Область техники.
Изобретение относится к области биотехнологии, а именно к способам получения рекомбинантной CAS13A-нуклеазы свободной от бактериальных эндотоксинов, пригодной для использования в системе CRISPR/Cas.
Уровень техники.
CRISPR-нуклеазы являются частью молекулярных систем CRISPR-Cas (CRISPR, clustered regularly interspaced short palindromic repeats; Cas, CRISPR associated protein), ответственных за иммунитет бактерий и архей к чужеродным генетическим элементам, таким как бактериофаги и плазмиды (Источник [1]: Van der Oost J., Westra E.R., Jackson R.N., Wiedenheft B. //Nat. Rev. Microbiol. 2014. V. 12, № 7. P. 479-492. Источник [2]. Koonin E.V., Makarova K.S., Zhang F. //Curr. Opin. Microbiol. 2017. V. 37, P. 67-78.).
Способность систем CRISPR-Cas селективно узнавать заданные последовательности ДНК и расщеплять их в определённых местах легла в основу наиболее широко распространённой на сегодня технологии направленного редактирования геномов, в первую очередь с использованием нуклеазы Cas9 и её генетически модифицированных вариантов (Источник [3]: Hsu P.D., Lander E.S., Zhang F. //Cell. 2014. V. 157, № 6, P. 1262-1278.). Селективность узнавания определяется формированием комплекса Cas-нуклеазы с направляющей РНК (нРНК, от англ. guide RNA, gRNA), которая содержит участок (спейсер), комплементарный участку таргетной ДНК (протоспейсер), и участок, ответственный за связывание с Cas-нуклеазой (повтор). При формировании гетеродуплекса «спейсер-протоспейсер» находящаяся в комплексе с нРНК Cas-нуклеаза расщепляет ДНК-мишень при наличии в непосредственной близости к протоспейсеру определённого нуклеотидного мотива (PAM, protospacer adjacent motif) (Источник [3]).
CRISPR-нуклеаза Cas13a (ранее известная как C2c2) принадлежит к VI типу CRISPR-Cas систем и обладает рядом особенностей, отличающих её от большей части CRISPR-нуклеаз, включая Cas9. Нуклеаза Cas13a является рибонуклеазой с функцией селективного узнавания таргетной молекулы РНК (через формирование комплементарного дуплекса спейсера нРНК с соответствующим участком РНК-мишени) с последующим её расщеплением, которое не зависит от наличия PAM (Источник [4]: Abudayyeh O.O., Gootenberg J.S., Konermann S., Joung J., Slaymaker I.M., Cox D.B., Shmakov S., Makarova K.S., Semenova E., Minakhin L., Severinov K., Regev A., Lander E.S., Koonin E.V., Zhang F. //Science. 2016. V. 353, № 6299. P. aaf5573. Источник [5]: Pickar-Oliver A., Gersbach C.A. //Nat. Rev. Mol. Cell. Biol. 2019. V. 20. № 8, P. 490-507). После активации и расщепления РНК-мишени нуклеаза теряет специфичность (активность только в отношении РНК-мишени) и приобретает так называемую «коллатеральную активность» – начинает расщеплять любые молекулы РНК. При этом коллатеральная активность Cas13a проявляется в условиях in vitro и при гетерологичной коэкспрессии Cas13a и нРНК в бактериальных клетках, но отсутствует при их экспрессии в эукариотических клетках (Источник [6]: Abudayyeh O.O., Gootenberg J.S., Essletzbichler P., Han S., Joung J., Belanto J.J., Verdine V., Cox D.B.T., Kellner M.J., Regev A., Lander E.S., Voytas D.F., Ting A.Y., Zhang F. //Nature. 2017. V. 550. № 7675. P. 280-284.Источник [7]: Cox D.B.T., Gootenberg J.S., Abudayyeh O.O., Franklin B., Kellner M.J., Joung J., Zhang F. // Science. 2017. V. 358. № 6366. P. 1019-1027.). Следует отметить, что последнее создаёт основу для развития технологии направленного редактирования транскриптома (ibid).
Локусы, кодирующие нуклеазу Cas13a, были впервые идентифицированы в геномах пяти бактериальных таксонов: α-протеобактерии, Bacilli, Clostridia, Fusobacteria и Bacteroidetes (Источник [8]: Shmakov S., Abudayyeh O.O., Makarova K.S., Wolf Y.I., Gootenberg J.S., Semenova E., Minakhin L., Joung J., Konermann S., Severinov K., Zhang F., Koonin E.V. //Mol. Cell. 2015. V. 60. № 3. P. 385-397.). В настоящее время в экспериментальных исследованиях наиболее интенсивно используются Cas13a-нуклеазы, кодируемые геномами бактерий рода Leptotrichia (Источник 4;6. Источник [9]: Tambe A., East-Seletsky A., Knott G.J., Doudna J.A., O'Connell M.R. //Cell Rep. 2018. V. 24. № 4. P. 1025-1036. Источник [10]: East-Seletsky A., O'Connell M.R., Knight S.C., Burstein D., Cate J.H., Tjian R., Doudna J.A. //Nature. 2016. V. 538. № 7624. P. 270-273. Источник [11]: Gootenberg J.S., Abudayyeh O.O., Lee J.W., Essletzbichler P., Dy A.J., Joung J., Verdine V., Donghia N., Daringer N.M., Freije C.A., Myhrvold C., Bhattacharyya R.P., Livny J., Regev A., Koonin E.V., Hung D.T., Sabeti P.C., Collins J.J., Zhang F. //Science. 2017. V. 356. № 6336. P. 438-442.). Экспериментальное тестирование 15 ортологов Сas13a показало, что Cas13а-нуклеаза обнаруженная у вида Leptotrichia wadei, обозначаемая LwaCas13a, наиболее эффективно подавляет рост клеток Escherichia coli (E. coli) при гетерологичной коэкспрессии Сas13a и нРНК (источник [6]). Нуклеотидная последовательность гена нуклеазы LwaCas13a включает ~3,5 тыс. пар оснований и кодирует полипептид длиной 1162 аминокислотных остатка (а.о.); молекулярный вес белка составляет ≈138 кДa. Коллатеральная активность LwaCas13 была положена в основу способа ультрачувствительной детекции бактерий и вирусов, получившего название SHERLOCK (источник [11], источник [12]: Gootenberg J.S., Abudayyeh O.O., Kellner M.J., Joung J., Collins J.J., Zhang F. //Science. 2018. V. 360. № 6387. P. 439-444. Источник [13]: Kellner M.J., Koob J.G., Gootenberg J.S., Abudayyeh O.O., Zhang F. //Nat. Protoc. 