ЗАЯВЛЕНИЕ ПРИОРИТЕТА
Данная заявка заявляет преимущество и приоритет по предварительной заявке США №62/235727, поданной 1 октября 2015 г., и предварительной заявке США №62/263313, поданной 4 декабря 2015 г., и предварительной заявке США №62/372574, поданной 9 августа 2016 г., каждая из которых включена в данный документ посредством ссылки в полном объеме.
ОБЛАСТЬ ТЕХНИКИ
Данное изобретение относится, помимо прочего, к композициям и способам, включая химерные белки, которые находят применение при лечении заболеваний, например, в иммунотерапии рака и аутоиммунных нарушений.
ОПИСАНИЕ ТЕКСТОВОГО ФАЙЛА, ПОДАННОГО В ЭЛЕКТРОННОМ ВИДЕ
Содержание текстового файла, поданного с этим документом в электронном виде, включено в данный документ посредством ссылки в полном объеме: машиночитаемая копия Перечня последовательностей (название файла: HTB-023PC-SequenceListing. txt; дата записи: 29 сентября 2016 г.; размер файла: 140 KB).
УРОВЕНЬ ТЕХНИКИ
Взаимодействие между раковыми клетками и иммунной системой является сложным и многогранным. См. de Visser et al., Nat. Rev. Cancer (2006) 6:24-37. В то время как у многих пациентов с раком, по-видимому, формируется противоопухолевый иммунный ответ, раковые клетки также развивают свои стратегии, направленные на уклонение от иммунного ответа и разрушение иммунной системы. В последнее время были разработаны иммунотерапевтические средства для лечения и профилактики рака и других заболеваний. Преимущество иммунотерапии заключается в том, что она обладает клеточной специфичностью, что не характерно для других методов лечения. Таким образом, способы повышения эффективности иммунотерапии могут быть клинически полезными. Достижения в определении механизмов и молекул, которые регулируют иммунные ответы, обеспечили новые терапевтические цели в лечении рака. Например, костимулирующие и коингибирующие молекулы играют решающую роль в регуляции Т-клеточных иммунных ответов. Однако, несмотря на впечатляющие ответы пациентов на лечение антителами, нацеленными на эти костимулирующие и коингибирующие молекулы, включая, например, анти-PD-1/PD-L1, у многих пациентов терапия с применением ингибиторов контрольной точки все еще является неэффективной. Поэтому, как и при применении большинства лекарственных средств для лечения рака, остается потребность в новых композициях и методах, которые могут повысить эффективность этих агентов.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Соответственно, в различных аспектах данного изобретения предлагаются композиции и способы, которые являются пригодными для применения в иммунотерапии рака, например, с целью управления или модификации иммунных сигналов для получения терапевтического эффекта. В различных вариантах осуществления с помощью данного изобретения обращают или подавляют иммунные ингибирующие сигналы, обеспечивая при этом активирующие или ко-стимулирующе сигналы в благоприятном контексте. Например, в одном аспекте данного изобретения предлагается химерный белок, содержащий: (а) первый внеклеточный домен трансмембранного белка типа I на или около N-конца, (b) второй внеклеточный домен трансмембранного белка типа II на или около С-конца и (с) линкер, при этом один из первого и второго внеклеточных доменов является иммуноингибирующим сигналом, и один из первого и второго внеклеточных доменов является иммуностимулирующим сигналом. Посредством связывания этих двух молекул в функциональной ориентации может быть достигнута координация между положительным и отрицательным сигналами. Например, в различных вариантах осуществления данного изобретения предлагается маскирование отрицательных иммунных сигналов и стимуляция положительных иммунных сигналов в одной конструкции. В различных вариантах осуществления данного изобретения предлагаются композиции, которые не являются антителами, или не являются композициями на основе антигенсвязывающих доменов, производных от антител (например, областей, определяющих комплементарность, CDR), а скорее обеспечивают прямое взаимодействие между рецептором и лигандом.
У пациентов с раком можно стимулировать иммунный ответ против опухолевых антигенов с целью активации собственной иммунной системы пациента для уничтожения опухолевых клеток. Тем не менее, некоторые раковые клетки разрабатывают свои стратегии, направленные на уклонение от иммунного ответа в процессе, известном как иммуноредактирование. Это может включать деактивацию специфических антигенов, деактивацию МНС I, активацию иммунорегуляторных поверхностных молекул (PD-L1, PD-L2, СЕАСАМ1, галектин-9, В7-Н3, В7-Н4, VISTA, CD47 и т.д.) или активацию растворимых иммуноингибирующих молекул (IDO, TGF-β, MICA и т.д.). В большинстве случаев, эти стратегии направлены на увеличение количества опухолевых клеток и реализуются в результате того, что при контакте инфильтрирующих опухоль иммунных клеток-киллеров с опухолевой клеткой, клетки-киллеры непосредственно ингибируются иммуносупресивными факторами и поэтому не могут уничтожить опухолевую клетку. Многие из иммуносупресивных лигандов, кооптированных опухолевыми клетками для подавления иммунного ответа, взаимодействуют с рецепторами, которые являются мембранными белками типа I. В некоторых вариантах осуществления данного изобретения химерный белок по данному изобретению содержит внеклеточный домен иммуноингибирующего агента, включая, без ограничения, один или большее количество из следующих элементов: TIM-3, BTLA, PD-1, CTLA-4, В7-Н4, PD-L1, PD-L2, В7-Н3, CD244, TIGIT, CD172a/SIRPα, VISTA/VSIG8, CD115, CD200, CD223, и TMIGD2. В некоторых вариантах осуществления данного изобретения химерный белок по данному изобретению содержит внеклеточный домен мембранного белка типа I, который обладает иммуноингибирующими свойствами. В различных вариантах осуществления данного изобретения химерный белок сконструирован таким образом, чтобы нарушать, блокировать, уменьшать и/или ингибировать передачу иммуноингибирующего сигнала, в качестве неограничивающего примера, связывание PD-1 с PD-L1 или PD-L2 и/или связывание CD172a с CD47 и/или связывание TIM-3 с одним или большим количеством элементов: галектином-9 и/или фосфатидилсерином.
Кроме того, в дополнение к подавлению иммуноингибирующей сигнализации часто является желательным улучшить передачу иммуностимулирующего сигнала для усиления иммунного ответа, например, для усиления противоопухолевого иммунного ответа у пациента. В некоторых вариантах осуществления данного изобретения химерный белок по данному изобретению содержит внеклеточный домен иммуностимулирующего сигнала, который, без ограничения, представляет собой один или большее количество из следующих элементов: лиганд ОХ-40, LIGHT (CD258), лиганд GITR, CD70, лиганд CD30, лиганд CD40, лиганд CD137, TRAIL и TL1A. В некоторых вариантах осуществления данного изобретения химерный белок по данному изобретению содержит внеклеточный домен мембранного белка типа II, который обладает иммуностимулирующими свойствами. В различных вариантах осуществления данного изобретения химерный белок сконструирован таким образом, чтобы усиливать, повышать и/или стимулировать передачу иммуностимулирующего сигнала, в качестве неограничивающего примера, связывание GITR с одним или большим количеством лигандов GITR, и/или связывание ОХ40 с OX40L, и/или CD40 с лигандом CD40.
В различных вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего рецептора и внеклеточный домен иммуностимулирующего лиганда, который может, без ограничений, доставлять иммунную стимуляцию в Т-клетку при маскировании иммуноингибирующих сигналов опухолевой клетки. В различных вариантах осуществления данного изобретения описанные химерные белки обеспечивают улучшенные иммунотерапевтические преимущества, эффективно вызывая замещение иммуноингибирующего сигнала на иммуностимулирующий сигнал. Например, конструкция химерного белка, содержащая (i) внеклеточный домен PD-1 и (ii) внеклеточный домен OX40L, способствует разрушение ингибирующего сигнала PD-L1/L2 и замену его стимулирующим OX40L. Соответственно, описанные химерные белки в некоторых вариантах осуществления данного изобретения могут применяться или находят свое применение в способах, связанных с ослаблением или устранением ингибирующего иммунного сигнала и/или усилением или активированием иммуностимулирующего сигнала. Указанные полезные характеристики усиливаются посредством подхода с применением одной конструкции описанных химерных белков. Например, замену сигнала можно осуществлять практически одновременно, и при этом замену сигнала проводят с учетом локальной цели в клинически значимом месте (например, в микроокружении опухоли). В других вариантах осуществления данного изобретения применяется тот же принципи и к другим конструкциям химерных белков, таким как, например, (i) внеклеточный домен PD-1 и (ii) внеклеточный домен GITRL; (i) внеклеточный домен BTLA и (ii) внеклеточный домен OX40L; (i) внеклеточный домен TIGIT и (ii) внеклеточный домен OX40L; (i) внеклеточный домен TMIGD2 и (ii) внеклеточный домен OX40L; (i) внеклеточный домен TIM3 и (ii) внеклеточный домен OX40L; и (i) внеклеточный домен CD172a или CD115 и (ii) внеклеточный домен CD40L; среди прочих.
Кроме того, в некоторых вариантах осуществления данного изобретения описанные химерные белки могут применяться или находят свое применение в способах, связанных с изменением баланса иммунных клеток в пользу иммунной атаки опухоли. Например, описанные химерные белки могут сдвигать соотношение иммунных клеток в клинически значимом месте в пользу клеток, которые могут уничтожить опухоль (например, Т-клетки, цитотоксические Т-лимфоциты, Т-хелперные клетки, клетки-естественные киллеры (NK), клетки-естественные киллеры Т (NKT), противоопухолевые макрофаги (например, макрофаги M1), В-клетки и дендритные клетки, в противовес клеткам, которые защищают опухоли (например, супрессорные клетки миелоидного происхождения (MDSC), регуляторные Т-клетки (Treg), ассоциированные с опухолью нейтрофилы (TAN), макрофаги М2 и ассоциированные с опухолью макрофаги (ТАМ)). В некоторых вариантах осуществления данного изобретения описанный химерный белок способен увеличивать соотношение эффекторных Т-клеток к регуляторным Т-клеткам.
В различных вариантах осуществления данного изобретения описанный химерный белок непредвиденно обеспечивает связывание компонентов внеклеточного домена с их соответствующими партнерами по связыванию с более низкими скоростями диссоциации (Kd or Koff) и, следовательно, помимо прочего, обеспечивает более длительное пребывание рецептора в лиганде и наоборот. Например, в некоторых вариантах осуществления данного изобретения это обеспечивает устойчивый эффект маскирования отрицательного сигнала. Кроме того, в некоторых вариантах осуществления данного изобретения это обеспечивает более длительный положительный сигнал, например, для обеспечения адекватной стимуляции эффекторной клетки (например, для пролиферации и/или высвобождения стимулирующих сигналов, таких как цитокины). Кроме того, этот стабильный синапс клеток (например, опухолевой клетки, несущей отрицательные сигналы, и Т-клетки, которая может атаковать опухоль) обеспечивает пространственную ориентацию в пользу уменьшения опухоли - например, позиционирование Т-клеток для атаки опухолевых клеток и/или стерическое предотвращение доставки отрицательных сигналов опухолевой клетки, включая отрицательные сигналы, выходящие за пределы тех, которые замаскированы химерным белком по данному изобретению. В других вариантах осуществления данного изобретения это обеспечивает более длительный целевой период полувыведения химерных белков (например, внутри опухоли) (t1/2) по сравнению с t1/2 в сыворотке. Такие характеристики могут иметь комбинированное преимущество, заключающееся в снижении уровня нецелевых токсинов, ассоциированных с системным распределением химерных белков.
Также, в различных аспектах, описанный химерный белок применяют в способе лечения рака, включающем введение эффективного количества фармацевтической композиции, содержащей химерный белок, нуждающемуся в этом пациенту. В других аспектах, описанный химерный белок применяют в способе лечения инфекций, включая, без ограничений, вирусные инфекции или инфекции, вызванные другими внутриклеточными патогенами. В еще одном аспекте, описанный химерный белок применяют в способе лечения аутоиммунных заболеваний.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фигуре 1 проиллюстрированы ориентации белков мембранных белков типа I (слева) и типа II (справа) в клеточной мембране. В мембранном белке типа I, изображенном на левой панели, аминоконец (обозначенный как "N") обращен во внеклеточную среду, а карбоксильный конец (обозначенный как "С") локализуется во внутриклеточной среде. Напротив, мембранный белок типа II, изображенный на правой панели, характеризуется обращением карбоксильного конца во внеклеточную среду и локализацией аминоконца во внутриклеточном пространстве.
На Фигуре 2 проиллюстрирована иммуноингибирующая и иммуномодулирующая сигнализация, которая имеет отношение к данному изобретению (из Mahoney, Nature Reviews Drug Discovery 2015:14; 561-585).
На Фигуре 3 приведена схематическая иллюстрация того, как можно сконструировать мембранный белок типа I и типа II (панель А) с удаленными трансмембранными и внутриклеточными доменами (панель В) и присоединенными с помощью линкерной последовательности (панель С) для получения одного слитого белка, в котором внеклеточные домены мембранных белков типа I и типа II обращены наружу в одном слитом белке (панель D). На панели С проиллюстрирована связь мембранного белка типа I и типа II путем удаления трансмембранных и внутриклеточных доменов каждого белка и соединения освобожденных внеклеточных доменов (ECD) из каждого белка с помощью линкерной последовательности. ECD в этом описании может включать в себя всю аминокислотную последовательность белка типа I или типа II, который обычно локализуется вне клеточной мембраны, или любой его части, которая сохраняет связывание с предполагаемым рецептором или лигандом. На панели С проиллюстрированы присоединенные внеклеточные домены в линейной конструкции, в которой внеклеточный домен мембранного белка типа I обращен к "левой" стороне конструкции, а внеклеточный домен мембранного белка типа II обращен к "правой" стороне конструкции.
На Фигуре 4 проиллюстрировано, что опухолевые клетки могут экспрессировать PD-L1 на поверхности клетки (панель А), которая может связываться с PD-1, экспрессируемым Т-клеткой (панель В). Это взаимодействие подавляет активацию Т-клеток. Слитый белок внеклеточного домена PD-1, присоединенный к внеклеточному домену OX40L, может связываться с PD-L1 на поверхности опухолевой клетки, предотвращая связывание с PD-1 на поверхности Т-клетки (панель С), Слитый белок может затем "свисать" с поверхности опухолевой клетки, а часть OX40L слитого белка может затем связываться с ОХ40, экспрессируемым на поверхности Т-клетки. Это будет приводить к замещению ингибирующего сигнала PD-L1 костимулирующим сигналом OX40L и тем самым усиливать противоопухолевую активность Т-клеток.
На Фигуре 5 проиллюстрирована экспрессия химерного PD-1-Fc мыши (m) и лиганда PD-1-Fc-OX40 (L) из клеток СНО-K1, обнаруженная с помощью анализа ELISA с захватом IgG мыши и определением анти-mIgG.
На Фигуре 6 проиллюстрированы результаты анализа ELISA, подтверждающие связывание mPD-1-Fc-OX40L с mOX40. На панели А приведено схематическое изображение метода ELISA, применяемого для обнаружения связывания mPD-1-Fc-OX40L с mOX40. Рекомбинантный mOX40, слитый с Fc (mOX40-hFc) человека, применяли для захвата mPD-1-Fc-OX40L в культуральной среде. Поликлональное антитело кролика к mPD-1 применяли для определения домена mPD-1 в химерном белке, с последующим определением с применением конъюгированного с пероксидазой хрена (HRP) поликлонального антитела к IgG (Н+L) кролика. На панели В проиллюстрированы результаты, в которых двукратные серийные разведения культуральной среды СНО-K1, содержащей белок mPD-1-Fc-OX40L, инкубировали со связанным на планшете mOX40-hFc, при этом связывание измеряли при показателе поглощения 450 нм. В качестве отрицательных контролей применяли белок mPD-1-Fc (связывание которого с рекомбинантным ОХ40 мыши не прогнозировалось), содержащий культуральную среду, а также отдельно взятую культуральную среду.
На Фигуре 7 проиллюстрированы результаты анализа ELISA, подтверждающие связывание mPD-1-Fc-OX40L с mPD-L1. На панели А приведено схематическое изображение метода ELISA, применяемого для обнаружения связывания mPD-1-Fc-OX40L с mPD-L1. Рекомбинантный mPD-L1, слитый с Fc (mPD-L1-hFc) человека, применяли для захвата химерного белка mPD-1-Fc-OX40L в культуральной среде. Для обнаружения связанных белков применяли конъюгированное с пероксидазой хрена (HRP) поликлональное антитело к IgG (Н+L) мыши. На панели В проиллюстрированы результаты, в которых двукратные серийные разведения культуральной среды СНО-K1, содержащей белок mPD-1-Fc-OX40L, инкубировали со связанным на планшете mPD-L1-hFc, при этом связывание измеряли при показателе поглощения 450 нм. В качестве положительного контроля применяли белок mPD-1-Fc, содержащий культуральную среду, а в качестве отрицательного контроля применяли отдельно взятую культуральную среду.
На Фигуре 8 проиллюстрировано, что внутриопухолевая доставка плазмидной ДНК in vivo, кодирующей PD-1-Fc-OX40L мыши (m), обуславливала увеличение количества антигенспецифических CD8+ Т-клеток. Обозначение "только ЭП" означает электропорацию в качестве отрицательного контроля. В этом эксперименте мышей C57BL/6 адоптивно трансфицировали овальбуминспецифическими CD8+ Т-клетками (ОТ-I) за 2 дня до инокуляции опухоли. Опухоли B16-F10-ova затем имплантировали в задний фланк каждой мыши в день 0. 7-дневные прижившиеся опухоли B16-F10-ova инъецировали плазмидной ДНК, кодирующей mPD1-Fc-OX40L, и сразу после этого электропорировали в дни 7 и 10 и сравнивали с контролем, предусматривающим только ЭП. Частоту клеток OT-I измеряли в указанные дни в периферической крови методом проточной цитометрии.
На Фигуре 9 проиллюстрировано, что внутриопухолевая доставка плазмидной ДНК in vivo, кодирующей mPD-1-Fc-OX40L, обуславливала регрессию опухоли в модели опухоли меланомы В16. F10-ova. Обозначение "только ЭП" означает электропорацию в качестве отрицательного контроля. На мышей воздействовали, как указано на Фигуре 9, и в указанные дни измеряли диаметр опухоли.
На Фигуре 10 проиллюстрированы результаты дополнительной характеристики mPD-1-Fc-OX40L. На панели А проиллюстрирован анализ вестерн-блоттинга с зондированием антителами для mPD-1 (левый гель), mFc (средняя панель) и mOX40L (правая панель), выполненный в восстанавливающих или невосстанавливающих условиях с дегликозилазой PNGазой F или без нее (на что указывают знаки "+" или "-" над каждым пятном). Белок мыши имел прогнозируемую молекулярную массу ~ 60 кДа в качестве мономерного белка. На панели В проиллюстрированы результаты функционального анализа ELISA, демонстрирующие связывание mPD-1-Fc-OX40L с mPD-L1 и mOX40. Для каждого набора гистограмм, столбцы представляют собой, слева направо, серийное разведение очищенного слитого белка mPD1-Fc-OX40L. На панели С проиллюстрированы результаты функционального анализа ELISA, демонстрирующие связывание mPD-1-Fc-OX40L с mFc (для каждой концентрации, OX40-His - левый столбец, a HVEM-His - правый столбец). На панели D проиллюстрировано связывание mPD-1-Fc-OX40L с активированными спленоцитами мыши, что обнаруживали на HLA I-A/I-E+PD-L1+(APC+PD-L1+) и CD4+OX40+ клетках (для каждого графика: популяции клеток, изображенные слева, представляют собой клетки APC-PD-L1- или CD4-OX40-, а популяции клеток, изображенные справа, представляют собой клетки APC+PD-L1+ или CD4+OX40+). На панели Е проиллюстрирована идентификация клеточных линий PD-L1низк (4Т1) и PD-L1выс (В16. F10) Ha панели F проиллюстрированы результаты анализа совместного культивирования спленоцитов/опухоли. IL2 ELISA проводили через 5 дней после первичного сбора клеток, выросших в культуре. Линейные диаграммы слева направо представляют +4Т1 клетки (-FP), +4Т1 клетки (+500 нг FP), +4Т1 клетки (+5 мкг FP), +В16 клетки (-FP), +В16 клетки (+500 нг FP), and +В16 клетки (+5 мкг FP).
На Фигуре 11 проиллюстрирована противоопухолевая эффективность mPD-1-Fc-OX40L. На панели А проиллюстрирована кинетика роста опухоли МС38 после применения указанных схем. Мышам Balb. с инокулировали в задний фланк 2,5×105 опухолевых клеток MC38-ova. В дни 5 и 8 мышам вводили препарат в соответствии с указанной группой лечения. Животным, предварительно получавшим анти-ОХ40, вводили 100 мкг OX86 mAb в каждый из двух дней, животным, предварительно получавшим анти-PD-L1, вводили 100 мкг 10F. 9G2 mAb в каждый из двух дней, животным, предварительно получавшим комбинацию анти-ОХ40 и анти-PD-L1, вводили 100 мкг каждого из OX86 и 10F. 9G2 в каждый из двух дней, а мышам, предварительно получавшим mPD1-Fc-OX40L, вводили в общей сложности 100 мкг mPD1-Fc-OX40L в каждый из двух дней. Площадь опухоли рассчитывали в указанные дни с учетом результатов измерения перпендикулярного диаметра опухоли с помощью электронного штангенциркуля. В день 40, мышам, в которых отмечалось полное отторжение существовавшей ранее опухоли (отсутствовала видимая или пальпируемая опухоль), вводили 2,5×105 МС38 родительских (не экспрессирующих ova) опухолевых клеток без какой-либо дополнительной обработки, при этом площадь опухоли рассчитывали, как указано выше. На панели В проиллюстрирована общая выживаемость для каждой группы лечения в течение эксперимента, которую определяли совокупным размером опухоли, превышающим 150 мм2 в соответствии с протоколами IACUC (на день 65; кривые сверху вниз: αOX40/αPD-L1, mPD1-Fc-OX40L, αOX40, αPD-L1 и без лечения). На панели С проиллюстрированы результаты анализа периферической крови на предмет соотношения CD4/CD8 (сверху) и процента клеток FOXP3+ Treg (снизу) для каждой указанной группы лечения (на обоих графиках, группы лечения слева направо: без лечения, αOX40, αPD-L1, αOX40/αPD-L1 и mPD1-Fc-OX40L). На панели D проиллюстрированы результаты анализ цитокинов сыворотки IFNγ, TNFα, IL4 и IL6. Для каждого набора данных, линейные графики слева направо представляют: без лечения, α-OX40, α-PD-L1, α-ОХ40/α-PD-L1 и mPD-1-Fc-OX40L (на четырех графиках, группы лечения слева вправо: без лечения, α-OX40, α-PD-L1, α-OX40/αPD-L1 и mPD1-Fc-OX40L). На панели Е проиллюстрирован средний размер опухоли для каждой группы лечения на день 13 эксперимента (на каждом графике, образцы слева направо: без лечения, mPD1-Fc-OX40L, mPD1-Fc-GITRL, mCD172a-Fc-CD40L, αОХ40, αPD-L1 и α GITR). На панели F проиллюстрирована процентная доля KSP тетрамер-специфических CD8+ Т-клеток, выделенных из опухоли (TIL) для каждой группы лечения на день 13 эксперимента (на каждом графике, образцы слева направо: без лечения, mPD1-Fc-OX40L, mPD1-Fc-GITRL, mCD172a-Fc-CD40L, αOX40, αPD-L1 и α GITR). На панели G проиллюстрирован фенотип CD8+ спленоцитов согласно хорошо изученным маркерам "иммунной памяти" на день 13 эксперимента для каждой группы лечения (на каждом графике, образцы слева направо: без лечения, mPD1-Fc-OX40L, mPD1-Fc-GITRL, mCD172a-Fc-CD40L, αOX40, αPD-L1 и α GITR). На панели H проиллюстрировано соотношение клеток CD4 к CD8 в периферической крови (левая панель), селезенке (средняя панель) и опухоли (правая панель) для каждой группы лечения на день 13 эксперимента (на каждом графике, образцы слева направо: без лечения, mPD1-Fc-OX40L, mPD1-Fc-GITRL, mCD172a-Fc-CD40L, αOX40, αPD-L1 и α GITR). На панели I проиллюстрирована схема того, как каждое животное подвергалось лечению в каждом эксперименте с применением модели опухоли толстой кишки СТ26. На панели J представлены репрезентативные участки проточной цитометрии, применяемые для расчета концентрации в сыворотке каждого указанного сывороточного цитокина, с применением набора матрицы гранул Legend Plex от компании BioLegend. Обозначается каждый указанный цитокин, включенный в панель, а средняя интенсивность флюоресценции каждого кластера гранул учитывают при расчете относительной концентрации каждого цитокина в сыворотке. На панели K представлен пример того, как можно применять анализ Legend Plex в качестве фармакодинамического биомаркера эффекта дозы для слитого белка PD1-Fc-OX40L. Продемонстрировано, что, применяя концентрацию IFNγ в качестве типовой, увеличение концентраций этого цитокина соответствует увеличению количества введений PD1-Fc-OX40L (на панели K продемонстрировано слева направо, без лечения, 40 мкг × 1, 40 мкг × 2, 100 мкг × 1, и 100 мкг × 2). На панели L проиллюстрирован кинетика роста опухоли СТ26 для каждой группы лечения.
На Фигуре 12, панель А, проиллюстрирована прогнозируемая третичная структура PD-1-Fc-OX40L человека, как определено с помощью анализа RaptorX. На панели В проиллюстрирована оценка иммуногенности PD-1-FC-OX40 человека с применением iTope, в алгоритме моделирования (ANTITOPE/ABZENA) in silico, с перекрестной ссылкой на запатентованную базу данных эпитопов Т-клеток.
На Фигуре 13 проиллюстрированы характеристики PD-1-Fc-OX40L человека (также называемого SL-279252).). На панели А проиллюстрированы пики элюирования белка А (OD450) из SL-279252, очищенные от стабильных (внутрилабораторных) или транзиторных трансфекционных (Thermo) препаратов. Результаты ELISA каждого пика элюирования накладываются на показания поглощения, чтобы показать, что белок SL279252 содержится в первом пике элюирования из колонки. На панели В проиллюстрирован анализ вестерн-блоттинга SL-279252, выполненный путем зондирования очищенного белка антителами анти-PD-1 человека (левый гель), анти-Fc (средний гель) и анти-OX40L (правый гель), в восстанавливающих или невосстанавливающих условиях с дегликозилазой PNGазой F или без нее Прогнозируемая молекулярная масса мономерного SL-279252 составляет 60,3 кДа. На панели С проиллюстрированы результаты функциональных анализов ELISA с применением захвата с помощью рекомбинантного hOX40 и обнаружением с помощью Gt-hOX40L/Gt-HRP по сравнению с рекомбинантным стандартом OX40L-Fc человека. На панели D проиллюстрированы результаты функциональных анализов ELISA, предназначенных для оценки функционального связывания для каждой стороны SL-279252 одновременно. В частности, рекомбинантный PD-L1 человека абсорбировали на планшете и применяли для захвата SL-279252. Захваченный белок затем обнаруживали с применением рекомбинантного hOX40-his/HRP анти-his кролика по отношению к HVEM-his, взятого в качестве отрицательного контроля для специфичности.
На Фигуре 14 проиллюстрированы результаты поверхностного плазмонного резонанса (SPR) и анализа полувыведения SL-279252. "Скорость ассоциации (Ka)", "скорость диссоциации (Kd)", и аффинность связывания (KD) определяли для SL-279252 при связывании с PD-L1 человека (панель A), PD-L2 человека (панель В), ОХ40 человека (панель C), FcγR1A человека (панель D) и FcRn (панель Е), по сравнению с соответствующими контролями. На панели F обобщены показатели скорости ассоциации (Ka), скорости диссоциации (Kd), и аффинности связывания (KD) для каждого проанализированного состояния. Связывание модифицированной конструкции SL-279252, содержащей отдельный лидерный пептид, а также мутации в области Fc для усиления связывания с FcRn, также анализировали при связывании с PD-L1 человека (панель G), PD-L2 человека (панель Н), ОХ40 человека (панель I), FcγR1A человека (панель J) и FcRn (панель K), по сравнению с соответствующими контролями. На панели L обобщены показатели скорости ассоциации (Ka), скорости диссоциации (Kd), и аффинности связывания (KD) для каждого проанализированного состояния. На панели М проиллюстрирован период полувыведения SL-279252 в сыворотке мышей C57BL/6 in vivo.
На панели N проиллюстрирован интратуморальный период полувыведения SL-279252 in vivo у мышей с ослабленным иммунитетом (NSG), которым были имплантированы PD-L1-положительные опухоли человека в один фланк (HeLa-PD-L1) и PD-L1-отрицательные опухоли человека в противоположный фланк (HeLa). В указанные дни две опухоли вырезали и рассекали на две части. Половину рассеченной на две части опухоли дезагрегировали и анализировали на предмет связывание SL-279252 с помощью проточной цитометрии с применением антител против OX40L человека. На панели О проиллюстрированы замороженные срезы из другой половины рассеченной на две части опухоли через 6 часов, 2 дня и 5 дней после введения одной инъекции SL-279252. Изображение на фигуре подтверждает персистенция SL-279252 в течение не менее 5 дней после введения.
На Фигуре 15 проиллюстрировано связывание SL-279252 с клетками in vitro. На панели А проиллюстрирована оценка родительских клеток Jurkat (клеточная популяция, изображенная слева) и Jurkat/hOX40 (клеточная популяция, изображенная справа) с помощью проточной цитометрии с применением антитела hOX40-APC. На панели В проиллюстрирована оценка родительских клеток СНО-K1 (клеточная популяция, изображенная слева) и CHO-K1/hPD-L1 (клеточная популяция, изображенная справа) с помощью проточной цитометрии с применением антитела hPD-L1-APC. На панели С проиллюстрирована оценка родительских клеток СНО-K1 (клеточная популяция, изображенная слева) и CHO-K1/hCD47 (клеточная популяция, изображенная справа) с помощью проточной цитометрии с применением антитела hCD47-APC. На панели D, возрастающие концентрации SL-279252 инкубировали с родительскими клетками СНО-K1 (левая панель) и CHO-K1/hPD-L1 (средняя панель) и детектировали с применением антитела против человеческого OX40L-APC. На правой панели проиллюстрирована кривая титрования для возрастающих концентраций SL-279252. На панели Е, возрастающие концентрации SL-279252 инкубировали с родительскими клетками Jurkat (левая панель) или клетками Jurkat/hOX40 (средняя панель) и детектировали с применением антитела против человеческого OX40L-APC. На правой панели проиллюстрирована кривая титрования для возрастающих концентраций SL-279252. На панели F, возрастающие концентрации hCD172a-Fc-OX40L инкубировали с родительскими клетками СНО-K1 (левая панель) или клетками CHO-K1-CD47 (средняя панель) и детектировали с применением антитела против человеческого OX40L-APC. На правой панели проиллюстрирована кривая титрования для возрастающих концентраций hCD172a-Fc-OX40L. На панели G проиллюстрировано связывание возрастающих концентраций hCD172a-Fc-OX40L с родительскими клетками Jurkat (левая панель) или клетками Jurkat-hOX40 (средняя панель). На правой панели проиллюстрирована кривая титрования для возрастающих концентраций hCD172a-Fc-OX40L. На панели Н, PD-L1низк человека (клетки РС3; клеточная популяция, изображенная слева) и PD-L1выс (НСС827; клеточная популяция, изображенная справа) идентифицировали с помощью проточной цитометрии. На панели I, возрастающие концентрации SL-279252 инкубировали с клетками РС3. На панели J, возрастающие концентрации SL-279252 инкубировали с клетками НСС827 в течение 2 часов. Клетки промывали и анализировали с помощью проточной цитометрии для связывания SL-279252 (антитело Fc-PE).
На Фигуре 16 проиллюстрированы функциональные характеристики SL-279252 in vitro. На панели А, экспрессию ОХ40 обнаруживали в Т-клетках человека, выделенных из МКПК, обработанных в течение 2 дней РМА/РНА/лономицином (ион.). На панели В, связывание SL-279252 оценивали в активированных CD4+ и CD8+ клетках (вторичный Fc-PE). На панели С представлено схематическое изображение анализа совместного культивирования Т-клеток/опухолей для выявления активации Т-клеток, а также временная шкала эксперимента. На панели D, среду для совместного культивирования оценивали с помощью IL2 ELISA через 6 дней после первичного выделения Т-клеток. Линейные диаграммы слева направо представляют +РС3 (-FP), +РС3 (+500 нг FP), +РС3 (+5 мкг FP), +НСС827 (-FP), +НСС827 (+500 нг FP) и +НСС827 (+5 мкг FP). На панели Е, совместно культивированные Т-клетки анализировали с помощью проточной цитометрии через 5 дней после первичного выделения на предмет пролиферации CD4+ и CD8+ клеток (Ki67) и через 7 дней после выделения на предмет экспрессии цитокинов в CD8+ клетках. Линейные диаграммы слева направо представляют +НСС827 (-FP), +НСС827 (+500 нг FP) и +НСС827 (+5 мкг FP).
На Фигуре 17, панель А, проиллюстрирован результат вестерн-блоттинга различных химерных белков, включая hCD172a-Fc-OX40L, hPD1-Fc-TL1A, hBTLA-Fc-OX40L, hTMIGD2-Fc-OX40L, hTM3-Fc-OX40L, mPD1-Fc-GITRL, mPD1-Fc-4-1BBL, mPD1-Fc-TL1A, mCD172a-Fc-CD40L. Каждый химерный белок зондировали антителами, специфичными для каждого связывающего конца и центрального домена Fc. Анализы ELISA проводили для подтверждения связывания различных химерных белков с ОХ40 человека. На панели В приведено схематическое изображение метода ELISA, применяемого для обнаружения связывания химерных белков с ОХ40 человека. На панели С проиллюстрированы результаты связывания PD1-Fc-OX40L человека с родительскими клетками Jurkat (левая панель, левая кривая) или с клетками Jurkat/OX40 (левая панель, правая кривая). Для демонстрации специфичности применяли два отрицательных контроля: PD1-Fc-TL1A человека (средняя панель) и PD1-Fc-OX40L собаки (правая панель). На панели D проиллюстрирована прогнозируемая третичная структура CD172a-Fc-OX40L человека, как определено с помощью анализа RaptorX. На панели Е проиллюстрирована типовая продукция и очистка PD1-Fc-OX40L человека (SL-279252), включая гель Кумасси (вверху слева), анти-IgG вестерн-блот (вверху справа), концентрацию элюированного белка (внизу слева) и профиль элюирования от аффинной хроматографии (внизу справа). На панели F проиллюстрирована типовая продукция и очистка CD172a-Fc-OX40L человека, включая гель Кумасси (вверху слева), анти-IgG вестерн-блот (вверху справа), концентрацию элюированного белка (внизу слева) и профиль элюирования от аффинной хроматографии (внизу справа). На панели G проиллюстрирована типовая продукция и очистка CD172a-Fc-CD40L мыши, включая параметры очистки (верхняя таблица) и анализ очищенного белка LabChip (нижняя панель). На панели Н проиллюстрирована типовая продукция и очистка TIGIT-Fc-OX40L человека, включая гель Кумасси (вверху слева), анти-IgG вестерн-блот (вверху справа), концентрацию элюированного белка (внизу слева) и профиль элюирования от аффинной хроматографии (внизу справа). На панели I проиллюстрирована аффинность связывания CD172a-Fc-OX40L человека с иммобилизованным рекомбинантным CD47. На панели J проиллюстрирована аффинность связывания CD172a-Fc-OX40L человека с иммобилизованным рекомбинантным ОХ40 человека. На панели K проиллюстрирована аффинность связывания CD172a-Fc-OX40L человека с иммобилизованным рекомбинантным FcγR1A. На панели L проиллюстрирована аффинность связывания CD172a-Fc-OX40L человека с иммобилизованным рекомбинантным FcRn человека. На панели М обобщены показатели скорости ассоциации (Ka), скорости диссоциации (Kd), и аффинности связывания (KD) для каждого проанализированного состояния. На панели N проиллюстрирована типовая продукция и очистка PD1-Fc-OX40L собаки, включая гель Кумасси (вверху слева), анти-IgG вестерн-блот (вверху справа), концентрацию элюированного белка (внизу слева) и профиль элюирования от аффинной хроматографии (внизу справа). На панели О проиллюстрирована типовая продукция и очистка PD1-Fc-OX40L мыши, включая гель Кумасси (вверху слева), анти-IgG вестерн-блот (вверху справа), концентрацию элюированного белка (внизу слева) и профиль элюирования от аффинной хроматографии (внизу справа). На панели Р проиллюстрирована типовая продукция и очистка PD1-Fc-GITRL мыши, включая гель Кумасси (вверху слева), анти-IgG вестерн-блот (вверху справа), концентрацию элюированного белка (внизу слева) и профиль элюирования от аффинной хроматографии (внизу справа). На панели Q проиллюстрирована типовая продукция и очистка PD1-Fc-41BBL мыши, включая гель Кумасси (вверху слева), анти-IgG вестерн-блот (вверху справа), концентрацию элюированного белка (внизу слева) и профиль элюирования от аффинной хроматографии (внизу справа). На панели R проиллюстрирована типовая продукция и очистка PD1-Fc-TL1A мыши, включая гель Кумасси (вверху слева), анти-IgG вестерн-блот (вверху справа), концентрацию элюированного белка (внизу слева) и профиль элюирования от аффинной хроматографии (внизу справа). На панели S проиллюстрирована типовая продукция и очистка CD115-Fc-CD40L мыши, включая гель Кумасси (вверху слева), анти-IgG вестерн-блот (вверху справа), концентрацию элюированного белка (внизу слева) и профиль элюирования от аффинной хроматографии (внизу справа). На панели Т проиллюстрирована типовая продукция и очистка PD1-Fc-GITRL человека, включая гель Кумасси (вверху слева), анти-IgG вестерн-блот (вверху справа), концентрацию элюированного белка (внизу слева) и профиль элюирования от аффинной хроматографии (внизу справа).
ПОДРОБНОЕ ОПИСАНИЕ
Данное изобретение основано, в частности, на открытии того, что химерные белки можно сконструировать из внеклеточных или эффекторных областей иммуномодулирующих трансмембранных белков посредством способа, в котором используется ориентация этих белков (например, тип I по сравнению с типом II) и, следовательно, который позволяет доставлять иммуностимулирующие и/или иммуноингибирующие сигналы, включая, например, маскирование иммуноингибирующего сигнала и замену его иммуномодулирующим сигналом при лечении рака.
Химерные белки
В одном аспекте данного изобретения предлагается химерный белок, содержащий: (а) первый внеклеточный домен трансмембранного белка типа I на или около N-конца, (b) второй внеклеточный домен трансмембранного белка типа II на или около С-конца и (с) линкер, при этом один из первого и второго внеклеточных доменов является иммуноингибирующим сигналом, и один из первого и второго внеклеточных доменов является иммуностимулирующим сигналом.
В некоторых вариантах осуществления данного изобретения химерный белок относится к рекомбинантному слитому белку, т.е. одному полипептиду, содержащему внеклеточные домены, описанные в данном документе (и, необязательно, линкер). Например, в различных вариантах осуществления данного изобретения химерный белок транслируется как единый элемент в клетке. В некоторых вариантах осуществления данного изобретения химерный белок относится к рекомбинантному белку из множества полипептидов, например, множества внеклеточных доменов, описанных в данном документе, которые являются связанными с целью получения единого элемента, например, in vitro (например, с одним или большим количеством синтетических линкеров, описанных в данном документе).
В некоторых вариантах осуществления данного изобретения внеклеточный домен относится к части трансмембранного белка, которая способна взаимодействовать с внеклеточной средой. В различных вариантах осуществления данного изобретения внеклеточный домен относится к части трансмембранного белка, которая является состоятельной для связывания с лигандом или рецептором и эффективной передачи сигнала в клетку. В различных вариантах осуществления данного изобретения внеклеточный домен представляет собой всю аминокислотную последовательность трансмембранного белка, которая является внешней по отношению к клетке или клеточной мембране. В различных вариантах осуществления данного изобретения внеклеточный домен представляет собой ту часть аминокислотной последовательности трансмембранного белка, которая является внешней по отношению к клетке или клеточной мембране, и необходима для передачи сигнала и/или связывания лиганда, что можно проанализировать с помощью способов, известных в данной области техники (например, анализов связывания лиганда и/или клеточной активации in vitro).
В некоторых вариантах осуществления данного изобретения иммуноингибирующий сигнал относится к сигналу, который уменьшает или устраняет иммунный ответ. Например, в контексте онкологии такие сигналы могут уменьшать или устранять противоопухолевый иммунитет. В нормальных физиологических условиях ингибирующий сигнал полезен для поддержания аутотолерантности (например, предотвращения аутоиммунных заболеваний), а также для защиты тканей от повреждения, когда иммунная система реагирует на патогенную инфекцию. Например, без ограничения, иммуноингибирующий сигнал можно идентифицировать путем обнаружения усиления клеточной пролиферации, повышения продукции цитокинов, активности клеточной гибели или фагоцитарной активности, когда такой ингибирующий сигнал блокируется. Конкретные примеры таких ингибирующих сигналов включают блокаду PD-1 PD-L1/L2 с применением опосредуемой антителом блокады или путем конкурентного ингибирования PD-L1/L2 с применением PD-1, содержащего слитые белки. Когда такой ингибирующий сигнал блокируется посредством ингибирования PD-L1/L2, это приводит к усилению гибели опухолевых клеток в результате активности Т-клеток, поскольку они больше не ингибируются PD-L1 или PD-L2. В другом примере ингибирующий сигнал может обеспечиваться CD47 и достигать макрофагов, экспрессирующих CD172a. Как правило, связывание CD47 с CD172a ингибирует способность макрофага фагоцитировать клетку-мишень, которая может быть восстановлена посредством блокады CD47 с помощью блокирующих антител или посредством конкурентного ингибирования CD47 с применением CD172a, содержащего слитые белки.
В некоторых вариантах осуществления данного изобретения иммуностимулирующий сигнал относится к сигналу, который усиливает иммунный ответ. Например, в контексте онкологии, такие сигналы могут повышать противоопухолевый иммунитет. Например, без ограничения, иммуностимулирующий сигнал можно идентифицировать путем путем обнаружения непосредственного стимулирования пролиферации, продукции цитокинов, активной гибели клеток или фагоцитарной активности лейкоцитов. Конкретные примеры включают в себя непосредственную стимуляцию рецепторов суперсемейства TNF, таких как ОХ40, 4-1ВВ или TNFRSF25, с применением либо антител-агонистов рецепторов, либо с применением слитых белков, кодирующих лиганды для таких рецепторов (OX40L, 4-1BBL, TL1A, соответственно). Стимуляция от любого из этих рецепторов может непосредственно стимулировать пролиферацию и продукцию цитокинов отдельных подмножеств Т-клеток. Другой пример включает является непосредственную стимуляцию иммуноингибирующей клетки посредством рецептора, который ингибирует активность такой иммуносупрессорной клетки. Это может включать, например, стимуляцию CD4+FoxP3+ регуляторных Т-клеток антителом-агонистом GITR или слитым белком GITRL, что может уменьшать способность этих регуляторных Т-клеток подавлять пролиферацию обычных CD4+ или CD8+ Т-клеток. В другом примере, это может включать стимуляцию CD40 на поверхности антигенпредставляющей клетки с применением антитела-агониста CD40 или слитого белка, содержащего CD40L, что обуславливает активацию антигенпредставляющих клеток, включая повышенную способность этих клеток представлять антиген в отношении соответствующих нативных костимулирующих молекул, в том числе в надсемействе В7 или TNF.
Как правило, мембранные белки состоят из внеклеточного домена, одного или ряда трансмембранных доменов и внутриклеточного домена. Не желая быть связанными какой-либо теорией, отметим, что внеклеточный домен мембранного белка является ответственным за взаимодействие с растворимым или связанным с мембраной рецептором или лигандом. Не желая быть связанными какой-либо теорией, отметим, что трансмембранный(е) домен(ы) является ответственным за локализацию белка в плазматической мембране. Не желая быть связанными какой-либо теорией, отметим, что внутриклеточный домен мембранного белка является ответственным за координацию взаимодействий с клеточными сигнальными молекулами с целью координации внутриклеточных ответов с внеклеточной средой (или наоборот). Существует два типа однопроходных мембранных белков: белки, которые имеют внеклеточный аминоконец и внутриклеточный карбоксильный конец (тип I), и белки, которые имеют внеклеточный карбоксильный конец и внутриклеточный аминоконец (тип II). Мембранные белки как типа I, так и типа II могут быть либо рецепторами, либо лигандами. В мембранных белках типа I аминоконец белка обращен во внеклеточную среду и, следовательно, содержит функциональные домены, которые являются ответственными за взаимодействие с другими партнерами по связыванию (либо лигандами, либо рецепторами) во внеклеточной среде (Фиг. 1, левое изображение). Для мембранных белков типа II, карбоксиконец белка обращен во внеклеточную среду и, следовательно, содержит функциональные домены, которые являются ответственными за взаимодействие с другими партнерами по связыванию (либо лигандами, либо рецепторами) во внеклеточной среде (Фиг. 1, правое изображение). Таким образом, эти два типа белков имеют противоположную ориентацию друг к другу.
Поскольку обращенные наружу домены мембранных белков типа I и типа II являются противоположными (Фиг. 1), можно связать внеклеточные домены мембранного белка типа I и типа II таким образом, что "обращенные наружу" домены будут также находится в противоположной ориентации друг к другу (Фиг. 3). Таким образом, полученная конструкция будет состоять из внеклеточного домена мембранного белка типа I на "левой" стороне молекулы, связанной с внеклеточным доменом мембранного белка типа II на "правой" стороне молекулы с помощью линкерной последовательности. Указанная конструкция может быть получена путем клонирования этих трех фрагментов (внеклеточный домен белка типа I с последующей линкерной последовательностью и последующим внеклеточным доменом белка типа II) в вектор (плазмиду, вирусную или другую), при этом аминоконец полной последовательности соответствовал "левой" стороне молекулы, содержащей белок типа I, а карбоксильный конец полной последовательности соответствовал "правой" стороне молекулы, содержащей белок типа II. Соответственно, в различных вариантах осуществления данного изобретения описанные химерные белки сконструированы в указанном качестве.
В некоторых вариантах осуществления данного изобретения внеклеточный домен можно применять с целью продуцирования растворимого белка для конкурентного ингибирования передачи сигнала лигандом этим рецептора. В некоторых вариантах осуществления данного изобретения внеклеточный можно применять с целью обеспечения искусственной передачи сигнала.
В некоторых вариантах осуществления данного изобретения внеклеточный домен трансмембранного белка типа I представляет собой иммуноингибирующий сигнал. В некоторых вариантах осуществления данного изобретения внеклеточный домен трансмембранного белка типа II представляет собой иммуностимулирующий сигнал.
В некоторых вариантах осуществления данного изобретения описанные химерные белки содержат внеклеточный домен трансмембранного белка типа I или его функциональный фрагмент. В некоторых вариантах осуществления данного изобретения описанные химерные белки содержат внеклеточный домен трансмембранного белка типа II или его функциональный фрагмент. В некоторых вариантах осуществления данного изобретения описанные химерные белки содержат внеклеточный домен трансмембранного белка типа I или его функциональный фрагмент, и внеклеточный домен трансмембранного белка типа II или его функциональный фрагмент.
В различных вариантах осуществления данного изобретения описанные химерные белки содержат внеклеточный домен трансмембранного белка типа I человека, как указано в ТАБЛИЦЕ 1, или его функциональный фрагмент. В различных вариантах осуществления данного изобретения описанные химерные белки содержат внеклеточный домен трансмембранного белка типа II человека, как указано в ТАБЛИЦЕ 2, или его функциональный фрагмент. В некоторых вариантах осуществления данного изобретения описанные химерные белки содержат внеклеточный домен трансмембранного белка типа I, как указано в ТАБЛИЦЕ 2, или его функциональный фрагмент, а также внеклеточный домен трансмембранного белка типа II, как указано в ТАБЛИЦЕ 2, или его функциональный фрагмент. ТАБЛИЦЫ 1 и 2 приведены в данном документе в другом месте.
В различных вариантах осуществления данного изобретения описанные химерные белки могут быть сконструированы для нацеливания на одну или большее количество молекул, которые находятся на лейкоцитах человека, включая, без ограничения, внеклеточные домены (где это применимо) SLAMF4, IL-2 R α, 4-1BB/TNFRSF9, IL-2 R β, ALCAM, В7-1, IL-4 R, В7-Н3, BLAME/SLAMFS, СЕАСАМ1, IL-6 R, IL-7 Rα, IL-10R α, IL-1 0 R β, IL-12 R β 1, IL-12 R β 2, CD2, IL-13 R α 1, IL-13, CD3, CD4, ILT2/CDS5j, ILT3/CDS5k, ILT4/CDS5d, ILT5/CDS5a, лютегрина α 4/CD49d, CDS, интегрина α E/CD103, CD6, интегрина α M/CD 11b, CDS, интегрина α X/CD11c, интегрина β 2/CD1S, KIR/CD15S, CD27/TNFRSF7, KIR2DL1, CD2S, KIR2DL3, CD30/TNFRSFS, KIR2DL4/CD15Sd, CD31/PECAM-1, KIR2DS4, CD40 лиганда/TNFSFS, LAG-3, CD43, LAIR1, CD45, LAIR2, CDS3, лейкотриена B4-R1, CDS4/SLAMF5, NCAM-L1, CD94, NKG2A, CD97, NKG2C, CD229/SLAMF3, NKG2D, CD2F-10/SLAMF9, NT-4, CD69, NTB-A/SLAMF6, общей γ цепи/IL-2 R γ, остеопонтина, CRACC/SLAMF7, PD-1, CRTAM, PSGL-1, CTLA-4, RANK/TNFRSF11A, CX3CR1, CX3CL1, L-селектина, SIRP β 1, SLAM, TCCR/WSX-1, DNAM-1, тимопоетин, EMMPRIN/CD147, TIM-1, EphB6, TIM-2, Fas/TNFRSF6, TIM-3, Fas-лиганд/TNFSF6, TIM-4, Fcγ RIII/CD16, TIM-6, TNFR1/TNFRSF1A, гранулизина, TNFRIII/TNFRSF1B, TRAIL R1/TNFRSF1OA, ICAM-1/CD54, TRAIL R2/TNFRSF10B, ICAM-2/CD102, TRAILR3/TNFRSF10C, IFN-γR1, TRAILR4/TNFRSF10D, IFN-γR2, TSLP, IL-1 R1 и TSLP R.
На активацию регуляторных Т-клеток весьма существенно влияют костимулирующие и коингибирующие сигналы. Два основных семейства костимулирующих молекул включают в себя семейство В7 и семейство фактора некроза опухоли (TNF). Эти молекулы связываются с рецепторами на Т-клетках, относящихся к семействам рецепторов CD28 или TNF, соответственно. Многие четко определенные коингибиторы и их рецепторы относятся к семействам В7 и CD28.
В различных вариантах осуществления данного изобретения описанные химерные белки сконструированы для нацеливания на одну или большее количество молекул, участвующих в ингибировании иммунной системы, включая, например: CTLA-4, PD-L1, PD-L2, PD-1, BTLA, HVEM, TIM3, GAL9, LAG3, VISTA/VSIG8, KIR, 2B4, TIGIT, CD160 (также называемого BY55), CHK1- и CHK2-киназы, A2aR, СЕАСАМ (например, СЕАСАМ-1, СЕАСАМ-3 и/или СЕАСАМ-5), и различные лиганды семейства В-7 (включая, но не ограничиваясь ими, В7-1, В7-2, B7-DC, В7-Н1, В7-Н2, В7-Н3, В7-Н4, В7-Н5, В7-Н6 и В7-Н7).
В различных вариантах осуществления данного изобретения the химерный белок по данному изобретению содержит внеклеточный домен иммуноингибирующего агента, включая, без ограничения, один или большее количество из следующих элементов: TIM-3, BTLA, PD-1, CTLA-4, CD244, CD 160, TIGIT, SIRPα/CD172a, 2В4, VISTA, VSIG8, LAG3, CD200 и TMIGD2.
В некоторых вариантах осуществления данного изобретения химерный белок по данному изобретению содержит внеклеточный домен мембранного белка типа I, который обладает иммуноингибирующими свойствами. В различных вариантах осуществления данного изобретения химерный белок сконструирован таким образом, чтобы нарушать, блокировать, уменьшать и/или ингибировать передачу иммуноингибирующего сигнала, в качестве неограничивающего примера, связывание PD-1 с PD-L1 или PD-L2 и/или связывание CD172a с CD47 и/или связывание TIM-3 с галектином-9 и/или фосфатидилсерином.
В некоторых вариантах осуществления данного изобретения химерный белок по данному изобретению содержит внеклеточный домен иммуностимулирующего сигнала, который, без ограничения, представляет собой один или большее количество из следующих элементов: лиганд ОХ-40 (OX-40L), LIGHT (CD258), лиганд GITR (GITRL), CD70, лиганд CD30, лиганд CD40 (CD40L), лиганд CD137, TRAIL и TL1A.
В различных вариантах осуществления данного изобретения химерный белок имитирует связывание ингибирующего сигнал лиганда с его родственным рецептором (например, PD-1 с PD-L1 или PD-L2; например, CD172a с CD47; например, CD115 с CSF1; например, TIM-3 с галектином-9 или фосфатидилсерином), но ингибирует передачу ингибирующего сигнала в иммунную клетку (например, Т-клетку, макрофаг или другой лейкоцит).
В различных вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего рецептора и внеклеточный домен иммуностимулирующего лиганда, который может, без ограничений, доставлять иммунную стимуляцию в Т-клетку при маскировании иммуноингибирующих сигналов опухолевой клетки. В различных вариантах осуществления данного изобретения химерный белок доставляет сигнал, который имеет конечный результат активации Т-клеток.
В некоторых вариантах осуществления данного изобретения химерный белок содержит иммуноингибирующий сигнал, который является ECD рецептора иммуноингибирующего сигнала, и действует на опухолевую клетку, которая несет родственный лиганд иммуноингибирующего сигнала. В некоторых вариантах осуществления данного изобретения химерный белок содержит иммуностимулирующий сигнал, который является ECD лиганда иммуностимулирующего сигнала, и действует на опухолевую клетку, которая несет родственный рецептор иммуностимулирующего сигнала. В некоторых вариантах осуществления данного изобретения химерный белок содержит как (i) иммуноингибирующий сигнал, который является рецептором иммуноингибирующего сигнала, и действует на опухолевую клетку, которая несет родственный лиганд иммуноингибирующего сигнала, и (ii) иммуностимулирующий сигнал, который является лигандом иммунного стимулирующего сигнала, и действует на Т-клетку, которая несет родственный рецептор иммуностимулирующего сигнала.
В некоторых вариантах осуществления данного изобретения химерный белок по данному изобретению содержит внеклеточный домен одного или большего количества иммуномодулирующих агентов, описанных в публикации Mahoney, Nature Reviews Drug Discovery 2015: 14; 561-585, все содержание которых включено в данное описание посредством ссылки. Например, со ссылкой на указанную Фиг. 2, химерный белок несет иммуноингибирующий сигнал (обозначаемый как "-"), который является рецептором пары (т.е. правая сторона фигуры), а опухолевая клетка несет лиганд, выбранный с левой стороны фигуры. В качестве дополнительного примера, со ссылкой на указанную Фиг. 2, химерный белок несет иммуностимулирующий сигнал (обозначаемый как "+"), который является лигандом пары (т.е. левая сторона фигуры), а опухолевая клетка несет рецептор, выбранный из правой стороны фигуры.
В некоторых вариантах осуществления данного изобретения химерный белок по данному изобретению содержит внеклеточный домен мембранного белка типа II, который обладает иммуностимулирующими свойствами. В различных вариантах осуществления данного изобретения химерный белок сконструирован таким образом, чтобы усиливать, повышать и/или стимулировать передачу иммуностимулирующего сигнала, в качестве неограничивающего примера, связывание GITR с одним или большим количеством лигандов GITR, и/или связывание ОХ40 с OX40L, и/или связывание CD40 с лигандом CD40.
В некоторых вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента PD-1 и спарен с иммуностимулирующим агентом, указанным ниже: PD-1/4-1BBL; PD-1/OX-40L; PD-1/LIGHT; PD-1/GITRL; PD-1/CD70; PD-1/CD30L; PD-1/CD40L; и PD-1/TL1A.
В одном варианте осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента PD-1 и спарен с иммуностимулирующим агентом OX-40L. В одном варианте осуществления данного изобретения химерный белок содержит аминокислотную последовательность SEQ ID NO: 22. В различных вариантах осуществления данного изобретения химерный белок связывается с PD-L1 или PD-L2 человека со значением <g idKD, составляющим от около 1 нМ до около 5 нМ, например, около 1 нМ, около 1,5 нМ, около 2 нМ, около 2,5 нМ, около 3 нМ, около 3,5 нМ, около 4 нМ, около 4,5 нМ или около 5 нМ. В различных вариантах осуществления данного изобретения химерный белок связывается с PD-L1 человека со значением KD, составляющим от около 5 нМ до около 15 нМ, например, около 5 нМ, около 5,5 нМ, около 6 нМ, около 6,5 нМ, около 7 нМ, около 7,5 нМ, около 8 нМ, около 8,5 нМ, около 9 нМ, около 9,5 нМ, около 10 нМ, около 10,5 нМ, около 11 нМ, около 11,5 нМ, около 12 нМ, около 12,5 нМ, около 13 нМ, около 13,5 нМ, около 14 нМ, около 14,5 нМ или около 15 нМ.
В различных вариантах осуществления данного изобретения химерный белок проявляет повышенную стабильность и период полувыведения белка. В некоторых вариантах осуществления настоящего изобретения химерный белок связывается с FcRn с высокой аффинностью. В различных вариантах осуществления данного изобретения химерный белок может связываться с FcRn с KD, составляющим от около 70 нМ до около 80 нМ. Например, химерный белок может связываться с FcRn со значением KD, составляющим около 70 нМ, около 71 нМ, около 72 нМ, около 73 нМ, около 74 нМ, около 75 нМ, около 76 нМ, около 77 нМ, около 78 нМ, около 79 нМ или около 80 нМ. В некоторых вариантах осуществления данного изобретения химерный белок по существу не связывается с другими рецепторами Fc (т.е., кроме FcRn) с эффекторной функцией.
В некоторых вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента PD-L1 или PD-L2 и спарен с иммуностимулирующим рецептором, указанным ниже: PD-L1/4-1ВВ; PD-L1/OX-40; PD-L1/HVEM; PD-L1/GITR; PD-L1/CD27; PD-L1/CD28; PD-L1/CD30; PD-L1/CD40 и PD-L1/CD137.
В некоторых вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента PD-L2 и спарен с иммуностимулирующим рецептором, указанным ниже: PD-L2/4-1ВВ; PD-L2/OX-40; PD-L2/HVEM; PD-L2/GITR; PD-L2/CD27; PD-L2/CD28; PD-L2/CD30; PD-L2/CD40 и PD-L2/CD137.
В некоторых вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента TIM-3 и спарен с иммуностимулирующим агентом, указанным ниже: TIM-3/ОХ-40L; TIM-3/LIGHT; TIM-3/GITRL; TIM-3/CD70; TIM-3/CD30L; TIM-3/CD40L; TIM-3/CD137L; TIM-3/TL1A; и TIM-3/OX40L.
В некоторых вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента BTLA и спарен с иммуностимулирующим агентом, указанным ниже: BTLA/ОХ-40L; BTLA/LIGHT; BTLA/GITRL; BTLA/CD70; BTLA/CD30L; BTLA/CD40L; BTLA/CD137L; BTLA/TL1A; и BTLA/OX40L.
В некоторых вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента CD172a/SIRPα и спарен с иммуностимулирующим агентом, указанным ниже: CD172a/ОХ-40L; CD172a/LIGHT; CD172a/CD70; CD172a/CD30L; CD172a/CD40L; CD172a/CD137L; CD172a/TL1A; и CD172a/OX40L.
В некоторых вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента CD115 и спарен с иммуностимулирующим агентом, указанным ниже: CD115/ОХ-40L; CD115/LIGHT; CD115/CD70; CD115/CD30L; CD115/CD40L; CD115/CD137L; CD115/TL1A; и CD115/OX40L.
В некоторых вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента TIGIT и спарен с иммуностимулирующим агентом, указанным ниже: TIGIT/ОХ-40L; TIGIT/LIGHT; TIGIT/GITRL; TIGIT/CD70; TIGIT/CD30L; TIGIT/CD40L; TIGIT/CD137L; TIGIT/TL1A; и TIGIT/OX40L.
В некоторых вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента TMIGD2 и спарен с иммуностимулирующим агентом, указанным ниже: TMIGD2/ОХ-40L; TMIGD2/LIGHT; TMIGD2/GITRL; TMIGD2/CD70; TMIGD2/CD30L; TMIGD2/CD40L; TMIGD2/CD137L; TMIGD2/TL1A; и TMIGD2/OX40L.
В некоторых вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента LAG3 и спарен с иммуностимулирующим агентом, указанным ниже: LAG3/ОХ-40L; LAG3/LIGHT; LAG3/GITRL; LAG3/CD70; LAG3/CD30L; LAG3/CD40L; LAG3/CD137L; LAG3/TL1A; и LAG3/OX40L.
В некоторых вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента VSIG8 и спарен с иммуностимулирующим агентом, указанным ниже: VSIG8/ОХ-40L; VSIG8/LIGHT; VSIG8/GITRL; VSIG8/CD70; VSIG8/CD30L; VSIG8/CD40L; VSIG8/CD137L; VSIG8/TL1A; и VSIG8/OX40L.
В некоторых вариантах осуществления данного изобретения химерный белок содержит внеклеточный домен иммуноингибирующего агента CD200 и спарен с иммуностимулирующим агентом, указанным ниже: CD200/ОХ-40L; CD200/LIGHT; CD200/GITRL; CD200/CD70; CD200/CD30L; CD200/CD40L; CD200/CD137L; CD200/TL1A; и CD200/OX40L.
В различных вариантах осуществления данного изобретения описанные химерные белки могут содержать варианты описанных в данном документе внеклеточных доменов, например последовательность, имеющую по меньшей мере около 60%, или по меньшей мере около 61%, или по меньшей мере около 62%, или по меньшей мере около 63%, или по меньшей мере около 64%, или по меньшей мере около 65%, или по меньшей мере около 66%, или по меньшей мере около 67%, или по меньшей мере около 68%, или по меньшей мере около 69%, или по меньшей мере около 70%, или по меньшей мере около 71%, или по меньшей мере около 72%, или по меньшей мере около 73%, или по меньшей мере около 74%, или по меньшей мере около 75%, или по меньшей мере около 76%, или по меньшей мере около 77%, или по меньшей мере около 78%, или по меньшей мере около 79%, или по меньшей мере около 80%, или по меньшей мере около 81%, или по меньшей мере около 82%, или по меньшей мере около 83%, или по меньшей мере около 84%, или по меньшей мере около 85%, или по меньшей мере около 86%, или по меньшей мере около 87%, или по меньшей мере около 88%, или по меньшей мере около 89%, или по меньшей мере около 90%, или по меньшей мере около 91%, или по меньшей мере около 92%, или по меньшей мере около 93%, или по меньшей мере около 94%, или по меньшей мере около 95%, или по меньшей мере около 96%, или по меньшей мере около 97%, или по меньшей мере около 98%, или по меньшей мере около 99%) идентичности последовательности с известной аминокислотной последовательностью или последовательностью нуклеиновой кислоты внеклеточных доменов, например, внеклеточных доменов человека, например, одной или большим количеством из SEQ ID NO: 1-15 в целом или относительно указанных доменов. В изобретение включены различные иллюстративные последовательности, как SEQ ID NO: 1-15, в которых внеклеточные домены показаны как подчеркнутые или выделенные жирным шрифтом, а линкер - обычным шрифтом. В различных вариантах осуществления данного изобретения линкер может быть заменен другим, описанным в данном документе.
В иллюстративном варианте осуществления данного изобретения химерный белок по данному изобретению содержит внеклеточный домен PD-1 и внеклеточный домен OX40L с применением домена шарнир-СН2-СН3 из последовательности антитела IgG4 человека. В этом варианте осуществления данного изобретения внеклеточный домен PD-1 подчеркнут, за ним следует домен с шарнир-СН2-СН3 IgG4 человека и короткий линкер (обычный текст), за которым следует внеклеточный домен OX40L (полужирный текст):


Эта последовательность кодирует белок с аминокислотной последовательностью:

Кроме того, эта аминокислотная последовательность, а также аминокислотные последовательности любого из внеклеточных доменов, описанных в данном документе (независимо от того, указаны ли они явно), также могут быть получены с помощью кодон-оптимизированных последовательностей нуклеиновой кислоты, таких как следующая последовательность, которая оптимизирована для экспрессия клеток китайского хомяка (СНО):


Другой вариант осуществления описанного химерного белка содержит внеклеточный домен PD-1 и внеклеточный домен костимулирующего лиганда, такого как TL1A, 4-1BBL, ICOSL, GITRL, CD27 или CD40L. Типовая последовательность, кодирующая внеклеточный домен PD-1 (подчеркнутый) -Fc (обычный текст) - внеклеточный домен TL1A (полужирный текст) представляет собой:


Эта нуклеотидная последовательность SEQ ID NO: 4 может быть кодон-оптимизированной с целью кодирования белка с аминокислотной последовательностью:
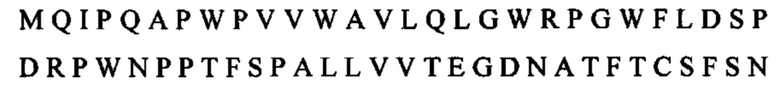

Существует много мембранных белков типа I, экспрессируемых опухолевыми клетками, которые могут быть замаскированы слитым белком, кодирующим внеклеточный домен родственного рецептора. Дополнительные примеры включают в себя слитый белок, кодирующий внеклеточный домен BTLA, связанный посредством Fc с OX40L. Такая конструкция может быть кодирована последовательностью нуклеиновой кислоты:


Эта нуклеотидная последовательность кодирует белок с аминокислотной последовательностью:
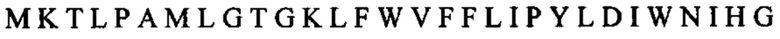

Другой пример может включать слитый белок, содержащий внеклеточный домен TIGIT, связанный посредством линкера Fc с OX40L.:


Эта последовательность может быть кодон-оптимизированной с целью кодирования белка с аминокислотной последовательностью:


Другой пример может включать слитый белок, содержащий внеклеточный домен TIM3, связанный посредством области Fc с OX40L человека:


Такая последовательность может быть кодон-оптимизированной с целью кодирования белка с аминокислотной последовательностью:


Другой пример может включать внеклеточный домен CD172a, присоединенный с помощью Fc-линкерной последовательности к внеклеточному домену OX40L человека:


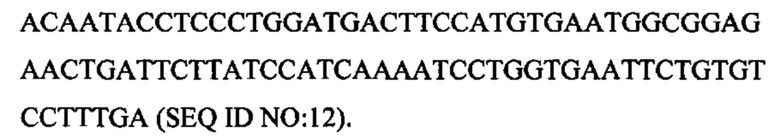
Такая последовательность может быть кодон-оптимизированной с целью кодирования белка с аминокислотной последовательностью:

Другой пример может включать внеклеточный домен TMIGD2, присоединенный с помощью Fc-линкерной последовательности к внеклеточному домену OX40L человека:
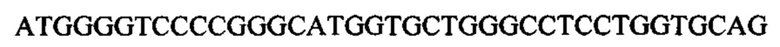

Такая последовательность может быть кодон-оптимизированной с целью кодирования белка с аминокислотной последовательностью:

