Изобретение относится к универсальному рекомбинантному вектору pBuc и плазмидной генетической конструкции pBuc-7В4, обеспечивающей синтез и секрецию полноразмерных рекомбинантных белков - моноклональных антител, в частности, антитела 7B4 против вируса Западного Нила, в клетках яичника китайского хомяка (CHO) и полученная с использованием указанного вектора pBuc и может быть использовано в медицине и биотехнологии. Данные белки будут обладать терапевтическим потенциалом. Также их можно будет использовать в научно-исследовательских целях.
Уровень техники
За последние три десятилетия моноклональные антитела (MAb) претерпели поразительную трансформацию из научных инструментов в мощные помощники для человека в медицинской практике. С тех пор, как MAb были впервые выведены на рынок в 90-х годах, было разработано множество стратегий для оптимизации их структуры и функций (Kennedy P. J. et al. Monoclonal antibodies: technologies for early discovery and engineering. Critical reviews in biotechnology. 2018; 38(3): 394-408). Первые MAb были получены с помощью гибридомной технологии. Исходно эта технология применялась только для получения мышиных антител. Позднее были созданы мыши, трансгенные по генам иммуноглобулинов человека, с помощью которых стало возможным получение полностью человеческих MAb. Несмотря на свой солидный возраст, гибридомная технология продолжает оставаться одним из основных методов получения MAb. Однако этот метод имеет некоторые существенные недостатки, главный из которых - низкая вероятность гибридизации клеток-партнеров (10–4–10–5). Вследствие этого получение гибридом к слабоиммуногенным белкам происходит с низкой эффективностью. Кроме того, использование мышей, трансгенных по генам Ig, строго лицензируется и требует больших финансовых затрат.
Альтернативным подходом к получению MAb стали различные комбинаторные методы: фаговый, ДНК-, РНК-, а также дрожжевой дисплеи. В основе комбинаторных методов лежит использование обширных библиотек генов вариабельных фрагментов иммуноглобулинов. Одним из распространенных дисплейных методов является метод фагового дисплея, с помощью которого получено несколько иммунотерапевтических MAb, в том числе адалимумаб, ингибирующий связывание фактора некроза опухоли со своим рецептором. Однако успехи данного подхода в сравнении с гибридомной технологией выглядят более чем скромными. Недостаток комбинаторных методов заключается в том, что получаемые антитела не проходят стадию селекции in vivo. Кроме того, при создании антител зачастую наблюдается нефизиологическое спаривание цепей Ig, то есть спаривание, которое никогда не встречается при естественном иммунном ответе. С этой точки зрения реагенты, полученные комбинаторными методами, можно рассматривать как антитело миметики, которые при длительном применении вызывают образование в организме человека нейтрализующих антител (Лушова А. А. и др. Новое поколение методов получения человеческих моноклональных антител. Молекулярная биология. 2017; 51(6): 899-906).
Поиски разработка новых антител получили толчок, когда технология моноклональных антител(антитело, полученное из одного клона B-клеток) была объединена с технологией рекомбинантной ДНК. Этот подход позволяет отбирать высоконейтрализующие антитела из очень большого пула антител (Kennedy P. J. et al. Monoclonal antibodies: technologies for early discovery and engineering. Critical reviews in biotechnology. 2018; 38(3): 394-408). Рекомбинантные технологии позволяют изменять биохимические и иммунохимические свойства антител, в том числе и изменять (увеличивать) аффинность антител. Несомненным достоинством рекомбинантных технологий является то, что с их помощью можно не только получать рекомбинантные аналоги уже существующих антител, но и создавать антитела, специфичные к различным антигенам, de novo (Альтшулер Е. П. и др. Получение рекомбинантных антител и способы увеличения их аффинности. Усп. биол. химии. 2010; 50: 203-258).
Благодаря технологии химеризации и гуманизации антител, и переходу к использованию полностью человеческих моноклональных антител, количество одобренных моноклональных антител, доступных на рынке для лечения различных заболеваний, резко возросло. По состоянию на декабрь 2019 года 79 терапевтических MAb были одобрены FDA США, но все еще существует значительный потенциал роста (Lu R. M. et al. Development of therapeutic antibodies for the treatment of diseases. Journal of biomedical science. 2020; 27(1); 1-30).Моноклональные антитела нашли широкое применение при лечении онкологических, аутоиммунных, инфекционных и некоторых других заболеваний.
Антитела представляют собой белки, продуцируемые В-лимфоцитами, которые обладают способностью связываться со своей молекулярной мишенью с высокой аффинностью и специфичностью. В классическом варианте антитело состоит из двух тяжелых цепей и двух легких цепей, и каждая из идентичных цепей содержит константную и вариабельную области. Четыре цепи соединены дисульфидными мостиками, образуют классическую Y-образную форму. В одном мономере антитела есть две антигенсвязывающие области (Fab)и одна константная область(Fc). Гибкая шарнирная область соединяет каждое из плеч Fab с фрагментом Fc, обеспечивая эффективное связывание даже с антигенами имеющих разные геометрические характеристики (Kennedy P. J. et al. Monoclonal antibodies: technologies for early discovery and engineering. Critical reviews in biotechnology. 2018; 38(3): 394-408).
Несмотря на высокие производственные затраты из-за дорогостоящей среды и требований к условиям культивирования, сегодня 60-70% фармацевтических рекомбинантных белков и все одобренные в настоящее время терапевтические антитела производятся при помощи линий клеток млекопитающих. Успех систем продукции клеток млекопитающих обусловлен их механизмом секреции и посттрансляционными модификациями, которые позволяют получать антитела, неотличимые от антител, вырабатываемых в организме человека. Более того, все еще продолжающийся прогресс в технологии культивирования клеток млекопитающих уже достиг уровней продукции антител около 5 г/л, что в 10 раз выше, чем они были несколько лет назад. Основными параметрами, ответственными за это развитие, являются улучшенное поколение линий клеток-продуцентов и длительное культивирование при очень высокой плотности клеток во время продукции (Schirrmann T. еt al. Production systems for recombinant antibodies. Front Biosci. 2008; 13(13): 4576-94). Тем не менее, линии клеток млекопитающих все еще находятся под критическим наблюдением в отношении неизвестного вирусного заражения и высвобождения факторов с онкогенным или патогенным потенциалом. Для снижения затрат, а также рисков заражения патогенами или агентами губчатой энцефалопатии крупного рогатого скота были разработаны среды без добавления сыворотки животных или компонентов сыворотки. Более того, все еще продолжаются исследования в области оптимизации процессов и генной инженерии производственных линий клеток в отношении гомогенности гликозилирования, а также улучшения метаболизма, снижения апоптоза и индуцибельной остановки клеточного цикла для продления продукции белка (Schirrmann T. еt al. Production systems for recombinant antibodies. Front Biosci. 2008; 13(13): 4576-94; Butler M. Animal cell cultures: recent achievements and perspectives in the production of biopharmaceuticals. Applied microbiology and biotechnology. 2005; 68(3): 283-291).
Важные достижения в области инженерии антител, сделанные за последнее десятилетие, повысили безопасность и эффективность терапевтических антител. Эти разработки, наряду с более глубоким пониманием иммуномодулирующих свойств антител, проложили путь для следующего поколения новых и улучшенных лекарств на основе антител для лечения заболеваний человека(Lu R. M. et al. Development of therapeutic antibodies for the treatment of diseases. Journal of biomedical science. 2020; 27(1); 1-30).В этой связи очень важное значение приобретает развитие и совершенствование различных методов продукции MAb.
Ближайшие аналоги
Известна американская патентная заявка, в которой описывается система экспрессионных векторов, обеспечивающая получение рекомбинантных белков в клетках млекопитающих (US 20110097798, МПК С12N15/63, опубл.28.04.2011 г.). В изобретении делается акцент на добавлении или модификации цис-регуляторных элементов ДНК экспрессирущих плазмид. Цис-регуляторные элементы предотвращают замалчивание встроенного в хромосому гена, что позволяет существенно увеличить его экспрессию. В данном случае авторы использовали АТ-богатые последовательности matrix attachment region (MAR), фланкирующие транскрибируемый ген. Также конструкции содержат CMV/SFH/CAG/CEFR/EFHT промоторы, FMDVIRES/GTXIRES, SV40 Ter/BGH Ter, различные маркеры для селекции (неомицин, зеоцин, пуромицин, гигромицин Б). Основной задачей авторов патента было сравнение эффективности сконструированных векторов для синтеза и секреции некоторых белков в различных клеточных линиях.
Однако использование невирусных плазмидных векторов не позволяет добиться высокой скорости доставки векторов к ядрам клеток-мишеней и интеграции трансгенов, что является недостатком данного изобретения.
Наиболее близким аналогом (прототипом) является универсальный вектор, описанный в работе (CN105255895, МПК C12N15/85, опубл. 20.01.2016 г.) «MAR транскрипционная регуляция и контрольный элемент для повышения уровня экспрессии белка в эукариотической клеточной линии и экспрессионная система для эукариотической клеточной линии». Предложенная авторами система экспрессии обеспечивает высокий выход белка, экспрессия является стабильной и длительной. Вектор создан на основе транспозазы SB, что позволяет легко интегрировать нужную последовательность в геном клетки. В качестве цис-регуляторного элемента выступает matrix attachment region - MAR2. Вектор содержит цитомегаловирусный промотр CMV, тяжелая и легкая цепи антитела соединены с помощью сайта внутренней посадки рибосомы или «IRES», который описывает последовательность, функционально обеспечивающую инициацию трансляции и позволяющую двум цистронам быть транслированными с одного транскрипта в животной клетке.
Недостатками данного изобретения являются отсутствие сильного лидерного пептида, обеспечивающего секрецию белка из клетки, применение не самых эффективных цис-регуляторных последовательностей (MAR2) и эукариотического промотора (CMV). Стоит отметить, что высокий выход целевого белка обеспечивается, скорее всего, условиями культивирования белка, а именно наработкой в суспензионной культуре.
Прототипом рекомбинантной плазмиды, обеспечивающей синтез полноразмерного антитела против вируса Западного Нила является рекомбинантная конструкция, описанная в патентной заявке США (US 20160024189, МПК А61К39/12, опубл. 28.01.2016 г.). Достоинством данного изобретения является использование сигнала полиаденилирования бычьего гормона роста (BGH poly A region), который представляет собой специализированную последовательность терминации для экспрессии белка в эукариотических клетках. Эта последовательность ДНК полезна для получения фармацевтических белков и пептидов в культуре, увеличения экспрессии генов при генной терапии и продукции белка у трансгенных животных (F.C. Rottman et al. Structure and Regulated Expression of Bovine Prolactin and Bovine Growth Hormone Genes. Advances in experimental medicine and biology. 1986; 205: 281-299).
Недостатками генетической конструкции в прототипе является наличие одинаковых цитомегаловирусных промоторов перед легкой и тяжелой цепью, отсутствие цис-регуляторных элементов. Это может привести к снижению целевого выхода антител.
Техническим результатом заявляемого изобретения является создание универсального интеграционного вектора pBuc и плазмидной генетической конструкции pBuc-7B4, обеспечивающая синтез и секрецию антитела 7B4 против вируса Западного Нила в клетках животных и полученная с использованием указанного вектора pBuc, который лишен недостатков прототипов.
Указанный технический результат достигается тем, что универсальный интеграционный вектор pBuc создания целевых плазмидных конструкций, обеспечивающих синтез и секрецию моноклональных антител, имеет нуклеотидную последовательность SEQ ID No. 1 и содержит в соответствии с физической и генетической картой, представленной на Фиг. 1, следующие элементы:
1. Участок начала репликации ori (координаты с 9663 по 10251 п.н.);
2.CHEF-1aпромотор (координаты с 1981 по 3477 п.н.), обеспечивает автономную репликацию в клетках млекопитающих.
3. GL последовательность (координаты с 4410 по 4460 п.н.), кодирующую лидерный пептид люциферазы Gaussia, обеспечивающей экспорт белка из клетки;
4. 176 последовательность (координаты с 3483 по 3545 п.н.), кодирующую гибридный лидерный пептид люциферазы (Cypridina noctiluca) и фиброина (Dendrolimus spectabilis), обеспечивающий экспорт белка из клетки;
5. Linker VL (координаты с 3546 по 4019 п.н.), синтезированную последовательность легкой цепи, фланкируемую уникальными сайтами рестрикции Bsa29I, AsuNHI;
6. EMCV IRES(координаты с 5936 по 6510 п.н.), участок внутренней посадки рибосомы;
7. Linker VH (координаты с 4464 по 4931 п.н.), синтезированную последовательность тяжелой цепи, фланкируемую уникальными сайтами рестрикции SphI, AsisI;
8. Human IgG1, (CL- СН1-СН2-СН3) (координаты с 4939 по 5915 п.н.) последовательность, кодирующая константную цепь антитела человека. Для увеличения периода полураспада антител в организме в данную последовательность были введены мутации (M252Y, S254T, T256E, E333A, H433K/N434F). Увеличения периода полураспада антител происходит за счет взаимодействия с неонатальными рецепторами;
9. SV40 poly(A) signal (координаты с 7188 по 7309 п.н.),последовательность, обеспечивающая стабилизацию мРНК-транскриптов;
10. PuroR (координаты с 6554 по 7153 п.н.), ген устойчивости к антибиотику пуромицину;
11. AmpR (координаты с 10422 по 11282 п.н.), ген устойчивости к антибиотику ампициллину;
12.UCOE,(координаты с 416 по 1964 п.н.) (координаты с 7319 по 8867 п.н.) последовательности, предотвращающие эпигенетическое «замалчивание» экспрессии трансгена.
