ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка заявляет приоритет по предварительной заявке США с серийным №62/087184, поданной 3 декабря 2014, которая включена в данный документ в полном объеме посредством ссылки по любому назначению.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящая заявка содержит Список последовательностей, представленный в электронном виде в формате ASCII и включенный в данный документ в полном объеме посредством ссылки. Указанная копия ASCII, созданная 20 ноября 2015, имеет название P32433-WO_SL.txt и размер, составляющий 190541 байт.
ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к антителам против тейхоевой кислоты клеточной стенки ("анти-WTA"), которые конъюгированны с антибиотиками группы рифамицина, и к использованию полученных конъюгатов антитело-антибиотик в лечении инфекций Staphylococcus.
УРОВЕНЬ ТЕХНИКИ
Staphylococcus aureus (S. aureus; SA) является основной причиной бактериальных инфекций у людей во всем мире и представляет собой серьезную проблему здравоохранения, как в больницах, так и в общественных местах. Однако, S. aureus не является исключительно патогеном и обычно колонизирует ноздри и кожу здоровых людей. Когда инфекция действительно имеет место, наиболее серьезные инфекции, такие как эндокардит, остеомиелит, некротизирующая пневмония и сепсис возникают после распространения бактерий в кровоток (Lowy, F.D. (1998) "Staphylococcus aureus infections" N Engl J Med 339, 520-532). За последние несколько десятилетий заражение S. aureus становится все труднее лечить из-за появления и быстрого распространения метициллин-резистентного S. aureus (MRSA), который устойчив ко всем известным бета-лактамным антибиотикам (Boucher, H.W., et al. (2009) "Bad bugs, no drugs: no ESKAPE! An update from the Infectious Diseases Society of America" Clin Infect Dis 48, 1-12). Инвазивные инфекции MRSA трудно поддаются лечению, смертность составляет ~ 20% и является основной причиной смерти, вызваннойинфекционными агентами, в США. Таким образом, немногими антибиотиками выбора для лечения инвазивных инфекций MRSA стали ванкомицин, линезолид и даптомицин (Boucher, H., Miller, L.G.& Razonable, R.R.(2010) "Serious infections caused by methicillin-resistant Staphylococcus aureus" Clin Infect Dis 51 Suppl 2, S183-197). Однако, о снижении восприимчивости к ванкомицину и перекрестной резистентности к линезолиду и даптомицину уже сообщалось в отношении клинических штаммов MRSA (Nannini, Е., Murray, В.Е. & Arias, С.А. (2010) "Resistance or decreased susceptibility to glycopeptides, daptomycin, and linezolid in methicillin-resistant Staphylococcus aureus" Curr Opin Pharmacol 10, 516-521). Co временем доза ванкомицина, необходимая для преодоления резистентности, приблизилась к уровням нефротоксичности. Таким образом, смертность и заболеваемость от инвазивных инфекций MRSA остаются высокими, несмотря на приминение этих антибиотиков.
Исследования показали, что S. aureus способен внедряться и выживать внутри клеток млекопитающих, включая фагоцитарные клетки, которые ответствечают за очищение от бактерий (Thwaites, G.E. & Gant, V.(2011) Are bloodstream leukocytes Trojan Horses for the metastasis of Staphylococcus aureus? Nat Rev Microbiol 9, 215-222; Rogers, D.E.,Tompsett, R. (1952) "The survival of staphylococci within human leukocytes" J.Exp.Med 95, 209-230; Gresham, H.D., et al.(2000) "Survival of Staphylococcus aureus inside neutrophils contributes to infection" J Immunol 164, 3713-3722); Kapral, F.A. & Shayegani, M.G. (1959) "Intracellular survival of staphylococci" J Exp Med 110, 123-138; Anwar, S., et al. (2009) "The rise and rise of Staphylococcus aureus: laughing in the face of granulocytes" Clin Exp Immunol 157, 216-224); Fraunholz, M. & Sinha, B.(2012) "Intracellular Staphylococcus aureus:live-in and let die" Front Cell Infect Microbiol 2, 43; Garzoni, C. & Kelley, W.L. (2011) "Return of the Trojan horse: intracellular phenotype switching and immune evasion by Staphylococcus aureus" EMBO Mol Med 3, 115-117). S. aureus поглощается фагоцитарными клетками-хозяевами, главным образом нейтрофилами и макрофагами, в течение нескольких минут после внутривенного инфицирования (Rogers, D.E. (1956) "Studies on Bacterimia: Mechanisms Relating to the Persistence of Bacteremia in Rabbits Following the Intravanous Injection of Staphylococci" JEM 103, 713). В то время как большинство бактерий эффективно уничтожаются этими клетками, неполное очищение от S. aureus переносимых кровью фагоцитов может позволить этим зараженным клеткам выступать в роли «троянского коня» для распространения бактерий от начального участка инфекции. Действительно, пациенты с нормальным числом нейтрофилов могут быть болеше склонны к диссеминированному заболеванию, чем пациенты со сниженным числом нейтрофилов (Thwaites, G.E. & Gant, V.(2011) выше). После доставки в ткани, S. aureus может поражать различные типы нефагоцитарных клеток, а внутриклеточный S. aureus в тканях ассоциируется с хроническими или рецидивирующими инфекциями. Кроме того, воздействие внутриклеточных бактерий на субоптимальные концентрации антибиотиков может способствовать появлению устойчивых к антибиотикам штаммов, что делает эту клиническую проблему более острой. В соответствии с этими наблюдениями, лечение пациентов с инвазивными инфекциями MRSA, такими как бактериемия или эндокардит, ванкомицином или даптомицином связано с частотой неудач более 50% (Kullar, R., Davis, S.L., Levine, D.P. & Rybak, M.J. Impact of vancomycin exposure on outcomes in patients with methicillin-resistant Staphylococcus aureus bacteremia: support for consensus guidelines suggested targets.C/w/са/ infectious diseases: an official publication of the Infectious Diseases Society of America 52, 975-981 (2011); Fowler, V.G., Jr. et al. Daptomycin versus standard therapy for bacteremia and endocarditis caused by Staphylococcus aureus. The New England journal of medicine 355, 653-665 (2006); Yoon, Y.K., Kim, J.Y., Park, D.W., Sohn, J.W. & Kim, M.J. Predictors of persistent methicillin-resistant Staphylococcus aureus bacteremia in patients treated with vancomycin. The Journal of antimicrobial chemotherapy 65:1015-1018 (2010)). Следовательно, более успешная антистафилококковая терапия должна включать элиминацию внутриклеточных бактерий.
Ансамицины представляют собой класс антибиотиков, включающий рифамицин, рифампин, рифампицин, рифабутин, рифапентин, рифалазил, ABI-1657 и их аналоги, которые ингибируют бактериальную РНК-полимеразу и обладают исключительной активностью против грамположительных и селективных грамотрицательных бактерий (Rothstein, D.M., et al (2003) Expert Opin.Invest.Drugs 12(2): 255-271; US 7342011; US 7271165).
Сообщается об иммунотерапии для профилактики и лечения S. aureus (включая MRS А) инфекций. US 2011/0262477 касается использования бактериальных адгезионных белков Еар, Emp и AdsA в качестве вакцин для стимуляции иммунного ответа против MRSA. WO 2000/071585 описывает изолированные моноклональные антитела, реагирующие на специфические изоляты штаммов S. aureus. В US 2011/0059085А1 предлагается стратегия на основе Ат, с использанием IgM Ат, специфичные для одного или нескольких капсульных антигенов SA, хотя фактические антитела не описаны.
Конъюгаты антитело-лекарственное средство (ADC), также известные как иммуноконъюгаты, являются целевыми химиотерапевтическими молекулами, которые сочетают в себе идеальные свойства как антител, так и цитотоксических лекарств, которые действуют путем нацеливания сильнодействующих цитотоксических лекарственных средств на антиген-экспрессирующие опухолевые клетки (Teicher, В.А. (2009) Curr.Cancer Drug Targets 9: 982-1004), тем самым увеличивая терапевтический индекс, максимизируя эффективность и сводя к минимуму побочную токсичность (Carter, P.J. and Senter P.D. (2008) The Cancer J..14(3): 154-169; Chari, R.V.(2008) Acc.Chem.Res. 41:98-107. ADC содержат целевое антитело, ковалентно связанное через линкерный блок с фрагментом цитотоксического лекарственного средства. Иммуноконъюгаты обеспечивают адресную доставку молекулы лекарственного вещества в опухоль, и внутриклеточное накопление в опухоли, тогда как системное введение неконъюгированных лекарственных веществ для достижения элиминации опухолевых клеток может привести к неприемлемым для нормальных клеток уровням tokch4hocth(Polakis Р. (2005) Curr. Opin. Pharmacol.5: 382-387).
Описаны неспецифические конъюгаты иммуноглобулин-антибиотик, которые связываются с поверхностью бактерий-мишеней через антибиотик для лечения сепсиса (США 5545721, США 6660267). Описаны антибиотик-конъюгированные антитела, которые имеют антигенсвязывающую часть, специфичную для бактериального антигена (такую как капсульный полисахарид SA), но не имеют константного участка, который реагирует с бактериальным Fc-связывающим белком, например, стафилококковым белком А (США 7569677).
Учитывая вызывающую опасения скорость сопротивления MRSA традиционным антибиотикам и, как следствие, смертность и заболеваемость от инвазивных инфекций MRSA, существует высокая неудовлетворенная потребность в новых терапевтических средствах для лечения инфекций S. aureus. Настоящее изобретение удовлетворяет эту потребность и предоставляет композиции и способы, которые преодолевают ограничения современных терапевтических композиций, а также предлагают дополнительные преимущества, которые будут очевидны из подробного описания ниже.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение предусматривает уникальное терапевтическое средство, которое включает элиминацию внутриклеточных бактерий. Данное изобретение демонстрирует, что такое терапевтическое действие является эффективным in vivo, когда обычные антибиотики, такие как ванкомицин, терпят неудачу.
Изобретение предусматривает композиции, называемые "конъюгаты антитело-антибиотик» или "ААС", содержащие антитело, конъюгированное с помощью ковалентного присоединения к одной или более молекулам антибиотика группы рифамицина.
Аспектом изобретения является соединение, конъюгат антитело-антибиотик, содержащее антитело против тейхоевой кислоты клеточной стенки (WTA), ковалентно присоединенное с помощью расщепляемого протеазой непептидного линкера к антибиотику группы рифамицина.
Иллюстративный вариант реализации изобретения представляет собой конъюгат антитело-антибиотик по п. 1, имеющий формулу:
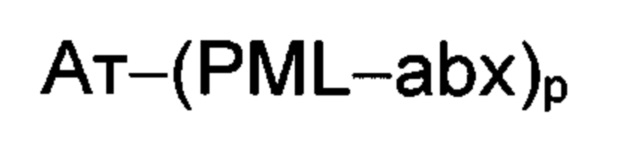
причем:
Ат представляет собой антитело против тейхоевой кислоты клеточной стенки; PML представляет собой расщепляемый протеазой непептидный линкер, имеющий формулу:
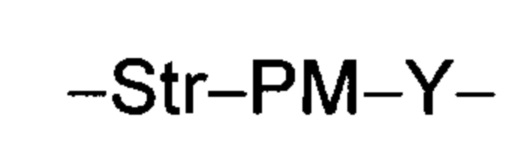
причем Str представляет собой растягивающую единицу; РМ представляет собой пептидомиметическую единицу, и Y представляет собой спейсерную единицу;
abx представляет собой антибиотик рифамицинового ряда; и
Р представляет собой целое число от 1 до 8. Соединения конъюгата антитело-антибиотик любого из предыдущих вариантов реализации изобретения могут содержать любой из описанных в данном документе Ат против тейхоевых кислот клеточной стенки (WTA). Эти антитела к WTA связываются со Staphylococcus aureus. В одном варианте реализации изобретения антитело представляет собой анти-WTAα моноклональное антитело. В иллюстративных анти-WTAα антителах, Ат представляет собой моноклональное антитело, содержащее легкую (L) цепь и тяжелую (Н) цепь, L-цепь, содержащую CDR L1, CDR L2, и CDR L3, и Н-цепь, содержащую, CDR H1, CDR Н2 и CDR Н3, причем CDR L1, CDR L2, и CDR L3 и CDR H1, CDR Н2 и CDR Н3 содержат аминокислотные последовательности CDR каждого из Ат 4461 (SEQ ID NO. 1-6), 4624 (SEQ ID NO. 7-12), 4399 (SEQ ID NO. 13-18), и 6267 (SEQ ID NO. 19-24) соответственно, как продемонстрировано в таблицах 1А и 1В.
В некоторых вариантах реализации изобретения, анти-WTA антитело содержит вариабельный участок тяжелой цепи (VH), где VH содержит по меньшей мере 95% идентичности последовательности по длине VH участка, выбранного из VH последовательности SEQ ID NO. 26, SEQ ID NO. 28, SEQ ID NO. 30, SEQ ID NO. 32 антител 4461, 4624, 4399, и 6267, соответственно. Антитела могут дополнительно содержать вариабельный участок L-цепи (VL), причем VL содержит по меньшей мере 95% идентичности последовательности по длине VL участка, выбранного из VL последовательности SEQ ID NO: 25, SEQ ID NO: 27, SEQ ID NO. 29, SEQ ID NO. 31 антител 4461, 4624, 4399 и 6267 соответственно.
В другом варианте реализации изобретения, конъюгат антитело-антибиотик по изобретению содержит анти-WTAβp моноклональное антитело. Иллюстративное анти-WTAβ антитело содержит легкую цепь и Н-цепь, L-цепь, содержащую CDR L1, CDR L2, и CDR L3, и Н-цепь, содержащую CDR H1, CDR Н2 и CDR Н3, причем CDR L1, CDR L2, и CDR L3 и CDR H1, CDR Н2 и CDR Н3 содержат аминокислотные последовательности соответствующих CDR каждого из Ат, продемонстрированных на Фиг. 12 (SEQ ID NO. 33-110).
Другое анти-WTAβ антитело, полезное для генерирования ААС по изобретению, содержит вариабельный участок L-цепи (VL), причем VL содержит по меньшей мере 95% идентичности последовательности по длине VL участка, выбранного из VL последовательности, соответствующей каждому из антител 6078, 6263, 4450, 6297, 6239, 6232, 6259, 6292, 4462, 6265, 6253, 4497 и 4487 соответственно, как продемонстрировано на Фиг. 15А-1, 15А-2, 15А-3 в положениях 1-107 по Кабат. Это антитело может дополнительно содержать вариабельный участок тяжелой цепи (VH), причем VH содержит по меньшей мере 95% идентичности последовательности по длине VH участка, выбранного из VH последовательности, соответствующей каждому из антител 6078, 6263, 4450, 6297, 6239, 6232, 6259, 6292, 4462, 6265, 6253, 4497, и 4487 соответственно, как продемонстрировано на Фиг. 15В-1 до 15В-6 в положениях 1-113 по Кабат.
В другом анти-WTAβ антителе, VL содержит последовательность SEQ ID NO. 111, и VH содержит последовательность SEQ ID NO. 112, причем X представляет собой Q или Е и X1 представляет собой М, I или V.
Изобретение предусматривает анти-WTAβ, пригодное для получения ААС по изобретению, причем легкая цепь антитела содержит сконструированный цистеин и содержит последовательность SEQ ID NO. 115, и Н-цепь содержит SEQ ID NO. 116, где X представляет собой М, I или V. В альтернативных спаривающихся L и Н цепях, легкая цепь антитела содержит последовательность SEQ ID NO. 113, и Н-цепь содержит сконструированный цистеин и содержит SEQ ID NO. 117, где X представляет собой М, I или V. Cys может быть сконструирован в каждой из L и Н цепей; в одном примере такого антитела WTAβ легкая цепь содержит сконструированный цистеин и содержит последовательность SEQ ID NO. 115, и Н-цепь содержит сконструированный цистеин и содержит SEQ ID NO. 117, где X представляет собой М, I или V.
Другое анти-WTAP антитело, пригодное для конъюгации, содержит VH и VL, причем VH содержит по меньшей мере 95% идентичности последовательности по длине VH SEQ ID NO. 156, и VL содержит по меньшей мере 95% идентичности последовательности по длине VL последовательности SEQ ID NO. 119. В конкретном варианте реализации изобретения, анти-WTAp антитело содержит VH, содержащую последовательность SEQ ID NO. 156, и VL, содержащую последовательность SEQ ID NO. 119.
Анти-WTAβ антитело по изобретению может содержать L-цепь, содержащую последовательность SEQ ID NO. 121, и Н-цепь, содержащую последовательность SEQ ID NO. 124. В другом примере, анти-WTAβ антитело содержит L-цепь, содержащую последовательность SEQ ID NO. 123, и Н-цепь, содержащую последовательность SEQ ID NO. 157 или SEQ ID NO. 124.
В других вариантах реализации изобретения, антитело содержит: I) CDR L-цепи и Н-цепи SEQ ID NO 99-104 или CDR L-цепи и Н-цепи SEQ ID NO. 33-38; или ii) VL SEQ ID NO. 119 или SEQ ID NO. 123 в паре с VH SEQ ID NO. 120 или SEQ ID NO. 156; или iii) VL SEQ ID NO. 111 в паре с VH SEQ ID NO. 112.
В некоторых вариантах реализации ААС по изобретению, антитело связывается с тем же эпитопом, что и антитело любого из предшествующих вариантов реализации изобретения.
Антитело по любому из предшествующих вариантов реализации изобретения может быть антигенсвязывающим фрагментом, не имеющим Fc-участка. В некоторых вариантах реализации изобретения антитело представляет собой F(ab) или F(ab ')2. В некоторых вариантах реализации изобретения, антитело дополнительно содержит константный участок тяжелой цепи и/или константный участок легкой цепи, причем константный участок тяжелой цепи и/или константный участок легкой цепи содержит одну или более аминокислот, которые замещены остатками цистеина. В некоторых вариантах реализации изобретения, константный участок тяжелой цепи содержит аминокислотную замену А118С и/или S400C, и/или константный участок легкой цепи содержит аминокислотную замену V205C, причем нумерационная система соответствует нумерации ЕС.
В некоторых вариантах реализации любого из антител, описанных выше, антитело не является изотипом IgM. В некоторых вариантах реализации любого из антител, описанных выше, антитело представляет собой изотип IgG (например, IgG1, IgG2, IgG3, IgG4), IgE, IgD или IgA (например, IgA1 или IgA2).
Иллюстративный вариант реализации изобретения представляет собой фармацевтическую композицию, содержащую соединение конъюгата антитело антибиотик и фармацевтически приемлемый носитель, глидант, разбавитель или вспомогательное вещество.
Анти-WTA-AAC по изобретению применимы в качестве антимикробных агентов, эффективных для лечения стафилококков человека и в ветеринарии, например S. aureus, S. saprophytics и S. simulans, а также Listeria, например Listeria monocytogenes. В конкретном аспекте ААС по изобретению полезны для лечения инфекций S. aureus. Таким образом, изобретение также обеспечивает способ лечения стафилококковой инфекции у человека или ветеринарного пациента, включающий введение пациенту терапевтически эффективного количества конъюгата антитело-антибиотик любого из предшествующих вариантов реализации изобретения. В одном варианте реализации изобретения, бактериальная инфекция представляет собой инфекцию Staphylococcus aureus. В некоторых вариантах реализации изобретения, у пациента была диагностирована инфекция S. aureus. В некоторых вариантах реализации, лечение бактериальной инфекции включает в себя уменьшение бактериальной нагрузки или количества. В одном варианте реализации изобретения, способ лечения представляет собой введение пациентам, у которых бактериальная инфекция, включая S. aureus, приводит к бактериемии. В конкретных вариантах реализации изобретения, способ используется для лечения стафилококкового эндокардита или остеомиелита. В одном варианте реализации изобретения, соединение конъюгата антитело-антибиотик вводят инфицированному пациенту в дозе в диапазоне от около 50 мг/кг до 100 мг/кг.
Также предложен способ уничтожения внутриклеточного S. aureus в клетках инфицированного S. aureus пациента, не убивая клетки-хозяева, путем введения соединения конъюгата анти-WTA-антибиотик любого из вышеуказанных вариантов реализации изобретения. Другой способ предусмотрен для уничтожения стойких стафилококковых бактериальных клеток (например, S. aureus) in vivo путем контактирования персистентных бактерий с ААС любого из предыдущих вариантов реализации изобретения.
В другом варианте реализации изобретения, способ лечения дополнительно включает в себя введение второго терапевтического средства. В следующем варианте реализации изобретения, вторым терапевтическим агентом является антибиотик, включающий в себя антибиотик против Staph aureus в целом или MRSA в частности.
В одном варианте реализации изобретения второй антибиотик, вводимый в комбинации с соединением конъюгата антитело-антибиотик по изобретению, выбирают из структурных классов: (i) аминогликозиды; (ii) бета-лактамы; (iii) макролиды/циьслические пептиды; (iv) тетрациклины; (V) фторхинолины/фторхинолоны; (Vi) и оксазолидиноны.
В одном варианте реализации изобретения, второй антибиотик, вводимый в комбинации с соединением конъюгата антитело-антибиотик по изобретению, выбирают из клиндамицина, новобиоцина, ретапамулина, даптомицина, GSK-2140944, CG-400549, ситафлоксацина, тейкопланина, триклозана, нафтиридона, радезолида, доксорубицина, ампициллина, ванкомицина, имипенема, дорипенема, гемцитабина, далбаванцина и азитромицина.
В некоторых вариантах реализации данного изобретения, бактериальная нагрузка у инфицированного пациента была снижена до не обнаруживаемого уровня после лечения. В одном варианте реализации изобретения, культура крови пациента является отрицательной после лечения по сравнению с положительной культурой крови перед лечением. В некоторых вариантах реализации данного изобретения, устойчивость к бактериям у субъекта является неопределяемой или низкой. В некоторых вариантах реализации данного изобретения, субъект не реагирует на лечение метициллином или ванкомицином.
Иллюстративный вариант реализации изобретения представляет собой процесс получения конъюгата антитело-антибиотик, включающий конъюгирование антибиотика рифамицинового ряда с антителом против тейхоевой кислоты клеточной стенки (WTA).
Иллюстративный вариант реализации изобретения представляет собой набор для лечения бактериальной инфекции, включающий:
А) фармацевтическую композицию, содержащую соединение конъюгата антитело-антибиотик, и фармацевтически приемлемый носитель, глидант, разбавитель или вспомогательное вещество; и
Б) инструкции по использованию.
Одним аспектом изобретения является промежуточный антибиотик-линкер, имеющий формулу II:
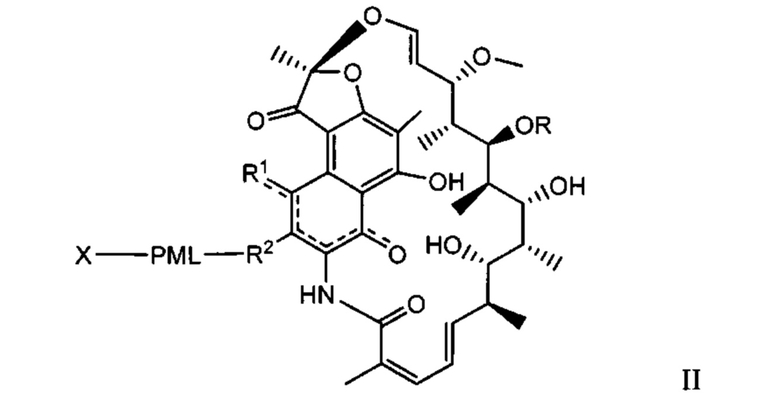
где:
пунктирные линии обозначают необязательную связь;
R представляет собой Н, С1-С12 алкил, или С(O)СН3;
R1 представляет собой ОН;
R2 представляет собой СН=N-(гетероциклил), причем гетероциклил необязательно замещен одной или более группами, независимо выбранными из С(O)СН3, С1-С12 алкил, С1-С12 гетероарил, С2-С20 гетероциклил, С6-С20 арил, и С3-С12 карбоциклил;
или R1 и R2 образуют пяти- или шестичленный конденсированный гетероарил или гетероциклил и необязательно образуют спиро или конденсированный шестичленный гетероарил, гетероциклил, арил или карбоциклическое кольцо, причем спиро или конденсированный шестичленный гетероарил, гетероциклил, арил или карбоциклическое кольцо необязательно замещено Н, F, Cl, Br, I, С1-С12 алкилом или ОН;
PML представляет собой расщепляемый протеазой непептидный линкер, присоединенный к R2 или конденсированный гетероарил или гетероциклил, образованный R1 и R2; и имеющий формулу:

причем Str представляет собой растягивающую единицу; РМ представляет собой пептидомиметическую единицу, и Y представляет собой спейсерную единицу; и
X представляет собой реакционноспособную функциональную группу, выбранную из малеимида, тиола, амино, бромида, бромацетамида, иодацетамида, п-толуолсульфоната, иодида, гидроксила, карбоксила, пиридилдисульфида и N-гидроксисукцинимида.
Следует понимать, что одна, несколько или все характеристики различных вариантов реализации изобретения, описанных в данном документе, могут быть объединены с образованием других вариантов реализации данного изобретения. Эти и другие аспекты изобретения будут понятны для специалиста в данной области техники.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1: Внутриклеточные хранилища MRS А защищены от ванкомицина in vivo и in vitro. На Фиг. 1А показана схема экспериментальной конструкции для создания свободных бактерий (планктонных) против внутриклеточных бактерий. Четыре когорты мышей заражали внутривенной инъекцией с приблизительно эквивалентными дозами жизнеспособных свободных бактерий или внутриклеточных бактерий, и отобранные группы были обработаны ванкомицином сразу же после инфицирования, а затем один раз в день (см. Пример 2). На Фиг. 1В и Фиг. 1С показаны бактериальные нагрузки в почках и головном мозге, соответственно, инфицированных мышей через 4 дня после заражения. Пунктирная линия указывает предел обнаружения для анализа. Фиг. 1D показывает, что MRSA защищен от ванкомицина при культивировании на монослое инфицированных клеток. (ND = не обнаружено). Фиг. 1Е и Фиг. 1F показывают, что MRSA способен расти в присутствии ванкомицина при культивировании на монослое инфицированных клеток. MRSA (свободные бактерии) высевали в среду, среду + ванкомицин, или среду + ванкомицин и высевали на монослой остеобластов MG63 (Фиг. 1Е) или клеток сосудов головного мозга человека (НВМЕС, Фиг. 1F). Внеклеточные бактерии (свободные бактерии) хорошо росли в среде исключительно, но уничтожались ванкомицином. В лунках, содержащих монослой клеток млекопитающих (внутриклеточный + ванко), часть бактерий была защищена от ванкомицина в течение первых 8 часов после инфицирования и была способна расширяться внутри внутриклеточного компартмента в течение 24 часов. Планки погрешностей показывают стандартное отклонение для трех лунок.
Фиг. 2 демонстрирует концепцию конъюгата антитело-антибиотик (ААС). В одном примере ААС состоит из антитела, направленного против эпитопа на поверхности S. aureus, связанного с мощным антибиотиком рифамицинового ряда (например, Рифалог) через линкер, который расщепляется лизосомальными протеазами.
На Фиг. 3 показан возможный механизм активации лекарственного средства для конъюгатов антитело-антибиотик (ААС). ААС связываются с внеклеточными бактериями через антигенсвязывающий домен (Fab) антитела и способствуют поглощению опсонизированных бактерий через Fc-опосредованный фагоцитоз. Линкер расщепляется лизосомальными протеазами, такими как катепсин В. После расщепления линкера, линкер гидролизуется, высвобождая свободный антибиотик внутри фаголизосомы. Свободный антибиотик убивает опсонизированные и фагоцитированные бактерии вместе с любыми ранее интернализованными бактериями, находящимися в одном и том же компартменте.
На Фиг. 4 показана клеточная стенка грамположительных бактерий, таких как S. Aureus, со схематическим изображением тейхоевых кислот клеточных стенок (WTA), липотейхоевой кислоты (LTA) и оболочек пептидогликана (PGN), которые стабилизируют клеточную мембрану и обеспечивают сайты прикрепления.
На Фиг. 5 показана химическая структура и гликозильные модификации тейхоевых кислот клеточных стенок (WTA), подробно описанные в разделе «Определения».
На Фиг. 6А и 6В приведены характеристики Ат после первичного скрининга библиотеки мАт, демонстрирующие позитивное связывание ИФА с препаратами клеточной стенки из USA300 или штамма Wood46 штаммов S. aureus, как описано в примере 3. Из Ат, которые связываются с WTA, 4 специфичны для WTA альфа и 13 специфически связываются с WTA бета.
На Фиг. 7А показано титрование меченого Alexa-488 анти-β-GlcNAC WTA или анти-α-GlcNAC WTA антитела на MRSA, выделенной непосредственно из инфицированных почек мыши. Анти-CMV-gD антитело служило в качестве контрольного изотипа антитела. Фиг. 7В демонстрирует, что антитело, используемое для генерации ААС, распознает эпитоп тейхоевой кислоты на клеточной стенке, который опосредуется гликозилтрансферазой TarS. Анализ FACS с использованием анти-β-GlcNAC WTA антитела или контрольным изотипом на ДТ USA300, USA300-TarM или USA300-TarS.
На Фиг. 8 продемонстрирован выбор мощного антибиотика рифамицинового ряда (рифалог) диметил pipBOR по его способности убивать нереплицирующиеся MRSA.
Фиг. 9: Анализ ингибирования роста, демонстрирующий, что интактный ТАС (форма ААС) не убивает планктонные бактерии, если антибиотик не выделяется при лечении катепсином В. ТАС инкубировали только в буфере (не окрашенные кружки) или обрабатывали катепсином В (окрашенные кружки). Интактный ТАС не был способен предотвращать рост бактерий после инкубации в течение ночи. Предварительная обработка ТАС катепсином В высвобождала достаточную антибиотическую активность для предотвращения роста бактерий при 0,6 мкг/мл ТАС, которая, как прогнозируется, содержит 0,006 мкг/мл антибиотика.
На Фиг. 10 продемонстрировано лечение зараженных S. aureus мышей анти-WTA-PML ААС, которое значительно уменьшало количество бактерий или уничтожало их в инфицированных органах по сравнению с голым антителом, как описано в примере 10. На Фиг. 10А схематично показан график времени эксперимента и моменты времени инъекции, как описано в примере 10. На Фиг. 10В показана обработка AAC (DAR2) из Таблицы 3, уменьшившей бактериальную нагрузку в почках примерно в 7000 раз. Фиг. 10С показывает, что лечение с AAC (DAR2) уменьшало бактериальную нагрузку в сердце примерно в 500 раз.
На Фиг. 11А представлено выравнивание аминокислотной последовательности вариабельных участков легкой цепи (VL) четырех человеческих анти-WTA-альфа антител 4461. 4624, 4399, 6267 (SEQ ID NO: 25, 27, 29 и 31, соответственно, в порядке появления). CDR-последовательности CDRL1, L2 и L3 в соответствии с нумерацией Кабата подчеркнуты. Фиг. 11В демонстрирует выравнивание аминокислотной последовательности вариабельных участков тяжелой цепи (VH) четырех человеческих анти-WTA-альфа антител по Фиг. 11А. CDR-последовательности CDR H1, Н2 и Н3 в соответствии с нумерацией согласно Кабат подчеркнуты (SEQ ID NOS 26, 28, 30 и 32, соответственно, в порядке появления).
На Фиг. 12 показаны CDR-последовательности L и Н цепей 13 человеческих анти-WTA бета-антител (SEQ ID NO: 33-110).
На Фиг. 13А-1 и 13А-2 показано выравнивание полной длины L-цепи (легкой цепи) анти-WTA-бета Ат 6078 (немодифицированное) и его вариантов v2, v3, v4 (SEQ ID NO 113, 113, 115, 113, 115, 113, 115 и 115, соответственно, в порядке появления). CDR-последовательности CDRL1, L2 и L3 в соответствии с нумерацией Кабат подчеркнуты. Боксы демонстрируют контактные остатки и CDR остатки в соответствии с нумерацией по Кабат и Хотиа. Варианты L-цепей, которые содержат сконструированный Cys, обозначены буквой С в черном боксе рядом с концом константного участка (в нумерации ЕС остаток №205 в этом случае). Название варианта, например, v2LC-Cys означает вариант 2, содержащий Cys, сконструированный в L-цепь. HCLC-Cys означает, что каждая из Н и L цепей содержит сконструированный Cys. Варианты 2, 3 и 4 имеют изменения в начале Н-цепи, как показано на Фиг. 13В.
На Фиг. 13В-1, 13В-2, 13В-3, 13В-4 показано выравнивание полной длины Н-цепи (тяжелой цепи) анти-WTA-бета Ат 6078 (немодифицированное) и его вариантов, v2, v3, v4 (SEQ ID NOS 114, 139-144 и 143, соответственно, в порядке появления), которые имеют изменения в начале Н-цепи. Варианты Н-цепей, которые содержат сконструированный Cys, обозначены С в черном боксе в начале константной области (в нумерации ЕС остаток №118 в этом случае).
На Фиг. 14А-1 и 14А-2 показано выравнивание полной длины L-цепи анти-WTA бета Ат 4497 (немодифицированное) и Cys-сконструированных L-цепей (SEQ ID NO 121, 123, 145 и 145 соответственно в порядке появления). CDR-последовательности CDRL1, L2 и L3 в соответствии с нумерацией согласно Кабат подчеркнуты. Боксы демонстрируют контактные остатки и CDR остатки в соответствии с нумерацией по Кабату и Хотиа. Варианты L-цепей, которые содержат сконструированный Cys, обозначены буквой С в пунктирном боксе рядом с концом константного участка (в нумерации ЕС остаток №205 в этом случае).
На Фиг. 14В-1, 14В-2, 14В-3 показано выравнивание полной длины Н-цепи анти-WTA-бета Ат 4497 (немодифицированное) и его варианта v8 с D, измененным на Е в положении CDR Н3 96, с или без сконструированного Cys (SEQ ID NOS 146-147, 157 и 147, соответственно, в порядке появления). Варианты Н-цепей, которые содержат сконструированный Cys, обозначены С в черном боксе в начале константной области (в нумерации ЕС остаток №118 в этом случае).
На Фиг. 15А-1, 15А-2, 15А-3 показано выравнивание аминокислотной последовательности полной длины легкой цепи тринадцати человеческих анти-WTA-бета антител (SEQ ID NOS 113, 158-167, 121 и 168, соответственно, в порядке появления). Вариабельный участок (VL) охватывает положения аминокислот по нумерации Кабат от 1 до 107. CDR-последовательности CDRL1, L2 и L3 в соответствии с нумерацией Кабата подчеркнуты.
На Фиг. 15В-1 до 15В-6 показано выравнивание аминокислотной последовательности полной длины легкой цепи тринадцати человеческих анти-WTA-бета антител по Фиг. 15А-1, 15А-2, 15А-3 (SEQ ID NOS 114, 169-176, 133-134, 138 и 127, соответственно, в порядке появления). Вариабельный участок (VH) охватывает положения аминокислот по нумерации Кабат 1-113. CDR-последовательности CDR H1, Н2 и Н3 в соответствии с нумерацией согласно Кабат подчеркнуты. Положение 118 Н-цепи в соответствии с нумерацией ЕС, отмеченное звездочкой, может быть изменено на Cys для конъюгации лекарственного средства. Остатки, выделенные черным цветом, могут быть заменены другими остатками, которые не влияют на связывание антигена, чтобы избежать дезамидирования, изомеризации аспарагиновой кислоты, окисления или N-связанного гликозилирования.
На Фиг. 16 показано сравнение Ат 4497 и его мутантов в выделенных положениях аминокислот и их относительной прочности связывания антигена, как было проверено с помощью ИФА. На Фиг. 16 представлены SEQ ID NOS 177, 177, 177, 178, 178, 179, 179, 180, 180 и 180, соответственно, в порядке появления.
На Фиг. 17 показано, что предварительная обработка 50 мг/кг свободными антителами неэффективна при модели внутривенной инфекции. Мышам Balb/c давали однократную дозу контроля растворителя (PBS) или 50 мг/кг антител путем внутривенной инъекции за 30 минут до инфицирования 2×107 КОЕ USA300. Группы лечения включали контрольный изотип антитела, которое не связывается с S. aureus (gD), антитело, направленное против бета-модификации тейхоевой кислоты клеточной стенки (4497), или антитело, направленное против альфа-модификации тейхоевой кислоты клеточной стенки (7578). Контрольные мыши получали два раза в день лечение с использованием внутрибрюшинной инъекции (Ванко) 110 мг/кг ванкомицина.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Далее будет приведено детальное описание некоторых вариантов реализации изобретения, примеры которых раскрыты с помощью сопутствующих структур и формул. Хотя изобретение будет описано в сочетании с пронумерованными вариантами реализации, включая способы, материалы и примеры, такое описание не является ограничивающим, и изобретение предназначено для охвата всех альтернатив, модификаций и эквивалентов, независимо от того, являются ли они общеизвестными или включены в данный документ. В случае если одна или более позиций используемой литературы, патентов, и подобных материалов отличается или противоречит настоящей заявке, включающей, но не ограничивающейся определенными условиями, сроком использования, описанными методами и т.п., настоящая заявка имеет преимущество. Если не указано иное, все технические и научные термины, используемые в данном документе, имеют то же значение, которое обычно подразумевается обычным специалистом в данной области техники, к которой относится настоящее изобретение. Специалисту в данной области техники будут понятны многие способы и материалы, аналогичные или эквивалентные описанным в данном документе, которые могут быть использованы при практическом осуществлении настоящего изобретения. Настоящее изобретение никаким образом не ограничено описанными способами и материалами.
Все публикации, патентные заявки, патенты, и другие ссылки, упомянутые в настоящем документе, включены посредством ссылки в полном объеме.
I. Общие методы
Способы и процедуры, описанные или упомянутые в данном документе, как правило, хорошо известны и обычно применяются с использованием обычной методики специалистами в данной области техники, такие как, например, широко используемые методики, описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual 3d edition (2001) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.; Current Protocols in Molecular Biology (F.M. Ausubel, et al. eds., (2003)); the series Methods in Enzymology (Academic Press, Inc.): PCR 2:A Practical Approach (M.J. MacPherson, B.D. Hames and G.R.
Taylor eds. (1995)), Harlow and Lane, eds. (1988) Antibodies, A Laboratory Manual, and Animal Cell Culture (R.I. Freshney, ed. (1987)); Oligonucleotide Synthesis (M.J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R.I. Freshney), ed., 1987); Introduction to Cell and Tissue Culture (J.P. Mather and P.E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J.B. Griffiths, and D.G. Newell, eds., 1993-8) J. Wiley and Sons; Handbook of Experimental Immunology (D.M. Weir and C.C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J.M. Miller and M.P. Calos, eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (J.E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C.A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: A Practical Approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal Antibodies: A Practical Approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using Antibodies: A Laboratory Manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M. Zanetti and J.D. Capra, eds., Harwood Academic Publishers, 1995); и Cancer: Principles and Practice of Oncology (V.T. DeVita et al., eds., J.B. Lippincott Company, 1993).
Номенклатура, используемая в данной заявке, основана на систематической номенклатуре IUPAC, если не указано иное. Если не указано иное, все технические и научные термины, используемые в данном документе, имеют такое же значение, которое обычно понимается средним специалистом в данной области техники, к которой относится настоящее изобретение, и согласуются с: Singleton et al (1994) Dictionary of Microbiology and Molecular Biology, 2nd Ed., J. Wiley & Sons, New York, NY; and Janeway, C., Travers, P., Walport, M., Shlomchik (2001) Immunobiology, 5th Ed.,Garland Publishing, New York.
II. ОПРЕДЕЛЕНИЯ
«Конъюгат антитело-антибиотик» или ААС представляет собой соединение, состоящее из антитела, которое химически связано с антибиотиком с помощью линкера. Антитело связывает антиген или эпитоп на поверхности бактерий, например, компонент клеточной стенки бактерий. Как используется в этом изобретении, линкер представляет собой расщепляемый протеазой непептидный линкер, который разработан для расщепления протеазами, включая катепсин В, лизосомальную протеазу, обнаруженную в большинстве типов клеток млекопитающих (Dubowchik et al., 2002) Bioconj. Chem. 13:855-869). Диаграмма ААС с его тремя компонентами показана на Фиг. 2. "THIOMAB™ Конъюгат с антибиотиками" или "ТАС" представляет собой форму ААС, в которой антитело химически конъюгировано с линкером-антибиотиком через один или более цистеинов, как правило, цистеин, который рекомбинантно сконструирован в антителе на специфическом сайте(ах) антитела таким образом, чтобы не мешать антигенсвязывающей функции.
Термин "тейхоевая кислота клеточной стенки" (WTA) означает анионные гликополимеры, которые ковалентно присоединены к пептидогликану посредством фосфодиэфирной связи с С6 гидроксилом Сахаров N-ацетилмураминовой кислоты. Хотя точная химическая структура может варьировать среди организмов, в одном варианте реализации WTA представляет собой рибитол тейхоевую кислоту с повторяющимися единицами 1,5-фосфодиэфирных связей D-рибитола и D-аланилового эфира в положении 2 и гликозильных заместителей в положении 4. Гликозильные группы могут быть N-ацетилглюкозаминилом α (альфа) или β (бета) как представлено в S. aureus. Гидроксилы на повторах фосфата алдитол/сахарного спирта были замещены катионными D-аланиновыми эфирами и моносахаридами, такими как N-ацетилглюкозамин. В одном аспекте гидроксильные заместители включают D-аланил и альфа (α) или бета (β) GlcNHAc. В одном конкретном аспекте, WTA включает соединение формулы:
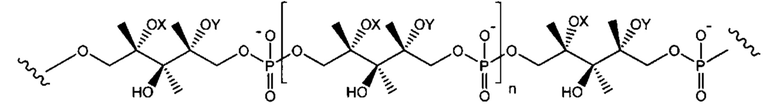
где волнистые линии указывают повторяющиеся звенья связывания или участки связывания полиальдиатола-Р или пептидогликана, где X представляет собой D-аланил или -Н; и Y представляет собой α (альфа)-GlcNHAc или β (бета)-GlcNHAc.
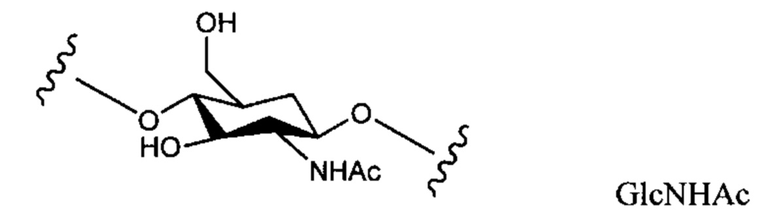
В S. aureus, WTA ковалентно связана с 6-ОН N-ацетилмураминовой кислоты (MurNAc) через дисахарид, состоящий из N-ацетилглюкозамина (GlcNAc) -1-Р и N-ацетилманнозамина (ManNAc), за которым следуют две или три единицы глицеринфосфатов. Фактический полимер WTA в таком случае состоит из 11-40 рибитол-фосфатных (Rbo-P) повторяющихся звеньев. Поэтапный синтез WTA сначала инициируется ферментом, называемым TagO, и штаммы S. aureus, у которых отсутствует ген TagO (путем искусственной делеции гена), не образуют никаких WTA. Повторяющиеся звенья могут быть дополнительно сшиты с D-аланином (D-Ala) в С2-ОН и/или с N-ацетилглюкозамином (GlcNAc) в положении С4-ОН через α- (альфа) или β- (бета) гликозидные связи. В зависимости от штамма S. aureus или фазы роста бактерий гликозидные связи могут представлять собой α-, β- или смесь двух аномеров.
Используемый в данном документе, термин «антитело WTA» относится к любому антителу, которое связывает WTA, независимо от того, является ли WTA альфа или WTA бета. Термины «альфа антитело против тейхоевой кислоты клеточной стенки» или «анти-WTA альфа антитело» или «анти-αWTA» или «антитело против α-GlcNac WTA» используются взаимозаменяемо для обозначения антитела, которое специфически связывает тейхоевую кислоту клеточной стенки (WTA) альфа. Точно так же термины «бета антитело против тейхоевой кислоты клеточной стенки» или «анти-WTA бета-антитело» или «анти-βТАА» или «антитело против β-GlcNac WTA» используются взаимозаменяемо для обозначения антитела, которое специфически связывается с тейхоевой кислотой клеточной стенки (WTA) бета.
Термин «антибиотик» (abx или Abx) включает любую молекулу, которая специфически ингибирует рост или уничтожение микроорганизмов, таких как бактерии, но не смертельна для хозяина в интервале концентрации и дозирования введения. В конкретном аспекте антибиотик не токсичен для хозяина при введенных концентрациях и интервалах дозирования. Антибиотики, эффективные против бактерий, можно широко классифицировать как бактерицидные (т.е., непосредственно убивает) или бактериостатические (т.е., предотвращают деление). Антибактерицидные антибиотики могут быть дополнительно классифицированы как антибиотики узкого спектра или широкого спектра действия. Антибиотик широкого спектра действия эффективен против широкого спектра бактерий, включающих как грамположительные и грамотрицательные бактерии, в отличие от антибиотика с узким спектром, который эффективен против меньшего диапазона или специфических семейств бактерий. Примеры антибиотиков включают: (i) аминогликозиды, например, амикацин, гентамицин, канамицин, неомицин, нетилмицин, стрептомицин, тобрамицин, паромицин, (ii) ансамицины, например, гельданамицин, гербимицин, (iii) карбацефемы, например, лоракарбеф, (iv), карбапенемы, например, эртапенум, дорипенем, имипенем/циластатин, меропенем, (v) цефалоспорины (первое поколение), например, цефадроксил, цефазолин, цефалотин, цефалексин, (vi) цефалоспорины (второе поколение), например цефлаклор, цефамандол, цефокситин, цефпрозил, цефуроксим, (vi) цефалоспорины (третье поколение), например цефиксим, цефдинир, цефдиторин, цефоперазон, цефотаксим, цефподоксим, цефтазидим, цефтибутен, цефтизоксим, цефтриаксон, (vii) цефалоспорины (четвертое поколение), например, цефепим, (viii), цефалоспорины (пятое поколение), например, цефтобипрол, (ix) гликопептиды, например, тейкопланин, ванкомицин, (X) макролиды, например, акситромицин, кларитромицин, диритромицин, эритромицин, рокситромицин, тролеандомицин, телитромицин, спектиномицин, (xi) монобактамы, например, акстреонам, (xii) пеницилины, например, амоксициллин, ампициллин, аклоциллин, карбенициллин, клоксациллин, диклоксациллин, флюклоксациллин, мезоциллин, метициллин, нафциллин, оксациллин, пенициллин, пеперациллин, тикарциллин, (xiii) антибиотические полипептиды, например, бацитрацин, колистин, полимиксин В, (Xiv) хинолоны, например, ципрофлоксацин, эноксацин, гатифлоксацин, левофлоксацин, лемефлоксацин, моксифлоксацин, норфлоксацин, орфлоксацин, тровафлоксацин, (xv) сульфонамиды, например, мафенид, пронтозил, сульфацетамид, сульфаметизол, сульфаниламид, сульфасалазин, сульфоксазол, триметоприм, триметоприм-сульфаметоксазол (TMP-SMX), (xvi) тетрациклины, например, демеклоциклин, доксициклин, миноциклин, окситетрациклин, тетрациклин и (xvii) другие, такие как арспенамин, хлорамфеникол, клиндамицин, линкомицин, этамбутол, фосфомицин, фузидиновая кислота, фуразолидон, изониазид, линезолид, метронидазол, мупироцин, нитрофурантоин, плаценсимицин, пиразинамид, квинупристин/дальфопристин, рифампицин/рифампицин или тинидазол.
Staphylococcus aureus также упоминается здесь как Staph А или S. aureus сокращенно. Термин «устойчивый к метициллину Staphylococcus aureus» (MRSA), альтернативно известный как устойчивый к множественным лекарствам Staphylococcus aureus или оксациллин-устойчивый Staphylococcus aureus (ORSA), относится к любому штамму Staphylococcus aureus, который устойчив к бета-лактамным антибиотикам, которые включают пенициллины (например, метициллин, диклоксациллин, нафциллин, оксациллин, и т.д.) и цефалоспорины. "Метициллинчувствительный Staphylococcus aureus" (MSSA) относится к любому штамму Staphylococcus aureus, который чувствителен к бета-лактамным антибиотикам.
Термины "анти-Staph антитело" и "антитело, которое связывается со Staph а" относятся к антителу, которое способно связывать антиген с Staphylococcus aureus ("S. aureus") с достаточной аффинностью, так что антитело может быть использовано в качестве диагностического и/или терапевтического агента для нацеливания на S. aureus. В одном варианте реализации изобретения, степень связывания антитела анти-Staph с неродственным белком, не являющимся Staph, составляет менее чем около 10% связывания антитела с MRSA при измерении, например, с помощью радиоиммуноанализа (РИА). В некоторых вариантах реализации изобретения антитело, связывающееся с Staph, характеризуется константой диссоциации (Kd), составляющей 1 ≤ мкм, ≤100 нМ, ≤10 нМ, ≤5 нМ, ≤4 нМ, ≤3 нМ, ≤2 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ, или ≤0,001 нМ (например, 10-8 M или менее, например от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М). В некоторых вариантах реализации изобретения анти-Staph антитело связывается с эпитопом Staph, который является консервативным среди Staph у различных видов.
Термин «минимальная ингибирующая концентрация" ("МИК") относится к самой низкой антимикробной концентрации, которая будет ингибировать видимый рост микроорганизма после инкубации в течение ночи. Известны анализы определения МИК. Один из способов описан в разделе «Примеры» ниже.
Термин «антитело» в настоящем документе используется в самом широком смысле и, конкретно, включает моноклональные антитела, поликлональные антитела, димеры, мультимеры, мультиспецифические антитела (например, биспецифические антитела), и их антигенсвязывающие фрагменты антител, (Miller et al (2003) J. of Immunology 170:4854-4861). Антитела могут быть мышиными, человеческими, гуманизированными, химерными или полученными от других видов. Антитело представляет собой белок, вырабатываемый иммунной системой, который способен распознавать и связываться со специфическим антигеном (Janeway, С., Travers, P., Walport, М., Shlomchik (2001) Immuno Biology, 5th Ed., Garland Publishing, New York). Целевой антиген обычно имеет многочисленные сайты связывания, также называемые эпитопами, которые распознаются CDR на нескольких антителах. Каждое антитело, которое специфически связывается с другим эпитопом, имеет различную структуру. Таким образом, один антиген может быть распознан и связан более чем одним соответствующим антителом. Антитело включает полноразмерную молекулу иммуноглобулина или иммунологически активную часть полноразмерной молекулы иммуноглобулина, т.е. молекулы, которая содержит сайт связывания антигена, который иммуноспецифически связывает антиген представляющей интерес мишени или ее части, такие мишени, включают, но не ограничиваются ими, раковые клетки или клетки, которые продуцируют аутоиммунные антитела, связанные с аутоиммунным заболеванием, инфицированные клетки или микроорганизмы, такие как бактерии. Раскрытый в данном документе иммуноглобулин (Ig) может быть любого изотипа, за исключением IgM (например, IgG, IgE, IgD, и IgA) и подкласса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2). Иммуноглобулин может быть получен из любых видов. В одном аспекте Ig имеет человеческое, мышиное или кроличье происхождение. В конкретном варианте реализации изобретения, Ig имеет человеческое происхождение.
"Класс" антитела относится к типу константного домена или константной области, содержащейся в его тяжелой цепи. Существует пять основных классов антител: IgA, IgD, IgE, IgG, и IgM, и некоторые из них могут быть дополнительно разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1, и IgA2. Константные домены тяжелой цепи, соответствующие различным классам иммуноглобулинов, называются α, δ, ε, γ, и μ, соответственно.
"Нативные антитела" относятся к природным молекулам иммуноглобулина с различной структурой. Например, нативные антитела IgG представляют собой гетеротетрамерные гликопротеины с молекулярной массой около 150000 дальтон, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, связанных дисульфидными связями. Каждая тяжелая цепь от N-конца к С-концу содержит вариабельную область (VH), также называемую вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, с последующими тремя константными доменами (CH1, СН2 и СН3). Аналогично, каждая легкая цепь от N-конца к С-концу содержит вариабельную область (VL), также называемую вариабельным легким доменом или вариабельным доменом легкой цепи, с последующим константным легким доменом (CL). В зависимости от аминокислотной последовательности константного домена легкую цепь антитела можно отнести к одному из двух типов, называемых каппа (κ) и лямбда (λ).
В данном контексте термины "полноразмерное антитело", "интактное антитело" и "целое антитело" являются взаимозаменяемыми и относятся к антителу, структура которого по существу аналогична структуре нативного антитела, или которое включает тяжелые цепи, содержащие Fc-область, описанную в настоящем документе.
"Фрагмент антитела" относится к молекуле, не являющейся интактным антителом, но содержащей фрагмент интактного антитела, связывающий антиген, с которым связывается интактное антитело. Примеры фрагментов антитела включают, но не ограничиваются ими, Fv, Fab, Fab', Fab'-SH, F(ab')2; диатела; линейные антитела; молекулы одноцепочечных антител (например, scFv); и полиспецифические антитела, образованные из фрагментов антител.
В данном контексте термин "моноклональное антитело" относится к антителу, полученному из популяции по существу однородных антител, т.е., отдельные антитела, составляющие популяцию, являются идентичными и/или связывают один и тот же эпитоп, за исключением возможных вариантных антител, например, содержащих природные мутации или возникающие во время продукции моноклонального антитела (например, естественные изменения в гликозилировании), причем такие варианты присутствуют в незначительных количествах. Одним из таких возможных вариантов антител IgG1 является расщепление С-концевого лизина (К) константного участка тяжелой цепи. В отличие от препаратов поликлональных антител, которые, как правило, содержат различные антитела против различных детерминант (эпитопов), каждое моноклональное антитело из препарата моноклонального антитела направлено против одной детерминанты антигена. Таким образом, модификатор "моноклональное" указывает на то, что антитело получают по существу из однородной популяции антител; его не следует интерпретировать как требование относительно продукции антитела с помощью какого-либо конкретного способа. Например, моноклональные антитела для использования согласно настоящему изобретению можно получить с помощью различных способов, включая, но не ограничиваясь ими, гибридомный способ, методики рекомбинантных ДНК, методики фагового дисплея и способы с использованием трансгенных животных, полностью или частично содержащие локусы человеческого иммуноглобулина, причем такие способы и другие типовые способы получения моноклональных антител описаны в настоящем документе. Кроме своей специфичности, моноклональные антитела обладают тем преимуществом, что они в процессе их синтеза не происходит загрязнения другими антителами.
Термин "химерное антитело" относится к антителу, в котором фрагмент тяжелой и/или легкой цепи происходит из конкретного источника или вида, в то время как остальная часть тяжелой и/или легкой цепи происходит из другого источника или вида.
"Антитело человека" представляет собой антитело, обладающее аминокислотной последовательностью, соответствующей последовательности антитела, продуцированного в организме человека или клетке человека, или происходящее из нечеловеческого источника, использующего репертуар антител человека или другие последовательности, кодирующие антитело человека. Из этого определения антитела человека, в частности, исключено гуманизированное антитело, содержащее антигенсвязывающие остатки нечеловеческого происхождения.
"Гуманизированное" антитело относится к химерному антителу, содержащему аминокислотные остатки из HVR нечеловеческого происхождения и аминокислотные остатки из FR человека. В некоторых вариантах реализации изобретения, гуманизированное антитело содержит по существу все из по меньшей мере одного, а обычно - двух вариабельных доменов, в которых все или по существу все HVR (например, CDR) соответствуют аналогичным участкам антитела нечеловеческого происхождения, и все или по существу все FR соответствуют последовательности антитела человека. Гуманизированное антитело необязательно может содержать по меньшей мере фрагмент константного участка антитела, происходящий из антитела человека. Термин "гуманизированная форма" антитела, например, антитела нечеловеческого происхождения, относится к антителу, подвергшемуся гуманизации.
Термин "вариабельный участок" или "вариабельный домен" относится к домену тяжелой или легкой цепи антитела, участвующему в связывании антитела с антигеном.
Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела обычно обладают аналогичной структурой, причем каждый домен содержит четыре консервативных каркасных участков (FR) и три гипервариабельных участка (HVR). (См., например, Kindt et al. Kuby Immunology, 6th ed., W.H. Freeman and Co., page 91 (2007). Для придания антигенсвязывающей специфичности может быть достаточно одного домена VH или VL. Кроме того, антитела, которые связываются с конкретным антигеном, можно выделить, используя VH или VL домен антитела, связывающийся с антигеном, для скрининга библиотеки последовательностей, комплементарных доменам VL или VH, соответственно. См., например, Portolano et al., J. Immunol. 150:880-887 (1993); Clarkson et al., Nature 352:624-628 (1991).
Термин "гипервариабельный участок," "HVR," или "HV," как используется в данном документе, относится к участкам вариабельного домена антитела, которые являются гипервариабельными по последовательности ("участки, определяющие комплементарность" или "CDR"), и/или образуют структурно определенные петли, и/или содержат остатки, контактирующие с антигеном ("контакты антигена"). Как правило, антитела включают шесть HVR; три в VH (H1, Н2, Н3), и три в VL (LI, L2, L3). В нативных антителах Н3 и L3 проявляют наибольшее многообразие из шести HVR, и, в частности, полагают, что Н3 играет уникальную роль в обеспечении высокой специфичности антител. См., например, Xu et al., Immunity 13:37-45 (2000); Johnson and Wu, in Methods in Molecular Biology 248:1-25 (Lo, ed., Human Press, Totowa, NJ, 2003). Действительно, природные антитела верблюдов, состоящие только из тяжелой цепи, являются функциональными и стабильными в отсутствие легкой цепи (Hamers-Casterman et al., (1993) Nature 363:446-448; Sheriff et al., (1996) Nature Struct. Biol. 3:733-736).
Существует ряд определений HVR, охватываемых в данном документе. Области, определяющие комплементарность, по Кабат (CDR) основаны на вариабельности последовательностей и являются наиболее часто используемыми (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Напротив, Хотиа обращает внимание на локализацию структурных петель (Chothia and Lesk, (1987) J. Mol. Biol. 196:901-917). Для антигенных контактов см. MacCallum et al. J. Mol. Biol. 262:732-745 (1996). АтМ HVR представляют собой компромисс между HVR по Кабат и структурными петлями по Хотиа, и используются программным обеспечением для моделирования антител АтМ от Oxford Molecular. «Контактные» HVR основаны на анализе доступных сложных кристаллических структур. Остатки из каждого из этих HVR указаны ниже.
KabatChothiaKabatChothia
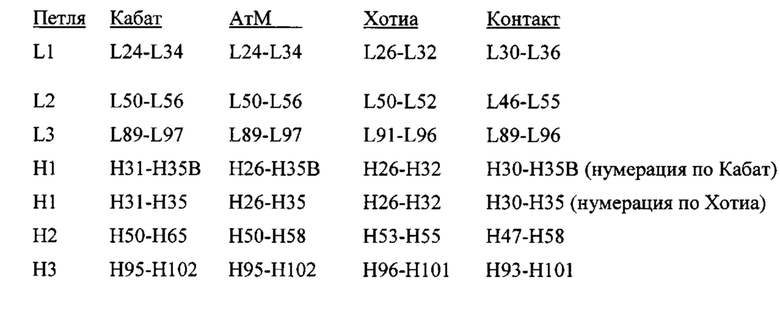
HVR могут содержать "расширенные HVR" такие как: 24-36 или 24-34 (L1), 46-56 или 50-56 (L2), и 89-97 или 89-96 (L3) в VL и 26-35 (H1), 50-65 или 49-65 (Н2) и 93-102, 94-102, или 95-102 (Н3) в VH. Если не указано иное, остатки HVR и другие остатки в вариабельном домене (например, FR-остатки) в данном документе нумеруют по Кабат и другие.
Выражения "нумерация остатков вариабельного домена по Кабат" или "нумерация положений аминокислот по Кабат" и их варианты относятся к системе нумерации, используемой для вариабельных доменов тяжелой или легкой цепи согласно представлению об антителах в публикации Кабат другие, ранее. При использовании этой системы нумерации фактическая линейная аминокислотная последовательность может содержать меньшее количество аминокислот или дополнительные аминокислоты, соответствующие укорачиванию или инсерции в FR или HVR вариабельного домена. Например, вариабельный домен тяжелой цепи может включать вставку одной аминокислоты (остаток 52а согласно Кабат) после остатка 52 Н2 и вставленные остатки (например, остатки 82а, 82b, и 82с, и т.д. согласно Кабат) после остатка 82 FR тяжелой цепи. Нумерацию остатков по Кабат можно определить для данного антитела путем выравнивания областей гомологии последовательности антитела со «стандартной» пронумерованной по Кабат последовательностью.
Термин "каркас" или "FR" относится к остаткам вариабельного домена, за исключением остатков гипервариабельной области (HVR). FR вариабельного домена обычно состоит из четырех доменов FR: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR в составе VH (или VL), как правило, определяются в следующей последовательности: FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
"Акцепторный каркас человека" для целей настоящего изобретения представляет собой каркас, содержащий аминокислотную последовательность каркаса вариабельного домена легкой цепи (VL) или каркаса вариабельного домена тяжелой цепи (VH), полученный из каркаса иммуноглобулина человека или консенсусной последовательности каркаса человека, как указано ниже. Акцепторный каркас человека, "полученный из" каркаса иммуноглобулина человека или консенсусной последовательности каркаса человека, может содержать совпадающую с ними аминокислотную последовательность или может содержать изменения аминокислотной последовательности. В некоторых вариантах реализации изобретения количество аминокислотных изменений составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. В некоторых вариантах реализации изобретения последовательность VL-акцепторного каркаса человека идентична последовательности VL-каркаса иммуноглобулина человека или консенсусной последовательности каркаса человека.
"Консенсусный каркас человека" является каркасом, представляющим наиболее распространенные аминокислотные остатки при отборе VL- или VH-каркасных последовательностей иммуноглобулина человека. Как правило, выбор последовательностей VL или VH иммуноглобулина человека проводят из подгруппы последовательностей вариабельных доменов. Как правило, подгруппа последовательностей представляет собой подгруппу по Кабати другие, Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), vols. 1-3. В одном из вариантов реализации VL подгруппа представляет собой подгруппу каппа I по Кабат и другие. В одном из вариантов реализации VH подгруппа представляет собой подгруппу III по Кабат и другие., выше.
В данном контексте термин "Fc-участок" применяется для обозначения С-концевого участка тяжелой цепи иммуноглобулина. Термин включает Fc-участки с нативными последовательностями и вариантные Fc-участки. Хотя границы Fc участка тяжелой цепи иммуноглобулина могут меняться, Fc участок тяжелой цепи IgG человека обычно определяют как область от аминокислотного остатка в положении Cys226 или от Pro230 до его карбокси-конца. С-концевой остаток лизина (остаток 447 по системе нумерации ЕС - также называемый индексом ЕС, как описано у Кабат et al., Sequences of Proteins of Immunological Interest, 5th Ed.Public Health Service, National Institutes of Health, Bethesda, MD, 1991) Fc-участок может быть удален, например, в процессе продуцирования или очистки антитела, или посредством рекомбинантного конструирования нуклеиновой кислоты, кодирующей тяжелую цепь антитела. Соответственно, композиция интактных антител может содержать популяции антител, у которых удалены все остатки K447, популяции антител, у которых не удален ни один остаток K447, и популяции антител, содержащих смесь антител, имеющих и не имеющих остаток K447. Термин "Fc-рецептор" или "FcR" также включает неонатальный рецептор, FcRn, который отвечает за перенос материнских IgG плоду. Guyer et al., J. Immunol. 117:587 (1976) and Kim et al., J. Immunol. 24:249 (1994). В данной области техники известны способы измерения связывания с FcRn (см., например, Ghetie and Ward, Immunol. Today 18:(12):592-8 (1997); Ghetie et al., Nature Biotechnology 15 (7):637-40 (1997); Hinton et al., J. Biol. Chem. 279(8):6213-6 (2004); WO 2004/92219 (Hinton et al.). Анализ связывания FcRn in vivo и период полувыведения из крови высокоафинных FcRn-связывающих полипептидов можно осуществить, например, на трансгенных мышах или на трансфецированных человеческих клеточных линиях, экспрессирующих человеческий FcRn, или на приматах, которым вводят полипептиды с вариантом Fc-участка. WO 2004/42072 (Presta) описывает варианты антител, которые улучшают или ухудшают связывание с FcR. См. также, например, Shields et al., J. Biol. Chem. 9(2):6591-6604 (2001).
Термин "антитело с созревшей аффинностью" относится к антителу с одним или более изменениями в одном или более гипервариабельньгх участках (HVR) по сравнению с исходным антителом, которое не содержит таких изменений, причем такие изменения приводят к повышению аффинности антитела к антигену.
Термин «эпитоп» относится к конкретному сайту молекулы антигена, с которым связывается антитело.
