Изобретение относится к универсальному интеграционному вектору pVEAL и рекомбинантной плазмиде pVEAL-15742, обеспечивающей синтез и секрецию scFv-Fc антител ADI-15742, способных нейтрализовать вирус Эбола в клетках млекопитающих с применением универсального вектора pVEAL и может быть использовано в молекулярной генетике и биотехнологии.
Уровень техники. Препараты на основе моноклональных антител являются наиболее перспективным инструментом экстренного противодействия вирусам вследствие их высокой специфичности и минимальным побочным эффектам. В настоящий момент по меньшей мере 570 антител по всему миру прошли клинические испытания (Lu R.М. et al. Development of therapeutic antibodies for the treatment of diseases. Journal of biomedical science. 2020; 27(1): 1-30; Wong G. et al. Post-exposure therapy of filovirus infections. Trends Microbiol. 2014; 22(8):456-63; Saphire E.O. An update on the use of antibodies against the filoviruses. Immunotherapy. 2013; 5(11): 1221- 33). Значительные технологические достижения сделали открытие и разработку методов лечения с помощью моноклональных антител более быстрыми и эффективными.
В настоящее время широко распространено получение антител формата scFv-Fc. Это одноцепочечные (single chain) антитела, состоящие из вариабельных фрагментов легкой и тяжелой цепей антитела и константной цепи. У scFv-Fc формата существует несколько преимуществ перед большими фрагментами или целыми антителами: меньшее количество сайтов расщепления для циркулирующих протеолитических ферментов и, следовательно, более высокая стабильность; scFv-Fc вариант подразумевает синтез одной цепи, что облегчает создание векторов и конструкций, упрощает схему получения и проверки, одноцепочечные антитела достигают цели быстрее, быстрее выводятся из организма (Huston J.S. et al. Protein engineering of single-chain Fv analogs and fusion proteins. Methods in enzymology. 1991, 203: 46-88; WO 1993/16185; US 5571894). Возможные аллергические реакции на неканоническую структуру несущественны в случае антител против инфекционных заболеваний. Такие антитела, в первую очередь должны обладать эффективностью  et al. High level transient production of recombinant antibodies and antibody fusion proteins in HEK293 cells. BMC biotechnology. 2013, 13(1): 52; Girgis M.D. et al. Targeting CEA in pancreas cancer xenografts with a mutated scFv-Fc antibody fragment. EJNMMI research. 2011, 1(1): 1-10).
et al. High level transient production of recombinant antibodies and antibody fusion proteins in HEK293 cells. BMC biotechnology. 2013, 13(1): 52; Girgis M.D. et al. Targeting CEA in pancreas cancer xenografts with a mutated scFv-Fc antibody fragment. EJNMMI research. 2011, 1(1): 1-10).
Одним из способов получения моноклональных scFv-Fc антител является встройка нуклеотидных последовательностей, кодирующих вариабельные последовательности тяжелой и легкой цепей антител, в интеграционные вектора. Эти вектора обеспечивают репликацию и экспрессию рекомбинантных генных конструкций в подходящем хозяине путем интеграции в хромосому данного хозяина. Интеграционный вектор включает в себя: рекомбинантные иммуноглобулиновые домены, точку начала репликации или автономно реплицирующуюся последовательность, доминантные маркерные последовательности, сигнальные последовательности, последовательности, контролирующие экспрессию и транскрипцию рекомбинантных генов, систему, отвечающую за интеграцию в геном (RU 2469093 С2, RU 2639519). Трансфекция линий клеток млекопитающих данными векторами и дальнейшее их культивирование позволяет клеткам синтезировать антитела непосредственно в культуральную жидкость (RU 2292353 С2).
В качестве интеграционного вектора могут выступать системы доставки на основе вирусов и невирусные ДНК-векторы. Системы доставки на основе вирусов имеют следующие недостатки: 1) препараты вирусов имеют риск заражения инфекционными агентами, включая репликационно-компетентный вирус; 2) вирусный вектор может вызывать нежелательные клеточные последствия, например острые иммунные, воспалительные или нейротоксические ответы; 3) эффективное генетическое размножение вирусных векторов может ограничивать размер терапевтического груза и может потребовать генетических мотивов для регуляции репликации вектора. Напротив, препараты невирусных ДНК-векторов на основе плазмид относительно недороги для очистки, в значительной степени неиммуногенны и не имеют жестких ограничений на последовательности, которые могут быть доставлены. Основные проблемы невирусных систем на основе плазмид заключаются в следующем: 1) низкие скорости доставки векторов к ядрам клеток-мишеней; 2) низкие скорости интеграции трансгенов, как правило, с сопутствующими плазмидными последовательностями, которые имеют склонность к подавлению экспрессии; 3) интеграция множественных копий трансгена, что также может подавлять их экспрессию (Hackett Р.В., et al. А transposon and transposase system for human application. Molecular therapy. 2010, 18(4): 674-683).
Указанные выше основные проблемы, связанные с невирусными векторами, можно преодолеть, используя систему на основе транспозазы SB (Sleeping Beauty). Эта система проста и не требует создания вирусных конструкций. Для обеспечения интеграции последовательности в геном необходимо наличие двух векторов, один должен содержать последовательность целевой встройки, фланкированной последовательностями IR/DR (L/R), второй вектор долен обеспечивать синтез транспозазы SB. По увеличению активности фермента выделяют транспозазы SB10, SB11, SB100 и т.д. (Hackett Р.В., et al. A transposon and transposase system for human application. Molecular therapy. 2010, 18(4): 674-683; Tipanee J. et al. Transposons: moving forward from preclinical studies to clinical trials. Human Gene Therapy. 2017, 28(11): 1087-1104). Достоинства систем на основе транспозазы: биобезопасность, легкость в работе, длина встраиваемого фрагмента не ограничена, стабильная экспрессия встраиваемого гена.
Ближайшие аналоги. В работе (Оксанич А.С. и др. Конструирование плазмидного вектора для получения химерных антител заданной специфичности в клетках млекопитающих. Журн. микробиол. 2017, 6:56-63) представлен универсальный плазмидный вектор, позволяющий получать полноразмерные химерные антитела различной специфичности различных классов и изотипов. Данный вектор создан на основе известного эукариотического плазмидного вектора pCI-neo и включает в себя сильный промотор CMV, гены устойчивости к антибиотикам, легкую и тяжелую цепи заданного антитела. Вектор прост в конструировании, для получения готовых препаратов антител используются доступные и недорогие методики. Однако конечный выход антитела невелик (4 мг/л).
В международной заявке WO 2007/081336 описан вектор для получения рекомбинантных белков с высоким уровнем секреции. Достоинством данного изобретения является тройная лидерная последовательность, CMV промотор в комбинации с регуляторным элементом elongation factor-1, что должно обеспечивать значительный уровень синтеза белка. Также вектор содержит PolyA, SV40 ori, SV40 PA, pUC Origin Site, гены устойчивости к антибиотикам ампициллин, неомицин. В описании изобретения отсутствуют данные о том, какой выход белка можно получить.
Недостатками приведенных выше изобретений является отсутствие цис-регуляторных элементов. При трансфекции клеток такими векторами последние хаотично встраиваются в хромосому, конформация участка встройки значительно влияет на транскрипцию гена и его экспрессию. Цис-регуляторные элементы позволяют увеличить экспрессию гена без воздействия на структуру хроматина в регионе встройки. К цис-регуляторным элементам относятся: locus control region (LCR), scaffold/matrix attachment region (S/MAR), insulator, ubiquitous chromatin open element (UCOE), anti-repressor (STAR element).
Наиболее близким аналогом (прототипом) является универсальный вектор, описанный в патенте Китая (CN 105255895, МПК C12N 15/85, опубл. 20.01.2016 г.). Авторы утверждают, что предложенная система экспрессии обеспечивает высокий выход белка, экспрессия является стабильной и длительной. Вектор создан на основе транспозазы SB, что позволяет легко интегрировать нужную последовательность в геном клетки. В качестве цис-регуляторного элемента выступает matrix attachment region - MAR2. Тяжелая и легкая цепи антитела соединены с помощью IRES. «Сайт внутренней посадки рибосомы» или «IRES» описывает последовательность, которая функционально обеспечивает инициацию трансляции и позволяет двум цистронам (открытым рамкам считывания) быть транслированными с одного транскрипта в животной клетке. Применение IRES элементов в векторных конструкциях описано ранее (Ramesh, N., et al. Nucl. Acids Res. 1996, 24: 2697-2700; Mosser, D.D. et al. BioTechniques. 1997, 22: 150-152). Для инициации транскрипции использовался цитогмегаловирусный промотр CMV.
Однако, известный универсальный вектор-прототип обеспечивает синтез только полноразмерных антител, которые, как упоминалось ранее, по многим параметрам уступают антителам формата scFv-Fc.
Прототипом рекомбинантной плазмиды, обеспечивающей синтез антитела против вируса Эбола, является конструкция, описанная в патенте США (US 10640550, МПК А61Р 31/14, опубл. 05.05.2020 г.). Вектор, сконструированный в данной работе, содержит промотор с энхансером, IRES, PolyA, гены устойчивости к антибиотикам ампициллин, неомицин. Возможна интеграция генетического материала в геном с помощью системы на основе траспозазы. Наработка антитела может осуществляться с помощью гибридомной технологии, либо путем трансфекции эукариотических клеток с последующим выходом синтезированных антител в культуральную жидкость.
Недостатками данной конструкции является отсутствие цис-регуляторных элементов, необходимость встройки полноразмерных легкой и тяжелой цепей антитела. В описании вектора не указано какой промотор использовался для инициации транскрипции, отсутствуют данные о выходе целевого белка, что не позволяет судить об эффективности данной конструкции.
Техническим результатом заявляемого изобретения является создание универсального интеграционного вектора pVEAL и плазмидной генетической конструкции pVEAL-15742, обеспечивающей синтез и секрецию целевого белка - scFv-Fc антитела ADI 15742 против вируса Эбола (Anna Z. Wec, et al. Antibodies from a Human Survivor Define Sites of Vulnerability for Broad Protection against Ebolaviruses. Cell. 2017, 169(5): 878-890) в клетках млекопитающих CHO-K1, полученного с использованием указанного универсального вектора pVEAL, который лишен недостатков прототипа.
Указанный технический результат достигается тем, что универсальный интеграционный вектор pVEAL служит для создания целевых плазмидных конструкций, обеспечивающих синтез и секрецию scFv-Fc моноклональных антител, имеет нуклеотидную последовательность SEQ ID No. 1 и содержит в соответствии с физической и генетической картой, представленной на Фиг. 1, следующие элементы:
- участок начала репликации ori (8616-9204);
- последовательности IR/DR (L/R), являющиеся частью системы на основе транспозазы SB100 (139-403, 7832-8127);
- последовательности UCOE, предотвращающих хромосомное «замалчивание» рекомбинантной экспрессионной кассеты (416 - 1549, 6272-7820);
- CMV Enhancer (2108-2487), CMV промотор (2488-2699) - сильный наиболее часто используемый промотор в генно-терапевтических конструкциях;
- Chimeric intron (2860-2992), увеличивающий эффективность CMV промотора;
- лидерный пептид VI9, обеспечивающий экспорт белка из клетки (3114-3164);
- место для встройки вариабельных фрагментов тяжелой и легкой цепей антитела, соединенных пептидным линкером, по сайтам гидролиза AfeI (3157), BamHI (3339);
- константную цепь антитела человека IgGl (СН1-СН2-СН3) (3348-4031) с мутациями M252Y, S254T, Т256Е, Е333А, H433K/N434F. Мутации увеличивают период полураспада антитела за счет взаимодействия с неонатальными рецепторами;
- участок внутренней посадки рибосомы EMCV IRES (4046-4620);
- последовательность Bleo, кодирующую фактор устойчивости к антибиотику блеомицину (4625-4995);
- последовательность SV40 poly(A) signal, необходимую для стабилизации мРНК-транскриптов (5042-5163);
- SV40 промотор (5921-6262), обеспечивающий автономную репликацию в клетках млекопитающих, комбинация с CMV промотором существенно увеличивает выход белка;
- ген устойчивости к антибиотику ампициллин AmpR и бактериальный промотор гена устойчивости к ампициллину, позволяющие проводить препаративную наработку плазмиды в E.coli (9375-10235).
Указанный технический результат достигается также тем, что плазмидная генетическая конструкция pVEAL-15742, обеспечивающая синтез и секрецию целевого белка - scFv-Fc антитела ADI 15742 против вируса Эбола, имеет нуклеотидную последовательность SEQ ID No. 2 и содержит в соответствии с физической и генетической картой, представленной на Фиг. 2, следующие элементы:
- участок начала репликации ori (8616-9204);
- последовательности IR/DR (L/R), являющиеся частью системы на основе транспозазы SB 100 (139-403, 8393-8688);
- последовательности UCOE, предотвращающих хромосомное «замалчивание» рекомбинантной экспрессионной кассеты (416 - 1549, 6833-8381);
- CMV Enhancer (2108-2487), CMV промотор (2488-2699) - сильный наиболее часто используемый промотор в генно-терапевтических конструкциях;
- Chimeric intron (2860-2992), увеличивающий эффективность CMV промотора;
- лидерный пептид VI9, обеспечивающий экспорт белка из клетки (3114-3164);
- ADI-15742-VH, последовательность, кодирующая вариабельный фрагмент тяжелой цепи антитела 15742 (3165-3533);
- ADI-15742-VK, последовательность, кодирующая вариабельный фрагмент легкой цепи антитела 15742 (3579-3903);
- константную цепь антитела человека IgG1 (СН1-СН2-СН3) (3909-4592) с мутациями M252Y, S254T, Т256Е, ЕЗЗЗА, H433K/N434F. Мутации увеличивают период полураспада антитела за счет взаимодействия с неонатальными рецепторами;
- участок внутренней посадки рибосомы EMCV IRES (4607-5181);
- последовательность Bleo, кодирующую фактор устойчивости к антибиотику блеомицину (5186-5556);
- последовательность SV40 poly(A) signal, необходимую для стабилизации мРНК-транскриптов (5603-5724);
- SV40 промотор (6482-6823), обеспечивающий автономную репликацию в клетках млекопитающих, комбинация с CMV промотором существенно увеличивает выход белка;
- ген устойчивости к антибиотику ампициллин AmpR и бактериальный промотор гена устойчивости к ампициллину, позволяющие проводить препаративную наработку плазмиды в E.coli (9936-10796).