13. P2A (координаты с 4353 по 4409 п.н.), пептидный линкер.
14.IR/DR(L)Lmut44,IR/DR(R)Rmut13Δ13(координаты с 139 по 403 п.н. и координаты с 8879 по 9174 п.н.), участки распознавания транспазазой.
Указанный технический результат достигается также тем, что плазмидная генетическая конструкция pBuc-7B4, обеспечивающая синтез и секрецию целевого белка – 7B4 против вируса Западного Нила, имеет нуклеотидную последовательность SEQ ID No. 2 и содержит в соответствии с физической и генетической картой, представленной на Фиг. 2, следующие элементы:
1. Участок начала репликации ori (координаты с 9663 по 10251 п.н.);
2.CHEF-1aпромотор (координаты с 1981 по 3477 п.н.), обеспечивает автономную репликацию в клетках млекопитающих.
3. GL последовательность (координаты с 4410 по 4460 п.н.), кодирующую лидерный пептид люциферазы Gaussia, обеспечивающей экспорт белка из клетки;
4. 176 последовательность (координаты с 3483 по 3545 п.н.), кодирующую гибридный лидерный пептид люциферазы (Cypridina noctiluca) и фиброина (Dendrolimus spectabilis), обеспечивающий экспорт белка из клетки;
5. 7B4-VH (координаты с 4308 по 4673 п.н.), последовательность, кодирующая тяжелую цепь антитела 7B4;
6. EMCV IRESatt (координаты с 5687 по 6284 п.н.), участок внутренней посадки рибосомы;
7. 7B4-VL (координаты с 3546 по 3863 п.н.), последовательность, кодирующая легкую цепь антитела 7B4;
8. Human IgG1, (CL- СН1-СН2-СН3) (координаты с 4939 по 5915 п.н.) последовательность, кодирующая константную цепь антитела человека. Для увеличения периода полураспада антител в организме в данную последовательность были введены мутации (M252Y, S254T, T256E, E333A, H433K/N434F). Увеличения периода полураспада антител происходит за счет взаимодействия с неонатальными рецепторами;
9. SV40 poly(A) signal(координаты с 7188 по 7309 п.н.), последовательность, обеспечивающая стабилизацию мРНК-транскриптов;
10. PuroR (координаты с 6554 по 7153 п.н.), ген устойчивости к антибиотику пуромицину;
11. AmpR (координаты с 10422 по 11282 п.н.), ген устойчивости к антибиотику ампициллину;
12.UCOE,(координаты с 416 по 1964 п.н.) (координаты с 7319 по 8867 п.н.) последовательности, предотвращающие эпигенетическое «замалчивание» экспрессии трансгена.
13. P2A (координаты с 4353 по 4409 п.н.), пептидный линкер.
14.IR/DR(L)Lmut44,IR/DR(R)Rmut13Δ13 (координаты с 139 по 403 п.н. и координаты с 8879 по 9174 п.н.), участки распознавания транспазазой.
Заявляемые плазмиды являются улучшенной версией прототипов т.к. в качестве системы интеграции используется транспозаза SB100, имеющая высокую активность фермента (Mates L etal. Molecular evolution of a novel hyperactive Sleeping Beauty transposase enables robust stable gene transfer in vertebrates. NatGenet. 2009, 41(6):753-61). Специфично связываясь с последовательностями IR/DR(L)Lmut44, IR/DR(R)Rmut13Δ13, SB100 обеспечивает вырезание и точную интеграцию целевой кассеты в участки генома позвоночных животных. Обычно специфическим сайтом интеграции для транспозона SB является пара оснований динуклеотида ТА. Использованием технологии транспозонов может обеспечить стабильный перенос генов с высокой эффективностью в клетки млекопитающих (до 95%) (Ruf S. et al. Large-scale analysis of the regulatory architecture of the mouse genome with a transposon-associated sensor. Naturegenetics. 201; 43(4): 379-386).
Указанный технический результат достигается наличием сильных цис-регуляторных элементов – универсальных элементов, открывающих хроматин (UCOE), предотвращающих хромосомное замалчивание (Neville JJ et al. Ubiquitous Chromatin-opening Elements (UCOEs): Applications in biomanufacturing and gene therapy. Biotechnol Adv. 2017, 35(5):557-564). UCOE широко применяется для получения рекомбинантного белка в линиях клеток млекопитающих (Bell A. C. et al. The protein CTCF is required for the enhancer blocking activity of vertebrate insulators. Cell. 1999; 98(3): 387-396).
Указанный технический результат достигается обеспечением высокого и стабильного выхода целевого продукта, благодаря включению в состав экспрессирующего вектора последовательности «сильного» гомологичного промотора фактора элонгацииCHEF-1a. По литературным данным этот промотор показывает более высокие уровни синтеза белка в клетка CHO в сравнении с другими используемыми промоторами (Wang W. et al. Impact of different promoters, promoter mutation, and an enhancer on recombinant protein expression in CHO cells. Sci Rep. 2017; 7(1): 1-10; Ebadat S. et al. Evaluating the efficiency of CHEF and CMV promoter with IRES and Furin/2A linker sequences for monoclonal antibody expression in CHO cells. PLoS One. 2017. 12(10): e0185967).
Указанный технический результат так же достигается уменьшением времени селекции до 3-х дней для получения стабильного штамма-продуцента антител, за счет включения в состав вектора нуклеотидной последовательности гена антибиотика пуромицина (PuroR).
Указанный технический результат так же достигается включением в состав экспрессионного вектора последовательности, кодирующей пептид P2A.P2A пептид обеспечивает строгую регуляцию соотношения синтезируемых тяжелой и легкой цепей антитела. Этот эффект достигается за счет того, что обе цепи антитела синтезируются в составе единого полипротеина, который расщепляется в С-концевой области P2A пептида.
Указанный технический результат так же достигается включением сайта протеолиза фуриновой протеазы. Включение этого сайта позволяет проводить удаление остатка аминокислотной последовательности P2A пептида.
Указанный технический результат так же достигается включением в состав экспрессионного вектора последовательности, кодирующей «ослабленный» вариант IRES перед последовательностью гена пуромицина (IRES satt). IRES функционирует как сайт посадки рибосом для внутренней инициации трансляции. Использование IRES satt позволяет уменьшить эффективность инициации синтеза пуромицина, поэтому после селекции выжившие клоны будут иметь повышенный уровень синтеза антитела.
Указанный технический результат так же достигается увеличением эффективности секреции белков за счет использования лидерных пептидов 176 иGL кодируемых участками вектора. Лидерный пептид 176 является гибридной сигнальной последовательностью люциферазы (Cypridina noctiluca) и фиброина (Dendrolimus spectabilis) и имеет состав MMRTLILAVLLVYFCATVHC. Лидерный пептид GL является сигнальной последовательностью люциферазы Gaussia и имеет состав MGVKVLFALICIAVAEA.
Изобретение поясняется графическими материалами, представленными на фиг. 1-3. На фиг. 1 изображена физическая и генетическая карта универсального рекомбинантного вектора pBUC. На фиг. 2 изображена физическая и генетическая карта плазмидной генетической конструкции pBuc-7В4. На фиг. 3 приведена электрофореграмма препарата трансгенной клеточной линии Cho-7B4, продуцента рекомбинантных антител 7B4, в 10% ПААГ.
Для лучшего понимания сущности предлагаемого изобретения ниже приведены примеры его осуществления. Все стандартные генно-инженерные и микробиологические манипуляции, а также амплификацию и секвенирование ДНК проводили по известным методикам (Маниатис Т., Фрич Э, Сэмбрук Дж. Молекулярное клонирование, М.: Мир, 1984; Клонирование ДНК. Методы. Под ред. Д. Гловера, Пер. с англ., Москва, Мир, 1988; Saiki R.K. et al. Science. 1988, 239(4839): 487-491; SangerF. etal. Proc. Nat. Acad. Sci. 1977, 74:5463-5467).
Пример 1. Конструирование рекомбинантного универсального вектора, обеспечивающего синтез антител в клетках млекопитающих.
Для конструирования вектора pBuc за основу брали акцепторный вектор pT2HB, содержащий кассету экспрессии, фланкированную двумя последовательностями с инвертированным / прямым повторами (IR/DR).
Интеграционный вектор pBuc был получен в несколько этапов. На первом этапе из вектора pT2HB, при помощи сайт-направленного мутагенеза был удален сайт гидролиза BamHI, получен вектор pT2HB2. Для этого были использованы праймеры pT2HB-Fи pT2HB-R (Таблица 1). ПЦР фрагмента PCR-pT2HB проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (плазмиды pT2HB), 10 пМ каждого праймера (pT2HB-F, pT2HB-R), 5 мкл 10хTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95ºС; 30 с – 95ºС, 30 с – 58ºС, 45 с – 72ºС (25 циклов); 120 с - 72ºС.
Реакционную смесь, содержащую фрагмент PCR-pT2HB, разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосу, соответствующую продукту амплификации - 709 п.н., визуализированную при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирку объемом 1,5 мл. Фрагмент выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Затем выделенный ПЦР-продукт PCR-pT2HB и исходную плазмиду pT2HB обрабатывали эндонуклеазами рестрикции Psp124BI и KpnI согласно протоколу фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам – 688 и 2859 п.н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable с генотипом F' proA+B+lacIq ∆(lacZ)M15 zzf::Tn10 (TetR) ∆(ara-leu) 7697 araD139 fhuA ∆lacX74 galK16 galE15 e14- Φ80dlacZ∆M15 recA1 relA1 endA1 nupG rpsL (StrR) rph spoT1 ∆(mrr-hsdRMS-mcrBC), предоставленные производителем “Invitrogen” (США). Для этого к 100 мкл клеток добавляли 10 мкл лигазной смеси, инкубировали на льду в течение 40 минут. После этого клетки подвергали «температурному шоку» при 420С в течение 40 сек. Охлаждали клетки на льду в течение 4 минут, затем добавляли по 200 мкл среды “SOС” (20 г/л триптона, 5 г/л дрожжевого экстракта, 0,5 г/л NaCl, 250 мМ КСl, 10 мМ MgCl2) и инкубировали при 300С в течение 60 минут. По окончании инкубации трансформированные клетки высевали на чашку Петри с твёрдой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов “PlasmidMiniKit”, “Qiagen” (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена плазмида pT2HB2.
Следующим шагом была встройка последовательностей UCOE-R и UCOE-L. Для получения последовательностей UCOE-R и UCOE-L использовали геномную ДНК человека. Для амплификации последовательности UCOE-R была проведена ПЦР с помощью праймеров (таблица 1).
ПЦР фрагмента UCOE-R проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (геномная ДНК человека), 10 пМ каждого праймера (UCOE-R-F, UCOE-R-R), 5 мкл 10хTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95ºС; 30 с – 95ºС, 30 с – 59ºС, 95 с – 72ºС (30 циклов); 120 с - 72ºС.
Реакционную смесь, содержащую фрагмент UCOE-R, разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосу, соответствующую продукту амплификации - 1572 п.н., визуализированную при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирку объемом 1,5 мл. Фрагмент выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Затем выделенный ПЦР-продукт UCOE-R обрабатывали эндонуклеазами рестрикции BseX3I и BamHI, плазмиду pT2HB2 обрабатывали эндонуклеазами рестрикции BseX3I и BglII согласно протоколу фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам – 1555 и 3529 п.н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable, как было описано выше. Трансформированные клетки высевали на чашку Петри с твёрдой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов “Plasmid Mini Kit”, “Qiagen” (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена плазмида pT2HB2-UCOE-R.
ПЦР фрагмента UCOE-L проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (геномная ДНК человека), 10 пМ каждого праймера (UCOE-L-F, UCOE-L-R), 5 мкл 10хTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95ºС; 30 с – 95ºС, 30 с – 59ºС, 97 с – 72ºС (30 циклов); 120 с - 72ºС.
Реакционную смесь, содержащую фрагмент UCOE-L, разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосу, соответствующую продукту амплификации - 1593 п.н., визуализированную при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирку объемом 1,5 мл. Фрагмент выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Затем выделенный ПЦР-продукт UCOE-Lи плазмиду pT2HB2-UCOE-R обрабатывали эндонуклеазами рестрикции HindIII и BseX3I согласно протоколу фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам – 1581 и 5051 п.н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable, как было описано выше. Трансформированные клетки высевали на чашку Петри с твёрдой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов “Plasmid Mini Kit”, “Qiagen” (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена плазмида pT2HB2-UCOE-RL.
