Перекрестная ссылка на родственные заявки
[0001] Данная заявка испрашивает преимущество по предварительной заявке на патент США №62/454476, поданной 3 февраля 2017 года, содержание которой тем самым включено посредством ссылки во всей своей полноте.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка содержит перечень последовательностей, который был подан в электронном виде в формате ASCII и тем самым включен посредством ссылки во всей своей полноте. Указанная ASCII-копия, созданная 10 января 2018 года, имеет название PAT057594-WO-PCT_SL.txt и размер 387054 байта.
Область техники, к которой относится изобретение
[0003] Настоящее изобретение относится к антителам к CCR7, фрагментам антител и их иммуноконъюгатам, а также путям их применения для лечения или предупреждения рака.
Предпосылки изобретения
[0004] CC‑хемокиновый рецептор 7 (CCR7) впервые был идентифицирован в 1993 году в качестве специфического для лимфоцитов рецептора (см., например, Birkenbach et al., J Virol. 1993 Apr;67(4):2209-20). Его экспрессия ограничена субпопуляциями иммунных клеток, таких как наивные T-клетки, T-клетки центральной памяти (Tcm), регуляторные T-клетки (Treg), наивные B-клетки, NK-клетки и зрелые антигенпрезентирующие дендритные клетки (DC). CCR7 регулирует хоминг иммунных клеток в лимфоидные органы и миграцию в пределах них и, таким образом, играет ключевую роль в балансе иммунитета и переносимости (см., например, Förster et al., Nat Rev Immunol. 2008 May;8(5):362-71).
[0005] CCR7 представляет собой родопсин-подобный рецептор, сопряженный с G-белком (GPCR), класса A с двумя лигандами, CCL21 и CCL19. Структура CCR7 еще не полностью выяснена, однако, были обнаружены некоторые мотивы, необходимые для активности рецептора (см., например, Legler et al., Int J Biochem Cell Biol. 2014 Jul 1).
CCR7 и рак
[0006] Также известно, что CCR7 (также называемый EBI1, BLR2, CC-CKR-7, CMKBR7, CD197 и CDw197) сверхэкспрессируется на целом ряде злокачественных опухолей, включая, среди прочего, B-клеточные злокачественные новообразования (например, CLL, MCL, лимфому Беркитта), T-клеточные злокачественные новообразования (например, ATLL), HNSCC, ESCC, карциному желудка, NSCLC, колоректальную карциному, рак поджелудочной железы, рак щитовидной железы, рак молочной железы и рак шейки матки. Например, сверхэкспрессия CCR7 на клетках колоректальной карциномы, ESCC, рака поджелудочной железы, HNSCC и рака желудка была ассоциирована с поздней стадией опухоли, метастазированием в лимфатические узлы и неблагоприятным прогнозом в отношении выживания (см., например, Malietzis et al., Journal of Surgical Oncology 2015;112:86-92; Irino et al., BMC Cancer 2014, 14:291; Guo et al., Oncology Letters 5: 1572-1578, 2013; Xia et al., Oral Dis. 2015 Jan;21(1):123-31; Du et al., Gastric Cancer. 2016 Mar 16).
[0007] Кроме того, было показано, например, что экспрессия CCR7 на клетках HNSCC играет роль в резистентности к химиотерапии (см., например, Wang et al., JNCI J Natl Cancer Inst (2008) 100 (7): 502-512.). При некоторых типах рака, таких как рак поджелудочной железы и носоглоточная карцинома (NPC), как известно, CCR7 стимулирует метастазирование раковых стволовых клеток и образование сфер (см., например, Zhang et al., PLOS ONE 11 (8); Lun et al., PLOS ONE 7(12)). Роль CCR7 в миграции клеток, инвазивности и EMT (эпителиально-мезенхимальном переходе) описана у различных типов рака, таких как рак молочной железы и поджелудочной железы in vitro и in vivo (см., например, Pang et al., Oncogene (2015), 1-13); Sperveslage et al., Int. J. Cancer: 131, E371-E381 (2012)). Ключевые пути, которые были описаны как необходимые для передачи сигнала с помощью CCR7, включают путь передачи сигнала p38/ERK1/2, опосредованный b-аррестином, и путь передачи сигнала Rho (см., например, Noor et al., J Neuroinflammation 2012 Apr 25; 9:77).
[0008] Известны многочисленные сопряженные с раком процессы, которые индуцируют экспрессию CCR7. Показано, что при HNSCC экспрессия CCR7 индуцируется факторами транскрипции NF-kB и AP1 за счет прямого связывания с сайтами в промоторе CCR7 (Mburu et al., J. Biol. Chem. 2012, 287:3581-3590). В частности, экспрессия CCR7 регулируется различными факторами в микроокружении опухоли. В данном контексте, известно, что экспрессия CCR7 индуцируется за счет пути b-дефензин 3/NF-kB в HNSCC (см., например, Mburu et al., Carcinogenesis vol.32 no.2 pp.168-174, 2010) и рецептора A эндотелина и индуцируемого гипоксией фактора-1 в клетках опухоли молочной железы (см., например, Wilson et al., Cancer Res 2006;66:11802-11807).
Конъюгаты антитела и лекарственного средства
[0009] Конъюгаты антитела и лекарственного средства ("ADC") нашли применение в местной доставке цитотоксических средств при лечении рака (см., например, Lambert, Curr. Opinion In Pharmacology 5:543-549, 2005). ADC обеспечивают возможность нацеленной доставки фрагмента, представляющего собой лекарственное средство, при которой может достигаться максимальная эффективность при минимальной токсичности. ADC включают антитело, выбранное за его способность связываться с клеткой, на которую нацелено терапевтическое вмешательство, соединенное с лекарственным средством, выбранным за его цитостатическую или цитотоксическую активность. Тем самым, при связывании антитела с целевой клеткой лекарственное средство доставляется в сайт, в котором требуется его терапевтический эффект.
[0010] Для применения в ADC было раскрыто множество антител, которые распознают и селективно связываются с целевыми клетками, например, раковыми клетками. Несмотря на масштабную работу над ADC, связывания антитела с конкретной мишенью, представляющей интерес, недостаточно для предсказания успеха в применениях ADC. Примеры факторов, которые могут воздействовать на терапевтическую эффективность ADC (кроме особенностей, присущих мишени), включают различные аспекты, которые необходимо модифицировать путем тонкой настройки, такие как оптимальная аффинность антитела как баланс между мишень-опосредованным распределением (TMDD) и управляющим эффективностью воздействием, оценка Fc-опосредованных функций (антителозависимая клеточноопосредованная цитотоксичность, ADCC), способ конъюгации (сайт-специфическая или нет), отношение молекул лекарственного средства/полезной нагрузки у такого конъюгата с каждым антителом ("DAR" или "соотношение лекарственного средства и антитела"), расщепляемость или стабильность линкера, стабильность ADC и склонность ADC к образованию агрегатов.
[0011] Остается потребность в антителах, способах прикрепления и цитотоксических полезных нагрузках с улучшенными свойствами для применения в качестве эффективных терапевтических композиций на основе ADC и способов, связанных с ними.
Краткое описание настоящего изобретения
[0012] В настоящей заявке раскрыты антитело или его антигенсвязывающий фрагмент, которые связываются с белком CCR7 человека, где антитело или его антигенсвязывающий фрагмент характеризуются сниженной или незначительной эффекторной функцией по сравнению с антителом дикого типа того же изотипа. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент характеризуются сниженным или незначительным уровнем активности антителозависимой клеточноопосредованной цитотоксичности (ADCC). В одном варианте осуществления антитело или его антигенсвязывающий фрагмент содержат "молчащую" область Fc. В некоторых вариантах осуществления антитело содержит в области Fc мутацию, выбранную из: D265A; P329A; P329G; N297A; D265A и P329A; D265A и N297A; L234 и L235A; P329A, L234A и L235A; а также P329G, L234A и L235A. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент характеризуются незначительной активностью цитолиза клеток. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент связываются с большей аффинностью с клетками, экспрессирующими более высокие уровни CCR7, чем с клетками, экспрессирующими более низкие уровни CCR7. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связываются с большей аффинностью с раковыми клетками, которые экспрессируют более высокие уровни CCR7, чем с нормальными клетками, которые экспрессируют более низкие уровни CCR7. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент в незначительной степени истощают нормальные гемопоэтические клетки, которые экспрессируют CCR7.
[0013] В одном варианте осуществления настоящей заявки раскрыты антитело или его антигенсвязывающий фрагмент, которые связывают CCR7, содержащие:
a. вариабельную область тяжелой цепи, которая содержит HCDR1 (определяющая комплементарность область 1 тяжелой цепи) под SEQ ID NO:1, HCDR2 (определяющая комплементарность область 2 тяжелой цепи) под SEQ ID NO:2 и HCDR3 (определяющая комплементарность область 3 тяжелой цепи) под SEQ ID NO:3; и вариабельную область легкой цепи, которая содержит LCDR1 (определяющая комплементарность область 1 легкой цепи) под SEQ ID NO:17, LCDR2 (определяющая комплементарность область 2 легкой цепи) под SEQ ID NO:18 и LCDR3 (определяющая комплементарность область 3 легкой цепи) под SEQ ID NO:19;
b. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:4, HCDR2 под SEQ ID NO:5 и HCDR3 под SEQ ID NO:6; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:20, LCDR2 под SEQ ID NO:21 и LCDR3 под SEQ ID NO:22;
c. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:7, HCDR2 под SEQ ID NO:8 и HCDR3 под SEQ ID NO:9; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:23, LCDR2 под SEQ ID NO:24 и LCDR3 под SEQ ID NO:25;
d. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:10, HCDR2 под SEQ ID NO:11 и HCDR3 под SEQ ID NO:12; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:26, LCDR2 под SEQ ID NO:27 и LCDR3 под SEQ ID NO:28;
e. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:33, HCDR2 под SEQ ID NO:34 и HCDR3 под SEQ ID NO:35; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:49, LCDR2 под SEQ ID NO:50 и LCDR3 под SEQ ID NO:51;
f. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:36, HCDR2 под SEQ ID NO:37 и HCDR3 под SEQ ID NO:38; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:52, LCDR2 под SEQ ID NO:53 и LCDR3 под SEQ ID NO:54;
g. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:39, HCDR2 под SEQ ID NO:40 и HCDR3 под SEQ ID NO:41; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:55, LCDR2 под SEQ ID NO:56 и LCDR3 под SEQ ID NO:57;
h. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:42, HCDR2 под SEQ ID NO:43 и HCDR3 под SEQ ID NO:44; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:58, LCDR2 под SEQ ID NO:59 и LCDR3 под SEQ ID NO:60;
i. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:65, HCDR2 под SEQ ID NO:66 и HCDR3 под SEQ ID NO:67; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:81, LCDR2 под SEQ ID NO:82 и LCDR3 под SEQ ID NO:83;
j. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:68, HCDR2 под SEQ ID NO:69 и HCDR3 под SEQ ID NO:70; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:84, LCDR2 под SEQ ID NO:85 и LCDR3 под SEQ ID NO:86;
k. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:71, HCDR2 под SEQ ID NO:72 и HCDR3 под SEQ ID NO:73; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:87, LCDR2 под SEQ ID NO:88 и LCDR3 под SEQ ID NO:89;
l. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:74, HCDR2 под SEQ ID NO:75 и HCDR3 под SEQ ID NO:76; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:90, LCDR2 под SEQ ID NO:91 и LCDR3 под SEQ ID NO:92;
m. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:596, HCDR2 под SEQ ID NO:597 и HCDR3 под SEQ ID NO:598; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:612, LCDR2 под SEQ ID NO:613 и LCDR3 под SEQ ID NO:614;
n. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:599, HCDR2 под SEQ ID NO:600 и HCDR3 под SEQ ID NO:601; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:615, LCDR2 под SEQ ID NO:616 и LCDR3 под SEQ ID NO:617;
o. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:602, HCDR2 под SEQ ID NO:603 и HCDR3 под SEQ ID NO:604; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:618, LCDR2 под SEQ ID NO:619 и LCDR3 под SEQ ID NO:620; или
p. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:605, HCDR2 под SEQ ID NO:606 и HCDR3 под SEQ ID NO:607; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:621, LCDR2 под SEQ ID NO:622 и LCDR3 под SEQ ID NO:623.
[0014] Антитело или его антигенсвязывающий фрагмент, которые связывают CCR7 по настоящей заявке, также могут содержать:
a. вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:13, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:29;
b. вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:45, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:61;
c. вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:77, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:93; или
d. вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:608, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:624.
[0015] В другом варианте осуществления антитело или его антигенсвязывающий фрагмент, которые связывают CCR7, содержат:
a. тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:15, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:31;
b. тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:47, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:63;
c. тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:79, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:95; или
d. тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:610, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:626.
[0016] Антитело или его антигенсвязывающий фрагмент, описанные в данном документе, могут содержать одну или несколько цистеиновых замен. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент содержат одну или несколько цистеиновых замен, выбранных из S152C, S375C или обеих S152C и S375C, в тяжелой цепи антитела или его антигенсвязывающего фрагмента, где нумерация положения соответствует системе EU. Антитело, раскрытое в данном документе, может представлять собой моноклональное антитело.
[0017] В настоящей заявке раскрыт конъюгат антитела и лекарственного средства, предусматривающий формулу:
Ab-(L-(D)m)n,
или ее фармацевтически приемлемую соль; где
Ab представляет собой антитело или его антигенсвязывающий антигенсвязывающий фрагмент, раскрытые в данном документе;
L представляет собой линкер;
D представляет собой фрагмент, представляющий собой лекарственное средство;
m составляет целое число от 1 до 8; и
n составляет целое число от 1 до 12.
В некоторых вариантах осуществления m составляет 1. В одном варианте осуществления n составляет от приблизительно 3 до приблизительно 4. В одном варианте осуществления линкер выбран из группы, состоящей из расщепляемого линкера, нерасщепляемого линкера, гидрофильного линкера, предварительно заряженного линкера и линкера на основе дикарбоновой кислоты.
[0018] В одном варианте осуществления линкер происходит из сшивающего реагента, выбранного из группы, состоящей из N-сукцинимидил-3-(2-пиридилдитио)пропионата (SPDP), N-сукцинимидил-4-(2-пиридилдитио)пентаноата (SPP), N-сукцинимидил-4-(2-пиридилдитио)бутаноата (SPDB), N-сукцинимидил-4-(2-пиридилдитио)-2-сульфо-бутаноата (сульфо-SPDB), N-сукцинимидилйодацетата (SIA), N-сукцинимидил(4-йодацетил)аминобензоата (SIAB), малеимид-PEG-NHS, N-сукцинимидил-4-(малеимидометил)циклогексанкарбоксилата (SMCC), N-сульфосукцинимидил-4-(малеимидометил)циклогексанкарбоксилата (сульфо-SMCC) и 2,5-диоксопирролидин-1-ил-17-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)-5,8,11,14-тетраоксо-4,7,10,13-тетраазагептадекан-1-оата (CX1-1).
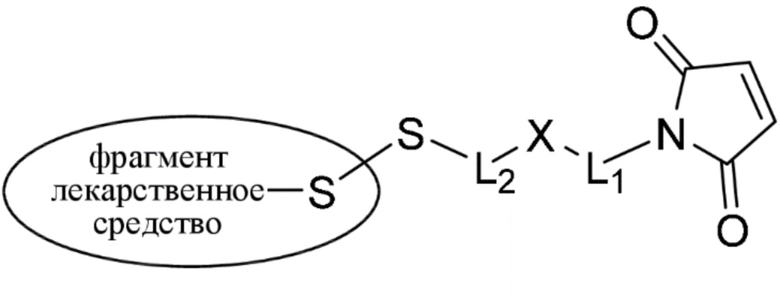
[0019] В других вариантах осуществления линкер имеет следующую формулу (IIA):
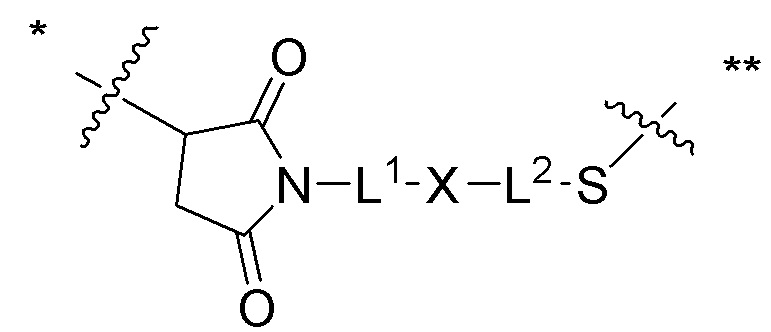 (IIA);
(IIA);
где * соединен с тиольной функциональной группой на антителе, и ** соединен с тиольной функциональной группой фрагмента, представляющего собой лекарственное средство; и где:
L1 представляет собой C1-6алкилен, при этом одна из метиленовых групп может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11;
X представляет собой -C(O)-NH-, -NHC(O)- или триазол; а алкилен является линейным или разветвленным.
[0020] В другом варианте осуществления линкер имеет следующую формулу:
где y составляет от 1 до 11; * соединен с тиольной функциональной группой на антителе, и ** соединен с тиольной функциональной группой фрагмента, представляющего собой лекарственное средство.
[0021] В одном варианте осуществления фрагмент, представляющий собой лекарственное средство, выбран из группы, состоящей из ингибитора V-АТФазы, проапоптического средства, ингибитора Bcl2, ингибитора MCL1, ингибитора HSP90, ингибитора IAP, ингибитора mTor, стабилизатора микротрубочек, дестабилизатора микротрубочек, ауристатина, аманитина, пирролобензодиазепина, ингибитора РНК-полимеразы, доластатина, майтанзиноида, MetAP (метионинаминопептидазы), ингибитора ядерного экспорта белков CRM1, ингибитора DPPIV, ингибиторов протеасомы, ингибиторов реакций переноса фосфорила в митохондриях, ингибитора синтеза белка, ингибитора киназ, ингибитора CDK2, ингибитора CDK9, ингибитора кинезина, ингибитора HDAC, ДНК-повреждающего средства, ДНК-алкилирующего средства, ДНК-интеркалятора, средства, связывающего малую бороздку ДНК, и ингибитора DHFR. В некоторых вариантах осуществления цитотоксическое средство представляет собой майтанзиноид, где майтанзиноид представляет собой N(2')-деацетил-N(2')-(3-меркапто-l-оксопропил)-майтанзин (DM1), N(2')-деацетил-N(2')-(4-меркапто-1-оксопентил)-майтанзин (DM3) или N(2')-деацетил-N2-(4-меркапто-4-метил-1-оксопентил)-майтанзин (DM4).
[0022] В одном варианте осуществления конъюгаты антитела и лекарственного средства, раскрытые в данном документе, предусматривают следующую формулу (VIII):
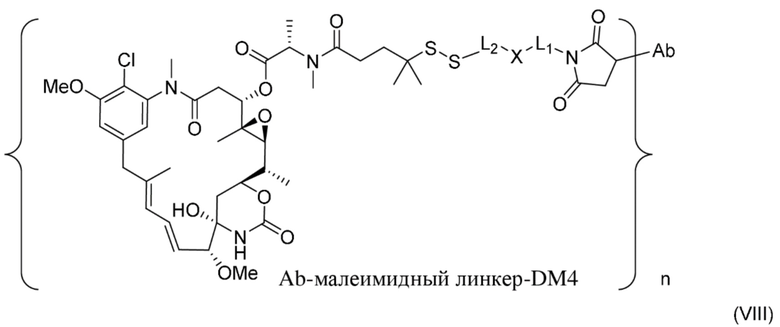
где L1 представляет собой C1-6алкилен, при этом одна метиленовая группа может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11;
X представляет собой -C(O)-NH-, -NHC(O)- или триазол; и алкилен является линейным или разветвленным; и где n составляет от приблизительно 3 до приблизительно 4; или ее фармацевтически приемлемую соль.
[0023] В одном варианте осуществления конъюгаты антитела и лекарственного средства, раскрытые в данном документе, имеют следующую формулу:
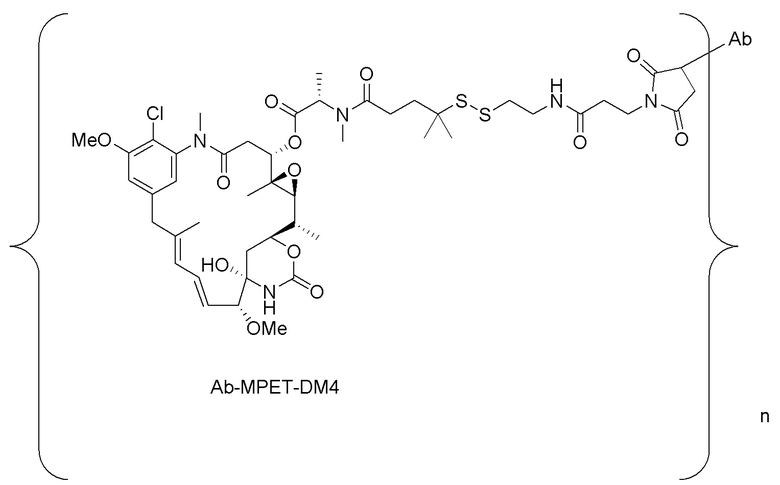
где n составляет от приблизительно 3 до приблизительно 4, и Ab представляет собой антитело, содержащее тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:47, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:63; или ее фармацевтически приемлемую соль.
[0024] В настоящей заявке также раскрыта фармацевтическая композиция, содержащая антитела или их антигенсвязывающие фрагменты, раскрытые в данном документе, и фармацевтически приемлемый носитель. В настоящей заявке также раскрыта фармацевтическая композиция, содержащая конъюгаты антитела и лекарственного средства, раскрытые в данном документе.
[0025] В настоящей заявке также раскрыты способы лечения или предупреждения рака у пациента, нуждающегося в этом, предусматривающие введение указанному пациенту конъюгатов антитела и лекарственного средства или фармацевтических композиций, раскрытых в данном документе, где рак экспрессирует CCR7.
[0026] В некоторых вариантах осуществления способов лечения или предупреждения рака конъюгат антитела и лекарственного средства или фармацевтическую композицию вводят пациенту в комбинации с одним или несколькими дополнительными терапевтическими соединениями. В одном варианте осуществления одно или несколько дополнительных терапевтических соединений выбраны из стандартного химиотерапевтического средства, костимулируюшей молекулы или ингибитора контрольных точек. В одном варианте осуществления костимулирующая молекула выбрана из агониста OX40, CD2, CD27, CDS, ICAM-1, LFA-1 (CD11a/CD18), ICOS (CD278), 4-1BB (CD137), GITR, CD30, CD40, BAFFR, HVEM, CD7, LIGHT, NKG2C, SLAMF7, NKp80, CD160, B7-H3, STING или лиганда CD83. В другом варианте осуществления ингибитор контрольных точек выбран из ингибитора PD-1, PD-L1, PD-L2, CTLA4, TIM3, LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и/или TGFR-бета.
[0027] В настоящей заявке также раскрыты конъюгаты антитела и лекарственного средства или фармацевтические композиции, раскрытые в данном документе, для применения в качестве лекарственного препарата. В одном варианте осуществления конъюгаты антитела и лекарственного средства или фармацевтические композиции, раскрытые в данном документе, предназначены для применения в лечении или предупреждении рака, экспрессирующего CCR7, у пациента, нуждающегося в этом.
[0028] В одном варианте осуществления настоящей заявки раскрыто применение антител или их антигенсвязывающих фрагментов, конъюгатов антитела и лекарственного средства или фармацевтических композиций, раскрытых в данном документе, для лечения или предупреждения рака, экспрессирующего CCR7, у пациента, нуждающегося в этом.
[0029] В одном варианте осуществления настоящей заявки раскрыто применение антител или их антигенсвязывающих фрагментов, конъюгатов антитела и лекарственного средства или фармацевтических композиций, раскрытых в данном документе, в изготовлении лекарственного препарата.
[0030] В одном варианте осуществления рак выбран из группы, состоящей из хронического лимфоцитарного лейкоза (CLL), периферических T-клеточных лимфом (PTCL), таких как T-клеточный лейкоз/лимфома взрослых (ATLL) и анапластическая крупноклеточная лимфома (ALCL), неходжкинской лимфомы (NHL), такой как лимфома из клеток мантийной зоны (MCL), лимфома Беркитта и диффузная В-крупноклеточная лимфома (DLBCL), карциномы желудка, немелкоклеточного рака легкого, мелкоклеточного рака легкого, рака головы и шеи, носоглоточной карциномы (NPC), рака пищевода, колоректальной карциномы, рака поджелудочной железы, рака щитовидной железы, рака молочной железы, почечноклеточного рака и рака шейки матки. В конкретных вариантах осуществления рак выбран из группы, состоящей из хронического лимфоцитарного лейкоза (CLL), периферических T-клеточных лимфом (PTCL), таких как T-клеточный лейкоз/лимфома взрослых (ATLL) и анапластическая крупноклеточная лимфома (ALCL), неходжкинской лимфомы (NHL), такой как лимфома из клеток мантийной зоны (MCL), лимфома Беркитта и диффузная В-крупноклеточная лимфома (DLBCL), и немелкоклеточного рака легкого.
[0031] В настоящей заявке также раскрыты нуклеиновые кислоты, которые кодируют антитела или антигенсвязывающие фрагменты, раскрытые в данном документе. В одном варианте осуществления нуклеиновая кислота содержит нуклеотидную последовательность под SEQ ID NO: 14, 16, 30, 32, 46, 48, 62, 64, 78, 80, 94, 96, 481, 483, 497 или 499. В данной заявке также раскрыты векторы, содержащие нуклеиновые кислоты, и клетки-хозяева, содержащие векторы или нуклеиновые кислоты. В данной заявке также раскрыт способ получения антител или антигенсвязывающих фрагментов, раскрытых в данном документе, предусматривающий культивирование клетки-хозяина и извлечения антитела из культуры клеток. В одном варианте осуществления способ извлечения антитела из культуры клеток предусматривает стадии:
a) удаления клеток и фильтрования культуры;
b) проведения очистки культуры с помощью аффинной хроматографии;
c) инактивации любых вирусов в культуре путем доведения pH до 3,4-3,6, затем повторного доведения pH до 5,8-6,2 и фильтрования культуры;
d) проведения очистки культуры с помощью катионообменной хроматографии и осуществления восстановления культуры на колонке;
e) осуществления анионообменной хроматографии в отношении культуры;
f) удаления вирусов с помощью нанофильтрации;
g) фильтрования культуры, содержащей антитело; и
h) получения очищенного антитела.
[0032] В еще одном варианте осуществления в данном документе раскрыт способ получения конъюгата антитела к CCR7 и лекарственного средства, предусматривающий:
(a) предварительное образование фрагмента, представляющего собой линкер-лекарственное средство, следующей формулы:
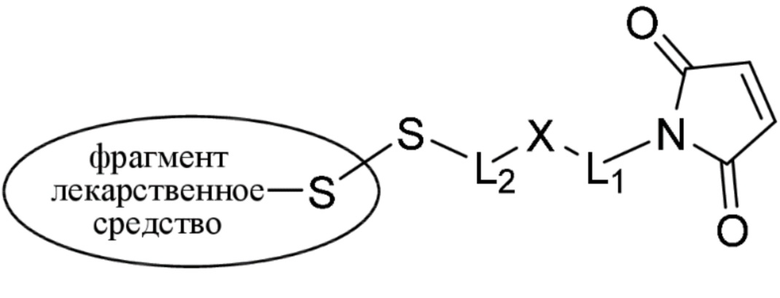
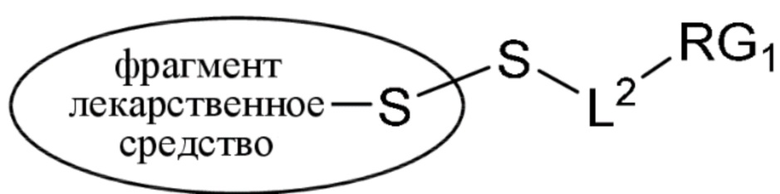
где:
фрагмент, представляющий собой лекарственное средство, представляет собой DM1, DM3 или DM4, и фрагмент, представляющий собой лекарственное средство, прикреплен к линкеру посредством своей тиольной функциональной группы;
L1 представляет собой C1-6алкилен, при этом одна из метиленовых групп может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11;
X представляет собой -C(O)-NH-, -NHC(O)- или триазол; а алкилен является линейным или разветвленным;
(b) конъюгирование указанного фрагмента, представляющего собой линкер-лекарственное средство, с антителом, извлеченным из культуры клеток, раскрытой в данном документе, с получением конъюгата антитела и лекарственного средства; и
(c) проведение очистки конъюгата антитела и лекарственного средства.
В одном варианте осуществления способ предусматривает:
(a) предварительное образование фрагмента, представляющего собой линкер-лекарственное средство, следующей формулы:
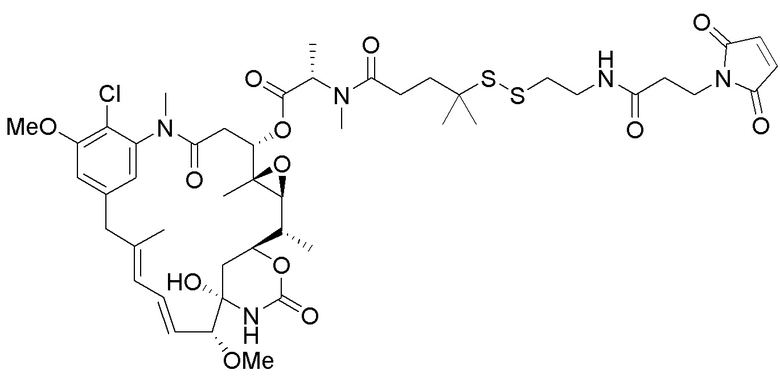 , и
, и
(b) конъюгирование указанного фрагмента, представляющего собой линкер-лекарственное средство, с антителом, извлеченным из культуры клеток, раскрытой в данном документе, с получением конъюгата антитела и лекарственного средства; и
(c) проведение очистки конъюгата антитела и лекарственного средства.
В другом варианте осуществления способ получения конъюгата антитела к CCR7 и лекарственного средства предусматривает:
(a) предварительное образование фрагмента, представляющего собой линкер-лекарственное средство, следующей формулы:
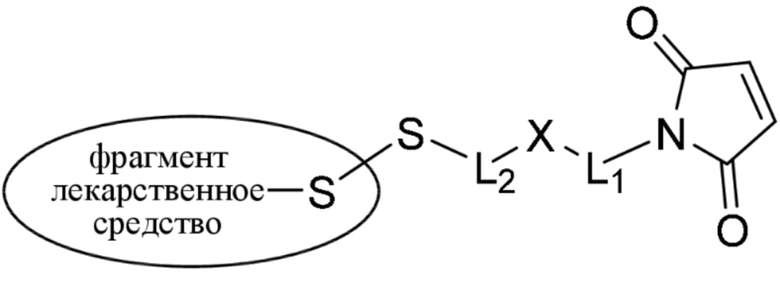
где:
фрагмент, представляющий собой лекарственное средство, представляет собой DM1, DM3 или DM4, и фрагмент, представляющий собой лекарственное средство, прикреплен к линкеру посредством своей тиольной функциональной группы;
L1 представляет собой C1-6алкилен, при этом одна из метиленовых групп может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11;
X представляет собой -C(O)-NH-, -NHC(O)- или триазол; а алкилен является линейным или разветвленным;
(b) конъюгирование указанного фрагмента, представляющего собой линкер-лекарственное средство, с антителом, раскрытым в данном документе, с получением конъюгата антитела и лекарственного средства; и
(c) проведение очистки конъюгата антитела и лекарственного средства.
В другом варианте осуществления способ получения конъюгата антитела к CCR7 и лекарственного средства предусматривает:
(a) предварительное образование фрагмента, представляющего собой линкер-лекарственное средство, следующей формулы:
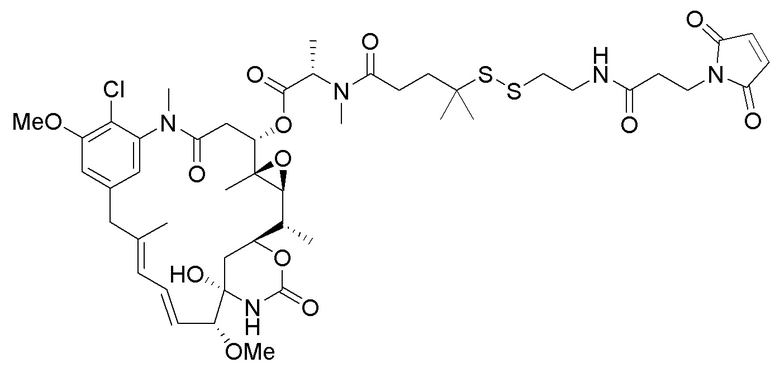
(b) конъюгирование указанного фрагмента, представляющего собой линкер-лекарственное средство, с антителом или его антигенсвязывающим фрагментом, раскрытыми в данном документе, с получением конъюгата антитела и лекарственного средства; и
(c) проведение очистки конъюгата антитела и лекарственного средства.
В другом варианте осуществления стадия предварительного образования указанного фрагмента, представляющего собой линкер-лекарственное средство, предусматривает:
a) осуществление реакции фрагмента, представляющего собой лекарственное средство, посредством его тиольной функциональной группы с:
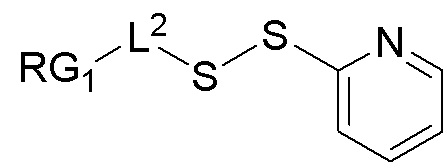 ;
;
с образованием:
 ;
;
b) осуществление реакции образованного
 с:
с:
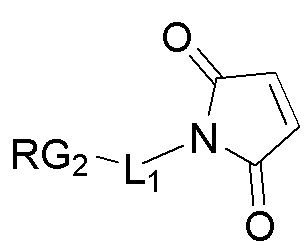 ;
;
с образованием фрагмента, представляющего собой линкер-лекарственное средство:
 ;
;
где:
L1 представляет собой C1-6алкилен, при этом одна из метиленовых групп может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11; и
X представляет собой -C(O)-NH-, -NHC(O)- или триазол;
где алкилен является линейным или разветвленным; и
RG1 и RG2 представляют собой 2 реакционноспособные группы, образующие группу X.
В другом варианте осуществления стадия предварительного образования указанного фрагмента, представляющего собой линкер-лекарственное средство, предусматривает:
a) осуществление реакции фрагмента, представляющего собой лекарственное средство, посредством его тиольной функциональной группы с:
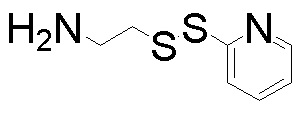 ;
;
с образованием:
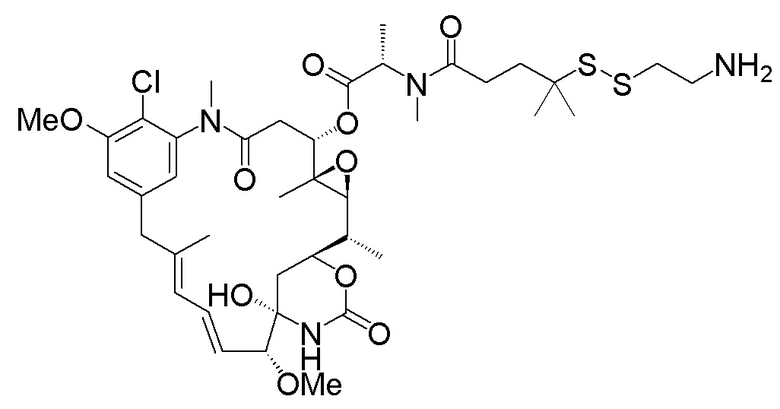 ;
;
b) осуществление реакции образованного
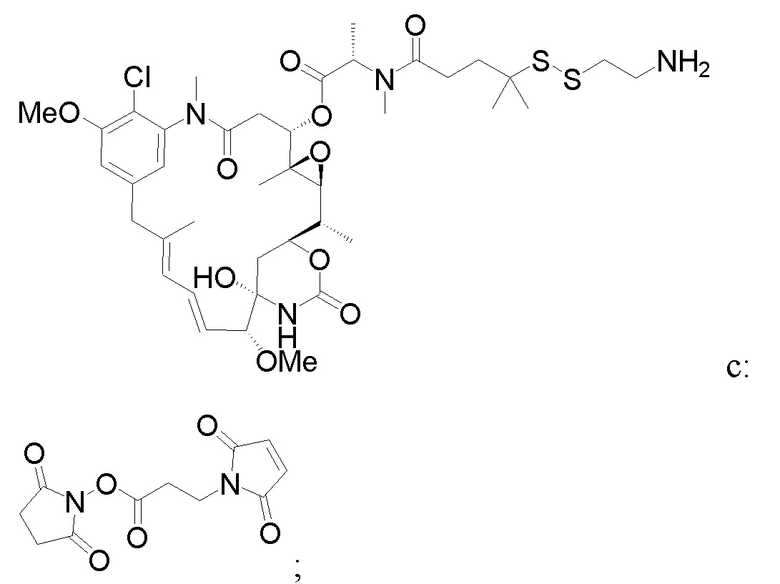
с образованием фрагмента, представляющего собой линкер-лекарственное средство:
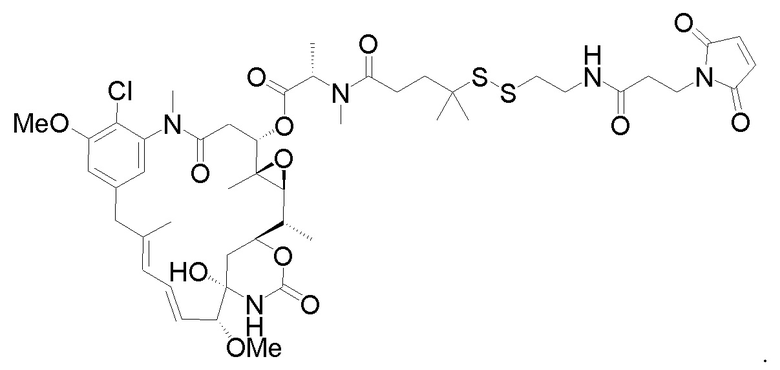
В некоторых вариантах осуществления конъюгат антитела и лекарственного средства, полученный согласно вышеуказанным способам, характеризуется средним DAR, измеренным с помощью УФ-спектрофотометра, составляющим от приблизительно 3 до приблизительно 4.
[0033] В другом варианте осуществления данной заявки раскрыт способ получения конъюгата антитела к CCR7 и лекарственного средства, предусматривающий:
(a) химическое соединение SMCC или MPET с фрагментом, представляющим собой лекарственное средство, DM-1 или DM-4, с образованием линкера-лекарственного средства;
(b) конъюгирование указанного линкера-лекарственного средства с антителом или его антигенсвязывающим фрагментом, раскрытыми в данном документе; и
(c) проведение очистки конъюгата антитела и лекарственного средства.
В одном варианте осуществления конъюгат антитела и лекарственного средства, полученный согласно данному способу, характеризуется средним DAR, измеренным с помощью УФ-спектрофотометра, составляющим от приблизительно 3 до приблизительно 4.
[0034] В настоящей заявке также раскрыт диагностический реагент, содержащий антитело или его антигенсвязывающий фрагмент, раскрытые в данном документе. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент являются меченными с помощью радиоактивной метки, флуорофора, хромофора, средства для визуализации или иона металла.
Краткое описание графических материалов
[0035] На фигуре 1 изображены экспериментальные данные по in vitro активности ADCC негуманизированного и гуманизированного антител к CCR7 антител в формате CysMab с применением суррогатного репортерного анализа ADCC.
[0036] На фигуре 2 изображены экспериментальные данные по in vitro активности ADCC у негуманизированных антител к CCR7, содержащих варианты Fc с мутацией DAPA, с применением суррогатного репортерного анализа ADCC.
[0037] На фигуре 3 изображены экспериментальные данные по связыванию с рекомбинантным hCCR7 антител к CCR7 в формате CysMab.DAPA с применением анализа на основе ELISA.
[0038] На фигуре 4A-C изображены экспериментальные данные по функциональности исходных антител к CCR7 с применением анализа с β-аррестином в режиме агониста (фигура 4A) и режиме антагониста (фигура 4B, фигура 4C).
[0039] На фигуре 5 изображены экспериментальные данные по конкуренции с лигандом CCR7 антител к CCR7 в формате CysMab.DAPA с применением FACS-анализа.
[0040] На фигуре 6 изображены экспериментальные данные по картированию эпитопов исходных антител к CCR7 с применением мутантных белков CCR7.
[0041] На фигуре 7A-B изображены экспериментальные данные по анализам с применением piggyback ADC (pgADC) исходных антител к CCR7 в комплексе с фрагментом, представляющим собой вторичное антитело, конъюгированное с полезной нагрузкой.
[0042] На фигуре 8 изображены экспериментальные данные по анализам цитолиза с применением "piggyback" ADC (pgADC) цитотоксических эффектов исходного Ab 121G12 в комплексе с фрагментом, представляющим собой вторичное антитело, конъюгированное с полезной нагрузкой, с применением клеточных линий, отрицательных по наличию мишени.
[0043] На фигуре 9 изображены графики, иллюстрирующие истощение CD4+ и CD8a+ T-клеток с помощью мышиного исходного Ab 121G12, характеризующегося перекрестной реактивностью к CCR7, в формате CysMab с Fc дикого типа, либо в виде антитела отдельно, либо конъюгированного с цитотоксином ауристатином, при этом от данных эффектов избавляются путем переключения на формат "молчащей" Fc с мутациями DAPA.
[0044] На фигуре 10 изображен график, иллюстрирующий дозозависимую эффективность конъюгатов антитела и лекарственного средства, 121G12.CysMab.DAPA.MPET.DM4 и 121G12.DAPA.sSPDB.DM4, в отношении ксенотрансплантатной модели множественной миеломы KE97.
[0045] На фигуре 11 изображен график, иллюстрирующий активность конъюгатов антитела и лекарственного средства, 121G12.CysMab.DAPA.MPET.DM4 и 121G12.sSPDB.DM4, на модели множественной миеломы KE97 с началом введения дозы при большей первоначальной опухолевой массе, чем на фиг. 10.
[0046] На фигуре 12 изображен график, иллюстрирующий in vivo активность конъюгата 121G12.CysMab.DAPA.MPET.DM4 на модели первичной немелкоклеточной опухоли легкого HLUX1934.
[0047] На фигуре 13 изображен график, иллюстрирующий активность конъюгированного исходного антитела 684E12.SMCC.DM1 на ксенотрансплантатной модели множественной миеломы KE97.
[0048] На фигуре 14A-B изображены фотографии IHC для выявления фосфо-гистона H3 (фигура 14A) и количественная оценка сигнала фосфо-гистона H3 (фигура 14B) для опухолей KE97 через 48 часов после обработки однократной дозой либо 121G12.CysMab.DAPA.MPET.DM4 при 2, 5 или 10 мг/кг, либо изотипического контроля, IgG1.CysMab.DAPA.MPET.DM4, при 10 мг/кг, демонстрирующие индукцию митотического блока (фосфо-гистон H3) после обработки с помощью ADC к CCR7.
[0049] На фигуре 15 изображен график, иллюстрирующий дозозависимую эффективность 121G12.CysMab.DAPA.MPET.DM4 в отношении ксенотрансплантатной модели ABC-DLBCL OCI-LY3.
[0050] На фигуре 16 изображен график, иллюстрирующий дозозависимую эффективность 121G12.CysMab.DAPA.MPET.DM4 в отношении ксенотрансплантатной модели GCB-DLBCL Toledo.
[0051] На фигуре 17 изображен график, иллюстрирующий эффективность 121G12.CysMab.DAPA.MPET.DM4 в отношении ксенотрансплантатной модели ALCL DEL.
[0052] На фигуре 18 изображен график, иллюстрирующий дозозависимую эффективность 121G12.CysMab.DAPA.MPET.DM4 в отношении модели NSCLC на основе полученного от пациента ксенотрансплантата HLUX1787.
Подробное описание изобретения
Определения
[0053] Если не указано иное, подразумевается, что следующие термины и фразы, используемые в данном документе, имеют следующие значения.
[0054] Термин "алкил" относится к одновалентной насыщенной углеводородной цепи, содержащей указанное число атомов углерода. Например, C1-6 алкил относится к алкильной группе, содержащей от 1 до 6 атомов углерода. Алкильная группа может быть прямой или разветвленной. Типичная разветвленная алкильная группа содержит одну, две или три ветви. Примеры алкильных групп включают без ограничения, метил, этил, пропил (н-пропил и изопропил), бутил (н-бутил, изобутил, втор-бутил и трет-бутил), пентил (н-пентил, изопентил и неопентил) и гексил. Термин "алкилен" обозначает двухвалентную форму "алкила".
[0055] Используемый в данном документе термин "антитело" относится к полипептиду из семейства иммуноглобулинов, который способен к связыванию соответствующего антигена нековалентным, обратимым и специфическим образом. Например, встречающееся в природе антитело IgG представляет собой тетрамер, содержащий по меньшей мере две тяжелые (H) цепи и две легкие (L) цепи, соединенные между собой посредством дисульфидных связей. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно обозначаемой в данном документе как VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно обозначаемой в данном документе как VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена, CL. Области VH и VL могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), которые чередуются с более консервативными областями, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, которые включают различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента.
[0056] Термин "антитело" предусматривает без ограничения моноклональные антитела, человеческие антитела, гуманизированные антитела, химерные антитела и антиидиотипические (анти-Id) антитела (включая, например, анти-Id антитела к антителам по настоящему изобретению). Антител могут относиться к любому изотипу/классу (например, IgG, IgE, IgM, IgD, IgA и IgY) или подклассу (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2).
[0057] "Определяющие комплементарность домены" или "определяющие комплементарность области" ("CDR") взаимозаменяемо относятся к гипервариабельным областям VL и VH. В цепях антитела CDR представляют собой сайт связывания белка-мишени, который обуславливает специфичность в отношении такого белка-мишени. В каждой человеческой VL или VH имеется по три CDR (CDR1-3, пронумерованные последовательно от N-конца), составляющие приблизительно 15-20% от вариабельных доменов. CDR являются структурно комплементарными эпитопу белка-мишени и, таким образом, непосредственно ответственны за специфичность связывания. Остальные отрезки VL или VH, так называемые каркасные области, проявляют меньшую изменчивость аминокислотной последовательности (Kuby, Immunology, 4th ed., Chapter 4. W.H. Freeman & Co., New York, 2000).
[0058] Положения CDR и каркасных областей могут быть определены с применением различных определений, широко известных из уровня техники, например, Kabat, Chothia, международной базы данных ImMunoGeneTics (IMGT) (во всемирной сети по адресу www.imgt.org/) и AbM (см., например, Johnson et al., Nucleic Acids Res., 29:205-206 (2001); Chothia and Lesk, J. Mol. Biol., 196:901-917 (1987); Chothia et al., Nature, 342:877-883 (1989); Chothia et al., J. Mol. Biol., 227:799-817 (1992); Al-Lazikani et al., J.Mol.Biol., 273:927-748 (1997)). Определения антигенсвязывающих активных центров также описаны в следующих источниках: Ruiz et al., Nucleic Acids Res., 28:219-221 (2000); и Lefranc, M.P., Nucleic Acids Res., 29:207-209 (2001); MacCallum et al., J. Mol. Biol., 262:732-745 (1996); и Martin et al., Proc. Natl. Acad. Sci. USA, 86:9268-9272 (1989); Martin et al., Methods Enzymol., 203:121-153 (1991); и Rees et al., в Sternberg M.J.E. (ed.), Protein Structure Prediction, Oxford University Press, Oxford, 141-172 (1996).
[0059] Как легкая, так и тяжелая цепи подразделяются на области структурной и функциональной гомологии. Термины "константный" и "вариабельный" применяются в функциональном смысле. В связи с этим следует понимать, что вариабельные домены из частей как легкой (VL), так и тяжелой (VH) цепей определяют распознавание антигена и специфичность в его отношении. Напротив, константные домены легкой цепи (CL) и тяжелой цепи (CH1, CH2 или CH3) придают важные биологические свойства, такие как секреция, перемещение через плаценту, связывание с рецептором Fc, связывание с комплементом и т. п. Принято, что номера доменов константной области увеличиваются по мере их удаления от антигенсвязывающего сайта или амино-конца антитела. N-конец представляет собой вариабельную область, а на C-конце находится константная область; домены CH3 и CL фактически содержат карбокси-концевые домены тяжелой и легкой цепи, соответственно.
[0060] Используемый в данном документе термин "антигенсвязывающий фрагмент" относится к одной или нескольким частям антитела, которые сохраняют способность специфически взаимодействовать (например, посредством связывания, стерического несоответствия, стабилизации/дестабилизации, пространственного распределения) с эпитопом антигена. Примеры связывающих фрагментов включают без ограничения одноцепочечные Fv (scFv), антитела верблюжьих, соединенные дисульфидными связями Fv (sdFv), Fab-фрагменты, F(ab')-фрагменты, одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; F(ab)2-фрагмент, двухвалентный фрагмент, содержащий два Fab-фрагмента, соединенные дисульфидным мостиком в шарнирной области; Fd-фрагмент, состоящий из доменов VH и CH1; Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела; dAb-фрагмент (Ward et al., Nature 341:544-546, 1989), который состоит из домена VH; а также выделенную определяющую комплементарность область (CDR) или другие эпитопсвязывающие фрагменты антитела.
[0061] Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, они могут быть соединены с применением рекомбинантных способов с помощью синтетического линкера, что обеспечивает их получение в виде одной белковой цепи, в которой области VL и VH соединяются в пару с образованием одновалентной молекулы (известной как одноцепочечный Fv ("scFv"); см., например, Bird et al., Science 242:423-426, 1988; и Huston et al., Proc. Natl. Acad. Sci. 85:5879-5883, 1988). Предусматривается, что такие одноцепочечные антитела также охватываются термином "антигенсвязывающий фрагмент". Такие антигенсвязывающие фрагменты получают с применением традиционных методик, известных специалистам в данной области техники, и фрагменты подвергают скринингу на применимость таким же способом, что и интактные антитела.
[0062] Антигенсвязывающие фрагменты также могут быть включены в состав однодоменных антител, максител, минител, однодоменных антител, интрател, диател, триател, тетрател, v-NAR и бис-scFv (см., например, Hollinger and Hudson, Nature Biotechnology 23:1126-1136, 2005). Антигенсвязывающие фрагменты можно прививать на остовы на основе полипептидов, таких как фибронектин типа III (Fn3) (см. патент США № 6703199, в котором описаны монотела на основе полипептида фибронектина).
[0063] Антигенсвязывающие фрагменты можно вводить в состав одноцепочечных молекул, содержащих пару тандемных сегментов Fv (VH-CH1-VH-CH1), которые вместе с комплементарными полипептидами легкой цепи образуют пару антигенсвязывающих областей (Zapata et al., Protein Eng. 8:1057-1062, 1995; и патент США № 5641870).
[0064] Используемый в данном документе термин "моноклональное антитело" или "композиция на основе моноклонального антитела" относится к полипептидам, включающим антитела и антигенсвязывающие фрагменты, которые характеризуются практически идентичной аминокислотной последовательностью или происходят из одного генетического источника. Данный термин также охватывает препараты молекул антител одного молекулярного состава. Композиция на основе моноклонального антитела проявляет одну специфичность и аффинность связывания в отношении конкретного эпитопа.
[0065] Используемый в данном документе термин "человеческое антитело" охватывает антитела, имеющие вариабельные области, в которых как каркасные области, так и CDR получены из последовательностей, происходящих от человека. Кроме того, если антитело содержит константную область, то константная область также происходит из таких человеческих последовательностей, например, последовательностей зародышевой линии человека, или мутантных вариантов последовательностей зародышевой линии человека, или антитела, содержащего консенсусные каркасные последовательности, полученные за счет анализа человеческих каркасных последовательностей, например, как описано в Knappik et al., J. Mol. Biol. 296:57-86, 2000). Также подразумеваются антитела, происходящие из человеческих последовательностей, где одна или несколько CDR были подвергнуты мутированию в процессе созревания аффинности или для целей изготовления/конъюгации с полезной нагрузкой. См. Kilpatrick et al., "Rapid development of affinity matured monoclonal antibodies using RIMMS", Hybridoma. 1997 Aug;16(4):381-9.
[0066] Человеческие антитела по настоящему изобретению могут включать аминокислотные остатки, которые не закодированы в человеческих последовательностях (например, мутации, введенные посредством случайного или сайт-специфического мутагенеза in vitro или за счет соматических мутаций in vivo, или консервативная замена, которая содействует стабильности или облегчает изготовление).
[0067] Используемый в данном документе термин "распознавать" относится к антителу или его антигенсвязывающему фрагменту, которые отыскивают свой эпитоп и взаимодействуют (например, связываются) с ним, независимо от того является ли эпитоп линейным или конформационным. Термин "эпитоп" относится к сайту на антигене, с которым специфически связываются антитело или антигенсвязывающий фрагмент по настоящему изобретению. Эпитопы могут быть образованы как смежными аминокислотами, так и несмежными аминокислотами, размещаемыми рядом за счет третичной укладки белка. Эпитопы, образуемые из смежных аминокислот, как правило, сохраняются при воздействии денатуририрующих растворителей, в то время как эпитопы, образуемые за счет третичной укладки, как правило, утрачиваются при обработке денатурирующими растворителями. Эпитоп, как правило, содержит по меньшей мере 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 аминокислот в уникальной пространственной конформации. Способы определения пространственной конформации эпитопов включают методики из уровня техники, например, рентгеноструктурную кристаллографию и 2-мерный ядерный магнитный резонанс (см., например, Epitope Mapping Protocols в Methods in Molecular Biology, Vol. 66, G. E. Morris, Ed. (1996)).
[0068] Используемый в данном документе термин "аффинность" относится к силе взаимодействия между антителом и антигеном в отдельных антигенных сайтах. В пределах каждого антигенного сайта вариабельная область "плеча" антитела взаимодействует с антигеном посредством слабых нековалентных сил в многочисленных сайтах; при этом, чем больше взаимодействий, тем сильнее аффинность.
[0069] Термин "выделенное антитело" относится к антителу, которое практически не содержит других антител с отличающейся антигенной специфичностью. Однако выделенное антитело, которое специфически связывается с одним антигеном, может характеризоваться перекрестной реактивностью в отношении других антигенов. Более того, выделенное антитело может практически не содержать другого клеточного материала и/или химических веществ.
[0070] Термин "соответствующая последовательность зародышевой линии человека" относится к последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность или подпоследовательность человеческой вариабельной области, которые обладают самой высокой установленной идентичностью аминокислотной последовательности с эталонной аминокислотной последовательностью или подпоследовательностью вариабельной области в сравнении со всеми другими всеми другими известными аминокислотными последовательностями вариабельной области, закодированными в последовательностях вариабельной области иммуноглобулина зародышевой линии человека. Соответствующая последовательность зародышевой линии человека также может относиться к аминокислотной последовательности или подпоследовательности человеческой вариабельной области, характеризующимся самой высокой идентичностью аминокислотной последовательности с эталонной аминокислотной последовательностью или подпоследовательностью вариабельной области в сравнении со всеми другими подвергнутыми оценке аминокислотными последовательностями вариабельной области. Соответствующей последовательностью зародышевой линии человека могут быть только каркасные области, только определяющие комплементарность области, каркасные и определяющие комплементарность области, вариабельный сегмент (как определено выше) или другие комбинации последовательностей или подпоследовательностей, которые составляют вариабельную область. Идентичность последовательности может быть определена с применением способов, описанных в данном документе, например, выравнивания двух последовательностей с применением BLAST, ALIGN или другого алгоритма выравнивания, известного из уровня техники. Соответствующая последовательность нуклеиновой кислоты или аминокислотная последовательность зародышевой линии человека может характеризоваться по меньшей мере приблизительно 90%, 91, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, или 100% идентичностью последовательности с эталонной последовательностью нуклеиновой кислоты или аминокислотной последовательностью вариабельной области. Соответствующие последовательности зародышевой линии человека могут быть определены, например, с помощью международной базы данных ImMunoGeneTics (IMGT), находящейся в открытом доступе (во всемирной сети по адресу www.imgt.org/) и V-base (во всемирной сети по адресу vbase.mrc-cpe.cam.ac.uk).
[0071] Фраза "специфически связывает" или "селективно связывает", когда она применяется в контексте описания взаимодействия между антигеном (например, белком) и антителом, фрагментом антитела или связывающим средством, происходящим из антитела, относится к реакции связывания, которая является определяющей для установления присутствия антигена в неоднородной популяции белков и других биологических веществ, например, в биологическом образце, например, крови, сыворотке, плазме или образце ткани. Таким образом, при некоторых обозначенных условиях проведения иммунологического анализа антитела или связывающие средства, характеризующиеся конкретной специфичностью связывания, связываются с конкретным антигеном в по меньшей мере два раза сильнее, чем фоновый уровень, и практически не связываются в значительном количестве с другими антигенами, присутствующими в образце. В одном варианте осуществления при обозначенных условиях иммунологического анализа антитело или связывающее средство с конкретной специфичностью связывания связывается с конкретным антигеном в по меньшей мере десять (10) раз сильнее относительно фонового уровня, и практически не связывается в значительном количестве с другими антигенами, присутствующими в образце. Специфическое связывание с антителом или связывающим средством в таких условиях может предусматривать то, что антитело или средство должно отбираться за его специфичность в отношении конкретного белка. При желании или необходимости данный отбор можно проводить путем отбрасывания антител, которые вступают в перекрестные реакции с молекулами от другого вида (например, мыши или крысы) или других подтипов. В качестве альтернативы в некоторых вариантах осуществления отбирают антитела или фрагменты антител, которые вступают в перекрестные реакции с некоторыми требуемыми молекулами.
[0072] Целый ряд форматов иммунологического анализа может применяться для отбора антител, характеризующихся специфической иммунной реактивностью в отношении конкретного белка. Например, твердофазные иммунологические анализы ELISA традиционно применяются для отбора антител, характеризующихся специфической реактивностью в отношении белка (см., например, Harlow & Lane, Using Antibodies, A Laboratory Manual (1998), где описаны форматы и условия иммунологического анализа, которые можно применять для определения специфической иммунной реактивности). Как правило, реакция специфического или селективного связывания будет приводить к сигналу, в по меньшей мере два раза превышающему фоновый уровень, и, более типично, в по меньшей мере 10-100 раз превышающему фоновый уровень.
[0073] Термин "равновесная константа диссоциации (KD [M])" относится к константе скорости диссоциации (kd [время-1]), поделенной на константу скорости ассоциации (ka [время-1, M-1]). Равновесные константы диссоциации могут быть измерены с применением любого способа, известного из уровня техники. Обычно антитела по настоящему изобретению будут характеризоваться равновесной константой диссоциации, составляющей менее приблизительно 10-7 или 10-8 M, например, менее приблизительно 10-9 M или 10-10 M, в некоторых вариантах осуществления менее приблизительно 10-11 M, 10-12 M или 10-13 M.
[0074] Термин "биодоступность" относится к системной доступности (т. е. уровням в крови/плазме) заданного количества лекарственного средства, вводимого пациенту. Биодоступность представляет собой абсолютный термин, обозначающий показатель как времени (скорости), в течение которого лекарственное средство достигает общего кровообращения из введенной лекарственной формы, так и общего количества (величины) лекарственного средства в нем.
[0075] Используемая в данном документе фраза "состоящий фактически из" относится к родам или видам активных фармацевтических средств, включенных в способ или композицию, а также любым вспомогательным веществам, не проявляющим активность в отношении намеченной цели применения способов или композиций. В некоторых вариантах осуществления фраза "состоящий фактически из" однозначно исключает включение одного или нескольких дополнительных активных средств, отличных от конъюгата антитела и лекарственного средства по настоящему изобретению. В некоторых вариантах осуществления фраза "состоящий фактически из" однозначно исключает включение одного или нескольких дополнительных активных средств, отличных от конъюгата антитела и лекарственного средства по настоящему изобретению и второго средства, вводимого совместно.
[0076] Термин "аминокислота" относится к встречающимся в природе, синтетическим и неприродным аминокислотам, а также аналогам аминокислот и миметикам аминокислот, которые функционируют подобно встречающимся в природе аминокислотам. Встречающиеся в природе аминокислоты представляют собой аминокислоты, закодированные в генетическом коде, а также такие аминокислоты, которые были впоследствии модифицированы, например, гидроксипролин, γ-карбоксиглутамат и O-фосфосерин. Аналоги аминокислот относятся к соединениям, которые характеризуются такой же основной химической структурой, что и встречающаяся в природе аминокислота, то есть имеют α-углерод, который связан с водородом, карбоксильной группой, аминогруппой и R-группой, например, к гомосерину, норлейцину, метионинсульфоксиду, метионинметилсульфонию. Такие аналоги имеют модифицированные R-группы (например, норлейцин) или модифицированные пептидные остовы, но сохраняют такую же основную химическую структуру, что и встречающаяся в природе аминокислота. Миметики аминокислот относятся к химическим соединениям, которые имеют структуру, отличающуюся от общей химической структуры аминокислоты, но которые функционируют подобно встречающейся в природе аминокислоте.
[0077] Термин "консервативно модифицированный вариант" применяется в отношении как аминокислотных последовательностей, так и последовательностей нуклеиновой кислоты. Применительно к конкретным последовательностям нуклеиновой кислоты, консервативно модифицированные варианты относятся к тем нуклеиновым кислотам, которые кодируют идентичные или фактически идентичные аминокислотные последовательности, или же, если нуклеиновая кислота не кодирует аминокислотную последовательность, к фактически идентичным последовательностям. Вследствие вырожденности генетического кода любой заданный белок кодируется большим количеством функционально идентичных нуклеиновых кислот. Например, все из кодонов GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в каждом положении, в котором кодоном задан аланин, кодон может быть изменен на любой из соответствующих описанных кодонов без изменения кодируемого полипептида. Такие вариации нуклеиновой кислоты являются "молчащими вариациями", которые представляют собой одну разновидность вариаций с консервативными модификациями. В данном документе каждая последовательность нуклеиновой кислоты, которая кодирует полипептид, также описывает каждую возможную молчащую вариацию нуклеиновой кислоты. Специалист в данной области будет осознавать, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который обычно является единственным кодоном для метионина, и TGG, который обычно является единственным кодоном для триптофана) может быть модифицирован с получением функционально идентичной молекулы. Соответственно, каждая молчащая вариация нуклеиновой кислоты, которая кодирует полипептид, неявно определена в каждой описанной последовательности.
[0078] В случае полипептидных последовательностей "консервативно модифицированные варианты" охватывают отдельные замены, делеции или добавления в полипептидной последовательности, которые приводят к замене аминокислоты на аналогичную по химическим свойствам аминокислоту. Таблицы консервативных замен, обеспечивающие функционально аналогичные аминокислоты, хорошо известны из уровня техники. Такие консервативно модифицированные варианты дополняют, а не исключают, полиморфные варианты, межвидовые гомологи и аллели по настоящему изобретению. Следующие восемь групп содержат аминокислоты, которые являются консервативными заменами друг для друга: 1) аланин (A), глицин (G); 2) аспарагиновая кислота (D), глутаминовая кислота (E); 3) аспарагин (N), глутамин (Q); 4) аргинин (R), лизин (K); 5) изолейцин (I), лейцин (L), метионин (M), валин (V); 6) фенилаланин (F), тирозин (Y), триптофан (W); 7) серин (S), треонин (T) и 8) цистеин (C), метионин (M) (см., например, Creighton, Proteins (1984)). В некоторых вариантах осуществления термин "консервативные модификации последовательности" используется для обозначения аминокислотных модификаций, которые не оказывают значительного влияния на характеристики связывания у антитела, содержащего аминокислотную последовательность, или не изменяют их.
[0079] Используемый в данном документе термин "оптимизированная" относится к нуклеотидной последовательности, кодирующей аминокислотную последовательность, которая была изменена с применением кодонов, предпочтительных в продуцирующих клетке или организме, обычно в эукариотической клетке, например, дрожжевой клетке, клетке Pichia, грибной клетке, клетке Trichoderma, клетке яичника китайского хомячка (CHO) или человеческой клетке. Оптимизированную нуклеотидную последовательность конструируют таким образом, чтобы полностью или насколько это возможно сохранить аминокислотную последовательность, изначально закодированную в первоначальной нуклеотидной последовательности, которая также известна как "исходная" последовательность.
[0080] Термины "процент идентичности" или "процентная идентичность", в контексте двух или более нуклеиновых кислот или полипептидных последовательностей, относятся к степени, в которой две или более последовательности или подпоследовательности являются одинаковыми. Две последовательности являются "идентичными", если они имеют одинаковую последовательность из аминокислот или нуклеотидов на протяжении области, подлежащей сравнению. Две последовательности являются "практически идентичными", если две последовательности имеют указанную процентную долю аминокислотных остатков или нуклеотидов, которые являются одинаковыми (т. е. 60% идентичность, необязательно 65%, 70%, 75%, 80%, 85%, 90%, 95% или 99% идентичность на протяжении указанной области или, если не указано, на протяжении всей последовательности), при сравнении и выравнивании для обеспечения максимального соответствия на протяжении окна сравнения или обозначенной области, как измерено с применением одного из следующих алгоритмов сравнения последовательности или посредством ручного выравнивания и визуального просмотра. Необязательно идентичность существует на протяжении области, длина которой составляет по меньшей мере приблизительно 30 нуклеотидов (или 10 аминокислот), или более предпочтительно на протяжении области, длина которой составляет 100-500 или 1000 или более нуклеотидов (или 20, 50, 200 или более аминокислот).
[0081] При сравнении последовательностей обычно одна последовательность выступает в качестве эталонной последовательности, с которой сравнивают тестируемые последовательности. При использовании алгоритма сравнения последовательностей тестируемую и эталонную последовательности вводят в компьютер, если необходимо, устанавливают координаты подпоследовательностей и устанавливают программные параметры алгоритма для анализа последовательностей. Могут применяться программные параметры по умолчанию или можно устанавливать альтернативные параметры. На основании программных параметров алгоритм сравнения последовательностей затем рассчитывает значения процента идентичности последовательностей для тестируемых последовательностей относительно эталонной последовательности.
