Область техники
Настоящее изобретение относится к моноклональному антителу, которое подавляет (ингибирует) иммуносупрессорные функции патогена, антиген-связывающему фрагменту указанного антитела и к гибридомам, вырабатывающим указанное антитело. В частности настоящее изобретение относится к моноклональному антителу, которое подавляет иммуносупрессорные функции, обусловленные веществом-иммуносупрессором, секретируемым или вырабатываемым патогеном, усиливая иммунную систему хозяина указанного патогена, и к антиген-связывающий фрагмент указанного антитела, и к гибридомам, вырабатывающим указанное антитело.
Уровень техники
Helicobacter pylori (Н. pylori) представляет собой грамотрицательную бактерию, которой инфицирована половина взрослого населения во всем мире. Хроническое воспаление, индуцируемое Н. pylori, может приводить к разным последствиям, включая язву желудка и двенадцатиперстной кишки и рак желудка, в зависимости от степени и распространенности гастрита, вызванного данным патогеном, не смотря на то, что в организме хозяина преимущественно активируется иммунный ответ слизистых оболочек, в основном опосредованный Th1, такого иммунного ответа недостаточно для создания защитного иммунитета от Н. pylori, что приводит к хроническим инфекциям и развитию патологий желудка у некоторых пациентов. Предыдущие исследования показали, что лизаты Н. pylori могут подавлять митоген-индуцированную пролиферацию Т-лимфоцитов, что указывает на то, что в лизате присутствуют определенные факторы, связанные с иммуносупрессорной активностью. Такие факторы снижают активность Т-лимфоцитов, независимо от генов вирулентности бактерий CagA и VacA. Для объяснения того, как Н. pylori прямо или косвенно подавляет опосредованный Т-лимфоцитами иммунитет, было предложено несколько механизмов: Н. pylori может подавлять пролиферацию Т-лимфоцитов и экспрессию TCR (рецепторов Т-лимфоцитов) посредством аргиназы, стимулировать выделение ингибирующего цитокина TGF-β, препятствовать зависимой от инвариантной цепи презентации антигена через VacA, подавлять функции DC через фосфорилирование CagA или подавлять фагоцитоз, осуществляемый профессиональными фагоцитами, через VirB7 и VirB11.
Несмотря на возможное участие перечисленных выше механизмов, в настоящее время главными регуляторными компонентами в подавлении активности Т-лимфоцитов и в поддержании равновесия между воспалением и выживанием бактерий считаются регуляторные Т-лимфоциты (Treg-лимфоциты). В 2003 году было обнаружено, что в вызываемой Н. pylori иммунопатологии и колонизации участвуют CD4+CD25+Т-лимфоциты. Дальнейшие исследования показали, что Treg-лимфоциты хозяина играют важную роль для защиты инфицированного Н. pylori хозяина от чрезмерного воспаления желудка и совокупности симптомов при болезни, но в то же время способствуют колонизации бактерий на слизистой оболочке желудка и двенадцатиперстной кишки. Кроме того, экспрессия В7-Н1 клетками эпителия желудка стимулирует развитие CD4+CD25+FoxP3+Treg-лимфоцитов после воздействия Н. pylori, что свидетельствует о том, что данный патоген стимулирует индукцию Treg-лимфоцитов хозяина. В последующих исследованиях изучали функции таких индуцированных Н. pylori Treg-лимфоцитов, и было показано, что они могут подавлять активность или вызывать толерантность специфичных к Н. pylori эффекторных Т-лимофоцитов. Кроме того, индуцированный Н. pylori гастрит ассоциирован с привлечением природных FoxP3+ Treg-лимфоцитов, которое коррелирует со степенью колонизации бактериями и экспрессии TGF-β1 (трансформирующего фактора роста-β1) в слизистой оболочке. В совокупности указанные результаты указывают на то, что реакции с участием Treg-лимфоцитов хозяина, в ответ на инфекцию Н. pylori, являются важными механизмами регуляции иммунного ответа на Н. pylori и вовлечены в патогенез заболеваний, связанных с Н. pylori.
Белок теплового шока 60 Н. pylori (HpHSP60) может индуцировать экспрессию провоспалительных цитокинов и TGF-β1 в моноцитах. Сообщалось, что HpHSP60 экспрессируется в клеточной стенке бактерий, ассоциирован с уреазой и может выступать в роли молекулы адгезии для клеток эпителия желудка. Кроме того, было показано, что введение антитела против HSP60 препятствует росту Н. pylori. Следовательно, HpHSP60 является не только необходимым фактором для выживания Н. pylori, но также важной молекулой, которая облегчает колонизацию в желудке человека. Однако многие исследования показали, что HpHSP60 выступает в роли сильного иммуногена, приводя к сильной индукции провоспалительных цитокинов, таких как ФНО-α (фактор некроза опухоли α), ИЛ-8 (интерлейкин-8) и ИЛ-6. Указанные цитокины приводят к воспалению в очаге инфекции, и такое HpHSP60-индуцированное воспаление может обладать свойством стимуляции процессов образования злокачественных опухолей, включая ангиогенез и метастазирование. HpHSP60 также является важным фактором вирулентности для инфекции Н. pylori в организме человека-хозяина.
Взятые в совокупности, данные о взаимосвязи между HpHSP60 и Treg-лимфоцитами, являются многообещающими и являются основанием для дальнейших исследований. Однако самые последние исследования сосредоточены на воспалении, вызываемом HpHSP60. Очень малое число исследований посвящено взаимосвязи между HpHSP60 и иммуносупрессорными реакциями, проявляемыми в организме хозяина.
В патенте США №6403099 раскрываются конъюгированные соединения, содержащие белок теплового шока и олигосахарид или полисахарид капсулы. Указанные соединения способны индуцировать образование антител к полисахариду. Указанный белок теплового шока включает белок теплового шока Н. pylori.
Краткое описание изобретения
Согласно настоящему изобретению определенные патогены способны подавлять иммунитет своего хозяина. Такие иммуносупрессорные функции в частности обеспечиваются иммуносупрессорными веществами, секретируемыми или вырабатываемыми указанными патогенами. Можно ингибировать иммунносупрессорные функции указанных патогенов, если выключить активность указанных иммуносупрессорных веществ. Когда указанная иммуносупрессорная активность ингибирована, активность иммунитета хозяина не подавляется. В результате, не ингибированная или усиленная иммунная активность хозяина может уменьшать количество патогенов или даже уничтожать их.
Авторы настоящего изобретения обнаружили новое моноклональное антитело, которое в существенной степени ингибирует иммуносупрессорные функции или вещества конкретных патогенов. На основании данного открытия предложено моноклональное антитело согласно настоящему изобретению, его антиген-связывающий фрагмент, гибридомы для выработки таких антител, а также способы их получения.
Следовательно, целью настоящего изобретения является обеспечение нового моноклонального антитела. Указанное моноклональное антитело в значительной степени подавляет иммуносупрессорные функции конкретных патогенов.
Еще одна задача настоящего исследования состоит в том, чтобы обеспечить антиген-связывающий фрагмент указанного моноклонального антитела, а также гибридомы, которые генерируют такое антитело.
Еще одна задача настоящего исследования состоит в том, чтобы обеспечить способы применения моноклонального антитела, его антиген-связывающего фрагмента и гибридом, которые генерируют такое антитело.
Согласно одному аспекту настоящего изобретения моноклональное антитело или его антиген-связывающий фрагмент соединяются с пептидом, содержащим аминокислотную последовательность MEKVGKDGVITVE (SEQ ID №1).
Также согласно настоящему изобретению предложены гибридомы, способные вырабатывать указанное моноклональное антитело или его антиген-связывающий фрагмент.
Моноклональное антитело согласно настоящему изобретению проявляет достоверное ингибирующее влияние на иммуносупрессорную активность конкретных патогенов и может применяться при блокировании иммуносупрессии, вызываемой патогенами. Согласно определенным вариантам реализации настоящего изобретения иммуносупрессорная функция патогенов обеспечивается иммуносупрессорным веществом, секретируемым или вырабатываемым указанными патогенами. Согласно таким вариантам реализации моноклональное антитело или его антиген-связывающий фрагмент согласно настоящему изобретению преимущественно приводят к уничтожению патогенов посредством подавления иммуносупрессорных функций указанных патогенов, и, следовательно, активации иммунных ответов хозяина на указанные патогены.
Согласно другому аспекту настоящего изобретения предложены способы получения моноклонального антитела или его антиген-связывающего фрагмента, и гибридомы, способные вырабатывать указанное антитело или указанные фрагменты. Указанный способ, включает: применение пептида, содержащего аминокислотную последовательность MEKVGKDGVITVE (SEQ ID №1), в качестве антигена, чтобы индуцировать в организме млекопитающего иммунный ответ на указанный антиген; получение иммунной клетки из организма такого млекопитающего, иммунизированного против указанных патогенов, и осуществление слияния полученной иммунной клетки с клеткой миеломы млекопитающих с получением гибридомы; клонирование полученной гибридомы с получением желаемой гибридомы. Способ согласно настоящему изобретению может также включать этапы: применения полученной гибридомы для получения антител; и сбор антител, выработанных указанной гибридомой.
Согласно предпочтительному варианту реализации настоящего изобретения указанная иммунная клетка включает клетку селезенки.
Моноклональное антитело согласно настоящему изобретению или его антиген-связывающий фрагмент можно применять непосредственно или в составе фармацевтической композиции, которая включает фармацевтически приемлемые вспомогательные вещества и др. Согласно одному варианту реализации настоящего изобретения предложена фармацевтическая композиция, и указанная композиция содержит моноклональное антитело или его антиген-связывающий фрагмент согласно настоящему изобретению. Согласно другому варианту реализации настоящего изобретения указанную фармацевтическую композицию применяют в качестве функционального ингибитора в отношении конкретных иммуносупрессорных веществ. Также согласно настоящему изобретению предложено применение моноклонального антитела согласно настоящему изобретению, включая его применение при получении фармацевтической композиции.