2019. V. 14. № 10. P. 2986-3012.). В методе SHERLOCK на первом этапе проводится изотермическая амплификация целевого фрагмента нуклеиновой кислоты, а затем детекция продуктов амплификации с применением LwaCas13a в комплексе с нРНК. Коллатеральная активность LwaCas13a приводит к деградации добавленных в реакционную смесь коротких молекул РНК, меченных флуорофором и тушителем (РНК-субстрат или РНК-репортеры), с последующей регистрацией флуоресцентного сигнала (источник [11-13]). Все реакции не требуют специализированного оборудования, а детекция может проводиться в формате тест-полосок (при замене флуорофора на биотиновую или пероксидазную метку), что открывает перспективы для полевой диагностики бактериальных и вирусных патогенов (Источник [14]: Myhrvold C., Freije C.A., Gootenberg J.S., Abudayyeh O.O., Metsky H.C., Durbin A.F., Kellner M.J., Tan A.L., Paul L.M., Parham L.A., Garcia K.F., Barnes K.G., Chak B., Mondini A., Nogueira M.L., Isern S., Michael S.F., Lorenzana I., Yozwiak N.L., MacInnis B.L., Bosch I., Gehrke L., Zhang F., Sabeti P.C. //Science. 2018. V. 360. № 6387. P. 444-448.). Cas13a нклеаза и аналогичные ей CRISPR-нуклеазы, чья коллатеральная активность проявляется при селективном узнавании заданных последовательностей в таргетных ДНК и РНК, рассматриваются сегодня как основа для создания нового поколения биосенсоров c чувствительностью детекции на уровне единичных ДНК/РНК молекул (Источник [15]: Li Y., Li S., Wang J., Liu G. //Trends Biotechnol. 2019. V. 37. № 7. P. 730-743.).
В настоящее время для получения рекомбинантной рибонуклеазы Cas-нуклеазы используют гетерологичную экспрессии в бактерий Escherichia coli. Для получения рекомбинантной рибонуклеазы Cas13a, требуемой чистоты, свободной от токсинов, используют трёхстадийную очистку, включающую последовательно стадии аффинной хроматографии (Strep-tag), ионнообменной хроматографии и эксклюзионной ВЭЖХ гель-фильтрации (Источники [11;12;13]). Экспрессия Cas13a-нуклеазы в клетках бактерии Escherichia coli отличается высоким выходом рекомбинантной рибонуклеазы Cas13a, обусловленной доступностью получения и воспроизведения бактерии Escherichia coli, однако фрагменты клеток бактерии Escherichia coli соединяются с получаемой Cas13a-нуклеазой и чистота получаемой Cas13a-нуклеазы недостаточна для применения, что требует выполнения дополнительной очистки Cas13a-нуклеазы от бактериальных экдотоксинов.
Недостаток известного способа получения Cas13a-нуклеазы заключается в технологической сложности осуществления способа, необходимостью использования трёхстадийной очистки для обеспечения достаточной чистоты конечного препарата Cas13a-нуклеазы от бактериальных эндотоксинов. Требуется техническое обеспечение (оборудование) для выполнения последовательной очистки и доочистки путем аффинной хроматографии (Strep-tag), ионнообменной хроматографии и эксклюзионной ВЭЖХ (гель-фильтрации), что делает практическое использование Cas13a-нуклеазы труднодоступным, ограничивает возможности для ее широкого (массового) применения.
Задачей изобретения является повышение доступности очищенного (в интервале 80-90%) рекомбинантного белка Cas13a-нуклеазы с функциональной коллатеральной РНКазной активностью, способного расщеплять любые молекулы РНК. Обеспечение получения очищенного рекомбинантного белка Cas13a-нуклеазы с коллатеральной РНКазной активностью, технологически простым способом, снижающим потребность в использовании технического обеспечения (различного оборудования) и не требующим многостадийной доочистки препарата от бактериальных эндотоксинов.
Авторы настоящего изобретения, в результате собственных исследований, пришли к решению, что задача получения рекомбинантной Cas13a-нуклеазы с функциональной активностью, способной расщеплять любые молекулы РНК, с достаточной степенью чистоты может быть успешно решена путем трансформации экспрессионного плазмидного вектора pET His6-TwinStrep-SUMO-LwaCas13a в клетки E.coli c последующим выделением и очисткой целевого белка Cas13a-нуклеазы путем использования аффинной или металл-хелатной хроматографии с использованием N-концевой полигистидиновой метки (6хHis-tag) и гравитационных хроматографических колонок.
Плазмидный вектор pET His6-TwinStrep-SUMO-LwaCas13a идентичен плазмиде pC013 His6-TwinStrep-SUMO-LwCas13a, депонированной в репозиторий ADDGENE (www.addgene.org/90097), которая была сконструирована для гетерологичной экспрессии рекомбинантного белка LwaCas13a (источник [11]). Конструкция данного плазмидного вектора предполагает индуцированную экспрессию с Т7-промотора, при этом в последовательность целевого белка с N-конца включены последовательно 6xHis-tag/(Twin)Strep-tag/SUMO. Это позволяет проводить очистку рекомбинантного белка в одну стадию путем хроматографии. При этом выход целевого белка максимален с 2 (второй) по 6 (шестую) фракцию элюата с хроматографической колонки (фиг.1).
Технический результат, заключается в получении очищенного рекомбинантного белка Cas13a-нуклеазы с функциональной коллатеральной РНКазной активностью, технологически простым способом. Упрощение процедуры получения и очистки рекомбинантного белка Cas13a-нуклеазы с функциональной коллатеральной РНКазной активностью делает Cas13a-нуклеазы более доступными для практического применения и проведения исследований, что позволяет расширить возможности по разработке и практическому использованию диагностических биосенсорных систем нового поколения, в технике, связанной с распознаванием и редактированием генов.
