ОБЛАСТЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ:
Настоящее изобретение относится к областям генофонда микроорганизмов и генной инженерии, а в частности к клонированию, анализу и исследованию функций биосинтетических генных кластеров антибиотиков в генной инженерии, а также к применению этих кластеров.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ:
Карримицин, также известный под наименованиями шэнцзимицин и биотекспирамицин - это 16-членный макролидный антибиотик, разработанный посредством синтетической биологической технологии [приводится в патентах под номерами ZL 971044406 и ZL 021487715]. Карримицин представляет собой спирамицин с мультиацилированным 4-гидроксилом, основными компонентами которого являются 4''-изовалерил-спирамицин III, II и I, причем доля компонента III составляет от 30% и более, компонента II - около 25%, а компонента I - не более 10%.
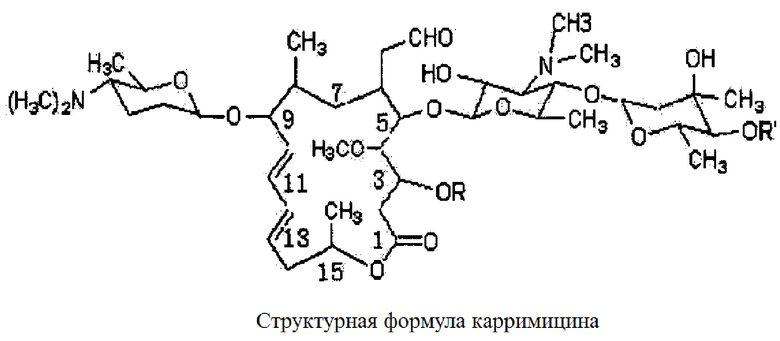
Изовалерил-спирамицин III: R=COCH2CH3 R'=COCH2CH(CH3)2
Изовалерил-спирамицин II: R=COCH3 R'=СОСН2СН(СН3)2
Изовалерил-спирамицин I: R=H R'=СОСН2СН(СН3)2
Карримицин оказывает относительно сильное воздействие на грампозитивные бактерии, а также обладает противобактериальным действием в отношении эритромицина, бактерий, резистентных к антибиотикам на основе бета-лактамов, гемофильной палочки, гонококка, легионеллы, бактероидов bacteroides fragillis и бактерий Clostridium perfringens. В частности, карримицин оказывает относительно сильное воздействие на микоплазмы пневмонии, хламидию трахоматис и хламидию пневмонии [Ю, Ланьсян и соавт. Sichuan Journal of Physiological Sciences; 1998, 20(3), патент №: 2003101224209] и обладает лучшими постантибиотическими и антибиотическими суб-МИК-эффектами. У карримицина отсутствует перекрестная лекарственная резистентность при использовании в комплексе с аналогичными препаратами. Согласно фармакокинетическим исследованиям карримицин обладает повышенной липофильностью и высоким внутриклеточным противобактериальным действием, высокой скоростью абсорбции при пероральном приеме, высокими показателями абсолютной биологической доступности, проницаемости тканей, широты распределения в тканях, времени периода полувыведения и длительности времени удерживания при введении in-vivo, а также показателями концентрации карримицина в тканях выше концентрации плазмы в крови [Сунь, Ливэнь и соавт. Chinese Pharmacological Bulletin 2000, 16 (6): 694-8; Чжун, Дафан и соавт. J chromatography В. 2003, 791: 45; Си, Сянгуо и соавт. Asian Journal of Drug Metabolism and Pharmacokinetics. 2003, 3 (2): 134; Си, Сянгуо и соавт. Chinese Chemical letter 2004, 15: 431; Си, Сянгуо и соавт. ActaPharmacologicaSinica, 2004, 25: 1396]. Согласно результатам фармакологических, токсикологических и завершенных трехэтапных клинических исследований карримицин применяется при лечении инфекций дыхательных путей, обладает выраженным эффектом лечения и коэффициентом побочных реакций, и в частности слабо воздействует на печень и обладает хорошими показателями безопасности [Линь, Футянь и соавт. Подборка документов восьмого всегосударственного симпозиума 1997 г., стр. 167; Чжао, Чуньянь и соавт. Chinese Journal of Antibiotics, 1998, 23 (4): 306; Сунь, Tao и соавт. Chinese Journal of Antibiotics, 2001, 26 (1): 49-51]. Карримицин - продукт непосредственной ферментации бактерии, полученной посредством генной инженерии, путем использования технологии рекомбинации генов. Процесс приготовления прост и удобен, и во время него легко предотвращается химическое загрязнение, а также осуществляется энергосбережение. Пероральные формы карримицина удобно принимать, и в день требуется всего один прием, что полезно для соблюдения пациентами лечения, а также удобно при включении в ряд базовых препаратов, входящих в медицинскую страховку.
Карримицин - продукт ферментации бактерий, полученных посредством генной инженерии (Streptomyces spiramyceticus WSJ-1), получаемый за счет клонирования экспрессии генов 4-изовалерил-трансферазы бактерий, вырабатывающих карбомицин, в бактериях, вырабатывающих спирамицин (Streptomyces spiramyceticus F21). Бактерии, вырабатывающие спирамицин (Streptomyces spiramyceticus F21), были извлечены нашей лабораторией из почвы в 1982 году в уезде Юнчан провинции Ганьсу, Китай. Морфологические, физиологические и биохимические характеристики, химический состав стенок клеток, нуклеотидная последовательность гена рРНК 16S и 5 белков конститутивного гена этих бактерий при анализе кладограммы не имеют ничего общего с известными зарубежными бактериями Streptomyces ambofaciens АТСС23877, вырабатывающими спирамицин, и спиромицетами. Таким образом, весьма вероятно, что настоящие бактерии, вырабатывающие спирамицин (Streptomyces spiramyceticus F21), являются новым видом стрептомицет [Даи, Цзяньлу и соавт. Journal of Microbiology, 2012, 39(4): 503-514].
Было выполнено секвенирование генных кластеров, относящихся к биосинтезу спирамицина у бактерий Streptomyces ambofaciens АТСС23877, вырабатывающих его [Каррей Ф. Microbiology, 2007, 153: 4111-4122], а также были составлены отчеты о последовательностях биосинтетических генных кластеров других макролидных антибиотиков, таких как авермектин, канавалиамицин, эритромицин, халкомицин, тилозин и медемицин [Икэда X. и соавт. Nat. Biotechnol. 2003, 21(5): 526-531, Хэйдок и соавт. Microbiology, 2005, 151, 3161-3169; Олийнык М. и соавт. Nat. Biotechnol. 2007, 25(4): 447-453; Уордс Л. и соавт. Antimicrob. Agents & Chemotherapy, 2004, 48(12): 4703-4712; Кандифф Э. и соавт. Antonie Van Leeuwenhoek, 2001, 79(3-4): 229-234; Мидох Наоки и соавт. Патент США 7070980]. Биосинтетические генные кластеры макролидных антибиотиков обладают полной длиной около 50-80 т.п.н. и общей характеристикой, заключающейся в том, что они состоят из поликетидсинтазы (ПКС) для считывания биосинтетической модульной структуры из 16-членного макролидного кольца, ферментов, связанных с единицей интенсификации синтеза поликетона, ферментов, отвечающих за модификацию разных групп радикалов лактонового кольца, а также включает гены синтеза и переноса гликозила, гены, связанные с функциями резистентности, регулирования и контроля, и т.д. Макролиды образуются посредством выполнения непрерывной реакции конденсации для катализа определенных простых молекул карбоновой кислоты с помощью ПКС, состоящей из модульных структур, способом, аналогичным используемому при биосинтезе жирных кислот. Каждый модуль отвечает только за одноэтапную реакцию конденсации в процессе формирования поликетонной цепочки, и такой модуль содержит как минимум один структурный домен бета-кетоацилсинтетазы (КС), один структурный домен ацилтрансферазы (AT) и один структурный домен ацилпереносящего белка (АПБ). Кроме того, этот модуль может дополнительно включать структурный домен бета-кетоацилредуктазы (КР), дегидразы (ДГ) и эфир-ацилредуктазы (ЭР), а также этими структурными доменами определяется степень редукции добавляемых единиц интенсификации. В то же время, действие структурного домена тиоэстеразы (ТЭ) также требует катализа циклического соединения и высвобождения поликетонных цепочек. В заключение, также требуется выполнить этапы модификации, такие как гидроксилирование, метилирование, метоксилирование и ацилирование, для образования различных структур макролидных антибиотиков. В целом, все макролиды связаны с гликозильными (или гликозиламинными) группами в разных количествах. Например, карримицин включает три гликозильные группы, а именно: форозамин, микаминозу и микарозу. Гликозильные группы задействуются ферментами, связанными с синтезом и переносом гликозила. Резистентные гены наделяют вырабатывающие бактерии способностью противостоять антибиотику собственной выработки и в целом относятся к белкам-переносчикам ABC. Гены, отвечающие за функции регуляции и контроля, участвуют в регуляции и контроле антибиотиков, производимых в результате собственного биосинтеза.
