ПРИОРИТЕТ
Настоящая заявка испрашивает приоритет согласно предварительной заявке США №62/463,997, поданной 27 февраля 2017 г., содержание которой полностью включено в настоящий документ посредством ссылки.
ВКЛЮЧЕНИЕ МАТЕРИАЛОВ, ПОДАННЫХ В ЭЛЕКТРОННОЙ ФОРМЕ, ПОСРЕДСТВОМ ССЫЛКИ
Настоящая заявка содержит перечень последовательностей. Он был подан в электронном виде через EFS-Web ав виде текстового файла ASCII под названием "SHK-002PC_SequenceListing_ST25". Размер файла с перечнем последовательностей составляет 92976 байт, и он был создан 27 февраля 2018 г.или приблизительно в эту дату. Перечень последовательностей полностью включен в настоящий документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится в том числе к химерным белкам, которые включают внеклеточный домен рецептора колониестимулирующего фактора 1 (CSF1R) и их применению в лечении заболеваний, таком как иммунотерапия рака и аутоиммунных нарушений.
УРОВЕНЬ ТЕХНИКИ
Недавние клинические данные демонстрируют выраженную реакцию пациента на агенты, оказывающие адресное воздействие на иммунные коингибиторные молекулы, включая, например, клинические исследования, приведшие к разрешению на применение YERVOY, KEYTRUDA и OPDIVO. Эти иммунотерапевтические агенты совместно характеризуются как ингибиторы ключевых этапов иммунного ответа, и, к сожалению, эти терапевтические средства обеспечивают клинический благоприятный эффект только у ~15-30% онкологических пациентов. Один потенциальный подход к улучшению частоты клинической реакции по расширенной популяции онкологических пациентов включает объединение терапевтического ингибитора ключевого этапа иммунного ответа с другим терапевтическим средством. Применение таких комбинаций нескольких отдельных терапевтических средств может привести к улучшенному клиническому благоприятному эффекту, однако разработка таких средств является трудоемкой. Кроме того, многие иммунотерапевтические средства характеризуются осложнениями в виде тяжелых побочных эффектов, которые существенно ограничивают терапевтическое окно при лечении пациентов.
Сохраняется потребность в новых способах и композициях, обеспечивающих получение эффективных иммунотерапевтических средств, в том числе объединении нескольких терапевтических механизмов в одном лекарственном средстве.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Соответственно, в настоящем изобретении предложены, в частности, композиции и способы, которые находят применение при лечении рака, например, путем преодоления множественных суппрессивных механизмов в микроокруженим опухоли, и стимуляции иммунологических противоопухолевых механизмов. Аналогичным образом, указанные композиции и способы находят применение при лечении воспалительных заболеваний. Например, в настоящем изобретении предложены, в частности, композиции и способы, позволяющие оказывать двойное адресное воздействие на суппрессивные миелоидные популяции, ингибируя сигнальный путь CSF1/CSF1R, и активировать антигенпрезентирующие клетки путем стимуляции сигнального пути CD40/CD40L. Такая одновременная блокада CSF1R и агонизм CD40 вызывают, в числе прочего, общее снижение количества иммуносуппрессивных клеток и сдвиг в сторону более воспалительного окружения опухоли и усиленного противоопухолевого эффекта.
В различных аспектах настоящего изобретения предложен гетерологичный химерный белок, содержащий: (а) первый домен, содержащий фрагмент рецептора колониестимулирующего фактора 1 (CSF1R), способный связывать лиганд CSF1R; (b) второй домен, содержащий фрагмент лиганда CD40 (CD40L), способный связывать рецептор CD40L; и (с) линкер, соединяющий первый и второй домены. В различных аспектах настоящего изобретения предложены способы лечения рака с применением этого гетерологичного химерного белка. В различных аспектах настоящего изобретения предложены способы лечения воспалительного заболевания с применением этого гетерологичного химерного белка.
В ряде вариантов реализации настоящего изобретения предложен рекомбинантный гибридный белок, содержащий общую структуру: N-конец (а) - (b) - (с) - С-конец, где (а) - первый домен, содержащий внеклеточный домен CSF1R, по меньшей мере на 95% идентичный аминокислотной последовательности SEQ ID NO: 2 и способный связывать лиганд CSF1R, (b) представляет собой линкер, связывающий первый домен и второй домен и содержащий шарнир-СН2-СН3 Fc-домен, происходящие из IgG4 человека (например, на 95% идентичные аминокислотной последовательности SEQ ID NO: 25, SEQ ID NO: 26, или SEQ ID NO: 27, а (с) представляет собой второй домен, содержащий внеклеточный домен лиганда CD40 (CD40L), по меньшей мере на 95% идентичный аминокислотной последовательности SEQ ID NO: 4 и способный связывать рецептор CD40L. В ряде вариантов реализации настоящего изобретения предложены способы лечения рака с применением этого гетерологичного химерного белка. В ряде вариантов реализации настоящего изобретения предложены способы лечения воспалительного заболевания с применением этого гетерологичного химерного белка.
Любой аспект или вариант реализации, описанные в настоящем документе, можно объединить с любым другим аспектом или вариантом реализации, раскрытым в настоящем документе.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1, безотносительно к теоретическим представлениям, показана схема механизма действия химерного белка CSF1R-Fc-CD40L. На фиг. 1В показан синапс, образованный химерным белком между опухолевой клеткой и Т-клеткой. На фиг. 1С показана прогнозируемая вторичная структура CSF1R-Fc-CD40L человека с указанием прогнозируемого формирования трех доменов в природном состоянии. Прогнозируемая молекулярная масса мономера химерного белка CSF1R-Fc-CD40L составляет приблизительно 105,4 кДа.
На фиг. 2 показано исследование трех доменов CSF1R-Fc-CD40L человека с использованием вестерн-блоттинга в невосстанавливающих условиях с нагреванием, восстанавливающих условиях с нагреванием и в восстанавливающих условиях с дегликозилированием и нагреванием (PNGa3a). Размер полос подтверждает, что прогнозируемая молекулярная масса мономера составляет приблизительно 105,4 кДа, и указывает, что в нативном состоянии он существует в виде гликозилированного димера. Как показано, дорожка 1, начиная с левого края каждого блота, представляет собой маркер молекулярной массы белков.
На фиг.3 показан функциональный твердофазный иммуноферментный анализ (твердофазный ИФА), демонстрирующий связывание CSF1R-Fc-CD40L человека с мишенями трех доменов по отдельности (Fc показан на верхнем левом, CSF1R - на верхнем правом график, a CD40L - на нижнем левом графике), а также одновременное связывание в рекомбинантными CSF1 и CD40, показанное на нижнем правом графике. На верхнем левом графике верхняя кривая представляет собой стандартный IgG, а нижняя кривая - hCSFR1-Fc-CD40L. На нижнем левом графике верхняя кривая представляет собой CD40L-FC, а нижняя кривая - hCSFR1-Fc-CD40L.
На фиг.4 показан клеточный анализ связывания in vitro, демонстрирующий способность химерного белка CSF1R(CD115)-Fc-CD40L человека связывать рецептор CD40, экспрессируемый клетками Jurkat (линия Т-клеток человека). Измеренная ЕС50 связывания составляет 77 нМ. "ARC" относится к химерному белку hCSF1R-Fc-CD40L.
На фиг.5A-5F показано сродство связывания CSF1R-Fc-CD40L, измеренное в системе Octet. Скорости ассоциации, скорости диссоциации и сродство (KD) определяли для CSF1R-Fc-CD40L человека к CD40-His (фиг.5А), доступного для приобретения одностороннего CD40L-Fc к CD40-His (фиг.5 В), доступного для приобретения антитела против CD40 к CD40-His (фиг.5С), hCSF1R-Fc-CD40L к CSF1-His (фиг.5D) и доступного для приобретения CSF1R-Fc к CSFl-His (фиг.5Е). CSF1R-Fc-CD40L человека связывал CD40 со сродством 4,83 нМ, a CSF1 - 646 пМ (фиг.5F). Термин "CSF1R-Fc-CD40L ARC" относится к химерному белку CSF1R-Fc-CD40L. На всех фигурах с фиг.5А по фиг.5Е кривые расположены в следующем порядке (сверху вниз): 100 мМ тестируемый агент, 33 мМ тестируемый агент, 11 мМ тестируемый агент и холостой раствор.
На фиг.6 показано исследование относительного сродства связывания CSF1R-Fc-CD40L человека с рекомбинантными CD40, CSF1 и ИЛ-34 человека посредством интерферометрии в биослое (Octet). Идентичное связывание наблюдали для двух лигандов CSF1R: CSF1 и ИЛ-34; таким образом, их кривые накладываются друг на друга. Таким образом, кривые расположены в следующем порядке: CD40-his наверху, a CSFl-his и ИЛ-34-his внизу и перекрываются.
На фиг.7А и 7В показано исследование CSF1R-Fc-CD40L мыши с использованием вестерн-блоттинга и функционального твердофазного ИФА связывания. На фиг.7А показано обнаружение всех трех доменов химерного белка mCSF1R-Fc-CD40L посредством вестерн-блоттинга в невосстанавливающих (дорожка 2), восстанавливающих (дорожка 3) и восстанавливающих условиях с обработкой PNGазой (дорожка 4). Восстановленная дегликозилированная форма белка мигрирует в положение прогнозируемой молекулярной массы приблизительно 105 кДа. На фиг.7В показан твердофазный ИФА, выполненный с целью обнаружения связывания CSF1R с рекомбинантным CSF1 (левый график), Fc с IgG (центральный график) и CD40L с rCD40 (правый график) с использованием способов обнаружения, схематично указанных над каждым графиком. CD115 представляет собой синоним CSF1R. На фиг.7В на левом графике mCD115-Fc-CD40L соответствует верхней кривой, а на среднем и правом графике - нижней кривой.
На фиг.8 показано связывание CSF1R-Fc-CD40L мыши in vitro с клетками СНО-К1, сверхэкспрессирующими CD40 мыши (верхняя кривая) по сравнению с исходной линией клеток СНО-K1, не экспрессирующей mCD40 (нижняя кривая). Измеренная ЕС50 связывания составляет 91,1 нМ.
На фиг.9 показаны данные анализа сигнального пути NF-κB/NIK in vitro с использованием химерного белка CSF1R-Fc-CD40L человека. Клетки U20S для анализа DiscoverX сигнального пути NIK культивировали с титрованием за счет доступного для приобретения одностороннего CD40L-Fc, одностороннего CSF1R-Fc или антитела против CD40 или химерного белка CSF1R-Fc-CD40L человека. Относительные люциферазные единицы (RLU) указывают на относительную активность сигнального пути NF-κB/NIK, активируемого после обработки согласно указанным схемам. Кривые обозначены следующим образом: в точке, соответствующей 0,01 мкг/мл на оси X, сверху вниз: CD40L-Fc, hCSFlR-Fc-CD40L, CSF1R-Fc и антитело против CD40.
На фиг.10А и фиг.10 В показаны функциональные данные об активности CSF1R-Fc-CD40L мыши in vivo. На фиг.10А показан анализ поглощения CSF1. Мышам, не несущим опухоли, вводили разовую дозу антитела против CD115 (CSF1R) в день 0. На 2 день мышам не вводили никаких агентов или вводили разовую дозу химерного белка CSF1R-Fc-CD40L. Сыворотку крови собирали на 2 день перед инъекцией химерного белка и на 3 день после обработки химерным белком. Сыворотку подвергали твердофазному ИФА для выявления CSF1 мыши; показано, что химерный белок CSF1R-Fc-CD40L мыши связывается с CSF1 и способствует его выведению. (На фиг.10В показана индукция ИЛ15Rα in vivo. Мышей с опухолями обрабатывали путем введения двух доз по 150 мкг mCSF1R-Fc-CD40L ARC на 5 и 7 дни после исходной инокуляции опухоли. На 13 день группу мышей умерщвляли, извлекали селезенки и лимфатические узлы и диссоциировали их для проточно-цитометрического анализа ИЛ15Rα. В соответствии с известными механизмами функции CD40L, мыши, обработанные химерным белком CSF1R-Fc-CD40L, демонстрировали увеличение уровня ИЛ15Rα в обоих тканевых компартментах. CD115 представляет собой синоним CSF1R. Верхняя кривая на графике на фиг.10А соответствует +αCD115, средняя кривая - +αCD115, а затем CD115-Fc-CD40L на 2 день, а нижняя кривая - отсутствию обработки. На фиг.10В (верхний и нижний графики) левые точки представляют собой контроль, а правые - CSF1R-Fc-CD40L.
На фиг.11А - фиг.11С показана противоопухолевая эффективность CSF1R-Fc-CD40L мыши в опухолях ободочной и прямой кишки СТ26. Мышам Balb/с инокулировали опухоли СТ26 в день 0. Через 4 дня роста опухоли, когда опухоли достигали диаметра 4-5 мм, мышей обрабатывали с применением контрольных антител или химерного белка mCSF1R-Fc-CD40L. Обработку повторно выполняли на 7 день. Вышеприведенная фигура включает: (фиг.11А) отдельные кривые роста опухолей для каждой экспериментальной группы, (FIG. 11А) общую выживаемость до 60 дня эксперимента и (фиг.11А) сводную таблицу исхода лечения для каждой группы. CD115 представляет собой синоним CSF1R. На фиг.11В в точке, соответствующей 35 дню, кривые представляют собой (сверху вниз): CD115-Fc-CD40L (150 мкг × 2), αCD115, αCD115/CD40, αCD40 (необработанные мыши не доживали до этого срока).
На фиг.12А - фиг.12Е показано иммунофенотипирование in vivo у мышей, несущих опухоли. Иммунофенотипирование у особей, несущих опухоли, также выполняли в каждой экспериментальной группе путем анализа спленоцитов, клеток лимфатических узлов и опухоль-инфильтрирующих лимфоцитов у мышей каждой группы на 13 день после инокуляции. На фиг.12А показаны результаты, демонстрирующие, что частота CD4+ и CD8+ Т-клеток в селезенке (но не лимфатических узлах или опухоли) была повышена у мышей, обработанных CSF1R-Fc-CD40L мыши, по сравнению с контрольными особями. На фиг.12В показано снижение доли CD4+CD25+ клеток в селезенке и опухоли, что может указывать на снижение количества иммунорегуляторных Т-клетокПримечательно, что несмотря на незначительное увеличение доли общих CD8+ клеток в опухоли (см. фиг.12С), обнаружено значительное увеличение доли CD8+ Т-клеток, специфичных по отношению к опухолевому антигену АН1 (при окрашивании тетрамером). Для определения потенциального доказательства активации рецептора CD40 анализировали индукцию CD19+ клеток (фиг.12D) и ИЛ-15Rα-положительных клеток (фиг.12Е). На всех фигурах с фиг.12А по фиг.12Е левые точки представляют собой контроль, а правые - CSF1R-Fc-CD40L.
На фиг.13А и фиг.13В показана безопасность CSF1R-Fc-CD40L мыши по сравнению с агонистическим антителом к CD40. Монотерапия с применением агонистического антитела к CD40 (клона FGK4.5) или комбинированная терапия с применением агонистического антитела к CD40 и антитела против CD115 (CSF1R) (клона AFS98) приводила к выраженной диарее и потере массы тела у мышей в ходе эксперимента. Эти данные показывают, что агонистическое антитело к CD40 инициировало воспалительный ответ в кишечнике, приводивший к диарее и потере массы тела, что значительно усугублялось за счет комбинации с блокадой CD115. Мыши в группе комбинации антител теряли >25% массы тела (см. фиг.13В), выглядели больными (фиг.13А), и в некоторых случаях этот воспалительный ответ приводил к смерти. Важно, что мыши, обработанные химерным белком CD115-Fc-CD40L мыши (другое название химерного белка mCSF1R-Fc-CD40L), выглядели здоровыми, у них не развивались признаки диареи или потери массы тела, и вели себя нормально (см. фотографии на левой панели). Эти данные соответствуют клиническим данным у людей, получавших агонистические антитела к CD40, и указывают на благоприятный профиль безопасности mCD115-Fc-CD40L. CD115 представляет собой синоним CSF1R. На фиг.13В столбики расположены в следующем порядке: отсутствие обработки, αCD115, αCD40, αCD115+αCD40, CD115-Fc-CD40L FP.
На фиг.14 показаны четыре потенциальные конфигурации типичных химерных белков (PD1-FC-OX40L).
На фиг.15 показаны вестерн-блоты химерных белков PD1-Fc-OX40L, прошедших электрофорез в ДСН-ПААГ в невосстанавливающих условиях, восстанавливающих условиях, и восстанавливающих условиях после обработки пептид-N-гликозидазой F (PNGазой F).
На фиг.16 показана хроматограмма химерных белков PD-1-Fc-OX40L, полученная с помощью эксклюзионной хроматографии (SEC).
На фиг.17 показаны гели электрофореза в ДСН-ПААГ и нативном ПААГ (без ДСН) химерных белков PD-1-Fc-OX40L, выполненных в невосстанавливающих условиях («-») или в восстанавливающих условиях («+»).
На фиг.18 показаны результаты электрофореза в нативном ПААГ (без SDS) химерных белков PD-1-No Fc-OX40L, линкер которых не содержит Fc-домена.
На фиг.19 показана, безотносительно к теоретическим представлениям, модель образования гексамера и конкатемеров из химерных белков согласно настоящему изобретению.
Фиг.20 представляет собой таблицу, в которой показаны соединяющие линкеры и Fc-линкеры, которые можно объединить в типичные модульные линкеры. Показанные типичные модульные линкеры можно объединять с любыми описанными в настоящем документе белками I типа и II типа и/или внеклеточными доменами описанных в настоящем документе белков I типа и II типа с образованием химерного белка согласно настоящему изобретению.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение основано, в частности, на открытии сконструированных химерных белков, содержащих первый домен, содержащий фрагмент рецептора колониестимулирующего фактора 1 (CSF1R), способный связывать лиганд CSF1R. В ряде вариантов реализации химерный белок дополнительно содержит второй домен, содержащий фрагмент лиганда CD40 (CD40L), способный связывать рецептор CD40L. В ряде вариантов реализации первый домен и второй домен соединены линкером. В ряде вариантов реализации химерный белок согласно настоящему изобретению обеспечивает иммуностимулирующий сигнал, например, способный активировать макрофаги и антигенпрезентирующие клетки, при этом представляя собой локализованную ловушку для ингибиторного сигнала, который в противном случае сдвигает равновесие в сторону иммуносупрессии (например, CSF1 или ИЛ-34). Таким образом, в различных вариантах реализации настоящего изобретения предложено эффективное средство для лечения онкологических и/или воспалительных заболеваний. Химерные белки
В ряде вариантов реализации настоящее изобретение относится к химерным белкам, конструкция которых содержит домен, например, внеклеточный домен иммуноингибиторного рецептора колониестимулирующего фактора 1 (CSF1R), также известного как рецептор макрофагального колониестимулирующего фактора (M-CSFR) и кластер дифференцировки 115 (CD115). Таким образом, в настоящем документе CSF1R и CD115 являются синонимами при упоминании по отдельности и/или в контексте химерного белка; таким образом, например, CSF1R-Fc-CD40L является тем же химерным белком, что и CD115-Fc-CD40L. CSF1R представляет собой однопроходный мембранный белок I типа, действующий как рецептор колониестимулирующего фактора 1 (CSF1). Кроме того, показано, что CSF1R является рецептором ИЛ-34. Связывание CSF1R с CSF1 или ИЛ-34 играет важную роль в выживании, пролиферации и дифференцировке кроветворных клеток-предшественников, особенно мононуклеарных фагоцитов, например, макрофагов и моноцитов. Кроме того, показано, что CSF1R связывается с CSF1 или ИЛ-34 в микроокружении опухоли. Связывание рецептора с этими лигандами индуцирует иммуносупрессию посредством, в числе прочего, индукции опухолеассоциированных макрофагов (ТАМ) и супрессорных клеток миелоидного происхождения (MDSC).
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит домен, например, внеклеточный домен CSF1R человека. CSF1R человека содержит аминокислотную последовательность SEQ ID NO: 1 (с аминокислотной последовательностью внеклеточного домена, содержащей аминокислоты 20 - 517).
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит внеклеточный домен CSF1R человека с аминокислотной последовательностью SEQ ID NO: 2. В ряде вариантов реализации химерные белки согласно настоящему изобретению содержат внеклеточный домен CSF1R, описанный в настоящем документе, или его вариант или функциональный фрагмент.Например, химерный белок может содержать последовательность внеклеточного домена CSF1R, представленную выше, или ее вариант или функциональный фрагмент, обладающий по меньшей мере приблизительно 60%, или по меньшей мере приблизительно 61%, или по меньшей мере приблизительно 62%, или по меньшей мере приблизительно 63%, или по меньшей мере приблизительно 64%, или по меньшей мере приблизительно 65%, или по меньшей мере приблизительно 66%, или по меньшей мере приблизительно 67%, или по меньшей мере приблизительно 68%, или по меньшей мере приблизительно 69%, или по меньшей мере приблизительно 70%, или по меньшей мере приблизительно 71%, или по меньшей мере приблизительно 72%, или по меньшей мере приблизительно 73%, или по меньшей мере приблизительно 74%, или по меньшей мере приблизительно 75%, или по меньшей мере приблизительно 76%, или по меньшей мере приблизительно 77%, или по меньшей мере приблизительно 78%, или по меньшей мере приблизительно 79%, или по меньшей мере приблизительно 80%, или по меньшей мере приблизительно 81%, или по меньшей мере приблизительно 82%, или по меньшей мере приблизительно 83%, или по меньшей мере приблизительно 84%, или по меньшей мере приблизительно 85%, или по меньшей мере приблизительно 86%, или по меньшей мере приблизительно 87%, или по меньшей мере приблизительно 88%, или по меньшей мере приблизительно 89%, или по меньшей мере приблизительно 90%, или по меньшей мере приблизительно 91%, или по меньшей мере приблизительно 92%, или по меньшей мере приблизительно 93%, или по меньшей мере приблизительно 94%, или по меньшей мере приблизительно 95%, или по меньшей мере приблизительно 96%, или по меньшей мере приблизительно 97%, или по меньшей мере приблизительно 98%, или по меньшей мере приблизительно 99% идентичностью последовательности по отношению к аминокислотной последовательности внеклеточного домена CSF1R, описанного в настоящем документе.
Структура CSF1R описана, например, в работе W.D. Тар, et al., "Structure-Guided Blockade of CSF1R Kinase in Tenosynovial Giant-Cell Tumor", N. Engl. J. Med. 2015 Jul 30; 373(5):428-37. Можно получить производные CSF1R на основе имеющихся структур CSF1R.
В ряде вариантов реализации химерные белки согласно настоящему изобретению могут содержать вариантный внеклеточный домен CSF1R человека, в котором сигнальный пептид (например, как представлено в SEQ ID NO: 1) замещен альтернативным сигнальным пептидом. В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит вариантный внеклеточный домен CSF1R, экспрессируемый с кДНК, прошедшей оптимизацию кодонов для экспрессии в клетках, продуцирующих белки, например, клетках яичника китайского хомяка (СНО) или почки эмбриона человека (HEK).
В ряде вариантов реализации внеклеточный домен CSF1R относится к фрагменту белка, способному взаимодействовать с внеклеточной средой. В ряде вариантов реализации внеклеточный домен CSF1R представляет собой полную аминокислотную последовательность белка, находящуюся вне клетки или клеточной мембраны. В ряде вариантов реализации внеклеточный домен CSF1R представляет собой фрагмент аминокислотной последовательности белка, который находится вне клетки или клеточной мембраны и необходим для передачи сигнала и/или связывания лиганда, что можно проанализировать с использованием способов, известных в данной области техники (например, анализов связывания лиганда и/или активации клеток in vitro).
В ряде вариантов реализации внеклеточный домен CSF1R относится к фрагменту белка, способному связываться с колониестимулирующим фактором 1 (CSF1). В ряде вариантов реализации химерный белок связывается с CSF1 человека с KD менее приблизительно 1 мкМ, приблизительно 900 нМ, приблизительно 800 нМ, приблизительно 700 нМ, приблизительно 600 нМ, приблизительно 500 нМ, приблизительно 400 нМ, приблизительно 300 нМ, приблизительно 200 нМ, приблизительно 150 нМ, приблизительно 130 нМ, приблизительно 100 нМ, приблизительно 90 нМ, приблизительно 80 нМ, приблизительно 70 нМ, приблизительно 60 нМ, приблизительно 55 нМ, приблизительно 50 нМ, приблизительно 45 нМ, приблизительно 40 нМ, приблизительно 35 нМ, приблизительно 30 нМ, приблизительно 25 нМ, приблизительно 20 нМ, приблизительно 15 нМ, приблизительно 10 нМ или приблизительно 5 нМ или приблизительно 1 нМ (согласно измерениям, например, с помощью поверхностного плазмонного резонанса или интерферометрии в биослое). В ряде вариантов реализации химерный белок связывается с CSF1 человека с KD менее приблизительно 1 нМ, приблизительно 900 пМ, приблизительно 800 пМ, приблизительно 700 пМ, приблизительно 600 пМ, приблизительно 500 пМ, приблизительно 400 пМ, приблизительно 300 пМ, приблизительно 200 пМ, приблизительно 100 пМ, приблизительно 90 пМ, приблизительно 80 пМ, приблизительно 70 пМ, приблизительно 60 пМ приблизительно 55 пМ приблизительно 50 пМ приблизительно 45 пМ, приблизительно 40 пМ, приблизительно 35 пМ, приблизительно 30 пМ, приблизительно 25 пМ, приблизительно 20 пМ, приблизительно 15 пМ или приблизительно 10 пМ или приблизительно 1 пМ (согласно измерениям, например, с помощью поверхностного плазмонного резонанса или интерферометрии в биослое).
В ряде вариантов реализации внеклеточный домен CSF1R относится к фрагменту белка, способному связываться с ИЛ-34. В ряде вариантов реализации химерный белок связывается с ИЛ-34 человека с KD менее приблизительно 1 мкМ, приблизительно 900 нМ, приблизительно 800 нМ, приблизительно 700 нМ, приблизительно 600 нМ, приблизительно 500 нМ, приблизительно 400 нМ, приблизительно 300 нМ, приблизительно 200 нМ, приблизительно 100 нМ, приблизительно 90 нМ, приблизительно 80 нМ, приблизительно 70 нМ, приблизительно 60 нМ, приблизительно 55 нМ, приблизительно 50 нМ, приблизительно 45 нМ, приблизительно 40 нМ, приблизительно 35 нМ, приблизительно 30 нМ, приблизительно 25 нМ, приблизительно 20 нМ, приблизительно 15 нМ, приблизительно 10 нМ или приблизительно 5 нМ или приблизительно 1 нМ (согласно измерениям, например, с помощью поверхностного плазмонного резонанса или интерферометрии в биослое). В ряде вариантов реализации химерный белок связывается с ИЛ-34 человека с KD менее приблизительно 1 нМ, приблизительно 900 пМ, приблизительно 800 пМ, приблизительно 700 пМ, приблизительно 600 пМ, приблизительно 500 пМ, приблизительно 400 пМ, приблизительно 300 пМ, приблизительно 200 пМ, приблизительно 100 пМ, приблизительно 90 пМ, приблизительно 80 пМ, приблизительно 70 пМ, приблизительно 60 пМ, приблизительно 55 пМ, приблизительно 50 пМ, приблизительно 45 пМ, приблизительно 40 пМ, приблизительно 35 пМ, приблизительно 30 пМ, приблизительно 25 пМ, приблизительно 20 пМ, приблизительно 15 пМ или приблизительно 10 пМ или приблизительно 1 пМ (согласно измерениям, например, с помощью поверхностного плазмонного резонанса или интерферометрии в биослое). В ряде вариантов реализации химерный белок связывается с CSF1 человека с KD, составляющей от приблизительно 100 пМ до приблизительно 600 пМ.
Указанный химерный белок содержит домен, например, внеклеточный домен лиганда иммуностимулятора CD40 (CD40L, также известного как CD154). CD40L является трансмембранным белком II типа, принадлежащим к надсемейству факторов некроза опухолей (ФИО). CD40 связывается с рецептором CD40 на макрофагах и антигенпрезентирующих клетках (АПК), в том числе антигенпрезентирующих В-клетках, что приводит к многим эффектам в зависимости от типа клетки-мишени. Кроме того, показано, что CD40L связывает интегрины α5β1 и αIIbβ3. CD40L действует как костимулятор и в особенности важен для субпопуляции Т-клеток, называемых фолликулярными Т-хелперными клетками (TFH-клетками). CD40L на TFH-клетках стимулирует созревание В-клеток и действует, активируя CD40 на поверхности В-клеток,тем самым облегчая обмен информацией между клетками.
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит домен, например, внеклеточный домен CD40L человека. CD40L человека содержит аминокислотную последовательность SEQ ID NO: 3 (с аминокислотной последовательностью внеклеточного домена, содержащей аминокислоты 47 - 261). В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит внеклеточный домен CD40L человека с аминокислотной последовательностью SEQ ID NO: 4. В ряде вариантов реализации химерные белки согласно настоящему изобретению могут содержать внеклеточный домен CD40L, описанный в настоящем документе, или его вариант или функциональный фрагмент.Например, химерный белок может содержать последовательность внеклеточного домена CD40L, представленную выше, или ее вариант или функциональный фрагмент, обладающий по меньшей мере приблизительно 60%, или по меньшей мере приблизительно 61%, или по меньшей мере приблизительно 62%, или по меньшей мере приблизительно 63%, или по меньшей мере приблизительно 64%, или по меньшей мере приблизительно 65%, или по меньшей мере приблизительно 66%, или по меньшей мере приблизительно 67%, или по меньшей мере приблизительно 68%, или по меньшей мере приблизительно 69%, или по меньшей мере приблизительно 70%, или по меньшей мере приблизительно 71%, или по меньшей мере приблизительно 72%, или по меньшей мере приблизительно 73%, или по меньшей мере приблизительно 74%, или по меньшей мере приблизительно 75%, или по меньшей мере приблизительно 76%, или по меньшей мере приблизительно 77%, или по меньшей мере приблизительно 78%, или по меньшей мере приблизительно 79%, или по меньшей мере приблизительно 80%, или по меньшей мере приблизительно 81%, или по меньшей мере приблизительно 82%, или по меньшей мере приблизительно 83%, или по меньшей мере приблизительно 84%, или по меньшей мере приблизительно 85%, или по меньшей мере приблизительно 86%, или по меньшей мере приблизительно 87%, или по меньшей мере приблизительно 88%, или по меньшей мере приблизительно 89%, или по меньшей мере приблизительно 90%, или по меньшей мере приблизительно 91%, или по меньшей мере приблизительно 92%, или по меньшей мере приблизительно 93%, или по меньшей мере приблизительно 94%, или по меньшей мере приблизительно 95%, или по меньшей мере приблизительно 96%, или по меньшей мере приблизительно 97%, или по меньшей мере приблизительно 98%, или по меньшей мере приблизительно 99% идентичностью последовательности по отношению к аминокислотной последовательности внеклеточного домена CD40L, описанного в настоящем документе.
Можно сконструировать производные CD40L на основании имеющихся структурных данных, в том числе описанных в статье Oganesyan V., et al., "Fibronectin type III domains engineered to bind CD40L: cloning, expression, purification, crystallization and preliminary X-ray diffraction analysis of two complexes", Acta Crystallogr Sect F Struct Biol Cryst Commun. 2013 Sep; 69(Pt 9): 1045-8.
В ряде вариантов реализации химерные белки согласно настоящему изобретению могут содержать вариантный внеклеточный домен CD40L, в котором сигнальный пептид (например, как представлено в SEQ ID NO: 3) замещен альтернативным сигнальным пептидом. В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит вариантный внеклеточный домен CD40L, экспрессируемый с кДНК, прошедшей оптимизацию кодонов для экспрессии в клетках, продуцирующих белки, например, клетках яичника китайского хомяка (СНО) или HEK.
В ряде вариантов реализации внеклеточный домен CD40L относится к фрагменту белка, способному взаимодействовать с внеклеточной средой. В ряде вариантов реализации внеклеточный домен CD40L представляет собой полную аминокислотную последовательность белка, находящуюся вне клетки или клеточной мембраны. В ряде вариантов реализации внеклеточный домен CD40L представляет собой фрагмент аминокислотной последовательности белка, который находится вне клетки или клеточной мембраны и необходим для передачи сигнала и/или связывания лиганда, что можно проанализировать с использованием способов, известных в данной области техники.
В ряде вариантов реализации внеклеточный домен CD40L относится к фрагменту белка, способному связываться с рецептором CD40. Аналогично другим членам надсемейства ФНО, мембраносвязанный CD40L существует в виде гомотримера. CD40L связывается с CD40 - представителем надсемейства рецепторов ФНО, который преимущественно экспрессируется на антигенпрезентирующих клетках, в том числе дендритных клетках (ДК), В-клетках и макрофагах. Взаимодействие CD40L/CD40 оказывает значительный эффект на дендритные клетки, В-клетки и эндотелиальные клетки среди многих клеток кроветворных и некроветворных компартментоа. Например, сигнальный путь CD40 индуцирует созревание ЛК и запускает активацию и дифференцировку Т-клеток. Сигнальный путь CD40 в В-клетках стимулирует образование зародышевых центров (GC), переключение изотипов иммуноглобулинов (Ig), соматические гипермутации (SHM) Ig, усиливая сродство к антигену и образование долгоживущих плазматических клеток и В-клеток памяти. Сигнальный путь CD40 также важен для выживания иммунных клеток.
В ряде вариантов реализации химерный белок согласно настоящему изобретению связывается с CD40 человека с KD менее приблизительно 1 мкМ, приблизительно 900 нМ, приблизительно 800 нМ, приблизительно 700 нМ, приблизительно 600 нМ, приблизительно 500 нМ, приблизительно 400 нМ, приблизительно 300 нМ, приблизительно 200 нМ, приблизительно 100 нМ, приблизительно 90 нМ, приблизительно 80 нМ, приблизительно 70 нМ, приблизительно 60 нМ, приблизительно 55 нМ, приблизительно 50 нМ, приблизительно 45 нМ, приблизительно 40 нМ, приблизительно 35 нМ, приблизительно 30 нМ, приблизительно 25 нМ, приблизительно 20 нМ, приблизительно 15 нМ, приблизительно 10 нМ или приблизительно 5 нМ или приблизительно 1 нМ (согласно измерениям, например, с помощью поверхностного плазмонного резонанса или интерферометрии в биослое). В ряде вариантов реализации химерный белок связывается с CD40 человека с KD менее приблизительно 1 нМ, приблизительно 900 пМ, приблизительно 800 пМ, приблизительно 700 пМ, приблизительно 600 пМ, приблизительно 500 пМ, приблизительно 400 пМ, приблизительно 300 пМ, приблизительно 200 пМ, приблизительно 100 пМ, приблизительно 90 пМ, приблизительно 80 пМ, приблизительно 70 пМ, приблизительно 60 пМ приблизительно 55 пМ приблизительно 50 пМ приблизительно 45 пМ, приблизительно 40 пМ, приблизительно 35 пМ, приблизительно 30 пМ, приблизительно 25 пМ, приблизительно 20 пМ, приблизительно 15 пМ или приблизительно 10 пМ или приблизительно 1 пМ (согласно измерениям, например, с помощью поверхностного плазмонного резонанса или интерферометрии в биослое). В ряде вариантов реализации химерный белок связывается с CD40 человека с KD, составляющей от приблизительно 300 пМ до приблизительно 700 пМ.
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит внеклеточный домен CSF1R (SEQ ID NO: 2).
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит внеклеточный домен CD40L (SEQ ID NO: 4).
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит внеклеточный домен OX40L (SEQ ID NO: 7).
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит внеклеточный домен CSF1R (SEQ ID NO: 2) и внеклеточный домен CD40L (SEQ ID NO: 4).
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит внеклеточный домен CSF1R (SEQ ID NO: 2) и внеклеточный домен OX40L (SEQ ID NO: 7).
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит домен шарнир-СН2-СН3 из последовательности антитела IgG4 человека (SEQ ID NO: 25, SEQ ID NO: 26 или SEQ ID NO: 27).
В ряде вариантов реализации химерный белок содержит модульный линкер, показанный на фиг.20.
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит внеклеточный домен CSF1R и внеклеточный домен CD40L с использованием домена шарнир-СН2-СН3 из последовательности антитела IgG4 человека в качестве линкера (эта химера CSF1R-Fc-CD40L представляет собой SEQ ID NO: 5).
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит внеклеточный домен CSF1R и внеклеточный домен OX40L с использованием домена шарнир-СН2-СНЗ из последовательности антитела IgG4 человека в качестве линкера (эта химера CSF1R-Fc-OX40L представляет собой SEQ ID NO: 8).
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит SEQ ID NO: 5, т.е. мономерный химерный белок CSF1R-Fc-CD40L (SL-115154) или его вариант или функциональный фрагмент.
В ряде вариантов реализации химерный белок может обладать по меньшей мере приблизительно 60%, или по меньшей мере приблизительно 61%, или по меньшей мере приблизительно 62%, или по меньшей мере приблизительно 63%, или по меньшей мере приблизительно 64%, или по меньшей мере приблизительно 65%, или по меньшей мере приблизительно 66%, или по меньшей мере приблизительно 67%, или по меньшей мере приблизительно 68%, или по меньшей мере приблизительно 69%, или по меньшей мере приблизительно 70%, или по меньшей мере приблизительно 71%, или по меньшей мере приблизительно 72%, или по меньшей мере приблизительно 73%, или по меньшей мере приблизительно 74%, или по меньшей мере приблизительно 75%, или по меньшей мере приблизительно 76%, или по меньшей мере приблизительно 77%, или по меньшей мере приблизительно 78%, или по меньшей мере приблизительно 79%, или по меньшей мере приблизительно 80%, или по меньшей мере приблизительно 81%, или по меньшей мере приблизительно 82%, или по меньшей мере приблизительно 83%, или по меньшей мере приблизительно 84%, или по меньшей мере приблизительно 85%, или по меньшей мере приблизительно 86%, или по меньшей мере приблизительно 87%, или по меньшей мере приблизительно 88%, или по меньшей мере приблизительно 89%, или по меньшей мере приблизительно 90%, или по меньшей мере приблизительно 91%, или по меньшей мере приблизительно 92%, или по меньшей мере приблизительно 93%, или по меньшей мере приблизительно 94%, или по меньшей мере приблизительно 95%, или по меньшей мере приблизительно 96%, или по меньшей мере приблизительно 97%, или по меньшей мере приблизительно 98%, или по меньшей мере приблизительно 99% идентичностью последовательности по отношению к аминокислотной последовательности любой из SEQ ID NO: 5 или 8.
В ряде вариантов реализации химерный белок согласно настоящему изобретению может содержать последовательность, содержащую одну или более из мутаций аминокислот по отношению к любой из последовательностей, описанных в настоящем документе. В ряде вариантов реализации химерный белок содержит последовательность, содержащую приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 100 или более аминокислотных мутаций по отношению к любой из аминокислотных последовательностей химерных белков, описанных в настоящем документе.
В ряде вариантов реализации одна или более аминокислотных мутаций могут быть независимо выбраны из замен, инсерций, делеций и укорачиваний.
В ряде вариантов реализации аминокислотные мутации представляют собой аминокислотные замены и могут включать консервативные и/или неконсервативные замены.
«Консервативные замены» можно выполнить, например, на основе сходства по полярности, заряду, размеру, растворимости, гидрофобности, гидрофильности и/или амфипатической природе участвующих аминокислотных остатков. 20 природных аминокислот можно разделить на следующие шесть стандартных групп аминокислот: (1) гидрофобные: Met, Ala, Val, Leu, He; (2) нейтральные гидрофильные: Cys, Ser, Thr; Asn, Gln; (3) кислые: Asp, Glu; (4) основные: His, Lys, Arg; (5) остатки, которые влияют на ориентацию цепи: Gly, Pro; и (6) ароматические: Trp, Tyr, Phe.
В контексте настоящего документа «консервативные замены» определяют как замену аминокислоты на другую аминокислоту, входящую в ту же группу из шести стандартных групп аминокислот, приведенных выше. Например, замена Asp на Glu сохраняет один отрицательный заряд в модифицированном таким образом полипептиде. Кроме того, глицин и пролин можно заменять друг на друга на основе их способности нарушать структуру α-спирали.
В контексте настоящего документа «неконсервативные замены» определяют как замену одной аминокислоты на другую аминокислоту, входящую в другую группу из шести стандартных групп аминокислот (1)-(6), приведенных выше.
В ряде вариантов реализации замены могут также включать неклассические аминокислоты (например, селенцистеин, пирролизин, N-формилметионин, β-аланин, ГАМК и δ-аминолевулиновую кислоту, 4-аминобензойную кислоту (ПАБК), D-изомеры стандартных аминокислот, 2,4-диаминомасляную кислоту, α-аминоизомасляную кислоту, 4-аминомасляную кислоту, Abu, 2-аминомасляную кислоту, γ-Abu, ε-Ahx, 6-аминогексановую кислоту, Aib, 2-аминоизомасляную кислоту, 3-аминопропионовую кислоту, орнитин, норлейцин, норвалин, гидроксипролин, саркозин, цитруллин, гомоцитруллин, цистеиновую кислоту, трет-бутилглицин, трет-бутилаланин, фенилглицин, циклогексилаланин, β-аланин, фтораминокислоты, сконструированные аминокислоты, например, β-метиламинокислоты, С-α-метиламинокислоты, N-α-метиламинокислоты и аналоги аминокислот в целом).