Технический результат достигается тем, что в способе получения рекомбинантной Cas13a-нуклеазы с коллатеральной активностью включающем трансформацию клеток бактерии Escherichia coli экспрессионным вектором, экспрессию и очистку целевого рекомбинантного белка Cas13a, очистку целевого рекомбинантного белка Cas13a-нуклеазы производят в одну стадию путем хроматографии.
Предполагается, что очистку целевого рекомбинантного белка Cas13a-нуклеазы производят с использованием последовательности аминокислотных остатков, добавленной генно-инженерными методами к C- или N-концу рекомбинантного белка Cas13a-нуклеазы. Предусмотрено, что при очистке проводят оценку функциональной активности белка Cas13a-нуклеазы.
Предполагается, что в экспрессионный вектор в последовательность гена белка добавляют последовательность нуклеотидов, кодирующую полигистидиновую метку His-tag на N- или С- конце белка, состоящую из шести или более аминокислотных остатков гистидина. Очистку целевого рекомбинантного белка Cas13a-нуклеазы производят в одну стадию путем аффинной хроматографии или металл-хелатной хроматографии. Предполагается, что применяют экспрессионный вектор pET His6-TwinStrep-SUMO-LwaCas13a который включает последовательность меток 6xHis-tag/(Twin)Strep-tag/SUMO.
Предусмотрено, что очистку рекомбинантного белка Cas13a-нуклеазы производят путем аффинной хроматографии на сефарозе с иммобилизованным Strep-Tactin.
Предусмотрено, что применяют аффинную хроматографию, основанную на специфическом взаимодействии добавленной последовательности с лигандом, иммобилизованном на твердофазном носителе.
Предусмотрено, что очистку целевого рекомбинантного белка Cas13a-нуклеазы производят с использованием полигистидиновой аминокислотной последовательности и матрицы в виде крупнопорового кремния, заряженного ионами никеля или кобальта в качестве хелатирующего лиганда.
Предполагается, что при очистке целевого рекомбинантного белка Cas13a-нуклеазы путем хроматографии, проводят оценку содержания целевого рекомбинантного белка Cas13a-нуклеазы с использованием гравитационных хроматографических колонок. Оценку целевого рекомбинантного белка Cas13a-нуклеазы во фракциях элюата с хроматографических колонок проводят с использованием гравитационных хроматографических колонок, содержащих хелатирующий лиганд, матрицу в виде крупнопорового кремния с заряженным Ni2+, обеспечивающих очистку белков с полигистидиновой меткой His-tag.
Предусмотрено, что бактерии Escherichia coli культивируют при 37°С, рост клеток контролируют измеряя оптическую плотность на 600 нм, при достижении D600 значения 0,6 в культуру добавляют индуктор экспрессии изопропил-β-D-1-тиогалактопиранозид до конечной концентрации 50 мкМ, экспрессию целевого рекомбинантного белка Cas13a-нуклеазы проводят течение 16 часов при 18°С, клетки бактерии осаждают центрифугированием, промывают фосфатно-солевым буфером.
Предусмотрено, что операции по получению целевого рекомбинантного белка Cas13a-нуклеазы проводят при +4°С, ресуспендируют клеточный осадок в колонке в буфере LEW, вносят лизоцим до конечной концентрации 1 мг/мл, фенилметансульфонил фторид до 1 мМ и ДНКаза до 1 мг/мл, инкубируют клетки бактерии Escherichia coli на льду в течение 30 минут, разрушают клетки бактерии Escherichia coli механическим способом с помощью френч-пресса, полученный гомогенат осветляют центрифугированием, наносят гомогенат на колонку, промывают колонку, связавшийся материал элюируют буфером, содержащим 250 мМ имидазола, последовательно собирают фракции объемом 0,5 мл, анализируют фракции путем электрофореза по Лэммли в 9% ПААГ с окраской красителем Кумасси G-250, оценивают относительное количество целевого рекомбинантного белка Cas13a-нуклеазы путем сопоставления площадей пиков, соответствующих белковым полосам на геле, после его денситометрии на гель-сканере, очищенный целевой белок Cas13a-нуклеазы переводят в буфер для хранения с использованием диализа.
Заявляемый в настоящем изобретении способ выгодно отличается от способов известных из уровня техники, возможностью получения достаточно чистого рекомбинантного белка Cas13a-нуклеазы с функциональной коллатеральной РНКазной путем одностадийной очистки.
Изобретение поясняется графическими материалами:
Фиг.1 –Фотография. ПААГ-электрофорез исходного гомогената клеток E. coli и фракций элюата при очистке целевого белка Cas13a-нуклеазы металл-хелатной хроматографией на колонках Protino Ni-TED 1000. 1 – гомогенат клеток, 2-6 – фракции элюата со второй по шестую. Целевой белок Cas13a-нуклеазы указан стрелкой. Слева показаны молекулярные массы в кДа.
Фиг.2 –График. Изменение относительной флуоресценции при инкубации флуоресцентно-меченного РНК-субстрата с целевым белком Cas13a-нуклеазы при 37ºС. Относительная флуоресценция вычислялась как отношение F/F0, F – флуоресценция в указанный момент времени, F0 – флуоресценция в начальный момент в отсутствии целевого белка Cas13a-нуклеазы. 1 – изменение флуоресценции в отсутствии РНК-мишени; 2, 3, 4, 5 и 6 – в присутствии 10, 25, 80, 250 и 800 нг суммарной РНК E. coli, соответственно. Показаны среднеарифметические значения и среднеквадратичные отклонения для трипликатных измерений.