На следующем этапе была проведена встройка фрагментов, кодирующих гены селективного антибиотика пуромицина (Puro-R), последовательностей EMCV IRES и SV40 poly(A) signal.
ПЦР фрагмента Puro-R проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (плазмиды pSBbi-Pur c геном пуромицина), 10 пМ каждого праймера (Pur-F и Pur-R) (таблица 1), 5 мкл 10хTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин – 95ºС, 30 с – 95ºС, 30 с – 58ºС, 40 с – 72ºС (30 циклов).
ПЦР фрагмента EMCV IRES проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (плазмиды pT2HB2-UCOE-RL-scFv-ADI-CH-IRES-Bleo41, кодирующую последовательность EMCV IRES), 10 пМ каждого праймера (IRES-RBD-F и IRES-RBD-R) (таблица 1), 5 мкл 10хTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95ºС, 30 с – 95ºС, 30 с – 58ºС, 40 с – 72ºС (30 циклов).
Далее проводили отжиг полученных ПЦР продуктов (IRES-Pur) с помощью праймеров IRES-RBD-F и Pur-R (таблица 1). Реакционная смесь объемом 50 мкл содержала 2 мкг каждого ПЦР продукта (Puro-R EMCV IRES), 10 пМ каждого праймера, 5 мкл 10хTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95ºС, 30 с – 95ºС, 30 с – 58ºС, 1 м 20 с – 72ºС (30 циклов).
ПЦР фрагмента SV40 poly(A) signal проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (плазмиды pT2HB2-UCOE-RL-scFv-ADI-CH-IRES-Bleo41, кодирующую последовательность SV40 poly(A) signal), 10 пМ каждого праймера (RBD-PA-F и RBD-PA-R) (таблица 1), 5 мкл 10хTaq буфера (фирмы Sibenzyme (Россия)), смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. ДНК-полимеразы Taq-SE фирмы Sibenzyme (Россия); реакцию осуществляли при следующих параметрах: 3 мин - 95ºС, 30 с – 95ºС, 30 с – 59ºС, 15 с – 72ºС (30 циклов).
Реакционные смеси, содержащие фрагменты IRES-Pur, SV40 poly(A) signal, разделяли электрофорезом в 1,5% агарозном геле и выявляли продукт реакции - единичный фрагмент размером 1240, 174 п.н. соответственно. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия). Затем выделенные ПЦР-продукты IRES-Pur, SV40 poly(A) signal обрабатывали эндонуклеазами рестрикции BamHI + FauNDI и FauNDI+BseX3I (Sibenzyme (Россия)) соответственно в соответствии с инструкцией производителя.
Предварительно наработанную и очищенную плазмиду pT2HB2-UCOE-RL обрабатывали эндонуклеазами рестрикции BamHI и BseX3I (Sibenzyme (Россия)).Реакцию гидролиза проводили в условиях, рекомендованных производителем. С целью очистки рестриционного вектора плазмидную ДНК наносили на 1%-ный агарозный гель и выделяли из геля с использованием набора «Gel Extraction Kit» фирмы Qiagen (Германия).
Реакцию лигирования проводили с использованием ДНК-лигазы бактериофага Т4 (Sibenzyme (Россия)). Реакция проводилась при +4°С в течение ночи. Полученной лигазной смесью трансформировали химически компетентные клетки E. Coli штамм NebStable, как описано ранее.
После трансформации клоны отбирали на агаризованнной среде LB, содержащей антибиотик ампициллин (50 мг/мл). Затем плазмидную ДНК выделяли из бактериальных клеток с помощью коммерческих наборов DNA mini kit фирмы «Qiagen» согласно рекомендациям производителя. Первичную структуру полученной плазмиды pLet проверяли секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск).
Далее в вектор pLet были встроены5 ПЦР-фрагментов. С помощью синтезированных последовательностей Linker VL, Linker VH в конструкцию были введены уникальные сайты рестрикции Bsa29I, AsuNHI и SphI, AsisI соответственно. В будущем по этим сайтам рестрикции можно будет клонировать тяжелые и легкие цепи любого антитела.
С помощью метода ПЦР был амплифицирован ген антибиотика пуромицина (PCR-Pur, 648 п.н.)., при этом был удален сайт BsmBI ПЦР проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (плазмиды pSBbi-Pur c геном пуромицина), 10 пкМ каждого праймера (BPur-F и Pur-R) (таблица 1), 10 мкл 5х Q5 реакционного буфера,10 мкл 5хQ5 High GC Enhancer, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 0.5 ед. Q5 High-Fidelity ДНК-полимеразы, реакцию осуществляли при следующих параметрах: 10 с – 980С, 20 с – 660С, 15 с – 720С (33 цикла).
С помощью метода ПЦР была амплифицирована последовательность EMCV IRES (PCR-BIRES, 642 п.н.). ПЦР проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл, содержащая 2 мкг ДНК (плазмидаpVEAL2cпоследовательностью EMCV IRES), 10 пкМ каждого праймера (BIRES-F и BIRES-R)(таблица 1),10 мкл 5х Q5 реакционного буфера,10 мкл 5хQ5 High GC Enhancer, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 0.5 ед. Q5 High-Fidelity ДНК-полимеразы, реакцию осуществляли при следующих параметрах: 10 с – 980С, 20 с – 700С, 15 с – 720С (33 цикла).
С помощью метода ПЦР была амплифицирована последовательность промотора CHEF-1a (PCR-CHEF, 1536 п.н.). ПЦР проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл, содержащая 2 мкг ДНК (геномной ДНК клеток СНО содержащей последовательность промотора CHEF-1a), 10 пкМ каждого праймера (TEF-Fи CHEF-R) (таблица 1), 10 мкл 5х Q5 реакционного буфера,10 мкл 5хQ5 High GC Enhancer, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 0.5 ед. Q5 High-Fidelity ДНК-полимеразы, реакцию осуществляли при следующих параметрах: 10 с – 980С, 20 с – 660С, 45 – 720С (33 цикла).
С помощью метода ПЦР была амплифицирована последовательность, кодирующая Linker VL, 176 последовательность, Furin, P2A пептид, (Linker VL, 943 п.н.). ПЦР проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл, содержащая 2 мкг ДНК (плазмида GL), 10 пкМ каждого праймера (VL-F и VL-R) (таблица 1), 10 мкл 5х Q5 реакционного буфера,10 мкл 5хQ5 High GC Enhancer, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 0.5 ед. Q5 High-Fidelity ДНК-полимеразы, реакцию осуществляли при следующих параметрах: 20с – 980С, 20 с – 630С, 30с – 720С (33 цикла).
Таблица 1 – Последовательности олигонуклеотидов, использованные для получения плазмиды pBuc
CGAGGAGACGCCGTGGCC-3’
CCATGaccgagtacaagcccac -3’
CTTATCATattatcatcgtgtttttcaaagga-3’
GAGTGAAGGTGCTGT-3’
GA-3’
CTGGCTGTGCTGCTGGTGTACTTCTGTGCC-3’
AGCAGACTAAAGTTGGTG-3’
С помощью метода ПЦР был амплифицирована последовательность, кодирующая Linker VH, Human IgG1 [CH1-CH2-CH3], GL (1562 п.н.). ПЦР проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл, содержащая 2 мкг ДНК (плазмида GH), 10 пкМ каждого праймера (VH-F и VH-R) (таблица 1), 10 мкл 5х Q5 реакционного буфера,10 мкл 5хQ5 High GC Enhancer, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 0.5 ед. Q5 High-Fidelity ДНК-полимеразы, реакцию осуществляли при следующих параметрах: 20с – 980С, 20 с – 650С, 45с – 720С (33 цикла).
Готовые ПЦР-продукты были выделены и очищены из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя. ПЦР-продукты Linker VH, Linker VL,PCR-BIRES обрабатывали эндонуклеазой рестрикции BsmBI. ПЦР-продукты PCR-Pur, PCR-CHEF обрабатывали общей эндонуклеазой рестрикции BsmBI, а также FauNDI и BamHI соответственно. Предварительно наработанную и очищенную плазмиду pLet обрабатывали эндонуклеазами рестрикции BamHI и FauNDI. Реакцию гидролиза проводили в условиях, рекомендованных производителем. С целью очистки рестрицированного вектора плазмидную ДНК наносили на 1%-ный агарозный гель и выделяли из геля с использованием набора «Gel Extraction Kit» фирмы Qiagen (Германия). Реакцию лигирования проводили с использованием ДНК-лигазы бактериофага Т4 («СибЭнзим», г. Новосибирск). Реакция проводилась при +4°С в течение ночи. Полученной лигазной смесью трансформировали компетентные клетки E. Coli штамм NebStable.
Первичную проверку на наличие вставки проводили при помощи ПЦР с колонии. Разделение продуктов амплификации проводили в 1%-м агарозном геле с последующим окрашиванием бромистым этидием (0,5 мкг/мл).
Положительные колонии, вносили в 5 мл среды LB с ампициллином (50 мкг/мл) и растили в течение ночи при 37°С при 170 об/мин. Затем плазмидную ДНК выделяли из бактериальных клеток с помощью коммерческих наборов DNAminikit фирмы «Qiagen» согласно рекомендациям производителя. Первичную структуру экспрессионного вектора подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена оригинальная плазмида pBuc, содержащая уникальные сайты рестрикции (Bsa29I, AsuNHI и SphI, AsisI), с помощью которых можно будет провести быструю замену цепей, для получения различных рекомбинантных антител в клетках млекопитающих.
Секвенирование плазмидной ДНК положительных клонов в районе встройки позволило отобрать клоны с отсутствием дефектов встраиваемых генов (вставки, делеции, замены), после чего из отобранных клонов была наработана и выделена плазмидная ДНК.
Пример 2. Конструирование рекомбинантнойплазмидыpBuc-7B4, обеспечивающей синтез и секрецию антитела 7B4 против вируса Западного Нила
Для получения рекомбинантной плазмиды pBuc-7В4 в универсальный вектор pBuc последовательно встраивались нуклеотидные последовательности, кодирующие легкую и тяжелую цепи антитела 7В4 против вируса Западного Нила. Последовательности праймеров, использованные для сборки плазмиды pBuc-7В4, представлены в таблице 2.
Таблица 2 – Последовательности олигонуклеотидов, использованные для получения плазмидыpBuc-7B4
С помощью ПЦР была амплифицирована последовательность, кодирующая легкую цепь антитела 7B4 (7B4-VL, 341 п.н.). ПЦР проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (плазмида pJet_7B4VL, содержащая последовательность легкой цепи 7B4), 10 пкМ каждого праймера (7В4L-F и 7В4L-R), 10 мкл 5х Q5 реакционного буфера,10 мкл 5хQ5 High GC Enhancer, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 0.5 ед. Q5 High-Fidelity ДНК-полимеразы, реакцию осуществляли при следующих параметрах: 20с – 980С, 20 с – 650С, 20с – 720С (33 цикла).
С помощью ПЦР была амплифицирована последовательность, кодирующая тяжелую цепь антитела 7В4 (7В4-VH, 381 п.н.). ПЦР проводили с использованием ПЦР-амплификатора «БИС» фирмы ООО БИС-Н (Россия). Реакционная смесь объемом 50 мкл содержала 2 мкг ДНК (плазмида pJet_7В4VH, содержащая последовательность тяжелой цепи 7B4), 10 пкМ каждого праймера (7В4H-F и 7В4H-R), 10 мкл 5х Q5 реакционного буфера,10 мкл 5хQ5 High GC Enhancer, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 0.5 ед. Q5 High-Fidelity ДНК-полимеразы, реакцию осуществляли при следующих параметрах: 20с – 980С, 20 с – 680С, 20с – 720С (33 цикла).
Реакционные смеси, содержащие фрагменты 7B4-VL, 7В4-VH, разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие продуктам амплификации – 341 и 381 п.н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Универсальный вектор pBuc и ПЦР-продукт 7B4-VL обрабатывали эндонуклеазами рестрикции AsuNHIи BSA29I в соответствии с протоколом фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам – 11494 и 323 п.н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable как было описано выше. Трансформированные клетки высевали на чашку Петри с твёрдой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов “Plasmid Mini Kit”, “Qiagen” (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена плазмида pBuc-7B4-VL.
Далее плазмиду pBuc-7B4-VL и ПЦР-продукт 7В4-VH обрабатывали эндонуклеазами рестрикции AsisSI, SphIв соответствии с протоколом фирмы-производителя (Sibenzyme (Россия)). Рестрицированные фрагменты разделяли электрофорезом в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК полосы, соответствующие необходимым фрагментам – 11361 и 363 п.н., визуализированные при помощи ультрафиолета (УФ), вырезали из агарозного геля, помещали в пробирки объемом 1,5 мл. Фрагменты выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия) в соответствии с инструкцией производителя.