Кроме того, можно внедрить мутации в нуклеотидные последовательности химерных белков со ссылкой на генетический код, в том числе с учетом вырожденности кодонов.
В ряде вариантов реализации химерный белок содержит линкер. В ряде вариантов реализации линкер содержит по меньшей мере один остаток цистеина, способный образовывать дисульфидную связь. Как описано в других местах настоящего документа, такой по меньшей мере один остаток цистеина, способный образовывать дисульфидную связь, отвечает, безотносительно к теоретическим представлениям, за поддержание надлежащего мультимерного состояния химерного белка и обеспечение эффективной продукции.
В ряде вариантов реализации химерный белок согласно настоящему изобретению содержит (а) первый домен, содержащий фрагмент рецептора колониестимулирующего фактора 1 (CSF1R), например, внеклеточный домен CSF1R, способный связывать лиганд CSF1R; (B) второй домен, содержащий фрагмент лиганда CD40 (CD40L), например, внеклеточный домен CD40L, способный связывать рецептор CD40L; и (с) линкер, соединяющий первый и второй домены.
В ряде вариантов реализации химерный белок представляет собой рекомбинантный гибридный белок, например, одиночный полипептид, содержащий внеклеточные домены, описанные в настоящем документе (и, необязательно, линкер). Например, в ряде вариантов реализации химерный белок транслируется в клетке как единое целое. В ряде вариантов реализации химерный белок относится к рекомбинантному белку из нескольких полипептидов, например, нескольких внеклеточных доменов, описанных в настоящем документе, связанных с образованием единого целого, например, in vitro (например, с помощью одного или более синтетических линкеров, описанных в настоящем документе). В ряде вариантов реализации химерный белок синтезируют химическим путем в виде одного полипептида, или можно отдельно синтезировать каждый домен химическим путем и затем объединить их. В ряде вариантов реализации фрагмент химерного белка транслируют, а другой фрагмент синтезируют химическим путем.
В ряде вариантов реализации химерные белки согласно настоящему изобретению могут представлять собой варианты, описанные в настоящем документе, например, химерные белки согласно настоящему изобретению могут содержать последовательность, обладающую по меньшей мере приблизительно 60%, или по меньшей мере приблизительно 61%, или по меньшей мере приблизительно 62%, или по меньшей мере приблизительно 63%, или по меньшей мере приблизительно 64%, или по меньшей мере приблизительно 65%, или по меньшей мере приблизительно 66%, или по меньшей мере приблизительно 67%, или по меньшей мере приблизительно 68%, или по меньшей мере приблизительно 69%, или по меньшей мере приблизительно 70%, или по меньшей мере приблизительно 71%, или по меньшей мере приблизительно 72%, или по меньшей мере приблизительно 73%, или по меньшей мере приблизительно 74%, или по меньшей мере приблизительно 75%, или по меньшей мере приблизительно 76%, или по меньшей мере приблизительно 77%, или по меньшей мере приблизительно 78%, или по меньшей мере приблизительно 79%, или по меньшей мере приблизительно 80%, или по меньшей мере приблизительно 81%, или по меньшей мере приблизительно 82%, или по меньшей мере приблизительно 83%, или по меньшей мере приблизительно 84%, или по меньшей мере приблизительно 85%, или по меньшей мере приблизительно 86%, или по меньшей мере приблизительно 87%, или по меньшей мере приблизительно 88%, или по меньшей мере приблизительно 89%, или по меньшей мере приблизительно 90%, или по меньшей мере приблизительно 91%, или по меньшей мере приблизительно 92%, или по меньшей мере приблизительно 93%, или по меньшей мере приблизительно 94%, или по меньшей мере приблизительно 95%, или по меньшей мере приблизительно 96%, или по меньшей мере приблизительно 97%, или по меньшей мере приблизительно 98%, или по меньшей мере приблизительно 99% идентичностью последовательности по отношению к аминокислотной последовательности указанных химерных белков, например, одной или более из любой из SEQ ID NO: 5 или 8.
В ряде вариантов реализации химерный белок содержит линкер. В ряде вариантов реализации линкер может происходить от встречающихся в природе многодоменных белков или представлять собой эмпирические линкеры, описанные, например, в источниках Chichili et al., (2013), Protein Sci. 22(2): 153-167, Chen et al., (2013), Adv Drug Deliv Rev. 65(10): 1357-1369, полностью включенных в настоящий документ посредством ссылки. В ряде вариантов реализации линкер можно сконструировать с использованием баз данных и компьютерных программ, например, описанных в источниках Chen et al., (2013), Adv Drug Deliv Rev. 65(10): 1357-1369 и Crasto et. al., (2000), Protein Eng. 13(5):309-312, полностью включенных в настоящий документ посредством ссылок.
В ряде вариантов реализации линкер представляет собой синтетический линкер, например, ПЭГ.
В ряде вариантов реализации линкер содержит полипептид. В ряде вариантов реализации длина полипептида составляет менее приблизительно 500 аминокислот, приблизительно 450 аминокислот, приблизительно 400 аминокислот, приблизительно 350 аминокислот, приблизительно 300 аминокислот, приблизительно 250 аминокислот, приблизительно 200 аминокислот, приблизительно 150 аминокислот или приблизительно 100 аминокислот.Например, длина линкера может составлять менее приблизительно 100, приблизительно 95, приблизительно 90, приблизительно 85, приблизительно 80, приблизительно 75, приблизительно 70, приблизительно 65, приблизительно 60, приблизительно 55, приблизительно 50, приблизительно 45, приблизительно 40, приблизительно 35, приблизительно 30, приблизительно 25, приблизительно 20, приблизительно 19, приблизительно 18, приблизительно 17, приблизительно 16, приблизительно 15, приблизительно 14, приблизительно 13, приблизительно 12, приблизительно 11, приблизительно 10, приблизительно 9, приблизительно 8, приблизительно 7, приблизительно 6, приблизительно 5, приблизительно 4, приблизительно 3 или приблизительно 2 аминокислот.В ряде вариантов реализации линкер является гибким. В одном варианте реализации линкер является жестким.
В ряде вариантов реализации линкер по существу состоит из остатков глицина и серина (например, содержит приблизительно 30%, или приблизительно 40%, или приблизительно 50%, или приблизительно 60%, или приблизительно 70%, или приблизительно 80%, или приблизительно 90%, или приблизительно 95%, или приблизительно 97%, или приблизительно 98%, или приблизительно 99%, или приблизительно 100% остатков глицина и серина).
В ряде вариантов реализации линкер представляет собой шарнирную область антитела (например, IgG, IgA, IgD и IgE, включая подклассы (например, IgG1, IgG2, IgG3 и IgG4, и IgA1 и IgA2)). Шарнирная область, присутствующая в антителах классов IgG, IgA, IgD и IgE, действует как гибкий спейсер, позволяя Fab-фрагменту свободно перемещаться в пространстве. В отличие от константных областей, шарнирные домены структурно разнообразны и различаются как по последовательности, так и по длине в иммуноглобулинах различных классов и подклассов. Например, длина и гибкость шарнирной области различается среди подклассов IgG. Шарнирная область IgG1 охватывает аминокислоты 216-231 и, поскольку она является гибкой в любых направлениях, Fab-фрагменты могут вращаться вокруг своих осей симметрии и перемещаться внутри сферы, центр которой находится в первом из двух дисульфидных мостиков между тяжелыми цепями. IgG2 содержит более короткий шарнир, чем IgG1; он состоит из 12 аминокислотных остатков и четырех дисульфидных мостиков. В шарнирной области IgG2 отсутствует остаток глицина, она является относительно короткой и содержит жесткую полипролиновую двойную спираль, стабилизированную дополнительными дисульфидными мостиками между тяжелыми цепями. Эти свойства ограничивают гибкость молекулы IgG2. IgG3 отличается от других подклассов своей уникальной увеличенной шарнирной областью (приблизительно в четыре раза более длинной, чем шарнир IgG1), содержащей 62 аминокислоты (включая 21 остаток пролина и 11 остатков цистеина) и образующей негибкую полипролиновую двойную спираль. В IgG3 Fab-фрагменты находятся относительно далеко от Fc-фрагмента, что придает молекуле большую гибкость. Удлиненный шарнир в IgG3 также отвечает за его более высокую молекулярную массу по сравнению с другими подклассами. Шарнирная область IgG4 короче, чем у IgG1, и ее гибкость является промежуточной между гибкостью IgG1 и IgG2. По имеющейся информации, гибкость шарнирных областей уменьшается в следующем порядке: IgG3>IgG1>IgG4>IgG2. В ряде вариантов реализации линкер может происходить от IgG4 человека и содержать одну или более из мутаций для усиления димеризации (включая S228P) или связывания FcRn.
Согласно кристаллографическим исследованиям, шарнирную область иммуноглобулина можно с функциональной точки зрения дополнительно разделить на три области: верхнюю шарнирную область, центральную область и нижнюю шарнирную область. См. источник Shin et al., 1992 Immunological Reviews 130:87. Верхняя шарнирная область включает аминокислоты от С-конца CH1 до первого остатка в шарнире, который ограничивает движение, обычно первого остатка цистеина, который образует межцепочечную дисульфидную связь между двумя тяжелыми цепями. Длина верхней шарнирной области коррелирует с сегментальной гибкостью антитела. Центральная шарнирная область содержит дисульфидные мостики между тяжелыми цепями, а нижняя шарнирная область присоединяется к N-концу домена CH2 и включает остатки в CH2. См. тот же источник. Центральная шарнирная область IgG1 человека дикого типа содержит последовательность СРРС (SEQ ID NO: 48), которая при димеризации путем образования дисульфидной связи образует циклический октапептид, который, как полагают, действует в качестве стержня, таким образом обеспечивая гибкость. В ряде вариантов реализации указанный линкер содержит одну или две или три из верхней шарнирной области, центральной шарнирной области и нижней шарнирной области любого антитела (например, IgG, IgA, IgD и IgE, включая подклассы (например, IgG1, IgG2, IgG3 и IgG4, а также IgA1 и IgA2)). Шарнирная область также может содержать один или более из сайтов гликозилирования, которые включают ряд структурно различающихся типов сайтов для присоединения углеводов. Например, IgA1 содержит пять сайтов гликозилирования в сегменте шарнирной области длиной 17 аминокислот, что придает полипептиду шарнирной области устойчивость к кишечным протеазам, что считается желательным свойством для секреторного иммуноглобулина. В ряде вариантов реализации линкер согласно настоящему изобретению содержит один или более из сайтов гликозилирования.
В ряде вариантов реализации линкер содержит Fc-домен антитела (например, IgG, IgA, IgD и IgE, включая подклассы (например, IgG1, IgG2, IgG3 и IgG4, а также IgA1 и IgA2)). В ряде вариантов реализации линкер содержит Fc-домен шарнир-СН2-СН3, полученный из антитела IgG4 человека. В ряде вариантов реализации линкер содержит Fc-домен шарнир-СН2-СН3, полученный из антитела IgG1 человека. В ряде вариантов реализации Fc-домен характеризуется повышенным сродством и усиленным связыванием с неонатальным Fc-рецептором (FcRn). В ряде вариантов реализации Fc-домен включает одну или более из мутаций, увеличивающих сродство и усиливающих связывание с FcRn. Безотносительно к теоретическим представлениям, считается, что повышенное сродство и усиленное связывание с FcRn повышает время полужизни указанных химерных белков in vivo.
В ряде вариантов реализации Fc-домен в линкере содержит одну или более из аминокислотных замен по аминокислотному остатку 250, 252, 254, 256, 308, 309, 311, 416, 428, 433 или 434 (в соответствии с системой нумерации согласно Kabat, описанной в источнике Kabat, et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991), специально включенном в настоящий документ посредством ссылки), или их эквиваленты. В одном варианте реализации аминокислотная замена по аминокислотному остатку 250 представляет собой замену на глутамин. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 252 представляет собой замену на тирозин, фенилаланин, триптофан или треонин. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 254 представляет собой замену на треонин. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 256 представляет собой замену на серии, аргинин, глутамин, глутаминовую кислоту, аспарагиновую кислоту или треонин. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 308 представляет собой замену на треонин. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 309 представляет собой замену на пролин. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 311 представляет собой замену на серии. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 385 представляет собой замену на аргинин, аспарагиновую кислоту, серии, треонин, гистидин, лизин, аланин или глицин. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 386 представляет собой замену на треонин, пролин, аспарагиновую кислоту, серии, лизин, аргинин, изолейцин или метионин. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 387 представляет собой замену на аргинин, пролин, гистидин, серии, треонин или аланин. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 389 представляет собой замену на пролин, серии или аспарагин. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 416 представляет собой замену на серии. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 428 представляет собой замену на лейцин. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 433 представляет собой замену на аргинин, серии, изолейцин, пролин или глутамин. В ряде вариантов реализации аминокислотная замена по аминокислотному остатку 434 представляет собой замену на гистидин, фенилаланин или тирозин.
В ряде вариантов реализации Fc-домен в линкере (например, содержащий константную область IgG) содержит одну или более из мутаций, например, замен по аминокислотному остатку 252, 254, 256, 433, 434 или 436 (в соответствии с системой нумерации согласно Kabat, описанной в источнике Kabat, et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991), специально включенном в настоящий документ посредством ссылки). В ряде вариантов реализации константная область IgG включает тройную мутацию M252Y/S254T/T256E или мутацию YTE. В одном варианте реализации константная область IgG включает тройную мутацию H433K/N434F/Y436H или мутацию KFH. В ряде вариантов реализации константная область IgG включает мутацию YTE в комбинации с мутацией KFH.
В ряде вариантов модифицированные гуманизированные антитела согласно настоящему изобретению содержат константную область IgG, содержащую одну или более из мутаций по аминокислотным остаткам 250, 253, 307, 310, 380, 428, 433, 434 и 435 (в соответствии с системой нумерации согласно Kabat, описанной в источнике Kabat, et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991), специально включенном в настоящий документ посредством ссылки). Типичные мутации включают T250Q, M428L, Т307А, Е380А, I253A, Н310А, M428L, H433K, N434A, N434F, N434S и Н435А. В ряде вариантов реализации константная область IgG содержит мутацию M428L/N434S или мутацию LS. В одном варианте реализации константная область IgG содержит мутацию T250Q/M428L или мутацию QL. В одном варианте реализации константная область IgG содержит мутацию N434A. В одном варианте реализации константная область IgG содержит мутацию T307A/E380A/N434A или мутацию AAA. В одном варианте реализации константная область IgG содержит мутацию I253A/H310A/H435A или мутацию IHH. В одном варианте реализации константная область IgG содержит мутацию H433K/N434F. В одном варианте реализации константная область IgG содержит мутацию M252Y/S254T/T256E в комбинации с мутацией H433K/N434F.
Дополнительные типичные мутации в константной области IgG описаны, например, в источниках Robbie, et al., Antimicrobial Agents and Chemotherapy (2013), 57(12):6147-6153, Dall'Acqua et al., JBC (2006), 281(33):23514-24, Dall'Acqua et al., Journal of Immunology (2002), 169:5171-80, Ко et al. Nature (2014) 514:642-645, Grevys et al. Journal of Immunology. (2015), 194(11):5497-508 и патенте США №7083784, полностью включенных в настоящий документ посредством ссылок.
В ряде вариантов реализации линкер содержит аминокислотную последовательность SEQ ID NO: 25 (см. таблицу ниже) или последовательность, идентичную ей по меньшей мере на 90%, или 93%, или 95%, или 97%, или 98%, или 99%. В ряде вариантов реализации мутации вносят в SEQ ID NO: 25 для увеличения стабильности и/или времени полужизни. Например, в ряде вариантов реализации Fc-домен в линкере содержит аминокислотную последовательность SEQ ID NO: 26 (см. таблицу ниже) или последовательность, идентичную ей по меньшей мере на 90%, или 93%, или 95%, или 97%, или 98%, или 99%. Примером Fc-стабилизирующего мутанта является S228P. Примерами мутантов Fc, увеличивающих время полужизни, являются T250Q, M428L, V308T, L309P и Q311S, и указанные линкеры могут содержать 1, или 2, или 3, или 4, или 5 из этих мутаций. Например, в ряде вариантов реализации Fc-домен в линкере содержит аминокислотную последовательность SEQ ID NO: 27 (см. таблицу ниже) или последовательность, идентичную ей по меньшей мере на 90%, или 93%, или 95%, или 97%, или 98%, или 99%.
Кроме того, один или более из соединяющих линкеров можно использовать для соединения Fc-домена в линкере (например, одной из SEQ ID NO: 25, SEQ ID NO: 26, SEQ ID NO: 27, или последовательности, идентичной им по меньшей мере на 90%, или 93%, или 95%, или 97%, или 98%, или 99%) и внеклеточных доменов. Например, любая из SEQ ID NO: 28, SEQ ID NO: 29, SEQ ID NO: 30, SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33 или их варианты могут соединять внеклеточный домен (ВКД), описанный в настоящем документе, и Fc-домен в линкере, описанный в настоящем документе. Любая из SEQ ID NO: 28-74 или их варианты необязательно располагаются между внеклеточным доменом, описанным в настоящем документе, и Fc-доменом, описанным в настоящем документе. В ряде вариантов реализации химерный белок содержит один соединяющий линкер, предшествующий Fc-домену, и второй соединяющий линкер, следующий за Fc-доменом; таким образом, химерный белок может содержать следующую структуру:
ВКД 1 (например, CSF1R) - соединяющий линкер 1 - Fc-домен - соединяющий линкер 2 - ВКД 2 (например, CD40L).
В ряде вариантов реализации первый и второй соединяющие линкеры могут быть разными или одинаковыми.
В ряде вариантов реализации первый и второй соединяющие линкеры могут быть выбраны из аминокислотных последовательностей SEQ ID NO: 25 - 74 и приведены в таблице 1 ниже:



В ряде вариантов реализации соединяющий линкер в основном содержит остатки глицина и серина (например, приблизительно 30%, или приблизительно 40%, или приблизительно 50%, или приблизительно 60%, или приблизительно 70%, или приблизительно 80%, или приблизительно 90%, или приблизительно 95%, или приблизительно 97%, или приблизительно 98%, или приблизительно 99%, или приблизительно 100% остатков глицина и серина). Например, в ряде вариантов реализации соединяющий линкер представляет собой (Gly4Ser)n, где п составляет от приблизительно 1 до приблизительно 8, например, 1, 2, 3, 4, 5, 6, 7 или 8 (SEQ ID NO: 49 - SEQ ID NO: 56, соответственно). В ряде вариантов реализации последовательность соединяющего линкера представляет собой GGSGGSGGGGSGGGGS (SEQ ID NO: 57). Дополнительные типичные соединяющие линкеры включают линкеры, содержащие последовательность LE, (Gly)8 (SEQ ID NO: 58), (Gly)6 (SEQ ID NO: 59), (EAAAK)n (n=1-3) (SEQ ID NO: 60 - SEQ ID NO: 62), A(EAAAK)nA (n=2-5) (SEQ ID NO: 63 - SEQ ID NO: 66), A(EAAAK)4ALEA(EAAAK)4A (SEQ ID NO: 67), PAPAP (SEQ ID NO: 68), KESGSVSSEQLAQFRSLD (SEQ ID NO: 69), GSAGSAAGSGEF (SEQ ID NO: 70) и (XP)n, где X обозначает любую аминокислоту, например, Ala, Lys или Glu, но не ограничиваются ими. В ряде вариантов реализации соединяющий линкер представляет собой GGS.
В ряде вариантов реализации соединяющий линкер представляет собой один или более из GGGSE (SEQ ID NO: 71), GSESG (SEQ ID NO: 72), GSEGS (SEQ ID NO: 73), GEGGSGEGSSGEGSSSEGGGSEGGGSEGGGSEGGS (SEQ ID NO: 74), и соединяющий линкер, состоящий из G, S и Е, случайным образом размещенных через каждые 4 аминокислоты.
В ряде вариантов реализации химерный белок содержит модульный линкер, показанный на фиг.20.
В ряде вариантов реализации линкер может быть функциональным. Например, линкер может выполнять функцию, состоящую в улучшении фолдинга и/или стабильности, улучшения экспрессии, улучшения фармакокинетики и/или улучшения биологической активности настоящего химерного белка, но не ограничиваться этим. В еще одном примере линкер может выполнять функцию, состоящую в обеспечении адресного воздействия химерного белка на конкретный тип клеток или конкретное местоположение.
В ряде вариантов реализации химерный белок демонстрирует повышенную стабильность и время полужизни белка. В ряде вариантов реализации химерный белок связывается с FcRn с высоким сродством. В ряде вариантов реализации химерный белок способен связываться с FcRn с KD, составляющей от приблизительно 1 нМ до приблизительно 80 нМ. Например, химерный белок способен связываться с FcRn с KD, составляющей приблизительно 1 нМ, приблизительно 2 нМ, приблизительно 3 нМ, приблизительно 4 нМ, приблизительно 5 нМ, приблизительно 6 нМ, приблизительно 7 нМ, приблизительно 8 нМ, приблизительно 9 нМ, приблизительно 10 нМ, приблизительно 15 нМ, приблизительно 20 нМ, приблизительно 25 нМ, приблизительно 30 нМ, приблизительно 35 нМ, приблизительно 40 нМ, приблизительно 45 нМ, приблизительно 50 нМ, приблизительно 55 нМ, приблизительно 60 нМ, приблизительно 65 нМ, приблизительно 70 нМ, приблизительно 71 нМ, приблизительно 72 нМ, приблизительно 73 нМ, приблизительно 74 нМ, приблизительно 75 нМ, приблизительно 76 нМ, приблизительно 77 нМ, приблизительно 78 нМ, приблизительно 79 нМ или приблизительно 80 нМ. В ряде вариантов реализации химерный белок способен связываться с FcRn с KD, составляющей приблизительно 9 нМ. В ряде вариантов реализации химерный белок по существу не связывается с другими Fc-рецепторами (т.е. отличающимися от FcRn) с эффекторной функцией.
В ряде вариантов реализации химерный белок с формулой ВКД 1 - соединяющий линкер 1 - Fc-домен - соединяющий линкер 2 - ВКД 2, где ВКД 1 представляет собой CSF1R, а ВКД 2 представляет собой CD40L, в настоящем документе может называться CSF1R-Fc-CD40L. В ряде вариантов реализации химерный белок не содержит один или оба соединяющие линкеры; такой химерный белок в настоящем документе также может называться CSF1R-Fc-CD40L.
В ряде вариантов реализации химерный белок представляет собой гибридный белок с формулой N-конец - (а) - (b) - (с) - С-конец, где (а) представляет собой CSF1R, (b) представляет собой линкер, содержащий по меньшей мере фрагмент Fc-домена, и (с) представляет собой CD40L, и может называться в настоящем документе CSF1R-Fc-CD40L.
В ряде вариантов реализации химерный белок оптимизирован для/специфичен по отношению к лигандам/рецепторам мыши; примером такого химерного белка является CSF1R-Fc-CD40L мыши, также называемый в настоящем документе mCSF1R-Fc-CD40L.
В ряде вариантов реализации химерный белок оптимизирован для/специфичен по отношению к лигандам/рецепторам человека; примером такого химерного белка является CSF1R-Fc-CD40L человека, также называемый в настоящем документе hCSF1R-Fc-CD40L.
Эти химерные белки могут не содержать один или оба соединяющие линкеры. Типичные соединяющие линкеры 1, Fc-домены и соединяющие линкеры 2 описаны выше в таблице 1; модульные линкеры, пригодные для образования химерных белков и содержащие конкретные соединяющие линкеры 1, Fc-домены и соединяющие линкеры 2, показаны на фиг.20. В ряде вариантов реализации химерный белок сконструирован так, что его мишенью является иммуноингибиторный сигнальный путь CSF1R/CSF1. В ряде вариантов реализации химерный белок сконструирован так, чтобы нарушать, блокировать, уменьшать и/или ингибировать передачу иммуноингибиторного сигнала, опосредованного связыванием CSF1 с CSF1R. В ряде вариантов реализации иммунноингибиторный сигнал относится к сигналу, который уменьшает или устраняет иммунный ответ. Например, в контексте онкологии такие сигналы могут уменьшать или устранять противоопухолевый иммунитет.В нормальных физиологических условиях ингибиторные сигналы используются для поддержания аутотолерантности (например, для предотвращения аутоиммунитета), а также для защиты тканей от повреждений при реакции иммунной системы на патогенную инфекцию. Например, в числе прочего, иммуноингибиторный сигнал можно выявить путем обнаружения усиления клеточной пролиферации, продукции цитокинов, активности по уничтожению клеток или фагоцитарной активности при блокировании такого ингибиторного сигнала.
В ряде вариантов реализации химерный белок согласно настоящему изобретению нарушает, блокирует, уменьшает и/или ингибирует передачу иммуноингибиторного сигнала, опосредованного связыванием CSF1 или ИЛ-34 с CSF1R. В ряде вариантов реализации химерный белок связывается с CSF1 или ИЛ-34 и блокирует их, тем самым нарушая, блокируя, уменьшая и/или ингибируя передачу ингибиторного сигнала к иммунной клетке (например, опухолеассоциированному макрофагу, антигенпрезентирующей клетке, миелоидной клетке или Т-клетке).
В ряде вариантов реализации химерные белки согласно настоящему изобретению могут осуществлять или находят применение в способах, включающих ингибирование или уменьшение связывания пар иммуноингибиторный рецептор/лиганд. CSF1R/CSF1 или CSFlR/ИЛ-34. В ряде вариантов реализации химерный белок согласно настоящему изобретению блокирует, уменьшает и/или ингибирует активацию CSF1R, например, снижая связывание CSF1R на иммунных клетках с CSF1 или ИЛ-34.
В ряде вариантов реализации мишенью указанного химерного белка является иммуностимулирующий сигнал, опосредованный связыванием CD40L с CD40. В одном варианте реализации химерный белок сконструирован таким образом, чтобы усиливать, увеличивать и/или стимулировать передачу иммуностимулирующего сигнала, опосредованного связыванием CD40L с CD40. В ряде вариантов реализации иммуностимулирующий сигнал относится к сигналу, который усиливает иммунный ответ.Например, в контексте онкологии такие сигналы могут усиливать противоопухолевый иммунитет.Например, в числе прочего, иммуностимулирующий сигнал можно выявить по прямой стимуляции пролиферации, продукции цитокинов, активности по уничтожению или фагоцитарной активности лейкоцитов, в том числе субпопуляций Т-клеток.
В ряде вариантов реализации химерный белок согласно настоящему изобретению усиливает, увеличивает и/или стимулирует передачу иммуностимулирующего сигнала, опосредованного связыванием CD40L с CD40. В ряде вариантов реализации химерный белок согласно настоящему изобретению, содержащий внеклеточный домен CD40L, действует на иммунную клетку (например, дендритную клетку, В-клетку, макрофаг, антигенпрезентирующую клетку или Т-клетку), экспрессирующую CD40, и усиливает, увеличивает и/или стимулирует передачу стимулирующего сигнала к иммунной клетке (например, дендритной клетке, В-клетке, макрофагу и Т-клетке).
В ряде вариантов реализации химерные белки согласно настоящему изобретению могут осуществлять или находят применение в способах, включающих стимуляцию или усиление связывания пары иммуностимулирующий рецептор/лиганд: CD40:CD40L. В ряде вариантов реализации химерный белок согласно настоящему изобретению увеличивает и/или стимулирует CD40 и/или связывание CD40 с одним или более лигандами CD40L.
В ряде вариантов реализации химерный белок содержит внеклеточный белка II типа, отличающегося от CD40L. Примеры белков II типа включают 4-1BBL, CD30L, FasL, GITRL, LIGHT, OX40L, TL1A и TRAIL. Настоящее изобретение дополнительно включает химерные белки и способы применения следующих химерных белков: CSF1R/4-1BBL, CSF1R/CD30L, CSFlR/FasL, CSF1R/GITRL, CSF1R/LIGHT, CSF1R/OX40L, CSF1R/TL1A и CSF1R/TRAIL. В ряде вариантов реализации химерный белок обладает общей структурой одного из белков CSF1R-Fc-4-1BBL, CSF1R-Fc-CD30L, CSF1R-Fc-FasL, CSF1R-Fc-GITRL, CSF1R-Fc-LIGHT, CSF1R-Fc-OX40L, CSF1R-Fc-TL1A и CSF1R-Fc-TRAIL.
Аминокислотные последовательности 4-1BBL, CD30L, FasL, GITRL, LIGHT, OX40L, TL1A и TRAIL содержат соответственно SEQ ID NO: 9, 11, 13, 15, 17, 6, 21 и 23.
В ряде вариантов реализации химерный белок содержит внеклеточный домен одного из 4-1BBL, CD30L, FasL, GITRL, LIGHT, OX40L, TL1A и TRAIL, которые содержат, соответственно, SEQ ID NO: 10, 12, 14, 16, 18, 7, 22 и 24. В ряде вариантов реализации химерные белки согласно настоящему изобретению могут содержать внеклеточный домен 4-1BBL, CD30L, FasL, GITRL, LIGHT, OX40L, TL1A или TRAIL, описанный в настоящем документе, или его вариант или функциональный фрагмент. Например, химерный белок может содержать последовательность внеклеточного домена 4-1BBL, CD30L, FasL, GITRL, LIGHT, OX40L, TL1A или TRAIL, представленную выше, или ее вариант или функциональный фрагмент, обладающий по меньшей мере приблизительно 60%, или по меньшей мере приблизительно 61%, или по меньшей мере приблизительно 62%, или по меньшей мере приблизительно 63%, или по меньшей мере приблизительно 64%, или по меньшей мере приблизительно 65%, или по меньшей мере приблизительно 66%, или по меньшей мере приблизительно 67%, или по меньшей мере приблизительно 68%, или по меньшей мере приблизительно 69%, или по меньшей мере приблизительно 70%, или по меньшей мере приблизительно 71%, или по меньшей мере приблизительно 72%, или по меньшей мере приблизительно 73%, или по меньшей мере приблизительно 74%, или по меньшей мере приблизительно 75%, или по меньшей мере приблизительно 76%, или по меньшей мере приблизительно 77%, или по меньшей мере приблизительно 78%, или по меньшей мере приблизительно 79%, или по меньшей мере приблизительно 80%, или по меньшей мере приблизительно 81%, или по меньшей мере приблизительно 82%, или по меньшей мере приблизительно 83%, или по меньшей мере приблизительно 84%, или по меньшей мере приблизительно 85%, или по меньшей мере приблизительно 86%, или по меньшей мере приблизительно 87%, или по меньшей мере приблизительно 88%, или по меньшей мере приблизительно 89%, или по меньшей мере приблизительно 90%, или по меньшей мере приблизительно 91%, или по меньшей мере приблизительно 92%, или по меньшей мере приблизительно 93%, или по меньшей мере приблизительно 94%, или по меньшей мере приблизительно 95%, или по меньшей мере приблизительно 96%, или по меньшей мере приблизительно 97%, или по меньшей мере приблизительно 98%, или по меньшей мере приблизительно 99% идентичностью последовательности по отношению к аминокислотной последовательности внеклеточного домена 4-1BBL, CD30L, FasL, GITRL, LIGHT, OX40L, TL1A, или TRAIL, описанного в настоящем документе.
В ряде вариантов реализации химерный белок согласно настоящему изобретению выполняет иммунную стимуляцию иммунной клетки (например, антигенпрезентирующей клетки), при этом представляя собой локализованную ловушку или блокатор иммуноингибиторного сигнала. В ряде вариантов реализации химерный белок обеспечивает передачу сигналов, в конечном итоге приводящую к активации иммунного ответа.
В ряде вариантов реализации химерные белки согласно настоящему изобретению могут осуществлять и могут применяться в способах, включающих стимулирование иммунной активации (например, против опухолей). В ряде вариантов реализации химерные белки согласно настоящему изобретению могут осуществлять и могут применяться в способах, включающих подавление иммунного ингибирования (например, позволяющего опухолям выживать). В ряде вариантов реализации химерные белки согласно настоящему изобретению обеспечивают улучшенную иммунную активацию и/или улучшенное подавление иммунного ингибирования из-за близости передаваемых сигналов, обеспечиваемой химерной природой конструктов.
В ряде вариантов реализации химерные белки согласно настоящему изобретению могут осуществлять или могут применяться в способах, включающих модуляцию амплитуды иммунного ответа, например, модуляцию уровня эффекторного ответа. В ряде вариантов реализации, например, при применении для лечения рака и/или воспалительного заболевания, химерные белки согласно настоящему изобретению изменяют степень иммунной стимуляции по сравнению с иммунным ингибированием с целью увеличения амплитуды Т-клеточного ответа, включая, без ограничений, стимуляцию повышенных уровней продукции цитокинов, пролиферации или потенциала уничтожения мишеней.
В ряде вариантов реализации химерные белки согласно настоящему изобретению могут осуществлять или находят применение в способах, включающих маскирование ингибиторного лиганда на поверхности опухолевой клетки и замену этого иммуноингибиторного лиганда иммуностимулирующим лигандом. Например, химерный белок согласно настоящему изобретению содержит (а) внеклеточный домен CSF1R и (b) внеклеточный домен CD40L, и позволяет нарушать ингибиторный сигнал CSF1/CSF1R и заменять его стимулирующим сигналом CD40L/CD40. Соответственно, химерные белки согласно настоящему изобретению в ряде вариантов реализации способны осуществлять или находят применение в способах, включающих уменьшение или устранение иммуноингибиторного сигнала и/или увеличение или активацию иммуностимулирующего сигнала. Например, опухоль, несущая ингибиторный сигнал (и таким образом уклоняющаяся от иммунного ответа), может быть заменена положительным сигналом, связывающимся с макрофагом или Т-клеткой, которая затем может атаковать опухолевую клетку. Соответственно, в ряде вариантов реализации иммуноингибиторный сигнал маскируется конструктами согласно настоящему изобретению и активируется иммуностимулирующий сигнал. Такие полезные свойства усиливаются подходом, используемым в химерных белках согласно настоящему изобретению, состоящим в использовании единого конструкта. Например, замену сигнала можно осуществлять почти одновременно, например, в одно и то же время, и замена сигнала приспособлена для локализации в области, имеющей клиническое значение (например, в микроокружении опухоли).
В ряде вариантов реализации химерные белки согласно настоящему изобретению способны осуществлять или находят применение в способах, включающих усиление, восстановление, способствование и/или стимуляцию иммунной модуляции. В ряде вариантов реализации химерные белки согласно настоящему изобретению, описанные в настоящем документе, восстанавливают, способствуют и/или стимулируют активность или активацию одной или более иммунных клеток против опухолевых клеток, включая: Т-клетки, цитотоксические Т-лимфоциты, Т-хелперные клетки, естественные киллеры (NK-клетки), Т-клетки, обладающие свойствами естественных киллеров (NKT), противоопухолевые макрофаги (например, макрофаги M1), В-клетки и дендритные клетки. В ряде вариантов реализации химерные белки согласно настоящему изобретению усиливают, восстанавливают, способствуют и/или стимулируют активность и/или активацию Т-клеток, включая, в качестве неограничивающего примера, активацию и/или стимуляцию одного или более из собственных Т-клеточных сигналов, включая сигнал, способствующий выживанию; аутокринный или паракринный сигнал роста; сигнал, опосредованный р38 MAPK, ERK, STAT, JAK, AKT или PI3K; антиапоптозный сигнал; и/или сигнал, способствующий и/или необходимый для одного или более из: продукции провоспалительных цитокинов или миграции Т-клеток или Т-клеточной инфильтрации опухоли.
В ряде вариантов реализации химерные белки согласно настоящему изобретению способны осуществлять или находят применение в способах, включающих увеличение количества одного или более из: Т-клеток (включая цитотоксические Т-лимфоциты, Т-хелперные клетки, Т-клетки, обладающие свойствами естественных киллеров (NKT), но не ограничиваясь ими), В клеток, естественных киллеров (NK-клеток), Т-клеток, обладающих свойствами естественных киллеров (NKT), дендритных клеток, моноцитов и макрофагов (например, одного или более из M1 и М2) в опухоли или в микроокружении опухоли. В ряде вариантов реализации химерный белок усиливает распознавание опухолевых антигенов CD8+ Т-клетками, в частности, Т-клетками, инфильтрирующими микроокружение опухоли. В ряде вариантов реализации химерный белок согласно настоящему изобретению индуцирует экспрессию CD19 и/или, увеличивает количество CD19-положительных клеток (например, CD19-положительных В-клеток). В одном варианте реализации химерный белок согласно настоящему изобретению индуцирует экспрессию ИЛ-15Rα и/или увеличивает количество ИЛ-15Rα-положительных клеток (например, ИЛ-15Rα-положительных дендритных клеток).
В ряде вариантов реализации химерные белки согласно настоящему изобретению способны осуществлять или находят применение в способах, включающих ингибирование и/или снижение рекрутирования иммуносупрессивных клеток (например, клеток-супрессоров миелоидного происхождения (MDSC), регуляторных Т-клеток (Treg), опухолеассоциированных нейтрофилов (TAN), макрофагов М2 и опухолеассоциированных макрофагов (ТАМ)), в особенности в опухоли и/или микроокружении опухоли (ТМЕ). В ряде вариантов реализации терапевтические средства согласно настоящему изобретению могут изменять соотношение макрофагов M1 и М2 в области опухоли и/или ТМЕ в пользу макрофагов M1.
В ряде вариантов реализации химерные белки согласно настоящему изобретению способны осуществлять и могут применяться в способах, включающих ингибирование и/или снижение инактивации Т-клеток и/или иммунной толерантности к опухоли, включающих введение субъекту эффективного количества химерного белка, описанного в настоящем документе. В ряде вариантов реализации химерные белки согласно настоящему изобретению способны увеличивать уровни различных цитокинов в сыворотке, включая один или более из ИФНγ, ФНОα, ИЛ-2, ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-9, ИЛ-10, ИЛ-13, ИЛ-17А, ИЛ-17Т и ИЛ-22, но не ограничиваясь ими. В ряде вариантов реализации химерные белки согласно настоящему изобретению способны повышать уровень ИЛ-2, ИЛ-4, ИЛ-5, ИЛ-10, ИЛ-13, ИЛ-17А, ИЛ-22, ФНОα или ИФНγ в сыворотке субъекта, получающего лечение. Обнаружение такого цитокинового ответа может обеспечить способ определения оптимальной схемы применения указанного химерного белка.
В ряде вариантов реализации химерные белки согласно настоящему изобретению ингибируют, блокируют и/или уменьшают гибель противоопухолевых CD8+ и/или CD4+ Т-клеток; или стимулируют, вызывают и/или увеличивают гибель проопухолевых Т-клеток. Истощение Т-клеток - это состояние дисфункции Т-клеток, характеризующееся прогрессирующей потерей пролиферативных и эффекторных функций, завершающееся клональной делецией. Соответственно, проопухолевая Т-клетка относится к состоянию дисфункции Т-клеток, которое возникает во время многих хронических инфекций, воспалительных заболеваний и рака. Эта дисфункция определяется слабыми пролиферативными и/или эффекторными функциями, устойчивой экспрессией ингибиторных рецепторов и состоянием транскрипции, отличным от такового у функциональных эффекторных Т-клеток или Т-клеток памяти. Истощение препятствует оптимальному контролю инфекции и опухолей. Примеры проопухолевых Т-клеток включают Treg, CD4+ и/или CD8+ Т-клетки, экспрессирующие один или более рецепторов, ингибирующих ключевые этапы иммунного ответа, Th2-клетки и Th17-клетки, но не ограничиваются ими. Рецепторы, ингибирующие ключевые этапы иммунного ответа, относятся к рецепторам, экспрессируемым на иммунных клетках, которые предотвращают или ингибируют неконтролируемые иммунные реакции. В противоположность этому, противоопухолевые CD8+ и/или CD4+ Т-клетки относятся к Т-клеткам, которые могут вызывать иммунный ответ против опухоли.
