Область техники
Изобретение относится к интегративному плазмидному вектору pVEAL2-S-RBD, обеспечивающему экспрессию и секрецию рекомбинантного рецептор-связывающего домена (RBD) коронавируса SARS-CoV-2 в клетках млекопитающих, штамму рекомбинантной клеточной линии СНО-K1-RBD и рекомбинантному белку RBD SARS-CoV-2, продуцируемому указанной клеточной линией СНО-K1-RBD и может быть использовано в области генной инженерии и биотехнологии. Рекомбинантный белок RBD может служить, в первую очередь, для создания вакцины против новой коронавирусной инфекции COVID-19, а также для создания диагностикумов.
Уровень техники
Пандемия COVID-19 вызвана новым высокопатогенным вирусом SARS-CoV-2, последствия заражения которым варьируют от бессимптомного течения и легких респираторных симптомов до летального исхода.
Вакцинопрофилактика считается наиболее эффективной мерой для предотвращения распространения вируса. На сегодняшний день во всем мире в разработке находятся более 190 вакцин-кандидатов против SARS-CoV-2 (https://www.who.int/who-documents-detail/draft-landscape-of-covid-19-candidate-vaccines). Привлекательной мишенью для создания вакцины является поверхностный гликопротеин S SARS-CoV-2. Гликопротеин S связывается с поверхностным белком ACE2 (ангиотензинпревращающий фермент 2) клеток-мишеней, опосредуя проникновение вируса в клетки (Gralinski & Menachery, 2020; Shang et al., 2020).
Эктодомен S белка включает два основных домена: рецептор-связывающий домен (RBD) и N-концевой домен (NTD) (Pallesen et al., 2017; Song et al., 2018; Zhou et al., 2019; Wrapp et al., 2020). Исследования показывают, что антитела нейтрализующие вирус в основном нацелены на домен RBD, к тому же он является одним из консервативных регионов (Ul Qamar et al., 2019; Gralinski & Menachery, 2020; Shang et al., 2020; Wrapp et al., 2020). Именно поэтому RBD рассматривается как одна из основных В-клеточных мишеней при создании вакцины против SARS-CoV-2 (https://pubmed.ncbi.nlm.nih.gov/32203189/). На данный момент 5 субъединичных вакцин на основе RBD находятся на этапе клинических испытаний и 8 на доклинических (https://www.who.int/who-documents-detail/draft-landscape-of-covid-19-candidate-vaccines).
Критическим фактором для разработки вакцины на основе RBD является выбор системы экспрессии. Наиболее перспективными являются клетки млекопитающих, поскольку позволяют получать целевой белок в наиболее близкой к нативной форме, т.е. прошедший необходимые пострансляционные модификации, что весьма важно для иммуногенности рекомбинантного белка. Так было показано, что на поверхности гликопротеина S SARS-CoV-2 имеется до двадцати мест гликозилирования, два из которых приходятся на регион RBD (Zeyu Sun, et al. // Mass spectrometry analysis of newly emerging coronavirus HCoV-19 spike S protein and human ACE2 reveals camouflaging glycans and unique post-translational modifications. // bioRxiv, 2020. DOI: 10.1101/2020.04.29.068098).
Известна заявка на изобретение Китая (CN 111217917, МПК А61K 39/215, опубл. 02.06.2020 г.), в которой описан способ получения слитых белков RBD-CTB и RBD-CRM197(A) для создания рекомбинантных субъединичных вакцин для лечения и профилактики COVID-19. Для этого в экспрессионный вектор pET9a встраивали нуклеотидные последовательности, кодирующие RBD соединенные линкером с адъювантами CTB или CRM197(A), и проводили трансформацию клеток Escherichia coli BL21 полученными конструкциями.
Недостатком данного изобретения является выбор системы экспрессии в клетах Escherichia coli BL21, поскольку известно, что у прокариот отсутствуют некоторые системы посттрансляционной модификации белков, например, гликозилирование, необходимое для получения корректной формы RBD, так же существует риск некорректного фолдинга молекул. Такие особенности могут критически снижать уровень иммуногенности итоговой вакцины.
Также известна заявка на изобретение Китая (CN 111254155, МПК А61K 39/215, опубл. 09.06.2020 г.), в которой варианты RBD для субъединичных вакцин против вируса SARS-CoV-2 получали с использованием растений-хозяев. Авторы с помощью транзиентной экспресии в Lactuca sativa (салат-латук) получали слитые белки CTB-RBD и RBD-Fc, используемые в вакцине для профилактики COVID-19.
Недостатком изобретения является способ выделения целевых белков, включающий многоступенчатую систему очистки, а так же особенности экспрессии связанные с некорректным гликозилированием и фолдингом белков в растительных системах экспрессии.
Наиболее близким аналогом (прототипом) является (патент РФ № 2723008, МПК C07K 14/165, C12N 15/50, опубл. 08.06.2020 г.)
- генетическая конструкция для экспрессии рекомбинантного белка RBD вируса SARS-CoV-2, включающая SEQ ID NO: 1;
- штамм клеток яичника китайского хомячка CHO-S-RBD, продуцент рекомбинантного белка RBD вируса SARS-CoV-2, содержащий генетическую конструкцию, включающую SEQ ID NO: 1;
- способ получения штамма клеток яичника китайского хомячка CHO-S-RBD, включающий: получение генетической конструкции, включающей SEQ ID NO: 1; введение указанной генетической конструкции в клетки путем липофекции; селекцию клеток на антибиотике Hygromycin B;
- рекомбинантный белок RBD вируса SARS-CoV-2 для выявления антител к SARS-CoV-2, имеющий SEQ ID NO: 2, являющийся продуктом штамма CHO-S-RBD;
- способ получения рекомбинантного белка RBD вируса SARS-CoV-2, включающий: культивирование штамма клеток яичника китайского хомячка CHO-S-RBD; хроматографическую очистку рекомбинантного белка RBD вируса SARS-CoV-2 из культуральной среды штамма клеток яичника китайского хомячка CHO-S-RBD; подтверждение получения рекомбинантного белка RBD вируса SARS-CoV-2, имеющего SEQ ID NO: 2.
Недостатком изобретения является использование относительно слабого лидерного пептида тяжелой цепи иммуноглобулина G человека, что может снизить выход целевого белка. Кроме того, негативное влияние на выход продукта может оказать тот факт, что авторы не проводили оптимизацию кодонного состава нуклеотидной последовательности, кодирующей RBD, для продуцента, а так же использовали вектор для эписомальной (внехромомсомной) экспрессии. Авторами показан довольно низкий уровень продукции 5-10 мг/л суспензионной культуры.
Таким образом, существует потребность в разработке стабильной клеточной линии-продуцента рекомбинантного RBD SARS-CoV-2.
Раскрытие изобретения
Техническим результатом заявленного изобретения является повышение выхода целевого рекомбинантного белка RBD SARS-CoV-2 за счет создания кодон-оптимизированной нуклеотидной последовательности, кодирующей белок RBD SARS-CoV-2 для экспрессии в клетках СНО, конструирования интегративного плазмидного вектора pVEAL2-RBD4, содержащего кодон-оптимизированный ген RBD и обеспечивающего его экспрессию в клетках млекопитающих, и разработки с помощью указанного вектора штамма-продуцента рекомбинантного белка RBD SARS-CoV-2 на основе рекомбинантного штамма клеточной линии яичников китайского хомячка СНО-К1.