Заявляемые плазмиды являются значительно усовершенствованными по сравнению с прототипами т.к. в качестве системы интеграции используется транспозаза SB 100, имеющая высокую активность фермента (Mates L et al. Molecular evolution of a novel hyperactive Sleeping Beauty transposase enables robust stable gene transfer in vertebrates. Nat Genet. 2009, 41(6):753-61), имеются сильные цис-регуляторные элементы UCOE, предотвращающие хромосомное замалчивание (Neville JJ et al. Ubiquitous Chromatin-opening Elements (UCOEs): Applications in biomanufacturing and gene therapy. Biotechnol Adv. 2017, 35(5):557-564), комбинация CMV промотора с энхансером, интроном А и промотром SV40 существенно увеличивает выход белка (RU 2639519), сильный лидерный пептид V19, увеличивающий секрецию рекомбинантного белка (WO 2008/148519 А2), связанная посредством EMCV IRES соэкспрессия селекционного маркера Bleo и последовательностей антител может улучшить селекционный процесс (RU 2639519), scFv-Fc формат антител облегчает создание конструкции, упрощает схему получения и проверки необходимого антитела.
Изобретение иллюстрируется следующими графическими фигурами. На фиг. 1 представлена физическая и генетическая карта универсального интеграционного вектора pVEAL. На фиг. 2 представлена физическая и генетическая карта интеграционного вектора pVEAL со вставкой scFv-Fc антитела ADI-15742 против вируса Эбола - pVEAL-15742. На фиг. 3 представлено определение минимальной концентрации блеомицина, необходимой для гибели нетрансфецированной линии клеток-хозяев. На фиг. 4 представлен анализ нейтрализующей активности моноклонального scFv-Fc антитела ADI 15742 против вируса Эбола. На фиг. 5 представлен анализ культуральной среды штамма-продуцента scFv-Fc антитела 15742. На фиг. 6 представлен анализ препарата scFv-Fc антитела 15742 после хроматографии на белке А.
Для лучшего понимания сущности предлагаемого изобретения ниже приведены примеры (1-5) его осуществления. Все стандартные генно-инженерные и микробиологические манипуляции, а также амплификацию и секвенирование ДНК проводили по известным методикам (Маниатис Т., Фрич Э, Сэмбрук Дж. Молекулярное клонирование, М.: Мир, 1984; Клонирование ДНК. Методы. Под ред. Д. Гловера, Пер. с англ., Москва, Мир, 1988; Saiki R.K. et al. Science. 1988, 239(4839):487-491; Sanger F. et al. Proc. Nat. Acad. Sci. 1977, 74:5463-5467).
Пример 1. Конструирование универсального интеграционного вектора pVEAL
Для проектирования вектора pVEAL-использовали акцепторный вектор рТ2НВ (Addgene plasmid # 26557). Интеграционный вектор был получен в несколько этапов. На первом этапе из вектора рТ2НВ при помощи сайт-направленного мутагенеза был удален сайт гидролиза BamHI. Для этого проводили ПЦР с использованием праймеров:
- 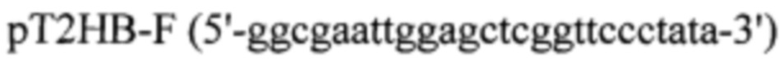 и
и
- 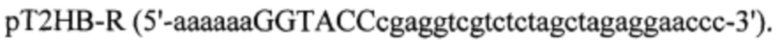
ПЦР фрагмента PCR-pT2HB проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (плазмиды рТ2НВ), 10 пМ каждого праймера (pT2HB-F, pT2HB-R), 5 мкл 10xTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95°С; 30 с - 95°С 30 с - 58°С, 45 с - 72°С (25 циклов); 120 с - 72°С.
Реакционную смесь, содержащую фрагмент PCR-pT2HB, разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосу, соответствующую продукту амплификации - 709 п.н., визуализированную при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирку объемом 1,5 мл. Фрагмент выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Затем выделенный ПНР-продукт PCR-pT2HB и исходную плазмиду рТ2НВ обрабатывали эндонуклеазами рестрикции Psp124BI и KpnI согласно протоколу фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам - 688 и 2859 п. н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable с генотипом F' proA+В+lacIq Δ(lacZ)M15 zzf::Tn10 (TetR) Δ(ara-leu) 7697 araD139 fhuA ΔlacX74 galK16 galE15 el4- Ф80dlacZΔM15 recA1 relA1 endA1 nupG rpsL (StrR) rph spoT1 Δ(mrr-hsdRMS-mcrBC), предоставленные производителем "Invitrogen" (США). Для этого к 100 мкл клеток добавляли 10 мкл лигазной смеси, инкубировали на льду в течение 40 минут. После этого клетки подвергали «температурному шоку» при 42°С в течение 40 сек. Охлаждали клетки на льду в течение 4 минут, затем добавляли по 200 мкл среды "SOC" (20 г/л триптона, 5 г/л дрожжевого экстракта, 0,5 г/л NaCl, 250 мМ KCl, 10 мМ MgCl2) и инкубировали при 30°С в течение 60 минут. По окончании инкубации трансформированные клетки высевали на чашку Петри с твердой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов "Plasmid Mini Kit", "Qiagen" (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена плазмида рТ2НВ2.
Следующим шагом была встройка последовательностей UCOE-R и UCOE-L. Для получения последовательностей UCOE-R и UCOE-L использовали геномную ДНК человека. Для амплификации последовательности UCOE-R была проведена ПЦР с помощью праймеров:
-  и
и
- 
ПЦР фрагмента UCOE-R проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (геномная ДНК человека), 10 пМ каждого праймера (UCOE-R-F, UCOE-R-R), 5 мкл 10xTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95°С; 30 с - 95°С, 30 с - 59°С, 95 с - 72°С (30 циклов); 120 с - 72°С.
Реакционную смесь, содержащую фрагмент UCOE-R, разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосу, соответствующую продукту амплификации - 1572 п.н., визуализированную при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирку объемом 1,5 мл. Фрагмент выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Затем выделенный ПЦР-продукт UCOE-R обрабатывали эндонуклеазами рестрикции BseX3I и BamHI, плазмиду рТ2НВ2 обрабатывали эндонуклеазами рестрикции BseX3I и BglII согласно протоколу фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам - 1555 и 3529 п. н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable как было описано выше. Трансформированные клетки высевали на чашку Петри с твердой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов "Plasmid Mini Kit", "Qiagen" (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена плазмида pT2HB2-UCOE-R.
Амплификация последовательности UCOE-L была проведена при помощи ПЦР с использованием праймеров:
-  и
и
- 
ПЦР фрагмента UCOE-L проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (геномная ДНК человека), 10 пМ каждого праймера (UCOE-L-F, UCOE-L-R), 5 мкл 10xTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95°С; 30 с - 95°С, 30 с - 59°С, 97 с - 72°С (30 циклов); 120 с - 72°С.
Реакционную смесь, содержащую фрагмент UCOE-L, разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосу, соответствующую продукту амплификации - 1593 п.н., визуализированную при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирку объемом 1,5 мл. Фрагмент выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Затем выделенный ПЦР-продукт UCOE-L и плазмиду pT2HB2-UCOE-R обрабатывали эндонуклеазами рестрикции HindIII и BseX3I согласно протоколу фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам - 1581 и 5051 п.н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable как было описано выше. Трансформированные клетки высевали на чашку Петри с твердой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов "Plasmid Mini Kit", "Qiagen" (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена плазмида pT2HB2-UCOE-RL.
Дальнейшее конструирование вектора заключалось во встройке последовательности V19-ADI (синтезирована по заказу ООО «ДНК-синтез»), кодирующей лидерный пептид V19 и уникальные сайты гидролиза AfeI и BamHI, по которым будет проводиться вставка цепей необходимого антитела и scFV-Fc фрагмента.
Лигирование последовательности V19-ADI с коммерческим вектором pJet (CloneJET PCR Cloning Kit, Thermo Scientific) проводили согласно инструкции производителя. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable как было описано выше. Трансформированные клетки высевали на чашку Петри с твердой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов "Plasmid Mini Kit", "Qiagen" (Германия) в соответствии с инструкциями производителя. Получена конструкция pJet-V19-ADI.
Далее вставку последовательности V19-ADI проводили в вектор pCI-neo-hEST2 (#1781). Этот вектор был выбран, потому что содержит в своем составе сильный эукариотический промотор CMV, а также набор генов устойчивости к антибиотикам, делающим его удобным для работы.
Вектор pCI-neo-hEST2 и pJet-V19-ADI обрабатывали эндонуклеазами рестрикции PspCI и SalI согласно протоколу фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% и 2% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам - 5467 и 182 п.н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable как было описано выше. Трансформированные клетки высевали на чашку Петри с твердой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов "Plasmid Mini Kit", "Qiagen" (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена плазмида pCI-neo-hEST2-V19-ADI.
Далее было проведено клонирование последовательности, кодирующей константную часть тяжелой цепи антитела человека (СН) Human IgG1 в вектор pCI-neo-hEST2-V19-ADI. Для амплификации последовательности, кодирующей константную часть тяжелой цепи антитела человека, использовали праймеры:
- 
 и
и
- 
В качестве ДНК матрицы использовали плазмиду pMAB-CH-100VH.
ПЦР фрагмента PCR-Human IgG1 проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (рМАВ-СН-100VH), 10 пМ каждого праймера (Hinge_Bspl3I_F, CH3_SalI_R), 5 мкл 10xTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95°С; 30 с - 95°С, 30 с - 60°С, 43 с - 72°С (30 циклов); 120 с - 72°С.
Реакционную смесь, содержащую фрагмент PCR-Human IgG1, разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосу, соответствующую продукту амплификации - 708 п. н., визуализированную при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирку объемом 1,5 мл. Фрагмент выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Затем выделенный ПЦР-продукт PCR-Human IgG1 и плазмиду pCI-neo-hEST2-V19-ADI обрабатывали эндонуклеазами рестрикции Bsp13I и SalI согласно протоколу фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам - 690 и 5635 п.н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable как было описано выше. Трансформированные клетки высевали на чашку Петри с твердой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов "Plasmid Mini Kit", "Qiagen" (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена плазмида pCI-neo-hEST2-V19-ADI-CH-21.
Далее проводилась встройка гена устойчивости к антибиотику блеомицину и последовательности EMCV IRES. Праймеры, используемые для амплификации:
- 
- 
ПЦР фрагмента PCR-Bleo-scFv проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (FUGW, Addgene plasmid # 14883), 10 пМ каждого праймера (Bleo-scFv-F, Bleo-scFv-R), 5 мкл 10xTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95°С; 30 с - 95°С, 30 с - 58°С, 25 с - 72°С (30 циклов); 120 с - 72°С.
Реакционную смесь, содержащую фрагмент PCR-Bleo-scFv, разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосу, соответствующую продукту амплификации - 400 п.н., визуализированную при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирку объемом 1,5 мл. Фрагмент выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Затем выделенный ПЦР-продукт PCR-Bleo-scFv и плазмиду pJet-EMCV-IRES-V17-100VH-CH-2018 обрабатывали эндонуклеазами рестрикции Msp20I и AsuNHI согласно протоколу фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам - 380 и 4578 п. н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable как было описано выше. Трансформированные клетки высевали на чашку Петри с твердой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов "Plasmid Mini Kit", "Qiagen" (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена плазмида pJet-IRES-Bleo.
ПЦР фрагмента EMCV-IRES-BleoR проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Праймеры, используемые для амплификации:
-  и
и
- 
Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (pJet-IRES-Bleo), 10 пМ каждого праймера (IRES-scFv-R и IRES-Bleo-scFv-R), 5 мкл 10xTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95°С; 30 с - 95°С, 30 с -58°С, 60 с - 72°С (30 циклов); 120 с - 72°С.
Реакционную смесь, содержащую фрагмент EMCV-IRES-BleoR, разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосу, соответствующую продукту амплификации - 984 п.н., визуализированную при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирку объемом 1,5 мл. Фрагмент выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Затем выделенный ПЦР-продукт EMCV-IRES-BleoR и плазмиду pCI-neo-hEST2-V19-ADI-CH-21 обрабатывали эндонуклеазами рестрикции XhoI и NotI согласно протоколу фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам - 965 и 6300 п.н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable как было описано выше. Трансформированные клетки высевали на чашку Петри с твердой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов "Plasmid Mini Kit", "Qiagen" (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена плазмида pCI-neo- hEST2-V19-ADI-CH-IRES-BLEO-21.