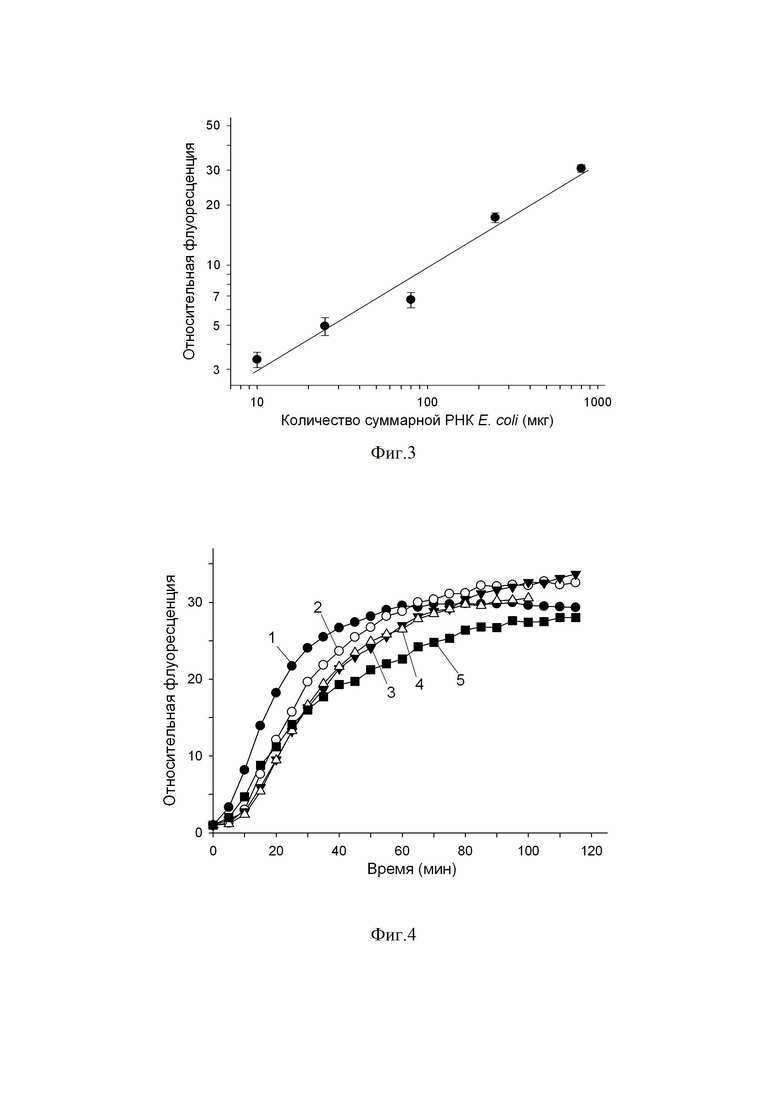
Фиг.3 – График. Изменение относительной флуоресценции (после 100 минутной инкубации флуоресцентно-меченного РНК-субстрата с целевым белком Cas13a-нуклеазы при 37ºС) как функция количества суммарной РНК E. coli.
Фиг.4 – График. Зависимость функциональной активности Cas13a-нуклеазы от времени хранения. 1 – кинетика относительной флуоресценции при инкубации флуоресцентно-меченного РНК-субстрата со свежеприготовленной Cas13a-нуклеазой; 2, 3, 4 и 5 – после её хранения при -80 ºС в течении 1, 2, 3,5 и 5 месяцев, соответственно. Количество суммарной РНК E. coli – 800 нг. Показаны среднеарифметические значения для трипликатных измерений; относительные среднеквадратичные отклонения не превышали 15%.
Фиг.5 – Таблица. Последовательности олигонуклеотидов, использованных в работе. Последовательность, комплементарная последовательности Т7-промотера, показана курсивом. Полужирным шрифтом выделена последовательность спейсера.
Осуществление изобретения.
Способ получения рекомбинантной Cas13a-нуклеазы с коллатеральной активностью включает трансформацию клеток бактерии Escherichia coli экспрессионным плазмидным вектором, экспрессию и очистку целевого рекомбинантного белка Cas13a-нуклеазы. Очистку целевого рекомбинантного белка Cas13a-нуклеазы производят в одну стадию путем хроматографии. Причем для очистки применяют (выбирают) аффинную хроматографию или металл-хелатную хроматографию.
Предполагается, что очистку целевого рекомбинантного белка Cas13a-нуклеазы производят с использованием последовательности аминокислотных остатков, добавленной генно-инженерными методами к C- или N-концу рекомбинантного белка Cas13a-нуклеазы.
В экспрессионный плазмидный вектор в последовательность гена белка предварительно добавляют последовательность нуклеотидов, кодирующую полигистидиновую метку His-tag на N- или С- конце белка, состоящую из шести или более аминокислотных остатков гистидина. Предполагается, что экспрессионный вектор pET His6-TwinStrep-SUMO-LwaCas13a включает последовательность меток 6xHis-tag/(Twin)Strep-tag/SUMO.
Очистку целевого рекомбинантного белка Cas13a-нуклеазы производят с использованием полигистидиновой аминокислотной последовательности и матрицы в виде крупнопорового кремния, заряженного ионами никеля или кобальта в качестве хелатирующего лиганда.
Предусмотрено, что производят очистку рекомбинантного белка с путем аффинной хроматографии на сефарозе с иммобилизованным Strep-Tactin. В одном варианте осуществления способа применяют аффинную хроматографию, основанную на специфическом взаимодействии добавленной последовательности с лигандом, иммобилизованном на твердофазном носителе.
При очистке целевого рекомбинантного белка Cas13a-нуклеазы путем хроматографии, проводят оценку содержания целевого рекомбинантного белка Cas13a-нуклеазы с использованием гравитационных хроматографических колонок. Оценку целевого рекомбинантного белка Cas13a-нуклеазы во фракциях элюата с хроматографических колонок проводят с использованием гравитационных хроматографических колонок, содержащих хелатирующий лиганд, матрицу в виде крупнопорового кремния с заряженным Ni2+, обеспечивающих очистку белков с полигистидиновой меткой His-tag. Выход целевого рекомбинантного белка Cas13a-нуклеазы максимален с второй по шестую фракцию элюата с хроматографической колонки.