В ряде вариантов реализации химерные белки согласно настоящему изобретению способны осуществлять и могут применяться в способах, включающих увеличение отношения эффекторных Т-клеток к регуляторным Т-клеткам. Примеры эффекторных Т-клеток включают ICOS+ эффекторные Т-клетки; цитотоксические Т-клетки (например, αβ TCR, CD3+, CD8+, CD45RO+); CD4+ эффекторные Т-клетки (например, αβ TCR, CD3+, CD4+, CCR7+, CD62L(высокий), ИЛ-7R/CD127+); CD8+ эффекторные Т-клетки (например, αβ TCR, CD3+, CD8+, CCR7+, CD62L(высокий), ИЛ-7R/CD127+); эффекторные Т-клетки памяти (например, CD62L(низкий), CD44+, TCR, CD3+, ИЛ-7R/CD127+, ИЛ-15R+, CCR7(низкий)); центральные Т-клетки памяти (например, CCR7+, CD62L+, CD27+; или CCR7(высокий), CD44+, CD62L(высокий), TCR, CD3+, ИЛ-7R/CD127+, ИЛ-15R+); CD62L+ эффекторные Т-клетки; CD8+ эффекторные Т-клетки памяти (ТЕМ), включая ранние эффекторные Т-клетки памяти (CD27+CD62L-) и поздние эффекторные Т-клетки памяти (CD27- CD62L-) (TemE и TemL, соответственно); CD127(+)CD25(низкий/-) эффекторные Т-клетки; CD127(-)CD25(-) эффекторные Т-клетки; CD8+ стволовые эффекторные клетки памяти (TSCM) (например, CD44(низкий)CD62L(высокий)CD122(высокий)sca(+)); ТН1 эффекторные Т-клетки (например, CXCR3+, CXCR6+ и CCR5+; или αβ TCR, CD3+, CD4+, ИЛ-12R+, ИФНγR+, CXCR3+), ТН2 эффекторные Т-клетки (например, CCR3+, CCR4+ и CCR8+; или αβ TCR, CD3+, CD4+, ИЛ-4R+, HH-33R+, CCR4+, HH-17RB+, CRTH2+); TH9 эффекторные Т-клетки (например, αβ TCR, CD3+, CD4+); TH17 эффекторные Т-клетки (например, αβ TCR, CD3+, CD4+, ИЛ-23R+, CCR6+, ИЛ-1R+); CD4+CD45RO+CCR7+ эффекторные Т-клетки, CD4+CD45RO+CCR7(-) эффекторные Т-клетки; и эффекторные Т-клетки, секретирующие ИЛ-2, ИЛ-4 и/или ИФН-γ. Примеры регуляторных Т-клеток включают ICOS+ регуляторные Т-клетки, CD4+CD25+FOXP3+ регуляторные Т-клетки, CD4+CD25+ регуляторные Т-клетки, CD4+CD25- регуляторные Т-клетки, CD4 CD25(высокий) регуляторные Т-клетки, TIM-3+PD-1+ регуляторные Т-клетки, ген-3 активации лимфоцитов (LAG-3)+ регуляторные Т-клетки, CTLA-4/CD152+ регуляторные Т-клетки, нейтропилин-1 (Nrp-1)+ регуляторные Т-клетки, CCR4+CCR8+ регуляторные Т-клетки, CD62L (L-селектин)+ регуляторные Т-клетки, CD45RB(низкий) регуляторные Т-клетки, CD127(низкий) регуляторные Т-клетки, LRRC32/GARP+ регуляторные Т-клетки, CD39+ регуляторные Т-клетки, GITR+ регуляторные Т-клетки, LAP+ регуляторные Т-клетки, 1В11+ регуляторные Т-клетки, BTLA+ регуляторные Т-клетки, регуляторные Т-клетки 1 типа (Tr1-клетки), Т-хелперы 3 типа (Th3-клетки), регуляторные клетки с фенотипом Т-клеток, обладающих свойствами естественных киллеров (NKTreg), CD8+ регуляторные Т-клетки, CD8+CD28- регуляторные Т-клетки и/или регуляторные Т-клетки, секретирующие ИЛ-10, ИЛ-35, ТФР-β, ФНО-α, галектин-1, ИФН-γ и/или МСР1.
В ряде вариантов реализации химерный белок согласно настоящему изобретению вызывает увеличение количества эффекторных Т-клеток (например, CD4+CD25- Т-клеток).
В ряде вариантов реализации химерный белок вызывает снижение количества регуляторных Т-клеток (например, CD4+CD25+ Т-клеток).
В ряде вариантов реализации химерный белок генерирует анамнестическую реакцию, которая может, например, предотвратить рецидив или защитить животное от повторной антигенной стимуляции. Таким образом, животное, получившее лечение химерным белком, впоследствии способно атаковать опухолевые клетки и/или предупреждать развитие опухолей при повторной антигенной стимуляции после первоначального лечения химерным белком. Соответственно, химерный белок согласно настоящему изобретению стимулирует как активное разрушение опухоли, так и иммунологическое распознавание опухолевых антигенов, которые необходимы для программирования анамнестической реакции, способной предотвратить рецидив.
В ряде вариантов реализации химерные белки согласно настоящему изобретению способны осуществлять и могут применяться в способах, включающих временную стимуляцию эффекторных иммунных клеток на срок не более приблизительно 12 часов, приблизительно 24 часов, приблизительно 48 часов, приблизительно 72 часов или приблизительно 96 часов, или приблизительно 1 недели, или приблизительно 2 недель. В ряде вариантов реализации химерные белки согласно настоящему изобретению способны осуществлять и могут применяться в способах, включающих временное истощение или ингибирование регуляторных или иммуносупрессорных клеток на срок не более приблизительно 12 часов, приблизительно 24 часов, приблизительно 48 часов, приблизительно 72 часов или приблизительно 96 часов, или приблизительно 1 недели, или приблизительно 2 недель. В ряде вариантов реализации временная стимуляция эффекторных Т-клеток и/или временное истощение или ингибирование иммуноингибиторных клеток происходит по существу в кровотоке пациента или в конкретной ткани/месте, включая лимфоидные ткани, например, костный мозг, лимфатический узел, селезенка, тимус, лимфоидную ткань, ассоциированную со слизистыми оболочками (MALT), нелимфоидные ткани или микроокружение опухоли.
В ряде вариантов реализации химерные белки согласно настоящему изобретению обладают преимуществами, включающими, в числе прочего, простоту использования и простоту производства. Это связано с тем, что два отдельных агента для иммунотерапии объединены в единый продукт, что позволяет использовать один производственный процесс вместо двух независимых производственных процессов. Кроме того, введение одного агента вместо двух отдельных агентов обеспечивает упрощенное введение и лучшее соблюдение пациентом режима лечения.
В ряде вариантов реализации химерный белок согласно настоящему изобретению может продуцироваться в клетке-хозяине млекопитающего в виде единой секретируемой и полностью функциональной полипептидной цепи.
В ряде вариантов реализации химерный белок согласно настоящему изобретению неожиданно обеспечивает связывание компонентов внеклеточного домена с соответствующими партнерами по связыванию с низкими скоростями диссоциации (Kd или Koff). В ряде вариантов реализации это обеспечивает неожиданно длительное взаимодействие рецептора с лигандом и наоборот.Такое действие обеспечивает устойчивый эффект маскирования отрицательного сигнала. Кроме того, в ряде вариантов реализации это обеспечивает более продолжительный положительный сигнальный эффект, например, позволяющий обеспечить адекватную стимуляцию эффекторной клетки для противоопухолевого действия. Например, химерный белок согласно настоящему изобретению, например, ввиду связывания с низкой скоростью диссоциации, обеспечивает достаточную передачу сигнала для обеспечения пролиферации иммунных клеток и позволяет провести противоопухолевую атаку. В качестве дополнительного примера, химерный белок согласно настоящему изобретению, например, ввиду связывания с низкой скоростью диссоциации, обеспечивает достаточную передачу сигнала для высвобождения стимулирующих сигналов, например, цитокинов.
Стабильный синапс клеток, стимулируемый настоящими агентами (например, опухолевой клетки, несущей отрицательные сигналы, и Т-клетки, способной атаковать опухоль), обеспечивает пространственную ориентацию, благоприятствующую уменьшению опухоли, например, размещение Т-клеток для атаки опухолевых клеток и/или стерическое предотвращение доставки отрицательных сигналов опухолевыми клетками, включая отрицательные сигналы, отличающиеся от сигналов, которые маскирует химерный белок согласно настоящему изобретению.
В ряде вариантов реализации химерный белок согласно настоящему изобретению обладает Kd (1/с) по отношению к CSF1 или ИЛ-34 человека, составляющей более приблизительно 2×106, приблизительно 2,5×106, приблизительно 3×106, приблизительно 3,5×106, приблизительно 4×106, приблизительно 4,5×106, приблизительно 5×106, приблизительно 5,5×106, приблизительно 6×106, приблизительно 6,5×106, приблизительно 7×106, приблизительно 7,5×106, приблизительно 8×106, приблизительно 8,5×106, приблизительно 9×106 или приблизительно 9,5×106 (согласно измерениям с помощью, например, поверхностного плазмонного резонанса или интерферометрии в биослое). В ряде вариантов реализации химерный белок связывается с CSF1 человека с KD, составляющей от приблизительно 100 пМ до приблизительно 600 пМ. В ряде вариантов реализации химерный белок связывается с CSF1 человека со скоростью ассоциации Ka (1/Мс), составляющей приблизительно 5,7×104, и отделяется от CSF1 человека со скоростью диссоциации Kd (1/с), составляющей приблизительно 7,3×10-6.
В ряде вариантов реализации химерный белок согласно настоящему изобретению обладает Kd (1/с) по отношению к CD40 человека, составляющей более приблизительно 2×106, приблизительно 2,5×106, приблизительно 3×106, приблизительно 3,5×106, приблизительно 4×106, приблизительно 4,5×106, приблизительно 5×106, приблизительно 5,5×106, приблизительно 6×106, приблизительно 6,5×106, приблизительно 7×106, приблизительно 7,5×106, приблизительно 8×106, приблизительно 8,5×106, приблизительно 9×106 или приблизительно 9,5×106 (согласно измерениям с помощью, например, поверхностного плазмонного резонанса или интерферометрии в биослое). В ряде вариантов реализации химерный белок связывается с CD40 человека со скоростью ассоциации Ка (1/Мс), составляющей приблизительно 1,3×104, и отделяется от CD40 человека со скоростью диссоциации Kd (1/с), составляющей приблизительно 6,7×10-6.
В ряде вариантов реализации это обеспечивает более длительное время полужизни 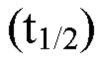 на мишени (например, внутри опухоли) по сравнению с
на мишени (например, внутри опухоли) по сравнению с  химерных белков в сыворотке. Такие свойства могут иметь совместное преимущество, заключающееся в снижении неспецифической токсичности, которая может быть связана с системным распределением химерных белков.
химерных белков в сыворотке. Такие свойства могут иметь совместное преимущество, заключающееся в снижении неспецифической токсичности, которая может быть связана с системным распределением химерных белков.
Действительно, сообщалось, что последовательная обработка антителами, блокирующими CSF1, и агонистическими антителами против CD40, например, индуцирует гепатотоксичность. См., например, Byrne et al. J. Immunology, 2016. Данные, описанные в настоящем документе (см., например, фиг.13), аналогичным образом продемонстрировали высокую токсичность этих двух антител при совместном введении мышам, которое вызывало смертельное воспаление кишечника и диарею. Как ни странно, в противоположность этому, обработка химерным белком CSF1R-Fc-CD40L блокирует CSF1R (что ингибирует передачу иммуноингибиторного сигнала) и активирует CD40L (что усиливает, увеличивает и/или стимулирует передачу иммуностимулирующего сигнала), не вызывая токсичности, возникавшей при совместной обработке антителами. Кроме того, в ряде вариантов реализации химерные белки согласно настоящему изобретению обеспечивают синергетическое терапевтическое действие (например, противоопухолевое действие), поскольку они позволяют улучшить сайт-специфическое взаимодействие двух иммунотерапевтических агентов. В ряде вариантов реализации химерные белки согласно настоящему изобретению обеспечивают синергетическое терапевтическое действие по сравнению с агонистическими антителами к CD40 и/или антагонистическими антителами против CSF1R. В ряде вариантов реализации химерные белки согласно настоящему изобретению обеспечивают возможность снижения неспецифической и/или системной токсичности.
В ряде вариантов реализации химерный белок согласно настоящему изобретению обладает усиленным профилем безопасности. В одном варианте реализации химерный белок согласно настоящему изобретению обладает ослабленным профилем токсичности. Например, введение химерного белка согласно настоящему изобретению может привести к ослаблению побочных эффектов, например, одного или более из диареи, воспаления (например, кишечника) или потери массы тела, наблюдавшихся при введении агонистических антител к CD40 и/или антагонистических антител против CD115. В ряде вариантов реализации химерные белки согласно настоящему изобретению обеспечивают улучшенную безопасность по сравнению с агонистическими антителами к CD40 и/или антагонистическими антителами против CSF115R без ущерба для эффективности.
В ряде вариантов реализации химерные белки согласно настоящему изобретению обеспечивают ослабление побочных эффектов, например, осложнений со стороны ЖКТ, по сравнению с существующими иммунотерапевтическими средствами, например, антителами к молекулам-ключевым элементам иммунного ответа, описанными в настоящем документе. Примеры осложнений со стороны ЖКТ включают боль в животе, потерю аппетита, аутоиммунные эффекты, запоры, судороги, обезвоживание, диарею, проблемы с питанием, усталость, метеоризм, образование жидкости в брюшной полости или асцит, дисбиоз желудочно-кишечного тракта (ЖКТ), мукозит ЖКТ, воспалительное заболевание кишечника, синдром раздраженного кишечника (СРК-Д и СРК-3), тошноту, боль, изменения стула или мочи, язвенный колит, рвоту, увеличение массы тела из-за удержания жидкости и/или слабость.
Заболевания, способы лечения и отбор пациентов
В ряде вариантов реализации настоящее изобретение относится к раковым заболеваниям и/или опухолям; например, лечению или профилактике раковых заболеваний и/или опухолей. Как описано в других частях настоящего документа, лечение рака может включать, в ряде вариантов реализации, модуляцию иммунной системы с помощью химерных белков согласно настоящему изобретению, благоприятствующую иммуностимуляции по сравнению с ингибированием иммунного ответа.
Раковые заболевания или опухоли относятся к неконтролируемому росту клеток и/или патологическому увеличению выживаемости клеток, и/или ингибированию апоптоза, что мешает нормальному функционированию органов и систем организма. Настоящее изобретение охватывает доброкачественные и злокачественные опухоли, полипы, гиперплазию, а также латентные опухоли или микрометастазы. Кроме того, настоящее изобретение охватывает клетки, характеризующиеся патологической пролиферацией, не сдерживаемой иммунной системой (например, клетки, инфицированные вирусом). Рак может представлять собой первичный или метастатический рак. Первичный рак может представлять собой область раковых клеток в месте возникновения, которая становится клинически обнаруживаемой, и может представлять собой первичную опухоль. Метастатический рак, напротив, может представлять собой распространение заболевания от одного органа или части к другому несмежному органу или части. Метастатический рак может быть вызван раковой клеткой, которая приобретает способность проникать и инфильтрировать окружающие нормальные ткани в локальной области, образуя новую опухоль, которая может представлять собой локальный метастаз. Рак также может быть вызван раковой клеткой, которая приобретает способность проникать через стенки лимфатических и/или кровеносных сосудов, после чего раковая клетка может циркулировать в кровотоке (тем самым являясь циркулирующей опухолевой клеткой) в другие места и ткани в организме. Рак может быть вызван таким процессом, как распространение в лимфатической или кровеносной системе. Рак также может быть вызван опухолевой клеткой, которая останавливается в другом месте, повторно проникает через сосуд или стенки, продолжает размножаться и в конечном итоге образует другую клинически обнаруживаемую опухоль. Рак может представлять собой эту новую опухоль, которая может представлять собой метастатическую (или вторичную) опухоль.
Рак может быть вызван метастазирующими опухолевыми клетками, которые могут представлять собой вторичную или метастатическую опухоль. Клетки опухоли могут быть такими же, как и в исходной опухоли. Например, если рак молочной железы или рак толстой кишки метастазирует в печень, вторичная опухоль, хотя и находится в печени, состоит из патологических клеток молочной железы или толстой кишки, а не патологических клеток печени. Таким образом, опухоль в печени может представлять собой метастатический рак молочной железы или метастатический рак толстой кишки, а не рак печени.
Рак может происходить из любой ткани. Рак может происходить из меланомы, толстой кишки, молочной железы или предстательной железы и, таким образом, может состоять из клеток, которые изначально представляли собой клетки кожи, толстой кишки, молочной железы или предстательной железы, соответственно. Рак также может представлять собой гематологическое злокачественное новообразование, которое может представлять собой лейкоз или лимфому. Рак может поражать такие ткани, как печень, легкое, мочевой пузырь или кишечник.
Примеры раковых заболеваний и/или опухолей согласно настоящему изобретению включают базальноклеточную карциному, рак желчевыводящих протоков; рак мочевого пузыря; рак кости; рак головного мозга и центральной нервной системы; рак молочной железы; рак брюшины; рак шейки матки; хориокарциному; рак толстой кишки и прямой кишки; рак соединительной ткани; рак пищеварительной системы; рак эндометрия; рак пищевода; рак глаза; рак головы и шеи; рак желудка (включая рак желудочно-кишечного тракта); глиобластому; карциному печени; гепатому; интраэпителиальное новообразование; рак почки; рак гортани; лейкоз; рак печени; рак легкого (например, мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого и плоскоклеточную карциному легкого); меланому; миелому; нейробластому; рак ротовой полости (губ, языка, рта и глотки); рак яичника; рак поджелудочной железы; рак предстательной железы; ретинобластому; рабдомиосаркому; рак прямой кишки; рак дыхательной системы; карциному слюнной железы; саркому; рак кожи; плоскоклеточный рак; рак желудка; рак яичка; рак щитовидной железы; рак матки или эндометрия; рак мочевыделительной системы; рак вульвы; лимфому, включая лимфому Ходжкина и неходжкинскую лимфому, а также В-клеточную лимфому (включая неходжкинскую лимфому (НХЛ) низкой степени злокачественности/фолликулярную НХЛ), мелкоклеточную лимфоцитарную (МЛ) НХЛ, НХЛ средней степени злокачественности/ фолликулярную НХЛ, диффузную НХЛ средней степени злокачественности; иммунобластную НХЛ высокой степени злокачественности, лимфобластную НХЛ высокой степени злокачественности, мелкоклеточную НХЛ с нерассеченными ядрами высокой степени злокачественности, НХЛ с массивным поражением, лимфому из клеток мантийной ткани, лимфому, связанную со СПИДом, и макроглобулинемию Вальденстрема, хронический лимфоцитарный лейкоз (ХЛЛ), острый лимфобластный лейкоз (ОЛЛ); волосатоклеточный лейкоз; хронический миелобластный лейкоз; а также другие карциномы и саркомы; и посттрансплантационное лимфопролиферативное расстройство (PTLD), а также патологическую пролиферацию сосудов, связанную с факоматозами, отек (например, связанный с опухолями головного мозга) и синдром Мейгса, но не ограничиваются ими.
В ряде вариантов реализации химерный белок применяют для лечения субъекта с невосприимчивым к лечению раком. В ряде вариантов реализации химерный белок применяют для лечения субъекта, невосприимчивого к одному или более иммуномодулирующим агентам. Например, в вариантах реализации химерный белок применяют для лечения субъекта, не реагирующего на лечение или даже демонстрирующего прогрессирование спустя приблизительно 12 недель лечения. Например, в вариантах реализации субъект невосприимчив к агенту PD-1, и/или PD-L1, и/или PD-L2, включая, например, пациентов, невосприимчивых к ниволумабу (ONO-4538/BMS-936558, MDX1106, OPDIVO, BRISTOL MYERS SQUIBB), пембролизумабу (KEYTRUDA, MERCK), пидилизумабу (CT-011, CURE TECH), MK-3475 (MERCK), BMS 936559 (BRISTOL MYERS SQUIBB), ибрутинибу (PHARMACYCLICS/ABBVIE), атезолизумабу (TECENTRIQ, GENENTECH) и/или MPDL3280A (ROCHE). Например, в ряде вариантов реализации субъект невосприимчив к агенту против CTLA-4, например, ипилимумабу (YERVOY) (например, у пациентов с меланомой). Соответственно, в вариантах реализации настоящего изобретения предложены способы лечения рака, позволяющие лечить пациентов, не реагирующих на различные виды терапии, включая монотерапию одним или более иммуномодулирующими агентами.
В ряде вариантов реализации способы согласно настоящему изобретению обеспечивают возможность лечения химерным белком пациента, невосприимчивого к дополнительному агенту, причем такие «дополнительные агенты» описаны в других частях настоящего документа, включая различные химиотерапевтические агенты, описанные в настоящем документе, но не ограничиваясь ими.
В ряде вариантов реализации химерные белки применяют для лечения, контроля или профилактики одного или более воспалительных заболеваний или состояний. Неограничивающие примеры воспалительных заболеваний включают обыкновенные угри, острое воспаление, аллергический ринит, астму, атеросклероз, атопический дерматит, аутоиммунное заболевание, аутоиммунные воспалительные заболевания, аутосомно-рецессивную спастическую атаксию, бронхоэктазию, целиакию, хронический холецистит, хроническое воспаление, хронический простатит, колит, дивертикулит, семейную эозинофилию (СЭ), гломерулонефрит, недостаточность глицеринкиназы, гнойный гидраденит, гиперчувствительность, воспаление, воспалительное заболевание кишечника, воспалительное заболевание органов таза, интерстициальный цистит, воспалительное заболевание гортани, синдром Ли, красный плоский лишай, синдром активации тучных клеток, мастоцитоз, воспалительное заболевание глаз, отит, боль, воспалительное заболевание органов таза, реперфузионное повреждение, респираторное заболевание, рестеноз, ревматизм, ревматоидный артрит, ринит, саркоидоз, септический шок, силикоз и другие пневмокониозы, отторжение трансплантата, туберкулез и васкулит.
В ряде вариантов реализации воспалительное заболевание представляет собой аутоиммунное заболевание или состояние, например, рассеянный склероз, сахарный диабет, волчанку, целиакию, болезнь Крона, язвенный колит, синдром Гийена-Барре, склеродермию, синдром Гудпасчера, гранулематоз Вегенера, аутоиммунную эпилепсию, энцефалит Расмуссена, первичный склероз желчных протоков, склерозирующий холангит, аутоиммунный гепатит, болезнь Аддисона, тиреоидит Хашимото, фибромиалгию, синдром Меньера; отторжение трансплантата (например, профилактику отторжения аллотрансплантата), пернициозную анемию, ревматоидный артрит, системную красную волчанку, дерматомиозит, синдром Шегрена, красную волчанку, рассеянный склероз, миастению гравис, синдром Рейтера, болезнь Грейвса и другие аутоиммунные заболевания.
В некоторых аспектах химерные агенты согласно настоящему изобретению применяют в способах активации антигенпрезентирующей клетки, например, за счет внеклеточного домена CD40L.
В некоторых аспектах химерные агенты согласно настоящему изобретению применяют в способах предупреждения передачи клетками иммуносупрессивного сигнала посредством внеклеточного домена CSF1R.
Комбинированная терапия и конъюгирование
В ряде вариантов реализации настоящего изобретения предложены химерные белки и способы, дополнительно включающим введение субъекту дополнительного агента. В ряде вариантов реализации настоящее изобретение относится к совместному введению и/или комбинированному составу. Любую из композиций, описанных в настоящем документе, можно включить в состав комбинированного состава и/или совместно вводить.
В ряде вариантов реализации любой химерный белок, описанный в настоящем документе, действует синергически при совместном введении с другим агентом, и его вводят в пониженных дозах по сравнению с дозами, обычно применяемыми при использовании таких агентов в качестве монотерапии. В ряде вариантов реализации, любой агент, упомянутый в настоящем документе, можно применять в комбинации с любым из химерных белков, описанных в настоящем документе.
В ряде вариантов реализации химерный белок, содержащий внеклеточный домен CSF1R, описанный в настоящем документе, вводят совместно с еще одним химерным белком. В ряде вариантов реализации химерный белок согласно настоящему изобретению, содержащий внеклеточный домен CSF1R, описанный в настоящем документе, вводят совместно с еще одним химерным белком, например, модулирующим адаптивный иммунный ответ.В ряде вариантов реализации химерные белки согласно настоящему изобретению, содержащие внеклеточный домен CSF1R, описанные в настоящем документе, вводят совместно с химерным белком, содержащим один или более из OX40L, PD-1, GITRL, 4-1BBL, SIRPα, TIM3, TIGIT, LIGHT или VSIG8. Безотносительно к теоретическим представлениям, считается, что комбинированная схема, включающая введение химерного белка согласно настоящему изобретению, индуцирующего врожденный иммунный ответ, и одного или более из других химерных белков, индуцирующих адаптивный иммунный ответ, может обеспечить синергические эффекты (например, синергические противоопухолевые эффекты).
В настоящем изобретении можно применять любой химерный белок, индуцирующий врожденный иммунный ответ.Например, химерный белок может представлять собой любой из химерных белков, описанных в заявке США 62/464002, индуцирующий адаптивный иммунный ответ.В таких вариантах реализации химерный белок содержит первый внеклеточный домен трансмембранного белка I Типа на N-конце или вблизи него и второй внеклеточный домен трансмембранного белка II типа на С-конце или вблизи него, причем один из первого и второго внеклеточных доменов обеспечивает иммуноингибиторный сигнал и один из первого и второго внеклеточных доменов обеспечивает иммуностимулирующий сигнал, как описано в заявке США 62/464002, полное содержание которой включено в настоящий документ посредством ссылки. В типичном варианте реализации химерный белок, индуцирующий адаптивный иммунный ответ, представляет собой химерный белок, содержащий внеклеточный домен PD-1 на N-конце и внеклеточный домен OX40L, GITRL или 4-1BBL на С-конце. В одном варианте реализации химерный белок, индуцирующий адаптивный иммунный ответ, представляет собой химерный белок, содержащий внеклеточный домен VSIG8 на N-конце и внеклеточный домен OX40L, GITRL или 4-1BBL на С-конце.
В ряде вариантов реализации химерный белок согласно настоящему изобретению, содержащий внеклеточный домен CSF1R, описанный в настоящем документе, вводят пациенту для стимуляции врожденного иммунного ответа, и впоследствии (например, через 1 день или через 2 дня или через 3 дня или через 4 дня или через 5 дней или через 6 дней или через 1 неделю или через 2 недели или через 3 недели или через 4 недели) после введения химерного белка, который индуцирует адаптивный иммунный ответ.
В ряде вариантов реализации, в том числе при при раке, но не ограничиваясь им, настоящее изобретение относится к применению химиотерапевтических агентов в качестве дополнительных агентов. Примеры химиотерапевтических агентов включают алкилирующие агенты, например, тиотепу и циклофосфамид CYTOXAN; алкилсульфонаты, например, бусульфан, импросульфан и пипосульфан; азиридины, например, бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилолмеламин; ацетогенины (например, буллатацин и буллатацинон); камптотецин (включая синтетический аналог топотекан); бриостатин; каллистатин; СС-1065 (включая его синтетические аналоги адозелезин, карзелезин и бизелезин); криптофицины (например, криптофицин-1 и криптофицин-8); доластатин; дуокармицин (включая синтетические аналоги KW-2189 и СВ 1-ТМ1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; производные азотистого иприта, например, хлорамбуцил, хлорнафазин, хлорфосфамид, эстрамустин, ифосфамид, мехлорэтамин, гидрохлорид оксида мехлорэтамина, мелфалан, новэмбихин, фенестерин, преднимустин, трофосфамид, урамустин; производные нитрозомочевины, например, кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимустин; антибиотики, например, ендииновые антибиотики (например, калихеамицин, в особенности калихеамицин гамма II и калихеамицин омега II (см, например, Agnew, Chem. Intl. Ed. Engl., 33:183-186 (1994)); динемицин, включая динемицин А; бифосфонаты, например, клодронат; эсперамицин; а также хромофор неокарциностатин и родственные хромопротеин-ендииновые антибиотики-хромофоры), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, каминомицин, карзинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин адриамицин (включая морфолиндоксорубицин, цианморфолиндоксорубицин, 2-пирролиндоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, например, митомицин С, микофеноловую кислоту, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, например, метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, например, деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, например, флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидина, например, анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, например, калустерон, дростанолон, эпитиостанол, мепитиостан, тестолактон; средства, угнетающие функции надпочечников, например, аминоглутетимид, митотан, трилостан; компенсатор фолиевой кислоты, например, фролиновую кислоту; ацеглатон; алдофосфамида гликозид; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатрексат; демеколцин; диазиквон; элформитин; эллиптиния ацетат; эпотилон; этоглуцид; нитрат галлия; гидроксимочевину; лентинан; лонидаинин; майтанзиноиды, например, майтанзин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; подофиллиновую кислоту; 2-этилгидразид; прокарбазин; полисахаридный комплекс ПСК (JHS Natural Products, Юджин, штат Орегон, США); разоксан; ризоксин; сизофуран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2',2''-трихлортриэтиламин; трихотецены (например, токсин Т-2, верракурин А, роридин А и ангуидин); уретан; виндезин; дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид ("Ara-С"); циклофосфамид; тиотепу; таксоиды, например, паклитаксел TAXOL (Bristol-Myers Squibb Oncology, Принстон, штат Нью-Джерси, США),абраксан - препарат из наночастиц паклитаксела, не содержащий кремофора и сконструированный на основе альбумина (American Pharmaceutical Partners, Schaumberg, 111.), и доцетаксел TAXOTERE (Rhone-Poulenc Rorer, Антони, Франция); хлоранбуцил; гемцитабин GEMZAR; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, например, цисплатин, оксалиплатин и карбоплатин; винбластин; платину; этопозид (VP-16); ифосфамид; митоксантрон; винкристин; винорелбин NAVELBINE; новантрон; тенипозид; эдатрексат; дауномицин; аминоптерин; кселоду; ибандронат; иринотекан (Camptosar, СРТ-11) (включая схему лечения иринотеканом с 5-FU и лейковорином); ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, например, ретиноевую кислоту; капецитабин; комбретастатин; лейковорин (LV); оксалиплатин, включая схему лечения оксалиплатином (FOLFOX); лапатиниб (TYKERB); ингибиторы PKC-α, Raf, H-Ras, EGFR (например, эрлотиниб (Tarceva)) и VEGF-A, снижающие пролиферацию клеток, и фармацевтически приемлемые соли, кислоты или производные любых перечисленных выше агентов, но не ограничиваются ими. Кроме того, способы лечения могут дополнительно включать применение излучений. Кроме того, способы лечения могут дополнительно включать применение фотодинамической терапии.
В ряде вариантов реализации, включая применение при раке, но не ограничиваясь им, дополнительный агент согласно настоящему изобретению представляет собой один или более из иммуномодуляторов, выбранных из агента, блокирующего, уменьшающего и/или ингибирующего PD-1 и PD-L1 или PD-L2 и/или связывание PD-1 с PD-L1 или PD-L2 (в качестве неограничивающего примера, один или более из ниволумаба (ONO-4538/BMS-936558, MDX1106, OPDIVO, BRISTOL MYERS SQUIBB), пембролизумаба (KEYTRUDA, Merck), MK-3475 (MERCK), BMS 936559 (BRISTOL MYER SQUIBB), атезолизумаба (TECENTRIQ, GENENTECH), MPDL3280A (ROCHE)), агента, увеличивающего и/или стимулирующего CD137 (4-1ВВ) и/или связывание CD137 (4-1ВВ) с одним или более из лигандов 4-1ВВ (в качестве неограничивающего примера, урелумаб (BMS-663513 и антитело против 4-1ВВ), и агента, блокирующего, уменьшающего и/или ингибирующего активность CTLA-4 и/или связывание CTLA-4 с одним или более из АР2М1, CD80, CD86, SHP-2 и PPP2R5A, и/или связывание ОХ40 с OX40L (в качестве неограничивающего примера, GBR 830 (GLENMARK), MEDI6469 (MEDFMMUNE).
В ряде вариантов реализации, включая применение при инфекционных заболеваниях, но не ограничиваясь им, настоящее изобретение относится к применению противоинфекционных средств в качестве дополнительных агентов. В ряде вариантов реализации противоинфекционное средство представляет собой противовирусный агент, включая абакавир, ацикловир, адефовир, ампренавир, атазанавир, цидофовир, дарунавир, делавирдин, диданозин, докозанол, эфавиренц, эльвитегравир, эмтрицитабин, энфувиртид, этравирин, фамцикловир и фоскарнет, но не ограничиваясь ими. В ряде вариантов реализации противоинфекционное средство представляет собой антибактериальный агент, включая цефалоспориновые антибиотики (цефалексин, цефуроксим, цефадроксил, цефазолин, цефалотин, цефаклор, цефамандол, цефокситин, цефпрозил и цефтобипрол); фторхинолоновые антибиотики (ципро, левахин, флоксин, текин, авелокс и норфлоксацин); тетрациклиновые антибиотики (тетрациклин, миноциклин, окситетрациклин и доксициклин); пенициллиновые антибиотики (амоксициллин, ампициллин, пенициллин V, диклоксациллин, карбенициллин, ванкомицин и метициллин); монобактамовые антибиотики (азтреонам); и карбапенемные антибиотики (эртапенем, дорипенем, имипенем/циластатин и меропенем), но не ограничиваясь ими. В ряде вариантов реализации противоинфекционные средства включают противомалярийные агенты (например, хлорохин, хинин, мефлохин, примахин, доксициклин, артеметер/люмефантрин, атовахон/прогуанил и сульфадоксин/пириметамин), метронидазол, тинидазол, ивермектин, пирантела памоат и альбендазол.
В ряде вариантов реализации, включая применение при аутоиммунных заболеваниях, но не ограничиваясь им, дополнительный агент представляет собой иммунодепрессант.В ряде вариантов реализации иммунодепрессант представляет собой противовоспалительный агент, например, стероидное противовоспалительное средство или нестероидное противовоспалительное средство (НПВС). Стероиды, в частности, кортикостероиды надпочечников и их синтетические аналоги, хорошо известны в данной области техники. Примеры кортикостероидов, подходящих для применения в настоящем изобретении, включают гидроксилтриамцинолон, альфа-метилдексаметазон, бета-метилбетаметазон, дипропионат беклометазона, бензоат беклометазона, дипропионат бетаметазона, валерат бетаметазона, валерат клобетазола, дезонид, дезоксиметазон, дексаметазон, диацетат дифлоразона, валерат дифлукортолона, флуадренолон, ацетонид флухлоролона, пивалат флуметазона, ацетонид флуоцинолона, флуоцинониды, бутиловый эфир флукортина, флуокортолон, ацетат флупреднидена (флупреднилидена), флурандренолон, галцинонид, ацетат гидрокортизона, бутират гидрокортизона, метилпреднизолон, ацетонид триамцинолона, кортизон, кортодоксон, флуцетонид, флудрокортизон, диацетат дифлуорозона, ацетонид флурадренолона, медризон, амцинафел, амцинафид, бетаметазон и равновесная смесь его сложных эфиров, хлорпреднизон, клокортелон, клесцинолон, дихлоризон, дифлупреднат, флуклоронид, флунизолид, фторметалон, флуперолон, флупреднизолон, гидрокортизон, мепреднизон, параметазон, преднизолон, преднизон, дипропионат беклометазона, но не ограничиваются ими. (НПВС), которые можно применять в настоящем изобретении, включают салициловую кислоту, ацетилсалициловую кислоту, метилсалицилат, салицилат гликоля, салицилмиды, бензил-2,5-диацетоксибензойную кислоту, ибупрофен, фулиндак, напроксен, кетопрофен, этофенамат, фенилбутазон и индометацин, но не ограничиваются ими. В ряде вариантов реализации иммунодепрессант может представлять собой цитостатики, например, алкилирующие агенты, антиметаболиты (например, азатиоприн, метотрексат), цитотоксические антибиотики, антитела (например, базиликсимаб, даклизумаб и муромонаб), антииммунофилины (например, циклоспорин, такролимус, сиролимус), интерфероны, опиоиды, ФНО-связывающие белки, микофенолаты и малые биологические агенты (например, финголимод, мириоцин).
В ряде вариантов реализации химерные белки (и/или дополнительные агенты), описанные в настоящем документе, включают производные, которые являются модифицированными, например, путем ковалентного присоединения к композиции молекулы любого типа таким образом, что ковалентное присоединение не препятствует проявлению активности композиции. Например, но не в качестве ограничения, производные включают композиции, модифицированные, в числе прочего, путем гликозилирования, липидизации, ацетилирования, пэгилирования, фосфорилирования, амидирования, модификации известными защитными/блокирующими группами, протеолитического расщепления, связывания с клеточным лигандом или другим белком, и т.д. Любую из множества химических модификаций можно выполнить известными способами, включая специфическое химическое расщепление, ацетилирование, формилирование, метаболический синтез туникамицина и т.д., но не ограничиваясь ими. Кроме того, производное может содержать одну или более из неклассических аминокислот. В других вариантах реализации химерные белки (и/или дополнительные агенты), описанные в настоящем документе, дополнительно содержат цитотоксический агент, в типичных вариантах реализации включающий токсин, химиотерапевтическое средство, радиоактивный изотоп и агент, вызывающий апоптоз или гибель клеток. Такие средства можно конъюгировать с композицией, описанной в настоящем документе.
Химерные белки (и/или дополнительные агенты), описанные в настоящем документе, можно, таким образом, подвергать посттрансляционной модификации с целью добавления эффекторных фрагментов, например, химических линкеров, обнаружимых групп, например, флуоресцентных красителей, ферментов, субстратов, биолюминесцентных материалов, радиоактивных материалов и хемилюминесцентных групп или функциональных фрагментов, например, стрептавидина, авидина, биотина, цитотоксина, цитотоксического агента и радиоактивных материалов.
Составы
Химерные белки (и/или дополнительные агенты), описанные в настоящем документе, могут содержать достаточно основную функциональную группу, способную вступать в реакцию с неорганической или органической кислотой, или карбоксильную группу, способную вступать в реакцию с неорганическим или органическим основанием с образованием фармацевтически приемлемой соли. Фармацевтически приемлемая соль добавления кислоты образуется из фармацевтически приемлемой кислоты, как хорошо известно в данной области техники. Такие соли включают фармацевтически приемлемые соли, перечисленные, например, в Journal of Pharmaceutical Science, 66, 2-19 (1977) и The Handbook of Pharmaceutical Salts; Properties, Selection, and Use. P.H. Stahl, и С.G. Wermuth (eds.),Verlag, Zurich (Switzerland) 2002, полностью включенных в настоящий документ посредством ссылки.
В ряде вариантов реализации композиции, описанные в настоящем документе, находятся в форме фармацевтически приемлемой соли.
Кроме того, любой химерный белок (и/или дополнительные агенты), описанный в настоящем документе, можно вводить субъекту в качестве компонента композиции, содержащей фармацевтически приемлемый наполнитель или носитель. Такие композиции могут необязательно содержать подходящее количество фармацевтически приемлемого вспомогательного соединения, обеспечивая форму для надлежащего введения. Фармацевтические вспомогательные вещества могут представлять собой жидкости, например, воду и масла, включая масла нефтяного, животного, растительного или синтетического происхождения, например, арахисовое масло, соевое масло, минеральное масло, кунжутное масло и т.п. Фармацевтические вспомогательные вещества могут представлять собой, например, физиологический раствор, аравийскую камедь, желатин, крахмальную пасту, тальк, кератин, коллоидный диоксид кремния, мочевину и т.п. Кроме того, можно применять вспомогательные, стабилизирующие, загущающие, смазывающие агенты и красители. В одном из вариантов реализации фармацевтически приемлемые вспомогательные вещества являются стерильными при введении субъекту. Вода является подходящим вспомогательным веществом при внутривенном введении любого агента, описанного в настоящем документе. Растворы хлорида натрия и водные растворы декстрозы и глицерина также можно применять в качестве жидких вспомогательных веществ, в частности, для растворов для инъекции. Подходящие фармацевтические вспомогательные вещества также включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое обезжиренное молоко, глицерин, пропиленгликоль, воду, этанол и т.п. Любой агент, описанный в настоящем документе, при необходимости может также содержать незначительные количества увлажнителей или эмульгаторов или рН-буферных агентов.
В ряде вариантов реализации композиции, описанные в настоящем документе, повторно суспендируют в буфере на основе хлорида натрия (включая TBS, PBS и т.п., но не ограничиваясь этим).
В ряде вариантов реализации химерные белки можно конъюгировать и/или присоединить к другому агенту с целью продления времени полужизни или улучшения фармакодинамических и фармакокинетических свойств иным образом. В ряде вариантов реализации химерные белки можно присоединить к или конъюгировать с одним или более из ПЭГ, XTEN (например, в виде rПЭГ (высвобождаемого ПЭГ)), полисиаловой кислоты (POLYXEN), альбумина (например, человеческого сывороточного альбумина или HAS), эластин-подобного белка (ELP), PAS, НАР, GLK, СТР, трансферрина и т.п. В ряде вариантов реализации каждый из отдельных химерных белков присоединяют к одному или более агентам, описанным в источнике BioDrugs (2015) 29:215-239, полностью включенном в настоящий документ посредством ссылки.
Введение, подбор дозы и схемы лечения
Настоящее изобретение включает описанный химерный белок (и/или дополнительные агенты) в различных составах. Любой химерный белок (и/или дополнительные агенты), описанный в настоящем документе, может принимать форму растворов, суспензий, эмульсий, капель, таблеток, пилюль, гранул, капсул, капсул, содержащих жидкости, порошков, составов с замедленным высвобождением, суппозиториев, эмульсий, аэрозолей, спреев, суспензий или любых других форм, подходящих для применения. Кроме того, можно использовать конструкты ДНК или РНК, кодирующие белковые последовательности. В одном варианте реализации композиция представлена в форме капсулы (см., например, патент США №5698155). Другие примеры подходящих фармацевтических вспомогательных веществ описаны в источнике Remington's Pharmaceutical Sciences 1447-1676 (Alfonso R. Gennaro eds., 19th ed. 1995), включенном в настоящий документ посредством ссылки.
