Изобретение принадлежит области биотехнологии и представляет собой частный случай условий обеспечивающих пассивную иммобилизацию труднорастворимых белков, полученных генно-инженерным способом (например, мембранных), на поверхности полистирола, способствующий в итоге формированию иммунных комплексов при постановке твердофазного иммуноферментного анализа (ИФА).
Иммуноферментный анализ прочно удерживает позиции в арсенале современных аналитических методов исследования благодаря высокой специфичности, чувствительности, универсальности, скорости и относительной простоты выполнения. Метод включает последовательное развитие иммунохимических взаимодействий, при этом начальные и последующие составляющие анализа иммобилизованы на твердой фазе, что позволяет легко удалять использованные реагенты, вводить новые и осуществлять накопление детектируемых продуктов в реакционных объемах растворов, кратно увеличивая тем самым чувствительность определения в широком динамическом диапазоне [EngvallE: 1977. Med Biol., 55 (4): 193-200, Kohl Т.О., Ascoli C.A: 2017(7). Cold Spring HarbProtoc, doi: 10.1101/pdb.prot093757, Kohl Т.О., Ascoli C.A: 2017(6). ColdSpringHarbProtoc., doi: 10.1101/pdb.prot093724.]. Базируясь поначалу на формате 96-луночного полистирольного микропланшета (далее в тексте - планшет) и отвечая возникающим задачам, физическая форма метода подверглась разнообразным изменениям трансформируясь в тест-полоски, мембраны биодатчиков, гелевые матрицы, микросуспензии частиц (полистирол, стекло, латекс, металл), при этом значительно увеличились масштабы тестов и автоматизация работ с жидкостями, чтением и обработкой результатов.
Хорошо известные разнообразные тесты для серодиагностики инфекций, радиоиммунологические, иммунодиффузия, иммунофлуоресценция, иммунопреципитация, агглютинация, гемолиз, нейтрализация и твердофазный иммуноферментный анализ (ИФА), включающие способы обнаружения антительных иммунных комплексов, этап усиления сигнала и детекцию различными методами, такими как оптические, по интенсивности цвета, флуоресценции, размеру частиц, альтернативно - путем измерения радиоактивности меток в иммунных комплексах. Диагностическим тестом, часто используемым в практике является иммуноферментный анализ (ИФА) с множеством вариантов проведения, типичным начальным шагом является иммобилизация антигенов или антител на твердой фазе, такой как лунки планшета [Патент РФ 2689143 С1], этап иммобилизации является критическим и, в целом, определяет чувствительность теста. На следующих стадиях возможно использование двух вариантов постановки анализа: так называемого теста с «захватом» или сэндвич-ИФА и конкурентнго ИФА (блокирующего, ингибирующего). При сэндвич-ИФА происходит последовательное введение компонентов тест-системы, вследствие иммунохимических взаимодействий молекулярные слои как бы наслаиваются один на другой. Во втором варианте, как полагают более специфичном, определенный компонент системы конкурирует за связывание с меченым антигеном или антителом и уменьшает максимальный регистрируемый сигнал, что фиксируется и выражается в виде доли ингибирования максимального сигнала ИФА. Естественным и необходимым условием является растворимость антигенов в рабочих буферах. В силу относительной простоты, доступности, универсальности и чувствительности ИФА широко используют не только в качестве метода лабораторной диагностики, но и в эпидемиологической практике, как инструмент массового обследования населения и отбора лиц для проведения вакцинации. В этом контексте критичным является чувствительность и специфичность ИФА-тестов.
Известно, что необходимой первичной составляющей ИФА является наличие подготовленной подложки с иммобилизованными компонентами иммунохимической системы, в частности белковыми антигенами. В настоящее время используют три разновидности иммобилизации белков: пассивную, ковалентную и аффинную. Пассивное связывание адекватно отвечает разнообразным задачам как исследований макромолекул, так и рутинных анализов и наиболее широко используется в практике. Степень пассивного связывания определяется соотношением гидрофильных и гидрофобных компартментов макромолекулы и соответствующим этому параметру характерам сорбирующих поверхностей полистирола, от гидрофобных до преимущественно гидрофильных. При оптимальном соотношении этих параметров количество сорбированного белка на поверхности полистирола может превосходить величину 600 нг/см2. Ковалентные поверхности для ИФА представляют собой химически модифицированный полистирол, введенные активные группы которого способны образовывать прочные ковалентные связи с NH2-, СООН- группами гаптенов, пептидов или 5'-производными олигонуклеотидов. Такие планшеты преимущественно используют для фиксации небольших молекул [Rasmussen S.R., Larsen M.R., Rasmussen S.E: 1991. AnalBiochem 198 (1): 138-142]. Различают псевдоковалентную иммобилизацию, когда на полистирольных поверхностях различными методами создают специальные подложки, на которые уже прикрепляют антигены. Известны примеры модификации полистирола с целью иммобилизации «нетипичных» белков, а именно глутаровым альдегидом [Morissette C., Goulet J., LamoureuxG: 1991. ApplEnvironMicrobiol. 57 (3): 836-842, GallD., NielsenK: 1994. JImmunoassay. 15 (3): 277-291], полилизином, поливинилбензил-лактониламидом [Suzuki N., Quesenberry M.S., Wang J.K., Lee R.T., Kobayashi K., Lee Y.C: 1997. AnalBiochem. 247 (2): 412-416] и 3-аминопропилртиэтоксисиланом [Kaur J., Boro R.C, Wangoo N., Singh K.R., Suri C.R: 2008. AnalChimActa, 607 (1): 92-99], дегидратацией проб, когда в лунки вносят раствор антигена в хлороформ-спиртовой смеси с последующим полным упариванием [Bantroch S.,  , Lam J.S: 1994. ClinDiagnLablmmunol. 1 (1): 55-62], малеимид-активированным БСА [Cuccuru M.A., Dessi D., RappelliP., FioriP.L: 2012. JImmunolMethods. 382 (1-2): 216-219]. Аффинные поверхности можно рассматривать как разновидность ковалентных, у которых функцию связывающих активных групп выполняют привитые аффинные лиганды. специфически взаимодействующие со стрептавидином, либо ионами двухвалентных металлов. Область применения аффинных планшетов ограничена анализом биотинилированных, либо генно-инженерных белков, первичные последовательности которых дополнены специальными якорными фрагментами, обеспечивающими высокоспецифичное взаимодействие с аффиными лигандами.
, Lam J.S: 1994. ClinDiagnLablmmunol. 1 (1): 55-62], малеимид-активированным БСА [Cuccuru M.A., Dessi D., RappelliP., FioriP.L: 2012. JImmunolMethods. 382 (1-2): 216-219]. Аффинные поверхности можно рассматривать как разновидность ковалентных, у которых функцию связывающих активных групп выполняют привитые аффинные лиганды. специфически взаимодействующие со стрептавидином, либо ионами двухвалентных металлов. Область применения аффинных планшетов ограничена анализом биотинилированных, либо генно-инженерных белков, первичные последовательности которых дополнены специальными якорными фрагментами, обеспечивающими высокоспецифичное взаимодействие с аффиными лигандами.
