Предшествующий уровень техники
Рак простаты (PCa) представляет собой третий самый обычный тип рака в Соединенных Штатах с более чем 200000 новых прогнозируемых случаев, которые будут диагностированы в этом году. У приблизительно 80% пациентов с РСа фенотип опухоли включает сверхэкспрессию антигена стволовой клетки простаты (PSCA). Кроме того, PSCA экспрессируется почти на 100% раковых заболеваний простаты с метастазами в кости, делая его теоретически привлекательной иммунотерапевтической мишенью. Недавние клинические испытания с CAR (химерный рецептор антигена), нацеленными на CD19, против злокачественных заболеваний B-клеток продемонстрировали впечатляющие результаты, тем не менее, повторение этого успеха с другими антигенами-мишенями остается неясным. Иммунотерапия против солидных опухолей налагает более сложный вызов, связанный с опухолью, из-за недостатка такой ограниченной экспрессии антигена (т.е. CD19 для злокачественных заболеваний В-клеток) и присутствия иммунодепрессивной микросреды, которая может значительно мешать эффективности CAR. Важно то, что имели место случаи токсичности на мишени, вне опухоли из-за низких уровней экспрессии антигена на нормальной ткани.
В то время как основные компоненты, необходимые для создания CAR, способного к связыванию с желательной мишенью, достаточно хорошо понятны, вызовом является конструирование CAR, который имеет качества, требующиеся для применения в безопасной и эффективной терапии. Например, важно избегать избыточной активности против нераковых клеток, которые экспрессируют низкий уровень мишени или вообще не экспрессируют мишень. Также важно избегать индуцирования высокого уровня продукции цитокинов, который может индуцировать нежелательные эффекты вне опухоли. Другие факторы, которые могут влиять на терапевтический потенциал, включают способность к репликации и продолжительность жизни Т-клеток, экспрессирующих CAR, и общую эффекторную функцию Т-клеток, экспрессирующих CAR, требующуюся для надежного противоопухолевого ответа, но не ограничиваются ими.
Краткое изложение сущности изобретения
В данном документе описаны химерные трансмембранные иммунорецепторы (химерные рецепторы антигена или «CAR»), которые содержат внеклеточный домен, трансмембранную область и внутриклеточный домен сигнализации. Внеклеточный домен включает scFv (одноцепочечный Fv), нацеленный против PSCA. Описанные в данном документе CAR являются полезными для лечения рака простаты и метастазов рака простаты в кости.
Помимо scFv, нацеленного на PSCA, внеклеточный домен включает спейсер, содержащий, например, часть домена Fc человеческого IgG4. Трансмембранная часть CAR включает, например, трансмембранный домен CD4, трансмембранный домен CD8, трансмембранный домен CD28, трансмембранный домен CD3 или трансмембранный домен 4IBB. Внутриклеточный домен сигнализации включает домен сигнализации из зета цепи комплекса человеческого CD3 (CD3ζ) и костимулирующий домен, например, костимулирующий домен ОХ40, CD28, CD28gg или 4-IBB (CD137). Внеклеточный домен обеспечивает то, что CAR, при экспрессии на поверхности Т-клетки, направляет активность Т-клетки на те клетки, которые экспрессируют PSCA. Такие клетки включают раковые клетки простаты. Включение костимулирующего домена последовательно с (но не обязательно непосредственно рядом с) CD3ζ во внутриклеточной области обеспечивает получение Т-клеткой костимулирующих сигналов. Т-клетки, например, специфичные для пациента, аутологичные Т-клетки, можно генетически модифицировать для экспрессии CAR, описанных в данном документе, и данные генетически модифицированные клетки могут быть размножены и использованы в ACT (адоптивная клеточная терапия). Можно использовать разные поднаборы Т-клеток. Кроме того, CAR могут быть экспрессированы в других иммунных клетках, таких как клетки NK (природные киллеры). При лечении пациента иммунной клеткой, экспрессирующей CAR, описанный в данном документе, данная клетка может быть аутологичной или аллогенной Т-клеткой. В некоторых случаях использованные клетки представляют собой центральные Т-клетки памяти (TCM) CD4+ и CD8+, которые представляют собой CD45RA-CD62L+, или клетки TCM/SCM/N (CD45RA+CD62L+), и применение таких клеток может улучшать долговременную жизнестойкость клеток после адоптивного переноса, по сравнению с применением других типов Т-клеток, специфичных для пациента. Важно то, что общая конструкция CAR избегает нежелательной активности против нераковых клеток, включая нераковые клетки, экспрессирующие только относительно низкий уровень PSCA.
scFv против PSCA может включать последовательность:
DIQLTQSPSTLSASVGDRVTITCSASSSVRFIHWYQQKPGKAPKRLIYDTSKLASGVPSR FSGSGSGTDFTLTISSLQPEDFATYYCQQWGSSPFTFGQGTKVEIKGSTSGGGSGGGS GGGGSSEVQLVEYGGGLVQPGGSLRLSCAASGFNIKDYYIHWVRQAPGKGLEWVAWI DPENGDTEFVPKFQGRATMSADTSKNTAYLQMNSLRAEDTAVYYCKTGGFWGQGTLV TVSS (SEQ ID NO: 38) или ее вариант, имеющий вплоть до 5 аминокислотных замен (например, консервативных замен).
В данном документе описана молекула нуклеиновой кислоты, кодирующая CAR, содержащий: scFv, направленный против PSCA (например, SEQ ID NO: 1), или его вариант, имеющий 1-5 (например, 1 или 2) модификаций аминокислот; трансмембранный домен, выбранный из: трансмембранного домена CD4 или его варианта, имеющего 1-10 (например, 1 или 2) модификаций аминокислот, трансмембранного домена CD8 или его варианта, имеющего 1-5 (например, 1 или 2) модификаций аминокислот, трансмембранного домена CD28 или его варианта, имеющего 1-5 (например, 1 или 2) модификаций аминокислот, и трансмембранного домена CD3ζ или его варианта, имеющего 1-10 (например, 1 или 2) модификаций аминокислот; костимулирующий домен; и домен сигнализации CD3ζ или его вариант, имеющий 1-5 (например, 1 или 2) модификаций аминокислот. Спейсерная область расположена между scFv и трансмембранным доменом. Спейсерная область, описанная более подробно ниже, может включать всю человеческую область Fc или ее часть.
В некоторых воплощениях молекула нуклеиновой кислоты экспрессирует полипептид, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 26-37; химерный рецептор антигена содержит аминокислотную последовательность, выбранную из SEQ ID NO: 26-37 с 1-5 (например, 1 или 2) модификациями аминокислот (например, заменами).
Также раскрыта популяция человеческих Т-клеток, трансдуцированных вектором, содержащим экспрессионную кассету, кодирующую химерный рецептор антигена, где химерный рецептор антигена содержит scFv, направленный на PSCA, который включает костимулирующий домен 4-IBB. В разных воплощениях популяция человеческих Т-клеток содержит вектор, экспрессирующий химерный рецептор антигена, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 26-37; данная популяция человеческих Т-клеток содержит центральные Т-клетки памяти (TCM) (например, по меньшей мере 20%, 30%, 40%, 50%, 60%, 70%, 80% данных клеток представляют собой клетки TCM; по меньшей мере 15%, 20%, 25%, 30%, 35% клеток TCM являются CD4+, и по меньшей мере 15%, 20%, 25%, 30%, 35% клеток TCM являются клетками CD8+).
Также описан способ лечения рака у пациента, включающий введение популяции аутологичных или аллогенных человеческих Т-клеток (например, аутологичных или аллогенных Т-клеток, содержащих Т-клетки, например, по меньшей мере 20%, 30%, 40%, 50%, 60%, 70%, 80% клеток представляют собой клетки TCM; по меньшей мере 15%, 20%, 25%, 30%, 35% клеток TCM являются CD4+, и по меньшей мере 15%, 20%, 25%, 30%, 35% клеток TCM являются клетками CD8+), трансдуцированных вектором, содержащим экспрессионную кассету, кодирующую химерный рецептор антигена, где химерный рецептор антигена содержит аминокислотную последовательность, выбранную из SEQ ID NO: 26-37. В разных воплощениях популяция человеческих Т-клеток содержит центральные Т-клетки памяти; рак представляет собой глиобластому; и трансдуцированные человеческие Т-клетки были получены способом, включающим получение Т-клеток от пациента, обработку Т-клеток для выделения центральных Т-клеток памяти и трансдуцирование по меньшей мере части центральных клеток памяти вирусным вектором, содержащим экспрессионную кассету, кодирующую химерный рецептор антигена, где химерный рецептор антигена содержит аминокислотную последовательность, выбранную из SEQ ID NO: 26-37.
Также описана молекула нуклеиновой кислоты, кодирующая полипептид, содержащий аминокислотную последовательность, которая является по меньшей мере на 95% идентичной аминокислотной последовательности, выбранной из SEQ ID NO: 26-37; молекула нуклеиновой кислоты, кодирующая полипептид, содержащий аминокислотную последовательность, которая является идентичной аминокислотной последовательности, выбранной из SEQ ID NO: 26-37, за исключением присутствия не более чем 5 аминокислотных замен, делеций или вставок; молекула нуклеиновой кислоты, кодирующая полипептид, содержащий аминокислотную последовательность, которая является идентичной аминокислотной последовательности, выбранной из SEQ ID NO: 26-37, за исключением присутствия не более чем 5 аминокислотных замен; и молекула нуклеиновой кислоты, кодирующая полипептид, содержащий аминокислотную последовательность, которая является идентичной аминокислотной последовательности, выбранной из SEQ ID NO: 26-37, за исключением присутствия не более чем 2 аминокислотных замен.
Т-клетки, экспрессирующие CAR, нацеленный на PSCA, могут быть полезными в лечении рака простаты, включая устойчивый к гормонам рак простаты и метастазы рака простаты, включающие метастазы в кости, печени и легком, а также других раковых заболеваний, которые экспрессируют PSCA, которые включают раковые заболевания поджелудочной железы, мочевого пузыря, толстой кишки и глиобластому (первичный, головного мозга), но не ограничиваются ими.
Таким образом, данное раскрытие включает способы лечения рака с использованием Т-клеток, экспрессирующих CAR, описанный в данном документе.
Данное раскрытие также включает молекулы нуклеиновой кислоты, которые кодируют любые из CAR, описанных в данном документе (например, векторы, которые включают последовательность нуклеиновой кислоты, кодирующую один из CAR), и выделенные Т-лимфоциты, которые экспрессируют любой из CAR, описанных в данном документе.
CAR, описанный в данном документе, может включать спейсерную область, расположенную между доменом, нацеленным на PSCA (т.е. scFv, распознающим PSCA, или его вариантом), и трансмембранным доменом. Можно использовать целый ряд разных спейсеров. Некоторые из них включают по меньшей мере часть человеческой области Fc, например, шарнирную часть человеческой области Fc или домен СН3 или его варианты. В Таблице 1 ниже предложены разные спейсеры, которые можно использовать в CAR, описанных в данном документе.
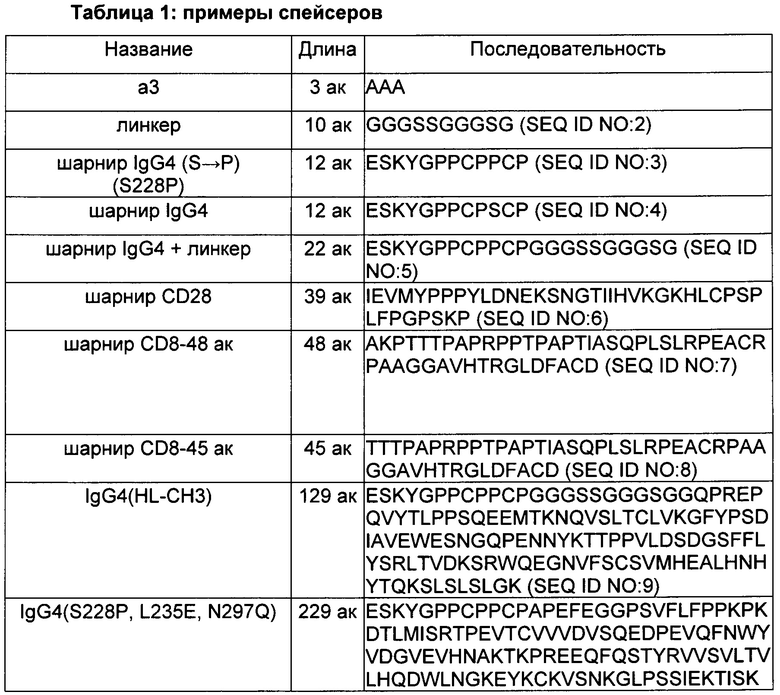

Некоторые спейсерные области включают всю шарнирную область иммуноглобулина (например, IgG1, IgG2, IgG3, IgG4) или ее часть, т.е. последовательность, которая попадает между доменами СН1 и СН2 иммуноглобулина, например, шарнир Fc IgG4 или шарнир CD8. Некоторые спейсерные области включают домен СН3 иммуноглобулина или и домен СН3, и домен СН2. Последовательности, происходящие из иммуноглобулина, могут включать одну или более чем одну модификацию аминокислот, например, 1, 2, 3, 4 или 5 замен, например, замен, которые уменьшают связывание вне мишени.
Термин «модификация аминокислот» относится к аминокислотной замене, вставке и/или делеции в последовательности белка или пептида. «Аминокислотная замена» или «замена» относится к замене аминокислоты в конкретном положении в последовательности родительского пептида или белка другой аминокислотой. Замену можно делать для изменения аминокислоты в полученном в результате белке неконсервативным образом (т.е. посредством замены кодона от аминокислоты, принадлежащей к группе аминокислот, имеющих конкретный размер или характеристику, до аминокислоты, принадлежащей к другой группе) или консервативным образом (т.е. посредством замены кодона от аминокислоты, принадлежащей к группе аминокислот, имеющих конкретный размер или характеристику, до аминокислоты, принадлежащей к той же самой группе). Такое консервативное изменение обычно приводит к меньшему изменению в структуре и функции образующегося белка. Следующее представляет собой примеры разных групп аминокислот: 1) аминокислоты с неполярными группами R: аланин, валин, лейцин, изолейцин, пролин, фенилаланин, триптофан, метионин; 2) аминокислоты с незаряженными полярными группами R: глицин, серии, треонин, цистеин, тирозин, аспарагин, глутамин; 3) аминокислоты с заряженными полярными группами R (отрицательно заряженными при рН 6,0): аспарагиновая кислота, глутаминовая кислота; 4) основные аминокислоты (положительно заряженные при рН 6,0): лизин, аргинин, гистидин (при рН 6,0). Другая группа может представлять собой те аминокислоты, которые имеют фенильные группы: фенилаланин, триптофан и тирозин.
В некоторых воплощениях спейсер происходит из IgG1, IgG2, IgG3 или IgG4 и включает один или более чем один аминокислотный остаток, замененный аминокислотным остатком, отличающимся от остатка, присутствующего в немодифицированном спейсере. Один или более чем один замененный аминокислотный остаток выбран из одного или более чем одного аминокислотного остатка в положениях 220, 226, 228, 229, 230, 233, 234, 235, 234, 237, 238, 239, 243, 247, 267, 268, 280, 290, 292, 297, 298, 299, 300, 305, 309, 218, 326, 330, 331, 332, 333, 334, 336, 339 или их комбинации, но не ограничивается ими. В данной схеме нумерации, описанной ниже более подробно, первая аминокислота в спейсере IgG4(L235E, N297Q) в Таблице 1 является 219-ой, и первая аминокислота в спейсере IgG4(HL-CH3) в Таблице 1 является 219-ой, также как и первая аминокислота в последовательности шарнира IgG и в последовательности шарнирного линкера (HL) IgG4 в Таблице 1.
В некоторых воплощениях модифицированный спейсер происходит из IgG1, IgG2, IgG3 или IgG4, который включает одну или более чем одну из следующих замен аминокислотных остатков: C220S, C226S, S228P, C229S, P230S, E233P, V234A, L234V, L234F, L234A, L235A, L235E, G236A, G237A, P238S, S239D, F243L, P247I, S267E, H268Q, S280H, K290S, K290E, K290N, R292P, N297A, N297Q, S298A, S298G, S298D, S298V, T299A, Y300L, V305I, V309L, E318A, K326A, K326W, K326E, L328F, A330L, A330S, A331S, P331S, I332E, E333A, E333S, E333S, K334A, A339D, A339Q, P396L или их комбинацию, но не ограничивается ими.
В некоторых воплощениях модифицированный спейсер происходит из области IgG4, которая включает один или более чем один аминокислотный остаток, замененный аминокислотным остатком, отличным от аминокислотного остатка, присутствующего в немодифицированной области. Один или более чем один замененный аминокислотный остаток выбран из одного или более чем одного аминокислотного остатка в положениях 220, 226, 228, 229, 230, 233, 234, 235, 234, 237, 238, 239, 243, 247, 267, 268, 280, 290, 292, 297, 298, 299, 300, 305, 309, 218, 326, 330, 331, 332, 333, 334, 336, 339 или их комбинации, но не ограничивается ими.
В некоторых воплощениях модифицированный спейсер происходит из области IgG4, которая включает одну или более чем одну из следующих замен аминокислотных остатков: 220S, 226S, 228P, 229S, 230S, 233P, 234A, 234V, 234F, 234A, 235A, 235E, 236A, 237A, 238S, 239D, 243L, 247I, 267E, 268Q, 280H, 290S, 290E, 290N, 292P, 297A, 297Q, 298A, 298G, 298D, 298V, 299A, 300L, 305I, 309L, 318A, 326A, 326W, 326E, 328F, 330L, 330S, 331S, 331S, 332E, 333A, 333S, 333S, 334A, 339D, 339Q, 396L или их комбинацию, но не ограничивается ими, где аминокислота в немодифицированном спейсере заменяется определенными выше аминокислотами в указанном положении.
Для положений аминокислот в иммуноглобулине, обсуждаемых в данном документе, нумерация осуществляется согласно индексу EU или схеме нумерации EU (Kabat et al. 1991 Sequences of Proteins of Immunological Interest, 5th Ed., United States Public Health Service, National Institutes of Health, Bethesda, тем самым целиком включена посредством ссылки). Индекс EU или индекс EU по Kabat или схема нумерации EU относится к нумерации антитела EU (Edelman et al. 1969 Proc Natl Acad Sci USA 63:78-85).
В изобретении можно использовать целый ряд трансмембранных доменов. Таблица 2 включает примеры подходящих трансмембранных доменов. При наличии спейсерного домена трансмембранный домен располагается ближе к карбоксильному концу относительно спейсерного домена.

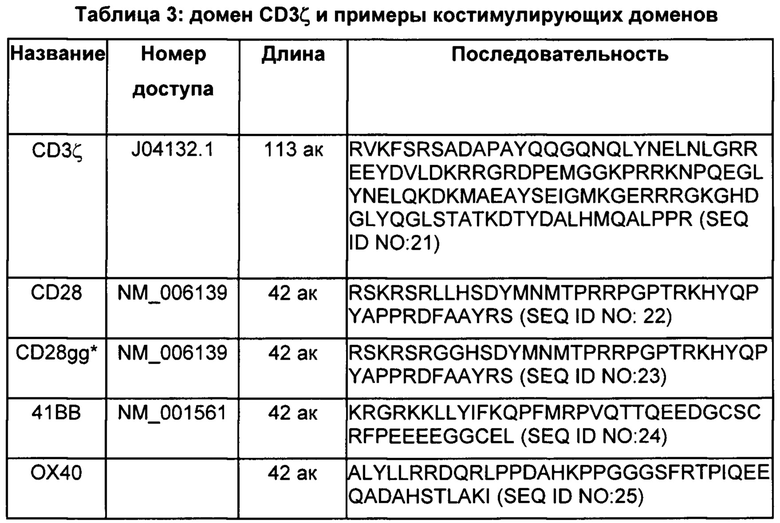
Многие из CAR, описанных в данном документе, включают один или более чем один (например, два) костимулирующих домена. Костимулирующий(щие) домен(ны) располагаются между трансмембранным доменом и доменом сигнализации CD3ζ. Таблица 3 включает примеры подходящих костимулирующих доменов, наряду с последовательностью домена сигнализации CD3ζ.