Последним шагом в получении целевого вектора pVEAL являлось объединение конструкций pT2HB2-UCOE-RL и pCI-neo- hEST2-V19-ADI-CH-IRES-BLEO-21. Плазмиду pT2HB2-UCOE-RL обрабатывали эндонуклеазами рестрикции BamHI и Mox20I, плазмиду pCI-neo-scFv-ADI-CH-IRES-BLEO-21 обрабатывали эндонуклеазами рестрикции BglII и HpaI.
Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам - 6617 и 4297 п.н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable как было описано выше. Трансформированные клетки высевали на чашку Петри с твердой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов "Plasmid Mini Kit", "Qiagen" (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск).
Встройка последовательностей вариабельных фрагментов тяжелой и легкой цепей любого антитела в универсальный вектор pVEAl проводится по уникальным сайтам гидролиза AfeI и BamHI.
Пример 2. Получение генетической конструкции pVEAL-15742, кодирующей scFv-Fc антитело ADI-15742, для трансфекции линии клеток СНО-K1.
Для подтверждения эффективности универсального интеграционного вектора pVEAL проводили встройку вариабельных последовательностей тяжелой и легкой цепей антитела ADI-15742 и связывающего их sc-Fv фрагмента.
Для начала были амплифицированы тяжелая и легкая цепи антитела ADI-15742. ПЦР фрагмента PCR-ADI-15742VH проводилась с синтезированной последовательности ADI-15742VH (ООО «ДНК-синтез») с помощью праймеров:
- 
 и
и
- 
ПЦР фрагмента PCR-ADI-15742VK с синтезированной последовательности ADI-15742VK (ООО «ДНК-синтез») с помощью праймеров:
- 
 и
и
- 
ПЦР фрагментов PCR-ADI-15742VH, PCR-ADI-15742VK проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционные смеси объемом 50 мкл содержали 2 мкг ДНК (ADI-15742VH/ ADI-15742VK), 10 пМ каждого праймера (V19-ADI-F, V19-ADI-L-R/ V19-ADI-L-F, V19-ADI-R), 5 мкл 10xTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95°С; 30 с - 95°С, 30 с -58°С, 30 с - 72°С (30 циклов); 120 с - 72°С.
Реакционные смеси, содержащие фрагменты PCR-ADI-15742VH, PCR-ADI-15742VK, разделяли электрофорезом в 1% агарозном геле е с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие продуктам амплификации - 487 и 384 п. н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее был проведен отжиг и последующая амплификация последовательностей PCR-ADI-15742VH, PCR-ADI-15742VK с использованием праймеров V19-ADI-F и V19-ADI-R при помощи ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК каждого фрагмента (PCR-ADI-15742VH, PCR-ADI-15742VK), 10 пМ каждого праймера (V19-ADI-F и V19-ADI-R), 5 мкл 10xTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95°С; 30 с - 95°С, 30 с -58°С, 51 с - 72°С (30 циклов); 120 с - 72°С.
Реакционную смесь, содержащую амплифицированный фрагмент scFv-ADI-15742, разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосу, соответствующую продукту амплификации - 851 п.н., визуализированную при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирку объемом 1,5 мл. Фрагмент выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Универсальный интеграционный вектор pVEAL и фрагмент scFv-ADI-15742 обрабатывали эндонуклеазами рестрикции AfeI и BamHI согласно протоколу фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам - 10732 и 743 п.н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable как было описано выше. Трансформированные клетки высевали на чашку Петри с твердой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов "Plasmid Mini Kit", "Qiagen" (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г.Новосибирск). В результате была получена плазмида pVEAL-15742.
Пример 3. Получение клонов-продуцентов scFv-Fc антитела 15742
Для получения клеток продуцентов были выбраны клетки СНО-K1 (клон К1 линии СНО, выделенной из яичника китайского хомячка). Использование этих клеток опирается на опыт их коммерческого использования для создания продуцентов терапевтических белков, в том числе антител. Линия поддерживалась в среде на основе DMEM/F12 с добавлением 10% термоинактивированной (30 мин при температуре 56°С) фетальной сыворотки крови коров (FBS), L-глутамина. По достижении монослоя клетки пересаживались.
Для получения стабильной клеточной линии, синтезирующей рекомбинантный белок, определяли минимальную концентрацию селективного антибиотика блеомицина, необходимую для гибели нетрансфецированной линии клеток-хозяев. Для определения IC50 и IC100 блеомицина клетки рассаживали в 12-луночный культуральный планшет со средой DMEM/F12 с добавлением 10% фетальной сыворотки крови коров (FBS) и L-глутамином, по достижении 90% конфлюэнтности монослоя производили замену культуральной среды на 2% DMEM/F12 с добавлением антибиотика блеомицина (50-700 мкг/мл). В течение 14 суток каждые 48-72 часов производили замену культуральной среды в лунках на 2% DMEM/F12 с добавлением блеомицина (50-700 мкг/мл).
При добавлении блеомицина чувствительные к нему клетки проявляли следующие морфологические изменения:
• Увеличение размера (аналогично воздействию цитомегаловируса, инфицирующего клетки)
• Аномальная форма клеток, включая появление длинных придатков
• Наличие больших пустых везикул в цитоплазме (пробой эндоплазматического ретикулума и аппарата Гольджи)
• Разрушение плазмы и ядерной мембраны (появление множества отверстий в этих мембранах). В конце концов, клетки полностью разрушатся, и останется только клеточный «мусор».
Указанные изменения проиллюстрированы на фиг. 3. На фиг. 3а представлена фотография клеток, не подвергшихся воздействию блеомицина, - положительный контроль К+. На фиг. 3б - 3и представлены фотографии клеток, подвергшихся воздействию блеомицина в количестве от 50 до 700 мкг/мл. На данных фигурах показано, что при увеличении концентрации антибиотика клетки существенно меняются морфологически. На фиг. 3д при концентрации блеомицина 300 мкг/мл видно, что примерно половина клеток сохраняют жизнеспособность и присущую им морфологию. На фиг. 3е, 3ж клетки практически полностью погибли, преобладают измененные и разрушенные клетки.
Таким образом, установлена полуингибирующая и ингибирующая дозы блеомицина для СНО-K1 (IC50=325 мкг/мл, IC100=475 мкг/мл).
Для встройки экспрессионой кассеты проводили котрансфекцию методом катионно-липидной трансфекции (Lipofectamine 3000, Invitrogen) клеток СНО-K1 двумя плазмидами. Первая содержала последовательность scFv-Fc антитела 15742, pVEAL-15742, вторая содержала последовательность транспозазы SB100, pCMV(CAT)T7-SB100. После трансфекции инкубировали клетки 96 часов в CO2-инкубаторе согласно рекомендациям производителя Lipofectamine 3000. Затем проводили замену среды на поддерживающую (с содержанием 2% фетальной бычьей сыворотки) с добавлением селективного антибиотика блеомицина (475 мкг/мл). Эту манипуляцию повторяли каждые 72-96 часов на протяжении 2-3 недель до тех пор, пока в отрицательном контроле клетки не погибли. Затем делали один пассаж на культуральных чашках и рассаживали в 96-луночные планшеты по одной клетке на лунку.
Получено 4 клона 15742 с нейтрализующим титром, определенным для IC90, (анализировали культуральную среду) от 1:8 до 1:32768. Реакцию вируснейтрализации проводили на псевдовирусной системе.
Пример 4. Анализ нейтрализующей активности scFv-Fc моноклонального антитела ADI15742 против вируса Эбола
Препараты антитела 15742 титровали с шагом 4, инкубировали 30 мин в CO2-инкубаторе со смесью псевдовирусных частиц вируса Эбола (50 мкл, физический титр 5×108 частиц) и переносили в 96-луночный плоскодонный планшет к суспензии культуры клеток HEK293T (50 мкл, 100 тыс/мл). Инкубировали 48 ч в CO2-инкубаторе. По истечении времени из лунок планшета удаляли ростовую среду, промывали 100 мкл PBS, добавляли 40 мкл 1×лизирующего буфера (Promega, США), инкубировали 10 минут при комнатной температуре, суспендировали и переносили по 35 мкл в оптический планшет, добавляли 35 мкл LAR (Promega, США), и считывали результаты при помощи люминометра (LuMate). Результаты анализа на примере одного из клонов представлены на фиг. 4. График, приведенный на фиг. 4, отражает зависимость нейтрализующей активности scFv-Fc антитела ADI 15742 против вируса Эбола, определенной для IC90, от степени разведения препарата. При разведении 1:8 препарат антитела обладает стопроцентной нейтрализующей активностью. При разведении до 1:32768 нейтрализующая активность составляет 75%.
Пример 5. Наработка и очистка scFv-Fc моноклонального антитела ADI15742
Каждый индивидуальный клон-продуцент рекомбинантного scFv-Fc антитела 15742 нарабатывался в культуральной среде DMEM/F12 с 10% содержанием фетальной телячьей сыворотки. Культивирование клонов-продуцентов проводилось в объеме 200 мл в роллерных бутылях при 37°С в течение 10 суток. Затем среда заменялась на поддерживающую, содержащую 2% фетальной телячьей сыворотки. Культивирование продолжали до тех пор, пока клетки сохраняли жизнеспособность. Далее полученную культуральную жидкость сливали, осаждали клеточный дебрис при помощи центрифугирования при 6000 g, 4°С в течение 15 мин.
Очистку рекомбинантных антител проводили при помощи аффинной хроматографии с использованием сорбента, содержащего на своей поверхности белок A (MabSelect SuRe, GE Healthcare, США), и хроматографической системы AKTAprime plus (GE Healthcare, США). Очистка включает в себя следующие стадии: уравновешивание хроматографической колонки, нанесение образца, промывка колонки, элюция белка.
Уравновешивание хроматографической колонки включает в себя промывание пятикратным объемом базового буфера (50 mM Трис-НСl, 150 мМ NaCl, рН 7,0-7,2). Скорость потока определялась временем контакта буфера с сорбентом, время контакта составляло не менее 4 мин.
Далее наносили культуральную жидкость, содержащую антитела, на колонку, время контакта образца с сорбентом составляло не менее 5 мин.
Отмывку колонки от несвязавшихся белков, а также от белков, обладающих низкой силой взаимодействия с сорбентом, проводили при помощи последовательной промывки хромоатографической колонки базовым буфером и буфером для промывки (20 мМ Трис-HCl, рН 6,8-7,0). Объемы базового и промывочного буферов составляли 3,5 объема колонки. Скорость потока определялась временем контакта буфера с сорбентом, время контакта составляло не менее 3,5 мин.
Элюцию проводили пятикратным объемом элюирующего буфера (100 mM Глицин-HCl, рН 3,4-3,5). Скорость потока определялась временем контакта буфера с сорбентом, время контакта составляло не менее 6 мин. Полученные образцы нейтрализовали до рН 7,0 при помощи буфера для титрования (1М Трис). Полученные препараты анализировали при помощи разделения в полиакриламидном геле (ПААГ) по стандартной методике (Фиг. 5). Препараты, нанесенные на ПААГ: дорожка 1 - исходная культуральная жидкость, содержащая наработанное антитело, до очистки на хроматографической колонке, 10 мкл; дорожка 2 - культуральная жидкость, прошедшая через хроматографическую колонку, и содержащая несвязавшиеся с сорбентом клеточные белки, 10 мкл; дорожка 3 - препарат после промывки колонки уравновешивающим буфером, содержит слабо связавшиеся с сорбентом клеточные белки, 10 мкл; дорожка 4 - препарат после промывки колонки буфером для промывки, содержит остатки несвязавшихся белков, 10 мкл; 5 - маркер длин белков 10-260 kDa; дорожка 6 - препарат антитела 15742 после очистки, 10 мкл. На представленной электрофореграмме видно, что полученное антитело очищено от посторонних белков, имеет массу порядка 50 kDa.
Препараты, содержащие рекомбинантное антитело, фильтровали от выпавших в осадок белков при помощи насадки на шприц с диаметром пор 0,22 мкм (ТРР, Швейцария). Элюаты концентрировались при помощи центрифугирования препаратов антитела в концентраторах (Sartorius AG, Германия) при 6000 g, 17°С. Концентрация антител определялась с помощью прибора для определения сверхмалых концентраций NanoPhotometer NP80 (Implen, Германия) согласно инструкции производителя. Контроль проводили при помощи разделения в ПААГ. Пример определения концентрации препарата очищенного scFv-Fc антитела 15742, полученного с одного из клонов-продуцентов, приведен на фиг. 6. Для определения приблизительной концентрации разводили препараты очищенного scFv-Fc антитела 15742 в 10, 20, 40, 80 раз и сравнивали с бычьим сывороточным альбумином (БСА) в концентрации от 0,025 до 1 мкг. Препараты, нанесенные на ПААГ: дорожка 1 - препарат очищенного антитела 15742 после концентрирования в 1-кратном разведении (выделение из 1 л культуральной среды), 10 мкл; дорожка 2 - препарат очищенного антитела 15742 до концентрирования в 10-кратном разведении (выделение из 1 л культуральной среды), 10 мкл; дорожка 3 - препарат очищенного антитела 15742 до концентрирования в 20-кратном разведении (выделение из 1 л культуральной среды), 10 мкл; дорожка 4 - препарат очищенного антитела 15742 до концентрирования в 40-кратном разведении (выделение из 1 л культуральной среды), 10 мкл; дорожка 5 - препарат очищенного антитела 15742 до концентрирования в 80-кратном разведении (выделение из 1 л культуральной среды), 10 мкл; дорожка 6-маркер длин белков 10-260 kDa; 7 - БСА - 1 мкг; 8 - БСА - 0,1 мкг; 9 - БСА - 0,05 мкг; 10 - БСА - 0,025 мкг. При сопоставлении концентраций scFv-Fc антитела 15742 и БСА сделан вывод, что концентрация целевого белка, полученная с одного из клонов-продуцентов, составляет примерно 150 мг/л.