[0082] Используемое в данном документе "окно сравнения" предусматривает ссылку на сегмент из любого количества смежных положений, выбранных из группы, состоящей из от 20 до 600, обычно от приблизительно 50 до приблизительно 200, чаще от приблизительно 100 до приблизительно 150, в котором последовательность можно сравнивать с эталонной последовательностью с таким же количеством смежных положений, после того как две последовательности подвергли оптимальному выравниванию. Способы выравнивания последовательностей для проведения сравнения хорошо известны из уровня техники. Оптимальное выравнивание последовательностей для сравнения можно проводить, например, с помощью алгоритма поиска локальной гомологии Смита-Уотермана, Adv. Appl. Math. 2:482c (1970), с помощью алгоритма выравнивания областей гомологии Нидлмана-Вунша, J. Mol. Biol. 48:443 (1970), с помощью способа поиска сходства Пирсона-Липмана, Proc. Natl. Acad. Sci. USA 85:2444 (1988), с помощью компьютерных реализаций таких алгоритмов (GAP, BESTFIT, FASTA и TFASTA в составе пакета программного обеспечения Wisconsin Genetics, Genetics Computer Group, 575 Science Dr., Мэдисон, Висконсин) или с помощью ручного выравнивания и визуального просмотра (см., например, Brent et al., Current Protocols in Molecular Biology, 2003).
[0083] Два примера алгоритмов, которые подходят для определения процента идентичности последовательностей и сходства последовательностей, представляют собой алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul et al., Nuc. Acids Res. 25:3389-3402, 1977; и Altschul et al., J. Mol. Biol. 215:403-410, 1990, соответственно. Программное обеспечение для осуществления анализов BLAST находится в открытом доступе от Национального центра биотехнологической информации. На первом этапе данный алгоритм предусматривает идентификацию пар последовательностей с высоким показателем сходства (HSP) за счет идентификации коротких слов длиной W в запрашиваемой последовательности, которые либо совпадают, либо удовлетворяют некоторому пороговому баллу T с положительным значением при выравнивании со словом такой же длины в последовательности из базы данных. T известен под названием пороговый балл соседних слов (Altschul et al., выше). Эти первоначальные совпадения соседних слов выступают в качестве "затравок" для начала поисков, предназначенных для нахождения более длинных HSP, содержащие их. Совпадения слов продлеваются в обоих направлениях вдоль каждой последовательности до тех пор, пока может увеличиваться суммарный балл выравнивания. В случае нуклеотидных последовательностей суммарные баллы рассчитывают с применением параметров M (вознаграждающий балл за пару совпадающих остатков; всегда >0) и N (штрафной балл за несовпадающие остатки; всегда <0). В случае аминокислотных последовательностей для подсчета суммарного балла применяют матрицу замен. Продление совпадений слов в каждом направлении останавливается, когда суммарный балл выравнивания уменьшается на величину X относительно своего максимального достигнутого значения; суммарный балл стремится к нулю или ниже вследствие накопления одного или нескольких выравниваний остатков с отрицательными баллами; или достигается конец любой из последовательностей. Параметры W, T и X алгоритма BLAST определяют чувствительность и скорость выравнивания. В программе BLASTN (для нуклеотидных последовательностей) в качестве параметров по умолчанию применяется длина слова (W), составляющая 11, ожидание (E), составляющее 10, M=5, N=-4 и сравнение обеих нитей. В случае аминокислотных последовательностей в программе BLASTP в качестве параметров по умолчанию применяется длина слова, составляющая 3, и ожидание (E), составляющее 10, и матрица замен BLOSUM62 (см. Henikoff and Henikoff (1989) Proc. Natl. Acad. Sci. USA 89:10915), выравнивания (B), составляющие 50, ожидание (E), составляющее 10, M=5, N=-4 и сравнение обеих нитей.
[0084] Алгоритм BLAST также осуществляет статистический анализ сходства между двумя последовательностями (см., например, Karlin and Altschul, Proc. Natl. Acad. Sci. USA 90:5873-5787, 1993). Одной мерой сходства, предусмотренной в алгоритме BLAST, является наименьшая суммарная вероятность (P (N)), которая указывает на вероятность, с которой совпадение между двумя нуклеотидными или аминокислотными последовательностями возникло случайно. Например, нуклеиновая кислота считается схожей с эталонной последовательностью, если наименьшая суммарная вероятность при сравнении тестируемой нуклеиновой кислоты с эталонной нуклеиновой кислотой составляет менее приблизительно 0,2, более предпочтительно менее приблизительно 0,01 и наиболее предпочтительно менее приблизительно 0,001.
[0085] Процентную идентичность между двумя аминокислотными последовательностями также можно определить с использованием алгоритма из E. Meyers и W. Miller (Comput. Appl. Biosci. 4:11-17, (1988) который был включен в программу ALIGN (версия 2.0), с применением таблицы весов замен остатков PAM120, штрафа за продление гэпа, составляющего 12, и штрафа за открытие гэпа, составляющего 4. Кроме того, процентная идентичность между двумя аминокислотными последовательностями может быть определена с применением алгоритма Нидлмана-Вунша, J. Mol. Biol. 48:444-453, (1970), который был включен в программу GAP в составе пакета программного обеспечения GCG (доступного на www.gcg.com), с применением либо матрицы BLOSUM62, либо матрицы PAM250, и также штрафа за открытие гэпа, составляющего 16, 14, 12, 10, 8, 6 или 4, и штрафа за продление гэпа, составляющего 1, 2, 3, 4, 5 или 6.
[0086] Помимо процента идентичности последовательностей, упомянутого выше, еще одним показателем того, что две последовательности нуклеиновой кислоты или полипептида являются практически идентичными, является то, что полипептид, кодируемый первой нуклеиновой кислотой, является иммунологически перекрестно реактивным с антителами, выработка которых индуцирована полипептидом, кодируемым второй нуклеиновой кислотой, как описано ниже. Таким образом, полипептид, как правило, является практически идентичным второму полипептиду, например, если два пептида отличаются только консервативными заменами. Другим показателем того, что две последовательности нуклеиновой кислоты являются практически идентичными, является то, что две молекулы или их комплементарные цепи гибридизируются друг с другом в жестких условиях, как описано ниже. Еще одним показателем того, что две последовательности нуклеиновой кислоты являются практически идентичными, является то, что для амплификации последовательности могут применяться одни и те же праймеры.
[0087] Термин "нуклеиновая кислота" используется в данном документе взаимозаменяемо с термином "полинуклеотид" и относится к дезоксирибонуклеотидам или рибонуклеотидам и полимерам на их основе в одно- или двухнитевой форме. Термин охватывает нуклеиновые кислоты, содержащие известные аналоги нуклеотидов или модифицированные остатки или связи в остове, которые являются синтетическими, встречающимися в природе и не встречающимися в природе, которые характеризуются свойствами связывания, подобными эталонной нуклеиновой кислоте, и которые метаболизируются подобно эталонным нуклеотидам. Примеры таких аналогов включают без ограничения фосфоротиоаты, фосфорамидаты, метилфосфонаты, хиральные метилфосфонаты, 2-O-метилрибонуклеотиды, пептидонуклеиновые кислоты (PNA).
[0088] Если не указано иное, конкретная последовательность нуклеиновой кислоты также в неявном виде охватывает ее консервативно модифицированные варианты (например, замены на основе вырожденных кодонов) и комплементарные последовательности, а также последовательность, указанную явным образом. А именно, как подробно описано ниже, замены на основе вырожденных кодонов можно проводить за счет создания последовательностей, в которых третье положение в одном или нескольких выбранных (или всех) кодонах заменено на любой из канонических нуклеозидов и/или остаток дезоксиинозина (Batzer et al., Nucleic Acid Res. 19:5081, 1991; Ohtsuka et al., J. Biol. Chem. 260:2605-2608, 1985; и Rossolini et al., Mol. Cell. Probes 8:91-98, 1994).
[0089] В контексте нуклеиновых кислот термин "функционально связанный" относится к функциональной взаимосвязи двух или более сегментов полинуклеотида (например, ДНК). Как правило, он относится к функциональной взаимосвязи регулирующей транскрипцию последовательности с транскрибируемой последовательностью. Например, промоторная или энхансерная последовательность является функционально связанной с кодирующей последовательностью, если она стимулирует или модулирует транскрипцию кодирующей последовательности в соответствующей клетке-хозяине или другой системе экспрессии. Обычно регулирующие транскрипцию промоторные последовательности, которые являются функционально связанными с транскрибируемой последовательностью, являются физически смежными с транскрибируемой последовательностью, т.е. они функционируют в цис-положении. Однако некоторые регулирующие транскрипцию последовательности, такие как энхансеры, не обязательно должны быть физически смежными или располагаться в непосредственной близости от кодирующих последовательностей, транскрипцию которых они усиливают.
[0090] Термины "полипептид" и "белок" используются в данном документе взаимозаменяемо для обозначения полимера из аминокислотных остатков. Термины применимы к полимерам из аминокислот, в которых один или несколько аминокислотных остатков представляют собой искусственный химический миметик соответствующей встречающейся в природе аминокислоты, а также к полимерам из встречающихся в природе аминокислот и полимеру из не встречающихся в природе аминокислот. Если не указано иное, конкретная полипептидная последовательность также в неявном виде охватывает ее консервативно модифицированные варианты.
[0091] Используемый в данном документе термин "конъюгат антитела и лекарственного средства" или "иммуноконъюгат" относится к связи антитела или его антигенсвязывающего фрагмента с другим средством, таким как химиотерапевтическое средство, токсин, иммунотерапевтическое средство, зонд для визуализации и т.п. Связь может представлять собой ковалентные связи или нековалентные взаимодействия, такие как посредством электростатических сил. Для образования конъюгата антитела и лекарственного средства могут использоваться различные линкеры, известные из уровня техники. Кроме того, конъюгат антитела и лекарственного средства может предусматриваться в форме слитого белка, который может экспрессироваться с полинуклеотида, кодирующего иммуноконъюгат. Используемый в данном документе "слитый белок" относится к белкам, созданным за счет соединения двух или более генов или фрагментов генов, которые изначально кодировали отдельные белки (в том числе пептиды и полипептиды). Трансляция слитого гена приводит к единому белку с функциональными свойствами, происходящими из каждого из исходных белков.
[0092] Термин "субъект" охватывает человека и отличных от человека животных. Отличные от человека животные охватывают всех позвоночных, например, млекопитающих и отличных от млекопитающих животных, как, например, отличных от человека приматов, овцу, собаку, корову, кур, амфибий и рептилий. За исключением случаев, когда это отмечается, термины "пациент" или "субъект" используются в данном документе взаимозаменяемо.
[0093] Используемый в данном документе термин "цитотоксин" или "цитотоксическое средство" относится к любому средству, которое является вредным для роста и пролиферации клеток и которое может функционировать с уменьшением, ингибированием или разрушением клетки или злокачественной опухоли.
[0094] Используемый в данном документе термин "противораковое средство" относится к любому средству, которое может использоваться для лечения или предупреждения нарушения пролиферации клеток, такого как рак, в том числе без ограничения к цитотоксическим средствам, химиотерапевтическим средствам, лучевой терапии и средствам для лучевой терапии, нацеливающим противораковым средствам и иммунотерапевтическим средствам.
[0095] Используемый в данном документе термин "фрагмент, представляющий собой лекарственное средство" или "полезная нагрузка" относится к химическому фрагменту, который конъюгирован с антителом или антигенсвязывающим фрагментом по настоящему изобретению, и может охватывать любое терапевтическое или диагностическое средство, например, противораковое, противовоспалительное, противоинфекционное (например, противогрибковое, антибактериальное, антипаразитарное, противовирусное) или анестетическое средство. Например, фрагмент, представляющий собой лекарственное средство, может представлять собой противораковое средство, такое как цитотоксин. В некоторых вариантах осуществления фрагмент, представляющий собой лекарственное средство, выбран из ингибитора V-АТФазы, ингибитора HSP90, ингибитора IAP, ингибитора mTor, стабилизатора микротрубочек, дестабилизатора микротрубочек, ауристатина, доластатина, майтанзиноида, MetAP (метионинаминопептидазы), ингибитора РНК-полимеразы, пирролобензодиазепина (PBD), аманитина, ингибитора ядерного экспорта белков CRM1, ингибитора DPPIV, ингибитора реакций переноса фосфорила в митохондриях, ингибитора синтеза белка, ингибитора киназ, ингибитора CDK2, ингибитора CDK9, ингибитора протеасомы, ингибитора кинезина, ингибитора HDAC, ДНК-повреждающего средства, ДНК-алкилирующего средства, ДНК-интеркалятора, средства, связывающего малую бороздку ДНК, и ингибитора DHFR. Способы прикрепления каждого из них к линкеру, совместимому с антителами и способом по настоящему изобретению, известны из уровня техники. См., например, Singh et al., (2009) Therapeutic Antibodies: Methods and Protocols, vol. 525, 445-457. Кроме того, полезная нагрузка может представлять собой зонд для биофизического обнаружения, флуорофор, спиновую метку, зонд для инфракрасного обнаружения, зонд для обнаружения аффинности, хелатор, зонд для спектроскопического обнаружения, радиоактивный зонд, липидную молекулу, полиэтиленгликоль, полимер, спиновую метку, ДНК, РНК, белок, пептид, поверхность, антитело, фрагмент антитела, наночастицу, квантовую точку, липосому, частицу PLGA, сахарид или полисахарид.
[0096] Термин "фрагмент, представляющий собой майтанзиноидное лекарственное средство", обозначает часть конъюгата антитела и лекарственного средства, которая имеет структуру майтанзиноидного соединения. Сначала майтанзин был выделен из куста Maytenus serrata, произрастающего в восточной Африке (патент США № 3896111). Впоследствии было установлено, что некоторые микроорганизмы также вырабатывают майтанзиноиды, такие как майтанзинол и сложные эфиры C-3-майтанзинола (патент США № 4151042). Сообщалось о синтетическом майтанзиноле и аналогах майтанзинола. См. патенты США №№ 4137230; 4248870; 4256746; 4260608; 4265814; 4294757; 4307016; 4308268; 4308269; 4309428; 4313946; 4315929; 4317821; 4322348; 4331598; 4361650; 4364866; 4424219; 4450254; 4362663 и 4371533, и Kawai et al., (1984) Chem. Pharm. Bull. 3441-3451), каждый из которых однозначно включен посредством ссылки. Примеры конкретных майтанзиноидов, применимых для конъюгации, включают DM1, DM3 и DM4.
[0097] "Опухоль" относится к росту и пролиферации неопластических клеток, неважно злокачественных или доброкачественных, а также всех предраковых и раковых клеток и тканей.
[0098] Термин "противоопухолевая активность" означает снижение скорости пролиферации, жизнеспособности или метастатической активности опухолевых клеток. Например, противоопухолевая активность может быть показана за счет падения скорости роста аномальных клеток, которое появляется во время терапии, или стабилизации или уменьшения размера опухоли, или более длительного выживания, обусловленного терапией, по сравнению с контролем без терапии. Такую активность можно оценить с применением принятых in vitro или in vivo моделей опухоли, в том числе без ограничения ксенотрансплантатных моделей, моделей с аллогенными трансплантатами, моделей MMTV и других известных моделей, известных из уровня техники для исследования противоопухолевой активности.
[0099] Термин "злокачественная опухоль" относится к недоброкачественной опухоли или раку. Используемый в данном документе термин "рак" охватывает злокачественную опухоль, которая характеризуется дерегулированным или неконтролируемым ростом клеток. Иллюстративные типы рака включают: карциномы, саркомы, лейкозы и лимфомы.
[00100] Термин "рак" охватывает первичные злокачественные опухоли (например, опухоли, клетки которых не мигрировали в локализации в организме субъекта, отличные от локализации исходной опухоли) и вторичные злокачественные опухоли (например, опухоли, развившиеся за счет метастазирования, миграции опухолевых клеток во вторичные локализации, которые отличны от локализации исходной опухоли).
[00101] Термин "CCR7" (также известный как BLR2, CC-CKR-7, CCR-7, CD197, CDw197, CMKBR7, EBI1 или хемокиновый рецептор 7 с C-C мотивом) относится к представителю семейства рецепторов, сопряженных с G белком. Последовательность нуклеиновой кислоты и аминокислотная последовательность CCR7 человек была опубликована в GenBank под следующими №№ доступа: NP_001829, NP_001288643, NP_001288645, NP_001288646, NP_001288647 (аминокислотные последовательности) и NM_001838, NM_001301714, NM_001301716, NM_001301717, NM_001301718 (нуклеотидные последовательности). Используемый в данном документе термин "CCR7" применяют для совместного обозначения всех встречающихся в природе изоформ белка CCR7 или его варианта.
[00102] Термин "вариант" относится к полипептиду, который имеет практически идентичную аминокислотную последовательность с эталонным полипептидом, или кодируется практически идентичной нуклеотидной последовательностью и может характеризоваться одной или несколькими активностями эталонного полипептида. Например, вариант может характеризоваться приблизительно 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более высокой идентичностью последовательности с эталонным полипептидом, при этом он сохраняет одну или несколько активностей эталонного полипептида.
[00103] Используемые в данном документе термины "лечить", "осуществление лечения" или "лечение" любого заболевания или нарушения в одном варианте осуществления относятся к уменьшению тяжести заболевания или нарушения (т.е. замедлению, или остановке, или снижению развития заболевания или по меньшей мере одного из его клинических симптомов). В другом варианте осуществления "лечить", "осуществление лечения" или "лечение" относятся к облегчению или уменьшению тяжести по меньшей мере одного физического параметра, в том числе таких, которые могут быть неявными для пациента. В еще одном варианте осуществления "лечить", "осуществление лечения" или "лечение" означают модулирование заболевания или нарушения либо физически (например, стабилизацию явного симптома), либо физиологически (например, стабилизацию физического параметра), либо с помощью того и другого.
[00104] Используемый в данном документе термин "предупреждать", "осуществление предупреждения" или "предупреждение" любого заболевания или нарушения относится к профилактическому лечению заболевания или нарушения или замедлению начала или прогрессирования заболевания или нарушения.
[00105] Термин "терапевтически приемлемое количество" или "терапевтически эффективная доза" взаимозаменяемо относится к количеству, достаточному для произведения требуемого результата (т.е. уменьшения размера опухоли, ингибирования роста опухоли, предотвращения метастазирования, ингибирования или предупреждения вирусной, бактериальной, грибковой или паразитарной инфекции). В некоторых вариантах осуществления терапевтически приемлемое количество не индуцирует или не вызывает нежелательные побочные эффекты. В некоторых вариантах осуществления терапевтически приемлемое количество индуцирует или вызывает побочные эффекты, но только такие, которые являются допустимыми с точки зрения медицинских работников, учитывая состояние пациента. Терапевтически приемлемое количество можно определять, вводя вначале низкую дозу, а затем постепенно увеличивать данную дозу до тех пор, пока не будет достигнут требуемый эффект. "Профилактически эффективная доза" и "терапевтически эффективная доза" молекул по настоящему изобретению может предупреждать проявление или, соответственно, приводить к уменьшению тяжести симптомов заболевания, в том числе симптомов, ассоциированных с раком.
[00106] Термин "совместное введение" относится к присутствию двух активных средств в крови индивидуума. Активные средства, которые вводятся совместно, могут доставляться одновременно или последовательно.
[00107] В настоящем изобретении представлены антитела, фрагменты антител (например, антигенсвязывающие фрагменты) и их конъюгаты с лекарственным средством, т.е. конъюгаты антитела и лекарственного средства или ADC, которые связываются с CCR7. В частности, в настоящем изобретении представлены антитела и фрагменты антител (например, антигенсвязывающие фрагменты), которые связываются с CCR7 и подвергаются интернализации после такого связывания. Антитела и фрагменты антител (например, антигенсвязывающие фрагменты) по настоящему изобретению могут применяться для получения конъюгатов антитела и лекарственного средства. Кроме того, в настоящем изобретении представлены конъюгаты антитела и лекарственного средства, которые характеризуются требуемыми фармакокинетическими характеристиками и другими требуемыми свойствами, и, таким образом, они могут применяться для лечения или предупреждения рака, экспрессирующего CCR7. В настоящем изобретении дополнительно предусмотрены фармацевтические композиции, содержащие конъюгаты антитела и лекарственного средства по настоящему изобретению; а также способы изготовления и применения таких фармацевтических композиций для лечения или предупреждения рака.
Конъюгаты антитела и лекарственного средства
[00108] В настоящем изобретении предусмотрены конъюгаты антитела и лекарственного средства, также называемые иммуноконъюгатами, где антитело, антигенсвязывающий фрагмент или его функциональный эквивалент, которые специфически связываются с CCR7, соединены с фрагментом, представляющим собой лекарственное средство. В одном аспекте антитела, антигенсвязывающие фрагменты или их функциональные эквиваленты по настоящему изобретению соединены посредством ковалентной связи за счет линкера с фрагментом, представляющим собой лекарственное средство, которое представляет собой противораковое средство. Конъюгаты антитела и лекарственного средства по настоящему изобретению могут доставлять эффективную дозу противоракового средства (например, цитотоксического средства) в ткани опухоли, экспрессирующей CCR7, вследствие чего может достигаться большая селективность (и более низкая эффективная доза).
[00109] В одном аспекте настоящего изобретения представлен иммуноконъюгат формулы (I):
Ab-(L-(D)m)n,
где Ab представляет собой CCR7-связывающее антитело, описанное в данном документе;
L представляет собой линкер;
D представляет собой фрагмент, представляющий собой лекарственное средство;
m составляет целое число от 1 до 8; и
n составляет целое число от 1 до 20. В одном варианте осуществления n составляет целое число от 1 до 10, от 2 до 8 или от 2 до 5. В конкретном варианте осуществления n составляет 2, 3 или 4. В некоторых вариантах осуществления m составляет 1; в других вариантах осуществления m составляет 2, 3 или 4.
[00110] Хотя в конкретной молекуле конъюгата отношение лекарственного средства к антителу имеет точное значение (например, n, умноженное на m в формуле (I)), известно, что зачастую значение будет представлять собой среднее значение, когда его применяют для описания образца, содержащего множество молекул, вследствие некоторой степени гетерогенности, обычно сопряженной со стадией конъюгации. Средняя нагрузка для образца иммуноконъюгата обозначается в данном документе как отношение лекарственного средства к антителу или "DAR". В некоторых вариантах осуществления, когда лекарственное средство представляет собой майтанзиноид, его называют "MAR." В некоторых вариантах осуществления DAR составляет от приблизительно 2 до приблизительно 6, и обычно составляет приблизительно 3, 3,5, 4, 4,5, 5, 5,5, 6, 6,5, 7,0, 7,5, 8,0. В некоторых вариантах осуществления по меньшей мере 50% образца по весу составляет соединение, характеризующееся средним DAR плюс или минус 2, и предпочтительно по меньшей мере 50% образца составляет конъюгат, который подразумевает среднее DAR плюс или минус 1. Варианты осуществления охватывают иммуноконъюгаты, у которых DAR составляет приблизительно 3,5, 3,6, 3,7, 3,8 или 3,9. В некоторых вариантах осуществления DAR, составляющее «приблизительно n», означает, что измеренное значение DAR находится в пределах 20% от n.
[00111] Настоящее изобретение также направлено на иммуноконъюгаты, содержащие антитела, фрагменты антител (например, антигенсвязывающие фрагменты) и их функциональные эквиваленты, раскрытые в данном документе, соединенные или конъюгированные с фрагментом, представляющим собой лекарственное средство. В одном варианте осуществления фрагмент, представляющий собой лекарственное средство D, представляет собой фрагмент, представляющий собой майтанзиноидное лекарственное средство, в том числе характеризующиеся структурой:
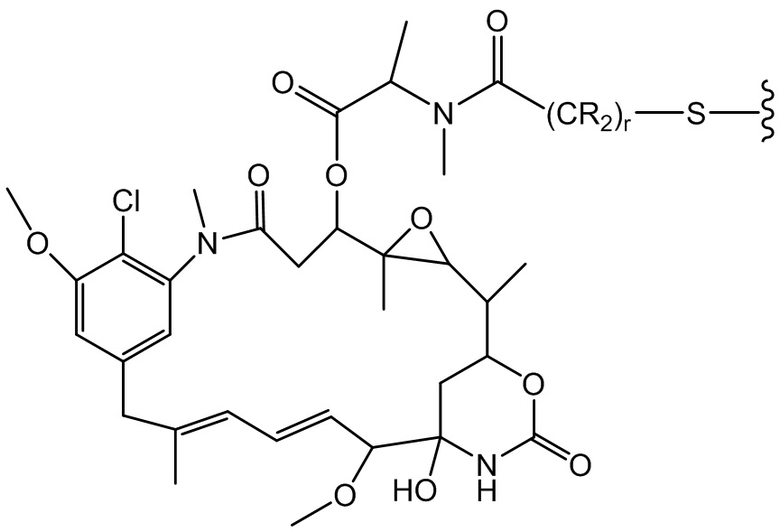
где волнистая линия указывает на ковалентную связь атома серы майтанзиноида с линкером конъюгат антитела и лекарственного средства. R в каждом случае независимо представляет собой H или C1-C6алкил. Алкиленовая цепь, прикрепляющая амидную группу к атому серы, может представлять собой метанил, этанил или пропил, т.е. r составляет 1, 2 или 3. (Патент США № 633410, патент США № 5208020, Chari et al. (1992) Cancer Res. 52;127-131, Lui et al. (1996) Proc. Natl. Acad. Sci. 93:8618-8623).
[00112] Все стереоизомеры фрагмента, представляющего собой майтанзиноидное лекарственное средство, предусмотрены для применения в иммуноконъюгатах по настоящему изобретению, т. е. любая комбинация R- и S-конфигураций по хиральным атомам углерода майтанзиноида. В одном варианте осуществления фрагмент, представляющий собой майтанзиноидное лекарственное средство, характеризуется следующей стереохимией.
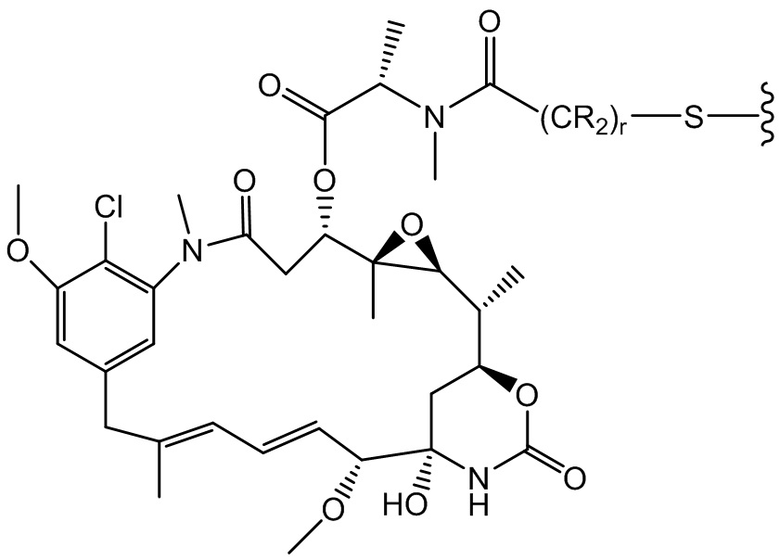
[00113] В одном варианте осуществления фрагмент, представляющий собой майтанзиноидное лекарственное средство, представляет собой N2'-деацетил-N2'-(3-меркапто-1-оксопропил)-майтанзин (также известный как DM1). DM1 представлен следующей структурной формулой.
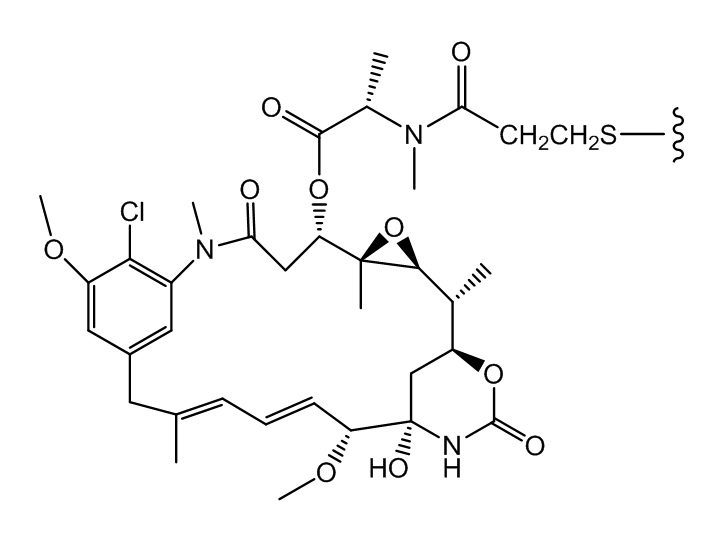
DM1
[00114] В другом варианте осуществления фрагмент, представляющий собой майтанзиноидное лекарственное средство, представляет собой N2'-деацетил-N2'-(4-меркапто-1-оксопентил)-майтанзин (также известный как DM3). DM3 представлен следующей структурной формулой.
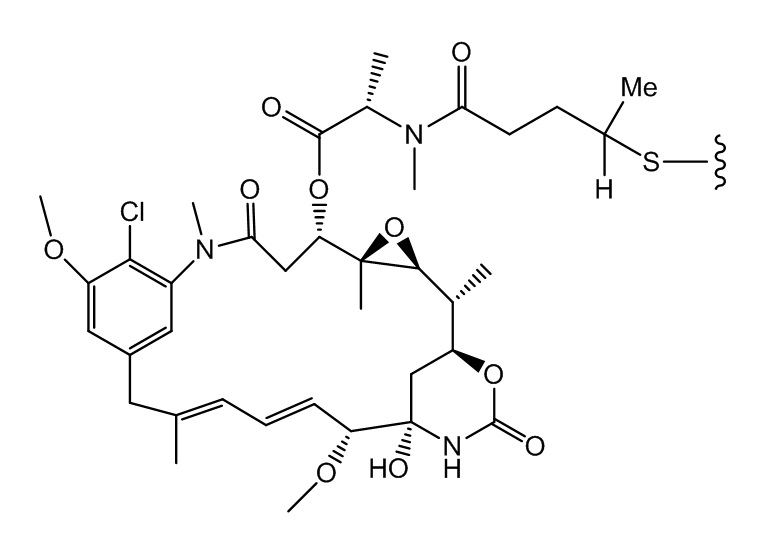
DM3
[00115] В другом варианте осуществления фрагмент, представляющий собой майтанзиноидное лекарственное средство, представляет собой N2'-деацетил-N2'-(4-метил-4-меркапто-1-оксопентил)-майтанзин (также известный как DM4). DM4 представлен следующей структурной формулой.
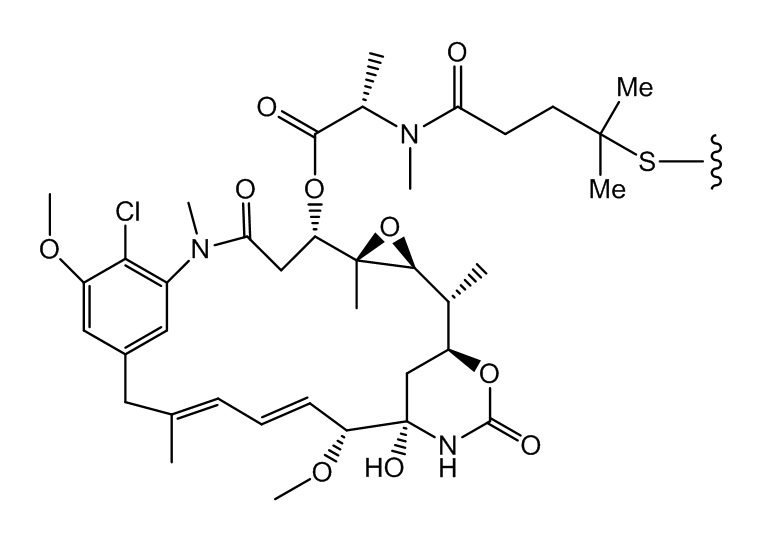
DM4
[00116] Фрагмент, представляющий собой лекарственное средство D, может быть соединен с антителом Ab с помощью линкера L. L представляет собой любой химический фрагмент, способный к соединению фрагмента, представляющего собой лекарственное средство, с антителом за счет ковалентных связей. Сшивающий реагент представляет собой бифункциональный или мультифункциональный реагент, который может использоваться для соединения фрагмента, представляющего собой лекарственное средство, и антитела с образованием конъюгатов антитела и лекарственного средства. Конъюгаты антитела и лекарственного средства могут быть получены с применением сшивающего реагента, имеющего реакционноспособные функциональные группы, способные к связывание как с фрагментом, представляющим собой лекарственное средство, так и с антителом. Например, цистеин, тиол или амин, например, на N-конце или боковой цепи аминокислоты, такой как лизин, в антителе могут образовать связь с функциональной группой сшивающего реагента. В качестве альтернативы конъюгаты антитела и лекарственного средства могут быть получены за счет предварительного образования фрагмента, представляющего собой линкер-лекарственное средство (или фрагмента, представляющего собой лекарственное средство-линкер, оба термина используются взаимозаменяемо), и осуществления реакции фрагмента, представляющего собой линкер-лекарственное средство, с антителом. В некоторых случаях фрагмент, представляющий собой линкер, собирается на лекарственном средстве постепенно с помощью нескольких соединяющих фрагментов до тех пор, пока не получится требуемый фрагмент, представляющий собой линкер-лекарственное средство,.
[00117] В одном варианте осуществления L представляет собой расщепляемый линкер. В другом варианте осуществления L представляет собой нерасщепляемый линкер. В некоторых вариантах осуществления L представляет собой кислотолабильный линкер, фотолабильный линкер, расщепляемый пептидазами линкер, расщепляемый эстеразами линкер, линкер с расщепляемой дисульфидной связью, гидрофильный линкер, предварительно заряженный линкер, расщепляемый гликозидазами линкер, расщепляемый фосфодиэстеразами линкер, расщепляемый фосфатазами линкер или линкер на основе дикарбоновой кислоты.
[00118] Подходящие сшивающие реагенты, которые образуют нерасщепляемый линкер между фрагментом, представляющим собой лекарственное средство, например майтанзиноид, и антителом, широко известны из уровня техники, и они могут образовывать нерасщепляемые линкеры, которые содержат атом серы (такие как SMCC), или линкеры, которые не содержат атом серы. Предпочтительные сшивающие реагенты, которые образуют нерасщепляемые линкеры между фрагментом, представляющим собой лекарственное средство, например майтанзиноид, и антителом, содержат фрагмент на основе малеимида или галогенацетила. В соответствии с настоящим изобретением про такие нерасщепляемые линкеры говорится, что они происходят из фрагментов на основе малеимида или галогенацетила.
[00119] Сшивающие реагенты, содержащие фрагмент на основе малеимида включают без ограничения N-сукцинимидил-4-(малеимидометил)циклогексанкарбоксилат (SMCC), сульфосукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (сульфо-SMCC), N-сукцинимидил-4-(малеимидометил)циклогексан-1-карбокси-(6-амидокапроат), который представляет собой "длинноцепочечный" аналог SMCC (LC-SMCC), N-сукцинимидильный сложный эфир κ-малеимидоундекановой кислоты (KMUA), N-сукцинимидильный сложный эфир γ-малеимидомасляной кислоты (GMBS), N-сукцинимидильный сложный эфир ε-малеимидокапроновой кислоты (EMCS), m-малеимидобензоил-N-гидроксисукцинимидный сложный эфир (MBS), N-(α-малеимидоацетокси)-сукцинимидный сложный эфир (AMSA), сукцинимидил-6-(β-малеимидопропионамидо)гексаноат (SMPH), N-сукцинимидил-4-(p-малеимидофенил)-бутират (SMPB), N-(-p-малеимидофенил)изоцианат (PMIP) и сшивающие реагенты на основе малеимида, содержащие полиэтиленгликольный спейсер, такие как MAL-PEG-NHS. Данные сшивающие реагенты образуют нерасщепляемые линкеры, происходящие из фрагментов на основе малеимида. Типичные структуры сшивающих реагентов на основе малеимида показаны ниже.
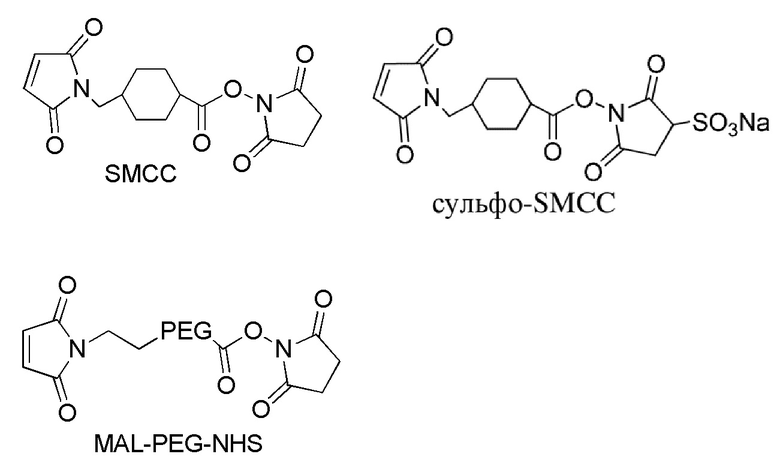
[00120] В другом варианте осуществления линкер L происходит из N-сукцинимидил-4-(малеимидометил)циклогексанкарбоксилата (SMCC), сульфосукцинимидил 4-(N-малеимидометил)циклогексан-1-карбоксилата (сульфо-SMCC) или MAL-PEG-NHS.
[00121] Сшивающие реагенты, содержащие фрагмент на основе галогенацетила, включают N-сукцинимидилйодацетат (SIA), N-сукцинимидил(4-йодацетил)аминобензоат (SIAB), N-сукцинимидилбромацетат (SBA) и N-сукцинимидил-3-(бромацетамидо)пропионат (SBAP). Данные сшивающие реагенты образуют нерасщепляемый линкер, происходящий из фрагментов на основе галогенацетила. Типичные структуры сшивающих реагентов на основе галогенацетила показаны ниже.
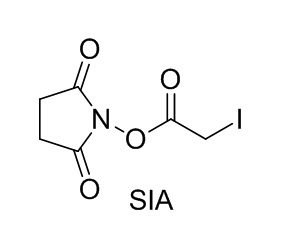 или
или 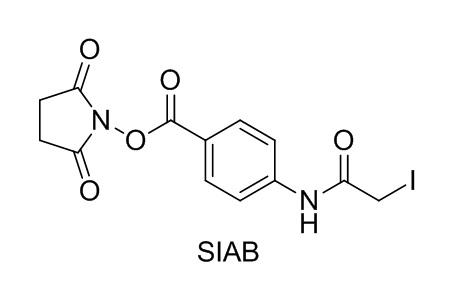
[00122] В одном варианте осуществления линкер L происходит из N-сукцинимидилйодацетата (SIA) или N-сукцинимидил(4-йодацетил)аминобензоата (SIAB).
[00123] Подходящие сшивающие реагенты, которые образуют расщепляемый линкер между фрагментом, представляющим собой лекарственное средство, например майтанзиноид, и антителом, широко известны из уровня техники. Линкеры, содержащие дисульфид, представляют собой линкеры, расщепляемые посредством дисульфидного обмена, который может происходить при физиологических условиях. В соответствии с настоящим изобретением про такие расщепляемые линкеры говорится, что они происходят из фрагментов на основе дисульфида. Подходящие дисульфидные сшивающие реагенты включают N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), N-сукцинимидил-4-(2-пиридилдитио)пентаноат (SPP), N-сукцинимидил-4-(2-пиридилдитио)бутаноат (SPDB) и N-сукцинимидил-4-(2-пиридилдитио)-2-сульфо-бутаноат (сульфо-SPDB), структуры которых показаны ниже. Данные дисульфидные сшивающие реагенты образуют расщепляемый линкер, происходящий из фрагментов на основе дисульфида.
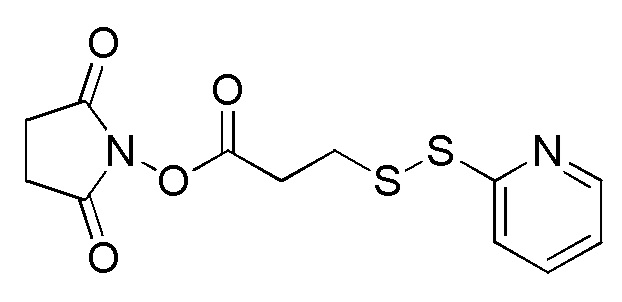
N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP),
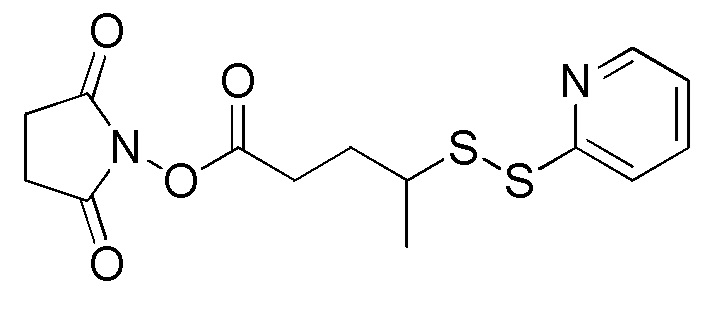
N-сукцинимидил-4-(2-пиридилдитио)пентаноат (SPP),
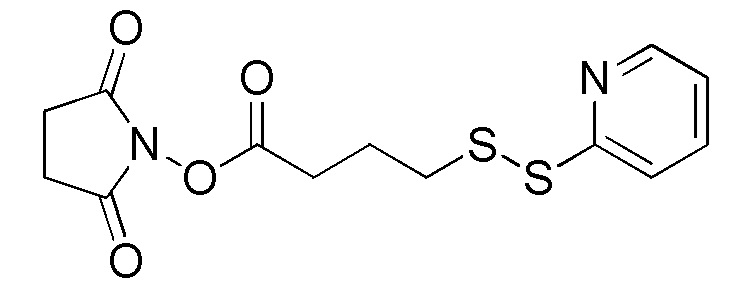
N-сукцинимидил-4-(2-пиридилдитио)бутаноат (SPDB) и
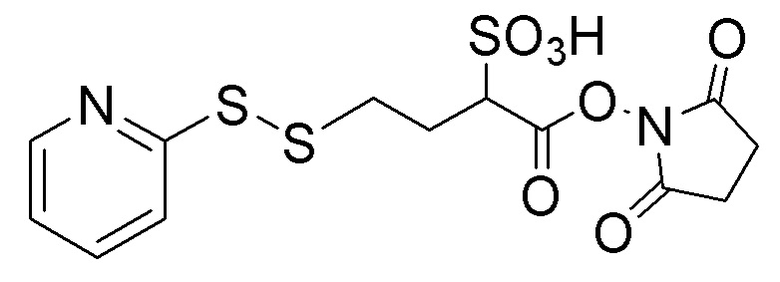
N-сукцинимидил-4-(2-пиридилдитио)-2-сульфо-бутаноат (сульфо-SPDB).
[00124] В одном варианте осуществления линкер L происходит из N-сукцинимидил-4-(2-пиридилдитио)бутаноата (SPDB).
[00125] Подходящие сшивающие реагенты, которые образуют заряженный линкер между фрагментом, представляющим собой лекарственное средство, например майтанзиноид, и антителом, известны как предварительно заряженные сшивающие реагенты. В одном варианте осуществления линкер L происходит из предварительно заряженного сшивающего реагента CX1-1. Структура CX1-1 приведена ниже.
2,5-диоксопирролидин-1-ил-17-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)-5,8,11,14-тетраоксо-4,7,10,13-тетраазагептадекан-1-оат (CX1-1)
[00126] Каждый из сшивающих реагентов, изображенных выше, на одном конце сшивающего реагента содержит NHS-сложный эфир, который реагирует с первичным амином антитела с образованием амидной связи, а на другом конце малеимидную группу или пиридинилдисульфидную группу, которая реагирует с сульфгидрилом фрагмента, представляющего собой майтанзиноидное лекарственное средство, с образованием тиоэфирной или дисульфидной связи.
[00127] В другом варианте осуществления подходящие сшивающие фрагменты, которые образуют расщепляемый линкер между фрагментом, представляющим собой лекарственное средство (например, майтанзиноид), и антителом представлены следующей формулой (II):
 (II);
(II);
где:
L1 представляет собой C1-6алкилен, при этом одна из метиленовых групп может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11; и
X представляет собой -C(O)-NH-, -NHC(O)- или триазол;
где алкилен является линейным или разветвленным.
В одном аспекте данного варианта осуществления y составляет 5, 7, 9 или 11. В другом аспекте данного варианта осуществления y составляет меньше 5.
В еще одном варианте осуществления подходящие сшивающие фрагменты в соответствии с формулой I выбраны из группы, состоящей из:
 ;
;
где y составляет от 1 до 11.
[00128] В случае сшивающих фрагментов, изображенных выше (т. е. MBT, MPET, MEPET; MMTBT, MPPT, MPBT), малеимидная группа обеспечивает возможность реакции с сульфгидрилом (или тиолом) цистеина в антителе, с образованием, тем самым, тиоэфирной связи; а тиольная функциональная группа сшивающего фрагмента соединяется с тиолом фрагмента, представляющего собой майтанзиноидное лекарственное средство, с образованием расщепляемой дисульфидной связи. В силу перекрестно-реакционноспособной природы соединяющего фрагмента формулы (I) (тиол и малеимид могут вступать в перекрестную реакцию), специалист в данной области техники будет осознавать, что соединяющий фрагмент должен собираться постепенно на фрагменте, представляющем собой лекарственное средство, как изображено на схеме 1.
[00129] В соответствии с вышеуказанным вариантом осуществления линкеры, полученные из сшивающих фрагментов (т.е. MBT, MPET, MEPET), могут быть изображены следующим образом:
(IIA);
где * соединен с тиольной функциональной группой на антителе, и ** соединен с тиольной функциональной группой фрагмента, представляющего собой лекарственное средство (например, фрагмента, представляющего собой майтанзиноидное лекарственное средство, DM1, DM3 или DM4).
[00130] В соответствии с вышеуказанным вариантом осуществления линкеры, полученные из сшивающих фрагментов (т.е. MBT, MPET, MEPET), могут быть изображены следующим образом:
где y составляет от 1 до 11; * соединен с тиольной функциональной группой на антителе, и ** соединен с тиольной функциональной группой фрагмента, представляющего собой лекарственное средство, (например, майтанзиноидное лекарственное средство, DM1, DM3 или DM4).
[00131] В предпочтительном варианте осуществления линкер имеет следующую формулу:
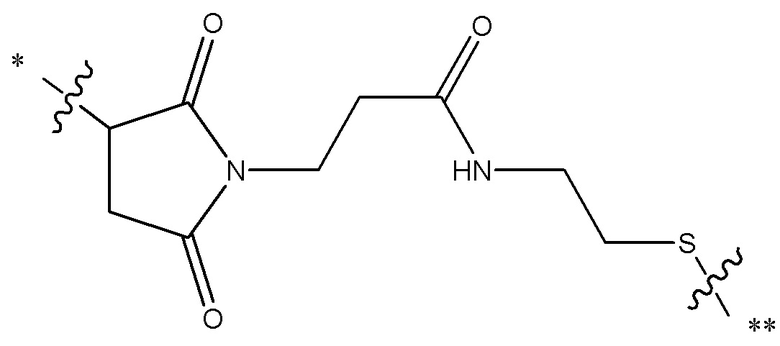 ;
;
где * соединен с тиольной функциональной группой на антителе, и ** соединен с тиольной функциональной группой майтанзиноидного лекарственного средства (DM1, DM3 или DM4).
[00132] В одном варианте осуществления настоящее изобретение относится к фрагменту, представляющему собой линкер-лекарственное средство, с формулой:
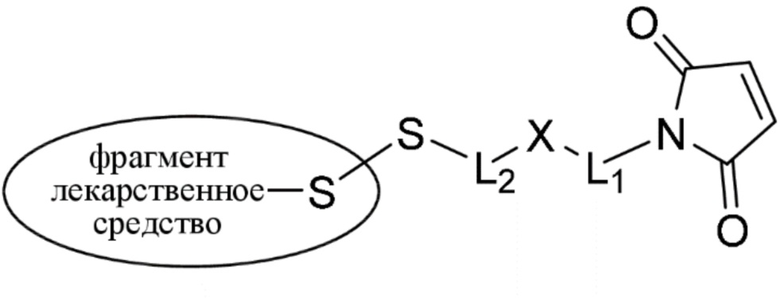 ; где
; где
L1 представляет собой C1-6алкилен, при этом одна из метиленовых групп может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11; и
X представляет собой -C(O)-NH-, -NHC(O)- или триазол;
где алкилен является линейным или разветвленным.
[00133] В другом варианте осуществления настоящее изобретение связано с постепенным образованием вышеуказанного конъюгата линкера и лекарственного средства, раскрытым на схеме 1 в данном документе.
[00134] В одном варианте осуществления настоящее изобретение относится к соединениям фрагмента, представляющего собой линкер-лекарственное средство, имеющим одну из следующих формул (III), (IV) и (V):
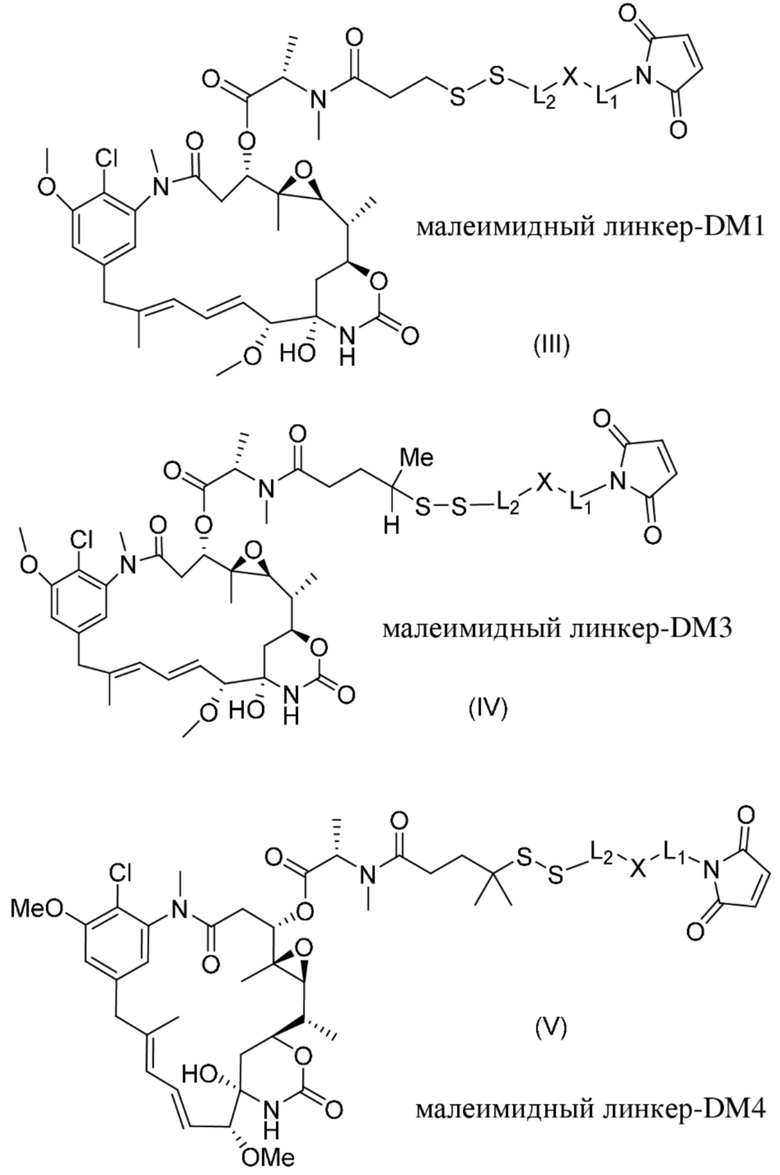
где L1 представляет собой C1-6алкилен, при этом одна метиленовая группа может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11; и
X представляет собой -C(O)-NH-, -NHC(O)- или триазол;
где алкилен является линейным или разветвленным.
[00135] В одном варианте осуществления настоящее изобретение относится к соединениям фрагмента, представляющего собой линкер-лекарственное средство, которые выбраны из следующих формул:
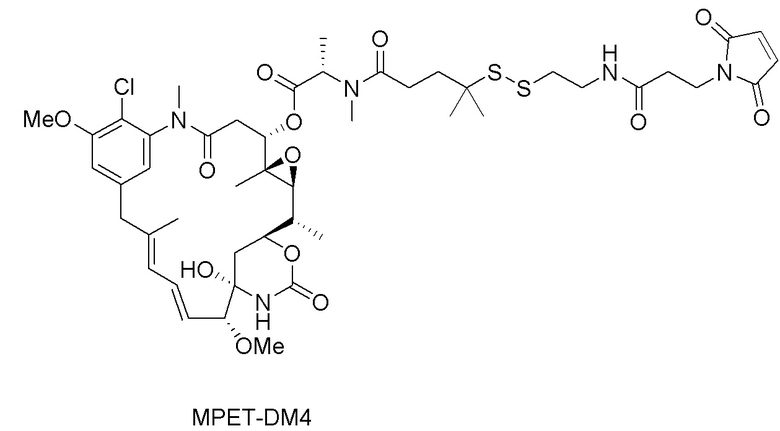 ;
;
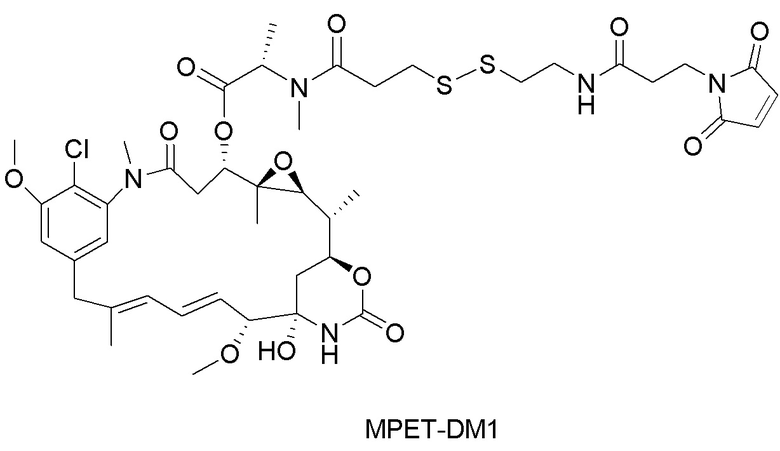 ;
;
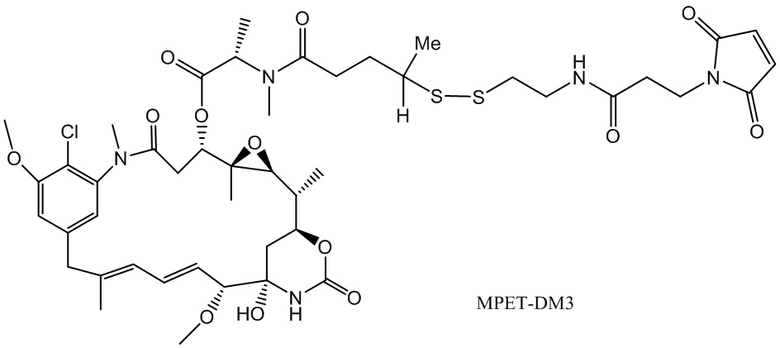 и
и
 .
.
[00136] В другом варианте осуществления настоящее изобретение относится к соединениям фрагмента, представляющего собой линкер-лекарственное средство, которые выбраны из следующих формул:
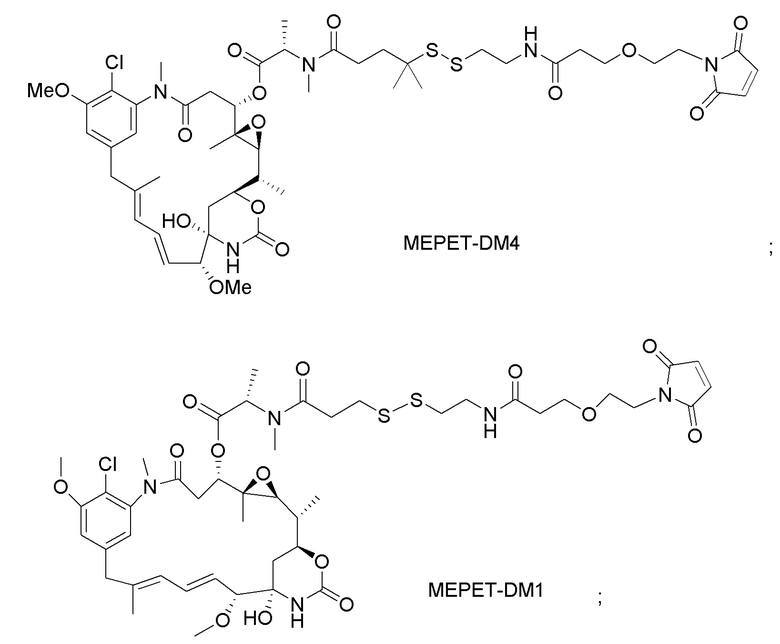
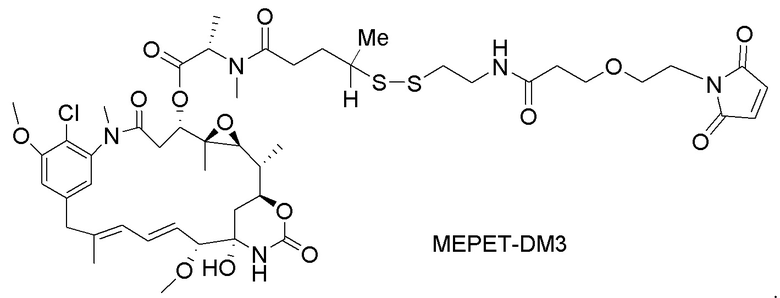
[00137] В другом варианте осуществления линкер-лекарственное средство по настоящему изобретению представлены любой из следующих формул:
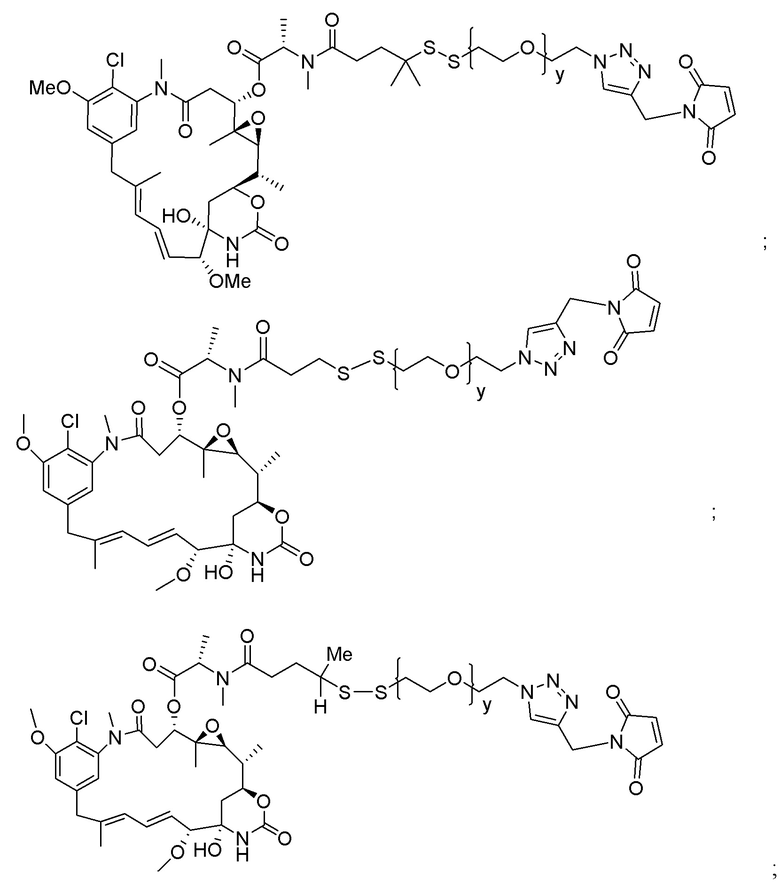 где y составляет от 1 до 11, предпочтительно от 1 до 5.
где y составляет от 1 до 11, предпочтительно от 1 до 5.
[00138] В одном варианте осуществления линкер-лекарственное средство по настоящему изобретению представлены любой из следующих структурных формул:
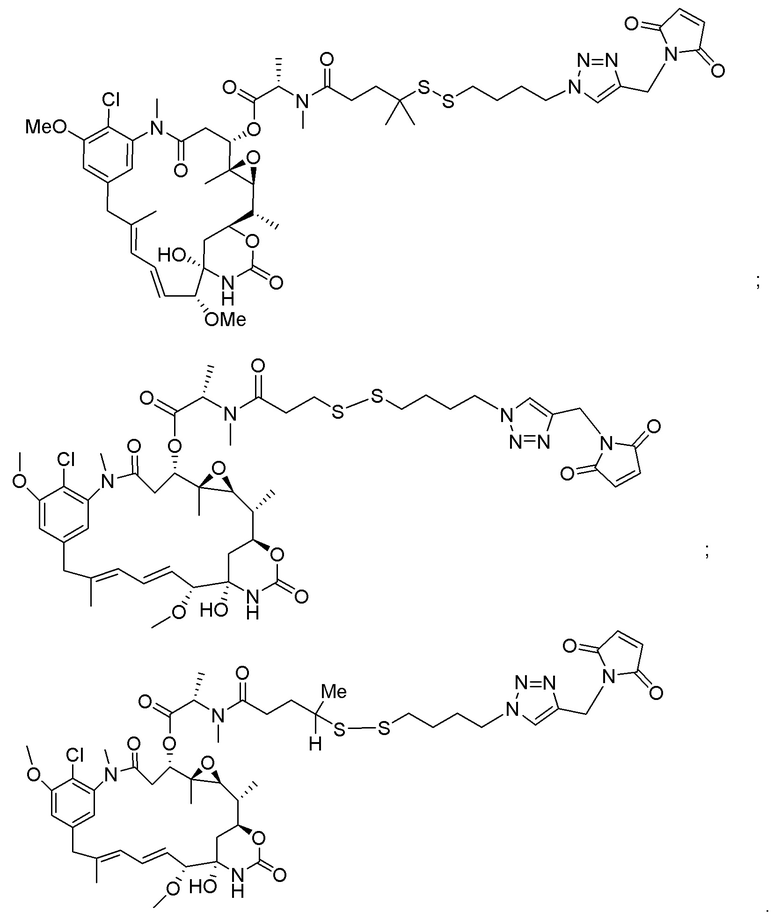
[00139] В одном варианте осуществления конъюгат по настоящему изобретению представлен любой из следующих структурных формул:
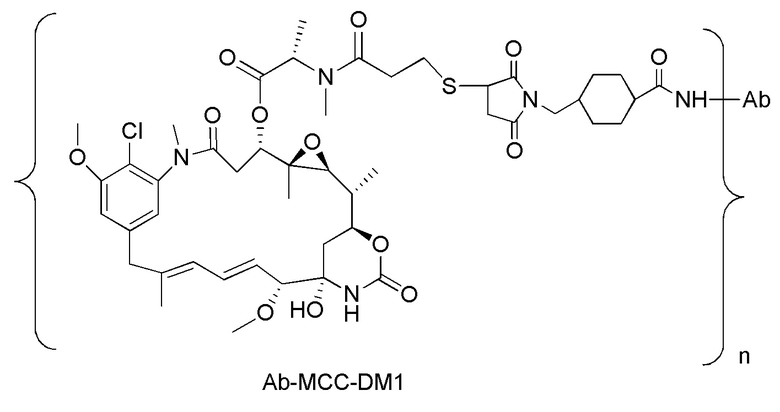
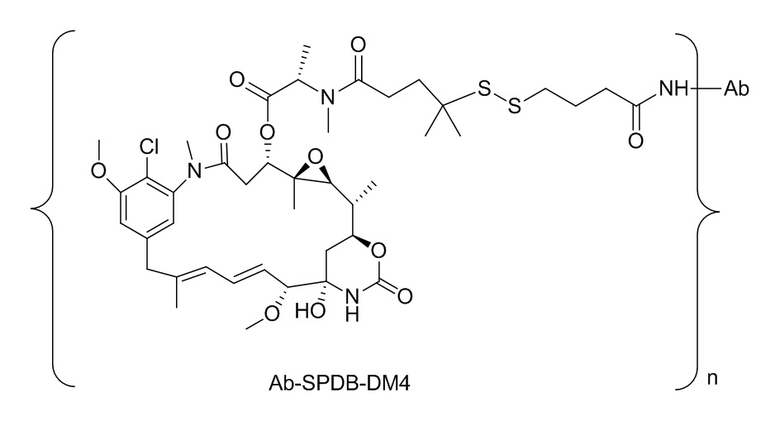 ,
,
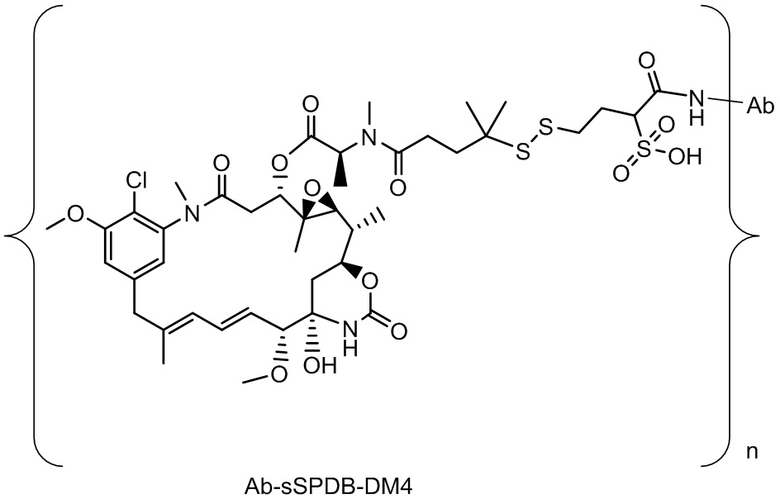
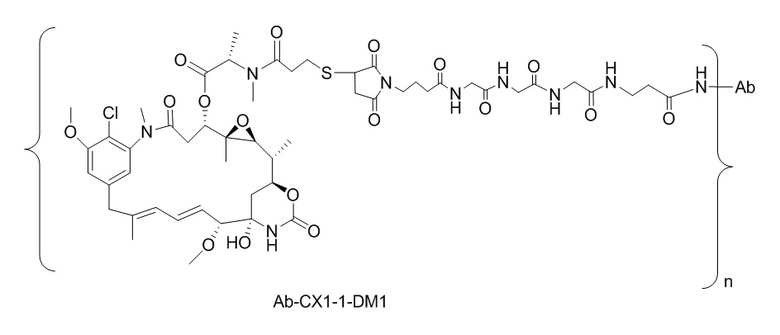
где:
Ab представляет собой антитело или его антигенсвязывающий фрагмент, которые специфически связывают CCR7;
n, который указывает на число групп линкер-лекарственное средство (L-D-), связанных с Ab за счет образования амидной связи с первичным амином на Ab, составляет целое число от 1 до 20. В одном варианте осуществления n составляет целое число от 1 до 10, от 2 до 8 или от 2 до 5. В конкретном варианте осуществления n составляет 3 или 4.