"Антитело, которое связывается с тем же эпитопом" в качестве эталонного антитела относится к антителу, которое блокирует связывание эталонного антитела с его антигеном в конкурентном анализе на 50% или более, и, наоборот, эталонное антитело блокирует связывание антитела с его антигеном в конкурентном анализе на 50% или более. Типовой конкурентный анализ приведен в данном документе.
Термин "свободное антитело" относится к антителу, не конъюгированному с гетерологичной группой (например, цитотоксической группой) или радиоактивной меткой. Свободное антитело может присутствовать в фармацевтической композиции.
Термин "эффекторные функции" относится к тем видам биологической активности, присущим Fc-участку антитела, которые изменяются зависимости от изотипа антитела. Примеры эффекторных функций антител включают: связывание Clq и комплемент-зависимую цитотоксичность (КЗЦ); связывание с рецептором Fc; антителозависимую клеточно-опосредованную цитотоксичность (АЗКЦ); фагоцитоз;
подавление рецепторов поверхности клетки (например, рецептора В-клеток); и активацию В-клеток.
"Антителозависимая клеточно-опосредованная цитотоксичность" или АЗКЦ относится к форме цитотоксичности, при которой секретируемый Ig, связанный с Fc-рецепторами (FcRs), присутствует в определенных цитотоксических клетках (например, естественных клетках-киллерах (NK), нейтрофилах и макрофагах), давая возможность этим цитотоксическим эффекторным клеткам специфически связываться с несущей антиген клеткой-мишенью и впоследствии убивать эту клетку-мишень цитотоксинами. Антитела являются "оружием" цитотоксических клеток и абсолютно необходимы для обеспечения такой гибели. Основные клетки, опосредующие АЗКЦ, NK-клетки, экспрессируют только Fcγ(гамма)RIII в то время как моноциты экспрессируют Fcγ(гамма)RI, Fcγ( гамма)RII и Fcγ( гамма)RIII. Сводная информация об экспрессии Fc на гемопоэтических клетках приведена в Таблице 3 на странице 464 публикации Ravetch and Kinet, Annu.Rev. Immunol. 9:457-92 (1991). Для оценки активности АЗКЦ представляющей интерес молекулы, может проводиться анализ АЗКЦ in vitro, такой как описано в патенте США №5500362 или США 5821337. Эффекторные клетки, пригодные для такого анализа, включают мононуклеары периферической крови (МКПК) и естественные киллеры (NK).B альтернативном или дополнительном варианте, АЗКЦ-активность молекулы, представляющей интерес, можно оценить in vivo, например, на животной модели, например, согласно описанию в публикации Clynes et al. PNAS USA 95:652-656 (1998).
«Фагоцитоз» относится к процессу, посредством которого патоген поглощается или интернализуется клеткой-хозяином (например, макрофагом или нейтрофилом). Фагоциты опосредуют фагоцитоз тремя путями: (i) прямые рецепторы клеточной поверхности (например, лектины, интегрины и рецепторы мутантов) (ii) комплемент, усиленный - с использованием рецепторов комплемента (включая CRI, рецептор для C3b, CR3 и CR4) для связывания и поглощения дополненных опсонизированных патогенов, и (iii) антитело, усиленное - с использованием Fc-рецепторов (включая FcγгаммаRI, FcγгаммаRIIA и FcγгаммаRIIIA) для связывания антител опсонизированных частиц, которые затем поглощаются и сливаются с лизосомами, и становятся фаголизосомами. В данном изобретении считается, что путь (iii) играет значительную роль в доставке терапевтических анти-MRSA средств ААС инфицированным лейкоцитам, например, нейтрофилам и макрофагам (Phagocytosis of Microbes: complexity in Action by D. Underhill and A. Ozinsky. (2002) Annual Review of Immunology, Vol 20:825).
"Комплементзависимая цитотоксичность" или "CDC" относится к лизису клетки-мишени в присутствии комплемента. Активация классического пути активации комплемента инициируется связыванием первого компонента системы комплемента (Clq) с антителами (соответствующего подкласса), которые связаны с распознаваемым ими антигеном. Для оценки активации комплемента, может быть выполнен анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996).
Углевод, присоединенный к Fc-участку, может быть изменен. Нативные антитела, продуцируемые клетками млекопитающих, обычно содержат разветвленный двухантенный олигосахарид, обычно присоединенный с помощью N-связи к Asn297 в СН2-домене Fc-участка. См., например, Wright et al. (1997) TIBTECH 15:26-32. Указанный олигосахарид может содержать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в "стебле" двухантенной олигосахаридной структуры. В некоторых вариантах реализации изобретения можно выполнить модификации олигосахарида в IgG с целью создания IgG с некоторыми дополнительными улучшенными свойствами. Например, предлагаются модификации антител, имеющие углеводную структуру, в которой отсутствует фукоза, присоединенная (прямо или косвенно) к Fc-участку. Такие модификации могут иметь улучшенную функцию АЗКЦ. См., например, US 2003/0157108 (Presta, L.); US 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Примеры публикаций, связанных с "дефукозилированными" или "фукозо дефицитными" модификациями антитела, включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; WO 2002/031140; Okazaki et al., J. Mol. Biol. 336:1239-1249 (2004); Yamane-Ohnuki et al. Biotech. Bioeng. 87:614 (2004). Примеры клеточных линий, способных продуцировать дефукозилированные антитела, включают клетки 13 СНО, дефектные по фукозилированию белков (Ripka et al. Arch. Biochem. Biophys. 249:533-545 (1986); публ. заявки патента США №2003/0157108 A1, Presta, L; и WO 2004/056312 A1, Adams et al., especially at Example 11), и клеточные линии с нокаутом, такие как ген альфа-1,6-фукозилтрансферазы, FUT8, нокаутные клетки СНО (см., например, Yamane-Ohnuki et al., Biotech. Bioeng. 87:614 (2004); Kanda, Y. et al, Biotechnol. Bioeng., 94(4):680-688 (2006); и WO 2003/085107).
"Выделенное антитело" представляет собой антитело, отделенное от компонента своего природного окружения. В некоторых вариантах реализации изобретения антитело является очищенным до более чем 95% или 99% чистоты, определяемой, например, посредством электрофореза (например, электрофореза в ДСН-ПААГ, изоэлектрического фокусирования (ИЭФ), капиллярного электрофореза) или хроматографии (например, ионообменной или обращенно-фазной ВЭЖХ). Обзор способов анализа чистоты антител см., например, Flatman et al., J.Chromatogr. В 848:79-87 (2007).
"Выделенная нуклеиновая кислота" относится к молекуле нуклеиновой кислоты, отделенной от компонента своего природного окружения. Выделенная нуклеиновая кислота содержит молекулу нуклеиновой кислоты, содержавшуюся в клетках, обычно содержащих указанную молекулу нуклеиновой кислоты, однако указанная молекула нуклеиновой кислоты присутствует вне хромосомы или в области хромосомы, отличающейся от ее естественного местоположения в хромосоме.
"Выделенная нуклеиновая кислота, кодирующая анти-WTA бета антитело" относится к одной или более молекулам нуклеиновой кислоты, кодирующим тяжелые и легкие цепи антитела, включая такую(ие) молекулу(ы) нуклеиновой кислоты в составе одного вектора или отдельных векторов и такую молекулу(ы) нуклеиновой кислоты, присутствующую в одном или более местах в клетке-хозяине.
В данном контексте термин "специфически связывающийся с" или "специфический для" относится к измеряемым и воспроизводимым взаимодействиям, таким как связывание между мишенью и антителом, что является определяющим фактором присутствия мишени в гетерогенной популяции молекул, включая биологические молекулы. Например, антитело, связывающееся с или специфически связывающееся с мишенью (которая может представлять собой эпитоп), представляет собой антитело, связывающее эту мишень с большим сродством, авидностью, более легко и/или на больший срок, чем оно связывается с другими мишенями. В одном варианте реализации изобретения, степень связывания антитела с мишенью, не связанной с WTA-бета, составляет менее чем около 10% связывания антитела с мишенью, как измерено, например, с помощью радиоиммуноанализа (РИА). В некоторых вариантах реализации изобретения, антитело, которое специфически связывается с WTA бета, имеет константу диссоциации (Kd) 1 ≤ мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, или ≤0,1 нМ. В некоторых вариантах реализации, антитело специфически связывается с эпитопом, консервативным для разных видов. В другом варианте реализации изобретения, специфическое связывание может включать, но необязательно требует, исключительное связывание.
"Аффинность связывания" обычно относится к интенсивности суммарных общих нековалентных взаимодействий между одиночным сайтом связывания молекулы (например, антитела) и ее партнера по связыванию (например, антигена). Если не указано иное, в данном контексте термин "аффинность связывания" относится к присущему молекуле сродству связывания, которое отражает взаимодействие между членами пары связывающихся компонентов (например, антителом и антигеном) при их соотношении 1:1. Сродство молекулы X к ее партнеру Y в целом можно выразить константой диссоциации (Kd). Аффинность можно измерить с помощью общепринятых в данной области техники способов, включая способы, описанные в настоящем документе. Низкоаффинные антитела в основном связывают антиген медленно и склонны легко диссоциировать, тогда как антитела с высокой аффинностью обычно связывают антиген быстрее и, как правило, остаются связанными дольше. В технике известно множество способов измерения аффинности связывания, любой из которых может быть использован для целей данного изобретения. Конкретные иллюстративные и типовые варианты реализации измерения аффинности связывания описаны ниже.
В одном варианте реализации, "Kd" или "значение Kd" в соответствии с настоящим изобретением измеряют с помощью анализа связывания антигена, меченного радиоактивной меткой (РИА), выполняемого с использованием Fab-версии исследуемого антитела и его антигена, как описано в следующем анализе. Аффинность связывания Fab с антигеном в растворе измеряют путем уравновешивания Fab с минимальной концентрацией (125I)-меченого антигена в присутствии серии разведений немеченого антигена, с последующим захватом связанного антигена с помощью планшета с иммобилизованным антителом против Fab (см., например, Chen et al., (1999) J. Mol. Biol. 293:865-881). Для создания условий для анализа микротитрационные планшеты (DYNEX Technologies, Inc.) покрывают в течение ночи улавливающим антителом к Fab (Cappel Labs) в концентрации 5 мкг/мл в 50 мМ растворе карбоната натрия (рН 9,6), а затем блокируют 2% (мас./об.) раствором бычьего сывороточного альбумина в PBS в течение двух-пяти часов при комнатной температуре (около 23°С). В неадсорбирующем планшете (Nunc #269620), 100 пМ или 26 пМ [125I]-антигена смешивают с серийными разведениями представляющего интерес Fab (например, в соответствии с оценкой антител анти-VEGF, Fab-12 в Presta et al., Cancer Res. 57:4593-4599 (1997)). Затем исследуемый Fab инкубируют в течение ночи; в то же время, инкубирование может продолжаться в течение более длительного периода (например, около 65 часов) для гарантии достижения равновесия. Затем смесь переносят на планшет для захвата и инкубируют при комнатной температуре (например, в течение часа). После инкубации раствор удаляют, а планшет промывают восемь раз поверхностно-активным веществом 0,1% TWEEN-20™ в PBS. После высушивания пластин, добавляют 150 мкл/на лунку сцинтиллятора (MICROSCINT-20™; Packard), и считают на гамма-счетчике TOPCOUNT™ (Packard) в течение десяти минут. Концентрации каждого Fab, обеспечивавшие связывание, меньшее или равное 20% от максимального, отбирают для применения в конкурентном анализе связывания.
Согласно еще одному варианту реализации, Kd измеряют с помощью анализа на основе поверхностного плазмонного резонанса с использованием прибора BIACORE®-2000 или BIACORE®-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C с чипами CM5 с иммобилизованным антигеном при ~ 10 единицах ответа (RU). Вкратце, чипы биосенсора с карбоксиметилированным декстраном (СМ5, BIAcore Inc.) активировали N-этил-N'-(3-диметиламинопропил)-карбодиимидгидрохлоридом (EDC) и N-гидроксисукцинимидом (NHS) согласно инструкциям поставщика. Антиген разбавляют 10 мМ раствором ацетата натрия, рН 4,8, до 5 мкг/мл (~0,2 мкМ) и вводят при скорости потока 5 мкл/минуту, до достижения около 10 единиц ответа (RU) связанного белка. После введения антигена вводят 1 М раствор этаноламина для блокирования непрореагировавших групп.Для измерений кинетики двукратные серийные разведения Fab (0,78 нМ до 500 нМ) вводят в PBS с 0,05% поверхностно-активным веществом TWEEN 20™ (PBST) при 25°С и скорости потока приблизительно 25 мкл/мин. Скорости ассоциации (kon) и диссоциации (koff) рассчитывали с помощью простейшей модели связывания Ленгмюра при соотношении один к одному (программное обеспечение BIAcore® Evaluation Software версии 3.2) путем одновременной аппроксимации сенсограмм ассоциации и диссоциации. Константа равновесной диссоциации (Kd) рассчитывается как отношение koff/kon. См., например, Chen et al., J. Mol. Biol. 293:865-881 (1999). Если скорость ассоциации согласно вышеописанному анализу поверхностного плазмонного резонанса превышает 106 М-1 s-1, то ее можно определить с помощью методики гашения флуоресценции, измеряющей увеличение или снижение интенсивности испускания флуоресценции (возбуждение = 295 нм; испускание = 340 нм, полоса пропускания 16 нм) при 25°С для 20 нМ раствора антител против антигена (в форме Fab) в PBS, рН 7,2, в присутствии возрастающих концентраций антигена, измеренных на спектрофотометре, например на спектрофотометре с устройством остановки потока (Aviv Instruments) или спектрофотометре SLM-AMINCO ™ 8000 серии (ThermoSpectronic) с перемешиваемой кюветой.
«Скорость передачи», «скорость ассоциации», «скорость ассоциации» или «kon» согласно данному изобретению также могут быть определены, как описано выше, с использованием системы BIACORE®-2000 или BIACORE®-3000 (BIAcore, Inc., Пискатуэй, Нью-Джерси).
Термины "клетка-хозяин", "линия клеток-хозяев" и "культура клеток-хозяев" применяются как взаимозаменяемые и относятся к клеткам, в которые введена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают "трансформанты" и "трансформированные клетки", которые включают первично трансформированные клетки и полученное от них потомство, независимо от количества пассажей. Потомство может не быть полностью идентичным исходной клетке по содержанию нуклеиновых кислот, и может содержать мутации. Настоящее описание включает мутантное потомство, обладающее той же функцией или биологической активностью, которую наблюдали во время скрининга или отбора из первоначально трансформированных клеток.
В данном документе термин "вектор" относится к молекуле нуклеиновой кислоты, способной обеспечивать репродукцию другой, связанной с ней нуклеиновой кислоты. Указанный термин включает вектор, находящийся в виде самореплицирующейся нуклеиновой кислоты, а также вектор, способный внедряться в геном клетки-хозяина, в которую его вводят.Некоторые векторы способны управлять экспрессией нуклеиновых кислот, с которыми они функционально связаны. Такие векторы в настоящем документе называют "экспрессирующими векторами".
"Процент (%) идентичности аминокислотной последовательности" по отношению к последовательностям эталонных полипептидов определяют как процентное содержание аминокислотных остатков в последовательности-кандидате, идентичных аминокислотным остаткам в конкретной последовательности эталонного полипептида после выравнивания последовательностей и, при необходимости, внедрения разрывов для достижения максимальной процентной идентичности последовательности, без учета консервативных замен как части идентичности последовательностей. Выравнивание для целей определения процентной идентичности аминокислотных последовательностей можно осуществить различными способами, известными специалистам в данной области техники, например, используя общедоступное компьютерное программное обеспечение, например, программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области техники могут определить соответствующие параметры выравнивания последовательностей, включая алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Однако для целей данного изобретения значение % идентичности аминокислотных последовательностей генерируется с использованием компьютерной программы сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 разработана компанией Genentech Inc., а исходный код представлен в документации пользователя в Бюро по охране авторских прав США, Вашингтон, округ Колумбия, 20559, где зарегистрирован под номером регистрации авторского права CIIIA TXU 510087. Программа ALIGN-2 находится в свободном доступе в Genentech, Inc., Саут-Сан-Франциско, Калифорния, или может быть скомпилирована из исходного кода. Для применения в операционной системе UNIX, включая цифровую версию UNIX V4.0D, программу ALIGN-2 следует компилировать. Все параметры сравнения последовательностей устанавливаются программой ALIGN-2 и остаются неизменными.
В случаях, когда ALIGN-2 используют для сравнения аминокислотных последовательностей, % идентичности аминокислотной последовательности данной аминокислотной последовательности А с, по сравнению с или относительно данной аминокислотной последовательности В (что можно иначе сформулировать так, что данная аминокислотная последовательность А характеризуется или включает определенный % идентичности аминокислотной последовательности с, по сравнению с или относительно данной аминокислотной последовательности В) рассчитывают следующим образом: 100-кратная доля X/Y, где X обозначает количество аминокислотных остатков, считающихся идентичными совпадениями при выравнивании А и В с помощью программы выравнивания последовательностей ALIGN-2, и где Y обозначает общее количество аминокислотных остатков в В. Следует принимать во внимание, что если длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, % идентичности аминокислотной последовательности А по отношению к В не равен % идентичности аминокислотной последовательности В по отношению к А. Если не указано иное, все используемые в данном документе процентные значения идентичности аминокислотных последовательностей получены, как описано.
Термин "антибиотик рифамицинового ряда" означает класс или группу антибиотиков, имеющих структуру или сходную структуру с рифамицином.
Термин "антибиотик рифалазилового ряда" означает класс или группу антибиотиков, имеющих структуру или сходную структуру с рифалазилом.
При указании числа заместителей термин «один или более» относится к диапазону от одного заместителя до максимально возможного числа замещений, то есть от замещения одного водорода до замещения всех атомов водорода заместителями. Термин «заместитель» означает атом или группу атомов, замещающих атом водорода на исходной молекуле. Термин «замещенный» означает, что указанная группа несет один или несколько заместителей. Когда любая группа может иметь несколько заместителей и предоставляется множество возможных заместителей, заместители выбирают независимо и они не обязательно должны быть одинаковыми. Термин «незамещенный» означает, что указанная группа не несет заместителей. Термин «необязательно замещенный» означает, что указанная группа является незамещенной или замещена одним или несколькими заместителями, независимо выбранными из группы возможных заместителей. При указании числа заместителей термин «один или более» означает от одного заместителя до максимально возможного числа замещений, то есть от замещения одного водорода до замещения всех атомов водорода заместителями.
Термин «алкил», в контексте данного документа, касается насыщенного линейного или разветвленного моновалентного углеводородного радикала, включающего от одного до двенадцати атомов углерода (С1-С12), где алкильный радикал может быть, возможно, независимо замещен одним или большим количеством заместителей, описанных ниже. В другом варианте реализации изобретения, алкильный радикал представляет собой от одного до восьми атомов углерода (C1-C8), или от одного до шести атомов углерода (C1-С6). Примеры алкильных групп включают, но не ограничиваются ими, метил (Me, -СН3), этил (Et, -СН2СН3), 1-пропил (n-Pr, n-пропил, -СН2СН2СН3), 2-пропил (i-Pr, i-пропил, -СН(СН3)2), 1-бутил (n-Bu, n-бутил, -СН2СН2СН2СН3), 2-метил-1-пропил (i-Bu, i-бутил, -СН2СН(СН3)2), 2-бутил (s-Bu, s-бутил, -СН(СН3)СН2СН3), 2-метил-2-пропил (t-Bu, t-t-butyl, -С(СН3)3), 1-пентил (n-пентил, -СН2СН2СН2СН2СН3), 2-пентил (-СН(СН3)СН2СН2СН3), 3-пентил (-СН(СН2СН3)2), 2-метил-2-бутил (-С(СН3)2СН2СН3), 3-метил-2-бутил (-СН(СН3)СН(СН3)2), 3-метил-1-бутил (-СН2СН2СН(СН3)2), 2-метил-1-бутил (-СН2СН(СН3)СН2СН3), 1-гексил (-СН2СН2СН2СН2СН2СН3), 2-гексил (-СН(СН3)СН2СН2СН2СН3), 3-гексил (-СН(СН2СН3)(СН2СН2СН3)), 2-метил-2-пентил (-С(СН3)2СН2СН2СН3), 3-метил-2-пентил (-СН(СН3)СН(СН3)СН2СН3), 4-метил-2-пентил (-СН(СН3)СН2СН(СН3)2), 3-метил-3-пентил (-С(СН3)(СН2СН3)2), 2-метил-3-пентил (-СН(СН2СН3)СН(СН3)2), 2,3-диметил-2-бутил (-С(СН3)2СН(СН3)2), 3,3-диметил-2-бутил (-СН(СН3)С(СН3)3, 1-гептил, 1-октил, и тому подобное.
Термин "алкилен", в контексте данного документа, касается насыщенного линейного или разветвленного двухвалентного углеводородного радикала, включающего от одного до двенадцати атомов углерода (С1-С12), где алкиленовый радикал может быть независимо замещен одним или более количеством заместителей, описанных ниже. В другом варианте реализации изобретения, алкиленовый радикал представляет собой от одного до восьми атомов углерода (C1-C8), или от одного до шести атомов углерода (C1-С6). Примеры алкиленовых групп включают, но не ограничиваются ими, метилен (-СН2-), этилен (-СН2СН2-), пропилен (-СН2СН2СН2-), и тому подобное.
Термин «алкенил» относится к линейному или разветвленному одновалентному углеводородному радикалу из двух-восьми углеродных атомов (С2-С8) с по меньшей мере одним сайтом ненасыщенности, то есть углерод-углерод, sp2 двойная связь, где алкинильный радикал может быть необязательно замещен независимо одним или более заместителей, описанных в данном документе, и включает радикалы, имеющие ориентации «цис» и «транс», или, альтернативно, ориентации «Е» и «Z». Примеры включают, но не ограничиваются ими, этиленил или винил (-СН=СН2), аллил (-СН2СН=СН2), и тому подобное.
Термин «алкенилен» относится к линейному или разветвленному двухвалентному углеводородному радикалу из двух-восьми углеродных атомов (С2-С8) с по меньшей мере одним сайтом ненасыщенности, то есть углерод-углерод, sp2 двойная связь, где алкениленовый радикал может быть необязательно замещен независимо одним или более заместителей, описанных в данном документе, и включает радикалы, имеющие ориентации «цис» и «транс», или, альтернативно, ориентации «Е» и «Z». Примеры включают, но не ограничиваются ими, этиленилен или винилен (-СН=СН-), аллил (-СН2СН=СН-), и тому подобное.
Термин «алкинил» относится к линейному или разветвленному одновалентному углеводородному радикалу из двух-восьми углеродных атомов (С2-С8) с по меньшей мере одним сайтом ненасыщенности, т.е. углерод-углерод, sp тройной связью, где алкинильный радикал может быть необязательно замещен независимо одним или более заместителями, описанными в данном документе. Примеры включают, но не ограничиваются ими, этинил (-С≡СН), пропинил (пропаргил, -CH2C≡СН), и тому подобное.
Термин «алкинилен» относится к линейному или разветвленному двухвалентному углеводородному радикалу из двух-восьми углеродных атомов (С2-С8) с по меньшей мере одним сайтом ненасыщенности, т.е. углерод-углерод, sp тройной связью, где алкиниленовый радикал может быть необязательно замещен независимо одним или более заместителями, описанными в данном документе. Примеры включают, но не ограничиваются ими, этинилен (-С≡С-), пропинилен (пропагилен, -СН2С≡С-), и тому подобное.
Термины "углеродное кольцо", "карбоциклил", "карбоциклическое кольцо» и «циклоалкил» относятся к одновалентному неароматическому, насыщенному или частично ненасыщенному кольцу, содержащему от 3 до 12 атомов (С3-С12) в качестве моноциклического кольца или от 7 до 12 атомов углерода в качестве бициклического кольца. Бициклические карбоциклы, содержащие от 7 до 12 атомов углерода, могут быть организованы, например, как бицикло[4,5], [5,5], [5,6] или [6,6] системы, а бициклические карбоциклы, содержащие 9 или 10 кольцевых атомов, могут быть организованы как бицикле- [5,6] или [6,6] системы, или как мостиковые системы, такие как бицикло[2.2.1]гептан, бицикло[2.2.2]октан и бицикло[3.2.2]нонан. В рамки указанного определения включены также спиро-фрагменты. Примеры моноциклических карбоциклов включают, но не ограничиваются ими, циклопропил, циклобутил, циклопентил, 1-циклопент-1-енил, 1-циклопент-2-енил, 1-циклопент-3-енил, циклогексил, 1-циклогекс-1-енил, 1-циклогекс-2-енил, 1-циклогекс-3-енил, циклогексадиенил, циклогептил, циклооктил, циклононил, циклодецил, циклоундецил, циклододецил и т.п. Карбоциклильные группы необязательно замещены независимо одним или несколькими заместителями, описанными в данном документе.
"Арил" означает одновалентный ароматический углеводородный радикал из 6-20 атомов углерода (С6-С20), полученный посредством удаления одного атома водорода от одного атома углерода исходной ароматической кольцевой системы. Некоторые арильные группы представлены в иллюстративных структурах как "Ar". Арил включает бициклические радикалы, содержащие ароматическое кольцо, конденсированное с насыщенным, частично ненасыщенным кольцом или с ароматическим карбоциклическим кольцом. Типичные арильные группы включают, но не ограничиваются ими, радикалы, полученные из бензола (фенил), замещенных бензолов, нафталина, антрацена, инденила, инданила, 1,2-дигидронафталина, 1,2,3,4-тетрагидронафталина и т.п. Арильные группы необязательно замещены независимо одним или более заместителями, описанными в данном документе.
"Арилен" означает двухвалентный ароматический углеводородный радикал с 6-20 атомами углерода (С6-С20), полученный удалением двух атомов водорода от двух атомов углерода исходной ароматической кольцевой системы. Некоторые ариленовые группы представлены в иллюстративных структурах как "Ar". Арилен включает бициклические радикалы, содержащие ароматическое кольцо, конденсированное с насыщенным, частично ненасыщенным кольцом или с ароматическим карбоциклическим кольцом. Типичные ариленовые группы включают, но не ограничиваются ими, радикалы, полученные из бензола (фенилена), замещенных бензолов, нафталина, антрацена, бифенилена, инденилена, инданилена, 1,2-дигидронафталина, 1,2,3,4-тетрагидронафталина и т.п. Ариленовые группы необязательно замещены одним или более заместителями, описанными в данном документе.
Термины "гетероцикл," "гетероциклил" и "гетероциклическое кольцо" использованы в данном документе взаимозаменяемо и относятся к насыщенному или частично ненасыщенному (т.е., содержащему одну или более двойных и/или тройных связей в кольце) карбоциклическому радикалу, содержащему от 3 до около 20 кольцевых атомов, в котором по меньшей мере один кольцевой атом представляет собой гетероатом, выбранный из азота, кислорода, фосфора и серы, а остальные кольцевые атомы представляют собой С, при этом один или более кольцевых атомов необязательно замещены независимо одним или более заместителями, описанными ниже. Гетероцикл может представлять собой моноцикл, содержащий от 3 до 7 кольцевых членов (от 2 до 6 атомов углерода и от 1 до 4 гетероатомов, выбранных из N, О, Р и S), или бицикл, содержащий от 7 до 10 кольцевых членов (от 4 до 9 атомов углерода и от 1 до 6 гетероатомов, выбранных из N, О, Р и S), например: бицикло [4,5], [5,5], [5,6] или [6,6] систему. Гетероциклы описаны в публикации Paquette, Leo A.; "Principles of Modern Heterocyclic Chemistry" (W.A. Benjamin, Нью-Йорк, 1968), в частности, в главах 1, 3, 4, 6, 7 и 9; "The Chemistry of Heterocyclic Compounds, A series of Monographs" (John Wiley & Sons, Нью-Йорк, с 1950 до настоящего времени), в частности, в томах 13, 14, 16, 19 и 28; и J. Am. Chem. Soc. (1960) 82:5566. "Гетероциклил" включает также радикалы, где гетероциклические радикалы слиты с насыщенным, частично ненасыщенным кольцом или ароматическим карбоциклическим или гетероциклическим кольцом. Примерами гетероциклических колец являются, не ограничиваясь ими, морфолин-4-ид, пиперидин-1-ил, пиперазинил, пиперазин-4-ил-2-он, пиперазин-4-ил-3-он, пирролидин-1-ил, тиоморфолин-4-ил, 8-диоксотиоморфолин-4-ил, азокан-1-ил, азетидин-1-ил, октагидропиридо[1,2-а]пиразин-2-ил, [1,4]диазепан-1-ил, пирролидинил, тетрагидрофуранил, дигидрофуранил, тетрагидротиенил, тетрагидропиранил, дигидропиранил, тетрагидротиопиранил, пиперидино, морфолино, тиоморфолино, тиоксанил, пиперазинил, гомопиперазинил, азетидинил, оксетанил, тиетанил, гомопиперидинил, оксепанил, тиепанил, оксазепанил, диазепинил, тиазепинил, 2-пирролинил, 3-пирролинил, индолинил, 2Н-пиранил, 4Н-пиранил, диоксанил, 1,3-диоксоланил, пиразолинил, дитианил, дитиоланил, дигидропиранил, дигидротиенил, дигидрофуранил, пиразолидинил, имидазолинил, имидазолидинил, 3-азабицикло[3.1.0]гексанил, 3-азабицикло[4.1.0]гептанил, азабицикло[2.2.2]гексанил, ЗН-индолил, хинолизинил и N-пиридилмочевины. В рамки указанного определения включены также спиро-фрагменты. Примеры гетероциклической группы, в которой 2 кольцевых атома замещены оксо-фрагментами (=O), представляют собой пиримидинонил и 1,1-диоксотиоморфолинил. Гетероциклические группы здесь необязательно независимо замещены одним или более количеством заместителей, описанных в данном документе.
Термин "гетероарил" относится к одновалентному ароматическому радикалу из 5-, 6-, или 7-членных колец и включает системы слитых колец (по меньшей мере одно из которых является ароматическим) и 5-20 атомов, содержащие один или более гетероатомов, независимо выбранных из азота, кислорода и серы. Примеры гетероарильных групп представляют собой пиридинил (включая, например, 2-гидроксипиридинил), имидазолил, имидазопиридинил, пиримидинил (включая, например, 4-гидроксипиримидинил), пиразолил, триазолил, пиразинил, тетразолил, фурил, тиенил, изоксазолил, тиазолил, оксадиазолил, оксазолил, изотиазолил, пирролил, хинолинил, изохинолинил, тетрагидроизохинолинил, индолил, бензимидазолил, бензофуранил, циннолинил, индазолил, индолизинил, фталазинил, пиридазинил, триазинил, изоиндолил, птеридинил, пуринил, оксадиазолил, триазолил, тиадиазолил, фуразанил, бензофуразанил, бензотиофенил, бензотиазолил, бензоксазолил, хиназолинил, хиноксалинил, нафтиридинил и фуропиридинил. Гетероарильные группы необязательно замещены независимо одним или более заместителями, описанными в данном документе.
Гетероциклическими или гетероарильными группами могут быть углерод (углерод-связанные) или связанный азот (азот-связанные), если это возможно. В качестве примера, но не ограничения, гетероциклы или гетероарилы, связанные через атом углерода, присоединены в положении 2, 3, 4, 5 или 6 пиридина, положении 3, 4, 5 или 6 пиридазина, положении 2, 4, 5 или 6 пиримидина, положении 2, 3, 5 или 6 пиразина, положении 2, 3, 4 или 5 фурана, тетрагидрофурана, тиофурана, тиофена, пиррола или тетрагидропиррола, положении 2, 4 или 5 оксазола, имидазола или тиазола, положении 3, 4 или 5 изоксазола, пиразола или изотиазола, положении 2 или 3 азиридина, положении 2, 3 или 4 азетидина, положении 2, 3, 4, 5, 6, 7 или 8 хинолина или положении 1, 3, 4, 5, 6, 7 или 8 изохинолина.
В качестве примера, но не ограничения, гетероциклы или гетероарилы, связанные через атом азота, присоединены в положении 1 азиридина, азетидина, пиррола, пирролидина, 2-пирролина, 3-пирролина, имидазола, имидазолидина, 2-имидазолина, 3-имидазолина, пиразола, пиразолина, 2-пиразолина, 3-пиразолина, пиперидина, пиперазина, индола, индолина, 1Н-индазола, положении 2 изоиндола или изоиндолина, положении 4 морфолина и положении 9 карбазола или β-карболина.
"Метаболит" представляет собой продукт, образованный посредством метаболизма в организме, определенного соединения или его соли. Метаболиты соединения могут быть идентифицированы с помощью стандартных методов, известных в данной области техники, и их активность определяют с помощью испытаний, таких как описаны в настоящем документе. Такие продукты могут быть образованы, например, в результате окисления, восстановления, гидролиза, амидирования, дезамидирования, эстерификации, деэстерификации, ферментативного расщепления и т.п. введенного соединения. Соответственно, настоящее изобретение включает метаболиты соединений, включая соединения, полученные способом, включающим контактирование соединения формулы I по данному изобретению с млекопитающим в течение периода времени, достаточного для получения его метаболического продукта.
Термин "фармацевтический состав" относится к препарату, который находится в такой форме, чтобы обеспечить эффективную биологическую активность активного ингредиента, содержащегося в нем, и не содержит дополнительных компонентов, являющихся неприемлемо токсичными для субъекта, которому должна быть введена композиция.
«Стерильная» композиция является асептической или свободной от всех живых микроорганизмов и их спор.
"Стабильная" композиция представляет собой композицию, в которой протеин в ней по существу сохраняет свою физическую и химическую стабильность и целостность при хранении. Различные аналитические методы для измерения стабильности белка доступны в данной области техники и рассматриваются в публикации Peptide and Protein Drug Delivery, 247-301, Vincent Lee Ed., Marcel Dekker, Inc., New York, New York, Pubs. (1991) и Jones, A. Adv. Drug Delivery Rev. 10:29-90 (1993). Стабильность может быть измерена при выбранной температуре в течение выбранного периода времени. Для быстрого скрининга препарат можно хранить при 40°С в течение от 2 недель до 1 месяца, после чего измеряют стабильность. Если композиция будет храниться при 2-8°С, в целом композиция должна быть стабильна при 30°С или 40°С в течение по меньшей мере 1 месяца и/или стабильна при 2-8°С в течение по меньшей мере 2 лет. Если композиция будет храниться при 30°С, в целом композиция должна быть стабильна в течение по меньшей мере 2 лет при 30°С и/или стабильна при 40°С в течение по меньшей мере 6 месяцев. Например, степень агрегации во время хранения может использоваться в качестве показателя стабильности белка. Таким образом, «стабильная» композиция может представлять собой композицию, в которой присутствует менее чем около 10% и предпочтительно менее чем около 5% белка в качестве агрегата в композиции. В других вариантах реализации изобретения, любое увеличение образования агрегатов во время хранения состава может быть определено.
"Изотоническая" композиция является той, которая имеет по существу такое же осмотическое давление, что и кровь человека. Изотонические композиции обычно имеют осмотическое давление от около 250 до 350 мОсм. Термин «гипотонический» описывает композицию с осмотическим давлением ниже, чем в крови человека. Соответственно, термин «гипертонический» используется для описания композиции с осмотическим давлением выше, чем у человеческой крови. Например, изотоничность может быть измерена с помощью осмометра давления пара и морозильного осмометра. Композиции по данному изобретению являются гипертоническими в результате добавления соли и/или буфера.
"Носители" включают фармацевтически приемлемые носители, вспомогательные вещества или стабилизаторы, которые являются нетоксичными для клетки или млекопитающего, подвергаемых их воздействию, в используемых дозировках и концентрациях. Часто физиологически приемлемый носитель представляет собой водный рН-буферный раствор. Примеры физиологически приемлемых носителей включают буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту; низкомолекулярные (менее чем около 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; спирты сахаров, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионогенные поверхностно-активные вещества, такие как TWEEN®, полиэтиленгликоль (PEG) и PLURONICS™.
"Фармацевтически приемлемый носитель" относится к ингредиенту в фармацевтическом составе, отличающемуся от активного ингредиента и являющемуся нетоксичным для субъекта. Фармацевтически приемлемый носитель включает буфер, вспомогательное вещество, стабилизатор или консервант, но не ограничивается ими. "Фармацевтически приемлемая кислота" включает неорганические и органические кислоты, которые являются нетоксичными при концентрации и способе их составления. Например, подходящие неорганические кислоты включают хлористоводородную, перхлорную, бромистоводородную, йодистоводородную, азотную, серную, сульфоновую, сульфиновую, сульфаниловую, фосфорную, угольную и т.д. Подходящие органические кислоты включают линейную и разветвленную алкильную, ароматическую, циклическую, циклоалифатическую, арилалифатическую, гетероциклическую, насыщенную, ненасыщенную, моно, ди- и трикарбоновую кислоту, включая, например, муравьиную, уксусную, 2-гидрокси уксусную, трифторуксусную, фенилуксусную, триметилуксусную, трет-бутил уксусную, антраниловую, пропановую, 2-гидроксипропановую, 2-оксопропановую, пропандионовую, циклопентанпропионовую, циклопентан пропионовую, 3-фенилпропионовую, бутановую, бутандиовую, бензойную, 3-(4-гидроксибензоил)бензойную, 2-ацетокси-бензойную, аскорбиновую, коричную, лаурилсульфоновую, стеариновую, муконовую, миндальную, янтарную, эмбоновую, фумаровую, яблочную, малеиновую, гидроксималеиновую, малоновую, молочную, лимонную, винную, гликолевую, глюконовую, глюконовую, пировиноградную, глиоксальную, щавелевую, мезиловую, салициловую, фталевую, пальмоидную, пальминовую, тиоциановую, метансульфоновую, этандисульфоновую, 1,2-этандисульфоновую, 2-гидроксиэтансульфоновую, бензолсульфоновую, 4-хоробензенесульфоновую, нафталин-2-сульфоновую, р-толуолсульфоновую, камфорсульфоновую, 4-метилбицикло[2.2.2]-окт-2-ен-1-карбоновую кислоту, глюкогептоновую кислоту, 4,4'-метиленбис-3-(гидрокси-2-ен-1-карбоновую кислоту), гидроксинафтойную кислоту.
"Фармацевтически приемлемые основания" включают неорганические и органические основания, которые являются нетоксичными по концентрации и способу их составления. Например, подходящие основания включают те, которые образованны из неорганического основания, образующего металлы, такие как литий, натрий, калий, магний, кальций, аммоний, железо, цинк, медь, марганец, алюминий, N-метилглюкамин, морфолин, пиперидин и органические нетоксичные основания, первичные, вторичные и третичные амины, замещенные амины, циклические амины и основные ионообменные смолы [например, N(R')4+ (где R' независимо представляет собой Н или С1-4 алкил, например аммоний, Трис)], например, изопропиламин, триметиламин, диэтиламин, триэтиламин, трипропиламин, этаноламин, 2-диэтиламиноэтанол, триметамин, дициклогексиламин, лизин, аргинин, гистидин, кофеин, прокаин, гидрабамин, холин, бетаин, этилендиамин, глюкозамин, метилглюкамин, теобромин, пурины, пиперазин, пиперидин, N-этилпиперидин, полиаминные смолы и тому подобное. В частности, предпочтительными органическими нетоксичными основаниями являются изопропиламин, диэтиламин, этаноламин, триметамин, дициклогексиламин, холин и кофеин.
Дополнительные фармацевтически приемлемые кислоты и основания, пригодные для использования в данном изобретении, включают те, которые получены из аминокислот, например гистидина, глицина, фенилаланина, аспарагиновой кислоты, глутаминовой кислоты, лизина и аспарагина.
"Фармацевтически приемлемые" буферы и соли включают соли, полученные путем добавления солей и вышеуказанных кислот и оснований. Конкретные буферы и/или соли включают гистидин, сукцинат и ацетат.
"Фармацевтически приемлемый сахар" представляет собой молекулу, которая в сочетании с представляющим интерес белком значительно предотвращает или уменьшает химическую и/или физическую нестабильность белка при хранении. Когда предполагается, что препарат лиофилизуют, а затем восстанавливают, «фармацевтически приемлемые сахара» также могут быть известны как «лиопротекторы». Иллюстративные сахара и их соответствующие сахарные спирты включают: аминокислоту, такую как глутамат натрия или гистидин; метиламин, такой как бетаин; лиотропную соль, такую как сульфат магния; полиол, такой как трехатомные спирты или спирты с более высоким молекулярным весом, например, глицерин, арабитол, ксилит, сорбит и маннит; пропиленгликоль; полиэтиленгликоль; PLURONICS®; и их комбинации. Дополнительные иллюстративные лиопротекторы включают глицерин и желатин, и сахара, меллибиозу, мелецитозу, рафинозу, маннотриозу и стахиозу. Примеры редуцирующих сахаров включают глюкозу, мальтозу, лактозу, мальтулозу, изо-мальтулозу и лактулозу. Примеры нередуцирующих сахаров включают нередуцирующие гликозиды полигидрокси соединений, выбранных из сахарных спиртов и других полиспиртов с прямой цепью. Предпочтительными сахарными спиртами являются моногликозиды, особенно те соединения, которые получают восстановлением дисахаридов, таких как лактоза, мальтоза, лактулоза и мальтулоза. Гликозидная боковая группа может быть глюкозидной или галактозидной. Дополнительными примерами сахарных спиртов являются глюцит, мальтит, лактит и изо-мальтулоза. Предпочтительными фармацевтически приемлемыми сахарами являются нередуцирующие сахара трегалоза или сахароза. Фармацевтически приемлемые сахара добавляют к композиции в «защитном количестве» (например, пре-лиофилизации), что означает, что белок по существу сохраняет свою физическую и химическую стабильность и целостность во время хранения (например, после восстановления и хранения).
Интересующий "разбавитель", как используется в данном документе, представляет собой тот, который является фармацевтически приемлемым (безопасным и нетоксичным для введения человеку) и полезен для приготовления жидкой композиции, такого как композиция, восстановленная после лиофилизации. Примеры разбавителей включают стерильную воду, бактериостатическую воду для инъекций (BWFI), рН-буферный раствор (например, фосфатно-солевой буфер), стерильный солевой раствор, раствор Рингера или раствор декстрозы. В альтернативном варианте реализации изобретения, разбавители могут включать водные растворы солей и/или буферы.
"Консервант" представляет собой соединение, которое может быть добавлено к композициям данного изобретения для снижения бактериальной активности. Добавление консерванта может, например, облегчать получение композиции многоразового использования (многократная доза). Примеры потенциальных консервантов включают хлорид октадецилдиметилбензиламмония, хлорид гексаметония, хлорид бензалкония (смесь хлоридов алкилбензилдиметиламмония, в которой алкильные группы являются длинноцепочечными соединениями) и хлорид бензетония. Другие типы консервантов включают ароматические спирты, такие как фенол, бутил и бензиловый спирт, алкилпарабены, такие как метил или пропилпарабен, катехол, резорцин, циклогексанол, 3-пентанол и м-крезол. Наиболее предпочтительным консервантом в контексте данного документа является бензиловый спирт.
"Индивидуум" или "субъект" представляет собой млекопитающее. Млекопитающие включают, но не ограничиваются ими, одомашненных животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и приматов, не являющихся людьми, таких как обезьяны), кроликов и грызунов (например, мышей и крыс). В некоторых вариантах реализации изобретения, индивидуум или субъект является человеком.
В настоящем описании термин «лечение» (и его грамматические варианты, такие как «лечить» или «проводить лечение») относится к клиническому вмешательству, предназначенному для изменения естественного течения заболевания у индивидуума, ткани или клетки, подвергающегося лечению в ходе клинического проявления патологии. Желаемые эффекты лечения включают в себя, но не ограничиваются ими, уменьшение скорости прогрессирования болезни, улучшение или ослабление болезненного состояния и ремиссию, или улучшение прогноза, все, поддающееся измерению специалистом в данной области, таким как врач. В одном из вариантов реализации изобретения, лечение может означать облегчение симптомов, уменьшение прямых или косвенных патологических последствий заболевания, снижение скорости прогрессирования заболевания, улучшение или временное улучшение течения заболевания, и ремиссию или улучшение прогноза. В некоторых вариантах реализации, ААС, ТАС согласно изобретению используются для замедления развития заболевания или замедления прогрессирования инфекционного заболевания, или снижения бактериальной нагрузки в кровотоке и/или в инфицированных тканях и органах.
Как используется в данном документе, "в сочетании с" относится к введению одного способа лечения в дополнение к другому способу лечения. Таким образом, «в сочетании с» относится к введению одного лечебного воздействия до, во время или после введения другого способа лечения индивидууму.
Термин «бактериемия» относится к присутствию бактерий в кровотоке, которые наиболее часто обнаруживается в культуре крови. Бактерии могут проникать в кровоток как тяжелое осложнение инфекции (например, пневмония или менингит), во время операции (особенно при вовлечении слизистых оболочек, таких как желудочно кишечный тракт), или из-за катетеров и других инородных тел, входящих в артерии или вены. Бактериемия может иметь несколько последствий. Иммунный ответ на бактерии может вызвать сепсис и септический шок, который приводит к относительно высокой смертности. Бактерии могут также использовать кровь для распространения в другие части тела, вызывая инфекции от исходного места инфекции. Примеры включают эндокардит или остеомиелит.
"Терапевтически эффективное количество" представляет собой минимальную концентрацию, необходимую для осуществления измеримого улучшения конкретного заболевания. В данном контексте терапевтически эффективное количество может варьироваться в зависимости от таких факторов, как течение заболевания, возраст, пол и масса тела пациента и способности антитела вызывать желаемый ответ у индивидуума. Терапевтически эффективное количество также является таким, при использовании которого терапевтически благоприятные эффекты превосходят любые токсические или пагубные эффекты антитела. В одном варианте реализации изобретения, терапевтически эффективное количество представляет собой количество, эффективное для снижения бактериемии при инфицировании in vivo. В одном аспекте, «терапевтически эффективное количество» представляет собой по меньшей мере такое количество, которое эффективно для снижения бактериальной нагрузки или колониеобразующих единиц (КОЕ), выделенных из образца пациента, такого как кровь, с по меньшей мере одним логарифмом по сравнению с предшествующим введением лекарственного средства. В более конкретном аспекте, сокращение составляет по меньшей мере 2 log. В другом аспекте, уменьшение составляет, по меньшей мере, 3, 4, 5 log. В еще одном аспекте, снижение достигает уровня, не детектируемого с использованием анализов, известных в данной области техники, включая анализы, приведенные в качестве примера. В другом варианте, терапевтически эффективное количество представляет собой количество ААС в одной или нескольких дозах, вводимых в течение периода лечения, которое приводит к негативной культуре крови (т.е. бактерии, являющиеся мишенью ААС, не вырастут) по сравнению с положительной культурой крови до или в начале лечения инфицированного пациента.
"Профилактически эффективное количество" относится к количеству, эффективному в дозах и в течение периодов времени, необходимых для достижения желательного профилактического результата. Поскольку профилактическую дозу используют для субъектов, находящихся до, на ранней стадии заболевания, или даже до воздействия условий, при которых повышен риск инфицирования, профилактически эффективное количество может быть меньше, чем терапевтически эффективное количество. В одном варианте реализации изобретения, профилактически эффективное количество представляет собой по меньшей мере количество, эффективное для снижения, предотвращения, возникновения или распространения инфекции от одной клетки к другой.
"Хроническое" введение относится к введению лекарственных(ого) средств(а) в непрерывном, а не в остром режиме, чтобы поддерживать первоначальный терапевтический эффект (активность) в течение длительного периода времени. "Прерывистое" введение это лечение, которое не выполняется последовательно без перерыва, а носит циклический характер.
Термин "вкладыш в упаковку" относится к инструкциям, в обычном порядке вкладываемым в упаковки терапевтических продуктов для продажи, которые содержат информацию о показаниях, применении, дозах, приеме, комбинированной терапии, противопоказаниях и/или предостережениях, касающихся применения таких терапевтических продуктов.
Термин «хиральный» относится к молекулам, которые обладают свойством не совпадать при наложении со своим зеркальным отображением, в то время как термин «ахиральный» относится к молекулам, которые при наложении совпадают со своим зеркальным отображением.
Термин "стереоизомеры" относится к соединениям, которые имеют одинаковый химический состав, но различаются расположением атомов или групп в пространстве.
"Диастереомер" относится к стереоизомеру с двумя или более центрами хиральности, молекулы которых не являются зеркальными отображениями друг друга. Диастереоизомеры имеют различные физические свойства, например, температуры плавления, температуры кипения, спектральные свойства и реакционную способность. Смеси диастереоизомеров можно разделить с помощью аналитических методик с высокой степенью разделения, таких как электрофорез и хроматография.
Термин "энантиомеры" относится к двум стереоизомерам соединения, которые являются зеркальными отображениями друг друга, не совпадающими при наложении.
Стереохимические определения и условные обозначения, используемые в настоящем документе, в целом соответствуют S.P. Parker, Ed., McGraw-Hill Dictionary of Chemical Terms (1984) McGraw-Hill Book Company, New York; and Eliel, E. and Wilen, S., Stereochemistry of Organic Compounds (1994) John Wiley & Sons, Inc., New York. Многие органические соединения существуют в оптически активных формах, т.е. они обладают способностью вращать плоскость плоскополяризованного света. В описании оптически активного соединения приставки D и L или R и S, используют для обозначения абсолютной конфигурации молекулы относительно ее хирального центра(-ов). Префиксы d и l или (+) и (-) используют для обозначения знака поворота плоскополяризованного света соединением, при этом префиксы (-) или 1 означают, что соединение является левовращающим. Соединение с приставкой (+) или d является правовращающим. Для данной химической структуры эти стереоизомеры являются одинаковыми, за исключением того, что они представляют собой зеркальные отражения друг друга. Конкретный стереоизомер также можно рассматривать в качестве энантиомера, при этом смесь таких изомеров часто называют энантиомерной смесью. Смесь энантиомеров 50:50 называют рацемической смесью или рацематом; указанная смесь может образовываться в случае, если в химической реакции или процессе отсутствуют стереоизбирательность или стереоспецифичность. Термины "рацемическая смесь" и "рацемат" относятся к эквимолярной смеси двух типов энантиомеров, не обладающей оптической активностью.
Термин "защитная группа" относится к заместителю, который обычно используют для блокирования или защиты конкретной функциональной группы, в то время как другие функциональные группы реагируют на соединение. Например, "аминозащитная группа" является заместителем, присоединенным к аминогруппе, который блокирует или защищает функциональную аминогруппу в соединении. Подходящие аминозащитные группы включают ацетил, трифторацетил, t-бутоксикарбонил (ВОС), бензилоксикарбонил (CBZ) и 9-флуоренилметиленоксикарбонил (Fmoc), но не ограничиваются ими. Общее описание защитных групп и их применение см. в T.W. Greene, Protective Groups in Organic Synthesis, John Wiley & Sons, New York, 1991 или более позднем издании этой книги.
В данном контексте термин "около" относится к обычному диапазону ошибок для соответствующего значения, хорошо известного специалисту в данной области техники. Упоминание термина «приблизительно» при значении или параметре в настоящем документе включает (и описывает) варианты реализации, направленные на само это значение или параметр.
В данном контексте и в прилагаемой формуле изобретения, формы единственного числа включают множественное число, если из контекста очевидно не следует иное. Например, ссылка на "антитело" представляет собой ссылку на от одного до нескольких антител, таких как молярные количества, и включает его эквиваленты, известные специалистам в данной области техники, и так далее.
III. КОМПОЗИЦИИ И СПОСОБЫ
КОНЪЮГАТЫ АНТИТЕЛО-АНТИБИОТИК (ААС)
Экспериментальные результаты данного документа являются убедительным свидетельством того, что терапия, направленная на устранение внутриклеточных бактерий, улучшит клинический успех. С этой целью данное изобретение предлагает уникальное терапевтическое средство, которое избирательно убивает организмы S. aureus, которые вторглись во внутриклеточные компартменты клеток-хозяев. Данное изобретение демонстрирует, что такое терапевтическое действие является эффективным в моделях in vivo, когда обычные антибиотики, такие как ванкомицин, терпят неудачу.
Изобретение обеспечивает антибактериальную терапию, которая направлена на предотвращение выхода антибиотиков благодаря нацеливанию на популяции бактерий, которые уклоняются от традиционной антибактериальной терапии. Новая антибактериальная терапия достигается с помощью конъюгата антитело-антибиотик (ААС), в котором антитело, специфичное для компонентов клеточной стенки, обнаруженных на S. aureus (включая MRSA), химически связано с мощным антибиотиком (производным рифамицина). Антибиотик присоединяется к антителу через расщепляемый протеазой непептидный линкер, который разработан для расщепления протеазами, включая катепсин В, лизосомальную протеазу, обнаруженную в большинстве типов клеток млекопитающих (Dubowchik et al (2002) Bioconj.Chem. 13:855-869). Диаграмма ААС с его тремя компонентами показана на Фиг. 2. Не ограничиваясь какой-либо одной теорией, один механизм действия ААС схематизируется на Фиг. 3. ААС действует как пролекарство, так как антибиотик неактивен (из-за большого размера антитела) до тех пор, пока линкер не расщеплен. Поскольку значительная часть S. aureus, обнаруживаемых в случае природной инфекции, поглощается клетками-хозяевами, главным образом нейтрофилами и макрофагами, в какой-то момент времени инфицирования хозяина, время, проведенное внутри клеток-хозяев, предоставляет бактерии существенную возможность для уклонения от активности антибиотиков. ААС изобретения предназначены для связывания с S. aureus и высвобождения антибиотика внутри фаголизосомы после того, как бактерии поглощаются клетками-хозяевами. Благодаря этому механизму ААС способны сконцентрировать активный антибиотик специально в месте, где S. aureus плохо поддаются воздействию обычных антибиотиков. Хотя изобретение не ограничено или не определено конкретным механизмом действия, ААС улучшают антибиотическую активность через три потенциальных механизма: (1) ААС доставляет антибиотик внутрь клеток млекопитающих, которые поглощают бактерии, тем самым повышая эффективность антибиотиков, которые слабо диффундируют в фаголизосомы, где изолированы бактерии; (2) ААС опсонизирует бактерии, тем самым увеличивая поглощение свободных бактерий фагоцитарными клетками и высвобождает антибиотик локально для уничтожения бактерий, в то время, когда они изолированы в фаголизосоме. Поскольку тысячи ААС могут связываться с одной бактерией, такая платформа высвобождает достаточное количество антибиотиков в этих внутриклеточных нишах для обеспечения максимального антимикробного уничтожения. Кроме того, поскольку большее количество бактерий высвобождается из ранее существовавших внутриклеточных резервуаров, высокая скорость такой терапии на основе антител обеспечивает немедленную «маркировку» этих бактерий, прежде чем они смогут уйти в соседние или отдаленные клетки, таким образом снижая дальнейшее распространение инфекции. (3) Благодаря связыванию антибиотика с антителом ААС увеличивает период полужизни антибиотиков in vivo (улучшенная фармакокинетика) по сравнению с антибиотиками, которые быстро выводятся из крови. Улучшенная фармакокинетика ААС обеспечивает доставку достаточного количества антибиотиков в области концентрации S. aureus, ограничивая общую дозу антибиотика, который необходимо вводить системно. Это свойство должно обеспечить долгосрочную терапию ААС, нацеленную на постоянную инфекцию с минимальными побочными эффектами антибиотиков.
Соединение конъюгата антитело-антибиотик, содержащее антитело против тейхоевой кислоты клеточной стенки (WTA), ковалентно присоединенное с помощью расщепляемого протеазой непептидного линкера к антибиотику типа рифамицина.
Иллюстративный вариант реализации изобретения представляет собой конъюгат антитело-антибиотик, имеющий формулу:
причем:
Ат представляет собой антитело против тейхоевой кислоты клеточной стенки; PML представляет собой расщепляемый протеазой непептидный линкер, имеющий формулу:

причем Str представляет собой растягивающую единицу; РМ представляет собой пептидомиметическую единицу, и Y представляет собой спейсерную единицу;
abx представляет собой антибиотик рифамицинового типа; и
Р представляет собой целое число от 1 до 8.
Антибиотик рифамицинового ряда может быть антибиотиком типа рифалазила.
Антибиотик рифамицинового ряда может содержать четвертичный амин, присоединенный к расщепляемому протеазой непептидному линкеру.
Иллюстративный вариант реализации конъюгата антитело-антибиотик имеет формулу I:
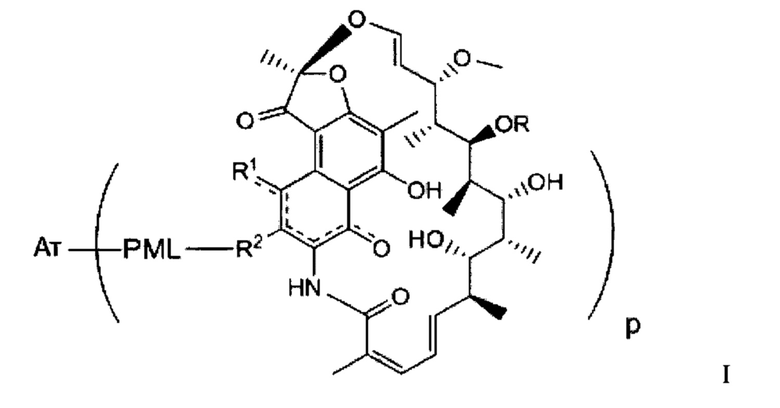
где:
пунктирные линии обозначают необязательную связь;
R представляет собой Н, С1-С12 алкил, или С(O)СН3;
R1 представляет собой ОН;
R2 представляет собой СН=N-(гетероциклил), причем гетероциклил необязательно замещен одной или более группами, независимо выбранными из С(O)СН3, С1-С12 алкил, С1-С12 гетероарил, С2-С20 гетероциклил, С6-С20 арил, и С3-С12 карбоциклил;
или R1 и R2 образуют пяти- или шестичленный конденсированный гетероарил или гетероциклил и необязательно образуют спиро или конденсированный шестичленный гетероарил, гетероциклил, арил или карбоциклическое кольцо, где спиро или конденсированный шестичленный гетероарил, гетероциклил, арил или карбоциклическое кольцо необязательно замещены Н, F, Cl, Br, I, С1-С12 алкилом или ОН;
PML представляет собой расщепляемый протеазой непептидный линкер, присоединенный к R2 или конденсированный гетероарил или гетероциклил, образованный R1 и R2; и
Ат представляет собой антитело против тейхоевой кислоты клеточной стенки (WTA).
Количество антибиотических фрагментов, которые могут быть конъюгированы через реактивный линкерный фрагмент с молекулой антитела, может быть ограничено числом свободных остатков цистеина, которые вводят описанными в данном документе способами. Иллюстративные ААС содержат антитела, которые имеют 1, 2, 3 или 4 сконструированные цистеиновые аминокислоты (Lyon, R. et al (2012) Methods in Enzym. 502:123-138).
АНТИТЕЛА ПРОТИВ ТЕЙХОЕВОЙ КИСЛОТЫ КЛЕТОЧНОЙ СТЕНКИ (WTA)
Некоторые анти-WTA Ат и конъюгированные анти-WTA антитела, которые связываются с WTA, экспрессируемыми рядом Gm+ бактерий, включая Staphylococcus aureus, раскрыты в данном документе. Анти-WTA антитела могут быть выбраны и получены способами, описанными в US 8283294; Meijer PJ et al (2006) J Mol Biol. 358(3):764-72; Lantto J, et al (2011) J Virol. 85 (4): 1820-33 и в примерах 3-4 ниже.
Клеточная стенка грамположительных бактерий состоит из толстого слоя оболочек из нескольких пептидогликанов (PGN), которые не только стабилизируют клеточную мембрану, но также обеспечивают множество участков, к которым могут быть присоединены другие молекулы (Фиг. 4). Основным классом этих гликопротеинов клеточной поверхности являются тейхоевые кислоты («ТА»), которые представляют собой богатые фосфатом молекулы, обнаруженные на многих гликано-связывающих белках (GPB). ТА бывают двух типов: (1) липотейхоевая кислота ("LTA"), которая прикреплена к плазматической мембране и простирается от клеточной поверхности до слоя пептидогликана; и (2) ТА клеточной стенки ("WTA"), которая ковалентно прикреплена к пептидогликану и проходит через клеточную стенку и за ее пределами (Фиг. 4). WTA может составлять 60% от общей массы GPB клеточной стенки. В результате он представляет высоко экспрессированный клеточный поверхностный антиген.
Химические структуры WTA различаются у разных организмов. В S. aureus, WTA ковалентно связана с 6-ОН N-ацетилмураминовой кислоты (MurNAc) через дисахарид, состоящий из N-ацетилглюкозамина (GlcNAc) -1-Р и N-ацетилманнозамина (ManNAc), за которым следуют две или три единицы глицеринфосфатов (Фиг. 5). Фактический полимер WTA состоит из около 11-40 повторяющихся единиц рибитолфосфата (Rbo-P). Поэтапный синтез WTA сначала инициируется ферментом, называемым TagO, и штаммы S. aureus, у которых отсутствует ген TagO (в следствие делеции гена), не образуют никаких WTA. Повторяющиеся звенья могут быть дополнительно сшиты с D-аланином (D-Ala) в С2-ОН и/или с N-ацетилглюкозамином (GlcNAc) в положении С4-ОН через α-(альфа) или β-(бета) гликозидные связи. В зависимости от штамма S. aureus или фазы роста бактерий гликозидные связи могут представлять собой α -, β - или смесь двух аномеров. Эти модификации сахара GlcNAc производятся двумя специфическим, выделенными из S. aureus гликозилтрансферазами (Gtfs): TarM Gtf опосредует образование α-гликозидных связей, тогда как TarS Gtfs опосредует образование β-(бета)гликозидных связей.
Учитывая убедительные доказательства того, что внутриклеточные хранилища MRSA защищены от антибиотиков, для предотвращения такого способа уклонения от
антибиотиков по данному изобретению были разработаны новые терапевтические композиции с использованием специфических антител S. aureus для присоединения антибиотика к бактериям таким образом, чтобы когда бактерии поглощаются или иным образом интернализуются клеткой-хозяином in vivo, они переносили антибиотик в клетку-хозяина.
Анти-WTA антитело ААС по настоящему изобретению может быть анти-WTAα антителом или анти-WTAβ антителом. Иллюстративные анти-WTA Ат, предусмотренные в описании, клонировали из В-клеток из S. aureus инфицированных пациентов (как показано в примерах ниже). В одном варианте реализации анти-WTA и анти-Staph aureus Ат являются человеческими моноклональными антителами. ААС или ТАС по данному изобретению включают химерные Ат и гуманизированные Ат, содержащие CDR данных WTA Ат.
Для терапевтического применения данного изобретения, WTA Ат, конъюгированные с антибиотиками для получения ААС, могут быть любого изотипа, за исключением IgM. В одном варианте реализации изобретения, WTA Ат являются изотипом IgG человека. В более конкретных вариантах реализации изобретения, WTA Ат представляют собой человеческий IgG1.
Во всем описании и фигурах Ат, обозначенные 4-значным числом (например, 4497) также могут упоминаться с предшествующим «S», например, S4497; оба названия относятся к одному и тому же антителу, которое представляет собой немодифицированную последовательность антитела дикого типа (ДТ). Варианты антитела обозначены с помощью «v» после номера антитела, например, 4497v8. Если не указано (например, по номеру варианта), указанные аминокислотные последовательности являются исходными, немодифицированными/неизмененными последовательностями. Эти Ат могут быть изменены на один или несколько остатков, например, для улучшения рК, стабильности, экспрессии, технологичности (например, как описано в примерах ниже), при сохранении, по существу, того же или улучшенного связывания с антигеном по сравнению с немодифицированным антителом дикого типа. Варианты настоящих WTA антител, имеющих консервативные аминокислотные замены, охватываются данным изобретением. Ниже, если не указано иное, нумерация CDR соответствует Кабат, а нумерация константных доменов соответствует нумерации ЕС.
Для конъюгации с получением ТАС соединения, антитела против WTA по изобретению могут содержать сконструированные Cys в одной или обеих L и Н цепях для конъюгации с промежуточным соединением линкер-антибиотик, как описано ниже.
На Фиг. 11А и Фиг. 11В показано выравнивание аминокислотной последовательности вариабельных участков легкой цепи (VL) и вариабельного участка тяжелой цепи (VH), соответственно, четырех человеческих анти-WTA-альфа антител. CDR-последовательности CDR L1, L2, L3 и CDR H1, Н2, Н3 в соответствии с нумерацией Кабата подчеркнуты.