С помощью информации в последовательностях генных кластеров и структурного анализа можно производить дополнительные манипуляции с генами в отношении вырабатывающих бактерий для получения новых и более эффективных антибиотиков. Например, новые макролидные антибиотики получают посредством изменения модульных структур синтеза ПКС макролидных антибиотиков путем манипуляций с генами, при этом изменяя лактоновое кольцо после модификации и заменяя или модифицируя гликозильные группы. Количество получаемых антибиотиков можно повысить за счет выполнения генетической операции на резистентных или регуляторных генах. [Уилкинсон Б. и соавт. Chem Biol. 2000, 7 (2): 111-117; Кальц Л. и соавт. Med Res Rev. 1999, 19 (6): 543-58; Гудман К.Д. Антимикробные агенты и химиотерапия, 2013, 57(2): 907-913; Вон В. и соавт. ProcNatlAcadSci USA, 2014, 111(15): 5688-93; Стратигопулос и соавт. MolMicrobiol. 2004, 54(5): 1326-34; Новакова Р. и соавт. Folia Microbiol. 2011, 56(3): 276-82].
СУЩНОСТЬ ИЗОБРЕТЕНИЯ:
Сущностью настоящего изобретения является биосинтетический сцепленный генный кластер карримицина. Биосинтетический генный кластер в сумме включает 44 открытые рамки считывания генов (ОРС). Полный размер нуклеотидной последовательности составляет 89 315 н.п. (посл. 1). Генный кластер включает 5 ОРС (10-14), кодирующих поликетидсинтазу, состоящих из 8 модулей и 37 структурных доменов; 9 ОРС (1, 4-6, 15 и 36-39), относящихся к единице интенсификации синтеза и модификации поликетона; 16 ОРС (9, 16-22, 24, 26, 28, 29, 33-35 и 41), относящихся к синтезу гликозила; и 6 ОРС (7, 8, 30-32 и 40), относящихся к переносу гликозила. Кроме того, генный кластер дополнительно включает 2 ОРС (3 и 25), относящихся к резистентности и 4 ОРС (2, 23, 27 и 42), вероятно, относящихся к регуляции и контролю. Нуклеотидные последовательности отдельно отобраны из группы, включающей: ОРС 1 (1-645), ОРС 2 (1 810-1 208), ОРС 3 (3 133-2 285), ОРС 4 (3 614-4 840), ОРС 5 (4 846-5 511), ОРС 6 (7 150-5 801), ОРС 7 (8 444-7 179), ОРС 8 (9 729-8 482), ОРС 9 (10 543-9 830), ОРС 10 (16 215-10 543), ОРС И (21 076-16 328), ОРС 12 (32 511-21 124), ОРС 13 (38 599-32 585), ОРС 14 (52 259-38 643), ОРС 15 (53 099-54 310), ОРС 16 (54 495-54 845), ОРС 17 (54 842-56 041), ОРС 18 (56 038-56 946), ОРС 19 (56 930-57 967), ОРС 20 (57 937-60 174), ОРС 21 (60 836-61 984), ОРС 22 (62 796-62 077), ОРС 23 (63 633-65 645), ОРС 24 (67 379-66 318), ОРС 25 (69 004-67 352), ОРС 26 (69 349-70 650), ОРС 27 (72 156-70 708), ОРС 28 (72 422-73 462), ОРС 29 (74 601-73 561), ОРС 30 (74 913-76 160), ОРС 31 (76 218-77 486), ОРС 32 (77 606-78 781), ОРС 33 (78 783-79 775), ОРС 34 (79 772-80 779), ОРС 35 (82 055-80 823), ОРС 36 (83 164-82 052), ОРС 37 (84 400-83 279), ОРС 38 (84 713-84 393), ОРС 39 (85 576-84 710), ОРС 40 (85 825-87042), ОРС 41 (87 094-87 702) и ОРС 42 (89 315-88 143) в посл. 1. Кроме того, генный кластер дополнительно включает ОРС 43 (866-60) и ОРС 44 (2 337-1 174) в посл. 2 экзогенных генов, расцепленных с посл. 1, и с полным размером 2 337 п.н.
Настоящим изобретением также предлагается последовательность аминокислот 4-фосфопантетеинил-трансферазы (ФПТ), и эта последовательность состоит из 214 аминокислот в посл. 3. Она называется IA-W1, а нуклеотидная последовательность кодирующего гена выбирается из оснований 1-645 в посл. 1.
Настоящим изобретением также предлагается фактор регуляции и контроля переноса последовательности аминокислот семейства TetR, и эта последовательность состоит из 200 аминокислот в посл. 4. Она называется IA-W2, а нуклеотидная последовательность кодирующего гена выбирается из оснований 1 810-1 208 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот рРНК-метилтрансферазы 23 S, и эта последовательность состоит из 282 аминокислот в посл. 5. Она называется IA-W3, а нуклеотидная последовательность кодирующего гена выбирается из оснований 3 133-2 285 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот 3-О-ацилтрасферазы, и эта последовательность состоит из 408 аминокислот в посл. 6. Она называется IA-W4, а нуклеотидная последовательность кодирующего гена выбирается из оснований 3 614-4 840 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот О-метилтрасферазы, и эта последовательность состоит из 221 аминокислот в посл. 7. Она называется IA-W5, а нуклеотидная последовательность кодирующего гена выбирается из оснований 4 846-5 511 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот кротонил-КоА-редуктазы, и эта последовательность состоит из 449 аминокислот в посл. 8. Она называется IA-W6, а нуклеотидная последовательность кодирующего гена выбирается из оснований 7 150-5 801 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот гликозилтрансферазы, и эта последовательность состоит из 421 аминокислот в посл. 9. Она называется IA-W7, а нуклеотидная последовательность кодирующего гена выбирается из оснований 8 444-7 179 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот акцессорного белка гликозилтрансферазы, и эта последовательность состоит из 415 аминокислот в посл. 10. Она называется IA-W8, а нуклеотидная последовательность кодирующего гена выбирается из оснований 9 729-8 482 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-аминогексозы-N-диметилтрансферазы, и эта последовательность состоит из 237 аминокислот в посл. 11. Она называется IA-W9, а нуклеотидная последовательность кодирующего гена выбирается из оснований 10 543-9 830 в посл. 1.
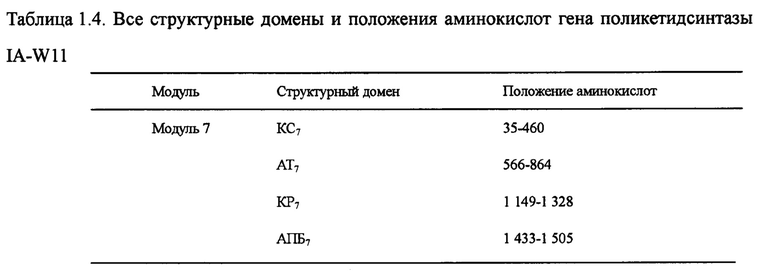
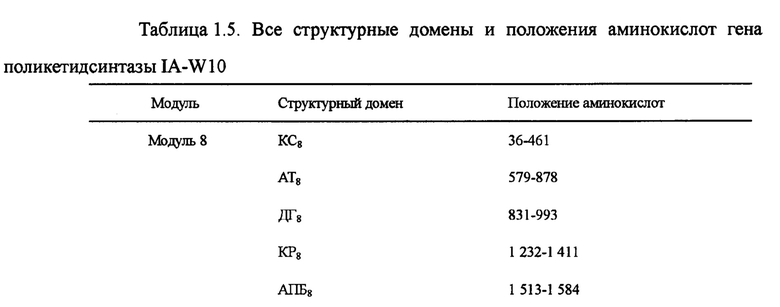
Настоящим изобретением также предлагается последовательность аминокислот, включающая структурный домен поликетидсинтазы, состоящий из кетосинтазы (КС)8-ацилтрансферазы (АТ)8-кеторедуктазы (КР)8-ацилпереносящего белка (АПБ)8-высвобождающей цепочку тиоэстеразы (ТЭ), и эта последовательность состоит из 1 890 аминокислот в посл. 12. Она называется IA-W10, а нуклеотидная последовательность кодирующего гена выбирается из оснований 16215-10543 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот, включающая структурный домен поликетидсинтазы, состоящий из КС7-АТ7-КР7-АПБ7, и эта последовательность состоит из 1 582 аминокислот в посл. 13. Она называется IA-W11, а нуклеотидная последовательность кодирующего гена выбирается из оснований 21 076-16 328 в посл. 1.
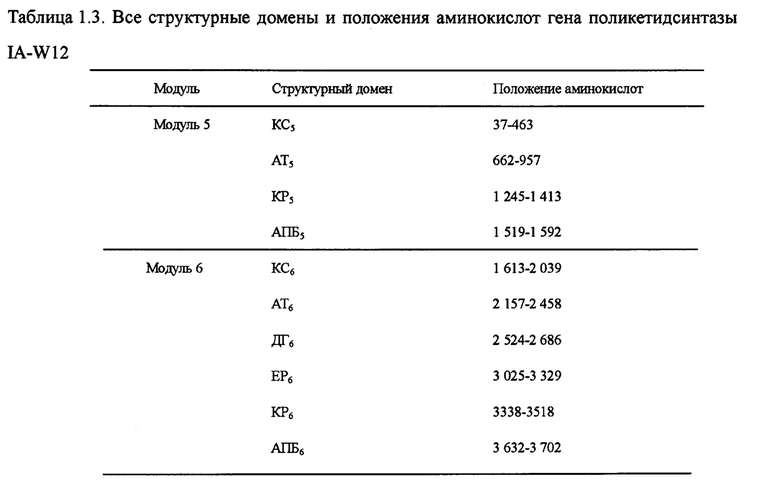
Настоящим изобретением также предлагается последовательность аминокислот, включающая структурный домен поликетидсинтазы, состоящий из КС5-АТ5-КР5-АПБ5-КС6-АТ6-ДГ6 (дегидразы)-ЕР6 (еноилредуктазы)-КР6-АПБ6, и эта последовательность состоит из 3 795 аминокислот в посл. 14. Она называется IA-W12, а нуклеотидная последовательность кодирующего гена выбирается из оснований 32 511-21 124 в посл. 1.