[00140] В другом варианте осуществления конъюгат по настоящему изобретению представлен любой из следующих формул (VI), (VII) и (VIII):
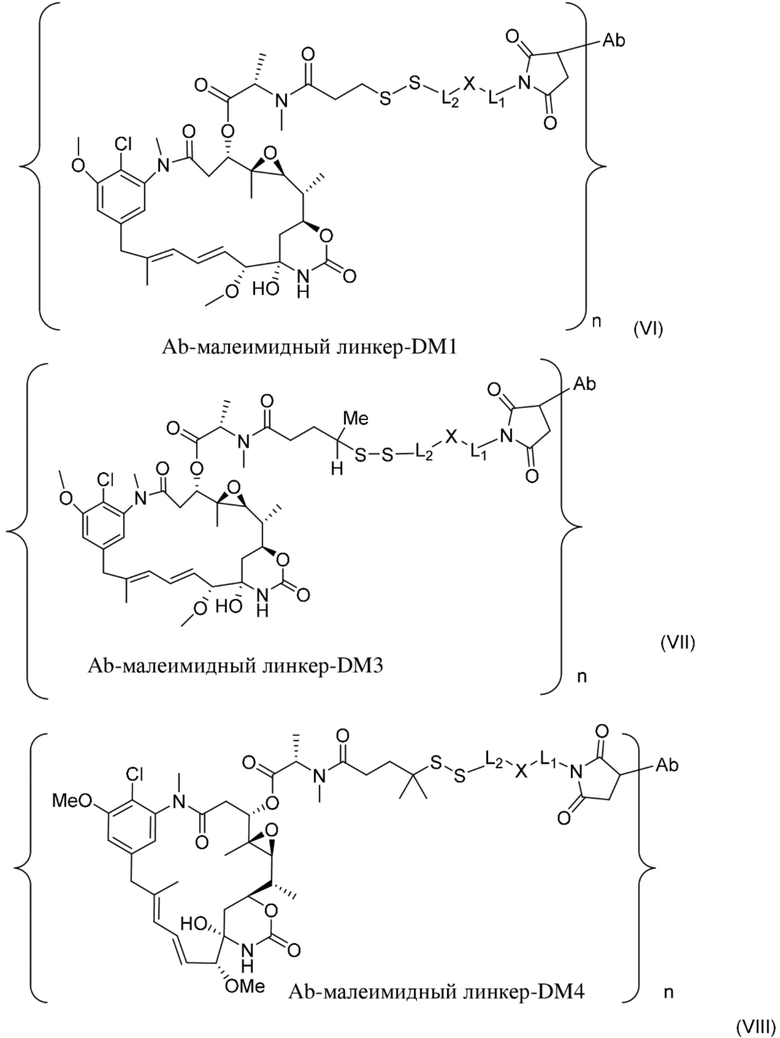 ;
;
где:
L1 представляет собой C1-6алкилен, при этом одна из метиленовых групп может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11; и
X представляет собой -C(O)-NH-, -NHC(O)- или триазол;
где алкилен является линейным или разветвленным; и
Ab представляет собой антитело или его антигенсвязывающий фрагмент;
n, который указывает на число групп линкер-лекарственное средство (L-D-), связанных с Ab за счет образования амидной связи с первичным амином на Ab, составляет целое число от 1 до 20. В одном варианте осуществления n составляет целое число от 1 до 10, от 2 до 8 или от 2 до 5. В конкретном варианте осуществления n составляет 3 или 4. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связываются с антигеном, который экспрессируется на опухолевых клетках. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент специфически связывают CCR7. В других вариантах осуществления антитело или его антигенсвязывающий фрагмент специфически связывают P-кадгерин, кадгерин 6, FGFR2 или FGFR4.
[00141] В другом варианте осуществления конъюгат по настоящему изобретению имеет формулу (VIA) или (VIB), соответствующую открытым формам сукцинимида конъюгата формулы (VI):

где:
L1 представляет собой C1-6алкилен, при этом одна из метиленовых групп может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11; и
X представляет собой -C(O)-NH-, -NHC(O)- или триазол;
где алкилен является линейным или разветвленным; и
Ab представляет собой антитело или антигенсвязывающий фрагмент;
n, который указывает на число групп линкер-лекарственное средство (L-D-), связанных с Ab за счет образования амидной связи с первичным амином на Ab, составляет целое число от 1 до 20. В одном варианте осуществления n составляет целое число от 1 до 10, от 2 до 8 или от 2 до 5. В конкретном варианте осуществления n составляет 3 или 4. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связываются с антигеном, который экспрессируется на опухолевых клетках. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент специфически связывают CCR7. В других вариантах осуществления антитело или его антигенсвязывающий фрагмент специфически связывают P-кадгерин, кадгерин 6, FGFR2 или FGFR4.
[00142] В другом варианте осуществления конъюгат по настоящему изобретению имеет формулу (VIIA) или (VIIB), соответствующую открытым формам сукцинимида конъюгата формулы (VII):
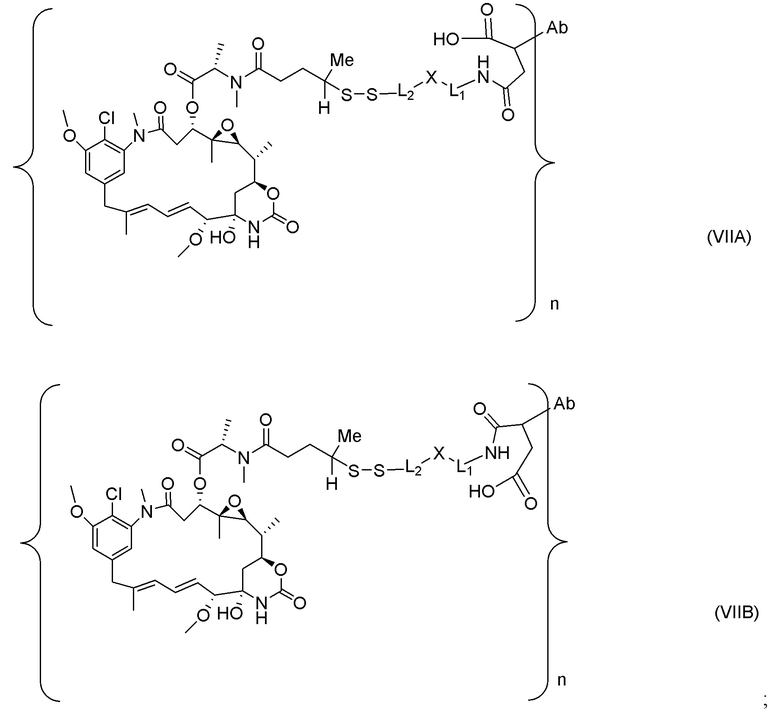
где:
L1 представляет собой C1-6алкилен, при этом одна из метиленовых групп может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11; и
X представляет собой -C(O)-NH-, -NHC(O)- или триазол;
где алкилен является линейным или разветвленным; и
Ab представляет собой антитело или его антигенсвязывающий фрагмент;
n, который указывает на число групп линкер-лекарственное средство (L-D), связанных с Ab за счет образования амидной связи с первичным амином на Ab, составляет целое число от 1 до 20. В одном варианте осуществления n составляет целое число от 1 до 10, от 2 до 8 или от 2 до 5. В конкретном варианте осуществления n составляет 3 или 4. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связываются с антигеном, который экспрессируется на опухолевых клетках. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент специфически связывают CCR7. В других вариантах осуществления антитело или его антигенсвязывающий фрагмент специфически связывают P-кадгерин, кадгерин 6, FGFR2 или FGFR4.
[00143] В другом варианте осуществления конъюгат по настоящему изобретению имеет формулу (VIIIA), (VIIIB), соответствующую открытым формам сукцинимида конъюгата формулы (VIII):
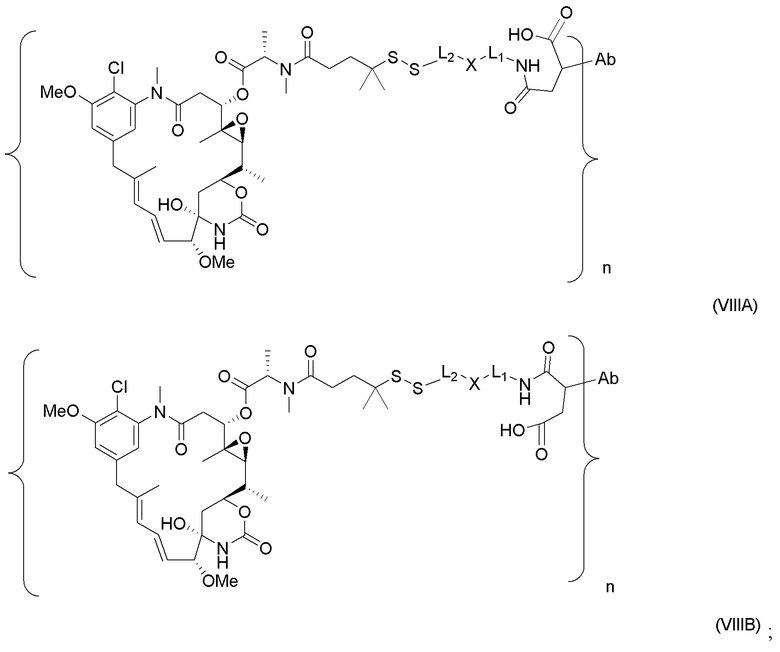
где:
L1 представляет собой C1-6алкилен, при этом одна из метиленовых групп может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11; и
X представляет собой -C(O)-NH-, -NHC(O)- или триазол;
где алкилен является линейным или разветвленным; и
Ab представляет собой антитело или его антигенсвязывающий фрагмент;
n, который указывает на число групп линкер-лекарственное средство (L-D-), связанных с Ab за счет образования амидной связи с первичным амином на Ab, составляет целое число от 1 до 20. В одном варианте осуществления n составляет целое число от 1 до 10, от 2 до 8 или от 2 до 5. В конкретном варианте осуществления n составляет 3 или 4. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связываются с антигеном, который экспрессируется на опухолевых клетках. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент специфически связывают CCR7. В других вариантах осуществления антитело или его антигенсвязывающий фрагмент специфически связывают P-кадгерин, кадгерин 6, FGFR2 или FGFR4.
[00144] В одном варианте осуществления каждый конъюгат антитела и лекарственного средства, раскрытый в данном документе, где фрагмент, представляющий собой линкер-лекарственное средство, присоединен к антителу с помощью сукцинимида, также может существовать в виде открытых форм сукцинимида, как в общем изображено в формулах (VIA), (VIB), (VIIA), (VIIB), (VIIIA) и (VIIIB).
[00145] В еще одном варианте осуществления конъюгат по настоящему изобретению представлен любой из следующих структурных формул:
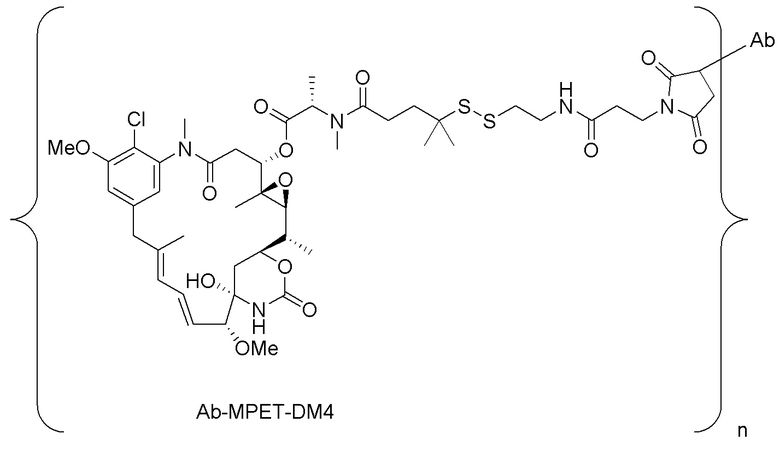
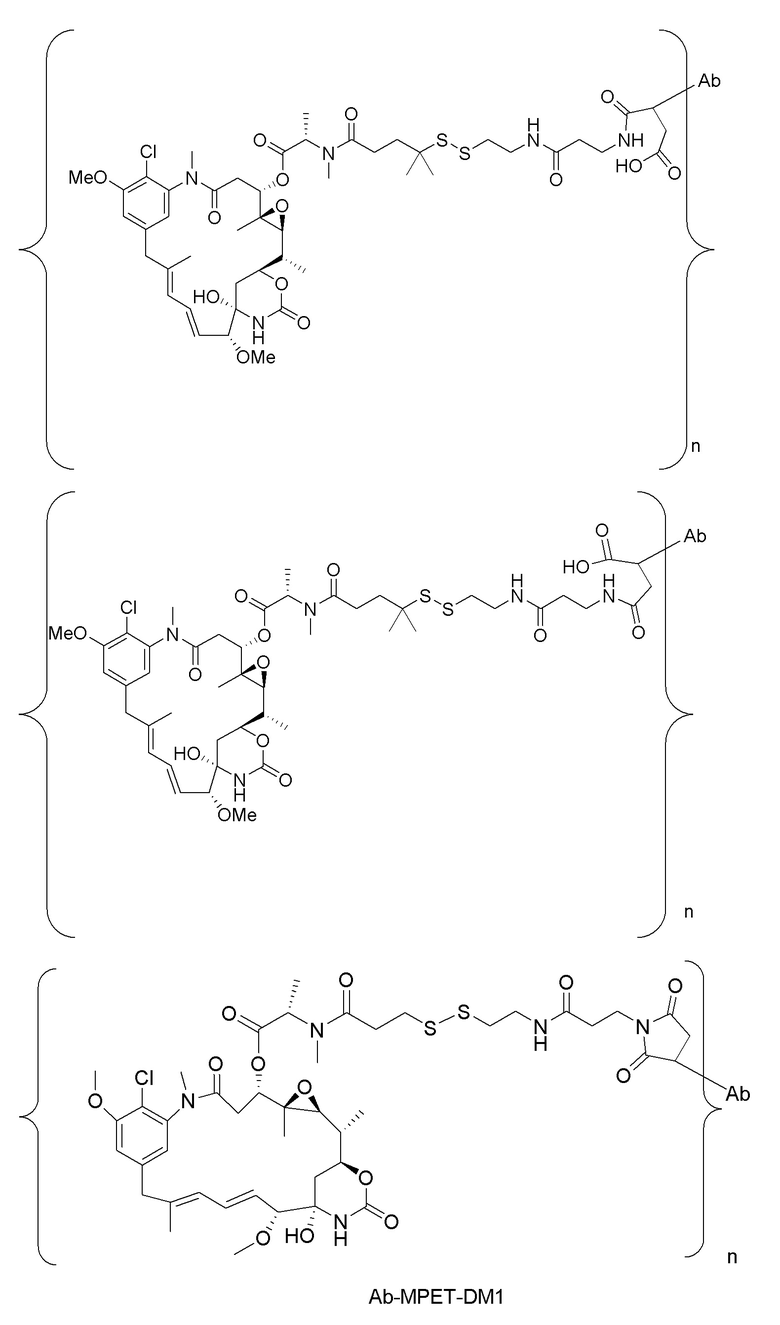
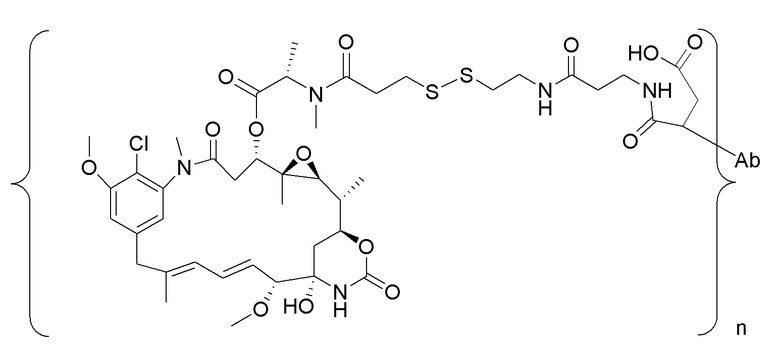
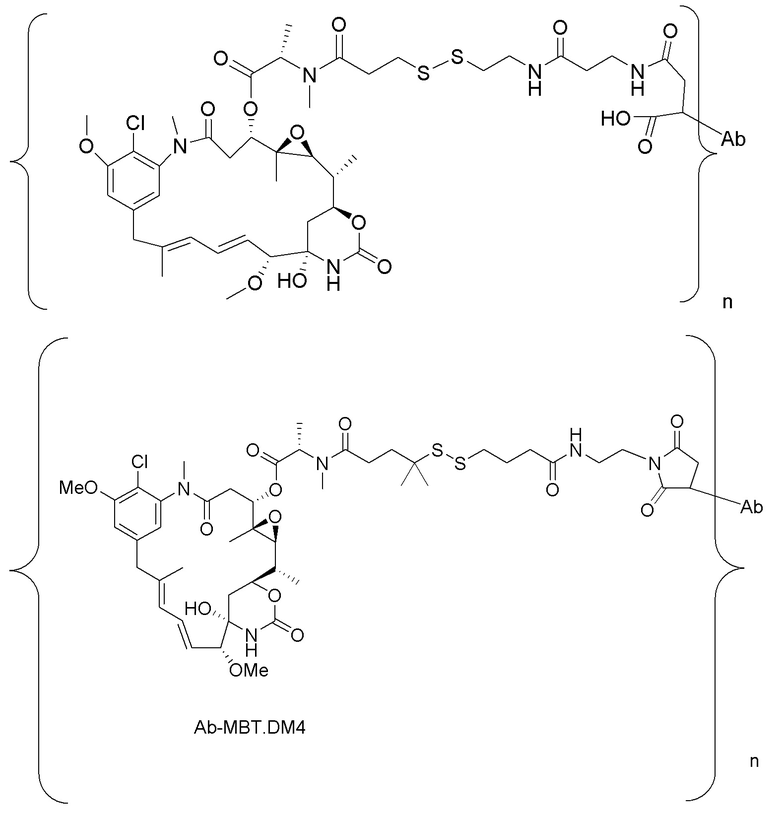
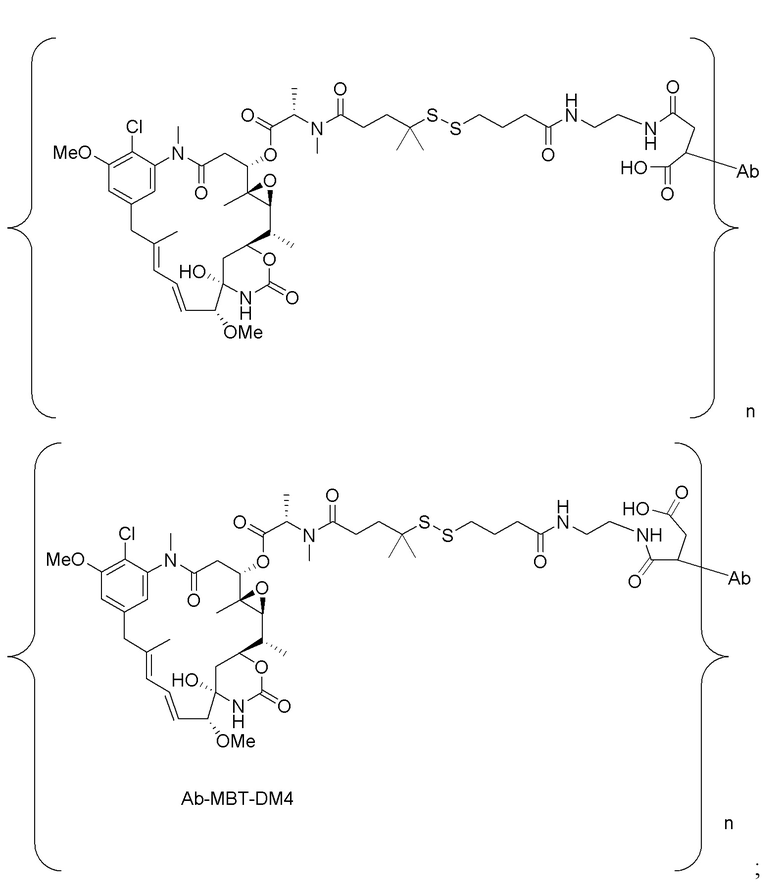
где:
Ab представляет собой антитело или его антигенсвязывающий фрагмент;
n, который указывает на число групп D-L, связанных с Ab за счет образования тиоэфирной связи с сульфгидрилом Ab, составляет целое число от 1 до 12, или от 1 до 8, или предпочтительно от 1 до 4. В конкретном варианте осуществления n составляет 3 или 4. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связываются с антигеном, который экспрессируется на опухолевых клетках. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент специфически связывают CCR7. В других вариантах осуществления антитело или его антигенсвязывающий фрагмент специфически связывают P-кадгерин, кадгерин 6, FGFR2 или FGFR4.
[00146] В другом варианте осуществления конъюгат по настоящему изобретению представлен любой из следующих структурных формул:
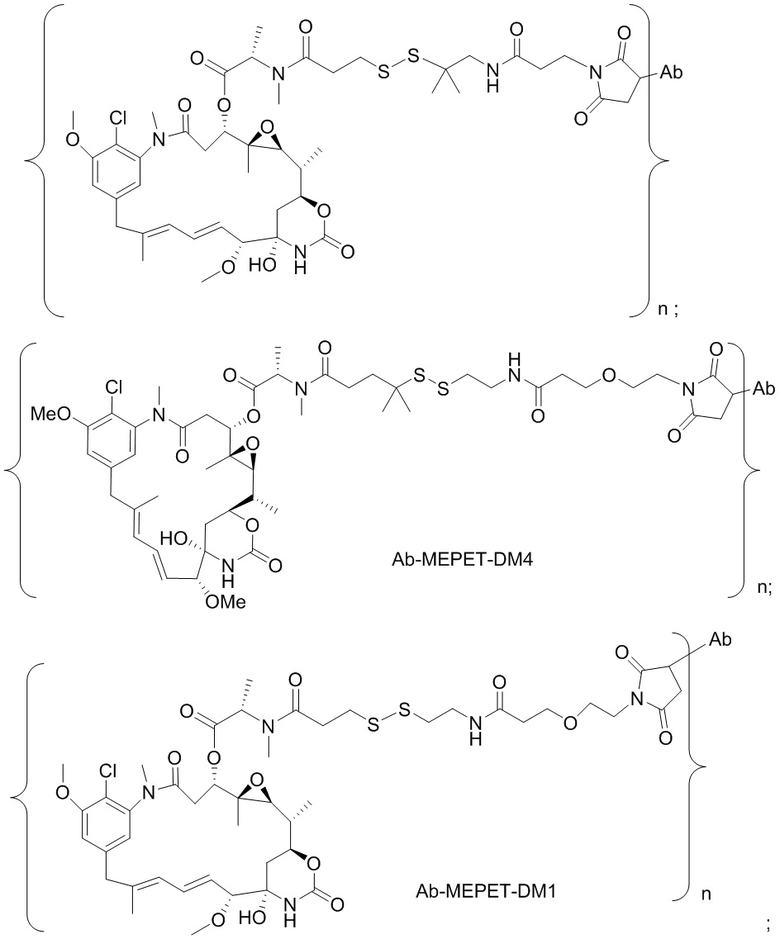 , и
, и
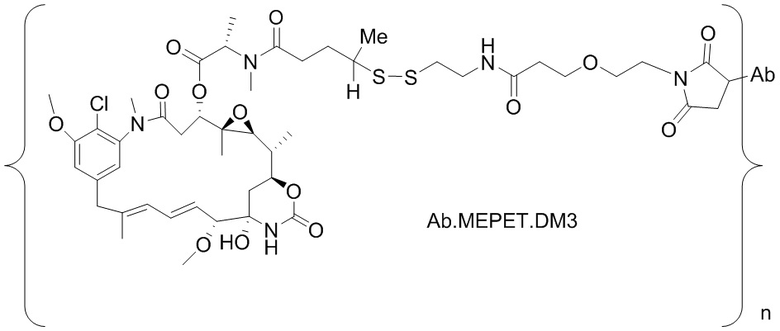 ;
;
а также соответствующими открытыми формами сукцинимида;
где:
Ab представляет собой антитело или его антигенсвязывающий фрагмент;
n, который указывает на число групп D-L, связанных с Ab за счет образования тиоэфирной связи с сульфгидрилом Ab, составляет целое число от 1 до 12, или от 1 до 8, или предпочтительно от 1 до 4. В конкретном варианте осуществления n составляет 3 или 4. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связываются с антигеном, который экспрессируется на опухолевых клетках. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент специфически связывают CCR7. В других вариантах осуществления антитело или его антигенсвязывающий фрагмент специфически связывают P-кадгерин, кадгерин 6, FGFR2 или FGFR4.
[00147] В другом варианте осуществления конъюгат по настоящему изобретению представлен любой из следующих структурных формул:
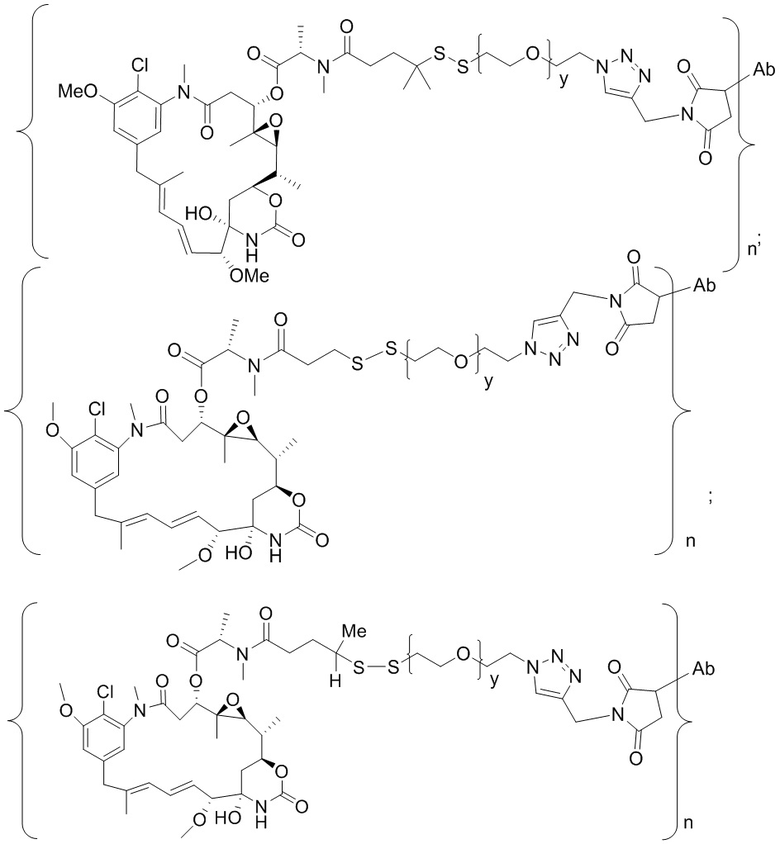 ;
;
а также соответствующими открытыми формами сукцинимида;
где:
y составляет от 1 до 11, предпочтительно от 1 до 5;
Ab представляет собой антитело или его антигенсвязывающий фрагмент;
n, который указывает на число групп D-L, связанных с Ab за счет образования тиоэфирной связи с сульфгидрилом Ab, составляет целое число от 1 до 12, или от 1 до 8, или предпочтительно от 1 до 4. В конкретном варианте осуществления n составляет 3 или 4. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связываются с антигеном, который экспрессируется на опухолевых клетках. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент специфически связывают CCR7. В других вариантах осуществления антитело или его антигенсвязывающий фрагмент специфически связывают P-кадгерин, кадгерин 6, FGFR2 или FGFR4.
[00148] В еще одном варианте осуществления конъюгат по настоящему изобретению представлен следующими формулами:
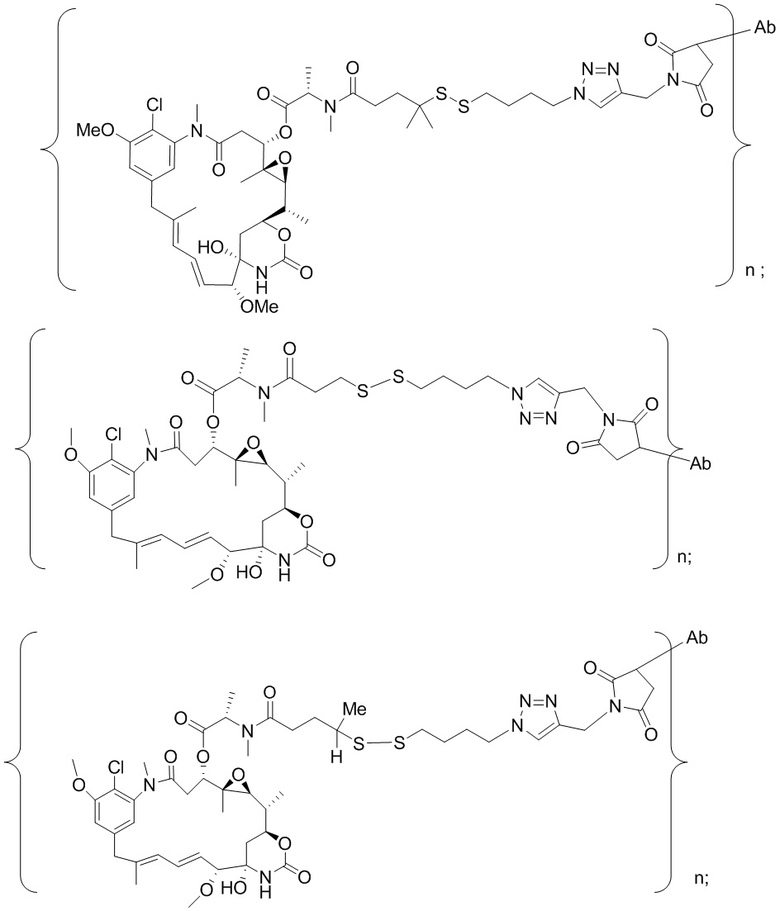
а также соответствующими открытыми формами сукцинимида;
где:
Ab представляет собой антитело или его антигенсвязывающий фрагмент;
n, который указывает на число групп D-L, связанных с Ab за счет образования тиоэфирной связи с сульфгидрилом Ab, составляет целое число от 1 до 12, или от 1 до 8, или предпочтительно от 1 до 4. В конкретном варианте осуществления n составляет 3 или 4. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связываются с антигеном, который экспрессируется на опухолевых клетках. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент специфически связывают CCR7. В других вариантах осуществления антитело или его антигенсвязывающий фрагмент специфически связывают P-кадгерин, кадгерин 6, FGFR2 или FGFR4.
[00149] В предпочтительном варианте осуществления конъюгат по настоящему изобретению представлен следующей формулой:
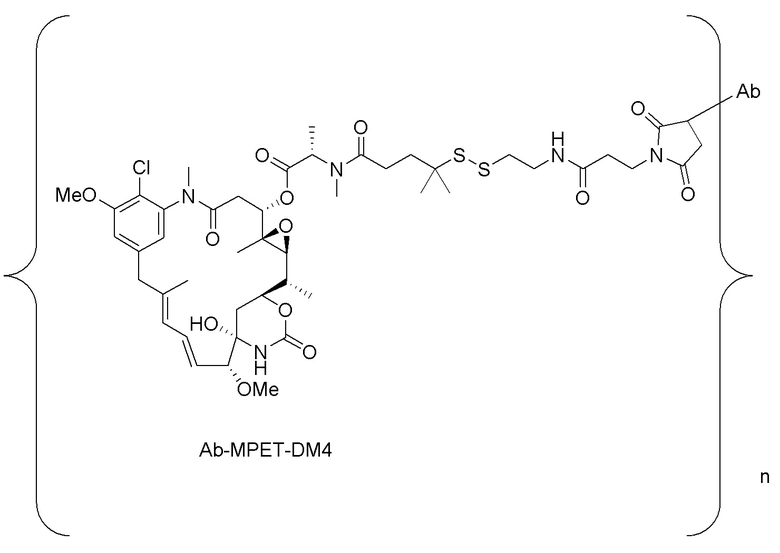
где:
Ab представляет собой антитело или его антигенсвязывающий фрагмент;
n, который указывает на число групп D-L, связанных с Ab за счет образования тиоэфирной связи с сульфгидрилом Ab, составляет целое число от 1 до 20. В некоторых вариантах осуществления n составляет целое число от 1 до 12. В некоторых вариантах осуществления n составляет целое число от 1 до 8. В некоторых вариантах осуществления n составляет целое число от 1 до 4. В конкретном варианте осуществления n составляет 3 или 4. В другом варианте осуществления среднее значение n составляет от приблизительно 3 до приблизительно 4. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связываются с антигеном, который экспрессируется на опухолевых клетках. В одном варианте осуществления антитело или его антигенсвязывающий фрагмент специфически связывают CCR7. В других вариантах осуществления антитело или его антигенсвязывающий фрагмент специфически связывают P-кадгерин, кадгерин 6, FGFR2 или FGFR4.
[00150] В одном варианте осуществления среднее молярное отношение лекарственного средства (например, DM1, DM3 или DM4) к антителу в конъюгате (т. е. среднее значение n, также известное как отношение майтанзиноида к антителу (MAR)) составляет от приблизительно 1 до приблизительно 10, от приблизительно 2 до приблизительно 8 (например, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4, 2,5, 2,6, 2,7, 2,8, 2,9, 3,0, 3,1, 3,2, 3,3, 3,4, 3,5, 3,6, 3,7, 3,8, 3,9, 4,0, 4,1, 4,2, 4,3, 4,4, 4,5, 4,6, 4,7, 4,8, 4,9, 5,0, 5,1, 5,2, 5,3, 5,4, 5,5, 5,6, 5,7, 5,8, 5,9, 6,0, 6,1, 6,2, 6,3, 6,4, 6,5, 6,6, 6,7, 6,8, 6,9, 7,0, 7,1, 7,2, 7,3, 7,4, 7,5, 7,6, 7,7, 7,8, 7,9, 8,0, или 8,1), от приблизительно 2,5 до приблизительно 7, от приблизительно 3 до приблизительно 5, от приблизительно 2,5 до приблизительно 4,5 (например, приблизительно 2,5, приблизительно 2,6, приблизительно 2,7, приблизительно 2,8, приблизительно 2,9, приблизительно 3,0, приблизительно 3,1, приблизительно 3,3, приблизительно 3,4, приблизительно 3,5, приблизительно 3,6, приблизительно 3,7, приблизительно 3,8, приблизительно 3,9, приблизительно 4,0, приблизительно 4,1, приблизительно 4,2, приблизительно 4,3, приблизительно 4,4, приблизительно 4,5), от приблизительно 3,0 до приблизительно 4,0, от приблизительно 3,2 до приблизительно 4,2 или от приблизительно 4,5 до 5,5 (например, приблизительно 4,5, приблизительно 4,6, приблизительно 4,7, приблизительно 4,8, приблизительно 4,9, приблизительно 5,0, приблизительно 5,1, приблизительно 5,2, приблизительно 5,3, приблизительно 5,4 или приблизительно 5,5).
[00151] В одном аспекте настоящего изобретения конъюгат по настоящему изобретению характеризуется в значительной степени высокой чистотой и имеет один или несколько из следующих признаков: (a) более приблизительно 90% (например, больше или равно приблизительно 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%), предпочтительно более приблизительно 95% разновидностей конъюгата являются мономерными, (b) уровень неконъюгированного линкера в препарате конъюгата составляет менее приблизительно 10% (например, меньше или равно приблизительно 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1% или 0%) (относительно общего количества линкера), (c) менее 10% разновидностей конъюгата являются сшитыми (например, меньше или равно приблизительно 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1% или 0%), (d) уровень свободного лекарственного средства (например, DM1, DM3 или DM4) в препарате конъюгата составляет менее приблизительно 2% (например, меньше или равно приблизительно 1,5%, 1,4%, 1,3%, 1,2%, 1,1%, 1,0%, 0,9%, 0,8%, 0,7%, 0,6%, 0,5%, 0,4%, 0,3%, 0,2%, 0,1%, или 0%) (моль/моль относительно общего количества цитотоксического средства); и/или (e) не происходит значительное повышение уровня свободного лекарственного средства (например, DM1, DM3 или DM4) после хранения (например, после приблизительно 1 недели, приблизительно 2 недель, приблизительно 3 недель, приблизительно 1 месяца, приблизительно 2 месяцев, приблизительно 3 месяцев, приблизительно 4 месяцев, приблизительно 5 месяцев, приблизительно 6 месяцев, приблизительно 1 года, приблизительно 2 лет, приблизительно 3 лет, приблизительно 4 лет или приблизительно 5 лет). "Значительное повышение" уровня свободного лекарственного средства (например, DM1, DM3 или DM4) означает, что после некоторого времени хранения (например, приблизительно 1 недели, приблизительно 2 недель, приблизительно 3 недель, приблизительно 1 месяца, приблизительно 2 месяцев, приблизительно 3 месяцев, приблизительно 4 месяцев, приблизительно 5 месяцев, приблизительно 6 месяцев, приблизительно 1 года, приблизительно 2 лет, приблизительно 3 лет, приблизительно 4 лет или приблизительно 5 лет) повышение уровня свободного лекарственного средства (например, DM1, DM3 или DM4) составляет менее приблизительно 0,1%, приблизительно 0,2%, приблизительно 0,3%, приблизительно 0,4%, приблизительно 0,5%, приблизительно 0,6%, приблизительно 0,7%, приблизительно 0,8%, приблизительно 0,9%, приблизительно 1,0%, приблизительно 1,1%, приблизительно 1,2%, приблизительно 1,3%, приблизительно 1,4%, приблизительно 1,5%, приблизительно 1,6%, приблизительно 1,7%, приблизительно 1,8%, приблизительно 1,9%, приблизительно 2,0%, приблизительно 2,2%, приблизительно 2,5%, приблизительно 2,7%, приблизительно 3,0%, приблизительно 3,2%, приблизительно 3,5%, приблизительно 3,7% или приблизительно 4,0%.
[00152] Применяемый в данном документе термин "неконъюгированный линкер" относится к антителу, которое ковалентно связано с линкером, происходящим из сшивающего реагента (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1), где антитело не присоединено ковалентной связью к лекарственному средству (например, DM1, DM3 или DM4) за счет линкера (т.е. "неконъюгированный линкер" может быть представлен как Ab-MCC, Ab-SPDB или Ab-CX1-1).
1. Фрагмент, представляющий собой лекарственное средство
[00153] В настоящем изобретении представлены иммуноконъюгаты, которые специфически связываются с CCR7. Конъюгаты антитела и лекарственного средства по настоящему изобретению содержат антитела к CCR7, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты, которые конъюгированы с фрагментом, представляющим собой лекарственное средство, например, противораковым средством, средством лечения аутоиммунных заболеваний, противовоспалительным средством, противогрибковым средством, антибактериальным средством, антипаразитическим средством, противовирусным средством или анестетическим средством. Антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению могут быть конъюгированы с несколькими идентичными или различными фрагментами, представляющими собой лекарственное средство, с применением любых способов, известных из уровня техники.
[00154] В некоторых вариантах осуществления фрагмент, представляющий собой лекарственное средство, из иммуноконъюгатов по настоящему изобретению, выбран из группы, состоящей из ингибитора V-АТФазы, проапоптического средства, ингибитора Bcl2, ингибитора MCL1, ингибитора HSP90, ингибитора IAP, ингибитора mTor, стабилизатора микротрубочек, дестабилизатора микротрубочек, ингибитора РНК-полимеразы, аманитина, пирролобензодиазепина, ауристатина, доластатина, майтанзиноида, MetAP (метионинаминопептидазы), ингибитора ядерного экспорта белков CRM1, ингибитора DPPIV, ингибитора Eg5, ингибиторов протеасомы, ингибитора реакций переноса фосфорила в митохондриях, ингибитора синтеза белка, ингибитора киназ, ингибитора CDK2, ингибитора CDK9, ингибитора кинезина, ингибитора HDAC, ДНК-повреждающего средства, ДНК-алкилирующего средства, ДНК-интеркалятора, средства, связывающего малую бороздку ДНК, и ингибитора DHFR.
[00155] В определенном варианте осуществления фрагмент, представляющий собой лекарственное средство, из иммуноконъюгатов по настоящему изобретению представляет собой ауристатин, раскрытый в PCT-публикациях под номерами: WO 2015/095301 и WO2015/189791, при этом обе заявки тем самым включены посредством ссылки. Неограничивающие примеры конструкций из фрагмента, представляющего собой ауристатиновое лекарственное средство, и линкера представляют собой: 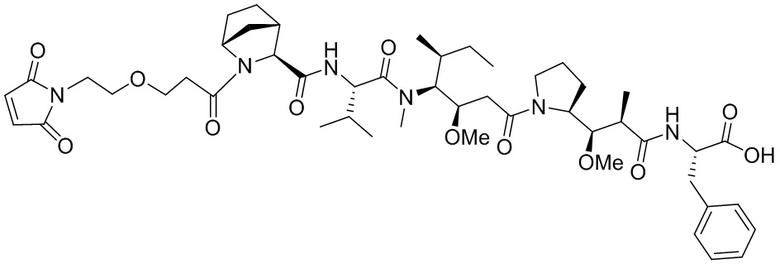
(которая представляет собой AURIX2, как раскрыто в настоящей заявке) и
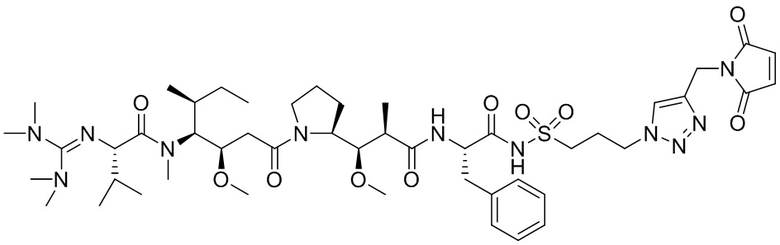
которая представляет собой AURIX1, как раскрыто в настоящей заявке).
[00156] В одном варианте осуществления фрагмент, представляющий собой лекарственное средство, из иммуноконъюгатов по настоящему изобретению представляет собой фрагмент, представляющий собой майтанзиноидное лекарственное средство, такое как без ограничения DM1, DM3 или DM4.
[00157] Кроме того, антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению могут быть конъюгированы с фрагментом, представляющим собой лекарственное средство, модифицирующее заданный биологический ответ. Фрагменты, представляющие собой лекарственные средства, не следует рассматривать как ограниченные классическими химическими терапевтическими средствами. Например, фрагмент, представляющий собой лекарственное средство, может представлять собой белок, пептид или полипептид, обладающие требуемой биологической активностью. Такие белки могут включать, например, токсин, такой как абрин, рицин A, экзотоксин Pseudomonas, холерный токсин или дифтерийный токсин; белок, такой как фактор некроза опухоли, α-интерферон, β-интерферон, фактор роста нервов, фактор роста тромбоцитов, тканевой активатор плазминогена, цитокин, апоптическое средство, антиангиогенное средство или модификатор биологического ответа, такой как, например, лимфокин.
[00158] В одном варианте осуществления антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению конъюгированы с фрагментом, представляющим собой лекарственное средство, таким как цитотоксин, лекарственное средство (например, иммуносупрессор) или радиотоксин. Примеры цитотоксинов включают без ограничения таксаны (см., например, международные (PCT) заявки на патент №№ WO 01/38318 и PCT/US03/02675), ДНК-алкилирующие средства (например, аналоги CC-1065), антрациклины, аналоги тубулизина, аналоги дуокармицина, ауристатин E, ауристатин F, майтанзиноиды и цитотоксические средства, содержащие фрагмент, представляющий собой реакционноспособный полиэтиленгликоль (см., например, Sasse et al., J. Antibiot. (Tokyo), 53, 879-85 (2000), Suzawa et al., Bioorg. Med. Chem., 8, 2175-84 (2000), Ichimura et al., J. Antibiot. (Tokyo), 44, 1045-53 (1991), Francisco et al., Blood (2003) (электронная публикация до печатной публикации), патенты США №№ 5475092, 6340701, 6372738 и 6436931, публикацию заявки на патент США № 2001/0036923 A1, находящиеся на рассмотрении заявки на патент США с порядковыми №№ 10/024290 и 10/116053, и международную (PCT) заявку на патент № WO 01/49698), таксон, цитохалазин B, грамицидин D, этидия бромид, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, t. колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин, а также их аналоги или гомологи. Терапевтические средства также включают, например, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил, декарбазин), разрушающие средства (например, мехлорэтамин, тиотепу, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин C и цис-дихлордиаминплатину (II) (DDP), цисплатин, антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (AMC)) и антимитотические средства (например, винкристин и винбластин). (См. например, US20090304721 от Seattle Genetics).
[00159] Другие примеры цитотоксинов, которые могут быть конъюгированы с антителами, фрагментами антител (антигенсвязывающими фрагментами) или функциональными эквивалентами по настоящему изобретению, включают дуокармицины, калихеамицины, майтанзины и ауристатины, а также их производные.
[00160] Различные типы цитотоксинов, линкеров и способы конъюгирования терапевтических средств с антителами известны из уровня техники, см., например, Saito et al., (2003) Adv. Drug Deliv. Rev. 55:199-215; Trail et al., (2003) Cancer Immunol. Immunother. 52:328-337; Payne, (2003) Cancer Cell 3:207-212; Allen, (2002) Nat. Rev. Cancer 2:750-763; Pastan and Kreitman, (2002) Curr. Opin. Investig. Drugs 3:1089-1091; Senter and Springer, (2001) Adv. Drug Deliv. Rev. 53:247-264.
[00161] Антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению также можно конъюгировать с радиоактивным изотопом для создания цитотоксических радиофармацевтических препаратов, называемых радиоиммуноконъюгаты. Примеры радиоактивных изотопов, которые можно конъюгировать с антителами для применения в диагностических или терапевтических целях, включают без ограничения йод-131, индий-111, иттрий-90 и лютеций-177. Способы получения радиоиммуноконъюгатов определены в уровне техники. Примеры радиоиммуноконъюгатов являются доступными коммерчески, в том числе ZevalinTM (DEC Pharmaceuticals) и BexxarTM (Corixa Pharmaceuticals), а также подобные способы можно использовать для получения радиоиммуноконъюгатов с применением антител по настоящему изобретению. В некоторых вариантах осуществления макроциклический хелатор представляет собой 1,4,7,10-тетраазациклододекан-N,N',N'',N'''-тетрауксусную кислоту (DOTA), которая может быть прикреплена к антителу через линкерную молекулу. Такие линкерные молекулы широко известны из уровня техники и описаны в Denardo et al., (1998) Clin Cancer Res. 4(10):2483-90; Peterson et al., (1999) Bioconjug. Chem. 10(4):553-7; и Zimmerman et al., (1999) Nucl. Med. Biol. 26(8):943-50, каждый из которых включен посредством ссылки во всей своей полноте.
[00162] Антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению также можно конъюгировать с гетерологичным белком или полипептидом (или их фрагментом, предпочтительно с полипептидом из по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90 или по меньшей мере 100 аминокислот) для создания слитых белков. В частности, в настоящем изобретении представлены слитые белки, содержащие фрагмент антитела (например, антигенсвязывающий фрагмент), описанный в данном документе (например, Fab-фрагмент, Fd-фрагмент, Fv-фрагмент, F(ab)2-фрагмент, домен VH, CDR VH, домен VL или CDR VL), и гетерологичный белок, полипептид или пептид.
[00163] Дополнительные слитые белки можно создавать за счет методик шаффлинга генов, шаффлинга мотивов, шаффлинга экзонов и/или шаффлинга кодонов (совокупно называемых "шаффлингом ДНК"). Шаффлинг ДНК можно использовать для изменения активности антител по настоящему изобретению или их фрагментов (например, получения антител или их фрагментов с более высокими уровнями аффинности и более низкими значениями скорости диссоциации). См., в целом, патенты США №№ 5605793, 5811238, 5830721, 5834252 и 5837458; Patten et al., (1997) Curr. Opinion Biotechnol. 8:724-33; Harayama, (1998) Trends Biotechnol. 16(2):76-82; Hansson et al., (1999) J. Mol. Biol. 287:265-76; и Lorenzo and Blasco, (1998) Biotechniques 24(2):308-313 (каждый из данных патентов и публикаций тем самым включен посредством ссылки во всей своей полноте). Антитела или их фрагменты, или кодируемые антитела или их фрагменты можно изменять путем воздействия на них случайного мутагенеза с помощью ПЦР с внесением ошибок, случайной вставки нуклеотидов или других способов до проведения рекомбинации. Полинуклеотид, кодирующий антитело или его фрагмент, которые специфически связываются с антигеном, может быть подвергнут рекомбинации с одним или несколькими компонентами, мотивами, секциями, частями, доменами, фрагментами и т. д. одной или нескольких гетерологичных молекул.
[00164] Более того, антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению могут быть конъюгированы с маркерными последовательностями, такими как пептид, для облегчения очистки. В предпочтительных вариантах осуществления маркерная аминокислотная последовательность представляет собой гексагистидиновый пептид (SEQ ID NO: 628), такой как метка, предусмотренная в векторе pQE (QIAGEN, Inc., 9259 Eton Avenue, Чатсворт, Калифорния, 91311), среди прочих, многие из которых являются коммерчески доступными. Как описано в Gentz et al., (1989) Proc. Natl. Acad. Sci. USA 86:821-824, например, гексагистидин (SEQ ID NO: 628) обеспечивает удобную очистку слитого белка. Другие пептидные метки, применимые для очистки, включают без ограничения гемагглютининовую ("HA") метку, которая соответствует эпитопу, происходящему из белка гемагглютинина вируса гриппа (Wilson et al., (1984) Cell 37:767), и метку "FLAG" (A. Einhauer et al., J. Biochem. Biophys. Methods 49: 455-465, 2001). Согласно настоящему изобретению для повышения эффективности антител или антигенсвязывающих фрагментов их также можно конъюгировать с проникающими в опухоль пептидами.
[00165] В других вариантах осуществления антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению конъюгируют с диагностическим или выявляемым средством. Такие иммуноконъюгаты могут быть применимыми для контроля или прогнозирования проявления, развития, прогрессирования и/или степени тяжести заболевания или нарушения в рамках процедуры клинического тестирования, такой как определение эффективности конкретной терапии. Такая диагностика и обнаружение могут осуществляться путем сопряжения антитела с выявляемыми веществами, в том числе без ограничения с различными ферментами, такими как без ограничения пероксидаза хрена, щелочная фосфатаза, бета-галактозидаза или ацетилхолинэстераза; простетическими группами, такими как без ограничения стрептавидин/биотин и авидин/биотин; флуоресцентными материалами, такими как без ограничения Alexa Fluor 350, Alexa Fluor 405, Alexa Fluor 430, Alexa Fluor 488, Alexa Fluor 500, Alexa Fluor 514, Alexa Fluor 532, Alexa Fluor 546, Alexa Fluor 555, Alexa Fluor 568, Alexa Fluor 594, Alexa Fluor 610, Alexa Fluor 633, Alexa Fluor 647, Alexa Fluor 660, Alexa Fluor 680, Alexa Fluor 700, Alexa Fluor 750, умбеллиферон, флуоресцеин, флуоресцеина изотиоцианат, родамин, дихлортриазиниламинфлуоресцеин, дансилхлорид или фикоэритрин; люминесцентными материалами, такими как без ограничения люминол; биолюминесцентными материалами, такими как без ограничения люцифераза, люциферин и экворин; радиоактивными материалами, такими как без ограничения йод (131I, 125I, 123I и 121I), углерод (14C), сера (35S), тритий (3H), индий (115In, 113In, 112In и 111In), технеций (99Tc), таллий (201Ti), галлий (68Ga, 67Ga), палладий (103Pd), молибден (99Mo), ксенон (133Xe), фтор (18F), 153Sm, 177Lu, 159Gd, 149Pm, 140La, 175Yb, 166Ho, 90Y, 47Sc, 186Re, 188Re, 142Pr, 105Rh, 97Ru, 68Ge, 57Co, 65Zn, 85Sr, 32P, 153Gd, 169Yb, 51Cr, 54Mn, 75Se, 64Cu, 113Sn и 117Sn; и позитронно-активными металлами с применением различных видов позитронной эмиссионной томографии, и нерадиоактивными ионами парамагнитных металлов.
[00166] Антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или функциональные эквиваленты по настоящему изобретению также могут быть прикреплены к твердым подложкам, которые особенно применимы для иммунологических анализов или очистки целевого антигена. Такие твердые подложки включают без ограничения стекло, целлюлозу, полиакриламид, нейлон, полистирол, поливинилхлорид или полипропилен.
2. Линкер
[00167] Используемый в данном документе "линкер" представляет собой любой химический фрагмент, который способен соединить антитело, фрагмент антитела (например, антигенсвязывающие фрагменты) или функциональный эквивалент с другим фрагментом, таким как фрагмент, представляющий собой лекарственное средство. Линкеры могут поддаваться расщеплению (расщепляемый линкер), такому как индуцированное кислотами расщепление, фотоиндуцированное расщепление, индуцированное пептидазами расщепление, индуцированное эстеразами расщепление, индуцированное гликозидазами расщепление, индуцированное фосфодиэстеразами расщепление, индуцированное фосфатазами расщепление и расщепление дисульфидной связи, в условиях, при которых соединение или антитело остаются активными. В качестве альтернативы линкеры могут быть в значительной степени устойчивыми к расщеплению (например, стабильный линкер или нерасщепляемый линкер). В некоторых аспектах линкер представляет собой предварительно заряженный линкер, гидрофильный линкер или линкер на основе дикарбоновой кислоты.
[00168] В одном аспекте линкер, применяемый в настоящем изобретении, происходит из сшивающего реагента, такого как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), N-сукцинимидил-4-(2-пиридилдитио)пентаноат (SPP), N-сукцинимидил-4-(2-пиридилдитио)бутаноат (SPDB), N-сукцинимидил-4-(2-пиридилдитио)-2-сульфо-бутаноат (сульфо-SPDB), N-сукцинимидилйодацетат (SIA), N-сукцинимидил-(4-йодацетил)аминобензоат (SIAB), малеимид-PEG-NHS, N-сукцинимидил-4-(малеимидометил)циклогексанкарбоксилат (SMCC), N-сульфосукцинимидил-4-(малеимидометил)циклогексанкарбоксилат (сульфо-SMCC) или 2,5-диоксопирролидин-1-ил-17-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)-5,8,11,14-тетраоксо-4,7,10,13-тетраазагептадекан-1-оат (CX1-1).
[00169] Нерасщепляемые линкеры представляют собой любой химический фрагмент, который способен соединить лекарственное средство, такое как майтанзиноид, с антителом с помощью устойчивой ковалентной связи и не попадает в категории, перечисленные выше для расщепляемых линкеров. Таким образом, нерасщепляемые линкеры являются в значительной степени устойчивыми к индуцированному кислотами расщеплению, фотоиндуцированному расщеплению, индуцированному пептидазами расщеплению, индуцированному эстеразами расщеплению и расщеплению дисульфидной связи. Кроме того, нерасщепляемый относится к способности химической связи в линкере или прикрепляемой к линкеру противостоять расщеплению, индуцированному кислотой, фотолабильным расщепляющим средством, пептидазой, эстеразой или химическим или физиологическим соединением, которое расщепляет дисульфидную связь, в условиях, при которых лекарственное средство, такое как майтанзиноид или антитело, не теряет свою активность.
[00170] Кислотолабильные линкеры представляют собой линкеры, расщепляемые при кислотном pH. Например, некоторые внутриклеточные компартменты, такие как эндосомы и лизосомы, имеют кислотный pH (pH 4-5) и обеспечивают условия, подходящие для расщепления кислотолабильных линкеров.
[00171] Фотолабильные линкеры представляют собой линкеры, которые применимы на поверхности тела и во многих полостях тела, которые доступны для воздействия света. Кроме того, инфракрасный свет может проникать внутрь ткани.
[00172] Некоторые линкеры могут расщепляться под действием пептидаз, т.е. являются расщепляемыми пептидазами линкерами. Только некоторые пептиды легко расщепляются внутри или за пределами клеток, см., например, Trout et al., 79 Proc. Natl. Acad. Sci. USA, 626-629 (1982) и Umemoto et al. 43 Int. J. Cancer, 677-684 (1989). Кроме того, пептиды состоят из α-аминокислот и пептидных связей, которые химически представляют собой амидные связи между карбоксильной группой одной аминокислоты и аминогруппой второй аминокислоты. Понятно, что другие амидные связи, такие как связь между карбоксильной группой и ε-аминогруппой лизина, не являются пептидными связями и считаются нерасщепляемыми.
[00173] Некоторые линкеры могут расщепляться под действием эстераз, т.е. являются расщепляемыми эстеразами линкерами. Опять-таки, только некоторые сложные эфиры могут расщепляться под действием эстераз, присутствующих внутри или за пределами клеток. Сложные эфиры образуются путем конденсации карбоновой кислоты и спирта. Простые сложные эфиры представляют собой сложные эфиры, полученные с помощью простых спиртов, таких как алифатические спирты, и небольших циклических и небольших ароматических спиртов.
[00174] Предварительно заряженные линкеры происходят из заряженных сшивающих реагентов, которые сохраняют свой заряд после введения в конъюгат антитела и лекарственного средства. Примеры предварительно заряженных линкеров можно найти в US 2009/0274713.
3. Конъюгация и получение ADC
[00175] Многочисленные способы конъюгирования линкеров-полезных нагрузок с антигенсвязывающим фрагментом известны из уровня техники (обзор приведен, например, в: Antibody-Drug Conjugate, Methods in Molecular Biology, Vol. 1045, Editor L. Ducry, Humana Press (2013)). Традиционно лекарственные средства конъюгируют с нативными лизиновыми или нативными цистеиновыми остатками антитела. Полученные препараты представляют собой сложные смеси. В последнее время для улучшения терапевтического индекса и гомогенности препаратов ADC используются способы сайт-специфической конъюгации (обзор приведен в: Panowski, S.; Bhakta, S.; Raab, H.; Polakis, P.; Junutula, J. R. mAbs 2014, 6, 34). Помимо применения гликоинжиниринга (Zhou, Q. et al. Bioconjugate chemistry 2014, 25, 510; Zhu, Z. et al. mAbs 2014, 6, 1190); некоторые более общеизвестные способы получения сайт-специфических ADC основаны на введении сконструированных цистеинов (Junutula, J. R. et al., Nature biotechnology 2008, 26, 925; Shinmi, D. et al., Bioconjugate chemistry 2016, 27, 1324), неканонических аминокислот (Tian, F. et al., Proceedings National Academy of Sciences USA 2014, 111, 1766; Axup, J. Y. et al., Proceedings National Academy of Sciences USA 2012, 109, 16101) или коротких пептидных последовательностей в остов антитела (Drake, P. M. et al., Bioconjugate chemistry 2014, 25, 1331; Strop, P. et al., Chemistry & biology 2013, 20, 161; Beerli, R. R. et al., PloS one 2015, 10, e0131177; Grunewald, J. et al., Bioconjugate chemistry 2015, 26, 2554). Данные способы обеспечивают контроль за стехиометрией и сайтом прикрепления цитотоксина, что приводит к лучшей фармакокинетике (PK), безопасности и профилям эффективности конъюгатов по сравнению с традиционно полученными ADC.
[00176] Конъюгаты по настоящему изобретению могут быть получены с помощью любых способов, известных из уровня техники, таких как описанные в патентах США №№ 7811572, 6411163, 7368565 и 8163888, публикациях заявок на патент США 2011/0003969, 2011/0166319, 2012/0253021 и 2012/0259100, и PCT-публикациях WO2014/124316 и WO2015/138615. Полное изложение идей данных патентов и публикаций заявок на патент включено в данный документ посредством ссылки.
Способ конъюгации с введенными путем конструирования цистеиновыми остатками антитела
[00177] Конъюгаты по настоящему изобретению могут быть получены с применением цистеиновых остатков, введенных путем конструирования в антитело, например, с помощью сайт-направленного мутагенеза. Такие сайт-специфические конъюгаты являются гомогенными и характеризуются улучшенными свойствами (Junutula JR, Raab H, Clark S, Bhakta S, Leipold DD, Weir S, Chen Y, Simpson M, Tsai SP, Dennis MS, Lu Y, Meng YG, Ng C, Yang J, Lee CC, Duenas E, Gorrell J, Katta V, Kim A, McDorman K, Flagella K, Venook R, Ross S, Spencer SD, Lee Wong W, Lowman HB, Vandlen R, Sliwkowski MX, Scheller RH, Polakis P, Mallet W. (2008) Nature Biotechnology 26:925-932.)
[00178] Поскольку введенные путем конструирования цистеины в антителах, экспрессируемых в клетках млекопитающих, во время биосинтеза модифицируются за счет аддуктов (дисульфидов), таких как глутатион (GSH) и/или цистеин (Chen et al. 2009), изначально экспрессированные введенные путем конструирования цистеиновые остатки в продукте не реагируют с реагентами, вступающими в реакцию с тиолами, такими как малеимидная или бром- или йод-ацетамидная группы. Для осуществления конъюгации полезной нагрузки с введенным путем конструирования цистеином после экспрессии глутатионовый или цистеиновый аддукты должны быть удалены путем восстановления данных дисульфидных аддуктов, что обычно вызывает также восстановление нативных дисульфидов в экспрессированном белке. Удаление аддуктов у введенных путем конструирования цистеинов можно осуществлять, вначале подвергая антитело воздействию восстановителя, например, дитиотреитола (DTT), TCEP, или восстановленного цистеина, с последующим проведением процедуры, которая обеспечивает возможность повторного окисления всех нативных дисульфидных связей антитела для восстановления и/или стабилизации функциональной структуры антитела.
[00179] Можно использовать несколько способов восстановления и повторного окисления антител с введенными путем конструирования Cys-остатками для получения конъюгатов антитела и лекарственного средства. Попытки следовать ранее описанным в литературе протоколам повторного окисления с применением высоких концентраций CuSO4 приводили к осаждению белка (Junutula JR, Raab H, Clark S, Bhakta S, Leipold DD, Weir S, Chen Y, Simpson M, Tsai SP, Dennis MS, Lu Y, Meng YG, Ng C, Yang J, Lee CC, Duenas E, Gorrell J, Katta V, Kim A, McDorman K, Flagella K, Venook R, Ross S, Spencer SD, Lee Wong W, Lowman HB, Vandlen R, Sliwkowski MX, Scheller RH, Polakis P, Mallet W. (2008) Nature Biotechnology 26:925). Авторы настоящего изобретения успешно подготовили и получили конъюгаты антитела и лекарственного средства с помощью нескольких различных способов восстановления и повторного окисления антитела.
[00180] В одном примере свежеполученный DTT добавляют к очищенным антителам с Cys-мутацией до конечной концентрации, составляющей 10 мМ. После инкубации с DTT при комнатной температуре в течение 1 часа смесь диализировали против PBS при 4°C в течение трех дней с ежедневной заменой буфера для удаления DTT и повторного окисления нативных дисульфидных связей антитела. Альтернативный способ заключается в удалении восстанавливающих реагентов путем пропускания через обессоливающую колонку, такую как Sephadex G-25, уравновешенную с помощью PBS. После того как белок полностью восстановился, к обессоленным образцам необязательно добавляют 1 мМ окисленного аскорбата (дигидроаскорбиновая кислота) и инкубации для повторного окисления проводят в течение 20-24 часов.
[00181] В другом иллюстративном способе освобождение введенных путем конструирования Cys-остатков осуществляют путем добавления полностью восстановленного цистеина при концентрации 20 мМ к антителам, связанным со смолой на основе сефарозы с белком A. Восстановление аддуктов на Cys достигается путем инкубации в течение примерно 30-60 минут при комнатной температуре, затем восстановитель быстро удаляют путем промывания смолы с помощью 50 объемов слоя PBS. Повторное окисление восстановленного антитела достигается инкубацией промытой взвеси при комнатной температуре с добавлением или без добавления 50-2000 нМ CuCl2 в качестве катализатора. За исключением применения сульфата меди, в примерах из данного документа применялся каждый из протоколов, описанных в данном документе, с получением аналогичных результатов. Повторное окисление восстанавливает внутрицепочечные дисульфиды, в то время как диализ, обессоливание или хроматография с белком A удаляет восстановитель, а также цистеины и глутатионы, изначально присоединенные к введенному(-ым) путем конструирования цистеину(-ам) антитела. Для отслеживания процесса повторного окисления обычно применяют HPLC-хроматографию с обращенной фазой: антитела загружают на колонку PLRP-S (4000 Å, 50 мм × 2,1 мм, Agilent), нагретую до 80°C и элюируют с применением линейного градиента 30-45% CH3CN в воде, содержащей 0,1% TFA, при 1,5 мл/мин и пике обнаружения при 215, 254 и 280 нм.
[00182] После повторного окисления антитело конъюгируют с предварительно образованным фрагментом, представляющим собой линкер-лекарственное средство. В качестве примера предварительно образованный фрагмент, представляющий собой линкер-лекарственное средство (такой как, например, MMTBT-DM4; MPET-DM4; MBT-DM4; MEPET-DM4, MPBT-DM1; и другие фрагменты, представляющие собой линкер-лекарственное средство, описанные в данном документе) добавляют к повторно окисленному антителу с Cys-мутацией из расчета 10 молярных эквивалентов относительно антитела в буфере PBS (pH 7,2). Инкубации проводят в течение 1 часа. Процесс конъюгации отслеживают с помощью HPLC с обращенной фазой, которая может отделить конъюгированные антитела от неконъюгированных. Реакционные смеси в процессе конъюгации анализируют на колонке PRLP-S (4000 Å, 50 мм × 2,1 мм, Agilent), нагретой до 80°C, а элюирование колонки проводят с помощью линейного градиента 30-60% ацетонитрила в воде, содержащей 0,1% TFA при расходе 1,5 мл/мин. Элюирование белков из колонки отслеживают при 280 нм, 254 нм и 215 нм.