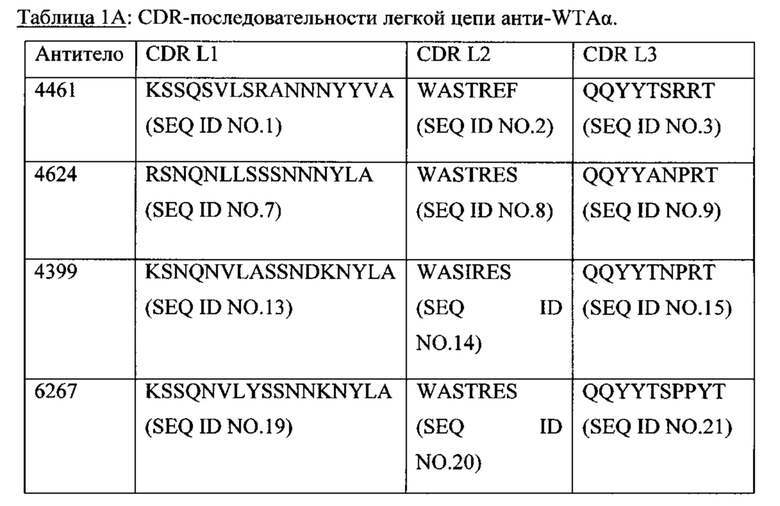
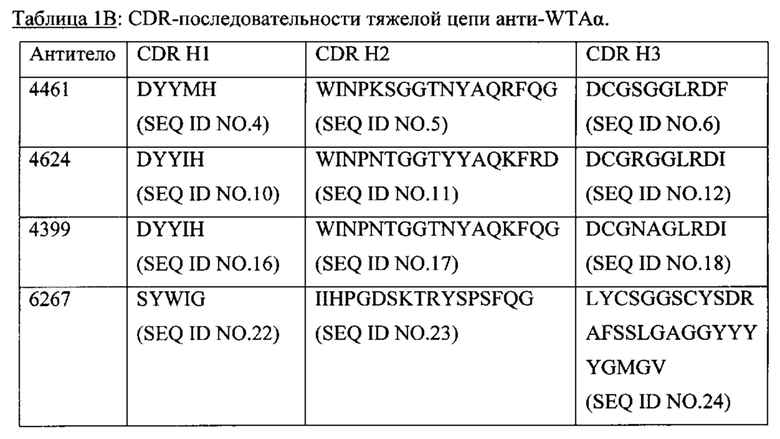
Последовательности каждой пары VL и VH являются следующими: 4461 Вариабельный участок легкой цепи
DIQMTQSPDSLAVSLGERATrNCKSSQSVLSRANNNYYVAWYQHKPGQPPKXLIYWAST REFGVPDRFSGSGSGTDFTLTINSLQAEDVAVYYCOOYYTSPvRTFGQGTKVEIK (SEQ ID NO. 25)
4461 Вариабельный участок тяжелой цепи
OVOLVOSGAEVRKPGASVKVSCKASGYSFTDYYMHWVROAPGOGLEWMGWrNPKSG
GmYAQRFQGRVTMTGDTSISAAYMDLASLTSDDTAVYYCVKDCGSGGLRDFWGOGT TVTVSS (SEQ ID NO. 26)
4624 Вариабельный участок легкой цепи
DIOMTOSPDSLSVSLGERATiNCRSNONLLSSSNNNYLAWYOQKPGQPLKLLIYWASTRESGVPDRFSGSGSGTDFTLTISSLOAEDVAVYYCOOYYANPRTFGOGTKVEIK (SEQ ID NO. 27)
4624 Вариабельный участок тяжелой цепи
OVOLOOSRVEVKRPGTSVKVSCKTSGYTFSDYYIHWVRLAPGQGLELMGWrNPNTGGTYYAQKFRDRVTMTRDTSIATAYLEMSSLTSDDTAVYYCAKDCGRGGLRDIWGPGTMVTVSS (SEQ ID NO. 28)
4399 Вариабельный участок легкой цепи
EIVLTOSPDSLAVSLGERATINCKSNONVLASSNDKNYLAWFOHKPGQPLKLLIYWASIRESGVPDRFSGSGSGTDFTLTISSLRAEDVAVYYCOOYYTNPRTFGQGTKVEFN (SEQ ID NO. 29)
4399 Вариабельный участок тяжелой цепи
EVOLVOSGAEVKKPGTSVKVSCKASGYTFTDYYIHWVRLAPGOGLELMGWrNPNTGGTNYAOKFOGRVTMTRDTSIATAYMELSSLTSDDTAVYYCAKDCGNAGLRDIWGQGTTVTVSS (SEQ ID NO. 30)
6267 Вариабельный участок легкой цепи
DIOLTOSPDSLAVSLGERATINCKSSONVLYSSNNKNYLAWYOQKPGQPPKLLIYWASTRESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCOOYYTSPPYTFGQGTKLEIE (SEQ ID NO. 31)
6267 Вариабельный участок тяжелой цепи
FVOT.VOSGAF.VKKPGEST.KTSCKGSGYSFTSYWIGWVROMPGKGLEWMGIIHPGDSKTRYSPSFOGOVTISADKSISTAYLOWNSLKASDTAMYYCARLYCSGGSCYSDRAFSSLGAGGYYYYGMGVWGOGTTVTVSS (SEQ ID NO. 32).
Для получения соединения AAC, данное изобретение предусматривает изолированное моноклональное антитело, которое связывает альфа тейхоевую кислоту клеточной стенки (WTAα), и содержит легкую цепь и Н-цепь, причем L-цепь, содержит CDR L1, L2, L3 и Н-цепь, содержит CDR H1, Н2, Н3, причем CDR L1, L2, L3 и H1, Н2, Н3 содержат аминокислотные последовательности CDR каждого из Ат 4461 (SEQ ID NO. 1-6), 4624 (SEQ ID NO. 7-12), 4399 (SEQ ID NO. 13-18), и 6267 (SEQ ID NO. 19-24) соответственно, как продемонстрировано в таблице 1А и таблице 1В выше.
В другом варианте изолированное моноклональное Ат, которое связывает WTAα, содержит вариабельный участок Н цепи (VH) и вариабельный участок L-цепи (VL), причем VH содержит, по меньшей мере, 95% идентичности последовательности по длине VH участка последовательности каждого из антител 4461, 4624, 4399, и 6267, соответственно. В еще одном конкретном аспекте, идентичность последовательности составляет 96%, 97%, 98%, 99% или 100%.
Данное изобретение также относится к ААС, содержащему анти-WTA бета-антитело из списка Ат, проиллюстрированного на Фиг. 12. В одном варианте реализации изобретения, изолированное анти-WTA бета моноклональное Ат содержит CDR L1, L2, L3 и H1, Н2, Н3, выбранные из группы, состоящей из CDR каждого из 13 Ат на Фиг. 12. В другом варианте реализации, изобретение предусматривает изолированное анти-WTA бета Ат, содержащее по меньшей мере 95% идентичности последовательности по длине V участка доменов каждого из 13 антител. В еще одном конкретном аспекте, идентичность последовательности составляет 96%, 97%, 98%, 99% или 100%.
Из 13 анти-WTA бета Ат 6078 и 4497 были модифицированы для создания вариантов i) имеющих сконструированный Cys в одной или обеих цепях L и Н для конъюгации с промежуточными соединениями линкер-антибиотик; и ii) отличающихся тем, что первый остаток в Н-цепи Q изменен на Е (v2), или первые два остатка QM были изменены на EI или EV (v3 и v4).
Фиг. 13А-1 и 13А-2 демонстрируют аминокислотную последовательность полной длины L-цепи анти-WTA бета Ат 6078 (немодифицированное) и его варианты, v2, v3, v4. Варианты L-цепей, которые содержат сконструированный Cys, обозначены буквой С в черном прямоугольнике в конце константной области (в нумерации ЕС остаток №205 в этом случае). Название варианта, например, v2LC-Cys означает вариант 2, содержащий Cys, сконструированный в L-цепь. HCLC-Cys означает, что обе цепи Н и L антитела содержат сконструированный Cys. На Фиг. 13В-1 до 13В-4 показано выравнивание полной длины Н-цепи анти-WTA бета Ат 6078 (немодифицированное) и его вариантов, v2, v3, v4, которые имеют изменения в первом или вторых 2 остатках Н-цепи. Варианты Н-цепей, которые содержат сконструированный Cys, обозначены буквой С в черном прямоугольнике в конце константной области (в нумерации ЕС остаток №118).
6078 Вариабельный участок легкой цепи (VL)
DIVMTQSPSILSASVGDRVTITCRASQTISGWLAWYQQKPAEAPKLLIYKASTLESGVPSRFSGSGSGTEFTLTISSLQPDDFGIYYCQQYKSYSFNFGQGTKVEIK (SEQ ID NO. 111)
6078 Вариабельный участок тяжелой цепи (VH)
XXiQLVQSGAEVKKPGASVKVSCEASGYTLTSYDINWVRQATGQGPEWMGWMNANSGNTGYAQKFQGRVTLTGDTSISTAYMELSSLRSEDTAVYYCARSSILVRGALGRYFDLWGRGTLVTVSS (SEQ ID NO. 112) где X представляет собой Q или Е; и X1 представляет собой М, I или V.
6078 Легкая цепь
DIVMTQSPSILSASVGDRVTITCRASQTISGWLAWYQQKPAEAPKLLIYKASTLESGVPSRFSGSGSGTEFTLTISSLQPDDFGIYYCQQYKSYSFNFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO. 113)
6078 Легкая цепь, сконструированная с использованием остатков цистеина
DIVMTQSPSILSASVGDRVTITCRASQTISGWLAWYQQKPAEAPKLLIYKASTLESGVPSRFSGSGSGTEFTLTISSLQPDDFGIYYCQQYKSYSFNFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPCTKSFNRGEC (SEQ ID NO. 115)
6078 Полноразмерная тяжелая цепь ДТ
QMQLVQSGAEVKKPGASVKVSCEASGYTLTSYDINWVRQATGQGPEWMGWMNANSGNTGYAQKFQGRVTLTGDTSISTAYMELSSLRSEDTAVYYCARSSILVRGALGRYFDLWGRGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG (SEQ ID NO. 114)
6078 Вариант (v2, v3 или v4) полноразмерной тяжелой цепи
EXQLVQSGAEVKKPGASVKVSCEASGYTLTSYDINWVRQATGQGPEWMGWMNANSGNTGYAQKFQGRVTLTGDTSISTAYMELSSLRSEDTAVYYCARSSILVRGALGRYFDLWGRGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGV HTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISbCAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALFINHYTQKSLSLSPG (SEQ ID NO. 116) где X может быть M, I или V.
6078 Вариант (v2, v3 или v4), Cys-сконструированной тяжелой цепи
EXQLVQSGAEVKKPGASVKVSCEASGYTLTSYDINWVRQATGQGPEWMGWMNANSGNTGYAQKFQGRVTLTGDTSISTAYMELSSLRSEDTAVYYCARSSILVRGALGRYFDLWGRGTLVTVSSCSTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG (SEQ ID NO. 117), где X представляет собой М, I или V.
В одном варианте реализации, изобретение предусматривает изолированное анти-WTA бета антитело, содержащее тяжелую и легкую цепь, причем тяжелая цепь содержит VH, имеющую, по меньшей мере, 95% идентичности последовательности с SEQ ID NO. 112. В дополнительном варианте реализации изобретения, это антитело дополнительно содержит VL, имеющую, по меньшей мере, 95% идентичности последовательности с SEQ ID NO. 111. В конкретном варианте реализации изобретения, анти-WTA бета антитело содержит легкую цепь и тяжелую цепь, причем L-цепь содержит VL SEQ ID NO. 111 и Н-цепь содержит VH SEQ ID NO. 112. В еще более конкретном варианте реализации изобретения, изолированное анти-WTA бета антитело содержит L-цепь SEQ ID NO. 113 и Н-цепь SEQ ID NO. 114.
6078 Cys-сконструированные варианты Н и L цепей могут быть спарены в любой из следующих комбинаций, чтобы сформировать полные Ат для конъюгирования с промежуточными продуктами линкер-Abx для создания анти-WTA ААС по изобретению. Немодифицированную L-цепь (SEQ ID NO. 113) можно спарить с Cys-сконструированным вариантом Н-цепи SEQ ID NO. 117; вариант может быть таким, в котором X представляет собой М, I или V. Cys-сконструированную L-цепь SEQ ID NO. 115 можно спарить с: Н-цепью SEQ ID NO: 114; вариантом Н-цепи SEQ ID NO. 116; или Cys-сконструированным вариантом Н-цепи SEQ ID NO. 117 (в этой версии обе цепи Н и L являются Cys-сконструированными). В конкретном варианте реализации изобретения, анти-WTA бета антитело и анти-WTA бета-ААС по изобретению содержат L-цепь SEQ ID NO. 115 и Н-цепь SEQ ID NO. 116.
На Фиг. 14А-1 и 14А-2 представлена полноразмерная L-цепь анти-WTA-бета Ат 4497 (немодифицированное) и его варианты v8. Варианты L-цепей, которые содержат спроектированный Cys, обозначены буквой С в черном боксе рядом с концом константного участка (в нумерации ЕС остаток №205). На Фиг. 14В-1, 14В-2, 14В-3 показано выравнивание полной длины Н-цепи анти-WTA бета-Ат 4497 (немодифицированное) и его варианта v8 с D, замененным на Е в положении CDR Н3 96, с или без сконструированного Cys. Варианты Н-цепей, которые содержат сконструированный Cys, обозначены С в черном боксе в начале константного участка CH1 (в нумерации ЕС остаток №118 в этом случае). Немодифицированный CDR Н3 представляет собой GDGGLDD (SEQ ID NO. 104); 4497v8 CDR Н3 представляет собой GEGGLDD (SEQ ID NO. 118).
4497 Вариабельный участок легкой цепи
DIQLTQSPDSLAVSLGERATINCKSSQSIFRTSRNKNLLNWYQQRPGQPPRLLIHWASTR KSGVPDRFSGSGFGTDFTLTITSLQAEDVAIYYCQQYFSPPYTFGQGTKLEIK (SEQ ID NO. 119)
4497 Вариабельный участок тяжелой цепи
EVQLVESGGGLVQPGGSLRLSCSASGFSFNSFWMHWVRQVPGKGLVWISFTNNEGTTTAYADSVRGRFIISRDNAKNTLYLEMNNLRGEDTAVYYCARGDGGLDDWGQGTLVTVSS(SEQ ID NO. 120)
4497.v8 Вариабельный участок тяжелой цепи
EVQLVESGGGLVQPGGSLRLSCSASGFSFNSFWMHWVRQVPGKGLVWISFTNNEGTTTAYADSVRGRFIISRDNAKNTLYLEMNNLRGEDTAVYYCARGEGGLDDWGQGTLVTVSS(SEQ ID NO. 156)
4497 Легкая цепь
DIQLTQSPDSLAVSLGERATINCKSSQSIFRTSRNKNLLNWYQQRPGQPPRLLIHWASTRKSGVPDRFSGSGFGTDFTLTITSLQAEDVAIYYCQQYFSPPYTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO. 121)
4497 v.8 Тяжелая цепь
EVQLVESGGGLVQPGGSLRLSCSASGFSFNSFWMHWVRQVPGKGLVWISFTNNEGTTTAYADSVRGRFIISRDNAKNTLYLEMNNLRGEDTAVYYCARGEGGLDDWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQS SGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG (SEQ ID NO. 122)
4497 - Cys Легкая цепь
DIQLTQSPDSLAVSLGERATINCKSSQSIFRTSRNKNLLNWYQQRPGQPPRLLIHWASTRKSGVPDRFSGSGFGTDFTLTITSLQAEDVAIYYCQQYFSPPYTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO. 123)
4497. v8 - Тяжелая цепь
EVQLVESGGGLVQPGGSLRLSCSASGFSFNSFWMHWVRQVPGKGLVWISFTNNEGTTTAYADSVRGRFIISRDNAKNTLYLEMNNLRGEDTAVYYCARGEGGLDDWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVFINAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG (SEQ ID NO. 157; то же, что и SEQ ID NO. 122).
4497.v8 - Cys Тяжелая цепь
EVQLVESGGGLVQPGGSLRLSCSASGFSFNSFWMHWVRQVPGKGLVWISFTNNEGTTTAYADSVRGRFIISRDNAKNTLYLEMNNLRGEDTAVYYCARGEGGLDDWGQGTLVTVSSCSTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG (SEQ ID NO. 124)
Другое изолированное анти-WTA бета антитело, предлагаемое в настоящем изобретении, содержит тяжелую и легкую цепь, причем тяжелая цепь содержит VH, имеющую, по меньшей мере, 95% идентичности последовательности с SEQ ID NO. 120. В дополнительном варианте реализации изобретения, это антитело дополнительно содержит VL, имеющую, по меньшей мере, 95% идентичности последовательности с SEQ ID NO. 119. В конкретном варианте реализации изобретения, анти-WTA бета антитело содержит легкую цепь и тяжелую цепь, причем L-цепь содержит VL SEQ ID NO. 119, и Н-цепь содержит VH SEQ ID NO. 120. В еще более конкретном варианте реализации изобретения, изолированное анти-WTA бета антитело содержит L-цепь SEQ ID NO. 121 и Н-цепь SEQ ID NO. 122.
4497 Cys-сконструированные варианты Н и L цепей могут быть спарены в любой из следующих комбинаций, чтобы сформировать полные Ат для конъюгирования с промежуточными продуктами линкер-Abx для создания анти-WTA ААС по изобретению. Немодифицированную L-цепь (SEQ ID NO. 121) можно спарить с Cys-сконструированным вариантом Н-цепи SEQ ID NO. 124. Cys-сконструированную L-цепь SEQ ID NO. 123 можно спарить с: вариантом Н-цепи SEQ ID NO. 157; или Cys-сконструированным вариантом Н-цепи SEQ ID NO. 124 (в этой версии обе цепи Н и L являются Cys-сконструированными). В конкретном варианте реализации изобретения, анти-WTA бета-антитело и анти-WTA бета-ААС по изобретению содержат L-цепь SEQ ID NO. 123.
Еще одним вариантом реализации является антитело, которое связывается с тем же эпитопом, что и каждый из анти-WTA альфа-Ат на Фиг. 11А и Фиг. 11В. Также представлено антитело, которое связывается с тем же эпитопом, что и каждое из анти-WTA бета-Ат Фиг. 12, Фиг. 13А и 13В и Фиг. 14А и 14В.
Связывание анти-WTA антител с WTA находится под влиянием аномерной ориентации модификаций GlcNAc-caxapa WTA. WTA модифицированы модификациями сахара N-ацетилглюкозамина (GlcNAc) в положении С4-ОН посредством α- или β-гликозидных связей, с помощью TarM гликозилтрансферазы или TarS гликозилтрансферазы, соответственно. Соответственно, препараты клеточной стенки из гликозилтрансферазо-мутантных штаммов с отсутствием TarM (ΔTarM), TarS (ΔTarS) или обоих TarM и TarS (ΔTarM/ΔTarS) подвергали иммуноблоттингу с помощью антител против WTA. WTA-антитело (S7574), специфичное к модификациям α-GlcNAc на WTA, не связывается с препаратом клеточной стенки из штамма ΔTarM (Meijer, P.J., et al. (2006) "Isolation of human antibody repertoires with preservation of the natural heavy and light chain pairing." Journal of molecular biology 358, 764-772). И наоборот, антитело WTA (S4462), специфичное к модификациям β-GlcNAc на WTA, не связывается с препаратом клеточной стенки из штамма ΔTarS. Как и ожидалось, оба эти антитела не связываются с препаратами клеточной стенки из делеционного штамма, лишенного как гликозилтрансфераз (ΔTarM/ΔTarS), так и штамма, лишенного каких-либо WTA (ΔTagO). Согласно такому анализу, антитела были охарактеризованы как мАт анти-α-GlcNAc WTA или как мАт анти-β-GlcNAc WTA, как указано в таблице на Фиг. 6А и 6В.
Цистеиновые аминокислоты могут быть сконструированы в реакционноспособных сайтах антитела и не образуют внутрицепочечных или межмолекулярных дисульфидных связей (Junutula, et al., 2008b Nature Biotech., 26(8):925-932; Dornan et al (2009) Blood 114(13):2721-2729; US 7521541; US 7723485; WO 2009/052249, Shen et al (2012) Nature Biotech., 30(2): 184-191; Junutula et al (2008) Jour of Immun. Methods 332:41-52). Сконструированные тиолы цистеина могут вступать в реакцию с линкерными реагентами или интермедиатами линкер-антибиотик по данному изобретению, которые имеют тиол реактивные электрофильные группы, такие как малеимид или альфа-галогеноамиды, с образованием ААС с модифицированными цистеином антителами (THIOMAB™ или thioMabs) и антибиотические (abx) фрагменты. Таким образом, местоположение антибиотической части может быть сконструировано, контролироваться и известно. Нагрузку антибиотика можно контролировать, поскольку сконструированные цистеиновые тиольные группы обычно реагируют с тиол-реактивными линкерными реагентами или промежуточными соединениями линкер-антибиотик с высоким выходом. Разработка анти-WTA антитела для введения аминокислоты цистеина путем замещения в одном сайте тяжелой или легкой цепи дает два новых цистеина на симметричном тетрамерном антителе. Может быть достигнута загрузка антибиотика около 2 и близкая гомогенность продукта конъюгации ААС.
В некоторых вариантах реализации изобретения, может быть желательно создать анти-WTA антитела, сконструированные с использованием цистеина, например, "thioMAb", в которых один или более остатков антитела замещены остатками цистеина. В конкретных вариантах реализации изобретения, замещенные остатки расположены в доступных сайтах антитела. В результате замещения указанных остатков цистеином в доступных участках антитела располагаются реакционноспособные тиольные группы, которые можно использовать для конъюгирования антитела с другими фрагментами, такими как фрагменты антибиотика, линкер-антибиотические фрагменты, с получением иммуноконъюгата, как дополнительно описано в данном документе. В некоторых вариантах реализации один или более из следующих остатков можно заменить на цистеин, включая V205 (нумерация по Кабат) легкой цепи; А118 (нумерация ЕС) тяжелой цепи; и S400 (нумерация ЕС) Fc-участка тяжелой цепи. Показана неограничивающая иллюстративная сконструированная с помощью цистеина тяжелая цепь А118С (SEQ ID NO: 149) и легкая цепь V205C (SEQ ID NO: 151) мутантов анти-WTA антитела, Анти-WTA антитела, сконструированные с использованием цистеина, можно получить, как описано (Junutula, et al., 2008b Nature Biotech., 26(8):925-932; US 7521541; US-2011/0301334).
В другом варианте реализации, изобретение предусматривает изолированное анти-WTA антитело для конъюгации с получением ААС, антитело, содержащее тяжелую цепь и легкую, причем тяжелая цепь содержит последовательность константного участка тяжелой цепи дикого типа или цистеин-сконструированный мутант (ThioMab), и легкая цепь содержит последовательность константного участка легкой цепи дикого типа или цистеин-сконструированный мутант (ThioMab). В одном аспекте тяжелая цепь имеет по меньшей мере 95% идентичности последовательности:
Константному участку тяжелой цепи (IgG1), дикого типа
ASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 148)
Константному участку тяжелой цепи (IgG1), A118C "ThioMab"
CSTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQ S SGL YS LS S V VT VP S S SLGTQT YICN VNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 149)
и легкая цепь имеет по меньшей мере 95% идентичности последовательности:
Константному участку легкой цепи (каппа), дикого типа
RTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC (SEQ ID NO: 150)
Константному участку легкой цепи (каппа), V205C "ThioMab"
RTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPCTKSFNRGEC (SEQ ID NO: 151)
ААС изобретения включают в себя цистеин-сконструированные анти-WTA антитела, где одна или несколько аминокислот дикого типа или родительского анти-WTA антитела заменены на аминокислоту цистеин. Любая форма антитела может быть сконструирована таким образом, т.е. мутирована. Например, родительский Fab-фрагмент антитела может быть сконструирован с образованием цистеин сконструированного Fab, упомянутого здесь как «ThioFab». Аналогичным образом родительское моноклональное антитело может быть сконструировано для образования «ThioFab». Следует отметить, что один сайт мутации дает один сконструированный цистеиновый остаток в ThioFab, в то время как мутация одного сайта дает два сконструированных остатка цистеина в ThioMab из-за димерной природы IgG-антитела. Мутанты с замещенными («сконструированными») остатками цистеина (Cys) оценивают на реакционную способность вновь введенных, сконструированных цистеиновых тиольных групп.
Антитела, описанные в данном документе, могут быть получены с использованием клеток-хозяев в культуре. Клетки-хозяева могут быть трансформированы векторами (векторами экспрессии или клонирования), содержащими одну или более нуклеиновых кислот, кодирующих описанные в данном документе антитела. Клетки могут быть культивированы в условиях, подходящих для получения антител, и антитела, продуцируемые клеткой, могут быть дополнительно очищены. Подходящие клетки для получения антител могут включать прокариотические, дрожжевые или высшие эукариотические клетки (например, млекопитающих). В некоторых вариантах реализации, используют клетку млекопитающего (человек или клетка млекопитающего, не являющегося человеком). В некоторых вариантах реализации, используют клетку из линии клеток яичника китайского хомячка (СНО).
Клетки млекопитающих могут быть культивированы, а размножение клеток млекопитающих в культуре (культура ткани) стало обычной процедурой. Примеры линий клеток-хозяев млекопитающих могут включать, не ограничиваясь только ими, линию клеток почки обезьяны CV1, трансформированную SV40 (COS-7, АТСС CRL 1651); эмбриональную линию клеток почки человека (клетки 293 или 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol. 36:59 (1977)); клетки почки детеныша хомяка (BHK, АТСС CCL 10); мышиные клетки Сертоли (ТМ4, Mather, Biol. Reprod. 23:243-251 (1980)); клетки почек обезьяны (CV1 АТСС CCL 70); клетки почек африканской зеленой мартышки (VERO-76, АТСС CRL-1587); клетки карциномы шейки матки человека (HELA, АТСС CCL 2); клетки почек собачьих (MDCK, АТСС CCL 34); клетки печени серой крысы (BRL 3А, АТСС CRL 1442); клетки легких человека (W138, АТСС CCL 75); клетки печени человека (Hep G2, НВ 8065); опухоль молочной железы мышей (ММТ 060562, АТСС CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci. 383:44-68 (1982)); клетки MRC 5; клетки FS4; и линию гепатомы человека (Hep G2). Другие подходящие линии клеток-хозяев млекопитающих включают клеточные линии миеломы, такие как NS0 и Sp2/0. Обзор по некоторым полученным из млекопитающих линиям клеток-хозяев, пригодных для получения антител, см., например, Yazaki and Wu, Methods in Molecular Biology, Vol. 248 (B.K.C.Lo, ed., Humana Press, Totowa, N.J., 2003), pp. 255-268.
В качестве хозяев также могут использоваться растительные клеточные культуры из хлопка, кукурузы, картофеля, сои, петунии, томатов, ряски (Leninaceae), люцерны (М. truncatula) и табака.
Подходящие прокариотические клетки для этой цели включают эубактерии, такие как грамотрицательные или грамположительные организмы, например, Enterobacteriaceae, такие как Escherichia, например, Е. coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например, Salmonella typhimurium, Serratia, например,Serratia marcescans, и Shigella, а также Bacilli, такие как В. subtilis и В. licheniformis (например, В. licheniformis 41P, описанную в DD 266,710 опубликованной 12 апреля 1989), Pseudomonas, такие как Р. aeruginosa, и Streptomyces. Одним из предпочтительных хозяев для клонирования является Е. coli 294 (АТСС 31,446), хотя можно применять другие штаммы, такие как Е. coli В, Е. coli X1776 (АТСС 31,537), и Е. coli W3110 (АТСС 27,325). Эти примеры являются иллюстративными, а не ограничивающими.
Помимо прокариот, эукариотические микроорганизмы, такие как мицелиальные грибы или дрожжи, являются подходящими хозяевами для клонирования или экспрессирования векторов, кодирующих антитело. Saccharomyces cerevisiae, или обычные хлебопекарные дрожжи, наиболее часто используются среди низших эукариотических микроорганизмов-хозяев. Тем не менее, ряд других родов, видов и штаммов общедоступны и полезны в данном документе, такие как Schizosaccharomyces pombe; Kluyveromyces хозяева, такие как, например, K. lactis, K. fragilis (АТСС 12,424), К. bulgaricus (АТСС 16,045), K. wickeramii (АТСС 24,178), K. waltii (АТСС 56,500), K. drosophilarum (АТСС 36,906), К. thermotolerans, и К. marxianus; тысячелистник (ЕР 402,226); Pichia pastoris (ЕР 183,070); Candida; Trichoderma reesia (ЕР 244,234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и нитчатые грибы, такие как, например, Neurospora, Penicillium, Tolypocladium, и Aspergillus хозяева, такие как A. nidulans и A. niger.
ФРАГМЕНТЫ АНТИБИОТИКОВ РИФАМИЦИНОВОГО РЯДА
Антибиотическая часть (abx) конъюгатов антитело-антибиотик (ААС) по изобретению представляет собой антибиотик или группу рифамицинового типа, которая обладает цитотоксическим или цитостатическим эффектом. Рифамицины представляют собой группу антибиотиков, которые получают либо естественным образом из бактерией, Nocardia mediterranei, Amycolatopsis mediterranei или искусственно. Они являются подклассом более крупного семейства ансамицина, который ингибирует бактериальную РНК-полимеразу (Fujii et al., 1995) Antimicrob.Agents Chemother. 39:1489-1492; Feklistov, et al (2008) Proc Natl Acad Sci USA, 105(39): 14820-5) и обладают потенциалом против грамположительных и селективных грамотрицательных бактерий. Рифамицины особенно эффективны против микобактерий и, следовательно, используются для лечения инфекций туберкулеза, проказы и микобактерий avium complex (MAC). Группа рифамицинового типа включает «классические» препараты рифамицина, а также производное рифамицина рифампицин (рифампицин, СА Рег. №13292-46-1), рифабутин (СА Рег. №72559-06-9; US 2011/0178001), рифапентин и рифалазил (СА Рег. №129791-92-0, Rothstein et al (2003) Expert Opin.Investig.Drugs 12(2):255-271; Fujii et al (1994) Antimicrob. Agents Chemother. 38:1118-1122. Многие антибиотики группы рифамицина обладают нежелательным свойством - развитием резистентности (Wichelhaus et al (2001) J. Antimicrob. Chemother. 47:153-156). Рифамицины впервые были выделены в 1957 году из ферментационной культуры Streptomyces mediterranei. Было обнаружено около семи рифамицинов, названных Рифамицин А, В, С, D, Е, S и SV (US 3150046). Рифамицин В был первым выпущен на рынок и использован при лечении лекарственно-устойчивого туберкулеза в 1960-х годах. Рифамицины используются для лечения многих заболеваний, наиболее значимыми из которых является туберкулез, связанный с ВИЧ. Из-за большого количества доступных аналогов и производных рифамицины широко используются для ликвидации патогенных бактерий, устойчивых к обычно используемым антибиотикам. Например, рифампицин известен своим мощным эффектом и способностью предотвращать лекарственную устойчивость. Он быстро убивает быстро делящиеся штаммы бактерий, а также «резистентные» клетки, которые остаются биологически неактивными в течение длительных периодов времени, что позволяет им уклоняться от антибиотической активности. Кроме того, и рифабутин, и рифапентин используются против приобретенного туберкулеза у ВИЧ-положительных пациентов.
Антибиотические фрагменты (abx) конъюгатов антитело-антибиотик формулы I представляют собой фрагменты типарифамицина, имеющие структуру:
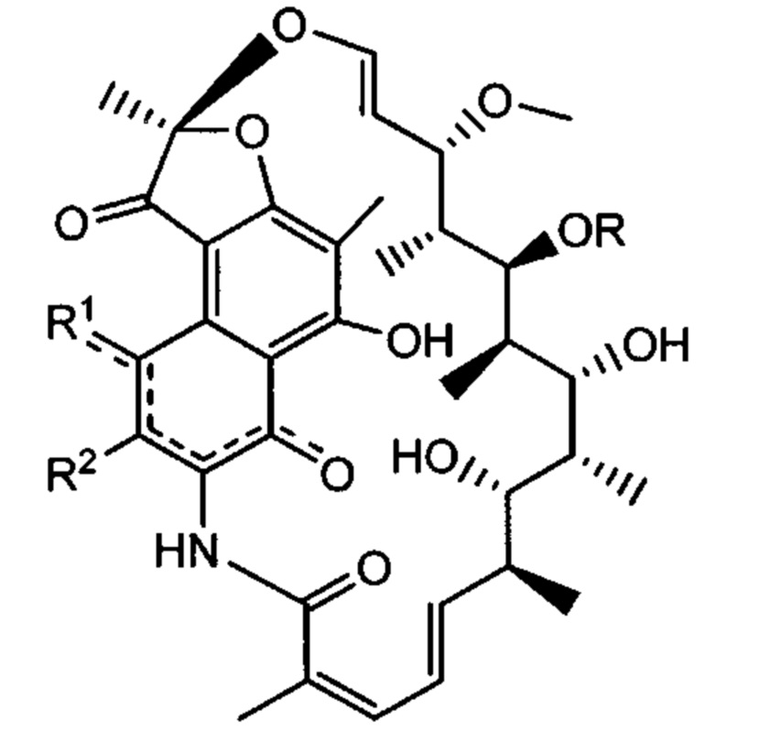
где:
пунктирные линии указывают на необязательную связь;
R представляет собой Н, С1-С12 алкил, или С(O)СН3;
R1 представляет собой ОН;
R2 представляет собой СН=N-(гетероциклил), причем гетероциклил необязательно замещен одной или более группами, независимо выбранными из С(O)СН3, С1-С12 алкил, С1-С12 гетероарил, С2-С20 гетероциклил, С6-С20 арил, и С3-С12 карбоциклил;
или R1 и R2 образуют пяти- или шестичленный конденсированный гетероарил или гетероциклил и необязательно образуют спиро или конденсированный шестичленный гетероарил, гетероциклил, арил или карбоциклическое кольцо, причем спиро или конденсированный шестичленный гетероарил, гетероциклил, арил или карбоциклическое кольцо необязательно замещены Н, F, Cl, Br, I, C1-C12 алкиом или ОН; и
где непептидный линкер PML ковалентно присоединен с R2.
Вариант реализации фрагмента типа рифамицина представляет собой:
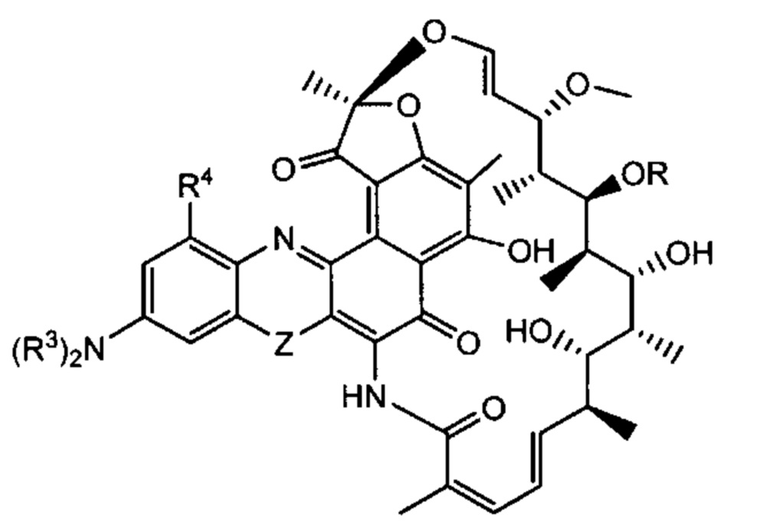
где R3 независимо выбран из Н и С1-С12 алкил; R4 выбран из Н, F, Cl, Br, I, С1-С12 алкила и ОН; и Z выбран из NH, N(C1-C12 алкил), О и S; и где непептидный линкер PML ковалентно присоединен к атому азота N(R3)2.
Вариант реализации фрагмента типа рифампицина представляет собой:
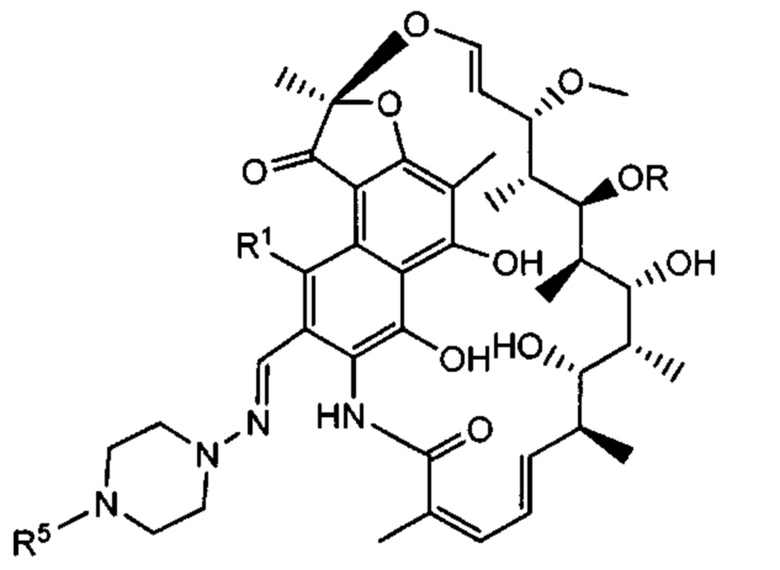
где
R5 выбран из Н и С1-С12 алкила; и где непептидный линкер PML ковалентно присоединен к атому азота NR5.
Вариант реализации фрагмента типа рифабутина представляет собой:
где R5 выбран из Н и С1-С12 алкила; и где непептидный линкер PML ковалентно присоединен к атому азота NR5.
Вариант реализации фрагмента бензоксазинорифамицинового типа представляет собой:
где R5 выбран из Н и С1-С12 алкила; и где непептидный линкер PML ковалентно присоединен к атому азота NR5.
Вариант реализации фрагмента бензоксазинорифамицинового типа, обозначаемом в данном документе как pipBOR, представляет собой:
где R3 независимо выбран из Н и С1-С12 алкила; и где непептидный линкер PML ковалентно присоединен к атому азота N(R3)2.
Вариант реализации фрагмента бензоксазинорифамицинового типа, обозначаемом в данном документе как диметил pipBOR, представляет собой:
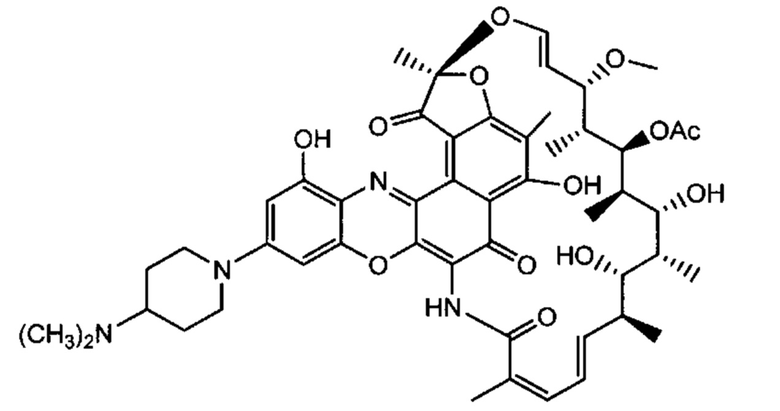
где непептидный линкер PML ковалентно присоединен к диметиламин атому азота.
Полусинтетическое производное рифамицина S или восстановленной формы натриевой соли рифамицина SV, можно превращать в антибиотики типа рифализин в несколько стадий, где R представляет собой Н или Ac, R3 независимо выбран из Н и С1-С12 алкила; R4 выбран из Н, F, Cl, Br, I, С1-С12 алкила, и ОН; и Z выбран из NH, N(C1-C12 алкил), О и S (см., например, Фиг. 23А и В, и Фиг. 25А и В в WO 2014/194247). Бензоксазино (Z = О), бензтиазино (Z = S), бенздиазино (Z = NH, N(C1-C12 алкил) рифамицины могут быть получены (US 7271165). Аналоги бензоксазинорифамицина (BOR), бензтиазинорифамицина (BTR) и бенздиазинорифамицина (BDR), которые содержат заместители, нумеруются согласно схеме нумерации, представленной в формуле А, в колонке 28 в US 7271165, который включен в качестве ссылки для этой цели. Под «25-О-деацетилом» рифамицин подразумевается аналог рифамицина, в котором ацетильная группа в положении 25 удалена. Аналоги, в которых это положение дополнительно дериватизировано, называются «25-O-деацетил-25-(заместитель)рифамицин», в котором номенклатура для дериватизирующей группы заменяется «заместитель» в полном названии соединения.
Фрагменты антибиотиков рифамицинового ряда могут быть синтезированы способами, аналогичными способам, раскрытым в US 4610919; US 4983602; US 5786349; US 5981522; US 4859661; US 7271165; US 2011/0178001; Seligson, et al., (2001) Anti-Cancer Drugs 12:305-13; Chem. Pharm. Bull., (1993) 41:148, и WO 2014/194247, каждая из которых включена сюда посредством ссылки). Фрагменты антибиотиков типа рифамицин можно скринировать на антимикробную активность путем измерения их минимальной ингибирующей концентрации (МИК) с использованием стандартных анализов МИК in vitro (Tomioka et al., (1993) Antimicrob. Agents Chemother. 37:67).
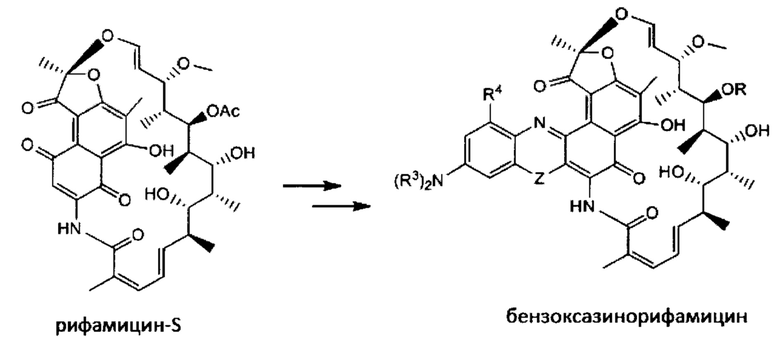
РАСЩЕПЛЯЕМЫЕ ПРОТЕАЗОЙ НЕПЕПТИДНЫЕ ЛИНКЕРЫ
«Расщепляемый протеазой непептидный линкер» (PML) представляет собой бифункциональный или многофункциональный фрагмент, который ковалентно присоединен к одному или более антибиотическим фрагментам (abx) и единице антитела (Ат) с образованием конъюгатов антитело-антибиотик (ААС) Формула I. Расщепляемые протеазой непептидные линкеры в ААС являются субстратами для расщепления внутриклеточными протеазами, в том числе в лизосоме. Протеазы включают различные катепсины и каспазы. Расщепление непептидного линкера ААС внутри клетки может высвобождать антибиотик рифамицинового ряда с антибактериальными эффектами.
Конъюгаты антитело-антибиотик (ААС) могут быть легко получены с использованием линкерного реагента или промежуточного соединения линкер-антибиотик, обладающего реакционной функциональностью для связывания с антибиотиком (abx) и с антителом (Ат). В одном иллюстративном варианте реализации изобретения, цистеин тиол сконструированного с использованием цистеина антитела (Ат) может образовывать связь с функциональной группой линкерного реагента, антибиотическим фрагментом промежуточного соединения антибиотик-линкер.
PML фрагмент ААС может содержать один аминокислотный остаток.
PML фрагмент ААС содержит пептидомиметическую единицу.
В одном аспекте, линкерный реагент или промежуточное соединение линкер-антибиотик имеет реакционноспособный сайт, который имеет электрофильную группу, которая реагирует с нуклеофильным цистеином, присутствующим на антителе. Цистеин тиол антитела может реагировать с электрофильной группой на линкерном реагенте или линкер-антибиотике, образуя ковалентную связь. Подходящие электрофильные группы включают, но не ограничиваются ими, малеимидные и галогенацитамидные группы.
Модифицированные цистеином антитела реагируют с линкерными реагентами или промежуточными соединениями линкер-антибиотик, с электрофильными функциональными группами, такими как малеимид или α-галокарбонил, в соответствии с методом конъюгации на стр. 766 Klussman, et al (2004), Bioconjugate Chemistry 15(4):765-773, и согласно протоколу примера 19.
В другом варианте реализации изобретения, реакционная группа линкерного реагента или промежуточного соединения линкер-антибиотик содержит тиол-реактивную функциональную группу, которая может образовывать связь со свободным тиолом цистеина антитела. Иллюстративные тиол-реактивные функциональные группы включают, но не ограничиваются ими, малеимид, α-галогенацетил, активированные сложные эфиры, такие как эфиры сукцинимидов, 4-нитрофениловые эфиры, пентафторфениловые сложные эфиры, тетрафторфениловые эфиры, ангидриды, хлорангидриды кислот, сульфонилхлориды, изоцианаты и изотиоцианаты.
В другом варианте реализации изобретения, линкерный реагент или промежуточное соединение линкер-антибиотик имеет реакционноспособную функциональную группу, которая имеет нуклеофильную группу, которая может реагировать с электрофильной группой, присутствующей на антителе. Подходящие электрофильные группы на антителе включают, но не ограничиваются ими, пиридилдисульфид, альдегид и кетоновые карбонильные группы. Гетероатом нуклеофильной группы линкерного реагента или промежуточного соединения линкер-антибиотик может вступать в реакцию с электрофильной группой на антителе и образовывать ковалентную связь с единицей антитела. Пригодные для этого нуклеофильные группы линкерного реагента или промежуточного соединения антибиотик-линкер включают, но не ограничиваются ими, гидразид, оксим, амино, тиол, гидразин, тиосемикарбазон, гидразинкарбоксилат и арилгидразид. Электрофильная группа на антителе обеспечивает удобный сайт для присоединения к линкерному реагенту или промежуточному соединению антибиотик-линкер.
Фрагмент PML может содержать один или несколько линкерных компонентов. Иллюстративные компоненты линкера включают одну аминокислоту, такую как цитруллин ("cit"), 6-малеимидкапроил ("МС"), малеимидпропаноил ("MP"), и п-аминобензилоксикарбонил ("РАВ"), N-сукцинимидил 4-(2-пиридилтио)пентаноат ("SPP"), и 4-(N-малеимидметил) циклогексан-1 карбоновую кислоту ("МСС"). В данной области техники известны различные компоненты линкера, некоторые из которых описаны ниже.
В другом варианте реализации изобретения, линкер может быть замещен группами, которые модулируют растворимость или реакционную способность. Например, заряженный заместитель, такой как сульфонат (-SO3-) или аммоний, может повысить водорастворимость реагента и облегчить реакцию связывания линкерного реагента с антителом или фрагментом антибиотика, или облегчить реакцию связывания Ат-L (промежуточное антитело-линкерное соединение) с abx, или abx-L (промежуточное антитело-линкерное соединение) с Ат, в зависимости от способа синтеза, используемого для получения ААС.
ААС данного изобретения явно рассматривает, но не ограничивается ими, препараты, полученные с помощью линкерных реагентов: ВМРЕО, BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC, сульфо-SMPB, SVSB (сукцинимидил-(4-винилсульфон)бензоат), и реагенты бисмалеимида, такие как DTME, BMB, BMDB, BMH, BMOE, BM(PEG)2, и BM(PEG)3. Бисмалеимидные реагенты позволяют присоединять тиоловые группы сконструированного цистеина в антителе к тиолсодержащему антибиотическому фрагменту, метке или линкерному промежуточному соединению последовательным или конвергентным образом. Другие функциональные группы, кроме малеимида, которые реагируют с тиольной группой сконструированного цистеина в антителе,, антибиотическим фрагментом или промежуточным соединением линкер-антибиотик, включают иодацетамид, бромацетамид, винилпиридин, дисульфид, пиридилдисульфид, изоцианат и изотиоцианат.
Полезные линкерные реагенты можно также получить из других коммерческих источников, такие как Molecular Biosciences Inc. (Boulder, CO), или синтезировать в соответствии с процедурами, описанными в Toki et al (2002) J.Org. Chem. 67:1866-1872; Dubowchik, et al. (1997) Tetrahedron Letters, 38:5257-60; Walker, M.A. (1995) J. Org. Chem. 60:5352-5355; Frisch et al (1996) Bioconjugate Chem. 7:180-186; US 6214345; WO 02/088172; US 2003130189; US 2003096743; WO 03/026577; WO 03/043583; и WO 04/032828.
В другом варианте реализации изобретения, фрагмент PML ААС содержит линкер дендритного типа для ковалентного присоединения более чем одного фрагмента антибиотика к антителу через разветвленный многофункциональный фрагмент линкера (Sun et al (2002) Bioorganic & Medicinal Chemistry Letters 12:2213-2215; Sun et al (2003) Bioorganic & Medicinal Chemistry 11:1761-1768). Дендритные линкеры могут увеличивать молярное соотношение антибиотика к антителу, т.е нагрузку, которая связана с эффективностью ААС. Таким образом, если сконструированное с цистеином антитело несет только одну реакционноспособную цистеиновую тиоловую группу, через дендритный линкер можно присоединить множество антибиотических фрагментов.
В некоторых вариантах реализации изобретения, ААС формулы I расщепляемый протеазой непептидный линкер PML имеет формулу:

причем Str представляет собой растягивающую единицу; РМ представляет собой пептидомиметическую единицу, и Y представляет собой спейсерную единицу; abx представляет собой антибиотик рифамицинового типа; и
Р представляет собой целое число от 1 до 8.
В одном варианте реализации изобретения, растягивающая единица "Str" имеет формулу:
где R6 выбирают из группы, состоящей из С1-С12 алкилена, С1-С12 алкилен -С(=O), С1-С12 алкилен-NH, (CH2CH2O)r, (CH2CH2O)r-С(=O), (CH2CH2O)r-CH2, и С1-С12 алкилен-NHC(=O)СН2СН(тиофен-3-ил), где г представляет собой целое число от 1 до 10.
Иллюстративные растягивающиеся блоки показаны ниже (где волнистая линия указывает на сайты ковалентного присоединения к антителу):
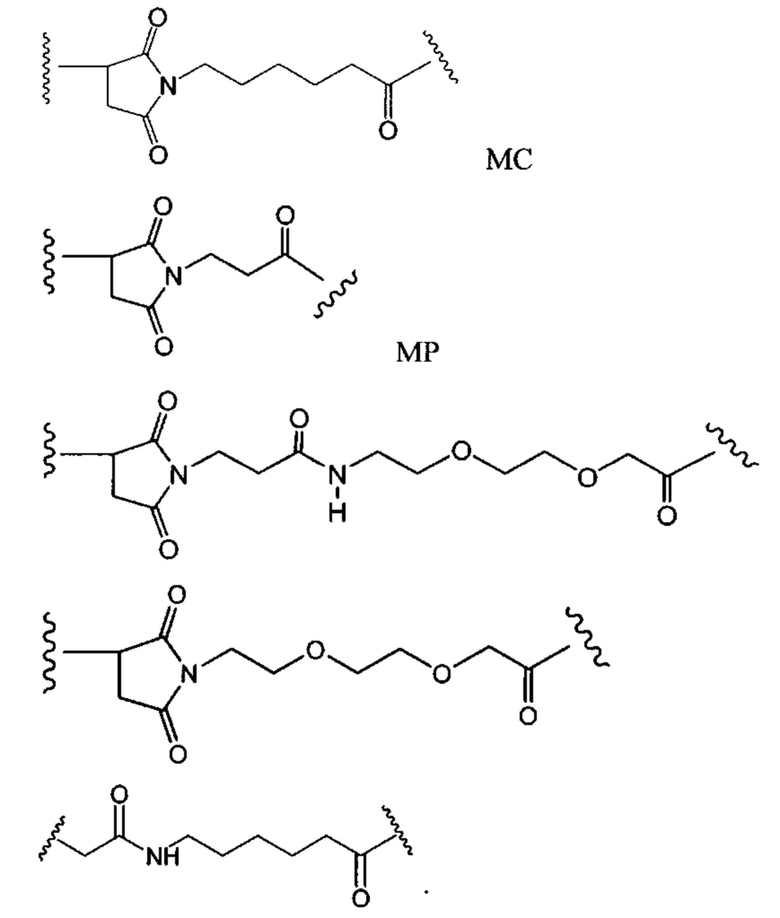
В одном варианте реализации изобретения, РМ имеет формулу:
где R7 и R8 вместе образуют С3-С7 циклоалкильное кольцо и
АА представляет собой боковую цепь аминокислоты, выбранную из Н, -СН3, -СН2(С6Н5), -CH2CH2CH2CH2NH2, -CH2CH2CH2NHC(NH)NH2, -СНСН(СН3)СН3, и -CH2CH2CH2NHC(O)NH2.
В одном варианте реализации изобретения, спейсерный блок Y содержит пара-аминобензил (РАВ) или пара-аминобензилоксикарбонил (РАВС).
Спейсерный блок позволяет высвобождать антибиотический фрагмент без отдельной стадии гидролиза. Спейсерный блок может быть «саморасщепляющимся» или «не саморасщепляющимся». В некоторых вариантах реализации изобретения, спейсерный блок линкера содержит п-аминобензиловый блок (РАВ). В одном таком варианте реализации изобретения, п-аминобензиловый спирт присоединен к аминокислотному блоку посредством амидной связи, карбамата, метилкарбамата или карбоната между п-аминобензильной группой и антибиотической частью (Hamann et al. (2005) Expert Opin.Ther.Patents (2005) 15:1087-1103). В одном варианте реализации изобретения, спейсерный блок представляет собой п-аминобензилоксикарбонил (РАВ).
В одном варианте реализации изобретения, антибиотик образует четвертичный амин, такой как диметиламинопиперидильная группа, когда он присоединен к РАВ-спейсерному блоку непептидного линкера PML. Примерами таких четвертичных аминов являются промежуточные соединения линкер-антибиотик (PLA) PLA-1-4 из таблицы 2. Четвертичная аминогруппа может модулировать расщепление антибиотического фрагмента для оптимизации антибактериальных эффектов ААС. В другом варианте реализации изобретения, антибиотик связан с РАВС-спейсерной единицей непептидного линкера PML, образуя карбаматную функциональную группу в ААС.Такая карбаматная функциональная группа может также оптимизировать антибактериальные эффекты ААС. Примерами РАВС карбаматных промежуточных соединений линкер-антибиотиков (PLA) являются PLA-5 и PLA-6 из Таблицы 2.
Другие примеры саморасщепляющихся спейсеров включают, но не ограничиваются ими, ароматические соединения, электронная структура которых аналогична РАВ-группе, например, производные 2-аминоимидазол-5-метанола (US 7375078; Hay et al. (1999) Bioorg.Med.Chem.Lett. 9:2237) и орто- или пара-аминобензилацетальдегиды. Можно использовать спейсеры, подвергающиеся циклизации при гидролизе амидной связи, например, замещенные и незамещенные амиды 4-аминомасляной кислоты (Rodrigues et al (1995) Chemistry Biology 2:223), соответственно замещенные бицикл[2.2.1] и бицикло[2.2.2] кольцевые системы (Storm et al (1972) J.Amer.Chem.Soc. 94:5815) и амиды 2-аминофенилпропионовой кислоты (Amsberry, et al (1990) J.Org.Chem. 55:5867). Ликвидация аминсодержащих лекарств, которые замещены у глицина (Kingsbury et al (1984) J. Med. Chem. 27:1447) также является примером самонесущих спейсеров, полезных в ААС.
Количество активного антибиотика, высвобождаемого при расщеплении ААС, можно измерить с помощью анализа высвобождения каспазы из примера 8.
ПРОМЕЖУТОЧНЫЕ СОЕДИНЕНИЯ ЛИНКЕР-АНТИБИОТИК ПОЛЕЗНЫЕ ДЛЯ ААС
Промежуточные соединения PML-линкер-антибиотик (PLA) формулы II и таблицы 2 получали путем связывания фрагмента антибиотика рифамицинового ряда с линкерным реагентом, примеры 11-21. Реагенты линкера получали способами, описанными в WO 2012/113847; US 7659241; US 7498298; US 20090111756; US 2009/0018086; US 6214345;
Dubowchik et al (2002) Bioconjugate Chem. 13(4):855-869
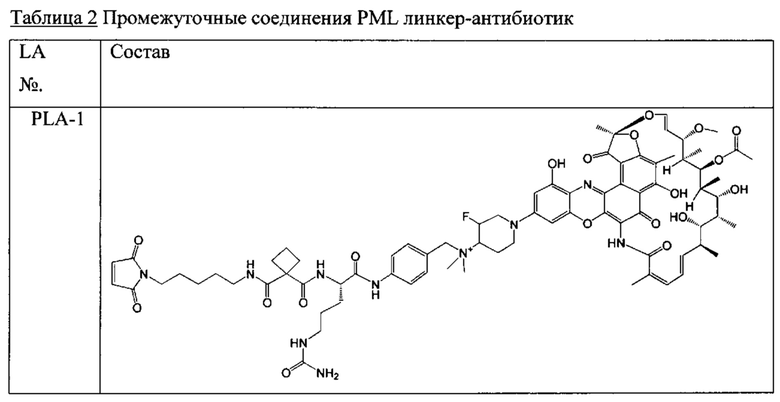
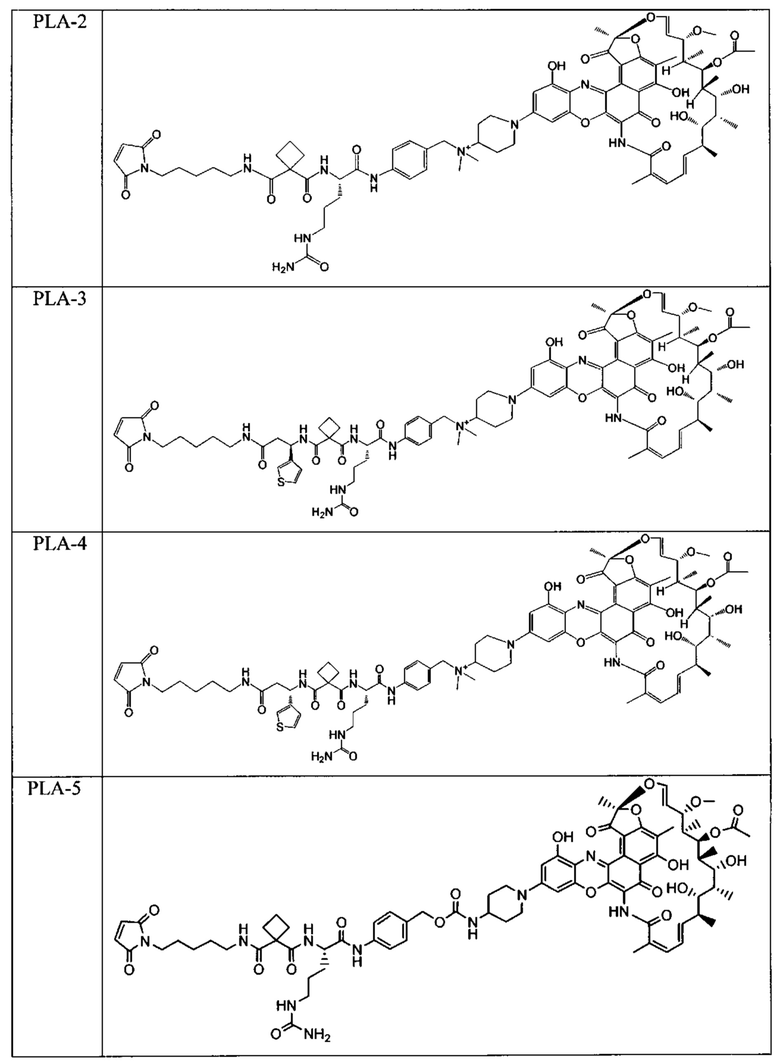
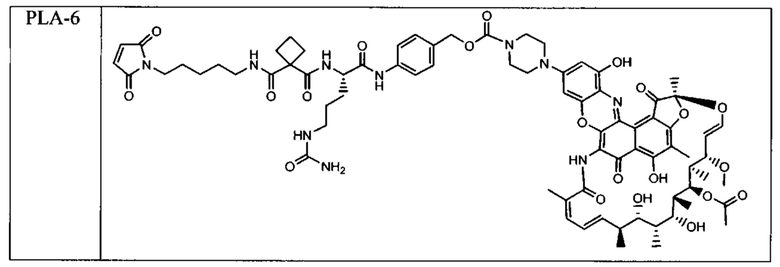
ВАРИАНТЫ РЕАЛИЗАЦИИ КОНЪЮГАТОВ АНТИБИОТИК-АНТИТЕЛО
Сконструированные с цистеином анти-WTA антитела были связаны через свободную тиоловую группу цистеина с производными рифамицина, названными pipBOR и другими, с помощью расщепляемого протеазой непептидного линкера с образованием конъюгатов антитело-антибиотик (ААС) из таблицы 3. Линкер предназначен для расщепления лизосомальными протеазами, включая катепсины В, D и другими. Образование промежуточного соединения линкер-антибиотик, состоящего из антибиотика и линкера PML, и других подробно описано в примерах 11-21. Линкер сконструирован таким образом, что расщепление амидной связи в РАВ-фрагменте отделяет антитело от антибиотика в активном состоянии.
ААС, названный «диметил pipBOR», идентичен ААС «pipBOR», за исключением диметилированной аминогруппы антибиотика и оксикарбонильной группы линкера.
На Фиг. 3 показан возможный механизм активации лекарственного средства для конъюгатов антитело-антибиотик (ААС). Активный антибиотик (АЬ) выделяется только после интернализации ААС внутрь клеток млекопитающих. Fab-часть антитела в ААС связывает S. aureus, тогда как Fc-часть ААС усиливает поглощение бактерий посредством опосредованного Fc-рецептором связывания с фагоцитирующими клетками, включая нейтрофилы и макрофаги. После интернализации в фаголизосому линкер может быть расщеплен лизосомальными протеазами, высвобождающими активный антибиотик внутрь фаголизосомы.
Вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает Формулу I:
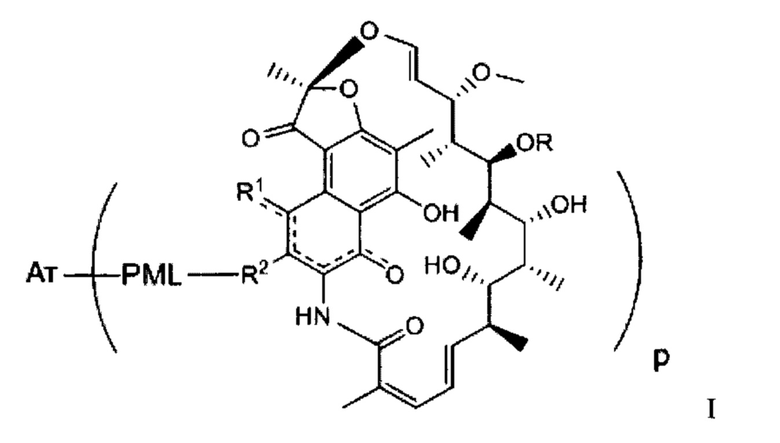
где:
пунктирные линии обозначают необязательную связь;
R представляет собой Н, С1-С12 алкил, или С(O)СН3;
R1 представляет собой ОН;
R2 представляет собой СН=N-(гетероциклил), причем гетероциклил необязательно замещен одной или более группами, независимо выбранными из С(O)СН3, С1-С12 алкил, С1-С12 гетероарил, С2-С20 гетероциклил, С6-С20 арил, и С3-С12 карбоциклил;
или R1 и R2 образуют пяти- или шестичленный конденсированный гетероарил или гетероциклил и необязательно образуют спиро или конденсированный шестичленный гетероарил, гетероциклил, арил или карбоциклическое кольцо, причем спиро или конденсированный шестичленный гетероарил, гетероциклил, арил или карбоциклическое кольцо необязательно замещены Н, F, Cl, Br, I, C1-C12 алкилом или ОН;
PML представляет собой расщепляемый протеазой непептидный линкер, присоединенный к R2 или конденсированный гетероарил или гетероциклил, образованный R1 и R2;
Ат представляет собой антитело против тейхоевой кислоты клеточной стенки (WTA); и
Р представляет собой целое число от 1 до 8.
Другой вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает формулу:
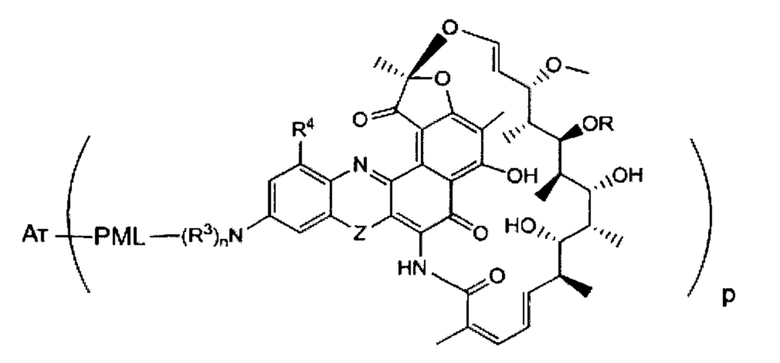
где
R3 независимо выбран из Н и C1-С12 алкила;
n равно 1 или 2;
R4 выбран из Н, F, Cl, Br, I, С1-С12 алкила, и ОН; и
Z выбран из NH, N(C1-C12 алкил), О и S.
Другой вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает формулу:
где
R5 выбран из Н и С1-С12 алкил; и
n равен 0 или 1.