В различных вариантах осуществления данного изобретения химерный белок может содержать аминокислотную последовательность, имеющую одну или большее количество аминокислотных мутаций по отношению к любой из описанных в данном документе белковых последовательностей. В некоторых вариантах осуществления данного изобретения одна или большее количество аминокислотных мутаций могут быть независимо выбраны из замен, вставок, делеций и усечений.
В некоторых вариантах осуществления данного изобретения аминокислотные мутации представляют собой аминокислотные замены и могут включать консервативные и/или неконсервативные замены.
"Консервативные замены" могут быть осуществлены, например, на основе сходства по полярности, заряду, размеру, растворимости, гидрофобности, гидрофильности и/или амфипатической природы применяемых аминокислотных остатков. 20 встречающихся в природе аминокислот можно сгруппировать в следующие шесть стандартных групп аминокислот: (1) гидрофобные: Met, Ala, Val, Leu, Ile; (2) нейтральные гидрофильные: Cys, Ser, Thr; Asn, Gln; (3) кислотные: Asp, Glu; (4) основные: His, Lys, Arg; (5) остатки, влияющие на ориентацию цепи: Gly, Pro; и (6) ароматические: Trp, Tyr, Phe.
В данном контексте "консервативные замены" определяются как обмен аминокислоты на другую аминокислоту, указанную в той же группе из шести стандартных групп аминокислот, приведенных выше. Например, обмен Asp на Glu сохраняет один отрицательный заряд в таком модифицированном полипептиде. Кроме того, глицин и пролин могут быть заменены друг на друга в зависимости от их способности разрушать α-спирали.
В данном контексте "неконсервативные замены" определяются как обмен аминокислоты на другую аминокислоту, указанную в разных группах из шести стандартных групп аминокислот от (1) до (6), приведенных выше.
В различных вариантах осуществления данного изобретения замены могут также включать неклассические аминокислоты (например, селеноцистеин, пирролизин, N-формилметионин β-аланин, ГАМК и δ-аминолевулиновую кислоту, 4-аминобензойную кислоту (РАВА), D-изомеры общих аминокислот, 2,4-диаминомасляную кислоту, α-аминомасляную кислоту, 4-аминомасляную кислоту, Abu, 2-аминоизомасляную кислоту, γ-Abu, ε-Ahx, 6-аминогексановую кислоту, Aib, 2-аминоизомасляную кислоту, 3-аминопропионовую кислоту, орнитин, норлейцин, норвалин, гидроксипролин, саркосем, цитруллин, гомоцитруллин, цистеиновую кислоту, трет-бутилглицин, трет-бутилаланин, фенилглицин, циклогексилаланин, β-аланин, фтороаминокислоты, разработанные аминокислоты, такие как β-метил аминокислоты, С α-метиламинокислоты, N α-метиламинокислоты и аналоги аминокислот в целом).
Могут также осуществить мутации с нуклеотидными последовательностями химерных белков со ссылкой на генетический код, в том числе с учетом вырождения кодонов.
В различных вариантах осуществления данного изобретения химерный белок содержит линкер. В различных вариантах осуществления данного изобретения линкер можно получить из встречающихся в природе многодоменных белков или линкер является эмпирическим линкером, как описано, например, в публикациях Chichili et al., (2013), Protein Sci. 22(2): 153-167, Chen et al., (2013), Adv Drug Deliv Rev. 65(10): 1357-1369, все содержание которых включено в данный документ посредством ссылки. В некоторых осуществления данного изобретения линкер может быть разработан с применением баз данных проектирования и компьютерных программ, таких как описанные в публикациях Chen et al., (2013), Adv Drug Deliv Rev. 65(10): 1357-1369 и Crasto et. al., (2000), Protein Eng. 13(5): 309-312, все содержание которых включено в данный документ посредством ссылки.
В некоторых вариантах осуществления данного изобретения, линкер является синтетическим линкером, таким как ПЭГ.
В других вариантах осуществления данного изобретения, линкер представляет собой полипептид. В некоторых вариантах осуществления данного изобретения линкер имеет длину менее чем около 500 аминокислот, около 450 аминокислот, около 400 аминокислот, около 350 аминокислот, около 300 аминокислот, около 250 аминокислот, около 200 аминокислот, около 150 аминокислот, или около 100 аминокислот. Например, линкер может имеет длину менее чем около 100, около 95, около 90, около 85, около 80, около 75, около 70, около 65, около 60, около 55, около 50, около 45, около 40, около 35, около 30, около 25, около 20, около 19, около 18, около 17, около 16, около 15, около 14, около 13, около 12, около 11, около 10, около 9, около 8, около 7, около 6, около 5, около 4, около 3 или около 2 аминокислот. В некоторых вариантах осуществления данного изобретения линкер является гибким. В другом варианте осуществления данного изобретения линкер является жестким.
В различных вариантах осуществления данного изобретения линкер по существу состоит из глициновых и сериновых остатков (например, около 30% или около 40%, или около 50%, или около 60%, или около 70%, или около 80%, или около 90%, или около 95%, или около 97% глицинов и серинов).
В различных вариантах осуществления данного изобретения линкер представляет собой шарнирную область антитела (например, IgG, IgA, IgD и IgE, включая подклассы (например, IgG1, IgG2, IgG3 и IgG4, а также IgA1 и IgA2)). Шарнирная область, обнаруженная в классах IgG, IgA, IgD и IgE, функционирует как гибкий спейсер, позволяющий части Fab свободно перемещаться в пространстве. В отличие от константных областей, шарнирные домены являются структурно разнообразными, варьируя как по последовательности, так и по длине среди классов и подклассов иммуноглобулина. Например, длина и гибкость шарнирной области варьируются между подклассами IgG. Шарнирная область IgG1 охватывает аминокислоты 216-231 и, поскольку она является свободно гибкой, фрагменты Fab могут вращаться вокруг своих осей симметрии и перемещаться внутри сферы, сосредоточенной на первом из двух дисульфидных мостиков между тяжелыми цепями. IgG2 имеет более короткий шарнир, чем IgG1, с 12 аминокислотными остатками и четырьмя дисульфидными мостиками. Шарнирная область IgG2 с отсутствующим глициновым остатком является относительно короткой и содержит жесткую двойную полипролиновую спираль, стабилизированную дополнительными дисульфидными мостиками между тяжелыми цепями. Эти свойства ограничивают гибкость молекулы IgG2. IgG3 отличается от других подклассов своей уникальной расширенной шарнирной областью (примерно в четыре раза длиннее шарнира IgG1), содержащей 62 аминокислоты (включая 21 пролин и 11 цистеинов), образуя негибкую полипролинную двойную спираль. В IgG3 фрагменты Fab относительно далеко располоджены от фрагмента Fc, придавая молекуле большую гибкость. Удлиненный шарнир в IgG3 также отвечает за его более высокую молекулярную массу по сравнению с другими подклассами. Шарнирная область IgG4 короче, чем IgG1, а параметры ее гибкости является промежуточными между IgG1 и IgG2. Сообщается, что гибкость шарнирных областей уменьшается в следующем порядке: IgG3>IgG1>IgG4>IgG2. В других вариантах осуществления данного изобретения линкер может быть производным из IgG4 человека и содержать одну или большее количество мутаций для усиления димеризации (включая S228P) или связывания FcRn.
Согласно кристаллографическим исследованиям, шарнирная область иммуноглобулина может быть дополнительно функционально подразделена на три области: верхнюю шарнирную область, сердцевинную область и нижнюю шарнирную область. См. Shin et al., 1992 Immunological Reviews 130:87. Верхняя шарнирная область включает аминокислоты от карбоксильного конца CH1 до первого остатка в шарнире, который ограничивает движение, как правило, до в первого остатка цистеина, который образует межцепочечную дисульфидную связь между двумя тяжелыми цепями. Длина верхней шарнирной области коррелирует с сегментной гибкостью антитела. Сердцевинная шарнирная область содержит дисульфидные мостики между тяжелыми цепями, а нижняя шарнирная область соединяется с аминоконцевым участком домена CH2 и включает остатки в CH2. Id. Сердцевинная шарнирная область IgG1 дикого типа человека содержит последовательность Cys-Pro-Pro-Cys, в результате димеризации которой путем образования дисульфидной связи образовывается циклический октапептид, который, как полагают, выступает в качестве стержня, тем самым придавая гибкость. В различных вариантах осуществления данного изобретения указанный линкер содержит одну или две или три области, выбранной из верхней шарнирной области, сердцевинной шарнирной области и нижней шарнирной области любого антитела (например, IgG, IgA, IgD и IgE, включая подклассы (например, IgG1, IgG2, IgG3 и IgG4, а также IgA1 и IgA2)). Шарнирная область может также содержать один или большее количество участков гликозилирования, которые включают в себя ряд участков, имеющих структурно различные типы, для прикрепления углеводов. Например, IgA1 содержит пять участков гликозилирования в 17-аминокислотном сегменте, что придает устойчивость полипептиду шарнирной области к действию кишечных протеаз и считается преимущественным свойством для секреторного иммуноглобулина. В различных вариантах осуществления данного изобретения линкер по данному изобретению содержит один или большее количество участков гликозилирования.
В различных вариантах осуществления данного изобретения линкер содержит домен Fc антитела (например, IgG, IgA, IgD и IgE, включая подклассы (например, IgG1, IgG2, IgG3 и IgG4, а также IgA1 и IgA2)). В различных вариантах осуществления данного изобретения линкер содержит домен Fc шарнир-СН2-СН3, производный от антитела IgG4 человека. В различных вариантах осуществления данного изобретения линкер содержит домен Fc шарнир-СН2-СН3, производный от антитела IgG1 человека. В некоторых вариантах осуществления данного изобретения домен Fc проявляет повышенную аффинность и усиленное связывание с неонатальным рецептором Fc (FcRn). В некоторых вариантах осуществления данного изобретения домен Fc включает одну или большее количество мутаций, которые повышают аффнность и усиливают связывание с FcRn. Не желая быть связанными какой-либо теорией, считается, что повышенная аффинность и усиленное связывание с FcRn увеличивает период полувыведения описанных химерных белков in vivo.
В некоторых вариантах осуществления данного изобретения линкер домена Fc содержит одну или большее количество аминокислотных замен в аминокислотных остатках 250, 252, 254, 256, 308, 309, 311, 428, 433 или 434 (в соответствии с нумерацией по Кабату) или их эквивалентах. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 250 представляет собой замену на глутамин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 252 представляет собой замену на тирозин, фенилаланин, триптофан или треонин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 254 представляет собой замену на треонин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 256 представляет собой замену на серин, аргинин, глутамин, глутаминовую кислоту, аспарагинову кислоту или треонин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 308 представляет собой замену на треонин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 309 представляет собой замену на пролин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 311 представляет собой замену на серин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 385 представляет собой замену на аргинин, аспарагиновую кислоту, серин, треонин, гистидин, лизин, аланин или глицин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 386 представляет собой замену на треонин, пролин, аспарагиновую кислоту, серин, лизин, аргинин, изолейцин или метионин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 387 представляет собой замену на аргинин, пролин, гистидин, серин, треонин или аланин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 389 представляет собой замену на пролин, серин или аспарагин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 428 представляет собой замену на лейцин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 433 представляет собой замену на аргинин, серин, изолейцин, пролин или глутамин. В одном варианте осуществления данного изобретения, аминокислотная замена в аминокислотном остатке 434 представляет собой замену на гистидин, фенилаланин или тирозин.
В некоторых вариантах осуществления данного изобретения линкер домена Fc (например, содержащий константную область IgG) содержит одну или большее количество мутаций, таких как замены в аминокислотных остатках 252, 254, 256, 433, 434 или 436 (в соответствии с нумерацией по Кабату). В одном варианте осуществления данного изобретения константная область IgG включает в себя тройную мутацию M252Y/S254T/T256E или мутацию YTE. В другом варианте осуществления данного изобретения константная область IgG включает в себя тройную мутацию H433K/N434F/Y436H или мутацию KFH. В дополнительном варианте осуществления данного изобретения константная область IgG включает в себя мутацию YTE и KFH в комбинации.
В некоторых вариантах осуществления данного изобретения модифицированные гуманизированные антитела по данному изобретению содержат константную область IgG, которая содержит одну или большее количество мутаций в аминокислотных остатках 250, 253, 307, 310, 380, 428, 433, 434 и 435. Типовые мутации включают T250Q, M428L, Т307А, Е380А, I253A, Н310А, M428L, H433K, N434A, N434F, N434S и Н435А. В одном варианте осуществления данного изобретения константная область IgG содержит мутацию M428L/N434S или мутацию LS. В другом варианте осуществления данного изобретения константная область IgG содержит мутацию T250Q/M428L или мутацию QL. В другом варианте осуществления данного изобретения константная область IgG содержит мутацию N434A. В другом варианте осуществления данного изобретения константная область IgG содержит мутацию T307A/E380A/N434A или мутацию AAA. В другом варианте осуществления данного изобретения константная область IgG содержит мутацию I253A/H310A/H435A или мутацию IHH. В другом варианте осуществления данного изобретения константная область IgG содержит мутацию H433K/N434F. В другом варианте осуществления данного изобретения константная область IgG содержит мутацию M252Y/S254T/T256E и мутацию H433K/N434F в комбинации.
Дополнительные типовые мутации в константной области IgG описаны, например, в публикациях Robbie, et al., Antimicrobial Agents and Chemotherapy (2013), 57(12): 6147-6153,  et al., JBC (2006), 281(33): 23514-24, et al. Journal of Immunology (2002), 169: 5171-80, Ko et al. Nature (2014) 514: 642-645, Grevys et al. Journal of Immunology. (2015), 194(11): 5497-508, и в патенте США №7083784, все содержание которых включено в данный документ посредством ссылки.
et al., JBC (2006), 281(33): 23514-24, et al. Journal of Immunology (2002), 169: 5171-80, Ko et al. Nature (2014) 514: 642-645, Grevys et al. Journal of Immunology. (2015), 194(11): 5497-508, и в патенте США №7083784, все содержание которых включено в данный документ посредством ссылки.
В некоторых вариантах осуществления настоящего изобретения линкер содержит аминокислотную последовательность SEQ ID NO: 70 или по меньшей мере идентичную ей на 90%, или 93%, или 95%, или 97%, или 98%, или 99%. В различных вариантах осуществления данного изобретения мутации осуществляют в SEQ ID No: 70 для повышения стабильности и/или увеличения периода полувыведения. Например, в некоторых вариантах осуществления настоящего изобретения линкер содержит аминокислотную последовательность SEQ ID NO: 71 или 72, или по меньшей мере идентичную ей на 90%, или 93%, или 95%, или 97%, или 98%, или 99%. Типовой стабилизирующий Fc мутант представляет собой S228P. Типовые мутанты, продлевающие период полувыведения Fc, представляют собой T250Q, M428L, V308T, L309P и Q311S, при этом описанные линкеры могут содержать 1 или 2 или 3 или 4 или 5 из этих мутантов.
Кроме того, один или большее количество соединительных линкеров могут применяться для соединения описанных линкеров IgG (например, одной или SEQ ID NO: 70, 71, или 71, или по меньшей мере идентичной им на 90%, или 93%, или 95%, или 97%, или 98%, или 99%) и внеклеточных доменов. Например, любая из SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или ее варианты могут связывать внеклеточный домен, как описано в данном документе, и линкер, как описано в данном документе. Необязательно, любая из SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или ее варианты являются смещенными между внеклеточным доменом, как описано в данном документе, и линкером, как описано в данном документе.
Дополнительные типовые линкеры включают, но не ограничиваются ими, линкеры, имеющие последовательность LE, GGGGS (SEQ ID NO: 23), (GGGGS)n (n=1-4), (Gly)8, (Gly)6, (EAAAK)n (n=1-3) (SEQ ID NO: 24), A(EAAAK)nA (n=2-5) (SEQ ID NO: 25), AEAAAKEAAAKA (SEQ ID NO: 26), A(EAAAK)4ALEA(EAAAK)4A (SEQ ID NO: 27), PAPAP (SEQ ID NO: 28), KESGSVSSEQLAQFRSLD (SEQ ID NO: 29), EGKSSGSGSESKST (SEQ ID NO: 30), GSAGSAAGSGEF (SEQ ID NO: 31), и (XP)n, с X, обозначающим любую аминокислоту, например, Ala, Lys или Glu.
В различных вариантах осуществления данного изобретения линкер может быть функциональным. Например, без ограничения, линкер может функционировать для улучшения фолдинга и/или стабильности, повышения уровня экспрессии, улучшения параметров фармакокинетики и/или повышения биологической активности описанного химерного белка. В другом примере линкер может функционировать для нацеливания химерного белка на конкретный тип или местоположение клетки.
В различных вариантах осуществления данного изобретения описанные химерные белки способны и могут применяться в способах, включающих стимулирование иммунной активации (например, против опухолей). В различных вариантах осуществления данного изобретения описанные химерные белки способны и могут применяться в способах, включающих подавление иммунного ингибирования (например, что позволяет опухолям выживать). В различных вариантах осуществления данного изобретения описанные химерные белки обеспечивают повышенную иммунную активацию и/или усиленное подавление иммунного ингибирования из-за близости сигнализации, что обеспечивается химерной природой конструкций.
В различных вариантах осуществления данного изобретения описанные химерные белки способны или могут применяться в способах, включающих модулирование амплитуды иммунного ответа, например, модулирование уровня эффекторного ответа. В некоторых вариантах осуществления данного изобретения, например, при применении для лечения рака описанные химерные белки изменяют степень иммунной стимуляции по сравнению с иммунным ингибированием для увеличения амплитуды ответа Т-клеток, включая, без ограничения, стимулирование повышенного уровня продукции цитокинов, пролиферации или потенциала уничтожения мишеней.
В различных вариантах осуществления данного изобретения описанные химерные белки, в некоторых вариантах осуществления, способны или находят применение в способах, включающих маскирование ингибирующего лиганда на поверхности опухолевой клетки и замену этого иммуноингибирующего лиганда иммуностимулирующим лигандом (см., например, Фиг. 4). Например, конструкция химерного белка, содержащая (i) внеклеточный домен PD-1 и (ii) внеклеточный домен OX40L, способствует разрушению ингибирующего сигнала PD-L1 и замену его стимулирующим OX40L. Соответственно, описанные химерные белки в некоторых вариантах осуществления данного изобретения могут применяться или находят свое применение в способах, связанных с ослаблением или устранением ингибирующего иммунного сигнала и/или усилением или активированием иммуностимулирующего сигнала. Например, опухолевая клетка, несущая ингибирующий сигнал (и, таким образом, уклоняющаяся от иммунного ответа), может быть заменена для связывания положительного сигнала на Т-клетке, которая затем может атаковать опухолевую клетку. Соответственно, в некоторых вариантах осуществления данного изобретения, ингибирующий иммунный сигнал маскируется описанными конструкциями, при этом стимулирующий иммунный сигнал активируется. Указанные полезные характеристики усиливаются посредством подхода с применением одной конструкции описанных химерных белков. Например, замену сигнала можно осуществлять практически одновременно, и при этом замену сигнала проводят с учетом локальной цели в клинически значимом месте (например, в микроокружении опухоли). В дополнительных вариантах осуществления данного изобретения применяется тот же принцип и к другим химерным белковым конструкциям, таким как, например, (i) внеклеточный домен PD-1 и (ii) внеклеточный домен GITRL; (i) внеклеточный домен BTLA и (ii) внеклеточный домен OX40L; (i) внеклеточный домен TIGIT и (ii) внеклеточный домен OX40L; (i) внеклеточный домен TIM3 и (ii) внеклеточный домен OX40L; и (i) внеклеточный домен CD172a и (ii) внеклеточный домен CD40L; и (i) внеклеточный домен CD115 и (ii) внеклеточный домен CD40L; и (i) внеклеточный домен TIM3 и (ii) внеклеточный домен OX40L; и (i) внеклеточный домен TIGIT и (ii) внеклеточный домен OX40L; среди прочих.
В различных вариантах осуществления данного изобретения описанные химерные белки могут применяться или находят свое применение в способах, включающих стимулирование или усиление связывание пар "иммуностимулирующий рецептор/лиганд". Типовые Т-клеточные костимулирующие рецепторы и их лиганды включают OX-40:OX40-L, CD27:CD70, CD30:CD30-L, CD40:CD40-L; CD137:CD137-L, HVEM:LIGHT, GITR:GITR-L, TNFRSF25:TL1A, DR5:TRAIL и BTLA:HVEM. В различных вариантах осуществления данного изобретения описанные химерные белки могут применяться или находят свое применение в способах, включающих ингибирование или уменьшение связывания пар "иммуномодулирующий рецептор/лиганд". Типовые Т-клеточные коингибирующие рецепторы и их лиганды включают, например, CTLA-4: CD80/CD86, PD-1: PD-L1/PD-L2, BTLA: HVEM, TIM-3: галектин-9/фосфатидилсерин, TIGIT/CD155 или CD112, VISTA/VSIG8, CD172a/CD47, B7H3R/B7H3, B7H4R/B7H4, CD244/CD48, TMIGD2/HHLA2, среди прочих.
В различных вариантах осуществления данного изобретения описанный химерный белок блокирует, уменьшает и/или ингибирует PD-1 и PD-L1 или PD-L2 и/или связывание PD-1 с PD-L1 или PD-L2. В различных вариантах осуществления данного изобретения описанный химерный белок блокирует, уменьшает и/или ингибирует активность CTLA-4 и/или связывание CTLA-4 с одним или большим количеством из элементов: АР2М1, CD80, CD86, SHP-2 и PPP2R5A. В различных вариантах осуществления данного изобретения описанный химерный белок повышает и/или стимулирует GITR и/или связывание GITR с одним или большим количеством лигандов GITR. В различных вариантах осуществления данного изобретения описанный химерный белок повышает и/или стимулирует ОХ40 и/или связывание ОХ40 с одним или большим количеством лигандов ОХ40.
В различных вариантах осуществления данного изобретения описанные химерные белки могут применяться или находят свое применение в способах, включающих усиление, восстановление, инициирование и/или стимулирование иммунной модуляции. В некоторых вариантах осуществления данного изобретения, химерные белки, описанные в данном документе, восстанавливают, инициируют и/или стимулируют активность или активацию одной или большего количества иммунных клеток против опухолевых клеток, включая, но не ограничиваясь ими: Т-клетки, цитотоксические Т-лимфоциты, Т-хелперные клетки, клетки-естественные киллеры (NK), клетки-естественные киллеры Т (NKT), противоопухолевые макрофаги ((например, M1 макрофаги), В-клетки и дендритные клетки. В некоторых вариантах осуществления данного изобретения описанные химерные белки восстанавливают, инициируют и/или стимулируют активность и/или активацию Т-клеток, включая, в качестве неограничивающего примера, активацию и/или стимуляцию одного или большего количества внутренних собственных сигналов Т-клеток, включая сигнал, способствующий выживанию; сигнал аутокринного или паракринного роста; а р38 MAPK-, ERK-, STAT-, JAK-, AKT- или PI3K-опосредованный сигнал; антиапоптотический сигнал; и/или сигнал, способствующий и/или необходимый для одного или большего количества процессов, выбранных из продукция провоспалительных цитокинов или миграция Т-клеток или инфильтрация опухоли Т- клетками.
В некоторых вариантах осуществления данного изобретения описанные химерные белки могут применяться или находят свое применение в способах, обуславливающих увеличение количества одного или более типов Т-клеток (включая, без ограничения, цитотоксические Т-лимфоциты, Т-хелперные клетки, клетки-естественные киллеры Т (NKT), В-клетки, клетки-естественные киллеры (NK), клетки-естественные киллеры Т (NKT), дендритные клетки, моноциты и макрофаги (например, один или более типов макрофагов, выбранных из M1 и М2) в опухоли или в микроокружении опухоли. В некоторых вариантах осуществления данного изобретения описанные химерные белки могут применяться или находят свое применение в способах, включающих ингибирование и/или обуславливающих снижение поступления иммуносупрессорных клеток (например, супрессорных клеток миелоидного происхождения (MDSC), регуляторных Т-клеток (Treg), ассоциированных с опухолью нейтрофилов (TAN), макрофагов М2 и ассоциированных с опухолью макрофагов (ТАМ)) в опухоль и/или микроокружение опухоли (МОО). В некоторых вариантах осуществления данного изобретения описанные препараты могут изменить соотношение макрофагов M1 к макрофагам М2 в области опухоли и/или в МОО в пользу макрофагов M1.
В различных вариантах осуществления данного изобретения описанные химерные белки могут применяться и находят свое применение в способах, обеспечивающих ингибирование и/или уменьшение Т-клеточной инактивации и/или иммунологической толерантности к опухоли, и включающих введение субъекту эффективного количества химерного белка, описанного в данном документе. В некоторых вариантах осуществления данного изобретения описанные химерные белки способны повышать уровни различных цитокинов в сыворотке, включая, но не ограничиваясь ими, один или большее количество из следующих элементов: IFNγ, TNFα, IL-2, IL-4, IL-5, IL-6, IL-9, IL-10, IL-13, IL-17A, IL-17F и IL-22. В некоторых вариантах осуществления данного изобретения описанные химерные белки способны усиливать функцию IL-2, IL-4, IL-5, IL-10, IL-13, IL-17A, IL-22, TNFα или IFNγ в сыворотке субъекта, получающего лечение (см., например, Фигуру 11, панель J). Обнаружение такого цитокинового ответа может обеспечить возможность определения оптимальной схемы введения доз для указанного химерного слитого белка (см., например, Фигуру 11, панель K).
В различных вариантах осуществления данного изобретения описанные химерные белки ингибируют, блокируют и/или снижают клеточную гибель противоопухолевой CD8+ и/или CD4+ Т-клетки; или стимулируют, индуцируют и/или повышают клеточную гибель гибель проопухолевой Т-клетки. Истощение Т-клеток представляет собой дисфункциональное состояние Т-клеток, характеризующееся прогрессирующей потерей пролиферативных и эффекторных функций, кульминацией которой является клональная делеция. Соответственно, проопухолевая Т-клетка представляет собой дисфункциональное состояние Т-клетки, которое возникает при многих хронических инфекциях и раке. Эта дисфункция определяется слабой пролиферативной и/или эффекторной активностью, устойчивой экспрессией ингибиторных рецепторов и транскрипционным состоянием, отличающимся от функциональных эффекторных клеток или Т-клеток памяти. Истощение препятствует оптимальному контролю за инфекцией и опухолями. Кроме того, противоопухолевая CD8+ и/или CD4+ Т-клетка относится к Т- клеткам, которые могут осуществлять иммунный ответ против опухоли. Типовые проопухолевые Т-клетки включают, но не ограничиваются ими, Tregs, CD4+ и/или CD8+ Т-клетки, экспрессирующие один или большее количество ингибирующих рецепторов контрольных точек, клетки Th2 и клетки Th17. Ингибирующие рецепторы контрольных точек относятся к рецепторам (например, CTLA-4, В7-Н3, В7-Н4, TIM-3), экспрессирующихся на иммунных клетках, которые предотвращают или ингибируют неконтролируемые иммунные ответы.
В различных вариантах осуществления данного изобретения описанные химерные белки способны и находят свое применение в способах, включающих увеличение соотношения эффекторных Т-клеток к регуляторным Т-клеткам. Типовые эффекторные Т-клетки включают ICOS+ эффекторные Т-клетки; цитотоксические Т-клетки (например, αβ TCR, CD3+, CD8+, CD45RO+); CD4+ эффекторные Т-клетки (например, αβ TCR, CD3+, CD4+, CCR7+, CD62Lhi, IL-7R/CD127+); CD8+ эффекторные Т-клетки (например, αβ TCR, CD3+, CD8+, CCR7+, CD62Lhi, IL-7R/CD127+); эффекторные Т-клетки памяти (например, CD62L(низк), CD44+, TCR, CD3+, IL-7R/CD127+, IL-15R+, CCR7(низк); центральные T-клетки памяти (например, CCR7+, CD62L+, CD27+; или CCR7hi, CD44+, CD62Lhi, TCR, CD3+, IL-7R/CD127+, IL-15R+); CD62L+ эффекторные Т-клетки; CD8+ эффекторные T-клетки памяти (ТЕМ), включая ранние эффекторные Т-клетки памяти (CD27+ CD62L-) и поздние эффекторные Т-клетки памяти (CD27- CD62L-) (TemE и TemL, соответственно); CD127(+)CD25(низк/-) эффекторные Т-клетки; CD127(-)CD25(-) эффекторные Т-клетки; CD8+ стволовые эффекторные клетки памяти (TSCM) (например, CD44(низк)CD62L(выс)CD122(выс)sca(+)); ТН1 эффекторные Т-клетки (например, CXCR3+, CXCR6+ и CCR5+; или αβ TCR, CD3+, CD4+, IL-12R+, IFNγR+, CXCR3+), TH2 эффекторные Т-клетки (например, CCR3+, CCR4+ и CCR8+; или αβ TCR, CD3+, CD4+, IL-4R+, IL-33R+, CCR4+, IL-17RB+, CRTH2+); TH9 эффекторные Т-клетки (например, αβ TCR, CD3+, CD4+); TH17 эффекторные Т-клетки (например, αβ TCR, CD3+, CD4+, IL-23R+, CCR6+, IL-1R+); CD4+CD45RO+CCR7+ эффекторные Т-клетки, CD4+CD45RO+CCR7(-) эффекторные Т-клетки; и эффекторные Т-клетки, секретирующие IL-2, IL-4 и/или IFN-γ. Типовые регуляторные Т-клетки включают ICOS+ регуляторные Т-клетки, CD4+CD25+FOXP3+ регуляторные Т-клетки, CD4+CD25+ регуляторные Т-клетки, CD4+CD25- регуляторные Т-клетки, CD4+CD25(выс) регуляторные Т-клетки, TIM-3+PD-1+ регуляторные Т-клетки, гена активации лимфоцитов-3 (LAG-3)+ регуляторные Т-клетки, CTLA-4/CD152+ регуляторные Т-клетки, нейропилина-1 (Nrp-1)+ регуляторные Т-клетки, CCR4+CCR8+ регуляторные Т-клетки, CD62L (L-селектин)+ регуляторные Т-клетки, CD45RB(низк) MK регуляторные Т-клетки, CD127(низк) регуляторные Т-клетки, LRRC32/GARP+ регуляторные Т-клетки, CD39+ регуляторные Т-клетки, GITR+ регуляторные Т-клетки, LAP+ регуляторные Т-клетки, 1В11+ регуляторные Т-клетки, BTLA+ регуляторные Т-клетки, type 1 регуляторные Т-клетки (клетки Tr1), Т-клетки -хелперы типа 3 (Th3), регуляторную клетку фенотипа Т-клетки натурального киллера (NKTreg), CD8+ регуляторные Т-клетки, CD8+CD28- регуляторные Т-клетки и/или регуляторные Т-клетки, секретирующие IL-10, IL-35, TGF-β, TNF-α, галектин-1, IFN-γ и/или МСР1.
В различных вариантах осуществления данного изобретения описанные химерные белки способны и находят свое применение в способах, включающих транзиентную стимуляцию эффекторных Т-клеток на протяжении периода времени не более чем около 12 часов, около 24 часов, около 48 часов, около 72 часов или около 96 часов или около 1 недели или около 2 недель. В различных вариантах осуществления данного изобретения описанные химерные белки способны и находят свое применение в способах, включающих транзиентное истощение или ингибирование регуляторных Т-клеток на протяжении периода времени не более чем около 12 часов, около 24 часов, около 48 часов, около 72 часов или около 96 часов или около 1 недели или около 2 недель. В различных вариантах осуществления данного изобретения транзиентная стимуляция эффекторных Т-клеток и/или транзиентное истощение или ингибирование регуляторных Т-клеток происходит, по существу, в кровотоке пациента или в определенной ткани/месте, включая лимфоидные ткани, такие как, например, костный мозг, лимфатический узел, селезенку, тимус, лимфоидную ткань, ассоциированную с слизистой оболочкой (MALT), нелимфоидные ткани или в микроокружении опухоли.
В различных вариантах осуществления данного изобретения описанные химерные белки обеспечивают преимущества, включая, без ограничения, простоту применения и простоту изготовления. Это объясняется тем, что два отдельных иммунотерапевтических агента объединяются в один продукт, который позволяет применять один производственный процесс вместо двух независимых производственных процессов. Кроме того, введение одного агента вместо двух отдельных агентов обеспечивает более легкое введение и более полную приверженность пациента к лечению. Кроме того, в отличие от, например, моноклональных антител, которые представляют собой крупные мультимерные белки, содержащие многочисленные дисульфидные связи и посттрансляционные модификации, такие как гликозилирование, данные химерные белки изготавливать легче и экономичнее.
В различных вариантах осуществления данного изобретения описанный химерный белок можно продуцировать в клетке-хозяине млекопитающего в виде секретируемой и полностью функциональной одиночной полипептидной цепи (см., например, Фигура 13, панель А, Фигура 17, панели Е-Н, Фигура 17, панели N-S).
В различных вариантах осуществления данного изобретения описанный химерный белок непредвиденно обеспечивает связывание компонентов внеклеточного домена с их соответствующими партнерами по связыванию с более низкими скоростями диссоциации (Kd или Koff). В некоторых вариантах осуществления данного изобретения это обеспечивает неожиданно длительное взаимодействие рецептора с лигандом и наоборот. Такое взаимодействие обеспечивает устойчивый эффект маскирования отрицательного сигнала (см., например, Фигура 14, Фигура 17, панели I-М). Кроме того, в некоторых вариантах осуществления данного изобретения, это обеспечивает более длительный положительный сигнал, например, чтобы дать возможность эффекторной клетке адекватно стимулировать противоопухолевый эффект. Например, описанный химерный белок, например, посредством связывания при низкой скорости диссоциации, обеспечивает эффективную передачу сигнала, тем самым обеспечивая пролиферацию Т- клеток и осуществление противоопухолевой атаки. В качестве дополнительного примера, описанный химерный белок, например, посредством связывания при низкой скорости диссоциации, обеспечивает эффективную передачу сигнала, тем самым обеспечивая высвобождение стимулирующих сигналов, таких как, например, цитокины Also. Этот стабильный синапс клеток, поддерживаемые описанными агентами (например, опухолевой клетки, несущей отрицательные сигналы, и Т-клетки, которая может атаковать опухоль) обеспечивает пространственную ориентацию в пользу уменьшения опухоли - например, позиционирование Т-клеток для атаки опухолевых клеток и/или стерическое предотвращение доставки отрицательных сигналов опухолевой клетки, включая отрицательные сигналы, выходящие за пределы тех, которые замаскированы химерным белком по данному изобретению.
В некоторых вариантах осуществления данного изобретения это обеспечивает более длительный целевой период полувыведения химерных белков (например, внутри опухоли) (t1/2) по сравнению с t1/2 в сыворотке. Такие характеристики могут иметь комбинированное преимущество, заключающееся в снижении уровня нецелевых токсинов, ассоциированных с системным распределением химерных белков (см., например, Фигура 14, панели М-О).
Дополнительно, в различных вариантах осуществления данного изобретения описанные химерные белки обеспечивают синергетический терапевтический эффект, поскольку позволяют усилить сайт-специфическое взаимодействие двух иммунотерапевтических агентов. В различных вариантах осуществления данного изобретения описанные химерные белки обеспечивают возможность снижения нецелевого воздействия и/или системной токсичности.
Заболевания; способы лечения и выбор пациентов
В различных вариантах осуществления настоящее изобретение относится к раковым заболеваниям и/или опухолям; например, лечению или предотвращению раковых заболеваний и/или опухолей. Как описано в данном документе в другом месте, лечение рака может включать в себя различные варианты, модулирующие иммунную систему с помощью описанных химерных белков, что обеспечивает преобладание стимуляции иммунной системы над ее ингибированием.
Раковые заболевания или опухоли характеризуются неконтролируемым ростом клеток и/или аномально повышенной выживаемостью клеток, и/или ингибированием апоптоза, что препятствует нормальному функционированию органов и систем организма. Данное определение включает доброкачественные и злокачественные раковые заболевания, полипы, гиперплазию, а также неактивные опухоли или микрометастазы. Кроме того, данное определение включает клетки с аномальной пролиферацией, которые не блокируются иммунной системой (, например, инфицированные вирусом клетки). Рак может быть первичным раком или метастатическим раком. Первичный рак может представлять собой участок раковых клеток в месте происхождения, которое становится клинически обнаруживаемым и может быть первичной опухолью. Напротив, метастатический рак может представлять собой распространение заболевание от одного органа или части к другому несмежному органу или его части. Метастатический рак может образовываться из раковой клетки, которая приобретает способность проникать и инфильтрировать окружающие нормальные ткани в локальной области, образуя новую опухоль, которая может быть местным метастазом. Рак также может образовываться из раковой клетки, которая приобретает способность проникать сквозь стенки лимфатических и/или кровеносных сосудов, после чего раковая клетка может циркулировать по кровотоку (тем самым, являясь циркулирующей опухолевой клеткой) к другим областям и тканям в организме. Рак может быть обусловлен процессом, таким как лимфатическое или гематогенное распространение. Рак может быть обусловлен опухолевой клеткой, которая поступает в другой участок, повторно проникает через сосуд или стенки, продолжает размножаться и в конечном итоге образует другую клинически обнаруживаемую опухоль. Рак может представлять собой эту новую опухоль, которая может быть метастатической (или вторичной) опухолью.
Рак может быть обусловлен опухолевыми клетками, которые являются метастазированными, и может быть вторичной или метастатической опухолью. Клетки опухоли могут быть подобны клеткам в исходной опухоли. Например, если рак молочной железы или рак толстой кишки метастазирует в печень, вторичная опухоль, обнаруживаемая в печени, состоит из раковых клеток молочной железы или толстой кишки, а не раковых клеток печени. Таким образом, опухоль в печени может быть метастатическим раком молочной железы или метастатическим раком толстой кишки, но не раком печени.
Рак может происходить из любой ткани. Рак может происходить из меланомы, толстой кишки, молочной железы или предстательной железы и, следовательно, может состоять из клеток, которые первоначально были клетками кожи, толстой кишки, молочной железы или предстательной железы, соответственно. Рак также может представлять собой гематологическую злокачественную опухоль, которая может быть лейкозом или лимфомой. Рак может проникать в ткань, такую как печень, легкие, мочевой пузырь или кишечник.
Репрезентативные виды раковых заболеваний и/или опухолей по данному изобретению выбирают из группы, состоящей из, но не ограничиваясь этим, базально-клеточной карциномы, рака желчных путей; рака мочевого пузыря; рака кости; рака головного мозга и центральной нервной системы; рака молочной железы; рака брюшной полости; рака шейки матки; хориокарциномы; рака толстой кишки и прямой кишки; рака соединительной ткани; рака пищеварительной системы; рака эндометрия; рака пищевода; рака глаз; рака головы и шеи; рака желудка (включая рак желудочно-кишечного тракта); глиобластомы; карциномы печени; гепатомы; интраэпителиального новообразования; рака почек или ренального рака; рака гортани; лейкоза; рака печени; рака легкого (например, мелкоклеточного рака легких, немелкоклеточного рака легких, аденокарциномы легких и плоскоклеточного рака легких); меланомы; миеломы; нейробластомы; рака ротовой полости полости (губ, языка, рта и глотки); рака яичников; рака поджелудочной железы; рака предстательной железы; ретинобластомы; рабдомиосаркомы; рака прямой кишки; рака дыхательной системы; рака слюнной железы; саркомы; рака кожи; плоскоклеточного рака; рака желудка; рака яичек; рака щитовидной железы; рака матки или рака эндометрия; рака мочеполовой системы; рака вульвы; лимфомы, включая лимфому Ходжкина и неходжкинскую лимфому, а также В-клеточной лимфомы (включая низкой степени злокачественности/фолликулярную неходжкинскую лимфому (НХЛ), малую лимфоцитарную (МЛ) НХЛ, средней степени злокачественности/фолликулярную НХЛ, средней степени злокачественности диффузную НХЛ; высшей степени злокачественности иммунобластную НХЛ, высшей степени злокачественности лимфобластную НХЛ, высшей степени злокачественности мелкоклеточную лимфому с нерассеченными ядрами НХЛ, НХЛ с массивным поражением, лимфому клеток мантийной ткани, лимфому связанную со СПИДом и макроглобулинемию Вальденстрема, хронический лимфолейкоз (ХЛЛ), острый лимфобластный лейкоз (ОЛЛ); волосатоклеточный лейкоз, хронический миелобластный лейкоз), а также других карцином и сарком, а также посттрансплантационного лимфопролиферативного синдрома (PTLD), а также патологической пролиферации сосудов, ассоциированной с факоматозами, эдему (такую как ассоциированую с опухолями головного мозга) и синдрома Мейгса.
В некоторых вариантах осуществления данного изобретения химерный белок применяют для лечения субъекта, который имеет устойчивый к лечению рак. В некоторых вариантах осуществления данного изобретения химерный белок применяют для лечения субъекта, который является устойчивым к лечению одним или большим количеством иммуномодулирующих агентов. Например, в некоторых вариантах осуществления данного изобретения химерный белок применяют для лечения субъекта, который не отвечает на лечение, или даже характеризуется прогрессированием заболевания, в результате приема лечения в течение 12 недель или около того. Например, в некоторых вариантах осуществления данного изобретения субъект является устойчивым к лечению агентом PD-1 и/или PD-L1 и/или PD-L2, включая, например, ниволумаб (ONO-4538/BMS-936558, MDX1106, ОПДИВО (OPDIVO), BRISTOL MYERS SQUIBB), пембролизумабом (КЕЙТРУДА (KEYTRUDA), MERCK), пидилизумабом (CT-011, CURE TECH), MK-3475 (MERCK), BMS 936559 (BRISTOL MYERS SQUIBB), ибрутинибом (PHARMACYCLICS/ABBVIE), атезолизумабом (ТЕЦЕНТРИК (TECENTRIQ), GENENTECH) и/или является устойчивым к лечению препаратом MPDL328OA (ROCHE). Например, в некоторых вариантах осуществления данного изобретения субъект является пациентом, является устойчивым к лечению агентом анти-CTLA-4, например, ипилимумабом (ЕРВОЙ (YERVOY)) (например, пациенты с меланомой). Соответственно, в различных вариантах осуществления данного изобретения предлагаются способы лечения рака, которые являются пригодными для пациентов, не отвечающих на различные виды терапии, включая монотерапию одним или большим количеством иммуномодулирующих агентов.
В некоторых вариантах осуществления данного изобретения описанные способы обеспечивают лечение с применением химерного белка у пациента, который является устойчивым к лечению дополнительным агентом, таким "дополнительные агенты", описанные в данном документе в другом месте, включая, без ограничения, различные химиотерапевтические агенты, описанные в данном документе.
В некоторых аспектах, описанные химерные агенты применяют для элиминации внутриклеточных патогенов. В некоторых аспектах, описанные химерные агенты применяют для лечения одной или большего количества инфекций. В некоторых вариантах осуществления данного изобретения описанные химерные белки применяют в способах лечения вирусных инфекций (включая, например, ВИЧ и ВГС), паразитарных инфекций (включая, например, малярию), и бактериальных инфекций. В различных вариантах осуществления данного изобретения инфекции индуцируют иммуносупрессию. Например, ВИЧ-инфекция часто приводит к иммуносупрессии у инфицированных субъектов. Соответственно, как описано в данном документе в другом месте, лечение таких инфекций может включать в себя различные варианты, модулирующие иммунную систему с помощью описанных химерных белков, что обеспечивает преобладание стимуляции иммунной системы над ее ингибированием. В альтернативном варианте, в данном изобретении предлагаются способы лечения инфекций, которые индуцируют иммуноактивацию. Например, кишечные гельминтные инфекции ассоциированы с хронической иммунной активацией. В этих вариантах осуществления данного изобретения лечение таких инфекций может включать в себя модулирование иммунной системы с помощью описанных химерных белков, что обеспечивает преобладание ингибирования иммунной системы над ее стимуляцией.
В различных вариантах осуществления данного изобретения предлагаются способы лечения вирусных инфекций, включая, без ограничения, острые или хронические вирусные инфекции, например, инфекции респираторного тракта, папилломатозные вирусные инфекции, инфекции, вызванные вирусом простого герпеса (ВПГ), вирусом иммунодефицита человека (ВИЧ), а также вирусные инфекции внутренних органов, например, инфицирование вирусами гепатита. В некоторых вариантах осуществления данного изобретения вирусная инфекция вызвана вирусом семейства Flaviviridae. В некоторых вариантах осуществления данного изобретения вирус семейства Flaviviridae выбирают из вируса желтой лихорадки, вируса лихорадки Западного Нила, вируса лихорадки Денге, вируса японского энцефалита, вируса энцефалита Сент-Луис и вируса гепатита С. В других вариантах осуществления данного изобретения вирусная инфекция вызвана вирусом семейства Picornaviridae, например, полиовирусом, риновирусом, коксакивирусом. В других вариантах осуществления данного изобретения вирусная инфекция вызвана представителем семейства Orthomyxoviridae, например, вирусом гриппа. В других вариантах осуществления данного изобретения вирусная инфекция вызвана представителем семейства Retroviridae, например, лентивирусом. В других вариантах осуществления данного изобретения вирусная инфекция вызвана представителем семейства Paramyxoviridae, например, респираторно-синцитиальным вирусом, вирусом парагриппа человека, рубулавирусом (например, вирусом эпидемического паротита), вирусом кори и метапневмовирусом человека. В других вариантах осуществления данного изобретения вирусная инфекция вызвана представителем семейства Bunyaviridae, например, хантавирусом. В других вариантах осуществления данного изобретения вирусная инфекция вызвана представителем семейства Reoviridae, например, ротавирусом.
В различных вариантах осуществления данного изобретения предлагаются способы лечения паразитарных инфекций, таких как протозойные инфекции или гельминтозы. В некоторых вариантах осуществления данного изобретения паразитарная инфекция вызывается протозойным паразитом. В некоторых вариантах осуществления настоящего изобретения протозойный паразит выбирают из кишечного протозойного паразита, тканевого протозойного паразита или кровяного протозойного паразита. Типовые протозойные паразиты включают, но не ограничиваются ими, Entamoeba hystolytica, Giardia lamblia, Cryptosporidium muris, Trypanosomatida gambiense, Trypanosomatida rhodesiense, Trypanosomatida crusi, Leishmania mexicana, Leishmania braziliensis, Leishmania tropica, Leishmania donovani, Toxoplasma gondii, Plasmodium vivax, Plasmodium ovale, Plasmodium malariae, Plasmodium falciparum, Trichomonas vaginalis и Histomonas meleagridis. В некоторых вариантах осуществления данного изобретения паразитарная инфекция вызывается гельминтным паразитом, таким как нематоды (например, Adenophorea). В некоторых вариантах осуществления данного изобретения паразита выбирают из Secementea (например, Trichuris trichiura, Ascaris lumbricoides, Enterobius vermicularis, Ancylostoma duodenale, Necator americanus, Strongyloides stercoralis, Wuchereria bancrofti, Dracunculus medinensis). В некоторых вариантах осуществления данного изобретения паразита выбирают из рода трематод (например, шистосомы, печеночной двуустки, кишечных сосальщиков и легочных сосальщиков). В некоторых вариантах осуществления данного изобретения паразита выбирают из: Schistosoma mansoni, Schistosoma haematobium, Schistosoma japonicum, Fasciola hepatica, Fasciola gigantica, Heterophyes heterophyes, Paragonimus westermani. В некоторых вариантах осуществления данного изобретения паразита выбирают из цестод (например, Taenia solium, Taenia saginata, Hymenolepis nana, Echinococcus granulosus).
В различных вариантах осуществления данного изобретения предлагаются способы лечения бактериальных инфекций. В различных вариантах осуществления данного изобретения бактериальная инфекция вызывается грамположительными бактериями, грамотрицательными бактериями, аэробными и/или анаэробными бактериями. В различных вариантах осуществления данного изобретения бактерию выбирают, но не ограничиваясь этим, из Staphylococcus, Lactobacillus, Streptococcus, Sarcina, Escherichia, Enterobacter, Klebsiella, Pseudomonas, Acinetobacter, Mycobacterium, Proteus, Campylobacter, Citrobacter, Nisseria, Baccillus, Bacteroides, Peptococcus, Clostridium, Salmonella, Shigella, Serratia, Haemophilus, Brucella и других организмов. В некоторых вариантах осуществления данного изобретения бактерию выбирают, но не ограничиваясь этим, из Pseudomonas aeruginosa, Pseudomonas fluorescens, Pseudomonas acidovorans, Pseudomonas alcaligenes, Pseudomonas putida, Stenotrophomonas maltophilia, Burkholderia cepacia, Aeromonas hydrophilia, Escherichia coli, Citrobacter freundii, Salmonella typhimurium, Salmonella typhi, Salmonella paratyphi, Salmonella enteritidis, Shigella dysenteriae, Shigella flexneri, Shigella sonnei, Enterobacter cloacae, Enterobacter aerogenes, Klebsiella pneumoniae, Klebsiella oxytoca, Serratia marcescens, Francisella tularensis, Morganella morganii, Proteus mirabilis, Proteus vulgaris, Providencia alcalifaciens, Providencia rettgeri, Providencia stuartii, Acinetobacter baumannii, Acinetobacter calcoaceticus, Acinetobacter haemolyticus, Yersinia enterocolitica, Yersinia pestis, Yersinia pseudotuberculosis, Yersinia intermedia, Bordetella pertussis, Bordetella parapertussis, Bordetella bronchiseptica, Haemophilus influenzae, Haemophilus parainfluenzae, Haemophilus haemolyticus, Haemophilus parahaemolyticus, Haemophilus ducreyi, Pasteurella multocida, Pasteurella haemolytica, Branhamella catarrhalis, Helicobacter pylori, Campylobacter fetus, Campylobacter jejuni, Campylobacter coli, Borrelia burgdorferi, Vibrio cholerae, Vibrio parahaemolyticus, Legionella pneumophila, Listeria monocytogenes, Neisseria gonorrhoeae, Neisseria meningitidis, Kingella, Moraxella, Gardnerella vaginalis, Bacteroides fragilis, Bacteroides distasonis, 3452A гомологичной группы Bacteroides, Bacteroides vulgatus, Bacteroides ovalus, Bacteroides thetaiotaomicron, Bacteroides uniformis, Bacteroides eggerthii, Bacteroides splanchnicus, Clostridium difficile, Mycobacterium tuberculosis, Mycobacterium avium, Mycobacterium intracellulare, Mycobacterium leprae, Corynebacterium diphtheriae, Corynebacterium ulcerans, Streptococcus pneumoniae, Streptococcus agalactiae, Streptococcus pyogenes, Enterococcus faecalis, Enterococcus faecium, Staphylococcus aureus, Staphylococcus epidermidis, Staphylococcus saprophyticus, Staphylococcus intermedius, Staphylococcus hyicussubsp. hyicus, Staphylococcus haemolyticus, Staphylococcus hominis, или Staphylococcus saccharolyticus.
В некоторых аспектах, описанные химерные агенты применяют для лечения одногог или большего количества аутоиммунных заболеваний или нарушений. В различных вариантах осуществления данного изобретения лечение аутоиммунного заболевания или нарушения может включать в себя модулирование иммунной системы с помощью описанных химерных белков, что обеспечивает преобладание ингибирования иммунной системы над ее стимуляцией. Типовые аутоиммунные заболевания или нарушения, поддающиеся лечению с помощью описанных химерных белков, включают те, в которых собственные антигены организма становятся мишенями для иммунного ответа, такие как, например, ревматоидный артрит, системная красная волчанка, сахарный диабет, анкилозирующий спондилоартрит, синдром Шегрена, воспалительные заболевания кишечника (например, язвенный колит, болезнь Крона), рассеянный склероз, саркоидоз, псориаз, болезнь Грейвса, тиреоидит Хашимото, псориаз, реакции гиперчувствительности (например, аллергии, сенная лихорадка, астма и острый отек, обусловленный реакцией гиперчувствительности I типа) и васкулит.
В еще одном аспекте данное изобретение направлено на способы лечения и предотвращения заболеваний и нарушений, опосредуемых Т-клетками, таких как, но не ограничиваясь ими, заболевания или нарушения, описанные в данном документе в другом месте, а также воспалительные заболевания или нарушения, болезнь "трансплантат против хозяина" (БТПХ), отторжение трансплантата и Т-клеточное пролиферативное нарушение. Конкретные примеры доменов ECD типа I, пригодных для применения в этом способе, включают в себя, но не ограничиваются: TNFRSF1b, BTNL2, PD-L1, PD-L2, CTLA-4, B7-Н3, В7-Н4, CD40, ОХ40, CD137, среди прочих.
В некоторых аспектах описанные химерные агенты применяют в способах активации Т- клетки, например, через внеклеточный домен, имеющий иммуностимулирующий сигнал.
В некоторых аспектах, описанные химерные агенты применяют для в способах предотвращения клеточной передачи иммуносупрессивного сигнала.
Комбинированные виды терапии и соединение
В некоторых вариантах осуществления данного изобретения предлагаются химерные белки и способы, которые дополнительно включают введение субъекту дополнительного агента. В некоторых вариантах реализации настоящее изобретение относится к совместному введению и/или совместному составлению. Любая из композиций, описанная в данном документе, может быть совместно составлена и/или совместно введена.
В некоторых вариантах осуществления данного изобретения любой химерный белок, описанный в данном документе, действует синергически при совместном введении с другим агентом, и вводится в дозах, которые являются ниже по сравнению с дозами, обычно применяемыми в случаях, когда такие агенты используются в качестве монотерапии. В различных вариантах осуществления данного изобретения, любой агент, упомянутый в данном документе, можно применять в комбинации с любым из химерных белков, описанных в данном документе.
В некоторых вариантах осуществления, включая, без ограничения, применение данного изобретения при раке, данное изобретение относится к химиотерапевтическим агентам в качестве дополнительных агентов. Примеры химиотерапевтических агентов включают, но не ограничиваются ими, алкилирующие агенты, такие как тиотепа и ЦИТОКСАНциклофосфамид; алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилолмеламин; ацетогенины (например, буллатацин и буллатацинон); камптотецин (включая синтетический аналог: топотекан); бриостатин; каллистатин; СС-1065 (включая его синтетические аналоги: адозелезин, карзелезин и бизелезин); криптофицины (например, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги: KW-2189 и СВ1-ТМ1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; нитроиприты, такие как хлорамбуцил, хлорнафазин, хлорфосфамид, эстрамустин, ифосфамид, мехлорэтамин, мехлорэтамина оксид гидрохлорид, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урациловый иприт; препараты нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихеамицин, особенно калихеамицин гамма 11 и калихеамицин омега 11 (см., например, Agnew, Chem. Intl. Ed. Engl., 33:183-186 (1994)); динемицин, включая динемицин А; бисфосфонаты, такие как клодронат; эсперамицин; а также неокарзиностатин хромофор и родственные хромопротеиновые энедииновые антибактериальные хромофоры), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, каминомицин, карзинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, АДРИАМИЦИН доксорубицин (включая морфолино-доксрубицин, цианоморфолино-доксорубицин, 2-пирролино-доклубицин и деоксидокрубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин С, микофенольная кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, келамицин, родорубицин, стрептогнин, стрептозоцин, туберцидин, убенимекс, циностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-ФУ); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; пуриновые аналоги, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; пиримидиновые аналоги, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флуксуридин; андрогены, такие как калустерон, пропионат дромоностанола, эпитиостанол, мепитиостан, тестолактон; агенты, угнетающие функции надпочечников, такие как миноглутетимид, митотан, трилостан; заменитель фолиевой кислоты, такой как фролиновая кислота; ацеглатон; альдофосфамидгликозид; аминолевулиновая кислота; энилурацил; амсакрин; бестрабуцил; бизантрен; эдатраксат; демеколцин; диазиквон; элформитин; эллиптиний ацетат; эпотилон; этоглюцид; нитрат галлия; гидроксимочевина; лентинан; лонидаинин; майтансиноиды, такие как майтансин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитрерин; пентостатин; фенамет; пирарубицин; лозоксантрон; подофиллиновая кислота; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK (JHS Natural Products, Юджин, штат Орегон); разоксан; ризоксин; сизофуран; спирогерманий; тенуазоновая кислота; триазиквон; 2,2', 2'-трихлортриэтиламин, трихотецены (например, токсин Т-2, верракурин А, роридин А и ангидин); уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид ("Ara-С"); циклофосфамид; тиотеп; таксоиды, например, ТАКСОЛ паклитаксел (Bristol-Myers Squibb Oncology, Принстон, штат Нью-Джерси), АБРАКСАН, не содержащий кремофора, альбуминовые нанокомпозиции паклитаксела (American Pharmaceutical Partners, Schaumberg, 111.) и ТАКСОТЕРдоцетаксел ( Rorer, Антони, Франция); хлоранбуцил; ГЕМЗАР гемцитабин; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин, оксалиплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митоксантрон; винкристин; НАВЕЛБИН винорелбин; новантрон; тенипозид; эдатрексат; дауномицин; аминоптерин; кселода; ибандронат; иринотекан (Камптозар, СРТ-11) (включая схему лечения иринотеканом с 5-ФУ и лейковорином); ингибитор топоизомеразы RFS 2000; дифлюорометилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин; комбретастатин; лейковорин (LV); оксалиплатин, включая схему лечения оксалиплатином (ФОЛФОКС); лапатиниб (ТИКЕРБ); ингибиторы PKC-α, Raf, H-Ras, EGFR (например, эрлотиниб (Тарцева)) и VEGF-A, которые уменьшают пролиферацию клеток, и фармацевтически приемлемые соли, кислоты или производные любого из вышеуказанных агентов. Кроме того способы лечения могут дополнительно включать применение лучевой терапии. Кроме того, способы лечения могут дополнительно включать применение фотодинамической терапии.
Rorer, Антони, Франция); хлоранбуцил; ГЕМЗАР гемцитабин; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин, оксалиплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митоксантрон; винкристин; НАВЕЛБИН винорелбин; новантрон; тенипозид; эдатрексат; дауномицин; аминоптерин; кселода; ибандронат; иринотекан (Камптозар, СРТ-11) (включая схему лечения иринотеканом с 5-ФУ и лейковорином); ингибитор топоизомеразы RFS 2000; дифлюорометилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин; комбретастатин; лейковорин (LV); оксалиплатин, включая схему лечения оксалиплатином (ФОЛФОКС); лапатиниб (ТИКЕРБ); ингибиторы PKC-α, Raf, H-Ras, EGFR (например, эрлотиниб (Тарцева)) и VEGF-A, которые уменьшают пролиферацию клеток, и фармацевтически приемлемые соли, кислоты или производные любого из вышеуказанных агентов. Кроме того способы лечения могут дополнительно включать применение лучевой терапии. Кроме того, способы лечения могут дополнительно включать применение фотодинамической терапии.
В различных вариантах осуществления, включая, без ограничения, применение данного изобретения при раке, указанным дополнительным агентом является один или большее количество иммуномодулирующих агентов, выбранных из агента, который блокирует, уменьшает и/или ингибирует PD-1 и PD-L1 или PD-L2 и/или связывание PD-1 с PD-L1 или PD-L2 (в качестве неограничивающего примера, один или большее количество из следующих агентов: ниволумаб (ONO-4538/BMS-936558, MDX1106, ОПДИВО (OPDIVO), BRISTOL MYERS SQUIBB), пембролизумаб (КЕЙТРУДА, Merck), пидилизумаб (СТ-011, CURE TECH), MK-3475 (MERCK), BMS 936559 (BRISTOL MYER SQUIBB) атезолизумаб (ТЕЦЕНТРИК, GENENTECH), MPDL328OA (ROCHE)), агента, который повышает и/или стимулирует CD137 (4-1ВВ) и/или связывание CD137 (4-1ВВ) с одним или большим количеством лигандов 4-1ВВ (в качестве неограничивающего примера, урелумаб (BMS-663513 и антитело анти-4-1ВВ), и агента, который блокирует, снижает и/или ингибирует активность CTLA-4 и/или связывание CTLA-4 с одним или большим количеством из следующих агентов: АР2М1, CD80, CD86, SHP-2 и PPP2R5A, и/или связывание ОХ40 с OX40L (в качестве неограничивающего примера, GBR 830 (GLENMARK), MEDI6469 (MEDIMMUNE).
В некоторых вариантах осуществления, включая, без ограничения, применение данного изобретения при инфекционных заболеваниях, данное изобретение относится к противоинфнекционным агентам в качестве дополнительных агентов. В некоторых вариантах осуществления данного изобретения противоинфекционный агент представляет собой противовирусный агент, включая, но не ограничиваясь ими, абакавир, ацикловир, адефовир, ампренавир, атазанавир, цидофовир, дарунавир, делавирдин, диданозин, докозанол, эфавиренц, эльвитрегравир, эмтрицитабин, энфувиртид, этравирин, фамцикловир и фоскарнет. В некоторых вариантах осуществления данного изобретения противоинфекционный агент представляет собой антибактериальный агент включая, но не ограничиваясь ими, цефалоксиновые антибиотики (цефалексин, цефуроксим, цефадроксил, цефазолин, цефалотин, цефаклор, цефамандол, цефокситин, цефпрозил и цефтобипрол); фторхинолоновые антибиотики (ципро, левахин, флоксин, техин, авелокс и норфлокс); тетрациклиновые антибиотики (тетрациклин, миноциклин, окситетрациклин и доксициклин); пенициллиновые антибиотики (амоксициллин, ампициллин, пенициллин V, диклоксациллин, карбенициллин, ванкомицин и метициллин); монобактамные антибиотики (азтреонам); и карбапенемовые антибиотики (эртапенем, дорипенем, имипенем/циластатин и меропенем). В некоторых вариантах осуществления данного изобретения противоинфекционные агенты включают противомалярийные агенты (например, хлорохин, хинин, мефлохин, примахин, доксициклин, артеметер/люмефантрин, атовакон/прогуанил и сульфадоксин/пириметамин), метронидазол, тинидазол, ивермектин, пирантел памоат и альбендазол.
В некоторых вариантах осуществления, включая, без ограничения, применение данного изобретения при аутоиммунных заболеваниях, дополнительный агент представляет собой иммуносупрессивный агент. В некоторых вариантах осуществления данного изобретения иммуносупрессивный агент представляет собой противовоспалительный агент, такой как стероидный противовоспалительный агент или нестероидный противовоспалительный агент (НПВП). Стероиды, особенно кортикостероиды надпочечников и их синтетические аналоги, хорошо известны в данной области техники. Примеры кортикостероидов, пригодных для применения в данном изобретении, включают, без ограничения, гидроксилтриамцинолон, альфа-метил дексаметазон, бета-метил бетаметазон, беклометазона дипропионат, бетаметазона бензоат, бетаметазона дипропионат, бетаметазона валерат, клобетазола валерат, дезонид, дезоксиметазон, дексаметазон, дифлоразона диацетат, дифторкотолона валерат, флуадренолон, флуклоролона ацетонид, флуметазона пивалат, флуозинолона ацетонид, флуоцинонид, флукортина бутилэфир, флуокортолон, флупреднидена (флупреднилидена) ацетат, флурандренолон, галцинонид, гидрокортизона ацетат, гидрокортизона бутират, метилпреднизолон, триамцинолона ацетонид, кортизон, кортодоксон, флуцетонид, флудрокортизон, дифлуорозона диацетат, флурадренолона ацетонид, медризон, амцинафел, амцинафид, бетаметазон и баланс его сложных эфиров, хлорпреднизон, хлокортелон, клесцинолон, дихлоризон, дифлупреднат, флукронид, флунизолид, фторметалон, флуперолон, флупреднизолон, гидрокортизон, мепреднизон, параметазон, преднизолон, преднизон, беклометазон дипропионат. (НПВП), которые можно применять в данном изобретении, включают, но не ограничиваются ими, салициловую кислоту, ацетилсалициловую кислоту, метилсалицилат, гликоль салицилат, салицилмиды, бензил-2,5-диацетоксибензойную кислоту, ибупрофен, фулиндак, напроксен, кетопрофен, этофенамат, фенилбутазон и индометацин. В некоторых вариантах осуществления данного изобртения иммуносупрессивный агент может представлять собой цитостатики, такие как алкилирующие агенты, антиметаболиты (например, азатиоприн, метотрексат), цитотоксические антибиотики, антитела (например, базиликсимаб, даклизумаб и муромонаб), антииммунофилины (например, циклоспорин, такролимус, сиролимус), инфероны, опиоиды, белки, связывающие TNF, микофеноляты и небольшие биологические агенты (например, фингомилод, мириоцин).
В некоторых вариантах осуществления данного изобретения химерные белки (и/или дополнительные агенты), описанные в данном документе, включают производные, которые являются модифицированными, т.е. путем ковалентного присоединения любого типа молекулы к композиции таким образом, что ковалентное присоединение не препятствует активности композиции. В качестве примера, но не в качестве ограничения, производные включают композиции, которые были модифицированы, в частности, гликозилированием, липидизацией, ацетилированием, пэгилированием, фосфорилированием, амидированием, дериватизацией известными защитными/блокирующими группами, протеолитическим расщеплением, связыванием с клеточным лигандом или другим белком, и т.д.. Любую из многочисленных химических модификаций можно осуществить с помощью известных методов, в том числе, но не ограничиваясь ими, специфическим химическим расщеплением, ацетилированием, формилированием, метаболическим синтезом турамицина и т.д. Кроме того, производное может содержать одну или большее количество неклассических аминокислот. В других вариантах осуществления данного изобретения химерные белки (и/или дополнительные агенты), описанные в данном документе, дополнительно содержат цитотоксический агент, содержащий в типовых вариантах осуществления, токсин, химиотерапевтический агент, радиоактивный изотоп, и агент, который вызывает апоптоз или гибель клетки. Такие агенты могут быть конъюгированы с композицией, описанной в данном документе.
Химерные белки (и/или дополнительные агенты), описанные в данном документе, могут быть, таким образом модифицированы посттрансляционно для добавления эффекторных фрагментов, таких как химические линкеры, детектируемые фрагменты, такие как, например, флуоресцентные красители, ферменты, субстраты, биолюминесцентные материалы, радиоактивные материалы, а также хемилюминесцентные фрагменты или функциональные фрагменты, такие как, например, стрептавидин, авидин, биотин, цитотоксин, цитотоксический агент и радиоактивные материалы.
Составы
Химерные белки (и/или дополнительные агенты), описанные в данном документе, могут обладать полноценной основной функциональной группой, которая может вступать в реакцию с неорганической или органической кислотой, или карбоксильную группу, которая может вступать в реакцию с неорганическим или органическим основанием с образованием фармацевтически приемлемой соли. Фармацевтически приемлемая кислотно-аддитивная соль образуется из фармацевтически приемлемой кислоты, как хорошо известно в данной области техники. Такие соли включают фармацевтически приемлемые соли, перечисленные, например, в Journal of Pharmaceutical Science, 66, 2-19 (1977) и в The Handbook of Pharmaceutical Salts; Properties, Selection, and Use. P.H. Stahl and C.G. Wermuth (eds.), Verlag, Zurich (Switzerland) 2002, содержание которых включено в данный документ посредством ссылки во всей их полноте.
В некоторых вариантах осуществления данного изобретения композиции, описанные в данном документе, представлены в форме фармацевтически приемлемой соли.
Кроме того, любой химерный белок (и/или дополнительные агенты), описанный в данном документе, можно вводить субъекту в качестве компонента композиции, которая содержит фармацевтически приемлемый носитель или несущую среду. Такие композиции могут необязательно содержать пригодное количество фармацевтически приемлемого вспомогательного вещества с тем, чтобы обеспечить форму для надлежащего введения. Фармацевтические вспомогательные вещества могут быть жидкостями, такими как вода и масла, включая, полученные из нефти, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и тому подобное. Фармацевтические вспомогательные вещества могут быть, например, солевым раствором, аравийской камедью, желатином, крахмальной пастой, тальком, кератином, коллоидным диоксидом кремния, мочевиной и тому подобное. Кроме того, могут применяться вспомогательные, стабилизирующие, загущающие, смазывающие агенты и красители. В одном из вариантов осуществления данного изобретения фармацевтически приемлемые вспомогательные вещества являются стерильными при введении субъекту. Вода является пригодным вспомогательным веществом в тех случаях, когда любой агент, описанный в данном документе, вводят внутривенно. Солевые растворы и водные растворы декстрозы и глицерина также можно применять в качестве жидких вспомогательных веществ, в частности, для инъекционных растворов. Пригодные фармацевтические вспомогательные вещества также включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое обезжиренное молоко, глицерин, пропилен, гликоль, воду, этанол и тому подобное. Любой агент, описанный в данном документе, если желательно, может также содержать незначительные количества смачивающих или эмульгирующих агентов или рН-буферные агенты.
В некоторых вариантах осуществления данного изобретения композиции, описанные в данном документе, являются ресуспендированными в солевом буфере (включая, без ограничения, TBS, PBS и тому подобное).
В различных вариантах осуществления данного изобретения химерные белки могут быть конъюгированными и/или слитыми с другим агентом, чтобы продлить период полувыведения или иным образом улучшить фармакодинамические и фармакокинетические свойства. В некоторых вариантах осуществления данного изобретения химерные белки могут быть слиты или конъюгированы с одним или большим количеством из следующих веществ: ПЭГ, XTEN (например, в виде rПЭГ), полисиаловой кислотой (ПОЛИКСЕН), альбумином (например, сывороточным альбумином человека или HAS), эластиноподобным белком (ELP), PAS, НАР, GLK, СТР, трансферрином и тому подобное. В различных вариантах осуществления данного изобретения каждый из отдельно взятых химерных белков является слитым с одним или большим количеством агентов, описанных в публикации BioDrugs (2015) 29: 215-239, все содержание которой включено в данный документ посредством ссылки.
Введение, дозы и схемы лечения
Данное изобретение включает в себя описанный химерный белок (и/или дополнительные агенты) в различных составах. Любой химерный белок (и/или дополнительные агенты), описанный в данном документе, может иметь форму растворов, суспензий, эмульсии, капель, таблеток, пилюль, гранул, капсул, капсул, содержащих жидкости, порошков, препаратов с замедленным высвобождением, суппозиториев, эмульсий, аэрозолей, спреев, суспензий или любую другую форму, подходящую для применения. Также могут применяться конструкции ДНК или РНК, кодирующие белковые последовательности. В одном варианте осуществления данного изобретения композиция представлена в форме капсулы (см., например, . , в патенте США №5698155). Другие примеры подходящих фармацевтических вспомогательных веществ описаны в публикации Remington's Pharmaceutical Sciences 1447-1676 (Alfonso R. Gennaro eds.,19th ed. 1995), которая включена в данный документ посредством ссылки.
При необходимости, составы, содержащие химерный белок (и/или дополнительные агенты), также могут включать солюбилизирующий агент. Кроме того, указанные агенты можно доставлены с помощью пригодного носителя или устройства для доставки, как известно в данной области техники. Комбинированные препараты, описанные в данном документе, могут быть совместно доставлены в одной доставке с помощью носителя или устройства для доставки. Композиции для введения могут необязательно включать местный анестетик, такой как, например, лидокаин, для того, чтобы уменьшить боль в месте инъекции.
Препараты, содержащие химерный белок (и/или дополнительные агенты) по настоящему изобретению могут быть представлены в виде единичных лекарственных форм и могут быть получены любым из способов, хорошо известных в области фармации. Такие способы, как правило, включают этап приведения терапевтических агентов в контакт с носителем, который состоит из одного или большего количества вспомогательных ингредиентов. Как правило, указанные составы получают путем равномерного и тщательного приведения терапевтического агента в контакт с жидким носителем, тонко измельченным твердым носителем, или обоими, а затем, при необходимости, приданием продукту формы, характерной для лекарственных форм желаемого состава (например, влажное или сухое гранулирование, порошковые смеси и т.д., с последующим таблетированием с помощью общепринятых способов, известных в данной области техники).
В одном варианте осуществления данного изобретения любой химерный белок (и/или дополнительный агент), описанный в данном документе, составляют в соответствии с общеизвестными процедурами в виде композиции, адаптированной для способа введения, описанного в данном документе.
Пути введения включают, например: внутрикожный, внутримышечный, внутрибрюшинный, внутривенный, подкожный, интраназальный, эпидуральный, пероральный, подъязычный, интраназальный, интрацеребральный, интравагинальный, трансдермальный, ректальный, путем ингаляции или местный в частности, в уши, нос, глаза или на кожу. В некоторых вариантах осуществления данного изобретения введение осуществляют перорально или с помощью парентеральной инъекции. В большинстве случаев введение приводит к высвобождению любого агента, описанного в данном документе, в кровоток.
Любой химерный белок (и/или дополнительные агенты), описанный в данном документе, можно вводить перорально. Такие химерные белки (и/или дополнительные агенты) можно вводить любым другим удобным путем, например, путем внутривенной инфузии или болюсной инъекции, путем абсорбции через эпителиальные или кожно-слизистые выстилающие ткани (например, слизистую оболочку полости рта, прямой кишки и слизистой оболочки кишечника и т.д.) и можно вводить вместе с другим биологически активным агентом. Введение может быть системным или местным. Известны различные системы доставки, например, инкапсулирование в липосомы, микрочастицы, микрокапсулы, капсулы и т.п., которые можно применять для введения.
В конкретных вариантах осуществления данного изобретения возможно будет желательным локальное введение в область, нуждающуюся в лечении. В одном варианте осуществления данного изобретения, например, при лечении рака, химерный белок (и/или дополнительные агенты) вводят в микроокружение опухоли (например, клетки, молекулы, внеклеточный матрикс и/или кровеносные сосуды, которые окружают и/или питают опухолевую клетку, включая, например, сосудистую сеть опухоли; инфильтрирующие опухоль лимфоциты; фибробластные ретикулярные клетки; эндотелиальные клетки-предшественники (ЕРС); ассоциированные с раком фибробласты; перициты; другие стромальные клетки; компоненты внеклеточного матрикса (ЕСМ); дендритные клетки; антигенпредставляющие клетки; Т-клетки; регуляторные Т-клетки; макрофаги; нейтрофилы и другие иммунные клетки, локализованные в непосредственной близости от опухоли) или лимфатические узлы и/или нацеливают на микроокружение опухоли или лимфатический узел. В различных вариантах осуществления данного изобретения, например, при лечении рака, химерный белок (и/или дополнительные агенты) вводят внутрь опухоли.
В различных вариантах осуществления данного изобретения описанный химерный белок проявляет двойной эффект, при котором наблюдается меньше побочных эффектов по сравнению с традиционной иммунотерапией (например, при лечении одним или большим количеством из следующих препаратов: ОПДИВО, КЕЙТРУДА, ЕРВОЙ и ТЕЦЕНТРИК). Например, описанные химерные белки уменьшают или предотвращают часто наблюдаемые нежелательные эффекты, связанные с патологией иммунной системы, которая поражает различные ткани и органы, включая кожу, желудочно-кишечный тракт, почки, периферическую и центральную нервную систему, печень, лимфатические узлы, глаза, поджелудочную железу и эндокринную систему; например, гипофизит, колит, гепатит, пневмонит, сыпь и ревматическая болезнь. Кроме того, описанное местное введение, например, внутриопухолевое, устраняет нежелательные явления, наблюдаемые при стандартном системном введении, например, при в/в инфузиях, которые применяются при традиционной иммунотерапии (например, при лечении одним или большим количеством из следующих препаратов: ОПДИВО, КЕЙТРУДА, ЕРВОЙ и ТЕЦЕНТРИК).
Лекарственные формы, пригодные для парентерального введения (например, внутривенного, внутримышечного, внутрибрюшинного, подкожного и внутрисуставного введения и инфузионного) включают, например, растворы, суспензии, дисперсии, эмульсии и тому подобное. Указанные лекарственные формы также могут быть изготовлены в виде стерильных твердых композиций (например, лиофилизированной композиции), которые могут быть растворены или суспендированы в стерильной среде для инъекций непосредственно перед применением. Указанные лекарственные формы могут содержать, например, суспендирующие или диспергирующие агенты, известные в данной области техники.
Дозы любого химерного белка (и/или дополнительных агентов), описанного в данном документе, а также схема введения может зависеть от различных параметров, включая, но не ограничиваясь этим, заболевание, подлежащую лечению, общее состояние здоровья субъекта, а также решение лечащего врача. Любой химерный белок, описанный в данном документе, можно вводить до (например, за 5 минут 15 минут 30 минут 45 минут, 1 час, 2 часа, 4 часа, 6 часов, 12 часов, 24 часа, 48 часов, 72 часа, 96 часов, 1 неделю, 2 недели, 3 недели, 4 недели, 5 недель, 6 недель, 8 недель или 12 недель), одновременно или после (например, по истечению 5 минут 15 минут 30 минут 45 минут, 1 час, 2 часа, 4 часа, 6 часов, 12 часов, 24 часа, 48 часов, 72 часа, 96 часов, 1 неделю, 2 недели, 3 недели, 4 недели, 5 недель, 6 недель, 8 недель или 12 недель) введения дополнительного агента субъекту, нуждающемуся в этом. В различных вариантах осуществления данного изобретения любой химерный белок и дополнительный агент, описанные в данном документе, вводят с интервалом в 1 минуту, с интервалом в 10 минут, с интервалом в 30 минут, с интервалом менее 1 часа, с интервалом 1 час, с интервалом от 1 часа до 2 часов, с интервалом от 2 часов до 3 часов, с интервалом от 3 часов до 4 часов, с интервалом от 4 часов до 5 часов, с интервалом от 5 часов до 6 часов, с интервалом от 6 часов до 7 часов, с интервалом от 7 часов до 8 часов, с интервалом от 8 часов до 9 часов, с интервалом от 9 часов до 10 часов, с интервалом от 10 часов до 11 часов, с интервалом от 11 часов до 12 часов, с интервалом не более 24 часов или с интервалом не более 48 часов.
Дозы любого химерного белка (и/или дополнительных агентов), описанного в данном документе, могут зависеть от нескольких факторов, включая тяжесть патологического состояния, независимо от того, подлежит это состояние лечению или предотвращению, а также возраст, вес и состояние здоровья субъекта, подлежащего лечению. Кроме того, на дозу для конкретного субъекта могут влиять фармакогеномные данные (эффект генотипа на фармакокинетический, фармакодинамический профили или профиль терапевтической эффективности). Кроме того, точные индивидуальные дозы могут быть скорректированы в некоторой степени в зависимости от целого ряда факторов, включая конкретную комбинацию вводимых агентов, время введения, способ введения, характеристики состава, скорость экскреции, конкретное заболевание, подлежащее лечению, тяжесть нарушения и анатомическая локализация нарушения. Можно предполагать некоторые изменения в дозе.
Для введения любого химерного белка (и/или дополнительных агентов), описанного в данном документе, путем парентеральной инъекции, доза обычно составляет от 0,1 мг до 250 мг в сутки, от 1 мг до 20 мг в сутки или от 3 мг до 5 мг в сутки. Инъекции можно осуществлять до четырех раз в сутки. В общем случае, при введении перорально или парентерально, доза любого агента, описанного в данном документе, как правило, составляет от 0,1 мг до 1500 мг в сутки, или от 0,5 мг до 10 мг в сутки, или от 0,5 мг до 5 мг в сутки. Можно вводить дозу до 3000 мг в сутки.
В другом варианте осуществления данного изобретения доставка может осуществляться в везикуле, в частности в липосоме (см. Langer, 1990, Science 249:1527-1533; Treat et al., in Liposomes in the Therapy of Infectious Disease and Cancer, Lopez-Berestein and Fidler (eds.),Liss, New York, pp.353-365 (1989).
Любой химерный белок (и/или дополнительные агенты), описанный в данном документе, можно вводить путем контролируемого высвобождения или замедленного высвобождения или с помощью устройств для доставки, которые хорошо известны обычным специалистам в данной области техники. Примеры включают, но не ограничиваются ими, те, которые описаны в патентах США №№3845770, 3916899; 3536809; 3598123; 4008719; 5674533; 5059595; 5591767; 5120548; 5073543; 5639476; 5354556; и 5733556, каждый из которых включен в данное описание посредством ссылки в полном объеме. Такие лекарственные формы могут быть пригодны для обеспечения контролируемого или замедленного высвобождения одного или большего количества активных ингредиентов с применением, например, гидропропилметил целлюлозы, других полимерных матриц, гелей, проницаемых мембран, осмотических систем, многослойных покрытий, микрочастиц, липосом, микросфер или их комбинации для того, чтобы обеспечить желаемый профиль высвобождения в различных пропорциях. Контролируемое или замедленное высвобождение активного ингредиента можно стимулировать различными условиями, в том числе, но не ограничиваясь этим, изменениями рН, изменениями температуры, стимуляцией с помощью соответствующей длины волны света, концентрацией или наличием ферментов, концентрацией или наличием воды, или другими физиологическими условиями или соединениями.
В другом варианте осуществления данного изобретения можно применять полимерные материалы (см. Medical Applications of Controlled Release, Langer and Wise (eds.), CRC Pres., Boca Raton, Florida (1974); Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen and Ball (eds.), Wiley, New York (1984); Ranger and Peppas, 1983, J. Macromol. Sci. Rev. Macromol. Chem. 23:61; см. также Levy et al., 1985, Science 228:190; During et al., 1989, Ann. Neurol. 25:351; Howard et al.,1989, J. Neurosurg. 71:105).
В другом варианте осуществления данного изобретения система контролированного высвобождения может быть помещена вблизи от целевой области, подлежащей лечению, при этом требуется только часть системной дозы (см. например, Goodson, в Medical Applications of Controlled Release, выше, vol. 2, pp. 115-138 (1984)). Можно применять другие системы с контролируемым высвобождением, обсуждаемые в обзоре Langer, 1990, Science 249:1527-1533.
Введение любого химерного белка (и или дополнительных агентов), описанного в данном документе, можно, независимо, осуществлять от одного до четырех раз в сутки, или от одного до четырех раз в месяц, или от одного до шести раз в год, или один раз в два, три, четыре или пять лет. Введение можно осуществлять на протяжении одного дня или одного месяца, двух месяцев, трех месяцев, шести месяцев, одного года, двух лет, трех лет и может быть даже на протяжении всей жизни субъекта.
Схему введения с применением любого химерного белка (и/или дополнительных агентов), описанного в данном документе, можно выбрать с учетом различных факторов, включая тип, вид, возраст, вес, пол и состояние здоровья субъекта; тяжесть патологического состояния, подлежащего лечению; путь введения; почечную или печеночную функцию у субъекта; фармакогеномный профиль индивидуума; и применяемое специфическое соединение по данному изобретению. Любой химерный белок (и/или дополнительные агенты), описанный в данном документе, можно вводить в виде одной суточной дозы или общую суточную дозу можно вводить в виде разделенных доз на два, три или четыре раза в сутки. Кроме того, любой химерный белок (и/или дополнительные агенты), описанный в данном документе, можно вводить непрерывно, а не периодически, в ходе всей схемы приема.
Клетки и нуклеиновые кислоты
В различных вариантах осуществления данного изобретения предлагается экспрессионный вектор, содержащий нуклеиновую кислоту, кодирующую описанный в данном документе химерный белок. В различных вариантах осуществления данного изобретения экспрессионный вектор содержит ДНК или РНК. В различных вариантах осуществления данного изобретения экспрессионный вектор представляет собой экспрессионный вектор млекопитающих.
Для экспрессии химерного белка можно применять как прокариотические, так и эукариотические векторы. Прокариотические векторы включают конструкции на основе последовательностей Е. coli (см., например, Makrides, Microbiol Rev 1996, 60:512-538). Неограничивающие примеры регуляторных областей, которые можно применять для экспрессии в Е. coli, включают lac, trp, lpp, phoA, recA, tac, Т3, T7 и λPL. Неограничивающие примеры прокариотических экспрессионных векторов могут включать серии вектора λgt, такие как λgt11 (Huynh et al., в "DNA Cloning Techniques, Vol.I:A Practical Approach," 1984, (D. Glover, ed.), pp. 49-78, IRL Press, Oxford), и серии вектора pET (Studier et al. Methods Enzymol 1990, 185:60-89). Однако протокариотические системы векторов-хозяев не могут выполнять большую часть посттрансляционного процессинга клеток млекопитающих. Таким образом, эукариотические системысистемы векторов-хозяев могут быть особенно пригодными. Для экспрессии химерных белков в клетках-хозяевах млекопитающих можно применять множество регуляторных областей. Например, можно применять ранние и поздние промоторы SV40, непосредственный ранний промотор цитомегаловируса (CMV) и промотор длинного концевого повтора вируса саркомы Рауса (RSV-LTR). Индуцируемые промоторы, которые могут быть пригодны для применения в клетках млекопитающих, включают, без ограничения, промоторы, ассоциированные с геном металлотионеина II, длинные концевые повторы вируса опухоли молочной железы мышей, отвечающей на лечение глюкокортикоидами (MMTV-LTR), ген β-интерферона и ген hsp70 (см. Williams et al., Cancer Res 1989, 49:2735-42; and Taylor et al. Mol Cell Biol 1990, 10:165-75). Для стимулирования экспрессии слитых белков в рекомбинантных клетках-хозяевах также могут быть полезными промоторы теплового шока или промоторы стресса.
В некоторых вариантах осуществления данного изобретения экспрессионные векторы по данному изобретению содержат нуклеиновую кислоту, кодирующую химерные белки (и/или дополнительные агенты), или их комплемент, функционально связанный с областью контроля экспрессии, или их комплемент, который является функциональным в клетке млекопитающего. Область контроля экспрессии способна стимулировать экспрессию функционально связанного блокирующего и/или стимулирующего агента, кодирующего нуклеиновую кислоту, в результате чего блокирующий и/или стимулирующий агент продуцируется в клетке человека, трансформированной указанным экспрессионным вектором.
Областями контроля экспрессии являются регуляторные полинуклеотиды (иногда называемые в данном документе "элементами"), такие как промоторы и энхансеры, которые влияют на экспрессию функционально связанной нуклеиновой кислоты. Область контроля экспрессии экспрессионного вектора согласно данному изобретению способна экспрессировать функционально связанную кодирующую нуклеиновую кислоту в клетке человека. В одном варианте осуществления данного изобретения клетка представляет собой клетку опухоли. В другом варианте осуществления данного изобретения клетка представляет собой неопухолевую клетку. В одном варианте осуществления данного изобретения область контроля экспрессии обеспечивает функционально связанной нуклеиновой кислоте экспрессию с возможностью регуляции. Сигнал (иногда называемый "стимулом") может увеличивать или уменьшать экспрессию нуклеиновой кислоты, функционально связанной с такой областью контроля экспрессии. Такие области контроля экспрессии, которые увеличивают экспрессию в ответ на сигнал, часто называются "индуцибельными". Такие области контроля экспрессии, которые уменьшают экспрессию в ответ на сигнал, часто называются "репрессируемыми". Как правило, увеличение или уменьшение количества таких элементов является пропорциональным количеству присутствующего сигнала; чем больше количество сигнала, тем больше увеличение или уменьшение экспрессии.
В одном варианте осуществления данного изобретения предполагается применение индуцибельных промоторов, способных к высокому уровню транзиентной экспрессии в ответ на сигнал. Например, в случаях непосредственной близости опухолевой клетки, клетка, трансформированная экспрессионным вектором для химерного белка (и/или дополнительных агентов), содержащим такую последовательность контроля экспрессии, индуцируется с целью транзиентного продуцирования высокого уровня агента путем воздействия соответствующего стимула на указанную трансформированную клетку. Типовые индуцибельные области контроля экспрессии включают те, которые содержат индуцибельный промотор, который стимулируется сигналом, таким как неизкомолекулярное химическое соединение. Конкретные примеры можно найти, например, в патенте США№№5989910, 5935934, 6015709 и 6004941, каждый из который включен в данный документ посредством ссылки в полном объеме.
Области контроля экспрессии и области контроля локуса включают полноразмерные промоторные последовательности, такие как нативные промоторные и энхансерные элементы, а также последовательности или полинуклеотидные варианты, которые сохраняют всю или часть полноразмерной или невариантной функции. В данном контексте термин "функциональный" и его грамматические варианты, при применении в отношении последовательности нуклеиновой кислоты, подпоследовательности или фрагмента, означает, что последовательность имеет одну или большее количество функций нативной последовательности нуклеиновой кислоты (например, невариантная или немодифицированная последовательность).
В данном контексте термин "функциональная связь" относится к физическому сопоставлению компонентов, определенных таким образом, что обеспечивает для них возможность функционировать по назначению. В примере элемента контроля экспрессии в функциональной связи с нуклеиновой кислотой, взаимосвязь является таковой, что элемент контроля экспрессии модулирует экспрессию нуклеиновой кислоты. Как правило, область контроля экспрессии, которая модулирует транскрипцию, размещается вблизи 5'-конца транскрибируемой нуклеиновой кислоты (то есть, "против хода транскрипции"). Области контроля экспрессии также могут быть расположены на 3'-конце транскрибируемой последовательности (то есть, "за ходом транскрипции") или внутри транскрипта (например, в интроне). Элементы контроля экспрессии могут быть расположены на некотором расстоянии от транскрибируемой последовательности (например, от 100 до 500, от 500 до 1000, от 2000 до 5000 или более нуклеотидов от нуклеиновой кислоты). Конкретным примером элемента контроля экспрессии является промотор, который обычно расположен на 5'-конце транскрибируемой последовательности. Другим примером элемента контроля экспрессии является энхансер, который может быть расположен на 5'- или 3'-конце транскрибируемой последовательности или внутри транскрибируемой последовательности.
Системы экспрессии с возможностью функционирования в клетках человека, хорошо известны в данной области техники и включают вирусные системы. Как правило, промотор с возможностью функционирования в клетке человека представляет собой любую последовательность ДНК, способную связывать РНК-полимеразу млекопитающих и инициировать транскрипцию кодирующей последовательности в мРНК по направлению к (3')-концу. Промотор будет иметь область инициации транскрипции, которая, как правило, расположена ближе к 5'-концу кодирующей последовательности, а ТАТА-бокс, как правило, расположен на 25-30 пар оснований от участка инициации транскрипции против ее хода. Считается, что ТАТА-бокс направляет РНК-полимеразу II для начала синтеза РНК на правильном участке. Как правило, промотор также содержит промоторный элемент (энхансерный элемент) перед участком инициации транскрипции ("левый элемент"), как правило, расположенный в пределах от 100 до 200 пар оснований "левее" ТАТА-бокса. "Левый" промоторный элемент определяет скорость, с которой инициируется транскрипция, и может функционировать в любой ориентации. Особенно часто в качестве промоторов применяют промоторы из вирусных генов млекопитающих, поскольку вирусные гены во многих случаях экспрессируются в высокой степени и имеют широкий диапазон хозяев. Примеры включают в себя ранний промотор SV40, промотор LTR опухоли молочной железы мыши, основной поздний промотор аденовируса, промотор вируса простого герпеса и промотор CMV.
Как правило, последовательности терминации транскрипции и полиаденилирования, распознаваемые клетками млекопитающих, являются регуляторными областями, расположенными 3' к стоп-кодону трансляции и, таким образом, вместе с промоторными элементами фланкируют кодирующую последовательность. 3'-конец зрелой мРНК образован специфическим для участка посттрансляционным расщеплением и полиаденилированием. Примеры терминатора транскрипции и сигналов полиаденилирования включают те, которые являются производными от SV40. Интроны также могут быть включены в конструкции экспрессии.
Существуют различные способы введения нуклеиновых кислот в жизнеспособные клетки. Методики, пригодные для переноса нуклеиновой кислоты в клетки млекопитающих in vitro, включают применение липосом, электропорацию, микроинъекцию, слияние клеток, системы на основе полимеров, DEAE-декстран, вирусную трансдукцию, метод осаждения фосфатом кальция и т.д. Для переноса гена in vivo может быть также применен ряд методов и реагентов, включая липосомы; устройства для доставки на основе природного полимера, такого как хитозан и желатин; для трансдукции in vivo являются также пригодными вирусные векторы. В некоторых ситуациях является желательным получение агента целенаправленного воздействия, такого как антитело или лиганд, специфичный для белка наружного слоя мембраны опухолевой клетки. В случаях применения липосом, для нацеливания и/или для облегчения поглощения можно применять белки, которые связываются с белком наружного слоя мембраны клетки, ассоциированным с эндоцитозом, например, капсидные белки или их фрагменты, тропные к клеткам конкретного типа, антитела к белкам, подвергающимся интернализации при клеточном цикле, белки, мишенью которых являются внутриклеточные структуры, и белки, удлиняющие внутриклеточный период полужизни. Методика рецептор-опосредованного эндоцитоза описана, например, в публикациях Wu et al., J. Biol. Chem. 262, 4429-4432 (1987); и Wagner et al., Proc. Natl. Acad. Sci. USA 87, 3410-3414 (1990).
При необходимости также можно применять агенты доставки генов, такие как, например, интеграционные последовательности. В данной области техники известны многочисленные интеграционные последовательности (см., например, Nunes-Duby et al., Nucleic Acids Res. 26:391-406, 1998; Sadwoski, J. Bacteriol., 165:341-357, 1986; Bestor, Cell, 122(3):322-325, 2005; Plasterk et al., TIG 15:326-332, 1999; Kootstra et al., Ann. Rev. Pharm. Toxicol., 43:413-439, 2003). К ним относятся рекомбиназы и транспозазы. Примеры включают Cre (Sternberg and Hamilton, J. Mol. Biol., 150:467-486, 1981), лямбда (Nash, Nature, 247, 543-545, 1974), FIp (Broach, et al., Cell, 29:227-234, 1982), R (Matsuzaki, et al., J. Bacteriology, 172:610-618, 1990), cpC31 (см., например, Groth et al., J. Mol. Biol. 335: 667-678, 2004), транспозоны "спящая красавица", транспозоны "семейства моряков" (Plasterk et al., выше) и компоненты для интеграции вирусов, таких как AAV, ретровирусов и антивирусов, имеющих компоненты, которые обеспечивают интеграцию вирусов, таких как LTR последовательности ретровирусов или лентивирусов и ITR-последовательности AAV (Kootstra et al., Ann. Rev. Pharm. Toxicol., 43:413-439, 2003). Кроме того, стратегии прямой и нацеленной генетической интеграции можно применять для вставки последовательностей нуклеиновых кислот, кодирующих химерные слитые белки, включая CRISPR/CAS9, "цинковые пальцы", TALEN и технологии редактирование генома мегануклеазой.
В одном аспекте данного изобретения предлагаются экспрессионные векторы для экспрессии химерных белков (и/или дополнительных агентов), которые являются вирусными векторами. Известны многие вирусные векторы, пригодные для генной терапии (см., например, Lundstrom, Trends Biotechnol., 21:1 17, 122, 2003. Типовые вирусные векторы включают те, которые выбраны из антивирусов (LV), ретровирусов (RV), аденовирусов (AV), аденоассоциированных вирусов (AAV) и α-вирусов, хотя можно применять и другие вирусные векторы. Для применения in vivo пригодны вирусные векторы, которые не интегрируются в геном хозяина, такие как α-вирусы и аденовирусы. Типовые типы α-вирусов включают вирус Синдбис, вирус венесуэльского энцефалита (VEE) и вирус леса Семлики (SFV). Для применения in vitro пригодны вирусные векторы, которые интегрируются в геном хозяина, такие как ретровирусы, AAV и антивирусы. В одном варианте осуществления данного изобретения предлагаются способы трансдуцирования клетки человека in vivo, включающие приведение в контакт солидной опухоли in vivo с вирусным вектором по данному изобретению.
В различных вариантах осуществления данного изобретения предлагается клетка-хозяин, содержащая экспрессионный вектор, который содержит описанный в данном документе химерный белок.
Для получения описанных химерных белков экспрессионные векторы можно вводить в клетки-хозяева. Например, клетки могут быть культивированы in vitro или генетически сконструированы. Пригодные клетки-хозяева млекопитающих включают, без ограничения, клетки, полученные от людей, обезьян и грызунов (см., например, Kriegler в "Gene Transfer and Expression:A Laboratory Manual," 1990, New York, Freeman & Co.). К ним относятся линии клеток почек обезьян, трансформированные SV40 (например COS-7, АТСС CRL 1651), линии эмбриональных клеток почек человека (например, 293, 293-EBNA, или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al. J Gen Virol 1977, 36:59); клетки почек детеныша хомяка (например, (ВНК, АТСС CCL 10); клетки яичника китайского хомяка-DHFR (например, СНО, Urlaub and Chasin, Proc Natl Acad Sci USA 1980, 77:4216); клетки DG44 СНО, клетки CHO-K1, клетки Сертоли мыши (Mather, Biol Reprod 1980, 23:243-251); фибробластные клетки мыши (например, NIH-3T3), клетки почек обезьян (например, CV1 АТСС CCL 70); клетки почек африканских зеленых обезьян, (например, VERO-76, АТСС CRL-1587); клетки карциномы шейки матки человека (например, HELA, АТСС CCL 2); клетки почки собаки (например, MDCK, АТСС CCL 34); клетки печени крыс буффало (например, BRL 3А, АТСС CRL 1442); клетки легких человека (например, W138, АТСС CCL 75); клетки печени человека (например, Hep G2, НВ 8065); и опухолевые клетки молочной железы мыши (например, ММТ 060562, АТСС CCL51). Иллюстративные типы раковых клеток для экспрессии слитых белков, описанных в данном документе, включают линию клеток фибробластов мыши, NIH3T3, линию клеток карциномы легкого Льюиса мыши, LLC, линию клеток мастоцитомы мыши, Р815, линию клеток лимфомы мыши, EL4 и ее овальбуминовый трансфектант, Е. G7, линию клеток меланомы мыши, B16F10, линию клеток фибросаркомы мыши, МС57, и линии клеток мелкоклеточной карциномы легкого человека, SCLC # 2 и SCLC # 7.
Клетки-хозяева можно получить от здоровых или пораженных субъектов, включая здоровых людей, пациентов с раком и пациентов с инфекционным заболеванием, из коллекций частных лабораторий, общедоступных коллекций культур, таких как Американская коллекция типовых культур, или от коммерческих поставщиков.
Клетки, которые можно применять для продукции описанных химерных белков in vitro, ex vivo и/или in vivo, включают, без ограничения, эпителиальные клетки, эндотелиальные клетки, кератиноциты, фибробласты, мышечные клетки, гепатоциты; клетки крови, такие как Т-лимфоциты, В-лимфоциты, моноциты, макрофаги, нейтрофилы, эозинофилы, мегакариоциты, гранулоциты; различные стволовые клетки или клетки-предшественники, в частности гемопоэтические стволовые клетки или клетки-предшественники (например, полученные из костного мозга), пуповинную кровь, периферическую кровь, фетальную печень и т.д. Выбор типа клетки зависит от типа опухоли или инфекционного заболевания, которое подлежит лечению или предотвращению, и может быть определен специалистом в данной области техники.
Субъекты и/или животные
В некоторых вариантах осуществления данного изобретения субъект и/или животное представляет собой млекопитающее, например, человека, мышь, крысу, морскую свинку, собаку, кошку, лошадь, корову, свинью, кролика, овцу или примата, не относящегося к человеку, такого как обезьяна, шимпанзе или бабуин. В других вариантах осуществления данного изобретения субъект и/или животное представляет собой не млекопитающее, такое как, например, данио рерио. В некоторых вариантах осуществления данного изобретения субъект и/или животное может содержать флуоресцентно-меченые клетки (например, меченые с помощью GFP). В некоторых вариантах осуществления данного изобретения субъект и/или животное представляет собой трансгенное животное, содержащее флуоресцентную клетку.
В некоторых вариантах осуществления данного изобретения субъектом и/или животным является человек. В некоторых вариантах осуществления данного изобретения человеком является человеческий ребенок. В других вариантах осуществления данного изобретения человеком является взрослый человек. В других вариантах осуществления данного изобретения человеком является пожилой человек. В других вариантах осуществления данного изобретения человеком может называться пациент.
В некоторых вариантах осуществления данного изобретения человек имеет возраст в диапазоне от около 0 месяцев до около 6 месяцев, от около 6 до около 12 месяцев, от около 6 до около 18 месяцев, от около 18 до около 36 месяцев, от около 1 до около 5 лет, от около 5 до около 10 лет, от около 10 до около 15 лет, от около 15 до около 20 лет, от около 20 до около 25 лет, от около 25 до около 30 лет, от около 30 до около 35 лет, от около 35 до около 40 лет, от около 40 до около 45 лет, от около 45 до около 50 лет, от около 50 до около 55 лет, из около от 55 до около 60 лет, от около 60 до около 65 лет, от около 65 до около 70 лет, от около 70 до около 75 лет, от около 75 до около 80 лет, от около 80 до около 85 лет, от около 85 до около 90 лет, от около 90 до около 95 лет или от около 95 до около 100 лет.
В других вариантах осуществления данного изобретения субъект представляет собой животное, не относящееся к человеку, и, следовательно, данное изобретение относится к применению в ветеринарии. В конкретном варианте осуществления данного изобретения, животное, не относящееся к человеку, представляет собой домашнее животное. В другом конкретном варианте осуществления данного изобретения, животное, не относящееся к человеку, представляет собой животное домашнего скота.
Наборы
Данное изобретение относится к наборам, которые могут упростить введение любого агента, описанного в данном документе. Типовой набор по данному изобретению содержит любую композицию, описанную в данном документе, в виде единичной лекарственной формы. В одном варианте осуществления данного изобретения единичная лекарственная форма представляет собой контейнер, такой как предварительно заполненный шприц, который может быть стерильным, содержащим любой агент, описанный в данном документе, и фармацевтически приемлемый носитель, растворитель, вспомогательное вещество или несущую среду. Набор может дополнительно содержать этикетку или напечатанные инструкции, дающие указания относительно применения любого агента, описанного в данном документе. Набор может также включать векорасширитель, местный анестетик и очищающий агент для места введения. Набор может также дополнительно включать один или большее количество дополнительных агентов, описанных в данном документе. В одном варианте осуществления данного изобретения набор содержит контейнер, содержащий эффективное количество композиции по данному изобретению и эффективное количество другой композиции, например, той, которая описана в данном документе. Данное изобретение будет дополнительно описано в следующих примерах, которые не ограничивают объем изобретения, описанный в Формуле изобретения.
ПРИМЕРЫ
Пример 1. Конструкция и характеристика конструкции PD-1-Fc-OX40L мыши
Создавали химерную конструкцию PD-1-Fc-OX40L мыши, после чего ее экспрессию в клетках СНО-K1 анализировали с помощью анализа ELISA с захватом IgG мыши (в данном случае, Fc получали из IgG1). В частности, клетки СНО-K1 стабильно нуклеофицировали векторами pVITRO2-GS-hygro или pcDNA3. 4, экспрессирующими либо внеклеточный домен (ECD) мыши PD-1, слитый с Fc (mPD-1-Fc), либо mPD-1-Fc, слитый с ECD OX40L (mPD-1-Fc-OX40L). С помощью предельного разведения выделяли антибиотикорезистентные одноклеточные клоны. Концентрацию каждого химерного белка, секретируемого в культуральную среду, определяли с помощью ELISA с захватом mIgG, как проиллюстрировано на Фигуре 5.
Анализы связывания проводили для характеристики способности PD-1-FC-OX40L мыши связываться с mOX40, а также с mPD-L1. На Фигуре 6, панель А, приведено схематическое изображение анализа ELISA, применяемого для обнаружения связывания mPD-1-Fc-OX40L мыши с mOX40. Рекомбинантный mOX40, слитый с Fc (mOX40-hFc) человека, специально применяли для захвата mPD-1-Fc-OX40L в культуральной среде. Поликлональное антитело кролика к mPD-1 применяли для определения домена mPD-1 в химерном белке, с последующим определением с применением конъюгированного с пероксидазой хрена (HRP) поликлонального антитела к IgG (Н+L) кролика. На Фигуре 6, панель В, проиллюстрировано, что PD-1-Fc-OX40L мыши эффективно связывается с ОХ40 по сравнению с отрицательным контролем mPD-1-Fc. На Фигуре 7, панель А, приведено схематическое изображение анализа ELISA, применяемого для обнаружения связывания mPD-1-Fc-OX40L мыши с mPD-L1. Рекомбинантный mPD-L1, слитый с Fc (mPD-L1-hFc) человека, специально применяли для захвата химерного белка mPD-1-Fc-OX40L в культуральной среде. Для обнаружения связанных белков применяли конъюгированное с пероксидазой хрена (HRP) поликлональное антитело к IgG (Н+L) мыши. На Фигуре 7, панель В, проиллюстрировано, что PPD-1-Fc-OX40L эффективно связывается с PD-L1 по сравнению с отрицательным контролем среды и положительным контролем с применением рекомбинантного PD1-Fc мыши.
Эксперименты проводили для характеристики активности PD-1-Fc-OX40L мыши при выявлении ответа Т-клеток и при лечении опухолей. Овальбуминовые антиген-специфические OT-I/EGFP, , CD8+ Т-клетки цыпленка (5×105) адоптивно трансфицировали мышам С57/BL6-альбино посредством инъекции в хвостовоую вену за 2 дня до инокуляции опухолевых клеток В16. F10-ova (5×105) в правый фланк мышей. После достижения опухолью диаметра 3-5 мм, PD-1-Fc-OX40L, экспрессирующий ДНК (50 мкг), электропорировали в опухоль с помощью заданного электрического импульса (1500 В/см), применяя при этом 8 импульсов при 100 мкС. Процент клеток CD8+ OT- I/EGFP в периферической крови определяли количественно с помощью анализа проточной цитометрии на протяжении заданного периода времени после электропорации. На Фигуре 8 проиллюстрировано, что внутриопухолевая доставка PD-1-Fc-OX40L мыши (m) in vivo обуславливала увеличение количества антигенспецифических CD8+ Т-клеток.
На Фигуре 9 проиллюстрировано, что внутриопухолевая доставка mPD-1-Fc-OX40L in vivo обуславливала регрессию опухоли в модели опухоли В16. F10-ova. Опухоли В16. F10-ova генерировали у мышей С57/В16-альбино, которых адоптивно трансфицировали клетками CD8+ OT-I/EGFP и электропорировали один раз с mPD-1-Fc-OX40L (50 мкг), экспрессирующим ДНК. Контрольные мыши не получали ДНК, но подвергались электропорации (только ЭП). Диаметры опухолей измеряли с помощью цифрового штангенциркуля на протяжении заданного периода времени после электропорации. На Фигуре 9 проиллюстрировано, что введение mPD-1-Fc-OX40L значительно уменьшает размер опухоли.
Пример 2. Дополнительная характеристика конструкции PD-1-Fc-OX40L мыши
Генерировали конструкцию mPD-1-Fc-OX40L, которая включала внеклеточный домен (ECD) PD-1 мыши, слитый с ECD OX40L посредством домена Fc шарнир-СН2-СН3, производного от IgG1 (mPD-1-Fc-OX40L). Конструкцию mPD-1-Fc-OX40L транзиентно экспрессировали в клетках 293 и очищали с помощью аффинной хроматографии с применением белка А. Для валидации обнаружения и связывания всех трех компонентов mPD-1-Fc-OX40L проводили анализ вестерн-блота и функциональный анализ ELISA (Фигура 10, панель А). Количественное определение mPD1-Fc-OX40L можно осуществить с помощью ELISA с захватом и обнаружением IgG мыши (Фигура 10, панель В). Продемонстрировано одновременное связывание mPD-1 и mOX40L с их партнерами mPD-L1 и mOX40, соответственно, путем захвата mPD-1-Fc-OX40L с mPD-L1-Fc и обнаружения его с mOX40-His, с последующим количественным хемилюминесцентным определением His-HRP (Фигура 8, панель С). Отмечено также, что существуют мономерные и димерные конформации mPD-1-Fc-OX40L.
Для оценки клеточного связывания mPD-1-Fc-OX40L ex vivo, первичные спленоциты мыши выделяли и активировали в течение 2 дней с помощью РМА/РНА/лономицина с целью усиления экспрессии ОХ40 и PD-L1. Затем активированные спленоциты обрабатывали 500 нг/мл mPD-1-Fc-OX40L и анализировали с помощью проточной цитометрии на предмет связывания (Fc-PE) (Фигура 8, панель D). Для выделения клеток, экспрессирующих PD-L1, спленоциты совместно окрашивали антителом, нацеленным на МНС II на антигенпредставляющих клетках (I-A/I-E). Для выделения клеток, экспрессирующих ОХ40, спленоциты совместно окрашивали CD4. mPD-1-Fc-OX40L в значительной мере связывался с популяциями спленоцитов PD-L1+ и ОХ40+, что указывает на то, что mPD-1-Fc-OX40L был создан и очищен надлежащим образом, чтобы связывать свои мишени с первичными производными клетками. Также оценивали активность связывания mPD1-Fc-OX40L с первичными линиями опухолевых клеток мыши, экспрессирующих PD-L1. Линию опухолевых клеток 4Т1 мыши идентифицировали как экспрессирующую низкие количества PD-L1, при том, что линия опухолевых клеток В16. F10 экспрессировала сравнительно высокие количества PD-L1. Продемонстрировано, что mPD1-Fc-OX40L связывает PD-L1-положительную линию опухолевых клеток В16. F10 в большей степени, чем линию опухолевых клеток 4Т1 с низким количеством PD-L1 (Фигура 10, панель Е).
Дополнительные функциональные активности mPD-1-Fc-OX40L были охарактеризованы с помощью анализа совместного культивирования активированных Т-клеток/опухоли. Во- первых, клетки PD-L1низк (4Т1) и PD-L1выс (В16. F10) мыши идентифицировали с помощью проточной цитометрии (Фигура 8, панель Е). Затем спленоциты мыши активировали в течение 2 дней гранулами CD3/CD28 и суб-насыщающей концентрации IL2. Через 2 дня активированные спленоциты совместно культивировали с облученными клетками 4Т1 или В16. F10 в присутствии или отсутствие mPD-1-Fc-OX40L. Через пять дней после первичного выделения спленоцитов культуральную среду собирали и анализировали на цитокина EL2 с помощью ELISA (Фигура 8, панель F). Обнаружено, что mPD-1-Fc-OX40L способен к значительной индукции секреции IL2, особенно в совместных культурах, содержащих опухолевые клетки с PD-L1выс. Не желая быть связанными какой-либо теорией, считается, что mPD-1-Fc-OX40L одновременно блокирует супрессивные эффекты PD-L1, одновременно активируя Т-клетки через сигнализацию OX40/OX40L, тем самым индуцируя секрецию IL2. В целом, эти данные свидетельствуют о том, что mPD-1-Fc-OX40L может обеспечить значительный противоопухолевый иммунитет в доклинических моделях.
Противоопухолевую активность mPD-1-Fc-OX40L анализировали с применением нескольких систем доклинических моделей опухолей. В частности, для оценки влияния mPD-1-Fc-OX40L на рост опухоли, общую выживаемость и индукцию ответа цитокинов в сыворотке после терапии, применяли модели колоректального рака мыши (СТ26 и МС38). Эти эксперименты проводили на равных с широко охарактеризованными антителами- агонистами ОХ40 (OX86) и антителами, блокирующими PD-L1 (10F. 9G2), которые применяли в виде монотерапии или в комбинации, в эквивалентной активной дозе mPD-1-Fc-OX40L, посредством внутрибрюшинной инъекции (2 дозы по 100 мкг каждая). Как проиллюстрировано на Фигуре 11, панель A, mPD-1-Fc-OX40L значительно уменьшал размер опухоли в модели МС38. Более конкретно, введение mPD-1-Fc-OX40L приводило к большей регрессии опухоли по сравнению с введением антитела-агониста ОХ40 и антитела, блокирующего PD-L1, вводимых отдельно или в комбинации. Важно отметить, что в каждой группе осуществляли повторную нагрузку родительской линией опухолевых клеток МС38 мышам, которые отторгали первичную опухоль. Эти данные продемонстрировали, что в отсутствие повторного лечения, мышам, получавшим mPD1-Fc-OX40L, удалось отторгнуть повторную нагрузку родительской опухолью в большей степени по сравнению с мышами любой другой группы лечения (Фигура 11, панели А и В). Кроме того, изготавливали и другие слитые конструкции, включая mPD1-Fc-GITRL и mPD1-Fc-41BBL, и применяли их у мышей с опухолью, как описано выше для mPD1-Fc- OX40L. Конструкции, содержащие как GITRL, так и 41BBL, приводили к уменьшению размера опухоли у пролеченных животных.
В дополнение к измерению размера опухоли также определяли фармакодинамический биомаркер - передачу сигналов mPD-1-Fc-OX40L in vivo. В частности, проводили анализ цитокинов сыворотки у мышей, получавших антитела анти-PD-L1 и анти-ОХ40, а также PD-1-Fc-OX40L Как проиллюстрировано на Фигуре 11, панели В и С, после лечения с применением mPD-1-Fc-OX40L регистрировали зависимый от дозы цитокиновый профиль, который был заметно похожий на цитокиновый профиль, наблюдаемый после комбинированного введения анти-PD-L1 и анти-ОХ40 и включающий повышенные уровни IFNγ, TNFα, IL-2, IL-4, IL-5, IL-6, IL-10, IL-17A и IL-22 (Фигура 11, панели С, D и J). Важно отметить, что обнаружение ответа цитокинов сыворотки после лечения с применением mPD1-Fc-OX40L было зависимым от дозы. В частности, лечение с применением одной или двух инъекций 40 мкг не приводило к детектируемому ответу цитокинов сыворотки, в то время как лечение с однократным применением 100 мкг приводило к промежуточному ответу цитокинов, а лечение с двухкратным применением 100 мкг приводило к более высокому ответу цитокинов (Фигура 11, панель K). Также продемонстрировано, что лечение мышей с применением mPD1-Fc-GITRL стимулировало специфический ответ цитокинов сыворотки.
В некоторых экспериментах мышей с опухолью МС38, умерщвляли на день 13 эксперимента для оценки клеточного иммунного ответа в опухоли, периферической крови и селезенке. На день 13 эксперимента, mPD1-Fc-OX40L, mPD1-Fc-GITRL и mCD172a-Fc- CD40L, как было показано, вызывают снижение темпов роста опухоли по сравнению с животными, не получавшими лечения, или животными, получавшими антитело-агонист ОХ40, антитело-агонист GITR или антитело, блокирующее PD-L1 (Фигура 11, панель Е). В соответствии с этими данными, было продемонстрировано, что у мышей, получавших mPD1-Fc-OX40L или mPD1-Fc-GITRL, на день 13 эксперимента увеличивалось количество инфильтрирующих опухоль лимфоцитов, специфичных к опухолевому антигену (TIL) (Фигура 11, панель F). Проанализировали фенотип памяти в CD8+ Т-клетках селезенки (Фигура 11, панель G), а также провели сравнение соотношения Т- клеток CD4/CD8 в зависимости от разных методов лечения (Фигура 11, панель Н).
Фармакодинамические биомаркеры передачи сигналов PD-1-Fc-OX40L in vivo также определяли с применением модели СТ26. В частности, проводили анализ цитокинов сыворотки у мышей, получавших антитела анти-PD-L1 и анти-ОХ40, отдельно или в комбинации, а также PD-1-Fc-OX40L. Как проиллюстрировано на Фигуре 11, панель D, после лечения с применением mPD-1-Fc-OX40L регистрировали цитокиновый профиль, который был заметно похожий на цитокиновый профиль, наблюдаемый после комбинированного введения антител анти-PD-L1 и анти-ОХ40. В частности, цитокиновый профиль состоял из повышенных уровней IFNγ, TNFα, IL-2, IL-4, IL-5, IL-6, IL -9, IL-10, IL-13, IL-17A, IL-17F и IL-22 (Фигура 11, панели J и K).
В соответствии с результатами, полученными из модели МС38, введение mPD-1-Fc- OX40L также значительно уменьшало размер опухоли в модели колоректального рака СТ26. Более конкретно, введение mPD-1-Fc-OX40L приводило к большей регрессии опухоли по сравнению с введением антитела-агониста ОХ40 и антитела, блокирующего PD-L1, вводимых отдельно или в комбинации (Фигура 11, панель L).. Кроме того, у мышей, получавших mPD-1-Fc-OX40L, наблюдали более длительный период выживания, по сравнению с мышами, которым вводили антитело-агонист ОХ40 и антитело, блокирующее PD-L1 (Фигура 11, панель L). Кроме того, было показано, что при применении других химерных слитых белковых конструкции, включая PD1-Fc-GITRL, PD1-Fc-41BBL и PD1-Fc-TL1A, отмечали замедление роста и регрессию опухоли в модели СТ26 (Фигура 11, панель L).
В целом эти данные наглядно демонстрируют, в частности, функциональную активность mPD-1-Fc-OX40L in vivo.
Пример 3. Конструкция и характеристика PD-1-Fc-OX40L человека Сконструировали PD-1-Fc-OX40L человека, содержащий PD-1 человека, связанный с OX40L через домен Fc шарнир-СН2-СН3, полученный из антитела иммуноглобулина 4 человека (IgG4). Эта конструкция получила название SL-279252.
Последовательность мРНК OX40L человека была следующей:




Аминокислотная последовательность OX40L человека была следующей (затененный - внеклеточный домен):

Последовательность нуклеиновой кислоты последовательности шарнир-СН2-СН3 из IgG4 человека была следующей:
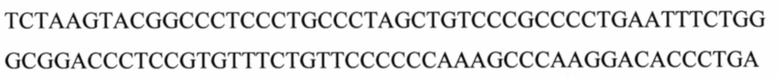

Последовательность кДНК PD-1 человека была следующей:

Последовательность нуклеиновой кислоты PD-1-Fc-OX40L человека была следующей:

Последовательность была кодон-оптимизированной относительно экспрессии клетками китайского хомяка (СНО) и выглядела следующим образом:

Соответственно, аминокислотная последовательность SL-279252 была следующей:

В альтернативном варианте SL-279252 может включать другие сигнальные пептиды, такие как соединения, полученные из коллагена V человека или тяжелой цепи IgG человека. В альтернативном варианте SL-279252 может включать одну или большее количество мутаций в домене Fc, с целью повышения стабильности или увеличения аффинности связывания с FcRn, например, как ранее описанные. Конструкцию PD-1-Fc-OX40L человека импортировали в программное обеспечение RaptorX для прогнозирования третичной структуры белка, чтобы обеспечить надлежащий фолдинг трех основных доменов (см. Фигуру 12, панель А). Третичные структуры каждого компонента (то есть, PD-1, Fc и OX40L) принимали свои собственные конформации в большей макромолекуле, что указывает на то, что PD-1-FC-OX40L может поддерживать связывающую способность и молекулярную функцию всех доменов. Затем иммуногенную возможность PD-1-Fc-OX40L оценивали с помощью алгоритма молекулярного моделирования в силиконе, перекрестно ссылающегося на базу данных эпитопов Т-клеток (ABZENA/ANTITOPE, Фигура 12, панель В). Несмотря на то, что все кодирующие последовательности были человеческими, отмечался минимальный потенциал для лидерных и линкерных последовательностей относительно получения иммунного ответа после лечения. Дальнейший анализ проводили с применением технологии прогнозирования антигена iTope в силиконе (ANTITOPE). На основе этого анализа спрогнозировано, что SL-279252 будет иметь "низкий риск" иммуногенности, поскольку не было обнаружено идентифицируемых Т-клеточных эпитопов. Соответственно, ожидалось, что SL-279252 будет обладать низкой иммуногенностью.
Затем синтезировали кодон-оптимизированную последовательность ДНК SL-279252, которую направленно клонировали в экспрессионные векторы pcDNA3. 4-гидро-mcs (THERMO FISHER) и pVITRO2-гидро-mcs (INVIVOGEN). После этого векторы либо транзиентно, либо стабильно трансфицировали в клетки СНО-К1 и 293Т, а культуральные супернатанты очищали с помощью стандартной аффинной хроматографии на агарозе с белком А. Анализы ELISA для выявления Fc/IgG человека на элюированных фракциях (из экспериментов с устойчивой трансфекцией) очищенного белка демонстрировали четкие пики, которые совпадали с первым основным пиком, обнаруженным при крупномасштабной очистке и полученным в экспериментах с транзиентной трансфекцией (Фигура 13, панель А), что указывает на то, что эффективного продуцирования SL-279252 можно достичь с помощью обычных методов очистки белка, таких как методы с применением белка А.
Для подтверждения того, что все три домена SL-279252 являются интактными и распознаваемыми с помощью анализа обнаружения белка, проводили анализ вестерн- блоттинга на выявление анти-PD-1, анти-Fc и анти-OX40L человека на очищенном слитом белке (Фигура 13, панель В). Обнаружение SL-279252 проводили с применением всех трех антител, и когда эксперимент с белком осуществляли в восстановительных условиях, SL- 279252 мигрировал при около 75 кДа. Около 50% невосстановленного белка "перегонялось" в качестве димера, что было потенциальным преимуществом, учитывая олигомеризацию in vivo, ассоциированную с передачей сигналов и функцией OX40/L. Прогнозируемая молекулярная масса SL-279252 составляла 60,3 кДа. Восстановленную фракцию SL-279252 обнаруживали с более высокой молекулярной массой, что, отметим, не желая быть связанными какой-либо теорией, может быть обусловлено гликозилированием. Это подтверждалось обработкой SL-279252 дегликозилазой белка, PNGазой F (Фигура 13, панель В). После дегликозилирования восстановленная фракция SL-279252 мигрировала точно при спрогнозированной молекулярной массе 60,3 кДа. Это свидетельствовало о том, что SL-279252 был со/ посттрансляционно модифицирован с помощью гликозилирования, что сыграла важную роль для правильного фолдинга и стабильности белков, а также адгезии "клетка-к-клетке" (Dalziel М, Dwek RA. Science 2014, Maverakis Е, Lebrilla СВ. J Autoimmun. 2015).
Затем проводили анализ с целью определения способности SL-279252 связываться со своими рецепторными/лигандными мишенями с применением рекомбинантных белков, иммобилизованных на планшете, в функциональных анализах ELISA. SL-279252 успешно захватывался рекомбинантным ОХ40 человека (Фигура 13, панель С) и обнаруживался с помощью антитела анти-OX40L человека/антикозьего антитела, конъюгированного с пероксидазой хрена (HRP). В связи с этим, захват SL-279252 с помощью ОХ40 человека с последующим обнаружением посредством двухэтапной инкубации с козьим анти-OX40L, а затем - антикозьим антителом с HRP, обуславливал эффективное обнаружение. С целью определения способности обоих концов SL-279252 одновременно связываться со своим соответствующим рецептором/лигандом, был разработан другой анализ ELISA, который захватывает SL-279252 с помощью абсорбированного на планшете PD-L1 человека и обнаруживает SL-279252 с помощью рекомбинантного OX40-his (Фигура 13, панель D). Этот анализ демонстрирует, что SL-279252 может одновременно связывать PD-L1 человека и ОХ40 человека.
Затем проводили анализ поверхностного плазмонного резонанса (ППР) для определения аффинности, с которой SL-279252 связывался с hPD-L1, hPD-L1, hOX40 и различными Fc- рецепторами человека (Фигура 14). В частности, меченые полигистидином версии рекомбинантного PD-L1 человека, PD-L2 и ОХ40 человека связывали с чипами ProteOn HTG трис-NTA (BIORAD). Затем SL-279252 пропускали через связанные лиганды в течение определенного периода времени, и для расчета аффинности связывания (KD) SL-279252 с каждым партнером генерировали относительный индекс "скорости ассоциации" (Ka) и "скорости диссоциации"(Kd). Рекомбинантные PD-1-Fc и OX40L-Fc человека применяли в качестве положительного контроля связывания. Эти контрольные образцы имеют относительно быструю "скорость ассоциации" и такую же быструю "скорость диссоциации", что приводит к низкой наномолярной аффинности связывания. В соответствии с этими результатами "скорость ассоциации" SL-279252 с PD-L1 человека была высокой, однако "скорость диссоциации" была намного ниже, фактически ~ в 20 раз медленнее, чем "скорость диссоциации" рекомбинантного PD-1-Fc, что указывает на то, что SL-279252 быстро и стабильно связывается и длительное время пребывает в соответствующей мишени (Фигура 14, панель А). Установлено, что значение KD связывания SL-279252 с PD-L1 человека составляет 2,08 нМ, что почти идентично наблюдаемому значению KD для ОПДИВО (OPDIVO) от компании BMS (~ 4 нМ). Значение KD связывания SL-279252 с PD-L2 человека было определено как 1,24 нМ (Фигура 14, панель В). SL-279252 связывался с высокой аффинностью к ОХ40 человека (246 нМ), опять же, с высокой "скоростью ассоциации" и низкой "скоростью диссоциации" (Фигура 14, панель С).
Для дополнительного определения молекулярных характеристик SL-279252 проводили ППР, анализируя аффинности связывания SL-279252 со связанными с чипом Fcγ-рецепторами FcγR1A и неонатальным рецептором FcRn. Продемонстрировано, что иммуноглобулин IgG1 человека связывается с самой высокой аффинностью с FcγR1A, а затем с FcRn, в дополнение к связыванию низкой степени с FcγR2b (Фигура 14, панель С и D). SL-279252 не связывался с FcγR1A или FcγR2B, но связывался с FcRn с аффинностью 73 нМ (Фигура 14, панель D и Е). Не желая быть связанными какой-либо теорией, отметим, что эта характеристика связывания может быть важной для слитого белка, поскольку FcRn участвует в рециклинге IgG на поверхности клетки, при этом избегая расщепления лизосомами и потенциально увеличивая период полувыведения SL-279252 in vivo. Сводные данные относительно аффинности связывания SL-279252 описаны в данном документе (Фигура 14, панель F).
Затем проводили анализ поверхностного плазмонного резонанса (ППР) для определения аффинности, с помощью которой анализировали связывание мутированной конструкции SL-279252 с hPD-L1, hPD-L1, hOX40 и различными рецепторами Fc человека, при этом указанная конструкция содержит лидерный пептид коллагена V и мутации области Fc для увеличения связывания с FcRn (и названа colPD1-FcRnOX40L) (Фигура 14). В частности, меченые полигистидином версии рекомбинантного PD-L1 человека, PD-L2 и ОХ40 человека связывали с чипами ProteOn HTG трис-NTA (BIORAD). Затем colPD1-FcRnOX40L пропускали через связанные лиганды в течение определенного периода времени, и для расчета аффинности связывания (KD) colPD1-FcRnOX40L с каждым партнером генерировали относительный индекс "скорости ассоциации" (Ka) и "скорости диссоциации" (Kd). Рекомбинантные PD-1-Fc и OX40L-Fc человека применяли в качестве положительного контроля связывания. Эти контрольные образцы имеют относительно быструю "скорость ассоциации" и такую же быструю "скорость диссоциации", что приводит к низкой наномолярной аффинности связывания. В соответствии с этими результатами, "скорость ассоциации" colPD1-FcRnOX40L для PD-L1 человека была высокой, однако "скорость диссоциации" была намного ниже, фактически ~ в 10 раз медленнее, чем "скорость диссоциации" рекомбинантного PD-1-Fc, что указывает на то, что colPD1-FcRnOX40L быстро и стабильно связывается и длительное время пребывает в соответствующей мишени (Фигура 14, панель G). Установлено, что значение Ко связывания colPD1-FcRnOX40L с PD-L1 человека, составляет 6,35 нМ, что почти идентично наблюдаемому значению KD для ОПДИВО (OPDIVO) от компании BMS (~ 4 нМ). Значение KD связывания colPD1-FcRnOX40L с PD-L2 человека было определено как 7,93 нМ (Фигура 14, панель Н). colPD1-FcRnOX40L связывался с высокой аффинностью к ОХ40 человека (9,61 нМ), опять же, с высокой "скоростью ассоциации" и низкой "скоростью диссоциации" (Фигура 14, панель I).
Для дополнительного определения молекулярных характеристик colPD1-FcRnOX40L проводили ППР, анализируя аффинности связывания colPD1-FcRnOX40L со связанными с чипом Fcγ рецепторами FcγR1A и неонатальным рецептором FcRn. Продемонстрировано, что иммуноглобулин IgG1 человека связывается с самой высокой аффинностью с FcγR1A, а затем с FcRn, в дополнение к связыванию низкой степени с FcγR2b (Фигура 14, панель J и K). colPD1-FcRnOX40L не связывался с FcγR1A или FcγR2B, но связывался с FcRn с аффинностью 2,51 нМ (Фигура 14, панель K). Не желая быть связанными какой-либо теорией, отметим, что эта характеристика связывания может быть важной для слитого белка, поскольку FcRn участвует в рециклинге IgG на поверхности клетки, при этом избегая расщепления лизосомами и потенциально увеличивая период полувыведения colPD1-FcRnOX40L in vivo. Сводные данные относительно аффинности связывания colPD1-FcRnOX40L описаны в данном документе (Фигура 14, панель L).
Кроме того, период полужизни очищенного SL-279252 in vivo анализировали на мышах C57BL/6 после введения им 200 мкг белка с помощью внутрибрюшинной инъекции. Затем у животных, получивших указанный белок, собирали кровь путем пункции сердца через 10 минут, 30 минут, 1 час, 3, 6, 12 и 24 часа и оставляли сгущаться в течение 2 часов при комнатной температуре. Затем анализировали сыворотку с помощью анализа ELISA, специфичного для IgG или OX40L человека, как описано выше. Как проиллюстрировано на Фигуре 14, панель М, период полувыведения SL-279252 в сыворотке мышей после однократной инъекции 200 мкг белка определялся в пределах от 7 до 15 часов. Предполагается, что конструкции, содержащие мутации для усиления связывания с FcRn, приведут к удлинению периода полувыведения in vivo.
Медленные скорости диссоциации, обнаруженные с помощью ППР, дают основания предположить, что SL-279252 может иметь более длительный период полувыведения из целевой области (то есть, из опухоли), по сравнению с периодом полувыведения из сыворотки. Чтобы исследовать этот вопрос, иммунокомпрометированным мышам NSG в один фланк имплантировали PD-L1-отрицательную опухоль HeLa (рак шейки матки человека), а в противоположный фланк имплантировали PD-L1-экспрессирующую опухоль HeLa. Мышам однократно вводили инъекции 200 мкг SL-279252, а отдельных мышей умерщвляли в определенные моменты времени. После умерщвления обе опухоли HeLa вырезали и рассекали на две части. Половину опухоли разделяли и анализировали на связывание SL-279252 с помощью проточной цитометрии. Этот анализ продемонстрировал, что SL-279252 специфически накапливается в PD-L1-позитивных, но не в PD-L1-негативных опухолях. Повышение концентрации SL-279252 в опухоли наблюдали в течение 48 часов после введения (Фигура 14, панель N). Кроме того, иммуногистохимический анализ другой половины каждой опухоли продемонстрировал, что существенное окрашивание OX40L человека определялось в течение 5 дней после лечения, наталкивая на предположение о том, что SL-279252 может обнаруживается в PD-L1-позитивных опухолях человека по меньшей мере в течение 5 дней после одного введения (Фигура 14, панель О).
Пример 4. Дополнительная функциональная характеристика PD-1-Fc-OX40L человека
Предыдущие данные продемонстрировали, что SL-279252 связывается с иммобилизованными мишенями с низкой наномолярной аффинностью и может быть обнаружен с помощью множества анализов с применением белка. С целью определения способности SL-279252 связывать свои мишени на поверхности живых клеток in vitro был проведен дополнительный анализ. Для оценки связывания SL-279252 с рецептором ОХ40 человека, сконструировали линию Т-клеток ОМЛ Jurkat человека для сверхэкспрессии ОХ40, создавая клетки Jurkat/hOX40 (верифицированные с помощью проточной цитометрии, Фигура 15, панель А). Для оценки связывания с PD-L1, линию клеток яичника китайского хомячка СНО-K1, которая не экспрессирует PD-L1 человека, трансфицировали для стабильной экспрессии PD-L1 человека (Фигура 15, панель В). Для оценки связывания с CD47 человека, клетки СНО-K1 трансфицировали для стабильной экспрессии CD47 человека (Фигура 15, панель С).
Клетки СНО-K1 или CHO-K1-PD-L1 затем обрабатывали повышающимся количеством SL-279252 и анализировали с помощью проточной цитометрии для определения домена OX40L человека с применением антител против OX40L-APC человека. SL-279252 не связывался с родительскими клетками СНО-K1, поскольку они не экспрессировали обнаруживаемые уровни PD-L1 человека. Однако почти вся популяция клеток СНО-K1-PD-L1 значительно изменилась, что указывает на то, что компонент PD1 человека в составе SL-279252 способен связывать свой рецептор на поверхности живых клеток (Фигура 15, панель D). Клетки Jurkat или Jurkat/OX40 затем обрабатывали повышающимся количеством SL-279252 и анализировали с помощью проточной цитометрии для определения домена OX40L человека с применением антител против OX40L-APC человека. SL-279252 не связывался с родительскими клетками Jurkat, поскольку они экспрессировали низкие уровни уровни ОХ40 человека. Однако почти вся популяция клеток Jurkat/OX40 значительно изменилась, что указывает на то, что компонент OX40L человека в составе SL-279252 способен связывать свой рецептор на поверхности живых клеток (Фигура 15, панель Е).
С целью исследования связывания другого химерного слитого белка, клетки CD172a-Fc-OX40L человека, СНО-K1 или CHO-K1-CD47 человека затем обрабатывали повышающимся количеством CD172a-Fc-OX40L и анализировали с помощью проточной цитометрии для определения домена OX40L человека с применением антител против OX40L-APC человека. CD172a-Fc-OX40L не связывался с родительскими клетками СНО-K1, поскольку они не экспрессировали обнаруживаемые уровни CD47 человека. Однако почти вся популяция клеток CHO-K1-PD-L1 значительно изменилась, что указывает на то, что компонент CD 172а человека в составе CD172a-Fc-OX40L способен связывать свой рецептор на поверхности живых клеток (Фигура 15, панель F). Клетки Jurkat или Jurkat/OX40 затем обрабатывали повышающимся количеством CD172a-Fc-OX40L и анализировали с помощью проточной цитометрии для определения домена OX40L человека с применением антител против OX40L-APC человека. CD172a-Fc-OX40L не связывался с родительскими клетками Jurkat, поскольку они экспрессировали низкие уровни ОХ40 человека. Однако почти вся популяция клеток Jurkat/OX40 значительно изменилась, что указывает на то, что компонент OX40L человека в составе CD172a-Fc-OX40L способен связывать свой рецептор на поверхности живых клеток (Фигура 15, панель G).
Кроме того, ряд линий опухолевых клеток человека подвергали скринингу на различные уровни эндогенной экспрессии PD-L1 человека с помощью проточной цитометрии. Идентифицировали линию клеток рака простаты (PC3) в виде PD-L1низк и линию клеток аденокарциномы легкого (НСС827) в виде PD-L1выс (Фигура 15, панель Н). Клетки РС3 и НСС827 инкубировали с повышающимся количеством SL-279252, а связывание обнаруживали с помощью проточной цитометрии. SL-279252 эффективно не связывался с клетками РС3 (PD-L1низк) (Фигура 15, панель I). Однако SL-279252 существенно связывался с клетками НСС827 (PD-L1выс) зависимым от концентрации образом (Фигура 15, панель J). Эти данные четко продемонстрировали, что SL-279252 может связываться как с ОХ40, так и с PD-L1 человека, экспрессирумыми на поверхности клетки, что дает убедительные доказательства функциональности SL-279252 относительно способности к двойному связыванию.
С целью подробного объяснения полученных результатов, были проведены эксперименты связывания SL-279252 с первичными Т-клетками, выделенными из мононуклеарных клеток периферической крови (МКПК), индуцированных в течение 2 дней ex vivo химической комбинацией, известной своей стимуляцией экспрессии OX40 (форбол 12-миристат 13-ацетат; РМА, фитогемагглютинин, РНА и лономицин). Как и ожидалось, после введения РМА/РНА/лон наблюдали значительное увеличение экспрессии OX40 на CD4+ и CD8+ Т-клетках (Фигура 16, панель А). Связывание SL-279252 с клетками CD4+ и CD8+ подтверждали с помощью способов, описанных выше (Фигура 16, панель В). Отмечено, что SL-279252 эффективно связывался с Т-клетками человека (как CD4+, так и CD8+).
Анализ активации Т-клеток/высвобождения IL2 применяли для оценки степени экспрессии PD-L1 на опухолевых клетках, ингибируемой секрецией Т-клетками противоопухолевого цитокина IL2, при совместном культивировании клеток (Фигура 16, панель С). Через 2 дня активированные Т-клетки высевали на облученные линии клеток рака, экспрессирующие PD-L1низ (PC3) и PD-L1выс (HCC827), в присутствии или отсутствии SL-279252. Различные данные активации Т-клеток оценивали в течение 1 недели после первичного выделения Т-клеток, включая секрецию IL2 (Фигура 16, панель D), пролиферации и экспрессию цитокинов (Фигура 16, панель Е). Исходные уровни секреции IL2 (в отсутствие SL-279252) были значительно выше в совместных культурах PD-L1низк PC3, чем с клетками PD-L1выс HCC827 через 6 дней после выделения Т-клеток, что указывает на то, что экспрессия опухоли PD-L1 прямо или косвенно подавляла дальнейшую активацию Т-клеток, что определялось секрецией IL2 (Фигура 16, панель D). Добавление SL-279252 к обоим совместным культурам PC3 и HCC827 повышало секрецию IL2 зависимым от концентрации образом. В частности, наблюдаемое увеличение IL-2 при совместном культивировании HCC827 (PD-L1выс) от исходного уровня (без SL-279252) до 5 мкг/мл SL-279252 составляло 1,92 раза по сравнению с 1,27 раза при совместном культивировании с клетками PC3 (PD-L1низк).
Кроме того, были проанализированы дополнительные характеристики активации Т-клеток, включая экспрессию маркера пролиферации Ki67 (Фигура 16, панель Е; верхняя часть). Совместная культура активированных Т-клеток с PD-L1выс HCC827 клетками ингибировала пролиферацию по сравнению с уровнем, наблюдаемым в отсутствие клеток HCC827 (черная линия). Добавление SL-279252 к совместной культуре усиливало окрашивание Ki67 как в CD4+, так и в CD8+ Т-клетках. Более того, активированные Т-клетки экспрессировали более высокие уровни цитокинов IFNγ и TNFα при совместном культивировании на клетках HCC827, чем при культивировании Т-клеток отдельно, возможно, из-за секреции других стимулирующих факторов опухолевыми клетками (Фигура 16, панель Е, снизу). Экспрессия этих цитокинов значительно повышалась после введения SL-279252.
В совокупности эти данные демонстрируют, среди прочего, что SL-279252 является сильно связанным со своими партнерами по связыванию PD-L1 и OX40 и способен отменить PD-L1-опосредованное ингибирование Т-клеток PD-L1-позитивными опухолевыми клетками человека in vitro.
Пример 5. Конструкция и характеристика дополнительных химерных белков
Были созданы дополнительные конструкции, которые включают: дополнительные конструкции PD-1-Fc-OX40L человека, а также hCD172a-Fc-OX40L человека, hPD1-Fc-TL1A, hBTLA-Fc-OX40L, hTMIGD2-Fc-OX40L, hTIM3-Fc-OX40L, mPD1-Fc-GITRL, mPD1-Fc-41BBL, mPD1-Fc-TL1A, mCD172a-Fc-CD40L, hTIGIT-Fc-OX40L и PD-1-Fc-OX40L собаки. Каждую из этих конструкций оптимизировали кодоном с целью экспрессии в клетках яичника китайского хомячка (СНО), трансфицировали в клетки СНО, и выбирали отдельные клоны с высоким уровнем экспрессии. Клоны с высоким уровнем экспрессии затем применяли для мелкомасштабного изготовления в перемешиваемых биореакторах в бессывороточной среде, а соответствующие химерные слитые белки очищали с применением колонок со смолой, связывающей белок А. На Фигуре 17, панель А, проиллюстрирован результат анализа вестерн-блоттинга различных химерных белков, включая hCD172a-Fc-OX40L, hPD1-Fc-TL1A, hBTLA-Fc-OX40L, hTMIGD2-Fc-OX40L, hTIM3-Fc-OX40L, mPD1-Fc-GITRL, mPD1-Fc-4-1BBL, mPD1-Fc-TL1A, mCD172a-Fc-CD40L, hTIGIT-Fc-OX40L.
Анализы связывания проводили для характеристики способности различных конструкций ECD-Fc-OX40L человека связываться с hOX40. Что касается hXECD-Fc-OX40L, то Х относится к ECD каждого белка, перечисленному в фигурной скобке слева (со ссылкой на Фигуру 17, панель В). На панели В Фигуры 17 приведено схематическое изображение метода ELISA, применяемого для обнаружения связывания hXECD-Fc-OX40L с hOX40. Рекомбинантный hOX40, слитый с Fc (hOX40-hFc) человека, применяли для захвата hXECD-Fc-OX40L в культуральной среде. Поскольку слитый белок hOX40, применяемый для захвата целевых слитых белков, также содержит область hIgG, блокирование проводили с применением антитела анти-hIgG, неконъюгированного с HRP, перед инкубацией с супернатантами культуры, содержащими целевые слитые белки. Поликлональное антитело кролика к hIgG применяли для определения домена hIgG в химерном белке, с последующим определением с применением конъюгированного с пероксидазой хрена (HRP) поликлонального антитела к IgG (Н+L) кролика.
Связывание SL-279252 с поверхностью клеток, экспрессируемых OX40 на клетках Jurkat, определяемое с помощью проточной цитометрии, сравнивали с двумя отрицательными контрольными белками, которые, как ожидается, не связываются с OX40 человека. Эти данные демонстрируют, что SL-279252 эффективно связывается с OX40 человека (левая панель), в то время как связывания ни pd1-fc-tl1a человека, ни PD1-Fc-OX40L собаки, с OX40 человека не наблюдали (Фигура 17, панель С).
Конструкцию CD172a-Fc-OX40L человека импортировали в программное обеспечение RaptorX для прогнозирования третичной структуры белка с целью определения указанной третичной структуры. Спрогнозированная третичная структура приведена на Фигуре 17, панель D.
Затем синтезировали кодон-оптимизированную последовательность ДНК нескольких химерных слитых белков, которую направленно клонировали в pVITRO2, pcDNA3. 4 и другие экспрессионные векторы. После этого векторы либо транзиентно, либо стабильно трансфицировали в клетки СНО и 293, и выбирали отдельные клоны с высоким уровнем экспрессии. Например, SL-279252 получали в результате транзиентного трансфицирования из клеток 293, очищали с помощью аффинной хроматографии с применением колонок с белком А и оценивали с помощью окрашивания красителем Кумасси, вестерн-блоттинга, а также проводили количественную оценку по сравнению со стандартом BCG (Фигура 17, панель Е).
В другом примере, CD172a-Fc-OX40L получали в результате транзиентного трансфицирования из клеток 293, очищали с помощью аффинной хроматографии с применением колонок с белком А и оценивали с помощью окрашивания красителем Кумасси, вестерн-блоттинга, а также проводили количественную оценку по сравнению со стандартом BCG (Фигура 17, панель F). В другом примере, CD172a-Fc-CD40L получали в результате транзиентного трансфицирования из клеток 293, очищали с помощью аффинной хроматографии с применением колонок с белком А и оценивали с помощью системы Perkin Elmer LabChip, а также проводили количественную оценку по сравнению со стандартом BCG (Фигура 17, панель G). В другом примере, TIGIT-Fc-OX40L человека получали в результате транзиентного трансфицирования из клеток 293, очищали с помощью аффинной хроматографии с применением колонок с белком А и оценивали с помощью окрашивания красителем Кумасси, вестерн-блоттинга, а также проводили количественную оценку по сравнению со стандартом BCG (Фигура 17, панель Н).
Аффинность связывания CD172a-Fc-OX40L человека с hCD47, hOX40 и различными Fc-рецепторами человека оценивали с помощью анализа поверхностного плазменного резонанса (ППР) (Фигура 17, панели I-М). В частности, меченые полигистидином версии рекомбинантного CD47 человека и OX40 человека связывали с чипами ProteOn HTG трис-NTA (BIORAD). Затем CD172a-Fc-OX40L пропускали через связанные лиганды в течение определенного периода времени, и для расчета аффинности связывания (KD) CD172a-Fc-OX40L с каждым партнером генерировали относительный индекс "скорости ассоциации" (Ka) и "скорости диссоциации"(Kd). Рекомбинантные CD47-Fc и OX40L-Fc человека применяли в качестве положительного контроля связывания. Эти контрольные образцы имеют относительно быструю "скорость ассоциации" и такую же быструю "скорость диссоциации", что приводит к низкой наномолярной аффинности связывания. В соответствии с этими результатами "скорость ассоциации" CD172a-Fc-OX40L с CD47 человека была высокой, однако "скорость диссоциации" была намного ниже, фактически ~ в 40 раз медленнее, чем "скорость диссоциации" рекомбинантного CD47-Fc, что указывает на то, что CD172a-Fc-OX40L быстро и стабильно связывается и длительное время пребывает в соответствующей мишени (Фигура 17, панель I). Значение KD связывания CD172a-Fc-OX40L с CD47 человека было определено как 3,59 нМ. CD172a-Fc-OX40L связывался с высокой аффинностью к OX40 человека (869 нМ), опять же, с высокой "скоростью ассоциации" и низкой "скоростью диссоциации" (Фигура 17, панель J).
Для дополнительного определения молекулярных характеристик CD172a-Fc-OX40L проводили ППР, анализируя аффинности связывания CD172a-Fc-OX40L со связанными с чипом Fcγ-рецепторами FcγR1A и неонатальным рецептором FcRn. Продемонстрировано, что иммуноглобулин IgG1 человека связывается с самой высокой аффинностью с FcγR1A, а затем с FcRn, в дополнение к связыванию низкой степени с FcγR2b (Фигура 17, панель K и L). CD172a-Fc-OX40L не связывался с FcγR1A или FcγR2B, но связывался с FcRn с аффинностью 790 нМ (Фигура 17, панель L). Не желая быть связанными какой-либо теорией, отметим, что эта характеристика связывания может быть важной для слитого белка, поскольку FcRn участвует в рециклинге IgG на поверхности клетки, при этом избегая расщепления лизосомами и потенциально увеличивая период полувыведения CD172a-Fc-OX40L in vivo. Сводные данные относительно аффинности связывания CD172a-Fc-OX40L описаны в данном документе (Фигура 17, панель М).
Затем синтезировали кодон-оптимизированную последовательность ДНК нескольких дополнительных химерных слитых белков, которую направленно клонировали в pVITRO2, pcDNA3. 4 и другие экспрессионные векторы. После этого векторы либо транзиентно, либо стабильно трансфицировали в клетки СНО и 293, и выбирали отдельные клоны с высоким уровнем экспрессии. Например, PD1-Fc-OX40L собаки получали в результате транзиентного трансфицирования из клеток 293, очищали с помощью аффинной хроматографии с применением колонок с белком А и оценивали с помощью окрашивания красителем Кумасси, вестерн-блоттинга, а также проводили количественную оценку по сравнению со стандартом BCG (Фигура 17, панель N). В другом примере, PD1-Fc-OX40L мыши получали в результате транзиентного трансфицирования из клеток 293, очищали с помощью аффинной хроматографии с применением колонок с белком А и оценивали с помощью окрашивания красителем Кумасси, вестерн-блоттинга, а также проводили количественную оценку по сравнению со стандартом BCG (Фигура 17, панель О). В другом примере, PD1-Fc-GITRL мыши получали в результате транзиентного трансфицирования из клеток 293, очищали с помощью аффинной хроматографии с применением колонок с белком А и оценивали с помощью окрашивания красителем Кумасси, вестерн-блоттинга, а также проводили количественную оценку по сравнению со стандартом BCG (Фигура 17, панель Р). В другом примере, PD1-Fc-41BBL мыши получали в результате транзиентного трансфицирования из клеток 293, очищали с помощью аффинной хроматографии с применением колонок с белком А и оценивали с помощью окрашивания красителем Кумасси, вестерн-блоттинга, а также проводили количественную оценку по сравнению со стандартом BCG (Фигура 17, панель Q). В другом примере, pdi-fc-tl1a мыши получали в результате транзиентного трансфицирования из клеток 293, очищали с помощью аффинной хроматографии с применением колонок с белком А и оценивали с помощью окрашивания красителем Кумасси, вестерн-блоттинга, а также проводили количественную оценку по сравнению со стандартом BCG (Фигура 17, панель R). В еще одном примере, CD115-Fc-CD40L получали в результате транзиентного трансфицирования из клеток 293, очищали с помощью аффинной хроматографии с применением колонок с белком А и оценивали с помощью окрашивания красителем Кумасси, вестерн-блоттинга, а также проводили количественную оценку по сравнению со стандартом BCG (Фигура 17, панель S).
Каждый очищенный белок характеризовали с помощью анализов ELISA с целью связывания с маркером, например, предполагаемым ингибирующим лигандом, а также предполагаемым костимулирующим рецептором. Например, для анализа связывания очищенного PD-1-Fc-OX40L человека, рекомбинантный PD-L1-Fc адсорбировали на микротитровальных планшетах и применяли для захвата PD-1-Fc-OX40L. Любой связанный PD-1-Fc-OX40L затем обнаруживали с применением рекомбинантного OX40-Fc человека, связанного с биотином, который в последствии обнаруживали в хромогенном анализе путем связывания со стрептравидином-HRP.
Кроме того, каждый очищенный белок характеризовали с помощью проточной цитометрии с целью связывания с предполагаемым ингибирующим лигандом, а также предполагаемым костимулирующим рецептором. Например, линии опухолевых клеток человека характеризовали на предмет эндогенной экспрессии PD-L1, уровень которой оказался особенно высоким на нескольких линиях опухолевых клеток меланомы человека. Продемонстрировано, что эти же линии опухолевых клеток являлись отрицательными относительно OX40L человека. После инкубации с PD-1-Fc-OX40L любой связанный химерный слитый белок обнаруживали с применением специфических антител OX40L человека. Аналогично, клетки Jurkat человека трансфицировали OX40 человека, при этом было продемонстрировано, что они оказались отрицательными относительно PD-L1 человека. После инкубации с химерными конструкциями PD-1-Fc-OX40L любой связанный комплекс обнаруживали с применением специфичных антител против PD-L1 человека. Создали серию скрининговых линий клеток с целью обнаружения специфического связывания клеточной поверхности каждого химерного слитого белка с его соответствующим рецептором/лигандом, в том числе: CHO-K1-CD47, CHO-K1-PD-L1, CHO-K1-HVEM, CHO-K1-HHLA2, CHO-K1-VISTA, CHO-K1-Gal9, HeLa-PD-L1, HeLa-CD47, HeLa-HVEM, HeLa-HHLA2, HeLa-VISTA, HeLa-Gal9.
Для определения функциональной активности каждого рецептора анализы пролиферации Т-клеток in vitro проводили в присутствии опухолевых клеток человека с наличием ингибирующего лиганда. Например, в опухолевые клетки меланомы человека, экспрессирующие PD-L1, вводили пептиды, специфичные для лизоцима куриного яйца (HEL), и инкубировали с HEL-специфическими Т-клетками человека, экспрессирующими рецептор OX40. Пролиферацию этих клеток контролировали в присутствии и в отсутствие конструкции PD-1-Fc-OX40L, при этом было обнаружено, что уровень пролиферации функционально зависел от присутствия химерных конструкций. В подобной системе применяют опухоли человека, экспрессирующие HVEM, CD47, галектин-9, рецепторы TIGIT или рецепторы TMIGD2.
В некоторых экспериментах PD-1-Fc-OX40L мыши или PD-1-Fc-TL1A мыши применяли для лечения опухолей мыши, о которых известно, что они являлись позитивными относительно PD-L1 мыши (включая меланому B16-F10, карциному толстой кишки МС38 и карциному толстой кишки СТ26). В этих системах прижившиеся опухоли обрабатывали очищенными химерными слитыми белками по сравнению со слитыми белками PD-1-Fc, моноклинальными антителами анти-PD-1 или анти-PD-L1 или моноклональными антителами анти-ОХ40 или анти-GITR. Как продемонстрировано в этих экспериментах, активность химерных конструкций приводит к усилению антигенспецифических Т-клеточных реакций и увеличению скорости отторжения опухоли по сравнению с отдельно применяемыми терапевтическими средствами. В некоторых экспериментах конструкции нуклеиновых кислот, кодирующие PD-1-Fc-OX40L или PD-1-fc-tl1a, непосредственно электропорировали в прижившиеся опухоли. Продемонстрировано, что в этих экспериментах химерные конструкции приводят к увеличению скорости отторжения опухоли, а также к увеличению пролиферации CD8+ Т-клеток, специфической для опухолевых антигенов и обнаруживаемой как в периферической крови, так и в пределах прижившихся опухолей.
С целью определения связывания очищенных химерных слитых белков с эксплантатами опухоли человека, получали свежезамороженные образцы опухоли человека и инкубировали их с каждым химерным слитым белком. Любой связанный слитый белок обнаруживали с помощью антитела против OX40L человека и контролировали при фоновом окрашивании путем отдельного окрашивания с помощью антитела против OX40L человека.
Для определения молекулярных характеристик каждого слитого белка очищенные химерные слитые белки характеризовали с помощью эксклюзионной хроматографии. Этот анализ был значимым, поскольку, например, известно, что OX40L ECD образует гомотример, в то время как область Fc, как известно, образует гомодимер, и в то время как рецептор, связывающийся с ингибирующим лигандом, может быть либо мономерным (например, PD-1) или образует гомомультимеры (например, TIM3). Таким образом, существует несколько возможных вариантов для отдельных видов, которые могут быть образованы этими химерными конструкциями. Также определяли дополнительные молекулярные характеристики относительно масс-спектрограммы, термостабильности, стабильности рН, физической стабильности, профиля заряда, гидрофобности, физической стабильности, буферной совместимости и растворимости до 100 мг/мл.
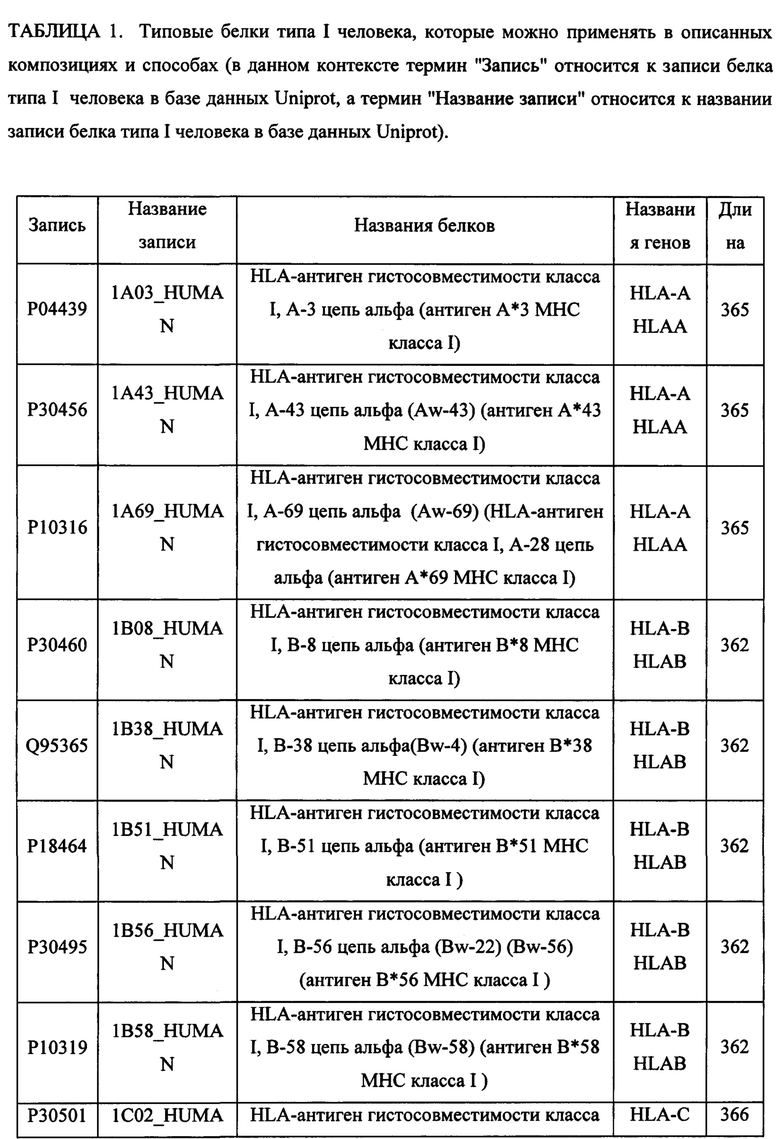
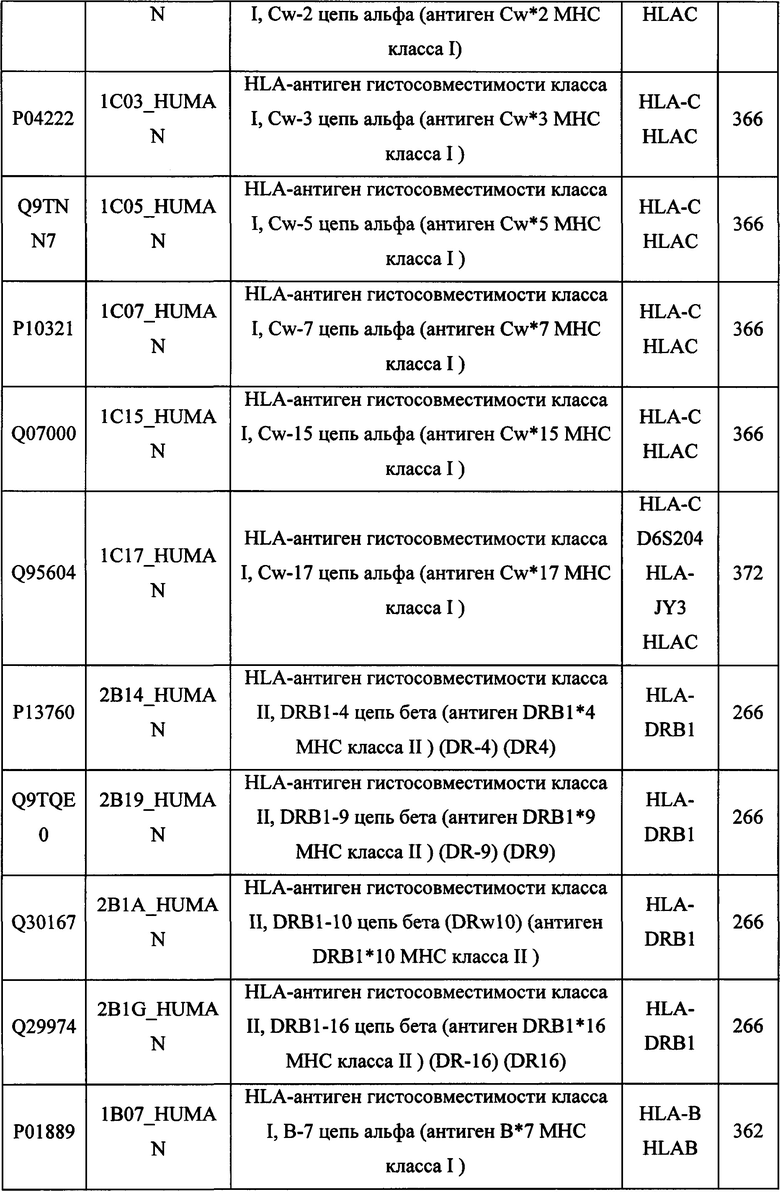
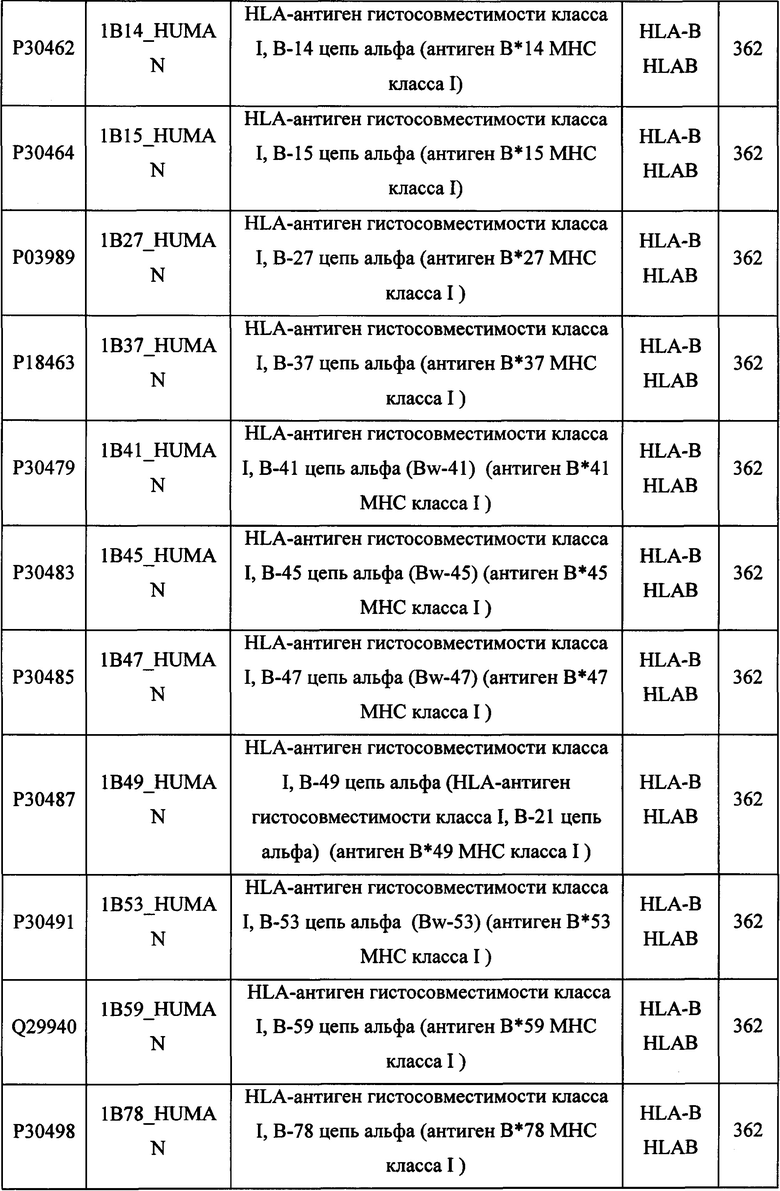
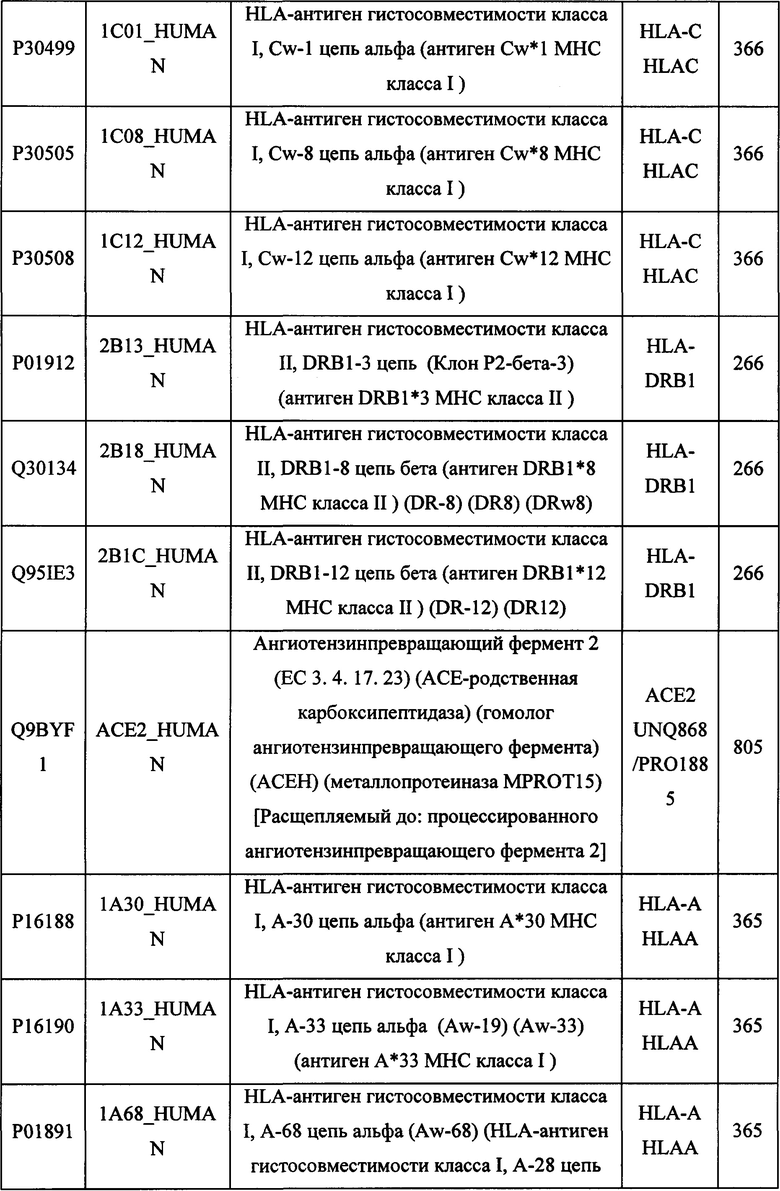

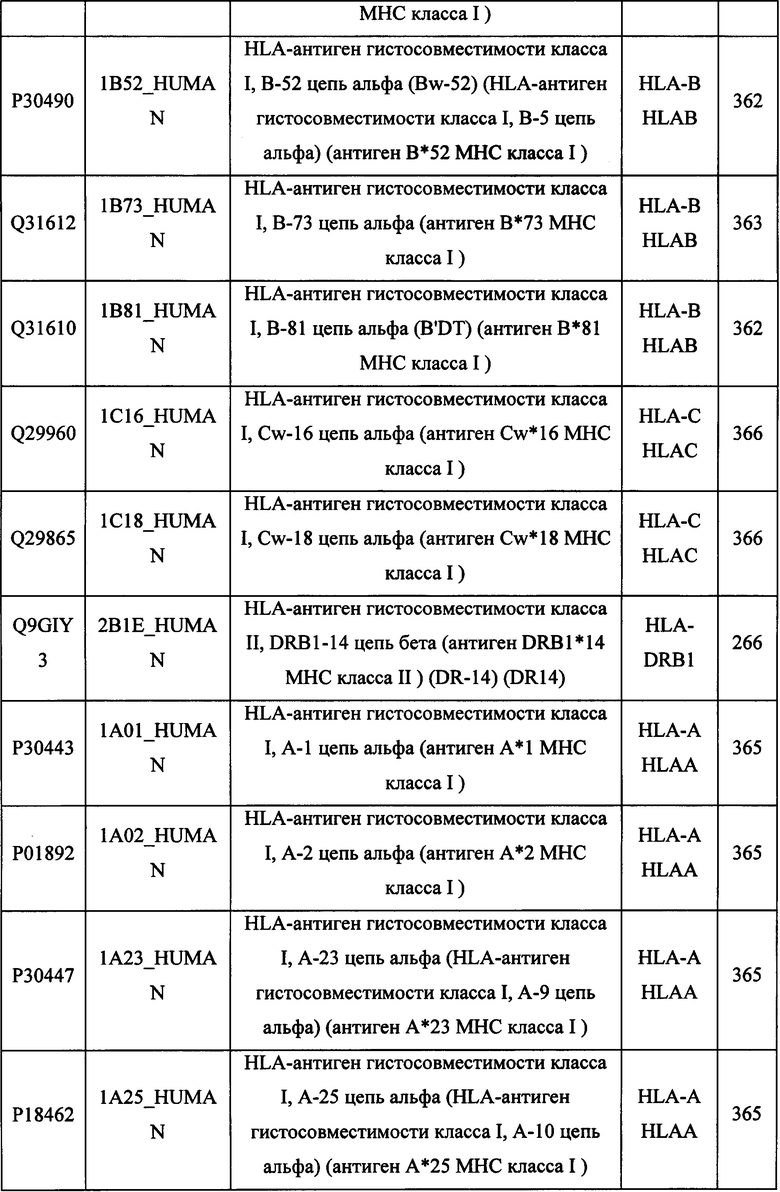
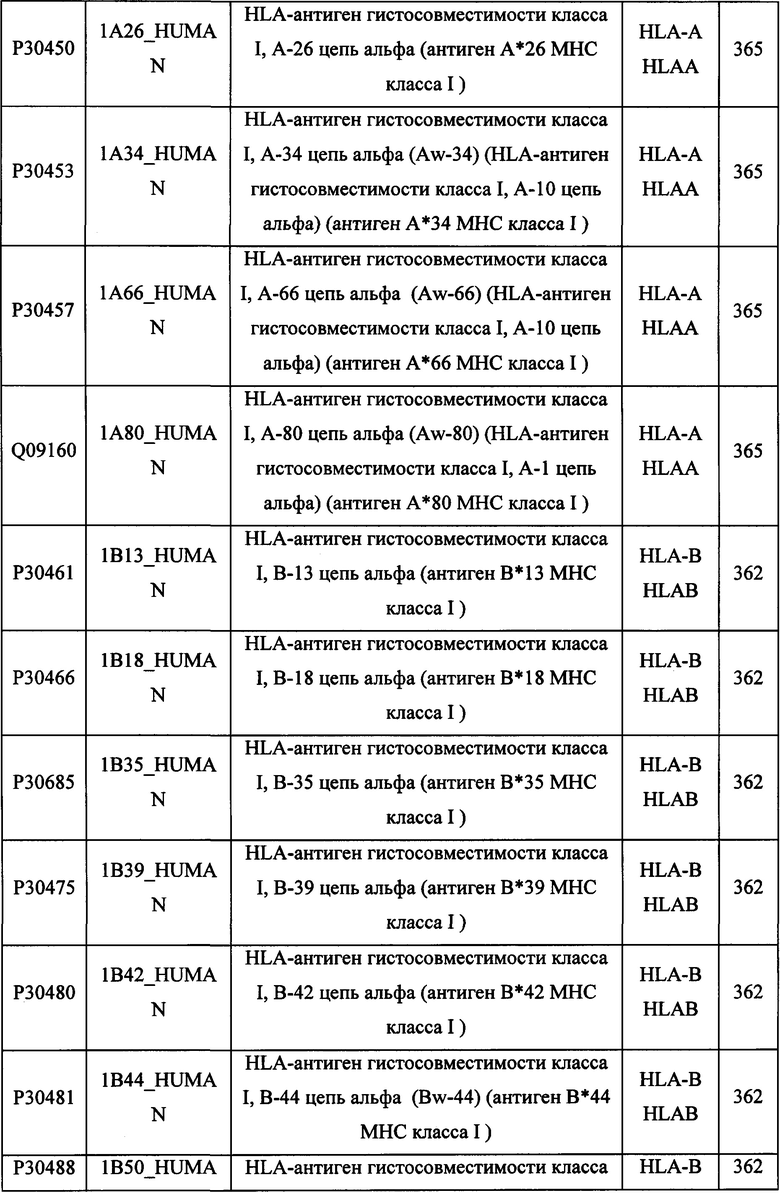
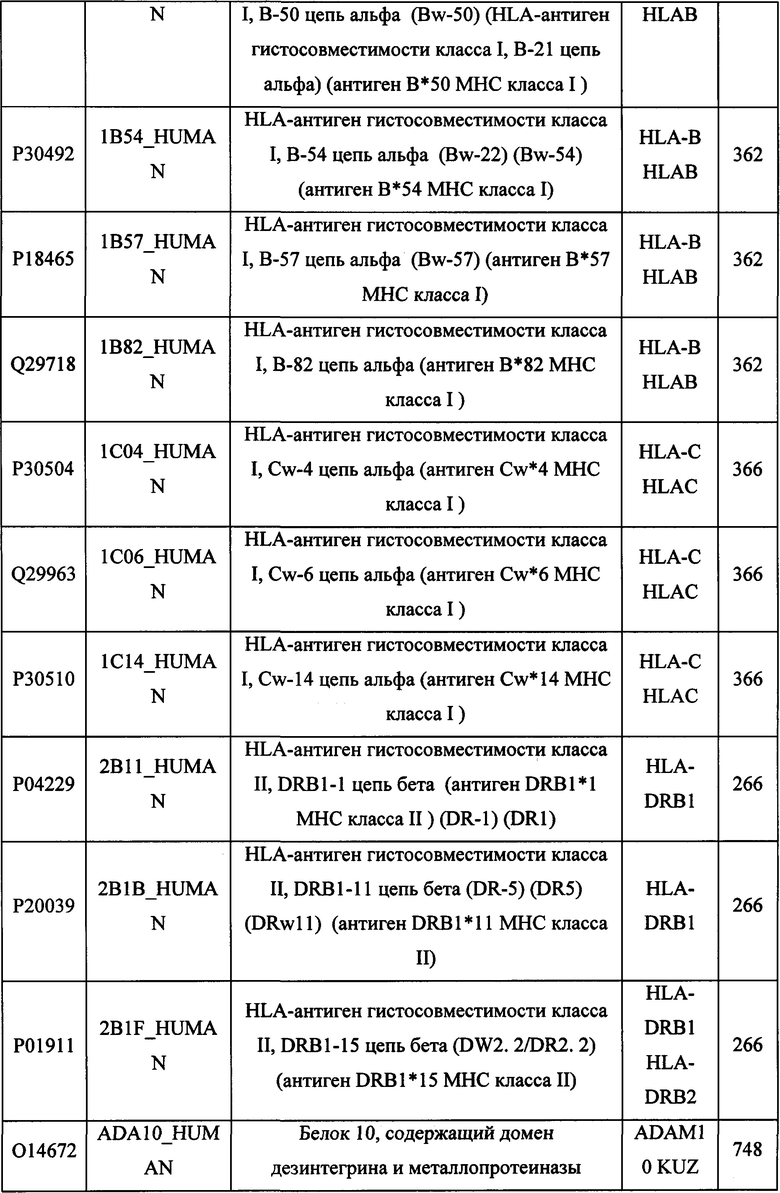