Краткое описание чертежей
На Фиг. 1 показаны результаты экспериментов по оценке влияния HpHSP60 на пролиферацию мононуклеарных клеток периферической крови (МКПК).
На Фиг. 2 показаны результаты экспериментов по оценке влияния HpHSP60 на пролиферацию Т-лимфоцитов.
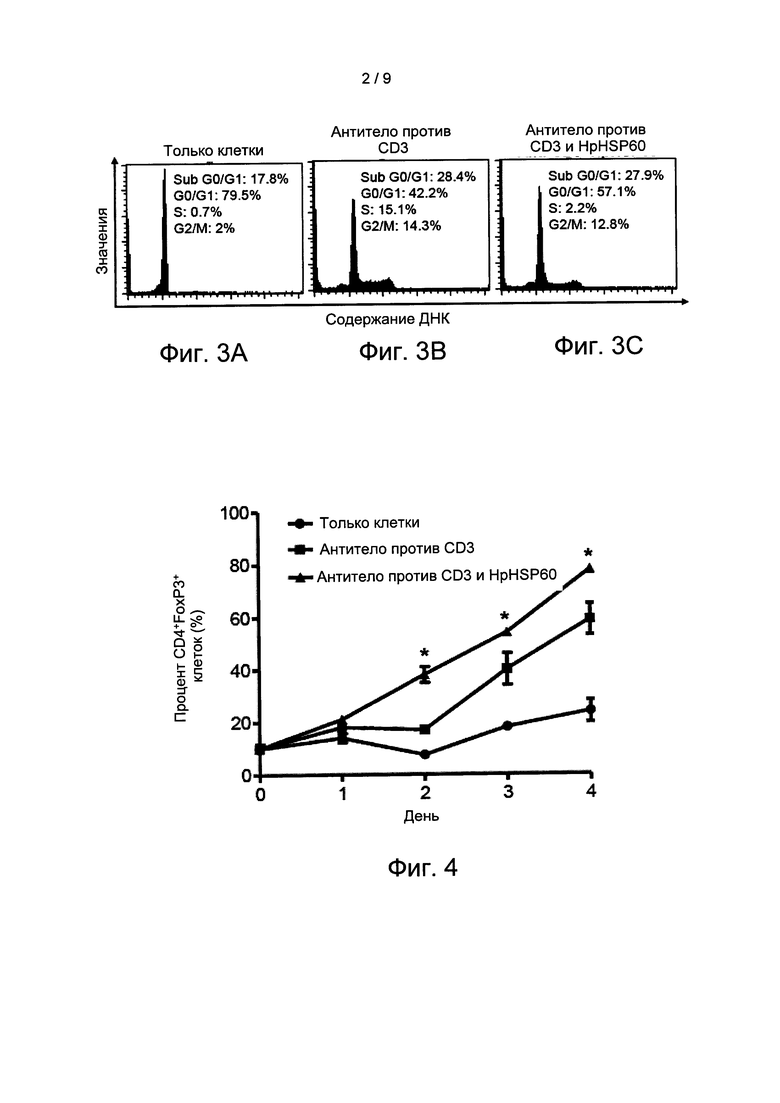
На Фиг. 3А-3С, соответственно, показаны результаты экспериментов по оценке влияния HpHSP60 на клеточные циклы МКПК.
На Фиг. 4 показаны результаты экспериментов по оценке индукции Treg-лимфоцитов in vitro белком HpHSP60.
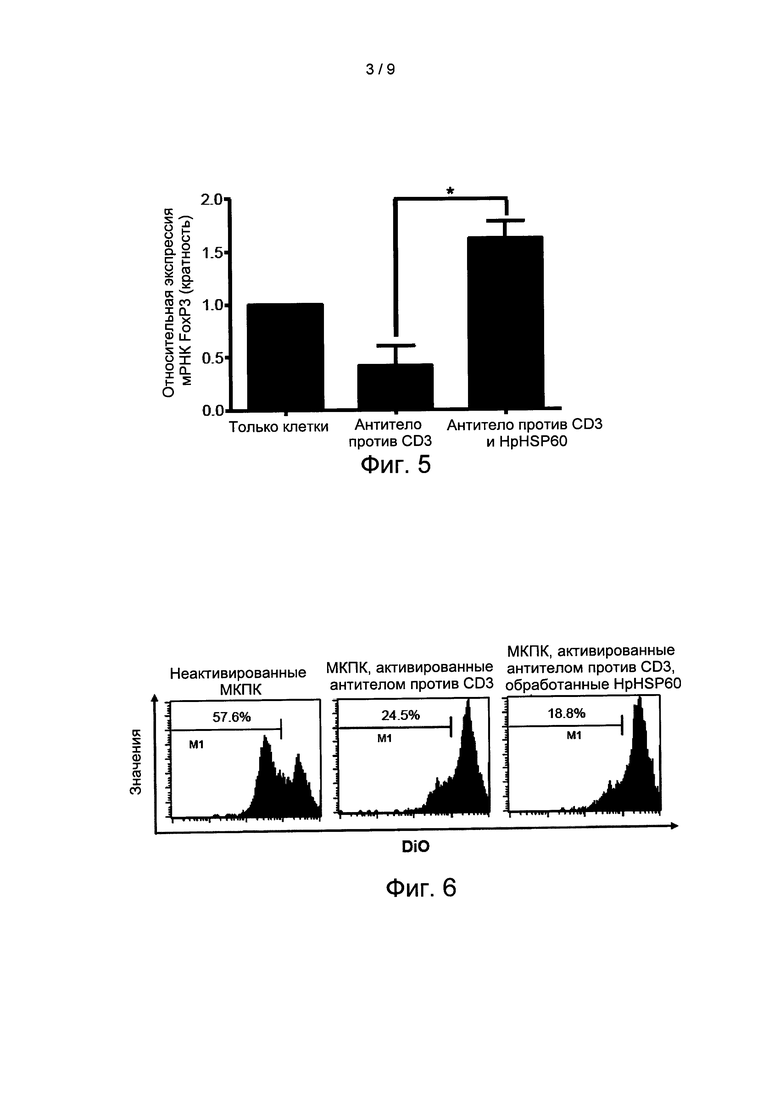
На Фиг. 5 показаны результаты экспериментов по оценке влияния HpHSP60 на пролиферацию Treg-лимфоцитов.
На Фиг. 6 показаны результаты экспериментов по оценке влияния HpHSP60-индуцированных Treg-лимфоцитов на пролиферацию Т-лимфоцитов.
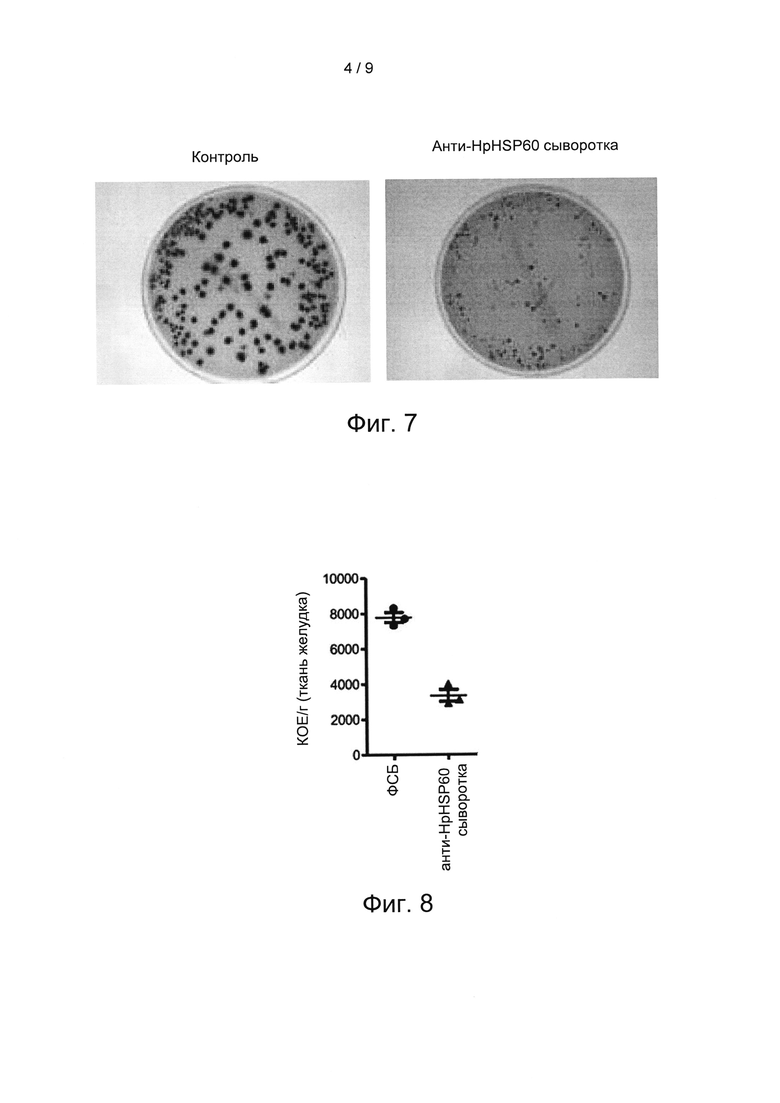
На Фиг. 7 показаны результаты экспериментов по оценке подавления роста Н. pylori in vivo вследствие блокирования иммуносупрессорных функций HpHSP60.
На Фиг. 8 показаны результаты другого эксперимента по оценке подавления роста Н. pylori in vivo вследствие блокирования иммуносупрессорных функций HpHSP60.
