее раскрытому в WO 2013/126006 как SEQ ID NO: 753.
Пример 2
Анализ стабильности модифицированных С5-связывающих полипептидов и соединений
Модифицированные С5-связывающие полипептиды и соединения были синтезированы и очищены по существу, как описано в WO 2013/126006.
В кратком изложении, ДНК, кодирующие С5-связывающие Z-варианты, были Е. coli. кодон-оптимизированными и синтезированными GeneArt, GmbH. Синтетические гены, представляющие новые С5-связывающие варианты Ζ субклонировали и экспрессировали в Е. coli.
Внутриклеточно экспресированные Z-варианты очищали с помощью обычных хроматографических методов. Гомогенизацию и осветление проводили посредством обработки ультразвуком с последующим центрифугированием и фильтрованием. Анионообменную хроматографию использовали в качестве стадии захвата. Дальнейшую очистку выполняли с помощью хроматографии гидрофобных взаимодействий. Очистку выполняли в кислых условиях (рН 5,5). Доочистку и замену буфера выполняли с помощью гель-фильтрационной хроматографии.
Очищенные белки готовили в 25 мМ NaP/125 мМ NaCl, рН 7,0, и подвергали ускоренному исследованию стабильности в течение 2 недель при 37°С. Стабильность анализировали по появлению новых вариантов после анализа стабильности посредством SDS-PAGE и обращенно-фазовой ВЭЖХ (RPC). В обоих анализах исходный образец и образец, подвергнутый исследованию стабильности, пропускали параллельно. Для SDS-PAGE в каждую лунку загружали 7,5 мкг белка. Пример полученного геля показан на Фиг. 4.
RPC проводили на Agilent 1100 ВЭЖХ, используя подвижную фазу А, состоящую из 0,1% трифторуксусной кислоты (TFA) в воде, и подвижную фазу В, состоящую из смеси 0,1% TFA/45% МеОН/45% изопропиламина (IPA)/10% воды. Пример полученной хроматограммы для SEQ ID NO: 5 показан на Фиг. 5. Результаты анализа стабильности обобщены в Таблице 1.
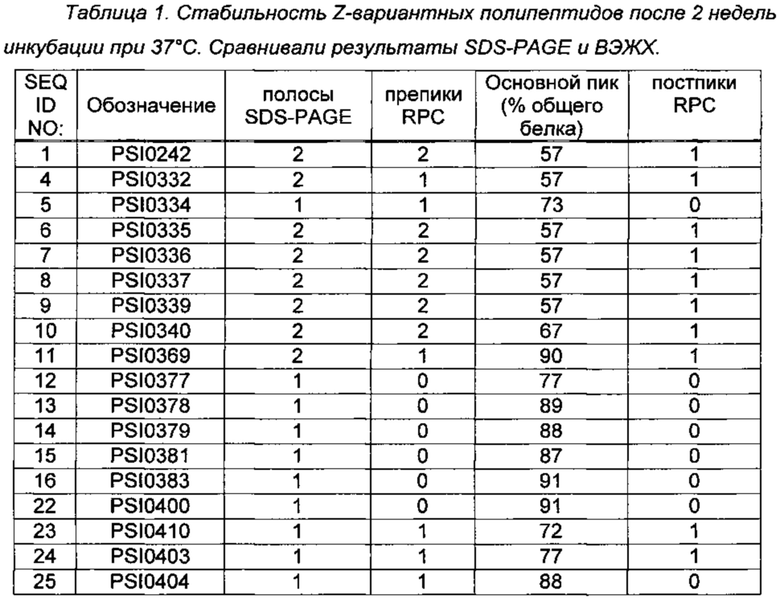
Из Таблицы 1 можно сделать вывод, что некоторые модифицированные С5-связывающие полипептиды или соединения обладают улучшенными свойствами, такими как повышенная стабильность по сравнению с PSI0242. Такие улучшенные С5-связывающие полипептиды или соединения включают PSI0334 (SEQ ID NO: 5), PSI0340 (SEQ ID NO: 10), PSI0369 (SEQ ID NO: 11), PSI0377 (SEQ ID NO: 12), PSI0378 (SEQ ID NO: 13), PSI0379 (SEQ ID NO: 14), PSI0381 (SEQ ID NO: 15), PSI0383 (SEQ ID NO: 16), PSI0400 (SEQ ID NO: 22), PSI0410 (SEQ ID NO: 23), PSI0403 (SEQ ID NO: 24) и PSI0404 (SEQ ID NO: 25). Общим у шести из указанных вариантов (SEQ ID NO: 5, 12, 13, 14, 16 и 22) является то, что аминокислотные остатки в положениях 52-53 были заменены с ND (cf. PSI0242) на SE. В SEQ ID NO: 15 соответствующая замена представляет собой ND на ES. В SEQ ID NO: 24 только аминокислотный остаток в положении 53 был заменен с D на Е, в то время как в SEQ ID NO: 25 аминокислотный остаток в положении 52 был заменен с N на S.
Пример 3
Связывание модифицированных соединений с человеческим С5
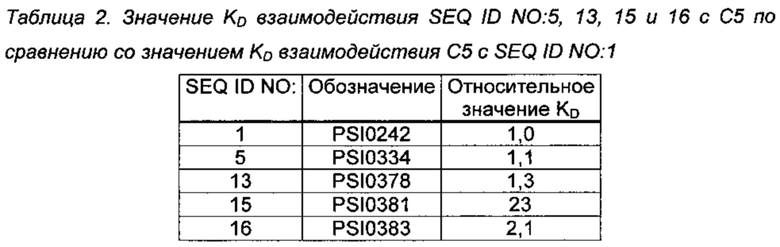
Человеческий сывороточный альбумин иммобилизовывали на тестовые биосенсоры (Dip and Read Biosensors) с реакционноспособным амином 2-го поколения (AR2G) (Pall Life sciences (ForteBio) каталожный номер 18-5092) посредством аминого сочетания. Загружали PSI0242 (SEQ ID NO: 1; 1 мкМ) и модифицированные С5-связывающие соединения (1 мкМ) в буфере для считывания (HBS-EP Buffer [10 мМ HEPES рН 7,4, 150 мМ NaCl, 3 мМ EDTA, 0,005% сурфактант Р20], GE Healthcare, каталожный номер BR100188), каждый на отдельный сенсор с HSA, в течение 120 секунд с последующей регистрацией базисной линии в течение 60 секунд в буфере для считывания перед воздействием человеческого С5 (Quidel, каталожный номер А403) в концентрациях, варьирующихся от 0,79 нМ до 25 нМ, в буфере для считывания с циклом регенерации и записью базисной линии между каждой концентрацией. Условия регенерации для сенсоров представляли собой 10 мМ глицин, рН 2 (три импульса по 30 секунд и подвижный буфер в течение 60 секунд). Из каждой спектрограммы вычитали исходные значения для аналогичной конструкции, содержащей альбумин-связывающий домен. (SEQ ID NO: 2), но без С5-связывающей способности. Данные анализировали согласно модели Ленгмюра 1:1 с использованием ForteBio Analysis 7.1 (программа Pall Life sciences (ForteBio) kinetics).
Относительное значение KD взаимодействия PSI0242 (SEQ ID NO; 1) с C5 показано в Таблице 2. KD PSI0242 (SEQ ID NO: 1) варьируется от 1 до 3 нМ в разных прогонах.
Результаты в Таблице 2 показывают, что С5-связывающие соединения по настоящему изобретению обладают способностью связываться с человеческим С5, сопоставимой с этой способностью у полипептида PSI0242 (SEQ ID NO: 1), раскрытого в WO 2013/126006.
Пример 4
Стабильность химически синтезированного С5-связывающего полипептида
Химически синтезированный PSI0400 (SEQ ID NO: 22) был заказан в ВАСНЕМ AG. Стабильность полипептида оценивали в соответствии с той же методикой, что и в Примере 2. Результаты анализа стабильности показаны в Таблице 3.
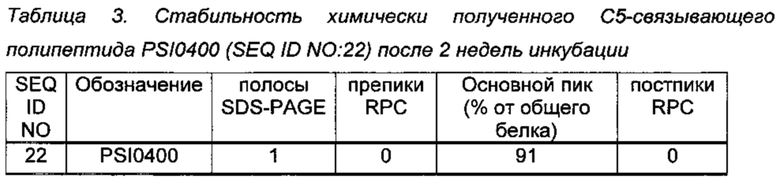
Стабильность PSI0400 была сравнима с тем же полипептидом, полученным в E. coli в Примере 2.
Целостность свертывания PSI0400 (SEQ ID NO: 22) сравнивали с рекомбинантным С5-связывающим полипептидом (PSI0257, SEQ ID NO: 26), полученным в соответствии с методами Примера 2, используя спектр кругового дихроизма (CD) в далекой УФ-области.
CD-спектр регистрировали с помощью спектрополяриметра J-720 CD (Jasco, Japan). Образцы разбавляли до концентрации белка 0,17 мг/мл, используя буфер Pi (5 мМ Na-K-PO4, рН 7,0). Сначала регистрировали CD-спектр буфера Pi, затем регистрировали спектры каждого из образцов и, наконец, еще раз буфер Pi. Если спектры двух буферов совпадали, первый зарегистрированный спектр использовали в качестве спектра буфера. Спектр буфера сглаживали, используя метода Савицкого-Голея с шириной свертки 25. Другие спектры сглаживали в соответствии с тем же методом с шириной свертки 15. Сглаженный спектр буфера затем вычитали из каждого из других сглаженных спектров. Программу CDNN использовали для оценки вторичной структуры белков, полученные оценки представлены в Таблице 4. Результаты показали, что ни две аминокислотные замены в положении 52 и 53, ни получение полипептида посредством химического синтеза не влияют на вторичную структуру химически синтезированного полипептида. Целостность вторичной структуры сравнивали с рекомбинантно полученным PSI0257 (SEQ ID NO: 26).

Пример 5
Связывание модифицированных Z-вариантов и полипептидов с человеческим С5
Аффинность связывания С5-связывающих соединений PSI0242 (SEQ ID NO: 1), PSI0340 (SEQ ID NO: 10), PSI0378 (SEQ ID NO: 13) и PSI0410 (SEQ ID NO: 23), а также С5-связывающего полипептида PSI0400 (SEQ ID NO: 22) с человеческим C5 анализировали с использованием прибора Biacore Т200 (GE Healthcare). Человеческий С5 (Quidel, каталожный номер А403) соединяли с СМ5 сенсорным чипом (900 RU), используя химию аминного сочетания в соответствии с протоколом производителя. Сочетание выполняли путем введения hC5 в концентрации 7,5 мкг/мл в 10 мМ Na-ацетатном буфере, рН 5 (GE Healthcare). Эталонную клетку обрабатывали теми же реагентами без введения человеческого С5. Связывание полипептида С5 и соединений с иммобилизованным hC5 изучали с помощью метода кинетики одного цикла, в котором пять концентраций образца, как правило 25, 12,5, 6,25, 3,12 и 1,56 нМ в буфере HBS-EP вводили один за другим при скорости потока 30 мкл/мин при 25°С в том же цикле без регенерации между введениями. Данные для эталонной клетки вычитали для компенсации изменений рефракционного индекса в массе. В большинстве случаев введение HBS-EP также было включено в качестве контроля, так что сенсограммы сравнивали с двумя контролями. Поверхности регенерировали в буфере HBS-EP. Кинетические константы рассчитывали из сенсограммы, используя модель Ленгмюра 1:1 для анализируемого вещества программного обеспечения Biacore Т200 Evaluation версия 1.0. Полученные значения KD взаимодействий представлены в Таблице 5.
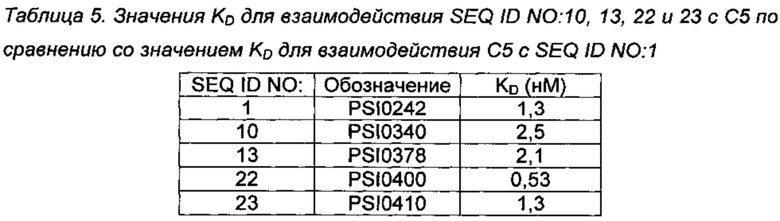
Представленные данные показывают, что аминокислотные замены, повышающие стабильность, не оказывает существенного негативного влияния на способность молекул связываться с С5 и, таким образом, не влияет на их биологическую активность.
Пример 6
Ингибирование гемолиза
Для изучения функционирования классического пути комплемента и его ингибирования С5-связывающими соединениями PSI0378 (SEQ ID NO: 13) и PSI0410 (SEQ ID NO: 23) и С5-связывающим полипептидом PSI0400 (SEQ ID NO: 22), получали бараньи эритроциты из свежей цельной бараньей крови в растворе Олсвера (Шведский национальный институт ветеринарии). Эритроциты затем обрабатывали кроличьей сывороткой против бараньих эритроцитов (Sigma) для получения сенсибилизированных антителами бараньих эритроцитов (ЕА). Весь процесс проводили в асептических условиях. Все другие реагенты были из коммерческих источников.
Анализ in vitro выполняли в 96-луночном (с лунками U-образной формы) микротитрационном планшете посредством последовательного добавления тестируемого белка, сывороточного комплемента и суспензии ЕА. Конечные концентрации всех агентов в общем реакционном объеме 50 мкл на лунку и при рН 7,3-7,4 составляли: 0,15 мМ CaCl2; 0.5 мМ MgCl2; 3 мМ NaN3; 138 мМ NaCl; 0,1% желатина; 1,8 мМ барбитала натрия; 3,1 мМ барбитуровой кислоты; 5 миллионов ЕА; сывороточный белок С5 комплемента в подходящем разведении и С5-связывающее соединение или полипептид в нужной концентрации.
С5-связывающие соединения и полипептид предварительно инкубировали с вышеуказанным сывороточным комплементом в течение 20 минут на льду перед началом реакции путем добавления суспензии ЕА. Гемолитической реакции давали возможность протекать при 37°С при перемешивании в течение 45 мин и затем при необходимости прекращали реакцию добавлением 100 мкл ледяного физиологического раствора, содержащего 0,02% Tween 20. Клетки центрифугировали на дно флакона и верхнюю часть, соответствующую 100 мкл супернатанта, переносили в прозрачный микропланшет, имеющий плоскодонные лунки с половинным объемом. Результаты реакции анализировали по оптической плотности, используя микротитрационный планшетный ридер при длине волны 415 нм. Контрольный образец (PSI0242, SEQ ID NO: 1) и носитель были включены в каждый планшет для определения значений для не ингибированных и полностью ингибированных реакций, соответственно. Эти величины использовали для расчета % ингибирования обусловленного комлементом гемолиза при любой данной концентрации образца. Эффективность ингибирования (значения IC 50) протестированных С5-связывающих соединений и полипептида определяли путем применения того же анализа в присутствии контролируемой концентрации человеческого С5, добавленного к обедненной по С5 сыворотке. Для сильнодействующих ингибиторов (от низкой наномолярной до субнаномолярной концентрации) конечную концентрацию С5 в реакционной смеси контролировали при 0,1 нМ, которую была при необходимости установлена посредством использования истощенной или дефицитной по С5 сыворотки. Результаты представлены ниже в Таблице 6.
