Область техники, к которой относится изобретение
Изобретение относится к области фармацевтики и биотехнологии, и может быть использовано для усиления роста новых сосудов в ишемизированных тканях. Изобретение представляет собой плазмидную бицистронную конструкцию, несущую ген hHGF фактора роста гепатоцитов человека и ген hVEGF165 фактора роста эндотелия сосудов. Заявляемая бицистронная конструкция может быть использована как лекарственное средство в медицинских целях для обеспечения стимуляции ангиогенеза, роста и ремоделирования сосудов, а также восстановления кровоснабжения в ишемизированных тканях, для лечения ряда заболеваний человека, обусловленных нарушением кровоснабжения тканей.
Уровень техники
Ишемическая болезнь сердца (ИБС) и критическая ишемия нижних конечностей (КИНК), наряду с ишемическими нарушениями мозгового кровообращения, в настоящее время относят к основным причинам инвалидизации и смертности населения.
Одним из перспективных способов лечения КИНК является «терапевтический ангиогенез» - стимуляция роста новых кровеносных сосудов с помощью генной терапии. Для индукции ангиогенеза в ишемизированной скелетной мускулатуре было предложено использовать плазмидные конструкции, несущие кДНК генов b-FGF, VEGF165, HGF и других факторов роста. Безопасность таких конструкций была показана во многих исследованиях [Pavel Igorevich Makarevich and Yelena Viktorovna Parfyonova (April 5th 2017). Therapeutic Angiogenesis: Foundations and Practical Application, Physiologic and Pathologic Angiogenesis, Dan Simionescu and Agneta Simionescu, IntechOpen, DOI: 10.5772/66411].
В 2011 году на территории РФ был зарегистрирован препарат "Неоваскулген", представляющий собой плазмиду, кодирующую фактор роста эндотелия сосудов (pCMV-VEGF165), его эффективность и безопасность в комплексной терапии пациентов с хронической ишемией нижних конечностей была доказана в ходе клинических исследований. Результат клинического исследования 2б-3 фазы с участием 100 пациентов показал значительное улучшение их функционального состояния, что выражалось достоверным увеличением дистанции безболевой ходьбы на 110%. Отсроченное наблюдение не выявило каких-либо побочных эффектов, связанных с применением препарата и показало, что терапевтический эффект после стандартного курса терапии сохраняется на протяжении трех лет. Селективный анализ эффективности терапевтического ангиогенеза у пациентов с фоновым СД (n=15), которые участвовали в исследовании, показал достоверное увеличении дистанции безболевой ходьбы на 202% через 6 месяцев после стандартного курса терапии.
Весной 2019 года Министерство здравоохранения, труда и социального обеспечения Японии (MHLW) разрешило применение генотерапевтического препарата Collatagene компании AnGes, основываясь на результатах успешно завершившихся клинических исследований. Активным компонентом препарата является плазмида с геном фактора роста гепатоцитов - HGF, который обладает ангиогенными свойствами, препарат переназначен для лечения критической ишемии нижних конечностей (КИНК). В ходе предыдущего плацебо-контролируемого рандомизированного исследования 2 фазы было показано, что его применение обеспечивает достоверное улучшение функционального состояния пациентов с КИНК, что выражается в снижении выраженности болевого синдрома и увеличении частоты заживления язвенных дефектов (Shigematsu Н., Yasuda К., Iwai Т., Sasajima Т., Ishimaru S., Ohashi Y., Yamaguchi Т., Ogihara Т., Morishita R. Randomized, double-blind, placebo-controlled clinical trial of hepatocyte growth factor plasmid for critical limb ischemia // Gene Ther. 2010. T. 17. №9. - С. 1152-1161). Китайскими учеными в ходе 1 фазы открытого клинического исследования с участием 21 пациента с КИНК на фоне облитерирующего атеросклероза была доказана безопасность плазмиды с генами двух изоформ HGF. Также были получены предварительные данные об эффективности, которые выражались в увеличение показателей транскутанного напряжения кислорода и лодыжечно-плечевого индекса (A phase I clinical study of naked DNA expressing two isoforms of hepatocyte growth factor to treat patients with critical limb ischemia / Y. Gu, J. Zhang, L. Guo, S. Cui // J. Gene Medicine. - 2011. - Vol. 13. P. 602-610).
Стоит учитывать, что ангиогенез - это многоэтапный процесс, и каждый этап находится под контролем различных цитокинов и факторов роста, поэтому применение только одного ангиогенного фактора (как в виде рекомбинантного белка, так и виде генетической конструкции) не позволяет добиться значительных результатов.
Известно использование в терапевтическом ангиогенезе комплексного введения нескольких плазмид, кодирующих разные ангиогенные факторы роста (АФР), в комбинации друг с другом.
Из уровня техники известно средство для стимуляции ангиогенеза в ишемизиро-ванной ткани, содержащее смесь в растворе 0,9%-ного NaCl плазмидных конструкций pC4W-hVEGFopt, несущую оптимизированный ген hVEGFopt фактора роста эндотелия сосудов, и pC4W-hHGFopt, несущую оптимизированный ген hHGFopt фактора роста гепатоцитов (патент РФ №2449799, 10.05.2012). Эффективный ангиогенез в ишемизированной ткани, достигается при использовании данного средства в массовом соотношении плазмидных конструкций pC4W-hVEGFopt и pC4W-hHGFopt от 1:3 до 3:1. Лучшие результаты достигались при их массовом соотношении 1:1.
Существенными недостатками введения смеси векторов является низкая вероятность котрансфекции клеток двумя плазмидами, различия экспрессии вводимых генов и снижение эффективности их совместного действия. Кроме того, при введении смеси плазмид не представляется возможным предсказать эффективность трансфекции клеток каждым из векторов, описать их фармакокинетику и конечную концентрацию кодируемых АФР, что значительно затрудняет фармацевтическую разработку лекарственных средств на основе комбинации двух и более генетических конструкций. Для ко-экспрессии двух и более генов разрабатываются мультицистронные конструкции, например, векторы, включающие внутренние сайты посадки рибосом (IRES). Плазмиды, несущие IRES, позволяют осуществлять одновременную экспрессию нескольких генов, однако, эффективность трансляции, обеспечиваемой IRES, обычно существенно уступает кэп-зависимой инициации, а конечное соотношение концентраций синтезируемых белков варьирует в зависимости от выбранного IRES. Группой польских ученых было проведено клиническое исследование плазмидной конструкции, несущей гены VEGFA и FGF2, разделенные последовательностью IRES вируса EMCV, для лечения ишемической болезни сердца [Kukula К, Chojnowska L, Dabrowski М, Witkowski A, Chmielak Z, Skwarek M, Kadziela J, Teresinska A, Malecki M, Janik P, et al. Intramyocardial plasmid-encoding human vascular endothelial growth factor A165/basic fibroblast growth factor therapy using percutaneous transcatheter approach in patients with refractory coronary artery disease (VIF-CAD) Am Heart J. 2011;161:581-589]. Введение плазмиды не привело к улучшению перфузии миокарда, однако, улучшились переносимость физической нагрузки и клинические проявления болезни.
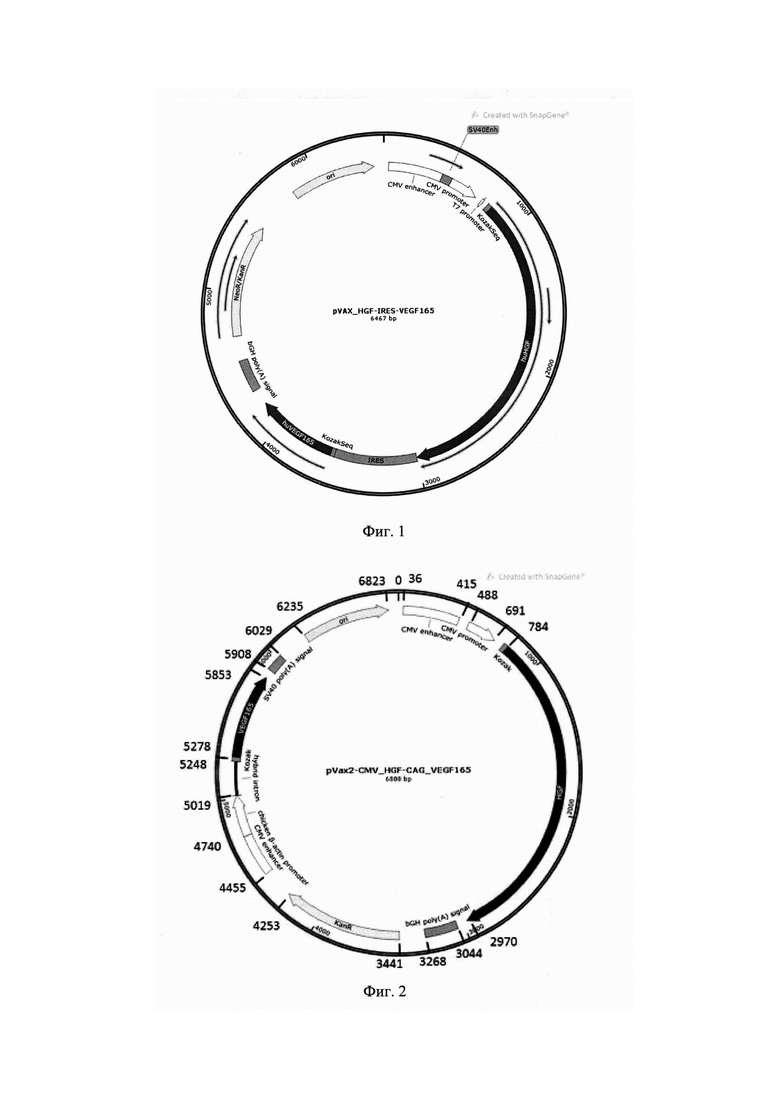
Наиболее близким к заявляемому решению является плазмидная конструкция, в которых гены факторов роста HGF и VEGF165 разделены сайтом внутренней посадки рибосом - IRES [Слободкина Е.А., Нимирицкий П.П., Долинкин А.О., Макаревич П.И., Парфенова Е.В. Разработка генотерапевтической плазмидной конструкции, кодирующей фактор роста гепатоцитов (HGF) и фактор роста эндотелия сосудов (VEGF165), Гены и Клетки - Материалы III Национального Конгресса по Регенеративной Медицине. Москва, 15-18 ноября 2017 года, серия 3, место издания Институт Стволовых Клеток Человека, Москва, Россия Москва, том 12, тезисы]. Общая схема данной плазмиды представлена на фиг. 1. Генно-инженерная конструкция, представляет собой экспрессионную векторную плазмиду, в которую клонированы вставки кДНК, кодирующие фактор роста эндотелия сосудов и фактор роста гепатоцитов, и содержит:
Основные элементы конструкции:
последовательность гена HGF
последовательность гена VEGF165
последовательность IRES (различные варианты)
Регуляторные элементы экспрессии и трансляции конструкции:
Энхансер 1 - CMV enhancer
Промотор pCMV (промотор цитомегаловируса)
Терминатор - bGH poly А (участок полиаденилирования гена бычьего гормона роста)
Регуляторные элементы экспрессии и трансляции конструкции в бактерии Е. coli:
Точка начала репликации (ori)
Селективный маркер для отбора трансформированных бактерий: - ген устойчивости к канамицину.