Другой вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает формулу:
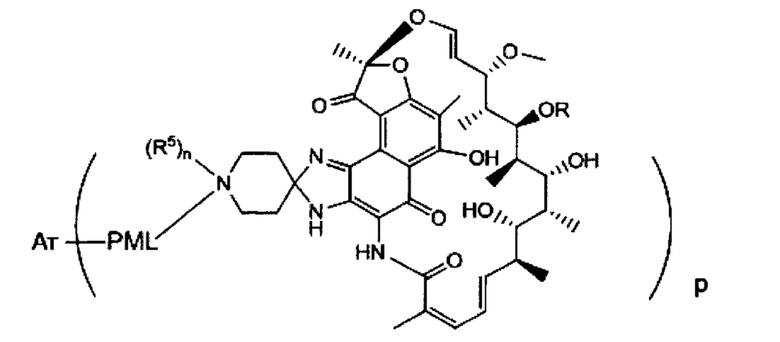
где
R5 выбран из Н и С1-С12 алкил; и
n равен 0 или 1.
Другой вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает формулу:
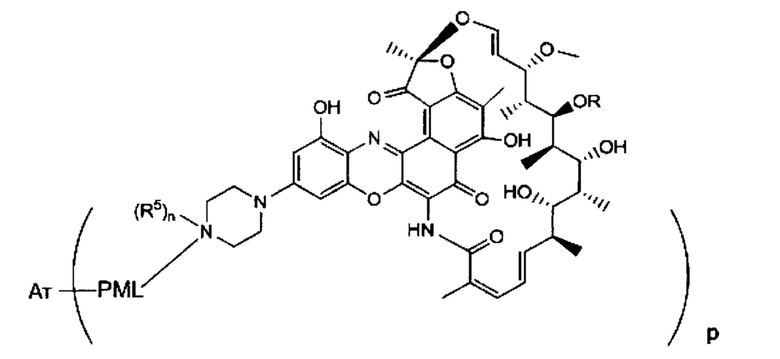
где
R5 независимо выбран из Н и С1-С12 алкила; и
n равен 0 или 1.
Другой вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает формулу:
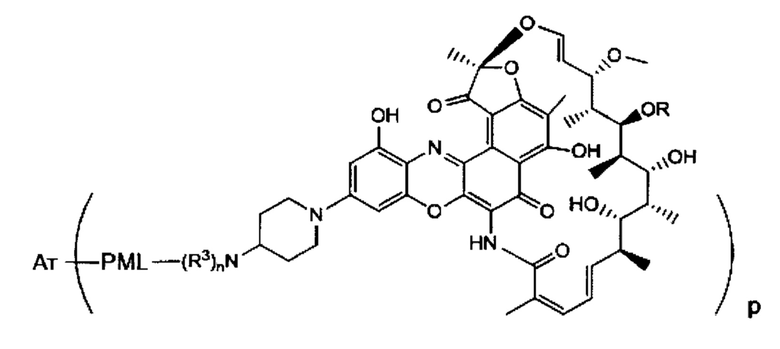
где
R3 независимо выбран из Н и С1-С12 алкила; и
n равно 1 или 2;
Другой вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает формулу:
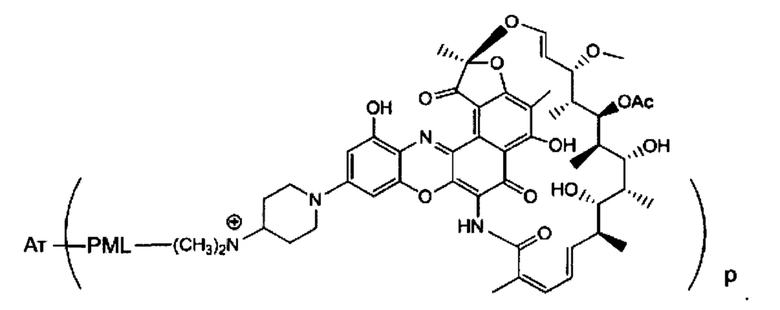
Другой вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает формулу:

Другой вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает формулу:
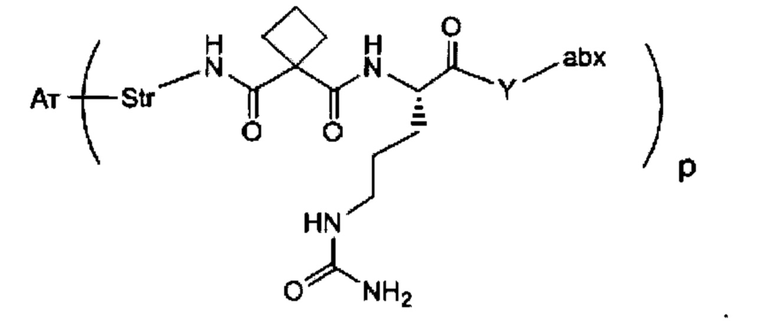
Другой вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает формулу:
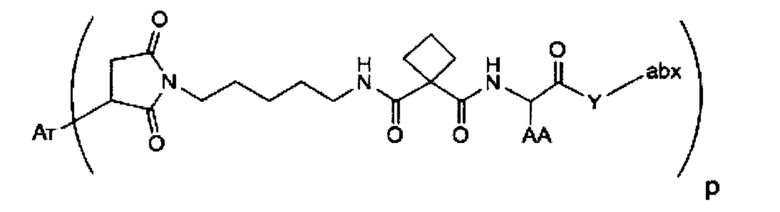
Другой вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает формулу:
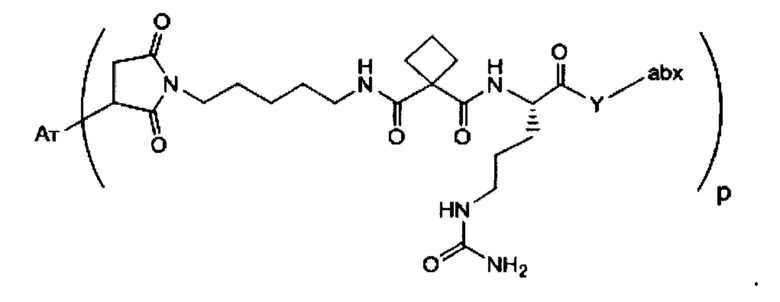
Другой вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает формулы:
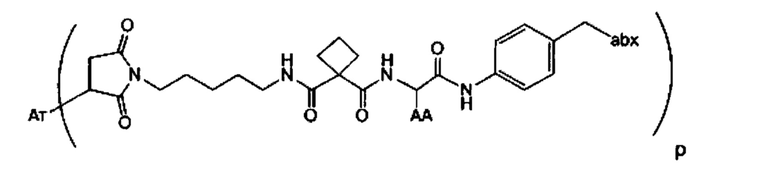
и
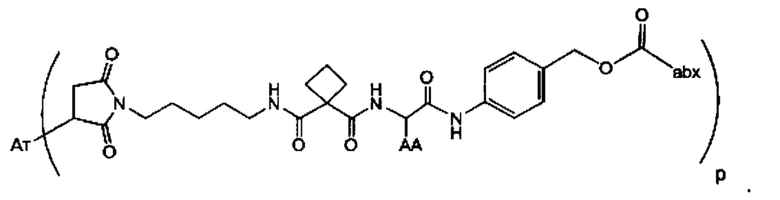
Другой вариант реализации соединений конъюгата антитело-антибиотик (ААС) по изобретению включает формулы:

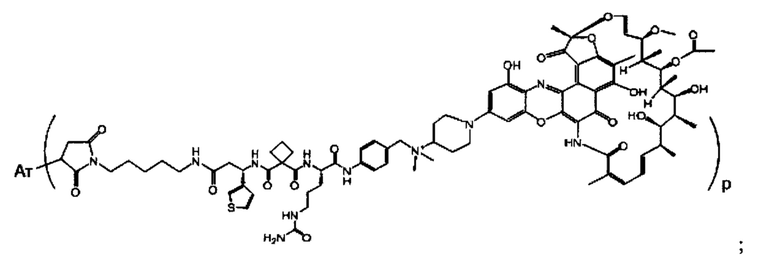
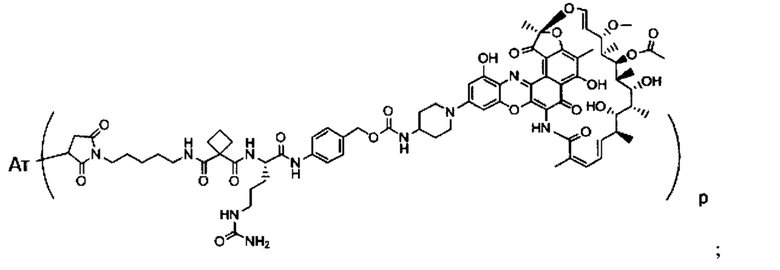
и
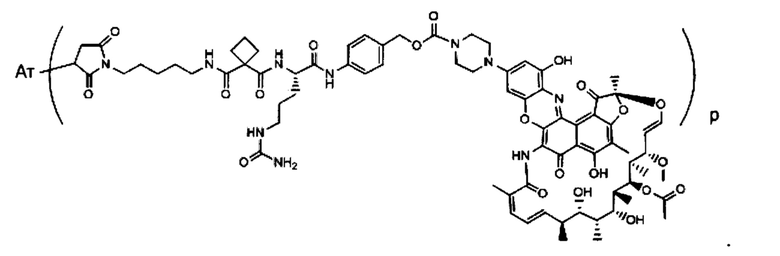
АНТИБИОТИЧЕСКАЯ НАГРУЗКА ААС
Нагрузка антибиотика представлена р, числом антибиотических (abx) фрагментов на антитело в молекуле формулы I. Нагрузка антибиотика может составлять от 1 до 20 фрагментов антибиотиков (D) на антитело. ААС формулы I включают коллекции или пул антител, конъюгированных с рядом антибиотических фрагментов, от 1 до 20. Среднее количество антибиотических фрагментов на антитело в препаратах ААС в результате реакций конъюгирования может быть охарактеризовано обычными способами, такими как масс-спектроскопия, ИФА-анализ и ВЭЖХ. Кроме того, можно определить количественное распределение ААС по отношению к р. В некоторых случаях путем таких средств, как обращенно-фазная ВЭЖХ или электрофорез, можно достичь отделения, очистки и оценки характеристик однородного ААС, в котором р представляет собой определенное значение, от ААС с другими значениями антибиотической нагрузки.
Для некоторых конъюгатов антитело-антибиотик р может быть ограничен количеством сайтов связывания на антителе. Например, если присоединяемая группа представляет собой тиоловую группу цистеина, как в иллюстративных вышеприведенных вариантах реализации, антитело может содержать только одну или несколько тиоловых групп цистеина, или может содержать только одну или несколько достаточно реакционноспособных тиоловых групп, к которым можно присоединить линкер. В некоторых вариантах реализации изобретения высокая антибиотическая нагрузка, например, р > 5, может привести к агрегации, нерастворимости, токсичности или потере способности некоторых конъюгатов антитело-антибиотик. В некоторых вариантах реализации изобретения, нагрузка антибиотика для ААС по настоящему изобретению составляет от 1 до около 8; от около 2 до около 6; от около 2 до около 4; или от около 3 до около 5; около 4; или около 2.
В некоторых вариантах реализации изобретения, количество фрагментов антибиотика, конъюгированных с антителом в ходе реакции конъюгирования, составляет менее теоретического максимального значения. Антитело может содержать, например, остатки лизина, которые не реагируют с промежуточным соединением антибиотик-линкер или линкерным реагентом, как обсуждается ниже. Как правило, антитела не содержат большого количества свободных и реакционноспособных тиоловых групп цистеина, которые могут связываться с фрагментом антибиотика; фактически, большинство тиоловых остатков цистеина в составе антител существуют в виде дисульфидных мостиков. В некоторых вариантах реализации, антитело можно восстановить с помощью восстанавливающего агента, например, дитиотреитола (ДТТ) или трикарбонилэтилфосфина (ТСЕР) при частично или полностью восстановительных условиях с целью получения реакционноспособных тиоловых групп цистеина. В некоторых вариантах реализации антитело подвергают воздействию денатурирующих условий с целью выявления реакционноспособных нуклеофильных групп, например, лизина или цистеина.
Нагрузка (отношение антибиотиков/антител, "AAR") ААС, также может упоминаться в данном документе, как отношение лекарственного средства к антителу (DAR), может контролироваться различными способами, например, с помощью: (i) ограничения молярного избытка промежуточного соединения антибиотика-линкера или линкерного реагента по отношению к антителу, (ii) ограничения времени или температуры реакции конъюгирования и (iii) посредством частичных или лимитирующих восстановительных условий для модификации тиоловых групп цистеина.
Следует понимать, что при реакции более чем одной нуклеофильной группы с промежуточным соединением антибиотик-линкер или линкерным реагентом, за которым следует реагент антибиотического фрагмента, полученный продукт представляет собой смесь ААС соединений с распределением одного или более антибиотических фрагментов, присоединенных к антителу. Среднее количество антибиотиков на антитело в смеси можно рассчитать с помощью двойного твердофазного ИФА антитела, специфичного по отношению к антителу и антибиотику. Отдельные молекулы ААС в смеси можно выявить с помощью масс-спектроскопии и разделить с помощью ВЭЖХ, например, гидрофобной хроматографии (см., например, McDonagh et al (2006) Prot. Engr. Design & Selection 19(7):299-307; Hamblett et al (2004) Clin.Cancer Res. 10:7063-7070; Hamblett, K.J., et al. "Effect of drug loading on the pharmacology, pharmacokinetics, and toxicity of an anti-CD30 antibody-drug conjugate," Abstract No.624, American Association for Cancer Research, 2004 Annual Meeting, March 27-31, 2004, Proceedings of the AACR, Volume 45, March 2004; Alley, S.C., et al. "Controlling the location of drug attachment in antibody-drug conjugates," Abstract No. 627, American Association for Cancer Research, 2004 Annual Meeting, March 27-31, 2004, Proceedings of the AACR, Volume 45, March 2004). В некоторых вариантах реализации, из конъюгационной смеси посредством электрофореза или хроматографии можно выделить однородный ААС с одинаковым значением нагрузки. Сконструированные с цистеином антитела по изобретению позволяют получать более гомогенные препараты, поскольку реакционный сайт на антителе в основном ограничен тиолом сконструированного цистеина. В одном варианте реализации изобретения, среднее количество антибиотических фрагментов на антитело находится в диапазоне от около 1 до около 20. В некоторых вариантах реализации изобретения, диапазон выбирается и регулируется от около 1 до 4.
СПОСОБЫ ПОЛУЧЕНИЯ КОНЪЮГАТОВ АНТИТЕЛО-АНТИБИОТИК
ААС формулы I можно получить различными способами, используя реакции органической химии, условия и реагенты, известные специалистам в данной области техники, включая: (1) реакцию нуклеофильной группы антитела с бивалентным линкерным реагентом для образования Ат-L через ковалентную связь, с последующей реакцией с фрагментом антибиотика (abx); и (2) реакцию нуклеофильной группы фрагмента антибиотика с двухвалентным линкерным реагентом для образования L-abx, через ковалентную связь с последующей реакцией с нуклеофильной группой антитела.
Иллюстративные способы получения ААС формулы I при помощи недавних методов, описаны в патенте США 7498298, содержание которого включено в данный документ посредством ссылки.
Примеры нуклеофильных групп антител включают: (i) N-концевые аминогруппы, (ii) аминогруппы боковых цепей, например, лизина, (iii) тиоловые группы боковых цепей, например, цистеина, и (iv) гидроксильные или аминогруппы углевода, если антитело является гликозилированным. Аминные, тиоловые и гидроксильные группы являются нуклеофильными и могут реагировать с образованием ковалентных связей с электрофильными группами на молекулах линкера и линкерных реагентов, включая: (i) активные эфиры, например, NHS-эфиры, HOBt-эфиры, галоформиаты и галогенангидриды; (ii) алкил- и бензилгалиды, например, галоацетамиды; и (iii) альдегиды, кетоны, карбоксильные и малеимидные группы. Некоторые антитела содержат восстанавливаемые межцепочечные дисульфидные связи, т.е. цистеиновые мостики. Антителам можно придать способность к реакции конъюгирования с линкерными реагентами путем обработки восстановителем, например, DTT (дитиотриэтолом) или трикарбонилэтилфосфином (ТСЕР), получая полностью или частично восстановленное антитело. Каждый цистеиновый мостик, таким образом, теоретически образует две реакционноспособные тиоловые нуклеофильные группы. В антитела можно внедрить дополнительные нуклеофильные группы путем модификации остатков лизина, например, посредством реакции остатков лизина с 2-иминтиоланом (реагентом Трота), приводящей к конверсии аминов в тиолы. Реакционноспособные тиоловые группы можно внедрить в антитело путем внедрения одного, двух, трех, четырех или более остатков цистеина (например, получения вариантов антител, включающих один или более аминокислотных остатков цистеина неприродного происхождения).
Конъюгаты антитело-антибиотик по изобретению также могут быть получены реакцией между электрофильной группой антитела, например, альдегидной или кетоновой карбонильной группой, с нуклеофильной группой линкерного реагента или антибиотиком. Пригодные для этого нуклеофильные группы линкерного реагента включают гидразид, оксим, аминогруппу, гидразин, тиосемикарбазон, гидразинкарбоксилат и арилгидразид, но не ограничиваются ими. В одном варианте реализации антитело модифицируют с целью внедрения электрофильных групп, которые способны реагировать с нуклеофильными заместителями в составе линкерного реагента или антибиотика. В еще одном варианте реализации можно окислить углеводы гликозилированных антител, например, периодатными окислителями, образуя альдегидные или кетоновые группы, которые могут реагировать с аминной группой линкерного реагента или фрагментами антибиотика.
Полученные группы иминового шиффова основания могут образовывать стабильную связь или восстанавливаться, например, боргидридными реагентами, образуя стабильные аминные связи. В одном варианте реализации, реакция углеводного фрагмента гликозилированного антитела с галактозооксидазой или мета-периодатом натрия могла приводить к образованию карбонильных (альдегидных и кетоновых) групп в составе антитела, которые могли реагировать с соответствующими группами на антибиотике (Hermanson, Bioconjugate Techniques). В еще одном варианте реализации, антитела, содержащие N-концевые остатки серина или треонина, могли реагировать с мета-периодатом натрия, что приводило к образованию альдегида вместо первой аминокислоты (Geoghegan & Stroh, (1992) Bioconjugate Chem. 3:138-146; US 5362852). Такой альдегид может реагировать с нуклеофильной группой антибиотического фрагмента или линкера.
Нуклеофильные группы на антибиотическом фрагменте включают, но не ограничиваются ими: аминные, тиоловые, гидроксильные, гидразидные, оксимные, гидразиновые, тиосемикарбазоновые, гидразинкарбоксилатные и арилгидразидные группы, способные реагировать с образованием ковалентных связей с электрофильными группами в составе молекул линкера и линкерных реагентов, включающими: (i) активные эфиры, например, NHS-эфиры, HOBt-эфиры, галоформиаты и галогенангидриды; (ii) алкил- и бензилгалиды, например, галоацетамиды; и (iii) альдегиды, кетоны, карбоксильные и малеимидные группы.
Конъюгаты антитело-антибиотик (ААС) в таблице 3 получали конъюгацией описанных анти-WTA антител и промежуточных соединений линкер-антибиотик из таблицы 2 и в соответствии с описанными способами в примере 7. ААС тестировали на эффективность с помощью анализа макрофагов in vitro (пример 9) и in vivo мышиной модели почек (пример 10).
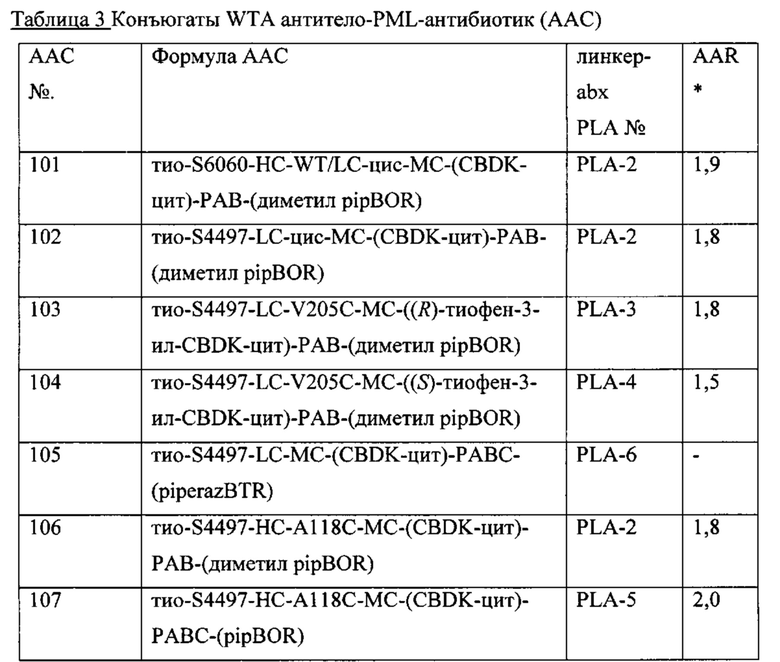
*AAR = среднее соотношение антибиотик/антитело
Дикий тип («ДТ»), мутантное антитело с модифицированным цистеином («тио»), легкая цепь («LC»), тяжелая цепь («НС»), 6-малеимидокапроил («МС»), малеимидопропаноил ("MP"), циклобутилдикето ("CBDK"), цитруллин ("cit"), цистеин ("cys"), п-аминобензил ("РАВ"), и п-аминобензилоксикарбонил ("РАВС").
СПОСОБЫ ЛЕЧЕНИЯ И ПРЕДОТВРАЩЕНИЯ ИНФЕКЦИЙ КОНЪЮГАТАМИ АНТИТЕЛО-АНТИБИОТИК
Анти-WTA-AAC по изобретению применимы в качестве антимикробных агентов, эффективных против Staphylococci человека и в ветеринарии, например, S. aureus, S. saprophyticus и S. simulans. В конкретном аспекте, ААС по изобретению полезны для лечения инфекций S. aureus.
После входа в кровоток, S. aureus может вызвать метастатическую инфекцию практически в любом органе. Вторичные инфекции встречаются примерно в одной трети случаев до начала терапии (Fowler et al., (2003) Arch. Intern. Med. 163: 2066-2072) и даже у 10% пациентов после начала терапии (Khatib et al., (2006) Scand. J. Infect. Dis., 38:7-14). Отличительными признаками инфекций являются большие резервуары гноя, разрушение тканей и образование абсцессов (все из которых содержат большое количество нейтрофилов). У около 40% пациентов развиваются осложнения, если бактериемия сохраняется после трех дней.
Предложенный механизм действия ААС описан выше (под заголовком «Конъюгаты антитело-антибиотик»). Конъюгаты анти-WTA антитело-антибиотик (ААС) по изобретению обладают значительными терапевтическими преимуществами для лечения внутриклеточных патогенов. ААС-линкер расщепляется путем воздействия на фаголизосомальные ферменты, высвобождая активный антибиотик. Из-за ограниченного пространства и относительно высокой концентрации местных антибиотиков (около 104 на бактерию), в результате фаголизосома больше не поддерживает выживание внутриклеточного патогена. Поскольку ААС по существу является неактивным пролекарством, терапевтический индекс антибиотика может быть увеличен относительно свободной (неконъюгированной) формы. Антитело обеспечивает специфическое нацеливание на патоген, тогда как расщепляемый линкер расщепляется в условиях, специфичных для внутриклеточного расположения патогена. Эффект может быть как непосредственно на опсонизированный возбудитель, так и другие патогены, которые локализируются в фаголизосоме. Антибиотикоустойчивость - это способность болезнетворного возбудителя противостоять уничтожению антибиотиками и другими противомикробными средствами, которая с точки зрения задействованного механизма отличается от множественной лекарственной устойчивости (Lewis K (2007). "Persister cells, dormancy and infectious disease". Nature Reviews Microbiology 5 (1):48-56. doi: 10.1038/nrmicro1557). Скорее, эта форма толерантности вызвана небольшой подгруппой микробных клеток, называемых устойчивыми организмами (Bigger JW (14 October 1944). "Treatment of staphylococcal infections with penicillin by intermittent sterilization".Lancet 244 (6320):497-500). Эти клетки не являются лекарственно устойчивыми в классическом смысле, но скорее являются спящими клетками, которые устойчивы к антибиотическому лечению, которое может убить их генетически идентичных родственников. Эта толерантность к антибиотикам индуцируется не- или чрезвычайно медленным разделяющим физиологическим состоянием. Когда противомикробное лечение не в состоянии уничтожить эти стойкие клетки, они становятся резервуаром для повторяющихся хронических инфекций. Конъюгаты антитело-антибиотик по изобретению обладают уникальным свойством убивать эти стойкие клетки и подавлять появление устойчивых к множественным лекарственным средствам бактериальных популяций.
В другом варианте реализации, анти-WTA-AAC по изобретению можно использовать для лечения инфекции независимо от внутриклеточного компартмента, в котором выживает патоген.
В другом варианте реализации изобретения, анти-WTA-AAC по изобретению можно также использовать для нацеливания на бактерии Staphylococci в планктонной форме или форме биопленки. Бактериальные инфекции, обработанные конъюгатами антитело-антибиотик (ААС) по изобретению, включают в себя лечение бактериальных легочных инфекций, таких как пневмония S. aureus, остеомиелит, рецидивирующий риносинусит, бактериальный эндокардит, бактериальные окулярные инфекции, такие как трахома и конъюнктивит, инфекции сердца, головного мозга или кожи, инфекции желудочно-кишечного тракта, такие как диарея путешественников, язвенный колит, синдром раздраженной толстой кишки (IBS), болезнь Крона и IBD (воспалительное заболевание кишечника) в общем, бактериальный менингит и абсцессы в любом органе, таком как мышцы, печень, мозговые оболочки или легкие. Бактериальные инфекции могут быть в других частях тела, таких как мочевой путь, кровоток, рана или место введения катетера. ААС по изобретению применимы для трудно поддающихся лечению инфекций, которые включают в себя биопленки, имплантаты или места связывания (например, остеомиелит и инфекции протезированных суставов) и высоко летальные инфекции, такие как больничная пневмония и бактериемия. Уязвимые группы пациентов, которые можно лечить для профилактики инфекции Staphylococcal aureus, включают пациентов с гемодиализом, пациентов с нарушенным иммунитетом, пациентов в отделениях интенсивной терапии и некоторых хирургических пациентов. В другом аспекте данное изобретение относится к способу уничтожения, лечения или предотвращения микробной инфекции у животного, предпочтительно млекопитающего и наиболее предпочтительно человека, который включает введение животному анти-WTA ААС или фармацевтической композиции ААС по изобретению. Изобретение дополнительно включает в себя лечение или профилактику заболеваний, связанных с или которые оппортунистически являются результатом таких микробных инфекций. Такие способы лечения или профилактики могут включать пероральное, местное, внутривенное, внутримышечное или подкожное введение композиции по изобретению. Например, перед операцией или введением катетера IV, при лечении интенсивной терапии, в трансплантационной медицине, с или после химиотерапии рака, или в других действиях, которые несут высокий риск инфицирования, ААС по изобретению можно вводить для предотвращения появления или распространения инфекции.
Бактериальная инфекция может быть вызвана бактериями с активной и неактивной формой, и ААС вводится в количестве и на протяжении времени, достаточных для лечения как активной, так и неактивной, скрытой формы бактериальной инфекции, продолжительность которой больше, чем необходимо для лечения активной формы бактериальной инфекции.
Анализ различных Gram + бактерий показал, что WTA бета экспрессируется на всех S. aureus, включая штаммы MRSA и MSSA, а также штаммы Staph, такие как S. saprophyticus и S. simulans. WTA альфа (Альфа-GLcNAc рибитол WTA) присутствует в большинстве, но не у всех S. aureus, а также присутствует на Listeria monocytogenes. WTA отсутствует на Gram- бактериях. Поэтому одним аспектом изобретения является способ лечения пациента, инфицированного одним или несколькими из S. aureus, S. saprophyticus или S. simulans, путем введения терапевтически эффективного количества анти-WTA бета-ААС по изобретению. Другим аспектом изобретения является способ лечения пациента, инфицированного S. aureus и/или Listeria monocytogenes, путем введения терапевтически эффективного количества анти-WTA альфа-ААС по изобретению. Изобретение также предусматривает способ профилактики инфекций одним или более из S. aureus или S. saprophyticus или S. simulans путем введения терапевтически эффективного количества анти-WTA бета-ААС по изобретению в условиях стационара, таких как хирургия, ожоговый пациент и трансплантация органов.
Для пациента, нуждающийся в лечении бактериальной инфекции, как это определено врачом в данной области техники, вид бактерий, которыми он/она инфицирован может, но не обязан, быть диагностирован. Так как у пациента с бактериальной инфекцией может очень быстро наступить ухудшение, в течение нескольких часов пациенту при поступлении в больницу может быть назначен анти-WTA-ААС по изобретению наряду с одним или более стандартами способами лечения Abx, таким как ванкомицин или ципрофлоксацин. Когда результаты диагностики становятся доступными и указывают на наличие, например, S. aureus при инфекции, пациент может продолжить лечение анти-WTA ААС. Поэтому в одном из вариантов реализации способа лечения бактериальной инфекции или, конкретно, инфекции S. aureus, пациенту вводят терапевтически эффективное количество анти-WTA бета-ААС. В способах лечения или профилактики по настоящему изобретению ААС по настоящему изобретению можно вводить в качестве единственного терапевтического агента или в сочетании с другими агентами, такими как описанные ниже. ААС по изобретению демонстрируют преимущество ванкомицина в лечении MRSA в доклинических моделях. Сравнение ААС с SOC может быть измерено, например, путем снижения смертности. Пациент, получающий лечение, оцениваться на реакцию на лечение ААС с помощью ряда измеряемых факторов. Примерами признаков и симптомов, которые могут использовать клиницисты для оценки улучшения состояния их пациентов, являются следующие: нормализация уровня лейкоцитов, если они повышены при диагностике, нормализация температуры тела, если повышена (лихорадка) во время диагностики, очистка культур крови, визуальное улучшение раны, включая уменьшение эритемы и дренирование гноя, снижение требований к искусственной вентиляции легких, такие, как снижение потребности в кислороде или снижение скорости вентиляции у пациента, который вентилируется, полное отключение искусственной вентиляции, если пациент вентилируется во время диагностики, использование меньшего количества лекарств для поддержания стабильного кровяного давления, если эти лекарства необходимы во время диагностики, нормализация лабораторных аномалий, которые свидетельствуют о недостаточности органов-мишеней, таких как повышенный уровень креатинина или печени, если они были аномальными во время постановки диагноза, и улучшение рентгенографии (например, рентгенография грудной клетки, которая ранее предлагала пневмонию с разрешением). У пациента в ОИТ эти факторы могут измеряться, по меньшей мере, ежедневно. Лихорадка контролируется так же, как и количество лейкоцитов, включая абсолютное количество нейтрофилов, а также доказательства того, что «левый сдвиг» (появление бластов, указывающих на увеличение производства нейтрофилов в ответ на активную инфекцию) исчез.
В контексте настоящих способов лечения данного изобретения считается, что пациент с бактериальной инфекцией поддается лечению, если наблюдается значительное измеримое улучшение, согласно оценке врача в данной области техники, по меньшей мере в двух или более из предшествующих факторов по сравнению со значениями, признаками или симптомами до или в начале лечения, или во время диагностики. В некоторых вариантах реализации изобретения, наблюдается измеримое улучшение в 3, 4, 5, 6 или более из вышеупомянутых факторов. Если в некоторых вариантах реализации изобретения, улучшение измеримых факторов составляет по меньшей мере на 50%, 60%, 70%, 80%, 90%, 95% или 100% по сравнению со значениями до воздействия. Обычно пациент может считаться полностью излеченным от бактериальной инфекции (например, инфекции S. aureus), если измеримые улучшения включают следующее: I) повторяющиеся культуры крови или ткани (обычно несколько), которые не показывают первоначально идентифицировавшие бактерии; ii) лихорадка нормализуется; iii) БКК нормализуются; и iv) доказательства того, что поражение конечного органа (сердце, легкие, печень, почки, сосудистый коллапс) полностью или частично разрешилось с учетом ранее существовавших сопутствующих заболеваний у пациента.
Схема приема
В любом из вышеприведенных аспектов при лечении инфицированного пациента дозировка ААС обычно составляет от около 0,001 до 1000 мг/кг/день. В одном варианте реализации изобретения, на пациента с бактериальной инфекцией воздействовали дозой ААК в диапазоне от около 1 до около 150 мг/кг, обычно от около 5 до около 150 мг/кг, более конкретно, от около 25 мг/кг до около 125 мг/кг, от 50 мг/кг до 125 мг/кг, еще более конкретно - от около 50 мг/кг до 100 мг/кг. ААС может назначаться ежедневно (например, разовая доза 5-50 мг/кг/день) или реже (например, разовая доза 5, 10, 25 или 50 мг/кг /неделя). Одна доза может быть разделена на 2 дня, например, 25 мг/кг на один день и 25 мг/кг на следующий день. Пациенту можно вводить дозу один раз каждые 3 дня (q3D), один раз в неделю, раз в две недели (qOW), в течение 1-8 недель. В одном варианте реализации изобретения, пациенту вводят ААС по изобретению через IV один раз в неделю в течение 2-6 недель вместе с стандартным лечением (SOC) бактериальной инфекции, такой как инфекция стафилококка. Продолжительность лечения будет определяться состоянием пациента или масштабом инфекции, например, продолжительность 2 недели для неосложненной бактериемии или 6 недель для бактериемии с эндокардитом.
В одном варианте реализации изобретения, ААС вводят в начальной дозе от 2,5 до 100 мг/кг в течение одного-семи дней с последующей поддерживающей дозой от 0,005 до 10 мг/кг один раз в течение одного-семи дней в течение одного месяца.
Способ приема
Для лечения бактериальных инфекций ААС по изобретению можно вводить в любой из предшествующих доз внутривенно (в.в.) или подкожно. В одном варианте реализации изобретения, WTA-AAC вводят внутривенно. В конкретном варианте реализации изобретения, WTA-AAC, вводимый внутривенно, представляет собой WTA-бета-ААС, конкретнее, при этом антитело WTA-бета представляет собой антитело, выбранное из группы Ат с аминокислотными последовательностями, как показанные на Фиг. 12, Фиг. 13А1 и А2 и Фиг. 13В1-В4, Фиг. 14А1-А2 и Фиг. 14В1-В3 и Фиг. 15А1-А3 и Фиг. 15В1-В6.
Комбинированная терапия
ААС можно вводить в сочетании с одним или более дополнительными, например, другими терапевтическими или профилактическими агентами, как это определено врачом, лечащим пациента.
В одном варианте реализации изобретения, второй антибиотик, вводимый в комбинации с соединением конъюгата антитело-антибиотик по изобретению, выбирают из структурных классов: (i) аминогликозиды; (ii) бета-лактамы; (iii) макролиды/циклические пептиды; (iv) тетрациклины; (v) фторхинолины/фторхинолоны; (vi) и оксазолидиноны. См.: Shaw, K. and Barbachyn, М. (2011) Ann. N.Y. Acad. Sci. 1241:48-70; Sutcliffe, J. (2011) Arm. N.Y. Acad. Sci. 1241:122-152.
В одном варианте реализации изобретения, второй антибиотик, вводимый в комбинации с соединением конъюгата антитело-антибиотик по изобретению, выбирают из клиндамицина, новобиоцина, ретапамулина, даптомицина, GSK-2140944, CG-400549, ситафлоксацина, тейкопланина, триклозана, нафтиридона, радезолида, доксорубицина, ампициллина, ванкомицина, имипенема, дорипенема, гемцитабина, далбаванцина и азитромицина.
Дополнительные примеры таких дополнительных терапевтических или профилактических агентов представляют собой противовоспалительные агенты (например, нестероидные противовоспалительные препараты (НПВП; например, детопрофен, диклофенак, дифлунизал, этодолак, фенопрофен, флурбипрофен, ибупрофен, индометацин, кетопрофен, меклофенамат, мефенамовая кислота, мелоксикам, набуметон, напроксен натрий, оксапрозин, пироксикам, сулиндак, толметин, целекоксиб, рофекоксиб, аспирин, холин салицилат, салсалат и натрий и магний салицилат) и стероиды (например, кортизон, дексаметазон, гидрокортизон, метилпреднизолон, преднизолон, преднизон, триамцинолон), антибактериальные агенты (например, азитромицин, кларитромицин, эритромицин, гатифлоксацин, левофлоксацин, амоксициллин, метронидазол, пенициллин G, пенициллин V, метициллин, оксациллин, клоксациллин, диклоксациллин, нафциллин, ампициллин, карбенициллин, тикарциллин, мезлоциллин, пиперациллин, азлоциллин, темоциллин, цепалотин, цефапирин, цефрадин, цефалоридин, цефазолин, цефамандол, цефуроксим, цефалексин, цефпрозил, цефаклор, лоракарбеф, цефокситин, цефматозол, цефотаксим, цефтизоксим, цефтриаксон, цефоперазон, цефтазидим, цефиксим, цефподоксим, цефтибутен, цефдинир, цефпиром, цефепим, BAL5788, BAL9141, имипенем, эртапенем, меропенем, азтреонам, клавуланат, сульбактам, тазобактам, стрептомицин, неомицин, канамицин, паромицин, гентамицин, тобрамицин, амикацин, нетилмицин, спектиномицин, сизомицин, дибекалин, изепамицин, тетрациклин, хлортетрациклин, демеклоциклин, миноциклин, окситетрациклин, метациклин, доксициклин, телитромицин, АВТ-773, линкомицин, клиндамицин, ванкомицин, оритаванцин, далбаванцин, тейкопланин, хинупристин и дальфопристин, сульфаниламид, пара-аминобензойная кислота, сульфадиазин, сульфизоксазол, сульфаметоксазол, сульфаталидин, линезолид, налидиксовая кислота, оксолиновая кислота, норфлоксацин, пефлоксацин, эноксацин, офлоксацин, ципрофлоксацин, темафлоксацин, ломефлоксацин, флероксацин, грепафлоксацин, спарфлоксацин, тровафлоксацин, клинафлоксацин, моксифлоксацин, гемифлоксацин, ситафлоксацин, даптомицин, гареноксацин, рамопланин, фаропенем, полимиксин, тигециклин, AZD2563, или триметоприм), антибактериальные антитела, включая антитела к тому же или другому антигену от ААС нацеленного на Ag, ингибиторы агрегации тромбоцитов (например, абциксимаб, аспирин, цилостазол, клопидогрель, дипиридамол, эптифибатид, тиклопидин или тирофибан), антикоагулянты (например, далтепарин, данапароид, эноксапарин, гепарин, тинзапарин или варфарин), жаропонижающие средства (например, ацетаминофен), или липидоснижающие агенты (например, холестирамин, колестипол, никотиновая кислота, гемфиброзил, пробукол, эзетимиб или статины, такие как аторвастатин, розувастатин, ловастатин симвастатин, правастатин, церивастатин и флувастатин). В одном варианте реализации изобретения, ААС по изобретению вводят в комбинации со стандартным лечением (SOC) для S. aureus (включая устойчивые к метициллину и чувствительные к метициллину штаммы). Обычно на MSSA, как правило, воздействуют нафциллином или оксациллином, также на MRSA обычно воздействуют ванкомицином или цефазолином.
Эти дополнительные агенты можно вводить в течение 14 дней, 7 дней, 1 дня, 12 часов или 1 часа после введения ААС или одновременно с ними. Дополнительные терапевтические агенты могут присутствовать в тех же или разных фармацевтических композициях, что и ААС.При наличии в различных фармацевтических композициях, могут быть использованы различные пути введения. Например, ААС можно вводить внутривенно или подкожно, в то время как второй агент можно вводить перорально.
ФАРМАЦЕВТИЧЕСКИЕ СОСТАВЫ
Данное изобретение также относится к фармацевтическим композициям, содержащим WTA-AAC, и к способам лечения бактериальной инфекции с использованием фармацевтических композиций, содержащих ААС. Такие композиции могут дополнительно содержать подходящие эксципиенты, такие как фармацевтически приемлемые эксципиенты (носители), включая буферы, кислоты, основания, сахара, разбавители, скользящие вещества, консерванты и т.п., которые хорошо известны в данной области техники и описанные в данном документе. Данные способы и композиции могут использоваться по отдельности или в сочетании с другими конвенционными способами и/или агентами для лечения инфекционных заболеваний. В некоторых вариантах реализации изобретения, фармацевтическая композиция включает 1) анти-WTAβ-AAC по изобретению и 2) фармацевтически приемлемый носитель. В некоторых вариантах реализации изобретения, фармацевтическая композиция включает 1) ААС по изобретению и необязательно 2) по меньшей мере один дополнительный терапевтический агент.
Фармацевтические композиции, содержащие ААС по изобретению, готовят для хранения путем смешивания ААС, имеющего желаемую степень чистоты, с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)) в виде водных растворов или лиофилизированных или других высушенных композиций. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов при используемых дозировках и концентрациях и включают буферы, такие как фосфатный, цитратный буфер, гистидин и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (менее чем около 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белка); и/или неионные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (ПЭГ). Фармацевтические составы, используемые для введения in vivo, как правило, являются стерильными, стерильность легко достигается путем фильтрации через стерильные фильтрующие мембраны.
Действующие вещества могут также заключаться в микрокапсулы, полученные, например, методиками коацервации или межфазной полимеризации, например, микрокапсулы из гидроксиметилцеллюлозы или желатина и полиметилметакрилатные микрокапсулы, соответственно, в коллоидные системы доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие методики описаны в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Можно получить препараты замедленного высвобождения. Подходящие препараты с замедленным высвобождением включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащих антитело или ААС по изобретению, причем матрицы находятся в форме профилированных изделий, например, пленок или микрокапсул. Примеры матриц с пролонгированным высвобождением включают полиэфиры, гидрогели (например, поли(2-метакрилат) или поли(виниловый спирт)), полилактиды (патент США №3773919), сополимеры L-глутаминовой кислоты и γ этил-L-глутамата, неразрушаемый этилвинилацетат, разрушаемые сополимеры молочной кислоты-гликолевой кислоты, такие как LUPRON DEPOT™ (инъекционные микросферы, состоящие из сополимера молочной кислоты-гликолевой кислоты и лейпролида ацетата), и поли-D-(-)-3-гидроксимасляную кислоту. В то время как полимеры, такие как этиленвинилацетат и молочная кислота-гликолевая кислота, обеспечивают возможность высвобождения молекулы дольше 100 суток, определенные гидрогели высвобождают белки в течении более коротких периодов времени. Если инкапсулированные антитела или ААС остаются в организме в течение длительного времени, то они могут денатурировать или агрегировать в результате воздействия влаги при 37°С, что приводит к потере биологической активности и возможным изменениям иммуногенности. Для стабилизации могут быть разработаны рациональные стратегии, зависящие от задействованного механизма. К примеру, если обнаружено, что механизм агрегации вызывает образование межмолекулярной связи S-S посредством тиодисульфидного обмена, то стабилизация может быть достигнута модификацией сульфгидрильных остатков, лиофилизацией из кислых растворов, регулированием содержания влаги, использованием подходящих добавок и разработкой специальных композиций с полимерными матрицами.
ААС может быть составлен в любой подходящей форме для доставки в клетку/ткань мишень. Например, ААС могут быть составлены в виде липосом, небольшого пузырька, состоящего из различных типов липидов, фосфолипидов и/или поверхностно-активного вещества, которые могут быть использованы для доставки лекарственного средства млекопитающему. Компоненты липосомы обычно расположены в двухслойном слое, подобном липидному расположению биологических мембран. Липосомы, содержащие антитело, получают способами, известными в данной области техники, такими как описанные в Epstein et al., (1985) Proc. Natl. Acad. Sci. USA 82:3688; Hwang et al., (1980) Proc. Natl Acad. Sci. USA 77:4030; US 4485045; US 4544545; WO 97/38731; US 5013556.
Особенно полезные липосомы могут быть получены методом обратного фазового испарения с липидной композицией, содержащей фосфатидилхолин, холестерин и ПЭГ-производный фосфатидилэтаноламин (ПЭГ-ПЭ). Липосомы экструдируют через фильтры с определенным размером пор, чтобы получить липосомы с требуемым диаметром.
МАТЕРИАЛЫ И МЕТОДЫ
Бактериальные штаммы:
Все эксперименты проводили с MRSA-USA300 NRS384, полученными из NARSA (http://www.narsa.net/control/member/repositories), если не указано иное.
Определение МИК для внеклеточных бактерий
МИК для внеклеточных бактерий определяли, производя серийные 2-кратные разведения антибиотика в триптическом соевом бульоне. Растворы антибиотика получали в четырех повторностях в 96-луночных планшетах для культивирования. MRSA (NRS384 штамм NRS384 из US A300) брали из экспоненциально растущей культуры и разводили до 1×104 КОЕ/мл. Бактерии культивировали в присутствии антибиотика в течение 18-24 часов при встряхивании при 37°С и определяли рост бактерий, считывая оптическую плотность (ОП) при 630 нМ. МИК был определен как доза антибиотика, которая ингибировала рост бактерий > 90%.
Определение МИК для внутриклеточных бактерий
Внутриклеточный МИК определяли у бактерий, которые секвестрировались внутри перитонеальных макрофагов мыши (см. ниже для генерации перитонеальных макрофагов мыши). Макрофаги высевали в 24-луночные культуральные чашки с плотностью 4x105 клеток/мл и инфицировали MRSA в соотношении 10-20 бактерий на макрофаг. Культуры макрофагов поддерживали в ростовых средах с добавлением 50 мкг/мл гентамицина (антибиотика, который активен только по отношению к внеклеточным бактериям) для ингибирования роста внеклеточных бактерий, и тестируемые антибиотики добавляли к среде для роста через 1 день после заражения. Выживание внутриклеточных бактерий оценивали через 24 часа после добавления антибиотиков. Макрофаги лизировали буферизованным солевым раствором Хэнкса, дополненным 0,1% альбумином бычьей сыворотки и 0,1% Тритоном-Х, и серийные разведения лизата готовили в фосфатном буферном солевом растворе, содержащем 0,05% Твин-20. Количество выживших внутриклеточных бактерий определяли путем нанесения на чашки триптического соевого агара с 5% дефибринированной овечьей кровью.
Выделение перитонеальных макрофагов:
Перитонеальные макрофаги выделяли из брюшины 6-8-недельных мышей Balb/c (Charles River Laboratories, Холлистер, Калифорния). Для увеличения выхода макрофагов, мышам предварительно вводили внутрибрюшинную инъекцию 1 мл тиогликолятной среды (Becton Dickinson). Тиогликолятную среду готовили в концентрации 4% в воде, стерилизовали автоклавированием и выдерживали в течение от 20 дней до 6 месяцев перед использованием. Перитонеальные макрофаги собирали через 4 дня после обработки тиогликолятом, промывая перитонеальную полость холодным фосфатным буферным раствором. Макрофаги высевали в модифицированную Дульбекко среду Игла (DMEM), дополненную 10% фетальной телячьей сывороткой, и 10 мМ HEPES, без антибиотиков, с плотностью 4×105 клеток/лунка в 24-луночных планшетах для культивирования. Макрофаги культивировали в течение ночи, чтобы обеспечить прилипание к чашке. Этот анализ также использовали для проверки внутриклеточного уничтожения в нефагоцитарных клетках. Линии клеток MG63 (CRL-1427) и А549 (CCL185) получали из АТСС и поддерживали в среде для культивирования тканей RPMI 1640, дополненной 10 мМ Hepes и 10% фетальной телячьей сыворотки (RPMI-10). Клетки HUVEC получали из Lonza и поддерживали в полной среде эндотелиальных клеток EGM (Lonza, Валкерсвилль, Мэриленд).
Инфекция макрофагов с опсонизированным MRSA:
Штамм USA300 MRSA (NRS384) был получен из репозитория NARSA (Шантильи, Вирджиния). В некоторых экспериментах использовали штамм Newman S. aureus (АТСС25904). Во всех экспериментах бактерии культивировали в триптическом соевом бульоне. Для оценки внутриклеточного уничтожения с помощью ААС, USA300 был взят из экспоненциально растущей культуры и промыт в НВ (сбалансированный солевой раствор Хэнкса с добавкой 10 мМ HEPES и 0,1% альбумина бычьей сыворотки). ААС или антитела разводили в НВ и инкубировали с бактериями в течение 1 часа для обеспечения связывания антител с бактериями (опсонизации), а опсонизированные бактерии использовали для инфицирования макрофагов в соотношении 10-20 бактерий на макрофаг (4×106 бактерий в 250 мкл НВ на лунку). Макрофаги предварительно промывали средой DMEM без сыворотки непосредственно перед заражением и инфицировали инкубацией при 37°С в инкубаторе с увлажненной тканевой культурой с 5% СО2, чтобы обеспечить фагоцитоз бактерий. Через 2 часа инфекционную смесь удаляли и заменяли нормальной питательной средой (DMEM, дополненной 10% фетальной сыворотки теленка, 10 мМ HEPES) и гентамицин добавляли при 50 мкг/мл для предотвращения роста внеклеточных бактерий. В конце инкубационного периода макрофаги промывали безсывороточной средой и клетки лизировали в НВ, дополненном 0,1% Тритона-Х (лизировали макрофаги без повреждения внутриклеточных бактерий). В некоторых экспериментах жизнеспособность макрофагов оценивали в конце периода культивирования путем детектирования высвобождения цитоплазматической лактатдегидрогеназы (LDH) в культуральный надосадок с использованием набора детектирования цитотоксичности LDH (продукт 11644793001, Roche Diagnostics Corp, Индианаполис, Индиана). Надосадки собирали и анализировали сразу в соответствии с инструкциями производителя. Серийные разведения лизата проводили в забуференном фосфатом физиологическом растворе, дополненном 0,05% Твин-20 (для разрушения агрегатов бактерий), и общее количество выживших внутриклеточных бактерий определяли путем нанесения на триптический соевый агар с 5% дефибринированной овечьей кровью.
Получение MRSA инфицированных перитонеальных клеток.
6-8-недельных самок мышей А/J (JAX™ Mice, Jackson Laboratories) инфицировали с 1×108 КОЕ штамма NRS384 в USA300 путем инъекции перитонеальной жидкости. Перитонеальную промывку собирали через день после заражения, и зараженные перитонеальные клетки обрабатывали 50 мкг/мл лизостафина, разбавленного в буфере Hepes, с добавлением 0,1% BSA (буфера НВ) в течение 30 минут при 37°С. Затем перитонеальные клетки промывали 2 раза в ледяном буфере НВ. Перитонеальные клетки разводили до 1×106 клеток/мл в тканевой культуральной среде RPMI 1640, дополненной 10 мМ Hepes и 10% фетальной телячьей сывороткой и 5 мкг/мл ванкомицина. Свободный MRSA от первичной инфекции хранили в течение ночи при 4°С в фосфатном буферном солевом растворе в качестве контроля внеклеточных бактерий, которые не подвергались уничтожению нейтрофилами.
Перенос инфекции из перитонеальных клеток в остеобласты:
Линию клеток остеобластов MG63 получали из АТСС (CRL-1427) и выдерживали в среде для культивирования ткани RPMI 1640, дополненной 10 мМ Hepes и 10% фетальной телячьей сыворотки (RPMI-10). Остеобласты высевали в 24-луночные планшеты для тканевой культуры и культивировали для получения непрерывного слоя. В день эксперимента остеобласты промывали один раз в RPMI (без добавок). MRSA или инфицированные перитонеальные клетки разбавляли в полном RPMI-10, и ванкомицин добавляли при 5 мкг/мл непосредственно перед инфицированием. Перитонеальные клетки добавляли к остеобластам при 1×106 перитонеальных клетках/мл. Образец клеток лизировали 0,1% Тритона-Х, чтобы определить фактическую концентрацию живых внутриклеточных бактерий во время инфекции. Фактический титр для всех инфекций определяли путем нанесения серийных разведений бактерий на триптическом соевом агаре с 5% дефибринированной овечьей кровью.
Остеобласты MG63 высевали в 4-х луночные стеклянные слайд-камеры и культивировали в среде для культивирования тканей RPMI 1640, дополненной 10 мМ Hepes и 10% фетальной телячьей сывороткой (RPMI-10), пока они не образовывали непрерывные слои. В день заражения лунки промывали безсывороточной средой и инфицировали суспензией зараженных перитонеальных клеток или штаммом US A3 00 MRSA, разведенным в полном RPMI-10, дополненном 5 мкг/мл ванкомицина. Через один день после заражения клетки промывали фосфатным буферным раствором (PBS) и фиксировали в течение 30 минут при комнатной температуре в PBS с 2% параформальдегидом. Лунки промывали 3 раза в PBS и пермеабилизировали PBS с 0,1% сапонина в течение 30 минут при комнатной температуре.
Модель передачи инфекции in vivo:
Запасы US A300 были подготовлены для заражения из активно растущих культур в триптическом соевом бульоне. Бактерии промывали 3 раза в фосфатном буферном растворе (PBS) и аликвоты замораживали при -80°С в PBS 25% глицерина. Инфекции внутриклеточных бактерий: Для этих экспериментов были выбраны мыши A/J, потому что они легко инфицировались относительно низкими дозами MRSA (2×106 КОЕ/мышь). 7-недельных самок мышей A/J получали из Jackson Lab и инфицировали перитонеальной инъекцией 5×107 КОЕ USA300. Мышей умерщвляли через 1 сутки после инфицирования, а брюшину промывали 5 мл холодного PBS. Перитонеальные промывки центрифугировали в течение 5 минут при 1000 об/мин при 4°С в настольной центрифуге. Клеточный осадок, содержащий перитонеальные клетки, собирали и клетки обрабатывали 50 мкг/мл лизостафина (Cell Sciences Inc.Canton MA, CRL 309C) в течение 20 минут при 37°С для уничтожения загрязняющих внеклеточных бактерий. Перитонеальные клетки промывали 3 раза в ледяном PBS для удаления лизостафина. Перитонеальные клетки донорских мышей объединяли, а реципиентным мышам вводили клетки, полученные от 5 доноров на каждого реципиента путем внутривенной инъекции в хвостовую вену. Чтобы определить количество живых внутриклеточных КОЕ, образец перитонеальных клеток лизировали в НВ (сбалансированный солевой раствор Хэнкса с добавкой 10 мМ HEPES и 0,1% альбумина бычьей сыворотки) с 0,1% Тритон-Х и серийные разведения лизата получали в PBS с 0,05% Твин-20. Инфекции свободных бактерий: Мышей A/J инфицировали различными дозами свободных бактерий, используя свежую аликвоту запасов глицерола, используемых для перитонеальных инъекций. Фактические дозы заражения были подтверждены нанесением КОЕ. Для данных, показанных на Фиг. 1А, фактическая доза инфицирования для внутриклеточных бактерий составляла 1,8×106 КОЕ/мышь, а фактическая доза инфицирования для свободных бактерий составляла 2,9×106 КОЕ/мышь. Отобранным мышам вводили однократную дозу 100 мг/кг ванкомицина путем внутривенной инъекции сразу после заражения.
Передача инфекции in vitro в нефагоцитарные клетки.
Генерация MRSA инфицированных перитонеальных клеток: 6-8-недельных самок мышей А/J (Jackson Lab) инфицировали с 1×108 КОЕ штамма NRS384 US A300 путем перитонеальной инъекции. Перитонеальную промывку собирали через день после заражения, и зараженные перитонеальные клетки обрабатывали 50 мкг/мл лизостафина, разбавленного в буфере Hepes, с добавлением 0,1% BSA (буфера НВ) в течение 30 минут при 37°С. Затем перитонеальные клетки промывали 2 раза в ледяном буфере НВ. Перитонеальные клетки разводили до 1×106 клеток/мл в тканевой культуральной среде RPMI 1640, дополненной 10 мМ Hepes и 10% фетальной телячьей сывороткой и 5 мкг/мл ванкомицина. Свободный MRSA от первичной инфекции хранили в течение ночи при 4°С в фосфатном буферном солевом растворе в качестве контроля внеклеточных бактерий, которые не подвергались уничтожению нейтрофилами.
Инфекция остеобластов или НВМЕС.Линию клеток MG63 получали из АТСС (CRL-1427) и выдерживали в среде для культивирования ткани RPMI 1640, дополненной 10 мМ Hepes и 10% фетальной телячьей сыворотки (RPMI-10). Клетки НВМЕС (Каталог # 1000) и среды ЕСМ (каталог # 1001) были получены в исследовательских лабораториях SciencCell (Карлсбад, Калифорния). Клетки высевали в 24-луночные планшеты для тканевой культуры и культивировали для получения непрерывного слоя. В день эксперимента, клетки промывали один раз в RPMI (без добавок). MRSA или инфицированные перитонеальные клетки разбавляли в полном RPMI-10, и ванкомицин добавляли при 5 мкг/мл непосредственно перед инфицированием. Перитонеальные клетки добавляли к остеобластам при 1×106 перитонеальных клетках/мл. Образец клеток лизировали 0,1% тритона-х, чтобы определить фактическую концентрацию живых внутриклеточных бактерий во время инфекции. Фактический титр для всех инфекций определяли путем нанесения серийных разведений бактерий на триптическом соевом агаре с 5% дефибринированной овечьей кровью.
Получение анти-S. aureus антител.
Человеческие IgG-антитела против анти-бета-GlcNAc WTA мАт клонировали из периферических В-клеток у пациентов после инфицирования S. aureus с использованием технологии обнаружения моноклональных антител, которая сохраняет родственное спаривание тяжелой и легкой цепей антитела38. Отдельные клоны антител экспрессировали трансфекцией клеток млекопитающих (Meijer, P.J., Nielsen, L.S., Lantto, J. & Jensen, A.(2009) Human antibody repertoires. Methods Mol Biol 525, 261-277, xiv; Meijer, P.J., et al. (2006) "Isolation of human antibody repertoires with preservation of the natural heavy and light chain pairing." Journal of molecular biology 358, 764-772). Надосадки, содержащие полноразмерные антитела IgGl, собирали через семь дней и использовали для скрининга связывания антигена с помощью ИФА. Эти антитела были положительными для связывания с препаратами клеточной стенки из USA300. Затем были получены антитела в 200-мл транзиентных трансфекциях и очищены хроматографией на белке A (MabSelect SuRe, GE Life Sciences, Пискатавей, Нью-Джерси) для дальнейшего тестирования. Выделение и использование этих антител было одобрено областной этической комиссией.
Конъюгация линкерного препарата с антителом.
Конструирование и производство THIOMAB варианта анти-WTA антитела (Ат) проводили следующим образом. Остаток цистеина был сконструирован в положении Val 205 легкой цепи Анти-WTA Ат, чтобы получить его THIOMAB™ вариант антитела с модифицированным цистеином. Это тио Анти-WTA конъюгировали с PML линкером-антибиотиком-интермедиатом из Таблицы 2. Антитело восстанавливали в присутствии пятидесятикратного молярного избытка DTT в течение ночи. Восстановительный агент и блоки цистеина и глутатиона очищали с использованием колонки HiTrap SP-HP (GE Healthcare). Антитело повторно окисляли в присутствии пятнадцатикратного молярного избытка дегидроаскорбиновой кислоты (МП Biomedical) в течение 2,5 часов. Образование межцепочечных дисульфидных связей контролировали LC/MS. Трехкратный молярный избыток промежуточного белка PML-линкера-антибиотика над белком инкубировали с THIOMAB в течение одного часа. Конъюгат лекарственного средства антитела очищали фильтрованием через 0,2 мкм фильтр SFCA (Millipore). Избыточный свободный линкерный препарат удаляли фильтрованием. Конъюгат заменяли буфером на 20 мМ гистидина ацетата с рН 5,5/240 мМ сахарозой путем диализа. Количество конъюгированных антибиотиков рифамицинового типа на мАт определяли количественно с помощью LC/MS-анализа в качестве соотношения антибиотик/антитело (AAR).Чистоту также оценивали с помощью вытеснительной хроматографии.
Масс-спектрометрический анализ
Анализ LC/MS проводили на 6530 квадрупольном/времяпролетном хромато-масс-спектрометре (Q-TOF) LC/MS (Agilent Technologies). Образцы хроматографировали на колонке PRLP-S, 1000 А, 8 мкм (50 мм ×2,1 мм, Agilent Technologies), нагретой до 80°С. Использовали линейный градиент от 30 до 60% В в течение 4,3 мин (растворитель 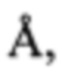 0,05% TFA в воде, растворитель В, 0,04% TFA в ацетонитриле) и элюент непосредственно ионизировали с использованием электрораспылительного источника. Данные собирали и деконволюционировали с использованием программного обеспечения для качественного анализа Agilent Mass Hunter. Перед анализом LC/MS, конъюгат антитело-лекарство обрабатывали лизил-эндопептидазой (Wako) в течение 30 минут при соотношении фермента к антителу 1:100 мас./мас., рН 8,0 и 37°С, чтобы получить Fab и Fc-часть для облегчения анализа. Соотношение антибиотиков и антител (AAR) (используемое взаимозаменяемо в данном документе с отношением лекарственное средство к антителу (DAR)) рассчитывали с использованием излишка Fab и Fab+1, вычисленного с помощью программного обеспечения MassHunter.
0,05% TFA в воде, растворитель В, 0,04% TFA в ацетонитриле) и элюент непосредственно ионизировали с использованием электрораспылительного источника. Данные собирали и деконволюционировали с использованием программного обеспечения для качественного анализа Agilent Mass Hunter. Перед анализом LC/MS, конъюгат антитело-лекарство обрабатывали лизил-эндопептидазой (Wako) в течение 30 минут при соотношении фермента к антителу 1:100 мас./мас., рН 8,0 и 37°С, чтобы получить Fab и Fc-часть для облегчения анализа. Соотношение антибиотиков и антител (AAR) (используемое взаимозаменяемо в данном документе с отношением лекарственное средство к антителу (DAR)) рассчитывали с использованием излишка Fab и Fab+1, вычисленного с помощью программного обеспечения MassHunter.
Анализ бактерий, выделенных от инфицированных мышей: мышей Balb/c инфицировали 1×107 КОЕ MRSA (USA300) путем внутривенной инъекции и почки собирали на 3-й день после инфицирования. Почки гомогенизировали, используя диссоциатор GentleMACS в объеме 5 мл на 2 почки, используя M-Tubes и программу RNA01.01 (Miltenyi Biotec, Оберн, Калифорния). Буфер для гомогенизации был: PBS + 0,1% Тритон-Х100, 10 мкг/мл ДНКазы (бычья поджелудочная железа II степени, Roche) и ингибиторы протеазы (полный коктейль ингибитора протеазы, Roche 11-836-153001). После гомогенизации образцы инкубировали при комнатной температуре в течение 10 минут, а затем разбавляли ледяным PBS и фильтровали через клеточный фильтр с ячейкой 40 мкМ. Гомогенаты ткани промывали 2 раза в ледяном PBS и затем суспендировали в объеме 0,5 мл на 2 почки в буфере НВ (сбалансированный солевой раствор Хэнкса с добавлением 10 мМ HEPES и 0,1% альбумина бычьей сыворотки). Клеточная суспензия снова фильтруется и 25 мкл бактериальной суспензии отбирается для каждой реакции окрашивания.
Проточная цитометрия для сравнения экспрессии анти-MRSA антител: Окрашивание антител для проточной цитометрии. Бактерии (1×107 выращенных in vitro бактерий или 25 мкл тканевого гомогената, описанные выше) суспендировали в НВ (выше) и блокировали инкубацией с 400 мкг/мл (микрограмм на миллилитр) мышиного IgG (SIGMA, 15381) в течение 1 часа. Флуоресцентно меченые антитела добавляли непосредственно к реакции блокирования и инкубировали при комнатной температуре еще 10-20 минут.Бактерии промывали 3 раза в буфере НВ и затем фиксировали в PBS 2% параформальдегида до анализа FACS. Тестируемые антитела (анти-βWTA: 4497, анти-αWTA: 7578 или контрольный изотип: gD) конъюгировали с А1еха-488, используя реакционноспособные реагенты амина (Invitrogen, сукцинимидилэфир Alexa Fluor 488, NHS-A488). Антитела в 50 мМ фосфате натрия подвергали взаимодействию с 5-10-кратным молярным избытком NHS-A488 в темноте в течение 2-3 часов при комнатной температуре. Этикетировочную смесь наносят на колонку GE Sepharose S200, уравновешенную в PBS для удаления избыточных реагентов из конъюгированного антитела. Количество молекул/антител А488 определяли с использованием УФ-метода, как описано изготовителем.
Для анализа бактерий в гомогенатах тканей использовали неконкурентное анти-5. aureus антитело (rF1 - Hazenbos, W.L., et al. (2013) Novel staphylococcal glycosyltransferases SdgA and SdgB mediate immunogenicity and protection of virulence-associated cell wall proteins. PLoS Pathog 9, e1003653), которое было конъюгировано с Alexa-647, чтобы отличить S. aureus от частиц подобного размера. Тестируемые антитела исследовали в диапазоне доз от 80 нг/мл до 50 мкг/мл. Проточную цитометрию проводили с использованием Beckton Dickson FACS ARIA (BD Biosciences, Сан-Хосе, Калифорния) и анализ проводили с использованием программного обеспечения для анализа FlowJo (Flow Jo LLC, Ашланд, Орегон).