[00183] В одном варианте осуществления примеры фрагмента, представляющего собой линкер-лекарственное средство, для конъюгации по цистеину могут быть получены в соответствии со схемами 1-3:
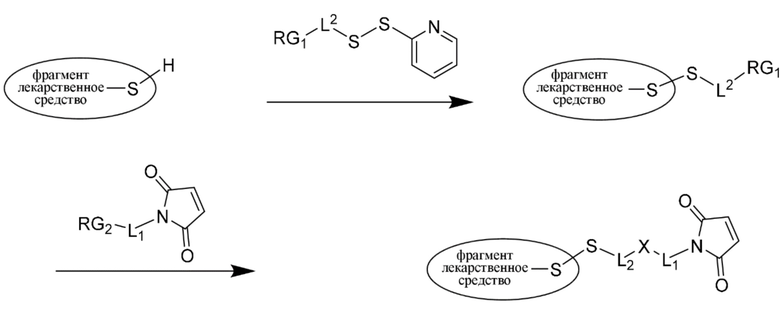
Схема 1.
где:
фрагмент, представляющий собой лекарственное средство, прикреплен к линкеру посредством тиольной функциональной группы;
L1 представляет собой C1-6алкилен, при этом одна из метиленовых групп может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11; и
X представляет собой -C(O)-NH-, -NHC(O)- или триазол;
где алкилен является линейным или разветвленным; и
RG1 и RG2 представляют собой 2 реакционноспособные группы, образующие группу X.
Осуществление реакции групп, которые образуют амид или триазол, широко известно из уровня техники.
Один пример предварительного образования фрагмента, представляющего собой линкер-лекарственное средство, представлен на схеме 2, где фрагмент, представляющий собой лекарственное средство, представляет собой DM4; RG1 представляет собой аминогруппу, а RG2 представляет собой активированную кислоту, которые приводят к образованию амидной связи (X):
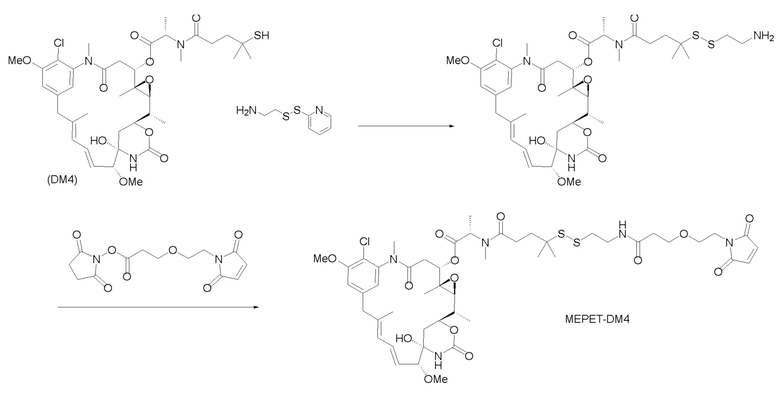
Схема 2.
Другой пример предварительного образования фрагмента, представляющего собой линкер-лекарственное средство, представлен на схеме 3, где фрагмент, представляющий собой лекарственное средство, представляет собой DM4; RG1 представляет собой азидную группу, а RG2 представляет собой алкиновую группу, которые приводят к образованию тетразола (X):
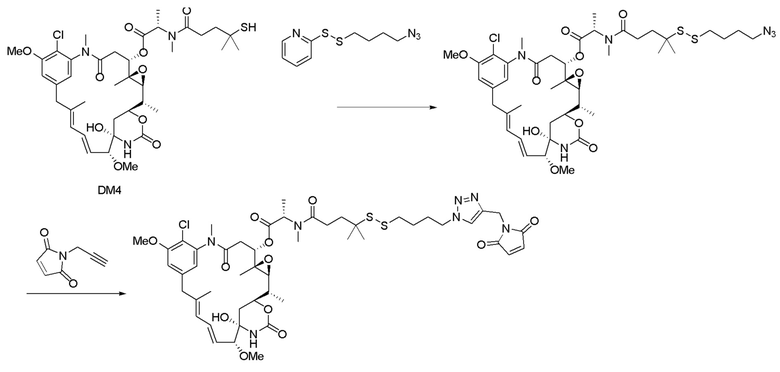
Схема 3.
[00184] Эффективность конъюгации различных фрагментов, представляющих собой лекарственное средство, имеющих присоединенный малеимид, с антителом с Cys-мутацией, варьирует в зависимости от растворимости применяемого фрагмента, представляющего собой лекарственное средство, однако, многие реакции приводят к более 90% конъюгата. Для оценки состояния агрегации полученные конъюгаты анализируют на колонке для эксклюзионной хроматографии (GE, Superdex200, 3,2/30) при расходе 0,1 мл/мин в PBS. Все конъюгаты являются преимущественно мономерными. Большая часть конъюгатов содержит менее 3% димерного и олигомерного материала, что указывает на то, что конъюгация фрагмента, представляющего собой лекарственное средство, имеющего присоединенный малеимид, с антителом с Cys-мутацией не вызывает агрегацию.
[00185] Иммуноконъюгаты также характеризуют с точки зрения средней нагрузки фрагмента, представляющего собой лекарственное средство, на фрагмент, представляющий собой антитело связывания, обычно называемой отношение лекарственного средства-к-антителу (DAR). Значение DAR экстраполируют, например, на основании данных LC-MS для восстановленных и дегликозилированных образцов. LC/MS обеспечивает возможность получения количественной оценки среднего числа молекул полезной нагрузки (фрагмент, представляющий собой лекарственное средство), прикрепленных к антителу в ADC. HPLC разделяет антитело на легкую и тяжелую цепи, а также разделяет тяжелую цепь (HC) и легкую цепь (LC) в соответствии с числом групп линкер-полезная нагрузка на цепь. Данные масс-спектров позволяют идентифицировать разновидности компонентов в смеси, например, LC, LC+1, LC+2, HC, HC+1, HC+2 и т.д. На основании средней нагрузки цепей LC и HC для ADC может быть рассчитано среднее DAR. DAR для данного образца иммуноконъюгата представляет собой среднее число молекул лекарственного средства (полезная нагрузка), прикрепленных к тетрамерному антителу, содержащему две легкие цепи и две тяжелые цепи.
Способ конъюгации с нативными цистеиновыми остатками антитела
[00186] Фрагменты, представляющие собой линкер-лекарственное средство, описанные в данном документе, также можно конъюгировать с нативными цистеиновыми остатками не подвергнутых конструированию антител с применением процедуры, которая предусматривает частичное восстановление антител (Doronina, S. O., Toki, B. E., Torgov, M. Y., Mendelsohn, B. A., Cerveny, C. G., Chace, D. F., DeBlanc, R. L., Gearing,R. P., Bovee, T. D., Siegall, C. B., Francisco, J. A., Wahl, A. F., Meyer, D. L., and Senter, P. D. (2003) Development of potent monoclonal antibody auristatin conjugates for cancer therapy. Nat. Biotechnol. 21, 778-784). Следующий протокол является неограничивающим примером того, как такие конъюгаты могут быть получены. Межцепочечные и внутрицепочечные дисульфидные связи антитела (обычно при концентрации 5-10 мг/мл) сначала частично восстанавливают в PBS, содержащем 2 мМ EDTA, путем добавления TCEP до конечной концентрации 10 мМ и инкубации смеси при 37°C в течение 1 часа. После обессоливания и добавления 1% вес/объем детергента PS-20 частично восстановленные антитела (1-2 мг/мл) подвергают реакции в течение ночи при 4°C с 0,5-1 мг соединения малеимидсодержащего линкера и полезной нагрузки на 10 мг антитела. Полученные конъюгаты очищают хроматографией с белком A посредством стандартных способов и проводят замену буфера на PBS, и их обычно анализируют с помощью масс-спектрометрии (MS), аналитической эксклюзионной хроматографии (AnSEC) и аналитической хроматографии гидрофобных взаимодействий (AnHIC) для определения отношения лекарственного средства-к-антителу, склонности к агрегации и гидрофобности, а также с помощью анализов активности.
Одностадийный способ для сшивания с лизиновыми остатками антитела
[00187] В одном варианте осуществления конъюгаты по настоящему изобретению могут быть получены с помощью одностадийного способа сшивания лекарственного средства с лизиновыми остатками на антителе. Способ предусматривает объединение антитела, лекарственного средства и сшивающего средства в значительной степени водной среде, необязательно содержащей один или несколько сорастворителей, при подходящем pH. В одном варианте осуществления способ предусматривает стадию приведения антитела по настоящему изобретению в контакт с лекарственным средством (например, DM1 или DM4) с образованием первой смеси, содержащей антитело и лекарственное средство, и затем приведение первой смеси, содержащей антитело и лекарственное средство, в контакт со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) в растворе, характеризующемся pH от приблизительно 4 до приблизительно 9, с получением смеси, содержащей (i) конъюгат (например, Ab-MCC-DM1, Ab-SPDB-DM4 или Ab-CX1-1-DM1), (ii) свободное лекарственное средство (например, DM1 или DM4) и (iii) побочные продукты реакции.
[00188] В одном варианте осуществления одностадийный способ предусматривает приведение антитела в контакт с лекарственным средством (например, DM1 или DM4), а затем со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) в растворе, характеризующемся pH приблизительно 6 или больше (например, от приблизительно 6 до приблизительно 9, от приблизительно 6 до приблизительно 7, от приблизительно 7 до приблизительно 9, от приблизительно 7 до приблизительно 8,5, от приблизительно 7,5 до приблизительно 8,5, от приблизительно 7,5 до приблизительно 8,0, от приблизительно 8,0 до приблизительно 9,0 или от приблизительно 8,5 до приблизительно 9,0). Например, способ по настоящему изобретению предусматривает приведение связывающего клетку средства в контакт с лекарственным средством (DM1 или DM4), а затем со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) в растворе, характеризующемся pH приблизительно 6,0, приблизительно 6,1, приблизительно 6,2, приблизительно 6,3, приблизительно 6,4, приблизительно 6,5, приблизительно 6,6, приблизительно 6,7, приблизительно 6,8, приблизительно 6,9, приблизительно 7,0, приблизительно 7,1, приблизительно 7,2, приблизительно 7,3, приблизительно 7,4, приблизительно 7,5, приблизительно 7,6, приблизительно 7,7, приблизительно 7,8, приблизительно 7,9, приблизительно 8,0, приблизительно 8,1, приблизительно 8,2, приблизительно 8,3, приблизительно 8,4, приблизительно 8,5, приблизительно 8,6, приблизительно 8,7, приблизительно 8,8, приблизительно 8,9, или приблизительно 9,0. В конкретном варианте осуществления способ по настоящему изобретению предусматривает приведение связывающего клетку средства в контакт с лекарственным средством (например, DM1 или DM4), а затем со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) в растворе, характеризующемся pH приблизительно 7,8 (например, pH от 7,6 до 8,0 или pH от 7,7 до 7,9).
[00189] Одностадийный способ (т.е. приведение антитела в контакт с лекарственным средством (например, DM1 или DM4), а затем со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) можно проводить при любой подходящей температуре, известной из уровня техники. Например, одностадийный способ может происходить при приблизительно 20°C или меньше (например, от приблизительно -10°C (при условии, что не допускается замерзание раствора, например, за счет присутствия органического растворителя, применяемого для растворения цитотоксического средства и бифункционального сшивающего реагента) до приблизительно 20°C, от приблизительно 0°C до приблизительно 18°C, от приблизительно 4°C до приблизительно 16°C), при комнатной температуре (например, от приблизительно 20°C до приблизительно 30°C или от приблизительно 20°C до приблизительно 25°C) или при повышенной температуре (например, от приблизительно 30°C до приблизительно 37°C). В одном варианте осуществления одностадийный способ происходит при температуре от приблизительно 16°C до приблизительно 24°C (например, приблизительно 16°C, приблизительно 17°C, приблизительно 18°C, приблизительно 19°C, приблизительно 20°C, приблизительно 21°C, приблизительно 22°C, приблизительно 23°C, приблизительно 24°C или приблизительно 25°C). В другом варианте осуществления одностадийный способ проводят при температуре приблизительно 15°C или меньше (например, от приблизительно -10°C до приблизительно 15°C или от приблизительно 0°C до приблизительно 15°C). Например, способ предусматривает приведение антитела в контакт с лекарственным средством (например, DM1 или DM4), а затем со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) при температуре приблизительно 15°C, приблизительно 14°C, приблизительно 13°C, приблизительно 12°C, приблизительно 11°C, приблизительно 10°C, приблизительно 9°C, приблизительно 8°C, приблизительно 7°C, приблизительно 6°C, приблизительно 5°C, приблизительно 4°C, приблизительно 3°C, приблизительно 2°C, приблизительно 1°C, приблизительно 0°C, приблизительно -1°C, приблизительно -2°C, приблизительно -3°C, приблизительно -4°C, приблизительно -5°C, приблизительно -6°C, приблизительно -7°C, приблизительно -8°C, приблизительно -9°C или приблизительно -10°C, при условии, что не допускается замерзание раствора, например, за счет присутствия органического(-их) растворителя(-ей), применяемого(-ых) для растворения сшивающего средства (например, SMCC, сульфо-SMCC, сульфо-SPDB, SPDB или CX1-1). В одном варианте осуществления способ предусматривает приведение антитела в контакт с лекарственным средством (например, DM1 или DM4), а затем со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) при температуре от приблизительно -10°C до приблизительно 15°C, от приблизительно 0°C до приблизительно 15°C, от приблизительно 0°C до приблизительно 10°C, от приблизительно 0°C до приблизительно 5°C, от приблизительно 5°C до приблизительно 15°C, от приблизительно 10°C до приблизительно 15°C или от приблизительно 5°C до приблизительно 10°C. В другом варианте осуществления способ предусматривает приведение антитела в контакт с лекарственным средством (например, DM1 или DM4), а затем со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) при температуре приблизительно 10°C (например, температуре от 8°C до 12°C или температуре от 9°C до 11°C).
[00190] В одном варианте осуществления описанное выше приведение в контакт осуществляют путем внесения антитела, затем приведения антитела в контакт с лекарственным средством (например, DM1 или DM4) с образованием первой смеси, содержащей антитело и лекарственное средство (например, DM1 или DM4), а затем приведения первой смеси, содержащей антитело и лекарственное средство (например, DM1 или DM4) в контакт со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1). Например, в одном варианте осуществления антитело вносится в реакционный сосуд, лекарственное средство (например, DM1 или DM4) добавляют в реакционный сосуд (тем самым приводя в контакт с антителом), а затем сшивающее средство (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) добавляют в смесь, содержащую антитело и лекарственное средство (например, DM1 или DM4) (тем самым приводя в контакт со смесью, содержащей антитело и лекарственное средство). В одном варианте осуществления антитело вносится в реакционный сосуд, а лекарственное средство (например, DM1 или DM4) добавляют в реакционный сосуд непосредственно после внесения антитела в сосуд. В другом варианте осуществления антитело вносится в реакционный сосуд, а лекарственное средство (например, DM1 или DM4) добавляют в реакционный сосуд через некоторый временной интервал после внесения антитела в сосуд (например, через приблизительно 5 минут, приблизительно 10 минут, приблизительно 20 минут, приблизительно 30 минут, приблизительно 40 минут, приблизительно 50 минут, приблизительно 1 час, приблизительно 1 день или больше после внесения связывающего клетку средства в емкость). Лекарственное средство (например, DM1 или DM4) можно добавлять быстро (т.е. за короткий временной интервал, такой как приблизительно 5 минут, приблизительно 10 минут) или медленно (как, например, с применением насоса).
[00191] Смесь, содержащую антитело и лекарственное средство (например, DM1 или DM4), затем можно привести в контакт со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1), либо непосредственно после приведения антитела в контакт с лекарственным средством (например, DM1 или DM4), либо немного позже (например, от приблизительно 5 минут до приблизительно 8 часов или больше) после приведения антитела в контакт с лекарственным средством (например, DM1 или DM4). Например, в одном варианте осуществления сшивающее средство (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) добавляют в смесь, содержащую антитело и лекарственное средство (например, DM1 или DM4), непосредственно после добавления лекарственного средства (например, DM1 или DM4) в реакционный сосуд, содержащий антитело. В качестве альтернативы смесь, содержащую антитело и лекарственное средство (например, DM1 или DM4), можно привести в контакт со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) через приблизительно 5 минут, приблизительно 10 минут, приблизительно 20 минут, приблизительно 30 минут, приблизительно 1 час, приблизительно 2 часа, приблизительно 3 часа, приблизительно 4 часа, приблизительно 5 часов, приблизительно 6 часов, приблизительно 7 часов, приблизительно 8 часов или больше после приведения антитела в контакт с лекарственным средством (например, DM1 или DM4).
[00192] После того, как смесь, содержащую антитело и лекарственное средство (например, DM1 или DM4), приводят в контакт со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1), обеспечивают возможность протекания реакции в течение приблизительно 1 часа, приблизительно 2 часов, приблизительно 3 часов, приблизительно 4 часов, приблизительно 5 часов, приблизительно 6 часов, приблизительно 7 часов, приблизительно 8 часов, приблизительно 9 часов, приблизительно 10 часов, приблизительно 11 часов, приблизительно 12 часов, приблизительно 13 часов, приблизительно 14 часов, приблизительно 15 часов, приблизительно 16 часов, приблизительно 17 часов, приблизительно 18 часов, приблизительно 19 часов, приблизительно 20 часов, приблизительно 21 часа, приблизительно 22 часов, приблизительно 23 часов, приблизительно 24 часов или больше (например, приблизительно 30 часов, приблизительно 35 часов, приблизительно 40 часов, приблизительно 45 часов или приблизительно 48 часов).
[00193] В одном варианте осуществления одностадийный способ дополнительно предусматривает стадию блокирования, чтобы заблокировать любое непрореагировавшее лекарственное средство (например, DM1 или DM4) и/или непрореагировавшее сшивающее средство (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1). Обычно стадию блокирования проводят до очистки конъюгата. В одном варианте осуществления смесь блокируют путем приведения смеси в контакт с блокирующим реагентом. Используемый в данном документе "блокирующий реагент" относится к реагенту, который реагирует со свободным лекарственным средством (например, DM1 или DM4) и/или сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1). В одном варианте осуществления можно применять малеимидные или галогенацетамидные блокирующие реагенты, такие как 4-малеимидомасляная кислота, 3-малеимидопропионовая кислота, N-этилмалеимид, йодацетамид или йодацетамидопропионовая кислота, чтобы гарантировать, что любая непрореагировавшая группа (такая как тиол) в лекарственном средстве (например, DM1 или DM4) заблокирована. Стадия блокирования может способствовать предупреждению димеризации лекарственного средства (например, DM1). Димеризованный DM1 может быть сложно удалить. После блокирования с помощью полярных, заряженных, блокирующих тиол реагентов (таких как 4-малеимидомасляная кислота или 3-малеимидопропионовая кислота) избыточный, непрореагировавший DM1 превращается в полярный, заряженный, водорастворимый аддукт, который можно легко отделить от конъюгата, соединенного ковалентной связью, во время стадии очистки. Также можно применять блокирование с помощью неполярных и нейтральных блокирующих тиол реагентов. В одном варианте осуществления смесь блокируют путем приведения смеси в контакт с блокирующим реагентом, который реагирует с непрореагировавшим сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1). Например, чтобы блокировать любой непрореагировавший SMCC, в смесь можно добавлять нуклеофилы. Предпочтительно нуклеофил представляет собой нуклеофил, содержащий аминогруппу, такой как лизин, таурин и гидроксиламин.
[00194] В предпочтительном варианте осуществления обеспечивают возможность протекания реакции (т.е. приведения антитела в контакт с лекарственным средством (например, DM1 или DM4), а затем со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1)) до ее завершения перед приведением смеси в контакт с блокирующим реагентом. В связи с этим блокирующий реагент добавляют в смесь через от приблизительно 1 часа до приблизительно 48 часов (например, через приблизительно 1 час, приблизительно 2 часа, приблизительно 3 часа, приблизительно 4 часа, приблизительно 5 часов, приблизительно 6 часов, приблизительно 7 часов, приблизительно 8 часов, приблизительно 9 часов, приблизительно 10 часов, приблизительно 11 часов, приблизительно 12 часов, приблизительно 13 часов, приблизительно 14 часов, приблизительно 15 часов, приблизительно 16 часов, приблизительно 17 часов, приблизительно 18 часов, приблизительно 19 часов, приблизительно 20 часов, приблизительно 21 час, приблизительно 22 часа, приблизительно 23 часа, приблизительно 24 часа или от приблизительно 25 часов до приблизительно 48 часов) после приведения смеси, содержащей антитело и лекарственное средство (например, DM1 или DM4), в контакт со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1).
[00195] В качестве альтернативы смесь блокируют путем снижения pH смеси до приблизительно 5,0 (например, 4,8, 4,9, 5,0, 5,1 или 5,2). В другом варианте осуществления смесь блокируют путем снижения pH до менее 6,0, менее 5,5, менее 5,0, менее 4,8, менее 4,6, менее 4,4, менее 4,2, менее 4,0. В качестве альтернативы pH снижают до значения от приблизительно 4,0 (например, 3,8, 3,9, 4,0, 4,1 или 4,2) до приблизительно 6,0 (например, 5,8, 5,9, 6,0, 6,1 или 6,2), от приблизительно 4,0 до приблизительно 5,0, от приблизительно 4,5 (например, 4,3, 4,4, 4,5, 4,6 или 4,7) до приблизительно 5,0. В одном варианте осуществления смесь блокируют путем снижения pH смеси до 4,8. В другом варианте осуществления смесь блокируют путем снижения pH смеси до 5,5.
[00196] В одном варианте осуществления одностадийный способ дополнительно предусматривает стадию удерживания для высвобождения нестабильно связавшихся линкеров из антитела. Стадия удерживания предусматривает удерживание смеси до очистки конъюгата (например, после стадии реакции, между стадией реакции и стадией блокирования или после стадии блокирования). Например, способ предусматривает (a) приведение антитела в контакт с лекарственным средством (например, DM1, DM3 или DM4) с образованием смеси, содержащей антитело и лекарственное средство (например, DM1, DM3 или DM4); а затем приведение смеси, содержащей антитело и лекарственное средство (например, DM1, DM3 или DM4), в контакт со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) в растворе, характеризующемся pH от приблизительно 4 до приблизительно 9, с получением смеси, содержащей (i) конъюгат (например, Ab-MCC-DM1, Ab-SPDB-DM4 или Ab-CX1-1-DM1), (ii) свободное лекарственное средство (например, DM1, DM3 или DM4) и (iii) побочные продукты реакции, (b) удержание смеси, полученной на стадии (a), до высвобождения нестабильно связавшихся линкеров из связывающего клетку средства и (c) проведение очистки смеси с получением очищенного конъюгата.
[00197] В другом варианте осуществления способ предусматривает (a) приведение антитела в контакт с лекарственным средством (например, DM1, DM3 или DM4) с образованием смеси, содержащей антитело и лекарственное средство (например, DM1, DM3 или DM4); и затем приведение смеси, содержащей антитело и лекарственное средство (например, DM1, DM3 или DM4), в контакт со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) в растворе, характеризующемся pH от приблизительно 4 до приблизительно 9, с получением смеси, содержащей (i) конъюгат, (ii) свободное лекарственное средство (например, DM1, DM3 или DM4) и (iii) побочные продукты реакции, (b) блокирование смеси, полученной на стадии (a), для блокирования любого непрореагировавшего лекарственного средства (например, DM1, DM3 или DM4) и/или непрореагировавшего сшивающего средства (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1), (c) удержание смеси, полученной на стадии (b), до высвобождения нестабильно связавшихся линкеров из связывающего клетку средства и (d) проведение очистки смеси с получением очищенного конъюгата (например, Ab-MCC-DM1, Ab-SPDB-DM4 или Ab-CX1-1-DM1).
[00198] В качестве альтернативы стадию удерживания можно проводить после очистки конъюгата, с последующей дополнительной стадией очистки.
[00199] В предпочтительном варианте осуществления обеспечивают возможность протекания реакции до завершения перед стадией удерживания. В связи с этим стадию удерживания проводят через от приблизительно 1 часа до приблизительно 48 часов (например, через приблизительно 1 час, приблизительно 2 часа, приблизительно 3 часа, приблизительно 4 часа, приблизительно 5 часов, приблизительно 6 часов, приблизительно 7 часов, приблизительно 8 часов, приблизительно 9 часов, приблизительно 10 часов, приблизительно 11 часов, приблизительно 12 часов, приблизительно 13 часов, приблизительно 14 часов, приблизительно 15 часов, приблизительно 16 часов, приблизительно 17 часов, приблизительно 18 часов, приблизительно 19 часов, приблизительно 20 часов, приблизительно 21 час, приблизительно 22 часа, приблизительно 23 часа, приблизительно 24 часа или от приблизительно 24 часов до приблизительно 48 часов) после приведения смеси, содержащей антитело и лекарственное средство (например, DM1, DM3 или DM4), в контакт со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1).
[00200] Стадия удерживания предусматривает поддержание раствора при подходящей температуре (например, от приблизительно 0°C до приблизительно 37°C) в течение подходящего периода времени (например, от приблизительно 1 часа до приблизительно 1 недели, от приблизительно 1 часа до приблизительно 24 часов, от приблизительно 1 часа до приблизительно 8 часов или от приблизительно 1 часа до приблизительно 4 часов) для высвобождения нестабильно связавшихся линкеров из антитела, при этом не происходит значительного высвобождения стабильно связавшихся линкеров из антитела. В одном варианте осуществления стадия удерживания предусматривает поддержание раствора при приблизительно 20°C или меньше (например, от приблизительно 0°C до приблизительно 18°C, от приблизительно 4°C до приблизительно 16°C), при комнатной температуре (например, от приблизительно 20°C до приблизительно 30°C или от приблизительно 20°C до приблизительно 25°C) или при повышенной температуре (например, от приблизительно 30°C до приблизительно 37°C). В одном варианте осуществления стадия удерживания предусматривает поддержание раствора при температуре от приблизительно 16°C до приблизительно 24°C (например, приблизительно 15°C, приблизительно 16°C, приблизительно 17°C, приблизительно 18°C, приблизительно 19°C, приблизительно 20°C, приблизительно 21°C, приблизительно 22°C, приблизительно 23°C, приблизительно 24°C или приблизительно 25°C). В другом варианте осуществления стадия удерживания предусматривает поддержание раствора при температуре от приблизительно 2°C до приблизительно 8°C (например, приблизительно 0°C, приблизительно 1°C, приблизительно 2°C, приблизительно 3°C, приблизительно 4°C, приблизительно 5°C, приблизительно 6°C, приблизительно 7°C, приблизительно 8°C, приблизительно 9°C или приблизительно 10°C). В другом варианте осуществления стадия удерживания предусматривает поддержание раствора при температуре приблизительно 37°C (например, приблизительно 34°C, приблизительно 35°C, приблизительно 36°C, приблизительно 37°C, приблизительно 38°C, приблизительно 39°C или приблизительно 40°C).
[00201] Продолжительность стадии удерживания зависит от температуры и pH, при которых проводят стадию удерживания. Например, продолжительность стадии удерживания может быть значительно уменьшена путем проведения стадии удерживания при повышенной температуре, при этом максимальная температура ограничивается стабильностью конъюгата связывающего клетку средства и цитотоксического средства. Стадия удерживания может предусматривать поддержание раствора в течение от приблизительно 1 часа до приблизительно 1 дня (например, приблизительно 1 часа, приблизительно 2 часов, приблизительно 3 часов, приблизительно 4 часов, приблизительно 5 часов, приблизительно 6 часов, приблизительно 7 часов, приблизительно 8 часов, приблизительно 9 часов, приблизительно 10 часов, приблизительно 12 часов, приблизительно 14 часов, приблизительно 16 часов, приблизительно 18 часов, приблизительно 20 часов, приблизительно 22 часов или приблизительно 24 часов), от приблизительно 10 часов до приблизительно 24 часов, от приблизительно 12 часов до приблизительно 24 часов, от приблизительно 14 часов до приблизительно 24 часов, от приблизительно 16 часов до приблизительно 24 часов, от приблизительно 18 часов до приблизительно 24 часов, от приблизительно 20 часов до приблизительно 24 часов, от приблизительно 5 часов до приблизительно 1 недели, от приблизительно 20 часов до приблизительно 1 недели, от приблизительно 12 часов до приблизительно 1 недели (например, приблизительно 12 часов, приблизительно 16 часов, приблизительно 20 часов, приблизительно 24 часов, приблизительно 2 дней, приблизительно 3 дней, приблизительно 4 дней, приблизительно 5 дней, приблизительно 6 дней или приблизительно 7 дней) или от приблизительно 1 дня до приблизительно 1 недели.
[00202] В одном варианте осуществления стадия удерживания предусматривает поддержание раствора при температуре от приблизительно 2°C до приблизительно 8°C в течение периода, составляющего от по меньшей мере приблизительно 12 часов вплоть до недели. В другом варианте осуществления стадия удерживания предусматривает поддержание раствора при температуре от приблизительно 2°C до приблизительно 8°C в течение ночи (например, от приблизительно 12 до приблизительно 24 часов, предпочтительно приблизительно 20 часов).
[00203] Значение pH для стадии удерживания предпочтительно составляет от приблизительно 4 до приблизительно 10. В одном варианте осуществления значение pH для стадии удерживания составляет приблизительно 4 или больше, но менее приблизительно 6 (например, от 4 до 5,9), или приблизительно 5 или больше, но менее приблизительно 6 (например, от 5 до 5,9). В другом варианте осуществления значения pH для стадии удерживания находятся в диапазоне от приблизительно 6 до приблизительно 10 (например, от приблизительно 6,5 до приблизительно 9, от приблизительно 6 до приблизительно 8). Например, значения pH для стадии удерживания могут составлять приблизительно 6, приблизительно 6,5, приблизительно 7, приблизительно 7,5, приблизительно 8, приблизительно 8,5, приблизительно 9, приблизительно 9,5, или приблизительно 10.
[00204] В конкретных вариантах осуществления стадия удерживания может предусматривать инкубирование смеси при 25°C при pH приблизительно 6-7,5 в течение от приблизительно 12 часов до приблизительно 1 недели, инкубирование смеси при 4°C при pH приблизительно 4,5-5,9 в течение от приблизительно 5 часов до приблизительно 5 дней или инкубирование смеси при 25°C при pH приблизительно 4,5-5,9 в течение от приблизительно 5 часов до приблизительно 1 дня.
[00205] Одностадийный способ необязательно может предусматривать добавление сахарозы на стадии реакции для повышения растворимости и извлечения конъюгатов. Желательно сахарозу добавляют при концентрации от приблизительно 0,1% (вес/объем) до приблизительно 20% (вес/объем) (например, приблизительно 0,1% (вес/объем), 1% (вес/объем), 5% (вес/объем), 10% (вес/объем), 15% (вес/объем) или 20% (вес/объем)). Предпочтительно сахарозу добавляют при концентрации от приблизительно 1% (вес/объем) до приблизительно 10% (вес/объем) (например, приблизительно 0,5% (вес/объем), приблизительно 1% (вес/объем), приблизительно 1,5% (вес/объем), приблизительно 2% (вес/объем), приблизительно 3% (вес/объем), приблизительно 4% (вес/объем), приблизительно 5% (вес/объем), приблизительно 6% (вес/объем), приблизительно 7% (вес/объем), приблизительно 8% (вес/объем), приблизительно 9% (вес/объем), приблизительно 10% (вес/объем) или приблизительно 11% (вес/объем)). Кроме того, стадия реакции также может предусматривать добавление буферного средства. Можно применять любое подходящее буферное средство, известное из уровня техники. Подходящие буферные средства включают, например, цитратный буфер, ацетатный буфер, сукцинатный буфер и фосфатный буфер. В одном варианте осуществления буферное средство выбрано из группы, состоящей из HEPPSO (N-(2-гидроксиэтил)пиперазин-N'-(2-гидроксипропансульфоновая кислота)), POPSO (дегидрат пиперазин-1,4-бис-(2-гидроксипропансульфоновой кислоты)), HEPES (4-(2-гидроксиэтил)пиперазин-1-этансульфоновая кислота), HEPPS (EPPS) (4-(2-гидроксиэтил)пиперазин-1-пропансульфоновая кислота), TES (N-[трис(гидроксиметил)метил]-2-аминоэтансульфоновая кислота) и их комбинации.
[00206] В одном варианте осуществления одностадийный способ может дополнительно предусматривать стадию проведения очистки смеси с получением очищенного конъюгата (например, Ab-MCC-DM1, Ab-SPDB-DM4 или Ab-CX1-1-DM1). Любые способы очистки, известные из уровня техники, можно применять для очистки конъюгатов по настоящему изобретению. В одном варианте осуществления конъюгаты по настоящему изобретению очищают с применением тангенциальной поточной фильтрации (TFF), неадсорбционной хроматографии, адсорбционной хроматографии, адсорбционной фильтрации, селективного осаждения или любого другого подходящего способа очистки, а также их комбинации. В другом варианте осуществления перед воздействием на конъюгаты способа очистки, описанного выше, конъюгаты вначале фильтруют через одну или несколько мембран из PVDF. В качестве альтернативы конъюгаты фильтруют через одну или несколько мембран из PVDF после воздействия на конъюгаты способа очистки, описанного выше. Например, в одном варианте осуществления конъюгаты фильтруют через одну или несколько мембран из PVDF, а затем очищают с применением тангенциальной поточной фильтрации. В качестве альтернативы конъюгаты очищают с применением тангенциальной поточной фильтрации, а затем фильтруют через одну или несколько мембран из PVDF.
[00207] Для очистки можно использовать любые подходящие TFF-системы, в том числе систему типа Pellicon (Millipore, Биллерика, Массачусетс), кассетную систему Sartocon (Sartorius AG, Эджвуд, Нью-Йорк) и систему типа Centrasette (Pall Corp., Ист-Хиллс, Нью-Йорк).
[00208] Для очистки можно использовать любую подходящую смолу для адсорбционной хроматографии. Предпочтительные смолы для адсорбционной хроматографии предусматривают хроматографию на гидроксиапатите, гидрофобную хроматографию с индуцированием заряда (HCIC), хроматографию гидрофобных взаимодействий (HIC), ионообменную хроматографию, ионообменную хроматографию с комбинированным режимом работы, аффинную хроматографию на иммобилизованных ионах металла (IMAC), хроматографию с лигандом красителем, аффинную хроматографию, хроматографию с обращенной фазой и их комбинации. Примеры подходящих гидроксиапатитных смол включают керамический гидроксиапатит (CHT типа I и типа II, Bio-Rad Laboratories, Геркулес, Калифорния), гидроксиапатит HA Ultrogel (Pall Corp., Ист-Хиллс, Нью-Йорк) и керамический фторапатит (CFT типа I и типа II, Bio-Rad Laboratories, Геркулес, Калифорния). Примером подходящей смолы для HCIC является смола MEP Hypercel (Pall Corp., Ист-Хиллс, Нью-Йорк). Примеры подходящих смол для HIC включают смолы на основе бутил-сефарозы, гексил-сефарозы, фенил-сефарозы и октил-сефарозы (все от GE Healthcare, Пискатавей, Нью-Джерси), а также метиловую смолу Macro-prep и трет-бутиловую смолу Macro-Prep (Biorad Laboratories, Геркулес, Калифорния). Примеры подходящих ионообменных смол включают смолы на основе SP-сефарозы, CM-сефарозы и Q-сефарозы (все от GE Healthcare, Пискатавей, Нью-Джерси) и смолу Unosphere S (Bio-Rad Laboratories, Геркулес, Калифорния). Примеры подходящих смолы для ионообменной хроматографии с комбинированным режимом работы включают смолу Bakerbond ABx (JT Baker, Филлипсбург, Нью-Джерси). Примеры подходящих смол для IMAC включают смолу Chelating Sepharose (GE Healthcare, Пискатавей, Нью-Джерси) и смолу Profinity IMAC (Bio-Rad Laboratories, Геркулес, Калифорния). Примеры подходящих смол для хроматографии с лигандом красителем включают смолу Blue Sepharose (GE Healthcare, Пискатавей, Нью-Джерси) и смолу Affi-gel Blue (Bio-Rad Laboratories, Геркулес, Калифорния). Примеры подходящих смол для аффинной хроматографии включают смолу на основе сефарозы с белком A (например, MabSelect, GE Healthcare, Пискатавей, Нью-Джерси) и смолы для аффинной хроматографии с лектинами, например, смолу на основе сефарозы с лектином чечевицы (GE Healthcare, Пискатавей, Нью-Джерси), где антитело несет подходящие сайты связывания лектина. Примеры подходящих смол для хроматографии с обращенной фазой включают смолы C4, C8 и C18 (Grace Vydac, Хесперия, Калифорния).
[00209] Для очистки можно использовать любую подходящую смолу для неадсорбционной хроматографии. Примеры подходящих смол для неадсорбционной хроматографии включают без ограничения смолы SEPHADEX™ G-25, G-50, G-100, SEPHACRYL™ (например, S-200 и S-300), смолы SUPERDEX™ (например, SUPERDEX™ 75 и SUPERDEX™ 200), смолы BIO-GEL® (например, P-6, P-10, P-30, P-60 и P-100) и другие, известные рядовым специалистам в данной области техники.
Двухстадийный способ и способ в одном реакционном сосуде для сшивания с лизиновыми остатками антитела
[00210] В одном варианте осуществления конъюгаты по настоящему изобретению могут быть получены, как описано в патенте США № 7811572 и публикации заявки на патент США № 2006/0182750. Способ предусматривает стадии: (a) приведения антитела по настоящему изобретению в контакт со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1) с присоединением с помощью ковалентной связи линкера (т.е. Ab-SMCC, Ab-SPDB или Ab-CX1-1) к антителу и тем самым получения первой смеси, содержащей антитело, имеющее связанный с ним линкер; (b) необязательно воздействия на первую смесь способа очистки с получением очищенной первой смеси из антитела, имеющего связанный с ним линкер; (c) конъюгирования лекарственного средства (например, DM1, DM3 или DM4) с антителом, имеющим связанный с ним линкер, в первой смеси путем осуществления реакции антитела, имеющего связанный с ним линкер, с лекарственным средством (например, DM1, DM3 или DM4) в растворе, характеризующемся pH от приблизительно 4 до приблизительно 9, с получением второй смеси, содержащей (i) конъюгат (например, Ab-MCC-DM1, Ab-SPDB-DM4 или Ab-CX1-1-DM1), (ii) свободное лекарственное средство (например, DM1, DM3 или DM4) и (iii) промежуточные продукты реакции; и (d) воздействия на вторую смесь способа очистки с очисткой конъюгата от других компонентов второй смеси. В качестве альтернативы стадия (b) очистки может не проводиться. На стадиях (b) и (d) могут применяться любые способы очистки, описанные в данном документе. В одном варианте осуществления на обеих стадиях (b) и (d) применяется TFF. В другом варианте осуществления TFF применяется на стадии (b), а адсорбционная хроматография (например, CHT) применяется на стадии (d).
Реагент для одностадийного способа и in-situ способ для сшивания с лизиновыми остатками антитела
[00211] В одном варианте осуществления конъюгаты по настоящему изобретению могут быть получены путем конъюгирования предварительно образованного соединения линкер-лекарственное средство (например, SMCC-DM1, сульфо-SMCC-DM1, SPDB-DM4 или CX1-1-DM1) с антителом по настоящему изобретению, как описано в патенте США № 6441163 и публикации заявки на патент США №№ 2011/0003969 и 2008/0145374, с последующей стадией очистки. Можно применять любые способы очистки, описанные в данном документе. Соединение линкер-лекарственное средство получают путем осуществления реакции лекарственного средства (например, DM1, DM3 или DM4) со сшивающим средством (например, SMCC, сульфо-SMCC, SPDB, сульфо-SPDB или CX1-1). Соединение линкер-лекарственное средство (например, SMCC-DM1, сульфо-SMCC-DM1, SPDB-DM4 или CX1-1-DM1) необязательно подвергают очистке перед конъюгированием с антителом.
Антитела к CCR7
[00212] В настоящем изобретении представлены антитела или фрагменты антител (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7. Антитела или фрагменты антител (например, антигенсвязывающие фрагменты) по настоящему изобретению включают без ограничения человеческие моноклональные антитела или их фрагменты, выделенные, как описано в примерах.
[00213] В некоторых вариантах осуществления настоящего изобретения представлены антитела или фрагменты антител (например, антигенсвязывающие фрагменты), которые специфически связывают CCR7, при этом указанные антитела или фрагменты антител (например, антигенсвязывающие фрагменты) содержат домен VH, имеющий аминокислотную последовательность под SEQ ID NO: 13, 45, 77 или 608. В некоторых вариантах осуществления настоящего изобретения также представлены антитела или фрагменты антител (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, при этом указанные антитела или фрагменты антител (например, антигенсвязывающие фрагменты) содержат CDR VH, имеющую аминокислотную последовательность любой из CDR VH, перечисленных в таблицах 1 и 4, ниже. В конкретных вариантах осуществления настоящего изобретения представлены антитела или фрагменты антител (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, при этом указанные антитела содержат (или в качестве альтернативы состоят из) одну, две, три, четыре, пять или больше CDR VH, имеющих аминокислотную последовательность любой из CDR VH, перечисленных в таблицах 1 и 4, ниже.
[00214] В настоящем изобретении представлены антитела или фрагменты антител (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, при этом указанные антитела или фрагменты антител (например, антигенсвязывающие фрагменты) содержат домен VL, имеющий аминокислотную последовательность под SEQ ID NO: 29, 61, 93 или 624. В настоящем изобретении также представлены антитела или фрагменты антител (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, при этом указанные антитела или фрагменты антител (например, антигенсвязывающие фрагменты) содержат CDR VL, имеющую аминокислотную последовательность любой из CDR VL, перечисленных в таблицах 1 и 4 ниже. В частности, в настоящем изобретении представлены антитела или фрагменты антител (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, при этом указанные антитела или фрагменты антител (например, антигенсвязывающие фрагменты) содержат (или в качестве альтернативы состоять из) одну, две, три или больше CDR VL, имеющих аминокислотную последовательность любой из CDR VL, перечисленных в таблицах 1 и 4, ниже.
[00215] Другие антитела или фрагменты антител (например, антигенсвязывающие фрагменты) по настоящему изобретению включают аминокислоты, которые были подвергнуты мутации, но при этом они характеризуются по меньшей мере 60, 70, 80, 90 или 95 процентной идентичностью в областях CDR с областями CDR, показанными в последовательностях, описанных в таблицах 1 и 4. В некоторых вариантах осуществления антитела содержат мутантные аминокислотные последовательности, в которых не более 1, 2, 3, 4 или 5 аминокислот были подвергнуты мутации в областях CDR в сравнении с областями CDR, показанными в последовательности, описанной в таблицах 1 и 4.
[00216] В настоящем изобретении также представлены последовательности нуклеиновой кислоты, которые кодируют VH, VL, полноразмерную тяжелую цепь и полноразмерную легкую цепь антител, которые специфически связываются с CCR7. Такие последовательности нуклеиновой кислоты могут быть оптимизированы для экспрессии в клетках млекопитающих.
[00217] На протяжении текста настоящей заявки может встречаться несоответствие между текстом описания и перечнем последовательностей, при этом текст описания имеет преимущественную силу.
Таблица 1. Примеры антител к CCR7 по настоящему изобретению
(DAPA, CysMab, мутации подчеркнуты)
(DAPA, CysMab, мутации подчеркнуты)
(DAPA, CysMab, мутации подчеркнуты)
[00218] Другие антитела по настоящему изобретению включают антитела, в которых аминокислоты или нуклеиновые кислоты, кодирующие аминокислоты, были подвергнуты мутации, но при этом они характеризуются по меньшей мере 60, 70, 80, 90 или 95 процентной идентичностью с последовательностями, описанными в таблицах 1 и 4. В некоторых вариантах осуществления 1, 2, 3, 4 или 5 аминокислот были подвергнуты мутации в вариабельных областях по сравнению с вариабельными областями, показанными в последовательности, описанной в таблицах 1 и 4, при этом они сохраняют практически такую же терапевтическую активность, что и антитела, перечисленные в таблицах 1 и 4.
[00219] Поскольку каждое из данных антител может связываться с CCR7 последовательности (аминокислотные последовательности и нуклеотидные последовательности, кодирующие аминокислотные последовательности) VH, VL, полноразмерной легкой цепи и полноразмерной тяжелой цепи можно подвергать "смешиванию и подбору" для создания других CCR7-связывающих антител по настоящему изобретению. Такие подвергнутые "смешиванию и подбору" CCR7-связывающие антитела можно тестировать с применением анализов связывания, известных из уровня техники (например, разновидностей ELISA и других анализов, описанных в разделе Примеры). Когда такие цепи подвергают смешиванию и подбору, последовательность VH из конкретной пары VH/VL следует заменить структурно подобной последовательностью VH. Аналогичным образом, последовательность полноразмерной тяжелой цепи из конкретной пары полноразмерная тяжелая цепь/полноразмерная легкая цепь следует заменить структурно подобной последовательностью полноразмерной тяжелой цепи. Аналогичным образом, последовательность VL из конкретной пары VH/VL следует заменить структурно подобной последовательностью VL. Аналогичным образом, последовательность полноразмерной легкой цепи из конкретной пары полноразмерная тяжелая цепь/полноразмерная легкая цепь следует заменить структурно подобной последовательностью полноразмерной легкой цепи. Соответственно, в одном аспекте настоящего изобретения представлены выделенные моноклональное антитело или его антигенсвязывающая область, имеющие: вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 13, 45, 77 и 608; и вариабельную область легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 29, 61, 93 и 624; где антитело специфически связывается с CCR7.
[00220] В другом аспекте настоящего изобретения представлены: (i) выделенное моноклональное антитело, имеющее: полноразмерную тяжелую цепь, содержащую аминокислотную последовательность, которая была оптимизирована для экспрессии в системе экспрессии, представляющей собой клетку млекопитающего, выбранную из группы, состоящей из SEQ ID NO: 15, 47, 79 и 610; и полноразмерную легкую цепь, содержащую аминокислотную последовательность, которая была оптимизирована для экспрессии в клетке млекопитающего, выбранную из группы, состоящей из SEQ ID NO: 31, 63, 95 и 626; или (ii) функциональный белок, содержащий его антигенсвязывающую часть.
[00221] В другом аспекте настоящего изобретения представлены CCR7-связывающие антитела, которые содержат CDR1, CDR2 и CDR3 тяжелой цепи и легкой цепи, описанные в таблицах 1 и 4, или их комбинации. Аминокислотные последовательности CDR1 VH антител показаны, например, под SEQ ID NO: 1, 4, 7, 10, 33, 36, 39, 42, 65, 68, 71 и 74. Аминокислотные последовательности CDR2 VH антител показаны, например, под SEQ ID NO: 2, 5, 8, 11, 34, 37, 40, 43, 66, 69, 72 и 75. Аминокислотные последовательности CDR3 VH антител показаны, например, под SEQ ID NO: 3, 6, 9, 12, 35, 38, 41, 44, 67, 70, 73 и 76. Аминокислотные последовательности CDR1 VL антител показаны, например, под SEQ ID NO: 17, 20, 23, 26, 49, 52, 55, 58, 81, 84, 87 и 90. Аминокислотные последовательности CDR2 VL антител показаны, например, под SEQ ID NO: 18, 21, 24, 27, 50, 53, 56, 59, 82, 85, 88 и 91. Аминокислотные последовательности CDR3 VL антител показаны, например, под SEQ ID NO: 19, 22, 25, 28, 51, 54, 57, 60, 83, 86, 89 и 92.
[00222] С учетом того, что каждое из данных антител может связываться с CCR7, и что специфичность связывания антигена в первую очередь обеспечивается областями CDR1, 2 и 3, последовательности CDR1, CDR2 и CDR3 VH и последовательности CDR1, CDR2 и CDR3 VL могут быть подвергнуты "смешиванию и подбору" (т.е. CDR из различных антител могут быть подвергнуты смешиванию и подбору. Такие подвергнутые "смешиванию и подбору" CCR7-связывающие антитела можно тестировать с применением анализов связывания, известных из уровня техники и описанных в Примерах (например, разновидностей ELISA). Когда последовательности CDR VH подвергают смешиванию и подбору, последовательность CDR1, CDR2 и/или CDR3 из конкретной последовательности VH следует заменить структурно подобной(-ыми) последовательностью(-ими) CDR. Аналогичным образом, когда последовательности CDR VL подвергают смешиванию и подбору, последовательность CDR1, CDR2 и/или CDR3 из конкретной последовательности VL следует заменить структурно подобной(-ыми) последовательностью(-ями) CDR. Рядовому специалисту в данной области техники будет совершенно очевидно, что новые последовательности VH и VL можно создавать путем замены одной или нескольких последовательностей областей CDR VH и/или VL структурно подобными последовательностями из последовательностей CDR, показанных в данном документе для моноклональных антител по настоящему изобретению.
[00223] Соответственно, в некоторых вариантах осуществления настоящего изобретения представлены выделенные моноклональное антитело или его антигенсвязывающая область, содержащие CDR1 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, 4, 7, 10, 33, 36, 39, 42, 65, 68, 71, 74, 596, 599, 602 и 605; CDR2 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, 5, 8, 11, 34, 37, 40, 43, 66, 69, 72, 75, 597, 600, 603 и 606; CDR3 тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 3, 6, 9, 12, 35, 38, 41, 44, 67, 70, 73, 76, 598, 601, 604 и 607; CDR1 легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 17, 20, 23, 26, 49, 52, 55, 58, 81, 84, 87, 90, 612, 615, 618 и 621; CDR2 легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 18, 21, 24, 27, 50, 53, 56, 59, 82, 85, 88, 91, 613, 616, 619 и 622; и CDR3 легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 19, 22, 25, 28, 51, 54, 57, 60, 83, 86, 89, 92, 614, 617, 620 и 623; где антитело специфически связывает CCR7.
[00224] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат CDR1 тяжелой цепи под SEQ ID NO:1, CDR2 тяжелой цепи под SEQ ID NO:2; CDR3 тяжелой цепи под SEQ ID NO:3; CDR1 легкой цепи под SEQ ID NO:17; CDR2 легкой цепи под SEQ ID NO:18 и CDR3 легкой цепи под SEQ ID NO:19.
[00225] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат CDR1 тяжелой цепи под SEQ ID NO:4, CDR2 тяжелой цепи под SEQ ID NO:5; CDR3 тяжелой цепи под SEQ ID NO:6; CDR1 легкой цепи под SEQ ID NO:20; CDR2 легкой цепи под SEQ ID NO:21 и CDR3 легкой цепи под SEQ ID NO:22.
[00226] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат CDR1 тяжелой цепи под SEQ ID NO:7, CDR2 тяжелой цепи под SEQ ID NO:8; CDR3 тяжелой цепи под SEQ ID NO:9; CDR1 легкой цепи под SEQ ID NO:23; CDR2 легкой цепи под SEQ ID NO:24 и CDR3 легкой цепи под SEQ ID NO:25.
[00227] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат CDR1 тяжелой цепи под SEQ ID NO:10, CDR2 тяжелой цепи под SEQ ID NO:11; CDR3 тяжелой цепи под SEQ ID NO:12; CDR1 легкой цепи под SEQ ID NO:26; CDR2 легкой цепи под SEQ ID NO:27 и CDR3 легкой цепи под SEQ ID NO:28.
[00228] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат CDR1 тяжелой цепи под SEQ ID NO:33, CDR2 тяжелой цепи под SEQ ID NO:34; CDR3 тяжелой цепи под SEQ ID NO:35; CDR1 легкой цепи под SEQ ID NO:49; CDR2 легкой цепи под SEQ ID NO:50 и CDR3 легкой цепи под SEQ ID NO:51.
[00229] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат CDR1 тяжелой цепи под SEQ ID NO:36, CDR2 тяжелой цепи под SEQ ID NO:37; CDR3 тяжелой цепи под SEQ ID NO:38; CDR1 легкой цепи под SEQ ID NO:52; CDR2 легкой цепи под SEQ ID NO:53 и CDR3 легкой цепи под SEQ ID NO:54.
[00230] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат CDR1 тяжелой цепи под SEQ ID NO:39, CDR2 тяжелой цепи под SEQ ID NO:40; CDR3 тяжелой цепи под SEQ ID NO:41; CDR1 легкой цепи под SEQ ID NO:55; CDR2 легкой цепи под SEQ ID NO:56 и CDR3 легкой цепи под SEQ ID NO:57.
[00231] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат CDR1 тяжелой цепи под SEQ ID NO:42, CDR2 тяжелой цепи под SEQ ID NO:43; CDR3 тяжелой цепи под SEQ ID NO:44; CDR1 легкой цепи под SEQ ID NO:58; CDR2 легкой цепи под SEQ ID NO:59 и CDR3 легкой цепи под SEQ ID NO:60.
[00232] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат CDR1 тяжелой цепи под SEQ ID NO:65, CDR2 тяжелой цепи под SEQ ID NO:66; CDR3 тяжелой цепи под SEQ ID NO:67; CDR1 легкой цепи под SEQ ID NO:81; CDR2 легкой цепи под SEQ ID NO:82 и CDR3 легкой цепи под SEQ ID NO:83.
[00233] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат CDR1 тяжелой цепи под SEQ ID NO:68, CDR2 тяжелой цепи под SEQ ID NO:69; CDR3 тяжелой цепи под SEQ ID NO:70; CDR1 легкой цепи под SEQ ID NO:84; CDR2 легкой цепи под SEQ ID NO:85 и CDR3 легкой цепи под SEQ ID NO:86.
[00234] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат CDR1 тяжелой цепи под SEQ ID NO:71, CDR2 тяжелой цепи под SEQ ID NO:72; CDR3 тяжелой цепи под SEQ ID NO:73; CDR1 легкой цепи под SEQ ID NO:87; CDR2 легкой цепи под SEQ ID NO:88 и CDR3 легкой цепи под SEQ ID NO:89.
[00235] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат CDR1 тяжелой цепи под SEQ ID NO:74, CDR2 тяжелой цепи под SEQ ID NO:75; CDR3 тяжелой цепи под SEQ ID NO:76; CDR1 легкой цепи под SEQ ID NO:90; CDR2 легкой цепи под SEQ ID NO:91 и CDR3 легкой цепи под SEQ ID NO:92.
[00236] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:596, HCDR2 под SEQ ID NO:597 и HCDR3 под SEQ ID NO:598; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:612, LCDR2 под SEQ ID NO:613 и LCDR3 под SEQ ID NO:614.
[00237] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:599, HCDR2 под SEQ ID NO:600 и HCDR3 под SEQ ID NO:601; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:615, LCDR2 под SEQ ID NO:616 и LCDR3 под SEQ ID NO:617.
[00238] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:602, HCDR2 под SEQ ID NO:603 и HCDR3 под SEQ ID NO:604; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:618, LCDR2 под SEQ ID NO:619 и LCDR3 под SEQ ID NO:620.
[00239] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:605, HCDR2 под SEQ ID NO:606 и HCDR3 под SEQ ID NO:607; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:621, LCDR2 под SEQ ID NO:622 и LCDR3 под SEQ ID NO:623.
[00240] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:13, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:29.
[00241] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:45, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:61.
[00242] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:77, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:93.
[00243] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:608, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:624.
[00244] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:15, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:31.
[00245] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:47, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:63.
[00246] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:79, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:95.
[00247] В конкретном варианте осуществления антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), которые специфически связываются с CCR7, содержат тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:610, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:626
[00248] В некоторых вариантах осуществления антитело, которое специфически связывается с CCR7, представляет собой антитело или фрагмент антитела (например, антигенсвязывающий фрагмент), которые описаны в таблицах 1 и 4.
1. Идентификация эпитопов и антител, которые связываются с одним и тем же эпитопом
[00249] В настоящем изобретении также представлены антитела и фрагменты антител (например, антигенсвязывающие фрагменты), которые специфически связываются с тем же эпитопом, что и антитела к CCR7, описанные в таблицах 1 и 4, или вступает в перекрестную конкуренцию с антителами, описанными в таблицах 1 и 4. Следовательно, дополнительные антитела и фрагменты антител (например, антигенсвязывающие фрагменты) можно идентифицировать на основе их способности к перекрестной конкуренции (например, к конкурентному ингибированию связывания, статистически значимым образом) с другими антителами по настоящему изобретению в анализах связывания CCR7, например, с помощью BIACORE или анализов, известных специалистам в данной области техники для измерения связывания. Способность тестируемого антитела ингибировать связывание антител и фрагментов антител (например, антигенсвязывающих фрагментов) по настоящему изобретению с CCR7 (например, CCR7 человека) демонстрирует, что тестируемое антитело может конкурировать с таким антителом или фрагментом антитела (например, антигенсвязывающими фрагментами) за связывание с CCR7; при этом, согласно неограничивающей теории, такое антитело может связываться с тем же или родственным (например, структурно подобным или пространственно близким или перекрывающимся) эпитопом на белке CCR7, что и антитело или фрагмент антитела (например, антигенсвязывающие фрагменты), с которым оно конкурирует. В некоторых вариантах осуществления антитела, которые связываются с тем же эпитопом на CCR7, что и антитела или фрагменты антител (например, антигенсвязывающие фрагменты), описанные в таблицах 1 и 4, представляют собой человеческие или гуманизированные моноклональные антитела. Такие человеческие или гуманизированные моноклональные антитела могут быть получены и выделены, как описано в данном документе.
2. Дополнительное изменение каркасной области в области Fc
[00250] Иммуноконъюгаты по настоящему изобретению могут содержать модифицированные антитела или их антигенсвязывающие фрагменты, которые дополнительно содержат модификации каркасных остатков в пределах VH и/или VL, например, для улучшения свойств антитела. В некоторых вариантах осуществления модификации каркасной области осуществляют для снижения иммуногенности антитела. Например, один подход заключается в том, что один или несколько каркасных остатков "мутируют к первоначальному виду" в направлении соответствующей последовательности зародышевой линии. Более конкретно, антитело, которое подверглось соматической мутации, может содержать каркасные остатки, которые отличаются от последовательности зародышевой линии, из которой происходит антитело. Такие остатки могут быть идентифицированы посредством сравнения каркасных последовательностей антитела с последовательностями зародышевой линии, из которых происходит антитело. Чтобы вернуть последовательности каркасной области в их конфигурацию зародышевой линии, соматические мутации можно вводить для "мутирования к первоначальному виду" в последовательности зародышевой линии, например, с помощью сайт-направленного мутагенеза. Подразумевается, что такие подвергнутые "мутации к первоначальному виду" антитела также охватываются настоящим изобретением.
[00251] Другой тип модификации каркасной области предусматривает мутацию одного или нескольких остатков в пределах каркасной области или даже в пределах одной или нескольких областей CDR, чтобы удалить T-клеточные эпитопы, со снижением тем самым потенциальной иммуногенности антитела. Данный подход также называют "деиммунизацией", и он более подробно описан в публикации заявки на патент США № 20030153043 от Carr et al.
[00252] В качестве дополнения или альтернативы модификациям, осуществляемым в пределах каркасных областей или областей CDR, антитела по настоящему изобретению можно конструировать таким образом, чтобы они включали модификации в пределах области Fc, как правило, для изменения одного или нескольких функциональных свойств антитела, таких как период полужизни в сыворотке крови, фиксация комплемента, связывание с Fc-рецептором и/или антигензависимая клеточная цитотоксичность (ADCC). Кроме того, антитело по настоящему изобретению может быть модифицировано химически (например, к антителу можно присоединять один или несколько химических фрагментов), или оно может быть модифицировано для изменения характера его гликозилирования, чтобы, опять-таки, изменить одно или несколько функциональных свойств антитела. Каждый из данных вариантов осуществления более подробно описан ниже.
[00253] В одном варианте осуществления шарнирную область CH1 модифицируют таким образом, что меняется число цистеиновых остатков в шарнирной области, например, увеличивается или уменьшается. Данный подход дополнительно описан в патенте США № 5677425 от Bodmer et al. Число цистеиновых остатков в шарнирной области CH1 изменяют, например, чтобы облегчить сборку легкой и тяжелой цепей, или чтобы увеличить или уменьшить стабильность антитела.
[00254] В некоторых вариантах осуществления антитело или фрагмент антитела, раскрытые в данном документе, содержат модифицированные или введенные путем конструирования аминокислотные остатки, например, один или несколько цистеиновых остатков, в качестве сайтов для конъюгации с фрагментом, представляющим собой лекарственное средство (Junutula JR, et al., Nat Biotechnol 2008, 26:925-932). В одном варианте осуществления настоящего изобретения представлены модифицированные антитело или фрагмент антитела, содержащие замену одной или нескольких аминокислот на цистеин в положениях, описанных в данном документе. Сайты для цистеиновой замены расположены в константных областях антитела или фрагмента антитела и, таким образом, применимы к множеству антител или фрагментов антител, и сайты выбирают для обеспечения стабильных и однородных конъюгатов. Модифицированные антитело или фрагмент могут иметь одну, две или более цистеиновых замен, и такие замены могут применяться в комбинации с другими способами модификации и конъюгации, описанными в данном документе. Способы для вставки цистеина в специфические местоположения антитела известны из уровня техники, см., например, Lyons et al, (1990) Protein Eng., 3:703-708, WO 2011/005481, WO2014/124316, WO 2015/138615. В некоторых вариантах осуществления модифицированное антитело содержит замену одной или нескольких аминокислот на цистеин в своей константной области в положении, выбранном из положений 117, 119, 121, 124, 139, 152, 153, 155, 157, 164, 169, 171, 174, 189, 191, 195, 197, 205, 207, 246, 258, 269, 274, 286, 288, 290, 292, 293, 320, 322, 326, 333, 334, 335, 337, 344, 355, 360, 375, 382, 390, 392, 398, 400 и 422 тяжелой цепи антитела, и где нумерация положений соответствует системе EU. В некоторых вариантах осуществления модифицированные антитело или фрагмент антитела содержат замену одной или нескольких аминокислот на цистеин в своей константной области в положении, выбранном из положений 107, 108, 109, 114, 129, 142, 143, 145, 152, 154, 156, 159, 161, 165, 168, 169, 170, 182, 183, 197, 199, и 203 легкой цепи антитела или фрагмента антитела, где нумерация положений соответствует системе EU, и где легкая цепь представляет собой человеческую каппа-легкую цепь. В некоторых вариантах осуществления модифицированные антитело или фрагмент такого антитела содержат комбинацию из замен двух или более аминокислот на цистеин в своих константных областях, где комбинации предусматривают замены в положениях 375 тяжелой цепи антитела, положении 152 тяжелой цепи антитела, положении 360 тяжелой цепи антитела или положении 107 легкой цепи антитела, и где нумерация положений соответствует системе EU. В некоторых вариантах осуществления модифицированные антитело или фрагмент такого антитела содержат замену одной аминокислоты на цистеин в своих константных областях, где замена находится в положении 375 тяжелой цепи антитела, положении 152 тяжелой цепи антитела, положении 360 тяжелой цепи антитела, положении 107 легкой цепи антитела, положении 165 легкой цепи антитела или положении 159 легкой цепи антитела, и где нумерация положений соответствует системе EU, и где легкая цепь представляет собой каппа-цепь. В конкретных вариантах осуществления модифицированные антитело или фрагмент такого антитела содержат комбинацию из замен двух аминокислот на цистеин в своих константных областях, где комбинации предусматривают замены в положениях 375 тяжелой цепи антитела и положении 152 тяжелой цепи антитела, где нумерация положений соответствует системе EU. В конкретных вариантах осуществления модифицированные антитело или фрагмент такого антитела содержат замену одной аминокислоты на цистеин в положении 360 тяжелой цепи антитела, где нумерация положений соответствует системе EU. В других конкретных вариантах осуществления модифицированные антитело или фрагмент такого антитела содержат замену одной аминокислоты на цистеин в положении 107 легкой цепи антитела, и где нумерация положений соответствует системе EU, и где легкая цепь представляет собой каппа-цепь.
[00255] В дополнительных вариантах осуществления антитела или фрагменты антител (например, антигенсвязывающий фрагмент), применимые в иммуноконъюгатах по настоящему изобретению, включают модифицированные или сконструированные антитела, такие как антитело, модифицированное для введения одной или нескольких других реакционноспособных аминокислот (отличных от цистеина), в том числе Pcl, пирролизина, пептидных меток (таких как метки S6, A1 и ybbR) и неприродных аминокислот, вместо по меньшей мере одной аминокислоты из нативной последовательности, тем самым обеспечивая реакционноспособный сайт на антителе или антигенсвязывающем фрагменте для конъюгации с фрагментом, представляющим собой лекарственное средство, или фрагментом, представляющим собой линкер-лекарственное средство, с комплементарной реакционной группой. Например, антитела или фрагменты антител могут быть модифицированы для введения Pcl или пирролизина (W. Ou, et al., (2011) PNAS 108 (26), 10437-10442; WO2014124258) или неприродных аминокислот (J.Y. Axup, et al., Proc Natl Acad Sci U S A, 109 (2012), pp. 16101-16106; обзор см. в C.C. Liu and P.G. Schultz (2010) Annu Rev Biochem 79, 413-444; C.H. Kim, et al., (2013) Curr Opin Chem Biol. 17, 412-419) в качестве сайтов для конъюгации с лекарственным средством. Аналогичным образом в антитело могут быть введены пептидные метки для способов ферментативной конъюгации (Strop P., et al., Chem Biol. 2013, 20(2):161-7; Rabuka D., Curr Opin Chem Biol. 2010 Dec;14(6):790-6; Rabuka D, et al., Nat Protoc. 2012, 7(6):1052-67). Одним другим примером является применение 4'-фосфопантетеинилтрансфераз (PPTаза) для конъюгации аналогов кофермента A (WO2013184514) и (Grünewald et al., (2015) Bioconjugate Chem. 26 (12), 2554-62). Способы конъюгации таких модифицированных или сконструированных антител с полезными нагрузками или комбинациями линкер-полезная нагрузка известны из уровня техники.
