ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к антителам к альфа-синуклеин αу и способу их применения для лечения синуклеинопатий. В частности, настоящее изобретение относится к антителам к человеческому альфа-синуклеину и их применению для лечения болезни Паркинсона.
УРОВЕНЬ ТЕХНИКИ
Альфа-синуклеин представляет собой небольшой растворимый белок длиной 140 аминокислот, существующий в совершенно разных формах. Альфа-синуклеин в основном обнаруживается в пресинаптических нервных окончаниях, и, хотя его точная функция неизвестна, исследователи полагают, что он играет центральную роль в многочисленных нейродегенеративных процессах.
В последние 15 лет было показано, что альфа-синуклеин играет ключевую роль в патогенезе всех форм болезни Паркинсона. Генетические мутации или увеличение числа копий гена альфа-синуклеина являются причиной развития семейной формы болезни Паркинсона с ранним началом (PD). Интересно, что в семьях с увеличенным количеством копий генных локусов патогенный эффект в бесспорно зависит от дозы гена. Дупликации генов являются причиной развития относительно ранней формы PD (~47 лет) с нормальным течением заболевания, в то время как трипликации гена связаны с очень ранним возрастом появления (~33 года) и очень быстрым течением заболевания. При всех формах болезни Паркинсона альфа-синуклеин является основным компонентом телец Леви, ключевым патологическим признаком заболевания.
Патология телец Леви распространяется в процессе заболевания, и предполагается, что альфа-синуклеин действует как прионоподобный белок с неправильной сборкой, приводящей к образованию токсичных олигомеров и агрегатов, которые могут распространяться от пораженных на непораженные нейроны (Olanow CW et al. Movement Disorders, Vol.28, No.1, 2013). Существующие в настоящее время методы лечения не способны остановить распространение заболевания и помогают только при лечении симптомов, связанных с прогрессирующей потерей зависимых от мотонейронов активностей. В 2014 году Tran H.T. et al. (Tran H.T. et al., Cell Reports 7, 2054-2065, 2014) показали, что внутрибрюшинное введение моноклонального антитела к неправильно упакованному альфа-синуклеину мышам, которым ранее была введена интрастриатальная инъекция предварительно сформированных фибрилл альфа-синуклеина, приводило к снижению выраженности патологии телец Леви, уменьшало потерю допаминергических нейронов черной субстанции и ослабляло моторные нарушения. Следовательно, все еще остается потребность в пассивной иммунотерапии, которая могла бы быть терапевтически эффективной при PD и других альфа-синуклеинопатиях.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение позволяет удовлетворить определенную выше потребность благодаря антителам к альфа-синуклеину согласно следующим вариантам осуществления.
Вариант осуществления 1: Антитело или его антигенсвязывающий фрагмент, которые связываются с альфа-синуклеином, где антитело содержит:
вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:44;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
вариабельную область тяжелой цепи, содержащую:
i. CDR-H1, содержащую SEQ ID NO:4;
ii. CDR-H2, содержащую SEQ ID NO:45, и
iii. CDR-H3, содержащую SEQ ID NO:46.
Вариант осуществления 2: Антитело или его антигенсвязывающий фрагмент согласно варианту осуществления 1, которое связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131; где эпитоп необязательно содержит A124 и G132.
Вариант осуществления 3: 3. Антитело или его антигенсвязывающий фрагмент по любому из п.1 или п.2, где антитело или антигенсвязывающий фрагмент предотвращают агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина.
Вариант осуществления 4: Антитело или его антигенсвязывающий фрагмент по любому из вариантов осуществления 1-3, где антитело или его антигенсвязывающий фрагмент способны связывать альфа-синуклеин в виде мономера и в фибриллах.
Вариант осуществления 5: Антитело или его антигенсвязывающий фрагмент по любому из предшествующих вариантов осуществления, которое имеет более высокое сродство связывания с альфа-синуклеином в фибриллах по сравнению с альфа-синуклеином в виде мономера, характеризующееся константой диссоциации (KD), по меньшей мере в 10 раз более высокой для мономерного альфа-синуклеина, чем для альфа-синуклеина в фибриллах.
Вариант осуществления 6: Антитело или его антигенсвязывающий фрагмент по любому из предшествующих вариантов осуществления с величиной (KD) к альфа-синуклеину в фибриллах 300 пМ или менее.
Вариант осуществления 7: Антитело или его антигенсвязывающий фрагмент по любому из предшествующих вариантов осуществления, которое не связывается с бета-синуклеином и/или гамма-синуклеином.
Вариант осуществления 8: Антитело или его антигенсвязывающий фрагмент по любому из предшествующих вариантов осуществления, представляющее собой химерное, гуманизированное или человеческое антитело.
Вариант осуществления 9: Антитело по любому из предшествующих вариантов осуществления, представляющее собой полноразмерное антитело.
Вариант осуществления 10: Антитело по варианту осуществления 9, где полноразмерное антитело выбирают из IgG1, IgG4 или IgG4P.
Вариант осуществления 11: Его антигенсвязывающий фрагмент по любому из вариантов осуществления 1-8, где антигенсвязывающий фрагмент выбирают из Fab, Fab', F(ab')2, scFv, dAb или VHH.
Вариант осуществления 12: Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, где антитело или его антигенсвязывающий фрагмент содержит:
a. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5, и CDR-H3, содержащую SEQ ID NO:6; и
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:31; или
c. легкую цепь, содержащую SEQ ID NO:17, и тяжелую цепь, содержащую SEQ ID NO:33.
Вариант осуществления 13: Антитело или его антигенсвязывающий фрагмент по любому из пп.1-11, где антитело или его антигенсвязывающий фрагмент содержит:
a. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5, и CDR-H3, содержащую SEQ ID NO:6; и
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:23; или
c. легкую цепь, содержащую SEQ ID NO:17, и тяжелую цепь, содержащую SEQ ID NO:25.
Вариант осуществления 14: Антитело или его антигенсвязывающий фрагмент по любому из пп.1-11, где антитело или его антигенсвязывающий фрагмент содержит:
a. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:8, и CDR-H3, содержащую SEQ ID NO:9; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:27 или 35; или
c. легкую цепь, содержащую SEQ ID NO:17, и тяжелую цепь, содержащую SEQ ID NO:29 или 37.
Вариант осуществления 15: Антитело или его антигенсвязывающий фрагмент по любому из пп.1-11, где антитело или его антигенсвязывающий фрагмент содержит:
a. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:7, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5, и CDR-H3, содержащую SEQ ID NO:6; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:19, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:23 или 31; или
c. легкую цепь, содержащую SEQ ID NO:21, и тяжелую цепь, содержащую SEQ ID NO:25 или 33.
Вариант осуществления 16: Антитело или его антигенсвязывающий фрагмент по любому из пп.1-11, где антитело или его антигенсвязывающий фрагмент содержит:
a. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:7, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:8, и CDR-H3, содержащую SEQ ID NO:9; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:19, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:27 или 35; или
c. легкую цепь, содержащую SEQ ID NO:21, и тяжелую цепь, содержащую SEQ ID NO:29 или 37.
Вариант осуществления 17: Антитело или его антигенсвязывающий фрагмент, которое:
a. конкурирует за связывание альфа-синуклеина с антителом или его антигенсвязывающим фрагментом по любому из предшествующих пунктов; и/или
b. перекрестно блокирует или является перекрестно блокированным в отношении связывания альфа-синуклеина антителом или его антигенсвязывающим фрагментом по любому из предшествующих пунктов; и/или
с. связывается с тем же эпитопом альфа-синуклеина, что и антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов; и/или
d. содержит вариабельную область тяжелой цепи, имеющую по меньшей мере 80% идентичности или сходства с последовательностью, приведенной в SEQ ID NO:23, SEQ ID NO:31, SEQ ID NO:27 или SEQ ID NO:35; и/или
е. содержит вариабельную область легкой цепи, имеющую по меньшей мере 80% идентичности или сходства с последовательностью, приведенной в SEQ ID NO:15 или SEQ ID NO:19.
Вариант осуществления 18: Выделенный полинуклеотид, кодирующий антитело или его антигенсвязывающий фрагмент по любому из вариантов осуществления 1-16.
Вариант осуществления 19: Выделенный полинуклеотид по варианту осуществления 18, кодирующий:
a. вариабельную область легкой цепи, где полинуклеотид:
i. является на по меньшей мере 90% идентичным SEQ ID NO:16 или SEQ ID NO:20; или
ii. содержит SEQ ID NO:16 или 20; или
iii. по существу состоит из SEQ ID NO:16 или SEQ ID NO:20;
b. вариабельную область тяжелой цепи, где полинуклеотид:
i. является на по меньшей мере 90% идентичным SEQ ID NO:24 или SEQ ID NO:28, или SEQ ID NO:32, или SEQ ID NO:36; или
ii. содержит SEQ ID NO:24 или SEQ ID NO:28, или SEQ ID NO:32, или SEQ ID NO:36; или
iii. по существу состоит из SEQ ID NO:24 или SEQ ID NO:28, или SEQ ID NO:32, или SEQ ID NO:36;
c. легкую цепь, где полинуклеотид:
i. является на по меньшей мере 90% идентичным SEQ ID NO:18 или SEQ ID NO:22; или
ii. содержит SEQ ID NO:18 или 22; или
iii. по существу состоит из SEQ ID NO:18 или SEQ ID NO:22;
d. тяжелую цепь, где полинуклеотид:
i. является на по меньшей мере 90% идентичным SEQ ID NO:26 или SEQ ID NO:30, или SEQ ID NO:34, или SEQ ID NO:38; или
ii. содержит SEQ ID NO:26 или SEQ ID NO:30, или SEQ ID NO:34, или SEQ ID NO:38; или
iii. по существу состоит из SEQ ID NO:26 или SEQ ID NO:30, или SEQ ID NO:34, или SEQ ID NO:38.
Вариант осуществления 20: Вектор клонирования или экспрессии, содержащий один или более полинуклеотидов по любому из вариантов осуществления 18 или 19.
Вариант осуществления 21: Клетка-хозяин, содержащая:
a. один или более полинуклеотидов по любому из вариантов осуществления 18 или 19; или
b. один или более векторов экспрессии по варианту осуществления 20.
Вариант осуществления 22: Способ получения антитела или его антигенсвязывающего фрагмента по любому из вариантов осуществления 1-17, включающий культивирование клетки-хозяина по варианту осуществления 21 в условиях, подходящих для продуцирования антитела или его антигенсвязывающего фрагмента, и выделение антитела или его антигенсвязывающего фрагмента.
Вариант осуществления 23: Фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент по любому из вариантов осуществления 1-17 и один или более фармацевтически приемлемых носителей, наполнителей или разбавителей.
Вариант осуществления 24: Антитело или его антигенсвязывающий фрагмент по любому из вариантов осуществления 1-17 или фармацевтическая композиция по варианту осуществления 23 для применения в терапии.
Вариант осуществления 25: Антитело или его антигенсвязывающий фрагмент по любому из вариантов осуществления 1-17 или фармацевтическая композиция по варианту осуществления 23 для применения для лечения одной или более синуклеинопатий.
Вариант осуществления 26: Антитело или его антигенсвязывающий фрагмент по п.25, где синуклеинопатии выбирают из болезни Паркинсона (PD) (включая идиопатические и наследственные формы болезни Паркинсона), деменции с тельцами Леви (DLB), болезни диффузных телец Леви (DLBD), варианта болезни Альцгеймера с тельцами Леви (LBVAD), комбинированной болезни Альцгеймера и Паркинсона, множественной системной атрофии (MSA) и нейродегенерации с накоплением железа в мозге 1-го типа (NBIA-1).
Вариант осуществления 27: Применение антитела или его антигенсвязывающего фрагмента по варианту осуществления 26, где синуклеинопатия представляет собой болезнь Паркинсона.
Вариант осуществления 28: Способ лечения синуклеинопатии у пациента, включающий введение указанному пациенту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из вариантов осуществления 1-17 или фармацевтической композиции по варианту осуществления 23.
Вариант осуществления 29: Способ по варианту осуществления 29, где синуклеинопатию выбирают из болезни Паркинсона (PD) (включая идиопатические и наследственные формы болезни Паркинсона), деменции с тельцами Леви (DLB), болезни диффузных телец Леви (DLBD), варианта болезни Альцгеймера с тельцами Леви (LBVAD), комбинированной болезни Альцгеймера и Паркинсона, множественной системной атрофии (MSA) и нейродегенерации с накоплением железа в мозге 1-го типа (NBIA-1), предпочтительно болезни Паркинсона.
Вариант осуществления 30: Антитело или его антигенсвязывающий фрагмент по любому из вариантов осуществления 1-16 для применения для диагностики альфа-синуклеинопатий, предпочтительно диагностики болезни Паркинсона.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1. (A) SDS-PAGE образцов экспрессии альфа-синуклеина. Альфа-синуклеин с His-меткой (1) и после удаления His-метки TEV-протеазой (2), эксклюзионная хроматография на Superdex 75 человеческого альфа-синуклеина, обработанного TEV-протеазой (3). Маркер молекулярной массы белка SeeBluePlus2 (Invitrogen) (M). (B) SDS-PAGE человеческого альфа-синуклеина, очищенного из супернатанта Expi293 в виде немеченого белка дикого типа. (4) Маркер молекулярной массы белка SeeBluePlus2 (Invitrogen) (M).
Фиг.2. (A) Анализ фибрилл с помощью JC-1 анализа мономера без флуоресценции и фибрилл с максимальной флуоресценцией при 540 нм. (B) Типичный пример спектра случайных спиралей мономерного человеческого альфа-синуклеина (длина волны 1646 см-1) и внутреннего β-листа, образованного в фибриллах рекомбинантного человеческого альфа-синуклеина (длина волны 1625-1630 см-1)
Фиг.3. ELISA анализ связывания. ELISA связывание кроличьего IgG1 6470 с мономером и фибриллами рекомбинантного человеческого альфа-синуклеина и пептидом PVDPDNEAYE человеческого альфа-синуклеина.
Фиг.4. (A) Вестерн-блот, показывающий связывание кроличьего IgG1 6470 с человеческим альфа-синуклеином и человеческим бета-синуклеином. 1 - человеческий альфа-синуклеин; 2 - человеческий альфа-синуклеин (rПептид); 3 - человеческий бета-синуклеин (rПептид); Маркер, MagicMark XP. (B) Изменения химического сдвига в ЯМР, показывающие предсказанный эпитоп 6470 на человеческом альфа-синуклеине.
Фиг.5. Ингибирование связывания IgG 6470 с иммобилизованным альфа-синуклеином (столбцы слева, мономер и правильные фибриллы, соответственно, каждого из протестированных пептидов).
Фиг.6. Схематическое представление Fab 6470 в комплексе с пептидом 123-132.
Фиг.7. Схематическое представление контактов тяжелой цепи Fab 6470 с пептидом 123-132. Пептидные остатки помечены непосредственно, вариабельные остатки тяжелой цепи 6470 помечены номером vH-остатка.
Фиг.8. Схематическое представление контактов легкой цепи Fab 6470 с пептидом 123-132. Пептидные остатки помечены непосредственно, остатки вариабельной области легкой цепи 6470 помечены номером vL-остатка.
Фиг.9. Гуманизация легкой цепи. 6470 означает последовательность вариабельной области легкой цепи кролика. 6470gL3 означает гуманизированный трансплантат вариабельной области легкой цепи 6470 с использованием человеческой зародышевой линии IGKV1-16 в качестве акцепторного каркаса. CDR показаны жирным шрифтом/подчеркнуты. Донорские остатки показаны жирным шрифтом/курсивом и закрашены: Q48 и Q72. Мутация в CDRL1 N33R показана жирным шрифтом/подчеркнута и заштрихована.
Фиг.10. Гуманизация тяжелой цепи. 6470 означает последовательность вариабельной области тяжелой цепи кролика. 6470gH23 и gH36 означают гуманизированные трансплантаты вариабельной области тяжелой цепи антитела 6470 с использованием человеческой зародышевой линии IGHV3-23 в качестве акцепторного каркаса. CDR показаны жирным шрифтом/подчеркнуты. Донорские остатки показаны жирным шрифтом/курсивом и закрашены: V24, Y47, I48, G49, S73, V78 и R97. Мутации S56N и N102H в CDRH2 и CDRH3, соответственно, показаны жирным шрифтом/подчеркнуты и заштрихованы.
Фиг.11. Напряжение на границе раздела воздух-жидкость. 6470 антитела и мутанты в трех заранее приготовленных буферах через 3 и 24 часа после встряхивания.
Фиг.12. Иммуногистохимия. Иммунореактивность в срезах головного мозга пациентов с PD (A-E) и без PD (F-H). (A-C) В височной коре пациентов с PD антитело 6470 окрашивало нейропил и структуры типа телец Леви (белые стрелки) в сером веществе. (D, E) антитело 6470 окрашивало структуры типа телец Леви (белые стрелки) в черной субстанции пациентов с PD. (F, G). В тканях кортикального слоя височной области без PD 6470 также окрашивало нейропиль, но не окрашивало структуры подобные тельцам Леви. (H) В черной субстанции индивидуума без PD не наблюдали никаких структур, подобных тельцам Леви; черные стрелки указывают на неспецифическую окраску. Масштабная полоска: 50 мкм.
Фиг.13. Анализ агрегации на основе клеток (клетки HEK). Антитела по настоящему изобретению способны ингибировать агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, с IC50 менее 5 нМ. Планки погрешности представляют стандартную ошибку измерения (SEM, N=3, n=9). В условных обозначениях FL в конце каждого названия антитела означает «полноразмерный».
Фиг.14. Анализ агрегации на основе клеток (первичные нейроны). Репрезентативное антитело по настоящему изобретению способно ингибировать агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина в первичных мышиных нейронах, экспрессирующих эндогенные уровни альфа-синуклеина, с IC50 менее 4 нМ. Планки погрешности представляют стандартную ошибку измерения (SEM, N=4, n=18).
Фиг.15. Изображения иммуногистохимического окрашивания патологии альфа-синуклеина (стрелки) в различных областях мозга мышей C57Bl/6J дикого типа (A) и мышей SNCA-OVX (B), которым инъецировали мышиный или человеческий PFF, соответственно.
Фиг.16. Количественная оценка патологического альфа-синуклеина в различных областях мозга мышей дикого типа C57Bl/6J (A: кора головного мозга; B: полосатое тело; C: миндалина и D: черная субстанция), которым инъецировали мышиный PFF.
Фиг.17. Фармакокинетические профили антител к альфа-синуклеину: А. антитело 6470 у мыши дикого типа; B. антитело 6470 и антитело сравнения у макак Cynomolgus.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение описано ниже со ссылкой на его конкретные неограничивающие аспекты и варианты осуществления и со ссылкой на некоторые чертежи и примеры.
Технические термины используются согласно их здравому смыслу, если не указано иное. Если конкретное значение передается определенными терминами, определения терминов даны в контексте, в котором они используются.
Когда термин «содержащий» используется в настоящем описании и формуле изобретения, он не исключает другие элементы. Для целей настоящего изобретения термин «состоящий из» следует рассматривать как предпочтительный вариант термина «содержащий».
При использовании единственного числа, также включено множественное число этого существительного, если не указано иное.
Используемые в настоящем описании термины «лечение», «лечащий» и т.п. относятся к получению нужного фармакологического и/или физиологического эффекта. Эффект может быть профилактическим в смысле частичного или полного предупреждения развития заболевания или его симптома и/или может быть терапевтическим в смысле частичного или полного излечения заболевания и/или неблагоприятного эффекта, связанного с этим заболеванием. Таким образом, лечение охватывает любое лечение заболевания у млекопитающего, особенно у человека, и включает: (а) профилактику возникновения заболевания у субъекта, который может быть предрасположен к заболеванию, но который еще не диагностирован как имеющий это заболевание; (b) ингибирование заболевания, т.е. купирование его развития; и (с) облегчение заболевания, т.е. вызов регрессии заболевания.
«Терапевтически эффективное количество» относится к количеству антитела к альфа-синуклеину или его антигенсвязывающему фрагменту, которое при введении млекопитающему или другому субъекту с целью лечения заболевания является достаточным для осуществления такого лечения заболевания. Терапевтически эффективное количество будет меняться в зависимости от антитела к альфа-синуклеину или его антигенсвязывающего фрагмента, заболевания и его тяжести, а также возраста, веса и т.д., субъекта, подлежащего лечению.
В контексте настоящего описания термин «выделенный» означает, что антитело, антигенсвязывающий фрагмент или полинуклеотид, в зависимости от обстоятельств, существует в физической среде, отличной от той, в которой оно может встречаться в природе. Настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которые связываются с альфа-синуклеином, где антитело содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:44;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:45, и
vi. CDR-H3, содержащую SEQ ID NO:46.
В SEQ ID NO:44 Хаа представляет собой аспарагин (Asn; N) или аргинин (Arg; R). Независимо от этого в SEQ ID NO:45 Xaa представляет собой серин (Ser; S) или аспарагин (Asn N), и в SEQ ID NO:46 Xaa представляет собой аспарагин (Asn N) или гистидин (His; H).
В одном из вариантов осуществления Xaa в SEQ ID NO:44 и 46 представляет собой аспарагин, и Xaa в SEQ ID NO:45 представляет собой серин.
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент, которые связываются с альфа-синуклеином, содержат:
а. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:1;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:5, и
vi. CDR-H3, содержащую SEQ ID NO:6.
Альфа-синуклеин (или альфа-син; а-синуклеин; а-син или любой другой известный синоним) относится к общему названию этого белка и включает, без ограничения, альтернативные варианты сплайсинга, мутанты и альфа-синуклеин других видов (мыши, обезьяны и т.д.). Если не указано иное, когда имеется в виду человеческий альфа-синуклеин или когда он указан в явном виде, такой альфа-синуклеин содержит последовательность, приведенную в SEQ ID NO:10 или в Uniprot P37840.
В контексте настоящего описания термин «антитело», как правило, относится к интактным (цельным) антителам, т.е. содержащим элементы двух тяжелых цепей и двух легких цепей. Антитело также может содержать дополнительные связывающие домены, например, как в молекуле DVD-Ig, описанной в WO 2007/024715, или в так называемом (FabFv)2Fc, описанном в WO 2011/030107. Таким образом, антитело в том виде, как используется в настоящем изобретении, включает двух-, трех- или четырехвалентные полноразмерные антитела.
Антигенсвязывающие фрагменты антител включают одноцепочечные антитела (т.е. полноразмерные тяжелую и легкую цепи); Fab, модифицированный Fab, Fab', модифицированный Fab', F(ab')2, Fv, Fab-Fv, Fab-dsFv, однодоменные антитела (например, VH или VL или VHH), scFv, би-, три- или тетравалентные антитела, Bis-scFv, диатела, тритела, триатела, тетратела и эпитоп-связывающие фрагменты любого из вышеперечисленного (см., например, Holliger and Hudson, 2005, Nature Biotech. 23(9):1126-1136; Adair and Lawson, 2005, Drug Design Reviews - Online 2(3), 209-217). Способы создания и изготовления этих фрагментов антител хорошо известны в данной области (см., например, Verma et al., 1998, Journal of Immunological Methods, 216, 165-181). Формат Fab-Fv был впервые описан в WO2009/040562, а его версия, стабилизированная дисульфидными связями, Fab-dsFv, впервые описана в WO2010/035012. Другие фрагменты антител, используемые в настоящем изобретении, включают фрагменты Fab и Fab', описанные в международных заявках на патент WO 2005/003169, WO 2005/003170 и WO 2005/003171. Поливалентные антитела могут иметь несколько специфичностей, например, могут быть биспецифическими, или могут быть моноспецифическими (см., например, WO 92/22583 и WO 05/113605). Одним из примеров последнего является три-Fab (или TFM), описанное в WO92/22583.
Альтернативный антигенсвязывающий фрагмент содержит Fab, связанный с двумя scFv или dsscFv, где каждый scFv или dsscFv связывается с одной и той же или разными мишенями (например, один scFv или dsscFv, связывающий терапевтическую мишень, и один scFv или dsscFv, который увеличивает период полураспада путем связывания, например, альбумина). Такие фрагменты антител описаны в публикации международной заявки на патент № WO 2015/197772, которая полностью включена в настоящее описание в виде ссылки и, в частности, упоминается в контексте обсуждения фрагментов антител.
Типичная молекула Fab' содержит пару тяжелой и легкой цепей, где тяжелая цепь содержит вариабельную область VH, константный домен CH1 и природную или модифицированную шарнирную область, а легкая цепь содержит вариабельную область VL и константный домен CL. Димеры Fab’ по настоящему изобретению образуют F(ab’)2, в котором димеризация может быть осуществлена, например, через шарнирную область.
Антитело или его антигенсвязывающий фрагмент по настоящему изобретению связывается с эпитопом альфа-синуклеина.
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент содержит:
а. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:44;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:45, и
vi. CDR-H3, содержащую SEQ ID NO:46
и связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
В SEQ ID NO:44 Хаа представляет собой аспарагин (Asn; N) или аргинин (Arg; R). Независимо от этого в SEQ ID NO:45 Xaa представляет собой серин (Ser; S) или аспарагин (Asn N), и в SEQ ID NO:46 Xaa представляет собой аспарагин (Asn N) или гистидин (His; H).
В одном из вариантов осуществления Xaa в SEQ ID NO:44 и 46 представляет собой аспарагин, и Xaa в SEQ ID NO:45 представляет собой серин.
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент, которое связывается с альфа-синуклеином, содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:1;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:5, и
vi. CDR-H3, содержащую SEQ ID NO:6
и связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
В контексте настоящего изобретения термин «эпитоп» используется взаимозаменяемо как для конформационных, так и для линейных эпитопов, где конформационный эпитоп состоит из прерывистых участков первичной аминокислотной последовательности антигена, а линейный эпитоп образован последовательностью непрерывно следующих друг за другом аминокислот.
Эпитоп может быть идентифицирован любым подходящим способом картирования эпитопов, известным в данной области, в комбинации с любым из антител, представленных в настоящем изобретении. Примеры таких способов включают скрининг пептидов различной длины, полученных из полноразмерного альфа-синуклеина, в отношении связывания с антителом или его фрагментом по настоящему изобретению и определение наименьшего фрагмента, который может специфически связываться с антителом, содержащим последовательность эпитопа, распознаваемого антителом. Пептиды альфа-синуклеина могут быть получены синтетически или путем протеолитического расщепления белка альфа-синуклеина. Пептиды, которые связываются с антителом, могут быть идентифицированы, например, с помощью масс-спектрометрического анализа. В другом примере для идентификации эпитопа, связанного с антителом по настоящему изобретению, могут быть использованы ЯМР-спектроскопия или рентгеновская кристаллография. Как правило, когда определение эпитопа выполняют методом рентгеновской кристаллографии, аминокислотные остатки антигена в пределах 4Å от CDR считаются аминокислотными остатками, входящими в состав эпитопа. После идентификации эпитоп может служить для получения фрагментов, которые связываются с антителом по настоящему изобретению, и, при необходимости, его можно использовать в качестве иммуногена для получения дополнительных антител, которые связываются с тем же эпитопом.
В одном из вариантов осуществления эпитоп антитела или его антигенсвязывающего фрагмента определяют с помощью рентгеновской кристаллографии с использованием пептида альфа-синуклеина, содержащего остатки 123-132 последовательности SEQ ID NO:10.
Предпочтительно антитело или его антигенсвязывающий фрагмент по настоящему изобретению предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина.
В этом конкретном контексте термин «предотвращать» (и его грамматические варианты) используется в настоящем описании взаимозаменяемо с термином «ингибировать» и указывает на действие, оказываемое антителом по настоящему изобретению на агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина. Это действие может быть профилактическим с точки зрения полного или частичного предотвращения агрегации; или полного или частичного уменьшения, т.е. блокирования, агрегации, которая уже началась после дальнейшего прогрессирования, или полного или частичного уменьшения вероятности возникновения дальнейшей агрегации; или полного или частичного купирования агрегации, которая уже произошла.
Не ограничиваясь какой-либо теорией, предполагается, что антитело или его антигенсвязывающий фрагмент по настоящему изобретению связывается с альфа-синуклеином:
i) в мономерной форме и предотвращает образование олигомеров и агрегатов альфа-синуклеина; и/или
ii) в олигомерной и фибриллярной форме и предотвращает распространение альфа-синуклеина от нейрона к нейрону и/или
iii) в олигомерной и/или фибриллярной форме и предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, предпочтительно эндогенную агрегацию альфа-синуклеина.
В контексте настоящего изобретения термин «фибриллы», «фибриллярная форма» или «в фибриллах», используемый в отношении альфа-синуклеина, предназначен для обозначения немономерных форм альфа-синуклеина, включая олигомеры альфа-синуклеина, которые могут представлять собой формы, распространяющиеся внутри и между структурами мозга.
Следовательно, в одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент связывается с альфа-синуклеином и содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:44;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:45, и
vi. CDR-H3, содержащую SEQ ID NO:46,
предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина. Предпочтительно, антитело или его антигенсвязывающий фрагмент связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
В SEQ ID NO:44 Хаа представляет собой аспарагин (Asn; N) или аргинин (Arg; R). Независимо от этого в SEQ ID NO:45 Xaa представляет собой серин (Ser; S) или аспарагин (Asn N), и в SEQ ID NO:46 Xaa представляет собой аспарагин (Asn N) или гистидин (His; H).
В одном из вариантов осуществления Xaa в SEQ ID NO:44 и 46 представляет собой аспарагин, и Xaa в SEQ ID NO:45 представляет собой серин.
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент, которое связывается с альфа-синуклеином, содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:1;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:5, и
vi. CDR-H3, содержащую SEQ ID NO:6,
и предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина. Предпочтительно, антитело или его антигенсвязывающий фрагмент связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению способно связывать альфа-синуклеин в виде мономера и фибрилл. В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент имеет более высокое сродство связывания альфа-синуклеина в фибриллах по сравнению с альфа-синуклеином в виде мономера. Это характеризуется константой диссоциации (KD), которая по меньшей мере в 10 раз является более высокой для мономерного альфа-синуклеина, чем для альфа-синуклеина в фибриллах.
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению имеет константу диссоциации (KD) для мономерного альфа-синуклеина менее 15 нМ. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению имеет константу диссоциации (KD) для альфа-синуклеина в фибриллах менее 10 нМ. В одном из предпочтительных вариантов осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению имеет константу диссоциации (KD) для альфа-синуклеина в фибриллах менее 300 пМ.
В контексте настоящего описания термин «KD» относится к константе диссоциации, которую получают из отношения Kd к Ka (то есть Kd/Ka), выраженную в виде молярной концентрации (M). Kd и Ka относятся к скорости диссоциации и скорости ассоциации, соответственно, конкретного взаимодействия антиген-антитело (или его антигенсвязывающего фрагмента). Значения KD для антител могут быть определены методами, хорошо известными в данной области. Один из способов определения KD антитела заключается в использовании поверхностного плазмонного резонанса, например, с помощью системы Biacore®, например, как описано в приведенных в настоящем описании примерах, с использованием выделенного природного или рекомбинантного альфа-синуклеина, его подходящего слитого белка/полипептида или его фибрилл. В одном из примеров сродство измеряют с использованием рекомбинантного человеческого альфа-синуклеина, как описано в приведенных в настоящем описании примерах. Для осуществления метода поверхностного плазмонного резонанса молекулы-мишени иммобилизуют на твердой фазе и подвергают воздействию лигандов в подвижной фазе, текущей вдоль проточной ячейки. Если происходит связывание лиганда с иммобилизованной мишенью, локальный показатель преломления света изменяется, что приводит к изменению угла SPR, которое можно отслеживать в реальном времени путем детектирования изменения интенсивности отраженного света. Для получения кажущейся константы отношения фаз ассоциации и диссоциации в реакции связывания можно проанализировать скорости изменения сигнала SPR. Отношение этих значений дает кажущуюся константу равновесия (сродство) (см., например, Wolff et al., Cancer Res. 53:2560-65 (1993)).
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению имеет более высокое сродство связывания (т.е. меньшую величину KD) для альфа-синуклеина в фибриллах по сравнению с альфа-синуклеином в виде мономера. Термин «сродство» относится к силе взаимодействия между антителом или его антигенсвязывающим фрагментом с альфа-синуклеином.
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению имеет IC50 блокирования агрегации альфа-синуклеина, индуцированной альфа-синуклеином в фибриллах, составляющую менее 10 нМ, предпочтительно антитело или его антигенсвязывающий фрагмент по настоящему изобретению имеет IC50 блокирования агрегации альфа-синуклеина, индуцированной альфа-синуклеином в фибриллах, составляющую менее 5 нМ. Примеры анализа агрегации на основе клеток раскрыты в примерах.
Используемый в настоящем описании термин IC50 относится к половине максимальной ингибирующей концентрации, которая является мерой эффективности вещества, такого как антитело, при ингибировании специфической биологической или биохимической функции, агрегации по настоящему изобретению, индуцированной альфа-синуклеином, предпочтительно альфа-синуклеином в фибриллах. IC50 представляет собой количественную меру, которая показывает, какое количество конкретного вещества необходимо для ингибирования наполовину заданного биологического процесса.
В одном из вариантов осуществления по результатам в in vitro анализов антитело или его антигенсвязывающий фрагмент по настоящему изобретению имеет IC50 блокирования агрегации альфа-синуклеина, индуцированной альфа-синуклеином в фибриллах, составляющую менее 10 нМ, предпочтительно антитело или его антигенсвязывающий фрагмент по настоящему изобретению имеет IC50 блокирования агрегации альфа-синуклеина, индуцированной альфа-синуклеином в фибриллах, составляющую менее 5 нМ.
Антитело или его антигенсвязывающий фрагмент по настоящему изобретению не связывается с бета-синуклеином и/или гамма-синуклеином и является специфическими к альфа-синуклеину.
Термин «специфический», используемый в настоящем описании, предназначен для обозначения антитела, которое распознает только тот антиген, по отношению к которому оно является специфическим, или для антитела, которое обладает значительно более высоким сродством связывания с антигеном, по отношению к которому оно является специфическим (например, альфа-синуклеином), по сравнению с уровнем связывания антигенов, по отношению к которым оно не является специфическим (гамма- и бета-синуклеин), например, обладает по меньшей мере в 5, 6, 7, 8, 9, 10 раз более высоким сродством связывания.
Антитела по настоящему изобретению могут быть получены любым подходящим способом, известным в данной области. Для получения антител, которые специфически распознают альфа-синуклеин, можно использовать слитые белки, включающие полипептид/белок альфа-синуклеин, клетки, (рекомбинантно или естественно) экспрессирующие полипептид. Полипептид может представлять собой «зрелый» полипептид или его биологически активный фрагмент или производное.
В одном из вариантов осуществления полипептид (т.е. антиген) представляет собой мономер человеческого альфа-синуклеина или его фрагмент, предпочтительно полученный, как описано в примерах ниже.
Полипептиды, используемые для иммунизации хозяина, могут быть получены хорошо известными в данной области способами из полученных методами генной инженерии клеток-хозяев, содержащих системы экспрессии, или они могут быть получены из природных биологических источников. В настоящей заявке термин «полипептиды» включает пептиды, полипептиды и белки. Они используются взаимозаменяемо, если не указано иное. В некоторых случаях полипептид альфа-синуклеина или его фрагмент может быть частью более крупного белка, такого как слитый белок, например, слитый с аффинной или аналогичной меткой.
Сконструированные антитела к полипептиду альфа-синуклеина могут быть получены в тех случаях, когда необходима иммунизация животного путем введения полипептидов животному, например, животному, предпочтительно не являющемуся человеком, согласно хорошо известным и рутинным протоколам, см., например, Handbook of Experimental Immunology, D. M. Weir (ed.), Vol 4, Blackwell Scientific Publishers, Oxford, England, 1986). Иммунизированы могут быть многие теплокровные животные, такие как кролики, мыши, крысы, овцы, коровы, верблюды или свиньи. Тем не менее, мыши, кролики, свиньи и крысы, как правило, являются наиболее подходящими.
Моноклональные антитела могут быть получены любым способом, известным в данной области, таким как метод гибридомы (Kohler & Milstein, 1975, Nature, 256:495-497), метод триомы, метод гибридомы человеческих В-клеток (Kozbor et al., 1983, Immunology Today, 4:72) и метод EBV-гибридомы (Cole et al., Monoclonal Antibodies and Cancer Therapy, pp77-96, Alan R Liss, Inc., 1985).
Антитела для применения по изобретению также могут быть получены с помощью методов генерации антител из одного лимфоцита путем клонирования и экспрессии кДНК вариабельной области иммуноглобулина, полученных из отдельных лимфоцитов, отобранных для продуцирования специфических антител, например, способами, описанными Babcook, J. et al., 1996, Proc., Natl. Акад. Sci. USA 93 (15):7843-7848l; WO92/02551; WO2004/051268 и WO2004/106377.