Анализ культуральной жидкости различных клонов-продуцентов показал, что наработка антител у них идет неравномерно. Содержание антител в препаратах после хроматографии и концентрирования варьируется в пределах от 35 мг до 150,6 мг с одного литра культуральной жидкости.
Список последовательностей:
<110> Федеральное бюджетное учреждение науки «Государственный научный центр
вирусологии и биотехнологии «Вектор» Федеральной службы по надзору в
сфере защиты прав потребителей и благополучия человека
(ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора)
<120> Универсальный интеграционный вектор pVEAL и рекомбинантная плазмида
pVEAL-15742, обеспечивающая синтез и секрецию scFv-Fc антитела ADI-15742
против вируса Эбола в клетках млекопитающих и полученная с использованием
вектора pVEAL
<160> SEQ ID NO 2
<210> SEQ ID NO: 1
<211> 10914
<212> PRT
<213> Artificial Sequence
<223> Нуклеотидная последовательность универсального интеграционного
вектора pVEAL
<400> 1
1 caaggcgatt aagttgggta acgccagggt tttcccagtc acgacgttgt aaaacgacgg
61 ccagtgagcg cgcgtaatac gactcactat agggcgaatt ggagctcggt tccctataca
121 gttgaagtcg gaagtttaca tacacttaag ttggagtcat taaaactcgt ttttcaacta
181 ctccacaaat ttcttgttaa caaacaatag ttttggcaag tcagttagga catctacttt
241 gtgcatgaca caagtcattt ttccaacaat tgtttacaga cagattattt cacttataat
301 tcactgtatc acaattccag tgggtcagaa gtttacatac actaagttga ctgtgccttt
361 aaacagcttg gaaaattcca gaaaatgatg tcatggcttt agaagcttca ttggcgccct
421 ccgcgcctac agctcaagcc acatccgaag ggggagggag ccgggagctg cgcgcggggc
481 cgccgggggg aggggtggca ccgcccacgc cgggcggcca cgaagggcgg ggcagcgggc
541 gcgcgcgcgg cggggggagg ggccggcgcc gcgcccgctg ggaattgggg ccctaggggg
601 agggcggagg cgccgacgac cgcggcactt accgttcgcg gcgtggcgcc cggtggtccc
661 caaggggagg gaagggggag gcggggcgag gacagtgacc ggagtctcct cagcggtggc
721 ttttctgctt ggcagcctca gcggctggcg ccaaaaccgg actccgccca cttcctcgcc
781 cgccggtgcg agggtgtgga atcctccaga cgctggggga gggggagttg ggagcttaaa
841 aactagtacc cctttgggac cactttcagc agcgaactct cctgtacacc aggggtcagt
901 tccacagacg cgggccaggg gtgggtcatt gcggcgtgaa caataatttg actagaagtt
961 gattcgggtg tttccggaag gggccgagtc aatccgccga gttggggcac ggaaaacaaa
1021 aagggaaggc tactaagatt tttctggcgg gggttatcat tggcgtaact gcagggacca
1081 cctcccgggt tgagggggct ggatctccag gctgcggatt aagcccctcc cgtcggcgtt
1141 aatttcaaac tgcgcgacgt ttctcacctg ccttcgccaa ggcaggggcc gggaccctat
1201 tccaagaggt agtaactagc aggactctag ccttccgcaa ttcattgagc gcatttacgg
1261 aagtaacgtc gggtactgtc tctggccgca agggtgggag gagtacgcat ttggcgtaag
1321 gtggggcgta gagccttccc gccattggcg gcggataggg cgtttacgcg acggcctgac
1381 gtagcggaag acgccttagt gggggggaag gttctagaaa agcggcggca gcggctctag
1441 cggcagtagc agcagcgccg ggtcccgtgc ggaggtgctc ctcgcagagt tgtttctcca
1501 gcagcggcag ttctcactac agcgccagga cgagtccggt tcgtgttcgt ccgcggagat
1561 ctctctcatc tcgctcggct gcgggaaatc gggctgaagc gactgagtcc gcgatggagg
1621 taacgggttt gaaatcaatg agttattgaa aagggcatgg cgaggccgtt ggcgcctcag
1681 tggaagtcgg ccagccgcct ccgtgggaga gaggcaggaa atcggaccaa ttcagtagca
1741 gtggggctta aggtttatga acggggtctt gagcggaggc ctgagcgtac aaacagcttc
1801 cccaccctca gcctcccggc gccatttccc ttcactgggg gtgggggatg gggagctttc
1861 acatggcgga cgctgccccg ctggggtgaa agtggggcgc ggaggcggga cttcttattc
1921 cctttctaaa gcacgctgct tcgggggcca cggcgtctcc tcggggatct tcaatattgg
1981 ccattagcca tattattcat tggttatata gcataaatca atattggcta ttggccattg
2041 catacgttgt atctatatca taatatgtac atttatattg gctcatgtcc aatatgaccg
2101 ccatgttggc attgattatt gactagttat taatagtaat caattacggg gtcattagtt
2161 catagcccat atatggagtt ccgcgttaca taacttacgg taaatggccc gcctggctga
2221 ccgcccaacg acccccgccc attgacgtca ataatgacgt atgttcccat agtaacgcca
2281 atagggactt tccattgacg tcaatgggtg gagtatttac ggtaaactgc ccacttggca
2341 gtacatcaag tgtatcatat gccaagtccg ccccctattg acgtcaatga cggtaaatgg
2401 cccgcctggc attatgccca gtacatgacc ttacgggact ttcctacttg gcagtacatc
2461 tacgtattag tcatcgctat taccatggtg atgcggtttt ggcagtacac caatgggcgt
2521 ggatagcggt ttgactcacg gggatttcca agtctccacc ccattgacgt caatgggagt
2581 ttgttttggc accaaaatca acgggacttt ccaaaatgtc gtaacaactg cgatcgcccg
2641 ccccgttgac gcaaatgggc ggtaggcgtg tacggtggga ggtctatata agcagagctc
2701 gtttagtgaa ccgtcagatc actagaagct ttattgcggt agtttatcac agttaaattg
2761 ctaacgcagt cagtgcttct gacacaacag tctcgaactt aagctgcagt gactctctta
2821 aggtagcctt gcagaagttg gtcgtgaggc actgggcagg taagtatcaa ggttacaaga
2881 caggtttaag gagaccaata gaaactgggc ttgtcgagac agagaagact cttgcgtttc
2941 tgataggcac ctattggtct tactgacatc cactttgcct ttctctccac aggtgtccac
3001 tcccagttca attacagctc ttaaggctag agtacttaat acgactcact ataggctagc
3061 ctcgagaatt cgtcctgctg cgcacgtggg aagccctggc cccggccgcc accatgatga
3121 ggcccatcgt gctggtgctg ctgttcgcca cctcagcgct ggccggcgag tgcgagtgcg
3181 agtgcgagtc gtacgtacgt acgtacgtac gtacgtatat atatatatat attatatata
3241 ttaatatatg gcgagtgcga gtgcgagtgc gagtcgtacg tacgtacgta cgtacgtacg
3301 tatatatata tatatattat atatattaat atatatatgg atccggagac aaaactcaca
3361 catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc ctcttccccc
3421 caaaacccaa ggacaccctc tacatcaccc gggaacctga ggtcacatgc gtggtggtgg
3481 acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc gtggaggtgc
3541 ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt gtggtcagcg
3601 tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc aaggtctcca
3661 acaaagccct cccagccccc atcgcgaaaa ccatctccaa agccaaaggg cagccccgag
3721 aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac caggtcagcc
3781 tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg gagagcaatg
3841 ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac ggctccttct
3901 tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac gtcttctcat
3961 gctccgtgat gcatgaggct ctgaagttcc actacacgca gaagagcctc tccctgtctc
4021 cgggtaaatg agtcgagcgg ttcccgcccc tctccctccc ccccccctaa cgttactggc
4081 cgaagccgct tggaataagg ccggtgtgcg tttgtctata tgttattttc caccatattg
4141 ccgtcttttg gcaatgtgag ggcccggaaa cctggccctg tcttcttgac gagcattcct
4201 aggggtcttt cccctctcgc caaaggaatg caaggtctgt tgaatgtcgt gaaggaagca
4261 gttcctctgg aagcttcttg aagacaaaca acgtctgtag cgaccctttg caggcagcgg
4321 aaccccccac ctggcgacag gtgcctctgc ggccaaaagc cacgtgtata agatacacct
4381 gcaaaggcgg cacaacccca gtgccacgtt gtgagttgga tagttgtgga aagagtcaaa
4441 tggctcacct caagcgtatt caacaagggg ctgaaggatg cccagaaggt accccattgt
4501 atgggatctg atctggggcc tcggtgcaca tgctttacat gtgtttagtc gaggttaaaa
4561 aacgtctagg ccccccgaac cacggggacg tggttttcct ttgaaaaaca cgatgataat
4621 atggccaagt tgaccagtgc cgttccggtg ctcaccgcgc gcgacgtcgc cggagcggtc
4681 gagttctgga ccgaccggct cgggttctcc cgggacttcg tggaggacga cttcgccggt
4741 gtggtccggg acgacgtgac cctgttcatc agcgcggtcc aggaccaggt ggtgccggac
4801 aacaccctgg cctgggtgtg ggtgcgcggc ctggacgagc tgtacgccga gtggtcggag