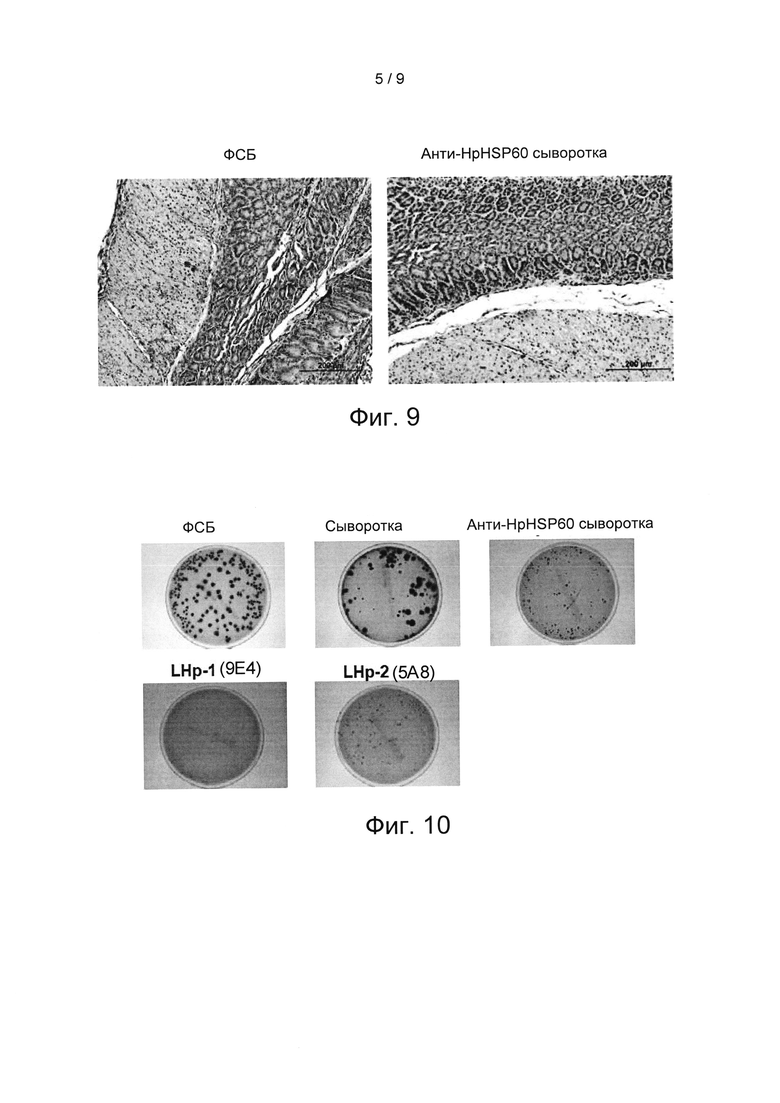
На Фиг. 9 показаны результаты экспериментов по оценке подавления Treg-лимфоцитов посредством блокирования иммуносупрессорных функций HpHSP60.
На Фиг. 10 показаны результаты экспериментов с активной последовательностью HpHSP60, которая индуцирует рост Treg-лимфоцитов.
На Фиг. 11 показана количественная оценка результатов экспериментов, показанных на Фиг. 10.
На Фиг. 12 показаны результаты экспериментов по изучению иммунологического механизма действия анти-HpHSP60 антител (антител против HpHSP60).
На Фиг. 13 показаны результаты экспериментов по оценке влияния анти-HpHSP60 антител на экспрессию в Treg-лимфоцитах в слизистой оболочке желудка мышей.
На Фиг. 14 показаны результаты экспериментов по оценке влияния анти-HpHSP60 антител на экспрессию ИЛ-10 в слизистой оболочке желудка мышей.
На Фиг. 15 показаны результаты экспериментов по распознаванию фрагментов HpHSP60 антителом LHP-1 (9Е4).
На Фиг. 16 показаны результаты дополнительных экспериментов по распознаванию фрагментов HpHSP60 антителом LHP-1 (9Е4).
Подробное описание изобретения
Не желая ограничивать настоящее изобретение какой-либо теорией, согласно настоящему изобретению определенные патогены способны подавлять иммунитет своих хозяев, секретируя или вырабатывая иммуносупрессорные вещества, которые обеспечивают пролиферацию или вызывают заболевание в организме хозяина. Примеры таких патогенов включают Н. pylori и другие сходные бактерии, такие как Helicobacter felis и Arcobacter suis. Авторы изобретения обнаружили, что белок теплового шока является одним из таких иммуносупрессорных веществ. Согласно вариантам реализации настоящего изобретения белок теплового шока 60 Н. pylori (HpHSP60) способен вступать в реакцию с моноцитами, стимулируя выработку иммуносупрессорных гормонов, таких как ИЛ-10 и TGF-β, и индуцируя пролиферацию Treg-лимфоцитов. Как следствие, иммунитет хозяина подавляется, и хозяин уже не способен сопротивляться хроническим инфекциям Н. pylori.
Согласно настоящему изобретению разработан новый способ усиления иммунитета хозяев. Для ингибирования активности иммуносупрессорных веществ применяют функциональный ингибитор, который блокирует функции иммуносупрессорных веществ. Таким образом, пролиферация Treg-лимфоцитов эффективно подавляется, и исключается иммуносупрессорный ответ хозяина.
Авторы изобретения обнаружили новое моноклональное антитело, которое в значительной степени подавляет иммуносупрессорные функции веществ конкретных патогенов. Таким образом, указанное моноклональное антитело или его антиген-связывающие фрагменты можно применять в качестве функционального ингибитора, который способен к идентификации иммуносупрессорного вещества или его фрагмента и блокированию функций такого иммуносупрессорного вещества.
Депонирование
Антитело LHP-1 (9Е4) было создано с применением в качестве антигена последовательности аминокислот в положениях 101-200 белка HpHSP60. Гибридомы, содержащие данное антитело - LHP-1 (9Е4) - были депонированы в Американской коллекции типовых культур (АТСС®), 10801 University Boulevard, Manassas, VA, 20110, USA, обозначение в ATCC: РТА-122900 (код доступа в АТСС: ). Дата депонирования 03.02.2016.
Моноклональные антитела и гибридомы
Моноклональное антитело или его антиген-связывающий фрагмент согласно настоящему изобретению связываются с пептидом, содержащим аминокислотную последовательность MEKVGKDGVITVE (SEQ ID №1), и способны эффективно ингибировать иммуносупрессорные реакции, вызываемые конкретными патогенами. Иммуносупрессорные функции патогенов обеспечиваются иммуносупрессорным веществом, секретируемым или вырабатываемым указанными патогенами. Моноклональное антитело или его фрагмент согласно настоящему изобретению преимущественно приводит к уничтожению патогенов, подавляя иммуносупрессорные функции указанных патогенов, и, следовательно, активируя иммунные ответы хозяина указанных патогенов. Моноклональное антитело согласно настоящему изобретению или его антиген-связывающий фрагмент проявляют достоверное функциональное подавление активности в отношении иммуносупрессорных функций, индуцируемых патогенами, с неожиданными результатами.
Согласно одному варианту реализации настоящего изобретения, указанное антитело или его антиген-связывающий фрагмент распознают пептид, содержащий аминокислотную последовательность, представленную MEKVGKDGVITVE (SEQ ID №1).
Антитело или его антиген-связывающий фрагмент согласно настоящему изобретению могут содержать тяжелую цепь и/или легкую цепь. Каждая легкая цепь и тяжелая цепь могут включать вариабельную область на своих N-концах и чередующиеся зоны, включающие 4 каркасных области (FR) и 3 участка, определяющих комплементарность, (CDR) в каждой из вариабельных областей.
Согласно одному варианту реализации настоящего изобретения антитело или его антиген-связывающий фрагмент могут включать в вариабельной области легкой цепи: CDR1, содержащий аминокислотную последовательность ASQSVDYDGDVFL (SEQ ID №2); CDR2, содержащий аминокислотную последовательность YAASN (SEQ ID №3); и CDR3, содержащий аминокислотную последовательность QSNEVPWT (SEQ ID №4). Согласно предпочтительному варианту реализации вариабельная область легкой цепи содержит аминокислотную последовательность, представленную в SEQ ID №5, т.е. положения 21-31 последовательности SEQ ID №6.
Согласно некоторым вариантам реализации настоящего изобретения, антитело или его антиген-связывающий фрагмент содержат в вариабельной области тяжелой цепи: CDR1, содержащий аминокислотную последовательность SGFTFSSFG (SEQ ID №7); CDR2, содержащий аминокислотную последовательность ISNGGS (SEQ ID №8); и CDR3, содержащий аминокислотную последовательность QGLRRRGAMDY (SEQ ID №9). Согласно предпочтительному варианту реализации вариабельная область тяжелой цепи содержит аминокислотную последовательность, представленную в SEQ ID №10, т.е. положения 20-139 последовательности SEQ ID №11.
Согласно еще одному предпочтительному варианту реализации настоящего изобретения, антитело или его антиген-связывающий фрагмент содержат вариабельные области легкой цепи и вариабельные области тяжелой цепи. Вариабельная область легкой цепи содержит: CDR1, содержащий аминокислотную последовательность ASQSVDYDGDVFL (SEQ ID №2), CDR2, содержащий аминокислотную последовательность YAASN (SEQ ID №3), и CDR3, содержащий аминокислотную последовательность QSNEVPWT (SEQ ID №4), а вариабельная область тяжелой цепи содержит: CDR1, содержащий аминокислотную последовательность SGFTFSSFG (SEQ ID №7), CDR2, содержащий аминокислотную последовательность ISNGGS (SEQ ID №8), и CDR3, содержащий аминокислотную последовательность QGLRRRGAMDY (SEQ ID №9).
Согласно вышеописанному варианту реализации более предпочтительным примером является антитело или его антиген-связывающий фрагмент, которые содержат вариабельную область легкой цепи и вариабельную область тяжелой цепи, причем указанная вариабельная область легкой цепи содержит аминокислотную последовательность, представленную положениями 21-131 последовательности SEQ ID №6, т.е. SEQ ID №5, а вариабельная область тяжелой цепи содержит последовательность, представленную положениями 20-139 последовательности SEQ ID №11, т.е. SEQ ID №10.
Согласно вариантам реализации настоящего изобретения, указанное моноклональное антитело преимущественно представляет собой химерное антитело, гуманизированное антитело или антитело человека.