Скрининг антител можно выполнять с помощью анализов измерения связывания с альфа-синуклеином и/или анализов измерения ингибирования образования фибрилл альфа-синуклеина в присутствии антитела или его фрагмента.
Антитело или его антигенсвязывающий фрагмент по настоящему изобретению содержит определяющие комплементарность области (CDR), три из тяжелой цепи и три из легкой цепи. Как правило, CDR находятся в каркасной области и вместе образуют вариабельную область. По соглашению, CDR в вариабельной области тяжелой цепи антитела или его антигенсвязывающего фрагмента обозначают как CDR-H1, CDR-H2 и CDR-H3, а в вариабельных областях легкой цепи - CDR-L1, CDR-L2 и CDR-L3. Они пронумерованы последовательно в направлении от N-конца к C-концу каждой цепи.
CDR обычно нумеруют в соответствии с системой, разработанной Kabat et al. Эта система изложена в Kabat et al., 1987, в Sequences of Proteins of Immunological Interest, US Depar™ent of Health and Human Services, NIH, USA (далее «Kabat et al. (см. выше)»). В настоящем описании используется эта система нумерации, если не указано иное.
Обозначения остатков по Кабат не всегда напрямую соответствуют линейной нумерации аминокислотных остатков. Фактическая линейная аминокислотная последовательность может содержать меньше или дополнительные аминокислотные остатки относительно строгой нумерации по Кабат, что соответствует укорочению или вставке в структурный компонент (будь то каркасная область или область, определяющая комплементарность (CDR)) основной структуры вариабельного домена. Правильная нумерация остатков по Кабат может быть определена для заданного антитела путем выравнивания гомологичных остатков в последовательности антитела относительно «стандартной» последовательности, пронумерованной по Кабат.
CDR вариабельного домена тяжелой цепи расположены в остатках 31-35 (CDR-H1), остатках 50-65 (CDR-H2) и остатках 95-102 (CDR-H3) согласно системе нумерации Кабат. Однако, согласно нумерации по Чотиа (Chothia, C. and Lesk, AMJ Mol. Biol., 196, 901-917 (1987)), петля, эквивалентная CDR-H1, простирается от остатка 26 до остатка 32. Таким образом, если не указано иное, в контексте настоящего описания «CDR-H1» относится к остаткам 26-35, согласно комбинированной системы нумерации по Кабат с топологическим определением петли по Чотиа.
CDR вариабельного домена легкой цепи расположены в остатках 24-34 (CDR-L1), остатках 50-56 (CDR-L2) и остатках 89-97 (CDR-L3) согласно системе нумерации Кабат.
В одном из предпочтительных вариантов осуществления антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3, и вариабельную область тяжелой цепи, содержащую CDR-H1, содержащую SEQ ID NO:4, CDR-H2, содержащую SEQ ID NO:5, и CDR-H3, содержащую SEQ ID NO:6.
Альтернативно, антитело или антигенсвязывающий фрагмент содержит вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1; CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи, содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:8, и CDR-H3, содержащую SEQ ID NO:9.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:7; CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи, содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5, и CDR-H3, содержащую SEQ ID NO:6.
В еще одном варианте осуществления антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:7; CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи, содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:8, и CDR-H3, содержащую SEQ ID NO:9.
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению могут содержать каркасные области животного, в котором было продуцировано антитело. Например, если антитело было продуцировано в организме кролика, оно будет содержать определенные выше CDR, и каркасные области кроличьего антитела, например антитело, содержащее вариабельную область легкой цепи в соответствии с SEQ ID NO:11 (нуклеотидная последовательность которой показана в SEQ ID NO:12) и вариабельной области тяжелой цепи в соответствии с SEQ ID NO:13 (нуклеотидная последовательность которой показана в SEQ ID NO:14).
В одном из вариантов осуществления антитело может представлять собой химерное, гуманизированное или человеческое антитело или его фрагмент.
Химерные антитела обычно получают с помощью методов рекомбинантной ДНК. ДНК может быть модифицирована путем замены кодирующей последовательности человеческих L- и H-цепей на соответствующие нечеловеческие (например, мышиные) H и L-константные области (Morrison; PNAS 81, 6851 (1984)).
Человеческие антитела содержат вариабельные области тяжелой или легкой цепей или полноразмерные тяжелые или легкие цепи, которые являются «продуктом» или «получены из» определенной последовательности зародышевой линии, если вариабельные области или полноразмерные цепи антитела получены из системы, в которой используются гены человеческого иммуноглобулина зародышевой линии. Такие системы включают иммунизацию трансгенной мыши, несущей гены человеческого иммуноглобулина, представляющим интерес антигеном или скрининг библиотеки генов человеческого иммуноглобулина, отображенной на фагах, с использованием представляющего интерес антигена. Человеческое антитело или его фрагмент, который является «продуктом» или «получен из» последовательности человеческого иммуноглобулина зародышевой лини, можно идентифицировать путем сравнения аминокислотной последовательности человеческого антитела с аминокислотными последовательностями человеческих иммуноглобулинов зародышевой линии и выбора последовательности человеческого иммуноглобулина зародышевой линии, которая наиболее близка к последовательности (т.е. имеет наибольший % идентичности) человеческого антитела. Человеческое антитело, которое является «продуктом» или «получено из» определенной последовательности человеческого иммуноглобулина зародышевой линии, может отличаться по аминокислотным остаткам от последовательности зародышевой линии, например, из-за встречающихся в природе соматических мутаций или преднамеренного введения сайт-направленной мутации. Однако выбранное человеческое антитело обычно имеет аминокислотную последовательность, которая на по меньшей мере 90% идентична аминокислотной последовательности, кодируемой геном человеческого иммуноглобулина зародышевой линии, и которая содержит аминокислотные остатки, определяющие человеческое антитело как являющееся человеческим при сравнении с аминокислотой последовательностью иммуноглобулина зародышевой линии других видов (например, мышиной последовательности зародышевой линии). В некоторых случаях человеческое антитело может иметь аминокислотную последовательность, которая на по меньшей мере 60%, 70%, 80%, 90% или на по меньшей мере 95%, или даже на по меньшей мере 96%, 97%, 98% или 99% идентична аминокислотной последовательности, кодируемой геном иммуноглобулина зародышевой линии. Как правило, человеческое антитело, полученное из конкретной последовательности человеческой зародышевой линии, будет иметь не более 10 аминокислотных отличий от аминокислотной последовательности, кодируемой геном человеческого иммуноглобулина зародышевой линии. В некоторых случаях человеческое антитело может демонстрировать не более 5 или даже не более 4, 3, 2 или 1 аминокислотного отличия от аминокислотной последовательности, кодируемой геном иммуноглобулина зародышевой линии.
Человеческие антитела могут быть получены несколькими способами, известными специалистам в данной области. Человеческие антитела могут быть получены методом гибридомы с использованием линий клеток человеческой миеломы или гетеромиеломы мыши-человека (Kozbor, J Immunol; (1984) 133:3001; Brodeur, Monoclonal Isolated Antibody Production Techniques and Applications, pp51-63, Marcel Dekker Inc, 1987). Альтернативные методы включают использование фаговых библиотек или трансгенных мышей, в каждом из которых используются репертуары человеческой вариабельной области (Winter G; (1994) Annu Rev Immunol 12:433-455, Green LL, (1999) J Immunol Methods 231:11-23).
В одном из предпочтительных вариантов осуществления настоящего изобретения антитело или его антигенсвязывающий фрагмент по изобретению является гуманизированными.
Следовательно, антитело или его антигенсвязывающий фрагмент связывается с альфа-синуклеином и содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:44;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:45, и
vi. CDR-H3, содержащую SEQ ID NO:46,
где антитело или его антигенсвязывающий фрагмент является гуманизированным. Предпочтительно, гуманизированное антитело или его антигенсвязывающий фрагмент предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и, более предпочтительно, связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
В SEQ ID NO:44 Хаа представляет собой аспарагин (Asn; N) или аргинин (Arg; R). Независимо от этого в SEQ ID NO:45 Xaa представляет собой серин (Ser; S) или аспарагин (Asn N), и в SEQ ID NO:46 Xaa представляет собой аспарагин (Asn N) или гистидин (His; H).
В одном из вариантов осуществления гуманизированное антитело или его антигенсвязывающий фрагмент связывается с альфа-синуклеином и содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:44;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:45, и
vi. CDR-H3, содержащую SEQ ID NO:46,
и предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где в SEQ ID NO:44 Xaa представляет собой аспарагин (Asn; N), в SEQ ID NO:45 Xaa представляет собой серин (Ser; S), и в SEQ ID NO:46 Xaa представляет собой аспарагин (Asn N).
В предпочтительном варианте осуществления гуманизированное антитело или его антигенсвязывающий фрагмент связывается с альфа-синуклеином и содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:1;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:5, и
vi. CDR-H3, содержащую SEQ ID NO:6,
и предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131.
В контексте настоящего описания термин «гуманизированное» антитело или его антигенсвязывающий фрагмент относится к антителу или его антигенсвязывающему фрагменту, в котором тяжелая и/или легкая цепь содержит одну или более CDR (включая, при необходимости, одну или более модифицированных CDR) из донорского антитела (например, антитела, не относящегося к человеческому, такому как мышиное или кроличье моноклональное антитело), привитую на каркас вариабельной области тяжелой и/или легкой цепи акцепторного антитела (например, человеческого антитела). Для обзора, см. Vaughan et al., Nature Biotechnology, 16, 535-539, 1998. В одном из вариантов осуществления вместо переноса всей CDR в каркас человеческого антитела переносят только один или более определяющих специфичность остатков из любой из описанных выше в настоящей заявке CDR (см., например, Kashmiri et al., 2005, Methods, 36, 25-34). В одном из вариантов осуществления в каркасную область человеческого антитела переносят только определяющие специфичность остатки из одной или более CDR, описанных выше в настоящей заявке. В другом варианте осуществления в каркасную область человеческого антитела переносят только определяющие специфичность остатки из каждой из описанных выше в настоящей заявке CDR.
Для прививки CDR можно использовать любую подходящую последовательность акцепторного каркасного участка вариабельной области с учетом класса/типа донорского антитела, из которого получены CDR, включая каркасные области мыши, примата и человека.
Соответственно, гуманизированное антитело по настоящему изобретению имеет вариабельный домен, содержащий человеческие акцепторные каркасные области, а также одну или более CDR, предоставленные, в частности, в настоящем описании. Таким образом, в одном из вариантов осуществления представлено блокирующее гуманизированное антитело, которое связывается с альфа-синуклеином, предпочтительно человеческим альфа-синуклеином, в котором вариабельный домен содержит акцепторные человеческие каркасные области и донорские нечеловеческие CDR.
Примерами человеческих каркасных областей, которые можно использовать в настоящем изобретении, являются KOL, NEWM, REI, EU, TUR, TEI, LAY и POM (Kabat et al., см. выше). Например, KOL и NEWM можно использовать для тяжелой цепи, REI можно использовать для легкой цепи, а EU, LAY и POM можно использовать как для тяжелой цепи, так и для легкой цепи. Альтернативно, можно использовать последовательности человеческой зародышевой линии, доступные по адресу: http://www.imgt.org/.
В гуманизированном антителе или его антигенсвязывающем фрагменте по настоящему изобретению акцепторные тяжелая и легкая цепи необязательно должны быть получены из одного и того же антитела и, при необходимости, могут содержать композитные цепи, имеющие каркасные области, полученные из разных цепей.
Подходящую каркасную область для легкой цепи гуманизированного антитела или его антигенсвязывающего фрагмента по настоящему изобретению получают из человеческой зародышевой линии IGKV1-16 JK4, имеющей SEQ ID NO:39, нуклеотидная последовательность которой приведена в SEQ ID NO:40.
Подходящую каркасную область для тяжелой цепи гуманизированного антитела или его антигенсвязывающего фрагмента по настоящему изобретению получают из человеческой зародышевой линии IGHV3-23 JH4, имеющей последовательность, приведенную в SEQ ID NO:41, нуклеотидная последовательность которой приведена в SEQ ID NO:42.
Соответственно, в одном из вариантов осуществления предложено гуманизированное антитело или его антигенсвязывающий фрагмент, включающее:
- последовательность, приведенную в SEQ ID NO:1 или SEQ ID NO:7 для CDR-L1, последовательность, приведенную в SEQ ID NO:2 для CDR-L2, и последовательность, приведенную в SEQ ID NO:3 для CDR-L3, где каркасную область легкой цепи получают из человеческой зародышевой линии IGKV1-16 JK4; и
- последовательность, приведенную в SEQ ID NO:4 для CDR-H1, последовательность, приведенную в SEQ ID NO:5 или SEQ ID NO:8 для CDR-H2, и последовательность, приведенную в SEQ ID NO:6 или SEQ ID NO:9 для CDR-H3, где каркасную область тяжелой цепи получают из человеческой зародышевой линии IGHV3-23 JH4.
В гуманизированном антителе или его антигенсвязывающем фрагменте по настоящему изобретению каркасные области могут не иметь точно такие же последовательности, как последовательности акцепторного антитела. Например, необычные остатки могут быть заменены остатками, более часто встречающимися в этом классе или типе акцепторной цепи. Альтернативно, выбранные остатки в акцепторных каркасных областях могут быть изменены таким образом, чтобы они соответствовали остаткам, найденным в том же положении в донорском антителе (см. Reichmann et al., 1998, Nature, 332, 323-324). Такие изменения должны быть сведены к минимуму, необходимому для восстановления сродства донорского антитела. Протокол для выбора остатков в акцепторных каркасных областях, которые, возможно, придется изменить, приведен в WO91/09967.
Таким образом, в одном из вариантов осуществления 1, 2, 3, 4, 5, 6, 7 или 8 остатков в каркасе заменены альтернативным аминокислотным остатком.
Соответственно, в одном из вариантов осуществления представлено гуманизированное антитело или его антигенсвязывающий фрагмент, в котором по меньшей мере остатки в каждом из положений 48 и 72 вариабельного домена легкой цепи (со ссылкой на SEQ ID NO:15 или 19) являются донорскими остатками, см., например, последовательности SEQ ID NO:15, 17, 19 и 21. Предпочтительно, остаток 48 вариабельного домена легкой цепи представляет собой глутамин, и/или остаток 72 вариабельного домена легкой цепи представляет собой глутамин.
Более предпочтительно оба остатка 48 и 72 в гуманизированной вариабельной области легкой цепи гуманизированного антитела или его антигенсвязывающего фрагмента по настоящему изобретению представляют собой глутамин.
В другом варианте осуществления предоставлено гуманизированное антитело или его антигенсвязывающий фрагмент, в котором по меньшей мере остатки в каждом из положений 24, 47, 48, 49, 73 и 97 (со ссылкой на SEQ ID NO:31 или 35) или 24, 47, 48, 49, 78 и 97 вариабельного домена тяжелой цепи (со ссылкой на SEQ ID NO:23 и 27) являются донорскими остатками, см., например, последовательности SEQ ID NO:23, 25, 27, 29, 31, 33, 35 и 37.
Предпочтительно остаток 24 вариабельного домена тяжелой цепи представляет собой валин, и/или остаток 47 вариабельного домена тяжелой цепи представляет собой тирозин, и/или остаток 48 вариабельного домена тяжелой цепи представляет собой изолейцин, и/или остаток 49 вариабельного домена тяжелой цепи представляет собой глицин, и/или остаток 97 вариабельного домена тяжелой цепи представляет собой аргинин, и/или остаток 73 вариабельного домена тяжелой цепи представляет собой серин, и/или остаток 78 вариабельного домена тяжелой цепи представляет собой валин.
Предпочтительно, в гуманизированной вариабельной области тяжелой цепи по настоящему изобретению остаток 24 представляет собой валин, остаток 47 представляет собой тирозин, остаток 48 представляет собой изолейцин, остаток 49 представляет собой глицин, остаток 73 представляет собой серин, и остаток 97 представляет собой аргинин. Также предпочтительно, в гуманизированной вариабельной области тяжелой цепи гуманизированного антитела или его антигенсвязывающего фрагмента по настоящему изобретению остаток 24 представляет собой валин, остаток 47 представляет собой тирозин, остаток 48 представляет собой изолейцин, остаток 49 представляет собой глицин, остаток 78 представляет собой валин, и остаток 97 представляет собой аргинин.
В одном из предпочтительных вариантов осуществления настоящего изобретения антитело или его антигенсвязывающий фрагмент связываются с альфа-синуклеином и содержит вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:31.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент содержит:
- вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:23; или
- вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:27 или 35; или
- вариабельную область легкой цепи, содержащую SEQ ID NO:19, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:23 или 31; или
- вариабельную область легкой цепи, содержащую SEQ ID NO:19, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:27 или 35.
В одном из вариантов осуществления изобретение относится к антителу или его антигенсвязывающему фрагменту, содержащему последовательность, которая на 80% аналогична или идентична последовательности, раскрытой в настоящем описании, например, на 85%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98% или 99% аналогична или идентична части или всей релевантной последовательности, например последовательности вариабельного домена, последовательности CDR или последовательности вариабельного домена, исключая CDR. В одном из вариантов осуществления соответствующая последовательность представляет собой SEQ ID NO:15. В одном из вариантов осуществления релевантная последовательность представляет собой SEQ ID NO:23 или SEQ ID NO:31.
В одном из вариантов осуществления настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которое связывается с человеческим альфа-синуклеином, содержащему легкую цепь, где вариабельный домен легкой цепи содержит последовательность, имеющую по меньшей мере 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98% или 99% идентичности или сходства с последовательностью, приведенной в SEQ ID NO:15 или SEQ ID NO:19, и/или вариабельный домен тяжелой цепи содержит последовательность, имеющую по меньшей мере 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98% или 99% идентичности или сходства с последовательностью приведенной в SEQ ID NO:31, SEQ ID NO:23, SEQ ID NO:27 или SEQ ID NO:35.
В одном из вариантов осуществления настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которое связывается с человеческим альфа-синуклеином, где антитело или его антигенсвязывающий фрагмент имеет вариабельный домен легкой цепи, который является на по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% аналогичным или идентичным последовательности, приведенной в SEQ ID NO:15, где антитело или его антигенсвязывающий фрагмент имеет последовательность, приведенную в SEQ ID NO:1 или SEQ ID NO:7 для CDR-L1, последовательность приведенную в SEQ ID NO:2 для CDR-L2, и последовательность, приведенную в SEQ ID NO:3 для CDR-L3.
В одном из вариантов осуществления настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которое связывается с человеческим альфа-синуклеином, где антитело или его антигенсвязывающий фрагмент имеет вариабельный домен тяжелой цепи, который является на по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% аналогичным или идентичным последовательности, приведенной в SEQ ID NO:31, и где антитело или его антигенсвязывающий фрагмент имеет последовательность, приведенную в SEQ ID NO:4 для CDR-H1, последовательность, приведенную в SEQ ID NO:5 или SEQ ID NO:8 для CDR-H2, и последовательность, приведенную в SEQ ID NO:6 или SEQ ID NO:9 для CDR-H3.
В контексте настоящего описания «идентичность» указывает на то, что в выровненных последовательностях в любом конкретном положении аминокислотные остатки являются идентичными между этими последовательностями. В контексте настоящего описания «сходство» указывает на то, что в выровненных последовательностях в любом конкретном положении находятся аминокислотные остатки схожего типа между этими последовательностями. Например, лейцин может быть заменен изолейцином или валином. Другие аминокислоты, которые часто могут заменять друг друга, включают, без ограничения:
- фенилаланин, тирозин и триптофан (аминокислоты, имеющие ароматические боковые цепи);
- лизин, аргинин и гистидин (аминокислоты, имеющие основные (щелочные) боковые цепи);
- аспартат и глутамат (аминокислоты, имеющие кислотные боковые цепи);
- аспарагин и глютамин (аминокислоты, имеющие амидные боковые цепи); и
- цистеин и метионин (аминокислоты, имеющие серосодержащие боковые цепи).
Степени идентичности и сходства могут быть легко вычислены (Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing. Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part 1, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987, Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991, the BLAST™ software available from NCBI (Altschul, S.F. et al., 1990, J. Mol. Biol. 215:403-410; Gish, W. & States, D.J. 1993, Nature Genet. 3:266-272. Madden, T.L. et al., 1996, Meth. Enzymol. 266:131-141; Altschul, S.F. et al., 1997, Nucleic Acids Res. 25:3389-3402; Zhang, J. & Madden, T.L. 1997, Genome Res. 7:649-656).
В одном из вариантов осуществления антигенсвязывающий фрагмент по настоящему изобретению может представлять собой, без ограничения, Fab, модифицированный Fab, Fab', модифицированный Fab', F(ab')2, Fv, однодоменные антитела (например, VH или VL или VHH), scFv, dsscFv, би-, три- или тетравалентные антитела, Bis-scFv, диатела, триатела, тетратела и эпитоп-связывающие фрагменты любого из вышеперечисленного (см., например, Holliger and Hudson, 2005, Nature Biotech. 23(9):1126-1136; Adair and Lawson, 2005, Drug Design Reviews - Online 2(3), 209-217). Способы создания и производства этих фрагментов антител хорошо известны в данной области (см., например, Verma et al., 1998, Journal of Immunological Methods, 216, 165-181). Другие фрагменты антител, используемые в настоящем изобретении, включают фрагменты Fab и Fab', описанные в WO2005/003169, WO2005/003170 и WO2005/003171. Поливалентные антитела могут иметь несколько специфичностей, например, могут быть биспецифическими или моноспецифическими (см., например, WO 92/22853, WO05/113605, WO2009/040562 и WO2010/035012).
Альтернативный антигенсвязывающий фрагмент содержит Fab, связанный с двумя scFv или dsscFv, где каждый из scFv или dsscFv связывается с одной и той же или разными мишенями (например, один scFv или dsscFv, связывающий терапевтическую мишень, и один scFv или dsscFv, который увеличивает период полураспада путем связывания, например, альбумина). Такие фрагменты антител описаны в публикации международной заявки на патент № WO 2015/197772, которая полностью включена в настоящее описание в виде ссылки и, в частности, в контексте обсуждения фрагментов антител.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению является частью альфа-синуклеин-связывающего слитого белка, который содержит, например, слитые антигенсвязывающие фрагменты по настоящему изобретению, например, в виде фрагмента Fab или Fab', и одно или два однодоменных антитела (dAb), непосредственно или опосредованно связанных с ними, например, как описано в документах WO2009/040562, WO2010035012, WO2011/030107, WO2011/061492 и WO2011/086091, каждый из которых включен в настоящее описание в виде ссылки. В одном из вариантов осуществления слитый белок содержит двухдоменные антитела, например, в виде спаренных вариабельного тяжелого (VH) и вариабельного легкого (VL) доменов, необязательно связанных дисульфидной связью.
В одном из вариантов осуществления элемент Fab или Fab' слитого белка имеет специфичность, такую же или аналогичную специфичности однодоменного антитела или антител. В одном из вариантов осуществления Fab или Fab' имеет специфичность к однодоменному антителу или антителам, другими словами, слитый белок является поливалентным. В одном из вариантов осуществления поливалентный слитый белок по настоящему изобретению имеет участок связывания альбумина, например, пара VH/VL этого белка обеспечивает участок связывания альбумина.
Домены константной области молекулы антитела по настоящему изобретению, если они присутствуют, могут быть выбраны с учетом предполагаемой функции молекулы антитела и, в частности, эффекторных функций, которые могут потребоваться. Например, домены константной области могут быть доменами человеческого IgA, IgD, IgE, IgG или IgM. В частности, можно использовать домены константной области человеческого IgG, особенно изотипов IgG1 и IgG3, если молекула антитела предназначена для терапевтического применения, и требуются эффекторные функции антитела. Альтернативно, изотипы IgG2 и IgG4 могут использоваться, если молекула антитела предназначена для терапевтических целей, и эффекторные функции антитела не требуются. Понятно, что также могут использоваться варианты последовательностей этих доменов константной области. Например, могут использоваться молекулы IgG4, в которых серин в положении 241 заменен пролином, описанный, например, в Angal et al. (Angal et al., Molecular Immunology, 1993, 30(1), 105-108), называемый в настоящем изобретении IgG4P. В одном из вариантов осуществления антитело представляет собой полноразмерное антитело, предпочтительно выбранное из IgG1 и IgG4 или IgG4P.
Следовательно, настоящее изобретение относится к полноразмерному гуманизированному антителу, которое связывается с альфа-синуклеином и содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:44;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:45, и
vi. CDR-H3, содержащую SEQ ID NO:46,
где гуманизированное антитело предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и предпочтительно связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E13, где эпитоп необязательно содержит A124 и G132, где антитело представляет собой изоформу IgG4P.
В SEQ ID NO:44 Хаа представляет собой аспарагин (Asn; N) или аргинин (Arg; R). Независимо от этого в SEQ ID NO:45 Xaa представляет собой серин (Ser; S) или аспарагин (Asn N), и в SEQ ID NO:46 Xaa представляет собой аспарагин (Asn N) или гистидин (His; H).
В одном из предпочтительных вариантов осуществления полноразмерное гуманизированное антитело, которое связывается с альфа-синуклеином, содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:44;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:45, и
vi. CDR-H3, содержащую SEQ ID NO:46,
и предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и предпочтительно связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132, где антитело представляет собой изоформу IgG4P, в которой в SEQ ID NO:44 Xaa представляет собой аспарагин (Asn; N), в SEQ ID NO:45 Xaa представляет собой серин (Ser; S) и в SEQ ID NO:46, Хаа представляет собой аспарагин (Asn N).
В наиболее предпочтительном варианте осуществления полноразмерное гуманизированное антитело, которое связывается с альфа-синуклеином, содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:1;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:5, и
vi. CDR-H3, содержащую SEQ ID NO:6,
и предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и предпочтительно связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
Специалисту в данной области также будет понятно, что антитела могут подвергаться различным пост-трансляционным модификациям. Тип и степень этих модификаций часто зависит от линии клеток-хозяев, используемых для экспрессии антитела, а также от условий культивирования. Такие модификации могут включать варианты гликозилирования, окисления метионина, образования дикетопиперазина, изомеризации аспартата и дезамидирования аспарагина. Частой модификацией является потеря карбоксиконцевого основного (щелочного) остатка (такого как лизин или аргинин) под действия карбоксипептидаз (как описано в Harris, RJ. Journal of Chromatography 705:129-134, 1995). Соответственно, С-терминальный лизин тяжелой цепи антитела может отсутствовать.
В одном из вариантов осуществления в процессе пост-трансляционных модификаций от молекулы антитела отщепляется С-концевая аминокислота.
В одном из вариантов осуществления в процессе пост-трансляционных модификаций от молекулы антитела отщепляется N-концевая аминокислота.
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи, приведенную в SEQ ID NO:15, и вариабельную область тяжелой цепи, выбранную из SEQ ID NO:23 или SEQ ID NO:31. Например, антитело может представлять собой полноразмерное антитело IgG4, содержащее вариабельную область легкой цепи, приведенную в SEQ ID NO:15, и вариабельную область тяжелой цепи, выбранную из SEQ ID NO:23 или SEQ ID NO:31. В другом варианте осуществления антитело представляет собой полноразмерное антитело IgG4, содержащее легкую цепь, приведенную в SEQ ID NO:17, и тяжелую цепь, выбранную из SEQ ID NO:25 или SEQ ID NO:33. В еще одном варианте осуществления антигенсвязывающий фрагмент представляет собой Fab', содержащий вариабельную область легкой цепи, приведенную в SEQ ID NO:15, и вариабельную область тяжелой цепи, выбранную из SEQ ID NO:23 или SEQ ID NO:31.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи, приведенную в SEQ ID NO:15, и вариабельную область тяжелой цепи, выбранную из SEQ ID NO:27 или SEQ ID NO:35. Например, антитело представляет собой полноразмерное антитело IgG4, содержащее вариабельную область легкой цепи, приведенную в SEQ ID NO:15, и вариабельную область тяжелой цепи, выбранную из SEQ ID NO:27 или SEQ ID NO:35. В другом варианте осуществления антитело представляет собой полноразмерное антитело IgG4, содержащее легкую цепь, приведенную в SEQ ID NO:17, и тяжелую цепь, выбранную из SEQ ID NO:29 или SEQ ID NO:37. В еще одном варианте осуществления антигенсвязывающий фрагмент представляет собой Fab', содержащий вариабельную область легкой цепи, приведенную в SEQ ID NO:15, и вариабельную область тяжелой цепи, выбранную из SEQ ID NO:27 или SEQ ID NO:35.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи, приведенную в SEQ ID NO:19, и вариабельную область тяжелой цепи, выбранную из SEQ ID NO:27 или SEQ ID NO:35. Например, антитело представляет собой полноразмерное антитело IgG4, содержащее вариабельную область легкой цепи, приведенную в SEQ ID NO:19, и вариабельную область тяжелой цепи, выбранную из SEQ ID NO:27 или SEQ ID NO:35. В другом варианте осуществления антитело представляет собой полноразмерное антитело IgG4, содержащее легкую цепь, приведенную в SEQ ID NO:21, и тяжелую цепь, выбранную из SEQ ID NO:29 или SEQ ID NO:37. В еще одном варианте осуществления антигенсвязывающий фрагмент представляет собой Fab', содержащий вариабельную область легкой цепи, приведенную в SEQ ID NO:19, и вариабельную область тяжелой цепи, выбранную из SEQ ID NO:27 или SEQ ID NO:35.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи, приведенную в SEQ ID NO:19, и вариабельную область тяжелой цепи, выбранную из SEQ ID NO:23 или SEQ ID NO:31. Например, антитело представляет собой полноразмерное антитело IgG4, содержащее вариабельную область легкой цепи, приведенную в SEQ ID NO:19, и вариабельную область тяжелой цепи, выбранную из SEQ ID NO:23 или SEQ ID NO:31. В другом варианте осуществления антитело представляет собой полноразмерное антитело IgG4, содержащее легкую цепь, приведенную в SEQ ID NO:21, и тяжелую цепь, выбранную из SEQ ID NO:25 или SEQ ID NO:33. В еще одном варианте осуществления антигенсвязывающий фрагмент представляет собой Fab', содержащий вариабельную область легкой цепи, приведенную в SEQ ID NO:21, и вариабельную область тяжелой цепи, выбранную из SEQ ID NO:25 или SEQ ID NO:33.
В предпочтительном варианте осуществления антитело связывается с альфа-синуклеином и представляет собой полноразмерное антитело IgG4, содержащее вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:31. Более предпочтительно, антитело предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и еще более предпочтительно антитело связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
В другом предпочтительном варианте осуществления антитело связывается с альфа-синуклеином и представляет собой полноразмерное антитело IgG4, содержащее легкую цепь, содержащую SEQ ID NO:17, и тяжелую цепь, содержащую SEQ ID NO:33. Более предпочтительно, антитело предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и еще более предпочтительно, антитело связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
Кроме того, настоящее изобретение также относится к антителу или его антигенсвязывающему фрагменту, которое конкурирует за связывание альфа-синуклеина с антителом или его антигенсвязывающим фрагментом по настоящему изобретению.
Следовательно, настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которое конкурирует за связывание альфа-синуклеина с антителами или антигенсвязывающими фрагментами по настоящему изобретению путем перекрестного блокирования или становится блокированным антителом или его антигенсвязывающим фрагментом по изобретению; и, в частности, антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи, содержащую SEQ ID NO:23, SEQ ID NO:31, SEQ ID NO:27 или SEQ ID NO:35, и вариабельную область легкой цепи, содержащую SEQ ID NO:15 или SEQ ID NO:19.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент конкурирует за связывание альфа-синуклеина с тем же эпитопом, что и антитело или его антигенсвязывающий фрагмент по настоящему изобретению, и, в частности, конкурирует с антителом или его антигенсвязывающим фрагментом с вариабельной областью тяжелой цепи, содержащей SEQ ID NO:23, SEQ ID NO:31, SEQ ID NO:27 или SEQ ID NO:35, и вариабельной области легкой цепи, содержащей SEQ ID NO:15 или SEQ ID NO:19, и за связывание с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, по меньшей мере, остатки M127, P128, S129, E130 и E131, предпочтительно остатки E123, Y125, E126, M127, P128, S129, E130 и E131.
В одном из вариантов осуществления такое антитело или его антигенсвязывающий фрагмент конкурирует с антителами или их фрагментами по настоящему изобретению и имеет вариабельную область тяжелой цепи, имеющую по меньшей мере 80% идентичности или сходства с последовательностью согласно SEQ ID NO:23, SEQ ID NO:31, SEQ ID NO:27 или SEQ ID NO:35; и/или имеет вариабельную область легкой цепи, имеющую по меньшей мере 80% идентичности или сходства с последовательностью согласно SEQ ID NO:15 или SEQ ID NO:19.
Конкурирующие антитела могут быть идентифицированы любым подходящим способом, известным в данной области, например, с помощью конкурентных анализов ELISA или BIAcore, в которых связывание перекрестно-блокирующего антитела с человеческим альфа синуклеином предотвращает связывание антитела по настоящему изобретению или наоборот. В таких анализах конкуренции можно использовать выделенный природный или рекомбинантный альфа синуклеин или подходящий слитый белок/полипептид. В одном из примеров конкуренцию измеряют с помощью рекомбинантного человеческого альфа-синуклеина (SEQ ID NO:10). В одном из примеров рекомбинантный человеческий альфа-синуклеин, меченный на N-конце или С-конце (например, слияние 6xHis-метки с сайтом распознавания TEV), используется в соответствии с примерами, приведенными в настоящем описании. В другом примере конкуренцию измеряют с помощью рекомбинантных фибрилл человеческого альфа-синуклеина.
В одном из вариантов осуществления конкурирующие антитела являются полностью человеческими или гуманизированными. В одном из вариантов осуществления конкурирующие антитела имеют сродство к человеческому альфа-синуклеину, составляющее 100 мкМ или менее, предпочтительно 50 мкМ или менее.
Биологические молекулы, такие как антитела или фрагменты, содержат кислотные и/или основные (щелочные) функциональные группы, придавая молекуле суммарный положительный или отрицательный заряд. Количество общего «наблюдаемого» заряда будет зависеть от абсолютной аминокислотной последовательности объекта, локального окружения заряженных групп в трехмерной структуре и условий окружающей среды молекулы. Изоэлектрическая точка (pI) - это pH, при котором конкретная молекула или ее поверхность в доступном растворителе не несет суммарного электрического заряда. В одном из примеров антитело к альфа-синуклеину или его антигенсвязывающий фрагмент по настоящему изобретению могут быть сконструированы таким образом, чтобы они имели подходящую изоэлектрическую точку. Благодаря этому можно получить антитела и/или фрагменты с более устойчивыми свойствами, в частности, с подходящими профилями растворимости и/или стабильности и/или улучшенными характеристиками очистки.
Таким образом, в одном из аспектов изобретение относится к гуманизированному антителу или его антигенсвязывающему фрагменту, которое связывается с альфа-синуклеином и которое сконструировано таким образом, что имеет изоэлектрическую точку, отличающуюся от точки первоначально идентифицированного антитела. Антитело может, например, быть сконструировано путем замены аминокислотного остатка, такого как замена кислотного аминокислотного остатка одним или более основными (щелочными) аминокислотными остатками. Альтернативно, могут быть введены основные аминокислотные остатки, или могут быть удалены кислотные аминокислотные остатки. Альтернативно, если молекула имеет неприемлемо высокое значение pI, то, при необходимости, могут быть введены кислотные остатки для снижения pI. Важно отметить, что в случае манипуляций с pI необходимо соблюдать осторожность для сохранения требуемой активности антитела или фрагмента. Таким образом, в одном из вариантов осуществления сконструированное антитело или его антигенсвязывающий фрагмент имеет такую же или по существу такую же активность, что и «немодифицированное» антитело или его фрагмент.
Для прогнозирования изоэлектрической точки антитела или фрагмента можно использовать такие программы, как **ExPASY http://www.expasy.ch/tools/pi_tool.h™l и http://www.iut-arles.up.univ-mrs.fr/w3bb/d_abim/compo-p.h™l.
Понятно, что сродство антител по настоящему изобретению может быть изменено любым подходящим способом, известным в данной области. Следовательно, настоящее изобретение также относится к вариантам молекул антител по настоящему изобретению, которые обладают улучшенным сродством к альфа-синуклеину, в частности, человеческому альфа-синуклеину. Такие варианты могут быть получены с помощью нескольких протоколов созревания сродства, включая мутирование CDR (Yang et al., J. Mol. Biol., 254, 392-403, 1995), перестановку цепей (Marks et al., Bio/Technology, 10, 779-783, 1992), использование штаммов-мутаторов E.coli (Low et al., J. Mol. Biol., 250, 359-368, 1996), перестановку в ДНК (Patten et al., Curr. Opin. Biotechnol., 8, 724-733, 1997), фаговый дисплей (Thompson et al., J. Mol. Biol., 256, 77-88, 1996) и ПЦР с имитацией полового размножения (Crameri et al., Nature, 391, 288-291, 1998). Обсуждение этих методов созревания сродства можно найти у Vaughan et al. (см. выше).