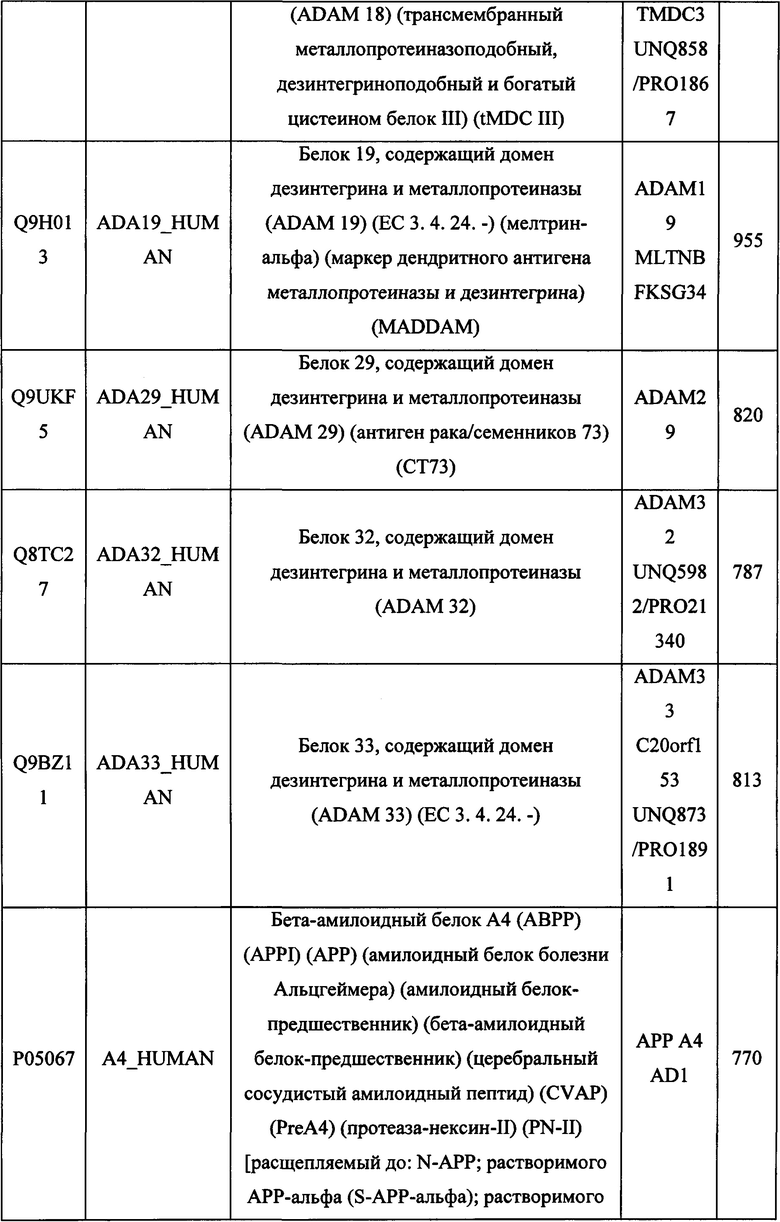
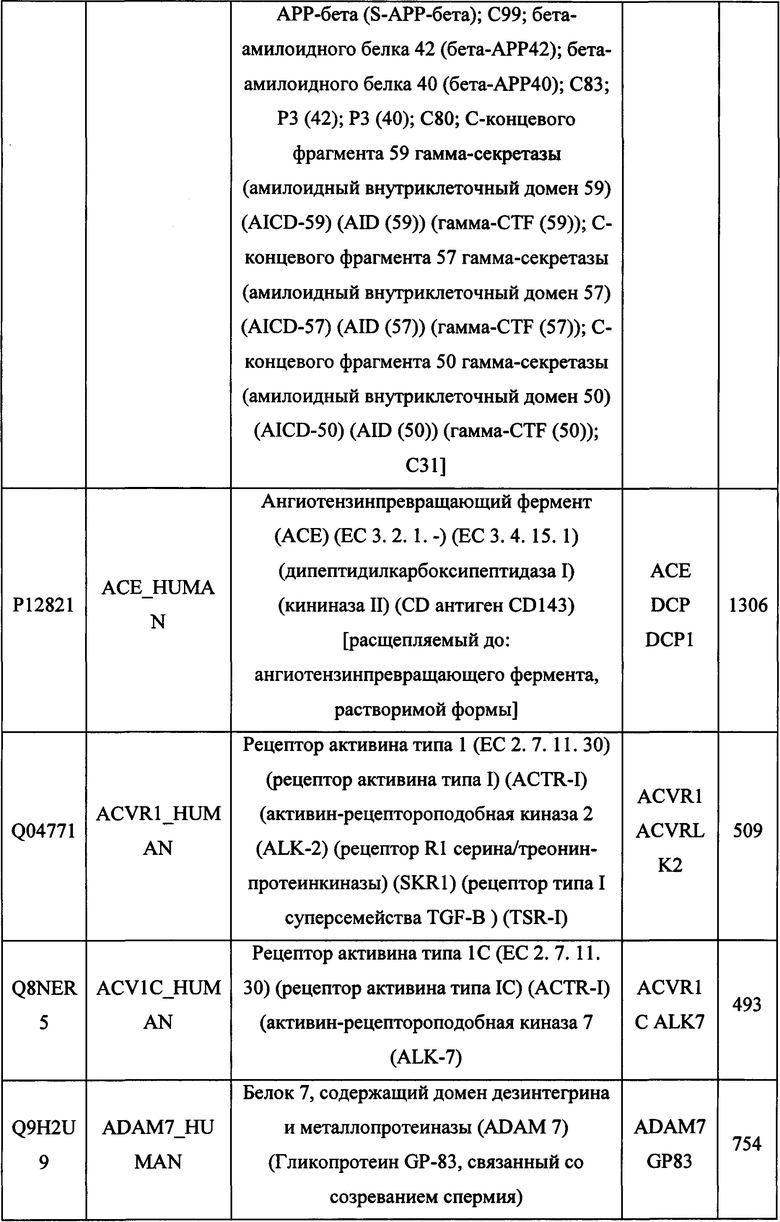

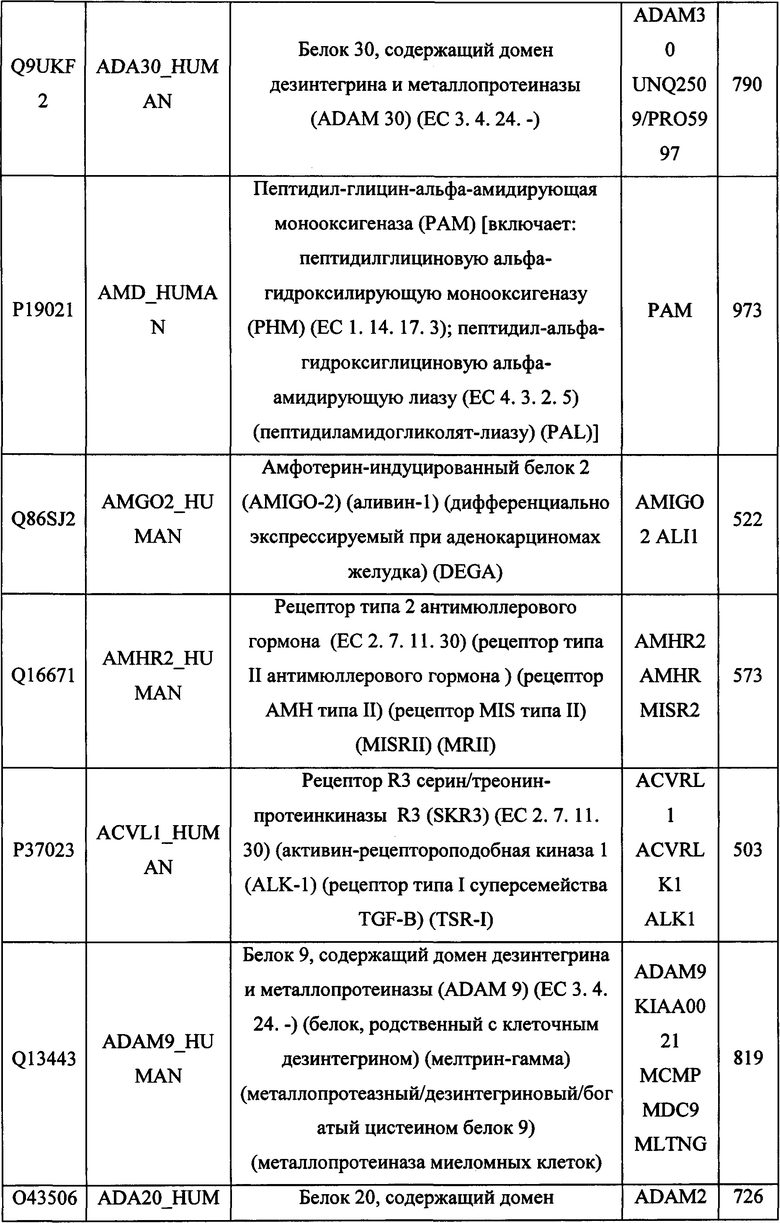
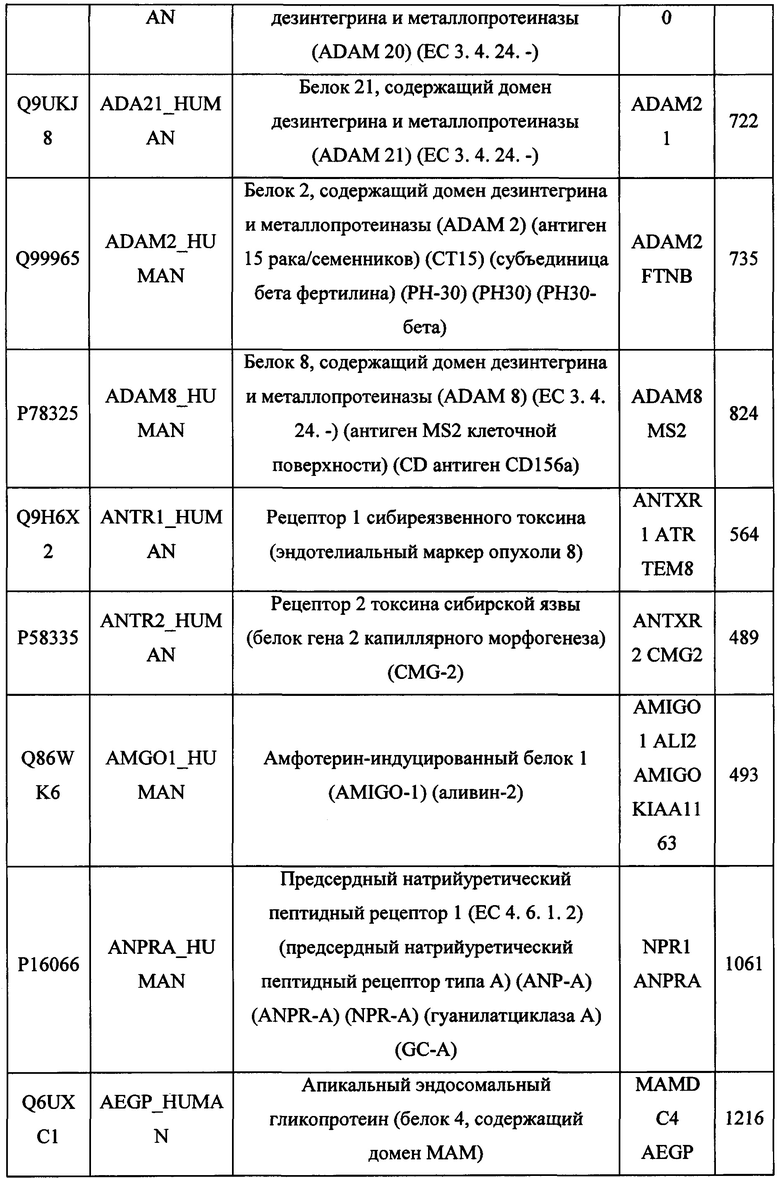
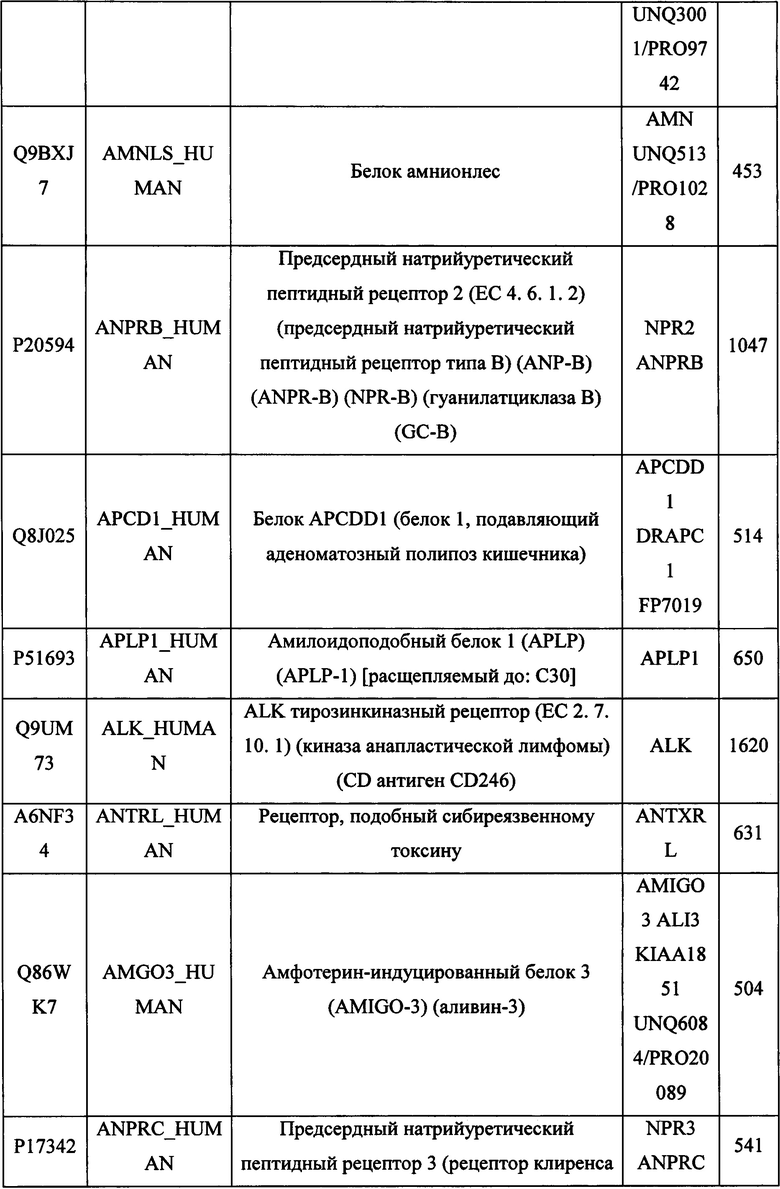
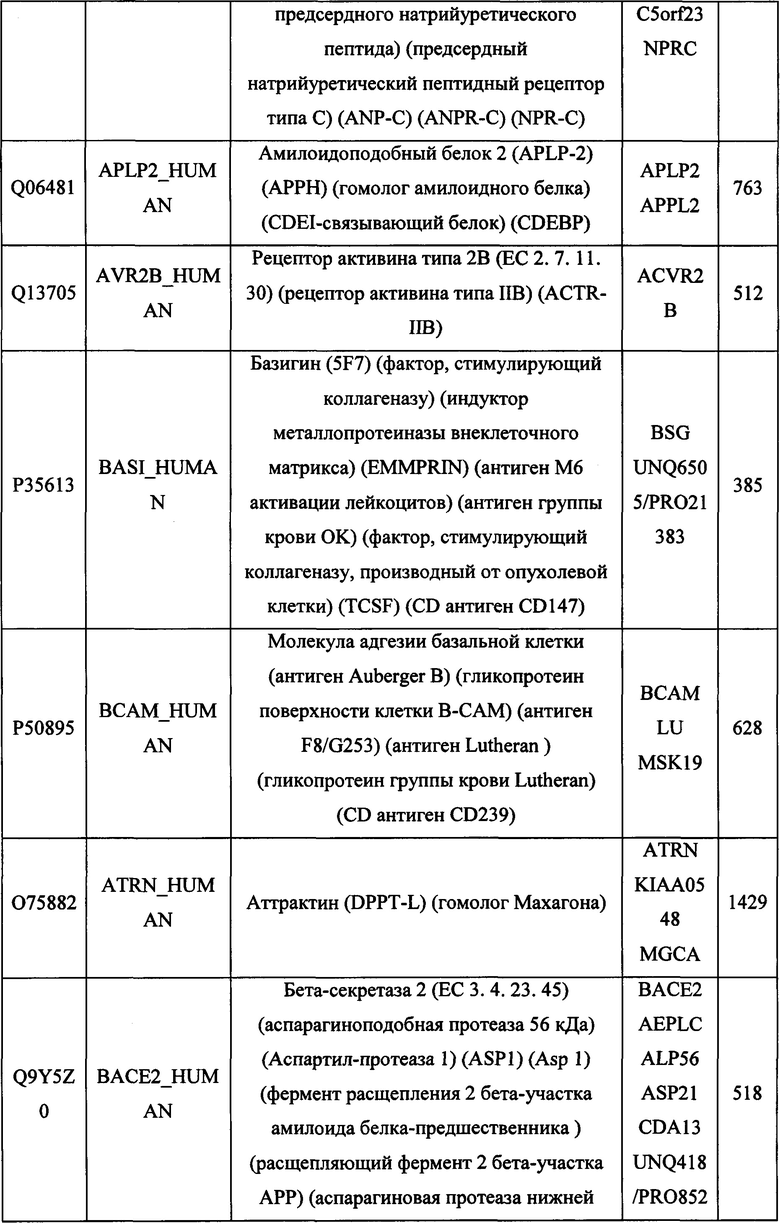
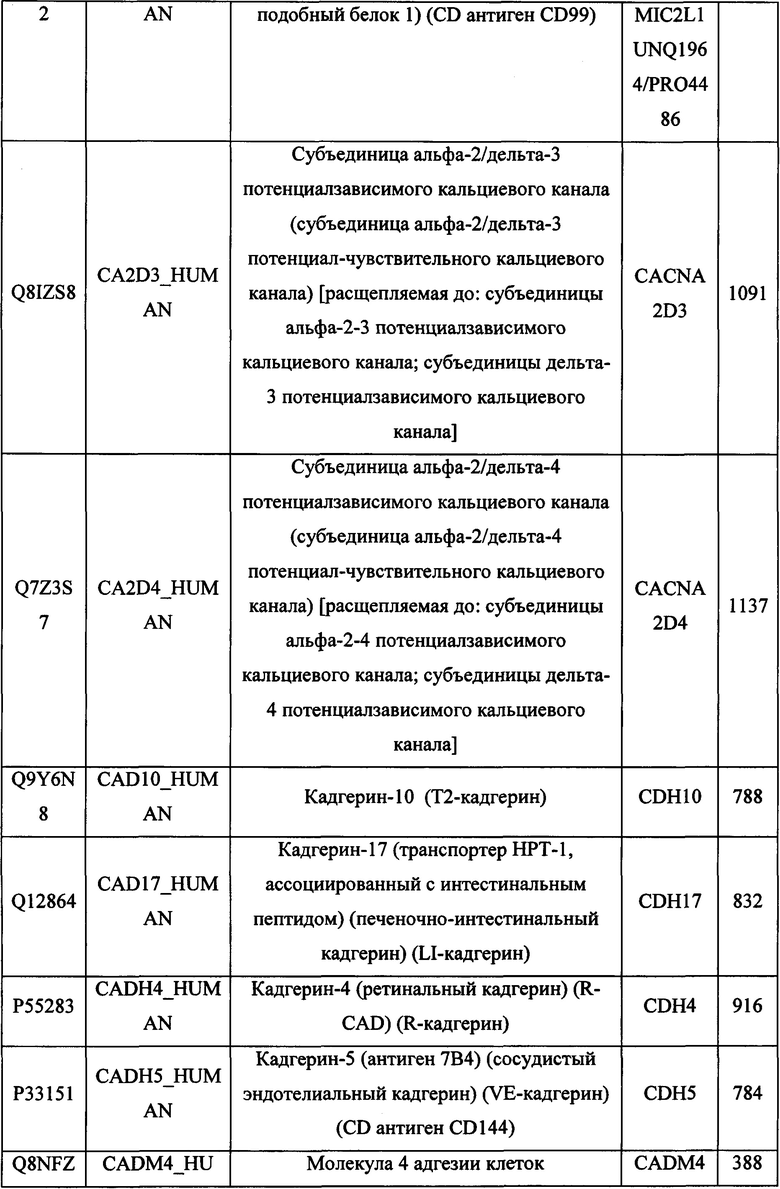
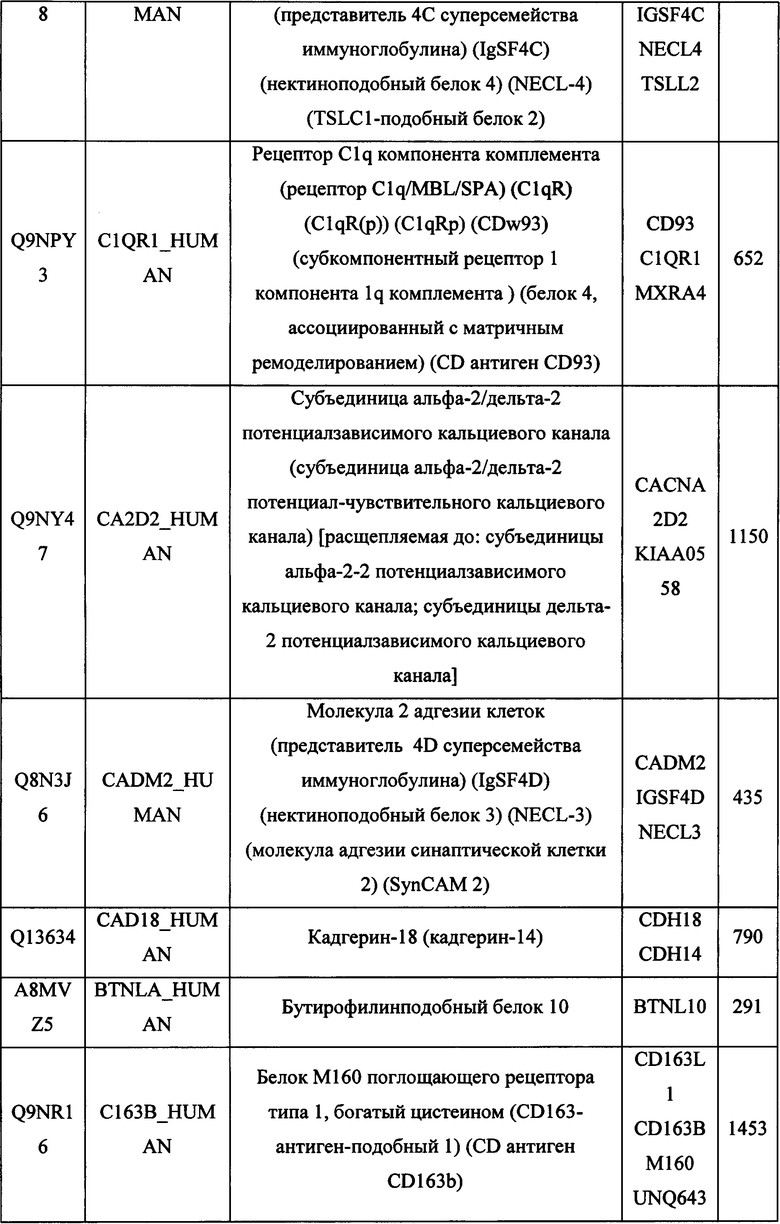
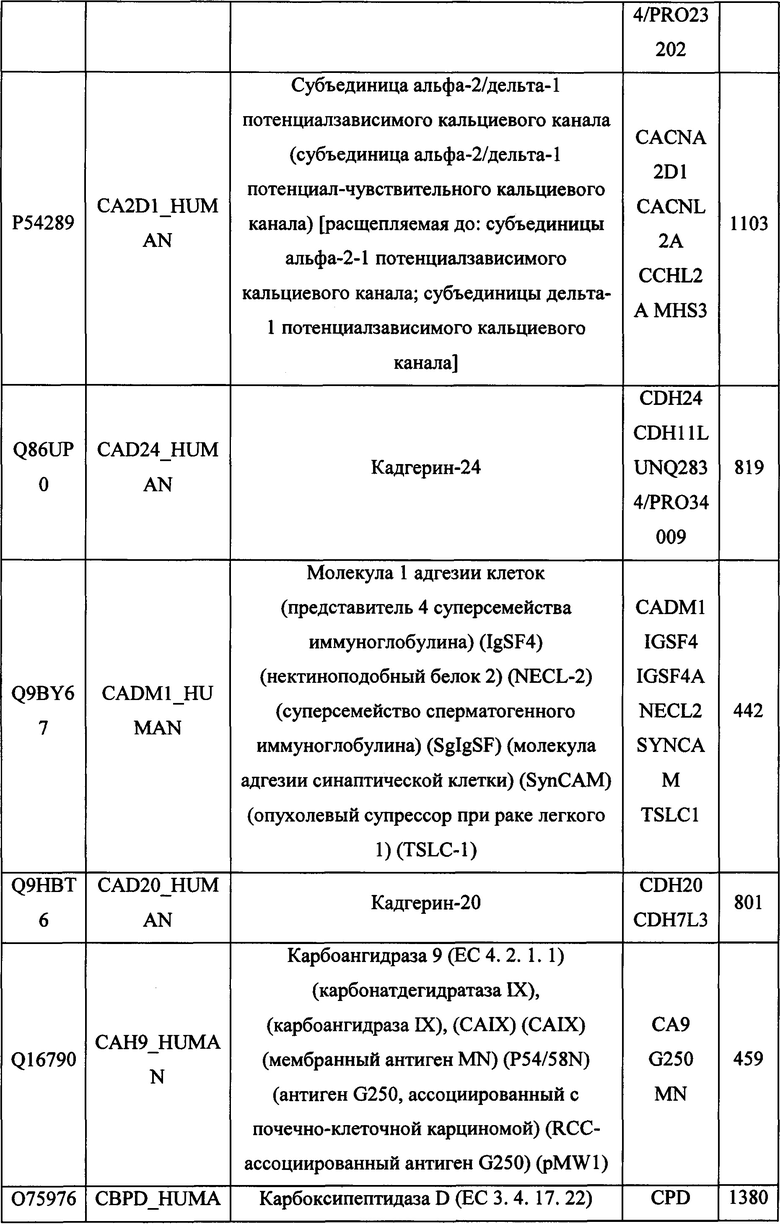
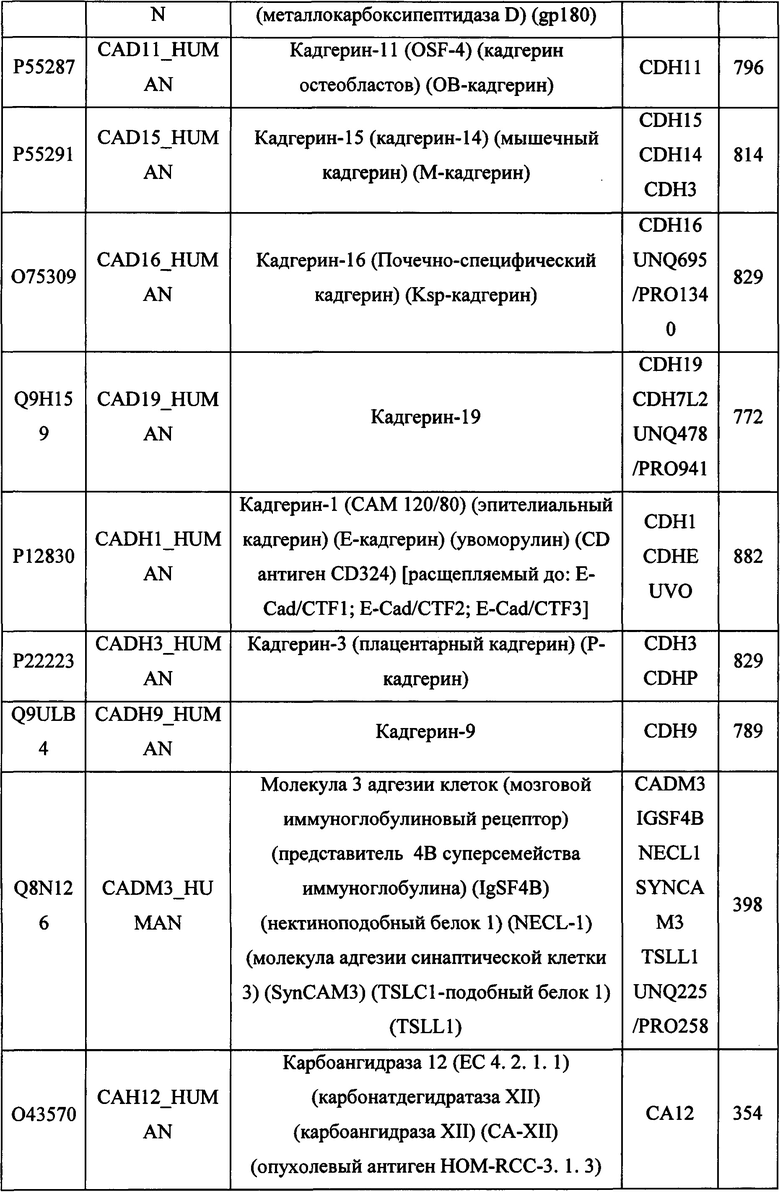
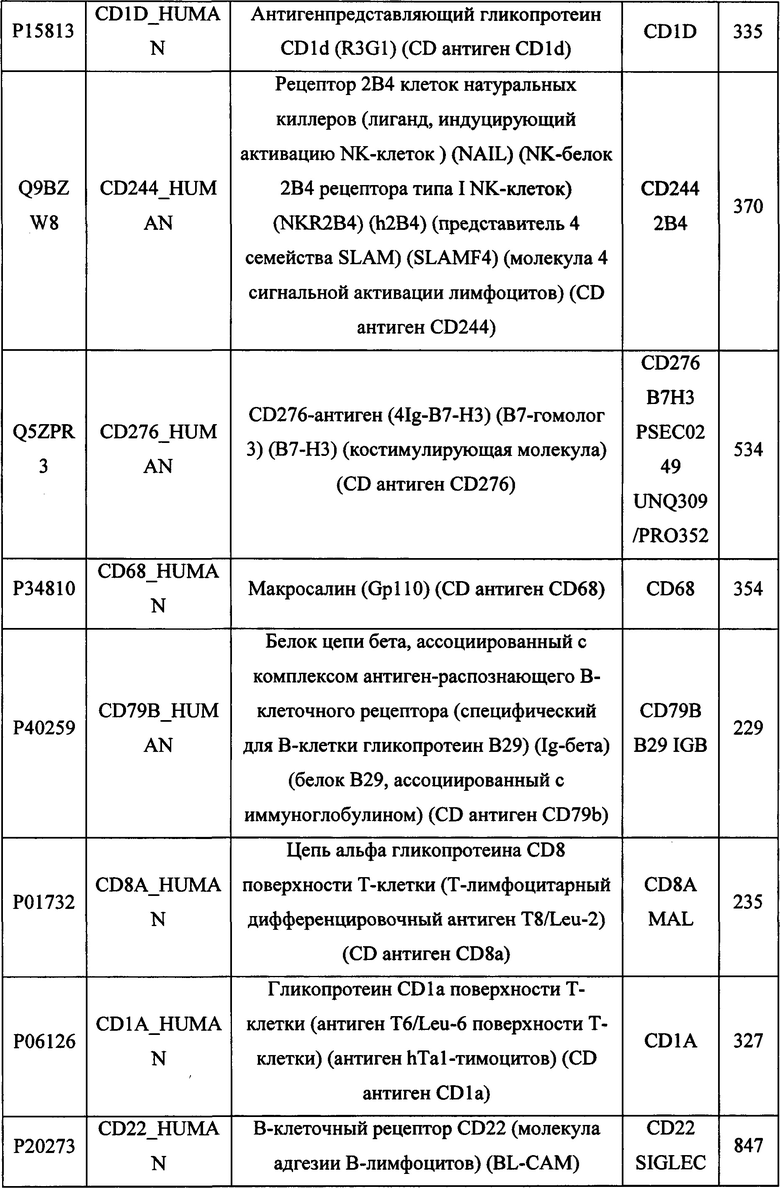
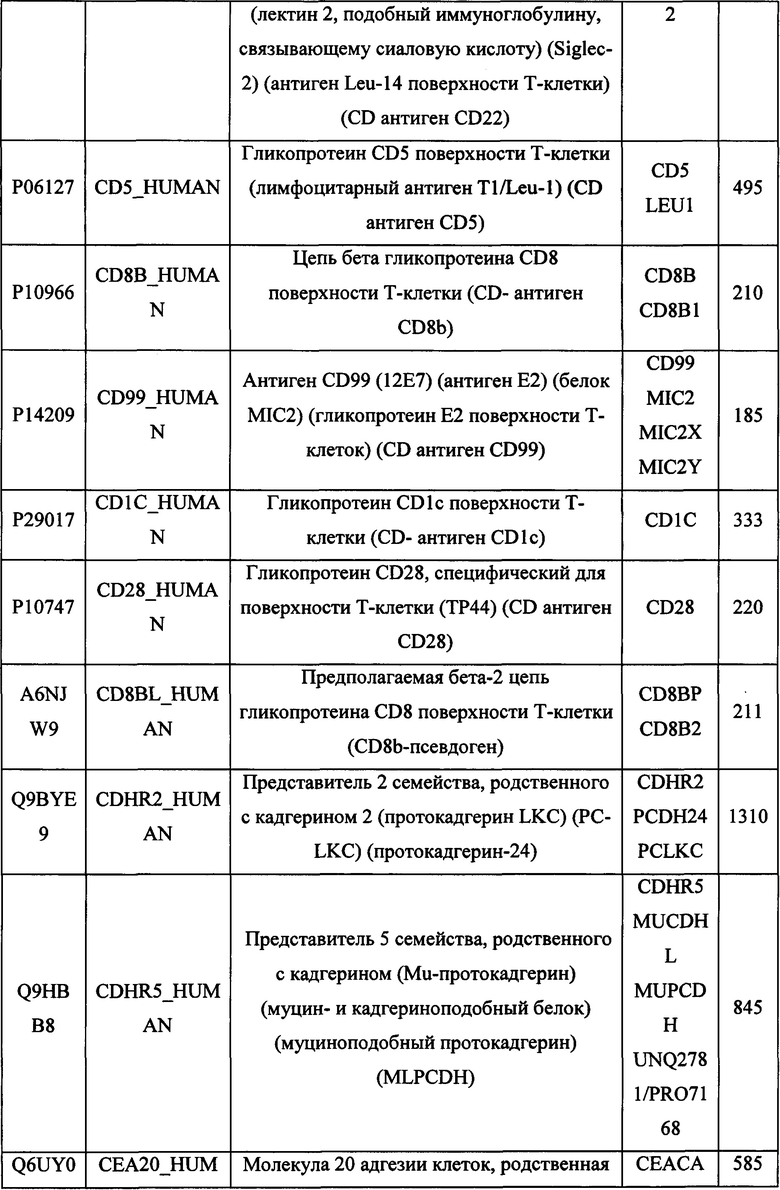
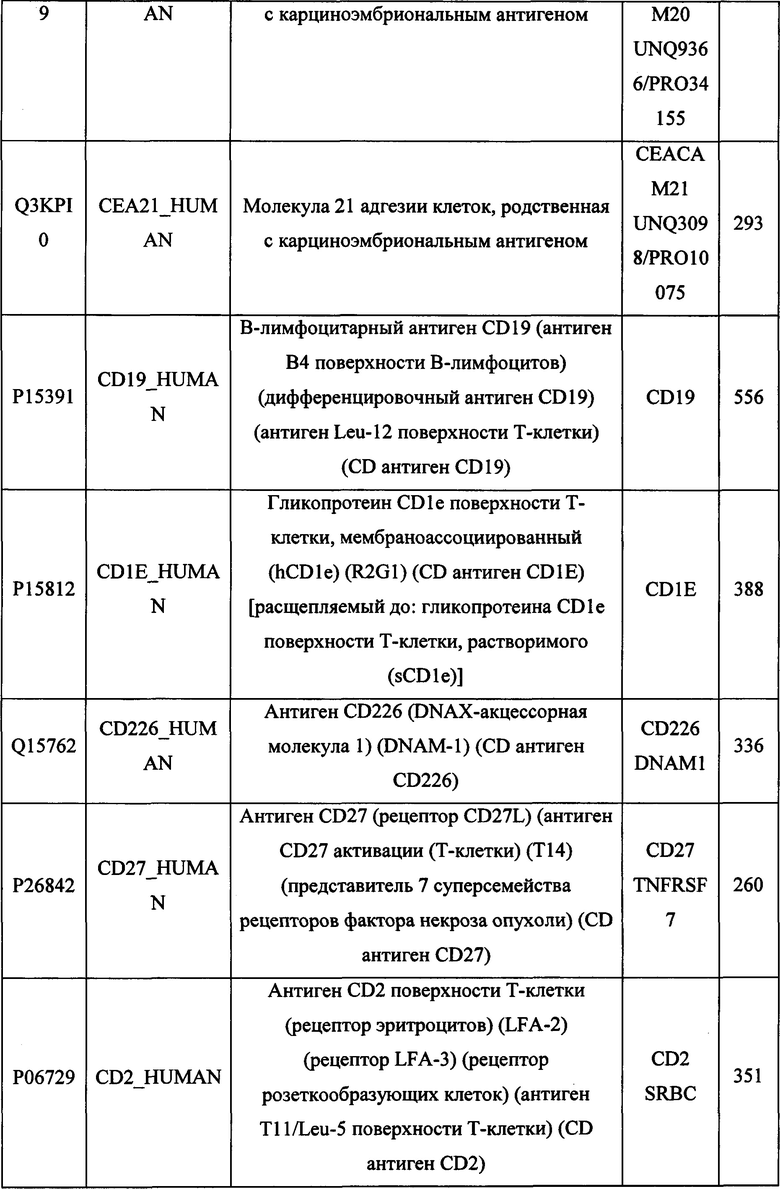
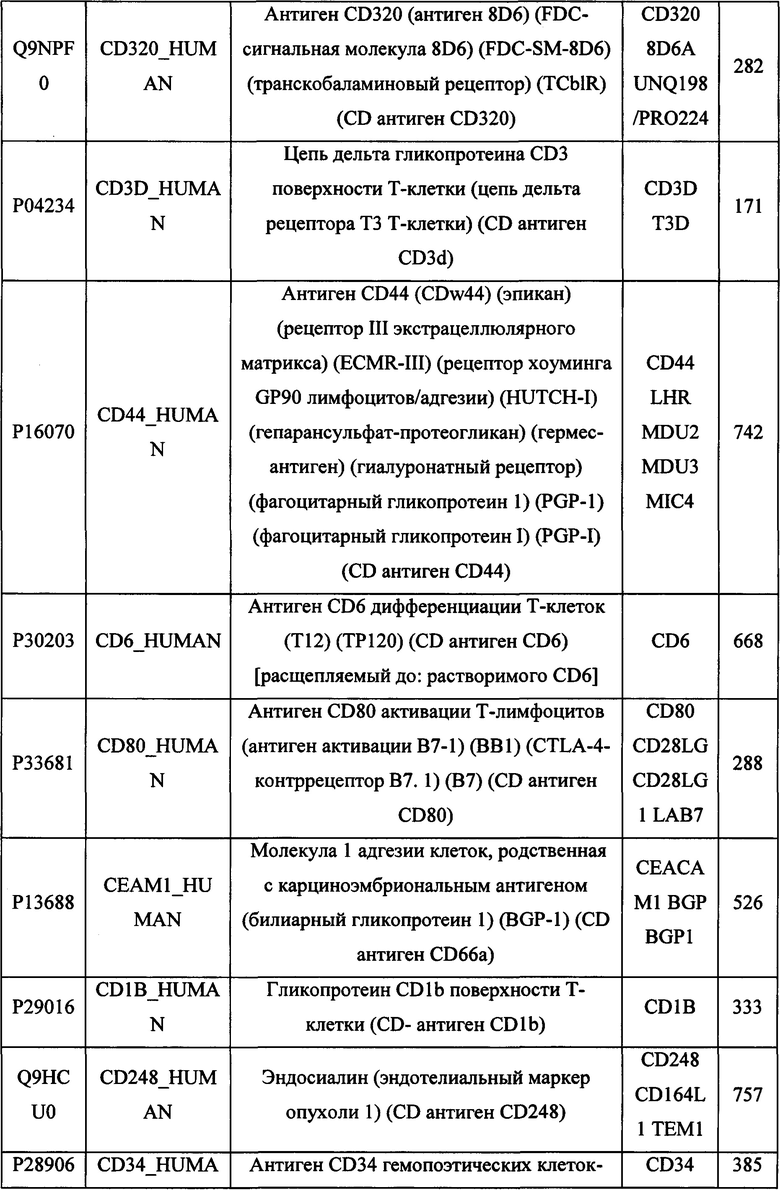



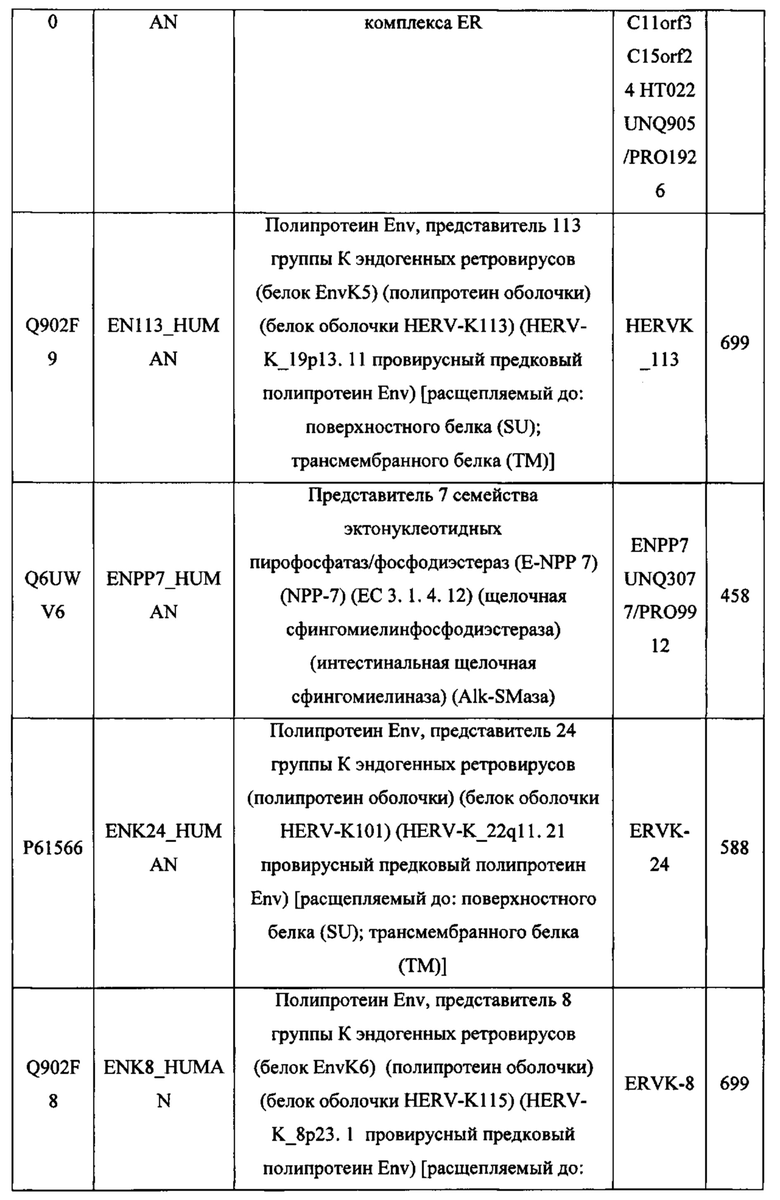
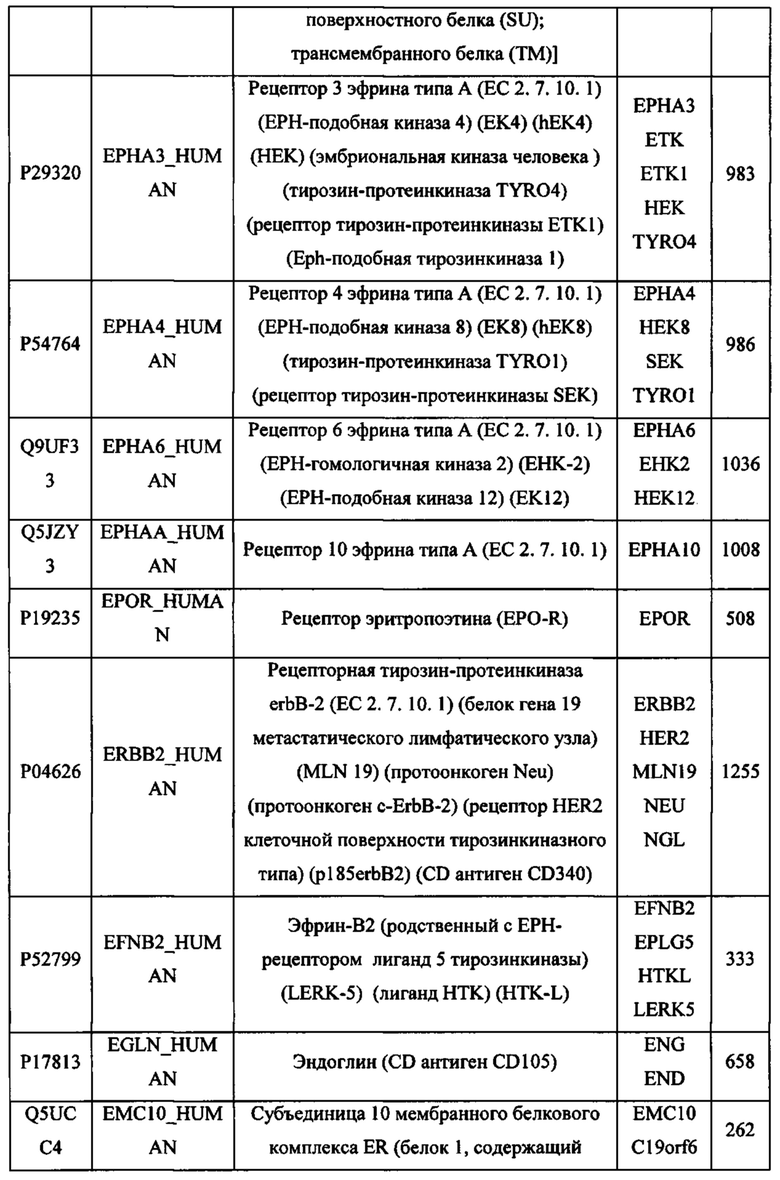
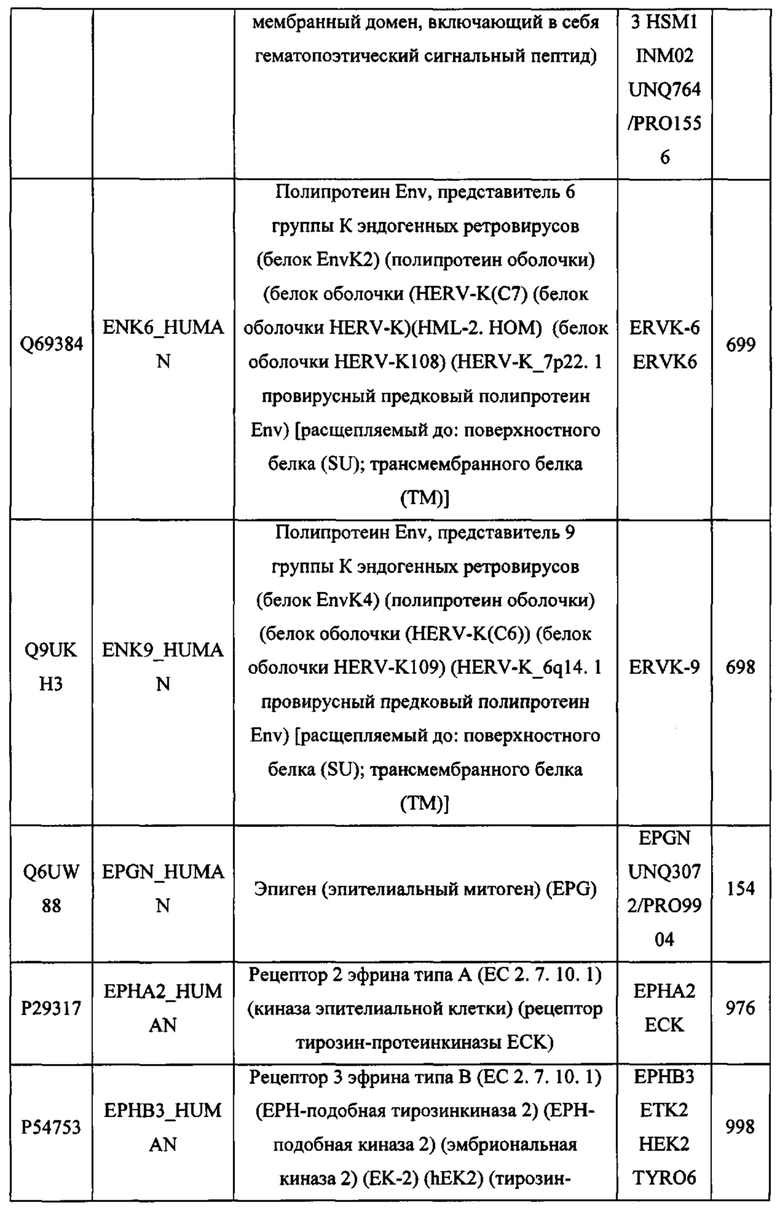
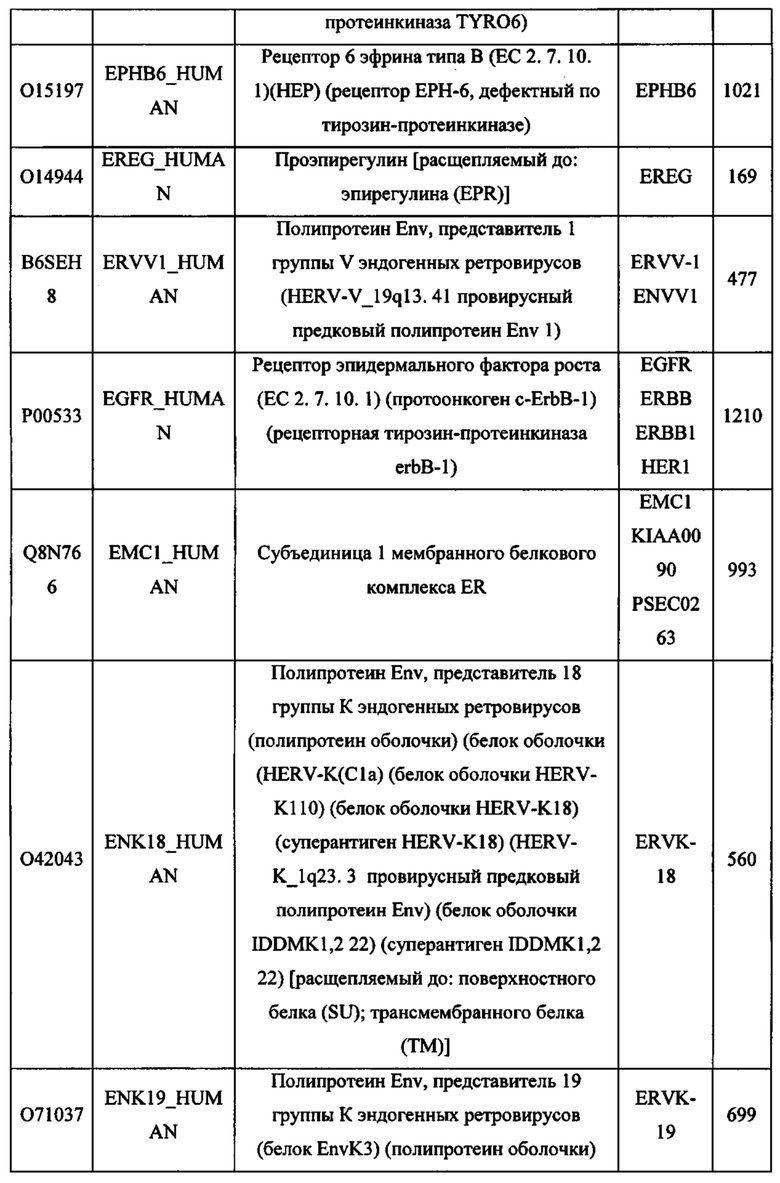
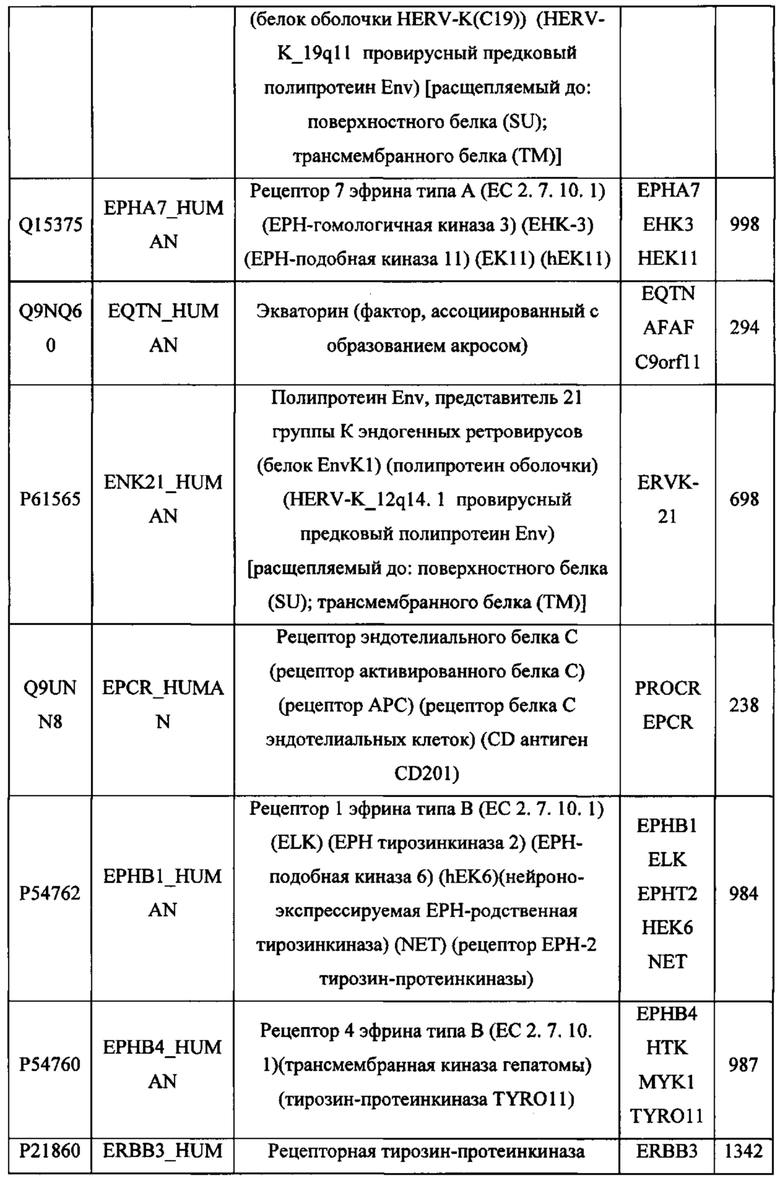
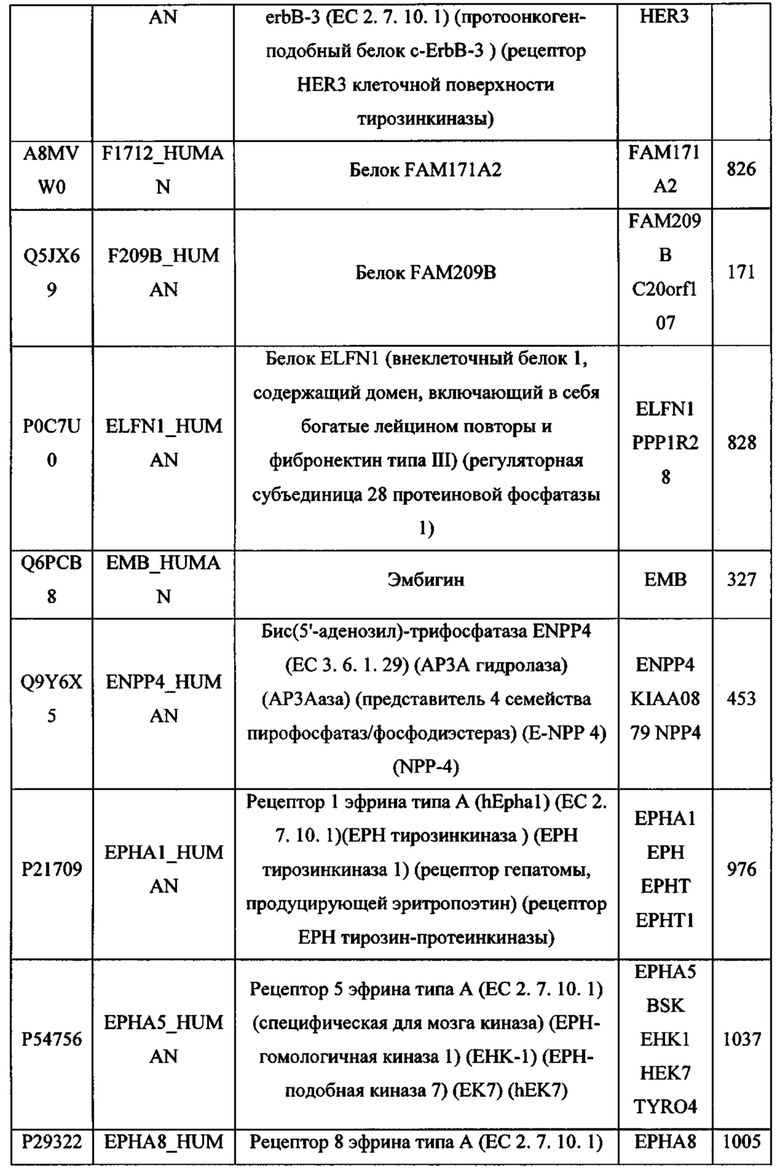
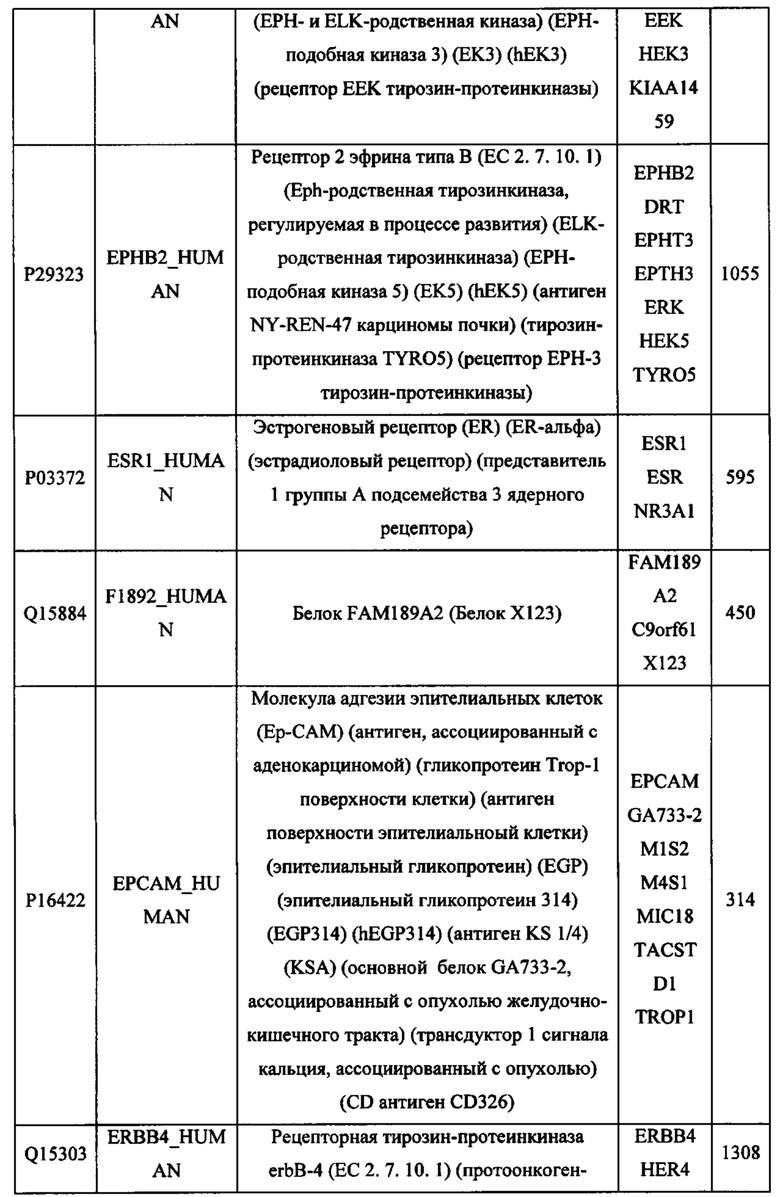
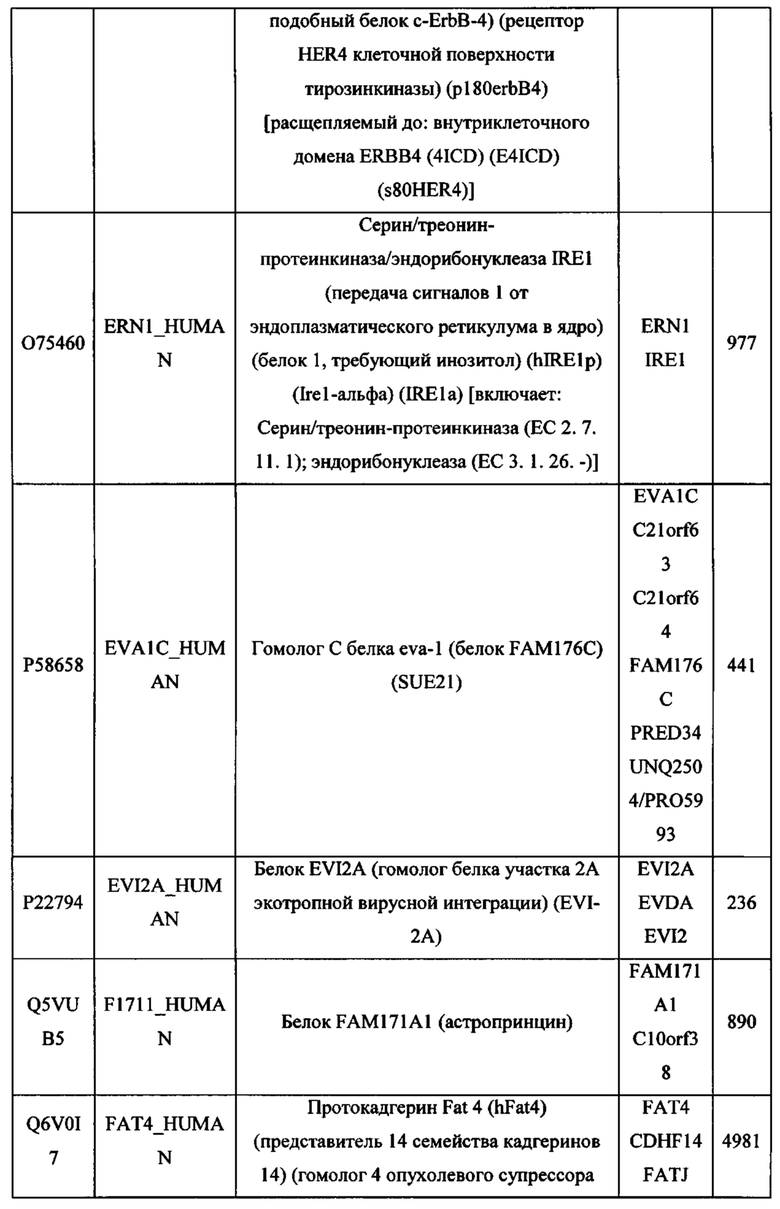

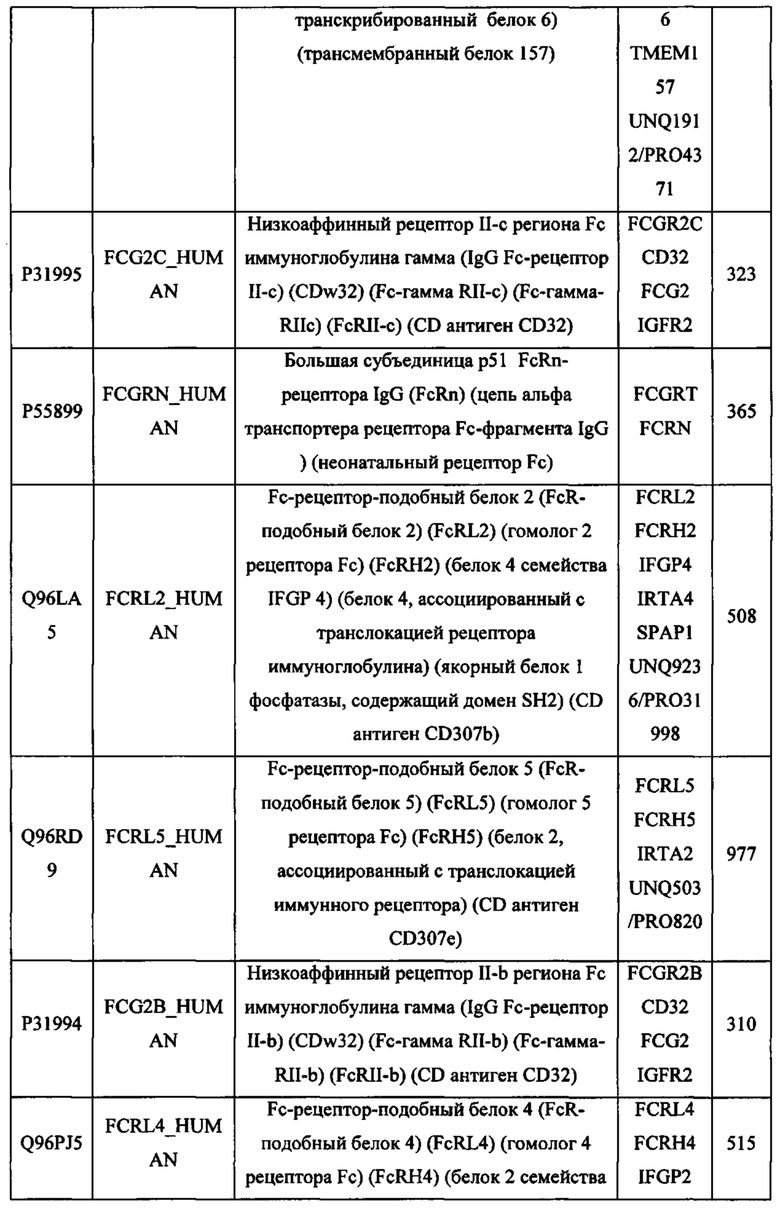
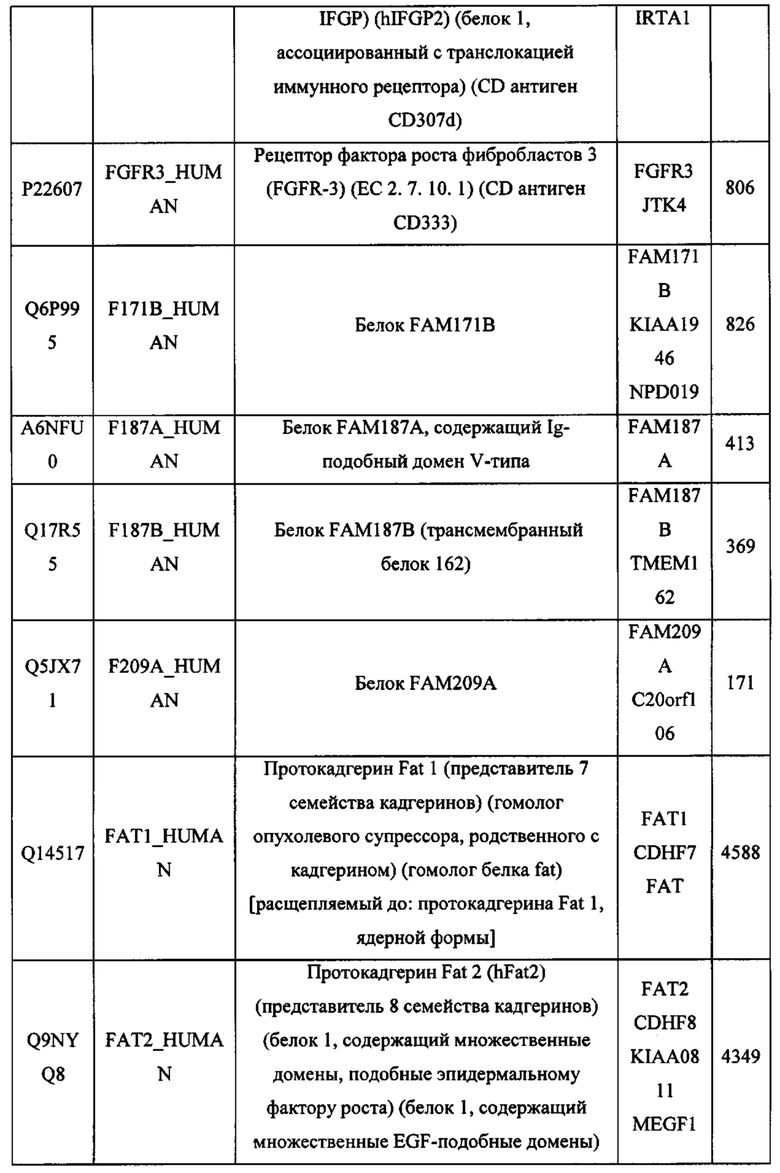
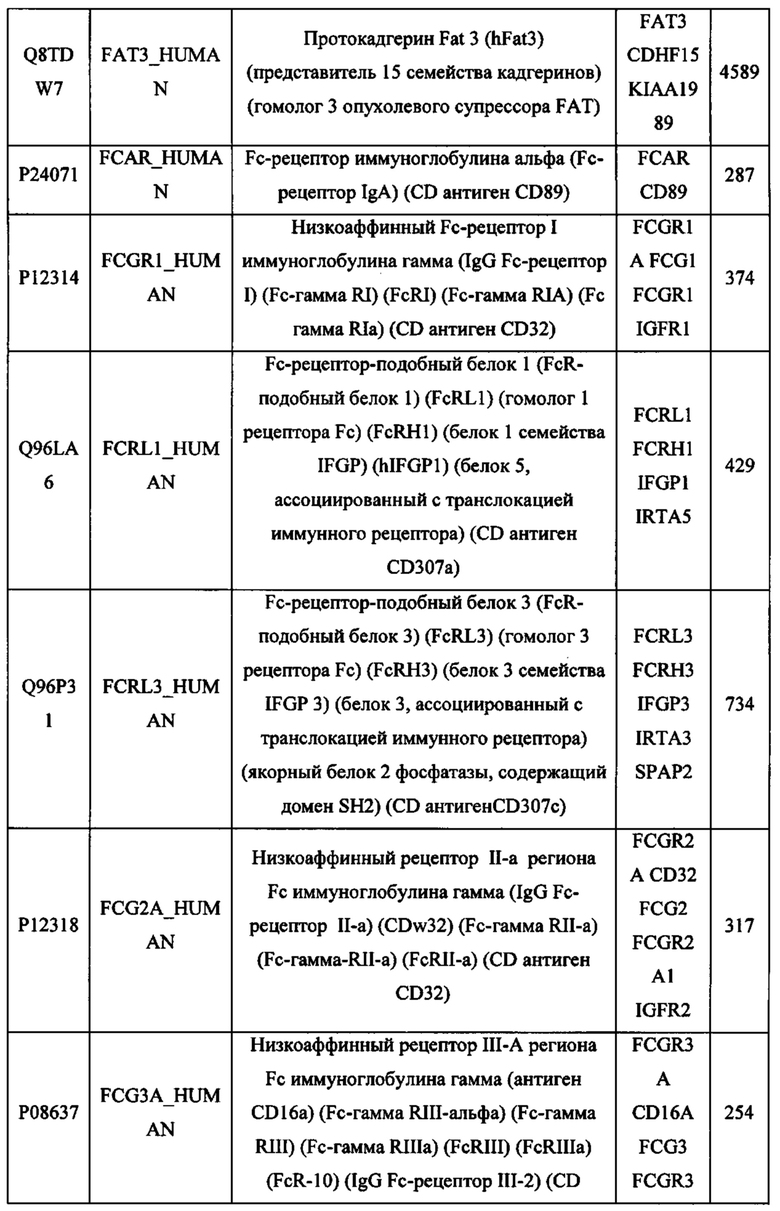

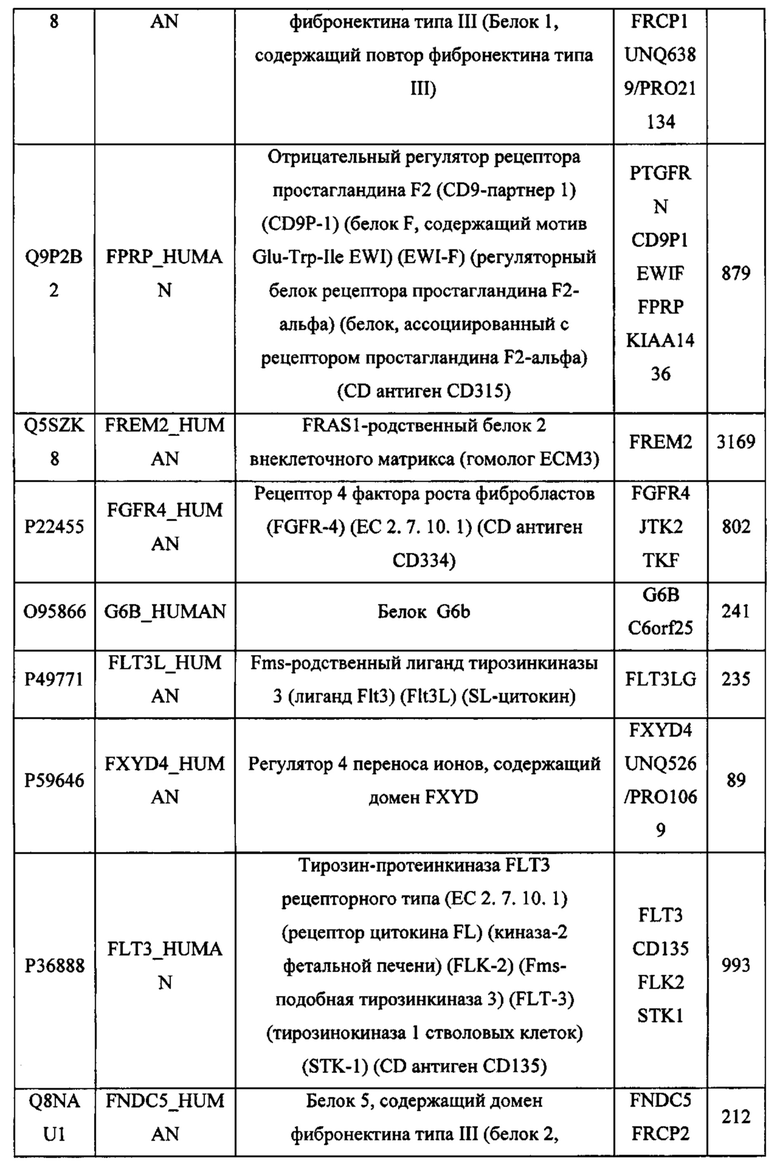

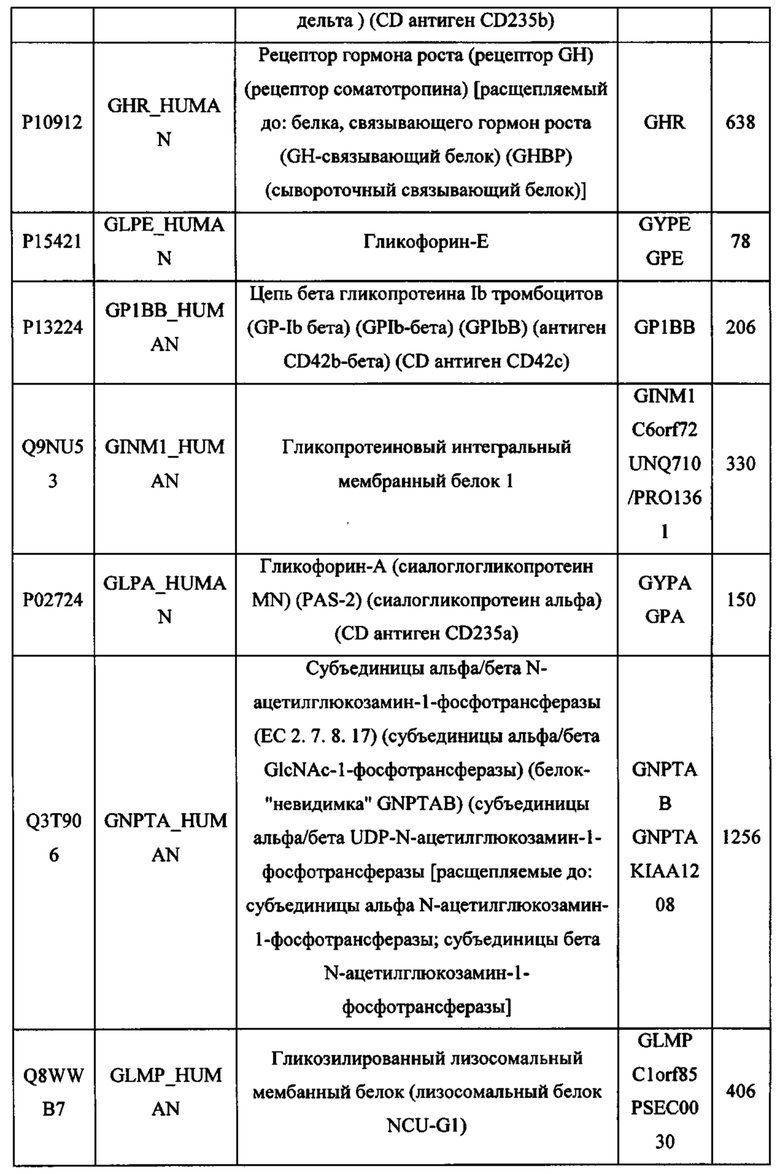
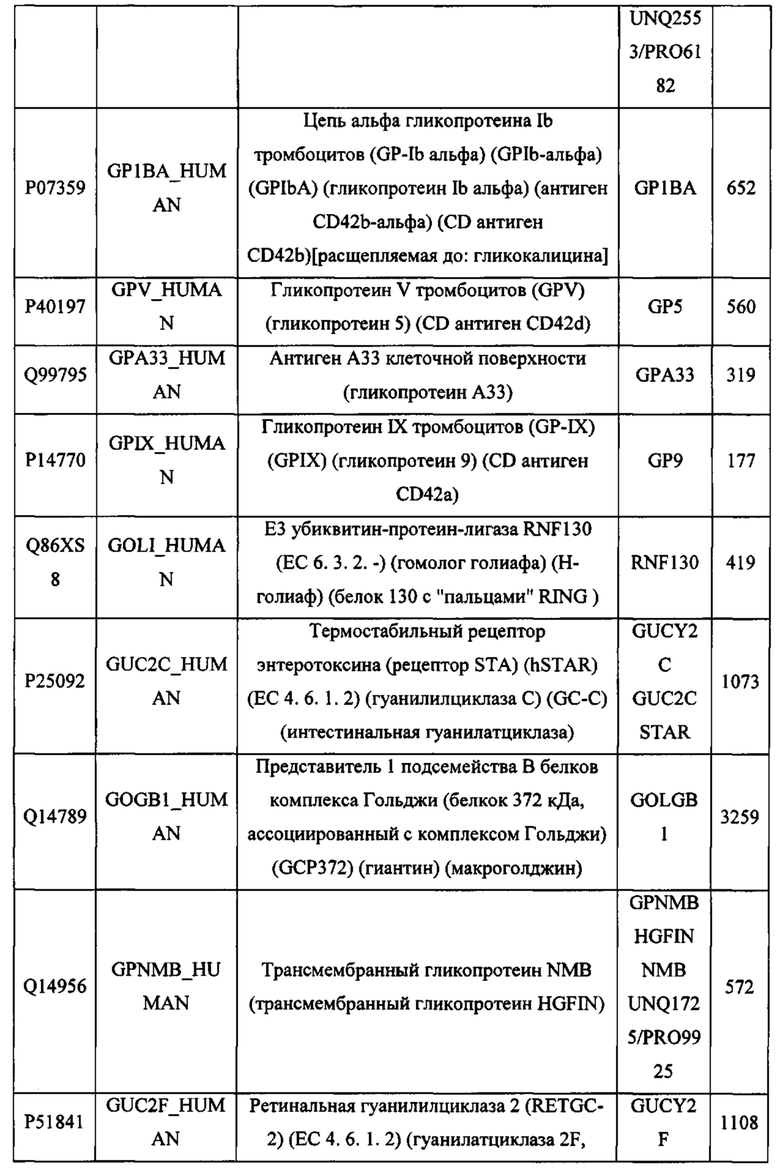
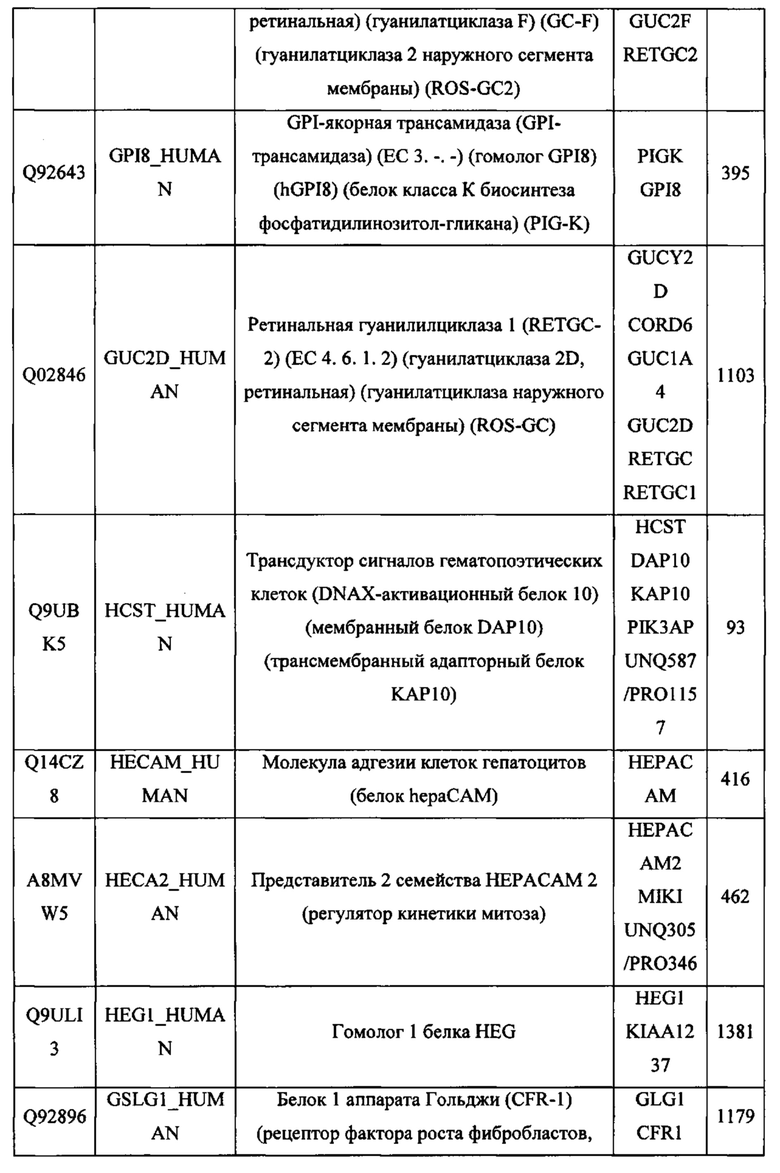

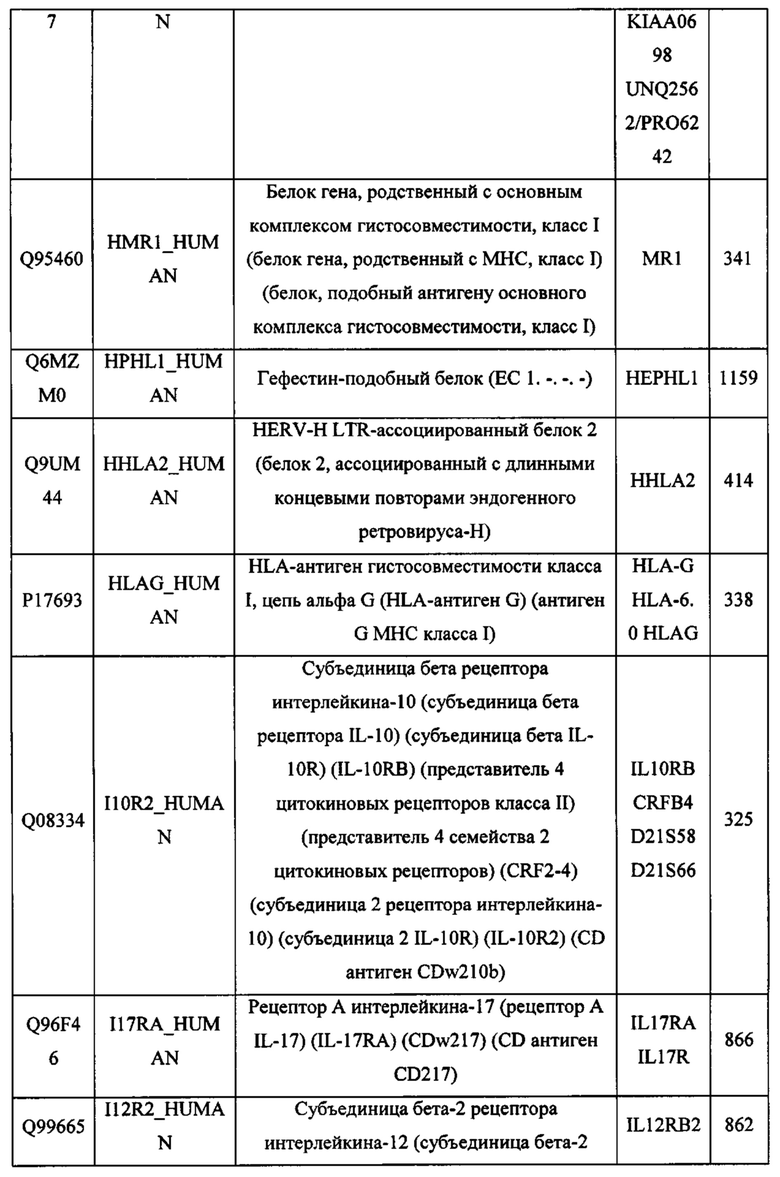
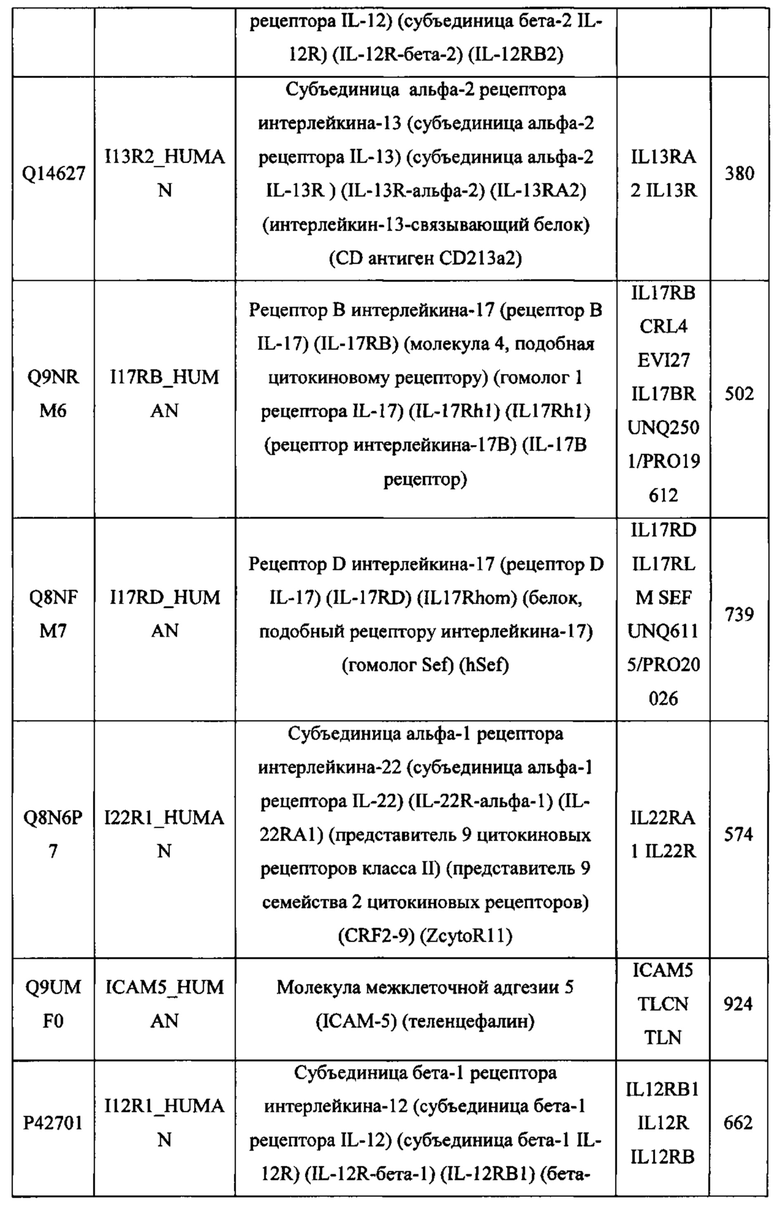
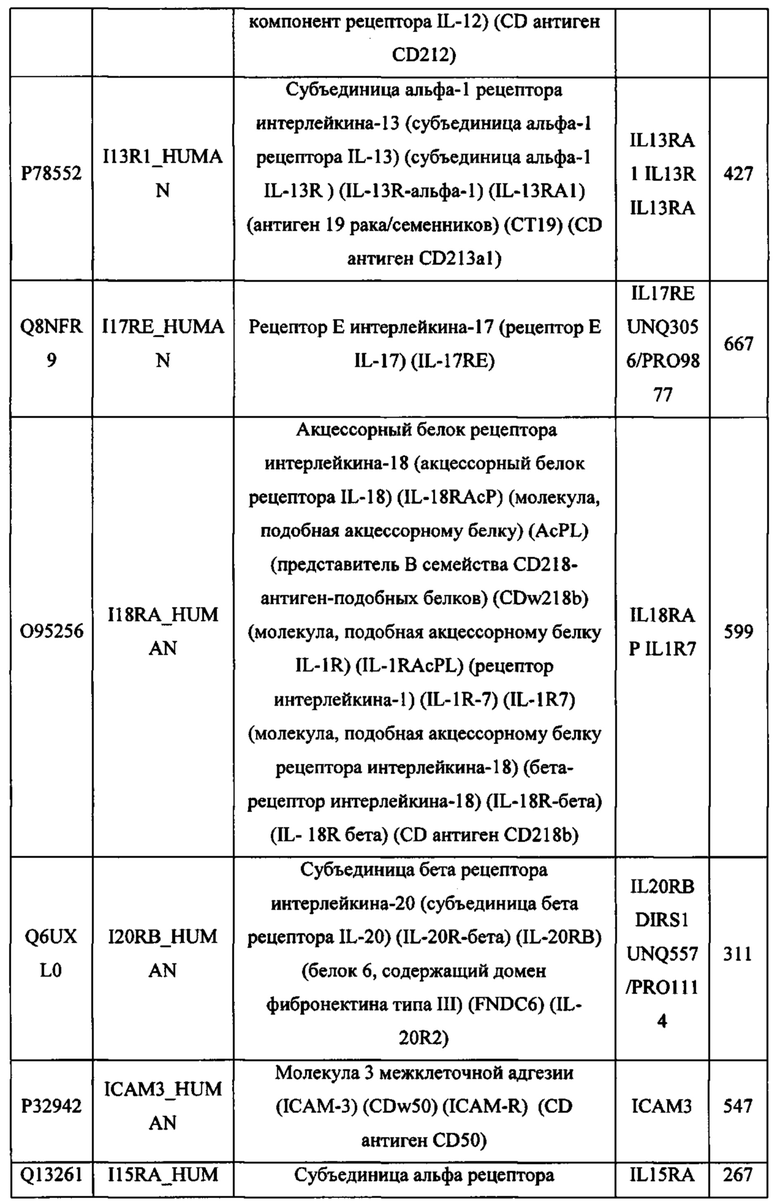
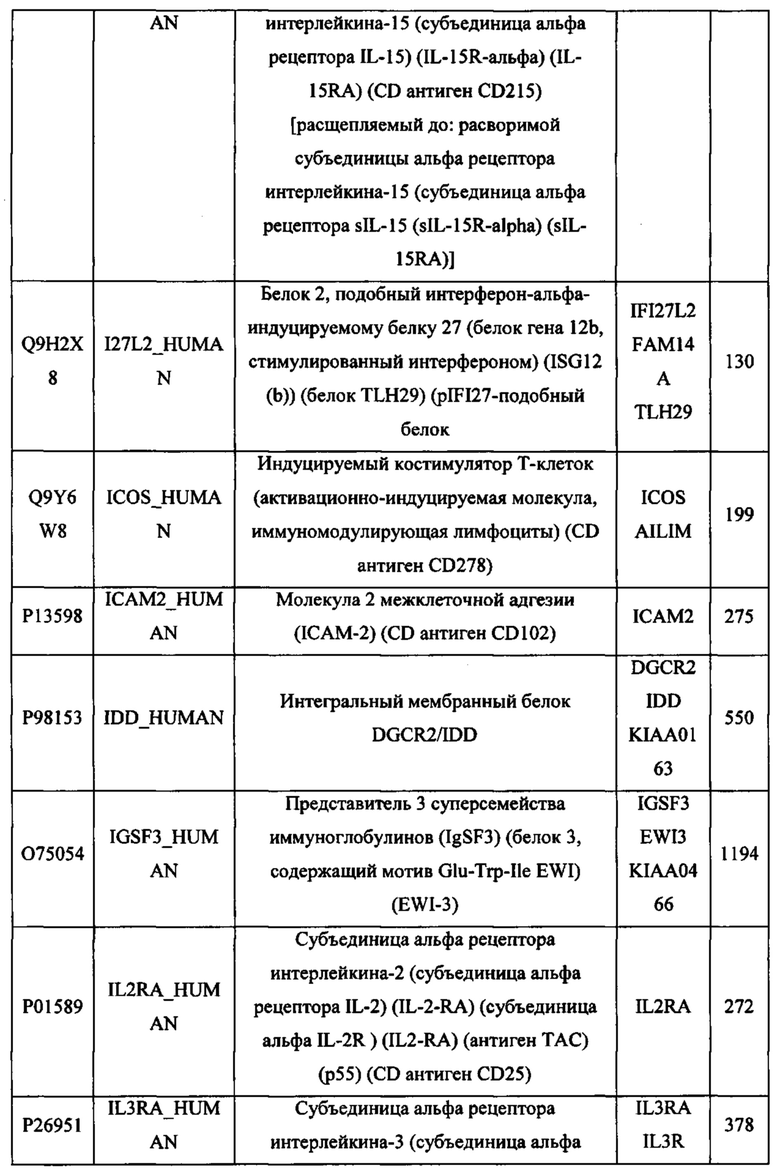
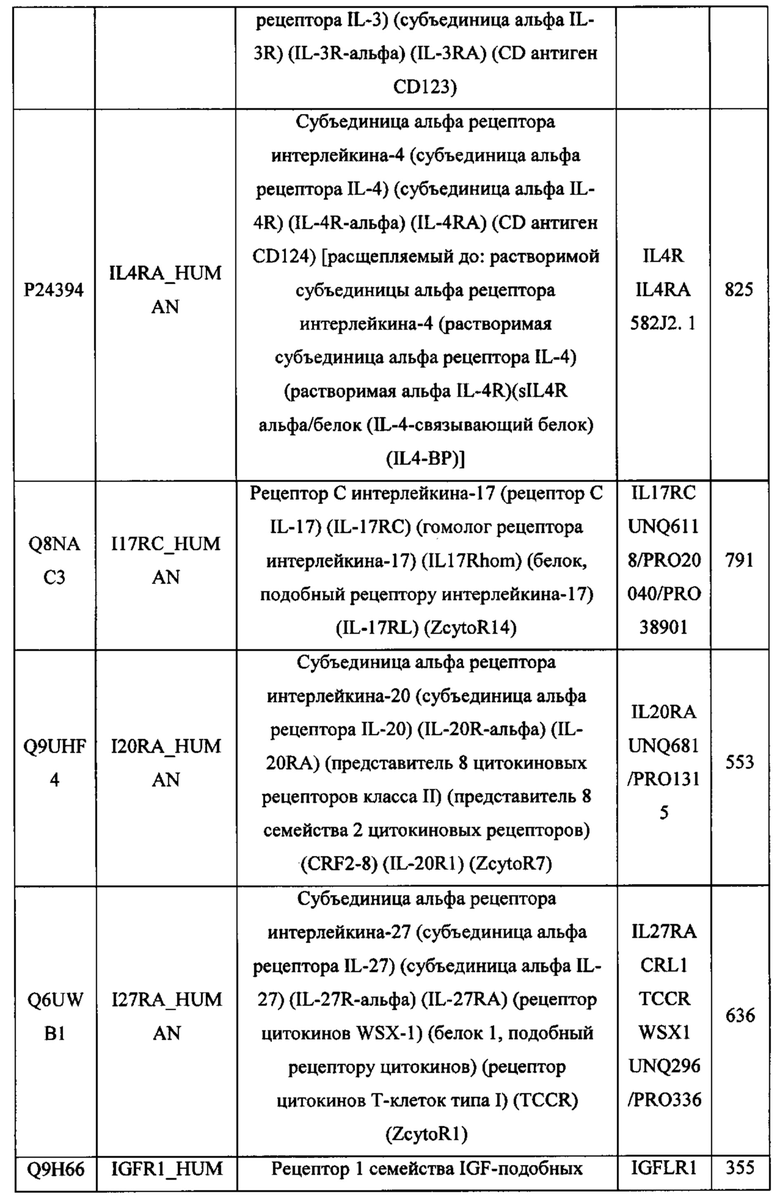
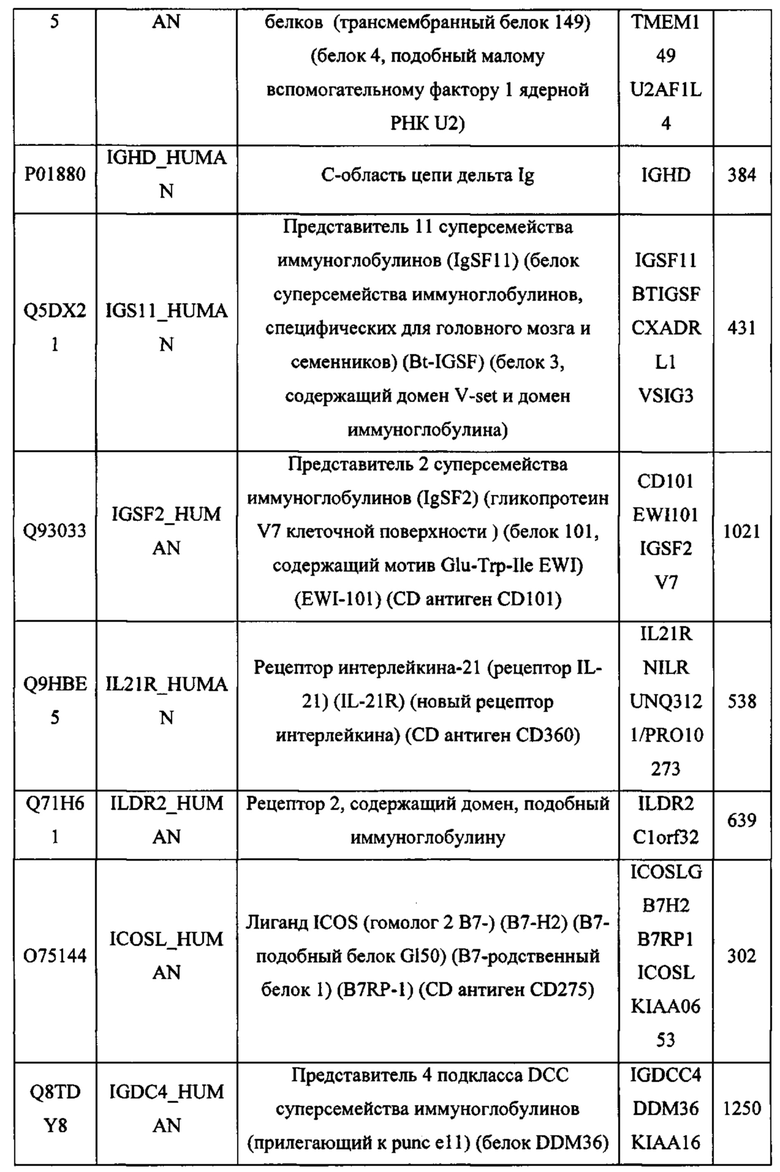
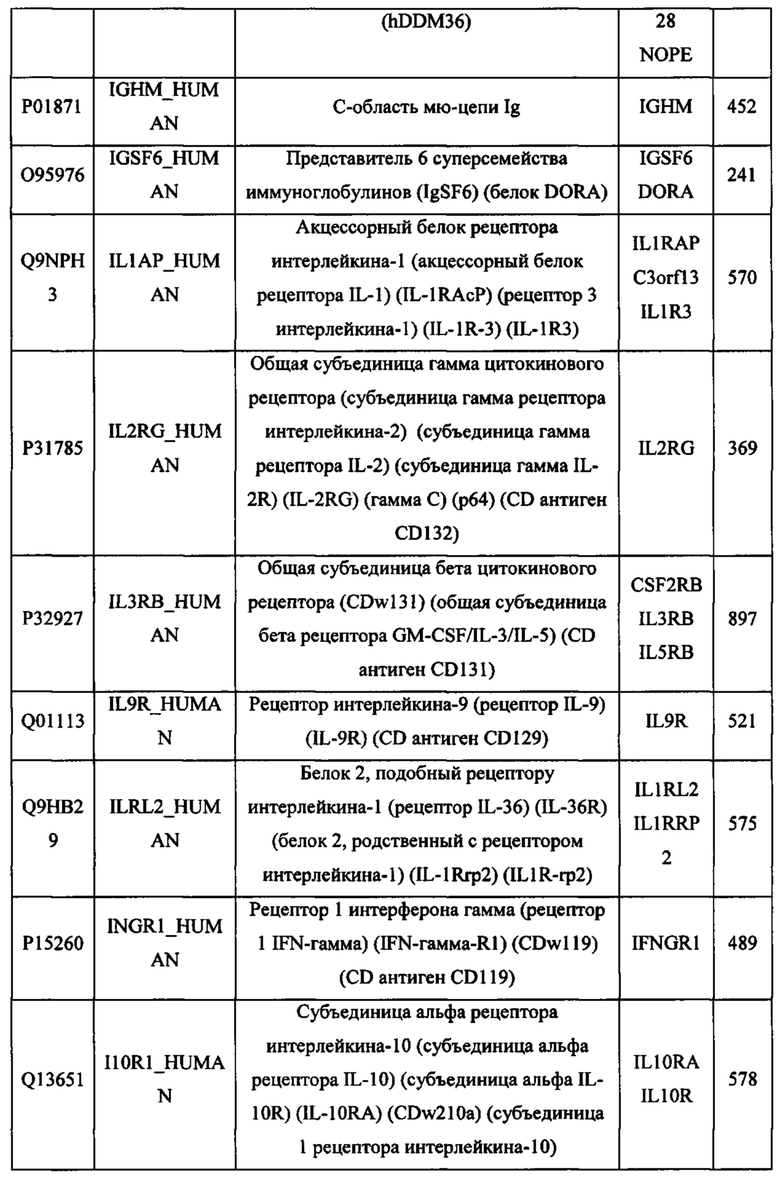
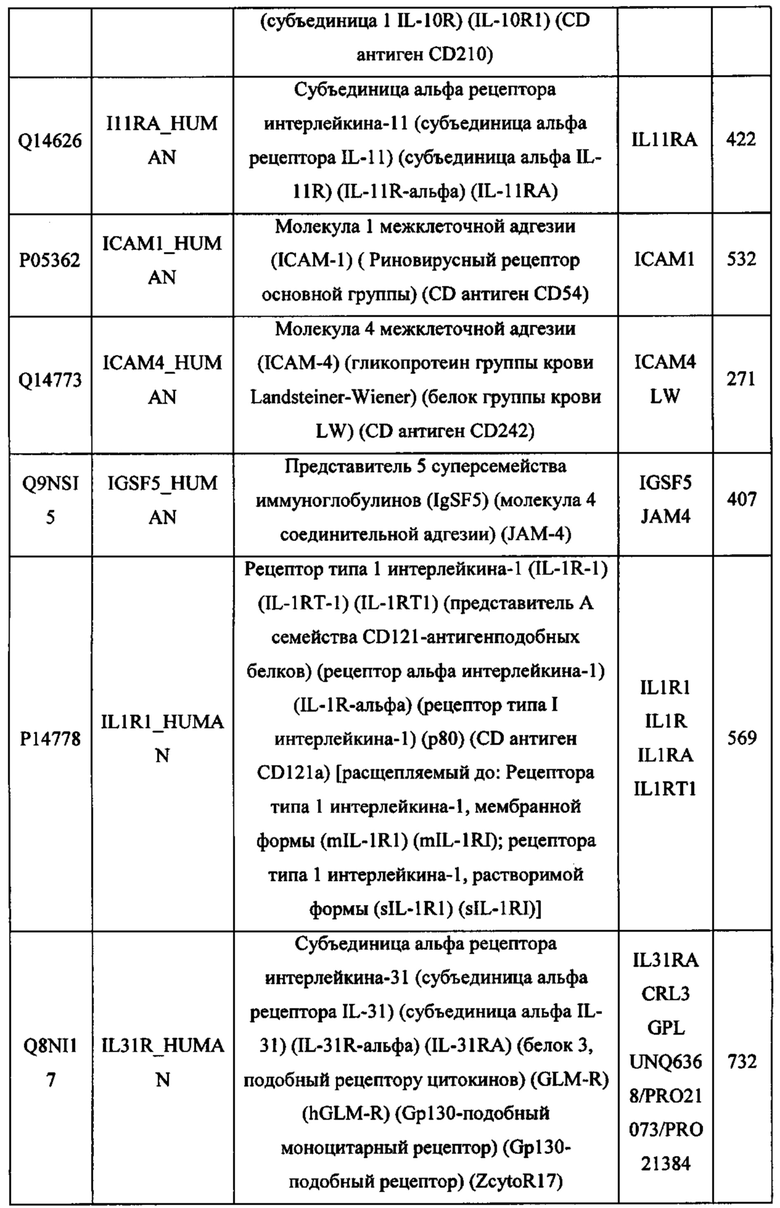
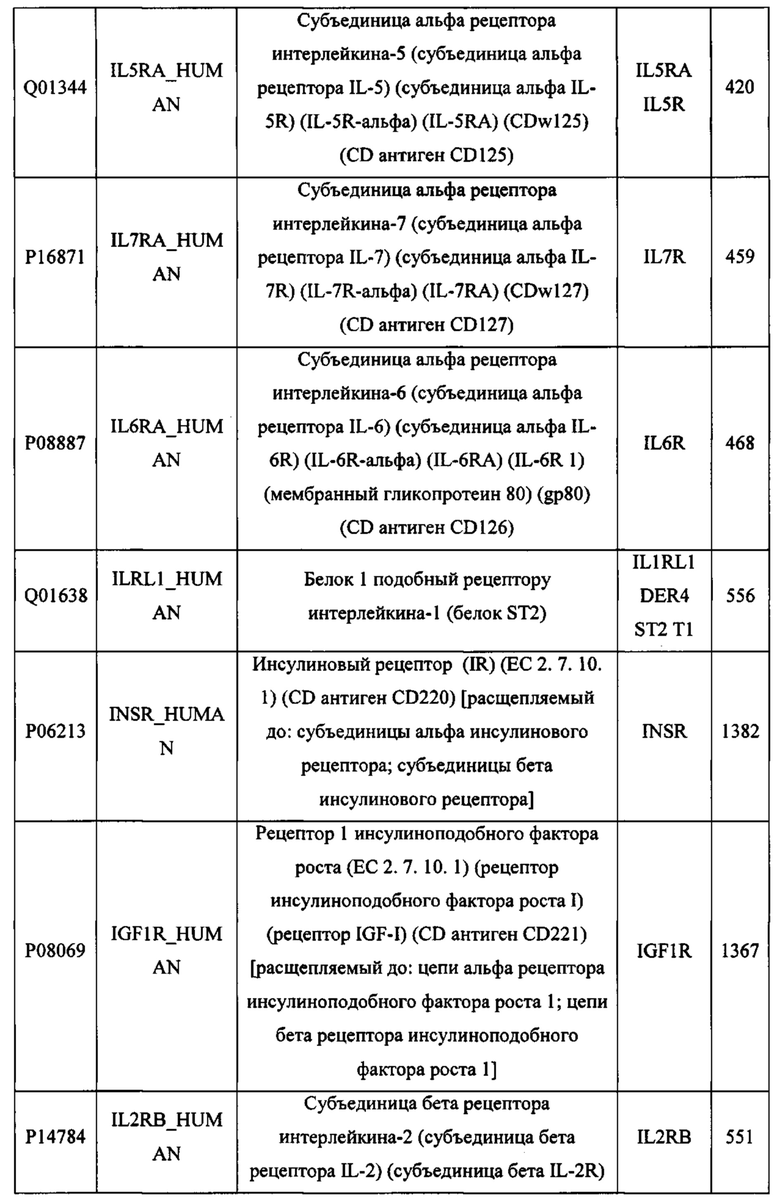