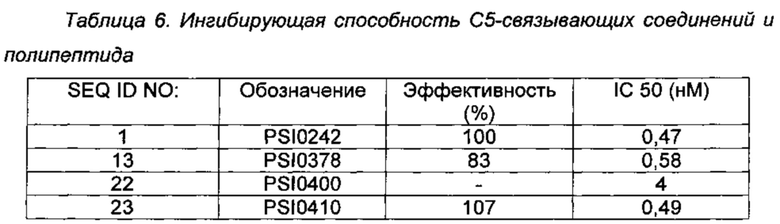
Результаты анализа гемолиза показывают, что улучшенные С5-связывающие соединения PSI0378 (SEQ ID NO: 13) и PSI0410 (SEQ ID NO: 23) не отличаются существенно от эталонного соединения PSI0242 (SEQ ID NO: 1) в отношении функции. С5-связывающий полипептид PSI0400 (SEQ ID NO: 22) является функциональным в этом анализе и поскольку он не содержит альбумин-связывающего домена, результаты не могут быть непосредственно сопоставлены с результатами эталонного соединения.
Пример 7
Связывание с человеческим альбумином
Для оценки аффинности С5-связывающих соединений к альбумину, использовали метод ELISA с человеческим альбумином, используя рекомбинантный человеческий альбумин в качестве покрытия (Novozymes) и имеющиеся в продаже антитела из Affibody АВ (первичные) и DakoCytomation (обнаруживающие). Стандарт для метода, полученный из PSI0242 (SEQ ID NO: 1) и содержащий С5-связывающий полипептид и альбумин-связывающий домен стрептококкового белка G, использовали для количественного определения образцов.
96-луночный микропланшет покрывали рекомбинантным человеческим альбумином. Затем планшет промывали фосфатно-солевым буфером, содержащим 0,05% Tween 20 (PBST) и блокировали в течение 1-2 часов 1%-ным казеином в PBS. После промывания планшета к нему добавляли стандарт, контроли метода, контрольный образец и тестируемые образцы. После инкубации в течение 2 часов несвязанное вещество удаляли промыванием. Анти-Affibody® IgG козы (Affibody АВ, каталожный номер 20.1000.01.0005) добавляли в лунки и планшет инкубировали в течение 1,5 часов, чтобы обеспечить связывание со связанными С5-связывающими соединениями. После промывания IgG HRP (DakoCytomation) кролика против козы оставляли связываться с козлиными антителами в течение 1 часа. После окончательной промывки определяли количество связанного HRP путем добавления ТМВ (3,3',5,5'-тетраметилбензидин) субстрата, который под воздействием фермента превращался в синий продукт. Добавление 1 М соляной кислоты через 30 минут останавливало реакцию, и цвет содержимого лунки менялся с синего на желтый. Поглощение при 450 нм измеряли фотометрически, используя поглощение при 650 нм в качестве эталонной длины волны. Интенсивность окраски была пропорциональна количеству PSI0242 (SEQ ID NO: 1) и концентрации образца определяли с помощью стандартной кривой.
Было показано, что С5-связывающие соединения, содержащие производное альбумин-связывающего домена из стрептококкового белка G (ABD), способны связываться с человеческим альбумином. Данные представлены в Таблице 7.

Интерпретация анализа заключается в том, что оба исследуемых С5-связывающих полипептида с улучшенной стабильностью сохраняют свою способность связываться с человеческим сывороточным альбумином.
Пример 8
Трехмесячный анализ стабильности С5-связывающих Z-вариантов и полипептидов
С5-связывающие варианты и полипептиды, которые показывали улучшенную стабильность по сравнению с PSI0242 в 2-недельном анализе стабильности при 37°С (Пример 2) подвергали более длительному 3-месячному анализу стабильности при 37°С. Параметры анализа стабильности и анализа посредством RPC (хроматография на обращенной фазе) были такими, как описано в Примере 2. Оценку стабильности выполняли путем измерения основного пика хроматограммы и расчета соответствующего процента от общего содержания белка. Данные из Примера 2 включены в Таблицу 8 ниже для упрощения интерпретации.
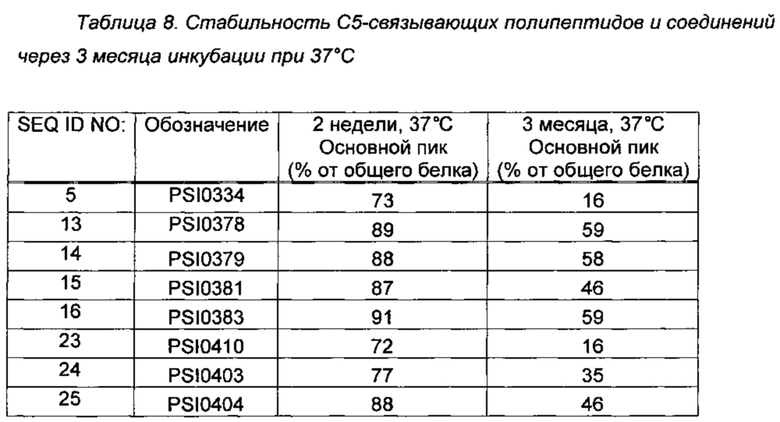
С5-связывающие соединения, содержащие аминокислотные замены ND на SE в положениях 52-53 (SEQ ID NO: 13, 14 и 16) по сравнению с PSI0242 показали более высокую долю белка в исходной форме через 3 месяца при 37°С, чем PSI0242 (SEQ ID NO: 1) через 2 недели при тех же условиях (см. Таблица 1). Другие исследованные соединения также демонстрировали повышенную стабильность по сравнению с PSI0242.
Пример 9
Получение, исследование стабильности и оценка связывания полипептидов с модифицированным каркасом со специфичностью к разным мишеням
Получение полипептидов с модифицированным каркасом со специфичностью к разным мишеням: Варианты полипептидов, содержащие новый каркас, описанный в данном описании изобретения, получают, взяв Z-вариантные полипептиды со специфичностью к разным мишеням и осуществив сайт-направленный мутагенез в выбранных положениях в каркасе. Новые молекулы альтернативно могут быть получены посредством химического синтеза всей молекулы или путем использования других методов молекулярной биологии, известных специалисту в данной области, для присоединения связывающего мотива Z-вариантного полипептида к новому каркасу.
Сравнительное исследование стабильности полипептидов с модифицированным каркасом со специфичностью к разным мишеням: Стабильность каждого нового полипептида, созданного, как описано выше, сравнивали со стабильностью исходного полипептида или другого сравнимого полипептида. Полипептиды подвергают воздействию различных условий, таких как приготовление в виде композиции в [25 мМ NaP, 125 мМ NaCl, рН 7,0] и инкубация при 37°С в течение 2 недель, как описано в Примере 2, и/или в течение 3 месяцев, как описано в Примере 8. Стабильность оценивают путем анализа появления новых вариантов, выполняя анализы SDS-PAGE и RPC, как описано в Примере 2.
Ожидается, что полипептиды с введенными модификациями в положения каркаса покажут улучшенную стабильность, аналогично результатам, представленными в Примере 2 и Примере 12.
Оценка связывания полипептидов с модифицированным каркасом:
Полипептиды, которые показали улучшенные свойства стабильности, дополнительно оценивали с точки зрения сохранения способности связываться с их мишенью после введения изменений в каркас. Изучение связывания проводили на биосенсорном приборе или любом другом приборе, известном специалисту в данной области, и измеряли взаимодействие между двумя или более молекулами. Например, молекулу-мишень или ее фрагмент иммобилизовывали на сенсорном чипе прибора и над чипом пропускали образец, содержащий тестируемый полипептид. Альтернативно, тестируемый полипептид иммобилизуют на сенсорном чипе прибора и образец, содержащий заданную мишень или ее фрагмент, пропускают над чипом. Аффинность связывания может быть протестирована в эксперименте, в котором образцы полипептида захватывают антителами, покрывающими ELISA планшеты, затем добавляют биотинилированную заданную мишень или ее фрагмент, а затем конъюгированный HRP стрептавидин. Добавляют субстрат ТМВ и измеряют поглощение при 450 нм, используя многолуночного планшетный ридер, такой как Victor3 (Perkin Elmer). Если необходимо количественное измерение, например, для определения значения ЕС50 (половина максимальной эффективной концентрации) взаимодействия, также можно использовать ELISA. Реакцию полипептида на серийное разведение заданной мишени или ее фрагмента измеряют, используя ELISA, как описано выше. Результаты, полученные в таких экспериментах, и значения ЕС50 могут быть рассчитаны из результатов с использованием, например, GraphPad Prism 5 и нелинейной регрессии. Если полипептид содержит альбумин-связывающий домен, влияние на связывание альбумина будет оцениваться аналогично описанному в Примере 3 или как описано в Примере 7.
Полипептиды, имеющие мутации каркаса, описанные в данном описании изобретения и, кроме того, аналогичную или улучшенную способность связывания со своей мишенью, считаются лучшими кандидатами для дальнейшей разработки, например, биофармацевтических продуктов.
Пример 10
Образование полипептидов с модифицированным каркасом и со специфичностью к четырем разным мишеням Варианты полипептидов, содержащих новый каркас, описанный в данном описании изобретения, были получены путем взятия Z-вариантных полипептидов со специфичностью к разным мишеням и выполнения сайт-направленного мутагенеза в выбранных положениях каркаса. Аминокислотные замены в положениях каркаса в полипептидных вариантах: Z02891 (SEQ ID NO: 27), нацеленном на рецептор 2 человеческого эпидермального фактора роста (HER2); Z15805 (SEQ ID NO: 30), нацеленном на бета-рецептор тромбоцитарного фактора роста (PDGF-RB); Z10103 (SEQ ID NO: 33), нацеленный на неонатальный Fc-рецептор (FcRn); и Z09782 (SEQ ID NO: 36), нацеленный на карбоангидразу IX (CAIX), указаны в Таблице 9.
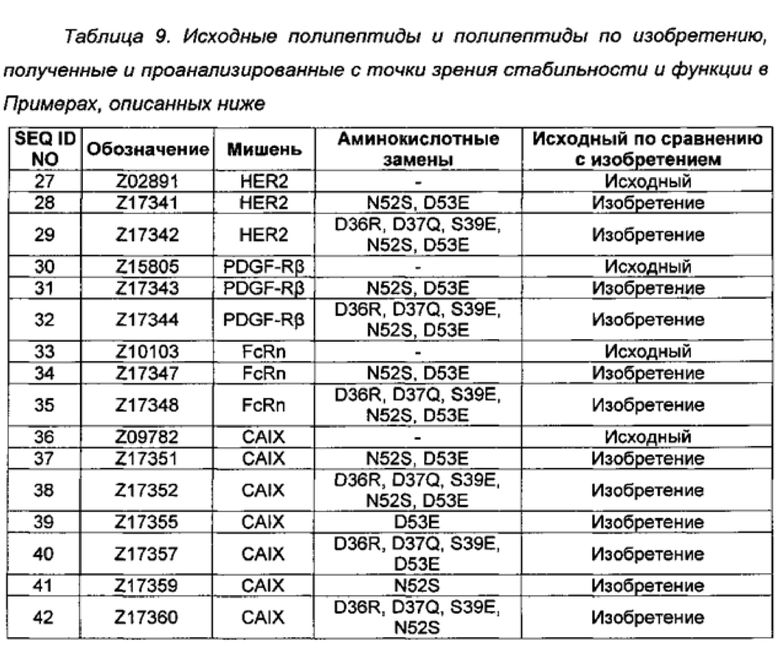
Все варианты клонировали с N-концевым 6×Гистидиновым тегом (His6) и получали конструкции, кодирующие полипептиды в формате MGSSHHHHHHLQ-[Z#####]. Мутации вводили в плазмиды полипептидов по изобретению, используя пары перекрывающихся олигонуклеотидных праймеров, кодирующих нужные аминокислотные замены и применяя общепринятие методы молекулярной биологии. Точные плазмидные последовательности подтверждали посредством ДНК-секвенирования.
Клетки Е. coli (штамм Т7Е2) (GeneBridge) трансформировали плазмидами, содержащими фрагменты генов, кодирующих исходные полипептиды и полипептиды по изобретению. Клетки культивировали при 37°С в среде TSB-YE с добавлением 50 мкг/мл канамицина, и затем индуцировали экспрессию белка путем добавления IPTG (изопропилтиогалактозид). Осажденные клетки разрушали, используя гомогенизатор FastPrep®-24 (Nordic Biolabs), и дебрис клеток удаляли центрифугированием. Каждый супернатант, содержащий Z-вариант в виде His6-меченного белка, очищали с помощью аффинной хроматографии с иммобилизованными ионами металла (IMAC), используя колонки His GraviTrap™ (GE Healthcare) в соответствии с инструкциями производителя. В очищенных Z-вариантах буфер меняли на фосфатно-солевой буферный раствор (PBS; 1,47 мМ KH2PO4, 8,1 мМ Na2HPO4, 137 мМ NaCl, 2,68 мМ KCl, рН 7,4), используя колонки для обессоливания PD-10 (GE Healthcare). Точную идентификацию каждого полипептида подтверждали посредством SDS-PAGE и ВЭЖХ-MS.
Пример 11
Спектроскопический анализ кругового дихроизма полипептидов с модифицированным каркасом
Анализ кругового дихроизма (CD) выполняли, чтобы определить температуры плавления (Tm) и оценить возможные изменения во вторичной структуре полипептидов по изобретению в результате аминокислотных замен.
Очищенные His6-меченные Z-варианты разбавляли до 0,5 мг/мл в PBS. Для каждого разведенного Z-варианта регистрировали спектр CD при 250-195 нм и при 20°С. Для определения Tm выполняли измерение переменной температуры (VTM). В ходе VTM измеряли поглощение при 221 нм, при этом температуру повышали от 20 до 90°С, с изменение температуры 5°С/мин. После VTM регистрировали второй спектр CD при 250-195 нм и при 20°С. Измерения CD выполняли на спектрополяриметре Jasco J-810 (Jasco Scandinavia АВ), используя кювету с длиной оптического пути 1 мм.
Tm каждого соответствующего полипептида, определенная из средней точки перехода на графике сигнала CD в зависимости от температуры, показана в Таблице 10. Все мутированные полипептиды продемонстрировали сохранение альфаспиральной структуры и обратимо или почти обратимо сворачивались даже после нагревания до 90°С. Выбранный набор спектров CD показан на Фиг. 7.