В рамках настоящего изобретения созревание сродства выполняли согласно IOTA (WO2014198951).
При необходимости, антитело или его антигенсвязывающий фрагмент по настоящему изобретению может быть конъюгировано с одной или более эффекторными молекулами. Понятно, что эффекторная молекула может содержать одну эффекторную молекулу или две, или более таких молекул, связанных с образованием единого фрагмента, который может быть присоединен к антителу или его антигенсвязывающему фрагменту по настоящему изобретению. Если необходимо получить фрагмент антитела, связанный с эффекторной молекулой, его можно получить с помощью стандартных химических процедур или процедур с рекомбинантными ДНК, в которых фрагмент антитела связывают с эффекторной молекулой либо непосредственно, либо через связывающий агент. Способы конъюгирования таких эффекторных молекул с антителами хорошо известны в данной области (см. Hellstrom et al., Controlled Drug Delivery, 2nd Ed., Robinson et al., Eds., 1987, pp. 623-53; Thorpe et al., 1982, Immunol. Rev., 62:119-58 и Dubowchik et al., 1999, Pharmacology and Therapeutics, 83, 67-123). Конкретные химические процедуры включают, например, процедуры, описанные в WO 93/06231, WO 92/22583, WO 89/00195, WO 89/01476 и WO 03/031581. Альтернативно, когда эффекторная молекула представляет собой белок или полипептид, связывание может быть достигнуто с помощью процедур с рекомбинантными ДНК, например, описанными в WO 86/01533 и EP0392745.
В контексте настоящего описания термин эффекторная молекула включает, например, противоопухолевые агенты, лекарственные вещества, токсины, биологически активные белки, например ферменты, другие антитела или фрагменты антител, синтетические или встречающиеся в природе полимеры, нуклеиновые кислоты и их фрагменты, например, ДНК, РНК и их фрагменты, радионуклиды, в частности, радиоактивный йод, радиоизотопы, хелатные металлы, наночастицы и репортерные группы, такие как флуоресцентные соединения или соединения, которые могут быть обнаружены с помощью ЯМР или ESR-спектроскопии.
Примеры эффекторных молекул могут включать цитотоксины или цитотоксические агенты, включая любой агент, который является вредным (например, убивает) для клетки. Примеры включают комбрестатины, доластатины, эпотилоны, стауроспорин, майтансиноиды, спонгистатины, ризоксин, галихондрины, роридины, гемиастерлины, таксол, цитохалазин В, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дигидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин и их аналоги или гомологи.
Эффекторные молекулы также включают, без ограничения, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацила декарбазин), алкилирующие агенты (например, мехлорэтамин, тиоэпа хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромоманнитол, стрептозотоцин, митомицин С и цис-дихлор-диамин платины (II) (DDP), цисплатин), антрациклины (например, даунорубицин (ранее дауномицин)) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин, антрамицин (AMC), калихеамицины или дуокармицины) и антимитотические агенты (например, винкристин и винбластин).
Другие эффекторные молекулы могут включать хелатные радионуклиды, такие как 111In и 90Y, Lu177, висмут213, калифорний252, иридий192 и вольфрам188/рений188; или лекарственные вещества, такие как, без ограничения, алкилфосфохолины, ингибиторы топоизомеразы I, таксоиды и сурамин.
Другие эффекторные молекулы включают белки, пептиды и ферменты. Представляющие интерес ферменты включают, без ограничения, протеолитические ферменты, гидролазы, лиазы, изомеразы, трансферазы. Представляющие интерес белки, полипептиды и пептиды включают, без ограничения, иммуноглобулины, токсины, такие как абрин, рицин А, экзотоксин Pseudomonas или дифтерийный токсин, белок, такой как инсулин, фактор некроза опухолей, α-интерферон, β-интерферон, фактор роста нервов, тромбоцитарный фактор роста или тканевой активатор плазминогена, тромботический агент или антиангиогенный агент, например ангиостатин или эндостатин, или модификатор биологического ответа, такой как лимфокин, интерлейкин-1 (IL-1), интерлейкин-2 (IL-2), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), гранулоцитарный колониестимулирующий фактор (G-CSF), фактор роста нервов (NGF) или другой фактор роста и иммуноглобулины.
Другие эффекторные молекулы могут включать детектируемые вещества, полезные, например, для диагностики. Примеры детектируемых веществ включают различные ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы, биолюминесцентные материалы, радиоактивные нуклиды, позитрон-испускающие металлы (для использования в позитронно-эмиссионной томографии) и нерадиоактивные ионы парамагнитных металлов. См. патент США № 4741900 касательно ионов металлов, которые могут быть конъюгированы с антителами для использования в качестве диагностируемых агентов. Подходящие ферменты включают пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу; подходящие простетические группы включают стрептавидин, авидин и биотин; подходящие флуоресцентные материалы включают умбеллиферон, флуоресцеин, изотиоцианат флуоресцеина, родамин, дихлортриазиниламина флуоресцеин, дансилхлорид и фикоэритрин; подходящие люминесцентные материалы включают люминол; подходящие биолюминесцентные материалы включают люциферазу, люциферин и экворин; и подходящие радиоактивные нуклиды включают 125I, 131I, 111In и 99Tc.
В другом примере эффекторная молекула может увеличивать период полувыведения антитела in vivo и/или уменьшать иммуногенность антитела и/или улучшать доставку антитела через эпителиальный барьер к иммунной системе. Примеры подходящих эффекторных молекул этого типа включают полимеры, альбумин, альбумин-связывающие белки или альбумин-связывающие соединения, такие как соединения, описанные в WO 05/117984.
Когда эффекторная молекула представляет собой полимер, она, как правило, может представлять собой синтетический или встречающийся в природе полимер, например необязательно замещенный полиалкиленовый, полиалкениленовый или полиоксиалкиленовый полимер с прямой или разветвленной цепью или разветвленный или неразветвленный полисахарид, например, гомо- или гетерополисахарид.
Конкретные необязательные заместители, которые могут присутствовать в вышеупомянутых синтетических полимерах, включают одну или более гидроксильные, метильные или метокси-группы.
Конкретные примеры синтетических полимеров включают необязательно замещенные с прямой или разветвленной цепью поли(этиленгликоль), поли(пропиленгликоль), поли(виниловый спирт) или их производные, особенно необязательно замещенный поли(этиленгликоль), такой как метоксиполи(этиленгликоль), или его производные.
Конкретные встречающиеся в природе полимеры включают лактозу, амилозу, декстран, гликоген или их производные.
В одном из вариантов осуществления полимер представляет собой альбумин или его фрагмент, такой как человеческий сывороточный альбумин или его фрагмент.
В контексте настоящего описания термин «производные» включает реакционноспособные производные, например, тиол-селективные реакционноспособные группы, такие как малеимиды и т.п. Реакционноспособная группа может быть связана с полимером непосредственно или через линкерный сегмент. Понятно, что остаток такой группы в некоторых случаях образует часть продукта в виде связующей группы между фрагментом антитела и полимером.
Размер полимера при необходимости может меняться, но обычно находится в диапазоне средней молекулярной массы от 500 Да до 50000 Да, например от 5000 до 40000 Да, например от 20000 до 40000 Да. Размер полимера, в частности, может быть выбран с учетом предполагаемого использования продукта, например, способности локализоваться в определенных тканях, таких как опухоли, или способности увеличивать период полувыведения из кровотока (обзор см. в Chapman, 2002, Advanced Drug Delivery Reviews, 54, 531-545). Таким образом, например, когда продукт предназначен для выведения из кровообращения и проникновения в ткани, например, для использования при лечении опухоли, предпочтительным может оказаться использование полимера с небольшой молекулярной массой, например, с молекулярной массой примерно 5000 Да. Для применений, в которых продукт остается в кровотоке, предпочтительным может оказаться использование полимера с более высокой молекулярной массой, например, молекулярной массой в диапазоне от 20000 Да до 40000 Да.
Подходящие полимеры включают полиалкиленовый полимер, такой как поли(этиленгликоль) или, особенно, метоксиполи(этиленгликоль) или его производное, и особенно с молекулярной массой в диапазоне от примерно 15000 Да до примерно 40000 Да.
В одном из примеров антитело или его антигенсвязывающий фрагмент по настоящему изобретению присоединено к фрагментам поли(этиленгликоля) (ПЭГ). В одном конкретном варианте осуществления антигенсвязывающий фрагмент по настоящему изобретению и молекулы ПЭГ могут быть присоединены через любую доступную аминокислотную боковую цепь или концевую аминокислотную функциональную группу, расположенную на фрагменте антитела, например любую свободную амино, имино, тиоловую, гидроксильную или карбоксильную группу. Такие аминокислоты могут встречаться в природе во фрагменте антитела или могут быть встроены во фрагмент с помощью методов рекомбинантной ДНК (см., например, US 5219,996; US 5667425; WO98/25971, WO2008/038024). В одном из примеров молекула антитела по настоящему изобретению представляет собой модифицированный фрагмент Fab, где модификация представляет собой добавление к С-концу его тяжелой цепи одной или более аминокислот для обеспечения возможности присоединения эффекторной молекулы. Соответственно, дополнительные аминокислоты образуют модифицированную шарнирную область, содержащую один или более остатков цистеина, к которым может быть присоединена эффекторная молекула. Для присоединения двух или более молекул ПЭГ могут быть использованы несколько участков.
Соответственно, молекулы ПЭГ ковалентно связывают через тиоловую группу, по меньшей мере, одного остатка цистеина, расположенного во фрагменте антитела. Каждая молекула полимера, присоединенная к модифицированному фрагменту антитела, может быть ковалентно связана с атомом серы остатка цистеина, расположенного на этом фрагменте. Ковалентная связь обычно представляет собой дисульфидную связь или, в частности, сероуглеродную связь. Когда в качестве точки прикрепления используют тиоловую группу, могут быть использованы соответствующим образом активированные эффекторные молекулы, например, могут быть использованы производные, селективные по отношению к тиолу, такие как малеимиды и производные цистеина. Активированный полимер можно использовать в качестве исходного материала при получении фрагментов антител, модифицированных полимером, как описано выше. Активированный полимер может быть любым полимером, содержащим тиоловую реакционноспособную группу, такую как t-галогенкарбоновая кислота или сложный эфир, например, йодацетамид, имид, например малеимид, винилсульфон или дисульфид. Такие исходные материалы являются коммерчески доступными (например, от компании Nektar, ранее известной как Shearwater Polymers Inc., Huntsville, AL, USA) или могут быть получены из коммерчески доступных исходных материалов с помощью обычных химических процедур. Конкретные молекулы ПЭГ включают 20К метокси-ПЭГ-амин (поставляемый компанией Nektar, ранее известной как Shearwater; Rapp Polymere; и компанией SunBio) и M-ПЭГ-SPA (поставляемый компанией Nektar, ранее известной как Shearwater).
В одном из вариантов осуществления антитело представляет собой модифицированный фрагмент Fab, фрагмент Fab' или диFab, который ПЭГилирован, т.е. имеет ковалентно связанный с ним ПЭГ (поли(этиленгликоль)), например, в соответствии со способом, раскрытым в ЕР 0948544 или ЕР1090037 [см. также "Poly(ethyleneglycol) Chemistry, Biotechnical and Biomedical Applications", 1992, J. Milton Harris (ed), Plenum Press, New York, "Poly(ethyleneglycol) Chemistry and Biological Applications", 1997, J. Milton Harris and S. Zalipsky (eds), American Chemical Society, Washington DC and "Bioconjugation Protein Coupling Techniques for the Biomedical Sciences", 1998, M. Aslam and A. Dent, Grove Publishers, New York; Chapman, A. 2002, Advanced Drug Delivery Reviews 2002, 54:531-545]. В одном из примеров ПЭГ присоединен к цистеину в шарнирной области. В одном из примеров фрагмент Fab, модифицированный ПЭГ, имеет малеимидную группу, ковалентно связанную с одной тиоловой группой в модифицированной шарнирной области. Остаток лизина может быть ковалентно связан с малеимидной группой, и к каждой из аминогрупп на остатке лизина может быть присоединен полимер метоксиполи(этиленгликоля) с молекулярной массой примерно 20000 Да. Таким образом, общая молекулярная масса ПЭГ, присоединенного к Fab-фрагменту, может составлять примерно 40000 Да.
Конкретные молекулы ПЭГ включают 2-[3-(N-малеимидо)пропионамидо]этиламид N,N'-бис(метоксиполи(этиленгликоля) ММ 20000), модифицированный лизином, также известный как PEG2MAL40K (доступный от Nektar, ранее Shearwater).
Альтернативные источники ПЭГ-линкеров включают NOF, поставляющую GL2-400MA3 (где m в приведенной ниже структуре равно 5) и GL2-400MA (где m равно 2) и n равно приблизительно 450:
Иными словами, каждый ПЭГ имеет размер, равный примерно 20000 Да.
Таким образом, в одном из вариантов осуществления ПЭГ представляет собой 2,3-бис(метилполиоксиэтилен-окси)-1-{[3-(6-малеимидо-1-оксогексил)амино]пропилокси}гексан (двухразветвленный ПЭГ, -СН2) 3NHCO(CH2)5-MAL, ММ 40000, известный как SUNBRIGHT GL2-400MA3.
Другие альтернативные ПЭГ-эффекторные молекулы следующего типа:
можно приобрести у Dr Reddy, NOF и Jenkem.
В одном из вариантов осуществления Fab или Fab' по настоящему изобретению конъюгированы с молекулой ПЭГ.
В одном из вариантов осуществления предложено антитело, которое является ПЕГилированным (например, с помощью раскрытого в настоящем описании ПЭГ), присоединенным через аминокислотный остаток цистеина в или около аминокислоты 226 цепи, например аминокислоты 226 тяжелой цепи (последовательная нумерация), например аминокислоты 223 в SEQ ID NO:33.
В одном из вариантов осуществления настоящее изобретение относится к молекуле Fab’-ПЭГ, содержащей один или более полимеров ПЭГ, например 1 или 2 полимера, таких как 40 кДа полимер или полимеры.
Молекулы Fab’-ПЭГ по настоящему изобретению могут быть особенно предпочтительными, поскольку они имеют период полураспада, независимый от Fc-фрагмента. В одном из вариантов осуществления предложен Fab', конъюгированный с полимером, таким как молекула ПЭГ, молекула крахмала или молекула альбумина. В одном из вариантов осуществления предложен scFv, конъюгированный с полимером, таким как молекула ПЭГ, молекула крахмала или молекула альбумина. В одном из вариантов осуществления Fab или Fab' по настоящему изобретению конъюгированы с человеческим сывороточным альбумином. В одном из вариантов осуществления антитело или фрагмент конъюгированы с молекулой крахмала, например, для увеличения периода полувыведения. Способы конъюгирования крахмала с белком описаны в патенте США № 801739, включенном в настоящее описание в качестве ссылки.
Настоящее изобретение также относится к выделенному полинуклеотиду, кодирующему антитело или его антигенсвязывающий фрагмент по настоящему изобретению. Выделенный полинуклеотид по настоящему изобретению может содержать синтетическую ДНК, например, полученную путем химической обработки, с помощью кДНК, геномной ДНК или любой их комбинации.
Для получения последовательностей ДНК, кодирующих антитело или его антигенсвязывающий фрагмент по настоящему изобретению, могут быть использованы стандартные методы молекулярной биологии. Требуемые последовательности ДНК могут быть синтезированы полностью или частично с помощью методов синтеза олигонуклеотидов. При необходимости можно использовать методы сайт-направленного мутагенеза и полимеразной цепной реакции (ПЦР).
В одном из вариантов осуществления выделенный полинуклеотид по изобретению кодирует:
a. вариабельную область легкой цепи, в которой полинуклеотид:
i. является на по меньшей мере 90% идентичным SEQ ID NO:16 или SEQ ID NO:20; или
ii. содержит SEQ ID NO:16 или 20; или
iii. по существу состоит из SEQ ID NO:16 или SEQ ID NO:20;
b. вариабельную область тяжелой цепи, в которой полинуклеотид:
i. является на по меньшей мере 90% идентичным SEQ ID NO:24 или SEQ ID NO:28, или SEQ ID NO:32, или SEQ ID NO:36; или
ii. содержит SEQ ID NO:24 или SEQ ID NO:28, или SEQ ID NO:32, или SEQ ID NO:36; или
iii. по существу состоит из SEQ ID NO:24 или SEQ ID NO:28, или SEQ ID NO:32, или SEQ ID NO:36;
c. легкую цепь, в которой полинуклеотид:
i. является на по меньшей мере 90% идентичным SEQ ID NO:18 или SEQ ID NO:22; или
ii. содержит SEQ ID NO:18 или 22; или
iii. по существу состоит из SEQ ID NO:18 или SEQ ID NO:22;
d. тяжелую цепь, в которой полинуклеотид:
i. является на по меньшей мере 90% идентичным SEQ ID NO:26 или SEQ ID NO:30, или SEQ ID NO:34, или SEQ ID NO:38; или
ii. содержит SEQ ID NO:26 или SEQ ID NO:30, или SEQ ID NO:34, или SEQ ID NO:38; или
iii. по существу состоит из SEQ ID NO:26 или SEQ ID NO:30, или SEQ ID NO:34, или SEQ ID NO:38;
e. вариабельную область легкой цепи, в которой полинуклеотид:
i. является на по меньшей мере 90% идентичным SEQ ID NO:12; или
ii. содержит SEQ ID NO:12; или
iii. по существу состоит из SEQ ID NO:12;
f. вариабельную область тяжелой цепи, в которой полинуклеотид:
i. является на по меньшей мере 90% идентичным SEQ ID NO:14; или
ii. содержит SEQ ID NO:14; или
iii. по существу состоит из SEQ ID NO:14.
В одном из вариантов осуществления настоящее изобретение относится к выделенному полинуклеотиду, кодирующему тяжелую цепь Fab’-фрагмента антитела или антитела IgG1 или IgG4 по настоящему изобретению, которая содержит последовательность, приведенную в SEQ ID NO:24, 28, 32 или 36. Также предоставлен выделенный полинуклеотид, кодирующий легкую цепь Fab'-фрагмента антитела или IgG1 или IgG4 антитела по настоящему изобретению, которая содержит последовательность, приведенную в SEQ ID NO:16 или 20.
В другом варианте осуществления настоящее изобретение относится к выделенному полинуклеотиду, кодирующему тяжелую цепь и легкую цепь антитела IgG4(P) по настоящему изобретению, в котором полинуклеотид, кодирующий тяжелую цепь, содержит последовательность, приведенную в SEQ ID NO:26, 30, 34 или 38, и полинуклеотид, кодирующий легкую цепь, содержит последовательность, приведенную в SEQ ID NO:18 или 22.
Настоящее изобретение также относится к вектору клонирования или экспрессии, содержащему один или более полинуклеотидов, раскрытых в настоящем описании. В одном примере вектор клонирования или экспрессии по настоящему изобретению содержит один или более выделенных полинуклеотидов, содержащих последовательность, выбранную из SEQ ID NO:16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36 или 38.
Общие способы, с помощью которых могут быть сконструированы векторы, способы трансфекции и способы культивирования, хорошо известны специалистам в данной области. В этой связи приведена ссылка на «Current Protocols in Molecular Biology», 1999, F. M. Ausubel (ed), Wiley Interscience, New York and the Maniatis Manual produced by Cold Spring Harbor Publishing.
Также предоставлена клетка-хозяин, содержащая одну или более выделенных полинуклеотидных последовательностей по изобретению или один или более векторов клонирования или экспрессии, содержащих одну или более выделенных полинуклеотидных последовательностей, кодирующих антитело по настоящему изобретению. Для экспрессии полинуклеотидных последовательностей, кодирующих антитело или его антигенсвязывающий фрагмент по настоящему изобретению, может быть использована любая подходящая система клетка-хозяин/вектор. Могут быть использованы бактериальные, например E.coli, и другие микробные системы, или могут быть использованы эукариотические, например млекопитающих, системы экспрессии клеток-хозяев. Подходящие клетки-хозяева млекопитающих включают клетки СНО, миеломы или гибридомы.
Подходящие типы клеток яичников китайского хомяка (клетки СНО), используемые в настоящем изобретении, могут включать клетки СНО и СНО-К1, включая клетки dhfr-СНО, такие как клетки СНО-DG44 и клетки СНО-DXB11, которые также можно использовать с DHFR-селективным маркером, или клетки CHOK1-SV, которые можно использовать с селективным маркером по глютамин-синтетазе. Другие типы клеток, используемые для экспрессии антител, включают лимфоцитарные клеточные линии, например клетки миеломы NSO и клетки SP2, клетки COS. Клетка-хозяин может быть стабильно трансформирована или трансфицирована выделенными полинуклеотидными последовательностями или векторами экспрессии по настоящему изобретению.
В одном из вариантов осуществления клетка-хозяин по настоящему изобретению представляет собой клетку CHO-DG44, стабильно трансфицированную векторами экспрессии, содержащими выделенные полинуклеотидные последовательности по настоящему изобретению, предпочтительно содержащие выделенные полинуклеотидные последовательности SEQ ID NO:18 и 26 или SEQ ID NO:18 и 34 или SEQ ID NO:18 и 30 или SEQ ID NO:18 и 38.
Настоящее изобретение также относится к способу получения антитела или его антигенсвязывающего фрагмента по настоящему изобретению, включающему культивирование клетки-хозяина по настоящему изобретению в условиях, подходящих для получения антитела или его антигенсвязывающего фрагмента по изобретению, и выделение антитела или его антигенсвязывающего фрагмента.
Антитело или его антигенсвязывающий фрагмент могут содержать только полипептид тяжелой или легкой цепи, и в этом случае для трансфекции клеток-хозяев необходимо использовать только кодирующую последовательность полипептида тяжелой цепи или легкой цепи. Для получения антител или их антигенсвязывающих фрагментов, содержащих как тяжелые, так и легкие цепи, клеточную линию можно трансфицировать двумя векторами: первым вектором, кодирующим полипептид легкой цепи, и вторым вектором, кодирующим полипептид тяжелой цепи. Альтернативно, можно использовать один вектор, который включает последовательности, кодирующие полипептиды легкой цепи и тяжелой цепи.
Таким образом, предложен способ культивирования клетки-хозяина и экспрессии антитела или его фрагмента, выделения последнего и, необязательно, его очистки с получением выделенного антитела или фрагмента. В одном из вариантов осуществления способ дополнительно включает этап конъюгирования эффекторной молекулы с выделенным антителом или фрагментом, например конъюгирование с полимером ПЭГ, в частности, как раскрыто в настоящем описании.
Таким образом, в одном из вариантов осуществления предоставлено очищенное антитело к альфа-синуклеину или его фрагмент, например гуманизированное антитело или его фрагмент, в частности антитело или его фрагмент по изобретению, по существу очищенные, в частности свободные или по существу свободные от эндотоксина и/или белка или ДНК клетки-хозяина.
По существу свободный от эндотоксина, как правило, относится к содержанию эндотоксина 1 МЕ на мг продукта антитела или менее, например 0,5 или 0,1 МЕ на мг продукта.
По существу свободный от белка или ДНК клетки-хозяина относится к содержанию белка и/или ДНК клетки-хозяина, равному 400 мкг на мг продукта антитела или менее, например 100 мкг на мг или менее, в частности 20 мкг на мг, в зависимости от ситуации.
Поскольку антитела по настоящему изобретению применимы для лечения, диагностики и/или профилактики патологического состояния, такого как альфа-синуклеинопатия, настоящее изобретение также относится к фармацевтической или диагностической композиции, содержащей антитело или его антигенсвязывающий фрагмент по настоящему изобретению в сочетании с одним или более из: фармацевтически приемлемого носителя, наполнителя или разбавителя.
Предпочтительно фармацевтическая или диагностическая композиция содержит гуманизированное антитело, которое связывается с альфа-синуклеином и содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:44;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:45, и
vi. CDR-H3, содержащую SEQ ID NO:46.
Более предпочтительно, фармацевтическая или диагностическая композиция содержит гуманизированное антитело, которое связывается с альфа-синуклеином и содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:1;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:5, и
vi. CDR-H3, содержащую SEQ ID NO:6.
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению является единственным активным ингредиентом. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению находится в комбинации с одним или более дополнительными активными ингредиентами. Альтернативно, фармацевтические композиции содержат антитело или его антигенсвязывающий фрагмент по настоящему изобретению, которое является единственным активным ингредиентом и которое можно вводить пациенту в виде отдельного средства в комбинации (например, одновременно, последовательно или отдельно) с другими агентами, лекарственными средствами или гормонами.
В другом варианте осуществления фармацевтическая композиция содержит антитело или его антигенсвязывающий фрагмент, содержащее вариабельную область легкой цепи SEQ ID NO:15 или 19 и содержащее вариабельную область тяжелой цепи SEQ ID NO:23, 27, 31 или 35, например, SEQ ID NO:15 и SEQ ID NO:23 или SEQ ID NO:15 и SEQ ID NO:31.
Предпочтительно настоящее изобретение относится к фармацевтической композиции, содержащей антитело или его антигенсвязывающий фрагмент, которое связывается с альфа-синуклеином и содержит вариабельную область легкой цепи SEQ ID NO:15 и вариабельную область тяжелой цепи SEQ ID NO:31.
Фармацевтические композиции по изобретению могут быть соответственно введены пациенту для определения необходимого терапевтически эффективного количества. В контексте настоящего описания термин «терапевтически эффективное количество» относится к количеству терапевтического агента, необходимому для лечения, улучшения или профилактики целевого заболевания или состояния или для проявления детектируемого терапевтического или профилактического эффекта. Для любого антитела терапевтически эффективное количество может быть изначально оценено с помощью анализа либо на клеточных культурах, либо на животных моделях, как правило, грызунов, кроликов, собак, свиней или приматов. Животную модель также можно использовать для определения подходящего диапазона концентраций и пути введения. Такая информация затем может быть использована для определения полезных доз и путей введения человеку.
Точное терапевтически эффективное количество для субъекта-человека будет зависеть от тяжести болезненного состояния, общего состояния здоровья субъекта, возраста, веса и пола субъекта, диеты, времени и частоты введений, комбинации(й) лекарственных веществ, аллергической реакции и толерантности/реакции на терапию. Это количество может быть определено путем обычных экспериментов, и его определение находится в пределах компетенции врача. Обычно терапевтически эффективное количество составляет от 0,01 до 500 мг/кг, например от 0,1 до 200 мг/кг, например 100 мг/кг. Фармацевтические композиции могут быть представлены в удобных единичных дозированных формах, содержащих заранее определенное количество активного агента по изобретению на дозу.
Фармацевтически приемлемые носители в терапевтических композициях могут дополнительно содержать жидкости, такие как вода, физиологический раствор, глицерин и этанол. Кроме того, в таких композициях могут присутствовать вспомогательные вещества, такие как смачивающие или эмульгирующие агенты или рН-буферные вещества. Такие носители позволяют получать фармацевтические композиции в виде таблеток, пилюль, драже, капсул, жидкостей, гелей, сиропов, кашицы и суспензий для приема пациентом.
Подходящие формы для введения включают формы, подходящие для парентерального введения, например путем инъекции или инфузии, например болюсной инъекции или непрерывной инфузии, для внутривенного, ингаляционного или подкожного введения. Если продукт предназначен для инъекции или инфузии, он может принимать форму суспензии, раствора или эмульсии в масляном или водном носителе и может содержать агенты, применяемые для получения соответствующей лекарственной формы, такие как суспендирующие, консервирующие, стабилизирующие и/или диспергирующие агенты. Альтернативно, антитело или его антигенсвязывающий фрагмент по изобретению могут быть в сухой форме, которую восстанавливают перед применением с помощью соответствующей стерильной жидкости. Также могут быть получены твердые формы, подходящие для растворения или суспендирования в жидких носителях перед инъекцией.
После получения лекарственной формы композиции по изобретению можно вводить непосредственно субъекту. Соответственно, настоящее изобретение относится к применению антитела или его антигенсвязывающего фрагмента по изобретению для изготовления лекарственного средства.
Подлежащие лечению субъекты могут быть животными. Предпочтительно, фармацевтические композиции по настоящему изобретению адаптированы для введения людям.
Следовательно, в другом аспекте настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту или фармацевтической композиции, содержащей антитело или его антигенсвязывающий фрагмент, для применения в терапии, где антитело или его антигенсвязывающий фрагмент связывается с альфа-синуклеином и содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:44;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:45, и
vi. CDR-H3, содержащую SEQ ID NO:46.
Предпочтительно, антитело или его антигенсвязывающий фрагмент является гуманизированным и предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и, более предпочтительно, связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
В предпочтительном варианте осуществления антитело или его антигенсвязывающий фрагмент или фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент, для применения в терапии, представляет собой антитело или его антигенсвязывающий фрагмент, которое связывается с альфа-синуклеином и содержит:
i. CDR-L1, содержащую SEQ ID NO:1;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:5, и
vi. CDR-H3, содержащую SEQ ID NO:6.
Предпочтительно, антитело или его антигенсвязывающий фрагмент является гуманизированным и предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и, более предпочтительно, связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
В частности, применение в терапии включает применение при лечении одной или более альфа-синуклеинопатий.
В еще одном аспекте настоящее изобретение относится к способу лечения одной или более синуклеинопатий у пациента, включающему введение указанному пациенту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по настоящему изобретению или фармацевтической композиции, содержащей антитело или его антигенсвязывающий фрагмент, где антитело или его антигенсвязывающий фрагмент связывается с альфа-синуклеином и содержит:
i. CDR-L1, содержащую SEQ ID NO:44;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
iv. CDR-H1, содержащую SEQ ID NO:4;
v. CDR-H2, содержащую SEQ ID NO:45, и
vi. CDR-H3, содержащую SEQ ID NO:46.
Предпочтительно, антитело или его антигенсвязывающий фрагмент является гуманизированным и предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и, более предпочтительно, связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
В одном из предпочтительных вариантов осуществления антитело или его антигенсвязывающий фрагмент или фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент, предназначены для применения при лечения одной или более альфа-синуклеинопатий, где антитело или его антигенсвязывающий фрагмент связывается с альфа синуклеином и соднржит:
а. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5, и CDR-H3, содержащую SEQ ID NO:6; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:31; или
c. легкую цепь, содержащую SEQ ID NO:17, и тяжелую цепь, содержащую SEQ ID NO:33.
Предпочтительно, это антитело или его антигенсвязывающий фрагмент является гуманизированным и предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и, более предпочтительно, связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
В другом предпочтительном варианте осуществления настоящее изобретение относится к способу лечения одной или более альфа-синуклеинопатий у пациента, включающему введение указанному пациенту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по настоящему изобретению или фармацевтической композиции, содержащей антитело или его антигенсвязывающий фрагмент, где антитело или его антигенсвязывающий фрагмент связывается с альфа-синуклеином и содержит:
а. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5, и CDR-H3, содержащую SEQ ID NO:6; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:31; или
c. легкую цепь, содержащую SEQ ID NO:17, и тяжелую цепь, содержащую SEQ ID NO:33.
Предпочтительно, антитело или его антигенсвязывающий фрагмент является гуманизированным и предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина, и, более предпочтительно, связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит A124 и G132.
Альтернативно, антитело или его антигенсвязывающий фрагмент или фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент, предназначено для применения в терапии или для лечения одной или более альфа-синуклеинопатий и представляет собой антитело или его антигенсвязывающий фрагмент содержащее:
а. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1 или SEQ ID NO:7; CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи, содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5 или SEQ ID NO:8, и CDR-H3, содержащую SEQ ID NO:6 или SEQ ID NO:9; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15 или 19, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:23 или SEQ ID NO:27 или SEQ ID NO:31 или SEQ ID NO:35; или
c. легкую цепь, содержащую SEQ ID NO:17 или SEQ ID NO:21, и тяжелую цепь, содержащую SEQ ID NO:25 или SEQ ID NO:29, или SEQ ID NO:33, или SEQ ID NO:37.
В другом варианте осуществления настоящего изобретения способ лечения одной или более альфа-синуклеинопатий у пациента включает введение указанному пациенту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по настоящему изобретению или фармацевтической композиции, содержащей антитело или его антигенсвязывающий фрагмент, где антитело или его антигенсвязывающий фрагмент связывается с альфа-синуклеином и содержит:
a. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1 или SEQ ID NO:7; CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи, содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5 или SEQ ID NO:8, и CDR-H3, содержащую SEQ ID NO:6 или SEQ ID NO:9; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15 или 19, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:23 или SEQ ID NO:27 или SEQ ID NO:31 или SEQ ID NO:35; или
c. легкую цепь, содержащую SEQ ID NO:17 или SEQ ID NO:21, и тяжелую цепь, содержащую SEQ ID NO:25 или SEQ ID NO:29, или SEQ ID NO:33, или SEQ ID NO:37.
Альфа-синуклеинопатии в соответствии с настоящим изобретением включают, без ограничения, болезнь Паркинсона (PD) (включая идиопатические и наследственные формы болезни Паркинсона), деменцию с тельцами Леви (DLB), болезнь диффузных телец Леви (DLBD), вариант болезни Альцгеймера с тельцами Леви (LBVAD), комбинированную болезнь Альцгеймера и Паркинсона, множественную системную атрофию (MSA) и нейродегенерацию с накоплением железа в мозге 1-го типа (NBIA-1). Предпочтительно альфа-синуклеинопатия представляет собой болезнь Паркинсона (PD).
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент или фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент, предназначены для применения при лечении болезни Паркинсона (PD) (включая идиопатические и наследственные формы болезни Паркинсона), деменции с тельцами Леви (DLB), болезни диффузных телец Леви (DLBD), варианта болезни Альцгеймера с тельцами Леви (LBVAD), комбинированной болезни Альцгеймера и Паркинсона, множественной системной атрофии (MSA) и нейродегенерации с накоплением железа в мозге 1-го типа (NBIA-1), предпочтительно болезни Паркинсона (PD), и представляет собой антитело или его антигенсвязывающий фрагмент, содержащее:
а. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5, и CDR-H3, содержащую SEQ ID NO:6; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:31; или
c. легкую цепь, содержащую SEQ ID NO:17, и тяжелую цепь, содержащую SEQ ID NO:33.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент или фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент, предназначены для применения при лечении болезни Паркинсона (PD) (включая идиопатические и наследственные формы болезни Паркинсона), деменции с тельцами Леви (DLB), болезни диффузных телец Леви (DLBD), варианта болезни Альцгеймера с тельцами Леви (LBVAD), комбинированной болезни Альцгеймера и Паркинсона, множественной системной атрофии (MSA) и нейродегенерации с накоплением железа в мозге 1-го типа (NBIA-1), предпочтительно болезни Паркинсона (PD), и представляет собой антитело или его антигенсвязывающий фрагмент, содержащее:
а. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1 или SEQ ID NO:7; CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи, содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5 или SEQ ID NO:8, и CDR-H3, содержащую SEQ ID NO:6 или SEQ ID NO:9; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15 или 19, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:23 или SEQ ID NO:27 или SEQ ID NO:31 или SEQ ID NO:35; или
c. легкую цепь, содержащую SEQ ID NO:17 или SEQ ID NO:21, и тяжелую цепь, содержащую SEQ ID NO:25 или SEQ ID NO:29, или SEQ ID NO:33 или SEQ ID NO:37.
В другом варианте осуществления предложен способ лечения болезни Паркинсона (PD) (включая идиопатические и наследственные формы болезни Паркинсона), деменции с тельцами Леви (DLB), болезни диффузных телец Леви (DLBD), варианта болезни Альцгеймера с тельцами Леви (LBVAD), комбинированной болезни Альцгеймера и Паркинсона, множественной системной атрофии (MSA) и нейродегенерации с накоплением железа в мозге 1-го типа (NBIA-1), предпочтительно болезни Паркинсона (PD), у пациента, включающий введение указанному пациенту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента или фармацевтической композиции, содержащей антитело или его антигенсвязывающий фрагмент, где антитело или его антигенсвязывающий фрагмент содержит:
a. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5, и CDR-H3, содержащую SEQ ID NO:6; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:31; или
c. легкую цепь, содержащую SEQ ID NO:17, и тяжелую цепь, содержащую SEQ ID NO:33.