CAR против PSCA, использованные в исследованиях, описанных в данном документе, представляют собой CAR, обобщенные в Таблице 4 (незрелые, включающие сигнальную последовательность GMCSFRa (рецептор а гранулоцитарно-макрофагального колониестимулирующего фактора), в которой указаны спейсерный домен и костимулирующий(щие) домен(ны) для каждого CAR. Все из них включают scFv против PSCA А11. Спейсер IgG4(HL-CH3) также называется спейсер IgG4ΔCH2. Зрелые последовательности (не имеющие сигнальной последовательности GMCSFRa) для SEQ ID NO: 26, 27, 28, 29, 30 и 31 представляют собой SEQ ID NO: 32, 33, 43, 35, 36 и 37.

Описание чертежей
ФИГ. 1: Схематическая диаграмма конструкций CAR с целым рядом спейсерных областей (более подробно описанных выше) и имеющих либо: трансмембранный домен CD28 и костимулирующий домен CD28; или трансмембранный домен CD4 и костимулирующий домен 4-IBB. В конструкциях использовались scFv МВ1 или scFv А11. Во всех конструкциях использовался цитолитический домен CD3ζ. Последовательность перескакивания рибосомы Т2А отделяет CAR от усеченного белка CD19 (CD19t), который используется для оценки экспрессии конструкции.
ФИГ. 2: измерение данных по экспрессии tCD19 и scFv (белок L) для разных конструкций на ФИГ. 1.
ФИГ. 3А-Д: характеризация in vitro двух разных Т-клеток с CAR против PSCA относительно линий клеток человеческого рака простаты. (А) Экспрессия PSCA в опухолевых клетках, генетически модифицированных для экспрессии PSCA (LCL, РС-3 и DU145). (Б-В) Дегрануляция CD107a и продукция IFNγ (интерферон-гамма) в Т-клетках с CAR после 5 ч сокультуры с опухолью-мишенью, измеренные проточной цитометрией. (Г-Д) Продукция IFNγ Т-клетками с CAR после 24 ч культуры с рекомбинантным белком PSCA или опухолями-мишенями, измеренная посредством ELISA (твердофазный иммуноферментный анализ).
ФИГ. 4А-Д: CAR против PSCA, содержащие костимулирующий домен 4-IBB, демонстрируют превосходную специфичность, пролиферацию и способность к умерщвлению опухолевых клеток. Умерщвление опухоли (А) и индукция PD-1 (Б) в Т-клетках с CAR PSCA(ΔCH2)28ζ или PSCA(ΔCH2)BBζ после 72 ч сокультуры с опухолями-мишенями (DU145, РС-3, DU145-PSCA и PC-3-PSCA), измеренные проточной цитометрией. (В) Умерщвление опухоли при соотношении эффектор:опухоль (Е:Т) от 0,25:1 до 4:1. (Г) Измеренная посредством CFSE (сукцинимидиловый эфир карбоксифлуоресцеина) пролиферация Т-клеток с CAR после 72 ч сокультуры с опухолями-мишенями. (Д) Кинетика умерщвления опухоли и индукции PD-1 в Т-клетках с CAR после 1-, 2- или 3-суточной сокультуры с опухолями-мишенями (DU145, слева; DU145-PSCA, справа).
ФИГ. 5А-Б: внеклеточный спейсер диктует функциональность in vitro CAR против PSCA. (А) Дегрануляция CD107a и продукция IFNγ в Т-клетках с CAR PSCA(EQ)BBζ, PSCA(ΔCH2)BBζ и PSCA(L)BBζ после 5 ч сокультуры с опухолями-мишенями (DU145 и DU145-PSCA), измеренные проточной цитометрией. (Б) IFNγ в Т-клетках с CAR после 24 ч культуры с рекомбинантным белком PSCA или опухолями-мишенями, измеренный посредством ELISA.
ФИГ. 6А-Г: Т-клетки с CAR против PSCA демонстрируют мощную противоопухолевую эффективность в ксенотрансплантатах рака простаты и ортотопических моделях. (А) Клетки PC-3-PSCA (2×106) подкожно инъецировали самцам мышей NSG, и, при достижении опухолями ~30-50 мм3, внутриопухолево инъецировали Tcm с CAR (5×106), и рост опухоли отслеживали посредством измерений штангенциркулем. (Б) Клетки DU145-PSCA (2×106) подкожно инъецировали самцам NSG, и клетки РВМС (одноядерные клетки периферической крови) с CAR (5×106) доставляли внутривенно. (В) Клетки PC-3-PSCA (2×105) инъецировали в большую берцовую кость самцов NSG, и внутривенно доставляли клетки РВМС с CAR (2×106 или 5×106). (Г) Стойкость Т-клеток с CAR в крови в 58 суток после инъекции опухоли в каждой группе.
ФИГ. 7А-7Г: Т-клетки с CAR против PSCA, содержащие костимулирующие домены CD28 или 4-IBB. (А) Диаграмма лентивирусной экспрессионной кассеты с CAR против PSCA, содержащим гуманизированный scFv (клон А11), нацеленный на PSCA, со 129-аминокислотным линкером на основе модифицированного человеческого Fc IgG4 (лишенного домена СН2 - ΔCH2), трансмембранным доменом (либо CD28, либо CD4), цитоплазматическим костимулирующим доменом CD28 или 4-IBB, и цитолитическим доменом CD3ζ. Усеченный несигнализирующий CD19 (CD19t) отделяется от CAR последовательностью перескакивания рибосомы Т2А для отслеживания клеток, экспрессирующих CAR. (Б) Имитацию (нетрансдуцированных), Т-клетки с CAR PSCA(ΔCH2)28ζ или PSCA(ΔCH2)BBζ оценивали посредством проточной цитометрии на экспрессию CD19t для выявления лентивирусной трансдукции CAR (наверху) или белка L для выявления scFv (внизу). (В) Кинетика размножения ex vivo для имитации и Т-клеток с CAR против PSCA на протяжении 25 суток в культуре. (Г) Экспрессия на поверхности клетки указанных маркеров клеточной поверхности Т-клеток с CAR против PSCA в конце размножения ex vivo, определенная проточной цитометрией. Все данные являются репрезентативными для по меньшей мере двух независимых экспериментов.
ФИГ. 8А-Ж: CAR против PSCA, содержащие костимулирующий домен 4-IBB, демонстрируют превосходное нацеливание на опухоль по сравнению с костимулированием CD28 в CAR против PSCA in vitro. (А) Гистограммы экспрессии PSCA в линиях клеток человеческого рака простаты, определенной посредством проточной цитометрии. Линии клеток DU145 и РС-3 лентивирусно трансдуцировали для сверхэкспрессии человеческого PSCA под контролем промотора EF1α (см. Материалы и методы). Линию клеток PC-3-PGK100p генерировали осуществлением экспрессии человеческого PSCA под контролем указанного мутантного промотора PGK (фосфоглицераткиназа). Клетки LAPC-9 эндогенно экспрессируют человеческий PSCA. (Б) Моментальные снимки анализа умерщвления опухоли, сравнивающего имитацию, Т-клетки с CAR PSCA(ΔCH2)28ζ или PSCA(ΔCH2)BBζ при соотношении эффектор:мишень 1:1, при анализе световой микроскопией, с последующей 3-суточной сокультурой с опухолевыми клетками РС-3 или PC-3-PSCA. (В) Аналогичный (Б) анализ умерщвления опухоли при оценке проточной цитометрией после 3-суточной сокультуры с указанными опухолями-мишенями. (Г) Репрезентативные зебра-графики экспрессии PD-1 в Т-клетках с CAR против PSCA после 3-суточной сокультуры с указанными опухолями-мишенями. (Д) Количественное измерение экспрессии PD-1 в Т-клетках CD8+ CAR+ после 3-суточной сокультуры с указанными опухолями-мишенями. (Е) Анализ умерщвления опухоли, сравнивающий Т-клетки с CAR против PSCA в 1, 2 или 3 суток сокультуры с DU145. Экспрессия PD-1 в Т-клетках по сравнению с Т-клетками, культивируемыми без опухолей-мишеней. (Ж) Анализ умерщвления опухоли с разными соотношениями эффектор : мишень при оценке проточной цитометрией после 3-суточной сокультуры с PC-3-PSCA или PC-3-PGK100p.Данные показаны в виде n, равного 2, на группу плюс/минус SD (стандартное отклонение). Все данные являются репрезентативными для по меньшей мере двух независимых экспериментов.
ФИГ. 9А-Е: CAR PSCA-BBζ демонстрируют антигензависимую продукцию цитокинов in vitro. (А) Продукция IFNγ, количественно измеренная ELISA в супернатантах от Т-клеток с CAR против PSCA, культивируемых в течение ночи с опухолевыми клетками DU145 или DU145-PSCA. (Б) То же самое, что и в (А) от Т-клеток с CAR против PSCA, культивируемых в течение ночи с опухолевыми клетками РС-3, PC-3-PGK100p или PC-3-PSCA. (В) Продукция IFNγ, количественно измеряемая ELISA в супернатантах от Т-клеток с CAR против PSCA, культивируемых в течение ночи на связанном с планшетом рекомбинантном человеческом PSCA при разных концентрациях белка. (Г) Репрезентативные зебра-графики, показывающие дегрануляцию CD107a Т-клетками с CAR против PSCA после 4-6 ч сокультуры с указанными опухолями-мишенями. (Д) Количественное измерение дегрануляции CD107a Т-клетками с CAR против PSCA из (Г). (Е) Репрезентативные зебра-графики экспрессии CD137 в имитации, Т-клетках с CAR PSCA(ΔCH2)28ζ или PSCA(ΔCH2)BBζ после 3-суточной сокультуры с указанными опухолями-мишеями. Данные показаны в виде n, равного 2, на группу плюс/минус SD. Все данные являются репрезентативными для по меньшей мере двух независимых экспериментов.
ФИГ. 10А-Е: надежная терапевтическая эффективность Т-клеток с CAR PSCA(ΔCH2)BBζ в моделях подкожных и ортотопических человеческих ксенотрансплантатов рака простаты с метастазами в кости. (А) Объем опухоли (мм3) у мышей NSG, несущих подкожные опухоли PC-3-PSCA (2,5×106), в сутки 0, обработанных имитацией или Т-клетками с CAR PSCA(ΔCH2)BBζ в указанных дозах, посредством инъекции в опухоль (в.о.) в сутки 34. N равно 4 мыши на группу. Данные являются репрезентативными для по меньшей мере двух независимых экспериментов. (Б) Мыши с большими опухолями (приблизительно 500 мм3), обработанные 5×106 Т-клеток имитации или Т-клеток с CAR посредством в.в. (внутривенной) инъекции в сутки 51. N равно 3 мыши на группу. Данные являются репрезентативными для по меньшей мере двух независимых экспериментов. (В) Иммуногистохимия опухолей PC-3-PSCA, отобранных через 11 суток после в.в. обработки Т-клетками, при окрашивании человеческим CD3 (верхние панели) и гранзимом В (нижние панели). (Г) Мыши, несущие опухоли в большеберцовой кости, с клетками РС-3 (дикого типа) (0,2×106) в правой задней лапе и клетками PC-3-PSCA (0,2×106) в левой задней лапе. В сутки 14 мышей обрабатывали 5×106 позитивных в отношении люциферазы светляка (~30%) Т-клеток имитации или Т-клеток с CAR PSCA(ΔCH2)BBζ посредством в.в. инъекции. Транспорт Т-клеток отслеживали в 4 часа, 1 сутки, 2 суток и 4 суток посредством неинвазивной оптической визуализации (Xenogen). Количественная оценка изображений потока, показывающая отношение PC-3-PSCA / РС-3 (дикого типа). N равно 4-6 мышей на группу. (Д) Мыши NSG, несущие внутрибольшеберцовые (левая задняя нога) PC-3-PSCA-eGFP-ffluc (0,2×106). Кинетику роста опухолей отслеживали посредством неинвазивной оптической визуализации (Xenogen). В сутки 14 мышам в.в. инъецировали 5×106 Т-клеток имитации или разные дозы Т-клеток с CAR PSCA(ΔCH2)BBζ. Показаны репрезентативные изображения потока для мышей в сутки 13 (предобработка) и сутки 33. (Е) Количественная оценка изображений потока (с интересующей областью (ROI) в месте инъекции опухоли) из групп только опухоли, Т-клеток имитации (5×106) и Т-клеток с CAR PSCA(ΔCH2)BBζ (5×106, 2,5×106, 1×106, 0,5×106). N равно 4 мыши на группу для групп CAR. Данные являются репрезентативными для по меньшей мере двух независимых экспериментов.
ФИГ. 11А-Г: надежная противоопухолевая эффективность Т-клеток с CAR PSCA(ΔCH2)BBζ по сравнению с Т-клетками с CAR PSCA(ΔCH2)28ζ в модели полученного от пациента ксенотрансплантата рака простаты с метастазами в кости. (А) Мыши NSG, несущие внутрибольшеберцовую (левая задняя лапа) LAPC-9-eGFP-ffluc (0,15×106). Кинетику роста опухолей отслеживали посредством неинвазивной оптической визуализации (Xenogen). В сутки 14 мышам в.в. инъецировали 5×106 Т-клеток имитации, Т-клеток с CAR PSCA(ΔCH2)28ζ или PSCA(ΔCH2)BBζ. Показаны репрезентативные изображения потока для мышей в указанные сутки. (Б) Количественная оценка изображений потока с ROI в большеберцовой кости (верхние панели) или от всего организма (нижние панели) из каждой группы обработки. (В) Уровни PSA, определенные ELISA, из сыворотки, отобранной у обработанных мышей (n равно 2-3 на группу) в сутки 76 после инъекции опухоли. (Г) Анализ проточной цитометрией периферической крови мышей в 24 и 76 суток после инъекции опухоли (n равно 2-3 на группу). Данные составлены из двух независимых экспериментов in vivo.
ФИГ. 12: фенотипы поверхности клеток популяций РВМС и TCM. (а) Исходные популяции РВМС и TCM анализировали проточной цитометрией на экспрессию CD4, CD8, CD45RA, CD62L, CCR7, CD14, CD25 и CD95. Показаны репрезентативные графики FACS (флуоресцентная сортировка клеток).
ФИГ. 13: анализ экспрессии мРНК PSCA в линиях опухолевых клеток. (а) кПЦР (количественная полимеразная цепная реакция) проводили на разных линиях раковых клеток простаты и не простаты для количественного измерения экспрессии PSCA. мРНК PSCA нормировали к мРНК GAPDH (глицеральдегид-3-фосфатдегидрогеназа).
ФИГ. 14А-В: сравнение CAR против PSCA, содержащих scFv МВ1 и содержащих scFv А11. (А) Диаграмма лентивирусной экспрессионной кассеты с CAR против PSCA, содержащим гуманизированный scFv МВ1 или А11, нацеленный на PSCA, со 129-аминокислотным линкером на основе модифицированного Fc человеческого IgG4 (не содержащего домена СН2, ΔСН2), трансмембранным доменом CD4, цитоплазматическим костимулирующим доменом 4-IBB и цитолитическим доменом CD3ζ. Усеченный несигнализрующий CD19 (CD19t) отделяется от CAR последовательностью перескакивания рибосомы Т2А для отслеживания клеток, экспрессирующих CAR. (Б) Т-клетки имитации (нетрансдуцированные), Т-клетки с CAR PSCA-MB1-(ΔCH2)BBζ или PSCA-A11-(ΔСН2)ВВζ, экспрессирующие CD19 для выявления лентивирусной трансдукции CAR (наверху) или белок L для выявления scFv (внизу) при определении проточной цитометрией. (В) Анализ умерщвления опухоли при оценке проточной цитометрией после 3-суточной сокультуры с указанными опухолями-мишенями.
ФИГ. 15: продукция цитокинов Т-клетками с CAR PSCA(ΔCH2)BBζ при трансдукции либо в РВМС, либо в TCM. Продукцию IFNγ количественно измеряли посредством ELISA в супернатантах от Т-клеток с CAR против PSCA, культивируемых на связанном с планшетом рекомбинантном человеческом PSCA, при разных концентрациях белка.
ФИГ. 16: активация и исчерпывающий фенотип Т-клеток имитации, Т-клеток с CAR PSCA(ΔCH2)28ζ и PSCA(ΔCH2)BBζ относительно связанного с планшетами OKT3. Экспрессия CD137 и PD1, определенная посредством проточной цитометрии в Т-клетках после 2-суточной инкубации с OKT3, связанным с планшетом (10 мкг/мл).
ФИГ. 17А-Г: лечение рецидивов PCSA-негативных опухолей с использованием Т-клеток с CAR, специфичным в отношении HER2 (человеческий рецептор эпидермального фактора роста 2). (А) Кинетика рецидивов опухолей у мышей, несущих PC-3-PSCA опухоль, обработанных PSCA(ΔCH2)BBζ. Каждая линия представляет индивидуальную мышь на группу. N равно 4 на группу. Данные являются репрезентативными для по меньшей мере двух независимых исследований. (Б) Иммуногистохимия опухолей PC-3-PSCA, отобранных из опухолей, обработанных имитацией (в первичный момент времени) или рецидивирующих опухолей, обработанных PSCA(ΔCH2)BBζ, окрашенных человеческим PSCA, CD3 и HER2. (В) Экспрессия HER2 в опухолевых клетках PC-3-PSCA, оцениваемая посредством проточной цитометрии. (Г) Объем опухоли (мм3) у мышей, несущих опухоли PC-3-PSCA, в.в. обработанных 5×106 Т-клеток имитации или Т-клеток с CAR PSCA(ΔCH2)BBζ (N равно 6 на группу), в сутки 24 («1-ая tx»). В сутки 81, когда мыши, обработанные Т-клетками с CAR, демонстрировали рецидив опухоли (50-100 мм3), мышам назначали вторую обработку («2-ая tx»), и они получали в.о. инъекции одних из: 5×106 Т-клеток имитации, Т-клеток с CAR PSCA(ΔCH2)BBζ или Т-клеток с CAR против HER2 (N равно 2 на группу).
ФИГ. 18: аминокислотная последовательность PSCAscFv-IgG4(HL-CH3)-CD4tm-4IBB-зета (SEQ ID NO:26).
ФИГ. 19: аминокислотная последовательность PSCAscFv-IgG4(EQ)-CD28tm-CD28gg-зета (SEQ ID NO:27).
ФИГ. 20: аминокислотная последовательность PSCAscFv-L-CD4tm-4IBB-зета (SEQ ID NO:28).
ФИГ. 21: аминокислотная последовательность PSCAscFv-IgG4(HL-CH3)-CD28tm-CD28gg-зета (SEQ ID NO:29).
ФИГ. 22: аминокислотная последовательность PSCAscFv-IgG4(EQ)-CD4tm-4IBB-зета (SEQ ID NO:30).
ФИГ. 23: аминокислотная последовательность PSCAscFv-L-CD28tm-4IBB-зета (SEQ ID NO:31).
Подробное описание изобретения
Ниже описаны структура, конструирование и характеризация разных химерных рецепторов антигена, нацеленных на PSCA. Химерный антиген (CAR) представляет собой рекомбинантную биомолекулу, которая содержит, как минимум, внеклеточный домен распознавания, трансмембранную область и внутриклеточный домен сигнализации. Термин «антиген», следовательно, не ограничивается молекулами, которые связываются антителами, но относится к любой молекуле, которая может специфично связываться с мишенью. Например, CAR может включать лиганд, который специфично связывается с рецептором поверхности клетки. Внеклеточный домен распознавания (также именуемый внеклеточным доменом или просто элементом распознавания, который он содержит) содержит элемент распознавания, который специфично связывается с молекулой, присутствующей на клеточной поверхности клетки-мишени. Трансмембранная область заякоривает CAR в мембране. Внутриклеточный домен сигнализации содержит домен сигнализации из зета цепи человеческого комплекса CD3 и, возможно, содержит один или более чем один костимулирующий домен сигнализации. CAR могут и связываться с антигеном, и трансдуцировать активацию Т-клеток, независимо от ограничения МНС (главный комплекс гистосовместимости). Таким образом, CAR представляют собой «универсальные» иммунорецепторы, которые могут лечить популяцию пациентов с антигенпозитивными опухолями, независимо от их генотипа HLA (человеческий лейкоцитарный антиген). Адоптивная иммунотерапия с использованием Т-лимфоцитов, которые экспрессируют опухолеспецифичные CAR, может быть мощной терапевтической стратегией для лечения рака.