Пример 12
Сравнительное исследование стабильности полипептидов с модифицированным каркасом со специфичностью к четырем разным мишеням
Для каждого нового полипептида, созданного, как описано в Примере 10, стабильность сравнивали со стабильностью исходного полипептида. Полипептиды, приготовленные в PBS, рН 7,4, разбавляли до 1 мг/мл, и 200 мкл аликвоты инкубировали при 37°С в течение 2 недель. Образцы, собранные до и после анализа стабильности, анализировали с помощью SDS-PAGE, используя 10% Bis-Tris NuPAGE гели (Invitrogen) и нанося 5 мкг белка в каждую лунку. Полученные окрашенные Кумасси синим гели показаны на Фиг. 8. Стабильность оценивали по появлению новых вариантов после инкубации при повышенной температуре и измененные варианты сравнивали с соответствующим исходным полипептидом.
Все полипептиды с модификациями, введенными в положения каркаса, представленные в Таблице 9, показали улучшенную стабильность по сравнению с соответствующим исходным полипептидом. В образцах исходных полипептидов вторая полоса была видна на геле чуть выше основной полосы. Соответствующая вторая полоса не была видна в образцах полипептидов по изобретению с заменой D53E и/или N52S. Это аналогично результатам, представленным в Примерах 2 и 4. Таким образом, стабилизирующий эффект, наблюдаемый для мутаций каркаса по изобретению, по-видимому, представляет собой общий эффект независимо от специфичности к мишени Z-варианта или полипептида, содержащего указанный Z-вариант. Полипептиды с заменами D53E и/или N52S, единственными или в комбинации с заменами D36R, D37Q и S39E, показали аналогичные профили на геле SDS-PAGE. Замена D53E, единственная или в комбинации с заменами D36R, D37Q и S39E, по-видимому, уменьшает количество видов с альтернативной конформацией, наблюдаемой как второй полоса на геле SDS-PAGE, но не может полностью предотвратить образование этих видов.
Пример 13
Оценка связывания полипептидов с модифицированным каркасом
Набор полипептидов, демонстрирующих свойства улучшенной стабильности в Примере 12, дополнительно оценивали с точки зрения сохранения свойств связывания с их мишенями после введения изменений в каркас, а также после проведения анализа стабильности, то есть инкубирования при 37°С в течение 2 недель. Сравнительные кинетические константы (kon и koff) и аффинности (KD) определяли, используя прибор Biacore 2000. Белки-мишени человеческий HER2-Fc (R&D Systems, каталожный номер 1129-ER-050), человеческий PDGF-Rβ (R&D Systems, каталожный номер 385-PR-100/CF), человеческий FcRn (Biorbyt, каталожный номер orb 84388) и человеческий CAIX (R&D Systems, каталожный номер 2188-СА) соответственно, были иммобилизованы на поверхности из слоя карбоксилированного декстрана чипов СМ5 (GE Healthcare). Иммобилизацию выполняли, используя химию аминного сочетания в соответствии с протоколом производителя и используя HBS-EP в качестве электродного буфера. Одну поверхность проточной ячейки на чипе активировали и деактивировали для использования в качестве контроля во время добавлений анализируемого вещества. Уровень иммобилизации целевого белка на соответствующей поверхности составлял приблизительно 850 RU для HER2, 2200 RU для PDGF-Rβ, 750 для FcRn и 580 RU для CAIX.
HBS-EP (HER2, PDGF-Rβ, CAIX) или буфер рН 6,0 Na2HPO4/лимонная кислота (126 мМ Na2HPO4, 37 мМ лимонная кислота) (FcRn) использовали в качестве электродного буфера и скорость потока составляла 30 мкл/мин в экспериментах по связыванию, выполняемых при 25°С, как описано ниже.
Z-варианты Z02891 (SEQ ID NO: 27), Z17341 (SEQ ID NO: 28) и Z17342 (SEQ ID NO: 29), нацеленные на HER2, разбавляли в электродном буфере до конечных концентраций 3,33, 10, 30 и 90 нМ и вводили в течение 5 минут, затем следовали 30 минут диссоциации в электродном буфере. Регенерацию посредством четырех импульсов со сменой 10 мМ HCl на 10 мМ NaOH, после чего следовало 5-минутное уравновешивание в электродном буфере, применяли после каждого введения анализируемого вещества.
Z-варианты Z15805 (SEQ ID NO: 30), Z17343 (SEQ ID NO: 31) и Z17344 (SEQ ID NO: 32), нацеленные на PDGF-Rβ, разбавляли в электродном буфере до конечных концентраций 6,67, 20, 60 и 180 нМ и вводили в течение 5 минут, затем следовали 20 минут диссоциации в электродном буфере. Регенерацию посредством трех импульсов 10 мМ NaOH с последующим 5-минутным уравновешиванием в электродном буфере применяли после каждого введения анализируемого вещества.
Z-варианты Z10103 (SEQ ID NO: 33), Z17347 (SEQ ID NO: 34) и Z17348 (SEQ ID NO: 35), нацеленные на FcRn, разбавляли в электродном буфере до конечных концентраций 3,33, 10 и 30 нМ и вводили в течение 3 минут, затем следовали 15 минут диссоциации в электродном буфере. Регенерацию посредством трех импульсов HBS-EP с последующим 10-минутным уравновешиванием в электродном буфере применяли после каждого введения анализируемого вещества.
Z-варианты Z09782 (SEQ ID NO: 36), Z17351 (SEQ ID NO: 37), Z17355 (SEQ ID NO: 39) и Z17359 (SEQ ID NO: 41), нацеленные на CAIX, разводили в электродном буфере до конечных концентраций 30, 90 и 270 нМ и вводили в течение 5 минут, затем следовали 15 минут диссоциации в электродном буфере. Регенерацию посредством трех импульсов 10 мМ глицина-HCl, рН 3,0, с последующим 5-минутным уравновешиванием в электродном буфере применяли после каждого введения анализируемого вещества.
Кинетические константы рассчитывали из сенсограмм, используя модель Ленгмюра 1:1 (HER2, FcRn, CAIX) или 1:1 модель связывания с переносом массы (PDGF-Rβ) программного обеспечения BiaEvaluation 4.1 (GE Healthcare). Кривые для контрольной поверхности вычитали из кривых для поверхностей с лигандом и данные для циклов с буфером вычитали из данных для циклов анализируемых образцов, чтобы скорректировать любой сдвиг сигнала.
Сравнительные кинетические константы для Z-вариантов, связывающихся со своей целевой молекулой, показаны в Таблице 11 и сенсограммы подгруппы анализируемых взаимодействий показаны на Фиг. 9. Данные показывают, что на аффинность лишь незначительно влияют замены ND на SE в положении 52-53, а для пары вариантов, Z17341 (SEQ ID NO: 28) и Z17343 (SEQ ID NO: 31), аффинность даже незначительно улучшается. Комбинация замен ND в SE в положениях 52-53 с заменами D36R, D37Q и S39E, например в Z17342 (SEQ ID NO: 29), Z17344 (SEQ ID NO: 32) и Z17348 (SEQ ID NO: 35), оказывает более отрицательное влияние на аффинность, главным образом из-за более высоких скоростей диссоциации, но тем не менее функциональные связующие вещества были получены с KD в диапазоне 10-9 М. Оцененные варианты также сохраняли способности к связыванию после 2 недель инкубации при 37°С.


ДЕТАЛИЗИРОВАННЫЙ ПЕРЕЧЕНЬ ВОПЛОЩЕНИЙ
1. Полипептид, содержащий аминокислотную последовательность, выбранную из:
1) EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q,
где каждый из Х2, Х3, Х4, Х6, Х7, Х10, Х11, Х17, X18, Х20, Х21, Х25 и Х28 независимо соответствует любому аминокислотному остатку; и
где, независимо друг от друга,
X16 выбран из N и Т;
Х26 выбран из K и S;
Х29Х30РХ32 выбран из DDPS и RQPE;
Х35 выбран из А и S;
Х36 выбран из Е и N;
Х39 выбран из А, С и S;
Х45 выбран из Е, N и S;
Х46 выбран из D, Е и S, при условии, что Х46 не является D, когда Х45 представляет собой N;
Х47 выбран из А и S; и
2) аминокислотной последовательности, которая обладает по меньшей мере 91%-ной идентичностью с последовательностью определенной в (1), при условии, что Х46 не является D, когда Х45 представляет собой N.
2. Полипептид по п. 1, где Х16 представляет собой Т.
3. Полипептид по п. 1 или 2, где Х26 представляет собой K.
4. Полипептид по любому из пп. 1-3, где Х29Х30РХ32 представляет собой DDPS.
5. Полипептид по пп. 1-3, где Х29Х30РХ32 представляет собой RQPE.
6. Полипептид по любому из пп. 1-5, где Х35 представляет собой S.
7. Полипептид по любому из пп. 1-6, где Х36 представляет собой Е.
8. Полипептид по любому из пп. 1-7, где Х39 представляет собой S.
9. Полипептид по любому из пп. 1-8, где Х45 выбран из Ε и S.
10. Полипептид по п. 9, где Х45 представляет собой Е.
11. Полипептид по п. 9, где Х45 представляет собой S.
12. Полипептид по любому из пп. 1-11, где Х46 выбран из Ε и S.
13. Полипептид по п. 12, где Х46 представляет собой Е.
14. Полипептид по п. 12, где Х46 представляет собой S.
15. Полипептид по п. 12, где Х46 представляет собой D.
16. Полипептид по любому из пп. 1-15, при условии, что Х46 не является D или Е, когда Х45 представляет собой N.
17. Полипептид по любому из пп. 1-16, где Х45Х46 выбран из ЕЕ, ES, SE и SS.
18. Полипептид по п. 17, где Х45Х46 выбран из ES и SE.
19. Полипептид по п. 18, где Х45Х46 представляет собой ES.
20. Полипептид по п. 18, где Х45Х46 представляет собой SE.
21. Полипептид по п. 18, где Х45Х46 представляет собой SD.
22. Полипептид по любому из пп. 1-21, где Х47 представляет собой S.
23. Полипептид по любому из пп. 1-22, содержащий дополнительные аминокислотные остатки.
24. Полипептид по п. 23, содержащий дополнительные аминокислотные остатки на С-конце указанного полипептида.
25. Полипептид по п. 24, где дополнительные аминокислотные остатки на С-конце указанного полипептида содержат АР.
26. Полипептид по п. 23, содержащий дополнительные аминокислотные остатки на N-конце указанного полипептида.
27. Полипептид по п. 26, где дополнительные аминокислотные остатки на N-конце указанного полипептида содержат AEAKYAK.
28. Полипептид по любому из пп. 23-27, где указанные дополнительные аминокислотные остатки добавлены с целью связывания, получения, очистки, стабилизации, сочетания или обнаружения полипептида.
29. Полипептид по любому из пп. 23-28, где указанные дополнительные аминокислотные остатки составляют один или более полипептидных доменов.
30. Полипептид по п. 29, где указанные один или более полипептидных доменов обладают функцией, выбранной из группы, включающей функцию связывания, ферментативную функцию, функцию хелатирования иона металла и флуоресцентную функцию или их сочетания.
31. Полипептид по любому из пп. 1-28, который содержит аминокислотную последовательность, выбранную из:
YAK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q АР; и
FNK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q АР,
где каждый Xy является таким, как он определен в любом из пп. 1-22.
32. Полипептид по п. 31, который содержит аминокислотную последовательность, выбранную из:
ADNNFNK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q APK;
ADNKFNK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q APK;
VDNKFNK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q APK;
VDAKYAK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q APK; и
AEAKYAK EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q APK;
где каждый Xy является таким, как он определен в любом из пп. 1-22.
33. Полипептид по любому из пп. 1-32, обладающий аффинностью к заданной мишени, где указанная мишень возможно выбрана из группы, состоящей из ABD, HER2, TNFα, EGFR, IGF1R, IgG, PDGFRβ, HER3, С5, FcRn, CAIX, амилоида β, CD4, IL8, IL6 и инсулина.
34. Слитый полипептид, содержащий полипептид по любому из пп. 1-33 в качестве фрагмента.
35. Полипептид или слитый полипептид по любому из пп. 1-34, дополнительно содержащий метку.
36. Полипептид или слитый полипептид по любому из пп. 1-35, дополнительно содержащий терапевтический агент.
37. Применение полипептида или слитого полипептида по любому из пп. 1-36 в качестве обнаруживающего реагента, реагента захвата или реагента разделения.
38. Полипептид или слитый полипептид по любому из пп. 1-36 для применения в терапии.
39. Полипептид или слитый полипептид по любому из пп. 1-36 для применения в качестве диагностического агента.
40. Полинуклеотид, кодирующий полипептид или слитый полипептид по любому из пп. 1-34.
41. Группа полипептидов-вариантов на основе общего каркаса, где каждый полипептид в группе содержит аминокислотную последовательность, выбранную из:
1) EX2X3X4AX6X7EIX10 X11LPNLX16X17X18QX20 X21AFIX25X26LX28X29X30 PX32QSX35X36LLX39E AKKLX45X46X47Q,
где каждый из Х2, Х3, Х4, Х6, Х7, Х10, Х11, Х17, Х18, Х20, Х21, Х25 и Х28 независимо соответствует любому аминокислотному остатку; и
где, независимо друг от друга,
Χ16 выбран из N и Т;
Х26 выбран из K и S;
Х29Х30РХ32 выбран из DDPS и RQPE;
Х35 выбран из А и S;
Х36 выбран из Ε и Ν;
Х39 выбран из А, С и S;
Х45 выбран из Ε, N и S;
Х46 выбран из D, Ε и S, при условии, что Х46 не является D, когда Х45 представляет собой Ν;
Х47 выбран из А и S; и
2) аминокислотной последовательности, которая обладает по меньшей мере 91%-ной идентичностью с последовательностью, определенной в (1), при условии, что Х46 не является D, когда Х45 представляет собой N.
42. Группа по п. 41, которая содержит по меньшей мере 1×104 уникальных полипептидных молекул.
43. Группа по п. 42, которая содержит по меньшей мере 1×106 уникальных полипептидных молекул.
44. Группа по п. 43, которая содержит по меньшей мере 1×108 уникальных полипептидных молекул.
45. Группа по п. 44, которая содержит по меньшей мере 1×1010 уникальных полипептидных молекул.
46. Группа по п. 45, которая содержит по меньшей мере 1×1012 уникальных полипептидных молекул.
47. Группа по п. 46, которая содержит по меньшей мере 1×1014 уникальных полипептидных молекул.
48. Группа полинуклеотидов, отличающаяся тем, что каждый его представитель кодирует представителя группы полипептидов по любому из пп. 41-47.