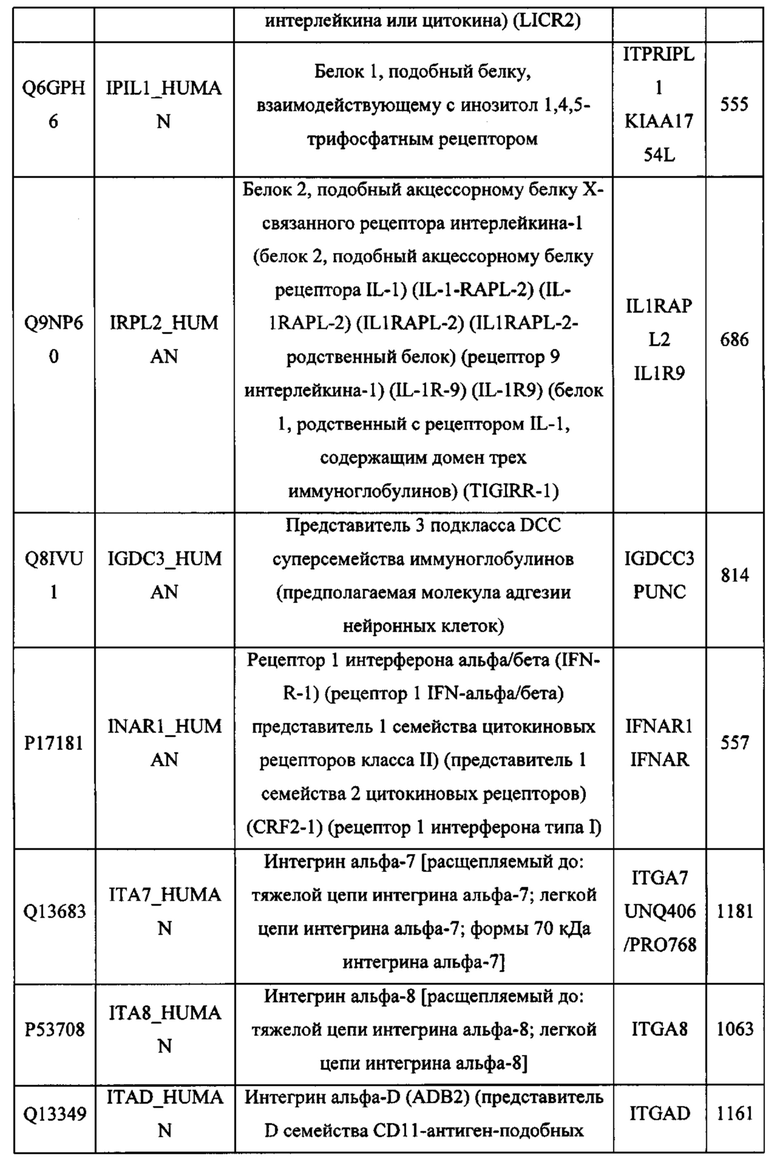
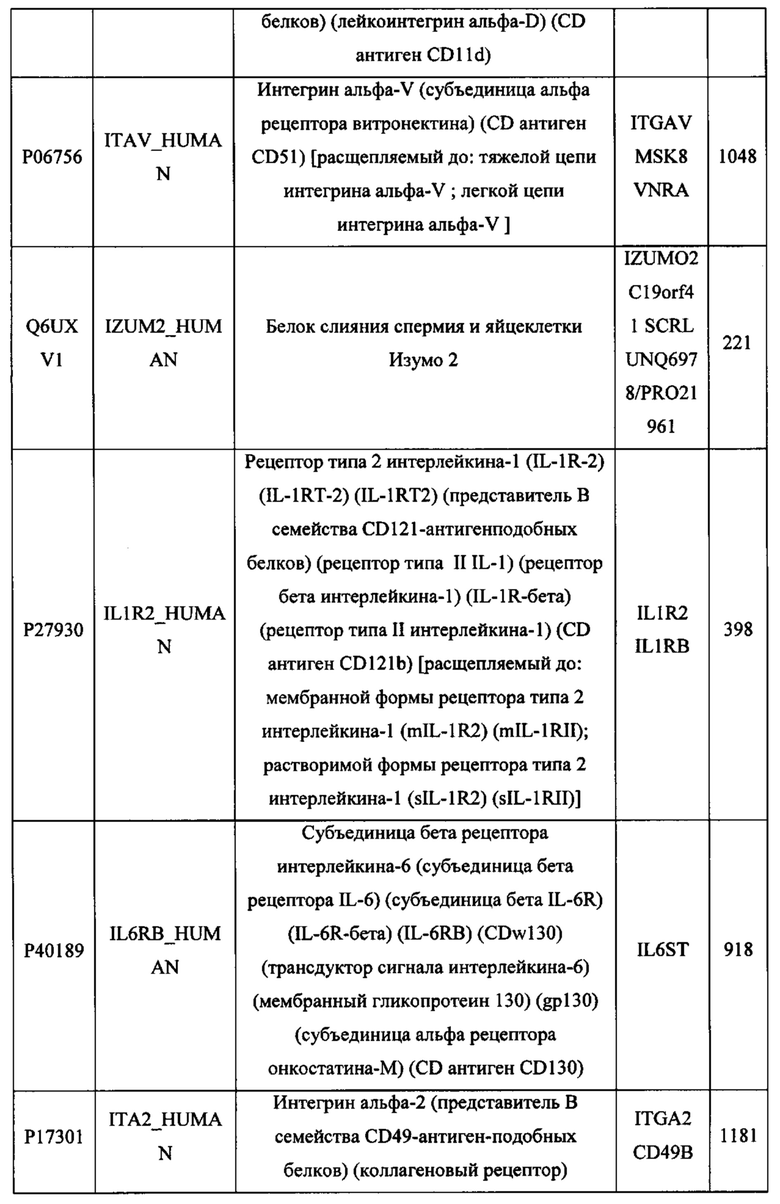
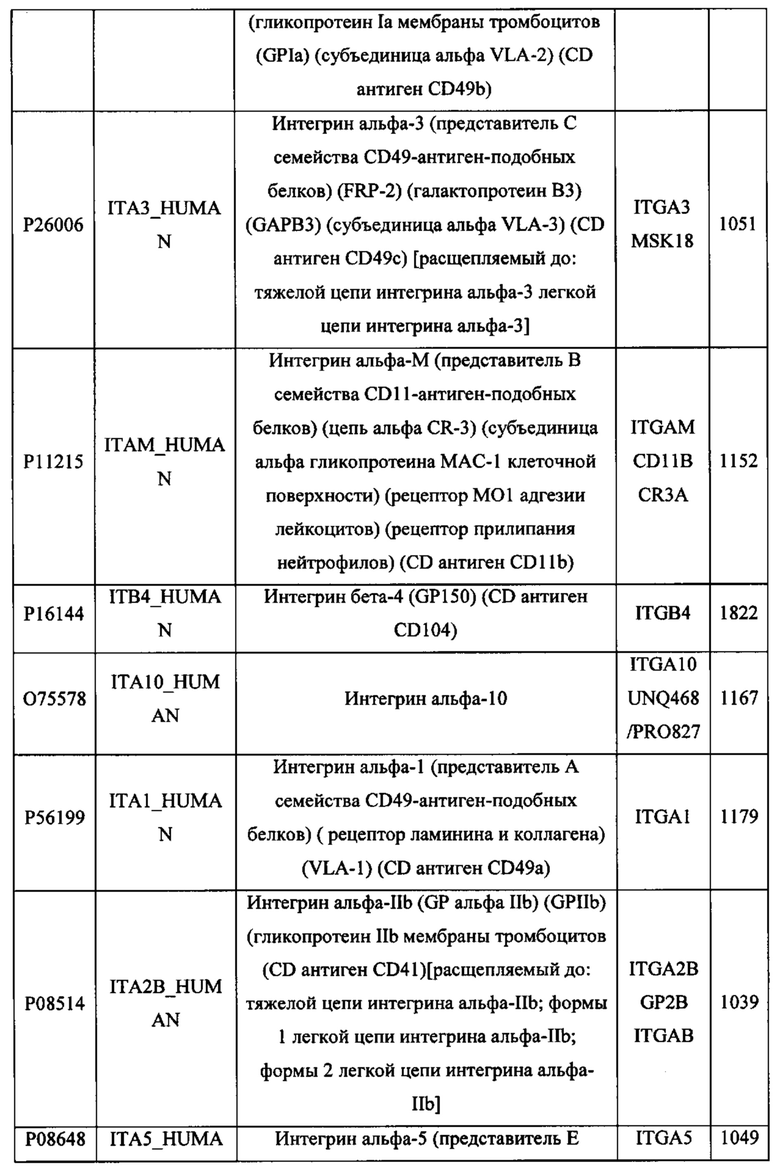
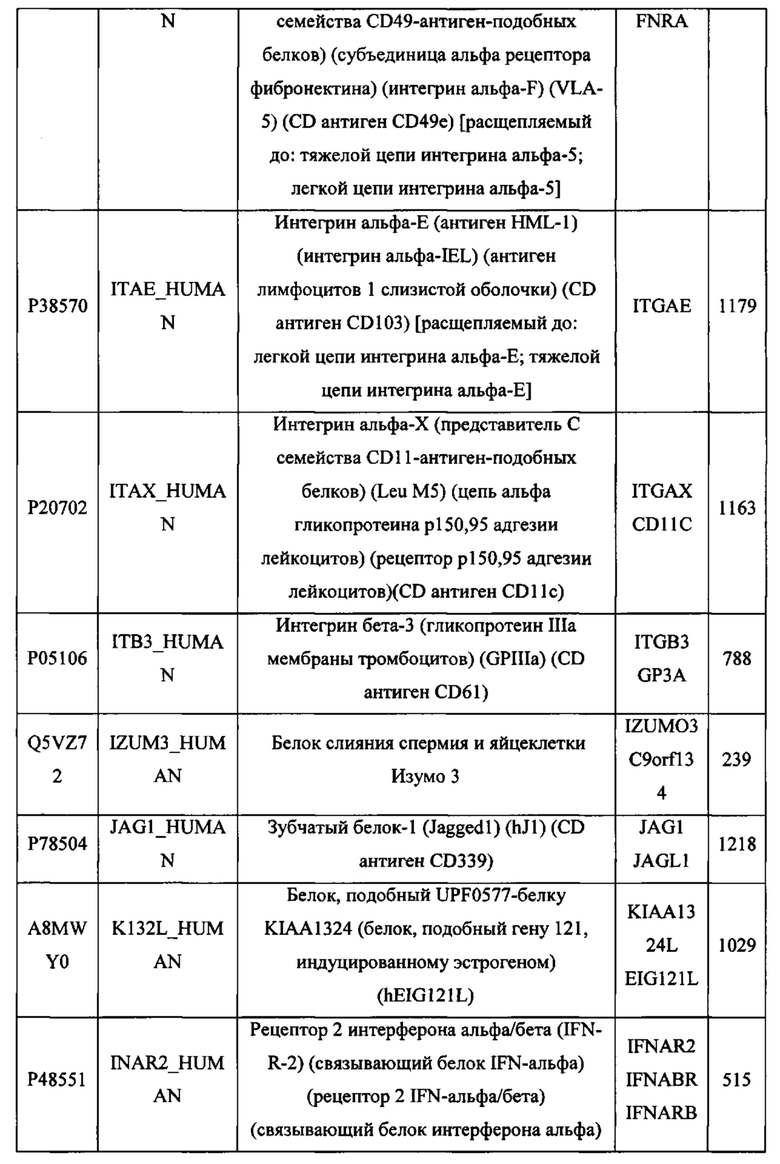
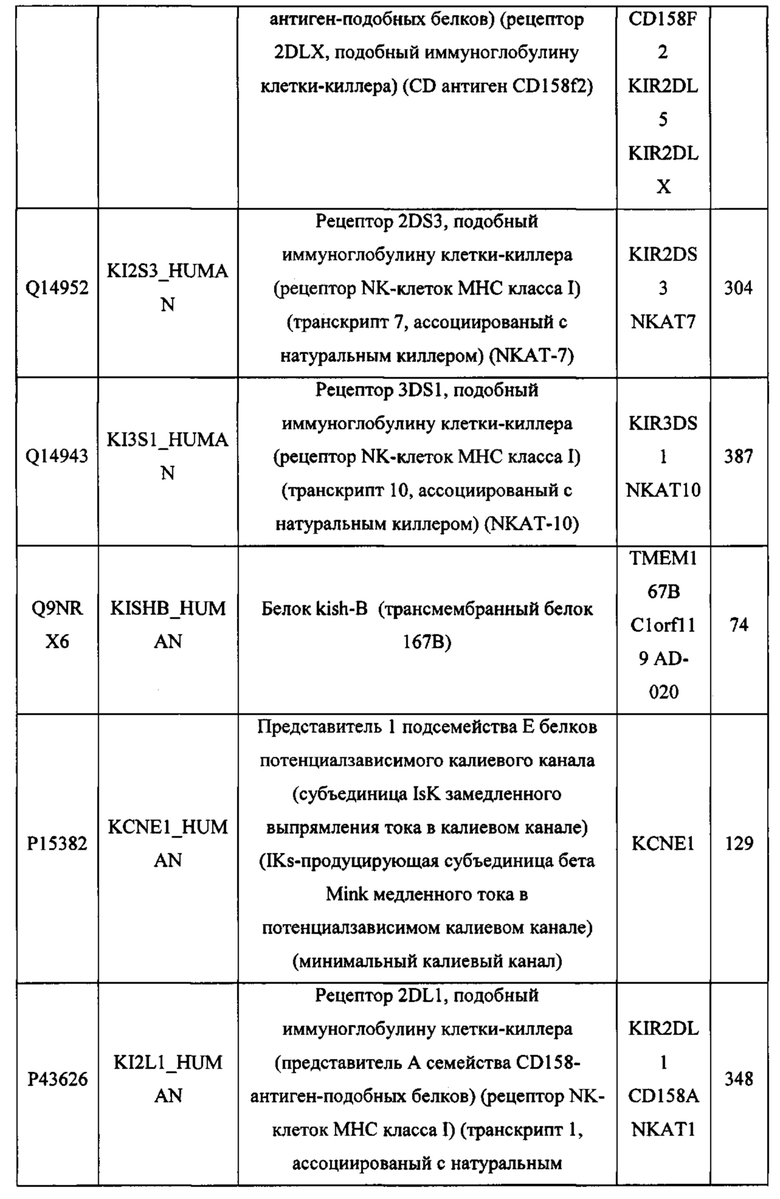
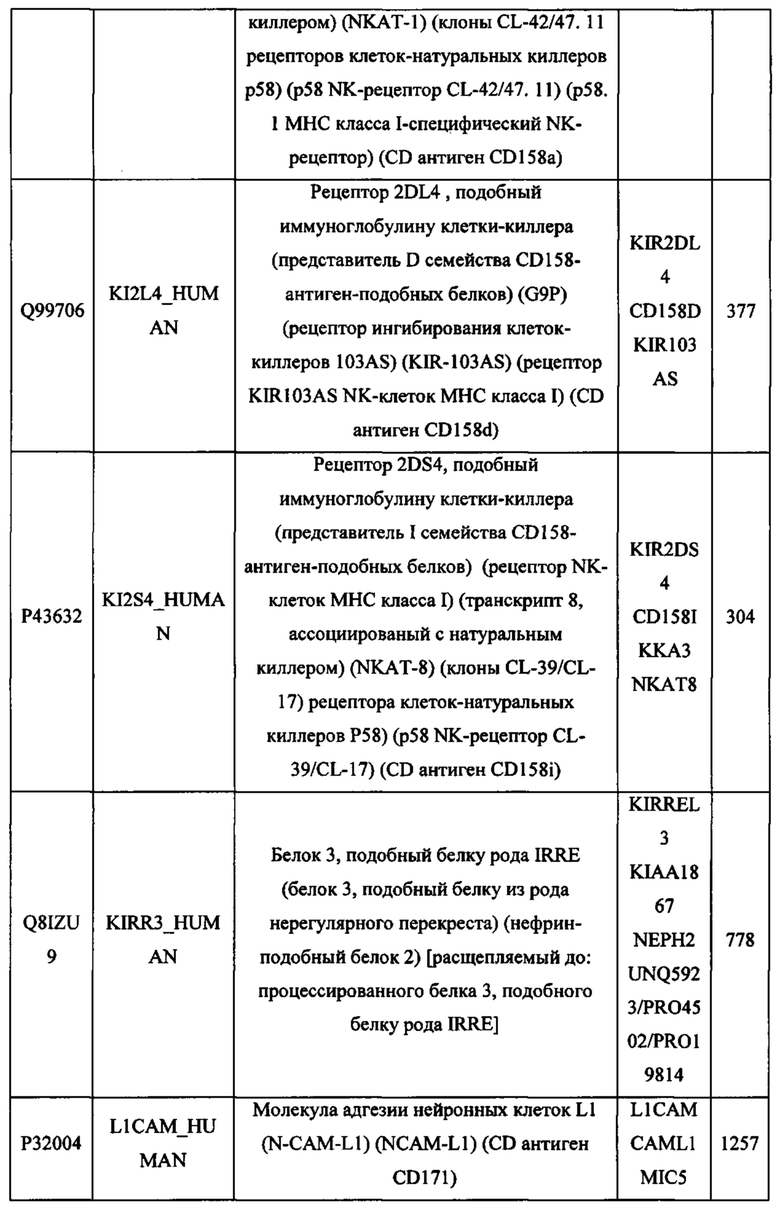
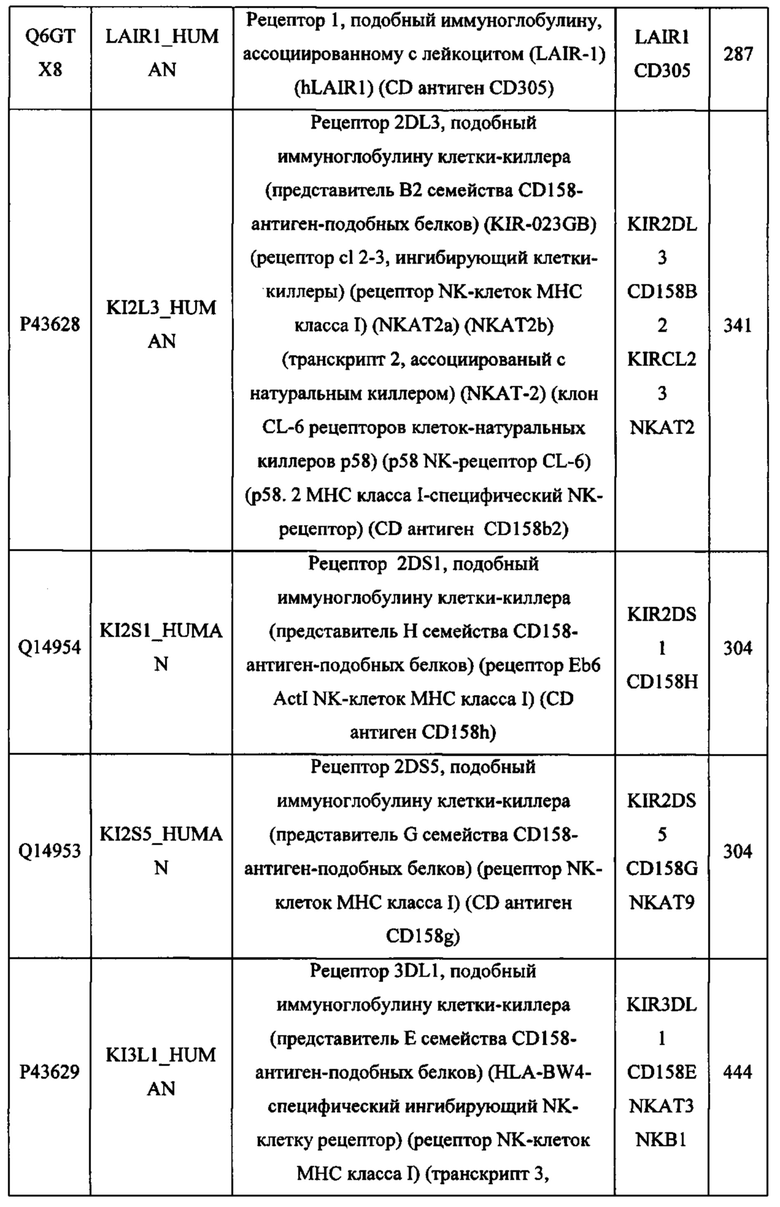
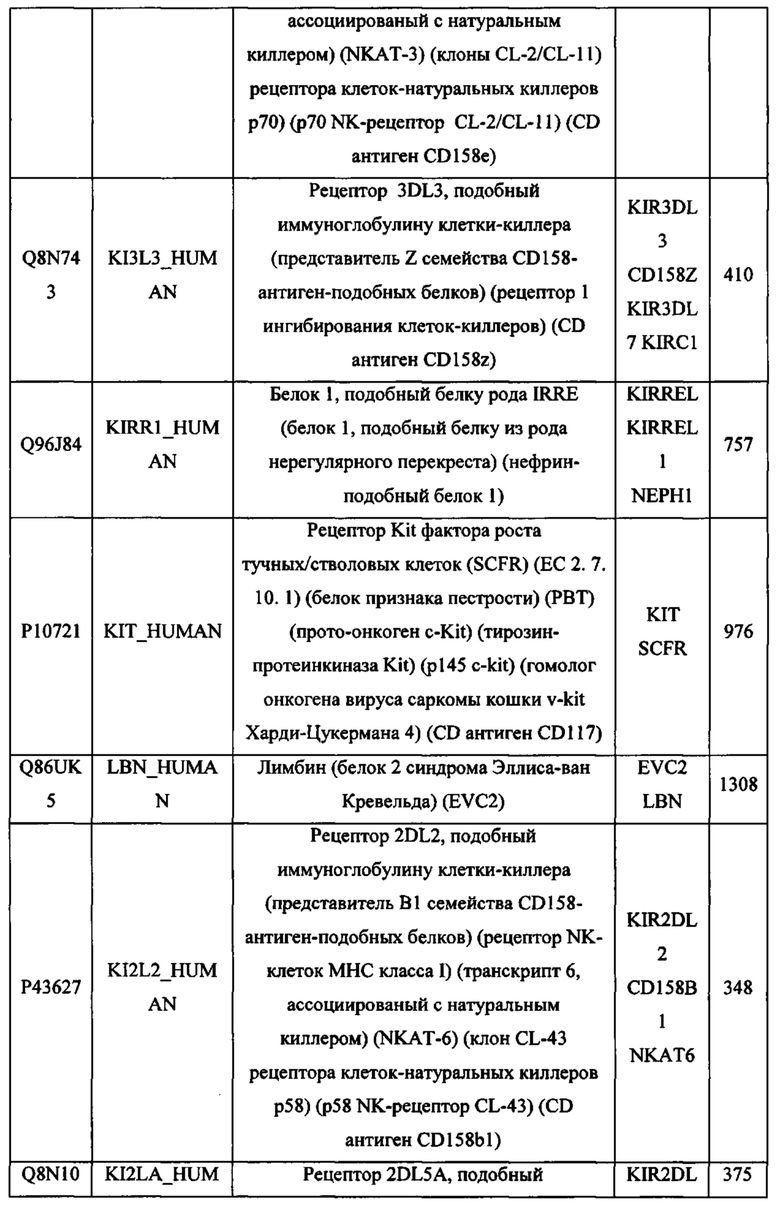
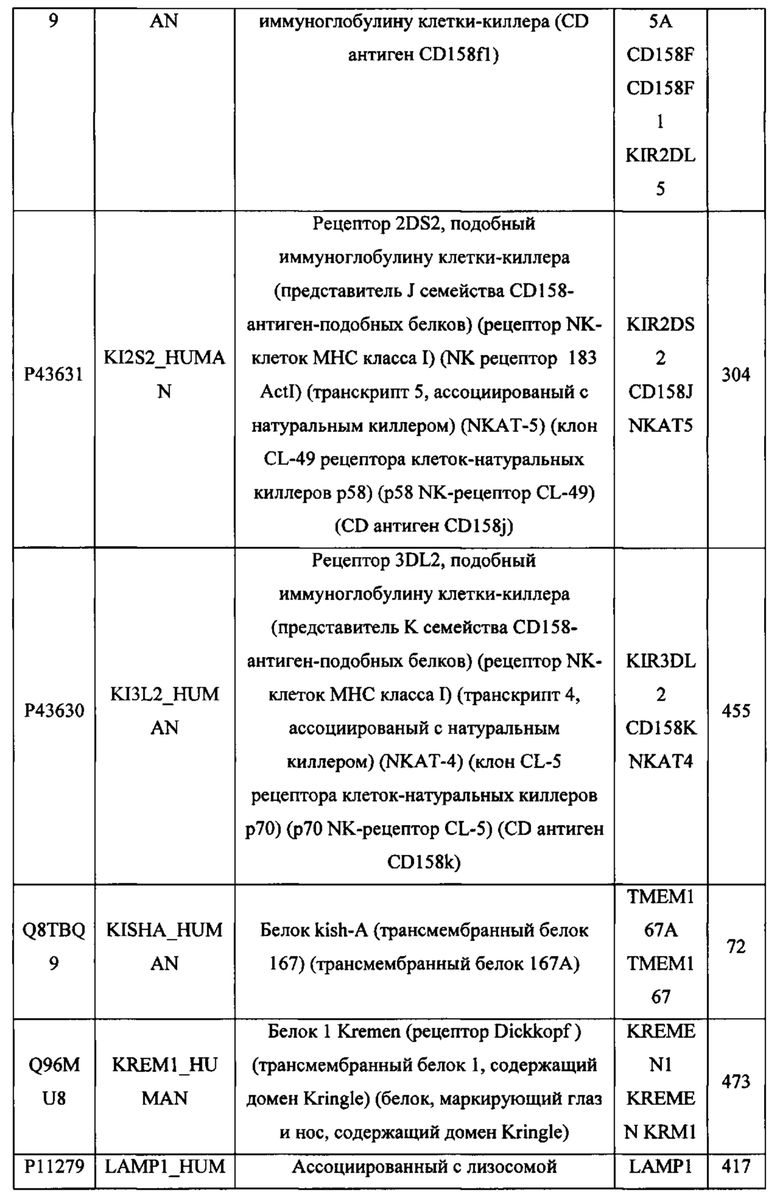
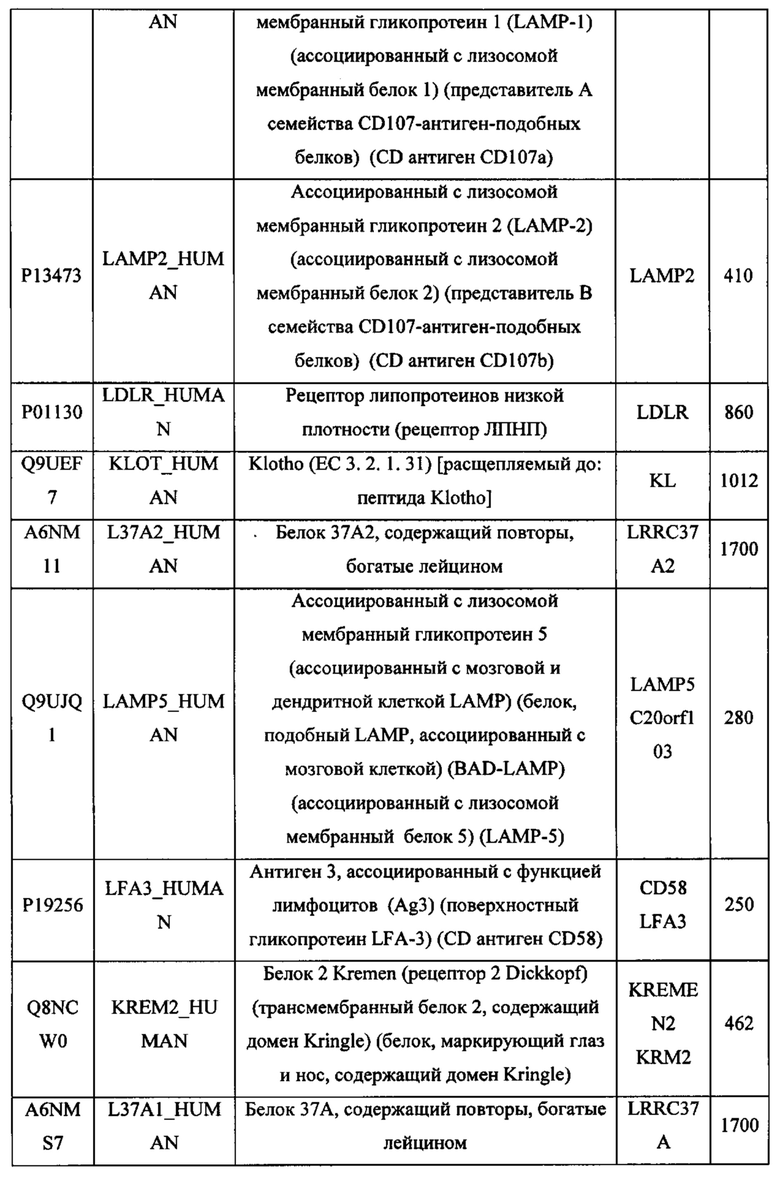
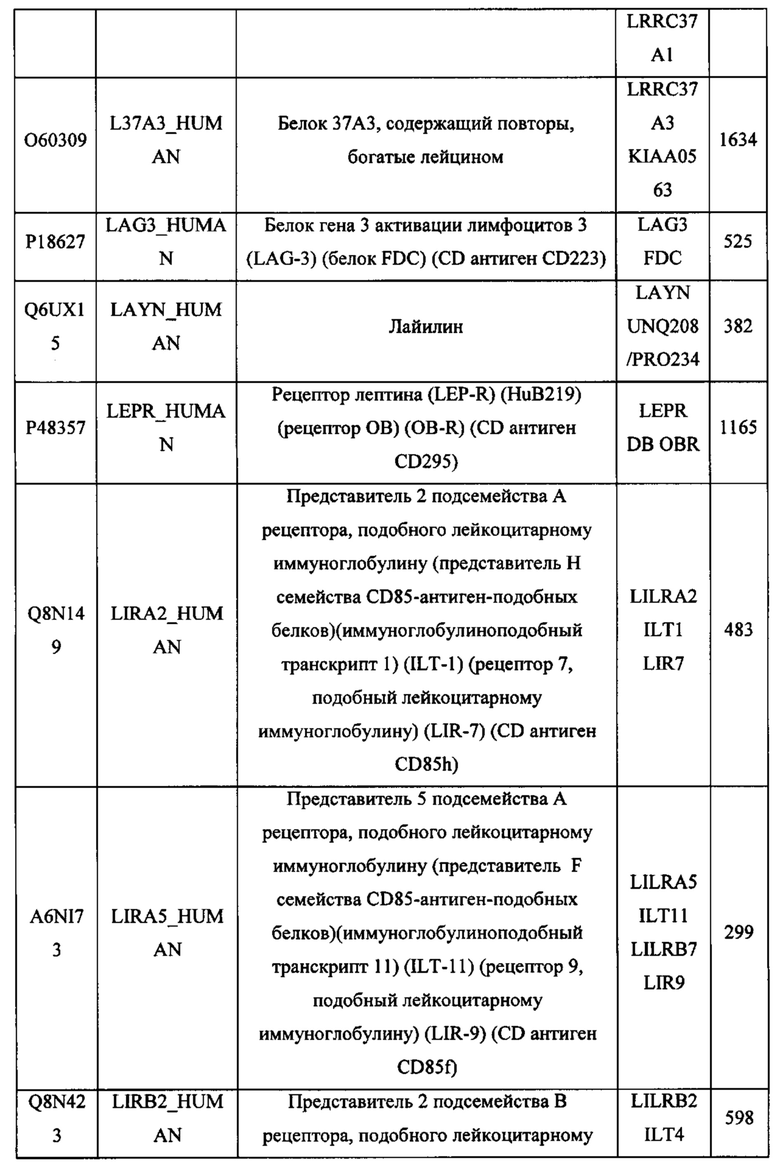
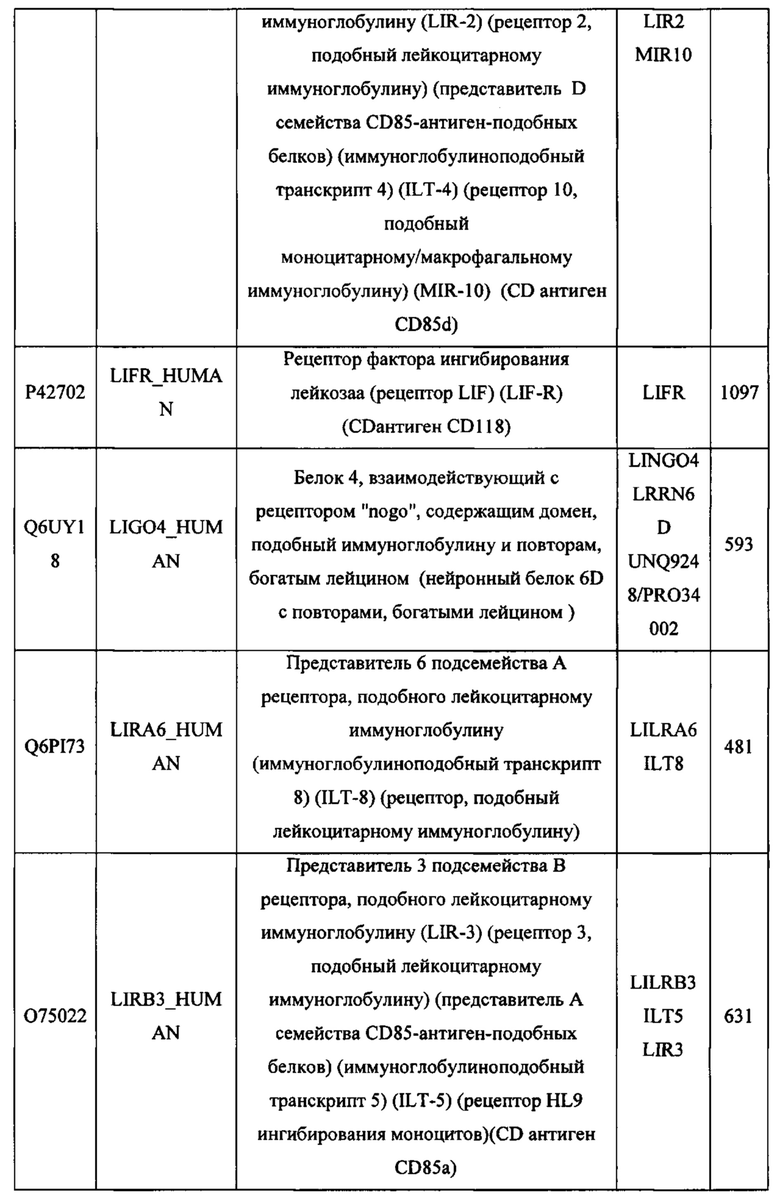

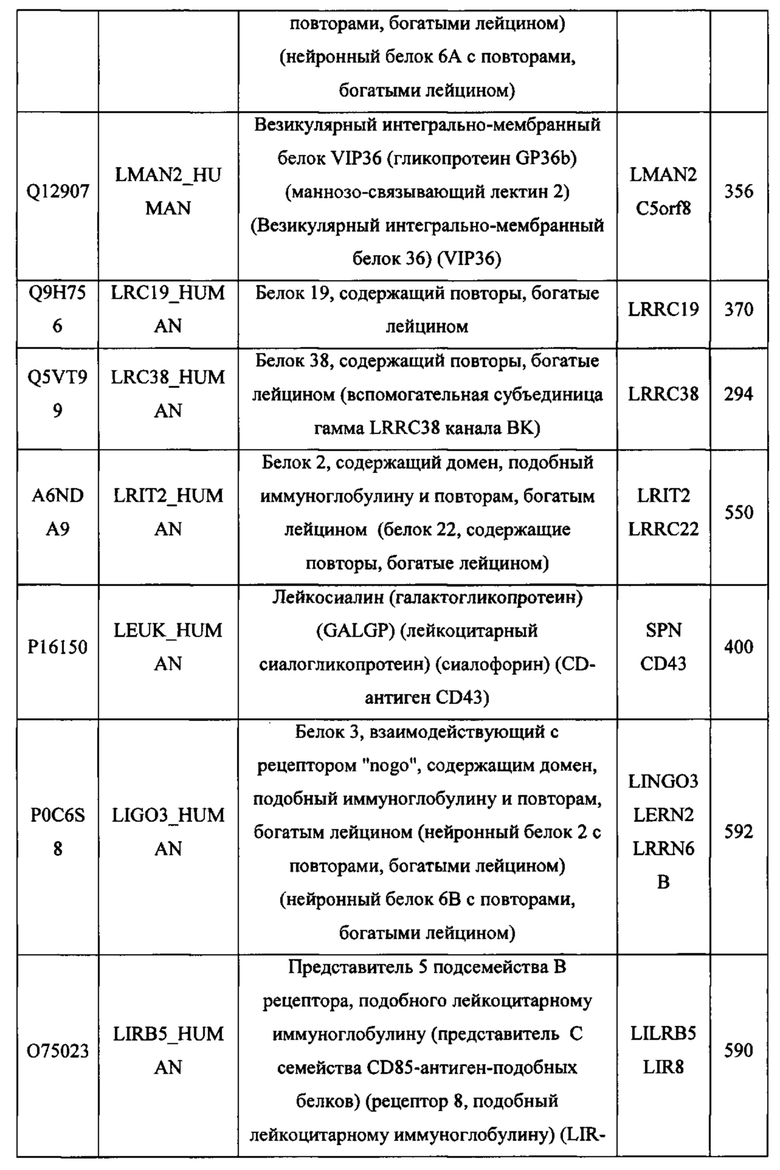
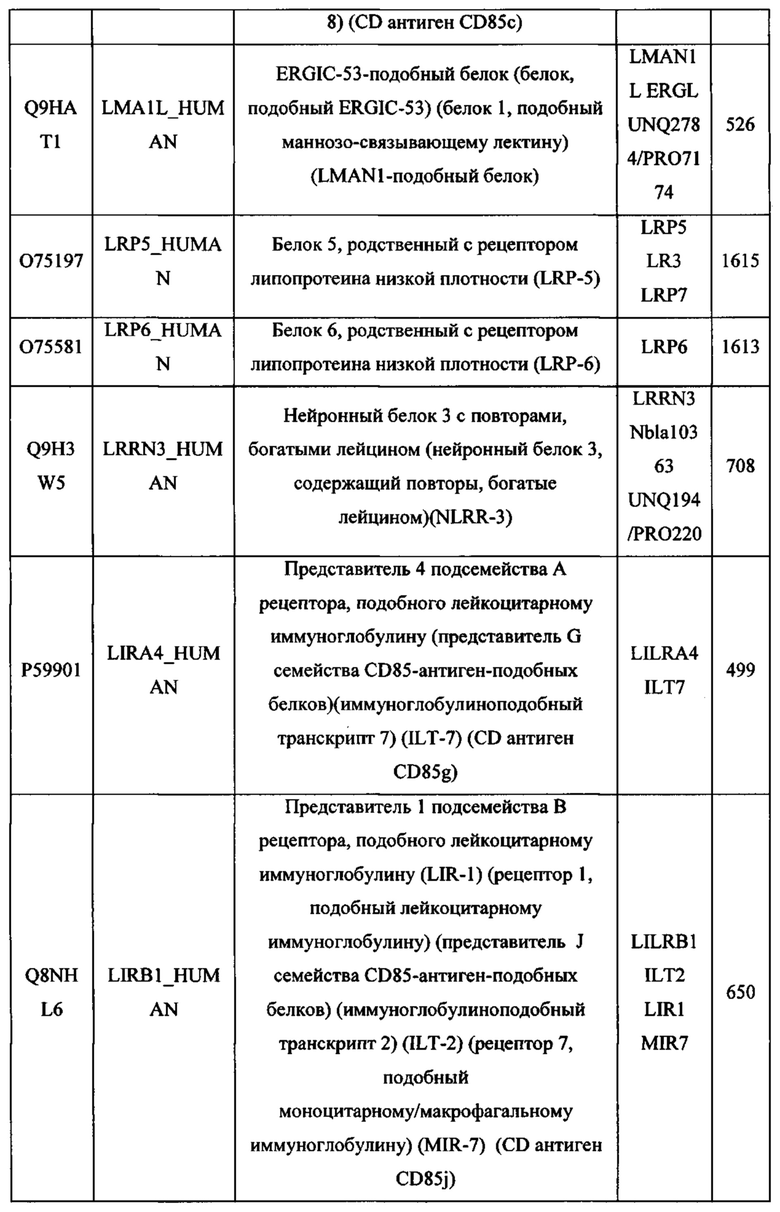
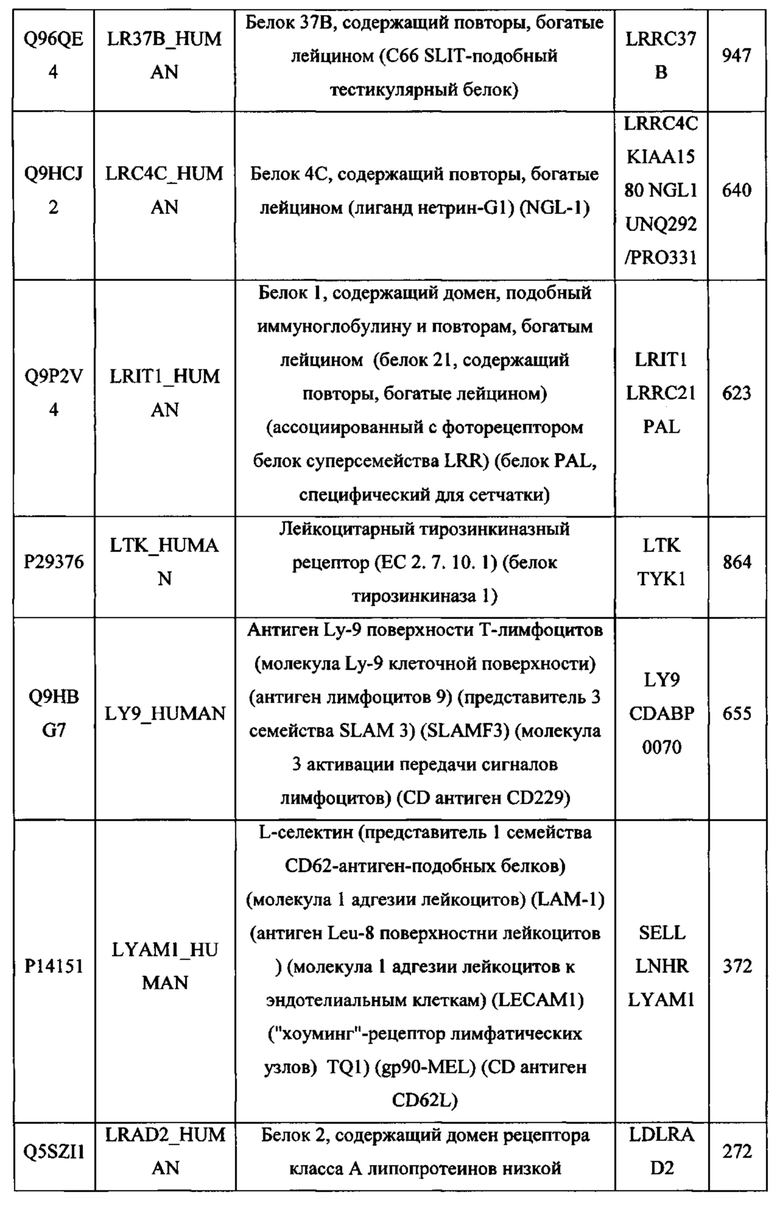
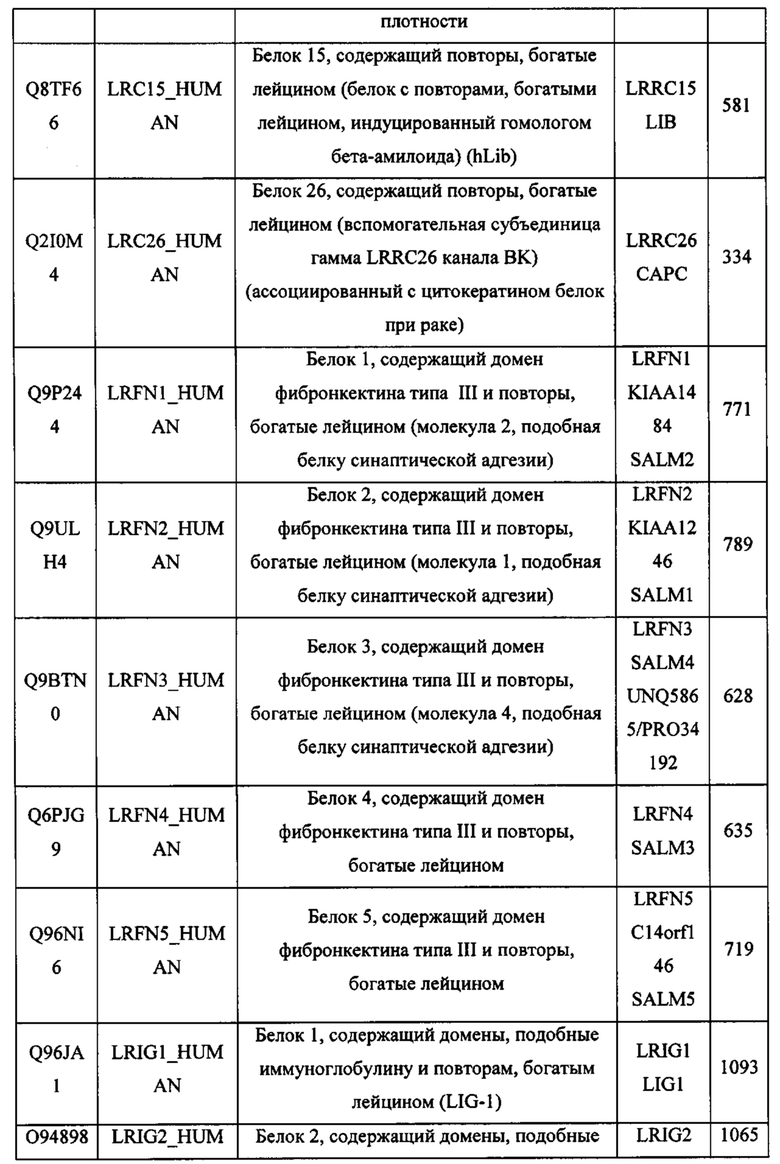
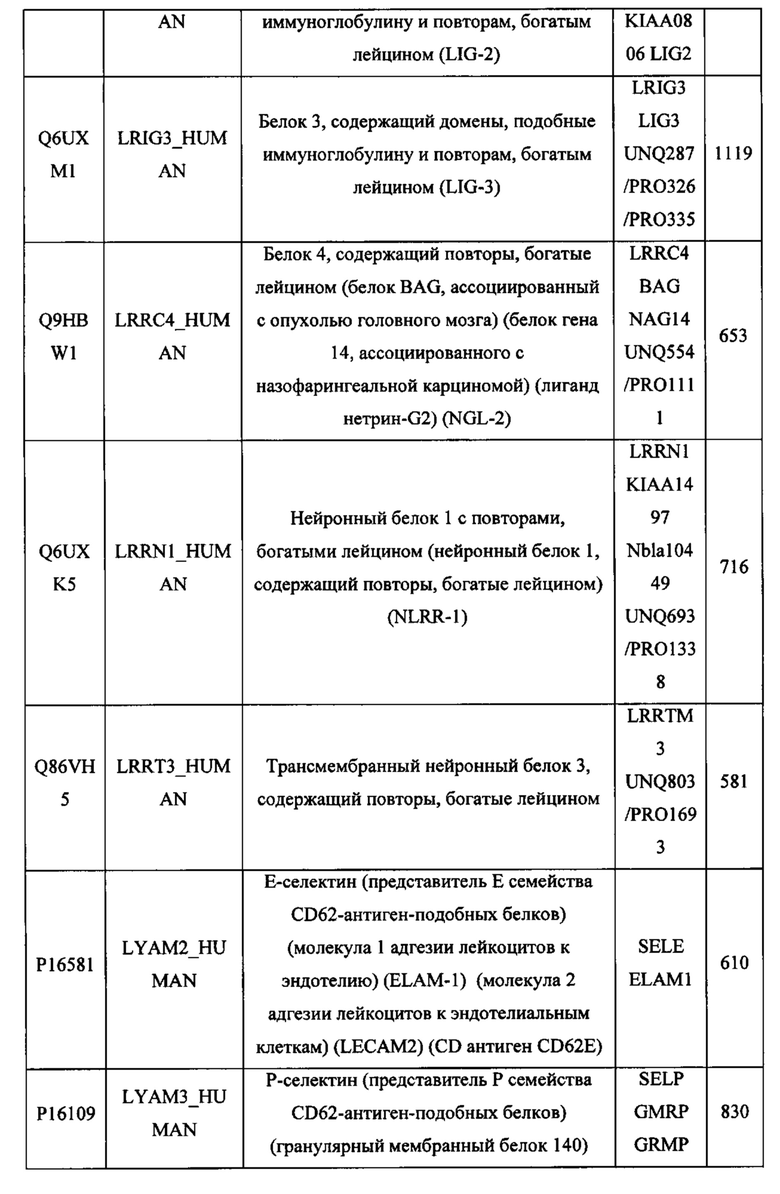
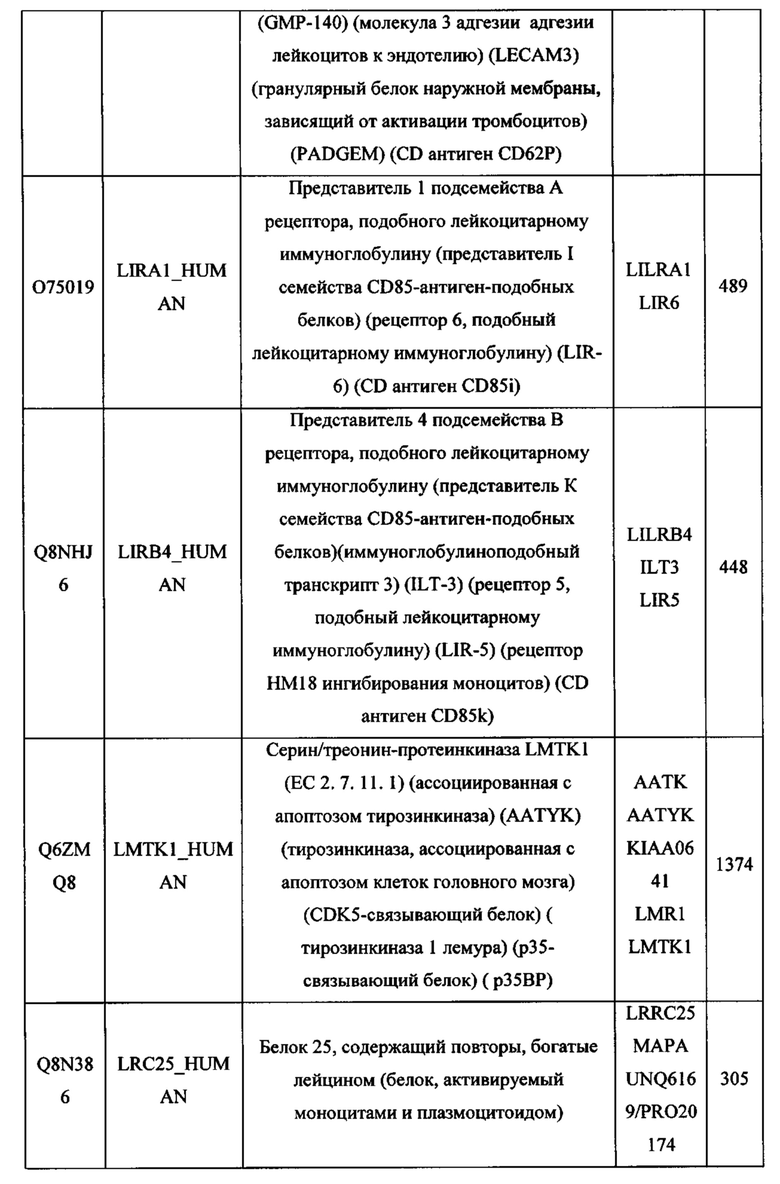
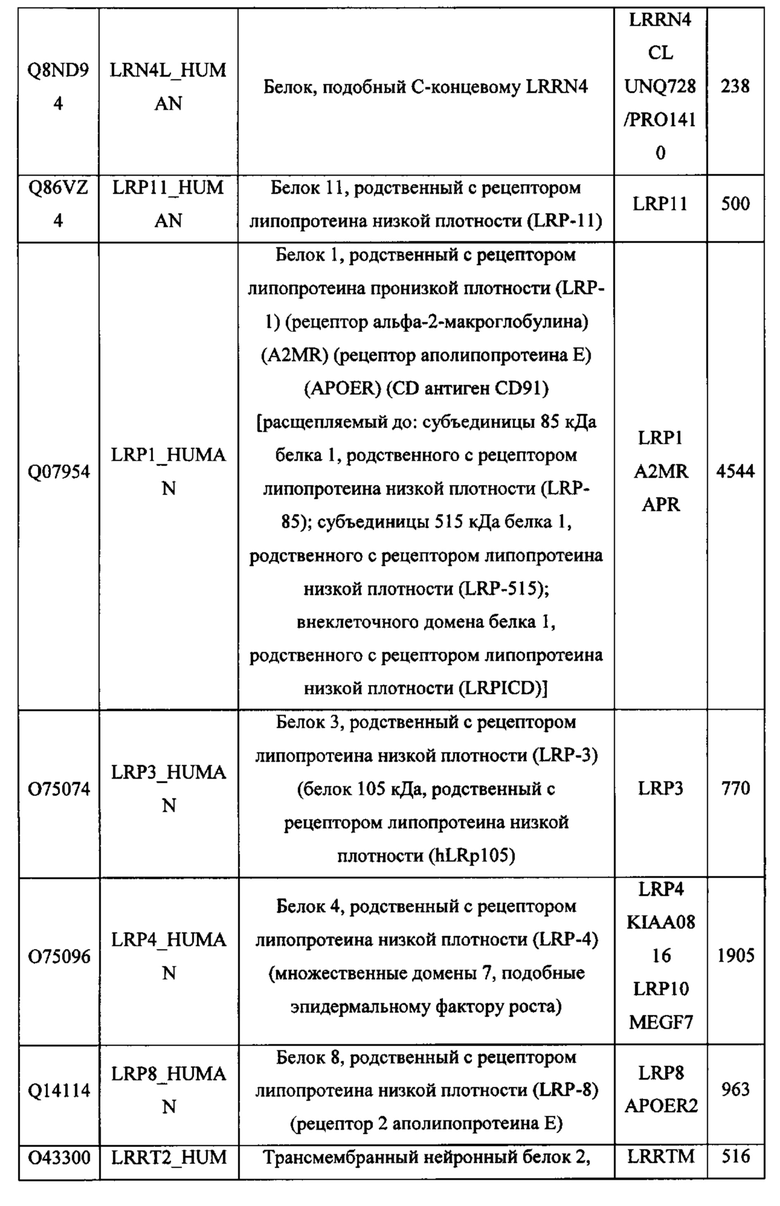
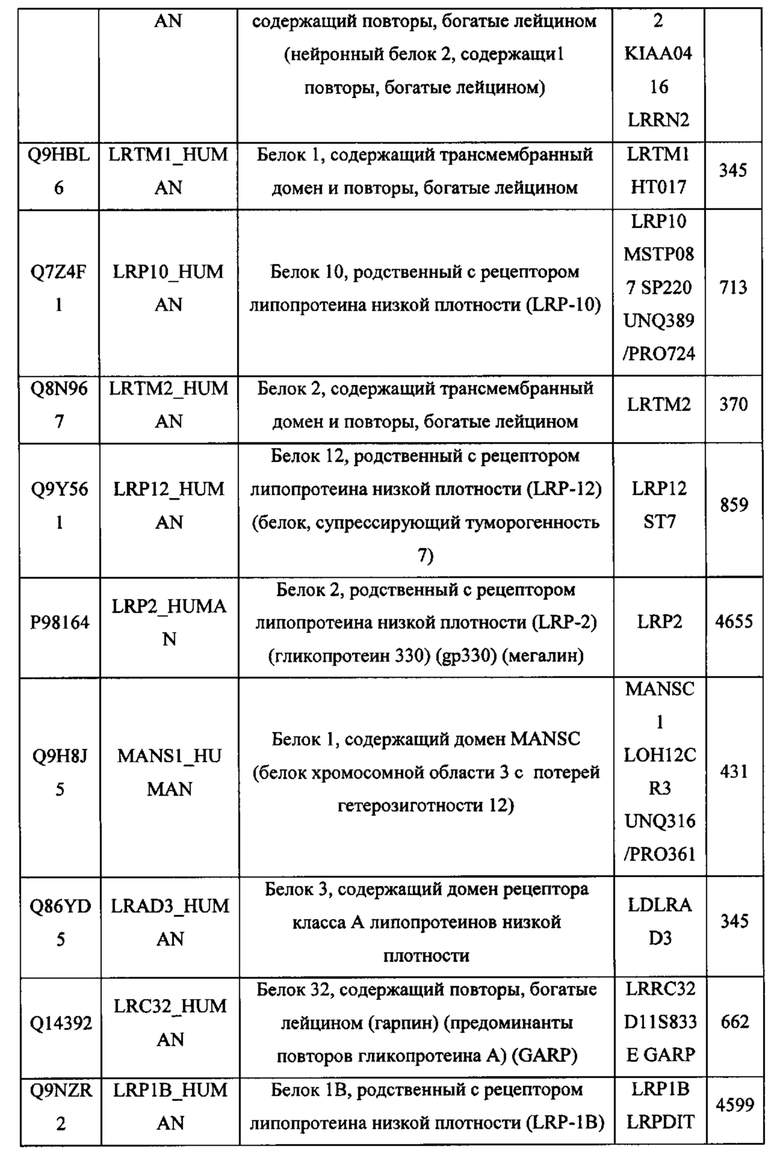
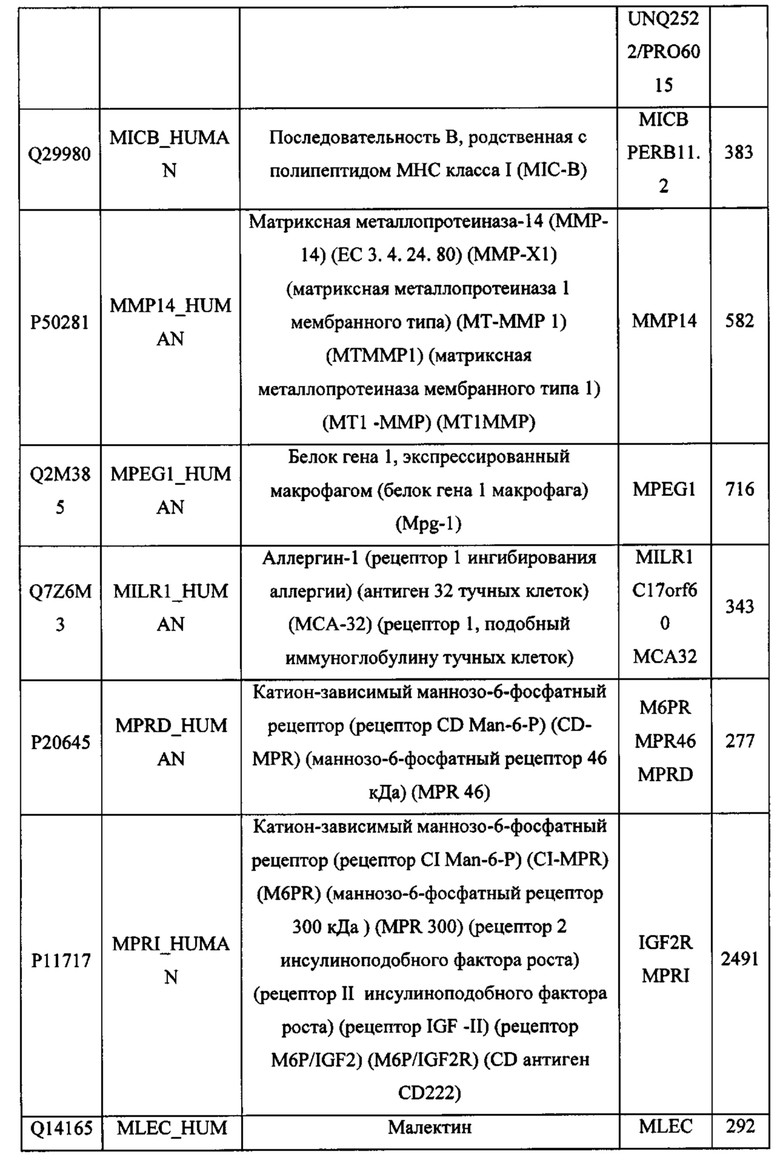
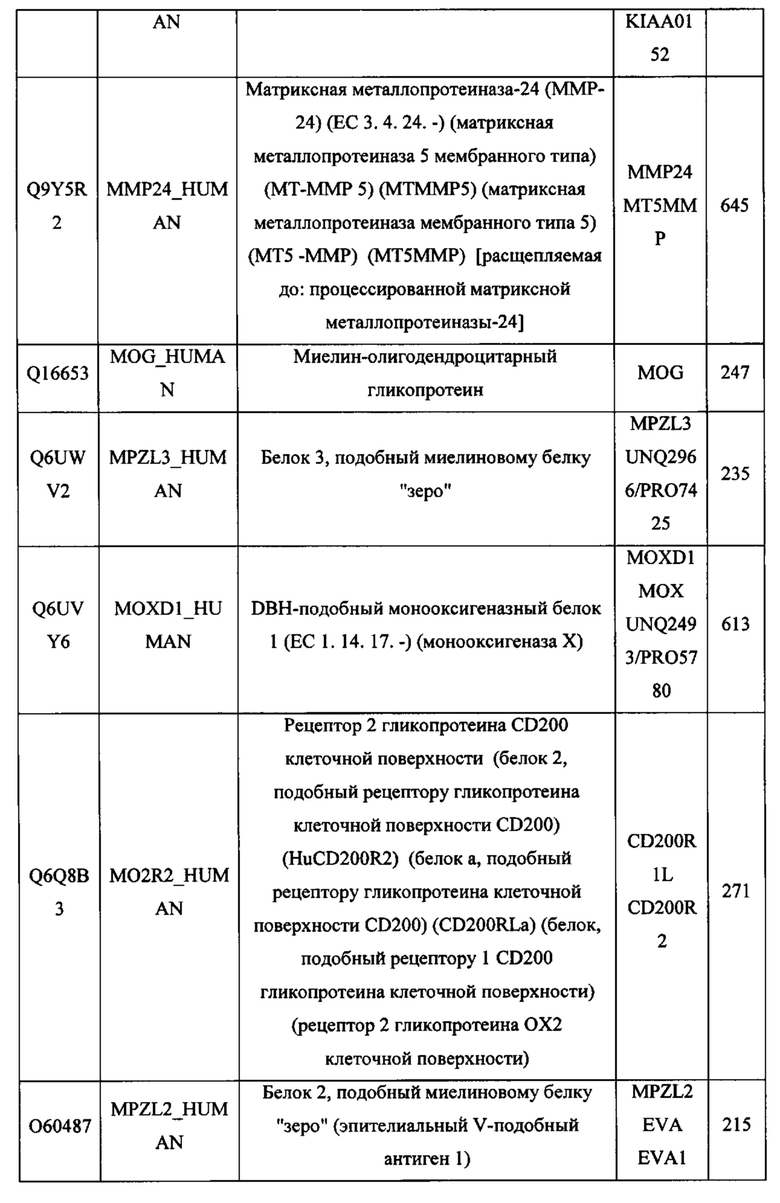
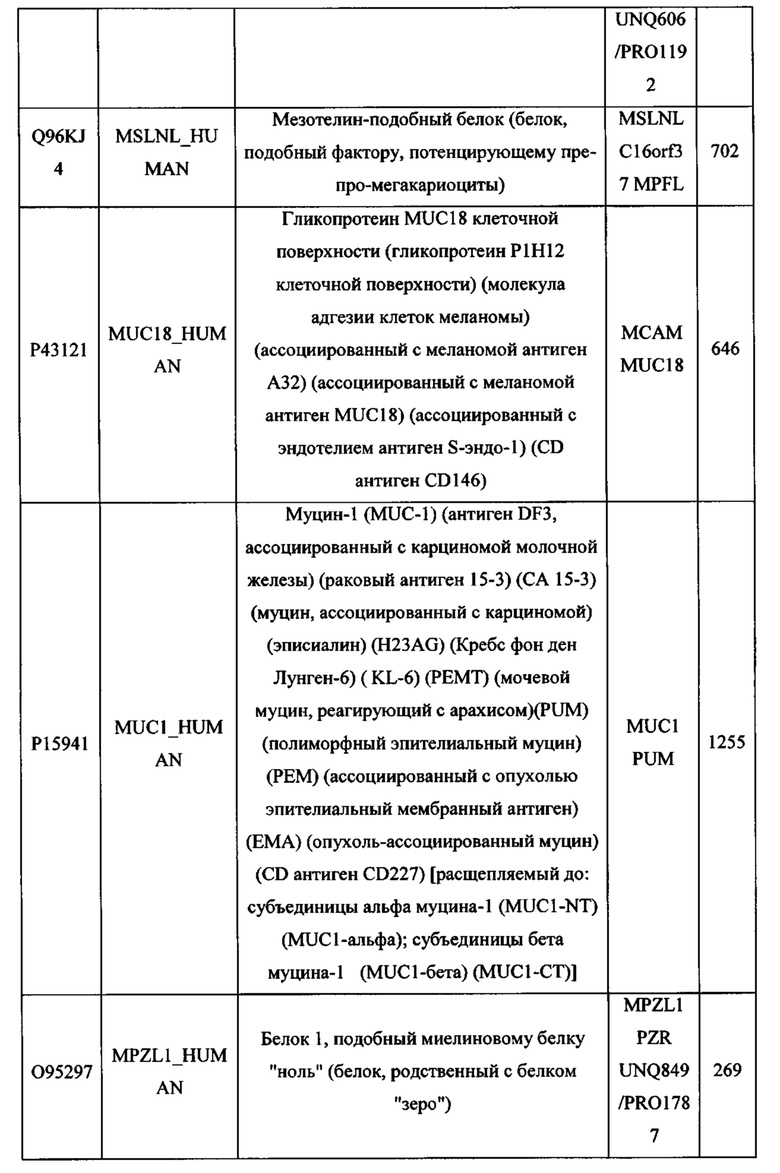
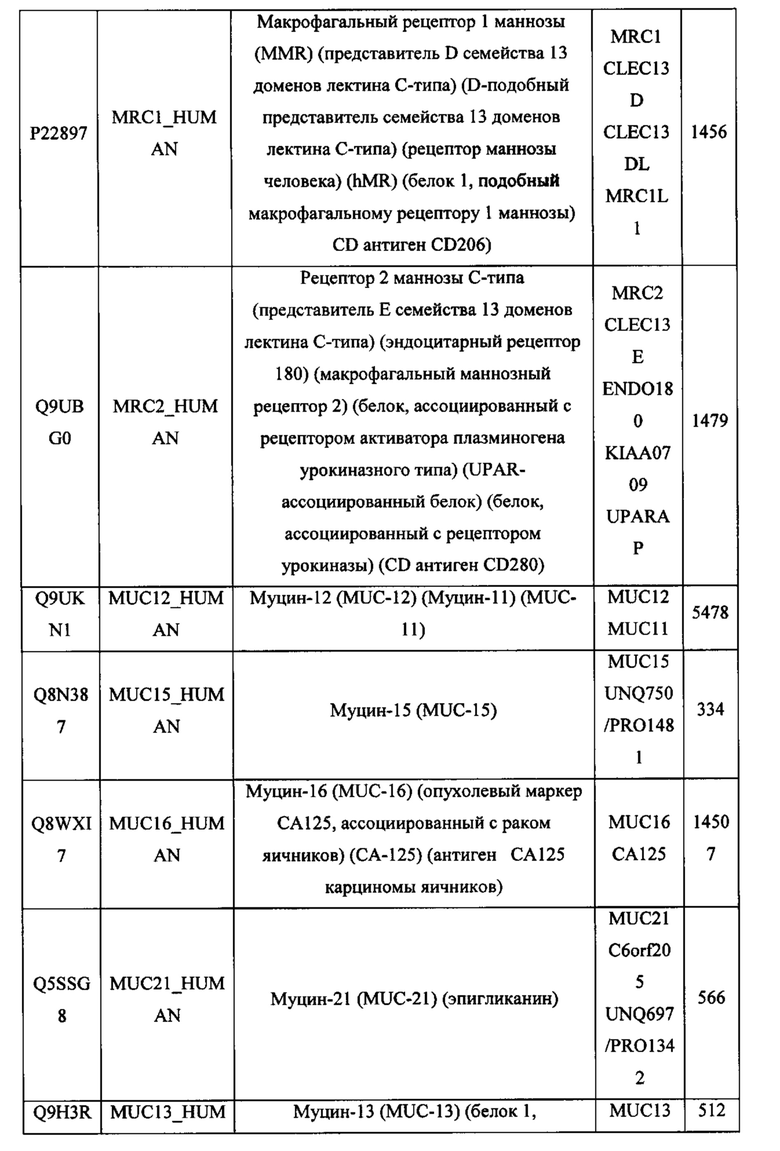
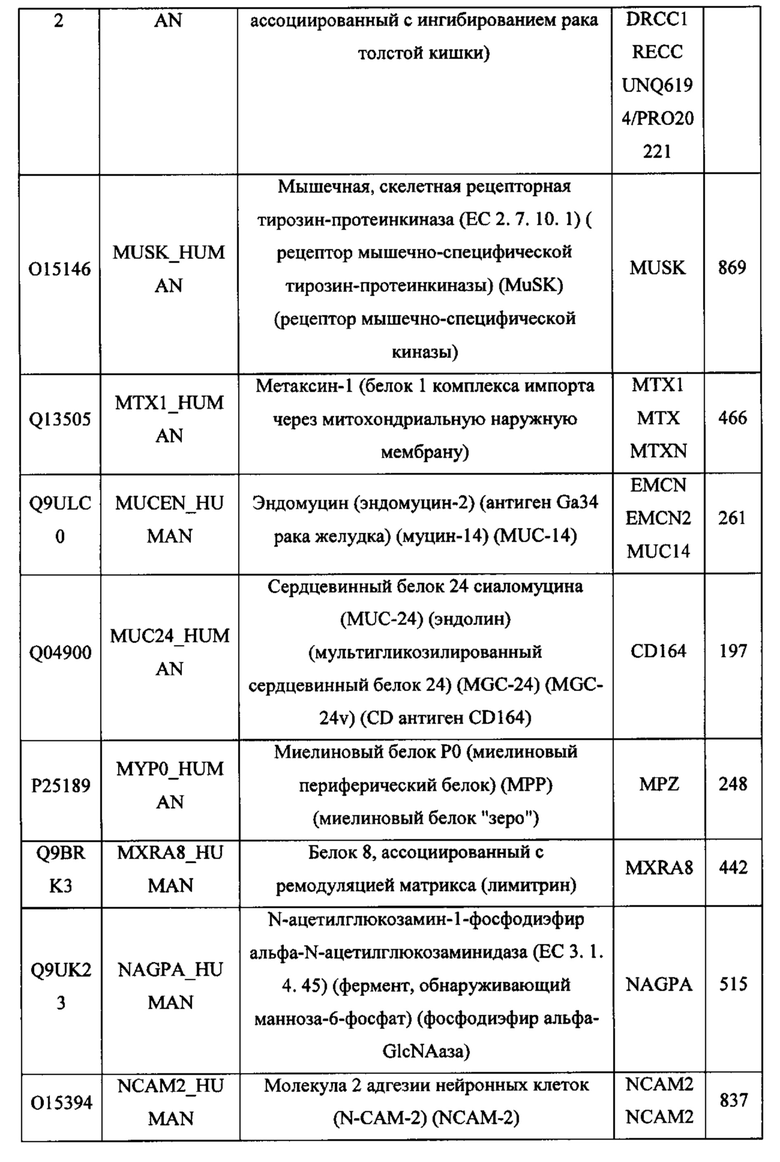
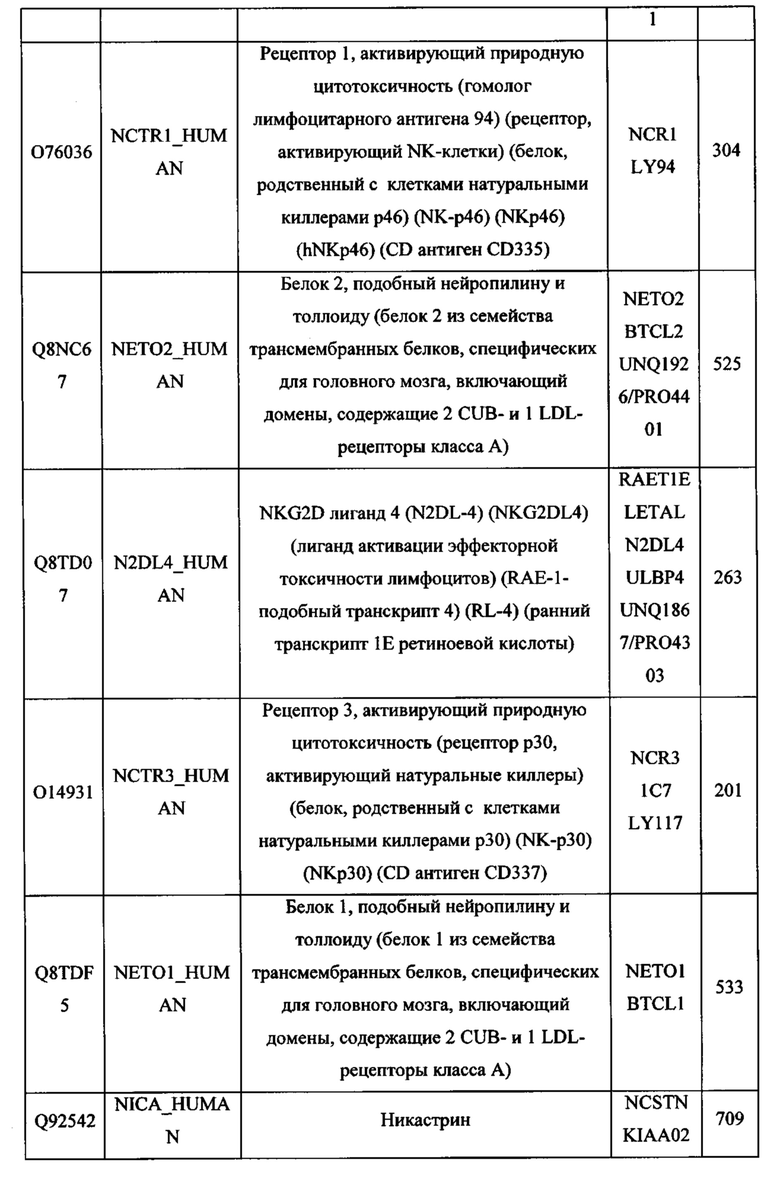

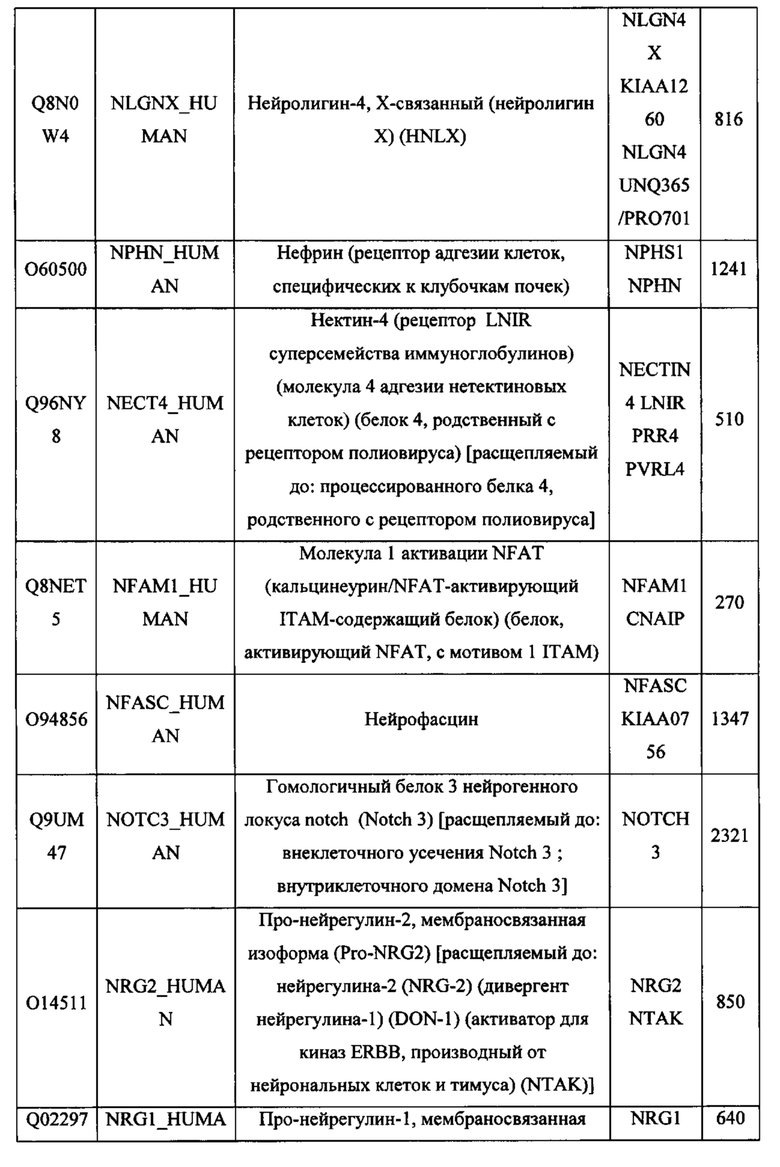
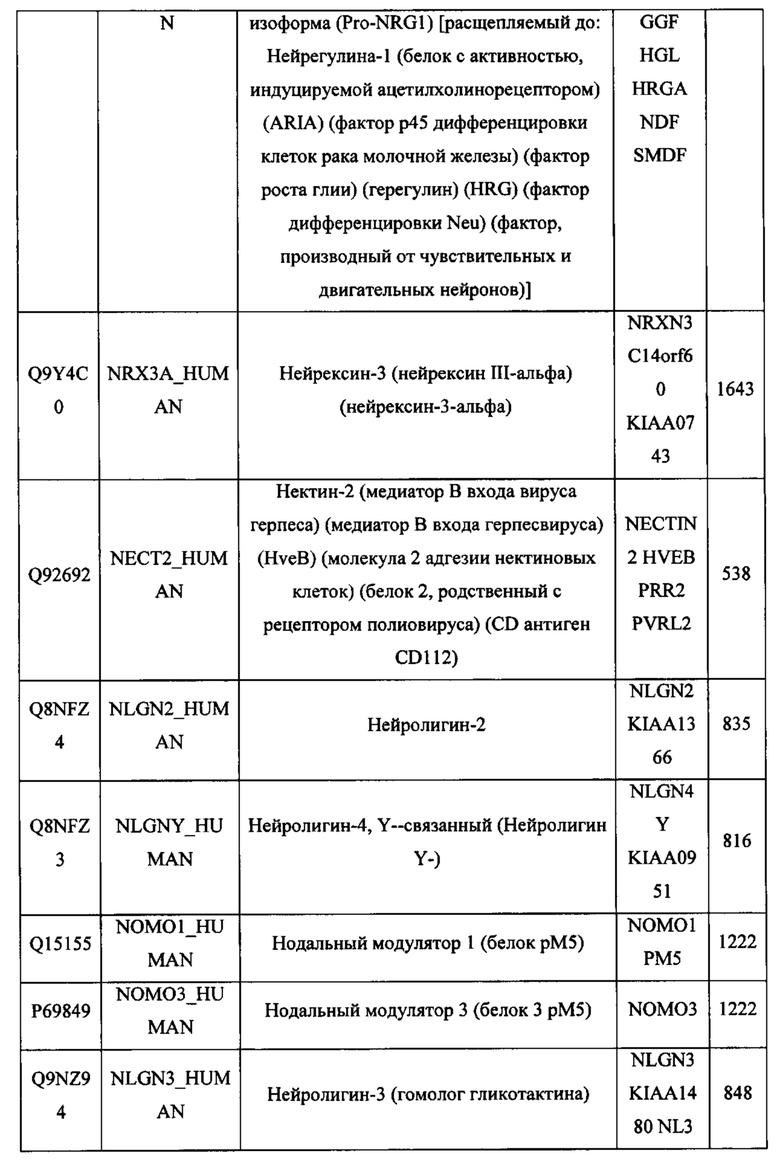

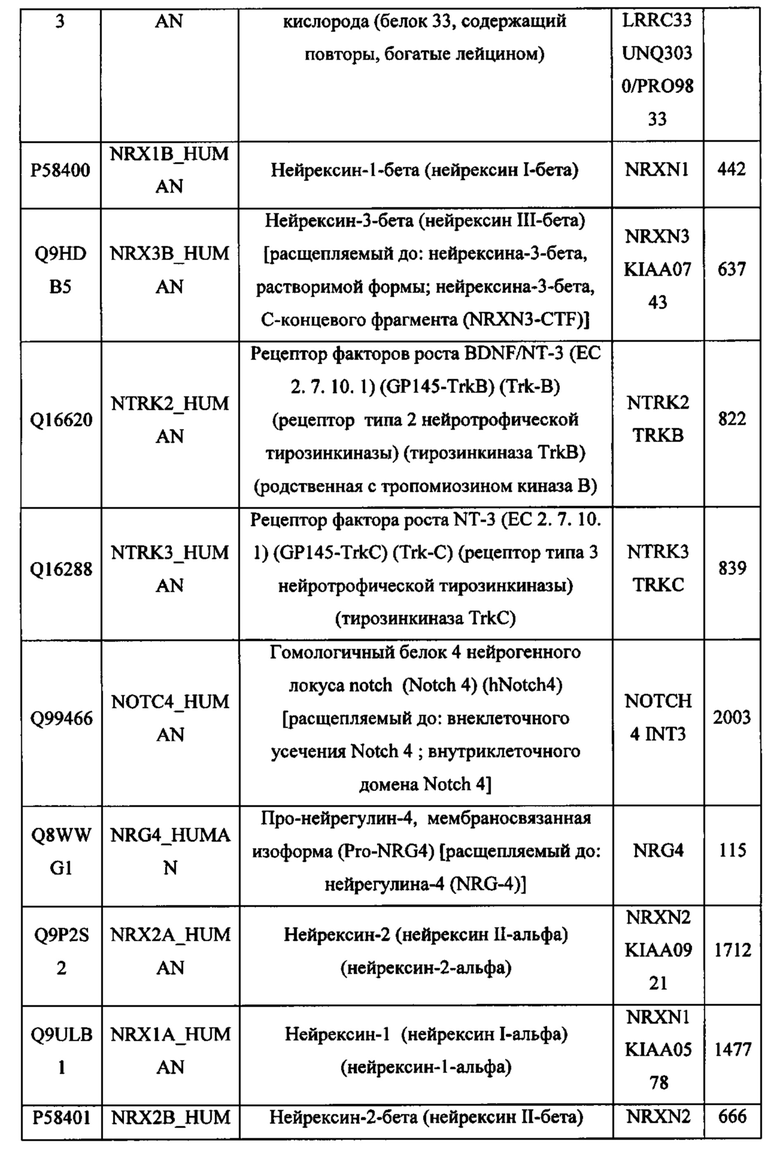
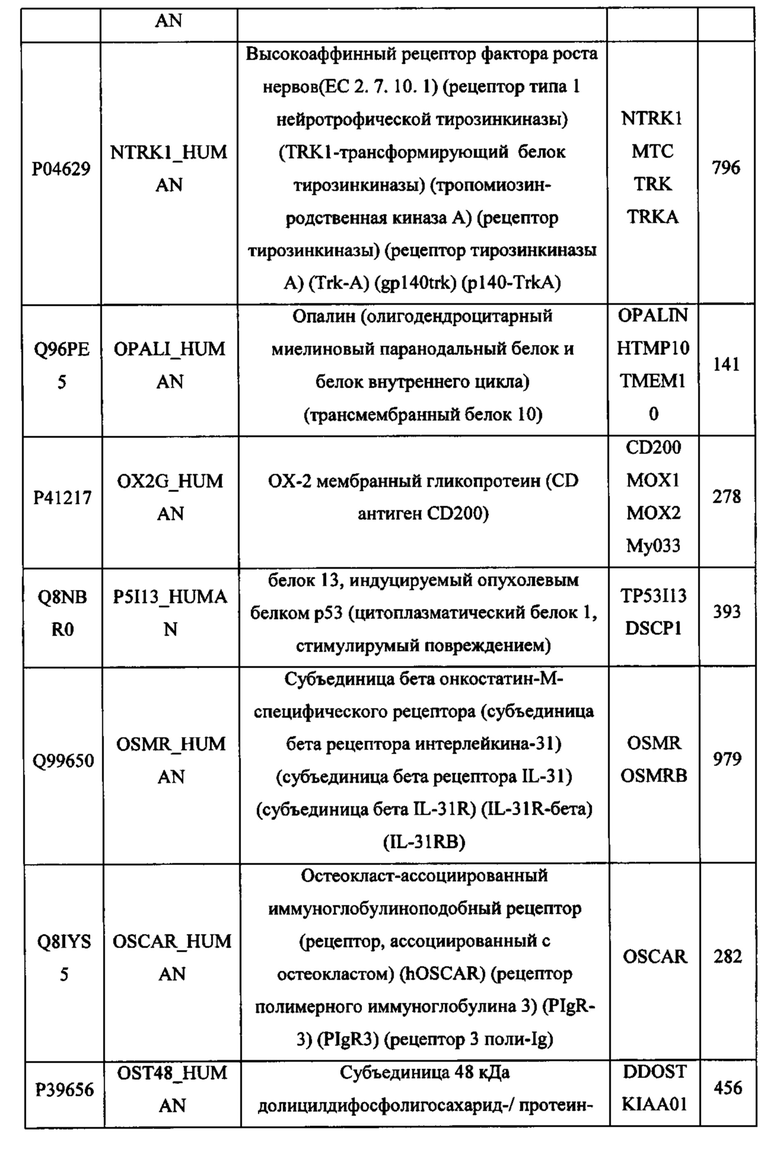
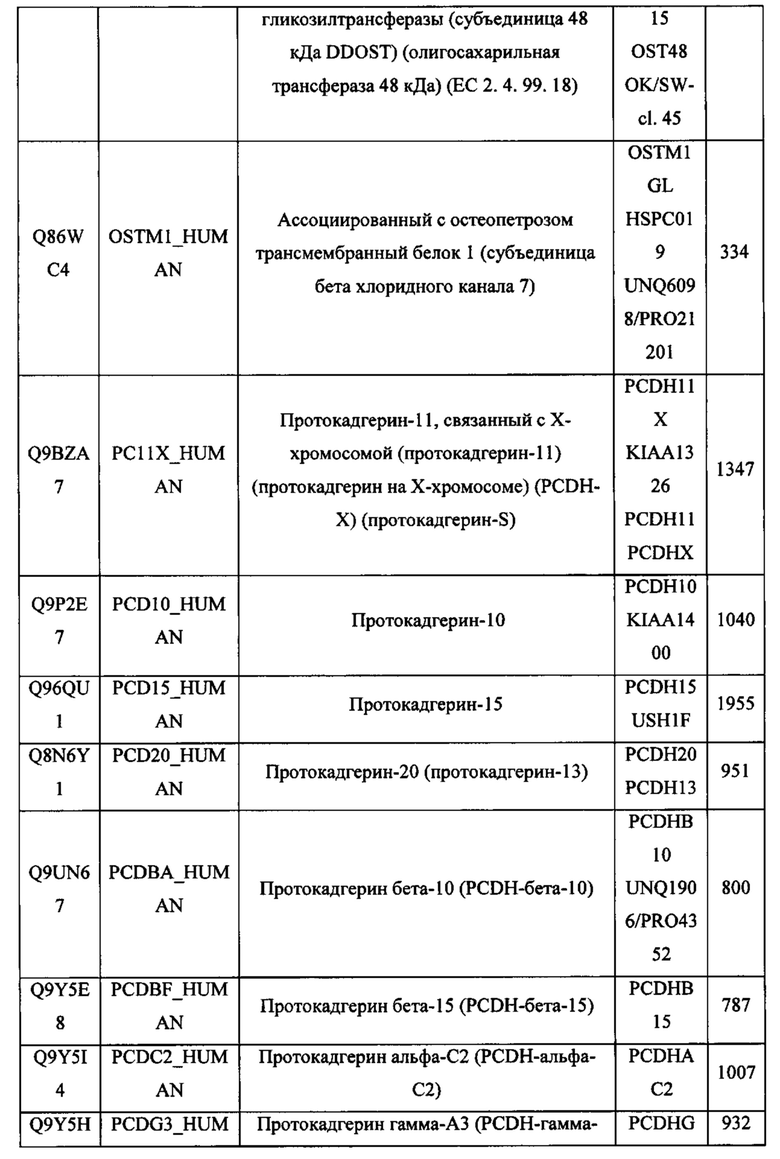
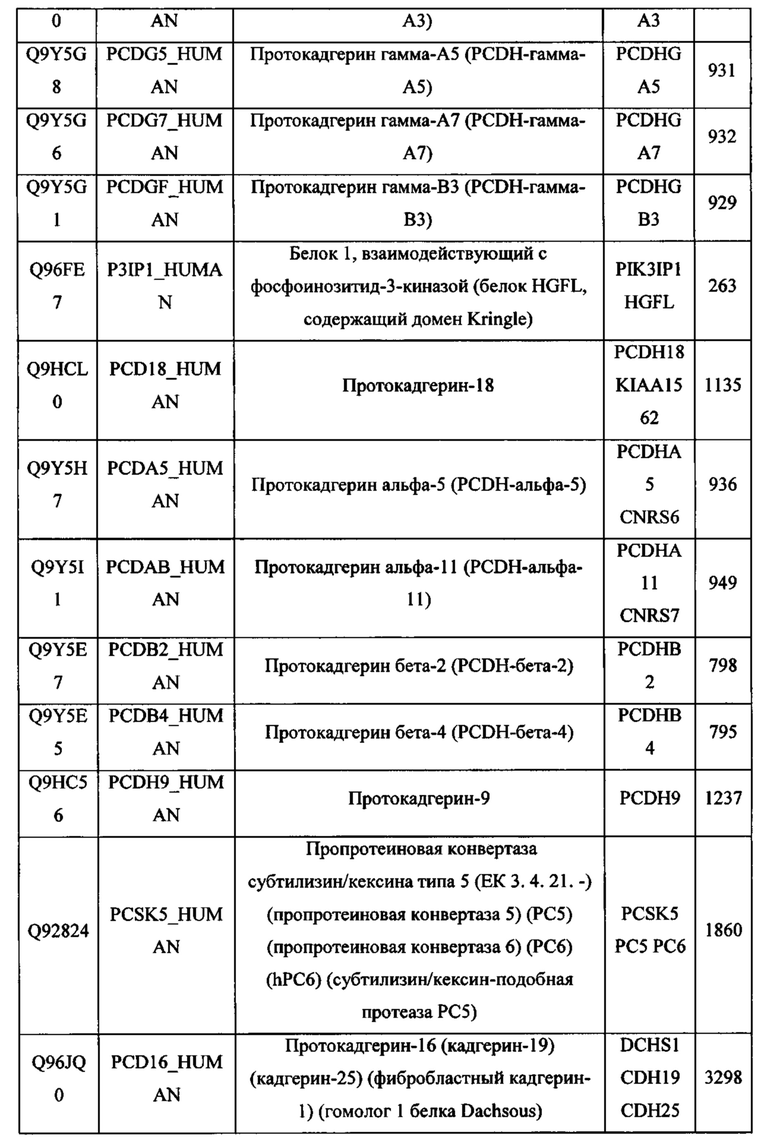
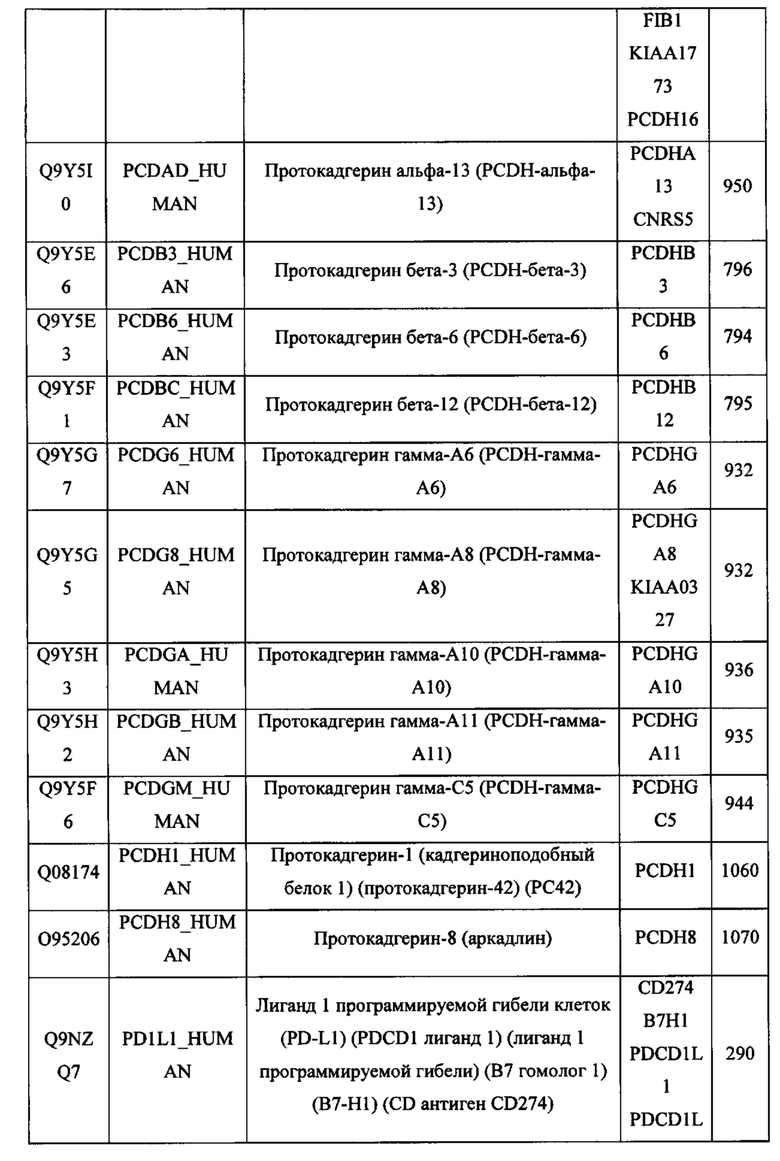
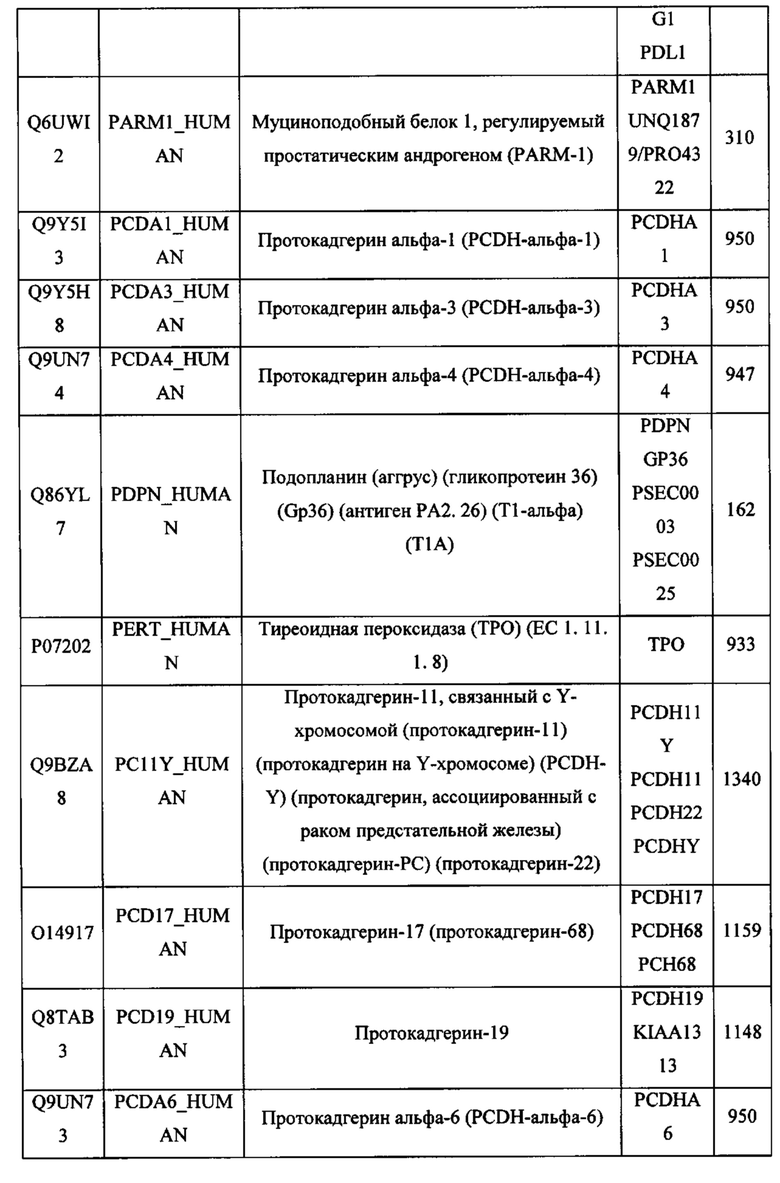