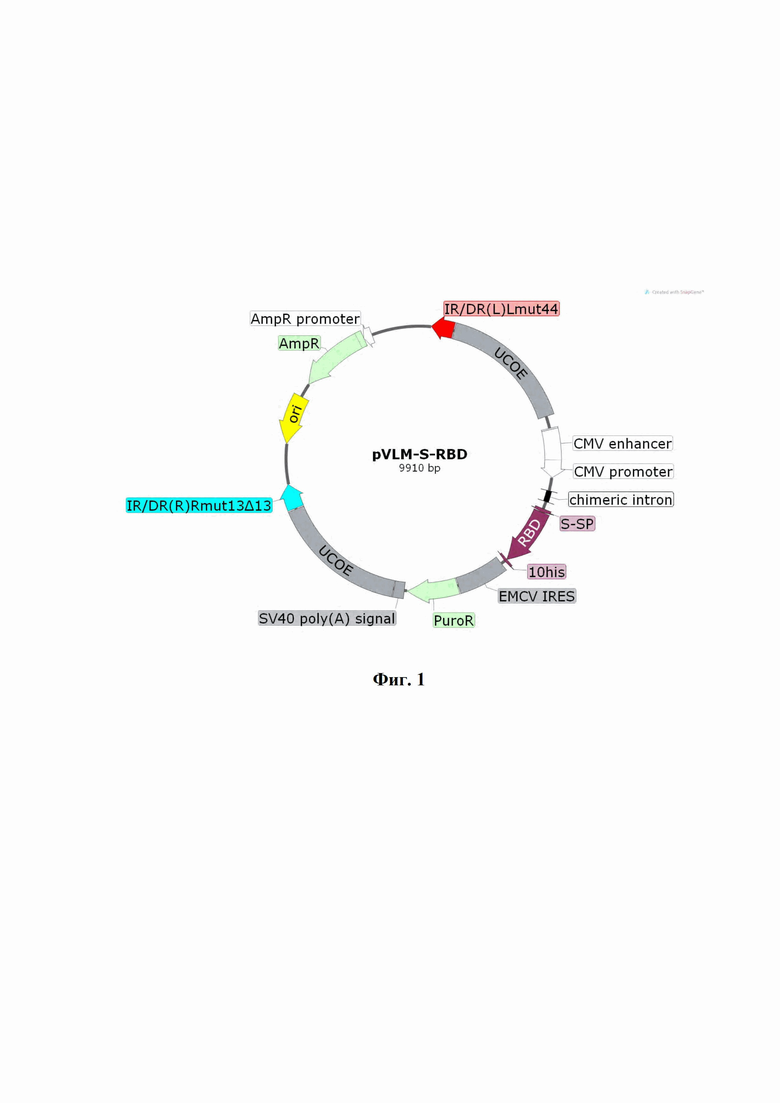
Технический результат достигается созданием интегративного плазмидного вектора pVEAL2-S-RBD, обеспечивающего экспрессию и секрецию белка RBD коронавируса SARS-CoV-2 в клетках млекопитающих, имеющего размер 9910 п.н., нуклеотидную последовательность SEQ ID No. 2 и содержащего в соответствии с физической и генетической картой, представленной на Фиг. 1, следующие элементы:
- участок начала репликации ori (7612-8200);
- последовательности прямых и инвертированных повторов IR/DR, содержащие сайты связывания с транспозазой SB100 (139-403, 6828-7123);
- последовательности UCOE, предотвращающие хромосомное «замалчивание» рекомбинантной экспрессионной кассеты (416 - 1964, 5268-6816);
- CMV энхансер (2115-2494), CMV промотора (2495-2706) - наиболее часто используемый промотор в генно-терапевтических конструкциях;
- Chimeric intron (2867-2999), химерный интрон, для усиления экспрессии целевого гена;
- лидерный пептид S-SP (3090-3134), обеспечивающий экспорт белка из клеток СНО-К1-RBD;
- кодон-оптимизированная нуклеотидная последовательность гена RBD (SEQ ID NO: 1) (3135-3836);
- последовательность 10хHis - полигистидиновый тэг для очистки рекомбинантного белка с помощью металл-хеллатной хроматографии (3837-3866);
- участок внутренней посадки рибосомы EMCV IRES (3916-4490);
- последовательность PuroR, кодирующая фактор устойчивости к антибиотику пуромицину (4503-5102);
- последовательность SV40 poly(A) signal для стабилизации мРНК-транскриптов за счет полиаденилирования (5137-5258);
- ген устойчивости к антибиотику ампициллину AmpR (8371-9231) и бактериальный промотор гена устойчивости к ампициллину (9232-9336), позволяющие проводить амплификацию плазмиды в E.coli.
Указанный технический результат достигается также созданием рекомбинантного штамма клеток яичника китайского хомячка СНО-K1-RBD - продуцента рекомбинантного белка RBD вируса SARS-CoV-2, содержащего интегративный плазмидный вектор pVEAL2-S-RBD по п. 1, имеющий нуклеотидную последовательность SEQ ID NO: 2, обеспечивающий экспрессию и секрецию белка RBD коронавируса SARS-CoV-2 и депонированный под номером 322 в коллекции культур клеток ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора (справка о депонировании прилагается).
Указанный технический результат достигается также получением рекомбинантного белка RBD коронавируса SARS-CoV-2, продуцируемого рекомбинантным штаммом клеток яичника китайского хомячка СНО-K1-RBD по п. 2 и имеющий аминокислотную последовательность SEQ ID NO: 3.
Заявляемая группа изобретений имеет существенные преимущества перед известными аналогами и прототипом. Оптимизацию кодонного состава нуклеотидной последовательности, имеющей улучшенный индекс адаптации кодонов CAI (англ. Codon Adaptation Index), обеспечивает повышение экспрессии рекомбинантного белка в клетках СНО путем увеличения эффективности трансляции. Интегративный вектор pVEAL2-RBD4 содержит оптимизированную нуклеотидную последовательность, кодирующую RBD, которая обеспечивает больший выход продукта в клетках линии СНО. UCOE-элементы, фланкирующие экспрессионную кассету указанного вектора, способствуют декомпактизации хроматина и, как следствие, усилению транскрипции гена RBD. Сигнальная последовательность играет решающую роль в секреции белка во внеклеточное пространство, поэтому в данном изобретении использована последовательность, обеспечивающая эффективный экспорт целевого белка из клеток, предотвращая токсическое воздействие рекомбинантного белка RBD на клетки продуцента.
Последовательность полигистидинового тэга 10×His введена для последующей очистки рекомбинантного белка RBD с помощью аффинной хроматографии. Добавление полигистидинового «хвоста» 10×His к одному из концов рекомбинантного белка представляет собой одну из известных генетических модификаций рекомбинантных белков, но свойства целевого белка могут значительно измениться в худшую сторону вплоть до потери его какого-либо свойства (например, потеря термостабильности белка, его агрегация и т.п.). При модификации заявляемого белка для исследователя неочевидно, что модифицированный белок будет сохранять свои специфические свойства: http://pushchinocity.ru/article/uchyonye-iz-g.puschino-vyyasnili-kak-vliyaet-poligistidinovyj-hvost-na-aktivnost-i-svojstva-endolizina-bakteriofaga-t5-81696.
Авторами установлено, что последовательность 10×His значительно повышает эффективность очистки рекомбинантного RBD в отличие от последовательности 6×His в прототипе. К тому же наличие 10×His на С-конце белка RBD не приводит к снижению иммуногенности, агрегации или иным изменениям белка в ходе хранения при +4°С в течение полугода.
Для разработки продуцента была выбрана клеточная линия СНО-К1, поскольку экспресcионная система клеток СНО позволяет получать наиболее корректную форму белка, обеспечивая все необходимые процессы посттрансляционной модификации. Для получения эффективной культуры-продуцента в заявленном изобретении проведен отбор наиболее продуктивных клонов культуры-продуцента CHO-K1-RBD. Это дополнительно увеличивает выход целевого продукта, поскольку поликлональная клеточная культура содержит клоны с разной способностью к продукции рекомбинантного белка.
Таким образом, заявляемая группа изобретений обеспечивает высокий выход рекомбинантного белка RBD SARS-CoV-2 в иммунологически корректной форме, что является хорошей платформой для создания иммунобиологических препаратов и вакцин. Кроме того, технические решения соотвествуют критериям «новизна» и «изобретательский уровень».
Осуществление изобретения
Описание фигур
Изобретение иллюстрируется следующими графическими фигурами.
На фиг. 1 изображена физическая и генетическая карта интегративного плазмидного pVEAL2-S-RBD.