Далее проводили лигирование фрагментов ДНК-лигазой фага Т4 (Sibenzyme (Россия)) в стандартном буферном растворе по методике производителя фермента. Полученной лигазной смесью трансформировали «компетентные» клетки E.coli штамма NEB Stable, как было описано выше. Трансформированные клетки высевали на чашку Петри с твёрдой питательной средой LB (среда LB с 1,5% агара), содержащей антибиотик (ампициллин, 50-100 мкг/мл). Чашку Петри помещали в термостат на 18 часов при 30°С. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона LB с добавлением ампициллина в концентрации 50 мкг/мл, наращивали 18 часов при 30°С и проводили выделение плазмидной ДНК при помощи наборов “Plasmid Mini Kit”, “Qiagen” (Германия) в соответствии с инструкциями производителя. Первичную структуру полученных плазмид подтверждали секвенированием. Секвенирование проводили по методу Сэнгера в ЦКП «Геномика» СО РАН (г. Новосибирск). В результате была получена плазмида pBuc-7B4.
Пример 3. Трансфекция клеток CHO-K1 плазмидой pBuc-7B4, получение и анализтрансгенной клеточной линии Cho-7B4, продуцента рекомбинантных антител 7B4
Для встройки экспрессионой кассеты проводили ко-трансфекцию методом катионно-липидной трансфекции (Lipofectamine 3000, Invitrogen) клеток CHO-K1 двумя плазмидами. Первая содержала последовательность антитела 7B4, pBuc-7B4, вторая содержала последовательность транспозазы SB100, pCMV(CAT)T7-SB100. После трансфекции инкубировали клетки 96 часов в СО2-инкубаторе согласно рекомендациям производителя. Через 3 дня в культуральную среду добавляли антибиотик пуромицин (InvivoGen, США) в конечной концентрации 10 мкг/мл, ген устойчивости к которому входит в состав вектора pBuc-7B4.Селекцию устойчивых клонов проводили в течение трех суток.
Отобранные клоны рассаживали в культуральные флаконы. И культивировали в среде DMEM/F12 с 10% содержанием фетальной телячьей сыворотки. Культивирование клонов–продуцентов проводилось при 37 °С в течение 10 суток. Затем среда заменялась на поддерживающую, содержащую 2% фетальной телячьей сыворотки. Культивирование продолжали до тех пор, пока клетки сохраняли жизнеспособность. Далее полученную культуральную жидкость сливали, осаждали клеточный дебрис при помощи центрифугирования при 6000 g, 4 °C в течение 15 мин. Полученную культуральную жидкость очищали с помощью аффинной хроматографии и оценивали продуктивность поликлона при помощи разделения в полиакриламидном геле (ПААГ).
Пример 4. Очистка антитела 7B4
Очистку рекомбинантного антитела 7B4проводили при помощи аффинной хроматографии с использованием сорбента, содержащего на своей поверхности белок А (MabSelect SuRe, GE Healthcare, США), и хроматографической системы AKTAprime plus (GE Healthcare, США). Очистка включает в себя следующие стадии: уравновешивание хроматографической колонки, нанесение образца, промывка колонки, элюция белка.
Уравновешивание хроматографической колонки включает в себя промывание пятикратным объемом базового буфера (50mM Трис-HCl, 150 мM NaCl, pH 7,0-7,2). Скорость потока определялась временем контакта буфера с сорбентом, время контакта составляло не менее 4 мин.
Далее наносили культуральную жидкость, содержащую антитела, на колонку, время контакта образца с сорбентом составляло не менее 5 мин.
Отмывку колонки от несвязавшихся белков, а также от белков, обладающих низкой силой взаимодействия с сорбентом, проводили при помощи последовательной промывки хромоатографической колонки базовым буфером и буфером для промывки (20 мM Трис-HCl, pH 6,8-7,0). Объемы базового и промывочного буферов составляли 3,5 объема колонки. Скорость потока определялась временем контакта буфера с сорбентом, время контакта составляло не менее 3,5 мин.
Элюцию проводили пятикратным объемом элюирующего буфера (100mM Глицин-HСl, pH 3,4-3,5). Скорость потока определялась временем контакта буфера с сорбентом, время контакта составляло не менее 6 мин. Полученные образцы нейтрализовали до рН 7,0 при помощи буфера для титрования (1M Трис).
На фиг.3 представлена электрофореграмма очищенного препаратарекомбинантного антитела 7В4 против вируса Западного Нила, полученного от трансгенной клеточной линии Cho-7B4,в ПААГ.1 –препаратантитела 7B4, нанесенный в количестве1 мкл; 2 - препаратантитела7B4,нанесенный в количестве0,5 мкл; 3 - препаратантитела7B4, нанесенный в количестве 0,25 мкл; 4 – БСА 10 мкг; 5 - БСА 5 мкг;6 - БСА 1 мкг;7 - БСА 0,5 мкг. Фиг. 3 подтверждает, что в клетках Cho-7B4 происходит синтез и секреция антитела 7B4 против вируса Западного Нила. Выход антитела составляет ≈ 12 мг/мл.
--->
ПРИЛОЖЕНИЕ
<110> Федеральное бюджетное учреждение науки «Государственный научный
центр вирусологии и биотехнологии «Вектор» Федеральной службы
по надзору в сфере защиты прав потребителей и благополучия
человека (ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора)
<120> Универсальный рекомбинантный вектор pBuc и плазмидная генетическая
конструкция pBuc-7В4 обеспечивающая синтез и секрецию антитела 7B4
против вируса Западного Нила и полученная с использованием
указанного универсального вектора рBuc.
<160> 2
<210> SEQ ID NO: 1
<211> 11961 п.н.
<212> DNA
<213> Artificial Sequence
<223> Нуклеотидная последовательность универсального интеграционного вектора
pBuc.
<400> 1
1 caaggcgatt aagttgggta acgccagggt tttcccagtc acgacgttgt aaaacgacgg
61 ccagtgagcg cgcgtaatac gactcactat agggcgaatt ggagctcggt tccctataca
121 gttgaagtcg gaagtttaca tacacttaag ttggagtcat taaaactcgt ttttcaacta
181 ctccacaaat ttcttgttaa caaacaatag ttttggcaag tcagttagga catctacttt
241 gtgcatgaca caagtcattt ttccaacaat tgtttacaga cagattattt cacttataat
301 tcactgtatc acaattccag tgggtcagaa gtttacatac actaagttga ctgtgccttt
361 aaacagcttg gaaaattcca gaaaatgatg tcatggcttt agaagcttca ttggcgccct
421 ccgcgcctac agctcaagcc acatccgaag ggggagggag ccgggagctg cgcgcggggc
481 cgccgggggg aggggtggca ccgcccacgc cgggcggcca cgaagggcgg ggcagcgggc
541 gcgcgcgcgg cggggggagg ggccggcgcc gcgcccgctg ggaattgggg ccctaggggg
601 agggcggagg cgccgacgac cgcggcactt accgttcgcg gcgtggcgcc cggtggtccc
661 caaggggagg gaagggggag gcggggcgag gacagtgacc ggagtctcct cagcggtggc
721 ttttctgctt ggcagcctca gcggctggcg ccaaaaccgg actccgccca cttcctcgcc
781 cgccggtgcg agggtgtgga atcctccaga cgctggggga gggggagttg ggagcttaaa
841 aactagtacc cctttgggac cactttcagc agcgaactct cctgtacacc aggggtcagt
901 tccacagacg cgggccaggg gtgggtcatt gcggcgtgaa caataatttg actagaagtt
961 gattcgggtg tttccggaag gggccgagtc aatccgccga gttggggcac ggaaaacaaa
1021 aagggaaggc tactaagatt tttctggcgg gggttatcat tggcgtaact gcagggacca
1081 cctcccgggt tgagggggct ggatctccag gctgcggatt aagcccctcc cgtcggcgtt
1141 aatttcaaac tgcgcgacgt ttctcacctg ccttcgccaa ggcaggggcc gggaccctat
1201 tccaagaggt agtaactagc aggactctag ccttccgcaa ttcattgagc gcatttacgg
1261 aagtaacgtc gggtactgtc tctggccgca agggtgggag gagtacgcat ttggcgtaag
1321 gtggggcgta gagccttccc gccattggcg gcggataggg cgtttacgcg acggcctgac
1381 gtagcggaag acgccttagt gggggggaag gttctagaaa agcggcggca gcggctctag
1441 cggcagtagc agcagcgccg ggtcccgtgc ggaggtgctc ctcgcagagt tgtttctcca
1501 gcagcggcag ttctcactac agcgccagga cgagtccggt tcgtgttcgt ccgcggagat
1561 ctctctcatc tcgctcggct gcgggaaatc gggctgaagc gactgagtcc gcgatggagg
1621 taacgggttt gaaatcaatg agttattgaa aagggcatgg cgaggccgtt ggcgcctcag
1681 tggaagtcgg ccagccgcct ccgtgggaga gaggcaggaa atcggaccaa ttcagtagca
1741 gtggggctta aggtttatga acggggtctt gagcggaggc ctgagcgtac aaacagcttc
1801 cccaccctca gcctcccggc gccatttccc ttcactgggg gtgggggatg gggagctttc
1861 acatggcgga cgctgccccg ctggggtgaa agtggggcgc ggaggcggga cttcttattc
1921 cctttctaaa gcacgctgct tcgggggcca cggcgtctcc tcggggatcc ctgggaaggg
1981 tccctaggat tgtgcacctc tcccgcgggg gacaagcagg ggatggcggg gctgacgtcg
2041 ggaggtggcc tccacgggaa gggacacccg gatctcgaca cagccttggc agtggagtca
2101 ggaagggtag gacagattct ggacgccctc ttggccagtc ctcaccgccc cacccccgat
2161 ggagccgaga gtaattcata caaaaggagg gatcgccttc gcccctggga atcccaggga
2221 ccgtcgctaa attctggccg gcctcccagc ccggaaccgc tgtgcccgcc cagcgcggcg
2281 ggaggagcct gcgcctaggg cggatcgcgg gtcggcggga gagcacaagc ccacagtccc
2341 cggcggtggg ggaggggcgc gctgagcggg ggcccgggag ccagcgcggg gcaaactggg
2401 aaagtggtgt cgtgtgctgg ctccgccctc ttcccgaggg tgggggagaa cggtataaaa
2461 gtgcggtagt cgcgttggac gttctttttc gcaacgggtt tgccgtcaga acgcaggtga
2521 gtggcgggtg tggcctccgc gggcccgggc tccctccttt gagcggggtc ggaccgccgt
2581 gcgggtgtcg tcggccgggc ttctctgcga gcgttcccgc cctggatggc gggctgtgcg
2641 ggagggcgag ggggggaggc ctggcggcgg ccccggagcc tcgcctcgtg tcgggcgtga
2701 ggcctagcgt ggcttccgcc ccgccgcgtg ccaccgcggc cgcgctttgc tgtctgcccg
2761 gctgccctcg attgcctgcc cgcggcccgg gccaacaaag ggagggcgtg gagctggctg
2821 gtagggagcc ccgtagtccg catgtcgggc agggagagcg gcagcagtcg ggggggggac
2881 cgggcccgcc cgtcccgcag cacatgtccg acgccgcctg gacgggtagc ggcctgtgtc
2941 ctgataaggc ggccgggcgg tgggttttag atgccgggtt caggtggccc cgggtcccgg
3001 cccggtctgg ccagtacccc gtagtggctt agctccgagg agggcgagcc cgcccgcccg
3061 gcaccagttg cgtgcgcgga aagatggccg ctcccgggcc ctgtagcaag gagctcaaaa
3121 tggaggacgc ggcagcccgg cggagcgggg cgggtgagtc acccacacaa aggaagaggg
3181 ccttgcccct cgccggccgc tgcttcctgt gaccccgtgg tgtaccggcc gcacttcagt
3241 caccccgggc gctctttcgg agcaccgctg gcctccgctg ggggagggga tctgtctaat
3301 ggcgttggag tttgctcaca tttggtgggt ggagactgta gccaggccag cctggccatg
3361 gaagtaattc ttggaatttg cccattttga gtttggagcg aagctgattg acaaagctgc
3421 ttagccgttc aaaggtattc ttcgaacttt ttttttaagg tgttgtgaaa accaccgtcg
3481 acatgatgcg gaccctgatc ctggctgtgc tgctggtgta cttctgtgcc accgtgcact
3541 gctccatcga ttcttatgaa ctgacccagg gcgacgaggc cgactactac tgccaagtgt
3601 gggactcctc ctccgactac ggcgacgagg ccgactacta ctgccaagtg tgggactcct
3661 cctccgacta cggcgacgag gccgactact actgccaagt gtgggactcc tcctccgact
3721 acggcgacga ggccgactac tactgcgagg ccgactacta ctgccaagtg tgggactcct
3781 cctccgacta cggcgacgag gccgactact actgccaagt gtgggactcc tcctccgact