[00256] В другом варианте осуществления для снижения биологического периода полужизни антитела подвергают мутации область Fc-шарнир антитела. Более конкретно, одну или несколько аминокислотных мутаций вводят в пограничную область между доменами CH2-CH3 фрагмента Fc-шарнир, вследствие чего антитело характеризуется нарушенным связыванием с белком A стафилококка (SpA) по сравнению со связыванием нативного домена Fc-шарнир с SpA. Данный подход более подробно описан в патенте США № 6165745 от Ward et al.
[00257] В других вариантах осуществления область Fc изменяют путем замены по меньшей мере одного аминокислотного остатка на отличный аминокислотный остаток для изменения эффекторных функций антитела. Например, одну или несколько аминокислот можно заменить на отличный аминокислотный остаток, вследствие чего антитело характеризуется измененной аффинностью в отношении эффекторного лиганда, но сохраняет способность к связыванию антигена исходного антитела. Эффекторный лиганд, аффинность в отношении которого изменяют, может представлять собой, например, Fc-рецептор или компонент C1 комплемента. Данный подход описан, например, в патентах США №№ 5624821 и 5648260, оба от Winter et al.
[00258] В другом варианте осуществления одну или несколько аминокислот, выбранных из аминокислотных остатков, можно заменить на отличный аминокислотный остаток, вследствие чего антитело характеризуется измененным связыванием C1q и/или сниженной или устраненной комплементзависимой цитотоксичностью (CDC). Данный подход описан, например, в патенте США № 6194551 от Idusogie et al.
[00259] В другом варианте осуществления один или несколько аминокислотных остатков изменяют, чтобы тем самым изменить способность антитела к фиксации комплемента. Данный подход описан, например, в PCT-публикации WO 94/29351 от Bodmer et al. Аллотипические аминокислотные остатки охватывают без ограничения константную область тяжелой цепи подклассов IgG1, IgG2 и IgG3, а также константную область легкой цепи изотипов каппа-цепи, как описано в Jefferis et al., MAbs. 1:332-338 (2009).
[00260] Комплексы на основе слитого белка с антителом, содержащим такие мутации, опосредуют сниженные антителозависимую клеточную цитотоксичность (ADCC) или комплементзависимую цитотоксичность (CDC) или их отсутствие. В некоторых вариантах осуществления аминокислотные остатки L234 и L235 в константной области IgG1 заменены на A234 и A235. В некоторых вариантах осуществления аминокислотный остаток N267 в константной области IgG1 заменен на A267. В некоторых вариантах осуществления аминокислотные остатки D265 и P329 в константной области IgG1 заменены на A265 и A329. В некоторых вариантах осуществления тяжелая цепь иммуноглобулина необязательно содержит мутацию или комбинацию мутаций, обусловливающих сниженную эффекторную функцию, выбранные из любой из D265A, P329A, P329G, N297A, D265A/P329A, D265A/N297A, L234/L235A, P329A/L234A/L235A и P329G/L234A/L235A. В конкретных вариантах осуществления иммуноконъюгат содержит тяжелую цепь иммуноглобулина, содержащую мутацию или комбинацию мутаций, обусловливающих сниженную эффекторную функцию, выбранных из любой из D265A, P329A, P329G, N297A, D265A/P329A, D265A/N297A, L234/L235A, P329A/L234A/L235A и P329G/L234A/L235A.
[00261] В другом варианте осуществления один или несколько аминокислотных остатков изменяют, чтобы тем самым изменить способность антитела к фиксации комплемента. Данный подход описан, например, в PCT-публикации WO 94/29351 от Bodmer et al. В конкретном варианте осуществления одна или несколько аминокислот антитела или его антигенсвязывающего фрагмента по настоящему изобретению заменены на один или несколько аллотипических аминокислотных остатков. Аллотипические аминокислотные остатки также включают без ограничения константную область тяжелой цепи подклассов IgG1, IgG2 и IgG3, а также константную область легкой цепи изотипа каппа-цепи, как описано в Jefferis et al., MAbs. 1:332-338 (2009).
[00262] В еще одном варианте осуществления модифицируют характер гликозилирования антитела. Например, можно получать aгликозилированное антитело (т. е. антитело, у которого отсутствует гликозилирование). Характер гликозилирования может быть изменен, например, для повышения аффинности антитела в отношении "антигена". Такие модификации углеводов можно осуществлять, например, посредством изменения одного или нескольких сайтов гликозилирования в пределах последовательности антитела. Например, можно произвести одну или несколько аминокислотных замен, которые приводят к устранению одного или нескольких сайтов гликозилирования в каркасной области вариабельной области, тем самым устраняя гликозилирования в этом сайте. Такое агликозилирование может повышать аффинность антитела в отношении антигена. Такой подход описан, например, патентах США №№ 5714350 и 6350861 от Co et al.
[00263] В другом варианте осуществления антитело модифицируют для увеличения его биологического периода полужизни. Возможны различные подходы. Например, можно вводить одну или несколько из следующих мутаций: T252L, T254S, T256F, как описано в патенте США № 6277375 от Ward. В качестве альтернативы для увеличения биологического периода полужизни антитело можно изменять в пределах области CH1 или CL таким образом, чтобы оно содержало эпитоп для связывания с рецептором реутилизации, взятый из двух петель домена CH2 из области Fc IgG, как описано в патентах США №№ 5869046 и 6121022 от Presta et al.
3. Выработка антител к CCR7
[00264] Антитела к CCR7 и фрагменты таких антител (например, антигенсвязывающие фрагменты) могут быть получены с помощью любых средств, известных из уровня техники, в том числе без ограничения рекомбинантной экспрессии, химического синтеза и ферментативного расщепления тетрамеров антител, при этом полноразмерные моноклональные антитела можно получать, например, с помощью гибридом или рекомбинантного продуцирования. Рекомбинантная экспрессия может осуществляться в любых подходящих клетках-хозяевах, известных из уровня техники, например, в клетках-хозяевах, представляющих собой клетки млекопитающих, бактериальных клетках-хозяевах, дрожжевых клетках-хозяевах, клетках-хозяевах, представляющих собой клетки насекомых, и т.д.
[00265] В настоящем изобретении дополнительно представлены полинуклеотиды, кодирующие антитела, описанные в данном документе, например, полинуклеотиды, кодирующие вариабельные области или сегменты тяжелой или легкой цепей, содержащие определяющие комплементарность области, описанные в данном документе. В некоторых вариантах осуществления полинуклеотид, кодирующий вариабельные области тяжелой цепи, характеризуется по меньшей мере 85%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью последовательности нуклеиновой кислоты с полинуклеотидом, выбранным из группы, состоящей из SEQ ID NO: 14, 46, 78 и 609. В некоторых вариантах осуществления полинуклеотид, кодирующий вариабельные области легкой цепи, характеризуется по меньшей мере 85%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью последовательности нуклеиновой кислоты с полинуклеотидом, выбранным из группы, состоящей из SEQ ID NO: 30, 62, 94 и 625.
[00266] В некоторых вариантах осуществления полинуклеотид, кодирующий тяжелую цепь, характеризуется по меньшей мере 85%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью последовательности нуклеиновой кислоты с полинуклеотидом под SEQ ID NO: 16, 48, 80 или 611. В некоторых вариантах осуществления полинуклеотид, кодирующий легкую цепь, характеризуется по меньшей мере 85%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичностью последовательности нуклеиновой кислоты с полинуклеотидом под SEQ ID NO: 32, 64, 96 или 627.
[00267] Полинуклеотиды по настоящему изобретению могут кодировать только последовательность вариабельной области антитела к CCR7. Они также могут кодировать как вариабельную область, так и константную область антитела. Некоторые из полинуклеотидных последовательностей кодируют полипептид, который содержит вариабельные области как тяжелой цепи, так и легкой цепи одного из иллюстративных мышиных антител к CCR7. Некоторые другие полинуклеотиды кодируют два полипептидных сегмента, которые в значительной степени идентичны вариабельным областям тяжелой цепи и легкой цепи, соответственно, одного из мышиных антител.
[00268] Полинуклеотидные последовательности можно получать с помощью твердофазного синтеза ДНК de novo или с помощью ПЦР-мутагенеза существующей последовательности (например, последовательностей, которые описаны в Примерах ниже), кодирующей антитело к CCR7 или его связывающий фрагмент. Прямой химический синтез нуклеиновых кислот можно осуществлять с помощью способов, известных из уровня техники, таких как фосфотриэфирный способ из Narang et al., Meth. Enzymol. 68:90, 1979; фосфодиэфирный способ из Brown et al., Meth. Enzymol. 68:109, 1979; диэтилфосфорамидитный способ из Beaucage et al., Tetra Lett., 22:1859, 1981; и способ с использованием твердой подложки из патента США № 4458066. Введение мутаций в полинуклеотидную последовательность с помощью ПЦР можно осуществлять, как описано, например, в PCR Technology: Principles and Applications for DNA Amplification, H.A. Erlich (Ed.), Freeman Press, NY, NY, 1992; PCR Protocols: A Guide to Methods and Applications, Innis et al. (Ed.), Academic Press, San Diego, CA, 1990; Mattila et al., Nucleic Acids Res. 19:967, 1991; и Eckert et al., PCR Methods and Applications 1:17, 1991.
[00269] Также в настоящем изобретении представлены векторы экспрессии и клетки-хозяева для продуцирования антитела к CCR7, описанных выше. Различные векторы экспрессии можно использовать для экспрессии полинуклеотидов, кодирующих цепи или связывающие фрагменты антитела к CCR7. Как вирусные, так и невирусные векторы экспрессии можно применять для продуцирования антител в клетке-хозяине, представляющей собой клетку млекопитающего. Невирусные векторы и системы включают плазмиды, эписомные векторы, как правило, с кассетой экспрессии для экспрессии белка или РНК, а также искусственные человеческие хромосомы (см., например, Harrington et al., Nat Genet. 15:345, 1997). Например, невирусные векторы, применимые для экспрессии полинуклеотидов и полипептидов, связывающих CCR7, в клетках млекопитающих (например, человеческих), включают pThioHis A, B и C, pcDNATM3.1/His, pEBVHis A, B и C (Invitrogen, Сан-Диего, Калифорния), векторы MPSV и многочисленные другие векторы, известные из уровня техники для экспрессии других белков. Применимые вирусные векторы включают векторы на основе ретровирусов, аденовирусов, аденоассоциированных вирусов, вирусов герпеса, векторы на основе SV40, папилломавируса, вируса Эпштейна-Барр HBP, векторы на основе вируса коровьей оспы и вируса леса Семлики (SFV). См. Brent et al., выше; Smith, Annu. Rev. Microbiol. 49:807, 1995; и Rosenfeld et al., Cell 68:143, 1992.
[00270] Выбор вектора экспрессии зависит от предполагаемых клеток-хозяев, в которых следует экспрессировать вектор. Как правило, векторы экспрессии содержать промотор и другие регуляторные последовательности (например, энхансеры), которые функционально связаны с полинуклеотидами, кодирующими цепь или фрагмент антитела к CCR7. В некоторых вариантах осуществления индуцируемый промотор используют для предотвращения экспрессии встроенных последовательностей в условиях, отличающихся от индуцирующих. Индуцируемые промоторы включают, например, арабинозный, lacZ, металлотионеиновый промотор или промотор белка теплового шока. Культуры трансформированных организмов можно размножать в неиндуцирующих условиях без смещения популяции в направлении кодирующих последовательностей, продукты экспрессии которых лучше переносятся клетками-хозяевами. Для эффективной экспрессии цепи или фрагмента антитела к CCR7 кроме промоторов также могут быть необходимы или требоваться другие регуляторные элементы. Такие элементы, как правило, включают кодон инициации трансляции ATG и смежный сайт связывания рибосомы или другие последовательности. Кроме того, эффективность экспрессии можно повысить за счет включения энхансеров, соответствующих применяемой клеточной системе (см., например, Scharf et al., Results Probl. Cell Differ. 20:125, 1994; и Bittner et al., Meth. Enzymol., 153:516, 1987). Например, для повышения экспрессии в клетках-хозяевах, представляющих собой клетки млекопитающих, можно применять энхансер SV40 или энхансер CMV.
[00271] В векторах экспрессии также может быть предусмотрено положение сигнальной последовательности секреции для образования слитого белка с полипептидами, кодируемыми вставленными последовательностями антитела к CCR7. Чаще вставляемые последовательности антитела к CCR7 соединяют с сигнальными последовательностями до включения в вектор. Векторы, подлежащие применению для получения последовательностей, кодирующих вариабельные домены легкой и тяжелой цепей антитела к CCR7, иногда также кодируют константные области или их части. Такие векторы обеспечивают возможность экспрессии вариабельных областей в виде слитых белков с константными областями, приводя тем самым к получению интактных антител или их фрагментов. Как правило, такие константные области являются человеческими.
[00272] Клетки-хозяева, несущие и экспрессирующие цепи антитела к CCR7, могут быть или прокариотическими, или эукариотическими. E. coli является одним прокариотическим хозяином, применимым для клонирования и экспрессии полинуклеотидов по настоящему изобретению. Другие подходящие для применения микробные хозяева включают бацилл, таких как Bacillus subtilis, и других энтеробактерий, таких как Salmonella, Serratia и различные виды Pseudomonas. Для этих прокариотических хозяев также можно создать векторы экспрессии, которые, как правило, содержат последовательности для управления экспрессией, совместимые с клеткой-хозяином (например, точку начала репликации). Кроме того, будет присутствовать любое число разнообразных хорошо известных промоторов, как, например, лактозная промоторная система, триптофановая (trp) промоторная система, бета-лактамазная промоторная система или промоторная система из фага лямбда. Как правило, промоторы управляют экспрессией, необязательно с помощью операторной последовательности, и имеют последовательности сайта связывания рибосомы и т.п. для инициации и завершения транскрипции и трансляции. Для экспрессии полипептидов, связывающих CCR7, по настоящему изобретению также можно использовать других микроорганизмов, таких как дрожжи. Также можно применять клетки насекомых в сочетании с бакуловирусными векторами.
[00273] В некоторых предпочтительных вариантах осуществления для экспрессии и продуцирования полипептидов, связывающих CCR7, по настоящему изобретению применяют клетки-хозяева, представляющие собой клетки млекопитающих. Например, они могут представлять собой либо линию клеток гибридомы, экспрессирующих эндогенные гены иммуноглобулинов (например, клоны гибридомы из клеток миеломы, как описано в Примерах), либо линию клеток млекопитающих, несущих экзогенный вектор экспрессии (например, клетки миеломы SP2/0, приведенные в качестве примера ниже). Они охватывают любые нормальные мортальные или нормальные или аномальные иммортальные клетки животных или человека. Например, был разработан ряд подходящих линий клеток-хозяев, способных секретировать интактные иммуноглобулины, в том числе линии клеток CHO, различные линии клеток Cos, клетки HeLa, линии клеток миеломы, трансформированные B-клетки и гибридомы. Применение тканевой культуры клеток млекопитающих для экспрессии полипептидов в общих чертах обсуждается, например, в Winnacker, From Genes to Clones, VCH Publishers, N.Y., N.Y., 1987. Векторы экспрессии для клеток-хозяев, представляющих собой клетки млекопитающих, могут включать последовательности для управления экспрессией, такие как точка начала репликации, промотор и энхансер (см., например, Queen et al., Immunol. Rev. 89:49-68, 1986), и необходимые сайты, несущие информацию для процессинга, такие как сайты связывания рибосом, сайты сплайсинга РНК, сайты полиаденилирования и последовательности терминатора транскрипции. Такие векторы экспрессии обычно содержат промоторы, происходящие из генов млекопитающих или из вирусов млекопитающих. Подходящие промоторы могут быть конститутивными, специфическими в отношении типа клеток, специфическими в отношении стадии развития и/или модулируемыми или регулируемыми. Применимые промоторы включают без ограничения металлотионеиновый промотор, конститутивный большой поздний промотор аденовируса, индуцируемый дексаметазоном промотор MMTV, промотор SV40, промотор polIII MRP, конститутивный промотор MPSV, индуцируемый тетрациклином промотор CMV (такой как немедленно-ранний промотор CMV человека), конститутивный промотор CMV и комбинации промотор-энхансер, известные из уровня техники.
[00274] Способы введения векторов экспрессии, содержащих представляющие интерес полинуклеотидные последовательности, варьируют в зависимости от типа клетки-хозяина. Например, трансфекцию с использованием хлорида кальция обычно используют для прокариотических клеток, тогда как обработку фосфатом кальция или электропорацию можно применять для других клеток-хозяев (см. в общих чертах Sambrook et al., выше). A Laboratory Manual, 4th ed.). Другие способы включают, например, электропорацию, обработку фосфатом кальция, трансформацию, опосредованную липосомами, инъекцию и микроинъекцию, баллистические способы, виросомы, иммунолипосомы, конъюгаты поликатион:нуклеиновая кислота, депротеинизированную ДНК, искусственные вирионы, слияние со структурным белком VP22 вируса герпеса (Elliot and O'Hare, Cell 88:223, 1997), усиленное средством поглощение ДНК и трансдукцию ex vivo. Чтобы обеспечить длительное получение рекомбинантных белков с высоким выходом, зачастую будет требоваться стабильная экспрессия. Например, линии клеток, которые стабильно экспрессируют цепи или связывающие фрагменты антитела к CCR7, можно получить с применением векторов экспрессии по настоящему изобретению, которые содержат вирусные точки начала репликации или эндогенные экспрессионные элементы и ген селектируемого маркера. После введения вектора клетки можно оставить расти в течение 1-2 дней в обогащенной среде перед их переносом на селективную среду. Задачей селектируемого маркера является придание устойчивости к факторам отбора, и его присутствие обеспечивает возможность роста клеток, которые успешно экспрессируют введенные последовательности, в селективных средах. Пролиферацию устойчивых стабильно трансфицированных клеток можно обеспечивать с применением методик тканевой культуры, соответствующих типу клеток.
Пути терапевтического применения
[00275] Антитела, фрагменты антител (например, антигенсвязывающие фрагменты) и конъюгаты антитела и лекарственного средства по настоящему изобретению применимы в виде разнообразных применений, в том числе без ограничения лечения или предупреждения рака, такого как типы солидного рака или гемобластозы. В некоторых вариантах осуществления антитела, фрагменты антител (например, антигенсвязывающие фрагменты) и конъюгаты антитела и лекарственного средства по настоящему изобретению применимы для ингибирования роста опухоли, индуцирования дифференцировки, уменьшения объема опухоли и/или снижения туморогенности опухоли. Способы применения могут представлять собой in vitro, ex vivo или in vivo способы.
[00276] В одном аспекте антитела, фрагменты антител (например, антигенсвязывающие фрагменты) и конъюгаты антитела и лекарственного средства по настоящему изобретению применимы для выявления присутствия CCR7 в биологическом образце. Используемый в данном документе термин "осуществление обнаружения" охватывает количественное или качественное обнаружение. В некоторых вариантах осуществления биологический образец содержит клетку или ткань. В некоторых вариантах осуществления такие ткани включают нормальные и/или раковые ткани, которые экспрессируют CCR7 на более высоких уровнях по сравнению с другими тканями.
[00277] В одном аспекте настоящего изобретения представлен способ обнаружения присутствия CCR7 в биологическом образце. В некоторых вариантах осуществления способ предусматривает приведение биологического образца в контакт с антителом к CCR7 в условиях, допускающих связывание антитела с антигеном, и выявление того, формируется ли комплекс между антителом и антигеном.
[00278] В одном аспекте настоящего изобретения представлен способ диагностики нарушения, ассоциированного с повышенной экспрессией CCR7. В некоторых вариантах осуществления способ предусматривает приведение тестируемой клетки в контакт с антителом к CCR7; определение уровня экспрессии (или количественно, или качественно) CCR7 на тестируемой клетке путем обнаружения связывания антитела к CCR7 с антигеном CCR7 и сравнение уровня экспрессии CCR7 в тестируемой клетке с уровнем экспрессии CCR7 на контрольной клетке (например, нормальной клетке, происходящей из той же ткани, что и тестируемая клетка, или клетке, которая экспрессирует CCR7 на уровнях, сравнимых с таковыми у нормальной клетки), где более высокий уровень экспрессии CCR7 на тестируемой клетке по сравнению с контрольной клеткой указывает на присутствие нарушения, ассоциированного с повышенной экспрессией CCR7. В некоторых вариантах осуществления тестируемую клетку получают от индивидуума с подозрением на наличие нарушения, ассоциированного с повышенной экспрессией CCR7. В некоторых вариантах осуществления нарушение представляет собой нарушение пролиферации клеток, такое как рак или опухоль. В некоторых вариантах осуществления способ предусматривает измерение числа копий гена CCR7 в тестируемой клетке.
[00279] В некоторых вариантах осуществления способ диагностики или обнаружения, такой как описываемые выше, предусматривает обнаружение связывания антитела к CCR7 с CCR7, экспрессируемым на поверхности клетки или в препарате мембраны, полученном из клетки, экспрессирующей CCR7 на своей поверхности. Иллюстративным анализом для обнаружения связывания антитела к CCR7 с CCR7, экспрессируемым на поверхности клетки, является "FACS"-анализ.
[00280] Для обнаружения связывания антитела к CCR7 с CCR7 могут применяться некоторые другие способы. Такие способы включают без ограничения анализы связывания антигена, которые широко известны из уровня техники, такие как вестерн-блоттинг, радиоиммунологические анализы, ELISA (твердофазный иммуноферментный анализ), иммунологические "сэндвич"-анализы, анализы иммунопреципитации, флуоресцентные иммунологические анализы, иммунологические анализы с белком A и иммуногистохимическое исследование (IHC).
[00281] В некоторых вариантах осуществления антитела к CCR7 являются меченными. Метки включают без ограничения метки или фрагменты, которые обнаруживают непосредственно (такие как флуоресцентные, хромофорные, электронноплотные, хемилюминесцентные и радиоактивные метки), а также фрагменты, такие как ферменты или лиганды, которые обнаруживают опосредованно, например, за счет ферментативной реакции или молекулярного взаимодействия.
[00282] В некоторых вариантах осуществления антитела к CCR7 являются иммобилизированными на нерастворимой матрице. Иммобилизация приводит к отделению антитела к CCR7 от любого белка CCR7, который остается свободным в растворе. Традиционно это осуществляют либо посредством перевода антитела к CCR7 в нерастворимую форму перед процедурой анализа в результате адсорбции на нерастворимой в воде матрице или поверхности (Bennich et al., патент США № 3720760) или соединения ковалентной связью (например, с применением сшивания глутаральдегидом), либо посредством перевода антитела к CCR7 в нерастворимую форму после образования комплекса между антителом к CCR7 и белком CCR7, например, посредством иммунного осаждения.
[00283] Любой из вышеизложенных вариантов осуществления диагностики или обнаружения может проводиться с применением иммуноконъюгата по настоящему изобретению вместо или в дополнение к антителу к CCR7.
[00284] В одном варианте осуществления настоящего изобретения представлен способ лечения или предупреждения заболевания, предусматривающий введение пациенту антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства по настоящему изобретению. В настоящем изобретении также представлено применение антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства по настоящему изобретению для лечения или предупреждения заболевания у пациента. В некоторых вариантах осуществления настоящего изобретения представлены антитела, фрагменты антител (например, антигенсвязывающие фрагменты) или конъюгаты антитела и лекарственного средства по настоящему изобретению для применения в лечении или предупреждении заболевания у пациента. В дополнительных вариантах осуществления настоящего изобретения представлено применение антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства по настоящему изобретению при изготовлении лекарственного препарата для лечения или предупреждения заболевания у пациента.
[00285] В некоторых вариантах осуществления заболевание, которое лечат с помощью антител, фрагментов антител (например, антигенсвязывающих фрагментов) и конъюгатов антитела и лекарственного средства по настоящему изобретению, представляет собой рак. В некоторых вариантах осуществления рак характеризуется наличием клеток, экспрессирующих CCR7, с которыми связываются антитела, фрагменты антител (например, антигенсвязывающие фрагменты) и конъюгаты антитела и лекарственного средства по настоящему изобретению. В некоторых вариантах осуществления рак характеризуется повышением экспрессии CCR7 по сравнению с уровнем экспрессии у здорового пациента. В некоторых вариантах осуществления экспрессия CCR7 может быть измерена по повышению РНК CCR7. В других вариантах осуществления рак характеризуется повышением числа копий ДНК CCR7. Специалистам в данной области техники известны другие способы измерения или определения уровней экспрессии CCR7. Примеры заболеваний, которые можно лечить и/или предупреждать, включают без ограничения, хронический лимфоцитарный лейкоз (CLL), периферические T-клеточные лимфомы (PTCL), такие как T-клеточный лейкоз/лимфома взрослых (ATLL) и анапластическая крупноклеточная лимфома (ALCL), неходжкинскую лимфому (NHL), такую как лимфома из клеток мантийной зоны (MCL), лимфома Беркитта и диффузная В-крупноклеточная лимфома (DLBCL), карциному желудка, немелкоклеточный рак легкого, мелкоклеточный рак легкого, рак головы и шеи, носоглоточную карциному (NPC), рак пищевода, колоректальную карциному, рак поджелудочной железы, рак щитовидной железы, рак молочной железы, почечноклеточный рак и рак шейки матки.
[00286] В настоящем изобретении представлены способы лечения или предупреждения рака, предусматривающие введение терапевтически эффективного количества антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства по настоящему изобретению. В некоторых вариантах осуществления рак представляет собой солидный рак. В некоторых вариантах осуществления субъектом является человек. В некоторых вариантах осуществления рак представляет собой устойчивый рак и/или рецидивирующий рак. В определенных аспектах, например, устойчивый рак является устойчивым к ингибиторам тирозинкиназы, ингибиторам EGFR, ингибиторам Her2, ингибиторам Her3, ингибиторам IGFR и ингибиторам Met.
[00287] В некоторых вариантах осуществления настоящего изобретения представлены способы ингибирования роста опухоли, предусматривающие введение субъекту терапевтически эффективного количества антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства по настоящему изобретению. В некоторых вариантах осуществления субъектом является человек. В некоторых вариантах осуществления субъект имеет опухоль или у него была удалена опухоль. В некоторых вариантах осуществления опухоль является устойчивой к другим ингибиторам тирозинкиназы, в том числе без ограничения к ингибиторам EGFR, ингибиторам Her2, ингибиторам Her3, ингибиторам IGFR и ингибиторам Met.
[00288] В некоторых вариантах осуществления опухоль экспрессирует CCR7, с которым связываются антитела к CCR7. В некоторых вариантах осуществления опухоль сверхэкспрессирует CCR7 человека. В некоторых вариантах осуществления опухоль характеризуется повышенным числом копий гена CCR7.
[00289] В настоящем изобретении также представлены способы отбора пациентов для лечения антителами, фрагментами антител (например, антигенсвязывающими фрагментами) или конъюгатами антитела и лекарственного средства по настоящему изобретению, предусматривающие введение терапевтически эффективного количества указанных антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства. В определенных аспектах способ предусматривает отбор пациентов с раком, устойчивым к ингибиторам тирозинкиназы. В определенных аспектах предполагается, что рак, устойчивый к ингибиторам тирозинкиназы, является устойчивым к ингибиторам EGFR, ингибиторам Her2, ингибиторам Her3, ингибиторам IGFR и/или ингибиторам Met. В определенных аспектах предполагается, что устойчивый рак представляет собой рак, устойчивый к Her2. В определенных аспектах предполагается, что рак представляет собой de novo устойчивый рак, а в еще аспектах предполагается, что рак представляет собой рецидивирующий рак. В определенных аспектах настоящего изобретения способы предусматривают отбор пациента с de novo устойчивым или рецидивирующим раком и измерение экспрессии CCR7. Предполагается, что в определенных аспектах рецидивирующий рак или опухоль изначально не были раком или опухолью, экспрессирующими CCR7, но становятся CCR7 положительным раком, который представляет собой устойчивые к тирозинкиназе или рецидивирующие рак или опухоль, после лечения ингибиторами тирозинкиназы.
[00290] В случае лечения или предупреждения заболеваний подходящая доза антител, фрагментов антител (например, антигенсвязывающих фрагментов) или конъюгатов антитела и лекарственного средства по настоящему изобретению зависит от различных факторов, таких как тип заболевания, подлежащего лечению, тяжесть и течение заболевания, ответная реакция заболевания, предыдущая терапия, клинический анамнез пациента и т. д. Антитело или средство могут вводиться один раз или на протяжении курса лечения, длящегося от нескольких дней до нескольких месяцев, или до осуществления излечения, или до достижения ослабления болезненного состояния (например, уменьшения размера опухоли). Оптимальные схемы приема можно рассчитать на основании измерений накопления лекарственного средства в организме пациента, и они будут варьироваться в зависимости от относительной активности индивидуальных антитела, фрагментов антитела (например, антигенсвязывающего фрагмента) или конъюгатов антитела и лекарственного средства. Лечащий врач может оценить частоту повторения введения дозы на основании измеренного времени удержания и концентраций лекарственного средства в физиологических жидкостях или тканях.
Комбинированная терапия
[00291] В определенных случаях антитело, фрагмент антитела (например, антигенсвязывающий фрагмент) или конъюгат антитела и лекарственного средства по настоящему изобретению объединяют с другими терапевтическими средствами, такими как другие противораковые средства, противоаллергические средства, средства против тошноты (или противорвотные средства), обезболивающие средства, цитопротекторные средства и их комбинации.
[00292] В одном варианте осуществления антитело, фрагмент антитела (например, антигенсвязывающий фрагмент) или конъюгат антитела и лекарственного средства по настоящему изобретению объединяют в фармацевтическом комбинированном составе со вторым соединением, характеризующимся противораковыми свойствами, или схема введения доз предусматривает комбинированную терапию с ним. Второе соединение фармацевтического комбинированного состава или схемы введения доз может характеризоваться видами активности, которые взаимно дополняют антитело или иммуноконъюгат из комбинации, вследствие чего они не оказывают неблагоприятного эффекта друг на друга. Например, антитело, фрагмент антитела (например, антигенсвязывающий фрагмент) или конъюгат антитела и лекарственного средства по настоящему изобретению могут вводиться в комбинации без ограничения с химиотерапевтическим средством, ингибитором тирозинкиназы, ингибитором пути передачи нисходящего сигнала с участием CCR7, ингибиторами IAP, ингибиторами Bcl2, ингибиторами Mcl1 и другими ингибиторами CCR7.
[00293] Используемый в данном документе термин "фармацевтическая комбинация" относится либо к фиксированной комбинации в форме единицы дозирования, либо к нефиксированной комбинации или набору из частей для комбинированного введения, где два или более терапевтических средств могут вводиться независимо в одно и то же время или по отдельности в пределах временных интервалов, в частности, когда такие временные интервалы обеспечивают возможность демонстрации кооперативного, например синергетического, эффекта партнеров по комбинации.
[00294] Термин "комбинированная терапия" относится к введению двух или более терапевтических средств для лечения или предупреждения терапевтического состояния или нарушения, описанного в настоящем изобретении. Такое введение охватывает совместное введение этих терапевтических средств практически одновременно, например, в одной капсуле, имеющей фиксированное отношение активных ингредиентов. В качестве альтернативы такое введение охватывает совместное введение в виде нескольких или отдельных контейнеров (например, капсулы, порошки и жидкости) для каждого активного ингредиента. Порошки и/или жидкости могут быть восстановлены или разбавлены до требуемой дозы перед введением. Кроме того, такое введение также охватывает применение каждого типа терапевтического средства последовательным образом, либо приблизительно в одно и то же время, либо в разное время. В любом случае схема лечения будет обеспечивать приносящие пользу эффекты комбинации лекарственных средств при лечении или предупреждении состояний или нарушений, описанных в данном документе.
[00295] Комбинированная терапия может обеспечивать "синергию" и оказаться "синергетической", т.e. эффект, достигаемый, когда активные ингредиенты применяются вместе, превышает сумму эффектов, которые являются результатом применения соединений по отдельности. Синергетический эффект может быть достигнут, когда активные ингредиенты: (1) составлены вместе и вводятся или доставляются одновременно в виде комбинированного состава с однократной дозой; (2) доставляются по очереди или параллельно как отдельные составы или (3) применяется какая-либо другая схема. При доставке в виде терапии с чередованием синергетический эффект может достигаться, когда соединения вводят или доставляют одно за другим, например, посредством различных инъекций в отдельных шприцах. В целом, во время терапии с чередованием эффективную дозу каждого активного ингредиента вводят одну за другой, т.е. сериями, в то время как при комбинированной терапии эффективную дозу двух или более активных ингредиентов вводят вместе.
[00296] Распространенные химиотерапевтические средства, рассматриваемые для применения в видах комбинированной терапии, включают анастрозол (Arimidex®), бикалутамид (Casodex®), блеомицина сульфат (Blenoxane®), бусульфан (Myleran®), бусульфан в виде инъекции (Busulfex®), капецитабин (Xeloda®), N4-пентоксикарбонил-5-дезокси-5-фторцитидин, карбоплатин (Paraplatin®), кармустин (BiCNU®), хлорамбуцил (Leukeran®), цисплатин (Platinol®), кладрибин (Leustatin®), циклофосфамид (Cytoxan® или Neosar®), цитарабин, цитозина арабинозид (Cytosar-U®), цитарабин в виде липосомной инъекции (DepoCyt®), дакарбазин (DTIC-Dome®), дактиномицин (актиномицин D, Cosmegan), даунорубицина гидрохлорид (Cerubidine®), даунорубицина цитрат в виде липосомной инъекции (DaunoXome®), дексаметазон, доцетаксел (Taxotere®), доксорубицина гидрохлорид (Adriamycin®, Rubex®), этопозид (Vepesid®), флударабина фосфат (Fludara®), 5-фторурацил (Adrucil®, Efudex®), флутамид (Eulexin®), тезацитибин, гемцитабин (дифтордезоксицитидин), гидроксимочевину (Hydrea®), идарубицин (Idamycin®), ифосфамид (IFEX®), иринотекан (Camptosar®), L-аспарагиназу (ELSPAR®), лейковорин кальция, мелфалан (Alkeran®), 6-меркаптопурин (Purinethol®), метотрексат (Folex®), митоксантрон (Novantrone®), милотарг, паклитаксел (Taxol®), феникс (иттрий 90/MX-DTPA), пентостатин, полифероспан 20 с имплантатом кармустина (Gliadel®), тамоксифена цитрат (Nolvadex®), тенипозид (Vumon®), 6-тиогуанин, тиотепу, тирапазамин (Tirazone®), топотекана гидрохлорид для инъекции (Hycamptin®), винбластин (Velban®), винкристин (Oncovin®) и винорелбин (Navelbine®), а также пеметрексед.
[00297] В одном аспекте настоящего изобретения представлен способ лечения или предупреждения рака путем введения субъекту, нуждающемуся в этом, конъюгата антитела и лекарственного средства по настоящему изобретению в комбинации с одним или несколькими ингибиторами тирозинкиназы, в том числе без ограничения ингибиторами BTK, ингибиторами EGFR, ингибиторами Her2, ингибиторами Her3, ингибиторами IGFR и ингибиторами Met.
[00298] Например, ингибиторы тирозинкиназы включают без ограничения ибрутиниб (PCI-32765); эрлотиниба гидрохлорид (Tarceva®); линифаниб (N-[4-(3-амино-1H-индазол-4-ил)фенил]-N'-(2-фтор-5-метилфенил)мочевина, также известный как ABT 869, доступный от Genentech); сунитиниба малат (Sutent®); босутиниб (4-[(2,4-дихлор-5-метоксифенил)амино]-6-метокси-7-[3-(4-метилпиперазин-1-ил)пропокси]хинолин-3-карбонитрил, также известный как SKI-606 и описанный в патенте США № 6780996); дазатиниб (Sprycel®); пазопаниб (Votrient®); сорафениб (Nexavar®); зактима (ZD6474) и иматиниб или иматиниба мезилат (Gilvec® и Gleevec®).
[00299] Ингибиторы рецептора эпидермального фактора роста рецептор (EGFR) ингибиторы включают без ограничения, эрлотиниба гидрохлорид (Tarceva®), гефитиниб (Iressa®); N-[4-[(3-хлор-4-фторфенил)амино]-7-[[(3''S'')-тетрагидро-3-фуранил]окси]-6-хиназолинил]-4(диметиламино)-2-бутенамид, Tovok®); вандетаниб (Caprelsa®); лапатиниб (Tykerb®); (3R,4R)-4-амино-1-((4-((3-метоксифенил)амино)пирроло[2,1-f][1,2,4]триазин-5-ил)метил)пиперидин-3-ол (BMS690514); канертиниба дигидрохлорид (CI-1033); 6-[4-[(4-этил-1-пиперазинил)метил]фенил]-N-[(1R)-1-фенилэтил]- 7H-пирроло[2,3-d]пиримидин-4-амин (AEE788, CAS 497839-62-0); мубритиниб (TAK165); пелитиниб (EKB569); афатиниб (BIBW2992); нератиниб (HKI-272); N-[4-[[1-[(3-фторфенил)метил]-1H-индазол-5-ил]амино]-5-метилпирроло[2,1-f][1,2,4]триазин-6-ил]-карбаминовую кислоту, (3S)-3-морфолинилметиловый сложный эфир (BMS599626); N-(3,4-дихлор-2-фторфенил)-6-метокси-7-[[(3aα,5β,6aα)-октагидро-2-метилциклопента[c]пиррол-5-ил]метокси]-4-хиназолинамин (XL647, CAS 781613-23-8) и 4-[4-[[(1R)-1-фенилэтил]амино]-7H-пирроло[2,3-d]пиримидин-6-ил]-фенол (PKI166, CAS 187724-61-4).
[00300] Антитела, связывающие EGFR, включают без ограничения, цетуксимаб (Erbitux®); панитумумаб (Vectibix®); матузумаб (EMD-72000); нимотузумаб (hR3); залутумумаб; TheraCIM h-R3; MDX0447 (CAS 339151-96-1) и ch806 (mAb-806, CAS 946414-09-1).
[00301] Ингибиторы рецептора эпидермального фактора роста 2 (рецептора Her2) человека (также известного как Neu, ErbB-2, CD340 или p185) включают без ограничения трастузумаб (Herceptin®); пертузумаб (Omnitarg®); трастузумаб эмтанзин (Kadcyla®); нератиниб (HKI-272, (2E)-N-[4-[[3-хлор-4-[(пиридин-2-ил)метокси]фенил]амино]-3-циано-7-этоксихинолин-6-ил]-4-(диметиламино)бут-2-енамид и описанный в PCT-публикации № WO 05/028443); лапатиниб или лапатиниба дитозилат (Tykerb®); (3R,4R)-4-амино-1-((4-((3-метоксифенил)амино)пирроло[2,1-f][1,2,4]триазин-5-ил)метил)пиперидин-3-ол (BMS690514); (2E)-N-[4-[(3-хлор-4-фторфенил)амино]-7-[[(3S)-тетрагидро-3-фуранил]окси]-6-хиназолинил]-4-(диметиламино)-2-бутенамид (BIBW-2992, CAS 850140-72-6); N-[4-[[1-[(3-фторфенил)метил]-1H-индазол-5-ил]амино]-5-метилпирроло[2,1-f][1,2,4]триазин-6-ил]-карбаминовую кислоту, (3S)-3-морфолинилметиловый сложный эфир (BMS 599626, CAS 714971-09-2); канертиниба дигидрохлорид (PD183805 или CI-1033) и N-(3,4-дихлор-2-фторфенил)-6-метокси-7-[[(3aα,5β,6aα)-октагидро-2-метилциклопента[c]пиррол-5-ил]метокси]-4-хиназолинамин (XL647, CAS 781613-23-8).
[00302] Ингибиторы Her3 включают без ограничения LJM716, MM-121, AMG-888, RG7116, REGN-1400, AV-203, MP-RM-1, MM-111 и MEHD-7945A.
[00303] Ингибиторы Met включают без ограничения кабозатиниб (XL184, CAS 849217-68-1); форетиниб (GSK1363089, ранее XL880, CAS 849217-64-7); тивантиниб (ARQ197, CAS 1000873-98-2); 1-(2-гидрокси-2-метилпропил)-N-(5-(7-метоксихинолин-4-илокси)пиридин-2-ил)-5-метил-3-оксо-2-фенил-2,3-дигидро-1H-пиразол-4-карбоксамид (AMG 458); кризотиниб (Xalkori®, PF-02341066); (3Z)-5-(2,3-дигидро-1H-индол-1-илсульфонил)-3-({3,5-диметил-4-[(4-метилпиперазин-1-ил)карбонил]-1H-пиррол-2-ил}метилен)-1,3-дигидро-2H-индол-2-он (SU11271); (3Z)-N-(3-хлорфенил)-3-({3,5-диметил-4-[(4-метилпиперазин-1-ил)карбонил]-1H-пиррол-2-ил}метилен)-N-метил-2-оксоиндолин-5-сульфонамид (SU11274); (3Z)-N-(3-хлорфенил)-3-{[3,5-диметил-4-(3-морфолин-4-илпропил)-1H-пиррол-2-ил]метилен}-N-метил-2-оксоиндолин-5-сульфонамид (SU11606); 6-[дифтор[6-(1-метил-1H-пиразол-4-ил)-1,2,4-триазоло[4,3-b]пиридазин-3-ил]метил]-хинолин (JNJ38877605, CAS 943540-75-8); 2-[4-[1-(хинолин-6-илметил)-1H-[1,2,3]триазоло[4,5-b]пиразин-6-ил]-1H-пиразол-1-ил]этанол (PF04217903, CAS 956905-27-4); N-((2R)-1,4-диоксан-2-илметил)-N-метил-N'-[3-(1-метил-1H-пиразол-4-ил)-5-оксо-5H-бензо[4,5]циклогепта[1,2-b]пиридин-7-ил]сульфамид (MK2461, CAS 917879-39-1); 6-[[6-(1-метил-1H-пиразол-4-ил)-1,2,4-триазоло[4,3-b]пиридазин-3-ил]тио]-хинолин (SGX523, CAS 1022150-57-7) и (3Z)-5-[[(2,6-дихлорфенил)метил]сульфонил]-3-[[3,5-диметил-4-[[(2R)-2-(1-пирролидинилметил)-1-пирролидинил]карбонил]-1H-пиррол-2-ил]метилен]-1,3-дигидро-2H-индол-2-он (PHA665752, CAS 477575-56-7).
[00304] Ингибиторы IGF1R включают без ограничения, BMS-754807, XL-228, OSI-906, GSK0904529A, A-928605, AXL1717, KW-2450, MK0646, AMG479, IMCA12, MEDI-573 и BI836845. Обзор см., например, в Yee, JNCI, 104; 975 (2012).
[00305] В другом аспекте настоящего изобретения представлен способ лечения или предупреждения рака путем введения субъекту, нуждающемуся в этом, конъюгата антитела и лекарственного средства по настоящему изобретению в комбинации с одним или несколькими ингибиторами пути передачи нисходящего сигнала с участием CCR7, в том числе без ограничения ингибиторами β-аррестина, ингибиторами GRK, ингибиторами MAPK, ингибиторами PI3K, ингибиторами JAK и т.д.
[00306] Например, ингибиторы фосфоинозитид 3-киназы (PI3K) включают без ограничения иделалисиб (Zydelig, GS-1101, Cal-101), 4-[2-(1H-индазол-4-ил)-6-[[4-(метилсульфонил)пиперазин-1-ил]метил]тиено[3,2-d]пиримидин-4-ил]морфолин (также известный как GDC 0941 и описанный в PCT-публикациях №№ WO 09/036082 и WO 09/055730); 2-метил-2-[4-[3-метил-2-оксо-8-(хинолин-3-ил)-2,3-дигидроимидазо[4,5-c]хинолин-1-ил]фенил]пропионитрил (также известный как BEZ 235 или NVP-BEZ 235 и описанный в PCT-цубликации № WO 06/122806); 4-(трифторметил)-5-(2,6-диморфолинoпиримидин-4-ил)пиридин-2-амин (также известный как BKM120 или NVP-BKM120 и описанный в PCT-публикации № WO2007/084786); тозасертиб (VX680 или MK-0457, CAS 639089-54-6); (5Z)-5-[[4-(4-пиридинил)-6-хинолинил]метилен]-2,4-тиазолидиндион (GSK1059615, CAS 958852-01-2); (1E,4S,4aR,5R,6aS,9aR)-5-(ацетилокси)-1-[(ди-2-пропениламино)метилен]-4,4a,5,6,6a,8,9,9a-октагидро-11-гидрокси-4-(метоксиметил)-4a,6a-диметил-циклопента[5,6]нафто[1,2-c]пиран-2,7,10(1H)-трион (PX866, CAS 502632-66-8) и 8-фенил-2-(морфолин-4-ил)-хромен-4-он (LY294002, CAS 154447-36-6).
[00307] В еще одном аспекте настоящего изобретения представлен способ лечения или предупреждения рака путем введения субъекту, нуждающемуся в этом, конъюгата антитела и лекарственного средства по настоящему изобретению в комбинации с одним или несколькими проапоптотическими средствами, в том числе без ограничения ингибиторами IAP, ингибиторами Bcl2, ингибиторами MCl1, средствами Trail, ингибиторами Chk.
[00308] Например, ингибиторы IAP включают без ограничения LCL161, GDC-0917, AEG-35156, AT406 и TL32711. Другие примеры ингибиторов IAP включают без ограничения раскрытые в WO04/005284, WO 04/007529, WO05/097791, WO 05/069894, WO 05/069888, WO 05/094818, US2006/0014700, US2006/0025347, WO 06/069063, WO 06/010118, WO 06/017295 и WO08/134679, все из которых включены в данный документ посредством ссылки.
[00309] Ингибиторы BCL-2 включают без ограничения венетоклакс (также известный как GDC-0199, ABT-199, RG7601); 4-[4-[[2-(4-хлорфенил)-5,5-диметил-1-циклогексен-1-ил]метил]-1-пиперазинил]-N-[[4-[[(1R)-3-(4-морфолинил)-1-[(фенилтио)метил]пропил]амино]-3-[(трифторметил)сульфонил]фенил]сульфонил]бензамид (также известный как ABT-263 и описанный в PCT-публикации № WO 09/155386); тетрокарцин A; антимицин; госсипол ((-)BL-193); обатоклакс; этил-2-амино-6-циклопентил-4-(1-циано-2-этокси-2-оксоэтил)-4H-хромон-3-карбоксилат (HA14-1); облимерсен (G3139, Genasense®); пептид Bak BH3; (-)-госсипол-уксусную кислоту (AT-101); 4-[4-[(4'-хлор[1,1'-бифенил]-2-ил)метил]-1-пиперазинил]-N-[[4-[[(1R)-3-(диметиламино)-1-[(фенилтио)метил]пропил]амино]-3-нитрофенил]сульфонил]-бензамид (ABT-737, CAS 852808-04-9) и навитоклакс (ABT-263, CAS 923564-51-6).
[00310] Агонисты проапоптических рецепторов (PARA), в том числе DR4 (TRAILR1) и DR5 (TRAILR2), включают без ограничения дуланермин (AMG-951, RhApo2L/TRAIL); мапатумумаб (HRS-ETR1, CAS 658052-09-6); лексатумумаб (HGS-ETR2, CAS 845816-02-6); апомаб (Apomab®); конатумумаб (AMG655, CAS 896731-82-1) и тигатузумаб (CS1008, CAS 946415-34-5, доступен от Daiichi Sankyo).
[00311] Ингибиторы киназы контрольных точек (CHK) включают без ограничения 7-гидроксистауроспорин (UCN-01); 6-бром-3-(1-метил-1H-пиразол-4-ил)-5-(3R)-3-пиперидинил-пиразолo[1,5-a]пиримидин-7-амин (SCH900776, CAS 891494-63-6); 5-(3-фторфенил)-3-уреидотиофен-2-карбоновой кислоты N-[(S)-пиперидин-3-ил]амид (AZD7762, CAS 860352-01-8); 4-[((3S)-1-азабицикло[2.2.2]окт-3-ил)амино]-3-(1H-бензимидазол-2-ил)-6-хлорхинолин-2(1H)-он (CHIR 124, CAS 405168-58-3); 7-аминодактиномицин (7-AAD), изогранулатимид, дебромгимениалдизин; N-[5-бром-4-метил-2-[(2S)-2-морфолинилметокси]-фенил]-N'-(5-метил-2-пиразинил)мочевину (LY2603618, CAS 911222-45-2); сульфорафан (CAS 4478-93-7, 4-метилсульфинилбутилизотиоцианат); 9,10,11,12-тетрагидро- 9,12-эпокси-1H-дииндолo[1,2,3-fg:3',2',1'-kl]пирроло[3,4-i][1,6]бензодиазоцин-1,3(2H)-дион (SB-218078, CAS 135897-06-2) и TAT-S216A (YGRKKRRQRRRLYRSPAMPENL (SEQ ID NO: 629)), а также CBP501 ((d-Bpa)sws(d-Phe-F5)(d-Cha)rrrqrr).
[00312] В дополнительном варианте осуществления настоящего изобретения представлен способ лечения или предупреждения рака путем введения субъекту, нуждающемуся в этом, конъюгата антитела и лекарственного средства по настоящему изобретению в комбинации с одним или несколькими иммуномодуляторами (например, одним или несколькими из активатора костимулирующей молекулы или ингибитора молекулы иммунных контрольных точек).
[00313] В некоторых вариантах осуществления иммуномодулятор представляет собой активатор костимулирующей молекулы. В одном варианте осуществления агонист костимулирующей молекулы выбран из агониста (например, агонистического антитела или его антигенсвязывающего фрагмента, или растворимого слитого белка) OX40, CD2, CD27, CDS, ICAM-1, LFA-1 (CD11a/CD18), ICOS (CD278), 4-1BB (CD137), GITR, CD30, CD40, BAFFR, HVEM, CD7, LIGHT, NKG2C, SLAMF7, NKp80, CD160, B7-H3, STING или лиганда CD83.
[00314] В некоторых вариантах осуществления иммуномодулятор представляет собой ингибитор молекулы иммунных контрольных точек. В одном варианте осуществления иммуномодулятор представляет собой ингибитор PD-1, PD-L1, PD-L2, CTLA4, TIM3, LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и/или TGFR-бета. В одном варианте осуществления ингибитор молекулы иммунных контрольных точек ингибирует PD-1, PD-L1, LAG-3, TIM-3 или CTLA4 или любую их комбинацию. Термин "ингибирование" или "ингибитор" подразумевает снижение определенного параметра, например, активности данной молекулы, например, ингибитора контрольных точек. Например, данный термин подразумевает ингибирование активности, например, активности PD-1 или PD-L1, на по меньшей мере 5%, 10%, 20%, 30%, 40%, 50% или больше. Таким образом, ингибирование не обязательно должно составлять 100%.
[00315] Ингибирование ингибирующей молекулы может происходить на уровне ДНК, РНК или белка. В некоторых вариантах осуществления для подавления экспрессии ингибирующей молекулы может применяться ингибирующая нуклеиновая кислота (например, dsRNA, siRNA или shRNA). В других вариантах осуществления ингибитор ингибирующего сигнала представляет собой полипептид, например, растворимый лиганд (например, Ig, связывающий PD-1, или Ig, связывающий CTLA-4) или антитело или его антигенсвязывающий фрагмент, которые связываются с ингибирующей молекулой; например, антитело или его фрагмент (также обозначаемые в данном документе как "молекула антитела"), которые связываются с PD-1, PD-L1, PD-L2, CTLA4, TIM3, LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и/или TGFR-бета или их комбинацией.
[00316] В одном варианте осуществления молекула антитела представляет собой полное антитело или его фрагмент (например, Fab, F(ab')2, Fv или одноцепочечный Fv-фрагмент (scFv)). В других вариантах осуществления молекула антитела имеет константную область тяжелой цепи (Fc), выбранную, например, из константных областей тяжелой цепи IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgD и IgE; в частности, выбранную, например, из константных областей тяжелой цепи IgG1, IgG2, IgG3 и IgG4, более конкретно, константной области тяжелой цепи IgG1 или IgG4 (например, IgG1 или IgG4 человека). В одном варианте осуществления константной область тяжелой цепи получена из IgG1 человека или IgG4 человека. В одном варианте осуществления константная область изменена, например, подвергнута мутации для модификации свойств молекулы антитела (например, повышения или снижения одного или нескольких из связывания с рецептором Fc, гликозилирования антител, числа цистеиновых остатков, функции эффекторных клеток или функции комплемента).
[00317] В некоторых вариантах осуществления молекула антитела находится в форме биспецифической или полиспецифической молекулы антитела. В одном варианте осуществления молекула биспецифического антитела характеризуется первой специфичностью связывания с PD-1 или PD-L1 и второй специфичностью связывания, например, второй специфичностью связывания с TIM-3, LAG-3 или PD-L2. В одном варианте осуществления молекула биспецифического антитела связывается с PD-1 или PD-L1 и TIM-3. В другом варианте осуществления молекула биспецифического антитела связывается с PD-1 или PD-L1 и LAG-3. В другом варианте осуществления молекула биспецифического антитела связывается с PD-1 и PD-L1. В еще одном варианте осуществления молекула биспецифического антитела связывается с PD-1 и PD-L2. В другом варианте осуществления молекула биспецифического антитела связывается с TIM-3 и LAG-3. Любая комбинация вышеуказанных молекул может быть составлена в молекулу полиспецифического антитела, например, триспецифическое антитело, которое предусматривает первую специфичность связывания с PD-1 или PD-1, а также вторую и третью специфичности связывания с двумя или более из TIM-3, LAG-3 или PD-L2.
[00318] В некоторых вариантах осуществления иммуномодулятор представляет собой ингибитор PD-1, например PD-1 человека. В другом варианте осуществления иммуномодулятор представляет собой ингибитор PD-L1, например PD-L1 человека. В одном варианте осуществления ингибитор PD-1 или PD-L1 представляет собой молекулу антитела к PD-1 или PD-L1. Ингибитор PD-1 или PD-L1 может вводиться отдельно или в комбинации с другими иммуномодуляторами, например, в комбинации с ингибитором LAG-3, TIM-3 или CTLA4. В иллюстративном варианте осуществления ингибитор PD-1 или PD-L1, например, молекулу антитела к PD-1 или PD-L1, вводят в комбинации с ингибитором LAG-3, например, молекулой антитела к LAG-3. В другом варианте осуществления ингибитор PD-1 или PD-L1, например, молекулу антитела к PD-1 или PD-L1, вводят в комбинации ингибитором TIM-3, например, молекулой антитела к TIM-3. В других вариантах осуществления ингибитор PD-1 или PD-L1, например, молекулу антитела к PD-1, вводят в комбинации с ингибитором LAG-3, например, молекулой антитела к LAG-3, и ингибитором TIM-3, например, молекулой антитела к TIM-3. Другие комбинации иммуномодуляторов с ингибитором PD-1 (например, с одним или несколькими из PD-L2, CTLA4, TIM3, LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и/или TGFR) также предусмотрены в пределах настоящего изобретения. Любая из молекул антитела, известная из уровня техники или раскрытая в данном документе, может использоваться в вышеуказанной комбинации ингибиторов молекулы контрольных точек.
[00319] В одном варианте осуществления ингибитор PD-1 представляет собой антитело к PD-1, выбранное из ниволумаба, пембролизумаба или пидилизумаба. В некоторых вариантах осуществления антитело к PD-1 представляет собой ниволумаб. К альтернативным названиям ниволумаба относятся MDX-1106, MDX-1106-04, ONO-4538 или BMS-936558. В некоторых вариантах осуществления антитело к PD-1 представляет собой ниволумаб (регистрационный номер CAS: 946414-94-4). Ниволумаб представляет собой полностью человеческое моноклональное антитело IgG4, которое специфически блокирует PD1. Ниволумаб (клон 5C4) и другие человеческие моноклональные антитела, которые специфически связываются с PD1, раскрыты в патенте США № 8008449 и PCT-публикации № WO2006/121168.
[00320] В других вариантах осуществления антитело к PD-1 представляет собой пембролизумаб. Пембролизумаб (торговое наименование KEYTRUDA, ранее ламбролизумаб, также известный как Merck 3745, MK-3475 или SCH-900475) представляет собой гуманизированное моноклональное антитело IgG4, которое связывается с PD1. Пембролизумаб раскрыт, например, в Hamid, O. et al. (2013) New England Journal of Medicine 369 (2): 134-44, PCT-публикации № WO2009/114335 и патенте США № 8354509.
[00321] В некоторых вариантах осуществления антитело к PD-1 представляет собой пидилизумаб. Пидилизумаб (CT-011; Cure Tech) представляет собой гуманизированное моноклональное антитело IgG1k, которое связывается с PD1. Пидилизумаб и другие гуманизированные моноклональные антитела к PD-1 раскрыты в PCT-публикации № WO2009/101611. Другие антитела к PD1 раскрыты в патенте США № 8609089, публикации заявки на патент США № 2010028330 и/или публикации заявки на патент США № 20120114649. Другие антитела к PD1 включают AMP 514 (Amplimmune).
[00322] В некоторых вариантах осуществления ингибитор PD-1 представляет собой PDR001 или любое другое антитело к PD-1, раскрытое в WO2015/112900.
[00323] В некоторых вариантах осуществления ингибитор PD-1 представляет собой иммуноадгезин (например, иммуноадгезин, содержащий внеклеточную или PD-1-связывающую часть PD-L1 или PD-L2, слитую с константной областью (например, областью Fc последовательности иммуноглобулина). В некоторых вариантах осуществления ингибитор PD-1 представляет собой AMP-224.
[00324] В некоторых вариантах осуществления ингибитор PD-L1 представляет собой антитело к PD-L1. В некоторых вариантах осуществления ингибитор на основе антитела к PD-L1 выбран из YW243.55.S70, MPDL3280A, MEDI-4736 или MDX-1105MSB-0010718C (также называемого A09-246-2), раскрытого, например, WO 2013/0179174, и имеет последовательность, раскрытую в данном документе (или последовательность практически идентичную или сходную с ней, например, последовательность, которая характеризуется по меньшей мере 85%, 90%, 95% или более высокой идентичностью с указанной последовательностью).
[00325] В одном варианте осуществления ингибитор PD-L1 представляет собой MDX-1105. MDX-1105, также известный как BMS-936559, представляет собой антитело к PD-L1, описанное в PCT-публикации № WO2007/005874.
[00326] В одном варианте осуществления ингибитор PD-L1 представляет собой YW243.55.S70. Антитело YW243.55.S70 представляет собой антитело к PD-L1, описанное в PCT-публикации № WO 2010/077634 (последовательности вариабельных областей тяжелой и легкой цепей показаны под SEQ ID No: 20 и 21, соответственно).
[00327] В одном варианте осуществления ингибитор PD-L1 представляет собой MDPL3280A (Genentech/Roche). MDPL3280A представляет собой человеческое моноклональное антитело IgG1 с оптимизированной Fc, которое связывается с PD-L1. MDPL3280A и другие человеческие моноклональные антитела к PD-L1 раскрыты в патенте США № 7943743 и публикации заявки на патент США № 20120039906.
[00328] В других вариантах осуществления ингибитор PD-L2 представляет собой AMP-224. AMP-224 представляет собой растворимый рецептор на основе слитого белка PD-L2 и Fc, который блокирует взаимодействие между PD1 и B7-H1 (B7-DCIg; Amplimmune; например, раскрытый в PCT-публикациях №№ WO2010/027827 и WO2011/066342).
[00329] В одном варианте осуществления ингибитор LAG-3 представляет собой молекулу антитела к LAG-3. В одном варианте осуществления ингибитор LAG-3 представляет собой BMS-986016.
Фармацевтические композиции
[00330] Для получения фармацевтических или стерильных композиций, содержащих иммуноконъюгаты, иммуноконъюгаты по настоящему изобретению смешивают с фармацевтически приемлемым носителем или вспомогательным веществом. Композиции могут дополнительно содержать один или несколько других терапевтических средств, которые подходят для лечения или предупреждения рака, экспрессирующего CCR7 (в том числе без ограничения хронического лимфоцитарного лейкоза (CLL), периферических T-клеточных лимфом (PTCL), таких как T-клеточный лейкоз/лимфома взрослых (ATLL) и анапластическая крупноклеточная лимфома (ALCL), неходжкинской лимфомы (NHL), такой как лимфома из клеток мантийной зоны (MCL), лимфома Беркитта и диффузная В-крупноклеточная лимфома (DLBCL), карциномы желудка, немелкоклеточного рака легкого, мелкоклеточного рака легкого, рака головы и шеи, носоглоточной карциномы (NPC), рака пищевода, колоректальной карциномы, рака поджелудочной железы, рака щитовидной железы, рака молочной железы, почечноклеточного рака и рака шейки матки).
[00331] Составы терапевтических и диагностических средств могут быть получены путем смешивания с физиологически приемлемыми носителями, вспомогательными веществами или стабилизаторами в форме, например, лиофилизированных порошков, взвесей, водных растворов, лосьонов или суспензий (см., например, Hardman et al., Goodman and Gilman's The Pharmacological Basis of Therapeutics, McGraw-Hill, New York, N.Y., 2001; Gennaro, Remington: The Science and Practice of Pharmacy, Lippincott, Williams, and Wilkins, New York, N.Y., 2000; Avis, et al. (eds.), Pharmaceutical Dosage Forms: Parenteral Medications, Marcel Dekker, NY, 1993; Lieberman, et al. (eds.), Pharmaceutical Dosage Forms: tablets, Marcel Dekker, NY, 1990; Lieberman, et al. (eds.) Pharmaceutical Dosage Forms: Disperse Systems, Marcel Dekker, NY, 1990; Weiner and Kotkoskie, Excipient Toxicity and Safety, Marcel Dekker, Inc., New York, N.Y., 2000).
[00332] Выбор схемы введения для терапевтического средства зависит от нескольких факторов, в том числе интенсивности метаболизма сыворотки или ткани для объекта, уровня симптомов, иммуногенности объекта и доступности клеток-мишеней в биологической матрице. В некоторых вариантах осуществления схема введения максимизирует количество терапевтического средства, доставляемого пациенту, в соответствии с приемлемым уровнем побочных эффектов. Соответственно, количество доставляемого биологического средства частично зависит от конкретного объекта и тяжести состояния, подлежащего лечению. Руководства по подбору подходящих доз антител, цитокинов и малых молекул являются доступными (см., например, Wawrzynczak, Antibody Therapy, Bios Scientific Pub. Ltd, Oxfordshire, UK, 1996; Kresina (ed.), Monoclonal Antibodies, Cytokines and Arthritis, Marcel Dekker, New York, N.Y., 1991; Bach (ed.), Monoclonal Antibodies and Peptide Therapy in Autoimmune Diseases, Marcel Dekker, New York, N.Y., 1993; Baert et al., New Engl. J. Med. 348:601-608, 2003; Milgrom et al., New Engl. J. Med. 341:1966-1973, 1999; Slamon et al., New Engl. J. Med. 344:783-792, 2001; Beniaminovitz et al., New Engl. J. Med. 342:613-619, 2000; Ghosh et al., New Engl. J. Med. 348:24-32, 2003; Lipsky et al., New Engl. J. Med. 343:1594-1602, 2000).
[00333] Определение подходящей дозы выполняет лечащий врач, например, с применением параметров или факторов, которые, как известно или ожидаемо из уровня техники, воздействуют на лечение или предупреждение или, как прогнозируют, воздействуют на лечение или предупреждение. Обычно начинают дозирование с количества, несколько меньшего, чем оптимальная доза, и после этого ее повышают небольшими шагами до тех пор, пока не достигается требуемый или оптимальный эффект по сравнению с любыми отрицательными побочными эффектами. Важные диагностические показатели включают, например, показатели, являющиеся симптомами воспаления, или уровень продуцируемых воспалительных цитокинов.
[00334] Фактические уровни дозы активных ингредиентов в фармацевтических композициях по настоящему изобретению могут варьироваться для того, чтобы получить количество активного ингредиента, которое является эффективным для достижения требуемого терапевтического ответа у конкретного пациента, эффективным в отношении композиции и способа введения, при этом не токсично для пациента. Выбранный уровень дозы будет зависеть от ряда фармакокинетических факторов, в том числе активности конкретных используемых композиций по настоящему изобретению или их сложного эфира, соли или амида, пути введения, времени введения, скорости выведения конкретного используемого соединения, длительности лечения, других лекарственных средств, соединений и/или материалов, применяемых в комбинации с конкретными используемыми композициями, возраста, пола, массы тела, состояния, общего состояния здоровья и анамнеза пациента, подлежащего лечению, и аналогичных факторов, известных в области техники медицины.
[00335] Композиции, содержащие антитела или их фрагменты по настоящему изобретению, могут вводиться в помощью непрерывной инфузии или с помощью доз с интервалами, составляющими, например, один день, одну неделю, или вводиться 1-7 раз в неделю, раз в две недели, раз в три недели, раз в четыре недели, раз в пять недель, раз в шесть недель, раз в семь недель или раз в восемь недель. Дозы могут вводиться внутривенно, подкожно, местно, перорально, назально, ректально, внутримышечно, внутрь мозга или посредством ингаляции. Специфический протокол введения доз является таким, который предусматривает максимальную дозу или частоту введения доз, позволяющие избежать значительных нежелательных побочных эффектов.
[00336] Для иммуноконъюгатов по настоящему изобретению доза, вводимая пациенту, может составлять от 0,0001 мг/кг до 100 мг/кг массы тела пациента. Доза может быть составлять от 0,0001 мг/кг до 30 мг/кг, от 0,0001 мг/кг до 20 мг/кг, от 0,0001 мг/кг и 10 мг/кг, от 0,0001 мг/кг до 5 мг/кг, от 0,0001 до 2 мг/кг, от 0,0001 до 1 мг/кг, от 0,0001 мг/кг до 0,75 мг/кг, от 0,0001 мг/кг до 0,5 мг/кг, от 0,0001 мг/кг до 0,25 мг/кг, от 0,0001 до 0,15 мг/кг, от 0,0001 до 0,10 мг/кг, от 0,001 до 0,5 мг/кг, от 0,01 до 0,25 мг/кг или от 0,01 до 0,10 мг/кг массы тела пациента. Доза антител или их фрагментов по настоящему изобретению может быть рассчитана с применением массы пациента в килограммах (кг), умноженной на подлежащую введению дозу в мг/кг.