Согласно предпочтительным вариантам реализации настоящего изобретения указанный антиген-связывающий фрагмент может представлять собой Fab, Fab', (Fab') 2, Fv или scFv. Изотип иммуноглобулина может быть IgG1, IgG2, IgG4, IgA, IgE или IgD.
Также согласно настоящему изобретению предложена гибридома для получения моноклонального антитела согласно настоящему изобретению, или его антиген-связывающего фрагмента. Согласно предпочтительным вариантам реализации настоящего изобретения указанной гибридомой является гибридома 9Е4.
Моноклональное антитело и его антиген-связывающий фрагмент, а также гибридому согласно настоящему изобретению можно получить в соответствии со следующими этапами: применение пептида, содержащего аминокислотную последовательность MEKVGKDGVITVE (SEQ ID №1), в качестве антигена, чтобы вызвать в организме млекопитающего иммунный ответ на антиген; получение плазмоцита (иммунной клетки) из организма млекопитающего, иммунизированного против указанных патогенов, и слияние полученной иммунной клетки с клеткой миеломы млекопитающих с получением гибридомы; клонирование полученной гибридомы и получение желаемой гибридомы. Способ согласно настоящему изобретению может дополнительно включать этапы: применения полученной гибридомы для получения антитела; и сбор антител, вырабатываемых указанной гибридомой.
Согласно описанному выше способу, способ иммунизации млекопитающего может включать любой способ введения, известный в данной области техники. Подходящие способы включают: внутрибрюшинную инъекцию, инъекцию в селезенку, внутримышечную инъекцию, подкожную инъекцию, внутрикожную инъекцию, пероральное введение, введение в слизистые оболочки, чрескожное введение и подобные. Среди них предпочтительными являются внутрибрюшинная инъекция и инъекция в селезенку. Интервалы между введением указанного антигена можно определить в зависимости от общего количества вводимого антигена, вида млекопитающего и других условий, например, введение несколько раз в месяц.
Указанное иммунизированное млекопитающее не ограничивается конкретным видом. Однако выбор следует делать с учетом таких условий, как совместимость клеток миеломы, применяемых при слиянии клеток. Подходящие млекопитающие включают мышей, крыс и хомяков. Среди них предпочтительными являются мыши.
Указанными иммунными клетками предпочтительно являются клетки селезенки, но это не составляет какого-либо технического ограничения.
Слияние иммунных клеток с клетками миеломы осуществляют при помощи любого из известных способов. Подходящие способы включают способ, предложенный Milstein et al. (Methods Enzymol., 73, 3-46, 1981). Указанный способ включает этапы: в присутствии ускорителя слияния смешивание иммунных клеток с клетками миеломы в питательной среде. В процессе слияния клеток соответствующим образом добавляют дополнительную питательную среду. Проводят разделение посредством многократного центрифугирования и получают гибридомы.
Подходящие питательные среды для применения при слиянии клеток включают: среду RPMI-1640, среду MEM (минимальная поддерживающая среды) и т.п. Указанные среды часто применяют при слиянии клеток. В процессе слияния при необходимости можно добавлять добавки, такие как сыворотка, например, фетальная телячья сыворотка.
Как правило, температура для слияния клеток предпочтительно составляет 25-37°C, более предпочтительно 30-37°C. Соотношение иммунных клеток и клеток миеломы предпочтительно составляет от приблизительно 1:1 до приблизительно 1:10.
Подходящие ускорители слияния включают: полиэтиленгликоль (ПЭГ), вирус Сендай и т.п. Среди них предпочтителен ПЭГ. Если применяют ПЭГ, его молекулярная масса может быть выбрана соответствующим образом, например, средняя молекулярная масса от приблизительно 1000 до приблизительно 6000. Кроме того, концентрация ПЭГ в среде может варьировать в диапазоне от приблизительно 30 до приблизительно 60% (масса/объем).
Согласно способу, описанному выше, можно отбирать гибридомы, с включением следующих этапов: Гибридомы, полученные путем слияния клеток, культивируют в питательной среде. Указанная среда предпочтительно является селективной средой, такой как среда HAT (среда с гипоксантином, аминоптерином и тимидином) и другие коммерческие среды. Для скрининга полученных гибридом применяют способ предельного разведения, например, с применением антительных значений пептида, включая аминокислотную последовательность MEKVGKDGVITVE (SEQ ID №. 1), в качестве индикатора. Время культивирования должно быть достаточно длительным, чтобы клетки, отличные от гибридом, успели погибнуть, и обычно оно составляет от нескольких дней до нескольких недель. Гибридомы, полученные на указанных этапах, можно пересевать в традиционную питательную среду, или длительное время хранить в жидком азоте.
Согласно предложенному в изобретении способу сбор моноклонального антитела или его антиген-связывающих фрагментов согласно настоящему изобретению включает этапы: применения известного способа культивирования гибридомы; и получения моноклональных антител из надосадочной жидкости их культуры. Другой способ включает этапы: введения гибридом млекопитающим, адаптированным к указанным гибридомам, для пролиферации гибридом; и получение моноклональных антител из асцита указанного млекопитающего. Среди применимых способов получение моноклональных антител из надосадочной жидкости культуры позволяет получить антитела более высокой степени чистоты, тогда как получение моноклональных антител из асцитов позволяет проводить массовое производство антител. Специалисты в данной области техники могут адекватно выбрать способ в зависимости от цели сбора.
Моноклональные антитела или их антиген-связывающий фрагмент, полученные при описанных выше этапах, можно подвергать дальнейшей очистке. Процесс очистки может быть основан на любом из известных способов, например, фракционировании солями, гель-фильтрации, аффинной хроматографии и т.п.
Предложенные в соответствии с настоящим изобретением моноклональные антитела и их антиген-связывающие фрагменты оказывают значимое функциональное ингибирующее действие на иммуносупрессорные функции патогенов. При применении, моноклональное антитело согласно настоящему изобретению или его антиген-связывающие фрагменты можно вводить напрямую или в составе фармацевтической композиции, которая содержит фармацевтически приемлемые вспомогательные вещества и др. Согласно настоящему изобретению указанная фармацевтическая композиция содержит эффективную дозу моноклонального антитела согласно настоящему изобретению или его антиген-связывающего фрагмента. Указанную фармацевтическую композицию можно применять в качестве функционального ингибитора в отношении иммуносупрессорных функций конкретных патогенов. Также согласно настоящему изобретению предложено применение моноклонального антитела согласно настоящему изобретению, включая его применение при получении фармацевтической композиции.
Фармацевтическая композиция согласно настоящему изобретению является композицией, содержащей функциональный ингибитор, который подавляет иммуносупрессорные функции, и ее получают посредством следующих этапов: Осуществление разведения или суспендирования моноклонального антитела или его антиген-связывающего фрагмента согласно настоящему изобретению в физиологическом растворе, дистиллированной воде или буферном растворе, таком как жидкий буфер для инъекций; и модулирование с целью получения композиции. Композиция с ингибитором иммуносупрессорных функций согласно настоящему изобретению может содержать другие вспомогательные вещества. Подходящие вспомогательные вещества включают: растворители, вещества, повышающие растворимость, консерванты, стабилизаторы, эмульгаторы, суспендирующие агенты, успокаивающие средства, изотоничные агенты, буферы, вспомогательные вещества, загустители, красители и традиционные основы, такие как разные рибосомы, полиаминокислотные основы, синтетические полимеры, природные полимеры и т.д.
Согласно настоящему изобретению предложен способ снижения подавления иммунитета, вызываемого белком теплового шока 60, секретируемым такими патогенами, как Н. pylori или другими похожими бактериями. После того как живому хозяину вводят моноклональное антитело согласно настоящему изобретению или его антиген-связывающий фрагмент, подавление иммунитета, вызванное белком теплового шока 60 указанных патогенов, уменьшается, и как следствие иммунная система живого хозяина активируется, и патогены уничтожаются.
Согласно способу, предложенному в соответствии с настоящим изобретением, моноклональное антитело согласно настоящему изобретению или его антиген-связывающий фрагмент можно вводить хозяину системным или местным образом. Способ введения включает любые известные способы, например, капельное введение, внутривенную инъекцию, внутримышечную инъекцию, подкожную инъекцию, внутрикожную инъекцию, пероральное введение, введение в слизистые оболочки, чрескожное введение и т.п.
Эффективное количество моноклонального антитела согласно настоящему изобретению или его антиген-связывающего фрагмента не является техническим ограничением. Специалисты в данной области техники могут адекватно определить его в зависимости от типа, природы, пола и возраста хозяина и т.п.
Ниже приведены конкретные примеры со ссылками на чертежи, чтобы проиллюстрировать моноклональное антитело, подавляющее иммуносупрессорные функции патогенов, его антиген-связывающий фрагмент и гибридомы, вырабатывающие такое антитело согласно настоящему изобретению. Однако следует понимать, что область настоящего изобретения не ограничивается ни одним из описанных вариантов реализации. Например, хотя в подробном описании механизм патогенеза, связанного с Н. pylori, и функциональный ингибитор HpHSP60 применены в качестве примеров для описания изобретения, белок теплового шока других бактерий, таких как Helicobacter felis (известная причина хронического энтерита) и Arcobacter suis (известная причина заболевания периодонта) также включает идентичный фрагмент HpHSP60, такой как HSP60 101-200. Следовательно, способ и применение данного изобретения также полезны в отношении патогенеза, связанного с этими и другими бактериями, и другими патогенами, которые имеют сходный патогенетический механизм.