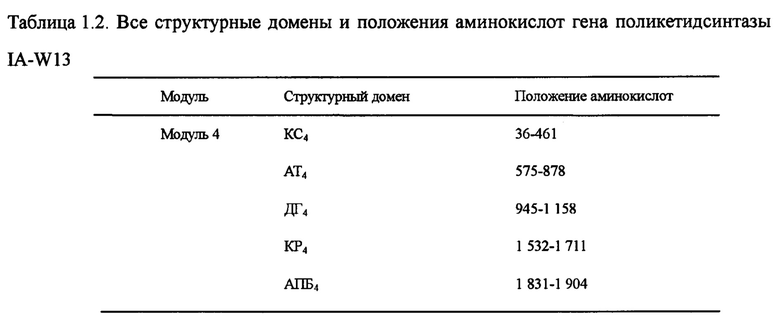
Настоящим изобретением также предлагается последовательность аминокислот, включающая структурный домен поликетидсинтазы, состоящий из КС4-АТ4-ДГ4-КР4-АПБ4, и эта последовательность состоит из 2 004 аминокислот в посл. 15. Она называется IA-W13, а нуклеотидная последовательность кодирующего гена выбирается из оснований 38 599-32 585 в посл. 1.
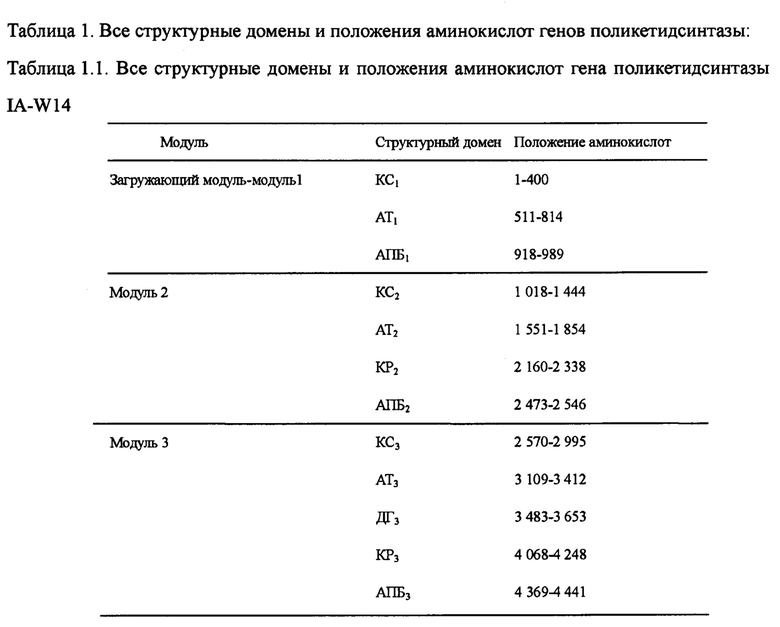
Настоящим изобретением также предлагается последовательность аминокислот, включающая структурный домен поликетидсинтазы, состоящий из КС1-АТ1-АПБ1-КС2-АТ2-КР2-АПИ2-КС3-АТ3-ДГ3-КР3-АПБ3, и эта последовательность состоит из 4 538 аминокислот в посл. 16. Она называется IA-W14, а нуклеотидная последовательность кодирующего гена выбирается из оснований 52 259-38 643 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот цитохром P-450-оксидазы, и эта последовательность состоит из 403 аминокислот в посл. 17. Она называется IA-W15, а нуклеотидная последовательность кодирующего гена выбирается из оснований 53 099-54 310 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-гексозы-изомеразы, и эта последовательность состоит из 116 аминокислот в посл. 18. Она называется IA-W16, а нуклеотидная последовательность кодирующего гена выбирается из оснований 54 495-54 845 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-гексозы-аминотрансферазы, и эта последовательность состоит из 399 аминокислот в посл. 19. Она называется IA-W17, а нуклеотидная последовательность кодирующего гена выбирается из оснований 54 842-56 041 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-глюкозы-синтазы, и эта последовательность состоит из 302 аминокислот в посл. 20. Она называется IA-W18, а нуклеотидная последовательность кодирующего гена выбирается из оснований 56 038-56 946 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-глюкозы-4, 6-дегидразы, и эта последовательность состоит из 345 аминокислот в посл. 21. Она называется IA-W19, а нуклеотидная последовательность кодирующего гена выбирается из оснований 56 930-57 967 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-гексозы-2, 3-дегидразы/тиоэстеразы, и эта последовательность состоит из 745 аминокислот в посл. 22. Она называется IA-W20, а нуклеотидная последовательность кодирующего гена выбирается из оснований 57937-60174 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-гексозы-аминотрансферазы, и эта последовательность состоит из 382 аминокислот в посл. 23. Она называется IA-W21, а нуклеотидная последовательность кодирующего гена выбирается из оснований 60 836-61 984 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-аминогексозы-N-диметилтрансферазы, и эта последовательность состоит из 239 аминокислот в посл. 24. Она называется IA-W22, а нуклеотидная последовательность кодирующего гена выбирается из оснований 62 796-62 077 в посл. 1.
Настоящим изобретением также предлагается фактор регуляции и контроля переноса последовательности аминокислот, и эта последовательность состоит из 670 аминокислот в посл. 25. Она называется IA-W23, а нуклеотидная последовательность кодирующего гена выбирается из оснований 63 633-65 645 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-аминогексозы-изомеразы, и эта последовательность состоит из 354 аминокислот в посл. 26. Она называется IA-W24, а нуклеотидная последовательность кодирующего гена выбирается из оснований 67 379-66 318 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот белка-переносчика ABC, и эта последовательность состоит из 550 аминокислот в посл. 27. Она называется IA-W25, а нуклеотидная последовательность кодирующего гена выбирается из оснований 69 004-67 352 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-гексозы-дегидразы, и эта последовательность состоит из 354 аминокислот в посл. 28. Она называется IA-W26, а нуклеотидная последовательность кодирующего гена выбирается из оснований 69 349-70 650 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот аналогичного ГТП-фермента, и эта последовательность состоит из 482 аминокислот в посл. 29. Она называется IA-W27, а нуклеотидная последовательность кодирующего гена выбирается из оснований 72 156-70 708 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-гексозы-изомеразы, и эта последовательность состоит из 346 аминокислот в посл. 30. Она называется IA-W28, а нуклеотидная последовательность кодирующего гена выбирается из оснований 72 422-73 462 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-гексозы-кеторедуктазы, и эта последовательность состоит из 346 аминокислот в посл. 31. Она называется IA-W29, а нуклеотидная последовательность кодирующего гена выбирается из оснований 74 601-73 561 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот акцессорного белка гликозилтрансферазы, и эта последовательность состоит из 415 аминокислот в посл. 32. Она называется IA-W30, а нуклеотидная последовательность кодирующего гена выбирается из оснований 74 913-76 160 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот гликозилтрансферазы, и эта последовательность состоит из 422 аминокислот в посл. 33. Она называется IA-W31, а нуклеотидная последовательность кодирующего гена выбирается из оснований 76 218-77 486 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот гликозилтрансферазы, и эта последовательность состоит из 391 аминокислот в посл. 34. Она называется IA-W32, а нуклеотидная последовательность кодирующего гена выбирается из оснований 77 606-78 781 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-гексозы-кеторедуктазы, и эта последовательность состоит из 330 аминокислот в посл. 14. Она называется IA-W33, а нуклеотидная последовательность кодирующего гена выбирается из оснований 78 783-79 775 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-гексозы-редуктазы, и эта последовательность состоит из 335 аминокислот в посл. 36. Она называется IA-W34, а нуклеотидная последовательность кодирующего гена выбирается из оснований 79 772-80 779 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-гексозы-метилтрансферазы, и эта последовательность состоит из 410 аминокислот в посл. 37. Она называется IA-W35, а нуклеотидная последовательность кодирующего гена выбирается из оснований 82 055-80 823 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот метокси-малонил-синтетазы, и эта последовательность состоит из 370 аминокислот в посл. 38. Она называется IA-W36, а нуклеотидная последовательность кодирующего гена выбирается из оснований 83 164-82 052 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот дегидрогеназы, и эта последовательность состоит из 373 аминокислот в посл. 39. Она называется IA-W37, а нуклеотидная последовательность кодирующего гена выбирается из оснований 84 400-83 279 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот ацилпереносящего белка, и эта последовательность состоит из 106 аминокислот в посл. 40. Она называется IA-W38, а нуклеотидная последовательность кодирующего гена выбирается из оснований 84 713-84 393 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот метокси-малонил-дегидрогеназы, и эта последовательность состоит из 288 аминокислот в посл. 41. Она называется IA-W39, а нуклеотидная последовательность кодирующего гена выбирается из оснований 85 576-84 710 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот гликозилтрансферазы, и эта последовательность состоит из 405 аминокислот в посл. 42. Она называется IA-W40, а нуклеотидная последовательность кодирующего гена выбирается из оснований 85 825-87 042 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот NDP-гексозы-изомеразы, и эта последовательность состоит из 202 аминокислот в посл. 43. Она называется IA-W41, а нуклеотидная последовательность кодирующего гена выбирается из оснований 87 094-87 702 в посл. 1.