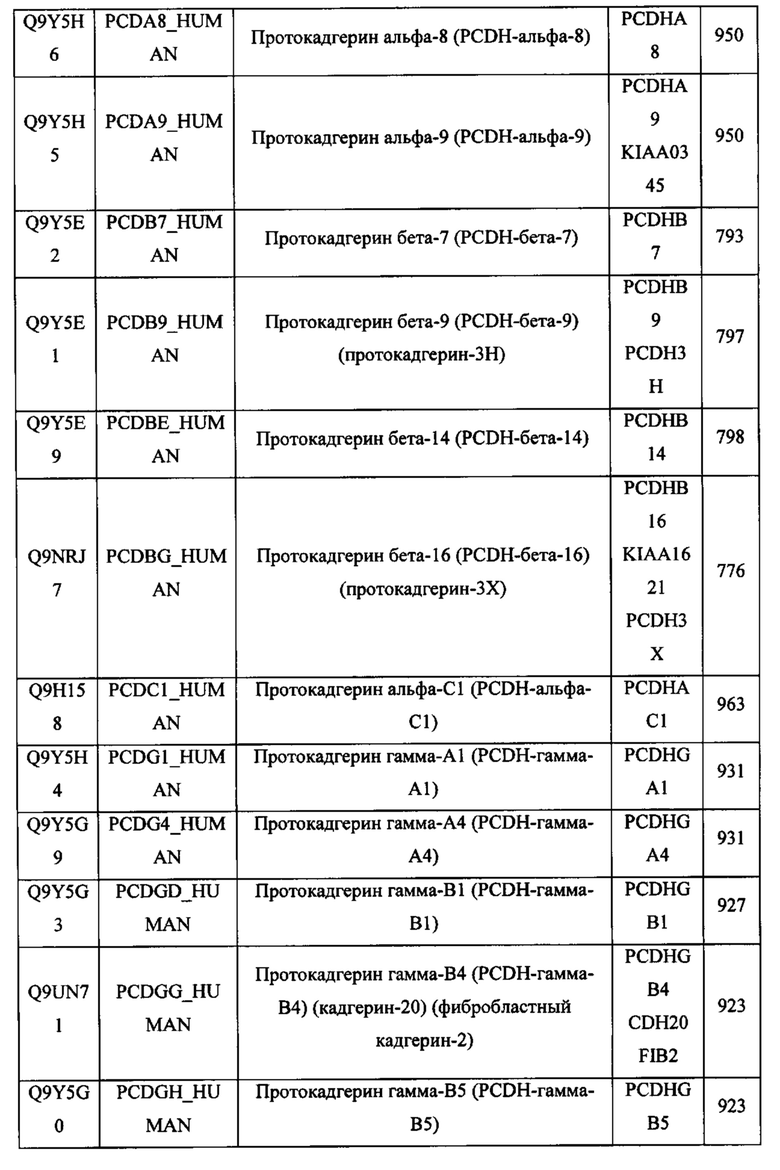
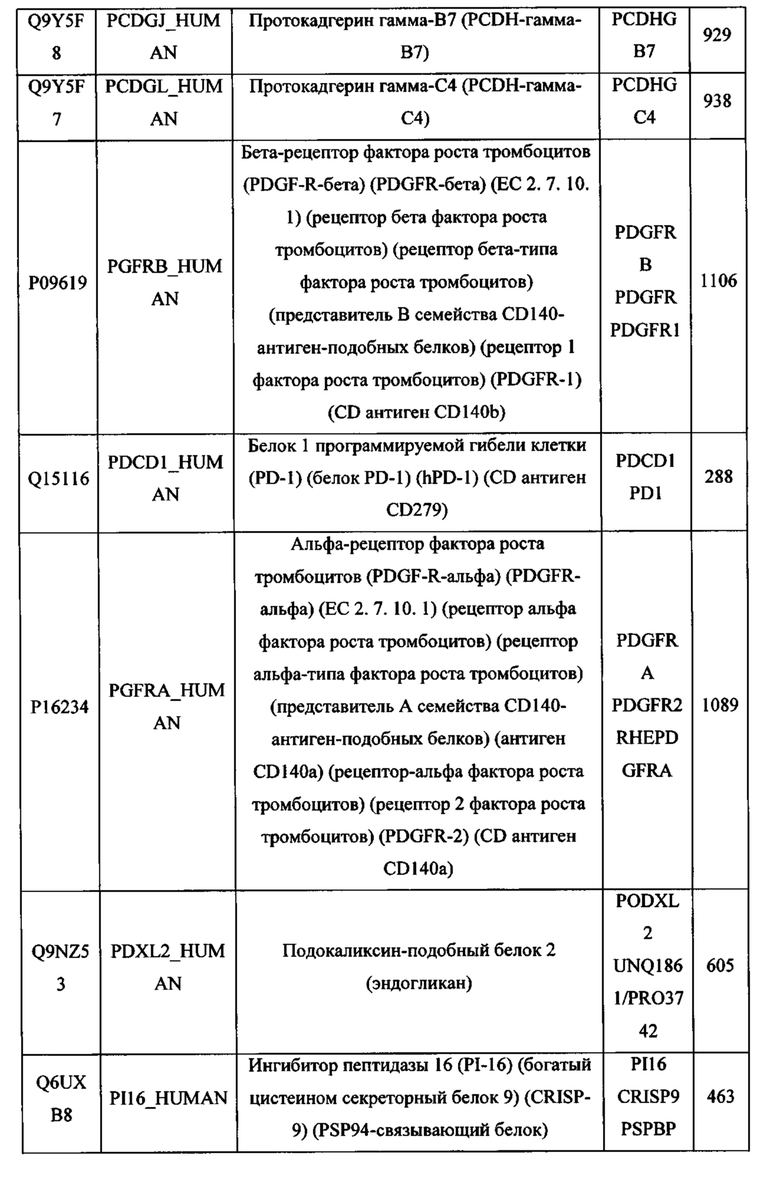
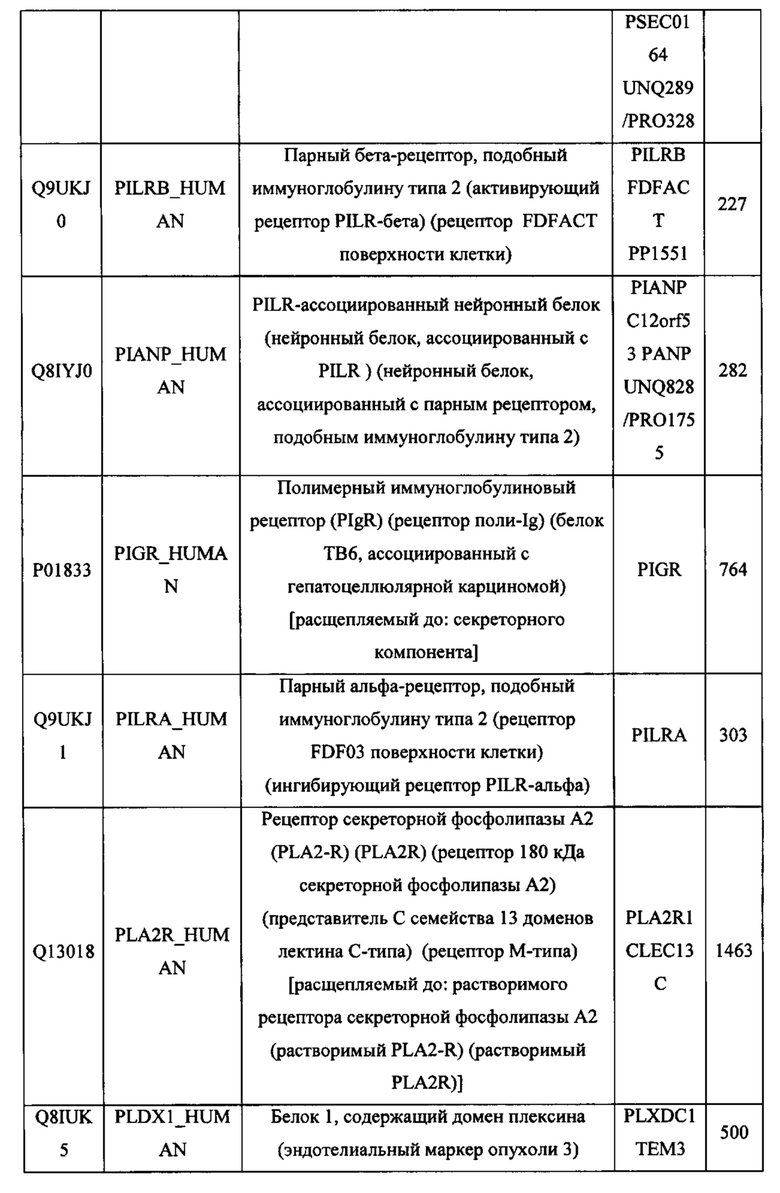
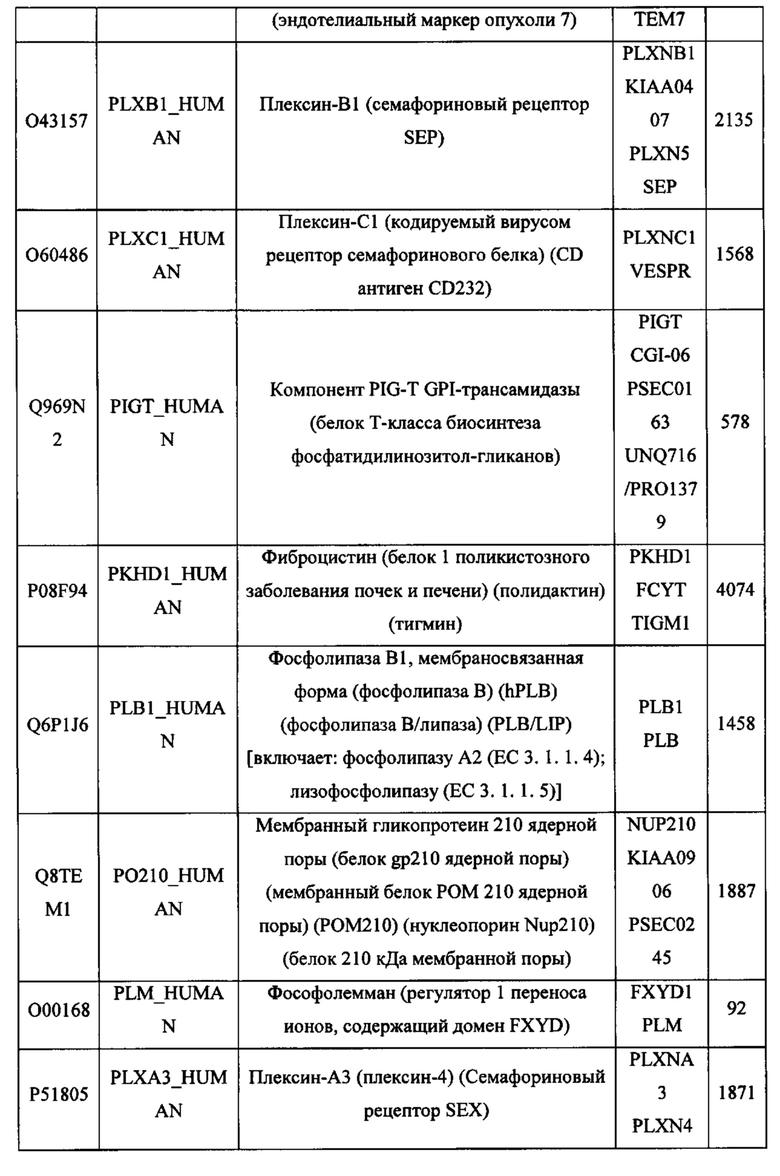
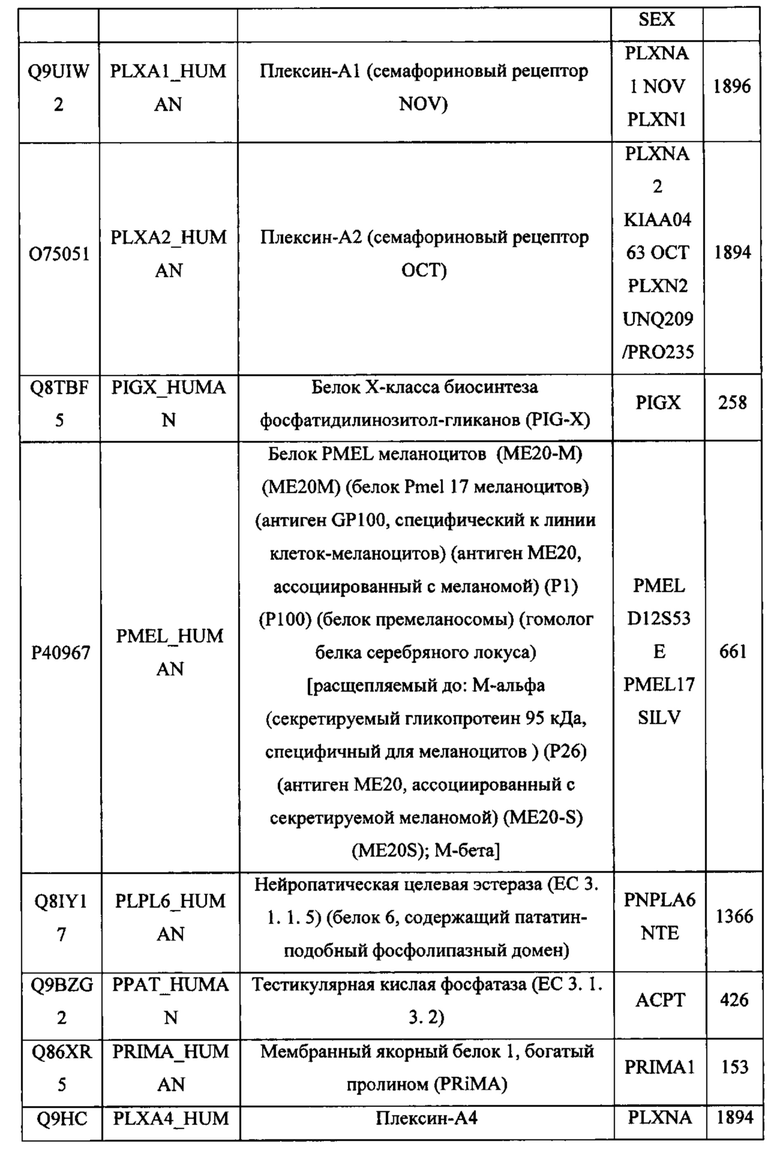
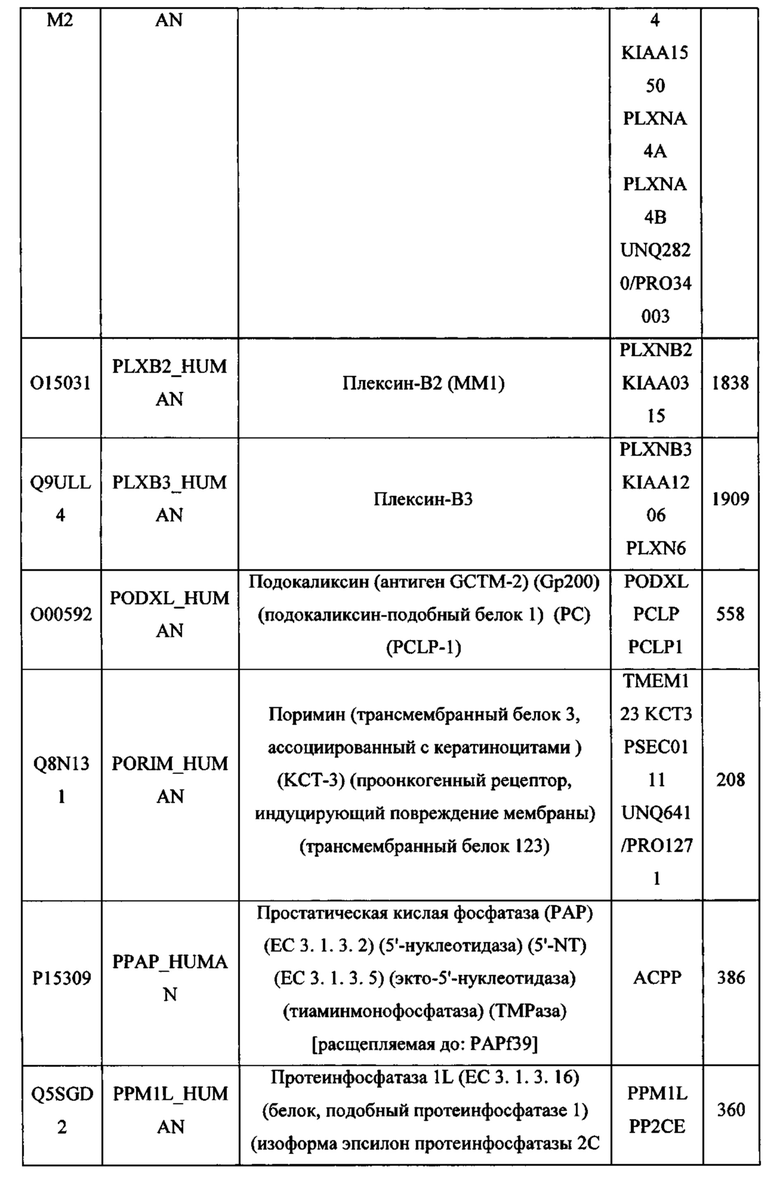
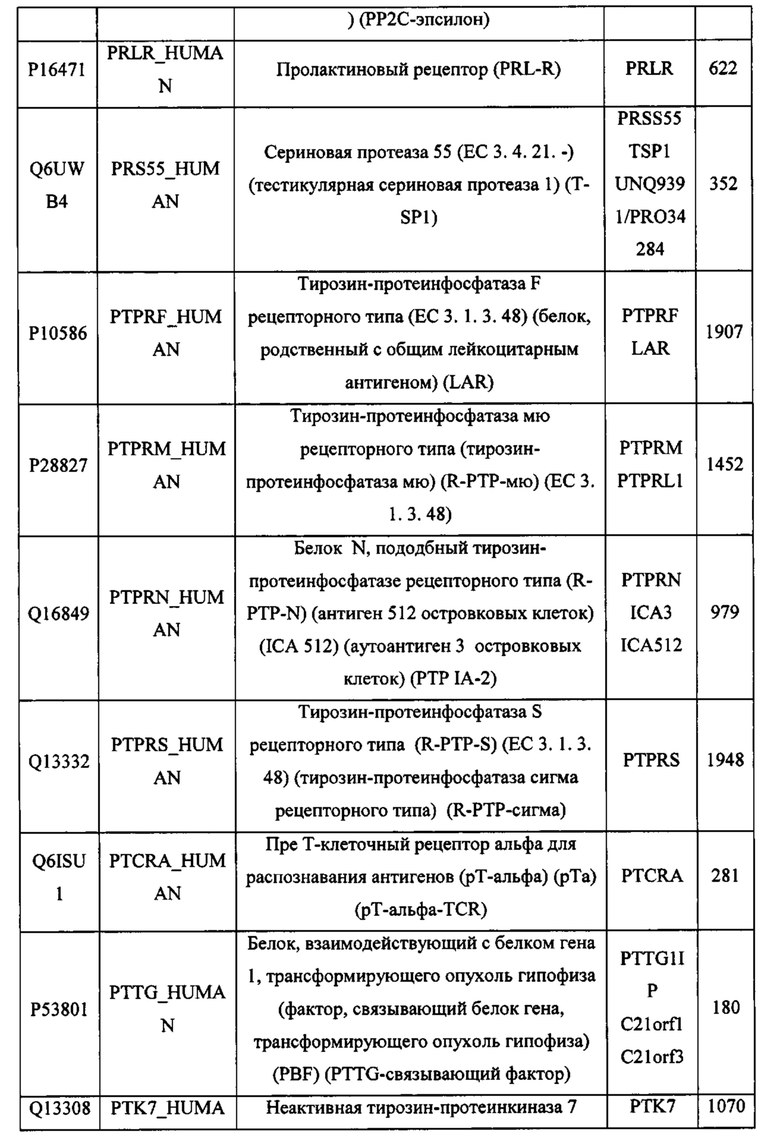
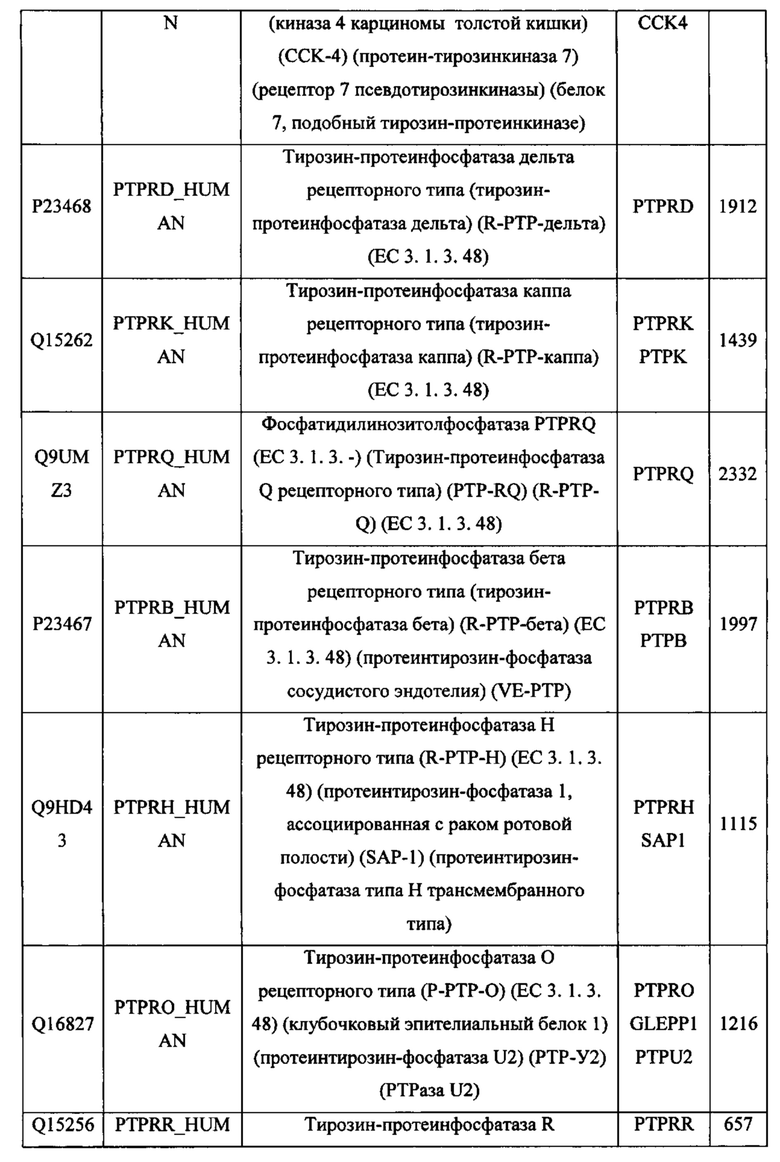
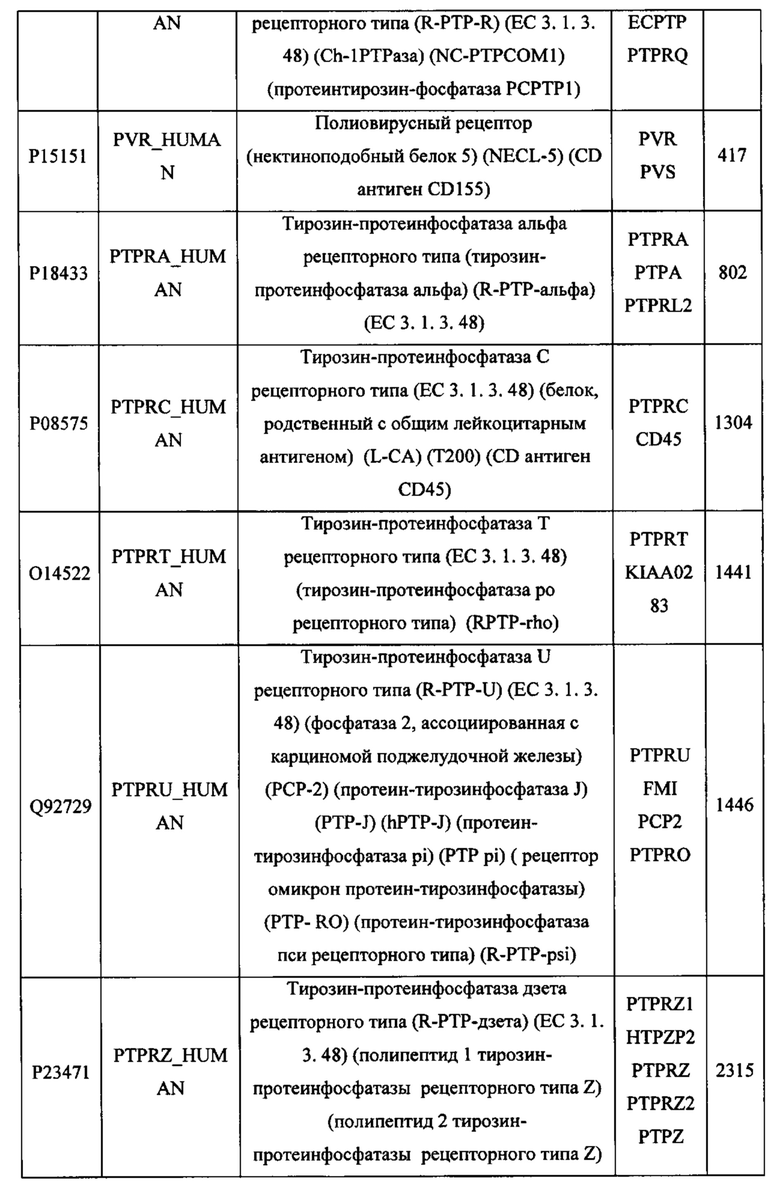
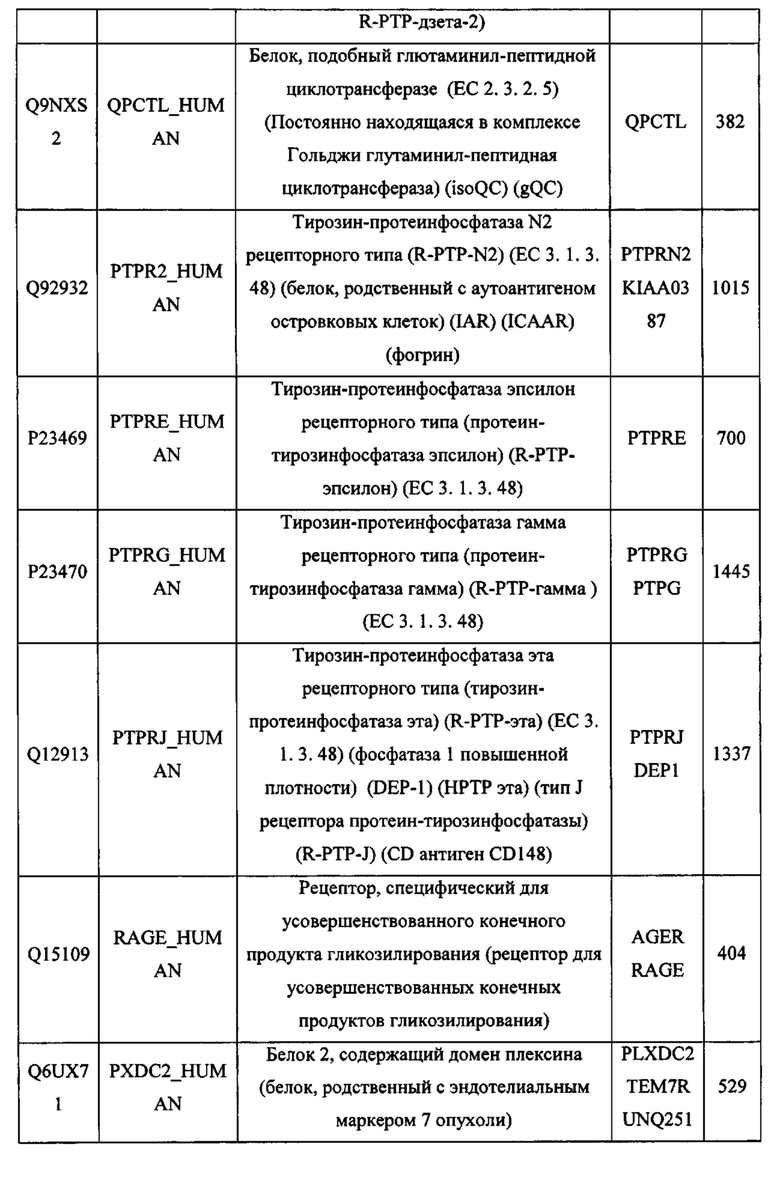
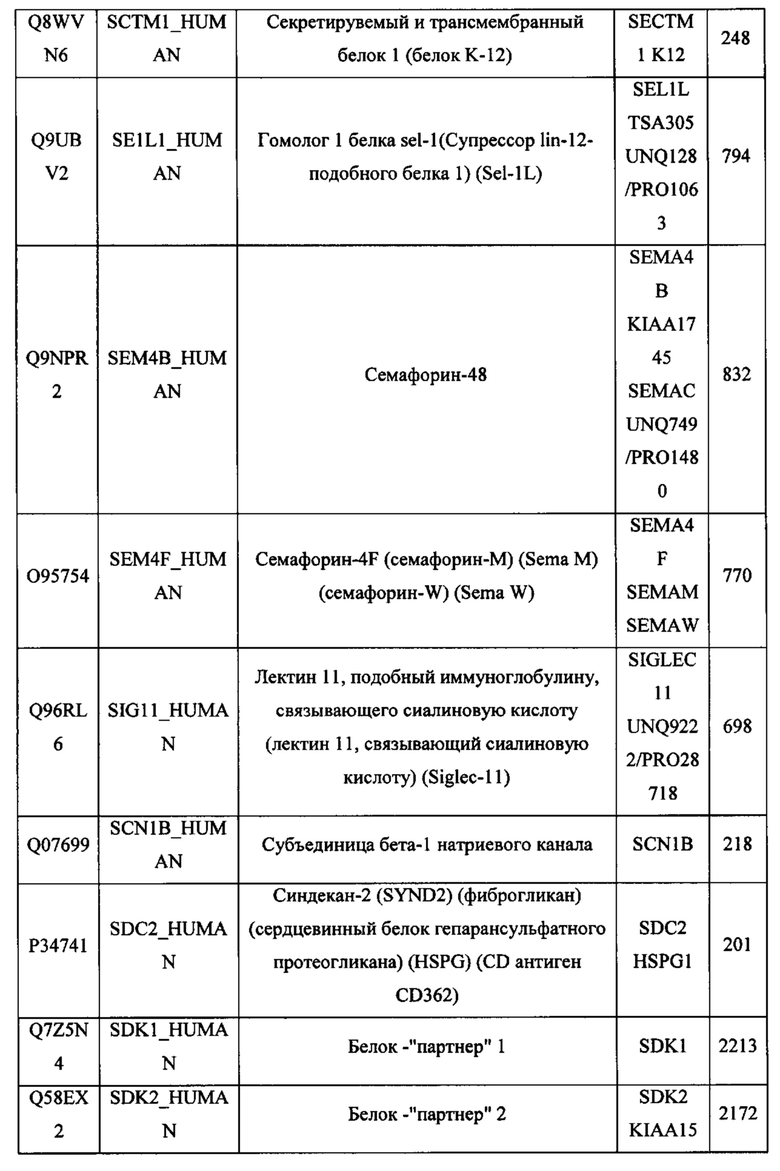
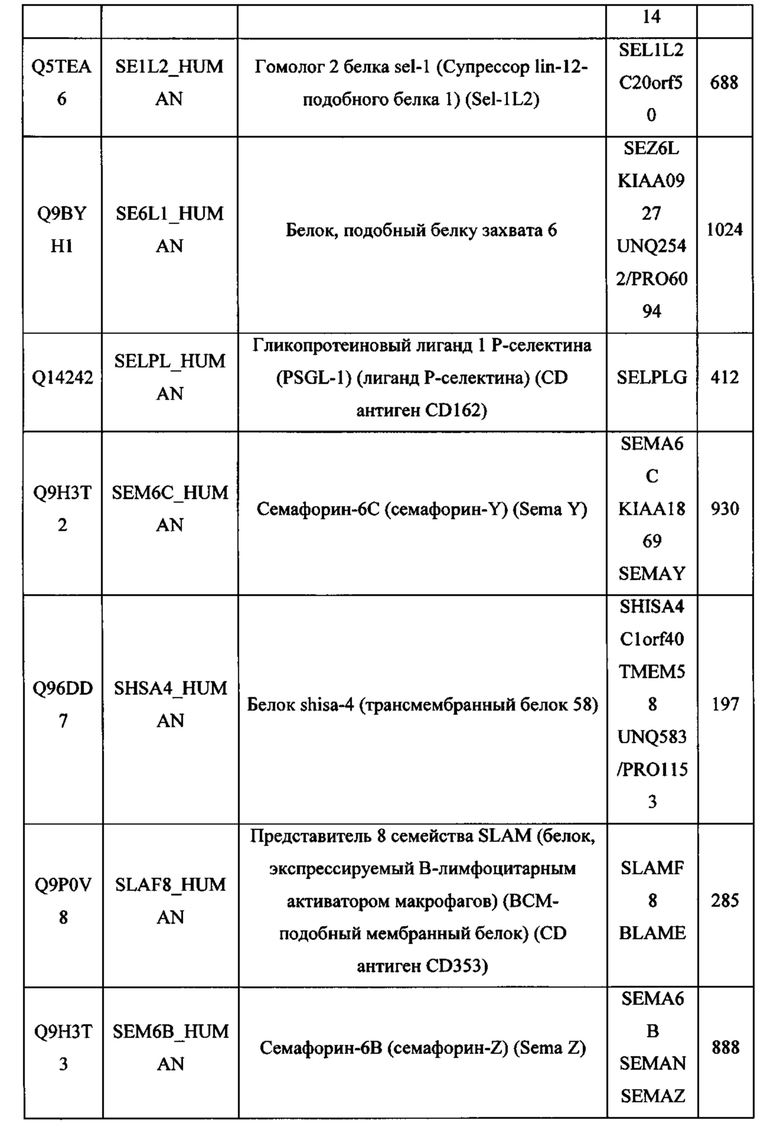
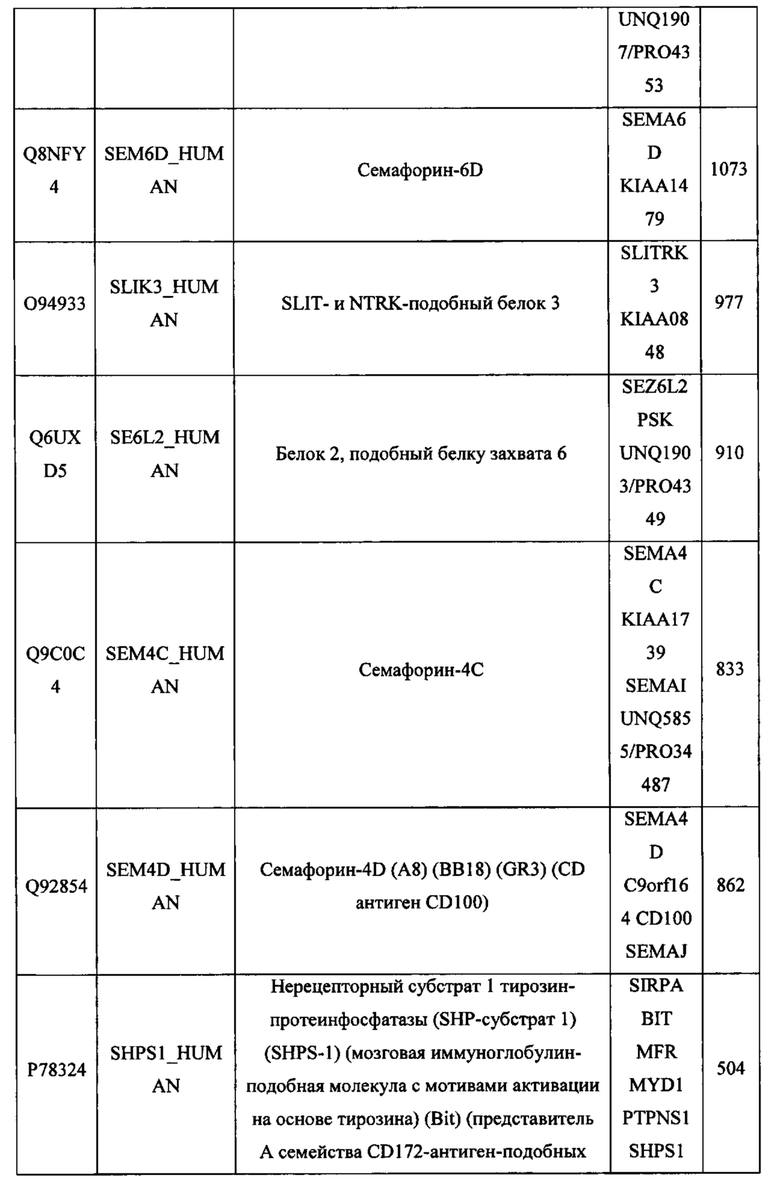
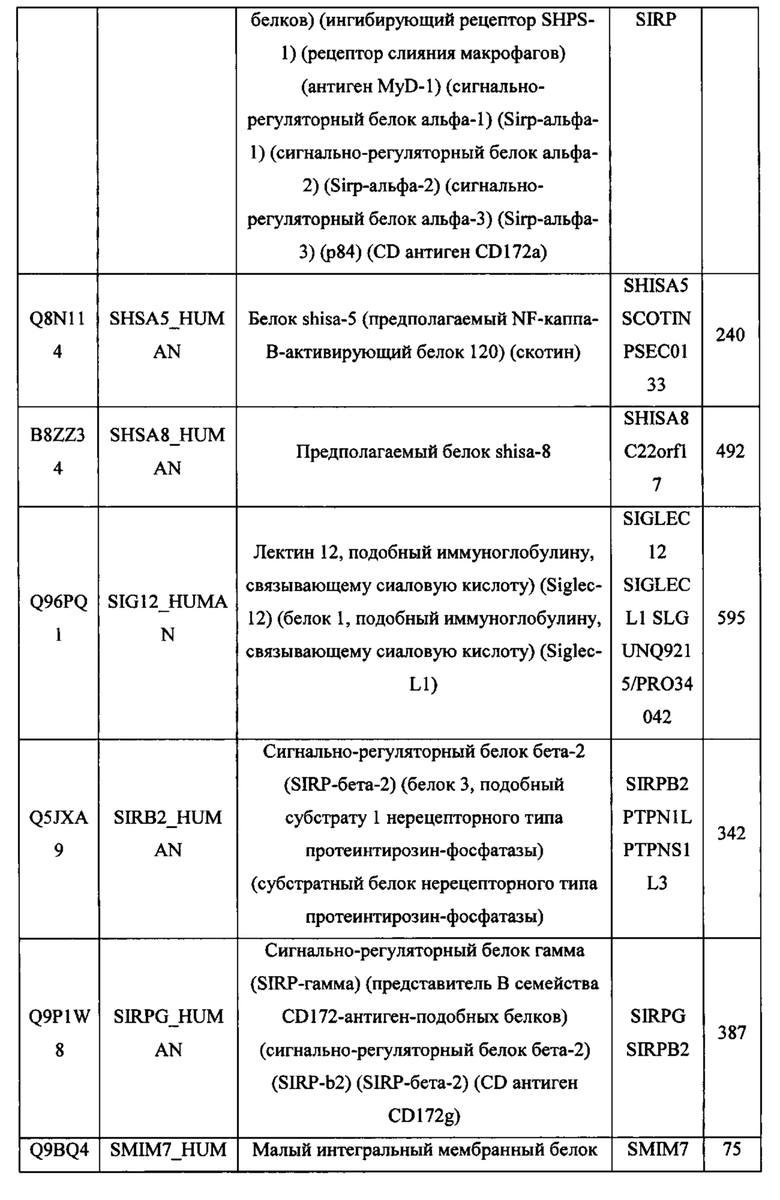
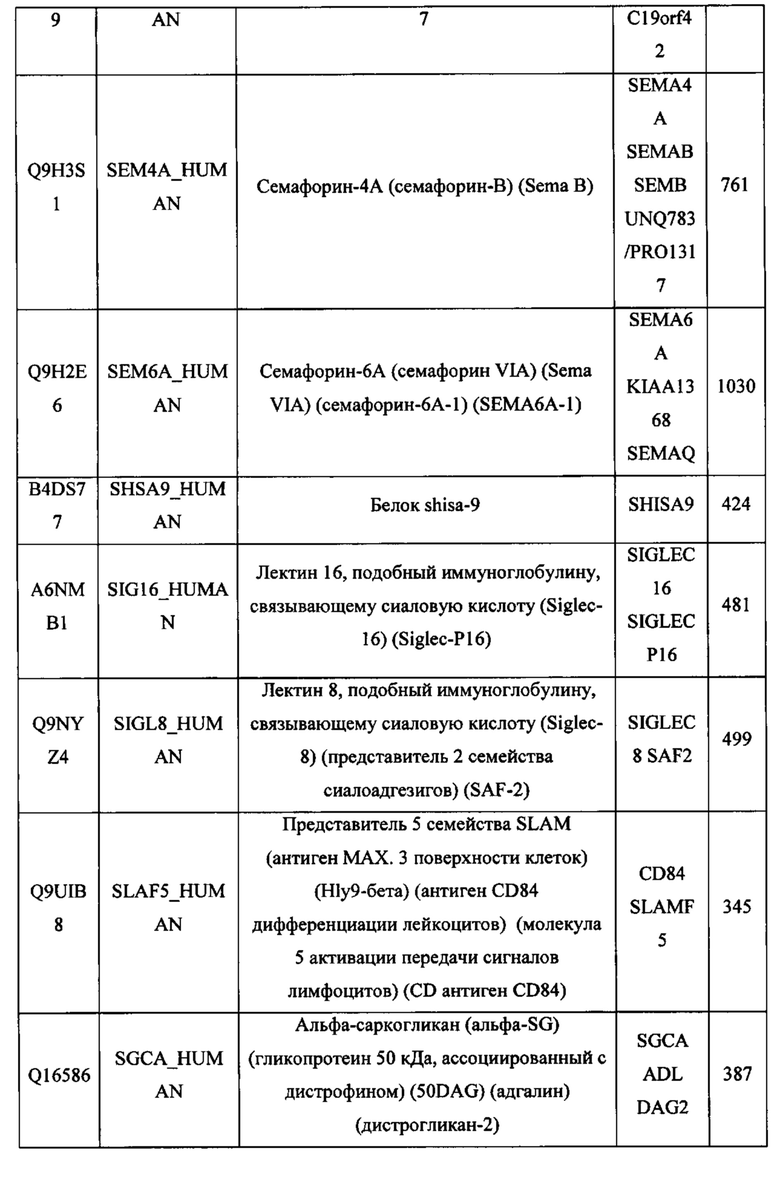
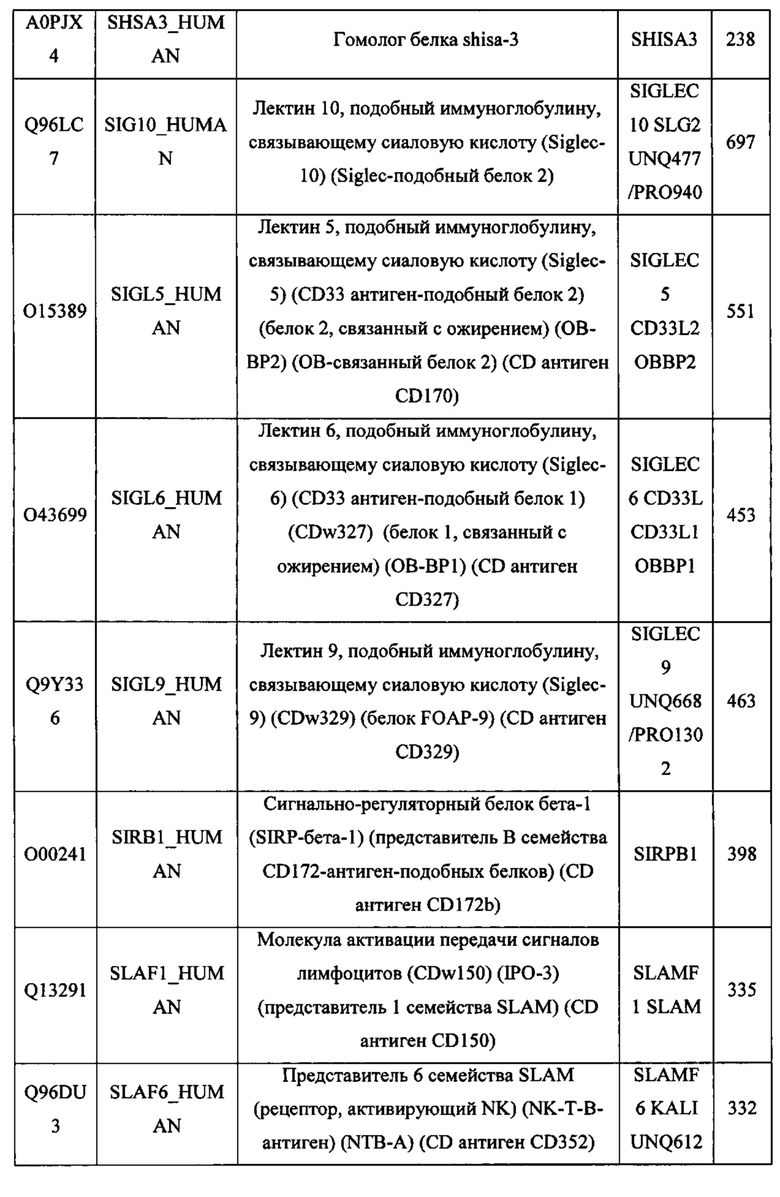
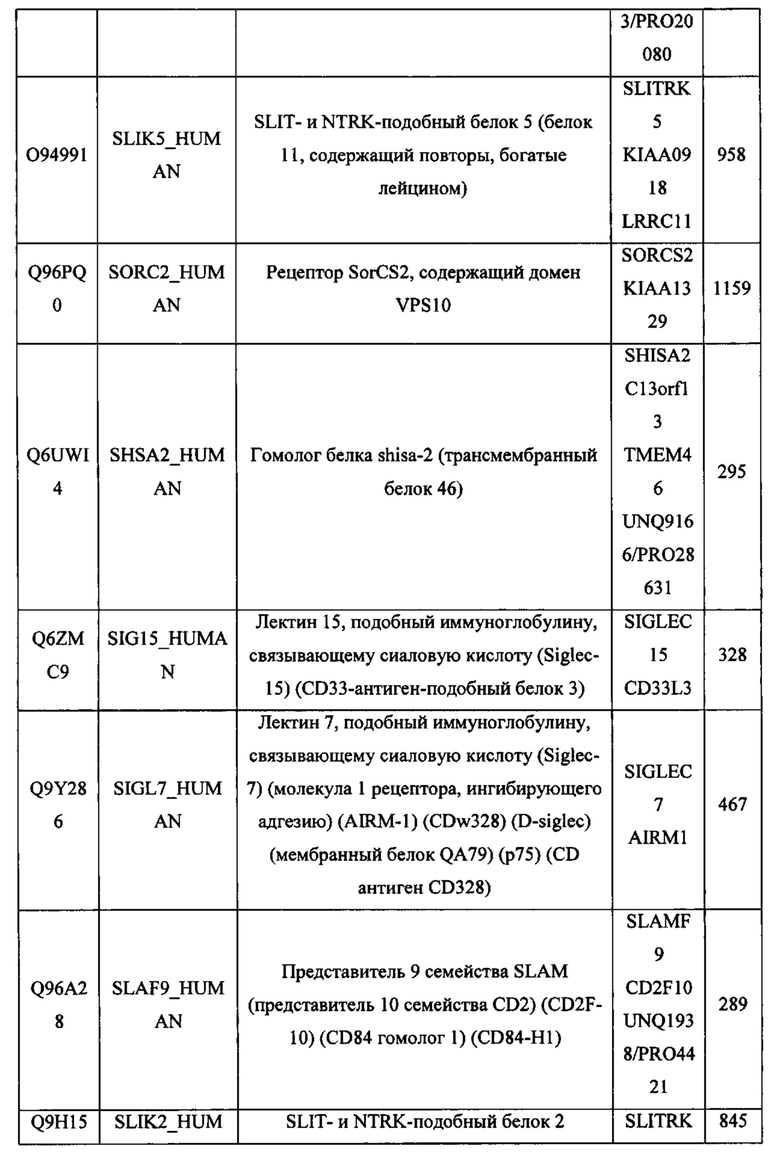
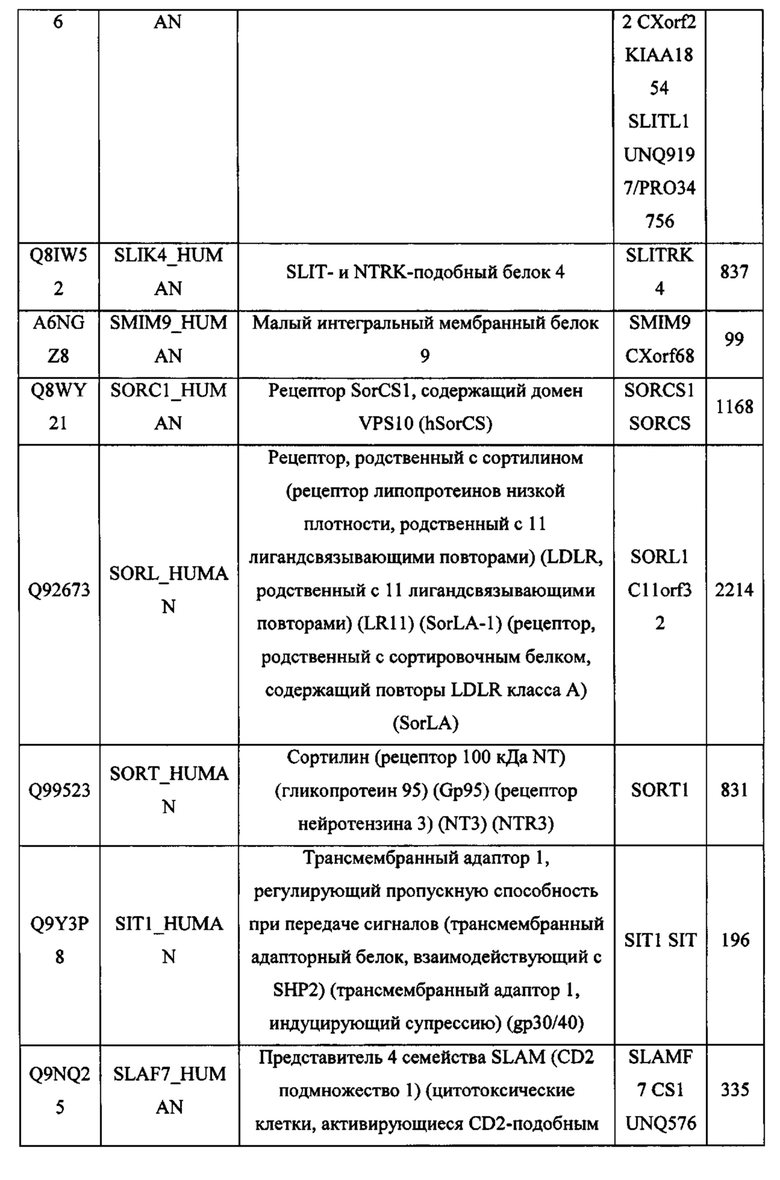
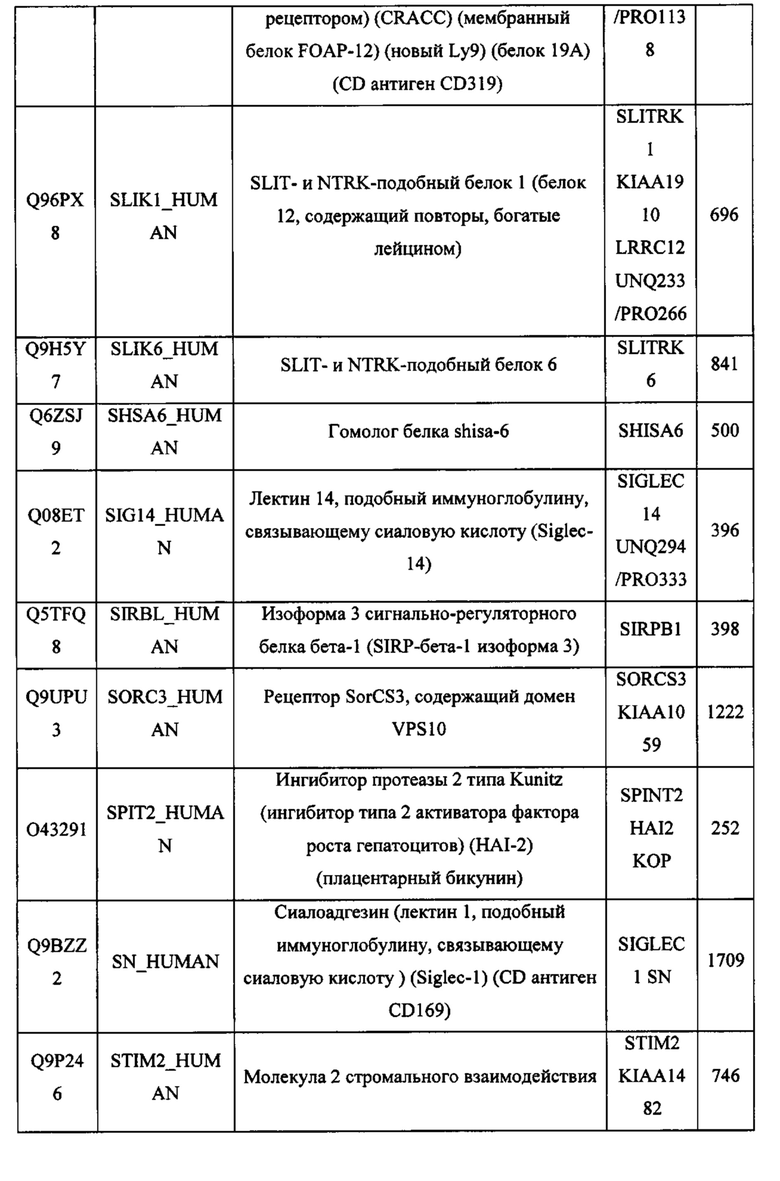
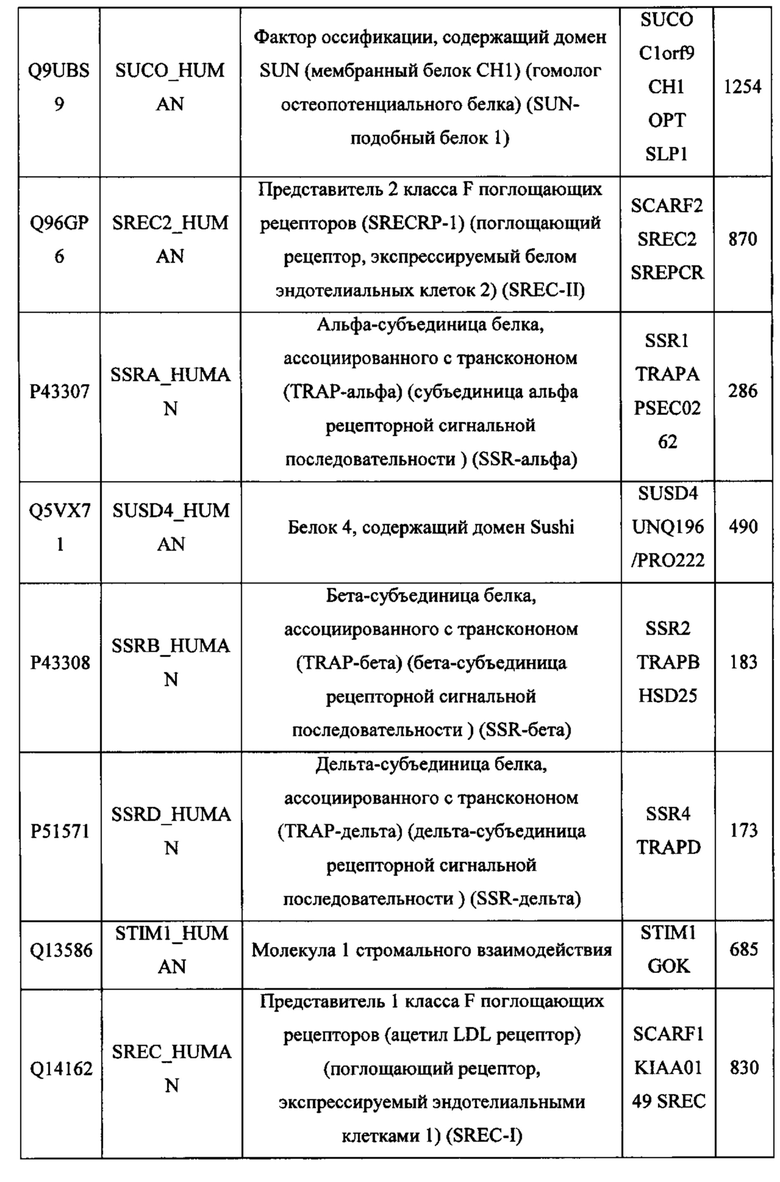

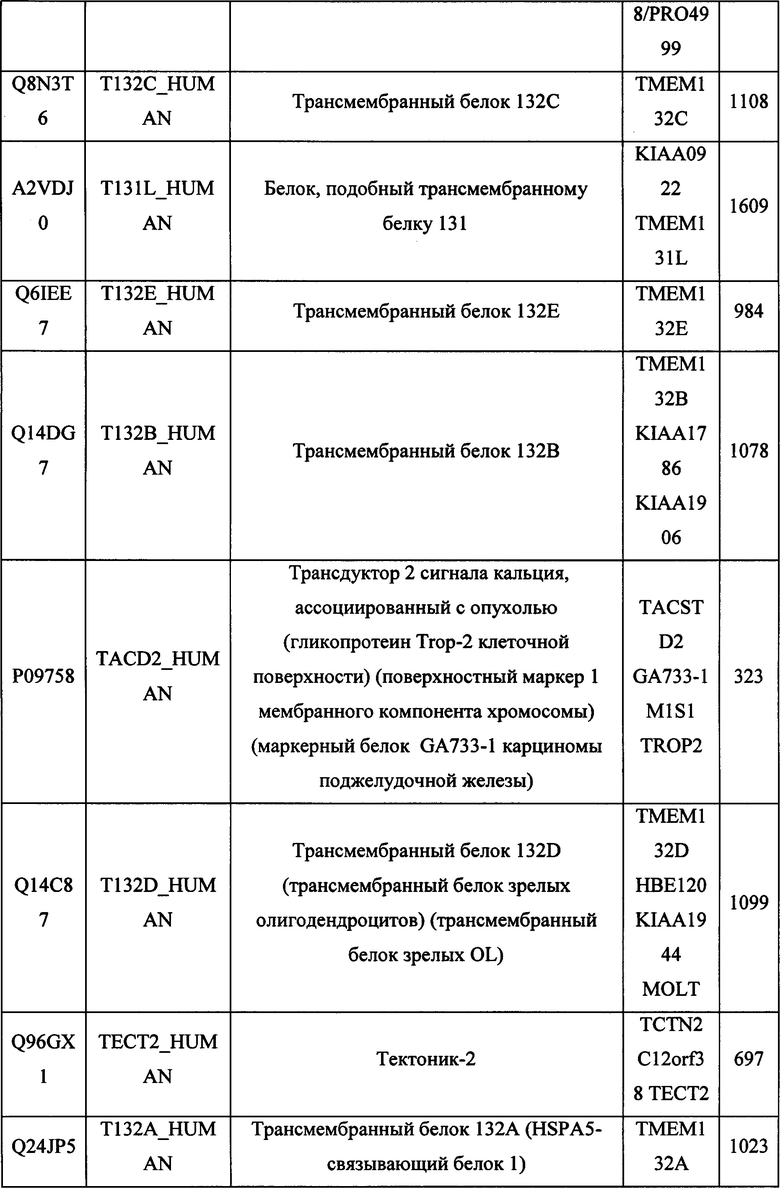

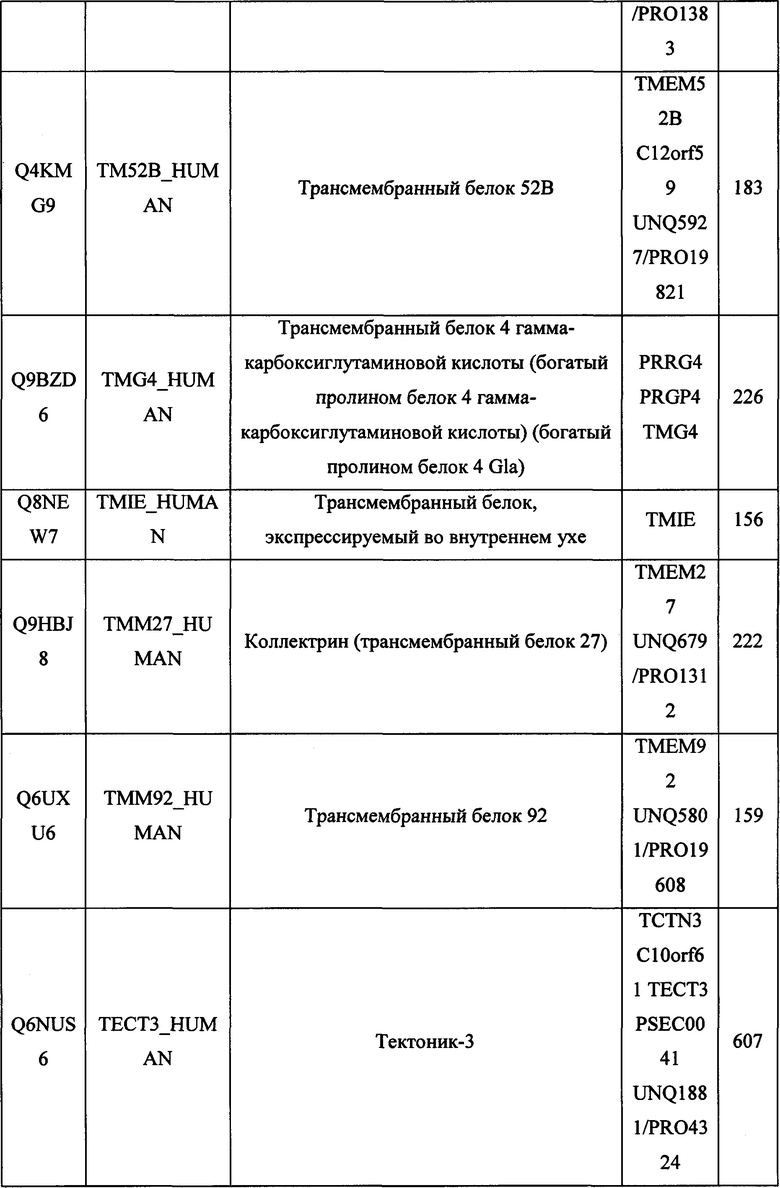
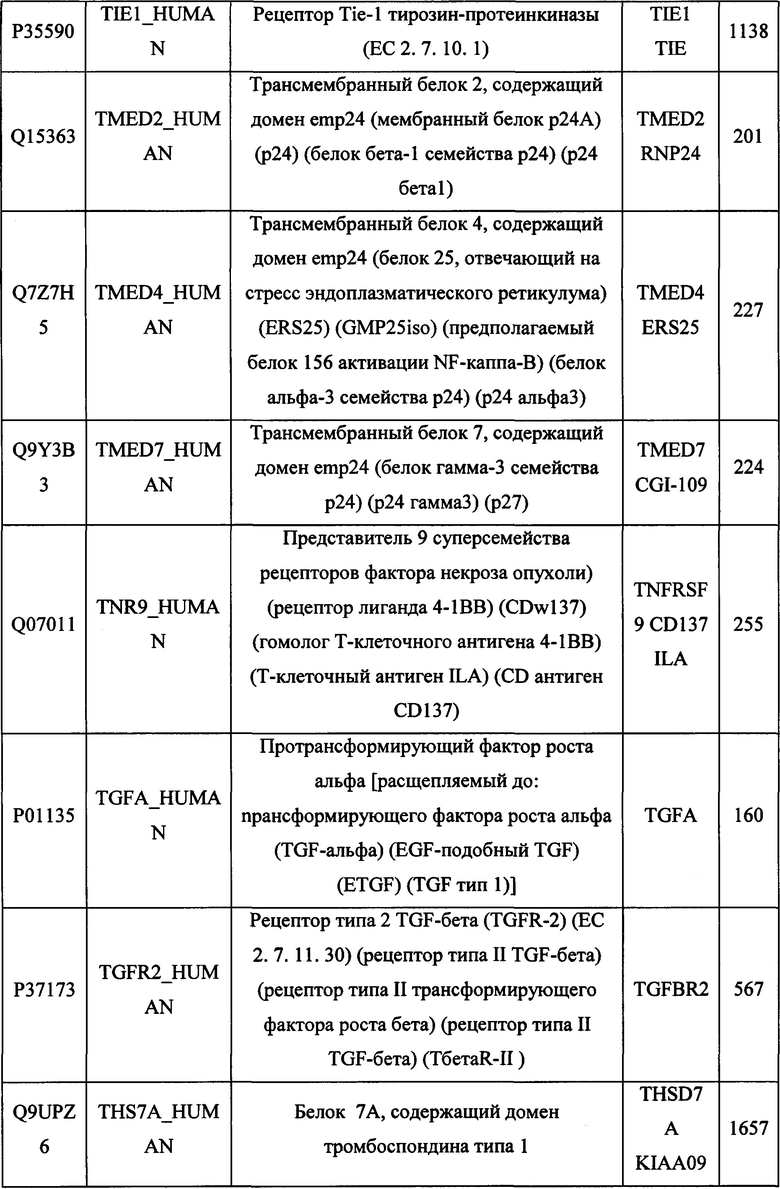

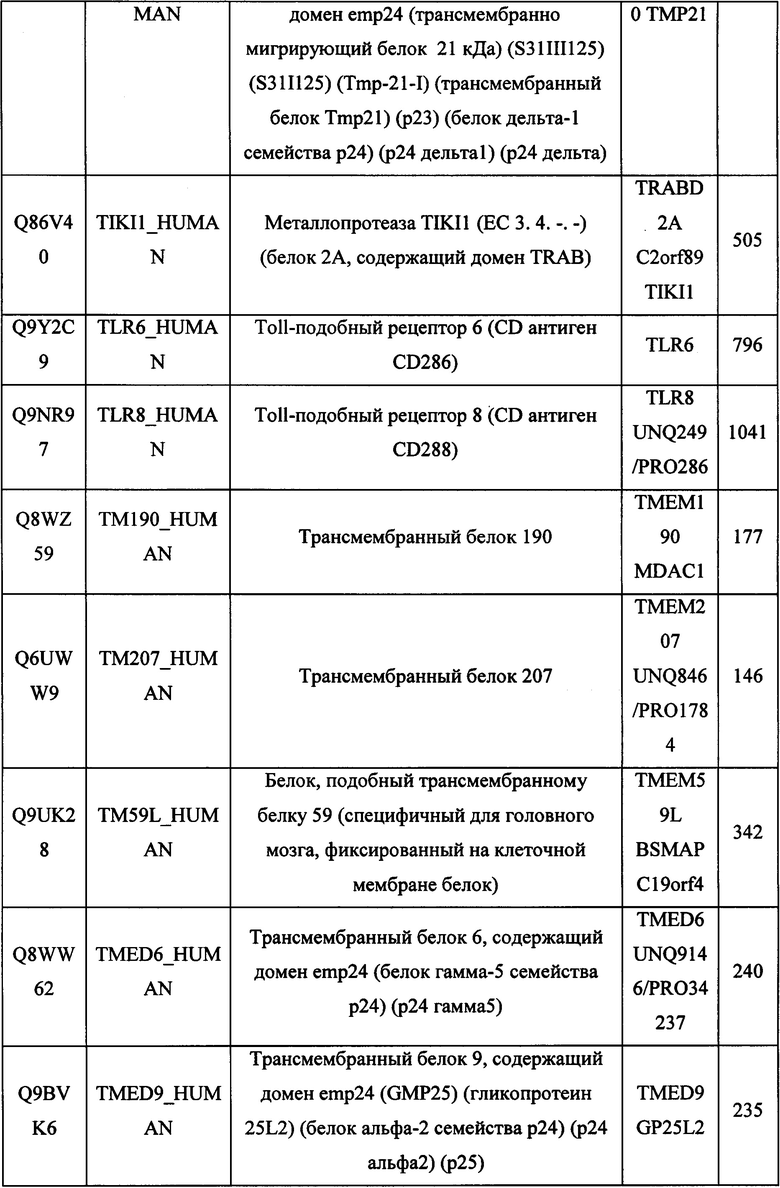
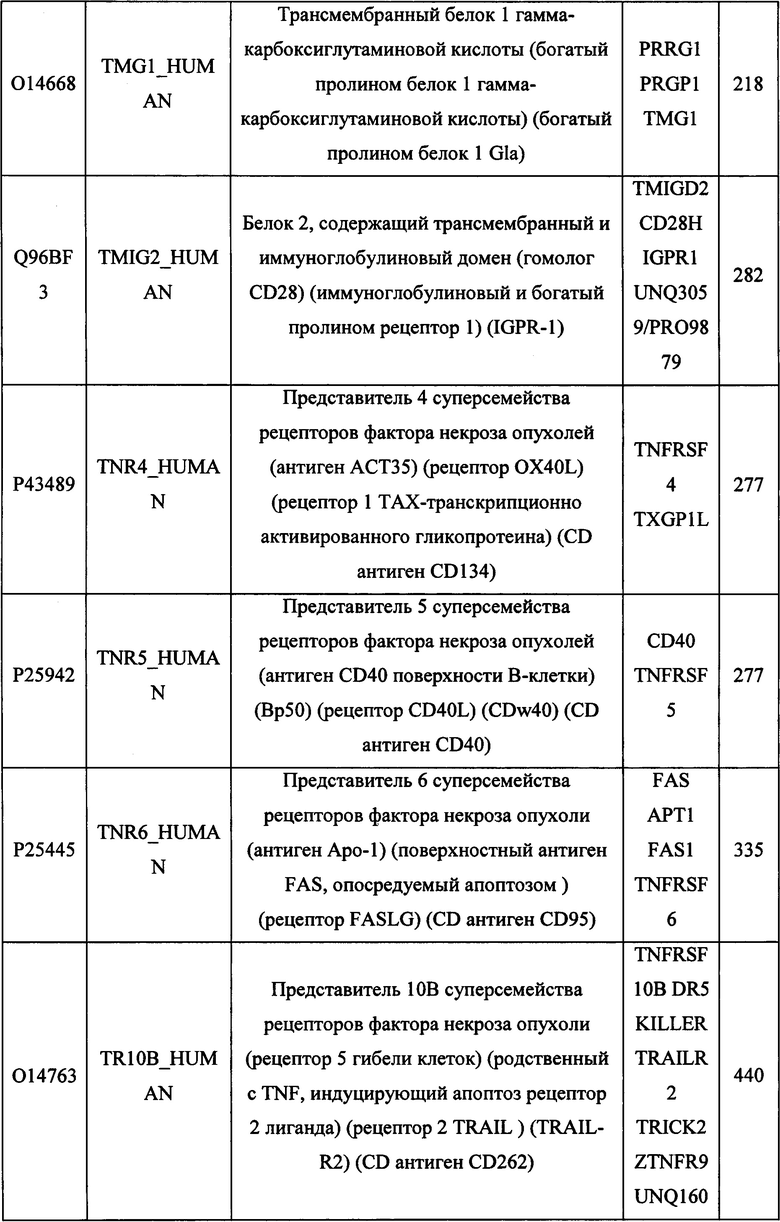
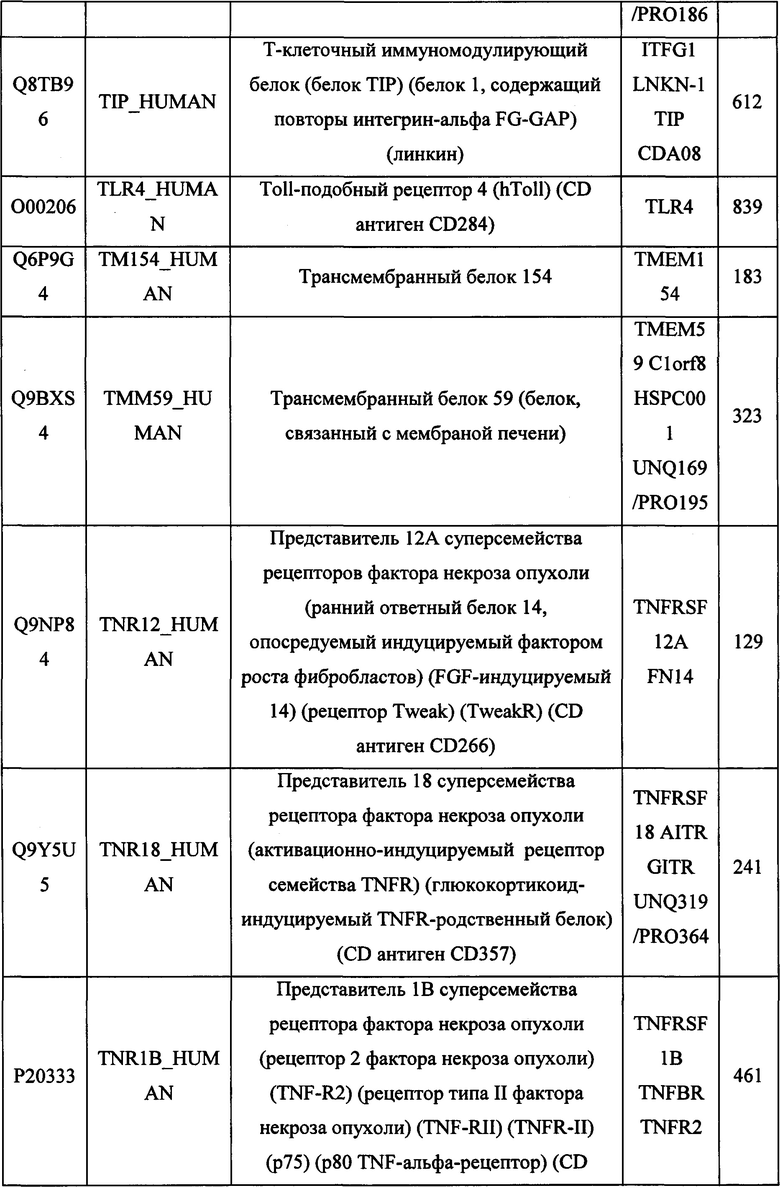

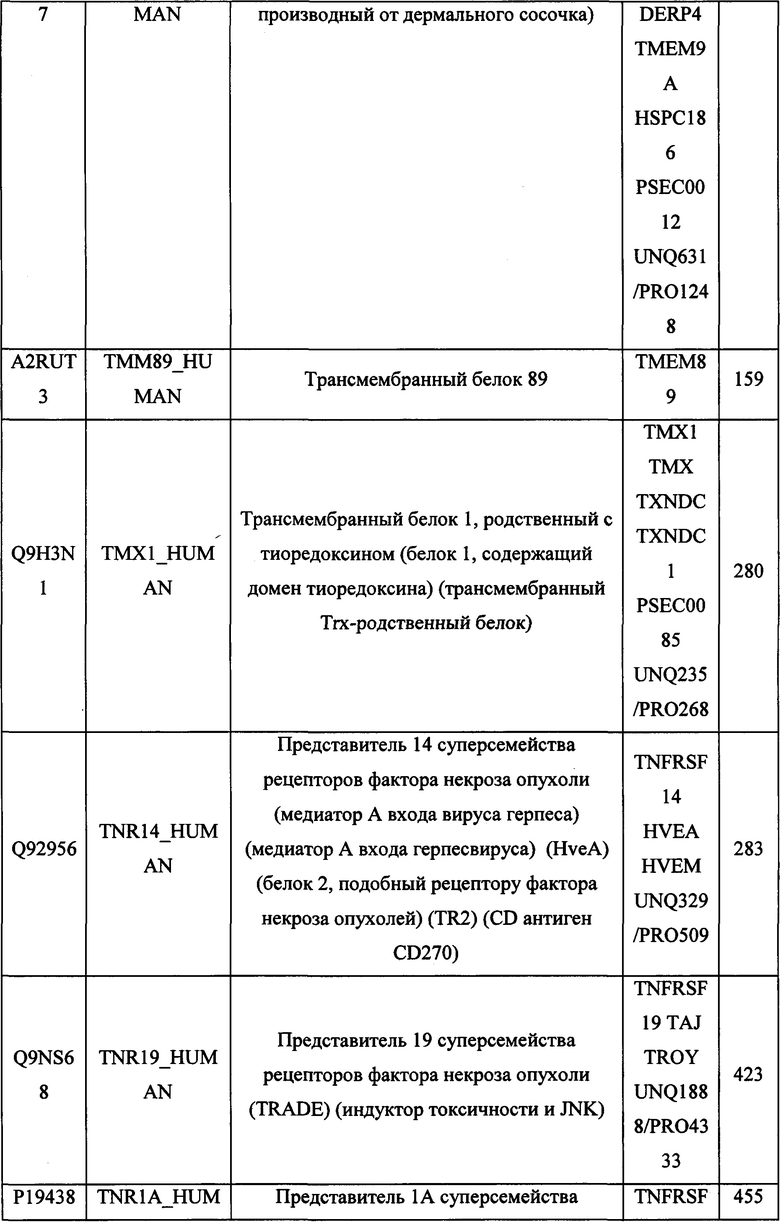
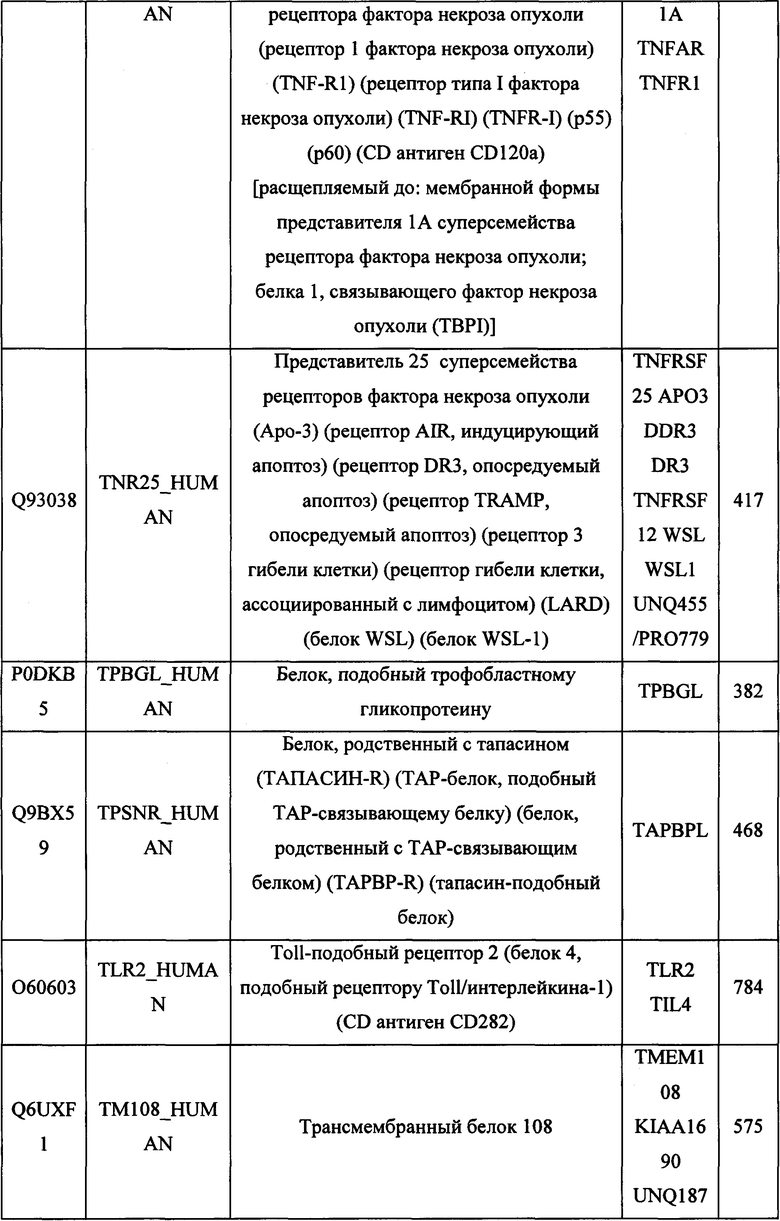
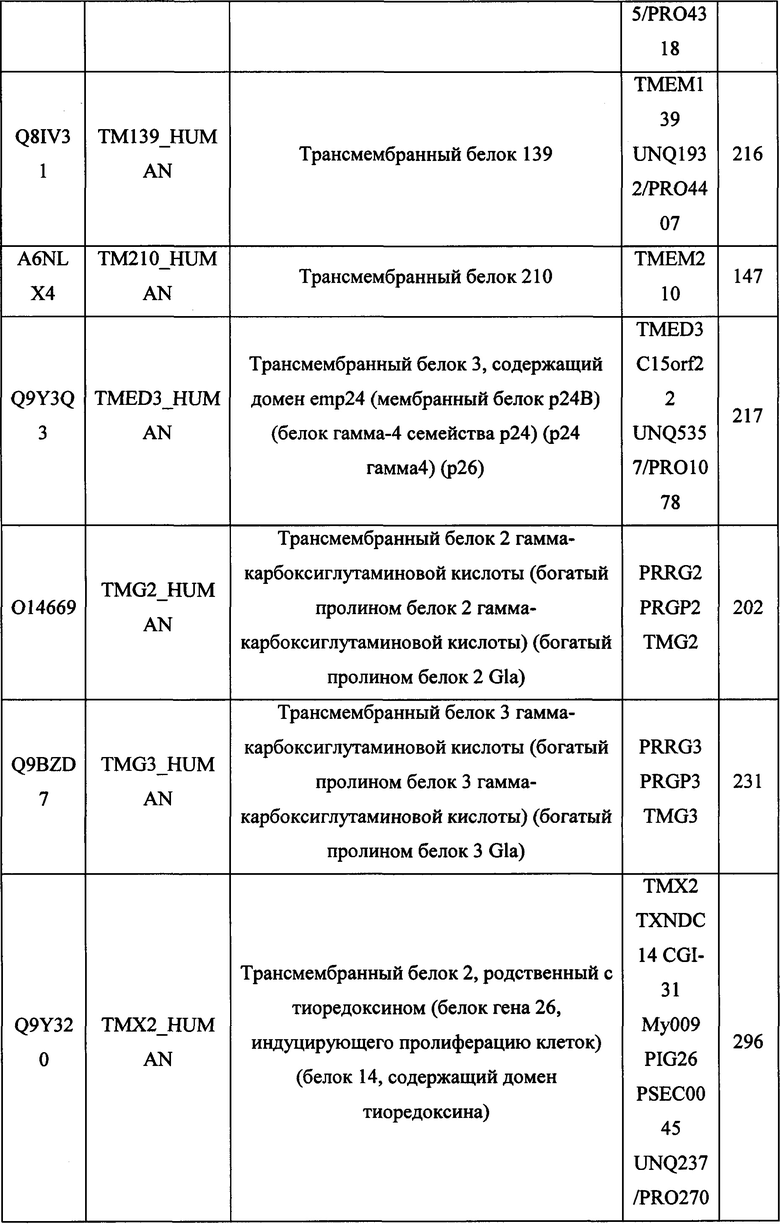
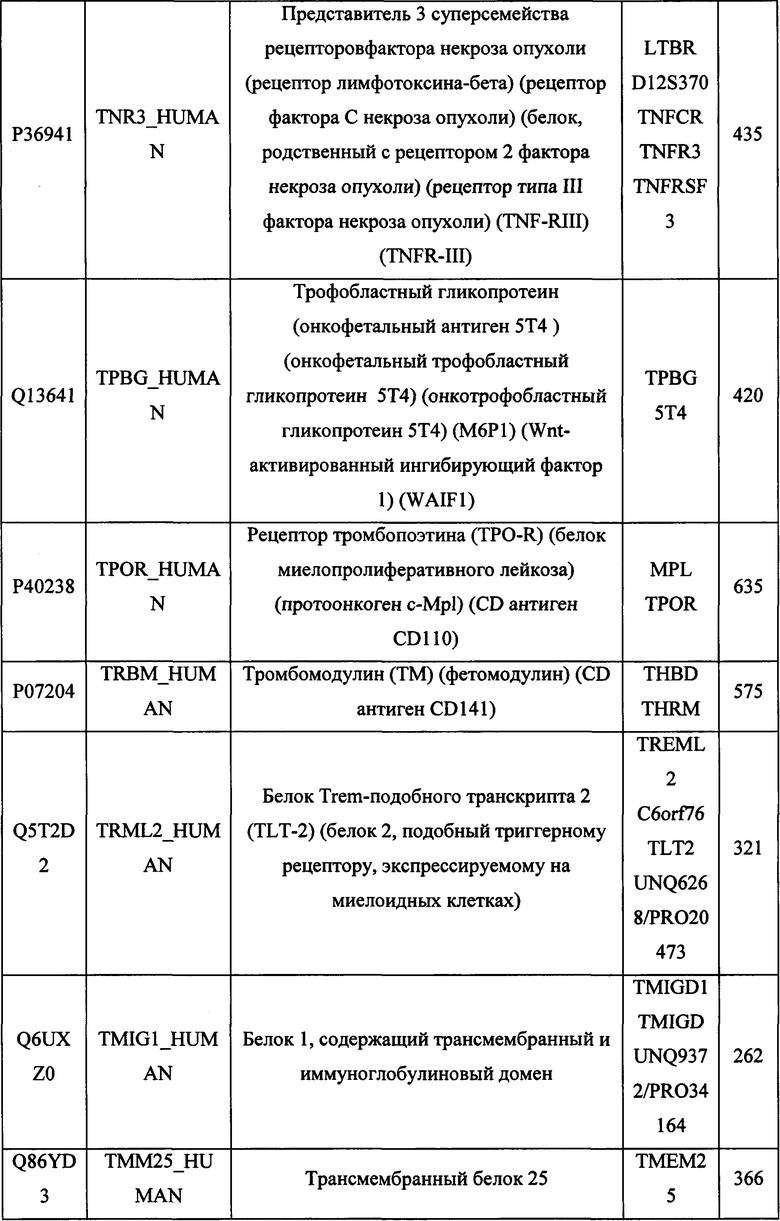
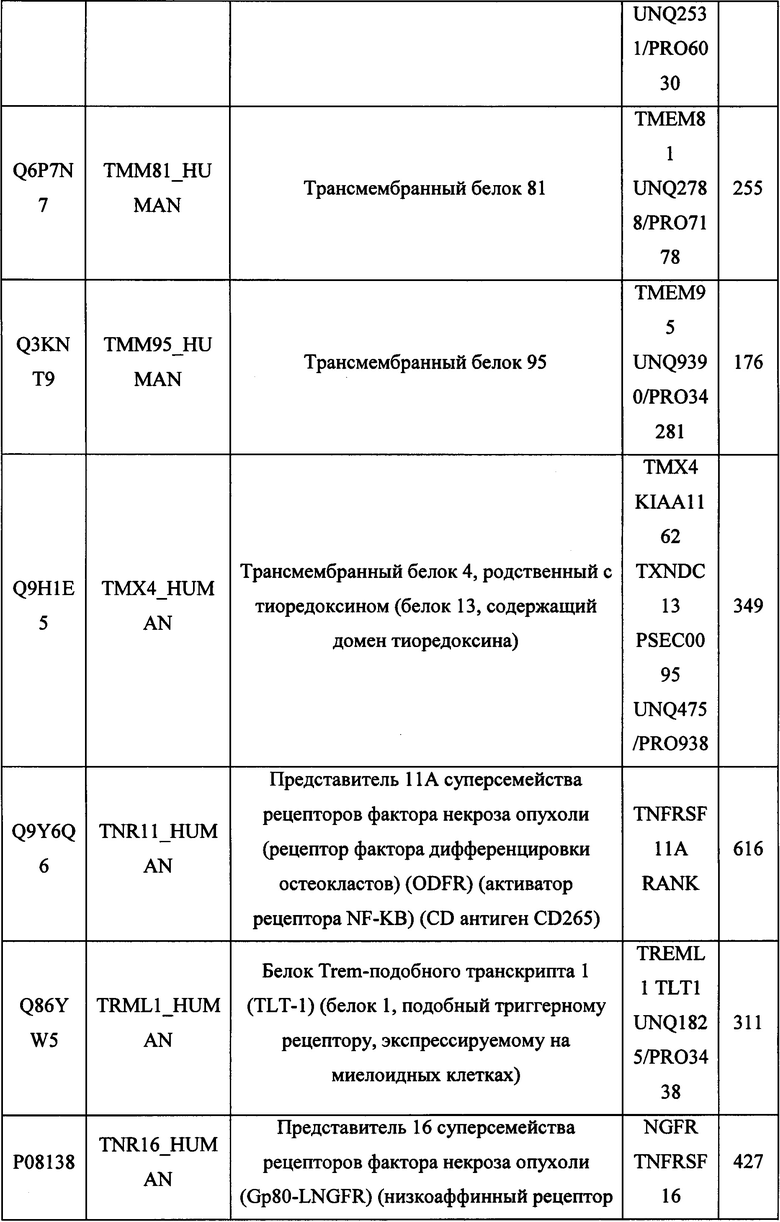
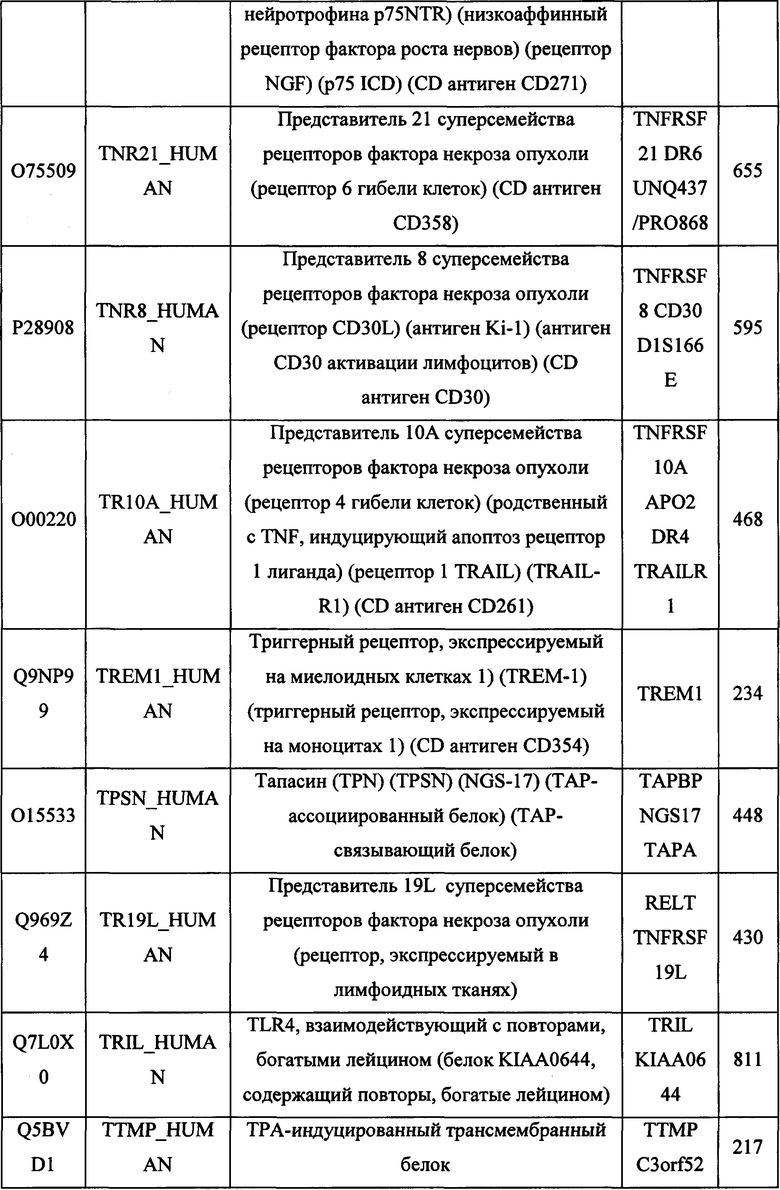
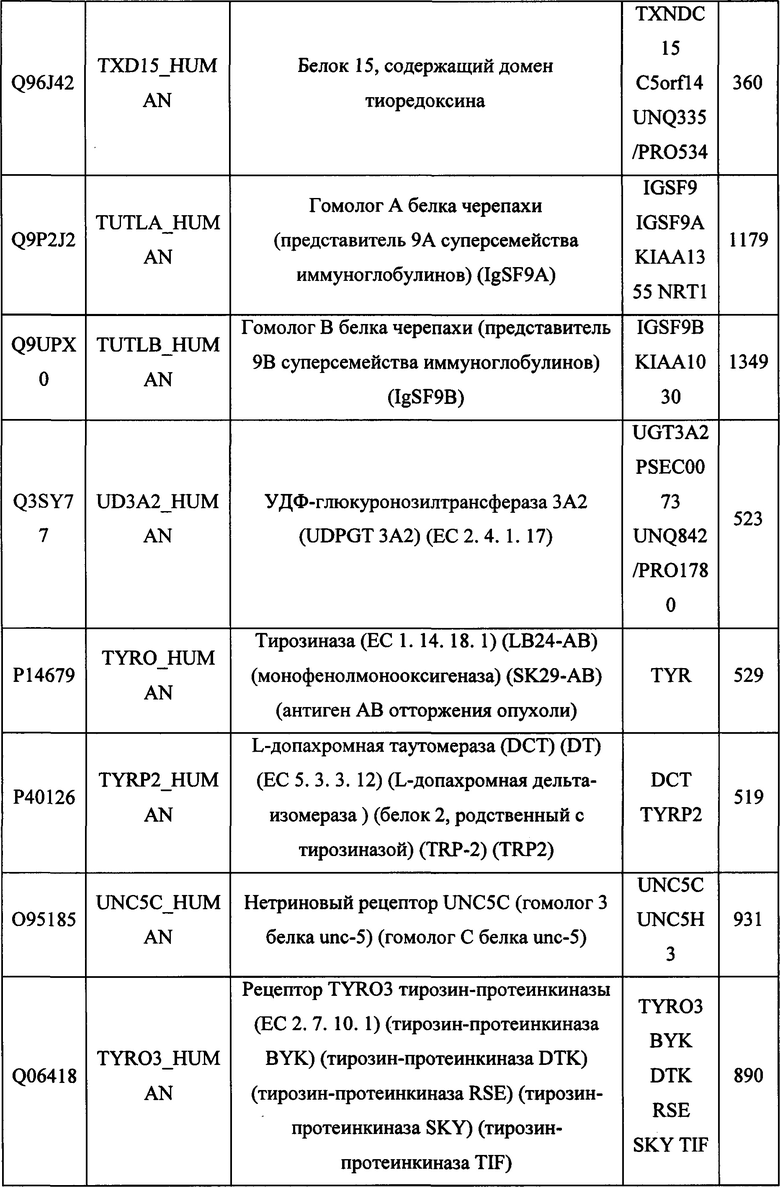
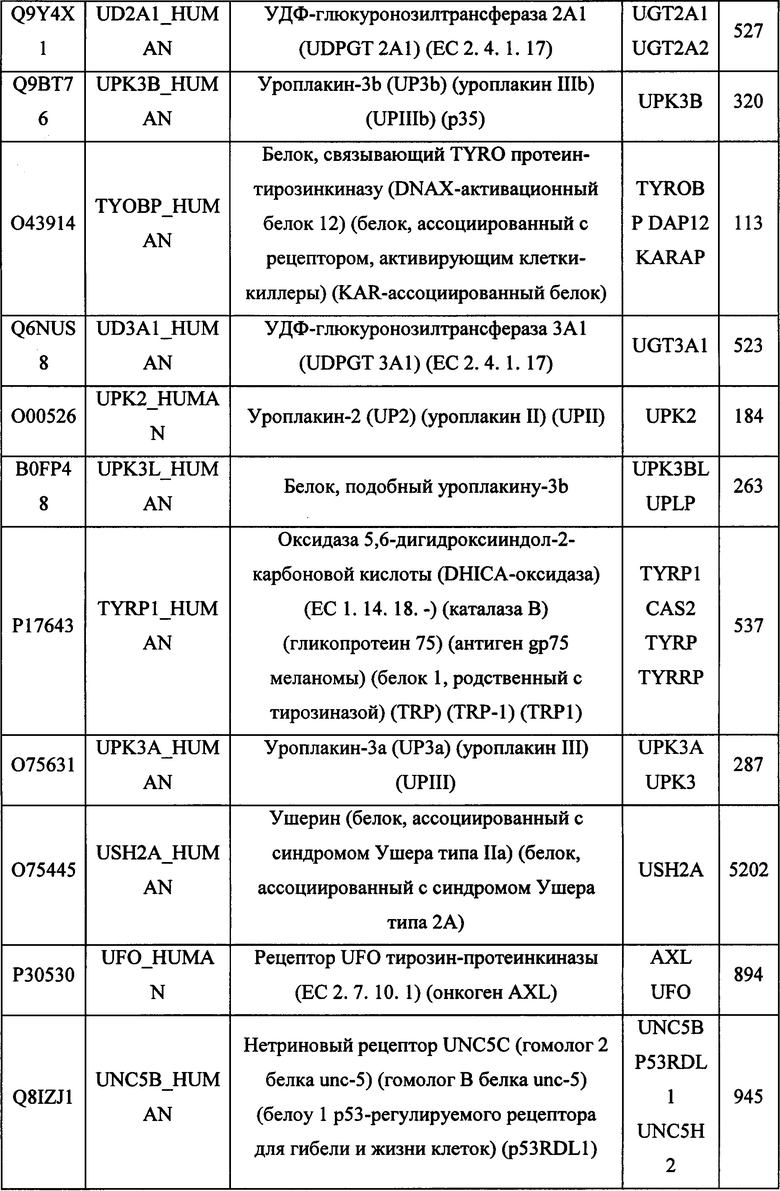
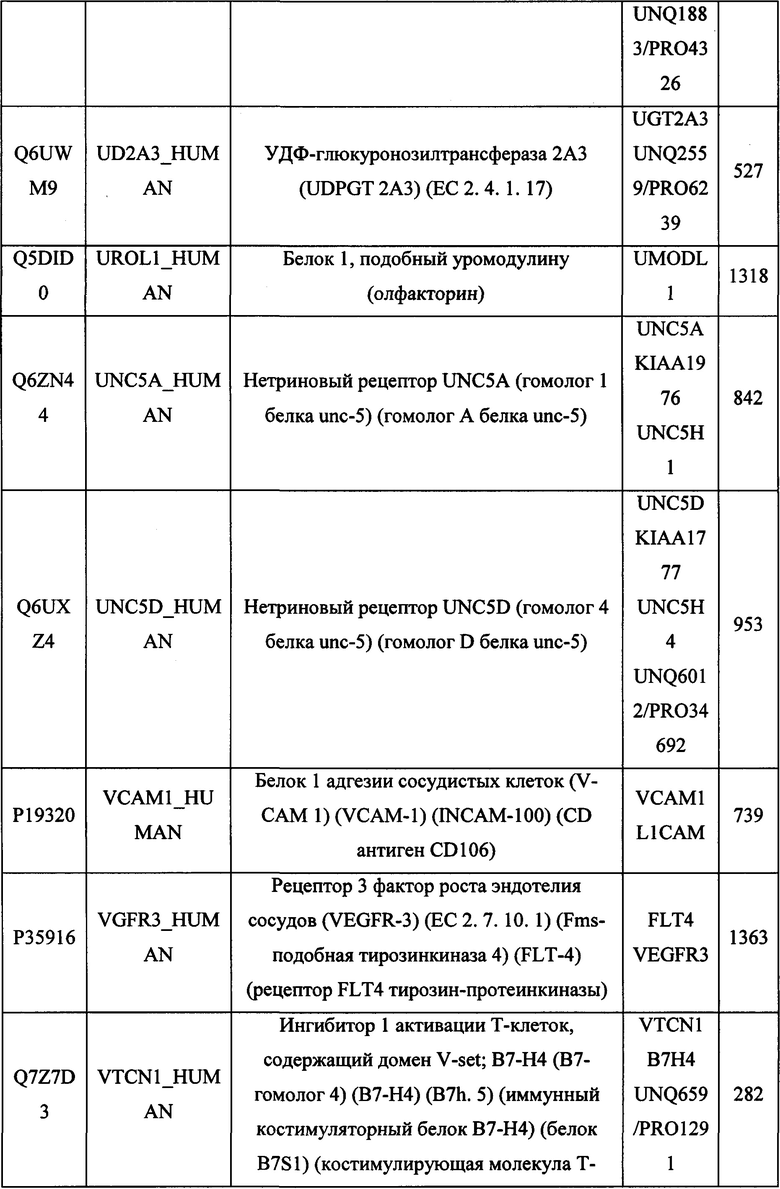

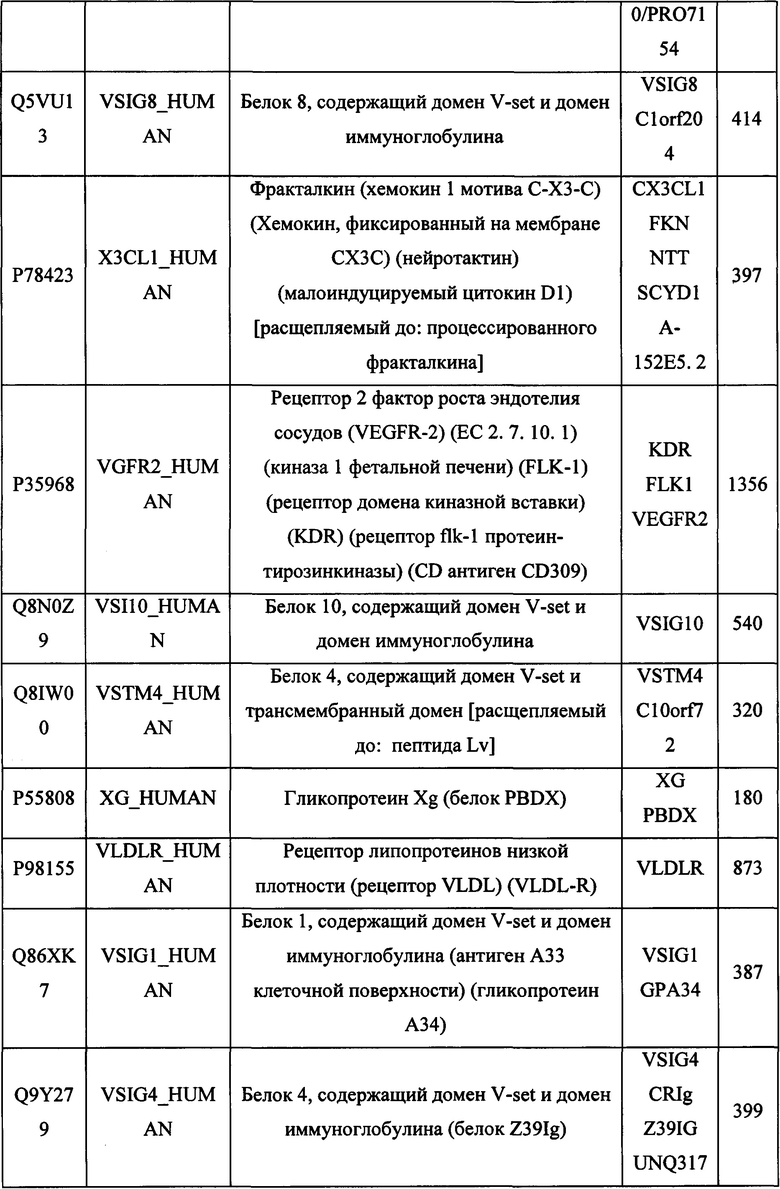
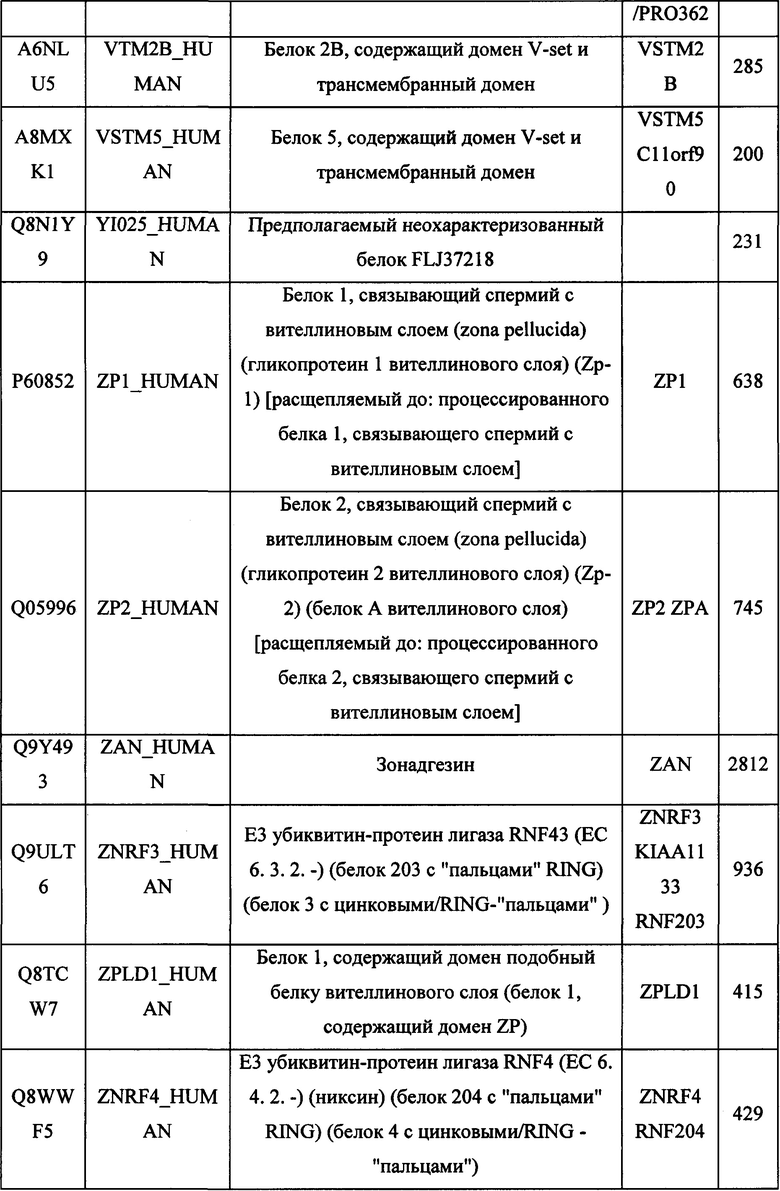
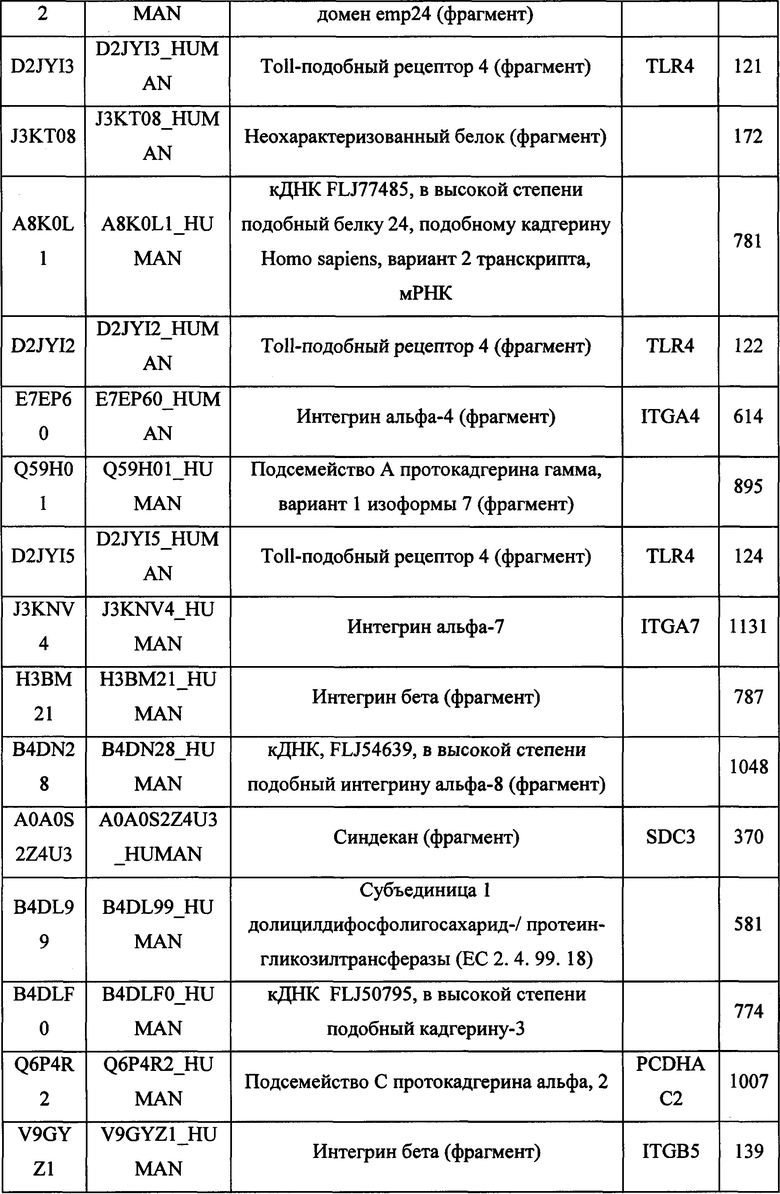
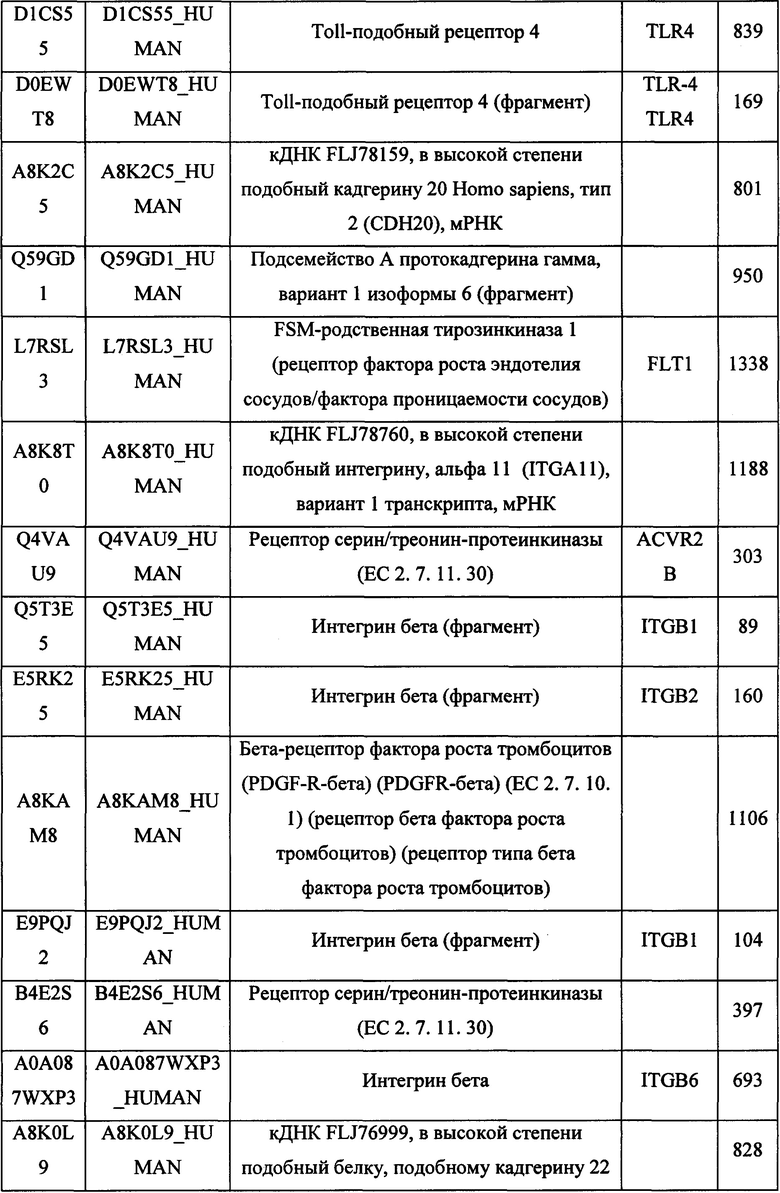
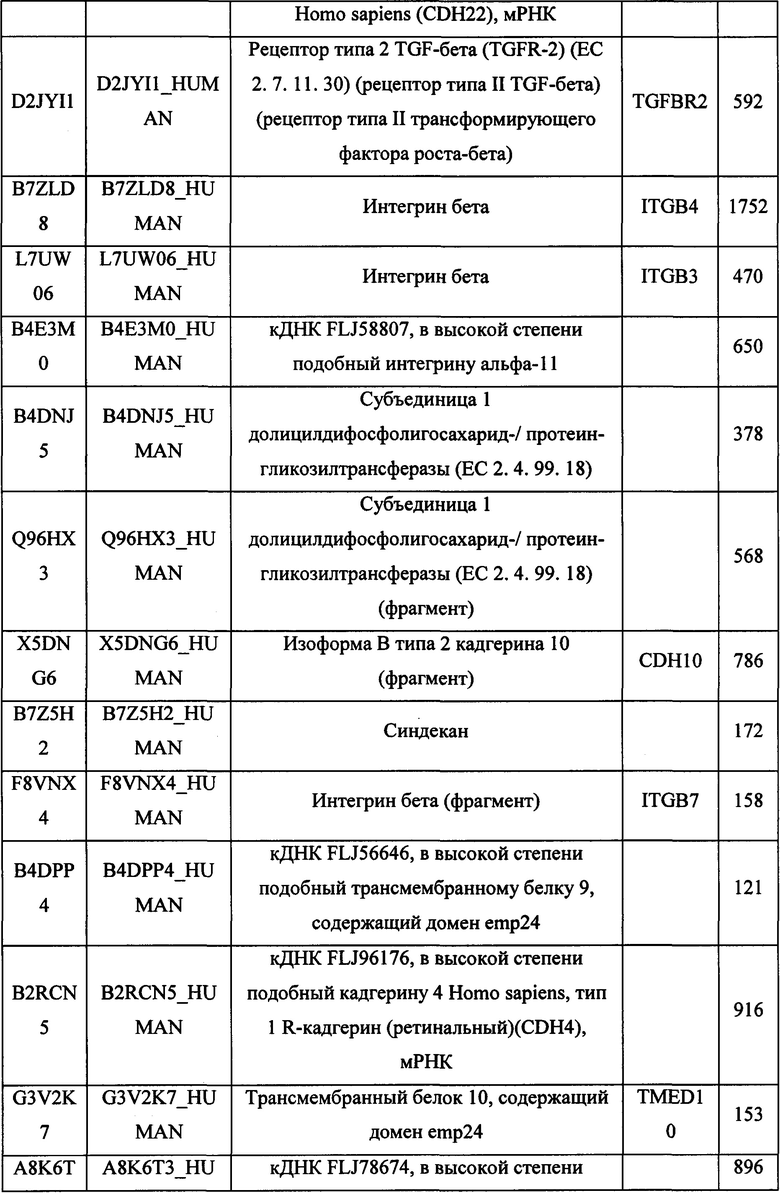
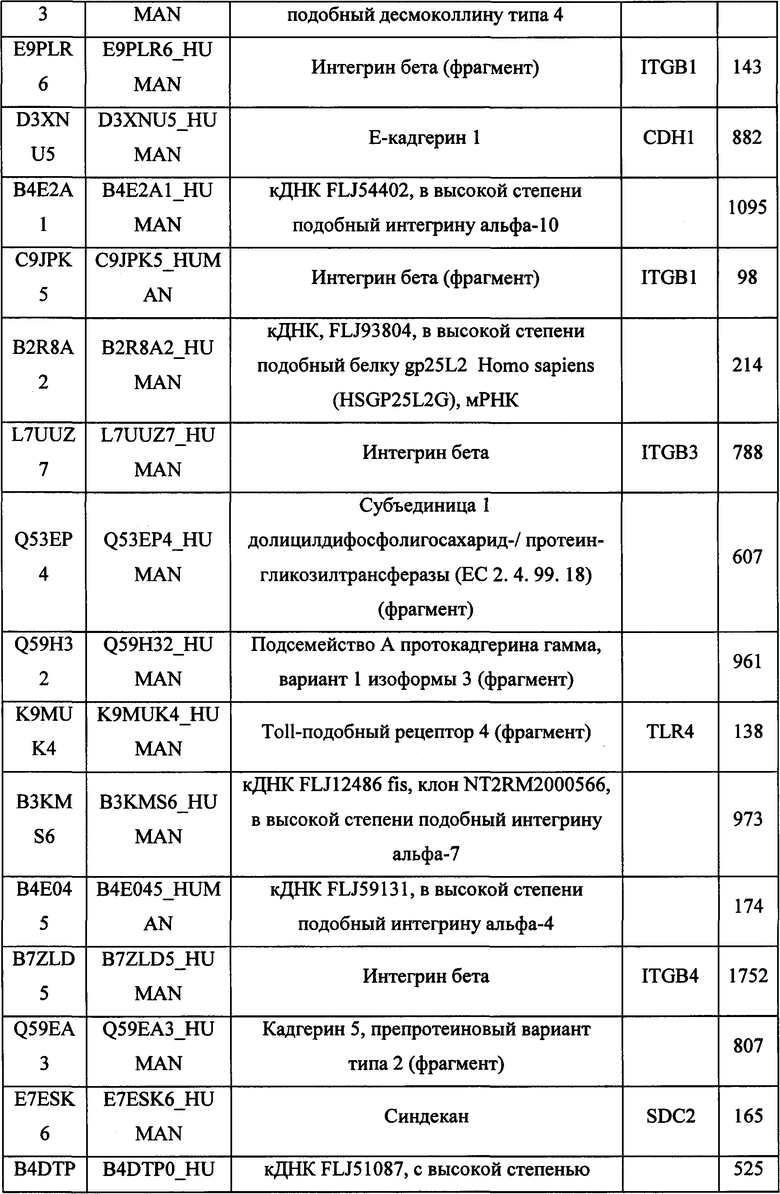


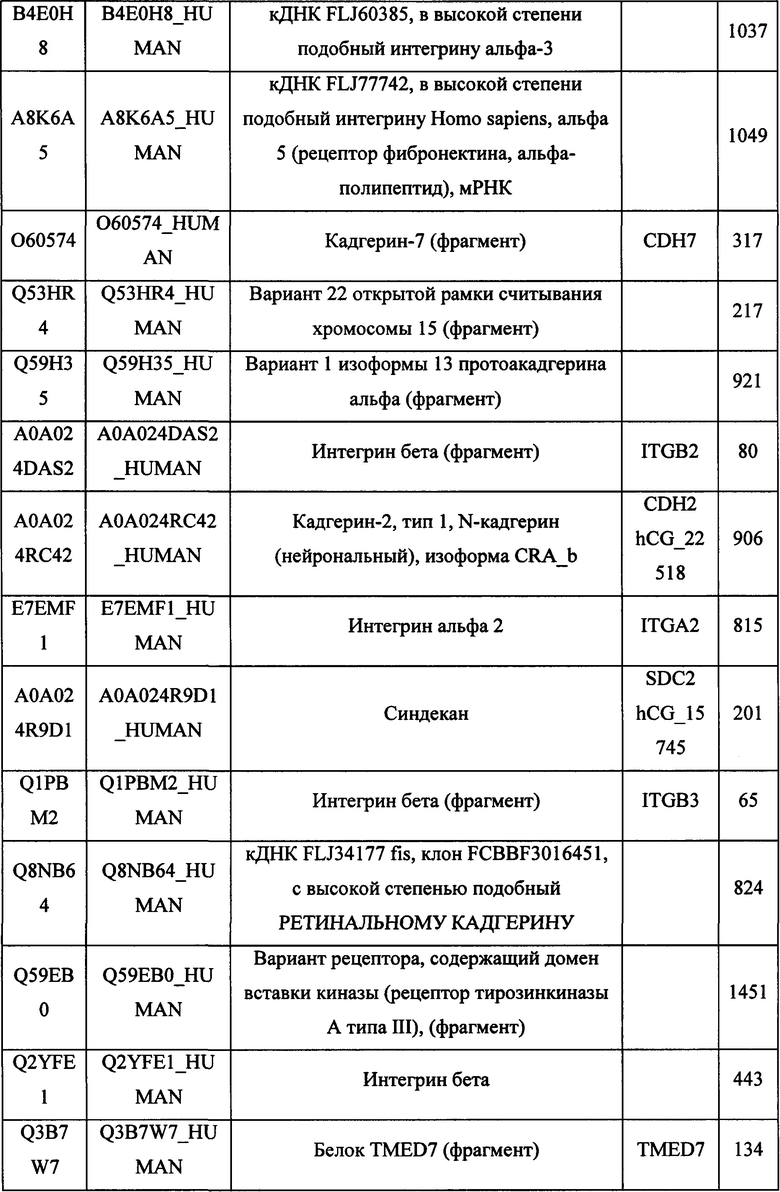
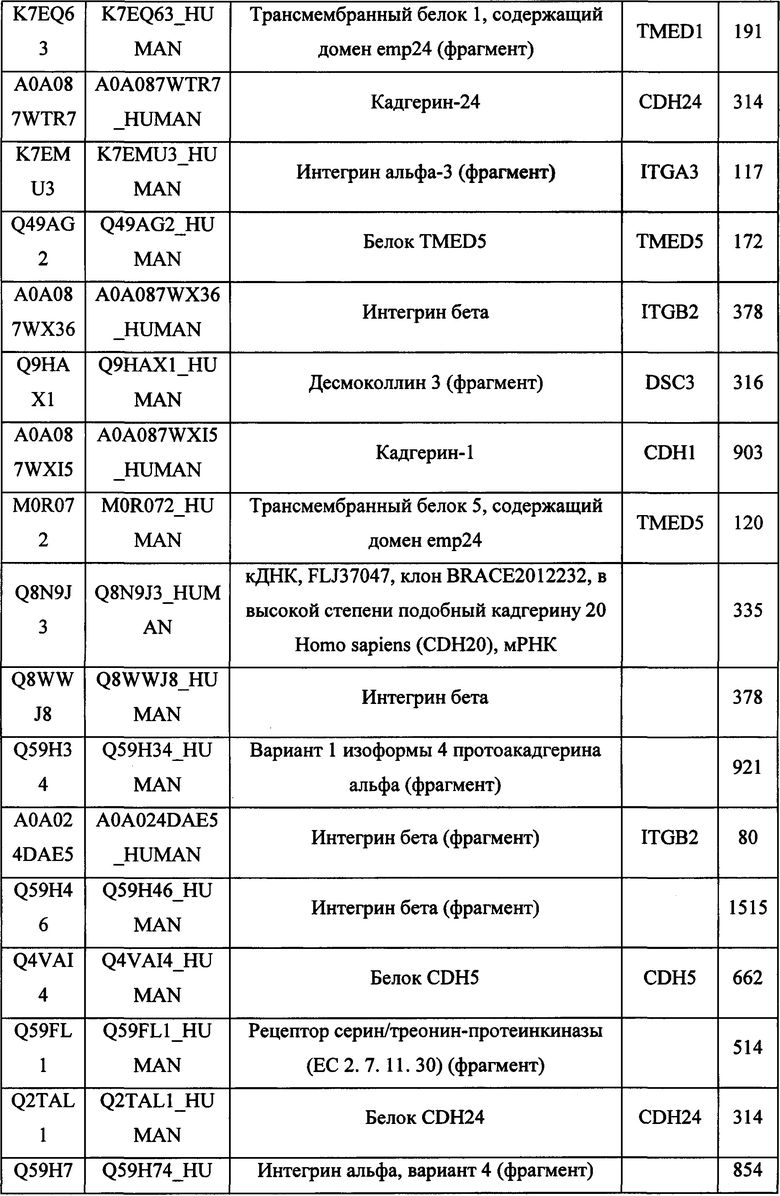


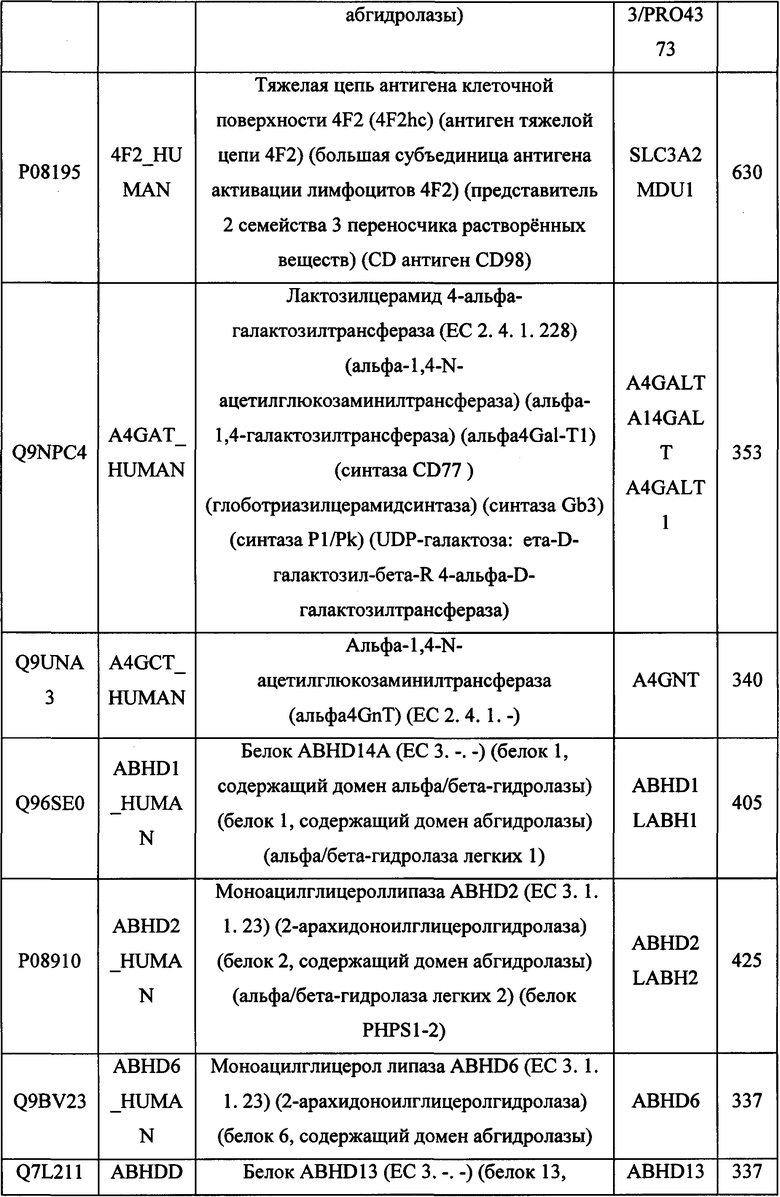
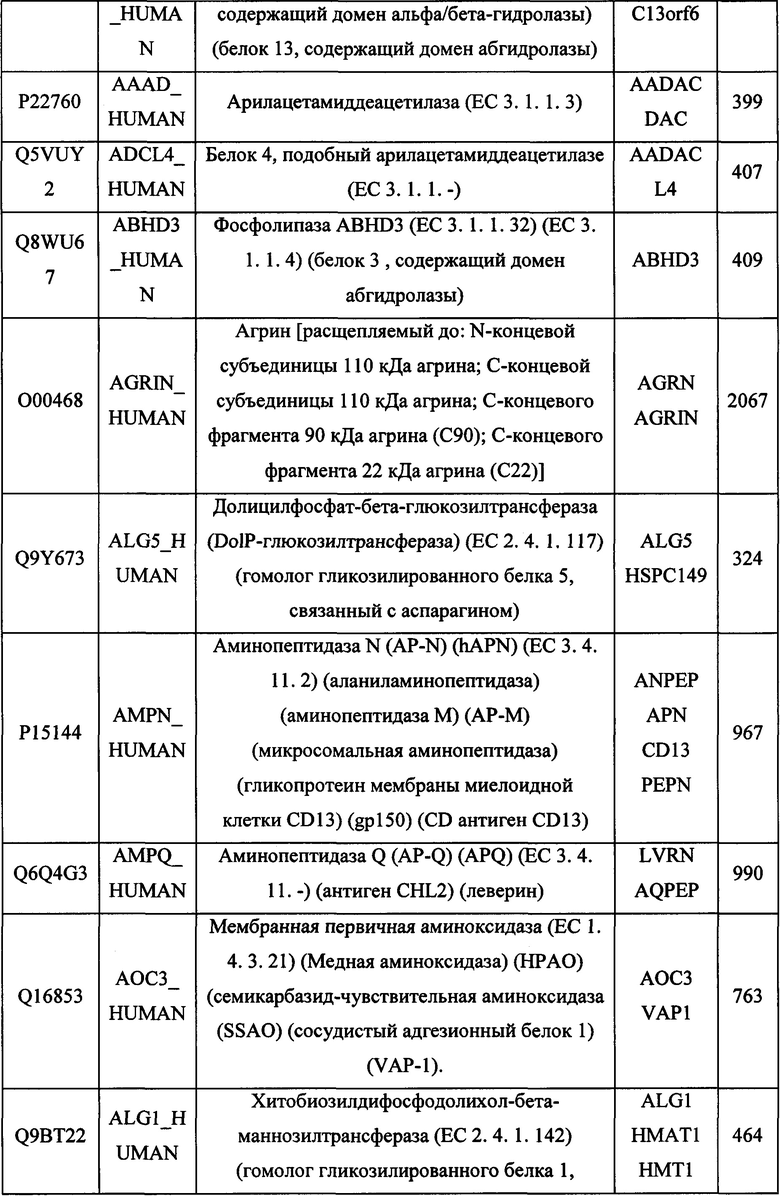

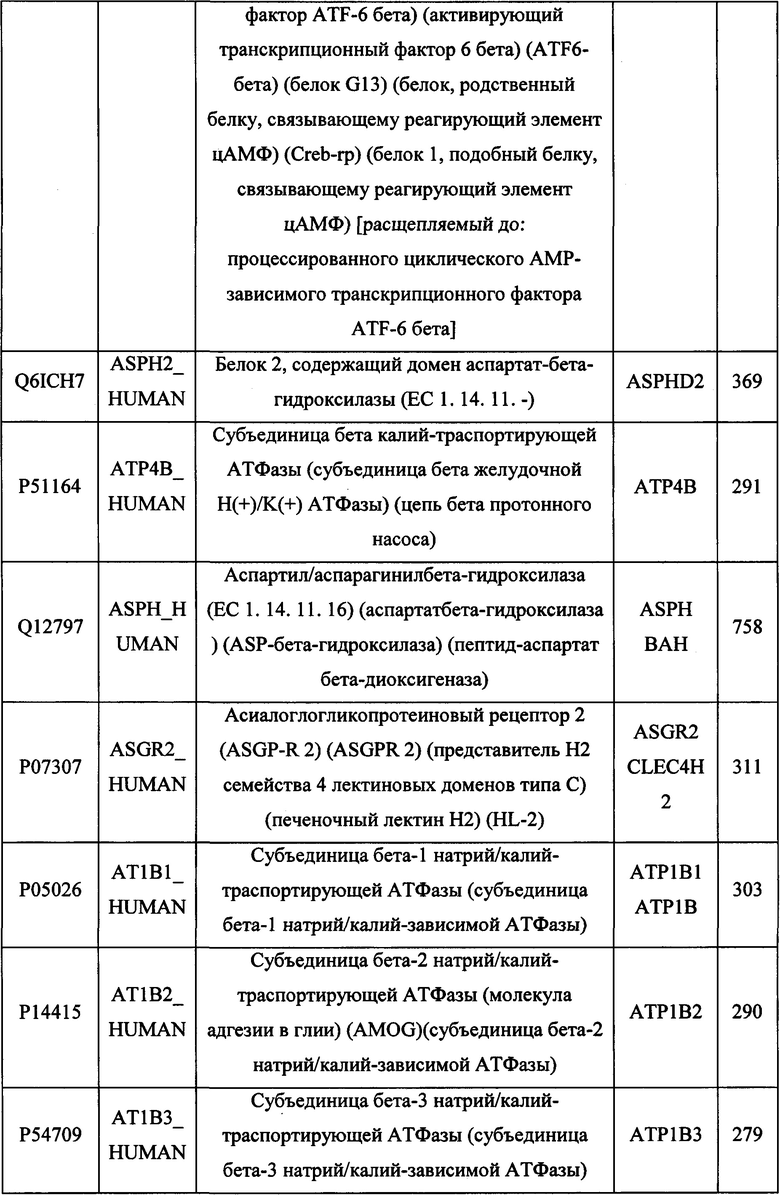

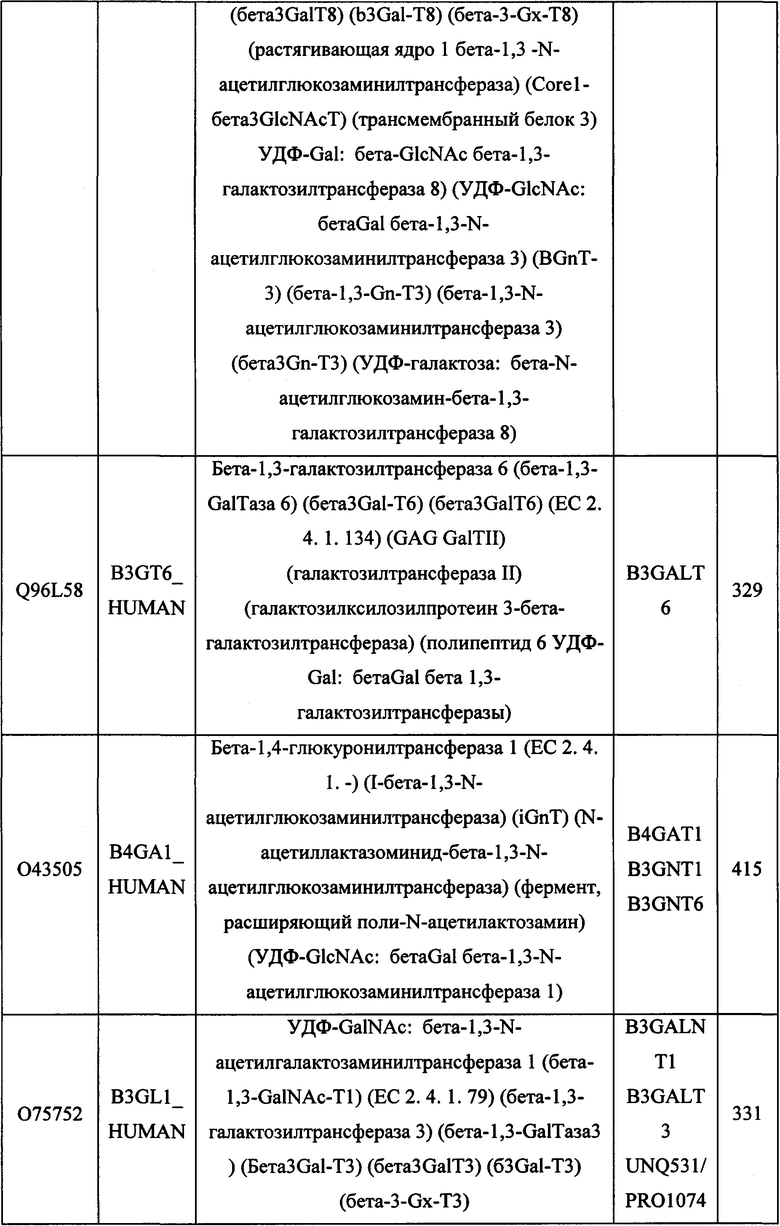
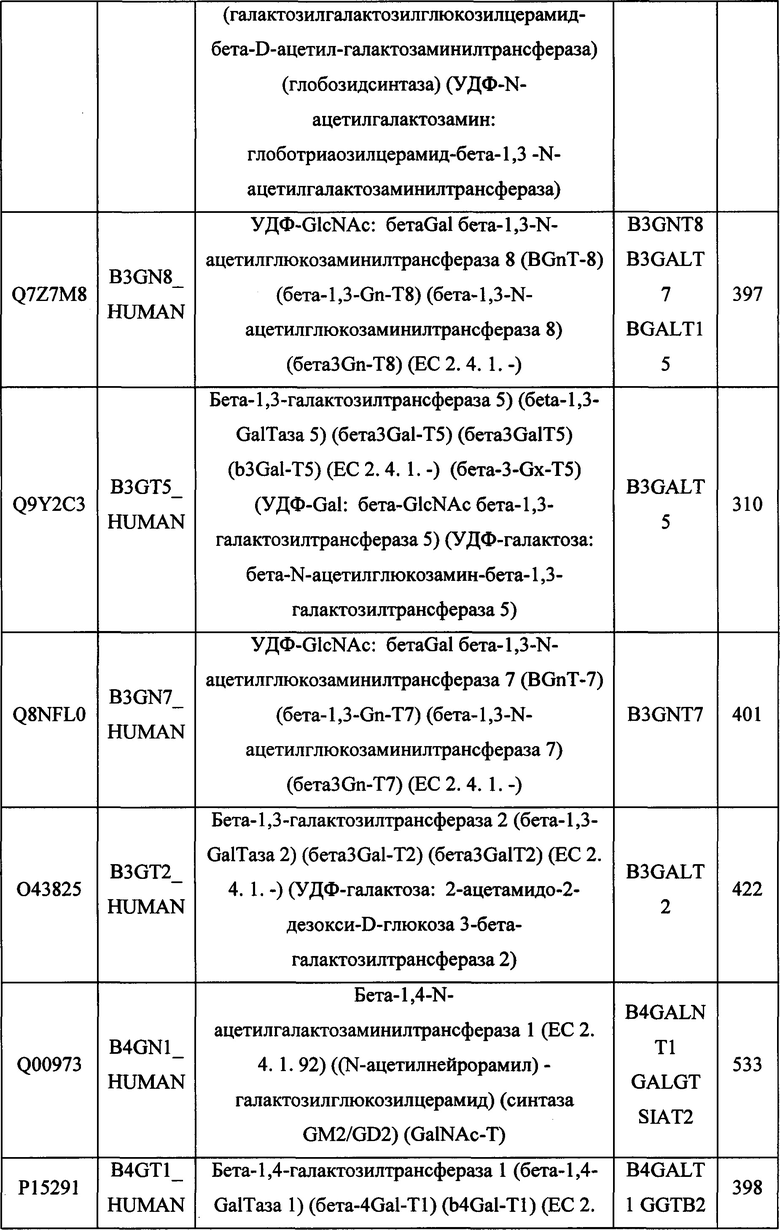
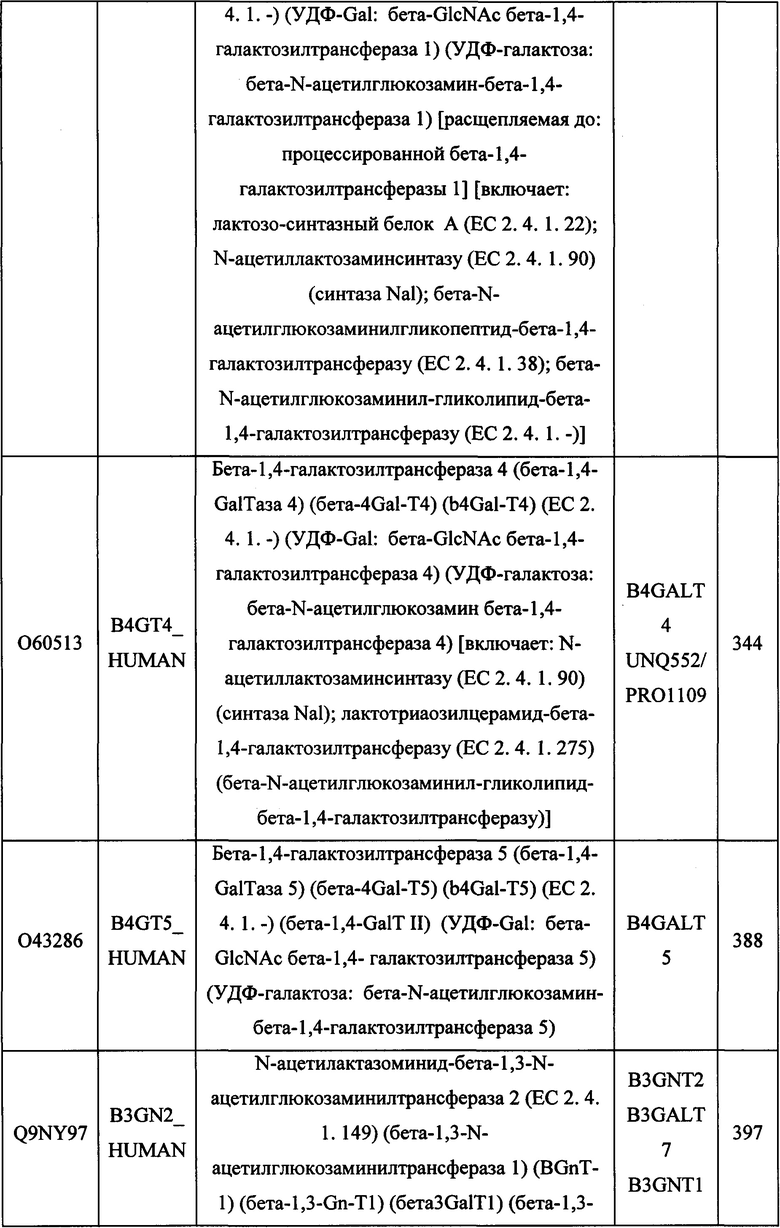