Вариант реализации 1: Культура клеток и выделение МКПК и Т-лимфоцитов
Мононуклеарные клетки периферической крови (МКПК) человека от здоровых доноров выделяли путем центрифугирования в градиенте плотности с применением фикол-пак плюс (GE Healthcare, Упсала, Швеция) и повторно суспендировали в RPMI-1640 с 10% инактивированной фетальной телячьей сыворотки и 1% пенициллином-стрептомицином. Для истощения моноцитов МКПК культивировали в 10-см чашках Петри с плотностью 106/мл в течение ночи для прикрепления моноцитов. Затем суспендированные клетки отбирали путем центрифугирования при скорости 1500 оборотов/мин в течение 15 мин. Все Т-лимфоциты выделяли из числа МКПК путем отрицательной селекции с применением сортирующего устройства для магнитных частиц (Miltenyi Biotec, Массачусетс, США). Если кратко, МКПК инкубировали со смесью биотин-конъюгированных антител, а затем с антителами на биотин, конъюгированными с микрогранулами, для магнитного истощения. Т-лимфоциты десорбировали в соответствии с протоколами производителя.
Вариант реализации 2: Действие HpHSP60 на пролиферацию МКПК
Пролиферацию МКПК, стимулированных анти-CD3 моноклональным антителом, обработанных HpHSP60, rGFP или прокипяченным HpHSP60 в разных дозах, отслеживали в пробе на пролиферацию клеток. Для измерения пролиферации клеток 0,2 мл клеток в концентрации 1×106 клеток/мл засевали в каждую лунку микропланшета на 96-лунок, в который предварительно наносили анти-CD3 моноклональное антитело. Пролиферацию клеток определяли посредством пробы МТТ (на основе 3-4,5-диметилтиазол-2-ил-2,5-дифенилтетразолия бромида) через 96 часов. Результаты показаны на Фиг. 1: результаты экспериментов по оценке влияния HpHSP60 на пролиферацию МКПК. Показанные на фигуре данные представлены как индекс пролиферации.
Индекс пролиферации клеток рассчитывали по следующей формуле: Индекс пролиферации (100%)=(OD595 клеток, обработанных анти-CD3+HpHSP60) / OD595 клеток, обработанных анти-CD3)* 100%. Результаты, которые значимо отличаются от необработанной группы, отмечены * (р<0,05) (n=15).
На Фиг. 1 символы  показывают, что после того, как HpHSP60 добавляют в МКПК, подавляется пролиферация Т-лимфоцитов. Символы
показывают, что после того, как HpHSP60 добавляют в МКПК, подавляется пролиферация Т-лимфоцитов. Символы  показывают, что rGFP (рекомбинантный зеленый флуоресцентный белок), который в данной экспериментальной системе был контрольным белком, не влияет на пролиферацию Т-лимфоцитов. Символами
показывают, что rGFP (рекомбинантный зеленый флуоресцентный белок), который в данной экспериментальной системе был контрольным белком, не влияет на пролиферацию Т-лимфоцитов. Символами  показан прокипяченный HpHSP60, который включает последовательность HpHSP60, но структура белка была нарушена. Результаты показывают, что прокипяченный HpHSP60 не влияет на пролиферацию Т-лимфоцитов.
показан прокипяченный HpHSP60, который включает последовательность HpHSP60, но структура белка была нарушена. Результаты показывают, что прокипяченный HpHSP60 не влияет на пролиферацию Т-лимфоцитов.
Вариант реализации 3: Влияние HpHSP60 на пролиферацию Т-лимфоцитов в МКПК
После обработки анти-CD3 моноклональным антителом, МКПК подвергали или не подвергали обработке HpHSP60 (20 нг). Т-лимфоциты или не Т-лимфоциты среди МКПК определяли путем окрашивания на маркер CD3 на поверхности клетки. Затем количество клеток рассчитывали посредством проточно-цитометрического анализа.
Для окрашивания на маркер CD3 на поверхности клетки собирали и окрашивали 1 мкг мышиного моноклонального антитела на CD3 IgG человека (ОКТЗ), а затем 0,5 мкг кроличьего вторичного антитела на IgG мыши-FITC («Biolegend», Калифорния, США). Для внутриклеточного окрашивания FoxP3 клетки собирали и окрашивали мышиным моноклональным антителом на CD4 человека-FITC («Biolegend», Калифорния, США), а затем фиксировали и пермеабилизировали, после чего проводили внутриклеточное окрашивание мышиным моноклональным антителом на FoxP3 человека-ПЭ («BD Biosciences)), Массачусетс, США) в соответствии с протоколом производителя. Для анализа клеточного цикла клетки собирали 72 часа спустя и 106 клеток фиксировали 70% ледяным этанолом. ДНК окрашивали ДНК-окрашивающим буфером (5% Тритон-Х, 0,1 мг/мл РНКазы А и 4 мкг/мл иодида пропидия) в течение 30 мин при комнатной температуре. Затем определяли изменения в содержании ДНК. Флуоресценцию анализировали при помощи проточного цитометра FACS (клеточный сортировщик с возбуждением флуоресценции) («Becton Dickinson», Гейдельберг, Германия) и программы CELLQuest Pro («Becton Dickinson», Гейдельберг, Германия). Результаты показаны на Фиг. 2: результаты экспериментов по оценке влияния HpHSP60 на пролиферацию Т-лимфоцитов.
На Фиг. 2 индекс пролиферации клеток рассчитывали по следующей формуле: индекс пролиферации (кратность)=(кол-во Т-лимфоцитов или не-Т-лимфоцитов в группе, обработанной анти-CD3/HpHSP60) / (кол-во Т-лимфоцитов или не-Т-лимфоцитов в необработанном контроле). Значимое различие показано как *(р<0,05) (n=4). Результаты показывают, что HpHSP60 способен подавлять пролиферацию Т-лимфоцитов. На данной фигуре символами  показаны Т-лимфоциты среди МКПК. Символами
показаны Т-лимфоциты среди МКПК. Символами  показаны не-Т-лимфоциты среди МКПК. Очевидно, что HpHSP60 подавляет пролиферацию именно Т-лимфоцитов.
показаны не-Т-лимфоциты среди МКПК. Очевидно, что HpHSP60 подавляет пролиферацию именно Т-лимфоцитов.
Вариант реализации 4: Влияние HpHSP60 на клеточный цикл
Определяли влияние HpHSP60 на клеточный цикл МКПК. Из продуктов МКПК, описанных в варианте реализации 3, получены сами МКПК, CD3-активированные МКПК и МКПК, обработанные анти-CB3 и HpHSP60, соответственно. Наблюдается процент клеток в фазах G1, G1, S и G2/M и выражается на гистограммах, как показано на Фиг. 3А-3С. На Фиг. 3А-3С, соответственно, показаны результаты экспериментов по оценке влияния HpHSP60 на клеточный цикл МКПК. Фигуры являются репрезентативными для трех повторов.
На Фиг. 3А-3С показано, что HpHSP60 подавляет пролиферацию Т-лимфоцитов, но не вызывает их гибель. На Фиг. 3А показано, что Т-лимфоциты без активации CD3 (только клетки) остаются в своей латентной фазе (G0/G). На Фиг. 3В показано, что после активации CD3 рост Т-лимфоцитов активировался, и образовывались типичные графики клеточного цикла. На Фигуре 3С не выявляется никакого значимого отличия от Фиг. 3В. МКПК, обработанные ани-CD3 и HpHSP60 (анти- CD3+HpHSP60), демонстрируют такое же отношение в субфазах G0/G1 (соответствующее гибели клеток), что и группа с анти-CD3. Результаты экспериментов показывают, что роль HpHSP60 состоит в том, чтобы подавлять рост Т-лимфоцитов, но не вызывать их гибель.
Вариант реализации 5: Результаты индукции Treg-лимфоцитов in vitro белком HpHSP60
Долю CD4+FoxP3+-клеток среди МКПК, обработанных HpHSP60 измеряли во времени. Значимое отличие от анти-CD3-контроля обозначено *(р<0.05) (n=5). Результаты показаны на Фиг. 4: результаты экспериментов с индукцией Treg-лимфоцитов in vitro белком HpHSP60.
Поскольку CD4 и FoxP3 являются маркерами Treg-лимфоцитов, можно идентифицировать влияние HpHSP60 на рост Т-лимоцитов на основании Фиг. 4. На данной фигуре « только клетка» отображает исходную кривую роста Т-лимфоцитов. «
только клетка» отображает исходную кривую роста Т-лимфоцитов. « анти-CD3» отображает кривую роста Т-лимфоцитов, активированных CD3. «
анти-CD3» отображает кривую роста Т-лимфоцитов, активированных CD3. « анти-CD3 + HpHSP60» показывает значительную пролиферацию Т-лимфоцитов. Результаты экспериментов показывают, что HpHSP60 способен усиливать пролиферацию Treg-лимфоцитов.
анти-CD3 + HpHSP60» показывает значительную пролиферацию Т-лимфоцитов. Результаты экспериментов показывают, что HpHSP60 способен усиливать пролиферацию Treg-лимфоцитов.
Вариант реализации 6: HpHSP60 усиливает пролиферацию Treg-лимфоцитов
После процедур согласно варианту реализации 5, клетки собирали через 72 часа для выделения тотальной ДНК. Для измерения экспрессии мРНК FoxP3 применяли ПЦР (полимеразную цепную реакцию) в реальном времени. Значимые отличия от анти-CD3 контроля показаны *(р<0,05) (n=4). Результаты показаны на Фиг. 5: результаты экспериментов по оценке влияния HpHSP60 на пролиферацию Treg-лимфоцитов.
Поскольку FoxP3 является маркером Treg-лимфоцитов, когда Treg-лимфоциты активируются, экспрессия FoxP3 увеличивается. Результаты анализа мРНК на Фиг. 5 показывают, что после добавления HpHSP60 экспрессия FoxP3 достоверно увеличивается. Данный эксперимент также подтверждает тот факт, что добавление HpHSP60 усиливает пролиферацию Treg-лимфоцитов.
Вариант реализации 7: Активность HpHSP60-индуцированных Treg-лимфоцитов в отношении пролиферации Т-лимфоцитов
Для измерения активности HpHSP60-индуцированных Treg-лимфоцитов в отношении пролиферации клеток применяли функциональный анализ. Результаты показаны на Фиг. 6. На Фиг. 6 показаны результаты экспериментов по оценке влияния HpHSP60-индуцированных Treg-лимфоцитов на пролиферацию Т-лимфоцитов. Цифры на гистограммах показывают процент пролиферативных клеток. Гистограмма является репрезентативной для трех повторов.