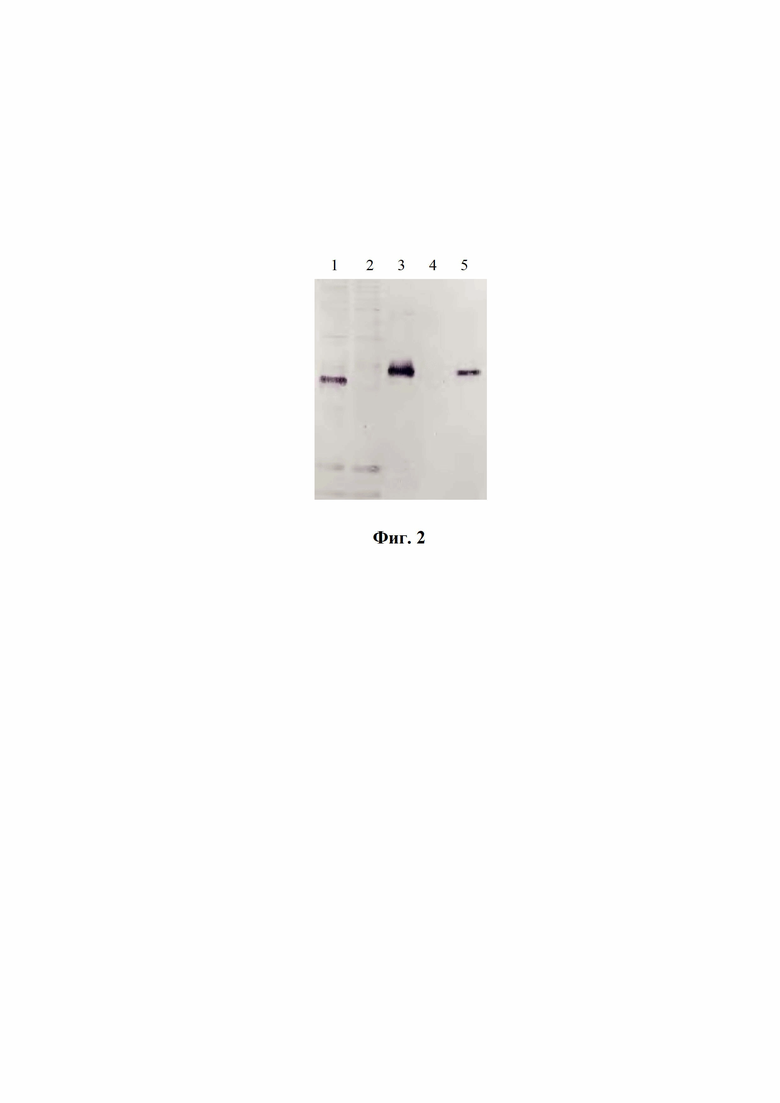
На фиг. 2 представлены результаты иммуноблоттинга белков при окрашивании моноклональными антителами к полигистидиновому тэгу.
На фиг. 3 представлен электрофорез белков в ДСН-ПААГ.
На фиг. 4 представлен анализ специфичности сывороток мышей, иммунизированных RBD, с помощью иммуноферментного анализа.
На фиг. 5 представлены результаты исследования нейтрализующей активности сывороток мышей, иммунизированных RBD, в реакции нейтрализации цитопатического действия (ЦПД) вируса на культуре клеток VERO E6.
Сведения, подтверждающие возможность осуществления изобретения. Ниже приведены примеры 1-5 конкретного осуществления изобретения.
Пример 1. Оптимизация кодонного состава нуклеотидной последовательности, кодирующей RBD SARS-CoV-2
Последовательность гена, кодирующего домен RBD (308V - 542N) белка S коронавируса SARS-CoV-2 (GenBank MN908947) извлекали из базы данных GenBank и проводили оптимизацию кодонного состава для клеток CHO при помощи инструмента GeneOptimizer (https://www.thermofisher.com/ru/en/home/life-science/cloning/gene-synthesis/geneart-gene-synthesis/geneoptimizer.html). Индекс адаптации кодонов CAI (англ. Codon Adaptation Index) был улучшен с 0,63 до 0,73.
Итоговую нуклеотидную последовательность (SEQ ID NO: 1) синтезировали на заказ в ООО «ДНК-синтез» в составе вектора pGH-RBD.
Пример 2. Конструирование интегративного плазмидного вектора pVEAL2-S-RBD для синтеза и секреции RBD SARS-CoV-2 в клетках млекопитающих
На основе вектора pVEAL2 получали конструкцию pVEAL2-S-RBD (фиг. 1), содержащую нуклеотидную последовательность RBD (SEQ ID NO: 1), последовательность 10хHis и лидерный пептид S-SP белка S SARS-CoV-2 (MFVFLVLLPLVSSQC).
Нуклеотидную последовательность сигнального пептида S-SP, 10хHis и сайтов рестрикции Sfr274I и SalI вводили в последовательность RBD одновременно с помощью ПЦР и последующего отжига продуктов амплификации. С использованием праймеров pVL-F и S-SP-R получали ПЦР-продукты на матрице вектора pVEAL2, а с использованием праймеров S-SPRBD-F и RBDH-R на матрице pGH-RBD (таблица 1).
Амплификацию фрагментов проводили по стандартному протоколу. Реакционные смеси, объемом 50 мкл, содержали 5-10 нг ДНК-матрицы; 0,2 мМ каждого из четырех дезоксинуклеозидтрифосфатов; 0,2 мкМ каждого прaймеров; 5 мкл SE-буфера для PfuSE ДНК полимеразы («СибЭнзим», г. Новосибирск); 1 мг/мл BSA; 50 ед/мл PfuSE ДНК полимеразы («СибЭнзим», г. Новосибирск).
Таблица 1. Нуклеотидные последовательности олигонуклеотидов
Температурный профиль реакции состоял из начальной денатурации при 95°С в течение 3 мин и 30 циклов (30 сек денатурации при 95°С, 20 сек отжига при 60°С и 1-2 мин элонгации при 72°С) с финальной элонгацией в течение 2 мин при 72°С.
Далее проводили отжиг соответствующих ПЦР-фрагментов и амплификацию полученных фрагментов с использованием праймеров pVL-F и RBDH-R согласно протоколу описаному выше. Затем проводили встройку в вектор pVEAL2 рестриктазно-лигазным методом по сайтам рестрикции Sfr274I и SalI. Подтверждение структуры полученного вектора проводили с помощью секвенирования.
Пример 3. Получение штамма рекомбинантной клеточной линии CHO-K1-RBD, продуцента рекомбинантного домена RBD SARS-CoV-2
Штамм рекомбинантной клеточной линии CHO-K1-RBD продуцент рекомбинантного домена RBD SARS-CoV-2 получен в 2020 г. на основе клеточной линии яичников китайского хомячка СНО-K1 с использованием разработанной конструкции pVEAL2-S-RBD. Клетки растили в инкубаторе при 5% содержания CO2, 80%-ной влажности. При достижении 80% плотности монослоя проводили трансфекцию клеток плазмидой pVEAL2-S-RBD с помощью Lipofectamine 3000 (ThermoFisher, США) в соответствии с инструкцией производителя. Для интеграции экспрессионной кассеты вектора pVEAL2-S-RBD в геном клеток совместно с плазмидой pVEAL2-S-RBD добавляли плазмиду pCMV(CAT)T7-SB100, кодирующую транспозазу SB100, в отношении 10:1. Через 3 дня в культуральную среду добавляли селективный антибиотик пуромицин (InvivoGen, США) в конечной концентрации 10 мкг/мл, ген устойчивости к которому входит в состав вектора.
Селекцию устойчивых клонов проводили в течение трех суток, затем поликлональную клеточную культуру рассевали в 96 луночном планшете в концентрации 1 клетка на лунку. Спустя две недели, анализировали наличие единичных колоний в лунках, отбирали культуральную жидкость и оценивали продуктивность клонов с помощью ИФА, из которых и получен заявляемый рекомбинантный штамм CHO-K1-RBD.
Штамм депонирован 19.01.2021 г. в коллекции ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора под номером 322.
Характеристика рекомбинантного штамма CHO-K1-RBD.
Морфология: веретеновидные и эпителиоподобные клетки с круглыми ядрами, содержащими от 1 до 2 ядрышек.
Способ культивирования: монослойный. Среда для культивирования: питательная среда DМЕМ/F-12 (1:1) - 90 %, сыворотка крови плодов коровы - 10 %.
Температура культивирования: 37°С. Посевная концентрация: 100 тыс. клеток в 1 мл. Метод снятия: 0,25 % трипсин (1/3) и 0,02 % версен (2/3).
Кратность рассева: 1:3. Частота пассирования: 3-4 суток.
Условия криоконсервации: питательная среда DМЕМ/F-12 (1:1) - 50 %, сыворотка крови плодов коровы - 40 %, ДМСО - 10 %.
Режим замораживания: при температуре 4°С - 1 ч, минус 80°С - 12 ч, минус 196°С.
Условия хранения: в криопробирках в количестве 5 шт. хранится в жидком азоте при температуре минус 196°С.
Номер пассажа в жидком азоте: 10. Жизнеспособность после криоконсервации: 85-90 %.
Маркерый признак: наличие в культуральной среде рекомбинантного белка RBD размером ~35 кДа, подтверждается с помощью белкового электрофореза и иммуноблотинга. Область применения: биотехнология, генетическая инженерия.
Пример 4. Получение рекомбинантного белка RBD с использованием штамма рекомбинантной клеточной линии CHO-K1-RBD.