Авторы изобретения генерировали широкий спектр CAR против PSCA и тестировали в нескольких анализах для идентификации CAR, имеющих подходящую активность и специфичность, не индуцируя избыточную продукцию цитокинов.
В некоторых случаях CAR, описанные в данном документе, могут быть продуцированы с использованием вектора, в котором после открытой рамки считывания CAR следует последовательность перескакивания рибосомы Т2А и усеченный CD19 (CD19t), у которого отсутствует цитоплазматический сигнализирующий хвост (усеченная аминокислота 323). При данной организации коэкспрессия CD19t дает инертный неиммуногенный поверхностный маркер, который обеспечивает точное измерение генномодифицированных клеток и делает возможной позитивную селекцию генномодифицированных клеток, а также эффективное отслеживание клеток и/или визуализацию терапевтических Т-клеток in vivo после адоптивного переноса. Коэкспрессия CD19t дает маркер для иммунологического нацеливания на трансдуцированные клетки in vivo с использованием клинически доступных антител и/или реактивов на основе иммунотоксина для избирательного устранения терапевтических клеток и функционирования, посредством этого, в качестве суицидного переключателя.
CAR, описанный в данном документе, может быть получен посредством любых способов, известных в данной области, хотя предпочтительно его получают с использованием методик генной инженерии. Нуклеиновые кислоты, кодирующие несколько областей химерного рецептора, могут быть получены и собраны в полную кодирующую последовательность посредством стандартных методик молекулярного клонирования, известных в данной области (скрининг геномных библиотек, ПЦР, лигирование с помощью праймера, сайтнаправленный мутагенез и т.д.), которые являются удобными. Образующуюся кодирующую область предпочтительно вставляют в экспрессионный вектор и используют для трансформации подходящей экспрессионной линии клеток-хозяев, предпочтительно линии клеток Т-лимфоцитов и, наиболее предпочтительно, аутологичной линии клеток Т-лимфоцитов.
Вектором для экспрессии CAR могут быть трансдуцированы разные поднаборы Т-клеток, выделенных из пациента, включая не подвергавшиеся селекции РВМС или Т-клетки, обогащенные CD3, или обогащенные CD3, или поднаборы Т-клеток памяти. Центральные Т-клетки памяти являются одним полезным поднабором Т-клеток. Центральные Т-клетки памяти могут быть выделены из одноядерных клеток периферической крови (РВМС) посредством отбора клеток CD45RO+/CD62L+с использованием, например, прибора CliniMACS® для иммуномагнитной селекции клеток, экспрессирующих желательные рецепторы. Клетки, обогащенные центральными Т-клетками памяти, можно активировать антителами против CD3/CD28, трансдуцировать, например, лентивирусным вектором SIN, который управляет экспрессией CAR, а также усеченным человеческим CD19 (CD19t) - неиммуногенным поверхностным маркером как для выявления in vivo, так и для потенциальной селекции ex vivo. Активированные/генетически модифицированные центральные Т-клетки памяти можно размножать in vitro с использованием IL-2/IL-15 и затем криоконсервировать.
Пример 1: конструирование CAR, нацеленного на PSCA
На ФИГ. 1 схематически проиллюстрированы элементы в открытой рамке считывания экспрессионного вектора, использованного для экспрессии разных CAR (верхняя панель), и образующийся CAR (нижняя панель). В данном CAR используется scFv МВ1, нацеленный на PSCA. scFv А11 не использовали, но он является подходящей альтернативой. Использовали три разных спейсера: IgG4(EQ), который включает область Fc IgG4, включающую области СН3, СН4 и шарнир, и имеет две аминокислотные замены, которые уменьшают связывание с нативными рецепторами Fc; IgG4(HL-CH3), который является аналогичным IgG4(EQ), но не имеет домена СН2 и имеет короткую линкерную последовательность, расположенную между шарнирной областью и областью СН3; и L, которая представляет собой короткую линкерную последовательность. Все три спейсера подробно описаны в Таблице 1. Использовали два альтернативных трансмембранных домена: CD4 и CD28, которые оба более подробно описаны в Таблице 2. Использовали два альтернативных костимулирующих домена: CD28gg - вариант костимулирующего домена CD28 и 4-IBB. Оба подробно описаны в Таблице 3. Все CAR включали цитоплазматический домен сигнализации CD3ζ, также описанный в Таблице 3. После кодирующих последовательностей CAR следовала последовательность перескакивания рибосомы Т2А и усеченная последовательность CD17 для обеспечения коэкспрессии поверхностного, некомпетентного к сигнализации, усеченного CD19 в качестве маркера.
Массу центральных Т-клеток памяти, которые включали клетки CD4+ и клетки CD8+, трансдуцировали лентивирусом, экспрессирующим один из шести разных CAR, проиллюстрированных в Таблице 4. Таким образом, CAR включал либо костимулирующий домен 4-IBB (и трансмембранный домен CD4), либо костимулирующий домен CD22gg (и трансмембранный домен CD28) и один из трех разных спейсерных доменов: IgG4(EQ), IgG4(HL-CH3) или L (обозначенные EQ, ΔСН2 или L на ФИГ. 2). Осуществляли FACS для измерения Т-клеток, экспрессирующих CD19 (CD19t) для выявления CAR, и белок L для выявления scFv для определения стабильности. Результаты данного анализа проиллюстрированы на ФИГ. 2.
Пример 2: клетки опухоли простаты, экспрессирующие PSCA Для экспрессии PSCA генетически модифицировали две разные линии опухолевых клеток рака простаты: РС-3 и DU145. На ФИГ. 3А приведены данные по экспрессии PSCA для родительских клеток и генетически модифицированных клеток, а также для клеток LCL.
Пример 3: продукция IFNγ разными Т-клетками, нацеленными на PSCA
На ФИГ. 3Б-Д приводятся данные по продукции IFNγ и данные по дегрануляции CD107a для двух разных CAR после 5 ч сокультуры с опухолью-мишенью (клетки DU145, клетки РС3, клетки DU145, трансфицированные экспрессионным вектором PSCA, или клетки РС3, трансфицированные экспрессионным вектором PSCA), измеренные посредством проточной цитометрии. На ФИГ. 4Г-Д приводятся данные по продукции IFNγ Т-клетками с CAR после 24 ч культуры с рекомбинантным белком PSCA или опухолями-мишенями, измеренные ELISA. Здесь также можно видеть то, что CAR с костимулирующим доменом 4-IBB продуцируют меньше IFNγ и меньшие уровни маркера дегрануляции, чем CAR с костимулирующим доменом CD28.
Данная оценка дегрануляции и внутриклеточной продукции IFNγ выявила то, что все CAR, которые включают костимулирующий домен CD22gg, демонстрируют неспецифичную активность против клеток DU145 дикого типа и клеток РС3 дикого типа, тогда как CAR, которые включают костимулирующий домен 4-IBB, демонстрируют значительно меньшую неспецифичную активность. Кроме того, CAR, которые включают костимулирующий домен CD22gg, продуцируют в целом больше цитокинов, чем те, которые включают костимулирующий домен 4-IBB.
Пример 4: умерщвление клеток разными CAR
Сравнение CAR, имеющих костимулирующий домен CD28, и CAR, имеющих костимулирующий домен 4-IBB (описанных на ФИГ. 3А), продемонстрировало то, что CAR против PSCA, содержащие костимулирующий домен 4-IBB, демонстрируют превосходные специфичность, пролиферацию и способность к умерщвлению опухолевых клеток. Результаты данного анализа показаны на ФИГ. 4А-Д. Умерщвление опухолей было более специфичным для CAR, имеющих костимулирующий домен 4-IBB, чем для CAR, имеющих костимулирующий домен CD28, что продемонстрировано меньшим умерщвлением клеток, не трансфицированных экспрессионной конструкцией PSCA (ФИГ. 4А). CAR, имеющие 4-IBB, также демонстрировали меньшие уровни индукции PD-1 (ФИГ. 4Б). Умерщвление и индукцию PD-1 измеряли после 72 ч сокультуры с опухолями-мишенями (DU145, РС-3, DU145-PSCA и PC-3-PSCA). На ФИГ. 4 В показаны результаты анализа умерщвления опухолей при использовании соотношений эффектор : опухоль (Е:Т) от 0,25:1 до 4:1. На ФИГ. 4Г показаны результаты анализа пролиферации Т-клеток с CAR после 72 ч сокультуры с опухолями-мишенями, и на ФИГ. 4Д показана кинетика умерщвления опухоли и индукции PD-1 в Т-клетках с CAR после 1, 2 или 3 суток сокультуры с опухолями-мишенями (DU145 - слева; DU145-PSCA - справа).
Пример 5: влияние спейсера на функцию CAR
Исследования, проиллюстрированные на ФИГ. 5А-Б показывают то, что спейсерная область может влиять на экспрессию CD107a (дегрануляция) и продукцию IFNγ. Все CAR здесь включают трансмембранный домен CD4 и костимулирующий домен 4-IBB.
Пример 6: противоопухолевая эффективность в ксенотрансплантате рака простаты и ортотопических моделях
Два типа Т-клеток с CAR против PSCA, описанных выше, демонстрируют мощную противоопухолевую эффективность в ксенотрансплантате рака простаты и ортотопических моделях. Клетки PC-3-PSCA (2×106) подкожно инъецировали самцам мышей NSG, и при достижении опухолями ~30-50 мм3 TCM с CAR (5×106) инъецировали в опухоль, и рост опухоли отслеживали посредством измерений штангенциркулем (ФИГ. 6А). Клетки DU145-PSCA (2×106) подкожно инъецировали самцам NSG, и клетки РВМС с CAR (5×106) доставляли внутривенно (ФИГ. 6Б). Для создания ортотопической модели клетки PC-3-PSCA (2×105) инъецировали у самцов NSG в большеберцовую кость, и клетки РВМС с CAR (2×106 или 5×106) доставляли внутривенно (ФИГ. 6 В). Оценивали стойкость Т-клеток с CAR в крови в 58 суток после инъекции опухоли в каждой группе из панели Б (ФИГ. 6Г).
Пример 7: CAR, нацеленный на PSCA, содержащий домен 4-IBB, демонстрирует превосходную селективность и уменьшает исчерпание Т-клеток по сравнению с доменом CD28
Сравнивали две конструкции CAR против PSCA, которые включают гуманизированный scFv против PSCA, полученный из 1G8 (клон А11) [Lepin et al. 2010 Eur J Nucl Med Mol Imaging 37:529), внеклеточный спейсер ΔСН2, цитолитический домен CD3ζ и домен для отслеживания клеток CD19t и отличаются только их костимулирующим доменом (4-IBB по сравнению с CD28) (ФИГ. 7А). В данном Примере и Примерах 8-11 описаны исследования с использованием CAR против PSCA, генетически модифицированных в Т-клетках, происходящих из РВМС, если не указано иное. Например, в некоторых исследованиях использовали центральные Т-клетки памяти (TCM), которые имеют отличный исходный фенотип клеточной поверхности Т-клетки (ФИГ. 12).
Оба CAR против PSCA стабильно экспрессировались (ФИГ. 7Б) при определении посредством выявления scFV и CD19t проточной цитометрией, хотя и с меньшими уровнями для PSCA(ΔCH2)BBζ по сравнению с PSCA(ΔCH2)28ζ. Данные Т-клетки с CAR демонстрировали сравнимую кинетику размножения Т-клеток ex vivo (ФИГ. 7В) и аналогичные фенотипы клеточной поверхности Т-клеток (ФИГ. 7Г).
Затем несколько линий клеток человеческого рака простаты, которые были стабильно генетически модифицированы для экспрессии человеческого гена PSCA под контролем промотора EF1α, использовали для определения способностей Т-клеток с CAR PSCA(ΔCH2)28ζ и PSCA(ΔCH2)BBζ к умерщвлению опухолей (ФИГ. 8А). Опухолевые клетки РС-3 также генетически модифицировали PSCA под управлением мутантного промотора PGK (фосфоглицераткиназа) (Frigault et al. 2015 Cancer Immunol Res 3:356) для получения линии клеток с низкой плотностью антигена (обозначенной PGK100p). LAPC-9 представляет собой ксенотрансплантат первичной опухоли, полученный от пациента с раком простаты с метастазами в кости (Craft et al. 1999 Cancer Res 59:503), который эндогенно экспрессирует PSCA. Т-клетки с CAR PSCA(ΔCH2)28ζ или PSCA(ΔCH2)BBζ сокультивировали с разными опухолями-мишенями. Визуализация клеток качественно продемонстрировала то, что оба CAR умерщвляли с аналогичной кинетикой (ФИГ. 8Б). В отдельном анализе умерщвления опухоли использовали проточную цитометрию для количественного измерения умерщвления опухоли посредством Т-клеток с CAR PSCA(ΔCH2)28ζ и PSCA(ΔCH2)BBζ. В то время как и Т-клетки с CAR PSCA(ΔCH2)28ζ, и Т-клетки с CAR PSCA(ΔCH2)BBζ умерщвляли опухолевые клетки, экспрессирующие PSCA, с аналогичной эффективностью, PSCA(ΔCH2)28ζ демонстрировал нацеливание на опухолевые клетки DU145 и РС-3 дикого типа, не экспрессирующие PSCA (ФИГ. 8В). На всех опухолях-мишенях проводили количественный ПЦР-анализ в реальном времени экспрессии PSCA, и он показал то, что в то время как экспрессия белка PSCA не выявлялась проточной цитометрией в клеках DU145 и РС-3 дикого типа, в данных линиях выявлялась экспрессия мРНК (ФИГ. 13), что вероятно способствовало нацеливанию CAR, содержащих CD28.
Исследовали влияние альтернативного scFv против PSCA - МВ1 [33] (ФИГ. 14А). В то время как оба CAR против PSCA, содержащих 4-IBB, на основе МВ1 и А11 экспресировались с аналогичной стабильностью (ФИГ. 14Б), CAR, содержащие scFv МВ1, демонстрировали значимое нацеливание на опухолевые клетки дикого типа по сравнению с CAR, содержащими scFv А11 (ФИГ. 14 В). Эти данные свидетельствуют о том, что домены нацеливания на антиген и костимулирующие домены согласованно работают для обеспечения селективности CAR в отношении опухоли, и что неселективность одного домена может перевешивать селективность, управляемую другим доменом.
Помимо усиленной селективности и отсутствия умерщвления дикого типа - опухолевых клеток, не экспрессирующих PSCA, Т-клетки с CAR PSCA(ΔCH2)BBζ демонстрировали меньше доказательств исчерпания по сравнению с Т-клетками с CAR PSCA(ΔCH2)28ζ, на что указывает пониженная экспрессия белка программируемой смерти-1 (PD-1) (ФИГ. 8Г). Различие в экспрессии PD-1 между PSCA(ΔCH2)28ζ и PSCA(ΔCH2)BBζ, главным образом, наблюдали в поднаборе CD8+ Т-клеток с CAR (ФИГ. 8Д). Кроме того, аналогичные тренды, хотя и менее надежные, наблюдали с другими маркерами исчерпывания, включающими LAG3 и TIM3 (данные не показаны).
Осуществляли анализ динамики умерщвления, в котором способность к умерщвлению PSCA(ΔCH2)28ζ и PSCA(ΔCH2)BBζ в одни, двое и трое суток сокультуры с опухолевыми клетками использовали для проверки кинетики экспрессии PD-1 (ФИГ. 8Е). Эти данные количественно подтвердили то, что PSCA(ΔCH2)28ζ и PSCA(ΔCH2)BBζ эквивалентно умерщвляли DU145-PSCA с течением времени, но что PSCA(ΔCH2)28ζ имел более высокую экспрессию PD-1.
В другом исследовании PSCA(ΔCH2)28ζ и PSCA(ΔCH2)BBζ сокультивировали относительно линии опухоли с низкой экспрессией PSCA (РС-3-PGK100p) и линии опухоли с высокой экспрессией PSCA (PC-3-PSCA) при разных соотношениях эффектор : мишень (Е:Т). Это исследование показало то, что при более низких соотношениях Е:Т PSCA(ΔCH2)BBζ был более селективным в отношении опухолевых клеток с высокой экспрессией PSCA по сравнению с PSCA(ΔCH2)28ζ (ФИГ. 8Ж). Аналогичные результаты наблюдали с использованием Т клеток с CAR против PSCA, происходящих либо из РВМС, либо TCM (данные не показаны). В совокупности эти данные свидетельствуют о том, что костимуляция 4-IBB обеспечивает мощное и селективное умерщвление опухолевых клеток с высокой экспрессией PSCA при минимизации активности против клеток с меньшей экспрессией PSCA, тогда как CAR, содержащие CD28, не имеют такой селективности нацеливания.
Пример 8: CAR против PSCA, содержащие 4-IBB, демонстрируют ослабленную, тем не менее селективную продукцию цитокинов по сравнению с CAR против PSCA, содержащими CD28
Для дальнейшего исследования различий между CAR против PSCA, содержащими CD28 и 4-IBB, провели исследования для сравнения соответствующей им активации Т-клеток и продукции цитокинов. Данные исследования выявили значимое ослабление продукции IFNγ Т-клетками с CAR PSCA(ΔCH2)BBζ по сравнению с Т-клетками с CAR PSCA(ΔCH2)28ζ после сокультивирования в течение ночи с опухолевыми клетками DU145-PSCA (ФИГ. 9А). Аналогичное ослабление продукции цитокинов наблюдали для CAR против PC-3-PSCA, содержащих 4-IBB. В то время как Т-клетки с CAR против PSCA, содержащие CD28, продуцировали эквивалентные уровни IFNγ относительно опухолевых клеток, экспрессирующих низкие и высокие уровни PSCA, Т-клетки с CAR, содержащие 4-IBB, продуцировали меньшие уровни IFNγ относительно опухолевых клеток, экспрессирующих низкие уровни PSCA (ФИГ. 9Б). Для исключения потенциальных не опосредованных CAR эффектов на продукцию цитокинов опухолевыми клетками проводили аналогичные измерения IFNγ посредством Т-клеток с CAR PSCA(ΔCH2)BBζ относительно рекомбинантного человеческого белка PSCA, связанного с планшетом. В то время как Т-клетки с CAR, содержащим CD28, демонстрировали насыщающий ответ против низких или высоких уровней PSCA, продукция IFNγ Т-клетками с CAR против PSCA, содержащими 4-IBB, была пропорциональной плотности антигена (ФИГ. 9 В). Аналогичные ответы цитокинов наблюдали независимо от поднабора Т-клеток, использованного для генерации Т-клеток с CAR против PSCA (ФИГ. 15).
CAR против PSCA, содержащие 4-IBB, демонстрировали легкое снижение дегрануляции CD107a по сравнению с CAR, содержащими CD28, относительно опухолевых клеток, экспрессирующих PSCA (ФИГ. 9Г и ФИГ. 9Д). Наблюдали значимое нацеливание PSCA(ΔCH2)28ζ на опухолевые клетки, не экспрессирующие PSCA, при измерении по экспрессии CD107a. Статус активации Т-клеток с CAR PSCA(ΔCH2)28ζ и PSCA(ΔCH2)BBζ был сравнимым при измерении по экспрессии 4-IBB (CD137) в 3-суточном анализе умерщвления опухоли (ФИГ. 9Е). Для того чтобы убедиться в том, что различия между Т-клетками с CAR против PSCA были обусловлены нацеливанием на антиген, а не собственным дефектом активности Т-клеток, авторы изобретения подтвердили аналогичную активацию (CD137) и исчерпание (PD-1) в Т-клетках, стимулированных антителом против человеческого CD3, связанным с планшетом - OKT3 (ФИГ. 16).