В другом варианте осуществления, способ лечения болезни Паркинсона (PD) (включая идиопатические и наследственные формы болезни Паркинсона), деменции с тельцами Леви (DLB), болезни диффузных телец Леви (DLBD), варианта болезни Альцгеймера с тельцами Леви (LBVAD), комбинированной болезни Альцгеймера и Паркинсона, множественной системной атрофии (MSA) и нейродегенерации с накоплением железа в мозге 1-го типа (NBIA-1), предпочтительно болезни Паркинсона (PD), у пациента, включает введение указанному пациенту терапевтически эффективного осуществления антитела или его антигенсвязывающего фрагмента или фармацевтической композиции, содержащей антитело или его антигенсвязывающий фрагмент, где антитело или его антигенсвязывающий фрагмент содержит:
a. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1 или SEQ ID NO:7; CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи, содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5 или SEQ ID NO:8, и CDR-H3, содержащую SEQ ID NO:6 или SEQ ID NO:9; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15 или 19, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:23 или SEQ ID NO:27 или SEQ ID NO:31 или SEQ ID NO:35; или
c. легкую цепь, содержащую SEQ ID NO:17 или SEQ ID NO:21, и тяжелую цепь, содержащую SEQ ID NO:25 или SEQ ID NO:29, или SEQ ID NO:33 или SEQ ID NO:37.
Альтернативно, изобретение также относится к применению антитела или его антигенсвязывающего фрагмента для производства лекарственного средства для лечения альфа-синуклеинопатии, где альфа-синуклеинопатия предпочтительно представляет собой болезнь Паркинсона (PD) (включая идиопатические и наследственные формы болезни Паркинсона), деменцию с тельцами Леви (DLB), болезнь диффузных телец Леви (DLBD), вариант болезни Альцгеймера с тельцами Леви (LBVAD), комбинированную болезнь Альцгеймера и Паркинсона, множественную системную атрофию (MSA) и нейродегенерацию с накоплением железа в мозге 1-го типа (NBIA-1), более предпочтительно болезнь Паркинсона (PD), где антитело или его антигенсвязывающий фрагмент содержит:
a. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5, и CDR-H3, содержащую SEQ ID NO:6; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:31; или
c. легкую цепь, содержащую SEQ ID NO:17, и тяжелую цепь, содержащую SEQ ID NO:33.
Частью настоящего изобретения также является применение антител к альфа-синуклеину или антигенсвязывающих фрагментов для применения в качестве диагностически активных агентов или в диагностических анализах, например, для диагностики альфа-синуклеинопатий, таких как болезнь Паркинсона (PD) (включая идиопатические и наследственные формы болезни Паркинсона), деменцию с тельцами Леви (DLB), болезнь диффузных телец Леви (DLBD), вариант болезни Альцгеймера с тельцами Леви (LBVAD), комбинированную болезнь Альцгеймера и Паркинсона, множественную системную атрофию (MSA) и нейродегенерацию с накоплением железа в мозге 1-го типа (NBIA-1).
Диагноз предпочтительно может быть выполнен на биологических образцах. «Биологический образец» охватывает множество типов образцов, получаемых от индивидуума, и может использоваться в диагностическом анализе или для мониторинга. Определение охватывает спинномозговую жидкость, кровь, такую как плазма и сыворотка, и другие жидкие образцы биологического происхождения, такие как моча и слюна, образцы твердой ткани, такие как образец биопсии или культуры ткани или клетки, полученные из нее, и их потомство. Определение также включает образцы, которыми манипулировали любым способом после их получения, такими как обработка реагентами, солюбилизация или обогащение в отношении определенных компонентов, таких как полинуклеотиды.
Диагностическое тестирование предпочтительно может выполняться на биологических образцах, которые не находятся в контакте с организмом человека или животного. Такое диагностическое тестирование также называется тестированием in vitro. Диагностическое тестирование in vitro может быть основано на методе in vitro обнаружения альфа-синуклеина в биологическом образце, который был получен от индивидуума, включающем этапы i) контактирования биологического образца с раскрытым в настоящем описании антителом к альфа-синуклеину или его антигенсвязывающим фрагментом; и ii) детектирование связывания антитела к альфа-синуклеину или его антигенсвязывающего фрагмента с альфа-синуклеином. Путем сравнения детектированного уровня альфа-синуклеина или наличия конкретной пост-трансляционно модифицированной формы альфа-синуклеина с подходящим контролем можно идентифицировать одну или более альфа-синуклеинопатий. Таким образом, такой способ детектирования можно использовать для определения, страдает ли субъект или имеет риск развития альфа-синуклеинопатии, включая определение стадии (тяжести) альфа-синуклеинопатии.
Следовательно, настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту для применения для диагностики альфа-синуклеинопатий, предпочтительно для диагностики или при наличии болезни Паркинсона, где антитело или его антигенсвязывающий фрагмент связывается с альфа-синуклеином и содержит:
a. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:44, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи, содержащую CDR-H1, содержащую SEQ ID NO:4, CDR-H2, содержащую SEQ ID NO:45 и CDR-H3, содержащую SEQ ID NO:46.
Предпочтительно антитело или его антигенсвязывающий фрагмент для применения для диагностики альфа-синуклеинопатий, предпочтительно для диагностики или при наличии болезни Паркинсона, связывается с альфа-синуклеином и содержит:
a. вариабельную область легкой цепи, содержащую CDR-L1, содержащую SEQ ID NO:1, CDR-L2, содержащую SEQ ID NO:2, и CDR-L3, содержащую SEQ ID NO:3; и вариабельную область тяжелой цепи содержащую CDR-H1, содержащую SEQ ID NO:4; CDR-H2, содержащую SEQ ID NO:5, и CDR-H3, содержащую SEQ ID NO:6; или
b. вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:31; или
c. легкую цепь, содержащую SEQ ID NO:17, и тяжелую цепь, содержащую SEQ ID NO:33.
Последовательности, включенные в настоящее изобретение, показаны в таблице 1:
P37840
VL
Н.п. VL
VL N33R
Н.п. N33R VL
Легкая цепь N33R
Н.п. легкой цепи N33R
VH
Н.п. VH
Тяжелая цепь
Н.п. тяжелой цепи
VH S56N N102H
Н.п. VH S56N N102H
Тяжелая цепь S56N N102H
Н.п. тяжелой цепи S56N N102H
VH
Н.п. VH
Тяжелая цепь
Н.п. тяжелой цепи
VH S56N N102H
Н.п. VH S56N N102H
Тяжелая цепь S56N N102H
Н.п. тяжелой цепи S56N N102H
X33
(X = N or R)
X56
(X = S or N)
X102
(X = N or H)
Ниже изобретение дополнительно описано с помощью примеров со ссылками на варианты осуществления, показанные на прилагаемых чертежах.
ПРИМЕРЫ
Пример 1: Экспрессия мономера и фибрилл человеческого альфа-синуклеина
Ген, кодирующий человеческий альфа-синуклеин, синтезировали и субклонировали в вектор pMH 10His TEV (содержащий промотор CMV) стандартными методами молекулярной биологии для создания вектора, сконструированного для получения альфа-синуклеина с N-концевой меткой 10His-TEV. Полученный вектор трансфицировали в клетки Expi293F с помощью системы экспрессии Expi293™ (Invitrogen) в соответствии с протоколами производителя. Белок альфа-синуклеин накапливали в культуральной среде, откуда затем извлекали с помощью аффинной хроматографии с иммобилизованным ионом металла на колонке HisTrap excel (GE Healthcare). Колонку промывали 25 мМ Трис-HCl, 300 мМ NaCl, pH 8,0, и белок элюировали ступенчатым градиентом 500 мМ имидазола в том же буфере. Метку 10His удаляли с помощью протеазы TEV. Затем образец концентрировали и обессоливали перед повторным введением расщепленного белка в колонку HisTrap excel и сбором расщепленного альфа-синуклеина в потоке. Альфа-синуклеин дополнительно очищали гель-фильтрацией на колонке HiLoad 26/600 Superdex 75 (GE Healthcare), и эндотоксин удаляли, пропуская через картридж Proteus NoEndo (Generon). SEC MALS подтвердил, что очищенный альфа-синуклеин является мономерным (фиг. 1А).
Дикий тип (немеченный) человеческого альфа-синуклеина также экспрессировали в клетках Expi293F. Белок извлекали из культуральной среды с помощью анионного обмена, используя колонку HiTrap Q (GE Healthcare). Колонку промывали 20 мМ Трис-HCl, рН 8,0, и белок элюировали градиентом хлорида натрия до 400 мМ. Фракции концентрировали и обессоливали, пропуская через колонку HiPrep 26/10 (GE Healthcare) и элюируя 20 мМ трис-HCl, рН 8,0. Затем белок очищали, используя колонку MonoQ 10/100GL, элюировали градиентом хлорида натрия до 400 мМ в 20 мМ Tris HCl, рН 8,0, с последующей гель-фильтрацией на колонке HiLoad 26/600 Superdex 75 (GE Healthcare) с элюцией в PBS pH 7,4 (рис. 1Б).
Этот мономер (немеченого) альфа-синуклеина дикого типа использовали для приготовления фибрилл альфа-синуклеина, полученных путем перемешивания очищенного рекомбинантного мономера альфа-синуклеина (9-10 мг/мл в PBS pH 7,4) со скоростью 1200 об/мин при 37°C при встряхивании инкубатора Vortemp56 (Labnet) непрерывно в течение 10 дней. Образование фибрилл оценивали с помощью анализа JC-1 (Lee et al., Biochem. J. 2009, 418, 311-323) и инфракрасной спектроскопии раствора с Фурье-преобразованием. Невключенный мономер в растворах фибрилл оценивали ультрацентрифугированием и пропусканием через мембрану с массой отсекаемых частиц 100 кДа с последующим гель-электрофорезом. В дальнейших исследованиях использовали только фибриллы с JC-1 ответом >15, низким количеством растворимого мономера (<5%) и FTIR-спектром с основным поглощением между 1625 и 1630 см-1 (фиг. 2). Приготовленные фибриллы хранили при -80°С.
Пример 2: Иммунизация и выделение антител
Выполняли несколько стратегий иммунизации с использованием различных веществ и иммуногенов. Антитело 6470 получали от самки новозеландского белого кролика (>2 кг), которую подкожно иммунизировали кроличьим Fc-слитым белком, содержащим остатки человеческого альфа-синуклеина 68-140, слитые с кроличьим Fc (SEQ ID NO:43).
Слитый белок, состоящий из альфа-синуклеина (68-140) и кроличьего Fc, экспрессировали для иммунизации в клетках Expi293F, используя систему экспрессии Expi293™ (Invitrogen) в соответствии с протоколом производителя. Белок очищали от супернатанта аффинной хроматографией на колонке MabSelectSure (GE Healthcare). Колонку уравновешивали буфером 50 мМ глицин/глицинат натрия, рН 8,8, и элюировали градиентом 0,1 М лимонной кислоты, рН 2,0, в том же буфере. Белковые фракции нейтрализовали 2М Трис HCl, pH 8,5, концентрировали и дополнительно очищали гель-фильтрацией на колонке HiLoad 26/600 Superdex 200 (GE Healthcare), уравновешенной и элюированной в PBS, pH 7,4. Кролики получили первичную иммунизацию, включающую 500 мкг слитого белка, эмульгированного в равном объеме полного адъюванта Фрейнда (CFA). Кроликам вводили 2 бустерные инъекции с интервалом в 21 день, используя неполный адъювант Фрейнда (IFA), и кровь брали из уха через 14 дней после иммунизации. Забор крови прекращали через 14 дней после последней бустерной инъекции одноклеточных суспензий селезенки, костного мозга и мононуклеарных клеток периферической крови, приготовленных и замороженных в 10% диметилсульфоксиде (ДМСО) в фетальной бычьей сыворотке (FCS) при -80°C.
В-клеточная культура
Культуры В-клеток получали способом, аналогичным описанному Tickle et al., 2015. J Biomol Screen: 20(4), 492-497. Вкратце, В-клетки, полученные из лимфатических узлов или спленоцитов иммунизированных животных, культивировали с плотностью примерно 2000-5000 клеток на лунку в 96-луночных планшетах для культуры ткани со штрих-кодом с 200 мкл/лунку средой RPMI 1640 (Gibco BRL), дополненной 10% FCS (Sigma Aldrich), 2% HEPES (Sigma Aldrich), 1% L-глутамина (Gibco BRL), 1% раствора пенициллина/стрептомицина (Gibco BRL), 0,1% β-меркаптоэтанолом (Gibco BRL), 1% супернатанта активированных человеческих PBMC (BSS), и облученными рентгеновским излучением мутантными клетками мышиной тимомы EL4 (5×104/лунку) в течение семи дней при 37°C в атмосфере 5% CO2. Культуры создавали, используя В-клетки, полученные от всех иммунизированных животных, и в целом было отобрано примерно 1,7×109 В-клеток.
6470, антитело по настоящему изобретению, получали из активированных В-клеток, полученных из лимфатических узлов, которые культивировали с плотностью приблизительно 5000 клеток на лунку. Лимфатический узел использовали дополнительно к спленоцитам, который, как оказалось, является альтернативным источником для получения образцов В-клеток и идентификации новых антител. Антитела с родственными последовательностями идентифицировали из В-клеток, полученных из лимфатического узла, но не из селезенки. У кролика, иммунизированного С-концевым белком человеческого альфа-синуклеина отбирали примерно 9,6×107 клеток.
Первичный скрининг
Наличие антител, специфических к человеческому альфа-синуклеину, в супернатантах В-клеточной культуры определяли с помощью гомогенного флуоресцентного анализа связывания с использованием сфер Superavidin™ (Bangs Laboratories), покрытых биотинилированным рекомбинантным полноразмерным человеческим альфа-синуклеином в качестве источника целевого антигена. Рекомбинантный человеческий альфа-синуклеин, раскрытый в настоящем описании, биотинилировали с помощью 3-кратного молярного избытка биотина. Низкий молярный избыток биотина использовали для того, чтобы избежать полной модификации всех семи остатков лизина, которые находятся в молекуле альфа-синуклеина. Мономер альфа-синуклеина инкубировали в течение ночи при 40°С с биотином, и свободный биотин удаляли на следующий день с помощью спин-обессоливающей колонки Zeba™. Скрининг включал перенос 10 мкл супернатанта из 96-луночных планшетов для тканевых культур со штрих-кодом в 384-луночные планшеты для анализа с черными стенками со штрих-кодом, содержащие биотинилированный мономер рекомбинантного человеческого альфа-синуклеина, иммобилизованный на сферах суперавидина (10 мкл/лунку) с помощью манипулятора для жидкости Agilent Bravo. Связывание выявляли с помощью конъюгата козий антикроличий IgG Fcγ-специфический Alexafluor647 (Jackson). Планшеты считывали на устройстве TTP Labtech Mirrorball для идентификации лунок, содержащих IgG, специфический к альфа-синуклеину.
Вторичный скрининг
После первичного скрининга положительные супернатанты объединяли в 96-луночных мастер-планшетах со штрих-кодом с помощью робота-манипулятора Beckman Coulter BiomekNXP, с B-клетками в планшетах для культивирования клеток, замороженных при -80°C. Затем мастер-планшеты подвергали скринингу в анализе ELISA с захватом стрептавидина, используя биотинилированный мономер рекомбинантного человеческого альфа-синуклеина или биотинилированные фибриллы рекомбинантного человеческого альфа-синуклеина, для идентификации лунок, которые демонстрировали связывание как с мономерным, так и фибриллярным рекомбинантным человеческим альфа-синуклеином, и для исключения любых лунок с ложноположительным результатом, демонстрирующих нецелевое связывание со сферами Superavidin™. Учитывая нерастворимую природу фибрилл, не подходили обычные протоколы покрытия ELISA, которые используются с белками в растворе. Было решено использовать минимальный протокол биотинилирования для сохранения фибриллярной структуры и для облегчения получения эффективного покрытия фибриллами планшета ELISA, который был предварительно покрыт стрептавидином.
Общий пул фибрилл биотинилированного альфа-синуклеина получали, как описано в настоящем описании, путем объединения биотинилированного мономера рекомбинантного альфа-синуклеина (как описано выше) с 50-кратным избытком немеченого рекомбинантного альфа-синуклеина в PBS. Образование фибрилл подтверждали анализом JC1 (Lee et al., Biochem. J. 2009, 418, 311-323).
Биотинилированный мономер или биотинилированные фибриллы в PBS захватывали на 384-луночных планшетах Maxisorp, покрытых стрептавидином, в карбонатном покрывающем буфере (dH2O + 0,16% Na2CO3 + 0,3% NaHCO3.) Планшеты блокировали 1% мас./об. ПЭГ/PBS и затем инкубировали с 10 мкл/лунку супернатанта культуры В-клеток (разбавленных 1:1 блокирующим буфером). В планшеты добавляли вторичные HRP-конъюгированные козьи антикроличьи IgG Fc антитела (Stratech Scientific Ltd/Jackson ImmunoResearch) с последующей визуализацией связывания с субстратом ™B (3,3',5,5'-тетраметилбензидин, от EMD Millipore; 10 мкл/лунку). Оптическую плотность измеряли при 630 нМ с помощью считывающего устройства для микропланшетов BioTek Synergy 2. Анализ первичного связывания выявил 640 искомых событий, и после скрининга ELISA было показано, что 491 из них связывается как с мономерным, так и с фибриллярным рекомбинантным человеческим альфа-синуклеином.
Супернатанты В-клеток, демонстрирующие самые сильные ELISA сигналы связывания с рекомбинантными фибриллами, отбирали для дальнейшего анализа с помощью поверхностного плазмонного резонанса для идентификации тех, которые демонстрируют самую лучшую скорость диссоциации от мономера рекомбинантного человеческого альфа-синуклеинова, фибрилл рекомбинантного человеческого альфа-синуклеина и фибрилл рекомбинантного мышиного альфа-синуклеина. Тестировали супернатанты из 80 разных В-клеток, девять лунок продемонстрировали значения скорости диссоциации (kd) от рекомбинантных человеческих фибрилл <1×10-5. Из них семь имели значения скорости диссоциации (kd) от рекомбинантных мышиных фибрилл менее чем 1×10-5, и два дали значения скорости диссоциации (kd) от рекомбинантного человеческого мономера менее чем 1×10-5. Все девять супернатантов были отобраны для извлечения вариабельной области.
Извлечение вариабельной области
Для извлечения генов вариабельных областей антител из отобранных представляющих интерес супернатантов, необходимо было выполнить этап деконволюции, чтобы обеспечить идентификацию антиген-специфических В-клеток в данной лунке, которая содержала гетерогенную популяцию В-клеток. Это было достигнуто с помощью метода флуоресцирующих фокусов (Clargo et al., 2014. MAbs: 6(1), 143-159). Вкратце, В-клетки, секретирующие иммуноглобулин из положительной лунки, смешивали со сферами стрептавидина (New England Biolabs), покрытыми биотинилированными фибриллами рекомбинантного человеческого альфа-синуклеина (полученными с помощью смеси 1:50, как описано выше), и конъюгатом козий анти-кроличий Fcγ-специфический FITC (Jackson) с конечным разведением 1:1200. После статической инкубации при 37°С в течение 1 часа антиген-специфические В-клетки можно было идентифицировать по наличию флуоресцентного гало, окружающего эту В-клетку. Несколько этих отдельных клонов В-клеток, идентифицированных с помощью микроскопа Olympus, затем отбирали с помощью микроманипулятора Eppendorf и помещали в пробирку для ПЦР.
Гены вариабельной области антитела извлекали из отдельных клеток с помощью ПЦР с обратной транскрипцией (ОТ) с использованием праймеров, специфических к вариабельной области тяжелой и легкой цепей. Два раунда ПЦР выполняли с помощью гнездовой 2° ПЦР, включающей сайты рестрикции на 3'- и 5'-концах, позволяющие клонировать вариабельную область в вектор экспрессии млекопитающих, экспрессирующий кроличий IgG (VH) или каппа-цепь (VL) кроличьего IgG. Гены антитела к альфа-синуклеину из 5 разных супернатантов успешно клонировали в векторы экспрессии. Конструкции тяжелой и легкой цепей котрансфицировали в клетки Expi-293 с использованием ExpiFectamine 293 (Invitrogen) для экспрессии рекомбинантного антитела в 125 мл колбе Эрленмейера объеме 30 мл. Через 5-7 дней экспрессии супернатанты собирали и очищали с помощью аффинной хроматографии.
ELISA скрининг содержащих транзиенты супернатантов
Затем очищенные антитела подвергали дальнейшему скринингу с помощью ELISA. Биотинилированные мономеры и фибриллы рекомбинантного человеческого альфа-синуклеина захватывали на 384-луночных планшетах Maxisorp (ThermoScientific/Nunc), покрытых стрептавидином, в карбонатном покрывающем буфере (dH2O + 0,16% Na2CO3 + 0,3% NaHCO3). Отдельные планшеты также покрывали биотинилированным пептидом, соответствующим остаткам 117-126 человеческого альфа-синуклеина, приведенного в SEQ ID NO:10 (пептид PVDPDNEAYE), для проверки, связаны ли транзиенты с этой или другой областью молекулы. Планшеты блокировали 1% мас./об. ПЭГ/PBS и затем инкубировали с несколькими разведениями очищенного временного супернатанта. Вторичные HRP-конъюгированные козьи антикроличьи IgG Fc антитела (Stratech Scientific Ltd/Jackson ImmunoResearch) добавляли в планшеты с последующей визуализацией связывания с субстратом ™B (3,3',5,5'-тетраметилбензидин, от EMD Millipore; 10 мкл/лунку). Оптическую плотность измеряли при 630 нМ с помощью считывающего устройства для микропланшетов BioTek Synergy 2. Данные для 6470 показаны на фиг. 3. Как видно, 6470 показывает связывание как с мономерным, так и с фибриллярным рекомбинантным человеческим альфа-синуклеином, но не дает связывания с пептидом 117-126.
Затем антитела (IgG) тестировали с помощью анализа агрегации на основе клеток, как описано далее в примере 7. Кинетику связывания всех антител, проявляющих активность в клеточном анализе, впоследствии определяли с помощью поверхностного плазмонного резонанса. Антитела тестировали в формате IgG и Fab для определения авидности (двухвалентного связывания) и сродства (моновалентного связывания), соответственно.
Пример 3: Характеристика антител
Кинетика по Biacore
Кинетику взаимодействия определяли методом поверхностного плазмонного резонанса на приборе Biacore T200. Каждый из трех разных лигандов, включая рекомбинантный полноразмерный мономер человеческого альфа-синуклеина, очищенные фибриллы рекомбинантного человеческого альфа-синуклеина и очищенные фибриллы рекомбинантного мышиного альфа-синуклеина, полученные согласно настоящему описанию, иммобилизовали в трех различных проточных ячейках на поверхности чипа СМ5 путем иммобилизации по аминогруппе. Три лиганда готовили в 10 мМ NaAc, pH 3,5, и иммобилизовали на поверхности отдельных проточных ячеек до достижения уровня иммобилизации примерно 30 единиц ответа (RU) для мономера альфа-синуклеина, примерно 40 RU для фибрилл человеческого альфа-синуклеина и примерно 300 RU для фибрилл мышиного альфа-синуклеина, соответственно, при скорости потока 10 мкл/мин. Буфер HBS-EP+ (GE Health Bio-Sciences AB) использовали в качестве рабочего буфера как для иммобилизации лиганда, так и для анализа кинетики. Затем измеряли связывание моноклонального кроличьего IgG1 6470 (содержащего SEQ ID NO:47 и 48) и моноклонального кроличьего Fab 6470 (содержащего SEQ ID NO:47 и 49) с тремя лигандами. Моноклональные антитела IgG или Fab вводили в 7 различных концентрациях от 800 нМ до 0,195 нМ, с каждой концентрацией в 3 проточные ячейки, с временем контакта 3 мин и временем диссоциации 30 мин при скорости потока 100 мкл/мин. Для регенерации поверхности вводили одну инъекцию 50 мМ HCl в течение 90 с со скоростью 10 мкл/мин и еще одну инъекцию 50 мМ HCl в течение 60 с со скоростью 10 мкл/мин. Данные анализировали с помощью программного обеспечения Biacore T200 (версия 3.0) с использованием модели двухвалентного аналита с предполагаемым отсутствием общего вклада (RI = 0) и глобальным Rmax для формата IgG, а также с использованием модели 1:1 с гибким общим вкладом (локальное значение RI) и глобальным значением Rmax.
Кинетические значения для связывания как IgG, так и Fab с иммобилизованными мишенями приведены в таблице 2. Формат IgG продемонстрировал явное селективное сродство к фибриллам человеческого альфа-синуклеина по сравнению со сродством к мономеру человеческого альфа-синуклеина, поскольку константа диссоциации KD для человеческих фибрилл ниже более чем в 10 раз.
(1/мс)
(1/с)
(нМ)
(1/мс)
(1/с)
(нМ)
(1/мс)
(1/с)
(нМ)
кроличий Fab
кроличий IgG1
Связывание с бета-синуклеином
Связывание антител к человеческому альфа-синуклеину с человеческим бета-синуклеином тестировали с помощью вестерн-блот анализа с использованием бета-синуклеина rPeptide. Один микрограмм синуклеина прогоняли в 4-12% бис/трис-геле и промокали на мембрану PVDF. Мембрану блокировали в PBS с 3% BSA и 0,1% Твин20. К блокированным блотам добавляли кроличьи антитела IgG1 6470 и инкубировали в течение 1 часа при комнатной температуре, промывали PBS, 0,1% Твин20 и инкубировали в течение 1 часа со конъюгатом вторичное антитело-HRP (конъюгат антикроличьих H+L с HRP, Bethyl, A120-101P). Блот тщательно промывали в PBS с 0,1% Твин20, PBS и водой. Хемилюминесценцию измеряли после добавления субстрата для вестерн-блоттинга ECL (Pierce). Как показано на фиг.4(А), кроличье IgG1 6470 (дорожка 3) не связывается с человеческим бета-синуклеином.
Картирование эпитопов
ЯМР
Человеческий альфа-синуклеин клонировали в вектор экспрессии pET28a так, чтобы белок экспрессировался без каких-либо меток. Конструкцию трансформировали в клетки E.coli BL21 (DE3) (Stratagene), и клетки выращивали в определенной среде с С13-меченной DL-глюкозой и N15-меченым сульфатом аммония в присутствии и в отсутствие оксида дейтерия (D2O). Экспрессию индуцировали при OD600нм=1 с 300 мМ IPTG, и культуру инкубировали при 30°C в течение 4 часов. Клетки осаждали и лизировали с помощью трех циклов замораживания-оттаивания в 100 мл буфера для лизиса (20 мМ Трис/HCl pH 8,0, 25 единиц бензоназы (Merck Millipore), полный коктейль без ингибиторов протеазы, без EDTA (2 таблетки, Roche) и 10 мг лизоцима (Sigma)). Лизат осветляли центрифугированием при 18000 об/мин, и очищенный лизат пропускали через 0,22 мкм фильтр (Stericup, Millipore). Стерильный лизат загружали в колонку MonoQ 10/100GL (GE Healthcare), уравновешенную 20 мМ Трис/HCl, рН 8,0, 5CV, и белок элюировали градиентом до 500 мМ NaCl в том же буфере. Дальнейшую очистку наиболее чистых фракций повторяли на колонке MonoQ 10/100GL после 5-кратного разбавления в 20 мМ Трис/HCl, рН 8,0. Самые чистые фракции объединяли, концентрировали с помощью центробежного концентратора MWCO 10 кДа (Centriprep, Millipore), очищали путем исключения по размеру на колонке HiLoad 26/600 Superdex 75 (GE Healthcare) и элюировали в 25 мМ натрий-фосфатном буфере, 100 мМ NaCl (рН 6,4). Фракции из колонки Superdex 75 объединяли и добавляли азид натрия (конечная концентрация 0,02%) и AEBSF (конечная концентрация 10 мкМ). Конечная концентрация белка составляла примерно 5 мг/мл.
Кроличий Fab 6470 (содержащий VL с SEQ ID NO:11 и VH с SEQ ID NO:13, а также содержащий SEQ ID NO:47 и 49) экспрессировали в CHO SXE в виде His-меченых объектов и очищали от супернатанта с помощью His-tag аффинной хроматографии, связывая белок с HisTrap Excel (GE Healthcare) из супернатанта и элюируя его с помощью 250 мМ имидазола в PBS. Пул элюции загружали на колонку HiTrap GammaBind Plus Sepharose (GE Healthcare), колонку промывали PBS, и белок элюировали 0,1 М глицин-HCl, рН 2,6, и рН доводили до 6 с помощью 0,75 М фосфата натрия, рН 9. В элюированном Fab-His белке заменяли буфер на буфер ЯМР (25 мМ фосфата натрия, рН 6,4, 100 мМ NaCl) на обессоливающей колонке HiPrep 26/10. Перед стерилизацией на 0,22 мкм фильтре Millex GV Fab-His белковые фракции концентрировали, и добавляли ингибиторы протеазы AEBSF (конечная концентрация 10 мкМ) и азид натрия (конечная концентрация 0,02%). Для кристаллографии концентрированный Fab-His 6470 очищали препаративной эксклюзионной хроматографией на колонке HiLoad 26/600 Superdex 75 (GE Healthcare) и элюировали 25 мМ фосфатом натрия, рН 6,4, 100 мМ NaCl. Чистоту конечных пулов проверяли методом UPLC-SEC на чистоту >99%. Конечные пулы пропускали через 0,22 мм фильтр Millex GV для стерилизации.
Определение остова α-синуклеина
Образцы ЯМР обычно имели объем 350 мкл с концентрацией белка 360 мкМ 13C/15N-меченого или 430 мкМ 2H/13C/15N-меченого человеческого α-синуклеина в 5 мм пробирках Shigemi. Условия буфера были следующими: 100 мМ NaCl, 25 мМ фосфата натрия, рН 6,4, 10 мкМ AEBSF, 0,02% NaN3. Все эксперименты регистрировали при 20°С либо на спектрометре Bruker AVIII с частотой 600 МГц, либо на спектрометре Bruker AVII с частотой 800 МГц, оснащенном зондами с криогенным охлаждением. Последовательные связи между ЯМР сигналами остатков в остове белка, HN(i)-N(i)-N(i±1) выполняли в 3D (H)N(CA)NNH эксперименте (Weisemann et al., 1993 3D Triple-resonance NMR techniques for the sequential assignment of NH and 15N resonances in 15N- and 13C-labelled proteins. J. Biomol. NMR 3), регистрировали со спектральной шириной 28, 28 и 10 ppm и временем сканирования 117 (F1) 117 (F2) и 140 (F3) мс в измерениях на ядрах 15N, 15N и 1H, соответственно, с 8 сканированиями на приращение и релаксационной задержкой 1,5 с. Использовали неоднородную выборку с плотностью выборки 10% (4000 из 40000 гиперкомплексных точек), что дало общее время сканирования 2,75 дня. Подтверждали последовательные связи, и типы остатков идентифицировали с помощью экспериментов TROSY-HNCA (Grzesiek and Bax, 1992 Improved 3D triple-resonance NMR techniques applied to a 31 kDa protein. J. Magn. Reson. 96, 432-440; Salzmann et al., 1998. TROSY in triple-resonance experiments: new perspectives for sequential NMR assignment of large proteins. Proc. Natl. Acad. Sci. USA. 95, 13585-90) и TROSY-HNCACB (Wittekind and Mueller, 1993 HNCACB, a High-Sensitivity 3D NMR Experiment to Correlate Amide-Proton and Nitrogen Resonances with the Alpha- and Beta-Carbon Resonances in Proteins. J. Magn. Reson. Ser. B 101, 201-205; Salzmann et al., 1999. TROSY-type Triple Resonance Experiments for Sequential NMR Assignment of Large Proteins. J. Am. Chem. Soc. 121, 844-848). В эксперименте TROSY-HNCA регистрировали спектры с шириной 23, 28, 10 ppm и временем сканирования 12,1 (F1), 21,7 (F2) и 100 (F3) мс в измерениях на ядрах 13C, 15N и 1H, соответственно (8 сканирований на приращение, релаксационная задержка 1,5 с, общее время сканирования 1 день), в то время как в эксперименте TROSY-HNCACB регистрировали спектры с шириной 56, 28 и 10 ppm и временем сканирования 8,2 (F1), 21,7 (F2) и 100 (F3) мс в измерениях на ядрах 13C, 15N и 1H, соответственно (8 сканирований на приращение, релаксационная задержка 1,5 с, общее время сканирования 1,7 дня). Расположение карбонильных групп в остове получали из спектра TROSY-HNCO (Grzesiek and Bax, 1992 Improved 3D triple-resonance NMR techniques applied to a 31 kDa protein. J. Magn. Reson. 96, 432-440; Salzmann et al., 1998. TROSY in triple-resonance experiments: new perspectives for sequential NMR assignment of large proteins. Proc. Natl. Acad. Sci. USA. 95, 13585-90), который записывали со спектральной шириной 10, 29, 10 ppm и временем сканирования 80 (F1), 21,7 (F2) и 150 (F3) мс в измерениях на ядрах 13C, 15N и 1H, соответственно (8 сканирований на приращение и релаксационной задержкой 1,5 с). Использовали неоднородную выборку с плотностью выборки 15% (1208 из 8050 гиперкомплексных точек), что дало общее время сканирования 19 часов. Спектры ЯМР обрабатывали, используя программное обеспечение NMRPipe (Delaglio et al., 1995 NMRPipe: a multidimensional spectral processing system based on UNIX pipes. J. Biomol. NMR 6, 277-93), с линейным предсказанием, используемым для однократного увеличения эффективного времени сканирования по ядрам азота. Данные неоднородной выборки восстанавливали с помощью гарвардского итерационного метода с мягким пороговым критерием (Hyberts et al., 2012), где данные восстанавливали до следующего числа Фурье, что приводило к увеличению косвенного времени сканирования на 60%. Анализ данных выполняли методом Sparky (Goddard and Kneller, D. G. SPARKY 3. In University of California, San Francisco), в результате чего определяли амидные протонные и азотные резонансы 133 остатков, что соответствует 99% остатков (исключая остатки пролина и N-концевого метионина).
Картирование связывающего сайта Fab 6470 осуществляли, используя 150 мкМ образца 2H/13C/15N-меченного человеческого альфа-синуклеина, содержащего 10% молярный избыток немеченого Fab 6470. Образцы готовили в том же буфере, как описано выше, для определения каркаса альфа-синуклеина. Изменения химического сдвига в спектрах 1H, 15N и 13C определяли путем сравнения спектра, полученного в эксперименте TROSY-HNCO (Grzesiek and Bax, 1992 Improved 3D triple-resonance NMR techniques applied to a 31 kDa protein. J. Magn. Reson. 96, 432-440; Salzmann et al., 1998. TROSY in triple-resonance experiments: new perspectives for sequential NMR assignment of large proteins. Proc. Natl. Acad. Sci. USA. 95, 13585-90), полученного для комплекса альфа-синуклеин/Fab, с эквивалентным контрольным спектром, полученным для свободного альфа-синуклеина. Контрольный спектр в эксперименте TROSY-HNCO со свободным альфа-синуклеином получали со спектральной шириной 10, 28 и 10 ppm и временем сканирования 80 (F1), 22 (F2) и 150 (F3) мс в измерениях на ядрах 13C, 15N и 1H, соответственно (16 сканов на приращение, релаксационная задержка 1,5 с). Использовали неоднородную выборку с плотностью выборки 25% (2013 из 8050 гиперкомплексных точек), что дало общее время сканирования 2,7 дня. Эксперимент TROSY-HNCO для комплекса альфа-синуклеин/Fab выполняли, используя спектральную ширину 10, 28 и 10 ppm и время сканирования 80 (F1), 21,7 (F2) и 80 (F3) мс в измерениях на ядрах 13C, 15N и 1H, соответственно (32 сканирования на приращение, релаксационная задержка 1,5 с). Использовали неоднородную выборку с плотностью выборки 25% (1119 из 4477 гиперкомплексных точек), что дало общее время сканирования 2,8 дня. Спектры ЯМР обрабатывали с помощью программы NMRPipe (Delaglio et al., 1995 NMRPipe: a multidimensional spectral processing system based on UNIX pipes. J. Biomol. NMR 6, 277-93) with reconstruction of the NUS data performed using mddnmr. Analysis of non-uniformly sampled spectra with Multi-Dimensional Decomposition. Prog. Nucl. Magn. Reson. Spectrosc., 59, p. 271-292). Эффективное время сканирования по ядрам азота 1-кратно увеличивали во время восстановления данных.
Изменения химического сдвига анализировали с помощью подхода минимального сдвига (Williamson et al., 1997 Mapping the binding site for matrix metalloproteinase on the N-terminal domain of the tissue inhibitor of metalloproteinases-2 by NMR chemical shift perturbation. Biochemistry 36, 13882-9), по существу, как было описано ранее (Veverka et al., 2008 Structural characterization of the interaction of mTOR with phosphatidic acid and a novel class of inhibitor: compelling evidence for a central role of the FRB domain in small molecule-mediated regulation of mTOR. Oncogene 27, 585-95), за исключением модификации уравнения, используемого для расчета изменения комбинированного химического сдвига (Δδ) с учетом химического сдвига карбонильного атома, в результате чего получаем следующее уравнение:
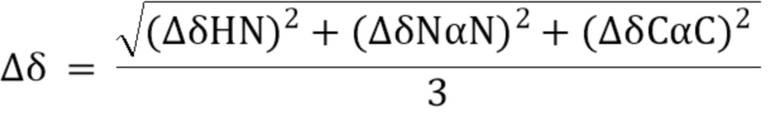
где ΔδHN, ΔδN и ΔδC - различия в химических сдвигах 1H, 15N и 13C, соответственно. αN и αC соответствуют коэффициентам масштабирования 0,2 и 0,35, соответственно, используемым для учета различий в диапазонах химических сдвигов протона амида, азота и карбонильного атома.