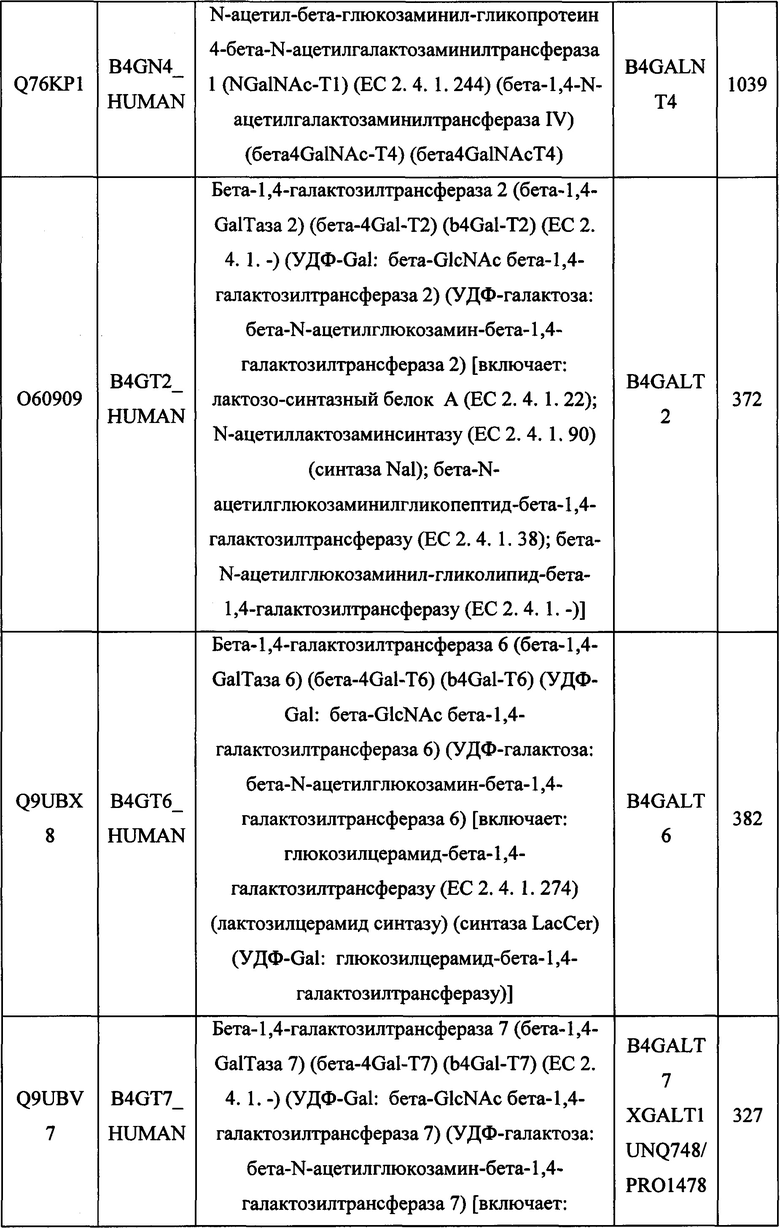

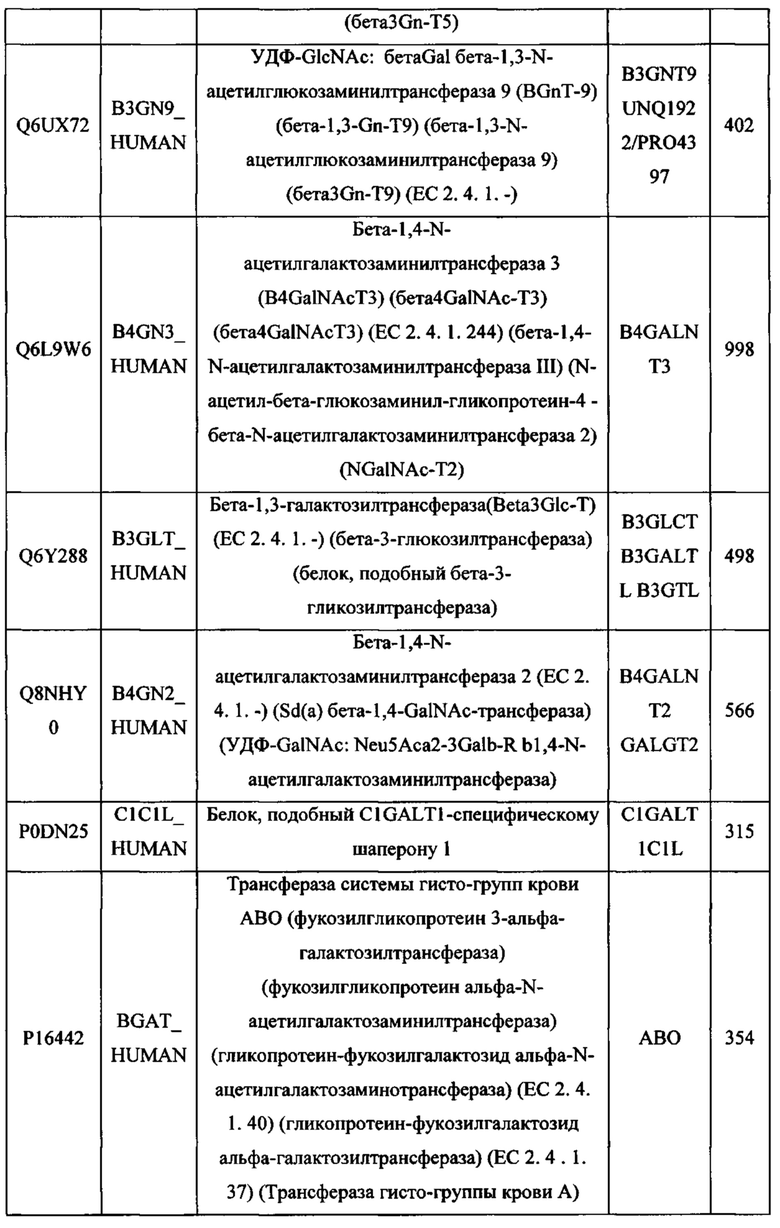
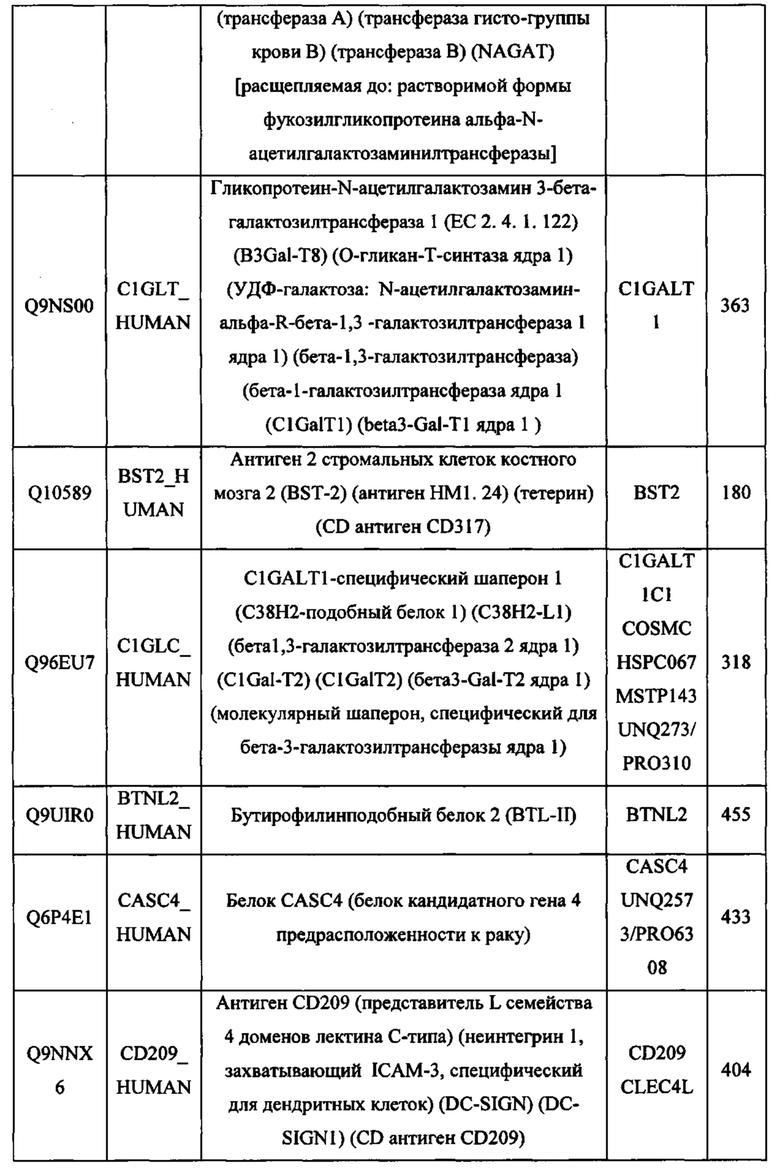
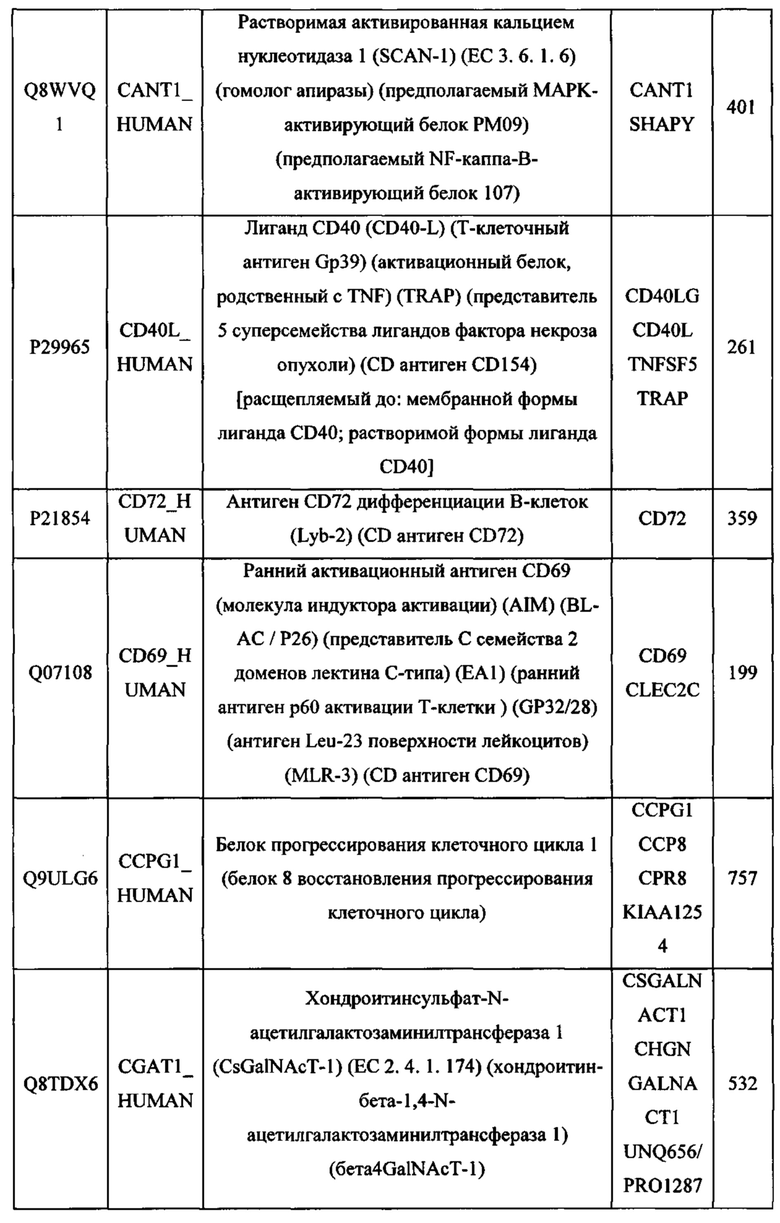
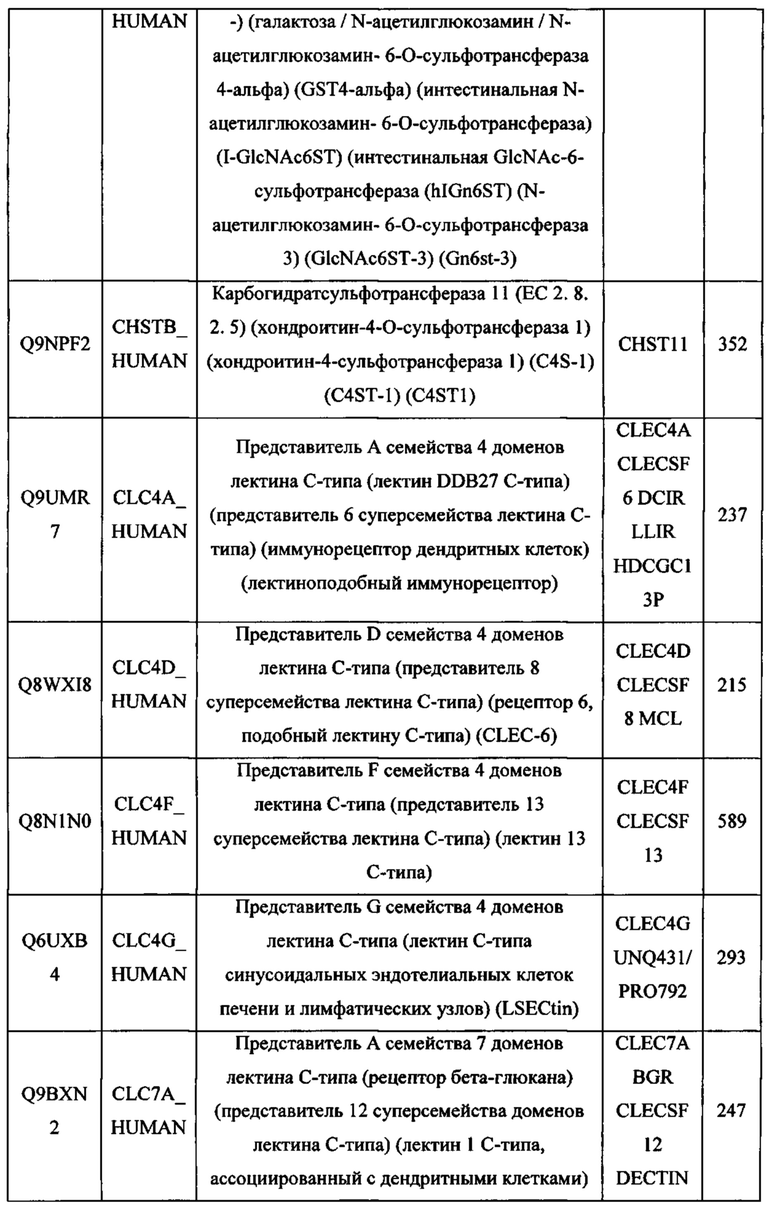
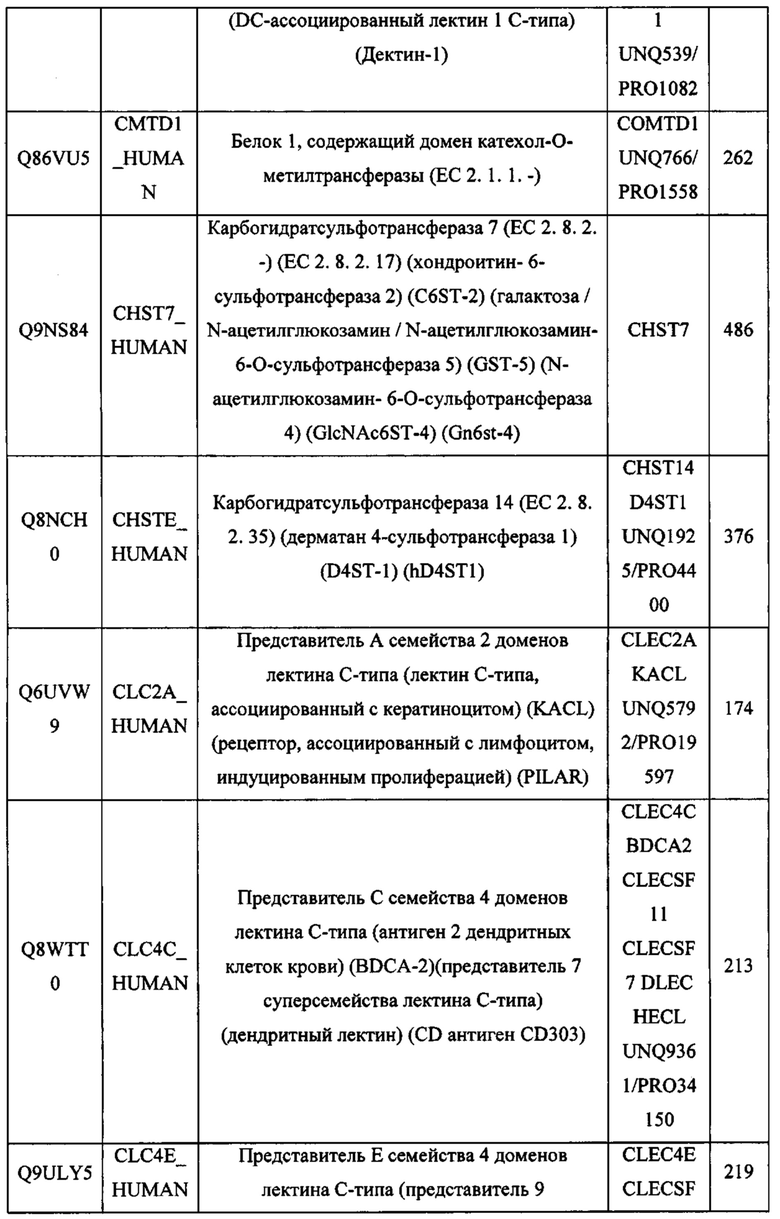
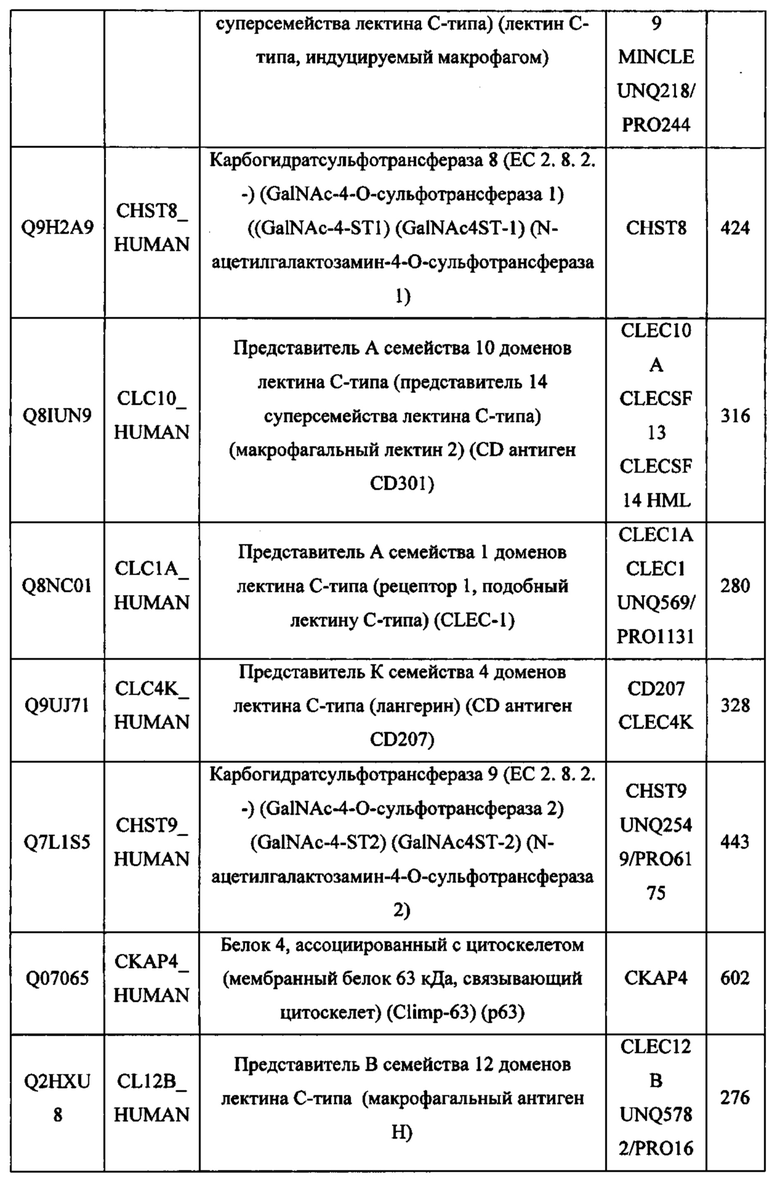
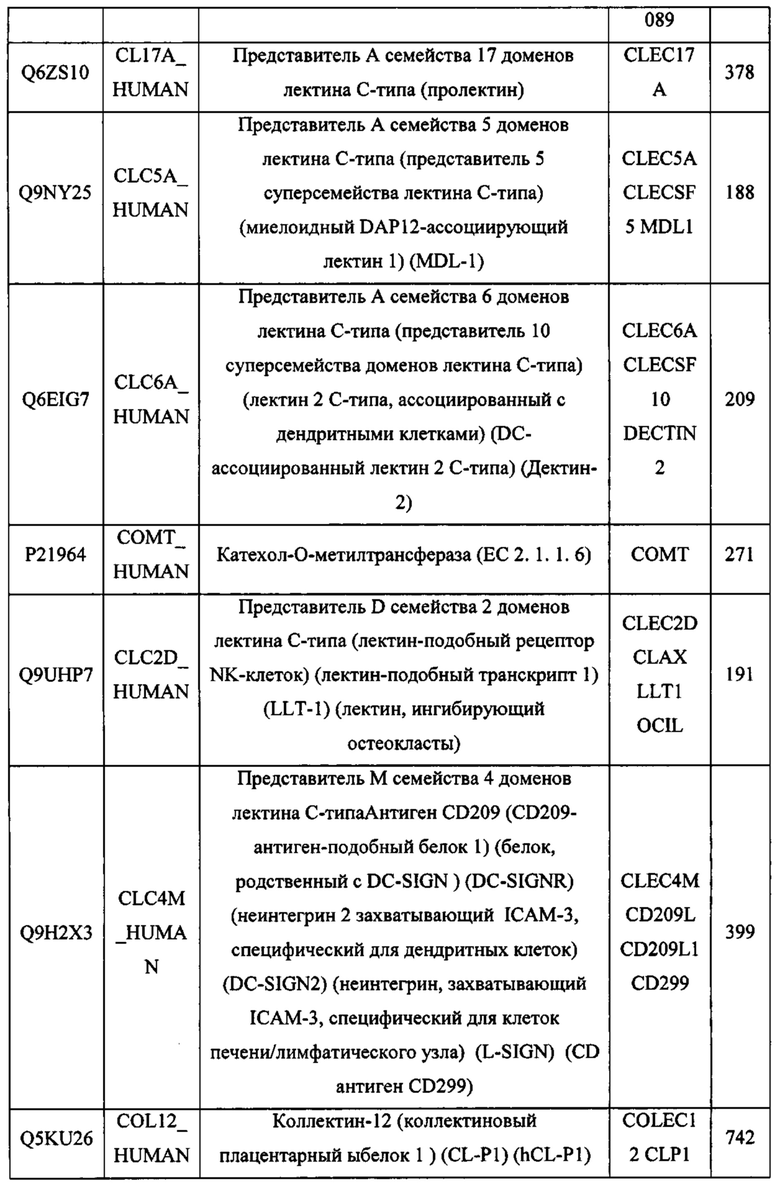
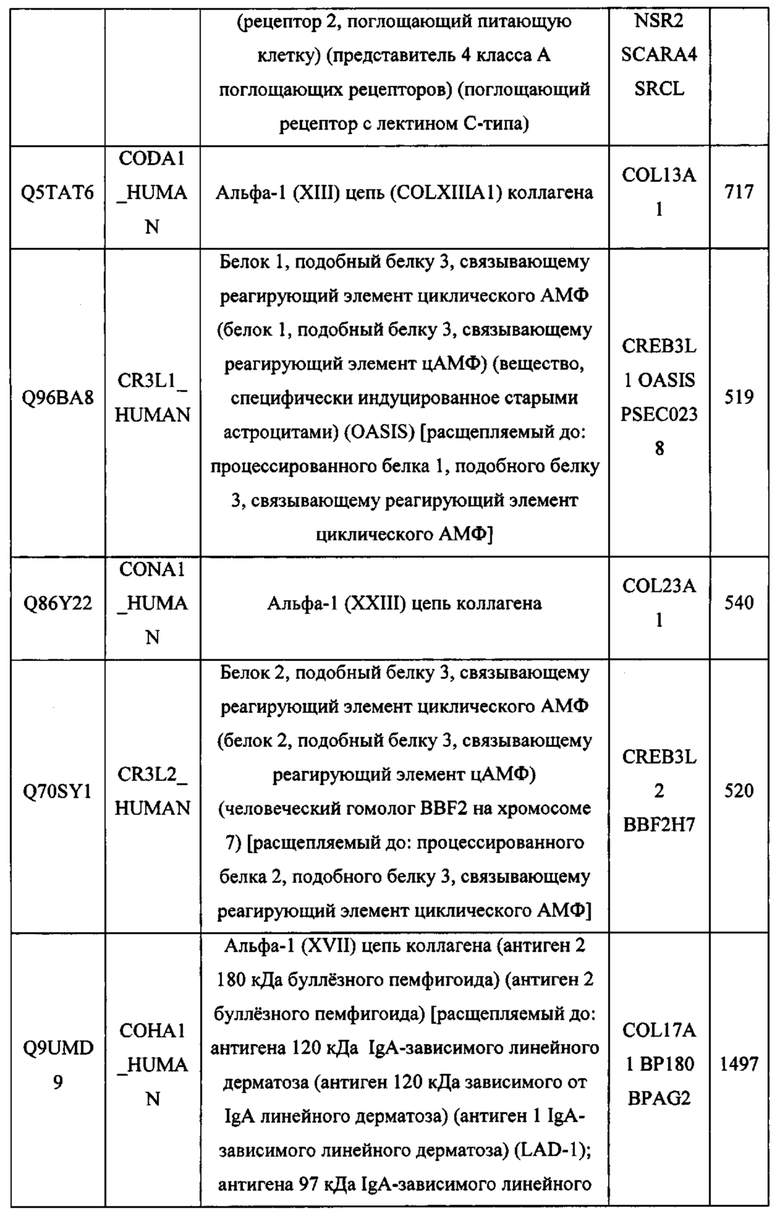
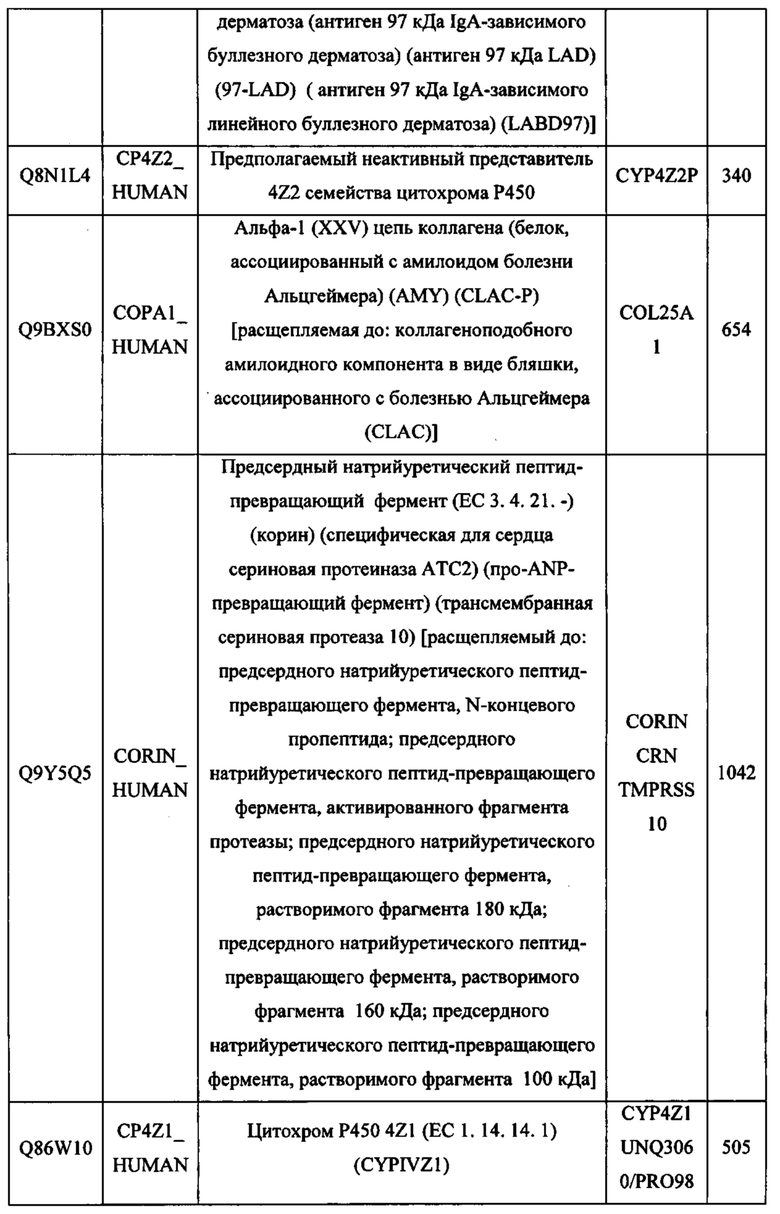
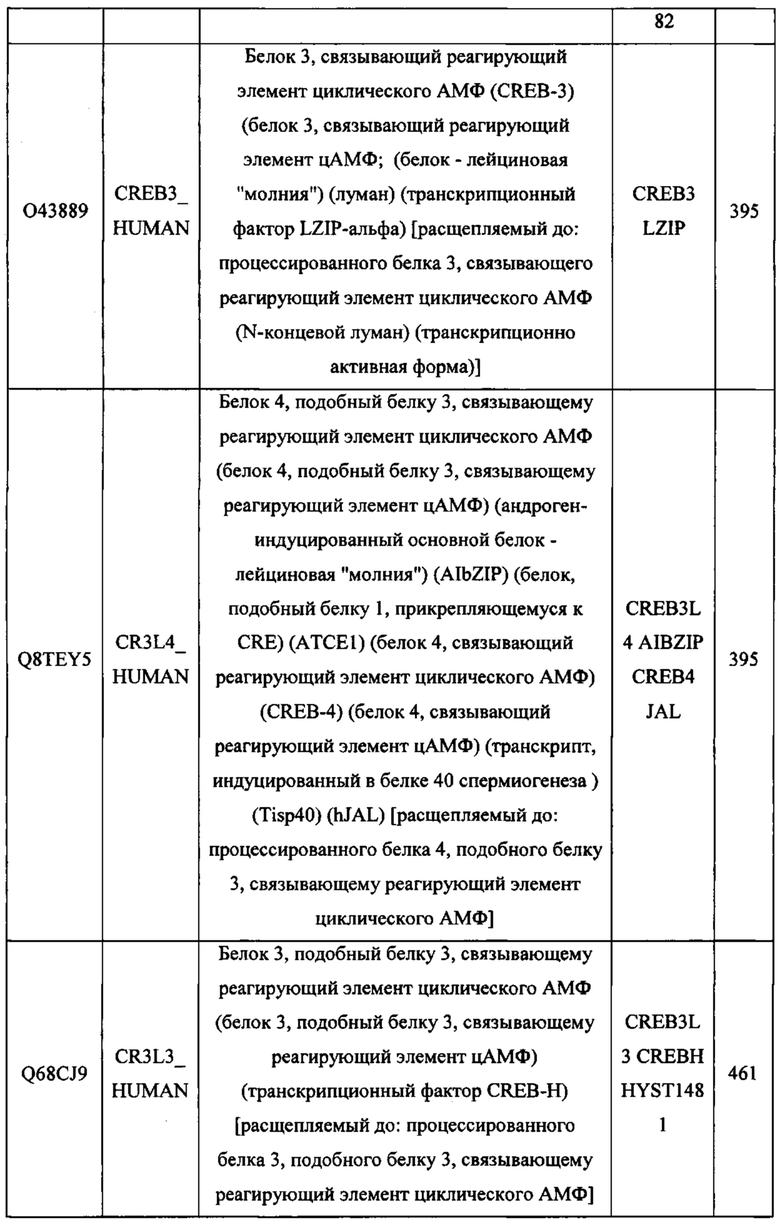
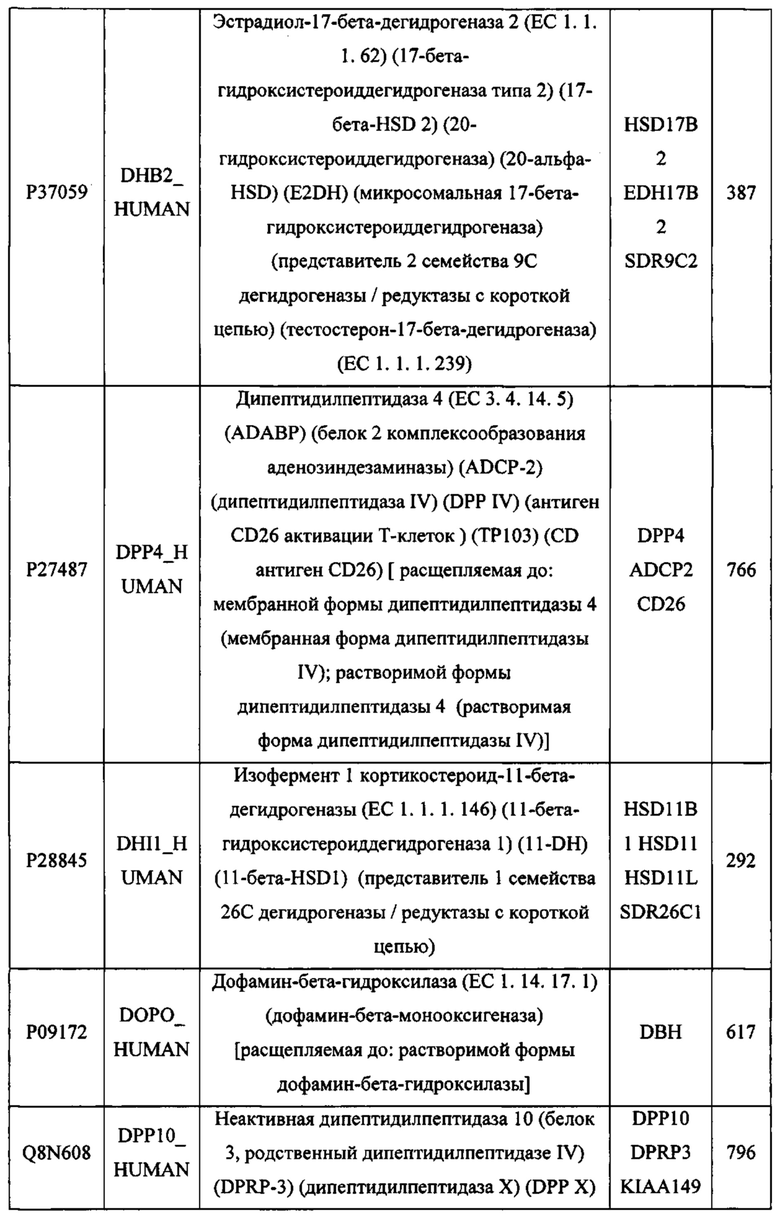
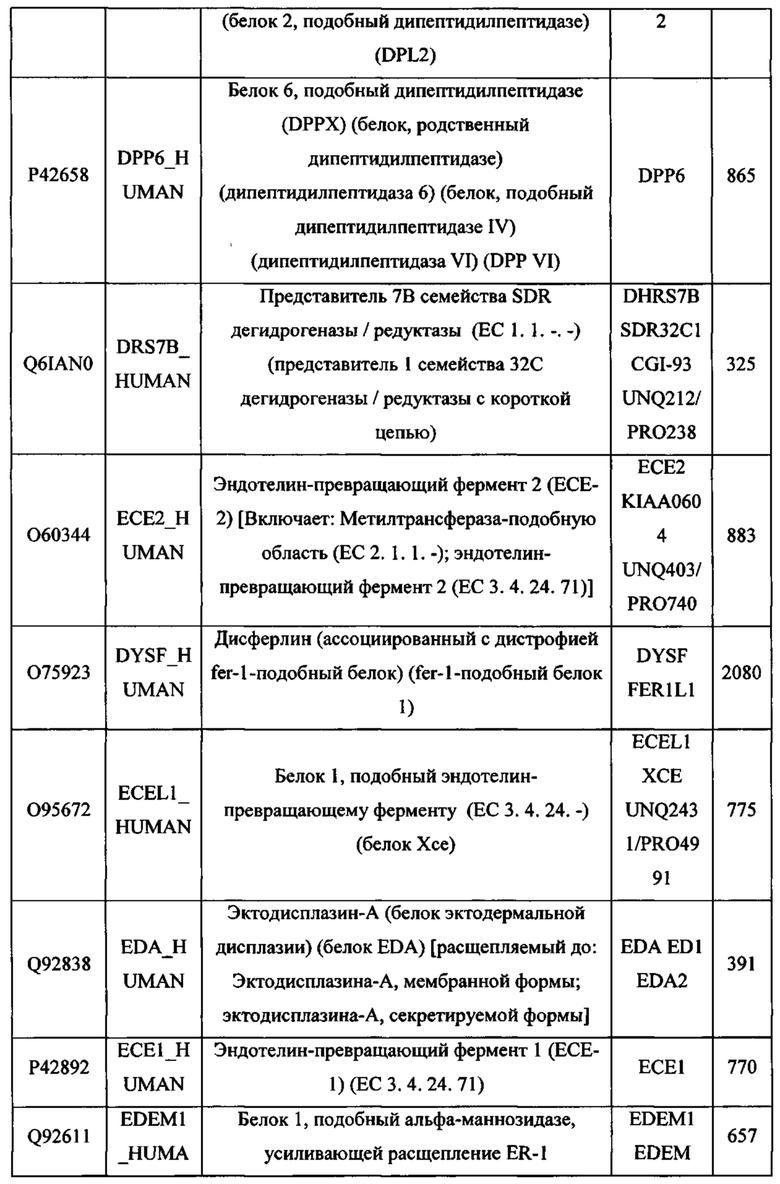
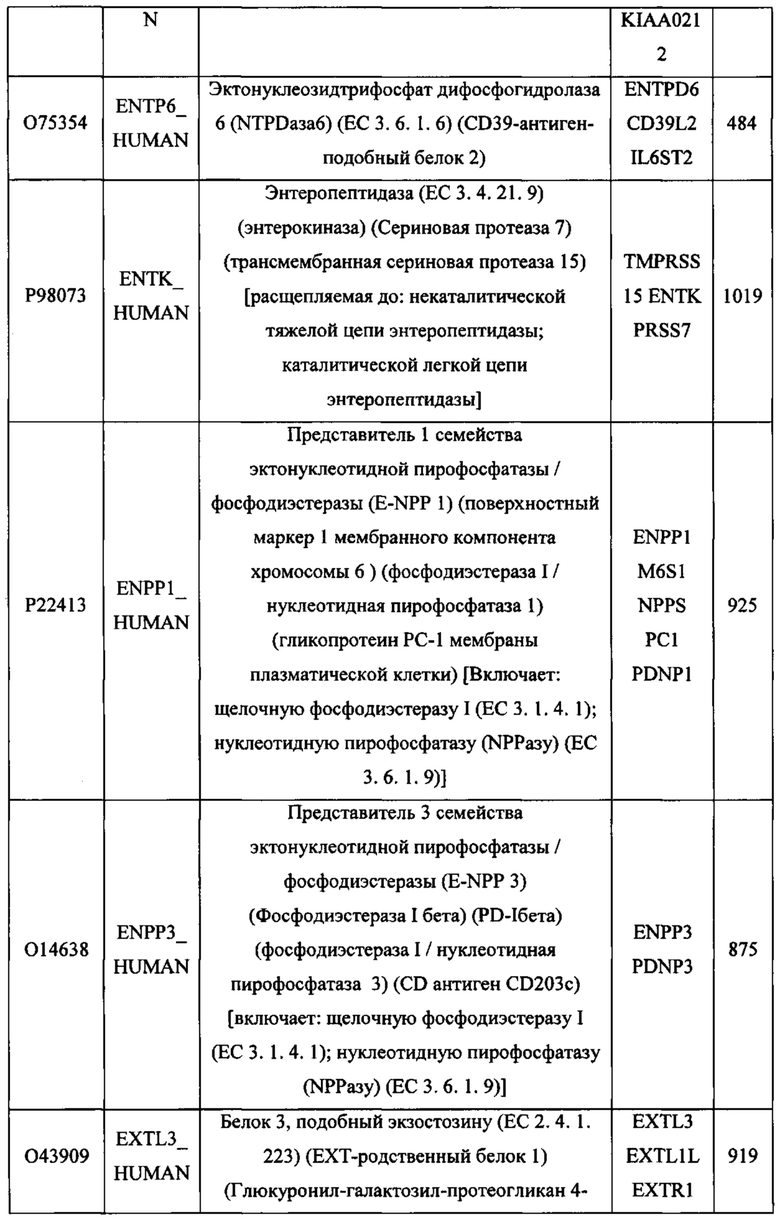
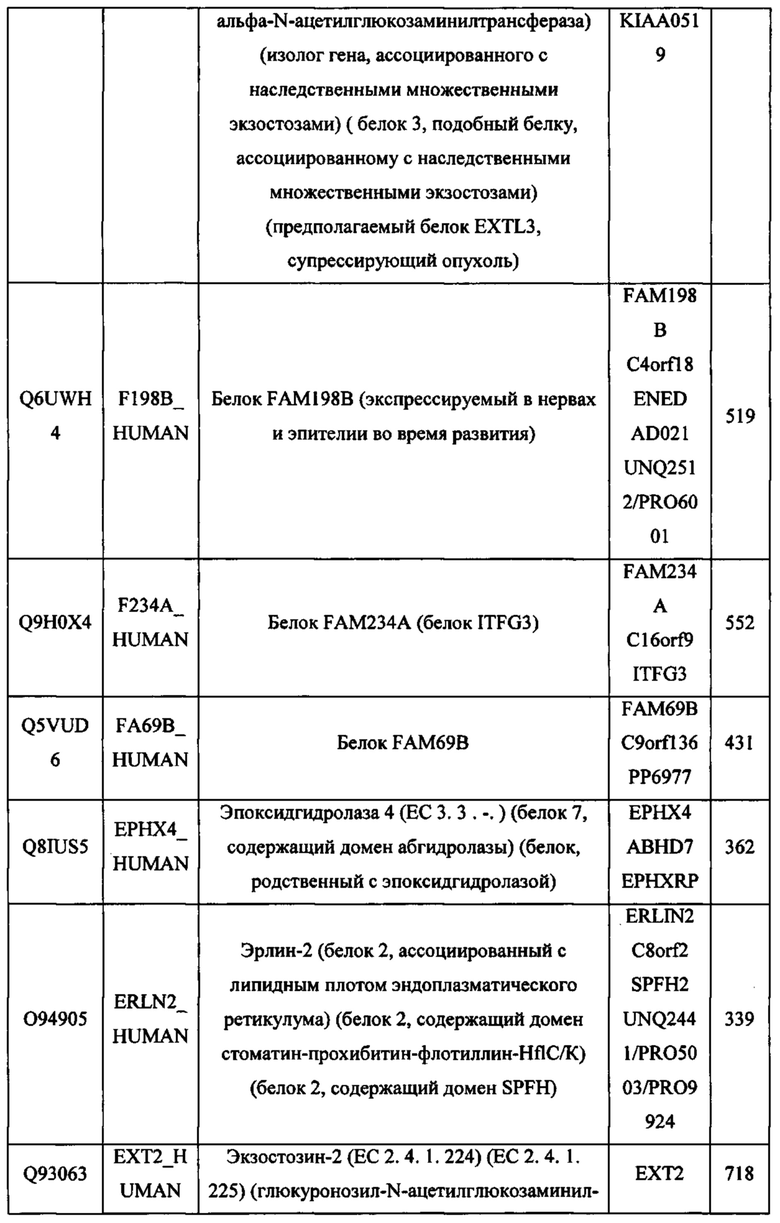
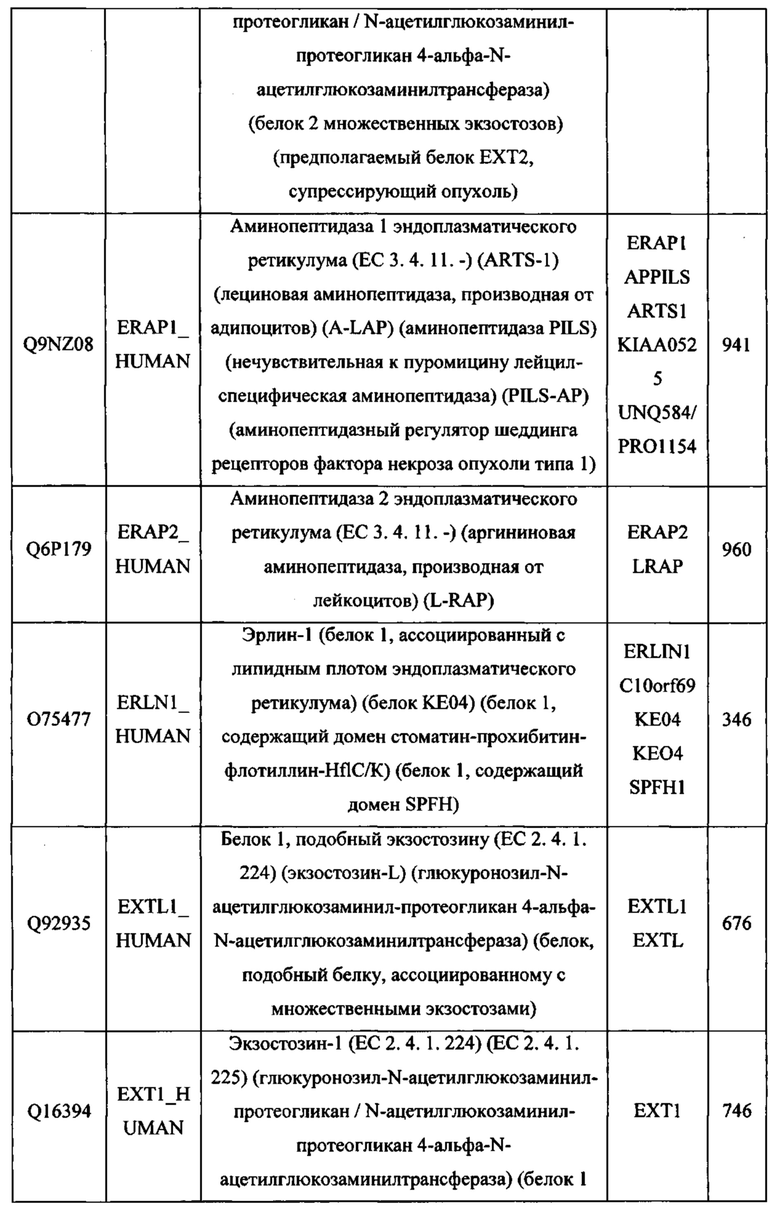
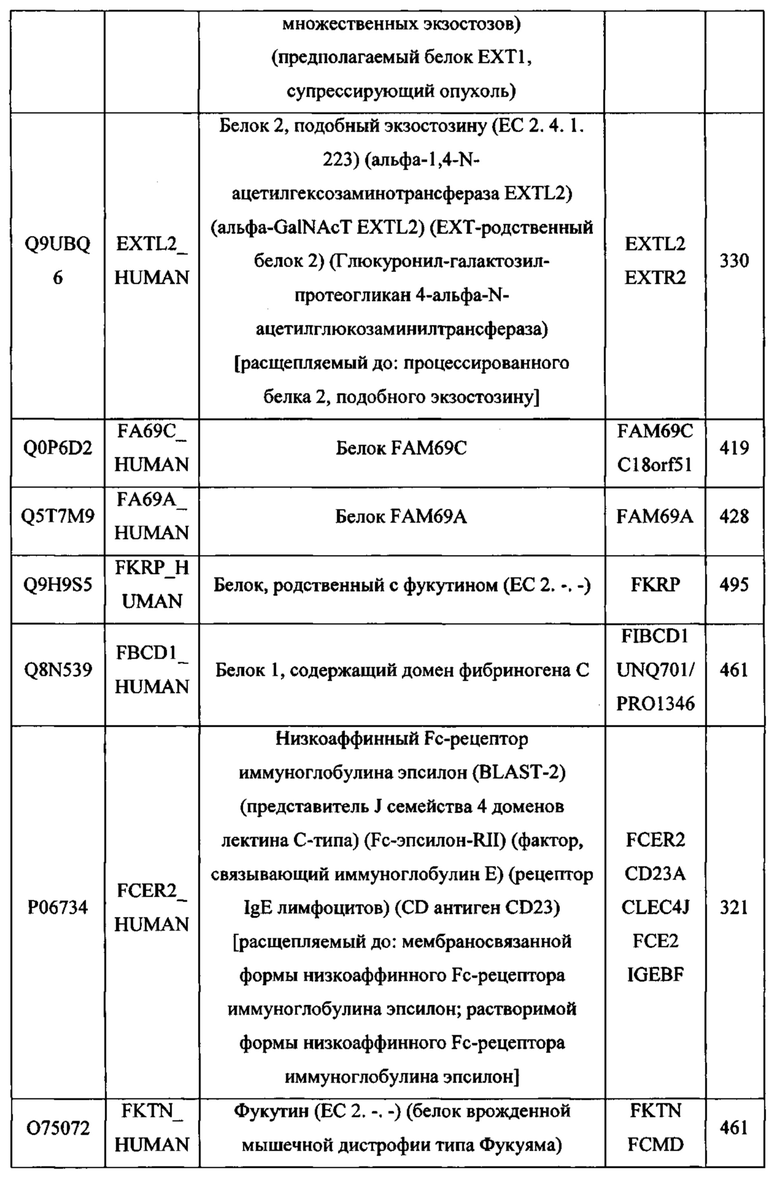
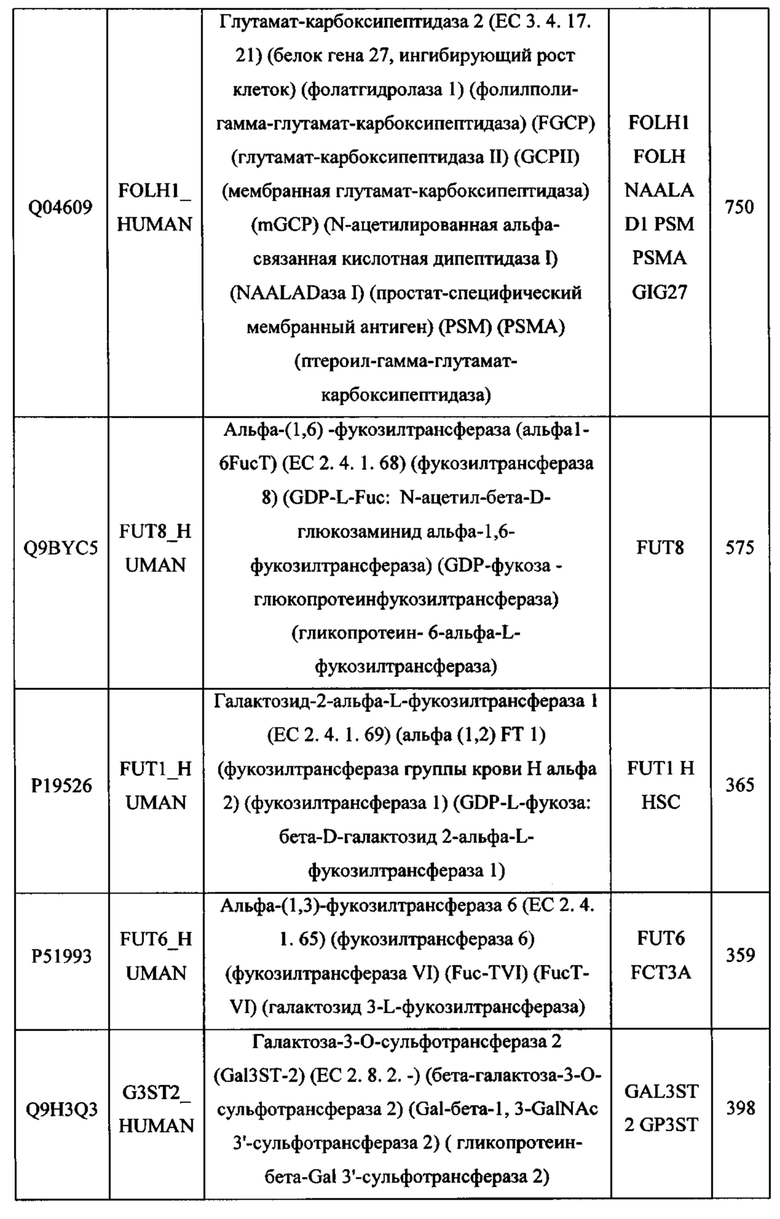
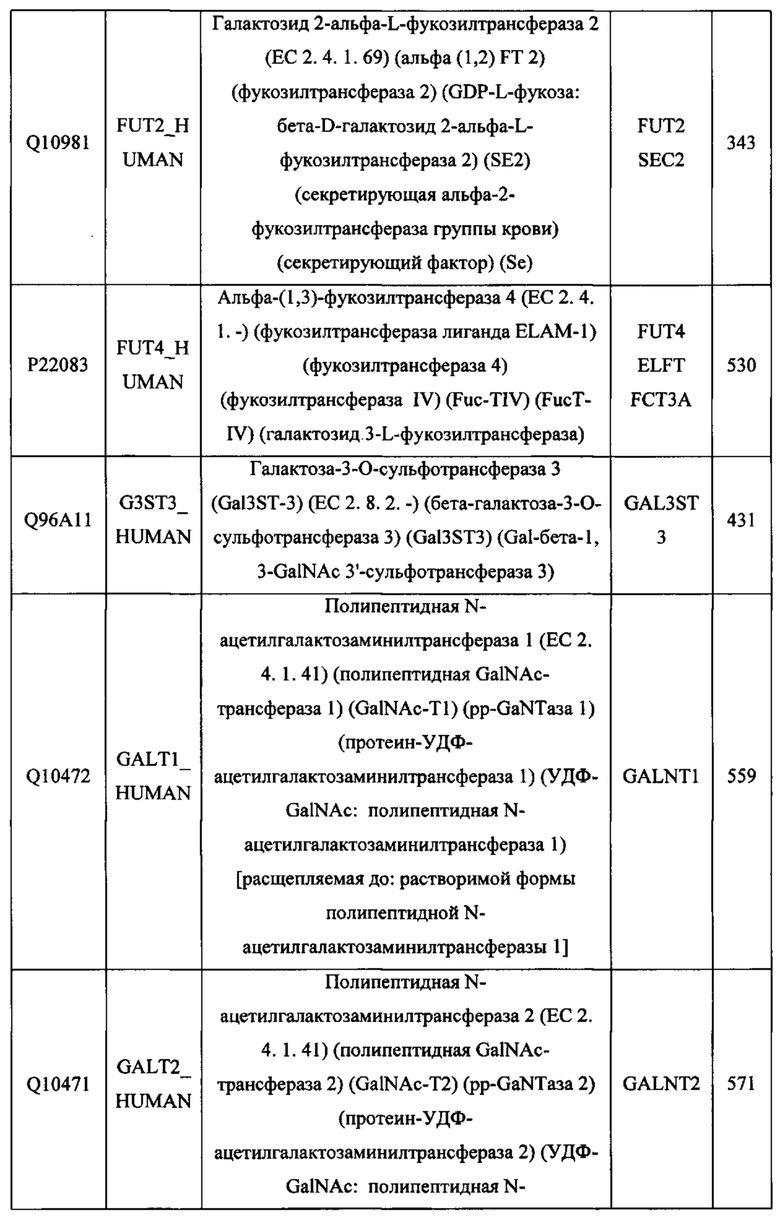

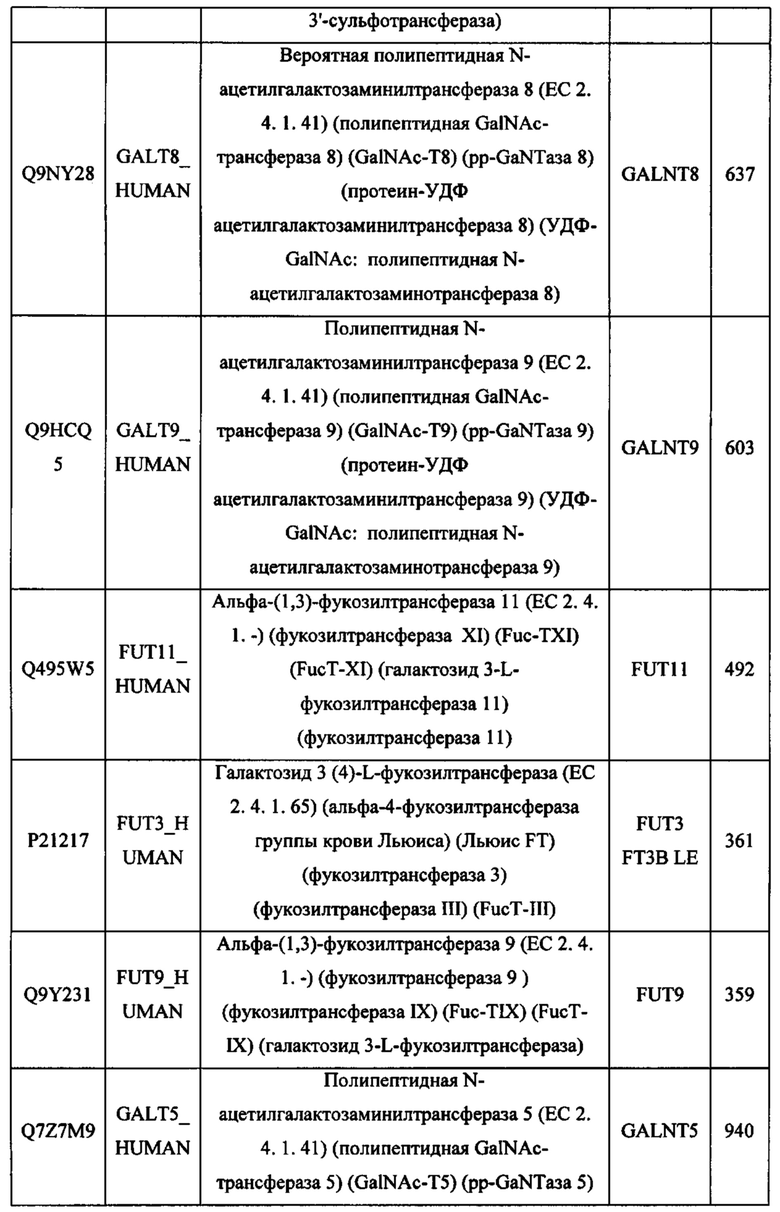
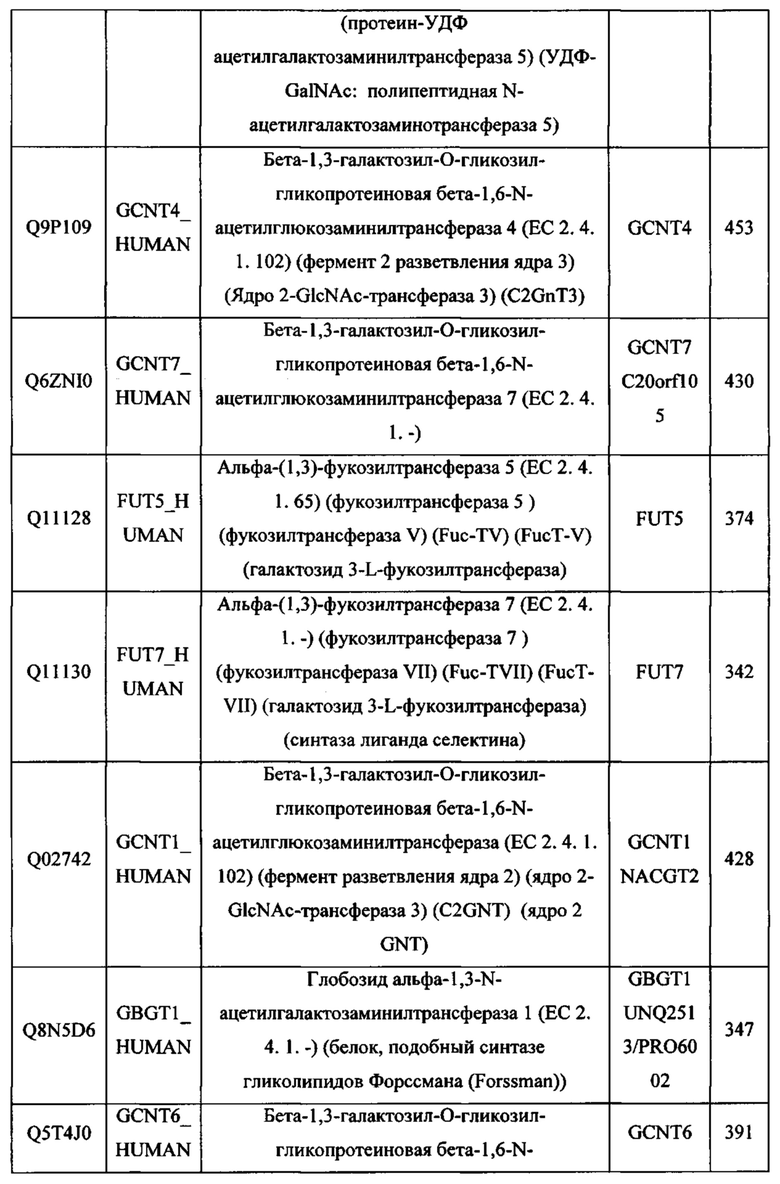
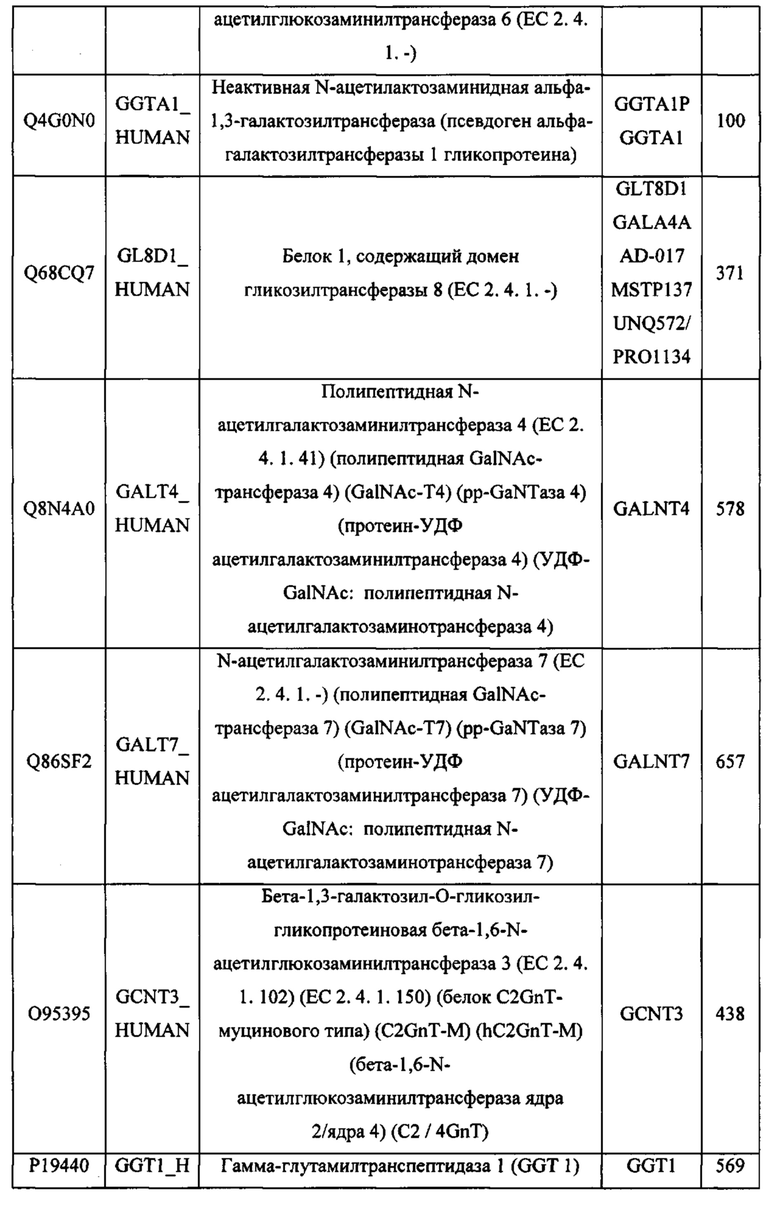
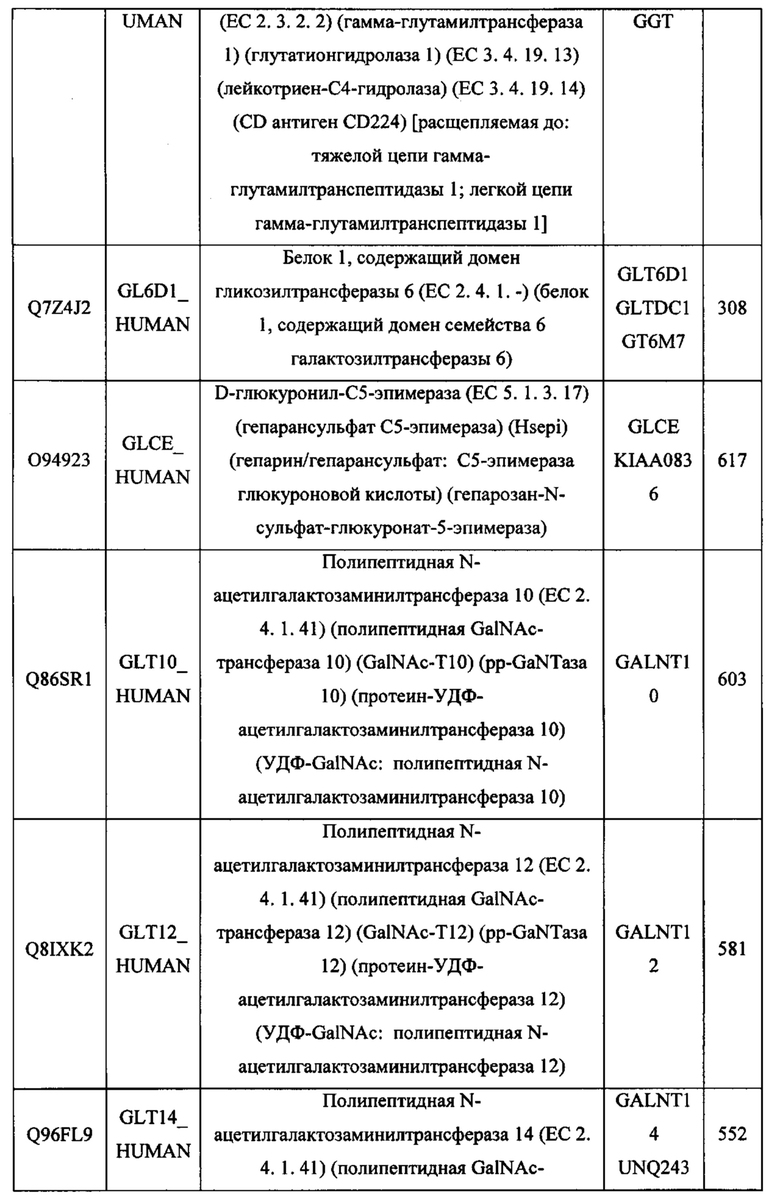
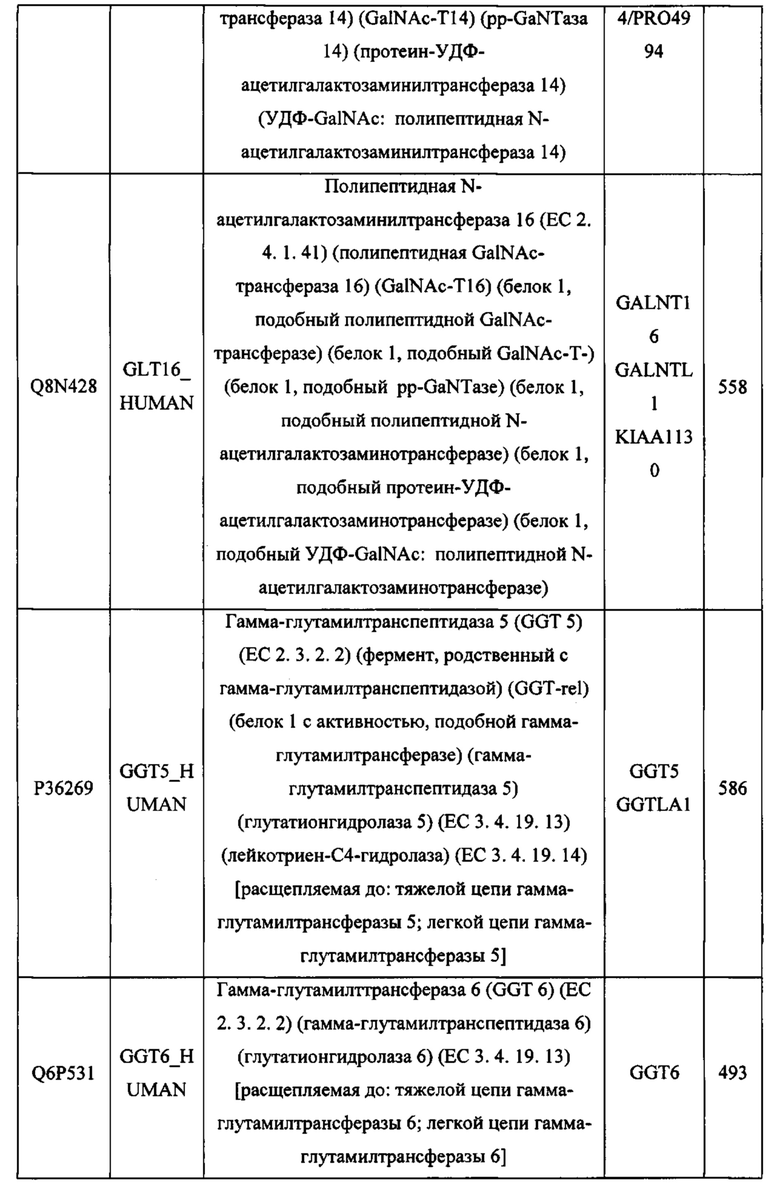
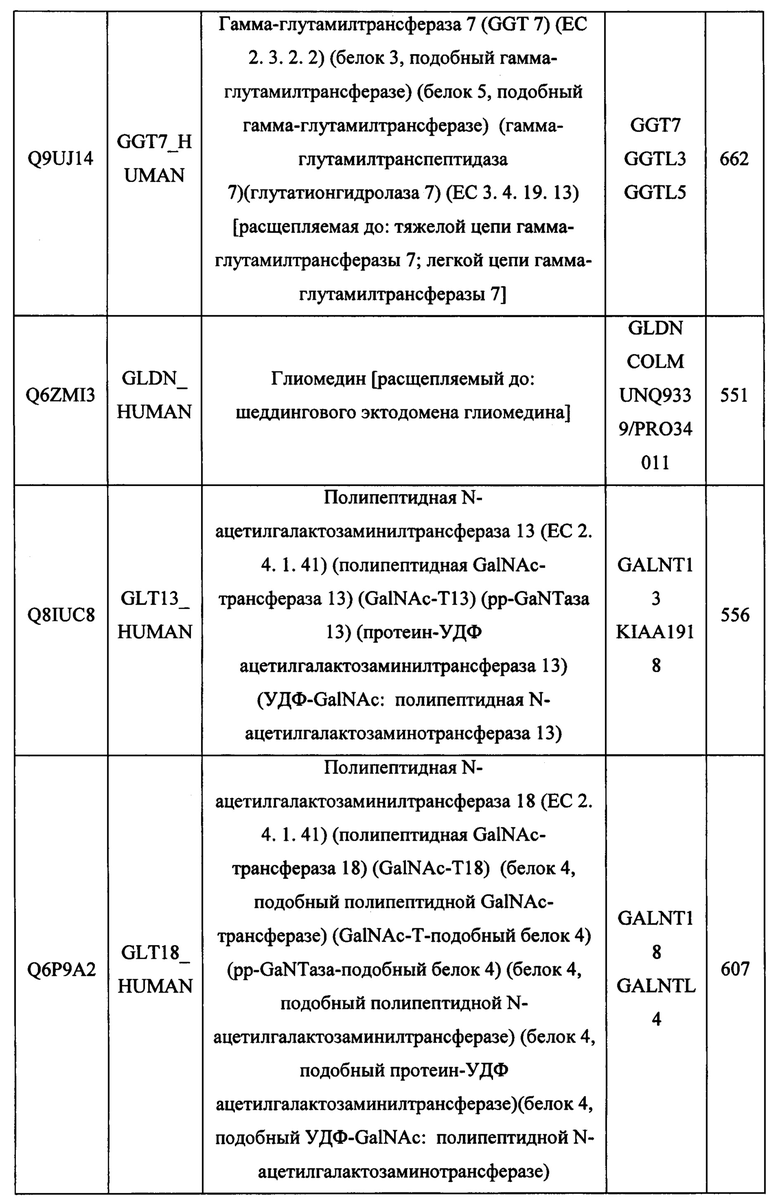
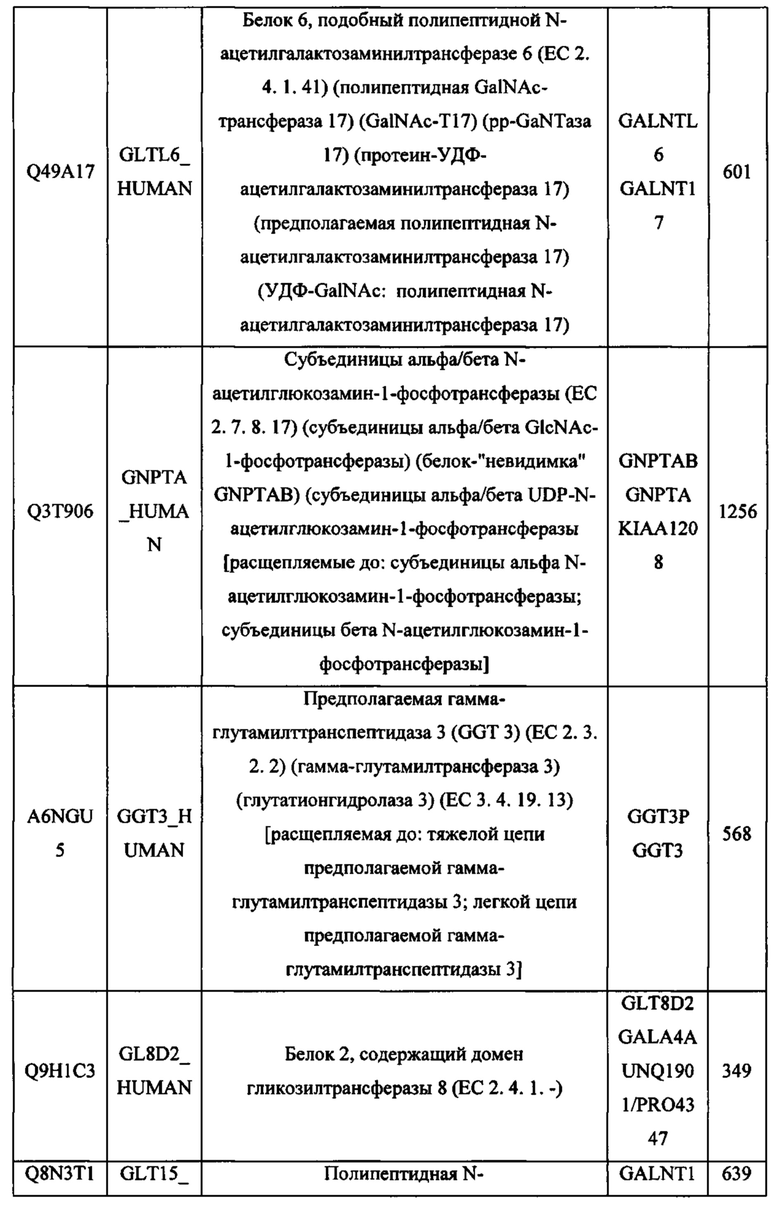
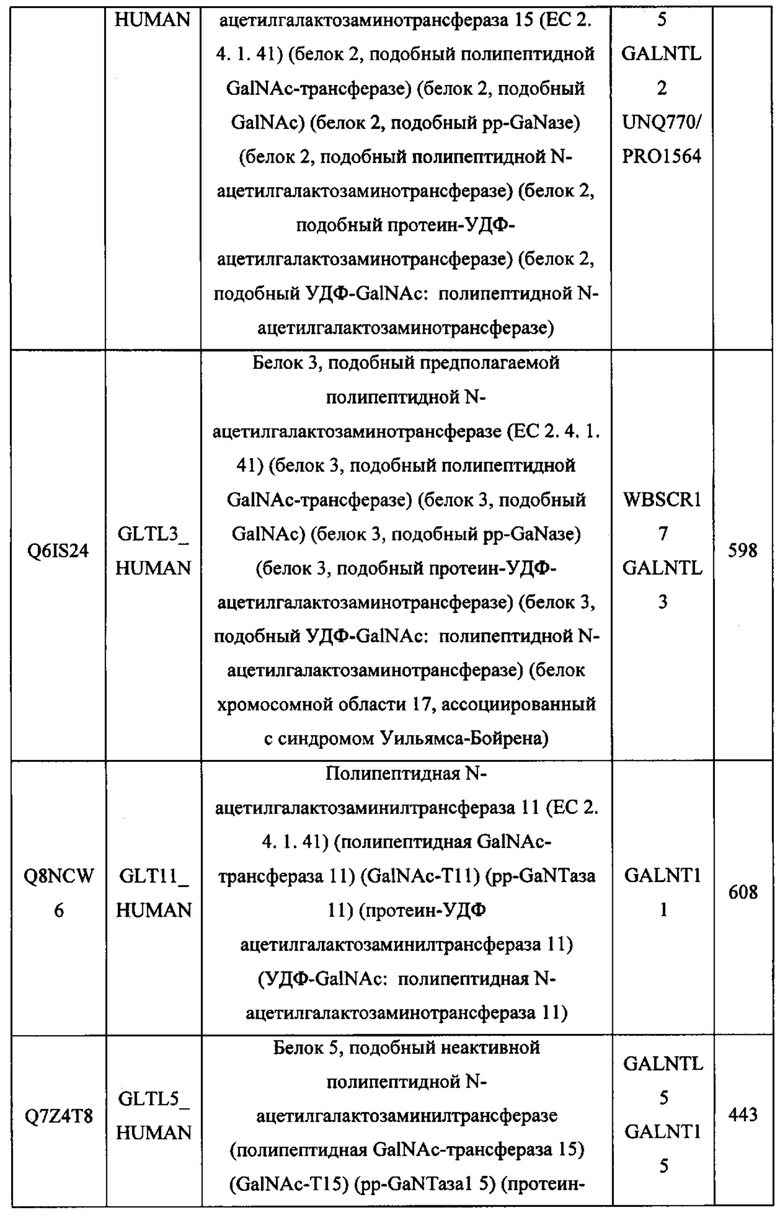
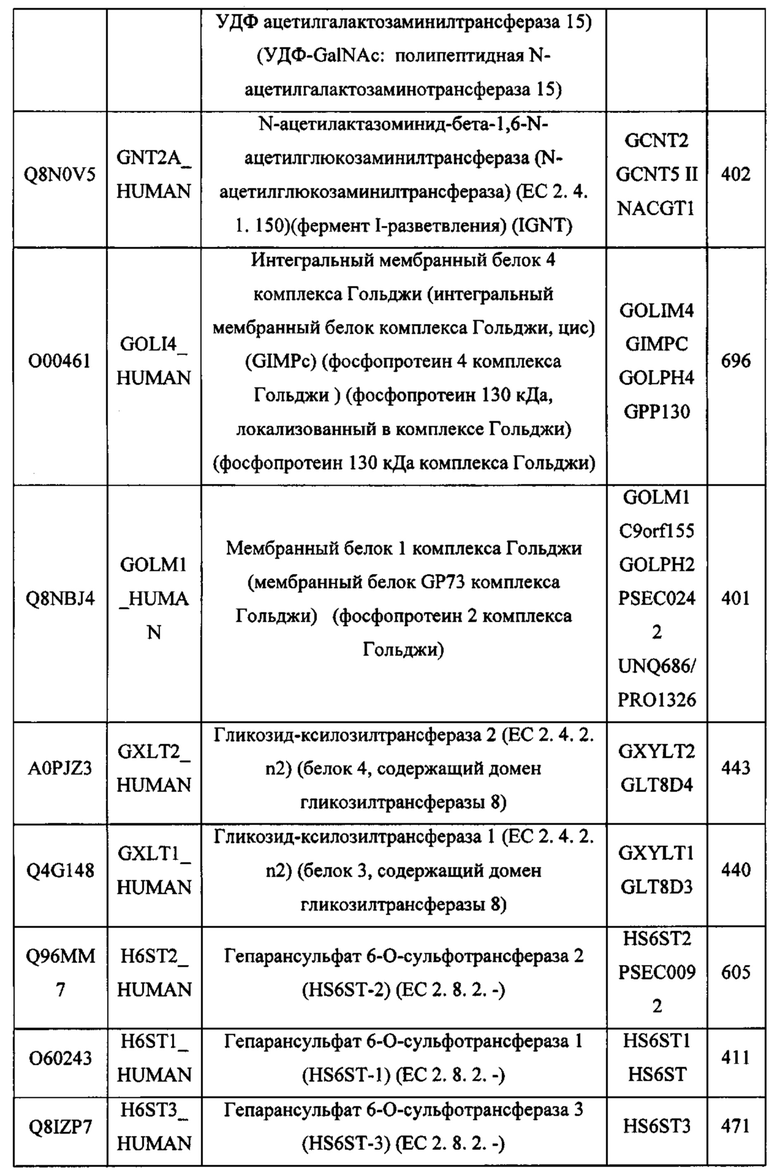
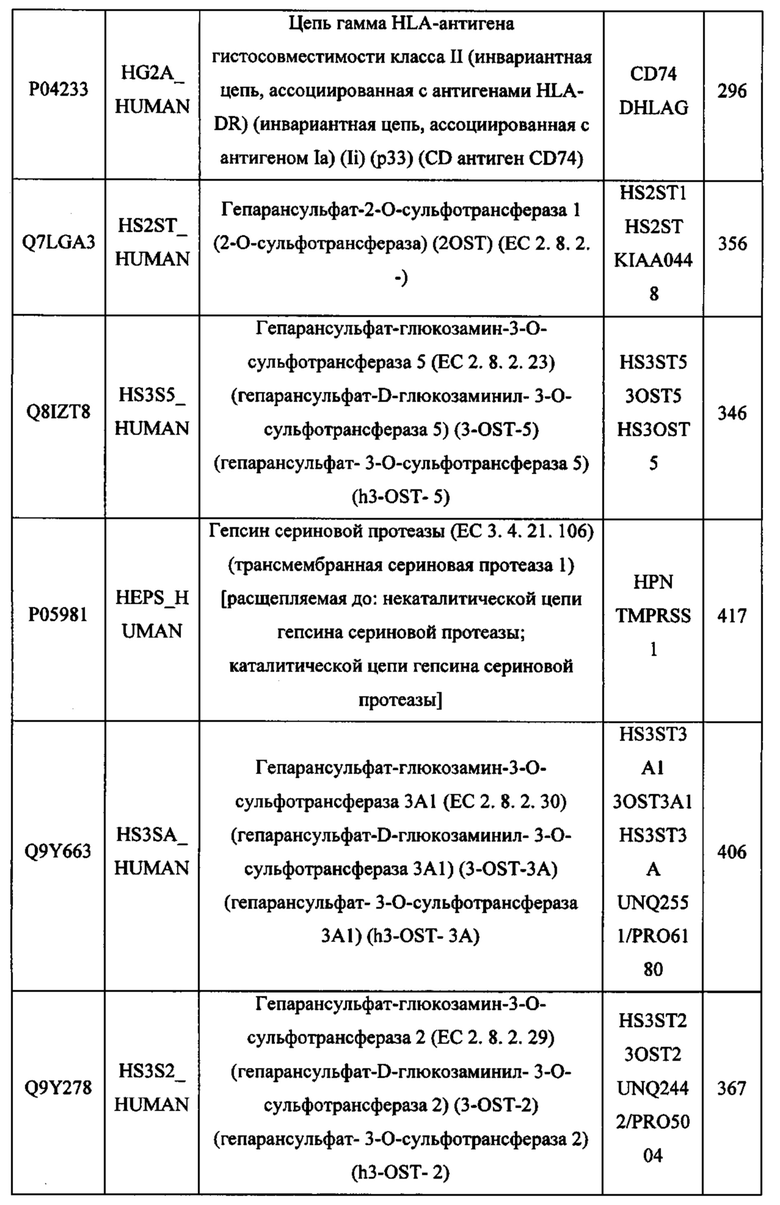

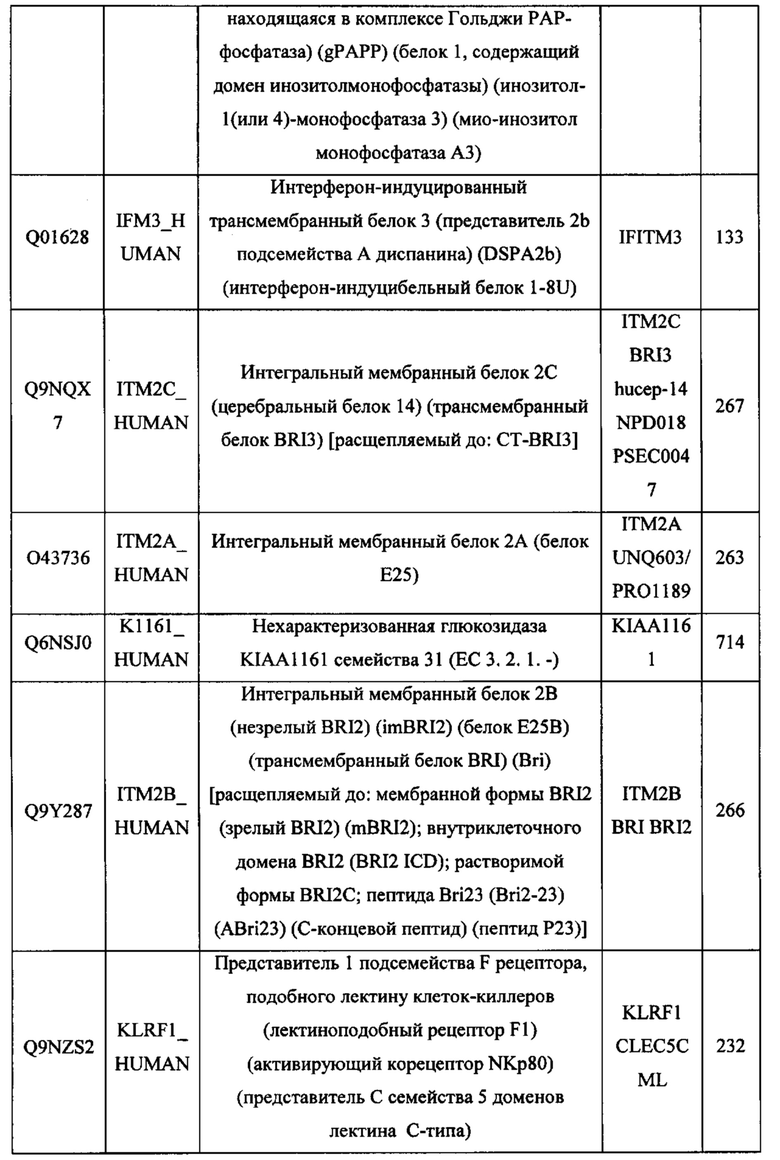
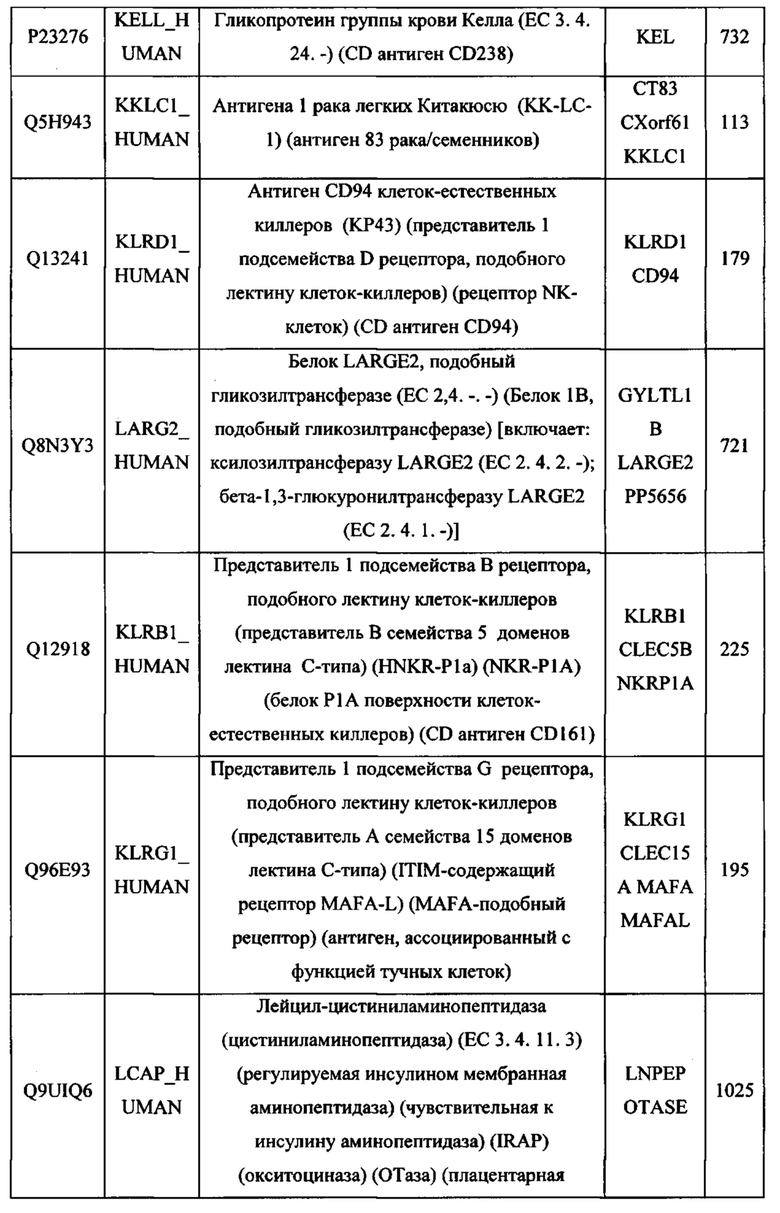
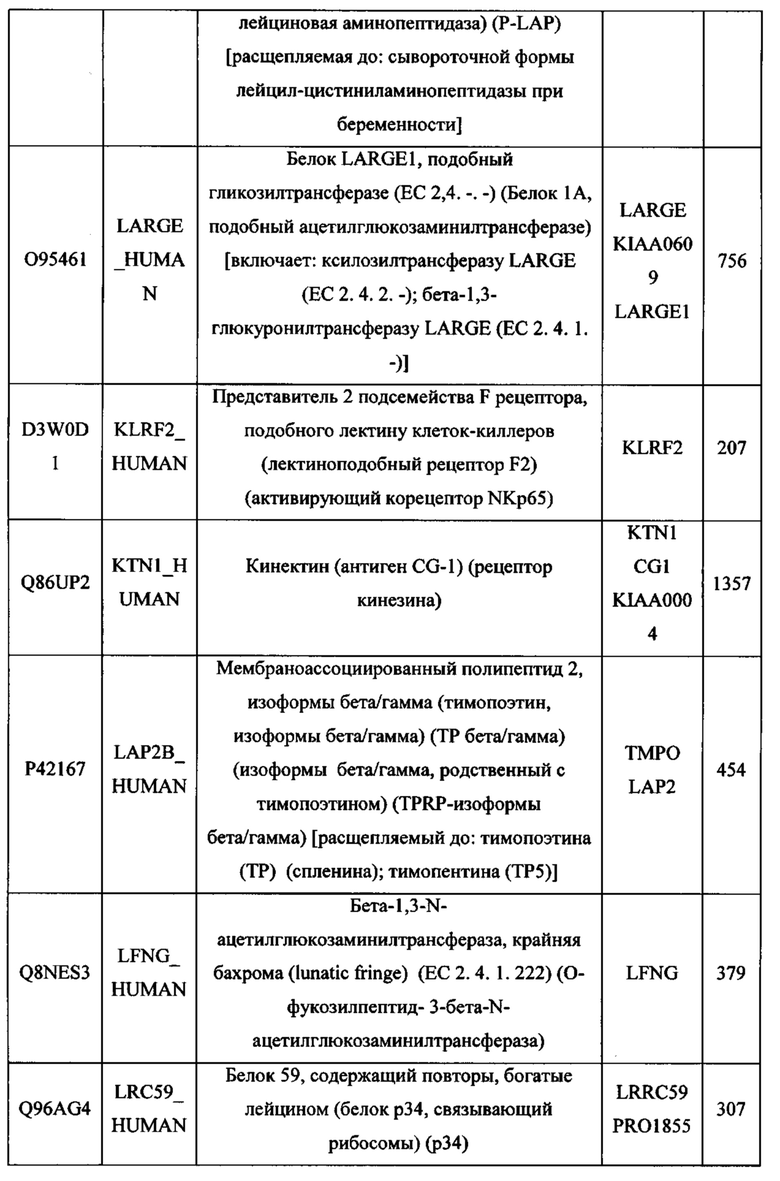

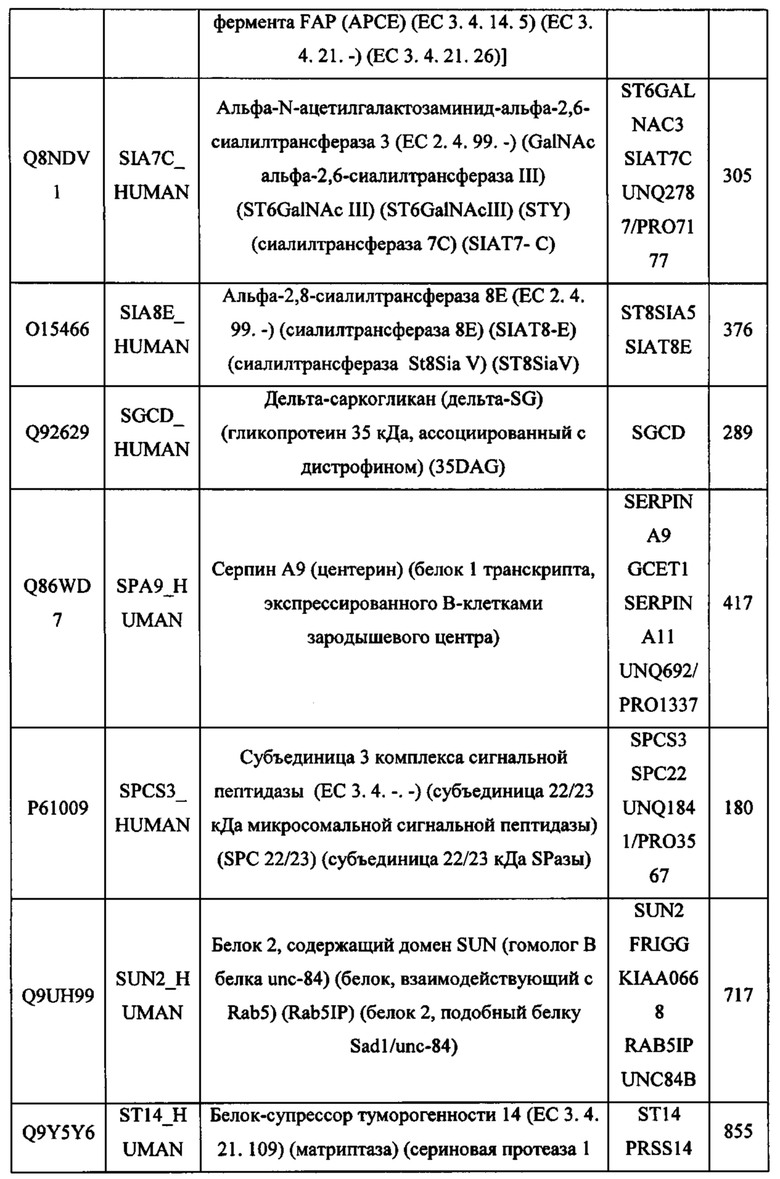
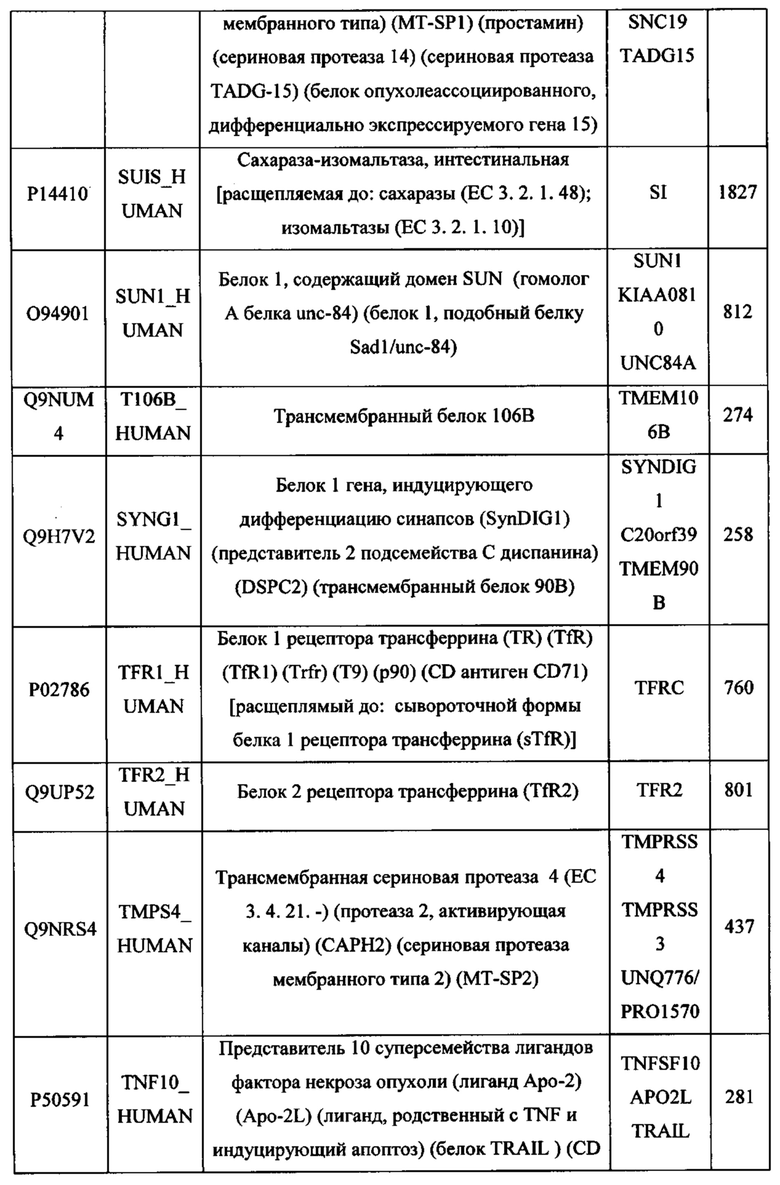
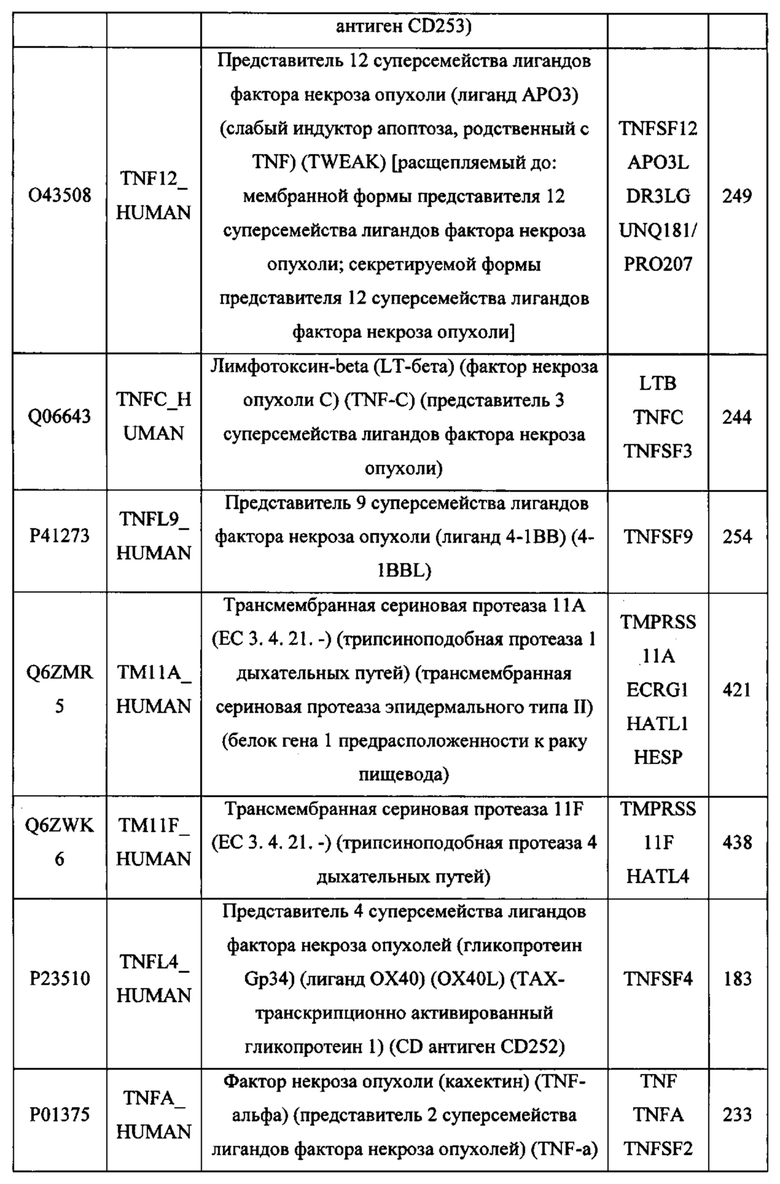
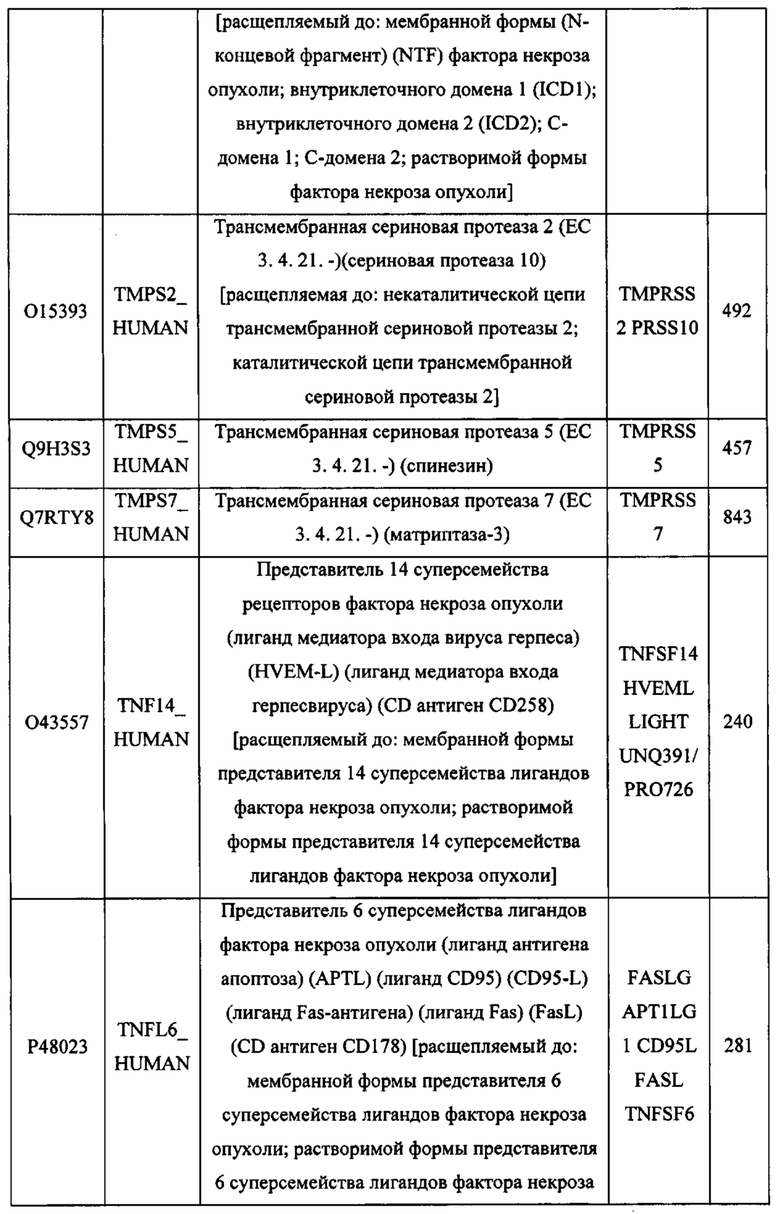
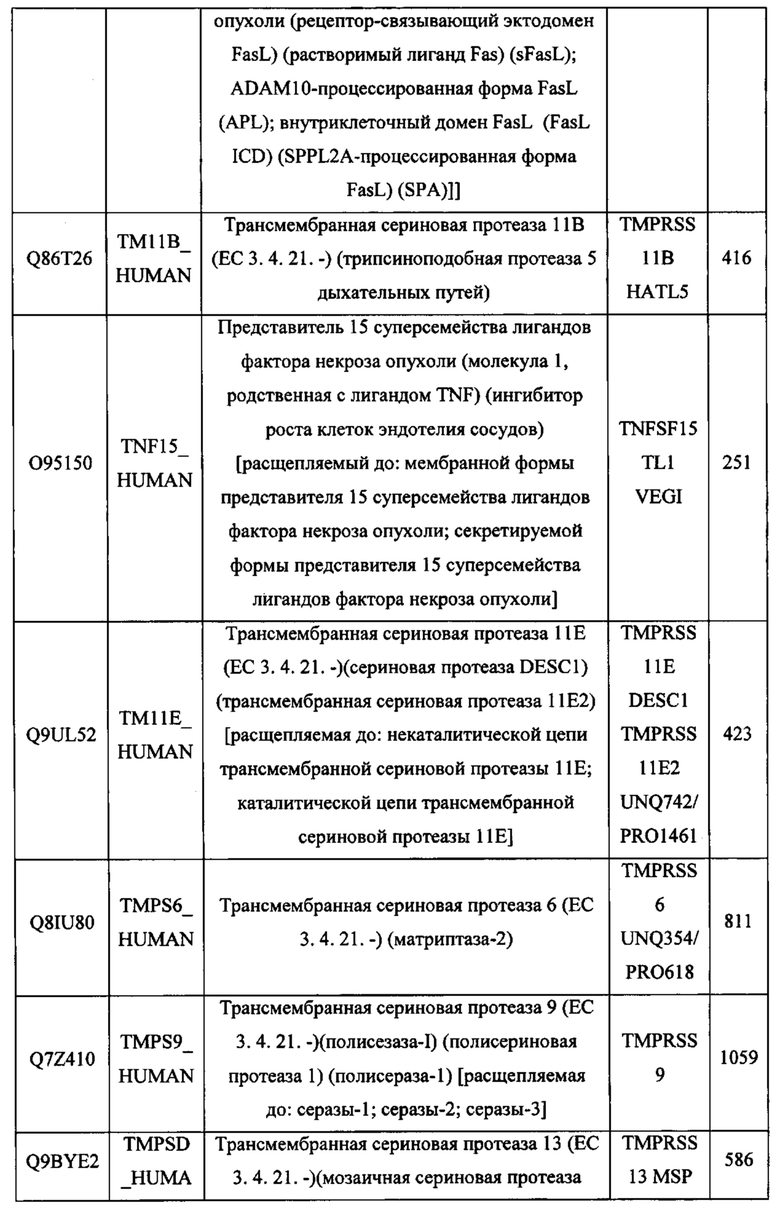




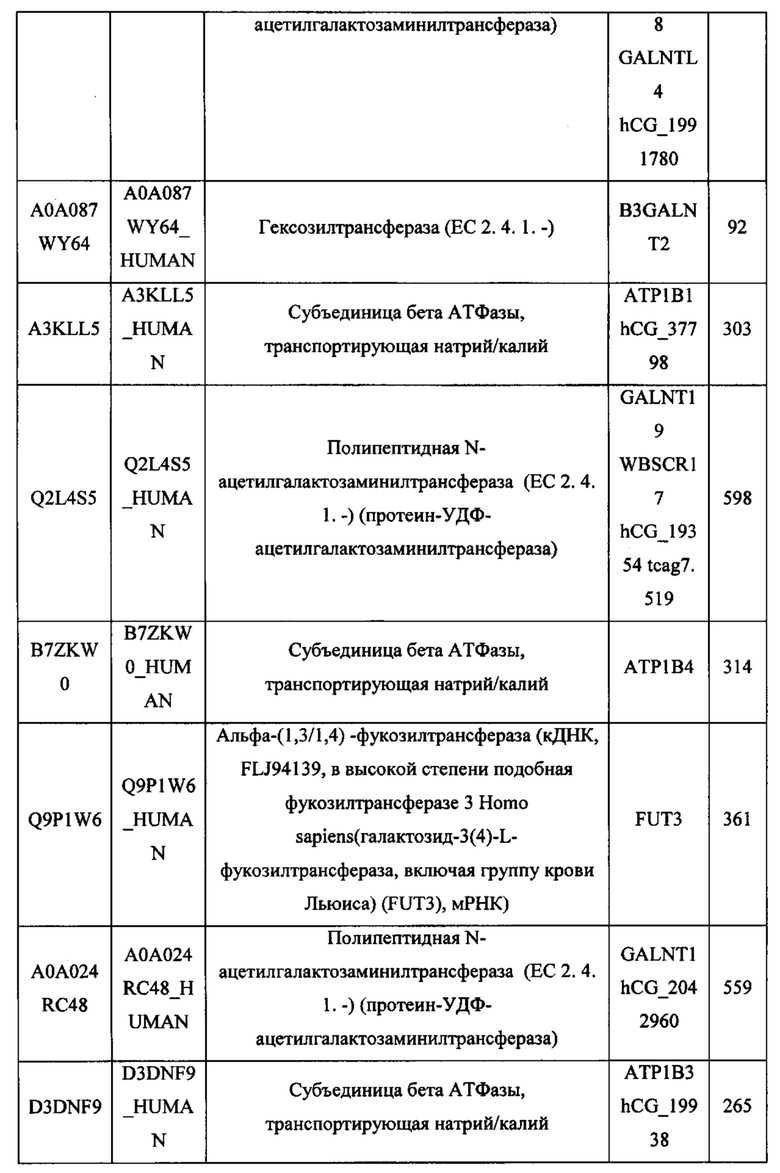
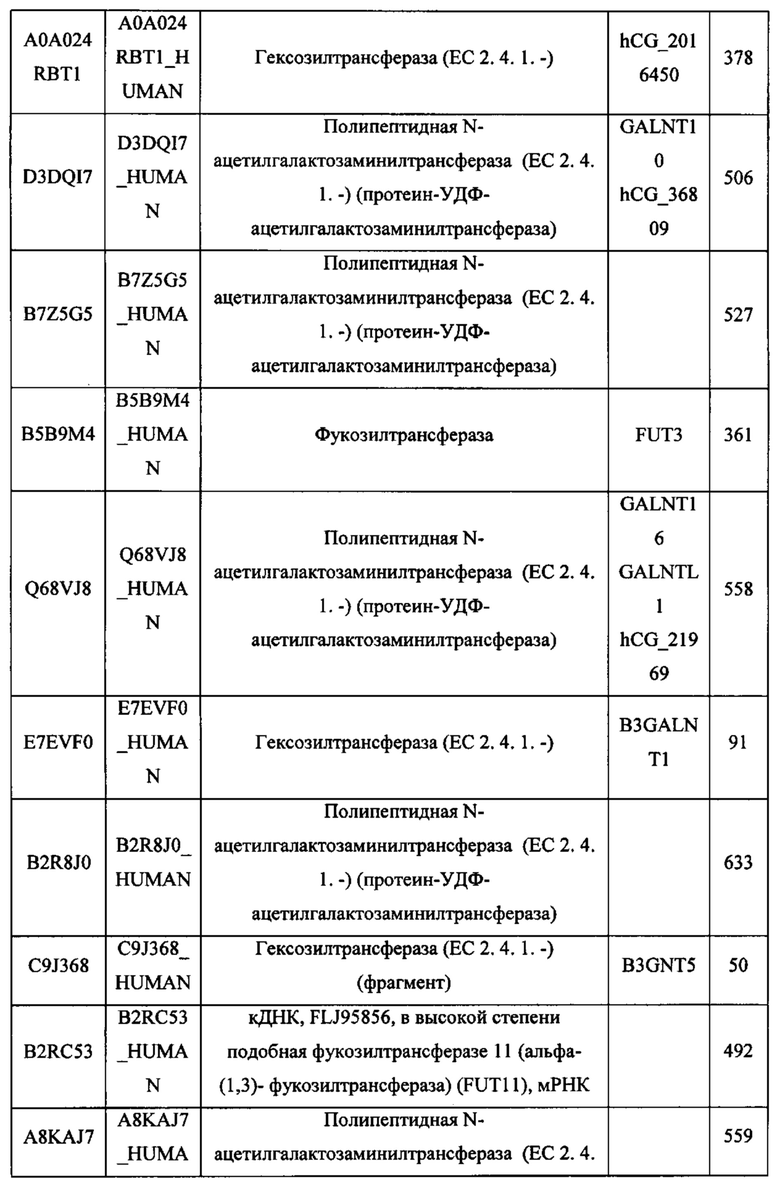
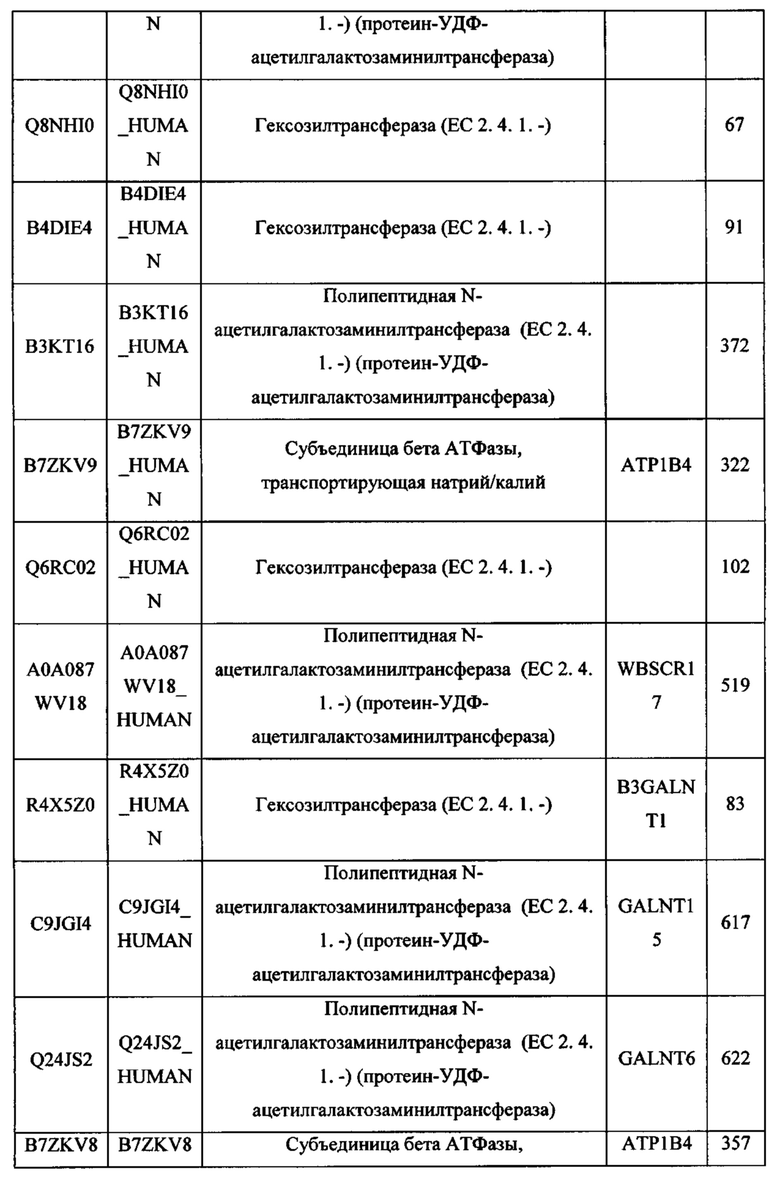
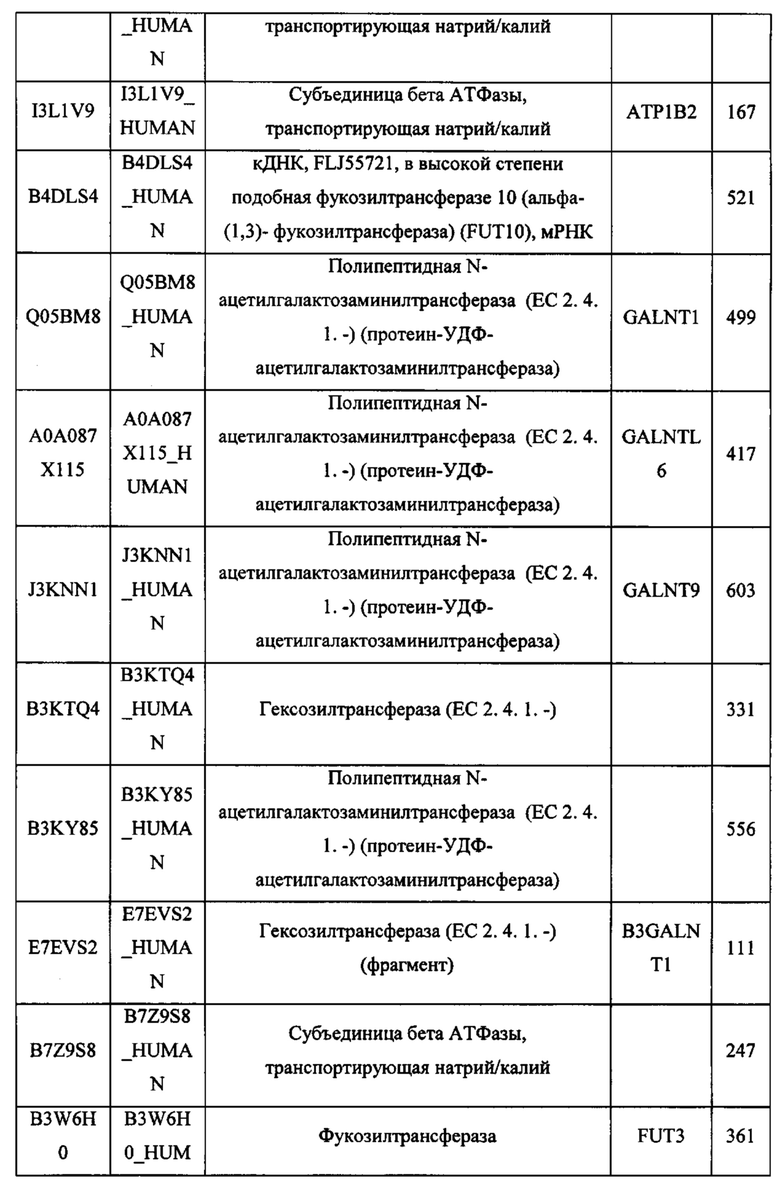
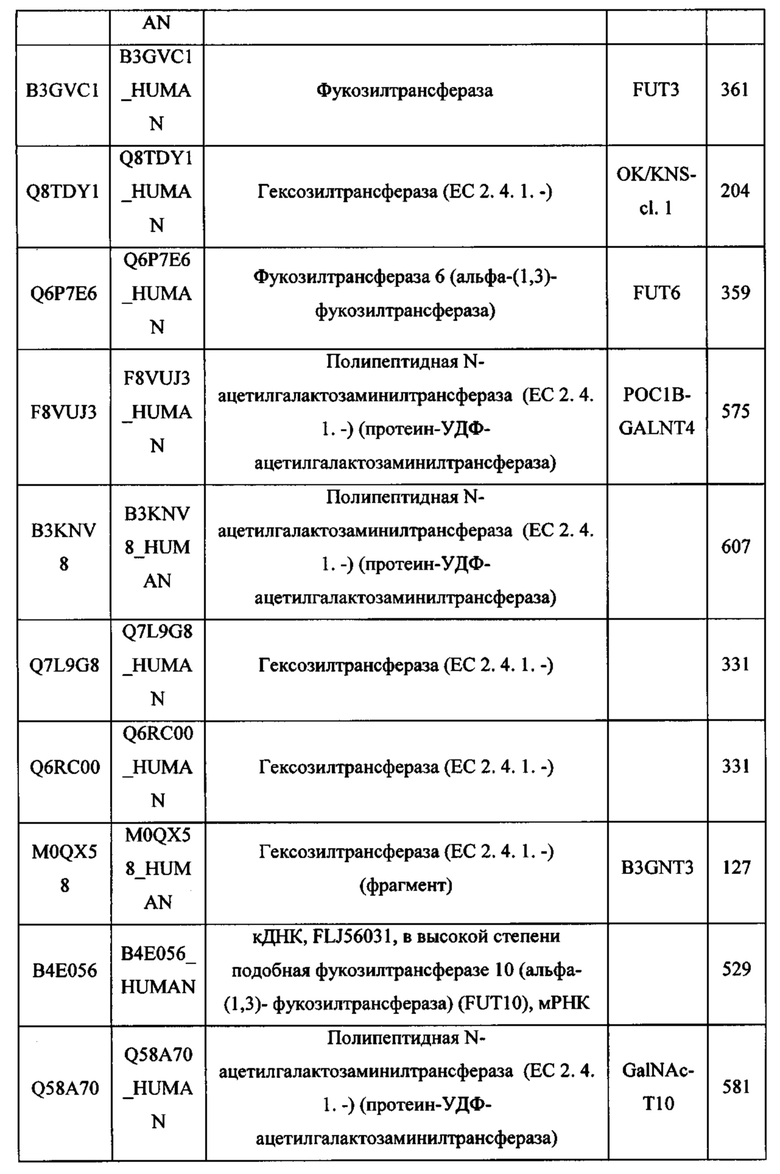
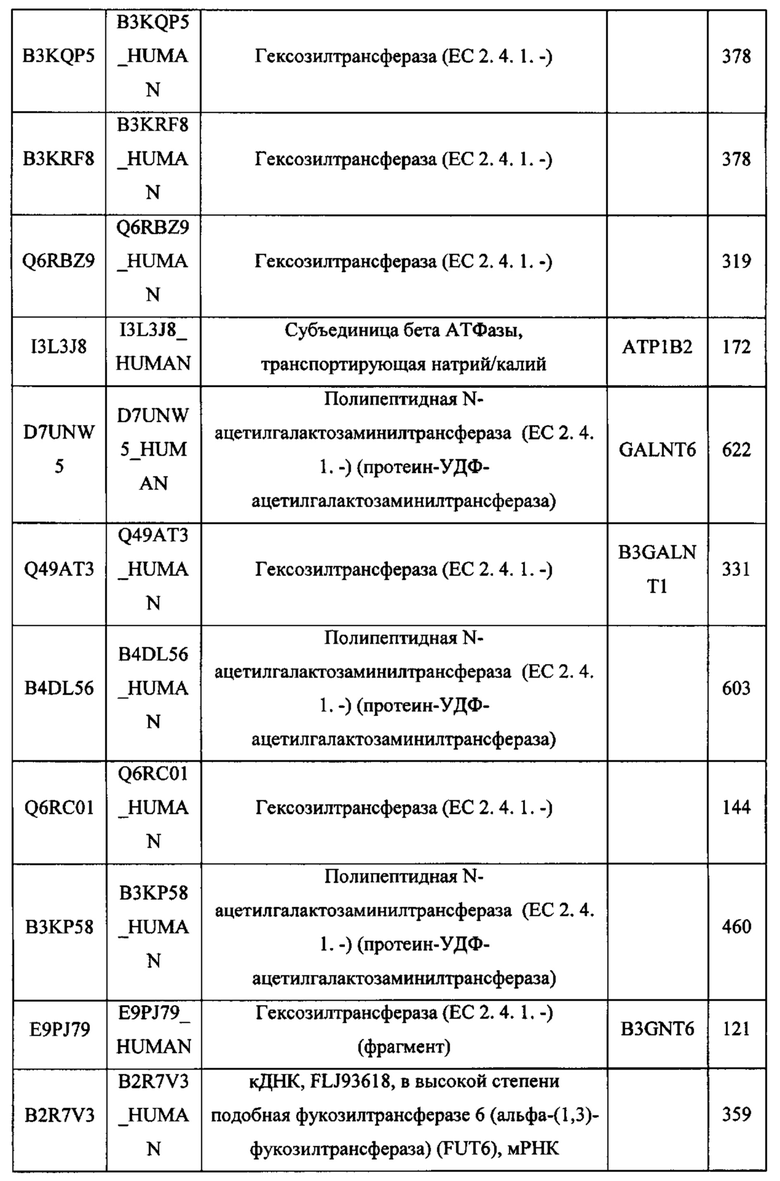
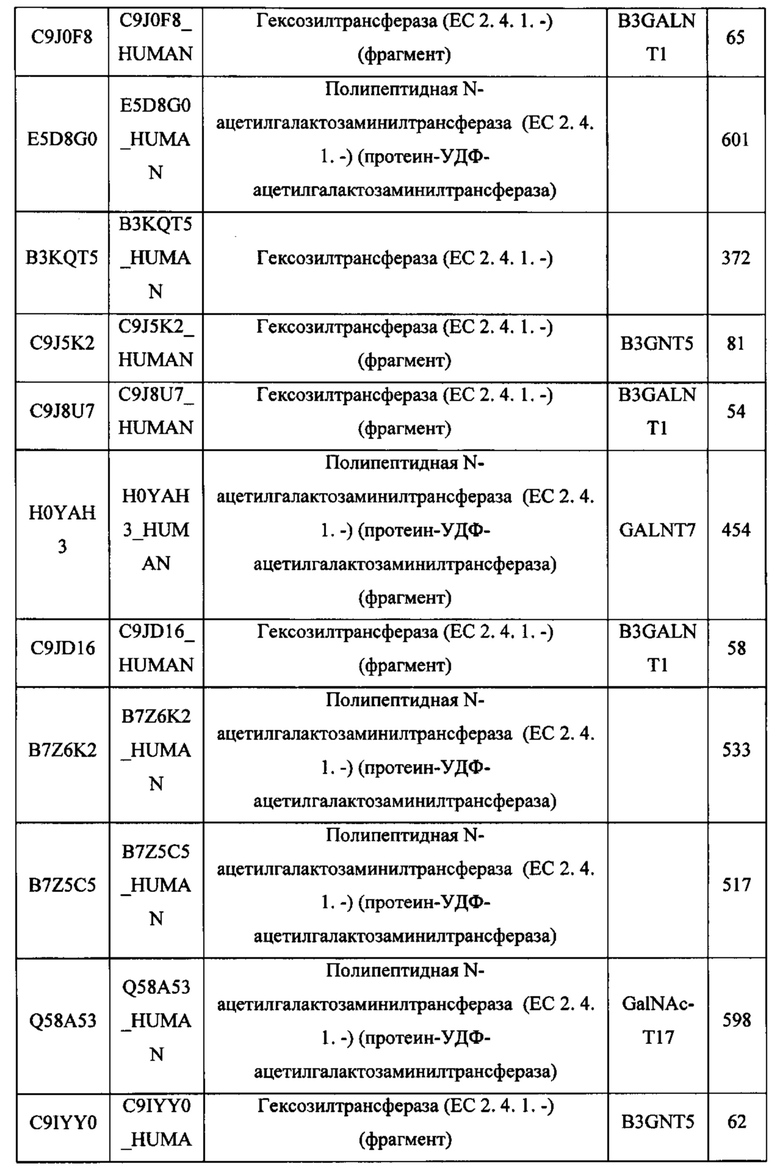