При необходимости составы, содержащие химерный белок (и/или дополнительные агенты), также могут включать солюбилизирующий агент.Кроме того, агенты можно доставлять с подходящим носителем или устройством для доставки, известными в данной области техники. Комбинированные терапевтические средства, описанные в настоящем документе, можно совместно доставлять в одном носителе или устройстве для доставки. Композиции для введения могут необязательно включать местный анестетик, например, лидокаин, для уменьшения боли в месте инъекции.
Составы, содержащие химерный белок (и/или дополнительные агенты) согласно настоящему изобретению, можно представить в удобных лекарственных формах и получать любым из способов, хорошо известных в области фармации. Такие способы обычно включают этап смешивания терапевтических агентов с носителем, состоящим из одного или более вспомогательных ингредиентов. Как правило, составы получают, равномерно и тщательно смешивая терапевтический агент с жидким носителем, тонкодисперсным твердым носителем или жидким и тонкодисперсным твердым носителями, а затем, при необходимости, выполняя формирование продукта в лекарственные формы желательного состава (например, влажных или сухих гранулированных порошков, порошковых смесей и т.д., с последующим изготовлением таблеток с помощью обычных способов, известных в данной области техники)
В одном из вариантов реализации любой химерный белок (и/или дополнительный агент), описанный в настоящем документе, включают в состав препарата в соответствии с общеизвестными процедурами в виде композиции, адаптированной для способа введения, описанного в настоящем документе.
Способы введения включают, например: внутрикожный, внутримышечный, внутрибрюшинный, внутривенный, подкожный, интраназальный, эпидуральный, пероральный, сублингвальный, интраназальный, интрацеребральный, интравагинальный, трансдермальный, ректальный, ингаляциооный или наружный, в частности, в уши, нос, глаза или на кожу. В ряде вариантов реализации введение осуществляют перорально или путем парентеральной инъекции. В некоторых случаях введение приводит к высвобождению агента, описанного в настоящем документе, в кровоток, или, в качестве альтернативы, агент вводят непосредственно в область активного заболевания.
Любой химерный белок (и/или дополнительные агенты), описанный в настоящем документе, можно вводить перорально. Такие химерные белки (и/или дополнительные агенты) можно также вводить любым другим удобным путем, например, путем внутривенного вливания или болюсной инъекции, путем всасывания через эпителиальные или кожно-слизистые выстилающие ткани (например, слизистую оболочку полости рта, прямой кишки и слизистую оболочку кишечника и т.д.), и можно вводить вместе с другим биологически активным агентом. Введение может быть системным или местным. Известны различные системы для доставки, например, инкапсулирование в липосомы, микрочастицы, микрокапсулы, капсулы и т.п., которые можно применять для введения.
В конкретных вариантах реализации может быть желательным местное введение в область, нуждающуюся в лечении. В одном из вариантов реализации, например, при лечении рака, химерный белок (и/или дополнительные агенты) вводят в микроокружение опухоли (например, клетки, молекулы, внеклеточный матрикс и/или кровеносные сосуды, окружающие и/или питающие опухолевую клетку, включая, например, сосудистую сеть опухоли; инфильтрирующие опухоль лимфоциты; фибробластные ретикулярные клетки; эндотелиальные клетки-предшественники (ЕРС); ассоциированные с раком фибробласты; перициты; другие стромальные клетки; компоненты внеклеточного матрикса (ЕСМ); дендритные клетки; антигенпрезентирующие клетки; Т-клетки; регуляторные Т-клетки; макрофаги; нейтрофилы; и другие иммунные клетки, расположенные в непосредственной близости от опухоли) или лимфатические узлы, и/или их мишенью является микроокружение опухоли или лимфатический узел. В ряде вариантов реализации, например, при лечении рака, химерный белок (и/или дополнительные агенты) вводят интратуморально.
В ряде вариантов реализации химерный белок согласно настоящему изобретению проявляет двойной эффект, при котором наблюдается меньше побочных действий по сравнению с традиционной иммунотерапией (например, при лечении одним или более из препаратов OPDIVO, KEYTRUDA, YERVOY и TECENTRIQ). Например, описанные химерные белки снижают или предупреждают часто наблюдаемые нежелательные явления, связанные с иммунной системой, которые поражают различные ткани и органы, включая кожу, желудочно-кишечный тракт, почки, периферическую и центральную нервную систему, печень, лимфатические узлы, глаза, поджелудочную железу и эндокринную систему; например, гипофизит, колит, гепатит, пневмонит, сыпь и ревматизм. Кроме того, местное введение согласно настоящему изобретению, например, интратуморально, устраняет нежелательные явления, наблюдаемые при стандартном системном введении, например, внутривенных вливаниях, которые применяются при обычной иммунотерапии (например, при лечении одним или более из препаратов OPDIVO, KEYTRUDA, YERVOY и TECENTRIQ).
Лекарственные формы, подходящие для парентерального введения (например, внутривенной, внутримышечной, внутрибрюшинной, подкожной и внутрисуставной инъекции и вливания) включают, например, растворы, суспензии, дисперсии, эмульсии и т.п. Их также можно изготовить в форме стерильных твердых композиций (например, лиофилизированной композиции), которые можно растворить или суспендировать в стерильной среде для инъекций непосредственно перед применением. Они могут содержать, например, суспендирующие или диспергирующие агенты, известные в данной области техники.
Введение любого химерного белка (и/или дополнительных агентов), описанного в настоящем документе, а также схема введения может зависеть от различных параметров, включая заболевание, подлежащее лечению, общее состояние здоровья субъекта, а также мнение лечащего врача, но не ограничиваясь ими.
Любой химерный белок, описанный в настоящем документе, можно вводить до (например, за 5 минут, 15 минут, 30 минут, 45 минут, 1 час, 2 часа, 4 часа, 6 часов, 12 часов, 24 часа, 48 часов, 72 часа, 96 часов, 1 неделю, 2 недели, 3 недели, 4 недели, 5 недель, 6 недель, 8 недель или 12 недель), одновременно или после (например, через 5 минут, 15 минут, 30 минут, 45 минут, 1 час, 2 часа, 4 часа, 6 часов, 12 часов, 24 часа, 48 часов, 72 часа, 96 часов, 1 неделю, 2 недели, 3 недели, 4 недели, 5 недель, 6 недель, 8 недель или 12 недель) введения дополнительного агента субъекту, нуждающемуся в этом. В ряде вариантов реализации любой химерный белок и дополнительный агент, описанные в настоящем документе, вводят с интервалом в 1 минуту, с интервалом в 10 минут, с интервалом в 30 минут, с интервалом менее 1 часа, с интервалом в 1 час, с интервалом от 1 часа до 2 часов, с интервалом от 2 часов до 3 часов, с интервалом от 3 часов до 4 часов, с интервалом от 4 часов до 5 часов, с интервалом от 5 часов до 6 часов, с интервалом от 6 часов до 7 часов, с интервалом от 7 часов до 8 часов, с интервалом от 8 часов до 9 часов, с интервалом от 9 часов до 10 часов, с интервалом от 10 часов до 11 часов, с интервалом от 11 часов до 12 часов, с интервалом в 1 день, с интервалом в 2 дня, с интервалом в 3 дня, с интервалом в 4 дня, с интервалом в 5 дней, с интервалом в 6 дней, с интервалом в 1 неделю, с интервалом в 2 недели, с интервалом в 3 недели или с интервалом в 4 недели.
В ряде вариантов реализации настоящее изобретение относится к совместному введению химерного белка согласно настоящему изобретению, содержащего внеклеточный домен рецептора колониестимулирующего фактора 1 (CSF1R), и другого химерного белка, индуцирующего адаптивный иммунный ответ.В таких вариантах реализации химерный белок согласно настоящему изобретению можно вводить до, одновременно или после введения химерного белка, индуцирующего адаптивный иммунный ответ.Например, химерные белки можно вводить с интервалом в 1 минуту, с интервалом в 10 минут, с интервалом в 30 минут, с интервалом менее 1 часа, с интервалом в 1 час, с интервалом от 1 часа до 2 часов, с интервалом от 2 часов до 3 часов, с интервалом от 3 часов до 4 часов, с интервалом от 4 часов до 5 часов, с интервалом от 5 часов до 6 часов, с интервалом от 6 часов до 7 часов, с интервалом от 7 часов до 8 часов, с интервалом от 8 часов до 9 часов, с интервалом от 9 часов до 10 часов, с интервалом от 10 часов до 11 часов, с интервалом от 11 часов до 12 часов, с интервалом в 1 день, с интервалом в 2 дня, с интервалом в 3 дня, с интервалом в 4 дня, с интервалом в 5 дней, с интервалом в 6 дней, с интервалом в 1 неделю, с интервалом в 2 недели, с интервалом в 3 недели или с интервалом в 4 недели. В типичном варианте реализации химерный белок согласно настоящему изобретению, содержащий внеклеточный домен CSF1R, и химерный белок, индуцирующий адаптивный иммунный ответ, вводят с интервалом в 1 неделю или вводят попеременно раз в неделю (т.е. химерный белок, содержащий внеклеточный домен CSF1R, вводят через 1 неделю после введения химерного белка, индуцирующего адаптивный иммунный ответ, и т.д.).
Дозировка любого химерного белка (и/или дополнительных агентов), описанного в настоящем документе, может зависеть от различных факторов, включая тяжесть состояния, того, подлежит ли состояние лечению или профилактике, а также возраста, массы тела и состояния здоровья пациента, подлежащего лечению. Кроме того, на используемую дозировку может влиять фармакогеномная (влияние генотипа на фармакокинетический, фармакодинамический профиль или профиль эффективности лекарственного средства) информация о конкретном субъекте. Кроме того, точные индивидуальные дозы можно в некоторой степени регулировать в зависимости от целого ряда факторов, включая конкретную комбинацию вводимых агентов, время введения, путь введения, природу состава, скорость выведения, конкретное заболевание, подлежащее лечению, тяжесть расстройства и анатомическую локализацию расстройства. Можно ожидать некоторых изменений дозировки. При введении любого химерного белка (и/или дополнительных агентов), описанного в настоящем документе, путем парентеральной инъекции дозировка может составлять от приблизительно 0,1 мг до приблизительно 250 мг в сутки, от приблизительно 1 мг до приблизительно 20 мг в сутки или от приблизительно 3 мг до приблизительно 5 мг в сутки. Обычно при пероральном или парентеральном введении дозировка любого агента, описанного в настоящем документе, может составлять от приблизительно 0,1 мг до приблизительно 1500 мг в сутки, или от приблизительно 0,5 мг до приблизительно 10 мг в сутки, или от приблизительно 0,5 мг до приблизительно 5 мг в сутки, или от приблизительно 200 до приблизительно 1200 мг в сутки (например, приблизительно 200 мг, приблизительно 300 мг, приблизительно 400 мг, приблизительно 500 мг, приблизительно 600 мг, приблизительно 700 мг, приблизительно 800 мг, приблизительно 900 мг, приблизительно 1000 мг, приблизительно 1100 мг, приблизительно 1200 мг в сутки).
В ряде вариантов реализации введение химерного белка (и/или дополнительных агентов), описанного в настоящем документе, осуществляют с помощью парентеральной инъекции в дозировке от приблизительно 0,1 мг до приблизительно 1500 мг на курс лечения, или от приблизительно 0,5 мг до приблизительно 10 мг на курс лечения, или от приблизительно 0,5 мг до приблизительно 5 мг на курс лечения, или от приблизительно 200 до приблизительно 1200 мг на курс лечения (например, приблизительно 200 мг, приблизительно 300 мг, приблизительно 400 мг, приблизительно 500 мг, приблизительно 600 мг, приблизительно 700 мг, приблизительно 800 мг, приблизительно 900 мг, приблизительно 1000 мг, приблизительно 1100 мг, приблизительно 1200 мг на курс лечения).
В ряде вариантов реализации подходящая дозировка химерного белка (и/или дополнительных агентов) составляет от приблизительно 0,01 мг/кг до приблизительно 100 мг/кг массы тела, или от приблизительно 0,01 мг/кг до приблизительно 10 мг/кг массы тела субъекта, например, приблизительно 0,01 мг/кг, приблизительно 0,02 мг/кг, приблизительно 0,03 мг/кг, приблизительно 0,04 мг/кг, приблизительно 0,05 мг/кг, приблизительно 0,06 мг/кг, приблизительно 0,07 мг/кг, приблизительно 0,08 мг/кг, приблизительно 0,09 мг/кг, приблизительно 0,1 мг/кг, приблизительно 0,2 мг/кг, приблизительно 0,3 мг/кг, приблизительно 0,4 мг/кг, приблизительно 0,5 мг/кг, приблизительно 0,6 мг/кг, приблизительно 0,7 мг/кг, приблизительно 0,8 мг/кг, приблизительно 0,9 мг/кг, приблизительно 1 мг/кг, приблизительно 1,1 мг/кг, приблизительно 1,2 мг/кг, приблизительно 1,3 мг/кг, приблизительно 1,4 мг/кг, приблизительно 1,5 мг/кг, приблизительно 1,6 мг/кг, приблизительно 1,7 мг/кг, приблизительно 1,8 мг/кг, 1,9 мг/кг, приблизительно 2 мг/кг, приблизительно 3 мг/кг, приблизительно 4 мг/кг, приблизительно 5 мг/кг, приблизительно 6 мг/кг, приблизительно 7 мг/кг, приблизительно 8 мг/кг, приблизительно 9 мг/кг, приблизительно 10 мг/кг массы тела, включая все промежуточные значения и диапазоны между указанными дозировками.
В еще одном варианте реализации можно осуществлять доставку в везикуле, в частности, в липосоме (см. источник Langer, 1990, Science 249:1527-1533; Treat et al., in Liposomes in therapy of Infectious Disease and Cancer, Lopez-Berestein and Fidler (eds.),Liss, New York, pp.353-365 (1989).
Любой химерный белок (и/или дополнительные агенты), описанный в настоящем документе, можно вводить с помощью средств с контролируемым высвобождением или замедленным высвобождением, или с помощью устройств для доставки, хорошо известных специалистам в данной области техники. Примеры включают средства, описанные в патентах США №3845770, 3916899, 3536809, 3598123, 4008719, 5674533, 5059595, 5591767, 5120548, 5073543, 5639476, 5354556 и 5733556, каждый из которых полностью включен в настоящий документ посредством ссылки, но не ограничиваются ими. Такие лекарственные формы можно применять для обеспечения контролируемого или замедленного высвобождения одного или более активных ингредиентов с помощью, например, гидропропилметилцеллюлозы, других полимерных матриксов, гелей, проницаемых мембран, осмотических систем, многослойных покрытий, микрочастиц, липосом, микросфер или их комбинации для обеспечения желательного профиля высвобождения в различных пропорциях. Контролируемое или замедленное высвобождение активного ингредиента можно стимулировать различными условиями, включая изменения рН, изменения температуры, стимуляцию с помощью света подходящей длины волны, концентрацию или доступность ферментов, концентрацию или доступность воды, или другие физиологические условия или соединения, но не ограничиваясь ими.
В одном варианте реализации можно применять полимерные материалы (см. источники Medical Applications of Controlled Release, Langer and Wise (eds.),CRC Pres., Boca Raton, Florida (1974); Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen and Ball (eds.),Wiley, New York (1984); Ranger and Peppas, 1983, J. Macromol. Sci. Rev. Macromol. Chem. 23:61; см. также Levy et al., 1985, Science 228:190; During et al., 1989, Ann. Neurol. 25:351; Howard et al., 1989, J. Neurosurg. 71:105).
В одном варианте реализации систему с контролируемым высвобождением можно разместить в непосредственной близости от области-мишени, подлежащей лечению, при этом требуется только часть системной дозы (см., например, Goodson, in Medical Applications of Controlled Release, supra, vol. 2, pp. 115-138 (1984)). Можно использовать другие системы с контролируемым высвобождением, обсуждаемые в обзоре Langer, 1990, Science 249:1527-1533.
Любой химерный белок (и/или дополнительные агенты), описанный в настоящем документе, можно независимым образом вводить от одного до четырех раз в сутки, или от одного до четырех раз в месяц, или от одного до шести раз в год, или один раз каждые два, три, четыре или пять лет.Введение можно выполнять на протяжении одного дня или одного месяца, двух месяцев, трех месяцев, шести месяцев, одного года, двух лет, трех лет, и даже на протяжении всей жизни субъекта.
Схему введения с использованием любого химерного белка (и/или дополнительных агентов), описанного в настоящем документе, можно выбрать в соответствии с различными факторами, включая тип, вид, возраст, массу тела, пол и медицинское состояние субъекта; тяжесть состояния, подлежащего лечению; путь введения; функцию печени или почек субъекта; фармакогеномный характер субъекта; и конкретное применяемое соединение согласно настоящему изобретению. Любой химерный белок (и/или дополнительные агенты), описанный в настоящем документе, можно вводить в виде разовой суточной дозы, или общую суточную дозу можно вводить в виде дробных доз два, три или четыре раза в сутки. Кроме того, любой химерный белок (и/или дополнительные агенты), описанный в настоящем документе, можно вводить непрерывно, а не периодически, в ходе всей схемы введения.
Клетки и нуклеиновые кислоты
В ряде вариантов реализации настоящего изобретения предложен экспрессирующий вектор, содержащий нуклеиновую кислоту, кодирующую химерный белок, описанный в настоящем документе. В ряде вариантов реализации указанный экспрессирующий вектор содержит ДНК или РНК. В ряде вариантов реализации экспрессирующий вектор представляет собой экспрессирующий вектор млекопитающего.
Для экспрессии химерного белка можно использовать как прокариотические, так и эукариотические векторы. Прокариотические векторы включают конструкты, основанные на последовательностях Е. coli (см., например, Makrides, Microbiol Rev 1996, 60:512-538). Неограничивающие примеры регуляторных областей, которые можно применять для экспрессии в Е. coli, включают lac, tкp, lpp, phoA, recA, tac, Т3, T7 и λPL. Неограничивающие примеры прокариотических экспрессирующих векторов могут включать серию векторов λgt, например, λgt11 (Huynh et al., в «DNA Cloning Techniques, Vol. A Practical Approach," 1984, (D. Glover, ed.), стр. 49-78, IRL Press, Oxford), и серию векторов pET (Studier et al., Methods Enzymol 1990, 185:60-89). Однако прокариотические системы хозяин-вектор не могут выполнять большую часть посттрансляционного процессинга клеток млекопитающих. Таким образом, эукариотические системы хозяин-вектор могут быть особенно пригодными. Для экспрессии химерных белков в клетках-хозяевах млекопитающих можно применять различные регуляторные области. Например, можно применять ранние и поздние промоторы SV40, предранний промотор цитомегаловируса (ЦМВ) и промотор длинного концевого повтора вируса саркомы Рауса (RSV-LTR). Индуцибельные промоторы, которые можно применять в клетках млекопитающих, включают, без ограничения, промоторы, связанные с геном металлотионеина II, длинные концевые повторы вируса опухоли молочной железы мышей, чувствительные к глюкокортикоидам (MMTV-LTR), ген β-интерферона и ген hsp70 (см. Williams et al., Cancer Res 1989, 49: 2735-42 и Taylor et al., Mol Cell Biol 1990, 10: 165-75). Промоторы теплового шока или стресс-промоторы также могут обеспечивать преимущество при управлении экспрессией химерных белков в рекомбинантных клетках-хозяевах.
В ряде вариантов реализации экспрессирующие векторы согласно настоящему изобретению содержат нуклеиновую кислоту, кодирующую химерные белки (и/или дополнительные агенты) или ее комплементарную цепь, функционально связанные с областью контроля экспрессии или ее комплементарной цепью, которая является функциональной в клетке млекопитающего. Область контроля экспрессии способна управлять экспрессией функционально связанной нуклеиновой кислоты, кодирующей блокирующий и/или стимулирующий агент, так что блокирующий и/или стимулирующий агент продуцируется в клетке человека, трансформированной экспрессирующим вектором.
Области контроля экспрессии представляют собой регуляторные полинуклеотиды (иногда называемые в настоящем документе элементами), например, промоторы и энхансеры, которые влияют на экспрессию функционально связанной нуклеиновой кислоты. Область контроля экспрессии экспрессирующего вектора согласно настоящему изобретению способна экспрессировать функционально связанную кодирующую нуклеиновую кислоту в человеческой клетке. В ряде вариантах реализации клетка представляет собой опухолевую клетку. В одном варианте реализации клетка представляет собой неопухолевую клетку. В ряде вариантов реализации область контроля экспрессии обеспечивает регулируемую экспрессию функционально связанной нуклеиновой кислоты. Сигнал (иногда называемый стимулом) может увеличивать или уменьшать экспрессию нуклеиновой кислоты, функционально связанной с такой областью контроля экспрессии. Такие области контроля экспрессии, которые увеличивают экспрессию в ответ на сигнал, часто называют индуцибельными. Такие области контроля экспрессии, которые уменьшают экспрессию в ответ на сигнал, часто называют репрессируемыми. Как правило, величина увеличения или уменьшения, обеспечиваемая такими элементами, пропорциональна величине присутствующего сигнала; чем больше уровень сигнала, тем больше увеличение или уменьшение экспрессии.
В ряде вариантов реализации настоящее изобретение предусматривает применение индуцибельных промоторов, способных временно вызывать высокий уровень экспрессии в ответ на сигнал. Например, в клетке, трансформированной экспрессирующим вектором химерного белка (и/или дополнительных агентов), содержащим такую последовательность контроля экспрессии, при нахождении в непосредственной близости от опухолевой клетки индуцируется временная продукция высокого уровня агента за счет воздействия соответствующего сигнала на трансформированную клетку. Примеры индуцибельных областей контроля экспрессии включают области, содержащие индуцибельный промотор, стимулируемый сигналом, например, низкомолекулярным химическим соединением. Конкретные примеры можно найти, например, в патентах США №5989910, 5935934, 6015709 и 6004941, каждый из которых полностью включен в настоящий документ посредством ссылки.
Области контроля экспрессии и области контроля локуса включают полноразмерные промоторные последовательности, например, нативные промоторные и энхансерные элементы, а также подпоследовательности или полинуклеотидные варианты, полностью или частично сохраняющие функцию полноразмерной или невариантной последовательности. В контексте настоящего документа термин «функциональный» и его грамматические варианты при использовании в отношении нуклеотидной последовательности, подпоследовательности или фрагмента означает, что указанная последовательность обладает одной или более функций наивной нуклеотидной последовательности (например, не вариантной или немодифицированной последовательности).
В контексте настоящего документа термин «функциональная связь» относится к физическому смежному расположению описываемых компонентов, позволяющему им функционировать назначенным образом. В примере элемента контроля экспрессии, находящегося в функциональной связи с нуклеиновой кислотой, указанная связь обеспечивает модулирование экспрессии нуклеиновой кислоты за счет элемента контроля. Как правило, область контроля экспрессии, модулирующая транскрипцию, располагается рядом с 5'-концом транскрибируемой нуклеиновой кислоты (т.е. "выше" нее). Области контроля экспрессии также могут располагаться на 3'-конце транскрибируемой последовательности (т.е. "ниже" нее) или внутри транскрипта (например, в интроне). Элементы контроля экспрессии могут располагаться на некотором расстоянии от транскрибируемой последовательности (например, от 100 до 500, от 500 до 1000, от 2000 до 5000 или более нуклеотидов от указанной нуклеиновой кислоты). Конкретным примером элемента контроля экспрессии является промотор, который обычно расположен на 5'-конце транскрибируемой последовательности. Еще одним примером элемента контроля экспрессии является энхансер, который может располагаться на 5'- или 3'-конце транскрибируемой последовательности или внутри транскрибированной последовательности.
Системы экспрессии, функционирующие в клетках человека, хорошо известны в данной области техники и включают вирусные системы. Обычно промотор, функционирующий в клетке человека, представляет собой любую последовательность ДНК, способную связывать РНК-полимеразу млекопитающих и инициировать нисходящую (в 3'-направлении) транскрипцию кодирующей последовательности в мРНК. Промотор может содержать область инициации транскрипции, которая обычно располагается проксимально к 5'-концу кодирующей последовательности, и, как правило, ТАТА-бокс, расположенный на 25-30 пар оснований выше сайта инициации транскрипции. Предполагается, что ТАТА-бокс направляет РНК-полимеразу II на начало синтеза РНК в нужном сайте. Промотор также обычно содержит вышерасположенный промоторный элемент (энхансерный элемент), обычно расположенный на 100-200 пар оснований выше ТАТА-бокса. Вышерасположенный промоторный элемент определяет скорость, с которой инициируется транскрипция, и может действовать в любой ориентации. Особенно подходящими являются промоторы из вирусных генов млекопитающих, поскольку вирусные гены часто характеризуются высокой экспрессией и имеют широкий спектр хозяев. Примеры включают ранний промотор SV40, промотор LTR вируса опухоли молочной железы мышей, основной поздний промотор аденовируса, промотор вируса простого герпеса и промотор ЦМВ.
Как правило, последовательности терминации транскрипции и полиаденилирования, распознаваемые клетками млекопитающих, представляют собой регуляторные области, расположенные в направлении 3' относительно стоп-кодона трансляции и, таким образом, вместе с промоторными элементами фланкируют кодирующую последовательность. 3'-конец зрелой мРНК образуется путем сайт-специфического посттрансляционного расщепления и полиаденилирования. Примеры терминатора транскрипции и сигнала полиаденилирования включают области, происходящие от SV40. Кроме того, экспрессирующие конструкты могут содержать интроны.
Существует множество методик введения нуклеиновых кислот в жизнеспособные клетки. Методикм, подходящие для переноса нуклеиновой кислоты в клетки млекопитающих in vitro, включают применение липосом, электропорацию, микроинъекцию, слияние клеток, системы на основе полимеров, ДЭАЭ-декстран, вирусную трансдукцию, способ осаждения с фосфатом кальция и т.д. Для переноса генов in vivo также можно использовать ряд методик и реагентов, включая липосомы; носители для доставки на основе природных полимеров, например, хитозана и желатина; для трансдукции in vivo также подходят вирусные векторы. В некоторых ситуациях желательно применять агент для адресной доставки, например, антитело или лиганд, специфичный по отношению к мембранному белку на поверхности опухолевой клетки. При использовании липосом для адресной доставки и/или облегчения поглощения можно применять белки, которые связываются с мембранным белком клеточной поверхности, ассоциированным с эндоцитозом, например, капсидные белки или их фрагменты, тропные по отношению к клеткам конкретного типа, антитела к белкам, которые подвергаются интернализации при циклическом обновлении мембраны, белки, оказывающие адресное воздействие на внутриклеточные структуры и увеличивающие время полужизни в клетке. Методика рецептор-опосредованного эндоцитоза, описана, например, в источнике Wu et al., J. Biol. Chem. 262, 4429-4432 (1987); и Wagner et al., Proc. Natl. Acad. Sci. USA 87, 3410-3414 (1990).
В соответствующих случаях также можно использовать агенты для доставки генов, например, интеграционные последовательности. В данной области техники известно множество интеграционных последовательностей (см., например, источники Nunes-Duby et al., Nucleic Acids Res. 26:391-406, 1998; Sadwoski, J. Bacterid., 165:341-357, 1986; Bestor, Cell, 122(3):322-325, 2005; Plasterk et al., TIG 15:326-332, 1999; Kootstra et al., Ann. Rev. Pharm. Toxicol., 43: 413-439, 2003). Они включают рекомбиназы и транспозазы. Примеры включают Cre (Sternberg and Hamilton, J. Mol. Biol., 150:467-486, 1981), лямбда (Nash, Nature, 247, 543-545, 1974), FIp (Broach, et al., Cell, 29:227-234, 1982), R (Matsuzaki, et al., J. Bacteriology, 172:610-618, 1990), cpC31 (см., например, Groth et al., J. Mol. Biol. 335:667-678, 2004), систему транспозонов sleeping beauty, транспозазы семейства mariner (Plasterk et al., см. выше) и компоненты для интеграции вирусов, например, AAV, ретеровирусы и антивирусы, содержащие компоненты, обеспечивающие интеграцию вируса, например, последовательности LTR ретровирусов или лентивируса и последовательности ITR AAV (Kootstra et al., Ann. Rev. Pharm. Toxicol., 43: 413-439, 2003). Кроме того, для вставки нуклеотидных последовательностей, кодирующих химерные белки, можно применять стратегии прямой и направленной генетической интеграции, включая технологии редактирования генов с использованием CRISPR/CAS9, доменов цинкового пальца, TALEN и мегануклеазы.
В различных аспектах настоящего изобретения предложены экспрессирующие векторы для экспрессии химерных белков (и/или дополнительных агентов), представляющие собой вирусные векторы. Известно большое количество вирусных векторов, подходящих для генной терапии (см., например, Lundstrom, Trends Biotechnol., 21:1 17, 122, 2003). Примеры вирусных векторов включают векторы, выбранные из антивирусов (LV), ретровирусов (RV), аденовирусов (AV), аденоассоциированных вирусов (AAV) и α-вирусов, хотя можно применять и другие вирусные векторы. Для применения in vivo подходят вирусные векторы, которые не интегрируются в геном хозяина, например, α-вирусы и аденовирусы. Примеры типов α-вирусов включают вирус Синдбис, вирус венесуэльского энцефалита лошадей (VEE) и вирус леса Семлики (SFV). Для применения in vitro подходят вирусные векторы, которые интегрируются в геном хозяина, например, ретровирусы, AAV и антивирусы. В одном из вариантов реализации настоящего изобретения предложены способы трансдукции клетки человека in vivo, включающие приведение солидной опухоли в контакт с вирусным вектором согласно настоящему изобретению in vivo.
В ряде вариантов реализации настоящего изобретения предложена клетка-хозяин, содержащая экспрессирующий вектор, содержащий химерный белок, описанный в настоящем документе.
Экспрессирующие векторы можно вводить в клетки-хозяева для продукции химерных белков согласно настоящему изобретению. Клетки можно, например, культивировать in vitro или сконструировать генетически. Подходящие клетки-хозяева млекопитающих включают, без ограничений, клетки, происходящие от человека, обезьян и грызунов (см., например, Kriegler в «Gene Transfer and Expression: A Laboratory Manual," 1990, New York, Freeman & Co.). Они включают линии клеток почки обезьяны, трансформированные SV40 (например, COS-7, АТСС CRL 1651); эмбриональные линии почек человека (например, клетки 293, 293-EBNA или 293, субклонированные для роста в суспензионной культуре, Graham et al., J Gen Virol 1977, 36:59); клетки почки детеныша хомяка (например, ВНК, АТСС CCL 10); клетки яичника китайского хомяка-DHFR (например, СНО, Urlaub and Chasin, Proc Natl Acad Sci USA 1980, 77:4216); клетки CHO DG44, клетки CHO-K1, клетки Сертоли мыши (Mather, Biol Reprod 1980, 23: 243-251); фибробласты мыши (например, NIH-3Т3), клетки почки обезьяны (например, CVI АТСС CCL 70); клетки почки африканской зеленой мартышки(например, VERO-76, АТСС CRL-1587); клетки карциномы шейки матки человека (например, HELA, АТСС CCL 2); клетки почки собаки (например, MDCK, АТСС CCL 34); клетки печени крысы линии Buffalo (например, BRL 3А, АТСС CRL 1442); клетки легких человека (например, W138, АТСС CCL 75); клетки печени человека (например, Hep G2, НВ 8065); и клетки опухоли молочной железы мыши (например, ММТ 060562, АТСС CCL51). Примеры типов раковых клеток для экспрессии химерных белков, описанных в настоящем документе, включают линию фибробластов мыши NIH3T3, линию клеток Льюисв (карциномы легкого мыши, LLC), линию клеток мастоцитомы мыши Р815, линию клеток лимфомы мыши EL4 и ее овальбуминовый трансфектант Е. G7, линию клеток меланомы мыши B16F10, линию клеток фибросаркомы мыши МС57, и линии клеток мелкоклеточной карциномы легкого человека SCLC#2 и SCLC#7.
Клетки-хозяева можно получить от нормальных или больных субъектов, в том числе здоровых людей, онкологических пациентов и пациентов с инфекционным заболеванием, из депозитов частных лабораторий, общедоступных коллекций культур, например, Американской коллекции типовых культур, или от коммерческих поставщиков.
Клетки, которые можно применять для продукции химерных белков согласно настоящему изобретению in vitro, ex vivo и/или in vivo, включают, без ограничений, эпителиальные клетки, эндотелиальные клетки, кератиноциты, фибробласты, мышечные клетки, гепатоциты; клетки крови, например, Т-лимфоциты, В-лимфоциты, моноциты, макрофаги, нейтрофилы, эозинофилы, мегакариоциты, гранулоциты; различные стволовые клетки или клетки-предшественники, в частности, кроветворные стволовые клетки или клетки-предшественники (например, полученные из костного мозга), пуповинную кровь, периферическую кровь, эмбриональную печень и т.д. Выбор типа клеток зависит от типа опухоли или инфекционного заболевания, подлежащего лечению или профилактике, и его может сделать специалист в данной области техники.
Субъекты и/или животные
В ряде вариантов реализации субъект и/или животное представляет собой млекопитающее, например, человека, мышь, крысу, морскую свинку, собаку, кошку, лошадь, корову, свинью, кролика, овцу или не являющегося человеком примата, например, мартышку, шимпанзе или павиана. В вариантах реализации субъект и/или животное представляет собой животное, не являющееся млекопитающим, например, данио-рерио. В ряде вариантов реализации субъект и/или животное может содержать клетки, меченые флуоресцентной меткой (например, GFP). В ряде вариантов реализации субъект и/или животное представляет собой трансгенное животное, содержащее флуоресцентную клетку.
В ряде вариантов реализации субъект и/или животное представляет собой человека. В ряде вариантов реализации человек представляет собой ребенка. В ряде вариантов реализации человек представляет собой взрослого человека. В ряде вариантов реализации человек представляет собой пожилого человека. В ряде вариантов реализации человек может называться пациентом.
В ряде вариантов реализации возраст человека составляет от приблизительно О месяцев до приблизительно 6 месяцев, от приблизительно 6 до приблизительно 12 месяцев, от приблизительно 6 до приблизительно 18 месяцев, от приблизительно 18 до приблизительно 36 месяцев, от приблизительно 1 до приблизительно 5 лет, от приблизительно 5 до приблизительно 10 лет, от приблизительно 10 до приблизительно 15 лет, от приблизительно 15 до приблизительно 20 лет, от приблизительно 20 до приблизительно 25 лет, от приблизительно 25 до приблизительно 30 лет, от приблизительно 30 до приблизительно 35 лет, от приблизительно 35 до приблизительно 40 лет, от приблизительно 40 до приблизительно 45 лет, от приблизительно 45 до приблизительно 50 лет, от приблизительно 50 до приблизительно 55 лет, от приблизительно 55 до приблизительно 60 лет, от приблизительно 60 до приблизительно 65 лет, от приблизительно 65 до приблизительно 70 лет, от приблизительно 70 до приблизительно 75 лет, от приблизительно 75 до приблизительно 80 лет, от приблизительно 80 до приблизительно 85 лет, от приблизительно 85 до приблизительно 90 лет, от приблизительно 90 до приблизительно 95 лет или от приблизительно 95 до приблизительно 100 лет.
В ряде вариантов реализации субъект представляет собой не являющееся человеком животное и, следовательно, изобретение относится к применению в ветеринарии. В конкретном варианте реализации не являющееся человеком животное представляет собой домашнее животное. В еще одном конкретном варианте реализации не являющееся человеком животное представляет собой сельскохозяйственное животное.
Наборы
В настоящем изобретении предложены наборы, которые могут упростить введение любого агента, описанного в настоящем документе. Типичный набор согласно настоящему изобретению содержит любую композицию, описанную в настоящем документе, в виде лекарственной формы. В одном варианте реализации лекарственная форма представляет собой контейнер, например, предварительно заполненный шприц, который может быть стерильным, содержащий любой агент, описанный в настоящем документе, и фармацевтически приемлемый носитель, разбавитель, вспомогательное вещество или среду-носитель. Набор может дополнительно содержать этикетку или напечатанные инструкции по применению любого агента, описанного в настоящем документе. Набор может также включать векорасширитель, местный анестетик и чистящее средство для места введения. Набор может также дополнительно содержать один или более дополнительных агентов, описанных в настоящем документе. В одном из вариантов осуществления набор содержит контейнер, содержащий эффективное количество композиции согласно настоящему изобретению и эффективное количество другой композиции, например, описанной в настоящем документе.
Любой аспект или вариант реализации, описанные в настоящем документе, можно объединить с любым другим аспектом или вариантом реализации, раскрытым в настоящем документе.
Изобретение дополнительно описано в следующих примерах, которые не ограничивают сущность изобретения, описанную в формуле изобретения.
ПРИМЕРЫ
Пример 1: Прогнозируемый механизм действия и структура мономерного химерного белка CSF1R-Fc-CD40L на основе прогноза in silico.
На фиг.1А показано схематическое представление ожидаемого механизма действия химерного белка CSF1R-Fc-CD40L. Домен CSF1R связывает CSF1 и/или ИЛ-34, обеспечивая «эффект поглощения» и предотвращает связывание CSF1 и/или ИЛ-34 с CSF1R на поверхности антигенпрезентирующих клеток, тем самым блокируя иммуноингибиторный сигнал. Одновременно домен CD40L химерного белка связывает CD40 на поверхности антигенпрезентирующих клеток, тем самым обеспечивая сигнал активации иммунного ответа. Совместное действие этих двух явлений усиливает иммунный ответ, блокируя ингибиторный сигнал (за счет ИЛ-34 и/или CSF1) и обеспечивая сигнал активации за счет CD40.
На фиг.1В показан синапс, образованный химерным белком между опухолевой клеткой и Т-клеткой.
На фиг.1С показан прогноз in silico структуры мономерного химерного белка CSF1R-Fc-CD40L (SL-115154), содержащего 947 аминокислотных остатков (SEQ ID NO: 5) при значении р 1,69×10-29. Прогнозируемая молекулярная масса мономерного белка составляет 105,4 кДа. Структура химерного белка представлена на Фиг.1А.
Конкретно, прогноз структуры выявил, что положения 33 аминокислот (3%) могут быть неупорядочены. Прогноз вторичной структуры всей последовательности химерного белка показал, что белок состоит из 2% α-спиралей (Н), 51% β-листов (Е) и 45% витков (С). Кроме того, для химерного белка рассчитали GDT (глобальный критерий расстояния) и uGDT (ненормированный GDT) для абсолютного глобального качества, получив значение uGDT (GDT), равное 738 (78). Прогноз доступности остатков белка для растворителя при трех состояниях показал, что 33% остатков были открытыми (Е), 46% -промежуточными (М), а 19% - закрытыми (В).
Пример 2: Исследование химерного белка CSF1R-Fc-CD40L.
Химерный белок CSF1R-Fc-CD40L человека (также в настоящем документе называемый CD115-Fc-CD40L) сконструировали, как описано выше в разделе «Подробное описание» и заявке США 62/464002, содержание которого полностью включено в настоящий документ посредством ссылки. Химерный белок исследовали, выполнив вестерн-блоттинг каждого отдельного домена химерного белка, т.е., с использованием антител против CSF1R, против Fc и против CD40L.
Вестерн-блоты указывали на присутствие олигомерной структуры (возможно, димера) с кажущейся молекулярной массой, равной приблизительно 240 кДа, в невосстановленных дорожках (фиг.2, дорожка 2 на каждом блоте), восстановленной до гликозилированной мономерной полосы в присутствии восстановителя β-меркаптоэтанола (фиг.2, дорожка 3 на каждом блоте). Как показано на фиг.2, дорожка 4 на каждом блоте, химерный белок подвергали анализу в виде мономера с прогнозируемой молекулярной массой, равной приблизительно 105 кДа, в присутствии как восстановителя (β-меркаптоэтанола), так и эндогликозидазы (PNGазы).
Пример 3: Исследование связывающей способности различных доменов химерного белка CSF1R-Fc-CD40L с помощью твердофазного ИФА
Для демонстрации сродства связывания различных доменов hCSF1R-Fc-CD40L (в настоящем документе также называемого CD115-Fc-CD40L) с соответствующими партнерами по связыванию (т.е. CSF1, hIgG или CD40) разработали варианты твердофазного иммуноферментного анализа (твердофазного ИФА). Конкретно, Fc-фрагмент химерного белка обнаруживали путем связывания с иммобилизованным на планшете IgG человека и обнаружения за счет антитела против IgG человека, конъюгированного с ПХ (верхний левый квадрант на фиг.3). Домен CSF1R химерного белка hCSF1R-Fc-CD40L обнаруживали путем связывания с иммобилизованным на планшете рекомбинантным белком CSF1 человека и обнаружения за счет антитела против IgG человека, конъюгированного с ПХ (верхний правый квадрант на фиг.3). Домен CD40L химерного белка обнаруживали путем связывания с иммобилизованным на планшете рекомбинантным белком CD40 человека и обнаружения за счет CD40-специфического антитела (нижний левый квадрант на фиг.3). Наконец, одновременное связывание как с CSF1, так и с CD40 демонстрировали с использованием двойного формата твердофазного ИФА, в котором рекомбинантный CD40 использовали для связывания CSF1R-Fc-CD40L, а рекомбинантный CSF1 использовали для обнаружения CSF1R-Fc-CD40L (нижний правый квадрант фиг.3).
Пример 4: Исследование сродства связывания химерного белка CSF1R-Fc-CD40L с клетками ex vivo
Анализ связывания с клетками выполнили для демонстрации сродства связывания различных доменов химерного белка mCSF1R-Fc-CD40L с соответствующими партнерами по связыванию на поверхности мембраны клеток млекопитающих.