Результаты экспериментов показывают, что когда число Treg-лимфоцитов возрастает, активность Т-клеток соответственно подавляется. Это доказывает, что когда добавляют HpHSP60 к МКПК, активность Т-лимфоцитов подавляется, из-за увеличения числа Treg-лимфоцитов.
Вариант реализации 8: Получение сыворотки с анти-HpHSP60 и моноклональных антител к HpHSP60
Мышей C3H/HeN приобретали в Национальном центре разведения и исследования лабораторных животных, Тайбей, Тайвань, и содержали в изоляторах без патогенов. Всю пищу, воду, клетки и подстилку стерилизовали перед применением. Самцам 5-недельного возраста в/в вводили HpHSP60, чтобы вызвать реакции иммунизации. После повторной бустерной дозы HpHSP60 у мышей отбирали кровь. Выделяли сыворотку, получая сыворотку, содержащую антитело против HpHSP60, которую называли «анти-HpHSP60 сыворотка». Продукты, получаемые на данном этапе, представляли собой поликлональное антитело.
Клетки селезенки мышей сливали с клетками миеломы мыши, и образовывалась гибридома. Впоследствии продукты подвергали скринингу путем иммуноферментного анализа (ELISA), чтобы выделить специфичные антитела.
Полученные линии клеток разводили и повторно распределяли в планшете для культивирования клеток на 96 лунок. Производили расчет, чтобы гарантировать, что каждая лунка содержит только одну клетку. После того, как клетки вырастали, образуя колонии, указанные колонии снова подвергали скринингу посредством ELISA, чтобы получить специфические антитела. Таким образом, получают моноклональные антитела.
Вариант реализации 9: Оценка уничтожения Н. pylori посредством блокирования HSP60 in vivo
Мышей С3Н/HeN приобретали в Национальном центре разведения и исследования лабораторных животных, Тайбей, Тайвань, и содержали в изоляторах без патогенов. Всю пищу, воду, клетки и подстилку стерилизовали перед применением. Самцам 5-недельного возраста в/в вводили 0,1 мл сыворотки HpHSP60, полученной согласно варианту реализации 8, перед заражением Н. pylori. Через 24 часа после введения антисыворотки мышей инфицировали 0,5 мл живого Н. pylori (штамм АТСС 15415, приблизительно 109 колониеобразующих единиц) в бульоне BHI через желудочный зонд два раза за 3-дневный период. После развития инфекции Н. pylori мышам затем в/в вводили 0,1 мл анти-HSP60 сыворотки раз в 3 дня.
Через 8 недель после заражения Н. pylori всех мышей умерщвляли в асептических условиях, и интактные желудки вскрывали по малой кривизне. Каждый желудок разрезали на два разных продольных образца, содержащих тело желудка и антральный отдел. Уничтожение Н. pylori анализировали путем повторного культивирования Н. pylori и иммуногистохимического окрашивания по экспрессии FoxP3.
Результаты показаны как среднее ± SEM (стандартная ошибка среднего). Статистическую значимость оценивали при помощи одностороннего t-критерия Стьюдента; р<0,05 считалось значимым. Результаты показаны на Фиг. 7, 8 и 9. Среди них на Фиг. 7 и 8, соответственно, показаны результаты нескольких экспериментов по подавлению роста Н. pylori in vivo вследствие блокирования иммуносупрессорных функций HpHSP60. На Фиг. 9 показаны результаты экспериментов по подавлению Treg-лимфоцитов вследствие блокирования иммуносупрессорных функций HpHSP60.
Фиг. 7 и 8 показывают, что анти-HpHSP сыворотка достоверно уменьшает повторное возникновение колоний H.pylori из лизата ткани желудка через 8 недель после заражения H.pylori. Чтобы определить механизм уменьшения колоний под действием антител, оценивали экспрессию Treg-лимфоцитов в ткани инфицированных H.pylori желудков. Фиг. 9 показывает, что лечение анти-HpHSP60 сывороткой достоверно уменьшает экспрессию Treg-лимфоцитов на слизистой оболочке желудка. Описанные результаты показывают, что хроническая инфекция Н. pylori коррелирует с HpHSP60, и что блокирование HpHSP60 может уменьшать колонизацию Н. pylori и генерирование Treg-лимфоцитов.
Вариант реализации 10: Положения Treg-индуицбельной последовательности в HpHSP60
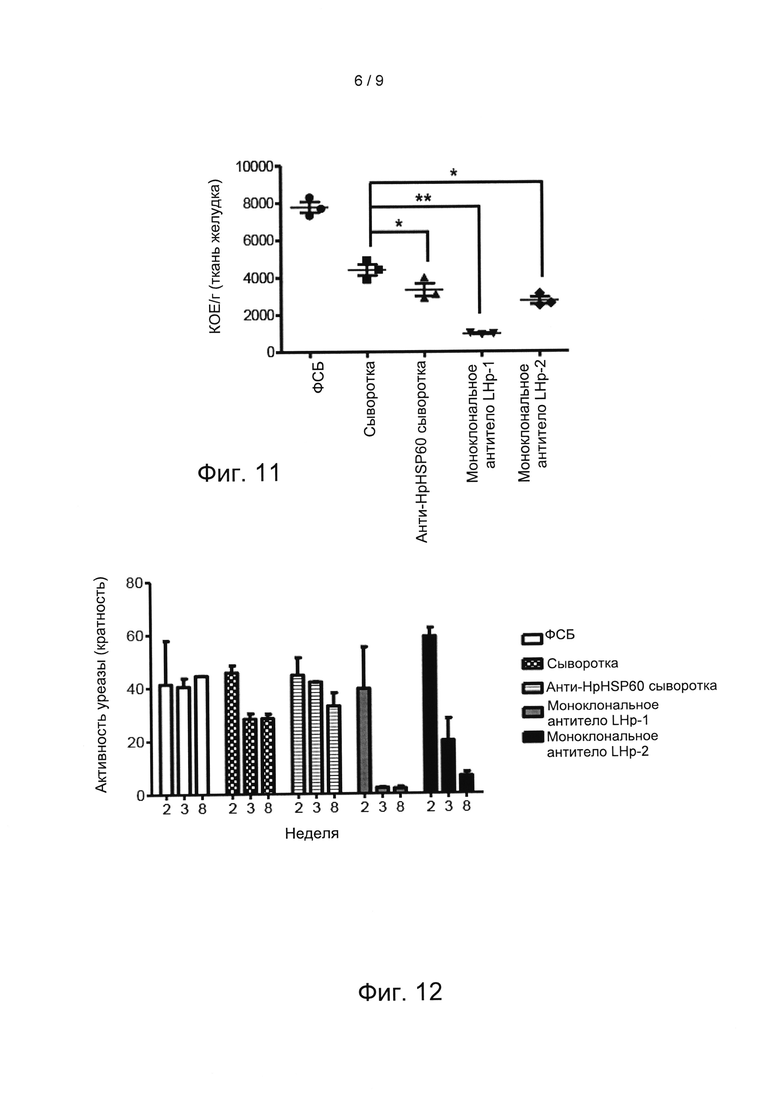
Чтобы локализовать положение активной последовательности в HpHSP60, которая индуцирует Treg-лимфоциты, получали анти- HpHSP60 моноклональные антитела, которые распознают полную последовательность или фрагменты HpHSP60. Применяют способ согласно варианту реализации 9. После 24-часового лечения анти- HpHSP60 сывороткой мышей инфицировали H. pylori, благодаря чему развивалась инфекция H. pylori. Затем мышам в/в вводили ОД мл ФСБ, сыворотку, анти-HSP60 сыворотку, моноклональное антитело LHP-1 (9Е4) и моноклональное антитело LHP-2 (5А8), соответственно, раз в 3 дня. Через 8 дней мышей умерщвляли. Стенку желудка измельчали и полученные гомогенаты желудка инкубировали в изолирующей сыворотке для H. pylori (агар EYE), чтобы подтвердить наличие H. pylori в желудке. Результаты показаны на Фигуре 10: результаты экспериментов по определению активной последовательности HpHSP60, которая индуцирует рост Treg-лимфоцитов.
Как показано на Фиг. 10, красные пятна на чашке Петри представляют собой колонии H. pylori. Данный эксперимент выявляет, что, хотя анти-HpHSP60 сыворотка подавляет рост H. pylori, антитело LHP-1 (9Е4) способно полностью уничтожать H. pylori.
Количество колоний (КОЕ) H. pylori определяли путем подсчета красных колоний на чашке с EYE. Значимые различия показаны *(р<0,05). Результаты показаны на Фиг. 11. На Фиг. 11 показана количественная оценка результатов экспериментов, показанных на Фин. 10. Как показано на Фиг. 11, H. pylori был полностью уничтожен после добавления антитела LHP-1 (9Е4).
Вариант реализации 11: Иммунологические механизмы анти- HpHSP60 антитела
Чтобы понять иммунологические механизмы действия анти-HpHSP60 антитела, измеряли активность желудочной уреазы мышей в соответствии с вариантом реализации 10 на 2-й, 3-й и 8-й неделе после заражения H. pylori. Активность уреазы нормировали на активность уреазы у контрольных мышей (без инфекции H. pylori). Результаты показаны на Фиг. 12: результаты экспериментов по изучению иммунологического механизма действия анти-HpHSP60 антител. Данные фигуры показывают, что антитело LHP-1 (9Е4) подавляет рост Н. pylori, или даже уничтожает Н. pylori, за счет ингибирования активности HpHSP60.
Антитело LHP-1 (9Е4) было сгенерировано при помощи аминокислотной последовательности положений 101-200 HpHSP60 в качестве антигена. Гибридомы, включая данное антитело - LHP-1 (9Е4) были депонированы в Американской коллекции типовых культур (АТСС®), Манассас, Виргиния, США Номер в АТСС: РТА-122900.