Настоящим изобретением также предлагается фактор регуляции и контроля переноса последовательности аминокислот белка, и эта последовательность состоит из 390 аминокислот в посл. 44. Она называется IA-W42, а нуклеотидная последовательность кодирующего гена выбирается из оснований 89 315-88 143 в посл. 1.
Настоящим изобретением также предлагается последовательность аминокислот экзогенной рРНК-метилазы 23 S (относящаяся к тиострептону и маркерному гену, резистентному к тиострептону), и эта последовательность состоит из 269 аминокислот в посл. 45. Она называется IA-W43, а нуклеотидная последовательность кодирующего гена выбирается из оснований 866-57 в посл. 2.
Настоящим изобретением также предлагается последовательность аминокислот экзогенной 4-микарозы-глюкозид-изовалерил-трансферазы, и эта последовательность состоит из 388 аминокислот в посл. 46. Она называется IA-W44, а нуклеотидная последовательность кодирующего гена выбирается из оснований 2337-1 171 в посл. 2.
На основании получения информации биосинтетического генного кластера карримицина и анализа возможных функций кодированных белков каждого гена посредством блокировки генов и гомологичного сопоставления, дополнительно приводятся все 44 гена биосинтетического генного кластера карримицина по настоящему изобретению, и этот генный кластер имеет структуру, изображенную на фиг. 1, а именно:
(1) пять генов поликетидсинтазы, включающих ОРС 10-14;
(2) девять генов, относящихся к единице интенсификации синтеза и модификации поликетона, включающих ОРС 1, ОРС 4-6, 15 и 36-39;
(3) шестнадцать генов, относящихся к синтезу гликозила, включающих ОРС 9, 16-22, 24, 26, 28, 29, 33-35 и 41;
(4) шесть генов, относящихся к переносу гликозила, включающих ОРС 7, 8, 30-32 и 40;
(5) два гена, относящихся к резистентности, включающих ОРС 3 и 25;
(6) 4 гена, относящихся к регуляции и контролю биосинтеза в целом, включающих ОРС 2, 23, 27 и 42; и
(7) два гена, включающих экзогенные маркерные гены, полученные посредством генной инженерии, ОРС 43 (тиострептон и ген, резистентный к тиострептону) и ген микарозы-4'-изовалерил-трансферазы ОРС 44, связанный с ОРС 43.
Пять генов поликетидсинтазы (ОРС 10-14) в посл. 1., комплементарные последовательности нуклеотидов и аминокислот которых очень важны для синтеза лактонового кольца карримицина. Пять генов поликетидсинтазы включают в себя 8 модулей и 37 структурных доменов, приведенных на фиг. 2 ОРС 14 включает 3 модуля: загружающий структурный домен 1, модуль 2 и модуль 3. В загружающем структурном домене KC1, AT1 и АПБ1 отвечают за начальный синтез лактонового кольца, а в качестве инициирующей единицы катализируется уксусная кислота. Модуль 2 включает структурные домены КС2, АТ2, КР2 и АПБ2, а модуль 3 - структурные домены КС3, АТ3, ДГ3, КР3 и АПБ3, и он отвечает за внедрение дополнительных 2 единиц интенсификации уксусной кислоты для окончательного образования каркасного участка с углеродной цепочкой С11-15 карримицина. ОРС 13 включает модуль 4, состоящий из КС4, АТ4, ДГ4, КР4 и АПБ4 и отвечающий за интенсификацию третьей единицы уксусной кислоты для окончательного образования каркасного участка с углеродной цепочкой С9-10 карримицина. ОРС 12 включает в себя модули 5 и 6, а модуль 5 состоит из структурного домена КС5-АТ5-КР5-АПБ5 и отвечает за внедрение единицы интенсификации пропионовой кислоты. Модуль 6 состоит из структурного домена КС6-АТ6-ДГ6-КР6-ЕР6-КР6-АПБ6 и отвечает за внедрение единицы интенсификации масляной кислоты для окончательного образования каркасного участка с углеродной цепочкой С5-С8 карримицина. ОРС 11 включает в себя модуль 7, состоящий из структурного домена КС7-АТ7-КР7-АПБ7, и отвечает за внедрение единицы интенсификации гликолевой кислоты для окончательного образования каркасного участка с углеродной цепочкой С3-С4 карримицина. ОРС 10 включает в себя модуль 8, состоящий из структурного домена КС8-АТ8-КР8-АПБ8-ТЭ, и отвечает за внедрение единицы интенсификации уксусной кислоты, а циклическое соединение и высвобождение углеродной цепочки выполняется за счет участия тиоэстеразы (ТЭ). На фиг. 2 приведена структурная схема генов поликетидсинтазы карримицина. В таблице 1 приведены все структурные домены и положения аминокислот генов поликетидсинтазы.
Нуклеотидные или комплементарные последовательности и их соответствующие последовательности аминокислот единицы интенсификации синтеза поликетона, а также гены ОРС 1, ОРС 4-6, 15 и 36-39, связанные с модификацией, представляют собой: IA-W1, кодирующий ФПТ-модифицированный поликетидсинтезированный ацилпереносящий белок (АПБ) для обеспечения возможности белку стать активным белком; IA-W4, кодирующий 3-О-ацилтрансферазу и отвечающий за ацилирование 3-позиционного гидроксила карримицина; IA-W5 и IA-W6, отдельно кодирующие О-метилазу и кротонил-КоА-редуктазу и отвечающие за обеспечение единиц интенсификации поликетида; IA-W15, кодирующий цитохром-Р450 - монооксидазу и отвечающий за окисление углеродной цепочки поликетида; а также IA-W36-39, отдельно кодирующие метокси-малонил-синтетазу, дегидрогеназу, ацилпереносящий белок и метокси-малонил-дегидрогеназу. Также все гены участвуют в синтезе и модификации единиц интенсификации поликетида.

Количество генов, относящихся к гликозильному синтезу карримицина, всего составляет 12, а именно: ОРС 9, 16-22, 24, 26, 28, 29, 33-35 и 41, причем ОРС 18, 19 и 28 кодируют ферменты синтеза, обезвоживания и изомеризации основных гликозильных единиц карримицина; ОРС 9, 20, 21, 24, 26 и 29 кодируют ферменты N-диметилирования, 2, 3-обезвоживания, аминирования, изомеризации, обезвоживания и кеторедукции NDP-гексозамина в синтезе форозамина; ОРС 16, 17 и 22 кодируют ферменты изомеризации, аминирования и N-диметилирования NDP-гексозамина микаминозы; а ОРС 33, 34, 35 и 41 кодируют ферменты кеторедукции, метилирования и изомеризации NDP-гексозамина микарозы.
Количество генов, относящихся к гликозильному переносу карримицина, всего составляет 6, а именно: ОРС 7, 8, 30-32 и 40, причем ОРС 7 кодирует гликозилазу микаминозы; ОРС 8 кодирует акцессорный белок гликозилации микаминозы; ОРС 31 и 32 кодируют гликозилазу форозамина; ОРС 30 кодирует акцессорный белок гликозилации форозамина; а ОРС 40 кодирует фермент гликозилации микарозы.
Количество генов, относящихся к резистентности карримицина, всего составляет 2, а именно: ОРС 3 и 25, причем ОРС 3 кодирует рРНК-метилазу 23S; а ОРС 25 кодирует белок-переносчик ABC. ОРС 3 и ОРС 25 наделяют бактерии, вырабатывающие карримицин, резистентностью к собственному антибиотику посредством механизма метилирования и накачки рибосомной РНК.
Количество генов карримицина, относящихся к регуляции и контролю биосинтеза, всего составляет 4, а именно: ОРС 2, 23, 27 и 42, причем ОРС 2 кодирует фактор ингибирования регуляции и контроля переноса последовательности аминокислот семейства TetR и, вероятно, участвует в отрицательной регуляции и контроле биосинтеза карримицина. ОРС 23 и 42 отдельно кодируют два фактора переноса положительной регуляции и контроля, и этот фактор переноса служит в качестве специального фактора положительной регуляции и контроля пути, а также используется для непосредственной регуляции и контроля биосинтеза карримицина. ОРС 27 кодирует ГТП-фермент и, вероятно, регулирует и контролирует биосинтез карримицина посредством функций регуляции и контроля клеток.
Экзогенные ОРС 43 и 44 относятся к биосинтезу карримицина, причем ОРС 43 кодирует ген рРНК-метилазы 23 S, относящийся к резистентности тиострептона. Этот ген связан с геном микарозы 4''-O-гидроксил-изовалерил-трансферазы ОРС 44, а экспрессия ОРС 43 может обеспечивать наличие идентифицирующего маркера для бактерии карримицина, полученной посредством генной инженерии.
Комплементарные последовательности согласно посл. 1 и посл. 2 по настоящему изобретению могут быть получены в любое время согласно принципу комплементарного спаривания оснований. Нуклеотидные последовательности или части нуклеотидных последовательностей согласно посл. 1 и посл. 2 могут быть получены посредством полимеразной цепной реакции (ПЦР) или расщепления ферментом соответствующей ДНК путем использования соответствующей рестрикционной эндонуклеазы или других соответствующих технологий. Гены, аналогичные генам биосинтеза карримицина, могут быть получены из других организмов посредством нуклеотидных последовательностей или их частей, предлагаемых в настоящем изобретении, путем применения метода полимеразной цепной реакции (ПНР) или метода Саузерн-гибридизации за счет использования ДНК, содержащей последовательности по настоящему изобретению, в качестве зонда.