Штамм культуры-продуцента CHO-K1-RBD, показавший наибольшую продуктивность, культивировали на роллерных установках и собирали культуральную среду. Наличие целевого белка определяли с помощью SDS-PAGE и иммуноблоттинга с использованием антител против гистидинового тэга (фиг. 2). На фиг. 2 представлены результаты иммуноблоттинга белков при окрашивании моноклональными антителами к полигистидиновому тэгу, где:
1 - образец клеток CHO-K1-RBD, продуцента RBD SARS-CoV-2;
2 - образец клеток CHO-К1;
3 - образец культуральной жидкости клеток CHO-K1-RBD;
4 - образец культуральной жидкости клеток CHO-К1;
5 - образец RBD, очищенного с помощью аффинной и ионообменной хроматографии.
Культуральную жидкость, содержащую RBD, центрифугировали и фильтровали через фильтровальные системы с размером пор 0,22 мкМ для удаления клеточного дебриса и очищали с помощью аффинной хроматографии на колонке с Ni-NTA (Qiagen, Германия) согласно инструкции производителя.
Затем проводили дополнительную очистку ионообменной хроматографией на соединенных последовательно колонках с катионообменным (SP-сефароза) и анионообменным (Q-сефароза) сорбентами, уравновешенными 20мМ Трис-HCl, рН 8,2. После нанесения белка колонки промывали 20мМ Трис-HCl, рН 8,2. Затем колонку с сорбентом SP-сефароза, на которую RBD не сорбируется в этих условиях (рН 8,2), отсоединяли, а белки, связавшиеся с сорбентом Q-сефароза, элюировали в линейном градиенте концентрации NaCI от 0 до 1 М в 20мМ Трис-HCl, рН 8,2.
Фракции элюатов анализировали при помощи электрофореза в денатурирующих условиях в 15% ДСН-ПААГ (фиг. 3). На фиг. 3 представлен электрофорез белков в ДСН-ПААГ, где:
М - маркер молекулярного веса;
1 - рекомбинантный RBD, полученный в клетках E.coli (~27 кДа);
2 - рекомбинантный RBD, полученный в клетках CНО-К1-RBD (~35 кДа).
Фракции, содержащие целевой белок размером ~35 кДа, диализовали против фосфатно-солевого буфера и подвергали стерилизующей фильтрации через фильтры с размером пор 0,22 мкМ. Для препаратов белка, полученных в трех независимых экспериментах, определяли концентрацию RBD на спектрофотометре Nanodrop 2000c. Выход рекомбинантного белка RBD SARS-CoV-2 после хроматографической очистки составил 50-100 мг с литра культуральной среды, что десятикратно превосходит выход белка RBD, представленный в прототипе, и является показателем эффективности группы заявленных изобретений.
Пример 5. Оценка иммуногенности рекомбинантного белка RBD SARS-CoV-2
Оценку иммуногенности полученного рекомбинантного RBD проводили в экспериментах по иммунизации мышей линии BALB/c (самки весом 16-18 г). Группы животных содержали в отдельных клетках (виварий ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора) в одинаковых условиях, на стандартном рационе со свободным доступом к пище и воде. Эксперименты были одобрены на заседании Биоэтической комиссии ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора (ГНЦ ВБ «Вектор»/10-09.2020, утвержден протоколом Биоэтической комиссии № 5 от 01.10.2020) и выполнены с соблюдением принципов гуманности, изложенных в директивах Европейского сообщества (86/609/ЕЕС) и Хельсинкской декларации.
Животные случайным образом были разделены на шесть групп (n=6). Препараты белков вводили внутрибрюшинно на 1-й и 14-й дни. Иммунизацию 1 группы проводили введением 80 мкг RBD, 2 группы - 80 мкг RBD в комбинации с неполным адъювантом Фрейнда (Sigma, США), 3 группы - 80 мкг RBD в комбинации с Al2O3 (Brenntag Biosector A/S, Denmark). Для сравнения иммуногенности 4 и 5 группам вводили RBD, полученный в прокариотической системе экспресии E.coli BL21 (proRBD) в количестве 80 мкг proRBD в комбинации с неполным адъювантом Фрейнда (Sigma, США) и Al2O3 (Brenntag Biosector A/S, Denmark) соответственно. Контрольной группе мышей вводили физиологический раствор. Через две недели после последней иммунизации, у животных проводили забор крови и получали сыворотки.
Специфическую активность полученных сывороток оценивали с помощью ИФА по стандартному протоколу. В качестве антигенов использовали препараты RBD, полученные в эу- и прокариотической системах экспрессии, а так же рекомбинантные тримеры белка S SARS-CoV-2, где домен RBD формируется в корректной форме. Антиегены в концентрации 1 мкг/мл сорбировали в лунках 96-луночного планшета в фосфатно-солевом буфере (PBS) (Greiner bio one, Германия) при 4°С в течение ночи. Затем отмывали буфером PBST (0.1% р-р Tween-20 в PBS) и блокировали 1 % раствором казеина в буфере PBST в течение 1 часа при комнатной температуре. После этого образцы сывороток вносили в трёхкратном последовательном разведении, начиная с 1:50, и инкубировали в течение 1 часа при 37°С. Затем добавляли антитела кролика против IgG мыши (разбавленные в 1% блокирующем растворе казеина 1:6000), конъюгированные с пероксидазой хрена (Sigma, США). После каждого этапа проводили отмывку несвязавшихся белков буфером PBST. Затем вносили раствор субстрата TMB (Amresco, США) и инкубировали в течение 10 минут, реакцию терминировали раствором 1H HCl и сразу измеряли оптическую плотность при длине волны 450 нм на ИФА-ридере (ChroMate Awareness technology inc., США). Графики были построены с помощью программы GraphPad Prism 6.0.
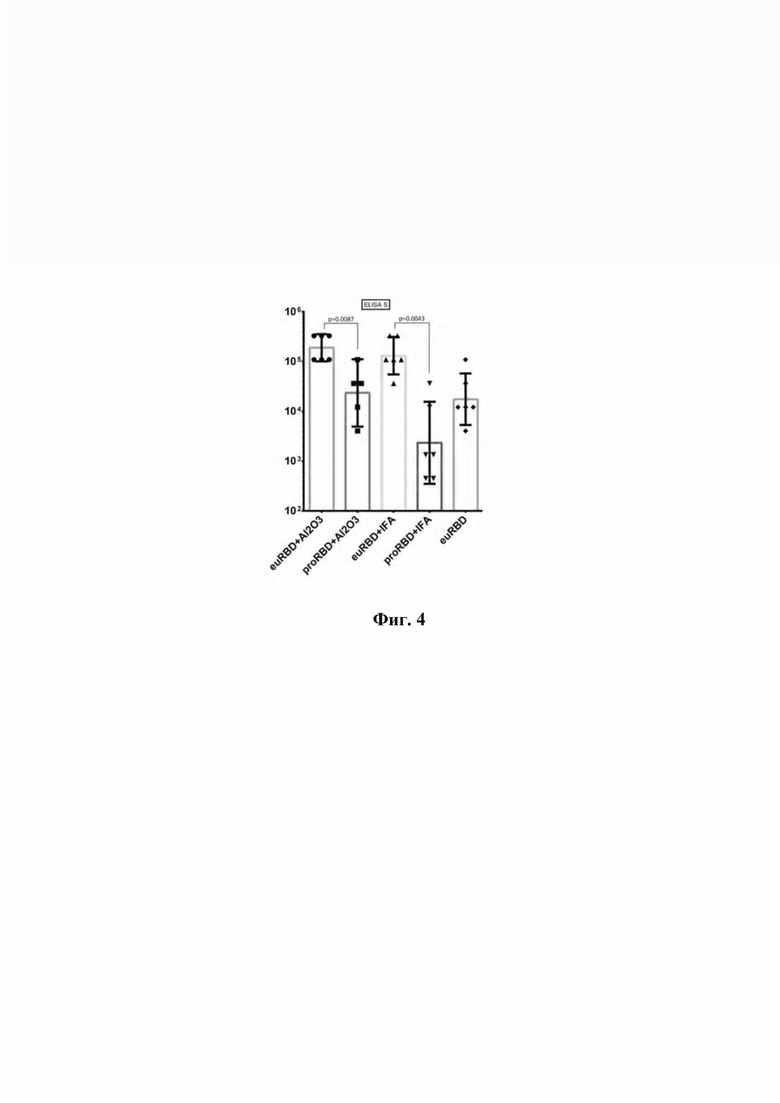
По результатам ИФА с использованием 3 указанных антигенов (фиг. 4) специфичность сывороток мышей, иммунизированных RBD, наработанным клетками CHO-K1-RBD, была выше, чем сывороток мышей, иммунизированных RBD, полученным в прокариотической системе экспрессии. На фиг. 4 представлен анализ специфичности сывороток мышей, иммунизированных RBD, с помощью иммуноферментного анализа, где:
- euRBD - RBD, полученный с помощью продуцента CHO-K1-RBD;
- proRBD - RBD, полученный с помощью продуцента E.coli BL21;
- Al2O3 - адъювант окись аллюминия;
- IFA - неполный адъювант Фрейнда.