Для идентификации участка связывания (эпитопов) Fab на альфа-синуклеине использовали гистограмму комбинированного минимального сдвига в зависимости от последовательности белка для выявления областей альфа-синуклеина, содержащих сильно искаженные сигналы. Если размер комбинированного изменения химического сдвига для отдельных аминокислот превышал пороговое значение среднего значения комбинированного изменения химического сдвига для всех аминокислот плюс одно стандартное отклонение от этого среднего, эти остатки отбирали для дальнейшей оценки в качестве возможных контактных остатков в связывающем участке Fab.
Сильно искаженные остатки идентифицировали как остатки с минимальным сдвигом, превышающем по меньшей мере среднее значение, вычисленное для всех сдвигов, плюс одно стандартное отклонение. Для идентификации остатков, связанных с Fab, применили четыре разные пороговые значения. Остатки, которые задействованы в связывающем участке, оценивали с возрастающим уровнем строгости как: остатки, минимальный сдвиг которых превышает среднее значение, вычисленное для всех сдвигов, плюс одно стандартное отклонение (составляющее >0,018925); остатки, минимальный сдвиг которых превышает среднее значение, вычисленное для всех сдвигов, плюс два стандартных отклонения (составляющее >0,032049); остатки, минимальный сдвиг которых превышает среднее значение, вычисленное для всех сдвигов, плюс три стандартных отклонения (составляющее >0,045174); остатки, минимальный сдвиг которых превышает среднее значение, вычисленное для всех сдвигов, плюс четыре стандартных отклонения (составляющее >0,058299). В этом анализе остатки пролина не могут быть идентифицированы, так как они не содержат амидного протона.
Следовательно, эпитоп для Fab 6470 определяется с возрастающим уровнем строгости как среднее значение, вычисленное для всех сдвигов, плюс одно стандартное отклонение: D121, N122, E123, A124, Y125, E126, M127, S129, E130, Y133, Q134, D135 и Y136; среднее значение, вычисленное для всех сдвигов, плюс два стандартных отклонения: E123, A124, Y125, E126, M127, S129, E130, D135 и Y136; среднее значение, вычисленное для всех сдвигов, плюс три стандартных отклонения: Y125, M127, S129 и D135; среднее значение, вычисленное для всех сдвигов, плюс четыре стандартных отклонения: M127, S129 и D135.
Как показано на фиг.4В, с помощью ЯМР-исследований было обнаружено, что антитело 6470 связывается по меньшей мере со следующими остатками (среднее значение +3SD) Y125, M127, S129 и D135 и, кроме того, также связывается со всеми следующими остатками (среднее значение +1SD) D121, N122, E123, A124, E126, E130, Y133, Q134 и Y136 человеческого альфа-синуклеина (SEQ ID NO:10).
Картирование пептида
Дальнейшую характеристику эпитопа, связанного с 6470, осуществляли с помощью коротких (обычно 9-мерных или 10-мерных) пептидов, характерных для и охватывающих С-концевую область человеческого альфа-синуклеина. Их использовали в конкурентном анализе поверхностного плазмонного резонанса для проверки, способны ли какие-либо из них ингибировать связывание антитела с мономерным альфа-синуклеином или предварительно образованными фибриллами альфа-синуклеина, иммобилизованными на чипе Biacore. Пептид, демонстрирующий максимальный уровень ингибирования, затем отбирали для изучения сокристаллизации с антителом для подтверждения точного эпитопа.
Пептиды приобретали в компании Peptide Protein Research Ltd., Bishop's Waltham, U.K., и синтезировали с помощью Fmoc твердофазного пептидного синтеза по методу Atherton и Sheppard (Ref: Atherton, E.; Sheppard, R.C. (1989). Solid Phase peptide synthesis: a practical approach. Oxford, England: IRL Press). N- и С-пептидные концы кэпировали ацетильной и амидной группами, соответственно, за исключением случая, когда пептиды представляли собой N-конец и С-конец α-синуклеина, при этом амино и карбоксильные группы, соответственно, оставались свободными. Исходные растворы пептидов готовили в ДМСО при 10 мМ. Полный список пептидов приведен в Таблице 3.
Рекомбинантный мономер человеческого альфа-синуклеина и предварительно сформированные фибриллы альфа-синуклеина иммобилизовали на чипе СМ5 с помощью инструмента Biacore 3000 (GE Healthcare). После активации поверхности карбоксиметилдекстрана путем инъекции 100 мкл свежей 1:1 (об./об.) смеси 50 мМ N-гидроксисукцимида и 200 мМ 1-этил-3-(3-диметиламинопропил)карбодиимида при скорости потока 10 мкл/мин HBS-EP (GE Healthcare) в качестве рабочего буфера добивались связывания путем введения 100 мкл мономера и фибрилл с концентрацией 5 мкМ в 10 мМ ацетате, рН 5,0, через отдельные проточные ячейки. Контрольную проточную ячейку активировали таким же образом, а затем поверхности всех проточных ячеек дезактивировали с помощью струи 50 мкл 1М этаноламина HCl, pH 8,5.
Пептидные растворы готовили в рабочем буфере при 100 мкМ, и контроль без пептида готовили в виде разведения 1:100, используя ДМСО в рабочем буфере. Раствор 6470 кроличьего Fab (содержащего SEQ ID NO:47 и 49) готовили в 50,5 нМ рабочем буфере до предварительной инкубации 198 мкл с 2 мкл пустого контроля или разбавленного пептида с получением конечной смеси 50 нМ Fab и 1 мкМ пептида или контроля. Сенсограммы регистрировали для каждого образца, вводя 30 мкл смеси со скоростью 10 мкл/мин, и регистрировали сигнал во временной точке за 5 секунд до окончания инъекции. В конце каждого цикла чип восстанавливали двумя 10 мкл инъекциями 40 мМ HCl и одной инъекцией 5 мМ NaOH. Контрольные циклы чередовали с пептидными циклами.
Степень ингибирования каждого пептида рассчитывали, как процент изменения единиц ответа, измеренных в точке записи сигнала, по сравнению со средним значением смежных контрольных циклов.
Уровень ингибирования каждого пептида альфа-синуклеина показан на фиг. 5. Значительное ингибирование связывания Fab 6470 либо с мономером альфа-синуклеина, либо с фибриллами наблюдали только для трех пептидов: AS121-130, AS123-132 и AS125-134, из них самые высокие уровни ингибирования наблюдали у AS123-132, которые составили 37% и 54% для связывания антитела с мономером и фибриллами, соответственно. Немного более низкие уровни ингибирования были получены для пептида AS125-134, 34% и 52%, соответственно, что указывает на то, что основной компонент эпитопа содержит остатки 125-132. Уровни ингибирования, обеспечиваемые пептидом AS121-130, были более низкими, 20% и 27%, соответственно, что позволило предположить, что общими остатками у всех трех пептидов являются 125-130, которые вносят основной вклад в эпитоп.
Поскольку эпитоп антитела 6470, по-видимому, содержит по меньшей мере последовательность YEMPSEEG, в исследованиях сокристаллизации с Fab 6470 изучали пептид AS123-132.
Рентгеновская кристаллография
Для получения комплексов 1 мл очищенного кроличьего Fab 6470 в концентрации примерно 10 мг/мл смешивали с пептидом альфа-синуклеина 123-132 (EAYEMPSEEG) в молярном соотношении Fab:пептид 1:2 и инкубировали в течение 1 часа при комнатной температуре. Условия, подходящие для роста кристаллов, определяли методом диффузии паров в сидячей капле с использованием коммерчески доступных экранов для кристаллизации (Qiagen). Для получения кристаллов дифракционного качества использовали метод диффузии паров висячих капель.
Для получения комплекса Fab 6470-пептид 123-132 1 мкл раствора белка смешивали с 1 мкл резервуарного раствора, содержащего 1,6 М сульфата аммония и 0,1 М буфера Hepes, рН 7,5. Кристаллы собирали и быстро замораживали в жидком азоте после быстрого прогона через раствор криопротектора, содержащий 1,6 М сульфата аммония, 0,1 М буфера Hepes, рН 7,5, и 20% глицерина. Кристаллы собирали и быстро замораживали в жидком азоте после быстрого прогона через раствор криопротектора, содержащий 0,2 М сульфата аммония и 35% (об./об.) полиэтиленгликоля 8000.
Дифракционные данные с монокристаллов Fab 6470-пептид 123-132 получали с разрешением до 2,9Å на линии луча i04-1 на приборе Diamond Synchrotron, Didcot, Oxfordshire, UK, и обрабатывали с помощью Mosflm, Aimless и Truncate. Структуру комплекса определяли методом молекулярного замещения с использованием программного обеспечения Phaser, используя координаты внутрилабораторного Fab в качестве поисковой модели.
Циклы уточнения и построения модели выполняли с помощью CNS (BBrunger et al., (2007) Nature Protocols 2, 2728-2733) и COOT (Emsley et al., (2004) Acta crystallographica. Section D, Biological crystallography 60, 2126-2132) до тех пор, пока вся статистика уточнения не сошлась для обеих моделей. Геометрию модели подтверждали с помощью программного обеспечения Molprobity43. Молекулярные визуализации получали с помощью Pymol44. Информацию об эпитопе, описанную ниже, получали путем рассмотрения атомов с разрешением 4Å на поверхности контакта Fab/пептид. Сбор даных и статистика уточнения приведена в таблице 4А и таблице 4В.
Таблица 4В
Значения в скобках относятся к оболочке, полученные при более высоком разрешении. Rsym = Σ⎪(I - <I>)⎪/Σ(I), где I - наблюдаемая интегральная интенсивность, <I> - средняя интегральная интенсивность, полученная из нескольких измерений, и суммирование производится по всем наблюдаемым отражениям. Rwork = Σ⎪⎪Fobs⎪ - k⎪Fcalc⎪⎪/Σ⎪Fobs⎪, где Fobs и Fcalc - наблюдаемые и рассчитанные структурные факторы соответственно. Rfree рассчитывается как Rwork с использованием 5% данных отражения, выбранных случайным образом и не учитываемых в расчетах уточнения.
Основные контактные участки между остатками тяжелой и легкой цепи и пептидом показаны в таблице 5.
Таким образом, эпитоп содержит остатки E123, Y125, E126, M127, P128, S129, E130 и E131. На фиг.6 показан Fab 6470 в комплексе с пептидом 123-132, а на фиг.7 и 8 показаны контакты между тяжелой цепью и легкой цепью Fab 64-132 и 6470, соответственно.
Пример 4: Гуманизация антитела и созревание сродства
Кроличье антитело 6470 гуманизировали путем прививки CDR из кроличьих V-областей на каркас V-области антитела человеческой зародышевой линии. Для восстановления активности антитела, в гуманизированной последовательности также сохраняли несколько остатков каркаса из кроличьих V-областей. Эти остатки отбирали с помощью протокола, описанного Adair et al. (1991) (WO91/09967). Выравнивание последовательностей V-областей кроличьих антител (донорских) с последовательностями V-областей человеческой зародышевой линии (акцепторной) показаны на фиг. 9 и 10 вместе с созданными гуманизированными последовательностями. CDR, привитые от донорской к акцепторной последовательности, определены по Kabat (Kabat et al., 1987), за исключением CDR-H1, где используется комбинированное определение по Чотиа/Кабат (см. Adair et al., WO91/09967).
Гены, кодирующие несколько вариантов последовательностей V-областей тяжелой и легкой цепей, разрабатывали и конструировали с помощью подхода автоматического синтеза DNA2.0 Inc. Другие варианты V-областей тяжелой и легкой цепей создавали путем модификации генов VH и VK с помощью олигонуклеотид-направленного мутагенеза, включая, в некоторых случаях, мутации в CDR. Для временной экспрессии в клетках млекопитающих гены гуманизированной V-области легкой цепи клонировали в вектор экспрессии pMhCK легкой цепи UCB, который содержит ДНК, кодирующую константную область человеческой Каппа-цепи (аллотип Km3). Гены гуманизированной V-области тяжелой цепи клонировали в вектор экспрессии тяжелой цепи человеческого гамма-4 UCB pMhγ4PFL, который содержит ДНК, кодирующую константную область тяжелой цепи человеческого гамма-4, с мутацией S241P, стабилизирующей шарнирную область (Angal et al., Mol. Immunol. 1993, 30(1):105-8). Химерное 6470, содержащее кроличьи V-области (SEQ ID NO:11 и 13) и человеческие константные области, также получали аналогичным образом и использованы в качестве антитела сравнения. Котрансфекция полученных векторов тяжелой и легкой цепей в суспензионные клетки Expi293™ дала экспрессию гуманизированных рекомбинантных антител в формате человеческого IgG4P.
Человеческую V-область IGKV1-16 плюс J-область JK4 (IMGT, http://www.imgt.org/) отбирали в качестве акцептора для CDR легкой цепи антитела 6470. Все каркасные остатки легкой цепи в трансплантате gL3 имели происхождение из гена человеческой зародышевой линии, за исключением остатков 48 и 72 (со ссылкой на SEQ ID NO:15), в котором были сохранены донорские остатки глутамина (Q48) и глутамина (Q72), соответственно. Сохранение остатков Q48 и Q72 было важным для сохранения полной активности гуманизированного антитела (фиг. 9 и таблица 6) для связывания фибрилл человеческого альфа-синуклеина.
варианты
Человеческую V-область IGHV3-23 плюс J-область JH4 (IMGT, http://www.imgt.org/) выбирали в качестве акцептора CDR тяжелой цепи антитела 6470. Как и во многих кроличьих антителах, ген VH антитела 6470 является более коротким, чем у выбранного человеческого акцептора. При выравнивании с человеческой акцепторной последовательностью в каркасе 1 VH-области антитела 6470 отсутствует N-концевой остаток, который сохраняется в гуманизированном антителе (фиг. 10). В каркасе 3 области VH кроличьего 6470 также отсутствуют два остатка (75 и 76) в петле между нитями D и E бета-листа: в гуманизированных трансплантатах зазор заполнен соответствующими остатками (лизин 75, K75; аспарагин 76, N76) из выбранной человеческой акцепторной последовательности (фиг. 10). Все остатки каркасной области тяжелой цепи в трансплантатах gH23 и gH36 происходят от гена человеческой зародышевой линии, за исключением одного или более остатков из группы, включающей остатки 24, 47, 48, 49, 73, 78 и 97 (со ссылкой на SEQ ID NO:23 и 31), в котором сохранены донорские остатки валина (V24), тирозина (Y47), изолейцина (I48), глицина (G49), серина (S73), валина (V78) и аргинина (R97), соответственно. Сохранение остатков V24, Y47, I48, G49 и R97 было важно для обеспечения полной активности гуманизированного антитела для связывания с фибриллами человеческого альфа-синуклеина.
Кроме того, гуманизированные гены VH клонировали в вектор экспрессии pMhFab10HIS человеческого Fab-HIS UCB, который содержит ДНК, кодирующую домен человеческой гамма-1 CH1-шарнирной области с С-концевой меткой из десяти гистидиновых остатков. Гистидиновая метка облегчает очистку экспрессированных Fab методом аффинной хроматографии. Котрансфекция полученных векторов тяжелой и легкой цепей в суспензионные клетки Expi293™ давала экспрессию гуманизированных рекомбинантных антител в форматах Fab-HIS.
Созревание сродства осуществляли в соответствии с методами IOTA, описанными в WO 2014198951. Интерфейс между Fab кроличьего 6470 и пептидом EAYEMPSEEG альфа-синуклеина (123-132) в комплексе, определенном с помощью рентгеновской кристаллографии, анализировали для выявления мутаций, которые потенциально могут улучшить сродство Fab кроличьего 6470 к белку альфа-синуклеина. IOTA является статистически потенциальным инструментом для определения вероятности встречаемости данного типа контактного атома в интерфейсе белка или сайта связывания.
Для оценки влияние этих мутаций на активность антител в отношении связывания мономера или фибрилл человеческого альфа-синуклеина, сначала изучили мутации на Fab кроличьего 6470 (Таблица 7А). Кинетику взаимодействия определяли методом поверхностного плазмонного резонанса на приборе Biacore T200, как описано в Примере 3. Остаток 33 в CDRL1 (со ссылкой на SEQ ID NO:11) мутировал из аспарагина (N) в аргинин (R) или лизин (K): мутация остатка 33 в аргинин привела к увеличению сродства к альфа-синуклеину (таблица 7A). Остаток 55 в CDRH2 мутировал из серина (S) в аспарагин (N), а остаток 99 в CDRH3 мутировал из аспарагина (N) в лизин (K) или в глютамин (Q), или в гистидин (H), или триптофан (W) (со ссылкой на SEQ ID NO:13), мутация остатка 55 в аспарагин и остатка 99 в гистидин привела к увеличению сродства к альфа-синуклеину (таблица 7А). Мутация аспарагина в CDRH3 (N99H) также удаляет потенциальный сайт дезамидирования.
Наконец, также были протестированы вновь идентифицированные мутации в ранее полученных полноразмерных гуманизированных антителах (таблица 6) и проверена их селективность в отношении человеческих фибрилл (таблица 7B). Кинетику взаимодействия определяли методом поверхностного плазмонного резонанса на приборе Biacore T200, как описано в Примере 3. Как показано в таблице 7B, мутации в положении 33 в легкой цепи (со ссылкой на SEQ ID NO:19) и 56 и 102 в тяжелой цепи (со ссылкой на SEQ ID NO:27 и 35) приводят к увеличению сродства к человеческим фибриллам, что является преимущественной характеристикой антител, которым необходимо преодолеть гематоэнцефалический барьер, чтобы связаться со своей мишенью. При систематическом введении антител, большое количество вводимого антитела может быть потеряно, потому что антитела имеют ограниченные системы для преодоления сложных физиологических барьеров.
Варианты цепей гуманизированных антител и их комбинации экспрессировали и оценивали в отношении их активности относительно исходного антитела, их биофизических свойств и пригодности для последующей обработки.
Пример 5: Характеристика гуманизированных антител
Биофизическую характеристику выполняли для шести гуманизированных антителах 64G IgG4P (таблица 8, последовательности приведены в таблице 1).
Все антитела подвергали скринингу на термостабильность (™), экспериментальное значение pI, гидрофобность, растворимость (анализ осаждения с помощью ПЭГ) и стабильность агрегации на границе раздела воздух/жидкость для определения, оказали ли мутации какое-либо влияние, в частности, на сродство, стабильность и способность к развитию.
Процесс скрининга также включал оценку химической стабильности (дезамидирование, склонность к изомеризации аспарагиновой кислоты), поскольку антитела содержат:
1. мотив Asn(102)S (дезамидирование) в CDR3 тяжелой цепи только gL3gH23 и gL3gH36;
2. мотив Asn(98)Asp(99) (дезамидирование) в CDR3 легкой цепи всех антител;
3. мотив Asn(32)Asn(33) (дезамидирование) в CDR1 легкой цепи всех мутантов, кроме N33;
4. мотив Asp(99)G (изомеризация Asp) в CDR3 легкой цепи всех антител.
Химическая нестабильность в этих участках может привести к неоднородности и иммуногенности продукта.
Измерение термостабильности (Тм)
Температуру плавления (™) или температуру в средней точке при развертывании белка определяли с помощью анализа Thermofluor. В этом методе флуоресцентный оранжевый краситель SYPRO® использовали для контроля процесса развертывания белка путем связывания с гидрофобными областями, которые становятся открытыми при повышении температуры.
Реакционная смесь содержала 5 мкл 30-кратного оранжевого красителя SYPRO® (Invitrogen™), разбавленного PBS из 5000X маточного раствора, и 45 мкл образца при 0,12 мг/мл (в PBS pH 7,4). Примерно 10 мкл смеси распределяли по 384-луночного планшета для оптическх измерений с четырьмя повторами и прогоняли на быстрой ПЦР-системе 7900HT (Applied Biosystems™). Нагревательное устройство системы ПЦР было настроено на температуру от 20°C до 99°C со скоростью увеличения 1,1°C/мин. Изменения флуоресценции в лунках контролировали с помощью прибора с зарядовой связью. Данные по увеличению интенсивности наносили на график, и точку перегиба наклона(ов) использовали для расчета ™, как описано ниже.
Все антитела имели два конформационных перехода при разворачивании глобулы. Первый можно отнести к ™ домена CH2. Второй можно отнести к среднему значению ™ разворачивающегося домена Fab и домена CH3, согласно литературным источникам (Garber E, Demarest SJ. Biochem Biophys Res Commun. 2007 Apr 13; 355(3):751-7). Результаты суммированы в таблице 9.
Термическая стабильность находится в пределах нормального диапазона (Heads et al “Relative stabilities of IgG1 and IgG4 Fab domains: influence of the light-heavy interchain disulfide bond architecture”. Protein Sci. 2012 Sep;21(9):1315-22), ожидаемого для молекул IgG4.
Экспериментальное значение pI
Экспериментальное значение pI антител 6470 получали с помощью системы cIEF iCE3™ с полностью капиллярным изображением (ProteinSimple).
Образцы готовили путем смешивания: 30 мкл образца (из исходного раствора 1 мг/мл в воде класса ВЭЖХ), 35 мкл 1% раствора метилцеллюлозы (Protein Simple), 4 мкл (pH 3-10) амфолитов (Pharmalyte), 0,5 мкл синтетических маркеров pI=4,65 и 0,5 мкл синтетических маркеров pI=9,77 (ProteinSimple), 12,5 мкл 8М раствора мочевины (Sigma-Aldrich®). Воду для ВЭЖХ использовали для доведения конечного объема до 100 мкл. Смесь встряхивали в течение короткого промежутка времени для обеспечения полного перемешивания и центрифугировали при 10000 об/мин в течение 3 минут для удаления пузырьков воздуха перед анализом. Образцы фокусировали в течение 1 минуты при 1,5 кВ, затем 5 минут при 3 кВ, и капиллярные изображения A280 получали с помощью программного обеспечения ProteinSimple. Полученные электрофореграммы сначала анализировали с помощью программного обеспечения iCE3, и определяли значения pI (линейная зависимость между маркерами pI). Затем откалиброванные электроферограммы интегрировали с помощью программного обеспечения Empower® (Waters).
Экспериментальное значение pI для всех антител 6470 находилось в диапазоне 8,4-9,2. Небольшое различие между молекулами имелось в соотношении заряженных частиц, однако для молекулы IgG4P это не является неожиданным. Все pI были высокими и, следовательно, могут быть использованы для производства антител.
Хроматография гидрофобного взаимодействия (HIC)
Хроматография гидрофобного взаимодействия (HIC) позвояет разделять молекулы в порядке увеличения гидрофобности. Молекулы связываются с гидрофобной стационарной фазой в присутствии высоких концентраций полярных солей и десорбируются в подвижную фазу при уменьшении концентрации соли. Более длительное время удерживания соответствует большей гидрофобности.
Образцы в количестве 2 мг/мл разбавляли 1:2 1,6 М сульфатом аммония и PBS (pH 7,4). 5 мкг (5 мкл) образца вводили в колонку Dionex ProPac™ HIC-10 (100 мм × 4,6 мм), последовательно соединенную с бинарным насосом ВЭЖХ Agilent 1200 с флуоресцентным детектором. Разделение контролировали по собственной флуоресценции (длины волн возбуждения и испускания, 280 нм и 340 нм, соответственно).
Используя буфер A (0,8 М 100 мМ фосфата фосфата аммония, pH 7,4) и буфер B (100 мМ фосфата, pH 7,4) анализировали образец с помощью градиентного элюирования следующим образом: (i) выдержка в течение 2 минут при 0% B, (ii) линейный градиент от 0 до 100% B за 30 минут (0,8 мл/мин) (iii) промывка колонки 100% B в течение 2 минут и повторное уравновешивание в 0% B в течение 10 минут перед следующей инъекцией образца. Температуру колонки поддерживали на уровне 20°С.
Стандарты, демонстрирующие низкую и высокую гидрофобность плюс контроль, также анализировали в той же последовательности прогонов для нормализации времени удерживания (таблица 11). Время удерживания (RT) образца нормализовали относительно стандартов низкой и высокой гидрофобности с помощью следующего уравнения:
[(Образец (RT) - низкий стандарт (RT)/Высокий стандарт (RT) - низкий стандарт (RT)] x 100.
Все антитела 6470 и мутанты показали одинаковые нормализованные времена удерживания и одинаковую низкую гидрофобность. Коммерчески доступные терапевтические антитела имеют тенденцию проявлять низкую гидрофобность (Jain et al «Biophysical properties of the clinical-stage antibody landscape», Proc Natl Acad Sci U S A. 2017 Jan 31;114(5):944-949). Низкая гидрофобность способствует стабильности (т.е. уменьшает агрегацию) во время производства.
Измерение растворимости с помощью анализа осаждения полиэтиленгликолем (ПЭГ).
Стабильность коллоидного раствора анализировали с помощью анализа осаждения полиэтиленгликолем (ПЭГ). ПЭГ использовали для количественного определения растворимости белка путем увеличения концентрации ПЭГ (вес/об.) и измерения количества белка, остающегося в растворе. Этот анализ служил для имитации эффекта высокой растворимости без использования традиционных методов концентрирования. Вызванное ПЭГ осаждение антител 6470 исследовали в присутствии 7-18% ПЭГ-3350 в PBS, рН 7,4, 50 мМ ацетат натрия/125 мМ хлорид натрия, рН 5,0 (ацетат, рН 5) и 20 мМ L-гистидина, 140 мМ NaCl, рН 6,0. Образцы заменяли буфером, где необходимо, с помощью диализа, и концентрацию доводили до 2 мг/мл. Для минимизации неравновесного осаждения подготовка образца состояла из смешивания растворов 2× белка и 2× ПЭГ в объемном соотношении 1:1. После перемешивания образцы инкубировали при 37°С в течение 30 минут для повторного растворения неравновесных агрегатов. После инкубации в течение ночи при 20°C образцы центрифугировали в течение 60 минут (4000 г). Аликвоты супернатанта переносили в половинный объем 96-луночных планшетов для оптических измерений и измеряли поглощение при 280 нм с помощью планшетного ридера BMG Labtech FLUOstar® Omega LVIS A280. Данные по концентрации наносили на график в зависимости от ПЭГ%, и рассчитанную среднюю точку (LogEC50), полученную путем нелинейного подбора кривой, с изменяющимся наклоном, получали как меру относительной растворимости коллоидного раствора образцов. В этом анализе более высокое значение LogEC50 соответствует более высокой стабильности коллоидного раствора.
Результаты (не показаны) показали, что с увеличением рН буфера стабильность коллоидного раствора уменьшалась у всех антител 6470. Кроме того, наблюдалась следующая тенденция: от более до менее растворимых: gL3gH23 и gL3gH36 > gL3gH23-S56N-N102H и gL3gH36-S56N-N102H > gL3-N33RgH23-S56N-N102H и gL3-N33RgH36-S56N-N102H.
Следовательно, мутации, введенные для созревания сродства, снижали стабильность коллоидного раствора молекул антител. Различий между трансплантатами gL3gH23 и gL3gH36 не наблюдалось.
Влияние напряжения на границе раздела воздух-жидкость (анализ агрегации)
Белки имеют тенденцию разворачиваться при контакте с поверхностью раздела воздух-жидкость, где гидрофобные поверхности представлены гидрофобной средой (воздух), а гидрофильные поверхности - гидрофильной средой (вода). Перемешивание растворов белка обеспечивает большую поверхность раздела воздух-жидкость, которая может стимулировать агрегацию. Этот анализ служит для имитации напряжений, которым подвергается молекула в процессе производства (например, ультрафильтрации), и для обеспечения жестких условий, чтобы попытаться различить разные молекулы антител.
Образцы в PBS pH 7,4, 50 мМ ацетат натрия/125 мМ хлорид натрия pH 5,0 (ацетат pH 5) и 20 мМ L-гистидина, 140 мМ NaCl, pH 6,0 подвергали воздействию вихревого потока на устройстве Eppendorf Thermomixer Comfort™. Буферы выбирали в виде обычных заранее приготовленных буферов. Перед встряхиванием концентрацию доводили до 1 мг/мл, используя соответствующие коэффициенты экстинкции (1,35 Ab 280 нм, 1 мг/мл, длина пути 1 см) и оптической плотности при 280 нм, 340 нм и 595 нм, полученные с помощью спектрофотометра Varian Cary® 50-Bio для установления нулевой точки считывания. Каждый образец суб-аликвотировали в 1,5 мл конические пробирки с крышкой типа Eppendorf® (4×250 мкл) и подвергали воздействию жестких условий для проверки устойчивости путем встряхивания при 1400 об/мин при 25°C в течение до 24 часов. Агрегацию, зависящую от времени (мутность), контролировали путем измерения образцов через 3 часа и 24 часа после встряхивания при 595 нм с помощью спектрофотометра Varian CaryТМ 50-Bio. Средние значения поглощения для каждого образца наносили на график в зависимости от времени.
Полученные результаты приведены на фиг. 11. Между антителами 6470 ни в одном из трех буферов не наблюдалось никаких различий через 24 часа, однако небольшое различие было выявлено в склонности к агрегации через 3 часа после встряхивания, которая, по-видимому, зависела от буфера.
Исследование воздействия экстремальных условий на дезамидирования/изомеризации Asp
Исследование в экстремальных условиях выполняли, используя антитела 6470 gL3gH23 и gL3gH36, для определения предрасположенности к дезамидированию/Asp-изомеризации четырех выявленных потенциальных последовательностей: Asn(102)S (мотив дезамидирования) в CDR3 тяжелой цепи; Asn(98)Asp(99) (мотив дезамидирования) в CDR3 легкой цепи; Asn(32)Asn(33) (мотив дезамидирования) в CDR1 легкой цепи и Asp(99)G (мотив изомеризации Asp) в CDR3 легкой цепи. Склонность/скорость дезамидирования и Asp-изомеризации не может быть предсказана, поскольку она зависит от первичной последовательности и трехмерной структуры, а также от свойств раствора (R C Stephenson and S Clarke (1989); K. Diepold et al (2012); Jasmin F. Sydow et al (2014); N.E. Robinson et al. (2004).
Также получали базовые уровни дезамидирования (не подвергнутые стрессовым условиям образцы) - низкие уровни указывают на низкую восприимчивость, но уровни могут изменяться в различных партиях/условиях производства.
Буфер в двух антителах 6470 заменяли буферами (i), которые, как известно, способствуют дезамидированию остатков Asn(N) (Tris pH 8/37°C), и (ii), которые, как известно, способствуют изомеризации Asp (ацетат, pH 5/37°C). Конечную концентрацию образца в каждом из буферов доводили до ~6,5 мг/мл и затем разделяли на две аликвоты, где одну хранили при 4°С и одну при 37°С в течение до 4 недель. Аликвоту немедленно удаляли (T0) и через 2 недели и 4 недели и хранили при -20°C.
Двухнедельные образцы оттаивали и анализировали путем триптического расщепления/масс-спектроскопии (МС) для анализа химической модификации следующим образом. Образцы подвергнутых стрессу белков восстанавливали с помощью TCEP и алкилировали хлоруксусной кислотой в буфере Tris-HCL, рН 8,0, содержащем 0,1% мас./об. моющего средства Rapigest. Добавляли трипсин (1:25 мас./мас.), и образцы расщепляли в течение ночи при комнатной температуре. Протеолиз останавливали добавлением муравьиной кислоты до 1% по объему, и образцы разбавляли до 0,5 мг/мл перед центрифугированием для удаления осажденного Rapigest™. Полученные в результате пептиды отделяли и анализировали на колонке Waters BEH C18, сопряженной с масс-спектрометром Thermo Fusion™, в которой используется зависящий от данных метод +ve-ion орбитрап-орбитрап с фрагментацией в результате диссоциации, вызванной столкновениями (CID). Данные ЖХ-МС анализировали с помощью программного обеспечения Thermo Xcalibur™ и PepfinderТМ.
Эксклюзионную ВЭЖХ и SDS PAGE также выполняли для контроля агрегации/деградации.
Результаты картирования пептида/масс-спектрометрии показали, что базовый уровень дезамидирования Asn во всех трех сайтах CDR составляет <1,5%, и что дезамидирование максимально увеличивается до ~6% через 2 недели при pH 8,0 и 37°C для сайта Asn(102)S в тяжелой цепи CDR3.
Модификация Asp(99) (образование сукцинимида) в CDR3 легкой цепи составило ~25% через 2 недели при рН 5,0 и 37°С.
Оценивали влияние химической модификации (дезамидирование в Asn(102) в CDR3 тяжелой цепи и образование промежуточного сукцинимида в Asp(99) в CDR3 легкой цепи) на сродство/авидность рекомбинантного полноразмерного мономера человеческого альфа-синуклеина и фибрилл очищенного рекомбинантного человеческого альфа-синуклеина. В исследовании использовали полностью дезамидированный продукт (Asn(102)Asp) и подверженный воздействию стрессовых условий материал (pH 5/2 неделя/37°C).
Пример 6: Иммуногистохимия
Иммуногистохимию выполняли на устройстве Asterand Bioscience (Royston, United Kingdoms). Криосрезы (10 мкм) сначала подвергали процедуре демаскировки антигена с помощью растворов для демаскировки целевого антитела Dako PT Link и EnVision FLEX Target Retrieval Solutions (pH 6) при 97°C в течение 20 минут с автоматическим нагреванием и охлаждением. Все последующие этапы инкубации проводили при комнатной температуре. Криосрезы сушили на воздухе в течение 30 минут, фиксировали в 4% параформальдегиде, приготовленном в 1X PBS, в течение 10 минут, промывали в промывочном буфере Dako EnVision™ FLEX (Dako) и затем загружали в Dako Autostainer Plus. Эндогенную активность пероксидазы блокировали путем инкубации срезов с блоком пероксидазы Dako (Dako) в течение 5 минут. Затем срезы дважды промывали 1X PBS перед инкубацией с белковым блоком Dako CSA II (Dako) в течение 10 минут. Раствор белкового блока удаляли воздушной струей, и срезы инкубировали в течение 30 минут с кроличьим IgG1 6470 (включающими SEQ ID NO:47 и 48), разбавленными (0,05 мкг/мл) в разбавителе антител Dako (Dako). После инкубации срезы дважды промывали 1X PBS, затем инкубировали с анти-кроличьим Dako Flex полимер-HRP субстратом (Dako) в течение 20 минут, дважды промывали и затем инкубировали с диаминобензидиновым субстратом (Dako) в течение 10 минут. Хромогенную реакцию останавливали, промывая предметные стекла дистиллированной водой. После хромогенеза срезы удаляли из Dako Autostainer Plus и вручную окрашивали гематоксилином, обезвоживали растворами этанола с увеличивающимися концентрациями, очищали тремя промывками ксилолом и покрывали средой для заливки DPX (Sigma-Aldrich). Цифровые изображения окрашенных срезов получали с помощью системы Aperio ScanScope AT Turbo (Leica Biosystems). Антитело 6470 тестировали на срезах мозга, полученных от пяти разных pS129-α-синуклеин-положительных и трех разных pS129-α-синуклеин-отрицательных доноров (1 срез/донор). Антитело 6470 окрашивало нейропили и структуры подобные тельцам Леви в височной коре и черной субстанции пациентов с PD (фиг. 12A-E). В тканях мозга без PD антитело 6470 окрашивало нейропили в височной коре, но в коре или черной субстанции не наблюдали никаких структур, подобных тельцам Леви (фиг. 12F-H). Эти наблюдения предполагают, что антитело 6470 связывается с нормальным α-синуклеином в нейропиле тканей мозга от пациентов с PD и без PD, в то время как оно связывается с патологическим α-синуклеином, присутствующим в тельцах Леви только у пациентов с PD.
Пример 7: Анализ агрегации на основе клеток
Клетки HEK Freestyle 293F (суспензионные клетки) засевали с плотностью 0,7×106 клеток/мл в среде для экспрессии Freestyle 293 (Invitrogen™) и культивировали до плотности 300×106 клеток/мл. Трансфекцию выполняли в соответствии с инструкциями производителя и быстро смешивали с 600 мкг pcDNA3.1(+), включающей ген альфа-синуклеина, в 20 мл среды OptiMEM, и при этом разводили 293Fectin в среде OptiMEM (Invitrogen™) и инкубировали в течение 5 минут при комнатной температуре. Разбавленную ДНК добавляли и инкубировали при комнатной температуре в течение 20 минут, а затем добавляли по каплям к клеткам (20 мл на колбу). Клетки инкубировали в течение 24 часов при 37°С, 125 об/мин, 8% СО2. Клетки либо использовали сразу, либо замораживали с плотностью 5 миллионов клеток/мл в FBS + 10% ДМСО.