Настоящим изобретением также предлагается способ получения как минимум части последовательности ДНК согласно посл. 1 и посл. 2 для построения рекомбинантного вектора.
Настоящим изобретением также предлагается способ блокирования генов биосинтеза карримицина, причем как минимум один из генов включает нуклеотидные последовательности согласно посл. 1.
Новые компоненты карримицина могут быть получены посредством блокирования одного или нескольких этапов биосинтеза карримицина путем использования клонированных генов или фрагментов ДНК нуклеотидных последовательностей или как минимум части этих последовательностей, раскрываемых в настоящем изобретении. Нуклеотидные последовательности включают фрагменты ДНК или гены, и их можно использовать для повышения количества получаемого карримицина или его компонентов.
Клонированная ДНК нуклеотидных последовательностей или как минимум их части, предлагаемая в настоящем изобретении, может быть использована для размещения большего количества клонотечных плазмид из геномной библиотеки. Эти клонотечные плазмиды включают как минимум часть последовательностей по настоящему изобретению, а также содержат ДНК регионов, прилегающих к клонотечным плазмидам, в геноме бактерий, вырабатывающих карримицин.
Нуклеотидные последовательности, предлагаемые в настоящем изобретении, могут модифицироваться или мутировать. Способы модификации или мутации включают в себя генную вставку или замещение, полимеразную цепную реакцию, ошибочно-опосредованную полимеразную цепную реакцию, сайт-специфическую мутацию, воссоединение разных последовательностей и мутацию под действием ультрафиолета или химического реагента.
Нуклеотидные последовательности, предлагаемые в настоящем изобретении, могут непосредственно изменяться (перестановка в ДНК) через разные части последовательностей или гомологические последовательности из других источников.
Фрагменты, структурные домены, модули, гены нуклеотидных последовательностей или как минимум их частей, предлагаемые в настоящем изобретении, могут быть использованы для построения библиотеки поликетидсинтаз или их компонентов, или библиотеки наборов. Новые поликетонные соединения получают посредством удаления или деактивации одного или нескольких структурных доменов поликетидсинтазы, модулей или генов тех же или других систем поликетидсинтазы или же путем удаления одного или нескольких структурных доменов, модулей или генов поликетидсинтазы.
Нуклеотидные последовательности генов-модификаторов и генов гликозильного синтеза и гликозилтрансферазы в настоящем изобретении обеспечивают способ получения компонентов карримицина посредством удаления, замещения или преобразования этих генов синтеза кликозила, переноса и модификации.
Фрагменты, структурные домены, модули, гены нуклеотидных последовательностей или как минимум их частей, предлагаемые в настоящем изобретении, могут быть использованы для повышения количества получаемого карримицина или его компонентов посредством удвоения количества.
Экспрессия клонированных генов нуклеотидных последовательностей или как минимум их части, предлагаемых в настоящем изобретении, может обеспечиваться в экзогенных хозяевах посредством соответствующих систем экспрессии для получения модифицированного карримицина или карримицина с повышенной биологической активностью или выходом. Эти экзогенные хозяева включают в себя стрептомицеты, кишечную палочку, бациллы, дрожжи, растения, животных и т.д.
Экспрессия генов или генных кластеров нуклеотидных последовательностей или как минимум их части, предлагаемых в настоящем изобретении, может обеспечиваться в гетерологичных хозяевах, а функции этих генов или генных кластеров в метаболических цепочках хозяев изучаются с помощью технологии ДНК-чипов.
Полипептиды аминокислотных последовательностей или как минимум их части, предлагаемые в настоящем изобретении, по-прежнему могут обладать биологической активностью, даже новой биологической активностью после исключения или замещения одной или нескольких аминокислот, или могут повышаться выход, динамические характеристики, или могут быть оптимизированы другие требуемые свойства белков. Новые белки или ферменты могут быть получены посредством соединения аминокислотных последовательностей по настоящему изобретению посредством соответствующей технологии удаления, после чего могут быть получены новые результаты или связанные с ними продукты.
Аминокислотные последовательности, предлагаемые в настоящем изобретении, могут быть использованы для отделения необходимых белков и применены для получения антител.
Аминокислотные последовательности, предлагаемые в настоящем изобретении, обеспечивают возможность прогнозирования трехмерной структуры поликетидсинтазы.
Гены, предлагаемые в настоящем изобретении, белки и их антитела также могут быть использованы для скрининга и разработки составов или белков для лекарственных средств, промышленности и сельского хозяйства.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ;
Фиг. 1: структура биосинтетического генного кластера карримицина.
Фиг.2: структура гена поликетидсинтазы карримицина.
Фиг. 3: схематическое изображение строения блокирующих рекомбинантных плазмид IA-W4, гена 3-О-ацилтрансферазы и двойного обмена.
Фиг. 4: схематическое изображение строения блокирующих рекомбинантных плазмид гена регуляции и контроля переноса IA-W42 и т.д.
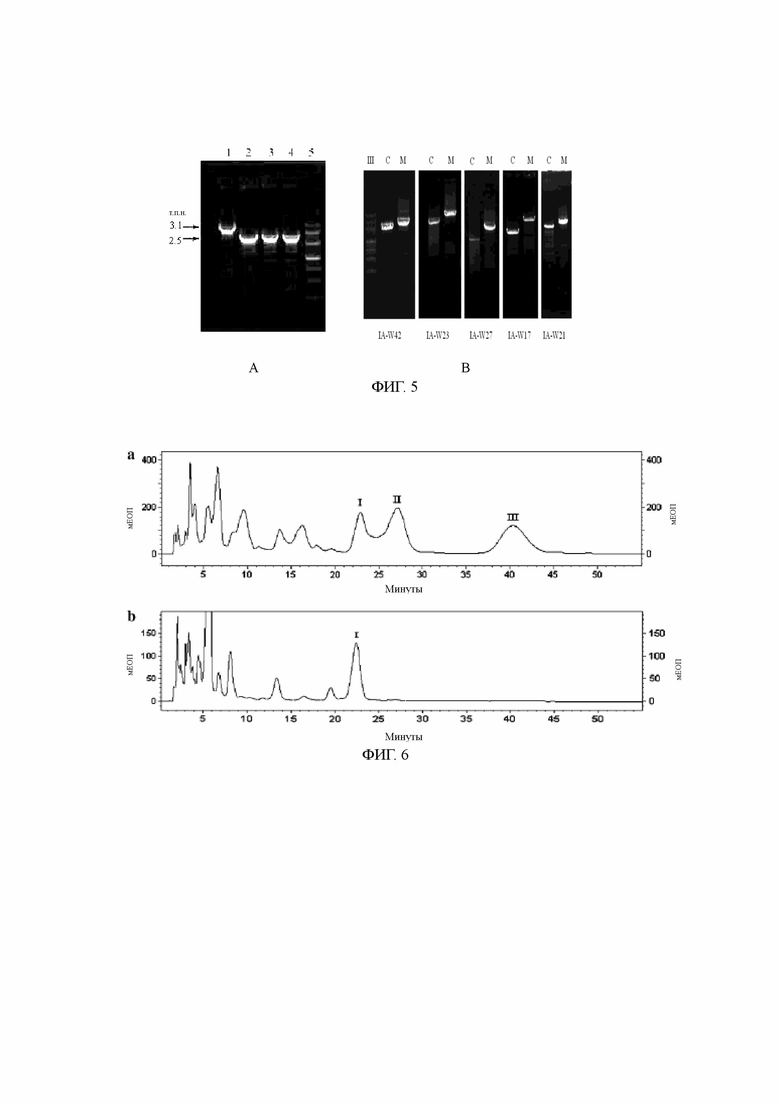
Фиг. 5А: верификация блокировки гена 3-О-ацилтрансферазы IA-W4 посредством ПЦР,
где: 1: исходный штамм; 2, 3, 4: генно-заблокированный мутант; и 5: ДНК-маркер III.
Фиг. 5В: верификация блокировки других генов посредством ПЦР,
где: III: ДНК-маркер III; С: исходный штамм; и М: генно-заблокированный мутант.
Фиг. 6: анализ продуктов ферментации генно-заблокированных мутантов 3-О-ацилтарнсферазы IA-W4 путем ВЭЖХ,
где: а: контроль карримицина; и b: ферментированный экстракт генно-заблокированного мутанта,
I, II и III - пики поглощения трех основных компонентов, т.е., изовалерил-спирамицина I, II и III карримицина соответственно.
Согласно настоящему изобретению мутантные штаммы получаются посредством опытов по блокировке генов. Опытным путем было доказано, что изменение компонента карримицина мутантных штаммов достигается за счет генной блокировки, или же карримицин более не вырабатывается. Таким образом, указывается, что полученная информация генного кластера относится к биосинтезу карримицина. Согласно настоящему изобретению экзогенный маркерный ген, резистентный к тиострептону (ОРС 43) и ген микарозы 4''-О-гидроксил-изовалерил-трансферазы (ОРС 44), связанный с ОРС 43, интегрируются в хромосомы бактерий, вырабатывающих карримицин, посредством генной гомологичной рекомбинации. При исследованиях в нашей лаборатории было доказано, что ОРС 43 и ОРС 44 важны для биосинтеза карримицина (Китайский журнал биотехнологий, том 15, выпуск 2, 1999, 171-176).