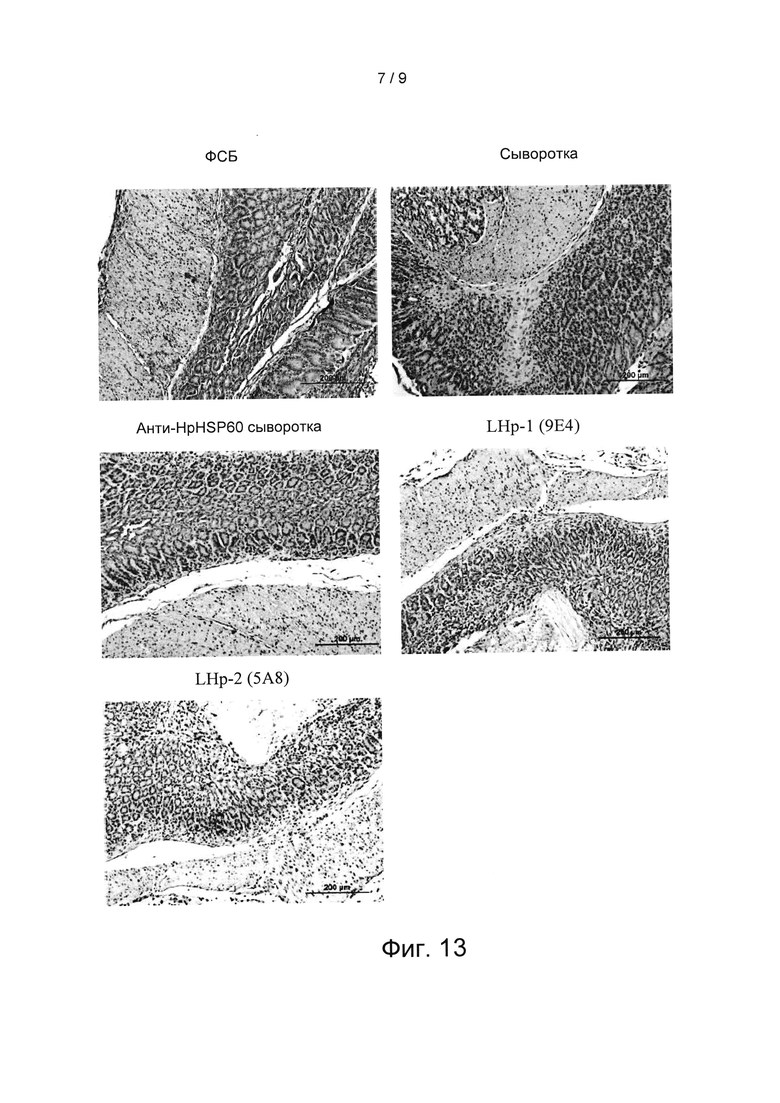
Вариант реализации 12: Оценка экспрессии Treg-лимфоцитов в слизистой оболочке желудка
Чтобы исследовать влияние анти- HpHSP60 антител на экспрессию Treg-лимфоцитов в слизистой оболочке желудка, желудки мышей, полученные во варианте реализации 10, фиксировали нейтральным буферным 10% формалином и заливали в парафин. Срезы толщиной пять микрометров окрашивали Г и Э (гематоксилином и эозином), затем проводили иммуногистохимическое окрашивание FoxP3. Результаты показаны на Фиг. 13. На Фиг. 13 показаны результаты экспериментов по оценке влияния анти- HpHSP60 антител на экспрессию Treg-лимфоцитов в слизистой оболочке желудка мышей. На данной фигуре все фотографии являются репрезентативными для мышей, умерщвленных на 8-ой неделе (200 мкм, начальное увеличение ×100). Результаты показывают, что экспрессии Treg в слизистой оболочке желудка мышей, которые получали анти- LHP-1 (9Е4) антитела, не наблюдалось.
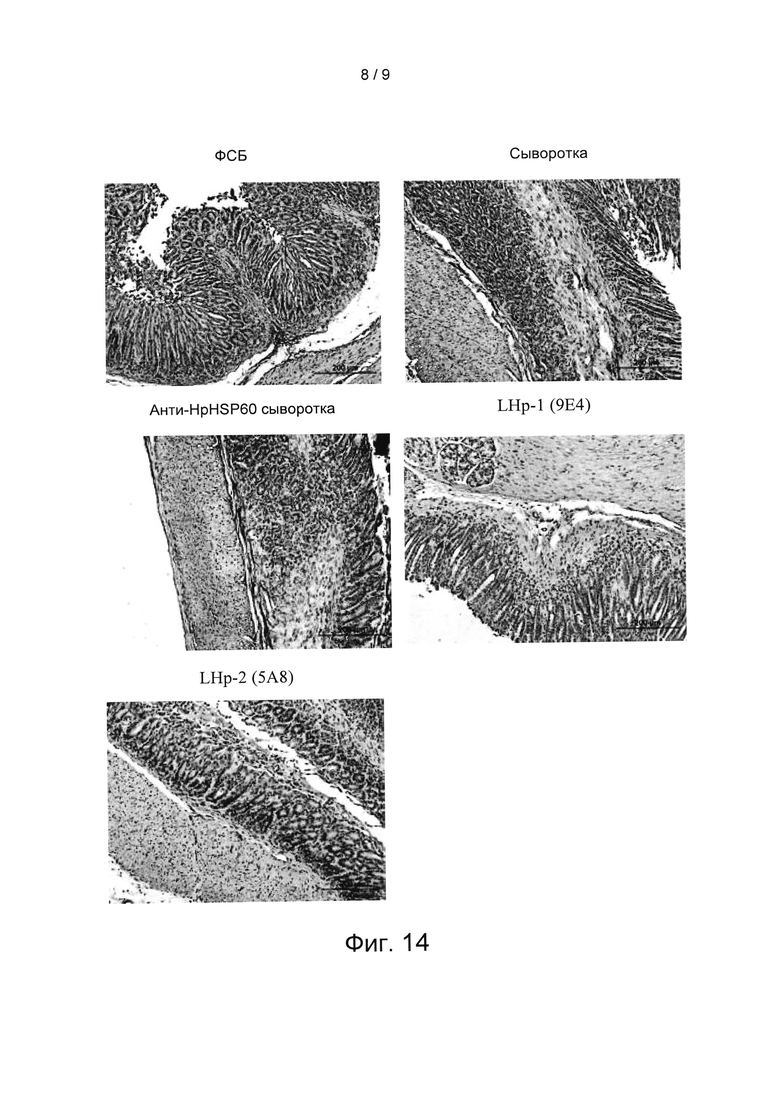
Вариант реализации 13: Влияние HpHSP60 на экспрессию ИЛ-10 в слизистой оболочке желудка.
Желудки мышей, полученные во варианте реализации 10, фиксировали нейтральным буферным 10% формалином и заливали в парафин. Срезы толщиной пять микрометров окрашивали Г и Э (гематоксилином и эозином), затем проводили иммуногистохимическое окрашивание на ИЛ-10. Результаты показаны на Фиг. 14. На Фиг. 14 показаны результаты экспериментов по оценке влияния анти- HpHSP60 антител на экспрессию ИЛ-10 в слизистой оболочке желудка мышей. На данной фигуре все фотографии являются репрезентативными для мышей, умерщвленных на 8-ой неделе (200 мкм начальное увеличение X100, 100 мкм начальное увеличение Х200). Результаты показывают, что в слизистой оболочке желудка мышей, которые получали анти- LHP-1 (9Е4) антитела, наблюдается экспрессия ИЛ-10.
Вариант реализации 14: Фрагменты HpHSP60, идентифицируемые антителом LHP-1 (9Е4)
Антитело LHP-1 (9Е4) применяют для идентификации фрагментов HpHSP60 разной длины, чтобы определять фрагменты HpHSP60, которые могут распознаваться антителом LHP-1 (9Е4). Результаты показаны на Фиг. 15. На Фиг. 15 показаны результаты экспериментов по распознаванию фрагментов HpHSP60 антителом LHP-1 (9Е4). На данной фигуре темные пятна отображают положительную идентификацию. Фрагменты, которые требуется идентифицировать, включают следующие, a IgK применяют в качестве положительного контроля, поскольку большинство моноклональных антител мыши каппа типа:
Целый полноразмерный HpHSP60, т.е. положения 1-547.
1-200 - фрагмент, включающий положения 1-200 HpHSP60.
101-200 - фрагмент, включающий положения 101-200 HpHSP60.
1-250 - фрагмент, включающий положения 1-250 HpHSP60.
200-300 - фрагмент, включающий положения 200-300 HpHSP60.
300-547 - фрагмент, включающий положения 300-547 HpHSP60.
Результаты показывают, что фрагмент HpHSP60, идентифицируемый антителом LHP-1 (9Е4), является последовательностью в положениях 101-200, и его аминокислотная последовательность следующая:

Вариант реализации 15: Дополнительное ограничение фрагмента HpHSP60, идентифицируемого антителом LHP-1 (9Е4)
В соответствии со способом, описанным в варианте реализации 14, антитело LHP-1 (9Е4) применяют для идентификации разных фрагментов с меньшей длиной. Результаты показаны на Фиг. 16. На Фиг. 16 показаны результаты дополнительных экспериментов по распознаванию фрагментов HpHSP60 антителом LHP-1 (9Е4). На данной фигуре темные пятна отображают положительную идентификацию. Δ [1] указывает положения 134-200 HpHSP60, с положительными результатами, тогда как Δ [5] указывает положения 101-168 HpHSP60, с отрицательными результатами. Следовательно, можно заключить, что фрагмент HpHSP60, идентифицируемый антителом LHP-1 (9Е4), включает аминокислотные положения 169-200 HpHSP60 и имеет следующую последовательность:
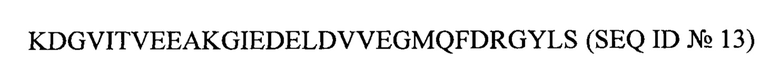
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Sagabio Co., Ltd.
