Изобретение относится к иммунологии и биотехнологии, а именно: к получению широконейтрализующего рекомбинантного антитела человека (рчАт) против вируса SARS-CoV-2, которое может быть использовано для разработки иммунобиологических лекарственных препаратов на основе антител, что расширяет возможности терапии/диагностики вирусной инфекции SARS-CoV-2 в медицине.
Объектом изобретения является рчАт класса IgG, имеющее аминокислотную последовательность SEQ ID NO:1 для вариабельного домена тяжелой цепи антитела и SEQ ID NO:2 для вариабельного домена легкой цепи антитела, специфически связывающееся с различными вариантами RBD домена белка S вируса SARS-CoV-2 (Wuhan-Hu-1, а также варианты Альфа, Гамма, Дельта и Омикрон) и обладающее широким спектром нейтрализующей активности.
Коронавирусная инфекция (COVID-19) - респираторное заболевание, вызываемое вирусом SARS-CoV-2, который появился в Китае в конце 2019 года и быстро распространился по всему миру, вызвав глобальную пандемию [Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. Nat Microbiol. 2020; 5:536-544. doi: 10.1038/s41564-020-0695-z; World Health Organization. Coronavirus disease 2019 (COVID-19). 2020; Situation Report-51]. Заболевание может протекать как в лёгкой, так и в тяжёлой форме, в том числе, приводя к смертельным исходам. Пациентам с легкими и умеренными симптомами COVID-19 назначается поддерживающая терапия с симптоматическим лечением, как, например, прием жаропонижающих средств от лихорадки, обезболивающие препараты, и наблюдение за признаками обострения, которое требует стационарного лечения. Риск тяжелого течения COVID-19 или смерти от заболевания угрожает любому человеку независимо от возраста. Тем не менее, тяжелая форма заболевания чаще всего развивается у людей, относящихся к группе риска: люди старше 60 лет, а также с хроническими сердечно-сосудистыми, респираторными, онкологическими заболеваниями, диабетом и др. В разгар пандемии вируснейтрализующие антитела были одобрены Всемирной организацией здравоохранения (ВОЗ) для лечения инфицированных SARS-CoV-2, если для пациента существовал высокий риск госпитализации или для лечения пациентов с тяжелым течением COVID-19. Нейтрализующие антитела появляются после заражения вирусом SARS-CoV-2 [Suthar M.S. et al, Rapid Generation of Neutralizing Antibody Responses in COVID-19 Patients. Cell Rep Med. 2020; 1(3):100040. doi: 10.1016/j.xcrm.2020.10004]) или вакцинации [Jackson L.A. et al, mRNA-1273 Study Group. An mRNA Vaccine against SARS-CoV-2 - Preliminary Report. N Engl J Med. 2020; 383(20):1920-1931. doi: 10.1056/NEJMoa2022483] сохраняются в сыворотке крови в течение нескольких месяцев и коррелируют с уровнем защиты от инфекции [Addetia, A. et al, Neutralizing Antibodies Correlate with Protection from SARS-CoV-2 in Humans during a Fishery Vessel Outbreak with a High Attack Rate. J Clin Microbiol. 