Однако данная плазмида не обеспечивает синтез достаточных количеств ангиогенных факторов роста ни in vitro, так, например, концентрации HGF и VEGF165 in vitro (в среде культивирования клеток линии НЕК293Т после их трансфекции) составляют не более 2,72 нМ и 1,6 нМ для HGF и VEGF165 соответственно. При этом соотношение концентраций HGF и VEGF165 непредсказуемо и в среднем составляет от 1:12 до 1:5 в зависимости от используемого IRES. Также данная плазмида не обеспечивает синтез достаточных количеств ангиогенных факторов роста ни in vitro ни при введении мышам – не более 0,72 нг/мл и вплоть до полного отсутствия. Следовательно, данный вектор не обладает параметрами, необходимыми для его использования в качестве экспрессионной конструкции для одновременной экспрессии генов фактора роста гепатоцитов и фактора роста эндотелия сосудов.
Технической проблемой является синтез двух генов в заданном соотношении, обеспечивающем эффективную стимуляцию ангиогенеза.
Раскрытие изобретения
Техническая проблема решается заявляемым плазмидным вектором, несущим гены ангиогенных факторов роста человека, и обеспечивающего синтез данных АФР in vivo в близком к эквимолярному соотношению.
Техническим результатом является создание плазмидной генетической конструкции, кодирующей кДНК фактора роста гепатоцитов (hHGF) и кДНК фактора роста эндотелия сосудов (hVEGF165) под независимыми промоторами, позволяющей осуществлять синтез данных АФР in vivo в соотношении молярных концентраций HGF/VEGF165 от
0,24:1 до 0,32:1 при обеспечении стабильного выхода белков. Полученный вектор оказался эффективен, и может быть использован для создания генотерапевтического препарата.
Для заявляемой генетической конструкции характерны следующие преимущества:
обеспечивает продукцию значительных количеств АФР: от 50 до 53 нМ и от 14 до 17 нМ для VEGF165 и HGF соответственно;
поскольку оба гена (VEGF165 и HGF) имеют собственные промоторы, энхансеры и терминаторы транскрипции, то экспрессия каждого из них происходит независимо и не влияет друг на друга.
Технический результат достигается созданием вектора, схема которого представлена на фиг.2, а именно генно-инженерной конструкцией для стимуляции ангиогенеза в ишемизированных тканях, включающей:
последовательность, кодирующую ген фактора роста гепатоцитов, находящуюся под контролем промотора цитомегаловируса - pCMV-HGF;
последовательность, кодирующую ген фактора роста эндотелия сосудов, находящуюся под контролем промотора гена β-актина цыпленка - pCAG-VEGF165.
При этом, в качестве последовательности, кодирующей ген фактора роста гепатоцитов, используют последовательность SEQ ID N0:1, а в качестве последовательности, кодирующей ген фактора роста эндотелия сосудов используют последовательность SEQ ID NО:2.
Генно-инженерная конструкция включает следующие последовательно расположенные элементы:
Энхансер 1 - CMV enhancer - 36 - 415 п.о.
Промотор 1 - pCMV (промотор цитомегаловируса)-488-691 п.о.
Ген hHGF - 784-2970 п.о.
Терминатор 1 - bGH poly А (участок полиаденилирования гена бычьего гормона роста) - 3044-3268 п.о.
Селективный маркер для отбора трансформированных бактерий - ген устойчивости к антибиотику - 3441-4253 п.о.
Энхансер 2 - CMV enhancer - 4455-4740 п.о.
Промотор 2 - pCAG (промотор гена β-актина цыпленка) - 4742-5019 п.о.
Гибридный интрон - 5019-5248 п.о.
Ген hVEGF165 - 5278-5853 п.о.
Терминатор 2 - SV40 poly А (участок полиаденилирования вируса SV40) - 5908-6029 п.о.
Точка начала репликации (ori) - 6235-6823 п.о.
Техническая проблема также решается применением заявляемой генно-инженерной конструкции в качестве действующего вещества генотерапевтического средства для стимуляции ангиогенеза в ишемизированных тканях.
Также техническая проблема решается средством для стимуляции ангиогенеза в ишемизированных тканях, включающее заявляемую плазмидную конструкцию и вспомогательные вещества для внутримышечного введения. При этом в качестве вспомогательных веществ использованы натрия хлорид или вода для инъекций.
Техническая проблема также решается способом стимуляции ангиогенеза в ишемизированных тканях, включающим внутримышечное введение лекарственного средства для стимуляции ангиогенеза в ишемизированных тканях, включающего заявляемую плазмидную конструкцию, в терапевтически эффективном количестве.
Краткое описание чертежей
Изобретение поясняется следующими чертежами.
На фиг. 1 представлена «карта» прототипа плазмидной генетической конструкции, кодирующей ген hHGF фактора роста гепатоцитов человека и ген hVEGF165 фактора роста эндотелия сосудов, разделенные последовательностью IRES.
На фиг. 2 представлена «карта» плазмидной генетической конструкции, кодирующей ген hHGF фактора роста гепатоцитов человека и ген hVEGF165 фактора роста эндотелия сосудов под двумя независимыми промоторами.
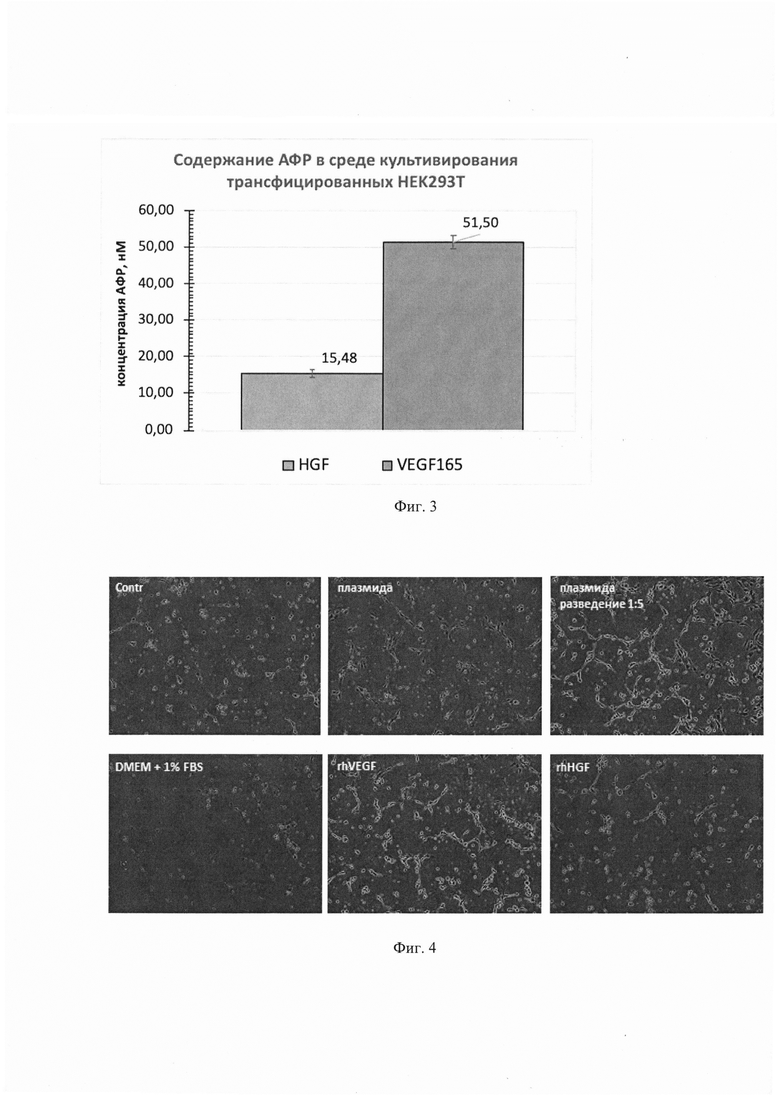
На фиг. 3. представлена диаграмма, демонстрирующая содержание HGF и VEGF165 в среде культивирования HEK293T после трансфекции кальций-фосфатным способом.
На фиг. 4. представлены фотографии, на которых показано формирование капилляроподобных структур клетками HUVEC на матригеле после их обработки экспериментальной и контрольными средами.
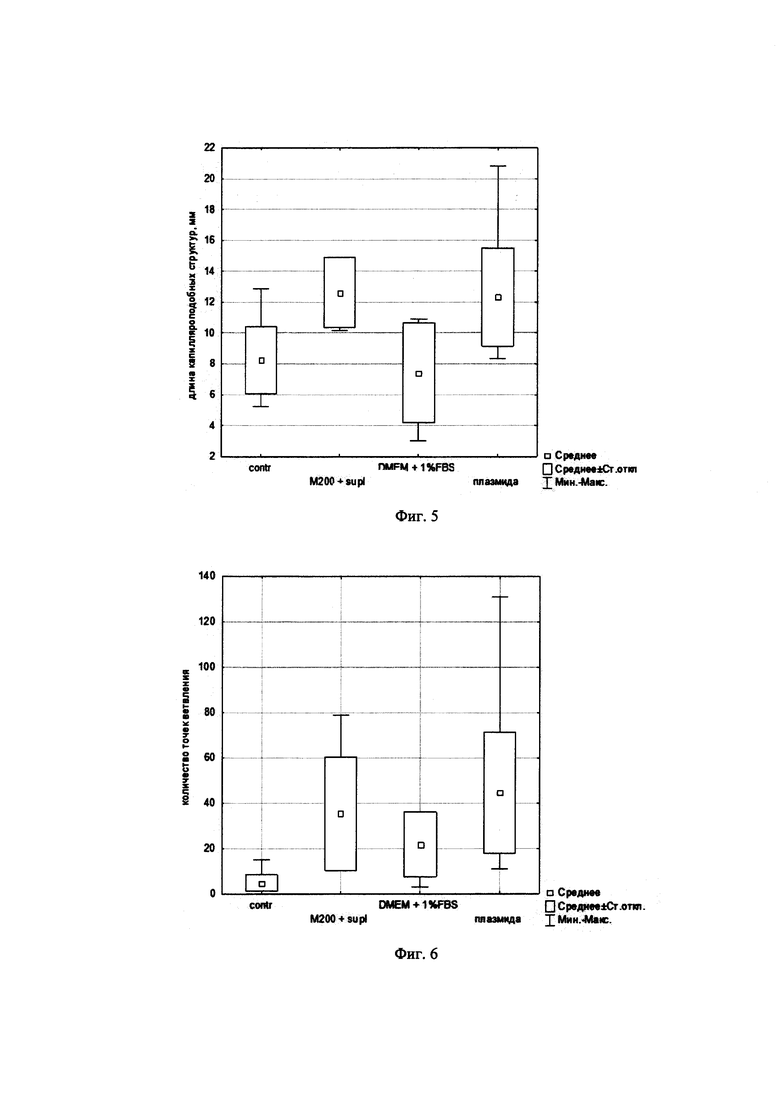
На фиг. 5 представлены графики, демонстрирующие общую длину капилляроподобных структур, сформированных клетками HUVEC на матригеле через 15 ч после обработки экспериментальной и контрольными средами.
На фиг. 6 представлены графики, демонстрирующие количество точек ветвления капилляроподобных структур, сформированных клетками HUVEC на матригеле через 15 ч после обработки экспериментальной и контрольными средами.
Осуществление изобретения
Заявляемая генно-инженерная конструкция представляет собой экспрессионную векторную плазмиду, в которую клонированы вставки кДНК, кодирующие фактор роста эндотелия сосудов и фактор роста гепатоцитов человека, а именно кольцевую ДНК длиной 6888 нуклеотидных пар.
Основные элементы конструкции:
участок pCMV-HGF
участок pCAG-VEGF 165
Расположение кДНК, кодирующих фактор роста эндотелия сосудов и фактор роста гепатоцитов человека, под независимыми промоторами, позволяет осуществлять синтез данных АФР in vivo в соотношении концентраций HGF/VEGF165 от 0,24:1 до 0,32:1 при обеспечении стабильного выхода белков.
Плазмидная конструкция несет оптимизированные гены VEGF165 (фактора роста эндотелия сосудов) и HGF (фактора роста гепатоцитов) человека, которые могут быть заменены на другие (например, нативные) последовательности данных генов.