Графики и статистическая обработка сделаны в программе GraphPad Prism. Изображены геометрические средние значения данных с 95% доверительным интервалом, статистический анализ проведён непараметрическим методом Манна-Уитни.
Анализ нейтрализующей активности сывороток мышей определяли в реакции нейтрализации цитопатического действия (ЦПД) вируса на культуре клеток. В работе использовали коронавирус SARS-Cov-2 штамм nCoV/Victoria/1/2020 (Государственная коллекция возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ БВ «Вектор» Роспотребнадзора, РФ), доза вируса составляла 100 ЦПД50. Культуру клеток Vero E6 растили в 96 луночных культуральных планшетах в ростовой питательной среде MEM с 10% фетальной бычьей сыворотки и добавлением антибиотиков (пенициллин 100 ед./мл, стрептомицин- 100 мкг/мл) до образования 100% монослоя.
Сыворотки мышей инактивировали при +56°С в течение 30 минут и готовили последовательные двукратные разведения на среде MEM начиная с разведения 1:10 (с 1:10 до 1:5120). Готовили смесь разведений сывороток и рабочего разведения вируса в равных объемах. Смесь инкубировали в течение 1 ч при комнатной температуре, затем наносили на монослой культуры клеток. Планшеты инкубировали в течение 4 сут при +37°С, 5%СО2. Затем проводили окрашивание клеток. Для этого в каждую лунку планшета вносили по 150 мкл 0,2% раствора генциан-виолета (1 г генциан виолета, 20 мл 96% этилового спирта, 120 мл 40% формалина и 350 мл раствора Хенкса). Через 30 мин удаляли жидкость из лунок и промывали водой. Оценку результатов проводили визуально, учитывая любое специфическое поражение культуры клеток в лунке как ЦПД. Титром сыворотки считали ее разведение, при котором регистрировали защиту в 50% лунок с культурой клеток от ЦПД вируса. Расчет титра нейтрализующих антител проводили по формуле Рида-Менча [23].
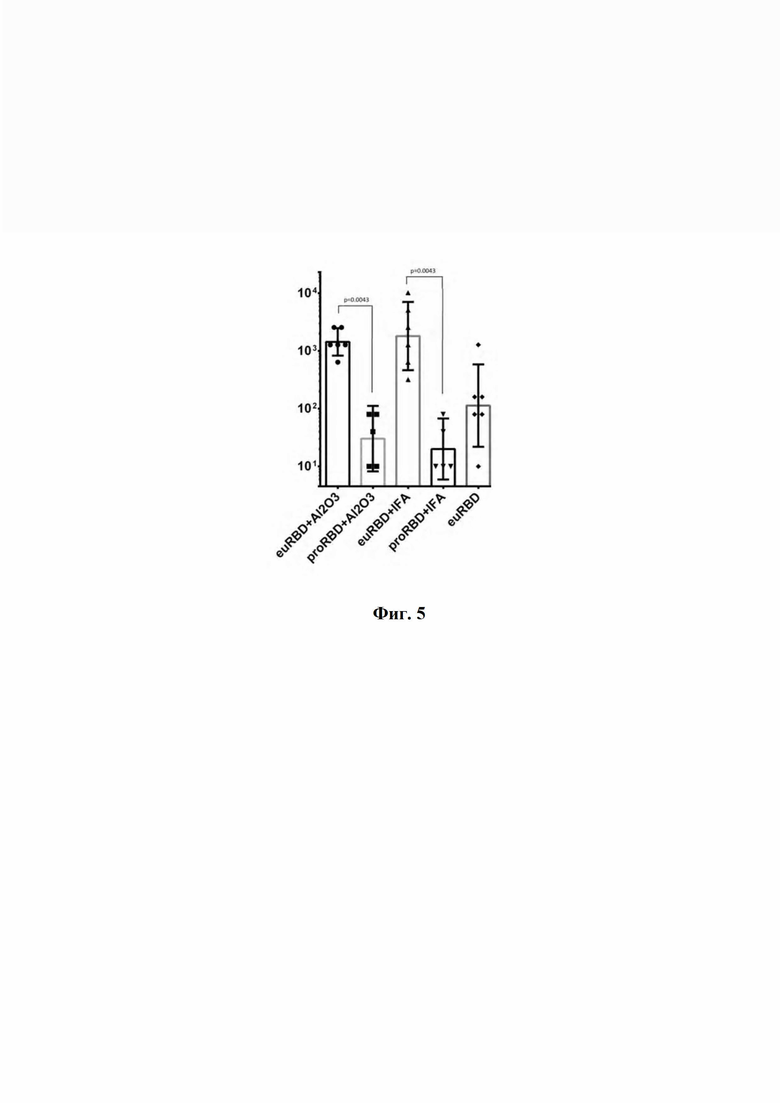
Результаты эксперимента (фиг. 5) подтверждают, что заявленный рекомбинантный RBD SARS-CoV-2 обеспечивает более эффективный гуморальный ответ, чем RBD, полученный в прокариотической системе экспресии. На фиг. 5 представлены результаты исследования нейтрализующей активности сывороток мышей, иммунизированных RBD, в реакции нейтрализации цитопатического действия (ЦПД) вируса на культуре клеток VERO E6, где:
- euRBD - RBD, полученный с помощью продуцента CHO-K1-RBD;
- proRBD - RBD, полученный с помощью продуцента E.coli BL21;
- Al2O3 - адъювант окись аллюминия;
- IFA - неполный адъювант Фрейнда.
Графики и статистическая обработка сделаны в программе GraphPad Prism. Изображены геометрические средние значения данных с 95% доверительным интервалом, статистический анализ проведён непараметрическим методом Манна-Уитни.
Таким образом, группа заявленных изобретений обеспечивает получение иммунологически корректной формы RBD SARS-CoV-2, который может быть использован как платформа для создания вакцины против COVID-19.
Технический результат заявляемого изобретения достигается за счет того, что выход рекомбинантного белка RBD SARS-CoV-2 после хроматографической очистки составляет 50-100 мг с литра культуральной среды, что десятикратно превосходит выход белка RBD, представленный в прототипе (продукция белка RBD SARS-CoV-2 в прототипе составляет от 5 мг/л до 10 мг/л).
--->
ПРИЛОЖЕНИЕ
<110> Федеральное бюджетное учреждение науки «Государственный научный центр
вирусологии и биотехнологии «Вектор» Федеральной службы по надзору в сфере защиты
прав потребителей и благополучия человека (ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора)
<120> Интегративный плазмидный вектор pVEAL2-S-RBD, обеспечивающий экспрессию и
секрецию рекомбинантного рецептор-связывающего домена (RBD) коронавируса SARS-CoV-2
в клетках млекопитающих, рекомбинантный штамм клеточной линии СНО-K1-RBD
и рекомбинантный белок RBD SARS-CoV-2, продуцируемый указанным штаммом клеточной
линии СНО-K1-RBD.
<160> 3
<210> SEQ ID NO: 1
<211> 702 п.н.
<212> DNA
<213> Artificial Sequence
<223> Кодон-оптимизированная нуклеотидная последовательность гена RBD.
<400> 1
1 gtggaaaagg gcatctacca gaccagcaac ttccgggtgc agcccaccga atccatcgtg
6 1 cggttcccca atatcaccaa tctgtgcccc ttcggcgagg tgttcaatgc caccagattc
121 gcctctgtgt acgcctggaa ccggaagcgg atcagcaatt gcgtggccga ctactccgtg
181 ctgtacaact ccgccagctt cagcaccttc aagtgctacg gcgtgtcccc taccaagctg
241 aacgacctgt gcttcacaaa cgtgtacgcc gacagcttcg tgatccgggg agatgaagtg
301 cggcagattg cccctggaca gacaggcaag atcgccgact acaactacaa gctgcccgac
361 gacttcaccg gctgtgtgat tgcctggaac agcaacaacc tggactccaa agtcggcggc
421 aactacaatt acctgtaccg gctgttccgg aagtccaatc tgaagccctt cgagcgggac
481 atctccaccg agatctatca ggccggcagc accccttgta acggcgtgga aggcttcaac
541 tgctacttcc cactgcagtc ctacggcttt cagcccacaa atggcgtggg ctatcagccc
601 tacagagtgg tggtgctgag cttcgaactg ctgcatgccc ctgccacagt gtgcggccct
661 aagaaaagca ccaatctcgt gaagaacaaa tgcgtgaact tc
<210> SEQ ID NO: 2
<211> 9910 п.н.