<120> МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, ПОДАВЛЯЮЩЕЕ ИММУНОСУПРЕССОРНЫЕ ФУНКЦИИ
ПАТОГЕНОВ, АНТИГЕН-СВЯЗЫВАЮЩИЙ ФРАГМЕНТ УКАЗАННОГО АНТИТЕЛА И
ГИБРИДОМЫ, ВЫРАБАТЫВАЮЩИЕ УКАЗАННОЕ АНТИТЕЛО
<130> SB3302P-EP
<160> 13
<210> 1
<211> 13
<212> PRT
<213> мыши
<400> 1
Met Glu Lys Val Gly Lys Asp Gly Val Ile Thr Val Glu
1 5 10
<210> 2
<211> 13
<212> PRT
<213> мыши
<400> 2
Ala Ser Gln Ser Val Asp Tyr Asp Gly Asp Val Phe Leu
1 5 10
<210> 3
<211> 5
<212> PRT
<213> мыши
<400> 3
Tyr Ala Ala Ser Asn
1 5
<210> 4
<211> 8
<212> PRT
<213> мыши
<400> 4
Gln Ser Asn Glu Val Phe Trp Thr
1 5
<210> 5
<211> 111
<212> PRT
<213> мыши
<400> 5
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Val Phe Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 6
<211> 131
<212> PRT
<213> мыши
<400> 6
Met Glu Thr Asp Thr Ile Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala
20 25 30
Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser
35 40 45
Val Asp Tyr Asp Gly Asp Val Phe Leu Asn Trp Tyr Gln Gln Lys Pro
50 55 60
Gly Gln Pro Pro Lys Leu Leu Ile Tyr Ala Ala Ser Asn Leu Glu Ser
65 70 75 80
Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Leu Asn Ile His Pro Val Glu Glu Asp Asp Ala Ala Thr Tyr Tyr Cys
100 105 110
Gln Gln Ser Asn Glu Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu
115 120 125
Glu Ile Lys
130
<210> 7
<211> 9
<212> PRT
<213> мыши
<400> 7
Ser Gly Phe Thr Phe Ser Ser Phe Gly
1 5
<210> 8
<211> 6
<212> PRT
<213> мыши
<400> 8
Ile Ser Asn Gly Gly Ser
1 5
<210> 9
<211> 11
<212> PRT
<213> мыши
<400> 9
Gln Gly Leu Arg Arg Arg Gly Ala Met Asp Tyr
1 5 10
<210> 10
<211> 120
<212> PRT
<213> мыши
<400> 10
Glu Val His Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asn Gly Gly Ser Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Thr Arg Gln Gly Leu Arg Arg Arg Gly Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 11
<211> 463
<212> PRT
<213> мыши
<400> 11
Met Asn Phe Gly Leu Ser Leu Ile Phe Leu Ala Leu Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val His Leu Val Glu Ser Gly Gly Asp Leu Val Lys
20 25 30
Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Ser Phe Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Asn Gly Gly Ser Tyr Thr Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Thr Arg Gln Gly Leu Arg Arg Arg Gly Ala Met Asp Tyr
115 120 125
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Lys Thr Thr Pro
130 135 140
Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser
145 150 155 160
Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val
165 170 175
Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe
180 185 190
Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr
195 200 205
Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala
210 215 220
His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp
225 230 235 240
Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val
245 250 255
Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr
260 265 270
Pro Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu
275 280 285
Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln
290 295 300
Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser
305 310 315 320
Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys
325 330 335
Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro
355 360 365
Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met
370 375 380
Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn
385 390 395 400
Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr
405 410 415
Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn
420 425 430
Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu
435 440 445
His Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
450 455 460
<210> 12
<211> 100
<212> PRT
<213> Helicobacter pylori
<400> 12
Glu Gly Leu Arg Asn Ile Thr Ala Gly Ala Asn Pro Ile Glu Val Lys
1 5 10 15
Arg Gly Met Asp Lys Ala Ala Glu Ala Ile Ile Asn Glu Leu Lys Lys
20 25 30
Ala Ser Lys Lys Val Gly Gly Lys Glu Glu Ile Thr Gln Val Ala Thr
35 40 45
Ile Ser Ala Asn Ser Asp His Asn Ile Gly Lys Leu Ile Ala Asp Ala
50 55 60
Met Glu Lys Val Gly Lys Asp Gly Val Ile Thr Val Glu Glu Ala Lys
65 70 75 80
Gly Ile Glu Asp Glu Leu Asp Val Val Glu Gly Met Gln Phe Asp Arg
85 90 95
Gly Tyr Leu Ser
100
<210> 13
<211> 31
<212> PRT
<213> Helicobacter pylori
<400> 13
Lys Asp Gly Val Ile Thr Val Glu Glu Ala Lys Gly Ile Glu Asp Glu
1 5 10 15
Leu Asp Val Val Glu Gly Met Gln Phe Asp Arg Gly Tyr Leu Ser
20 25 30
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, НАПРАВЛЕННЫЕ НА CD127 | 2015 |
|
RU2734076C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА, КОТОРЫЕ РАСПОЗНАЮТ АЛЬФА-СИНУКЛЕИН | 2012 |
|
RU2743738C2 |
| АНТИТЕЛО ПРОТИВ TNFR2 И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2793165C1 |
| АНТИ-ГАЛЕКТИН-9 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2840501C1 |
| АНТИТЕЛА В7-Н4 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2809243C2 |
| АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ ИНТЕРЛЕЙКИНУ-2 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2745451C1 |
| АНТИТЕЛО ПРОТИВ GARP | 2016 |
|
RU2769379C2 |
| АНТИ-HLA-A2 АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2782276C2 |
| АНТИТЕЛА К ЦЕРАМИДУ | 2015 |
|
RU2717651C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |

Изобретение относится к области биотехнологии, в частности к моноклональному антителу, его фрагменту, а также содержащей их фармацевтической композиции. Также раскрыта гибридома для получения вышеуказанного моноклонального антитела или антиген-связывающего фрагмента. Изобретение позволяет эффективно осуществлять функциональное ингибирование иммуносупрессорных функций патогена, где указанный патоген представляет собой Helicobacter pylori. 5 н. и 7 з.п. ф-лы, 16 ил., 15 пр.
1. Моноклональное антитело, которое связывается с пептидом, содержащим аминокислотную последовательность, представленную в SEQ ID № 1;
при этом указанное антитело содержит:
в вариабельной области легкой цепи: CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID № 2; CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID № 3; и CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID № 4; и
в вариабельной области тяжелой цепи: CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID № 7; CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID № 8; и CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID № 9.
2. Моноклональное антитело по п.1, распознающее пептид, содержащий аминокислотную последовательность, представленную в SEQ ID № 1.
3. Моноклональное антитело по п.1 или 2, содержащее в вариабельной области легкой цепи аминокислотную последовательность, представленную положениями 21-131 последовательности SEQ ID № 6.
4. Моноклональное антитело по п.1 или 2, содержащее в вариабельной области тяжелой цепи аминокислотную последовательность, представленную положениями 20-139 последовательности SEQ ID № 11.
5. Моноклональное антитело по п.1 или 2, отличающееся тем, что указанное антитело представляет собой антитело, выбранное из группы, состоящей из химерного антитела, гуманизированного антитела и антитела человека.
6. Моноклональное антитело по п.1 или 2, отличающееся тем, что указанное антитело продуцируется гибридомой с номером депонирования в ATCC: PTA-122900.
7. Моноклональное антитело по п.1 или 2, включающее изотип иммуноглобулина, выбранный из группы, состоящей из IgG1, IgG2, IgG4, IgA, IgE и IgD.
8. Антиген-связывающий фрагмент моноклонального антитела по любому из пп. 1-7, который связывается с пептидом, содержащим аминокислотную последовательность, представленную в SEQ ID № 1, при этом указанный антиген-связывающий фрагмент содержит:
CDR1, CDR2 и CDR3 вариабельной области легкой цепи, содержащие аминокислотные последовательности, представленные в SEQ ID № 2, SEQ ID № 3 и SEQ ID № 4 соответственно; и
CDR1, CDR2 и CDR3 вариабельной области тяжелой цепи, содержащие аминокислотные последовательности, представленные в SEQ ID № 7, SEQ ID № 8 и SEQ ID № 9 соответственно.
9. Антиген-связывающий фрагмент по п. 8, отличающийся тем, что антиген-связывающий фрагмент выбран из группы, состоящей из Fab, Fab ', (Fab') 2, Fv и scFv.
10. Гибридома для получения моноклонального антитела по любому из пп. 1-7 или антиген-связывающего фрагмента по п. 8 или 9, где указанная гибридома характеризуется номером депонирования в ATCC: PTA-122900.
11. Применение моноклонального антитела по любому из пп. 1-7 или антиген-связывающего фрагмента по п. 8 или п.9 в качестве лекарственного средства для функционального ингибирования иммуносупрессорных функций патогена, где указанный патоген представляет собой Helicobacter pylori.
12. Фармацевтическая композиция, содержащая эффективную дозу моноклонального антитела по любому из пп. 1-7 или антиген-связывающего фрагмента по п. 8 или 9 для функционального ингибирования иммуносупрессорных функций патогена, где указанный патоген представляет собой Helicobacter pylori.
| HIROYUKI YAMAGUCHI et al., Growth Inhibition of Helicobacter pylori by Monoclonal Antibody to Heat-Shock Protein 60, Microbiol | |||
| Immunol., 1997, Vol | |||
| Механический грохот | 1922 |
|
SU41A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Многоцилиндровый объемный водомер двойного действия с вращающимися цилиндрами | 1923 |
|
SU909A1 |
| SHIGERU KAMIYA et al., Immune response to heat shock protein of Helicobacter pylori -a candidate as a vaccine component, Keio J Med, 2002, Vol.51 Suppl 2, pp | |||
| Пишущая машина для тюркско-арабского шрифта | 1922 |
|
SU24A1 |
| TOMOYUKI | |||