Регуляторные элементы экспрессии и трансляции конструкции:
Энхансер 1 - CMV enhancer - 380 п.о.
Промотор 1 - pCMV (промотор цитомегаловируса) - 204 п.о.
Терминатор 1 - bGH poly А (участок полиаденилирования гена бычьего гормона роста) - 225 п.о.
Энхансер 2 - CMV enhancer - 286 п.о.
Промотор 2 - pCAG (промотор гена (β-актина цыпленка) - 278 п.о.
Г ибридный (химерный) интрон - 229 п.о.
Терминатор 2 - SV40 poly А (участок полиаденилирования вируса SV40) - 122 п.о.
Регуляторные элементы экспрессии и трансляции конструкции в бактерии Е. coli:
Точка начала репликации (ori) - 589 п.о.
Селективный маркер для отбора трансформированных бактерий: - ген устойчивости к канамицину - 813 п.о. (может быть заменен на другой ген устойчивости к антибиотикам или селективный маркер).
Синтез плазмиды осуществляли с помощью стандартной технологии и оборудования, применяемых для решения подобных задач в генной инженерии (Watson J.D., Gilman М, Witkowski J., Zoller M. - Recombinant DNA, Scientific American Books, New York, 1992). В качестве вектора для синтеза заявляемой плазмиды могут быть использованы любые коммерческие векторы, для которых характерна высококопийная репликация в Е. coli (более
150 копий на клетку), относительно небольшой размер (3-6 тыс. п.о.) и высокий уровень экспрессии клонируемого гена в клетках млекопитающих.
В качестве генов, кодирующих VEGF165 и HGF могут быть использованы как ко-дон-оптимизированные последовательности генов данных АФР с последовательностями SEQ ID NO: 1 и SEQ ID NO: 2, так и нативные гены.
Для трансформации и наработки плазмидной ДНК могут быть использованы компетентные штаммы Escherichia coli, например, Е. coli DH-5α.
Основные элементы конструкции:
Регуляторные элементы экспрессии и трансляции конструкции и гены VEGF165 (фактора роста эндотелия сосудов) и HGF (фактора роста гепатоцитов) человека:
 Энхансер 1 - CMV enhancer
Энхансер 1 - CMV enhancer
Промотор 1 - pCMV (промотор цитомегаловируса)
Последовательность гена HGF
Терминатор 1 - bGH poly А (участок полиаденилирования гена бычьего гормона роста)
Энхансер 2 - CMV enhancer
Промотор 2 - pCAG (промотор гена β-актина цыпленка)
Последовательность гена VEGF165
Гибридный (химерный) интрон
Терминатор 2 - SV40 poly А (участок полиаденилирования вируса SV40)
Регуляторные элементы экспрессии и трансляции конструкции в бактерии Е. coli:
Точка начала репликации (ori)
Селективный маркер для отбора трансформированных бактерий, выбранный из группы, включающей, но, не ограничиваясь ими, устойчивые к антибиотикам маркеры, такие как ампициллин, канамицин, неомицин и тому подобное.
Ниже представлено более подробное описание заявляемого изобретения. Настоящее изобретение может подвергаться различным изменениям и модификациям, понятным специалисту на основе прочтения данного описания. Такие изменения не ограничивают объем притязаний. Например, могут изменяться стратегия сборки генетической конструкции (используемые праймеры, сайты клонирования и т.д.), методы наращивания, выделения и очистки плазмид, способ трансфекции, методы оценки концентраций АФР и др.
Все используемые реагенты являются коммерчески доступными, все процедуры, если не оговорено особо, осуществляли при комнатной температуре или температуре окружающей среды, то есть в диапазоне от 18 до 25°С.
Фармацевтическая композиция на основе изобретения может вводиться парентерально, внутримышечно или любым другим способом, обеспечивающим возможность доставки соединения и/или композиции к клеткам/тканям. Например, фармацевтическую композицию можно вводить субъекту в виде внутримышечных инъекций. Для этих целей необходимо использовать стерильные растворы заявляемой композиции в фармацевтически приемлемом растворителе (см. ниже). Стерильность полученных композиций обеспечивается технологией и процессом их производства или получения в эксперименте.
Термин «фармацевтически приемлемый» относится к нетоксическому материалу, который не взаимодействует с действием активного компонента фармацевтической композиции. "Фармацевтически приемлемый носитель" относится к биосовместимому раствору, который в достаточной степени имеет такие характеристики, как стерильность, рН, изотоничность, стабильность и подобные, и может включать любые растворители, разбавители, включая стерильный физиологический раствор, раствор хлорида натрия для инъекций, раствор Рингера для инъекций, раствор декстрозы для инъекций, раствор декстрозы и хлорида натрия для инъекций, содержащий лактат раствор Рингера для инъекций и другие водные буферные растворы, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические вещества и подобные. Фармацевтически приемлемый носитель также может содержать стабилизаторы, консерванты, антиоксиданты или другие добавки, которые хорошо известны специалистам в данной области, или другой наполнитель, известный из уровня техники.
Концентрация плазмидного вектора для высокоэффективной экспрессии генов VEGF165 и HGF человека в носителе должна быть достаточной для обеспечения терапевтического ангиогенеза. Например, при применении раствора на основе 0,9% хлорида натрия терапевтическая доза может составлять приблизительно от 25 до 1000 мкг на кг веса пациента.
Введение рекомендуется осуществлять путем внутримышечной инъекции в место, максимально близкое к ишемизированному участку. Введение производится после стандартной обработки кожи, с соблюдением правил асептики, дробно через несколько вколов так, чтобы весь массив мышц пораженного сегмента был инфильтрирован раствором. Введение может осуществляться как однократно (разово), так и несколько раз в зависимости от показаний, который оценивает лечащий врач на основании специфических параметров пациента, таких как возраст, масса тела, пол, степень повреждения ткани и т.п.
1. Создание плазмидного вектора для высокоэффективной экспрессии генов VEGF165 и HGF человека.
Синтез генов и плазмид осуществляли с помощью стандартной технологии и оборудования, применяемых для решения подобных задач в генной инженерии [Sambrook J et al., Molecular Cloning - A laboratory Manual, Cold Spring Harbor Laboratory Press, New York, (1989)], [Watson, J.D., Oilman, M., Witkowski, J., Zoller, M. Recombinant DNA, Scientific American Books, New York, 1992].
Синтез плазмиды осуществляли на основе видоизмененной системы pVAX, преимуществом которой является факт разрешения ее к использованию FDA США, т.е. данные о безопасности входящих в ее состав компонентов. В качестве генов, кодирующих VEGF165 и HGF использовали последовательности SEQ ID NO: 1 и SEQ ID NO: 2. Для повышения уровня продукции HGF и VEGF165 в трансфицированных клетках была оптимизирована природная последовательность кДНК этих генов. Среди триплетов нуклеотидов, кодирующих аминокислоты белковых последовательностей природных генов HGF и VEGF165, были выявлены наиболее редко встречаемые последовательности в генах человека. Выявленные редкие триплеты, а также соседние с ними триплеты были заменены на триплеты, кодирующие ту же аминокислоту, но при этом наиболее часто встречающиеся в генах человека. После модификации все аминокислоты природных генов HGF и VEGF165 остались неизмененными.
Использованный дизайн плазмидного вектора, с одной стороны, позволил без существенного увеличения размера плазмиды вместить в нее кДНК двух факторов роста, а с другой, позволил обеспечить близкое к оптимальному соотношение продуцируемых белков, т.е. высокую продукцию рекомбинантного human-BDNF и умеренную продукцию human-uPA, различающуюся в среднем на порядок.
Ниже представлен пример реализации изобретения, который не ограничивает настоящее изобретение, а лишь демонстрирует возможность его осуществления с достижением технического результата.
Пример
Используемый экспрессионный вектор pVax2 был создан из коммерческого вектора pVax1 (ThermoFisher) и был взят как основной каркасный вектор для конструирования заявляемого изобретения. Модификация заключалась во вставке SV40 энхансера между CMV промотором и CMV энхансером pVax1 и замене гена устойчивости к канамицину KanR.
1. На первом этапе была проведена рестирикция pVax2 CeiI (в буфере W+BSA) и вставка фосфорилированного дуплекса с получением конструкции pVax2-VEGFIns.

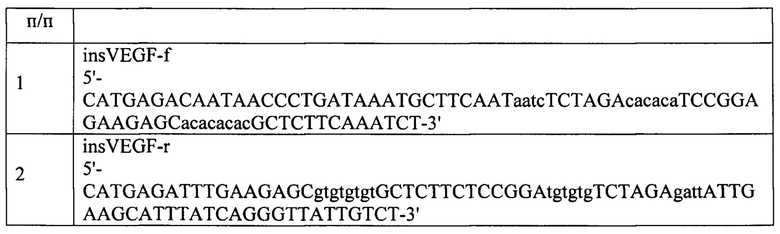
2. На втором этапе конструкция pVax2-VEGFIns подвергалась рестрикция AsuNHI+Pmel (CutSmart) с последующей вставкой оптимизированного hHGF, собранного из синтетических олигонуклеотидов и рестрицированного AsuNHI (CutSmart), с получением конструкции pVax2-CMV_hHGF-VEGFIns. Олигонуклеотиды для сборки hHGFopt приведены в таблице 2 и 3.





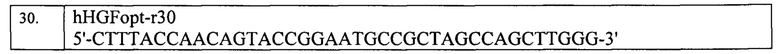
3. На третьем этапе была осуществлена последовательная рестрикция конструкции pVax2-hHGF-VEGFIns рестриктазами XbaI (CutSmart) и Bsp13I (2K), а затем вставка CAG-промотора, амплифицированного из вектора рХ458 с помощью праймеров, приведенных в таблице 4, и последовательно рестрицированного Xba1 (CutSmart) и Bsp13I (2K). В итоге была получена конструкция pVax2-CMV_hHGF-CAG_VEGFIns.

Затем, на четвертом этапе, была проведена рестрикция вектора pVax2-CMV_hHGF-CAG VEGFIns PciSI (В), и осуществлена вставка hVEGF165, собранного из синтетических олигонуклеотидов и рестрицированного PciSI (В) с получением заявляемой плазмиды pVax2-CMV_hHGF-CAG_hVEGF165. Олигонуклеотиды для сборки hVEGF165 приведены в таблицах 5 и 6.



Затем полученный продукт трансформировали в Е. coli DH5a и трансформанты отбирали на основании устойчивости к канамицину. Плазмидную ДНК, выделенную приблизительно из 10 таких колоний, анализировали на наличие кДНК HGF и VEGF165 ре-стрикционным расщеплением, используя различные ферменты рестрикции. Одну такую плазмиду секвенировали, используя автоматический ДНК секвенатор (ABI), и она была определена, как имеющая правильную интеграцию и последовательность кДНК HGF и VEGF165 в векторе pVax2.
Схема полученной плазмиды представлена на фиг. 2.
Таким образом, генно-инженерная конструкция, представляющая собой экспрессионную векторную плазмиду (кольцевую ДНК длиной 6888 нуклеотидных пар), в которую клонированы вставки кДНК, кодирующие фактор роста эндотелия сосудов и фактор роста гепатоцитов человека (несет оптимизированные гены VEGF165 (фактора роста эндотелия сосудов) и HGF (фактора роста гепатоцитов) человека), содержит основные элементы конструкции:
 участок pCMV-HGF 488-2970 п.о.
участок pCMV-HGF 488-2970 п.о.
участок pCAG-VEGF165 4742-5853 п.о.
Регуляторные элементы экспрессии и трансляции конструкции:
Энхансер 1 - CMV enhancer - 36-415 п.о.