49. Комбинация группы полипептидов по любому из пп. 41-47 с группой полинуклеотидов по п. 48, где каждый представитель указанной группы полипептидов физически или пространственно ассоциирован с полинуклеотидом, кодирующим этого представителя, посредством средства взаимосвязи генотип-фенотип.
50. Комбинация по п. 49, где указанное средство взаимосвязи генотип-фенотип включает систему фагового дисплея.
51. Комбинация по п. 49, где указанное средство взаимосвязи генотип-фенотип включает систему выбора на основе дисплея на клеточной поверхности.
52. Комбинация по п. 51, где указанная система дисплея клеточной поверхности содержит прокариотические клетки.
53. Комбинация по п. 52, где указанные прокариотические клетки представляют собой грамположительные клетки.
54. Комбинация по п. 51, где указанная система дисплея на клеточной поверхности включает эукариотические клетки.
55. Комбинация по п. 54, где указанные эукариотические клетки представляют собой клетки дрожжей.
56. Комбинация по п. 49, где указанное средство взаимосвязи генотип-фенотип включают бесклеточную дисплейную систему.
57. Комбинация по п. 56, где указанная бесклеточная дисплейная система содержит рибосомную дисплейную систему.
58. Комбинация по п. 56, где указанная бесклеточная дисплейная система включает дисплейную систему in vitro компартментализации.
59. Комбинация по п. 56, где указанная бесклеточная дисплейная система включает систему cis дисплея.
60. Комбинация по п. 56, где бесклеточная дисплейная система включает микрогранулированную дисплейную систему.
61. Комбинация по п. 49, где указанное средство взаимосвязи генотип-фенотип включает недисплейную систему.
62. Комбинация по п. 61, где указанная недисплейная система представляет комплементационный анализ фрагмента белка.
63. Способ выбора искомого полипептида, обладающего аффинностью к заданной мишени, из группы полипептидов, включающий стадии:
(а) предоставления группы полипептидов по любому из пп. 41-47;
(б) приведения этой группы полипептидов в контакт с заданной мишенью в условиях, которые обеспечивают возможность специфического взаимодействия между мишенью и по меньшей мере одним искомым полипептидом, обладающим аффинностью к этой мишени; и
(в) выбор, на основе указанного специфического взаимодействия, по меньшей мере одного искомого полипептида из остальной группы полипептидов.
64. Способ по п. 63, где стадия (а) включает подготовительные стадии предоставления группы полинуклеотидов по п. 48 и экспрессию указанной группы полинуклеотидов с получением указанной группы полипептидов.
65. Способ по п. 64, где каждый представитель указанной группы полипептидов физически или пространственно ассоциирован с полинуклеотидом, кодирующим этот представитель, посредством средства взаимосвязи генотип-фенотип.
66. Способ по п. 65, где указанное средство взаимосвязи генотип-фенотип является таким, как определено в любом из пп. 50-62.
67. Способ выделения полинуклеотида, кодирующего искомый полипептид, обладающий аффинностью к заданной мишени, включающий стадии:
- выбора указанного искомого полипептида и кодирующего его полинуклеотида из группы полипептидов с использованием способа по п. 63; и
- выделение таким образом отделенного полинуклеотида, кодирующего искомый полипептид.
68. Способ идентификации искомого полипептида, обладающего аффинностью к заданной мишени, включающий стадии:
- выделения полинуклеотида, кодирующего указанный искомый полипептид, с использованием способа по п. 67; и
- секвенирование полинуклеотида для установления посредством дедукции аминокислотной последовательности указанного искомого полипептида.
69. Способ выделения и идентификации искомого полипептида, обладающего аффинностью к заданной мишени, из группы полипептидов, включающий стадии:
(а) синтеза каждого представителя группы полипептидов по любому из пп. 41-47 на отдельном носителе или грануле;
(б) выбора или обогащения носителей или гранул на основании взаимодействия полипептида с заданной мишенью; и
(в) идентификации полипептида посредством метода характеристики белка.
70. Способ по п. 69, где метод характеристики белка, использованный на стадии (в), представляет собой масс-спектрометрический анализ.
71. Способ получения искомого полипептида, обладающего аффинностью к заданной мишени, включающий стадии:
- выделения и идентификации искомого полипептида с использованием способа по п. 68 или выбора и идентификации искомого полипептида с использованием способа по любому из пп. 69 и 70; и
- получение указанного искомого полипептида.
72. Способ по п. 71, где указанное получение выполняют, используя химический синтез искомого полипептида de novo.
73. Способ по п. 71, где указанное получение выполняют, используя рекомбинантную экспрессию полинуклеотида, кодирующего искомый полипептид.
74. Способ получения искомого полипептида, обладающего аффинностью к заданной мишени, включающий стадии:
(а1) выделения полинуклеотида, кодирующего указанный искомый полипептид, используя способ по п. 68; или
(а2) обратная трансляция полипептида, идентифицированного с использованием способа выбора и идентификации по любому из пп. 69 и 70; и
(б) после (а1) или (а2), экспрессия выделенного таким образом полинуклеотида с получением указанного искомого полипептида.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> AFFIBODY AB
<120> C5-связывающие полипептиды
<130> 21071642
<150> EP13182022.7
<151> 2013-08-28
<160> 54
<170> PatentIn version 3.5
<210> 1
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 1
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys Val Asp Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 2
<211> 46
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный альбумин-связывающий полипептид
<400> 2
Leu Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly
1 5 10 15
Val Ser Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu
20 25 30
Gly Val Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
35 40 45
<210> 3
<211> 1676
<212> PRT
<213> HOMO SAPIENS
<400> 3
Met Gly Leu Leu Gly Ile Leu Cys Phe Leu Ile Phe Leu Gly Lys Thr
1 5 10 15
Trp Gly Gln Glu Gln Thr Tyr Val Ile Ser Ala Pro Lys Ile Phe Arg
20 25 30
Val Gly Ala Ser Glu Asn Ile Val Ile Gln Val Tyr Gly Tyr Thr Glu
35 40 45
Ala Phe Asp Ala Thr Ile Ser Ile Lys Ser Tyr Pro Asp Lys Lys Phe
50 55 60
Ser Tyr Ser Ser Gly His Val His Leu Ser Ser Glu Asn Lys Phe Gln
65 70 75 80
Asn Ser Ala Ile Leu Thr Ile Gln Pro Lys Gln Leu Pro Gly Gly Gln
85 90 95
Asn Pro Val Ser Tyr Val Tyr Leu Glu Val Val Ser Lys His Phe Ser
100 105 110
Lys Ser Lys Arg Met Pro Ile Thr Tyr Asp Asn Gly Phe Leu Phe Ile
115 120 125
His Thr Asp Lys Pro Val Tyr Thr Pro Asp Gln Ser Val Lys Val Arg
130 135 140
Val Tyr Ser Leu Asn Asp Asp Leu Lys Pro Ala Lys Arg Glu Thr Val
145 150 155 160
Leu Thr Phe Ile Asp Pro Glu Gly Ser Glu Val Asp Met Val Glu Glu
165 170 175
Ile Asp His Ile Gly Ile Ile Ser Phe Pro Asp Phe Lys Ile Pro Ser
180 185 190
Asn Pro Arg Tyr Gly Met Trp Thr Ile Lys Ala Lys Tyr Lys Glu Asp
195 200 205
Phe Ser Thr Thr Gly Thr Ala Tyr Phe Glu Val Lys Glu Tyr Val Leu
210 215 220
Pro His Phe Ser Val Ser Ile Glu Pro Glu Tyr Asn Phe Ile Gly Tyr
225 230 235 240
Lys Asn Phe Lys Asn Phe Glu Ile Thr Ile Lys Ala Arg Tyr Phe Tyr
245 250 255
Asn Lys Val Val Thr Glu Ala Asp Val Tyr Ile Thr Phe Gly Ile Arg
260 265 270
Glu Asp Leu Lys Asp Asp Gln Lys Glu Met Met Gln Thr Ala Met Gln
275 280 285