Пример 9: Т-клетки с CAR PSCA(ΔCH2)BBζ демонстрируют надежную терапевтическую эффективность в подкожных моделях рака простаты
В данном исследовании мышей, несущих подкожные опухоли PC-3-PSCA, обрабатывали одной внутриопухолевой инъекцией 5×106 Т-клеток с CAR PSCA(ΔCH2)BBζ. Наблюдали полную регрессию опухоли в пределах двух недель после внутриопухолевой инъекции Т-клеток. Несмотря на то, что регрессия опухоли была очевидной в течение более чем 30 суток, опухоли в конечном счете давали рецидив у большинства животных с аналогичной кинетикой относительно первичной опухоли (ФИГ. 17А), что авторы изобретения будут рассматривать ниже. Для установления того, была ли достижимой в данной модели солидной опухоли системная терапия Т-клетками с CAR, внутривенно доставляли разные дозы Т-клеток с CAR PSCA(ΔCH2)BBζ. В то время как 5×106 Т-клеток с CAR против PSCA демонстрировали полную регрессию опухолей, аналогичную, хотя и отложенную терапевтическую эффективность, наблюдали с дозой CAR всего-лишь 0,25×106 (ФИГ. 10А). Для экстраполяции данных результатов на большую опухолевую нагрузку большие опухоли PC-3-PSCA (~500 мм3) обрабатывали одной внутривенной инъекцией 5×106 Т-клеток с CAR PSCA(ΔCH2)BBζ. В данном случае наблюдали быструю регрессию опухоли (ФИГ. 10Б). Значимую инфильтрацию опухоли человеческими Т-клетками наблюдали через 11 суток после инфузии Т-клеток с CAR (ФИГ. 10В, верхняя панель), которые также экспрессировали гранзим В (ФИГ. 10Г, нижняя панель) - маркер активности Т-клеток. Опухоли из мышей, обработанных имитацией, демонстрировали очень мало человеческих Т-клеток или низкую экспрессию гранзима В в тот же самый момент времени.
Рецидив после одной терапии Т-клетками с антигенспецифичным CAR возможно является ожидаемым явлением, принимая во внимание гетерогенный антигенный профиль солидных опухолей, но механизмы, лежащие в основе устойчивости/рецидивов, все еще исследуются. Для лучшего понимания отложенных рецидивов опухоли, которые наблюдали на ФИГ. 10А, использовали иммуногистохимию для оценки длительного присутствия антигена на опухолевых клетках и сохранения Т-клеток с CAR против PSCA. Интересно то, что в то время как опухоли, обработанные имитацией, были высокопозитивными в отношении PSCA, опухоли, которые давали рецидив после обработки Т-клетками с CAR PSCA(ΔCH2)BBζ, были PSCA-негативными (ФИГ. 17Б, верхняя панель). Однако в тех же самых дающих рецидив опухолях было много человеческих Т-клеток (ФИГ. 17Б, средняя панель), даже несмотря на то, что данные опухоли отбирали по меньшей мере через 2 месяца после инфузии Т-клеток с CAR. Клетки РС-3 также экспрессируют HER2 in vitro (ФИГ. 17В), и было подтверждено то, что и обработанные имитацией, и обработанные PSCA(ΔCH2)BBζ рецидивирующие опухоли экспрессировали HER2 с эквивалентными уровнями in vivo (ФИГ. 17Б, нижняя панель). Для определения того, были ли опухоли PSCA-негативными и все еще чувствительными к терапии Т-клетками с CAR, рецидивирующие опухоли обрабатывали внутриопухолевой инъекцией либо имитации, либо направленных на PSCA, либо направленных на HER2 Т-клеток с CAR. Несмотря на то, что рецидивирующие PSCA-негативные опухоли не отвечали на CAR против PSCA, они были чувствительными к обработке Т-клетками с CAR против HER2 (ФИГ. 17Г).
Пример 10: Т-клетки с CAR PSCA(ΔCH2)BBζ транспортируются в кость и демонстрируют противоопухолевую эффективность при раке простаты с метастазами в кости
Одним из главных препятствий для клеточной иммунотерапии является иммунодепрессивное микроокружение, которое может препятствовать эффективному транспорту и выживанию Т-клеток в солидных опухолях. Для прямой оценки транспорта и антигензависимого размножения Т-клеток с CAR в опухолях простаты с метастазами в кости Т-клетки с CAR PSCA(ΔCH2)BBζ, меченные люциферазой светляка, в.в. инъецировали мышам, несущим в большеберцовой кости опухоли РС-3 дикого типа (в анатомически правой большеберцовой кости) и PC-3-PSCA (в анатомически левой большеберцовой кости). Интересно то, что в то время как имитация и Т-клетки с CAR против PSCA демонстрировали одинаковый ранний транспорт в обе опухоли (через 4 часа после инфузии Т-клеток), Т-клетки с CAR против PSCA преимущественно обнаруживались в опухолях, экспрессирующих PSCA, в сутки 1 после инъекции Т-клеток, и их уровень увеличивался на протяжении 4 суток кинетической визуализации (ФИГ. 10Г), указывая на антигензависимый транспорт и/или пролиферацию Т-клеток с CAR в PSCA-позитивных опухолях. Затем провели исследование, в котором опухолевые клетки PC-3-PSCA инъецировали в пространство внутри большеберцовой кости. В сутки 14 после приживления опухоли данных мышей, несущих опухоль, внутривенно обрабатывали уменьшающимися дозами Т-клеток с CAR PSCA(ΔCH2)BBζ (от 0,5×106 до 5×106) (ФИГ. 10Д). Значительное большинство мышей, обработанных либо 5×106, либо 2,5×106 Т-клеток с CAR, демонстрировало полную регрессию опухоли, тогда как мыши, обработанные либо 1×106, либо 0,5×106 Т-клеток с CAR, имели более гетерогенный терапевтический ответ (ФИГ. 10Е). Клиническая релевантность данной модели является очевидной при сравнении эффективных доз из ортотопических исследований с дозами, используемыми в подкожной модели, где наблюдали полную регрессию с дозами Т-клеток с CAR всего-лишь 0,25×106. Вероятно то, что расхождение в общей терапии, наблюдаемое в данных моделях, обусловлено различиями в инфильтрации и выживании Т-клеток с CAR в данных микроокружениях опухолей.
Пример 11: костимулирование 4-IBB обеспечивает превосходную стойкость и надежные противоопухолевые ответы CAR против PSCA в клинически релевантной модели рака простаты с метастазами в кости
Описанные выше исследования расширяли с использованием полученного от пациента ксенотрансплантата опухоли - рака простаты с метастазами в кости, экспрессирующего PSCA - LAPC-9. В сутки 14 после приживления опухоли мыши, обработанные одной в.в. инъекцией 5×106 Т-клеток с CAR PSCA(ΔCH2)BBζ, демонстрировали почти полную регрессию опухолей в месте опухоли внутри большеберцовой кости (ФИГ. 11А). Несмотря на то, что на опухоли внутри большеберцовой кости осуществлялось эффективное нацеливание, опухоли LAPC-9 распространялись в другие места в организме, что было особенно очевидно обнаружено в разных лимфатических узлах (подмышечных и паховых) и тимусе, что подтверждается иммуногистохимией (данные не показаны). Несмотря на то, что они, по-видимому, росли в течение нескольких недель после исходной регрессии опухоли в кости, они, в конечном счете, устранялись Т-клетками с CAR PSCA(ΔCH2)BBζ.
Основываясь на требовании наличия устойчивых Т-клеток для полной противоопухолевой активности CAR против PSCA, было проведено исследование для сравнения CAR против PSCA, содержащих либо костимулирующий домен CD28, либо 4-IBB. В то время как и Т-клетки с CAR PSCA(ΔCH2)28ζ и Т-клетки с CAR PSCA(ΔCH2)BBζ демонстрировали кардинальную регрессию метастазов в кости, мыши, получающие CAR против PSCA, содержащие CD28, демонстрировали рецидив в месте первичной опухоли, а также метастатическое заболевание, тогда как мыши, обработанные CAR против PSCA, содержащим 4-IBB, демонстрировали полные противоопухолевые ответы (ФИГ. 11А и ФИГ. 11Б). Рецидив опухоли у мышей, обработанных Т-клетками с CAR PSCA(ΔCH2)28ζ, подтверждали посредством количественного измерения уровней PSA (специфический антиген простаты) в крови в сутки 76 после обработки Т-клетками с CAR (ФИГ. 10В). Т-клетки с CAR количественно измеряли в крови обработанных животных, и, в то время как Т-клетки с CAR наблюдали в обеих группах в сутки 24 после инъекции опухоли, Т-клетки с CAR PSCA(ΔCH2)BBζ были значительно более многочисленными в сутки 76, указывая на большую устойчивость (ФИГ. 10Г). В целом данные исследования демонстрируют мощную и надежную противоопухолевую эффективность Т-клеток с CAR PSCA(ΔCH2)BBζ во многих опухолевых системах, включая ортотопические модели метастазов в кости рака простаты.
Пример 12: конструирование и структура epHIV7, используемого для экспрессии CAR
Плазмида pHIV7 представляет собой родительскую плазмиду, из которой были получены разные экспрессионные векторы CAR в Исследовательской лаборатории терапевтических средств на основе Т-клеток (TCTRL) в City of Hope (СОН). Вектор epHIV7, использованный для экспрессии CAR, получали из вектора pHIV7. Важно то, что в данном векторе для управления экспрессией CAR используется человеческий промотор EF1. И 5'-, и 3'-последовательности данного вектора были получены из pv653RSN, который был ранее получен из провируса НХВс2. Вытесняемые последовательности полиуринового тракта ДНК (сРРТ) были получены из штамма pNL4-3 ВИЧ-1 (вирус иммунодефицита человека-1) из репозитория реактивов NIH AIDS (Национальный институт здравоохранения по СПИД). Последовательность посттранскрипционного регуляторного элемента лесного сурка (WPRE) была описана ранее.
Конструирование pHIV7 проводили следующим образом. Вкратце, pv653RSN, содержащую 653 п.о. из gag-pol плюс 5'- и 3'-длинные концевые повторы (LTR) с расположенным между ними геном SL3-неомицинфосфотрансферазы (Neo), субклонировали в pBluescript следующим образом: на стадии 1 из последовательностей от 5' LTR до элемента, отвечающего на rev (RRE), делали p5' HIV-1 51, и затем 5' LTR модифицировали посредством удаления последовательностей выше TATA бокса и лигировали сначала с энхансером CMV и затем с репликатором SV40 (p5'HIV-2). На стадии 2 после клонирования 3'LTR в pBluescript с получением p3'HIV-1 делали делецию 400 п.о. в 3'LTR энхансере/промоторе для удаления цис-регуляторных элементов в U3 HIV и образования p3'HIV-2. На стадии 3 фрагменты, выделенные из p5'HIV-3 и p3'HIV-2, лигировали с получением pHIV-3. На стадии 4 p3'HIV-2 дополнительно модифицировали посредством удаления дополнительных расположенных выше последовательностей HIV с получением р3' HIV-3, и фрагмент из 600 п.о. BamHI-Sall, содержащий WPRE, добавляли к p3'HIV-3 с получением p3'HIV-4. На стадии 5 уменьшали размер RRE pHIV-3 посредством ПЦР и лигировали с 5' фрагментом из pHIV-3 (не показан) и с p3'HIV-4 с получением pHIV-6. На Стадии 6 фрагмент BgIII-BamHI из 190 п.о., содержащий вытесняемую последовательность ДНК сРРТ из pNL4-3 HIV-1 (55), амплифицировали из pNL4-3 и помещали между последовательностями RRE и WPRE в pHIV6 с получением pHIV-7. Данную родительскую плазмиду - pHIV7-GFP (GFP - зеленый флуоресцентный белок) использовали для упаковки родительского вектора с использованием четырехплазмидной системы.
Упаковочный сигнал - пси, ψ - требуется для эффективной упаковки вирусного генома в вектор. RRE и WPRE усиливают транспорт РНК-транскрипта и экспрессию трансгена. Было продемонстрировано то, что вытесняемая последовательность в комбинации с WPRE усиливает эффективность трансдукции лентивирусного вектора в клетках млекопитающих.
Хелперные функции, требующиеся для продукции вирусного вектора, разделяются на три отдельные плазмиды для уменьшения вероятности образования компетентного к репликации лентивируса посредством рекомбинации: 1) pCgp кодирует белок gag/pol, требующийся для сборки вирусного вектора; 2) pCMV-Rev2 кодирует белок Rev, который действует на последовательность RRE для помощи в транспортировке вирусного генома для эффективной упаковки; и 3) pCMV-G кодирует гликопротеин вируса везикулостоматита (VSV), который требуется для инфективности вирусного вектора.
Имеется минимальная гомология последовательности ДНК между геномом вектора, кодируемого pHIV7, и хелперными плазмидами. Области гомологии включают область упаковочного сигнала из приблизительно 600 нуклеотидов, расположенную в последовательности gag/pol хелперной плазмиды pCgp; последовательность промотора CMV во всех трех хелперных плазмидах и последовательность RRE в хелперной плазмиде pCgp.Весьма маловероятно то, что из-за гомологии в данных областях мог бы быть генерирован компетентный к репликации рекомбинантный вирус, так как это потребовало бы многочисленных событий рекомбинации. Кроме того, у любых образующихся рекомбинантов отсутствовали бы функциональные последовательности LTR и tat, требующиеся для лентивирусной репликации.
Промотор CMV был заменен промотором EF1α-HTLV (EF1p), и новая плазмида была названа epHIV7. EF1p имел 563 п.о. и был введен в epHIV7 с использованием Nrul и Nhel, затем промотор CMV был вырезан.
Из данной системы был удален лентивирусный геном, исключая gag/pol и rev, которые необходимы для патогенности вируса дикого типа и требуются для продуктивной инфекции клеток-мишеней. Кроме того, конструкция вектора CLRX-IgG4Fc(EQ)-CD28-zeta-T2ACD19t_epHIV7 не содержит интактного промотора 3'LTR, таким образом, полученная в результате экспрессируемая и обратно транскрибированная ДНК провирусного генома в клетках-мишенях будет иметь неактивные LTR. В результате данной конструкции последовательности, происходящие из ВИЧ-1, не будут транскрибироваться из провируса, и только терапевтические последовательности будут экспрессироваться от соответствующих им промоторов. Ожидается то, что удаление промоторной активности LTR в векторе SIN значительно уменьшает возможность непреднамеренной активации генов хозяина.
Пример 13: получение векторов для трансдукции Т-клеток
Для каждой плазмиды, экспрессирующей CAR, был получен семенной банк, который используется для инокуляции ферментера с получением достаточных количеств плазмидной ДНК. Данную плазмидную ДНК тестировали на идентичность, стерильность и присутствие эндотоксина перед ее применением в производстве лентивирусного вектора.
Вкратце, клетки размножали от рабочей клетки 293Т (WCB), которая была протестирована для подтверждения стерильности и отсутствия вирусного загрязнения. Сосуд с клетками 293Т от WCB 293Т оттаивали. Клетки выращивали и размножали до наличия достаточного числа клеток для посева в подходящее число 10-слойных клеточных фабрик (CF) для продукции вектора и поддержания линии клеток. Для продукции можно использовать одну линию клеток.
Лентивирусный вектор продуцировали в подпартиях из вплоть до 10 CF. Две подпартии можно продуцировать в ту же самую неделю, приводя к продукции приблизительно 20 л лентивирусного супернатанта/неделю. Материал, продуцированный из всех подпартий, объединяли на протяжении последующей фазы переработки для того, чтобы произвести одну партию продукта. Клетки 293Т высевали в CF в среду 293Т (DMEM с 10% FBS (фетальная телячья сыворотка). Фабрики помещали в инкубатор при 37°С и горизонтально выравнивали для получения равномерного распределения клеток на всех слоях CF. Через двое суток клетки трансфицировали четырьмя лентивирусными плазмидами, описанными выше, с использованием способа с CaPO4, который включает смесь Tris:EDTA, 2 М CaCl2, 2× HBS и четырех ДНК плазмид. В сутки 3 после трансфекции супернатант, содержащий секретированные лентивирусные векторы, отбирали, очищали и концентрировали. После удаления супернатанта из CF из каждой CF отбирали клетки конца продуцирования. Клетки из каждой фабрики трипсинизировали и отбирали центрифугированием. Клетки ресуспендировали в среде для замораживания и криоконсервировали. Данные клетки позднее использовали для тестирования на лентивирус, компетентный в отношении репликации (RCL).
Для очистки и приготовления векторов в виде препарата неочищенный супернатант осветляли мембранной фильтрацией для удаления клеточных обломков. ДНК клетки-хозяина и остаточную плазмидную ДНК деградировали посредством расщепления эндонуклеазой (Бензоназа®). Вирусный супернатант осветляли от клеточных обломков с использованием 0,45 мкм фильтра. Осветленный супернатант отбирали в предварительно взвешенный контейнер, в который добавляют Бензоназу® (конечная концентрация 50 U/мл). Эндонуклеазное расщепление остаточной плазмидной ДНК и геномной ДНК хозяина проводили при 37°С в течение 6 ч. Для удаления остаточных низкомолекулярных компонентов из неочищенного супернатанта использовали исходное концентрирование супернатанта, обработанного эндонуклеазой, посредством ультрафильтрации с тангенциальным потоком (TFF), концентрируя вирус ~20 раз. Осветленный, обработанный эндонуклеазой вирусный супернатант циркулировал через картридж с полыми волокнами с NMWCO (номинальный порог отсечения молекулярной массы) 500 кДа при скорости тока, подобранной для поддержания скорости сдвига ~4000 с-1 или меньше при максимизации скорости потока. Во время процесса концентрирования запускали диафильтрацию супернатанта, обработанного нуклеазой, для поддержания эффективности картриджа. С использованием 4%-ной лактозы в PBS (фосфатно-солевой буферный раствор) в качестве диафильтрационного буфера устанавливали скорость замены пермеата 80%. Вирусный супернатант доводили до целевого объема, представляющего 20-кратное концентрирование неочищенного супернатанта, и диафильтрацию продолжали в течение 4 дополнительных смен объема со скоростью замены пермеата 100%.
Дальнейшее концентрирование вирусного продукта осуществляли с применением методики высокоскоростного центрифугирования. Каждую подпартию лентивируса осаждали с использованием центрифуги Sorvall RC-26 plus при 6000 об./мин (6088 RCF (относительная сила центрифугирования) при 6°С в течение 16-20 ч. Вирусный осадок из каждой подпартии затем растворяли в 50 мл объеме 4%-ной лактозы в PBS. Растворенный осадок в данном буфере представляет собой конечную композицию вирусного препарата. Весь процесс концентрирования вектора приводил приблизительно к 200-кратному уменьшению объема. После завершения получения всех подпартий материал затем помещали при -80°С, тогда как образцы из каждой подпартии тестировали на стерильность. После подтверждения стерильности образца подпартии быстро оттаивали при 37°С с частым встряхиванием. Материал затем объединяли и вручную аликвотировали в боксе биологической безопасности Класса II Типа А/В3 в помещении для работы с вирусными векторами. Использовали конфигурацию заполнения 1 мл концентрированного лентивируса в стерильных криососудах класса 6 USP (Фармакопея США) с наружной резьбой и О-кольцом. Системы качества (QS) Центра развития прикладной технологии (CATD) в СОН выпустили все материалы согласно политике и стандартным технологическим операциям для CBG и в соответствии с надлежащими производственными практиками (cGMP).
Для обеспечения чистоты препарата лентивирусного вектора его тестировали на остаточные загрязнения ДНК хозяина и перенос остаточной ДНК хозяина и плазмид. Наряду с другими тестами идентичность вектора оценивали посредством ПЦР-ОТ (полимеразная цепная реакция, сопряженная с обратной транскрипцией) для обеспечения того, что присутствует правильный вектор. Для вектора, предназначенного для применения в данном исследовании, удовлетворялись все критерии выпуска.
Пример 14: получение Т-клеток, подходящих для экспрессии CAR, нацеленных на PSCA
Т-лимфоциты получают от пациента посредством лейкафереза, и подходящий поднабор аллогенных или аутологичных Т-клеток, например, центральных Т-клеток памяти (TCM) генетически изменяется для экспрессии CAR, затем вводится обратно пациенту посредством любых клинически приемлемых способов для достижения противораковой терапии.
Подходящие TCM могут быть получены следующим образом. Продукты афереза, полученные от участников исследования, подписавших информированное согласие, разделяли на фиколе, промывали и инкубировали в течение ночи. Клетки затем обедняли в отношении популяций моноцитов, регуляторных Т-клеток и наивных Т-клеток с использованием реактивов против CD14, против CD25 и против CD45RA уровня качества GMP (Miltenyi Biotec) и прибора для разделения CliniMACS™. После обеднения негативная фракция клеток обогащается в отношении клеток TCM CD62L+ с использованием DREG56-биотина (клинический уровень качества СОН) и микрошариков против биотина (Miltenyi Biotec) на приборе для разделения CliniMACS™.