Промотор 1 - pCMV (промотор цитомегаловируса) - 488-691 п.о.
Терминатор 1 - bGH poly А (участок полиаденилирования гена бычьего гормона роста) - 3044-3268 п.о.
Энхансер 2 - CMV enhancer - 4455-4720 п.о.
Промотор 2 - pCAG (промотор гена β-актина цыпленка) - 4742-5019 п.о.
Гибридный интрон - 5020-5248 п.о.
Терминатор 2 - SV40 poly А (участок полиаденилирования вируса SV40) -5908-6029 п.о.
Регуляторные элементы экспрессии и трансляции конструкции в бактерии Е. coli:
Точка начала репликации (ori) - 6235-6823 п.о.
Селективный маркер для отбора трансформированных бактерий: - ген устойчивости к канамицину - 3441-4253 п.о. (может быть заменен на другой ген устойчивости к антибиотикам или селективный маркер).
Для амплификации всех полученных плазмидных ДНК использовались бактерии Е. coli штамма DH-5α. После трансформации бактерии культивировались в течение ночи в селективной среде LB с канамицином в концентрации 100 мкг/мл. После окончания инкубации бактерии осаждали центрифугированием и использовали для выделения плазмидной ДНК на колонке (Qiagen, США).
1. Приготовление заявляемой генетической конструкции
Для трансформации и наработки плазмидной ДНК (пДНК) использовали штамм Е. coli DH-5α. Исходный штамм Е. coli DH-5α является музейным и хранится во Всероссийской Коллекции Промышленных Микроорганизмов. Трансформированный штамм Е. coli DH-5α-CMV-HGF_CAG-VEGF165 отличается от исходного штамма Е. coli DH-5α устойчивостью к антибиотику канамицину, обеспечиваемой введенной в состав штамма плазмиды «CMV-HGF_CAG-VEGF165». Штамм-продуцент DH-5α-CMV-HGF_CAG-VEGF165 для наработки плазмидной ДНК получали трансформацией исходного компетентного штамма Е. coli DH-5α соответствующей плазмидой, с последующим отбором рекомбинантных клонов на среде LB с канамицином при 37°С. Трансформацию компетентных клеток Е. coli штамма DH5α проводили следующим образом. В эппендорф, содержащий 1 нг пДНК добавляли суспензию размороженных на льду бактерий (объем культуры 30 мкл), после чего проводили трансформацию методом «теплового шока». Для этого пробирку помещали в термостат, нагретый до 42°С на 45 сек, после чего на 5 мин переносили в лед. После этого к полученной культуре добавляли стерильную среду LB (500 мкл) и инкубировали на термошейкере в течение 60 мин (500 об/мин, 37°С). После этого 150 мкл полученной суспензии переносили в ламинарном бактериологическом боксе на заранее подготовленные чашки Петри диаметром 100 мм залитые LB-агаром, содержащим селектирующий агент - канамицин (100 мкг/мл). Контролем трансформации служила проба без внесения пДНК, в которой адекватным считалось отсутствие бактериального роста в LB-агаре с канамицином. После этого суспензию распределяли по поверхности LB-arapa с помощью прожженной проволочной петли и полученную культуру переносили в инкубатор на 15-17 ч при температуре 37°С. По окончании инкубации одну колонию бактерий с помощью стерильного шпателя переносили в жидкую культуру малого объема (15-20 мл, соотношение объема жидкости и воздуха не менее 1:4-1:5) на среде LB, содержащей 100 мкг/мл канамицина. После этого колбу с инокулированной средой помещали в термошейкер на 18 часов (250 об/мин, 37°С). Далее полученную культуру переносили в жидкую среду LB со 100 мкг/мл канамицина объемом 500 мл в соотношении 1:500, после чего ее инкубировали в аналогичных условиях в течение 15-18 ч для получения большого объема бактериальных клеток, содержащих пДНК. Полученную культуру бактерий в среде LB (объемом от 500 мл до 3 л в зависимости от количества пДНК) осаждали центрифугированием (12000 g, 15 мин), после чего жидкую фазу удаляли, а осадок использовали для выделения пДНК. Лизис, очистку от эндотоксинов и выделение плазмиды проводили с использованием наборов компании Qiagen (Midi или Maxi EndoFree kit) согласно протоколу производителя
2. Трансфекция клеток линии HEK293T
Трансфекция клеток линии HEK293T проводилась кальций-фосфатным методом, однако, возможно использование и другого метода (например, протокол трансфекции с помощью Lipofectamine 2000). Трансфекция проводилась в 6-луночном планшете по достижении клетками 80-90% конфлюэнтного монослоя. Для этого 2 мкг пДНК смешивали с раствором 0,3 М CaCl2 и по каплям добавляли к 2xHBS (рН=7,10), состоящему из 280 мМ NaCl; 50 мМ HEPES; 1,5 мМ Na2HPO4. После этого полученную смесь инкубировали 1 мин и по каплям добавляли к среде культивирования HEK293T (DMEM/10% ФБС, объем 2 мл/лунке). Через 15-16 часов клетки отмывали и к ним добавляли бессывороточную DMEM, которую через 48 ч собирали, откручивали от клеточного дебриса и использовали для оценки продукции VEGF165 и HGF. Концентрации VEGF165 и HGF в среде культивирования клеток HEK293T через 48 ч после трансфекции составили 51,50 и 15,48 нМ соответственно.
Соотношение молярных концентраций HGF7VEGF165 равнялось 0,3. Полученные данные представлены на фиг. 3
3. Тестирование биологической активности плазмид путем оценки формирования капилляроподобных структур клетками HUVEC на матригеле (tube assay) - модель ангиогенеза in vitro
Клетки HUVEC 3-4 пассажа высаживали на 48-луночный планшет, покрытый слоем Матригеля (BD Biosciences). Перед началом эксперимента клетки депривировали в бессывороточной среде для культивирования клеток эндотелия (М200) в течение 4 ч. Затем на клетки наносили среду, собранную с трансфицированных клеток HEK293T (общий объем среды 2000 мкл). В качестве положительного контроля использовали клетки, к которым добавлялась полная среда культивирования (M200+supl). Планшет с клетками помещали в систему прижизненного анализа клеток - IncuCyte®, клетки фотографировали, в режиме фазового контраста со 100х увеличением каждые 2 часа. Длину трубочек и число точек ветвления оценивали с использованием программного пакета ImageJ. Результат представлен на фиг. 4-6.
4. Инъекция плазмид в скелетные мышцы мышей с последующей низковольтовой электропорацией
Для внутримышечного введения плазмид использовался стерильный раствор пДНК в 0,9% NaCl, который вводился в m. tibialis anterior однократно. Животным вводилось по 100 мкг пДНК, растворенной в 30-50 мкл физиологического раствора (2-3 мг/мл). Чрескожно игла инсулинового шприца вводилась параллельно продольной оси мышцы, после чего раствор медленно вводился в толщу мышцы во избежание разрыва перимизия. Для повышения эффективности трансфекции был использован метод низковольтовой электропорации, описанный Schertzer и Lynch, с минимальной модификацией: было опущено введение гиалуронидазы, описанное в оригинальном методе. Непосредственно после инъекции плазмид на кожу, прилегающую к m. tibialis anterior, накладывались пластинчатые электроды, смазанные проводящим гелем. Затем с использованием генератора импульсных токов постоянной фор мы марки BTX-Harvard apparatus ЕСМ 830 (Harvard apparatus, США) проводилась электропорация тремя импульсами с напряжением 80 В/см расстояния между электродами, частотой 1 Гц и продолжительностью 20 мсек. Затем полярность электродов менялась и подавалось еще 3 импульса с аналогичными приведенным характеристиками. Поле проведения электропорации поверхность кожи очищалась 70% этиловым спиртом.
5. Опыты ex vivo на модели эксплантной культуры скелетной мышцы
Образцы т. tibialis anterior мыши были выделены на 2 день после введения пДНК. Подготовку эксплантной культуры на матригеле выполняли по методу Jang et al. (Jang HS, Kim HJ, Kim JM, Lee YS, Kim KL, et al. (2004) A novel ex vivo angiogenesis assay based on electroporation-mediated delivery of naked plasmid DNA to skeletal muscle. Mol Ther 9: 464-474.). Образцы скелетной мышцы высаживали на матригель и затем культивировали в полной среде DMEM с 4,5 г/л D-глюкозы (Life Technologies, США) и 2% ФБС (Gibco, США) при стандартных условиях. На 3 и 7 сутки культивирования эксплантов отбирали образцы культуральной среды для оценки содержания VEGF165 и HGF человека методом ИФА. Концентрации VEGF165 и HGF, продуцированных мышиными мышечными эксплантами после инъекции, составили 0,81 и 0,35 нг/мл соответственно на 3 день и 0,82 и 0,49 нг/мл на 7 день. Соотношение молярных концентраций HGF/VEGF165 равнялось 0,24 на 3 день и 0,32 на 7 день.
6. Сравнительная оценка продукции VEGF165 и HGF клетки НЕК29 разными плазмидами
На первом этапе на основе вектора pVAX нами были созданы прототипы заявляемой генетической конструкции, а именно плазмиды, несущие различные варианты IRES для осуществления одновременной экспрессии генов факторов роста HGF и VEGF165, характеризующиеся общей схемой строения: HGF-IRES -VEGF165. Для оценки продукции VEGF165 и HGF клетки НЕК293Т трансфицировали кальций фосфатным способом, концентрации VEGF165 и HGF в клеточной среде оценивались методом ИФА (результаты представлены в таблице 7).
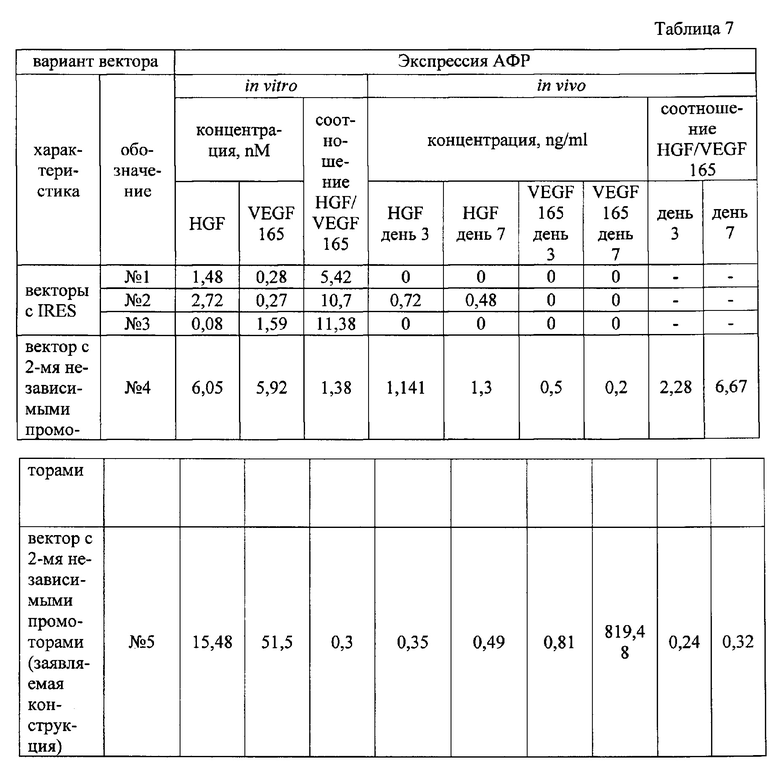
Плазмида №3 - конструкция HGF-IRES_FGF1-VEGF165 отличается от конструкции №1 тем, что включает последовательность IRES эукариотического белка FGF1. Молярные концентрации HGF и VEGF165 составили 0,08 и 1,59 нМ соответственно. Соотношение HGF/VEGF165 равнялось 11,38.