<212> DNA
<213> Artificial Sequence
<223> Нуклеотидная последовательность вектора pVEAL2-S-RBD.
<400> 2
1 caaggcgatt aagttgggta acgccagggt tttcccagtc acgacgttgt aaaacgacgg
61 ccagtgagcg cgcgtaatac gactcactat agggcgaatt ggagctcggt tccctataca
121 gttgaagtcg gaagtttaca tacacttaag ttggagtcat taaaactcgt ttttcaacta
181 ctccacaaat ttcttgttaa caaacaatag ttttggcaag tcagttagga catctacttt
241 gtgcatgaca caagtcattt ttccaacaat tgtttacaga cagattattt cacttataat
301 tcactgtatc acaattccag tgggtcagaa gtttacatac actaagttga ctgtgccttt
361 aaacagcttg gaaaattcca gaaaatgatg tcatggcttt agaagcttca ttggcgccct
421 ccgcgcctac agctcaagcc acatccgaag ggggagggag ccgggagctg cgcgcggggc
481 cgccgggggg aggggtggca ccgcccacgc cgggcggcca cgaagggcgg ggcagcgggc
541 gcgcgcgcgg cggggggagg ggccggcgcc gcgcccgctg ggaattgggg ccctaggggg
601 agggcggagg cgccgacgac cgcggcactt accgttcgcg gcgtggcgcc cggtggtccc
661 caaggggagg gaagggggag gcggggcgag gacagtgacc ggagtctcct cagcggtggc
721 ttttctgctt ggcagcctca gcggctggcg ccaaaaccgg actccgccca cttcctcgcc
781 cgccggtgcg agggtgtgga atcctccaga cgctggggga gggggagttg ggagcttaaa
841 aactagtacc cctttgggac cactttcagc agcgaactct cctgtacacc aggggtcagt
901 tccacagacg cgggccaggg gtgggtcatt gcggcgtgaa caataatttg actagaagtt
961 gattcgggtg tttccggaag gggccgagtc aatccgccga gttggggcac ggaaaacaaa
1021 aagggaaggc tactaagatt tttctggcgg gggttatcat tggcgtaact gcagggacca
1081 cctcccgggt tgagggggct ggatctccag gctgcggatt aagcccctcc cgtcggcgtt
1141 aatttcaaac tgcgcgacgt ttctcacctg ccttcgccaa ggcaggggcc gggaccctat
1201 tccaagaggt agtaactagc aggactctag ccttccgcaa ttcattgagc gcatttacgg
1261 aagtaacgtc gggtactgtc tctggccgca agggtgggag gagtacgcat ttggcgtaag
1321 gtggggcgta gagccttccc gccattggcg gcggataggg cgtttacgcg acggcctgac
1381 gtagcggaag acgccttagt gggggggaag gttctagaaa agcggcggca gcggctctag
1441 cggcagtagc agcagcgccg ggtcccgtgc ggaggtgctc ctcgcagagt tgtttctcca
1501 gcagcggcag ttctcactac agcgccagga cgagtccggt tcgtgttcgt ccgcggagat
1561 ctctctcatc tcgctcggct gcgggaaatc gggctgaagc gactgagtcc gcgatggagg
1621 taacgggttt gaaatcaatg agttattgaa aagggcatgg cgaggccgtt ggcgcctcag
1681 tggaagtcgg ccagccgcct ccgtgggaga gaggcaggaa atcggaccaa ttcagtagca
1741 gtggggctta aggtttatga acggggtctt gagcggaggc ctgagcgtac aaacagcttc
1801 cccaccctca gcctcccggc gccatttccc ttcactgggg gtgggggatg gggagctttc
1861 acatggcgga cgctgccccg ctggggtgaa agtggggcgc ggaggcggga cttcttattc
1921 cctttctaaa gcacgctgct tcgggggcca cggcgtctcc tcggggatcc cagatcttca
1981 atattggcca ttagccatat tattcattgg ttatatagca taaatcaata ttggctattg
2041 gccattgcat acgttgtatc tatatcataa tatgtacatt tatattggct catgtccaat
2101 atgaccgcca tgttggcatt gattattgac tagttattaa tagtaatcaa ttacggggtc
2161 attagttcat agcccatata tggagttccg cgttacataa cttacggtaa atggcccgcc
2221 tggctgaccg cccaacgacc cccgcccatt gacgtcaata atgacgtatg ttcccatagt
2281 aacgccaata gggactttcc attgacgtca atgggtggag tatttacggt aaactgccca
2341 cttggcagta catcaagtgt atcatatgcc aagtccgccc cctattgacg tcaatgacgg
2401 taaatggccc gcctggcatt atgcccagta catgacctta cgggactttc ctacttggca
2461 gtacatctac gtattagtca tcgctattac catggtgatg cggttttggc agtacaccaa
2521 tgggcgtgga tagcggtttg actcacgggg atttccaagt ctccacccca ttgacgtcaa
2581 tgggagtttg ttttggcacc aaaatcaacg ggactttcca aaatgtcgta acaactgcga
2641 tcgcccgccc cgttgacgca aatgggcggt aggcgtgtac ggtgggaggt ctatataagc
2701 agagctcgtt tagtgaaccg tcagatcact agaagcttta ttgcggtagt ttatcacagt
2761 taaattgcta acgcagtcag tgcttctgac acaacagtct cgaacttaag ctgcagtgac
2821 tctcttaagg tagccttgca gaagttggtc gtgaggcact gggcaggtaa gtatcaaggt
2881 tacaagacag gtttaaggag accaatagaa actgggcttg tcgagacaga gaagactctt
2941 gcgtttctga taggcaccta ttggtcttac tgacatccac tttgcctttc tctccacagg
3001 tgtccactcc cagttcaatt acagctctta aggctagagt acttaatacg actcactata
3061 ggctagcctc gagaattcgc tggctagcca tgtttgtttt tcttgtttta ttgccactag
3121 tctctagtca gtgtgtggaa aagggcatct accagaccag caacttccgg gtgcagccca
3181 ccgaatccat cgtgcggttc cccaatatca ccaatctgtg ccccttcggc gaggtgttca
3241 atgccaccag attcgcctct gtgtacgcct ggaaccggaa gcggatcagc aattgcgtgg
3301 ccgactactc cgtgctgtac aactccgcca gcttcagcac cttcaagtgc tacggcgtgt
3361 cccctaccaa gctgaacgac ctgtgcttca caaacgtgta cgccgacagc ttcgtgatcc
3421 ggggagatga agtgcggcag attgcccctg gacagacagg caagatcgcc gactacaact
3481 acaagctgcc cgacgacttc accggctgtg tgattgcctg gaacagcaac aacctggact
3541 ccaaagtcgg cggcaactac aattacctgt accggctgtt ccggaagtcc aatctgaagc
3601 ccttcgagcg ggacatctcc accgagatct atcaggccgg cagcacccct tgtaacggcg
3661 tggaaggctt caactgctac ttcccactgc agtcctacgg ctttcagccc acaaatggcg
3721 tgggctatca gccctacaga gtggtggtgc tgagcttcga actgctgcat gcccctgcca
3781 cagtgtgcgg ccctaagaaa agcaccaatc tcgtgaagaa caaatgcgtg aacttccatc
3841 accaccatca tcaccatcac caccactaag cttaagttta aaccgctgat cagcctcgtc
3901 gaccgagcgg ttcccgcccc tctccctccc ccccccctaa cgttactggc cgaagccgct
3961 tggaataagg ccggtgtgcg tttgtctata tgttattttc caccatattg ccgtcttttg
4021 gcaatgtgag ggcccggaaa cctggccctg tcttcttgac gagcattcct aggggtcttt
4081 cccctctcgc caaaggaatg caaggtctgt tgaatgtcgt gaaggaagca gttcctctgg
4141 aagcttcttg aagacaaaca acgtctgtag cgaccctttg caggcagcgg aaccccccac
4201 ctggcgacag gtgcctctgc ggccaaaagc cacgtgtata agatacacct gcaaaggcgg
4261 cacaacccca gtgccacgtt gtgagttgga tagttgtgga aagagtcaaa tggctcacct
4321 caagcgtatt caacaagggg ctgaaggatg cccagaaggt accccattgt atgggatctg
4381 atctggggcc tcggtgcaca tgctttacat gtgtttagtc gaggttaaaa aacgtctagg
4441 ccccccgaac cacggggacg tggttttcct ttgaaaaaca cgatgataat atggccacaa
4501 ccatgaccga gtacaagccc acggtgcgcc tcgccacccg cgacgacgtc cccagggccg
4561 tacgcaccct cgccgccgcg ttcgccgact accccgccac gcgccacacc gtcgatccgg
4621 accgccacat cgagcgggtc accgagctgc aagaactctt cctcacgcgc gtcgggctcg
4681 acatcggcaa ggtgtgggtc gcggacgacg gcgccgcggt ggcggtctgg accacgccgg
4741 agagcgtcga agcgggggcg gtgttcgccg agatcggccc gcgcatggcc gagttgagcg
4801 gttcccggct ggccgcgcag caacagatgg aaggcctcct ggcgccgcac cggcccaagg
4861 agcccgcgtg gttcctggcc accgtcggcg tctcgcccga ccaccagggc aagggtctgg