Для пассивного связывания макромолекул с различными типами поверхностей пластика необходимо, чтобы белок растворялся в буферах, которые контактируют с поверхностью, так как именно в водной среде возможно проявление сил, ведущих к образованию ионных, координационных, гидрофобных и вандерваальсовских связей [Nieto A.,  A., Moreno C.,
A., Moreno C.,  М., VivesJ: 1986. AnnlnstPasteurlmmunol, 137C (2): 161-172]. В настоящее время в подавляющем числе работ используют пассивное связывание и иммуноферментный анализ начинают с продолжительной инкубации белков, растворенных в 0,1 М карбонат-бикарбонатном буфере, рН 9,6 в течение 18-20 ч при температуре 4-6°C. При этих условиях происходит пассивное насыщение поверхности полистирольных лунок макромолекулами. Показано также, что сорбция происходит и при нейтральных значениях рН в фосфатно-солевой буферной системе [Chang L.S., Lin J., Chang C.С: 1994. BiochemMolBiolInt, 34 (2): 403-408], при этом необходимым условием является растворимость белка. Между тем, на практике в области генноинженерных исследований, многие рекомбинантные белки в процессе роста микробных клеток накапливаются в составе телец включения и не способны в этой форме переходить в буферы воспроизводящие физиологические условия.
М., VivesJ: 1986. AnnlnstPasteurlmmunol, 137C (2): 161-172]. В настоящее время в подавляющем числе работ используют пассивное связывание и иммуноферментный анализ начинают с продолжительной инкубации белков, растворенных в 0,1 М карбонат-бикарбонатном буфере, рН 9,6 в течение 18-20 ч при температуре 4-6°C. При этих условиях происходит пассивное насыщение поверхности полистирольных лунок макромолекулами. Показано также, что сорбция происходит и при нейтральных значениях рН в фосфатно-солевой буферной системе [Chang L.S., Lin J., Chang C.С: 1994. BiochemMolBiolInt, 34 (2): 403-408], при этом необходимым условием является растворимость белка. Между тем, на практике в области генноинженерных исследований, многие рекомбинантные белки в процессе роста микробных клеток накапливаются в составе телец включения и не способны в этой форме переходить в буферы воспроизводящие физиологические условия.
Широко распространены протоколы получения генно-инженерных белков под контролем сильных промоторов в клетках кишечной палочки, что позволяет наработать биомассу с высоким содержанием целевого белка, достигающего 30-40%. высокий выход ценного целевого продукта как раз является биотехнологической целью этого подхода [П.Х. Копылов П.Х., Светоч Т.Э., Иванов С.А., Комбарова Т.И., Перовская О.Н., Титарева Г.М., Анисимов А.П: 2019. Прикладная биохимия и микробиология, 55 (5): 471-480]. Сильные промоторы вызывают дисбаланс биосинтеза и структурообразования гетерологичных полипептидов, так что даже небольшие гидрофильные новосинтезированные белки приобретают разупорядоченную структуру и сбрасываются микробной клеткой в тельца включения, нерастворимые впоследствии в буферных системах воспроизводящие условия, близкие к физиологическим (обычно, 10 мМ фосфатно-солевой буфер, рН 7,2). Сложности, обусловленные растворимостью, возникающие, например, при очистке рекомбинантных белков аффинной хроматографией, снимаются применением сильных денатурирующих агентов (концентрированные растворы мочевины или хлористого гуанидина), которые разупорядочевают структуру белков телец включения и растворяют их, но не подавляют полностью координационные взаимодействия встроенной в полипептидную цепь полигистидиновой последовательности и аффинной матрицы. Однако, восстановление устойчивых и растворимых при физиологических условиях молекулярных форм белков, пригодных для пассивной иммобилизации в лунки планшетов, обусловлено предварительным сложением нативной третичной структуры (ренатурацией) полипептидов при удалении сильных денатурирующих веществ.
Известно, что вопросы ренатурации белков являются одними из самых актуальных и сложных в рамках современных биотехнологических наук. Способы ренатурации белков обладают одним базовым сходством, заключающимся в том, что происходит удаление тем, или иным образом, концентрированного раствора денатурирующего агента, что побуждает образование, восстановление нативной, биологически-активной структуры полипептидов, определяемой их первичной аминокислотной последовательностью [Burgess R.R: 2009. MethodsEnzymol, 463: 259-282]. Сложение нативной структуры белка последовательно проходит через астрономическое число промежуточных конформационных вариантов с очень высокой вероятностью неправильного завершения укладки полипептидной цепи, при этом флуктуации гидрофобных участков усиливают межмолекулярные взаимодействия, вызывая агрегацию макромолекул и образование нерастворимых осадков. Для получения растворимых белков с нативной конформацией существует множество протоколов ренатурации с использованием самых различных экспериментальных приемов, самым простым из которых является разведение ренатурационных растворов различными буферными растворами [Cabrita L.D., Bottomley S.P: 2004. BiotechnolAnnuRev, 10: 31-50]. Известны альтернативные методы перевода белков в растворы с низкой ионной силой диализом или ультрафильтрацией [Yoshii H., Furuta T., Yonehara T., ItoD., Linko Y.Y., LinkoP: 2000. BiosciBiotechnolBiochem, 64 (6): 1159-1165]. Все они в разной степени противостоят агрегации, но часто сопровождаются низким выходом и сильным разбавлением конечных растворов белка. Разработан ряд методических подходов, имеющих целью обойти концентрационный барьер агрегации ренатурирующих белков путем их иммобилизации на твердой фазе [LiM., PoliakovA., Danielson U.H., Su Z., Janson J.C: 2003. BiotechnolLett, 25 (20): 1729-1734]. Подобным образом для нескольких классов белков восстановление третичной структуры и функциональности возможно только при наличии соответствующих смежных структурных компонентов, например, липополисахаридов, детергентов [Korhonen T.K., Haiko J., Laakkonen L.,  2013. FrontCelllnfectMicrobiol, 3: 35. doi: 10.3389/fcimb.2013.00035.]. Растворимые формы таких белковых комплексов были получены, однако сопутствующие детергенты конкурируют с полипептидами при пассивной сорбции и практически блокируют поверхность полистирольных лунок при концентрациях обуславливающих мицеллообразование (CMC) [Gardas A., LewartowskaA: 1988. JImmunolMethods, 106 (2): 251-255]. Кроме способа удержания полипептидных цепей координационными или ковалентными «пришивками», сохраняющимися в окружении жестких денатурантов в лунках планшетов, исследователями в пептидных библиотеках были найдены додекапептиды с высоким специфическим сродством к полистирольной поверхности [Kumada Y., Tokunaga Y., Imanaka H., Imamura K., Sakiyama T., Katoh S., NakanishiK: 2006. BiotechnolProg, 22 (2): 401-405], которое сохранялось в растворах мочевины в интервале концентраций до 2 М [Kumada Y., Hamasaki K., Nakagawa A., Sasaki Е., Shirai Т., Okumura М., Inoue М., Kishimoto М: 2013. JImmunolMethods, 400-401: 70-77]. Иммобилизованные при этих условиях фрагменты антител, содержащие соответствующие додекапептиды восстанавливали структуру и обретали специфическую связывающую активность.