Для анализа связывания с клетками сконструировали иммортализованные линии клеток, стабильно экспрессирующие CD40 (Jurkat/CD40). Возрастающие концентрации химерного белка mPD-1-Fc-LIGHT инкубировали со сверхэкспрессирующей линией клеток (Jurkat/CD40) в течение 2 часов. Клетки собирали, промывали и окрашивали антителами для обнаружения связывания химерного белка с помощью проточной цитометрии.
Как показано на фиг.4, химерный белок CSF1R-Fc-CD40L связывался с CD40 на поверхности клеток в зависимости от концентрации и со сродством в области низких нМ. Конкретно, как показано на фиг.4, анализ связывания с клетками продемонстрировал, что CSF1R-Fc-CD40L связывался с CD40 со сродством приблизительно 77 нМ (в соответствии с расчетом ЕС50).
Пример 5: Исследование сродства связывания химерного белка CSF1R-Fc-CD40L с помощью поверхностного плазмонного резонанса (ПНР) и интерферометрии поверхности биослоя
Сродство связывания различных доменов химерного белка hCSF1R-Fc-CD40L измеряли с помощью поверхностного плазмонного резонанса (ПНР) на системе BioRad ProteOn XPR 360. Конкретно, сродство химерного белка по отношению к CSF1 и CD40 определяли и сравнивали с рекомбинантными контрольными белками; результаты показаны ниже в таблице:
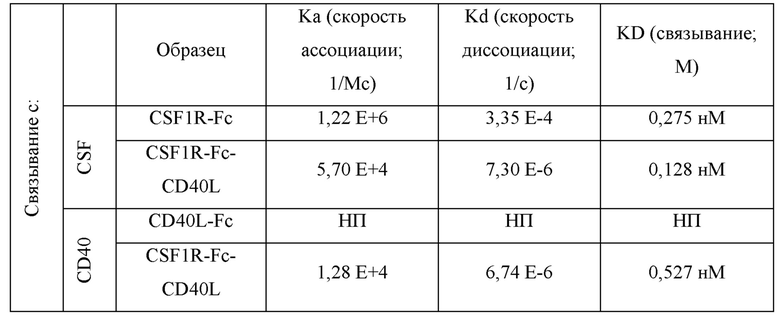
Установлено, что химерный белок hCSF1R-Fc-CD40L связывался с CSF1 и CD40 с высоким сродством. В частности, отмечено, что скорость диссоциации химерного белка hCSF1R-Fc-CD40L была значительно более медленной, чем у контрольных белков (т.е. CSF1R-Fc и CD40L-Fc). Например, скорость диссоциации химерного белка от CSF1 была в 45,9 раз более медленной, чем у белка CSF1R-Fc.
Кроме того, сродство связывания каждого домена CSF1R-Fc-CD40L измеряли с использованием системы Octet на основе интерферометрии поверхности биослоя (фиг.5А - фиг.5F). Эти результаты дополнительно подтвердили высокое сродство связывания химерного белка CSF1R-Fc-CD40L с каждым партнером по связыванию.
Пример 6. Сродство связывания с обоими лигандами CSF1R
Сообщалось, что CSF1R связывается с двумя лигандами: CSF1 и ИЛ-34. Таким образом, было желательно продемонстрировать, что CSF1R-Fc-CD40L способен связываться как с CSF1, так и с ИЛ-34. Это протестировали с использованием интерферометрии поверхности биослоя (Octet), результаты которой показаны на фиг.6.
Связывание CSF1R-Fc-CD40L с CSF1 и ИЛ-34 не отличалось друг от друга; таким образом, кривые виртуально накладывались друг на друга.
Пример 7. Исследование химерного белка CSF1R-Fc-CD40L мыши.
Химерный белок CSF1R-Fc-CD40L мыши (также в настоящем документе называемый mCSFlR-Fc-CD40L) сконструировали, как описано выше в разделе «Подробное описание» и заявке США 62/464002, содержание которого полностью включено в настоящий документ посредством ссылки. Химерный белок исследовали, выполнив вестерн-блоттинг каждого отдельного домена химерного белка, т.е., с использованием антител α-CSFIR, α-Fc и α-CD40L.
Вестерн-блоты указывали на присутствие олигомерной структуры (возможно, димера) с кажущейся молекулярной массой, равной приблизительно 240 кДа, в невосстановленных дорожках (фиг.7А, дорожка 2 на каждом блоте), восстановленной до гликозилированной мономерной полосы в присутствии восстановителя β-меркаптоэтанола (фиг.7А, дорожка 3 на каждом блоте). Как показано на фиг.7А, дорожка 4 на каждом блоте, химерный белок подвергали анализу в виде мономера с прогнозируемой молекулярной массой, равной приблизительно 105 кДа, в присутствии как восстановителя (β-меркаптоэтанола), так и эндогликозидазы (PNGазы).
Для демонстрации сродства связывания различных доменов mCSF1R-Fc-CD40L с соответствующими партнерами по связыванию (т.е. CSF1, mIgG или CD40) разработали варианты твердофазного иммуноферментного анализа (твердофазного ИФА). Конкретно, Fc-фрагмент химерного белка обнаруживали путем связывания с иммобилизованным на планшете IgG мыши и обнаружения за счет антитела против IgG мыши, конъюгированного с ПХ (средний график на фиг.7В). Домен CSF1R химерного белка mCSF1R-Fc-CD40L обнаруживали путем связывания с иммобилизованным на планшете рекомбинантным белком CSF1 мыши и обнаружения за счет антитела против IgG мыши, конъюгированного с ПХ (левый график на фиг.7В). Домен CD40L химерного белка обнаруживали путем связывания с иммобилизованным на планшете рекомбинантным белком CD40 мыши и обнаружения за счет CD40L-специфического антитела (правый график на фиг.7В).
Как показано на фиг.7В, различные домены химерного белка hCSF1R-Fc-CD40L эффективно взаимодействовали с соответствующими партнерами по связыванию с высоким сродством. В то же время обнаружено, что использование центральной области Fc в твердофазном ИФА для обнаружения химерных белков дает тенденцию к недооценке фактического содержания белка в образце. Таким образом, данный анализ позволял обнаружить низкий уровень химерного белка hCSF1R-Fc-CD40L по сравнению со стандартом.
Пример 8. Исследование сродства связывания химерного белка CSF1R-Fc-CD40L мыши с клетками ex vivo
Анализ связывания с клетками выполнили для демонстрации сродства связывания различных доменов химерного белка mCSF1R-Fc-CD40L с соответствующими партнерами по связыванию на поверхности мембраны клеток млекопитающих.
Для анализа связывания с клетками сконструировали иммортализованные линии клеток, стабильно экспрессирующие CD40 (CHOK1/CD40). Возрастающие концентрации химерного белка CSF1R-Fc-CD40L мыши инкубировали со сверхэкспрессирующей линией клеток (CHOK1/CD40) в течение 2 часов. Клетки собирали, промывали и окрашивали антителами для обнаружения связывания химерного белка с помощью проточной цитометрии.
Как показано на фиг.8, химерный белок CSF1R-Fc-CD40L мыши связывался с CD40 на поверхности клеток в зависимости от концентрации и со сродством в области низких нМ. Конкретно, как показано на фиг.8, анализ связывания с клетками продемонстрировал, что CSF1R-Fc-CD40L связывался с CD40 со сродством приблизительно 91,1 нМ (в соответствии с расчетом ЕС50). В качестве отрицательного контроля, обнаруживаемое связывание с исходной (не экспрессирующей CD40) линии клеток CHOK1 отсутствовало.
Пример 9. Индукция сигнального пути CD40 in vitro
CD40 человека представляет собой гомотримерный рецептор, который при активации индуцирует сигнальный каскад, включающий активацию как NF-κВ, так и NIK. На фиг.9 показаны типичные данные анализа сигнального пути NF-κB/NIK in vitro с использованием химерного белка CSF1R-Fc-CD40L человека. Клетки U20S для анализа DiscoverX сигнального пути NIK культивировали с титрованием за счет доступного для приобретения одностороннего CD40L-Fc, одностороннего CSF1R-Fc или агонистического антитела к CD40 или химерного белка CSF1R-Fc-CD40L человека. Относительные люциферазные единицы (RLU) указывают на относительную активность сигнального пути NF-κB/NIK, активируемого после обработки согласно указанным схемам. Показано, что hCSF1R-Fc-CD40L эффективно активирует сигнальный путь за счет NF-κВ и NIK в степени, сопоставимой с химерным белком CD40L-Fc. Агонистическое антитело к CD40 не стимулировало активацию CD40 в данном анализе, поскольку для этого антитела требуется перекрестное связывание Fc-рецептора для облегчения надлежащей кластеризаци рецептора CD40.
Пример 10: Функциональный анализ химерного белка CSF1R-Fc-CD40L.
Установлено, что CSF1R (также известный как CD115) является независимым ключевым элементом иммунного ответа из-за его роли в связывании с CSF1 и/или ИЛ-34 в микроокружении опухоли. Как показано на фиг.1А, связывание CSF1R с любым из этих двух лигандов стимулирует иммуносуппрессию посредством различных механизмов, в том числе индукции супрессорных клеток миелоидного происхождения. Безотносительно к теоретическим представлениям, считается, что химерный белок CSF1R-Fc-CD40L может одновременно действовать как цитокиновая ловушка для CSF11/ИЛ-34 и стимулирует макрофаги и антигенпрезентирующие клетки за счет CSF1R, тем самым обеспечивая мощный противоопухолевый иммунитет.
Для исследования функциональной активности химерного белка mCSF1R-Fc-CD40L разработаны два функциональных анализа.
Первый анализ представляет собой анализ перехвата/поглощения in vivo для оценки способности химерного белка mCSF1R-Fc-CD40L связывать растворимый CSF1 и снижать его уровень в сыворотке. Конкретно, мышам, не несущим опухоли, вводили разовую дозу антитела против CSF1R (также известного как антитело против CD115) в день 0. На 2 день мышам не вводили никаких агентов или вводили разовую дозу химерного белка CSF1R-Fc-CD40L. Сыворотку крови собирали на 2 день перед инъекцией химерного белка и на 3 день после обработки химерным белком. Сыворотку подвергали твердофазному ИФА для выявления CSF1 мыши. Как показано на фиг.10А, химерный белок mCSF1R-Fc-CD40L был способен связывать растворимый CSF1 и снижать его уровень в сыворотке, таким образом устраняя его обнаружение при твердофазном ИФА.
Второй анализ включал иммунологическое профилирование мышей, несущих опухоли, in vivo через 13 дней после обработки химерным белком mCSF1R-Fc-CD40L. Конкретно, уровень ИЛ15Rα+ клеток в селезенке и лимфатических узлах анализировали в качестве показателя иммунной активации химерным белком (в частности, CD40L-фрагментом химерного белка). Мышей с опухолями обрабатывали путем введения двух доз по 150 мкг химерного белка mCSF1R-Fc-CD40L на 5 и 7 дни после исходной инокуляции опухоли. На 13 день группу мышей умерщвляли, извлекали селезенки и лимфатические узлы и диссоциировали их для проточно-цитометрического анализа ИЛ15Rα. Уровень ИЛ15Rα+ - клеток в селезенке и лимфатических узлах определяли, как показано на фиг.10 В. В соответствии с известными механизмами функции CD40L, мыши, обработанные химерным белком, демонстрировали увеличение уровня ИЛ15Rα в селезенке и лимфатических узлах по сравнению с необработанными мышами, что явным образом указывало на стимуляцию иммунной активации химерным белком за счет пути CD40/CD40L.
Пример 11: Исследование противоопухолевой активности химерного белка CSF1R-Fc-CD40L in vivo
Противоопухолевую активность химерного белка mCSF1R-Fc-CD40L in vivo анализировали с использованием модели опухоли ободочной и прямой кишки СТ26 у мыши.
В одной серии экспериментов мышам Balb/c инокулировали опухолевые клетки СТ26 в день 0 и/или подвергали их повторной антигенной стимуляции путем второй инокуляции опухолевых клеток СТ26 на 30 день. Через 5 дней роста опухоли, когда опухоли достигали диаметра 4-5 мм, мышей обрабатывали с применением агонистических антител к CD40, блокирующих антител против CSF1R (CD115), комбинации этих двух антител или химерного белка mCSF1R-Fc-CD40L. Обработку повторно выполняли на 7 день.
Рост опухоли для каждой экспериментальной группы оценивали, как показано на фиг.11А. В частности, у необработанных мышей быстро развивались опухоли. Обработка с применением агонистических антител к CD40, блокирующих антител против CSF1R (CD115) или комбинации этих двух антител, по-видимому, немного задерживала развитие опухолей. В отличие от этого, обработка мышей химерным белком mCD115-Fc-CD40L в значительной степени предотвращала и/или замедляла развитие опухолей. Вышеприведенные данные указывают, что обработка химерным белком CSF1R(CD115)-Fc-CD40L вызывала эффект иммунной памяти in vivo. Таким образом, обработанное животное впоследствии могло атаковать опухолевые клетки и/или предупреждать развитие опухолей при повторной антигенной стимуляции после первоначальной обработки химерным белком.
Кроме того, оценивали общий процент выживаемости мышей спустя 50 дней после инокуляции опухоли. Все необработанные мыши погибли в течение 30 дней после инокуляции опухоли. Другие группы мышей, обработанных с применением агонистических антител к CD40, блокирующих антител против CSF1R (CD115) или комбинации этих двух антител, характеризовались более продолжительной выживаемостью, однако до срока 50 дней после инокуляции опухолей дожили менее 25% этих мышей. Важно, что более 70% мышей, обработанных с применением химерного белка mCD115-Fc-CD40L, остались в живых спустя 50 дней после инокуляции опухоли, как показано на фиг.11В. Как показано на фиг.11С, обработка с применением химерного белка приводила к значительно более выраженному отторжению опухоли, чем обработка с применением агонистических антител к CD40, блокирующих антител CSF1R (CD115) или комбинации этих двух антител.
Пример 12. Иммунофенотипирование популяций лимфоцитов у мышей, несущих опухоли.
Кроме того, выполняли иммунофенотипирование путем анализа спленоцитов, клеток лимфатических узлов и опухоль-инфильтрирующих лимфоцитов на 13 день после инокуляции. Как показано на фиг.12А, показаны результаты, демонстрирующие, что мыши, обработанные химерным белком mCD115-Fc-CD40L, демонстрировали повышенную частоту как CD4+, так и CD8+ Т-клеток в селезенке, но не лимфатических узлах или опухоли, по сравнению с необработанными мышами. Кроме того, мыши, обработанные химерным белком, демонстрировали снижение доли CD4+CD25+ клеток в селезенке и опухоли, что может указывать на снижение количества регуляторных Т-клеток за счет химерного белка (фиг.12В). Примечательно, что несмотря на незначительное увеличение доли общих CD8+ клеток в опухоли (см. фиг.12А), у мышей, обработанных химерным белком mCD115-Fc-CD40L, обнаружено значительное увеличение доли CD8+ Т-клеток, специфичных по отношению к опухолевому антигену АН1 (при окрашивании тетрамером) (фиг.12С), что указывало на то, что химерный белок усиливал распознавание опухоли CD8+ Т-клетками.
Для оценки активации рецептора CD40 химерным белком mCD115-Fc-CD40L анализировали индукцию CD19+ клеток и ИЛ-15Rα-положительных клеток химерным белком. Как показано на фиг.12D, у мышей, обработанных химерным белком, наблюдали значительное увеличение количества CD19+ -клеток в спленоцитах. Это увеличение количества CD19+ -клеток не наблюдали в лимфатических узлах или опухолевых клетках. Кроме того, в спленоцитах мышей, обработанных химерным белком, имело место значительное увеличение количества ИЛ-15Rα положительных клеток (фиг.12Е). Это увеличение также не наблюдалось в лимфатических узлах или опухолевых клетках.
Пример 13. Пониженная токсичность CSF1R-Fc-CD40L по сравнению с антителами против CSF1R and CD40.
Исследование in vivo неожиданно продемонстрировало, что химерный белок CSF115R-Fc-CD40L обладал расширенным профилем безопасности. Конкретно, обнаружено, что у мышей, обработанных с применением агонистического антитела к CD40 и комбинированной терапии с использованием антител против CD40 и CD115, развивалась выраженная диарея и потеря массы тела в ходе эксперимента. У мышей, обработанных агонистическим антителом к CD40, начинался воспалительный ответ в кишечнике, приводивший к диарее и потере массы тела, что значительно усугублялось за счет комбинированного лечения с блокадой CD115. Мыши в группе комбинации антител (против (CD115 и CD40) теряли >25% массы тела (см. фиг.13В), выглядели больными, и в некоторых случаях этот воспалительный ответ приводил к смерти (см. фиг.13А). В отличие от этого, мыши, обработанные химерным белком mCD115-Fc-CD40L, выглядели здоровыми, у них не развивались признаки диареи или потери массы тела, и вели себя нормально (фиг.13А и фиг.13В).
Совместно эти данные показывают, что обработка с применением химерного белка mCD115-Fc-CD40L приводила к значительно более высокой частоте полного отторжения опухоли, чем обработка с применением агонистических антител к CD40, блокирующих антител CD115 по отдельности или в комбинации агонистических антител к CD40 и блокирующих антител CD115. Далее, обработка химерным белком обеспечивала расширенный профиль безопасности по сравнению с обработкой антителами, которые были высокотоксичными при совместном введении мышам и вызывали смертельное воспаление кишечника и диарею.
Пример 14: Исследование вклада Fc-домена в линкере в функциональность химерных белков
В этом примере проанализировали вклад Fc-домена в линкере в функциональность химерных белков согласно настоящему изобретению. PD-1-Fc-OX40L использовали в качестве модели Fc-содержащих химерных белков. Таким образом, представленные ниже данные относятся к химерным белкам согласно настоящему изобретению.
В своем нативном состоянии PD-1 существует в виде мономера, в то время как OX40L имеют тенденцию к димеризации из-за электростатических взаимодействий между доменами OX40L; Fc-домены связываются друг с другом через дисульфидные связи. Совместно несколько межмолекулярных взаимодействий могут вносить вклад в четвертичную структуру PD-1-Fc-OX40L. Существует по меньшей мере четыре потенциальные конфигурации PD-1-Fc-OX40L, в которых химерный белок существует в виде мономера, димера, тримера или гексамера. См. фиг.14.
Существование мономерной и димерной конфигураций химерного белка проверяли, подвергая химерные белки воздействию восстанавливающих и невосстанавливающих условий и затем проводя электрофорез белков в ДСН-ПААГ. В невосстанавливающих условиях (восстановление: «-») химерный белок мигрировал при электрофорезе в ДСН-ПААГ на приблизительно 200 кДа. Вестерн-блоты зондировали антителами против PD-1, Fc или OX40L, соответственно, в левом, среднем и правом блотах, показанных на фиг.15. Поскольку предполагаемая молекулярная масса мономерной формы химерного белка составляет 57,6 кДа, ожидали, что разновидность с молекулярной массой 200 кДа является по меньшей мере димером. Однако в восстанавливающих условиях (восстановление: «+»), восстанавливающих дисульфидные связи (например, между доменами Fc), химерный белок мигрировал в ДСН-ПААГ на приблизительно 100 кДа. Поскольку разновидность с молекулярной массой 100 кДа была тяжелее, чем ожидалось, было спрогнозировано, что избыточная масса была обусловлена гликозилированием. Наконец, химерные белки обрабатывали пептид-N-гликозидазой F (PNGаза F «+») и выполняли электрофорез в ДСП-ПААГ в восстанавливающих условиях. В этих условиях химерный белок мигрировал на приблизительно 57,6 кДа. Эти данные свидетельствуют о том, что химерный белок гликозилирован и существует в природе по меньшей мере в виде димера; димеризация, по-видимому, обусловлена дисульфидной связью между Fc-доменами.
Способы с использованием геля ДСН-ПААГ не позволяют точно предсказать молекулярную массу белков с большим зарядом и/или высокой молекулярной массой. Таким образом, затем химерные белки исследовали с помощью эксклюзионной хроматографии (SEC). В отличие от электрофореза в ДСН-ПААГ, при котором отрицательно заряженный ДСН ослабляет зарядовые взаимодействия между пептидами, при SEC не используются детергенты или восстановители. При проведении SEC химерного белка PD-1-Fc-OX40L ни один из пиков не составлял приблизительно 200 кДа. Это говорит о том, что в нативной форме химерный белок не существует в виде димера. Вместо этого был обнаружен пик, соответствующий размеру более 670 кДа. См. фиг.16. Эти и предыдущие данные указывают, что химерный белок PD-1-Fc-OX40L в своем нативном состоянии существует в виде гексамера.
Как показано выше, при проведении электрофореза в ДСН-ПААГ в невосстанавливающих или в восстанавливающих условиях ДСН в образце и/или рабочем буфере преобразует гексамерный химерный белок PD-1-Fc-OX40L в преобладающую форму димера или мономера, соответственно, в отсутствие и в присутствии восстановителя. См. фиг.17 (гель слева). При проведении нативного электрофореза в ПААГ без ДСН и восстановителя химерный белок существует в виде гексамера. Однако при проведении нативного электрофореза в ПААГ в присутствии восстановителя (восстанавливающего дисульфидные связи) химерный белок мигрировал как более тяжелая молекула, чем ожидалось; как показано на фиг.17 (гель справа, дорожка 2), где химерный белок не мигрировал из загрузочной лунки в существенной степени. Эти данные свидетельствуют о том, что химерный белок олигомеризовался в белок более высокого порядка. Таким образом, в химерных белках дисульфидная связь, по-видимому, важна для контроля олигомеризации более высокого порядка.
Для дальнейшего подтверждения этого были сконструированы химерные белки без Fc-домена, например, «PD-1-без Fc-OX40L». Такие химерные белки не содержат дисульфидной связи, образуемой между Fc-доменами в химерных белках, описанных ранее. Как показано на фиг.18, при нативном электрофорезе в ПААГ химерных белков без Fc-доменов ни один из белков практически не мигрировал из своей загрузочной лунки (дорожки №1-4 с возрастающей концентрацией PDl-No Fc-OX40L); это, опять же, указывает на то, что химерные белки «без Fc» образовали конкатемероподобный комплекс, содержащий множество белков. Таким образом, отсутствие Fc-домена в химерном белке приводит к образованию белковых агрегатов. Эти данные показывают, что дисульфидная связь, например, между Fc-доменами на разных химерных белках, стабилизирует химерные белки и гарантирует, что каждый из них существует в виде гексамера, а не в виде белка/конкатемера более высокого порядка. Другими словами, неожиданно оказалось, что Fc-домен упорядочивает химерные белковые комплексы. Дорожки с 1 по 4 соответственно включают 2,5 мкг PD-1-без Fc-OX40L, 5 мкг PD-1-без FC-OX40L, 7,5 мкг PD-1-без Fc-OX40L и 10 мкг PD-1-без Fc-OX40L.
На фиг.19 показана модель, обобщающая вышеупомянутые данные и показывающая, как гексамер и конкатамеры образуются из химерных белков согласно настоящему изобретению. Типичный химерный белок (PD-1-Fc-OX40L) естественным образом превращается в гексамер (вследствие электростатического взаимодействия между доменами OX40L и димеризации с помощью Fc-доменов). Однако в отсутствие контролирующих эффектов дисульфидной связи между Fc-доменами в восстанавливающих условиях в белке PD-1-Fc-OX40L и из-за отсутствия Fc-доменов в PD-1-без Fc-OX40L эти последние химерные белки образуют конкатемеры.
Кроме того, сконструировали химерные белки, в которых Fc-домен (описанный в настоящем документе) заменили на фиколин (в котором отсутствуют остатки цистеина, необходимые для образования дисульфидной связи между химерными белками). Как и в случае с химерными белками без Fc и химерными белками, содержащими Fc и проанализированными с помощью нативного электрофореза в ПААГ в присутствии восстановителя (в обоих указанных случаях белки образовали агрегаты, которые не мигрировали в гель), химерные белки, содержащие фиколин, по-видимому, также образовывали структуры более высокого порядка, которые не мигрировали в гель. Эти данные подтверждают вывод о том, что дисульфидная связь важна для надлежащего фолдинга и функционирования химерных белков согласно настоящему изобретению.
Наконец, получали химерные белки с использованием спиральных Fc-доменов (CCDFc). При функциональной оценке было получено очень мало очищенного белка.
Соответственно, включение Fc-домена в линкер химерного белка (который способен образовывать дисульфидные связи между химерными белками) помогает избежать образования нерастворимых и, вероятно, нефункциональных конкатемеров и/или агрегатов белка.
Пример 15: Продукция дополнительных CSFlR-содержащих химерных белков, содержащих внеклеточные домены других белков II типа
В данном примере описаны дополнительные химерные белки согласно настоящему изобретению. Такие дополнительные химерные белки получали аналогично получению химерного белка CSF1R-Fc-CD40L, например, как описано выше в разделе «Подробное описание» и заявке США 62/464002, содержание которой полностью включено в настоящий документ посредством ссылки.
Эти дополнительные химерные белки имели общую формулу: ВКД 1 -соединяющий линкер 1 - Fc-домен - соединяющий линкер 2 - ВКД 2, где ВКД 1 представляет собой внеклеточный домен CSF1R, а ВКД 2 - внеклеточный домен белка II типа, отличного от CD40L. Примеры белков II типа включают 4-1BBL, CD30L, FasL, GITRL, LIGHT, OX40L, TL1A и TRAIL. Эти химерные белки могут не содержать один или оба соединяющие линкеры.
Эти химерные белки могут не содержать один или оба соединяющие линкеры. Типичные соединяющие линкеры 1, Fc-домены и соединяющие линкеры 2 описаны выше в таблице 1; модульные линкеры, пригодные для образования химерных белков и содержащие конкретные соединяющие линкеры 1, Fc-домены и соединяющие линкеры 2, показаны на фиг.20.
В качестве альтернативы, дополнительные химерные белки представляли собой гибридные белки со следующей общей формулой: N-конец - (а) - (b) - (с) - С-конец, где (а) представляет собой CSF1R, (b) представляет собой линкер, содержащий по меньшей мере часть Fc-домена, и (с) представляет собой внеклеточный домен белка II типа, отличного от CD40L. Примеры белков II типа включают 4-1BBL, CD30L, FasL, GITRL, LIGHT, OX40L, TL1A и TRAIL.
Аминокислотные последовательности 4-1BBL, CD30L, FasL, GITRL, LIGHT, OX40L, TL1A и TRAIL содержат соответственно SEQ ID NO: 9, 11, 13, 15, 17, 6, 21 и 23. Аминокислотная последовательность внеклеточного домена 4-1BBL, CD30L, FasL, GITRL, LIGHT, OX40L, TL1A и TRAIL содержит соответственно SEQ ID NO: 10, 12, 14, 16, 18, 7, 22 и 24. Аминокислотная последовательность CSF1R содержит SEQ ID NO: 1, а внеклеточный домен CSF1R содержит SEQ ID NO: 2. Химерные белки могут содержать вариант вышеупомянутых последовательностей, например, последовательность, по меньшей мере приблизительно на 95% идентичную вышеупомянутой последовательности.
Примеры линкеров описаны выше в таблице 1; модульные линкеры, подходящие для образования химерных белков и содержащие конкретные соединяющие линкеры 1, Fc-домены и соединяющие линкеры 2, показаны на фиг.20.
Соответственно, настоящее изобретение дополнительно включает следующие дополнительные химерные белки и способы с использованием дополнительных химерных белков (например, при лечении рака и/или лечении воспалительного заболевания): CSF1R-Fc-4-1BBL, CSF1R-Fc-CD30L, CSF1R-Fc-FasL, CSF1R-Fc-GITRL, CSF1R-Fc-LIGHT, CSF1R-Fc-OX40L, CSF1R-Fc-TL1A и CSF1R-Fc-TRAIL.
Дополнительные химерные белки исследовали, как описано выше для CSF1R-Fc-CD40L в примерах 1-13, но с реагентами (например, партнерами по связыванию, рекомбинантными клетками-мишенями и типами раковых клеток/опухолей), специфичными для дополнительных химерных белков, а не с реагентами, необходимыми для исследования CSF1R-Fc-CD40L. Таким образом, используя CSF1R-Fc-4-1BBL в качестве примера, можно исследовать CSF1R-Fc-4-1BBL аналогично примеру 2 с использованием антител против CSF1R, Fc и 4-1BBL, а не антител против CSF1R, Fc и CD40L, необходимых для CSF1R-Fc-CD40L.
Как и в случае химерных белков CSF1R-Fc-OX40L, дополнительные химерные белки могут быть эффективны при лечении рака и/или лечении воспалительного заболевания за счет блокирования CSF1R (что ингибирует передачу иммуноингибирующего сигнала) и усиления, увеличения и/или стимуляции передачи иммуностимулирующего сигнала посредством активации рецептора/лиганда одного из 4-1BBL, CD30L, FasL, GITRL, LIGHT, OX40L, TL1A и TRAIL. Кроме того, дополнительные химерные белки могут быть эффективны при лечении рака и/или воспалительного заболевания, при этом не вызывая токсичности, обусловленной терапевтическими средствами, включающими несколько антител, например, CSF1- или ИЛ-34-блокирующее антитело и агонистическое антитело к рецептору/лиганду одного из 4-1BBL, CD30L, FasL, GITRL, LIGHT, OX40L, TL1A и TRAIL.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ШАТТАК ЛАБС, ИНК.