3841 acggcgacga ggccgactac tactgccaag tgtgggactc ctcctccgac tacggcgacg
3901 aggccgacta ctactgccaa gtgtgggact cctcctccga ctacggcgac gaggccgact
3961 actactgcca agtgtgggac tcctcctccg actaccctcc tactgtgttg ggagctagcc
4021 gtacggtggc tgcaccatct gtcttcatct tcccgccatc tgatgagcag ttgaaatctg
4081 gaactgcctc tgttgtgtgc ctgctgaata acttctatcc cagagaggcc aaagtacagt
4141 ggaaggtgga taacgccctc caatcgggta actcccagga gagtgtcaca gagcaggaca
4201 gcaaggacag cacctacagc ctcagcagca ccctgacgct gagcaaagca gactacgaga
4261 aacacaaagt ctacgcctgc gaagtcaccc atcagggcct gagctcgccc gtcacaaaga
4321 gcttcaacag gggagagtgt agacggaaga gagccaccaa ctttagtctg ctgaagcagg
4381 ccggcgacgt ggaagagaat cctggaccta tgggagtgaa ggtgctgttc gccctgatct
4441 gtattgccgt ggccgaagct tctgcatgcg gccagggcac aacagtggcc cagaaagagt
4501 gcgccagaga taccgtggcc cagaaagagt gcgccagaga taccgtggcc cagaaagagt
4561 gcgccagaga taccgtgggc ccagaaagag tgcgccagag ataccgtggc ccagaaagag
4621 tgcgccagag ataccgtggc ccagaaagag tgcgccagag ataccgtgcc cagaaagagt
4681 gcgccagaga taccgtggcc cagaaagagt gcgccagaga taccgtggcc cagaaagagt
4741 gcgccagaga taccgtggcc cagaaagagt gcgccagaga taccgtggcc cagaaagagt
4801 gcgccagaga taccgtggcc cagaaagagt gcgccagaga taccgtggcc cagaaagagt
4861 gcgccagaga taccgtggcc cagaaagagt gcgccagaga taccgtgaca gtttctagcg
4921 cgatcgcagc aaagggccca agcgtcttcc ccctggcacc ctcctccaag agcacctctg
4981 ggggcacagc ggccctgggc tgcctggtca aggactactt ccccgaaccg gtgacggtgt
5041 cgtggaactc aggcgccctg accagcggcg tgcacacctt cccggctgtc ctacagtcct
5101 caggactcta ctccctcagc agcgtggtga ccgtgccctc cagcagcttg ggcacccaga
5161 cctacatctg caacgtgaat cacaagccca gcaacaccaa ggtggacaag aaagttgagc
5221 ccaaatcttg tgacaaaact cacacatgcc caccgtgccc agcacctgaa ctcctggggg
5281 gaccgtcagt cttcctcttc cccccaaaac ccaaggacac cctctacatc acccgggaac
5341 ctgaggtcac atgcgtggtg gtggacgtga gccacgaaga ccctgaggtc aagttcaact
5401 ggtacgtgga cggcgtggag gtgcataatg ccaagacaaa gccgcgggag gagcagtaca
5461 acagcacgta ccgtgtggtc agcgtcctca ccgtcctgca ccaggactgg ctgaatggca
5521 aggagtacaa gtgcaaggtc tccaacaaag ccctcccagc ccccatcgcg aaaaccatct
5581 ccaaagccaa agggcagccc cgagaaccac aggtgtacac cctgccccca tcccgggagg
5641 agatgaccaa gaaccaggtc agcctgacct gcctggtcaa aggcttctat cccagcgaca
5701 tcgccgtgga gtgggagagc aatgggcagc cggagaacaa ctacaagacc acgcctcccg
5761 tgctggactc cgacggctcc ttcttcctct acagcaagct caccgtggac aagagcaggt
5821 ggcagcaggg gaacgtcttc tcatgctccg tgatgcatga ggctctgaag ttccactaca
5881 cgcagaagag cctctccctg tctccgggta aatgacgagc ctgcaggcgg ttcccgcccc
5941 tctccctccc ccccccctaa cgttactggc cgaagccgct tggaataagg ccggtgtgcg
6001 tttgtctata tgttattttc caccatattg ccgtcttttg gcaatgtgag ggcccggaaa
6061 cctggccctg tcttcttgac gagcattcct aggggtcttt cccctctcgc caaaggaatg
6121 caaggtctgt tgaatgtcgt gaaggaagca gttcctctgg aagcttcttg aagacaaaca
6181 acgtctgtag cgaccctttg caggcagcgg aaccccccac ctggcgacag gtgcctctgc
6241 ggccaaaagc cacgtgtata agatacacct gcaaaggcgg cacaacccca gtgccacgtt
6301 gtgagttgga tagttgtgga aagagtcaaa tggctcacct caagcgtatt caacaagggg
6361 ctgaaggatg cccagaaggt accccattgt atgggatctg atctggggcc tcggtgcaca
6421 tgctttacat gtgtttagtc gaggttaaaa aacgtctagg ccccccgaac cacggggacg
6481 tggttttcct ttgaaaaaca cgatgataat atgataagct tgccacaacc ccgggagatg
6541 aggatcgaat tccatgaccg agtacaagcc cacggtgcgc ctcgccaccc gcgacgacgt
6601 ccccagggcc gtacgcaccc tcgccgccgc gttcgccgac taccccgcca cgcgccacac
6661 cgtcgatccg gaccgccaca tcgagcgggt caccgagctg caagaactct tcctcacgcg
6721 cgtcgggctc gacatcggca aggtgtgggt cgcggacgac ggcgccgcgg tggcggtctg
6781 gaccacgccg gagagcgtcg aagcgggggc ggtgttcgcc gagatcggcc cgcgcatggc
6841 cgagttgagc ggttcccggc tggccgcgca gcaacagatg gaaggcctcc tggcgccgca
6901 ccggcccaag gagcccgcgt ggttcctggc caccgtcggc gtttcgcccg accaccaggg
6961 caagggtctg ggcagcgccg tcgtgctccc cggagtggag gcggccgagc gcgccggggt
7021 gcccgccttc ctggagacct ccgcgccccg caacctcccc ttctacgagc ggctcggctt
7081 caccgtcacc gccgacgtcg aggtgcccga aggaccgcgc acctggtgca tgacccgcaa
7141 gcccggtgcc tgattcgcat atgggttaat gcttcgagca gacatgataa gatacattga
7201 tgagtttgga caaaccacaa ctagaatgca gtgaaaaaaa tgctttattt gtgaaatttg
7261 tgatgctatt gctttatttg taaccattat aagctgcaat aaacaagttc cacggccggc
7321 cctccgcgcc tacagctcaa gccacatccg aagggggagg gagccgggag ctgcgcgcgg
7381 ggccgccggg gggaggggtg gcaccgccca cgccgggcgg ccacgaaggg cggggcagcg
7441 ggcgcgcgcg cggcgggggg aggggccggc gccgcgcccg ctgggaattg gggccctagg
7501 gggagggcgg aggcgccgac gaccgcggca cttaccgttc gcggcgtggc gcccggtggt
7561 ccccaagggg agggaagggg gaggcggggc gaggacagtg accggagtct cctcagcggt
7621 ggcttttctg cttggcagcc tcagcggctg gcgccaaaac cggactccgc ccacttcctc
7681 gcccgccggt gcgagggtgt ggaatcctcc agacgctggg ggagggggag ttgggagctt
7741 aaaaactagt acccctttgg gaccactttc agcagcgaac tctcctgtac accaggggtc
7801 agttccacag acgcgggcca ggggtgggtc attgcggcgt gaacaataat ttgactagaa
7861 gttgattcgg gtgtttccgg aaggggccga gtcaatccgc cgagttgggg cacggaaaac
7921 aaaaagggaa ggctactaag atttttctgg cgggggttat cattggcgta actgcaggga
7981 ccacctcccg ggttgagggg gctggatctc caggctgcgg attaagcccc tcccgtcggc
8041 gttaatttca aactgcgcga cgtttctcac ctgccttcgc caaggcaggg gccgggaccc
8101 tattccaaga ggtagtaact agcaggactc tagccttccg caattcattg agcgcattta
8161 cggaagtaac gtcgggtact gtctctggcc gcaagggtgg gaggagtacg catttggcgt
8221 aaggtggggc gtagagcctt cccgccattg gcggcggata gggcgtttac gcgacggcct
8281 gacgtagcgg aagacgcctt agtggggggg aaggttctag aaaagcggcg gcagcggctc
8341 tagcggcagt agcagcagcg ccgggtcccg tgcggaggtg ctcctcgcag agttgtttct
8401 ccagcagcgg cagttctcac tacagcgcca ggacgagtcc ggttcgtgtt cgtccgcgga
8461 gatctctctc atctcgctcg gctgcgggaa atcgggctga agcgactgag tccgcgatgg
8521 aggtaacggg tttgaaatca atgagttatt gaaaagggca tggcgaggcc gttggcgcct
8581 cagtggaagt cggccagccg cctccgtggg agagaggcag gaaatcggac caattcagta
8641 gcagtggggc ttaaggttta tgaacggggt cttgagcgga ggcctgagcg tacaaacagc
8701 ttccccaccc tcagcctccc ggcgccattt cccttcactg ggggtggggg atggggagct
8761 ttcacatggc ggacgctgcc ccgctggggt gaaagtgggg cgcggaggcg ggacttctta
8821 ttccctttct aaagcacgct gcttcggggg ccacggcgtc tcctcgggga tctagcttgt
8881 ggaaggctac tcgaaatgtt tgacccaagt taaacaattt aaaggcaatg ctaccaaata
8941 ctaattgagt gtatgtaaac ttctgaccca ctgggaatgt gatgaaagaa ataaaagctg
9001 aaatgaatca ttctctctac tattattctg atatttcaca ttcttaaaat aaagtggtga
9061 tcctaactga cctaagacag ggaattttta ctaggattaa atgtcaggaa ttgtgaaaaa
9121 gtgagtttaa atgtatttgg ctaaggtgta tgtaaacttc cgacttcaac tgtatagggt
9181 tcctctagct agagacgacc tcgggtaccc agcttttgtt ccctttagtg agggttaatt
9241 gcgcgcttgg cgtaatcatg gtcatagctg tttcctgtgt gaaattgtta tccgctcaca
9301 attccacaca acatacgagc cggaagcata aagtgtaaag cctggggtgc ctaatgagtg
9361 agctaactca cattaattgc gttgcgctca ctgcccgctt tccagtcggg aaacctgtcg
9421 tgccagctgc attaatgaat cggccaacgc gcggggagag gcggtttgcg tattgggcgc
9481 tcttccgctt cctcgctcac tgactcgctg cgctcggtcg ttcggctgcg gcgagcggta
9541 tcagctcact caaaggcggt aatacggtta tccacagaat caggggataa cgcaggaaag
9601 aacatgtgag caaaaggcca gcaaaaggcc aggaaccgta aaaaggccgc gttgctggcg
9661 tttttccata ggctccgccc ccctgacgag catcacaaaa atcgacgctc aagtcagagg
9721 tggcgaaacc cgacaggact ataaagatac caggcgtttc cccctggaag ctccctcgtg
9781 cgctctcctg ttccgaccct gccgcttacc ggatacctgt ccgcctttct cccttcggga
9841 agcgtggcgc tttctcatag ctcacgctgt aggtatctca gttcggtgta ggtcgttcgc
9901 tccaagctgg gctgtgtgca cgaacccccc gttcagcccg accgctgcgc cttatccggt
9961 aactatcgtc ttgagtccaa cccggtaaga cacgacttat cgccactggc agcagccact
10021 ggtaacagga ttagcagagc gaggtatgta ggcggtgcta cagagttctt gaagtggtgg
10081 cctaactacg gctacactag aaggacagta tttggtatct gcgctctgct gaagccagtt
10141 accttcggaa aaagagttgg tagctcttga tccggcaaac aaaccaccgc tggtagcggt
10201 ggtttttttg tttgcaagca gcagattacg cgcagaaaaa aaggatctca agaagatcct
10261 ttgatctttt ctacggggtc tgacgctcag tggaacgaaa actcacgtta agggattttg
10321 gtcatgagat tatcaaaaag gatcttcacc tagatccttt taaattaaaa atgaagtttt
10381 aaatcaatct aaagtatata tgagtaaact tggtctgaca gttaccaatg cttaatcagt
10441 gaggcaccta tctcagcgat ctgtctattt cgttcatcca tagttgcctg actccccgtc
10501 gtgtagataa ctacgatacg ggagggctta ccatctggcc ccagtgctgc aatgataccg
10561 cgagacccac gctcaccggc tccagattta tcagcaataa accagccagc cggaagggcc
10621 gagcgcagaa gtggtcctgc aactttatcc gcctccatcc agtctattaa ttgttgccgg
10681 gaagctagag taagtagttc gccagttaat agtttgcgca acgttgttgc cattgctaca
10741 ggcatcgtgg tgtcacgctc gtcgtttggt atggcttcat tcagctccgg ttcccaacga
10801 tcaaggcgag ttacatgatc ccccatgttg tgcaaaaaag cggttagctc cttcggtcct
10861 ccgatcgttg tcagaagtaa gttggccgca gtgttatcac tcatggttat ggcagcactg
10921 cataattctc ttactgtcat gccatccgta agatgctttt ctgtgactgg tgagtactca
10981 accaagtcat tctgagaata gtgtatgcgg cgaccgagtt gctcttgccc ggcgtcaata
11041 cgggataata ccgcgccaca tagcagaact ttaaaagtgc tcatcattgg aaaacgttct
11101 tcggggcgaa aactctcaag gatcttaccg ctgttgagat ccagttcgat gtaacccact
11161 cgtgcaccca actgatcttc agcatctttt actttcacca gcgtttctgg gtgagcaaaa
11221 acaggaaggc aaaatgccgc aaaaaaggga ataagggcga cacggaaatg ttgaatactc
11281 atactcttcc tttttcaata ttattgaagc atttatcagg gttattgtct catgagcgga
11341 tacatatttg aatgtattta gaaaaataaa caaatagggg ttccgcgcac atttccccga
11401 aaagtgccac ctgacgcgcc ctgtagcggc gcattaagcg cggcgggtgt ggtggttacg
11461 cgcagcgtga ccgctacact tgccagcgcc ctagcgcccg ctcctttcgc tttcttccct
11521 tcctttctcg ccacgttcgc cggctttccc cgtcaagctc taaatcgggg gctcccttta
11581 gggttccgat ttagtgcttt acggcacctc gaccccaaaa aacttgatta gggtgatggt
11641 tcacgtagtg ggccatcgcc ctgatagacg gtttttcgcc ctttgacgtt ggagtccacg
11701 ttctttaata gtggactctt gttccaaact ggaacaacac tcaaccctat ctcggtctat
11761 tcttttgatt tataagggat tttgccgatt tcggcctatt ggttaaaaaa tgagctgatt
11821 taacaaaaat ttaacgcgaa ttttaacaaa atattaacgc ttacaatttc cattcgccat
11881 tcaggctgcg caactgttgg gaagggcgat cggtgcgggc ctcttcgcta ttacgccagc
11941 tggcgaaagg gggatgtgct g
<210> SEQ ID NO: 2
<211> 11724 п.н.