Время уничтожения для свободных антибиотиков на нереплицирующихся бактериях: S. aureus (USA300) отбирали из стационарной фазы культуры в течение ночи, промывали один раз в фосфатно-солевом буфере (PBS) и суспендировали в 1×107 КОЕ/мл в PBS без антибиотика или с 1×10-6 М антибиотика в объеме 10 мл в 50 мл пробирке для центрифугирования из полипропилена. Бактерии инкубировали при 37°С в течение ночи при встряхивании. В каждый момент времени три образца по 1 мл удаляли из каждой культуры и центрифугировали для сбора бактерий. Бактерии один раз промывали PBS для удаления антибиотика, и общее количество выживших бактерий определяли путем нанесения серийных разведений бактерий на чашки с агаром.
Уничтожение устойчивых клеток свободными антибиотиками: S. aureus (USA300) отбирали из стационарной фазы культуры в течение ночи, промывали один раз в триптическом соевом бульоне (TSB) и затем доводили до конечной концентрации 1×107 КОЕ/мл в общем объеме 10 мл TSB или TSB с ципрофлоксацином (0,05 мМ). Культуры инкубировали при встряхивании при 37°С в течение 6 часов и затем добавляли второй антибиотик, либо рифампицин (1 мкг/мл), либо другой антибиотик рифамицинового ряда (1 мкг/мл). В указанные моменты времени образцы удаляли из каждой культуры, один раз промывали PBS для удаления антибиотика и ресуспендировали в PBS. Общее количество выживших бактерий определяли путем нанесения серийных разведений бактерий на чашки с агаром. В последний момент времени собирали и наносили остатки каждой культуры.
Анализ высвобождения катепсина для ААС: Для количественного определения количества активного антибиотика, выделенного из ААС после обработки катепсином В, ААС разбавляли до 200 мкг/мл в катепсиновом буфере (20 мМ ацетата натрия, 1 мМ ЭДТА, 5 мМ L-цистеина, рН 5). Катепсин В (из бычьей селезенки, SIGMA С7800) добавляли при 10 мкг/мл и образцы инкубировали в течение 1 часа при 37°С. В качестве контроля ААС инкубировали в буфере отдельно. Реакцию останавливали добавлением 9 объемов бактериальных ростовых сред, триптического соевого бульона с рН 7,4 (TSB). Для оценки общего высвобождения активного антибиотика серийные разведения реакционной смеси были выполнены в четырех повторностях в TSB в 96-луночных планшетах, и MRSA (USA300) добавляли в каждую лунку при конечной плотности 2×103 КОЕ/мл. Культуры инкубировали в течение ночи при 37°С при встряхивании, и рост бактерий измеряли путем считывания оптической плотности при 630 нм с использованием планшет-ридера.
Синтез FRET конъюгата антитела S4497 для фаголизосомального процессинга.
Малеимидный FRET пептид был синтезирован и конъюгирован с S4497, сконструированным с цистеином, THIOMAB™ антителом. Пара FRET использовала тетраметилродамин (TAMRA) и флуоресцеин (Fischer, R., et al (2010) Bioconjug Chem 21, 64-73). Малеимидный FRET-пептид синтезировали по стандарту Fmoc твердофазной химии с использованием пептидного синтезатора PS3 (Protein Technologies, Inc). Вкратце, 0,1 ммоль амидной смолы Ринка использовали для получения С-концевого карбоксамида. Fmoc-Lys(Mtt)-OH на N- и С-концевых остатках использовали для удаления Mtt (монометокситритил) группы на смоле и проведения дополнительной реакции в боковой цепи для присоединения TAMRA и флуоресцеина. Пептидомиметическую единицу CBDK-cit добавляли между парой FRET в качестве расщепляемого катепсином спейсера. Необработанный малеимидный FRET-пептид или малеимидокапроил-K(ТАМРА)-С-CBDK-cit -K(флуоресцеин), отщепленный от смолы, подвергали дальнейшей очистке с помощью ВЭЖХ с обращенной фазой с колонкой Jupiter 5 мкм С4 (5 мкм, 10 мм × 250 мм, Phenomenex). Наш зонд FRET позволяет контролировать не только внутриклеточный перенос конъюгата антител, но также и процессинг линкера в фаголизосоме. Конъюгат интактного антитела флуоресцирует только красным цветом в следствие передачи энергии резонанса флуоресценции от донора. Однако при расщеплении субстрата пептида FRET в фаголизосоме ожидается появление зеленой флуоресценции от донора.
Видеомикроскопия для обнаружения расщепления линкера внутри макрофагов
Перитонеальные макрофаги мыши были высеяны на слайд-камере (Ibidi, Verona, WI, каталог 80826) в полных средах, как описано для анализа внутриклеточного уничтожения макрофагов. USA300 был помечен Cell Tracker Violet (Invitrogen C10094) при 100 мкг/мл в PBS 0,1% BSA путем инкубации в течение 30 минут при 37°С. Меченые бактерии опсонизировали зондом 4497-FRET путем инкубации в течение 1 часа в буфере НВ. Макрофаги промывали один раз непосредственно перед добавлением опсонизированных бактерий, и бактерии добавляли к клеткам при 1×107 бактерии/мл. Для контроля без фагоцитоза, макрофаги предварительно обрабатывали 60 нМ латрункулина A (Calbiochem) в течение 30 минут до и во время фагоцитоза. Слайды помещались в микроскоп сразу после добавления бактерий в клетки, и кинофрагменты были получены с помощью конфокального микроскопа Leica SP5, оборудованного камерой с СО2-инкубатором и контроллерами температуры от Ludin. Изображения снимались каждую минуту в течение 30 минут с использованием Plan АРО CS 40Х, N.A:1,25, масляная иммерсионная линза и лазерные линии 488 нм и 543 нм для возбуждения соответственно alexa 488 и TAMRA. Фазовые изображения также регистрировались с использованием лазерной линии 543 нм.
Количественная оценка высвобожденного антибиотика внутри макрофагов с помощью масс-спектрометрии.
Перитонеальные макрофаги мыши были инфицированы в 24-луночных планшетах для тканевой культуры, как описано ниже для внутриклеточного анализа уничтожения с MRSA, опсонизированной ААС, при 100 мкг/мл в НВ. После завершения фагоцитоза клетки промывали и в лунки добавляли 250 мкл полной среды+гентамицин и клетки инкубировали в течение 1 часа или 3 часов. В каждый момент времени надосадок и клеточные фракции собирали и добавляли ацетонитрил (ACN) до конечной концентрации 75 % и инкубировали в течение 30 минут. Экстракты клеток и надосадков лиофилизировали выпариванием в атмосфере N2 (TurboVap) и восстанавливали в 100 мкл 50% ACN, фильтровали и анализировали в системе Ab Sciex QTRAP 6500 LC/MS/MS.
In vitro анализ внутриклеточного уничтожения.
Типы нефагоцитарных клеток: Линии клеток MG63 (CRL-1427) и А549 (CCL185) получали из АТСС и поддерживали в среде для культивирования тканей RPMI 1640, дополненной 10 мМ Hepes и 10% фетальной телячьей сыворотки (RPMI-10). Клетки HUVEC получали из Lonza и поддерживали в полной среде эндотелиальных клеток EGM (Lonza, Валкерсвилль, Мэриленд). Клетки НВМЕС (Каталог # 1000) и среды ЕСМ (каталог # 1001) были получены в исследовательских лабораториях SciencCell (Карлсбад, Калифорния).
Мышиные макрофаги: Перитонеальные макрофаги выделяли из брюшины 6-8-недельных мышей Balb/c (Charles River Laboratories, Холлистер, Калифорния). Для увеличения выхода макрофагов, мышам предварительно вводили внутрибрюшинную инъекцию 1 мл тиогликолятной среды (Becton Dickinson). Тиогликолятную среду готовили в концентрации 4% в воде, стерилизовали автоклавированием и выдерживали в течение от 20 дней до 6 месяцев перед использованием. Перитонеальные макрофаги собирали через 4 дня после обработки тиогликолятом, промывая перитонеальную полость холодным фосфатным буферным раствором. Макрофаги высевали в модифицированную Дульбекко среду Игла (DMEM), дополненную 10% фетальной телячьей сывороткой, и 10 мМ HEPES, без антибиотиков, с плотностью 4×105 клеток/лунка в 24-луночных планшетах для культивирования. Макрофагов культивировали в течение ночи, чтобы обеспечить прилипание к чашке.
Человеческие макрофаги М2: CD14+ Моноциты очищали из нормальной крови человека с использованием набора для выделения моноцитов II (Miltenyi, Cat 130-091-153) и высевали при 1,5×105 клеток/см2 на чашках для культуры тканей, предварительно покрытых фетальной телячьей сывороткой (FCS), и культивировали в средах RPMI 1640 с 20% FCS + 100 нг/мл rhM-CSF. Среду обновляли в 1-й день и в 7 день, среду меняли до 5% сыворотки + 20 нг/мл IL-4. Макрофаги использовались через 18 часов.
Протокол анализа: Во всех экспериментах бактерии культивировали в триптическом соевом бульоне. Для оценки внутриклеточного уничтожения конъюгатами антибиотик-антитело (ААС), US A300 был взят из экспоненциально растущей культуры и промыт в НВ (сбалансированный солевой раствор Хэнкса с добавкой 10 мМ HEPES и 0,1% альбумина бычьей сыворотки). ААС или антитела разводили в НВ и инкубировали с бактериями в течение 1 часа для обеспечения связывания антител с бактериями (опсонизации), а опсонизированные бактерии использовали для инфицирования макрофагов в соотношении 10-20 бактерий на макрофаг (4×106 бактерий в 250 мкл НВ на лунку). Макрофаги предварительно промывали средой DMEM без сыворотки непосредственно перед заражением и инфицировали инкубацией при 37°С в инкубаторе с увлажненной тканевой культурой с 5% СО2, чтобы обеспечить фагоцитоз бактерий. Через 2 часа инфекционную смесь удаляли и заменяли нормальной питательной средой (DMEM, дополненной 10% фетальной сыворотки теленка, 10 мМ HEPES) и гентамицин добавляли при 50 мкг/мл для предотвращения роста внеклеточных бактерий. В конце инкубационного периода макрофаги промывали безсывороточной средой и клетки лизировали в НВ, дополненном 0,1% Тритона-Х (лизировали макрофаги без повреждения внутриклеточных бактерий). Серийные разведения лизата проводили в забуференном фосфатом физиологическом растворе, дополненном 0,05% Твин-20 (для разрушения агрегатов бактерий), и общее количество выживших внутриклеточных бактерий определяли путем нанесения на триптический соевый агар с 5% дефибринированной овечьей кровью.
ПРИМЕРЫ
Пример 1 Внутриклеточный MRSA защищен от традиционных антибиотиков
Для подтверждения гипотезы о том, что клетки млекопитающих обеспечивают защитную нишу для S. aureus в присутствии антибактериальной терапии было проведено сравнение эффективности трех основных антибиотиков, которые в настоящее время используются в качестве стандарта лечения (SOC) для инвазивных инфекций MRSA (ванкомицин, даптомицин и линезолид) против внеклеточных планктонных бактерий в сравнении с бактериями, секвестрованными внутри мышиных макрофагов (таблица 4).
Для внеклеточных бактерий, MRSA культивировали в течение ночи в триптическом соевом бульоне и определяли МИК как минимальную дозу антибиотика, которая предотвращала рост. Для внутриклеточных бактерий мышиные перитонеальные макрофаги инфицировали MRSA и культивировали в присутствии гентамицина для уничтожения внеклеточных бактерий. Тестируемые антибиотики добавляли в культуральную среду на следующий день после инфицирования, и общее количество выживших внутриклеточных бактерий определяли через 24 часа. Ожидаемые концентрации сыворотки для клинически значимых антибиотиков были зарегистрированы в Antimicrobial Agents, Andre Bryskier. ASM Press, Washington DC (2005).
Этот анализ с высоковирулентным внебольничным штаммом MRSA USA300 показал, что хотя внеклеточный MRSA очень восприимчив к ингибированию роста низкими концентрациями ванкомицина, даптомицина и линезолида в жидкой культуре, все три антибиотика не смогли уничтожить тот же самый штамм MRSA, изолированный внутри макрофагов, на который воздействовали клинически достижимыми концентрациям антибиотиков. Даже в отношении рифампицина, который считается относительно эффективным для удаления внутриклеточных патогенов (Vandenbroek, P.V. (1989) Antimicrobial Drugs, Microorganisms, and Phagocytes. Reviews of Infectious Diseases 11, 213-245), потребовалась в 6000 раз более высокая доза для устранения внутриклеточного MRSA по сравнению с дозой, необходимой для ингибирования роста (МИК) планктонных бактерий (Таблица 1), в соответствии с другими исследованиями, показывающими, что большинство существующих антибиотиков неэффективны при уничтожении внутриклеточного S. aureus как in vitro, так и in vivo (Sandberg, A., Hessler, J.H., Skov, R.L., Blom, J. & Frimodt-Moller, N. (2009) "Intracellular activity of antibiotics against Staphylococcus aureus in a mouse peritonitis model" Antimicrob Agents Chemother 53, 1874-1883).
Пример 2 Распространение инфекции с внутриклеточным MRSA
Эти эксперименты сравнивали вирулентность внутриклеточных бактерий против эквивалентной дозы свободноживущих планктонных бактерий и определяли, могут ли внутриклеточные бактерии создавать инфекцию в присутствии ванкомицина in vivo. Четыре когорты мышей заражали внутривенной инъекцией с приблизительно эквивалентными дозами S. aureus жизнеспособных свободных бактерий (2,9 × 106), взятых непосредственно из культуры бульонов или внутриклеточных бактерий (1,8 × 106), изолированных внутри макрофагов хозяина и нейтрофилов, образовавшихся в следствие перитонеальной инфекцией донорских мышей (Фиг. 1А), на отобранные группы воздействовали ванкомицином сразу же после инфицирования, а затем один раз в день. Мышей обследовали через 4 дня после инфицирования на предмет бактериальной колонизации в почках, орган, который последовательно колонизировали S. aureus у мышей. В трех независимых экспериментах было обнаружено эквивалентная или более высокая бактериальная нагрузка в почках мышей, инфицированных внутриклеточными бактериями, по сравнению с теми, которых инфицировали эквивалентной дозой планктонных бактерий (Фиг. 1 В). Неожиданно было обнаружено, что инфицирование внутриклеточными бактериями приводит к более плотной колонизации мозга, органа, который не эффективно колонизируется после заражения планктонными бактериями в этой модели (Фиг. 1С). Более того, внутриклеточные бактерии, но не планктонные бактерии, смогли создать инфекцию вопреки терапии ванкомицином в этой модели (Фиг. 1В, Фиг. 1С)
Дальнейший анализ in vitro более точно определил степень, до которой внутриклеточная выживаемость способствует уклонению от антибиотиков. С этой целью остеобласты MG63 инфицировали либо планктонным MRSA, или внутриклеточным MRSA, в присутствии ванкомицина.
Инфекция остеобластов или НВМЕС. Линию клеток MG63 получали из АТСС (CRL-1427) и выдерживали в среде для культивирования ткани RPMI 1640, дополненной 10 мМ Hepes и 10% фетальной телячьей сыворотки (RPMI-10). Клетки НВМЕС (каталог # 1000) и среды ЕСМ (каталог # 1001) были получены в исследовательских лабораториях SciencCell (Карлсбад, Калифорния). Клетки высевали в 24-луночные планшеты для тканевой культуры и культивировали для получения непрерывного слоя. В день эксперимента клетки промывали один раз в RPMI (без добавок). MRSA или инфицированные перитонеальные клетки разбавляли в полном RPMI-10, и ванкомицин добавляли из ресчета 5 мкг/мл непосредственно перед инфицированием. Перитонеальные клетки добавляли к остеобластам в концентрации 1x106 перитонеальных кл./мл. Образец клеток лизировали 0,1% Тритона-Х, чтобы определить фактическую концентрацию живых внутриклеточных бактерий во время инфекции. Фактический титр для всех инфекций определяли путем нанесения серийных разведений бактерий на триптическом соевом агаре с 5% дефибринированной овечьей кровью.
MRSA (свободные бактерии) высевали в среду, среду + ванкомицин или среду + ванкомицин, помещенный на монослой остеобластов MG63 (Фиг. 1Е) или на клетоки микрососудистых эндотелиальных клеток мозга человека (НВМЕС, Фиг. 1F). Планшеты центрифугировали для ускорения контакта бактерий с монослоем. В каждый момент времени культуральный надосадок собирали для извлечения внеклеточных бактерий или прикрепленные к субстрату клетки лизировали для высвобождения внутриклеточных бактерий.
Планктонные бактерии, подвергшиеся воздействию только ванкомицина, были эффективно уничтожены. Выжившие бактерии не восстанавливались после одного дня культивирования (Фиг. 1D). Когда такое же количество планктонных бактерий было высеяно на остеобласты MG63, на следующий день после инфицирования обнаруживалось небольшое количество выживших бактерий (примерно 0,06% от исходного), связанное с клетками MG63, которые были защищены от ванкомицина путем инвазии остеобластов.
MRSA, которые были секвестрированы внутри перитонеальных клеток, продемонстрировали резкое увеличение, как выживаемости, так и эффективности инфекции в присутствии ванкомицина. Около 15% внутриклеточного MRSA в лейкоцитах выжили в условиях идентичных тем, в которых ванкомицин стерилизовал культуры планктонных бактерий. Внутриклеточные бактерии также были способны инфицировать монослой остеобластов MG63 в присутствии ванкомицина, что привело к удвоению количества бактерий, восстановившихся за один день после воздействия ванкомицином (Фиг. 1D). Более того, количество внутриклеточных S. aureus увеличилось почти в 10 раз за 24-часовой период в клетках MG63 (Фиг. 1Е), первичных эндотелиальных клетках головного мозга человека (Фиг. 1F) и А549 бронхиальных эпителиальных клетках (не показаны) при постоянном воздействии концентрации ванкомицина, убивающего свободные живые бактерии. Несмотря на то, что они защищены от уничтожения антибиотиками, рост бактерий не наблюдался в культурах инфицированных перитонеальных макрофагов и нейтрофилов (не показаны). Вместе эти данные подтверждают, что внутриклеточные резервуары MRSA в миелоидных клетках могут способствовать распространению инфекции на новые сайты, даже при наличии активного лечения антибиотиками, и внутриклеточный рост может происходить в эндотелиальных и эпителиальных клетках даже в условиях постоянной антибактериальной терапии.
Для разработки реагента, который специфически уничтожает внутриклеточный S. aureus, антитела и компоненты антибиотика были тщательно отобраны и оптимизированы для максимальной эффективности. Приведенные ниже примеры демонстрируют эксперименты и результаты, приводящие к выбору антител против бета-тейхоевой кислоты клеточной стенки (анти-WTAP) и к определенным антибиотикам типа рифамицина, которые должны конъюгироваться с образованием ААС.
Пример 3 Выбор моноклонального анти-S.aureus антитела
Количество антибиотика, доставляемого ААС и, следовательно, его конечная эффективность, ограничено числом сайтов связывания антител на поверхности бактерии. Таким образом, было важно выбрать антитело, которое связывается с очень широко распространенным антигеном, который стабильно экспрессируется на MRSA на всех стадиях инфекции in vivo. В качестве начального шага, был клонирован и очищен от В-клеток перечень из более чем 40 анти-S. aureus антител, полученных из периферической крови пациентов, восстанавливающиеся от различных инфекций S. aureus и скриниронан на связывание с MRSA, выделенной непосредственно из почек инфицированных мышей. Получение, скрининг и отбор антител
Сокращения: MRSA (метициллин-резистентный S. aureus); MSSA (метициллин-чувствительный S. aureus); VISA (ванкомицин промежуточно-устойчивый S. aureus); LTA (липотейхоевая кислота); TSB (триптический соевый бульон); CWP (подготовка клеточной стенки).
IgG-антитела человека клонировали из периферических В-клеток пациентов после инфекции S. aureus, с использованием технологии Symplex™ (Symphogen, Ленгби, Дания), которая сохраняет родственное спаривание тяжелой и легкой цепей антитела, как описано в США 8283294:"Method for cloning cognate antibodies"; Meijer PJ et al. Journal of Molecular Biology 358:764-772 (2006); и Lantto J et al. J Virol. 85(4):1820-33 (Feb 2011); Plasma and memory cells were used as genetic source for the recombinant full-length IgG repertoires. Отдельные клоны антител экспрессировали трансфекцией клеток млекопитающих как описано в Meijer PJ, et al. Methods in Molecular Biology 525:261-277, xiv.(2009). Надосадки, содержащие полноразмерные антитела IgG1, собирали через семь дней и использовали для скрининга связывания антигена с помощью непрямого ИФА при первичном скрининге. Была создана библиотека мАт, показывающих положительное связывание ИФА с препаратами клеточной стенки из US A300 или штаммом Wood46 S. aureus штамма. Затем были получены антитела в 200-мл транзиентных трансфекциях и очищены хроматографией на белке A (MabSelect SuRe, GE Life Sciences, Пискатавей, Нью-Джерси) для дальнейшего тестирования. Для крупномасштабного производства антител их продуцировали в клетках СНО. Векторы, кодирующие VL и VH, трансфецировали в клетки СНО; IgG очищали от культуральной среды с помощью аффинной хроматографии с белком А.

CW#1 и CW#3 всегда смешивались при изготовлении покрытия ИФА
На Фиг. 6 суммируется первичный скрининг антител с помощью ИФА. Все (за исключением 4569) были выделаны при скрининге с приготовленной смесью клеточной стенки USA300 (среда без железа: TSB в соотношении 96:4). Все GlcNAc бета (за исключением 6259), SDR и PGN (4479) мАт также были положительными для PGN и WTA при первичном скрининге. Все GlcNAc альфа были обнаружены исключительно путем скрининга на связывание со смесью USA300 CW. 4569 (специфичный для LTA) был обнаружен путем скрининга на Wood46 CWP.
Самый высокий уровень связывания антител был обнаружен с человеческим IgGi, который распознает β-О-связанные модификации сахара GlcNAc на WTA (Таблица 6). Меньшее связывание было достигнуто с моноклональными антителами, распознающими а-О-связанный GlcNAc; контрольный изотип антитела против белка gD цитомегаловируса (CMV) проявлял некоторую минимальную реактивность благодаря белку А, экспрессированому на in-vivo-полученном S. aureus (Фиг. 7А). Антигенную специфичность антител определяли генетическими методами, так что антитела против модификаций сахара α- или β-GlcNAc на WTA не связывались со штаммами & aureus, лишенными соответствующих гликозилтрансфераз (как показано на Фиг. 7В). В соответствии со степенью связывания антител с in vivo-полученным MRSA, ААС, полученные с анти-β-GlcNAc WTA антителами, показали высокую эффективность по сравнению с антителами, полученными с анти-α-GlcNAc WTA антителами.
Выбор анти-WTA мАт из библиотеки с использованием ex vivo проточной цитометрии
Каждое мАт в этой библиотеке запрашивалось по трем критериям: (1) относительная интенсивность связывания мАт с поверхностью MRSA, как показатель высокой экспрессии соответствующего родственного антигена, который будет способствовать высокой доставке антибиотиков; (2) последовательность связывания мАт с MRSA, выделенной из большого разнообразия инфицированных тканей, как указание на стабильную экспрессию родственного антигена на поверхности MRSA in vivo во время инфекций; и (3) способность к связыванию мАт с группой клинических штаммов S. aureus, как показатель сохранения экспрессии родственного поверхностного антигена. С этой целью проточную цитометрию использовали для тестирования всех этих предварительно отобранных в библиотеке надосадков культуры мАт для реактивности с S. aureus из различных инфицированных тканей и из разных штаммов S. aureus.
Все мАт в библиотеке анализировали на их способность связывать MRSA с инфицированными почками, селезенкой, печенью и легкими у мышей, которые были инфицированы с MRSA USA300; и в сердцах или почках от кроликов, которые были инфицированы USA300 COL в модели кроличьего эндокардита. Способность антитела узнавать S. aureus из различных инфицированных тканей повышает вероятность того, что терапевтическое антитело будет активным в широком спектре различных клинических инфекций с S. aureus. Бактерии анализировали сразу после сбора органов, то есть без субкультуры, для предотвращения фенотипических изменений, вызванных культуральными условиями in vitro. Несколько поверхностных антигенов S. aureus, будучи экспрессированными в культуре in vitro, теряют экспрессию в инфицированных тканях. Антитела, направленные против таких антигенов, вряд ли будут полезны для лечения инфекций. Во время анализа этой библиотеки мАт на различных зараженных тканях это наблюдение было подтверждено для значительного числа антител, которые показали значительное связывание с бактериям S. aureus из культуры, но отсутствие связывания с бактериями из всех тестируемых инфицированных тканей. Некоторые антитела связываются с бактериями из некоторых, но не всех, тестируемых инфицированных тканей. Поэтому были отобраны антитела, которые были способны распознавать бактерии из всех протестированных условий инфекции. Оценивали следующие параметры: (1) относительную интенсивность флуоресценции, как меру излишка антигена; (2) количество органов, окрашенных положительно, как меру устойчивости экспрессии антигена; и (3) способность к связыванию мАт с группой клинических штаммов S. aureus как показатель сохранения экспрессии родственного поверхностного антигена. Интенсивность флуоресценции тестируемых антител определялась как относительно к контрольному изотипу антитела, которое направлено против не релевантного антигена, например, IgG1 мАт анти-герпес вируса gD:5237 (см. ниже). МАт против WTA-бета не только показали самый высокий избыток антигена, но также показали очень последовательное связывание с MRSA из всех инфицированных тканей, которые были протестированы и указаны выше.
Кроме того, оценивали способность этих мАт связываться со следующими штаммами S. aureus, и они были культивированы in vitro в TSB:USA300 (MRSA), USA400 (MRSA), COL (MRSA), MRSA252 (MRSA), Wood46 (MSSA), Розенбах (MSSA), Ньюман (MSSA), и Mu50 (VISA). Было обнаружено, что анти-WTA бета-мАт, но не анти-WTA альфа-мАт, являются реактивными со всеми этими штаммами. Анализ связывания с различными штаммами показал, что WTA-бета более консервативен, чем WTA-альфа, и поэтому лучше подходит для ААС.
Пример 4 Характеристика антител со специфичностью к тейхоевым кислотам клеточной стенки
Подтверждение специфичности WTA к Ат
Препараты клеточной стенки (CWP) из дикого типа штамма S. aureus (ДТ) и мутантного штамма S. aureus у которого отсутствует WTA (ATagO; штамм WTA-ноль), были получены путем инкубирования 40 мг гранулированных штаммов S. aureus с 1 мл 10 мМ Трис-HCl (рН 7,4), дополненного 30% рафинозой, 100 мкг/мл лизостафина (Cell Sciences, Кантон, Массачусетс), и коктейлем ингибитора протеазы без ЭДТА (Roche, Плезатон, Калифорния), в течение 30 мин при 37°С. Лизаты центрифугировали при 11600 × g в течение 5 мин и собирали надосадки, содержащие компоненты клеточной стенки. Для иммуноблот-анализа белки разделяли на 4-12% трис-глициновый гель и переносили на нитроцеллюлозную мембрану (Invitrogen, Карлсбадт, Калифорния) с последующим блоттингом с указанными тест-антителами против WTA или с контрольными антителами против PGN и LTA.
Иммуноблотинг показывает, что антитела против WTA связываются с препаратами клеточной стенки из ДТ S. aureus, но не с препаратами клеточной стенки из штамма ATagO без WTA. Контрольные антитела против пептидогликана (анти-PGN) и липотейхоевой кислоты (анти-LTA) хорошо связываются с препаратами клеточной стенки. Эти данные указывают на специфичность тест-антител против WTA.
i) Проточная цитометрия для определения степени связывания мАт с поверхностью MRSA
Экспрессию поверхностных антигенов на целых бактериях из инфицированных тканей анализировали способом проточной цитометрией, используя следующий протокол. Для окрашивания антител бактериями из зараженных тканей мышей 6-8-недельным самкам мышей С57 В1/6 (Charles River, Вилмингтон, Массачусетс) внутривенно вводили с 108 КОЕ лог-фазы USA300 в PBS. Органы мыши собирали через два дня после заражения. Инфекционный эндокардит кроликов (ИЭ) был создан, как описано ранее у Tattevin P. et al. Antimicrobial agents and chemotherapy 54:610-613 (2010). Кроликам внутривенно вводили 5×107 КОЕ стационарной фазы MRSA штамма COL, а сердечную вегетацию собирали спустя восемнадцать часов. Лечение ванкомицином в дозе 30 мг/кг проводили внутривенно дважды в день. 18 ч после инфицирования с 7×107 КОЕ стационарной фазы.
Чтобы лизировать мышиные или кроличьи клетки, ткани гомогенизировали в пробирках М (Miltenyi, Аубурн, Калифорния) с использованием мягкого диссоциатора клеток MACS (Miltenyi) с последующей инкубацией в течение 10 мин при RT в PBS, содержащем 0,1% Тритон-Х100 (Thermo), 10 мкг/мл DNAsel (Roche) и коктейль ингибитора полной мини-протеазы (Roche). Суспензии пропускали через 40-микронный фильтр (BD) и промывали HBSS без фенольного красного, дополненного 0,1% свободным белком IgG (Sigma) и 10 мМ Hepes, рН 7,4 (буфер НВ). Затем бактериальные суспензии инкубировали с 300 мкг/мл кроличьего IgG (Sigma) в буфере НВ в течение 1 часа при комнатной температуре (RT) для блокирования неспецифического связывания IgG. Бактерии окрашивали с 2 мкг/мл первичных антител, включая rF1 или изотип контроля IgG1 мАт анти-герпес вируса gD:5237 (Nakamura GR et al., J Virol 67:6179-6191 (1993)), а затем с флуоресцентными антителами против человеческого IgG (Jackson Immunoresearch, Вест Гров, Пенсильвания). Для того чтобы дифференцировать бактерии от остатков органов мыши или кроликов, было выполнено двойное окрашивание с использованием 20 мкг/мл пептидогликана мышиного мАт 702 анти-5. aureus (Abeam, Кембридж, Массачусетс) и меченое флуорохромом анти-мышиное IgG вторичное антитело (Jackson Immunoresearch). Бактерии промывали и анализировали с помощью FACSCalibur (BD). Во время проточного цитометрического анализа, бактерии подвергали стробированию для положительного окрашивания с помощью мАт 702 из двух участков флуоресценции.
ii) Измерение аффинности связывания с S. aureus и плотности антигена на MRSA
В таблице 6 показан анализ равновесного связывания антител MRSA, связывающихся с штаммом Newman-ΔSPA, и плотности антигена на бактерии.
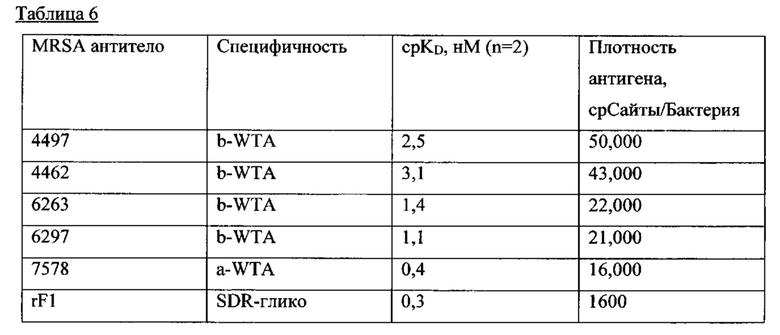
Kd и плотность антигена были получены с использованием анализа связывания радиолигандных клеток в следующих условиях анализа: DMEM + 2,5% мышиный связывающий сывороточный буфер; связывание раствора в течение 2 часов при комнатной температуре (RT); и использование 400000 бактерий/лунку.
Ат 6263 является 6078-подобным, так как последовательности очень похожи. За исключением второго остатка (R против G) в CDR Н3 все другие CDR последовательности L и Н цепи идентичны.
Пример 5 Аминокислотные модификации анти-WTA антител
Таким образом, участок VH каждого из aHTH-WTA-бета Ат клонировали и связывали с константным участком гамма-1 человеческой Н-цепи, a VL связывали с константным участком каппа для экспрессии Ат в качестве IgGl. Последовательности дикого типа были изменены в определенных положениях для улучшения стабильности антитела при сохранении связывания антигена, как описано ниже. Затем были получены сконструированные с цистеином Ат (ThioMabs, также упоминаемый как THIOMAB™).
i. Связывание вариабельных участков с константными участками
Участки VH WTA бета Ат, идентифицированные из вышеприведенной библиотеки антител человека, связывались с константными участками человека γ1, для получения полноразмерных IgG1 Ат.Цепи L были каппа L-цепями.
ii. Формирование вариантов стабильности
WTA Ат на Фиг. 12, (см., в частности, Фиг. 13А, 13В, 14А, 14В) были сконструированы для улучшения определенных свойств (чтобы избежать дезамидирования, изомеризации аспарагиновой кислоты, окисления или N-связанного гликозилирования) и протестированы на сохранение связывания антигена, а также химическую стабильность после замены аминокислот. Одноцепочечную ДНК клонов, кодирующую тяжелую или легкую цепь, очищали от фаговых частиц M13K07, выращенных в клетках Е. coli CJ236, с использованием набора QIAprep Spin М13 (Qiagen).
5'фосфорилированные синтетические олигонуклеотиды с последовательностями:
5'-CCCAGACTGCACCAGCTGGATCTCTGAATGTACTCCAGTTGC-3' (SEQ ID NO. 152)
5'-CCAGACTGCACCAGCTGCACCTCTGAATGTACTCCAGTTGC-3' (SEQ ID NO. 153) 5'СС AGGGTTCCCTGGCCCCAWTMGTCAAGTCCASCWKCACCTCTTGCACAGTAATAGACAGC-3' (SEQ ID NO. 154); и
5'-CCTGGCCCCAGTCGTCAAGTCCTCCTTCACCTCTTGCACAGTAATAGACAGC-3' (SEQ ID NO. 155) (коды ИЮПАК)
использовали для мутации клонов, кодирующих антитела, путем мутагенеза, направленного на сайт олигонуклеотида, как описано с помощью сайт-специфического мутагенеза, следуя методологии, описанной в Kunkel, Т.А. (1985). Rapid and efficient site-specific mutagenesis without phenotypic selection. Proceedings of the National Academy of Sciences USA 82(2):488-492. Мутантную ДНК использовали для трансформации клетки Е. coli XL1-Blue (Agilent Technologies) и высевали на чашки Luria Broth, содержащие 50 мкг/мл карбенициллина. Колонии собирали индивидуально и выращивали в жидкой среде Luria Broth, содержащей 50 мкг/мл карбенициллина. ДНК Miniprep секвенировали для подтверждения наличия мутаций.
Для Ат 6078 вторая аминокислота в VH, met (met-2), подвержена окислению. Поэтому met-2 мутировали в Ile или Val, чтобы избежать окисления остатка. Так как изменение met-2 может влиять на аффинность связывания, мутанты были протестированы на связывание со Staph CWP с помощью ИФА.
Было обнаружено, что мотивы CDR НЗ "DG" или "DD" подвержены трансформации в изо-аспарагиновую кислоту. Ат 4497 содержит DG в положениях 96 и 97 CDR Н3 (см. Фиг. 16) и было изменено для стабильности. CDR Н3, как правило, критичен для связывания антигена, поэтому несколько мутантов были протестированы на связывание с антигеном и химическую стабильность. Мутант D96E (v8) сохраняет связывание с антигеном, сходным с Ат 4497 дикого типа (Фиг. 16), и является стабильным и не образует изо-аспарагиновую кислоту.
Staph CWP ИФА
Для анализа 6078 мутантов антител, обработанный лизостафином препарат клеточной линии USA300 ΔSPA S. aureus (ДТ), состоящий из 1Х109 бактерий/мл, разбавляли 1/100 в 0,05 карбоната натрия с рН 9,6 и наносили на 384-луночные планшеты для ИФА (Nunc; Нептун, Нью-Джерси) во время инкубации в течение ночи при 4°С. Планшеты промывали PBS плюс 0,05% Твин-20 и блокировали во время 2-часовой инкубации с PBS плюс 0,5% бычьего сывороточного альбумина (BSA). Эту и все последующие инкубации проводили при комнатной температуре с легким перемешиванием. Пробы антител разбавляли в буфере для образца/стандартного разбавления (PBS, 0,5% BSA, 0,05% Твин 20, 0,25% CHAPS, 5 мМ ЭДТА, 0,35 М NaCl, 15 м.д. Proclin (рН 7,4)), добавляли к промытым планшетам и инкубировали в течение 1,5-2 часов. Границы чашки анти-S. aureus антител были обнаружены во время 1-часовой инкубации с конъюгированным пероксидазой фрагментом анти-человеческого IgG(Fc) F(ab')2 козла (Jackson ImmunoResearch; Вест Гров, Пенсильвания), разбавленным до 40 нг/мл в буфере для анализа (PBS, 0,5% BSA, 15 м.д. Proclin, 0,05% Твин 20). После окончательной промывки добавляли тетраметилбензидин (KPL, Гейтерсбург, Мэриленд), окрашивали в течение 5-10 минут и реакцию останавливали 1 М фосфорной кислотой. Планшеты считывали при 450 нм с эталонным значением 620 нм с использованием микропланшетного ридера.
iii. Создание мутантов с Cys-конструкцией (ThioMabs)
Полноразмерные ThioMab получали путем введения цистеина в Н-цепь (в СН1) или в L-цепь (Сκ) в заданном положении, как описано ранее и описано ниже, чтобы обеспечить конъюгацию антитела с промежуточным соединением линкер-антибиотик (Цистеиновые аминокислоты могут быть сконструированы в реакционноспособных сайтах в тяжелой цепи (НС) или легкой цепи (LC) антитела и не образуют внутрицепочечных или межмолекулярных дисульфидных связей) (Junutula, et al., 2008b Nature Biotech., 26(8):925-932; Dornan et al (2009) Blood 114(13):2721-2729; США 7521541; СЩА 7723485; WO 2011/156328; WO 2009/052249, Shen et al (2012) Nature Biotech., 30(2):184-191; Junutula et al (2008) Jour of Immun. Methods 332:41-52). Затем цепи H и L клонировали в отдельные плазмиды, а плазмиды, кодирующие Н и L, совместно трансфецировали в клетки 293, где они экспрессировались и собирались в интактные Ат. Как Н, так и L-цепи также могут быть клонированы в одну и ту же экспрессионную плазмиду. IgG1, имеющий 2 сконструированных Cys, по одному в каждой из Н-цепей; или 2 сконструированных Cys, по одному в каждой из цепочек L; или комбинацию сконструированного Cys в каждой из цепочек Н и L (HCLCCys), приводящую к 4 сконструированным Cys на тетрамер антитела, были получены путем экспрессии желаемой комбинации цепей cys мутанта и цепей дикого типа.
На Фиг. 13А и 13В показаны 6078 ДТ и мутантное Ат с комбинацией НС Cys и LC Cys. 6078 мутанты также тестировались на их способность связывать белок А с дефицитом US300 Staph А из ночной культуры. По результатам анализа FACS (данные не показаны) мутантное Ат связывало USA300 аналогично 6078 ДТ (неизмененным) антителам; изменения аминокислот у мутантов не влияли на связывание со Staph A. gD использовали в качестве неспецифического отрицательного контрольного антитела.
Пример 6: Выбор антибиотика
Антибиотики рифамицинового ряда были выбраны благодаря их высокому потенциалу, неизменной бактерицидной активности при низком фаголизосомальном рН, способности противостоять внутриклеточным влияниям и легкости, с которой они могут быть соединены с расщепляемым протеазой непептидным (PML) линкером, пригодным для конъюгации с анти-WTA антителом. Поскольку внутрифагоцитарные бактерии реплицировались наиболее медленно, для оптимизации антибиотика также требовалось, чтобы он был способен уничтожать нереплицирующийся MRSA при высвобождении из ААС.
MRSA собирали из стационарной фазы культуры и суспендировали в забуференном фосфатным буферном растворе, не содержащем антибиотик или 1×106 М рифампицина или рифамицинового производного (Рифалог) и инкубировали при 37°С. В указанные моменты времени образец культуры собирали и центрифугировали для удаления антибиотика, и общее число выживших бактерий определяли с помощью плакирования.
Интересно, что добавление антибиотика рифамицинового ряда (рифалог), но не рифампицина, привело к более чем 1000-кратному уменьшению числа жизнеспособных не реплицирующихся бактерий после инкубации в течение ночи в минимальном фосфатно-солевом буфере (PBS) (см. Фиг. 8). Аналогичным образом антибиотик рифамицинового ряда был способен уничтожать классически определенные устойчивые клетки, бактерии, которые входят в длительный период состояния покоя для выживание под действием антибиотиков (например, ципрофлоксацина) в растущих культурах (данные не показаны). Добавление рифампицина не влияло на их жизнеспособность, что согласуется с предыдущими наблюдениями (Cordon, В.Р., et al. (2013) Nature 503, 365-370). Напротив, добавление антибиотика рифамицинового ряда (рифалог) приводило к уничтожению устойчивых клеток до уровня ниже предела обнаружения. Эти результаты позволяют предположить, что антибиотики рифамицинового ряда обладают замечательной способностью убивать неактивные, не делящиеся клетки.
Пример 7: Получение анти-WTA антитело-антибиотик конъюгатов
Конъюгаты антитело-антибиотик против тейхоевой кислоты клеточной стенки (ААС) Таблицы 3 получали путем конъюгирования анти-WTA антитела с PML промежуточным соединением линкер-антибиотик, включая приведенные в Таблице 2. Перед конъюгацией анти-WTA антитела частично восстанавливали с помощью ТСЕР с использованием стандартных способов в соответствии с методикой, описанной в WO 2004/010957, рекомендации которого включены для справки в контексте данного документа. Частично восстановленные антитела конъюгировали с промежуточным соединением линкер-антибиотик с использованием стандартных способов в соответствии с методикой, описанной, например, в Doronina et al. (2003) Nat. Biotechnol. 21:778-784 и США 2005/0238649 A1. Вкратце, частично восстановленные антитела были объединены с промежуточным соединением линкер-антибиотик, чтобы обеспечить конъюгацию промежуточного соединения линкер-антибиотик с восстановленными остатками цистеина антитела. Реакции конъюгации гасили, и очищали ААС. Определяли нагрузку антибиотика (среднее количество антибиотических фрагментов на антитело) для каждого ААС, нагрузкасоставляла от около 1 до около 2 для антител против тейхоевой кислоты клеточной стенки, сконструированных с помощью единственного сайта цистеинового мутанта.
Восстановление/окисление ThioMab для конъюгации: Полноразмерные моноклональные антитела, сконструированные с использованием цистеина (ThioMabs - Junutula, et al., 2008b Nature Biotech., 26(8):925-932; Dornan et al (2009) Blood 114(13):2721-2729; США 7521541; США 7723485; WO 2009/052249, Shen et al (2012) Nature Biotech., 30(2):184-191; Junutula et al (2008) Jour of Immun. Methods 332:41-52), экспрессированные в клетках CHO, восстанавливали с около 20-40-кратным избытком ТСЕР (трис(2-карбоксиэтил)фосфина гидрохлорида или DTT (дитиотреитола) в 50 мМ Трис рН 7,5 с 2 мМ ЭДТА в течение 3 часов при 37°С или в течение ночи при комнатной температуре. (Getz et al (1999) Anal. Biochem. Vol 273:73-80; Soltec Ventures, Beverly, MA). Восстановленный ThioMab разводили и загружали в колонку HiTrap S в 10 мМ ацетата натрия, рН 5, и элюировали PBS, содержащим 0,3 М хлорида натрия. Альтернативно, антитело подкисляли добавлением 1/20 той объема 10% уксусной кислоты, разбавляли 10 мМ сукцинатом рН 5, загружали в колонку и затем промывали 10 объемами колонки янтарного буфера. Колонку элюировали 50 мМ Трис рН 7,5, 2 мМ ЭДТА.
Элюированный восстановленный ThioMab обрабатывали 15-кратным молярным избытком DHAA (дегидроаскорбиновой кислоты) или 200 нМ водного сульфата меди (CuSO4). Окисление межцепочечных дисульфидных связей завершалось примерно через три часа или более. Окисление окружающим воздухом также было эффективным. Повторно окисленное антитело диализуют в 20 мМ сукцината натрия рН 5, 150 мМ NaCl, 2 мМ ЭДТА и хранят замороженными при -20°С.
Конъюгация Thio-Mabs с промежуточным соединением линкер-антибиотик: Деблокированные, повторно окисленные тио-антитела (ThioMab) подвергали взаимодействию с 6-8-кратным молярным избытком промежуточного соединения линкер-антибиотик из Таблицы 2 (из стока ДМСО при концентрации 20 мМ) в 50 мМ Трис, рН 8, пока реакция была полной (16-24 часа), согласно анализу LC-MS реакционной смеси.
Неочищенные конъюгаты антитело-антибиотик (ААС) затем наносили на катионообменную колонку после разбавления 20 мМ сукцинатом натрия, рН 5. Колонку промывали по меньшей мере 10 объемами колонки 20 мМ сукцината натрия, рН 5, и антитело элюировали с PBS. ААС составляли в 20 мМ His/ацетат, рН 5, с 240 мМ сахарозы, используя гель-фильтрационные колонки. ААС характеризовали с помощью УФ-спектроскопии для определения концентрации белка, аналитической SEC (гель-фильтрационная хроматография) для агрегационного анализа и LC-MS до и после лечения с эндопептидазой лизина С.
Гель-фильтрационную хроматографию проводили с использованием колонки Shodex KW802.5 в 0,2 М фосфате калия с рН 6,2 с 0,25 мМ хлорида калия и 15% IPA со скоростью потока 0,75 мл/мин. Агрегационное состояние ААС определяли путем интегрирования поглощения элюированной пиковой области при 280 нм.
Анализ LC-MS проводили с использованием прибора Agilent QTOF 6520 ESI. В качестве примера, ААС, полученный с использованием этого реаганта, обрабатывали 1:500 мас./мас. эндопротеиназой Lys С (Promega) в Трис, рН 7,5, в течение 30 мин при 37°С. Полученные фрагменты расщепления загружали на 1000А, 8 мкм колонку PLRP-S, нагретую до 80°С, и элюировали с градиентом от 30% В до 40% В за 5 минут. Подвижная фаза А: H2O с 0,05% TFA. Подвижная фаза В: ацетонитрил с 0,04% TFA. Скорость потока: 0,5 мл/мин. Элюирование белка контролировали с помощью УФ-поглощения при 280 нм до электрораспылительной ионизации и МС-анализа. Обычно достигалось хроматографическое разрешение неконъюгированного Fc-фрагмента, остаточного неконъюгированного Fab и антибиотика-Fab. Полученные m/z спектры деконволировали с использованием программного обеспечения Mass Hunter ™ (Agilent Technologies) для вычисления массы фрагментов антитела.
Пример 8: Расщепление и вьщеление антибиотика
Антибиотики рифамицинового ряда тестировались на способность непосредственно уничтожать внеклеточные бактерии при их соединении с мАт анти-β WTA в формате ААС. ААС инкубировали в буфере отдельно или обрабатывали катепсином В. Серийные разведения полученной реакции добавляли в лунки, содержащие MRSA в триптическом соевом бульоне, и культивировали в течение ночи, чтобы идентифицировать лунки, содержащие достаточный активный антибиотик для предотвращения роста.
Как и было предсказано, растущие планктонные бактерии не пострадали от инкубации в течение ночи с интактным анти-MRSA ААС, если только ААС не был предварительно обработан катепсином В для высвобождения активного антибиотика (Фиг. 9). Предварительная обработка ААС катепсином В высвобождала достаточную антибиотическую активность для предотвращения роста бактерий при 0,6 мкг/мл ААС, которая, как прогнозируется, содержит 0,006 мкг/мл антибиотика.
Чтобы проверить, освобождается ли антибиотик из ААС только после интернализации ААС-опсонизированных бактерий в клетки, расщепление линкера можно исследовать зондом на основе флуоресцентного резонансного переноса энергии (FRET), состоящим из того же анти-MRSA антитела, конъюгированного с двумя молекулами красителя, используя тот же линкер, что и в ААС.MRSA опсонизировался конъюгатом FRET и добавлялся к культурам макрофагов. Поглощение бактерий и расщепление зонда контролируется с помощью видеомикроскопии. Линкер расщепляется в течение нескольких минут после поглощения бактерий макрофагами, которые визуализируются высвобождением зонда А488, аналогично высвобождению антибиотика типа рифамицина на ААС. С другой стороны, линкер оставался неповрежденным, когда бактерии не усваивались из-за обработки макрофагов латрункулином А, ингибитором фагоцитоза. Для подтверждения того, что свободный антибиотик действительно высвобождается внутри макрофагов после поглощения MRSA, покрытого действительными ААС также может быть использован масс-спектрометрический анализ.
Пример 9 Потенциал анти-WTAβ PML ААС in vitro
Анти-WTAβ-CBDK ААС эффективно уничтожает S. aureus при интернализации первичными человеческими и мышиными макрофагами и нескоторыми клеточными линиями человека in vitro. Анализ макрофагов in vitro.
S. aureus (штамм US A300 NRS384) инкубировали с различными дозами (100 мкг/мл, 10 мкг/мл, 1 мкг/мл или 0,1 мкг/мл) анти-WTAβ антитела 4497, At4497-CBDK-диметил pipBOR ААС нагруженному 2 молекулами антибиотика на антитело (DAR2) или с aHTH-WTA-CBDK-диметил pipBOR ААС нагруженному 4 молекулами антибиотика на антитело (DAR4) в течение 1 часа для обеспечения связывания антитела с бактериями. Полученные опсонизированные бактерии наносили на мышиные макрофаги и инкубировали при 37°С для обеспечения фагоцитоза. Через 2 часа инфекционную смесь удаляли и заменяли нормальной питательной средой, дополненной 50 мкг/мл гентамицина, для уничтожения любых оставшихся внеклеточных бактерий. Общее количество выживших внутриклеточных бактерий определяли через 2 дня путем нанесения серийных разведений лизатов макрофагов на триптические соевые агаровые пластины.
Пример 10 Эффективность in vivo анти-WTAβ-PML ААС
Лечение инфекции S. aureus у мышей снижает бактериальную нагрузку в органах на несколько порядков.
Чтобы определить, является ли терапевтическое направление специфическим для внутриклеточного S. aureus, и будет ли эффективностым во время инфекции, WTA-PML ААС были протестированы на модели внутривенной инфекции мыши. Этот пример демонстрирует, что WTA-PML ААС эффективны для значительного уменьшения или уничтожении внутриклеточных инфекций S. aureus, в модели внутривенной инфекции у мышей.
Модель перитонита. 7-недельных самок мышей А/J (Jackson Laboratories) заражали перитонеальной инъекцией с 5×107 КОЕ US A300. Мышей умерщвляли через 2 дня после заражения, а брюшину промывали 5 мл холодного фосфатного буферного раствора (PBS). Почки гомогенизировали в 5 мл PBS, как описано ниже для модели внутривенной инфекции. Перитонеальные промывки центрифугировали в течение 5 минут при 1000 об/мин при 4°С в настольной центрифуге. Надосадочную жидкость собирали как внеклеточные бактерии, а осадок клеток, содержащий перитонеальные клетки, собирали в качестве внутриклеточной фракции. Клетки обрабатывали 50 мкг/мл лизостафина в течение 20 минут при 37°С для уничтожения загрязняющих внеклеточных бактерий. Перитонеальные клетки промывали 3 раза в ледяном PBS для удаления лизостафина перед анализом. Чтобы посчитать количество внутриклеточных КОЕ, перитонеальные клетки лизировали в НВ (сбалансированный солевой раствор Хэнкса с добавкой 10 мМ HEPES и 0,1% альбумина бычьей сыворотки) с 0,1% Тритон-Х и серийные разведения лизата получали в PBS с 0,05% твин-20.
Модель внутривенной инфекции у мышей
Чтобы считаться клинически значимым, ААС должен быть в состоянии устранить уже установленную внутриклеточную инфекцию. Чтобы оценить это, лечение было отложено до 24 часов после начала бактериемии, время, когда лечение ванкомицином было минимально эффективным. Внутриклеточная инфекция быстро обосновывается в нейтрофилах; по крайней мере, в одной модели бактериемии 95% бактерий в крови попадают внутрь нейтрофилов в течение 15 минут7, что, по-видимому, объясняет снижение эффективности ванкомицина. В этих условиях лечение одной дозой ААС было эффективным и доказало превосходство над лечением эквивалентной дозой свободного антибиотика рифамицинового ряда.
S. aureus является обычным колонизатором кожи и слизистых оболочек человека. Предварительный анализ нескольких источников человеческой сыворотки, включая IGIV-GammaGard, препарат объединенного иммуноглобулина от ~ 10000 человек, показал, что человеческая сыворотка содержит приблизительно 300 мкг/мл анти-S. aureus антител, из которых ~ 70% направлены на модификации GlcNAc WTA. Сыворотка мыши не показывает детктируемых уровней анти-S1 aureus антитела. Чтобы определить, могут ли эндогенные анти-WTA антитела, обнаруженные в нормальной человеческой сыворотке, конкурировать за связывание с ААС, СВ17. SCID (Charles River Laboratories, Холлистер, Калифорния) мышей были реконстркированны с помощью иммуноглобулина GammaGard S/D IGIV (ASD Healthcare, Брукс, Кентукки) с использованием режима дозирования, оптимизированного для достижения постоянного уровня, по меньшей мере, 10 мг/мл человеческого IgG в сыворотке. IGIV вводили с начальной внутривенной дозой 30 мг на мышь, с последующей второй дозой 15 мг/мышь путем внутрибрюшинной (внутрибрюшинной) инъекции через 6 часов, и последующими ежедневными дозами 15 мг на мышь путем внутрибрюшинной инъекции в течение 3 последовательных дней. Такие мыши были одинаково восприимчивы к инфекции MRSA по сравнению с необработанными контролями.
Мышей (n = 8 для каждого из антител или ААС) инфицировали путем внутривенной инъекции через 4 часа после первой дозы IGIV с 1×107 КОЕ MRSA (штамм US A300 NRS384), разведенным в фосфатном буферном солевом растворе. Инфицированных мышей обрабатывали 50 мг/кг нативного антитела S4497 или S4497 ААС из таблицы 3. Мышам давали однократную дозу ААС в течение 24 часов после внутривенной инъекции, умерщвляли на 4 день после инфицирования, а почки и сердца собирали в 5 мл фосфатного буферного солевого раствора. Образцы тканей гомогенизировали с использованием GentleMACS Dissociator ™ (Miltenyi Biotec, Аубурн, Калифорния). Общее количество бактерий, полученных из органа, определяли путем нанесения серийных разведений гомогената ткани в PBS 0,05% Твин-20 на триптическом соевом агаре с 5% дефибринированной овечьей крови.
Как показывают результаты на Фиг. 10, несмотря на присутствие потенциально конкурирующих антител, одна доза анти-β-GlcNAc WTA ААС (S4497-AAC) значительно снижает или уничтожает бактериальные градации в инфицированных органах по сравнению с нативным антителом. Фиг. 10 В показывает, что обработка ААС (DAR2) уменьшала бактериальную нагрузку в почках примерно в 7000 раз. Фиг. 10С показывает, что обработка ААС (DAR2) из таблицы 3 уменьшала бактериальную нагрузку в сердце примерно в 500 раз. Лечение только нативным антибиотиком, диметил pipBOR (при равной молярной концентрации диметил pipBOR в ААС) в модели in vivo инфекции не было эффективным по сравнению с диметил pipBOR, конъюгированным с анти-WTA антителом в качестве ААС.
Неконъюгированные (свободные) анти-WTA антитела не эффективны in vivo На Фиг. 17 показано, что предварительная обработка 50 мг/кг свободным антителом неэффективна при модели внутривенной инфекции. Мышам Balb/c путем внутривенной инъекции давали однократную дозу контроля, растворителя (PBS), или 50 мг/кг антител за 30 минут до инфицирования 2×107 КОЕ USA300. Группы лечения включали контрольный изотип антитела, которое не связывается с S. aureus (gD), антитело, направленное против бета-модификации тейхоевой кислоты клеточной стенки (4497), или антитело, направленное против альфа-модификации тейхоевой кислоты клеточной стенки (7578). Контрольные мыши получали два раза в день лечение с использованием внутрибрюшинной инъекции (Ванко) 110 мг/кг ванкомицина.
Пример 11 Пиперидил бензоксазино рифамицин (pipBOR) 5
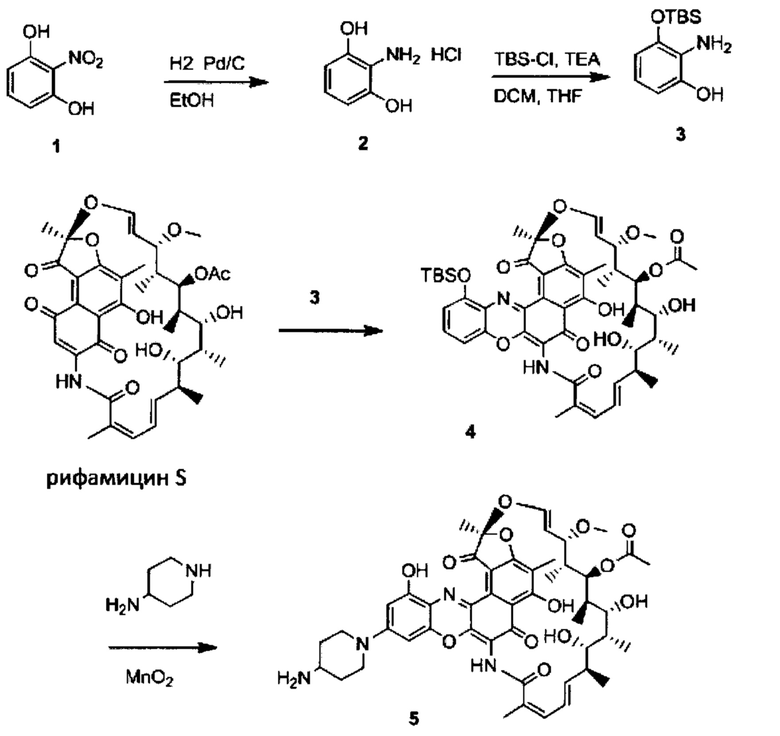
2-Нитробензол-1,3-диол 1 гидрировали в газообразном водороде с использованием катализатора палладий/углерод в этанольном растворителе с получением 2-аминобензол-1,3-диол 2, выделенном в виде гидрохлоридной соли. Монозащита 2 с трет-бутилдиметилсилилхлоридом и триэтиламином в дихлорметане/тетрагидрофуране дала 2-амино-3-(трет-бутилдиметилсилилокси)фенол 3. Рифамицин S (ChemShuttle Inc., Fremont, СА, США 7342011; США 7271165; США 7547692) подвергали взаимодействию с 3 путем окислительной конденсации с оксидом марганца или газообразным кислородом в толуоле при комнатной температуре с получением защищенного TBS бензоксазино-рифамицина 4. LCMS (ESI):M+H+ = 915,41. Реакция 4 с пиперидин-4-амином и оксидом марганца дала пиперидилбензоксазино рифамицин (pipBOR) 5. LCMS (ESI):M+H+ = 899,40
Пример 12 Диметил pipBOR 6
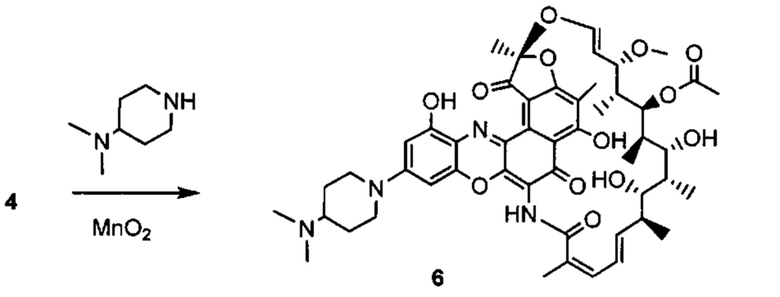
Реакция N,N-диметилпиперидин-4-амина с TBS-защищенным бензоксазино-рифамицином 4 дала диметилпиперидилбензоксазинорифамицин (диметил pipBOR) 6
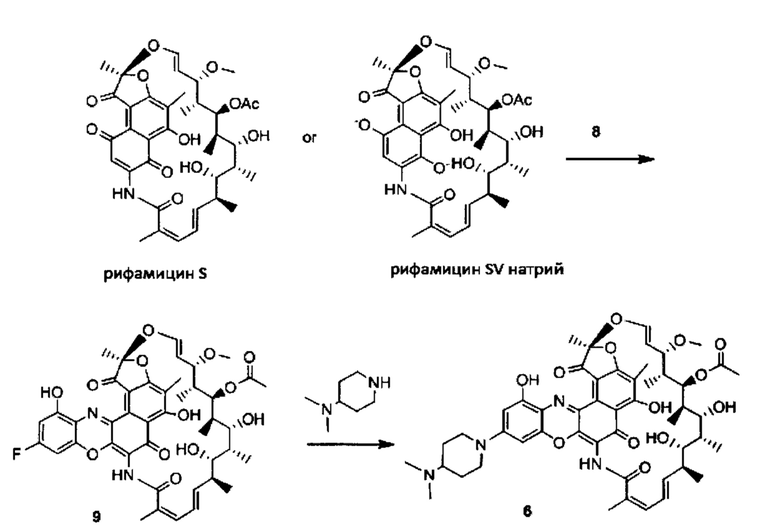
Альтернативно, (5-фтор-2-нитро-1,3-фенилен)бис(окси)бис(метилен) дибензол 7 гидрировали в газообразном водороде с использованием катализатора палладий/углерод в растворителе тетрагидрофуран/метанол до удаления бензильных групп, получая 2-амино-5-фторбензол-1,3-диол 8. LCMS (ESI):M+H+ = 144,04. Коммерчески доступный Рифамицин Били Рифамицин SV натрий (ChemShuttle Inc., Fremont, СА) подвергали взаимодействию с 2-амино-5-фторбензол-1,3-диолом 8 путем окислительной конденсации в воздухе или цианиде железа калия в этилацетате при 60°С с получением фторбензоксазино рифамицина 9. Смещение фтора с помощью N,N-диметилпиперидин-4-амина дало диметил pipBOR 6.LCMS (ESI):M+H+ = 927,43
Пример 13 (S)-N-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентил)-N-(1-(4-(гидроксиметил)фениламино)-1-оксо-5-уреидопентан-2-ил)циклобутан-1,1-дикарбоксамид 10
Шаг 1:Получение 1-(5-аминопентил)-1Н-пиррол-2,5-дион гидрохлорида 10а
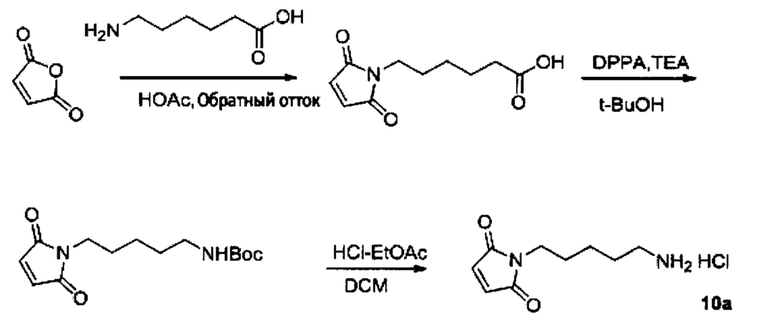
Малеиновый ангидрид, фуран-2,5-дион (150 г, 1,53 моль) добавляли к перемешиваемому раствору 6-аминогексановой кислоты (201 г, 1,53 моль) в НОАс (1000 мл). После перемешивания смеси при комнатной температуре в течение 2 ч, кипятили с обратным холодильником в течение 8 ч. Органические растворители удаляли при пониженном давлении и остаток экстрагировали с EtOAc (500 мл × 3), промывали Н2О. Объединенные органические слои сушат над Na2SO4 и концентрируют, получая неочищенный продукт. Его промывают петролейным эфиром с получением 6-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)гексановой кислоты в виде белог о твердого вещества (250 г, 77,4%). DPPA (130 г, 473 ммоль) и TEA (47,9 г, 473 ммоль) добавляли к раствору 6-(2,5-д иоксо-2,5-дигидро-1Н-пиррол-1-ил) гексановой кислоты (100 г, 473 ммоль) в t-BuOH (200 мл). Смесь кипятили с обратным холодильником в течение 8 ч под действием N2. Смесь концентрируют и остаток очищают колоночной хроматографией на силикагеле (РЕ:EtOAc = 3:1), получая трет-бутил 5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентилкарбамат (13 г, 10%). К раствору трет-бутил 5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентилкарбамата (28 г, 992 ммоль)в безводном EtOAc (30 мл) добавляли по каплям HCl/EtOAc (50 мл). После перемешивания смеси при комнатной температуре в течение 5 ч, фильтруют и твердое вещество сушат, получая 1-(5-α минопентил)-1Н-пиррол-2,5-дион гидрохлорид 10а (16 г, 73,7%). 1H NMR (400 МГц, ДМСО-d6): δ 8,02 (с, 2Н), 6,99 (с, 2Н), 3,37-3,34 (м, 2Н), 2,71-2,64 (м, 2Н), 1,56-1,43 (м, 4Н), 1,23-1,20 (м, 2Н).