<120> ХИМЕРНЫЕ БЕЛКИ НА ОСНОВЕ CSF1R
<130> SHK-002PC
<150> US 62/463 997
<151> 27.02.2017
<160> 80
<170> PatentIn версия 3.5
<210> 1
<211> 953
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 1
Ile Pro Val Ile Glu Pro Ser Val Pro Glu Leu Val Val Lys Pro Gly
1 5 10 15
Ala Thr Val Thr Leu Arg Cys Val Gly Asn Gly Ser Val Glu Trp Asp
20 25 30
Gly Pro Pro Ser Pro His Trp Thr Leu Tyr Ser Asp Gly Ser Ser Ser
35 40 45
Ile Leu Ser Thr Asn Asn Ala Thr Phe Gln Asn Thr Gly Thr Tyr Arg
50 55 60
Cys Thr Glu Pro Gly Asp Pro Leu Gly Gly Ser Ala Ala Ile His Leu
65 70 75 80
Tyr Val Lys Asp Pro Ala Arg Pro Trp Asn Val Leu Ala Gln Glu Val
85 90 95
Val Val Phe Glu Asp Gln Asp Ala Leu Leu Pro Cys Leu Leu Thr Asp
100 105 110
Pro Val Leu Glu Ala Gly Val Ser Leu Val Arg Val Arg Gly Arg Pro
115 120 125
Leu Met Arg His Thr Asn Tyr Ser Phe Ser Pro Trp His Gly Phe Thr
130 135 140
Ile His Arg Ala Lys Phe Ile Gln Ser Gln Asp Tyr Gln Cys Ser Ala
145 150 155 160
Leu Met Gly Gly Arg Lys Val Met Ser Ile Ser Ile Arg Leu Lys Val
165 170 175
Gln Lys Val Ile Pro Gly Pro Pro Ala Leu Thr Leu Val Pro Ala Glu
180 185 190
Leu Val Arg Ile Arg Gly Glu Ala Ala Gln Ile Val Cys Ser Ala Ser
195 200 205
Ser Val Asp Val Asn Phe Asp Val Phe Leu Gln His Asn Asn Thr Lys
210 215 220
Leu Ala Ile Pro Gln Gln Ser Asp Phe His Asn Asn Arg Tyr Gln Lys
225 230 235 240
Val Leu Thr Leu Asn Leu Asp Gln Val Asp Phe Gln His Ala Gly Asn
245 250 255
Tyr Ser Cys Val Ala Ser Asn Val Gln Gly Lys His Ser Thr Ser Met
260 265 270
Phe Phe Arg Val Val Glu Ser Ala Tyr Leu Asn Leu Ser Ser Glu Gln
275 280 285
Asn Leu Ile Gln Glu Val Thr Val Gly Glu Gly Leu Asn Leu Lys Val
290 295 300
Met Val Glu Ala Tyr Pro Gly Leu Gln Gly Phe Asn Trp Thr Tyr Leu
305 310 315 320
Gly Pro Phe Ser Asp His Gln Pro Glu Pro Lys Leu Ala Asn Ala Thr
325 330 335
Thr Lys Asp Thr Tyr Arg His Thr Phe Thr Leu Ser Leu Pro Arg Leu
340 345 350
Lys Pro Ser Glu Ala Gly Arg Tyr Ser Phe Leu Ala Arg Asn Pro Gly
355 360 365
Gly Trp Arg Ala Leu Thr Phe Glu Leu Thr Leu Arg Tyr Pro Pro Glu
370 375 380
Val Ser Val Ile Trp Thr Phe Ile Asn Gly Ser Gly Thr Leu Leu Cys
385 390 395 400
Ala Ala Ser Gly Tyr Pro Gln Pro Asn Val Thr Trp Leu Gln Cys Ser
405 410 415
Gly His Thr Asp Arg Cys Asp Glu Ala Gln Val Leu Gln Val Trp Asp
420 425 430
Asp Pro Tyr Pro Glu Val Leu Ser Gln Glu Pro Phe His Lys Val Thr
435 440 445
Val Gln Ser Leu Leu Thr Val Glu Thr Leu Glu His Asn Gln Thr Tyr
450 455 460
Glu Cys Arg Ala His Asn Ser Val Gly Ser Gly Ser Trp Ala Phe Ile
465 470 475 480
Pro Ile Ser Ala Gly Ala His Thr His Pro Pro Asp Glu Phe Leu Phe
485 490 495
Thr Pro Val Val Val Ala Cys Met Ser Ile Met Ala Leu Leu Leu Leu
500 505 510
Leu Leu Leu Leu Leu Leu Tyr Lys Tyr Lys Gln Lys Pro Lys Tyr Gln
515 520 525
Val Arg Trp Lys Ile Ile Glu Ser Tyr Glu Gly Asn Ser Tyr Thr Phe
530 535 540
Ile Asp Pro Thr Gln Leu Pro Tyr Asn Glu Lys Trp Glu Phe Pro Arg
545 550 555 560
Asn Asn Leu Gln Phe Gly Lys Thr Leu Gly Ala Gly Ala Phe Gly Lys
565 570 575
Val Val Glu Ala Thr Ala Phe Gly Leu Gly Lys Glu Asp Ala Val Leu
580 585 590
Lys Val Ala Val Lys Met Leu Lys Ser Thr Ala His Ala Asp Glu Lys
595 600 605
Glu Ala Leu Met Ser Glu Leu Lys Ile Met Ser His Leu Gly Gln His
610 615 620
Glu Asn Ile Val Asn Leu Leu Gly Ala Cys Thr His Gly Gly Pro Val
625 630 635 640
Leu Val Ile Thr Glu Tyr Cys Cys Tyr Gly Asp Leu Leu Asn Phe Leu
645 650 655
Arg Arg Lys Ala Glu Ala Met Leu Gly Pro Ser Leu Ser Pro Gly Gln
660 665 670
Asp Pro Glu Gly Gly Val Asp Tyr Lys Asn Ile His Leu Glu Lys Lys
675 680 685
Tyr Val Arg Arg Asp Ser Gly Phe Ser Ser Gln Gly Val Asp Thr Tyr
690 695 700
Val Glu Met Arg Pro Val Ser Thr Ser Ser Asn Asp Ser Phe Ser Glu
705 710 715 720
Gln Asp Leu Asp Lys Glu Asp Gly Arg Pro Leu Glu Leu Arg Asp Leu
725 730 735
Leu His Phe Ser Ser Gln Val Ala Gln Gly Met Ala Phe Leu Ala Ser
740 745 750
Lys Asn Cys Ile His Arg Asp Val Ala Ala Arg Asn Val Leu Leu Thr
755 760 765
Asn Gly His Val Ala Lys Ile Gly Asp Phe Gly Leu Ala Arg Asp Ile
770 775 780
Met Asn Asp Ser Asn Tyr Ile Val Lys Gly Asn Ala Arg Leu Pro Val
785 790 795 800
Lys Trp Met Ala Pro Glu Ser Ile Phe Asp Cys Val Tyr Thr Val Gln
805 810 815
Ser Asp Val Trp Ser Tyr Gly Ile Leu Leu Trp Glu Ile Phe Ser Leu
820 825 830
Gly Leu Asn Pro Tyr Pro Gly Ile Leu Val Asn Ser Lys Phe Tyr Lys
835 840 845
Leu Val Lys Asp Gly Tyr Gln Met Ala Gln Pro Ala Phe Ala Pro Lys
850 855 860
Asn Ile Tyr Ser Ile Met Gln Ala Cys Trp Ala Leu Glu Pro Thr His
865 870 875 880
Arg Pro Thr Phe Gln Gln Ile Cys Ser Phe Leu Gln Glu Gln Ala Gln
885 890 895
Glu Asp Arg Arg Glu Arg Asp Tyr Thr Asn Leu Pro Ser Ser Ser Arg
900 905 910
Ser Gly Gly Ser Gly Ser Ser Ser Ser Glu Leu Glu Glu Glu Ser Ser
915 920 925
Ser Glu His Leu Thr Cys Cys Glu Gln Gly Asp Ile Ala Gln Pro Leu
930 935 940
Leu Gln Pro Asn Asn Tyr Gln Phe Cys
945 950
<210> 2
<211> 498
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 2
Ile Pro Val Ile Glu Pro Ser Val Pro Glu Leu Val Val Lys Pro Gly
1 5 10 15
Ala Thr Val Thr Leu Arg Cys Val Gly Asn Gly Ser Val Glu Trp Asp
20 25 30
Gly Pro Pro Ser Pro His Trp Thr Leu Tyr Ser Asp Gly Ser Ser Ser
35 40 45
Ile Leu Ser Thr Asn Asn Ala Thr Phe Gln Asn Thr Gly Thr Tyr Arg
50 55 60
Cys Thr Glu Pro Gly Asp Pro Leu Gly Gly Ser Ala Ala Ile His Leu
65 70 75 80
Tyr Val Lys Asp Pro Ala Arg Pro Trp Asn Val Leu Ala Gln Glu Val
85 90 95
Val Val Phe Glu Asp Gln Asp Ala Leu Leu Pro Cys Leu Leu Thr Asp
100 105 110
Pro Val Leu Glu Ala Gly Val Ser Leu Val Arg Val Arg Gly Arg Pro
115 120 125
Leu Met Arg His Thr Asn Tyr Ser Phe Ser Pro Trp His Gly Phe Thr
130 135 140
Ile His Arg Ala Lys Phe Ile Gln Ser Gln Asp Tyr Gln Cys Ser Ala
145 150 155 160
Leu Met Gly Gly Arg Lys Val Met Ser Ile Ser Ile Arg Leu Lys Val
165 170 175
Gln Lys Val Ile Pro Gly Pro Pro Ala Leu Thr Leu Val Pro Ala Glu
180 185 190
Leu Val Arg Ile Arg Gly Glu Ala Ala Gln Ile Val Cys Ser Ala Ser
195 200 205
Ser Val Asp Val Asn Phe Asp Val Phe Leu Gln His Asn Asn Thr Lys
210 215 220
Leu Ala Ile Pro Gln Gln Ser Asp Phe His Asn Asn Arg Tyr Gln Lys
225 230 235 240
Val Leu Thr Leu Asn Leu Asp Gln Val Asp Phe Gln His Ala Gly Asn
245 250 255
Tyr Ser Cys Val Ala Ser Asn Val Gln Gly Lys His Ser Thr Ser Met
260 265 270
Phe Phe Arg Val Val Glu Ser Ala Tyr Leu Asn Leu Ser Ser Glu Gln
275 280 285
Asn Leu Ile Gln Glu Val Thr Val Gly Glu Gly Leu Asn Leu Lys Val
290 295 300
Met Val Glu Ala Tyr Pro Gly Leu Gln Gly Phe Asn Trp Thr Tyr Leu
305 310 315 320
Gly Pro Phe Ser Asp His Gln Pro Glu Pro Lys Leu Ala Asn Ala Thr
325 330 335
Thr Lys Asp Thr Tyr Arg His Thr Phe Thr Leu Ser Leu Pro Arg Leu
340 345 350
Lys Pro Ser Glu Ala Gly Arg Tyr Ser Phe Leu Ala Arg Asn Pro Gly
355 360 365
Gly Trp Arg Ala Leu Thr Phe Glu Leu Thr Leu Arg Tyr Pro Pro Glu
370 375 380
Val Ser Val Ile Trp Thr Phe Ile Asn Gly Ser Gly Thr Leu Leu Cys
385 390 395 400
Ala Ala Ser Gly Tyr Pro Gln Pro Asn Val Thr Trp Leu Gln Cys Ser
405 410 415
Gly His Thr Asp Arg Cys Asp Glu Ala Gln Val Leu Gln Val Trp Asp
420 425 430
Asp Pro Tyr Pro Glu Val Leu Ser Gln Glu Pro Phe His Lys Val Thr
435 440 445
Val Gln Ser Leu Leu Thr Val Glu Thr Leu Glu His Asn Gln Thr Tyr
450 455 460
Glu Cys Arg Ala His Asn Ser Val Gly Ser Gly Ser Trp Ala Phe Ile
465 470 475 480
Pro Ile Ser Ala Gly Ala His Thr His Pro Pro Asp Glu Phe Leu Phe
485 490 495
Thr Pro
<210> 3
<211> 261
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 3
Met Ile Glu Thr Tyr Asn Gln Thr Ser Pro Arg Ser Ala Ala Thr Gly
1 5 10 15
Leu Pro Ile Ser Met Lys Ile Phe Met Tyr Leu Leu Thr Val Phe Leu
20 25 30
Ile Thr Gln Met Ile Gly Ser Ala Leu Phe Ala Val Tyr Leu His Arg
35 40 45
Arg Leu Asp Lys Ile Glu Asp Glu Arg Asn Leu His Glu Asp Phe Val
50 55 60
Phe Met Lys Thr Ile Gln Arg Cys Asn Thr Gly Glu Arg Ser Leu Ser
65 70 75 80
Leu Leu Asn Cys Glu Glu Ile Lys Ser Gln Phe Glu Gly Phe Val Lys
85 90 95
Asp Ile Met Leu Asn Lys Glu Glu Thr Lys Lys Glu Asn Ser Phe Glu
100 105 110
Met Gln Lys Gly Asp Gln Asn Pro Gln Ile Ala Ala His Val Ile Ser
115 120 125
Glu Ala Ser Ser Lys Thr Thr Ser Val Leu Gln Trp Ala Glu Lys Gly
130 135 140
Tyr Tyr Thr Met Ser Asn Asn Leu Val Thr Leu Glu Asn Gly Lys Gln
145 150 155 160
Leu Thr Val Lys Arg Gln Gly Leu Tyr Tyr Ile Tyr Ala Gln Val Thr
165 170 175
Phe Cys Ser Asn Arg Glu Ala Ser Ser Gln Ala Pro Phe Ile Ala Ser
180 185 190
Leu Cys Leu Lys Ser Pro Gly Arg Phe Glu Arg Ile Leu Leu Arg Ala
195 200 205
Ala Asn Thr His Ser Ser Ala Lys Pro Cys Gly Gln Gln Ser Ile His
210 215 220
Leu Gly Gly Val Phe Glu Leu Gln Pro Gly Ala Ser Val Phe Val Asn
225 230 235 240
Val Thr Asp Pro Ser Gln Val Ser His Gly Thr Gly Phe Thr Ser Phe
245 250 255
Gly Leu Leu Lys Leu
260
<210> 4
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 4
His Arg Arg Leu Asp Lys Ile Glu Asp Glu Arg Asn Leu His Glu Asp
1 5 10 15
Phe Val Phe Met Lys Thr Ile Gln Arg Cys Asn Thr Gly Glu Arg Ser
20 25 30
Leu Ser Leu Leu Asn Cys Glu Glu Ile Lys Ser Gln Phe Glu Gly Phe
35 40 45
Val Lys Asp Ile Met Leu Asn Lys Glu Glu Thr Lys Lys Glu Asn Ser
50 55 60
Phe Glu Met Gln Lys Gly Asp Gln Asn Pro Gln Ile Ala Ala His Val
65 70 75 80
Ile Ser Glu Ala Ser Ser Lys Thr Thr Ser Val Leu Gln Trp Ala Glu
85 90 95
Lys Gly Tyr Tyr Thr Met Ser Asn Asn Leu Val Thr Leu Glu Asn Gly
100 105 110
Lys Gln Leu Thr Val Lys Arg Gln Gly Leu Tyr Tyr Ile Tyr Ala Gln
115 120 125
Val Thr Phe Cys Ser Asn Arg Glu Ala Ser Ser Gln Ala Pro Phe Ile
130 135 140
Ala Ser Leu Cys Leu Lys Ser Pro Gly Arg Phe Glu Arg Ile Leu Leu
145 150 155 160
Arg Ala Ala Asn Thr His Ser Ser Ala Lys Pro Cys Gly Gln Gln Ser
165 170 175
Ile His Leu Gly Gly Val Phe Glu Leu Gln Pro Gly Ala Ser Val Phe
180 185 190
Val Asn Val Thr Asp Pro Ser Gln Val Ser His Gly Thr Gly Phe Thr
195 200 205
Ser Phe Gly Leu Leu Lys Leu
210 215
<210> 5
<211> 947
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 5
Ile Pro Val Ile Glu Pro Ser Val Pro Glu Leu Val Val Lys Pro Gly
1 5 10 15
Ala Thr Val Thr Leu Arg Cys Val Gly Asn Gly Ser Val Glu Trp Asp
20 25 30
Gly Pro Pro Ser Pro His Trp Thr Leu Tyr Ser Asp Gly Ser Ser Ser
35 40 45
Ile Leu Ser Thr Asn Asn Ala Thr Phe Gln Asn Thr Gly Thr Tyr Arg
50 55 60
Cys Thr Glu Pro Gly Asp Pro Leu Gly Gly Ser Ala Ala Ile His Leu
65 70 75 80
Tyr Val Lys Asp Pro Ala Arg Pro Trp Asn Val Leu Ala Gln Glu Val
85 90 95
Val Val Phe Glu Asp Gln Asp Ala Leu Leu Pro Cys Leu Leu Thr Asp
100 105 110
Pro Val Leu Glu Ala Gly Val Ser Leu Val Arg Val Arg Gly Arg Pro
115 120 125
Leu Met Arg His Thr Asn Tyr Ser Phe Ser Pro Trp His Gly Phe Thr
130 135 140
Ile His Arg Ala Lys Phe Ile Gln Ser Gln Asp Tyr Gln Cys Ser Ala
145 150 155 160
Leu Met Gly Gly Arg Lys Val Met Ser Ile Ser Ile Arg Leu Lys Val
165 170 175
Gln Lys Val Ile Pro Gly Pro Pro Ala Leu Thr Leu Val Pro Ala Glu
180 185 190
Leu Val Arg Ile Arg Gly Glu Ala Ala Gln Ile Val Cys Ser Ala Ser
195 200 205
Ser Val Asp Val Asn Phe Asp Val Phe Leu Gln His Asn Asn Thr Lys
210 215 220
Leu Ala Ile Pro Gln Gln Ser Asp Phe His Asn Asn Arg Tyr Gln Lys
225 230 235 240
Val Leu Thr Leu Asn Leu Asp Gln Val Asp Phe Gln His Ala Gly Asn
245 250 255
Tyr Ser Cys Val Ala Ser Asn Val Gln Gly Lys His Ser Thr Ser Met
260 265 270
Phe Phe Arg Val Val Glu Ser Ala Tyr Leu Asn Leu Ser Ser Glu Gln
275 280 285
Asn Leu Ile Gln Glu Val Thr Val Gly Glu Gly Leu Asn Leu Lys Val
290 295 300
Met Val Glu Ala Tyr Pro Gly Leu Gln Gly Phe Asn Trp Thr Tyr Leu
305 310 315 320
Gly Pro Phe Ser Asp His Gln Pro Glu Pro Lys Leu Ala Asn Ala Thr
325 330 335
Thr Lys Asp Thr Tyr Arg His Thr Phe Thr Leu Ser Leu Pro Arg Leu
340 345 350
Lys Pro Ser Glu Ala Gly Arg Tyr Ser Phe Leu Ala Arg Asn Pro Gly
355 360 365
Gly Trp Arg Ala Leu Thr Phe Glu Leu Thr Leu Arg Tyr Pro Pro Glu
370 375 380
Val Ser Val Ile Trp Thr Phe Ile Asn Gly Ser Gly Thr Leu Leu Cys
385 390 395 400
Ala Ala Ser Gly Tyr Pro Gln Pro Asn Val Thr Trp Leu Gln Cys Ser
405 410 415
Gly His Thr Asp Arg Cys Asp Glu Ala Gln Val Leu Gln Val Trp Asp
420 425 430
Asp Pro Tyr Pro Glu Val Leu Ser Gln Glu Pro Phe His Lys Val Thr
435 440 445
Val Gln Ser Leu Leu Thr Val Glu Thr Leu Glu His Asn Gln Thr Tyr
450 455 460
Glu Cys Arg Ala His Asn Ser Val Gly Ser Gly Ser Trp Ala Phe Ile
465 470 475 480
Pro Ile Ser Ala Gly Ala His Thr His Pro Pro Asp Glu Phe Leu Phe
485 490 495
Thr Pro Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
500 505 510
Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
515 520 525
Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
530 535 540
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
545 550 555 560
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
565 570 575
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
580 585 590
Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys Gly Leu Pro
595 600 605
Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln Pro Arg Glu
610 615 620
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
625 630 635 640
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
645 650 655
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
660 665 670
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg
675 680 685
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
690 695 700
Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
705 710 715 720
Ser Leu Ser Leu Gly Lys Ile Glu Gly Arg Met Asp His Arg Arg Leu
725 730 735
Asp Lys Ile Glu Asp Glu Arg Asn Leu His Glu Asp Phe Val Phe Met
740 745 750
Lys Thr Ile Gln Arg Cys Asn Thr Gly Glu Arg Ser Leu Ser Leu Leu
755 760 765
Asn Cys Glu Glu Ile Lys Ser Gln Phe Glu Gly Phe Val Lys Asp Ile
770 775 780
Met Leu Asn Lys Glu Glu Thr Lys Lys Glu Asn Ser Phe Glu Met Gln
785 790 795 800
Lys Gly Asp Gln Asn Pro Gln Ile Ala Ala His Val Ile Ser Glu Ala
805 810 815
Ser Ser Lys Thr Thr Ser Val Leu Gln Trp Ala Glu Lys Gly Tyr Tyr
820 825 830
Thr Met Ser Asn Asn Leu Val Thr Leu Glu Asn Gly Lys Gln Leu Thr
835 840 845
Val Lys Arg Gln Gly Leu Tyr Tyr Ile Tyr Ala Gln Val Thr Phe Cys
850 855 860
Ser Asn Arg Glu Ala Ser Ser Gln Ala Pro Phe Ile Ala Ser Leu Cys
865 870 875 880
Leu Lys Ser Pro Gly Arg Phe Glu Arg Ile Leu Leu Arg Ala Ala Asn
885 890 895
Thr His Ser Ser Ala Lys Pro Cys Gly Gln Gln Ser Ile His Leu Gly
900 905 910
Gly Val Phe Glu Leu Gln Pro Gly Ala Ser Val Phe Val Asn Val Thr
915 920 925
Asp Pro Ser Gln Val Ser His Gly Thr Gly Phe Thr Ser Phe Gly Leu
930 935 940
Leu Lys Leu
945
<210> 6
<211> 183
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 6
Met Glu Arg Val Gln Pro Leu Glu Glu Asn Val Gly Asn Ala Ala Arg
1 5 10 15
Pro Arg Phe Glu Arg Asn Lys Leu Leu Leu Val Ala Ser Val Ile Gln
20 25 30
Gly Leu Gly Leu Leu Leu Cys Phe Thr Tyr Ile Cys Leu His Phe Ser
35 40 45
Ala Leu Gln Val Ser His Arg Tyr Pro Arg Ile Gln Ser Ile Lys Val
50 55 60
Gln Phe Thr Glu Tyr Lys Lys Glu Lys Gly Phe Ile Leu Thr Ser Gln
65 70 75 80
Lys Glu Asp Glu Ile Met Lys Val Gln Asn Asn Ser Val Ile Ile Asn
85 90 95
Cys Asp Gly Phe Tyr Leu Ile Ser Leu Lys Gly Tyr Phe Ser Gln Glu
100 105 110
Val Asn Ile Ser Leu His Tyr Gln Lys Asp Glu Glu Pro Leu Phe Gln
115 120 125
Leu Lys Lys Val Arg Ser Val Asn Ser Leu Met Val Ala Ser Leu Thr
130 135 140
Tyr Lys Asp Lys Val Tyr Leu Asn Val Thr Thr Asp Asn Thr Ser Leu
145 150 155 160
Asp Asp Phe His Val Asn Gly Gly Glu Leu Ile Leu Ile His Gln Asn
165 170 175
Pro Gly Glu Phe Cys Val Leu
180
<210> 7
<211> 133
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 7
Gln Val Ser His Arg Tyr Pro Arg Ile Gln Ser Ile Lys Val Gln Phe
1 5 10 15
Thr Glu Tyr Lys Lys Glu Lys Gly Phe Ile Leu Thr Ser Gln Lys Glu
20 25 30
Asp Glu Ile Met Lys Val Gln Asn Asn Ser Val Ile Ile Asn Cys Asp
35 40 45
Gly Phe Tyr Leu Ile Ser Leu Lys Gly Tyr Phe Ser Gln Glu Val Asn
50 55 60
Ile Ser Leu His Tyr Gln Lys Asp Glu Glu Pro Leu Phe Gln Leu Lys
65 70 75 80
Lys Val Arg Ser Val Asn Ser Leu Met Val Ala Ser Leu Thr Tyr Lys
85 90 95
Asp Lys Val Tyr Leu Asn Val Thr Thr Asp Asn Thr Ser Leu Asp Asp
100 105 110
Phe His Val Asn Gly Gly Glu Leu Ile Leu Ile His Gln Asn Pro Gly
115 120 125
Glu Phe Cys Val Leu
130
<210> 8
<211> 865
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 8
Ile Pro Val Ile Glu Pro Ser Val Pro Glu Leu Val Val Lys Pro Gly
1 5 10 15
Ala Thr Val Thr Leu Arg Cys Val Gly Asn Gly Ser Val Glu Trp Asp
20 25 30
Gly Pro Pro Ser Pro His Trp Thr Leu Tyr Ser Asp Gly Ser Ser Ser
35 40 45
Ile Leu Ser Thr Asn Asn Ala Thr Phe Gln Asn Thr Gly Thr Tyr Arg
50 55 60
Cys Thr Glu Pro Gly Asp Pro Leu Gly Gly Ser Ala Ala Ile His Leu
65 70 75 80
Tyr Val Lys Asp Pro Ala Arg Pro Trp Asn Val Leu Ala Gln Glu Val
85 90 95
Val Val Phe Glu Asp Gln Asp Ala Leu Leu Pro Cys Leu Leu Thr Asp
100 105 110
Pro Val Leu Glu Ala Gly Val Ser Leu Val Arg Val Arg Gly Arg Pro
115 120 125
Leu Met Arg His Thr Asn Tyr Ser Phe Ser Pro Trp His Gly Phe Thr
130 135 140
Ile His Arg Ala Lys Phe Ile Gln Ser Gln Asp Tyr Gln Cys Ser Ala
145 150 155 160
Leu Met Gly Gly Arg Lys Val Met Ser Ile Ser Ile Arg Leu Lys Val
165 170 175
Gln Lys Val Ile Pro Gly Pro Pro Ala Leu Thr Leu Val Pro Ala Glu
180 185 190
Leu Val Arg Ile Arg Gly Glu Ala Ala Gln Ile Val Cys Ser Ala Ser
195 200 205
Ser Val Asp Val Asn Phe Asp Val Phe Leu Gln His Asn Asn Thr Lys
210 215 220
Leu Ala Ile Pro Gln Gln Ser Asp Phe His Asn Asn Arg Tyr Gln Lys
225 230 235 240
Val Leu Thr Leu Asn Leu Asp Gln Val Asp Phe Gln His Ala Gly Asn
245 250 255
Tyr Ser Cys Val Ala Ser Asn Val Gln Gly Lys His Ser Thr Ser Met
260 265 270
Phe Phe Arg Val Val Glu Ser Ala Tyr Leu Asn Leu Ser Ser Glu Gln
275 280 285
Asn Leu Ile Gln Glu Val Thr Val Gly Glu Gly Leu Asn Leu Lys Val
290 295 300
Met Val Glu Ala Tyr Pro Gly Leu Gln Gly Phe Asn Trp Thr Tyr Leu
305 310 315 320
Gly Pro Phe Ser Asp His Gln Pro Glu Pro Lys Leu Ala Asn Ala Thr
325 330 335
Thr Lys Asp Thr Tyr Arg His Thr Phe Thr Leu Ser Leu Pro Arg Leu
340 345 350
Lys Pro Ser Glu Ala Gly Arg Tyr Ser Phe Leu Ala Arg Asn Pro Gly
355 360 365
Gly Trp Arg Ala Leu Thr Phe Glu Leu Thr Leu Arg Tyr Pro Pro Glu
370 375 380
Val Ser Val Ile Trp Thr Phe Ile Asn Gly Ser Gly Thr Leu Leu Cys
385 390 395 400
Ala Ala Ser Gly Tyr Pro Gln Pro Asn Val Thr Trp Leu Gln Cys Ser
405 410 415
Gly His Thr Asp Arg Cys Asp Glu Ala Gln Val Leu Gln Val Trp Asp
420 425 430
Asp Pro Tyr Pro Glu Val Leu Ser Gln Glu Pro Phe His Lys Val Thr
435 440 445
Val Gln Ser Leu Leu Thr Val Glu Thr Leu Glu His Asn Gln Thr Tyr
450 455 460
Glu Cys Arg Ala His Asn Ser Val Gly Ser Gly Ser Trp Ala Phe Ile
465 470 475 480
Pro Ile Ser Ala Gly Ala His Thr His Pro Pro Asp Glu Phe Leu Phe
485 490 495
Thr Pro Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
500 505 510
Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
515 520 525
Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
530 535 540
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
545 550 555 560
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
565 570 575
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
580 585 590
Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys Gly Leu Pro
595 600 605
Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln Pro Arg Glu
610 615 620
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
625 630 635 640
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
645 650 655
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
660 665 670
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg
675 680 685
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
690 695 700
Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
705 710 715 720
Ser Leu Ser Leu Gly Lys Ile Glu Gly Arg Met Asp Gln Val Ser His
725 730 735
Arg Tyr Pro Arg Ile Gln Ser Ile Lys Val Gln Phe Thr Glu Tyr Lys
740 745 750
Lys Glu Lys Gly Phe Ile Leu Thr Ser Gln Lys Glu Asp Glu Ile Met
755 760 765
Lys Val Gln Asn Asn Ser Val Ile Ile Asn Cys Asp Gly Phe Tyr Leu
770 775 780
Ile Ser Leu Lys Gly Tyr Phe Ser Gln Glu Val Asn Ile Ser Leu His
785 790 795 800
Tyr Gln Lys Asp Glu Glu Pro Leu Phe Gln Leu Lys Lys Val Arg Ser
805 810 815
Val Asn Ser Leu Met Val Ala Ser Leu Thr Tyr Lys Asp Lys Val Tyr
820 825 830
Leu Asn Val Thr Thr Asp Asn Thr Ser Leu Asp Asp Phe His Val Asn
835 840 845
Gly Gly Glu Leu Ile Leu Ile His Gln Asn Pro Gly Glu Phe Cys Val
850 855 860
Leu
865
<210> 9
<211> 254
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 9
Met Glu Tyr Ala Ser Asp Ala Ser Leu Asp Pro Glu Ala Pro Trp Pro
1 5 10 15
Pro Ala Pro Arg Ala Arg Ala Cys Arg Val Leu Pro Trp Ala Leu Val
20 25 30
Ala Gly Leu Leu Leu Leu Leu Leu Leu Ala Ala Ala Cys Ala Val Phe
35 40 45
Leu Ala Cys Pro Trp Ala Val Ser Gly Ala Arg Ala Ser Pro Gly Ser
50 55 60
Ala Ala Ser Pro Arg Leu Arg Glu Gly Pro Glu Leu Ser Pro Asp Asp
65 70 75 80
Pro Ala Gly Leu Leu Asp Leu Arg Gln Gly Met Phe Ala Gln Leu Val
85 90 95
Ala Gln Asn Val Leu Leu Ile Asp Gly Pro Leu Ser Trp Tyr Ser Asp
100 105 110
Pro Gly Leu Ala Gly Val Ser Leu Thr Gly Gly Leu Ser Tyr Lys Glu
115 120 125
Asp Thr Lys Glu Leu Val Val Ala Lys Ala Gly Val Tyr Tyr Val Phe
130 135 140
Phe Gln Leu Glu Leu Arg Arg Val Val Ala Gly Glu Gly Ser Gly Ser
145 150 155 160
Val Ser Leu Ala Leu His Leu Gln Pro Leu Arg Ser Ala Ala Gly Ala
165 170 175
Ala Ala Leu Ala Leu Thr Val Asp Leu Pro Pro Ala Ser Ser Glu Ala
180 185 190
Arg Asn Ser Ala Phe Gly Phe Gln Gly Arg Leu Leu His Leu Ser Ala
195 200 205
Gly Gln Arg Leu Gly Val His Leu His Thr Glu Ala Arg Ala Arg His
210 215 220
Ala Trp Gln Leu Thr Gln Gly Ala Thr Val Leu Gly Leu Phe Arg Val
225 230 235 240
Thr Pro Glu Ile Pro Ala Gly Leu Pro Ser Pro Arg Ser Glu
245 250
<210> 10
<211> 205
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 10
Ala Cys Pro Trp Ala Val Ser Gly Ala Arg Ala Ser Pro Gly Ser Ala
1 5 10 15
Ala Ser Pro Arg Leu Arg Glu Gly Pro Glu Leu Ser Pro Asp Asp Pro
20 25 30
Ala Gly Leu Leu Asp Leu Arg Gln Gly Met Phe Ala Gln Leu Val Ala
35 40 45
Gln Asn Val Leu Leu Ile Asp Gly Pro Leu Ser Trp Tyr Ser Asp Pro
50 55 60
Gly Leu Ala Gly Val Ser Leu Thr Gly Gly Leu Ser Tyr Lys Glu Asp
65 70 75 80
Thr Lys Glu Leu Val Val Ala Lys Ala Gly Val Tyr Tyr Val Phe Phe
85 90 95
Gln Leu Glu Leu Arg Arg Val Val Ala Gly Glu Gly Ser Gly Ser Val
100 105 110
Ser Leu Ala Leu His Leu Gln Pro Leu Arg Ser Ala Ala Gly Ala Ala
115 120 125
Ala Leu Ala Leu Thr Val Asp Leu Pro Pro Ala Ser Ser Glu Ala Arg
130 135 140
Asn Ser Ala Phe Gly Phe Gln Gly Arg Leu Leu His Leu Ser Ala Gly
145 150 155 160
Gln Arg Leu Gly Val His Leu His Thr Glu Ala Arg Ala Arg His Ala
165 170 175
Trp Gln Leu Thr Gln Gly Ala Thr Val Leu Gly Leu Phe Arg Val Thr
180 185 190
Pro Glu Ile Pro Ala Gly Leu Pro Ser Pro Arg Ser Glu
195 200 205
<210> 11
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 11
Met Asp Pro Gly Leu Gln Gln Ala Leu Asn Gly Met Ala Pro Pro Gly
1 5 10 15
Asp Thr Ala Met His Val Pro Ala Gly Ser Val Ala Ser His Leu Gly
20 25 30
Thr Thr Ser Arg Ser Tyr Phe Tyr Leu Thr Thr Ala Thr Leu Ala Leu
35 40 45
Cys Leu Val Phe Thr Val Ala Thr Ile Met Val Leu Val Val Gln Arg
50 55 60
Thr Asp Ser Ile Pro Asn Ser Pro Asp Asn Val Pro Leu Lys Gly Gly
65 70 75 80
Asn Cys Ser Glu Asp Leu Leu Cys Ile Leu Lys Arg Ala Pro Phe Lys
85 90 95
Lys Ser Trp Ala Tyr Leu Gln Val Ala Lys His Leu Asn Lys Thr Lys
100 105 110
Leu Ser Trp Asn Lys Asp Gly Ile Leu His Gly Val Arg Tyr Gln Asp
115 120 125
Gly Asn Leu Val Ile Gln Phe Pro Gly Leu Tyr Phe Ile Ile Cys Gln
130 135 140
Leu Gln Phe Leu Val Gln Cys Pro Asn Asn Ser Val Asp Leu Lys Leu
145 150 155 160
Glu Leu Leu Ile Asn Lys His Ile Lys Lys Gln Ala Leu Val Thr Val
165 170 175
Cys Glu Ser Gly Met Gln Thr Lys His Val Tyr Gln Asn Leu Ser Gln
180 185 190
Phe Leu Leu Asp Tyr Leu Gln Val Asn Thr Thr Ile Ser Val Asn Val
195 200 205
Asp Thr Phe Gln Tyr Ile Asp Thr Ser Thr Phe Pro Leu Glu Asn Val
210 215 220
Leu Ser Ile Phe Leu Tyr Ser Asn Ser Asp
225 230
<210> 12
<211> 172
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 12
Gln Arg Thr Asp Ser Ile Pro Asn Ser Pro Asp Asn Val Pro Leu Lys
1 5 10 15
Gly Gly Asn Cys Ser Glu Asp Leu Leu Cys Ile Leu Lys Arg Ala Pro