[00337] Дозы иммуноконъюгатов по настоящему изобретению могут быть повторными, а введения могут быть разделены менее чем 1 днем, по меньшей мере 1 днем, 2 днями, 3 днями, 5 днями, 10 днями, 15 днями, 30 днями, 45 днями, 2 месяцами, 75 днями, 3 месяцами, 4 месяцами, 5 месяцами или по меньшей мере 6 месяцами. В некоторых вариантах осуществления иммуноконъюгаты по настоящему изобретению можно давать два раза в неделю, раз в неделю, раз в две недели, раз в три недели, раз в четыре недели или менее часто. В конкретном варианте осуществления дозы иммуноконъюгатов по настоящему изобретению вводятся повторно через каждые 2 недели.
[00338] Эффективное количество для конкретного пациента может варьироваться в зависимости от таких факторов, как подлежащее лечению состояние, общее состояние здоровья пациента, способ, путь и доза введения и тяжесть побочных эффектов (см., например, Maynard et al., A Handbook of SOPs for Good Clinical Practice, Interpharm Press, Boca Raton, Fla., 1996; Dent, Good Laboratory and Good Clinical Practice, Urch Publ., London, UK, 2001).
[00339] Путем введения может быть, например, местное нанесение или нанесение на кожу, инъекция или инфузия посредством подкожного, внутривенного, внутрибрюшинного, внутримозгового, внутримышечного, внутриглазного, внутриартериального, внутриспинномозгового, внутриочагового введения, или системы замедленного высвобождения или имплантат (см., например, Sidman et al., Biopolymers 22:547-556, 1983; Langer et al., J. Biomed. Mater. Res. 15:167-277, 1981; Langer, Chem. Tech. 12:98-105, 1982; Epstein et al., Proc. Natl. Acad. Sci. USA 82:3688-3692, 1985; Hwang et al., Proc. Natl. Acad. Sci. USA 77:4030-4034, 1980; патенты США №№ 6350466 и 6316024). При необходимости композиция может также содержать солюбилизирующее средство и местный анестетик, такой как лидокаин, для облегчения боли в месте инъекции, или их оба. Кроме того, также можно использовать ингаляционное введение, например, за счет применения ингалятора или распылителя и состава со средством в виде аэрозоля. См., например, патенты США №№ 6019968, 5985320, 5985309, 5934272, 5874064, 5855913, 5290540 и 4880078; и публикации согласно PCT №№ WO 92/19244, WO 97/32572, WO 97/44013, WO 98/31346 и WO 99/66903, каждый из которых включен в данный документ посредством ссылки во всей своей полноте.
[00340] Композицию по настоящему изобретению также вводят с помощью одного или нескольких путей введения с применением одного или нескольких из целого ряда способов, известных из уровня техники. Как будет понятно специалисту в данной области техники, путь и/или способ введения будут варьироваться в зависимости от необходимых результатов. Выбранные пути введения иммуноконъюгатов по настоящему изобретению включают внутривенный, внутримышечный, внутрикожный, внутрибрюшинный, подкожный пути введения, введение в спинной мозг или другие парентеральные пути введения, например, с помощью инъекции или инфузии. Парентеральное введение может представлять способы введения, отличные от энтерального и местного введения, обычно производимые с помощью инъекции, и оно охватывает без ограничения внутривенные, внутримышечные, внутриартериальные, интратекальные, внутрикапсульные, интраорбитальные, внутрисердечные, внутрикожные, внутрибрюшинные, транстрахеальные, подкожные, субкутикулярные, внутрисуставные, подкапсулярные, субарахноидальные, интраспинальные, эпидуральные и интрастернальные инъекцию и инфузию. В качестве альтернативы композицию по настоящему изобретению можно вводить посредством отличного от парентерального пути, такого как местный, эпидермальный пути введения или введение через слизистые оболочки, например, интраназально, перорально, вагинально, ректально, сублингвально или местно. В одном варианте осуществления иммуноконъюгаты по настоящему изобретению вводят с помощью инфузии. В другом варианте осуществления иммуноконъюгаты по настоящему изобретению вводят подкожно.
[00341] Если иммуноконъюгаты по настоящему изобретению вводятся в системе контролируемого высвобождения или замедленного высвобождения, для осуществления контролируемого или замедленного высвобождения может применяться насос (см. Langer, выше; Sefton, CRC Crit. Ref Biomed. Eng. 14:20, 1987; Buchwald et al., Surgery 88:507, 1980; Saudek et al., N. Engl. J. Med. 321:574, 1989). Для осуществления контролируемого или замедленного высвобождения видов терапии по настоящему изобретению могут применяться полимерные материалы (см., например, Medical Applications of Controlled Release, Langer and Wise (eds.), CRC Pres., Boca Raton, Fla., 1974; Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen and Ball (eds.), Wiley, New York, 1984; Ranger and Peppas, J. Macromol. Sci. Rev. Macromol. Chem. 23:61, 1983; см. также Levy et al., Science 228:190, 1985; During et al., Ann. Neurol. 25:351, 1989; Howard et al., J. Neurosurg. 7 1:105, 1989; патент США № 5679377; патент США № 5916597; патент США № 5912015; патент США № 5989463; патент США № 5128326; PCT-публикацию № WO 99/15154 и PCT-публикацию № WO 99/20253. Примеры полимеров, используемых в составах с замедленным высвобождением, включают без ограничения поли(2-гидроксиэтилметакрилат), поли(метилметакрилат), поли(акриловую кислоту), сополимер(этилена и винилацетата), поли(метакриловую кислоту), полигликолиды (PLG), полиангидриды, поли(N-винилпирролидон), поли(виниловый спирт), полиакриламид, поли(этиленгликоль), полилактиды (PLA), сополимеры(лактида и гликолидов) (PLGA) и сложные полиортоэфиры. В одном варианте осуществления полимер, используемый в составе с замедленным высвобождением, является инертным, не содержит выщелачиваемых примесей, стабильный при хранении, стерильный и биологически разлагаемый. Систему контролируемого или замедленного высвобождения можно поместить вблизи профилактической или терапевтической мишени, при этом требуется только часть системной дозы (см., например, Goodson, в Medical Applications of Controlled Release, выше, vol. 2, pp. 115-138, 1984).
[00342] Системы контролируемого высвобождения рассмотрены в обзоре Langer, Science 249:1527-1533, 1990). Любую методику, известную специалисту в данной области техники, можно применять для получения составов с замедленным высвобождением, содержащих один или несколько иммуноконъюгатов по настоящему изобретению. См., например, патент США № 4526938, PCT-публикацию WO 91/05548, PCT-публикацию WO 96/20698, Ning et al., Radiotherapy & Oncology 39:179-189, 1996; Song et al., PDA Journal of Pharmaceutical Science & Technology 50:372-397, 1995; Cleek et al., Pro. Int'l. Symp. Control. Rel. Bioact. Mater. 24:853-854, 1997; и Lam et al., Proc. Int'l. Symp. Control Rel. Bioact. Mater. 24:759-760, 1997, каждый из которых включен в данный документ посредством ссылки во всей своей полноте.
[00343] Если иммуноконъюгаты по настоящему изобретению вводятся местно, они могут быть составлены в форме мази, крема, трансдермального пластыря, лосьона, геля, спрея, аэрозоля, раствора, эмульсии или другой формы, широко известной специалисту в данной области техники. См., например, Remington's Pharmaceutical Sciences and Introduction to Pharmaceutical Dosage Forms, 19th ed., Mack Pub. Co., Easton, Pa. (1995). В случае нераспыляемых местных лекарственных форм обычно используются вязкие-полутвердые или твердые формы, содержащие носитель или одно или несколько вспомогательных веществ, совместимые с местным нанесением и, в некоторых случаях, характеризующиеся динамической вязкостью больше, чем у воды. Подходящие составы включают без ограничения растворы, суспензии, эмульсии, кремы, мази, порошки, линименты, бальзамы и т. п., которые при необходимости стерилизуют или смешивают со вспомогательными средствами (например, консервантами, стабилизаторами, смачивающими средствами, буферами или солями) для воздействия на различные свойства, такие как, например, осмотическое давление. Другие подходящие местные лекарственные формы охватывают распыляемые аэрозольные препараты, где активный ингредиент, в некоторых случаях в комбинации с твердым или жидким инертным носителем, упакован в смеси с летучим веществом под давлением (например, газообразным пропеллентом, таким как фреон) или в бутылке-пульверизаторе. При необходимости в фармацевтические композиции и лекарственные формы также можно добавлять увлажнители или увлажняющие средства. Примеры таких дополнительных ингредиентов широко известны из уровня техники.
[00344] Если композиции, содержащие иммуноконъюгаты, вводят интраназально, они могут быть составлены в форме аэрозоля, спрея, тумана или в форме капель. В частности, в целях удобства профилактические или терапевтические средства для применения согласно настоящему изобретению можно доставлять в форме спрея-аэрозоля из пакетов под давлением или с помощью небулайзера с применением подходящего пропеллента (например, дихлордифторметана, трихлорфторметана, дихлортетрафторэтана, диоксида углерода или другого подходящего газа). В случае аэрозоля под давлением единица дозирования может определяться за счет обеспечения клапана для доставки отмеренного количества. Капсулы и картриджи (например, состоящие из желатина) для применения в ингаляторе или инсуффляторе можно составлять содержащими порошковую смесь соединения и подходящей порошковой основы, такой как лактоза или крахмал.
[00345] Способы совместного введения или лечения с применением второго терапевтического средства, например, цитокина, стероида, химиотерапевтического средства, антибиотика, или облучения, известны из уровня техники (см., например, Hardman et al., (eds.) (2001) Goodman and Gilman's The Pharmacological Basis of Therapeutics, 10th ed., McGraw-Hill, New York, N.Y.; Poole and Peterson (eds.) (2001) Pharmacotherapeutics for Advanced Practice:A Practical Approach, Lippincott, Williams & Wilkins, Phila., Pa.; Chabner and Longo (eds.) (2001) Cancer Chemotherapy and Biotherapy, Lippincott, Williams & Wilkins, Phila., Pa.). Эффективное количество терапевтического средства может снижать симптомы на по меньшей мере 10%; по меньшей мере 20%; по меньшей мере приблизительно 30%; по меньшей мере 40% или по меньшей мере 50%.
[00346] Дополнительные виды терапии (например, профилактические или терапевтические средства), которые можно применять в комбинации с иммуноконъюгатами по настоящему изобретению, можно применять с интервалом менее 5 минут, с интервалом менее 30 минут, с интервалом 1 час, с интервалом приблизительно 1 час, с интервалом от приблизительно 1 до приблизительно 2 часов, с интервалом от приблизительно 2 часов до приблизительно 3 часов, с интервалом от приблизительно 3 часов до приблизительно 4 часов, с интервалом от приблизительно 4 часов до приблизительно 5 часов, с интервалом от приблизительно 5 часов до приблизительно 6 часов, с интервалом от приблизительно 6 часов до приблизительно 7 часов, с интервалом от приблизительно 7 часов до приблизительно 8 часов, с интервалом от приблизительно 8 часов до приблизительно 9 часов, с интервалом от приблизительно 9 часов до приблизительно 10 часов, с интервалом от приблизительно 10 часов до приблизительно 11 часов, с интервалом от приблизительно 11 часов до приблизительно 12 часов, с интервалом от приблизительно 12 часов до 18 часов, с интервалом от 18 часов до 24 часов, с интервалом от 24 часов до 36 часов, с интервалом от 36 часов до 48 часов, с интервалом от 48 часов до 52 часов, с интервалом от 52 часов до 60 часов, с интервалом от 60 часов до 72 часов, с интервалом от 72 часов до 84 часов, с интервалом от 84 часов до 96 часов или с интервалом от 96 часов до 120 часов от введения иммуноконъюгатов по настоящему изобретению. Два или более видов терапии могут применяться во время одного и того же визита пациента.
[00347] В некоторых вариантах осуществления иммуноконъюгаты по настоящему изобретению могут составляться, чтобы гарантировать точное распределение in vivo. Например, гематоэнцефалический барьер (BBB) не пропускает множество высокогидрофильных соединений. Чтобы гарантировать то, что терапевтические соединения по настоящему изобретению пересекают BBB (при необходимости), их можно составлять, например, в липосомах. Способы изготовления липосомы, см., например, в патентах США №№ 4522811; 5374548 и 5399331. Липосомы могут содержать один или несколько фрагментов, которые селективно транспортируются в специфические клетки или органы, с улучшением тем самым нацеленной доставки лекарственного средства (см., например, Ranade, (1989) J. Clin. Pharmacol. 29:685). Иллюстративные нацеливающие фрагменты включают фолат или биотин (см., например, патент США № 5416016 от Low et al.); маннозиды (Umezawa et al., (1988) Biochem. Biophys. Res. Commun. 153:1038); антитела (Bloeman et al., (1995) FEBS Lett. 357:140; Owais et al., (1995) Antimicrob. Agents Chemother. 39:180); рецептор поверхностно-активного белка A (Briscoe et al., (1995) Am. J. Physiol. 1233:134); p 120 (Schreier et al., (1994) J. Biol. Chem. 269:9090); см. также K. Keinanen; M. L. Laukkanen (1994) FEBS Lett. 346:123; J. J. Killion; I. J. Fidler (1994) Immunomethods 4:273.
[00348] В настоящем изобретении предусмотрены протоколы для введения субъекту, нуждающемуся в этом, фармацевтической композиции, содержащей иммуноконъюгаты по настоящему изобретению, отдельно или в комбинации с другими видами терапии. Виды терапии (например, профилактические или терапевтические средства) из видов комбинированной терапии по настоящему изобретению могут применяться в отношении субъекта одновременно или последовательно. Терапия (например, профилактические или терапевтические средства) из видов комбинированной терапии по настоящему изобретению также можно вводить циклически. Циклическая терапия предусматривает введение первого средства терапии (например, первого профилактического или терапевтического средства) в течение некоторого периода времени, а затем введение второго средства терапии (например, второго профилактического или терапевтического средства) в течение некоторого периода времени и повторение такого последовательного введения, т. е. цикла, с целью снижения развития устойчивости к одному из видов терапии (например, средствам), для недопущения или снижения побочных эффектов одного из видов терапии (например, средств) и/или для улучшения эффективности видов терапии.
[00349] Виды терапии (например, профилактические или терапевтические средства) из видов комбинированной терапии по настоящему изобретению могут применяться в отношении субъекта одновременно.
[00350] Термин "одновременно" не ограничивается применением видов терапии (например, профилактических или терапевтических средств) точно в одно и то же время, а скорее означает, что фармацевтическую композицию, содержащую антитела или их фрагменты по настоящему изобретению, вводят субъекту последовательно и в течение временного интервала таким образом, чтобы конъюгаты антитела и лекарственного средства по настоящему изобретению могли действовать вместе с другим(-и) видом(-ами) терапии для обеспечения большей пользы, чем если бы их вводили иным способом. Например, каждый вид терапии может применяться в отношении субъекта в одно и то же время или последовательно в любом порядке в различные моменты времени; однако, если их не применяют в одно и то же время, их следует применять достаточно близко по времени, чтобы обеспечить требуемый терапевтический или профилактический эффект. Каждый вид терапии можно применять в отношении субъекта по отдельности, в любой подходящей форме и с помощью любого подходящего пути. В различных вариантах осуществления виды терапии (например, профилактические или терапевтические средства) применяют в отношении субъекта с интервалом менее 5 минут, с интервалом менее 15 минут, с интервалом менее 30 минут, с интервалом менее 1 часа, с интервалом приблизительно 1 час, с интервалом от приблизительно 1 часа до приблизительно 2 часов, с интервалом от приблизительно 2 часов до приблизительно 3 часов, с интервалом от приблизительно 3 часов до приблизительно 4 часов, с интервалом от приблизительно 4 часов до приблизительно 5 часов, с интервалом от приблизительно 5 часов до приблизительно 6 часов, с интервалом от приблизительно 6 часов до приблизительно 7 часов, с интервалом от приблизительно 7 часов до приблизительно 8 часов, с интервалом от приблизительно 8 часов до приблизительно 9 часов, с интервалом от приблизительно 9 часов до приблизительно 10 часов, с интервалом от приблизительно 10 часов до приблизительно 11 часов, с интервалом от приблизительно 11 часов до приблизительно 12 часов, с интервалом 24 часа, с интервалом 48 часов, с интервалом 72 часа или с интервалом 1 неделя. В других вариантах осуществления два или более видов терапии (например, профилактические или терапевтические средства) применяются во время одного и того же визита пациента.
[00351] Профилактические или терапевтические средства из видов комбинированной терапии могут вводиться субъекту в одной и той же фармацевтической композиции. В качестве альтернативы профилактические или терапевтические средства из видов комбинированной терапии могут вводиться субъекту одновременно в отдельных фармацевтических композициях. Профилактические или терапевтические средства могут вводиться субъекту с помощью одного и того же или различных путей введения. Профилактические или терапевтические средства из видов комбинированной терапии могут вводиться субъекту в одной и той же фармацевтической композиции. В качестве альтернативы профилактические или терапевтические средства из видов комбинированной терапии могут вводиться субъекту одновременно в отдельных фармацевтических композициях. Профилактические или терапевтические средства могут вводиться субъекту с помощью одного и того же или различных путей введения.
Примеры
Пример 1. Получение антител к CCR7
Получение конструкций для экспрессии CCR7 человека, крысы, мыши и яванского макака
[00352] Полноразмерные гены CCR7 человека, яванского макака и мыши синтезировали на основе аминокислотных последовательностей из баз данных GenBank или Uniprot (SEQ ID NO: 97, SEQ ID NO: 99, SEQ ID NO: 101). кДНК-матрицу CCR7 крысы получали посредством генного синтеза на основе сведений об аминокислотной последовательности, полученной с применением мРНК, выделенной из различных тканей крысы (SEQ ID NO: 103). Все синтезированные фрагменты ДНК клонировали в подходящие векторы экспрессии.
Таблица 2. Сведения об аминокислотной и нуклеотидной последовательности CCR7
Получение линий клеток, стабильно экспрессирующих CCR7
[00353] Линии клеток, стабильно экспрессирующих CCR7, получали с применением ретровирусной трансдукции. Клетки 293T подвергали совместной трансфекции ретровирусным вектором экспрессии CCR7 и упаковывающим вектором pCL-Eco или pCL-10A1 (Novus, США, № по кат. NBP2-29540 или NBP2-2952) с применением реагента трансфекции Fugene 6 (Promega, США, № по кат. E2692), следуя рекомендациям производителя. Клетки инкубировали при 37°C в увлажненном CO2-инкубаторе и вирусный супернатант собирали через 48 часов после трансфекции. Клетки NIH/3T3 и 300.19 выращивали до практически конфлюэнтного монослоя. У клеток удаляли ростовую среду и добавляли вирусный супернатант в присутствии 8 мкг полибрена/мл (конечная концентрация) (EMD Millipore, № по кат. TR-1003-G). После инкубации в течение 3-6 часов при 37°C добавляли свежую среду. Затем клетки культивировали в условиях, подходящих для отбора, с получением линий клеток, стабильно экспрессирующих CCR7.
Получение, экспрессия и очистка вирусоподобных частиц (VLP)
[00354] Клетки HEK293T или NIH/3T3 поддерживали в DMEM с 10% FBS. Для получения VLP клеткам меняли среду на DMEM с 4% FBS, затем подвергали совместной трансфекции плазмидой экспрессии CCR7 и ретровирусной плазмидой экспрессии Gag при соотношении 3:2 в мкг. Через сорок восемь часов после трансфекции клеточный супернатант собирали и просветляли центрифугированием при 2500 × g в течение 5 мин в настольной центрифуге и держали на льду. VLP очищали ультрацентрифугированием при 100000 × g через 20% сахарозную подушку в 38-мл центрифужных пробирках Beckman Ultra-Clear (№ по каталогу 344058) на Ti-роторе Beckman Coulter SW 32 на ультрацентрифуге Sorvall RC 6+. Полученные осадки ресуспендировали в 300 мкл холодного стерильный PBS и подвергали количественной оценке с применением BCA-анализа (Pierce, № по каталогу 23225).
Создание иммуногенного остова CCR7 на основании структуры
[00355] Члены семейства рецепторов, сопряженных с G-белком, представляют собой мембранные белки, которые содержат семь трансмембранных спиральных областей (TM1…TM7), каждая из которых соединена связывающими последовательностями варьирующей длины. Амино-конец белка расположен на внешней стороне клеточной поверхности, это означает, что 4 области белка потенциально находятся на поверхности клетки, амино-конец (N-конец) и 3 области внеклеточных петель (EC1, EC2 и EC3). Таким образом, такие области доступны в качестве антигенов для антител.
[00356] Предполагалось, что комбинацию из одного или нескольких таких 4 элементов, можно будет вставить в остов растворимого белка, чтобы он структурно приблизился к области CCR7, находящейся за пределами клетки.
[00357] Для определения оптимальных внеклеточных областей CCR7 строили модель с применением кристаллической структуры близкого гомолога CXCR4 (Wu et al., "Structures of the CXCR4 chemokine GPCR with small-molecule и cyclic peptide antagonists". (2010) Science 330: 1066-1071), применяя комбинирование структуры CXCR4 записями в базе данных структуры белков (3ODU, 3OE0, 3OE6, 3OE8, 3OE9) и программное обеспечение для моделирования. На основании модели сделали вывод, что аминокислоты, которые расположены между связывающими трансмембранными спиральными областями, находятся на поверхности белка. Данные области идентифицированы в таблице 3 ниже.
Таблица 3. Сведения об аминокислотной и нуклеотидной последовательности иммуногенного остова CCR7
(NP_001829.1| хемокиновый рецептор типа 7 с мотивом C-C)
Внеклеточные области выделены жирным шрифтом.
Области вставки или их производные обозначены жирным шрифтом и подчеркнуты.
Вставленные последовательности подчеркнуты и обозначены жирным шрифтом
[00358] Малый размер петель EC1 и EC3 и пространственное отделение EC3 от остальных трех областей делает приоритетным применение N-конца и петли EC2 в качестве эпитопов-кандидатов.
[00359] Моделирование слияния последовательности N-конца с кристаллической структурой мышиного Fab с последовательностью EC2, вставленной в различные области петель Fab, как, например, в каркасную область 1, CDR-H3 или CDR3-H1, показало, что если степень гибкости для данных двух последовательностей является допустимой, то они могут представлять собой обоснованные приближения для структуры данных областей в CCR7.
Создание иммуногенного остова для иммунизации мышей
[00360] Привитые конструкции для иммунизации мышей создавали путем слияния N-концевой последовательности из таблицы 3 с N-концом тяжелой цепи остова мышиного Fab (обозначенного как MGFTX1) и прививания модифицированного варианта последовательности EC2 с мутацией цистеинового остатка в положении 24 на серин (таблица 3 и подчеркнутые+обозначенные жирным шрифтом последовательности в SEQ ID NO: 113) в CDR3 из остова MGFTX1, затем продуцировали обе тяжелую и легкую цепи из цепей иммуноглобулин для создания конечных белковых конструкций. Таким образом, конструкции, обозначенные как FabCCR7M1, содержали мышиную каркасную область, которая должна характеризоваться иммунной толерантностью при объединении с человеческой последовательностью, на которую был направлен иммунный ответ мыши.
[00361] Последовательность N-конца напрямую сливали с тяжелой цепью остова MGFTX1. Точки для вставки EC2 выбирали так, чтобы она находилась посередине петли CDR, исходя из доступных данных для структурной или гомологичной модели. Белки с привитым Fab получали с применением стандартных методик молекулярной биологии путем использования рекомбинантной ДНК, кодирующей соответствующие последовательности.
[00362] Например, синтезировали вариабельную область каждого антитела, содержащего EC2, вставленную в CDR3 тяжелой цепи. ДНК, кодирующую вариабельную область, амплифицировали с помощью ПЦР и полученный фрагмент субклонировали в вектор, содержащий или константную область легкой цепи, или константную область и область Fc тяжелой цепи. Таким способом получали белки FabCCR7M1 с соответствующим слиянием N-конца и вставкой EC2 в H3. Полученные конструкции показаны в таблице 3. Трансфекции с применением подходящих комбинаций векторов, содержащих тяжелую и легкую цепь, приводят к экспрессии рекомбинантного Fab, содержащего одну молекулу N-конца и одну молекулу EC2.
[00363] Выбор того, какую CDR отбирают для прививания, определяется, исходя из параметров наружного расположения петли и близости к N-концу. В настоящее время программное обеспечение для моделирования только частично применимо для предсказания того, какая CDR и какое местоположение в пределах CDR будут обеспечивать требуемые параметры, что связано с гибкостью петель после проведения слияния и прививания. Структура остова MGFTX1 в комбинации с EC2_C24S (таблица 3, SEQ ID NO: 110) показала, что последовательность находится снаружи и является гибкой, о чем свидетельствует отсутствие электронной плотности у большей части последовательности.
[00364] Подводя итог, точку вставки в CDR выбирали на основе структуры, исходя из предположения, что прививание в CDR будет обеспечивать некоторый уровень структурного подобия с нативным антигеном.
Создание гибридомы
[00365] Трансгенных мышей Bcl-2 (линия C57BL/6-Tgn (bcl-2) 22 WEHI) иммунизировали антигеном с применением процедуры, которая называется "Повторная иммунизация в несколько сайтов" (RIMMS) (McIntyre GD. Hybridoma 1997). Вкратце, мышам инъецировали по 1-3 мкг иммуногена CCR7 в 8 специфических сайтов, близких к периферическим лимфатическим узлам (PLN). Данную процедуру повторяли 8 раз в течение 12-дневного периода. В день 12 собирали образцы крови и титр сывороточных антител анализировали с помощью FACS. В некоторых случаях мышей BALB/c и/или C57Bl/6 иммунизировали с помощью клеток NIH3T3 или 300.19, стабильно свехэкспрессирующих CCR7 человека (SEQ ID NO: 97). Животным инъецировали подкожно 5×106 клеток в PBS один раз в месяц в течение 3 месяцев с последующей внутривенной бустер-инъекцией 25 мкг VLP, экспрессирующих CCR7 человека. Через два дня после бустер-инъекции собирали образцы крови и титр сывороточных антител анализировали с помощью FACS. Селезенки и объединенные PLN извлекали у мышей с высокими титрами. Для сбора лимфоцитов селезенки и PLN промывали дважды с помощью DMEM, а затем разделяли на клетки за счет пропускания через сито с размером ячеек 70 микрон (Falcon, № 352350). Полученные лимфоциты промывали еще 2 раза перед проведением слияния в среде Cytofusion (среда для электропорации BTXpress Cytofusion®, № по кат. 47001).
[00366] Для проведения слияния клетки миеломы F0 смешивали с лимфоцитами при соотношении 1:4. Смесь клеток центрифугировали, суспендировали в среде Cytofusion и затем вносили в камеру для электрослияния (коаксиальная камера 9ML от Harvard Apparatus, изделие № 470020). Электрослияние проводили с соответствии с инструкциями производителя с применением системы для получения гибридом Hybrimune модели CEEF-50B (Cyto Pulse Sciences, Inc). Слитые клетки оставляли для восстановления в течение 5 мин в камере, разбавляли 1/10 в среде для слияния без HAT (DMEM+20% FBS, 1% пенициллина/стрептомицина/глутамина, 1X NEAA, 0,5X HFCS) и помещали при 37°C на один час. Добавляли 4X среду HAT (DMEM+20% FBS, 1% пенициллина/стрептомицина/глутамина, 1X NEAA, 4X HAT, 0,5X HFCS) с получением 1X раствора и плотность доводили до 1,67×104 клеток/мл. Клетки высевали в 384-луночные планшеты из расчета 60 мкл/лунка.
Скрининг с помощью FACS
[00367] Через десять дней после слияния планшеты с гибридомами подвергали скринингу на присутствие CCR7-специфических антител с применением проточной цитометрии для подтверждения специфического связывания антител-кандидатов с линиями клеток, стабильно сверхэкспрессирующими или экспрессирующими эндогенный CCR7. Клетки тщательно ополаскивали PBS и обрабатывали аккутазой (Millipore, № SCR005) для отделения от ростовых планшетов и ресуспендировали в холодном PBS. Клетки метили биотином с флуоресцентным красителем согласно инструкциям производителя (набор для биотинилирования клеточной поверхности FluoReporter, Thermo Fisher Scientific, № по кат. F-20650; стрептавидин PE-Cy7, ThermoFisher Scientific, № по кат. SA1012). Клетки ресуспендировали из расчета примерно 1×106 клеток/мл в буфере FACS (1X DPBS, 3% FBS, 5 мМ EDTA, 0,1% азида натрия). В 384-луночный планшет предварительно вносили по 20 мкл супернатанта гибридом и добавляли 20 мкл клеточной суспензии. Клетки инкубировали в течение 1 часа при 4°C, промывали дважды с помощью холодного буфера FACS и ресуспендировали в 20 мкл вторичного антитела, разведенного 1:400 в буфере FACS (конъюгированный с аллофикоцианином F(ab')2 козы к IgG человека, Fcγ-специфический; Jackson Immunoresearch, № по кат. 109-136-098). После дополнительной инкубации в течение 45 мин при 4°C, клетки промывали дважды с помощью буфера FACS и ресуспендировали в 20 мкл буфера FACS+2 мкг/мл пропидия йодида (Sigma Aldrich, № по кат. P4864-10ML). Геометрическое среднее интенсивности флуоресценции рассчитывали на живых одиночных клетках с применением программного обеспечения FlowJo™.
Очистка антител
[00368] Получали химерные антитела, содержащие мышиные вариабельные области и человеческие константные области. Кроме того, разрабатывали химерные варианты, содержащие цистеиновые мутации (например, цистеины в положении K36°C или в положениях E152C и S375C тяжелой цепи) для конъюгации с фрагментом, представляющим собой лекарственное средство, и получения ADC, описанных более подробно в данном документе. В случае каждой из отобранных гибридом, mAb121G12, mAb506E15, mAb674J13 и mAb684E12, получали ДНК-последовательности вариабельных областей (VH и VL) антител, производимых гибридомами, для создания оптимизированных последовательностей (например, гуманизации, придания предпочтительных характеристик). ДНК вариабельной области из мышиных моноклональных антител амплифицировали с помощью RACE на основе РНК, полученной из каждой отобранной линии клеток гибридомы, с применением стандартных способов. Полипептидные последовательности каждой из мышиных вариабельных областей тяжелой/легкой цепей показаны под SEQ ID NO: 128/SEQ ID NO: 144, SEQ ID NO: 160/SEQ ID NO: 176, SEQ ID NO: 192/SEQ ID NO: 208 и SEQ ID NO: 224/SEQ ID NO: 240, соответственно, для каждой из гибридом 674J13, 121G12, 506E15 и 684E12. Соответствующие производные нуклеотидные последовательности вариабельных областей тяжелой/легкой цепей для каждой из гибридом показаны под SEQ ID NO: 129/SEQ ID NO: 145, SEQ ID NO: 161/SEQ ID NO: 177, SEQ ID NO: 193/SEQ ID NO: 209 и SEQ ID NO: 225/SEQ ID NO: 241. Для получения химерных антител ДНК-последовательности, кодирующие домен VL и VH антитела из гибридомы субклонировали в векторы экспрессии, содержащие соответствующие последовательности человеческой тяжелой цепи дикого типа, или тяжелой цепи с введенным путем конструирования Cys, или тяжелой цепи с мутацией D265A/P329A (DAPA) и последовательность константной области человеческой легкой цепи (IgG1, каппа-цепь).
Гуманизация антител
[00369] Конструкции вариабельных областей подвергали разработке для гуманизации и оптимизации последовательностей (например, удаления посттрансляционных модификаций, нежелательных сайтов и т.д.), для включения цистеиновых мутаций (например, цистеинов в положении K36°C или в положениях E152C и S375C тяжелой цепи), предназначенных для конъюгации фрагмента, представляющего собой лекарственное средство, и получения ADC, описанных более подробно в данном документе; а также для модификации с помощью мутаций эффекторной области Fc (например, мутаций D265A/P329A в области Fc), чтобы обеспечить конструкции, характеризующиеся сниженной эффекторной функцией Fc, и обеспечения их комбинаций.
[00370] ДНК-последовательности, кодирующие гуманизированные домены VL и VH, заказывали в GeneArt (Life Technologies Inc., Регенсбург, Германия), включая оптимизацию кодонов для Cricetulus griseus. Последовательности, кодирующие домены VL и VH, субклонировали из векторов, взятых из GeneArt, в векторы экспрессии, подходящие для продуцирования белка в клетках млекопитающих. Тяжелые и легкие цепи клонировали в отдельные векторы экспрессии для обеспечения возможности совместной трансфекции.
[00371] Рекомбинантные антитела (IgG1, каппа-цепь) получали за счет совместной трансфекции векторов в экспрессирующие клетки Freestyle™ 293 (Invitrogen, США) с применением PEI (полиэтиленимин, MW 25000, линейный, Polysciences, США, № по кат. 23966) в качестве реагента для трансфекции. Исходный раствор PEI готовили растворением 1 г PEI в 900 мл воды, подходящей для культивирования клеток, при комнатной температуре (RT). Для облегчения растворения PEI раствор подкисляли добавлением HCl до pH 3-5 с последующей нейтрализацией с помощью NaOH с получением конечного pH, составляющего 7,05. Наконец, объем доводили до 1 л и раствор стерилизовали фильтрацией через фильтр с размером пор 0,22 мкм, разделяли на аликвоты и замораживали при -80°C до дальнейшего применения.
[00372] Клетки Freestyle™ 293 (Gibco™, ThermoFisher scientific, USA, № по кат. R79007) культивировали в среде Freestyle™ 293 (Gibco™, ThermoFisher scientific, США, № по кат. 12338018) во встряхиваемых колбах (Corning, Тьюксбери, Массачусетс) на орбитальном шейкере (100-120 об/мин) при 37°C в увлажненном инкубаторе при 5% CO2. В случае временных трансфекций клетки выращивали до плотности примерно 3×106 клеток/мл, а затем 1 мкг стерилизованной фильтрацией ДНК/мл культуры (0,5 мкг тяжелой цепи+0,5 мкг легкой цепи) добавляли в раствор 2 мкг PEI/1 мкг ДНК в OptiMem (ThermoFisher Scientific, США, #11058021) и инкубировали при RT в течение 8 минут. Смесь добавляли к клеткам Freestyle™ 293 по каплям с аккуратным перемешиванием круговыми движениями. После трансфекции клетки культивировали в течение одной-двух недель до проведения очистки антитела от супернатанта. Чтобы создать стабильные линии клеток для продуцирования антител, векторы трансфицировали совместно с помощью нуклеофекции (Nucleofector™ 96-well shuttle™; Lonza) в клетки CHO с применением рекомендаций производителя и их культивировали в условиях отбора в течение не более четырех недель во встряхиваемых колбах. Клетки собирали центрифугированием, а супернатант извлекали для очистки антитела. Антитело очищали с применением колонок с белком A, белком G или MabSelect SuRe (GE Healthcare Life Sciences). Перед загрузкой супернатанта смолу уравновешивали с помощью PBS. После связывания образца колонку промывали с помощью PBS и антитело элюировали с помощью буфера для элюирования IgG Thermo (Pierce), pH 2,8 (№ по кат. 21004). Элюированные фракции нейтрализовали с помощью буфера на основе дегидрата трехосновного цитрата натрия, pH 8,5 (Sigma Aldrich, № по кат. S4641-1Kg), и затем диализировали в течение ночи в PBS, pH 7,2.
Краткое описание антител
[00373] В таблице 4 изложены соответствующие сведения о последовательности исходных и гуманизированных антител к CCR7, происходящих из мышиных гибридомных антител. На всем протяжении данной заявки при описании антител термины "гибридомное" и "исходное" используются взаимозаменяемо и относятся к Ab, которые происходят из гибридомы.
Таблица 4. Сведения об аминокислотной и нуклеотидной последовательности для гибридомных и гуманизированных антител к CCR7
(Fc WT)
(Fc WT)
(Fc WT)
(CysMab, мутации подчеркнуты)
(DAPA, CysMab, мутации подчеркнуты)
(CysMab, мутации подчеркнуты)
(CysMab, мутации подчеркнуты)
(DAPA, CysMab, мутации подчеркнуты)
(CysMab, мутации подчеркнуты)
(DAPA, CysMab, мутации подчеркнуты)
(CysMab, мутации подчеркнуты)
(CysMab, мутации подчеркнуты)
(DAPA, CysMab, мутации подчеркнуты)
(DAPA, мутации подчеркнуты)
(DAPA, мутации подчеркнуты)
(DAPA, мутации подчеркнуты)
(DAPA, CysMab, мутации подчеркнуты)
Пример 2. In vitro оценка активности ADCC, индуцированной антителом
[00374] Способность антител-кандидатов опосредовать ADCC оценивали с применением суррогатного репортерного анализа ADCC. В качестве клеток-мишеней использовали клетки JVM2, экспрессирующие CCR7 и CD20. Клетки JVM2 промывали и ресуспендировали из расчета 8×104 клеток/мл клеток/мл. Эффекторными клетками в данном анализе была линия клеток Jurkat, стабильно экспрессирующих CD16V158 и NFAT-зависимый люциферазный репортер (Jurkat-V158); экспрессия люциферазы является заменителем канонической передачи сигнала ADCC через CD16. Вкратце, клетки Jurkat-V158, выращенные в суспензионной культуре, центрифугировали для удаления истощенной среды, осадок ресуспендировали в среде для анализа и доводили до 1,6×106 клеток/мл клеток/мл. Смешивали равные объемы эффекторных клеток и клеток-мишеней с получением основной смеси клеток, в которой отношение клеток-мишеней к эффекторным клеткам составляет 1:5 или 1:20. Титр антитела разбавляли в среде для анализа при конечной наибольшей концентрации, составляющей 50 мкг/мл в лунке для анализа. 12,5 мкл раствора Ab добавляли в 384-луночный круглодонный планшет и затем добавляли 12,5 мкл основной смеси клеток. Антитело и клетки тщательно смешивали за счет пипетирования и инкубировали в течение 4 часов при 37°C в 5% CO2. После инкубации в каждую лунку добавляли по 15 мкл субстрата Bright Glo (Promega № G7572) и встряхивали в течение 5 мин при RT при 1050 об/мин. Люминесцентный сигнал считывали на планшет-ридере Envision (Perkin Elmer).
[00375] В качестве контроля включали антитело, нацеливающееся на CD20, и оно показывало значительную передачу сигнала с помощью NFAT, измеренную по активности люциферазы. Аналогично, антитела-кандидаты к CCR7 индуцировали значительную активность ADCC (фигура 1).
[00376] В таблице ниже обобщены типичные результаты для антител различных форматов, включенных в анализ ADCC с применением клеток JVM2 и Jurkat-V158.
Таблица 5. Активность ADCC у негуманизированных и гуманизированных антител к CCR7
[00377] Анализ ADCC проводили с негуманизированным антителом 506E15 в качестве типичного антитела к CCR7, способного опосредовать ADCC, на различных линиях клеток с разным числом CCR7-рецепторов для определения минимальной плотности рецепторов, необходимой для активности ADCC, и наличие надлежащей границы безопасности по сравнению с нормальными CCR7+ T-клетками. В таблице ниже обобщены некоторые из данных.
Таблица 6. Активность ADCC у негуманизированного антитела 506E15
[00378] Данные показывают, что даже очень низких уровней CCR7-рецепторов, сравнимых с уровнями CCR7-рецепторов на нормальных T клетках, было достаточно для обеспечения значительной активности ADCC. ADCC также оценивали с применением анализа жизнеспособности при совместном культивировании NK-клеток с CCR7+ раковыми клетками и получали аналогичные результаты (не обсуждаются в данном документе).
[00379] Это согласуется с описанным ниже наблюдением, согласно которому отмечали истощение T-клеток in vivo и обнаружили, что оно основано на механизме ADCC. Эти результаты демонстрируют, что механизм ADCC делает CCR7 неприменимым с точки зрения безопасности. В связи с этим, всех кандидатов переключали в формат с "молчащей" Fc (DAPA) для улучшения общей безопасности с точки зрения мишени.
[00380] Репортерный in vitro анализ ADCC повторяли, и он подтвердил отсутствие активности ADCC у негуманизированных антител к CCR7, содержащих варианты Fc с мутацией DAPA (фигура 2).
Пример 3. Определение биохимических характеристик антител
Аффинность антител к CCR7 в отношении CCR7
[00381] Аффинность различных антител и ADC в отношении CCR7 и его видовых ортологов определяли с применением FACS. Очищенные IgG титровали, чтобы определить значения EC50 для связывания с CCR7, экспрессированным на поверхности клеток.
[00382] Для этого CCR7-положительные клетки собирали (адгезивные клетки отделяли с помощью аккутазы), промывали дважды с помощью буфера FACS (PBS/3% FCS/0,02% азида натрия) и разбавляли до примерно 2×106 клеток/мл в буфере FACS. Все последующие стадии проводили на льду, чтобы предотвратить интернализацию рецептора. По 100 мкл клеточной суспензии/лунка переносили в 96-луночные планшеты с U-образным дном (Falcon). 2×105 клеток/лунка инкубировали с последовательным разведением представляющего интерес антитела к CCR7, при этом концентрация антитела варьировала в диапазоне нескольких порядков, начиная от самой высокой 100 нМ, в течение 60 минут при 4°C с аккуратным встряхиванием. После инкубации клетки центрифугировали (1200 об/мин, 2 мин, 4°C) и трижды промывали буфером FACS. Антитело обнаружения, конъюгированное с флуорофором APC антитело к hFc-гамма (Jackson Immunoresearch), добавляли из расчета 1:400 и образцы инкубировали в течение 1 ч на льду в темноте с аккуратным встряхиванием. После финальной промывки клетки ресуспендировали в 100 мкл буфера FACS, содержащего 0,2 мкг/мл DAPI, с последующим считыванием на устройстве для проточной цитометрии (анализатор клеток BD LSRFortessa; № по кат. 647177). Среднюю интенсивность флуоресценции (MFI) живых одиночных клеток рассчитывали в Flowjo 10.0.8 и результаты экспортировали в Graphpad Prism6 для определения EC50.
[00383] Селективность оценивали путем измерения наблюдаемых аффинностей связывания с изогенной парой клеток, сконструированных для сверхэкспрессии CCR7, а также с линиями клеток, экспрессирующих паралоги CCR7, например, CCR9, CCR6, CXCR4 и CCR8. Все антитела к CCR7 связывались специфически образом только с клетками, экспрессирующими CCR7, как показано в таблице 7 ниже.
Таблица 7. Связывание различных антител к CCR7 с клетками, экспрессирующими CCR7
(CCR7-/ CCR9+/ CCR6+)
[00384] В аналогичном эксперименте антитела тестировали на перекрестную реактивность с применением наборов сконструированных изогенно-совпадающих линий клеток (серия NIH3T3) и CD4+ T клеток, которые очищали из нескольких партий PBMC от здоровых доноров, а также яванских макаков, крыс Вистар и мышей CD-1. Обнаружили, что все антитела специфически связывают CCR7 человека и яванского макака при аналогичных наблюдаемых аффинностях, как показано в таблицах 8 и 9 ниже. Только негуманизированное антитело 121G12 характеризуется перекрестной реактивностью в отношении молекулы грызунов.
Таблица 8. Перекрестная реактивность различных негуманизированных антител к CCR7
Таблица 9. Перекрестная реактивность различных гуманизированных антитела к CCR7
[00385] Чтобы определить влияние плотности рецепторов на наблюдаемую аффинность, и, следовательно, вклада авидности в связывание антитела с клеткой, проводили титриметрические эксперименты FACS на линиях раковых клеток, экспрессирующих CCR7 человека на различных уровнях экспрессии, и нормальных CCR7-положительных T клетках, происходящих из PBMC. Количественное определение рецепторов проводили с помощью FACS с применением микросфер от Bangs Laboratories в качестве стандартов подсчета импульсов и следовали инструкциям производителя. Иллюстративные результаты показаны в таблице 10 ниже.
Таблица 10. Вклад авидности в наблюдаемую аффинность в зависимости от плотности рецепторов
[00386] Для всех антител к CCR7 показан значительный вклад авидности в наблюдаемую аффинность, и сила связывания снижалась в зависимости от плотности рецепторов. В частности, для 121G12 показано значительно более слабое связывание на клетках с низким уровнем экспрессии CCR7, представленных нормальными CD4+ T клетками, по сравнению с раковыми клетками DEL, являющимися представителями клеток, для которых показано его применение.
[00387] Относительно низкая аффинность конкретного 121G12, которое продемонстрировало самый сильный эффект авидности среди всех антител к CCR7, является оптимальной для использования отличия в плотности рецепторов между нормальными и раковыми клетками в качестве метода для смещения связывания антитела в направлении раковых клеток.
Связывание с рекомбинантным hCCR7 в ELISA
[00388] Связывание и аффинность также оценивали в анализе на основе ELISA с применением рекомбинантного CCR7 (Origene, № TP306614). 384-луночные планшеты Maxisorp™ (Thermo Nunc) покрывали с помощью 3,5 мкг/мл рекомбинантного CCR7, разведенного в PBS. После блокирования с помощью 3% BSA (бычий сывороточный альбумин) в PBS в течение 1 часа при комнатной температуре, промывки планшетов 3x с помощью PBS-T (0,01% Tween 20 в PBS), добавляли последовательные разведения первичных антител и инкубировали в течение 1 часа при комнатной температуре. Планшеты промывали вновь и связавшиеся антитела обнаруживали посредством инкубации с 1:5000 антителом к hFc-гамма, конъюгированным с пероксидазой хрена (HRP; Jackson Immunoresearch, № по кат. 115-035-098), в течение 1 часа при комнатной температуре с последующей промывкой помощью PBS-T и последующего добавления субстрата для пероксидазы SureBlue (KPL, № 52-00-03). Спустя 15 мин регистрировали поглощение при 650 нм и анализировали в GraphPad Prism6.
[00389] Все протестированные антитела к CCR7 способны к связыванию рекомбинантного hCCR7 (таблица 11; фигура 3).
Таблица 11. Аффинность связывания гуманизированных антител к CCR7 с рекомбинантным hCCR7
Связывание с двойными FabGraft
[00390] FabGraft ELISA проводили для оценки связывания с эпитопом минимальной протяженности, который содержит N-конец и EC2 из CCR7. Вкратце, 384-луночные планшеты Maxisorp™ (Thermo Nunc) покрывали с помощью 5 мкг/мл FabGraft. Во всем остальном следовали общей инструкции для протокола ELISA, как описано выше. Все антитела к CCR7 способны к связыванию с двойными FabGraft, как показано в таблице 12 ниже.
Таблица 12. Аффинность связывания гуманизированных антител к CCR7 с FabGraft
Оценка pH-зависимости в ELISA с применением VLP
[00391] Известно, что CCL19, связавшийся с CCR7, подвергается интернализации как комплекс рецептор-лиганд. Однако в то время как CCR7 рециклируется обратно на поверхность клетки, CCL19 сортируется в лизосому для разложения, что показывает противоположную "судьбу" CCR7 и его лиганда, подвергнутых эндоцитозу (Otero et al., J Immunol 2006; 177:2314-2323). Для приводящего к нужному эффекту ADC к CCR7 предпочтительно, чтобы антитело вело себя аналогично лиганду, например, подвергалось быстрой интернализации, но не рециклировалось обратно вместе с CCR7. Чтобы достичь этого, авторы настоящего изобретения обеспечивали отбор pH-зависимых антител, которые будут проявлять более слабое связывание с CCR7 в условиях низкого pH.
[00392] Для оценки pH-зависимости антител к CCR7 проводили ELISA с применением вирусоподобных частиц (VLP), экспрессирующих CCR7. Вкратце, 384-луночные планшеты Maxisorp™ (Thermo Nunc) покрывали с помощью 25 мкг/мл VLP. Первичные антитела инкубировали либо в буфере с pH 5,8 (1:1; dH2O:0,1 M цитратный буфер, 150 мМ NaCl) или буфере с pH 7,4. Во всем остальном следовали общей инструкции для протокола ELISA, как описано выше.
[00393] При использовании целого ряда гуманизированных вариантов для каждого кандидата сравнение связывания антител при нейтральном (7,4) и кислом (5,8) pH показало, что все антитела-кандидаты к CCR7 характеризуются улучшенной аффинностью при pH 7,4 (таблица 13). Нижеприведенные объекты были выбраны на основании их самой большой pH-зависимости, в числе других признаков.
Таблица 13. pH-зависимость связывания гуманизированных антител к CCR7 с VLP, экспрессирующими CCR7
Анализ с b-аррестином
[00394] Чтобы определить функциональность антител к CCR7 проводили анализ с β-аррестином с применением набора для обнаружения импульса PathHunter от DiscoverX (№ 93-0247) либо в режиме агониста для оценки агонистической функции, либо в режиме антагониста для оценки антагонистической функции.
[00395] В случае режима агониста CHO-flpin-hCCR7 (разработанная DiscoverX линия клеток, экспрессирующих hCCR7 с меткой ProLink, β-аррестин-EA) высевали из расчета 8×104 клеток/лунка в 20 мкл/лунка ростовой среды (среда F-12 Хэма/Glutamax; Invitrogen+10% FBS+0,5 мг/мл G418+0,2 мг/мл гигромицина B; Invitrogen+5 мкг/мл бластицидина; Gibco) с 100 нг/мл Dox в 384-луночные планшеты, покрытые металлическими крышками, инкубировали при 37°C, 5% CO2 в течение ночи. На следующий день последовательное разведение 5x рабочего раствора в 1x буфере для анализа (20 мМ HEPES/0,1% BSA/1x HBSS, pH 7,4) выполняли для тестируемых антител или положительного контроля с применением лиганда hCCL19 (R&D, 361/MI-025/CF). В каждую лунку добавляли по 5 мкл 5x рабочего раствора антител или лиганда, проводили быстрое центрифугирование и инкубировали при 37°C/5% CO2 в течение 2 ч. После инкубации в каждую лунку добавляли по 25 мкл реагента для обнаружения, инкубировали при комнатной температуре в темноте при встряхивании в течение 20 мин. Затем измеряли сигнал люминесценции на приборе Envision, чтобы определить активность фермента. Наконец, активность фермента анализировали с применением Excel.
[00396] В режиме антагониста клетки CHO-flpin-hCCR7 высевали, как описано выше. На следующий день 6x рабочий раствор (0,5 мкМ x 6=3,0 мкМ) в 1x буфере для анализа получали для каждого тестируемого антитела или MAB197 положительного контроля (эталонное антитело; лиганд-антагонист от R&D) или hIgG отрицательного контроля. В каждую лунку добавляли по 5 мкл 6x рабочего раствора антител или контролей, проводили быстрое центрифугирование и инкубировали при 37°C/5% CO2 в течение 30 мин. Во время инкубация выполняли последовательные разведения 6x рабочего раствора hCCL19 в 1x буфере для анализа. После инкубации в каждую лунку добавляли по 5 мкл 6x рабочего раствора hCCL19, проводили быстрое центрифугирование и инкубировали при 37°C/5% CO2 в течение 90 мин. После инкубации в каждую лунку добавляли по 25 мкл реагента для обнаружения, инкубировали при комнатной температуре в темноте при встряхивании в течение 20 мин. Затем измеряли сигнал люминесценции на приборе Envision, чтобы определить активность фермента, и анализировали с помощью Excel.
[00397] Ни одно из исходных антител к CCR7 не продемонстрировало активность в модели агониста (фигура 4A). Однако при анализе в формате антагониста 506E15 и 121G12 идентифицировали как сильных антагонистов, например, антител, блокирующих связывание с лигандом (фигуры 4B, 4C). 674J12 представляет собой нейтральное антитело, не блокирующее связывание с лигандом. 684E12 представляет собой слабого антагониста.
Анализ конкурентного связывания в FACS с применением лиганда
[00398] Чтобы подтвердить то, что антитело конкурирует с лигандом CCR7, проводили FACS-анализ в присутствии избыточной концентрации лиганда. FACS-анализ проводили, как описано выше. В протокол вносили некоторые изменения, например, CCL19 поддерживали при постоянной концентрации 1 мкМ, в то время как концентрация первичного антитела, которое одновременно применяли к клеткам DEL, варьировала в диапазоне нескольких порядков, начиная с самой высокой концентрации 100 нМ. После 30-минутного периода инкубации в ледяном буфере FACS клетки промывали, в течение 15 мин применяли вторичное антитело к hFc, конъюгированное с PE, и MFI определяли, как описано выше.
[00399] На фигуре 5 показано, что на гуманизированное CysMab.DAPA 674J13 не влияло присутствие избытка CCL19, что подтверждает его нейтральную функциональность. На аффинность связывания гуманизированных CysMab.DAPA 121G12 и 506E15 действительно сильно влияло присутствие избытка лиганда.
Способность к интернализации в различных линиях клеток, характеризующихся диапазоном плотности рецепторов
[00400] Другим аспектом приводящего к нужному эффекту ADC к CCR7 является гарантия оптимального использования дифференциального распределение экспрессии CCR7. В качестве представителя нормальных клеток, экспрессирующих CCR7, CD4+ T-клетки выделяли из PBMC здоровых доноров. Кроме того, отобрали целый ряд CCR7-положительных линий раковых клеток, проявляющих диапазон плотностей рецепторов. Авторы настоящего изобретения специально выбрали антитела с более слабой наблюдаемой аффинностью связывания при FACS с CCR7+ T-клетками относительно CCR7+ раковых клеток. Кроме того, в данном документе описан анализ pHrodo, в котором флуорофорная метка, активируемая низким pH, на антителах к CCR7 используется для оценки того, как измеренный по флуоресценции захват антитела клетками коррелирует с плотностью рецепторов. Предпочтительно свести к минимуму захват антитела нормальными клетками, чтобы получить максимальное терапевтическое окно.
[00401] Вкратце, мечение антитела к CCR7 в формате CysMab с помощью малеимида-pHrodo (ThermoFisher) проводили следуя инструкциям производителя с получением объектов с DAR=4 (отношение лекарственного средства, например, флуорофора, к антителу). FACS-анализ проводили, как описано выше. В протокол вносили некоторые изменения, например, чтобы обеспечить возможность интернализации, первичное антитело инкубировали из расчета 5 мкг/мл с клетками при 37°C в среде для культивирования в течение 6 ч, затем промывали с помощью ледяного буфера FACS, содержащего азид натрия, для прекращения реакции.
[00402] В таблице 14 ниже обобщена способность к интернализации у трех антител с применением целого ряда из линий клеток. В качестве контроля применяли ненацеленное антитело, меченное с помощью pHrodo, и было установлено, что сигнал фонового шума составляет до 400 MFI, например, опосредованный нецелевым взаимодействием захват конъюгата антитела. Данные показывают, что для всех трех антител к CCR7 требуются уровни рецептора CCR7 в диапазоне, который является типичным для большинства CCR7+ линий раковых клеток, например более 20000 рецепторов, чтобы эффективно подвергаться интернализации и накапливать конъюгированное вещества, в то же время не попадать в нормальные CD4+ T-клетки.
Таблица 14. Способность к интернализации у различных антител к CCR7
DAPA
DAPA
DAPA
Эпитоп-специфическая сортировка с применением системы Octet Red96
[00403] Эпитоп-специфическую сортировку исходных антител к hCCR7 проводили с применением системы Octet Red96 (ForteBio, США), которая проводит биослойную интерферометрию (BLI). Иммуногенный остов CCR7 биотинилировали с помощью AviTag™ с использованием биотинлигазы BirA в соответствии с рекомендациями производителя (Avidity, LLC, США № по кат. BirA500). Биотинилированный иммуногенный остов загружали из расчета 1,5 мкг/мл на предварительно уравновешенные стрептавидиновые сенсоры (ForteBio, США). Затем сенсоры переносили в раствор, содержащий 100 нМ антитела A в 1X буфере для кинетического анализа (ForteBio, США). Сенсоры быстро промывали в 1X буфере для кинетического анализа и переносили во второй раствор, содержащий 100 нМ антитела-конкурента. Кинетику связывания определяли на основании необработанных данных с применением программного обеспечения для анализа системы Octet Red96 (версия 6.3, ForteBio, США). Антитела тестировали во всех попарных комбинациях в качестве как антитела A, так и антитела-конкурента.
Таблица 15. Результаты сортировки антител
Картирование эпитопов с применением мутаций CCR7
[00404] Дополнительное картирование эпитопов проводили с использованием линий клеток с мутантным CCR7. Создавали линии клеток NIH/3T3, экспрессирующих варианты CCR7 человека с мутациями. Мутации вводили в специфические положения для замены остатка CCR7 человека на соответствующий остаток CCR7 мыши. Введенные мутации охватывали D35E, F44Y, L47V, S49F, D198G, R201K, S202N, S204G, Q206D, A207T, M208L, I213V, T214S, E215A и H216Q. Конструкции плазмид с мутантным CCR7 создавали посредством сайт-направленного мутагенеза и вводили в клетки NIH/3T3 с получением линий клеток со стабильной экспрессией. Специфическое связывание антител-кандидатов с каждой линией клеток с мутантным CCR7 оценивали посредством проточной цитометрии. Клетки тщательно ополаскивали PBS и обрабатывали аккутазой (Millipore, № SCR005) для отделения от ростовых планшетов и ресуспендировали из расчета примерно 1×105 клеток/90 мкл в 1x буфере FACS (2% FBS+0,1% NaN3 в PBS). 10 мкл 10x раствора антитела в буфере FACS предварительно вносили в 96-луночный планшет с U-образным дном и добавляли 90 мкл клеточной суспензии. Клетки инкубировали в течение 30 минут при 4°C, промывали 1x с помощью холодного PBS и ресуспендировали в 100 мкл вторичного антитела, разведенного 1:500 в 1x буфере FACS (конъюгированный с аллофикоцианином F(ab')2 козы к IgG человека, Fcγ-специфический; Jackson Immunoresearch, № по кат. 109-136-098). После дополнительной инкубации в течение 15 мин при 4°C, клетки промывали дважды с помощью PBS и ресуспендировали в 100 мкл 1x буфера FACS+4 мкг/мл пропидия йодида (Life Technologies, № по кат. P3566). Геометрическое среднее интенсивности флуоресценции рассчитывали на живых одиночных клетках с применением FlowJo и наносили на график как % от геометрического среднего интенсивности флуоресценции при CCR7 WT.
[00405] Измеренные на основе EC50 показатели аффинности в отношении CCR7 с точковыми мутациями показали, что все протестированные антитела характеризуются различными профилями связывания, что свидетельствует о различном использовании конформационного эпитопа (фигура 6). Из всех показанных антител 506E15 характеризуется характером связывания, наиболее сходным с таковым MAB197, например, для обоих показано значимое резкое снижение аффинности связывания, когда остатки F44 или L47 на N-конце CCR7 мутированы, и отсутствие снижения, когда мутированы M208 или I213 в петле EC2. По-видимому, 121G12 и 674J13 используют различный набор критических точек контакта в пределах пространства конформационного эпитопа, поскольку они отличаются от MAB197 по меньшей мере 4 из 8 протестированных точковых мутаций.
Пример 4. Создание и определение характеристик конъюгатов антитела к CCR7 и лекарственного средства
В нижеприведенном разделе, мкл используется для обозначения микролитра. Аналогичным образом мкМ используется для микромоль/л; а мкм используется для обозначения микрометра.
[00406] Пример 4A. Получение конъюгата антитела и лекарственного средства 121G12.CysMab.DAPA.MPET.DM4
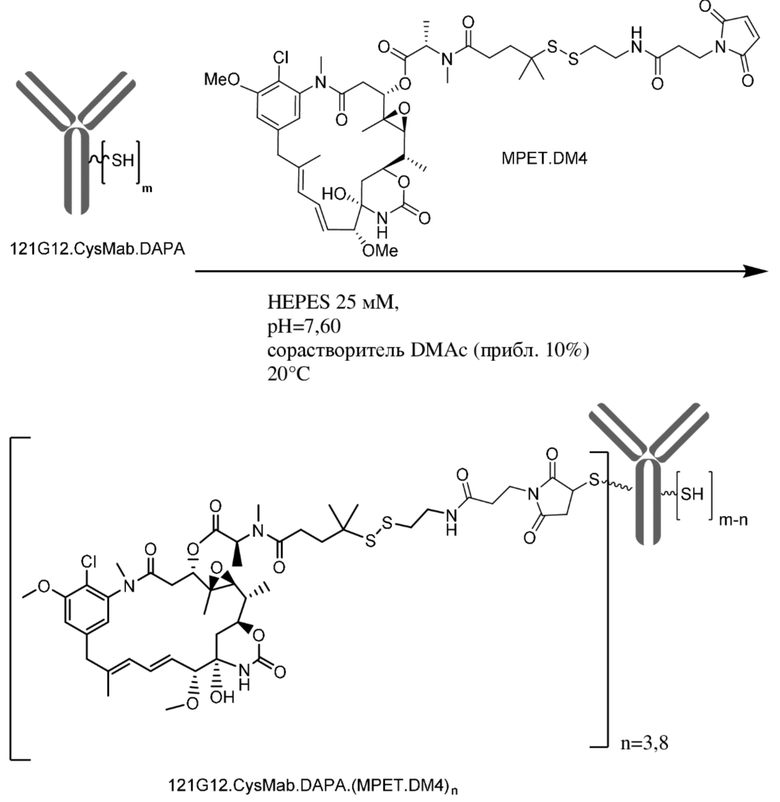
[00407] Конъюгация очищенных 121G12.CysMab.DAPA и MPET.DM4
[00408] Исходным материалом служило антитело 121G12.CysMab.DAPA при 127 мг/мл (для 10 мг/мл раствора IgG коэффициент экстинкции при OD280 составляет 13,7) в буфере на основе 10 мМ гидрохлорида гистидина. К 7,9 мл антитела (1003 мг) добавляли 16 мл 0,5 M фосфата натрия, pH 8 (Teknova S1280), проверяли, чтобы pH составлял >7, затем антитело абсорбировали на 100,3 мл смолы RMP с белком A (GE Healthcare 1-223BPO/I) в течение 25 минут с аккуратным перемешиванием круговыми движениями при комнатной температуре. Смолу, загруженную из расчета 10 мг Ab/мл слоя, промывали с помощью 15 объемов слоя 1x буфера PBS (Hyclone SH30256.02) посредством вакуум-фильтрации через фильтрующее устройство с размером пор 0,2 мкм (Nalgene 567-0020), помещаемое на горлышко бутылки, затем ресуспендировали в 100,3 мл 1x PBS с получением 50% взвеси.
[00409] К взвеси добавляли 4329 мкл 0,5 M цистеина (Sigma G121-03), составленного в 0,5 M фосфате, pH 8, в который был добавлен NaOH (Alfa Aesar A16037) при соотношении 13,6 г/л. Взвесь изредка перемешивали путем вращения сосуда при комнатной температуре в течение 30 минут, затем промывали посредством вакуум-фильтрации через фильтрующее устройство с размером пор 0,2 мкм, помещаемое на горлышко бутылки, с помощью 50 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления. Промытую смолу ресуспендировали в 100,3 мл 1x PBS (50% взвесь) и добавляли 1003 мкл 100 мкМ CuCl2 (Aldrich 751944) (суммарно 500 нМ Cu2+) для инициирования повторного окисления. Повторное окисление антитела тестировали путем изъятия 30-мкл аликвоты взвеси, добавления 1 мкл 20 мМ исходного раствора эталонного малеимида (пример 3, страница 110 в WO2015/095301), который, как известно, сдвигает пик антитела при RPLC, смешивания в течение 1 минуты, центрифугирования при 7000 × g в течение 10 секунд, удаления супернатанта, добавления 60 мкл буфера для элюирования IgG Thermo (Thermo Scientific 21009), центрифугирования при 14000 × g в течение 10 секунд, отбора образцов супернатанта и анализа продуктов с помощью RPLC следующим образом: 2-мкл образец вводили в нагретую (80°C) колонку 4,6×50 мм Agilent PLRP-S (частицы 5 мкм, размер пор 4000 Å), уравновешенную 0,1% трифторуксусной кислотой в 29,5% CH3CN/в воде (Millipore TX1280P-1, Burdick and Jackson 407-4) при расходе 1,5 мл/мин. Колонку элюировали с помощью 5-минутного градиента до 44,5% CH3CN/в воде, поддерживали данную концентрацию в течение 1,9 минуты и пики определяли при 280 нм. Оптимальное время для конъюгации определяется как время, при котором максимального значения достигает пик основного продукта, сокращены до минимума пики с более поздним элюированием, и более не возрастают пики с более ранним элюированием.
[00410] Когда анализ RPLC показывал, что повторное окисление было оптимальным (в данном случае 60 минут), добавляли 3010 мкл 20 мМ исходного раствора MPET.DM4 в DMSO и взвесь изредка аккуратно перемешивали путем вращения сосуда при комнатной температуре в течение 30 минут. Затем взвесь промывали с помощью 20 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления.
[00411] Затем взвесь переносили в колонку с пористым стеклянным фильтром (Pierce 7375021), предварительно элюировали с помощью 0,5 объема слоя буфера для элюирования IgG Thermo (отбрасывали), затем элюировали с помощью 2 объемов слоя того же буфера. Весь элюат (201 мл) нейтрализовали с помощью 20,1 мл 0,5 M фосфата натрия, pH 8, затем концентрировали до 60 мл с применением центрифужных концентраторов (Amicon UFC905024) при 3000 × g. Затем концентрат наносили на буферообменные колонки 24 x PD-10 (GE Healthcare 17-0851-01), уравновешенные 1x PBS, при загрузке 2,5 мл концентрата и элюировании с помощью 3,5 мл 1x PBS согласно рекомендациям изготовителя.
[00412] Для исследований стабильности материал объединяли с партией, полученной идентичным образом, для обеспечения 2 граммов исходного материала. Объединенный материал интенсивно диализировали (колба Slidealyzer, Thermo Scientific 87762) против буфера на основе 10 мМ хлорида гистидина, pH 5 (гистидин от JTBaker 2080-05), концентрировали до ~30 мг/мл, затем добавляли сахарозу (Millipore 1.00892.1003) и Tween 20 (JTBaker 4116-04) до 240 мМ и 0,02% (объем/объем), соответственно. Для избытка материала, который не требовался для исследований стабильности, проводили обратную замену буфера на 1x PBS. Образцы разделяли на аликвоты, подвергали быстрому замораживанию с помощью жидкого азота и хранили при -80°C. Конечные концентрации составляли 23,9 мг/мл для материала, составленного для тестирования стабильности, и 18,9 мг/мл для материала, составленного в 1x PBS.