Шаг 2: Получение (S)-1-(1-(4-(гидроксиметил)фениламино)-1-оксо-5-уреидопентан-2-илкарбамоил)циклобутанкарбоновой кислоты 10b

К смеси (S)-2-амино-5-уреидопентановой кислоты 10 г (17,50 г, 0,10 моль) в смеси диоксана и H2O (50 мл/75 мл) добавляли K2CO3 (34,55 г, 0,25 моль). Fmoc-Cl (30,96 г, 0,12 моль) медленно добавляли при 0°С. Реакционную смесь нагревали до комнатной температуры более 2 ч. Органический растворитель удаляли при пониженном давлении и водную суспензию доводили до рН=3 с помощью 6 М раствором HCl и экстрагировали с EtOAc (100 мл × 3). Органический слой сушат над Na2SO4, фильтровали и концентрировали при пониженном давлении с получением (S)-2-((((9Н-флуорен-9-ил)метокси)карбонил)амино)-5-уреидопентановую кислоты 10f (38,0 г, 95,6%). 10f является коммерчески доступным.
К раствору 10f (4 г, 10 ммоль) в смеси DCM и МеОН (100 мл / 50 мл) добавляли (4-аминофенил)метанол (1,6 г, 13 ммоль, 1,3 экв.) и 2-этокси-1-этоксикарбонил-1,2-дигидрохинолина, EEDQ, Sigma-Aldrich CAS Per. №16357-59-8 (3,2 г, 13 ммоль, 1,3 экв.). После перемешивания смеси при комнатной температуре в течение 16 ч при N2, его концентрировали, получая коричневое твердое вещество. Добавляли МТВЕ (200 мл) и перемешивали при 15°С в течение 2 часов. Твердое вещество собирали фильтрованием, промывали МТВЕ (50 мл × 2) и получали (S)-(9Н-флуорен-9-ил)метил(1-((4-(гидроксиметил)фенил)амино)-1-оксо-5-уреидопентан-2-ил)карбамат 10е в виде оранжевого твердого вещества (4,2 г, 84%). LCMS (ESI): m/z 503,0 [М+1].
К перемешиваемому раствору 10е (4,2 г, 8,3 ммоль) в сухом DMF (20 мл) добавляли пиперидин (1,65 мл, 17 ммоль, 2 экв.) по капле при комнатной температуре. Смесь перемешивали при комнатной температуре в течении 30 мин, до образования твердого осадока. Добавляли сухой DCM (50 мл), и смесь сразу становилась прозрачной. Смесь перемешивали при комнатной температуре в течении 30 мин, и LCMS показывал, что 10е был израсходован. Ее концентрировали досуха при пониженном давлении (чтобы не оставался пиперидин), и остаток распределяли между EtOAc и Н2О (50 мл/20 мл). Водную фазу промывали с EtOAc (50 мл × 2) и концентрировали с получением (S)-2-амино-N-(4-(гидроксиметил) фенил)-5-уреидопентанамида 10d в виде маслянистого остатка (2,2 г, 94%) (содержащего небольшое количество DMF).
Коммерчески доступная 1,1-циклобутандикарбоновая кислота, 1,1-диэтиловый эфир (CAS Рег. №3779-29-1) превращали путем ограниченного омыления водным основанием в половину кислоты/сложного эфира 1,1-циклобутандикарбоновой кислоты, и 1-этиловый эфир (CAS Рег. №54450-84-9) и активацию с реагентом сочетания, таким как TBTU (O-(Бензотриазол-1-ил)-N,N,N',N'-тетраметилурония тетрафторборат, называемый также: N,N,N',N'-Тетраметил-O-(бензотриазол-1-ил)уронийтетрафторборат, CAS № 125700-67-6, Sigma-Aldrich B-2903) и N-гидроксисукцинимид с эфиром NHS, 1-(2,5-диоксопирролидин-1-ил)1-этилциклобутан-1,1-дикарбоксилат.
К раствору 1-(2,5-диоксопирролидин-1-ил)-1-этилциклобутан-1,1-дикарбоксилата (8 г, 29,7 ммоль) в DME (50 мл) добавляли раствор 10d (6,0 г, 21,4 ммоль) и NaHCO3 (7,48 г, 89,0 ммоль)в воде (30 мл). После перемешивания смеси при комнатной температуре в течение 16 ч, его концентрировали досуха пр и пониженном давлении и остаток очищали к олоночной хроматографией (DCM:MeOH = 10:1) с получением (S)-этил-1-((1-(4-(гидроксиметил)фенил)-2-оксо-6-уреидогексан-3 ил) карбамоил)циклобутан карбоксилат 10с в виде белого твердого вещества (6,4 г, 68,7%). LCMS (ESI): m/z 435.0 [М+1]
К перемешиваемому раствору 10 с (6,4 г, 14,7 ммоль) в смеси THF и МеОН (20 мл/10 мл) добавляли раствор LiOH⋅H2O (1,2 г, 28,6 ммоль) в H2O (20 мл) при комнатной температуре. После перемешивания смеси при комнатной температуре в течение 16 ч, растворитель удаляли при пониженном давлении, полученный остаток очищали препаративной ВЭЖХ с получением (S)-1-(1-(4-(гидроксиметил)фениламино)-1-оксо-5-уреидопентан-2-илкарбамоил) циклобутанкарбоновой кислоты 10b (3,5 г, выход: 58,5%). LCMS (ESI): m/z 406,9 [М+1]. 1Н NMR (400 МГц, метанол-d4) δ 8,86 (d, J=8,4 Гц, 2 Н), 8,51 (d, J=8,4 Гц, 2 Н), 5,88-5,85 (м, 1 Н), 5,78 (с, 2 Н), 4,54-4,49 (м, 3 Н), 4,38-4,32 (м, 1 Н), 3,86-3,75 (м, 1 Н), 3,84-3,80 (м, 2 Н), 3,28-3,21 (м, 1 Н), 3,30-3,24 (м, 1 Н), 3,00-2,80 (м, 1 Н), 2,37-2,28 (м, 2 Н).
Шаг 3: Получение S)-N-(5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентил)-N-(1-(4-(гидроксиметил)фениламино)-1-оксо-5-уреидопентан-2-ил)циклобутан-1,1-дикарбоксамид 10

Диизопропилэтиламин, DIPEA (1,59 г, 12,3 ммоль) и бис(2-оксо-3-оксазолидинил)фосфиновый хлорид, ВОР-Cl (CAS Рег. №68641-49-6, Sigma-Aldrich, 692 мг, 2,71 ммоль) добавляли к раствору (S)-1-(1-(4-(гидроксиметил)фениламино)-1-оксо-5-уреидопентан-2-илкарбамоил)циклобутанкарбоновой кислоты 10b (1 г, 2,46 ммоль) в DMF (10 мл) при 0°С, с последующим 1-(5-аминопентил)-1Н-пиррол-2,5-дион гидрохлорид 10а (592 мг, 2,71 ммоль).Смесь перемешивают при 0°С в течение 0,5 часа. Реакционную смесь гасили раствором лимонной кислоты (10 мл), экстрагировали DCM/MeOH (10:1). Органический слой сушат и концентрируют, а остаток очищают колоночной хроматографией на силикагеле (DCM:MeOH = 10:1) с получением S)-N-(5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентил)-N-(1-(4-(гидроксиметил)фениламино)-1-оксо-5-уреидопентан-2-ил)циклобутан-1,1-дикарбоксамид 10 (1,0 г, 71%), также называемый MC-CBDK-cit-PAB-OH. LCMS (ESI):M+H+ = 571,28. 1Н NMR (400 МГц, ДМСО-d6): δ 10,00 (с, 1Н), 7,82-7,77 (м, 2Н), 7,53 (d, J=8,4 Гц, 2 Н), 7,19 (d, J=8,4 Гц, 2 Н), 6,96 (с, 2Н), 5,95 (t, J=6,4 Гц, 1Н), 5,39 (с, 2Н), 5,08 (t, J=5,6 Гц, 1H), 4,40-4,35 (м, 3Н), 4,09 (d, J=4,8 Гц, 1 Н), 3,01 (d, J=3,2 Гц, 2 Н), 3,05-2,72 (м, 4Н), 2,68-2,58 (м, 3Н), 2,40-2,36 (м, 4Н), 1,72-1,70 (м, 3Н), 1,44-1,42 (м, 1Н), 1,40-1,23 (м, 6Н), 1,21-1,16 (м, 4Н).
Пример 14 (S)-N-(1-(4-(хлорметил)фениламино)-1-оксо-5-уреидопентан-2-ил)-N-(5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентил) циклобутан-1,1-дикарбоксамид 11
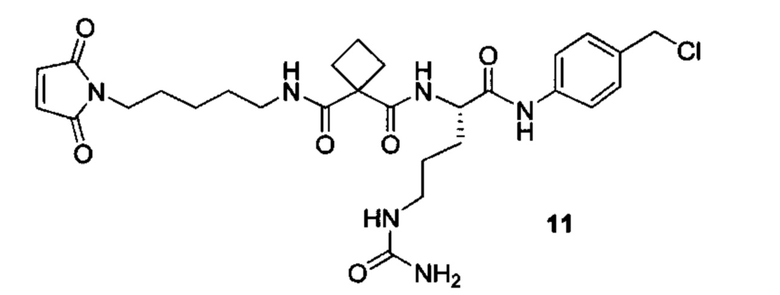
Раствор (S)-N-(5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентил)-N-(1-(4-(гидроксиметил)фениламино)-1-оксо-5-уреидопентан-2-ил)циклобутан-1,1-дикарбоксамид 10 (2,0 г, 3,5 ммоль) в N,N-диметилформамиде, DMF или N-метилпирролидоне, NMP (50 мл) обрабатывали тионилхлоридом, SOCl2 (1,25 г, 10,5 ммоль) порциями по каплям при 0°С. Реакция оставалась желтой. Реакцию контролировали с помощью LC/MS с показателем конверсии > 90%. После того, как реакционну ю смесь перемешивали при 20°С в течение 30 мин или несколько часов, ее разбавляли водой (50 мл) и экстрагировали с EtOAc (50 мл ×3). Органич еский слой сушили, концентрировали и очищали флеш-колонкой (DCM:MeOH = 20:1) до образования 11, также называемой MC-CBDK-cit-PAB-Cl как серое твердое вещество. LCMS:(5-95, АВ, 1,5 мин), 0,696 мин, m/z=589,0 [М+1]+.
Пример 15 (S)-4-(2-(1-(5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентилкарбамоил)циклобутанкарбоксамидо)-5-уреидопентанамидо)бензил-4-нитрофенилкарбонат 12
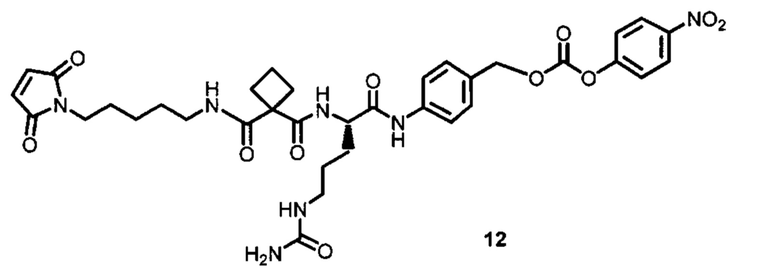
К раствору (S)-N-(5-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пентил)-N-(1-(4-(гидроксиметил)фениламино)-1-оксо-5-уреидопентан-2-ил)циклобутан-1,1-дикарбоксамида 10 в безводном DMF добавляют диизопропилэтиламин (DIEA), затем следует PNP карбонат (бис(4-нитрофенил)карбонат). Реакцио нный раствор перемешивали при комнатной т емпературе (комн. т.) в течение 4 часов и смесь очищали препаративной ВЭЖХ с получением 12. LCMS (ESI):M+H+ = 736,29.
Пример 16 Получение МС-(CBDK-cit)-РАВ-(диметил, фтор pipBOR) - PLA-1
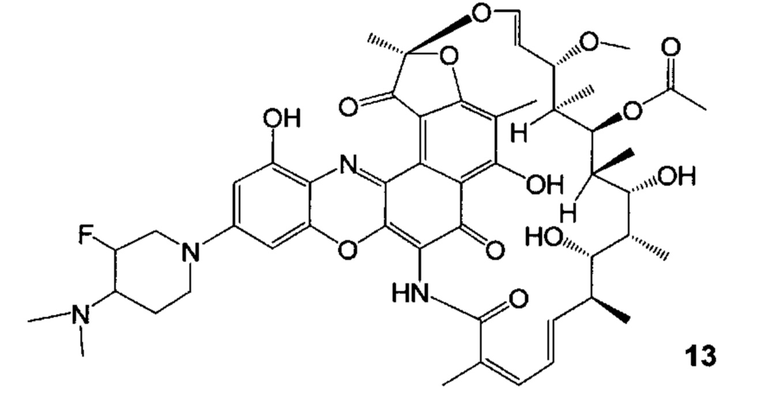
Следуя методике для PLA-2, (S)-N-(1-(4-(хлорметил)фениламино)-1-оксо-5-уреидопентан-2-ил)-N-(5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентил)циклобутан-1,1-дикарбоксамид 11 и фторированное производное рифамицина, диметилфтор pipBOR 13 (LCMS (ESI): М+Н+ = 945,43) подвергали взаимодействию с образованием MC-(CBDK-cit)-РАВ-(диметил, фтор pipBOR) - PLA-1, Таблица 2. LCMS (ESI):М+Н+ = 1499,7
Пример 17 Получение МС-(CBDK-cit)-РАВ-(диметил pipBOR) - PLA-2
(S)-N-(1-(4-(хлорметил)фениламино)-1-оксо-5-уреидопентан-2-ил)-N-(5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентил)циклобутан-1,1-дикарбоксамид 11 (0,035 ммоль) в DMF охлаждали до 0°С и добавляли диметил pipBOR 6, (10 мг, 0,011 ммоль). Смесь разбавляли еще 0,5 мл DMF. Перемешивали на открытом воздухе в течение 30 минут. Добавляли N,N-диизопропилэтиламин (DIEA, 10 мкл, 0,05 ммоль) и реакционную смесь перемешивали в течение ночи на открытом воздухе. С помощью LC/MS наблюдали 50% желаемого продукта. Добавляли дополнительное 0,2 экв. N,N-диизопропилэтиламинного основания, в то время как реакционную смесь перемешивали на открытом воздухе в течение еще 6 часов до тех пор, пока реакция не перестала прогрессировать. Реакционную смесь разбавляли с DMF и очищали на ВЭЖХ (20-60% ACN/HCOOH в⋅H2O), чтобы получить МС-(CBDK-cit)-РАВ-(диметил pipBOR) - PLA-2, Таблица 2. LCMS (ESI):M+H+ = 1481,8, выход 31%.
Пример 18 Получение МС-((R)-тиофен-3-ил-CBDK-cit)-РАВ-(диметил pipBOR) (PLA-3)
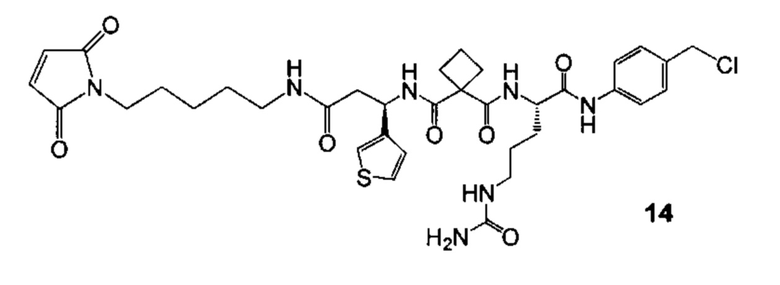
Следуя методике для PLA-2, (N-((S)-1-(4-(хлорметил)фениламино)-1-оксо-5-уреидопентан-2-ил)-N-((R)-3-(5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентиламино)-3-оксо-1-(тиофен-3-ил)пропил)циклобутан-1,1-дикарбоксамид 14 (LCMS (ESI): М+Н+ = 742,3) и диметил pipBOR 6 подвергали взаимодействию с получением МС-((R)-тиофен-3-ил-CBDK-cit)-РАВ-(диметил pipBOR) (PLA-3, Таблица 2). LCMS (ESI):M+H+ = 1633,9
Пример 19 Получение МС-((S)-тиофен-3-ил-CBDK-cit)-РАВ-(диметил pipBOR) (PLA-4)
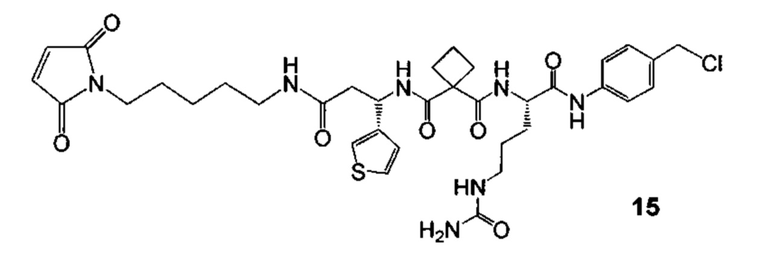
Следуя методике для PLA-2, (N-((R)-1-(4-(хлорметил)фениламино)-1-оксо-5-уреидопентан-2-ил)-N-((R)-3-(5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентиламино)-3-оксо-1-(тиофен-3-ил)пропил)циклобутан-1,1-дикарбоксамид 15 (LCMS (ESI): М+Н+ = 742,3) и диметил pipBOR 6 подвергали взаимодействию с получением МС-((R)-тиофен-3-ил-CBDK-cit)-РАВ-(диметил pipBOR) (PLA-4, Таблица 2). LCMS (ESI):M+H+ = 1633,9
Пример 20 Получение MC-(CBDK-cit)-PABC-(pipBOR) (PLA-5)
Пиперидил бензоксазино рифамицин (pipBOR) 5 (15 мг, 0,0167 ммоль) и затем (S)-4-(2-(1-(5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентилкарбамоил)циклобутанкарбоксамидо)-5-уреидопентанамидо)бензил-4-нитрофенилкарбонат 12 (12 мг, 0,0167 ммоль) взвешивали во флаконе. Добавляли диметилформамид, DMF (0,3 мл), затем диизопропилэтиламин, DIEA (0,006 мл, 0,0334 ммоль) и реакционную смесь перемешивали при комнатной температуре в течение 2 часов. Реакционный раствор непосредственно очищают с помощью ВЭЖХ (от 30 до 70% MeCN/вода + 1% муравьиная кислота) с получением МС - (CBDK-cit)-PABC-(pipBOR) (PLA-5, Таблица 2). LCMS (ESI):M+H+ = 1496,5
Пример 21 Получение MC-(CBDK-cit)-PABC-(piperazBTR) (PLA-6)
Следуя процедурам PLA-5, производное пиперидинового рифамицина, piperazBOR 16 (LCMS (ESI):M+H+ = 885,4) и (S)-4-(2-(1-(5-(2,5-диоксо-2,5-дигидро-1Н-пиррол-1-ил)пентилкарбамоил)циклобутанкарбоксамидо)-5-уреидопентанамидо)бензил-4-нитрофенилкарбонат 12 подвергали взаимодействию с получением MC-(CBDK-cit)-PABC-(piperazBTR) (PLA-6. Таблица 2). LCMS (ESI):M+H+ = 1482,5
Хотя для ясности понимания некоторые детали настоящего исследования описаны посредством иллюстраций и примеров, не следует считать, что описание и примеры ограничивают рамки настоящего изобретения. Все патенты, патентные заявки, и ссылки, цитируемые в описании, включены посредством ссылки в полном объеме.
--->
Перечень последовательностей
<110> ДЖЕНЕНТЕК, ИНК.p
<120> КОНЪЮГАТЫ АНТИТЕЛА К STAPHYLOCOCCUS AUREUS С РИФАМИЦИНОМ И ИХ ПРИМЕНЕНИЕ
<130> P32433-WO
<140>
<141>
<150> 62/087184
<151> 2014-12-03
<160> 180
<170> PatentIn версия 3.5
<210> 1
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид
peptide"
<400> 1
Lys Ser Ser Gln Ser Val Leu Ser Arg Ala Asn Asn Asn Tyr Tyr Val
1 5 10 15
Ala
<210> 2
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 2
Trp Ala Ser Thr Arg Glu Phe
1 5
<210> 3
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 3
Gln Gln Tyr Tyr Thr Ser Arg Arg Thr
1 5
<210> 4
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 4
Asp Tyr Tyr Met His
1 5
<210> 5
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 5
Trp Ile Asn Pro Lys Ser Gly Gly Thr Asn Tyr Ala Gln Arg Phe Gln
1 5 10 15
Gly
<210> 6
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 6
Asp Cys Gly Ser Gly Gly Leu Arg Asp Phe
1 5 10
<210> 7
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 7
Arg Ser Asn Gln Asn Leu Leu Ser Ser Ser Asn Asn Asn Tyr Leu Ala
1 5 10 15
<210> 8
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 8
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 9
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 9
Gln Gln Tyr Tyr Ala Asn Pro Arg Thr
1 5
<210> 10
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 10
Asp Tyr Tyr Ile His
1 5
<210> 11
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 11
Trp Ile Asn Pro Asn Thr Gly Gly Thr Tyr Tyr Ala Gln Lys Phe Arg
1 5 10 15
Asp
<210> 12
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 12
Asp Cys Gly Arg Gly Gly Leu Arg Asp Ile
1 5 10
<210> 13
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 13
Lys Ser Asn Gln Asn Val Leu Ala Ser Ser Asn Asp Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 14
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 14
Trp Ala Ser Ile Arg Glu Ser
1 5
<210> 15
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 15
Gln Gln Tyr Tyr Thr Asn Pro Arg Thr
1 5
<210> 16
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 16
Asp Tyr Tyr Ile His
1 5
<210> 17
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 17
Trp Ile Asn Pro Asn Thr Gly Gly Thr Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 18
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 18
Asp Cys Gly Asn Ala Gly Leu Arg Asp Ile
1 5 10
<210> 19
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 19
Lys Ser Ser Gln Asn Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 20
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 20
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 21
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 21
Gln Gln Tyr Tyr Thr Ser Pro Pro Tyr Thr
1 5 10
<210> 22
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 22
Ser Tyr Trp Ile Gly
1 5
<210> 23
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 23
Ile Ile His Pro Gly Asp Ser Lys Thr Arg Tyr Ser Pro Ser Phe Gln
1 5 10 15
Gly
<210> 24
<211> 29
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 24
Leu Tyr Cys Ser Gly Gly Ser Cys Tyr Ser Asp Arg Ala Phe Ser Ser
1 5 10 15
Leu Gly Ala Gly Gly Tyr Tyr Tyr Tyr Gly Met Gly Val
20 25
<210> 25
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 25
Asp Ile Gln Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Ser Arg
20 25 30
Ala Asn Asn Asn Tyr Tyr Val Ala Trp Tyr Gln His Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Phe Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Thr Ser Arg Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
100 105 110
Lys
<210> 26
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 26
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Arg Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Lys Ser Gly Gly Thr Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Gly Asp Thr Ser Ile Ser Ala Ala Tyr
65 70 75 80
Met Asp Leu Ala Ser Leu Thr Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Lys Asp Cys Gly Ser Gly Gly Leu Arg Asp Phe Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 27
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 27
Asp Ile Gln Met Thr Gln Ser Pro Asp Ser Leu Ser Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Arg Ser Asn Gln Asn Leu Leu Ser Ser
20 25 30
Ser Asn Asn Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro
35 40 45
Leu Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Tyr
85 90 95
Tyr Ala Asn Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 28
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 28
Gln Val Gln Leu Gln Gln Ser Arg Val Glu Val Lys Arg Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Thr Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile His Trp Val Arg Leu Ala Pro Gly Gln Gly Leu Glu Leu Met
35 40 45
Gly Trp Ile Asn Pro Asn Thr Gly Gly Thr Tyr Tyr Ala Gln Lys Phe
50 55 60
Arg Asp Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ala Thr Ala Tyr
65 70 75 80
Leu Glu Met Ser Ser Leu Thr Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Cys Gly Arg Gly Gly Leu Arg Asp Ile Trp Gly Pro Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 29
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 29
Glu Ile Val Leu Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Asn Gln Asn Val Leu Ala Ser
20 25 30
Ser Asn Asp Lys Asn Tyr Leu Ala Trp Phe Gln His Lys Pro Gly Gln
35 40 45
Pro Leu Lys Leu Leu Ile Tyr Trp Ala Ser Ile Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Arg Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Thr Asn Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Phe
100 105 110
Asn
<210> 30
<211> 119
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 30
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Ile His Trp Val Arg Leu Ala Pro Gly Gln Gly Leu Glu Leu Met
35 40 45
Gly Trp Ile Asn Pro Asn Thr Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ala Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Cys Gly Asn Ala Gly Leu Arg Asp Ile Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 31
<211> 114
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 31
Asp Ile Gln Leu Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Asn Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Thr Ser Pro Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu
100 105 110
Ile Glu
<210> 32
<211> 138
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 32
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile His Pro Gly Asp Ser Lys Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Asn Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Leu Tyr Cys Ser Gly Gly Ser Cys Tyr Ser Asp Arg Ala Phe
100 105 110
Ser Ser Leu Gly Ala Gly Gly Tyr Tyr Tyr Tyr Gly Met Gly Val Trp
115 120 125
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
130 135
<210> 33
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 33
Arg Ala Ser Gln Thr Ile Ser Gly Trp Leu Ala
1 5 10
<210> 34
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 34
Lys Ala Ser Thr Leu Glu Ser
1 5
<210> 35
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 35
Gln Gln Tyr Lys Ser Tyr Ser Phe Asn
1 5
<210> 36
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 36
Ser Tyr Asp Ile Asn
1 5
<210> 37
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 37
Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 38
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 38
Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp Leu
1 5 10 15
<210> 39
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 39
Arg Ala Ser Gln Thr Ile Ser Gly Trp Leu Ala
1 5 10
<210> 40
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 40
Lys Ala Ser Thr Leu Glu Ser
1 5
<210> 41
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 41
Gln Gln Tyr Lys Ser Tyr Ser Phe Asn
1 5
<210> 42
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 42
Ser Tyr Asp Ile Asn
1 5
<210> 43
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 43
Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 44
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 44
Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp Leu
1 5 10 15
<210> 45
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 45
Arg Ala Ser Gln Phe Val Ser Arg Thr Ser Leu Ala
1 5 10
<210> 46
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 46
Glu Thr Ser Ser Arg Ala Thr
1 5
<210> 47
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 47
His Lys Tyr Gly Ser Gly Pro Arg Thr
1 5
<210> 48
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 48
Asn Tyr Asp Phe Ile
1 5
<210> 49
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 49
Trp Met Asn Pro Asn Ser Tyr Asn Thr Gly Tyr Gly Gln Lys Phe Gln
1 5 10 15
Gly
<210> 50
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 50
Ala Val Arg Gly Gln Leu Leu Ser Glu Tyr
1 5 10
<210> 51
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 51
Arg Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala
1 5 10
<210> 52
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 52
Asp Ala Ser Ser Arg Ala Thr
1 5
<210> 53
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 53
Gln Lys Tyr Gly Ser Thr Pro Arg Pro
1 5
<210> 54
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 54
Ser Tyr Asp Ile Asn
1 5
<210> 55
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 55
Trp Met Asn Pro Asn Ser Gly Asn Thr Asn Tyr Ala Gln Arg Phe Gln
1 5 10 15
Gly
<210> 56
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 56
Glu Arg Trp Ser Lys Asp Thr Gly His Tyr Tyr Tyr Tyr Gly Met Asp
1 5 10 15
Val
<210> 57
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 57
Arg Ala Ser Leu Asp Ile Thr Asn His Leu Ala
1 5 10
<210> 58
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 58
Glu Ala Ser Ile Leu Gln Ser
1 5
<210> 59
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 59
Glu Lys Cys Asn Ser Thr Pro Arg Thr
1 5
<210> 60
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 60
Asn Tyr Asp Ile Asn
1 5
<210> 61
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 61
Trp Met Asn Pro Ser Ser Gly Arg Thr Gly Tyr Ala Pro Lys Phe Arg
1 5 10 15
Gly
<210> 62
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 62
Gly Gly Gly Tyr Tyr Asp Ser Ser Gly Asn Tyr His Ile Ser Gly Leu
1 5 10 15
Asp Val
<210> 63
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 63
Arg Ala Ser Gln Ser Val Gly Ala Ile Tyr Leu Ala
1 5 10
<210> 64
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 64
Gly Val Ser Asn Arg Ala Thr
1 5
<210> 65
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 65
Gln Leu Tyr Thr Ser Ser Arg Ala Leu Thr
1 5 10
<210> 66
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 66
Ala Tyr Ala Met Asn
1 5
<210> 67
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 67
Ser Ile Thr Lys Asn Ser Asp Ser Leu Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 68
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 68
Leu Ala Ala Arg Ile Met Ala Thr Asp Tyr
1 5 10
<210> 69
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 69
Arg Ala Ser Gln Gly Ile Arg Asn Gly Leu Gly
1 5 10
<210> 70
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 70
Pro Ala Ser Thr Leu Glu Ser
1 5
<210> 71
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 71
Leu Gln Asp His Asn Tyr Pro Pro Thr
1 5
<210> 72
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 72
Tyr Tyr Ser Met Ile
1 5
<210> 73
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 73
Ser Ile Asp Ser Ser Ser Arg Tyr Leu Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 74
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 74
Asp Gly Asp Asp Ile Leu Ser Val Tyr Arg Gly Ser Gly Arg Pro Phe
1 5 10 15
Asp Tyr
<210> 75
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 75
Arg Ala Ser Gln Gly Ile Arg Asn Gly Leu Gly
1 5 10
<210> 76
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 76
Pro Ala Ser Thr Leu Glu Ser
1 5
<210> 77
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 77
Leu Gln Asp His Asn Tyr Pro Pro Ser
1 5
<210> 78
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 78
Tyr Tyr Ser Met Ile
1 5
<210> 79
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 79
Ser Ile Asp Ser Ser Ser Arg Tyr Arg Tyr Tyr Thr Asp Ser Val Lys
1 5 10 15
Gly
<210> 80
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 80
Asp Gly Asp Asp Ile Leu Ser Val Tyr Gln Gly Ser Gly Arg Pro Phe
1 5 10 15
Asp Tyr
<210> 81
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 81
Arg Ala Ser Gln Ser Val Arg Thr Asn Val Ala
1 5 10
<210> 82
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 82
Gly Ala Ser Thr Arg Ala Ser
1 5
<210> 83
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 83
Leu Gln Tyr Asn Thr Trp Pro Arg Thr
1 5
<210> 84
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 84
Thr Asn Asp Met Ser
1 5
<210> 85
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 85
Thr Ile Ile Gly Ile Asp Asp Thr Thr His Tyr Ala Asp Ser Val Arg
1 5 10 15
Gly
<210> 86
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 86
Asn Ser Gly Ile Tyr Ser Phe
1 5
<210> 87
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 87
Arg Ala Ser Gln Asp Ile Gly Ser Ser Leu Ala
1 5 10
<210> 88
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 88
Ala Thr Ser Thr Leu Gln Ser
1 5
<210> 89
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 89
Gln Gln Leu Asn Asn Tyr Val His Ser
1 5
<210> 90
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 90
Asp Tyr Ala Met Gly
1 5
<210> 91