In vivo продукция белков VEGF165 и HGF оценивалась на модели эксплантной культуры скелетных мышц, выделенных после инъекции с электропорации векторов по пункту 1 (100 нг/мл) мышам. Образцы культуральной среды для оценки содержания VEGF165 и HGF методом ИФА отбирались на 3 и 7 сутки культивирования эксплантов.
Для плазмиды №1 концентрации HGF и VEGF165, продуцированных мышиными мышечными эксплантами, не детектировались методом ИФА.
Для плазмиды №2 концентрации HGF в среде культивирования мышечных эксплантов составили 0,72 нг/мл и 0,48 нг/мл на 3 и 7 дни соответственно. Концентрация белка VEGF165 была ниже порога чувствительности используемого метода детекции.
Для плазмиды №3 концентрации HGF и VEGF165 также не детектировались методом ИФА.
Таким образом, векторы №№1, 2, 3 не дали достаточной для оценки методом ИФА продукции АФР. Из полученных данных можно сделать вывод о том, что синтезируемые количества белков VEGF165 и HGF ниже констант диссоциации данных АФР с их рецепторами и недостаточны для проявления их биологического действия.
В качестве альтернативного варианта мультицистронного вектора была создана и протестирована следующая генетическая конструкция:
№4 - вектор, в котором гены белков VEGF165 и HGF имеют собственные независимые промоторы: цитомегаловируса (CMV) и куриного бета-актина (CAG) для генов HGF и VEGF165 соответственно - «CMV_CAG». Молярные концентрации HGF и VEGF165 составили 6,05 и 5,92 нМ соответственно. Соотношение HGF/VEGF165 равнялось 1,38.
Конструкция №4 была протестирована in vivo на модели эксплантной культуры скелетных мышц мышей, описанной в пункте 2. Продукция белков VEGF165 и HGF оценивалась методом ИФА.
После инъекции и электропорации мышам вектора №4 концентрации HGF в среде культивирования мышечных эксплантов составили 1,14 нг/мл и 1,3 нг/мл на 3 и 7 дни соответственно. Концентрации VEGF165 составили 0,5 нг/мл и 0,2 нг/мл на 3 и 7 дни соответственно. Соотношение HGF/VEGF165 равнялось 2,28 на 3 день и 6,67 на 7 день культивирования эксплантов.
Таким образом, только вектор, включающий два отдельных промотора для независимой экспрессии обоих АФР дает продукцию белков HGF и VEGF165 in vitro и in vivo.
На основе вектора №4, сконструирована заявляемая генетическая конструкция. Вектор представляет собой аналогичную плазмиде №4 конструкцию, отличающуюся тем, что в ней последовательностью гена HGF заменена на оптимизированную. Концентрации белков HGF и VEGF165 в среде культивирования трансфицированных плазмидой клеток HEK293T составили 15,48 и 51,5 нМ соответственно. Соотношение HGF/VEGF165 равнялось 0,3.
Как следует из описанного выше, единственным вектором, позволяющим достигать оптимальных концентраций и соотношения АФР является заявляемая конструкция с двумя отдельными промоторами для каждого из генов: CMV для гена HGF и CAG для гена VEGF165. Использование двух отдельных кассет с собственными энхансерами, промоторами и терминаторами транскрипции для каждого из генов обеспечивает независимую экспрессию каждого из них. Результаты экспериментов с вектором, выбранном в качестве прототипа - плазмидой №4, а также с заявляемой конструкцией показали активность обоих промоторов как in vitro в клетках человека, так и in vivo в эксплантной культуре мышц, полученной после инъекции плазмиды мышам. Однако только использование заявляемого вектора обеспечивает продукцию белков HGF и VEGF165 в близком к эквимолярному соотношению с небольшим преобладанием «активатора ангиогенеза» - фактора роста эндотелия сосудов. Все варианты векторов, основанные на отличных способах реализации, не позволяют достичь технического результата и не могут быть использованы для одновременной экспрессии in vivo двух ангиогенных факторов роста (АФР) с одной генетической конструкции в достаточных количествах и в оптимальном соотношении.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Открытое общество с ограниченной ответственностью «Генная и клеточная терапия» (ООО «Генная и клеточная терапия»)
<120> ГЕННО-ИНЖЕНЕРНАЯ КОНСТРУКЦИЯ ДЛЯ СТИМУЛЯЦИИ АНГИОГЕНЕЗА
<160> SEQ ID NО:1
<210> 1
<211> 2187
<212> ДНК
<213> Кодон-оптимизированная последовательность Homo sapiens (human)
<223> Нуклеотидная последовательность гена фактор роста гепатоцитов (hHGF)
<400> 1
1 ATGTGGGTGA CCAAGCTGCT GCCAGCCCTG CTGCTGCAGC ATGTCCTCCT
51 GCATCTCCTC CTGCTCCCCA TCGCCATCCC CTATGCAGAG GGACAAAGGA
101 AAAGAAGAAA TACAATTCAT GAATTCAAAA AATCAGCAAA GACCACCCTG
151 ATCAAGATCG ACCCCGCCCT GAAGATCAAG ACCAAAAAAG TGAATACTGC
201 AGACCAGTGT GCCAATAGAT GTACCAGGAA CAAGGGCCTG CCATTCACTT
251 GCAAGGCTTT TGTTTTTGAT AAAGCAAGAA AACAATGCCT CTGGTTCCCC
301 TTCAATAGCA TGTCAAGTGG AGTGAAAAAA GAATTTGGCC ATGAATTTGA
351 CCTCTATGAA AACAAAGACT ACATTAGAAA CTGCATCATT GGTAAAGGAC
401 GCAGCTACAA GGGAACAGTA TCTATCACTA AGAGTGGCAT CAAATGTCAG
451 CCCTGGAGTT CCATGATACC ACACGAACAC AGCTTTCTGC CTTCCAGCTA
501 CCGGGGCAAG GACCTGCAGG AGAACTACTG CCGGAACCCT CGAGGGGAAG
551 AAGGGGGACC CTGGTGTTTC ACAAGCAATC CAGAGGTACG CTACGAAGTC
601 TGTGACATTC CTCAGTGTTC AGAAGTTGAA TGCATGACCT GCAATGGGGA
651 GAGTTATCGA GGTCTCATGG ATCATACAGA ATCAGGCAAG ATTTGTCAGC
701 GCTGGGATCA TCAGACACCA CACCGGCACA AATTCTTGCC TGAAAGATAT
751 CCCGACAAGG GCTTTGATGA TAATTATTGC CGCAATCCCG ATGGCCAGCC
801 GAGGCCATGG TGCTATACTC TTGACCCTCA CACCCGCTGG GAGTACTGTG
851 CAATTAAAAC ATGCGCTGAC AATACTATGA ATGACACTGA TGTTCCTTTG
901 GAAACAACTG AATGCATCCA AGGTCAAGGA GAAGGCTACA GGGGCACTGT
951 CAATACCATT TGGAATGGAA TTCCATGTCA GCGTTGGGAT TCTCAGTATC
1001 CTCACGAGCA TGACATGACT CCTGAAAATT TCAAGTGCAA GGACCTACGA
1051 GAAAATTACT GCCGAAATCC AGATGGGTCT GAATCACCCT GGTGTTTTAC
1101 CACTGATCCA AACATCCGAG TTGGCTACTG CTCCCAAATT CCAAACTGTG
1151 ATATGTCACA TGGACAAGAT TGTTATCGTG GGAATGGCAA AAATTATATG
1201 GGCAACTTAT CCCAAACAAG ATCTGGACTA ACATGTTCAA TGTGGGACAA
1251 GAACATGGAA GACTTACATC GTCATATCTT CTGGGAACCA GATGCAAGTA
1301 AGCTGAATGA GAATTACTGC CGAAATCCAG ATGATGATGC TCATGGACCC
1351 TGGTGCTACA CGGGAAATCC ACTCATTCCT TGGGATTATT GCCCTATTTC
1401 TCGTTGTGAA GGTGATACCA CACCTACAAT AGTCAATTTA GACCATCCCG
1451 TAATATCTTG TGCCAAAACG AAACAATTGC GAGTTGTAAA TGGGATTCCA
1501 ACACGAACAA ACATAGGATG GATGGTTAGT TTGAGATACA GAAATAAACA
1551 TATCTGCGGA GGATCATTGA TAAAGGAGAG TTGGGTTCTT ACTGCACGAC
1601 AGTGTTTCCC TTCTCGAGAC TTGAAAGATT ATGAAGCTTG GCTTGGAATT
1651 CATGATGTCC ACGGAAGAGG AGATGAGAAA TGCAAACAGG TTCTCAATGT
1701 TTCCCAGCTG GTATATGGCC CTGAAGGATC AGATCTGGTT TTAATGAAGC
1751 TTGCCAGGCC TGCTGTCCTG GATGATTTTG TTAGTACGAT TGATTTACCT
1801 AATTATGGAT GCACAATTCC TGAAAAGACC AGTTGCAGTG TTTATGGCTG
1851 GGGCTACACT GGACTGATCA ACTACGACGG CCTGCTGCGG GTGGCCCACC
1901 TGTACATCAT GGGAAATGAG AAATGCAGCC AGCATCATCG AGGGAAGGTG
1951 ACTCTGAATG AGTCTGAAAT ATGTGCTGGG GCTGAAAAGA TTGGATCAGG
2001 ACCATGTGAG GGGGATTATG GTGGCCCACT TGTTTGTGAG CAACATAAAA
2051 TGAGAATGGT TCTTGGTGTC ATTGTTCCTG GTCGTGGATG TGCCATTCCA
2101 AATCGTCCTG GTATTTTTGT CCGAGTAGCA TATTATGCAA AATGGATCCA
2151 CAAGATCATC CTGACCTACA AGGTGCCACA GTCCTGA
<160> SEQ ID NО:2
<210> 2
<211> 576
<212> ДНК
<213> Кодон-оптимизированная последовательность Homo sapiens (human)
<223> Нуклеотидная последовательность гена фактора роста эндотелия сосудов (hVEGF165)
<400> 2
1 ATGAACTTTC TGCTGAGCTG GGTCCACTGG TCACTGGCAC TGCTGCTGTA
51 TCTGCATCAC GCAAAATGGT CACAGGCCGC ACCTATGGCC GAGGGCGGAG
101 GGCAGAACCA CCATGAAGTG GTCAAGTTCA TGGACGTGTA CCAGAGATCA
151 TATTGTCACC CTATCGAGAC ACTGGTCGAC ATTTTCCAGG AATACCCAGA
201 TGAGATCGAA TATATCTTCA AGCCAAGCTG CGTGCCACTG ATGCGGTGCG
251 GCGGATGCTG TAACGATGAG GGACTGGAAT GCGTCCCCAC CGAGGAATCT
301 AATATCACAA TGCAGATCAT GCGAATTAAG CCTCACCAGG GGCAGCATAT
351 TGGCGAGATG AGTTTCCTGC AGCACAACAA ATGCGAGTGT AGGCCAAAGA
401 AAGACCGAGC TCGACAGGAG AATCCATGTG GACCTTGCTC TGAAAGGAGA
451 AAGCATCTGT TTGTGCAGGA CCCCCAGACT TGCAAGTGTA GCTGCAAAAA
501 TACCGATTCC AGGTGTAAGG CACGGCAGCT GGAACTGAAT GAGCGGACCT
551 GTCGGTGTGA TAAGCCAAGA AGATAA
<160> SEQ ID NО:3
<210> 3
<211> 229
<212> ДНК
<213> искусственная последовательность: hybrid between chicken β-actin (CBA) and minute virus of mice (MMV) introns