В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 32 или последовательность, идентичную ей по меньшей мере на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 33 или последовательность, идентичную ей по меньшей мере на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 34 или последовательность, идентичную ей по меньшей мере на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 35 или последовательность, идентичную ей по меньшей мере на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 36, или последовательность, идентичную ей по меньшей мере на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 37, или последовательность, идентичную ей по меньшей мере на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 38, или последовательность, идентичную ей по меньшей мере на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 39, или последовательность, идентичную ей по меньшей мере на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 40, или последовательность, идентичную ей по меньшей мере на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 41, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 42, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 43, или последовательность, идентичную ей по меньшей мере на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 44, или последовательность, идентичную ей по меньшей мере на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (ii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 45, или последовательность, идентичную ей по меньшей мере на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 46 или по меньшей мере идентичную ей на 90%, или 93%, или 95%, или 97%, или 98%, или 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 47 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 48 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 49 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 50, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 51, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 52, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 53, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 54, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит: (i) аминокислотную последовательность SEQ ID NO: 55, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и (ii) одну из аминокислотных последовательностей SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58, SEQ ID NO: 59, SEQ ID NO: 60, SEQ ID NO: 61, SEQ ID NO: 62, SEQ ID NO: 63, SEQ ID NO: 64, SEQ ID NO: 65, SEQ ID NO: 66, SEQ ID NO: 67, SEQ ID NO: 68, и SEQ ID NO: 69 или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, и, необязательно (iii) линкер Ig, выбранный из аминокислотных последовательностей SEQ ID NO: 70, SEQ ID NO: 71, и SEQ ID NO: 72, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%, при этом, также необязательно, одна или большее количество из (i) и (ii) связаны с (iii) посредством (iv) соединительного линкера SEQ ID NO: 73, SEQ ID NO: 74, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, или его вариантов. В различных вариантах осуществления данного изобретения линкеры (iii) или (iv) могут быть замещены аминокислотной последовательностью SEQ ID NO: 23-31, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит аминокислотную последовательность SEQ ID NO: 2, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит аминокислотную последовательность SEQ ID NO: 5, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит аминокислотную последовательность SEQ ID NO: 7, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит аминокислотную последовательность SEQ ID NO: 9, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит аминокислотную последовательность SEQ ID NO: 11, или по меньшей мере идентичной ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит аминокислотную последовательность SEQ ID NO: 13, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит аминокислотную последовательность SEQ ID NO: 15, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
В различных вариантах осуществления данного изобретения настоящий химерный белок содержит аминокислотную последовательность SEQ ID NO: 22, или по меньшей мере идентичную ей на 90%, или на 93%, или на 95%, или на 97%, или на 98%, или на 99%.
ЭКВИВАЛЕНТЫ
В то время как данное изобретение было описано в связи с конкретными вариантами реализации, следует понимать, что возможны дополнительные модификации и данная заявка охватывает любые вариации, применения или адаптации данного изобретения, следуя, в общем, принципам изобретения и включая такие отклонения от настоящего описания, которые приходят от известной или обычной практики в данной области техники, к которой относится изобретение, и которые могут быть применены к важным признакам, изложенным ранее в данном документе и следующим образом в объеме прилагаемой формулы изобретения.
Специалистам в данной области техники будет понятно или они смогут установить, используя не более чем рутинные эксперименты, многочисленные эквиваленты конкретных вариантов осуществления, описанных конкретно в данном документе. Такие эквиваленты охватываются объемом следующей формулы изобретения.
ВКЛЮЧЕНИЕ ПОСРЕДСТВОМ ССЫЛКИ
Все патенты и публикации, упомянутые в данном документе, включены в данный документ посредством ссылки во всей их полноте.
Публикации, рассмотренные в данном документе, приведены исключительно для их описания до даты подачи настоящей заявки. Информацию в настоящем документе не следует толковать как признание того, что данное изобретение не имеет права датировать такую публикацию более ранним числом из-за действия более раннего изобретения.
Используемые в данном документе заголовки представлены исключительно для упорядочения структуры документа и никоим образом не ограничивают данное изобретение. Содержание каждого отдельного раздела может быть в равной степени применимо ко всем разделам.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Хит Байолоджикс, ИНК.
<120> КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ СОЕДИНЕНИЯ ВНЕКЛЕТОЧНЫХ ДОМЕНОВ
ТИПА I И ТИПА II В КАЧЕСТВЕ ГЕТЕРОЛОГИЧНЫХ ХИМЕРНЫХ БЕЛКОВ
<130> HTB-023PC
<150> US 62/235,727
<151> 2015-10-01
<150> US 62/263,313
<151> 2015-12-04
<150> US 62/372,574
<151> 2016-08-09
<160> 78
<170> PatentIn версия 3.5
<210> 1
<211> 1605
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 1
atgcagatcc cacaggcgcc ctggccagtc gtctgggcgg tgctacaact gggctggcgg 60
ccaggatggt tcttagactc cccagacagg ccctggaacc cccccacctt ctccccagcc 120
ctgctcgtgg tgaccgaagg ggacaacgcc accttcacct gcagcttctc caacacatcg 180
gagagcttcg tgctaaactg gtaccgcatg agccccagca accagacgga caagctggcc 240
gccttccccg aggaccgcag ccagcccggc caggactgcc gcttccgtgt cacacaactg 300
cccaacgggc gtgacttcca catgagcgtg gtcagggccc ggcgcaatga cagcggcacc 360
tacctctgtg gggccatctc cctggccccc aaggcgcaga tcaaagagag cctgcgggca 420
gagctcaggg tgacagagag aagggcagaa gtgcccacag cccaccccag cccctcaccc 480
aggccagccg gccagttcca atctaagtac ggccctccct gccctagctg tcccgcccct 540
gaatttctgg gcggaccctc cgtgtttctg ttccccccaa agcccaagga caccctgatg 600
atcagccgga cccccgaagt gacctgtgtg gtggtggatg tgtcccagga agatcccgag 660
gtgcagttca attggtacgt ggacggggtg gaagtgcaca acgccaagac caagcccaga 720
gaggaacagt tcaacagcac ctaccgggtg gtgtctgtgc tgaccgtgct gcaccaggat 780
tggctgagcg gcaaagagta caagtgcaag gtgtccagca agggcctgcc cagcagcatc 840
gaaaagacca tcagcaacgc caccggccag cccagggaac cccaggtgta cacactgccc 900
cctagccagg aagagatgac caagaaccag gtgtccctga catgcctcgt gaagggcttc 960
tacccctccg atatcgccgt ggaatgggag agcaacggcc agccagagaa caactacaag 1020
accacccccc cagtgctgga cagcgacggc tcattcttcc tgtactcccg gctgacagtg 1080
gacaagagca gctggcagga aggcaacgtg ttcagctgca gcgtgatgca cgaagccctg 1140
cacaaccact acacccagaa gtccctgagc ctgtccctgg gcaaaataga gggacgaatg 1200
gaccaggtat cacatcggta tcctcgaatt caaagtatca aagtacaatt taccgaatat 1260
aagaaggaga aaggtttcat cctcacttcc caaaaggagg atgaaatcat gaaggtgcag 1320
aacaactcag tcatcatcaa ctgtgatggg ttttatctca tctccctgaa gggctacttc 1380
tcccaggaag tcaacattag ccttcattac cagaaggatg aggagcccct cttccaactg 1440
aagaaggtca ggtctgtcaa ctccttgatg gtggcctctc tgacttacaa agacaaagtc 1500
tacttgaatg tgaccactga caatacctcc ctggatgact tccatgtgaa tggcggagaa 1560
ctgattctta tccatcaaaa tcctggtgaa ttctgtgtcc tttga 1605
<210> 2
<211> 534
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 2
Met Gln Ile Pro Gln Ala Pro Trp Pro Val Val Trp Ala Val Leu Gln
1 5 10 15
Leu Gly Trp Arg Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro Trp
20 25 30
Asn Pro Pro Thr Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly Asp
35 40 45
Asn Ala Thr Phe Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe Val
50 55 60
Leu Asn Trp Tyr Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu Ala
65 70 75 80
Ala Phe Pro Glu Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe Arg
85 90 95
Val Thr Gln Leu Pro Asn Gly Arg Asp Phe His Met Ser Val Val Arg
100 105 110
Ala Arg Arg Asn Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser Leu
115 120 125
Ala Pro Lys Ala Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg Val
130 135 140
Thr Glu Arg Arg Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser Pro
145 150 155 160
Arg Pro Ala Gly Gln Phe Gln Ser Lys Tyr Gly Pro Pro Cys Pro Ser
165 170 175
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
180 185 190
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
195 200 205
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
210 215 220
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
225 230 235 240
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
245 250 255
Leu His Gln Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser
260 265 270
Ser Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr
275 280 285
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
290 295 300
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
305 310 315 320
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
325 330 335
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
340 345 350
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Ser Trp Gln Glu Gly
355 360 365
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
370 375 380
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys Ile Glu Gly Arg Met
385 390 395 400
Asp Gln Val Ser His Arg Tyr Pro Arg Ile Gln Ser Ile Lys Val Gln
405 410 415
Phe Thr Glu Tyr Lys Lys Glu Lys Gly Phe Ile Leu Thr Ser Gln Lys
420 425 430
Glu Asp Glu Ile Met Lys Val Gln Asn Asn Ser Val Ile Ile Asn Cys
435 440 445
Asp Gly Phe Tyr Leu Ile Ser Leu Lys Gly Tyr Phe Ser Gln Glu Val
450 455 460
Asn Ile Ser Leu His Tyr Gln Lys Asp Glu Glu Pro Leu Phe Gln Leu
465 470 475 480
Lys Lys Val Arg Ser Val Asn Ser Leu Met Val Ala Ser Leu Thr Tyr
485 490 495
Lys Asp Lys Val Tyr Leu Asn Val Thr Thr Asp Asn Thr Ser Leu Asp
500 505 510
Asp Phe His Val Asn Gly Gly Glu Leu Ile Leu Ile His Gln Asn Pro
515 520 525
Gly Glu Phe Cys Val Leu
530
<210> 3
<211> 1782
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 3
atgcagattc ctcaggcccc ttggcctgtc gtgtgggctg tgctgcagct gggatggcgg 60
cctggctggt ttctggacag ccccgacaga ccctggaacc cccctacatt ttcccctgcc 120
ctgctggtcg tgaccgaggg cgacaatgcc accttcacct gtagcttcag caacaccagc 180
gagagcttcg tgctgaactg gtacagaatg agccccagca accagaccga caagctggcc 240
gccttccccg aggatagatc tcagcccggc caggactgcc ggttcagagt gacccagctg 300
cccaacggcc gggacttcca catgtctgtc gtgcgggcca gacggaacga cagcggcaca 360
tatctgtgcg gcgccatcag cctggccccc aaggcccaga tcaaagagag cctgagagcc 420
gagctgagag tgaccgagag aagggccgaa gtgcctaccg cccaccctag cccatctcca 480
agacctgccg gccagttcca gtctaagtac ggccctcctt gccccagctg tcccgcccct 540
gaatttctgg gcggacccag cgtgttcctg ttccccccaa agcccaagga caccctgatg 600
atcagccgga cccccgaagt gacctgcgtg gtggtggatg tgtcccagga agatcccgag 660
gtgcagttca attggtacgt ggacggcgtg gaagtgcaca acgccaagac caagcccaga 720
gaggaacagt tcaacagcac ctaccgggtg gtgtccgtgc tgaccgtgct gcaccaggat 780
tggctgagcg gcaaagagta caagtgcaag gtgtccagca agggcctgcc cagcagcatc 840
gagaaaacca tcagcaacgc caccggccag cccagggaac cccaggtgta cacactgccc 900
cctagccagg aagagatgac caagaaccag gtgtccctga cctgtctcgt gaagggcttc 960
tacccctccg atatcgccgt ggaatgggag agcaacggcc agcctgagaa caactacaag 1020
accacccccc cagtgctgga cagcgacggc tcatttttcc tgtactccag actgaccgtg 1080
gacaagagca gctggcagga aggcaacgtg ttcagctgct ccgtgatgca cgaggccctg 1140
cacaaccact acacccagaa gtccctgtct ctgagcctgg gcaagatcga gggccggatg 1200
gatagagccc agggcgaagc ctgcgtgcag ttccaggctc tgaagggcca ggaattcgcc 1260
cccagccacc agcaggtgta cgcccctctg agagctgacg gcgacaagcc tagagcccac 1320
ctgacagtcg tgcggcagac ccctacccag cacttcaaga atcagttccc agccctgcac 1380
tgggagcacg agctgggcct ggccttcacc aagaacagaa tgaactacac caacaagttt 1440
ctgctgatcc ccgagagcgg cgactacttc atctacagcc aagtgacctt ccggggcatg 1500
accagcgagt gcagcgagat cagacaggcc ggcagaccta acaagcccga cagcatcacc 1560
gtcgtgatca ccaaagtgac cgacagctac cccgagccca cacagctgct gatgggcacc 1620
aagagcgtgt gcgaagtggg cagcaactgg ttccagccca tctacctggg cgccatgttc 1680
agtctgcaag agggcgataa gctgatggtc aacgtgtccg acatctccct ggtggattac 1740
accaaagagg acaagacctt cttcggcgcc tttctgctct ga 1782
<210> 4
<211> 1782
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 4
atgcagatcc cacaggcgcc ctggccagtc gtctgggcgg tgctacaact gggctggcgg 60
ccaggatggt tcttagactc cccagacagg ccctggaacc cccccacctt ctccccagcc 120
ctgctcgtgg tgaccgaagg ggacaacgcc accttcacct gcagcttctc caacacatcg 180
gagagcttcg tgctaaactg gtaccgcatg agccccagca accagacgga caagctggcc 240
gccttccccg aggaccgcag ccagcccggc caggactgcc gcttccgtgt cacacaactg 300
cccaacgggc gtgacttcca catgagcgtg gtcagggccc ggcgcaatga cagcggcacc 360
tacctctgtg gggccatctc cctggccccc aaggcgcaga tcaaagagag cctgcgggca 420
gagctcaggg tgacagagag aagggcagaa gtgcccacag cccaccccag cccctcaccc 480
aggccagccg gccagttcca atctaagtac ggccctccct gccctagctg tcccgcccct 540
gaatttctgg gcggaccctc cgtgtttctg ttccccccaa agcccaagga caccctgatg 600
atcagccgga cccccgaagt gacctgtgtg gtggtggatg tgtcccagga agatcccgag 660
gtgcagttca attggtacgt ggacggggtg gaagtgcaca acgccaagac caagcccaga 720
gaggaacagt tcaacagcac ctaccgggtg gtgtctgtgc tgaccgtgct gcaccaggat 780
tggctgagcg gcaaagagta caagtgcaag gtgtccagca agggcctgcc cagcagcatc 840
gaaaagacca tcagcaacgc caccggccag cccagggaac cccaggtgta cacactgccc 900
cctagccagg aagagatgac caagaaccag gtgtccctga catgcctcgt gaagggcttc 960
tacccctccg atatcgccgt ggaatgggag agcaacggcc agccagagaa caactacaag 1020
accacccccc cagtgctgga cagcgacggc tcattcttcc tgtactcccg gctgacagtg 1080
gacaagagca gctggcagga aggcaacgtg ttcagctgca gcgtgatgca cgaagccctg 1140
cacaaccact acacccagaa gtccctgagc ctgtccctgg gcaaaataga gggacgaatg 1200
gaccgggccc agggagaggc ctgtgtgcag ttccaggctc taaaaggaca ggagtttgca 1260
ccttcacatc agcaagttta tgcacctctt agagcagacg gagataagcc aagggcacac 1320
ctgacagttg tgagacaaac tcccacacag cactttaaaa atcagttccc agctctgcac 1380
tgggaacatg aactaggcct ggccttcacc aagaaccgaa tgaactatac caacaaattc 1440
ctgctgatcc cagagtcggg agactacttc atttactccc aggtcacatt ccgtgggatg 1500
acctctgagt gcagtgaaat cagacaagca ggccgaccaa acaagccaga ctccatcact 1560
gtggtcatca ccaaggtaac agacagctac cctgagccaa cccagctcct catggggacc 1620
aagtctgtat gcgaagtagg tagcaactgg ttccagccca tctacctcgg agccatgttc 1680
tccttgcaag aaggggacaa gctaatggtg aacgtcagtg acatctcttt ggtggattac 1740
acaaaagaag ataaaacctt ctttggagcc ttcttactat ag 1782
<210> 5
<211> 593
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 5
Met Gln Ile Pro Gln Ala Pro Trp Pro Val Val Trp Ala Val Leu Gln
1 5 10 15
Leu Gly Trp Arg Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro Trp
20 25 30
Asn Pro Pro Thr Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly Asp
35 40 45
Asn Ala Thr Phe Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe Val
50 55 60
Leu Asn Trp Tyr Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu Ala
65 70 75 80
Ala Phe Pro Glu Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe Arg
85 90 95
Val Thr Gln Leu Pro Asn Gly Arg Asp Phe His Met Ser Val Val Arg
100 105 110
Ala Arg Arg Asn Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser Leu
115 120 125
Ala Pro Lys Ala Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg Val
130 135 140
Thr Glu Arg Arg Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser Pro
145 150 155 160
Arg Pro Ala Gly Gln Phe Gln Ser Lys Tyr Gly Pro Pro Cys Pro Ser
165 170 175
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
180 185 190
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
195 200 205
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
210 215 220
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
225 230 235 240
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
245 250 255
Leu His Gln Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser
260 265 270
Ser Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr
275 280 285
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
290 295 300
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
305 310 315 320
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
325 330 335
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
340 345 350
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Ser Trp Gln Glu Gly
355 360 365
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
370 375 380
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys Ile Glu Gly Arg Met
385 390 395 400
Asp Arg Ala Gln Gly Glu Ala Cys Val Gln Phe Gln Ala Leu Lys Gly
405 410 415
Gln Glu Phe Ala Pro Ser His Gln Gln Val Tyr Ala Pro Leu Arg Ala
420 425 430
Asp Gly Asp Lys Pro Arg Ala His Leu Thr Val Val Arg Gln Thr Pro
435 440 445
Thr Gln His Phe Lys Asn Gln Phe Pro Ala Leu His Trp Glu His Glu
450 455 460
Leu Gly Leu Ala Phe Thr Lys Asn Arg Met Asn Tyr Thr Asn Lys Phe
465 470 475 480
Leu Leu Ile Pro Glu Ser Gly Asp Tyr Phe Ile Tyr Ser Gln Val Thr
485 490 495
Phe Arg Gly Met Thr Ser Glu Cys Ser Glu Ile Arg Gln Ala Gly Arg
500 505 510
Pro Asn Lys Pro Asp Ser Ile Thr Val Val Ile Thr Lys Val Thr Asp
515 520 525
Ser Tyr Pro Glu Pro Thr Gln Leu Leu Met Gly Thr Lys Ser Val Cys
530 535 540
Glu Val Gly Ser Asn Trp Phe Gln Pro Ile Tyr Leu Gly Ala Met Phe
545 550 555 560
Ser Leu Gln Glu Gly Asp Lys Leu Met Val Asn Val Ser Asp Ile Ser
565 570 575
Leu Val Asp Tyr Thr Lys Glu Asp Lys Thr Phe Phe Gly Ala Phe Leu
580 585 590
Leu
<210> 6
<211> 1554
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 6
atgaagacat tgcctgccat gcttggaact gggaaattat tttgggtctt cttcttaatc 60
ccatatctgg acatctggaa catccatggg aaagaatcat gtgatgtaca gctttatata 120
aagagacaat ctgaacactc catcttagca ggagatccct ttgaactaga atgccctgtg 180
aaatactgtg ctaacaggcc tcatgtgact tggtgcaagc tcaatggaac aacatgtgta 240
aaacttgaag atagacaaac aagttggaag gaagagaaga acatttcatt tttcattcta 300
cattttgaac cagtgcttcc taatgacaat gggtcatacc gctgttctgc aaattttcag 360
tctaatctca ttgaaagcca ctcaacaact ctttatgtga cagatgtaaa aagtgcctca 420
gaacgaccct ccaaggacga aatggcaagc tctaagtacg gccctccctg ccctagctgt 480
cccgcccctg aatttctggg cggaccctcc gtgtttctgt tccccccaaa gcccaaggac 540
accctgatga tcagccggac ccccgaagtg acctgtgtgg tggtggatgt gtcccaggaa 600
gatcccgagg tgcagttcaa ttggtacgtg gacggggtgg aagtgcacaa cgccaagacc 660
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtctgtgct gaccgtgctg 720
caccaggatt ggctgagcgg caaagagtac aagtgcaagg tgtccagcaa gggcctgccc 780
agcagcatcg aaaagaccat cagcaacgcc accggccagc ccagggaacc ccaggtgtac 840
acactgcccc ctagccagga agagatgacc aagaaccagg tgtccctgac atgcctcgtg 900
aagggcttct acccctccga tatcgccgtg gaatgggaga gcaacggcca gccagagaac 960
aactacaaga ccaccccccc agtgctggac agcgacggct cattcttcct gtactcccgg 1020
ctgacagtgg acaagagcag ctggcaggaa ggcaacgtgt tcagctgcag cgtgatgcac 1080
gaagccctgc acaaccacta cacccagaag tccctgagcc tgtccctggg caaaatagag 1140
ggacgaatgg accaggtatc acatcggtat cctcgaattc aaagtatcaa agtacaattt 1200
accgaatata agaaggagaa aggtttcatc ctcacttccc aaaaggagga tgaaatcatg 1260
aaggtgcaga acaactcagt catcatcaac tgtgatgggt tttatctcat ctccctgaag 1320
ggctacttct cccaggaagt caacattagc cttcattacc agaaggatga ggagcccctc 1380
ttccaactga agaaggtcag gtctgtcaac tccttgatgg tggcctctct gacttacaaa 1440
gacaaagtct acttgaatgt gaccactgac aatacctccc tggatgactt ccatgtgaat 1500
ggcggagaac tgattcttat ccatcaaaat cctggtgaat tctgtgtcct ttga 1554
<210> 7
<211> 517
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 7
Met Lys Thr Leu Pro Ala Met Leu Gly Thr Gly Lys Leu Phe Trp Val
1 5 10 15
Phe Phe Leu Ile Pro Tyr Leu Asp Ile Trp Asn Ile His Gly Lys Glu
20 25 30
Ser Cys Asp Val Gln Leu Tyr Ile Lys Arg Gln Ser Glu His Ser Ile
35 40 45
Leu Ala Gly Asp Pro Phe Glu Leu Glu Cys Pro Val Lys Tyr Cys Ala
50 55 60
Asn Arg Pro His Val Thr Trp Cys Lys Leu Asn Gly Thr Thr Cys Val
65 70 75 80
Lys Leu Glu Asp Arg Gln Thr Ser Trp Lys Glu Glu Lys Asn Ile Ser
85 90 95
Phe Phe Ile Leu His Phe Glu Pro Val Leu Pro Asn Asp Asn Gly Ser
100 105 110
Tyr Arg Cys Ser Ala Asn Phe Gln Ser Asn Leu Ile Glu Ser His Ser
115 120 125
Thr Thr Leu Tyr Val Thr Asp Val Lys Ser Ala Ser Glu Arg Pro Ser
130 135 140
Lys Asp Glu Met Ala Ser Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys
145 150 155 160
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
165 170 175
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
180 185 190
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
195 200 205
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
210 215 220
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
225 230 235 240
His Gln Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser
245 250 255
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly
260 265 270
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
275 280 285
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
290 295 300
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
305 310 315 320
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
325 330 335
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Ser Trp Gln Glu Gly Asn
340 345 350
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
355 360 365
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys Ile Glu Gly Arg Met Asp
370 375 380
Gln Val Ser His Arg Tyr Pro Arg Ile Gln Ser Ile Lys Val Gln Phe
385 390 395 400
Thr Glu Tyr Lys Lys Glu Lys Gly Phe Ile Leu Thr Ser Gln Lys Glu
405 410 415
Asp Glu Ile Met Lys Val Gln Asn Asn Ser Val Ile Ile Asn Cys Asp
420 425 430
Gly Phe Tyr Leu Ile Ser Leu Lys Gly Tyr Phe Ser Gln Glu Val Asn
435 440 445
Ile Ser Leu His Tyr Gln Lys Asp Glu Glu Pro Leu Phe Gln Leu Lys
450 455 460
Lys Val Arg Ser Val Asn Ser Leu Met Val Ala Ser Leu Thr Tyr Lys
465 470 475 480
Asp Lys Val Tyr Leu Asn Val Thr Thr Asp Asn Thr Ser Leu Asp Asp
485 490 495
Phe His Val Asn Gly Gly Glu Leu Ile Leu Ile His Gln Asn Pro Gly
500 505 510
Glu Phe Cys Val Leu
515
<210> 8
<211> 1527
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 8
atgcgctggt gtctcctcct gatctgggcc caggggctga ggcaggctcc cctcgcctca 60
ggaatgatga caggcacaat agaaacaacg gggaacattt ctgcagagaa aggtggctct 120
atcatcttac aatgtcacct ctcctccacc acggcacaag tgacccaggt caactgggag 180
cagcaggacc agcttctggc catttgtaat gctgacttgg ggtggcacat ctccccatcc 240
ttcaaggatc gagtggcccc aggtcccggc ctgggcctca ccctccagtc gctgaccgtg 300
aacgatacag gggagtactt ctgcatctat cacacctacc ctgatgggac gtacactggg 360
agaatcttcc tggaggtcct agaaagctca gtggctgagc acggtgccag gttccagatt 420
ccatctaagt acggccctcc ctgccctagc tgtcccgccc ctgaatttct gggcggaccc 480
tccgtgtttc tgttcccccc aaagcccaag gacaccctga tgatcagccg gacccccgaa 540
gtgacctgtg tggtggtgga tgtgtcccag gaagatcccg aggtgcagtt caattggtac 600
gtggacgggg tggaagtgca caacgccaag accaagccca gagaggaaca gttcaacagc 660
acctaccggg tggtgtctgt gctgaccgtg ctgcaccagg attggctgag cggcaaagag 720
tacaagtgca aggtgtccag caagggcctg cccagcagca tcgaaaagac catcagcaac 780
gccaccggcc agcccaggga accccaggtg tacacactgc cccctagcca ggaagagatg 840
accaagaacc aggtgtccct gacatgcctc gtgaagggct tctacccctc cgatatcgcc 900
gtggaatggg agagcaacgg ccagccagag aacaactaca agaccacccc cccagtgctg 960
gacagcgacg gctcattctt cctgtactcc cggctgacag tggacaagag cagctggcag 1020
gaaggcaacg tgttcagctg cagcgtgatg cacgaagccc tgcacaacca ctacacccag 1080
aagtccctga gcctgtccct gggcaaaata gagggacgaa tggaccaggt atcacatcgg 1140
tatcctcgaa ttcaaagtat caaagtacaa tttaccgaat ataagaagga gaaaggtttc 1200
atcctcactt cccaaaagga ggatgaaatc atgaaggtgc agaacaactc agtcatcatc 1260
aactgtgatg ggttttatct catctccctg aagggctact tctcccagga agtcaacatt 1320
agccttcatt accagaagga tgaggagccc ctcttccaac tgaagaaggt caggtctgtc 1380
aactccttga tggtggcctc tctgacttac aaagacaaag tctacttgaa tgtgaccact 1440
gacaatacct ccctggatga cttccatgtg aatggcggag aactgattct tatccatcaa 1500
aatcctggtg aattctgtgt cctttga 1527
<210> 9
<211> 508
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 9
Met Arg Trp Cys Leu Leu Leu Ile Trp Ala Gln Gly Leu Arg Gln Ala
1 5 10 15
Pro Leu Ala Ser Gly Met Met Thr Gly Thr Ile Glu Thr Thr Gly Asn
20 25 30
Ile Ser Ala Glu Lys Gly Gly Ser Ile Ile Leu Gln Cys His Leu Ser
35 40 45
Ser Thr Thr Ala Gln Val Thr Gln Val Asn Trp Glu Gln Gln Asp Gln
50 55 60
Leu Leu Ala Ile Cys Asn Ala Asp Leu Gly Trp His Ile Ser Pro Ser
65 70 75 80
Phe Lys Asp Arg Val Ala Pro Gly Pro Gly Leu Gly Leu Thr Leu Gln
85 90 95
Ser Leu Thr Val Asn Asp Thr Gly Glu Tyr Phe Cys Ile Tyr His Thr
100 105 110
Tyr Pro Asp Gly Thr Tyr Thr Gly Arg Ile Phe Leu Glu Val Leu Glu
115 120 125
Ser Ser Val Ala Glu His Gly Ala Arg Phe Gln Ile Pro Ser Lys Tyr
130 135 140
Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
145 150 155 160
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
165 170 175
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
180 185 190
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
195 200 205
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
210 215 220
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Ser Gly Lys Glu
225 230 235 240
Tyr Lys Cys Lys Val Ser Ser Lys Gly Leu Pro Ser Ser Ile Glu Lys
245 250 255
Thr Ile Ser Asn Ala Thr Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
260 265 270
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
275 280 285
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
290 295 300
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
305 310 315 320
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
325 330 335
Ser Ser Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
340 345 350
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
355 360 365
Lys Ile Glu Gly Arg Met Asp Gln Val Ser His Arg Tyr Pro Arg Ile
370 375 380
Gln Ser Ile Lys Val Gln Phe Thr Glu Tyr Lys Lys Glu Lys Gly Phe
385 390 395 400
Ile Leu Thr Ser Gln Lys Glu Asp Glu Ile Met Lys Val Gln Asn Asn
405 410 415
Ser Val Ile Ile Asn Cys Asp Gly Phe Tyr Leu Ile Ser Leu Lys Gly
420 425 430
Tyr Phe Ser Gln Glu Val Asn Ile Ser Leu His Tyr Gln Lys Asp Glu
435 440 445
Glu Pro Leu Phe Gln Leu Lys Lys Val Arg Ser Val Asn Ser Leu Met
450 455 460
Val Ala Ser Leu Thr Tyr Lys Asp Lys Val Tyr Leu Asn Val Thr Thr
465 470 475 480
Asp Asn Thr Ser Leu Asp Asp Phe His Val Asn Gly Gly Glu Leu Ile
485 490 495
Leu Ile His Gln Asn Pro Gly Glu Phe Cys Val Leu
500 505
<210> 10
<211> 1710
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 10
atgttttcac atcttccctt tgactgtgtc ctgctgctgc tgctgctact acttacaagg 60
tcctcagaag tggaatacag agcggaggtc ggtcagaatg cctatctgcc ctgcttctac 120
accccagccg ccccagggaa cctcgtgccc gtctgctggg gcaaaggagc ctgtcctgtg 180
tttgaatgtg gcaacgtggt gctcaggact gatgaaaggg atgtgaatta ttggacatcc 240
agatactggc taaatgggga tttccgcaaa ggagatgtgt ccctgaccat agagaatgtg 300
actctagcag acagtgggat ctactgctgc cggatccaaa tcccaggcat aatgaatgat 360
gaaaaattta acctgaagtt ggtcatcaaa ccagccaagg tcacccctgc accgactcgg 420
cagagagact tcactgcagc ctttccaagg atgcttacca ccaggggaca tggcccagca 480
gagacacaga cactggggag cctccctgat ataaatctaa cacaaatatc cacattggcc 540
aatgagttac gggactctag attggccaat gacttacggg actctggagc aaccatcaga 600
ataggctcta agtacggccc tccctgccct agctgtcccg cccctgaatt tctgggcgga 660
ccctccgtgt ttctgttccc cccaaagccc aaggacaccc tgatgatcag ccggaccccc 720
gaagtgacct gtgtggtggt ggatgtgtcc caggaagatc ccgaggtgca gttcaattgg 780
tacgtggacg gggtggaagt gcacaacgcc aagaccaagc ccagagagga acagttcaac 840
agcacctacc gggtggtgtc tgtgctgacc gtgctgcacc aggattggct gagcggcaaa 900
gagtacaagt gcaaggtgtc cagcaagggc ctgcccagca gcatcgaaaa gaccatcagc 960
aacgccaccg gccagcccag ggaaccccag gtgtacacac tgccccctag ccaggaagag 1020
atgaccaaga accaggtgtc cctgacatgc ctcgtgaagg gcttctaccc ctccgatatc 1080
gccgtggaat gggagagcaa cggccagcca gagaacaact acaagaccac ccccccagtg 1140
ctggacagcg acggctcatt cttcctgtac tcccggctga cagtggacaa gagcagctgg 1200
caggaaggca acgtgttcag ctgcagcgtg atgcacgaag ccctgcacaa ccactacacc 1260
cagaagtccc tgagcctgtc cctgggcaaa atagagggac gaatggacca ggtatcacat 1320
cggtatcctc gaattcaaag tatcaaagta caatttaccg aatataagaa ggagaaaggt 1380
ttcatcctca cttcccaaaa ggaggatgaa atcatgaagg tgcagaacaa ctcagtcatc 1440
atcaactgtg atgggtttta tctcatctcc ctgaagggct acttctccca ggaagtcaac 1500
attagccttc attaccagaa ggatgaggag cccctcttcc aactgaagaa ggtcaggtct 1560
gtcaactcct tgatggtggc ctctctgact tacaaagaca aagtctactt gaatgtgacc 1620
actgacaata cctccctgga tgacttccat gtgaatggcg gagaactgat tcttatccat 1680
caaaatcctg gtgaattctg tgtcctttga 1710
<210> 11
<211> 569
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 11
Met Phe Ser His Leu Pro Phe Asp Cys Val Leu Leu Leu Leu Leu Leu
1 5 10 15
Leu Leu Thr Arg Ser Ser Glu Val Glu Tyr Arg Ala Glu Val Gly Gln
20 25 30
Asn Ala Tyr Leu Pro Cys Phe Tyr Thr Pro Ala Ala Pro Gly Asn Leu
35 40 45
Val Pro Val Cys Trp Gly Lys Gly Ala Cys Pro Val Phe Glu Cys Gly
50 55 60
Asn Val Val Leu Arg Thr Asp Glu Arg Asp Val Asn Tyr Trp Thr Ser
65 70 75 80
Arg Tyr Trp Leu Asn Gly Asp Phe Arg Lys Gly Asp Val Ser Leu Thr
85 90 95
Ile Glu Asn Val Thr Leu Ala Asp Ser Gly Ile Tyr Cys Cys Arg Ile
100 105 110
Gln Ile Pro Gly Ile Met Asn Asp Glu Lys Phe Asn Leu Lys Leu Val
115 120 125
Ile Lys Pro Ala Lys Val Thr Pro Ala Pro Thr Arg Gln Arg Asp Phe
130 135 140
Thr Ala Ala Phe Pro Arg Met Leu Thr Thr Arg Gly His Gly Pro Ala
145 150 155 160
Glu Thr Gln Thr Leu Gly Ser Leu Pro Asp Ile Asn Leu Thr Gln Ile
165 170 175
Ser Thr Leu Ala Asn Glu Leu Arg Asp Ser Arg Leu Ala Asn Asp Leu
180 185 190
Arg Asp Ser Gly Ala Thr Ile Arg Ile Gly Ser Lys Tyr Gly Pro Pro
195 200 205
Cys Pro Ser Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
210 215 220
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
225 230 235 240
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
245 250 255
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
260 265 270
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val
275 280 285
Leu Thr Val Leu His Gln Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys
290 295 300
Lys Val Ser Ser Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
305 310 315 320
Asn Ala Thr Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
325 330 335
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
340 345 350
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
355 360 365
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
370 375 380
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Ser Trp
385 390 395 400
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
405 410 415
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys Ile Glu
420 425 430
Gly Arg Met Asp Gln Val Ser His Arg Tyr Pro Arg Ile Gln Ser Ile
435 440 445
Lys Val Gln Phe Thr Glu Tyr Lys Lys Glu Lys Gly Phe Ile Leu Thr
450 455 460
Ser Gln Lys Glu Asp Glu Ile Met Lys Val Gln Asn Asn Ser Val Ile
465 470 475 480
Ile Asn Cys Asp Gly Phe Tyr Leu Ile Ser Leu Lys Gly Tyr Phe Ser
485 490 495
Gln Glu Val Asn Ile Ser Leu His Tyr Gln Lys Asp Glu Glu Pro Leu
500 505 510
Phe Gln Leu Lys Lys Val Arg Ser Val Asn Ser Leu Met Val Ala Ser
515 520 525
Leu Thr Tyr Lys Asp Lys Val Tyr Leu Asn Val Thr Thr Asp Asn Thr
530 535 540
Ser Leu Asp Asp Phe His Val Asn Gly Gly Glu Leu Ile Leu Ile His
545 550 555 560
Gln Asn Pro Gly Glu Phe Cys Val Leu
565
<210> 12
<211> 2223
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 12
atggagcccg ccggcccggc ccccggccgc ctcgggccgc tgctctgcct gctgctcgcc 60
gcgtcctgcg cctggtcagg agtggcgggt gaggaggagc tgcaggtgat tcagcctgac 120
aagtccgtgt tggttgcagc tggagagaca gccactctgc gctgcactgc gacctctctg 180
atccctgtgg ggcccatcca gtggttcaga ggagctggac caggccggga attaatctac 240
aatcaaaaag aaggccactt cccccgggta acaactgttt cagacctcac aaagagaaac 300
aacatggact tttccatccg catcggtaac atcaccccag cagatgccgg cacctactac 360
tgtgtgaagt tccggaaagg gagccccgat gacgtggagt ttaagtctgg agcaggcact 420
gagctgtctg tgcgcgccaa accctctgcc cccgtggtat cgggccctgc ggcgagggcc 480
acacctcagc acacagtgag cttcacctgc gagtcccacg gcttctcacc cagagacatc 540
accctgaaat ggttcaaaaa tgggaatgag ctctcagact tccagaccaa cgtggacccc 600
gtaggagaga gcgtgtccta cagcatccac agcacagcca aggtggtgct gacccgcgag 660
gacgttcact ctcaagtcat ctgcgaggtg gcccacgtca ccttgcaggg ggaccctctt 720
cgtgggactg ccaacttgtc tgagaccatc cgagttccac ccaccttgga ggttactcaa 780
cagcccgtga gggcagagaa ccaggtgaat gtcacctgcc aggtgaggaa gttctacccc 840
cagagactac agctgacctg gttggagaat ggaaacgtgt cccggacaga aacggcctca 900
accgttacag agaacaagga tggtacctac aactggatga gctggctcct ggtgaatgta 960
tctgcccaca gggatgatgt gaagctcacc tgccaggtgg agcatgacgg gcagccagcg 1020
gtcagcaaaa gccatgacct gaaggtctca gcccacccga aggagcaggg ctcaaatacc 1080
gccgctgaga acactggatc taatgaacgg aacatctatt ctaagtacgg ccctccctgc 1140
cctagctgtc ccgcccctga atttctgggc ggaccctccg tgtttctgtt ccccccaaag 1200
cccaaggaca ccctgatgat cagccggacc cccgaagtga cctgtgtggt ggtggatgtg 1260
tcccaggaag atcccgaggt gcagttcaat tggtacgtgg acggggtgga agtgcacaac 1320
gccaagacca agcccagaga ggaacagttc aacagcacct accgggtggt gtctgtgctg 1380
accgtgctgc accaggattg gctgagcggc aaagagtaca agtgcaaggt gtccagcaag 1440
ggcctgccca gcagcatcga aaagaccatc agcaacgcca ccggccagcc cagggaaccc 1500
caggtgtaca cactgccccc tagccaggaa gagatgacca agaaccaggt gtccctgaca 1560
tgcctcgtga agggcttcta cccctccgat atcgccgtgg aatgggagag caacggccag 1620
ccagagaaca actacaagac caccccccca gtgctggaca gcgacggctc attcttcctg 1680
tactcccggc tgacagtgga caagagcagc tggcaggaag gcaacgtgtt cagctgcagc 1740
gtgatgcacg aagccctgca caaccactac acccagaagt ccctgagcct gtccctgggc 1800
aaaatagagg gacgaatgga ccaggtatca catcggtatc ctcgaattca aagtatcaaa 1860
gtacaattta ccgaatataa gaaggagaaa ggtttcatcc tcacttccca aaaggaggat 1920
gaaatcatga aggtgcagaa caactcagtc atcatcaact gtgatgggtt ttatctcatc 1980
tccctgaagg gctacttctc ccaggaagtc aacattagcc ttcattacca gaaggatgag 2040
gagcccctct tccaactgaa gaaggtcagg tctgtcaact ccttgatggt ggcctctctg 2100
acttacaaag acaaagtcta cttgaatgtg accactgaca atacctccct ggatgacttc 2160
catgtgaatg gcggagaact gattcttatc catcaaaatc ctggtgaatt ctgtgtcctt 2220
tga 2223
<210> 13
<211> 740
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 13
Met Glu Pro Ala Gly Pro Ala Pro Gly Arg Leu Gly Pro Leu Leu Cys
1 5 10 15
Leu Leu Leu Ala Ala Ser Cys Ala Trp Ser Gly Val Ala Gly Glu Glu
20 25 30
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
35 40 45
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro Val Gly
50 55 60
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
65 70 75 80
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
85 90 95
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
100 105 110
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
115 120 125
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
130 135 140
Arg Ala Lys Pro Ser Ala Pro Val Val Ser Gly Pro Ala Ala Arg Ala
145 150 155 160
Thr Pro Gln His Thr Val Ser Phe Thr Cys Glu Ser His Gly Phe Ser
165 170 175
Pro Arg Asp Ile Thr Leu Lys Trp Phe Lys Asn Gly Asn Glu Leu Ser
180 185 190
Asp Phe Gln Thr Asn Val Asp Pro Val Gly Glu Ser Val Ser Tyr Ser
195 200 205
Ile His Ser Thr Ala Lys Val Val Leu Thr Arg Glu Asp Val His Ser
210 215 220
Gln Val Ile Cys Glu Val Ala His Val Thr Leu Gln Gly Asp Pro Leu
225 230 235 240
Arg Gly Thr Ala Asn Leu Ser Glu Thr Ile Arg Val Pro Pro Thr Leu
245 250 255
Glu Val Thr Gln Gln Pro Val Arg Ala Glu Asn Gln Val Asn Val Thr
260 265 270
Cys Gln Val Arg Lys Phe Tyr Pro Gln Arg Leu Gln Leu Thr Trp Leu
275 280 285
Glu Asn Gly Asn Val Ser Arg Thr Glu Thr Ala Ser Thr Val Thr Glu
290 295 300
Asn Lys Asp Gly Thr Tyr Asn Trp Met Ser Trp Leu Leu Val Asn Val
305 310 315 320
Ser Ala His Arg Asp Asp Val Lys Leu Thr Cys Gln Val Glu His Asp
325 330 335
Gly Gln Pro Ala Val Ser Lys Ser His Asp Leu Lys Val Ser Ala His
340 345 350
Pro Lys Glu Gln Gly Ser Asn Thr Ala Ala Glu Asn Thr Gly Ser Asn
355 360 365
Glu Arg Asn Ile Tyr Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro
370 375 380
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
385 390 395 400
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
405 410 415
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
420 425 430
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
435 440 445
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
450 455 460
Gln Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys
465 470 475 480
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln
485 490 495
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
500 505 510
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
515 520 525
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
530 535 540
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
545 550 555 560
Tyr Ser Arg Leu Thr Val Asp Lys Ser Ser Trp Gln Glu Gly Asn Val
565 570 575
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
580 585 590
Lys Ser Leu Ser Leu Ser Leu Gly Lys Ile Glu Gly Arg Met Asp Gln
595 600 605
Val Ser His Arg Tyr Pro Arg Ile Gln Ser Ile Lys Val Gln Phe Thr
610 615 620
Glu Tyr Lys Lys Glu Lys Gly Phe Ile Leu Thr Ser Gln Lys Glu Asp
625 630 635 640
Glu Ile Met Lys Val Gln Asn Asn Ser Val Ile Ile Asn Cys Asp Gly
645 650 655
Phe Tyr Leu Ile Ser Leu Lys Gly Tyr Phe Ser Gln Glu Val Asn Ile
660 665 670
Ser Leu His Tyr Gln Lys Asp Glu Glu Pro Leu Phe Gln Leu Lys Lys
675 680 685
Val Arg Ser Val Asn Ser Leu Met Val Ala Ser Leu Thr Tyr Lys Asp
690 695 700
Lys Val Tyr Leu Asn Val Thr Thr Asp Asn Thr Ser Leu Asp Asp Phe
705 710 715 720
His Val Asn Gly Gly Glu Leu Ile Leu Ile His Gln Asn Pro Gly Glu
725 730 735
Phe Cys Val Leu
740
<210> 14
<211> 1554
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 14
atggggtccc cgggcatggt gctgggcctc ctggtgcaga tctgggccct gcaagaagcc 60
tcaagcctga gcgtgcagca ggggcccaac ttgctgcagg tgaggcaggg cagtcaggcg 120
accctggtct gccaggtgga ccaggccaca gcctgggaac ggctccgtgt taagtggaca 180
aaggatgggg ccatcctgtg tcaaccgtac atcaccaacg gcagcctcag cctgggggtc 240
tgcgggcccc agggacggct ctcctggcag gcacccagcc atctcaccct gcagctggac 300
cctgtgagcc tcaaccacag cggggcgtac gtgtgctggg cggccgtaga gattcctgag 360
ttggaggagg ctgagggcaa cataacaagg ctctttgtgg acccagatga ccccacacag 420
aacagaaacc ggatcgcaag cttcccagga tctaagtacg gccctccctg ccctagctgt 480
cccgcccctg aatttctggg cggaccctcc gtgtttctgt tccccccaaa gcccaaggac 540
accctgatga tcagccggac ccccgaagtg acctgtgtgg tggtggatgt gtcccaggaa 600
gatcccgagg tgcagttcaa ttggtacgtg gacggggtgg aagtgcacaa cgccaagacc 660
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtctgtgct gaccgtgctg 720
caccaggatt ggctgagcgg caaagagtac aagtgcaagg tgtccagcaa gggcctgccc 780
agcagcatcg aaaagaccat cagcaacgcc accggccagc ccagggaacc ccaggtgtac 840
acactgcccc ctagccagga agagatgacc aagaaccagg tgtccctgac atgcctcgtg 900
aagggcttct acccctccga tatcgccgtg gaatgggaga gcaacggcca gccagagaac 960
aactacaaga ccaccccccc agtgctggac agcgacggct cattcttcct gtactcccgg 1020
ctgacagtgg acaagagcag ctggcaggaa ggcaacgtgt tcagctgcag cgtgatgcac 1080
gaagccctgc acaaccacta cacccagaag tccctgagcc tgtccctggg caaaatagag 1140
ggacgaatgg accaggtatc acatcggtat cctcgaattc aaagtatcaa agtacaattt 1200
accgaatata agaaggagaa aggtttcatc ctcacttccc aaaaggagga tgaaatcatg 1260
aaggtgcaga acaactcagt catcatcaac tgtgatgggt tttatctcat ctccctgaag 1320
ggctacttct cccaggaagt caacattagc cttcattacc agaaggatga ggagcccctc 1380
ttccaactga agaaggtcag gtctgtcaac tccttgatgg tggcctctct gacttacaaa 1440
gacaaagtct acttgaatgt gaccactgac aatacctccc tggatgactt ccatgtgaat 1500
ggcggagaac tgattcttat ccatcaaaat cctggtgaat tctgtgtcct ttga 1554
<210> 15
<211> 517
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 15
Met Gly Ser Pro Gly Met Val Leu Gly Leu Leu Val Gln Ile Trp Ala
1 5 10 15
Leu Gln Glu Ala Ser Ser Leu Ser Val Gln Gln Gly Pro Asn Leu Leu
20 25 30
Gln Val Arg Gln Gly Ser Gln Ala Thr Leu Val Cys Gln Val Asp Gln
35 40 45
Ala Thr Ala Trp Glu Arg Leu Arg Val Lys Trp Thr Lys Asp Gly Ala
50 55 60
Ile Leu Cys Gln Pro Tyr Ile Thr Asn Gly Ser Leu Ser Leu Gly Val
65 70 75 80
Cys Gly Pro Gln Gly Arg Leu Ser Trp Gln Ala Pro Ser His Leu Thr
85 90 95
Leu Gln Leu Asp Pro Val Ser Leu Asn His Ser Gly Ala Tyr Val Cys
100 105 110
Trp Ala Ala Val Glu Ile Pro Glu Leu Glu Glu Ala Glu Gly Asn Ile
115 120 125
Thr Arg Leu Phe Val Asp Pro Asp Asp Pro Thr Gln Asn Arg Asn Arg
130 135 140
Ile Ala Ser Phe Pro Gly Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys
145 150 155 160
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
165 170 175
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
180 185 190
Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp
195 200 205
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
210 215 220
Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
225 230 235 240
His Gln Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser
245 250 255
Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly
260 265 270
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu
275 280 285
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
290 295 300
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
305 310 315 320
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
325 330 335
Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Ser Trp Gln Glu Gly Asn
340 345 350
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
355 360 365
Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys Ile Glu Gly Arg Met Asp
370 375 380
Gln Val Ser His Arg Tyr Pro Arg Ile Gln Ser Ile Lys Val Gln Phe
385 390 395 400
Thr Glu Tyr Lys Lys Glu Lys Gly Phe Ile Leu Thr Ser Gln Lys Glu
405 410 415
Asp Glu Ile Met Lys Val Gln Asn Asn Ser Val Ile Ile Asn Cys Asp
420 425 430
Gly Phe Tyr Leu Ile Ser Leu Lys Gly Tyr Phe Ser Gln Glu Val Asn
435 440 445
Ile Ser Leu His Tyr Gln Lys Asp Glu Glu Pro Leu Phe Gln Leu Lys
450 455 460
Lys Val Arg Ser Val Asn Ser Leu Met Val Ala Ser Leu Thr Tyr Lys
465 470 475 480
Asp Lys Val Tyr Leu Asn Val Thr Thr Asp Asn Thr Ser Leu Asp Asp
485 490 495
Phe His Val Asn Gly Gly Glu Leu Ile Leu Ile His Gln Asn Pro Gly
500 505 510
Glu Phe Cys Val Leu
515
<210> 16
<211> 3493
<212> ДНК
<213> Homo sapiens
<400> 16
tcaatcgcct tttatctctg gccctgggac ctttgcctat tttctgattg ataggctttg 60
ttttgtcttt acctccttct ttctggggaa aacttcagtt ttatcgcacg ttcccctttt 120
ccatatcttc atcttccctc tacccagatt gtgaagatgg aaagggtcca acccctggaa 180
gagaatgtgg gaaatgcagc caggccaaga ttcgagagga acaagctatt gctggtggcc 240
tctgtaattc agggactggg gctgctcctg tgcttcacct acatctgcct gcacttctct 300
gctcttcagg tatcacatcg gtatcctcga attcaaagta tcaaagtaca atttaccgaa 360
tataagaagg agaaaggttt catcctcact tcccaaaagg aggatgaaat catgaaggtg 420
cagaacaact cagtcatcat caactgtgat gggttttatc tcatctccct gaagggctac 480
ttctcccagg aagtcaacat tagccttcat taccagaagg atgaggagcc cctcttccaa 540
ctgaagaagg tcaggtctgt caactccttg atggtggcct ctctgactta caaagacaaa 600
gtctacttga atgtgaccac tgacaatacc tccctggatg acttccatgt gaatggcgga 660
gaactgattc ttatccatca aaatcctggt gaattctgtg tcctttgagg ggctgatggc 720
aatatctaaa accaggcacc agcatgaaca ccaagctggg ggtggacagg gcatggattc 780
ttcattgcaa gtgaaggagc ctcccagctc agccacgtgg gatgtgacaa gaagcagatc 840
ctggccctcc cgcccccacc cctcagggat atttaaaact tattttatat accagttaat 900
cttatttatc cttatatttt ctaaattgcc tagccgtcac accccaagat tgccttgagc 960
ctactaggca cctttgtgag aaagaaaaaa tagatgcctc ttcttcaaga tgcattgttt 1020
ctattggtca ggcaattgtc ataataaact tatgtcattg aaaacggtac ctgactacca 1080
tttgctggaa atttgacatg tgtgtggcat tatcaaaatg aagaggagca aggagtgaag 1140
gagtggggtt atgaatctgc caaaggtggt atgaaccaac ccctggaagc caaagcggcc 1200
tctccaaggt taaattgatt gcagtttgca tattgcctaa atttaaactt tctcatttgg 1260
tgggggttca aaagaagaat cagcttgtga aaaatcagga cttgaagaga gccgtctaag 1320
aaataccacg tgcttttttt ctttaccatt ttgctttccc agcctccaaa catagttaat 1380
agaaatttcc cttcaaagaa ctgtctgggg atgtgatgct ttgaaaaatc taatcagtga 1440
cttaagagag attttcttgt atacagggag agtgagataa cttattgtga agggttagct 1500
ttactgtaca ggatagcagg gaactggaca tctcagggta aaagtcagta cggattttaa 1560
tagcctgggg aggaaaacac attctttgcc acagacaggc aaagcaacac atgctcatcc 1620
tcctgcctat gctgagatac gcactcagct ccatgtcttg tacacacaga aacattgctg 1680
gtttcaagaa atgaggtgat cctattatca aattcaatct gatgtcaaat agcactaaga 1740
agttattgtg ccttatgaaa aataatgatc tctgtctaga aataccatag accatatata 1800
gtctcacatt gataattgaa actagaaggg tctataatca gcctatgcca gggcttcaat 1860
ggaatagtat ccccttatgt ttagttgaaa tgtcccctta acttgatata atgtgttatg 1920
cttatggcgc tgtggacaat ctgatttttc atgtcaactt tccagatgat ttgtaacttc 1980
tctgtgccaa accttttata aacataaatt tttgagatat gtattttaaa attgtagcac 2040
atgtttccct gacattttca atagaggata caacatcaca gaatctttct ggatgattct 2100
gtgttatcaa ggaattgtac tgtgctacaa ttatctctag aatctccaga aaggtggagg 2160
gctgttcgcc cttacactaa atggtctcag ttggattttt ttttcctgtt ttctatttcc 2220
tcttaagtac accttcaact atattcccat ccctctattt taatctgtta tgaaggaagg 2280
taaataaaaa tgctaaatag aagaaattgt aggtaaggta agaggaatca agttctgagt 2340
ggctgccaag gcactcacag aatcataatc atggctaaat atttatggag ggcctactgt 2400
ggaccaggca ctgggctaaa tacttacatt tacaagaatc attctgagac agatattcaa 2460
tgatatctgg cttcactact cagaagattg tgtgtgtgtt tgtgtgtgtg tgtgtgtgtg 2520
tatttcactt tttgttattg accatgttct gcaaaattgc agttactcag tgagtgatat 2580
ccgaaaaagt aaacgtttat gactataggt aatatttaag aaaatgcatg gttcattttt 2640
aagtttggaa tttttatcta tatttctcac agatgtgcag tgcacatgca ggcctaagta 2700
tatgttgtgt gtgttgtttg tctttgatgt catggtcccc tctcttaggt gctcactcgc 2760
tttgggtgca cctggcctgc tcttcccatg ttggcctctg caaccacaca gggatatttc 2820
tgctatgcac cagcctcact ccaccttcct tccatcaaaa atatgtgtgt gtgtctcagt 2880
ccctgtaagt catgtccttc acagggagaa ttaacccttc gatatacatg gcagagtttt 2940
gtgggaaaag aattgaatga aaagtcagga gatcagaatt ttaaatttga cttagccact 3000
aactagccat gtaaccttgg gaaagtcatt tcccatttct gggtcttgct tttctttctg 3060
ttaaatgaga ggaatgttaa atatctaaca gtttagaatc ttatgcttac agtgttatct 3120
gtgaatgcac atattaaatg tctatgttct tgttgctatg agtcaaggag tgtaaccttc 3180
tcctttacta tgttgaatgt atttttttct ggacaagctt acatcttcct cagccatctt 3240
tgtgagtcct tcaagagcag ttatcaattg ttagttagat attttctatt tagagaatgc 3300
ttaagggatt ccaatcccga tccaaatcat aatttgttct taagtatact gggcaggtcc 3360
cctattttaa gtcataattt tgtatttagt gctttcctgg ctctcagaga gtattaatat 3420
tgatattaat aatatagtta atagtaatat tgctatttac atggaaacaa ataaaagatc 3480
tcagaattca cta 3493
<210> 17
<211> 183
<212> PRT
<213> Homo sapiens
<400> 17
Met Glu Arg Val Gln Pro Leu Glu Glu Asn Val Gly Asn Ala Ala Arg
1 5 10 15
Pro Arg Phe Glu Arg Asn Lys Leu Leu Leu Val Ala Ser Val Ile Gln
20 25 30
Gly Leu Gly Leu Leu Leu Cys Phe Thr Tyr Ile Cys Leu His Phe Ser
35 40 45
Ala Leu Gln Val Ser His Arg Tyr Pro Arg Ile Gln Ser Ile Lys Val
50 55 60
Gln Phe Thr Glu Tyr Lys Lys Glu Lys Gly Phe Ile Leu Thr Ser Gln
65 70 75 80
Lys Glu Asp Glu Ile Met Lys Val Gln Asn Asn Ser Val Ile Ile Asn
85 90 95
Cys Asp Gly Phe Tyr Leu Ile Ser Leu Lys Gly Tyr Phe Ser Gln Glu
100 105 110
Val Asn Ile Ser Leu His Tyr Gln Lys Asp Glu Glu Pro Leu Phe Gln
115 120 125
Leu Lys Lys Val Arg Ser Val Asn Ser Leu Met Val Ala Ser Leu Thr
130 135 140
Tyr Lys Asp Lys Val Tyr Leu Asn Val Thr Thr Asp Asn Thr Ser Leu
145 150 155 160
Asp Asp Phe His Val Asn Gly Gly Glu Leu Ile Leu Ile His Gln Asn
165 170 175
Pro Gly Glu Phe Cys Val Leu
180
<210> 18
<211> 684
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 18
tctaagtacg gccctccctg ccctagctgt cccgcccctg aatttctggg cggaccctcc 60
gtgtttctgt tccccccaaa gcccaaggac accctgatga tcagccggac ccccgaagtg 120
acctgtgtgg tggtggatgt gtcccaggaa gatcccgagg tgcagttcaa ttggtacgtg 180
gacggggtgg aagtgcacaa cgccaagacc aagcccagag aggaacagtt caacagcacc 240
taccgggtgg tgtctgtgct gaccgtgctg caccaggatt ggctgagcgg caaagagtac 300
aagtgcaagg tgtccagcaa gggcctgccc agcagcatcg aaaagaccat cagcaacgcc 360
accggccagc ccagggaacc ccaggtgtac acactgcccc ctagccagga agagatgacc 420
aagaaccagg tgtccctgac atgcctcgtg aagggcttct acccctccga tatcgccgtg 480
gaatgggaga gcaacggcca gccagagaac aactacaaga ccaccccccc agtgctggac 540
agcgacggct cattcttcct gtactcccgg ctgacagtgg acaagagcag ctggcaggaa 600
ggcaacgtgt tcagctgcag cgtgatgcac gaagccctgc acaaccacta cacccagaag 660
tccctgagcc tgtccctggg caaa 684
<210> 19
<211> 867
<212> ДНК
<213> Homo sapiens
<400> 19
atgcagatcc cacaggcgcc ctggccagtc gtctgggcgg tgctacaact gggctggcgg 60
ccaggatggt tcttagactc cccagacagg ccctggaacc cccccacctt ctccccagcc 120
ctgctcgtgg tgaccgaagg ggacaacgcc accttcacct gcagcttctc caacacatcg 180
gagagcttcg tgctaaactg gtaccgcatg agccccagca accagacgga caagctggcc 240
gccttccccg aggaccgcag ccagcccggc caggactgcc gcttccgtgt cacacaactg 300
cccaacgggc gtgacttcca catgagcgtg gtcagggccc ggcgcaatga cagcggcacc 360
tacctctgtg gggccatctc cctggccccc aaggcgcaga tcaaagagag cctgcgggca 420
gagctcaggg tgacagagag aagggcagaa gtgcccacag cccaccccag cccctcaccc 480
aggccagccg gccagttcca aaccctggtg gttggtgtcg tgggcggcct gctgggcagc 540
ctggtgctgc tagtctgggt cctggccgtc atctgctccc gggccgcacg agggacaata 600
ggagccaggc gcaccggcca gcccctgaag gaggacccct cagccgtgcc tgtgttctct 660
gtggactatg gggagctgga tttccagtgg cgagagaaga ccccggagcc ccccgtgccc 720
tgtgtccctg agcagacgga gtatgccacc attgtctttc ctagcggaat gggcacctca 780
tcccccgccc gcaggggctc agctgacggc cctcggagtg cccagccact gaggcctgag 840
gatggacact gctcttggcc cctctga 867
<210> 20
<211> 1623
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 20
gtcgacgcca ccatgcagat cccacaggcg ccctggccag tcgtctgggc ggtgctacaa 60
ctgggctggc ggccaggatg gttcttagac tccccagaca ggccctggaa cccccccacc 120
ttctccccag ccctgctcgt ggtgaccgaa ggggacaacg ccaccttcac ctgcagcttc 180
tccaacacat cggagagctt cgtgctaaac tggtaccgca tgagccccag caaccagacg 240
gacaagctgg ccgccttccc cgaggaccgc agccagcccg gccaggactg ccgcttccgt 300
gtcacacaac tgcccaacgg gcgtgacttc cacatgagcg tggtcagggc ccggcgcaat 360
gacagcggca cctacctctg tggggccatc tccctggccc ccaaggcgca gatcaaagag 420
agcctgcggg cagagctcag ggtgacagag agaagggcag aagtgcccac agcccacccc 480
agcccctcac ccaggccagc cggccagttc caatctaagt acggccctcc ctgccctagc 540
tgtcccgccc ctgaatttct gggcggaccc tccgtgtttc tgttcccccc aaagcccaag 600
gacaccctga tgatcagccg gacccccgaa gtgacctgtg tggtggtgga tgtgtcccag 660
gaagatcccg aggtgcagtt caattggtac gtggacgggg tggaagtgca caacgccaag 720
accaagccca gagaggaaca gttcaacagc acctaccggg tggtgtctgt gctgaccgtg 780
ctgcaccagg attggctgag cggcaaagag tacaagtgca aggtgtccag caagggcctg 840
cccagcagca tcgaaaagac catcagcaac gccaccggcc agcccaggga accccaggtg 900
tacacactgc cccctagcca ggaagagatg accaagaacc aggtgtccct gacatgcctc 960
gtgaagggct tctacccctc cgatatcgcc gtggaatggg agagcaacgg ccagccagag 1020
aacaactaca agaccacccc cccagtgctg gacagcgacg gctcattctt cctgtactcc 1080
cggctgacag tggacaagag cagctggcag gaaggcaacg tgttcagctg cagcgtgatg 1140
cacgaagccc tgcacaacca ctacacccag aagtccctga gcctgtccct gggcaaaata 1200
gagggacgaa tggaccaggt atcacatcgg tatcctcgaa ttcaaagtat caaagtacaa 1260
tttaccgaat ataagaagga gaaaggtttc atcctcactt cccaaaagga ggatgaaatc 1320
atgaaggtgc agaacaactc agtcatcatc aactgtgatg ggttttatct catctccctg 1380
aagggctact tctcccagga agtcaacatt agccttcatt accagaagga tgaggagccc 1440
ctcttccaac tgaagaaggt caggtctgtc aactccttga tggtggcctc tctgacttac 1500
aaagacaaag tctacttgaa tgtgaccact gacaatacct ccctggatga cttccatgtg 1560
aatggcggag aactgattct tatccatcaa aatcctggtg aattctgtgt cctttgagtc 1620
gac 1623
<210> 21
<211> 1637
<212> ДНК
<213> Homo sapiens
<400> 21
caccggcgag atctgccacc atgcagatcc ctcaggcccc ctggcctgtc gtgtgggctg 60
tgctgcagct gggatggcgg cctggctggt tcctggactc tcctgacaga ccctggaacc 120
cccccacctt tagccctgct ctgctggtcg tgaccgaggg cgacaacgcc accttcacct 180
gttccttcag caacacctcc gagtccttcg tgctgaactg gtacagaatg tcccccagca 240
accagaccga caagctggcc gccttccccg aggatagatc ccagcctgga caggactgcc 300
ggttcagagt gacccagctg cccaacggcc gggacttcca catgtctgtc gtgcgggcca 360
gacggaacga ctccggcaca tatctgtgcg gcgccatctc cctggccccc aaggctcaga 420
tcaaagagtc tctgcgggcc gagctgagag tgaccgagag aagggctgag gtgccaaccg 480
cccaccctag cccatctcca agacctgccg gccagttcca gtctaagtac ggccctcctt 540
gccctagctg ccctgcccct gaatttctgg gcggaccctc cgtgttcctg ttccccccaa 600
agcccaagga caccctgatg atctcccgga cccccgaagt gacctgcgtg gtggtggatg 660
tgtcccagga agatcccgag gtgcagttca attggtacgt ggacggcgtg gaagtgcaca 720
acgccaagac caagcccaga gaggaacagt tcaactccac ctaccgggtg gtgtccgtgc 780
tgaccgtgct gcaccaggat tggctgtccg gcaaagagta caagtgcaag gtgtcctcca 840
agggcctgcc ctccagcatc gaaaagacca tctctaacgc caccggccag ccccgggaac 900
cccaggtgta cacactgcct ccaagccagg aagagatgac caagaaccag gtgtccctga 960
cctgtctcgt gaagggcttc tacccctccg atatcgccgt ggaatgggag tccaacggcc 1020
agcctgagaa caactacaag accacccccc ctgtgctgga ctccgacggc tccttcttcc 1080
tgtactcccg cctgaccgtg gacaagtcct cctggcagga aggcaacgtg ttctcctgct 1140
ccgtgatgca cgaggccctg cacaaccact acacccagaa gtccctgtcc ctgtctctgg 1200
gcaagatcga gggccggatg gatcaggtgt cacacagata cccccggatc cagtccatca 1260
aagtgcagtt taccgagtac aagaaagaga agggattcat cctgacctcc cagaaagagg 1320
acgagatcat gaaggtgcag aacaactccg tgatcatcaa ctgcgacggg ttctacctga 1380
tctccctgaa gggctacttc agtcaggaag tgaacatcag cctgcactac cagaaggacg 1440
aggaacccct gttccagctg aagaaagtgc ggagcgtgaa ctccctgatg gtggcctctc 1500
tgacctacaa ggacaaggtg tacctgaacg tgaccaccga caatacctcc ctggacgact 1560
tccacgtgaa cggcggcgag ctgatcctga tccaccagaa ccctggcgag ttctgcgtgc 1620
tgtgactcga ggctagc 1637
<210> 22
<211> 534
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 22
Met Gln Ile Pro Gln Ala Pro Trp Pro Val Val Trp Ala Val Leu Gln
1 5 10 15
Leu Gly Trp Arg Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro Trp
20 25 30
Asn Pro Pro Thr Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly Asp
35 40 45
Asn Ala Thr Phe Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe Val
50 55 60
Leu Asn Trp Tyr Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu Ala
65 70 75 80
Ala Phe Pro Glu Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe Arg
85 90 95
Val Thr Gln Leu Pro Asn Gly Arg Asp Phe His Met Ser Val Val Arg
100 105 110
Ala Arg Arg Asn Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser Leu
115 120 125
Ala Pro Lys Ala Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg Val
130 135 140
Thr Glu Arg Arg Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser Pro
145 150 155 160
Arg Pro Ala Gly Gln Phe Gln Ser Lys Tyr Gly Pro Pro Cys Pro Ser
165 170 175
Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
180 185 190
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
195 200 205
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
210 215 220
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
225 230 235 240
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
245 250 255
Leu His Gln Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser
260 265 270
Ser Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr
275 280 285
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
290 295 300
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
305 310 315 320
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
325 330 335
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
340 345 350
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Ser Trp Gln Glu Gly
355 360 365
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
370 375 380
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys Ile Glu Gly Arg Met
385 390 395 400
Asp Gln Val Ser His Arg Tyr Pro Arg Ile Gln Ser Ile Lys Val Gln
405 410 415
Phe Thr Glu Tyr Lys Lys Glu Lys Gly Phe Ile Leu Thr Ser Gln Lys
420 425 430
Glu Asp Glu Ile Met Lys Val Gln Asn Asn Ser Val Ile Ile Asn Cys
435 440 445
Asp Gly Phe Tyr Leu Ile Ser Leu Lys Gly Tyr Phe Ser Gln Glu Val
450 455 460
Asn Ile Ser Leu His Tyr Gln Lys Asp Glu Glu Pro Leu Phe Gln Leu
465 470 475 480
Lys Lys Val Arg Ser Val Asn Ser Leu Met Val Ala Ser Leu Thr Tyr
485 490 495
Lys Asp Lys Val Tyr Leu Asn Val Thr Thr Asp Asn Thr Ser Leu Asp
500 505 510
Asp Phe His Val Asn Gly Gly Glu Leu Ile Leu Ile His Gln Asn Pro
515 520 525
Gly Glu Phe Cys Val Leu
530
<210> 23
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 23
Gly Gly Gly Gly Ser
1 5
<210> 24
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 24
Glu Ala Ala Ala Lys
1 5
<210> 25
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 25
Ala Glu Ala Ala Ala Lys Ala
1 5
<210> 26
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 26
Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Ala
1 5 10
<210> 27
<211> 46
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 27
Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys
1 5 10 15
Glu Ala Ala Ala Lys Ala Leu Glu Ala Glu Ala Ala Ala Lys Glu Ala
20 25 30
Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Ala
35 40 45
<210> 28
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 28
Pro Ala Pro Ala Pro
1 5
<210> 29
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 29
Lys Glu Ser Gly Ser Val Ser Ser Glu Gln Leu Ala Gln Phe Arg Ser
1 5 10 15
Leu Asp
<210> 30
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 30
Glu Gly Lys Ser Ser Gly Ser Gly Ser Glu Ser Lys Ser Thr
1 5 10
<210> 31
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 31
Gly Ser Ala Gly Ser Ala Ala Gly Ser Gly Glu Phe
1 5 10
<210> 32
<211> 143
<212> PRT
<213> Homo sapiens
<400> 32
Leu Asp Ser Pro Asp Arg Pro Trp Asn Pro Pro Thr Phe Ser Pro Ala
1 5 10 15
Leu Leu Val Val Thr Glu Gly Asp Asn Ala Thr Phe Thr Cys Ser Phe
20 25 30
Ser Asn Thr Ser Glu Ser Phe Val Leu Asn Trp Tyr Arg Met Ser Pro
35 40 45
Ser Asn Gln Thr Asp Lys Leu Ala Ala Phe Pro Glu Asp Arg Ser Gln
50 55 60
Pro Gly Gln Asp Cys Arg Phe Arg Val Thr Gln Leu Pro Asn Gly Arg
65 70 75 80
Asp Phe His Met Ser Val Val Arg Ala Arg Arg Asn Asp Ser Gly Thr
85 90 95
Tyr Leu Cys Gly Ala Ile Ser Leu Ala Pro Lys Ala Gln Ile Lys Glu
100 105 110
Ser Leu Arg Ala Glu Leu Arg Val Thr Glu Arg Arg Ala Glu Val Pro
115 120 125
Thr Ala His Pro Ser Pro Ser Pro Arg Pro Ala Gly Gln Phe Gln
130 135 140
<210> 33
<211> 343
<212> PRT
<213> Homo sapiens
<400> 33
Glu Glu Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala
1 5 10 15
Ala Gly Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro
20 25 30
Val Gly Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu
35 40 45
Ile Tyr Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser
50 55 60
Asp Leu Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn
65 70 75 80
Ile Thr Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys
85 90 95
Gly Ser Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu
100 105 110
Ser Val Arg Ala Lys Pro Ser Ala Pro Val Val Ser Gly Pro Ala Ala
115 120 125
Arg Ala Thr Pro Gln His Thr Val Ser Phe Thr Cys Glu Ser His Gly
130 135 140
Phe Ser Pro Arg Asp Ile Thr Leu Lys Trp Phe Lys Asn Gly Asn Glu
145 150 155 160
Leu Ser Asp Phe Gln Thr Asn Val Asp Pro Val Gly Glu Ser Val Ser
165 170 175
Tyr Ser Ile His Ser Thr Ala Lys Val Val Leu Thr Arg Glu Asp Val
180 185 190
His Ser Gln Val Ile Cys Glu Val Ala His Val Thr Leu Gln Gly Asp
195 200 205
Pro Leu Arg Gly Thr Ala Asn Leu Ser Glu Thr Ile Arg Val Pro Pro
210 215 220
Thr Leu Glu Val Thr Gln Gln Pro Val Arg Ala Glu Asn Gln Val Asn
225 230 235 240
Val Thr Cys Gln Val Arg Lys Phe Tyr Pro Gln Arg Leu Gln Leu Thr
245 250 255
Trp Leu Glu Asn Gly Asn Val Ser Arg Thr Glu Thr Ala Ser Thr Val
260 265 270
Thr Glu Asn Lys Asp Gly Thr Tyr Asn Trp Met Ser Trp Leu Leu Val
275 280 285
Asn Val Ser Ala His Arg Asp Asp Val Lys Leu Thr Cys Gln Val Glu
290 295 300
His Asp Gly Gln Pro Ala Val Ser Lys Ser His Asp Leu Lys Val Ser
305 310 315 320
Ala His Pro Lys Glu Gln Gly Ser Asn Thr Ala Ala Glu Asn Thr Gly
325 330 335
Ser Asn Glu Arg Asn Ile Tyr
340
<210> 34
<211> 127
<212> PRT
<213> Homo sapiens
<400> 34
Lys Glu Ser Cys Asp Val Gln Leu Tyr Ile Lys Arg Gln Ser Glu His
1 5 10 15
Ser Ile Leu Ala Gly Asp Pro Phe Glu Leu Glu Cys Pro Val Lys Tyr
20 25 30
Cys Ala Asn Arg Pro His Val Thr Trp Cys Lys Leu Asn Gly Thr Thr
35 40 45
Cys Val Lys Leu Glu Asp Arg Gln Thr Ser Trp Lys Glu Glu Lys Asn
50 55 60
Ile Ser Phe Phe Ile Leu His Phe Glu Pro Val Leu Pro Asn Asp Asn
65 70 75 80
Gly Ser Tyr Arg Cys Ser Ala Asn Phe Gln Ser Asn Leu Ile Glu Ser
85 90 95
His Ser Thr Thr Leu Tyr Val Thr Asp Val Lys Ser Ala Ser Glu Arg
100 105 110
Pro Ser Lys Asp Glu Met Ala Ser Arg Pro Trp Leu Leu Tyr Arg
115 120 125
<210> 35
<211> 120
<212> PRT
<213> Homo sapiens
<400> 35
Met Met Thr Gly Thr Ile Glu Thr Thr Gly Asn Ile Ser Ala Glu Lys
1 5 10 15
Gly Gly Ser Ile Ile Leu Gln Cys His Leu Ser Ser Thr Thr Ala Gln
20 25 30
Val Thr Gln Val Asn Trp Glu Gln Gln Asp Gln Leu Leu Ala Ile Cys
35 40 45
Asn Ala Asp Leu Gly Trp His Ile Ser Pro Ser Phe Lys Asp Arg Val
50 55 60
Ala Pro Gly Pro Gly Leu Gly Leu Thr Leu Gln Ser Leu Thr Val Asn
65 70 75 80
Asp Thr Gly Glu Tyr Phe Cys Ile Tyr His Thr Tyr Pro Asp Gly Thr
85 90 95
Tyr Thr Gly Arg Ile Phe Leu Glu Val Leu Glu Ser Ser Val Ala Glu
100 105 110
His Gly Ala Arg Phe Gln Ile Pro
115 120
<210> 36
<211> 181
<212> PRT
<213> Homo sapiens
<400> 36
Ser Glu Val Glu Tyr Arg Ala Glu Val Gly Gln Asn Ala Tyr Leu Pro
1 5 10 15
Cys Phe Tyr Thr Pro Ala Ala Pro Gly Asn Leu Val Pro Val Cys Trp
20 25 30
Gly Lys Gly Ala Cys Pro Val Phe Glu Cys Gly Asn Val Val Leu Arg
35 40 45
Thr Asp Glu Arg Asp Val Asn Tyr Trp Thr Ser Arg Tyr Trp Leu Asn
50 55 60
Gly Asp Phe Arg Lys Gly Asp Val Ser Leu Thr Ile Glu Asn Val Thr
65 70 75 80
Leu Ala Asp Ser Gly Ile Tyr Cys Cys Arg Ile Gln Ile Pro Gly Ile
85 90 95
Met Asn Asp Glu Lys Phe Asn Leu Lys Leu Val Ile Lys Pro Ala Lys
100 105 110
Val Thr Pro Ala Pro Thr Arg Gln Arg Asp Phe Thr Ala Ala Phe Pro
115 120 125
Arg Met Leu Thr Thr Arg Gly His Gly Pro Ala Glu Thr Gln Thr Leu
130 135 140
Gly Ser Leu Pro Asp Ile Asn Leu Thr Gln Ile Ser Thr Leu Ala Asn
145 150 155 160
Glu Leu Arg Asp Ser Arg Leu Ala Asn Asp Leu Arg Asp Ser Gly Ala
165 170 175
Thr Ile Arg Ile Gly
180
<210> 37
<211> 202
<212> PRT
<213> Homo sapiens
<400> 37
Gln Val Gln Val Val Thr Gln Asp Glu Arg Glu Gln Leu Tyr Thr Pro
1 5 10 15
Ala Ser Leu Lys Cys Ser Leu Gln Asn Ala Gln Glu Ala Leu Ile Val
20 25 30
Thr Trp Gln Lys Lys Lys Ala Val Ser Pro Glu Asn Met Val Thr Phe
35 40 45
Ser Glu Asn His Gly Val Val Ile Gln Pro Ala Tyr Lys Asp Lys Ile
50 55 60
Asn Ile Thr Gln Leu Gly Leu Gln Asn Ser Thr Ile Thr Phe Trp Asn
65 70 75 80
Ile Thr Leu Glu Asp Glu Gly Cys Tyr Met Cys Leu Phe Asn Thr Phe
85 90 95
Gly Phe Gly Lys Ile Ser Gly Thr Ala Cys Leu Thr Val Tyr Val Gln
100 105 110
Pro Ile Val Ser Leu His Tyr Lys Phe Ser Glu Asp His Leu Asn Ile
115 120 125
Thr Cys Ser Ala Thr Ala Arg Pro Ala Pro Met Val Phe Trp Lys Val
130 135 140
Pro Arg Ser Gly Ile Glu Asn Ser Thr Val Thr Leu Ser His Pro Asn
145 150 155 160
Gly Thr Thr Ser Val Thr Ser Ile Leu His Ile Lys Asp Pro Lys Asn
165 170 175
Gln Val Gly Lys Glu Val Ile Cys Gln Val Leu His Leu Gly Thr Val
180 185 190
Thr Asp Phe Lys Gln Thr Val Asn Lys Gly
195 200
<210> 38
<211> 128
<212> PRT
<213> Homo sapiens
<400> 38
Leu Ser Val Gln Gln Gly Pro Asn Leu Leu Gln Val Arg Gln Gly Ser
1 5 10 15
Gln Ala Thr Leu Val Cys Gln Val Asp Gln Ala Thr Ala Trp Glu Arg
20 25 30
Leu Arg Val Lys Trp Thr Lys Asp Gly Ala Ile Leu Cys Gln Pro Tyr
35 40 45
Ile Thr Asn Gly Ser Leu Ser Leu Gly Val Cys Gly Pro Gln Gly Arg
50 55 60
Leu Ser Trp Gln Ala Pro Ser His Leu Thr Leu Gln Leu Asp Pro Val
65 70 75 80
Ser Leu Asn His Ser Gly Ala Tyr Val Cys Trp Ala Ala Val Glu Ile
85 90 95
Pro Glu Leu Glu Glu Ala Glu Gly Asn Ile Thr Arg Leu Phe Val Asp
100 105 110
Pro Asp Asp Pro Thr Gln Asn Arg Asn Arg Ile Ala Ser Phe Pro Gly
115 120 125
<210> 39
<211> 162
<212> PRT
<213> Homo sapiens
<400> 39
Phe Lys Val Ala Thr Pro Tyr Ser Leu Tyr Val Cys Pro Glu Gly Gln
1 5 10 15
Asn Val Thr Leu Thr Cys Arg Leu Leu Gly Pro Val Asp Lys Gly His
20 25 30
Asp Val Thr Phe Tyr Lys Thr Trp Tyr Arg Ser Ser Arg Gly Glu Val
35 40 45
Gln Thr Cys Ser Glu Arg Arg Pro Ile Arg Asn Leu Thr Phe Gln Asp
50 55 60
Leu His Leu His His Gly Gly His Gln Ala Ala Asn Thr Ser His Asp
65 70 75 80
Leu Ala Gln Arg His Gly Leu Glu Ser Ala Ser Asp His His Gly Asn
85 90 95
Phe Ser Ile Thr Met Arg Asn Leu Thr Leu Leu Asp Ser Gly Leu Tyr
100 105 110
Cys Cys Leu Val Val Glu Ile Arg His His His Ser Glu His Arg Val
115 120 125
His Gly Ala Met Glu Leu Gln Val Gln Thr Gly Lys Asp Ala Pro Ser
130 135 140
Asn Cys Val Val Tyr Pro Ser Ser Ser Gln Asp Ser Glu Asn Ile Thr
145 150 155 160
Ala Ala
<210> 40
<211> 242
<212> PRT
<213> Homo sapiens
<400> 40
Val Arg Ile Asn Gly Asp Gly Gln Glu Val Leu Tyr Leu Ala Glu Gly
1 5 10 15
Asp Asn Val Arg Leu Gly Cys Pro Tyr Val Leu Asp Pro Glu Asp Tyr
20 25 30
Gly Pro Asn Gly Leu Asp Ile Glu Trp Met Gln Val Asn Ser Asp Pro
35 40 45
Ala His His Arg Glu Asn Val Phe Leu Ser Tyr Gln Asp Lys Arg Ile
50 55 60
Asn His Gly Ser Leu Pro His Leu Gln Gln Arg Val Arg Phe Ala Ala
65 70 75 80
Ser Asp Pro Ser Gln Tyr Asp Ala Ser Ile Asn Leu Met Asn Leu Gln
85 90 95
Val Ser Asp Thr Ala Thr Tyr Glu Cys Arg Val Lys Lys Thr Thr Met
100 105 110
Ala Thr Arg Lys Val Ile Val Thr Val Gln Ala Arg Pro Ala Val Pro
115 120 125
Met Cys Trp Thr Glu Gly His Met Thr Tyr Gly Asn Asp Val Val Leu
130 135 140
Lys Cys Tyr Ala Ser Gly Gly Ser Gln Pro Leu Ser Tyr Lys Trp Ala
145 150 155 160
Lys Ile Ser Gly His His Tyr Pro Tyr Arg Ala Gly Ser Tyr Thr Ser
165 170 175
Gln His Ser Tyr His Ser Glu Leu Ser Tyr Gln Glu Ser Phe His Ser
180 185 190
Ser Ile Asn Gln Gly Leu Asn Asn Gly Asp Leu Val Leu Lys Asp Ile
195 200 205
Ser Arg Ala Asp Asp Gly Leu Tyr Gln Cys Thr Val Ala Asn Asn Val
210 215 220
Gly Tyr Ser Val Cys Val Val Glu Val Lys Val Ser Asp Ser Arg Arg
225 230 235 240
Ile Gly
<210> 41
<211> 427
<212> PRT
<213> Homo sapiens
<400> 41
Glu Asp Phe Arg Val Ile Gly Pro Ala His Pro Ile Leu Ala Gly Val
1 5 10 15
Gly Glu Asp Ala Leu Leu Thr Cys Gln Leu Leu Pro Lys Arg Thr Thr
20 25 30
Met His Val Glu Val Arg Trp Tyr Arg Ser Glu Pro Ser Thr Pro Val
35 40 45
Phe Val His Arg Asp Gly Val Glu Val Thr Glu Met Gln Met Glu Glu
50 55 60
Tyr Arg Gly Trp Val Glu Trp Ile Glu Asn Gly Ile Ala Lys Gly Asn
65 70 75 80
Val Ala Leu Lys Ile His Asn Ile Gln Pro Ser Asp Asn Gly Gln Tyr
85 90 95
Trp Cys His Phe Gln Asp Gly Asn Tyr Cys Gly Glu Thr Ser Leu Leu
100 105 110
Leu Lys Val Ala Gly Leu Gly Ser Ala Pro Ser Ile His Met Glu Gly
115 120 125
Pro Gly Glu Ser Gly Val Gln Leu Val Cys Thr Ala Arg Gly Trp Phe
130 135 140
Pro Glu Pro Gln Val Tyr Trp Glu Asp Ile Arg Gly Glu Lys Leu Leu
145 150 155 160
Ala Val Ser Glu His Arg Ile Gln Asp Lys Asp Gly Leu Phe Tyr Ala
165 170 175
Glu Ala Thr Leu Val Val Arg Asn Ala Ser Ala Glu Ser Val Ser Cys
180 185 190
Leu Val His Asn Pro Val Leu Thr Glu Glu Lys Gly Ser Val Ile Ser
195 200 205
Leu Pro Glu Lys Leu Gln Thr Glu Leu Ala Ser Leu Lys Val Asn Gly
210 215 220
Pro Ser Gln Pro Ile Leu Val Arg Val Gly Glu Asp Ile Gln Leu Thr
225 230 235 240
Cys Tyr Leu Ser Pro Lys Ala Asn Ala Gln Ser Met Glu Val Arg Trp
245 250 255
Asp Arg Ser His Arg Tyr Pro Ala Val His Val Tyr Met Asp Gly Asp
260 265 270
His Val Ala Gly Glu Gln Met Ala Glu Tyr Arg Gly Arg Thr Val Leu
275 280 285
Val Ser Asp Ala Ile Asp Glu Gly Arg Leu Thr Leu Gln Ile Leu Ser
290 295 300
Ala Arg Pro Ser Asp Asp Gly Gln Tyr Arg Cys Leu Phe Glu Lys Asp
305 310 315 320
Asp Val Tyr Gln Glu Ala Ser Leu Asp Leu Lys Val Val Ser Leu Gly
325 330 335
Ser Ser Pro Leu Ile Thr Val Glu Gly Gln Glu Asp Gly Glu Met Gln
340 345 350
Pro Met Cys Ser Ser Asp Gly Trp Phe Pro Gln Pro His Val Pro Trp
355 360 365
Arg Asp Met Glu Gly Lys Thr Ile Pro Ser Ser Ser Gln Ala Leu Thr
370 375 380
Gln Gly Ser His Gly Leu Phe His Val Gln Thr Leu Leu Arg Val Thr
385 390 395 400
Asn Ile Ser Ala Val Asp Val Thr Cys Ser Ile Ser Ile Pro Phe Leu
405 410 415
Gly Glu Glu Lys Ile Ala Thr Phe Ser Leu Ser
420 425
<210> 42
<211> 235
<212> PRT
<213> Homo sapiens
<400> 42
Leu Pro Ala Gln Val Ala Phe Thr Pro Tyr Ala Pro Glu Pro Gly Ser
1 5 10 15
Thr Cys Arg Leu Arg Glu Tyr Tyr Asp Gln Thr Ala Gln Met Cys Cys
20 25 30
Ser Lys Cys Ser Pro Gly Gln His Ala Lys Val Phe Cys Thr Lys Thr
35 40 45
Ser Asp Thr Val Cys Asp Ser Cys Glu Asp Ser Thr Tyr Thr Gln Leu
50 55 60
Trp Asn Trp Val Pro Glu Cys Leu Ser Cys Gly Ser Arg Cys Ser Ser
65 70 75 80
Asp Gln Val Glu Thr Gln Ala Cys Thr Arg Glu Gln Asn Arg Ile Cys
85 90 95
Thr Cys Arg Pro Gly Trp Tyr Cys Ala Leu Ser Lys Gln Glu Gly Cys
100 105 110
Arg Leu Cys Ala Pro Leu Arg Lys Cys Arg Pro Gly Phe Gly Val Ala
115 120 125
Arg Pro Gly Thr Glu Thr Ser Asp Val Val Cys Lys Pro Cys Ala Pro
130 135 140
Gly Thr Phe Ser Asn Thr Thr Ser Ser Thr Asp Ile Cys Arg Pro His
145 150 155 160
Gln Ile Cys Asn Val Val Ala Ile Pro Gly Asn Ala Ser Met Asp Ala
165 170 175
Val Cys Thr Ser Thr Ser Pro Thr Arg Ser Met Ala Pro Gly Ala Val
180 185 190
His Leu Pro Gln Pro Val Ser Thr Arg Ser Gln His Thr Gln Pro Thr
195 200 205
Pro Glu Pro Ser Thr Ala Pro Ser Thr Ser Phe Leu Leu Pro Met Gly
210 215 220
Pro Ser Pro Pro Ala Glu Gly Ser Thr Gly Asp
225 230 235
<210> 43
<211> 438
<212> PRT
<213> Homo sapiens
<400> 43
Leu Glu Val Gln Val Pro Glu Asp Pro Val Val Ala Leu Val Gly Thr
1 5 10 15
Asp Ala Thr Leu Cys Cys Ser Phe Ser Pro Glu Pro Gly Phe Ser Leu
20 25 30
Ala Gln Leu Asn Leu Ile Trp Gln Leu Thr Asp Thr Lys Gln Leu Val
35 40 45
His Ser Phe Ala Glu Gly Gln Asp Gln Gly Ser Ala Tyr Ala Asn Arg
50 55 60
Thr Ala Leu Phe Pro Asp Leu Leu Ala Gln Gly Asn Ala Ser Leu Arg
65 70 75 80
Leu Gln Arg Val Arg Val Ala Asp Glu Gly Ser Phe Thr Cys Phe Val
85 90 95
Ser Ile Arg Asp Phe Gly Ser Ala Ala Val Ser Leu Gln Val Ala Ala
100 105 110
Pro Tyr Ser Lys Pro Ser Met Thr Leu Glu Pro Asn Lys Asp Leu Arg
115 120 125
Pro Gly Asp Thr Val Thr Ile Thr Cys Ser Ser Tyr Gln Gly Tyr Pro
130 135 140
Glu Ala Glu Val Phe Trp Gln Asp Gly Gln Gly Val Pro Leu Thr Gly
145 150 155 160
Asn Val Thr Thr Ser Gln Met Ala Asn Glu Gln Gly Leu Phe Asp Val
165 170 175
His Ser Ile Leu Arg Val Val Leu Gly Ala Asn Gly Thr Tyr Ser Cys
180 185 190
Leu Val Arg Asn Pro Val Leu Gln Gln Asp Ala His Ser Ser Val Thr
195 200 205
Ile Thr Pro Gln Arg Ser Pro Thr Gly Ala Val Glu Val Gln Val Pro
210 215 220
Glu Asp Pro Val Val Ala Leu Val Gly Thr Asp Ala Thr Leu Arg Cys
225 230 235 240
Ser Phe Ser Pro Glu Pro Gly Phe Ser Leu Ala Gln Leu Asn Leu Ile
245 250 255
Trp Gln Leu Thr Asp Thr Lys Gln Leu Val His Ser Phe Thr Glu Gly
260 265 270
Arg Asp Gln Gly Ser Ala Tyr Ala Asn Arg Thr Ala Leu Phe Pro Asp
275 280 285
Leu Leu Ala Gln Gly Asn Ala Ser Leu Arg Leu Gln Arg Val Arg Val
290 295 300
Ala Asp Glu Gly Ser Phe Thr Cys Phe Val Ser Ile Arg Asp Phe Gly
305 310 315 320
Ser Ala Ala Val Ser Leu Gln Val Ala Ala Pro Tyr Ser Lys Pro Ser
325 330 335
Met Thr Leu Glu Pro Asn Lys Asp Leu Arg Pro Gly Asp Thr Val Thr
340 345 350
Ile Thr Cys Ser Ser Tyr Arg Gly Tyr Pro Glu Ala Glu Val Phe Trp
355 360 365
Gln Asp Gly Gln Gly Val Pro Leu Thr Gly Asn Val Thr Thr Ser Gln
370 375 380
Met Ala Asn Glu Gln Gly Leu Phe Asp Val His Ser Val Leu Arg Val
385 390 395 400
Val Leu Gly Ala Asn Gly Thr Tyr Ser Cys Leu Val Arg Asn Pro Val
405 410 415
Leu Gln Gln Asp Ala His Gly Ser Val Thr Ile Thr Gly Gln Pro Met
420 425 430
Thr Phe Pro Pro Glu Ala
435
<210> 44
<211> 208
<212> PRT
<213> Homo sapiens
<400> 44
Cys Gln Gly Ser Ala Asp His Val Val Ser Ile Ser Gly Val Pro Leu
1 5 10 15
Gln Leu Gln Pro Asn Ser Ile Gln Thr Lys Val Asp Ser Ile Ala Trp
20 25 30
Lys Lys Leu Leu Pro Ser Gln Asn Gly Phe His His Ile Leu Lys Trp
35 40 45
Glu Asn Gly Ser Leu Pro Ser Asn Thr Ser Asn Asp Arg Phe Ser Phe
50 55 60
Ile Val Lys Asn Leu Ser Leu Leu Ile Lys Ala Ala Gln Gln Gln Asp
65 70 75 80
Ser Gly Leu Tyr Cys Leu Glu Val Thr Ser Ile Ser Gly Lys Val Gln
85 90 95
Thr Ala Thr Phe Gln Val Phe Val Phe Glu Ser Leu Leu Pro Asp Lys
100 105 110
Val Glu Lys Pro Arg Leu Gln Gly Gln Gly Lys Ile Leu Asp Arg Gly
115 120 125
Arg Cys Gln Val Ala Leu Ser Cys Leu Val Ser Arg Asp Gly Asn Val
130 135 140
Ser Tyr Ala Trp Tyr Arg Gly Ser Lys Leu Ile Gln Thr Ala Gly Asn
145 150 155 160
Leu Thr Tyr Leu Asp Glu Glu Val Asp Ile Asn Gly Thr His Thr Tyr
165 170 175
Thr Cys Asn Val Ser Asn Pro Val Ser Trp Glu Ser His Thr Leu Asn
180 185 190
Leu Thr Gln Asp Cys Gln Asn Ala His Gln Glu Phe Arg Phe Trp Pro
195 200 205
<210> 45
<211> 422
<212> PRT
<213> Homo sapiens
<400> 45
Val Pro Val Val Trp Ala Gln Glu Gly Ala Pro Ala Gln Leu Pro Cys
1 5 10 15
Ser Pro Thr Ile Pro Leu Gln Asp Leu Ser Leu Leu Arg Arg Ala Gly
20 25 30
Val Thr Trp Gln His Gln Pro Asp Ser Gly Pro Pro Ala Ala Ala Pro
35 40 45
Gly His Pro Leu Ala Pro Gly Pro His Pro Ala Ala Pro Ser Ser Trp
50 55 60
Gly Pro Arg Pro Arg Arg Tyr Thr Val Leu Ser Val Gly Pro Gly Gly
65 70 75 80
Leu Arg Ser Gly Arg Leu Pro Leu Gln Pro Arg Val Gln Leu Asp Glu
85 90 95
Arg Gly Arg Gln Arg Gly Asp Phe Ser Leu Trp Leu Arg Pro Ala Arg
100 105 110
Arg Ala Asp Ala Gly Glu Tyr Arg Ala Ala Val His Leu Arg Asp Arg
115 120 125
Ala Leu Ser Cys Arg Leu Arg Leu Arg Leu Gly Gln Ala Ser Met Thr
130 135 140
Ala Ser Pro Pro Gly Ser Leu Arg Ala Ser Asp Trp Val Ile Leu Asn
145 150 155 160
Cys Ser Phe Ser Arg Pro Asp Arg Pro Ala Ser Val His Trp Phe Arg
165 170 175
Asn Arg Gly Gln Gly Arg Val Pro Val Arg Glu Ser Pro His His His
180 185 190
Leu Ala Glu Ser Phe Leu Phe Leu Pro Gln Val Ser Pro Met Asp Ser
195 200 205
Gly Pro Trp Gly Cys Ile Leu Thr Tyr Arg Asp Gly Phe Asn Val Ser
210 215 220
Ile Met Tyr Asn Leu Thr Val Leu Gly Leu Glu Pro Pro Thr Pro Leu
225 230 235 240
Thr Val Tyr Ala Gly Ala Gly Ser Arg Val Gly Leu Pro Cys Arg Leu
245 250 255
Pro Ala Gly Val Gly Thr Arg Ser Phe Leu Thr Ala Lys Trp Thr Pro
260 265 270
Pro Gly Gly Gly Pro Asp Leu Leu Val Thr Gly Asp Asn Gly Asp Phe
275 280 285
Thr Leu Arg Leu Glu Asp Val Ser Gln Ala Gln Ala Gly Thr Tyr Thr
290 295 300
Cys His Ile His Leu Gln Glu Gln Gln Leu Asn Ala Thr Val Thr Leu
305 310 315 320
Ala Ile Ile Thr Val Thr Pro Lys Ser Phe Gly Ser Pro Gly Ser Leu
325 330 335
Gly Lys Leu Leu Cys Glu Val Thr Pro Val Ser Gly Gln Glu Arg Phe
340 345 350
Val Trp Ser Ser Leu Asp Thr Pro Ser Gln Arg Ser Phe Ser Gly Pro
355 360 365
Trp Leu Glu Ala Gln Glu Ala Gln Leu Leu Ser Gln Pro Trp Gln Cys
370 375 380
Gln Leu Tyr Gln Gly Glu Arg Leu Leu Gly Ala Ala Val Tyr Phe Thr
385 390 395 400
Glu Leu Ser Ser Pro Gly Ala Gln Arg Ser Gly Arg Ala Pro Gly Ala
405 410 415
Leu Pro Ala Gly His Leu
420
<210> 46
<211> 498
<212> PRT
<213> Homo sapiens
<400> 46
Ile Pro Val Ile Glu Pro Ser Val Pro Glu Leu Val Val Lys Pro Gly
1 5 10 15
Ala Thr Val Thr Leu Arg Cys Val Gly Asn Gly Ser Val Glu Trp Asp
20 25 30
Gly Pro Pro Ser Pro His Trp Thr Leu Tyr Ser Asp Gly Ser Ser Ser
35 40 45
Ile Leu Ser Thr Asn Asn Ala Thr Phe Gln Asn Thr Gly Thr Tyr Arg
50 55 60
Cys Thr Glu Pro Gly Asp Pro Leu Gly Gly Ser Ala Ala Ile His Leu
65 70 75 80
Tyr Val Lys Asp Pro Ala Arg Pro Trp Asn Val Leu Ala Gln Glu Val
85 90 95
Val Val Phe Glu Asp Gln Asp Ala Leu Leu Pro Cys Leu Leu Thr Asp
100 105 110
Pro Val Leu Glu Ala Gly Val Ser Leu Val Arg Val Arg Gly Arg Pro
115 120 125
Leu Met Arg His Thr Asn Tyr Ser Phe Ser Pro Trp His Gly Phe Thr
130 135 140
Ile His Arg Ala Lys Phe Ile Gln Ser Gln Asp Tyr Gln Cys Ser Ala
145 150 155 160
Leu Met Gly Gly Arg Lys Val Met Ser Ile Ser Ile Arg Leu Lys Val
165 170 175
Gln Lys Val Ile Pro Gly Pro Pro Ala Leu Thr Leu Val Pro Ala Glu
180 185 190
Leu Val Arg Ile Arg Gly Glu Ala Ala Gln Ile Val Cys Ser Ala Ser
195 200 205
Ser Val Asp Val Asn Phe Asp Val Phe Leu Gln His Asn Asn Thr Lys
210 215 220
Leu Ala Ile Pro Gln Gln Ser Asp Phe His Asn Asn Arg Tyr Gln Lys
225 230 235 240
Val Leu Thr Leu Asn Leu Asp Gln Val Asp Phe Gln His Ala Gly Asn
245 250 255
Tyr Ser Cys Val Ala Ser Asn Val Gln Gly Lys His Ser Thr Ser Met
260 265 270
Phe Phe Arg Val Val Glu Ser Ala Tyr Leu Asn Leu Ser Ser Glu Gln
275 280 285
Asn Leu Ile Gln Glu Val Thr Val Gly Glu Gly Leu Asn Leu Lys Val
290 295 300
Met Val Glu Ala Tyr Pro Gly Leu Gln Gly Phe Asn Trp Thr Tyr Leu
305 310 315 320
Gly Pro Phe Ser Asp His Gln Pro Glu Pro Lys Leu Ala Asn Ala Thr
325 330 335
Thr Lys Asp Thr Tyr Arg His Thr Phe Thr Leu Ser Leu Pro Arg Leu
340 345 350
Lys Pro Ser Glu Ala Gly Arg Tyr Ser Phe Leu Ala Arg Asn Pro Gly
355 360 365
Gly Trp Arg Ala Leu Thr Phe Glu Leu Thr Leu Arg Tyr Pro Pro Glu
370 375 380
Val Ser Val Ile Trp Thr Phe Ile Asn Gly Ser Gly Thr Leu Leu Cys
385 390 395 400
Ala Ala Ser Gly Tyr Pro Gln Pro Asn Val Thr Trp Leu Gln Cys Ser
405 410 415
Gly His Thr Asp Arg Cys Asp Glu Ala Gln Val Leu Gln Val Trp Asp
420 425 430
Asp Pro Tyr Pro Glu Val Leu Ser Gln Glu Pro Phe His Lys Val Thr
435 440 445
Val Gln Ser Leu Leu Thr Val Glu Thr Leu Glu His Asn Gln Thr Tyr
450 455 460
Glu Cys Arg Ala His Asn Ser Val Gly Ser Gly Ser Trp Ala Phe Ile
465 470 475 480
Pro Ile Ser Ala Gly Ala His Thr His Pro Pro Asp Glu Phe Leu Phe
485 490 495
Thr Pro
<210> 47
<211> 93
<212> PRT
<213> Homo sapiens
<400> 47
Leu Gln Cys Phe Cys His Leu Cys Thr Lys Asp Asn Phe Thr Cys Val
1 5 10 15
Thr Asp Gly Leu Cys Phe Val Ser Val Thr Glu Thr Thr Asp Lys Val
20 25 30
Ile His Asn Ser Met Cys Ile Ala Glu Ile Asp Leu Ile Pro Arg Asp
35 40 45
Arg Pro Phe Val Cys Ala Pro Ser Ser Lys Thr Gly Ser Val Thr Thr
50 55 60
Thr Tyr Cys Cys Asn Gln Asp His Cys Asn Lys Ile Glu Leu Pro Thr
65 70 75 80
Thr Val Lys Ser Ser Pro Gly Leu Gly Pro Val Glu Leu
85 90
<210> 48
<211> 214
<212> PRT
<213> Homo sapiens
<400> 48
His Gly Thr Glu Leu Pro Ser Pro Pro Ser Val Trp Phe Glu Ala Glu
1 5 10 15
Phe Phe His His Ile Leu His Trp Thr Pro Ile Pro Asn Gln Ser Glu
20 25 30
Ser Thr Cys Tyr Glu Val Ala Leu Leu Arg Tyr Gly Ile Glu Ser Trp
35 40 45
Asn Ser Ile Ser Asn Cys Ser Gln Thr Leu Ser Tyr Asp Leu Thr Ala
50 55 60
Val Thr Leu Asp Leu Tyr His Ser Asn Gly Tyr Arg Ala Arg Val Arg
65 70 75 80
Ala Val Asp Gly Ser Arg His Ser Asn Trp Thr Val Thr Asn Thr Arg
85 90 95
Phe Ser Val Asp Glu Val Thr Leu Thr Val Gly Ser Val Asn Leu Glu
100 105 110
Ile His Asn Gly Phe Ile Leu Gly Lys Ile Gln Leu Pro Arg Pro Lys
115 120 125
Met Ala Pro Ala Asn Asp Thr Tyr Glu Ser Ile Phe Ser His Phe Arg
130 135 140
Glu Tyr Glu Ile Ala Ile Arg Lys Val Pro Gly Asn Phe Thr Phe Thr
145 150 155 160
His Lys Lys Val Lys His Glu Asn Phe Ser Leu Leu Thr Ser Gly Glu
165 170 175
Val Gly Glu Phe Cys Val Gln Val Lys Pro Ser Val Ala Ser Arg Ser
180 185 190
Asn Lys Gly Met Trp Ser Lys Glu Glu Cys Ile Ser Leu Thr Arg Gln
195 200 205
Tyr Phe Thr Val Thr Asn
210
<210> 49
<211> 173
<212> PRT
<213> Homo sapiens
<400> 49
Glu Pro Pro Thr Ala Cys Arg Glu Lys Gln Tyr Leu Ile Asn Ser Gln
1 5 10 15
Cys Cys Ser Leu Cys Gln Pro Gly Gln Lys Leu Val Ser Asp Cys Thr
20 25 30
Glu Phe Thr Glu Thr Glu Cys Leu Pro Cys Gly Glu Ser Glu Phe Leu
35 40 45
Asp Thr Trp Asn Arg Glu Thr His Cys His Gln His Lys Tyr Cys Asp
50 55 60
Pro Asn Leu Gly Leu Arg Val Gln Gln Lys Gly Thr Ser Glu Thr Asp
65 70 75 80
Thr Ile Cys Thr Cys Glu Glu Gly Trp His Cys Thr Ser Glu Ala Cys
85 90 95
Glu Ser Cys Val Leu His Arg Ser Cys Ser Pro Gly Phe Gly Val Lys
100 105 110
Gln Ile Ala Thr Gly Val Ser Asp Thr Ile Cys Glu Pro Cys Pro Val
115 120 125
Gly Phe Phe Ser Asn Val Ser Ser Ala Phe Glu Lys Cys His Pro Trp
130 135 140
Thr Ser Cys Glu Thr Lys Asp Leu Val Val Gln Gln Ala Gly Thr Asn
145 150 155 160
Lys Thr Asp Val Val Cys Gly Pro Gln Asp Arg Leu Arg
165 170
<210> 50
<211> 186
<212> PRT
<213> Homo sapiens
<400> 50
Leu His Cys Val Gly Asp Thr Tyr Pro Ser Asn Asp Arg Cys Cys His
1 5 10 15
Glu Cys Arg Pro Gly Asn Gly Met Val Ser Arg Cys Ser Arg Ser Gln
20 25 30
Asn Thr Val Cys Arg Pro Cys Gly Pro Gly Phe Tyr Asn Asp Val Val
35 40 45
Ser Ser Lys Pro Cys Lys Pro Cys Thr Trp Cys Asn Leu Arg Ser Gly
50 55 60
Ser Glu Arg Lys Gln Leu Cys Thr Ala Thr Gln Asp Thr Val Cys Arg
65 70 75 80
Cys Arg Ala Gly Thr Gln Pro Leu Asp Ser Tyr Lys Pro Gly Val Asp
85 90 95
Cys Ala Pro Cys Pro Pro Gly His Phe Ser Pro Gly Asp Asn Gln Ala
100 105 110
Cys Lys Pro Trp Thr Asn Cys Thr Leu Ala Gly Lys His Thr Leu Gln
115 120 125
Pro Ala Ser Asn Ser Ser Asp Ala Ile Cys Glu Asp Arg Asp Pro Pro
130 135 140
Ala Thr Gln Pro Gln Glu Thr Gln Gly Pro Pro Ala Arg Pro Ile Thr
145 150 155 160
Val Gln Pro Thr Glu Ala Trp Pro Arg Thr Ser Gln Gly Pro Ser Thr
165 170 175
Arg Pro Val Glu Val Pro Gly Gly Arg Ala
180 185
<210> 51
<211> 163
<212> PRT
<213> Homo sapiens
<400> 51
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln
<210> 52
<211> 126
<212> PRT
<213> Homo sapiens
<400> 52
Lys Ala Met His Val Ala Gln Pro Ala Val Val Leu Ala Ser Ser Arg
1 5 10 15
Gly Ile Ala Ser Phe Val Cys Glu Tyr Ala Ser Pro Gly Lys Ala Thr
20 25 30
Glu Val Arg Val Thr Val Leu Arg Gln Ala Asp Ser Gln Val Thr Glu
35 40 45
Val Cys Ala Ala Thr Tyr Met Met Gly Asn Glu Leu Thr Phe Leu Asp
50 55 60
Asp Ser Ile Cys Thr Gly Thr Ser Ser Gly Asn Gln Val Asn Leu Thr
65 70 75 80
Ile Gln Gly Leu Arg Ala Met Asp Thr Gly Leu Tyr Ile Cys Lys Val
85 90 95
Glu Leu Met Tyr Pro Pro Pro Tyr Tyr Leu Gly Ile Gly Asn Gly Thr
100 105 110
Gln Ile Tyr Val Ile Asp Pro Glu Pro Cys Pro Asp Ser Asp
115 120 125
<210> 53
<211> 220
<212> PRT
<213> Homo sapiens
<400> 53
Phe Thr Val Thr Val Pro Lys Asp Leu Tyr Val Val Glu Tyr Gly Ser
1 5 10 15
Asn Met Thr Ile Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asp Leu
20 25 30
Ala Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Asn Ile Ile Gln
35 40 45
Phe Val His Gly Glu Glu Asp Leu Lys Val Gln His Ser Ser Tyr Arg
50 55 60
Gln Arg Ala Arg Leu Leu Lys Asp Gln Leu Ser Leu Gly Asn Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr Arg Cys
85 90 95
Met Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Val Lys Val
100 105 110
Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val Asp Pro
115 120 125
Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr Pro Lys
130 135 140
Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser Gly Lys
145 150 155 160
Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Phe Asn Val Thr
165 170 175
Ser Thr Leu Arg Ile Asn Thr Thr Thr Asn Glu Ile Phe Tyr Cys Thr
180 185 190
Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu Val Ile
195 200 205
Pro Glu Leu Pro Leu Ala His Pro Pro Asn Glu Arg
210 215 220
<210> 54
<211> 201
<212> PRT
<213> Homo sapiens
<400> 54
Leu Phe Thr Val Thr Val Pro Lys Glu Leu Tyr Ile Ile Glu His Gly
1 5 10 15
Ser Asn Val Thr Leu Glu Cys Asn Phe Asp Thr Gly Ser His Val Asn
20 25 30
Leu Gly Ala Ile Thr Ala Ser Leu Gln Lys Val Glu Asn Asp Thr Ser
35 40 45
Pro His Arg Glu Arg Ala Thr Leu Leu Glu Glu Gln Leu Pro Leu Gly
50 55 60
Lys Ala Ser Phe His Ile Pro Gln Val Gln Val Arg Asp Glu Gly Gln
65 70 75 80
Tyr Gln Cys Ile Ile Ile Tyr Gly Val Ala Trp Asp Tyr Lys Tyr Leu
85 90 95
Thr Leu Lys Val Lys Ala Ser Tyr Arg Lys Ile Asn Thr His Ile Leu
100 105 110
Lys Val Pro Glu Thr Asp Glu Val Glu Leu Thr Cys Gln Ala Thr Gly
115 120 125
Tyr Pro Leu Ala Glu Val Ser Trp Pro Asn Val Ser Val Pro Ala Asn
130 135 140
Thr Ser His Ser Arg Thr Pro Glu Gly Leu Tyr Gln Val Thr Ser Val
145 150 155 160
Leu Arg Leu Lys Pro Pro Pro Gly Arg Asn Phe Ser Cys Val Phe Trp
165 170 175
Asn Thr His Val Arg Glu Leu Thr Leu Ala Ser Ile Asp Leu Gln Ser
180 185 190
Gln Met Glu Pro Arg Thr His Pro Thr
195 200
<210> 55
<211> 235
<212> PRT
<213> Homo sapiens
<400> 55
Leu Ile Ile Gly Phe Gly Ile Ser Gly Arg His Ser Ile Thr Val Thr
1 5 10 15
Thr Val Ala Ser Ala Gly Asn Ile Gly Glu Asp Gly Ile Leu Ser Cys
20 25 30
Thr Phe Glu Pro Asp Ile Lys Leu Ser Asp Ile Val Ile Gln Trp Leu
35 40 45
Lys Glu Gly Val Leu Gly Leu Val His Glu Phe Lys Glu Gly Lys Asp
50 55 60
Glu Leu Ser Glu Gln Asp Glu Met Phe Arg Gly Arg Thr Ala Val Phe
65 70 75 80
Ala Asp Gln Val Ile Val Gly Asn Ala Ser Leu Arg Leu Lys Asn Val
85 90 95
Gln Leu Thr Asp Ala Gly Thr Tyr Lys Cys Tyr Ile Ile Thr Ser Lys
100 105 110
Gly Lys Gly Asn Ala Asn Leu Glu Tyr Lys Thr Gly Ala Phe Ser Met
115 120 125
Pro Glu Val Asn Val Asp Tyr Asn Ala Ser Ser Glu Thr Leu Arg Cys
130 135 140
Glu Ala Pro Arg Trp Phe Pro Gln Pro Thr Val Val Trp Ala Ser Gln
145 150 155 160
Val Asp Gln Gly Ala Asn Phe Ser Glu Val Ser Asn Thr Ser Phe Glu
165 170 175
Leu Asn Ser Glu Asn Val Thr Met Lys Val Val Ser Val Leu Tyr Asn
180 185 190
Val Thr Ile Asn Asn Thr Tyr Ser Cys Met Ile Glu Asn Asp Ile Ala
195 200 205
Lys Ala Thr Gly Asp Ile Lys Val Thr Glu Ser Glu Ile Lys Arg Arg
210 215 220
Ser His Leu Gln Leu Leu Asn Ser Lys Ala Ser
225 230 235
<210> 56
<211> 133
<212> PRT
<213> Homo sapiens
<400> 56
Gln Val Ser His Arg Tyr Pro Arg Ile Gln Ser Ile Lys Val Gln Phe
1 5 10 15
Thr Glu Tyr Lys Lys Glu Lys Gly Phe Ile Leu Thr Ser Gln Lys Glu
20 25 30
Asp Glu Ile Met Lys Val Gln Asn Asn Ser Val Ile Ile Asn Cys Asp
35 40 45
Gly Phe Tyr Leu Ile Ser Leu Lys Gly Tyr Phe Ser Gln Glu Val Asn
50 55 60
Ile Ser Leu His Tyr Gln Lys Asp Glu Glu Pro Leu Phe Gln Leu Lys
65 70 75 80
Lys Val Arg Ser Val Asn Ser Leu Met Val Ala Ser Leu Thr Tyr Lys
85 90 95
Asp Lys Val Tyr Leu Asn Val Thr Thr Asp Asn Thr Ser Leu Asp Asp
100 105 110
Phe His Val Asn Gly Gly Glu Leu Ile Leu Ile His Gln Asn Pro Gly
115 120 125
Glu Phe Cys Val Leu
130
<210> 57
<211> 132
<212> PRT
<213> Homo sapiens
<400> 57
Thr Ser Leu Lys Pro Thr Ala Ile Glu Ser Cys Met Val Lys Phe Glu
1 5 10 15
Leu Ser Ser Ser Lys Trp His Met Thr Ser Pro Lys Pro His Cys Val
20 25 30
Asn Thr Thr Ser Asp Gly Lys Leu Lys Ile Leu Gln Ser Gly Thr Tyr
35 40 45
Leu Ile Tyr Gly Gln Val Ile Pro Val Asp Lys Lys Tyr Ile Lys Asp
50 55 60
Asn Ala Pro Phe Val Val Gln Ile Tyr Lys Lys Asn Asp Val Leu Gln
65 70 75 80
Thr Leu Met Asn Asp Phe Gln Ile Leu Pro Ile Gly Gly Val Tyr Glu
85 90 95
Leu His Ala Gly Asp Asn Ile Tyr Leu Lys Phe Asn Ser Lys Asp His
100 105 110
Ile Gln Lys Thr Asn Thr Tyr Trp Gly Ile Ile Leu Met Pro Asp Leu
115 120 125
Pro Phe Ile Ser
130
<210> 58
<211> 205
<212> PRT
<213> Homo sapiens
<400> 58
Ala Cys Pro Trp Ala Val Ser Gly Ala Arg Ala Ser Pro Gly Ser Ala
1 5 10 15
Ala Ser Pro Arg Leu Arg Glu Gly Pro Glu Leu Ser Pro Asp Asp Pro
20 25 30
Ala Gly Leu Leu Asp Leu Arg Gln Gly Met Phe Ala Gln Leu Val Ala
35 40 45
Gln Asn Val Leu Leu Ile Asp Gly Pro Leu Ser Trp Tyr Ser Asp Pro
50 55 60
Gly Leu Ala Gly Val Ser Leu Thr Gly Gly Leu Ser Tyr Lys Glu Asp
65 70 75 80
Thr Lys Glu Leu Val Val Ala Lys Ala Gly Val Tyr Tyr Val Phe Phe
85 90 95
Gln Leu Glu Leu Arg Arg Val Val Ala Gly Glu Gly Ser Gly Ser Val
100 105 110
Ser Leu Ala Leu His Leu Gln Pro Leu Arg Ser Ala Ala Gly Ala Ala
115 120 125
Ala Leu Ala Leu Thr Val Asp Leu Pro Pro Ala Ser Ser Glu Ala Arg
130 135 140
Asn Ser Ala Phe Gly Phe Gln Gly Arg Leu Leu His Leu Ser Ala Gly
145 150 155 160
Gln Arg Leu Gly Val His Leu His Thr Glu Ala Arg Ala Arg His Ala
165 170 175
Trp Gln Leu Thr Gln Gly Ala Thr Val Leu Gly Leu Phe Arg Val Thr
180 185 190
Pro Glu Ile Pro Ala Gly Leu Pro Ser Pro Arg Ser Glu
195 200 205
<210> 59
<211> 195
<212> PRT
<213> Homo sapiens
<400> 59
Ser Gln Leu Arg Ala Gln Gly Glu Ala Cys Val Gln Phe Gln Ala Leu
1 5 10 15
Lys Gly Gln Glu Phe Ala Pro Ser His Gln Gln Val Tyr Ala Pro Leu
20 25 30
Arg Ala Asp Gly Asp Lys Pro Arg Ala His Leu Thr Val Val Arg Gln
35 40 45
Thr Pro Thr Gln His Phe Lys Asn Gln Phe Pro Ala Leu His Trp Glu
50 55 60
His Glu Leu Gly Leu Ala Phe Thr Lys Asn Arg Met Asn Tyr Thr Asn
65 70 75 80
Lys Phe Leu Leu Ile Pro Glu Ser Gly Asp Tyr Phe Ile Tyr Ser Gln
85 90 95
Val Thr Phe Arg Gly Met Thr Ser Glu Cys Ser Glu Ile Arg Gln Ala
100 105 110
Gly Arg Pro Asn Lys Pro Asp Ser Ile Thr Val Val Ile Thr Lys Val
115 120 125
Thr Asp Ser Tyr Pro Glu Pro Thr Gln Leu Leu Met Gly Thr Lys Ser
130 135 140
Val Cys Glu Val Gly Ser Asn Trp Phe Gln Pro Ile Tyr Leu Gly Ala
145 150 155 160
Met Phe Ser Leu Gln Glu Gly Asp Lys Leu Met Val Asn Val Ser Asp
165 170 175
Ile Ser Leu Val Asp Tyr Thr Lys Glu Asp Lys Thr Phe Phe Gly Ala
180 185 190
Phe Leu Leu
195
<210> 60
<211> 215
<212> PRT
<213> Homo sapiens
<400> 60
His Arg Arg Leu Asp Lys Ile Glu Asp Glu Arg Asn Leu His Glu Asp
1 5 10 15
Phe Val Phe Met Lys Thr Ile Gln Arg Cys Asn Thr Gly Glu Arg Ser
20 25 30
Leu Ser Leu Leu Asn Cys Glu Glu Ile Lys Ser Gln Phe Glu Gly Phe
35 40 45
Val Lys Asp Ile Met Leu Asn Lys Glu Glu Thr Lys Lys Glu Asn Ser
50 55 60
Phe Glu Met Gln Lys Gly Asp Gln Asn Pro Gln Ile Ala Ala His Val
65 70 75 80
Ile Ser Glu Ala Ser Ser Lys Thr Thr Ser Val Leu Gln Trp Ala Glu
85 90 95
Lys Gly Tyr Tyr Thr Met Ser Asn Asn Leu Val Thr Leu Glu Asn Gly
100 105 110
Lys Gln Leu Thr Val Lys Arg Gln Gly Leu Tyr Tyr Ile Tyr Ala Gln
115 120 125
Val Thr Phe Cys Ser Asn Arg Glu Ala Ser Ser Gln Ala Pro Phe Ile
130 135 140
Ala Ser Leu Cys Leu Lys Ser Pro Gly Arg Phe Glu Arg Ile Leu Leu
145 150 155 160
Arg Ala Ala Asn Thr His Ser Ser Ala Lys Pro Cys Gly Gln Gln Ser
165 170 175
Ile His Leu Gly Gly Val Phe Glu Leu Gln Pro Gly Ala Ser Val Phe
180 185 190
Val Asn Val Thr Asp Pro Ser Gln Val Ser His Gly Thr Gly Phe Thr
195 200 205
Ser Phe Gly Leu Leu Lys Leu
210 215
<210> 61
<211> 172
<212> PRT
<213> Homo sapiens
<400> 61
Gln Arg Thr Asp Ser Ile Pro Asn Ser Pro Asp Asn Val Pro Leu Lys
1 5 10 15
Gly Gly Asn Cys Ser Glu Asp Leu Leu Cys Ile Leu Lys Arg Ala Pro
20 25 30
Phe Lys Lys Ser Trp Ala Tyr Leu Gln Val Ala Lys His Leu Asn Lys
35 40 45
Thr Lys Leu Ser Trp Asn Lys Asp Gly Ile Leu His Gly Val Arg Tyr
50 55 60
Gln Asp Gly Asn Leu Val Ile Gln Phe Pro Gly Leu Tyr Phe Ile Ile
65 70 75 80
Cys Gln Leu Gln Phe Leu Val Gln Cys Pro Asn Asn Ser Val Asp Leu
85 90 95
Lys Leu Glu Leu Leu Ile Asn Lys His Ile Lys Lys Gln Ala Leu Val
100 105 110
Thr Val Cys Glu Ser Gly Met Gln Thr Lys His Val Tyr Gln Asn Leu
115 120 125
Ser Gln Phe Leu Leu Asp Tyr Leu Gln Val Asn Thr Thr Ile Ser Val
130 135 140
Asn Val Asp Thr Phe Gln Tyr Ile Asp Thr Ser Thr Phe Pro Leu Glu
145 150 155 160
Asn Val Leu Ser Ile Phe Leu Tyr Ser Asn Ser Asp
165 170
<210> 62
<211> 243
<212> PRT
<213> Homo sapiens
<400> 62
Thr Asn Glu Leu Lys Gln Met Gln Asp Lys Tyr Ser Lys Ser Gly Ile
1 5 10 15
Ala Cys Phe Leu Lys Glu Asp Asp Ser Tyr Trp Asp Pro Asn Asp Glu
20 25 30
Glu Ser Met Asn Ser Pro Cys Trp Gln Val Lys Trp Gln Leu Arg Gln
35 40 45
Leu Val Arg Lys Met Ile Leu Arg Thr Ser Glu Glu Thr Ile Ser Thr
50 55 60
Val Gln Glu Lys Gln Gln Asn Ile Ser Pro Leu Val Arg Glu Arg Gly
65 70 75 80
Pro Gln Arg Val Ala Ala His Ile Thr Gly Thr Arg Gly Arg Ser Asn
85 90 95
Thr Leu Ser Ser Pro Asn Ser Lys Asn Glu Lys Ala Leu Gly Arg Lys
100 105 110
Ile Asn Ser Trp Glu Ser Ser Arg Ser Gly His Ser Phe Leu Ser Asn
115 120 125
Leu His Leu Arg Asn Gly Glu Leu Val Ile His Glu Lys Gly Phe Tyr
130 135 140
Tyr Ile Tyr Ser Gln Thr Tyr Phe Arg Phe Gln Glu Glu Ile Lys Glu
145 150 155 160
Asn Thr Lys Asn Asp Lys Gln Met Val Gln Tyr Ile Tyr Lys Tyr Thr
165 170 175
Ser Tyr Pro Asp Pro Ile Leu Leu Met Lys Ser Ala Arg Asn Ser Cys
180 185 190
Trp Ser Lys Asp Ala Glu Tyr Gly Leu Tyr Ser Ile Tyr Gln Gly Gly
195 200 205
Ile Phe Glu Leu Lys Glu Asn Asp Arg Ile Phe Val Ser Val Thr Asn
210 215 220
Glu His Leu Ile Asp Met Asp His Glu Ala Ser Phe Phe Gly Ala Phe
225 230 235 240
Leu Val Gly
<210> 63
<211> 155
<212> PRT
<213> Homo sapiens
<400> 63
Gln Arg Phe Ala Gln Ala Gln Gln Gln Leu Pro Leu Glu Ser Leu Gly
1 5 10 15
Trp Asp Val Ala Glu Leu Gln Leu Asn His Thr Gly Pro Gln Gln Asp
20 25 30
Pro Arg Leu Tyr Trp Gln Gly Gly Pro Ala Leu Gly Arg Ser Phe Leu
35 40 45
His Gly Pro Glu Leu Asp Lys Gly Gln Leu Arg Ile His Arg Asp Gly
50 55 60
Ile Tyr Met Val His Ile Gln Val Thr Leu Ala Ile Cys Ser Ser Thr
65 70 75 80
Thr Ala Ser Arg His His Pro Thr Thr Leu Ala Val Gly Ile Cys Ser
85 90 95
Pro Ala Ser Arg Ser Ile Ser Leu Leu Arg Leu Ser Phe His Gln Gly
100 105 110
Cys Thr Ile Ala Ser Gln Arg Leu Thr Pro Leu Ala Arg Gly Asp Thr
115 120 125
Leu Cys Thr Asn Leu Thr Gly Thr Leu Leu Pro Ser Arg Asn Thr Asp
130 135 140
Glu Thr Phe Phe Gly Val Gln Trp Val Arg Pro
145 150 155
<210> 64
<211> 0
<212> PRT
<213> Homo sapiens
<400> 64
000
<210> 65
<211> 0
<212> PRT
<213> Homo sapiens
<400> 65
000
<210> 66
<211> 0
<212> PRT
<213> Homo sapiens
<400> 66
000
<210> 67
<211> 0
<212> PRT
<213> Homo sapiens
<400> 67
000
<210> 68
<211> 0
<212> PRT
<213> Homo sapiens
<400> 68
000
<210> 69
<211> 0
<212> PRT
<213> Homo sapiens
<400> 69
000
<210> 70
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 70
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Arg Leu Thr Val Asp Lys Ser Ser Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 71
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 71
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Arg Leu Thr Val Asp Lys Ser Ser Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 72
<211> 217
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 72
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Thr Pro His
65 70 75 80
Ser Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Arg Leu Thr Val Asp Lys Ser Ser Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 73
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 73
Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro
1 5 10
<210> 74
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 74
Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 75
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 75
Ser Lys Tyr Gly Pro Pro
1 5
<210> 76
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 76
Ile Glu Gly Arg Met Asp
1 5
<210> 77
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 77
Gly Gly Gly Val Pro Arg Asp Cys Gly
1 5
<210> 78
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 78
Ile Glu Gly Arg Met Asp Gly Gly Gly Gly Ala Gly Gly Gly Gly
1 5 10 15
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЕ БЕЛКИ НА ОСНОВЕ TIGIT И LIGHT | 2018 |
|
RU2775490C2 |
| ХИМЕРНЫЕ БЕЛКИ НА ОСНОВЕ CSF1R | 2018 |
|
RU2792239C2 |
| АНТИТЕЛА К PD-1 И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2761640C2 |
| АНТИ-ОХ40 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2753493C2 |
| ВЫСОКОАФФИННЫЕ АНТИ-PD-1 И АНТИ-LAG-3 АНТИТЕЛА И ПОЛУЧЕННЫЕ ИЗ НИХ БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ | 2019 |
|
RU2782381C2 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| АНТИТЕЛО, НАЦЕЛЕННОЕ НА ВСМА, И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2799655C2 |
| ВАРИАНТЫ FC-ОБЛАСТИ С ИЗМЕНЕННЫМ СВЯЗЫВАНИЕМ С НЕОНАТАЛЬНЫМ FC-РЕЦЕПТОРОМ (FCRN) ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2830231C2 |
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2766199C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ОСНОВЕ АНТИТЕЛА К PD-L1 И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2766590C2 |
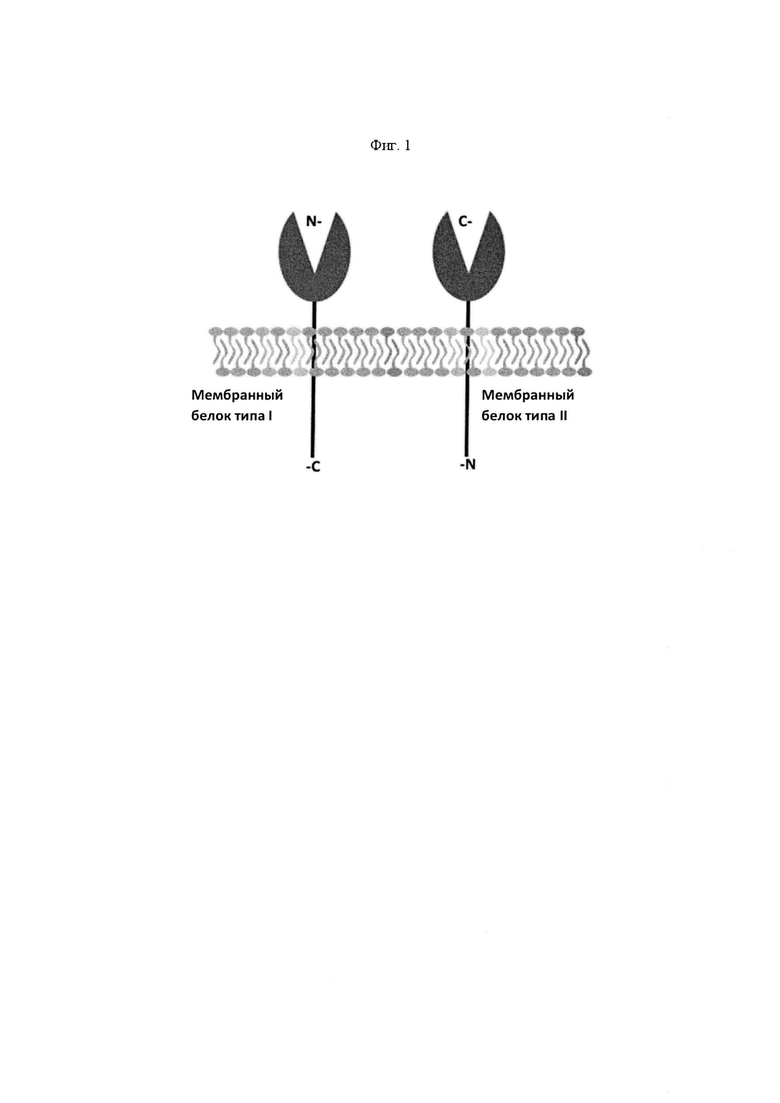
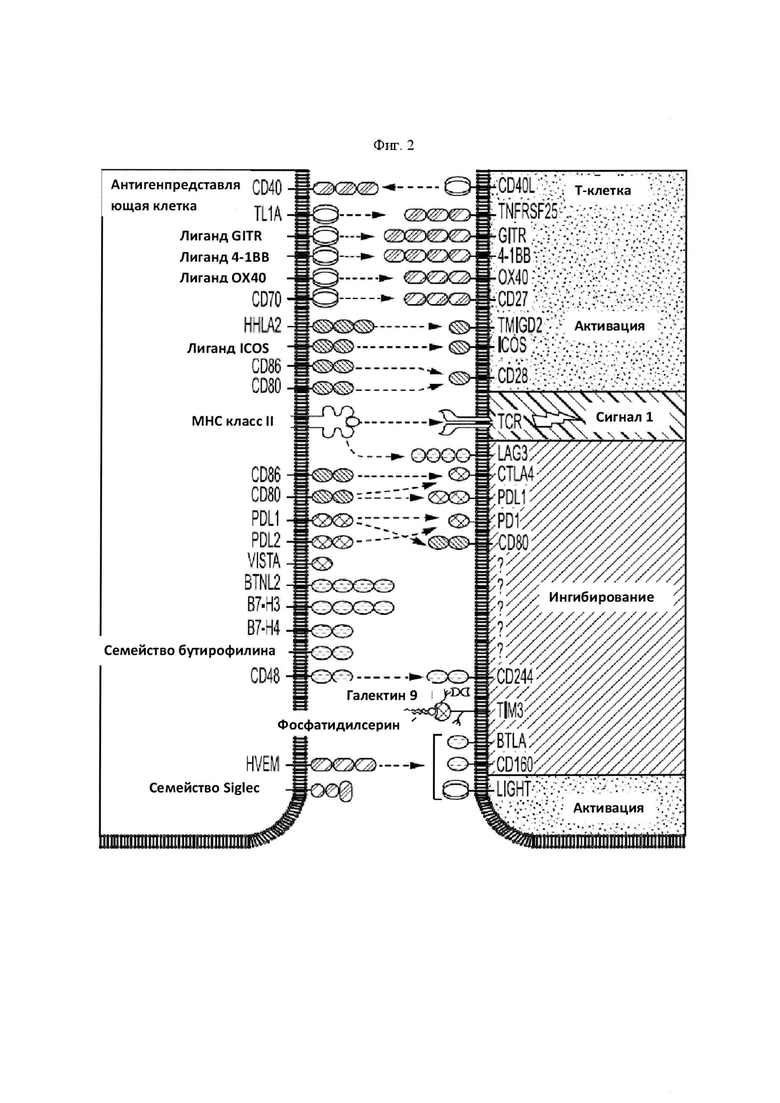
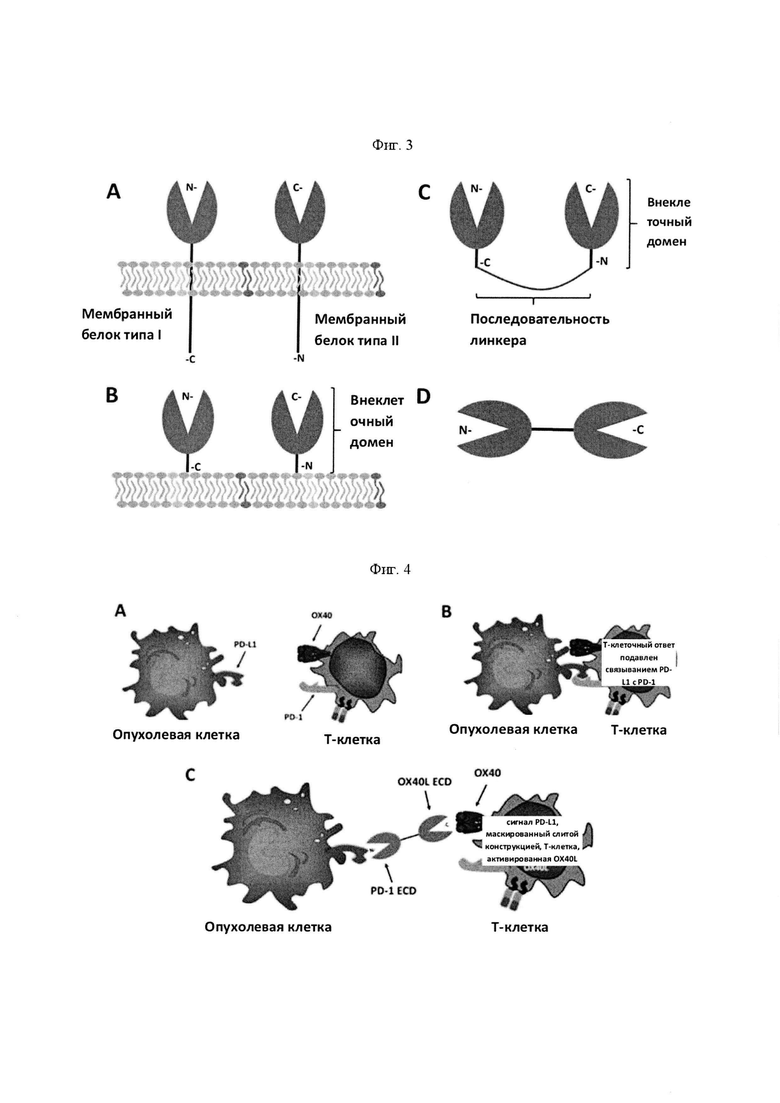
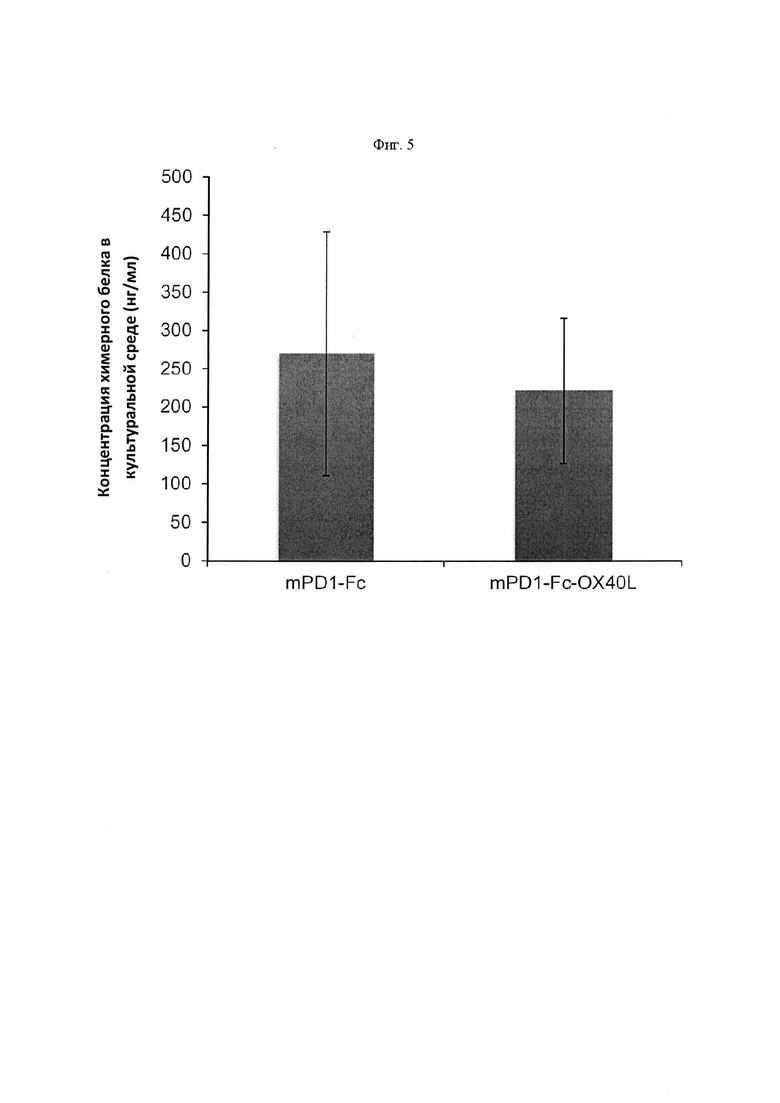
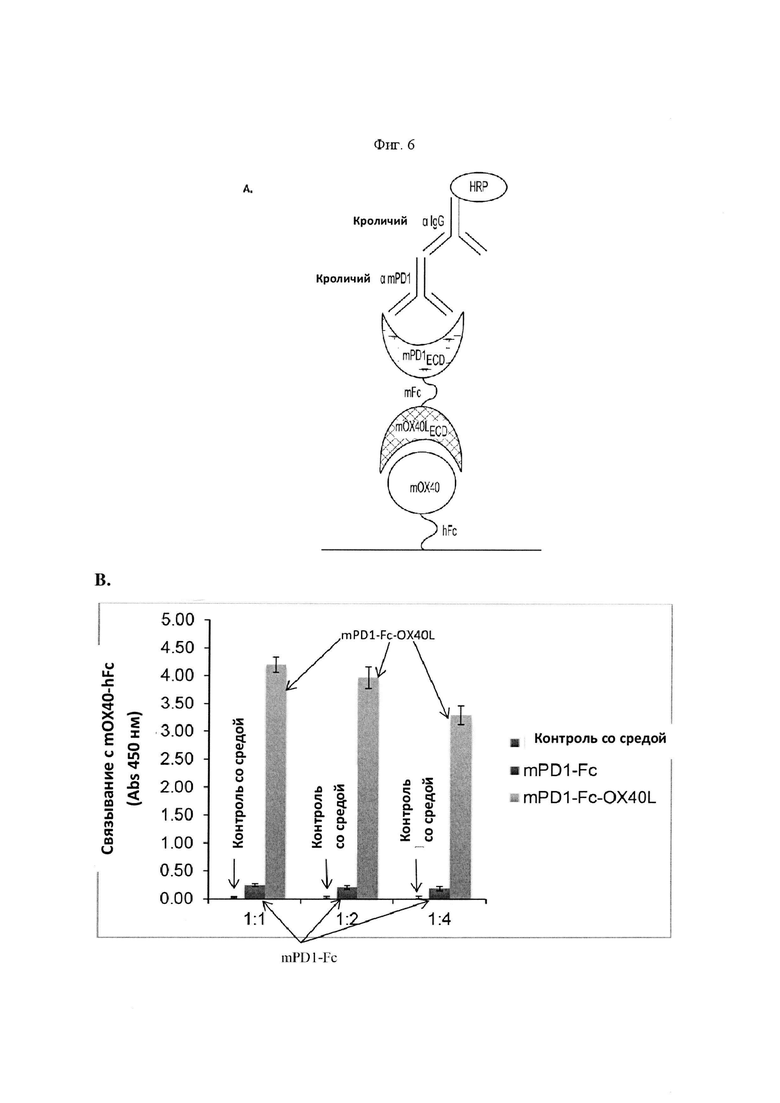