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 91
Val Val Thr Gly His Ser Tyr Arg Thr His Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 92
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 92
Arg Ile Trp Ser Tyr Gly Asp Asp Ser Phe Asp Val
1 5 10
<210> 93
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 93
Arg Ala Ser Gln Ser Ile Gly Asp Arg Leu Ala
1 5 10
<210> 94
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 94
Trp Ala Ser Asn Leu Glu Gly
1 5
<210> 95
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 95
Gln Gln Tyr Lys Ser Gln Trp Ser
1 5
<210> 96
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 96
Ser Tyr Ala Met Asn
1 5
<210> 97
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 97
Tyr Ile Ser Ser Ile Glu Thr Ile Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 98
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 98
Asp Arg Leu Val Asp Val Pro Leu Ser Ser Pro Asn Ser
1 5 10
<210> 99
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 99
Lys Ser Ser Gln Ser Ile Phe Arg Thr Ser Arg Asn Lys Asn Leu Leu
1 5 10 15
Asn
<210> 100
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 100
Trp Ala Ser Thr Arg Lys Ser
1 5
<210> 101
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 101
Gln Gln Tyr Phe Ser Pro Pro Tyr Thr
1 5
<210> 102
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 102
Ser Phe Trp Met His
1 5
<210> 103
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 103
Phe Thr Asn Asn Glu Gly Thr Thr Thr Ala Tyr Ala Asp Ser Val Arg
1 5 10 15
Gly
<210> 104
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 104
Gly Asp Gly Gly Leu Asp Asp
1 5
<210> 105
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 105
Arg Ala Ser Gln Phe Thr Asn His Tyr Leu Asn
1 5 10
<210> 106
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 106
Val Ala Ser Asn Leu Gln Ser
1 5
<210> 107
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 107
Gln Gln Ser Tyr Arg Thr Pro Tyr Thr
1 5
<210> 108
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 108
Ser Gly Tyr Tyr Asn
1 5
<210> 109
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 109
Tyr Ile Leu Ser Gly Ala His Thr Asp Ile Lys Ala Ser Leu Gly Ser
1 5 10 15
<210> 110
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид”
<400> 110
Ser Gly Val Tyr Ser Lys Tyr Ser Leu Asp Val
1 5 10
<210> 111
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 111
Asp Ile Val Met Thr Gln Ser Pro Ser Ile Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Thr Ile Ser Gly Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Ala Glu Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Gly Ile Tyr Tyr Cys Gln Gln Tyr Lys Ser Tyr Ser Phe
85 90 95
Asn Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 112
<211> 124
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<220>
<221> ВАРИАНТ
<222> (1)..(1)
<223> /замена="Glu"
<220>
<221> ВАРИАНТ
<222> (2)..(2)
<223> /замена="Ile" или "Val"
<220>
<221> MISC_FEATURE
<222> (1)..(124)
<223> /note=" Вариант остатков, приведенный в последовательности, не имеет предпочтения по сравнению с аннотациями для вариантов позиций"
<400> 112
Gln Met Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Leu Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Gly Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp
100 105 110
Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 113
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 113
Asp Ile Val Met Thr Gln Ser Pro Ser Ile Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Thr Ile Ser Gly Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Ala Glu Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Gly Ile Tyr Tyr Cys Gln Gln Tyr Lys Ser Tyr Ser Phe
85 90 95
Asn Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 114
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 114
Gln Met Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Leu Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Gly Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp
100 105 110
Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly
450
<210> 115
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 115
Asp Ile Val Met Thr Gln Ser Pro Ser Ile Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Thr Ile Ser Gly Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Ala Glu Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Gly Ile Tyr Tyr Cys Gln Gln Tyr Lys Ser Tyr Ser Phe
85 90 95
Asn Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Cys Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 116
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<220>
<221> ВАРИАНТ
<222> (2)..(2)
<223> /замена="Ile" или "Val"
<220>
<221> MISC_FEATURE
<222> (1)..(453)
<223> /note=" Вариант остатков, приведенный в последовательности, не имеет предпочтения по сравнению с аннотациями для вариантов позиций"
<400> 116
Glu Met Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Leu Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Gly Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp
100 105 110
Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly
450
<210> 117
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<220>
<221> ВАРИАНТ
<222> (2)..(2)
<223> /замена="Ile" или "Val"
<220>
<221> MISC_FEATURE
<222> (1)..(453)
<223> /note=" Вариант остатков, приведенный в последовательности, не имеет предпочтения по сравнению с аннотациями для вариантов позиций"
<400> 117
Glu Met Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Leu Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Gly Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp
100 105 110
Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Cys Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly
450
<210> 118
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 118
Gly Glu Gly Gly Leu Asp Asp
1 5
<210> 119
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 119
Asp Ile Gln Leu Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Ile Phe Arg Thr
20 25 30
Ser Arg Asn Lys Asn Leu Leu Asn Trp Tyr Gln Gln Arg Pro Gly Gln
35 40 45
Pro Pro Arg Leu Leu Ile His Trp Ala Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Phe Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Thr Ser Leu Gln Ala Glu Asp Val Ala Ile Tyr Tyr Cys Gln Gln
85 90 95
Tyr Phe Ser Pro Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 120
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 120
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Ser Phe Asn Ser Phe
20 25 30
Trp Met His Trp Val Arg Gln Val Pro Gly Lys Gly Leu Val Trp Ile
35 40 45
Ser Phe Thr Asn Asn Glu Gly Thr Thr Thr Ala Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Asn Asn Leu Arg Gly Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Asp Gly Gly Leu Asp Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 121
<211> 220
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 121
Asp Ile Gln Leu Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Ile Phe Arg Thr
20 25 30
Ser Arg Asn Lys Asn Leu Leu Asn Trp Tyr Gln Gln Arg Pro Gly Gln
35 40 45
Pro Pro Arg Leu Leu Ile His Trp Ala Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Phe Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Thr Ser Leu Gln Ala Glu Asp Val Ala Ile Tyr Tyr Cys Gln Gln
85 90 95
Tyr Phe Ser Pro Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 122
<211> 445
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 122
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Ser Phe Asn Ser Phe
20 25 30
Trp Met His Trp Val Arg Gln Val Pro Gly Lys Gly Leu Val Trp Ile
35 40 45
Ser Phe Thr Asn Asn Glu Gly Thr Thr Thr Ala Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Asn Asn Leu Arg Gly Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Glu Gly Gly Leu Asp Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 123
<211> 220
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 123
Asp Ile Gln Leu Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Ile Phe Arg Thr
20 25 30
Ser Arg Asn Lys Asn Leu Leu Asn Trp Tyr Gln Gln Arg Pro Gly Gln
35 40 45
Pro Pro Arg Leu Leu Ile His Trp Ala Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Phe Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Thr Ser Leu Gln Ala Glu Asp Val Ala Ile Tyr Tyr Cys Gln Gln
85 90 95
Tyr Phe Ser Pro Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 124
<211> 444
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 124
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Ser Phe Asn Ser Phe
20 25 30
Trp Met His Trp Val Arg Gln Val Pro Gly Lys Gly Leu Val Trp Ile
35 40 45
Ser Phe Thr Asn Asn Glu Gly Thr Thr Thr Ala Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Asn Asn Leu Arg Gly Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Glu Gly Gly Leu Asp Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Cys Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 125
<400> 125
000
<210> 126
<400> 126
000
<210> 127
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 127
Gln Met Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Ser Cys Ser Val Ser Gly Ala Ser Ala Ser Ser Gly
20 25 30
Tyr Tyr Asn Trp Val Arg Gln Thr Pro Gly Gly Gly Leu Glu Trp Ile
35 40 45
Ala Tyr Ile Leu Ser Gly Ala His Thr Asp Ile Lys Ala Ser Leu Gly
50 55 60
Ser Arg Val Ala Val Ser Val Asp Thr Ser Lys Asn Gln Val Thr Leu
65 70 75 80
Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Ser Gly Val Tyr Ser Lys Tyr Ser Leu Asp Val Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 128
<400> 128
000
<210> 129
<400> 129
000
<210> 130
<400> 130
000
<210> 131
<400> 131
000
<210> 132
<400> 132
000
<210> 133
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 133
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Ala Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ser Val Val Thr Gly His Ser Tyr Arg Thr His Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ile Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Arg Ile Trp Ser Tyr Gly Asp Asp Ser Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly
450
<210> 134
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 134
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ile Glu Thr Ile Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Asp Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Arg Leu Val Asp Val Pro Leu Ser Ser Pro Asn Ser Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly
450
<210> 135
<400> 135
000
<210> 136
<400> 136
000
<210> 137
<400> 137
000
<210> 138
<211> 445
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 138
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Ser Phe Asn Ser Phe
20 25 30
Trp Met His Trp Val Arg Gln Val Pro Gly Lys Gly Leu Val Trp Ile
35 40 45
Ser Phe Thr Asn Asn Glu Gly Thr Thr Thr Ala Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Asn Asn Leu Arg Gly Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Asp Gly Gly Leu Asp Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 139
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 139
Glu Met Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Leu Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Gly Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp
100 105 110
Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Cys Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly
450
<210> 140
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 140
Glu Met Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Leu Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Gly Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp
100 105 110
Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly
450
<210> 141
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 141
Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Leu Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Gly Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp
100 105 110
Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Cys Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly
450
<210> 142
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 142
Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Leu Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Gly Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp
100 105 110
Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly
450
<210> 143
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 143
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Leu Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Gly Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp
100 105 110
Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Cys Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly
450
<210> 144
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 144
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Leu Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Gly Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp
100 105 110
Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly
450
<210> 145
<211> 220
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 145
Asp Ile Gln Leu Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Ile Phe Arg Thr
20 25 30
Ser Arg Asn Lys Asn Leu Leu Asn Trp Tyr Gln Gln Arg Pro Gly Gln
35 40 45
Pro Pro Arg Leu Leu Ile His Trp Ala Ser Thr Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Phe Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Thr Ser Leu Gln Ala Glu Asp Val Ala Ile Tyr Tyr Cys Gln Gln
85 90 95
Tyr Phe Ser Pro Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Cys Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 146
<211> 445
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 146
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Ser Phe Asn Ser Phe
20 25 30
Trp Met His Trp Val Arg Gln Val Pro Gly Lys Gly Leu Val Trp Ile
35 40 45
Ser Phe Thr Asn Asn Glu Gly Thr Thr Thr Ala Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Asn Asn Leu Arg Gly Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Asp Gly Gly Leu Asp Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 147
<211> 445
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 147
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Ser Phe Asn Ser Phe
20 25 30
Trp Met His Trp Val Arg Gln Val Pro Gly Lys Gly Leu Val Trp Ile
35 40 45
Ser Phe Thr Asn Asn Glu Gly Thr Thr Thr Ala Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Asn Asn Leu Arg Gly Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Glu Gly Gly Leu Asp Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Cys Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 148
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 148
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 149
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 149
Cys Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 150
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 150
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 151
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 151
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Cys Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 152
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический олигонуклеотид»
<400> 152
cccagactgc accagctgga tctctgaatg tactccagtt gc 42
<210> 153
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический олигонуклеотид»
<400> 153
ccagactgca ccagctgcac ctctgaatgt actccagttg c 41
<210> 154
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический олигонуклеотид»
<400> 154
ccagggttcc ctggccccaw tmgtcaagtc cascwkcacc tcttgcacag taatagacag 60
c 61
<210> 155
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический олигонуклеотид»
<400> 155
cctggcccca gtcgtcaagt cctccttcac ctcttgcaca gtaatagaca gc 52
<210> 156
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид
<400> 156
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Ser Phe Asn Ser Phe
20 25 30
Trp Met His Trp Val Arg Gln Val Pro Gly Lys Gly Leu Val Trp Ile
35 40 45
Ser Phe Thr Asn Asn Glu Gly Thr Thr Thr Ala Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Asn Asn Leu Arg Gly Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Glu Gly Gly Leu Asp Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 157
<211> 445
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 157
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Ser Phe Asn Ser Phe
20 25 30
Trp Met His Trp Val Arg Gln Val Pro Gly Lys Gly Leu Val Trp Ile
35 40 45
Ser Phe Thr Asn Asn Glu Gly Thr Thr Thr Ala Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Asn Asn Leu Arg Gly Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Glu Gly Gly Leu Asp Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 158
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 158
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ile Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Thr Ile Ser Gly Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Ala Glu Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Gly Ile Tyr Tyr Cys Gln Gln Tyr Lys Ser Tyr Ser Phe
85 90 95
Asn Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 159
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 159
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Phe Val Ser Arg Thr
20 25 30
Ser Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Arg Leu Leu
35 40 45
Ile Tyr Glu Thr Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Met Tyr Tyr Cys His Lys Tyr Gly Ser Gly Pro
85 90 95
Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Val Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 160
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 160
Glu Thr Thr Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Lys Val Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Lys Tyr Gly Ser Thr Pro
85 90 95
Arg Pro Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 161
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 161
Asp Val Val Met Thr Gln Ser Ser Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Leu Asp Ile Thr Asn His
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Glu Leu Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Ile Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Glu Lys Cys Asn Ser Thr Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 162
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 162
Glu Ile Val Met Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Ala Ile
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Glu Pro Gly Arg Ala Pro Thr Leu Leu
35 40 45
Phe Tyr Gly Val Ser Asn Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Cys Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Leu Tyr Thr Ser Ser Arg
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 163
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 163
Glu Ile Val Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Arg Asn Gly
20 25 30
Leu Gly Trp Tyr Gln Gln Thr Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Pro Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Asp Arg Asp Phe Thr Leu Thr Ile Thr Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp His Asn Tyr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 164
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 164
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Arg Asn Gly
20 25 30
Leu Gly Trp Tyr Gln Gln Ile Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Pro Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Asp Arg Asp Phe Thr Leu Thr Ile Thr Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp His Asn Tyr Pro Pro
85 90 95
Ser Phe Ser Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 165
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 165
Asp Ile Gln Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Thr Val Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Arg Thr Asn
20 25 30
Val Ala Trp Tyr Arg His Lys Ala Gly Gln Ala Pro Met Ile Leu Val
35 40 45
Ser Gly Ala Ser Thr Arg Ala Ser Gly Ala Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Glu Phe Thr Leu Thr Ile Thr Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Leu Gln Tyr Asn Thr Trp Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Val Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 166
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 166
Asp Val Val Met Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Leu Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Arg Pro Gly Lys Ala Pro Asn Leu Leu Ile
35 40 45
Tyr Ala Thr Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Phe Gly Thr Glu Phe Thr Leu Thr Ile Ser Thr Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Leu Asn Asn Tyr Val His
85 90 95
Ser Phe Gly Pro Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 167
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 167
Glu Thr Thr Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Asp Arg
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Tyr Trp Ala Ser Asn Leu Glu Gly Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Thr Gly Ser Gly Thr Glu Phe Ala Leu Thr Ile Ser Gly Leu Gln Pro
65 70 75 80
Asp Asp Leu Ala Thr Tyr Tyr Cys Gln Gln Tyr Lys Ser Gln Trp Ser
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 168
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 168
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Phe Thr Asn His Tyr
20 25 30
Leu Asn Trp Tyr Gln His Lys Pro Gly Arg Ala Pro Lys Leu Met Ile
35 40 45
Ser Val Ala Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Thr Gly
50 55 60
Ser Glu Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Gly Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Arg Thr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Ser Arg Leu Glu Met Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 169
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 169
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Leu Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Met Asn Ala Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Gly Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Ser Ile Leu Val Arg Gly Ala Leu Gly Arg Tyr Phe Asp
100 105 110
Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly
450
<210> 170
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 170
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Ile Ile Ile Asn Tyr
20 25 30
Asp Phe Ile Trp Val Arg Gln Ala Thr Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Met Asn Pro Asn Ser Tyr Asn Thr Gly Tyr Gly Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Trp Asp Ser Ser Met Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Val Arg Gly Gln Leu Leu Ser Glu Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 171
<211> 455
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 171
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Arg Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Met Asn Pro Asn Ser Gly Asn Thr Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Met Thr Lys Asn Thr Ser Ile Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Glu Arg Trp Ser Lys Asp Thr Gly His Tyr Tyr Tyr Tyr Gly
100 105 110
Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 172
<211> 456
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 172
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Glu Ala Ser Gly Tyr Thr Val Ser Asn Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Met Asn Pro Ser Ser Gly Arg Thr Gly Tyr Ala Pro Lys Phe
50 55 60
Arg Gly Arg Val Thr Met Thr Arg Ser Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Gly Tyr Tyr Asp Ser Ser Gly Asn Tyr His Ile Ser
100 105 110
Gly Leu Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala
115 120 125
Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser
130 135 140
Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe
145 150 155 160
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly
165 170 175
Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
180 185 190
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
195 200 205
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
210 215 220
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
225 230 235 240
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
245 250 255
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
260 265 270
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
275 280 285
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
290 295 300
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
305 310 315 320
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
325 330 335
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
340 345 350
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
355 360 365
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
370 375 380
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
385 390 395 400
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
405 410 415
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
420 425 430
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