2020; 58(11):e02107-20. doi: 10.1128/JCM.02107-20]. По мере эволюции вируса SARS-CoV-2 под давлением отбора за счет естественного или индуцированного вакцинацией иммунитета, появляются его новые варианты, характеризующиеся повышенной трансмиссивностью, изменением течения заболевания или риском повторного инфицирования из-за изменения антигенного состава. Появление мутантных вариантов SARS-CoV-2 привело к снижению эффективности существующих нейтрализующих антител и ограничивает их использование на сегодняшний день. Поскольку мутации в значительной степени изменяют силу и/или кинетику связывания вируса с ACE2 рецептором, мутации, возникающие в рецептор-связывающем домене (RBD) S белка, представляют особый интерес [Amoutzias G.D. et al, The Remarkable Evolutionary Plasticity of Coronaviruses by Mutation and Recombination: Insights for the COVID-19 Pandemic and the Future Evolutionary Paths of SARS-CoV-2. Viruses. 2022; 14(1):78. doi: 10.3390/v14010078]. Альфа вариант коронавируса SARS-CoV-2 (линия B.1.1.7) был впервые обнаружен в начале декабря 2020 года во время пандемии COVID-19 в Великобритании, мутация N501Y в RBD привела к увеличению аффинности связывания вируса с клетками организма человека в 7 раз [Davies N.G. et al. Estimated Transmissibility and Impact of SARS-CoV-2 Lineage B.1.1.7 in England. Science. 2021; 372:eabg3055. doi: 10.1126/science.abg3055; Supasa P. et al. Reduced Neutralization of SARS-CoV-2 B.1.1.7 Variant by Convalescent and Vaccine Sera. Cell. 2021; 184:2201-11. doi: 10.1016/j.cell.2021.02.033]. Гамма вариант (P1) был идентифицирован в ноябре 2020 года Бразилии и вызвал резкую вспышку заболеваемости, данный вируса SARS-CoV-2 имеет 12 мутаций в S белке, включая K417T, E484K и N501Y в RBD [Wink P.L. et al. Genomic Surveillance of SARS-CoV-2 Lineages Indicates Early Circulation of P.1 (Gamma) Variant of Concern in Southern Brazil. Microbiol Spectr. 2022;10(1):e0151121. doi: 10.1128/spectrum.01511-21; Dejnirattisai W. et al. Antibody evasion by the P.1 strain of SARS-CoV-2. Cell. 2021; 184(11):2939-2954.e9. doi: 10.1016/j.cell.2021.03.055]. Дельта вариант SARS-CoV-2 (B.1.617.2) c мутациями L452R, T478K, ±E484Q в RBD был впервые идентифицирован в декабре 2020 года в Индии, его повышенная контагиозность привела к полному вытеснению исходного уханьского варианта (Wuhan-Hu-1) [Zhang L. et al. Analysis of SARS-CoV-2 variants B.1.617: host tropism, proteolytic activation, cell-cell fusion, and neutralization sensitivity. Emerg Microbes Infect. 2022; 11(1):1024-1036. doi: 10.1080/22221751]. Омикрон (B.1.1.529) впервые был идентифицирован в ноябре 2021 в Южно-Африканской Республике и Ботсване, содержал беспрецедентное количество мутаций, сконцентрированных в гене S белка (более 30 мутаций, в том числе 15 - в RBD), незамедлительно вытеснил вариант дельта и на сегодняшний день является преобладающим в России и во всем мире, продолжая мутировать.
Обширная мутационная нагрузка на основной антиген снизила активность известных терапевтических нейтрализующих антител, что привело к увеличению доз или пересмотру вариантов терапии больных с тяжелой формой течения заболевания [Hoffmann M. et al. SARS-CoV-2 Variant B. 1.617 Is Resistant to Bamlanivimab and Evades Antibodies Induced by Infection and Vaccination. Cell Rep. 2021; 36:109415. doi: 10.1016/j.celrep.2021.109415; Cao Y. et al. Omicron escapes the majority of existing SARS-CoV-2 neutralizing antibodies. Nature. 2022; 602:657-663. doi: 10.1038/s41586-021-04385-3].