Asn Thr Met Leu Ile Asn Gly Ile Ala Gln Val Thr Phe Asp Ser Glu
290 295 300
Thr Ala Val Lys Glu Leu Ser Tyr Tyr Ser Leu Glu Asp Leu Asn Asn
305 310 315 320
Lys Tyr Leu Tyr Ile Ala Val Thr Val Ile Glu Ser Thr Gly Gly Phe
325 330 335
Ser Glu Glu Ala Glu Ile Pro Gly Ile Lys Tyr Val Leu Ser Pro Tyr
340 345 350
Lys Leu Asn Leu Val Ala Thr Pro Leu Phe Leu Lys Pro Gly Ile Pro
355 360 365
Tyr Pro Ile Lys Val Gln Val Lys Asp Ser Leu Asp Gln Leu Val Gly
370 375 380
Gly Val Pro Val Thr Leu Asn Ala Gln Thr Ile Asp Val Asn Gln Glu
385 390 395 400
Thr Ser Asp Leu Asp Pro Ser Lys Ser Val Thr Arg Val Asp Asp Gly
405 410 415
Val Ala Ser Phe Val Leu Asn Leu Pro Ser Gly Val Thr Val Leu Glu
420 425 430
Phe Asn Val Lys Thr Asp Ala Pro Asp Leu Pro Glu Glu Asn Gln Ala
435 440 445
Arg Glu Gly Tyr Arg Ala Ile Ala Tyr Ser Ser Leu Ser Gln Ser Tyr
450 455 460
Leu Tyr Ile Asp Trp Thr Asp Asn His Lys Ala Leu Leu Val Gly Glu
465 470 475 480
His Leu Asn Ile Ile Val Thr Pro Lys Ser Pro Tyr Ile Asp Lys Ile
485 490 495
Thr His Tyr Asn Tyr Leu Ile Leu Ser Lys Gly Lys Ile Ile His Phe
500 505 510
Gly Thr Arg Glu Lys Phe Ser Asp Ala Ser Tyr Gln Ser Ile Asn Ile
515 520 525
Pro Val Thr Gln Asn Met Val Pro Ser Ser Arg Leu Leu Val Tyr Tyr
530 535 540
Ile Val Thr Gly Glu Gln Thr Ala Glu Leu Val Ser Asp Ser Val Trp
545 550 555 560
Leu Asn Ile Glu Glu Lys Cys Gly Asn Gln Leu Gln Val His Leu Ser
565 570 575
Pro Asp Ala Asp Ala Tyr Ser Pro Gly Gln Thr Val Ser Leu Asn Met
580 585 590
Ala Thr Gly Met Asp Ser Trp Val Ala Leu Ala Ala Val Asp Ser Ala
595 600 605
Val Tyr Gly Val Gln Arg Gly Ala Lys Lys Pro Leu Glu Arg Val Phe
610 615 620
Gln Phe Leu Glu Lys Ser Asp Leu Gly Cys Gly Ala Gly Gly Gly Leu
625 630 635 640
Asn Asn Ala Asn Val Phe His Leu Ala Gly Leu Thr Phe Leu Thr Asn
645 650 655
Ala Asn Ala Asp Asp Ser Gln Glu Asn Asp Glu Pro Cys Lys Glu Ile
660 665 670
Leu Arg Pro Arg Arg Thr Leu Gln Lys Lys Ile Glu Glu Ile Ala Ala
675 680 685
Lys Tyr Lys His Ser Val Val Lys Lys Cys Cys Tyr Asp Gly Ala Cys
690 695 700
Val Asn Asn Asp Glu Thr Cys Glu Gln Arg Ala Ala Arg Ile Ser Leu
705 710 715 720
Gly Pro Arg Cys Ile Lys Ala Phe Thr Glu Cys Cys Val Val Ala Ser
725 730 735
Gln Leu Arg Ala Asn Ile Ser His Lys Asp Met Gln Leu Gly Arg Leu
740 745 750
His Met Lys Thr Leu Leu Pro Val Ser Lys Pro Glu Ile Arg Ser Tyr
755 760 765
Phe Pro Glu Ser Trp Leu Trp Glu Val His Leu Val Pro Arg Arg Lys
770 775 780
Gln Leu Gln Phe Ala Leu Pro Asp Ser Leu Thr Thr Trp Glu Ile Gln
785 790 795 800
Gly Val Gly Ile Ser Asn Thr Gly Ile Cys Val Ala Asp Thr Val Lys
805 810 815
Ala Lys Val Phe Lys Asp Val Phe Leu Glu Met Asn Ile Pro Tyr Ser
820 825 830
Val Val Arg Gly Glu Gln Ile Gln Leu Lys Gly Thr Val Tyr Asn Tyr
835 840 845
Arg Thr Ser Gly Met Gln Phe Cys Val Lys Met Ser Ala Val Glu Gly
850 855 860
Ile Cys Thr Ser Glu Ser Pro Val Ile Asp His Gln Gly Thr Lys Ser
865 870 875 880
Ser Lys Cys Val Arg Gln Lys Val Glu Gly Ser Ser Ser His Leu Val
885 890 895
Thr Phe Thr Val Leu Pro Leu Glu Ile Gly Leu His Asn Ile Asn Phe
900 905 910
Ser Leu Glu Thr Trp Phe Gly Lys Glu Ile Leu Val Lys Thr Leu Arg
915 920 925
Val Val Pro Glu Gly Val Lys Arg Glu Ser Tyr Ser Gly Val Thr Leu
930 935 940
Asp Pro Arg Gly Ile Tyr Gly Thr Ile Ser Arg Arg Lys Glu Phe Pro
945 950 955 960
Tyr Arg Ile Pro Leu Asp Leu Val Pro Lys Thr Glu Ile Lys Arg Ile
965 970 975
Leu Ser Val Lys Gly Leu Leu Val Gly Glu Ile Leu Ser Ala Val Leu
980 985 990
Ser Gln Glu Gly Ile Asn Ile Leu Thr His Leu Pro Lys Gly Ser Ala
995 1000 1005
Glu Ala Glu Leu Met Ser Val Val Pro Val Phe Tyr Val Phe His
1010 1015 1020
Tyr Leu Glu Thr Gly Asn His Trp Asn Ile Phe His Ser Asp Pro
1025 1030 1035
Leu Ile Glu Lys Gln Lys Leu Lys Lys Lys Leu Lys Glu Gly Met
1040 1045 1050
Leu Ser Ile Met Ser Tyr Arg Asn Ala Asp Tyr Ser Tyr Ser Val
1055 1060 1065
Trp Lys Gly Gly Ser Ala Ser Thr Trp Leu Thr Ala Phe Ala Leu
1070 1075 1080
Arg Val Leu Gly Gln Val Asn Lys Tyr Val Glu Gln Asn Gln Asn
1085 1090 1095
Ser Ile Cys Asn Ser Leu Leu Trp Leu Val Glu Asn Tyr Gln Leu
1100 1105 1110
Asp Asn Gly Ser Phe Lys Glu Asn Ser Gln Tyr Gln Pro Ile Lys
1115 1120 1125
Leu Gln Gly Thr Leu Pro Val Glu Ala Arg Glu Asn Ser Leu Tyr
1130 1135 1140
Leu Thr Ala Phe Thr Val Ile Gly Ile Arg Lys Ala Phe Asp Ile
1145 1150 1155
Cys Pro Leu Val Lys Ile Asp Thr Ala Leu Ile Lys Ala Asp Asn
1160 1165 1170
Phe Leu Leu Glu Asn Thr Leu Pro Ala Gln Ser Thr Phe Thr Leu
1175 1180 1185
Ala Ile Ser Ala Tyr Ala Leu Ser Leu Gly Asp Lys Thr His Pro
1190 1195 1200
Gln Phe Arg Ser Ile Val Ser Ala Leu Lys Arg Glu Ala Leu Val
1205 1210 1215
Lys Gly Asn Pro Pro Ile Tyr Arg Phe Trp Lys Asp Asn Leu Gln
1220 1225 1230
His Lys Asp Ser Ser Val Pro Asn Thr Gly Thr Ala Arg Met Val
1235 1240 1245
Glu Thr Thr Ala Tyr Ala Leu Leu Thr Ser Leu Asn Leu Lys Asp
1250 1255 1260
Ile Asn Tyr Val Asn Pro Val Ile Lys Trp Leu Ser Glu Glu Gln
1265 1270 1275
Arg Tyr Gly Gly Gly Phe Tyr Ser Thr Gln Asp Thr Ile Asn Ala
1280 1285 1290
Ile Glu Gly Leu Thr Glu Tyr Ser Leu Leu Val Lys Gln Leu Arg
1295 1300 1305
Leu Ser Met Asp Ile Asp Val Ser Tyr Lys His Lys Gly Ala Leu
1310 1315 1320
His Asn Tyr Lys Met Thr Asp Lys Asn Phe Leu Gly Arg Pro Val
1325 1330 1335
Glu Val Leu Leu Asn Asp Asp Leu Ile Val Ser Thr Gly Phe Gly
1340 1345 1350
Ser Gly Leu Ala Thr Val His Val Thr Thr Val Val His Lys Thr
1355 1360 1365
Ser Thr Ser Glu Glu Val Cys Ser Phe Tyr Leu Lys Ile Asp Thr
1370 1375 1380
Gln Asp Ile Glu Ala Ser His Tyr Arg Gly Tyr Gly Asn Ser Asp
1385 1390 1395
Tyr Lys Arg Ile Val Ala Cys Ala Ser Tyr Lys Pro Ser Arg Glu
1400 1405 1410
Glu Ser Ser Ser Gly Ser Ser His Ala Val Met Asp Ile Ser Leu
1415 1420 1425
Pro Thr Gly Ile Ser Ala Asn Glu Glu Asp Leu Lys Ala Leu Val
1430 1435 1440
Glu Gly Val Asp Gln Leu Phe Thr Asp Tyr Gln Ile Lys Asp Gly
1445 1450 1455
His Val Ile Leu Gln Leu Asn Ser Ile Pro Ser Ser Asp Phe Leu
1460 1465 1470
Cys Val Arg Phe Arg Ile Phe Glu Leu Phe Glu Val Gly Phe Leu
1475 1480 1485
Ser Pro Ala Thr Phe Thr Val Tyr Glu Tyr His Arg Pro Asp Lys
1490 1495 1500
Gln Cys Thr Met Phe Tyr Ser Thr Ser Asn Ile Lys Ile Gln Lys
1505 1510 1515
Val Cys Glu Gly Ala Ala Cys Lys Cys Val Glu Ala Asp Cys Gly
1520 1525 1530
Gln Met Gln Glu Glu Leu Asp Leu Thr Ile Ser Ala Glu Thr Arg
1535 1540 1545
Lys Gln Thr Ala Cys Lys Pro Glu Ile Ala Tyr Ala Tyr Lys Val
1550 1555 1560
Ser Ile Thr Ser Ile Thr Val Glu Asn Val Phe Val Lys Tyr Lys
1565 1570 1575
Ala Thr Leu Leu Asp Ile Tyr Lys Thr Gly Glu Ala Val Ala Glu
1580 1585 1590
Lys Asp Ser Glu Ile Thr Phe Ile Lys Lys Val Thr Cys Thr Asn
1595 1600 1605
Ala Glu Leu Val Lys Gly Arg Gln Tyr Leu Ile Met Gly Lys Glu
1610 1615 1620
Ala Leu Gln Ile Lys Tyr Asn Phe Ser Phe Arg Tyr Ile Tyr Pro
1625 1630 1635
Leu Asp Ser Leu Thr Trp Ile Glu Tyr Trp Pro Arg Asp Thr Thr
1640 1645 1650
Cys Ser Ser Cys Gln Ala Phe Leu Ala Asn Leu Asp Glu Phe Ala
1655 1660 1665
Glu Asp Ile Phe Leu Asn Gly Cys
1670 1675
<210> 4
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 4
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Arg Gln Pro Glu Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys Val Asp Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 5
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 5
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys Val Asp Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 6
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 6