435 440 445
Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 173
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 173
Gln Ile Thr Leu Lys Glu Ser Gly Gly Gly Leu Ile Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Pro Phe Ser Ala Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Arg Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Thr Lys Asn Ser Asp Ser Leu Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Gly Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Val Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Leu Ala Ala Arg Ile Met Ala Thr Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 174
<211> 456
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 174
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Asp Tyr Tyr
20 25 30
Ser Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Asp Ser Ser Ser Arg Tyr Leu Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Gln Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Ser Gly Leu Arg Val Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Asp Asp Ile Leu Ser Val Tyr Arg Gly Ser Gly Arg
100 105 110
Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala
115 120 125
Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser
130 135 140
Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe
145 150 155 160
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly
165 170 175
Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
180 185 190
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
195 200 205
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
210 215 220
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
225 230 235 240
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
245 250 255
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
260 265 270
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
275 280 285
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
290 295 300
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
305 310 315 320
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
325 330 335
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
340 345 350
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
355 360 365
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
370 375 380
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
385 390 395 400
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
405 410 415
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
420 425 430
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
435 440 445
Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 175
<211> 456
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 175
Gln Val Gln Leu Gln Gln Ser Gly Gly Gly Leu Val Asn Pro Gly Glu
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Asn Tyr Tyr
20 25 30
Ser Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Asp Ser Ser Ser Arg Tyr Arg Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Gln Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Ser Ala Leu Arg Val Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Asp Asp Ile Leu Ser Val Tyr Gln Gly Ser Gly Arg
100 105 110
Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala
115 120 125
Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser
130 135 140
Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe
145 150 155 160
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly
165 170 175
Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
180 185 190
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
195 200 205
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
210 215 220
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
225 230 235 240
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
245 250 255
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
260 265 270
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
275 280 285
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
290 295 300
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
305 310 315 320
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
325 330 335
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
340 345 350
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
355 360 365
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
370 375 380
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
385 390 395 400
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
405 410 415
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
420 425 430
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
435 440 445
Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 176
<211> 445
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический полипептид»
<400> 176
Gln Val Gln Leu Gln Gln Ser Gly Gly Gly Leu Glu Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Ile Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Asn
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ser Thr Ile Ile Gly Ile Asp Asp Thr Thr His Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Thr Val Ser Arg Asp Thr Ser Lys Asn Met Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Val Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Lys Asn Ser Gly Ile Tyr Ser Phe Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 177
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 177
Ala Arg Gly Asp Gly Gly Leu
1 5
<210> 178
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 178
Ala Arg Gly Asp Ala Gly Leu
1 5
<210> 179
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 179
Ala Arg Gly Glu Gly Gly Leu
1 5
<210> 180
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /note=«Описание искусственной последовательности: синтетический пептид»
<400> 180
Ala Arg Gly Ala Gly Gly Leu
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ И СВЯЗЫВАЮЩИЕ АГЕНТЫ, НАЦЕЛЕННЫЕ НА DLL3 | 2020 |
|
RU2822366C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ НА ОСНОВЕ АКТИВИРУЮЩИХ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИХ АНТИГЕНСВЯЗЫВАЮЩИХ МОЛЕКУЛ ПРОТИВ CD3 И ФОЛАТНОГО РЕЦЕПТОРА 1 (FOLR1) И АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХСЯ С ОСЬЮ PD-1 | 2015 |
|
RU2753902C2 |
| Антитела против CXCR2 и их применение | 2019 |
|
RU2807067C2 |
| CD3-СВЯЗЫВАЮЩИЙ ДОМЕН | 2015 |
|
RU2742691C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР КЛЕТОК | 2015 |
|
RU2751362C2 |
| МОДИФИЦИРОВАННАЯ J-ЦЕПЬ | 2015 |
|
RU2761118C2 |
| ВЫДЕЛЕННОЕ АНТИТЕЛО (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛА, ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА, ЭКСПРЕССИОННАЯ КАССЕТА (ВАРИАНТЫ), ПЛАЗМИДА (ВАРИАНТЫ), КЛЕТКА-ХОЗЯИН, ФАРМАЦЕВТИЧЕСКИЙ ПРЕПАРАТ, НАБОР, СПОСОБ ЛЕЧЕНИЯ БОЛЬНОГО, ПОДВЕРЖЕННОГО РИСКУ ИЛИ СТРАДАЮЩЕГО ОТ ИНФЕКЦИИ E.COLI, И СПОСОБ ДИАГНОСТИКИ ОПРЕДЕЛЕНИЯ ИНФЕКЦИЙ E.COLI | 2014 |
|
RU2724530C2 |
| НОВЫЕ КОНСТРУКЦИИ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА, СОДЕРЖАЩИЕ ДОМЕНЫ TNFR2 | 2019 |
|
RU2808254C2 |
| ГИБРИДНЫЕ БЕЛКИ ВАРИАНТА sPD-1—FC | 2019 |
|
RU2785993C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2015 |
|
RU2743464C2 |
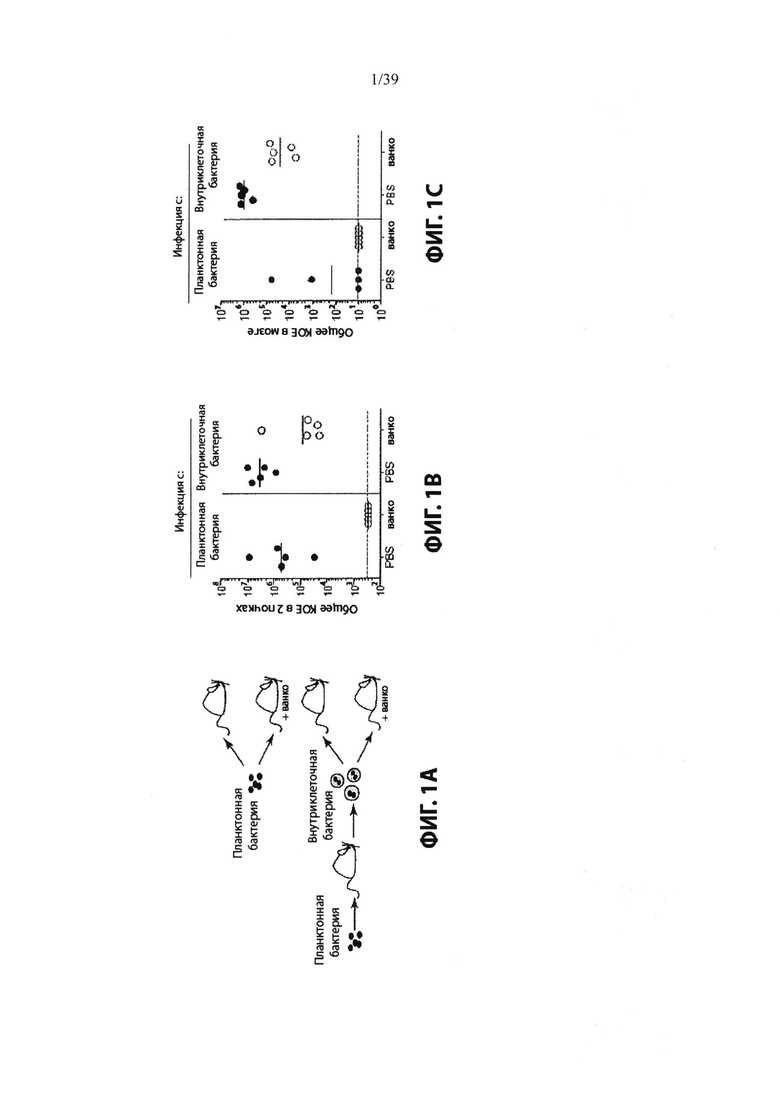
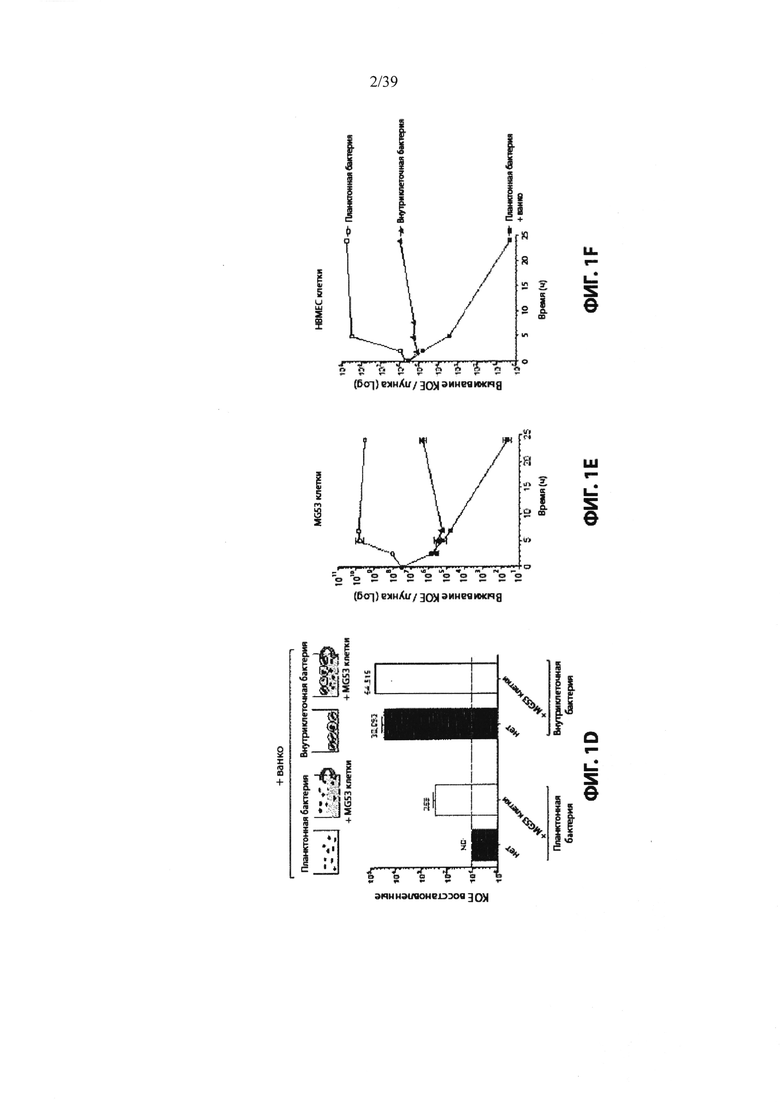
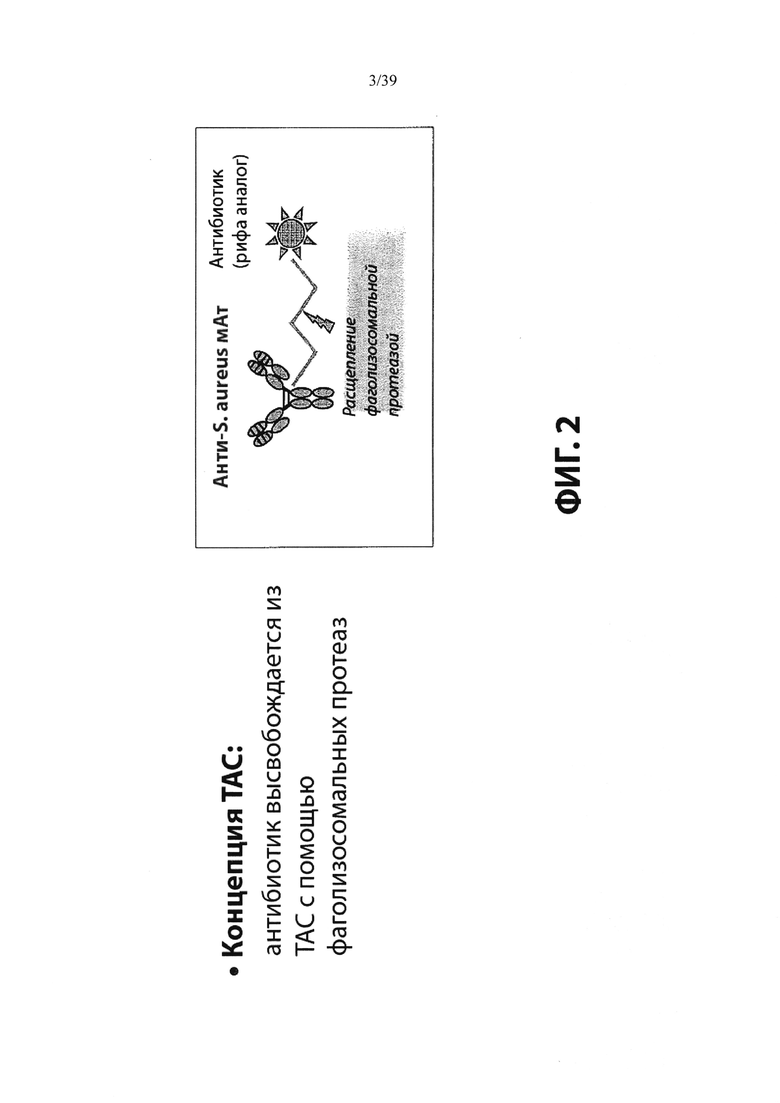
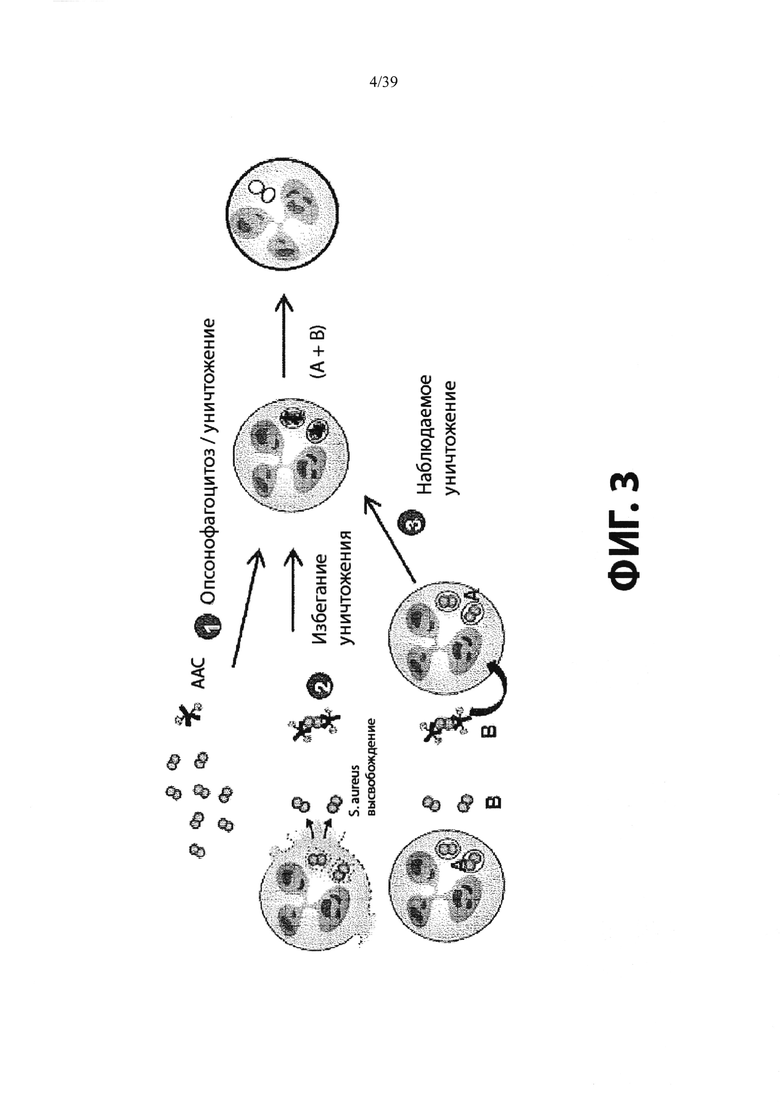

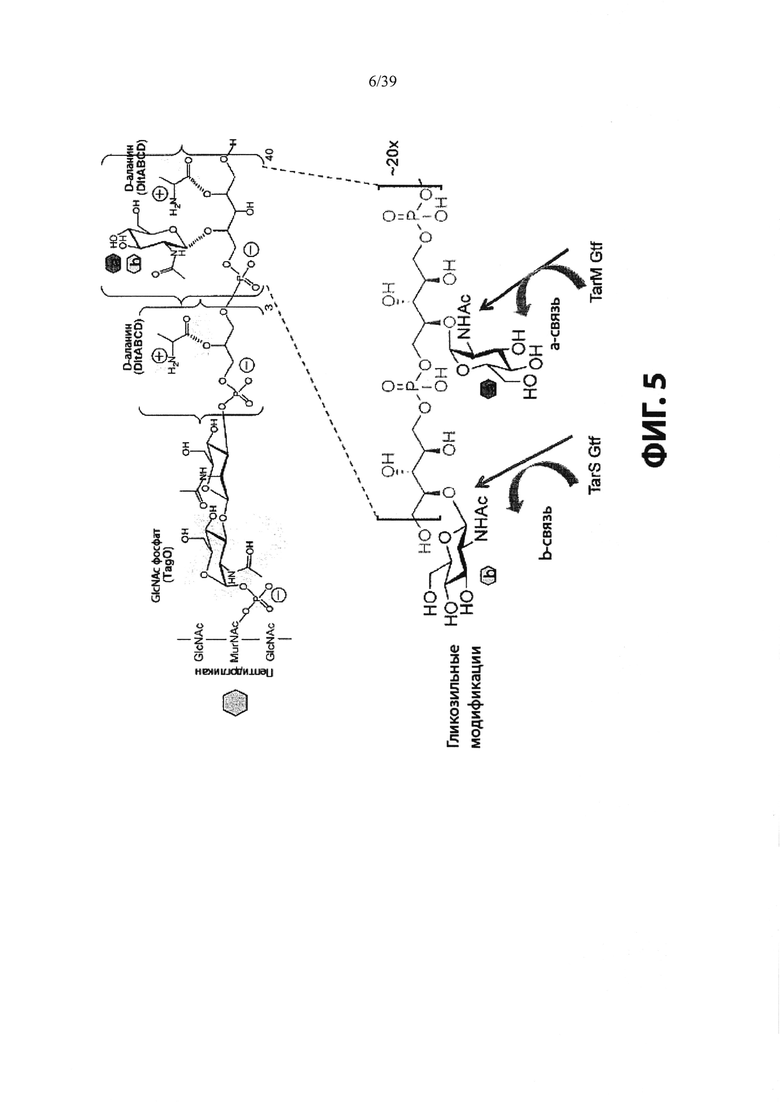
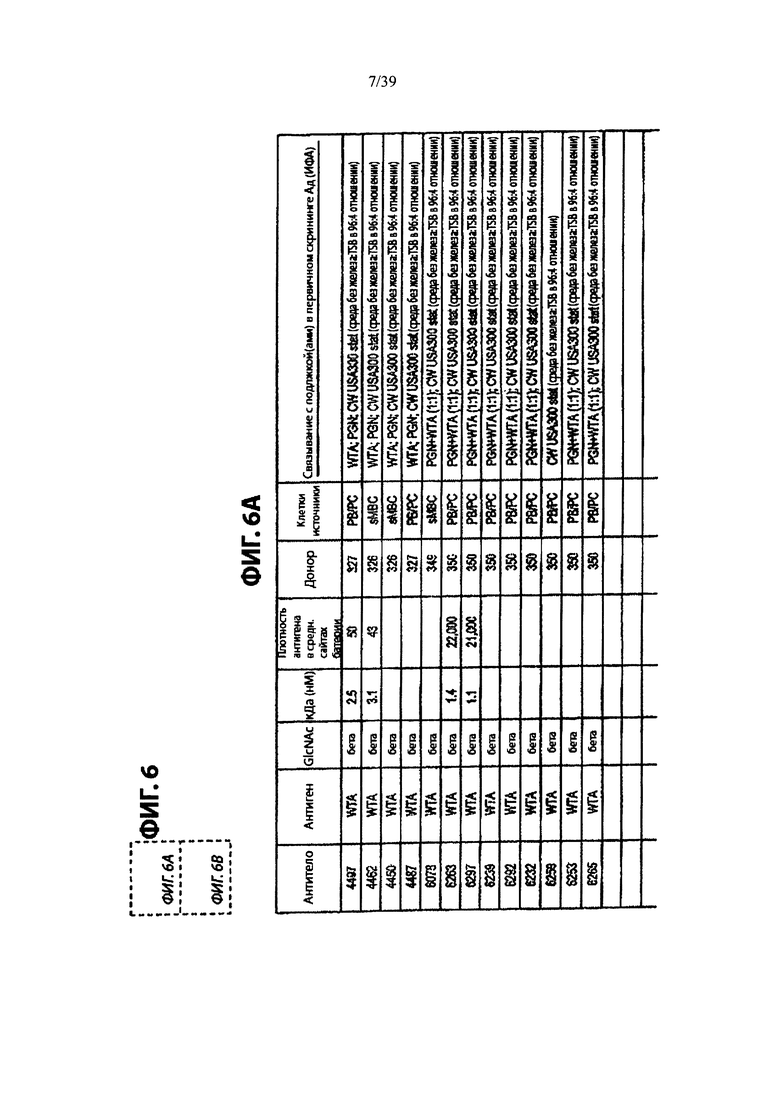
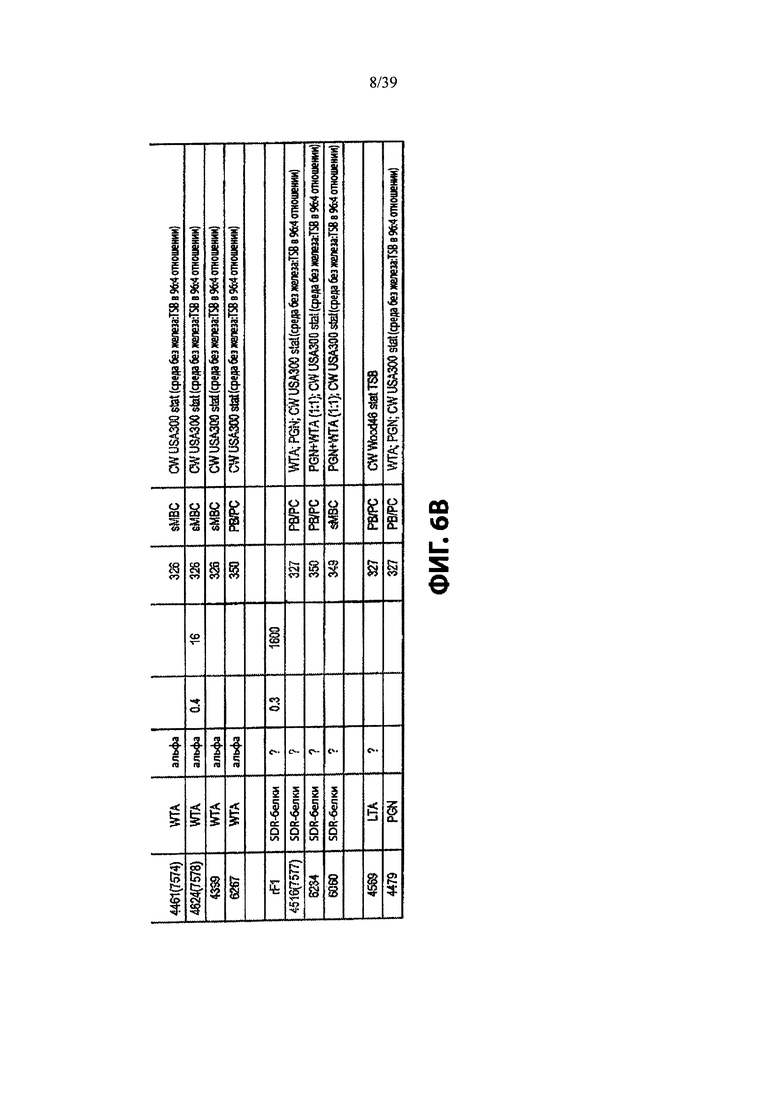
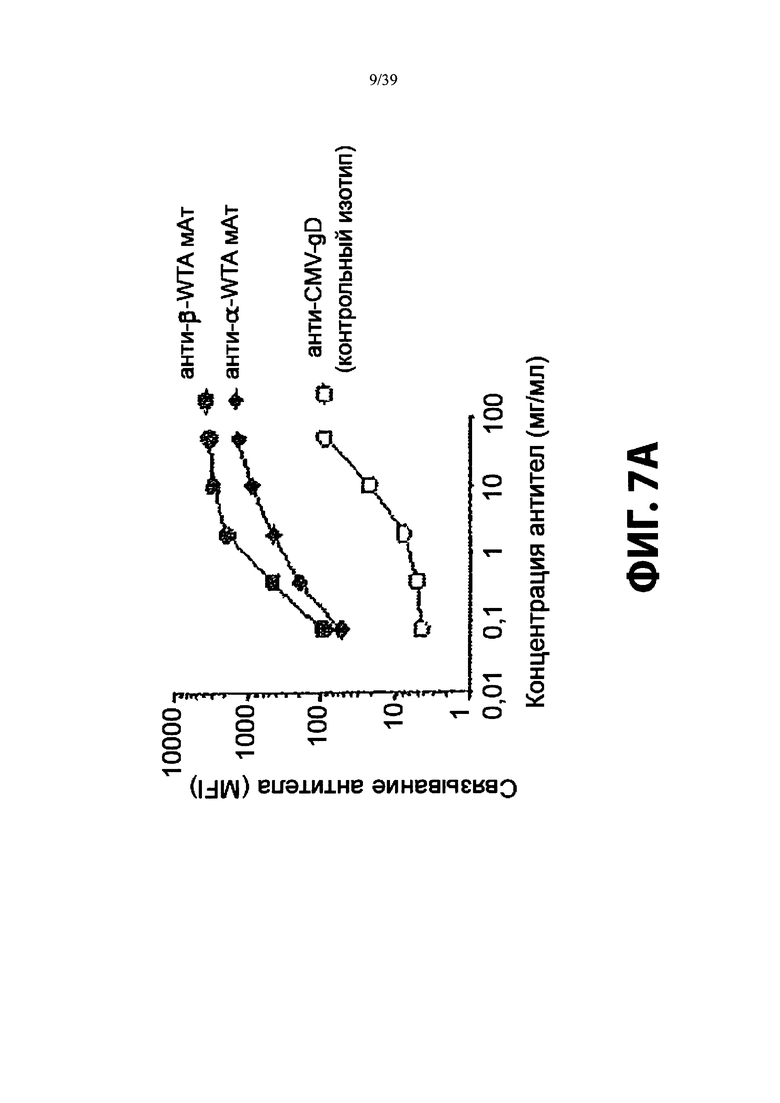
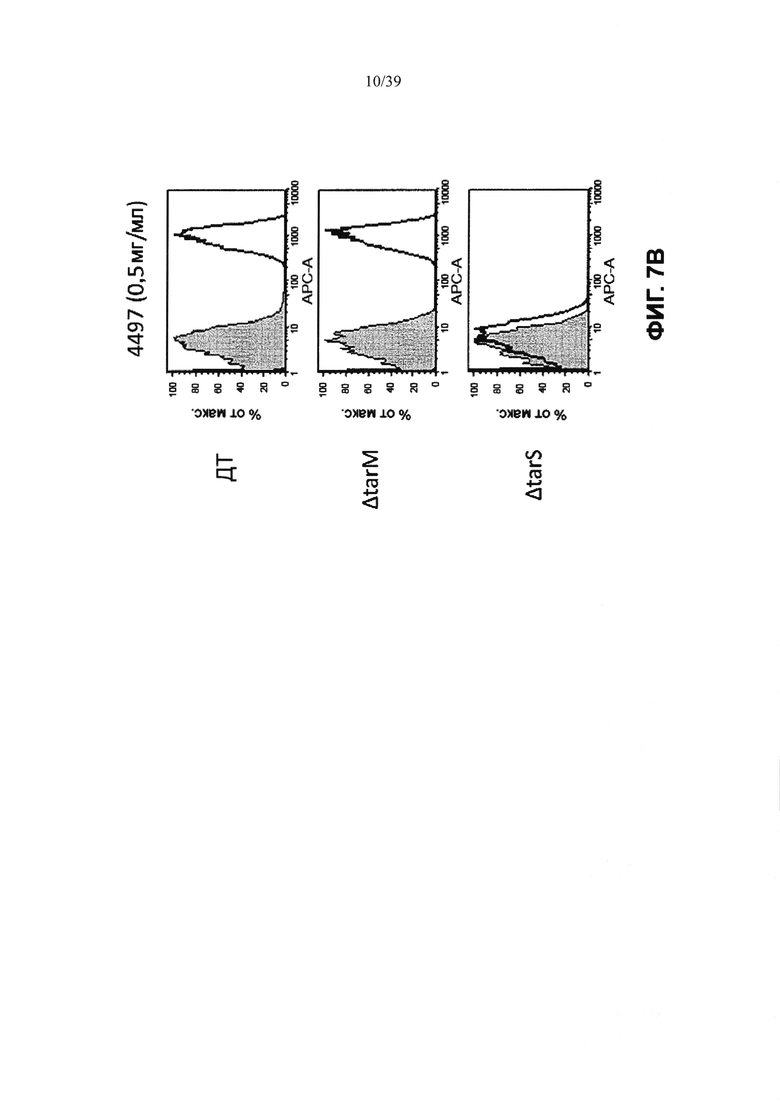
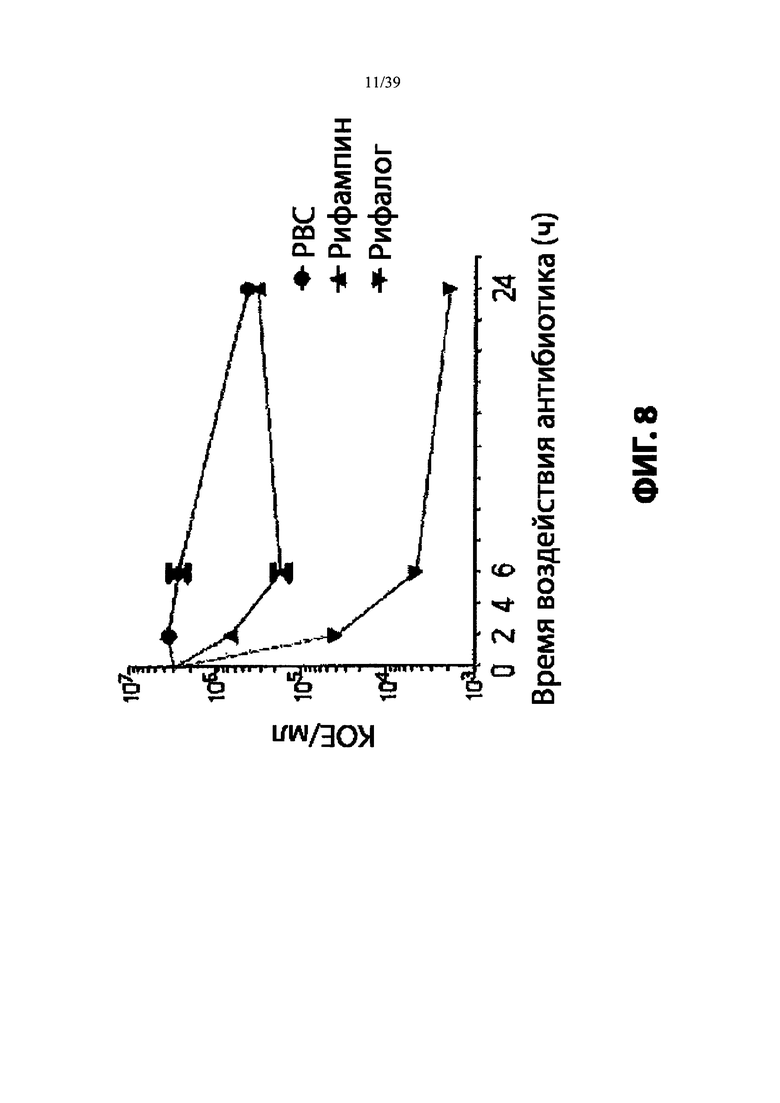
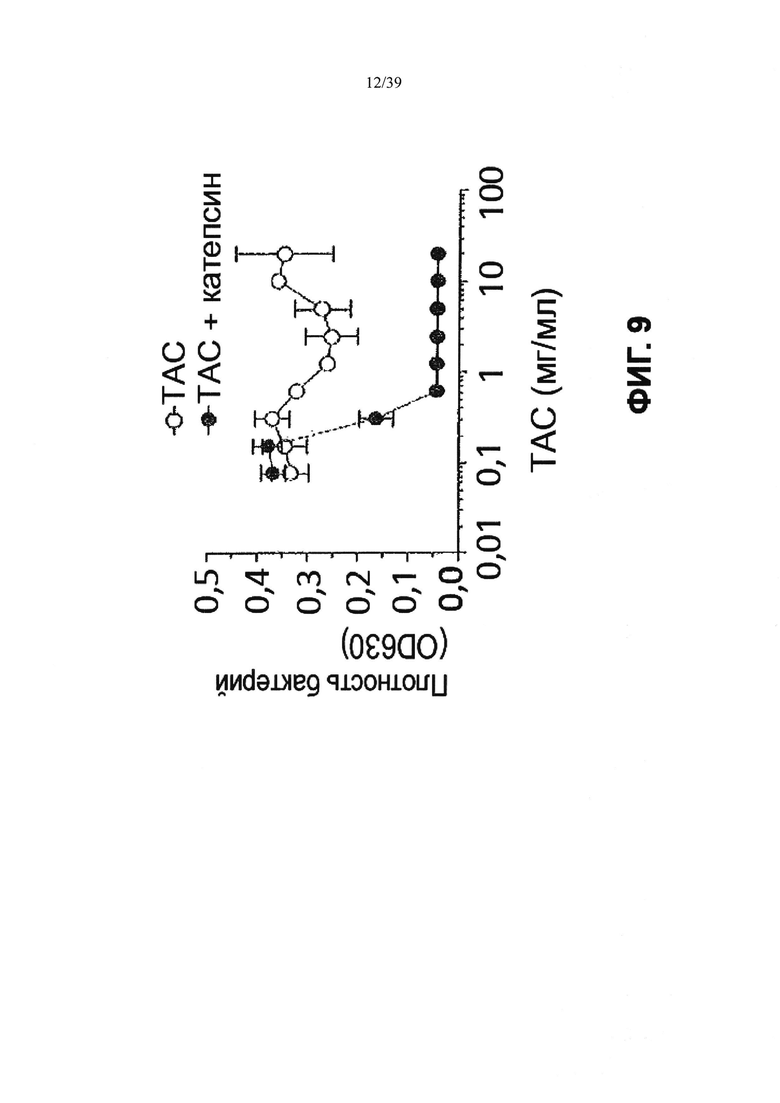
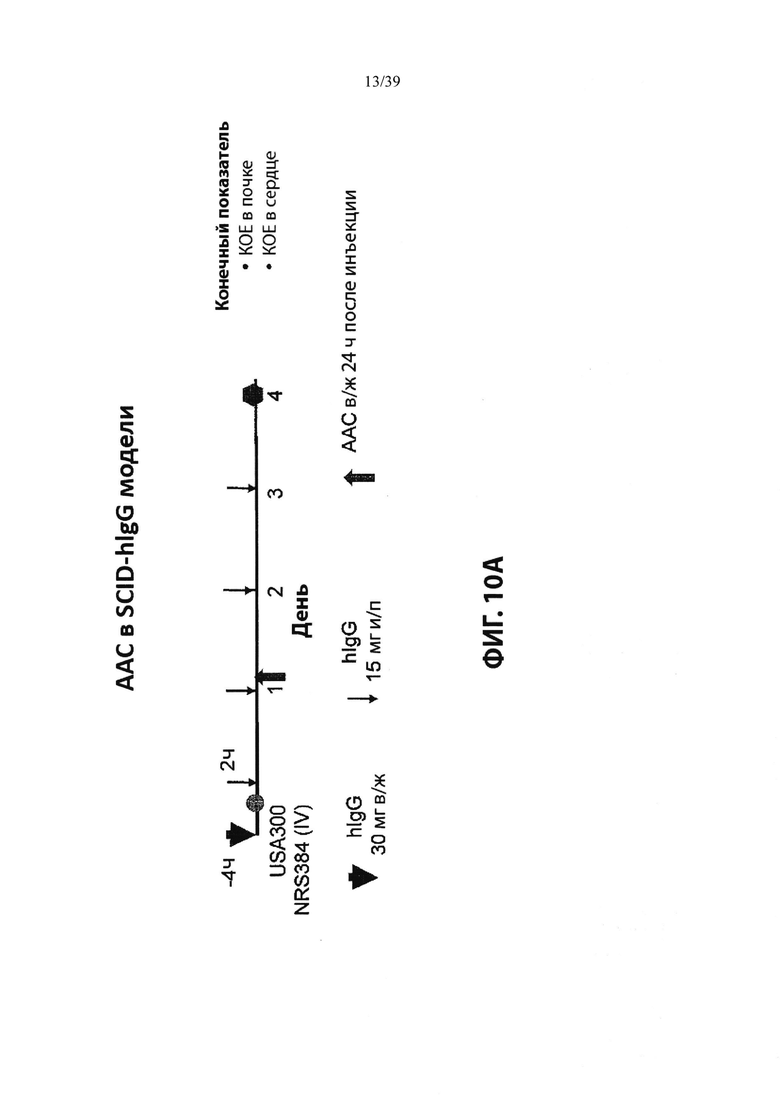
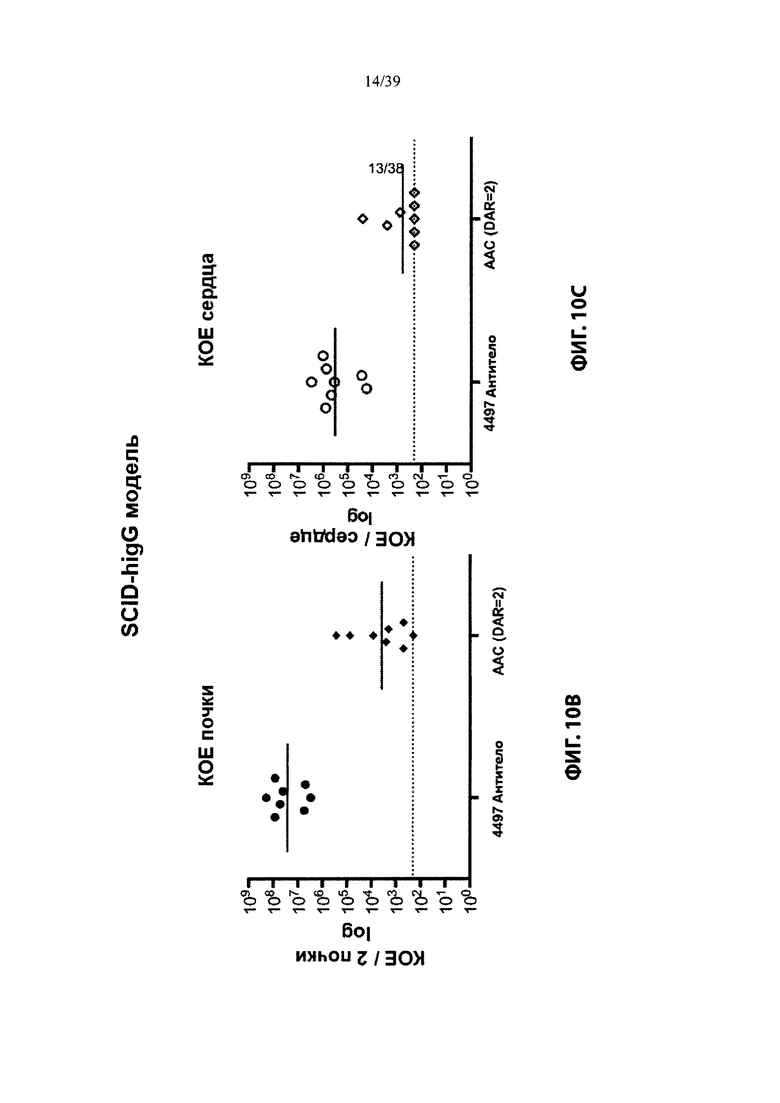
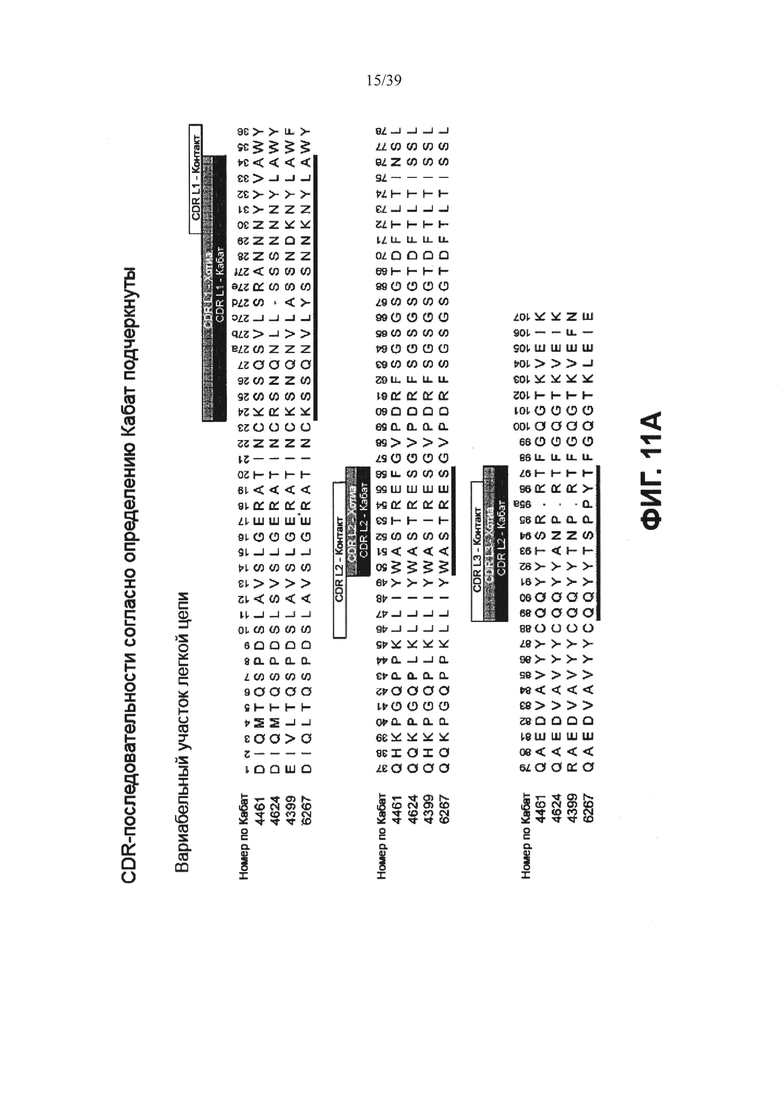
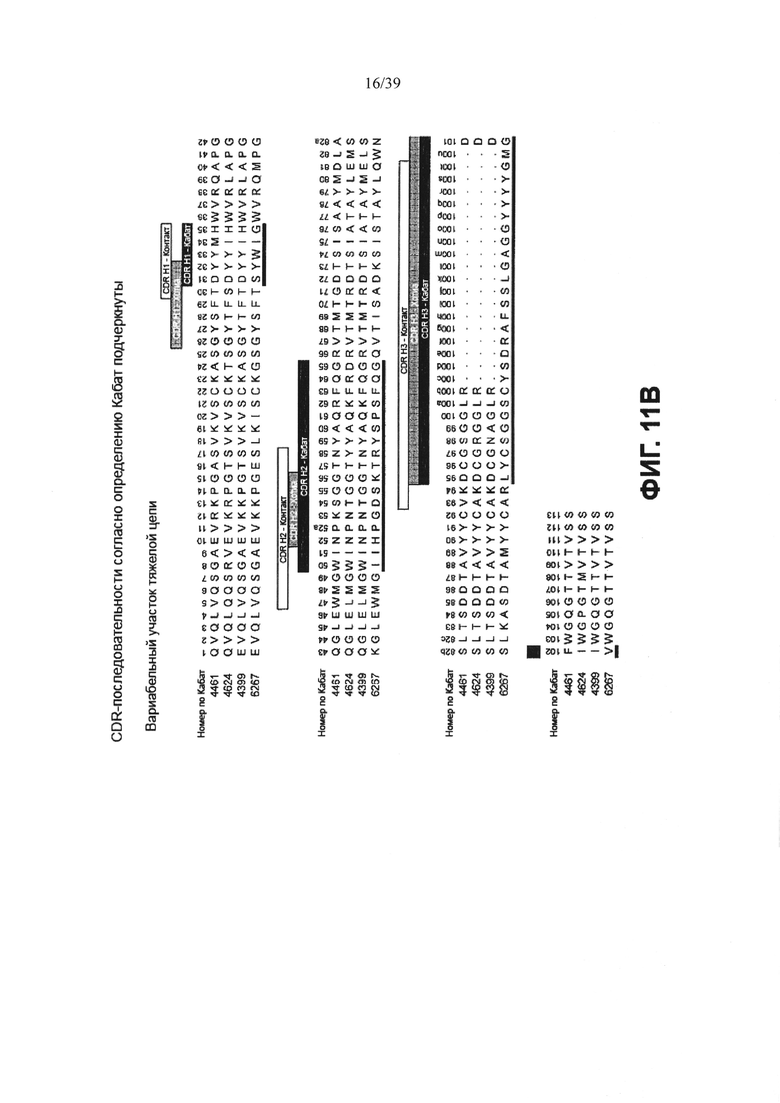
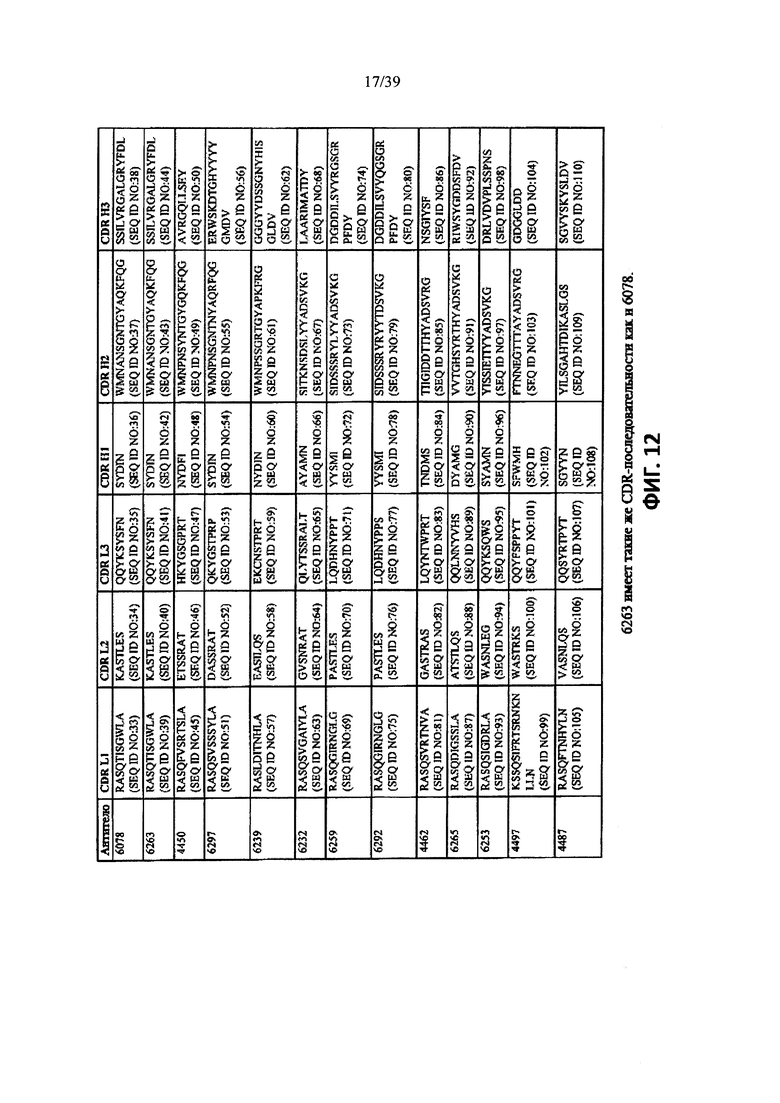
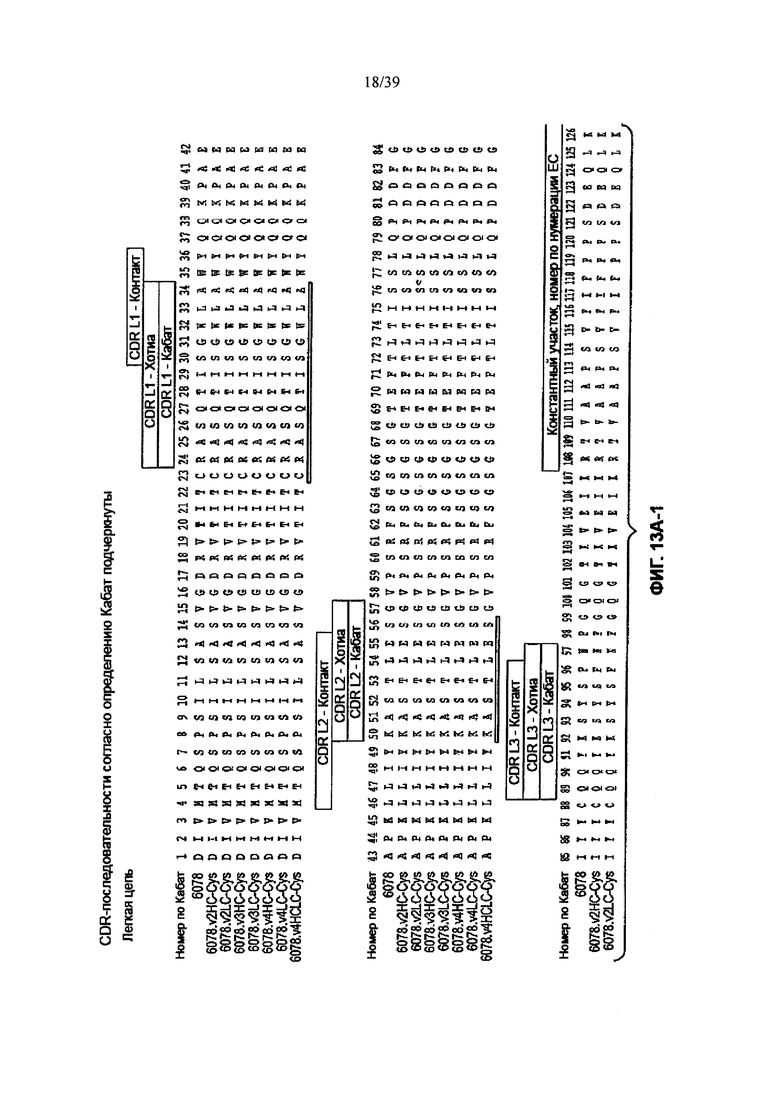
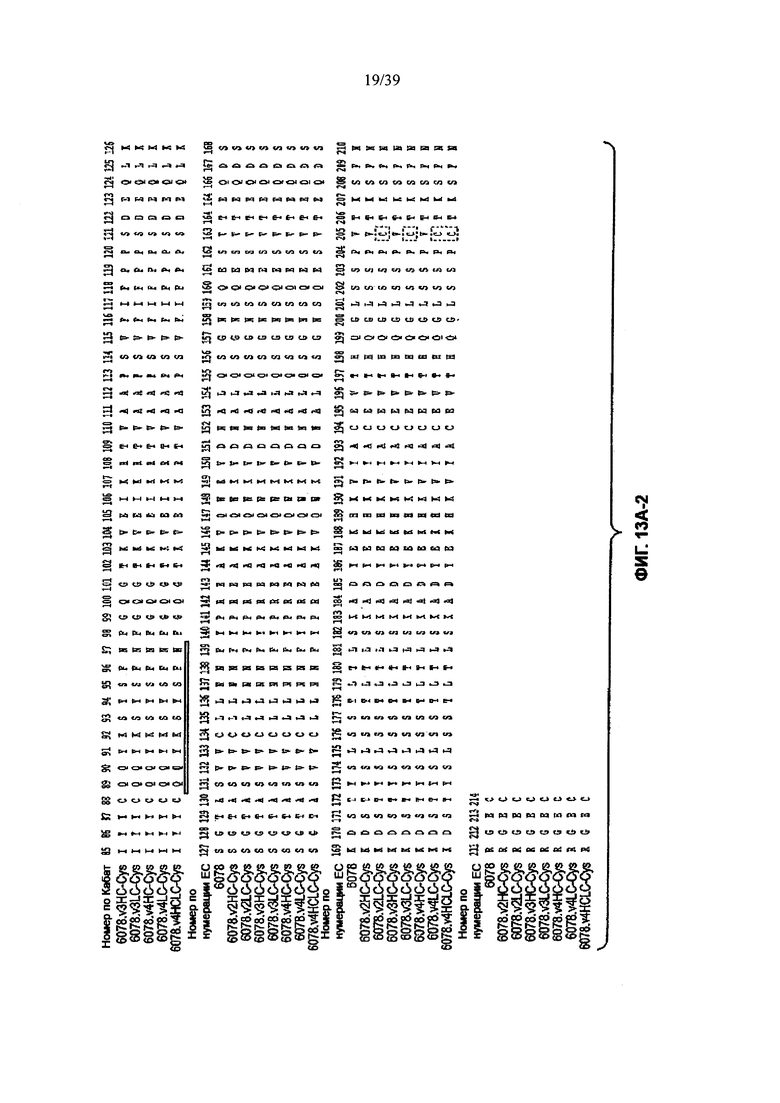
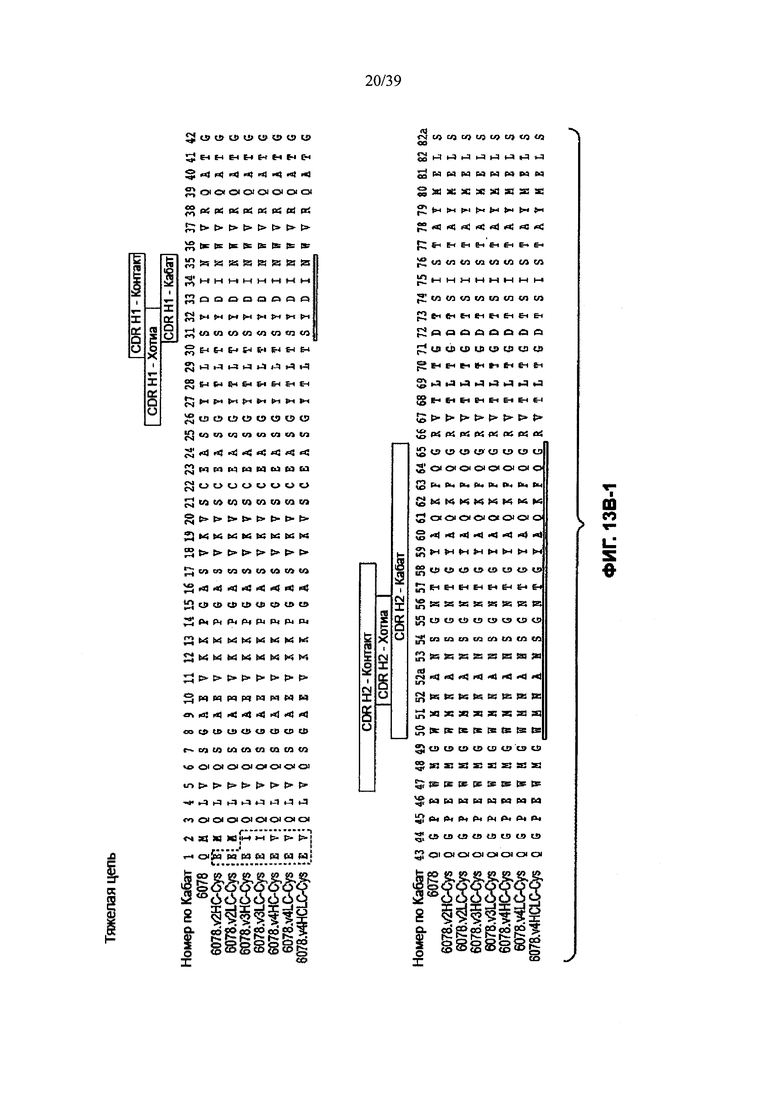
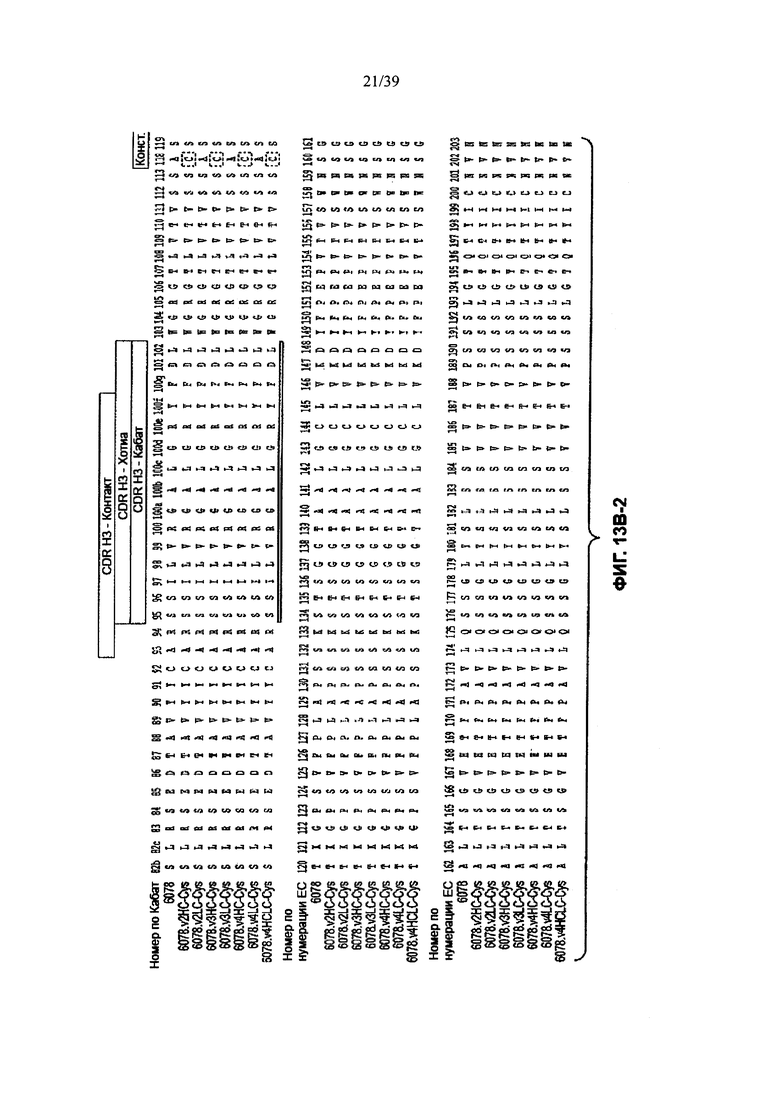
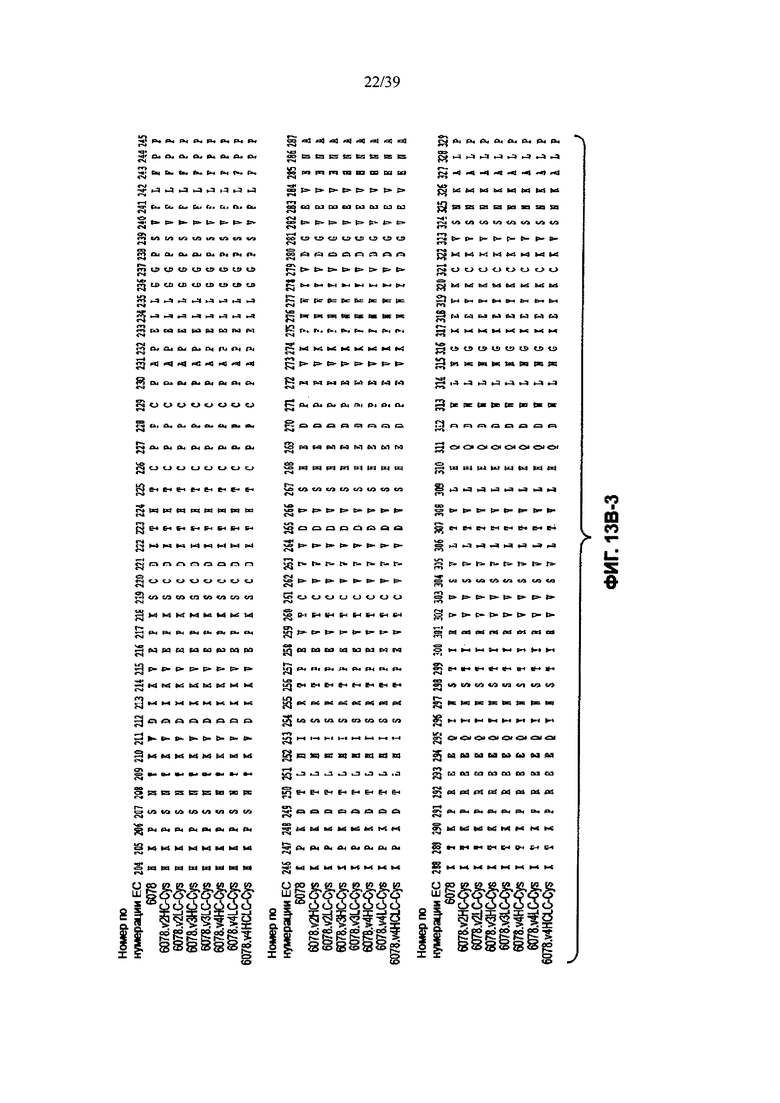
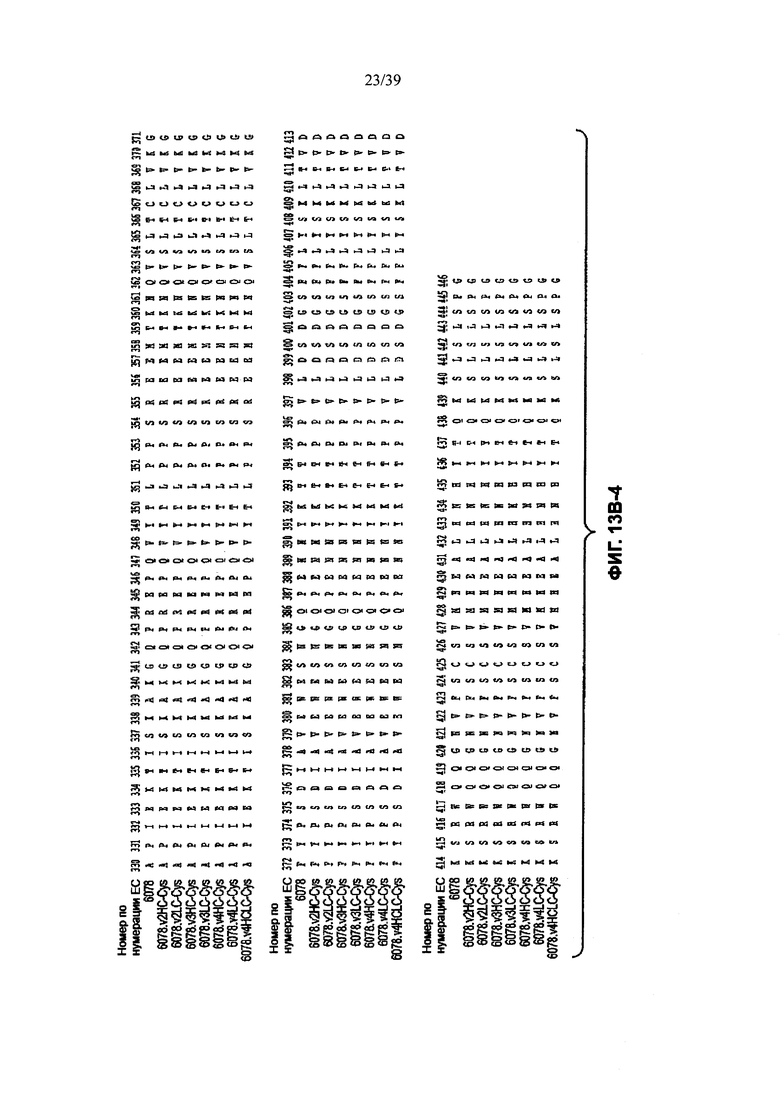
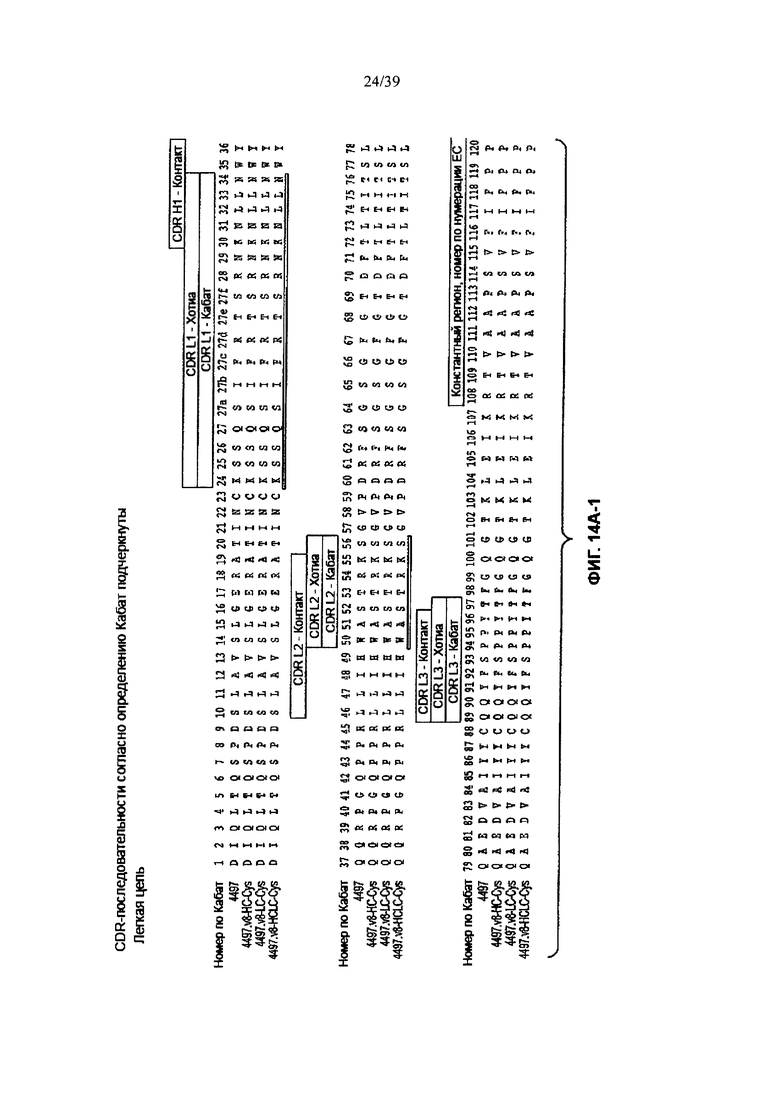
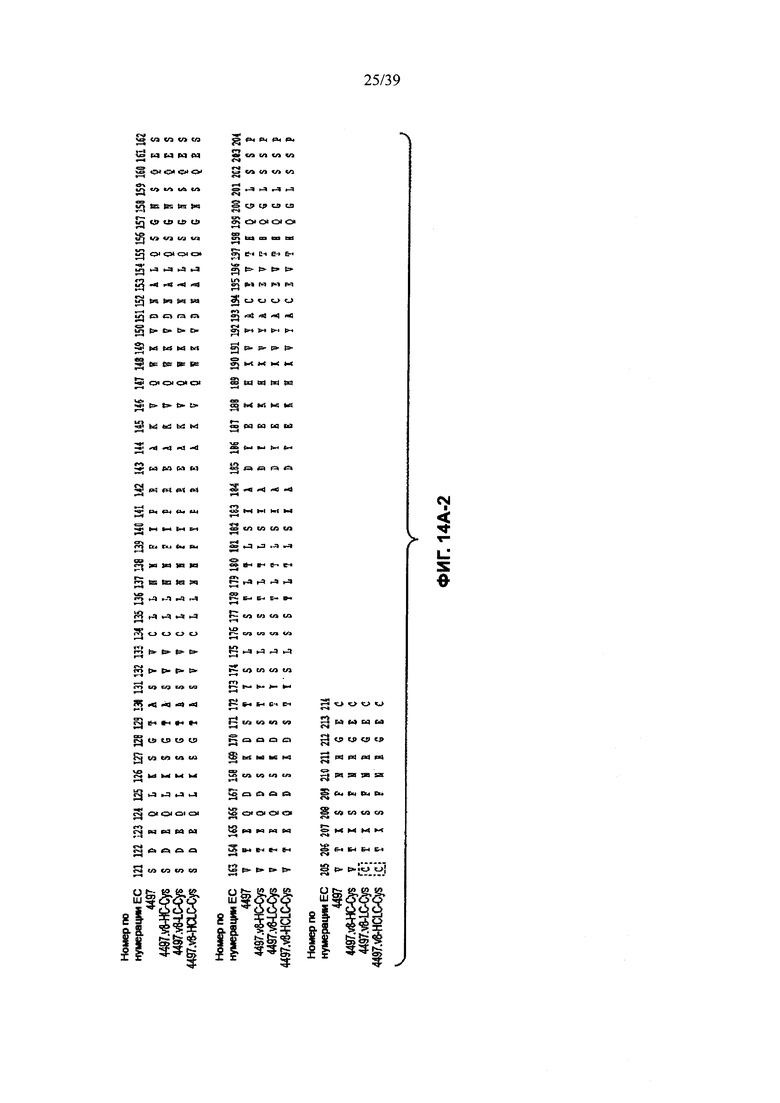
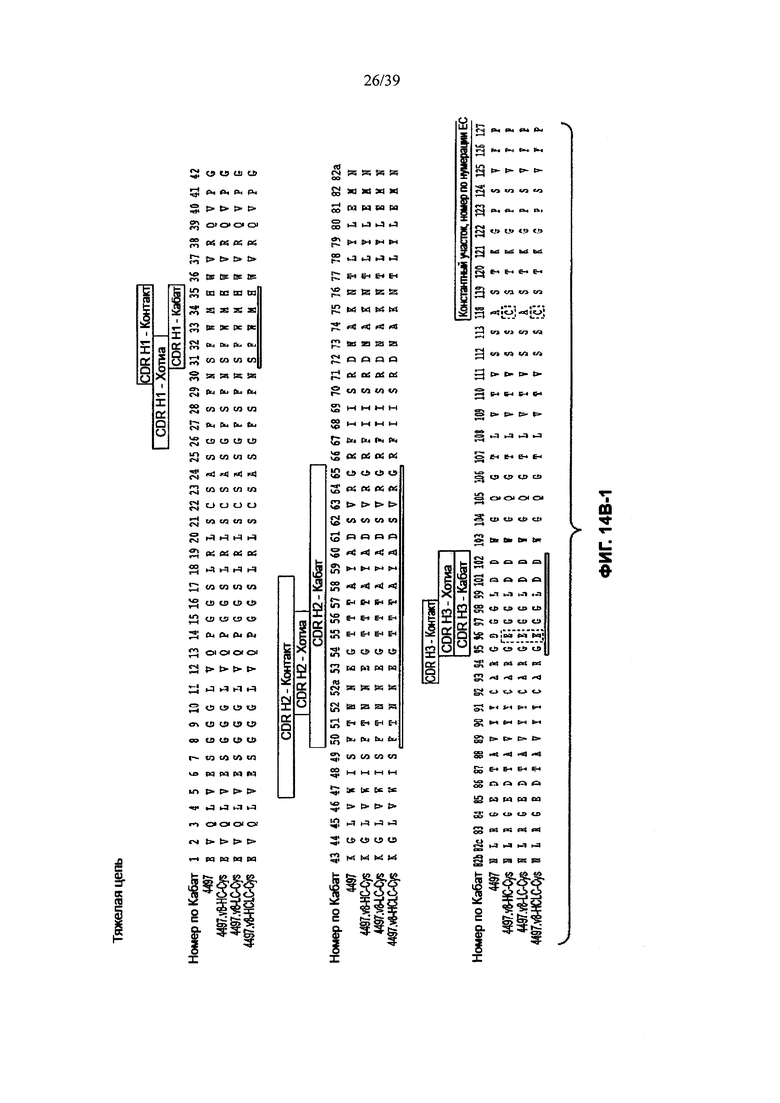
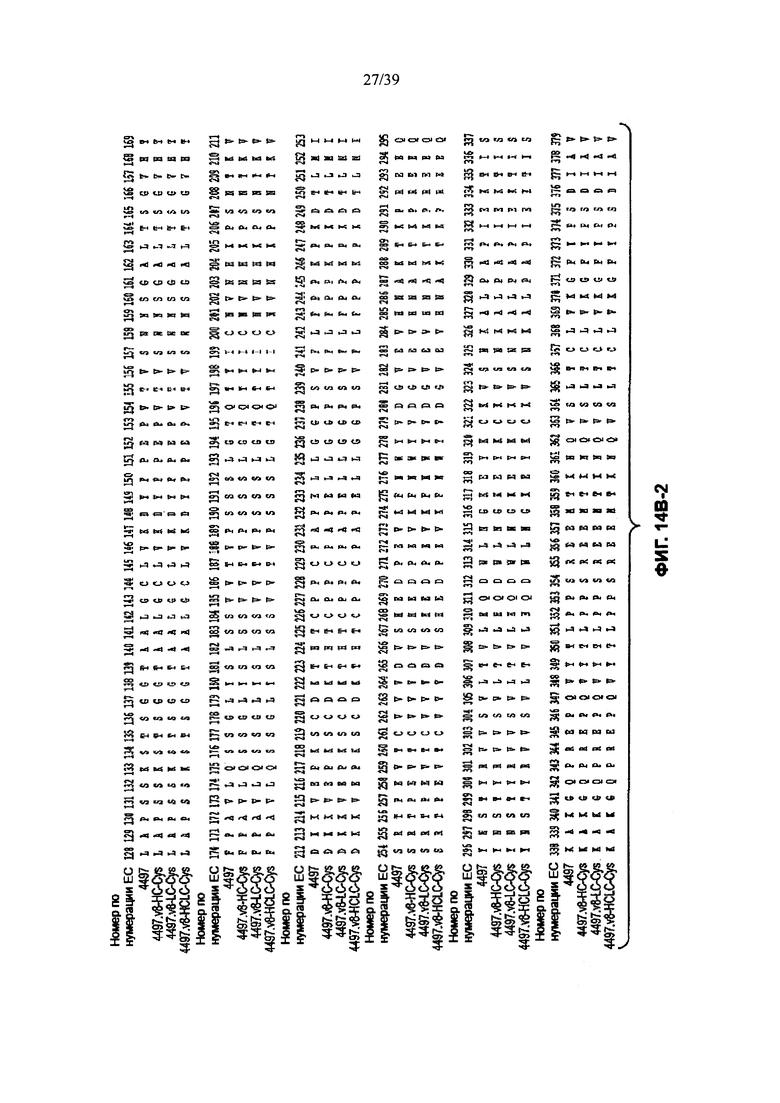
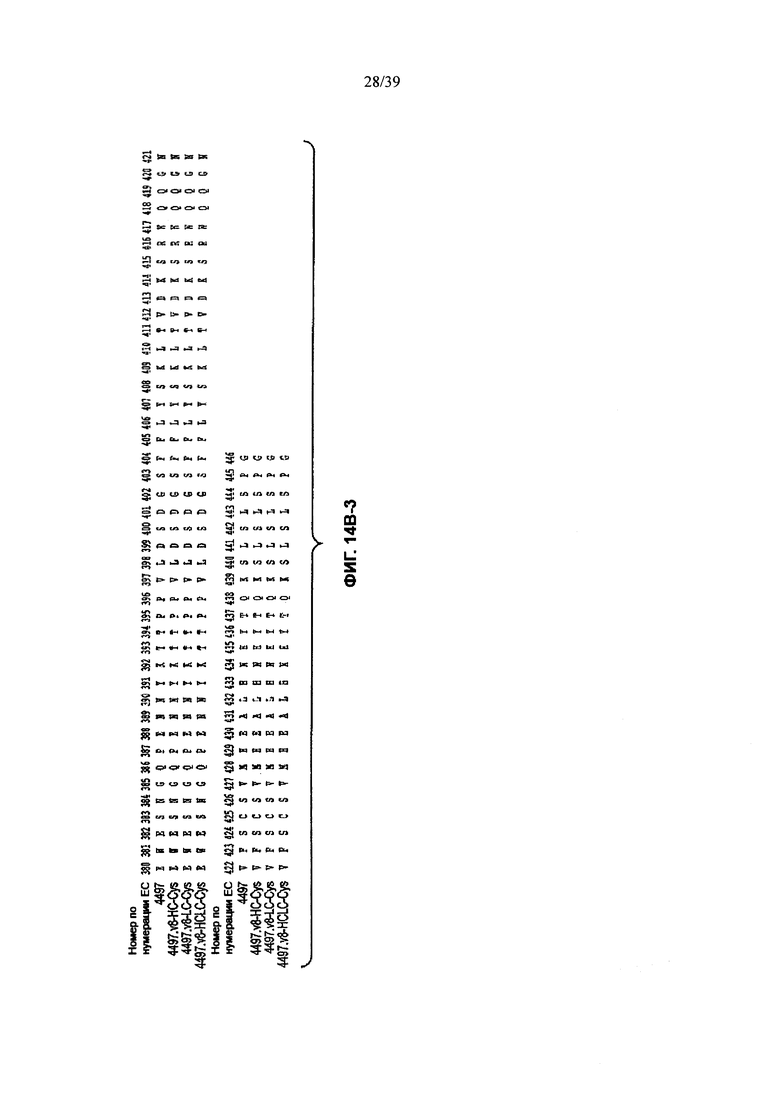
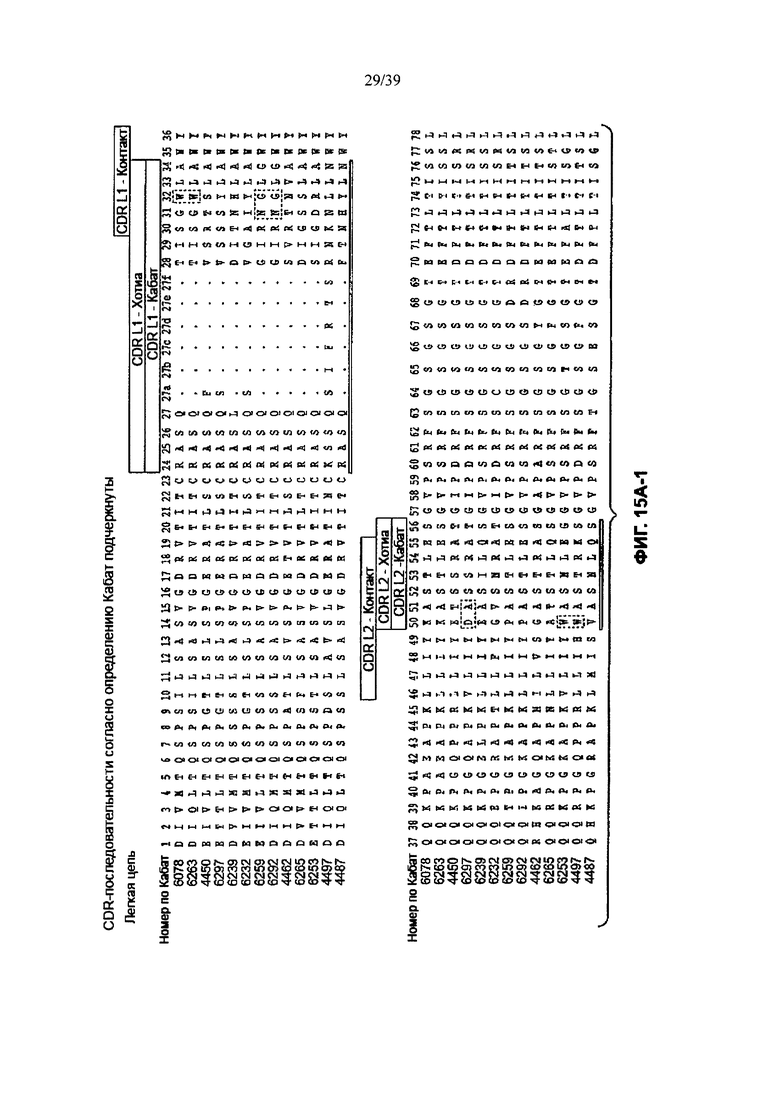
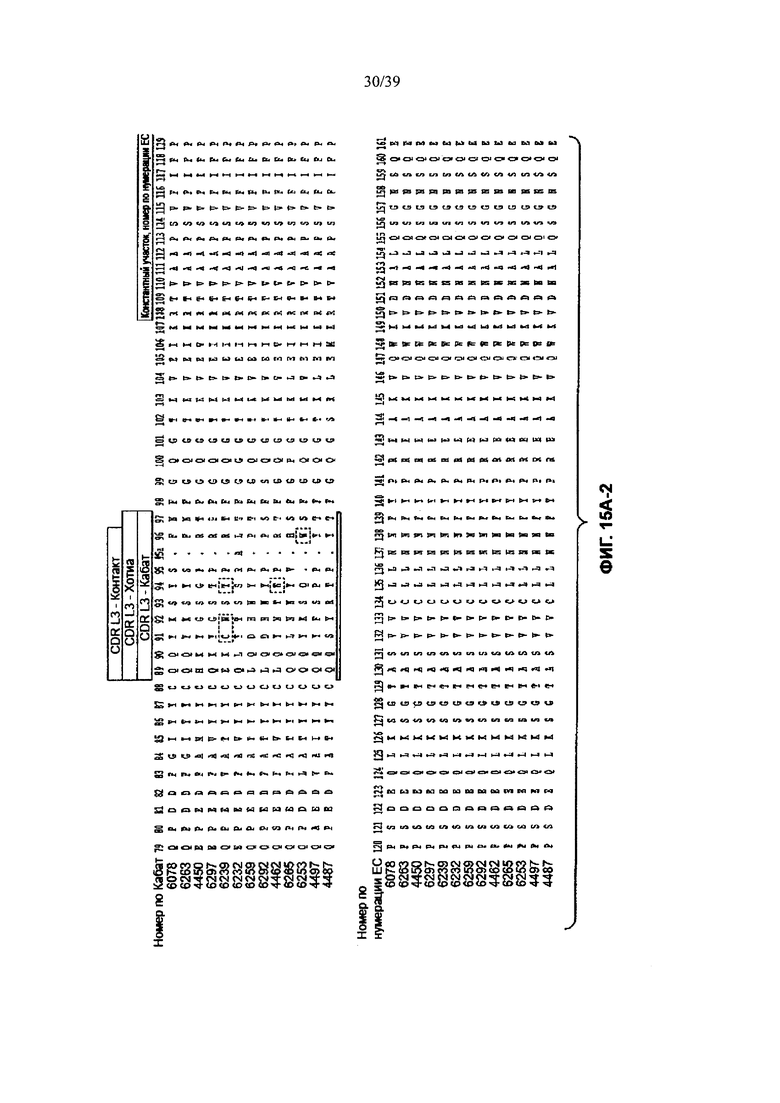
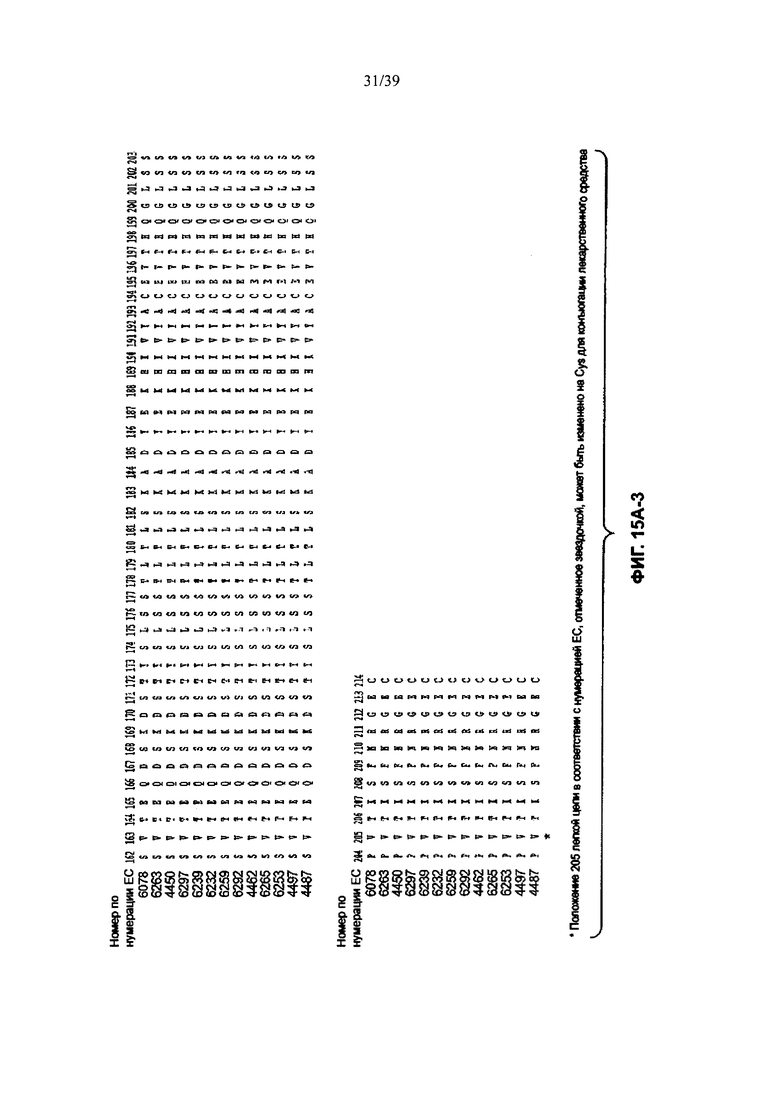
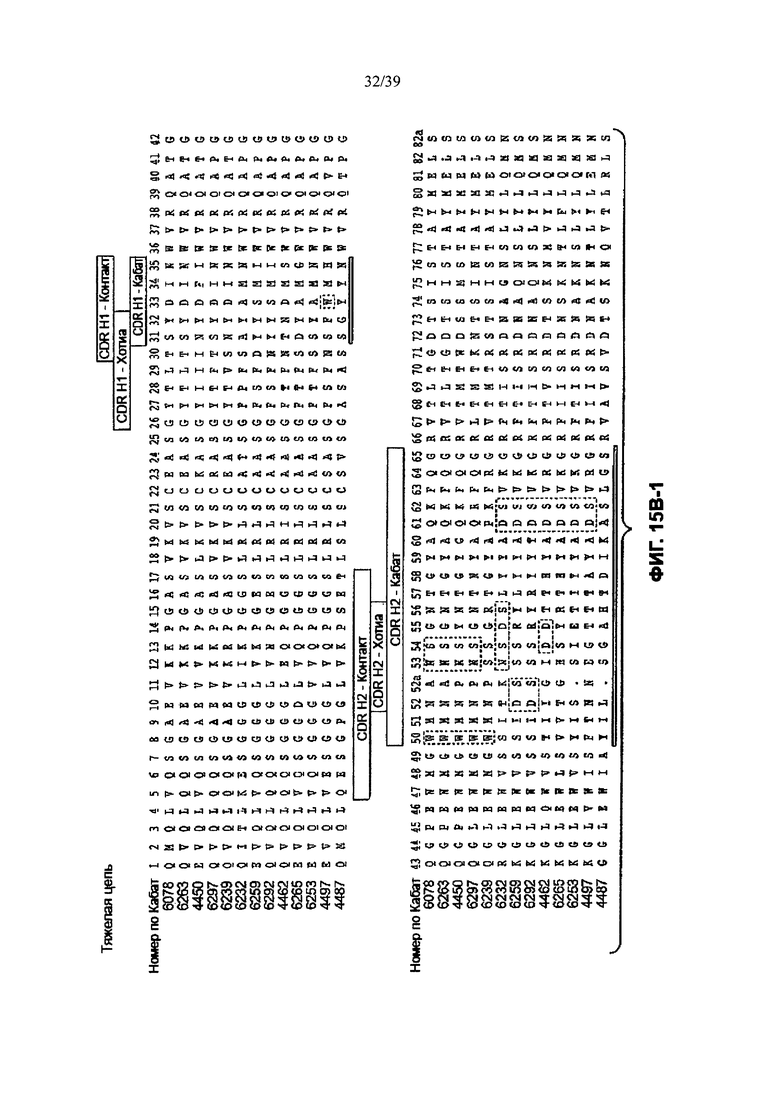
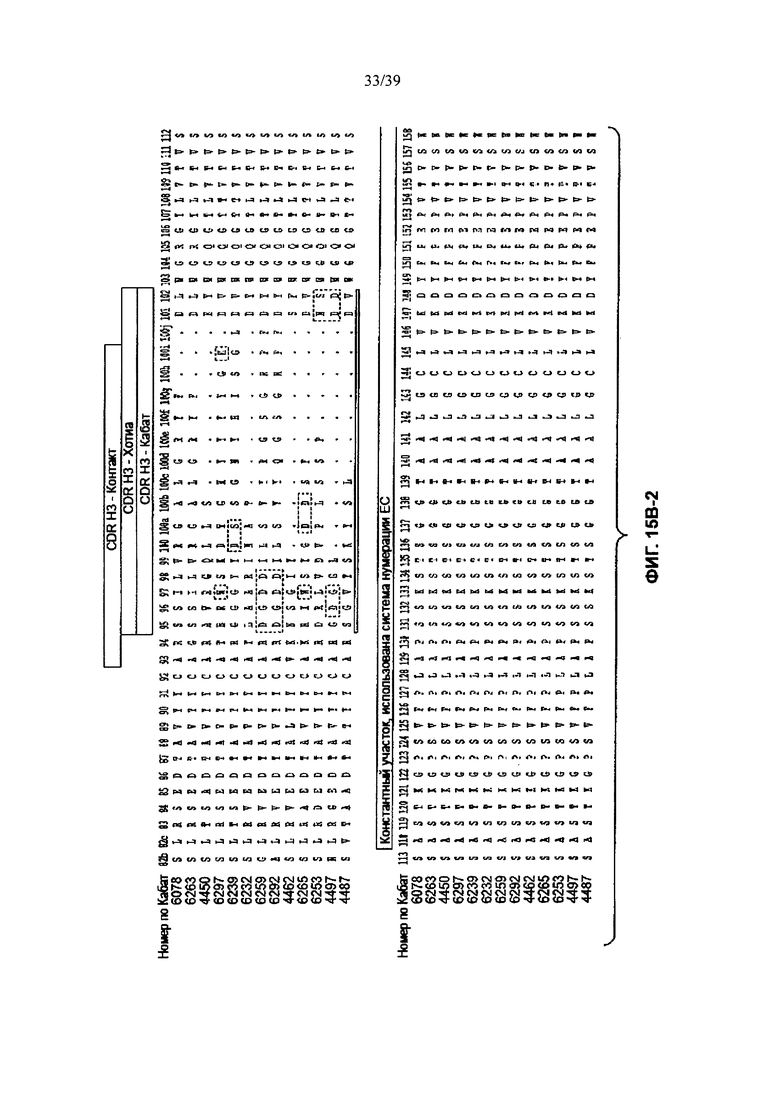
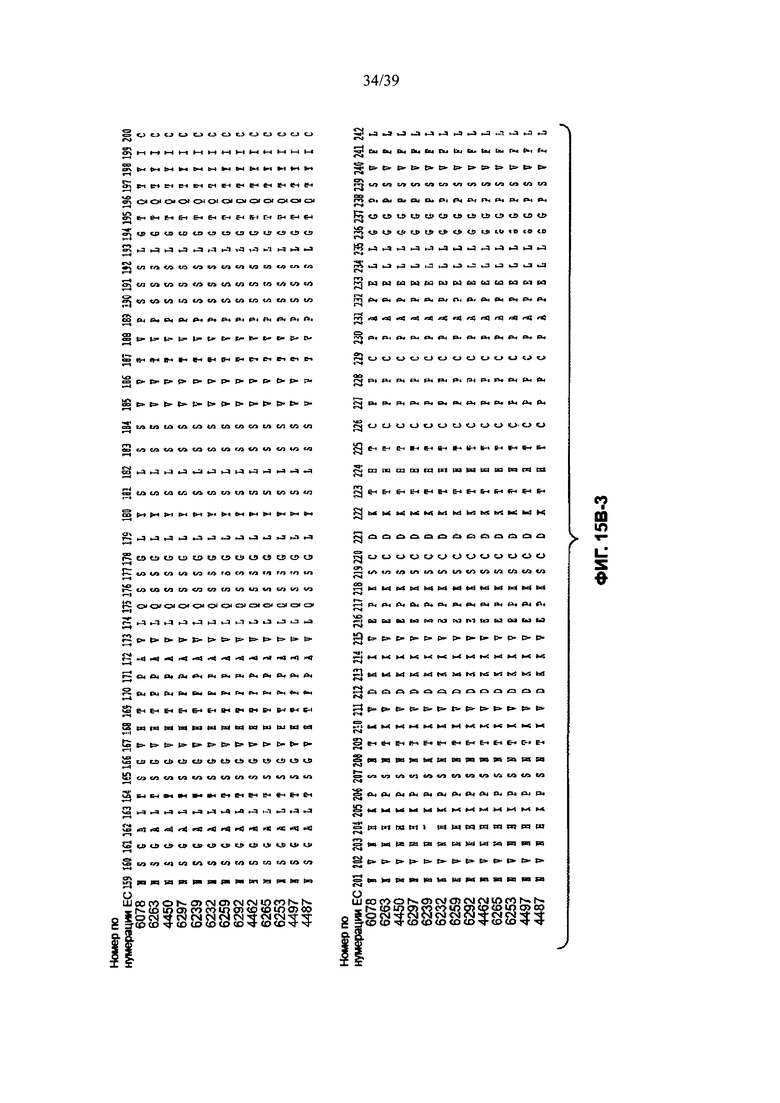
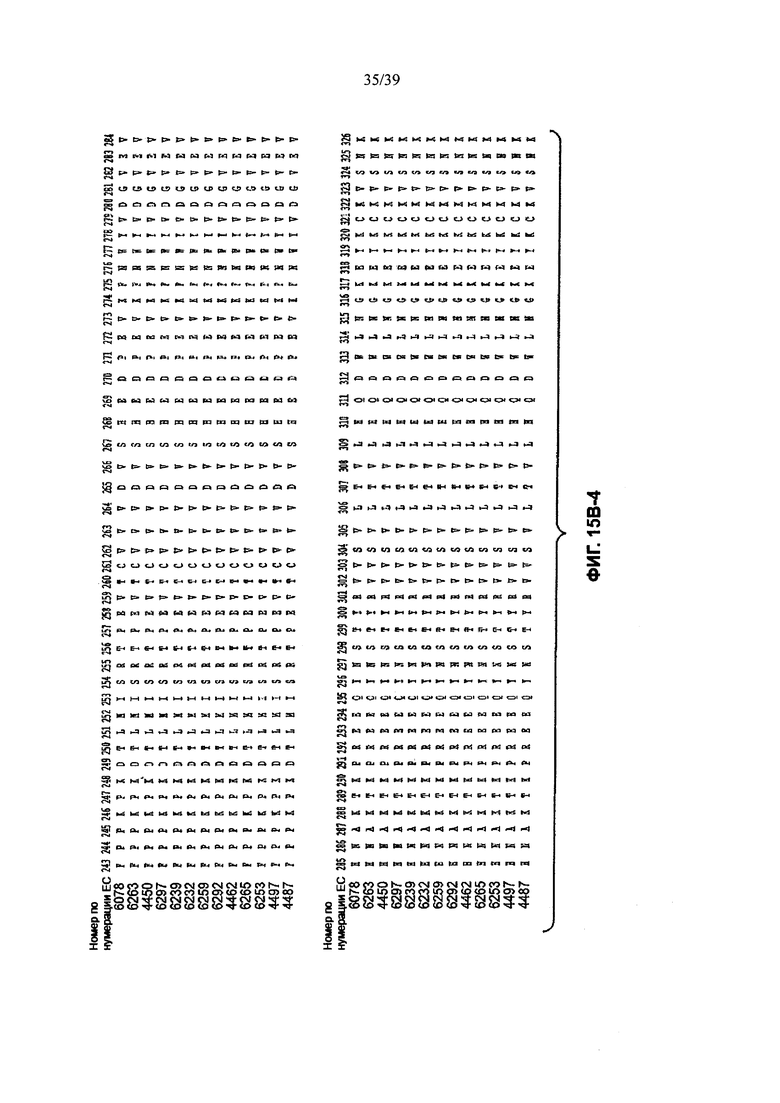
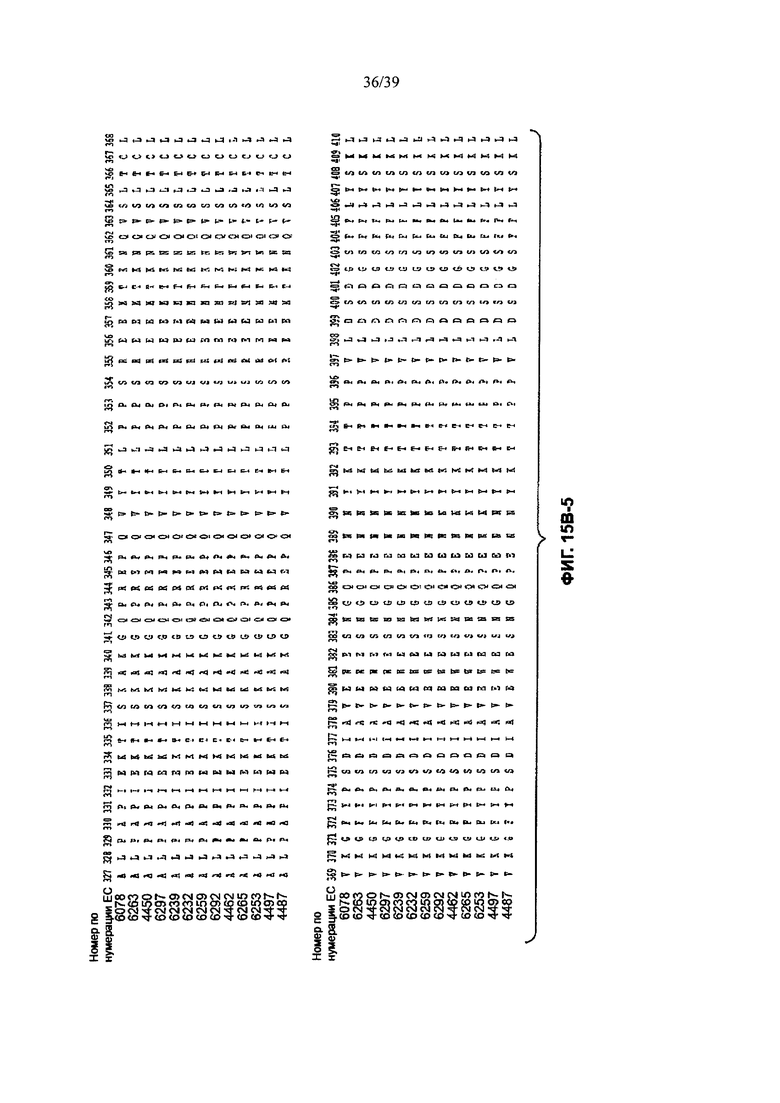
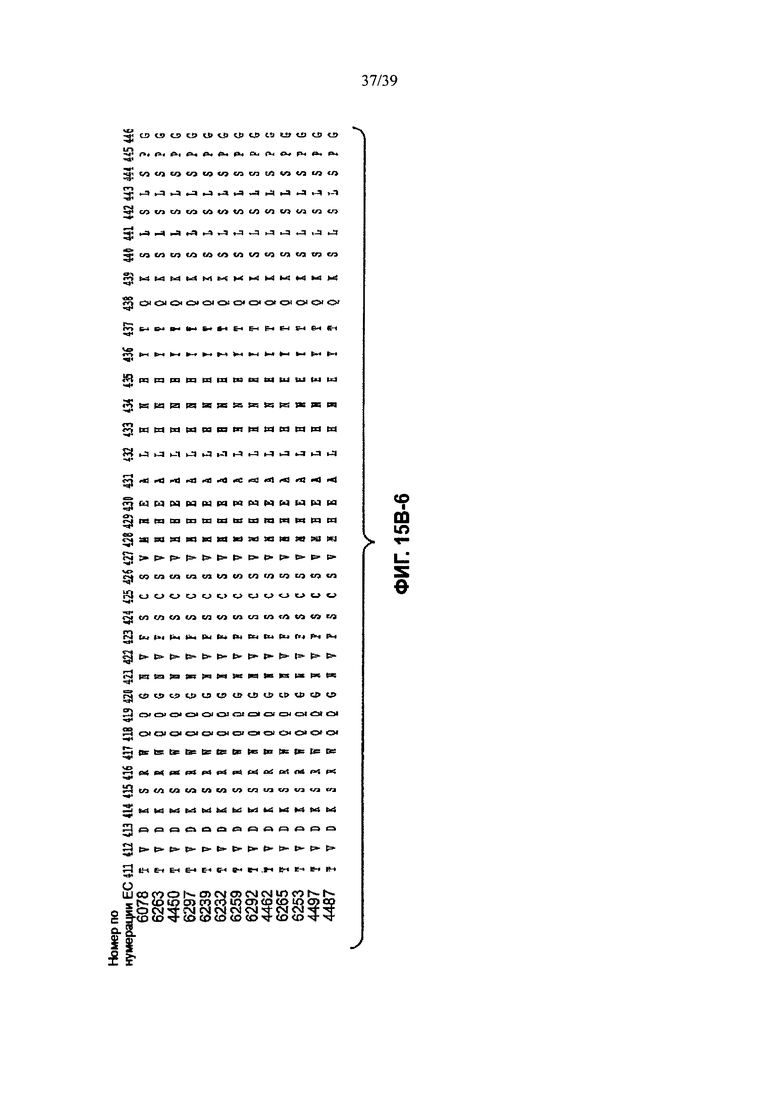
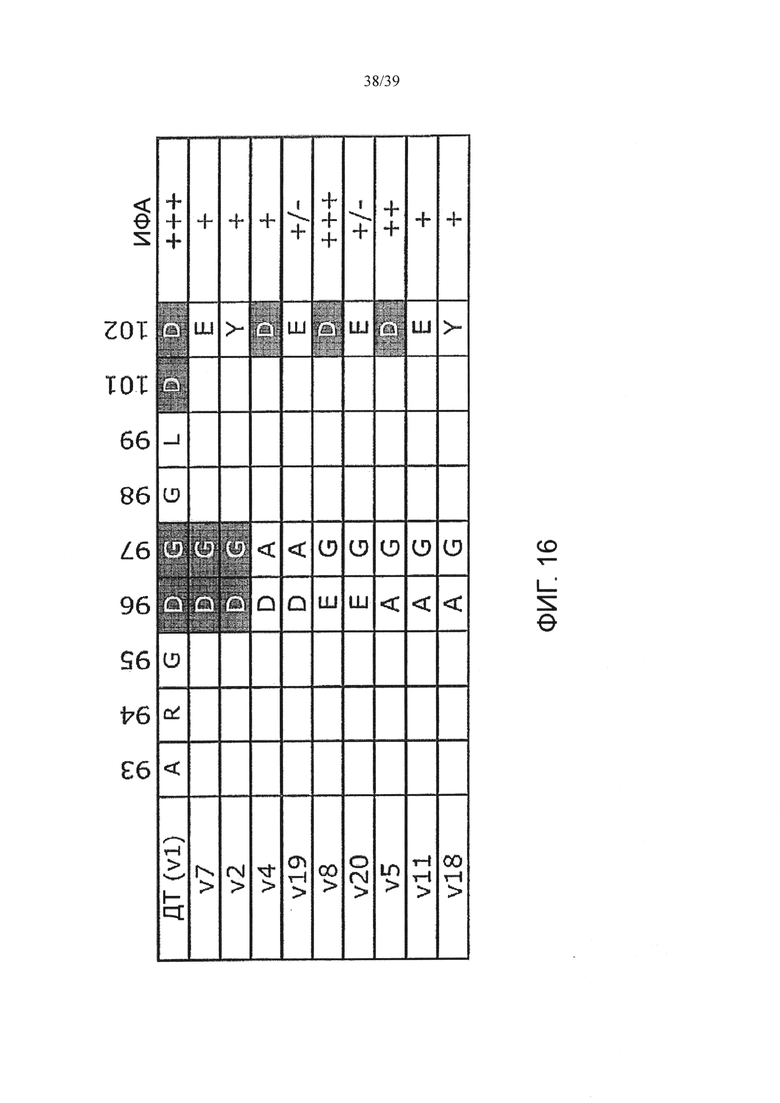
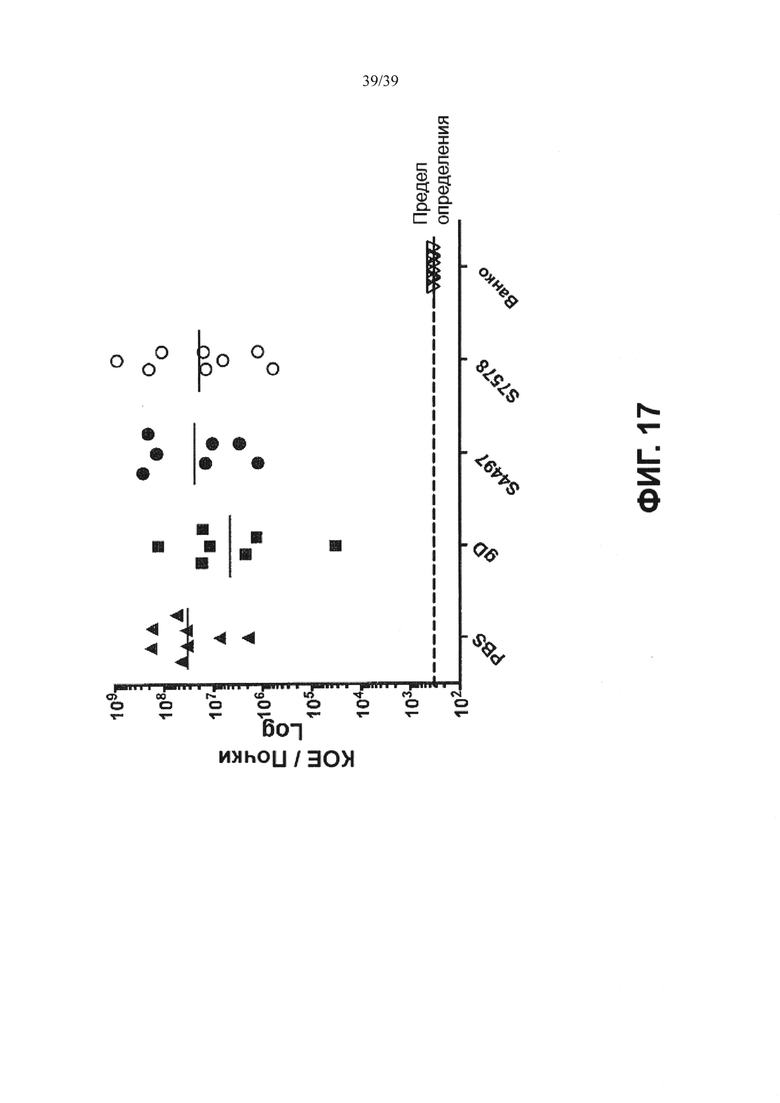
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая соединение конъюгата антитело-антибиотик для лечения бактериальной инфекции, вызванной одним или более из Staphylococcus aureus, Staphylococcus saprophytics или Staphylococcus simulans, содержащее WTA (тейхоевая кислота клеточной стенки) β антитело, фармацевтическую композицию для лечения бактериальной инфекции, вызванной одним или более из Staphylococcus aureus, Staphylococcus saprophyticus или Staphylococcus simulans, способ лечения бактериальной инфекции у пациента, инфицированного одним или более из Staphylococcus aureus, Staphylococcus saprophyticus или Staphylococcus simulans, способ уничтожения внутриклеточного Staphylococcus aureus в клетках пациента, инфицированного Staphylococcus aureus, без уничтожения клеток-хозяев, способ получения соединения конъюгата антитело-антибиотик, набор для лечения инфекции, вызванной Staphylococcus aureus, Staphylococcus saprophyticus и/или Staphylococcus simulans, и промежуточное соединение антибиотик-линкер для получения конъюгата антитело-антибиотик. Изобретение расширяет арсенал средств для лечения бактериальной инфекции, вызванной одним или более из Staphylococcus aureus, Staphylococcus saprophytics или Staphylococcus simulans. 7 н. и 22 з.п. ф-лы, 17 ил., 6 табл., 21 пр.
1. Соединение конъюгата антитело-антибиотик для лечения бактериальной инфекции, вызванной одним или более из Staphylococcus aureus, Staphylococcus saprophytics или Staphylococcus simulans, содержащее WTA (тейхоевая кислота клеточной стенки) β антитело, ковалентно присоединенное с помощью расщепляемого протеазой непептидного линкера к антибиотику типа рифамицина и имеющее формулу:
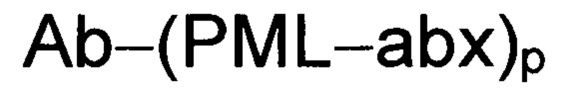
где:
Аb представляет собой WTAβ антитело,
где WTAβ антитело содержит легкую цепь (L) и тяжелую цепь (Н), причем: L-цепь содержит CDR L1, содержащий последовательность KSSQSIFRTSRNKNLLN (SEQ ID NO: 99), CDR L2, содержащий последовательность WASTRKS (SEQ ID NO: 100), и CDR L3, содержащий последовательность QQYFSPPYT (SEQ ID NO: 101); и Н-цепь содержит CDR H1, содержащий последовательность SFWMH (SEQ ID NO: 102), CDR Н2, содержащий последовательность FTNNEGTTTAYADSVRG (SEQ ID NO: 103), и CDR H3, содержащий последовательность GEGGLDD (SEQ ID NO: 118), или
где WTAβ антитело содержит легкую цепь (L) и тяжелую цепь (Н), причем: L-цепь содержит CDR L1, содержащий последовательность RASQTISGWLA (SEQ ID NO: 33), CDR L2, содержащий последовательность KASTLES (SEQ ID NO: 34), и CDR L3, содержащий последовательность QQYKSYSFN (SEQ ID NO: 35); и Н-цепь содержит CDR H1, содержащий последовательность SYDIN (SEQ ID NO: 36), CDR Н2, содержащий последовательность WMNANSGNTGYAQKFQG (SEQ ID NO: 37), и CDR H3, содержащий последовательность SSILVRGALGRYFDL (SEQ ID NO: 38), или
где WTAβ антитело содержит легкую цепь (L) и тяжелую цепь (Н), причем:
(а) L-цепь содержит CDR L1, содержащий последовательность RASQTISGWLA (SEQ ID NO: 39), CDR L2, содержащий последовательность KASTLES (SEQ ID NO: 40), и CDR L3 содержащий последовательность QQYKSYSFN (SEQ ID NO: 41); и Н-цепь содержит CDR H1, содержащий последовательность SYDIN (SEQ ID NO: 42), CDR H2, содержащий последовательность WMNANSGNTGYAQKFQG (SEQ ID NO: 43), и CDR Н3, содержащий последовательность SSILVRGALGRYFDL (SEQ ID NO: 44);
(b) L-цепь содержит CDR L1, содержащий последовательность RASQFVSRTSLA (SEQ ID NO: 45), CDR L2, содержащий последовательность ETSSRAT (SEQ ID NO: 46), и CDR L3, содержащий последовательность HKYGSGPRT (SEQ ID NO: 47); и Н-цепь содержит CDR H1, содержащий последовательность NYDFI (SEQ ID NO: 48), CDR H2, содержащий последовательность WMNPNSYNTGYGQKFQG (SEQ ID NO: 49), и CDR H3, содержащий последовательность AVRGQLLSEY (SEQ ID NO: 50);
(c) L-цепь содержит CDR L1, содержащий последовательность RASQSVSSSYLA (SEQ ID NO: 51), CDR L2, содержащий последовательность DASSRAT (SEQ ID NO: 52), и CDR L3, содержащий последовательность QKYGSTPRP (SEQ ID NO: 53); и Н-цепь содержит CDR H1, содержащий последовательность SYDIN (SEQ ID NO: 54), CDR H2, содержащий последовательность WMNPNSGNTNYAQRFQG (SEQ ID NO: 55), и CDR H3, содержащий последовательность ERWSKDTGHYYYYGMDV (SEQ ID NO: 56);
(d) L-цепь содержит CDR L1, содержащий последовательность RASLDITNHLA (SEQ ID NO: 57), CDR L2, содержащий последовательность EASILQS (SEQ ID NO: 58), и CDR L3, содержащий последовательность EKCNSTPRT (SEQ ID NO: 59); и Н-цепь содержит CDR H1, содержащий последовательность NYDIN (SEQ ID NO: 60), CDR H2, содержащий последовательность WMNPSSGRTGYAPKFRG (SEQ ID NO: 61), и CDR H3, содержащий последовательность GGGYYDSSGNYH1SGLDV (SEQ ID NO: 62);
(e) L-цепь содержит CDR L1, содержащий последовательность RASQSVGAIYLA (SEQ ID NO: 63), CDR L2, содержащий последовательность GVSNRAT (SEQ ID NO: 64), и CDR L3, содержащий последовательность QLYTSSRALT (SEQ ID NO: 65); и Н-цепь содержит CDR H1, содержащий последовательность AYAMN (SEQ ID NO: 66), CDR H2, содержащий последовательность SITKNSDSLYYADSVKG (SEQ ID NO: 67), и CDR H3, содержащий последовательность LAARIMATDY (SEQ ID NO: 68);
(f) L-цепь содержит CDR L1, содержащий последовательность RASQGIRNGLG (SEQ ID NO: 69), CDR L2, содержащий последовательность PASTLES (SEQ ID NO: 70), и CDR L3, содержащий последовательность LQDHNYPPT (SEQ ID NO: 71); и Н-цепь содержит CDR H1, содержащий последовательность YYSMI (SEQ ID NO: 72), CDR H2, содержащий последовательность SIDSSSRYLYYADSVKG (SEQ ID NO: 73), и CDR H3, содержащий последовательность DGDDILSVYRGSGRPFDY (SEQ ID NO: 74);
(g) L-цепь содержит CDR L1, содержащий последовательность RASQGIRNGLG (SEQ ID NO: 75), CDR L2, содержащий последовательность PASTLES (SEQ ID NO: 76), и CDR L3, содержащий последовательность LQDHNYPPS (SEQ ID NO: 77); и Н-цепь содержит CDR H1, содержащий последовательность YYSMI (SEQ ID NO: 78), CDR Н2, содержащий последовательность SIDSSSRYRYYTDSVKG (SEQ ID NO: 79), и CDR H3, содержащий последовательность DGDDILSVYQGSGRPFDY (SEQ ID NO: 80);
(h) L-цепь содержит CDR L1, содержащий последовательность RASQSVRTNVA (SEQ ID NO: 81), CDR L2, содержащий последовательность GASTRAS (SEQ ID NO: 82), и CDR L3, содержащий последовательность LQYNTWPRT (SEQ ID NO: 83); и Н-цепь содержит CDR H1, содержащий последовательность TNDMS (SEQ ID NO: 84), CDR H2, содержащий последовательность TIIGIDDTTHYADSVRG (SEQ ID NO: 85), и CDR H3, содержащий последовательность NSGIYSF (SEQ ID NO: 86);
(i) L-цепь содержит CDR L1, содержащий последовательность RASQDIGSSLA (SEQ ID NO: 87), CDR L2, содержащий последовательность ATSTLQS (SEQ ID NO: 88), и CDR L3, содержащий последовательность QQLNNYVHS (SEQ ID NO: 89); и Н-цепь содержит CDR H1, содержащий последовательность DYAMG (SEQ ID NO: 90), CDR H2, содержащий последовательность VVTGHSYRTHYADSVKG (SEQ ID NO: 91), и CDR H3, содержащий последовательность RIWSYGDDSFDV (SEQ ID NO: 92);
(j) L-цепь содержит CDR L1, содержащий последовательность RASQSIGDRLA (SEQ ID NO: 93), CDR L2, содержащий последовательность WASNLEG (SEQ ID NO: 94), и CDR L3, содержащий последовательность QQYKSQWS (SEQ ID NO: 95); и Н-цепь содержит CDR H1, содержащий последовательность SYAMN (SEQ ID NO: 96), CDR H2, содержащий последовательность YISSIETIYYADSVKG (SEQ ID NO: 97), и CDR H3, содержащий последовательность DRLVDVPLSSPNS (SEQ ID NO: 98);
(k) L-цепь содержит CDR L1, содержащий последовательность RASQFTNHYLN (SEQ ID NO: 105), CDR L2, содержащий последовательность VASNLQS (SEQ ID NO: 106), и CDR L3, содержащий последовательность QQSYRTPYT (SEQ ID NO: 107); и Н-цепь содержит CDR H1, содержащий последовательность SGYYN (SEQ ID NO: 108), CDR H2, содержащий последовательность YILSGAHTDIKASLGS (SEQ ID NO: 109), и CDR H3, содержащий последовательность SGVYSKYSLDV (SEQ ID NO: 110);
PML представляет собой расщепляемый протеазой непептидный линкер, имеющий формулу:

где Str представляет собой растягивающую единицу; РМ представляет собой пептидомиметическую единицу и Y представляет собой спейсерную единицу; abx представляет собой антибиотик рифамицинового типа и Р представляет собой целое число от 1 до 8, где Str имеет формулу:
где R6 выбирают из группы, состоящей из С1-С12 алкилена, С1-С12 алкилен-С(=O), C1-С12 алкилен-NH, (СН2СН2O)r, (СН2СН2O)r-С(=O), (СН2СН2O)r-СН2 и С1-С12 алкилен-NНС(=O)СН2СН(тиофен-3-ил), где r представляет собой целое число от 1 до 10;
где Y содержит пара-аминобензил (РАВ) или пара-аминобензилоксикарбонил (РАВС);
и
где РМ имеет формулу:
где R7 и R8 вместе образуют С3-С7 циклоалкильное кольцо и
АА представляет собой боковую цепь аминокислоты, выбранную из Н, -СН3, -СН2(С6Н5), -СН2СН2СН2СН2NH2, -СН2СН2СН2NHC(NH)NH2, -СНСН(СН3)СН3 и -СН2СН2СН2NHC(O)NH2.
2. Соединение конъюгата антитело-антибиотик по п. 1, где антибиотик рифамицинового типа представляет собой антибиотик типа рифалазила.
3. Соединение конъюгата антитело-антибиотик по п. 1, где антибиотик типа рифамицина содержит катион аммония, присоединенный к расщепляемому протеазой непептидному линкеру.
4. Соединение конъюгата антитело-антибиотик по п. 1, имеющее формулу:
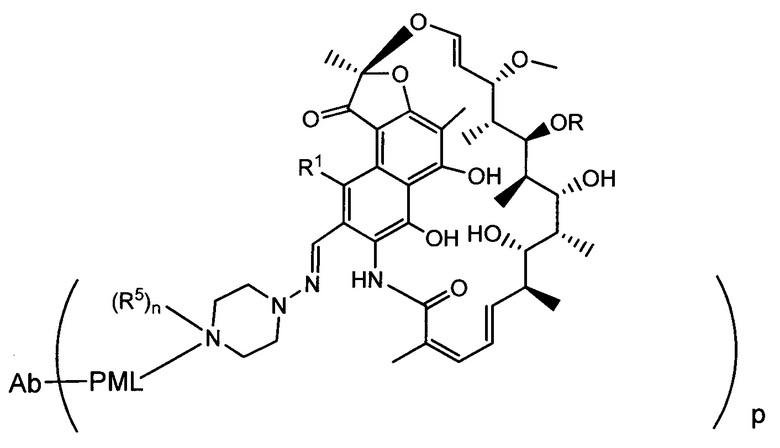
где
R5 выбран из Н и С1-С12 алкила; и
n равен 0 или 1.
5. Соединение конъюгата антитело-антибиотик по п. 1, имеющее формулу:
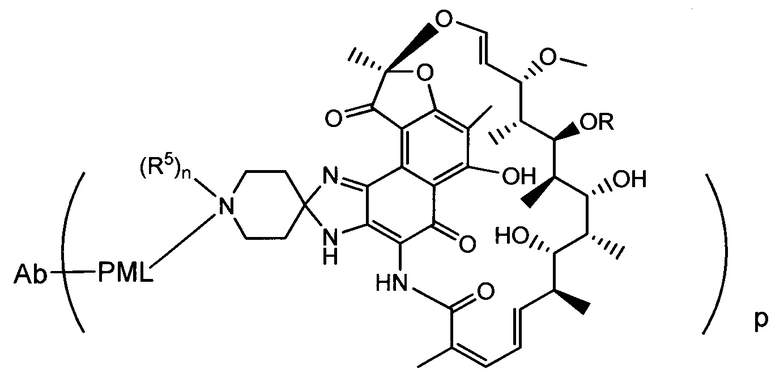
где
R5 выбран из Н и С1-С12 алкила; и
n равен 0 или 1.
6. Соединение конъюгата антитело-антибиотик по п. 1, имеющее формулу:
где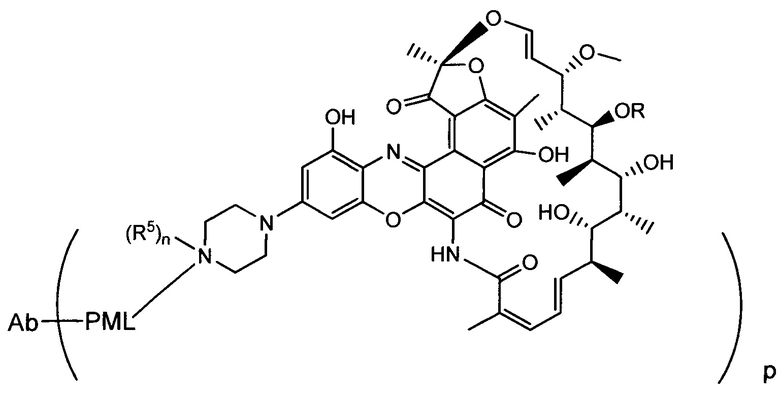
R5 независимо выбран из Н и C1-C12 алкила; и
n равен 0 или 1.
7. Соединение конъюгата антитело-антибиотик по п. 1, имеющее формулу:
где 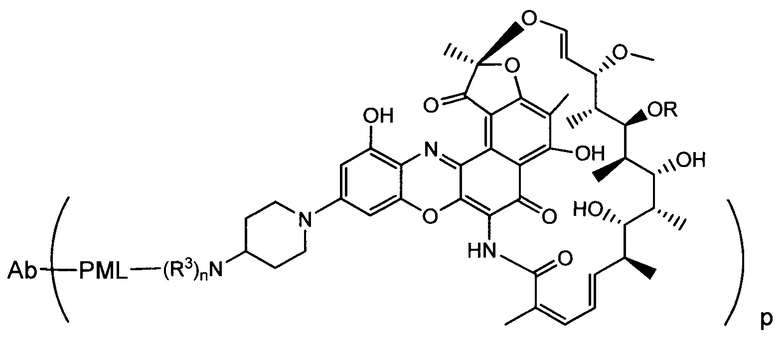
R3 независимо выбран из Н и C1-C12 алкила; и
n равно 1 или 2.
8. Соединение конъюгата антитело-антибиотик по п. 7, имеющее формулу:
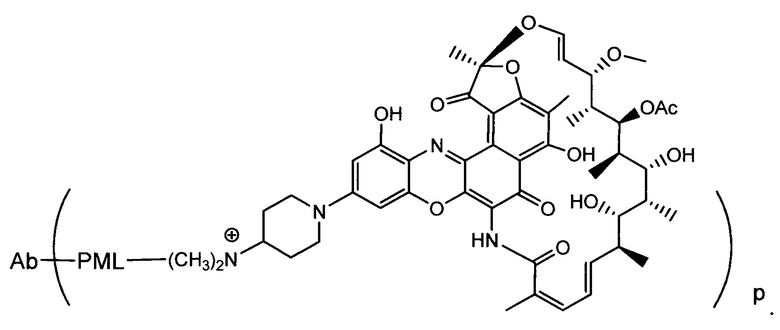
9. Соединение конъюгата антитело-антибиотик по п. 1, где R6 представляет собой (СН2)5.
10. Соединение конъюгата антитело-антибиотик по п. 1, где Y содержит пара-аминобензил или пара-аминобензилоксикарбонил.
11. Соединение конъюгата антитело-антибиотик по п. 1, выбранное из формул:
i)
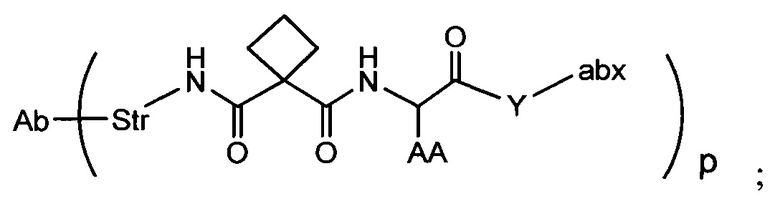
ii)
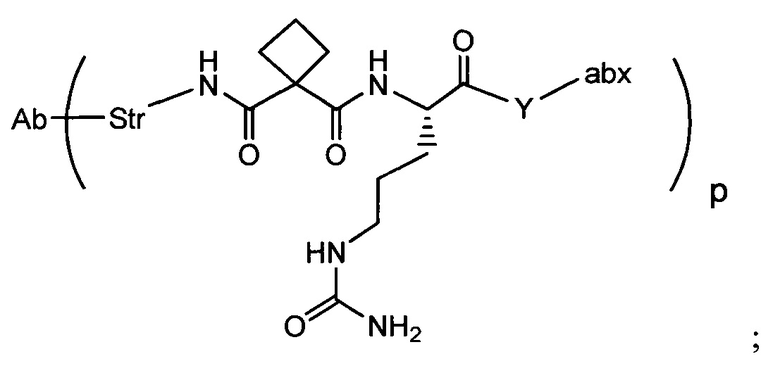
iii)
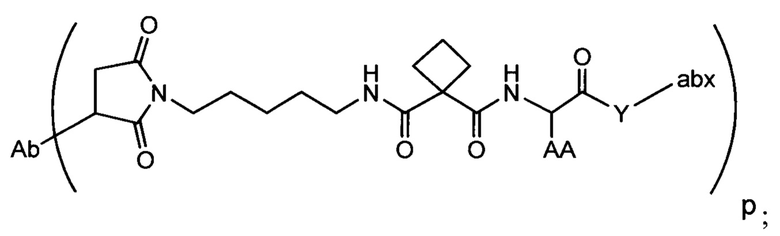
iv)
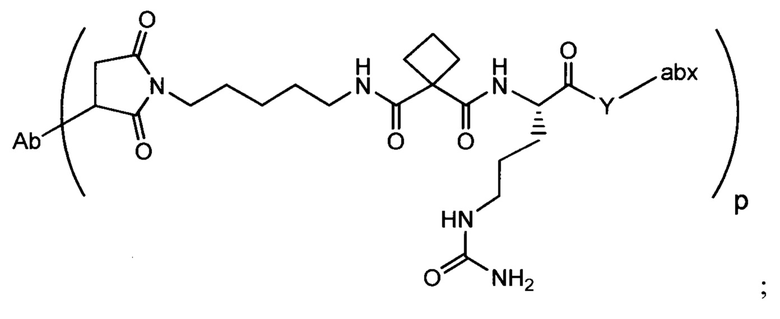
v)
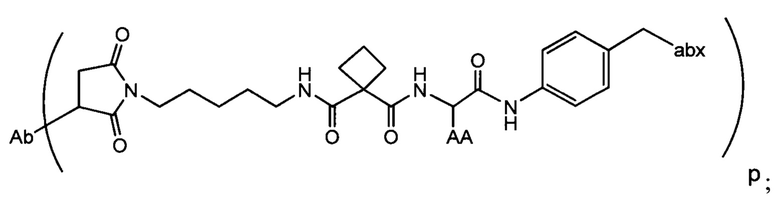
vi)
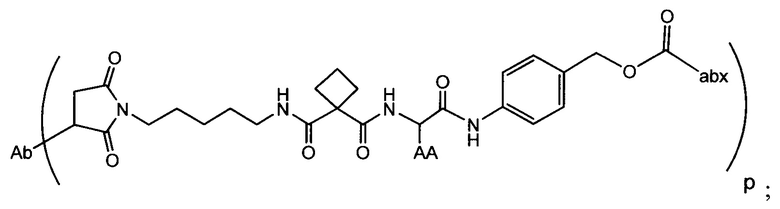
vii)
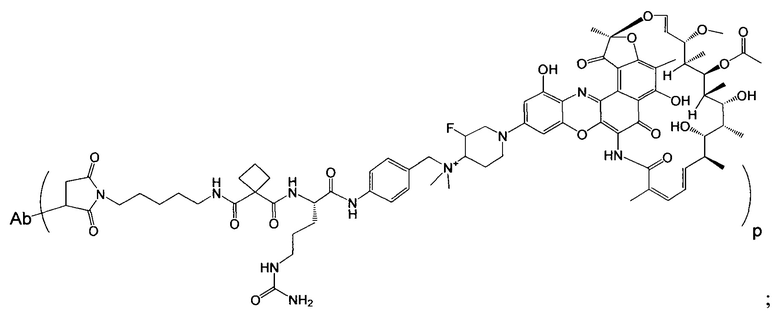
viii)
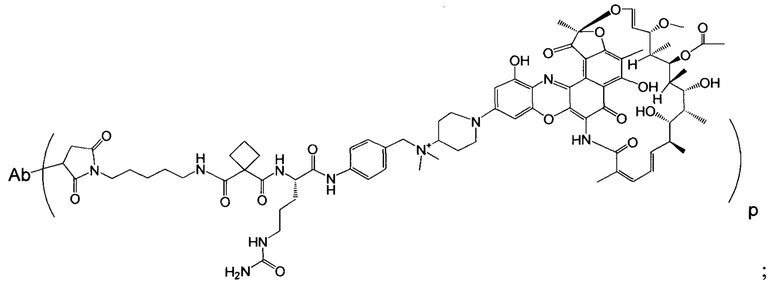
ix)
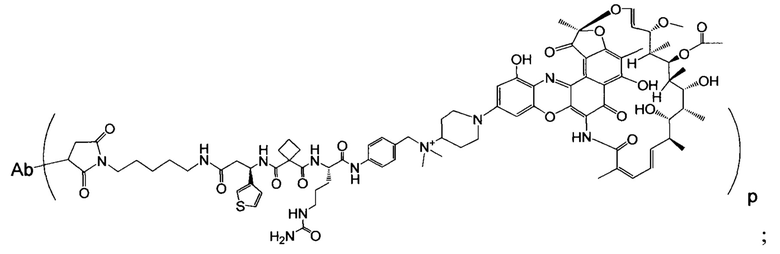
x)
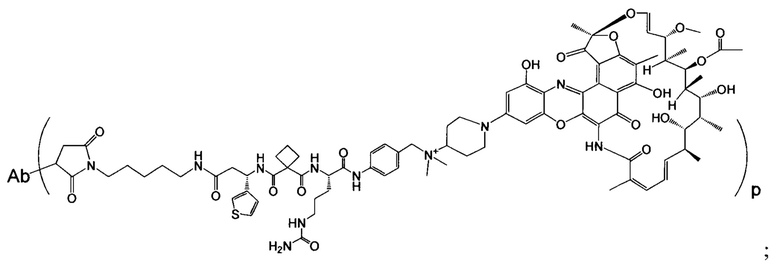
xi)
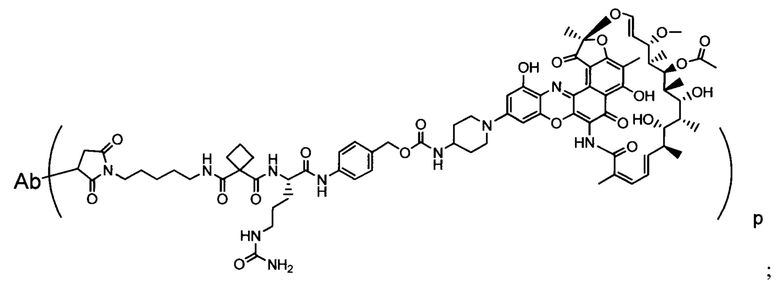
и
xii)
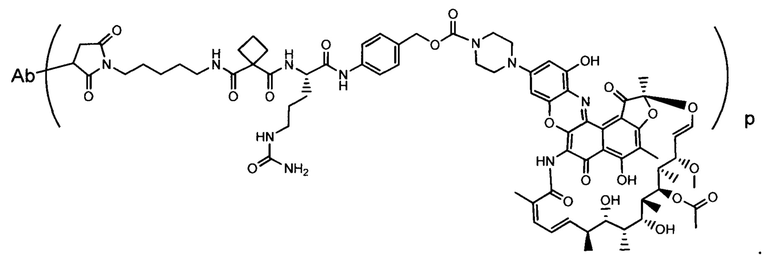
12. Соединение конъюгата антитело-антибиотик по п. 1, где антитело представляет собой WTAβ моноклональное антитело.
13. Соединение конъюгата антитело-антибиотик по п. 1, где WTAβ антитело содержит:
(а) вариабельную область (VH) тяжелой цепи, где VH содержит аминокислотную последовательность, имеющую SEQ ID NO: 156, и/или
(b) вариабельную область (VL) легкой цепи, где VL содержит аминокислотную последовательность, имеющую SEQ ID NO: 119.
14. Соединение конъюгата антитело-антибиотик по п. 1, где WTAβ антитело содержит:
(a) вариабельную область (VH) тяжелой цепи, где VH содержит аминокислотную последовательность, имеющую SEQ ID NO: 112, и/или
(b) вариабельную область (VL) легкой цепи, где VL содержит аминокислотную последовательность, имеющую SEQ ID NO: 111.
15. Соединение конъюгата антитело-антибиотик по любому из предшествующих пунктов, где WTAβ антитело связывается со Staphylococcus aureus.
16. Соединение конъюгата антитело-антибиотик по любому из предшествующих пунктов, где антитело представляет собой F(ab) или F(ab')2.
17. Фармацевтическая композиция для лечения бактериальной инфекции, вызванной одним или более из Staphylococcus aureus, Staphylococcus saprophyticus или Staphylococcus simulans, содержащая соединение конъюгата антитело-антибиотик, содержащее терапевтически эффективное количество WTAβ антитела, ковалентно присоединенного с помощью расщепляемого протеазой непептидного линкера к антибиотику типа рифамицина, по п. 1 и фармацевтически приемлемый носитель, глидант, разбавитель или вспомогательное вещество.
18. Способ лечения бактериальной инфекции у пациента, инфицированного одним или более из Staphylococcus aureus, Staphylococcus saprophyticus или Staphylococcus simulans, включающий введение пациенту терапевтически эффективного количества соединения конъюгата антитело-антибиотик по п. 1.
19. Способ по п. 18, где пациент инфицирован бактериями типа Staphylococcal.
20. Способ по п. 19, где пациент инфицирован Staphylococcus aureus.
21. Способ по п. 18, где соединение конъюгата антитело-антибиотик вводят пациенту в дозе в диапазоне от около 50 мг/кг до 100 мг/кг.
22. Способ уничтожения внутриклеточного Staphylococcus aureus в клетках пациента, инфицированного Staphylococcus aureus, без уничтожения клеток-хозяев путем введения соединения конъюгата антитело-антибиотик по п. 1.
23. Способ получения соединения конъюгата антитело-антибиотик по п. 1, включающий конъюгирование антибиотика рифамицинового типа с WTAβ антителом.
24. Набор для лечения инфекции, вызванной Staphylococcus aureus, Staphylococcus saprophyticus и/или Staphylococcus simulans, содержащий:
а) фармацевтическую композицию по п. 17; и
б) инструкции по использованию.
25. Промежуточное соединение антибиотик-линкер для получения конъюгата антитело-антибиотик, имеющее формулу II:
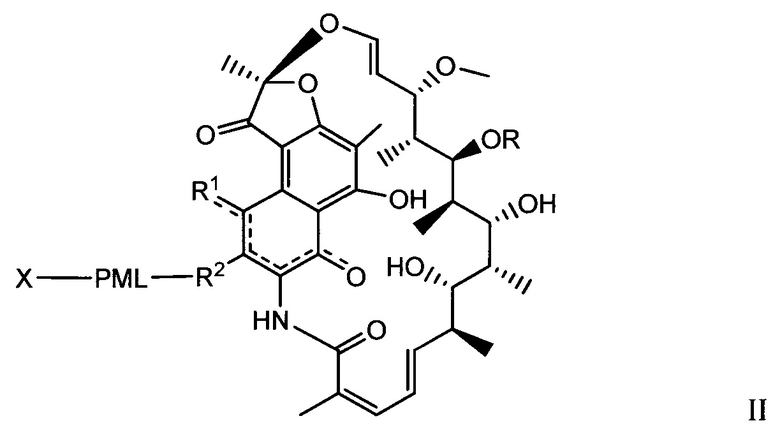
где:
пунктирные линии обозначают необязательную связь;
R представляет собой Н, C1-С12 алкил или С(O)СН3;
R1 представляет собой ОН; и
R2 представляет собой СН=N-(гетероциклил), причем гетероциклил замещен группой, независимо выбранной из С(O)СН3, C1-С12 алкил, C1-С12 гетероарил, С2-С20 гетероциклил, С6-С20 арил и С3-С12 карбоциклил;
или R1 и R2 образуют пяти- или шестичленный конденсированный гетероарил или гетероциклил и необязательно образуют спиро или конденсированный шестичленный гетероарил, гетероциклил, арил или карбоциклическое кольцо, где спиро или конденсированный шестичленный гетероарил, гетероциклил, арил или карбоциклическое кольцо необязательно замещены Н, F, Cl, Br, I, C1-C2 алкилом или ОН;
PML представляет собой расщепляемый протеазой непептидный линкер, присоединенный к R2 или конденсированный гетероарил или гетероциклил, образованный R1 и R2, и имеющий формулу:
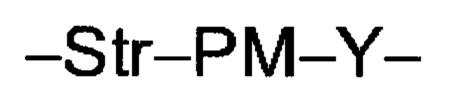
где Str представляет собой растягивающую единицу; РМ представляет собой пептидомиметическую единицу и Y представляет собой спейсерную единицу; и
X представляет собой реакционноспособную функциональную группу, выбранную из малеимида, тиола, амино, бромида, бромацетамида, иодацетамида, п-толуолсульфоната, иодида, гидроксила, карбоксила, пиридилдисульфида и N-гидроксисукцинимида,
где РМ имеет формулу:
где R7 и R8 вместе образуют С3-С7 циклоалкильное кольцо и
АА представляет собой боковую цепь аминокислоты, выбранную из Н, -СН3, -СН2(С6Н5), -СН2СН2СН2СН2NH2, -СН2СН2СН2NHC(NH)NH2, -СНСН(СН3)СН3 и -СН2СН2СН2NHC(O)NH2,
где Str имеет формулу:
где R6 выбирают из группы, состоящей из С1-С12 алкилена, С1-С12 алкилен-С(=O), C1-С12 алкилен-NH, (СН2СН2O)r, (СН2СН2O)r-С(=O), (СН2СН2O)r-СН2 и С1-С12 алкилен-NНС(=O)СН2СН(тиофен-3-ил), где r представляет собой целое число от 1 до 10; и
при этом Y содержит пара-аминобензил (РАВ) или пара-аминобензилоксикарбонил (РАВС).
26. Промежуточное соединение антибиотик-линкер по п. 25, где X является
 или
или 
27. Промежуточное соединение антибиотик-линкер по п. 25, имеющее формулу:
где 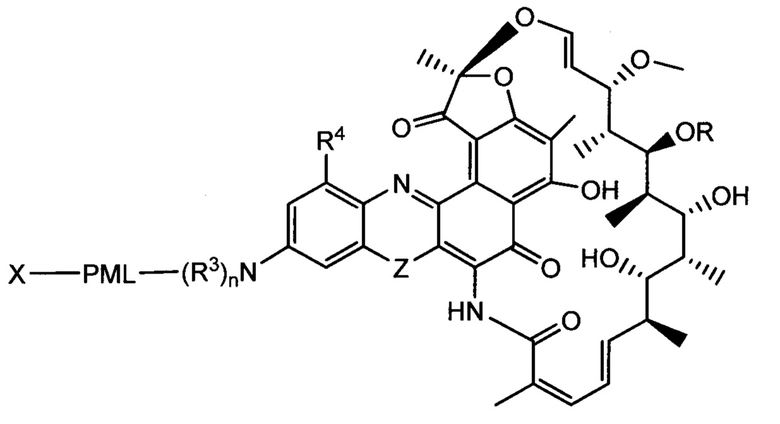
R3 независимо выбран из Н и C1-C12 алкила;
n равно 1 или 2;
R4 выбран из Н, F, Cl, Br, I, С1-С12 алкила и ОН; и
Z выбран из NH, N(C1-C12 алкил), О и S.
28. Промежуточное соединение антибиотик-линкер по п. 25, имеющее формулу:
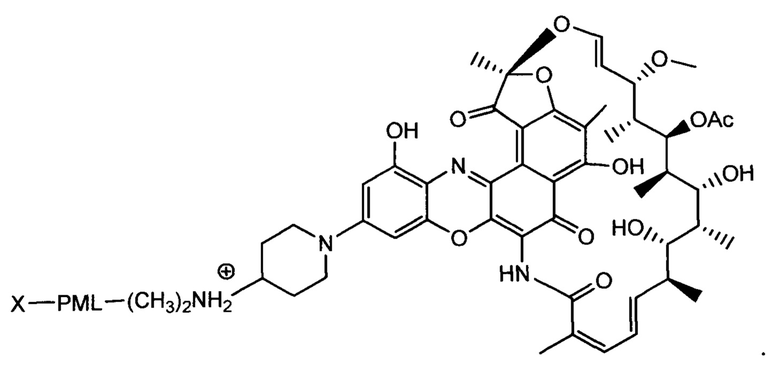
29. Промежуточное соединение антибиотик-линкер по п. 25, выбранное из формул:
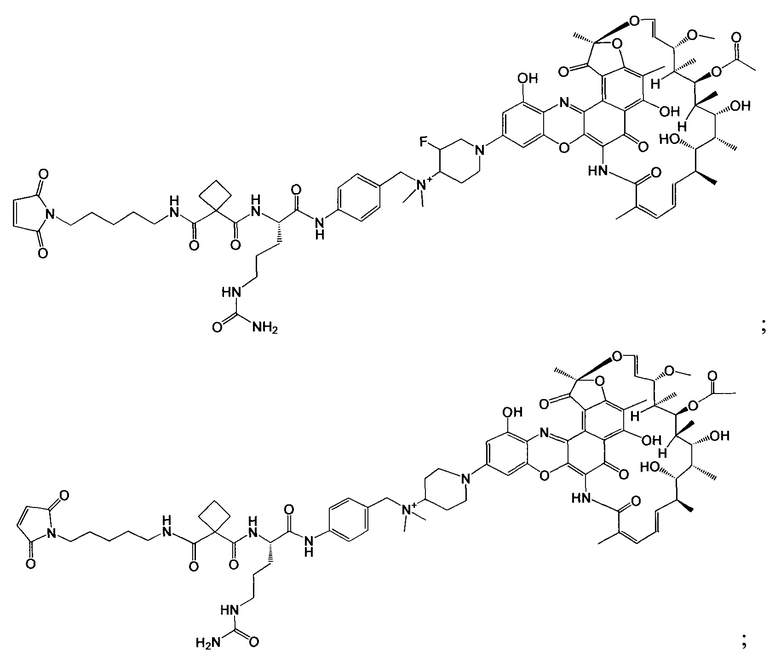
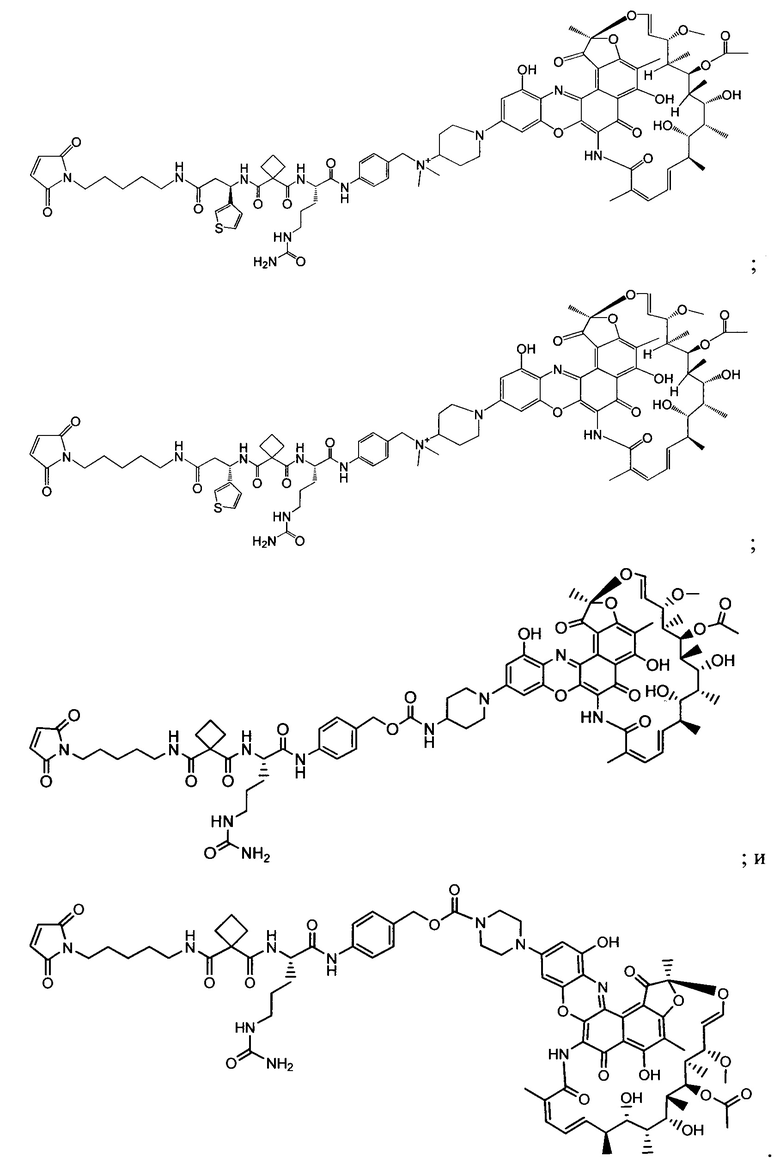
| WO 2011008092 A4, 20.01.2011 | |||
| WO 2014080251 A1, 30.05.2014 | |||
| WO 2005081711 A2, 09.09.2005 | |||
| WO 2007096703 A2, 30.08.2007 | |||
| RU 2015151046 A, 30.06.2017 | |||
| WO 2007121484 A2, 25.10.2007. |