ПОДРОБНОЕ ОПИСАНИЕ;
Варианты осуществления, приведенные ниже, используются исключительно для содействия специалистам в настоящей области техники для лучшего понимания настоящего изобретения и никоим образом не ограничивают его.
<Вариант осуществления 1> Извлечение полной ДНК бактерий (S. spiramyceticus), вырабатывающих карримицин
Формула культуральной среды R2YE (г/100 мл):

добавляются 0,2 мл раствора следового элемента, а также дистиллированная вода до получения объема 100 мл с рН 6,5
Раствор следового элемента (г/100 мл):

Выполняется стерилизация 15 фунтов в течение 20 мин при температуре 121°С
Было произведено выращивание S. spiramyceticus в 25 мл культуральной среды R2YE, произведено выращивание на вибростенде в течение 48 часов при температуре 28°С. Затем был выполнен пересев в 100 мл культуральной среды R2YE, произведено выращивание на вибростенде в течение 24 часов при температуре 28°С. Затем после центрифугирования в течение 10-15 мин на скорости 5000 об/мин был собран таллом (около 10 г). В основном, операция выполнялась согласно техническим требованиям к изделию от научной компании UPTECHTMlife. К таллому были добавлены 50 мл раствора ЭДТК для промывки с вибрацией, после чего раствор подвергся центрифугированию, и был удален супернатант. Мицелий был суспендирован в 25 мл раствора лизоцима (10 мг/мл, приготовлен из 10 ммоль Трис-HCl с рН 8,0, 2 ммоль ЭДТК и TritonX-100 1,2% посредством добавления 0,5 мл рибонуклеазы из 100 мг/мл), а также было произведено выращивание в течение около 1-2 ч при температуре 37°С до пропускания клетками света. Затем были добавлены 2,5 мл раствора протеиназы К и проведено выращивание в течение 30 мин при температуре 55°С. После этого были добавлены 20 мл раствора додецилсульфата натрия 10% и проведено выращивание в течение 10 мин при температуре 70°С. Был добавлен равный объем безводного этанола и проведено полное встряхивание. Затем раствор был помещен в колонну выделения ДНК и подвержен центрифугированию в течение 1 мин на скорости 12000 об/мин. После этого для промывки колонны были добавлены 50 мл раствора с содержанием протеиназы и проведено центрифугирование в течение 1 мин на скорости 12000 об/мин при комнатной температуре. После этого колонна была дважды промыта 50 мл раствора для ополаскивания, и было проведено центрифугирование в течение 1 мин на скорости 12000 об/мин. Затем были добавлены 5-10 мл элюента ТЭ и помещены в условия комнатной температуры на 2-5 мин, после чего было проведено центрифугирование в течение 1 мин при температуре 12000 об/мин. Раствор был собран, и полная ДНК была постановлена на хранение при температуре -20°С.
<Вариант осуществления 2> Верификация функций информации генов в посл. 1 посредством блокировки гена.
Выполнена блокировка генов, таких как IA-W1, IA-W42, с двух концов генных кластеров, и для получения мутантных штаммов отобраны IA-W4, 17, 21, 23 и 27. Опытным путем установлено, что способность этих заблокированных штаммов к вырабатыванию карримицина изменилась или была утрачена. Таким образом, полученная из генного кластера информация важна для выработки карримицина. Строение праймеров выполняется согласно кодирующим генам, приведенным выше, и их выше- и нижележащим последовательностям, и они помещаются в соответствующих сайтах расщепления ферментами. Последовательности праймеров приведены в таблице 2.


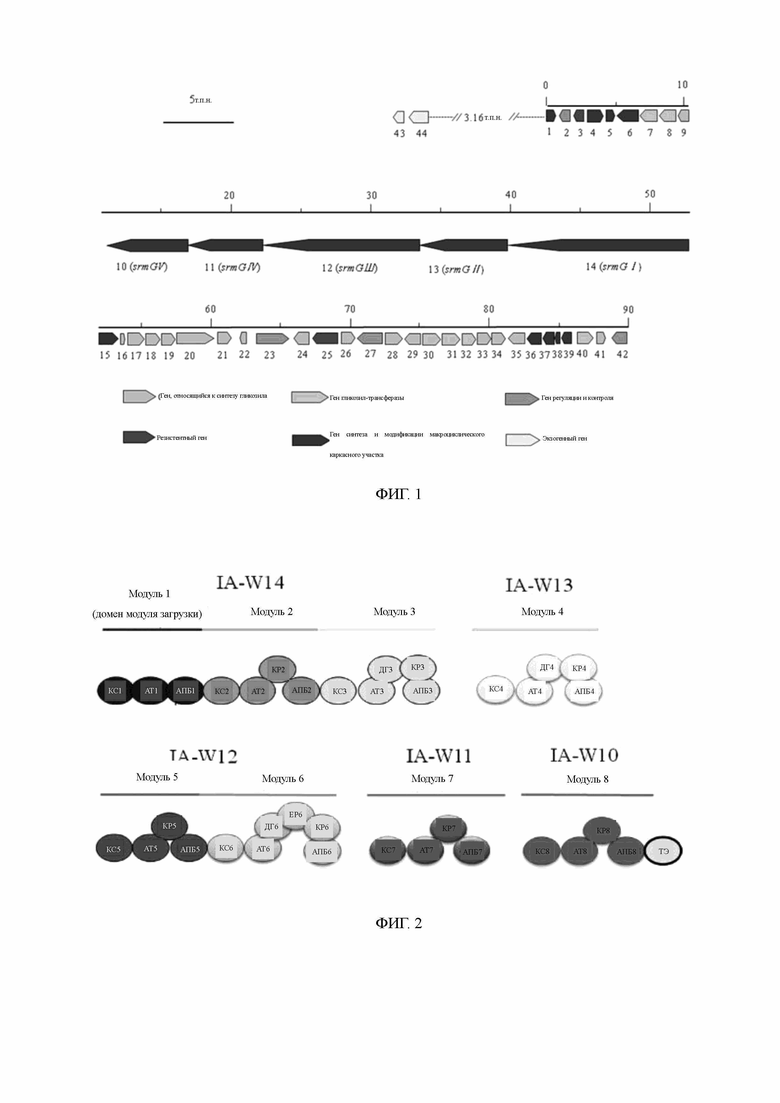
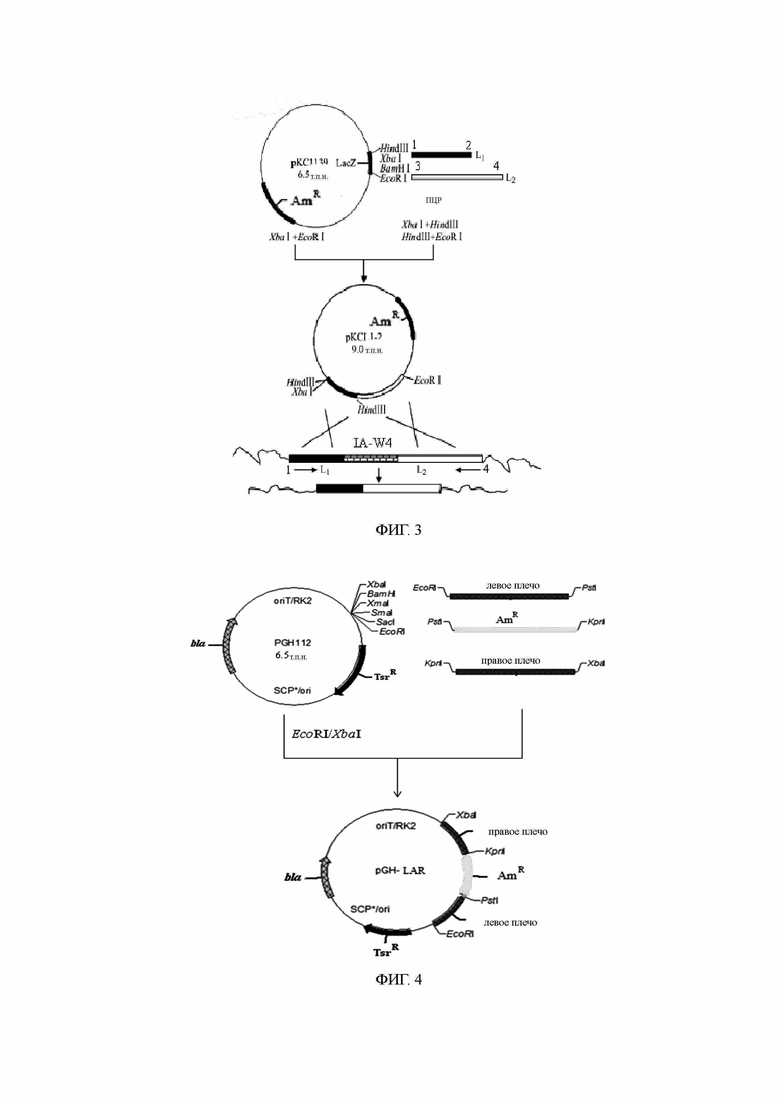
Соответствующие фрагменты гомологичных генов отдельно получаются посредством амплификации ПЦР, а рекомбинантная плазмида, содержащая гомологичные гены, была получена путем использования соответствующих сайтов расщепления ферментами, встраивания маркерных генов с резистентностью, прошедших скрининг (апрамицин, или Am) и связывания этих генов с температурочувствительным вектором рКС1139 [Бирман М. и соавт. Ген. 1992; 116(1): 43-9] или вектором кишечной палочки/стрептомицеты pGH112 [Youbao Biology Company]. Рекомбинантная плазмида была преобразована с помощью протопластов и перенесена в бактерии, вырабатывающие карримицин. После выращивания одиночные колонии были изолированы для получения штамма с гомологичным фрагментом блокирующего гена двойного обмена. Схематическое изображение строения блокирующих рекомбинантных плазмид IA-W4, гена 3-О-ацилтрансферазы и двойного обмена приведено на фиг. 3. Схематическое изображение блокирующих рекомбинантных плазмид генов, таких как ген регуляции и контроля переноса IA-W42, приведено на фиг. 4.