Моноклональные человеческие антитела, ранее разрешенные Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) с экстренным решением для применения (EUA) - казиривимаб и имдевимаб (Ronapreve (REGN10933 и REGN10987), Regeneron, США), бамланивимаб и этесевимаб (LY-CoV555 и LY-CoV016, Eli Lilly, США), вводимые в виде коктейля, а также регданвимаб (CT-P59, Celltrion, Южная Корея) в экспериментах in vitro с использованием псевдовирусной системы и живых вирусов показали полную потерю нейтрализующей активности препаратов против варианта Омикрон и их использование было ограничено [VanBlargan L. A. et al. An infectious SARS-CoV-2 B.1.1.529 Omicron virus escapes neutralization by therapeutic monoclonal antibodies. Nat. Med. 2022; 28:490-495. doi: 10.1038/s41591-021-01678-y; Tatham, L. et al. Lack of Ronapreve (REGN-CoV; casirivimab and imdevimab) virological efficacy against the SARS-CoV-2 Omicron variant (B.1.1.529) in K18-hACE2 mice. bioRxiv. 2022; doi: org/10.1101/2022.01.23.477397]. Эффективность сотровимаба (S309, VIR-7831, GlaxoSmithKline, Великобритания) и бебтеловимаба (LY-CoV1404, Eli Lilly, США) в отношении различных вариантов Омикрона сохранялась [Zhou T. et al. Structural basis for potent antibody neutralization of SARS-CoV-2 variants including B.1.1.529. Science. 2022; 376:eabn8897. doi: 10.1126/science.abn889]. Необходимо отметить, что применение сотровимаба до недавнего времени было разрешено на территории России [https://grls.rosminzdrav.ru/PandemicPermissions.aspx?TorgName=сотровимаб], однако дальнейшее исследование эффективности препарата показало, что его использование стимулирует эволюцию вируса SARS-CoV-2 у пациентов с ослабленным иммунитетом [Rockett R. et al. Resistance mutations in SARS-CoV-2 delta variant after sotrovimab use. N Engl J Med. 2022; 386:1477-1479. doi: 10.1056/NEJMc2120219; Destras G. et al. Sotrovimab drives SARS-CoV-2 omicron variant evolution in immunocompromised patients. Lancet Microbe. 2022; 3(8):e559. doi: 10.1016/S2666-5247(22)00120-3], что привело к пересмотру решения о его использовании [WHO, Therapeutics and COVID-19: living guideline, 16/09/2022]. Бебтеловимаб также больше не разрешен FDA к применению в США из-за отсутствия активности в отношении циркулирующих вариантов [FDA, Fact sheet for healthcare providers: emergency use authorization for bebtelovimab, ID: 5072477, 30/11/2022]. Таким образом, в настоящее время в мире нет моноклональных антител, разрешенных для лечения острой инфекции.
Лекарственный препарат на основе комбинации моноклональных антител тиксагевимаб и цилгавимаб (Эвушелд (AZD8895+AZD1061), AstraZeneca, Великобритания) одобрен для доконтактной профилактики COVID-19 у лиц c ослабленным иммунитетом (иммунодефицит или иммуносупрессивная терапия) или лицам, для которых вакцинация не рекомендована по причине тяжелой побочной реакции на вакцину или ее компоненты в анамнезе [Levin MJ, et al. Intramuscular AZD7442 (Tixagevimab-Cilgavimab) for Prevention of Covid-19. N Engl J Med. 2022.; 386:2188-2200. doi:10.1056/NEJMoa2116620; FDA, Fact sheet for healthcare providers: emergency use authorization for evusheld, ID: 5098106, 5/12/2022]. На территории России препарат также включен в «Реестр решений об обращении лекарственных препаратов во время пандемии» [https://grls.rosminzdrav.ru/PandemicPermissions.aspx?TorgName=эвушелд]. AZD8895 и AZD1061 сохраняют эффективность против Омикрона, несмотря на снижение нейтрализующей активности к его различным субвариантам [Otiniano A. et al. Tixagevimab/cilgavimab for Omicron SARS-CoV-2 infection in patients with haematologic diseases. Bone Marrow Transplant. 2022; doi: 10.1038/s41409-022-01894-1].
Комбинации антител к разными эпитопам S белка обеспечивают их синергетическое действие, а также снижают вероятность появления мутаций, придающих вирусу устойчивость против антител [Zost S.J. et al. Potently neutralizing and protective human antibodies against SARS-CoV-2. Nature. 2020; 584(7821):443-449. doi: 10.1038/s41586-020-2548-6; Yang, Y., Du, L. Neutralizing antibodies and their cocktails against SARS-CoV-2 Omicron and other circulating variants. Cell. Mol. Immunol. 2022; 19:962-964. doi: 10.1038/s41423-022-00890-1]. Таким образом, в связи с продолжающимся появлением новых вариантов вируса SARS-CoV-2, кодирующих все большее количество аминокислотных замен в S белке, требуется постоянный мониторинг эффективности действия уже существующих препаратов, а также поиск и расширение панели моноклональных антител, эффективных для лечения и профилактики COVID-19.