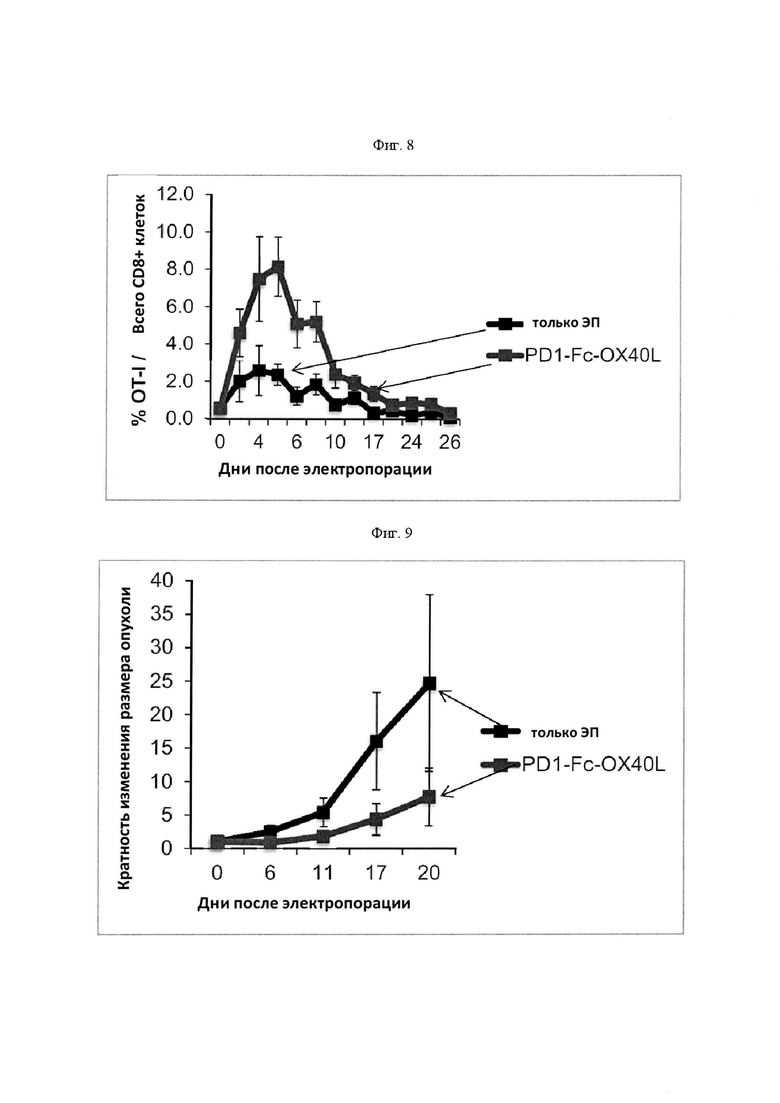
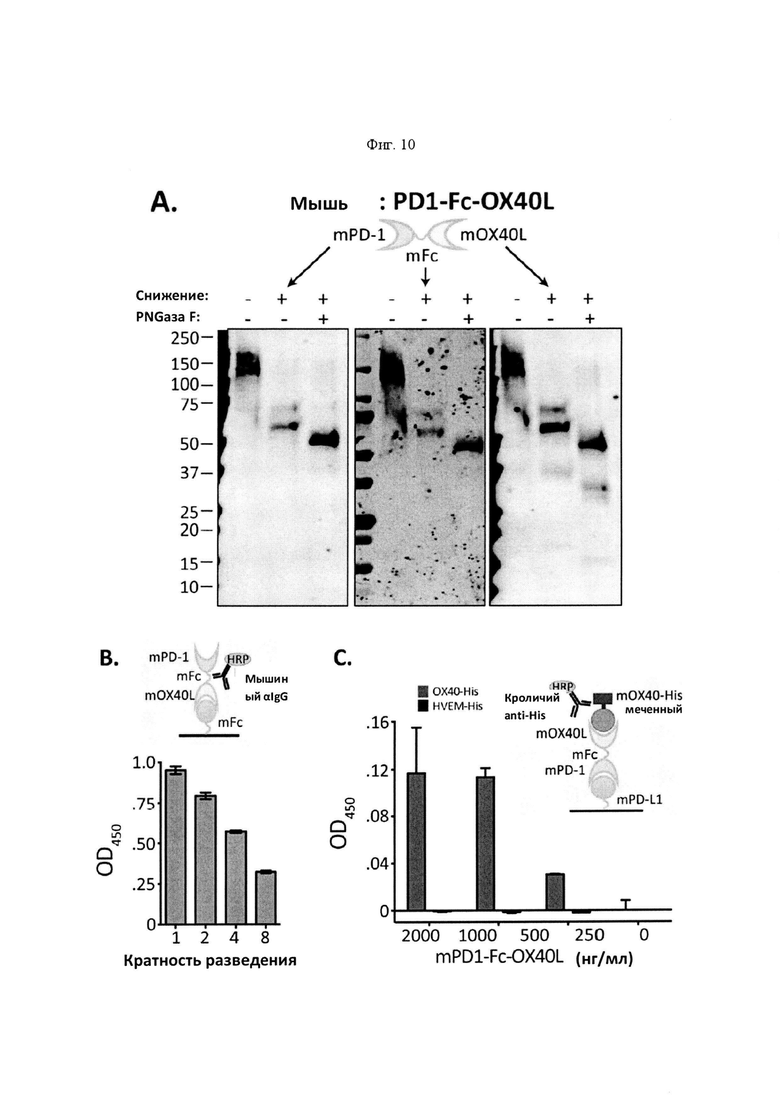
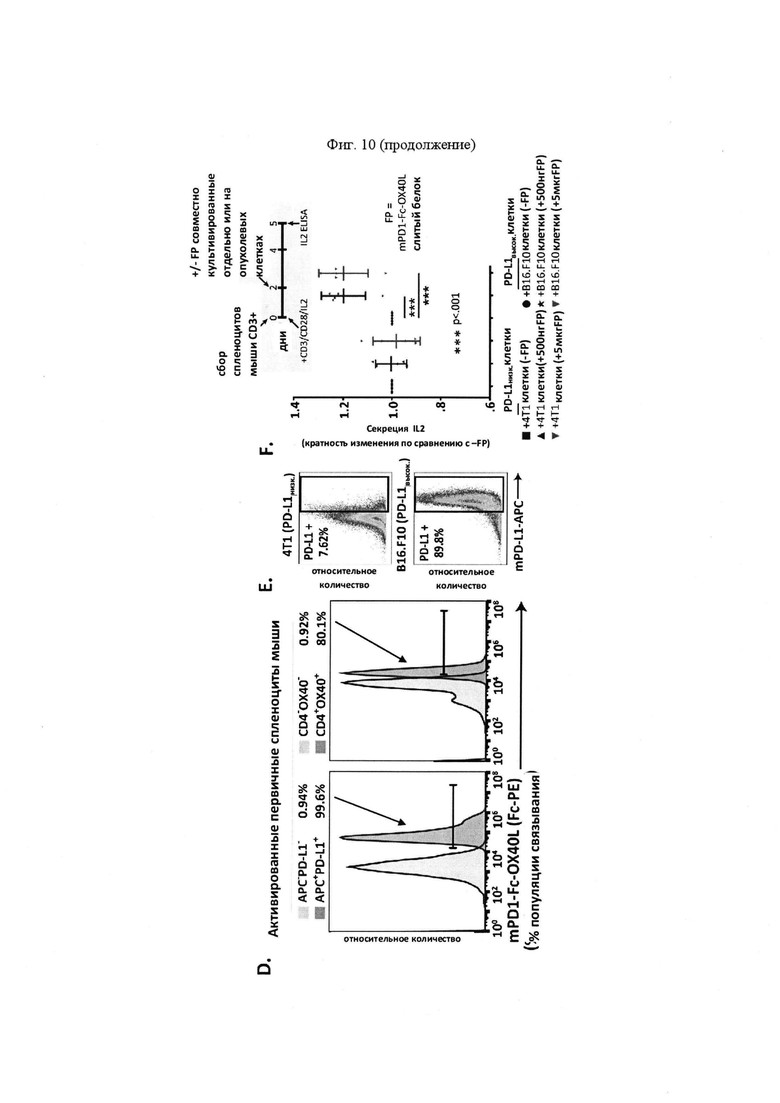
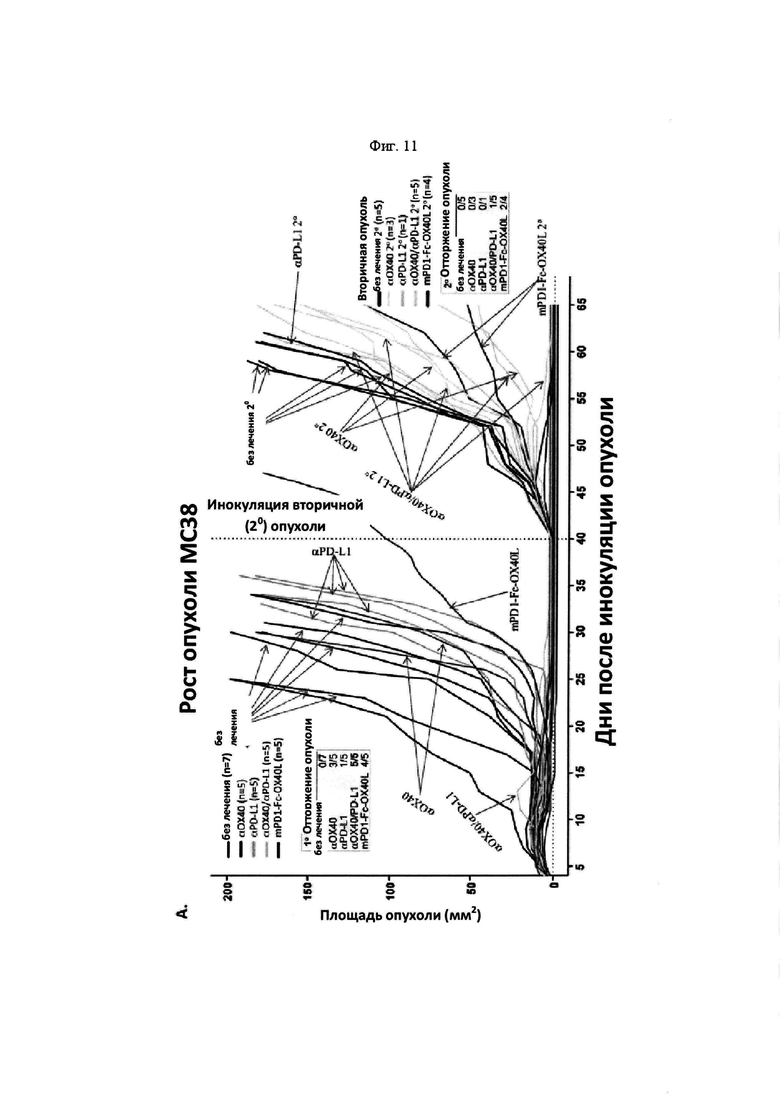
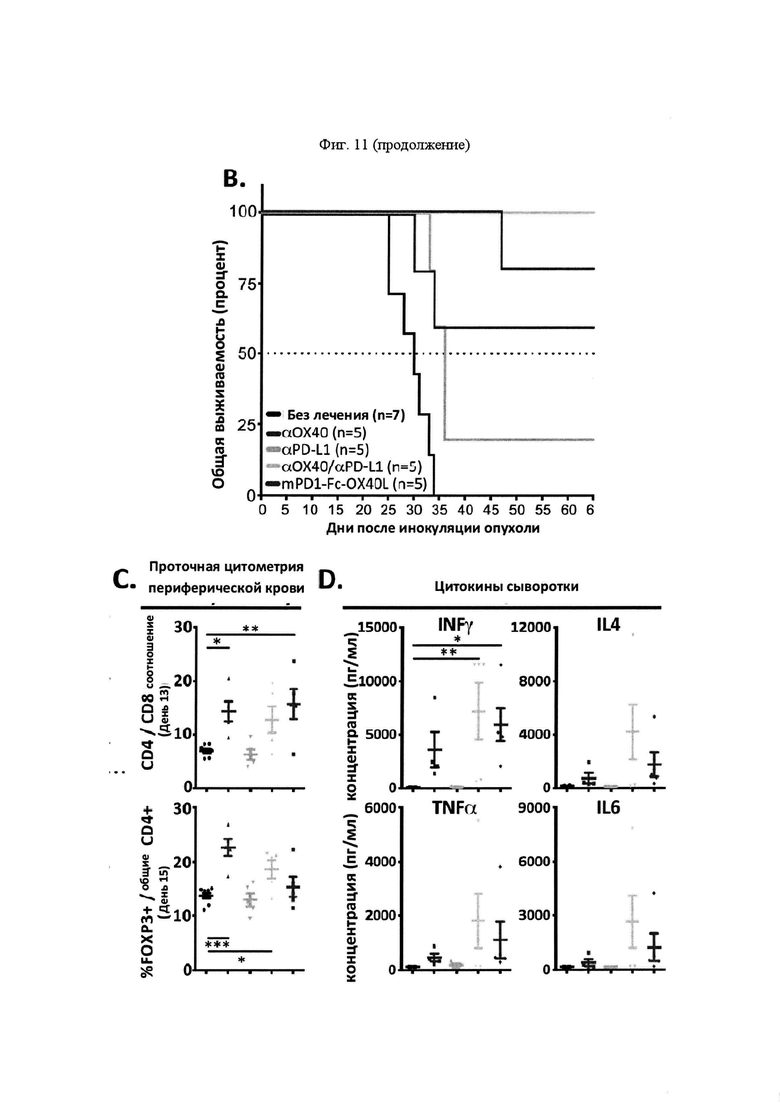
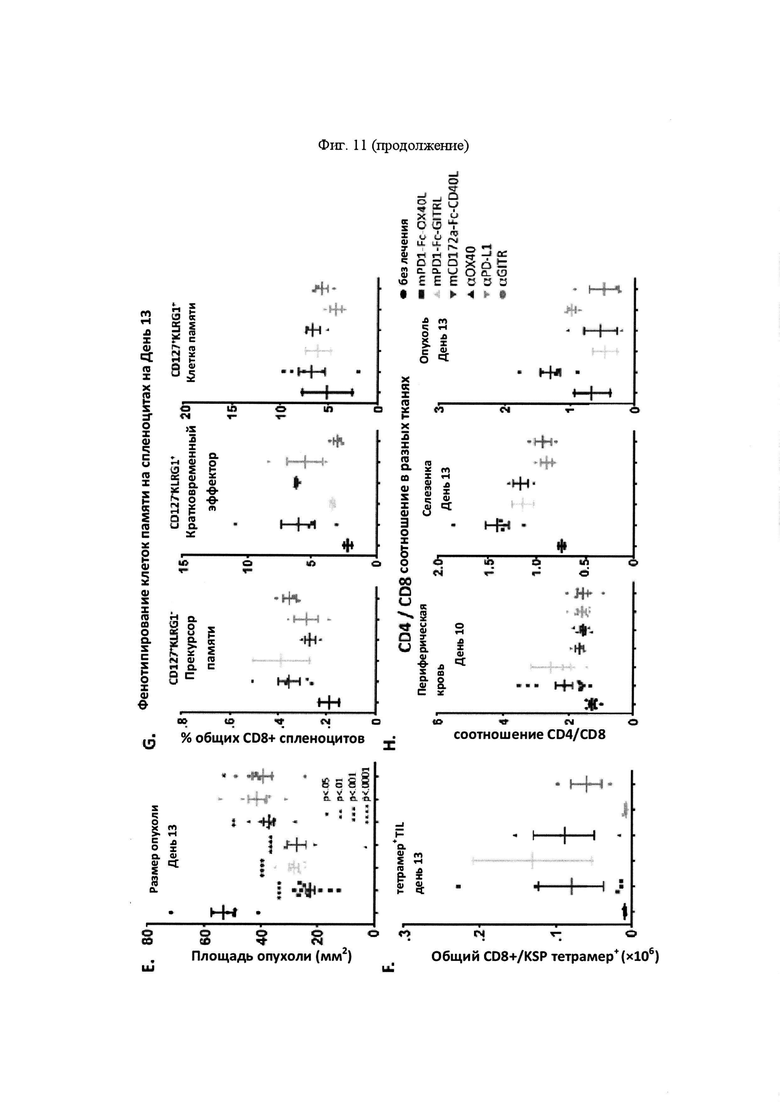
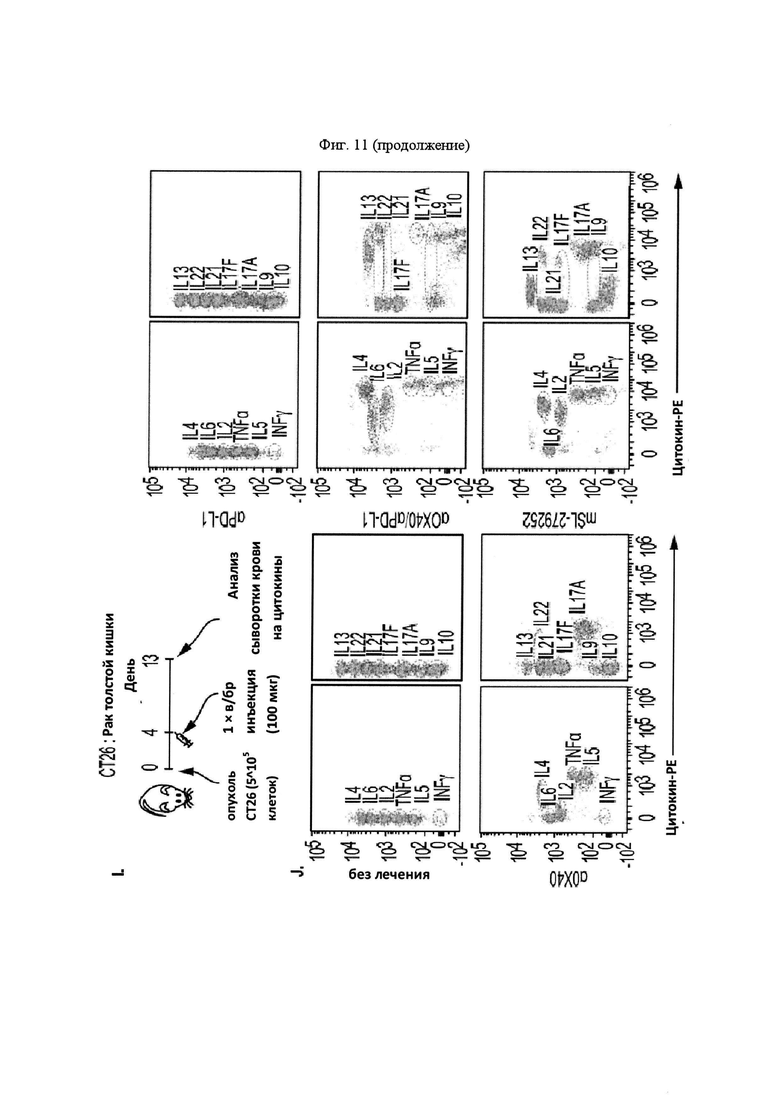
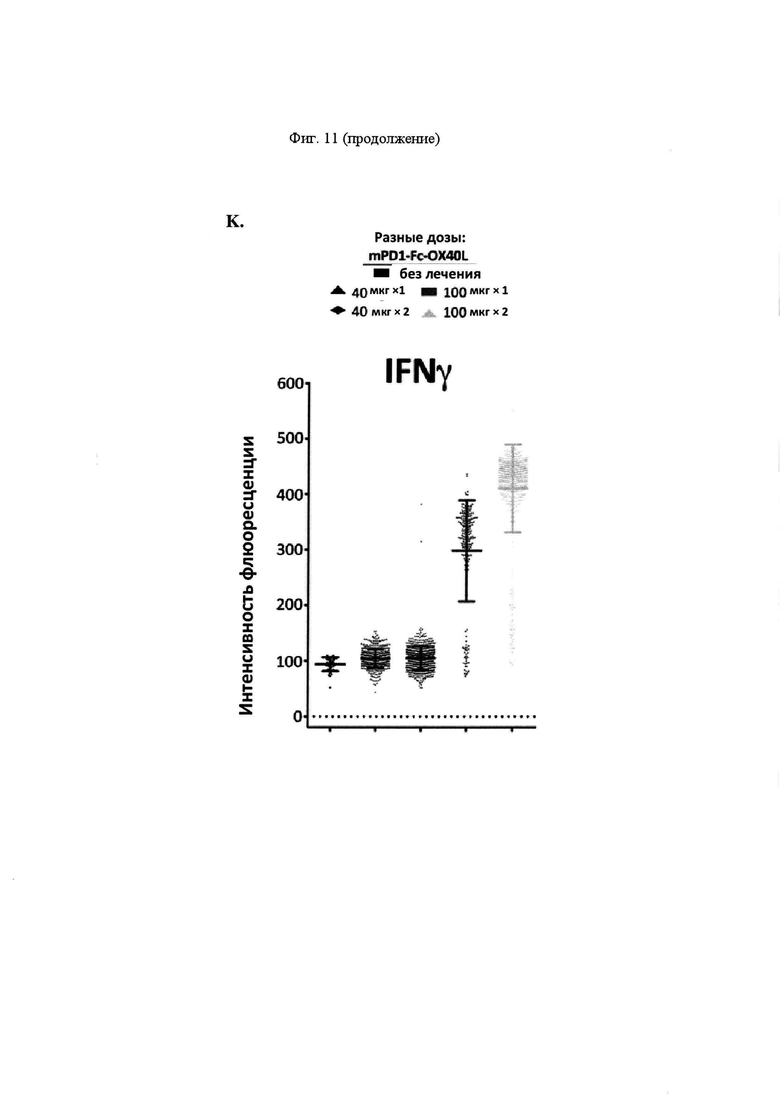


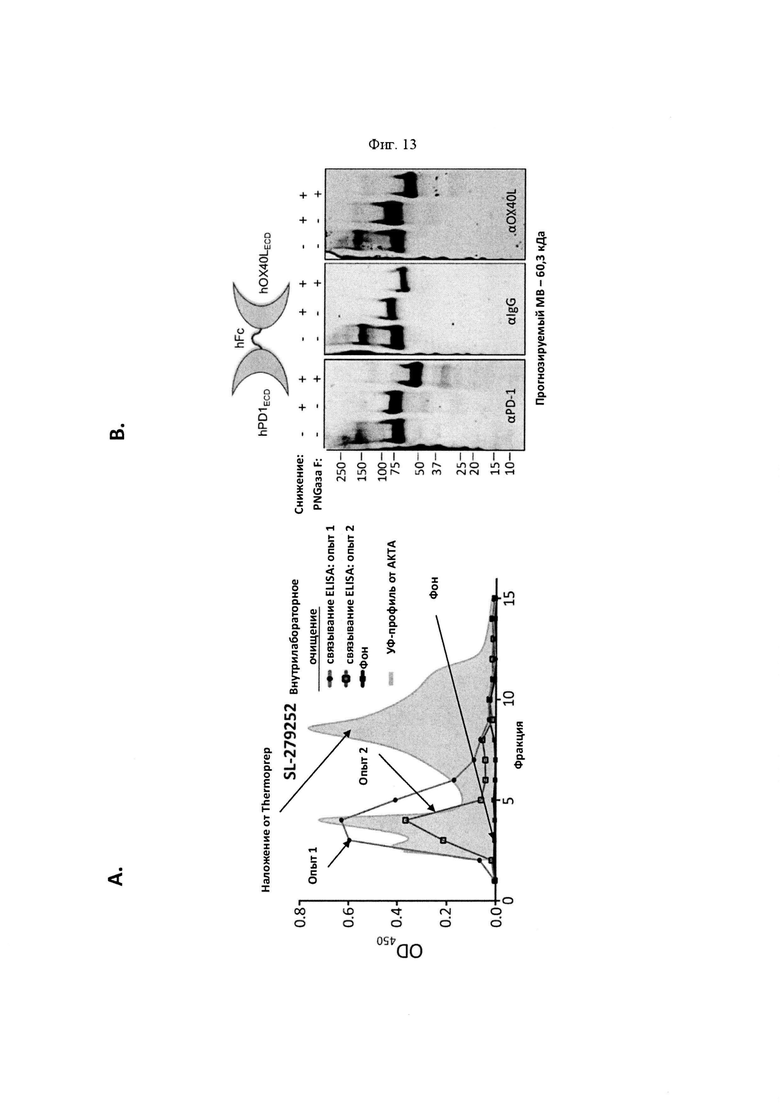
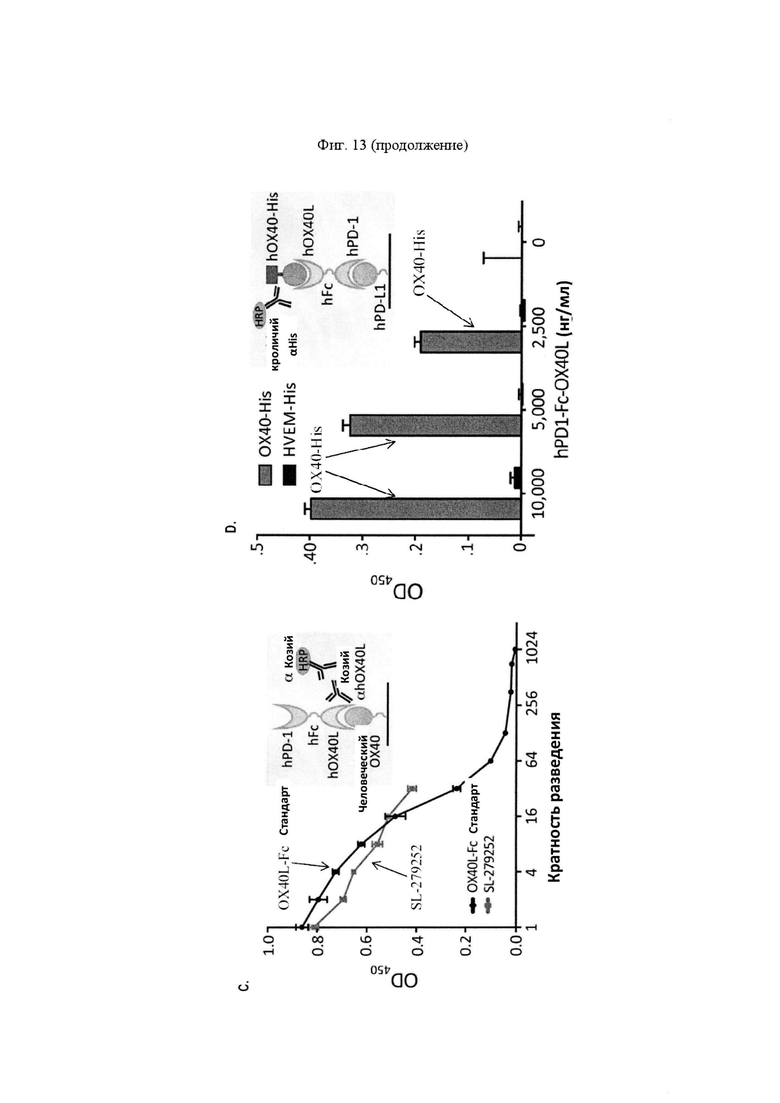
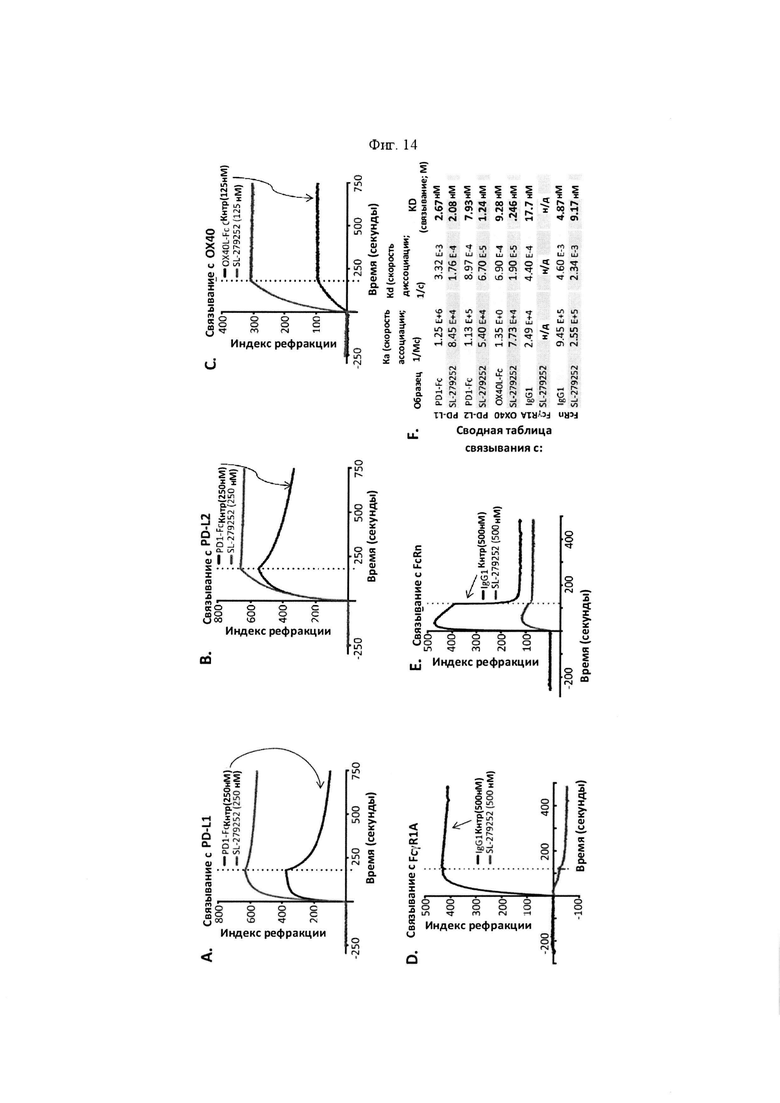
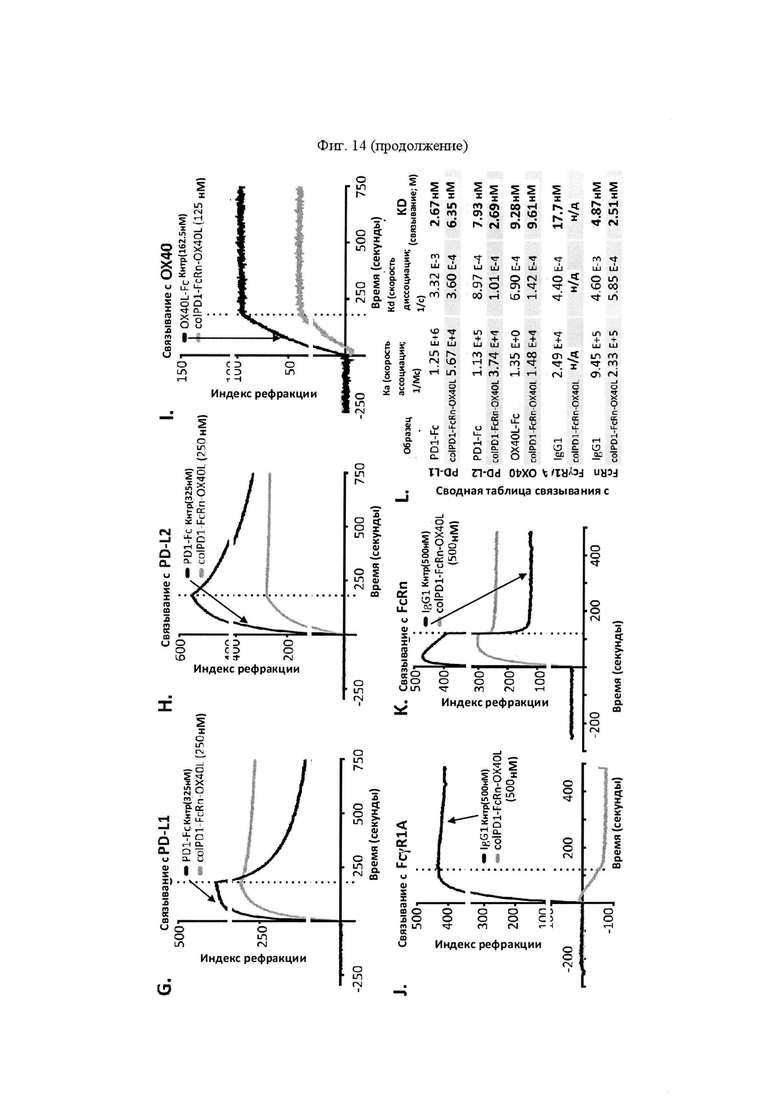
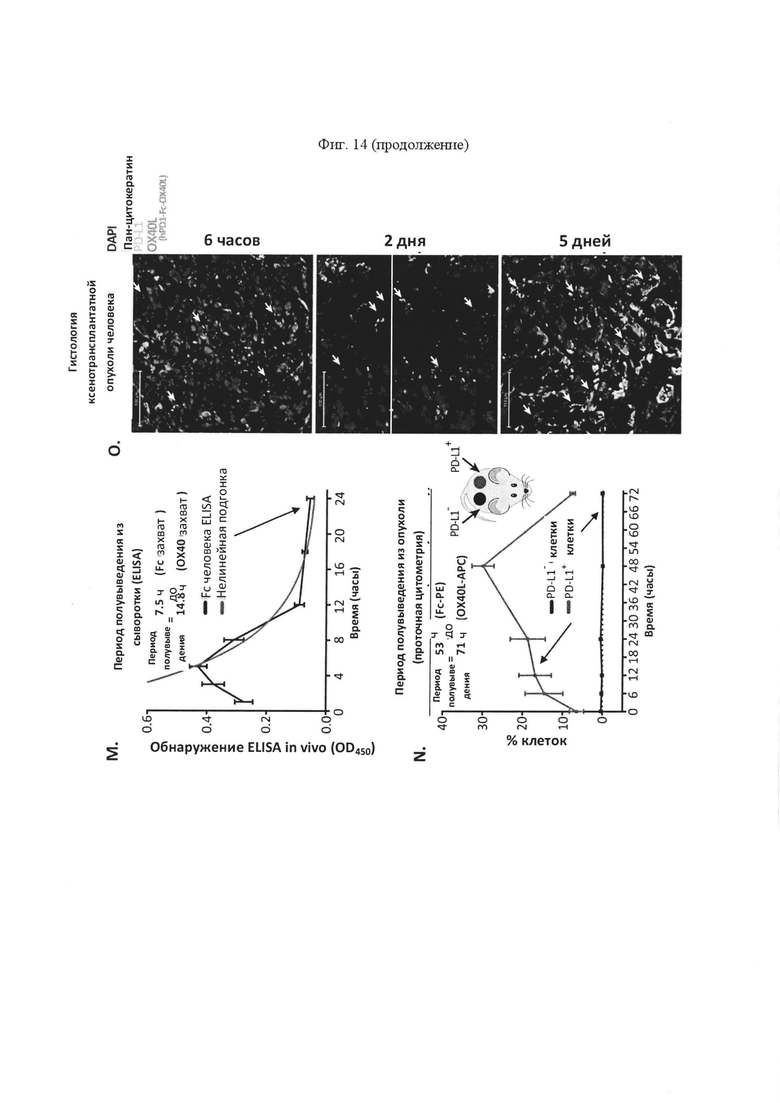



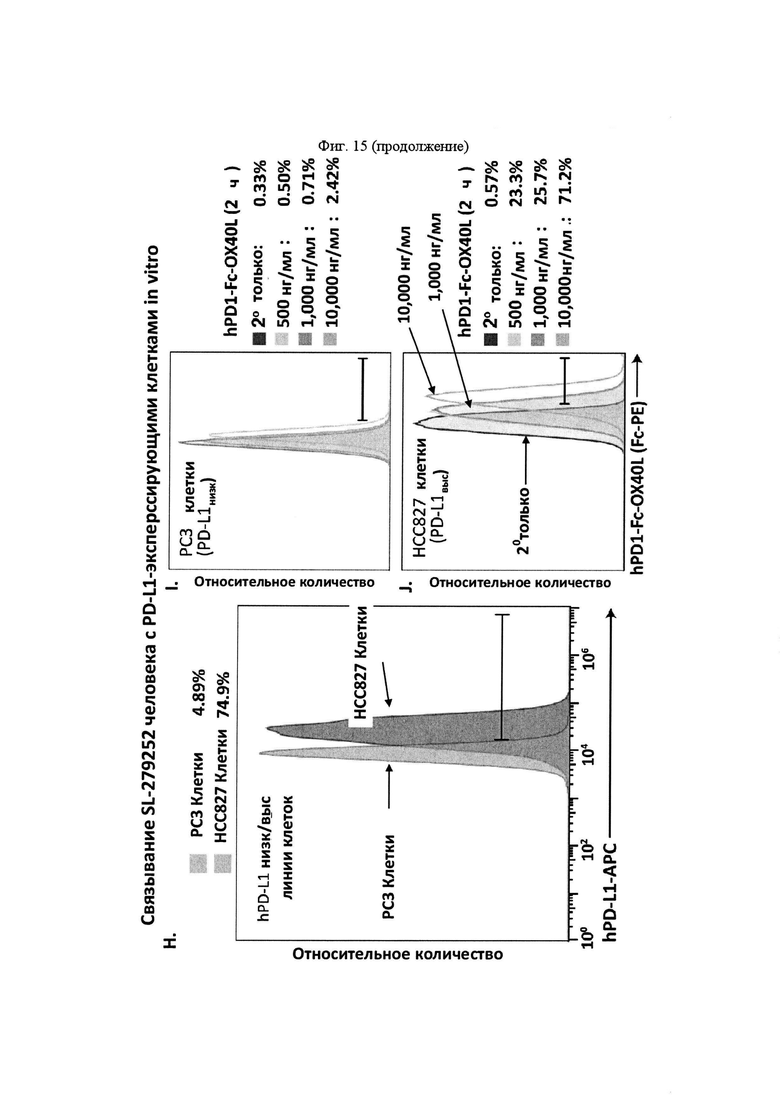
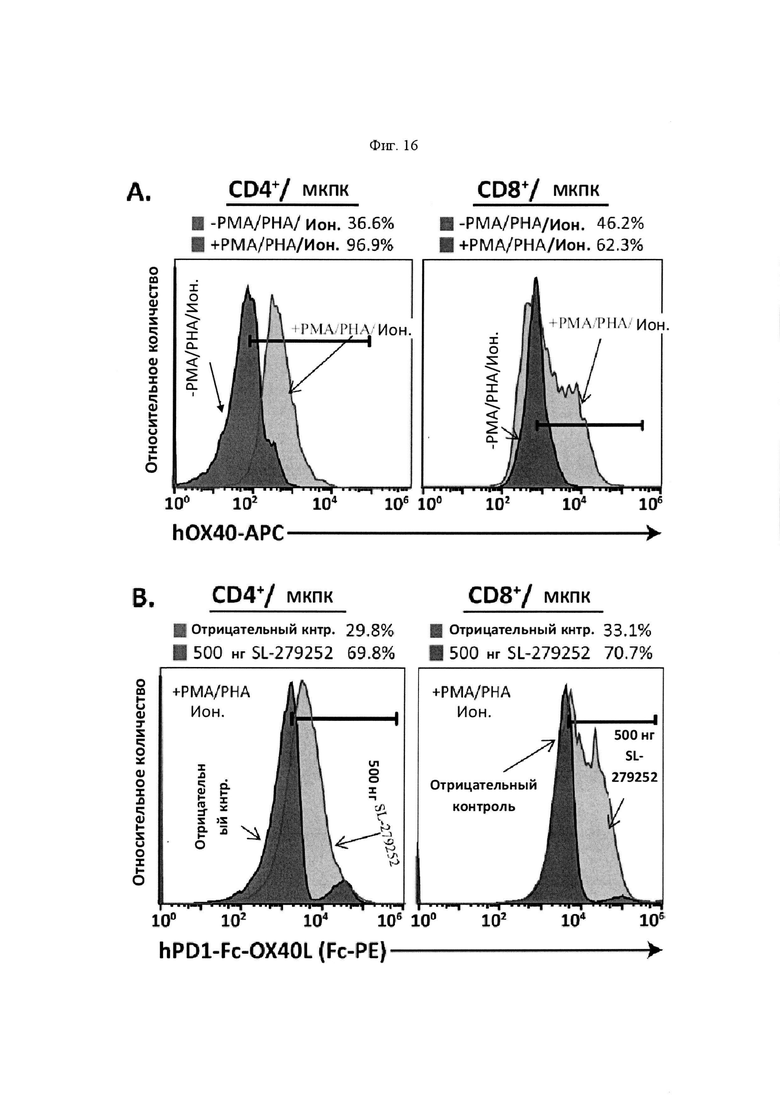
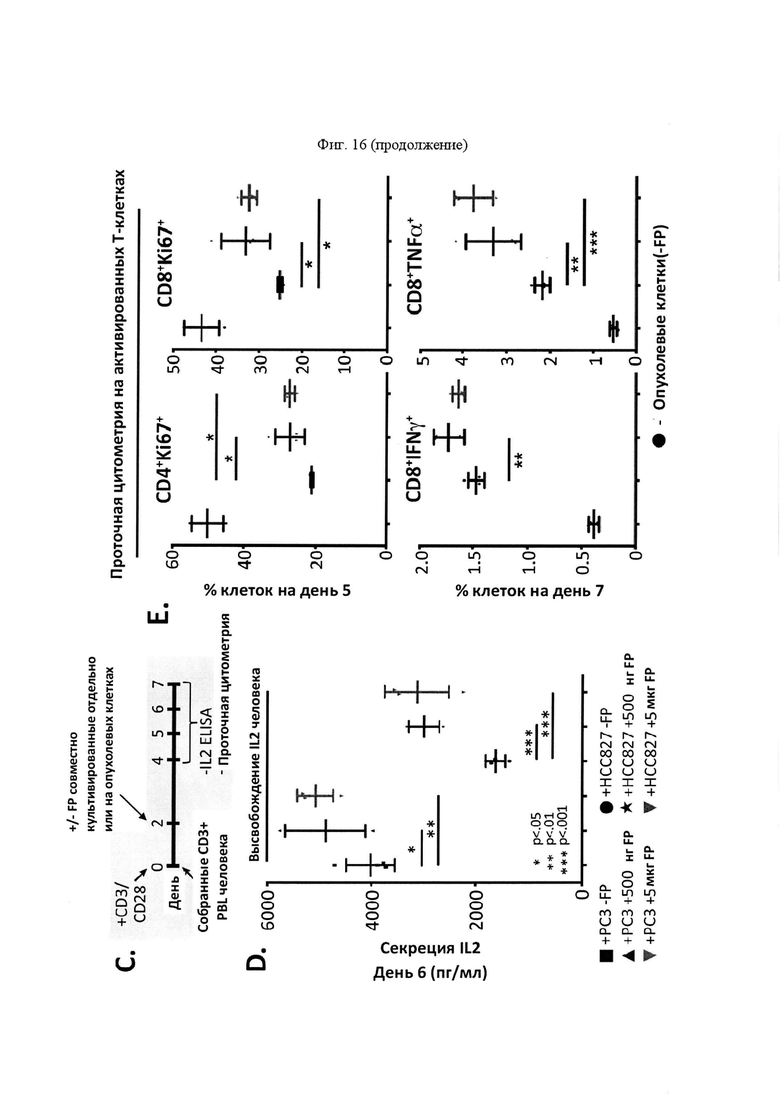
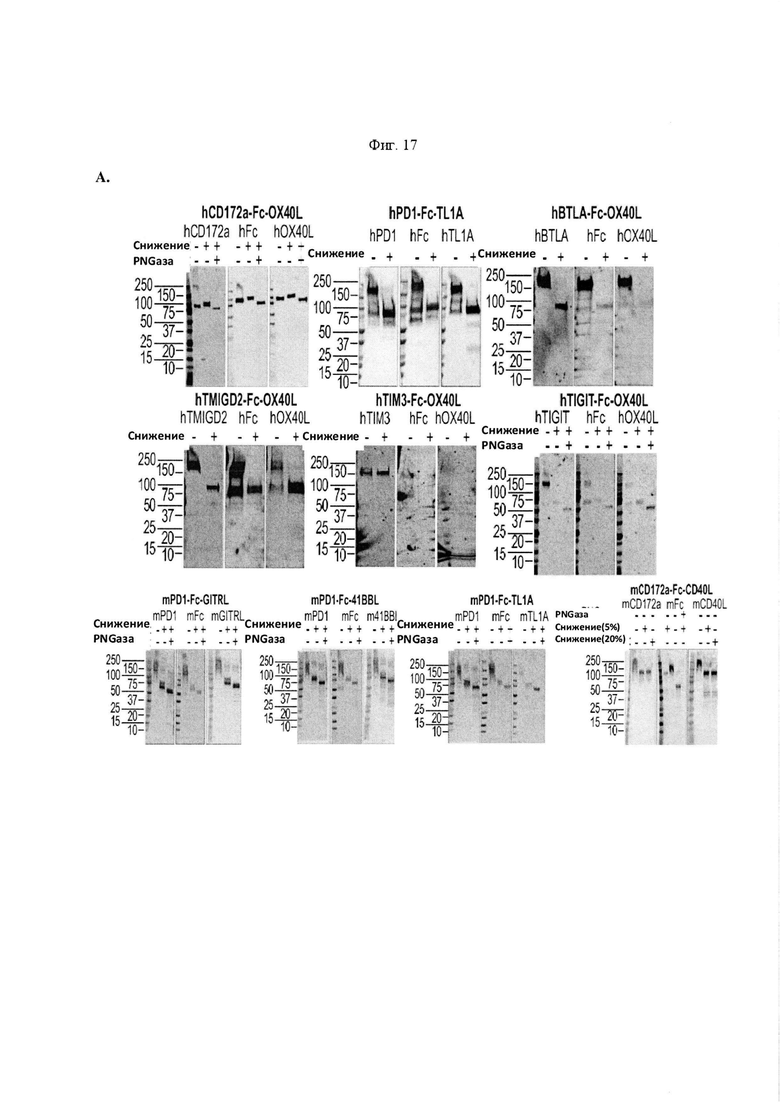


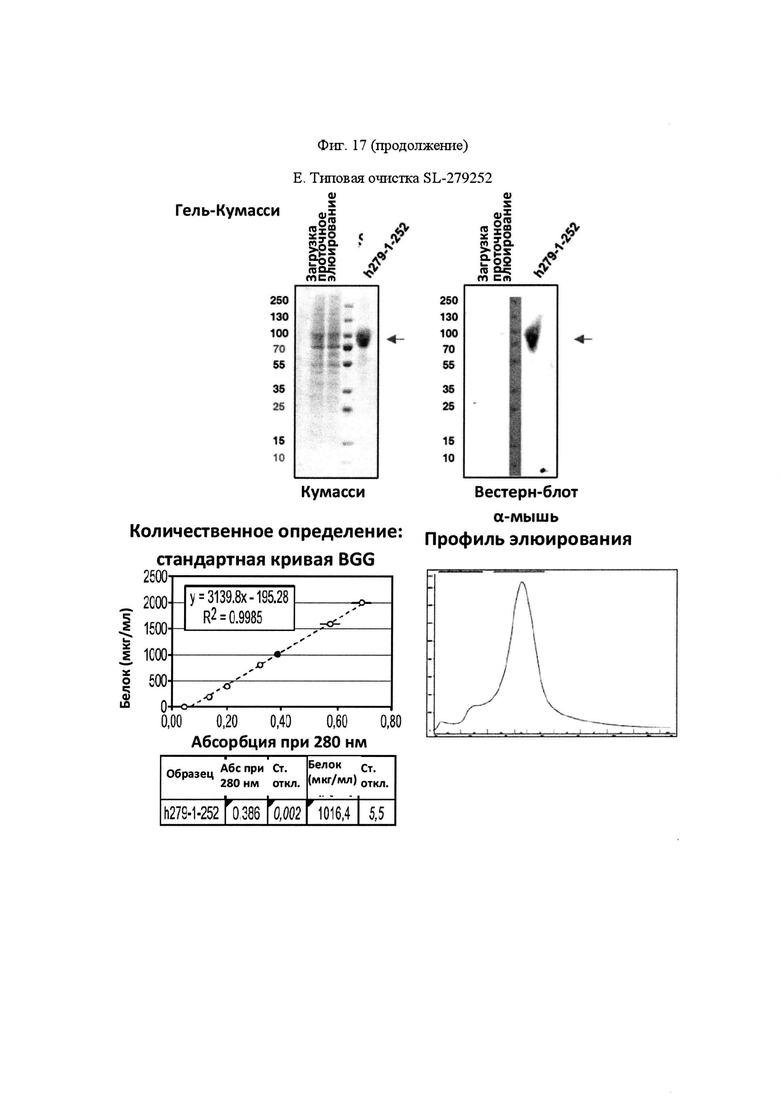
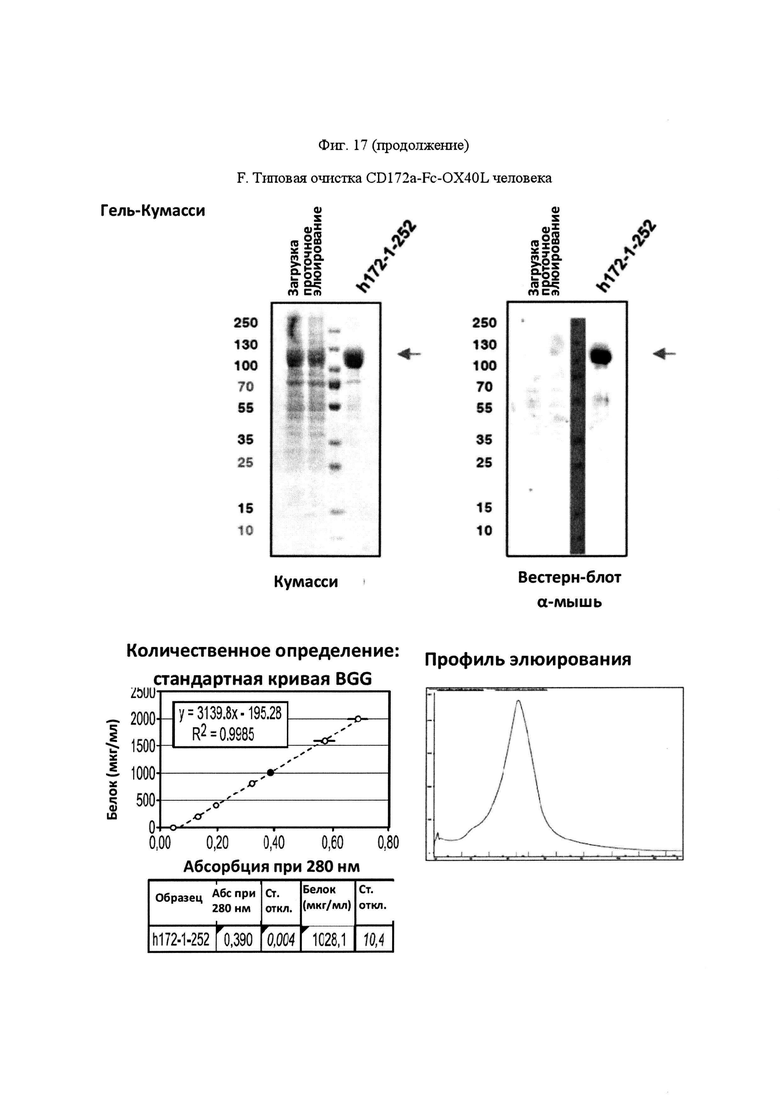

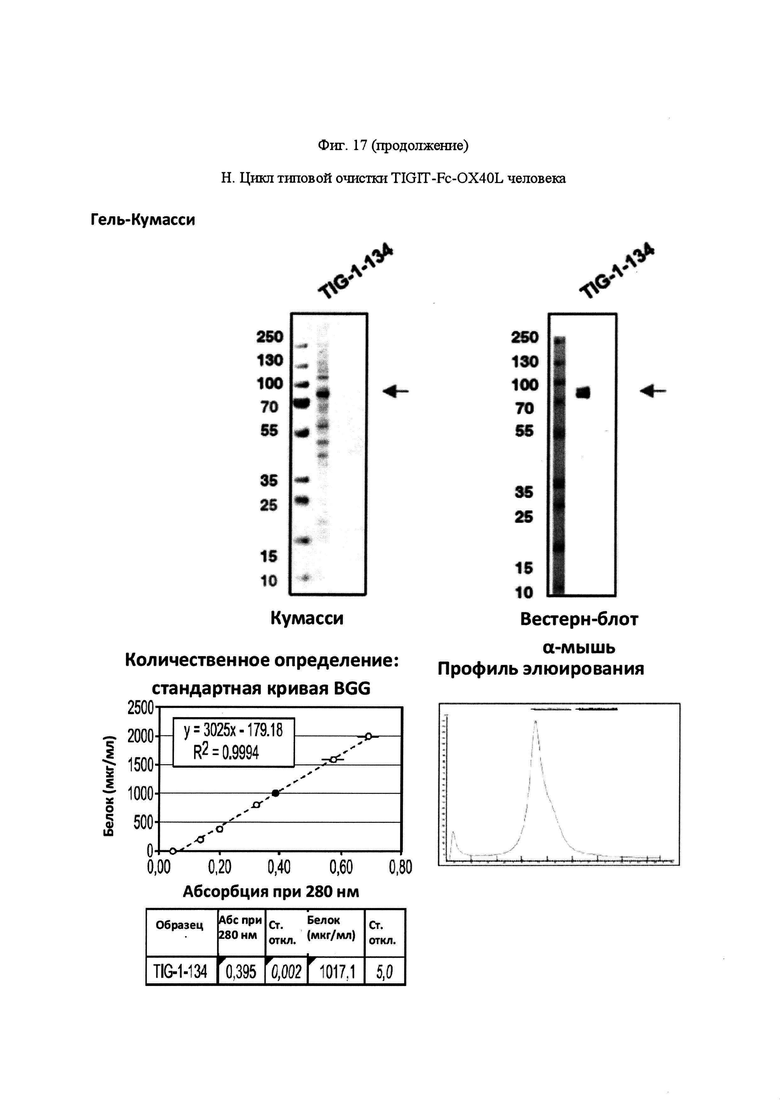
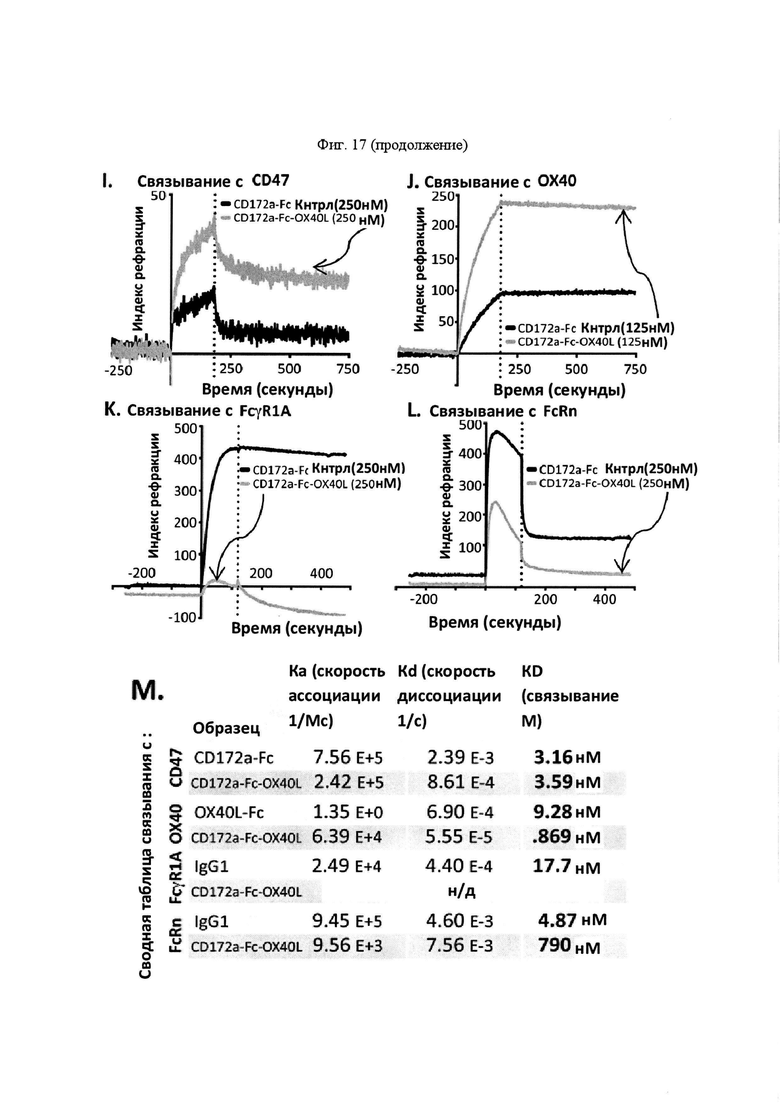
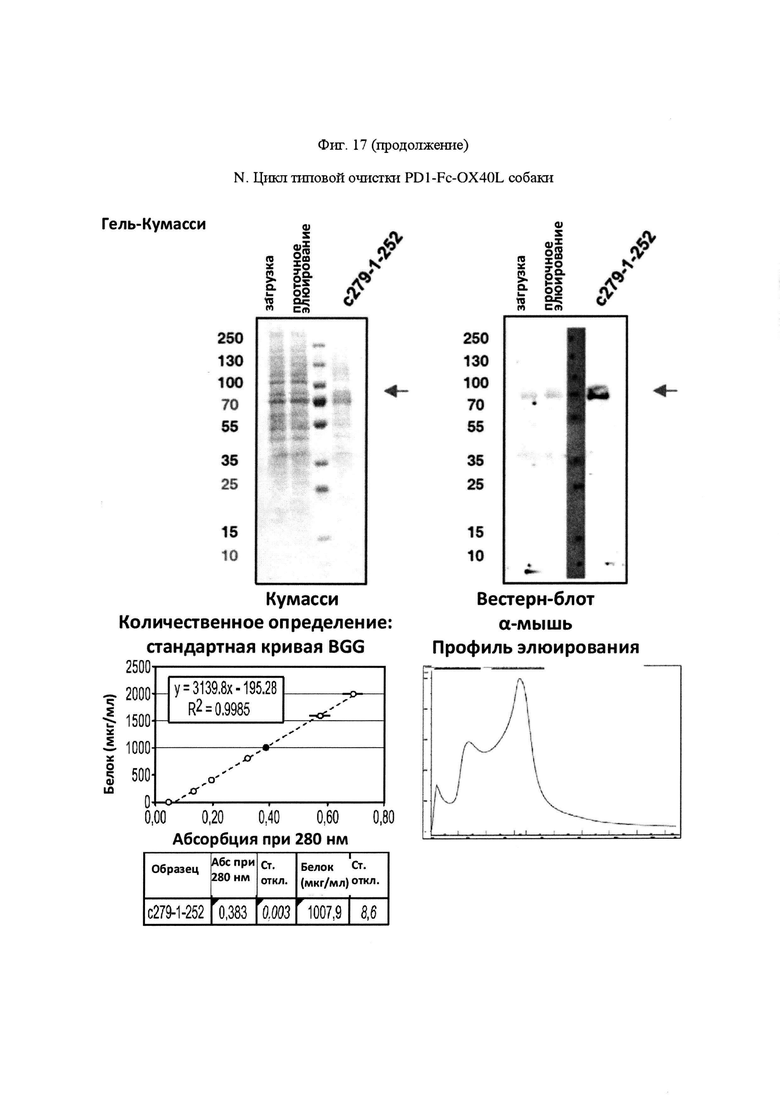
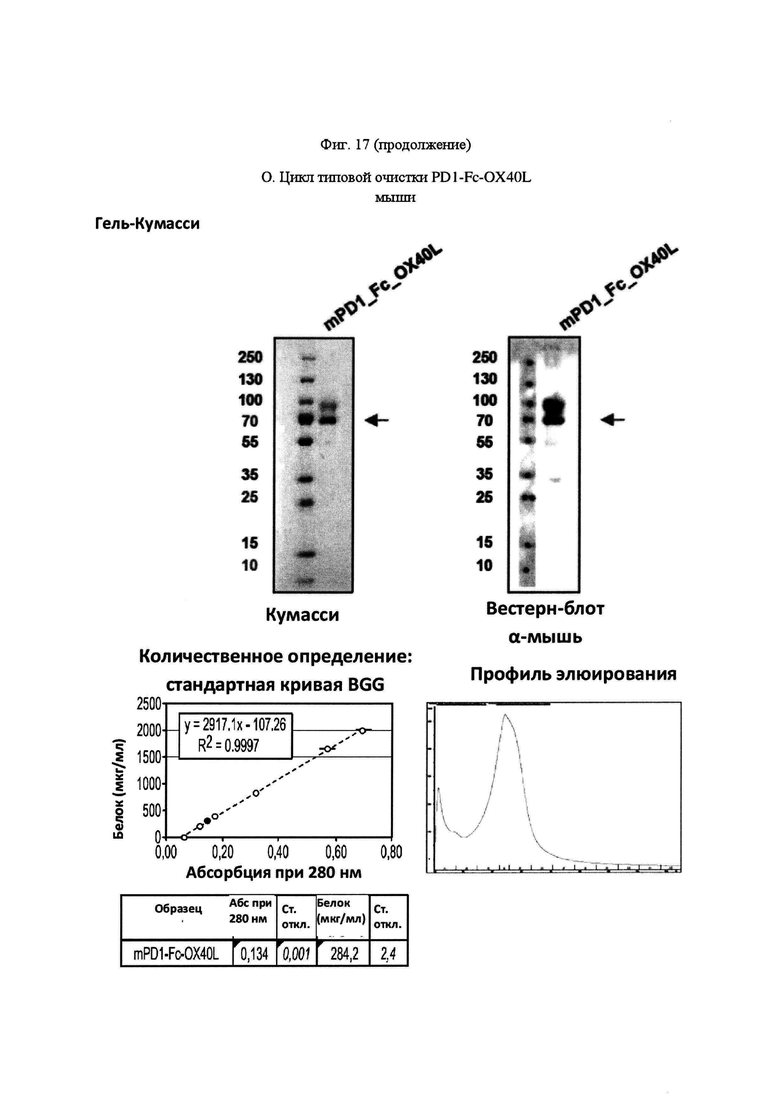
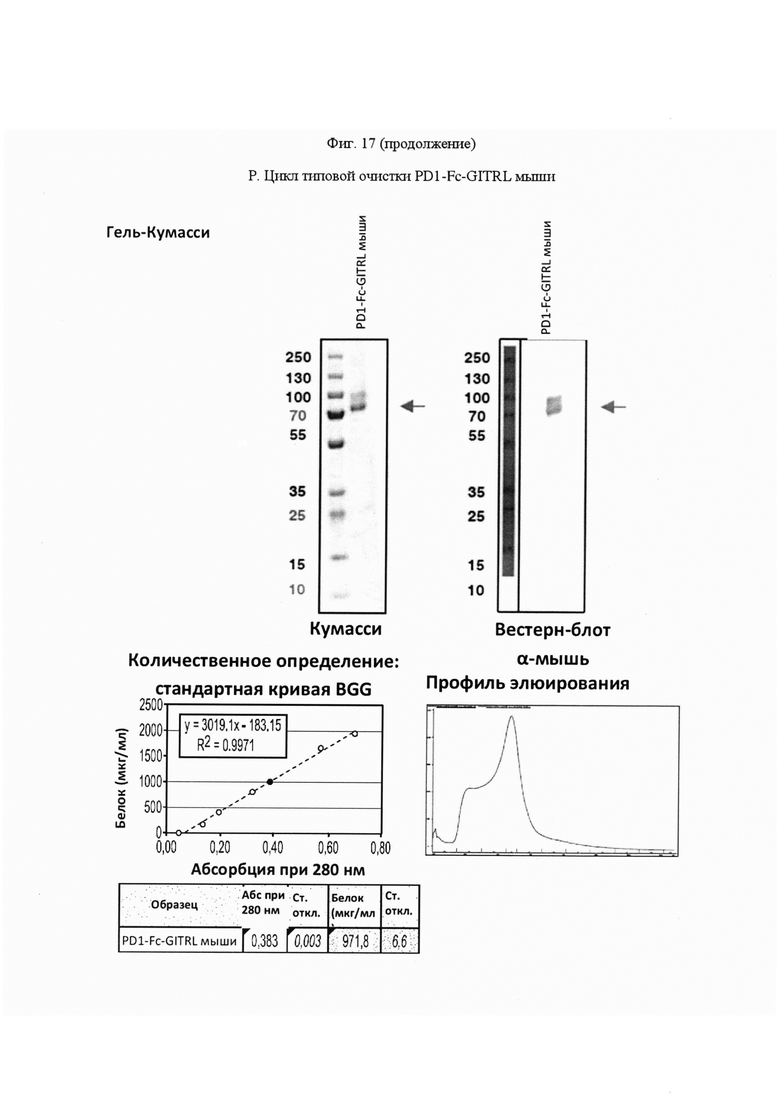
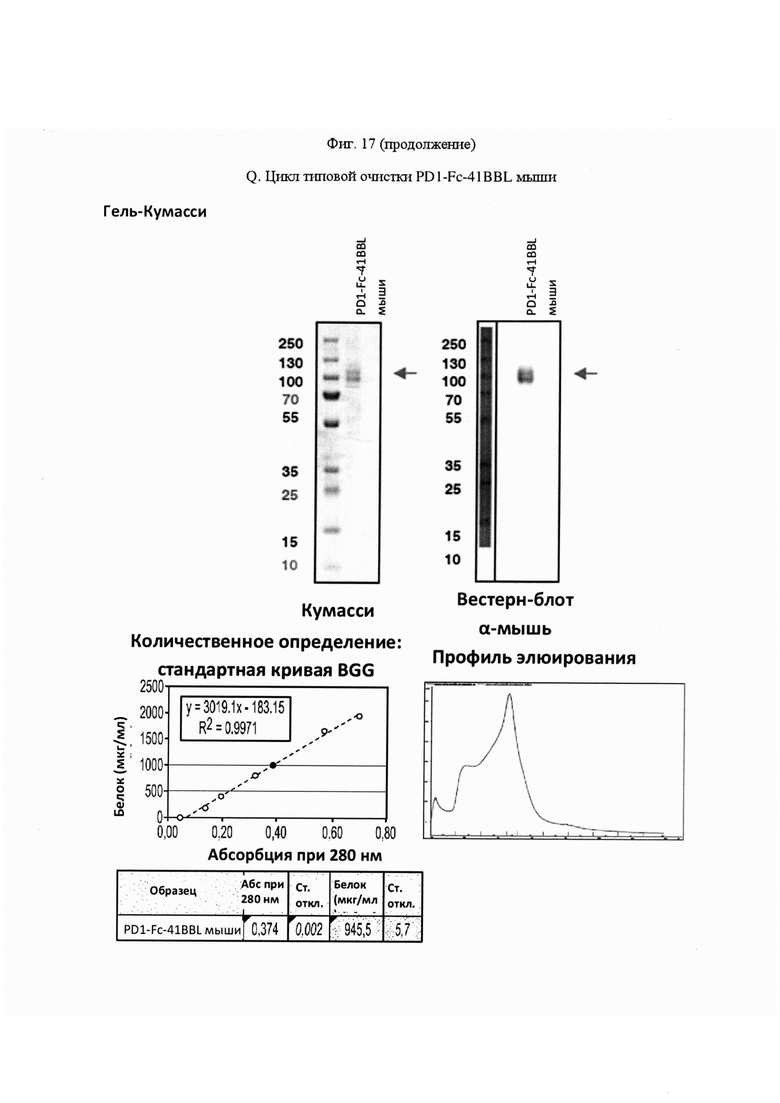
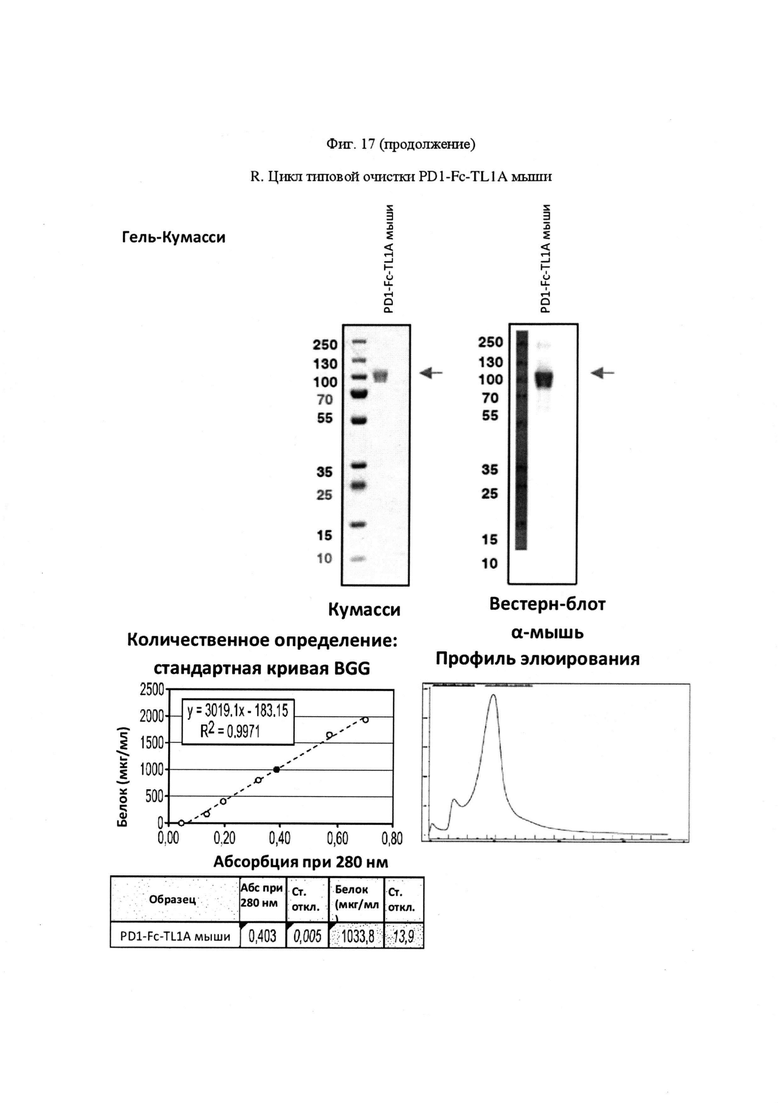
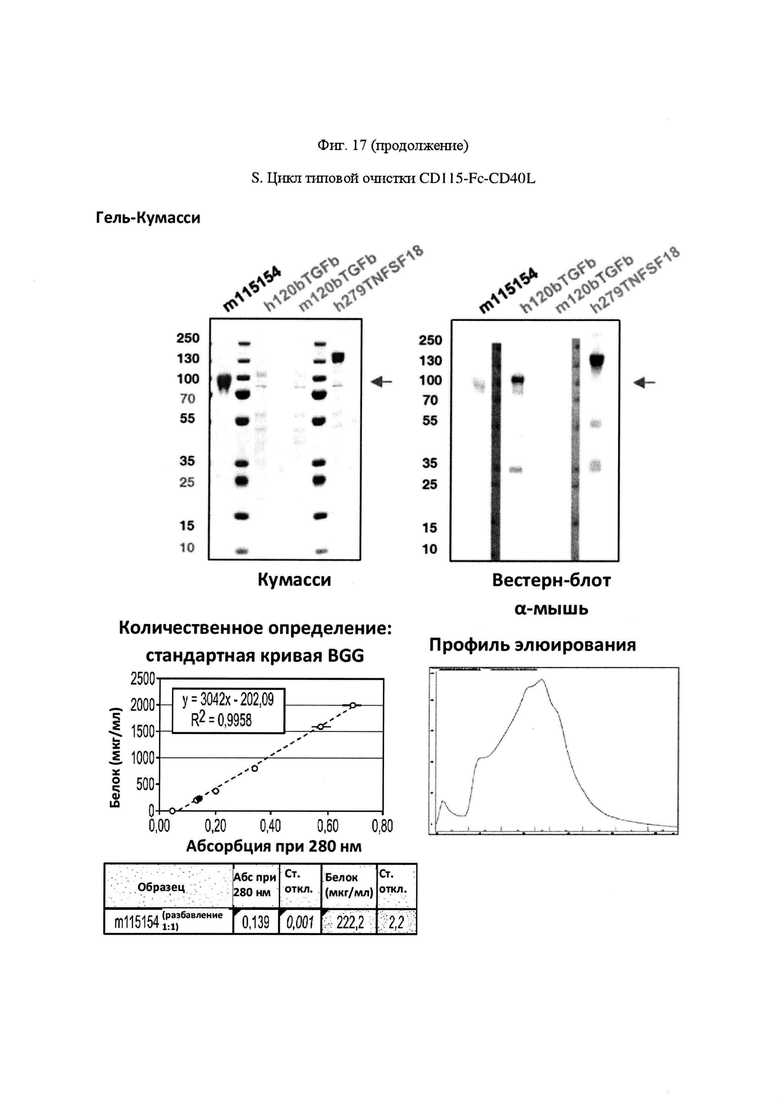
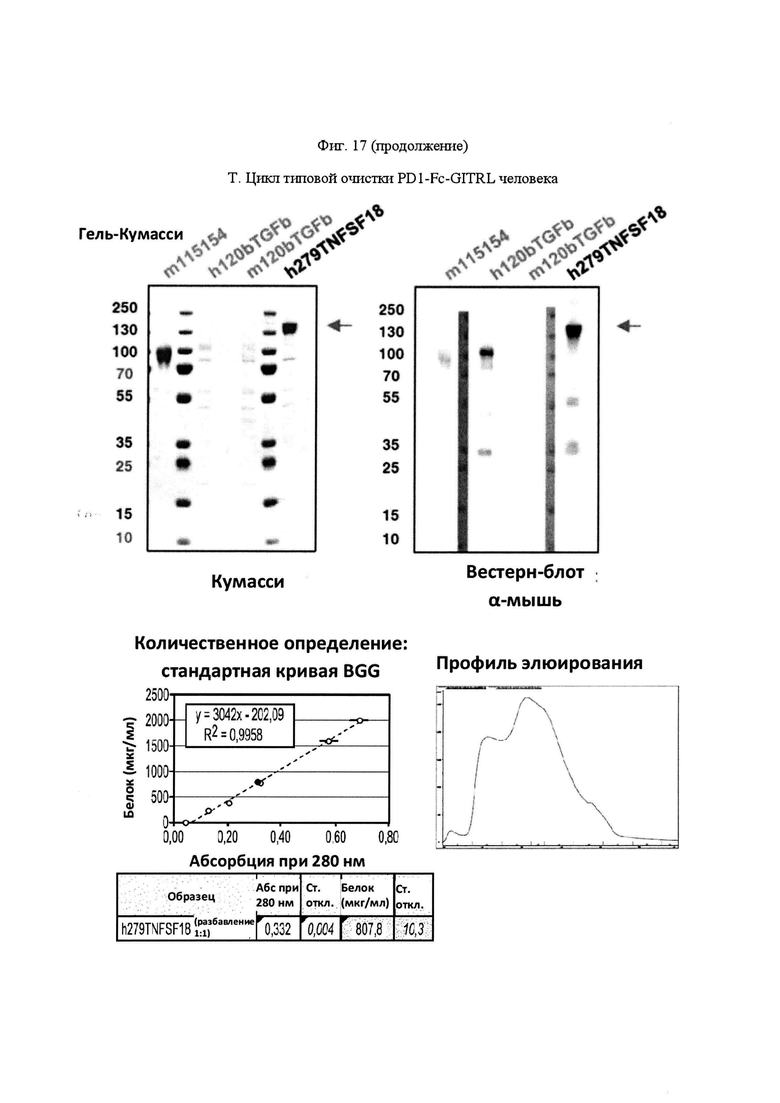
Изобретение относится к области биотехнологии, конкретно к гетерологичным химерным белкам, и может быть использовано в медицине для иммунотерапии рака. Сконструирован белок, включающий: (а) первый внеклеточный домен трансмембранного белка типа I на N-конце, (b) второй внеклеточный домен трансмембранного белка типа II на С-конце и (c) линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4, и соединяющий указанные первый и второй внеклеточные домены. При этом трансмембранный белок типа I и трансмембранный белок типа II представляют собой соответственно: PD-1 и лиганд OX40 (OX40L); PD-1 и лиганд 4-1BB (4-1BBL); PD-1 и лиганд GITR (GITRL); PD-1 и TL1A; TIM3 и OX40L; CD172a/SIRPα и OX40L; CD172a/SIRPα и лиганд CD40 (CD40L); TIGIT и OX40L; BTLA и OX40L; TMIGD2 и OX40L; или CD115(CSF1R) и CD40L. Изобретение обеспечивает эффективное модулирование иммунных сигналов при иммунотерапии рака. 17 н. и 25 з.п. ф-лы, 17 ил., 2 табл., 5 пр.
1. Гетерологичный химерный белок для иммунотерапии рака, содержащий:
(а) первый внеклеточный домен трансмембранного белка типа I на N-конце,
(b) второй внеклеточный домен трансмембранного белка типа II на С-конце; и
(c) линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4, и соединяющий указанные первый и второй внеклеточные домены;
при этом трансмембранный белок типа I и трансмембранный белок типа II представляют собой соответственно: PD-1 и лиганд OX40 (OX40L); PD-1 и лиганд 4-1BB (4-1BBL); PD-1 и лиганд GITR (GITRL); PD-1 и TL1A; TIM3 и OX40L; CD172a/SIRPα и OX40L; CD172a/SIRPα и лиганд CD40 (CD40L); TIGIT и OX40L; BTLA и OX40L; TMIGD2 и OX40L; или CD115(CSF1R) и CD40L.
2. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок содержит (i) внеклеточный домен PD-1 и (ii) внеклеточный домен OX40L.
3. Гетерологичный химерный белок по п. 2, отличающийся тем, что указанный химерный белок содержит аминокислотную последовательность, имеющую по меньшей мере 98% идентичности с последовательностью SEQ ID NO: 2.
4. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок содержит (i) внеклеточный домен PD-1 и (ii) внеклеточный домен GITRL.
5. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок содержит (i) внеклеточный домен BTLA и (ii) внеклеточный домен OX40L.
6. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок содержит (i) внеклеточный домен TIGIT и (ii) внеклеточный домен OX40L.
7. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок содержит (i) внеклеточный домен TIM3 и (ii) внеклеточный домен OX40L.
8. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок содержит (i) внеклеточный домен CD172a/SIPRα и (ii) внеклеточный домен CD40L.
9. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок содержит (i) внеклеточный домен PD-1 и (ii) внеклеточный домен 4-1BBL.
10. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок содержит (i) внеклеточный домен PD-1 и (ii) внеклеточный домен TL1A.
11. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок содержит (i) внеклеточный домен CD172a/SIPRα и (ii) внеклеточный домен OX40L.
12. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок содержит (i) внеклеточный домен TMIGD2 и (ii) внеклеточный домен OX40L.
13. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок содержит (i) внеклеточный домен CD115(CSF1R) и (ii) внеклеточный домен CD40L.
14. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок представляет собой рекомбинантный слитый белок.
15. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок способен увеличивать соотношение эффекторных Т-клеток к регуляторным Т-клеткам.
16. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок способен увеличивать и/или предотвращать уменьшение субпопуляции цитотоксических Т-клеток; эффекторных T-клеток памяти; центральных T-клеток памяти; CD8+ стволовых клеток эффекторных клеток памяти; TH1 эффекторных Т-клеток; TH2 эффекторных Т-клеток; TH9 эффекторных Т-клеток; TH17 эффекторных Т-клеток; и/или эффекторных Т-клеток, секретирующих IL-2, IL-4 и/или IFN-γ.
17. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок вызывает цитокиновый ответ в периферической крови животного, который можно применять в качестве метода определения подходящей дозы указанного химерного белка.
18. Гетерологичный химерный белок по п. 1, отличающийся тем, что линкер содержит домен Fc шарнир-CH2-CH3, происходящий от IgG1, необязательно IgG1 человека.
19. Гетерологичный химерный белок по п. 1, отличающийся тем, что линкер содержит домен Fc шарнир-CH2-CH3, происходящий от IgG4, необязательно IgG4 человека.
20. Гетерологичный химерный белок по п. 1, отличающийся тем, что указанный химерный белок можно продуцировать в клетке-хозяине млекопитающего в виде секретируемой и полностью функциональной одиночной полипептидной цепи.
21. Фармацевтическая композиция для иммунотерапии рака, содержащая эффективное количество гетерологичного химерного белка по любому из вышеуказанных пунктов и фармацевтически приемлемое вспомогательное вещество.
22. Экспрессионный вектор, содержащий нуклеиновую кислоту, кодирующую гетерологичный химерный белок по любому из пп. 1-20.
23. Экспрессионный вектор по п. 22, отличающийся тем, что указанный экспрессионный вектор представляет собой экспрессионный вектор млекопитающих.
24. Экспрессионный вектор по п. 23, отличающийся тем, что указанный экспрессионный вектор содержит ДНК или РНК.
25. Клетка-хозяин для экспресии гетерологичного химерного белка по любому из пп. 1-20, содержащая экспрессионный вектор по любому из пп. 22-24.
26. Вирусный вектор для экспрессии гетерологичного химерного белка по любому из пп. 1-20, содержащий последовательность нуклеиновой кислоты, кодирующую гетерологичный химерный белок по любому из пп. 1-20.
27. Способ лечения рака, включающий введение эффективного количества фармацевтической композиции по п. 21 субъекту, нуждающемуся в этом.
28. Способ иммунотерапии рака, включающий введение эффективного количества фармацевтической композиции по п. 21 субъекту, нуждающемуся в этом.
29. Способ модулирования иммунных сигналов для иммунотерапии рака, включающий введение эффективного количества фармацевтической композиции по п. 21 субъекту, нуждающемуся в этом.
30. Способ по любому из пп. 27-29, отличающийся тем, что Т-клетки субъекта активируются указанным первым внеклеточным доменом, имеющим иммуностимулирующий сигнал.
31. Способ по любому из пп. 27-30, отличающийся тем, что субъект имеет опухоль и предотвращена передача иммуносупрессивного сигнала, активированного указанным вторым внеклеточным доменом, обладающим иммуноингибирующим сигналом, от одной или большего количества опухолевых клеток.
32. Гетерологичный химерный белок для иммунотерапии рака, содержащий:
(i) аминокислотную последовательность SEQ ID NO: 32 или последовательность, идентичную ей по меньшей мере на 98%; аминокислотную последовательность SEQ ID NO: 56 или последовательность, идентичную ей по меньшей мере на 98%; и линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4;
(ii) аминокислотную последовательность SEQ ID NO: 32 или последовательность, идентичную ей по меньшей мере на 98%; аминокислотную последовательность SEQ ID NO: 58 или последовательность, идентичную ей по меньшей мере на 98%; и линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4;
(iii) аминокислотную последовательность SEQ ID NO: 32 или последовательность, идентичную ей по меньшей мере на 98%; аминокислотную последовательность SEQ ID NO: 57 или последовательность, идентичную ей по меньшей мере на 98%; и линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4;
(iv) аминокислотную последовательность SEQ ID NO: 32 или последовательность, идентичную ей по меньшей мере на 98%; аминокислотную последовательность SEQ ID NO: 59 или последовательность, идентичную ей по меньшей мере на 98%; и линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4;
(v) аминокислотную последовательность SEQ ID NO: 36 или последовательность, идентичную ей по меньшей мере на 98%; аминокислотную последовательность SEQ ID NO: 56 или последовательность, идентичную ей по меньшей мере на 98%; и линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4;
(vi) аминокислотную последовательность SEQ ID NO: 33 или последовательность, идентичную ей по меньшей мере на 98%; аминокислотную последовательность SEQ ID NO: 56 или последовательность, идентичную ей по меньшей мере на 98%; и линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4;
(vii) аминокислотную последовательность SEQ ID NO: 33 или последовательность, идентичную ей по меньшей мере на 98%; аминокислотную последовательность SEQ ID NO: 60 или последовательность, идентичную ей по меньшей мере на 98%; и линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4;
(viii) аминокислотную последовательность SEQ ID NO: 35 или последовательность, идентичную ей по меньшей мере на 98%; аминокислотную последовательность SEQ ID NO: 56 или последовательность, идентичную ей по меньшей мере на 98%; и линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4;
(ix) аминокислотную последовательность SEQ ID NO: 34 или последовательность, идентичную ей по меньшей мере на 98%; аминокислотную последовательность SEQ ID NO: 56 или последовательность, идентичную ей по меньшей мере на 98%; и линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4;
(x) аминокислотную последовательность SEQ ID NO: 38 или последовательность, идентичную ей по меньшей мере на 98%; аминокислотную последовательность SEQ ID NO: 56 или последовательность, идентичную ей по меньшей мере на 98%; и линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4; или
(xi) аминокислотную последовательность SEQ ID NO: 46 или последовательность, идентичную ей по меньшей мере на 98%; аминокислотную последовательность SEQ ID NO: 60 или последовательность, идентичную ей по меньшей мере на 98%; и линкер, содержащий домен Fc шарнир-CH2-CH3, происходящий от IgG1 или IgG4.
33. Гетерологичный химерный белок по п. 32, где линкер содержит аминокислотную последовательность одной из SEQ ID NO: 71, 72 или идентичную ей по меньшей мере на 98%.
34. Гетерологичный химерный белок по п. 32, дополнительно содержащий один или несколько соединительных линкеров, имеющих аминокислотную последовательность одной из SEQ ID NO: 23-31 и 73-78, при этом один или несколько соединительных линкеров примыкают к указанному линкеру.
35. Гетерологичный химерный белок для иммунотерапии рака, содержащий аминокислотную последовательность SEQ ID NO: 2 или последовательность, идентичную ей по меньшей мере на 98%.
36. Гетерологичный химерный белок для иммунотерапии рака, содержащий аминокислотную последовательность SEQ ID NO: 5 или последовательность, идентичную ей по меньшей мере на 98%.
37. Гетерологичный химерный белок для иммунотерапии рака, содержащий аминокислотную последовательность SEQ ID NO: 7 или последовательность, идентичную ей по меньшей мере на 98%.
38. Гетерологичный химерный белок для иммунотерапии рака, содержащий аминокислотную последовательность SEQ ID NO: 9 или последовательность, идентичную ей по меньшей мере на 98%.
39. Гетерологичный химерный белок для иммунотерапии рака, содержащий аминокислотную последовательность SEQ ID NO: 11 или последовательность, идентичную ей по меньшей мере на 98%.
40. Гетерологичный химерный белок для иммунотерапии рака, содержащий аминокислотную последовательность SEQ ID NO: 13 или последовательность, идентичную ей по меньшей мере на 98%.
41. Гетерологичный химерный белок для иммунотерапии рака, содержащий аминокислотную последовательность SEQ ID NO: 15 или последовательность, идентичную ей по меньшей мере на 98%.
42. Фармацевтическая композиция для иммунотерапии рака, содержащая эффективное количество гетерологичного химерного белка по любому из пп. 35-41 и фармацевтически приемлемое вспомогательное вещество.
| US 2015183881 A1, 02.07.2015 | |||
| WO 2014121093 A1, 07.08.2014 | |||
| US 20130287802 A1, 31.10.2013 | |||
| ALI S.A | |||
| et al., Anti-tumour therapeutic efficacy of OX40L in murine tumourmodel,Vaccine, 2004, v.9, n.22(27-28), p.3585-3594, c.3586-3587 | |||
| US 2015190506 A1, 09.07.2015 | |||
| MAHONEY K.M | |||
| et al., Combination Cancer Immunotherapy and New Immunomodulatory |