На первом этапе осуществляют трансформацию клеток бактерии Escherichia coli экспрессионным плазмидным вектором. Для бактериальной экспрессии белка Cas13a-нуклеазы используют плазмидный вектор pET His6-TwinStrep-SUMO-LwaCas13a, представленный компанией «Novoprolabs» (КНР; Кат. №V10159). Согласно спецификации поставщика, плазмидный вектор идентичен плазмиде pC013 His6-TwinStrep-SUMO-LwCas13a, депонированной в репозиторий ADDGENE (www.addgene.org/90097), который сконструирован гетерологичной экспрессии рекомбинантного белка LwaCas13a (Источник [11]). Конструкция данного плазмидного вектора pET His6-TwinStrep-SUMO-LwaCas13a предполагает индуцированную экспрессию с Т7-промотора, при этом в последовательность целевого белка с N-конца включены последовательно 6xHis-tag/(Twin)Strep-tag/SUMO. Это позволяет проводить очистку рекомбинантного белка белка Cas13a-нуклеазы в одну стадию путем применения либо металл-хелатной хроматографии, либо аффинной хроматографии на сефарозе с иммобилизованным Strep-Tactin.
Экспрессионный плазмидный вектор pET His6-TwinStrep-SUMO-LwaCas13a трансформируют в штамм бактерии E. coli например Rosetta™ 2(DE3)pLysS Singles™ Competent Cells («Merck», Германия). Ночная культура (среда LB с добавлением ампициллина и хлорамфеникола в стандартных концентрациях) в количестве 2 мл вносят в 300 мл среды Terrific Broth (содержит 12 г/л триптона, 2 г/л пептона, 24 г/л дрожжевого экстракта, 5 г/л глицерина, 12,5 г/л K2HPO4 и 2,3 г/л KH2PO4), культивируют при 37°С. Рост клеток контролируют измерением оптической плотности на 600 нм (D600). При достижении D600 значения 0,6 в культуру добавляют индуктор экспрессии изопропил-β-D-1-тиогалактопиранозид (ИПТГ) до конечной концентрации 50 мкМ.
Производят экспрессию целевого белка Cas13a-нуклеазы в течение 16 часов при 18˚С. Клетки бактерии осаждают центрифугированием (4000 g, 15 мин, 4°С), промывают фосфатно-солевым буфером (Phosphate Buffered Saline, «Sigma-Aldrich», США) и используют для получения целевого белка Cas13a-нуклеазы. Операции по получению целевого белка Cas13a проводят при +4°С.
Производят очистку целевого рекомбинантного белка Cas13a-нуклеазы. Очистку рекомбинантного белка Cas13a-нуклеазы осуществляют на хромотографических колонках, например Protino Ni-TED 1000 («Macherey-Nagel», Германия) в соответствии с протоколом производителя. Клеточный осадок ресуспендируют в буфере LEW (50 мМ NaH2PO4, 300 мМ NaCl, pH 8.0) из набора к колонкам Protino Ni-TED 1000. Дополнительно в буфер вносят лизоцим (например «Sigma-Aldrich», США) до конечной концентрации 1 мг/мл, фенилметансульфонил фторид (phenylmethanesulfonyl fluoride, «Sigma-Aldrich», США) до 1 мМ и ДНКаза (DNase I, «Roche», Швейцария) до 1 мг/мл. После инкубации на льду в течение 30 минут клетки разрушают механическим способом с помощью френч-пресса («Thermo Electron Corporation», США). Полученный гомогенат осветляют центрифугированием – 20 000 g, 40 мин, 4°С. Осветлённый гомогенат наносят на колонку. После промывки колонки, связавшийся материал элюируют буфером, содержащим 250 мМ имидазола, последовательно собирая фракции объемом 0,5 мл. Фракции анализируют методом электрофореза по Лэммли в 9% ПААГ с окраской красителем Кумасси G-250 («Bio-Rad», США).
Относительное количество целевого рекомбинантного белка Cas13a-нуклеазы оценивают сопоставлением площадей пиков, соответствующих белковым полосам на геле, после его денситометрии на гель-сканере ImageScanner III («GE Healthcare», США). Очищенный аффинной хроматографией целевой белок Cas13a-нуклеазы переводят в буфер для хранения (600 мМ NaCl, 50 мМ Tris-HCl, pH 7,5, 5% глицерин, 2 мМ DTT) с помощью диализа. Диализ проводят с использованием мембран SnakeSkin Dialysis Tubing 7000 MWCO («Thermo Fisher Scientific», США). Концентрации суммарного и целевого белка измеряют методом Брэдфорда, используя растворы БСА в качестве стандартов. Аликвоты раствора целевого белка хранят на -80°С.
Для масс-спектрометрической идентификации целевого рекомбинантного белка Cas13a-нуклеазы представляющие интерес белковые полосы вырезают из полиакриламидного геля после ПААГ-электрофореза препаратов очищенного белка и содержащийся в них материал подвергают трипсинолизу в геле в соответствии с протоколом, описанным в (Источник [16] Shevchenko A., Tomas H., Havlis J., Olsen J.V., Mann M. //Nat. Protoc. 2006. V. 1. № 6. P. 2856-2860.). Полученную смесь пептидов анализируют на времяпролетном MALDI-TOF/TOF масс-спектрометре Bruker Ultraflex II («Bruker Daltonics», Германия) и методом тандемной хромато-масс-спектрометрии (LC-ESI-MS/MS) на масс-спектрометре Orbitrap Q-Exactive («Thermo Fisher Scientific», США), сочленённом с хроматографической системой Agilent HPLC 1100 Series (“Agilent Technologies”, США). В последнем случае используют аналитическую колонку Zorbax 300SB-C18 (“Agilent Technologies”, США) с обращенной фазой.