<212> DNA
<213> Artificial Sequence
<223> Нуклеотидная последовательность плазмидной генетической конструкции
pBuc-7В4, обеспечивающей синтез и секрецию антитела 7B4 против
вируса Западного Нила
<400> 2
1 caaggcgatt aagttgggta acgccagggt tttcccagtc acgacgttgt aaaacgacgg
61 ccagtgagcg cgcgtaatac gactcactat agggcgaatt ggagctcggt tccctataca
121 gttgaagtcg gaagtttaca tacacttaag ttggagtcat taaaactcgt ttttcaacta
181 ctccacaaat ttcttgttaa caaacaatag ttttggcaag tcagttagga catctacttt
241 gtgcatgaca caagtcattt ttccaacaat tgtttacaga cagattattt cacttataat
301 tcactgtatc acaattccag tgggtcagaa gtttacatac actaagttga ctgtgccttt
361 aaacagcttg gaaaattcca gaaaatgatg tcatggcttt agaagcttca ttggcgccct
421 ccgcgcctac agctcaagcc acatccgaag ggggagggag ccgggagctg cgcgcggggc
481 cgccgggggg aggggtggca ccgcccacgc cgggcggcca cgaagggcgg ggcagcgggc
541 gcgcgcgcgg cggggggagg ggccggcgcc gcgcccgctg ggaattgggg ccctaggggg
601 agggcggagg cgccgacgac cgcggcactt accgttcgcg gcgtggcgcc cggtggtccc
661 caaggggagg gaagggggag gcggggcgag gacagtgacc ggagtctcct cagcggtggc
721 ttttctgctt ggcagcctca gcggctggcg ccaaaaccgg actccgccca cttcctcgcc
781 cgccggtgcg agggtgtgga atcctccaga cgctggggga gggggagttg ggagcttaaa
841 aactagtacc cctttgggac cactttcagc agcgaactct cctgtacacc aggggtcagt
901 tccacagacg cgggccaggg gtgggtcatt gcggcgtgaa caataatttg actagaagtt
961 gattcgggtg tttccggaag gggccgagtc aatccgccga gttggggcac ggaaaacaaa
1021 aagggaaggc tactaagatt tttctggcgg gggttatcat tggcgtaact gcagggacca
1081cctcccgggt tgagggggct ggatctccag gctgcggatt aagcccctcc cgtcggcgtt
1141 aatttcaaac tgcgcgacgt ttctcacctg ccttcgccaa ggcaggggcc gggaccctat
1201 tccaagaggt agtaactagc aggactctag ccttccgcaa ttcattgagc gcatttacgg
1261 aagtaacgtc gggtactgtc tctggccgca agggtgggag gagtacgcat ttggcgtaag
1321 gtggggcgta gagccttccc gccattggcg gcggataggg cgtttacgcg acggcctgac
1381 gtagcggaag acgccttagt gggggggaag gttctagaaa agcggcggca gcggctctag
1441 cggcagtagc agcagcgccg ggtcccgtgc ggaggtgctc ctcgcagagt tgtttctcca
1501 gcagcggcag ttctcactac agcgccagga cgagtccggt tcgtgttcgt ccgcggagat
1561 ctctctcatc tcgctcggct gcgggaaatc gggctgaagc gactgagtcc gcgatggagg
1621 taacgggttt gaaatcaatg agttattgaa aagggcatgg cgaggccgtt ggcgcctcag
1681 tggaagtcgg ccagccgcct ccgtgggaga gaggcaggaa atcggaccaa ttcagtagca
1741 gtggggctta aggtttatga acggggtctt gagcggaggc ctgagcgtac aaacagcttc
1801 cccaccctca gcctcccggc gccatttccc ttcactgggg gtgggggatg gggagctttc
1861 acatggcgga cgctgccccg ctggggtgaa agtggggcgc ggaggcggga cttcttattc
1921 cctttctaaa gcacgctgct tcgggggcca cggcgtctcc tcggggatcc ctgggaaggg
1981 tccctaggat tgtgcacctc tcccgcgggg gacaagcagg ggatggcggg gctgacgtcg
2041 ggaggtggcc tccacgggaa gggacacccg gatctcgaca cagccttggc agtggagtca
2101 ggaagggtag gacagattct ggacgccctc ttggccagtc ctcaccgccc cacccccgat
2161 ggagccgaga gtaattcata caaaaggagg gatcgccttc gcccctggga atcccaggga
2221 ccgtcgctaa attctggccg gcctcccagc ccggaaccgc tgtgcccgcc cagcgcggcg
2281 ggaggagcct gcgcctaggg cggatcgcgg gtcggcggga gagcacaagc ccacagtccc
2341 cggcggtggg ggaggggcgc gctgagcggg ggcccgggag ccagcgcggg gcaaactggg
2401 aaagtggtgt cgtgtgctgg ctccgccctc ttcccgaggg tgggggagaa cggtataaaa
2461 gtgcggtagt cgcgttggac gttctttttc gcaacgggtt tgccgtcaga acgcaggtga
2521 gtggcgggtg tggcctccgc gggcccgggc tccctccttt gagcggggtc ggaccgccgt
2581 gcgggtgtcg tcggccgggc ttctctgcga gcgttcccgc cctggatggc gggctgtgcg
2641 ggagggcgag ggggggaggc ctggcggcgg ccccggagcc tcgcctcgtg tcgggcgtga
2701 ggcctagcgt ggcttccgcc ccgccgcgtg ccaccgcggc cgcgctttgc tgtctgcccg
2761 gctgccctcg attgcctgcc cgcggcccgg gccaacaaag ggagggcgtg gagctggctg
2821 gtagggagcc ccgtagtccg catgtcgggc agggagagcg gcagcagtcg ggggggggac
2881 cgggcccgcc cgtcccgcag cacatgtccg acgccgcctg gacgggtagc ggcctgtgtc
2941 ctgataaggc ggccgggcgg tgggttttag atgccgggtt caggtggccc cgggtcccgg
3001 cccggtctgg ccagtacccc gtagtggctt agctccgagg agggcgagcc cgcccgcccg
3061 gcaccagttg cgtgcgcgga aagatggccg ctcccgggcc ctgtagcaag gagctcaaaa
3121 tggaggacgc ggcagcccgg cggagcgggg cgggtgagtc acccacacaa aggaagaggg
3181 ccttgcccct cgccggccgc tgcttcctgt gaccccgtgg tgtaccggcc gcacttcagt
3241 caccccgggc gctctttcgg agcaccgctg gcctccgctg ggggagggga tctgtctaat
3301 ggcgttggag tttgctcaca tttggtgggt ggagactgta gccaggccag cctggccatg
3361 gaagtaattc ttggaatttg cccattttga gtttggagcg aagctgattg acaaagctgc
3421 ttagccgttc aaaggtattc ttcgaacttt ttttttaagg tgttgtgaaa accaccgtcg
3481 acatgatgcg gaccctgatc ctggctgtgc tgctggtgta cttctgtgcc accgtgcact
3541 gctccatcga tatcgtgatg acccaggccc acaagttcat gagcaccagc gtgggcgacc
3601 gcgtgagcat cacctgcaag gccagccagg acgtgagcac cgccgtggcc tggtaccagc
3661 agaagcccgg ccagagcccc aaggtgctga tctactgggc cagcacccgc cacaccggcg
3721 tgcccgaccg cttcaccggc agcggcagcg gcaccgacta caccctgacc atcagcagcg
3781 tgcaggccga ggacctggcc ctgtactact gccagcagca ctacagcaac cccctgacct
3841 tcggcgccgg caccaagctg gagctgaagg ctagccgtac ggtggctgca ccatctgtct
3901 tcatcttccc gccatctgat gagcagttga aatctggaac tgcctctgtt gtgtgcctgc
3961 tgaataactt ctatcccaga gaggccaaag tacagtggaa ggtggataac gccctccaat
4021 cgggtaactc ccaggagagt gtcacagagc aggacagcaa ggacagcacc tacagcctca
4081 gcagcaccct gacgctgagc aaagcagact acgagaaaca caaagtctac gcctgcgaag
4141 tcacccatca gggcctgagc tcgcccgtca caaagagctt caacagggga gagtgtagac
4201 ggaagagagc caccaacttt agtctgctga agcaggccgg cgacgtggaa gagaatcctg
4261 gacctatggg agtgaaggtg ctgttcgccc tgatctgtat tgccgtggcc gaagcttctg
4321 catgccaggt gcagctgcag gagagcggcg ccgagctggt gaagcccggc gccagcgtga
4381 agctgagctg caaggccagc ggctacatct tcaccaacta cgacatcaac tgggtgcgcc
4441 agcgccccga gcagggcctg gagtggatcg gctggatctt ccccgaggac ggccgcacca
4501 agaacaacga gaagttcaag ggcaaggcca ccctgaccac cgacaagagc agcagcaccg
4561 cctacatgca gctgagccgc ctgaccagcg aggacagcgc cgtgtacttc tgcgcccgcg
4621 gcggcggcta cggcaactac ttcgactact ggggccaggg caccctggtg accgtgagca
4681 gcgcgatcgc agcaaagggc ccaagcgtct tccccctggc accctcctcc aagagcacct
4741 ctgggggcac agcggccctg ggctgcctgg tcaaggacta cttccccgaa ccggtgacgg
4801 tgtcgtggaa ctcaggcgcc ctgaccagcg gcgtgcacac cttcccggct gtcctacagt
4861 cctcaggact ctactccctc agcagcgtgg tgaccgtgcc ctccagcagc ttgggcaccc
4921 agacctacat ctgcaacgtg aatcacaagc ccagcaacac caaggtggac aagaaagttg
4981 agcccaaatc ttgtgacaaa actcacacat gcccaccgtg cccagcacct gaactcctgg
5041 ggggaccgtc agtcttcctc ttccccccaa aacccaagga caccctctac atcacccggg
5101 aacctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag gtcaagttca
5161 actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg gaggagcagt
5221 acaacagcac gtaccgtgtg gtcagcgtcc tcaccgtcct gcaccaggac tggctgaatg
5281 gcaaggagta caagtgcaag gtctccaaca aagccctccc agcccccatc gcgaaaacca
5341 tctccaaagc caaagggcag ccccgagaac cacaggtgta caccctgccc ccatcccggg