20 25 30
Phe Lys Lys Ser Trp Ala Tyr Leu Gln Val Ala Lys His Leu Asn Lys
35 40 45
Thr Lys Leu Ser Trp Asn Lys Asp Gly Ile Leu His Gly Val Arg Tyr
50 55 60
Gln Asp Gly Asn Leu Val Ile Gln Phe Pro Gly Leu Tyr Phe Ile Ile
65 70 75 80
Cys Gln Leu Gln Phe Leu Val Gln Cys Pro Asn Asn Ser Val Asp Leu
85 90 95
Lys Leu Glu Leu Leu Ile Asn Lys His Ile Lys Lys Gln Ala Leu Val
100 105 110
Thr Val Cys Glu Ser Gly Met Gln Thr Lys His Val Tyr Gln Asn Leu
115 120 125
Ser Gln Phe Leu Leu Asp Tyr Leu Gln Val Asn Thr Thr Ile Ser Val
130 135 140
Asn Val Asp Thr Phe Gln Tyr Ile Asp Thr Ser Thr Phe Pro Leu Glu
145 150 155 160
Asn Val Leu Ser Ile Phe Leu Tyr Ser Asn Ser Asp
165 170
<210> 13
<211> 281
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 13
Met Gln Gln Pro Phe Asn Tyr Pro Tyr Pro Gln Ile Tyr Trp Val Asp
1 5 10 15
Ser Ser Ala Ser Ser Pro Trp Ala Pro Pro Gly Thr Val Leu Pro Cys
20 25 30
Pro Thr Ser Val Pro Arg Arg Pro Gly Gln Arg Arg Pro Pro Pro Pro
35 40 45
Pro Pro Pro Pro Pro Leu Pro Pro Pro Pro Pro Pro Pro Pro Leu Pro
50 55 60
Pro Leu Pro Leu Pro Pro Leu Lys Lys Arg Gly Asn His Ser Thr Gly
65 70 75 80
Leu Cys Leu Leu Val Met Phe Phe Met Val Leu Val Ala Leu Val Gly
85 90 95
Leu Gly Leu Gly Met Phe Gln Leu Phe His Leu Gln Lys Glu Leu Ala
100 105 110
Glu Leu Arg Glu Ser Thr Ser Gln Met His Thr Ala Ser Ser Leu Glu
115 120 125
Lys Gln Ile Gly His Pro Ser Pro Pro Pro Glu Lys Lys Glu Leu Arg
130 135 140
Lys Val Ala His Leu Thr Gly Lys Ser Asn Ser Arg Ser Met Pro Leu
145 150 155 160
Glu Trp Glu Asp Thr Tyr Gly Ile Val Leu Leu Ser Gly Val Lys Tyr
165 170 175
Lys Lys Gly Gly Leu Val Ile Asn Glu Thr Gly Leu Tyr Phe Val Tyr
180 185 190
Ser Lys Val Tyr Phe Arg Gly Gln Ser Cys Asn Asn Leu Pro Leu Ser
195 200 205
His Lys Val Tyr Met Arg Asn Ser Lys Tyr Pro Gln Asp Leu Val Met
210 215 220
Met Glu Gly Lys Met Met Ser Tyr Cys Thr Thr Gly Gln Met Trp Ala
225 230 235 240
Arg Ser Ser Tyr Leu Gly Ala Val Phe Asn Leu Thr Ser Ala Asp His
245 250 255
Leu Tyr Val Asn Val Ser Glu Leu Ser Leu Val Asn Phe Glu Glu Ser
260 265 270
Gln Thr Phe Phe Gly Leu Tyr Lys Leu
275 280
<210> 14
<211> 179
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 14
Gln Leu Phe His Leu Gln Lys Glu Leu Ala Glu Leu Arg Glu Ser Thr
1 5 10 15
Ser Gln Met His Thr Ala Ser Ser Leu Glu Lys Gln Ile Gly His Pro
20 25 30
Ser Pro Pro Pro Glu Lys Lys Glu Leu Arg Lys Val Ala His Leu Thr
35 40 45
Gly Lys Ser Asn Ser Arg Ser Met Pro Leu Glu Trp Glu Asp Thr Tyr
50 55 60
Gly Ile Val Leu Leu Ser Gly Val Lys Tyr Lys Lys Gly Gly Leu Val
65 70 75 80
Ile Asn Glu Thr Gly Leu Tyr Phe Val Tyr Ser Lys Val Tyr Phe Arg
85 90 95
Gly Gln Ser Cys Asn Asn Leu Pro Leu Ser His Lys Val Tyr Met Arg
100 105 110
Asn Ser Lys Tyr Pro Gln Asp Leu Val Met Met Glu Gly Lys Met Met
115 120 125
Ser Tyr Cys Thr Thr Gly Gln Met Trp Ala Arg Ser Ser Tyr Leu Gly
130 135 140
Ala Val Phe Asn Leu Thr Ser Ala Asp His Leu Tyr Val Asn Val Ser
145 150 155 160
Glu Leu Ser Leu Val Asn Phe Glu Glu Ser Gln Thr Phe Phe Gly Leu
165 170 175
Tyr Lys Leu
<210> 15
<211> 199
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 15
Met Thr Leu His Pro Ser Pro Ile Thr Cys Glu Phe Leu Phe Ser Thr
1 5 10 15
Ala Leu Ile Ser Pro Lys Met Cys Leu Ser His Leu Glu Asn Met Pro
20 25 30
Leu Ser His Ser Arg Thr Gln Gly Ala Gln Arg Ser Ser Trp Lys Leu
35 40 45
Trp Leu Phe Cys Ser Ile Val Met Leu Leu Phe Leu Cys Ser Phe Ser
50 55 60
Trp Leu Ile Phe Ile Phe Leu Gln Leu Glu Thr Ala Lys Glu Pro Cys
65 70 75 80
Met Ala Lys Phe Gly Pro Leu Pro Ser Lys Trp Gln Met Ala Ser Ser
85 90 95
Glu Pro Pro Cys Val Asn Lys Val Ser Asp Trp Lys Leu Glu Ile Leu
100 105 110
Gln Asn Gly Leu Tyr Leu Ile Tyr Gly Gln Val Ala Pro Asn Ala Asn
115 120 125
Tyr Asn Asp Val Ala Pro Phe Glu Val Arg Leu Tyr Lys Asn Lys Asp
130 135 140
Met Ile Gln Thr Leu Thr Asn Lys Ser Lys Ile Gln Asn Val Gly Gly
145 150 155 160
Thr Tyr Glu Leu His Val Gly Asp Thr Ile Asp Leu Ile Phe Asn Ser
165 170 175
Glu His Gln Val Leu Lys Asn Asn Thr Tyr Trp Gly Ile Ile Leu Leu
180 185 190
Ala Asn Pro Gln Phe Ile Ser
195
<210> 16
<211> 128
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 16
Gln Leu Glu Thr Ala Lys Glu Pro Cys Met Ala Lys Phe Gly Pro Leu
1 5 10 15
Pro Ser Lys Trp Gln Met Ala Ser Ser Glu Pro Pro Cys Val Asn Lys
20 25 30
Val Ser Asp Trp Lys Leu Glu Ile Leu Gln Asn Gly Leu Tyr Leu Ile
35 40 45
Tyr Gly Gln Val Ala Pro Asn Ala Asn Tyr Asn Asp Val Ala Pro Phe
50 55 60
Glu Val Arg Leu Tyr Lys Asn Lys Asp Met Ile Gln Thr Leu Thr Asn
65 70 75 80
Lys Ser Lys Ile Gln Asn Val Gly Gly Thr Tyr Glu Leu His Val Gly
85 90 95
Asp Thr Ile Asp Leu Ile Phe Asn Ser Glu His Gln Val Leu Lys Asn
100 105 110
Asn Thr Tyr Trp Gly Ile Ile Leu Leu Ala Asn Pro Gln Phe Ile Ser
115 120 125
<210> 17
<211> 240
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 17
Met Glu Glu Ser Val Val Arg Pro Ser Val Phe Val Val Asp Gly Gln
1 5 10 15
Thr Asp Ile Pro Phe Thr Arg Leu Gly Arg Ser His Arg Arg Gln Ser
20 25 30
Cys Ser Val Ala Arg Val Gly Leu Gly Leu Leu Leu Leu Leu Met Gly
35 40 45
Ala Gly Leu Ala Val Gln Gly Trp Phe Leu Leu Gln Leu His Trp Arg
50 55 60
Leu Gly Glu Met Val Thr Arg Leu Pro Asp Gly Pro Ala Gly Ser Trp
65 70 75 80
Glu Gln Leu Ile Gln Glu Arg Arg Ser His Glu Val Asn Pro Ala Ala
85 90 95
His Leu Thr Gly Ala Asn Ser Ser Leu Thr Gly Ser Gly Gly Pro Leu
100 105 110
Leu Trp Glu Thr Gln Leu Gly Leu Ala Phe Leu Arg Gly Leu Ser Tyr
115 120 125
His Asp Gly Ala Leu Val Val Thr Lys Ala Gly Tyr Tyr Tyr Ile Tyr
130 135 140
Ser Lys Val Gln Leu Gly Gly Val Gly Cys Pro Leu Gly Leu Ala Ser
145 150 155 160
Thr Ile Thr His Gly Leu Tyr Lys Arg Thr Pro Arg Tyr Pro Glu Glu
165 170 175
Leu Glu Leu Leu Val Ser Gln Gln Ser Pro Cys Gly Arg Ala Thr Ser
180 185 190
Ser Ser Arg Val Trp Trp Asp Ser Ser Phe Leu Gly Gly Val Val His
195 200 205
Leu Glu Ala Gly Glu Lys Val Val Val Arg Val Leu Asp Glu Arg Leu
210 215 220
Val Arg Leu Arg Asp Gly Thr Arg Ser Tyr Phe Gly Ala Phe Met Val
225 230 235 240
<210> 18
<211> 182
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 18
Leu Gln Leu His Trp Arg Leu Gly Glu Met Val Thr Arg Leu Pro Asp
1 5 10 15
Gly Pro Ala Gly Ser Trp Glu Gln Leu Ile Gln Glu Arg Arg Ser His
20 25 30
Glu Val Asn Pro Ala Ala His Leu Thr Gly Ala Asn Ser Ser Leu Thr
35 40 45
Gly Ser Gly Gly Pro Leu Leu Trp Glu Thr Gln Leu Gly Leu Ala Phe
50 55 60
Leu Arg Gly Leu Ser Tyr His Asp Gly Ala Leu Val Val Thr Lys Ala
65 70 75 80
Gly Tyr Tyr Tyr Ile Tyr Ser Lys Val Gln Leu Gly Gly Val Gly Cys
85 90 95
Pro Leu Gly Leu Ala Ser Thr Ile Thr His Gly Leu Tyr Lys Arg Thr
100 105 110
Pro Arg Tyr Pro Glu Glu Leu Glu Leu Leu Val Ser Gln Gln Ser Pro
115 120 125
Cys Gly Arg Ala Thr Ser Ser Ser Arg Val Trp Trp Asp Ser Ser Phe
130 135 140
Leu Gly Gly Val Val His Leu Glu Ala Gly Glu Lys Val Val Val Arg
145 150 155 160
Val Leu Asp Glu Arg Leu Val Arg Leu Arg Asp Gly Thr Arg Ser Tyr
165 170 175
Phe Gly Ala Phe Met Val
180
<210> 19
<400> 19
000
<210> 20
<400> 20
000
<210> 21
<211> 251
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 21
Met Ala Glu Asp Leu Gly Leu Ser Phe Gly Glu Thr Ala Ser Val Glu
1 5 10 15
Met Leu Pro Glu His Gly Ser Cys Arg Pro Lys Ala Arg Ser Ser Ser
20 25 30
Ala Arg Trp Ala Leu Thr Cys Cys Leu Val Leu Leu Pro Phe Leu Ala
35 40 45
Gly Leu Thr Thr Tyr Leu Leu Val Ser Gln Leu Arg Ala Gln Gly Glu
50 55 60
Ala Cys Val Gln Phe Gln Ala Leu Lys Gly Gln Glu Phe Ala Pro Ser
65 70 75 80
His Gln Gln Val Tyr Ala Pro Leu Arg Ala Asp Gly Asp Lys Pro Arg
85 90 95
Ala His Leu Thr Val Val Arg Gln Thr Pro Thr Gln His Phe Lys Asn
100 105 110
Gln Phe Pro Ala Leu His Trp Glu His Glu Leu Gly Leu Ala Phe Thr
115 120 125
Lys Asn Arg Met Asn Tyr Thr Asn Lys Phe Leu Leu Ile Pro Glu Ser
130 135 140
Gly Asp Tyr Phe Ile Tyr Ser Gln Val Thr Phe Arg Gly Met Thr Ser
145 150 155 160
Glu Cys Ser Glu Ile Arg Gln Ala Gly Arg Pro Asn Lys Pro Asp Ser
165 170 175
Ile Thr Val Val Ile Thr Lys Val Thr Asp Ser Tyr Pro Glu Pro Thr
180 185 190
Gln Leu Leu Met Gly Thr Lys Ser Val Cys Glu Val Gly Ser Asn Trp
195 200 205
Phe Gln Pro Ile Tyr Leu Gly Ala Met Phe Ser Leu Gln Glu Gly Asp
210 215 220
Lys Leu Met Val Asn Val Ser Asp Ile Ser Leu Val Asp Tyr Thr Lys
225 230 235 240
Glu Asp Lys Thr Phe Phe Gly Ala Phe Leu Leu
245 250
<210> 22
<211> 192
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 22
Arg Ala Gln Gly Glu Ala Cys Val Gln Phe Gln Ala Leu Lys Gly Gln
1 5 10 15
Glu Phe Ala Pro Ser His Gln Gln Val Tyr Ala Pro Leu Arg Ala Asp
20 25 30
Gly Asp Lys Pro Arg Ala His Leu Thr Val Val Arg Gln Thr Pro Thr
35 40 45
Gln His Phe Lys Asn Gln Phe Pro Ala Leu His Trp Glu His Glu Leu
50 55 60
Gly Leu Ala Phe Thr Lys Asn Arg Met Asn Tyr Thr Asn Lys Phe Leu
65 70 75 80
Leu Ile Pro Glu Ser Gly Asp Tyr Phe Ile Tyr Ser Gln Val Thr Phe
85 90 95
Arg Gly Met Thr Ser Glu Cys Ser Glu Ile Arg Gln Ala Gly Arg Pro
100 105 110
Asn Lys Pro Asp Ser Ile Thr Val Val Ile Thr Lys Val Thr Asp Ser
115 120 125
Tyr Pro Glu Pro Thr Gln Leu Leu Met Gly Thr Lys Ser Val Cys Glu
130 135 140
Val Gly Ser Asn Trp Phe Gln Pro Ile Tyr Leu Gly Ala Met Phe Ser
145 150 155 160
Leu Gln Glu Gly Asp Lys Leu Met Val Asn Val Ser Asp Ile Ser Leu
165 170 175
Val Asp Tyr Thr Lys Glu Asp Lys Thr Phe Phe Gly Ala Phe Leu Leu
180 185 190
<210> 23
<211> 281
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 23
Met Ala Met Met Glu Val Gln Gly Gly Pro Ser Leu Gly Gln Thr Cys
1 5 10 15
Val Leu Ile Val Ile Phe Thr Val Leu Leu Gln Ser Leu Cys Val Ala
20 25 30
Val Thr Tyr Val Tyr Phe Thr Asn Glu Leu Lys Gln Met Gln Asp Lys
35 40 45
Tyr Ser Lys Ser Gly Ile Ala Cys Phe Leu Lys Glu Asp Asp Ser Tyr
50 55 60
Trp Asp Pro Asn Asp Glu Glu Ser Met Asn Ser Pro Cys Trp Gln Val
65 70 75 80
Lys Trp Gln Leu Arg Gln Leu Val Arg Lys Met Ile Leu Arg Thr Ser
85 90 95
Glu Glu Thr Ile Ser Thr Val Gln Glu Lys Gln Gln Asn Ile Ser Pro
100 105 110
Leu Val Arg Glu Arg Gly Pro Gln Arg Val Ala Ala His Ile Thr Gly
115 120 125
Thr Arg Gly Arg Ser Asn Thr Leu Ser Ser Pro Asn Ser Lys Asn Glu
130 135 140
Lys Ala Leu Gly Arg Lys Ile Asn Ser Trp Glu Ser Ser Arg Ser Gly
145 150 155 160
His Ser Phe Leu Ser Asn Leu His Leu Arg Asn Gly Glu Leu Val Ile
165 170 175
His Glu Lys Gly Phe Tyr Tyr Ile Tyr Ser Gln Thr Tyr Phe Arg Phe
180 185 190
Gln Glu Glu Ile Lys Glu Asn Thr Lys Asn Asp Lys Gln Met Val Gln
195 200 205
Tyr Ile Tyr Lys Tyr Thr Ser Tyr Pro Asp Pro Ile Leu Leu Met Lys
210 215 220
Ser Ala Arg Asn Ser Cys Trp Ser Lys Asp Ala Glu Tyr Gly Leu Tyr
225 230 235 240
Ser Ile Tyr Gln Gly Gly Ile Phe Glu Leu Lys Glu Asn Asp Arg Ile
245 250 255
Phe Val Ser Val Thr Asn Glu His Leu Ile Asp Met Asp His Glu Ala
260 265 270
Ser Phe Phe Gly Ala Phe Leu Val Gly
275 280
<210> 24
<211> 243
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 24
Thr Asn Glu Leu Lys Gln Met Gln Asp Lys Tyr Ser Lys Ser Gly Ile
1 5 10 15
Ala Cys Phe Leu Lys Glu Asp Asp Ser Tyr Trp Asp Pro Asn Asp Glu
20 25 30
Glu Ser Met Asn Ser Pro Cys Trp Gln Val Lys Trp Gln Leu Arg Gln
35 40 45
Leu Val Arg Lys Met Ile Leu Arg Thr Ser Glu Glu Thr Ile Ser Thr
50 55 60
Val Gln Glu Lys Gln Gln Asn Ile Ser Pro Leu Val Arg Glu Arg Gly
65 70 75 80
Pro Gln Arg Val Ala Ala His Ile Thr Gly Thr Arg Gly Arg Ser Asn
85 90 95
Thr Leu Ser Ser Pro Asn Ser Lys Asn Glu Lys Ala Leu Gly Arg Lys
100 105 110
Ile Asn Ser Trp Glu Ser Ser Arg Ser Gly His Ser Phe Leu Ser Asn
115 120 125
Leu His Leu Arg Asn Gly Glu Leu Val Ile His Glu Lys Gly Phe Tyr
130 135 140
Tyr Ile Tyr Ser Gln Thr Tyr Phe Arg Phe Gln Glu Glu Ile Lys Glu
145 150 155 160
Asn Thr Lys Asn Asp Lys Gln Met Val Gln Tyr Ile Tyr Lys Tyr Thr
165 170 175
Ser Tyr Pro Asp Pro Ile Leu Leu Met Lys Ser Ala Arg Asn Ser Cys
180 185 190
Trp Ser Lys Asp Ala Glu Tyr Gly Leu Tyr Ser Ile Tyr Gln Gly Gly
195 200 205
Ile Phe Glu Leu Lys Glu Asn Asp Arg Ile Phe Val Ser Val Thr Asn
210 215 220
Glu His Leu Ile Asp Met Asp His Glu Ala Ser Phe Phe Gly Ala Phe
225 230 235 240
Leu Val Gly
<210> 25
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 25
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Arg Leu Thr Val Asp Lys Ser Ser Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 26
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 26
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Thr Pro His
65 70 75 80
Ser Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Arg Leu Thr Val Asp Lys Ser Ser Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 27
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 27
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Ser Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 28
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 28
Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro
1 5 10
<210> 29
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 29
Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 30
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 30
Ser Lys Tyr Gly Pro Pro
1 5
<210> 31
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 31
Ile Glu Gly Arg Met Asp
1 5
<210> 32
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 32
Gly Gly Gly Val Pro Arg Asp Cys Gly
1 5
<210> 33
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 33
Ile Glu Gly Arg Met Asp Gly Gly Gly Gly Ala Gly Gly Gly Gly
1 5 10 15
<210> 34
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 34
Gly Gly Gly Ser Gly Gly Gly Ser
1 5
<210> 35
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 35
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
1 5 10
<210> 36
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 36
Glu Gly Lys Ser Ser Gly Ser Gly Ser Glu Ser Lys Ser Thr
1 5 10
<210> 37
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 37
Gly Gly Ser Gly
1
<210> 38
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 38
Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly
1 5 10
<210> 39
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 39
Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys
1 5 10 15
<210> 40
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 40
Glu Ala Ala Ala Arg Glu Ala Ala Ala Arg Glu Ala Ala Ala Arg Glu
1 5 10 15
Ala Ala Ala Arg
20
<210> 41
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 41
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala
1 5 10 15
Ser
<210> 42
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 42
Gly Gly Gly Gly Ala Gly Gly Gly Gly
1 5
<210> 43
<400> 43
000
<210> 44
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 44
Gly Ser Gly Ser Gly Ser
1 5
<210> 45
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 45
Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser
1 5 10
<210> 46
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 46
Gly Gly Gly Gly Ser Ala Ser
1 5
<210> 47
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 47
Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro
1 5 10 15
Ala Pro Ala Pro
20
<210> 48
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 48
Cys Pro Pro Cys
1
<210> 49
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 49
Gly Gly Gly Gly Ser
1 5
<210> 50
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 50
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 51
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 51
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 52
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 52
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 53
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 53
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 54
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 54
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25 30
<210> 55
<211> 35
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 55
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser
35
<210> 56
<211> 40
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 56
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser Gly Gly Gly Gly Ser
35 40
<210> 57
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 57
Gly Gly Ser Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 58
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 58
Gly Gly Gly Gly Gly Gly Gly Gly
1 5
<210> 59
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 59
Gly Gly Gly Gly Gly Gly
1 5
<210> 60
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 60
Glu Ala Ala Ala Lys
1 5
<210> 61
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 61
Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys
1 5 10
<210> 62
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 62
Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys
1 5 10 15
<210> 63
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 63
Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Ala
1 5 10
<210> 64
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 64
Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys
1 5 10 15
Ala
<210> 65
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 65
Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys
1 5 10 15
Glu Ala Ala Ala Lys Ala
20
<210> 66
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 66
Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys
1 5 10 15
Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Ala
20 25
<210> 67
<211> 46
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 67
Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys
1 5 10 15
Glu Ala Ala Ala Lys Ala Leu Glu Ala Glu Ala Ala Ala Lys Glu Ala
20 25 30
Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Ala
35 40 45
<210> 68
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 68
Pro Ala Pro Ala Pro
1 5
<210> 69
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 69
Lys Glu Ser Gly Ser Val Ser Ser Glu Gln Leu Ala Gln Phe Arg Ser
1 5 10 15
Leu Asp
<210> 70
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 70
Gly Ser Ala Gly Ser Ala Ala Gly Ser Gly Glu Phe
1 5 10
<210> 71
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 71
Gly Gly Gly Ser Glu
1 5
<210> 72
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 72
Gly Ser Glu Ser Gly
1 5
<210> 73
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 73
Gly Ser Glu Gly Ser
1 5
<210> 74
<211> 35
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 74
Gly Glu Gly Gly Ser Gly Glu Gly Ser Ser Gly Glu Gly Ser Ser Ser
1 5 10 15
Glu Gly Gly Gly Ser Glu Gly Gly Gly Ser Glu Gly Gly Gly Ser Glu
20 25 30
Gly Gly Ser
35
<210> 75
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 75
Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe Leu
1 5 10 15
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
20 25 30
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
35 40 45
Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
50 55 60
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
65 70 75 80
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Ser
85 90 95
Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys Gly Leu Pro Ser Ser
100 105 110
Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln Pro Arg Glu Pro Gln
115 120 125
Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val
130 135 140
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
145 150 155 160
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
165 170 175
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr
180 185 190
Val Asp Lys Ser Ser Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val
195 200 205
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
210 215 220
Ser Leu Gly Lys Ile Glu Gly Arg Met Asp
225 230
<210> 76
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 76
Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe Leu
1 5 10 15
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu
20 25 30
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
35 40 45
Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
50 55 60
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
65 70 75 80
Tyr Arg Val Val Ser Val Leu Thr Thr Pro His Ser Asp Trp Leu Ser
85 90 95
Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys Gly Leu Pro Ser Ser
100 105 110
Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln Pro Arg Glu Pro Gln
115 120 125
Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val
130 135 140
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
145 150 155 160
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
165 170 175
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr
180 185 190
Val Asp Lys Ser Ser Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val
195 200 205
Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
210 215 220
Ser Leu Gly Lys Ile Glu Gly Arg Met Asp
225 230
<210> 77
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 77
Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe Leu
1 5 10 15
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu
20 25 30
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
35 40 45
Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
50 55 60
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
65 70 75 80
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Ser
85 90 95
Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys Gly Leu Pro Ser Ser
100 105 110
Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln Pro Arg Glu Pro Gln
115 120 125
Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val
130 135 140
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
145 150 155 160
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
165 170 175
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr
180 185 190
Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val
195 200 205
Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
210 215 220
Ser Leu Gly Lys Ile Glu Gly Arg Met Asp
225 230
<210> 78
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 78
Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu
1 5 10 15
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
20 25 30
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
35 40 45
Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
50 55 60
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
65 70 75 80
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Ser
85 90 95
Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys Gly Leu Pro Ser Ser
100 105 110
Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln Pro Arg Glu Pro Gln
115 120 125
Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val
130 135 140
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
145 150 155 160
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
165 170 175
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr
180 185 190
Val Asp Lys Ser Ser Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val
195 200 205
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
210 215 220
Ser Leu Gly Lys Ile Glu Gly Arg Met Asp
225 230
<210> 79
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 79
Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu
1 5 10 15
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu
20 25 30
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
35 40 45
Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
50 55 60
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
65 70 75 80
Tyr Arg Val Val Ser Val Leu Thr Thr Pro His Ser Asp Trp Leu Ser
85 90 95
Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys Gly Leu Pro Ser Ser
100 105 110
Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln Pro Arg Glu Pro Gln
115 120 125
Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val
130 135 140
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
145 150 155 160
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
165 170 175
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr
180 185 190
Val Asp Lys Ser Ser Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val