Спектры пептидных масс, полученные методом MALDI-TOF масс-спектрометрии, сравнивают с теоретически ожидаемыми спектрами из базы данных NCBI при помощи веб-платформы Mascot Peptide Mass Fingerprint (“Matrix Science”, Великобритания), используют следующие параметры: погрешность определения массы (mass tolerance) моноизотопного, пропионамидирование цистеина считали фиксированной модификацией, окисление метионина – вариабельной, допускали один пропуск сайта трипсинолиза. В случае LC-ESI-MS/MS, масс-спектры обрабатывают с помощью программных продуктов SearchGUI (v. 3.3.16) (Источник [17]: Barsnes H., Vaudel M. Searchgui: //J. Proteome Res. 2018. V. 17. № 7. P. 2552-2555.) и PeptideShaker (v. 1.16.42) (Источник [18]: Vaudel M., Burkhart J.M., Zahedi R.P., Oveland E., Berven F.S., Sickmann A., Martens L., Barsnes H. //Nat. Biotechnol. 2015. V. 33. № 1. P. 22-24.). Поиск проводили тремя поисковыми алгоритмами (X! Tandem, MS-GF+ и OMSSA) (Источник [19]: Bjornson R.D., Carriero N.J., Colangelo C., Shifman M., Cheung K.H., Miller P.L., Williams K. //Proteome Res. 2008. V. 7, № 1. P. 293-299. Источник [20]: Kim S., Pevzner P.A. //Nat. Commun. 2014. V. 5. P. 5277. Источник [21]: Geer L.Y., Markey S.P., Kowalak J.A., Wagner L., Xu M., Maynard D.M., Yang X., Shi W., Bryant S.H. //J. Proteome Res. 2004. V. 3. № 5. P. 958-964.) в кастомизированных библиотеках белковых последовательностей, включающих последовательности белков E. coli K12 (Swiss-Prot, release 09-2019, 4533 белковые последовательности) и лабораторных белков-контаминантов cRAP (Swiss-Prot, release 01-2012, 115 последовательностей), а также последовательности белков L. wadei (штамм F0279; UniProt Proteome ID UP000016626, 2277 белков) и Leptotrichia buccalis (UniProt Proteome ID UP000001910; 2218 белков). При обработке результатов хромато-масс-спектрометрического анализа устанавливают следующие параметры поиска: погрешность определения массы пептидного иона – 10 ppm, фрагментного иона – 0,05 Да, один пропуск сайта трипсинолиза, фиксированная модификация – карбамидометилирование цистеина, вариабельная – окисление метионина. Белок считали идентифицированным, если удавалось детектировать не менее двух протеотипических пептидов, на каждый из которых приходилось не меньше 2 PSM (т.е. фактов совпадения теоретического спектра со спектром экспериментальным). Отсечение по ложноположительным идентификациям как для пептидов, так и для белков было установлено на уровне 1%.
Оценку функциональной активности выделенного белка проводят следующим образом. Для синтеза направляющей (нРНК) используют ДНК-матрицу D8R, содержащую на 3’-конце участок, комплементарный последовательности промотора ДНК-зависимой РНК-полимеразы фага Т7, и олигонуклеотид T7F с последовательностью Т7-промотора (фиг.5). ДНК-олигонуклеотиды были синтезированы компанией «Синтол» (Россия). Эквимолярную смесь олигонуклеотидов D8R и T7F отжигают для формирования дуплекса и использовали для синтеза in vitro нРНК с помощью набора TranscriptAid T7 High Yield Transcription Kit («Thermo Fisher Scientific», США) согласно инструкции производителя. Очистку нРНК от примесей проводят смесью фенол/хлороформ/изоамиловый спирт 25:24:1 («Acros Organics», Бельгия) с последующим осаждением этанолом. Осадок растворяют в воде, свободной от нуклеаз, и концентрацию нРНК определяют на спектрофотометре NanoDrop 1000 («Thermo Fisher Scientific», США). Аликвоты раствора нРНК хранили на -80ºС. Последовательность нРНК представлена в таблице (фиг.5)
В качестве мишени для оценки эндонуклеазной активности полученного рекомбинантного белка Cas13a-нуклеазы используют рибосомную РНК 16S E. coli штамма XL1 Blue. Суммарную РНК E. coli выделяют с помощью реагента ExtractRNA («Евроген», Россия) согласно инструкции производителя. Реакционная смесь для оценки коллатеральной активности включает буфер (40 мМ Трис-HCl pH 7,3; 60 мМ NaCl; 6 мМ MgCl2), 45 нМ рекомбинантного белка, 22,5 нМ нРНК, 125 нМ субстрата из набора RNase Alert v2 («Thermo Fisher Scientific», США), 2 μL ингибитора РНКаз (Murine RNase Inhibitor, «New England Biolabs», Великобритания), 100 нг референсной РНК (Universal Human Reference RNA, “Agilent Technologies”, США) в качестве фона и различное количество суммарной РНК E. coli. Перед добавлением суммарной РНК E. coli, референсной РНК и субстрата из набора RNase Alert v2, смесь инкубировали в течение 15 мин при 37°C. После их добавления, реакционную смесь инкубируют 1,5-2 часа при 37°C. Флуоресценцию, возникающую при деградации субстрата из набора RNase Alert v2 для оценки РНКазной активности, детектируют с помощью планшетного ридера Infinite M200 PRO («TECAN», Швейцария), используя длины волн 490 нм и 520 нм для возбуждения и эмиссии, соответственно.
Выход рекомбинантного белка Cas13a-нуклеазы при элюции с колонки максимален в третьей фракции (фиг. 1), при этом в первой и восьмой фракциях целевой белок полностью отсутствует. Существенное количество целевого рекомбинантного белка Cas13a-нуклеазы также присутствовует в четвёртой фракции. Материал третьей и четвертой фракций был объединён и использован для тестирования функциональной активности рекомбинантного белка как Cas13a-нуклеазы. Чистота препаратов рекомбинантного белка Cas13a-нуклеазы, полученных объединением третьей и четвёртой фракций элюата, лежит в интервале 80-90%.
В качестве мишени для проверки функциональной активности очищенного белка используют рибосомную РНК 16S штамма XL1 Blue E. coli. Такой выбор определяется доступностью данного штамма и простотой получения суммарной РНК, в которой рибосомальная РНК 16S представляет одну из доминирующих компонент. Последовательности возможных нРНК были подобраны с помощью программы CRISPR-RT (Источник [22]: Zhu H., Richmond E., Liang C. //Bioinformatics. 2018. V. 34. № 1. P. 117-119.), исходя из того, что полученный рекомбинантный белок представляет рибонуклеазу LwaCas13a. Последующий анализ вторичной структуры предложенных программой CRISPR-RT последовательностей нРНК с помощью программы Mfold (Источник [23]: Zuker M. //Nucleic Acids Res. 2003. V. 31. № 13. P. 3406-3415.) показал, что среди них присутствует значительное число последовательностей, у которых с высокой вероятностью могут образовываться нежелательные дуплексные участки между последовательностями спейсера и повтора. Такие дуплексы могут препятствовать формированию последовательностью повтора характерной шпилечной структуры, ответственной за взаимодействие с Cas13a (источник [11]). В результате проведённого анализа, последовательность нРНК, представленная в табл. (фиг.5), была выбрана как наиболее оптимальная. Последовательность спейсера данной нРНК комплементарна участку с 12 по 40 нуклеотид молекулы 16S РНК E. coli (протоспейсер).