<223> Нуклеотидная последовательность гибридного (химерного) интрона
<400> 3
1 ggagtcgctg cgacgctgcc ttcgccccgt gccccgctcc gccgccgcct
51 cgcgccgccc gccccggctc tgactgaccg cgttactccc acaggtgagc
101 gggcgggacg gcccttctcc tccgggctgt aattagctga gcaagaggta
151 agggtttaag ggatggttgg ttggtggggt attaatgttt aattacctgg
201 agcacctgcc tgaaatcact ttttttcag
<160> SEQ ID NО:4
<210> 4
<211> 82
<212> ДНК
<213> искусственная последовательность insVEGF-f
<223> Олигонуклеотид для вставки фосфорилированного дуплекса
<400> 4
1 CATGAGACAA TAACCCTGAT AAATGCTTCA ATaatcTCTA GAcacacaTC
51 CGGAGAAGAG CacacacacG CTCTTCAAAT CT
<160> SEQ ID NО:5
<210> 5
<211> 82
<212> ДНК
<213> искусственная последовательность insVEGF-r
<223> Олигонуклеотид для вставки фосфорилированного дуплекса
<400> 5
1 CATGAGATTT GAAGAGCgtg tgtgtGCTCT TCTCCGGAtg tgtgTCTAGA
51 gattATTGAA GCATTTATCA GGGTTATTGT CT
<160> SEQ ID NО:6
<210> 6
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f1
<223> Олигонуклеотид для сборки hHGFopt
<400> 6
1 CCCAAGCTGG CTAGCGGCAT TCCGGTACTG TTGGTAAAGC CACCATGTGG
51 GTGACCAAGC TGCTGCCAGC CCTGCTGCT
<160> SEQ ID NО:7
<210> 7
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f2
<223> Олигонуклеотид для сборки hHGFopt
<400> 7
1 GCAGCATGTC CTCCTGCATC TCCTCCTGCT CCCCATCGCC ATCCCCTATG
51 CAGAGGGACA AAGGAAAAGA AGAAATACA
<160> SEQ ID NО:8
<210> 8
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f3
<223> Олигонуклеотид для сборки hHGFopt
<400> 8
1 ATTCATGAAT TCAAAAAATC AGCAAAGACC ACCCTGATCA AGATCGACCC
51 CGCCCTGAAG ATCAAGACCA AAAAAGTGA
<160> SEQ ID NО:9
<210> 9
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f4
<223> Олигонуклеотид для сборки hHGFopt
<400> 9
1 ATACTGCAGA CCAGTGTGCC AATAGATGTA CCAGGAACAA GGGCCTGCCA
51 TTCACTTGCA AGGCTTTTGT TTTTGATAA
<160> SEQ ID NО:10
<210> 10
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f5
<223> Олигонуклеотид для сборки hHGFopt
<400> 10
1 AGCAAGAAAA CAATGCCTCT GGTTCCCCTT CAATAGCATG TCAAGTGGAG
51 TGAAAAAAGA ATTTGGCCAT GAATTTGAC
<160> SEQ ID NО:11
<210> 11
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f6
<223> Олигонуклеотид для сборки hHGFopt
<400> 11
1 CTCTATGAAA ACAAAGACTA CATTAGAAAC TGCATCATTG GTAAAGGACG
51 CAGCTACAAG GGAACAGTAT CTATCACTA
<160> SEQ ID NО:12
<210> 12
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f7
<223> Олигонуклеотид для сборки hHGFopt
<400> 12
1 AGAGTGGCAT CAAATGTCAG CCCTGGAGTT CCATGATACC ACACGAACAC
51 AGCTTTCTGC CTTCCAGCTA CCGGGGCAA
<160> SEQ ID NО:13
<210> 13
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f8
<223> Олигонуклеотид для сборки hHGFopt
<400> 13
1 GGACCTGCAG GAGAACTACT GCCGGAACCC TCGAGGGGAA GAAGGGGGAC
51 CCTGGTGTTT CACAAGCAAT CCAGAGGTA
<160> SEQ ID NО:14
<210> 14
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f9
<223> Олигонуклеотид для сборки hHGFopt
<400> 14
1 CGCTACGAAG TCTGTGACAT TCCTCAGTGT TCAGAAGTTG AATGCATGAC
51 CTGCAATGGG GAGAGTTATC GAGGTCTCA
<160> SEQ ID NО:15
<210> 15
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f10
<223> Олигонуклеотид для сборки hHGFopt
<400> 15
1 TGGATCATAC AGAATCAGGC AAGATTTGTC AGCGCTGGGA TCATCAGACA
51 CCACACCGGC ACAAATTCTT GCCTGAAAG
<160> SEQ ID NО:16
<210> 16
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f11
<223> Олигонуклеотид для сборки hHGFopt
<400> 16
1 ATATCCCGAC AAGGGCTTTG ATGATAATTA TTGCCGCAAT CCCGATGGCC
51 AGCCGAGGCC ATGGTGCTAT ACTCTTGAC
<160> SEQ ID NО:17
<210> 17
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f12
<223> Олигонуклеотид для сборки hHGFopt
<400> 17
1 CCTCACACCC GCTGGGAGTA CTGTGCAATT AAAACATGCG CTGACAATAC
51 TATGAATGAC ACTGATGTTC CTTTGGAAA
<160> SEQ ID NО:18
<210> 18
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f13
<223> Олигонуклеотид для сборки hHGFopt
<400> 18
1 CAACTGAATG CATCCAAGGT CAAGGAGAAG GCTACAGGGG CACTGTCAAT
51 ACCATTTGGA ATGGAATTCC ATGTCAGCG
<160> SEQ ID NО:19
<210> 19
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f14
<223> Олигонуклеотид для сборки hHGFopt
<400> 19
1 TTGGGATTCT CAGTATCCTC ACGAGCATGA CATGACTCCT GAAAATTTCA
51 AGTGCAAGGA CCTACGAGAA AATTACTGC
<160> SEQ ID NО:20
<210> 20
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f15
<223> Олигонуклеотид для сборки hHGFopt
<400> 20
1 CGAAATCCAG ATGGGTCTGA ATCACCCTGG TGTTTTACCA CTGATCCAAA
51 CATCCGAGTT GGCTACTGCT CCCAAATTC
<160> SEQ ID NО:21
<210> 21
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f16
<223> Олигонуклеотид для сборки hHGFopt
<400> 21
1 CAAACTGTGA TATGTCACAT GGACAAGATT GTTATCGTGG GAATGGCAAA
51 AATTATATGG GCAACTTATC CCAAACAAG
<160> SEQ ID NО:22
<210> 22
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f17
<223> Олигонуклеотид для сборки hHGFopt
<400> 22
1 ATCTGGACTA ACATGTTCAA TGTGGGACAA GAACATGGAA GACTTACATC
51 GTCATATCTT CTGGGAACCA GATGCAAGT
<160> SEQ ID NО:23
<210> 23
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f18
<223> Олигонуклеотид для сборки hHGFopt
<400> 23
1 AAGCTGAATG AGAATTACTG CCGAAATCCA GATGATGATG CTCATGGACC
51 CTGGTGCTAC ACGGGAAATC CACTCATTC
<160> SEQ ID NО:24
<210> 24
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f19
<223> Олигонуклеотид для сборки hHGFopt
<400> 24
1 CTTGGGATTA TTGCCCTATT TCTCGTTGTG AAGGTGATAC CACACCTACA
51 ATAGTCAATT TAGACCATCC CGTAATATC
<160> SEQ ID NО:25
<210> 25
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f20
<223> Олигонуклеотид для сборки hHGFopt
<400> 25
1 TTGTGCCAAA ACGAAACAAT TGCGAGTTGT AAATGGGATT CCAACACGAA
51 CAAACATAGG ATGGATGGTT AGTTTGAGA
<160> SEQ ID NО:26
<210> 26
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-f21
<223> Олигонуклеотид для сборки hHGFopt
<400> 26
1 TACAGAAATA AACATATCTG CGGAGGATCA TTGATAAAGG AGAGTTGGGT
51 TCTTACTGCA CGACAGTGTT TCCCTTCTC
<160> SEQ ID NО:27
<210> 27
<211> 78
<212> ДНК
<213> искусственная последовательность hHGFopt-f22
<223> Олигонуклеотид для сборки hHGFopt
<400> 27
1 GAGACTTGAA AGATTATGAA GCTTGGCTTG GAATTCATGA TGTCCACGGA
51 AGAGGAGATG AGAAATGCAA ACAGGTTC
<160> SEQ ID NО:28
<210> 28
<211> 78
<212> ДНК
<213> искусственная последовательность hHGFopt-f23
<223> Олигонуклеотид для сборки hHGFopt
<400> 28
1 TCAATGTTTC CCAGCTGGTA TATGGCCCTG AAGGATCAGA TCTGGTTTTA
51 ATGAAGCTTG CCAGGCCTGC TGTCCTGG
<160> SEQ ID NО:29
<210> 29
<211> 78
<212> ДНК
<213> искусственная последовательность hHGFopt-f24
<223> Олигонуклеотид для сборки hHGFopt
<400> 29
1 ATGATTTTGT TAGTACGATT GATTTACCTA ATTATGGATG CACAATTCCT
51 GAAAAGACCA GTTGCAGTGT TTATGGCT
<160> SEQ ID NО:30
<210> 30
<211> 78
<212> ДНК
<213> искусственная последовательность hHGFopt-f25
<223> Олигонуклеотид для сборки hHGFopt
<400> 30
1 GGGGCTACAC TGGACTGATC AACTACGACG GCCTGCTGCG GGTGGCCCAC
51 CTGTACATCA TGGGAAATGA GAAATGCA
<160> SEQ ID NО:31
<210> 31
<211> 78
<212> ДНК
<213> искусственная последовательность hHGFopt-f26
<223> Олигонуклеотид для сборки hHGFopt
<400> 31
1 GCCAGCATCA TCGAGGGAAG GTGACTCTGA ATGAGTCTGA AATATGTGCT
51 GGGGCTGAAA AGATTGGATC AGGACCAT
<160> SEQ ID NО:32
<210> 32
<211> 78
<212> ДНК
<213> искусственная последовательность hHGFopt-f27
<223> Олигонуклеотид для сборки hHGFopt
<400> 32
1 GTGAGGGGGA TTATGGTGGC CCACTTGTTT GTGAGCAACA TAAAATGAGA
51 ATGGTTCTTG GTGTCATTGT TCCTGGTC
<160> SEQ ID NО:33
<210> 33
<211> 80
<212> ДНК
<213> искусственная последовательность hHGFopt-f28
<223> Олигонуклеотид для сборки hHGFopt
<400> 33
1 GTGGATGTGC CATTCCAAAT CGTCCTGG TATTTTTGTC CGAGTAGCAT
51 ATTATGCAAA ATGGATCCAC AAGATCATCC
<160> SEQ ID NО:34
<210> 34
<211> 77
<212> ДНК
<213> искусственная последовательность hHGFopt-f29
<223> Олигонуклеотид для сборки hHGFopt
<400> 34
1 TGACCTACAA GGTGCCACAG TCCTGACTTA AGCTTGGTAC CGAGCTCGGA
51 TCCGCCCCTC TCCCTCCCCC CCCCTAA
<160> SEQ ID NО:35
<210> 35
<211> 39
<212> ДНК
<213> искусственная последовательность hHGFopt-r1
<223> Олигонуклеотид для сборки hHGFopt
<400> 35
1 TTAGGGGGGG GGGAGGGAGA GGGGCGGATC CGAGCTCGG
<160> SEQ ID NО:36
<210> 36
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r2
<223> Олигонуклеотид для сборки hHGFopt
<400> 36
1 TACCAAGCTT AAGTCAGGAC TGTGGCACCT TGTAGGTCAG GATGATCTTG
51 TGGATCCATT TTGCATAATA TGCTACTCG