Полная ДНК блокированных штаммов и полная ДНК исходных штаммов были подвержены верификации ПЦР путем отдельного применения соответствующих праймеров, как показано на фиг. 5А и 5В. Как показано на фиг. 5А, 613 п.о. в кодирующем гене ОРС 4 мутантного штамма были удалены при заблокированном гене IA-W4. Результат верификации ПЦР представлен на фиг. 5В. В сравнении с исходными штаммами длина продуктов ПЦР возрастала по мере того, как маркерные гены с резистентностью вводились в мутантный штамм при заблокированных кодирующих генах, связанных с ними.
Как установлено при опытах по ферментации и при определении продуктов путем ВЭЖХ, заблокированные мутантные штаммы IA-W4 более не вырабатывали 4-изовалерил-спирамицин III и II, и как основной компонент в них стал преобладать 4-изовалерил-спирамицин I (фиг. 6). Было установлено, что ген 3-О-ацетилтрансферазы IA-W4 в генной информации в посл. 1, предлагаемый в настоящем изобретении, участвует в биосинтезе карримицина. Ввиду блокировки этого гена мутантные штаммы утратили функцию ацилирования 3-позиционного гидроксила карримицина из лактонового кольца последнего.
Как было установлено при опытах по ферментации с другими штаммами с заблокированными генами, по противобактериальному действию и определению продуктов путем ВЭЖХ, заблокированные штаммы более не вырабатывали активированный карримицин. Было установлено, что генный кластер в посл. 1, предлагаемый в настоящем изобретении, участвует в биосинтезе карримицина.
<Вариант осуществления 3> Скрининг штаммов генного переноса и блокировка штаммов бактерий, вырабатывающих карримицин.
3.1 Подготовка протопласта:
Нововыращенные на скошенном агаре споры бактерий, вырабатывающих карримицин, были помещены в жидкую культуральную среду R2YE и подверглись взбалтыванию в течение 48 ч при температуре 28°С на скорости 220 об/мин. Со скоростью посева 10% культуральная жидкость помещалась в свежую жидкую культуральную среду R2YE с содержанием 0,5% глицина и подвергалась взбалтыванию в течение 20 ч при температуре 28°С. 10 мл бактериального раствора были помещены в центрифужную пробирку и подвергнуты центрифугированию на скорости 3000 об/мин для сбора мицелия. Мицелий был промыт фосфатным буфером.

Была выполнена стерилизация 15 фунтов в течение 30 мин при температуре 121°С.
После двукратной промывки мицелий был суспендирован в фосфатном буфере соответствующего объема, был добавлен раствор фосфатного буфера с лизоцимом (окончательная концентрация составила 2 мг/мл), равномерно перемешан и выдержан на водяной бане в течение 30-45 мин при температуре 37°С со встряхиванием каждые 10-15 мин. Условия образования протопласта контролировались с помощью фазоконтрастного микроскопа 10×40. Ферментолиз был прекращен, когда микроскопическое исследование показало, что большая часть мицелия образовала протопласт. После фильтрования через поглощающую хлопчатобумажную ткань фильтрат был подвергнут двукратной промывке фосфатным буфером с центрифугированием. В завершение протопласт был суспендирован в 1 мл фосфатного буфера, и суспензия была отдельно помещена в пробирки Эппендорф объемом 100 мкл/трубка и законсервированы при температуре -70°С для дальнейшего использования.
3.2 Трансформация протопласта плазмидной ДНК:
Были отобраны 100 мкл протопласта и помещены в раствор плазмидной ДНК 10 мкл, а стенка пробирки опрокидывалась для получения однородной смеси. Были быстро добавлены 400 мкл фосфатного буфера с содержанием 25% ПЭГ-1000 (продукция компании Britain Koch-Light), были выполнены продувка-всасывание и однородное перемешивание, и состав был помещен в условия комнатной температуры на 5 мин. Плоская плашка обезвоженного R2YE была покрыта 200 мкл смеси, были проведены выращивание в течение 20 ч при температуре 28°С, заливка 50 мг/мл стерильной воды с тиострептоном, выращивание в течение 5-7 дней при температуре 28°С и отбор трансформантов.
3.3 Скрининг мутантных штаммов с заблокированным геном
Трансформанты были помещены в культуральную среду, содержащую 50 мкг/мл тиострептона
Брикетированный порошок

Для подготовки использовалась деионизированная вода; в течение 30 мин выполнялась стерилизация 15 фунтов при температуре 121°С при естественном значении рН.
В течение 5-7 дней проводилось выращивание при температуре 28°С, и 4-5 генерация была перенесена в недозированную культуральную среду. Были отделены моноспоры. Моноспоры были отдельно и соответствующим образом подвергнуты скринингу в культуральной среде с содержанием апрамицина (50 мкг/мл Am) для отсеивания штаммов с заблокированными генами, выращенных в апрамицине, а не тиострептоне. Были отобраны заблокированные штаммы с устойчивой экспрессией маркера, а ДНК заблокированных штаммов была извлечена из геномов. Была проведена амплификация ПЦР путем применения соответствующих праймеров по варианту осуществления 2, и по размеру продуктов и путем секвенирования ДНК была выполнена оценка правильности блокировки генов.
<Вариант осуществления 4> Ферментация бактерий, вырабатывающих карримицин, и штаммов с заблокированным геном, определение активности продуктов
4.1 Ферментация
культуральная среда на скошенном агаре (г/л):


Для подготовки использовалась деионизированная вода; в течение 30 мин выполнялась стерилизация 15 фунтов при температуре 121°С при естественном значении рН.
Штаммы выращивались в культуральной среде со скошенным агаром в течение 10-12 дней при температуре 28°С. После выращивания путем пропитывания они были посеяны в треугольной колбе объемом 100 мл, содержащей 30 мл культуральной среды для ферментации, и выращены в течение 96-120 ч со встряхиванием при температуре 28°С.
Культуральная среда для ферментации (г/л):

Для подготовки использовалась деионизированная вода; в течение 30 мин выполнялась стерилизация 15 фунтов при температуре 121°С.
4.2 Определение активности продукта ферментации:
Сбраживаемая среда была подвергнута центрифугированию, был отобран и разбавлен супернатант, после чего было проведено определение путем отбора сенной палочки в качестве бактерий для определения с опорой на микробиологическую пробу на ацетилспирамицин (II), 2005 г., <Фармакопея Китайской Народной Республики>. Определение проводилось путем использования метода цилиндрической плашки с методом калибровочной кривой.
4.3 Извлечение и идентификация продукта ферментации:
Сбраживаемая среда была подвергнута центрифугированию в течение 15 мин на скорости 3000 об/мин при комнатной температуре, а рН супернатанта регулировался до показателя 8,5 посредством одномолярного раствора NaOH. После этого супернатант был извлечен с помощью 1/2 объема этилацетата. Была отобрана и подвергнута сушке обдувом фаза этилацетата на плоском зеркале, после чего высушенное вещество было растворено в хроматографически чистом метаноле, а затем были введены 10-20 мкл образца после фильтрации. Измерительные приборы для хроматографии: жидкостный хроматограф Shimadzu LC-10ATvp и диодно-матричный детектор; хроматографическая колонна: Кромасил Cig (4,5×150 мм, 5 мкм); потоковая фаза: СН3ОН/1% NaH2PO4 (55:45); длина волны детектирования: 231 нм; скорость потока: 1 мл/мин; температура колонны: 25°С. Продукты ферментации мутантных штаммов определяются путем отбора эталонного продукта карримицина в качестве контрольного образца (приобретен в Национальном институте по контролю фармацевтических и биологических продуктов).
Гены и белки, используемые в настоящем изобретении, представлены в таблице последовательностей.
Изобретение относится к биотехнологии и представляет собой биосинтетический генный кластер карримицина. Биосинтетический генный кластер включает 44 открытые рамки считывания генов (ОРС): 5 ОРС (ОРС 10-14), кодирующих поликетидсинтазу; 9 ОРС (ОРС 1, 4-6, 15 и 36-39), относящихся к единице интенсификации синтеза и модификации поликетона; 16 ОРС (ОРС 9, 16-22, 24, 26, 28, 29, 33-35 и 41), относящихся к синтезу гликозила; 6 ОРС (ОРС 7, 8, 30-32 и 40), относящихся к переносу гликозила; 2 ОРС (ОРС 3 и 25), относящихся к резистентности; 4 ОРС (ОРС 2, 23, 27 и 42), вероятно, относящихся к регуляции; ОРС маркерного гена, резистентного к тиостептрону (ОРС 43); и ОРС гена 4''-микарозы-глюкозид-изовалерил-трансферазы (ОРС 44). 9 з.п. ф-лы, 6 ил., 2 табл., 4 пр.