Если клетки были предварительно заморожены, криопробирки оттаивали и клетки ресуспендировали в среде Freestyle 293, центрифугировали при 500g в течение 5 минут, супернатант сливали, и осадок ресуспендировали в среде Freestyle 293 (Life Technologies™), содержащей Pen/Strep (Invitrogen™) с плотностью 2×106 клеток/мл. В 384-луночный планшет (Grainer™) добавляли 20 мкл клеточной суспензии (всего около 40000 клеток/лунку). В каждую лунку добавляли 150 нМ фибрилл человеческого альфа-синуклеина (полученных, как раскрыто в настоящем описании в Примере 1), а затем антитела (6470 gL3gH23; 6470 gL3gH36; 6470 gL3N33gH23 S56N N102; 6470 gL3N33gH36 S56N N102; 6470 gL3gH23 S56N N102; 6470 gL3gH36 S56N N102; все в виде полноразмерного IgG4P, последовательности приведены в таблице 1) в PBS для тестирования (в различных концентрациях). Планшеты инкубировали при 37°C, 5% CO2, 95% влажности в инкубаторе для клеточных культур в течение 2 дней.
В конце второго дня среду удаляли из всех лунок, и планшет промывали, оставляя по 20 мкл на лунку. Примерно 50 мкл PBS добавляли в каждую лунку, и планшеты центрифугировали при 500 g в течение 5 минут. Супернатант аспирировали из всех лунок с помощью устройства для промывки планшетов, оставляя по 20 мкл среды в каждой лунке. Добавляли Versene (Lonza™) (50 мкл/лунку), и планшеты центрифугировали при 500 g в течение 5 минут, супернатант аспирировали, оставляя только по 20 мкл среды на лунку. В каждую лунку добавляли 20 мкл 8% формальдегида (16% водный раствор, Life Technologies™) + 2% Тритон Х-100 (VWR™) в PBS. Планшеты инкубировали при комнатной температуре в течение 15 минут, и после этого добавляли 50 мкл буфера FACS, состоящего из HBSS (VWR™ без кальция и магния) + 2% FBS + 2 мМ EDTA (Life Technologies™). Планшеты центрифугировали при 2000 g в течение 1 минуты, и супернатант аспирировали, оставляя только 20 мкл среды в каждой лунке. В каждую лунку дополнительно добавляли 20 мкл буфера FACS с антителом к альфа-синуклеину pSer129 (AbCam™), разведенным 1:300. Планшеты инкубировали в течение 1 часа при комнатной температуре, а затем в каждую лунку добавляли 50 мкл буфера FACS перед повторным центрифугированием при 2000 g в течение 1 минуты. Супернатант удаляли перед добавлением в каждую лунку вторичного конъюгированного с Alexafluor647 анти-кроличьего антитела (Life Technologies™), разведенного 1:500, и DAPI (Life Technologies™). Планшеты инкубировали в течение 1 часа при комнатной температуре в темноте, а затем добавляли 50 мкл буфера FACS, и планшеты центрифугировали при 2000 g в течение 1 минуты. После промывки добавляли больше буфера FACS, и планшеты были готовы для помещения в проточный цитометр (BD FACS Canto II) для считывания.
Данные FACS анализировали с помощью программного обеспечения FlowJo. Во-первых, живые единичные клетки подвергали стробированию с помощью прямого и бокового рассеяния. Во-вторых, стробировали события DAPI+, и их количество использовали в качестве меры количества живых, ядросодержащих единичных клеток. Наконец, меченные фосфосерином 129 альфа-синуклеин-положительные (pSer129+) клетки подвергали стробированию. Процент pSer129+ клеток относительно всех DAPI+ клеток использовали в качестве меры агрегации. Данные нормализовали относительно лунок, обработанных только фибриллами и без антител, и выражали в процентах.
Результаты приведены на фиг. 13, где показана способность тестируемых антител ингибировать индуцированную фибриллами альфа-синуклеина агрегацию на клетках, экспрессирующих альфа-синуклеин. Эти данные подтверждают, что антитела по настоящему изобретению способны блокировать индуцированную фибриллами альфа-синуклеина агрегацию с IC50 менее 5 нМ.
Планки погрешности представляют стандартную ошибку измерения (SEM, N=3, n=9).
Пример 8: Анализ агрегации первичных нейронов
Гиппокампы из мышиных эмбрионов E17 иссекали в диссекционном буфере (HBSS без кальция и без магния, 0,6% D-(+)-глюкозы, 20 мМ Hepes). Затем диссекционный буфер удаляли и заменяли раствором для диссоциации (HBSS без кальция и без магния, 0,6% D-(+)-глюкозы, 20 мМ HEPES, 40 ед/мл папаина, 1 мг/мл ДНКазы, 1 мМ L-цистеина, 0,5 мм ЭДТА). После 30 минут инкубации при 37°С буфер для диссоциации удаляли, и гиппокампы промывали 3 раза средой для посева (среда Neurobasal™, добавка 2% B27, 1 мМ GlutaMAX, 2,5% FBS, 50 ед./мл пенициллин-стрептомицин). Сгустки ткани разбивали с помощью пипетки объемом 1 мл с получением суспензии единичных клеток. Клетки разводили до соответствующей концентрации в среде для посева. Примерно 15000 клеток высевали в каждую лунку 384-луночного планшета, покрытого PDL. Затем клетки хранили в инкубаторе для клеточных культур при 37°С, 5% СО2, 95% влажности.
На следующий день 80% среды заменяли средой для посева без FBS [среда Neurobasal™, добавка 2% B27, 1 мМ GlutaMAX, 50 ед/мл пенициллин-стрептомицин). Через семь дней после посева среду удаляли, оставляя по 20 мкл в каждой лунке. В каждую лунку добавляли 100 нМ фибрилл человеческого альфа-синуклеина (полученных, как раскрыто в настоящем описании в примере 1), а затем антитело 6470 (6470gL3gH36 hIgG4P; VR6470 на фиг.14, содержащее SEQ ID NO:17 и SEQ ID NO:33) в PBS для тестирования (в различных концентрациях). Планшет инкубировали при 37°C, 5% CO2, 95% влажности в инкубаторе для клеточных культур в течение дополнительных 7 дней. Через четырнадцать дней после посева среду аспирировали из всех лунок, оставляя 20 мкл на лунку. Каждую лунку промывали 80 мкл физиологического раствора Дульбекко с фосфатным буфером (DPBS). DPBS удаляли, и клетки инкубировали в 40 мкл буфера для фиксации (DPBS с 4% параформальдегидом) на лунку в течение 15 минут. Затем буфер для фиксации удаляли, и клетки снова промывали 80 мкл DPBS. DPBS удаляли и заменяли 40 мкл буфера для пермеабилизации (DPBS с 0,1% Тритон X-100) на лунку. Через 10 минут буфер для пермеабилизации удаляли, и клетки инкубировали в течение 1 часа в 40 мкл блокирующего буфера (PBS с 1% BSA и 0,1% Тритон X-100) на лунку. Затем блокирующий буфер удаляли и заменяли раствором первичного антитела с концентрацией 40 мкл/лунку (блокирующий буфер с 0,3%-ным меченным фосфосерином 129 кроличьим анти-антителом к альфа-синуклеину (AbCam™ ab51253). Раствор антитела инкубировали на клетках в течение 1 часа, затем трижды промывали (90 мкл/каждую, PBS). После последней промывки PBS удаляли и заменяли на 40 мкл раствора вторичного антитела (0,1% анти-кроличьего антитела, конъюгированного с AlexaFluor647, в PBS с 0,2% антитела к бета-III-тубулину, конъюгированного с AlexaFluor488). Раствор вторичного антитела инкубировали в лунках в течение 1 часа, затем удаляли и заменяли 40 мкл PBS, содержащего 0,3% CellMask Blue™. После 5 минут инкубации лунки промывали 3 раза 80 мкл PBS, затем заполняли 50 мкл PBS на лунку перед тем, как планшет запечатывали прозрачной пластиковой пленкой.
Планшеты обрабатывали на устройстве для визуализации планшетов Arrayscan (ThermoFisher Scientific™). Изображения анализировали с помощью программного обеспечения HCS Scan™ того же производителя. Плотность нейронов контролировали с помощью сигнала бета-III-тубулина. Зоны разреженного расположения клеток или зоны с поврежденны со слоем поврежденных нейрональных клеток, определяемые по значительному уменьшению поверхности с сигналом бета-III-тубулина, исключали. Наконец, для количественной оценки патологической агрегации альфа-синуклеина использовали поверхность с сигналом pSer129 альфа-синуклеина на одну зону.
Считается, что фосфорилирование альфа-синуклеина по S129 играет важную роль в контроле нормальных функций альфа-синуклеина, а также в регуляции его агрегации, образования LB и нейротоксичности. В нормальных условиях только небольшая доля альфа-синуклеина конститутивно фосфорилирована по S129 в мозге (Fujiwara H, et al. (2002) Nat Cell Biol, 4, 160-164), тогда как значительное накопление pS129 наблюдается в мозге пациентов, страдающих синуклеинопатиями (Kahle PJ, et al. (2000) Ann NY Acad Sci, 920, 33-41); Okochi M, et al. (2000) J Biol Chem, 275, 390-397); Anderson JP, et al. (2006) J Biol Chem, 281, 29739-29752).
Данные нормализовали относительно лунок, обработанных только фибриллами без антител, и выражали в процентах. Как показано на фиг. 14, 6470gL3gH36 (обозначен как VR6470) ингибирует агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина на первичных нейронах мышей, экспрессирующих эндогенные уровни альфа-синуклеина. Планки погрешности представляют стандартную ошибку измерения (SEM, N=4, n=18). Эти данные подтверждают, что 6470gL3gH36 способен блокировать индуцированную фибриллами альфа-синуклеина агрегацию в первичных мышиных нейронах с IC50 ниже, чем 4 нМ.
Пример 9: Оценка эффективности VR6470 in vivo
Антитело, содержащее вариабельную область SEQ ID NO:17 и SEQ ID NO:33 и константную область IgG1 мыши (назван в этом примере VR6470) тестировали на мышах-самцах дикого типа C57Bl/6J (Janvier, Франция) и в трансгенной модели нокаутной по α-синуклеину мыши, экспрессирующей человеческий альфа-синуклеин (далее упрминаемой как SNCA-OVX; Charles River, France).
Мышам C57Bl/6J и SNCA-OVX инъецировали указанное антитело и предварительно сформированные мышиные фибриллы (PFF) (полученные, как раскрыто в настоящем описании в примере 1). Антитело отрицательного контроля (101,4) и носитель также инъецировали вместе с антителом сравнения к альфа-синуклеину (C-терминальное Ab сравнения), которое связывается с последними девятью С-концевыми остатками альфа-синуклеина. Такое антитело сравнения (которое имеет CDR, отличные от антител по настоящему изобретению), показало сопоставимые характеристики связывания с антителами по настоящему изобретению. Антитело сравнения обладает более высоким сродством к альфа-синуклеину, чем антитела по настоящему изобретению, и аналогичными биофизическими свойствами. Оно также одинаково эффективно в предотвращении агрегации альфа-синуклеина в анализах на основе клеток HEK в соответствии с примером 8 (таблица 11).
Антитела предварительно инкубировали с PFF в течение 30 минут на шейкере при комнатной температуре перед непосредственным введением в мозг животных. Смеси антитело/PFF готовили в PBS с соотношением 1 мкг PFF/10 мкг антител. PBS (pH 7,4) использовали в качестве раствора носителя. Одну инъекцию вводили за 24 часа до комбинированного интрацеребрального введения.
Затем антитела вводили мышам внутрибрюшинно в дозе 30 мг/кг. Вторую внутрибрюшинную инъекцию вводили через 7 дней после первой, и затем использовали такую же схему лечения (одну внутрибрюшинную инъекцию в неделю в дозе 30 мг/кг с объемом введения 10 мл/кг) в течение 4 недель, получая, в общей сложности, 4 инъекции мышам-самцам дикого типа C57Bl/6J, и в течение 11 недель, получая, в общей сложности, 12 инъекций мышам SNCA-OVX. В обоих экспериментах мышей случайным образом распределяли по группам лечения лекарственными средствами, эксперимент выполняли слепым способом в отношении лечения.
Эксперименты на животных выполняли в соответствии с руководящими принципами Европейской директивы 2010/63/EU и бельгийского законодательства. Этический комитет по экспериментам на животных UCB Biopharma SPRL (LA1220040 и LA2220363) утвердил протокол эксперимента (ASYN-IC-PARKINSON-MO). На момент операции мыши весили от 25 до 30 г и были 17-недельного возраста. Мышей содержали в клетках (4 мыши на клетку, Macrolon типа 2). Их содержали в условиях суточного цикла 12:12 день/ночь, и свет включали в 06:00. Температуру поддерживали на уровне 20-21°С, а влажность составляла приблизительно 40%. Все животные имели свободный доступ к стандартным пищевым гранулам и воде перед распределением в экспериментальные группы. До и после операции предоставляли обогащенный корм и надлежащие условия содержания (Enviro-dri, Pharma Serv). За здоровьем животных ежедневно следил персонал по уходу за животными. Для минимизации страданиий животных были приложены все усилия. Животных умерщвляли под наркозом.
Операцию проводили под общим наркозом, используя смесь 50 мг/кг кетамина (Nimatek, Eurovet Animal Health B.V.) и 0,5 мг/кг медетомидина (Domitor, Orion Corporation), которую вводили внутрибрюшинно. Кроме того, для поддержания животных в бодрствующем состоянии давали 2,5 мг/кг атипамезола (Antisedan, Orion Corporation). Очищенные рекомбинантные PFF оттаивали и обрабатывали ультразвуком при комнатной температуре (Qsonica 500 - 20 кГц; мощность 65%, 30 импульсов продолжительностью 1 сек (ВКЛ.) с 1 сек перерывом (ВЫКЛ.) в течение одной минуты). Затем PFF предварительно смешивали с антителами в течение 30 минут и встряхивали при комнатной температуре в течение 30 минут перед инъекцией в мозг. Раствор (2 мкл) вводили со скоростью 0,2 мкл/мин, и иглу оставляли на месте еще на 2,5 минуты до ее медленного втягивания. Инъекции осуществляли в одностороннем порядке в правый стриатум в следующих координатах: AP = +0,20 мм, ML = -2,00 мм, DV = -3,20 мм.
После анестезии мышей перфузировали транскардиальной перфузией раствором 0,9% PBS, охлажденным льдом, содержащим 10 ед/мл гепарина, в течение 9 минут со скоростью потока 6 мл/мин через левый желудочек. Правое предсердие надрезали как маршрут оттока. Затем животных перфузировали охлажденным на льду 4% параформальдегидом в PBS в течение 15 минут со скоростью потока 6 мл/мин. Мозг фиксировали в течение ночи в PBS, содержащем 4% параформальдегида, при 4°C (день 0). На следующее утро (день +1) 4% параформальдегид удаляли, а мозг промывали холодным PBS и инкубировали в течение ночи. На следующий день (день +2) мозг промывали PBS в течение минимум 1 часа и переносили в PBS, содержащий 15% сахарозы, и хранили при 4°C до отправки.
Срезы мозга выполняли в Neuroscience Associates (TN, США). Сначала мозг обрабатывали в течение ночи 20% глицерином и 2% диметилсульфоксидом для предотвращения образования артефактов во время заморозки и помещали в желатиновую матрицу согласно методу MultiBrain®. После отверждения блоки быстро замораживали погружением в изопентан, охлажденный до -70°C измельченным сухим льдом, и устанавливали на стадии заморозки санного микротома AO860. Срезы блоков толщиной 40 мкм выполняли с помощью MultiBrain® в корональной плоскости. Все срезы собирали последовательно в 24 контейнера на блок, которые заполняли раствором Antigen Preserve (49% PBS pH 7,0, 50% этиленгликоль, 1% поливинилпирролидон). Неокрашенные срезы сразу помещали на хранение при -20°С.
Свободно плавающие срезы окрашивали иммунохимией с помощью антитела к альфа-синуклеину pSer129 (мышиный анти-альфа-синуклеин (pSer129), биотин - (Wako - 010-26481)), разведенного в соотношении 1:30000. Во всех инкубационных растворах из блокирующей сыворотки далее использовали буферный раствор Tris (TBS) с Тритон X-100 в качестве носителя; все промывки осуществляли TBS. Эндогенную пероксидазную активность блокировали обработкой перекисью водорода 0,9%, а неспецифическое связывание блокировали 1,26% цельной нормальной сывороткой. После промывки срезы окрашивали первичным антителом в течение ночи при комнатной температуре. Раствор носителя содержал 0,3% Тритон X-100 для пермеабилизации. После промывки срезы инкубировали с комплексом авидин-биотин-HRP (набор Vectastain Elite ABC, Vector Laboratories, Burlingame, CA) в течение одного часа при комнатной температуре. После промывки срезы обрабатывали диаминобензидин тетрагидрохлоридом (DAB) и 0,0015% перекисью водорода для создания видимого продукта реакции, закрепленного на желатинизированных (суббедовых) предметных стеклах, высушенных на воздухе, слегка окрашенных тионином, обезвоженных в спиртах, очищенных в ксилоле и покрытых Permount.
Флуоресцентную иммуногистохимию для p62/SQS™1 (известно, что p62 коагрегирует в тельцах Леви у человека) и Amytracker (для агрегатов белка в целом) выполняли на свободноплавающих срезах мозга. Было показано, что VR6470 уменьшает количество агрегированных белков, окрашенных Amytracker, и совместно локализуется с pS129. Это указывает на то, что антитело VR6470 не только уменьшает количество фосфосинуклеина, но также уменьшает количество агрегатов синуклеина (данные не показаны).
Количество сигнала pSer129 альфа-синуклеина на одно поле зрения сигнала pSer129 альфа-синуклеина использовали для количественной оценки патологической агрегации альфа-синуклеина на ипсилатеральной стороне полосатого тела, коры, базолатеральных комплексах миндалины и черной субстанции. Представляющие интерес области (ROI) очерчивали вручную, и автоматическое количественное определение сигнала pSer129 альфа-синуклеина в различных областях мозга выполняли с помощью программного обеспечения VisioPharm 6 (VisioPharm). Для количественной оценки сигнала pSer129 альфа-синуклеина использовали линейный байесовский алгоритм, который обеспечивает значение площади сигнала (площадь метки в мкм2). Площадь метки отражает величину патологии pSer129 альфа-синуклеина, которая охватывает различные области мозга. Все количественные оценки выполняли слепым методом до конца статистического анализа.
Анализ данных выполняли для % площади метки (т.е. отношение между площадью сигнала pSer129 в мкм2 и площадью представляющей интерес области в мкм2). Площадь метки в % оценивали повторно для нескольких срезов мозга, расположенных ростро-каудально (стриатум: 13-14 срезов от Bregma +1,1 до -0,94; кора: 13-14 срезов от +1,1 до -0,94; базолатеральная миндалина: 6-10 срезов от -0,58 до -2,06; черная субстанция: 6-8 срезов от -2,54 до -3,88), а AUC вычисляли отдельно для каждого испытуемого.
Для статистического анализа использовали односторонний ANOVA. ANOVA сопровождалось множеством попарных сравнений между средними без какого-либо учета множественности сравнеий (** р<0,01 и * р<0,05). Данные подвергали логарифмическому преобразованию для соответствия критериям нормальности и гомоскедастичности. Графики представляют геометрические средние непреобразованных данных.
Как показано на фиг.15А и 15В, антитело 6470 (содержащее вариабельные области SEQ ID NO:17 и 33 и константную область IgG1 мыши) заметно уменьшало патологию альфа-синуклеина (т.е. сигнал pSer129 альфа-синуклеина) по сравнению с тремя контрольными группами в четырех различных ипсилатеральных областях мозга, включая стриатум, кору головного мозга, миндалину и черную субстанцию, через один месяц после введения мышиных PFF мышам-самцам дикого типа C57Bl/6J (фиг. 15A) и через три месяца после введения человеческих PFF мышам-самцам SNCA-OVX (фиг. 15B).
На фиг. 16 показано количественное определение альфа-синуклеина, фосфорилированного по Ser129 (AUC% площади маркера) в ипсилатеральной коре, полосатом телеце, миндалине и черной субстанции у мышей дикого типа C57Bl/6J, соответственно. Антитело отрицательного контроля и С-концевое антитело сравнения не уменьшали патологию альфа-синуклеина по сравнению с группой, получавшей носитель. В отличие от этого, антитело 6470 значительно уменьшало уровень патологии (т.е. сигнал pSer129альфа-синуклеина) в коре, стриатуме, миндалине и черной субстанции (р<0,01) по сравнению с тремя контрольными группами мышей C57Bl/6J, которым вводили мышиные PFF. Среди протестированных мышей C57Bl/6J дикого типа группа, получавшая 6470, показала значительное уменьшение уровней pSer129 альфа-синуклеина в четырех различных структурах, три из которых - дистальные области от места инъекции (кора, черная субстанция и миндалина).
У мышей SNCA OVX, которым инъецировали человеческие PFF, антитело 6470 значительно снижало уровень патологии в коре и стриатуме по сравнению с мышами, которые получали носитель, антитело отрицательного контроля (101.4) и С-концевое антитело сравнения. У мышей SNCA-OVX 6470 значительное уменьшение уровня pSer129 наблюдалось по меньшей мере в двух различных структурах мозга (кора и стриатум), где по меньшей мере одна (кора головного мозга) расположена дистальнее места инъекции.
Эти результаты подтверждают, что антитела, содержащие структурные признаки по настоящему изобретению способны предотвращать in vivo появление альфа-синуклеина, фосфорилированного по Ser129.
Кроме того, результаты показали, что не все антитела, которые связываются с альфа-синуклеином в С-концевой области, являются эффективными in vivo. Антитело сравнения, которое связывается с самим С-концом альфа-синуклеина с высоким сродством и которое эффективно предотвращает агрегацию альфа-синуклеина в клеточных анализах, не смогло предотвратить фосфорилирование Ser129 in vivo.
Следовательно, антитело по настоящему изобретению можно использовать для лечения альфа-синуклеинопатий, например, для которых характерно увеличение фосфорилирования Ser129, включая болезнь Паркинсона (PD) (включая идиопатические и наследственные формы болезни Паркинсона), деменцию с тельцами Леви (DLB), болезнь диффузных телец Леви (DLBD), вариант болезни Альцгеймера с тельцами Леви (LBVAD), комбинированную болезнь Альцгеймера и Паркинсона, множественную системную атрофию (MSA) и нейродегенерацию с накоплением железа в мозге 1-го типа (NBIA-1).
Пример 10: Фармакокинетика антитела 6470 у мыши
Самцам мышей C57/Bl6 (n=3 на лекарство) внутривенно вводили в виде разовой дозы 2 мг/кг антитела 6470gL3gH36 IgG4P (содержащего SEQ ID NO:17 и 33; на фиг. 17 и далее обозначаемое как 6470).
Образцы крови брали (через 0,083, 1, 4, 8, 24, 72, 120, 168 и 336 часов после инъекции) из хвостовой вены и оставляли для коагуляции при комнатной температуре. Сыворотку выделяли после центрифугирования, которую затем замораживали и хранили до выполнения анализа. Количественную оценку 6470 выполняли методом LC-MS/MS. Образцы сыворотки, полученные в ходе исследования, размораживали, и выполняли количественную оценку по калибровочной кривой, полученной с помощью антитела 6470 или антитела сравнения, добавляемого в различных концентрациях в контрольную мышиную сыворотку. Перед инъекцией образцов в систему LC-MS/MS сыворотку денатурировали, восстанавливали и алкилировали с помощью ацетонитрила (VWR, Великобритания), TCEP-Трис (2-карбоксиэтил) гидрохлорида фосфина (Sigma, Великобритания) и йодацетамида (Sigma, Великобритания), соответственно. Затем алкилированные образцы восстанавливали в 100 мМ бикарбонатном буфере аммония (Sigma, Великобритания) и расщепляли в течение ночи ферментом трипсин (Promega, UK) при 37°C. Расщепление останавливали добавлением к образцам муравьиной кислоты для понижения рН, а затем обессоливали, используя планшет Waters HLB SPE. Полученный элюент упаривали с помощью вакуумного испарителя. После того, как образцы были полностью высушены, их восстанавливали 95/5 смесью вода/ацетонитрил, содержащей 0,1% муравьиной кислоты, и вводили в систему LC-MS/MS. Анализ ЖХ-МС/МС выполняли с помощью Prominence системы для ВЭЖХ (компании Schimadzu), соединенной с масс-спектрометром Triple Quad 6500. Расщепленный образец впрыскивали автосамплером в колонку для высокоэффективной жидкостной хроматографии с обращенной фазой (пептидная колонка Phenomenex Aeris C18 100×2.1 мм, 2,6 мкм), которую поддерживали при 50°C. В течение 6 минут применяли линейный градиент 5-70% ацетонитрила в 0,1% муравьиной кислоте, а затем увеличивали до 95% ацетонитрила в 0,1% муравьиной кислоте в течение 0,8 минут со скоростью потока 0,6 мл/мин. Масс-спектрометр устанавливали на выполнение анализа мониторинга множественных реакций для обнаружения множественных переходов пептидов 6470 или 5811 с временем выдержки 50 миллисекунд на переход. Анализ данных выполняли с помощью программного обеспечения Analyst 1.6.
Эти данные демонстрируют, что у мышей антитело 6470 показало очень хорошие фармакокинетические свойства (таблица 12 и фиг. 17А), что отражено низкими измеренными значениями клиренса. Они, по всей видимости, превосходят типичные диапазоны, указанные для препаратов человеческого IgG, вводимых мышам (3-16 мл/сутки/кг; Deng et al 2011, mabs 3:161-66).
Фармакокинетические свойства антитела 6470 также исследовали на обезьянах Cynomolgus и сравнивали с антителом предшествующего уровня техники. Самцам обезьян Cynomolgus (n=3 или n=6 на препарат) внутривенно вводили однократную дозу 2 или 3 мг/кг антитела 6470gL3gH36 IgG4P (6470) и другого антитела сравнения к альфа-синуклеину (IgG1 антитело к альфа-синуклеину, которое связывается с альфа-синуклеином по аминокислотам 118-126; WO2013/063516).
Образцы крови брали в различные моменты времени (0,083, 1, 3, 6, 24, 48, 96, 168, 240, 336, 504, 576, 672 часа после инъекций) и оставляли для коагуляции при комнатной температуре. Сыворотку выделяли после центрифугирования, которую затем замораживали и хранили до выполнения анализа. Образцы размораживали и анализировали с помощью LC/ESI MS/MS. Для 6470 использовали метод, описанный в этом примере выше, с количественным определением, выполненным путем получения стандартной кривой в сыворотке Cynomolgus. Для антитела сравнения в качестве внутреннего стандарта использовали лошадиный миоглобин, а количественную оценку выполняли путем сравнения сигналов с внутренним стандартным сигналом. Для этого образцы смешивали с внутренним стандартом. Затем образцы денатурировали, алкилировали и далее подвергали ферментативному расщеплению в течение ночи (трипсин). После расщепления образцы разбавляли, и сигнатурные пептиды всех аналитов подвергали анализу ЖХ-МС/МС. Образцы готовили только один раз и вводили дважды (один раз для каждого метода).
Профили концентрации в зависимости от времени анализировали с помощью Pharsight Phoenix 6, используя некомпартментальный анализ для определения фармакокинетических параметров клиренса и периода полувыведения у каждого отдельного животного. Значения среднего и стандартного отклонения приведены для каждой молекулы.
Как показано на фиг. 17B и в таблице 12, у обезьян Cynomolgus антитело 6470 также демонстрирует превосходные фармакокинетические свойства, с низким значением клиренса. Как и у мышей, его фармакокинетические характеристики, по всей видимости, превосходят типичный диапазон, указанный для препаратов человеческого IgG, вводимых обезьянам Cynomolgus (5-12 мл/сутки/кг; Deng et al 2011, mabs 3:161-66).
Быстрый клиренс антител сравнения, наблюдаемый у Cynomolgus, согласуется с опубликованными данными, полученными у людей (JAMA Neurology 2018, 75, 10:1206-14). Антитело 6470 превосходит антитело сравнения, демонстрирующее неудовлетворительные, атипичные фармакокинетические характеристики и параметры, как по экспозиции, так и по клиренсу.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЮСБ Биофарма СРЛ
<120> Антитела
<130> PF0130-WO
<150> GB1720975.0
<151> 2017-12-15
<160> 49
<170> PatentIn version 3.5
<210> 1
<211> 13
<212> Белок
<213> Oryctolagus cuniculus
<400> 1
Gln Ala Ser Gln Ser Val Tyr Lys Asn Asn Tyr Leu Ala
1. 5 10
<210> 2
<211> 7
<212> Белок
<213> Oryctolagus cuniculus
<400> 2
Gly Ala Ser Thr Leu Ala Ser
1. 5
<210> 3
<211> 12
<212> Белок
<213> Oryctolagus cuniculus
<400> 3
Ala Gly Tyr Lys Gly Gly Arg Asn Asp Gly Phe Ala
1. 5 10
<210> 4
<211> 10
<212> Белок
<213> Oryctolagus cuniculus
<400> 4
Gly Ile Asp Leu Ser Ser His Asp Met Tyr
1. 5 10
<210> 5
<211> 16
<212> Белок
<213> Oryctolagus cuniculus
<400> 5
Ala Ile Tyr Ala Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1. 5 10 15
<210> 6
<211> 9
<212> Белок
<213> Oryctolagus cuniculus
<400> 6
Ile His Tyr Gly Asn Ser Gly Gly Leu
1. 5
<210> 7
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR-L1 N33R
<400> 7
Gln Ala Ser Gln Ser Val Tyr Lys Asn Arg Tyr Leu Ala
1. 5 10
<210> 8
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR-H2 S56N
<400> 8
Ala Ile Tyr Ala Ser Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1. 5 10 15
<210> 9
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR-H3 N102H
<400> 9
Ile His Tyr Gly His Ser Gly Gly Leu
1. 5
<210> 10
<211> 140
<212> Белок
<213> Homo sapiens
<400> 10
Met Asp Val Phe Met Lys Gly Leu Ser Lys Ala Lys Glu Gly Val Val
1. 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Ala Glu Ala Ala Gly Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Lys Glu Gly Val
35 40 45
Val His Gly Val Ala Thr Val Ala Glu Lys Thr Lys Glu Gln Val Thr
50 55 60
Asn Val Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Val Lys
85 90 95
Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile
100 105 110
Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro
115 120 125
Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
130 135 140
<210> 11
<211> 112
<212> Белок
<213> Oryctolagus cuniculus
<400> 11
Ala Ile Val Met Thr Gln Thr Pro Ser Ser Lys Ser Val Ala Val Gly
1. 5 10 15
Asp Thr Val Thr Ile Asn Cys Gln Ala Ser Gln Ser Val Tyr Lys Asn
20 25 30
Asn Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Gln
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Val
65 70 75 80
Val Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Gly Gly
85 90 95
Arg Asn Asp Gly Phe Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 12
<211> 336
<212> ДНК
<213> Oryctolagus cuniculus
<400> 12
gccatcgtga tgacccagac tccatcttcc aagtctgtcg ctgtgggaga cacagtcacc 60
atcaattgcc aggccagtca gagtgtttat aagaacaact acttagcctg gtttcaacag 120
aaaccagggc agcctcccaa acaactgatc tatggtgcgt ccactctggc atctggggtc 180
ccatcgcggt tcaaaggcag tggatctggg acacagttca ctctcaccat cagcgatgtg 240
gtgtgtgacg atgctgccac ttactactgt gcaggatata aaggtggtcg taatgatggt 300
tttgctttcg gcggagggac cgaggtggtg gtcaaa 336
<210> 13
<211> 114
<212> Белок
<213> Oryctolagus cuniculus
<400> 13
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1. 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser His Asp
20 25 30
Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile Gly
35 40 45
Ala Ile Tyr Ala Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Met Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Ile His
85 90 95
Tyr Gly Asn Ser Gly Gly Leu Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser
<210> 14
<211> 342
<212> ДНК
<213> Oryctolagus cuniculus
<400> 14
cagtcggtgg aggagtccgg gggtcgcctg gtcacgcctg ggacacccct gacactcacc 60
tgcacagtct ctggaatcga cctcagtagc cacgacatgt attgggtccg ccaggctcca 120
gggaaggggc tggaatacat tggagccatt tatgctagtg gtagcacata ctacgcgagc 180
tgggcgaaag gccgattcac catctccaag acctcgacca cggtggatct gaaaatgacc 240
agtctgacaa ccgaggacac ggccacctat ttctgtgcca gaattcatta tggtaatagt 300
ggtgggttgt ggggccaagg caccctggtc accgtctcga gt 342
<210> 15
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 gL3 VL
<400> 15
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Val Tyr Lys Asn
20 25 30
Asn Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Gln
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Gly Gly
85 90 95
Arg Asn Asp Gly Phe Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 16
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 6470 gL3 VL нукл.
<400> 16
gacattcaga tgacccagtc cccttcatca ctgtccgcga gcgtgggcga cagagtgacc 60
attacgtgcc aagccagcca gtccgtgtac aagaacaact acctggcctg gttccagcaa 120
aagcccggga aggcgccaaa acagcttatc tacggtgcat ccactctcgc ctcgggagtg 180
ccgagccgct tctcgggatc tgggtccgga actcagttca ccctgactat ctcgtccctg 240
caacccgagg atttcgccac ctactactgc gccggctata agggaggacg gaacgacggc 300
ttcgcttttg gtggaggcac caaggtcgaa atcaag 336
<210> 17
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 gL3 Легкая цепь
<400> 17
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Val Tyr Lys Asn
20 25 30
Asn Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Gln
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Gly Gly
85 90 95
Arg Asn Asp Gly Phe Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 18
<211> 657
<212> ДНК
<213> Искусственная последовательностьe
<220>
<223> 6470 gL3 Легкая цепь нукл.
<400> 18
gacattcaga tgacccagtc cccttcatca ctgtccgcga gcgtgggcga cagagtgacc 60
attacgtgcc aagccagcca gtccgtgtac aagaacaact acctggcctg gttccagcaa 120
aagcccggga aggcgccaaa acagcttatc tacggtgcat ccactctcgc ctcgggagtg 180
ccgagccgct tctcgggatc tgggtccgga actcagttca ccctgactat ctcgtccctg 240
caacccgagg atttcgccac ctactactgc gccggctata agggaggacg gaacgacggc 300
ttcgcttttg gtggaggcac caaggtcgaa atcaagcgta cggtggccgc tccctccgtg 360
ttcatcttcc caccctccga cgagcagctg aagtccggca ccgcctccgt cgtgtgcctg 420
ctgaacaact tctacccccg cgaggccaag gtgcagtgga aggtggacaa cgccctgcag 480
tccggcaact cccaggaatc cgtcaccgag caggactcca aggacagcac ctactccctg 540
tcctccaccc tgaccctgtc caaggccgac tacgagaagc acaaggtgta cgcctgcgaa 600
gtgacccacc agggcctgtc cagccccgtg accaagtcct tcaaccgggg cgagtgc 657
<210> 19
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 gL3 VL N33R
<400> 19
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Val Tyr Lys Asn
20 25 30
Arg Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Gln
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Gly Gly
85 90 95
Arg Asn Asp Gly Phe Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 20
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 6470 gL3 VL N33R нукл.
<400> 20
gacattcaga tgacccagtc cccttcatca ctgtccgcga gcgtgggcga cagagtgacc 60
attacgtgcc aagccagcca gtccgtgtac aagaaccgtt acctggcctg gttccagcaa 120
aagcccggga aggcgccaaa acagcttatc tacggtgcat ccactctcgc ctcgggagtg 180
ccgagccgct tctcgggatc tgggtccgga actcagttca ccctgactat ctcgtccctg 240
caacccgagg atttcgccac ctactactgc gccggctata agggaggacg gaacgacggc 300
ttcgcttttg gtggaggcac caaggtcgaa atcaag 336
<210> 21
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 gL3 Легкая цепь N33R
<400> 21
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Val Tyr Lys Asn
20 25 30
Arg Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Gln
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Gly Gly
85 90 95
Arg Asn Asp Gly Phe Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 22
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 6470 gL3 Легкая цепь N33R нукл.