<160> SEQ ID NО:37
<210> 37
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r3
<223> Олигонуклеотид для сборки hHGFopt
<400> 37
1 GACAAAAATA CCAGGACGAT TTGGAATGGC ACATCCACGA CCAGGAACAA
51 TGACACCAAG AACCATTCTC ATTTTATGT
<160> SEQ ID NО:38
<210> 38
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r4
<223> Олигонуклеотид для сборки hHGFopt
<400> 38
1 TGCTCACAAA CAAGTGGGCC ACCATAATCC CCCTCACATG GTCCTGATCC
51 AATCTTTTCA GCCCCAGCAC ATATTTCAG
<160> SEQ ID NО:39
<210> 39
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r5
<223> Олигонуклеотид для сборки hHGFopt
<400> 39
1 ACTCATTCAG AGTCACCTTC CCTCGATGAT GCTGGCTGCA TTTCTCATTT
51 CCCATGATGT ACAGGTGGGC CACCCGCAG
<160> SEQ ID NО:40
<210> 40
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r6
<223> Олигонуклеотид для сборки hHGFopt
<400> 40
1 CAGGCCGTCG TAGTTGATCA GTCCAGTGTA GCCCCAGCCA TAAACACTGC
51 AACTGGTCTT TTCAGGAATT GTGCATCCA
<160> SEQ ID NО:41
<210> 41
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r7
<223> Олигонуклеотид для сборки hHGFopt
<400> 41
1 TAATTAGGTA AATCAATCGT ACTAACAAAA TCATCCAGGA CAGCAGGCCT
51 GGCAAGCTTC ATTAAAACCA GATCTGATC
<160> SEQ ID NО:42
<210> 42
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r8
<223> Олигонуклеотид для сборки hHGFopt
<400> 42
1 CTTCAGGGCC ATATACCAGC TGGGAAACAT TGAGAACCTG TTTGCATTTC
51 TCATCTCCTC TTCCGTGGAC ATCATGAAT
<160> SEQ ID NО:43
<210> 43
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r9
<223> Олигонуклеотид для сборки hHGFopt
<400> 43
1 TCCAAGCCAA GCTTCATAAT CTTTCAAGTC TCGAGAAGGG AAACACTGTC
51 GTGCAGTAAG AACCCAACTC TCCTTTATC
<160> SEQ ID NО:44
<210> 44
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r10
<223> Олигонуклеотид для сборки hHGFopt
<400> 44
1 AATGATCCTC CGCAGATATG TTTATTTCTG TATCTCAAAC TAACCATCCA
51 TCCTATGTTT GTTCGTGTTG GAATCCCAT
<160> SEQ ID NО:45
<210> 45
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r11
<223> Олигонуклеотид для сборки hHGFopt
<400> 45
1 TTACAACTCG CAATTGTTTC GTTTTGGCAC AAGATATTAC GGGATGGTCT
51 AAATTGACTA TTGTAGGTGT GGTATCACC
<160> SEQ ID NО:46
<210> 46
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r12
<223> Олигонуклеотид для сборки hHGFopt
<400> 46
1 TTCACAACGA GAAATAGGGC AATAATCCCA AGGAATGAGT GGATTTCCCG
51 TGTAGCACCA GGGTCCATGA GCATCATCA
<160> SEQ ID NО:47
<210> 47
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r13
<223> Олигонуклеотид для сборки hHGFopt
<400> 47
1 TCTGGATTTC GGCAGTAATT CTCATTCAGC TTACTTGCAT CTGGTTCCCA
51 GAAGATATGA CGATGTAAGT CTTCCATGT
<160> SEQ ID NО:48
<210> 48
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r14
<223> Олигонуклеотид для сборки hHGFopt
<400> 48
1 TCTTGTCCCA CATTGAACAT GTTAGTCCAG ATCTTGTTTG GGATAAGTTG
51 CCCATATAAT TTTTGCCATT CCCACGATA
<160> SEQ ID NО:49
<210> 49
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r15
<223> Олигонуклеотид для сборки hHGFopt
<400> 49
1 ACAATCTTGT CCATGTGACA TATCACAGTT TGGAATTTGG GAGCAGTAGC
51 CAACTCGGAT GTTTGGATCA GTGGTAAAA
<160> SEQ ID NО:50
<210> 50
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r16
<223> Олигонуклеотид для сборки hHGFopt
<400> 50
1 CACCAGGGTG ATTCAGACCC ATCTGGATTT CGGCAGTAAT TTTCTCGTAG
51 GTCCTTGCAC TTGAAATTTT CAGGAGTCA
<160> SEQ ID NО:51
<210> 51
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r17
<223> Олигонуклеотид для сборки hHGFopt
<400> 51
1 TGTCATGCTC GTGAGGATAC TGAGAATCCC AACGCTGACA TGGAATTCCA
51 TTCCAAATGG TATTGACAGT GCCCCTGTA
<160> SEQ ID NО:52
<210> 52
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r18
<223> Олигонуклеотид для сборки hHGFopt
<400> 52
1 GCCTTCTCCT TGACCTTGGA TGCATTCAGT TGTTTCCAAA GGAACATCAG
51 TGTCATTCAT AGTATTGTCA GCGCATGTT
<160> SEQ ID NО:53
<210> 53
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r19
<223> Олигонуклеотид для сборки hHGFopt
<400> 53
1 TTAATTGCAC AGTACTCCCA GCGGGTGTGA GGGTCAAGAG TATAGCACCA
51 TGGCCTCGGC TGGCCATCGG GATTGCGGC
<160> SEQ ID NО:54
<210> 54
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r20
<223> Олигонуклеотид для сборки hHGFopt
<400> 54
1 AATAATTATC ATCAAAGCCC TTGTCGGGAT ATCTTTCAGG CAAGAATTTG
51 TGCCGGTGTG GTGTCTGATG ATCCCAGCG
<160> SEQ ID NО:55
<210> 55
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r21
<223> Олигонуклеотид для сборки hHGFopt
<400> 55
1 CTGACAAATC TTGCCTGATT CTGTATGATC CATGAGACCT CGATAACTCT
51 CCCCATTGCA GGTCATGCAT TCAACTTCT
<160> SEQ ID NО:56
<210> 56
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r22
<223> Олигонуклеотид для сборки hHGFopt
<400> 56
1 GAACACTGAG GAATGTCACA GACTTCGTAG CGTACCTCTG GATTGCTTGT
51 GAAACACCAG GGTCCCCCTT CTTCCCCTC
<160> SEQ ID NО:57
<210> 57
<211> 79
<212> ДНК
<213> искусственная последовательность hHGFopt-r23
<223> Олигонуклеотид для сборки hHGFopt
<400> 57
1 GAGGGTTCCG GCAGTAGTTC TCCTGCAGGT CCTTGCCCCG GTAGCTGGAA
51 GGCAGAAAGC TGTGTTCGTG TGGTATCAT
<160> SEQ ID NО:58
<210> 58
<211> 78
<212> ДНК
<213> искусственная последовательность hHGFopt-r24
<223> Олигонуклеотид для сборки hHGFopt
<400> 58
1 GGAACTCCAG GGCTGACATT TGATGCCACT CTTAGTGATA GATACTGTTC
51 CCTTGTAGCT GCGTCCTTTA CCAATGAT
<160> SEQ ID NО:59
<210> 59
<211> 78
<212> ДНК
<213> искусственная последовательность hHGFopt-r25
<223> Олигонуклеотид для сборки hHGFopt
<400> 59
1 GCAGTTTCTA ATGTAGTCTT TGTTTTCATA GAGGTCAAAT TCATGGCCAA
51 ATTCTTTTTT CACTCCACTT GACATGCT
<160> SEQ ID NО:60
<210> 60
<211> 78
<212> ДНК
<213> искусственная последовательность hHGFopt-r26
<223> Олигонуклеотид для сборки hHGFopt
<400> 60
1 ATTGAAGGGG AACCAGAGGC ATTGTTTTCT TGCTTTATCA AAAACAAAAG
51 CCTTGCAAGT GAATGGCAGG CCCTTGTT
<160> SEQ ID NО:61
<210> 61
<211> 78
<212> ДНК
<213> искусственная последовательность hHGFopt-r27
<223> Олигонуклеотид для сборки hHGFopt
<400> 61
1 CCTGGTACAT CTATTGGCAC ACTGGTCTGC AGTATTCACT TTTTTGGTCT
51 TGATCTTCAG GGCGGGGTCG ATCTTGAT
<160> SEQ ID NО:62
<210> 62
<211> 78
<212> ДНК
<213> искусственная последовательность hHGFopt-r28
<223> Олигонуклеотид для сборки hHGFopt
<400> 62
1 CAGGGTGGTC TTTGCTGATT TTTTGAATTC ATGAATTGTA TTTCTTCTTT
51 TCCTTTGTCC CTCTGCATAG GGGATGGC
<160> SEQ ID NО:63
<210> 63
<211> 77
<212> ДНК
<213> искусственная последовательность hHGFopt-r29
<223> Олигонуклеотид для сборки hHGFopt
<400> 63
1 GATGGGGAGC AGGAGGAGAT GCAGGAGGAC ATGCTGCAGC AGCAGGGCTG
51 GCAGCAGCTT GGTCACCCAC ATGGTGG
<160> SEQ ID NО:64
<210> 64
<211> 39
<212> ДНК
<213> искусственная последовательность hHGFopt-r30
<223> Олигонуклеотид для сборки hHGFopt
<400> 64
1 CTTTACCAAC AGTACCGGAA TGCCGCTAGC CAGCTTGGG
<160> SEQ ID NО:65
<210> 65
<211> 55
<212> ДНК
<213> искусственная последовательность
<223> Праймер для амплификации CAG-промотора
<400> 65
1 CCACCGtcta gaggtacccg ttacataact tacggtaaat ggcccgcctg
51 gctga
<160> SEQ ID NО:66
<210> 66
<211> 74
<212> ДНК
<213> искусственная последовательность
<223> Праймер для амплификации CAG-промотора
<400> 66
1 CCACCGTCCG GAATGCCctg aaaaaaagtg atttcaggca ggtgctccag
51 gtaattaaac attaataccc cacc
<160> SEQ ID NО:67
<210> 67
<211> 78
<212> ДНК
<213> искусственная последовательность hVEGF165-f1
<223> Олигонуклеотид для сборки hVEGF165
<400> 67
1 CCACCAGCTC TTCCCGGTAC TGTTGGTAAA GCCACCATGA ACTTTCTGCT
51 GAGCTGGGTC CACTGGTCAC TGGCACTG
<160> SEQ ID NО:68
<210> 68
<211> 78
<212> ДНК
<213> искусственная последовательность hVEGF165-f2
<223> Олигонуклеотид для сборки hVEGF165
<400> 68
1 CTGCTGTATC TGCATCACGC AAAATGGTCA CAGGCCGCAC CTATGGCCGA
51 GGGCGGAGGG CAGAACCACC ATGAAGTG
<160> SEQ ID NО:69
<210> 69
<211> 78
<212> ДНК
<213> искусственная последовательность hVEGF165-f3
<223> Олигонуклеотид для сборки hVEGF165
<400> 69
1 GTCAAGTTCA TGGACGTGTA CCAGAGATCA TATTGTCACC CTATCGAGAC
51 ACTGGTCGAC ATTTTCCAGG AATACCCA
<160> SEQ ID NО:70
<210> 70
<211> 78
<212> ДНК
<213> искусственная последовательность hVEGF165-f4
<223> Олигонуклеотид для сборки hVEGF165
<400> 70
1 GATGAGATCG AATATATCTT CAAGCCAAGC TGCGTGCCAC TGATGCGGTG
51 CGGCGGATGC TGTAACGATG AGGGACTG
<160> SEQ ID NО:71
<210> 71
<211> 78
<212> ДНК
<213> искусственная последовательность hVEGF165-f5
<223> Олигонуклеотид для сборки hVEGF165