[00413] Анализ полученных образцов выглядит следующим образом.
[00414] Методы анализа. Концентрацию определяли на основании того, что для 10 мг/мл раствора IgG коэффициент экстинкции при OD280 составляет 13,7 Пироген определяли с применением анализа Kinetic QCL (Lonza Walkersville 50-650H) со считыванием на планшет-ридере TECAN Safire. Процент агрегатов определяли с помощью аналитической эксклюзионной хроматографии на предколонке Shodex KW-G (Thomson Instrument Company, № по кат. 6960955) и колонке KW-803 (TIC, № по кат. 6960940), уравновешенных подвижной фазой [20 мМ Tris, ~pH 7,65 (приготовленный с помощью 10 мМ Tris, pH 7,4, 10 мМ Tris, pH 8), 200 мМ NaCl, 0,02% азида натрия], с получением данных при 280 нм. Для определения DAR готовили аликвоту образца, которую получали разбавлением образца до 2 мг/мл в 1x PBS, дегликозилированием образца с помощью PNGазы F (получена в лаборатории) в течение 10 минут при 50°C, удалением PNGазы F за счет связывание с белком A, промывкой с помощью 1x PBS и элюированием с помощью 1% муравьиной кислоты. Затем образец вводили в колонку 2,1×50 мм PLRP-S (частицы 8 мкм, размер пор 1000 Å), уравновешенную 0,1% муравьиной кислотой в 20% CH3CN/в воде (Invitrogen) при расходе 0,5 мл/мин. Колонку промывали с помощью 20% CH3CN/в воде в течение 3 минут, затем элюировали с помощью 0,1-минутного градиента до 0,1% муравьиной кислоты в 90% CH3CN/в воде, при этом данную концентрацию поддерживали в течение 1,9 минуты. Данные масс-спектров получали на устройстве Agilent 1260, и деконволюцию проводили с помощью программного обеспечения для качественного анализа MassHunter версии B.05.00 в диапазоне 110-180 кДа. Площади пиков, соответствующих различным рассчитанным состояниям DAR, взвешивали в соответствии с DAR каждого пика, затем суммировали и взвешенную площадь пика DAR4 делили на сумму всех взвешенных пиков с получением значения DAR.
[00415] Получение линкера-полезной нагрузки MPET.DM4
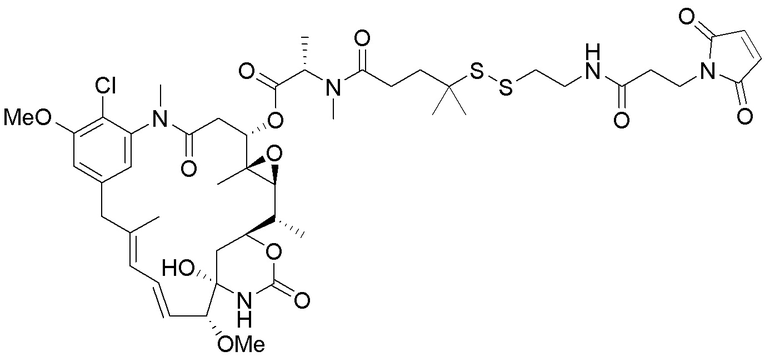
[00416] Аналитические способы
Если не указано иное, в Получении промежуточных соединений и Примерах применяли следующие способы HPLC и HPLC/MS.
Анализ LC/MS осуществляли на системе Agilent 1200sl/6140.
Колонка: Waters Acquity HSS T3 C18, 50×2,0, 1,8 мкм
Подвижная фаза: A) H2O+0,05% TFA; B: ацетонитрил+0,035% TFA
Параметры работы насоса:
Обнаружение: Диодно-матричный УФ-детектор при 190 нм - 400 нм
Сканирование при MS: 200-1350 amu
ELSD: 60°C
Параметры MS:
[00417] (14S,16S,32S,33S,2R,4S,10E,12E,14R)-86-хлор-14-гидрокси-85,14-диметокси-33,2,7,10-тетраметил-12,6-диоксо-7-аза-1(6,4)-оксазинана-3(2,3)-оксиранa-8(1,3)-бензолaциклотетрадекафан-10,12-диен-4-ил N-(4-((2-(3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропанамидо)этил)дисульфанил)-4-метилпентаноил)-N-метил-L-аланинат
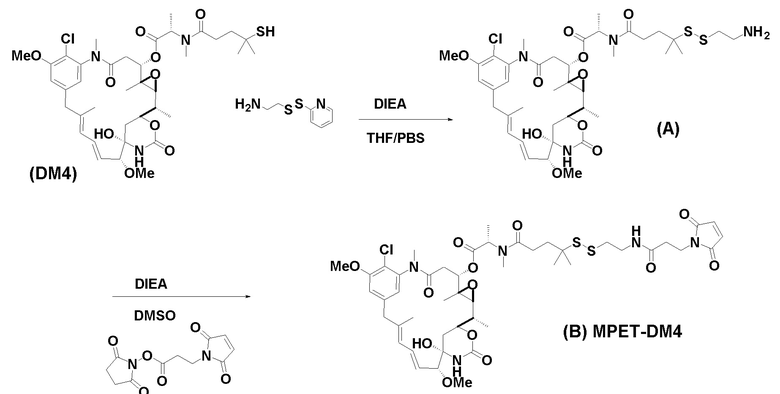
[00418] Стадия 1. Получение (14S,16S,32S,33S,2R,4S,10E,12E,14R)-86-хлор-14-гидрокси-85,14-диметокси-33,2,7,10-тетраметил-12,6-диоксо-7-аза-1(6,4)-оксазинана-3(2,3)-оксиранa-8(1,3)-бензолaциклотетрадекафан-10,12-диен-4-ил N-(4-((2-аминоэтил)дисульфанил)-4-метилпентаноил)-N-метил-L-аланината
[00419] К DM4 (480 мг, 0,62 ммоль), растворенному в буфере PBS (10,5 мл) и безводном THF (21 мл), добавляли 2-(пиридин-2-илдисульфанил)этан-1-амин (151 мг, 0,68 ммоль) и DIEA (0,27 мл, 1,54 ммоль) при комнатной температуре. Реакционную смесь перемешивали при комнатной температуре в течение 30 мин и концентрировали под вакуумом. Водный остаток разбавляли с помощью CH3CN (1 мл) и H2O (2 мл) и очищали посредством ISCO с обращенной фазой, элюировали с помощью 10-60% ацетонитрила в H2O, содержащей 0,05% TFA. Фракции, содержащие требуемый продукт, лиофилизировали с получением требуемого продукта (555 мг, выход 93%). 1H ЯМР (400 МГц, MeOD-d4) δ ppm 0,83 (s, 3 H) 1,21 (d, J=5,0 Гц, 3 H) 1,25 (s, 3 H) 1,28 (s, 3 H) 1,30 (d, J=5,0 Гц, 3 H) 1,45-1,55 (m, 3 H) 1,67 (s, 3 H) 1,84-1,88 (m, 1 H) 1,95-2,01 (m, 1 H) 2,14 (dd, J=5,0 и 15,0 Гц, 1 H) 2,37-2,43 (m, 1 H) 2,53-2,59 (m, 1 H) 2,64 (dd, J=10,0 и 15,0 Гц, 1 H) 2,82-2,89 (m, 5 H) 2,91 (d, J=10,0 Гц, 1 H) 3,16 (dd, J=5,0 и 10,0 Гц, 2 H) 3,20 (s, 3 H) 3,23 (d, J=10,0 Гц, 1 H) 3,35 (s, 3 H) 3,55 (d, J=5,0 Гц, 1 H) 3,58 (d, J=10,0 Гц, 1 H) 4,15-4,20 (m, 1 H) 4,64 (dd, J=5,0 и 10,0 Гц, 1 H) 5,43 (q, J=5,0 Гц, 2 H) 5,66 (dd, J=10,0 и 15,0 Гц, 1 H) ) 6,58 (dd, J=10,0 и 15,0 Гц, 1 H) 6,65 (d, J=10,0 Гц, 1 H) 6,66 (s, 1 H) 7,11 (bs, 1H) 7,28 (bs, 1H); MS масса/заряд 855,3 (M+H), Время удерживания 0,988 минуты.
[00420] Стадия 2. Получение (14S,16S,32S,33S,2R,4S,10E,12E,14R)-86-хлор-14-гидрокси-85,14-диметокси-33,2,7,10-тетраметил-12,6-диоксо-7-аза-1(6,4)-оксазинана-3(2,3)-оксиранa-8(1,3)-бензолaциклотетрадекафан-10,12-диен-4-ил N-(4-((2-(3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропанамидо)этил)дисульфанил)-4-метилпентаноил)-N-метил-L-аланината
[00421] К (14S,16S,32S,33S,2R,4S,10E,12E,14R)-86-хлор-14-гидрокси-85,14-диметокси-33,2,7,10-тетраметил-12,6-диоксо-7-аза-1(6,4)-оксазинана-3(2,3)-оксиранa-8(1,3)-бензолaциклотетрадекафан-10,12-диен-4-ил-N-(4-((2-аминоэтил)дисульфанил)-4-метилпентаноил)-N-метил-L-аланинату (555 мг, 0,57 ммоль), растворенному в безводном DMSO (7 мл), добавляли 2,5-диоксопирролидин-1-ил-3-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)пропаноат (171 мг, 0,63 ммоль) и DIEA (249 мл, 1,43 ммоль) при комнатной температуре. Реакционную смесь перемешивали при комнатной температуре в течение 15 мин и нейтрализовали с применением TFA. Смесь охлаждали до 0°C с помощью бани со льдом, с последующим добавлением CH3CN (2 мл) и H2O (7 мл), а затем очищали посредством ISCO с обращенной фазой, при этом элюировали с помощью 10-70% ацетонитрила в H2O, содержащей 0,05% TFA. Фракции, содержащие требуемый продукт, лиофилизировали с получением требуемого продукта (430 мг, выход 66%). 1H ЯМР (400 МГц, CDCl3) δ ppm 0,81 (s, 3 H) 1,23 (s, 3 H) 1,24 (s, 3 H) 1,25 (s, 1 H) 1,28 (d, J=5,0 Гц, 3 H) 1,31 (d, J=5,0 Гц, 3 H) 1,43-1,49 (m, 1 H) 1,61 (d, J=15,0 Гц, 1 H) 1,64 (s, 3 H) 1,81-1,87 (m, 1 H) 1,94-2,01 (m, 1 H) 2,19 (dd, J=5,0 и 15,0 Гц, 1 H) 2,30-2,36 (m, 1 H) 2,54 (t, J=5,0 Гц, 2 H) 2,61 (dd, J=10,0 и 15,0 Гц, 1 H) 2,70 (t, J=5,0 Гц, 2 H) 2,88 (s, 3 H) 3,00 (d, J=10,0 Гц, 1 H) 3,13 (d, J=10,0 Гц, 1 H) 3,21 (s, 3 H) 3,55 (s, 3 H) 3,45 (q, J=5,0 Гц, 2 H) 3,49 (d, J=5,0 Гц, 1 H) 3,62 (d, J=10,0 Гц, 1 H) 3,83 (t, J=5,0 Гц, 1 H) 3,98 (s, 3 H) 4,32 (m, 1 H) 4,80 (dd, J=5,0 и 10,0 Гц, 1 H) 5,28 (d, J=5,0 Гц, 1 H) 5,66 (dd, J=10,0 и 15,0 Гц, 1 H) ) 6,22 (bs, 1 H) 6,42 (dd, J=10,0 и 15,0 Гц, 1 H) 6,50 (s, 1 H) 6,63 (s, 1 H) 6,66 (d, J=10,0 Гц, 1 H) 6,70 (s, 2H) 6,83(s, 1H); MS масса/заряд 988,3 (M+H-H2O), Время удерживания 1,145 минуты.
[00422] Пример 4B. Получение конъюгата антитела и лекарственного средства 121G12.DAPA.sSPDB.DM4
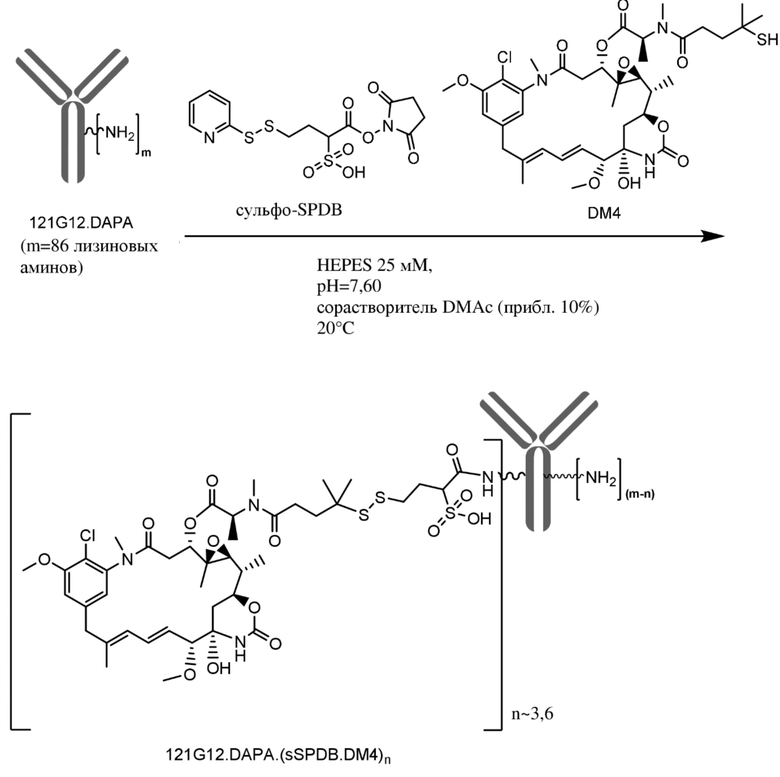
[00423] К перемешиваемому раствору 25 мМ буфера HEPES, pH 7,6 (3 мл; стерильный) и диметилацетамида (DMAc; 0,12 мл) при 22°C добавляли раствор (1,695 мл) 121G12.DAPA; MW ~145546 г/моль; 62 мг (0,426 мкмоль)) в буфере на основе фосфата калия (10 мМ, pH6; стерильный). Добавляли майтанзиноид DM4 (2,42 мг (3,101 мкмоль), растворенный в 0,242 мл DMAc. Добавляли линкер сульфо-SPDB (0,970 мг (2,386 мкмоль, скорректирован для анализа), растворенный в 0,970 мл DMAc. Спустя 18 ч реакционную смесь анализировали на завершенность реакции с помощью SEC-UV и HPLC.
[00424] Реакционную смесь очищали от низкомолекулярных побочных продуктов и проводили замену буфера путем фильтрации через мембранные ячейки Amicon; отсекаемая молекулярная масса 30 кДа, с применением 10 мМ буфера PBS, pH 7,4 (стерильный) для промывки. Полученные после применения Amicon ультраконцентраты объединяли и разбавляли до 10 мг/мл (УФ) с получением 2,9 мл раствора конъюгата антитела и лекарственного средства, 121G12.DAPA.sSPDB-DM4, в 10 мМ буфере PBS, pH 7,4 (извлечение белка 49%).
[00425] С помощью SEC-UV определили, что соотношение лекарственного средства и антитела составляет n= 3,6, а мономерная чистота составляет 98,7%. Уровень эндотоксинов составлял 0,14 ЕЭ/мг (тест BET Endosafe).
[00426] Пример 4C. Получение конъюгата антитела и лекарственного средства 684E12.SMCC.DM1
(n=3,6)
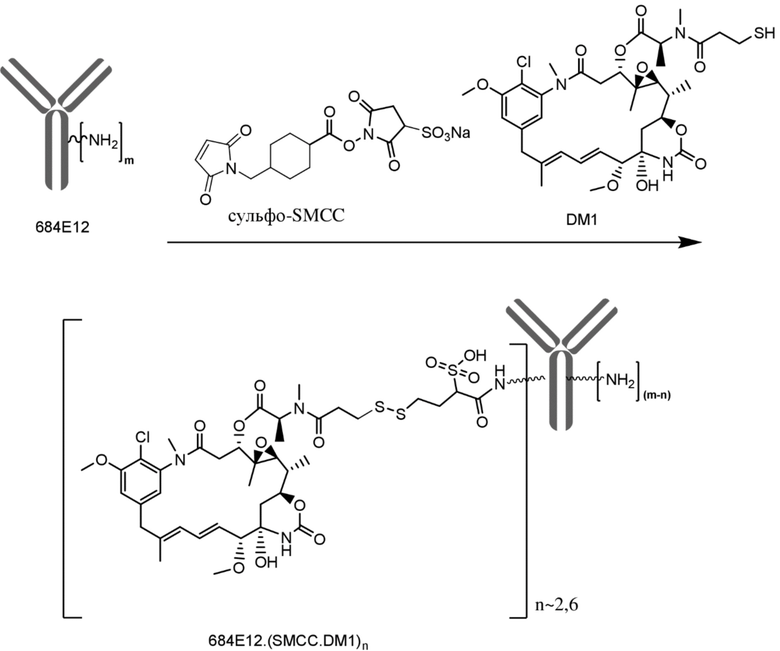
[00427] К раствору антитела (исходное 684E12) (7,1 мг/мл, 3,4 мл, приблизительно 47 мкМ, PBS, pH 7,4) добавляли 100 мкл 2 мМ DM1 (0,17 мг) в DMA и 50 мкл 4 мМ сульфо-SMCC (0,15 мг) в DMA и смесь инкубировали и аккуратно перемешивали при 4°C в течение ночи. После инкубации реакционную смесь очищали путем обессоливания на обессоливающей колонке HiPrep 26/10 (GE Healthcare) с применением PBS, pH 7,4, в качестве подвижного буфера, и стерилизовали фильтрацией. Очищенный конъюгат анализировали с помощью MALDI-MS, и, согласно оценкам, DAR составило 2,6. Аналитическая SEC показала, что в образце присутствует 3,7% агрегация (или 96,3% мономеров), а тестирование LAL (PTS, Charles River Laboratories) определило, что показатель эндотоксинов составляет 0,36 ЕЭ/мг.
[00428] Пример 4D. Получение конъюгата антитела и лекарственного средства 506E15.AURIX1
Исходным материалом служило антитело 506E15.CysMab (Fc WT) при 18,8 мг/мл (для 10 мг/мл раствора IgG коэффициент экстинкции при OD280 составляет 13,7) в 1x забуференном фосфатом солевом растворе (1x PBS). 1,76 мл антитела абсорбировали на 3 мл смолы RMP с белком A (GE Healthcare 1-223BPO/I) и к полученной взвеси добавляли 240 мкл 0,5 M цистеина (Sigma G121-03), составленного в 0,5 M фосфате, pH 8, в который был добавлен NaOH (Alfa Aesar A16037) при соотношении 13,6 г/л. Взвесь изредка перемешивали путем вращения сосуда при комнатной температуре в течение 30 минут, затем промывали посредством вакуум-фильтрации через фильтрующее устройство с размером пор 0,2 мкм, помещаемое на горлышко бутылки, с помощью 50 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления. Промытую смолу ресуспендировали в 2 мл 1x PBS (50% взвесь) и добавляли 60 мкл 100 мкМ CuCl2 (Aldrich 751944) (суммарно 100 нМ Cu2+) для инициирования повторного окисления. Спустя 420 минут добавляли дополнительные 90 мкл 100 мкМ CuCl2 для дополнительного ускорения повторного окисления. Повторное окисление антитела тестировали путем изъятия 30-мкл аликвоты взвеси, добавления 1 мкл 20 мМ исходного раствора эталонного малеимида (пример 3, страница 110 в WO2015/095301), который, как известно, сдвигает пик антитела при RPLC, смешивания в течение 1 минуты, центрифугирования при 7000 × g в течение 10 секунд, удаления супернатанта, добавления 60 мкл буфера для элюирования IgG Thermo (Thermo Scientific 21009), центрифугирования при 14000 × g в течение 10 секунд, отбора образцов супернатанта и анализа продуктов с помощью RPLC следующим образом: 2-мкл образец вводили в нагретую (80°C) колонку 4,6×50 мм Agilent PLRP-S (частицы 5 мкм, размер пор 4000 Å), уравновешенную 0,1% трифторуксусной кислотой в 29,5% CH3CN/в воде (Millipore TX1280P-1, Burdick and Jackson 407-4) при расходе 1,5 мл/мин. Колонку элюировали с помощью 5-минутного градиента до 44,5% CH3CN/в воде, поддерживали данную концентрацию в течение 1,9 минуты и пики определяли при 280 нм. Оптимальное время для конъюгации определяется как время, при котором максимального значения достигает пик основного продукта, сокращены до минимума пики с более поздним элюированием, и более не возрастают пики с более ранним элюированием.
[00429] Когда анализ RPLC показывал, что повторное окисление было оптимальным (в данном случае 565 минут), добавляли 220 мкл 20 мМ исходного раствора AURIX1 в DMSO и взвесь изредка аккуратно перемешивали путем вращения сосуда при комнатной температуре в течение 70 минут. Затем взвесь промывали с помощью 20 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления.
[00430] Затем взвесь переносили в колонку с пористым стеклянным фильтром (Pierce 7375021), предварительно элюировали с помощью 0,5 объема слоя буфера для элюирования IgG Thermo (отбрасывали), затем элюировали с помощью 2 объемов слоя того же буфера. Весь элюат (6 мл) нейтрализовали с помощью 0,6 мл 0,5 M фосфата натрия, pH 8, концентрировали до 2,5 мл с применением центрифужного концентратора (Amicon UFC905024) при 3000 × g, наносили на буферообменную колонку PD-10 (GE Healthcare 17-0851-01), уравновешенную 1x PBS, при загрузке 2,5 мл концентрата и элюировании с помощью 3,5 мл 1x PBS согласно рекомендациям изготовителя. Выход составил 22 мг (66%).
[00431] Пример 4E. Получение конъюгата антитела и лекарственного средства 506E15.CysMab.DAPA.AURIX2
Исходным материалом служило антитело 506E15.CysMab.DAPA при 10 мг/мл (для 10 мг/мл раствора IgG коэффициент экстинкции при OD280 составляет 13,7) в 1x забуференном фосфатом солевом растворе (1x PBS). 2,0 мл антитела абсорбировали на 2 мл смолы RMP с белком A (GE Healthcare 1-223BPO/I) и к полученной взвеси добавляли 160 мкл 0,5 M цистеина (Sigma G121-03), составленного в 0,5 M фосфате, pH 8, в который был добавлен NaOH (Alfa Aesar A16037) при соотношении 13,6 г/л. Взвесь изредка перемешивали путем вращения сосуда при комнатной температуре в течение 30 минут, затем промывали посредством вакуум-фильтрации через фильтрующее устройство с размером пор 0,2 мкм, помещаемое на горлышко бутылки, с помощью 50 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления. Промытую смолу ресуспендировали в 2 мл 1x PBS (50% взвесь) и добавляли 10 мкл 100 мкМ CuCl2 (Aldrich 751944) (суммарно 250 нМ Cu2+) для инициирования повторного окисления. Повторное окисление антитела тестировали путем изъятия 30-мкл аликвоты взвеси, добавления 1 мкл 20 мМ исходного раствора эталонного малеимида (пример 3, страница 110 в WO2015/095301), который, как известно, сдвигает пик антитела при RPLC, смешивания в течение 1 минуты, центрифугирования при 7000 × g в течение 10 секунд, удаления супернатанта, добавления 60 мкл буфера для элюирования IgG Thermo (Thermo Scientific 21009), центрифугирования при 14000 × g в течение 10 секунд, отбора образцов супернатанта и анализа продуктов с помощью RPLC следующим образом: 2-мкл образец вводили в нагретую (80°C) колонку 4,6×50 мм Agilent PLRP-S (частицы 5 мкм, размер пор 4000 Å), уравновешенную 0,1% трифторуксусной кислотой в 29,5% CH3CN/в воде (Millipore TX1280P-1, Burdick and Jackson 407-4) при расходе 1,5 мл/мин. Колонку элюировали с помощью 5-минутного градиента до 44,5% CH3CN/в воде, поддерживали данную концентрацию в течение 1,9 минуты и пики определяли при 280 нм. Оптимальное время для конъюгации определяется как время, при котором максимального значения достигает пик основного продукта, сокращены до минимума пики с более поздним элюированием, и более не возрастают пики с более ранним элюированием.
[00432] Когда анализ RPLC показывал, что повторное окисление было оптимальным (в данном случае 295 минут), добавляли 80 мкл 20 мМ исходного раствора AURIX2 в DMSO и взвесь изредка аккуратно перемешивали путем вращения сосуда при комнатной температуре в течение 85 минут. Затем взвесь промывали с помощью 20 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления.
[00433] Затем взвесь переносили в колонку с пористым стеклянным фильтром (Pierce 7375021), предварительно элюировали с помощью 0,5 объема слоя буфера для элюирования IgG Thermo (отбрасывали), затем элюировали с помощью 2 объемов слоя того же буфера. Весь элюат (4 мл) нейтрализовали с помощью 0,4 мл 0,5 M фосфата натрия, pH 8, наносили на 2 буферообменные колонки PD-10 (GE Healthcare 17-0851-01), уравновешенные 1x PBS, при загрузке 2,5 мл элюата и элюировании с помощью 3,5 мл 1x PBS согласно рекомендациям изготовителя. Выход составил 13,3 мг (67%).
[00434] Пример 4F. Получение конъюгата антитела и лекарственного средства 674J13.CysMab.AURIX1
[00435] Исходным материалом служило антитело 674J13.CysMab (Fc WT) при 9 мг/мл (для 10 мг/мл раствора IgG коэффициент экстинкции при OD280 составляет 13,7) в 1x забуференном фосфатом солевом растворе (1x PBS). К 57,6 мл антитела добавляли DTT до 200 мМ (Invitrogen 15508-013) и раствор инкубировали в течение 75 минут для сильного восстановления Ab. Затем восстановленные Ab наносили на буферообменные колонки PD-10 (GE Healthcare 17-0851-01), уравновешенные 1x PBS, при загрузке 2,5 мл концентрата и элюировании с помощью 3,5 мл 1x PBS согласно рекомендациям изготовителя. Элюаты из колонок PD10 объединяли, затем вновь наносили на свежие колонки PD10 для более полного удаления DTT. Обратите внимание, что в отдельных экспериментах колонки PD10 являются более эффективными в удалении DTT, чем можно было бы наблюдать при использовании только механизма эксклюзионной хроматографии, при условии, что колонки используют только один раз.
[00436] Повторное окисление антитела тестировали путем изъятия 30-мкл аликвоты взвеси, добавления 1 мкл 20 мМ исходного раствора AURIX1, который, как известно, сдвигает пик антитела при RPLC, смешивания в течение 1 минуты, центрифугирования при 7000 × g в течение 10 секунд, удаления супернатанта, добавления 60 мкл буфера для элюирования IgG Thermo (Thermo Scientific 21009), центрифугирования при 14000 × g в течение 10 секунд, отбора образцов супернатанта и анализа продуктов с помощью RPLC следующим образом: 2-мкл образец вводили в нагретую (80°C) колонку 4,6×50 мм Agilent PLRP-S (частицы 5 мкм, размер пор 4000 Å), уравновешенную 0,1% трифторуксусной кислотой в 29,5% CH3CN/в воде (Millipore TX1280P-1, Burdick and Jackson 407-4) при расходе 1,5 мл/мин. Колонку элюировали с помощью 5-минутного градиента до 44,5% CH3CN/в воде, поддерживали данную концентрацию в течение 1,9 минуты и пики определяли при 280 нм. Оптимальное время для конъюгации определяется как время, при котором максимального значения достигает пик основного продукта, сокращены до минимума пики с более поздним элюированием, и более не возрастают пики с более ранним элюированием.
[00437] Когда анализ RPLC показывал, что повторное окисление было оптимальным (в данном случае 180 минут), добавляли 290 мкл 20 мМ исходного раствора AURIX1 в DMSO вместе с 6 мл смолы RMP с белком A (GE Healthcare 1-223BPO/I). Взвесь изредка перемешивали путем вращения сосуда при комнатной температуре в течение 40 минут. Затем взвесь промывали с помощью 20 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления.
[00438] Затем взвесь переносили в колонку с пористым стеклянным фильтром (Pierce 7375021), предварительно элюировали с помощью 0,5 объема слоя буфера для элюирования IgG Thermo (отбрасывали), затем элюировали с помощью 2 объемов слоя того же буфера. Весь элюат (11,5 мл) нейтрализовали с помощью 1,2 мл 0,5 M фосфата натрия, pH 8, концентрировали до 2,5 мл с применением центрифужного концентратора (Amicon UFC905024) при 3000 × g, наносили на буферообменную колонку PD-10 (GE Healthcare 17-0851-01), уравновешенную 1x PBS, при загрузке 2,5 мл концентрата и элюировании с помощью 3,5 мл 1x PBS согласно рекомендациям изготовителя. Выход составил 26 мг (41%).
[00439] Пример 4G. Получение конъюгата антитела и лекарственного средства 674J13.CysMab.DAPA.AURIX2
[00440] Исходным материалом служило антитело 674J13.CysMab.DAR4.DAPA при 31,7 мг/мл (для 10 мг/мл раствора IgG коэффициент экстинкции при OD280 составляет 13,7) в 1x забуференном фосфатом солевом растворе (1x PBS). 9,5 мл антитела абсорбировали на 30,1 мл смолы RMP с белком A (GE Healthcare 1-223BPO/I) и к полученной взвеси добавляли 1800 мг DTT (Invitrogen 15508-013) для сильного восстановления Ab (суммарно 200 мМ DTT). Взвесь перемешивали путем вращения сосуда при комнатной температуре в течение 20 минут, затем промывали посредством вакуум-фильтрации через фильтрующее устройство с размером пор 0,2 мкм, помещаемое на горлышко бутылки, с помощью 50 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления. Промытую смолу ресуспендировали в 2 мл 1x PBS и перемешивали путем вращения сосуда при комнатной температуре, при этом не добавляли медь (она ускоряла повторное окисление слишком сильно). Повторное окисление антитела тестировали путем изъятия 30-мкл аликвоты взвеси, добавления 1 мкл 20 мМ исходного раствора эталонного малеимида (пример 3, страница 110 в WO2015/095301), который, как известно, сдвигает пик антитела при RPLC, смешивания в течение 1 минуты, центрифугирования при 7000 × g в течение 10 секунд, удаления супернатанта, добавления 60 мкл буфера для элюирования IgG Thermo (Thermo Scientific 21009), центрифугирования при 14000 × g в течение 10 секунд, отбора образцов супернатанта и анализа продуктов с помощью RPLC следующим образом: 2-мкл образец вводили в нагретую (80°C) колонку 4,6×50 мм Agilent PLRP-S (частицы 5 мкм, размер пор 4000 Å), уравновешенную 0,1% трифторуксусной кислотой в 29,5% CH3CN/в воде (Millipore TX1280P-1, Burdick and Jackson 407-4) при расходе 1,5 мл/мин. Колонку элюировали с помощью 5-минутного градиента до 44,5% CH3CN/в воде, поддерживали данную концентрацию в течение 1,9 минуты и пики определяли при 280 нм. Оптимальное время для конъюгации определяется как время, при котором максимального значения достигает пик основного продукта, сокращены до минимума пики с более поздним элюированием, и более не возрастают пики с более ранним элюированием.
[00441] Когда анализ RPLC показывал, что повторное окисление было оптимальным (в данном случае 80 минут), добавляли 903 мкл 20 мМ исходного раствора AURIX2 в DMSO и взвесь изредка аккуратно перемешивали путем вращения сосуда при комнатной температуре в течение 50 минут. Затем взвесь промывали с помощью 20 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления.
[00442] Затем взвесь переносили в колонку с пористым стеклянным фильтром (Pierce 7375021), предварительно элюировали с помощью 0,5 объема слоя буфера для элюирования IgG Thermo (отбрасывали), затем элюировали с помощью 2 объемов слоя того же буфера. Весь элюат (60,2 мл) нейтрализовали с помощью 6,0 мл 0,5 M фосфата натрия, pH 8, концентрировали до 17,5 мл с применением центрифужного концентратора (Amicon UFC905024) при 3000 × g, наносили на 7 буферообменных колонок PD-10 (GE Healthcare 17-0851-01), уравновешенных 1x PBS, при загрузке 2,5 мл концентрата и элюировании с помощью 3,5 мл 1x PBS согласно рекомендациям изготовителя. Выход составил 243 мг (81%)
[00443] Пример 4H. Получение конъюгата антитела и лекарственного средства 121G12.CysMab.DAPA.AURIX1
[00444] Исходным материалом служило антитело 121G12.CysMab.DAPA при 16,7 мг/мл (для 10 мг/мл раствора IgG коэффициент экстинкции при OD280 составляет 13,7) в 1x забуференном фосфатом солевом растворе (1x PBS). 3,6 мл антитела абсорбировали на 6 мл смолы RMP с белком A (GE Healthcare 1-223BPO/I) и полученную взвесь перемешивали путем вращения сосуда при комнатной температуре в течение 125 минут, затем добавляли 544 мкл 0,5 M цистеина (Sigma G121-03), составленного в 0,5 M фосфате, pH 8, в который был добавлен NaOH (Alfa Aesar A16037) при соотношении 13,6 г/л. Взвесь изредка перемешивали путем вращения сосуда при комнатной температуре в течение 60 минут, затем промывали посредством вакуум-фильтрации через фильтрующее устройство с размером пор 0,2 мкм, помещаемое на горлышко бутылки, с помощью 50 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления. Промытую смолу ресуспендировали в 2 мл 1x PBS (50% взвесь) и добавляли 16 мкл 100 мкМ CuCl2 (Aldrich 751944) (суммарно 250 нМ Cu2+) для инициирования повторного окисления. Повторное окисление антитела тестировали путем изъятия 30-мкл аликвоты взвеси, добавления 1 мкл 20 мМ исходного раствора AURIX1, который, как известно, сдвигает пик антитела при RPLC, смешивания в течение 1 минуты, центрифугирования при 7000 × g в течение 10 секунд, удаления супернатанта, добавления 60 мкл буфера для элюирования IgG Thermo (Thermo Scientific 21009), центрифугирования при 14000 × g в течение 10 секунд, отбора образцов супернатанта и анализа продуктов с помощью RPLC следующим образом: 2-мкл образец вводили в нагретую (80°C) колонку 4,6×50 мм Agilent PLRP-S (частицы 5 мкм, размер пор 4000 Å), уравновешенную 0,1% трифторуксусной кислотой в 29,5% CH3CN/в воде (Millipore TX1280P-1, Burdick and Jackson 407-4) при расходе 1,5 мл/мин. Колонку элюировали с помощью 5-минутного градиента до 44,5% CH3CN/в воде, поддерживали данную концентрацию в течение 1,9 минуты и пики определяли при 280 нм. Оптимальное время для конъюгации определяется как время, при котором максимального значения достигает пик основного продукта, сокращены до минимума пики с более поздним элюированием, и более не возрастают пики с более ранним элюированием.
[00445] Когда анализ RPLC показывал, что повторное окисление было оптимальным (в данном случае 170 минут), добавляли 67 мкл 20 мМ исходного раствора AURIX1 в DMSO и взвесь изредка аккуратно перемешивали путем вращения сосуда при комнатной температуре в течение 120 минут. Затем взвесь промывали с помощью 20 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления.
[00446] Затем взвесь переносили в колонку с пористым стеклянным фильтром (Pierce 7375021), предварительно элюировали с помощью 0,5 объема слоя буфера для элюирования IgG Thermo (отбрасывали), затем элюировали с помощью 2 объемов слоя того же буфера. Весь элюат (15 мл) нейтрализовали с помощью 1,5 мл 0,5 M фосфата натрия, pH 8, концентрировали до 5 мл с применением центрифужного концентратора (Amicon UFC905024) при 3000 x g, наносили на 2 буферообменные колонки PD-10 (GE Healthcare 17-0851-01), уравновешенные 1x PBS, при загрузке 2,5 мл элюата и элюировании с помощью 3,5 мл 1x PBS согласно рекомендациям изготовителя. Выход составил 54 мг (92%).
[00447] Пример 4I. Получение конъюгата антитела и лекарственного средства 121G12.CysMab.AURIX1
Исходным материалом служило антитело 121G12.CysMab (Fc WT) при 12,5 мг/мл (для 10 мг/мл раствора IgG коэффициент экстинкции при OD280 составляет 13,7) в 1x забуференном фосфатом солевом растворе (1xPBS). 4,8 мл антитела абсорбировали на 6 мл смолы RMP с белком A (GE Healthcare 1-223BPO/I) и полученную взвесь перемешивали путем вращения сосуда при комнатной температуре в течение 125 минут, затем добавляли 592 мкл 0,5 M цистеина (Sigma G121-03), составленного в 0,5 M фосфате, pH 8, в который был добавлен NaOH (Alfa Aesar A16037) при соотношении 13,6 г/л. Взвесь изредка перемешивали путем вращения сосуда при комнатной температуре в течение 60 минут, затем промывали посредством вакуум-фильтрации через фильтрующее устройство с размером пор 0,2 мкм, помещаемое на горлышко бутылки, с помощью 50 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления. Промытую смолу ресуспендировали в 2 мл 1x PBS (50% взвесь) и добавляли 16 мкл 100 мкМ CuCl2 (Aldrich 751944) (суммарно 250 нМ Cu2+) для инициирования повторного окисления. Повторное окисление антитела тестировали путем изъятия 30-мкл аликвоты взвеси, добавления 1 мкл 20 мМ исходного раствора AURIX1, который, как известно, сдвигает пик антитела при RPLC, смешивания в течение 1 минуты, центрифугирования при 7000 × g в течение 10 секунд, удаления супернатанта, добавления 60 мкл буфера для элюирования IgG Thermo (Thermo Scientific 21009), центрифугирования при 14000 × g в течение 10 секунд, отбора образцов супернатанта и анализа продуктов с помощью RPLC следующим образом: 2-мкл образец вводили в нагретую (80°C) колонку 4,6×50 мм Agilent PLRP-S (частицы 5 мкм, размер пор 4000 Å), уравновешенную 0,1% трифторуксусной кислотой в 29,5% CH3CN/в воде (Millipore TX1280P-1, Burdick and Jackson 407-4) при расходе 1,5 мл/мин. Колонку элюировали с помощью 5-минутного градиента до 44,5% CH3CN/в воде, поддерживали данную концентрацию в течение 1,9 минуты и пики определяли при 280 нм. Оптимальное время для конъюгации определяется как время, при котором максимального значения достигает пик основного продукта, сокращены до минимума пики с более поздним элюированием, и более не возрастают пики с более ранним элюированием.
[00448] Когда анализ RPLC показывал, что повторное окисление было оптимальным (в данном случае 160 минут), добавляли 67 мкл 20 мМ исходного раствора AURIX1 в DMSO и взвесь изредка аккуратно перемешивали путем вращения сосуда при комнатной температуре в течение 120 минут. Затем взвесь промывали с помощью 20 объемов слоя 1x PBS за по меньшей мере 10 циклов фильтрации и добавления.
[00449] Затем взвесь переносили в колонку с пористым стеклянным фильтром (Pierce 7375021), предварительно элюировали с помощью 0,5 объема слоя буфера для элюирования IgG Thermo (отбрасывали), затем элюировали с помощью 2 объемов слоя того же буфера. Весь элюат (15 мл) нейтрализовали с помощью 1,5 мл 0,5 M фосфата натрия, pH 8, концентрировали до 5 мл с применением центрифужного концентратора (Amicon UFC905024) при 3000 x g, наносили на 2 буферообменные колонки PD-10 (GE Healthcare 17-0851-01), уравновешенные 1x PBS, при загрузке 2,5 мл элюата и элюировании с помощью 3,5 мл 1x PBS согласно рекомендациям изготовителя. Выход составил 52 мг (89%).
Методы анализа. Концентрацию определяли на основании того, что для 10 мг/мл раствора IgG коэффициент экстинкции при OD280 составляет 13,7 Пироген определяли с применением анализа Kinetic QCL (Lonza Walkersville 50-650H) со считыванием на планшет-ридере TECAN Safire. Процент агрегатов определяли с помощью аналитической эксклюзионной хроматографии на предколонке Shodex KW-G (Thomson Instrument Company, № по кат. 6960955) и колонке KW-803 (TIC, № по кат. 6960940), уравновешенных подвижной фазой [20 мМ Tris, ~pH 7,65 (приготовленный с помощью 10 мМ Tris, pH 7,4, 10 мМ Tris, pH 8), 200 мМ NaCl, 0,02% азида натрия], с получением данных при 280 нм. Для определения DAR получали аликвоту образца посредством разбавления образца до 2 мг/мл в 1x PBS, дегликозилирования образца с помощью PNGазы F (получена в лаборатории) в течение 10 минут при 50°C, удаления PNGазы F за счет связывание с белком A, промывки с помощью 1x PBS и элюирования с помощью 1% муравьиной кислоты. Образец восстанавливали добавлением 1/4 объема 5 M ацетата аммония, pH 5,0, содержащего 0,5 M TCEP, и инкубирования при комнатной температуре в течение 30 минут. Затем образец вводили в колонку 2,1×50 мм PLRP-S (частицы 8 мкм, размер пор 1000 Å), уравновешенную 0,1% муравьиной кислотой в 20% CH3CN/в воде (Invitrogen) при расходе 0,5 мл/мин. Колонку промывали с помощью 20% CH3CN/в воде в течение 3 минут, затем элюировали с помощью 0,1-минутного градиента до 0,1% муравьиной кислоты в 90% CH3CN/в воде, при этом данную концентрацию поддерживали в течение 1,9 минуты. Данные масс-спектров получали на устройстве Agilent 1260, и деконволюцию проводили с помощью программного обеспечения для качественного анализа MassHunter версии B.05.00 в диапазоне 15-60 кДа. Площади пиков, соответствующих различным рассчитанным состояниям DAR, взвешивали в соответствии с DAR каждого пика, затем суммировали и взвешенную площадь пика DAR4 делили на сумму всех взвешенных пиков с получением значения DAR.
Анализ полученного образца выглядит следующим образом.
AURIX2
AURIX1
AURIX2
AURIX1
AURIX1
AURIX1
LC 0,03
[00450] Пример 4J. Получение дополнительных конъюгатов с применением других антител CysMab
[00451] Способы, описанные в примере 4A, также применяли для получения конъюгатов MPET.DM4 с другими антителами с цистеинами, введенными путем конструирования.
[00452] Способы применяют для получения конъюгатов Ab.CysMab.MPET.DM4, связывающих P-кадгерин,с применением антител NOV169N31Q(E152C-S375C), NEG0012(E152C-S375C), NEG0013(E152C-S375C), NEG0016(E152C-S375C), NEG0064(E152C-S375C), NEG0067(E152C-S375C), NOV169N31Q(K360C(HC)-K107C(LC)), NEG0012(K360C(HC)-K107C(LC)), NEG0013(K360C(HC)-K107C(LC)), NEG0016(K360C(HC)-K107C(LC)), NEG0064(K360C(HC)-K107C(LC)) и NEG0067(K360C(HC)-K107C(LC)), раскрытых в PCT-публикации № WO2016/203432.
Пример 5. In vitro определение характеристик ADC
[00453] Характеристики конъюгатов антитела и лекарственного средства (ADC) определяли с помощью различных функциональных и аналитических методов. По результатам оценки с помощью FACS ADC сохраняли связывание с белком-мишенью CCR7 на клетках. В случае всех ADC геометрическое среднее интенсивности флуоресценции в анализе связывания FACS находилось в пределах 20% от значения для неконъюгированного антитела. По результатам аналитической SEC было показано, что >95% материала ADC характеризовались требуемой молекулярной массой; в случаях, где это не наблюдалось для исходных продуктов реакции, применение препаративной SEC позволяло достичь необходимой спецификации. Соотношение лекарственного средства и антитела (DAR) оценивали с помощью LCMS образца дегликозилированного восстановленного антитела, при суммировании встречаемости различных разновидностей DAR и уравновешивания числа молекул лекарственного средства на каждой разновидности DAR (например, один ион с DAR2 считался как 2, один ион с DAR1 считался как 1). Конъюгаты, содержащие константные области, содержащие две цистеиновые мутации, характеризовались по меньшей мере DAR 3,4 или выше и наиболее обычно DAR 3,8 или выше. Для всех зарегистрированных конъюгатов данные были согласованы.
Пример 6. Ингибирование пролиферации клеток/анализ жизнеспособности клеток
[00454] Выше авторы настоящего изобретения показали, что конъюгированные с флуорофором варианты всех трех антител к CCR7 могли приводить к интернализации CCR7 и эффективному накоплению конъюгированного флуорофора в компартментах клеток с низким pH для целого набора линий клеток. В данном примере авторы настоящего изобретения показывают способность антитела к интернализации и накоплению конъюгированного вещества внутри клетки в режиме, при котором конъюгированное вещество представляет собой токсичную полезную нагрузку.
[00455] В режиме "piggyback" ADC (pgADC) цитотоксические эффекты антител к CCR7 в комплексе с фрагментом вторичного антитела, конъюгированного с полезной нагрузкой, изучали путем оценки жизнеспособности клеток после четырех дней обработки. Получали трехкратные разбавления CCR7-специфических IgG и смешивали их с постоянным количеством Fab-фрагмента, сопряженного с полезной нагрузкой. Конечная концентрация Fab-фрагмента, сопряженного с полезной нагрузкой, составляла 0,5 мкг/мл. Реагент Fab представляет собой направленный на Fc мыши Fab, конъюгированный или с MMAF, или с сапорином (Advanced Targeting Systems, Fab-Zap). После предварительной инкубации в течение 30 мин при комнатной температуре по 10 мкл/лунка комплекса антитело-полезная нагрузка добавляли в 384-луночные планшеты с белым дном в трех повторностях. Соответствующие CCR7(+) клетки высевали таким образом, чтобы их плотность составляла менее 1×106/мл в случае клеток из суспензионных культур и достигалась 80% конфлюэнтность в случае адгезивных клеток. Клетки собирали (адгезивные клетки отделяли с помощью аккутазы) и ресуспендировали до примерно 2×104 клеток/мл. Клетки добавляли в 384-луночные планшеты поверх комплекса антитело-полезная нагрузка (20 мкл/лунка). Планшет инкубировали в течение четырех дней при 37°C и 5% CO2. Затем готовили раствор CellTiter-Glo (CellTiter-Glo® люминесцентный анализ жизнеспособности клеток; Promega, № G7571) и добавляли к клеткам из расчета 20 мкл/лунка. Только жизнеспособные клетки продуцируют АТФ, необходимый для люциферазной реакции (предоставленной CellTiter-Glo), которая приводит к люминесценции. В соответствии с этим жизнеспособность клеток определяли по сигналу люминесценции, который измеряли после 10 мин инкубации при 22°C и 400 об/мин с помощью ридера для нескольких меток Envision 2104. Значения IC50 рассчитывали с применением программного обеспечения Graphpad Prism.
[00456] На фигуре 7A и фигуре 7B показано, что все четыре антитела к CCR7 способны к зависимому от концентрации цитолизу клеток при использовании CCR7+ KE97 клеток в анализе формата "piggyback" с применением MMAF-конъюгированного реагента. В таблице ниже обобщены результаты экспериментов с применением MMAF или сапорина в качестве инструмента-полезной нагрузки при формате "piggy-back".
Таблица 16. IC50 и AMAX антител к CCR7 в цитотоксическом анализе с применением pgADC
[00457] Специфичность цитолиза под действием pgADC оценивали с применением линий клеток, отрицательный по мишени. На фигуре 8 показан пример с использованием реагента FabZap. Специфическое CCR7-зависимое повышение активности 121G12 pgADC наблюдали в случае клеток KE97 и NIH3T3.hCCR7 в отличие от CCR7-отрицательных исходных клеток NIH3T3 или контрольного антитела mIgG.
Пример 7. Воздействие перекрестно реактивных для мыши неконъюгированных или конъюгированных с AURIX1 антител к CCR7 на нормальные гемопоэтические клетки мыши in vivo
[00458] Экспрессия в нормальных тканях в пределах вида ограничена клетками гемопоэтического происхождения, включая CD4+ и CD8+ T клетки в крови и лимфоидных органах, что делает ADC, нацеливающийся на CCR7, потенциально небезопасным, особенно в формате Fc дикого типа, который мог бы приводить к ADCC и истощению лимфоидных клеток.
[00459] Чтобы определить воздействие нацеливания ADC на CCR7, находящийся на нормальных гемопоэтических клетках in vivo, на здоровых самках мышей CD-1 возраста 6-8 недель оценивали перекрестно реактивное для мыши исходное Ab 121G12 как в неконъюгированном состоянии, так и конъюгированное с AURIX1, при этом формат Fc был дикого типа или "молчащий" (DAPA). Мышей обрабатывали путем введения однократной IV инъекции, содержащей исходное 121G12.Cys-Mab.Fc дикого типа.hIgG1 (121G12.wt.Fc), исходное 121G12.Cys-Mab.DAPA.hIgG1 (121G12.DAPA.Fc), исходное 121G12.Cys-Mab.Fc дикого типа.hIgG1.AURIX1 (121G12.wt.Fc.AURIX1) или исходное 121G12.Cys-Mab.DAPA.hIgG1.AURIX1 (121G12.DAPA.Fc.AURIX1) при конечной дозе, составляющей 10 мг/кг. Все дозы корректировали в соответствии с индивидуальной массой тела мышей.
[00460] В день 25 после обработки селезенки извлекали и разделяли на суспензию из отдельных клеток с применением gentleMACS Dissociator (Miltenyi Biotec Inc, Сан-Диего, Калифорния). Затем 1 миллион клеток в каждом образце окрашивали с помощью смеси Ab, которая содержала крысиное антитело к CD8a мыши BUV737, клон 53-6.7 (1:100) (BD Biosciences, Сан-Хосе, Калифорния, № по кат. 564297), и крысиное антитело к CD4 мыши BV510, клон RM4-5 (1:200) (BD Biosciences, Сан-Хосе, Калифорния, № по кат. 563106), чтобы определить воздействие отдельных обработок на CD4+ и CD8a+ T-клетки. Образцы инкубировали при 4°C в течение 30 мин, промывали в ледяном забуференном фосфатом солевом растворе HyClone (Hyclone Laboratories, Логан, Юта) и оценивали на клеточном анализаторе BD LSRFortessaTM (BD Biosciences, Сан-Хосе, Калифорния). Подсчет общего количества спленоцитов использовали для определения истощения CD4+ или CD8a+ T-клеток. T-критерий использовали для определения значимости различий между группами.
[00461] Как показано в таблице 17 и на фигуре 9, сильное сокращение числа CD4+ (FC 0,5-0,6) и CD8a+ T-клеток (FC 0,3-0,5) в селезенке наблюдали на день 3 обработки с помощью Ab 121G12.wt.FC или 121G12.wt.Fc.AURIX1 при 10 мг/кг, это позволяет предположить, что воздействие в виде истощения T-клеток не зависело от присутствия полезной нагрузки AURIX1. От таких эффектов избавляются путем получения "молчащей" Fc за счет введения мутаций DAPA. Как 121G12.DAPA.Fc, так и 121G12.DAPA.Fc.AURIX1 не оказывали воздействия на популяции T-клеток в сравнении с группой без обработки. Эти данные указывают на то, что ADC к CCR7 могут быть небезопасными с точки зрения истощения T-клеток, что можно предотвратить путем получения "молчащей" Fc за счет мутаций DAPA.
Таблица 17. Воздействие антитела 121G12 на популяции CD4+ и CD8a+ T-клеток у мышей CD-1
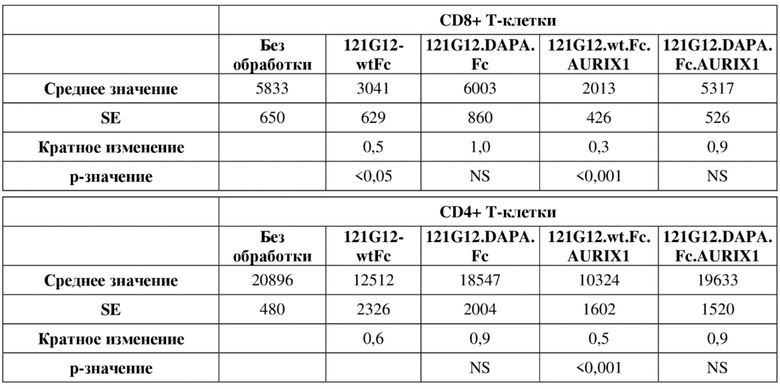
Эксперимент оценивали в день 25 после обработки. Кратное изменение (FC)=среднее количество спленоцитов в день 25 для указанной группы обработки/среднее количество спленоцитов для контрольной группы без обработки в день 3. T-критерий использовали для определения значимости в сравнении с группой без обработки (*p<0,05, ***p<0,001; NS=не значимо).
Пример 8. Активность ADC к CCR7 в формате прямых конъюгатов
[00462] Цитотоксические эффекты после связывания антител (ADC), напрямую конъюгированных с различными полезными нагрузками, и их интернализацию в CCR7(+) клетки исследовали путем оценки жизнеспособности клеток после четырех дней обработки. Трехкратные разбавления ADC вносили в 384-луночные планшеты с белым дном в трех повторностях (10 мкл/мл). Соответствующие CCR7(+) клетки высевали таким образом, чтобы их плотность составляла менее 1×106/мл в случае клеток из суспензионных культур и достигалась 80% конфлюэнтность в случае адгезивных клеток. Клетки собирали (адгезивные клетки отделяли с помощью аккутазы) и ресуспендировали до примерно 2×104 клеток/мл. Клетки добавляли в 384-луночные планшеты поверх антитела (20 мкл/лунка). Планшеты инкубировали в течение четырех дней при 37°C и 5% CO2. Затем готовили раствор CellTiter-Glo (CellTiter-Glo® люминесцентный анализ жизнеспособности клеток; Promega, № G7571) и добавляли к клеткам из расчета 20 мкл/лунка. Жизнеспособность клеток определяли по сигналу люминесценции, который измеряли после 10 мин инкубации при 22°C и 400 об/мин, с помощью ридера Envision. Значения IC50 рассчитывали с применением программного обеспечения Graphpad Prism.
[00463] В таблице ниже показаны примеры эффектов в отношении жизнеспособности клеток, измеренных для антител CysMab к CCR7 в формате Fc дикого типа или "молчащей" (DAPA) Fc, конъюгированных с AURIX1 или AURIX2.
Таблица 18. IC50 для ADC к CCR7 в цитотоксическом анализе
CysMab. AURIX1
CysMab.DAPA.
AURIX2
CysMab.DAPA.AURIX2
[00464] Чтобы оценить ADC на специфичность в отношении мишени и зависимость от уровня рецепторов, активность ADC тестировали на линиях клеток с различными уровнями CCR7-рецепторов. В таблице ниже показан пример гуманизированного антитела 674J13 в формате CysMab.DAPA и конъюгированного с AURIX2. Линии клеток отбирали на основании сходной чувствительности к полезной нагрузке.
Таблица 19. IC50 гуманизированного антитела 674J13 в формате DAPA и конъюгированного с AURIX2
[00465] Как видно из эксперимента pHrodo, для активности ADC требуется более высокая степень числа рецепторов, чем для механизма ADCC. Точное граничное значение числа рецепторов зависит от различных параметров (например, авидности антитела, активности полезной нагрузки), а данные, показанные в данном примере, описывают общие принципы. Применение зависимого от авидности антитела к CCR7 в формате DAPA сдвигает активность ADC в направлении раковых клеток, а не нормальных CCR7+ PBMC, которые в данном примере представлены линиями раковых клеток с менее 2000 CCR7-рецепторов.
Пример 9. Сайт-специфическое введение MPET.DM4 в ADC
[00466] ADC, конъюгированные с DM4, являются общепринятыми в области техники ADC. В данном примере авторы настоящего изобретения описывают создание и применение сайт-специфически конъюгированного MPET.DM4 с использованием варианта CysMab антител, что обеспечивает преимущество получения воспроизводимо однородной партии ADC с контролируемым DAR, в которой DAR (отношение лекарственного средства к антителу) составляет приблизительно 4. Было описано, что конъюгаты, полученные без использования сайт-специфической конъюгации, зачастую содержат значительные популяции с высоким DAR, которые связывали с нежелательными биофизическими особенностями, в том числе повышенной гидрофобностью и, следовательно, более быстрым клиренсом, неудовлетворительным PK-профилем и повышенной токсичностью. Ниже авторы настоящего изобретения показывают различные in vitro и in vivo оценки ADC к CCR7 на основе MPET.DM4 в сравнении с его аналогом на основе sSPDB.DM4.
Пример 10. Аффинность связывания ADC с MPET.DM4 в сравнении с ADC с sSPDB.DM4 в анализе FACS
[00467] Другим потенциальным благоприятным эффектом сайт-специфической конъюгации в сравнении с отсутствием сайт-специфической конъюгации могла быть потенциальная интерференция сайтов конъюгации с лизином, которые структурно расположены вблизи важных CSD-остатков. Можно ожидать, что конъюгация полезной нагрузки с такими лизиновыми сайтами будет воздействовать на аффинность связывания ADC. Чтобы протестировать аффинности связывания ADC с применением описанных в данном примере CysMab, конъюгированных с MPET.DM4, в сравнении с антителами, конъюгированными с sSPDB.DM4 через эндогенный лизин, аффинность связывания определяли с помощью FACS, как описано выше. В таблице ниже обобщены некоторые типичные данные аффинности, которые показывают умеренное снижение аффинности связывания sSPDB.DM4 ADC в сравнении с MPET.DM4 ADC.
Таблица 20. Аффинность связывания ADC к CCR7
CysMab.DAR4, неконъюгированное
CysMab.DAPA.
MPET.DM4
(DAR 3,8)
DAPA.sSPDB.DM4 (DAR3,9)
Пример 11. In vitro активность ADC с MPET.DM4 в сравнении с ADC с sSPDB.DM4
[00468] Цитотоксические эффекты ADC 121G12.CysMab.DAPA.MPET.DM4 и 121G12.DAPA.sSPDB.DM4 (DAR3,9) оценивали в in vitro анализе жизнеспособности, как описано выше. В таблице ниже показана аналогичная, слегка улучшенная активность конъюгата с MPET.DM4 по сравнению с конъюгатом с sSPDB.DM4. Это может быть следствием лучше сохраненной аффинности или других факторов.
Таблица 21. In vitro цитотоксическая активность ADC к CCR7
[00469] ADC 121G12.CysMab.DAPA.MPET.DM4 дополнительно тестировали на ряде линий раковых клеток, охватывающих различные показания, где он достигал значительного цитолиза клеток, который коррелировал с плотностью рецепторов.
Таблица 22. In vitro цитотоксическая активность ADC к CCR7 у линий клеток
Пример 12. Дозозависимая in vivo эффективность 121G12.CysMab.DAPA.MPET.DM4 и 121G12.DAPA.sSPDB.DM4 в отношении ксенотрансплантатной модели множественной миеломы KE97 у мышей SCID-Beige
[00470] Чтобы продемонстрировать нацеленную противоопухолевую активность 121G12.CysMab.DAPA.MPET.DM4 и 121G12.DAPA.sSPDB.DM4 in vivo, ксенотрансплантатную модель KE97 приживляли самкам мышей SCID-beige посредством подкожной инъекции 3×106 клеток в правый бок каждой мыши. После того как опухоли достигали примерно 135 мм3, мышей рандомизировали в соответствии с объемом опухоли в группы обработки (n=8 на группу). Мышей обрабатывали путем IV введения одного из 121G12.CysMab.DAPA.MPET.DM4 (DAR4) при конечной дозе, составляющей 0,5, 2 или 5 мг/кг, 121G12.DAPA.sSPDB.DM4 (DAR 3,9) из расчета 0,5, 2 или 5 мг/кг или неспецифического изотипического контрольного IgG1.CysMab.DAPA.MPET.DM4 из расчета 5 мг/кг. Все дозы корректировали в соответствии с индивидуальной массой тела мышей.
[00471] Все тестируемые средства хорошо переносились на протяжении исследования, и выраженных клинических симптомов токсичности или потери массы тела не наблюдали ни в одной из групп обработки (таблица 23).
Таблица 23. Дозозависимая эффективность ADC к CCR7 в отношении ксенотрансплантатной модели KE97
чество)
Однократная доза
Однократная доза
Однократная доза
Однократная доза
Однократная доза
Однократная доза
Однократная доза
Эксперимент оценивали в день 9 после обработки (день 23 после имплантации), * p<0,001 в сравнении с контрольной группой без обработки (однофакторный ANOVA/критерий Тьюки для множественных сравнений). % ΔT/ΔC=100 ΔT/ΔC, где: ΔT=средний объем опухоли в обработанной лекарственным средством группе в D23 исследования - средний объем опухоли в обработанной лекарственным средством группе в начальный день при введении дозы; ΔC=средний объем опухоли в контрольной группе в D23 исследования - средний объем опухоли в контрольной группе в начальный день введения дозы D14. % регрессии=(1-T конечный/T начальный)×100, где T конечный представляет собой средний объем опухоли в D23, а T начальный определяется как объем опухоль в D14 после имплантации. Δ массы тела (%)=(средняя масса тела в D23-средняя масса тела в D14) *100/средняя масса тела в D14 обработки.
[00472] Незначительную противоопухолевую эффективность наблюдали после обработки неспецифическим изотипическим контрольным IgG1.CysMab.DAPA.MPET.DM4 из расчета 5 мг/кг. Обработка 121G12.CysMab.DAPA.MPET.DM4 приводила к дозозависимой противоопухолевой эффективности со значением ΔT/ΔC, составляющим 78,62% (0,5 мг/кг), в тоже время дозы 2 и 5 мг/кг приводили к средней регрессии на 33% и 54%, соответственно, ко D9 после введения первой дозы (D23 после имплантации). Обработка 121G12.DAPA.sSPDB.DM4 также продемонстрировала дозозависимую противоопухолевую эффективность со значениями ΔT/ΔC, составляющими 85,38% (0,5 мг/кг) и 13,77% (2 мг/кг), в то же время доза 5 мг/кг приводила к средней регрессии, составляющей 33%, ко D9 после введения первой дозы (D23 после имплантации). Ко D23-D25 после имплантации мышей контрольных групп и групп обработки дозой 0,5 мг/кг умерщвляли, а оставшиеся группы обрабатывали второй дозой в D28 после имплантации одного из 121G12.CysMab.DAPA.MPET.DM4 из расчета 2 или 5 мг/кг или 121G12.DAPA.sSPDB.DM4 из расчета 2 или 5 мг/кг. Устойчивую регрессию опухоли наблюдали в случае введения 121G12.CysMab.DAPA.MPET.DM4 из расчета 2 и 5 мг/кг, а также в случае 121G12.DAPA.sSPDB.DM4 из расчета 5 мг/кг вплоть до конца исследования в D42 после имплантации. В случае 121G12.DAPA.sSPDB.DM4 из расчета 2 мг/кг реакция была более гетерогенной, причем у примерно 25% мышей проявлялась устойчивая регрессия опухоли, в то же время у остальной группы показаны или стабилизация заболевания, или прогрессирование опухоли (фигура 10, таблица 23).
Пример 13. Оценка эффективности 121G12.CysMab.DAPA.MPET.DM4 в сравнении с 121G12.DAPA.sSPDB.DM4 в отношении модели множественной миеломы KE97 при больших начальных объемах опухоли
[00473] In vivo модель c более высокими требованиями обеспечивали с применением линии клеток множественной миеломы KE97, чтобы провести дополнительное различие в противоопухолевой эффективности, обусловленной разными расщепляемыми линкерами, путем сравнения эффективности 121G12.CysMab.DAPA.MPET.DM4 121G12.DAPA.sSPDB.DM4 (DAR3,9) в отношении опухолей с большими начальными объемами опухоли при введении первой дозы. Ксенотрансплантатную модель KE97 приживляли самкам мышей SCID-beige посредством подкожной инъекции 3×106 клеток в правый бок каждой мыши. После того как опухоли достигали примерно 450 мм3, мышей рандомизировали в соответствии с объемом опухоли в две группы обработки (n=8). Мышей обрабатывали путем IV введения 2 мг/кг одного из 121G12.CysMab.DAPA.MPET.DM4 или 121G12.DAPA.sSPDB.DM4.
[00474] Незначительную противоопухолевую эффективность наблюдали после обработки 121G12.DAPA.sSPDB.DM4 из расчета 2 мг/кг. Обработка 121G12.CysMab.DAPA.MPET.DM4 приводила к частичной регрессии/продолжительной остановке роста у 75% (6 из 8) мышей с обработкой однократной дозой (фигура 11). 121G12.CysMab.DAPA.MPET.DM4 сильно превосходило 121G12.DAPA.sSPDB.DM4 в данной модели (в D25 средний объем опухоли составлял 524,83 ± 143,20 в сравнении с 1337,13 ± 35,13, соответственно, p<0,001; непарный T-критерий).
Пример 14. In vivo эффективность 121G12.CysMab.DAPA.MPET.DM4 в отношении модели опухоли на основе полученного от пациента первичного немелкоклеточного рака легкого HLUX1934
[00475] Противоопухолевую активность 121G12.CysMab.DAPA.MPET.DM4 оценивали на ксенотрансплантатной модели первичного немелкоклеточного рака легкого HLUX1934, экспрессирующего CCR7. Самкам бестимусных "голых" мышей имплантировали подкожно фрагменты опухоли в правый бок каждой мыши. После того как опухоли достигали примерно 100 мм3, мышей рандомизировали в соответствии с объемом опухоли в группы обработки (n=8). Мышей обрабатывали путем IV введения одного из 121G12.CysMab.DAPA.MPET.DM4 (DAR4) из расчета 10 мг/кг или неспецифического изотипического контрольного IgG1.CysMab.DAPA.MPET.DM4 из расчета 10 мг/кг. Вторую дозу каждого антитела вводили спустя 2 недели. Все дозы корректировали в соответствии с индивидуальной массой тела мышей.