2013. FrontCelllnfectMicrobiol, 3: 35. doi: 10.3389/fcimb.2013.00035.]. Растворимые формы таких белковых комплексов были получены, однако сопутствующие детергенты конкурируют с полипептидами при пассивной сорбции и практически блокируют поверхность полистирольных лунок при концентрациях обуславливающих мицеллообразование (CMC) [Gardas A., LewartowskaA: 1988. JImmunolMethods, 106 (2): 251-255]. Кроме способа удержания полипептидных цепей координационными или ковалентными «пришивками», сохраняющимися в окружении жестких денатурантов в лунках планшетов, исследователями в пептидных библиотеках были найдены додекапептиды с высоким специфическим сродством к полистирольной поверхности [Kumada Y., Tokunaga Y., Imanaka H., Imamura K., Sakiyama T., Katoh S., NakanishiK: 2006. BiotechnolProg, 22 (2): 401-405], которое сохранялось в растворах мочевины в интервале концентраций до 2 М [Kumada Y., Hamasaki K., Nakagawa A., Sasaki Е., Shirai Т., Okumura М., Inoue М., Kishimoto М: 2013. JImmunolMethods, 400-401: 70-77]. Иммобилизованные при этих условиях фрагменты антител, содержащие соответствующие додекапептиды восстанавливали структуру и обретали специфическую связывающую активность.
Представленные выше многочисленные и разнообразные способы связывания белковых антигенов и полистирольных поверхностей лунок планшетов направлены в первую очередь на модификацию твердой фазы, а именно, полистирольных поверхностей таким образом, чтобы между растворенными антигенами и модифицированным полистиролом возникли различные типы связей, обеспечивающие пассивную, ковалентную или аффинную иммобилизацию.
Технический результат предлагаемого изобретения прежде всего состоит в том, что оно делает возможным сенсибилизировать полистирольные планшеты белковыми антигенами, нерастворимыми ранее в стандартных буферных системах, используемых на первом этапе обработки и позволяет осуществлять постановку ELISA для большого числа труднорастворимых белковых антигенов, повышая чувствительность и специфичность теста.
Достижение технического результата показано на полипептидах чумного микроба не являющихся гомологами по первичным аминокислотным последовательностям, клонированным в клетках кишечной палочки, которые после выделения аффинной хроматографией образовали осадки, нерастворимые в буферных растворах, используемых для пассивной иммобилизации белков в лунках полистирольных планшетов. Используют полипептиды чумного микроба: субстрат связывающий белок ABC транспортера метионина (MetQ), белок наружной мембраны (OmpA), фруктозо-бисфосфатальдолаза (Fba), passenger-доменбелка-автотранспортера YapM, для которых результаты, полученные предлагаемым способом, идентичны.
Подготовительные этапы для получения технического результата состоят в том, что лизированные бактериальные культуры, продуцировавшие один из представленных здесь чумных полипептидов преимущественно в составе телец включения, центрифугируют в течение 20 мин при 15000×g и 10°C, полученный осадок переосаждают еще раз в тех же условиях. Растворимую часть удаляют, а осадок длительное время растворяют в двадцатикратном (вес/объем) избытке буферного раствора, содержащего 8 М мочевину, 5 мМ имидазол и 20 мМ Трис-HCl, рН 8, после чего снижают концентрацию мочевины до 6 М и перемешивают еще 2 часа. Падосадочную часть осторожно отделяют после центрифугирования и используют для выделения белка. Выделение полипептидов, несущих последовательность His6 проводят аффинной хроматографией носителем AF-ChelateToyopearl 650М, насыщенным ионами Ni++. Полученные фракции очищенных белков анализируют электрофорезом и диализуют значимые фракции против 20 мМ, рН 7,4 фосфатного буферного раствора в течение 18 час при 10°C. Образовавшийся осадки целевых белков Ме10 собирают центрифугированием, тщательно суспензируют в 5 мл фосфатного буфера, содержащего 5% трегалозы, и подвергают быстрой заморозке в низкотемпературном морозильнике по аликвотам. В аликвотах суспензии после размораживания определяют концентрацию белка при помощи ДСН электрофореза в восстанавливающих условиях. Количественное маркирование геля проводят при помощи бычьего сывороточного альбумина (БСА), внося соответственно по 0,2, 0,4, 0,6, 0,8, 1,0 и 1,4 мкг в разные лунки, при этом, пробы анализируемых очищенных белков разводят таким образом, чтобы количество белка, нагруженное в лунки геля, попало в интервал от 0,2 до 1,4 мкг. Гели окрашивают Кумасси ярко голубым G-250, концентрации белков в исходных суспензиях вычисляют по числовым изображениям гелей, используя значения площадей отдельных пиков БСА. искомых белков на электрофореграммах, степень разведения образцов и количество внесенных в лунки полипептидов.
Технический результат непосредственно достигается тем, что перед этапом сенсибилизации планшетов предварительно полностью растворяют гомогенную суспензию препаратов целевых белков в 8 М мочевине таким образом, чтобы в 10 мкл полученного раствора содержалось 800 нг белка. Сенсибилизацию планшетов проводят внесением в каждую лунку 10 мкл таких растворов с последующим 10-кратным разбавлением в лунках 0,1 М раствором карбонат-бикарбонатного буфера, рН 9,6, после чего планшеты герметизируют специальной пленкой и инкубируют 18 час при 6°C. Инкубационные растворы удаляют и промывают 4 раза сенсибилизированные планшеты 10 мМ фосфатно-солевым буфером, рН 7,4, содержащим 0,05% твин-20 (ФСБТ), после чего вносят в лунки по 120 мкл 1% раствора БСА в ФСБТ, герметизируют и инкубируют 3 час при 37°C. На последней стадии сенсибилизированные планшеты промывают 3 раза ФСБТ, удаляют буфер, герметизируют, замораживают и хранят при -20°C. Полученные таким образом планшеты несут на поверхности лунок пассивно иммобилизованные белковые антигены, обладающие присущими им иммунохимическими свойствами, способные связываться со специфическими антителами, выработанными животными (мыши) в ответ на иммунизацию этими антигенами, что позволяет определять титры специфических антител в сыворотках крови мышей отражающих протективные свойства исследуемых антигенов. Таким же образом была проведена пассивная иммобилизация всех перечисленных белковых антигенов чумного микроба, значительно отличающихся первичными аминокислотными последовательностями, но труднорастворимых в физиологических буферах.
Изобретение иллюстрируется следующими фигурами:
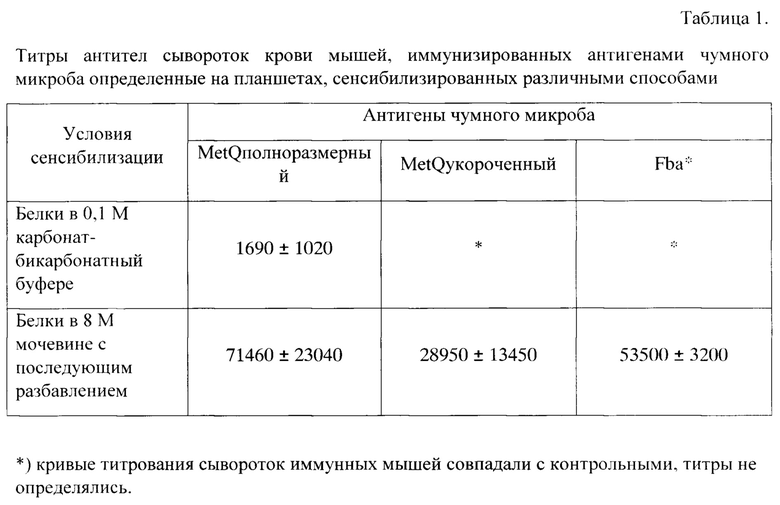
Фиг. 1. Электрофорез и иммуноблот препаратов различных белков на этапе подготовки клеточных лизатов.