1. Биосинтетический генный кластер карримицина, состоящий из 44 генов, включающих:
1) пять генов поликетидсинтазы, включающих ОРС 10-14 (выбирается из оснований 16215-10543 в последовательности № 1), 11 (выбирается из оснований 21076-16328 в последовательности № 1), 12 (выбирается из оснований 32511-21124 в последовательности 1), 13 (выбирается из оснований 38599-32585 в последовательности № 1), 14 (выбирается из оснований 52259-38643 в последовательности № 1);
2) девять генов, относящихся к единице интенсификации синтеза и модификации поликетона, включающих ОРС 1 (выбирается из оснований 1-645 в последовательности № 1), 4 (выбирается из оснований 3614-4840 в последовательности № 1), 5 (выбирается из оснований 4846-5511 в последовательности № 1), 6 (выбирается из оснований 7150-5801 в последовательности № 1), 15 (выбирается из оснований 53099-54310 в последовательности № 1) и 36 (выбирается из оснований 83164-82052 в последовательности № 1), 37 (выбирается из оснований 84400-83279 в последовательности № 1), 38 (выбирается из оснований 84713-84393 в последовательности № 1), 39 (выбирается из оснований 85576-84710 в последовательности № 1);
3) шестнадцать генов, относящихся к синтезу гликозила, включающих ОРС 9 (выбирается из оснований 1-645 в последовательности № 1), 16 (выбирается из оснований 54495-54845 в последовательности № 1), 17 (выбирается из оснований 54842-56041 в последовательности № 1), 18 (выбирается из оснований 56038-56946 в последовательности № 1), 19 (выбирается из оснований 56930-57967 в последовательности № 1), 21 (выбирается из оснований 60836-61984 в последовательности № 1), 22 (выбирается из оснований 62796-62077 в последовательности № 1), 24 (выбирается из оснований 67379-66318 в последовательности № 1), 26 (выбирается из оснований 69349-70650 в последовательности № 1), 28 (выбирается из оснований 72422-73462 в последовательности № 1), 29 (выбирается из оснований 74601-73561 в последовательности № 1), 33 (выбирается из оснований 78783-79775 в последовательности № 1), 34 (выбирается из оснований 79772-80779 в последовательности № 1), 35 (выбирается из оснований 82055-80823 в последовательности № 1) и 41 (выбирается из оснований 87094-87702 в последовательности № 1);
4) шесть генов, относящихся к переносу гликозила, включающих ОРС 7 (выбирается из оснований 8444-7179 в последовательности № 1), 8 (выбирается из оснований 9729-8482 в последовательности № 1), 30 (выбирается из оснований 74913-76160 в последовательности № 1), 31 (выбирается из оснований 76218-77486 в последовательности № 1), 32 (выбирается из оснований 77606-78781 в последовательности № 1) и 40 (выбирается из оснований 85825-87042 в последовательности № 1);
5) два гена, относящихся к резистентности, включающих ОРС 3 (выбирается из оснований 3133-2285 в последовательности № 1) и 25 (выбирается из оснований 69004-67352 в последовательности № 1);
6) четыре гена, относящихся к регуляции биосинтеза, включающих ОРС 2 (выбирается из оснований 1810-1208 в последовательности № 1), 23 (выбирается из оснований 63633-65645 в последовательности № 1), 27 (выбирается из оснований 72156-70708 в последовательности № 1) и 42 (выбирается из оснований 89315-88143 в последовательности № 1); и
7) два гена, включающих экзогенные маркерные гены, полученные посредством генной инженерии, ОРС 43 (выбирается из оснований 866-60 в последовательности № 2) и ген микарозы-4̋-О-изовалерил-трансферазы ОРС 44 (выбирается из оснований 2337-1174 в последовательности № 2), связанный с ОРС 43.
2. Биосинтетический генный кластер по п. 1, отличающийся тем, что пять генов поликетидсинтазы кодируют фермент биосинтеза поликетида, и этот фермент катализирует синтез 16-членного лактонового кольца карримицина, при этом аминокислотные последовательности, соответствующие нуклеотидным или комплементарным последовательностям 5 генов поликетидсинтазы ОРС 10-14 (выбирается из оснований 16215-10543 в последовательности № 1), 11 (выбирается из оснований 21076-16328 в последовательности № 1), 12 (выбирается из оснований 32511-21124 в последовательности № 1), 13 (выбирается из оснований 38599-32585 в последовательности 1), 14 (выбирается из оснований 52259-38643 в последовательности № 1), включают в себя IA-W10 в последовательности 12, IA-W11, определенную в последовательности № 13, IA-W12, определенную в последовательности № 14, IA-W13, определенную в последовательности № 15, и IA-W14, определенную в последовательности № 16.
3. Биосинтетический генный кластер по п. 1 или 2, отличающийся тем, что пять поликетидсинтаз включают в себя аминокислотные последовательности, соответствующие нуклеотидным или комплементарным последовательностям структурных доменов кетосинтазы (КС), ацилтрансферазы (АТ), кеторедуктазы (КР), дегидразы (ДГ), еноилредуктазы (ЕР), ацилпереносящего белка (АПБ) и тиоэстеразы.
4. Биосинтетический генный кластер по п. 3, отличающийся тем, что пять поликетидсинтаз включают в себя модули или структурные домены.
5. Биосинтетический генный кластер по п. 1, отличающийся тем, что аминокислотные последовательности, соответствующие нуклеотидным или комплементарным последовательностям генов, относящихся к единице интенсификации синтеза и модификации поликетона, содержащие ОРС 1, 4-6, 15 и 36-39, включают в себя IA-W1, определенную в последовательности № 3, IA-W4, определенную в последовательности № 6, IA-W5, определенную в последовательности № 7, IA-W6, определенную в последовательности № 8, IA-W15, определенную в последовательности № 17, IA-W36, определенную в последовательности № 38, IA-W37, определенную в последовательности № 39, IA-W38, определенную в последовательности № 40, и IA-W39, определенную в последовательности № 41.
6. Биосинтетический генный кластер по п. 1, отличающийся тем, что аминокислотные последовательности, соответствующие нуклеотидным или комплементарным последовательностям генов, относящихся к синтезу гликозила, содержащие ОРС 9, 16-22, 24, 26, 28, 29, 33-35 и 41, включают в себя IA-W9, определенную в последовательности № 11, IA-W16, определенную в последовательности № 18, IA-W17, определенную в последовательности № 19, IA-W18, определенную в последовательности № 20, IA-W19, определенную в последовательности № 21, IA-W20, определенную в последовательности № 22, IA-W21, определенную в последовательности № 23, IA-W22, определенную в последовательности № 24, IA-W24, определенную в последовательности № 26, IA-W26, определенную в последовательности № 28, IA-W28, определенную в последовательности № 30, IA-W29, определенную в последовательности № 31, IA-W33, определенную в последовательности № 35, IA-W34, определенную в последовательности № 36, IA-W35, определенную в последовательности № 37, и IA-W41, определенную в последовательности № 43.
7. Биосинтетический генный кластер по п. 1, отличающийся тем, что аминокислотные последовательности, соответствующие нуклеотидным или комплементарным последовательностям генов, относящихся к переносу гликозила, содержащие ОРС 7, 8, 30-32 и 40, включают в себя IA-W7, определенную в последовательности № 9, IA-W8, определенную в последовательности № 10, IA-W30, определенную в последовательности № 32, IA-W31, определенную в последовательности № 33, IA-W32, определенную в последовательности № 34, и IA-W40, определенную в последовательности № 42.
8. Биосинтетический генный кластер по п. 1, отличающийся тем, что аминокислотные последовательности, соответствующие нуклеотидным или комплементарным последовательностям генов, относящихся к резистентности, содержащие ОРС 3 и 25, включают в себя IA-W3, определенную в последовательности № 5, и IA-W25, определенную в последовательности № 27.
9. Биосинтетический генный кластер по п. 1, отличающийся тем, что аминокислотные последовательности, соответствующие нуклеотидным или комплементарным последовательностям генов, относящихся к регуляции биосинтеза, содержащие ОРС 2, 23, 27 и 42, включают в себя IA-W2, определенную в последовательности № 4, IA-W23, определенную в последовательности № 25, IA-W27, определенную в последовательности № 29, и IA-W42, определенную в последовательности № 44.
10. Биосинтетический генный кластер по п. 1, отличающийся тем, что аминокислотные последовательности, соответствующие нуклеотидным или комплементарным последовательностям экзогенных маркерных генов, полученных методом генной инженерии, содержащие ОРС 43 и ОРС 44, связанный с ОРС 43, включают в себя IA-W43, определенную в последовательности № 45, и IA-W44, определенную в последовательности № 46.
| SHANG, Guangdong et al | |||
| Construction of a Stable Bioengineered Strain of Biotechmycin, CHINESE JOURNAL OF BIOTECHNOLOGY, vol | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Аппарат для передачи изображений на расстояние | 1920 |
|
SU171A1 |
| ПЕРЕПЕЛОВ А.В | |||
| Способ смешанной растительной и животной проклейки бумаги | 1922 |
|
SU49A1 |