4861 gtcgtgtcca cgaacttccg ggacgcctcc gggccggcca tgaccgagat cggcgagcag
4921 ccgtgggggc gggagttcgc cctgcgcgac ccggccggca actgcgtgca cttcgtggcc
4981 gaggagcagg actgagcggc cgcttccctt tagtgagggt taatgcttcg agcagacatg
5041 ataagataca ttgatgagtt tggacaaacc acaactagaa tgcagtgaaa aaaatgcttt
5101 atttgtgaaa tttgtgatgc tattgcttta tttgtaacca ttataagctg caataaacaa
5161 gttaacaaca acaattgcat tcattttatg tttcaggttc agggggagat gtgggaggtt
5221 ttttaaagca agtaaaacct ctacaaatgt ggtaaaatcc gataaggatc gatccgggct
5281 ggcgtaatag cgaagaggcc cgcaccgatc gcccttccca acagttgcgc agcctgaatg
5341 gcgaatggac gcgccctgta gcggcgcatt aagcgcggcg ggtgtggtgg ttacgcgcag
5401 cgtgaccgct acacttgcca gcgccctagc gcccgctcct ttcgctttct tcccttcctt
5461 tctcgccacg ttcgccggct ttccccgtca agctctaaat cgggggctcc ctttagggtt
5521 ccgatttagt gctttacggc acctcgaccc caaaaaactt gattagggtg atggttcacg
5581 tagtgggcca tcgccctgat agacggtttt tcgccctttg acgttggagt ccacgttctt
5641 taatagtgga ctcttgttcc aaactggaac aacactcaac cctatctcgg tctattcttt
5701 tgatttataa gggattttgc cgatttcggc ctattggtta aaaaatgagc tgatttaaca
5761 aaaatttaac gcgaatttta acaaaatatt aacgcttaca atttcctgat gcggtatttt
5821 ctccttacgc atctgtgcgg tatttcacac cgcatacgcg gatctgcgca gcaccatggc
5881 ctgaaataac ctctgaaaga ggaacttggt taggtacctt ctgaggcgga aagaaccagc
5941 tgtggaatgt gtgtcagtta gggtgtggaa agtccccagg ctccccagca ggcagaagta
6001 tgcaaagcat gcatctcaat tagtcagcaa ccaggtgtgg aaagtcccca ggctccccag
6061 caggcagaag tatgcaaagc atgcatctca attagtcagc aaccatagtc ccgcccctaa
6121 ctccgcccat cccgccccta actccgccca gttccgccca ttctccgccc catggctgac
6181 taattttttt tatttatgca gaggccgagg ccgcctcggc ctctgagcta ttccagaagt
6241 agtgaggagg cttttttgga ggccacggcc ggccctccgc gcctacagct caagccacat
6301 ccgaaggggg agggagccgg gagctgcgcg cggggccgcc ggggggaggg gtggcaccgc
6361 ccacgccggg cggccacgaa gggcggggca gcgggcgcgc gcgcggcggg gggaggggcc
6421 ggcgccgcgc ccgctgggaa ttggggccct agggggaggg cggaggcgcc gacgaccgcg
6481 gcacttaccg ttcgcggcgt ggcgcccggt ggtccccaag gggagggaag ggggaggcgg
6541 ggcgaggaca gtgaccggag tctcctcagc ggtggctttt ctgcttggca gcctcagcgg
6601 ctggcgccaa aaccggactc cgcccacttc ctcgcccgcc ggtgcgaggg tgtggaatcc
6661 tccagacgct gggggagggg gagttgggag cttaaaaact agtacccctt tgggaccact
6721 ttcagcagcg aactctcctg tacaccaggg gtcagttcca cagacgcggg ccaggggtgg
6781 gtcattgcgg cgtgaacaat aatttgacta gaagttgatt cgggtgtttc cggaaggggc
6841 cgagtcaatc cgccgagttg gggcacggaa aacaaaaagg gaaggctact aagatttttc
6901 tggcgggggt tatcattggc gtaactgcag ggaccacctc ccgggttgag ggggctggat
6961 ctccaggctg cggattaagc ccctcccgtc ggcgttaatt tcaaactgcg cgacgtttct
7021 cacctgcctt cgccaaggca ggggccggga ccctattcca agaggtagta actagcagga
7081 ctctagcctt ccgcaattca ttgagcgcat ttacggaagt aacgtcgggt actgtctctg
7141 gccgcaaggg tgggaggagt acgcatttgg cgtaaggtgg ggcgtagagc cttcccgcca
7201 ttggcggcgg atagggcgtt tacgcgacgg cctgacgtag cggaagacgc cttagtgggg
7261 gggaaggttc tagaaaagcg gcggcagcgg ctctagcggc agtagcagca gcgccgggtc
7321 ccgtgcggag gtgctcctcg cagagttgtt tctccagcag cggcagttct cactacagcg
7381 ccaggacgag tccggttcgt gttcgtccgc ggagatctct ctcatctcgc tcggctgcgg
7441 gaaatcgggc tgaagcgact gagtccgcga tggaggtaac gggtttgaaa tcaatgagtt
7501 attgaaaagg gcatggcgag gccgttggcg cctcagtgga agtcggccag ccgcctccgt
7561 gggagagagg caggaaatcg gaccaattca gtagcagtgg ggcttaaggt ttatgaacgg
7621 ggtcttgagc ggaggcctga gcgtacaaac agcttcccca ccctcagcct cccggcgcca
7681 tttcccttca ctgggggtgg gggatgggga gctttcacat ggcggacgct gccccgctgg
7741 ggtgaaagtg gggcgcggag gcgggacttc ttattccctt tctaaagcac gctgcttcgg
7801 gggccacggc gtctcctcgg ggatctagct tgtggaaggc tactcgaaat gtttgaccca
7861 agttaaacaa tttaaaggca atgctaccaa atactaattg agtgtatgta aacttctgac
7921 ccactgggaa tgtgatgaaa gaaataaaag ctgaaatgaa tcattctctc tactattatt
7981 ctgatatttc acattcttaa aataaagtgg tgatcctaac tgacctaaga cagggaattt
8041 ttactaggat taaatgtcag gaattgtgaa aaagtgagtt taaatgtatt tggctaaggt
8101 gtatgtaaac ttccgacttc aactgtatag ggttcctcta gctagagacg acctcgggta
8161 cccagctttt gttcccttta gtgagggtta attgcgcgct tggcgtaatc atggtcatag
8221 ctgtttcctg tgtgaaattg ttatccgctc acaattccac acaacatacg agccggaagc
8281 ataaagtgta aagcctgggg tgcctaatga gtgagctaac tcacattaat tgcgttgcgc
8341 tcactgcccg ctttccagtc gggaaacctg tcgtgccagc tgcattaatg aatcggccaa
8401 cgcgcgggga gaggcggttt gcgtattggg cgctcttccg cttcctcgct cactgactcg
8461 ctgcgctcgg tcgttcggct gcggcgagcg gtatcagctc actcaaaggc ggtaatacgg
8521 ttatccacag aatcagggga taacgcagga aagaacatgt gagcaaaagg ccagcaaaag
8581 gccaggaacc gtaaaaaggc cgcgttgctg gcgtttttcc ataggctccg cccccctgac
8641 gagcatcaca aaaatcgacg ctcaagtcag aggtggcgaa acccgacagg actataaaga
8701 taccaggcgt ttccccctgg aagctccctc gtgcgctctc ctgttccgac cctgccgctt
8761 accggatacc tgtccgcctt tctcccttcg ggaagcgtgg cgctttctca tagctcacgc
8821 tgtaggtatc tcagttcggt gtaggtcgtt cgctccaagc tgggctgtgt gcacgaaccc
8881 cccgttcagc ccgaccgctg cgccttatcc ggtaactatc gtcttgagtc caacccggta
8941 agacacgact tatcgccact ggcagcagcc actggtaaca ggattagcag agcgaggtat
9001 gtaggcggtg ctacagagtt cttgaagtgg tggcctaact acggctacac tagaaggaca
9061 gtatttggta tctgcgctct gctgaagcca gttaccttcg gaaaaagagt tggtagctct
9121 tgatccggca aacaaaccac cgctggtagc ggtggttttt ttgtttgcaa gcagcagatt
9181 acgcgcagaa aaaaaggatc tcaagaagat cctttgatct tttctacggg gtctgacgct
9241 cagtggaacg aaaactcacg ttaagggatt ttggtcatga gattatcaaa aaggatcttc
9301 acctagatcc ttttaaatta aaaatgaagt tttaaatcaa tctaaagtat atatgagtaa
9361 acttggtctg acagttacca atgcttaatc agtgaggcac ctatctcagc gatctgtcta
9421 tttcgttcat ccatagttgc ctgactcccc gtcgtgtaga taactacgat acgggagggc
9481 ttaccatctg gccccagtgc tgcaatgata ccgcgagacc cacgctcacc ggctccagat
9541 ttatcagcaa taaaccagcc agccggaagg gccgagcgca gaagtggtcc tgcaacttta
9601 tccgcctcca tccagtctat taattgttgc cgggaagcta gagtaagtag ttcgccagtt
9661 aatagtttgc gcaacgttgt tgccattgct acaggcatcg tggtgtcacg ctcgtcgttt
9721 ggtatggctt cattcagctc cggttcccaa cgatcaaggc gagttacatg atcccccatg
9781 tgtgcaaaa aagcggttag ctccttcggt cctccgatcg ttgtcagaag taagttggcc
9841 gcagtgttat cactcatggt tatggcagca ctgcataatt ctcttactgt catgccatcc
9901 gtaagatgct tttctgtgac tggtgagtac tcaaccaagt cattctgaga atagtgtatg
9961 cggcgaccga gttgctcttg cccggcgtca atacgggata ataccgcgcc acatagcaga
10021 actttaaaag tgctcatcat tggaaaacgt tcttcggggc gaaaactctc aaggatctta
10081 ccgctgttga gatccagttc gatgtaaccc actcgtgcac ccaactgatc ttcagcatct
10141 tttactttca ccagcgtttc tgggtgagca aaaacaggaa ggcaaaatgc cgcaaaaaag
10201 ggaataaggg cgacacggaa atgttgaata ctcatactct tcctttttca atattattga
10261 agcatttatc agggttattg tctcatgagc ggatacatat ttgaatgtat ttagaaaaat
10321 aaacaaatag gggttccgcg cacatttccc cgaaaagtgc cacctgacgc gccctgtagc
10381 ggcgcattaa gcgcggcggg tgtggtggtt acgcgcagcg tgaccgctac acttgccagc
10441 gccctagcgc ccgctccttt cgctttcttc ccttcctttc tcgccacgtt cgccggcttt