Оценка функциональной рибонуклеазной активности полученного рекомбинантного белка, проведённая с помощью набора для детекции рибонуклеазной активности RNase Alert v2, показала, что в отсутствии РНК-мишени белок не проявляет рибонуклеазной активности (фиг. 2). Однако при добавлении РНК-мишени наблюдается развитие флуоресцентного сигнала, связанное с расщеплением РНК-репортеров в результате появившейся рибонуклеазной активности. Таким образом, полученный рекомбинантный белок обладает мишень-зависимой коллатеральной рибонуклеазной активностью, типичной для Cas13a-нуклеаз. В серии последовательных разведений суммарной РНК E. coli была показана зависимость развития флуоресцентного сигнала от концентрации мишени, которая в логарифмических координатах может быть хорошо аппроксимирована линейной функцией (фиг. 3).
Экспрессия функционально активного рекомбинантного белка Cas13a-нуклеазы проводилась при 18°С. Тестировали температуры культивирования клеток после индукции экспрессии (от 18°С до 37°С) и их влияние на уровень экспрессии и функциональную активность целевого белка. Было установлено, что повышение температуры до 37°С приводит к тому, что целевой белок практически весь переходит в клетках в нерастворимую форму. Инкубация в температурном диапазоне от 25°С до 30°С позволяет получать Cas13a-нуклеазу в растворимой форме, однако при этом полученный рекомбинантный белок проявляет неспецифическую рибонуклеазную активность. Это проявляется в том, что рекомбинантная Cas13a-нуклеаза расщепляет РНК-репортеры в отсутствии РНК-мишени. Культивирование при 18°С приводит к относительно невысокому уровню экспрессии целевого белка (количество чистого белка, которое получается в этих условиях, составляет ~1 мг на 100 мл среды), оно обеспечивает экспрессию функционально активной Cas13a-нуклеазы.
Таким образом, показано, что рекомбинантный белок Cas13a-нуклеазы с мишень-зависимой коллатеральной РНКазной активностью может быть получен после гетерологичной экспрессии в E. coli с помощью простой одностадийной очистки хроматографией с использованием N-концевой полигистидиновой метки. Это расширяет возможности исследования Cas13a-нуклеаз как основы для разработки диагностических биосенсорных систем нового поколения и их последующего практического использования.
Способ обеспечивает получение очищенного рекомбинантного белка Cas13a-нуклеазы с функциональной коллатеральной РНКазной активностью, технологически простым способом. Упрощение процедуры получения и очистки рекомбинантного белка Cas13a-нуклеазы с функциональной коллатеральной РНКазной активностью делает Cas13a-нуклеазы более доступными для практического применения и исследований, что позволяет расширить возможности по разработке и практическому использованию диагностических биосенсорных систем нового поколения, в технике, связанной с распознаванием и редактированием генов. Способ выгодно отличается от известных из уровня техники способов возможностью получения достаточно чистого рекомбинантного белка Cas13a-нуклеазы с функциональной коллатеральной РНКазной с использованием простой одностадийной очистки путем металл-хелатной хроматографии или аффинной хроматографии, основанной на специфическом взаимодействии добавленной последовательности с лигандом, иммобилизованном на твердофазном носителе.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ И ОЧИСТКИ ФУНКЦИОНАЛЬНО АКТИВНОГО РЕКОМБИНАНТНОГО БЕЛКА NSP1 SARS-COV-2 И СПОСОБ ИДЕНТИФИКАЦИИ ПОТЕНЦИАЛЬНЫХ ЛЕКАРСТВЕННЫХ СОЕДИНЕНИЙ ПРОТИВ COVID-19, НАПРАВЛЕННЫХ НА ИНГИБИРОВАНИЕ NSP1 | 2021 |
|
RU2822761C2 |
| Способ получения препарата рекомбинантной нуклеазы CAS, по существу, свободного от бактериальных эндотоксинов, полученный данным способом препарат и содержащий его набор для использования в системе CRISPR/Cas | 2019 |
|
RU2707542C1 |
| СИСТЕМА РЕДАКТИРОВАНИЯ ГЕНОМНОЙ ДНК ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ НА ОСНОВЕ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ, КОДИРУЮЩЕЙ БЕЛОК SUCAS9NLS | 2022 |
|
RU2804422C1 |
| НУКЛЕАЗА CPF1 ИЗ БАКТЕРИИ Ruminococcus bromii, МОЛЕКУЛА ДНК ИЛИ РНК, КОДИРУЮЩАЯ НУКЛЕАЗУ, ВЕКТОР, СОДЕРЖАЩИЙ УКАЗАННУЮ МОЛЕКУЛУ ДНК, СИСТЕМА CRISPR/CPF1, СОДЕРЖАЩАЯ УКАЗАННУЮ НУКЛЕАЗУ И НАПРАВЛЯЮЩУЮ РНК, КЛЕТКА-ХОЗЯИН ДЛЯ ПОЛУЧЕНИЯ НУКЛЕАЗЫ CPF1, СПОСОБ ПОЛУЧЕНИЯ НУКЛЕАЗЫ CPF1 И ЕЕ ПРИМЕНЕНИЕ | 2022 |
|
RU2816876C1 |
| Рекомбинантная плазмидная ДНК pCSBrz4M, кодирующая полипептид со свойствами сладкого белка браззеина, и рекомбинантный штамм E. coli - продуцент полипептида со свойствами сладкого белка браззеина | 2022 |
|
RU2799530C1 |
| Средство обнаружения нуклеиновых кислот на основе ScCas12a белка из бактерии Sedimentisphaera cyanobacteriorum | 2023 |
|
RU2820345C1 |
| ШТАММ ESCHERICHIA COLI BL21(DE3)PLYSS/PET15B-HISCPF1 - ПРОДУЦЕНТ РНК-НАПРАВЛЯЕМОЙ ЭНДОНУКЛЕАЗЫ CRISPR/CPF1 | 2021 |
|
RU2774120C1 |
| Рекомбинантный слитый белок, специфически связывающий и нейтрализующий вирус бешенства, способ его получения и применение | 2023 |
|
RU2829507C1 |
| Тест-система для детекции целевой ДНК или РНК с использованием короткого белка-аргонавта в комплексе с нуклеазой и флуоресцентного репортера | 2023 |
|
RU2821993C1 |
| Новая компактная нуклеаза CAS9 II типа из Anoxybacillus flavithermus | 2020 |
|
RU2749307C1 |


Изобретение относится к способу получения рекомбинантной CAS13A-нуклеазы, свободной от бактериальных эндотоксинов, пригодной для использования в системе CRISPR/Cas. Технический результат заключается в получении очищенного рекомбинантного белка Cas13a-нуклеазы с функциональной коллатеральной РНКазной активностью, технологически простым способом. 13 з.п. ф-лы, 5 ил.