4921 gcagcgccgt cgtgctcccc ggagtggagg cggccgagcg cgccggggtg cccgccttcc
4981 tggagacctc cgcgccccgc aacctcccct tctacgagcg gctcggcttc accgtcaccg
5041 ccgacgtcga ggtgcccgaa ggaccgcgca cctggtgcat gacccgcaag cccggtgcct
5101 gattcgcata tgggttaatg cttcgagcag acatgataag atacattgat gagtttggac
5161 aaaccacaac tagaatgcag tgaaaaaaat gctttatttg tgaaatttgt gatgctattg
5221 ctttatttgt aaccattata agctgcaata aacaagttcc acggccggcc ctccgcgcct
5281 acagctcaag ccacatccga agggggaggg agccgggagc tgcgcgcggg gccgccgggg
5341 ggaggggtgg caccgcccac gccgggcggc cacgaagggc ggggcagcgg gcgcgcgcgc
5401 ggcgggggga ggggccggcg ccgcgcccgc tgggaattgg ggccctaggg ggagggcgga
5461 ggcgccgacg accgcggcac ttaccgttcg cggcgtggcg cccggtggtc cccaagggga
5521 gggaaggggg aggcggggcg aggacagtga ccggagtctc ctcagcggtg gcttttctgc
5581 ttggcagcct cagcggctgg cgccaaaacc ggactccgcc cacttcctcg cccgccggtg
5641 cgagggtgtg gaatcctcca gacgctgggg gagggggagt tgggagctta aaaactagta
5701 cccctttggg accactttca gcagcgaact ctcctgtaca ccaggggtca gttccacaga
5761 cgcgggccag gggtgggtca ttgcggcgtg aacaataatt tgactagaag ttgattcggg
5821 tgtttccgga aggggccgag tcaatccgcc gagttggggc acggaaaaca aaaagggaag
5881 gctactaaga tttttctggc gggggttatc attggcgtaa ctgcagggac cacctcccgg
5941 gttgaggggg ctggatctcc aggctgcgga ttaagcccct cccgtcggcg ttaatttcaa
6001 actgcgcgac gtttctcacc tgccttcgcc aaggcagggg ccgggaccct attccaagag
6061 gtagtaacta gcaggactct agccttccgc aattcattga gcgcatttac ggaagtaacg
6121 tcgggtactg tctctggccg caagggtggg aggagtacgc atttggcgta aggtggggcg
6181 tagagccttc ccgccattgg cggcggatag ggcgtttacg cgacggcctg acgtagcgga
6241 agacgcctta gtggggggga aggttctaga aaagcggcgg cagcggctct agcggcagta
6301 gcagcagcgc cgggtcccgt gcggaggtgc tcctcgcaga gttgtttctc cagcagcggc
6361 agttctcact acagcgccag gacgagtccg gttcgtgttc gtccgcggag atctctctca
6421 tctcgctcgg ctgcgggaaa tcgggctgaa gcgactgagt ccgcgatgga ggtaacgggt
6481 ttgaaatcaa tgagttattg aaaagggcat ggcgaggccg ttggcgcctc agtggaagtc
6541 ggccagccgc ctccgtggga gagaggcagg aaatcggacc aattcagtag cagtggggct
6601 taaggtttat gaacggggtc ttgagcggag gcctgagcgt acaaacagct tccccaccct
6661 cagcctcccg gcgccatttc ccttcactgg gggtggggga tggggagctt tcacatggcg
6721 gacgctgccc cgctggggtg aaagtggggc gcggaggcgg gacttcttat tccctttcta
6781 aagcacgctg cttcgggggc cacggcgtct cctcggggat ctagcttgtg gaaggctact
6841 cgaaatgttt gacccaagtt aaacaattta aaggcaatgc taccaaatac taattgagtg
6901 tatgtaaact tctgacccac tgggaatgtg atgaaagaaa taaaagctga aatgaatcat
6961 tctctctact attattctga tatttcacat tcttaaaata aagtggtgat cctaactgac
7021 ctaagacagg gaatttttac taggattaaa tgtcaggaat tgtgaaaaag tgagtttaaa
7081 tgtatttggc taaggtgtat gtaaacttcc gacttcaact gtatagggtt cctctagcta
7141 gagacgacct cgggtaccca gcttttgttc cctttagtga gggttaattg cgcgcttggc
7201 gtaatcatgg tcatagctgt ttcctgtgtg aaattgttat ccgctcacaa ttccacacaa
7261 catacgagcc ggaagcataa agtgtaaagc ctggggtgcc taatgagtga gctaactcac
7321 attaattgcg ttgcgctcac tgcccgcttt ccagtcggga aacctgtcgt gccagctgca
7381 ttaatgaatc ggccaacgcg cggggagagg cggtttgcgt attgggcgct cttccgcttc
7441 ctcgctcact gactcgctgc gctcggtcgt tcggctgcgg cgagcggtat cagctcactc
7501 aaaggcggta atacggttat ccacagaatc aggggataac gcaggaaaga acatgtgagc
7561 aaaaggccag caaaaggcca ggaaccgtaa aaaggccgcg ttgctggcgt ttttccatag
7621 gctccgcccc cctgacgagc atcacaaaaa tcgacgctca agtcagaggt ggcgaaaccc
7681 gacaggacta taaagatacc aggcgtttcc ccctggaagc tccctcgtgc gctctcctgt
7741 tccgaccctg ccgcttaccg gatacctgtc cgcctttctc ccttcgggaa gcgtggcgct
7801 ttctcatagc tcacgctgta ggtatctcag ttcggtgtag gtcgttcgct ccaagctggg
7861 ctgtgtgcac gaaccccccg ttcagcccga ccgctgcgcc ttatccggta actatcgtct
7921 tgagtccaac ccggtaagac acgacttatc gccactggca gcagccactg gtaacaggat
7981 tagcagagcg aggtatgtag gcggtgctac agagttcttg aagtggtggc ctaactacgg
8041 ctacactaga aggacagtat ttggtatctg cgctctgctg aagccagtta ccttcggaaa
8101 aagagttggt agctcttgat ccggcaaaca aaccaccgct ggtagcggtg gtttttttgt
8161 ttgcaagcag cagattacgc gcagaaaaaa aggatctcaa gaagatcctt tgatcttttc
8221 tacggggtct gacgctcagt ggaacgaaaa ctcacgttaa gggattttgg tcatgagatt
8281 atcaaaaagg atcttcacct agatcctttt aaattaaaaa tgaagtttta aatcaatcta
8341 aagtatatat gagtaaactt ggtctgacag ttaccaatgc ttaatcagtg aggcacctat
8401 ctcagcgatc tgtctatttc gttcatccat agttgcctga ctccccgtcg tgtagataac
8461 tacgatacgg gagggcttac catctggccc cagtgctgca atgataccgc gagacccacg
8521 ctcaccggct ccagatttat cagcaataaa ccagccagcc ggaagggccg agcgcagaag
8581 tggtcctgca actttatccg cctccatcca gtctattaat tgttgccggg aagctagagt
8641 aagtagttcg ccagttaata gtttgcgcaa cgttgttgcc attgctacag gcatcgtggt
8701 gtcacgctcg tcgtttggta tggcttcatt cagctccggt tcccaacgat caaggcgagt
8761 tacatgatcc cccatgttgt gcaaaaaagc ggttagctcc ttcggtcctc cgatcgttgt
8821 cagaagtaag ttggccgcag tgttatcact catggttatg gcagcactgc ataattctct
8881 tactgtcatg ccatccgtaa gatgcttttc tgtgactggt gagtactcaa ccaagtcatt
8941 ctgagaatag tgtatgcggc gaccgagttg ctcttgcccg gcgtcaatac gggataatac
9001 cgcgccacat agcagaactt taaaagtgct catcattgga aaacgttctt cggggcgaaa
9061 actctcaagg atcttaccgc tgttgagatc cagttcgatg taacccactc gtgcacccaa
9121 ctgatcttca gcatctttta ctttcaccag cgtttctggg tgagcaaaaa caggaaggca
9181 aaatgccgca aaaaagggaa taagggcgac acggaaatgt tgaatactca tactcttcct
9241 ttttcaatat tattgaagca tttatcaggg ttattgtctc atgagcggat acatatttga
9301 atgtatttag aaaaataaac aaataggggt tccgcgcaca tttccccgaa aagtgccacc
9361 tgacgcgccc tgtagcggcg cattaagcgc ggcgggtgtg gtggttacgc gcagcgtgac
9421 cgctacactt gccagcgccc tagcgcccgc tcctttcgct ttcttccctt cctttctcgc
9481 cacgttcgcc ggctttcccc gtcaagctct aaatcggggg ctccctttag ggttccgatt
9541 tagtgcttta cggcacctcg accccaaaaa acttgattag ggtgatggtt cacgtagtgg
9601 gccatcgccc tgatagacgg tttttcgccc tttgacgttg gagtccacgt tctttaatag
9661 tggactcttg ttccaaactg gaacaacact caaccctatc tcggtctatt cttttgattt
9721 ataagggatt ttgccgattt cggcctattg gttaaaaaat gagctgattt aacaaaaatt
9781 taacgcgaat tttaacaaaa tattaacgct tacaatttcc attcgccatt caggctgcgc
9841 aactgttggg aagggcgatc ggtgcgggcc tcttcgctat tacgccagct ggcgaaaggg
9901 ggatgtgctg
<210> SEQ ID NO: 3
<211> 244 а.о.