10501 ccccgtcaag ctctaaatcg ggggctccct ttagggttcc gatttagtgc tttacggcac
10561 ctcgacccca aaaaacttga ttagggtgat ggttcacgta gtgggccatc gccctgatag
10621 acggtttttc gccctttgac gttggagtcc acgttcttta atagtggact cttgttccaa
10681 actggaacaa cactcaaccc tatctcggtc tattcttttg atttataagg gattttgccg
10741 atttcggcct attggttaaa aaatgagctg atttaacaaa aatttaacgc gaattttaac
10801 aaaatattaa cgcttacaat ttccattcgc cattcaggct gcgcaactgt tgggaagggc
10861 gatcggtgcg ggcctcttcg ctattacgcc agctggcgaa agggggatgt gctg
<210> SEQ ID NO: 2
<211> 11475
<212> PRT
<213> Artificial Sequence
<223> Нуклеотидная последовательность рекомбинантной плазмиды pVEAL-15742,
обеспечивающая синтез и секрецию scFv-Fc антитела ADI-15742 против
вируса Эбола.
<400> 2
1 caaggcgatt aagttgggta acgccagggt tttcccagtc acgacgttgt aaaacgacgg
61 ccagtgagcg cgcgtaatac gactcactat agggcgaatt ggagctcggt tccctataca
121 gttgaagtcg gaagtttaca tacacttaag ttggagtcat taaaactcgt ttttcaacta
181 ctccacaaat ttcttgttaa caaacaatag ttttggcaag tcagttagga catctacttt
241 gtgcatgaca caagtcattt ttccaacaat tgtttacaga cagattattt cacttataat
301 tcactgtatc acaattccag tgggtcagaa gtttacatac actaagttga ctgtgccttt
361 aaacagcttg gaaaattcca gaaaatgatg tcatggcttt agaagcttca ttggcgccct
421 ccgcgcctac agctcaagcc acatccgaag ggggagggag ccgggagctg cgcgcggggc
481 cgccgggggg aggggtggca ccgcccacgc cgggcggcca cgaagggcgg ggcagcgggc
541 gcgcgcgcgg cggggggagg ggccggcgcc gcgcccgctg ggaattgggg ccctaggggg
601 agggcggagg cgccgacgac cgcggcactt accgttcgcg gcgtggcgcc cggtggtccc
661 caaggggagg gaagggggag gcggggcgag gacagtgacc ggagtctcct cagcggtggc
721 ttttctgctt ggcagcctca gcggctggcg ccaaaaccgg actccgccca cttcctcgcc
781 cgccggtgcg agggtgtgga atcctccaga cgctggggga gggggagttg ggagcttaaa
841 aactagtacc cctttgggac cactttcagc agcgaactct cctgtacacc aggggtcagt
901 tccacagacg cgggccaggg gtgggtcatt gcggcgtgaa caataatttg actagaagtt
961 gattcgggtg tttccggaag gggccgagtc aatccgccga gttggggcac ggaaaacaaa
1021 aagggaaggc tactaagatt tttctggcgg gggttatcat tggcgtaact gcagggacca
1081 cctcccgggt tgagggggct ggatctccag gctgcggatt aagcccctcc cgtcggcgtt
1141 aatttcaaac tgcgcgacgt ttctcacctg ccttcgccaa ggcaggggcc gggaccctat
1201 tccaagaggt agtaactagc aggactctag ccttccgcaa ttcattgagc gcatttacgg
1261 aagtaacgtc gggtactgtc tctggccgca agggtgggag gagtacgcat ttggcgtaag
1321 gtggggcgta gagccttccc gccattggcg gcggataggg cgtttacgcg acggcctgac
1381 gtagcggaag acgccttagt gggggggaag gttctagaaa agcggcggca gcggctctag
1441 cggcagtagc agcagcgccg ggtcccgtgc ggaggtgctc ctcgcagagt tgtttctcca
1501 gcagcggcag ttctcactac agcgccagga cgagtccggt tcgtgttcgt ccgcggagat
1561 ctctctcatc tcgctcggct gcgggaaatc gggctgaagc gactgagtcc gcgatggagg
1621 taacgggttt gaaatcaatg agttattgaa aagggcatgg cgaggccgtt ggcgcctcag
1681 tggaagtcgg ccagccgcct ccgtgggaga gaggcaggaa atcggaccaa ttcagtagca
1741 gtggggctta aggtttatga acggggtctt gagcggaggc ctgagcgtac aaacagcttc
1801 cccaccctca gcctcccggc gccatttccc ttcactgggg gtgggggatg gggagctttc
1861 acatggcgga cgctgccccg ctggggtgaa agtggggcgc ggaggcggga cttcttattc
1921 cctttctaaa gcacgctgct tcgggggcca cggcgtctcc tcggggatct tcaatattgg
1981 ccattagcca tattattcat tggttatata gcataaatca atattggcta ttggccattg
2041 catacgttgt atctatatca taatatgtac atttatattg gctcatgtcc aatatgaccg
2101 ccatgttggc attgattatt gactagttat taatagtaat caattacggg gtcattagtt
2161 catagcccat atatggagtt ccgcgttaca taacttacgg taaatggccc gcctggctga
2221 ccgcccaacg acccccgccc attgacgtca ataatgacgt atgttcccat agtaacgcca
2281 atagggactt tccattgacg tcaatgggtg gagtatttac ggtaaactgc ccacttggca
2341 gtacatcaag tgtatcatat gccaagtccg ccccctattg acgtcaatga cggtaaatgg
2401 cccgcctggc attatgccca gtacatgacc ttacgggact ttcctacttg gcagtacatc
2461 tacgtattag tcatcgctat taccatggtg atgcggtttt ggcagtacac caatgggcgt
2521 ggatagcggt ttgactcacg gggatttcca agtctccacc ccattgacgt caatgggagt
2581 ttgttttggc accaaaatca acgggacttt ccaaaatgtc gtaacaactg cgatcgcccg
2641 ccccgttgac gcaaatgggc ggtaggcgtg tacggtggga ggtctatata agcagagctc
2701 gtttagtgaa ccgtcagatc actagaagct ttattgcggt agtttatcac agttaaattg
2761 ctaacgcagt cagtgcttct gacacaacag tctcgaactt aagctgcagt gactctctta
2821 aggtagcctt gcagaagttg gtcgtgaggc actgggcagg taagtatcaa ggttacaaga
2881 caggtttaag gagaccaata gaaactgggc ttgtcgagac agagaagact cttgcgtttc
2941 tgataggcac ctattggtct tactgacatc cactttgcct ttctctccac aggtgtccac
3001 tcccagttca attacagctc ttaaggctag agtacttaat acgactcact ataggctagc
3061 ctcgagaatt cgtcctgctg cgcacgtggg aagccctggc cccggccgcc accatgatga
3121 ggcccatcgt gctggtgctg ctgttcgcca cctcagcgct ggccggcgag gtgcagctgg
3181 tggagtccgg gggaggcttg gtacagccgg gggggtccct gagactctcc tgtgcagcct
3241 ctggattcac ctttagcagc tatgccatga gctgggtccg ccaggctcca gggaaggggc
3301 tggagtgggt ctccgaaatt agcggtcttg gtggtagcac atactacgca gactccgcga
3361 agggccggtt caccatctcc agagacaatt ccaagagcac cctgtatctg caaatgaaca
3421 gcctgagagc cgaagacacg gccgtatatt actgtgcgaa agatcatcgc gtttgggcac
3481 ctggatatta ctttgaccac tggggccagg gaaccctggt cactgtgagc tctggtggtg
3541 gtggtagcgg cggcggcggc tctggtggtg gtggttccga tattgtgctg acacagtctc
3601 cttccaccct gtctgcatct gtcggagaca gagtcaccat cacttgccgg gccagtcaga
3661 gtattagtag ctggttggcc tggtatcagc agaaaccagg gaaagcccct aaactcctga
3721 tctatgatgc ctccagtttg gaaagtgggg tcccatcaag gttcagcggc agtggatctg
3781 ggacagagtt cactctcacc atcagcagcc tgcagcctga tgattttgca acttatttct
3841 gccaacagta taataggtcc cccactttcg gcggagggac caaggtggaa atcaagggtg
3901 gatccggaga caaaactcac acatgcccac cgtgcccagc acctgaactc ctggggggac
3961 cgtcagtctt cctcttcccc ccaaaaccca aggacaccct ctacatcacc cgggaacctg
4021 aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag ttcaactggt
4081 acgtggacgg cgtggaggtg cataatgcca agacaaagcc gcgggaggag cagtacaaca
4141 gcacgtaccg tgtggtcagc gtcctcaccg tcctgcacca ggactggctg aatggcaagg
4201 agtacaagtg caaggtctcc aacaaagccc tcccagcccc catcgcgaaa accatctcca
4261 aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc cgggaggaga
4321 tgaccaagaa ccaggtcagc ctgacctgcc tggtcaaagg cttctatccc agcgacatcg
4381 ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg cctcccgtgc
4441 tggactccga cggctccttc ttcctctaca gcaagctcac cgtggacaag agcaggtggc
4501 agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgaagttc cactacacgc
4561 agaagagcct ctccctgtct ccgggtaaat gagtcgagcg gttcccgccc ctctccctcc
4621 ccccccccta acgttactgg ccgaagccgc ttggaataag gccggtgtgc gtttgtctat
4681 atgttatttt ccaccatatt gccgtctttt ggcaatgtga gggcccggaa acctggccct
4741 gtcttcttga cgagcattcc taggggtctt tcccctctcg ccaaaggaat gcaaggtctg
4801 ttgaatgtcg tgaaggaagc agttcctctg gaagcttctt gaagacaaac aacgtctgta
4861 gcgacccttt gcaggcagcg gaacccccca cctggcgaca ggtgcctctg cggccaaaag
4921 ccacgtgtat aagatacacc tgcaaaggcg gcacaacccc agtgccacgt tgtgagttgg
4981 atagttgtgg aaagagtcaa atggctcacc tcaagcgtat tcaacaaggg gctgaaggat