195 200 205
Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
210 215 220
Ser Leu Gly Lys Ile Glu Gly Arg Met Asp
225 230
<210> 80
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 80
Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu
1 5 10 15
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu
20 25 30
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
35 40 45
Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
50 55 60
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
65 70 75 80
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Ser
85 90 95
Gly Lys Glu Tyr Lys Cys Lys Val Ser Ser Lys Gly Leu Pro Ser Ser
100 105 110
Ile Glu Lys Thr Ile Ser Asn Ala Thr Gly Gln Pro Arg Glu Pro Gln
115 120 125
Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val
130 135 140
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
145 150 155 160
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
165 170 175
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr
180 185 190
Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val
195 200 205
Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
210 215 220
Ser Leu Gly Lys Ile Glu Gly Arg Met Asp
225 230
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЕ БЕЛКИ НА ОСНОВЕ TIGIT И LIGHT | 2018 |
|
RU2775490C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ СОЕДИНЕНИЯ ВНЕКЛЕТОЧНЫХ ДОМЕНОВ ТИПА I И ТИПА II В КАЧЕСТВЕ ГЕТЕРОЛОГИЧНЫХ ХИМЕРНЫХ БЕЛКОВ | 2016 |
|
RU2766200C1 |
| НОВЫЙ МОДИФИЦИРОВАННЫЙ СЛИТЫЙ БЕЛОК FC-ФРАГМЕНТА ИММУНОГЛОБУЛИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2800919C2 |
| Двухвалентное биспецифическое химерное антитело, включающее гетеродимер на основе MHC или MHC-подобных белков | 2021 |
|
RU2820683C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ПАТОЛОГИЧЕСКИХ СОСТОЯНИЙ АНТИТЕЛАМИ, КОТОРЫЕ СВЯЗЫВАЮТСЯ С РЕЦЕПТОРОМ КОЛОНИЕСТИМУЛИРУЮЩЕГО ФАКТОРА 1 (CSF1R) | 2013 |
|
RU2718751C2 |
| АНТИТЕЛА К CD40L И СПОСОБЫ ЛЕЧЕНИЯ СВЯЗАННЫХ С CD40L ЗАБОЛЕВАНИЙ И НАРУШЕНИЙ | 2016 |
|
RU2727646C2 |
| CD40L-СПЕЦИФИЧНЫЕ КАРКАСНЫЕ СТРУКТУРЫ, ПРОИСХОДЯЩИЕ ИЗ TN3, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2826189C2 |
| СПОСОБ ПОВЫШЕНИЯ ИММУНОГЕННОСТИ БЕЛКОВОГО/ПЕПТИДНОГО АНТИГЕНА | 2021 |
|
RU2830627C1 |
| АНТИТЕЛО К CD40, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2779128C2 |
| УНИВЕРСАЛЬНЫЙ АНТИТЕЛООПОСРЕДОВАННЫЙ БИОСЕНСОР | 2016 |
|
RU2746486C2 |
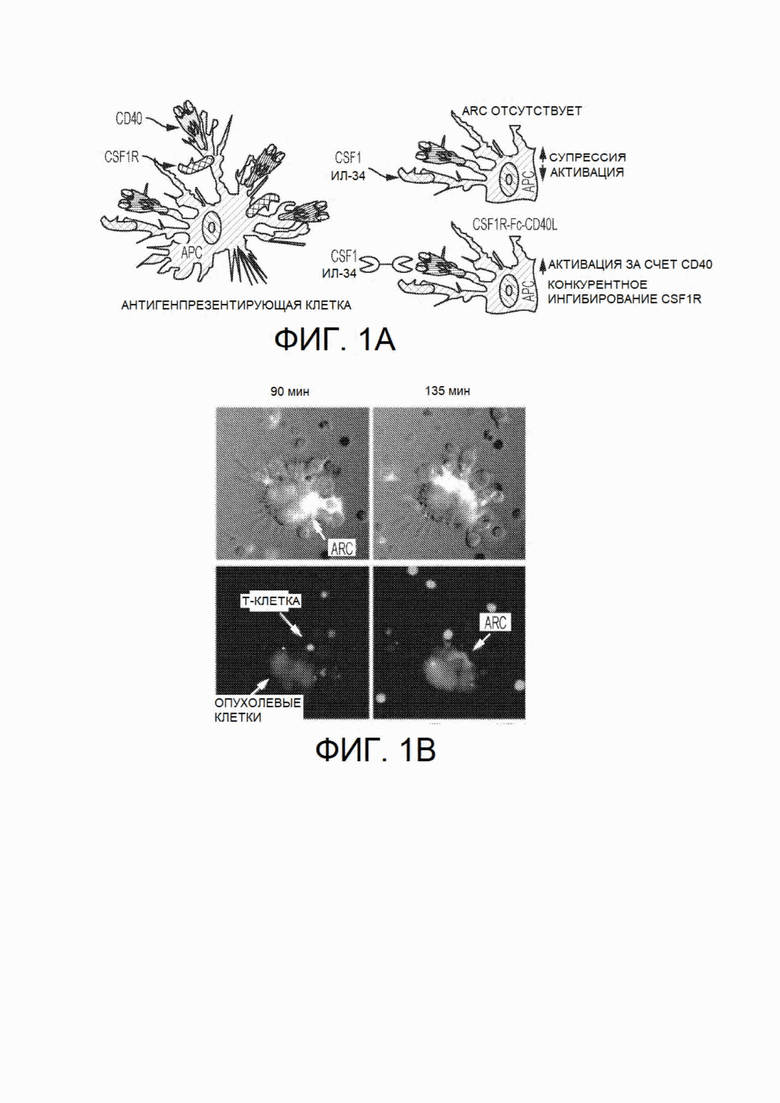
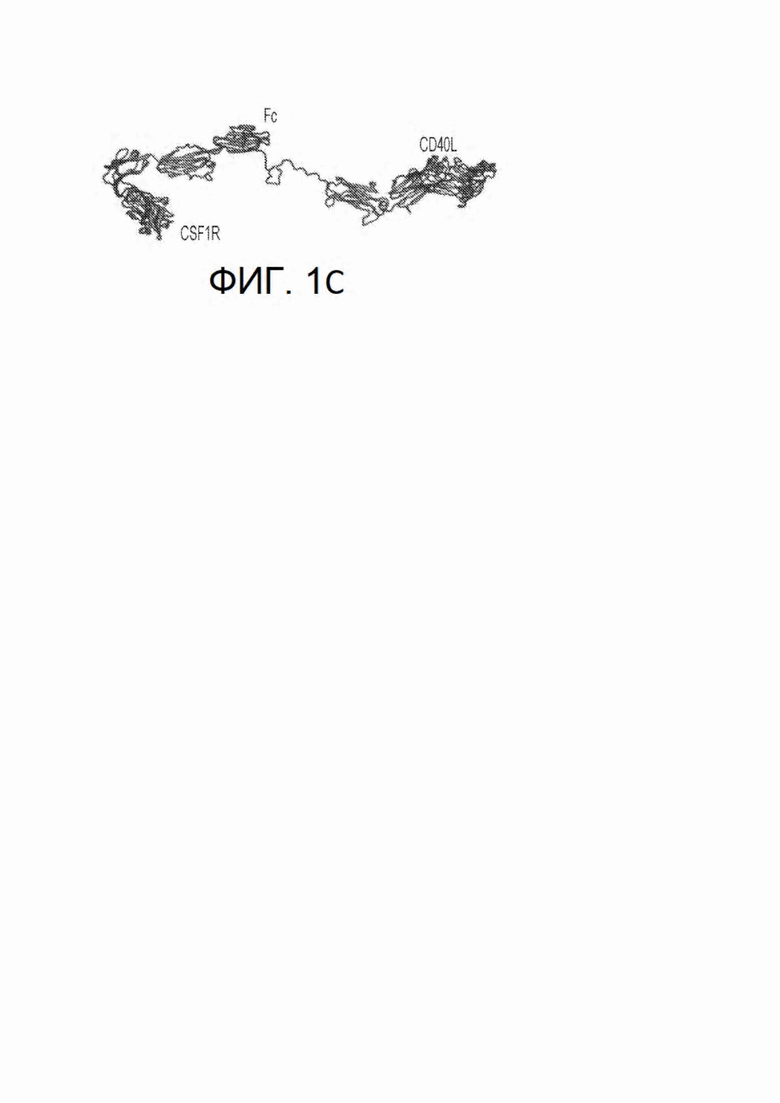
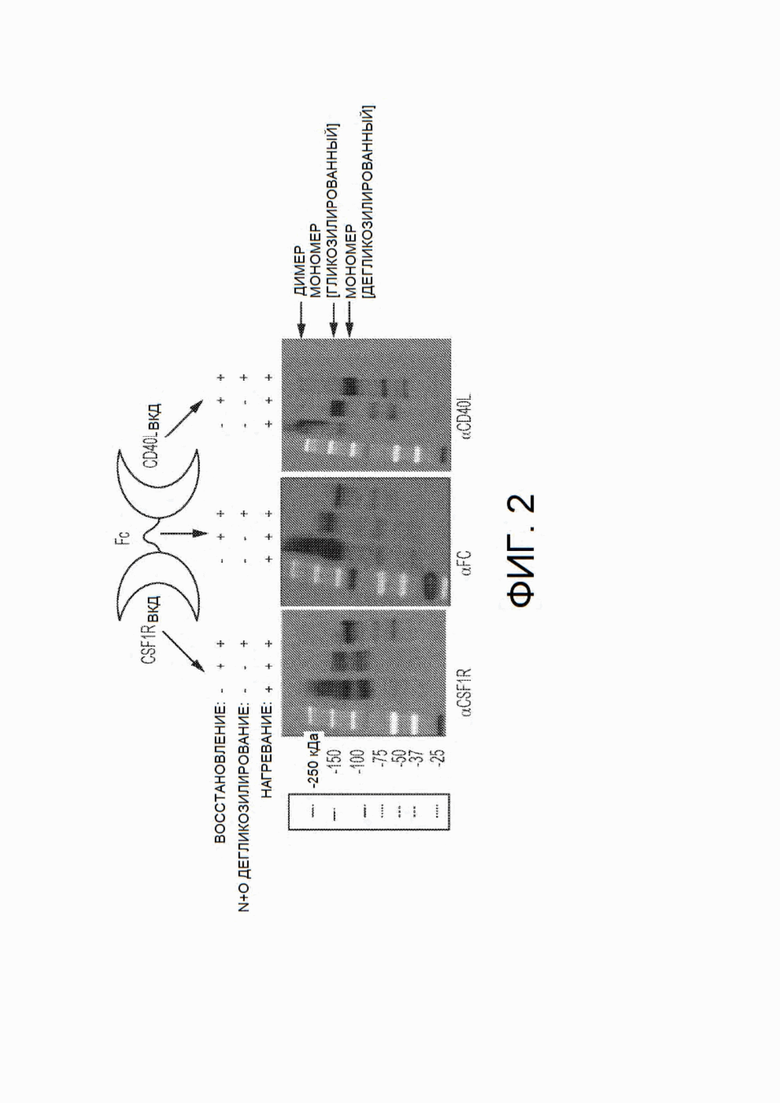
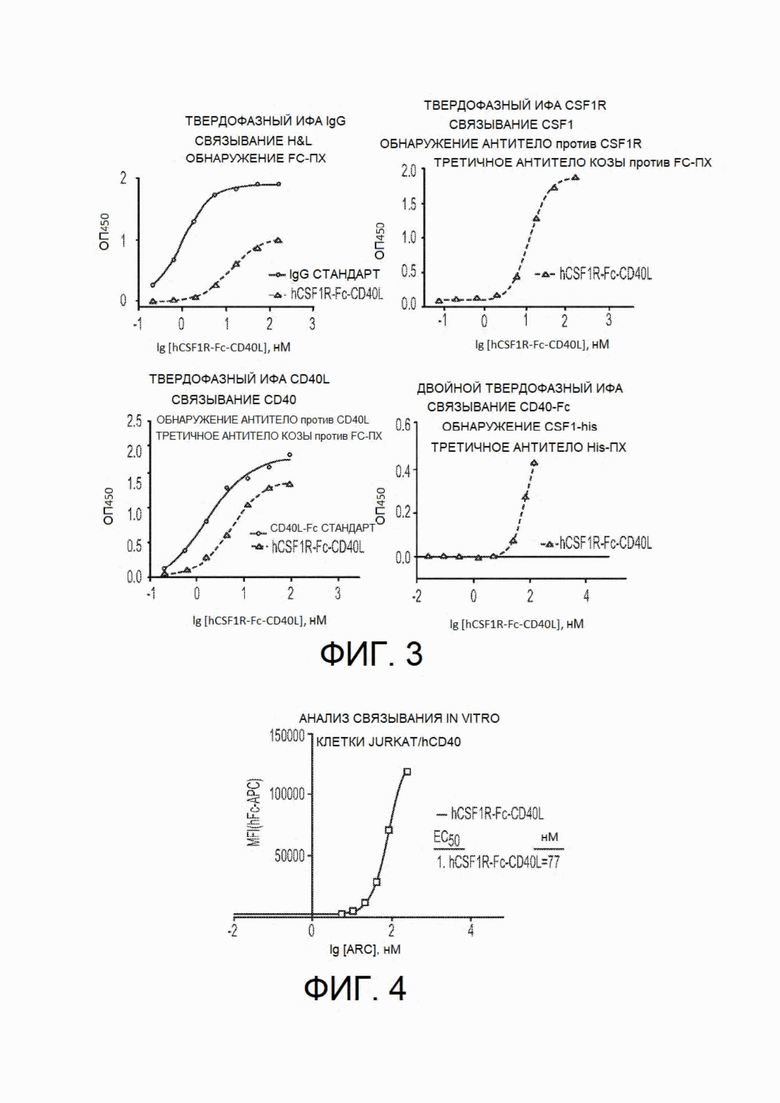
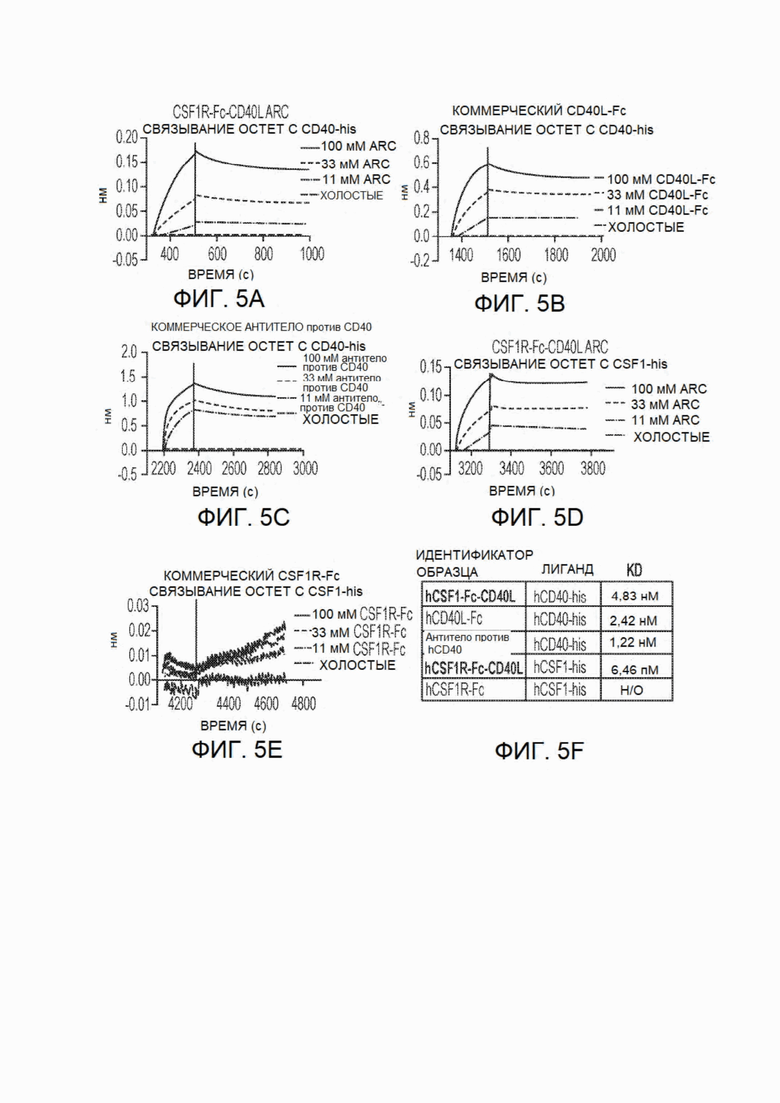
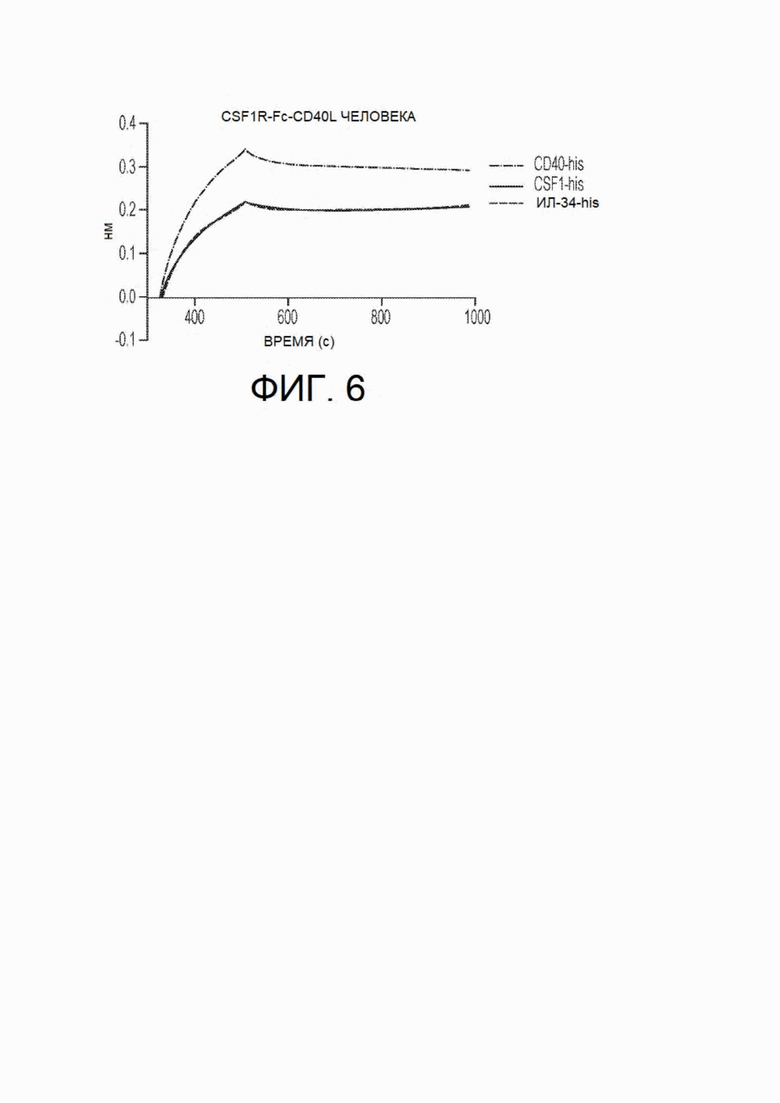
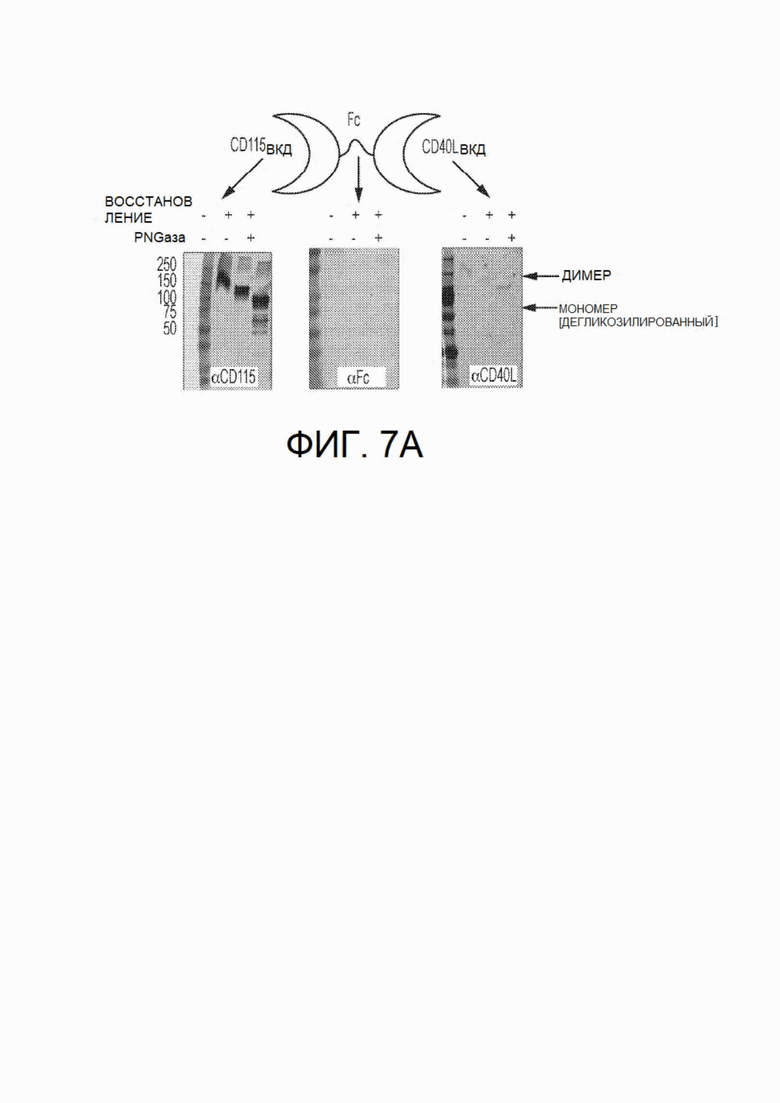
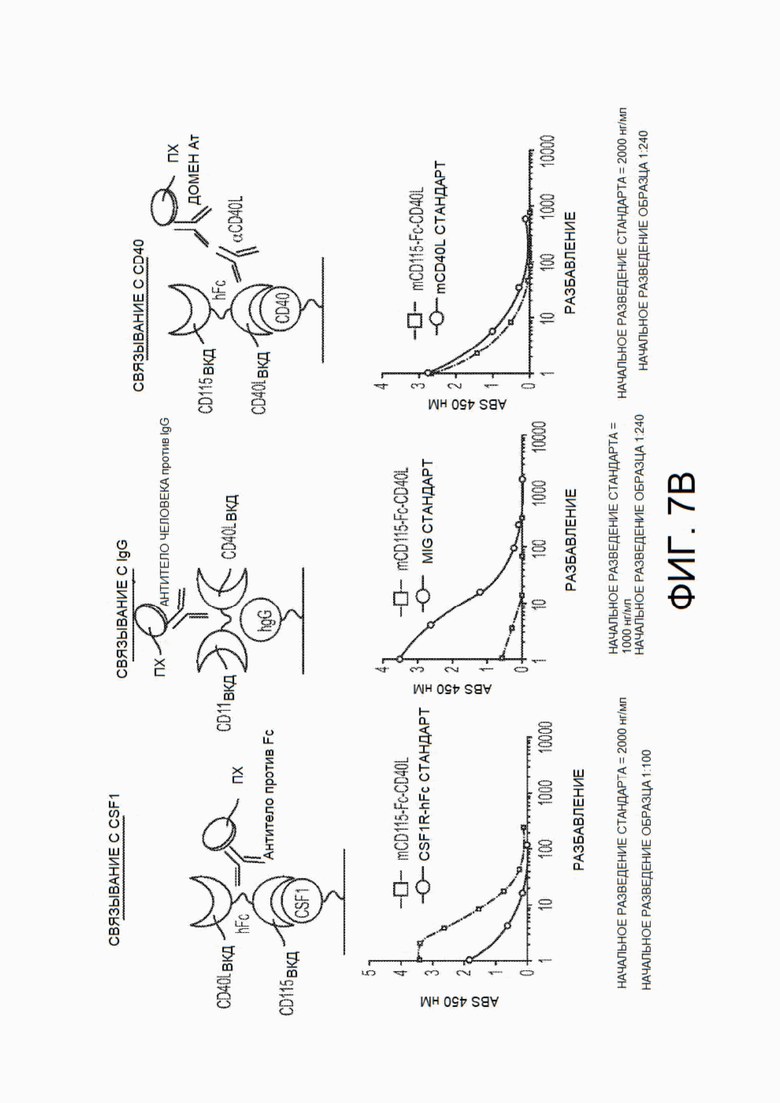
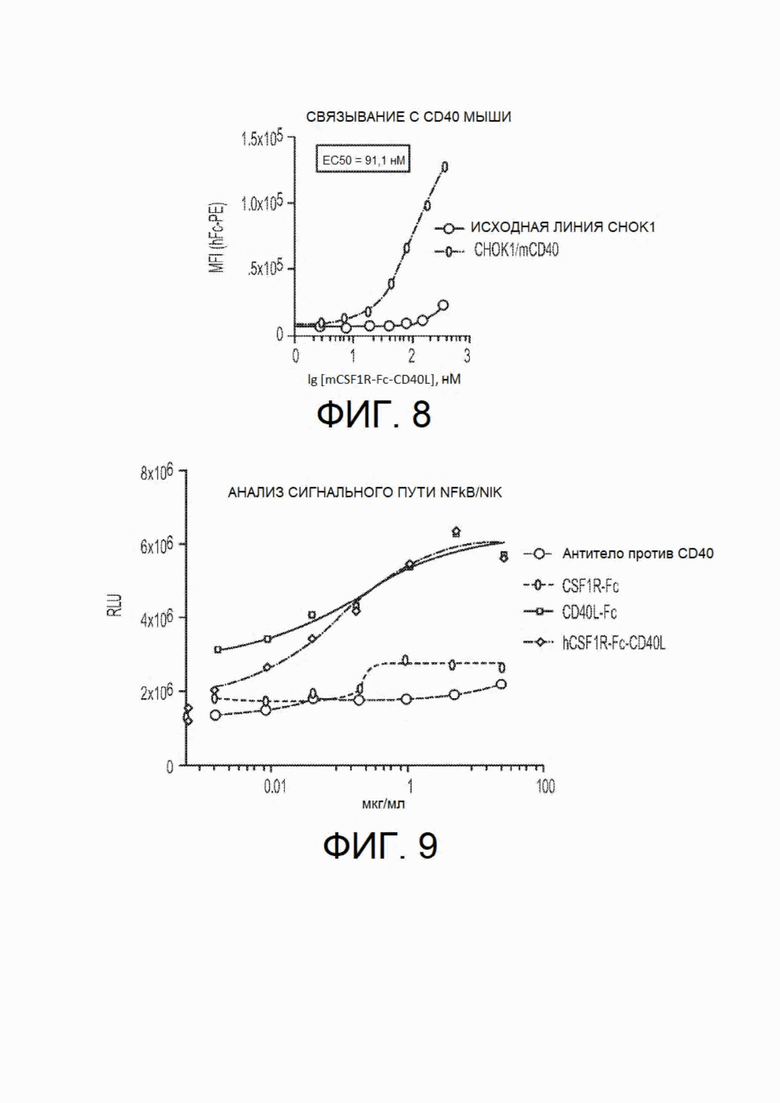
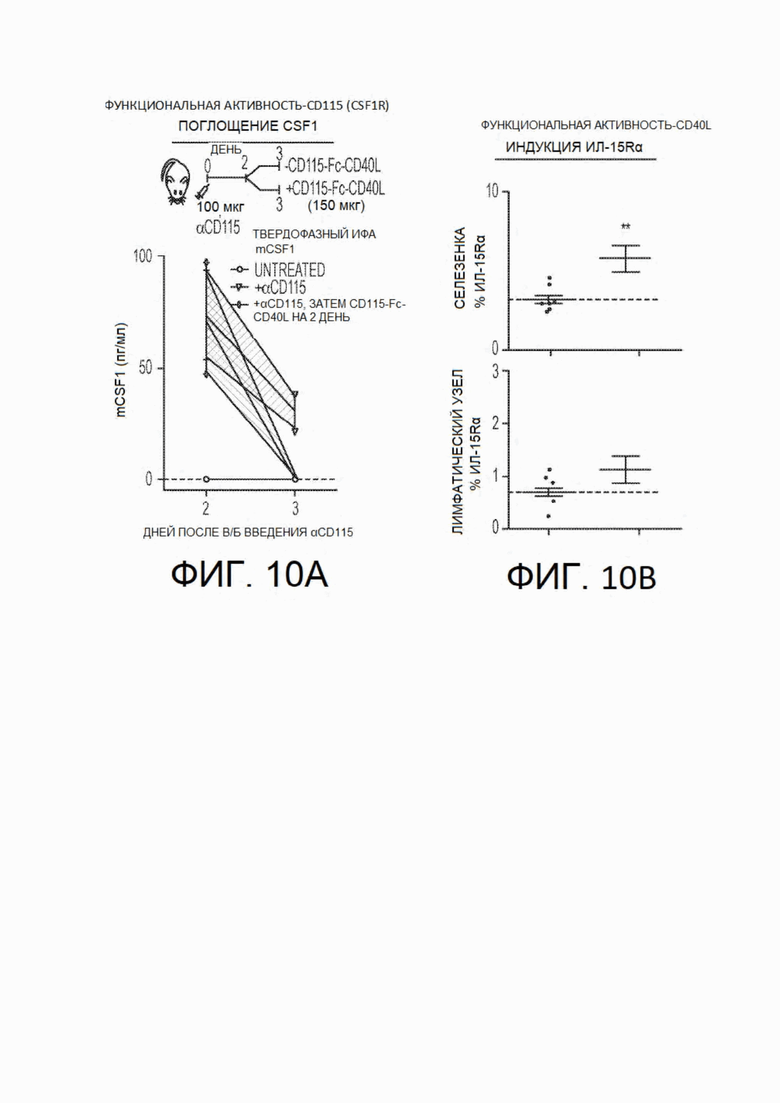
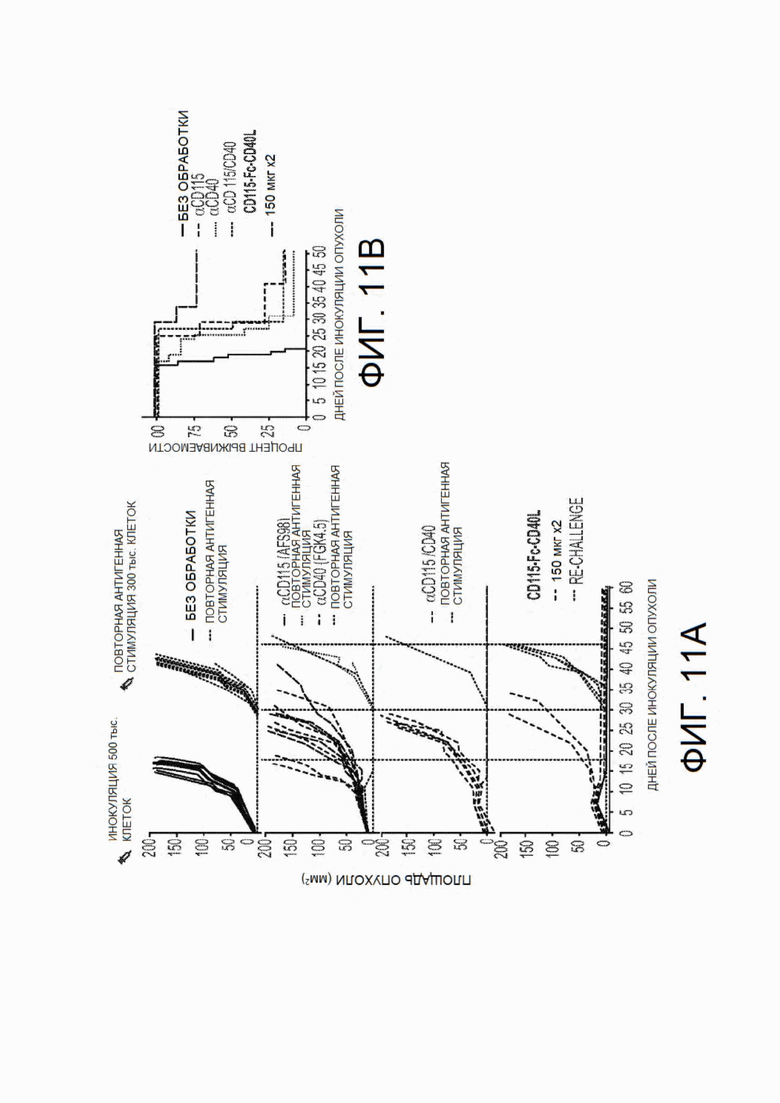
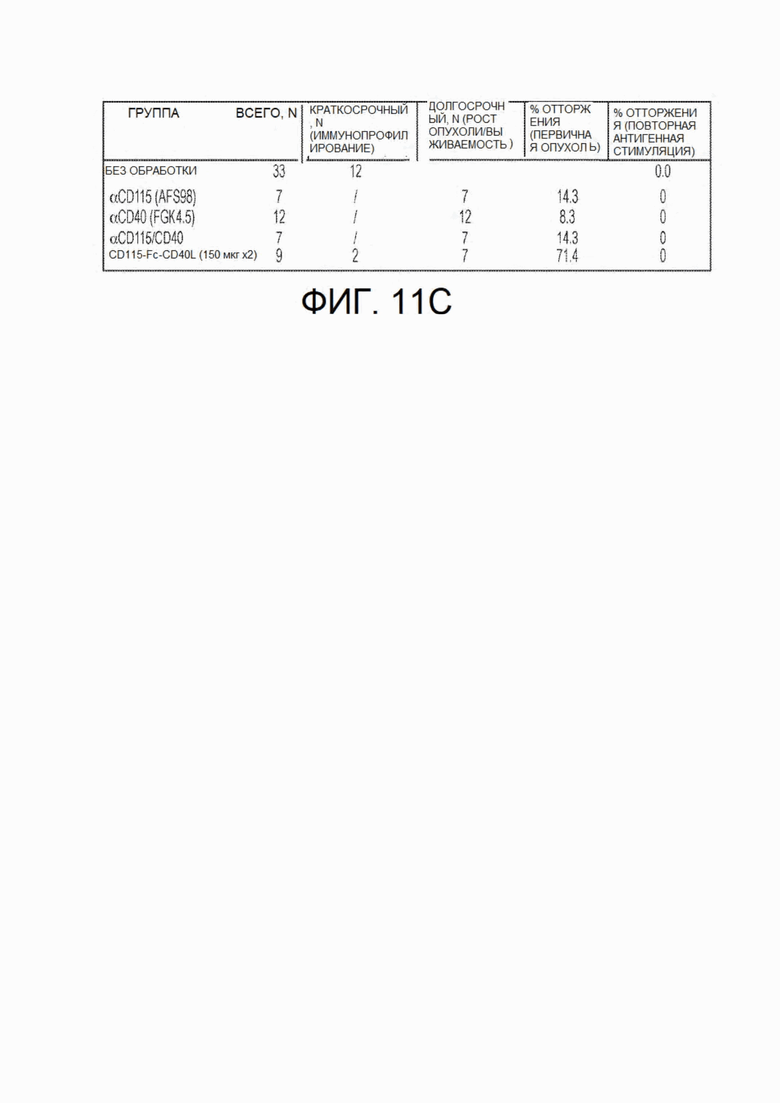
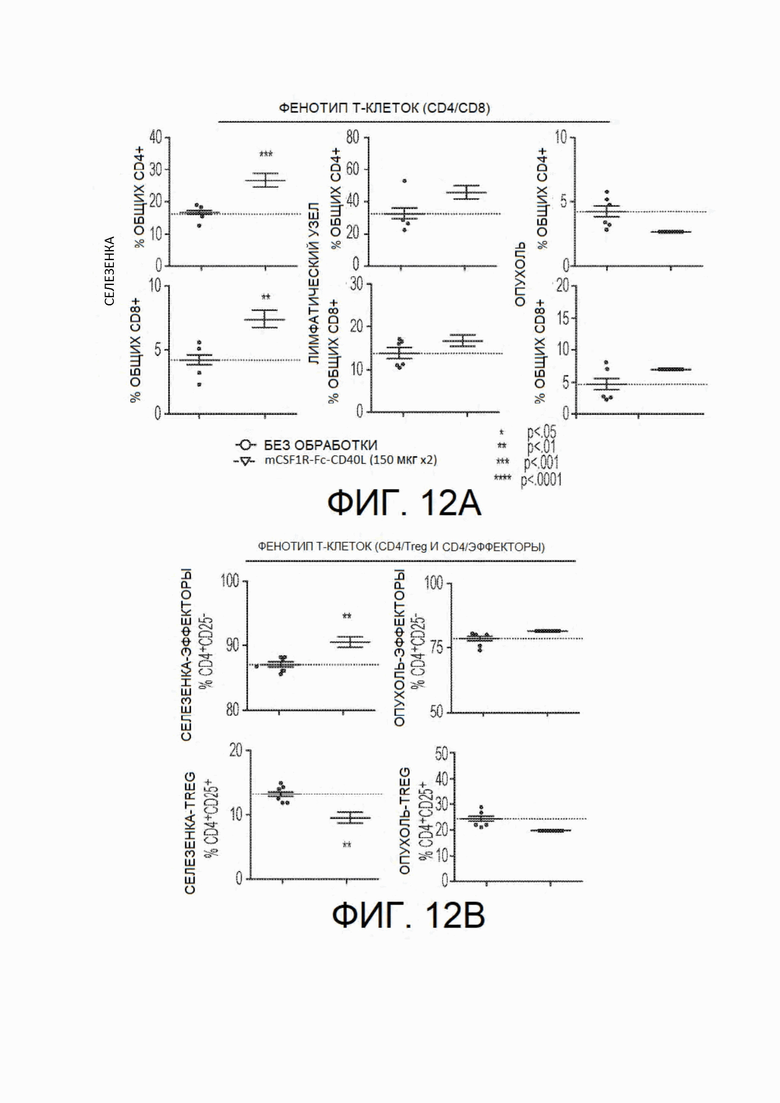
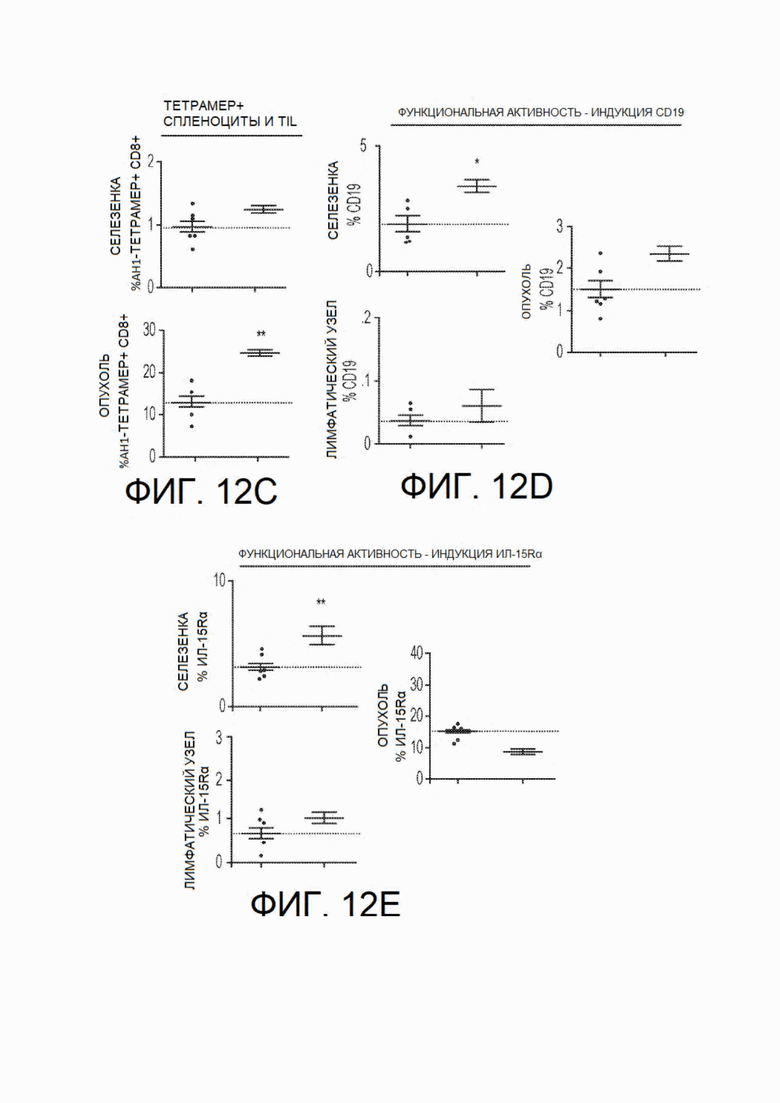
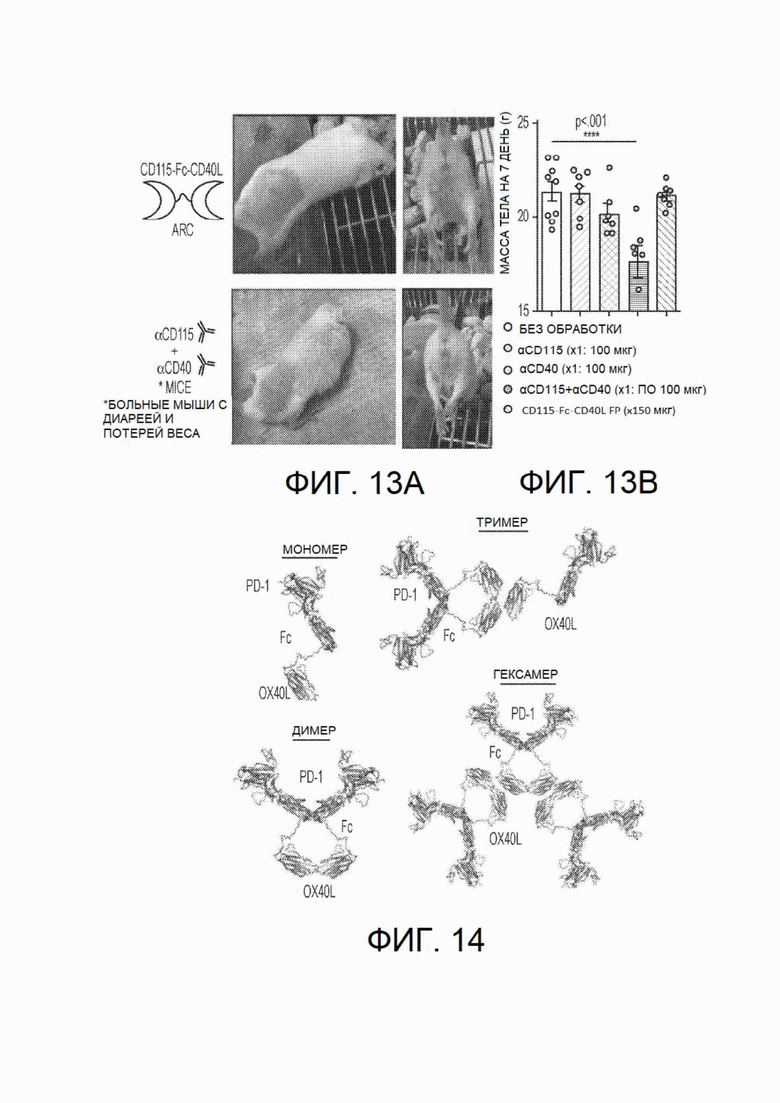
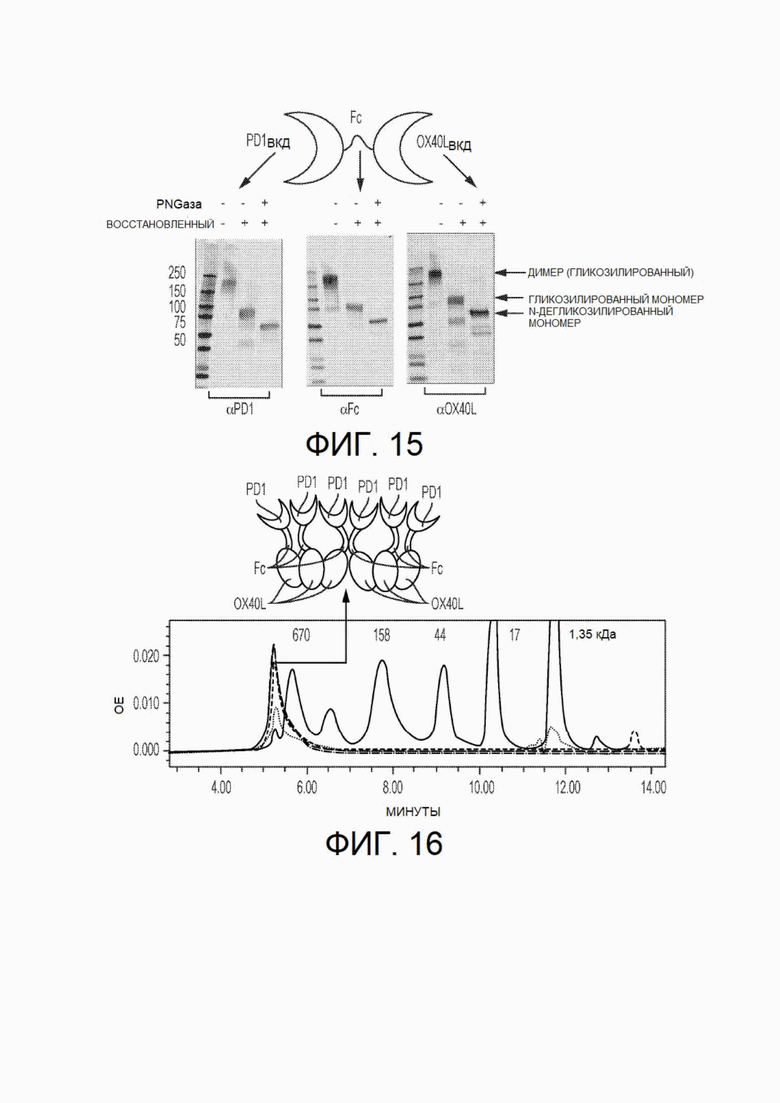
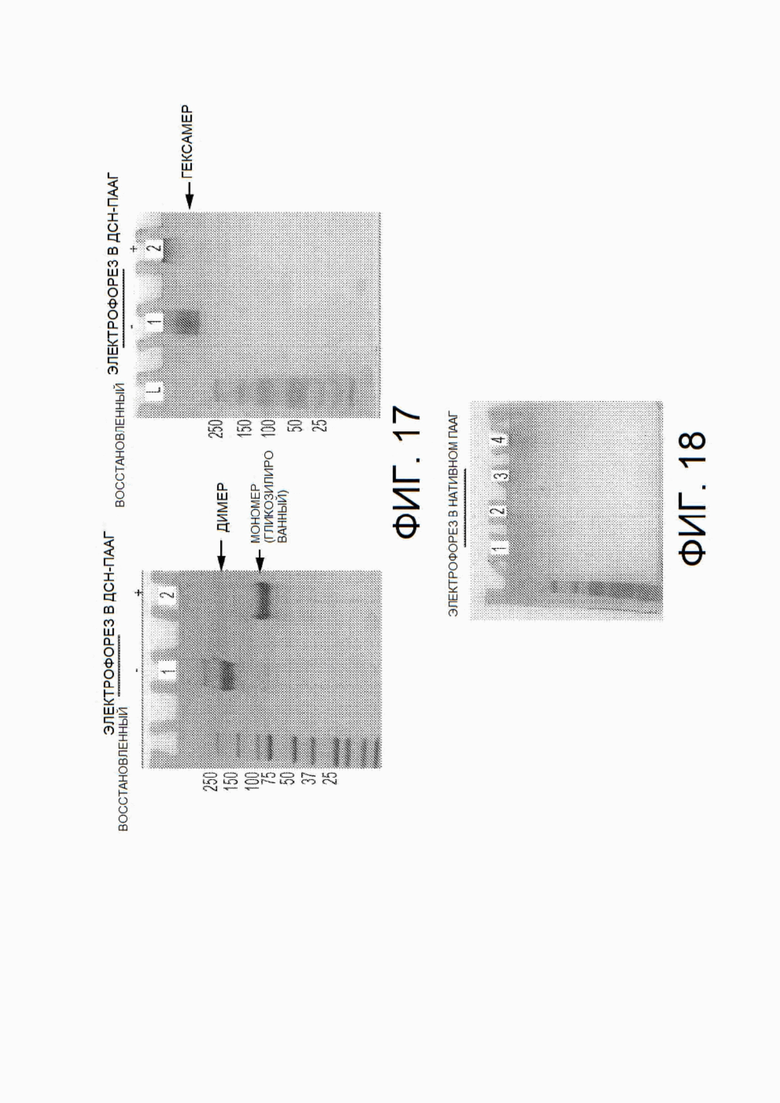
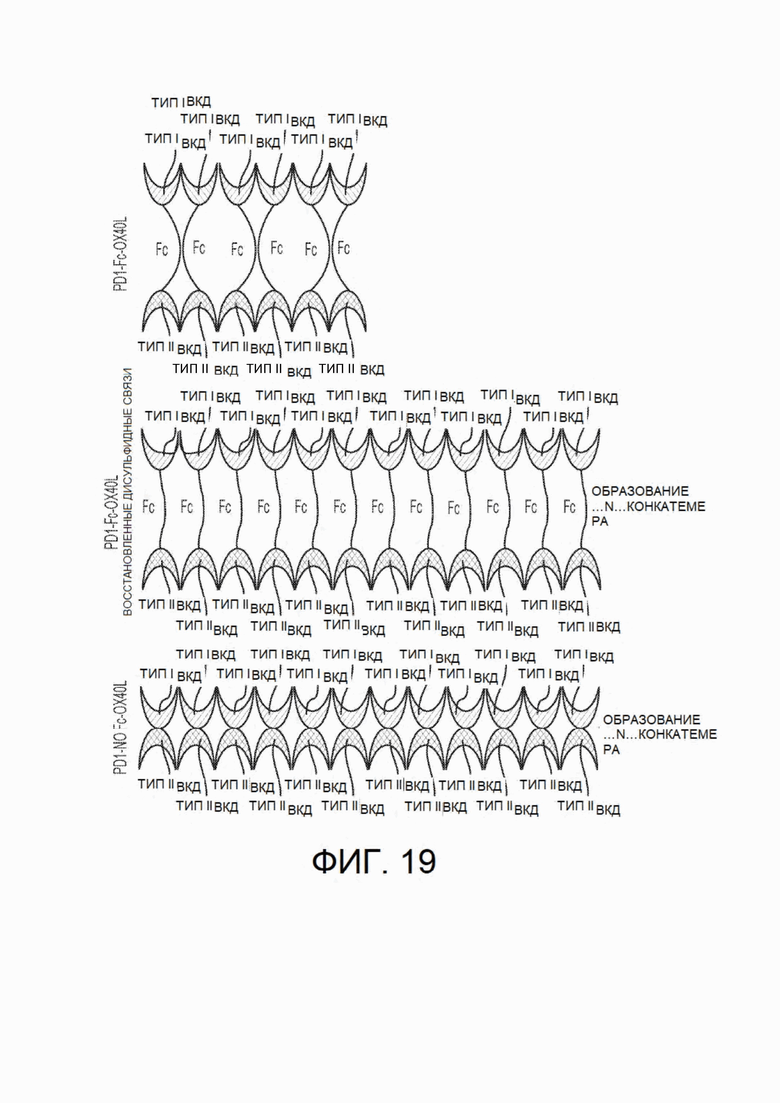
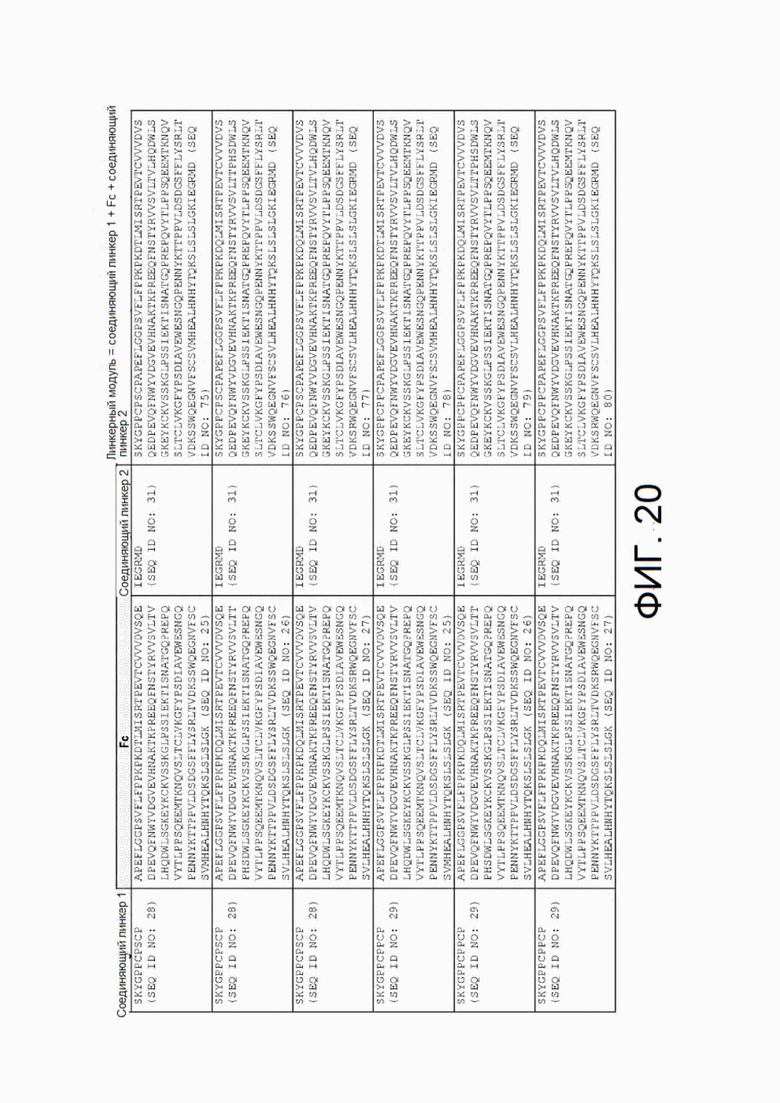
Изобретение относится к области биотехнологии, в частности к гетерологичным химерным белкам, содержащим внеклеточный домен рецептора колониестимулирующего фактора 1 (CSF1R), и их применению для одновременного маскирования иммуноингибиторного сигнала и активации иммуностимулирующего сигнала при лечении рака; к нуклеиновой кислоте, кодирующей упомянутые гетерологичные химерные белки; к клетке-хозяину, экспрессирующей упомянутые гетерологичные химерные белки; к фармацевтической композиции, содержащей терапевтически эффективное количество упомянутого гетерологичного химерного белка, и ее применению для модулирования иммунного ответа и лечения рака; к рекомбинантному гибридному белку для одновременного маскирования иммуноингибиторного сигнала и активации иммуностимулирующего сигнала и его применению в качестве лекарственного средства при лечении рака. Изобретение представляет собой эффективное иммунотерапевтическое средство, объединяющее несколько терапевтических механизмов. 13 н. и 53 з.п. ф-лы, 20 ил., 2 табл., 15 пр.
1. Гетерологичный химерный белок для одновременного маскирования иммуноингибиторного сигнала и активации иммуностимулирующего сигнала, содержащий общую структуру:
N-конец – (a) – (b) – (c) – C-конец,
где:
(а) представляет собой первый домен, содержащий внеклеточный домен рецептора колониестимулирующего фактора 1 (CSF1R), способный связывать лиганд CSF1R, и содержащий аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 2 или обладает по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами;
(b) представляет собой линкер, соединяющий первый домен и второй домен и содержащий шарнир-CH2-CH3-Fc-домен и один или более соединяющих линкеров, независимо выбранных из SEQ ID NO: 28-74, и
(с) представляет собой второй домен, содержащий внеклеточный домен лиганда CD40 (CD40L), способный связывать CD40, и содержащий аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 4 или обладает по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами.
2. Гетерологичный химерный белок по п. 1, отличающийся тем, что первый домен по существу полностью содержит внеклеточный домен CSF1R, а второй домен по существу полностью содержит внеклеточный домен CD40L.
3. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок может ингибировать иммуносупрессивный сигнал.
4. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок способен:
(а) ослаблять или устранять иммуноингибиторный сигнал при связывании фрагмента CSF1R со своим лигандом и/или
(b) увеличивать или активировать иммуностимулирующий сигнал при связывании фрагмента CD40L со своим рецептором, где рецептор CD40L представляет собой CD40.
5. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что лиганд CSF1R представляет собой CSF1 или ИЛ-34.
6. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок способен одновременно связывать лиганд CSF1R и рецептор CD40L, причем лиганд CSF1R представляет собой CSF1 или ИЛ-34, а рецептор CD40L представляет собой CD40.
7. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок способен одновременно связывать рекомбинантный CD40 человека и CSF1 человека in vitro.
8. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок вызывает истощение CSF1 и/или ИЛ-34, необязательно в сыворотке.
9. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок демонстрирует расширенный профиль безопасности и/или сокращенный профиль токсичности по сравнению с агонистическими антителами к CD40 и/или антагонистическими антителами против CSF1R.
10. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок демонстрирует усиленное противоопухолевое действие по сравнению с агонистическими антителами к CD40 и/или антагонистическими антителами против CSF1R.
11. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок способен увеличивать или предотвращать снижение численности субпопуляции CD4+ и/или CD8+ Т-клеток.
12. Гетерологичный химерный белок по любому из пп. 1 или 2, отличающийся тем, что указанный химерный белок способен усиливать активность Т-клеток по уничтожению опухоли.
13. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок способен обеспечивать долгосрочное иммуномодулирующее действие.
14. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок способен вызывать активацию антигенпрезентирующих клеток.
15. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок способен усиливать способность антигенпрезентирующих клеток презентировать антиген.
16. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок сдвигает соотношение иммунных клеток в пользу клеток, способных уничтожать опухоль, и против клеток, защищающих опухоль.
17. Гетерологичный химерный белок по п. 16, отличающийся тем, что клетка, способная уничтожать опухоль, выбрана из Т-клеток, цитотоксических Т-лимфоцитов, Т-хелперных клеток, естественных киллеров (NK-клеток), T-клеток, обладающих свойствами естественных киллеров (NKT), противоопухолевых макрофагов (например, макрофагов M1), B-клеток и дендритных клеток, при этом клетка, защищающая опухоль, выбрана из супрессорных клеток миелоидного происхождения (MDSC), регуляторных Т-клеток (Treg), опухоль-ассоциированных нейтрофилов (TAN), макрофагов М2 и опухоль-ассоциированных макрофагов (TAM).
18. Гетерологичный химерный белок по п. 17, отличающийся тем, что указанный химерный белок стимулирует противоопухолевые макрофаги и антигенпрезентирующие клетки, избегая индукции MDSC за счет ингибирования CSF1 и/или ИЛ-34.
19. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок увеличивает отношение эффекторных Т-клеток к регуляторным Т-клеткам.
20. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок обеспечивает долгосрочный маскирующий эффект иммуноингибиторных сигналов.
21. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок обеспечивает более длительное время полужизни (t1/2) на мишени (например, внутри опухоли) по сравнению с t1/2 химерных белков в сыворотке.
22. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок снижает токсичность по сравнению с лечением антителами против CSF1R и CD40.
23. Гетерологичный химерный белок по п. 1, отличающийся тем, что шарнир-CH2-CH3-Fc-домен происходит от IgG4.
24. Гетерологичный химерный белок по п. 23, где шарнир-CH2-CH3-Fc-домен происходит от IgG4 человека.
25. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок экспрессируется в клетке-хозяине млекопитающего в виде секретируемой и функциональной одиночной полипептидной цепи.
26. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что внеклеточный домен CSF1R содержит аминокислотную последовательность, которая по меньшей мере на 98% идентична аминокислотной последовательности SEQ ID NO: 2.
27. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что внеклеточный домен CD40L содержит аминокислотную последовательность, которая по меньшей мере на 98% идентична аминокислотной последовательности SEQ ID NO: 4.
28. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что линкер содержит аминокислотную последовательность, по меньшей мере на 97% идентичную аминокислотной последовательности SEQ ID NO: 25, SEQ ID NO: 26 или SEQ ID NO: 27.
29. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что линкер содержит два или более соединяющих линкеров, каждый из которых независимо выбран из SEQ ID NO: 28-74.
30. Гетерологичный химерный белок по п. 29, отличающийся тем, что один соединяющий линкер является N-концевым относительно шарнир-CH2-CH3-Fc-домена, а другой соединяющий линкер является C-концевым относительно шарнир-CH2-CH3-Fc-домена.
31. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок представляет собой рекомбинантный гибридный белок.
32. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что указанный химерный белок способен создавать стабильный синапс между клетками.
33. Гетерологичный химерный белок по п. 32, отличающийся тем, что стабильный синапс между клетками обеспечивает пространственную ориентацию, благоприятствующую уменьшению опухоли.
34. Гетерологичный химерный белок по п. 32 или 33, отличающийся тем, что пространственная ориентация приводит Т-клетки в положение для атаки опухолевых клеток и/или стерически предотвращает доставку опухолевыми клетками отрицательных сигналов, включая отрицательные сигналы помимо тех, которые маскирует химерный белок согласно настоящему изобретению.
35. Гетерологичный химерный белок по п. 1 или 2, отличающийся тем, что связывание одного из или обоих внеклеточных доменов с его соответствующим партнером по связыванию происходит с низкими скоростями диссоциации (Koff), что обеспечивает длительное взаимодействие рецептора и его лиганда.
36. Гетерологичный химерный белок по п. 35, отличающийся тем, что длительное взаимодействие обеспечивает более длительный положительный сигнальный эффект.
37. Гетерологичный химерный белок по п. 36, отличающийся тем, что более длительный положительный сигнальный эффект обеспечивает адекватную стимуляцию эффекторной клетки для противоопухолевого действия.
38. Гетерологичный химерный белок по п. 35 или 36, отличающийся тем, что длительное взаимодействие обеспечивает пролиферацию Т-клеток и позволяет выполнять противоопухолевую атаку.
39. Гетерологичный химерный белок по п. 35 или 36, отличающийся тем, что длительное взаимодействие обеспечивает достаточную передачу сигнала для обеспечения высвобождения стимулирующих сигналов.
40. Гетерологичный химерный белок по п. 39, отличающийся тем, что стимулирующий сигнал представляет собой цитокин.
41. Нуклеиновая кислота, кодирующая химерный белок по п. 1 или 2.
42. Клетка-хозяин, содержащая экспрессирующий вектор, содержащий нуклеиновую кислоту по п. 41, где клетка-хозяин не происходит из человеческих эмбриональных клеток.
43. Фармацевтическая композиция для одновременного маскирования иммуноингибиторного сигнала и активации иммуностимулирующего сигнала, содержащая терапевтически эффективное количество гетерологичного химерного белка по п. 1 или 2.
44. Применение фармацевтической композиции по п. 43 для лечения рака у нуждающегося в этом субъекта.
45. Применение фармацевтической композиции по п. 43 для модулирования иммунного ответа пациента у нуждающегося в этом субъекта, где модулирование включает:
усиление, увеличение и/или стимуляцию передачи иммуностимулирующего сигнала; и/или
нарушение, блокирование, уменьшение и/или ингибирование передачи иммуноингибиторного сигнала.
46. Применение по п. 44 или 45, отличающееся тем, что Т-клетки пациента активируются.
47. Применение по п. 44 или 45, отличающееся тем, что у пациента имеется опухоль и предотвращается передача иммуносупрессивного сигнала одной или более опухолевыми клетками.
48. Применение фармацевтической композиции для лечения рака у нуждающегося в этом субъекта, причем фармацевтическая композиция содержит гетерологичный химерный белок для одновременного маскирования иммуноингибиторного сигнала и активации иммуностимулирующего сигнала, содержащий общую структуру:
N-конец – (a) – (b) – (c) – C-конец,
где:
(а) первый домен, содержащий внеклеточный домен рецептора колониестимулирующего фактора 1 (CSF1R), способный связывать лиганд CSF1R, и содержащий аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 2 или обладает по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами;
(b) представляет собой линкер, соединяющий первый домен и второй домен и содержащий шарнир-CH2-CH3-Fc-домен и один или более соединяющих линкеров, независимо выбранных из SEQ ID NO: 28-74, и
(с) представляет собой второй домен, содержащий внеклеточный домен лиганда CD40 (CD40L), способный связывать CD40, и содержащий аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 4 или обладает по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами.
49. Применение по п. 48, отличающееся тем, что Т-клетки субъекта активируются при связывании вторым доменом гетерологичного химерного белка, и:
(а) при связывании одной или более опухолевых клеток первым доменом гетерологичного химерного белка предотвращается передача ими иммуносупрессивного сигнала,
(b) достигается количественно измеримый цитокиновый ответ в периферической крови субъекта и/или
(c) снижается рост опухоли у нуждающегося в этом субъекта по сравнению с субъектом, получавшим лечение блокирующими антителами против CD40 и/или блокирующими антителами против CSF1 или ИЛ-34.
50. Применение по п. 48 или 49, отличающееся тем, что оно ингибирует сигнальный путь CSF1R и ингибирует популяции супрессивных миелоидных клеток.
51. Применение по п. 48, отличающееся тем, что оно стимулирует сигнальный путь CD40 и активирует антигенпрезентирующие клетки.
52. Применение по п. 48, отличающееся тем, что оно уменьшает количество или активность опухоль-ассоциированных макрофагов (TAM) по сравнению с субъектами, не получавшими лечения, или субъектами, подвергавшимися адресному воздействию на CD40/CD40L и CSF1/CSF1R.
53. Применение по п. 48, отличающееся тем, что оно уменьшает количество или активность опухоль-ассоциированных макрофагов (TAM) в микроокружении опухоли (ТМЕ) по сравнению с субъектами, не получавшими лечения, или субъектами, подвергавшимися адресному воздействию на CD40/CD40L и CSF1/CSF1R.
54. Применение по п. 48, отличающееся тем, что оно уменьшает количество или активность регуляторных Т-клеток (Treg) по сравнению с субъектами, не получавшими лечения, или субъектами, подвергавшимися адресному воздействию на CD40/CD40L и CSF1/CSF1R.
55. Применение по п. 48, отличающееся тем, что оно уменьшает количество или активность ИЛ-10 и/или ИЛ-4 по сравнению с субъектами, не получавшими лечения, или субъектами, подвергавшимися адресному воздействию на CD40/CD40L и CSF1/CSF1R.
56. Применение по п. 48, отличающееся тем, что оно усиливает созревание и дифференцировку провоспалительных макрофагов и дендритных клеток по сравнению с субъектами, не получавшими лечения, или субъектами, подвергавшимися адресному воздействию на CD40/CD40L и CSF1/CSF1R.
57. Применение по п. 48, отличающееся тем, что оно усиливает примирование эффекторных Т-клеток в дренирующих лимфатических узлах субъекта по сравнению с субъектами, не получавшими лечения, или субъектами, подвергавшимися адресному воздействию на CD40/CD40L и CSF1/CSF1R.
58. Применение по п. 48, отличающееся тем, что оно вызывает общее снижение количества иммуносупрессивных клеток и сдвиг в сторону более воспалительного окружения опухоли по сравнению с субъектами, не получавшими лечения, или субъектами, подвергавшимися адресному воздействию на CD40/CD40L и CSF1/CSF1R.
59. Применение химерного белка в качестве лекарственного средства для одновременного маскирования иммуноингибиторного сигнала и активации иммуностимулирующего сигнала, где химерный белок содержит общую структуру:
N-конец – (a) – (b) – (c) – C-конец,
где:
(а) представляет собой первый домен, содержащий внеклеточный домен рецептора колониестимулирующего фактора 1 (CSF1R), способный связывать лиганд CSF1R, и содержащий аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 2 или обладает по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами;
(b) представляет собой линкер, соединяющий первый домен и второй домен и содержащий шарнир-CH2-CH3-Fc-домен и один или более соединяющих линкеров, независимо выбранных из SEQ ID NO: 28-74, и
(с) представляет собой второй домен, содержащий внеклеточный домен лиганда CD40 (CD40L), способный связывать CD40, и содержащий аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 4 или обладает по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами.
60. Применение химерного белка при лечении рака, где химерный белок содержит общую структуру:
N-конец – (a) – (b) – (c) – C-конец,
где:
(а) представляет собой первый домен, содержащий внеклеточный домен рецептора колониестимулирующего фактора 1 (CSF1R), способный связывать лиганд CSF1R, и содержащий аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 2 или обладает по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами;
(b) представляет собой линкер, соединяющий первый домен и второй домен и содержащий шарнир-CH2-CH3-Fc-домен и один или более соединяющих линкеров, независимо выбранных из SEQ ID NO: 28-74, и
(с) представляет собой второй домен, содержащий внеклеточный домен лиганда CD40 (CD40L), способный связывать CD40, и содержащий аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 4 или обладает по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами.
61. Применение химерного белка по п. 1 при производстве лекарственного средства для лечения рака.
62. Рекомбинантный гибридный белок для одновременного маскирования иммуноингибиторного сигнала и активации иммуностимулирующего сигнала, содержащий общую структуру:
N-конец – (a) – (b) – (c) – C-конец,
где:
(а) представляет собой первый домен, содержащий внеклеточный домен CSF1R, который способен связывать лиганд CSF1R и имеет аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 2, или ее вариант, обладающий по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами,
(b) представляет собой линкер, связывающий первый домен и второй домен и содержащий шарнир-CH2-CH3-Fc-домен, происходящий из IgG4 человека, и, необязательно, последовательность соединяющего линкера из SEQ ID NO: 28-74, и
(c) представляет собой второй домен, содержащий внеклеточный домен лиганда CD40 (CD40L), который способен связывать рецептор CD40 и имеет аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 4, или ее вариант, обладающий по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами.
63. Рекомбинантный гибридный белок по п. 62, отличающийся тем, что линкер содержит последовательность, по меньшей мере на 97% идентичную аминокислотной последовательности SEQ ID NO: 25, SEQ ID NO: 26 или SEQ ID NO: 27.
64. Применение рекомбинантного гибридного белка в качестве лекарственного средства для одновременного маскирования иммуноингибиторного сигнала и активации иммуностимулирующего сигнала, где белок содержит общую структуру:
N-конец – (a) – (b) – (c) – C-конец,
где:
(а) представляет собой первый домен, содержащий внеклеточный домен CSF1R, который способен связывать лиганд CSF1R и имеет аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 2, или ее вариант, обладающий по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами,
(b) представляет собой линкер, связывающий первый домен и второй домен и содержащий шарнир-CH2-CH3-Fc-домен, происходящий из IgG4 человека, и, необязательно, последовательность соединяющего линкера из SEQ ID NO: 28-74, и
(c) представляет собой второй домен, содержащий внеклеточный домен лиганда CD40 (CD40L), который способен связывать рецептор CD40 и имеет аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 4, или ее вариант, обладающий по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами.
65. Применение рекомбинантного гибридного белка при лечении рака, содержащего общую структуру:
N-конец – (a) – (b) – (c) – C-конец,
где:
(а) представляет собой первый домен, содержащий внеклеточный домен CSF1R, который способен связывать лиганд CSF1R и имеет аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 2, или ее вариант, обладающий по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами и,
(b) представляет собой линкер, связывающий первый домен и второй домен и содержащий шарнир-CH2-CH3-Fc-домен, происходящий из IgG4 человека, и, необязательно, последовательность соединяющего линкера из SEQ ID NO: 28-74, и
(c) представляет собой второй домен, содержащий внеклеточный домен лиганда CD40 (CD40L), который способен связывать рецептор CD40 и имеет аминокислотную последовательность, которая идентична аминокислотной последовательности SEQ ID NO: 4, или ее вариант, обладающий по меньшей мере 97% идентичностью указанной последовательности с консервативными аминокислотными заменами.
66. Применение рекомбинантного гибридного белка по п. 62 при производстве лекарственного средства для лечения рака.
| WO2010062401 A2, 03.06.2021 | |||
| SHEN J | |||
| et al., Single variable domain-IgG fusion: a novel recombinant approach to Fc domain-containing bispecific antibodies | |||
| Journal of Biological Chemistry | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| V | |||
| ПАРОПЕРЕГРЕВАТЕЛЬ ДЛЯ ТРУБЧАТЫХ ПАРОВЫХ КОТЛОВ С ЭЛЕМЕНТАМИ, СОСТОЯЩИМИ ИЗ ДВУХ ПЕТЕЛЬ, ВВОДИМЫХ В ПРОГАРНЫЕ ТРУБЫ КОТЛА | 1916 |
|
SU281A1 |
| N | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Pp | |||
| Висячий замок | 1926 |
|
SU10706A1 |
| COURTNEIDGE S | |||
| A | |||
| et al., Activation of Src family kinases by colony stimulating factor‐1, and their association with its | |||