После обогащения клетки TCM готовят в полной X-Vivo15 плюс 50 IU (международные единицы)/мл IL-2 и 0,5 нг/мл IL-15 и переносят в тефлоновый мешок для культуры клеток, где они стимулируются шариками Dynal ClinEx™ Vivo CD3/CD28. Вплоть до пяти суток после стимуляции клетки трансдуцируются лентивирусным вектором, экспрессирующим желательный CAR при множественности заражения (MOI) от 1,0 до 0,3. Культуры поддерживаются в течение вплоть до 42 суток с добавлением полной X-Vivo15 и цитокинов IL-2 и IL-15, как требуется для размножения клеток (поддерживая плотность клеток от 3×105 до 2×106 жизнеспособных клеток/мл и осуществляя дополнение цитокинами каждый понедельник, среду и пятницу культуры). При данных условиях клетки типично размножаются до приблизительно 109 клеток в пределах 21 суток. В конце периода культуры клетки отбирают, дважды промывают и готовят в виде препарата в криоконсервирующей среде клинического уровня качества (Cryostore CS5, BioLife Solutions).
В сутки инфузии Т-клеток криоконсервированный и выпущенный продукт оттаивают, промывают и готовят в виде препарата для повторной инфузии. Криоконсервированные сосуды, содержащие выпущенный клеточный продукт, удаляют из хранилища с жидким азотом, оттаивают, охлаждают и промывают промывочным буфером на основе PBS/2% человеческого сывороточного альбумина (HSA). После центрифугирования супернатант удаляют, и клетки ресуспендируют в инфузионном разбавителе на основе нормального физиологического раствора, не содержащего консервантов (PFNS)/2% HSA. Образцы удаляют для контроля качества.
Методики, использованные в Примерах 7-11
Линии клеток: линии клеток человеческого метастатического рака простаты DU145 (АТСС НТВ-81) и РС-3 (АТСС CRL-1435) культивировали в RPMI-1640 (Lonza), содержащей 10% фетальной телячьей сыворотки (FBS, Hyclone) и 1× антибиотик-противогрибковое средство (Gibco), содержащее 100 U/мл пенициллина, 100 мкг/мл стрептомицина и 0,25 мкг/мл фунгизона (полная RPMI). Линию клеток человеческой фибросаркомы - НТ1080 (АТСС CCL-121) и линию клеток человеческой эмбриональной почки - 293Т (АТСС CRL-3216) культивировали в среде Игла, модифицированной по Дульбекко (DMEM, Life Technologies), содержащей 10% FBS, 1× антибиотик-противогрибковое средство, 25 мМ HEPES (Irvine Scientific) и 2 мМ L-глутамин (Fisher Scientific) (полная DMEM). Ксенотрансплантат человеческого рака простаты LAPC-9 (любезный подарок Др. Роберта Райтера, UCLA) культивировали в среде Дульбекко, модифицированной по Искову (IMDM, Irvine Scientific), содержащей 20% FBS и 1× антибиотик-противогрибковое средство (полная IMDM). Клетки LAPC-9 серийно пассировали в самцах мышей NOD.Cg-Prkdcscid IL2rgtm1Wjl/SzJ (NSG), и получали одноклеточные суспензии, как было описано ранее (Craft et al. 1999 Cancer Res 59:5030). Вкратце, ткань опухоли отбирали, измельчали в чашке Петри и расщепляли 1% проназой Е (Roche). После промывки полной IMDM одноклеточные суспензии фильтровали через 40 мкм клеточное сито (Falcon), вновь промывали и немедленно замораживали. Лимфобластоидную линию клеток, трансформированную EBV (вирус Эпштейна-Барр) (LCL), и клетки LCL, содержащие связанный с мембраной агонист OKT3 на основе scFv, специфичного в отношении CD3 эпсилон (LCL-OKT3 (Wang et al. 2011 Blood 117:1888)), культивировали в полной RPMI. Все клетки культивировали при 37°С с 5% CO2. Проводили аутентификацию клеток DU145 и РС-3 посредством профилирования STR (короткие концевые повторы) и подтверждали то, что они являются негативными в отношении микоплазм (DDC Medical, ОН).
Конструкции ДНК и продукция лентивируса: опухолевые клетки DU145 и РС-3 генетически модифицировали для экспрессии PSCA посредством трансдукции лентивирусом epHIV7, несущим человеческий ген PSCA (№доступа: NM_005672.4) под контролем промотора EF1α. Клетки PSCA+ окрашивали мышиным антителом против человеческого PSCA (1G8), как описано ниже (см. раздел «Внутриклеточное/внеклеточное окрашивание и проточная цитометрия»), и затем сортировали FACS (флуоресцентная сортировка клеток) с использованием сортировщика клеток BD FACSAria™ - сделанного по специальному заказу исследовательского продукта (SORP). Для получения опухолевых клеток с низкой экспрессией PSCA ген PSCA помещали под контроль мутировавших версий промотора PGK, как описано ранее (Frigault et al. 2015 Cancer Immunol Res 3:356). Последовательность scFv A11 (Lepin et al. 2010 Eur J Nucl Med Mol Imaging 37:529) была любезно предоставлена докторами Анной By и Робертом Райтером (UCLA). Последовательность scFv МВ1 была опубликована ранее (Feldmann et al. 2012 J Immunol 189:3249). Конструкции CAR с усеченным геном CD19 (CD19t), отделенным последовательностью перескакивания рибосомы Т2А, клонировали в лентивирусный каркас epHIV7. Нацеленный на антиген домен включал либо scFv А11, либо scFv МВ1. Внеклеточный спейсерный домен включал 129-аминокислотную версию спейсера средней длины на основе Fc IgG4 с делецией СН2 (ΔСН2) (Jonnalagadda et al. 2015 Mol Ther 23:757). Внутриклеточный костимулирующий домен сигнализации содержал либо CD28 с трансмембранным доменом CD28, либо 4-IBB с трансмембранным доменом CD4. Цитолитический домен CD3ζ был описан ранее (Cooper et al. 2003 Blood 101:1637).
Лентивирус получали посевом клеток 293Т в колбы для культуры тканей Т-225 за 1 сутки до трансфекции упаковочными плазмидами и плазмидой на основе лентивирусного каркаса с желательным CAR. Супернатанты собирали через 3-4 суток, фильтровали и центрифугировали для удаления клеточных обломков, и инкубировали с 2 мМ магнием и 25 U/мл эндонуклеазы Бензоназы® (EMD Millipore) для удаления загрязняющих нуклеиновых кислот. Супернатанты объединяли и концентрировали посредством высокоскоростного центрифугирования (6080 g) в течение ночи при 4°С. Лентивирусные осадки затем ресуспендировали в растворе фосфато-солевого буфера (PBS)-лактозы (4 г лактозы на 100 мл PBS), аликвотировали и хранили при -80°С до последующего применения. Литры лентивируса, определенного по экспрессии CD19t, количественно измеряли с использованием клеток НТ1080.
Выделение Т-клеток, лентивирусная трансдукция и размножение ex vivo: продукты лейкафереза были получены от участников исследования, подписавших информированное согласие (здоровые доноры), согласно протоколам, одобренным внутренней контрольной комиссией (IRB) City of Норе (СОН). В сутки лейкафереза были выделены одноядерные клетки периферической крови (РВМС) посредством центрифугирования в градиенте плотности над Ficoll-Paque (GE Healthcare), с последующими многократными промывками в PBS/EDTA (Miltenyi Biotec). Клетки отдыхали в течение ночи при комнатной температуре (RT) на ротаторе, и затем их промывали и ресуспендировали в полной X-VIVO. Для исследований с использованием общих РВМС клетки немедленно замораживали в криоконсервирующих средах CryoStor® CS5 (BioLife Solutions). Вплоть до 5×109 РВМС инкубировали с микрошариками против CD14, против CD25 и против CD45RA (Miltenyi Biotec) в течение 30 мин при RT и магнитно удаляли с использованием системы CliniMACS® (Miltenyi Biotec) согласно протоколу изготовителя. Обедненные РВМС затем обогащали центральными Т-клетками памяти (TCM) посредством инкубирования с биотинилированным антителом против CD62L (произведенным Центром биомедицины и гететики в City of Норе) в течение 30 мин при RT и затем с микрошариками против биотина (Miltenyi Biotec) в течение дополнительных 30 мин при RT. TCM затем магнитно обогащали с использованием системы autoMACS® (Miltenyi Biotec) согласно протоколу изготовителя. Для исследований с использованием TCM клетки немедленно замораживали, как описано выше. Чистоту и фенотип РВМС и TCM подтверждали проточной цитометрией.
Свежеоттаявшие РВМС или TCM один раз промывали и культивировали в Х-VIVO-15 (Lonza) с 10% FBS (полная Х-VIVO), содержащей 100 U/мл рекомбинантного человеческого IL-2 (rhIL-2, Novartis Oncology) и 0,5 нг/мл рекомбинантного человеческого IL-15 (rhIL-15, CellGenix). Для лентивирусной трансдукции CAR Т-клетки культивировали с CD3/CD28 Dynabeads® (Life Technologies), понтамина сульфатом (АРР Pharmaceuticals), смесью цитокинов (как указано выше) и желательным лентивирусом при разных MOI либо в сутки, либо в сутки после стимуляции шариками. Центрифужную инокуляцию проводили центрифугированием при 2000 об./мин в течение 30 мин при 32°С без тормозов. Клетки затем культивировали в и подпитывали свежей полной X-VIVO, содержащей цитокины, каждые 2-3 суток. После 7-9 суток шарики магнитно удаляли, и клетки дополнительно размножали в полной X-VIVO, содержащей цитокины, для достижения желательного выхода клеток. Т-клетки с CAR подвергали позитивной селекции на CD19t с использованием набора I или II для позитивного обогащения CD19 EasySep™ (StemCell Technologies) согласно протоколу изготовителя. После дальнейшего размножения клетки замораживали до функциональных анализов in vitro и применения в моделях опухолей in vivo. Чистоту и фенотип Т-клеток с CAR подтверждали проточной цитометрией.
Внутриклеточное/внеклеточное окрашивание и проточная цитометрия: для анализа проточной цитометрией клетки ресуспендировали в буфере FACS (сбалансированный солевой раствор Хэнкса без Са2+, Mg2+ или фенолового красного (HBSS-/-, Life Technologies), содержащем 2% FBS и 1× антибиотик-противогрибковое средство). Для окрашивания PSCA мышиное антитело против человеческого PSCA (1G8) было любезно предоставлено Др. Робертом Райтером, UCLA. Для выявления scFv CAR использовали биотинилированный белок-L (GenScript USA), как было описано ранее [35]. Клетки инкубировали с первичными антителами в течение 30 минут при 4°С в темноте перед переходом ко вторичному окрашиванию. Для внеклеточного и вторичного окрашивания клетки дважды промывали перед 30 мин инкубацией при 4°С в темноте с антителами, конъюгированными с флуоресцеина изотиоцианатом (FITC), фикоэритрином (РЕ), комплексом перидинин-хлорофилл-белок (PerCP), PerCP-Су5.5, РЕ-Су7, аллофикоцианином (АРС) и АРС-Су7 (или APC-eFluor780) (против CD3, CD4, CD8, CD14, CD19, CD25, специфичных в отношении мышиного или человеческого CD45, CD45RA, CD45RO, CD62L, CD95, CD107a, CD137, LAG3 (CD223), PD-1 (CD279), TIM3 (CD366), CCR7, IFNγ, козы против мышиного Ig, и стрептавидина), приобретенными у BioLegend, eBioscience, BD Biosciences или Fisher Scientific. Жизнеспособность клеток определяли с использованием 4',6-диамидино-2-фенилиндола (DAPI, Sigma). Для внутриклеточного окрашивания клетки фиксировали, пермеабилизировали и обрабатывали набором для апоптоза РЕ Active-Caspase-3 (BD Biosciences) согласно протоколу изготовителя. Клетки затем инкубировали с антителами, конъюгированными с флуорофором, в течение 30 минут при 4°С в темноте и дважды промывали перед ресуспендированием в буфере для FACS и получением данных на MACSQuant Analyzer 10 (Miltenyi Biotec). Данные анализировали с использованием программы FlowJo (v10, TreeStar).
Функциональные анализы Т-клеток in vitro: для дегрануляции и анализов внутриклеточных цитокинов Т-клетки с CAR и опухоли-мишени сокультивировали при разных соотношениях эффектор : мишень в полной X-VIVO без экзогенных цитокинов в круглодонных 96-луночных обработанных планшетах для культуры ткани (Corning). FITC-CD107a добавляли в культуры, и после инкубирования в течение 4-6 ч при 37°С клетки фиксировали и пермеабилизировали перед анализом проточной цитометрией, как описано выше. Для анализов умерщвления опухолей Т-клетки с CAR и опухоли-мишени сокультивировали при разных соотношениях эффектор:мишень в полной X-VIVO без экзогенных цитокинов в 96-луночных планшетах в течение 1-5 суток и анализировали проточной цитометрией, как описано выше. Умерщвление опухолей Т-клетками с CAR рассчитывали сравнением числа CD45-негативных клеток по отношению к числу, наблюдаемому с Т-клетками имитации.
ELISA и мультиплексные анализы цитокинов: высокоаффинные 96-луночные плоскодонные планшеты (Corning) покрывали разными концентрациями рекомбинантного человеческого белка PSCA (аминокислоты 23-95; Abnova) в течение ночи в 1× PBS при 4°С. Лунки дважды промывали 1× PBS, блокировали 10% FBS в течение 1 ч и вновь промывали. В покрытые лунки добавляли Т-клетки с CAR (5×103 в 200 мкл). Когда это было определено, опухоли-мишени (5×103) инкубировали с Т-клетками в непокрытых лунках (конечный объем 200 мкл). После инкубации в течение ночи при 37°С супернатанты отбирали и обрабатывали согласно протоколу изготовителя для набора ELISA для человеческого IFNγ Ready-SET-GO!® (eBioscience). Считывание планшетов осуществляли при 450 нм с использованием многометочного счетчика Wallac Victor3 1420 (Perkin-Elmer) и программы Wallac 1420 Workstation. В качестве альтернативы, супернатанты анализировали на многие цитокины с использованием мультиплексного набора шариков для иммуноанализа (Invitrogen) согласно протоколу изготовителя. ELISA человеческого PSA/KLK3 (Abcam) на мышиной сыворотке проводили согласно протоколу изготовителя.
Количественная ПЦР: клетки опухоли (посеянные в концентрации 0,25×106/мл) культивировали в течение одних суток до выделения РНК. РНК экстрагировали с использованием очистки на колонке набора RNeasy® Mini (Qiagen). кДНК получали с использованием системы для синтеза первой нити Superscript™ IV (Invitrogen). РНК-праймеры получали с использованием анализов экспрессии генов TaqMan® специфичных в отношении либо PSCA (Hs04166224_g1, Life Technologies), либо GAPDH (глицеральдегид-3-фосфатдегидрогеназа) (Hs02758991_g1, Life Technologies). кПЦР проводили на системе для ПЦР в реальном времени ViiA™ 7 (Thermo Fisher). Наборы праймеров подтверждали с использованием стандартной кривой, охватывающей определенный динамический интервал, с одним пиком кривой плавления. Экспрессию генов-мишеней нормировали к GAPDH.
Исследования опухолей in vivo: все эксперименты на животных проводили согласно протоколам, одобренным Институциональным комитетом по уходу за и применению животных City of Норе. Для исследований подкожных опухолей клетки РС-3 и DU145 (2,5×106) готовили в HBSS-/- и подкожно инъецировали в левую выбритую часть брюха самцов мышей NSG. Рост опухолей отслеживали 3 раза в неделю посредством измерения штангенциркулем. Как только объемы опухолей достигали 50-500 мм3, Т-клетки с CAR готовили в PBS и инъецировали либо внутриопухолево (в.о.), либо внутривенно (в.в.). Как только опухоли достигали 15 мм в диаметре, мышей умерщвляли, и опухоли отбирали и обрабатывали для иммуногистохимии, как описано ниже. При рецидиве подкожных опухолей мышей обрабатывали в.о. инъекцией либо CAR против PSCA, либо CAR против HER2. У мышей, анестезированных изофлураном, отбирали периферическую кровь посредством ретроорбитального (RO) спускания крови через гепаринизированные капиллярные трубки (Chase Scientific) и в полистирольные пробирки, содержащие раствор гепарина/PBS (1000 единиц/мл, Sagent Pharmaceuticals). Отбирали приблизительно 150 мкл крови на мышь. Кровь лизировали 1× буфером для лизиса эритроцитов (Sigma) согласно протоколу изготовителя и затем промывали, окрашивали и анализировали проточной цитометрией, как описано выше.
Для ортотопических исследований опухолей внутри большеберцовой кости LAPC-9 и PC-3-PSCA трансдуцировали лентивирусом, несущим усиленный зеленый флуоресцентный белок (eGFP)/люциферазу светляка (ffluc), для обеспечения неинвазивной оптической визуализации (Xenogen) сразу после имплантации мышам (что приводит к линиям, названным LAPC-9-eGFP-ffluc и PC-3-PSCA-eGFP-ffluc). Вкратце, данные линии инкубировали с полибреном (4 мг/мл, Sigma) и лентивирусом eGFP-ffluc (см. выше), с последующей сортировкой клеток на клетки GFP+ с использованием сортировщика клеток BD FACSAria™ SORP. Свежеотсортированные клетки LAPC-9-eGFP-ffluc серийно пассировали в мышах NSG, как описано выше. Клетки PC-3-PSCA-eGFP-ffluc (2×105) или LAPC-9-eGFP-ffluc (1,5×105) получали так же, как и в подкожных моделях. Перед инъекцией опухоли мышей анестезировали внутрибрюшинной инъекцией (в.б.) кетамина/ксилазина в газообразном изофлуране. Опухолевые клетки (в 30 мкл HBSS-/-) инъецировали в пространство внутри большеберцовой кости задней лапы мыши. Через 14 суток мышам в.в. инъецировали Т-клетки с CAR. Рост опухоли отслеживали посредством оптической визуализации один раз в две недели (IVIS, Xenogen), и сигналы потока анализировали с использованием программы Living Image (Xenogen). Для визуализации мышам в.б. инъецировали 150 мкл калиевой соли D-люциферина (Perkin Elmer), суспендированной в PBS, в дозировке 4,29 мг/мышь.
Для исследований транспорта Т-клеток мышам имплантировали в пространство правой большеберцовой кости клетки РС-3 дикого типа (2×105), и в пространство левой большеберцовой кости - клетки PC-3-PSCA (2×105). Через 14 суток мышам в.в. инъецировали 5×106 Т-клеток имитации или Т-клеток с CAR PSCA(ΔCH2)BBζ, которые были сотрансдуцированы с лентивирусом, несущим eGFP-ffluc. Т-клетки были обогащенными в отношении CAR, и посредством проточной цитометрии было определено то, что приблизительно 30% являются eGFP+. Транспорт Т-клеток отслеживали посредством неинвазивной оптической визуализации (Xenogen) в 4 ч, 1 сутки, 2 суток и 4 суток после инфузии Т-клеток. Сигналы потока анализировали, как описано выше.
Иммуногистохимия: опухолевую ткань фиксировали вплоть до 3 суток в 4% параформальдегида (Boston BioProducts) и хранили в 70%-ном этаноле до дальнейшей обработки. Гистологию осуществляли в Центре патологии City of Норе. Вкратце, залитые в парафин срезы (10 мкм) окрашивали мышиным антителом против человеческого CD3 (DAKO), мышиным антителом против человеческого PSCA (Abcam), крысиным антителом против человеческого HER2 (DAKO) и крысиным антителом против человеческого гранзима-В (eBioscience). Изображения получали с использованием цифрового сканера предметных стекол Nanozoomer 2.0НТ и ассоциированной программы NDP.view2 (Hamamatzu).