5041 gcccagaagg taccccattg tatgggatct gatctggggc ctcggtgcac atgctttaca
5101 tgtgtttagt cgaggttaaa aaacgtctag gccccccgaa ccacggggac gtggttttcc
5161 tttgaaaaac acgatgataa tatggccaag ttgaccagtg ccgttccggt gctcaccgcg
5221 cgcgacgtcg ccggagcggt cgagttctgg accgaccggc tcgggttctc ccgggacttc
5281 gtggaggacg acttcgccgg tgtggtccgg gacgacgtga ccctgttcat cagcgcggtc
5341 caggaccagg tggtgccgga caacaccctg gcctgggtgt gggtgcgcgg cctggacgag
5401 ctgtacgccg agtggtcgga ggtcgtgtcc acgaacttcc gggacgcctc cgggccggcc
5461 atgaccgaga tcggcgagca gccgtggggg cgggagttcg ccctgcgcga cccggccggc
5521 aactgcgtgc acttcgtggc cgaggagcag gactgagcgg ccgcttccct ttagtgaggg
5581 ttaatgcttc gagcagacat gataagatac attgatgagt ttggacaaac cacaactaga
5641 atgcagtgaa aaaaatgctt tatttgtgaa atttgtgatg ctattgcttt atttgtaacc
5701 attataagct gcaataaaca agttaacaac aacaattgca ttcattttat gtttcaggtt
5761 cagggggaga tgtgggaggt tttttaaagc aagtaaaacc tctacaaatg tggtaaaatc
5821 cgataaggat cgatccgggc tggcgtaata gcgaagaggc ccgcaccgat cgcccttccc
5881 aacagttgcg cagcctgaat ggcgaatgga cgcgccctgt agcggcgcat taagcgcggc
5941 gggtgtggtg gttacgcgca gcgtgaccgc tacacttgcc agcgccctag cgcccgctcc
6001 tttcgctttc ttcccttcct ttctcgccac gttcgccggc tttccccgtc aagctctaaa
6061 tcgggggctc cctttagggt tccgatttag tgctttacgg cacctcgacc ccaaaaaact
6121 tgattagggt gatggttcac gtagtgggcc atcgccctga tagacggttt ttcgcccttt
6181 gacgttggag tccacgttct ttaatagtgg actcttgttc caaactggaa caacactcaa
6241 ccctatctcg gtctattctt ttgatttata agggattttg ccgatttcgg cctattggtt
6301 aaaaaatgag ctgatttaac aaaaatttaa cgcgaatttt aacaaaatat taacgcttac
6361 aatttcctga tgcggtattt tctccttacg catctgtgcg gtatttcaca ccgcatacgc
6421 ggatctgcgc agcaccatgg cctgaaataa cctctgaaag aggaacttgg ttaggtacct
6481 tctgaggcgg aaagaaccag ctgtggaatg tgtgtcagtt agggtgtgga aagtccccag
6541 gctccccagc aggcagaagt atgcaaagca tgcatctcaa ttagtcagca accaggtgtg
6601 gaaagtcccc aggctcccca gcaggcagaa gtatgcaaag catgcatctc aattagtcag
6661 caaccatagt cccgccccta actccgccca tcccgcccct aactccgccc agttccgccc
6721 attctccgcc ccatggctga ctaatttttt ttatttatgc agaggccgag gccgcctcgg
6781 cctctgagct attccagaag tagtgaggag gcttttttgg aggccacggc cggccctccg
6841 cgcctacagc tcaagccaca tccgaagggg gagggagccg ggagctgcgc gcggggccgc
6901 cggggggagg ggtggcaccg cccacgccgg gcggccacga agggcggggc agcgggcgcg
6961 cgcgcggcgg ggggaggggc cggcgccgcg cccgctggga attggggccc tagggggagg
7021 gcggaggcgc cgacgaccgc ggcacttacc gttcgcggcg tggcgcccgg tggtccccaa
7081 ggggagggaa gggggaggcg gggcgaggac agtgaccgga gtctcctcag cggtggcttt
7141 tctgcttggc agcctcagcg gctggcgcca aaaccggact ccgcccactt cctcgcccgc
7201 cggtgcgagg gtgtggaatc ctccagacgc tgggggaggg ggagttggga gcttaaaaac
7261 tagtacccct ttgggaccac tttcagcagc gaactctcct gtacaccagg ggtcagttcc
7321 acagacgcgg gccaggggtg ggtcattgcg gcgtgaacaa taatttgact agaagttgat
7381 tcgggtgttt ccggaagggg ccgagtcaat ccgccgagtt ggggcacgga aaacaaaaag
7441 ggaaggctac taagattttt ctggcggggg ttatcattgg cgtaactgca gggaccacct
7501 cccgggttga gggggctgga tctccaggct gcggattaag cccctcccgt cggcgttaat
7561 ttcaaactgc gcgacgtttc tcacctgcct tcgccaaggc aggggccggg accctattcc
7621 aagaggtagt aactagcagg actctagcct tccgcaattc attgagcgca tttacggaag
7681 taacgtcggg tactgtctct ggccgcaagg gtgggaggag tacgcatttg gcgtaaggtg
7741 gggcgtagag ccttcccgcc attggcggcg gatagggcgt ttacgcgacg gcctgacgta
7801 gcggaagacg ccttagtggg ggggaaggtt ctagaaaagc ggcggcagcg gctctagcgg
7861 cagtagcagc agcgccgggt cccgtgcgga ggtgctcctc gcagagttgt ttctccagca
7921 gcggcagttc tcactacagc gccaggacga gtccggttcg tgttcgtccg cggagatctc
7981 tctcatctcg ctcggctgcg ggaaatcggg ctgaagcgac tgagtccgcg atggaggtaa
8041 cgggtttgaa atcaatgagt tattgaaaag ggcatggcga ggccgttggc gcctcagtgg
8101 aagtcggcca gccgcctccg tgggagagag gcaggaaatc ggaccaattc agtagcagtg
8161 gggcttaagg tttatgaacg gggtcttgag cggaggcctg agcgtacaaa cagcttcccc
8221 accctcagcc tcccggcgcc atttcccttc actgggggtg ggggatgggg agctttcaca
8281 tggcggacgc tgccccgctg gggtgaaagt ggggcgcgga ggcgggactt cttattccct
8341 ttctaaagca cgctgcttcg ggggccacgg cgtctcctcg gggatctagc ttgtggaagg
8401 ctactcgaaa tgtttgaccc aagttaaaca atttaaaggc aatgctacca aatactaatt
8461 gagtgtatgt aaacttctga cccactggga atgtgatgaa agaaataaaa gctgaaatga
8521 atcattctct ctactattat tctgatattt cacattctta aaataaagtg gtgatcctaa
8581 ctgacctaag acagggaatt tttactagga ttaaatgtca ggaattgtga aaaagtgagt
8641 ttaaatgtat ttggctaagg tgtatgtaaa cttccgactt caactgtata gggttcctct
8701 agctagagac gacctcgggt acccagcttt tgttcccttt agtgagggtt aattgcgcgc
8761 ttggcgtaat catggtcata gctgtttcct gtgtgaaatt gttatccgct cacaattcca
8821 cacaacatac gagccggaag cataaagtgt aaagcctggg gtgcctaatg agtgagctaa
8881 ctcacattaa ttgcgttgcg ctcactgccc gctttccagt cgggaaacct gtcgtgccag
8941 ctgcattaat gaatcggcca acgcgcgggg agaggcggtt tgcgtattgg gcgctcttcc
9001 gcttcctcgc tcactgactc gctgcgctcg gtcgttcggc tgcggcgagc ggtatcagct
9061 cactcaaagg cggtaatacg gttatccaca gaatcagggg ataacgcagg aaagaacatg
9121 tgagcaaaag gccagcaaaa ggccaggaac cgtaaaaagg ccgcgttgct ggcgtttttc
9181 cataggctcc gcccccctga cgagcatcac aaaaatcgac gctcaagtca gaggtggcga
9241 aacccgacag gactataaag ataccaggcg tttccccctg gaagctccct cgtgcgctct
9301 cctgttccga ccctgccgct taccggatac ctgtccgcct ttctcccttc gggaagcgtg
9361 gcgctttctc atagctcacg ctgtaggtat ctcagttcgg tgtaggtcgt tcgctccaag
9421 ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct gcgccttatc cggtaactat
9481 cgtcttgagt ccaacccggt aagacacgac ttatcgccac tggcagcagc cactggtaac
9541 aggattagca gagcgaggta tgtaggcggt gctacagagt tcttgaagtg gtggcctaac
9601 tacggctaca ctagaaggac agtatttggt atctgcgctc tgctgaagcc agttaccttc
9661 ggaaaaagag ttggtagctc ttgatccggc aaacaaacca ccgctggtag cggtggtttt
9721 tttgtttgca agcagcagat tacgcgcaga aaaaaaggat ctcaagaaga tcctttgatc
9781 ttttctacgg ggtctgacgc tcagtggaac gaaaactcac gttaagggat tttggtcatg
9841 agattatcaa aaaggatctt cacctagatc cttttaaatt aaaaatgaag ttttaaatca
9901 atctaaagta tatatgagta aacttggtct gacagttacc aatgcttaat cagtgaggca
9961 cctatctcag cgatctgtct atttcgttca tccatagttg cctgactccc cgtcgtgtag
10021 ataactacga tacgggaggg cttaccatct ggccccagtg ctgcaatgat accgcgagac
10081 ccacgctcac cggctccaga tttatcagca ataaaccagc cagccggaag ggccgagcgc
10141 agaagtggtc ctgcaacttt atccgcctcc atccagtcta ttaattgttg ccgggaagct
10201 agagtaagta gttcgccagt taatagtttg cgcaacgttg ttgccattgc tacaggcatc
10261 gtggtgtcac gctcgtcgtt tggtatggct tcattcagct ccggttccca acgatcaagg
10321 cgagttacat gatcccccat gttgtgcaaa aaagcggtta gctccttcgg tcctccgatc
10381 gttgtcagaa gtaagttggc cgcagtgtta tcactcatgg ttatggcagc actgcataat
10441 tctcttactg tcatgccatc cgtaagatgc ttttctgtga ctggtgagta ctcaaccaag
10501 tcattctgag aatagtgtat gcggcgaccg agttgctctt gcccggcgtc aatacgggat
10561 aataccgcgc cacatagcag aactttaaaa gtgctcatca ttggaaaacg ttcttcgggg
10621 cgaaaactct caaggatctt accgctgttg agatccagtt cgatgtaacc cactcgtgca