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys Val Asp Gly Ser Leu Ala
50 55 60
Glu Ala Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asn Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Leu His Ile Leu Ala Ala Leu Pro
100 105
<210> 7
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 7
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Ser Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys Val Asp Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 8
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 8
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Glu Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys Val Asp Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 9
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 9
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Ala
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys Val Asp Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 10
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 10
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Glu Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys Val Asp Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 11
<211> 103
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 11
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Leu Ala Glu Ala Lys Glu Ala
50 55 60
Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser Asp Phe Tyr Lys Arg
65 70 75 80
Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val Glu Ala Leu Lys Asp
85 90 95
Ala Ile Leu Ala Ala Leu Pro
100
<210> 12
<211> 57
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 12
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro
50 55
<210> 13
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 13
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys Val Glu Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 14
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 14
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys Val Ala Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 15
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 15
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Glu Ser Ser Gln Ala Pro Lys Val Glu Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 16
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 16
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Arg Gln Pro Glu Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys Val Glu Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 17
<211> 57
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 17
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Glu Ser Ser Gln Ala Pro
50 55
<210> 18
<211> 57
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 18
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Arg Gln Pro Glu Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro
50 55
<210> 19
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 19
Glu Val Leu Glu Ala Trp Asp Glu Ile Asp Arg Leu Pro Asn Leu Thr
1 5 10 15
Ile Glu Gln Trp Leu Ala Phe Ile Asn Lys Leu Asp Asp Asp Pro Ser
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Ser Glu Ser Gln
35 40 45
<210> 20
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 20
Glu Val Leu Glu Ala Trp Asp Glu Ile Asp Arg Leu Pro Asn Leu Thr
1 5 10 15
Ile Glu Gln Trp Leu Ala Phe Ile Asn Lys Leu Asp Asp Asp Pro Ser
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Glu Ser Ser Gln
35 40 45
<210> 21
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 21
Glu Val Leu Glu Ala Trp Asp Glu Ile Asp Arg Leu Pro Asn Leu Thr
1 5 10 15
Ile Glu Gln Trp Leu Ala Phe Ile Asn Lys Leu Asp Arg Gln Pro Glu
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Ser Glu Ser Gln
35 40 45
<210> 22
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 22
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys
50 55
<210> 23
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 23
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys Val Glu Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 24
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 24
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Glu Ser Gln Ala Pro Lys Val Glu Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 25
<211> 108
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 25
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Asp Ser Gln Ala Pro Lys Val Glu Gly Ser Leu Ala
50 55 60
Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly Val Ser
65 70 75 80
Asp Phe Tyr Lys Arg Leu Ile Asp Lys Ala Lys Thr Val Glu Gly Val
85 90 95
Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
100 105
<210> 26
<211> 62
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный С5-связывающий Z-вариант
<400> 26
Ala Glu Ala Lys Tyr Ala Lys Glu Val Leu Glu Ala Trp Asp Glu Ile
1 5 10 15
Asp Arg Leu Pro Asn Leu Thr Ile Glu Gln Trp Leu Ala Phe Ile Asn
20 25 30
Lys Leu Asp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys Val Asp Gly Ser
50 55 60
<210> 27
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный HER2-связывающий Z-вариант
<400> 27
Ala Glu Ala Lys Tyr Ala Lys Glu Met Arg Asn Ala Tyr Trp Glu Ile
1 5 10 15
Ala Leu Leu Pro Asn Leu Thr Asn Gln Gln Lys Arg Ala Phe Ile Arg
20 25 30
Lys Leu Tyr Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys
50 55
<210> 28
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный HER2-связывающий Z-вариант
<400> 28
Ala Glu Ala Lys Tyr Ala Lys Glu Met Arg Asn Ala Tyr Trp Glu Ile
1 5 10 15
Ala Leu Leu Pro Asn Leu Thr Asn Gln Gln Lys Arg Ala Phe Ile Arg
20 25 30
Lys Leu Tyr Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys
50 55
<210> 29
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный HER2-связывающий Z-вариант
<400> 29
Ala Glu Ala Lys Tyr Ala Lys Glu Met Arg Asn Ala Tyr Trp Glu Ile
1 5 10 15
Ala Leu Leu Pro Asn Leu Thr Asn Gln Gln Lys Arg Ala Phe Ile Arg
20 25 30
Lys Leu Tyr Arg Gln Pro Glu Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys
50 55
<210> 30
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный PDGFR-бета-связывающий Z-вариант
<400> 30
Ala Glu Ala Lys Tyr Ala Lys Glu Leu Ile Glu Ala Ala Ala Glu Ile
1 5 10 15
Asp Ala Leu Pro Asn Leu Thr Arg Arg Gln Trp Asn Ala Phe Ile Lys
20 25 30
Lys Leu Val Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys
50 55
<210> 31
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный PDGFR-бета-связывающий Z-вариант
<400> 31
Ala Glu Ala Lys Tyr Ala Lys Glu Leu Ile Glu Ala Ala Ala Glu Ile
1 5 10 15
Asp Ala Leu Pro Asn Leu Thr Arg Arg Gln Trp Asn Ala Phe Ile Lys
20 25 30
Lys Leu Val Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys
50 55
<210> 32
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный PDGFR-бета-связывающий Z-вариант
<400> 32
Ala Glu Ala Lys Tyr Ala Lys Glu Leu Ile Glu Ala Ala Ala Glu Ile
1 5 10 15
Asp Ala Leu Pro Asn Leu Thr Arg Arg Gln Trp Asn Ala Phe Ile Lys
20 25 30
Lys Leu Val Arg Gln Pro Glu Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys
50 55
<210> 33
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный FcRn-связывающий Z-вариант
<400> 33
Ala Glu Ala Lys Tyr Ala Lys Glu Gln Asp Ala Ala Ala His Glu Ile
1 5 10 15
Arg Trp Leu Pro Asn Leu Thr Phe Asp Gln Arg Val Ala Phe Ile His
20 25 30
Lys Leu Ala Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys
50 55
<210> 34
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный FcRn-связывающий Z-вариант
<400> 34