Технической задачей изобретения является расширения арсенала антител, обладающих широким спектром нейтрализующей активности против вариантов вируса SARS-CoV-2.
Технический результат заключается в создании антитела Т11, которое эффективно связывает варианты RBD домена S белка вируса SARS-CoV-2, нейтрализуют вирус SARS-CoV-2 дикого типа (Wuhan-Hu-1), а также штаммы Дельта (B.1.617.2) и Омикрон (B.1.1.529) и может быть использовано для разработки иммунобиологических лекарственных препаратов на основе антител для терапии вирусной инфекции SARS-CoV-2.
Указанный технический результат достигается:
созданием рекомбинантных плазмидных ДНК pFuse/T11/CH и pFuse/T11/CL, кодирующих, соответственно, гены тяжелой и легкой цепи рекомбинантного антитела Т11, и состоящих из плазмиды pFuse, которая содержит высокоэффективный синтетический промотор hEF/HTLV, сигнальную последовательность интерлейкина-2 (IL-2 signal), обеспечивающую эффективную секрецию продукта экспрессии генов в культуральную среду, нуклеотидную последовательность гена вариабельного домена тяжелой цепи в единой рамке считывания с геном константного домена тяжелой цепи (CH) иммуноглобулина человека IgG1 (SEQ ID NO:3) или нуклеотидную последовательность гена вариабельного домена легкой цепи в единой рамке считывания с геном константного домена лямбда легкой цепи (CL) иммуноглобулина человека (SEQ ID NO:4), сигнальную последовательность полиаденилирования вируса SV40; нуклеотидную последовательность гена BleoR, продукт которого обеспечивает устойчивость трансформантов к антибиотику зеоцин, нуклеотидную последовательность точки начала репликации плазмидной ДНК (ori) для ее наработки в бактериальных клетках E. coli;
получением полноразмерного антитела Т11, предусматривающим котрансфекцию рекомбинантными плазмидными ДНК pFuse/T11/CH и pFuse/T11/CL клеток линии HEK293F, культивирование клеток в условиях, при которых продуцируется антитело Т11, имеющее аминокислотную последовательность SEQ ID NO:1 для вариабельного домена тяжелой цепи антитела и SEQ ID NO:2 для вариабельного домена легкой цепи антитела;
выделением и хроматографической очисткой рекомбинантного антитела Т11.
Изобретение иллюстрируют следующими примерами.
ПРИМЕР 1
Создание генетических конструкций pFuse/T11/CH и pFuse/T11/CL
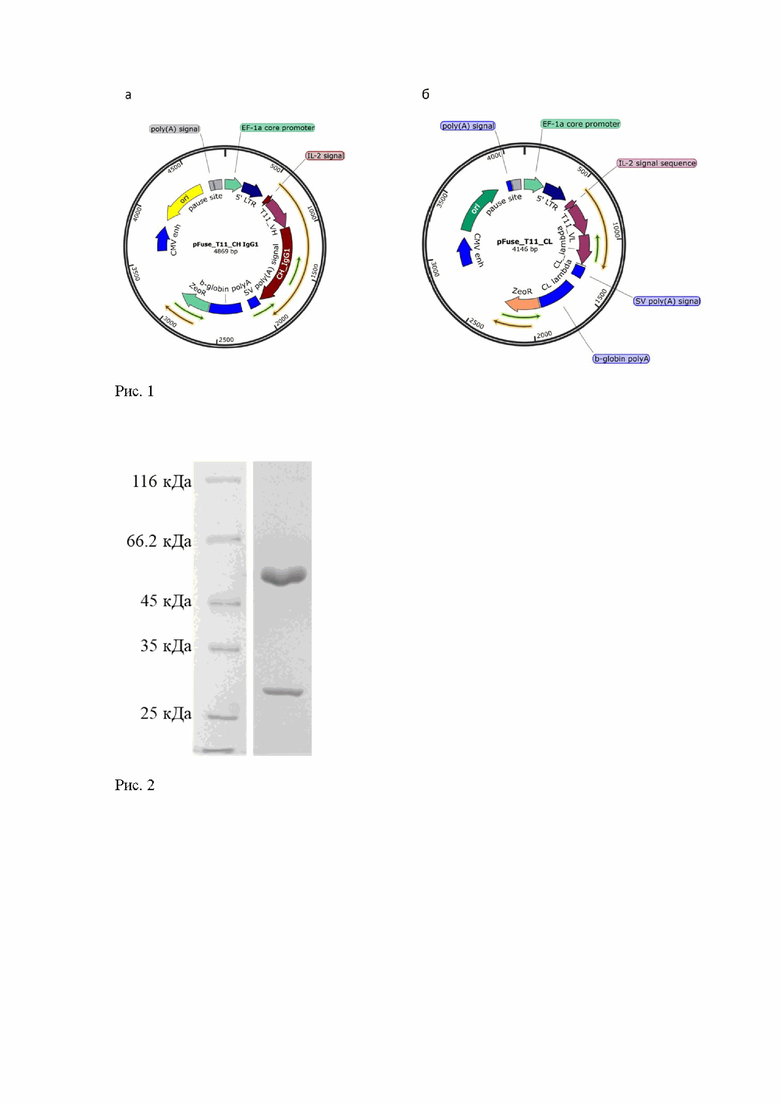