<400> 71
1 GAATGCGTCC CCACCGAGGA ATCTAATATC ACAATGCAGA TCATGCGAAT
51 TAAGCCTCAC CAGGGGCAGC ATATTGGC
<160> SEQ ID NО:72
<210> 72
<211> 78
<212> ДНК
<213> искусственная последовательность hVEGF165-f6
<223> Олигонуклеотид для сборки hVEGF165
<400> 72
1 GAGATGAGTT TCCTGCAGCA CAACAAATGC GAGTGTAGGC CAAAGAAAGA
51 CCGAGCTCGA CAGGAGAATC CATGTGGA
<160> SEQ ID NО:73
<210> 73
<211> 78
<212> ДНК
<213> искусственная последовательность hVEGF165-f7
<223> Олигонуклеотид для сборки hVEGF165
<400> 73
1 CCTTGCTCTG AAAGGAGAAA GCATCTGTTT GTGCAGGACC CCCAGACTTG
51 CAAGTGTAGC TGCAAAAATA CCGATTCC
<160> SEQ ID NО:74
<210> 74
<211> 77
<212> ДНК
<213> искусственная последовательность hVEGF165-f8
<223> Олигонуклеотид для сборки hVEGF165
<400> 74
1 AGGTGTAAGG CACGGCAGCT GGAACTGAAT GAGCGGACCT GTCGGTGTGA
51 TAAGCCAAGA AGATAAGTTT AAACCCG
<160> SEQ ID NО:75
<210> 75
<211> 77
<212> ДНК
<213> искусственная последовательность hVEGF165-f9
<223> Олигонуклеотид для сборки hVEGF165
<400> 75
1 CTGATCAGCC TCGACTTCGA GCAGACGCTT CGAGCAGACA TGATAAGATA
51 CATTGATGAG TTTGGACAAA CCACAAC
<160> SEQ ID NО:76
<210> 76
<211> 77
<212> ДНК
<213> искусственная последовательность hVEGF165-f10
<223> Олигонуклеотид для сборки hVEGF165
<400> 76
1 TAGAATGCAG TGAAAAAAAT GCTTTATTTG TGAAATTTGT GATGCTATTG
51 CTTTATTTGT AACCATTATA AGCTGCA
<160> SEQ ID NО:77
<210> 77
<211> 77
<212> ДНК
<213> искусственная последовательность hVEGF165-f11
<223> Олигонуклеотид для сборки hVEGF165
<400> 77
1 ATAAACAAGT TAACAACAAC AATTGCATTC ATTTTATGTT TCAGGTTCAG
51 GGGGAGATGT GGGAGGTTTT TTAAAGC
<160> SEQ ID NО:78
<210> 78
<211> 76
<212> ДНК
<213> искусственная последовательность hVEGF165-f12
<223> Олигонуклеотид для сборки hVEGF165
<400> 78
1 AACACGTGCT AAAACTTCAT TTTTAATTTA AAAGGATCTA GGTGAAGATC
51 CTTTTTGATA ATAGAAGAGC TGGTGG
<160> SEQ ID NО:79
<210> 79
<211> 39
<212> ДНК
<213> искусственная последовательность hVEGF165-r1
<223> Олигонуклеотид для сборки hVEGF165
<400> 79
1 CCACCAGCTC TTCTATTATC AAAAAGGATC TTCACCTAG
<160> SEQ ID NО:80
<210> 80
<211> 78
<212> ДНК
<213> искусственная последовательность hVEGF165-r2
<223> Олигонуклеотид для сборки hVEGF165
<400> 80
1 ATCCTTTTAA ATTAAAAATG AAGTTTTAGC ACGTGTTGCT TTAAAAAACC
51 TCCCACATCT CCCCCTGAAC CTGAAACA
<160> SEQ ID NО:81
<210> 81
<211> 78
<212> ДНК
<213> искусственная последовательность hVEGF165-r3
<223> Олигонуклеотид для сборки hVEGF165
<400> 81
1 TAAAATGAAT GCAATTGTTG TTGTTAACTT GTTTATTGCA GCTTATAATG
51 GTTACAAATA AAGCAATAGC ATCACAAA
<160> SEQ ID NО:82
<210> 82
<211> 78
<212> ДНК
<213> искусственная последовательность hVEGF165-r4
<223> Олигонуклеотид для сборки hVEGF165
<400> 82
1 TTTCACAAAT AAAGCATTTT TTTCACTGCA TTCTAGTTGT GGTTTGTCCA
51 AACTCATCAA TGTATCTTAT CATGTCTG
<160> SEQ ID NО:830
<210> 83
<211> 78
<212> ДНК
<213> искусственная последовательность hVEGF165-r5
<223> Олигонуклеотид для сборки hVEGF165
<400> 83
1 CTCGAAGCGT CTGCTCGAAG TCGAGGCTGA TCAGCGGGTT TAAACTTATC
51 TTCTTGGCTT ATCACACCGA CAGGTCCG
<160> SEQ ID NО:84
<210> 84
<211> 76
<212> ДНК
<213> искусственная последовательность hVEGF165-r6
<223> Олигонуклеотид для сборки hVEGF165
<400> 84
1 CTCATTCAGTTC CAGCTGCCGT GCCTTACACC TGGAATCGGT ATTTTTGCAG
51 CTACACTTGC AAGTCTGGGG GTCCTG
<160> SEQ ID NО:85
<210> 85
<211> 77
<212> ДНК
<213> искусственная последовательность hVEGF165-r7
<223> Олигонуклеотид для сборки hVEGF165
<400> 85
1 CACAAACAGA TGCTTTCTCC TTTCAGAGCA AGGTCCACAT GGATTCTCCT
51 GTCGAGCTCG GTCTTTCTTT GGCCTAC
<160> SEQ ID NО:86
<210> 86
<211> 77
<212> ДНК
<213> искусственная последовательность hVEGF165-r8
<223> Олигонуклеотид для сборки hVEGF165
<400> 86
1 ACTCGCATTT GTTGTGCTGC AGGAAACTCA TCTCGCCAAT ATGCTGCCCC
51 TGGTGAGGCT TAATTCGCAT GATCTGC
<160> SEQ ID NО:87
<210> 87
<211> 77
<212> ДНК
<213> искусственная последовательность hVEGF165-r9
<223> Олигонуклеотид для сборки hVEGF165
<400> 87
1 ATTGTGATAT TAGATTCCTC GGTGGGGACG CATTCCAGTC CCTCATCGTT
51 ACAGCATCCG CCGCACCGCA TCAGTGG
<160> SEQ ID NО:88
<210> 88
<211> 77
<212> ДНК
<213> искусственная последовательность hVEGF165-r10
<223> Олигонуклеотид для сборки hVEGF165
<400> 88
1 CACGCAGCTT GGCTTGAAGA TATATTCGAT CTCATCTGGG TATTCCTGGA
51 AAATGTCGAC CAGTGTCTCG ATAGGGT
<160> SEQ ID NО:89
<210> 89
<211> 77
<212> ДНК
<213> искусственная последовательность hVEGF165-r11
<223> Олигонуклеотид для сборки hVEGF165
<400> 89
1 GACAATATGA TCTCTGGTAC ACGTCCATGA ACTTGACCAC TTCATGGTGG
51 TTCTGCCCTC CGCCCTCGGC CATAGGT
<160> SEQ ID NО:90
<210> 90
<211> 77
<212> ДНК
<213> искусственная последовательность hVEGF165-r12
<223> Олигонуклеотид для сборки hVEGF165
<400> 90
1 GCGGCCTGTG ACCATTTTGC GTGATGCAGA TACAGCAGCA GTGCCAGTGA
51 CCAGTGGACC CAGCTCAGCA GAAAGTT
<160> SEQ ID NО:91
<210> 91
<211> 39
<212> ДНК
<213> искусственная последовательность hVEGF165-r13
<223> Олигонуклеотид для сборки hVEGF165
<400> 91
1 CATGGTGGCT TTACCAACAG TACCGGGAAG AGCTGGTGG
<---
Изобретение относится к области биотехнологии, конкретно, к генно-инженерным конструкциям для стимуляции ангиогенеза, и может быть использовано в медицине для лечения заболеваний, обусловленных нарушением кровоснабжения тканей. Предложена плазмидная бицистронная конструкция, несущая ген hHGF фактора роста гепатоцитов человека и ген hVEGF165 фактора роста эндотелия сосудов, с двумя отдельными промоторами для каждого из генов: промотора цитомегаловируса - pCMV для гена HGF и промотора гена β-актина цыпленка – pCAG для гена VEGF165. Бицистронная конструкция может быть использована в качестве лекарственного средства для обеспечения стимуляции ангиогенеза, роста и ремоделирования сосудов, а также восстановления кровоснабжения в ишемизированных тканях. Изобретение позволяет достигать оптимальных концентраций и соотношения ангиогенных факторов роста (АФР). 4 н. и 2 з.п. ф-лы, 7 табл., 1 пр., 6 ил.
1. Генно-инженерная конструкция для стимуляции ангиогенеза в ишемизированных тканях, включающая:
- последовательность нуклеотидов SEQ ID NO: 1, кодирующую ген фактора роста гепатоцитов, находящуюся под контролем промотора цитомегаловируса - pCMV-HGF;
- последовательность нуклеотдов SEQ ID NO: 2, кодирующую ген фактора роста эндотелия сосудов, находящуюся под контролем промотора гена β-актина цыпленка - pCAG-VEGF165.
2. Генно-инженерная конструкция по п. 1, характеризующаяся тем, что ее конструкция включает следующие последовательно расположенные элементы:
Энхансер 1 - CMV enhancer- 36-415 п.о.,
Промотор 1 - pCMV (промотор цитомегаловируса) - 488-691 п.о.,
Ген hHGF - 784-2970 п.о.,
Терминатор 1 - bGH poly А (участок полиаденилирования гена бычьего гормона роста) - 3044-3268 п.о.,
Селективный маркер для отбора трансформированных бактерий - ген устойчивости к антибиотику - 3441-4253 п.о.,
Энхансер 2 - CMV enhancer - 4455-4740 п.о.,
Промотор 2 - pCAG (промотор гена β-актина цыпленка) - 4742-5019 п.о.,
Гибридный интрон - 5019-5248 п.о.,
Ген hVEGF165 - 5278-5853 п.о.,
Терминатор 2 - SV40 poly А (участок полиаденилирования вируса SV40) - 5908-6029 п.о.,
Точка начала репликации (ori) - 6235-6823 п.о.
3. Применение генно-инженерной конструкции по п. 1 в качестве действующего вещества генотерапевтического средства для стимуляции ангиогенеза в ишемизированных тканях.
4. Средство для стимуляции ангиогенеза в ишемизированных тканях, включающее плазмидную конструкцию по п. 1 и вспомогательные вещества для внутримышечного введения.
5. Средство по п. 4, характеризующееся тем, что в качестве вспомогательных веществ использованы натрия хлорид или вода для инъекций.
6. Способ стимуляции ангиогенеза в ишемизированных тканях, включающий внутримышечное введение лекарственного средства по п. 4 в терапевтически эффективном количестве.
| СЛОБОДКИНА Е.А | |||
| и др., Разработка генотерапевтической плазмидной конструкции, кодирующей фактор роста гепатоцитов (HGF) и фактор роста эндотелия сосудов (VEGF165), Материалы III Национального конгресса по регенеративной медицине, Москва, 15-18 ноября 2017 г., Гены и Клетки, 2017, т.XII, н.3, с.227 | |||
| MASAKII.et al., Angiogenic gene therapy for |