Статистический анализ: данные представлены как среднее плюс/минус SEM (стандартная ошибка среднего), если не утверждается иное. Статистические сравнения между группами проводили с использованием непарного двухстороннего t-критерия Стьюдента для расчета значения р. *р меньше 0,05, **р меньше 0,01, ***р меньше 0,001; н.з. - не значимо.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> СИТИ ОФ ХОУП
<120> ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНА, НАЦЕЛЕННЫЕ НА PSCA
<130> 40056-0025WO1
<140> PCT/US2016/055761
<141> 2016-10-06
<150> US 62/238,062
<151> 2015-10-06
<160> 38
<170> PatentIn version 3.5
<210> 1
<211> 236
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> scFv, направленный против PSCA
<400> 1
Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Arg Phe Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Gly Ser Ser Pro Phe Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Ser Thr Ser Gly Gly
100 105 110
Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Glu Val Gln Leu
115 120 125
Val Glu Tyr Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Tyr Tyr Ile His Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Trp Ile Asp
165 170 175
Pro Glu Asn Gly Asp Thr Glu Phe Val Pro Lys Phe Gln Gly Arg Ala
180 185 190
Thr Met Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn
195 200 205
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Lys Thr Gly Gly
210 215 220
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
225 230 235
<210> 2
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Область линкера - основана на человеческой
<400> 2
gggssgggsg 10
<210> 3
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> мутантная последовательность - основана на шарнире IgG4 человека
<400> 3
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 4
<211> 12
<212> ПРТ
<213> Homo sapiens
<400> 4
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro
1 5 10
<210> 5
<211> 20
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность шарнира IgG4 человека и линкера
<400> 5
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Gly Gly Gly Ser
1 5 10 15
Ser Gly Gly Gly
20
<210> 6
<211> 39
<212> ПРТ
<213> Homo sapiens
<400> 6
Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu Lys Ser Asn
1 5 10 15
Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro Ser Pro Leu
20 25 30
Phe Pro Gly Pro Ser Lys Pro
35
<210> 7
<211> 48
<212> ПРТ
<213> Homo sapiens
<400> 7
Ala Lys Pro Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro
1 5 10 15
Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro
20 25 30
Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp
35 40 45
<210> 8
<211> 45
<212> ПРТ
<213> Homo sapiens
<400> 8
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp
35 40 45
<210> 9
<211> 129
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Слияние шарнир-линкер-CH3 /IgG4 HL-CH3
<400> 9
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Gly Gly Gly Ser
1 5 10 15
Ser Gly Gly Gly Ser Gly Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
20 25 30
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
35 40 45
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
50 55 60
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
65 70 75 80
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
85 90 95
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
100 105 110
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
115 120 125
Lys
<210> 10
<211> 229
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Слияние с мутациями
<400> 10
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Gln Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 11
<211> 0
<212> ПРТ
<213> Несуществующая
<400> 11
000
<210> 12
<211> 107
<212> ПРТ
<213> Homo sapiens
<400> 12
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
1 5 10 15
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
100 105
<210> 13
<211> 21
<212> ПРТ
<213> Homo sapiens
<400> 13
Leu Cys Tyr Leu Leu Asp Gly Ile Leu Phe Ile Tyr Gly Val Ile Leu
1 5 10 15
Thr Ala Leu Phe Leu
20
<210> 14
<211> 27
<212> ПРТ
<213> Homo sapiens
<400> 14
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
1 5 10 15
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val
20 25
<210> 15
<211> 28
<212> ПРТ
<213> Homo sapiens
<400> 15
Met Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser
1 5 10 15
Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val
20 25
<210> 16
<211> 22
<212> ПРТ
<213> Homo sapiens
<400> 16
Met Ala Leu Ile Val Leu Gly Gly Val Ala Gly Leu Leu Leu Phe Ile
1 5 10 15
Gly Leu Gly Ile Phe Phe
20
<210> 17
<211> 21
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> трансмембранный домен, CD8tm
<400> 17
Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu
1 5 10 15
Ser Leu Val Ile Thr
20
<210> 18
<211> 23
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> трансмембранный домен, CD8tm2
<400> 18
Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu
1 5 10 15
Ser Leu Val Ile Thr Leu Tyr
20
<210> 19
<211> 24
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> трансмембранный домен, CD8tm3
<400> 19
Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu
1 5 10 15
Ser Leu Val Ile Thr Leu Tyr Cys
20
<210> 20
<211> 27
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> трансмембранный домен, 41BB
<400> 20
Ile Ile Ser Phe Phe Leu Ala Leu Thr Ser Thr Ala Leu Leu Phe Leu
1 5 10 15
Leu Phe Phe Leu Thr Leu Arg Phe Ser Val Val
20 25
<210> 21
<211> 112
<212> ПРТ
<213> Homo sapiens
<400> 21
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 22
<211> 41
<212> ПРТ
<213> Homo sapiens
<400> 22
Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr
1 5 10 15
Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro
20 25 30
Pro Arg Asp Phe Ala Ala Tyr Arg Ser
35 40
<210> 23
<211> 41
<212> ПРТ
<213> Homo sapiens
<400> 23
Arg Ser Lys Arg Ser Arg Gly Gly His Ser Asp Tyr Met Asn Met Thr
1 5 10 15
Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro
20 25 30
Pro Arg Asp Phe Ala Ala Tyr Arg Ser
35 40
<210> 24
<211> 42
<212> ПРТ
<213> Homo sapiens
<400> 24
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
1 5 10 15
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
20 25 30
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
35 40
<210> 25
<211> 42
<212> ПРТ
<213> Homo sapiens
<400> 25
Ala Leu Tyr Leu Leu Arg Arg Asp Gln Arg Leu Pro Pro Asp Ala His
1 5 10 15
Lys Pro Pro Gly Gly Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln
20 25 30
Ala Asp Ala His Ser Thr Leu Ala Lys Ile
35 40
<210> 26
<211> 566
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность химерного белка, Фигура 18
<400> 26
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Ser Ala Ser
35 40 45
Ser Ser Val Arg Phe Ile His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
50 55 60
Pro Lys Arg Leu Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
85 90 95
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp
100 105 110
Gly Ser Ser Pro Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
115 120 125
Gly Ser Thr Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Ser Glu Val Gln Leu Val Glu Tyr Gly Gly Gly Leu Val Gln Pro
145 150 155 160
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys
165 170 175
Asp Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
180 185 190
Trp Val Ala Trp Ile Asp Pro Glu Asn Gly Asp Thr Glu Phe Val Pro
195 200 205
Lys Phe Gln Gly Arg Ala Thr Met Ser Ala Asp Thr Ser Lys Asn Thr
210 215 220
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
225 230 235 240
Tyr Cys Lys Thr Gly Gly Phe Trp Gly Gln Gly Thr Leu Val Thr Val
245 250 255
Ser Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Gly Gly
260 265 270
Gly Ser Ser Gly Gly Gly Ser Gly Gly Gln Pro Arg Glu Pro Gln Val
275 280 285
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
290 295 300
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
305 310 315 320
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
325 330 335
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
340 345 350
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
355 360 365
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
370 375 380
Leu Gly Lys Met Ala Leu Ile Val Leu Gly Gly Val Ala Gly Leu Leu
385 390 395 400
Leu Phe Ile Gly Leu Gly Ile Phe Phe Lys Arg Gly Arg Lys Lys Leu
405 410 415
Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln
420 425 430
Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly
435 440 445
Cys Glu Leu Gly Gly Gly Arg Val Lys Phe Ser Arg Ser Ala Asp Ala
450 455 460
Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu
465 470 475 480
Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp
485 490 495
Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu
500 505 510
Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile
515 520 525
Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr
530 535 540
Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met
545 550 555 560
Gln Ala Leu Pro Pro Arg
565
<210> 27
<211> 671
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> химерный белок PSCAscFv-IgG4(EQ)-CD28tm-CD28gg-зета, Фигура 19
<400> 27
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Ser Ala Ser
35 40 45
Ser Ser Val Arg Phe Ile His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
50 55 60
Pro Lys Arg Leu Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
85 90 95
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp
100 105 110
Gly Ser Ser Pro Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
115 120 125
Gly Ser Thr Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Ser Glu Val Gln Leu Val Glu Tyr Gly Gly Gly Leu Val Gln Pro
145 150 155 160
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys
165 170 175
Asp Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
180 185 190
Trp Val Ala Trp Ile Asp Pro Glu Asn Gly Asp Thr Glu Phe Val Pro
195 200 205
Lys Phe Gln Gly Arg Ala Thr Met Ser Ala Asp Thr Ser Lys Asn Thr
210 215 220
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
225 230 235 240
Tyr Cys Lys Thr Gly Gly Phe Trp Gly Gln Gly Thr Leu Val Thr Val
245 250 255
Ser Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
260 265 270
Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
275 280 285
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
290 295 300
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
305 310 315 320
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
325 330 335
Gln Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
340 345 350
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
355 360 365
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
370 375 380
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
385 390 395 400
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
405 410 415
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
420 425 430
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
435 440 445
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
450 455 460
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
465 470 475 480
Leu Ser Leu Ser Leu Gly Lys Met Phe Trp Val Leu Val Val Val Gly
485 490 495
Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile
500 505 510
Phe Trp Val Arg Ser Lys Arg Ser Arg Gly Gly His Ser Asp Tyr Met
515 520 525
Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro
530 535 540
Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Gly Gly Gly Arg
545 550 555 560
Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln
565 570 575
Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp
580 585 590
Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro
595 600 605
Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp
610 615 620
Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg
625 630 635 640
Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr
645 650 655
Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
660 665 670
<210> 28
<211> 447
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность химерного белка, Фигура 20
<400> 28
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Ser Ala Ser
35 40 45
Ser Ser Val Arg Phe Ile His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
50 55 60
Pro Lys Arg Leu Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
85 90 95
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp
100 105 110
Gly Ser Ser Pro Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
115 120 125
Gly Ser Thr Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Ser Glu Val Gln Leu Val Glu Tyr Gly Gly Gly Leu Val Gln Pro
145 150 155 160
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys
165 170 175
Asp Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
180 185 190
Trp Val Ala Trp Ile Asp Pro Glu Asn Gly Asp Thr Glu Phe Val Pro
195 200 205
Lys Phe Gln Gly Arg Ala Thr Met Ser Ala Asp Thr Ser Lys Asn Thr
210 215 220
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
225 230 235 240
Tyr Cys Lys Thr Gly Gly Phe Trp Gly Gln Gly Thr Leu Val Thr Val