<212> DNA
<213> Artificial Sequence
<223> Аминокислотная последовательность белка RBD.
<400> 3
1 VEKGIYQTSN FRVQPTESIV RFPNITNLCP FGEVFNATRF ASVYAWNRKR ISNCVADYSV
61 LYNSASFSTF KCYGVSPTKL NDLCFTNVYA DSFVIRGDEV RQIAPGQTGK IADYNYKLPD
121 DFTGCVIAWN SNNLDSKVGG NYNYLYRLFR KSNLKPFERD ISTEIYQAGS TPCNGVEGFN
181 CYFPLQSYGF QPTNGVGYQP YRVVVLSFEL LHAPATVCGP KKSTNLVKNK CVNFHHHHHH
241 HHHH
<---

Изобретение относится к генной инженерии, биотехнологии и медицине. Описан интегративный плазмидный вектор pVEAL2-S-RBD, обеспечивающий экспрессию и секрецию белка RBD коронавируса SARS-CoV-2 в клетках млекопитающих, имеющий размер 9910 п.н., нуклеотидную последовательность SEQ ID No. 2 и содержащий элементы в соответствии с физической и генетической картой, представленной на Фиг. 1. Также создан рекомбинантный штамм клеточной линии яичника китайского хомячка СНО-K1-RBD - продуцент рекомбинантного белка RBD вируса SARS-CoV-2, содержащего интегративный плазмидный вектор pVEAL2-S-RBD по п. 1, имеющий нуклеотидную последовательность SEQ ID NO: 2, обеспечивающий экспрессию и секрецию белка RBD коронавируса SARS-CoV-2 и депонированный под номером 322 в коллекции культур клеток ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора (справка о депонировании прилагается). Также получен рекомбинантный белок RBD коронавируса SARS-CoV-2, продуцируемый рекомбинантным штаммом клеток яичника китайского хомячка СНО-K1-RBD по п. 2 и имеющий аминокислотную последовательность SEQ ID NO: 3. Техническим результатом является повышение выхода целевого рекомбинантного белка RBD SARS-CoV-2. Изобретение может быть использовано для создания вакцин. 3 н.п. ф-лы, 5 ил., 1 табл., 5 пр.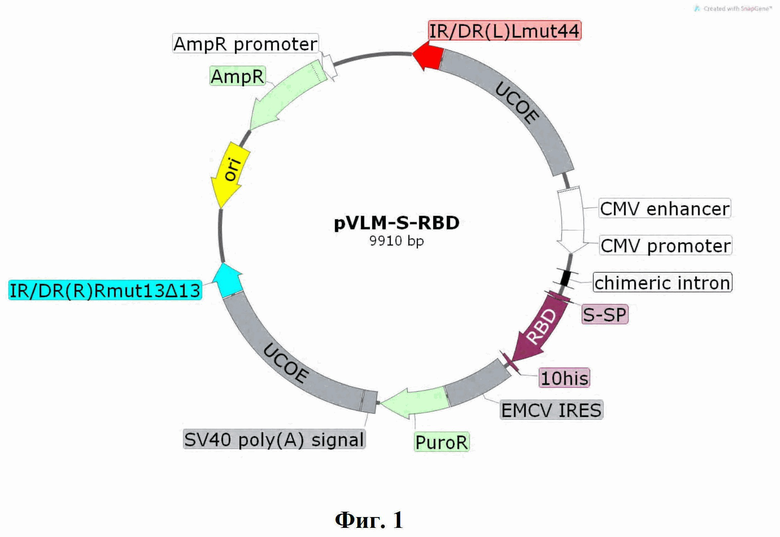
1. Интегративный плазмидный вектор pVEAL2-S-RBD, обеспечивающий экспрессию и секрецию белка RBD коронавируса SARS-CoV-2 в клетках млекопитающих, имеющий размер 9910 п.н., нуклеотидную последовательность SEQ ID NO: 2 и содержащий в соответствии с физической и генетической картой, представленной на Фиг. 1, следующие элементы:
- участок начала репликации ori (7612-8200);
- последовательности прямых и инвертированных повторов IR/DR, содержащие сайты связывания с транспозазой SB100 (139-403, 6828-7123);
- последовательности UCOE, предотвращающие хромосомное «замалчивание» рекомбинантной экспрессионной кассеты (416-1964, 5268-6816);
- CMV энхансер (2115-2494), CMV промотора (2495-2706);
- наиболее часто используемый промотор в генно-терапевтических конструкциях;
- Chimeric intron (2867-2999), химерный интрон, для усиления экспрессии целевого гена;
- лидерный пептид S-SP (3090-3134), обеспечивающий экспорт белка из клеток СНО-К1-RBD;
- кодон-оптимизированная нуклеотидная последовательность гена RBD (SEQ ID NO: 1) (3135-3836);
- последовательность 10хHis - полигистидиновый тэг для очистки рекомбинантного белка с помощью металл-хеллатной хроматографии (3837-3866);
- участок внутренней посадки рибосомы EMCV IRES (3916-4490);
- последовательность PuroR, кодирующая фактор устойчивости к антибиотику пуромицину (4503-5102);
- последовательность SV40 poly(A) signal для стабилизации мРНК-транскриптов за счет полиаденилирования (5137-5258);
- ген устойчивости к антибиотику ампициллину AmpR (8371-9231) и бактериальный промотор гена устойчивости к ампициллину (9232-9336), позволяющие проводить амплификацию плазмиды в E.coli.
2. Рекомбинантный штамм клеточной линии яичника китайского хомячка СНО-K1-RBD продуцента рекомбинантного белка RBD вируса SARS-CoV-2, содержащий интегративный плазмидный вектор pVEAL2-S-RBD по п. 1, имеющий нуклеотидную последовательность SEQ ID NO: 2, обеспечивающий экспрессию и секрецию белка RBD коронавируса SARS-CoV-2 и депонированный под номером 322 в коллекции культур клеток ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора.
3. Рекомбинантный белок RBD коронавируса SARS-CoV-2, продуцируемый рекомбинантным штаммом клеточной линии яичника китайского хомячка СНО-K1-RBD по п. 2, имеющий аминокислотную последовательность SEQ ID NO: 3 и предназначенный для создания иммунобиологических препаратов.
| Пептидные иммуногены и вакцинная композиция против коронавирусной инфекции COVID-19 с использованием пептидных иммуногенов | 2020 |
|
RU2738081C1 |
| МУТАНТНЫЕ ШТАММЫ MYCOPLASMA HYOPNEUMONIAE | 2013 |
|
RU2689671C2 |
| КОНЪЮГАТ ДЛЯ ИММУНИЗАЦИИ И ВАКЦИНАЦИИ И СПОСОБ ПОВЫШЕНИЯ ИММУНОГЕННОСТИ | 2008 |
|
RU2378015C2 |
| CN 111458504 A, 28.07.2020 | |||
| CN 111647053 A, 11.09.2020 | |||
| CN 111533790 A, 14.08.2020 | |||
| CN 111848753 A, 30.10.2020. | |||