5401 aggagatgac caagaaccag gtcagcctga cctgcctggt caaaggcttc tatcccagcg
5461 acatcgccgt ggagtgggag agcaatgggc agccggagaa caactacaag accacgcctc
5521 ccgtgctgga ctccgacggc tccttcttcc tctacagcaa gctcaccgtg gacaagagca
5581 ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggctctg aagttccact
5641 acacgcagaa gagcctctcc ctgtctccgg gtaaatgacg agcctgcagg cggttcccgc
5701 ccctctccct cccccccccc taacgttact ggccgaagcc gcttggaata aggccggtgt
5761 gcgtttgtct atatgttatt ttccaccata ttgccgtctt ttggcaatgt gagggcccgg
5821 aaacctggcc ctgtcttctt gacgagcatt cctaggggtc tttcccctct cgccaaagga
5881 atgcaaggtc tgttgaatgt cgtgaaggaa gcagttcctc tggaagcttc ttgaagacaa
5941 acaacgtctg tagcgaccct ttgcaggcag cggaaccccc cacctggcga caggtgcctc
6001 tgcggccaaa agccacgtgt ataagataca cctgcaaagg cggcacaacc ccagtgccac
6061 gttgtgagtt ggatagttgt ggaaagagtc aaatggctca cctcaagcgt attcaacaag
6121 gggctgaagg atgcccagaa ggtaccccat tgtatgggat ctgatctggg gcctcggtgc
6181 acatgcttta catgtgttta gtcgaggtta aaaaacgtct aggccccccg aaccacgggg
6241 acgtggtttt cctttgaaaa acacgatgat aatatgataa gcttgccaca accccgggag
6301 atgaggatcg aattccatga ccgagtacaa gcccacggtg cgcctcgcca cccgcgacga
6361 cgtccccagg gccgtacgca ccctcgccgc cgcgttcgcc gactaccccg ccacgcgcca
6421 caccgtcgat ccggaccgcc acatcgagcg ggtcaccgag ctgcaagaac tcttcctcac
6481 gcgcgtcggg ctcgacatcg gcaaggtgtg ggtcgcggac gacggcgccg cggtggcggt
6541 ctggaccacg ccggagagcg tcgaagcggg ggcggtgttc gccgagatcg gcccgcgcat
6601 ggccgagttg agcggttccc ggctggccgc gcagcaacag atggaaggcc tcctggcgcc
6661 gcaccggccc aaggagcccg cgtggttcct ggccaccgtc ggcgtttcgc ccgaccacca
6721 gggcaagggt ctgggcagcg ccgtcgtgct ccccggagtg gaggcggccg agcgcgccgg
6781 ggtgcccgcc ttcctggaga cctccgcgcc ccgcaacctc cccttctacg agcggctcgg
6841 cttcaccgtc accgccgacg tcgaggtgcc cgaaggaccg cgcacctggt gcatgacccg
6901 caagcccggt gcctgattcg catatgggtt aatgcttcga gcagacatga taagatacat
6961 tgatgagttt ggacaaacca caactagaat gcagtgaaaa aaatgcttta tttgtgaaat
7021 ttgtgatgct attgctttat ttgtaaccat tataagctgc aataaacaag ttccacggcc
7081 ggccctccgc gcctacagct caagccacat ccgaaggggg agggagccgg gagctgcgcg
7141 cggggccgcc ggggggaggg gtggcaccgc ccacgccggg cggccacgaa gggcggggca
7201 gcgggcgcgc gcgcggcggg gggaggggcc ggcgccgcgc ccgctgggaa ttggggccct
7261 agggggaggg cggaggcgcc gacgaccgcg gcacttaccg ttcgcggcgt ggcgcccggt
7321 ggtccccaag gggagggaag ggggaggcgg ggcgaggaca gtgaccggag tctcctcagc
7381 ggtggctttt ctgcttggca gcctcagcgg ctggcgccaa aaccggactc cgcccacttc
7441 ctcgcccgcc ggtgcgaggg tgtggaatcc tccagacgct gggggagggg gagttgggag
7501 cttaaaaact agtacccctt tgggaccact ttcagcagcg aactctcctg tacaccaggg
7561 gtcagttcca cagacgcggg ccaggggtgg gtcattgcgg cgtgaacaat aatttgacta
7621 gaagttgatt cgggtgtttc cggaaggggc cgagtcaatc cgccgagttg gggcacggaa
7681 aacaaaaagg gaaggctact aagatttttc tggcgggggt tatcattggc gtaactgcag
7741 ggaccacctc ccgggttgag ggggctggat ctccaggctg cggattaagc ccctcccgtc
7801 ggcgttaatt tcaaactgcg cgacgtttct cacctgcctt cgccaaggca ggggccggga
7861 ccctattcca agaggtagta actagcagga ctctagcctt ccgcaattca ttgagcgcat
7921 ttacggaagt aacgtcgggt actgtctctg gccgcaaggg tgggaggagt acgcatttgg
7981 cgtaaggtgg ggcgtagagc cttcccgcca ttggcggcgg atagggcgtt tacgcgacgg
8041 cctgacgtag cggaagacgc cttagtgggg gggaaggttc tagaaaagcg gcggcagcgg
8101 ctctagcggc agtagcagca gcgccgggtc ccgtgcggag gtgctcctcg cagagttgtt
8161 tctccagcag cggcagttct cactacagcg ccaggacgag tccggttcgt gttcgtccgc
8221 ggagatctct ctcatctcgc tcggctgcgg gaaatcgggc tgaagcgact gagtccgcga
8281 tggaggtaac gggtttgaaa tcaatgagtt attgaaaagg gcatggcgag gccgttggcg
8341 cctcagtgga agtcggccag ccgcctccgt gggagagagg caggaaatcg gaccaattca
8401 gtagcagtgg ggcttaaggt ttatgaacgg ggtcttgagc ggaggcctga gcgtacaaac
8461 agcttcccca ccctcagcct cccggcgcca tttcccttca ctgggggtgg gggatgggga
8521 gctttcacat ggcggacgct gccccgctgg ggtgaaagtg gggcgcggag gcgggacttc
8581 ttattccctt tctaaagcac gctgcttcgg gggccacggc gtctcctcgg ggatctagct
8641 tgtggaaggc tactcgaaat gtttgaccca agttaaacaa tttaaaggca atgctaccaa
8701 atactaattg agtgtatgta aacttctgac ccactgggaa tgtgatgaaa gaaataaaag
8761 ctgaaatgaa tcattctctc tactattatt ctgatatttc acattcttaa aataaagtgg
8821 tgatcctaac tgacctaaga cagggaattt ttactaggat taaatgtcag gaattgtgaa
8881 aaagtgagtt taaatgtatt tggctaaggt gtatgtaaac ttccgacttc aactgtatag
8941 ggttcctcta gctagagacg acctcgggta cccagctttt gttcccttta gtgagggtta
9001 attgcgcgct tggcgtaatc atggtcatag ctgtttcctg tgtgaaattg ttatccgctc
9061 acaattccac acaacatacg agccggaagc ataaagtgta aagcctgggg tgcctaatga
9121 gtgagctaac tcacattaat tgcgttgcgc tcactgcccg ctttccagtc gggaaacctg
9181 tcgtgccagc tgcattaatg aatcggccaa cgcgcgggga gaggcggttt gcgtattggg
9241 cgctcttccg cttcctcgct cactgactcg ctgcgctcgg tcgttcggct gcggcgagcg
9301 gtatcagctc actcaaaggc ggtaatacgg ttatccacag aatcagggga taacgcagga
9361 aagaacatgt gagcaaaagg ccagcaaaag gccaggaacc gtaaaaaggc cgcgttgctg
9421 gcgtttttcc ataggctccg cccccctgac gagcatcaca aaaatcgacg ctcaagtcag
9481 aggtggcgaa acccgacagg actataaaga taccaggcgt ttccccctgg aagctccctc
9541 gtgcgctctc ctgttccgac cctgccgctt accggatacc tgtccgcctt tctcccttcg
9601 ggaagcgtgg cgctttctca tagctcacgc tgtaggtatc tcagttcggt gtaggtcgtt
9661 cgctccaagc tgggctgtgt gcacgaaccc cccgttcagc ccgaccgctg cgccttatcc
9721 ggtaactatc gtcttgagtc caacccggta agacacgact tatcgccact ggcagcagcc
9781 actggtaaca ggattagcag agcgaggtat gtaggcggtg ctacagagtt cttgaagtgg
9841 tggcctaact acggctacac tagaaggaca gtatttggta tctgcgctct gctgaagcca
9901 gttaccttcg gaaaaagagt tggtagctct tgatccggca aacaaaccac cgctggtagc
9961 ggtggttttt ttgtttgcaa gcagcagatt acgcgcagaa aaaaaggatc tcaagaagat
10021 cctttgatct tttctacggg gtctgacgct cagtggaacg aaaactcacg ttaagggatt
10081 ttggtcatga gattatcaaa aaggatcttc acctagatcc ttttaaatta aaaatgaagt
10141 tttaaatcaa tctaaagtat atatgagtaa acttggtctg acagttacca atgcttaatc
10201 agtgaggcac ctatctcagc gatctgtcta tttcgttcat ccatagttgc ctgactcccc
10261 gtcgtgtaga taactacgat acgggagggc ttaccatctg gccccagtgc tgcaatgata
10321 ccgcgagacc cacgctcacc ggctccagat ttatcagcaa taaaccagcc agccggaagg
10381 gccgagcgca gaagtggtcc tgcaacttta tccgcctcca tccagtctat taattgttgc
10441 cgggaagcta gagtaagtag ttcgccagtt aatagtttgc gcaacgttgt tgccattgct
10501 acaggcatcg tggtgtcacg ctcgtcgttt ggtatggctt cattcagctc cggttcccaa
10561 cgatcaaggc gagttacatg atcccccatg ttgtgcaaaa aagcggttag ctccttcggt
10621 cctccgatcg ttgtcagaag taagttggcc gcagtgttat cactcatggt tatggcagca
10681 ctgcataatt ctcttactgt catgccatcc gtaagatgct tttctgtgac tggtgagtac
10741 tcaaccaagt cattctgaga atagtgtatg cggcgaccga gttgctcttg cccggcgtca
10801 atacgggata ataccgcgcc acatagcaga actttaaaag tgctcatcat tggaaaacgt
10861 tcttcggggc gaaaactctc aaggatctta ccgctgttga gatccagttc gatgtaaccc