<400> 22
gacattcaga tgacccagtc cccttcatca ctgtccgcga gcgtgggcga cagagtgacc 60
attacgtgcc aagccagcca gtccgtgtac aagaaccgtt acctggcctg gttccagcaa 120
aagcccggga aggcgccaaa acagcttatc tacggtgcat ccactctcgc ctcgggagtg 180
ccgagccgct tctcgggatc tgggtccgga actcagttca ccctgactat ctcgtccctg 240
caacccgagg atttcgccac ctactactgc gccggctata agggaggacg gaacgacggc 300
ttcgcttttg gtggaggcac caaggtcgaa atcaagcgta cggtggccgc tccctccgtg 360
ttcatcttcc caccctccga cgagcagctg aagtccggca ccgcctccgt cgtgtgcctg 420
ctgaacaact tctacccccg cgaggccaag gtgcagtgga aggtggacaa cgccctgcag 480
tccggcaact cccaggaatc cgtcaccgag caggactcca aggacagcac ctactccctg 540
tcctccaccc tgaccctgtc caaggccgac tacgagaagc acaaggtgta cgcctgcgaa 600
gtgacccacc agggcctgtc cagccccgtg accaagtcct tcaaccgggg cgagtgc 657
<210> 23
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 gH23 VH
<400> 23
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Ser His
20 25 30
Asp Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Ala Ile Tyr Ala Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ile His Tyr Gly Asn Ser Gly Gly Leu Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 24
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 6479gH23 VH нукл.
<400> 24
gaggttcagc tgctggagtc tggaggcggg cttgtccagc ctggagggag cctgcgtctc 60
tcttgtgcag taagcggcat cgacctgtcc agccacgaca tgtattgggt acgtcaggca 120
ccgggtaaag gtctggaata catcggcgcc atttatgcta gtggtagcac atactacgcg 180
agctgggcga aaggccgttt caccatctcc cgtgacaact ctaaaaacac cgtgtacctg 240
cagatgaact ctctgcgtgc ggaagacact gcggtttact attgcgcgcg tattcattat 300
ggtaatagtg gtgggttgtg gggtcagggt actctggtta ccgtctcgag c 351
<210> 25
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 gH23 Тяжелая цепь
<400> 25
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Ser His
20 25 30
Asp Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Ala Ile Tyr Ala Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ile His Tyr Gly Asn Ser Gly Gly Leu Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 26
<211> 1332
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 6470gH23 Тяжелая цепь нукл.
<400> 26
gaggttcagc tgctggagtc tggaggcggg cttgtccagc ctggagggag cctgcgtctc 60
tcttgtgcag taagcggcat cgacctgtcc agccacgaca tgtattgggt acgtcaggca 120
ccgggtaaag gtctggaata catcggcgcc atttatgcta gtggtagcac atactacgcg 180
agctgggcga aaggccgttt caccatctcc cgtgacaact ctaaaaacac cgtgtacctg 240
cagatgaact ctctgcgtgc ggaagacact gcggtttact attgcgcgcg tattcattat 300
ggtaatagtg gtgggttgtg gggtcagggt actctggtta ccgtctcgag cgcttctaca 360
aagggcccct ccgtgttccc tctggcccct tgctcccggt ccacctccga gtctaccgcc 420
gctctgggct gcctggtcaa ggactacttc cccgagcccg tgacagtgtc ctggaactct 480
ggcgccctga cctccggcgt gcacaccttc cctgccgtgc tgcagtcctc cggcctgtac 540
tccctgtcct ccgtcgtgac cgtgccctcc tccagcctgg gcaccaagac ctacacctgt 600
aacgtggacc acaagccctc caacaccaag gtggacaagc gggtggaatc taagtacggc 660
cctccctgcc ccccctgccc tgcccctgaa tttctgggcg gaccttccgt gttcctgttc 720
cccccaaagc ccaaggacac cctgatgatc tcccggaccc ccgaagtgac ctgcgtggtg 780
gtggacgtgt cccaggaaga tcccgaggtc cagttcaatt ggtacgtgga cggcgtggaa 840
gtgcacaatg ccaagaccaa gcccagagag gaacagttca actccaccta ccgggtggtg 900
tccgtgctga ccgtgctgca ccaggactgg ctgaacggca aagagtacaa gtgcaaggtg 960
tccaacaagg gcctgccctc cagcatcgaa aagaccatct ccaaggccaa gggccagccc 1020
cgcgagcccc aggtgtacac cctgccccct agccaggaag agatgaccaa gaaccaggtg 1080
tccctgacct gtctggtcaa gggcttctac ccctccgaca ttgccgtgga atgggagtcc 1140
aacggccagc ccgagaacaa ctacaagacc accccccctg tgctggacag cgacggctcc 1200
ttcttcctgt actctcggct gaccgtggac aagtcccggt ggcaggaagg caacgtcttc 1260
tcctgctccg tgatgcacga ggccctgcac aaccactaca cccagaagtc cctgtccctg 1320
agcctgggca ag 1332
<210> 27
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 gH23 VH S56N N102H
<400> 27
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Ser His
20 25 30
Asp Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Ala Ile Tyr Ala Ser Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ile His Tyr Gly His Ser Gly Gly Leu Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 28
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 6470 gH23 VH S56N N102H нукл.
<400> 28
gaggttcagc tgctggagtc tggaggcggg cttgtccagc ctggagggag cctgcgtctc 60
tcttgtgcag taagcggcat cgacctgtcc agccacgaca tgtattgggt acgtcaggca 120
ccgggtaaag gtctggaata catcggcgcc atttatgcta gtggtaatac atactacgcg 180
agctgggcga aaggccgttt caccatctcc cgtgacaact ctaaaaacac cgtgtacctg 240
cagatgaact ctctgcgtgc ggaagacact gcggtttact attgcgcgcg tattcattat 300
ggtcacagtg gtgggttgtg gggtcagggt actctggtta ccgtctcgag c 351
<210> 29
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 gH23 Тяжелая цепь S56N N102H
<400> 29
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Ser His
20 25 30
Asp Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Ala Ile Tyr Ala Ser Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ile His Tyr Gly His Ser Gly Gly Leu Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 30
<211> 1332
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 6470 gH23 Тяжелая цепь S56N N102H нукл.
<400> 30
gaggttcagc tgctggagtc tggaggcggg cttgtccagc ctggagggag cctgcgtctc 60
tcttgtgcag taagcggcat cgacctgtcc agccacgaca tgtattgggt acgtcaggca 120
ccgggtaaag gtctggaata catcggcgcc atttatgcta gtggtaatac atactacgcg 180
agctgggcga aaggccgttt caccatctcc cgtgacaact ctaaaaacac cgtgtacctg 240
cagatgaact ctctgcgtgc ggaagacact gcggtttact attgcgcgcg tattcattat 300
ggtcacagtg gtgggttgtg gggtcagggt actctggtta ccgtctcgag cgcttctaca 360
aagggcccct ccgtgttccc tctggcccct tgctcccggt ccacctccga gtctaccgcc 420
gctctgggct gcctggtcaa ggactacttc cccgagcccg tgacagtgtc ctggaactct 480
ggcgccctga cctccggcgt gcacaccttc cctgccgtgc tgcagtcctc cggcctgtac 540
tccctgtcct ccgtcgtgac cgtgccctcc tccagcctgg gcaccaagac ctacacctgt 600
aacgtggacc acaagccctc caacaccaag gtggacaagc gggtggaatc taagtacggc 660
cctccctgcc ccccctgccc tgcccctgaa tttctgggcg gaccttccgt gttcctgttc 720
cccccaaagc ccaaggacac cctgatgatc tcccggaccc ccgaagtgac ctgcgtggtg 780
gtggacgtgt cccaggaaga tcccgaggtc cagttcaatt ggtacgtgga cggcgtggaa 840
gtgcacaatg ccaagaccaa gcccagagag gaacagttca actccaccta ccgggtggtg 900
tccgtgctga ccgtgctgca ccaggactgg ctgaacggca aagagtacaa gtgcaaggtg 960
tccaacaagg gcctgccctc cagcatcgaa aagaccatct ccaaggccaa gggccagccc 1020
cgcgagcccc aggtgtacac cctgccccct agccaggaag agatgaccaa gaaccaggtg 1080
tccctgacct gtctggtcaa gggcttctac ccctccgaca ttgccgtgga atgggagtcc 1140
aacggccagc ccgagaacaa ctacaagacc accccccctg tgctggacag cgacggctcc 1200
ttcttcctgt actctcggct gaccgtggac aagtcccggt ggcaggaagg caacgtcttc 1260
tcctgctccg tgatgcacga ggccctgcac aaccactaca cccagaagtc cctgtccctg 1320
agcctgggca ag 1332
<210> 31
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 gH36 VH
<400> 31
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Ser His
20 25 30
Asp Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Ala Ile Tyr Ala Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ile His Tyr Gly Asn Ser Gly Gly Leu Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 32
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 6470 gH36 VH нукл.
<400> 32
gaggttcagc tgctggagtc tggaggcggg cttgtccagc ctggagggag cctgcgtctc 60
tcttgtgcag taagcggcat cgacctgtcc agccacgaca tgtattgggt acgtcaggca 120
ccgggtaaag gtctggaata catcggcgcc atttatgcta gtggtagcac atactacgcg 180
agctgggcga aaggccgttt caccatctcc cgtgactcca gcaaaaacac cctgtacctg 240
cagatgaact ctctgcgtgc ggaagacact gcggtttact attgcgcgcg tattcattat 300
ggtaatagtg gtgggttgtg gggtcagggt actctggtta ccgtctcgag c 351
<210> 33
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 gH36 Тяжелая цепь
<400> 33
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Ser His
20 25 30
Asp Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Ala Ile Tyr Ala Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ile His Tyr Gly Asn Ser Gly Gly Leu Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 34
<211> 1332
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 6470 gH36 Тяжелая цепь нукл.
<400> 34
gaggttcagc tgctggagtc tggaggcggg cttgtccagc ctggagggag cctgcgtctc 60
tcttgtgcag taagcggcat cgacctgtcc agccacgaca tgtattgggt acgtcaggca 120
ccgggtaaag gtctggaata catcggcgcc atttatgcta gtggtagcac atactacgcg 180
agctgggcga aaggccgttt caccatctcc cgtgactcca gcaaaaacac cctgtacctg 240
cagatgaact ctctgcgtgc ggaagacact gcggtttact attgcgcgcg tattcattat 300
ggtaatagtg gtgggttgtg gggtcagggt actctggtta ccgtctcgag cgcttctaca 360
aagggcccct ccgtgttccc tctggcccct tgctcccggt ccacctccga gtctaccgcc 420
gctctgggct gcctggtcaa ggactacttc cccgagcccg tgacagtgtc ctggaactct 480
ggcgccctga cctccggcgt gcacaccttc cctgccgtgc tgcagtcctc cggcctgtac 540
tccctgtcct ccgtcgtgac cgtgccctcc tccagcctgg gcaccaagac ctacacctgt 600
aacgtggacc acaagccctc caacaccaag gtggacaagc gggtggaatc taagtacggc 660
cctccctgcc ccccctgccc tgcccctgaa tttctgggcg gaccttccgt gttcctgttc 720
cccccaaagc ccaaggacac cctgatgatc tcccggaccc ccgaagtgac ctgcgtggtg 780
gtggacgtgt cccaggaaga tcccgaggtc cagttcaatt ggtacgtgga cggcgtggaa 840
gtgcacaatg ccaagaccaa gcccagagag gaacagttca actccaccta ccgggtggtg 900
tccgtgctga ccgtgctgca ccaggactgg ctgaacggca aagagtacaa gtgcaaggtg 960
tccaacaagg gcctgccctc cagcatcgaa aagaccatct ccaaggccaa gggccagccc 1020
cgcgagcccc aggtgtacac cctgccccct agccaggaag agatgaccaa gaaccaggtg 1080
tccctgacct gtctggtcaa gggcttctac ccctccgaca ttgccgtgga atgggagtcc 1140
aacggccagc ccgagaacaa ctacaagacc accccccctg tgctggacag cgacggctcc 1200
ttcttcctgt actctcggct gaccgtggac aagtcccggt ggcaggaagg caacgtcttc 1260
tcctgctccg tgatgcacga ggccctgcac aaccactaca cccagaagtc cctgtccctg 1320
agcctgggca ag 1332
<210> 35
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 gH36 VH S56N N102H
<400> 35
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Ser His
20 25 30
Asp Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Ala Ile Tyr Ala Ser Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ile His Tyr Gly His Ser Gly Gly Leu Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 36
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 6470 gH36 VH S56N N102H нукл.
<400> 36
gaggttcagc tgctggagtc tggaggcggg cttgtccagc ctggagggag cctgcgtctc 60
tcttgtgcag taagcggcat cgacctgtcc agccacgaca tgtattgggt acgtcaggca 120
ccgggtaaag gtctggaata catcggcgcc atttatgcta gtggtaatac atactacgcg 180
agctgggcga aaggccgttt caccatctcc cgtgactcca gcaaaaacac cctgtacctg 240
cagatgaact ctctgcgtgc ggaagacact gcggtttact attgcgcgcg tattcattat 300
ggtcacagtg gtgggttgtg gggtcagggt actctggtta ccgtctcgag c 351
<210> 37
<211> 444
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 gH36 Тяжелая цепь S56N N102H
<400> 37
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Ser His
20 25 30
Asp Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Ala Ile Tyr Ala Ser Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ile His Tyr Gly His Ser Gly Gly Leu Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 38
<211> 1332
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 6470 gH36 Тяжелая цепь S56N N102H нукл.
<400> 38
gaggttcagc tgctggagtc tggaggcggg cttgtccagc ctggagggag cctgcgtctc 60
tcttgtgcag taagcggcat cgacctgtcc agccacgaca tgtattgggt acgtcaggca 120
ccgggtaaag gtctggaata catcggcgcc atttatgcta gtggtaatac atactacgcg 180
agctgggcga aaggccgttt caccatctcc cgtgactcca gcaaaaacac cctgtacctg 240
cagatgaact ctctgcgtgc ggaagacact gcggtttact attgcgcgcg tattcattat 300
ggtcacagtg gtgggttgtg gggtcagggt actctggtta ccgtctcgag cgcttctaca 360
aagggcccct ccgtgttccc tctggcccct tgctcccggt ccacctccga gtctaccgcc 420
gctctgggct gcctggtcaa ggactacttc cccgagcccg tgacagtgtc ctggaactct 480
ggcgccctga cctccggcgt gcacaccttc cctgccgtgc tgcagtcctc cggcctgtac 540
tccctgtcct ccgtcgtgac cgtgccctcc tccagcctgg gcaccaagac ctacacctgt 600
aacgtggacc acaagccctc caacaccaag gtggacaagc gggtggaatc taagtacggc 660
cctccctgcc ccccctgccc tgcccctgaa tttctgggcg gaccttccgt gttcctgttc 720
cccccaaagc ccaaggacac cctgatgatc tcccggaccc ccgaagtgac ctgcgtggtg 780
gtggacgtgt cccaggaaga tcccgaggtc cagttcaatt ggtacgtgga cggcgtggaa 840
gtgcacaatg ccaagaccaa gcccagagag gaacagttca actccaccta ccgggtggtg 900
tccgtgctga ccgtgctgca ccaggactgg ctgaacggca aagagtacaa gtgcaaggtg 960
tccaacaagg gcctgccctc cagcatcgaa aagaccatct ccaaggccaa gggccagccc 1020
cgcgagcccc aggtgtacac cctgccccct agccaggaag agatgaccaa gaaccaggtg 1080
tccctgacct gtctggtcaa gggcttctac ccctccgaca ttgccgtgga atgggagtcc 1140
aacggccagc ccgagaacaa ctacaagacc accccccctg tgctggacag cgacggctcc 1200
ttcttcctgt actctcggct gaccgtggac aagtcccggt ggcaggaagg caacgtcttc 1260
tcctgctccg tgatgcacga ggccctgcac aaccactaca cccagaagtc cctgtccctg 1320
agcctgggca ag 1332
<210> 39
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Человеческий IGKV1-16 JK4 акцепторный каркас
<400> 39
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Ser Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 40
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Человеческая IGKV1-16 JK4 акцепторная каркасная нукл.
<400> 40
gacatccaga tgacccagtc tccatcctca ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgtc gggcgagtca gggcattagc aattatttag cctggtttca gcagaaacca 120
gggaaagccc ctaagtccct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcagcag cctgcagcct 240
gaagattttg caacttatta ctgccaacag tataatagtt accctctcac tttcggcgga 300
gggaccaagg tggagatcaa a 321
<210> 41
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> Человеческий IGHV3-23 JH4 акцепторы каркас
<400> 41
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser
<210> 42
<211> 339
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Человеческая IGHV3-23 JH4 акцепторная каркасная нукл.
<400> 42
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtat attactgtgc gaaatacttt 300
gactactggg gccaaggaac cctggtcacc gtctcctca 339
<210> 43
<211> 306
<212> Белок
<213> Искусственная последовательность
<220>
<223> Кроличий Fc - человеческий 68-140 a-syn
<400> 43
Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys Thr Val Glu
1. 5 10 15
Gly Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Val Lys Lys Asp Gln
20 25 30
Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile Leu Glu Asp
35 40 45
Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu
50 55 60
Gly Tyr Gln Asp Tyr Glu Pro Glu Ala Val Glu Lys Thr Val Ala Pro
65 70 75 80
Ser Thr Cys Ser Lys Pro Thr Cys Pro Pro Pro Glu Leu Leu Gly Gly
85 90 95
Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
100 105 110
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Asp
115 120 125
Asp Pro Glu Val Gln Phe Thr Trp Tyr Ile Asn Asn Glu Gln Val Arg
130 135 140