10681 cccaactgat cttcagcatc ttttactttc accagcgttt ctgggtgagc aaaaacagga
10741 aggcaaaatg ccgcaaaaaa gggaataagg gcgacacgga aatgttgaat actcatactc
10801 ttcctttttc aatattattg aagcatttat cagggttatt gtctcatgag cggatacata
10861 tttgaatgta tttagaaaaa taaacaaata ggggttccgc gcacatttcc ccgaaaagtg
10921 ccacctgacg cgccctgtag cggcgcatta agcgcggcgg gtgtggtggt tacgcgcagc
10981 gtgaccgcta cacttgccag cgccctagcg cccgctcctt tcgctttctt cccttccttt
11041 ctcgccacgt tcgccggctt tccccgtcaa gctctaaatc gggggctccc tttagggttc
11101 cgatttagtg ctttacggca cctcgacccc aaaaaacttg attagggtga tggttcacgt
11161 agtgggccat cgccctgata gacggttttt cgccctttga cgttggagtc cacgttcttt
11221 aatagtggac tcttgttcca aactggaaca acactcaacc ctatctcggt ctattctttt
11281 gatttataag ggattttgcc gatttcggcc tattggttaa aaaatgagct gatttaacaa
11341 aaatttaacg cgaattttaa caaaatatta acgcttacaa tttccattcg ccattcaggc
11401 tgcgcaactg ttgggaaggg cgatcggtgc gggcctcttc gctattacgc cagctggcga
11461 aagggggatg tgctg
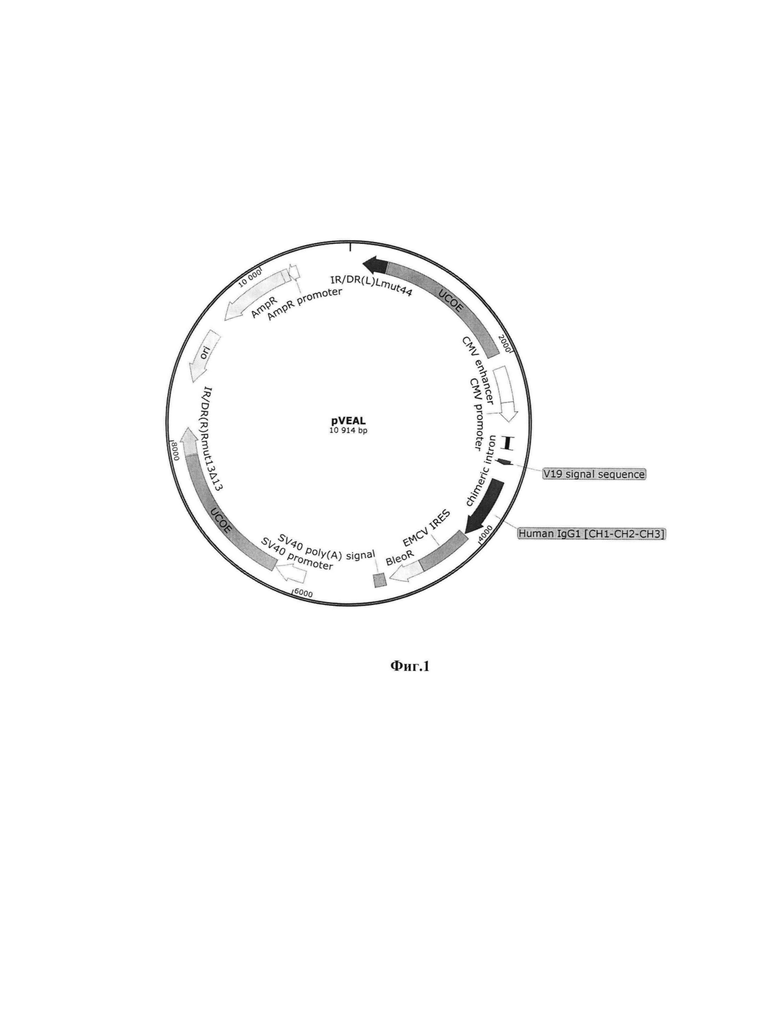
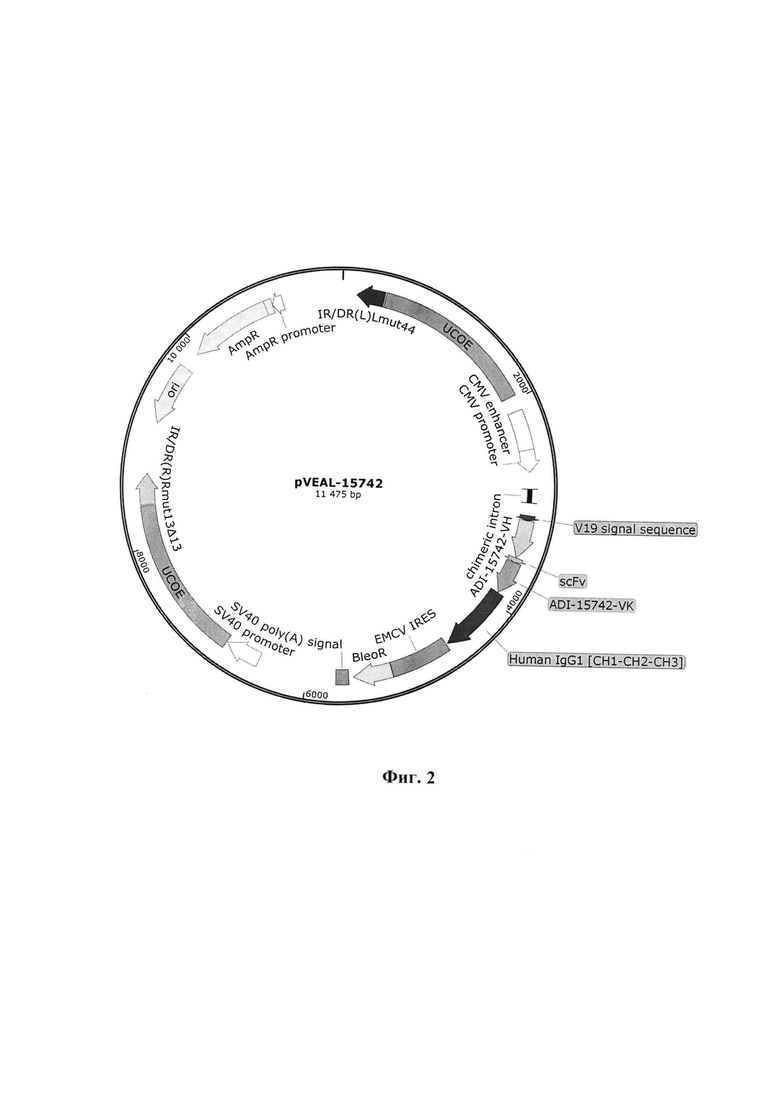
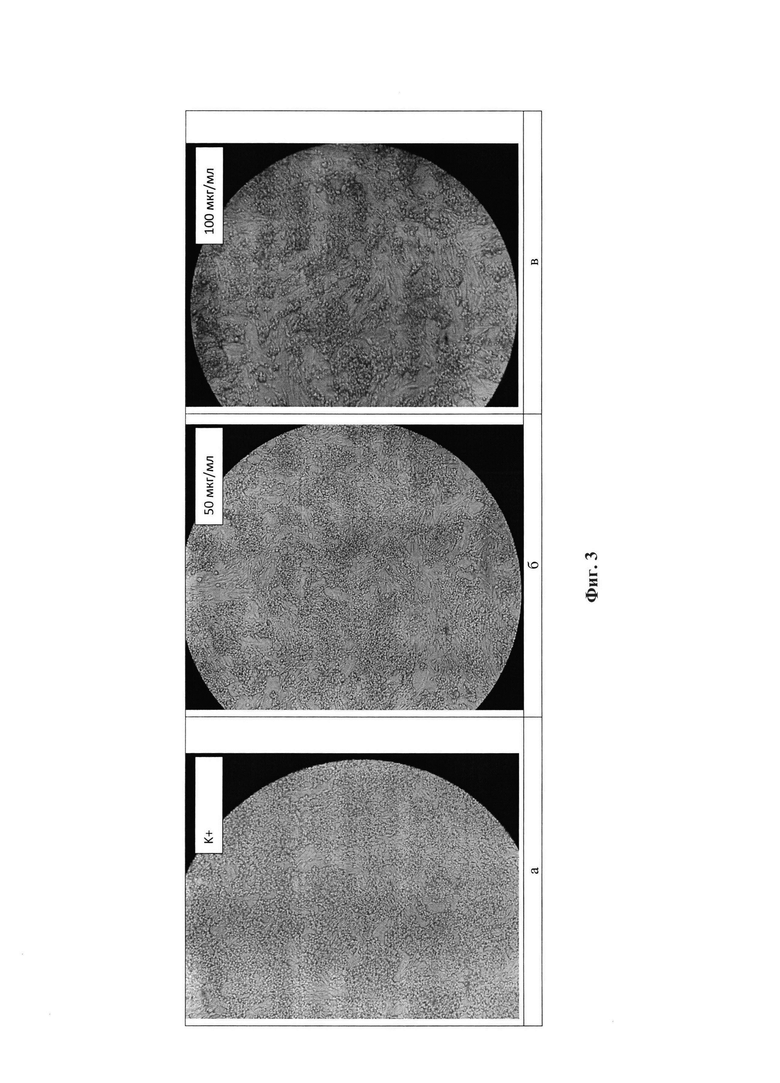
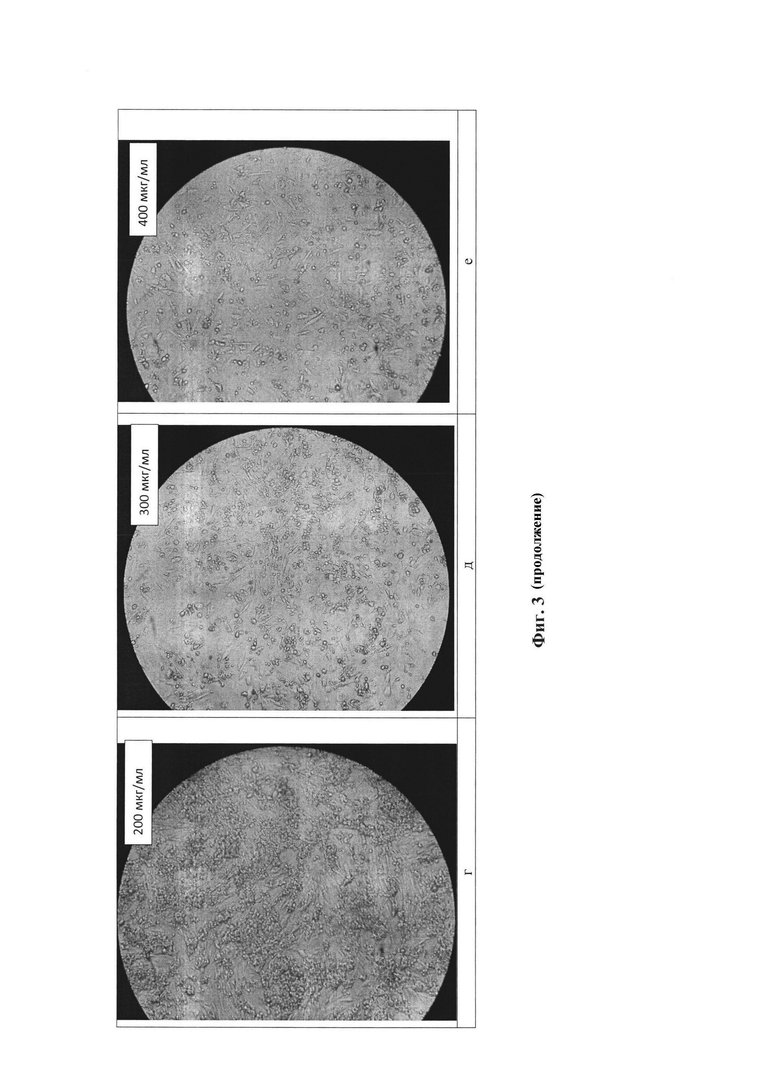
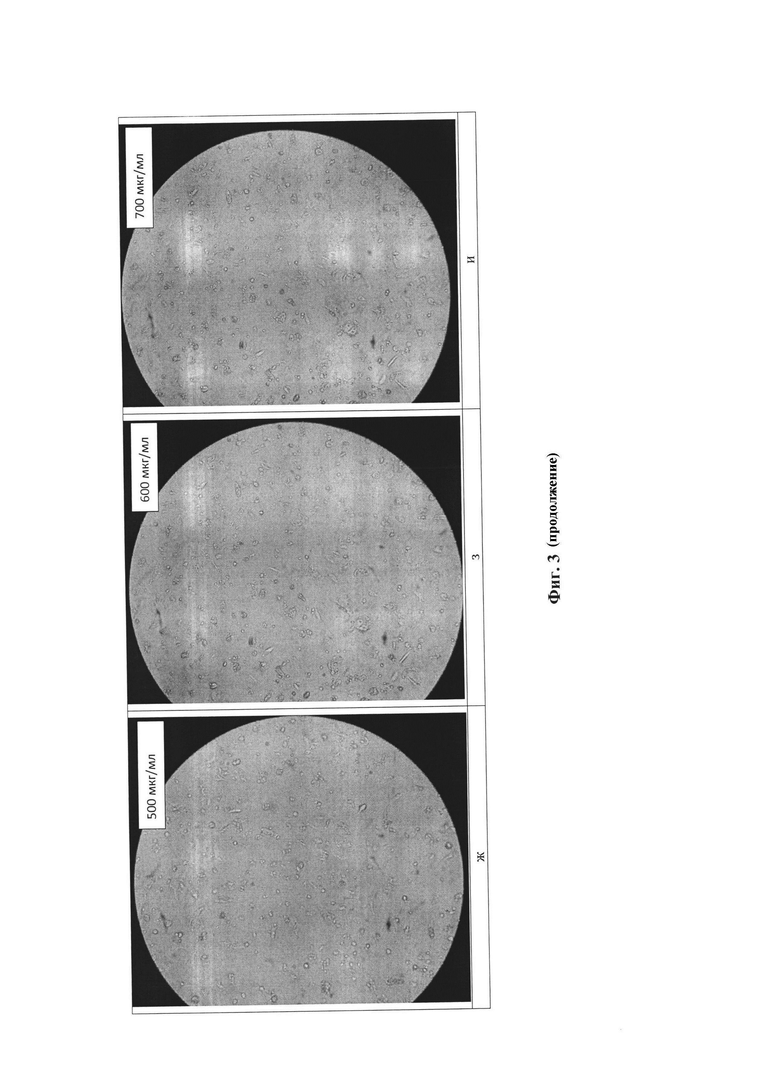
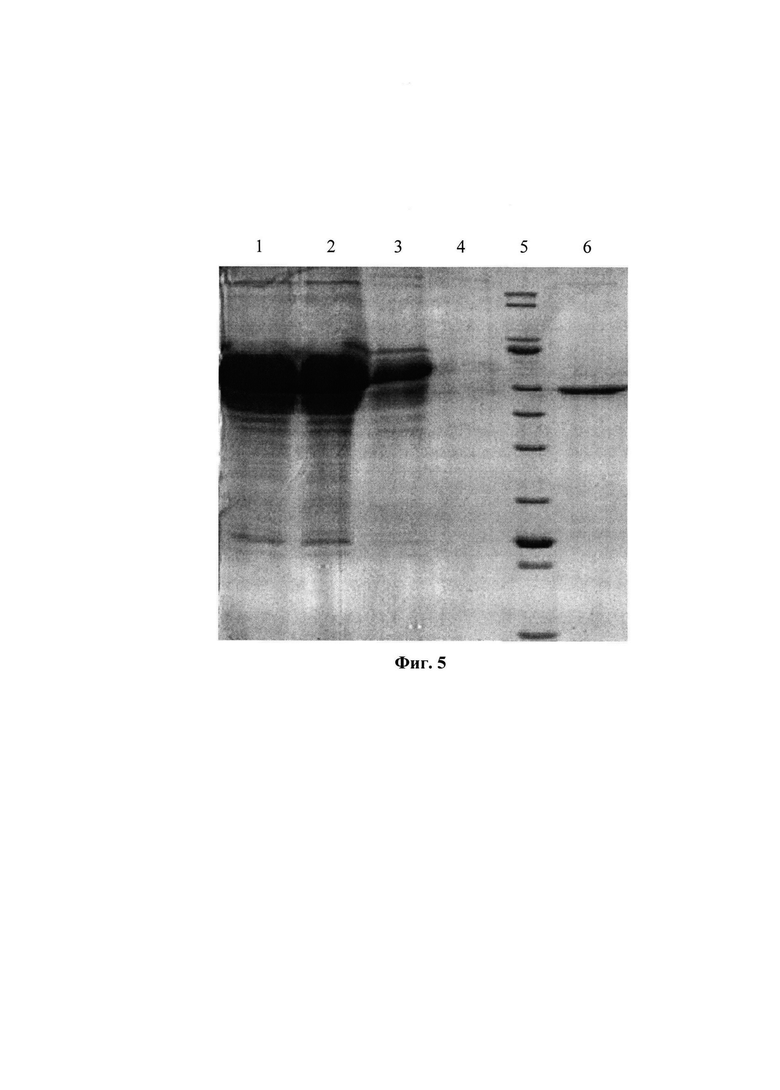
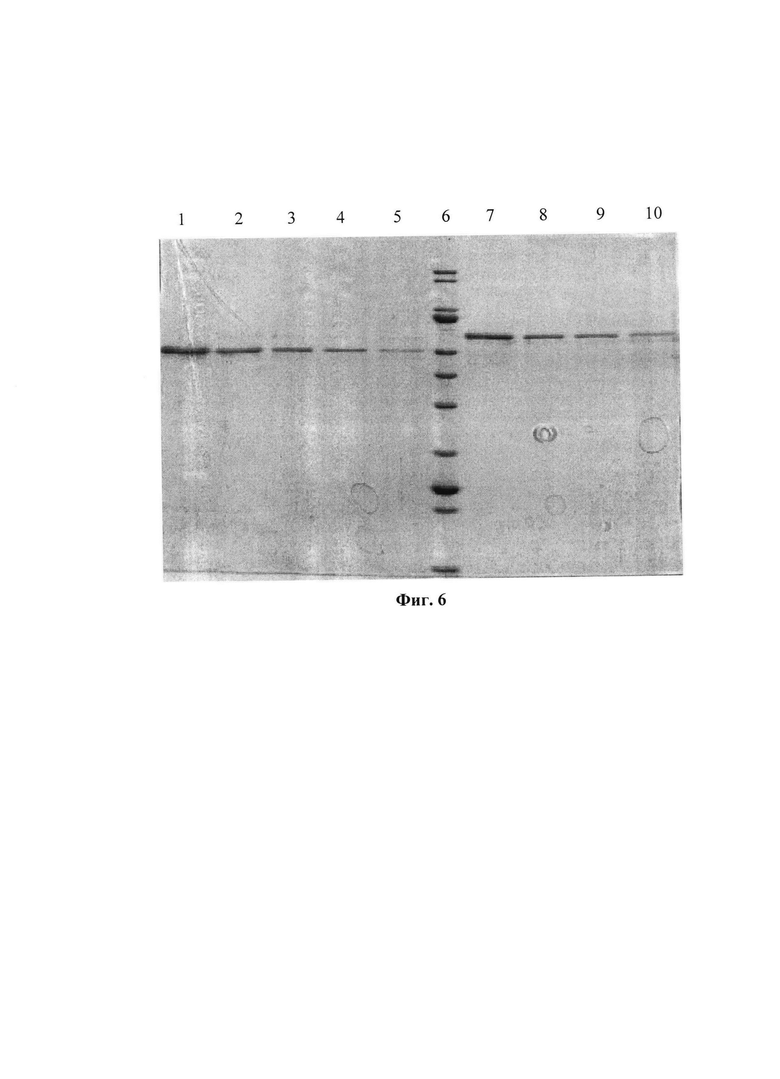
Изобретение относится к биотехнологии. Описан универсальный интеграционный вектор pVEAL и рекомбинантая плазмида pVEAL-15742, обеспечивающая синтез и секрецию scFv-Fc антител ADI-15742, способных нейтрализовать вирус Эбола в клетках млекопитающих. Универсальный интеграционный вектор pVEAL служит для создания целевых плазмидных конструкций, обеспечивающих синтез и секрецию scFv-Fc моноклональных антител, имеет нуклеотидную последовательность SEQ ID No. 1 и содержит элементы в соответствии с физической и генетической картой, представленной на Фиг. 1. Плазмидная генетическая конструкция pVEAL-15742, обеспечивающая синтез и секрецию целевого белка scFv-Fc антитела ADI 15742 против вируса Эбола, имеет нуклеотидную последовательность SEQ ID No. 2 и содержит элементы в соответствии с физической и генетической картой, представленной на Фиг. 2. Техническим результатом заявляемого изобретения является создание универсального интеграционного вектора pVEAL и плазмидной генетической конструкции pVEAL-15742, обеспечивающей синтез и секрецию целевого белка scFv-Fc антитела ADI 15742 против вируса Эбола в клетках млекопитающих СНО-K1, полученного с использованием указанного универсального вектора pVEAL. 2 н.п. ф-лы, 6 ил., 5 пр.
1. Универсальный интеграционный вектор pVEAL, служащий для создания плазмидной генетической конструкции pVEAL-15742, имеющий нуклеотидную последовательность SEQ ID No. 1 и содержащий в соответствии с физической и генетической картой, представленной на Фиг. 1, следующие элементы:
- участок начала репликации ori (8616-9204);
- последовательности IR/DR (L/R), являющиеся частью системы на основе транспозазы SB100 (139-403, 7832-8127);
- последовательности UCOE, предотвращающие хромосомное «замалчивание» рекомбинантной экспрессионной кассеты (416 - 1549, 6272-7820);
- CMV Enhancer (2108-2487), CMV промотор (2488-2699) - сильный наиболее часто используемый промотор в генно-терапевтических конструкциях;
- Chimeric intron (2860-2992), увеличивающий эффективность CMV промотора;
- лидерный пептид V19, обеспечивающий экспорт белка из клетки (3114-3164);
- место для встройки вариабельных фрагментов тяжелой и легкой цепей антитела, соединенных пептидным линкером, по сайтам гидролиза AfeI (3157), ВаmHI (3339);
- константную цепь антитела человека IgG1 (СН1-СН2-СН3) (3348-4031) с мутациями M252Y, S254T, Т256Е, Е333А, H433K/N434F, которые увеличивают период полураспада антитела за счет взаимодействия с неонатальными рецепторами;
- участок внутренней посадки рибосомы EMCV IRES (4046-4620);
- последовательность Bleo, кодирующую фактор устойчивости к антибиотику блеомицину (4625-4995);
- последовательность SV40 poly(A) signal, необходимую для стабилизации мРНК-транскриптов (5042-5163);
- SV40 промотор (5921-6262), обеспечивающий автономную репликацию в клетках млекопитающих, комбинация с CMV промотором существенно увеличивает выход белка;
- ген устойчивости к антибиотику ампициллин AmpR и бактериальный промотор гена устойчивости к ампициллину, позволяющие проводить препаративную наработку плазмиды в E.coli (9375-10235).
2. Плазмидная генетическая конструкция pVEAL-15742, обеспечивающая синтез и секрецию целевого белка scFv-Fc антитела ADI 15742 против вируса Эбола в клетках млекопитающих, имеющая нуклеотидную последовательность SEQ ID No. 2 и содержащая в соответствии с физической и генетической картой, представленной на Фиг. 2, следующие элементы:
- участок начала репликации ori (8616-9204);
- последовательности IR/DR (L/R), являющиеся частью системы на основе транспозазы SB100 (139-403, 8393-8688);
- последовательности UCOE, предотвращающие хромосомное «замалчивание» рекомбинантной экспрессионной кассеты (416 - 1549, 6833-8381);
- CMV Enhancer (2108-2487), CMV промотор (2488-2699) - сильный наиболее часто используемый промотор в генно-терапевтических конструкциях;
- Chimeric intron (2860-2992), увеличивающий эффективность CMV промотора;
- лидерный пептид V19, обеспечивающий экспорт белка из клетки (3114-3164);
- ADI-15742-VH, последовательность, кодирующая вариабельный фрагмент тяжелой цепи антитела 15742 (3165-3533);
- ADI-15742-VK, последовательность, кодирующая вариабельный фрагмент легкой цепи антитела 15742 (3579-3903);
- константную цепь антитела человека IgG1 (СН1-СН2-СН3) (3909-4592) с мутациями M252Y, S254T, Т256Е, Е333А, H433K/N434F, которые увеличивают период полураспада антитела за счет взаимодействия с неонатальными рецепторами;
- участок внутренней посадки рибосомы EMCV IRES (4607-5181);
- последовательность Bleo, кодирующую фактор устойчивости к антибиотику блеомицину (5186-5556);
- последовательность SV40 poly(A) signal, необходимую для стабилизации мРНК-транскриптов (5603-5724);
- SV40 промотор (6482-6823), обеспечивающий автономную репликацию в клетках млекопитающих, комбинация с CMV промотором существенно увеличивает выход белка;
- ген устойчивости к антибиотику ампициллин AmpR и бактериальный промотор гена устойчивости к ампициллину, позволяющие проводить препаративную наработку плазмиды в E.coli (9936-10796).
| Искусственные гены, кодирующие белки-иммуногены EV.CTL и EV.Th, рекомбинантные плазмидные ДНК pEV.CTL и pEV.Th, обеспечивающие экспрессию искусственных генов, и искусственные Т-клеточные полиэпитопные белки-иммуногены EV.CTL и EV.Th, содержащие эпитопы антигенов вируса Эбола, используемые для создания вакцины против вируса Эбола | 2018 |
|
RU2713723C1 |
| WO 2019025984 A1, 07.02.2019 | |||
| WO 2020132382 A1, 25.06.2020. | |||