Ala Glu Ala Lys Tyr Ala Lys Glu Gln Asp Ala Ala Ala His Glu Ile
1 5 10 15
Arg Trp Leu Pro Asn Leu Thr Phe Asp Gln Arg Val Ala Phe Ile His
20 25 30
Lys Leu Ala Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys
50 55
<210> 35
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный FcRn-связывающий Z-вариант
<400> 35
Ala Glu Ala Lys Tyr Ala Lys Glu Gln Asp Ala Ala Ala His Glu Ile
1 5 10 15
Arg Trp Leu Pro Asn Leu Thr Phe Asp Gln Arg Val Ala Phe Ile His
20 25 30
Lys Leu Ala Arg Gln Pro Glu Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys
50 55
<210> 36
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 36
Ala Glu Ala Lys Tyr Ala Lys Glu Asn Leu Phe Ala Gly Trp Glu Ile
1 5 10 15
Ser Asp Leu Pro Asn Leu Thr Asp Tyr Gln Arg Asn Ala Phe Ile Tyr
20 25 30
Lys Leu Trp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Asp Ser Gln Ala Pro Lys
50 55
<210> 37
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 37
Ala Glu Ala Lys Tyr Ala Lys Glu Asn Leu Phe Ala Gly Trp Glu Ile
1 5 10 15
Ser Asp Leu Pro Asn Leu Thr Asp Tyr Gln Arg Asn Ala Phe Ile Tyr
20 25 30
Lys Leu Trp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys
50 55
<210> 38
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 38
Ala Glu Ala Lys Tyr Ala Lys Glu Asn Leu Phe Ala Gly Trp Glu Ile
1 5 10 15
Ser Asp Leu Pro Asn Leu Thr Asp Tyr Gln Arg Asn Ala Phe Ile Tyr
20 25 30
Lys Leu Trp Arg Gln Pro Glu Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Glu Ser Gln Ala Pro Lys
50 55
<210> 39
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 39
Ala Glu Ala Lys Tyr Ala Lys Glu Asn Leu Phe Ala Gly Trp Glu Ile
1 5 10 15
Ser Asp Leu Pro Asn Leu Thr Asp Tyr Gln Arg Asn Ala Phe Ile Tyr
20 25 30
Lys Leu Trp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Glu Ser Gln Ala Pro Lys
50 55
<210> 40
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 40
Ala Glu Ala Lys Tyr Ala Lys Glu Asn Leu Phe Ala Gly Trp Glu Ile
1 5 10 15
Ser Asp Leu Pro Asn Leu Thr Asp Tyr Gln Arg Asn Ala Phe Ile Tyr
20 25 30
Lys Leu Trp Arg Gln Pro Glu Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Asn Glu Ser Gln Ala Pro Lys
50 55
<210> 41
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 41
Ala Glu Ala Lys Tyr Ala Lys Glu Asn Leu Phe Ala Gly Trp Glu Ile
1 5 10 15
Ser Asp Leu Pro Asn Leu Thr Asp Tyr Gln Arg Asn Ala Phe Ile Tyr
20 25 30
Lys Leu Trp Asp Asp Pro Ser Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Asp Ser Gln Ala Pro Lys
50 55
<210> 42
<211> 58
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 42
Ala Glu Ala Lys Tyr Ala Lys Glu Asn Leu Phe Ala Gly Trp Glu Ile
1 5 10 15
Ser Asp Leu Pro Asn Leu Thr Asp Tyr Gln Arg Asn Ala Phe Ile Tyr
20 25 30
Lys Leu Trp Arg Gln Pro Glu Gln Ser Ser Glu Leu Leu Ser Glu Ala
35 40 45
Lys Lys Leu Ser Asp Ser Gln Ala Pro Lys
50 55
<210> 43
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный HER2-связывающий Z-вариант
<400> 43
Glu Met Arg Asn Ala Tyr Trp Glu Ile Ala Leu Leu Pro Asn Leu Thr
1 5 10 15
Asn Gln Gln Lys Arg Ala Phe Ile Arg Lys Leu Tyr Asp Asp Pro Ser
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Ser Glu Ser Gln
35 40 45
<210> 44
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный HER2-связывающий Z-вариант
<400> 44
Glu Met Arg Asn Ala Tyr Trp Glu Ile Ala Leu Leu Pro Asn Leu Thr
1 5 10 15
Asn Gln Gln Lys Arg Ala Phe Ile Arg Lys Leu Tyr Arg Gln Pro Glu
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Ser Glu Ser Gln
35 40 45
<210> 45
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный PDGFR-бета-связывающий Z-вариант
<400> 45
Glu Leu Ile Glu Ala Ala Ala Glu Ile Asp Ala Leu Pro Asn Leu Thr
1 5 10 15
Arg Arg Gln Trp Asn Ala Phe Ile Lys Lys Leu Val Asp Asp Pro Ser
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Ser Glu Ser Gln
35 40 45
<210> 46
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный PDGFR-бета-связывающий Z-вариант
<400> 46
Glu Leu Ile Glu Ala Ala Ala Glu Ile Asp Ala Leu Pro Asn Leu Thr
1 5 10 15
Arg Arg Gln Trp Asn Ala Phe Ile Lys Lys Leu Val Arg Gln Pro Glu
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Ser Glu Ser Gln
35 40 45
<210> 47
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный FcRn-связывающий Z-вариант
<400> 47
Glu Gln Asp Ala Ala Ala His Glu Ile Arg Trp Leu Pro Asn Leu Thr
1 5 10 15
Phe Asp Gln Arg Val Ala Phe Ile His Lys Leu Ala Asp Asp Pro Ser
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Ser Glu Ser Gln
35 40 45
<210> 48
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный FcRn-связывающий Z-вариант
<400> 48
Glu Gln Asp Ala Ala Ala His Glu Ile Arg Trp Leu Pro Asn Leu Thr
1 5 10 15
Phe Asp Gln Arg Val Ala Phe Ile His Lys Leu Ala Arg Gln Pro Glu
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Ser Glu Ser Gln
35 40 45
<210> 49
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 49
Glu Asn Leu Phe Ala Gly Trp Glu Ile Ser Asp Leu Pro Asn Leu Thr
1 5 10 15
Asp Tyr Gln Arg Asn Ala Phe Ile Tyr Lys Leu Trp Asp Asp Pro Ser
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Ser Glu Ser Gln
35 40 45
<210> 50
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 50
Glu Asn Leu Phe Ala Gly Trp Glu Ile Ser Asp Leu Pro Asn Leu Thr
1 5 10 15
Asp Tyr Gln Arg Asn Ala Phe Ile Tyr Lys Leu Trp Arg Gln Pro Glu
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Ser Glu Ser Gln
35 40 45
<210> 51
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 51
Glu Asn Leu Phe Ala Gly Trp Glu Ile Ser Asp Leu Pro Asn Leu Thr
1 5 10 15
Asp Tyr Gln Arg Asn Ala Phe Ile Tyr Lys Leu Trp Asp Asp Pro Ser
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Asn Glu Ser Gln
35 40 45
<210> 52
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 52
Glu Asn Leu Phe Ala Gly Trp Glu Ile Ser Asp Leu Pro Asn Leu Thr
1 5 10 15
Asp Tyr Gln Arg Asn Ala Phe Ile Tyr Lys Leu Trp Arg Gln Pro Glu
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Asn Glu Ser Gln
35 40 45
<210> 53
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 53
Glu Asn Leu Phe Ala Gly Trp Glu Ile Ser Asp Leu Pro Asn Leu Thr
1 5 10 15
Asp Tyr Gln Arg Asn Ala Phe Ile Tyr Lys Leu Trp Asp Asp Pro Ser
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Ser Asp Ser Gln
35 40 45
<210> 54
<211> 48
<212> PRT
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<223> Сконструированный CAIX-связывающий Z-вариант
<400> 54
Glu Asn Leu Phe Ala Gly Trp Glu Ile Ser Asp Leu Pro Asn Leu Thr
1 5 10 15
Asp Tyr Gln Arg Asn Ala Phe Ile Tyr Lys Leu Trp Arg Gln Pro Glu
20 25 30
Gln Ser Ser Glu Leu Leu Ser Glu Ala Lys Lys Leu Ser Asp Ser Gln
35 40 45
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКРУТИРУЮЩИЕ Т-КЛЕТКИ ПОЛИПЕПТИДЫ, СПОСОБНЫЕ СВЯЗЫВАТЬ CD123 И TCR АЛЬФА/БЕТА | 2017 |
|
RU2775063C2 |
| УЛУЧШЕННЫЕ ОДИНОЧНЫЕ ВАРИАБЕЛЬНЫЕ ДОМЕНЫ ИММУНОГЛОБУЛИНА, СВЯЗЫВАЮЩИЕСЯ С СЫВОРОТОЧНЫМ АЛЬБУМИНОМ | 2017 |
|
RU2765384C2 |
| УСОВЕРШЕНСТВОВАННЫЕ АГЕНТЫ, СВЯЗЫВАЮЩИЕ СЫВОРОТОЧНЫЙ АЛЬБУМИН | 2018 |
|
RU2797270C2 |
| УСОВЕРШЕНСТВОВАННЫЕ СВЯЗЫВАЮЩИЕСЯ С СЫВОРОТОЧНЫМ АЛЬБУМИНОМ ВЕЩЕСТВА | 2018 |
|
RU2789495C2 |
| БИСПЕЦИФИЧЕСКИЕ 2+1 КОНТОРСТЕЛА | 2018 |
|
RU2797305C2 |
| ПОЛИПЕПТИД, СОДЕРЖАЩИЙ АНТИГЕНСВЯЗЫВАЮЩИЙ ДОМЕН И ТРАНСПОРТИРУЮЩИЙ СЕГМЕНТ | 2017 |
|
RU2827545C2 |
| ИММУНОГЛОБУЛИНЫ, СВЯЗЫВАЮЩИЕ АГГРЕКАН | 2018 |
|
RU2771818C2 |
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С РАКОВЫМИ КЛЕТКАМИ И ОБЕСПЕЧИВАЮТ НАПРАВЛЕННЫЙ ПЕРЕНОС РАДИОНУКЛИДОВ К УКАЗАННЫМ КЛЕТКАМ | 2020 |
|
RU2833191C2 |