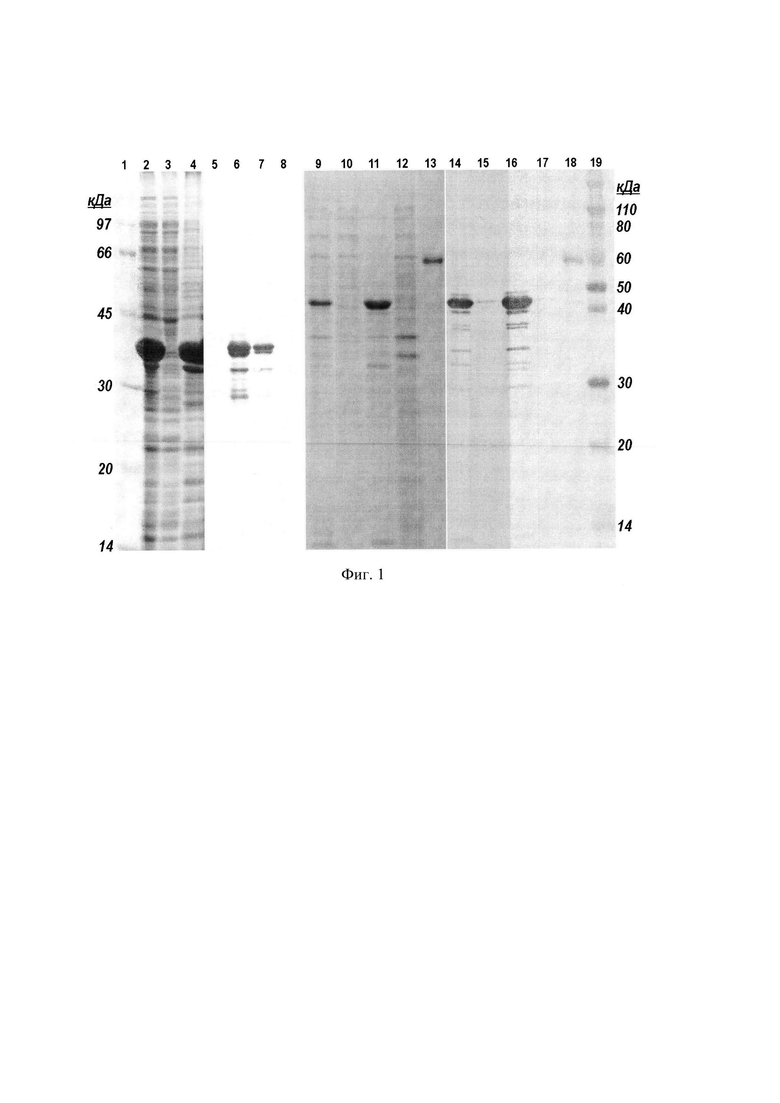
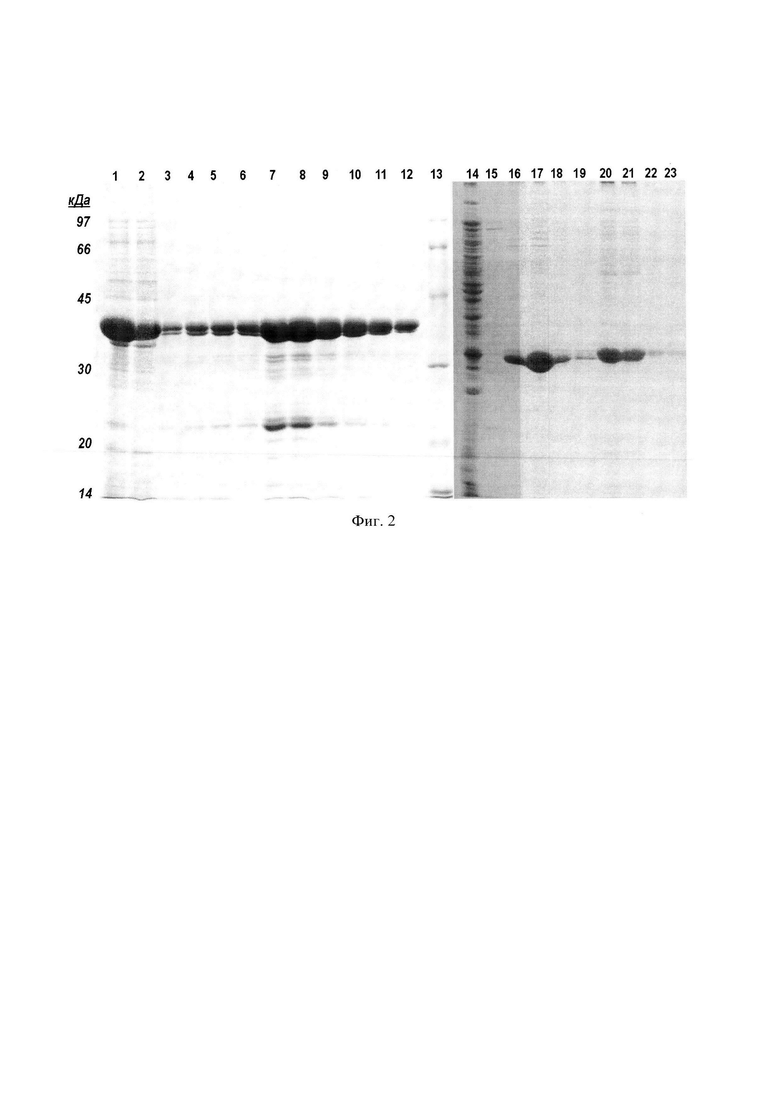
Фиг. 2. Электрофорез хроматографических фракций, полученных в результате аффинной хроматографии белков на колонке с AF-ChelateToyopearl 650М, нагруженной ионами Ni2+.