Антитело Т11 отобрано из библиотеки одноцепочечных антител человека, получившего полный курс вакцинации препаратом PfizerBioNTech с использованием дрожжевого дисплея. Последовательность гена, кодирующего вариабельный домен тяжелой цепи (VH), амплифицируют с помощью ПЦР с использованием олигонуклеотидов 5’-tcacgaattcgcaggtgcagctggtggagtc-3’ и 5’-ggtgcttgctgaggagacggtgacc-3’ и далее клонируют в вектор pFuse-IgG1 с использованием эндонуклеаз рестрикции EcoRI и BsmBI с получением генетической конструкции pFuse/T11/CH (Рис. 1а). Последовательность гена, кодирующего вариабельный домен легкой цепи (VL), амплифицируют с помощью ПЦР с использованием олигонуклеотидов 5’- tcacgaattcgcaggttgtgctgactcag-3’ и 5’-gctgacctaggacggtcagcttg-3’ и далее клонируют в вектор pFuse-Lambda с использованием эндонуклеаз рестрикции EcoRI и AvrII, с получением генетической конструкции pFuse/T11/CL (Рис. 1б).
ПРИМЕР 2
Получение полноразмерного рекомбинантного антитела Т11
Полученные генетические конструкциии pFuse/T11/CH и pFuse/T11/CL используют для временной трансфекций клеток линии HEK-293 с использованием среды FreeStyle 293 и полиэтиленимина (PEI) [N. Portolano et al. J. Vis. Exp. 2014]. Далее клетки культивируют на орбитальном шейкере, вращающемся со скоростью 130 об/мин, в СО2-инкубаторе, содержащем влажную атмосферу с 8% CO2 в воздухе при температуре 37°C в течение 7-10 дней для наработки белка. При показателе жизнеспособности клеток менее 70% культуральную жидкость последовательно центрифугируют при скорости 900 об/мин и в течение 10 минут, а затем при скорости 13000 об/мин и в течение 15 минут и используют для выделения белка. Рекомбинантное антитело Т11 очищают с использованием аффинной хроматографии на колонке HiTrap Protein G HP в соответствии с инструкцией производителя и гель-фильтрации на колонке Superdex 200 (GE Healthcare, USA) в фосфатно-солевом буфере. Чистоту полученного препарата рчАТ Т11 анализируют электрофоретически (Рис.2).
ПРИМЕР 3
Определение аффинности взаимодействия антитела Т11 с RBD WT и его вариантами - альфа, гамма, дельта, омикрон
Константы диссоциации комплекса антитела Т11 с RBDWT (Wuhan-Hu-1), а также с вариантами RBD Альфа (B.1.1.7), Гамма (P1), Дельта (B.1.617.2) и Омикрон (B.1.1.529) определяют c использованием метода поверхностного плазмонного резонанса. Для иммобилизации антигена RBD используют стандартные чипы BiaCore CM5. Иммобилизацию проводят по методике производителя, концентрации вариантов RBD варируют в диапазоне 1-10 мкг/мл. Для оценки связывания готовят разведения антитела Т11 в диапазоне концентраций 1 - 16 нМ. Эксперимент проводят на приборе BiaCore T200 в условиях одного цикла, где время ассоциации составляет 120 секунд, время диссоциации - 3600 секунд. Анализ кинетики проводят с использованием встроенного ПО, результаты представлены в Таблице 1.