10921 actcgtgcac ccaactgatc ttcagcatct tttactttca ccagcgtttc tgggtgagca
10981 aaaacaggaa ggcaaaatgc cgcaaaaaag ggaataaggg cgacacggaa atgttgaata
11041 ctcatactct tcctttttca atattattga agcatttatc agggttattg tctcatgagc
11101 ggatacatat ttgaatgtat ttagaaaaat aaacaaatag gggttccgcg cacatttccc
11161 cgaaaagtgc cacctgacgc gccctgtagc ggcgcattaa gcgcggcggg tgtggtggtt
11221 acgcgcagcg tgaccgctac acttgccagc gccctagcgc ccgctccttt cgctttcttc
11281 ccttcctttc tcgccacgtt cgccggcttt ccccgtcaag ctctaaatcg ggggctccct
11341 ttagggttcc gatttagtgc tttacggcac ctcgacccca aaaaacttga ttagggtgat
11401 ggttcacgta gtgggccatc gccctgatag acggtttttc gccctttgac gttggagtcc
11461 acgttcttta atagtggact cttgttccaa actggaacaa cactcaaccc tatctcggtc
11521 tattcttttg atttataagg gattttgccg atttcggcct attggttaaa aaatgagctg
11581 atttaacaaa aatttaacgc gaattttaac aaaatattaa cgcttacaat ttccattcgc
11641 cattcaggct gcgcaactgt tgggaagggc gatcggtgcg ggcctcttcg ctattacgcc
11701 agctggcgaa agggggatgt gctg
<---
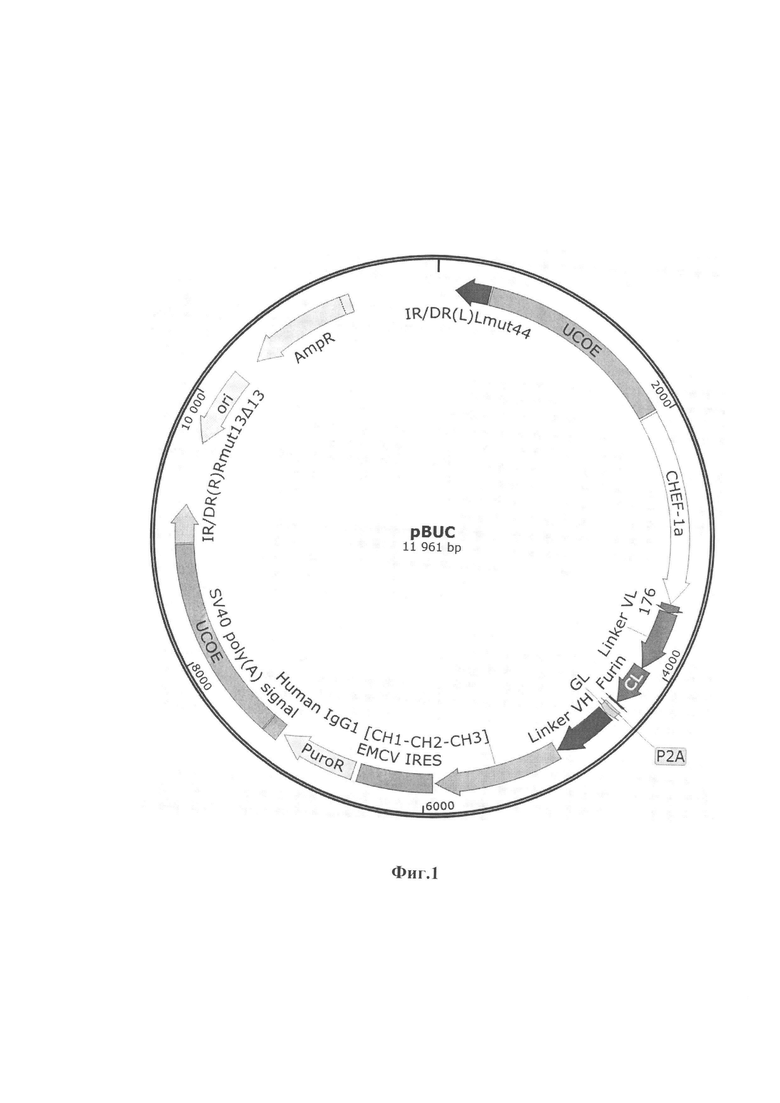
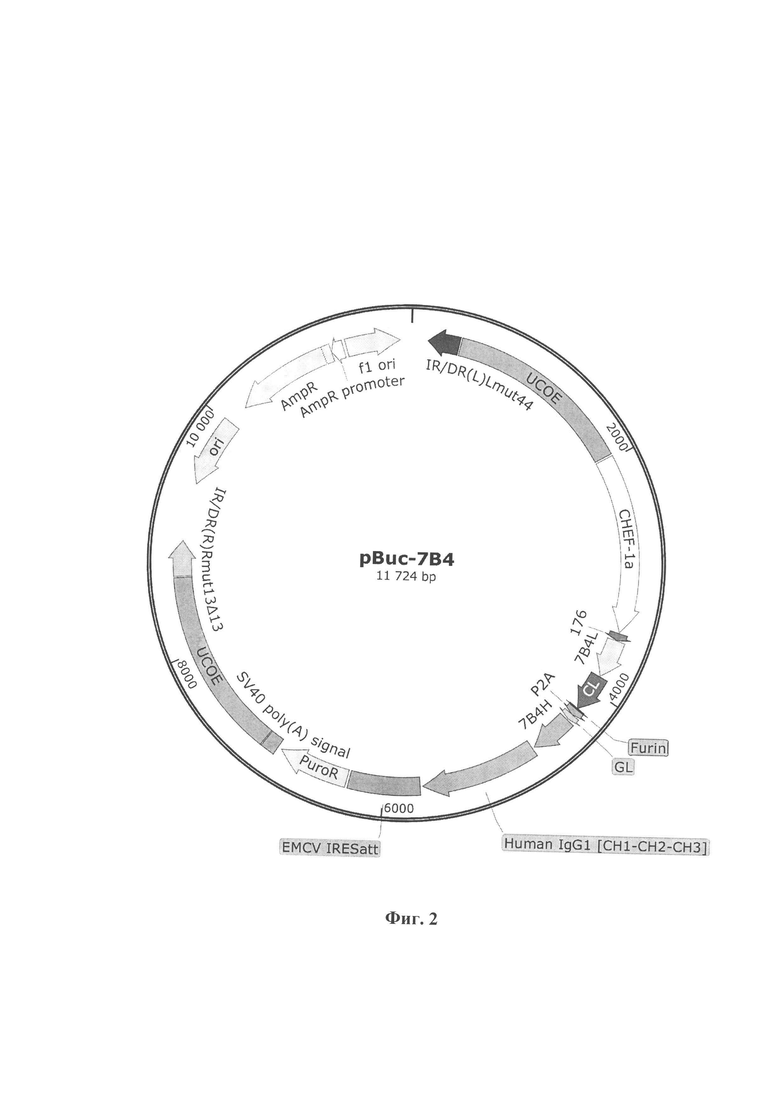
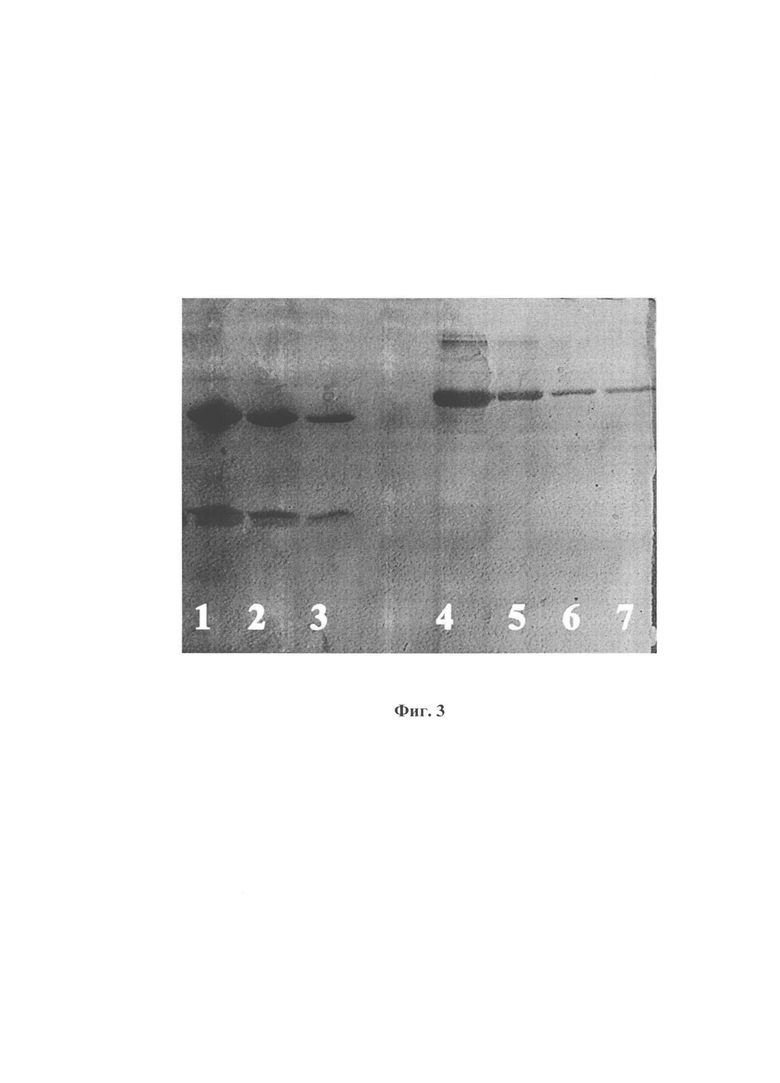
Изобретение относится к биотехнологии, в частности к универсальному рекомбинантному вектору pBuc и плазмидной генетической конструкции pBuc-7В4, обеспечивающей синтез и секрецию полноразмерных рекомбинантных белков моноклональных антител, в частности антитела 7В4 против вируса Западного Нила, в клетках яичника китайского хомяка (СНО) и полученной с использованием указанного вектора pBuc, и может быть использовано в медицине и биотехнологии. Данные белки будут обладать терапевтическим потенциалом. Также их можно будет использовать в научно-исследовательских целях. Изобретение позволяет создать универсальный интеграционный вектор pBuc и плазмидную генетическую конструкцию pBuc-7В4, обеспечивающую синтез и секрецию антитела 7В4 против вируса Западного Нила в клетках животных и полученную с использованием указанного вектора pBuc. 2 н.п. ф-лы, 3 ил., 2 табл., 4 пр.
1. Универсальный интеграционный вектор pBuc, служащий для создания целевых плазмидных конструкций, обеспечивающих синтез и секрецию моноклональных антител, имеет нуклеотидную последовательность SEQ ID No. 1 длиной 11961 п.н. и содержит в соответствии с физической и генетической картой, представленной на Фиг. 1, следующие элементы:
- участок начала репликации ori (координаты с 9663 по 10251 п.н.);
- CHEF-1а промотор (координаты с 1981 по 3477 п.н.), обеспечивающий автономную репликацию в клетках млекопитающих;
- GL последовательность (координаты с 4410 по 4460 п.н.), кодирующую лидерный пептид люциферазы Gaussia, обеспечивающей экспорт белка из клетки;
- 176 последовательность (координаты с 3483 по 3545 п.н.), кодирующую гибридный лидерный пептид люциферазы (Cypridina noctiluca) и фиброина (Dendrolimus spectabilis), обеспечивающий экспорт белка из клетки;
- Linker VL (координаты с 3546 по 4019 п.н.), синтезированную последовательность легкой цепи, фланкируемую уникальными сайтами рестрикции Bsa29I, AsuNHI;
- EMCV IRES (координаты с 5936 по 6510 п.н.), участок внутренней посадки рибосомы;
- Linker VH (координаты с 4464 по 4931 п.н.), синтезированную последовательность тяжелой цепи, фланкируемую уникальными сайтами рестрикции SphI, AsisI;
- Human IgG1 (CL- CHI- CH2- СН3) (координаты с 4939 по 5915 п.н.), последовательность, кодирующая константную цепь антитела человека с мутациями (M252Y, S254T, Т256Е, Е333А, H433K/N434F) для увеличения периода полураспада антител за счет взаимодействия с неонатальными рецепторами;
- SV40 poly(A) signal (координаты с 7188 по 7309 п.н.), последовательность, обеспечивающая стабилизацию мРНК-транскриптов;
- PuroR (координаты с 6554 по 7153 п.н.), ген устойчивости к антибиотику пуромицину;
- AmpR (координаты с 10422 по 11282 п.н.), ген устойчивости к антибиотику ампициллину;
- UCOE (координаты с 416 по 1964 п.н.) (координаты с 7319 по 8867 п.н), последовательности, предотвращающие эпигенетическое «замалчивание» экспрессии трансгена;
- Р2А (координаты с 4353 по 4409 п.н), пептидный линкер;
- IR/DR(L)Lmut44, IR/DR(R)Rmutl3Δ13 (координаты с 139 по 403 п.н. и координаты с 8879 по 9174 п.н.), участки распознавания транспазазой.
2. Плазмидная генетическая конструкция pBuc-7В4, обеспечивающая синтез и секрецию целевого белка - 7В4 против вируса Западного Нила, имеет нуклеотидную последовательность SEQ ID No. 2 длиной 11724 п.н. и содержит в соответствии с физической и генетической картой, представленной на Фиг. 2, следующие элементы:
- участок начала репликации ori (координаты с 9663 по 10251 п.н.);
- CHEF-1а промотор (координаты с 1981 по 3477 п.н.), обеспечивающий автономную репликацию в клетках млекопитающих;
- GL последовательность (координаты с 4410 по 4460 п.н.), кодирующую лидерный пептид люциферазы Gaussia, обеспечивающей экспорт белка из клетки;
- 176 последовательность (координаты с 3483 по 3545 п.н.), кодирующую гибридный лидерный пептид люциферазы Cypridina noctiluca и фиброина Dendrolimus spectabilis, обеспечивающий экспорт белка из клетки;
- 7B4-VH (координаты с 4308 по 4673 п.н.), последовательность, кодирующая тяжелую цепь антитела 7В4;
- EMCV IRESatt (координаты с 5687 по 6284 п.н.), участок внутренней посадки рибосомы;
- 7B4-VL (координаты с 3546 по 3863 п.н.), последовательность, кодирующая легкую цепь антитела 7В4;
- Human IgG1 (CL- CHI- CH2- СН3) (координаты с 4939 по 5915 п.н.), последовательность, кодирующая константную цепь антитела человека с мутациями (M252Y, S254T, Т256Е, Е333А, H433K/N434F) для увеличения периода полураспада антител в организме за счет взаимодействия с неонатальными рецепторами;
- SV40 poly(A) signal (координаты с 7188 по 7309 п.н.), последовательность, обеспечивающая стабилизацию мРНК-транскриптов;
- PuroR (координаты с 6554 по 7153 п.н.), ген устойчивости к антибиотику пуромицину;
- AmpR (координаты с 10422 по 11282 п.н.), ген устойчивости к антибиотику ампициллину;
- UCOE (координаты с 416 по 1964 п.н.) (координаты с 7319 по 8867 п.н), последовательности, предотвращающие эпигенетическое «замалчивание» экспрессии трансгена;
- Р2А (координаты с 4353 по 4409 п.н.), пептидный линкер;
- IR/DR(L)Lmut44, IR/DR(R)Rmutl3Δ13 (координаты с 139 по 403 п.н. и координаты с 8879 по 9174 п.н.), участки распознавания транспазазой.
| CN 105255895 A, 20.01.2016 | |||
| US 20110097798 A1, 28.04.2011 | |||
| US 20160024189 A1, 28.01.2016 | |||
| Лушова А.А | |||
| и др | |||
| Новое поколение методов получения человеческих моноклональных антител | |||
| Молекулярная биология | |||
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |