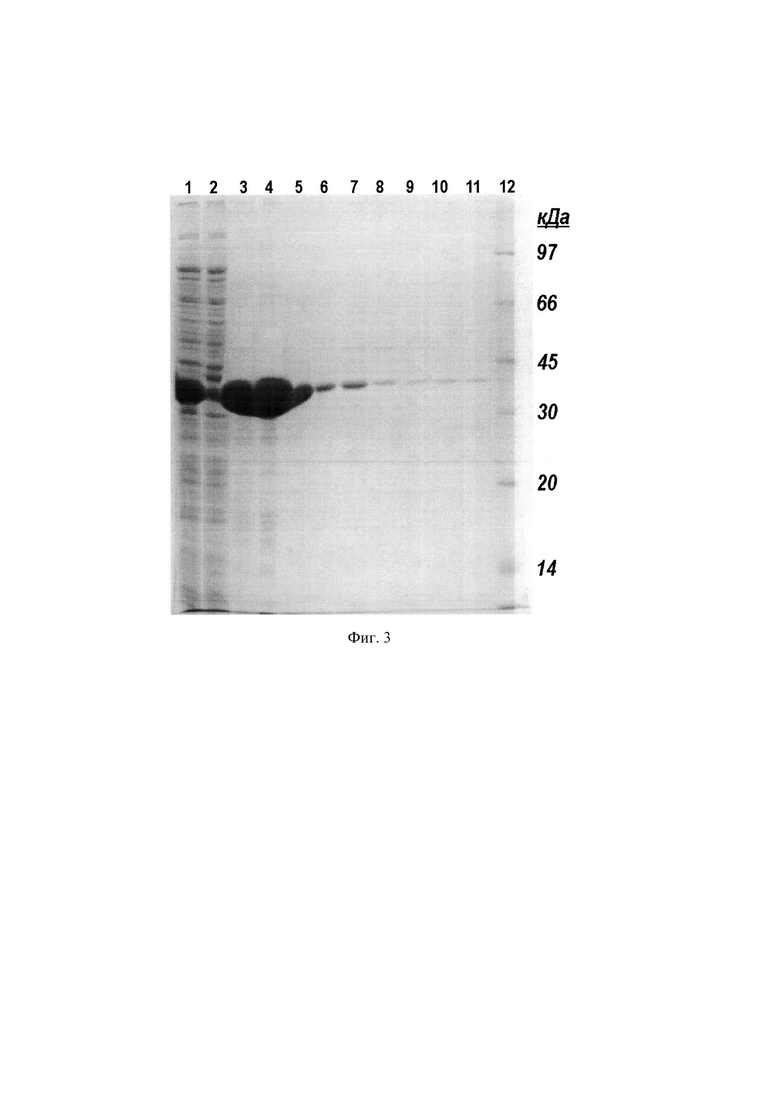
Фиг. 3. Электрофорез хроматографических фракций, полученных в результате аффинной хроматографии белка Fba на колонке с AF-ChelateToyopearl 650М, нагруженной ионами Ni2+.
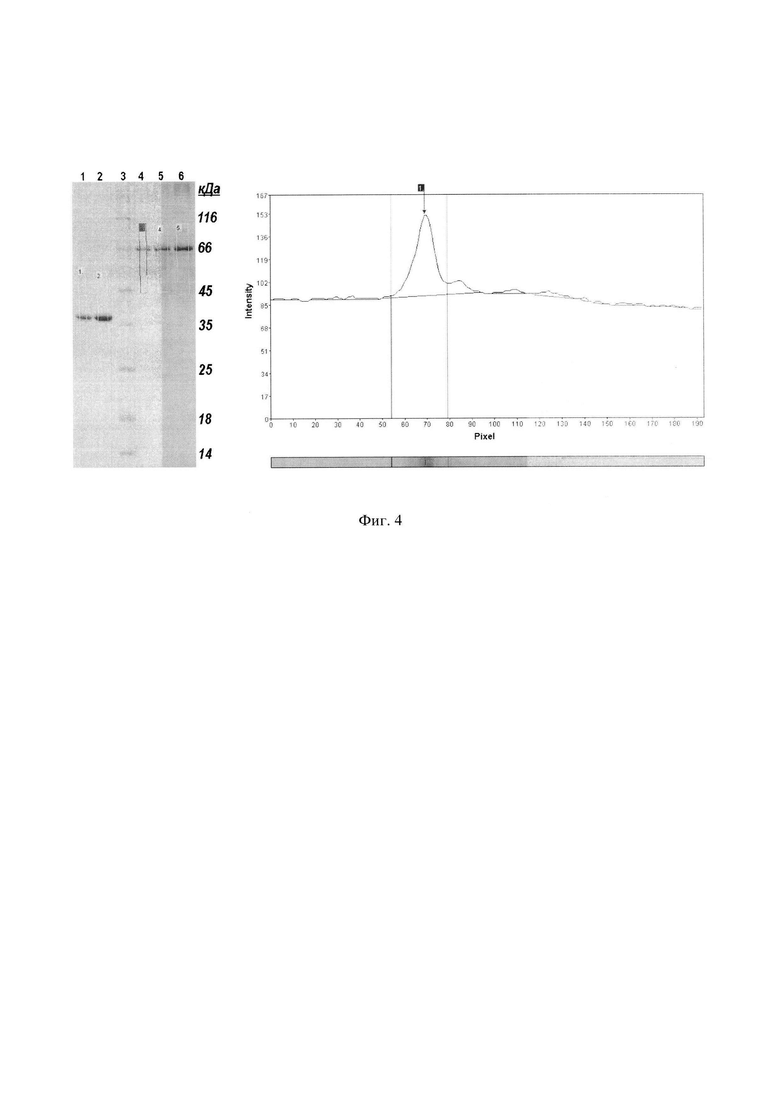
Фиг. 4. Анализ цифрового изображения геля с белком Fba программой «GelAnalyzer®».
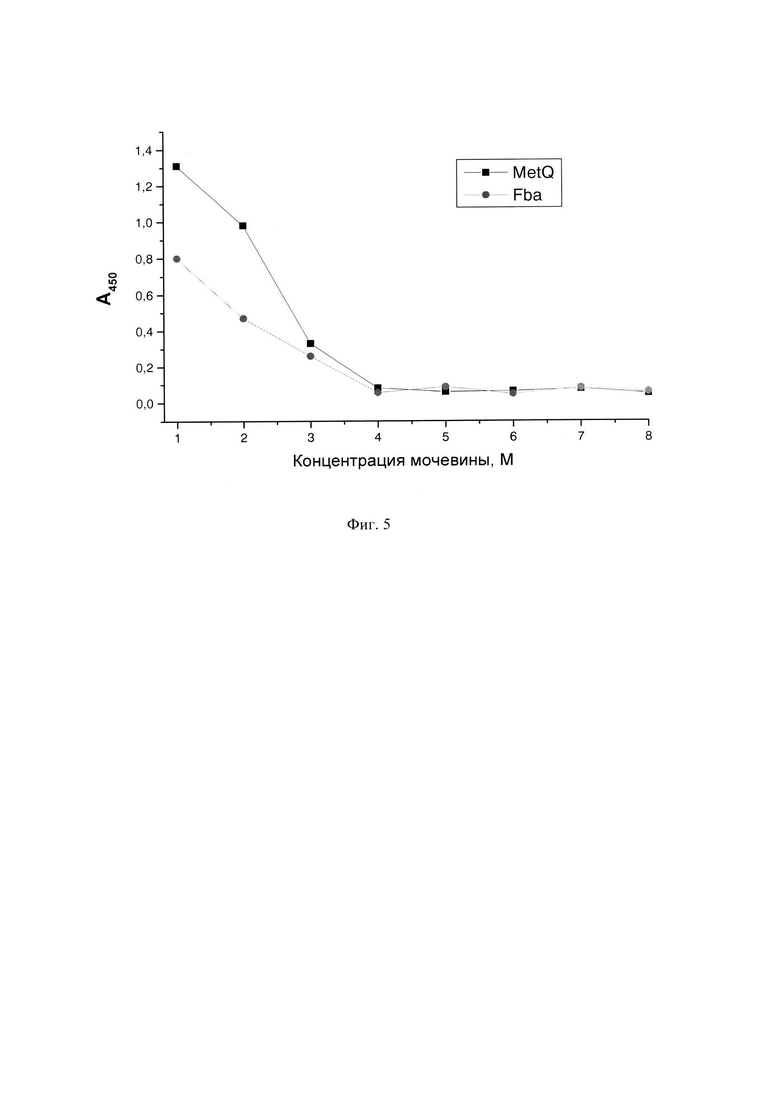
Фиг. 5. Определения пассивного связывания антигенов в растворах мочевины.
Пример 1. Подготовка лизатов клеток кишечной палочки для выделения белковых антигенов.
Для выделения белков чумного микроба, клонированных в составе вектора pET32b(+) в клетках штамма-продуцента Е. coli BL21(DE3), используют культуры, продуцирующие соответствующие целевые белки после индукции ИПТГ. Этапы подготовки материала для аффинной хроматографии одинаковы для всех целевых белков, приведенных в этой работе. На фиг. 1 в качестве примера представлены результаты подготовки лизатов, содержащие белки OmpA и YapM Y. pestis. Осадки влажной биомассы (10-12 г) суспензируют в десятикратном избытке (вес/объем) буферном растворе, содержащем 5 мМ имидазол и 20 мМ Трис-HCl, рН 8,0. Лизис клеток проводят в стакане, который охлаждают непрерывно на ледяной бане, наконечником ультразвукового дезинтегратора Bandelin (ЕС) при мощности 50% от максимальной циклами по 15 сек с равновеликими паузами до тех пор, пока величина затраченной энергии не составит 60 кДж. Клеточные лизаты центрифугируют в течение 20 мин при 15000×g и 10°C и отделяют растворимую часть от осадка, фракции хранят в замороженном состоянии до появления результатов электрофореза и иммуноблота. Каждую фракцию анализируют при помощи электрофореза в полиакриламидном геле в восстанавливающих условиях (ДСН-ПААГ), подтверждая присутствие целевых белков во фракциях осадка, то есть в тельцах включения. (Фиг. 1. Числами в верхней части рисунка обозначены линии: 1, 2, 3, 4 - электрофорез белка OmpA; 9, 10, 11, 12, 13 - электрофорез белка YapM; 5, 6, 7, 8 - иммуноблот белка OmpA; 14, 15, 16, 17, 18, 19 - иммуноблот белка YapM. Клеточные лизаты, линии 2, 5, 6, 9, 14, 17; надосадочные фракции, линии 3, 8, 10, 15; осадки, линии 4, 7, 11, 18. Линии: 5, 12, 17 - отрицательный контроль; 13, 18 - положительный контроль; 1,19 - маркеры молекулярных масс.) Осадки растворяют при непрерывном перемешивании в двадцатикратном (вес/объем) избытке буферного раствора, содержащего 8 М мочевину, 5 мМ имидазол, 500 мМ натрий хлористый и 20 мМ Трис-HCl, рН 8,0, в течение 20 ч на магнитной мешалке при 10°C, после чего разбавляют растворы этим же буфером, но без мочевины, таким образом, чтобы уменьшить ее концентрацию до 6 М. После разбавления раствор перемешивают еще 2 ч и центрифугируют 30 мин при 18000×g и 20°C, прозрачную надосадочную часть осторожно отделяют и используют для выделения белков MetQ, OmpA, Fba и YapM.
Пример 2. Очистка белковых антигенов аффинной хроматографией из подготовленных фракций лизатов клеток кишечной палочки.
Используют хроматографический носитель AF-ChelateToyopearl 650М, стационарная фаза которого объемом 25 мл упакована в колонку ХК-26 и предварительно насыщена ионами Ni2+. Колонку уравновешивают буферным раствором, содержащим 6 М мочевину, 5 мМ имидазол, 500 мМ натрий хлористый и 20 мМ Трис-HCl, рН 8,0 и пропускают прозрачную надосадочную фракцию центрифугата, полученную, как описано в примере 1. Такие же фракции, подготовленные из лизатов других штаммов Е. coli несущие, соответственно, белки MetQ, OmpA, Fba и YapM используют для их выделения аффинной хроматографией в одинаковых условиях. Колонку промывают и элюируют белки буферными растворами с увеличивающейся концентрацией имидазола, сначала буфер содержит 20 мМ имидазол и удаляет примеси с умеренным сродством к ионам Ni2+, затем элюируют целевые белки буферным раствором, содержащим 300 мМ имидазол. В некоторых случаях целевые белки элюируют вместе с буферным раствором, содержащим 20 мМ имидазол. Для белка MetQ наблюдали элюцию изоформ, различающихся сродством к аффинной матрице, причем, изоформы обладали различными молекулярными массами. (Фиг. 2. Белки, линии: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 - OmpA; 14, 15, 16, 17, 18, 19, 20, 21, 22, 23 - MetQ. Надосадочные фракции центрифугата, нанесенные на колонку, линии: 1, 14; фракции, прошедшие через колонку, линии: 2, 15; фракции элюированные 20 мМ имидазолом, линии: 3, 4, 5, 6, 16, 17, 18, 19; фракции элюированные 300 мМ имидазолом, линии: 7, 8, 9, 10, 11, 12, 20, 21, 22, 23. Маркеры молекулярных масс, линия 13). Белок OmpA элюируют преимущественно буферным раствором, содержащим 300 мМ имидазола, в то время, как белок Fba - 20 мМ имидазола. (Фиг. 3. Надосадочная фракция центрифугата, нанесенная на колонку, линия 1; фракция, прошедшая через колонку, линия 2; фракции элюированные 20 мМ имидазолом, линии: 3, 4, 5, 6; фракции элюированные 300 мМ имидазолом, линии: 7, 8, 9, 10, 11. Маркеры молекулярных масс, линия 12). Элюаты получают фракциями по 6 мл и анализируют их ДСН-ПААГ.
Пример 3. Замена буфера диализом и определение содержания целевых белков в суспензиях полученных препаратов.
Фракции, содержащие очищенные белки объединяют и диализуют против 100-кратного избытка 20 мМ фосфатного буферного раствора, рН 7,4 в течение 18 ч при 10°C. Буферный раствор меняют три раза. При диализе по мере уменьшения концентрации мочевины целевые белки теряют растворимость и образуют осадки, которые собирают центрифугированием проб в течение 3,5 мин при 15000×g и 10°C. К осадкам прибавляют по 5 мл фосфатного буфера, содержащего 5% трегалозы и тщательно суспензируют, после чего подвергают аликвоты проб быстрой заморозке с использованием низкотемпературного морозильного оборудования (хранение) или жидкого азота. Определение концентрации белка в пробах проводят после разморажвания единичных образцов; к суспензиям добавляют эквивалентные объемы «буферного раствора для приготовления электрофоретических образцов (буфер Лэммли)», перемешивают и выдерживают пробы при 95°C в течение 10 мин на водяной бане. Охлажденные до комнатной температуры пробы центрифугируют 15 мин при 16000×g и 20°C и для электрофореза используют образцы, осадки которых полностью растворены в буфере Лэммли. Пробы разводят таким образом, чтобы количество белка, нагружаемого в лунки концентрирующего геля, находилось в интервале от 0,3 мкг до 1,2 мкг. Каждый гель несет калибровочные пробы альбумина, количество которого лежит в этом же интервале нагруженного количества белка. Электрофорез проводят в режиме постоянного напряжения 200 в и прекращают, когда полоса лидерного красителя достигает нижнего края геля, после чего проводят окраску Кумасси ярко-голубым G-250 в течение 50 мин на качалке со скоростью вращения 60 об/мин. Отмывку избытков красителя также проводят на качалке при температуре 50°C 7,5% раствором уксусной кислоты, содержащей 5% этанола, с трехкратной заменой раствора, после чего гель фотографируют в проходящем свете с разрешением 300 dpi. Цифровые изображения гелей обрабатывают программами обработки гелей, например, «GelAnalyzer®», или эквивалентными. При анализе цифровых изображений следуют инструкциям разработчика программы. После маркирования искомых полос, построения базовых линий и определения площадей пиков, получают числовые параметры, характеризующие отдельные белковые полосы. Концентрацию белков в исходных образцах вычисляют по результатам оцифровки электрофоретических полос, с учетом примененных разведений и рабочих объемов анализируемых проб. (Фиг. 4. Линии на электрофореграмме: 1, 2 - диализованный белок Fba; 4, 5, 6 - БСА, соответственно, в линиях по 1,4 мкг, 0,6 мкг и 0,8 мкг; 3 - маркеры молекулярных масс; прямоугольниками выделены анализируемые белки. Денситограммой представлен анализируемый фрагмент линии 4, выделенный красным прямоугольником; синяя линия денситограммы - базовая линия).
Пример 4. Сенсибилизация планшетов антигенами MetQ, OmpA и Fba.
Перед сенсибилизацией планшетов, суспензии исследуемых антигенов продолжительное время инкубируют в концентрированных растворах мочевины или хлористого гуанидина, создающих жесткие условия растворения белков, сопровождающееся денатурацией полипептидной цепи, что обеспечивает исчерпывающее растворение белков. На практике, к определенному объему суспензии белка, размороженной после хранения в низкотемпературном морозильнике, добавляют постепенно при перемешивании расчетное количество мочевины таким образом, чтобы ее конечная концентрация составила 8 М, растворение реактива ведут при непрерывном перемешивании и комнатной температуре в течение 2 ч. Если белок не растворяется в мочевине, его растворение начинают с добавления хлористого гуанидина до конечной концентрации соли 7 М. С учетом количества белка в суспензии, определенного ранее по результатам электрофореза, из полученного раствора, или некоторой его части готовят сенсибилизирующий раствор для нанесения в лунки планшетов, путем его разбавления раствором 8М мочевины таким образом, чтобы в 10 мкл полученного сенсибилизирующего раствора содержалось 800 нг белка. Сенсибилизацию планшетов (пассивное связывание антигена) начинают внесением в лунки 10 мкл раствора антигенов в 8 М мочевине имея ввиду, что в этих условиях пассивная иммобилизация растворенных антигенов на поверхностях полистирольных лунок блокирована. Пассивное связывание белков запускается разбавлением содержимого лунок 90 мкл 0,1 М раствора карбонат-бикарбонатного буфера, рН 9,6, после чего планшеты герметизируют специальной пленкой и инкубируют 18 ч при 6°C для завершения процесса. Планшеты приводят в рабочее состояние удалением сенсибилизирующего раствора антигенов и четырехкратной промывкой 10 мМ фосфатно-солевым буфером, рН 7,4, содержащим 0,05% твин-20 (ФСБТ). После этого вносят в лунки по 120 мкл 1% раствора БСА в ФСБТ. герметизируют и инкубируют 3 ч при 37°C. На последнем этапе сенсибилизированные планшеты промывают 3 раза ФСБТ, удаляют буфер, герметизируют пленкой, замораживают и хранят пригодные для работы планшеты при -20°C.
Для определения пассивного связывания антигенов в присутствии концентрированных растворов мочевины готовили специальные планшеты. Растворы белка в 8 М мочевине разводили таким образом, чтобы конечные сенсибилизирующие растворы содержали одинаковое количество белка (8 мкг в 1000 мкл таких растворов), но различные концентрации мочевины, соответственно от 7 М до 1 М. Приготовленные разведения сразу же наносили по 100 мкл (800 нг) в лунки планшетов и инкубировали, как описано выше, включая этапы промывки, блокировки БСА, повторной промывки и хранения при -20°C.
Лунки сенсибилизировали белковыми антигенами в обычных условиях, используемых для их пассивной сорбции. Использованные антигены нерастворимы в стандартных буферных растворах, используемыми на этапе начальной обработки планшетов. Для этого, к размороженной суспензии, содержащей определенное количество белка добавляли 10 мМ фосфатно-солевой буферный раствор, рН 7,4, или 0,1 М карбонат-бикарбонатный буферный раствор, рН 9,6 и перемешивали полученные суспензии на магнитной мешалке при комнатной температуре в течение 2 ч. Объемы буферных растворов рассчитывали исходя из того, что 1000 мкл такого раствора должно содержать 8 мкг белка. По истечении 2 час полученными растворами сенсибилизировали планшеты, внося по 100 мкл в каждую лунку и инкубировали в течение 18 ч при 6°C. После инкубации проводили этапы промывки ФСБТ, блокировки БСА, повторной промывки ФСБТ, замораживали и хранили при -20°C. (Фиг. 5. Планшеты сенсибилизировали добавлением в лунки по 800 нг белков MetQ и Fba в растворах мочевины различных концентраций, обозначенных на оси абсцисс. По оси ординат отложены значения А450. измеренные после обработки планшетов суммарными сыворотками 5 мышей, иммунизированных, соответственно, белками MetQ и Fba, разведенными 1:1000 и специфическими антимышиными антителами, конъюгированными с пероксидазой хрена).
Пример 5. Определение титров специфических антител в сыворотке крови мышей против антигенов использованных для пассивной иммобилизации планшетов.
Для иммунизации мышей с целью получения «антительного» ответа белки сорбировали стерильно на геле гидроокиси алюминия и иммунизировали группы (n=5) мышей подкожно препаратом, в 0,2 мл суспензии которого находилось 10 мкг белка. Контрольные группы иммунизировали гелем гидроокиси алюминия без антигенов. Повторную иммунизацию проводили через 21 день, так же, как и забор крови. Для определения титров антител используют твердофазный иммуноферментный анализ на предварительно сенсибилизированных планшетах, хранившихся при температуре -20°C. Серию последовательных разведений каждой антисыворотки, состоящей из 8 ступеней начинают с пробы разведенной 1:1000 с последующим шагом 2,5 до пробы с разведением 1:610350, таким же образом разводят и контрольные сыворотки используя ФСБТ как для разведения антисывороток, так и в дальнейшем для конъюгированных антивидовых антител. После низкотемпературной камеры планшеты выдерживают при комнатной температуре 5 мин, удаляют пленку и вносят в лунки мультиканальной пипеткой по 100 мкл каждого разведения антисывороток и контролей. Планшеты заклеивают и инкубируют в течение 18 ч при 6°C, после чего удаляют пленку и промывают 4 раза ФСБТ, после чего вносят в лунки немедленно по 100 мкл раствора разведенных (1:6000) антимышиных антител, конъюгированных с пероксидазой хрена и инкубируют заклеенные планшеты в течение 1,5 ч при 37°C. Промывку и фотометрирование планшетов при длине волны 450 нм проводят роботизированной системой для ИФА «EvolisTwinPlus», фирмы Bio-Rad. На последнем этапе в лунки промытых планшетов вносят по 100 мкл раствора субстрата. Развитие цветной реакции происходит за счет донорно-акцепторной пары: перекись водорода - о-фенилендиамин в составе субстратного раствора. Реакцию останавливают через 15 мин добавлением в лунки по 100 мкл 1 М раствора серной кислоты. Строят кривые изменения оптической плотности растворов в лунках от степени разведения антисывороток и вычисляют значения титров, которые определяют, как величину наибольшего разведения антисыворотки, измеренная оптическая плотность которой в два раза превышает оптическую плотность раствора соответствующего контроля. Титры специфических антител наглядно представляют обратными величинами разведений антисывороток
Табл. 1.
Эффективность достигнутого технического результата изобретения проявляется прежде всего в том, что делает ненужным необходимый ранее сложный эмпирический этап ренатурации белков, в результате которого исследователи получали интересующие их объекты в растворимой форме. Исчезла необходимость воспроизводить или создавать методы ренатурации белков, цель которых, в данном случае заключалась бы в получении растворимого белкового продукта, пригодного для иммобилизации в лунки полистирольных планшетов. Кроме того, технический результат изобретения не предполагает использовать ковалентную, псевдоковалентную и аффинную иммобилизацию генноинженерных белков.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ВЫДЕЛЕНИЯ РЕНАТУРИРОВАННОГО БЕЛКА G ИЗ МАРКИРОВАННОГО ПОЛИАКРИЛАМИДНОГО ГЕЛЯ | 2016 |
|
RU2646103C2 |
| Рекомбинантная плазмидная ДНК pMBP-PUUV_N, обеспечивающая синтез и секрецию слитого белка MBP-PUUV_N в клетках E. coli, штамм Escherichia coli KRX/pMBP-PUUV_N, продуцирующий указанный белок, используемый для получения рекомбинантного белка PUUV_N, применяемого для диагностики геморрагической лихорадки с почечным синдромом | 2023 |
|
RU2838919C1 |
| Способ диагностики риккетсиозов группы клещевой пятнистой лихорадки, иммуноферментная диагностическая тест-система для его осуществления | 2019 |
|
RU2726484C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ПОВЫШЕННОГО УРОВНЯ АУТОАНТИТЕЛ К ПЛАЗМИНОГЕНУ ЧЕЛОВЕКА И ПРОДУКТАМ ЕГО ДЕГРАДАЦИИ И ДИАГНОСТИЧЕСКАЯ ТЕСТ-СИСТЕМА ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2014 |
|
RU2597783C2 |
| Способ дифференциации вирулентных штаммов Listeria monocytogenes от авирулентных штаммов с использованием поликлональных антител против факторов патогенности интерналина А (InlA) и интерналина В (InlB). | 2023 |
|
RU2808590C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ СОДЕРЖАНИЯ ПРОДУКТОВ ПРОТЕОЛИЗА В ПЛАЗМЕ КРОВИ И ДИАГНОСТИЧЕСКАЯ ТЕСТ-СИСТЕМА ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2015 |
|
RU2597782C1 |
| Способ получения рекомбинантного человеческого глиального фибриллярного кислого белка и используемый в нём штамм-продуцент | 2022 |
|
RU2811427C1 |
| НАБОР ДЛЯ КОЛИЧЕСТВЕННОГО ОПРЕДЕЛЕНИЯ АВЕРМЕКТИНОВ МЕТОДОМ ОДНОСТАДИЙНОГО КОНКУРЕНТНОГО ИММУНОФЕРМЕНТНОГО АНАЛИЗА | 2009 |
|
RU2416094C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ТРЕХ ГИБРИДНЫХ БЕЛКОВ ОБОЛОЧКИ ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА, ОПРЕДЕЛЯЮЩИХ ПРИНАДЛЕЖНОСТЬ К СИБИРСКОМУ (DBD2-D3S), ЕВРОПЕЙСКОМУ (DBD2-D3E) И ДАЛЬНЕВОСТОЧНОМУ (DBD2-D3D) ПОДТИПАМ ВИРУСА; РЕКОМБИНАНТНЫЕ ПЛАЗМИДЫ pDBD2-D3S, pDBD2-D3E И pDBD2-D3D; ШТАММЫ-ПРОДУЦЕНТЫ Escherichia coli M15 [pREP4]; ХИМЕРНЫЕ БЕЛКИ И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2560588C1 |
| Мозаичный рекомбинантный полипептид, содержащий фрагменты белков вируса гепатита Е 1 и 3 генотипов в одной полипептидной цепи, предназначенный для использования в тест-системах, применяемых в серодиагностике гепатита Е | 2020 |
|
RU2754791C1 |
Изобретение относится к области медицины и фармацевтики, а именно к способу сенсибилизации планшета для иммуноферментного анализа нерастворимыми белковыми антигенами чумного микроба, включающему разрушение бактериальной массы ультразвуком с последующим разделением субфракций, проведение аффинной хроматографии белков, выделение и растворение осадка очищенных целевых белков с денатурацией мочевиной и определение их концентрации в растворах, отличающемуся тем, что раствор денатурированного в мочевине очищенного белка MetQ, OmpA, Fba и YapM вносят в лунки планшета и проводят последующее десятикратное разбавление в лунках раствором карбонат-бикарбонатного буфера, сопровождающееся ренатурацией белков и их пассивной иммобилизацией в лунках полистирольных планшетов. Технический результат заключается в возможности сенсибилизировать полистирольные планшеты белковыми антигенами, нерастворимыми в стандартных буферных системах, используемых на первом этапе обработки, и осуществлять постановку ELISA для таких белковых антигенов, повышая чувствительность и специфичность теста. 5 ил., 1 табл., 5 пр.
Способ сенсибилизации планшета для иммуноферментного анализа нерастворимыми белковыми антигенами чумного микроба, включающий разрушение бактериальной массы ультразвуком с последующим разделением субфракций центрифугированием, переосаждением, ресуспензированием, растворением осадка, проведение аффинной хроматографии белков, выделение целевых белков, нерастворимых в буферных растворах, применяемых для сенсибилизации полистирольных планшетов, растворение осадка очищенных целевых белков с денатурацией 8 М мочевиной и определение их концентрации в растворах после ДСН-электрофореза в восстанавливающих условиях, отличающийся тем, что 10 мкл раствора денатурированного в 8 М мочевине каждого очищенного белка MetQ, OmpA, Fba и YapM с концентрацией 0,08 мг/мл вносят в лунки планшета и проводят последующее десятикратное разбавление в лунках 0,1 М раствором карбонат-бикарбонатного буфера, рН 9,6, сопровождающееся ренатурацией белков и их одновременной пассивной иммобилизацией в лунках полистирольных планшетов в течение 18 часов при температуре 6°C.
| Короткозамкнутый ротор асинхронного двигателя | 1929 |
|
SU19696A1 |
| МОДИФИЦИРОВАННЫЕ СУПЕРСПИРАЛЬНЫЕ БЕЛКИ С УЛУЧШЕННЫМИ СВОЙСТВАМИ | 2013 |
|
RU2677799C2 |
| HNASKO R | |||
| et al | |||
| A rapid method to improve protein detection by indirect ELISA // Biochemical and biophysical research communications | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| - V | |||
| Устройство анодов катодных ламп | 1923 |
|
SU410A1 |
| - No | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| - P | |||
| СТЕРЕОСКОП | 1921 |
|
SU726A1 |
| ТВЕРДОФАЗНЫЙ ИММУНОФЕРМЕНТНЫЙ АНАЛИЗ (ELISA) ДЛЯ ФАКТОРА РОСТА ЭНДОТЕЛИЯ СОСУДОВ (VEGF) | 2007 |
|
RU2517301C2 |