245 250 255
Ser Ser Gly Gly Gly Ser Ser Gly Gly Gly Ser Gly Met Ala Leu Ile
260 265 270
Val Leu Gly Gly Val Ala Gly Leu Leu Leu Phe Ile Gly Leu Gly Ile
275 280 285
Phe Phe Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro
290 295 300
Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys
305 310 315 320
Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Gly Gly Gly Arg
325 330 335
Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln
340 345 350
Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp
355 360 365
Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro
370 375 380
Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp
385 390 395 400
Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg
405 410 415
Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr
420 425 430
Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
435 440 445
<210> 29
<211> 571
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность химерного белка, Фигура 21
<400> 29
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Ser Ala Ser
35 40 45
Ser Ser Val Arg Phe Ile His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
50 55 60
Pro Lys Arg Leu Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
85 90 95
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp
100 105 110
Gly Ser Ser Pro Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
115 120 125
Gly Ser Thr Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Ser Glu Val Gln Leu Val Glu Tyr Gly Gly Gly Leu Val Gln Pro
145 150 155 160
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys
165 170 175
Asp Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
180 185 190
Trp Val Ala Trp Ile Asp Pro Glu Asn Gly Asp Thr Glu Phe Val Pro
195 200 205
Lys Phe Gln Gly Arg Ala Thr Met Ser Ala Asp Thr Ser Lys Asn Thr
210 215 220
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
225 230 235 240
Tyr Cys Lys Thr Gly Gly Phe Trp Gly Gln Gly Thr Leu Val Thr Val
245 250 255
Ser Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Gly Gly
260 265 270
Gly Ser Ser Gly Gly Gly Ser Gly Gly Gln Pro Arg Glu Pro Gln Val
275 280 285
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
290 295 300
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
305 310 315 320
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
325 330 335
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
340 345 350
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
355 360 365
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
370 375 380
Leu Gly Lys Met Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala
385 390 395 400
Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg
405 410 415
Ser Lys Arg Ser Arg Gly Gly His Ser Asp Tyr Met Asn Met Thr Pro
420 425 430
Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro
435 440 445
Arg Asp Phe Ala Ala Tyr Arg Ser Gly Gly Gly Arg Val Lys Phe Ser
450 455 460
Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr
465 470 475 480
Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys
485 490 495
Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn
500 505 510
Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu
515 520 525
Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly
530 535 540
His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr
545 550 555 560
Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
565 570
<210> 30
<211> 666
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> химерный белок PSCAscFv-IgG4(EQ)-CD4tm-4IBB-зета, Фигура 22
<400> 30
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Ser Ala Ser
35 40 45
Ser Ser Val Arg Phe Ile His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
50 55 60
Pro Lys Arg Leu Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
85 90 95
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp
100 105 110
Gly Ser Ser Pro Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
115 120 125
Gly Ser Thr Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Ser Glu Val Gln Leu Val Glu Tyr Gly Gly Gly Leu Val Gln Pro
145 150 155 160
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys
165 170 175
Asp Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
180 185 190
Trp Val Ala Trp Ile Asp Pro Glu Asn Gly Asp Thr Glu Phe Val Pro
195 200 205
Lys Phe Gln Gly Arg Ala Thr Met Ser Ala Asp Thr Ser Lys Asn Thr
210 215 220
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
225 230 235 240
Tyr Cys Lys Thr Gly Gly Phe Trp Gly Gln Gly Thr Leu Val Thr Val
245 250 255
Ser Ser Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
260 265 270
Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
275 280 285
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
290 295 300
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
305 310 315 320
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
325 330 335
Gln Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
340 345 350
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
355 360 365
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
370 375 380
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
385 390 395 400
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
405 410 415
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
420 425 430
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
435 440 445
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
450 455 460
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
465 470 475 480
Leu Ser Leu Ser Leu Gly Lys Met Ala Leu Ile Val Leu Gly Gly Val
485 490 495
Ala Gly Leu Leu Leu Phe Ile Gly Leu Gly Ile Phe Phe Lys Arg Gly
500 505 510
Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val
515 520 525
Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu
530 535 540
Glu Glu Gly Gly Cys Glu Leu Gly Gly Gly Arg Val Lys Phe Ser Arg
545 550 555 560
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
565 570 575
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
580 585 590
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
595 600 605
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
610 615 620
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
625 630 635 640
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
645 650 655
Ala Leu His Met Gln Ala Leu Pro Pro Arg
660 665
<210> 31
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> химерный белок PSCAscFv-L-CD28tm-4IBB-зета, Фигура 23
<400> 31
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Ser Ala Ser
35 40 45
Ser Ser Val Arg Phe Ile His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
50 55 60
Pro Lys Arg Leu Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
85 90 95
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp
100 105 110
Gly Ser Ser Pro Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
115 120 125
Gly Ser Thr Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Ser Glu Val Gln Leu Val Glu Tyr Gly Gly Gly Leu Val Gln Pro
145 150 155 160
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys
165 170 175
Asp Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
180 185 190
Trp Val Ala Trp Ile Asp Pro Glu Asn Gly Asp Thr Glu Phe Val Pro
195 200 205
Lys Phe Gln Gly Arg Ala Thr Met Ser Ala Asp Thr Ser Lys Asn Thr
210 215 220
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
225 230 235 240
Tyr Cys Lys Thr Gly Gly Phe Trp Gly Gln Gly Thr Leu Val Thr Val
245 250 255
Ser Ser Gly Gly Gly Ser Ser Gly Gly Gly Ser Gly Met Phe Trp Val
260 265 270
Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr
275 280 285
Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Gly Gly
290 295 300
His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg
305 310 315 320
Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg
325 330 335
Ser Gly Gly Gly Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala
340 345 350
Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg
355 360 365
Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu
370 375 380
Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn
385 390 395 400
Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
405 410 415
Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly
420 425 430
Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala
435 440 445
Leu Pro Pro Arg
450
<210> 32
<211> 544
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность химерного белка, Фигура 18 без сигнальной
<400> 32
Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Arg Phe Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Gly Ser Ser Pro Phe Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Ser Thr Ser Gly Gly
100 105 110
Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Glu Val Gln Leu
115 120 125
Val Glu Tyr Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Tyr Tyr Ile His Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Trp Ile Asp
165 170 175
Pro Glu Asn Gly Asp Thr Glu Phe Val Pro Lys Phe Gln Gly Arg Ala
180 185 190
Thr Met Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn
195 200 205
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Lys Thr Gly Gly
210 215 220
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Glu Ser Lys Tyr
225 230 235 240
Gly Pro Pro Cys Pro Pro Cys Pro Gly Gly Gly Ser Ser Gly Gly Gly
245 250 255
Ser Gly Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
260 265 270
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
275 280 285
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
290 295 300
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
305 310 315 320
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
325 330 335
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
340 345 350
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys Met Ala Leu
355 360 365
Ile Val Leu Gly Gly Val Ala Gly Leu Leu Leu Phe Ile Gly Leu Gly
370 375 380
Ile Phe Phe Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln
385 390 395 400
Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser
405 410 415
Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Gly Gly Gly
420 425 430
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
435 440 445
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
450 455 460
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
465 470 475 480
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
485 490 495
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
500 505 510
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
515 520 525
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
530 535 540
<210> 33
<211> 649
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> химерный белок PSCAscFv-IgG4(EQ)-CD28tm-CD28gg-зета, Фигура 19
без сигнальной
<400> 33
Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Arg Phe Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Gly Ser Ser Pro Phe Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Ser Thr Ser Gly Gly
100 105 110
Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Glu Val Gln Leu
115 120 125
Val Glu Tyr Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Tyr Tyr Ile His Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Trp Ile Asp
165 170 175
Pro Glu Asn Gly Asp Thr Glu Phe Val Pro Lys Phe Gln Gly Arg Ala
180 185 190
Thr Met Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn
195 200 205
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Lys Thr Gly Gly
210 215 220
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Glu Ser Lys Tyr
225 230 235 240
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Gly Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
275 280 285
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Gln Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
370 375 380
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
450 455 460
Lys Met Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr
465 470 475 480
Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys
485 490 495
Arg Ser Arg Gly Gly His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg
500 505 510
Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp
515 520 525
Phe Ala Ala Tyr Arg Ser Gly Gly Gly Arg Val Lys Phe Ser Arg Ser
530 535 540
Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu
545 550 555 560
Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg
565 570 575
Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln
580 585 590
Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr
595 600 605
Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp
610 615 620
Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala
625 630 635 640
Leu His Met Gln Ala Leu Pro Pro Arg
645
<210> 34
<211> 447
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность химерного белка, Фигура 20 без сигнальной
<400> 34
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Ser Ala Ser
35 40 45
Ser Ser Val Arg Phe Ile His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
50 55 60
Pro Lys Arg Leu Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