1. Способ получения рекомбинантной Cas13a-нуклеазы с коллатеральной активностью, включающий трансформацию клеток бактерии Escherichia coli экспрессионным вектором, экспрессию и очистку целевого рекомбинантного белка Cas13a, отличающийся тем, что трансформацию клеток бактерии Escherichia coli осуществляют экспрессионным вектором pET His6-TwinStrep-SUMO-LwaCas13a, очистку целевого рекомбинантного белка Cas13a-нуклеазы производят в одну стадию путем хроматографии, с использованием полигистидиновой метки His-tag и гравитационных хроматографических колонок.
2. Способ по п. 1, отличающийся тем, что очистку целевого рекомбинантного белка Cas13a-нуклеазы производят в одну стадию путем аффинной хроматографии.
3. Способ по п. 1, отличающийся тем, что очистку целевого рекомбинантного белка Cas13a-нуклеазы производят в одну стадию путем металл-хелатной хроматографии.
4. Способ по п. 1, отличающийся тем, что в экспрессионный вектор в последовательность гена белка добавлена последовательность нуклеотидов, кодирующая полигистидиновую метку His-tag на N- или С-конце белка, состоящая из шести или более аминокислотных остатков гистидина.
5. Способ по п. 3, отличающийся тем, что очистку целевого рекомбинантного белка Cas13a-нуклеазы производят с использованием полигистидиновой аминокислотной последовательности и матрицы в виде крупнопорового кремния, заряженного ионами никеля или кобальта в качестве хелатирующего лиганда.
6. Способ по п. 1, отличающийся тем, что очистку целевого рекомбинантного белка Cas13a-нуклеазы путем хроматографии производят с использованием последовательности аминокислотных остатков, добавленной генно-инженерными методами к C- или N-концу рекомбинантного белка.
7. Способ по п. 2, отличающийся тем, что применяют аффинную хроматографию, основанную на специфическом взаимодействии добавленной последовательности с лигандом, иммобилизованным на твердофазном носителе.
8. Способ по п. 1, отличающийся тем, что при очистке целевого рекомбинантного белка Cas13a-нуклеазы путем хроматографии проводят оценку содержания целевого рекомбинантного белка Cas13a-нуклеазы с использованием гравитационных хроматографических колонок.
9. Способ по п. 1, отличающийся тем, что экспрессионный вектор pET His6-TwinStrep-SUMO-LwaCas13a включает последовательность меток 6xHis-tag/(Twin)Strep-tag/SUMO.
10. Способ по п. 8, отличающийся тем, что оценку целевого рекомбинантного белка Cas13a-нуклеазы во фракциях элюата с хроматографических колонок проводят с использованием гравитационных хроматографических колонок, содержащих хелатирующий лиганд, матрицу в виде крупнопорового кремния с заряженным Ni2+, обеспечивающих очистку белков с полигистидиновой меткой His-tag.
11. Способ по п. 2, отличающийся тем, что производят очистку рекомбинантного белка путем аффинной хроматографии на сефарозе с иммобилизованным Strep-Tactin.
12. Способ по п. 1, отличающийся тем, что бактерии Escherichia coli с экспрессионным вектором культивируют при 37°С, рост клеток контролируют, измеряя оптическую плотность на 600 нм, при достижении D600 значения 0,6 в культуру добавляют индуктор экспрессии изопропил-β-D-1-тиогалактопиранозид до конечной концентрации 50 мкМ, экспрессию целевого белка проводят в течение 16 часов при 18°С, клетки осаждают центрифугированием, промывают фосфатно-солевым буфером.
13. Способ по п. 1, отличающийся тем, что операции по получению целевого белка Cas13a-нуклеазы проводят при +4°С, ресуспендируют клеточный осадок в буфере LEW, вносят лизоцим до конечной концентрации 1 мг/мл, фенилметансульфонил фторид до 1 мМ и ДНКазу до 1 мг/мл, инкубируют клетки бактерии на льду в течение 30 минут, разрушают клетки бактерии механическим способом с помощью френч-пресса, полученный гомогенат осветляют центрифугированием, наносят гомогенат на колонку, промывают колонку, связавшийся материал элюируют буфером, содержащим 250 мМ имидазола, последовательно собирают фракции объемом 0,5 мл, анализируют фракции путем электрофореза по Лэммли в 9% ПААГ с окраской красителем Кумасси G-250, оценивают относительное количество целевого белка Cas13а путем сопоставления площадей пиков, соответствующих белковым полосам на геле, после его денситометрии на гель-сканере, очищенный целевой белок переводят в буфер для хранения с использованием диализа.
14. Способ по п. 1, отличающийся тем, что при очистке проводят оценку функциональной активности белка Cas13a-нуклеазы.
| САВИНОВА А.С | |||
| и др., Cas13a: очистка и использование для обнаружения вирусной РНК, Вестник Российского государственного медицинского университета, 2018, н.2, стр.22-27 | |||
| GOOTENBERG J.S | |||
| et al., Nucleic acid detection with CRISPR-Cas13a/C2c2, Science, 2017, Vol.356, N6336, pp | |||
| Способ обогащения кислородных руд путем взбалтывания пены | 1911 |
|
SU438A1 |
| KOONIN E.V | |||
| et al., Diversity, classification and evolution of | |||