ПРИМЕР 4
Определение вируснейтрализующей активности антитела Т11
Для определения вируснейтрализующей активности антитела Т11 используют псевдотипированные вирусные частицы, несущих на поверхности S-белок SARS-CoV2 дикого типа (Wuhan-Hu-1), варианта Дельта (B.1.617.2) или Омикрон (B.1.1.529). Клетки линии HEK293T-ACE2 рассевают в 96-луночный планшет и растят в течение 18-24 часов до достижения конфлюентности 50-80%. В стерильном планшете проводят разведения антитела Т11 в среде (DMEM+10%FBS) в диапазоне концентраций 0.003-100 нМ, смешивают с 50 мкл псевдовирусов и добавляют в 96-луночный планшет с клетками HEK293T-ACE2, содержащими по 50 кондиционной среды. Через 72 часа инкубации клетки лизируют и используют для оценки люминесценции по методике производителя Luciferase Assay System Bright Glo (Promega) и прибора VarioScan (Thermo). Обработку данных осуществляют с использованием ПО Origin Pro. Рассчитанные IC50 (концентрации полумаксимального ингибирования) антитела Т11 в отношении вируса SARS-CoV-2 дикого типа (Wuhan-Hu-1) и Дельта (B.1.617.2) и Омикрон (B.1.1.529) представлена в таблице 2.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="Широко
нейтрализующее антитело против SARS-CoV-2 full.xml"
softwareName="WIPO Sequence" softwareVersion="2.2.0"
productionDate="2022-12-28">
<ApplicantFileReference>11</ApplicantFileReference>
<ApplicantName languageCode="ru">ИБХ РАН</ApplicantName>
<ApplicantNameLatin>IBCh RAS</ApplicantNameLatin>
<InventionTitle languageCode="ru">Широко нейтрализующее антитело
против SARS-CoV-2</InventionTitle>
<SequenceTotalQuantity>4</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>127</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..127</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>QVQLVESGAEVKKPGSSVKVSCKAPGGTFSSYAISWVRQAPGQGLEWMG
GIIPIFGTPNYAQKFQGRVTITADESTSTAYMELSSLRSEDTAVYYCARGYCGGDCYSLELVWYFDLWGR
GTLVTVSS</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>111</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..111</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>QVVLTQPPSVSGAPGQRVTISCTGSSSNIGAGYDVQWYQQLPGTAPKLL
IYGNSNRPSGVPDRFSGSKSGTSASLAITGLQAEDEADYYCQSYDSSLSGWVFGGGTKLTVL</INSDSe
q_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>1377</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1377</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>caggtgcagctggtggagtctggggctgaggtgaagaagcctgggtcct
cggtgaaggtctcctgcaaggctcctggaggcaccttcagcagctatgctatcagctgggtgcgacaggc
ccctggacaagggcttgagtggatgggagggatcatccctatctttggtacaccaaactacgcacagaag
ttccagggcagagtcacgattaccgcggacgaatccacgagcacagcctacatggagctgagcagcctga
gatctgaggacacggccgtgtattactgtgcgaggggatattgtggtggtgattgctattcattagaact
cgtctggtacttcgatctctggggccgtggcaccctggtcaccgtctcctcagcaagcaccaagggccca
tcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtca
aggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacacctt
cccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgactgtgccctctagcagcttg
ggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagc
ccccgaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtc
agtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtg
gtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcata
atgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcct
gcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatc
gagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccggg
atgagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgt
ggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggc
tccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgct
ccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaatga</
INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>654</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..654</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q8">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>caggttgtgctgactcagccgccctcagtgtctggggccccagggcaga
gggtcaccatctcctgcactgggagcagctccaacatcggggcaggttatgatgtacagtggtaccagca
gcttccaggaacagcccccaaactcctcatctatggtaacagcaatcggccctcaggggtccctgaccga
ttctctggctccaagtctggcacctcagcctccctggccatcactgggctccaggctgaggatgaggctg
attattactgccagtcctatgacagcagcctgagtggttgggtgttcggcggagggaccaagctgaccgt
cctaggtcagcccaaggctgccccctcggtcactctgttcccgccctcctctgaggagcttcaagccaac
aaggccacactggtgtgtctcataagtgacttctacccgggagccgtgacagtggcctggaaggcagata
gcagccccgtcaaggcgggagtggagaccaccacaccctccaaacaaagcaacaacaagtacgcggccag
cagctacctgagcctgacgcctgagcagtggaagtcccacaaaagctacagctgccaggtcacgcatgaa
gggagcaccgtggagaagacagtggcccctacagaatgttcatag</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Интегративный плазмидный вектор pVEAL3-RBDdel, обеспечивающий синтез и секрецию рекомбинантного белка рецепторсвязывающего домена RBDdelta коронавируса SARS-CoV-2 в клетках млекопитающих, рекомбинантный штамм клеточной линии CHO-K1- RBDdelta и рекомбинантный белок RBDdelta SARS-CoV-2, продуцируемый штаммом клеточной линии | 2023 |
|
RU2816175C1 |
| ПОЛИПЕПТИДНЫЙ МОДУЛЬ ДЛЯ СВЯЗЫВАНИЯ КОНСЕРВАТИВНОГО ЭПИТОПА РЕЦЕПТОР-СВЯЗЫВАЮЩЕГО ДОМЕНА БЕЛКА SPIKE КОРОНАВИРУСА SARS-COV-2 | 2022 |
|
RU2809183C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АКТИВНОСТИ НЕЙТРАЛИЗУЮЩИХ АНТИТЕЛ К SARS-CoV-2 В СЫВОРОТКЕ ИЛИ ПЛАЗМЕ КРОВИ ЛЮДЕЙ, ПЕРЕНЕСШИХ COVID-19 ИЛИ ПРИВИТЫХ ВАКЦИНАМИ ДЛЯ ПРОФИЛАКТИКИ НОВОЙ КОРОНАВИРУСНОЙ ИНФЕКЦИИ COVID-19, С ИСПОЛЬЗОВАНИЕМ НАБОРА РЕАГЕНТОВ ДЛЯ ИММУНОФЕРМЕНТНОГО АНАЛИЗА, СОДЕРЖАЩЕГО РЕКОМБИНАНТНЫЙ РЕЦЕПТОР-СВЯЗЫВАЮЩИЙ ДОМЕН (RBD) ПОВЕРХНОСТНОГО ГЛИКОПРОТЕИНА S КОРОНАВИРУСА SARS-COV-2 И РЕКОМБИНАНТНЫЙ ЧЕЛОВЕЧЕСКИЙ РЕЦЕПТОР АСЕ2 | 2021 |
|
RU2784655C1 |
| Плазмидная генетическая конструкция pET21a_SKP, обеспечивающая экспрессию в прокариотической системе E.coli рекомбинантного белка SKP, и рекомбинантный белок SKP, обладающий широконейтрализующими свойствами однодоменного наноантитела против SARS-CoV-2 | 2024 |
|
RU2839376C1 |
| Тяжелоцепочечные моноклональные антитела, специфически связывающиеся с S белком вируса SARS-CoV-2, и способ их применения для терапии заболеваний, вызываемых различными вариантами вируса SARS-CoV-2 | 2024 |
|
RU2836313C1 |
| Моноклональное антитело iC2 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817697C1 |
| Моноклональное антитело к RBD фрагменту в составе S белка вируса SARS-CoV-2 | 2020 |
|
RU2744274C1 |
| Рекомбинантная плазмида pVBL-RBDomik, обеспечивающая синтез и секрецию рекомбинантного рецептор-связывающего домена (RBD) коронавируса SARS-CoV-2 линии B.1.1.529 в клетках млекопитающих. | 2021 |
|
RU2772905C1 |
| Иммунобиологическое средство и способ его использования для индукции специфического иммунитета против вирусов SARS-CoV-2 вариант B.1.617.2 (Delta) и SARS-CoV-2 вариант B.1.1.529 (Omicron) (варианты) | 2022 |
|
RU2779634C1 |
| Моноклональное антитело iC1 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817696C1 |
Изобретение относится к области биотехнологии. Описано рекомбинантное антитело человека T11 класса IgG против вируса SARS-CoV-2, обладающее свойством селективно связываться с RBD доменом S белка вируса SARS-CoV-2 и нейтрализовать активность вируса SARS-CoV-2 и способ его получения. Антитело содержит вариабельный домен тяжелой цепи (VH) с последовательностью аминокислот, представленных на SEQ ID NO: 1 и вариабельный домен легкой цепи (VL) с последовательностью аминокислот, представленных на SEQ ID NO: 2. Изобретение расширяет арсенал антител, обладающих нейтрализующей активностью против вируса SARS-CoV-2. 2 н.п. ф-лы, 2 ил., 4 пр., 2 табл.
1. Рекомбинантное антитело человека T11 класса IgG против вируса SARS-CoV-2, содержащее вариабельный домен тяжелой цепи с последовательностью аминокислот, представленных на SEQ ID NO: 1 и вариабельный домен легкой цепи с последовательностью аминокислот, представленных на SEQ ID NO: 2, обладающее свойством селективно связываться с RBD доменом S белка вируса SARS-CoV-2 и нейтрализовать активность вируса SARS-CoV-2.
2. Способ получения полноразмерного рекомбинантного антитела человека Т11 по п.1, предусматривающий котрансфекцию рекомбинантными плазмидными ДНК pFuse/T11/CH и pFuse/T11/CL клеток линии HEK293F, при этом плазмидная ДНК pFuse/T11/CH содержит нуклеотидную последовательность гена вариабельного домена тяжелой цепи в единой рамке считывания с геном константного домена тяжелой цепи иммуноглобулина человека IgG1, представленную на SEQ ID NO:3, плазмидная ДНК pFuse/T11/CL содержит нуклеотидную последовательность гена вариабельного домена легкой цепи в единой рамке считывания с геном константного домена лямбда легкой цепи иммуноглобулина человека, представленную на SEQ ID NO:4, наработку рекомбинантного антитела Т11 и его хроматографическую очистку из культуральной среды.
| Иммунобиологическое средство и способ его использования для индукции специфического иммунитета против вируса тяжелого острого респираторного синдрома SARS-CoV-2 (варианты) | 2020 |
|
RU2720614C1 |
| Моноклональное антитело к RBD фрагменту в составе S белка вируса SARS-CoV-2 | 2020 |
|
RU2744274C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АКТИВНОСТИ НЕЙТРАЛИЗУЮЩИХ АНТИТЕЛ К SARS-CoV-2 В СЫВОРОТКЕ ИЛИ ПЛАЗМЕ КРОВИ ЛЮДЕЙ, ПЕРЕНЕСШИХ COVID-19 ИЛИ ПРИВИТЫХ ВАКЦИНАМИ ДЛЯ ПРОФИЛАКТИКИ НОВОЙ КОРОНАВИРУСНОЙ ИНФЕКЦИИ COVID-19, С ИСПОЛЬЗОВАНИЕМ НАБОРА РЕАГЕНТОВ ДЛЯ ИММУНОФЕРМЕНТНОГО АНАЛИЗА, СОДЕРЖАЩЕГО РЕКОМБИНАНТНЫЙ РЕЦЕПТОР-СВЯЗЫВАЮЩИЙ ДОМЕН (RBD) ПОВЕРХНОСТНОГО ГЛИКОПРОТЕИНА S КОРОНАВИРУСА SARS-COV-2 И РЕКОМБИНАНТНЫЙ ЧЕЛОВЕЧЕСКИЙ РЕЦЕПТОР АСЕ2 | 2021 |
|
RU2784655C1 |
| US 11312760 B1 26.04.2022 | |||
| WO 2021222935 A2 04.11.2021. | |||