85 90 95
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp
100 105 110
Gly Ser Ser Pro Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
115 120 125
Gly Ser Thr Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Ser Glu Val Gln Leu Val Glu Tyr Gly Gly Gly Leu Val Gln Pro
145 150 155 160
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys
165 170 175
Asp Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
180 185 190
Trp Val Ala Trp Ile Asp Pro Glu Asn Gly Asp Thr Glu Phe Val Pro
195 200 205
Lys Phe Gln Gly Arg Ala Thr Met Ser Ala Asp Thr Ser Lys Asn Thr
210 215 220
Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
225 230 235 240
Tyr Cys Lys Thr Gly Gly Phe Trp Gly Gln Gly Thr Leu Val Thr Val
245 250 255
Ser Ser Gly Gly Gly Ser Ser Gly Gly Gly Ser Gly Met Ala Leu Ile
260 265 270
Val Leu Gly Gly Val Ala Gly Leu Leu Leu Phe Ile Gly Leu Gly Ile
275 280 285
Phe Phe Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro
290 295 300
Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys
305 310 315 320
Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Gly Gly Gly Arg
325 330 335
Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln
340 345 350
Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp
355 360 365
Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro
370 375 380
Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp
385 390 395 400
Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg
405 410 415
Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr
420 425 430
Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
435 440 445
<210> 35
<211> 549
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> последовательность химерного белка, Фигура 21 без сигнальной
<400> 35
Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Arg Phe Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Gly Ser Ser Pro Phe Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Ser Thr Ser Gly Gly
100 105 110
Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Glu Val Gln Leu
115 120 125
Val Glu Tyr Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Tyr Tyr Ile His Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Trp Ile Asp
165 170 175
Pro Glu Asn Gly Asp Thr Glu Phe Val Pro Lys Phe Gln Gly Arg Ala
180 185 190
Thr Met Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn
195 200 205
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Lys Thr Gly Gly
210 215 220
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Glu Ser Lys Tyr
225 230 235 240
Gly Pro Pro Cys Pro Pro Cys Pro Gly Gly Gly Ser Ser Gly Gly Gly
245 250 255
Ser Gly Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
260 265 270
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
275 280 285
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
290 295 300
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
305 310 315 320
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
325 330 335
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
340 345 350
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys Met Phe Trp
355 360 365
Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val
370 375 380
Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Gly
385 390 395 400
Gly His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr
405 410 415
Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr
420 425 430
Arg Ser Gly Gly Gly Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro
435 440 445
Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly
450 455 460
Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro
465 470 475 480
Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr
485 490 495
Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly
500 505 510
Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln
515 520 525
Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln
530 535 540
Ala Leu Pro Pro Arg
545
<210> 36
<211> 644
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> химерный белок PSCAscFv-IgG4(EQ)-CD4tm-4IBB-зета, Фигура 22
без сигнальной
<400> 36
Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Arg Phe Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Gly Ser Ser Pro Phe Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Ser Thr Ser Gly Gly
100 105 110
Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Glu Val Gln Leu
115 120 125
Val Glu Tyr Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Tyr Tyr Ile His Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Trp Ile Asp
165 170 175
Pro Glu Asn Gly Asp Thr Glu Phe Val Pro Lys Phe Gln Gly Arg Ala
180 185 190
Thr Met Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn
195 200 205
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Lys Thr Gly Gly
210 215 220
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Glu Ser Lys Tyr
225 230 235 240
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Gly Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
275 280 285
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Gln Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
370 375 380
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
450 455 460
Lys Met Ala Leu Ile Val Leu Gly Gly Val Ala Gly Leu Leu Leu Phe
465 470 475 480
Ile Gly Leu Gly Ile Phe Phe Lys Arg Gly Arg Lys Lys Leu Leu Tyr
485 490 495
Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu
500 505 510
Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu
515 520 525
Leu Gly Gly Gly Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala
530 535 540
Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg
545 550 555 560
Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu
565 570 575
Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn
580 585 590
Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
595 600 605
Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly
610 615 620
Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala
625 630 635 640
Leu Pro Pro Arg
<210> 37
<211> 430
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> химерный белок PSCAscFv-L-CD28tm-4IBB-зета, Фигура 23 без
сигнальной
<400> 37
Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Arg Phe Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Gly Ser Ser Pro Phe Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Ser Thr Ser Gly Gly
100 105 110
Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Glu Val Gln Leu
115 120 125
Val Glu Tyr Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Tyr Tyr Ile His Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Trp Ile Asp
165 170 175
Pro Glu Asn Gly Asp Thr Glu Phe Val Pro Lys Phe Gln Gly Arg Ala
180 185 190
Thr Met Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn
195 200 205
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Lys Thr Gly Gly
210 215 220
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Ser
225 230 235 240
Ser Gly Gly Gly Ser Gly Met Phe Trp Val Leu Val Val Val Gly Gly
245 250 255
Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe
260 265 270
Trp Val Arg Ser Lys Arg Ser Arg Gly Gly His Ser Asp Tyr Met Asn
275 280 285
Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr
290 295 300
Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Gly Gly Gly Arg Val
305 310 315 320
Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn
325 330 335
Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val
340 345 350
Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg
355 360 365
Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys
370 375 380
Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg
385 390 395 400
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys
405 410 415
Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
420 425 430
<210> 38
<211> 236
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> scFv, направленный против PSCA
<400> 38
Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Arg Phe Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Gly Ser Ser Pro Phe Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Gly Ser Thr Ser Gly Gly
100 105 110
Gly Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Glu Val Gln Leu
115 120 125
Val Glu Tyr Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Tyr Tyr Ile His Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Trp Ile Asp
165 170 175
Pro Glu Asn Gly Asp Thr Glu Phe Val Pro Lys Phe Gln Gly Arg Ala
180 185 190
Thr Met Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn
195 200 205
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Lys Thr Gly Gly
210 215 220
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
225 230 235
<---


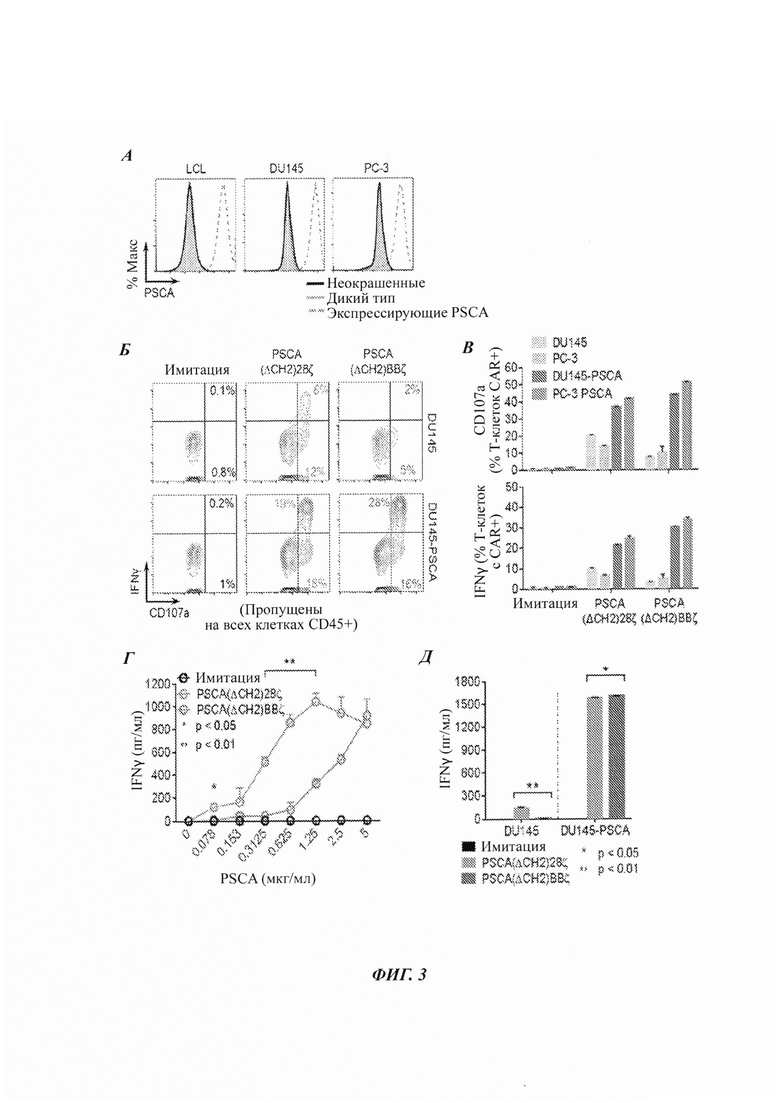
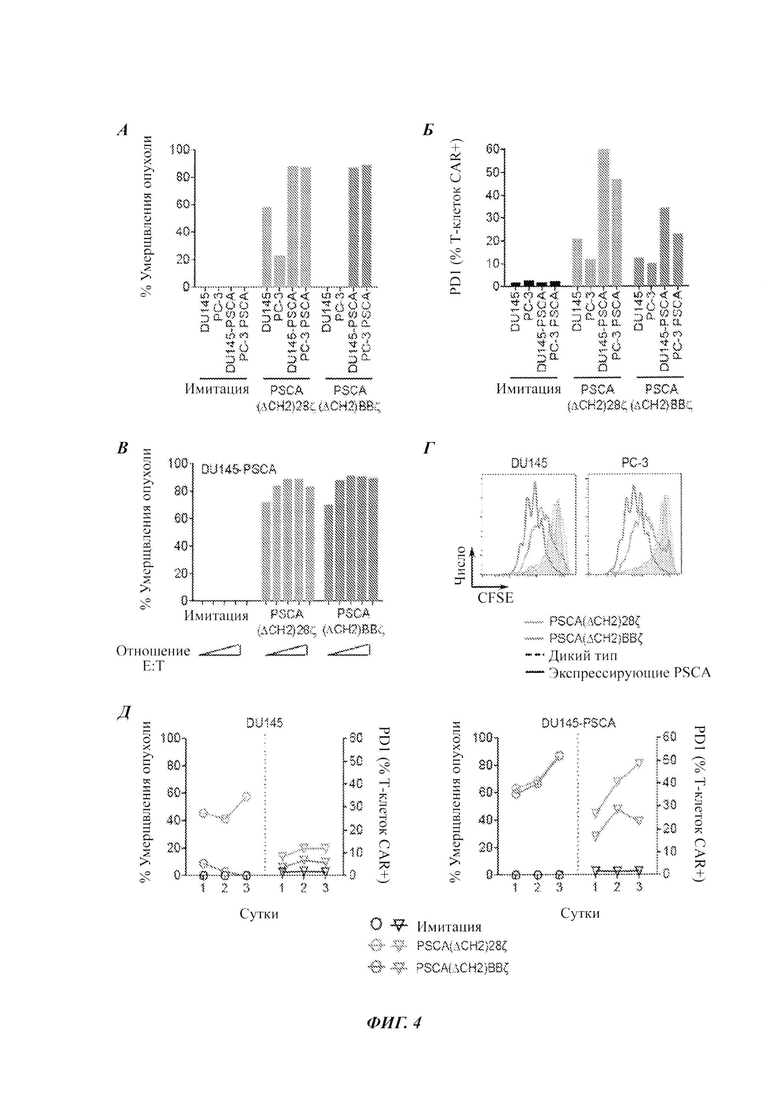
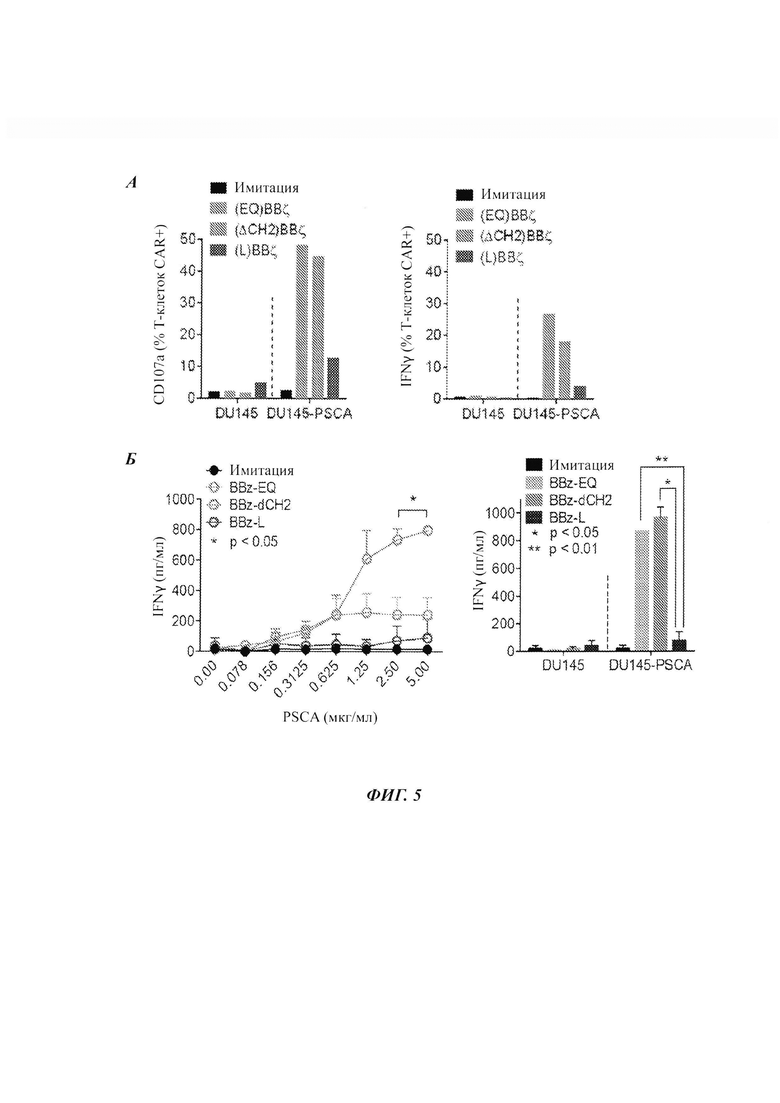
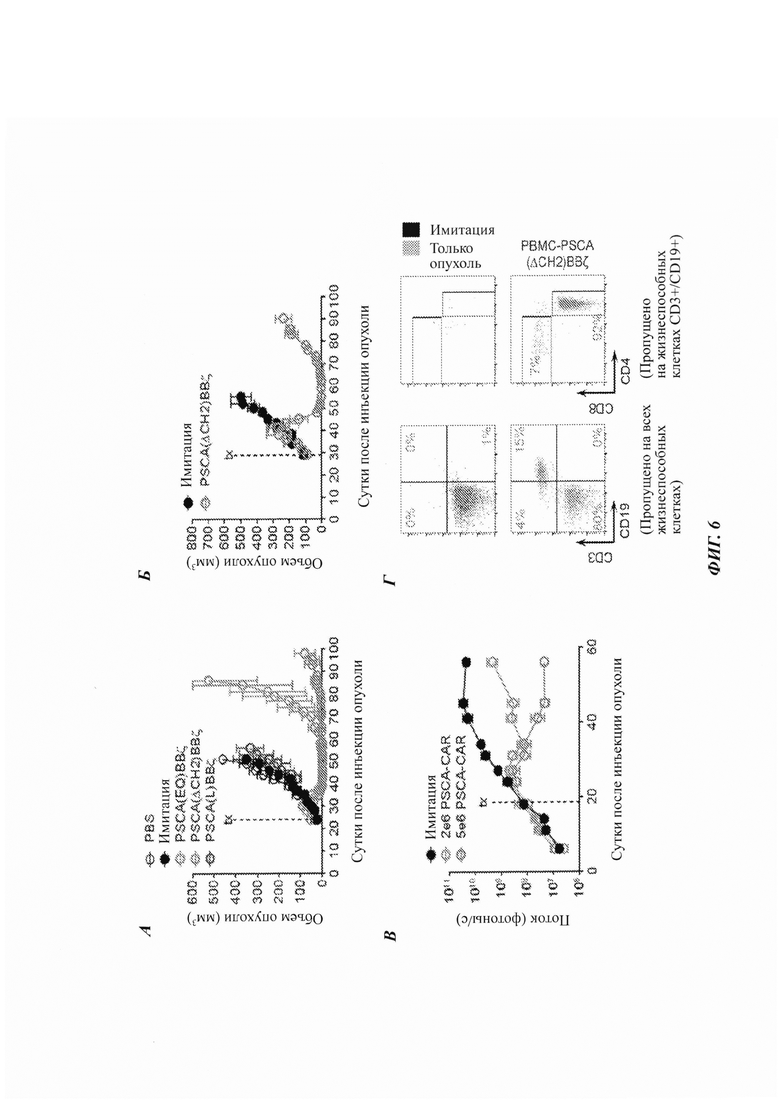
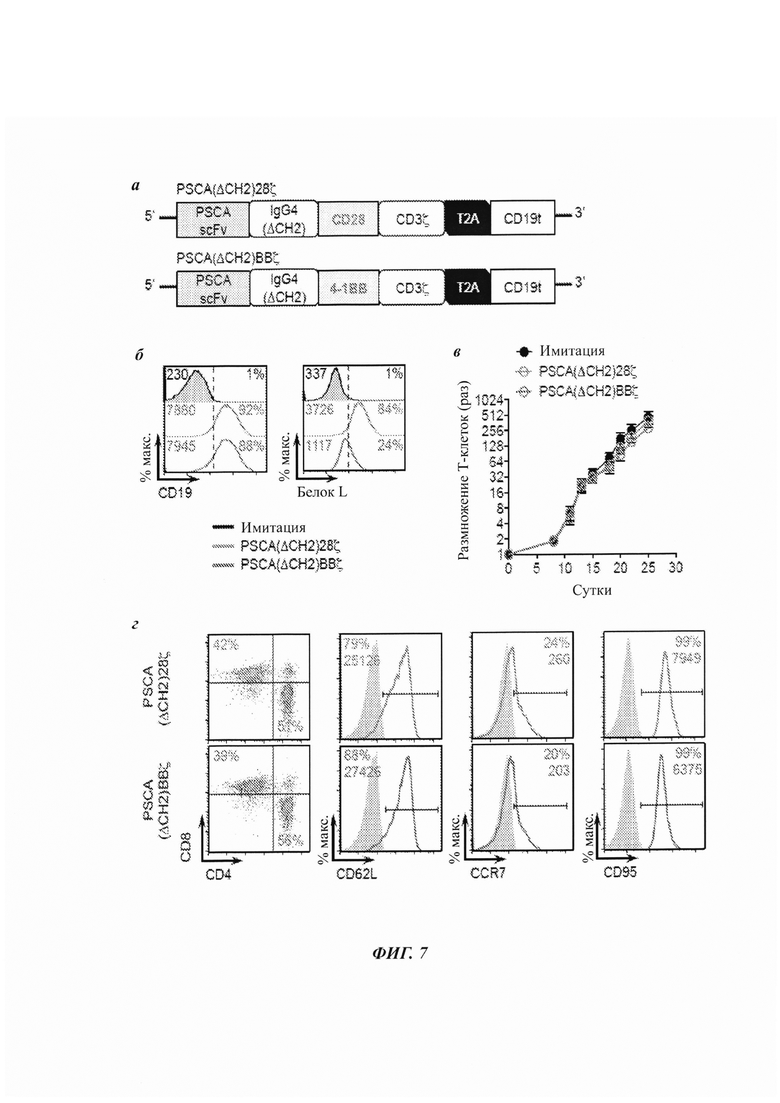
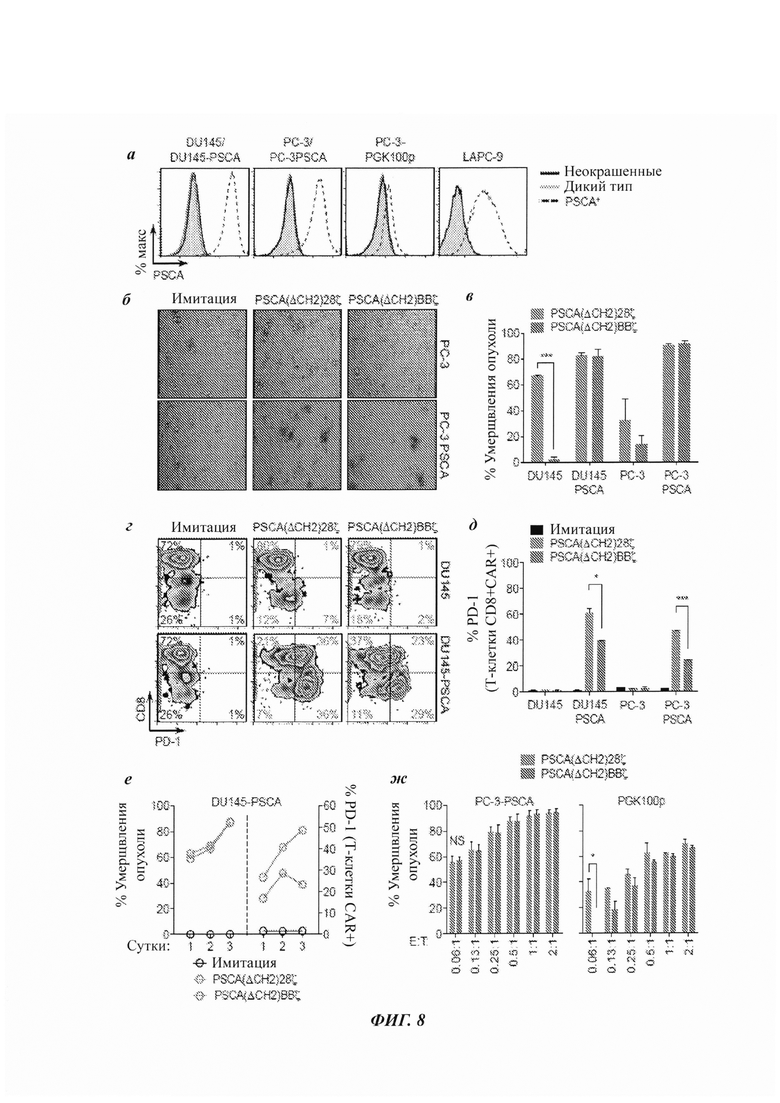

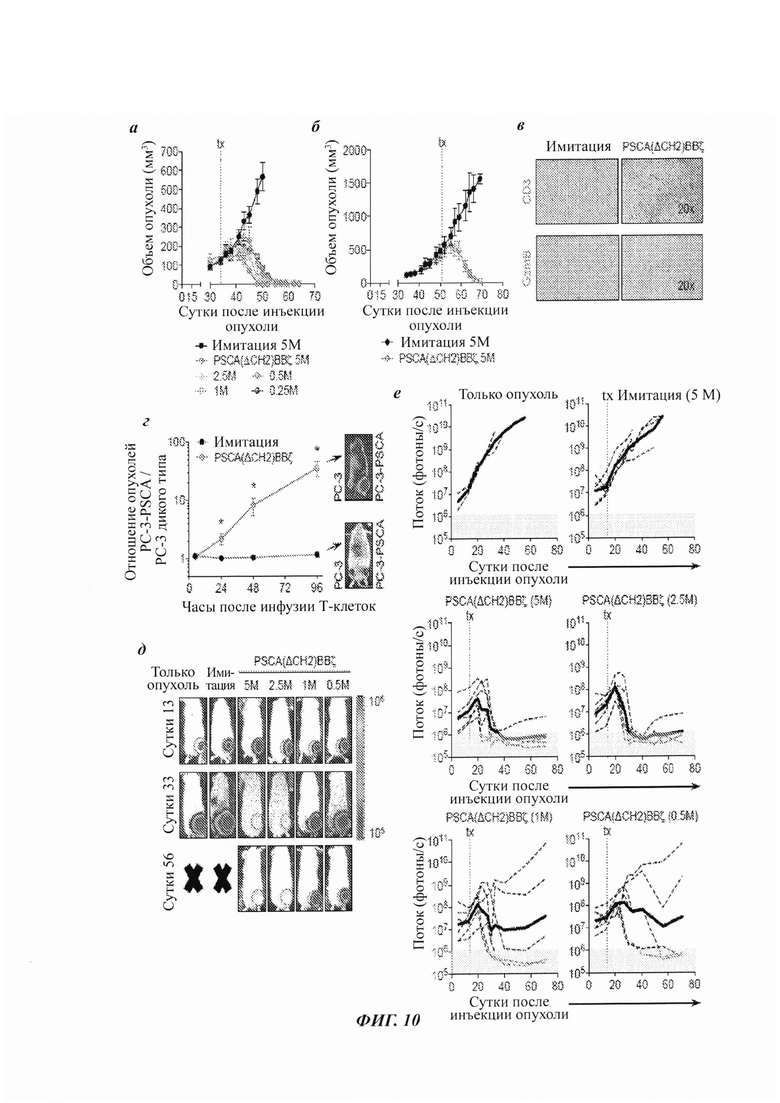
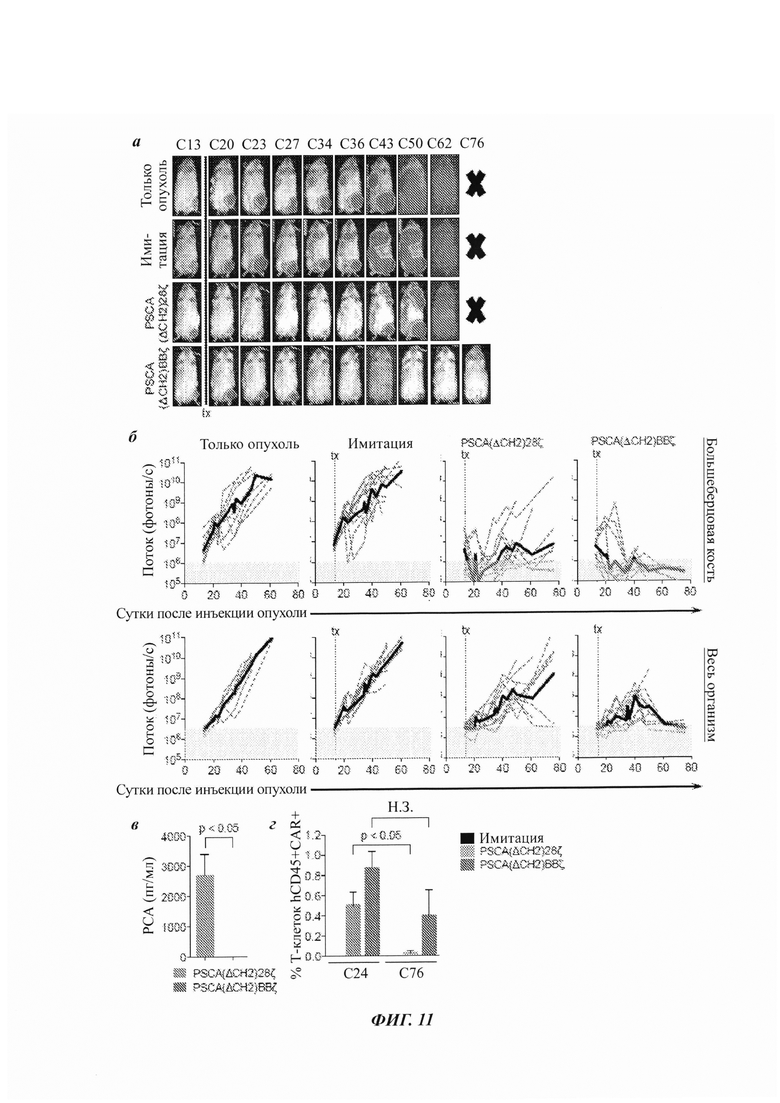
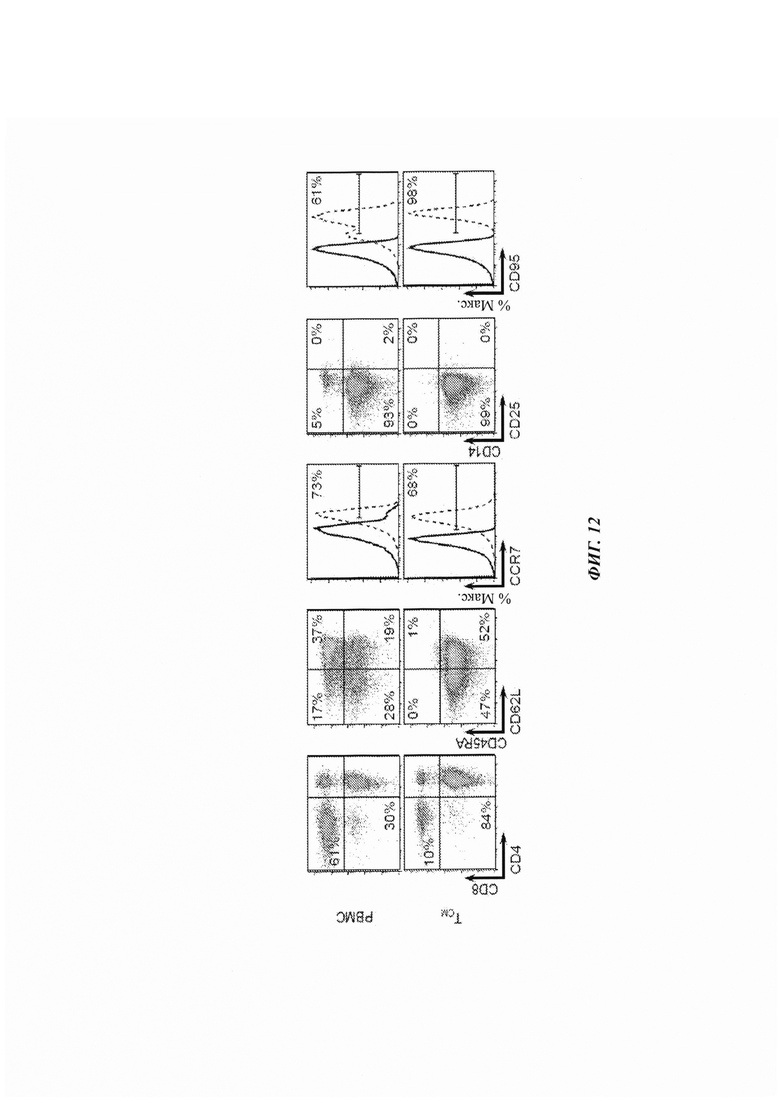


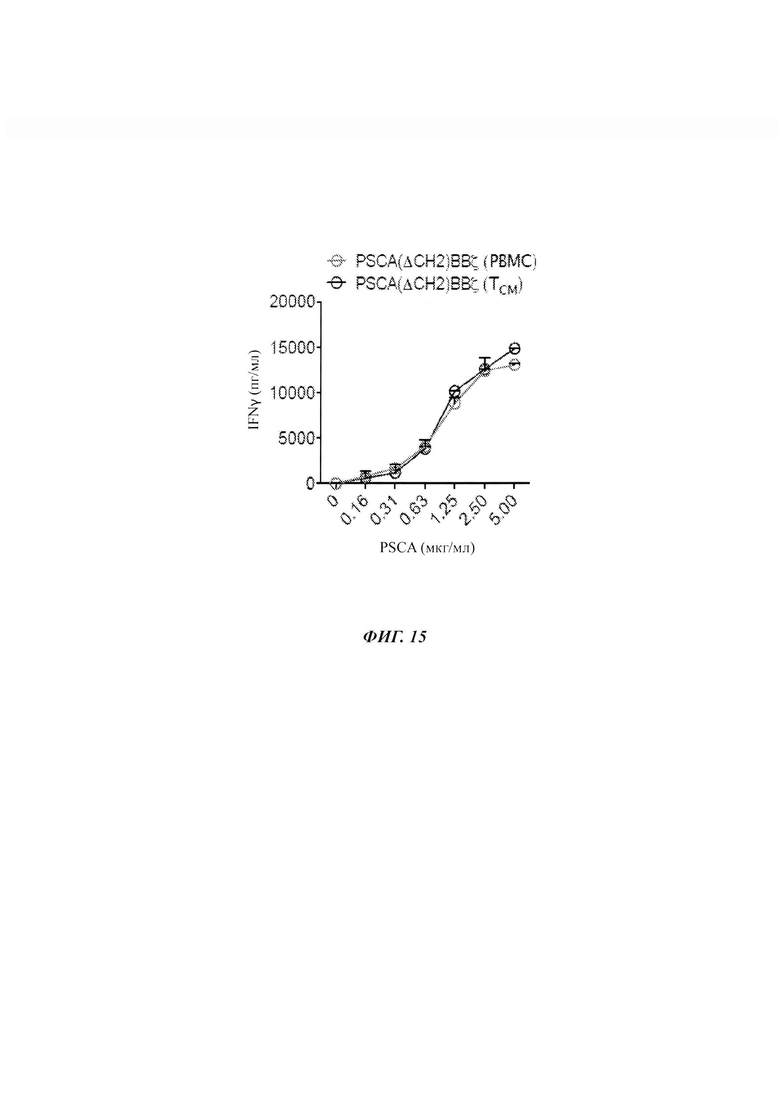
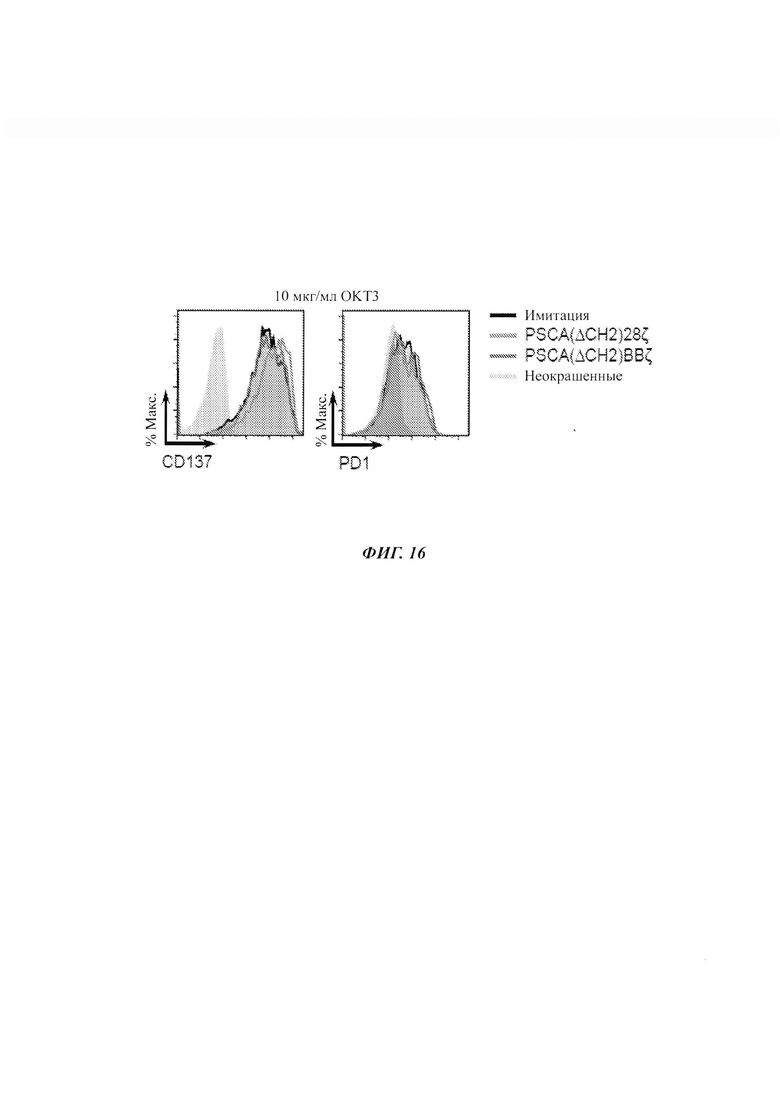
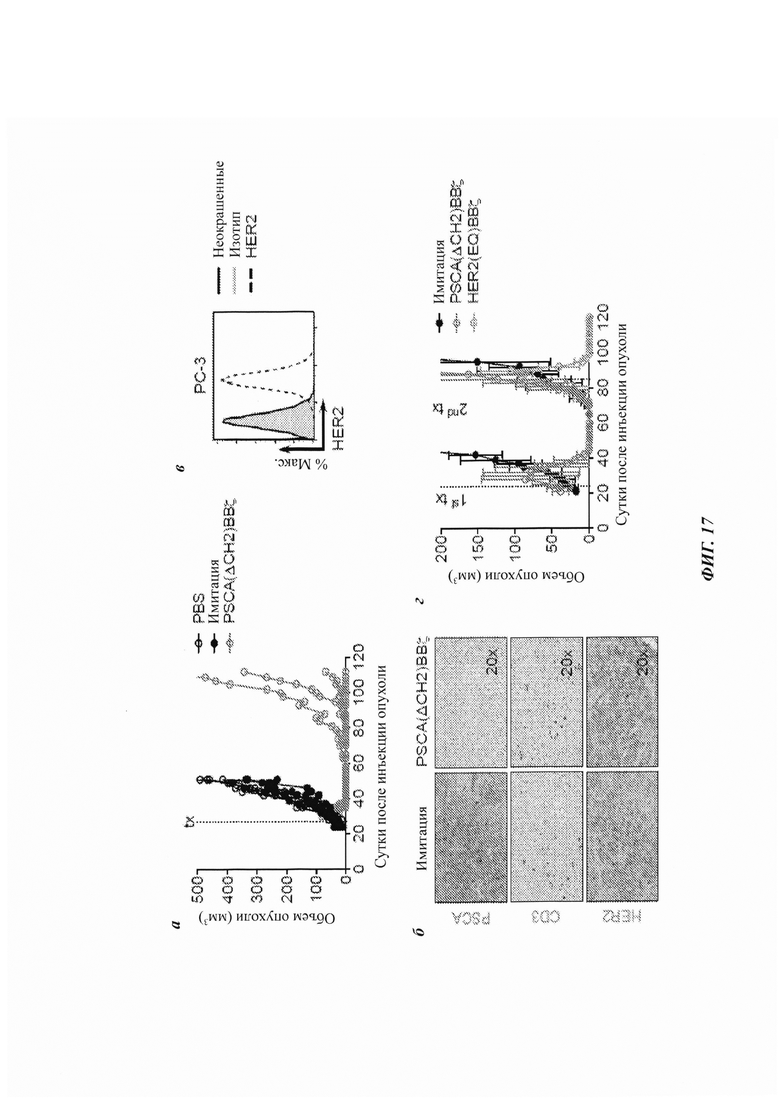






Изобретение относится к области биотехнологии. Описана группа изобретений, включающая молекулу нуклеиновой кислоты, кодирующую химерный рецептор антигена, нацеленный на PSCA (антиген стволовой клетки простаты), популяцию человеческих Т-клеток для лечения рака, экспрессирующего PSCA, композицию, содержащую популяцию аутологичных или аллогенных человеческих Т-клеток, для применения в способе лечения рака, экспрессирующего PSCA у пациента и химерный рецептор антигена, нацеленный на PSCA. В одном из вариантов реализации молекула нуклеиновой кислоты экспрессирует полипептид, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 26, 28, 30, 32, 34 и 36. Изобретение расширяет арсенал средств, кодирующих химерный рецептор антигена, нацеленный на PSCA. 4 н. и 10 з.п. ф-лы, 23 ил., 4 табл., 14 пр.
1. Молекула нуклеиновой кислоты, кодирующая химерный рецептор антигена, нацеленный на PSCA (антиген стволовой клетки простаты), где указанная молекула нуклеиновой кислоты экспрессирует полипептид, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 26, 28, 30, 32, 34 и 36.
2. Молекула нуклеиновой кислоты по п. 1, где указанная молекула нуклеиновой кислоты экспрессирует полипептид, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 26 и 32.
3. Молекула нуклеиновой кислоты по п. 1, где указанная молекула нуклеиновой кислоты экспрессирует полипептид, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 28 и 34.
4. Молекула нуклеиновой кислоты по п. 1, где указанная молекула нуклеиновой кислоты экспрессирует полипептид, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 30 и 36.
5. Популяция человеческих Т-клеток для лечения рака, экспрессирующего PSCA, содержащих вектор, экспрессирующий химерный рецептор антигена, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 26, 28, 30, 32, 34 и 36.
6. Популяция человеческих Т-клеток по п. 5, где указанная молекула нуклеиновой кислоты экспрессирует полипептид, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 26 и 32.
7. Популяция человеческих Т-клеток по п. 5, где указанный вектор экспрессирует химерный рецептор антигена, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 28 и 34.
8. Популяция человеческих Т-клеток по п. 5, где указанный вектор экспрессирует химерный рецептор антигена, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 30 и 36.
9. Популяция CD4+ и СD8+ человеческих Т-клеток по любому из пп. 5-8, где Т-клетки состоят из популяции центральных Т-клеток памяти.
10. Композиция, содержащая популяцию аутологичных или аллогенных человеческих Т-клеток, трансдуцированных вектором, содержащим экспрессионную кассету, кодирующую химерный рецептор антигена, где химерный рецептор антигена содержит аминокислотную последовательность, выбранную из SEQ ID NO: 26, 28, 30, 32, 34 и 36, для применения в способе лечения рака, экспрессирующего PSCA, у пациента, где способ включает введение указанной композиции.
11. Химерный рецептор антигена, нацеленный на PSCA, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 26, 28, 30, 32, 34 и 36.
12. Химерный рецептор антигена по п. 11, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 26 и 32.
13. Химерный рецептор антигена по п. 11, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 28 и 34.
14. Химерный рецептор антигена по п. 11, содержащий аминокислотную последовательность, выбранную из SEQ ID NO: 30 и 36.
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| CARTELLIERI M et | |||
| al | |||
| "A novel ex vivo isolation and expansion procedure for chimeric antigen receptor engrafted human T cells", PLOS ONE, 2014, Vol | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| HILLERDAL V et | |||
| al | |||
| "Systemic treatment with CAR-engineered T cells against PSCA delays subcutaneous tumor growth and prolongs survival of mice", | |||