Thr Ala Arg Pro Pro Leu Arg Glu Gln Gln Phe Asn Ser Thr Ile Arg
145 150 155 160
Val Val Ser Thr Leu Pro Ile Ala His Gln Asp Trp Leu Arg Gly Lys
165 170 175
Glu Phe Lys Cys Lys Val His Asn Lys Ala Leu Pro Ala Pro Ile Glu
180 185 190
Lys Thr Ile Ser Lys Ala Arg Gly Gln Pro Leu Glu Pro Lys Val Tyr
195 200 205
Thr Met Gly Pro Pro Arg Glu Glu Leu Ser Ser Arg Ser Val Ser Leu
210 215 220
Thr Cys Met Ile Asn Gly Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp
225 230 235 240
Glu Lys Asn Gly Lys Ala Glu Asp Asn Tyr Lys Thr Thr Pro Ala Val
245 250 255
Leu Asp Ser Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Pro
260 265 270
Thr Ser Glu Trp Gln Arg Gly Asp Val Phe Thr Cys Ser Val Met His
275 280 285
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Ile Ser Arg Ser Pro
290 295 300
Gly Lys
305
<210> 44
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR-L1 X33
<220>
<221> MISC_FEATURE
<222> (10)..(10)
<223> Xaa представляет собой Asn (N) или Arg (R)
<400> 44
Gln Ala Ser Gln Ser Val Tyr Lys Asn Xaa Tyr Leu Ala
1. 5 10
<210> 45
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR-H2 X56
<220>
<221> MISC_FEATURE
<222> (7)..(7)
<223> Xaa представляет собой Ser (S) или Asn (N)
<400> 45
Ala Ile Tyr Ala Ser Gly Xaa Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1. 5 10 15
<210> 46
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR-H3 X102
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> Xaa представляет собой Asn (N) или His (H)
<400> 46
Ile His Tyr Gly Xaa Ser Gly Gly Leu
1. 5
<210> 47
<211> 216
<212> Белок
<213> Oryctolagus cuniculus
<400> 47
Ala Ile Val Met Thr Gln Thr Pro Ser Ser Lys Ser Val Ala Val Gly
1. 5 10 15
Asp Thr Val Thr Ile Asn Cys Gln Ala Ser Gln Ser Val Tyr Lys Asn
20 25 30
Asn Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Gln
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Val
65 70 75 80
Val Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Gly Gly
85 90 95
Arg Asn Asp Gly Phe Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
Arg Thr Pro Val Ala Pro Thr Val Leu Ile Phe Pro Pro Ala Ala Asp
115 120 125
Gln Val Ala Thr Gly Thr Val Thr Ile Val Cys Val Ala Asn Lys Tyr
130 135 140
Phe Pro Asp Val Thr Val Thr Trp Glu Val Asp Gly Thr Thr Gln Thr
145 150 155 160
Thr Gly Ile Glu Asn Ser Lys Thr Pro Gln Asn Ser Ala Asp Cys Thr
165 170 175
Tyr Asn Leu Ser Ser Thr Leu Thr Leu Thr Ser Thr Gln Tyr Asn Ser
180 185 190
His Lys Glu Tyr Thr Cys Lys Val Thr Gln Gly Thr Thr Ser Val Val
195 200 205
Gln Ser Phe Asn Arg Gly Asp Cys
210 215
<210> 48
<211> 437
<212> Белок
<213> Oryctolagus cuniculus
<400> 48
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1. 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser His Asp
20 25 30
Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile Gly
35 40 45
Ala Ile Tyr Ala Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Met Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Ile His
85 90 95
Tyr Gly Asn Ser Gly Gly Leu Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser Gly Gln Pro Lys Ala Pro Ser Val Phe Pro Leu Ala Pro Cys
115 120 125
Cys Gly Asp Thr Pro Ser Ser Thr Val Thr Leu Gly Cys Leu Val Lys
130 135 140
Gly Tyr Leu Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Thr Leu
145 150 155 160
Thr Asn Gly Val Arg Thr Phe Pro Ser Val Arg Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Ser Val Thr Ser Ser Ser Gln Pro Val
180 185 190
Thr Cys Asn Val Ala His Pro Ala Thr Asn Thr Lys Val Asp Lys Thr
195 200 205
Val Ala Pro Ser Thr Cys Ser Lys Pro Thr Cys Pro Pro Pro Glu Leu
210 215 220
Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Thr
225 230 235 240
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
245 250 255
Ser Gln Asp Asp Pro Glu Val Gln Phe Thr Trp Tyr Ile Asn Asn Glu
260 265 270
Gln Val Arg Thr Ala Arg Pro Pro Leu Arg Glu Gln Gln Phe Asn Ser
275 280 285
Thr Ile Arg Val Val Ser Thr Leu Pro Ile Ala His Gln Asp Trp Leu
290 295 300
Arg Gly Lys Glu Phe Lys Cys Lys Val His Asn Lys Ala Leu Pro Ala
305 310 315 320
Pro Ile Glu Lys Thr Ile Ser Lys Ala Arg Gly Gln Pro Leu Glu Pro
325 330 335
Lys Val Tyr Thr Met Gly Pro Pro Arg Glu Glu Leu Ser Ser Arg Ser
340 345 350
Val Ser Leu Thr Cys Met Ile Asn Gly Phe Tyr Pro Ser Asp Ile Ser
355 360 365
Val Glu Trp Glu Lys Asn Gly Lys Ala Glu Asp Asn Tyr Lys Thr Thr
370 375 380
Pro Ala Val Leu Asp Ser Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu
385 390 395 400
Ser Val Pro Thr Ser Glu Trp Gln Arg Gly Asp Val Phe Thr Cys Ser
405 410 415
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Ile Ser
420 425 430
Arg Ser Pro Gly Lys
435
<210> 49
<211> 227
<212> Белок
<213> Искусственная последовательность
<220>
<223> 6470 кроличья Fab-His тяжелая цепь
<400> 49
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1. 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser His Asp
20 25 30
Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile Gly
35 40 45
Ala Ile Tyr Ala Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Met Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Ile His
85 90 95
Tyr Gly Asn Ser Gly Gly Leu Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser Gly Gln Pro Lys Ala Pro Ser Val Phe Pro Leu Ala Pro Cys
115 120 125
Cys Gly Asp Thr Pro Ser Ser Thr Val Thr Leu Gly Cys Leu Val Lys
130 135 140
Gly Tyr Leu Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Thr Leu
145 150 155 160
Thr Asn Gly Val Arg Thr Phe Pro Ser Val Arg Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Ser Val Thr Ser Ser Ser Gln Pro Val
180 185 190
Thr Cys Asn Val Ala His Pro Ala Thr Asn Thr Lys Val Asp Lys Thr
195 200 205
Val Ala Pro Ser Thr Cys Ser Lys Pro His His His His His His His
210 215 220
His His His
225 22
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Моноклональные антитела к альфа-синуклеину для предотвращения агрегации тау-белка | 2017 |
|
RU2760334C2 |
| АНТИТЕЛА | 2018 |
|
RU2798399C2 |
| СРЕДСТВА, ПУТИ ПРИМЕНЕНИЯ И СПОСОБЫ ЛЕЧЕНИЯ СИНУКЛЕОПАТИИ | 2016 |
|
RU2765303C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА, КОТОРЫЕ РАСПОЗНАЮТ АЛЬФА-СИНУКЛЕИН | 2012 |
|
RU2743738C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА СВЕРТЫВАНИЯ XI | 2017 |
|
RU2757314C2 |
| АНТИТЕЛО К АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 ЧЕЛОВЕКА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2807060C1 |
| КОМБИНАЦИИ АНТИТЕЛ К STAPHYLOCOCCUS AUREUS | 2019 |
|
RU2804815C2 |
| СПОСОБЫ ОБНАРУЖЕНИЯ ФОЛАТНОГО РЕЦЕПТОРА 1 В ОБРАЗЦЕ ПАЦИЕНТА | 2018 |
|
RU2759410C2 |
| АНТИТЕЛО, НАПРАВЛЕННОЕ ПРОТИВ ФАКТОРА СЛИПАНИЯ А (ClfA) S. aureus | 2019 |
|
RU2818805C2 |
| ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2018 |
|
RU2810094C2 |
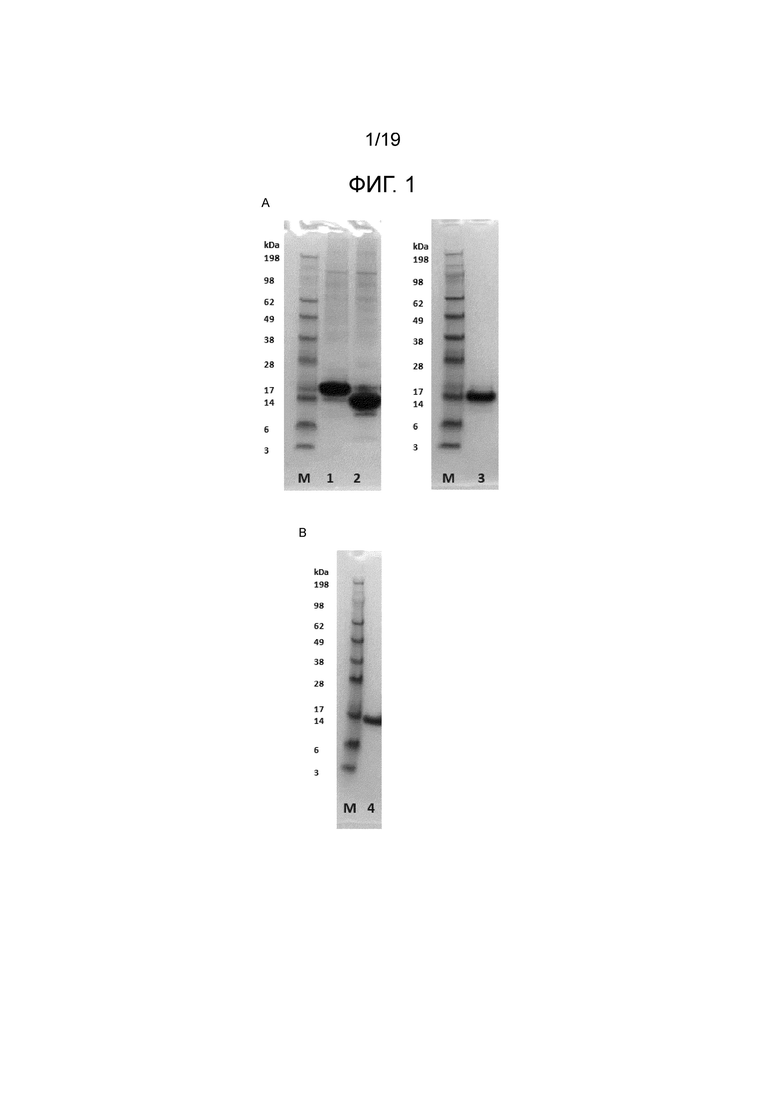
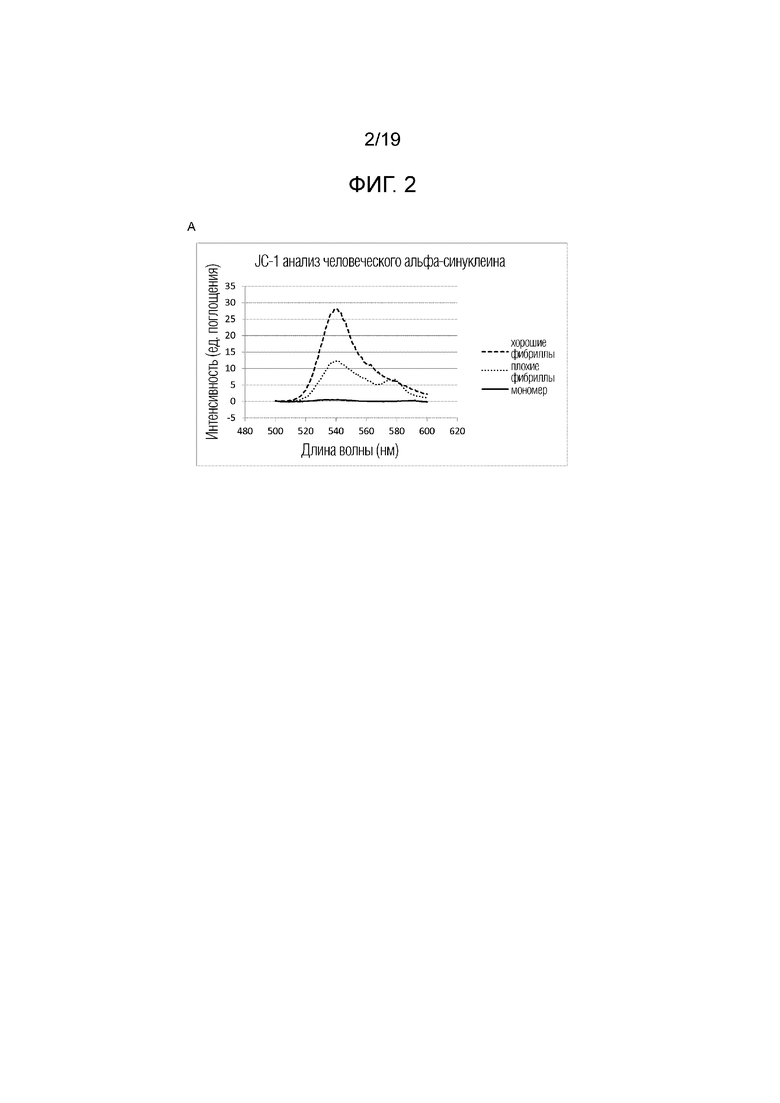
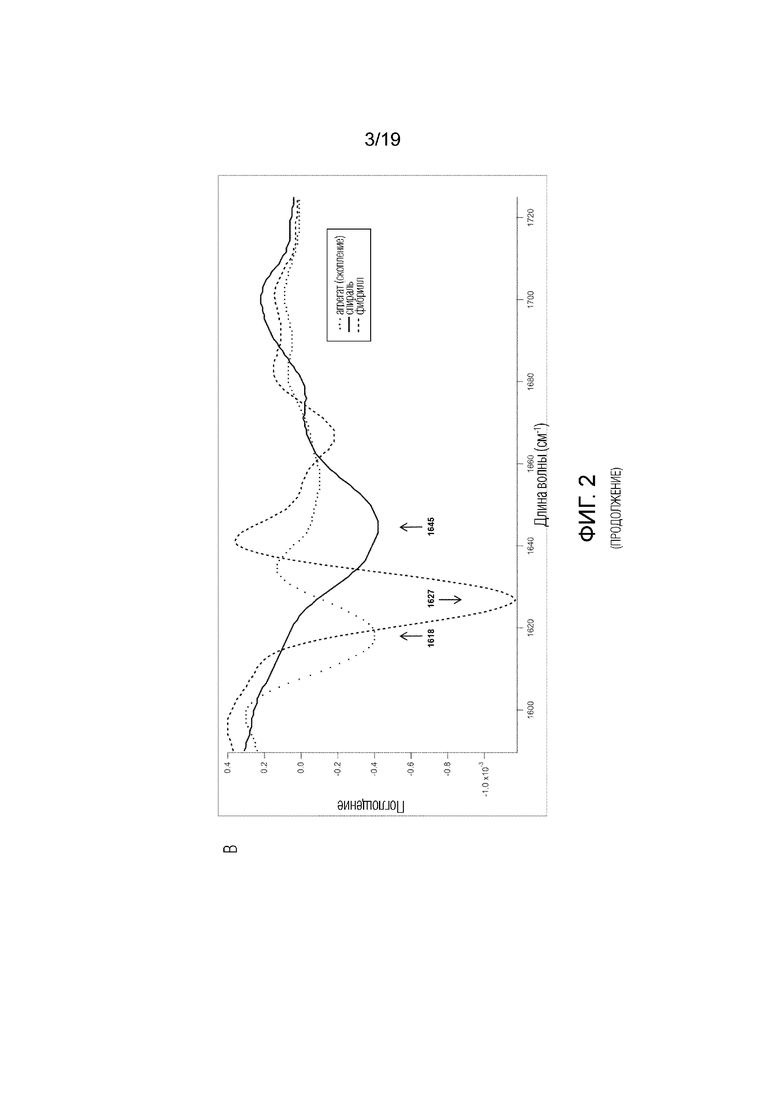
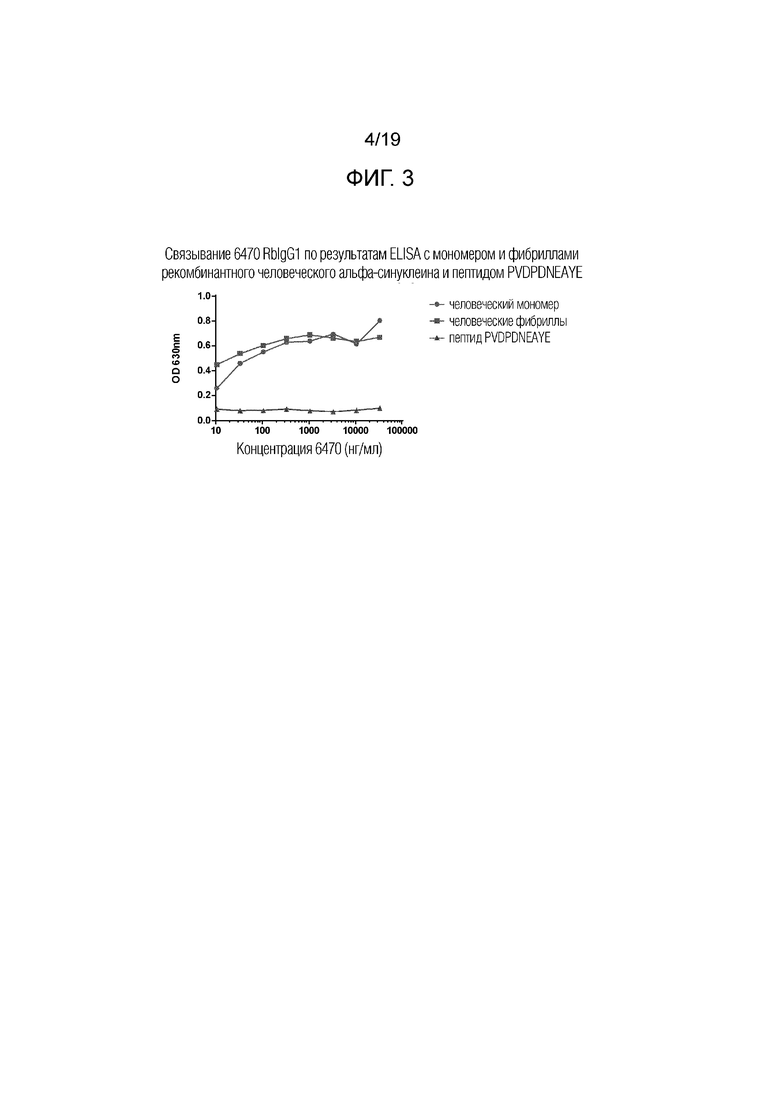
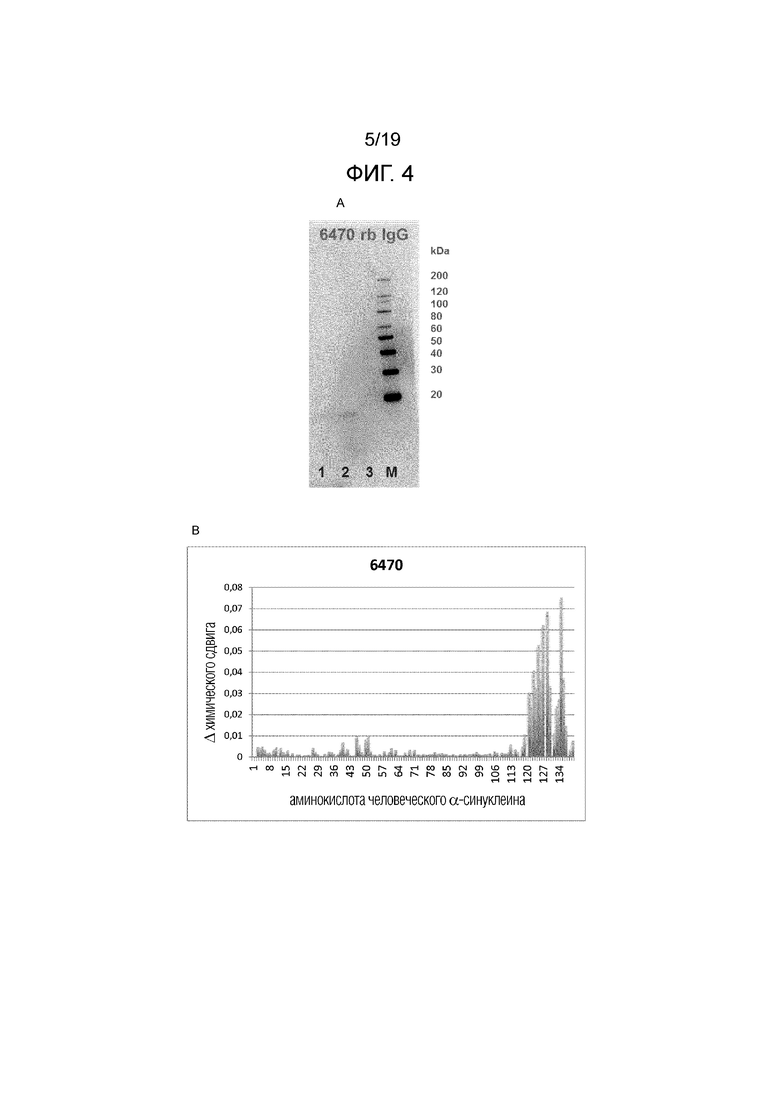
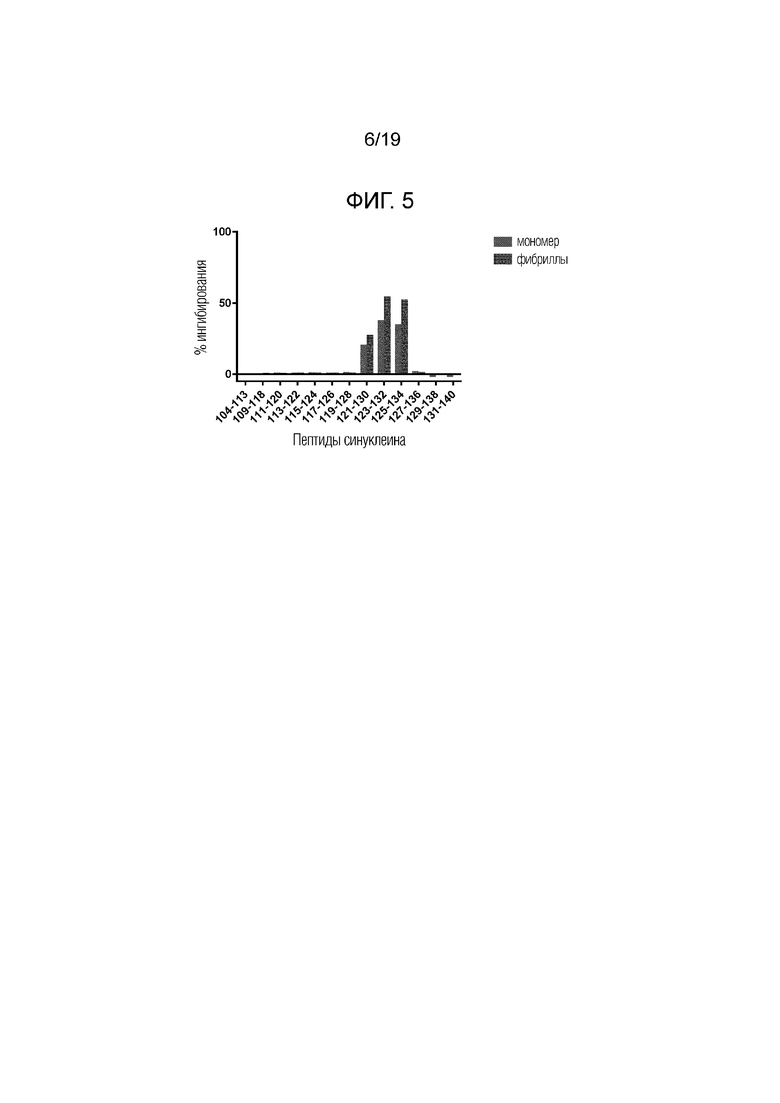

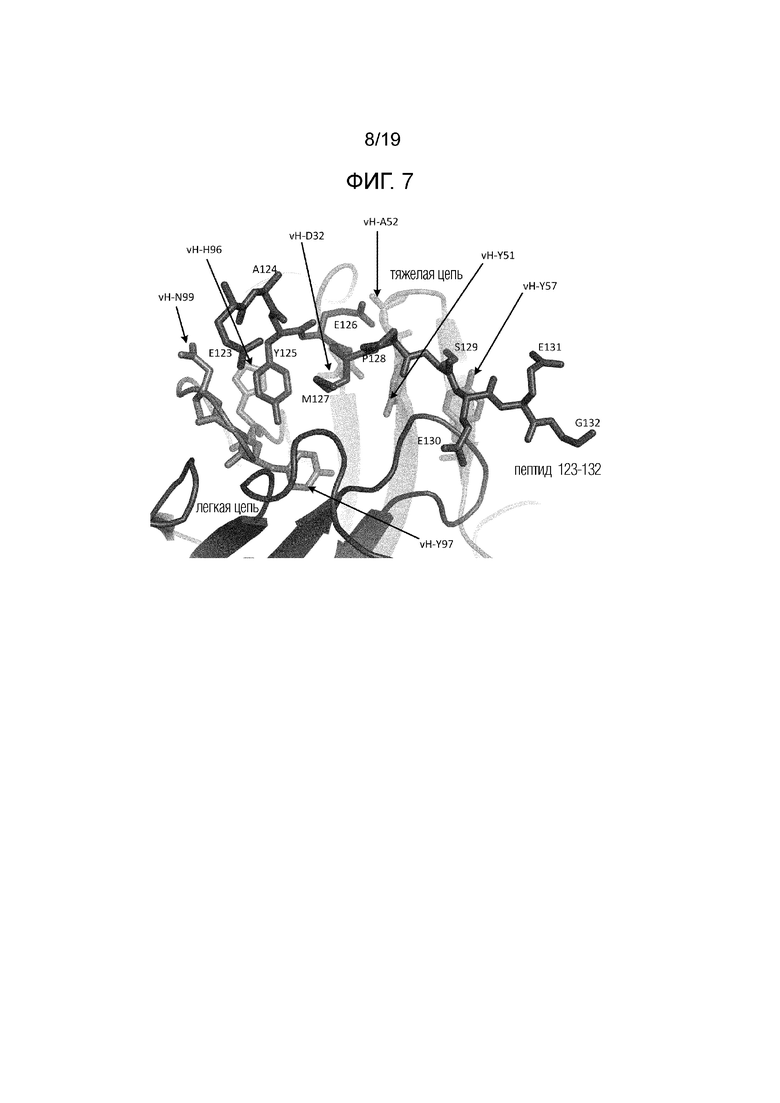
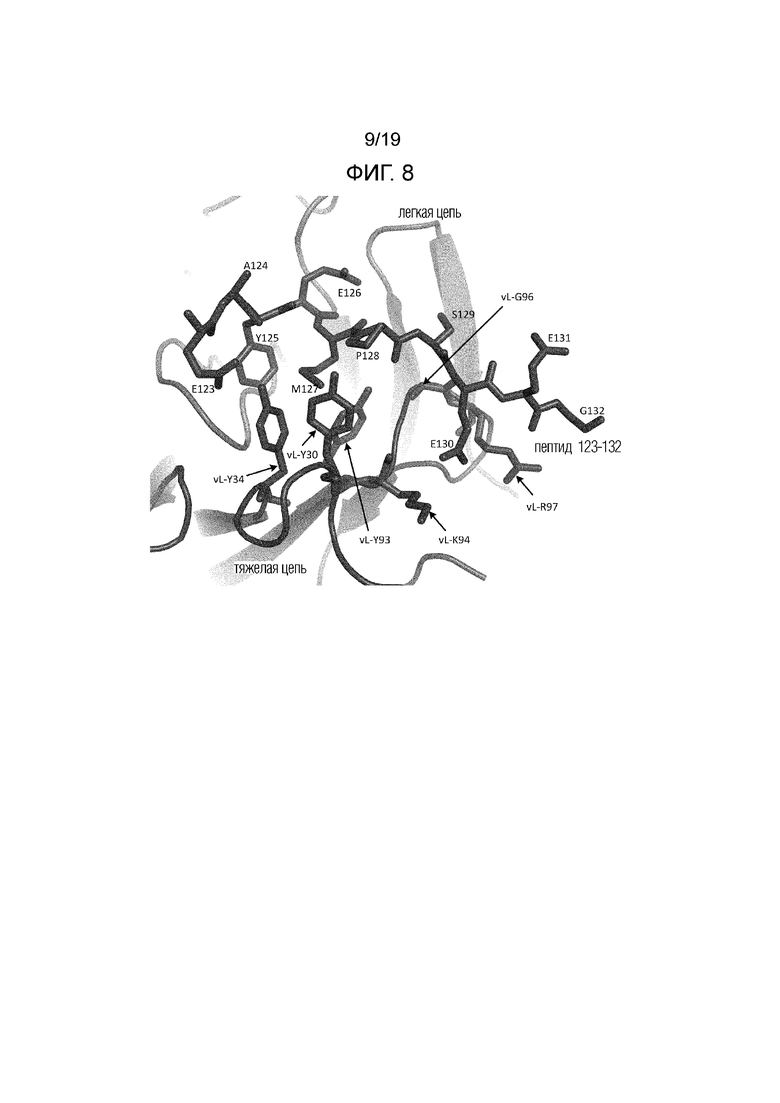
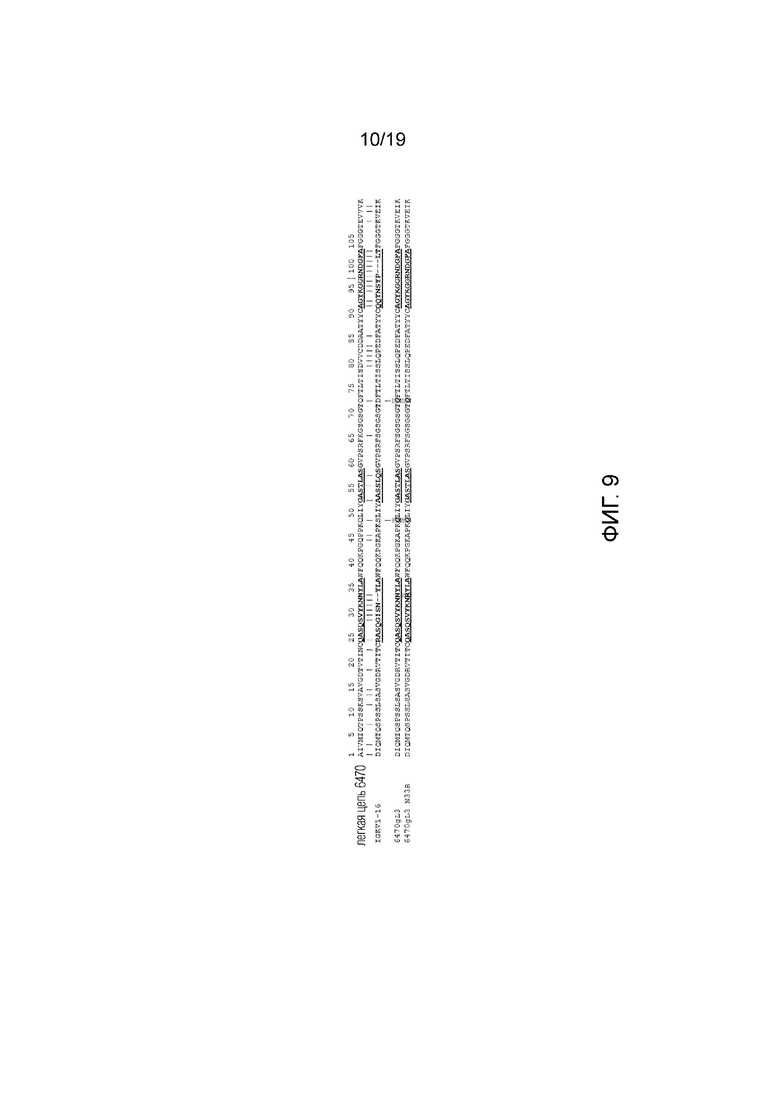


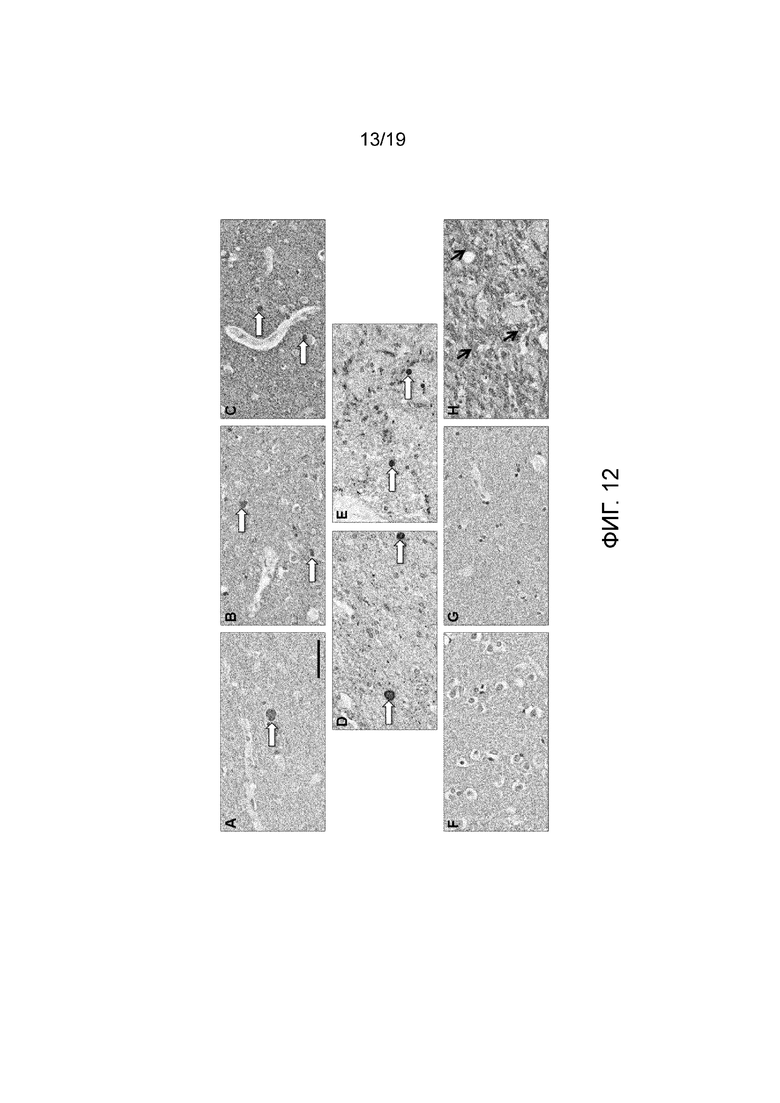
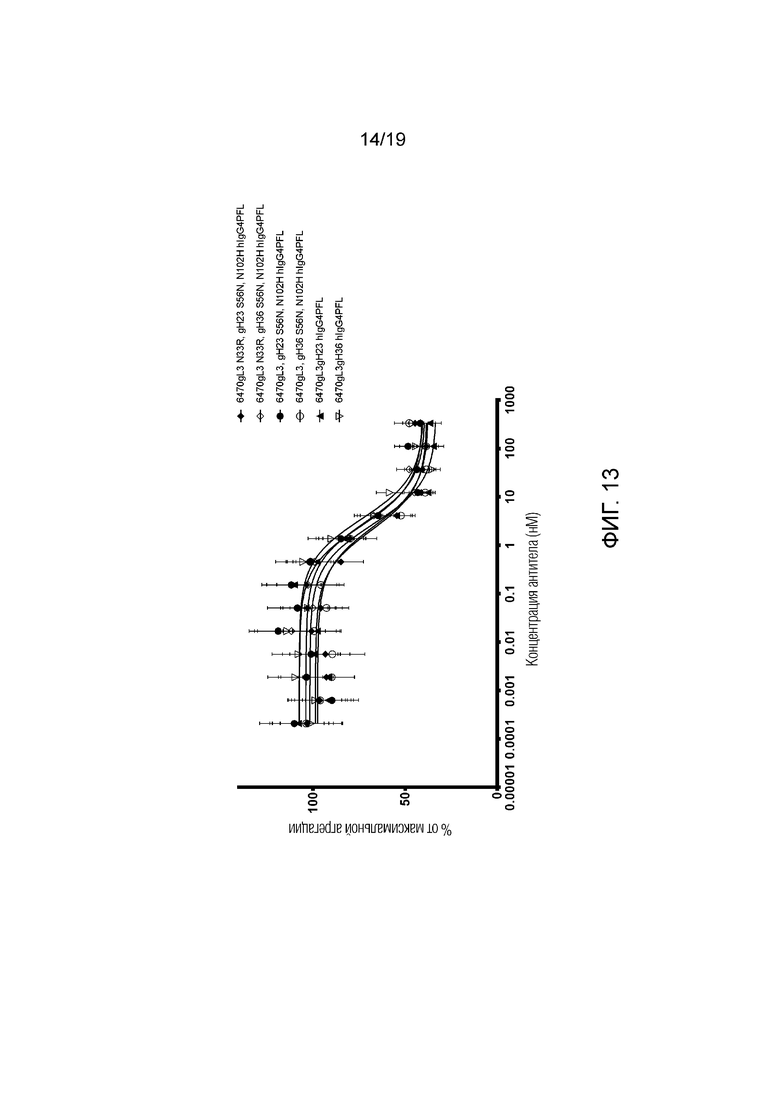
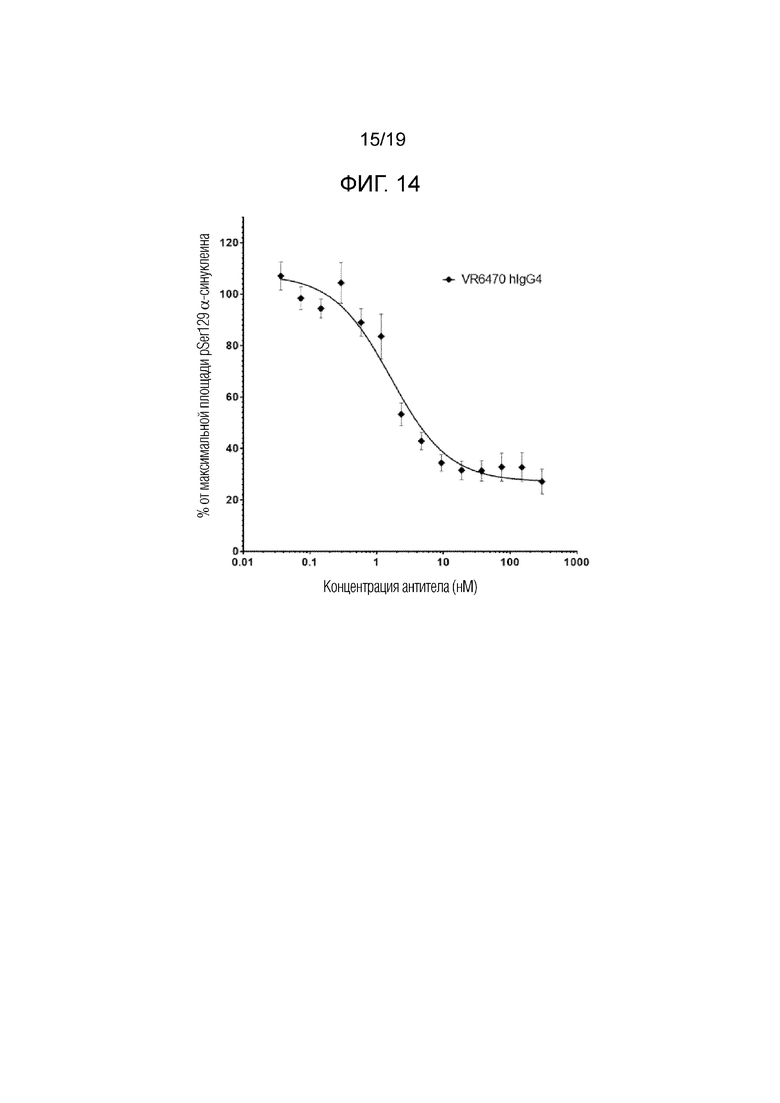

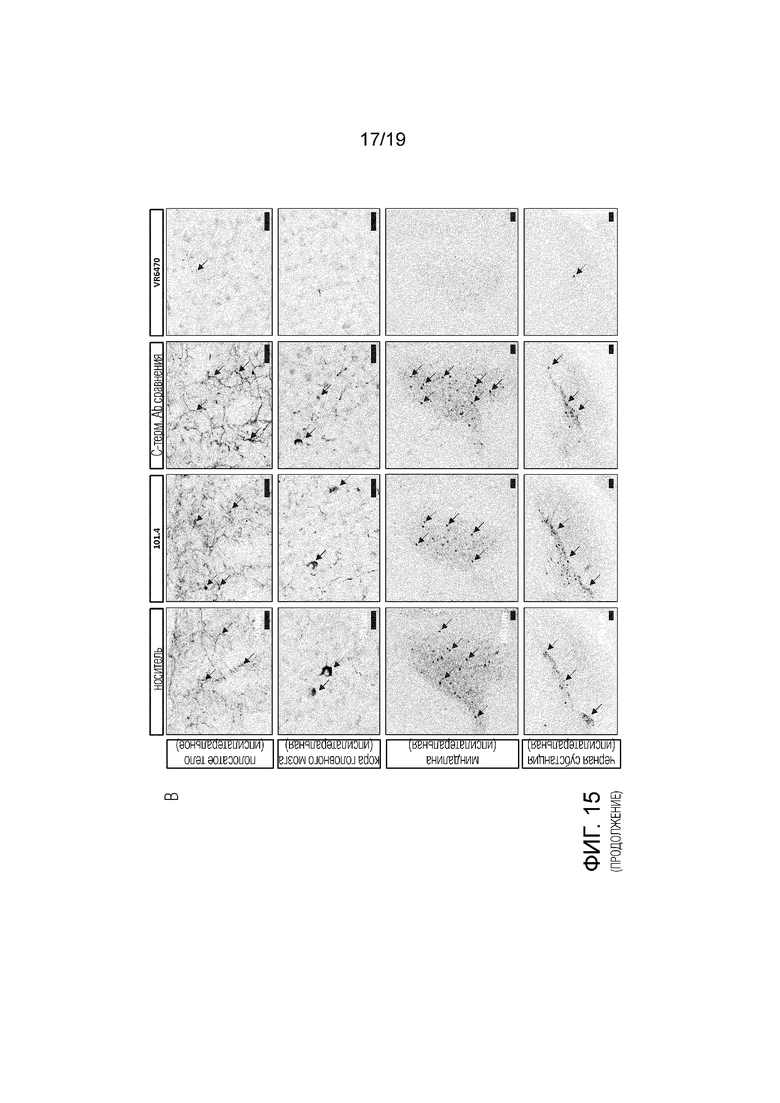
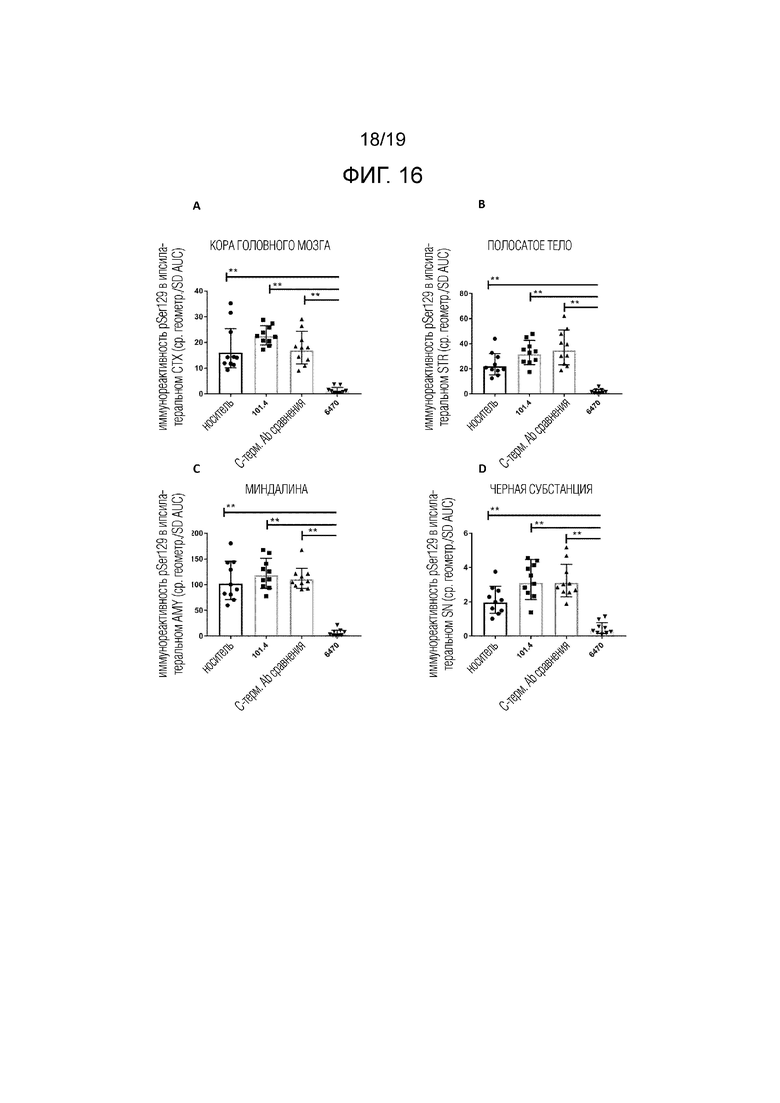
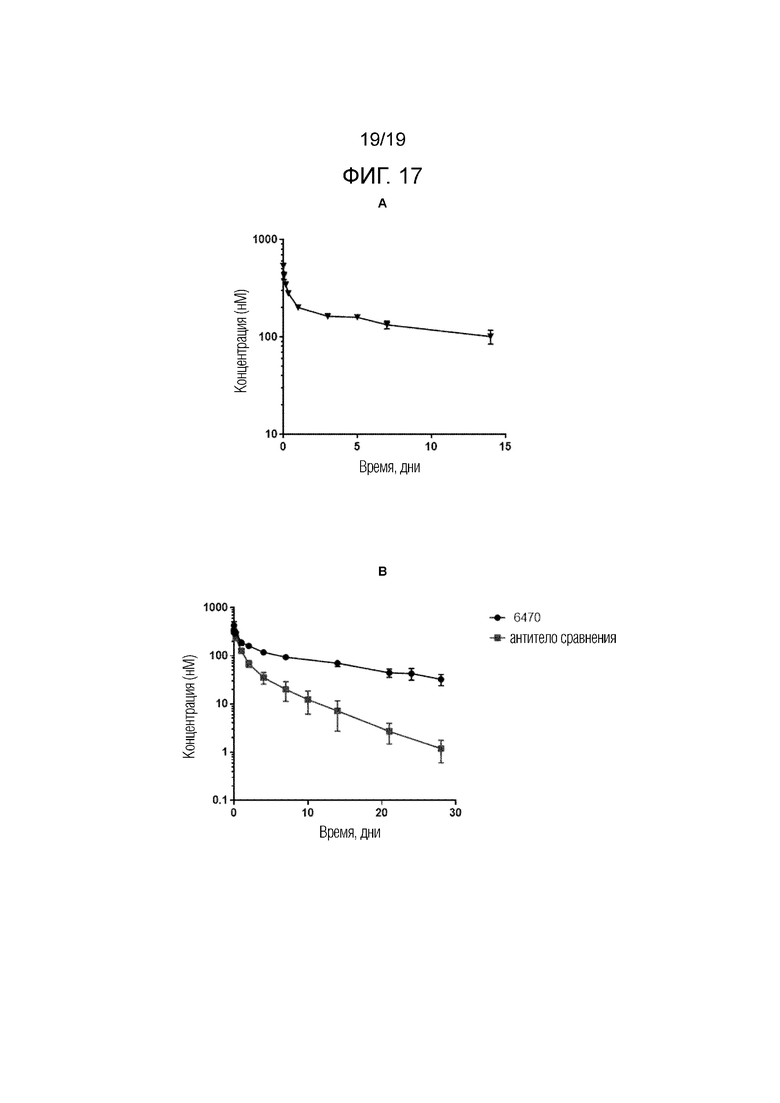
Группа изобретений относится к биотехнологии. Представлены антитело или его антигенсвязывающий фрагмент, которые связываются с альфа-синуклеином. При этом антитело или антигенсвязывающий фрагмент способны связываться с альфа-синуклеином как в виде мономера, так и в виде фибрилл и предотвращать агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина. Также представлены: выделенный полинуклеотид, кодирующий антитело или его антигенсвязывающий фрагмент, векторы клонирования и экспрессии, клетка-хозяин, способ получения антитела или его антигенсвязывающего фрагмента, фармацевтическая композиция для лечения болезни Паркинсона и способ лечения болезни Паркинсона. Изобретения можно применять для лечения синуклеинопатий, включая болезнь Паркинсона. 8 н. и 19 з.п. ф-лы, 17 ил., 12 табл., 10 пр.
1. Антитело или его антигенсвязывающий фрагмент, которое связывается с альфа-синуклеином, где антитело или антигенсвязывающий фрагмент содержит:
a. вариабельную область легкой цепи, содержащую:
i. CDR-L1, содержащую SEQ ID NO:1;
ii. CDR-L2, содержащую SEQ ID NO:2, и
iii. CDR-L3, содержащую SEQ ID NO:3; и
b. вариабельную область тяжелой цепи, содержащую:
i. CDR-H1, содержащую SEQ ID NO:4;
ii. CDR-H2, содержащую SEQ ID NO:5, и
iii. CDR-H3, содержащую SEQ ID NO:6.
2. Антитело или его антигенсвязывающий фрагмент по п.1, которое связывается с эпитопом альфа-синуклеина, содержащим, со ссылкой на SEQ ID NO:10, остатки E123, Y125, E126, M127, P128, S129, E130 и E131, где эпитоп необязательно содержит А124 и G132.
3. Антитело или его антигенсвязывающий фрагмент по любому из пп.1 или 2, где антитело или антигенсвязывающий фрагмент предотвращает агрегацию альфа-синуклеина, индуцированную фибриллами альфа-синуклеина.
4. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-3, где антитело или антигенсвязывающий фрагмент способно связываться с альфа-синуклеином как в виде мономера, так и в виде фибрилл.
5. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, которое имеет более высокое сродство связывания с альфа-синуклеином в виде фибрилл по сравнению с альфа-синуклеином в виде мономера, характеризующееся константой диссоциации (KD), по меньшей мере в 10 раз более высокой для мономерного альфа-синуклеина, чем для альфа-синуклеина в фибриллах.
6. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, которое имеет (KD) к альфа-синуклеину в фибриллах 300 пМ или менее.
7. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, которое не связывается с бета-синуклеином и/или гамма-синуклеином.
8. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, где антитело представляет собой химерное или гуманизированное антитело.
9. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, где антитело является полноразмерным антителом.
10. Антитело по п.9, где полноразмерное антитело выбрано из IgG1, IgG4 или IgGP.
11. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-8, где антигенсвязывающий фрагмент выбран из Fab, Fab', F(ab')2, scFv или dAb.
12. Антитело или его антигенсвязывающий фрагмент по любому из предшествующих пунктов, где антитело или его антигенсвязывающий фрагмент содержит:
а. вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:31; или
b. легкую цепь, содержащую SEQ ID NO:17, и тяжелую цепь, содержащую SEQ ID NO:33.
13. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-11, где антитело или его антигенсвязывающий фрагмент содержит:
a. вариабельную область легкой цепи, содержащую SEQ ID NO:15, и вариабельную область тяжелой цепи, содержащую SEQ ID NO:23; или
b. легкую цепь, содержащую SEQ ID NO:17, и тяжелую цепь, содержащую SEQ ID NO:25.
14. Выделенный полинуклеотид, кодирующий антитело или его антигенсвязывающий фрагмент по любому из пп.1-13.
15. Выделенный полинуклеотид по п.14, где полинуклеотид кодирует:
a. вариабельную область легкой цепи, где полинуклеотид:
i. по меньшей мере на 90% идентичен SEQ ID NO:16; или
ii. содержит SEQ ID NO:16; или
iii. по существу состоит из SEQ ID NO:16;
b. вариабельную область тяжелой цепи, где полинуклеотид:
i. по меньшей мере 90% на идентичен SEQ ID NO:24 или SEQ ID NO:32; или
ii. содержит SEQ ID NO:24 или SEQ ID NO:32; или
iii. по существу состоит из SEQ ID NO:24 или SEQ ID NO:32;
c. легкую цепь, где полинуклеотид:
i. по меньшей мере на 90% идентичен SEQ ID NO:18; или
ii. содержит SEQ ID NO:18; или
iii. по существу состоит из SEQ ID NO:18;
d. тяжелую цепь, где полинуклеотид:
i. по меньшей мере на 90% идентичен SEQ ID NO:26 или SEQ ID NO:34; или
ii. содержит SEQ ID NO:26 или SEQ ID NO:34; или
iii. по существу состоит из SEQ ID NO:26 или SEQ ID NO:34.
16. Вектор клонирования, содержащий один или более полинуклеотидов по любому из пп.14 или 15.
17. Вектор экспрессии, содержащий один или более полинуклеотидов по любому из пп.14 или 15.
18. Клетка-хозяин для получения антитела или антигенсвязывающего фрагмента по любому из пп.1-13, содержащая:
а. один или более полинуклеотидов по любому из пп.14 или 15 или
b. один или более векторов экспрессии по п.17.
19. Способ получения антитела или его антигенсвязывающего фрагмента по любому из пп.1-13, включающий культивирование клетки-хозяина по п.18 в подходящих условиях для получения антитела или его антигенсвязывающего фрагмента и выделение антитела или его антигенсвязывающего фрагмента.
20. Фармацевтическая композиция для лечения болезни Паркинсона, содержащая эффективное количество антитела или его антигенсвязывающего фрагмента по любому из пп.1-13 и один или более фармацевтически приемлемых носителей, наполнителей или разбавителей.
21. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-13 для применения в лечении одной или более синуклеинопатий.
22. Антитело или его антигенсвязывающий фрагмент для применения по п.21, где синуклеинопатия выбрана из болезни Паркинсона (PD) (включая идиопатические и наследственные формы болезни Паркинсона), деменции с тельцами Леви (DLB), болезни диффузных телец Леви (DLBD), варианта болезни Альцгеймера с тельцами Леви (LBVAD), комбинированной болезни Альцгеймера и Паркинсона, множественной системной атрофии (MSA) и нейродегенерации с накоплением железа в мозге 1-го типа (NBIA-1).
23. Антитело или его антигенсвязывающий фрагмент для применения по п.22, где синуклеинопатия представляет собой болезнь Паркинсона.
24. Способ лечения болезни Паркинсона (PD) у пациента, включающий введение указанному пациенту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.1-13 или фармацевтической композиции по п.20.
25. Способ по п.24, где болезнь Паркинсона представляет собой идиопатическую и наследственную формы болезни Паркинсона.
26. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-13 для применения для диагностики альфа-синуклеинопатии.
27. Антитело или антигенсвязывающий фрагмент для применения по п.26 для применения в диагностике болезни Паркинсона.
| WO 2013063516 A1, 02.05.2013 | |||
| W0 2014058924 A2, 01.09.2011 | |||
| WO 2012177972 A1, 28.08.2014 | |||
| VERED KUNIK et al.: "Structural Consensus among Antibodies Defines the Antigen Binding Site", PLOS COMPUTATIONAL BIOLOGY,vol | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С ПРОТОФИБРИЛЛАМИ, И ИХ ПРИМЕНЕНИЕ В МЕТОДАХ ТЕРАПИИ И ДИАГНОСТИКИ БОЛЕЗНИ ПАРКИНСОНА, ДЕМЕНЦИИ, АССОЦИИРОВАННОЙ С ОБРАЗОВАНИЕМ ДИФФУЗНЫХ ТЕЛЕЦ ЛЕВИ, И ДРУГИХ АЛЬФА-СИНУКЛЕИНОПАТИЙ | 2011 |
|
RU2555526C2 |