| ПОЛИПЕПТИД, ВКЛЮЧАЮЩИЙ АНТИГЕНСВЯЗЫВАЮЩИЙ ДОМЕН И ТРАНСПОРТИРУЮЩИЙ СЕГМЕНТ | 2018 |
|
RU2815452C2 |
| Новые полипептиды | 2015 |
|
RU2725442C2 |

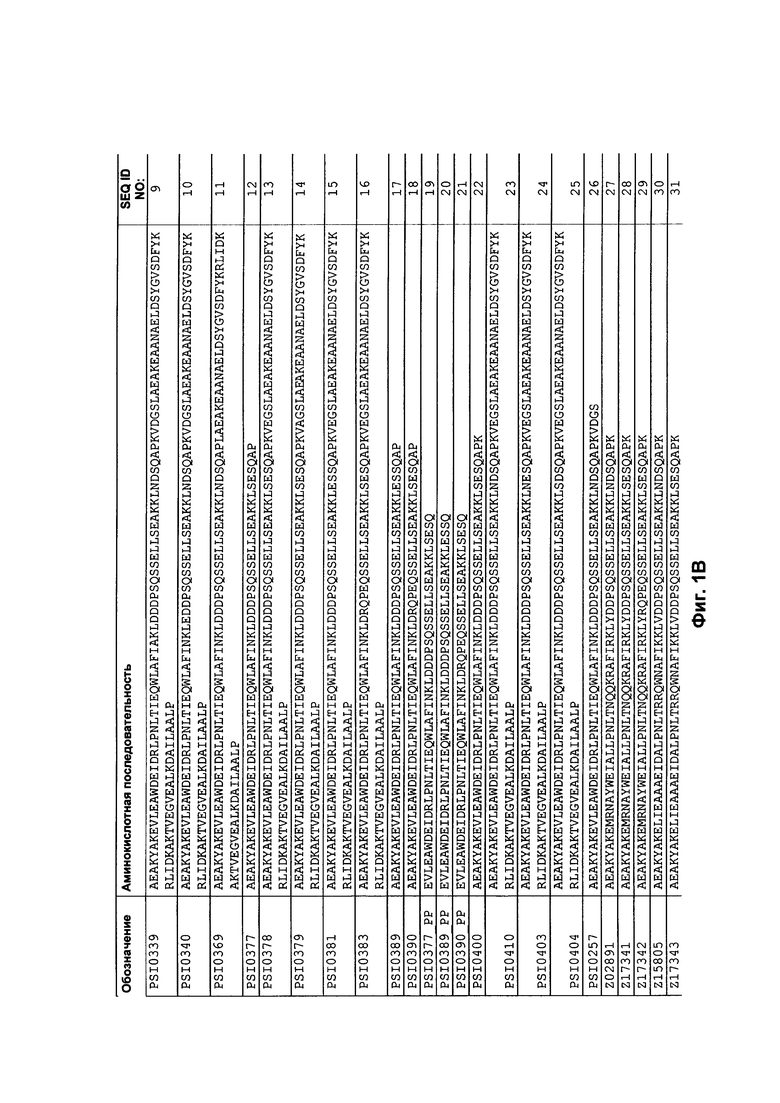


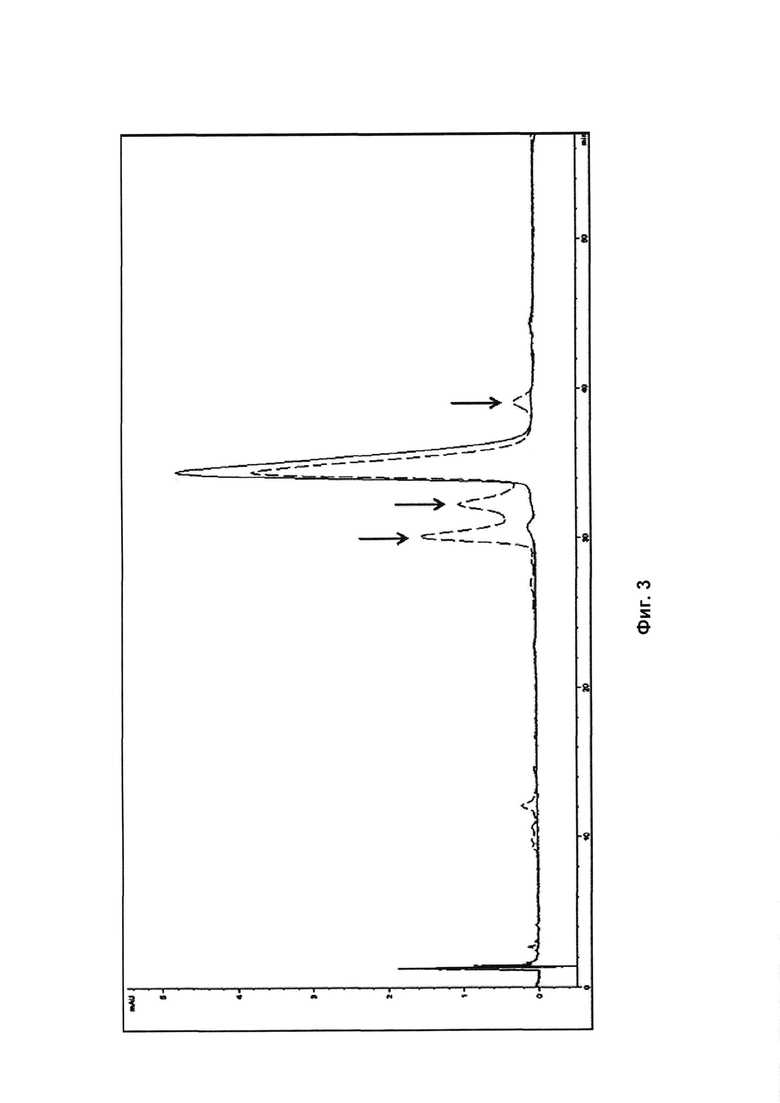
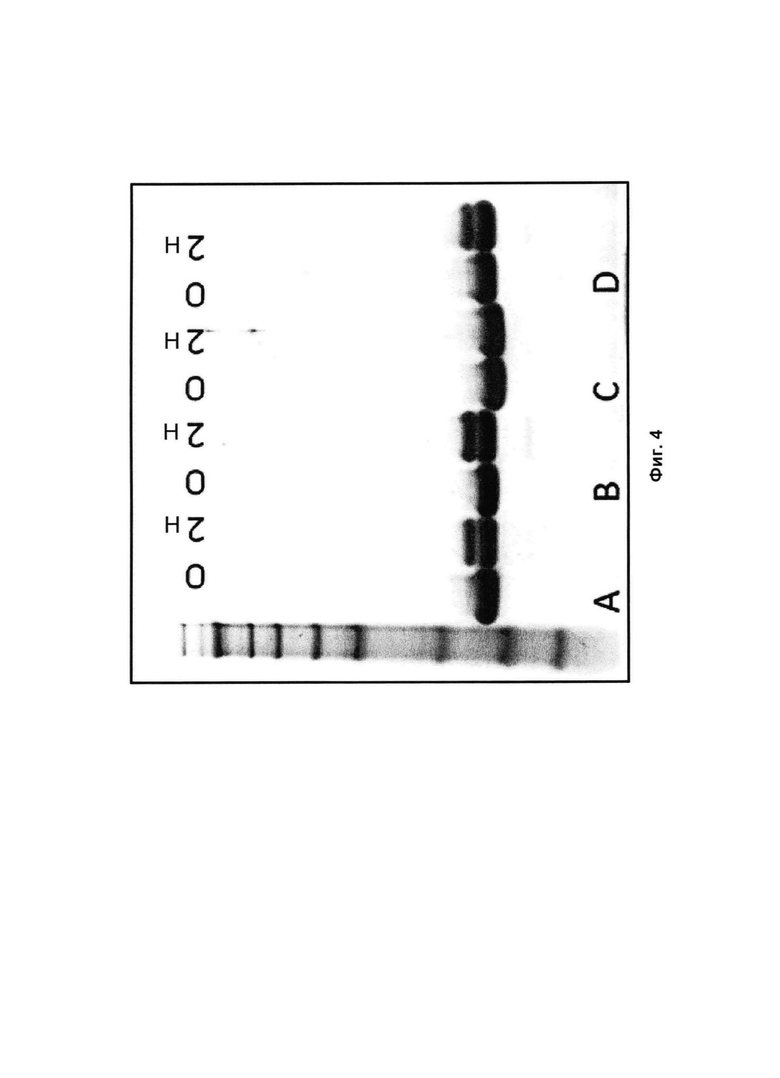



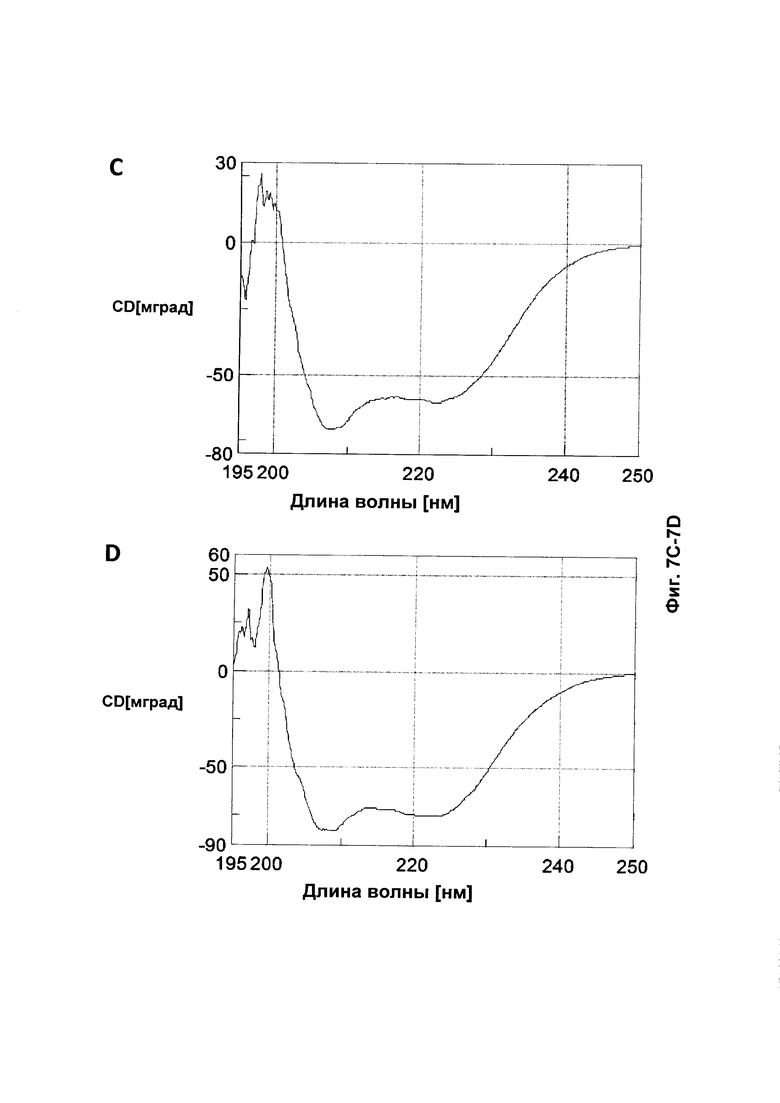
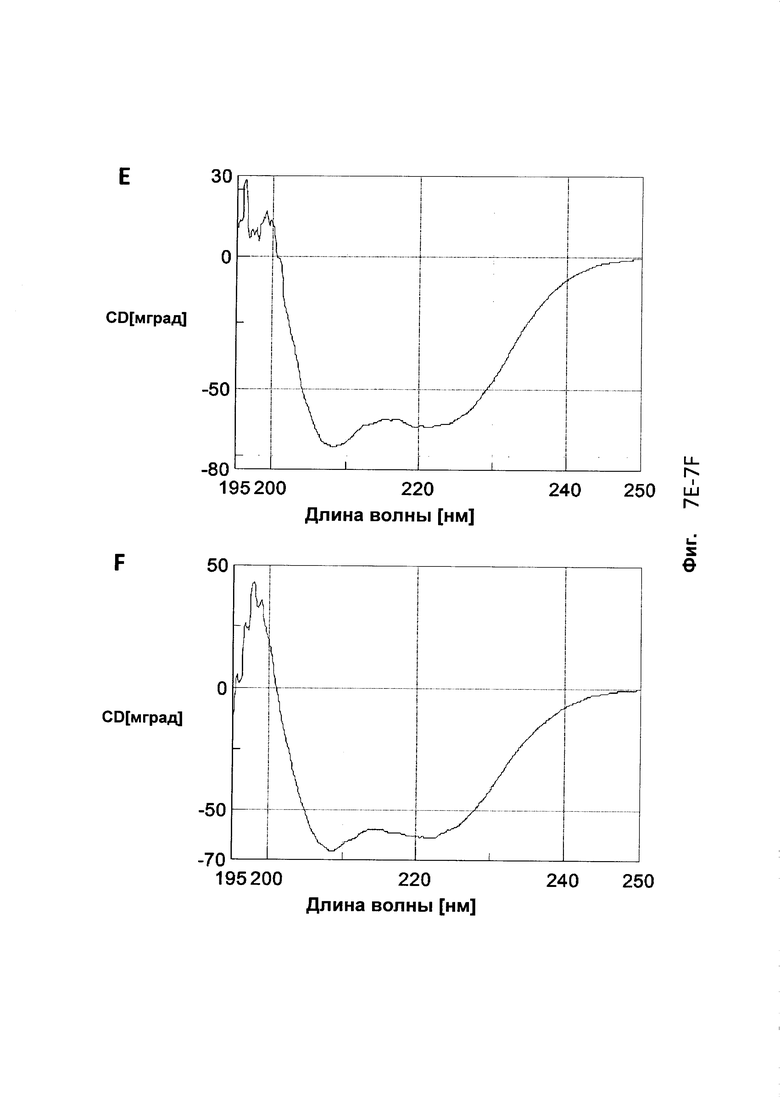
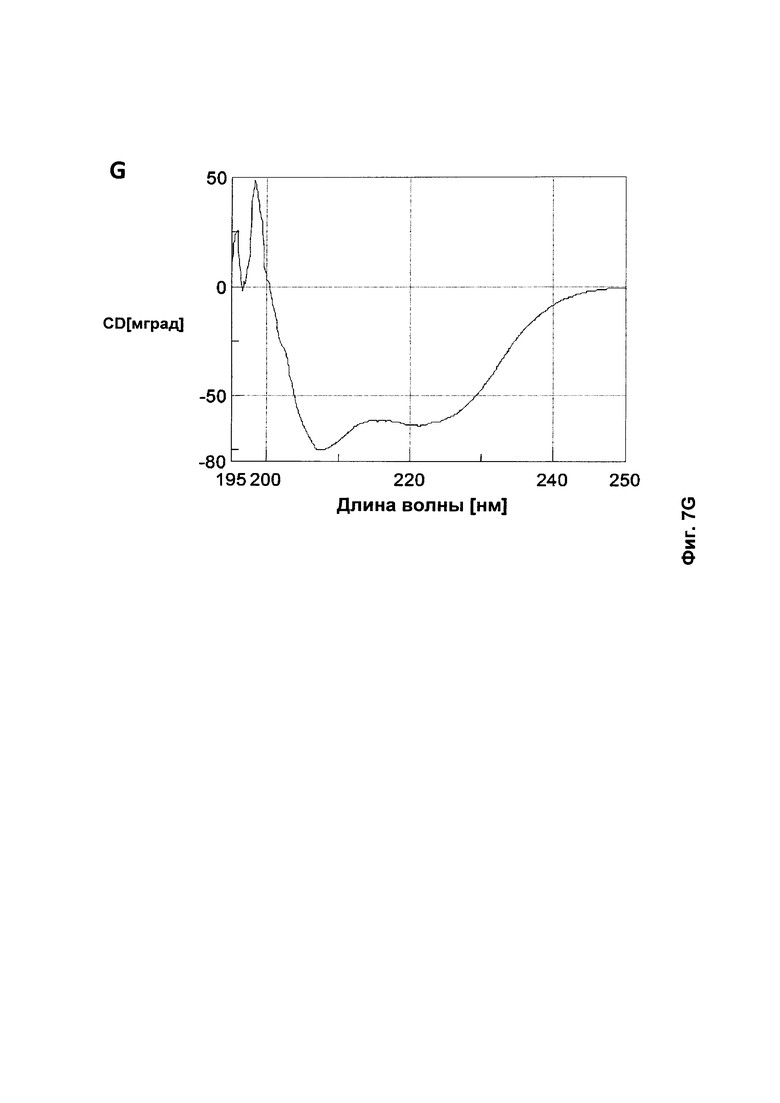

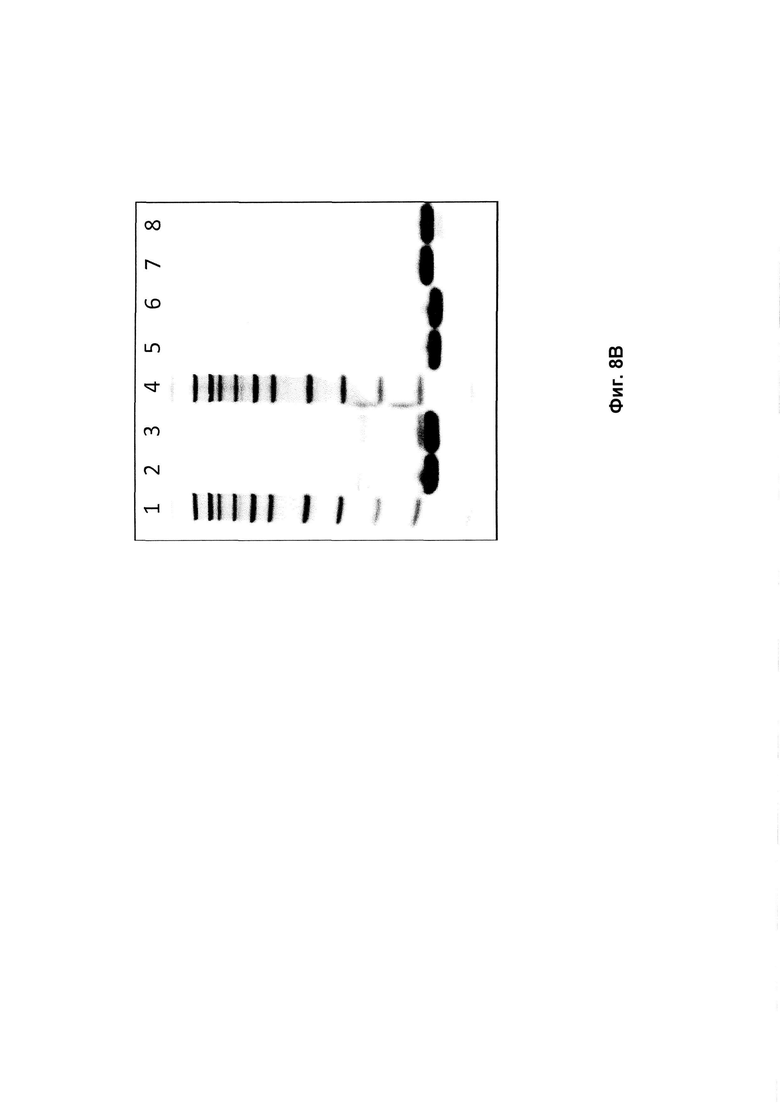
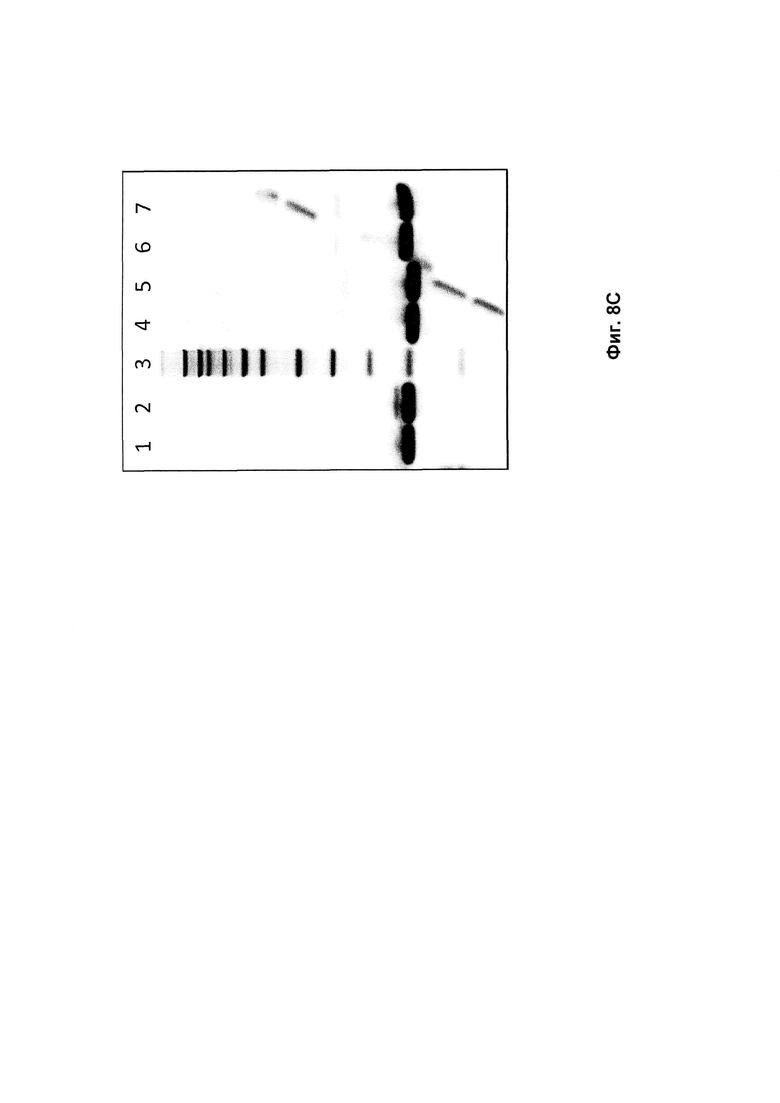

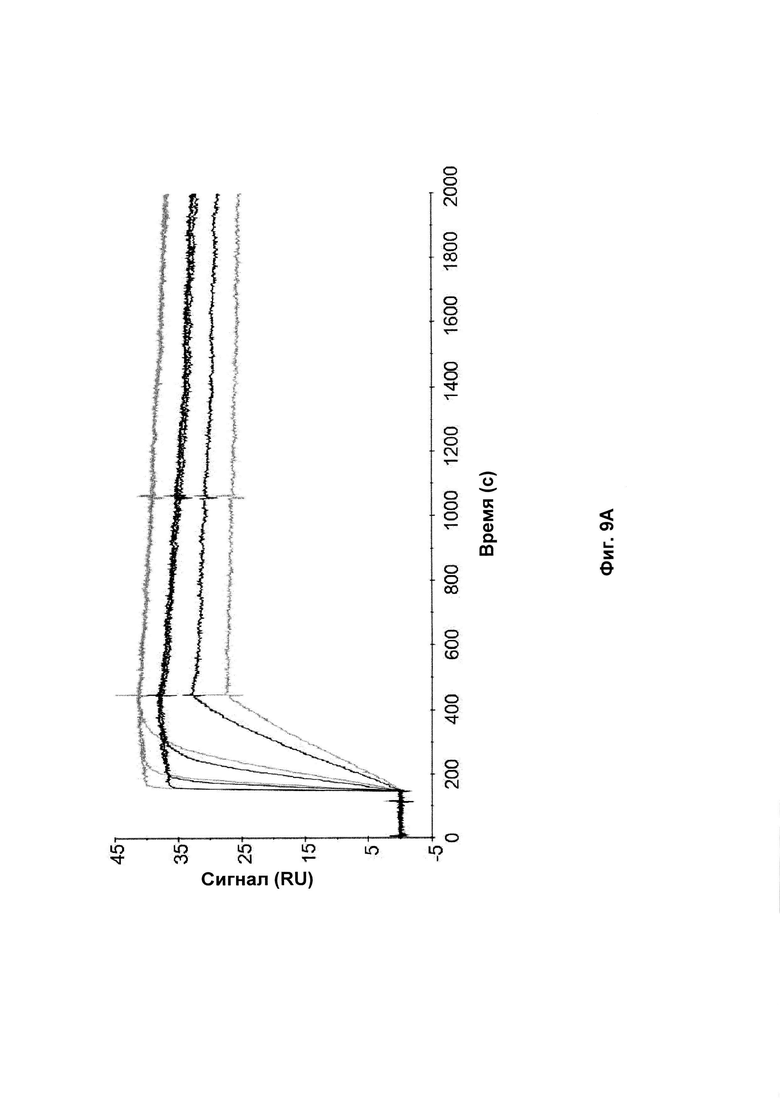
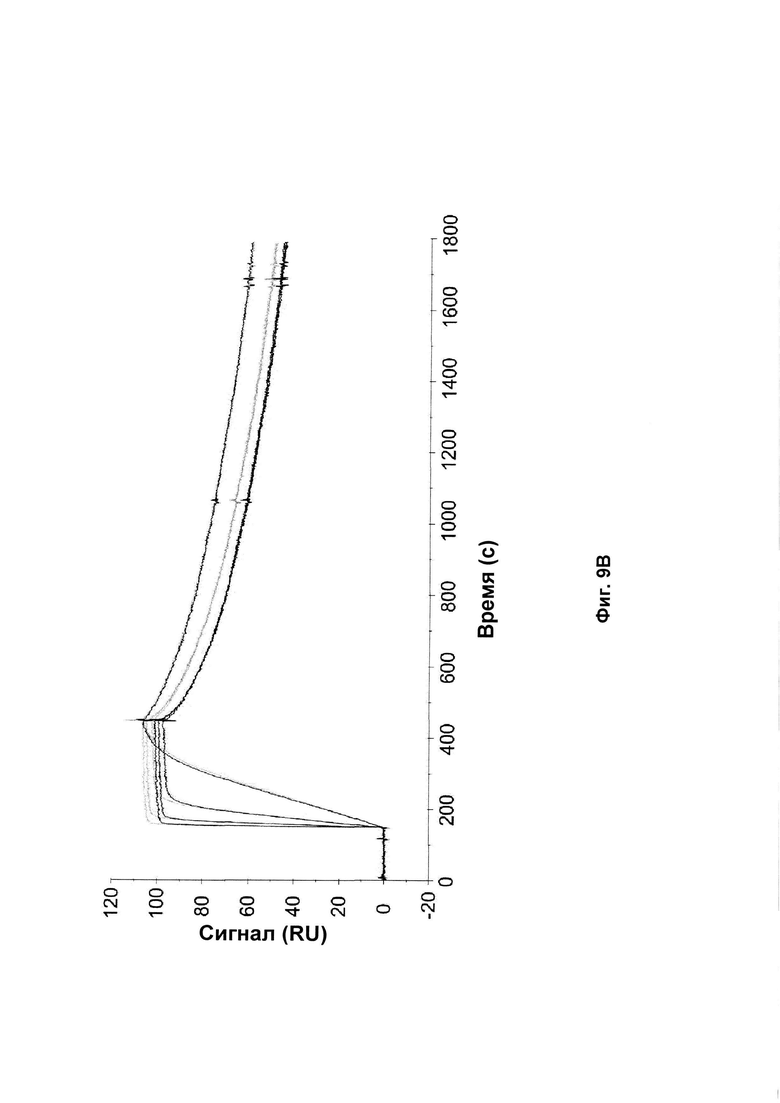

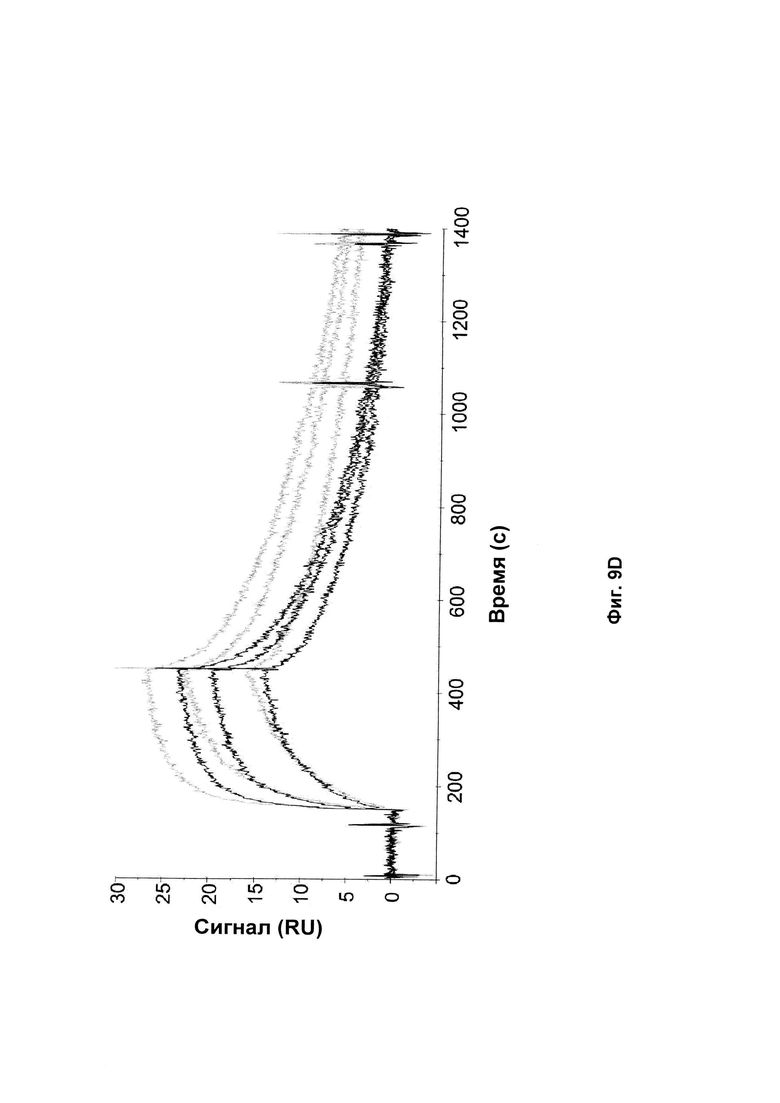
Изобретение относится к области биотехнологии, конкретно к полипептиду, содержащему последовательность EX2X3X4AX6
X7EIX10Х11LPNLX16X17X18QX20X21AFIX25X26
LX28X29X30PX32QSX35X36LLX39EAKKLX45X46X47
Q, и обладающему повышенной стабильностью. Настоящее изобретение также относится к группам полипептидов-вариантов на основе общего каркаса, где каждый полипептид в группе содержит аминокислотную последовательность EX2X3X4AX6X7EIX10Х11LPNLX16X17X18QX20X21
AFIX25X26LX28X29X30PX32QSX35X36LLX39
EAKKLX45X46X47Q, и к способам отбора нужного полипептида, обладающего аффинностью к заданной мишени. 11 н. и 6 з.п. ф-лы, 9 ил., 11 табл., 13 пр.
1. Полипептид, содержащий аминокислотную последовательность, выбранную из:
1) EX2X3X4AX6X7EIX10X11LPNLX16X17X18QX20X21AFIX25X26LX28X29X30PX32QSX35X36LLX39EAKKLX45X46X47Q,
где каждый из X2, X3, X4, X6, X7, X10, X11, X17, X18, X20, X21, X25 и X28
независимо соответствует любому аминокислотному остатку; и где, независимо друг от друга,
X16 выбран из N и T;
X26 выбран из K и S;
X29X30PX32 выбран из DDPS и RQPE;
X35 выбран из A и S;
X36 выбран из E и N;
X39 выбран из A, C и S;
X45 выбран из E и S;
X46 выбран из D, E и S;
X47 выбран из A и S; и
2) аминокислотной последовательности, которая обладает по меньшей мере 91%-ной идентичностью с последовательностью, определенной в (1), при условии, что X46 не является D или E, когда X45 представляет собой N, где указанный полипептид обладает повышенной стабильностью.
2. Полипептид по п. 1, где X45 представляет собой S.
3. Полипептид по п. 1, где X45X46 выбран из ES и SE.
4. Полипептид по п. 3, где X45X46 представляет собой SE.
5. Полипептид по любому из пп. 1-4, который содержит аминокислотную последовательность, выбранную из:
YAKEX2X3X4AX6X7EIX10X11LPNLX16X17X18QX20X21AFIX25X26LX28X29X30PX32QSX35X36LLX39EAKKLX45X46X47QAP; и
FNKEX2X3X4AX6X7EIX10X11LPNLX16X17X18QX20X21AFIX25X26LX28X29X30PX32QSX35X36LLX39EAKKLX45X46X47QAP,
где каждый Xy такой, как он определен в любом из пп. 1-4.
6. Слитый полипептид, содержащий полипептид по любому из пп. 1-5 в качестве группировки и вторую группировку, содержащую дополнительные аминокислотные остатки, где указанный слитый полипептид обладает повышенной стабильностью, и где указанные дополнительные аминокислотные остатки составляют один или более полипептидных доменов, имеющих функцию пролонгирования времени полувыведения.
7. Полинуклеотид, кодирующий полипептид по любому из пп. 1-5.
8. Полинуклеотид, кодирующий слитый полипептид по п. 6.
9. Библиотека полипептидов-вариантов для выбора искомого полипептида, на основе общего каркаса, где каждый полипептид в библиотеке содержит аминокислотную последовательность, выбранную из:
1) EX2X3X4AX6X7EIX10X11LPNLX16X17X18QX20X21AFIX25X26LX28X29X30PX32QSX35X36LLX39EAKKLX45X46X47Q,
где каждый из X2, X3, X4, X6, X7, X10, X11, X17, X18, X20, X21, X25 и X28
независимо соответствует любому аминокислотному остатку; и где, независимо друг от друга,
X16 выбран из N и T;
X26 выбран из K и S;
X29X30PX32 выбран из DDPS и RQPE;
X35 выбран из A и S;
X36 выбран из E и N;
X39 выбран из A, C и S;
X45 выбран из E и S;
X46 выбран из D, E и S;
X47 выбран из A и S; и
2) аминокислотной последовательности, которая обладает по меньшей мере 91%-ной идентичностью с последовательностью, определенной в (1), при условии, что X46 не является D или E, когда X45 представляет собой N, где указанный полипептид обладает повышенной стабильностью.
10. Библиотека по п. 9, которая содержит по меньшей мере 1×104 уникальных полипептидных молекул.
11. Библиотека полинуклеотидов для получения библиотеки полипептидов-вариантов по п. 9, отличающаяся тем, что каждый ее представитель кодирует представителя библиотеки полипептидов по любому из пп. 9-10.
12. Комбинация библиотеки полипептидов по любому из пп. 9-10 и библиотеки полинуклеотидов по п. 11 для ассоциирования полипептида с кодирующим его полинуклеотидом, где каждый представитель указанной библиотеки полипептидов физически или пространственно ассоциирован с полинуклеотидом, кодирующим этого представителя, при помощи системы дисплея или структурного анализа комплементации фрагментов белка (PCA).
13. Комбинация по п. 12, где указанная система дисплея включает систему фагового дисплея.
14. Способ выбора искомого полипептида, обладающего аффинностью к заданной мишени, из библиотеки полипептидов, включающий стадии:
(а) предоставления библиотеки полипептидов по любому из пп. 9-10;
(б) приведения этой библиотеки полипептидов в контакт с заданной мишенью в условиях, которые обеспечивают возможность специфического взаимодействия между этой мишенью и по меньшей мере одним искомым полипептидом, обладающим аффинностью к этой мишени; и
(в) выбора, на основе указанного специфического взаимодействия, по меньшей мере одного искомого полипептида из остальной библиотеки полипептидов.
15. Способ выделения полинуклеотида, кодирующего искомый полипептид, обладающий аффинностью к заданной мишени, включающий стадии:
- предоставления комбинации по любому из пп. 12-13;
- приведения этой комбинации в контакт с заданной мишенью в условиях, которые обеспечивают возможность специфического взаимодействия между этой мишенью и по меньшей мере одним искомым полипептидом в указанной комбинации, обладающим аффинностью к этой мишени;
- выбора, на основе указанного специфического взаимодействия, по меньшей мере одного искомого полипептида из остальной комбинации; и
- выделения таким образом отделенного полинуклеотида, кодирующего искомый полипептид.
16. Способ идентификации искомого полипептида, обладающего аффинностью к заданной мишени, включающий стадии:
- выделения полинуклеотида, кодирующего указанный искомый полипептид, с использованием способа по п. 15; и
- секвенирования полинуклеотида для установления аминокислотной последовательности указанного искомого полипептида.
17. Способ отбора и идентификации искомого полипептида,
обладающего аффинностью к заданной мишени, из библиотеки полипептидов, включающий стадии:
(а) синтеза каждого представителя библиотеки полипептидов по любому из пп. 9-10 на отдельном носителе или грануле;
(б) выбора или обогащения носителей или гранул на основании взаимодействия полипептида с заданной мишенью; и
(в) идентификации полипептида посредством метода анализа белка.
| WO 2005075507, 18.08.2005 | |||
| WO 2011056124, 12.05.2011 | |||
| GOUDA H | |||
| et al., Three-dimensional solution structure of the B domain of staphylococcal protein A: comparisons of the solution and crystal structures, Biochemistry, 1992, V | |||
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |
| Приспособление к шкивам для надевания ремня | 1928 |
|
SU9665A1 |
| SERGEEVA A | |||
| et al., Display technologies: application for the discovery of drug and gene delivery | |||