[00476] Неспецифическое изотипическое контрольное IgG1.CysMab.DAPA.MPET.DM4 из расчета 10 мг/кг, по-видимому, приводило к небольшой задержке роста опухоли по сравнению с группой без обработки (значение ΔT/ΔC составляло 53,07%), потенциально вследствие неспецифического связывания антитела с нецелевой мишенью в модели HLUX1934. Обработка 121G12.CysMab.DAPA.MPET.DM4 приводила к более выраженной эффективности, которая была устойчивой при введении второй дозы. Обработка 121G12.CysMab.DAPA.MPET.DM4 при дозе 10 мг/кг хорошо переносилась, при этом отсутствовала явная потеря массы тела, а значение ΔT/ΔC составляло 18,83% в D34 после имплантации (фигура 12, таблица 24).
Таблица 24. Эффективность 121G12.CysMab.DAPA.MPET.DM4 в отношении модели полученного от пациента NSCLC HLUX1934. Эксперимент оценивали в день 34 после имплантации (D23 после обработки)
* p<0,001 в сравнении с контрольной группой без обработки (однофакторный ANOVA/критерий Тьюки для множественных сравнений). % ΔT/ΔC=100 ΔT/ΔC, где: ΔT=средний объем опухоли в обработанной лекарственным средством группе в D34 исследования - средний объем опухоли в обработанной лекарственным средством группе в начальный день при введении дозы; ΔC=средний объем опухоли в контрольной группе в D34 исследования - средний объем опухоли в контрольной группе в начальный день введения дозы. Δ массы тела (%)=(средняя масса тела в D34-средняя масса тела в D11) *100/средняя масса тела в D11 обработки. ** В группе без обработки мышей умерщвляли из-за избыточной опухолевой массы в D25-D27 после обработки.
Пример 15. In vivo эффективность 684E12.SMCC.DM1 в отношении ксенотрансплантатной модели множественной миеломы KE97 у мышей SCID-Beige
[00477] Чтобы продемонстрировать нацеленную противоопухолевую активность 684E12.SMCC.DM1 in vivo, ксенотрансплантатную модель KE97 приживляли самкам мышей SCID-beige посредством подкожной инъекции 3×106 клеток в правый бок каждой мыши. После того как опухоли достигали примерно 200 мм3, мышей рандомизировали в соответствии с объемом опухоли в группы обработки (n=8 на группу). Мышей обрабатывали путем IV введения одного из исходного 684E12.SMCC.DM1 (DAR2,6) при конечной дозе 2 или 6 мг/кг или неспецифического изотипического контрольного IgG1.SMCC.DM1 из расчета 6 мг/кг. Все дозы корректировали в соответствии с индивидуальной массой тела мышей.
[00478] Все тестируемые средства хорошо переносились на протяжении исследования, и выраженных клинических симптомов токсичности или потери массы тела не наблюдали ни в одной из групп обработки. Незначительную противоопухолевую эффективность наблюдали после обработки неспецифическим изотипическим контрольным IgG1.SMCC.DM1 или исходным 684E12.SMCC.DM1 из расчета 2 мг/кг. Обработка исходным 684E12.SMCC.DM1 из расчета 6 мг/кг приводила к значению ΔT/ΔC, составляющему 6,72% в D11 после введения дозы (p<0,0001, однофакторный ANOVA/критерий Тьюки для множественных сравнений) (фигура 13).
Пример 16. In vivo целевая модуляция фармакодинамических маркеров под действием 121G12.CysMab.DAPA.MPET.DM4 в модели опухоли KE97
[00479] Накопление опухолевых клеток, положительных по маркеру фосфо-гистон H3, после обработки 121G12.CysMab.DAPA.MPET.DM4 применяли для оценки способности ADC к CCR7 индуцировать остановку на стадии G2/M in vivo.
[00480] Проводили исследование, при котором ксенотрансплантатную модель KE97 приживляли самкам мышей SCID-beige посредством подкожной инъекции 3×106 клеток в правый бок каждой мыши. После того как опухоли достигали примерно 140 мм3, мышей рандомизировали в соответствии с объемом опухоли в группы обработки (n=3 на группу). Мышей обрабатывали путем однократного IV введения одного из 121G12.CysMab.DAPA.MPET.DM4 при конечной дозе 2, 5 или 10 мг/кг или неспецифического изотипического контрольного IgG1.CysMab.DAPA.MPET.DM4 из расчета 10 мг/кг. Все дозы корректировали в соответствии с индивидуальной массой тела мышей. Через 48 часов после обработки опухоли собирали для оценки уровней фосфо-гистона H3 с помощью иммунного гистохимического окрашивания, описанного ниже.
[00481] Чтобы измерить накопление ядер, положительных по фосфо-гистону H3, с помощью иммуногистохимического исследования, поликлональное антитело кролика, нацеливающееся на остатки, окружающие фосфорилированный серин 10 гистона H3 человека, получали от Ventana Medical Systems (Тусон, Аризона, № по кат. 760-4591). Протокол IHC предусматривал нагревание и умеренное воздействие (32 мин) реагента для демаскирования антигена Ventana Discovery Cell Conditioner 1. Образцы инкубировали в течение 60 мин при комнатной температуре с первичным антителом (предварительно разведенным согласно рекомендациям производителя). Последующую инкубацию в течение 12 мин проводили с конъюгированным HRP вторичным Ab к кролику OmniMap (Ventana, Тусон, Аризона, № по кат. 760-4311) (предварительно разведенным согласно рекомендациям производителя).
[00482] Хотя на фигуре 14A на препаратах, полученных у представителей группы без обработки и группы изотипического контрольного IgG1.CysMab.DAPA.MPET.DM4, показаны редкие опухолевые клетки, положительные по фосфо-гистону H3, устойчивое дозозависимое повышение иммунного окрашивания фосфо-гистона H3 обнаружено через 48 часов после введения 121G12.CysMab.DAPA.MPET.DM4. Количественное определение сигнала выполняли с применением MatLab (MathWorks, Натик, Массачусетс), где общую площадь сигнала фосфо-гистона H3 (мкм2) нормализовали по общей площади ядер (мкм2), что давало значения относительного показателя фосфо-гистона H3 (%), показанные ниже для каждой из групп обработки. Данные на фигуре 14B указывают на то, что 121G12.CysMab.DAPA.MPET.DM4 способен вызывать сильно выраженную остановку на стадии G2/M на ксенотрансплантатах опухоли, что соответствует ожидаемому механизму действия полезной нагрузки.
Пример 17. Способ получения антитела 121G12.CysMab.DAPA
[00483] В данном примере описан способ получения антитела к CCR7, 121G12.CysMab.DAPA, с помощью культуры клеток, где Ab экспрессируется с вектора, который кодирует Ab. После того как Ab экспрессировался в культуре клеток, Ab очищали из культуры клеток следующим образом.
[00484] Первая стадия способа очистки промежуточного вещества лекарственной субстанции антитела 121G12.CysMab.DAPA заключалась в удалении клеток с помощью поточной глубокой фильтрации, с последующей фильтрацией через фильтр с размером пор 0,2 мкм.
[00485] Вторая стадия заключается в стадии аффинной жидкостной хроматографии с белком A. В зависимости от общего количества нерасфасованного продукта данную стадию проводят за несколько циклов. Каждый цикл допускает максимальную загрузку, составляющую примерно 20 г/л объема колонки. Элюирование проводят с помощью 50 мМ уксусной кислоты с примерным pH 3,0. Рабочая температура составляет 18-28°C. Все элюаты объединяют и хранят при 2-8°C до проведения стадии инактивации вируса.
[00486] Третья стадия представляет собой инактивацию вируса с помощью "обработки при низком pH". Температуру раствора промежуточного вещества из стадии 2 доводят до 18-28°C и корректируют pH до 3,5 (диапазон 3,4-3,6). Затем раствор промежуточного вещества продукта выдерживают для инактивации вируса в течение 70 минут (диапазон 60-90 минут). После завершения времени выдерживания раствор доводят до pH 6,0 (диапазон 5,8-6,2). В конце стадии раствор подвергают глубокой поточной фильтрации с помощью фильтра с размером пор 0,2 мкм и хранят при 2-8°C.
[00487] Четвертая стадия представляет собой катионообменную хроматографию в режиме связывание/элюирование, которая предусматривает интегрированное восстановление на колонке. В зависимости от титра данную стадию проводят за несколько циклов. Каждый цикл допускает загрузку, составляющую примерно 30 г/л объема колонки. Колонку уравновешивают с помощью буфера A, содержащего 20 мМ сукцинат натрия, pH 6,0. Восстановление на колонке проводят с применением 20 мМ фосфата натрия, 1 мМ EDTA, 7 мМ L-цистеина, pH 7,1, в качестве буфера для восстановления. Буфер для восстановления удаляют с помощью буфера A и элюирование проводят с линейным градиентом от 10% до 90% с помощью буфера A и буфера B, содержащего 10 мМ сукцината натрия, 300 мМ хлорида натрия, pH 6,0. Элюаты и объединенные элюаты можно хранить при 2-8°C до стадии анионообменной хроматографии на мультимодальных разделительных матрицах.
[00488] Пятая стадия способа представляет собой анионообменную хроматографию в режиме "с непрерывным потоком". В зависимости от титра данную стадию проводят за несколько циклов. Каждый цикл допускает максимальную загрузку, составляющую примерно 350 г/л объема колонки. Рабочая температура составляет 18-28°C. Уравновешивание проводят с помощью 20 мМ сукцината натрия, 119 мМ хлорида натрия, pH 6,0. Конечный продукт фильтрации хранят при 2-8°C перед стадией удаления вируса.
[00489] Вирусная фильтрация, шестая стадия, состоит из префильтрации с помощью фильтра с размером пор 0,1 мкм, с последующей вирусной фильтрацией с помощью нанофильтра Planova 20N. Температуру раствора промежуточного вещества из стадия 5 доводят до 18-28°C перед вирусной фильтрацией. Рабочая температура составляет 18-28°C. После нанофильтрации промежуточное вещество хранят при 2-8°C или 18-28°C.
[00490] Седьмая стадия, ультрафильтрация/диафильтрация, состоит из стадии концентрирования до примерно 70 г/л с последующей стадией 1ой диафильтрации с помощью 10 мМ фосфата калия, pH 6,0. При диафильтрации планируется получить коэффициент обмена, составляющий по меньшей мере 7, с последующим разбавлением до примерно 50 г/л. Конечное промежуточное вещество лекарственной субстанции пропускают через фильтр с размером пор 0,2 мкм и хранят при 2-8°C.
[00491] На восьмой и последней стадии конечным нерасфасованным промежуточным соединением лекарственной субстанции заполняют в виде аликвот подходящие контейнеры и хранят при температуре ниже -60°C после замораживания.
Таблица 25. Блок-схема способа очистки и восстановления
(MabSelect SuRE)
и
восстановление на колонке
(Fractogel EMD SO3 (M))
(Capto adhere)
(Planova 20N)
Пример 18. Дозозависимая in vivo эффективность 121G12.CysMab.DAPA.MPET.DM4 в отношении ксенотрансплантатной модели ABC-DLBCL OCI-LY3 у мышей NSG.
[00492] Чтобы продемонстрировать нацеленную противоопухолевую активность 121G12.CysMab.DAPA.MPET.DM4 in vivo на модели ABC-DLBCL, ксенотрансплантатную модель OCI-LY3 приживляли самкам мышей NSG посредством подкожной инъекции 10×106 клеток в правый бок каждой мыши. После того как опухоли достигали примерно 140 мм3, мышей рандомизировали в соответствии с объемом опухоли в группы обработки (n=6 на группу). Мышей обрабатывали путем IV введения одного из 121G12.CysMab.DAPA.MPET.DM4 (DAR4) при конечной дозе 0,5, 1 или 2 мг/кг или неспецифического изотипического контрольного hIgG1.CysMab.DAPA.MPET.DM4 из расчета 2 мг/кг в день 1 и день 15 исследования. Все дозы корректировали в соответствии с индивидуальной массой тела мышей. Все тестируемые средства хорошо переносились на протяжении исследования, и выраженных клинических симптомов токсичности или потери массы тела не наблюдали ни в одной из групп обработки (таблица 26).
[00493] Незначительную противоопухолевую эффективность наблюдали после обработки неспецифическим изотипическим контрольным hIgG1.CysMab.DAPA.MPET.DM4 из расчета 2 мг/кг. Обработка 121G12.CysMab.DAPA.MPET.DM4 приводила к дозозависимой противоопухолевой эффективности со значением ΔT/ΔC, составляющим 74,6% (0,5 мг/кг) и 10,7% (1 мг/кг), в то же время доза 2 мг/кг приводила к средней регрессии, составляющей 65,9%, ко дню 28 исследования. У 3 из 6 мышей в группе 121G12.CysMab.DAPA.MPET.DM4 из расчета 2 мг/кг проявлялась полная регрессия (фигура 15).
Таблица 26. Дозозависимая эффективность ADC к CCR7 в отношении ксенотрансплантатной модели OCI-LY3 в день 28 обработки.
общее количество)
при введении дозы в D1 и D15
при введении дозы в D1 и D15
при введении дозы в D1 и D15
при введении дозы в D1 и D15
Эксперимент оценивали в день 28 после обработки, ** p<0,005 в сравнении с контрольной группой без обработки (однофакторный ANOVA/критерий Тьюки для множественных сравнений). % ΔT/ΔC=100 ΔT/ΔC, где: ΔT=средний объем опухоли в обработанной лекарственным средством группе в D28 исследования - средний объем опухоли в обработанной лекарственным средством группе в начальный день при введении дозы; ΔC=средний объем опухоли в контрольной группе в D28 исследования - средний объем опухоли в контрольной группе в начальный день введения дозы D1. % регрессии=(1- T конечный/T начальный) x100 рассчитывали, если ΔT<0, где T конечный представляет собой средний объем опухоли в D28, а T начальный определяется как объем опухоли в D1 обработки. Δ массы тела (%)=(средняя масса тела в D28-средняя масса тела в D1) *100/средняя масса тела в D1 обработки.
Пример 19. Дозозависимая in vivo эффективность 121G12.CysMab.DAPA.MPET.DM4 в отношении ксенотрансплантатной модели GCB-DLBCL Toledo у мышей SCID-bg.
[00494] Чтобы продемонстрировать нацеленную противоопухолевую активность 121G12.CysMab.DAPA.MPET.DM4 in vivo на модели GCB-DLBCL, ксенотрансплантатную модель Toledo приживляли самкам мышей Scid-bg посредством подкожной инъекции 3×106 клеток в правый бок каждой мыши. После того как опухоли достигали примерно 100 мм3, мышей рандомизировали в соответствии с объемом опухоли в группы обработки (n=4 на группу). Мышей обрабатывали путем IV введения одного из 121G12.CysMab.DAPA.MPET.DM4 (DAR4) при конечной дозе 2 или 5 мг/кг или неспецифического изотипического контрольного hIgG1.CysMab.DAPA.MPET.DM4 из расчета 5 мг/кг в день 1 и день 15 исследования. Все дозы корректировали в соответствии с индивидуальной массой тела мышей. Все тестируемые средства хорошо переносились на протяжении исследования, и выраженных клинических симптомов токсичности или потери массы тела не наблюдали ни в одной из групп обработки (таблица 27).
[00495] Незначительную противоопухолевую эффективность наблюдали после обработки неспецифическим изотипическим контрольным hIgG1.CysMab.DAPA.MPET.DM4 из расчета 5 мг/кг. Обработка 121G12.CysMab.DAPA.MPET.DM4 приводила к дозозависимой противоопухолевой эффективности со значением ΔT/ΔC, составляющим 52,7% (2 мг/кг) и 20,6% (5 мг/кг) (фигура 16, таблица 27).
Таблица 27. Дозозависимая эффективность ADC к CCR7 в отношении ксенотрансплантатной модели Toledo в день 20 обработки.
при введении дозы в D1 и D15
при введении дозы в D1 и D15
при введении дозы в D1 и D15
Эксперимент оценивали в день 20 после обработки, * p<0,05, ** p<0,005 в сравнении с контрольной группой без обработки (однофакторный ANOVA/критерий Тьюки для множественных сравнений). % ΔT/ΔC=100 ΔT/ΔC, где: ΔT=средний объем опухоли в обработанной лекарственным средством группе в D20 исследования - средний объем опухоли в обработанной лекарственным средством группе в начальный день при введении дозы; ΔC=средний объем опухоли в контрольной группе в D20 исследования - средний объем опухоли в контрольной группе в начальный день введения дозы D1. Δ массы тела (%)=(средняя масса тела в D20-средняя масса тела в D1) *100/средняя масса тела в D1 обработки.
Пример 20. In vivo эффективность 121G12.CysMab.DAPA.MPET.DM4 на ксенотрансплантатной модели DEL ALCL у мышей SCID-bg.
[00496] Чтобы продемонстрировать нацеленную противоопухолевую активность 121G12.CysMab.DAPA.MPET.DM4 in vivo на модели CCR7-положительной ALCL, ксенотрансплантатную модель DEL приживляли самкам мышей Scid-bg посредством подкожной инъекции 3×106 клеток в правый бок каждой мыши. После того как опухоли достигали примерно 100 мм3, мышей рандомизировали в соответствии с объемом опухоли в группы обработки (n=4 на группу). Мышей обрабатывали путем IV введения одного из 121G12.CysMab.DAPA.MPET.DM4 (DAR4) при конечной дозе 2 мг/кг или неспецифического изотипического контрольного изотип.MPET.DM4 из расчета 2 мг/кг в день 1 и день 15 исследования. Все дозы корректировали в соответствии с индивидуальной массой тела мышей.
[00497] Незначительную противоопухолевую эффективность наблюдали после обработки неспецифическим изотипическим контрольным hIgG1.CysMab.DAPA.MPET.DM4 из расчета 2 мг/кг. Обработка 121G12.CysMab.DAPA.MPET.DM4 из расчета 2 мг/кг приводила к средней регрессии, составляющей 40,2%, ко дню 14 исследования после введения однократной дозы (p<0,01). Мышам вводили вторую дозу в день 15 и за ними наблюдали в течение еще трех недель. Одно животное-исключение не проявило реакцию на обработку второй дозой, и его пришлось подвергнуть умерщвлению в день 28 из-за опухолевой массы. Еще одно животное продемонстрировало медленное прогрессирование заболевания и его также исключили в день 28 из последующего наблюдения за экспрессией мишени. У двух из четырех мышей продолжали наблюдать устойчивое воздействие на рост опухоли (фигура 17). Все обработки хорошо переносились с отсутствием явной потери массы тела.
Пример 21. In vivo эффективность 121G12.CysMab.DAPA.MPET.DM4 в отношении модели опухоли на основе полученного от пациента первичного немелкоклеточного рака легкого HLUX1787
[00498] Противоопухолевую активность 121G12.CysMab.DAPA.MPET.DM4 оценивали на ксенотрансплантатной модели первичного немелкоклеточного рака легкого HLUX1787, экспрессирующего CCR7. Самкам NSG мышей имплантировали подкожно фрагменты опухоли в правый бок каждой мыши. После того как опухоли достигали примерно 150 мм3, мышей рандомизировали в соответствии с объемом опухоли в группы обработки (n=6 на группу). Мышей обрабатывали путем IV введения одного из 121G12.CysMab.DAPA.MPET.DM4 (DAR4) при 0,5, 2 или 5 мг/кг в день 1, а вторую дозу вводили спустя 2 недели в день 15. Все дозы корректировали в соответствии с индивидуальной массой тела мышей.
[00499] Незначительную противоопухолевую эффективность наблюдали в случае низких доз конъюгированного Ab, однако частичную регрессию или стабилизацию заболевания наблюдали в случае обработки 121G12.CysMab.DAPA.MPET.DM4 из расчета 5 мг/кг. Устойчивую эффективность воздействия на опухоли наблюдали через две недели после введения второй дозы, что привело к значению ΔT/ΔC, составляющему 1,3% в D30 обработки (p<0,001; однофакторный ANOVA/критерий Тьюки для множественных сравнений) (фигура 18). Все обработки хорошо переносились с отсутствием явной потери массы тела.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> NOVARTIS AG
<120> КОНЪЮГАТЫ АНТИТЕЛА К CCR7 И ЛЕКАРСТВЕННОГО СРЕДСТВА
<130> PAT057594-WO-PCT
<140>
<141>
<150> 62/454,476
<151> 2017-02-03
<160> 629
<170> PatentIn версия 3.5
<210> 1
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 1
Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
1. 5 10
<210> 2
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 2
Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 3
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 3
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 4
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 4
Ser Tyr Ala Met Ser
1. 5
<210> 5
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 5
Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 6
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 6
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 7
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 7
Gly Phe Thr Phe Ser Ser Tyr
1. 5
<210> 8
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 8
Ser Ser Gly Gly Ser Phe
1. 5
<210> 9
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 9
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 10
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 10
Gly Phe Thr Phe Ser Ser Tyr Ala
1. 5
<210> 11
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 11
Ile Ser Ser Gly Gly Ser Phe Thr
1. 5
<210> 12
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 12
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 13
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 13
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 14
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 14
gaagtgcagc tggtggaatc tggcggcgga ctggtcaagc ctggcggctc cctgagactg 60
tcttgcgccg cctccggctt caccttctcc agctacgcca tgtcctggat ccggcaggcc 120
cctggcaagg gactggagtg ggtggccacc atctcctccg gcggcagctt cacctactac 180
cccgactccg tgaagggccg gttcaccatc tcccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgtgc cagacgggcc 300
tccaccgtcg tgggcaccga tttcgatgtg tggggccagg gcacaaccgt gaccgtgtcc 360
tcc 363
<210> 15
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 15
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 16
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 16
gaagtgcagc tggtggaatc tggcggcgga ctggtcaagc ctggcggctc cctgagactg 60
tcttgcgccg cctccggctt caccttctcc agctacgcca tgtcctggat ccggcaggcc 120
cctggcaagg gactggagtg ggtggccacc atctcctccg gcggcagctt cacctactac 180
cccgactccg tgaagggccg gttcaccatc tcccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgtgc cagacgggcc 300
tccaccgtcg tgggcaccga tttcgatgtg tggggccagg gcacaaccgt gaccgtgtcc 360
tccgcctcca ccaagggacc ctccgtgttc cctctggccc cttccagcaa gtccacctct 420
ggcggcaccg ccgctctggg ctgcctggtc aaggactact tcccctgccc tgtgacagtg 480
tcctggaact ccggcgctct gacctccggc gtgcacacct tccctgccgt gctgcagtcc 540
tccggcctgt actccctgtc ctccgtcgtg accgtgcctt cctccagcct gggcacccag 600
acctacatct gcaacgtgaa ccacaagccc tccaacacca aagtggacaa gcgggtggaa 660
cccaagtcct gcgacaagac ccacacctgt cctccctgcc ctgcccctga gctgctggga 720
ggcccttccg tgttcctgtt ccctccaaag cccaaggaca ccctgatgat ctcccggacc 780
cctgaagtga cctgcgtggt ggtggccgtg tcccacgagg atcccgaagt gaagttcaat 840
tggtacgtgg acggcgtgga agtgcacaat gccaagacca agcccagaga ggaacagtac 900
aactccacct accgggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 960
aaagagtaca agtgcaaagt gtccaacaag gccctggccg ctcccatcga aaagaccatc 1020
tccaaggcca agggccagcc cagagagccc caagtgtaca cactgcctcc cagccgggaa 1080
gagatgacca agaaccaagt gtccctgacc tgcctcgtga agggcttcta cccctgcgat 1140
atcgccgtgg agtgggagtc caacggccag cccgagaaca actacaagac cacccctccc 1200
gtgctggaca gcgacggctc attcttcctg tactccaagc tgaccgtgga caagtcccgg 1260
tggcagcagg gcaacgtgtt ctcctgctcc gtgatgcacg aggccctgca caaccactac 1320
acccagaagt ccctgtccct gagccccggc aag 1353
<210> 17
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 17
Arg Ala Ser Gln Asp Ile Gly Ser Ser Leu Asn
1. 5 10
<210> 18
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 18
Ala Thr Ser Ser Leu Asp Ser
1. 5
<210> 19
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 19
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 20
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 20
Arg Ala Ser Gln Asp Ile Gly Ser Ser Leu Asn
1. 5 10
<210> 21
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 21
Ala Thr Ser Ser Leu Asp Ser
1. 5
<210> 22
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 22
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 23
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 23
Ser Gln Asp Ile Gly Ser Ser
1. 5
<210> 24
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 24
Ala Thr Ser
1
<210> 25
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 25
Tyr Ala Ser Ser Pro Pro
1. 5
<210> 26
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 26
Gln Asp Ile Gly Ser Ser
1. 5
<210> 27
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 27
Ala Thr Ser
1
<210> 28
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 28
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 29
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 29
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Leu Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Asn Trp Leu Gln Gln Lys Pro Gly Lys Ala Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Val Val Tyr Tyr Cys Leu Gln Tyr Ala Ser Ser Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 30
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 30
gacatccaga tgacccagag cccctccagc ctgtccgcct ccgtgggcga tagagtgacc 60
ctgacctgcc gggcctccca ggacatcggc tcctccctga actggctgca gcagaagccc 120
ggcaaggcca tcaagcggct gatctacgcc acctcctccc tggactccgg cgtgccctcc 180
cggttctctg gctccagatc cggcaccgac tacaccctga ccatctccag cctgcagccc 240
gaggacttcg tggtgtacta ctgcctgcag tacgcctcca gccctcccac cttcggcgga 300
ggcaccaagc tggaaatcaa g 321
<210> 31
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 31
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Leu Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Asn Trp Leu Gln Gln Lys Pro Gly Lys Ala Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Val Val Tyr Tyr Cys Leu Gln Tyr Ala Ser Ser Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 32
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 32
gacatccaga tgacccagag cccctccagc ctgtccgcct ccgtgggcga tagagtgacc 60
ctgacctgcc gggcctccca ggacatcggc tcctccctga actggctgca gcagaagccc 120
ggcaaggcca tcaagcggct gatctacgcc acctcctccc tggactccgg cgtgccctcc 180
cggttctctg gctccagatc cggcaccgac tacaccctga ccatctccag cctgcagccc 240
gaggacttcg tggtgtacta ctgcctgcag tacgcctcca gccctcccac cttcggcgga 300
ggcaccaagc tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag cggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 33
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 33
Gly Phe Thr Phe Ser Thr Tyr Ala Met Ser
1. 5 10
<210> 34
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 34
Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val Lys
1. 5 10 15
Gly
<210> 35
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 35
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 36
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 36
Thr Tyr Ala Met Ser
1. 5
<210> 37
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 37
Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val Lys
1. 5 10 15
Gly
<210> 38
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 38
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 39
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 39
Gly Phe Thr Phe Ser Thr Tyr
1. 5
<210> 40
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 40
Ser Asp Ala Gly Ser Tyr
1. 5
<210> 41
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 41
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 42
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 42
Gly Phe Thr Phe Ser Thr Tyr Ala
1. 5
<210> 43
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 43
Ile Ser Asp Ala Gly Ser Tyr Ser
1. 5
<210> 44
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 44
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10 15
<210> 45
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 45
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 46
<211> 366
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 46
gaagtgcagc tggtggaatc tggcggcgga ctggtcaagc ctggcggctc cctgagactg 60
tcttgcgccg cctccggctt caccttctcc acctacgcca tgtcctgggt ccgacaggcc 120
cctggaaagg gcctggagtg ggtggccacc atctccgacg ccggctccta ctcctactac 180
cccgacaacg tgaagggccg gttcaccatc tcccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgcgc cagacggggc 300
tccagatacg aagagtacta cgtgatggac tactggggcc agggcacaac cgtgaccgtg 360
tcctcc 366
<210> 47
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 47
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 48
<211> 1356
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 48
gaagtgcagc tggtggaatc tggcggcgga ctggtcaagc ctggcggctc cctgagactg 60
tcttgcgccg cctccggctt caccttctcc acctacgcca tgtcctgggt ccgacaggcc 120
cctggaaagg gcctggagtg ggtggccacc atctccgacg ccggctccta ctcctactac 180
cccgacaacg tgaagggccg gttcaccatc tcccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgcgc cagacggggc 300
tccagatacg aagagtacta cgtgatggac tactggggcc agggcacaac cgtgaccgtg 360
tcctccgcct ccaccaaggg accctccgtg ttccctctgg ccccttccag caagtccacc 420
tctggcggca ccgccgctct gggctgcctg gtcaaggact acttcccctg ccctgtgaca 480
gtgtcctgga actccggcgc tctgacctcc ggcgtgcaca ccttccctgc cgtgctgcag 540
tcctccggcc tgtactccct gtcctccgtc gtgaccgtgc cttcctccag cctgggcacc 600
cagacctaca tctgcaacgt gaaccacaag ccctccaaca ccaaagtgga caagcgggtg 660
gaacccaagt cctgcgacaa gacccacacc tgtcctccct gccctgcccc tgagctgctg 720
ggaggccctt ccgtgttcct gttccctcca aagcccaagg acaccctgat gatctcccgg 780
acccctgaag tgacctgcgt ggtggtggcc gtgtcccacg aggatcccga agtgaagttc 840
aattggtacg tggacggcgt ggaagtgcac aatgccaaga ccaagcccag agaggaacag 900
tacaactcca cctaccgggt ggtgtccgtg ctgaccgtgc tgcaccagga ctggctgaac 960
ggcaaagagt acaagtgcaa agtgtccaac aaggccctgg ccgctcccat cgaaaagacc 1020
atctccaagg ccaagggcca gcccagagag ccccaagtgt acacactgcc tcccagccgg 1080
gaagagatga ccaagaacca agtgtccctg acctgcctcg tgaagggctt ctacccctgc 1140
gatatcgccg tggagtggga gtccaacggc cagcccgaga acaactacaa gaccacccct 1200
cccgtgctgg actccgacgg ctcattcttc ctgtactcca agctgaccgt ggacaagtcc 1260
cggtggcagc agggcaacgt gttctcctgc tccgtgatgc acgaggccct gcacaaccac 1320
tacacccaga agtccctgtc cctgagcccc ggcaag 1356
<210> 49
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 49
Arg Ala Ser Gln Ser Ile Ser Asn Asn Leu His
1. 5 10
<210> 50
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 50
Tyr Ala Ser Gln Ser Ile Ser
1. 5
<210> 51
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 51
Gln Gln Ser Ser Ser Trp Leu Thr
1. 5
<210> 52
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 52
Arg Ala Ser Gln Ser Ile Ser Asn Asn Leu His
1. 5 10
<210> 53
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 53
Tyr Ala Ser Gln Ser Ile Ser
1. 5
<210> 54
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 54
Gln Gln Ser Ser Ser Trp Leu Thr
1. 5
<210> 55
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 55
Ser Gln Ser Ile Ser Asn Asn
1. 5
<210> 56
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 56
Tyr Ala Ser
1
<210> 57
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 57
Ser Ser Ser Trp Leu
1. 5
<210> 58
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 58
Gln Ser Ile Ser Asn Asn
1. 5
<210> 59
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 59
Tyr Ala Ser
1
<210> 60
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 60
Gln Gln Ser Ser Ser Trp Leu Thr
1. 5
<210> 61
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 61
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1. 5 10 15
Glu Arg Val Thr Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asn Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Ile Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Val Glu Pro
65 70 75 80
Glu Asp Phe Gly Val Tyr Phe Cys Gln Gln Ser Ser Ser Trp Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 62
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 62
gagatcgtgc tgacacagtc ccctgccacc ctgtctgtgt ctcccggcga gagagtgacc 60
ctgtcctgcc gggcctccca gtccatctcc aacaacctgc actggtatca gcagaagccc 120
ggccaggccc ctcggctgct gattaagtac gcctcccaga gcatctccgg catccctgcc 180
agattctccg gctccggcag cggcaccgac ttcaccctga ccatctccag cgtggaaccc 240
gaggacttcg gcgtgtactt ctgccagcag tcctcctcct ggctgacctt cggccagggc 300
accaagctgg aaatcaag 318
<210> 63
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 63
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1. 5 10 15
Glu Arg Val Thr Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asn Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Ile Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Val Glu Pro
65 70 75 80
Glu Asp Phe Gly Val Tyr Phe Cys Gln Gln Ser Ser Ser Trp Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 64
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 64
gagatcgtgc tgacacagtc ccctgccacc ctgtctgtgt ctcccggcga gagagtgacc 60
ctgtcctgcc gggcctccca gtccatctcc aacaacctgc actggtatca gcagaagccc 120
ggccaggccc ctcggctgct gattaagtac gcctcccaga gcatctccgg catccctgcc 180
agattctccg gctccggcag cggcaccgac ttcaccctga ccatctccag cgtggaaccc 240
gaggacttcg gcgtgtactt ctgccagcag tcctcctcct ggctgacctt cggccagggc 300
accaagctgg aaatcaagcg tacggtggcc gctcccagcg tgttcatctt cccccccagc 360
gacgagcagc tgaagagcgg caccgccagc gtggtgtgcc tgctgaacaa cttctacccc 420
cgggaggcca aggtgcagtg gaaggtggac aacgccctgc agagcggcaa cagccaggag 480
agcgtcaccg agcaggacag caaggactcc acctacagcc tgagcagcac cctgaccctg 540
agcaaggccg actacgagaa gcataaggtg tacgcctgcg aggtgaccca ccagggcctg 600
tccagccccg tgaccaagag cttcaacagg ggcgagtgc 639
<210> 65
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 65
Gly Tyr Ser Ile Thr Ser Gly Tyr Ser Trp His
1. 5 10
<210> 66
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 66
His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1. 5 10 15
<210> 67
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 67
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 68
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 68
Ser Gly Tyr Ser Trp His
1. 5
<210> 69
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 69
His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1. 5 10 15
<210> 70
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 70
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 71
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 71
Gly Tyr Ser Ile Thr Ser Gly Tyr
1. 5
<210> 72
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 72
His Ser Ser Gly Ser
1. 5
<210> 73
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 73
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 74
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 74
Gly Tyr Ser Ile Thr Ser Gly Tyr Ser
1. 5
<210> 75
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 75
Ile His Ser Ser Gly Ser Thr
1. 5
<210> 76
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 76
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr
1. 5 10
<210> 77
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 77
Asp Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp His Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp
35 40 45
Met Ala His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 78
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 78
gacgtgcagc tgcaggaatc tggccctggc ctggtcaagc cctcccagac cctgtccctg 60
acctgcaccg tgtccggcta ctctatcacc tccggctaca gctggcactg gatccggcag 120
caccccggca agggcctgga atggatggcc cacatccact cctccggctc caccaactac 180
aaccccagcc tgaagtcccg gatcaccatc tcccgggaca cctccaagaa ccagttctcc 240
ctgaagctgt cctccgtgac cgccgctgac accgccgtgt actactgtgc cagaggcggc 300
gtgcaggcct tcgcttattg gggccaggga accctggtca ccgtgtcctc c 351
<210> 79
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 79
Asp Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp His Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp
35 40 45
Met Ala His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Ala Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 80
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 80
gacgtgcagc tgcaggaatc tggccctggc ctggtcaagc cctcccagac cctgtccctg 60
acctgcaccg tgtccggcta ctctatcacc tccggctaca gctggcactg gatccggcag 120
caccccggca agggcctgga atggatggcc cacatccact cctccggctc caccaactac 180
aaccccagcc tgaagtcccg gatcaccatc tcccgggaca cctccaagaa ccagttctcc 240
ctgaagctgt cctccgtgac cgccgctgac accgccgtgt actactgtgc cagaggcggc 300
gtgcaggcct tcgcttattg gggccaggga accctggtca ccgtgtcctc cgccagcacc 360
aagggaccct ccgtgttccc tctggcccct tccagcaagt ccacctctgg cggcaccgcc 420
gctctgggct gcctcgtgaa ggactacttc ccctgccccg tgaccgtgtc ctggaactcc 480
ggcgctctga cctccggcgt gcacaccttc cctgccgtgc tgcagtcctc cggcctgtac 540
tccctgtcca gcgtcgtgac cgtgccctcc agctctctgg gcacccagac ctacatctgc 600
aacgtgaacc acaagccctc caacaccaaa gtggacaagc gggtggaacc caagtcctgc 660
gacaagaccc acacctgtcc tccctgccct gcccctgagc tgctgggagg cccttccgtg 720
ttcctgttcc ctccaaagcc caaggacacc ctgatgatct cccggacccc tgaagtgacc 780
tgcgtggtgg tggccgtgtc ccacgaggat cccgaagtga agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagtacaa ctccacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaagtgt ccaacaaggc cctggccgct cccatcgaaa agaccatctc caaggccaag 1020
ggccagccca gagagcccca agtgtacaca ctgcctccca gccgggaaga gatgaccaag 1080
aatcaagtgt ccctgacctg tctggtcaag ggcttctacc cctgcgatat cgccgtggag 1140
tgggagtcca acggccagcc cgagaacaac tacaagacca cccctcccgt gctggactcc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agtcccggtg gcagcagggc 1260
aacgtgttct cctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgagcctgt cccctggcaa g 1341
<210> 81
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 81
Ser Ala Ser Ser Ser Val Ile Tyr Met His
1. 5 10
<210> 82
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 82
Asp Thr Ser Lys Leu Ala Ser
1. 5
<210> 83
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 83
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 84
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 84
Ser Ala Ser Ser Ser Val Ile Tyr Met His
1. 5 10
<210> 85
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 85
Asp Thr Ser Lys Leu Ala Ser
1. 5
<210> 86
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 86
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 87
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 87
Ser Ser Ser Val Ile Tyr
1. 5
<210> 88
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 88
Asp Thr Ser
1
<210> 89
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 89
Trp Ser Ser Asn Pro Leu
1. 5
<210> 90
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 90
Ser Ser Val Ile Tyr
1. 5
<210> 91
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 91
Asp Thr Ser
1
<210> 92
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 92
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 93
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 93
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Ala Ser Pro Gly
1. 5 10 15
Glu Arg Val Thr Met Ser Cys Ser Ala Ser Ser Ser Val Ile Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Met Glu Pro Glu
65 70 75 80
Asp Ala Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 94
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 94
gagatcgtgc tgacacagtc ccctgccacc ctgtccgcct ctccaggcga gcgcgtgaca 60
atgtcctgct ccgcctcctc ctccgtgatc tacatgcact ggtatcagca gaagcccggc 120
caggcccctc ggcggtggat ctacgatacc tccaagctgg cctccggcgt gcccgccaga 180
ttctccggct ctggctctgg caccgactac accctgacca tctccagcat ggaacccgag 240
gacgccgccg tgtactactg ccagcagtgg tcctccaacc ctctgacctt cggccagggc 300
accaagctgg aaatcaag 318
<210> 95
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 95
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Ala Ser Pro Gly
1. 5 10 15
Glu Arg Val Thr Met Ser Cys Ser Ala Ser Ser Ser Val Ile Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Met Glu Pro Glu
65 70 75 80
Asp Ala Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 96
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 96
gagatcgtgc tgacacagtc ccctgccacc ctgtccgcct ctccaggcga gcgcgtgaca 60
atgtcctgct ccgcctcctc ctccgtgatc tacatgcact ggtatcagca gaagcccggc 120
caggcccctc ggcggtggat ctacgatacc tccaagctgg cctccggcgt gcccgccaga 180
ttctccggct ctggctctgg caccgactac accctgacca tctccagcat ggaacccgag 240
gacgccgccg tgtactactg ccagcagtgg tcctccaacc ctctgacctt cggccagggc 300
accaagctgg aaatcaagcg tacggtggcc gctcccagcg tgttcatctt cccccccagc 360
gacgagcagc tgaagagcgg caccgccagc gtggtgtgcc tgctgaacaa cttctacccc 420
cgggaggcca aggtgcagtg gaaggtggac aacgccctgc agagcggcaa cagccaggag 480
agcgtcaccg agcaggacag caaggactcc acctacagcc tgagcagcac cctgaccctg 540
agcaaggccg actacgagaa gcataaggtg tacgcctgcg aggtgaccca ccagggcctg 600
tccagccccg tgaccaagag cttcaacagg ggcgagtgc 639
<210> 97
<211> 378
<212> БЕЛОК
<213> Homo sapiens
<400> 97
Met Asp Leu Gly Lys Pro Met Lys Ser Val Leu Val Val Ala Leu Leu
1. 5 10 15
Val Ile Phe Gln Val Cys Leu Cys Gln Asp Glu Val Thr Asp Asp Tyr
20 25 30
Ile Gly Asp Asn Thr Thr Val Asp Tyr Thr Leu Phe Glu Ser Leu Cys
35 40 45
Ser Lys Lys Asp Val Arg Asn Phe Lys Ala Trp Phe Leu Pro Ile Met
50 55 60
Tyr Ser Ile Ile Cys Phe Val Gly Leu Leu Gly Asn Gly Leu Val Val
65 70 75 80
Leu Thr Tyr Ile Tyr Phe Lys Arg Leu Lys Thr Met Thr Asp Thr Tyr
85 90 95
Leu Leu Asn Leu Ala Val Ala Asp Ile Leu Phe Leu Leu Thr Leu Pro
100 105 110
Phe Trp Ala Tyr Ser Ala Ala Lys Ser Trp Val Phe Gly Val His Phe
115 120 125
Cys Lys Leu Ile Phe Ala Ile Tyr Lys Met Ser Phe Phe Ser Gly Met
130 135 140
Leu Leu Leu Leu Cys Ile Ser Ile Asp Arg Tyr Val Ala Ile Val Gln
145 150 155 160
Ala Val Ser Ala His Arg His Arg Ala Arg Val Leu Leu Ile Ser Lys
165 170 175
Leu Ser Cys Val Gly Ile Trp Ile Leu Ala Thr Val Leu Ser Ile Pro
180 185 190
Glu Leu Leu Tyr Ser Asp Leu Gln Arg Ser Ser Ser Glu Gln Ala Met
195 200 205
Arg Cys Ser Leu Ile Thr Glu His Val Glu Ala Phe Ile Thr Ile Gln
210 215 220
Val Ala Gln Met Val Ile Gly Phe Leu Val Pro Leu Leu Ala Met Ser
225 230 235 240
Phe Cys Tyr Leu Val Ile Ile Arg Thr Leu Leu Gln Ala Arg Asn Phe
245 250 255
Glu Arg Asn Lys Ala Ile Lys Val Ile Ile Ala Val Val Val Val Phe
260 265 270
Ile Val Phe Gln Leu Pro Tyr Asn Gly Val Val Leu Ala Gln Thr Val
275 280 285
Ala Asn Phe Asn Ile Thr Ser Ser Thr Cys Glu Leu Ser Lys Gln Leu
290 295 300
Asn Ile Ala Tyr Asp Val Thr Tyr Ser Leu Ala Cys Val Arg Cys Cys
305 310 315 320
Val Asn Pro Phe Leu Tyr Ala Phe Ile Gly Val Lys Phe Arg Asn Asp
325 330 335
Leu Phe Lys Leu Phe Lys Asp Leu Gly Cys Leu Ser Gln Glu Gln Leu
340 345 350
Arg Gln Trp Ser Ser Cys Arg His Ile Arg Arg Ser Ser Met Ser Val
355 360 365
Glu Ala Glu Thr Thr Thr Thr Phe Ser Pro
370 375
<210> 98
<211> 1134
<212> ДНК
<213> Homo sapiens
<400> 98
atggacctgg ggaaaccaat gaaaagcgtg ctggtggtgg ctctccttgt cattttccag 60
gtatgcctgt gtcaagatga ggtcacggac gattacatcg gagacaacac cacagtggac 120
tacactttgt tcgagtcttt gtgctccaag aaggacgtgc ggaactttaa agcctggttc 180
ctccctatca tgtactccat catttgtttc gtgggcctac tgggcaatgg gctggtcgtg 240
ttgacctata tctatttcaa gaggctcaag accatgaccg atacctacct gctcaacctg 300
gcggtggcag acatcctctt cctcctgacc cttcccttct gggcctacag cgcggccaag 360
tcctgggtct tcggtgtcca cttttgcaag ctcatctttg ccatctacaa gatgagcttc 420
ttcagtggca tgctcctact tctttgcatc agcattgacc gctacgtggc catcgtccag 480
gctgtctcag ctcaccgcca ccgtgcccgc gtccttctca tcagcaagct gtcctgtgtg 540
ggcatctgga tactagccac agtgctctcc atcccagagc tcctgtacag tgacctccag 600
aggagcagca gtgagcaagc gatgcgatgc tctctcatca cagagcatgt ggaggccttt 660
atcaccatcc aggtggccca gatggtgatc ggctttctgg tccccctgct ggccatgagc 720
ttctgttacc ttgtcatcat ccgcaccctg ctccaggcac gcaactttga gcgcaacaag 780
gccatcaagg tgatcatcgc tgtggtcgtg gtcttcatag tcttccagct gccctacaat 840
ggggtggtcc tggcccagac ggtggccaac ttcaacatca ccagtagcac ctgtgagctc 900
agtaagcaac tcaacatcgc ctacgacgtc acctacagcc tggcctgcgt ccgctgctgc 960
gtcaaccctt tcttgtacgc cttcatcggc gtcaagttcc gcaacgatct cttcaagctc 1020
ttcaaggacc tgggctgcct cagccaggag cagctccggc agtggtcttc ctgtcggcac 1080
atccggcgct cctccatgag tgtggaggcc gagaccacca ccaccttctc ccca 1134
<210> 99
<211> 378
<212> БЕЛОК
<213> Macaca fascicularis
<400> 99
Met Asp Leu Gly Lys Pro Met Lys Ser Val Leu Val Val Ala Leu Leu
1. 5 10 15
Val Ile Phe Gln Val Cys Leu Cys Gln Asp Glu Val Thr Asp Asp Tyr
20 25 30
Ile Gly Asp Asn Thr Thr Val Asp Tyr Thr Leu Phe Glu Ser Leu Cys
35 40 45
Ser Lys Lys Asp Val Arg Asn Phe Lys Ala Trp Phe Leu Pro Ile Met
50 55 60
Tyr Ser Ile Ile Cys Phe Val Gly Leu Leu Gly Asn Gly Leu Val Val
65 70 75 80
Leu Thr Tyr Ile Tyr Phe Lys Arg Leu Lys Thr Met Thr Asp Thr Tyr
85 90 95
Leu Leu Asn Leu Ala Val Ala Asp Ile Leu Phe Leu Leu Thr Leu Pro
100 105 110
Phe Trp Ala Tyr Ser Ala Ala Lys Ser Trp Val Phe Gly Val His Phe
115 120 125
Cys Lys Leu Ile Phe Ala Ile Tyr Lys Met Ser Phe Phe Ser Gly Met
130 135 140
Leu Leu Leu Leu Cys Ile Ser Ile Asp Arg Tyr Val Ala Ile Val Gln
145 150 155 160
Ala Val Ser Ala His Arg His Arg Ala Arg Val Leu Leu Ile Ser Lys
165 170 175
Leu Ser Cys Val Gly Ile Trp Ile Leu Ala Thr Val Leu Ser Ile Pro
180 185 190
Glu Leu Leu Tyr Ser Gly Leu Gln Arg Ser Ser Ser Glu Gln Ala Met
195 200 205
Arg Cys Ser Leu Ile Thr Glu His Val Glu Ala Phe Ile Thr Ile Gln
210 215 220
Val Ala Gln Met Val Ile Gly Phe Leu Val Pro Leu Leu Ala Met Ser
225 230 235 240
Phe Cys Tyr Leu Val Ile Ile Arg Thr Leu Leu Gln Ala Arg Asn Phe
245 250 255
Glu Arg Asn Lys Ala Ile Lys Val Ile Ile Ala Val Val Val Val Phe
260 265 270
Ile Val Phe Gln Leu Pro Tyr Asn Gly Val Val Leu Ala Gln Thr Val
275 280 285
Ala Asn Phe Asn Ile Thr Ser Ser Thr Cys Glu Leu Ser Lys Gln Leu
290 295 300
Asn Ile Ala Tyr Asp Val Thr Tyr Ser Leu Ala Cys Val Arg Cys Cys
305 310 315 320
Val Asn Pro Phe Leu Tyr Ala Phe Ile Gly Val Lys Phe Arg Asn Asp
325 330 335
Leu Phe Lys Leu Phe Lys Asp Leu Gly Cys Leu Ser Gln Glu Gln Leu
340 345 350
Arg Gln Trp Ser Ser Cys Arg His Ile Arg Arg Ser Ser Met Ser Val
355 360 365
Glu Ala Glu Thr Thr Thr Thr Phe Ser Pro
370 375
<210> 100
<211> 1134
<212> ДНК
<213> Macaca fascicularis
<400> 100
atggacctgg ggaaaccaat gaaaagcgtg ctggtggtgg ctctccttgt cattttccag 60
gtatgcctgt gtcaagatga ggtcacggac gattacatcg gagacaacac cacagtggac 120
tacactttgt tcgagtcttt gtgctccaag aaggacgtgc ggaactttaa agcctggttc 180
ctccctatca tgtactccat catttgtttc gtgggcctac tgggcaatgg gctggtcgtg 240
ttgacctata tctatttcaa gaggctcaag accatgaccg atacctacct gctcaacctg 300
gcggtggcag acatcctctt cctcctgacc cttcccttct gggcctacag cgcggccaag 360
tcctgggtct tcggtgtcca cttttgcaag ctcatctttg ccatctacaa gatgagcttc 420
ttcagtggca tgctcctact tctttgcatc agcattgacc gctacgtggc catcgtccag 480
gctgtctcag ctcaccgcca ccgtgcccgc gtccttctca tcagcaagct gtcctgtgtg 540
ggcatctgga tactagccac agtgctctcc atcccagagc tcctgtacag tggcctccag 600
aggagcagca gtgagcaagc gatgcgatgc tctctcatca cagagcatgt ggaggccttt 660
atcaccatcc aggtggccca gatggtgatc ggctttctgg tccccctgct ggccatgagc 720
ttctgttacc ttgtcatcat ccgcaccctg ctccaggcac gcaactttga gcgcaacaag 780
gccatcaagg tgatcatcgc tgtggtcgtg gtcttcatag tcttccagct gccctacaat 840
ggggtggtcc tggcccagac ggtggccaac ttcaacatca ccagtagcac ctgtgagctc 900
agtaagcaac tcaacatcgc ctacgacgtc acctacagcc tggcctgcgt ccgctgctgc 960
gtcaaccctt tcttgtacgc cttcatcggc gtcaagttcc gcaacgatct cttcaagctc 1020
ttcaaggacc tgggctgcct cagccaggag cagctccggc agtggtcttc ctgtcggcac 1080
atccggcgct cctccatgag tgtggaggcc gagaccacca ccaccttctc ccca 1134
<210> 101
<211> 378
<212> БЕЛОК
<213> Mus sp.
<400> 101
Met Asp Pro Gly Lys Pro Arg Lys Asn Val Leu Val Val Ala Leu Leu
1. 5 10 15
Val Ile Phe Gln Val Cys Phe Cys Gln Asp Glu Val Thr Asp Asp Tyr
20 25 30
Ile Gly Glu Asn Thr Thr Val Asp Tyr Thr Leu Tyr Glu Ser Val Cys
35 40 45
Phe Lys Lys Asp Val Arg Asn Phe Lys Ala Trp Phe Leu Pro Leu Met
50 55 60
Tyr Ser Val Ile Cys Phe Val Gly Leu Leu Gly Asn Gly Leu Val Ile
65 70 75 80
Leu Thr Tyr Ile Tyr Phe Lys Arg Leu Lys Thr Met Thr Asp Thr Tyr
85 90 95
Leu Leu Asn Leu Ala Val Ala Asp Ile Leu Phe Leu Leu Ile Leu Pro
100 105 110
Phe Trp Ala Tyr Ser Glu Ala Lys Ser Trp Ile Phe Gly Val Tyr Leu
115 120 125
Cys Lys Gly Ile Phe Gly Ile Tyr Lys Leu Ser Phe Phe Ser Gly Met
130 135 140
Leu Leu Leu Leu Cys Ile Ser Ile Asp Arg Tyr Val Ala Ile Val Gln
145 150 155 160
Ala Val Ser Ala His Arg His Arg Ala Arg Val Leu Leu Ile Ser Lys
165 170 175
Leu Ser Cys Val Gly Ile Trp Met Leu Ala Leu Phe Leu Ser Ile Pro
180 185 190
Glu Leu Leu Tyr Ser Gly Leu Gln Lys Asn Ser Gly Glu Asp Thr Leu
195 200 205
Arg Cys Ser Leu Val Ser Ala Gln Val Glu Ala Leu Ile Thr Ile Gln
210 215 220
Val Ala Gln Met Val Phe Gly Phe Leu Val Pro Met Leu Ala Met Ser
225 230 235 240
Phe Cys Tyr Leu Ile Ile Ile Arg Thr Leu Leu Gln Ala Arg Asn Phe
245 250 255
Glu Arg Asn Lys Ala Ile Lys Val Ile Ile Ala Val Val Val Val Phe
260 265 270
Ile Val Phe Gln Leu Pro Tyr Asn Gly Val Val Leu Ala Gln Thr Val
275 280 285
Ala Asn Phe Asn Ile Thr Asn Ser Ser Cys Glu Thr Ser Lys Gln Leu
290 295 300
Asn Ile Ala Tyr Asp Val Thr Tyr Ser Leu Ala Ser Val Arg Cys Cys
305 310 315 320
Val Asn Pro Phe Leu Tyr Ala Phe Ile Gly Val Lys Phe Arg Ser Asp
325 330 335
Leu Phe Lys Leu Phe Lys Asp Leu Gly Cys Leu Ser Gln Glu Arg Leu
340 345 350
Arg His Trp Ser Ser Cys Arg His Val Arg Asn Ala Ser Val Ser Met
355 360 365
Glu Ala Glu Thr Thr Thr Thr Phe Ser Pro
370 375
<210> 102
<211> 1134
<212> ДНК
<213> Mus sp.
<400> 102
atggacccag ggaaacccag gaaaaacgtg ctggtggtgg ctctccttgt cattttccag 60
gtgtgcttct gccaagatga ggtcaccgat gactacatcg gcgagaatac cacggtggac 120
tacaccctgt acgagtcggt gtgcttcaag aaggatgtgc ggaactttaa ggcctggttc 180
ctgcctctca tgtattctgt catctgcttc gtgggcctgc tcggcaacgg gctggtgata 240
ctgacgtaca tctatttcaa gaggctcaag accatgacgg atacctacct gctcaacctg 300
gccgtggcag acatcctttt cctcctgatt cttcccttct gggcctacag cgaagccaag 360
tcctggatct ttggcgtcta cctgtgtaag ggcatctttg gcatctataa gttaagcttc 420
ttcagcggga tgctgctgct cctatgcatc agcattgacc gctacgtagc catcgtccag 480
gccgtgtcgg ctcatcgcca ccgcgcccgc gtgcttctca tcagcaagct gtcctgtgtg 540
ggcatctgga tgctggccct cttcctctcc atcccggagc tgctctacag cggcctccag 600
aagaacagcg gcgaggacac gctgagatgc tcactggtca gtgcccaagt ggaggccttg 660
atcaccatcc aagtggccca gatggttttt gggttcctag tgcctatgct ggctatgagt 720
ttctgctacc tcattatcat ccgtaccttg ctccaggcac gcaactttga gcggaacaag 780
gccatcaagg tgatcattgc cgtggtggta gtcttcatag tcttccagct gccctacaat 840
ggggtggtcc tggctcagac ggtggccaac ttcaacatca ccaatagcag ctgcgaaacc 900
agcaagcagc tcaacattgc ctatgacgtc acctacagcc tggcctccgt ccgctgctgc 960
gtcaaccctt tcttgtatgc cttcatcggc gtcaagttcc gcagcgacct cttcaagctc 1020
ttcaaggact tgggctgtct cagccaggaa cggctccggc actggtcttc ctgccggcat 1080
gtacggaacg cgtcggtgag catggaggcg gagaccacca caaccttctc cccg 1134
<210> 103
<211> 378
<212> БЕЛОК
<213> Rattus sp.
<400> 103
Met Asp Leu Gly Lys Pro Thr Lys Asn Val Leu Val Val Ala Leu Leu
1. 5 10 15
Val Ile Phe Gln Val Cys Phe Cys Gln Asp Glu Val Thr Asp Asp Tyr
20 25 30
Ile Gly Glu Asn Thr Thr Val Asp Tyr Thr Leu Tyr Glu Ser Val Cys
35 40 45
Phe Lys Lys Asp Val Arg Asn Phe Lys Ala Trp Phe Leu Pro Leu Met
50 55 60
Tyr Ser Val Ile Cys Phe Val Gly Leu Leu Gly Asn Gly Leu Val Val
65 70 75 80
Leu Thr Tyr Ile Tyr Phe Lys Arg Leu Lys Thr Met Thr Asp Thr Tyr
85 90 95
Leu Leu Asn Leu Ala Val Ala Asp Ile Leu Phe Leu Met Ile Leu Pro
100 105 110
Phe Trp Ala Tyr Ser Glu Ala Lys Ser Trp Ile Phe Gly Ala Tyr Leu
115 120 125
Cys Lys Ser Ile Phe Gly Ile Tyr Lys Leu Ser Phe Phe Ser Gly Met
130 135 140
Leu Leu Leu Leu Cys Ile Ser Ile Asp Arg Tyr Val Ala Ile Val Gln
145 150 155 160
Ala Val Ser Ala His Arg His Arg Ala Arg Val Leu Leu Ile Ser Lys
165 170 175
Leu Ser Cys Ile Gly Ile Trp Thr Leu Ala Phe Phe Leu Ser Ile Pro
180 185 190
Glu Leu Leu Tyr Ser Gly Leu Gln Lys Asn Ser Gly Glu Asp Thr Trp
195 200 205
Arg Cys Ser Leu Val Ser Ala Gln Val Glu Ala Leu Ile Ala Ile Gln
210 215 220
Val Ala Gln Met Val Val Gly Phe Val Leu Pro Met Leu Ala Met Ser
225 230 235 240
Phe Cys Tyr Leu Val Ile Ile Arg Thr Leu Leu Gln Ala Arg Asn Phe
245 250 255
Glu Arg Asn Lys Ala Ile Lys Val Ile Ile Ala Val Val Val Val Phe
260 265 270
Val Val Phe Gln Leu Pro Tyr Asn Gly Val Val Leu Ala Gln Thr Val
275 280 285
Ala Asn Phe Asn Ile Thr Asn Ser Ser Cys Glu Ala Ser Lys Gln Leu
290 295 300
Asn Ile Ala Tyr Asp Val Thr Tyr Ser Leu Ala Ser Val Arg Cys Cys
305 310 315 320
Val Asn Pro Phe Leu Tyr Ala Phe Ile Gly Val Lys Phe Arg Ser Asp
325 330 335
Leu Phe Lys Leu Phe Lys Asp Leu Gly Cys Leu Ser Gln Glu Arg Leu
340 345 350
Arg Gln Trp Ser Ser Cys Arg His Val Arg His Thr Ser Val Ser Met
355 360 365
Glu Ala Glu Thr Thr Thr Thr Phe Ser Pro
370 375
<210> 104
<211> 1134
<212> ДНК
<213> Rattus sp.
<400> 104
atggacctgg ggaagcccac gaaaaacgtg ctggtggtgg ctctcctggt cattttccag 60
gtgtgcttct gccaagatga ggtcacagac gactacatcg gcgagaacac caccgtggac 120
tacaccctgt atgagtcggt gtgcttcaag aaggatgtgc ggaactttaa ggcctggttc 180
ctccctctca tgtactcagt catttgcttc gtgggcctgc taggcaatgg gctggtggtg 240
ctgacataca tctatttcaa gagactgaag accatgacgg atacctacct gctcaacctg 300
gccgtggcag acatcctctt cctcatgatc cttcccttct gggcctacag cgaagccaag 360
tcctggatct ttggtgccta cctgtgtaag agcatctttg gcatctacaa gttaagcttc 420
ttcagcggga tgttgctgct cctgtgtatc agcattgacc gctatgtggc catcgtccag 480
gccgtgtcag cccaccggca ccgcgcccgc gtgcttctca tcagcaagct gtcctgtata 540
ggcatctgga cgctggcctt tttcctttct atccctgagc tgctctacag cggcctccag 600
aagaacagcg gcgaggacac gtggagatgc tccctggtca gtgcccaagt ggaggccttg 660
atcgccatcc aagtggccca gatggttgtt gggtttgtac tgcctatgct ggctatgagt 720
ttctgctacc tggttatcat ccgcactctg ctccaggcgc gaaacttcga gcggaacaag 780
gccatcaagg tgatcatcgc tgtggtcgta gtgttcgtcg tcttccagct gccctacaat 840
ggggtggtcc tggcccagac cgtggccaat ttcaacatca ccaatagcag ctgcgaagcc 900
agcaagcagc tcaacattgc ctatgacgtc acctacagcc tggcctccgt ccgctgctgt 960
gtcaaccctt tcttgtatgc cttcatcggc gtcaagttcc gcagcgacct cttcaagctc 1020
ttcaaggact tgggctgcct cagccaggaa cggctccggc agtggtcttc ctgccgccat 1080
gtacggcaca cgtccgtgag catggaggcg gagactacca ccaccttctc cccg 1134
<210> 105
<211> 378
<212> БЕЛОК
<213> Homo sapiens
<400> 105
Met Asp Leu Gly Lys Pro Met Lys Ser Val Leu Val Val Ala Leu Leu
1. 5 10 15
Val Ile Phe Gln Val Cys Leu Cys Gln Asp Glu Val Thr Asp Asp Tyr
20 25 30
Ile Gly Asp Asn Thr Thr Val Asp Tyr Thr Leu Phe Glu Ser Leu Cys
35 40 45
Ser Lys Lys Asp Val Arg Asn Phe Lys Ala Trp Phe Leu Pro Ile Met
50 55 60
Tyr Ser Ile Ile Cys Phe Val Gly Leu Leu Gly Asn Gly Leu Val Val
65 70 75 80
Leu Thr Tyr Ile Tyr Phe Lys Arg Leu Lys Thr Met Thr Asp Thr Tyr
85 90 95
Leu Leu Asn Leu Ala Val Ala Asp Ile Leu Phe Leu Leu Thr Leu Pro
100 105 110
Phe Trp Ala Tyr Ser Ala Ala Lys Ser Trp Val Phe Gly Val His Phe
115 120 125
Cys Lys Leu Ile Phe Ala Ile Tyr Lys Met Ser Phe Phe Ser Gly Met
130 135 140
Leu Leu Leu Leu Cys Ile Ser Ile Asp Arg Tyr Val Ala Ile Val Gln
145 150 155 160
Ala Val Ser Ala His Arg His Arg Ala Arg Val Leu Leu Ile Ser Lys
165 170 175
Leu Ser Cys Val Gly Ile Trp Ile Leu Ala Thr Val Leu Ser Ile Pro
180 185 190
Glu Leu Leu Tyr Ser Asp Leu Gln Arg Ser Ser Ser Glu Gln Ala Met
195 200 205
Arg Cys Ser Leu Ile Thr Glu His Val Glu Ala Phe Ile Thr Ile Gln
210 215 220
Val Ala Gln Met Val Ile Gly Phe Leu Val Pro Leu Leu Ala Met Ser
225 230 235 240
Phe Cys Tyr Leu Val Ile Ile Arg Thr Leu Leu Gln Ala Arg Asn Phe
245 250 255
Glu Arg Asn Lys Ala Ile Lys Val Ile Ile Ala Val Val Val Val Phe
260 265 270
Ile Val Phe Gln Leu Pro Tyr Asn Gly Val Val Leu Ala Gln Thr Val
275 280 285
Ala Asn Phe Asn Ile Thr Ser Ser Thr Cys Glu Leu Ser Lys Gln Leu
290 295 300
Asn Ile Ala Tyr Asp Val Thr Tyr Ser Leu Ala Cys Val Arg Cys Cys
305 310 315 320
Val Asn Pro Phe Leu Tyr Ala Phe Ile Gly Val Lys Phe Arg Asn Asp
325 330 335
Leu Phe Lys Leu Phe Lys Asp Leu Gly Cys Leu Ser Gln Glu Gln Leu
340 345 350
Arg Gln Trp Ser Ser Cys Arg His Ile Arg Arg Ser Ser Met Ser Val
355 360 365
Glu Ala Glu Thr Thr Thr Thr Phe Ser Pro
370 375
<210> 106
<211> 30
<212> БЕЛОК
<213> Homo sapiens
<400> 106
Gln Asp Glu Val Thr Asp Asp Tyr Ile Gly Asp Asn Thr Thr Val Asp
1. 5 10 15
Tyr Thr Leu Phe Glu Ser Leu Cys Ser Lys Lys Asp Val Arg
20 25 30
<210> 107
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 107
Lys Ser Trp Val Phe Gly Val His
1. 5
<210> 108
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 108
Tyr Ser Asp Leu Gln Arg Ser Ser Ser Glu Gln Ala Met Arg Cys Ser
1. 5 10 15
Leu Ile Thr
<210> 109
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 109
Phe Asn Ile Thr Ser Ser Thr
1. 5
<210> 110
<211> 19
<212> БЕЛОК
<213> Homo sapiens
<400> 110
Tyr Ser Asp Leu Gln Arg Ser Ser Ser Glu Gln Ala Met Arg Ser Ser
1. 5 10 15
Leu Ile Thr
<210> 111
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 111
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met Leu Trp Val Arg Gln Ala Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Ser Thr Ile Tyr Tyr Ala Asp Arg Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Thr Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ser Thr Gly Thr Phe Ala Tyr Trp Gly Gln Gly Thr Pro Val Thr Val
100 105 110
Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly
115 120 125
Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys
130 135 140
Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu
145 150 155 160
Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr
165 170 175
Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Glu
180 185 190
Thr Val Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp
195 200 205
Lys Lys Ile Val Pro Arg Asp Cys
210 215
<210> 112
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 112
Asp Val Val Met Thr Gln Asn Pro Leu Ser Leu Pro Val Ser Leu Gly
1. 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Ile Tyr Asn
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Arg Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
115 120 125
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
130 135 140
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
145 150 155 160
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
180 185 190
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
195 200 205
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 113
<211> 265
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 113
Gln Asp Glu Val Thr Asp Asp Tyr Ile Gly Asp Asn Thr Thr Val Asp
1. 5 10 15
Tyr Thr Leu Phe Glu Ser Leu Cys Ser Lys Lys Asp Val Arg Glu Val
20 25 30
Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly Ser Leu
35 40 45
Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr Gly Met
50 55 60
Leu Trp Val Arg Gln Ala Pro Glu Lys Gly Leu Glu Trp Ile Ala Tyr
65 70 75 80
Ile Ser Ser Gly Ser Ser Thr Ile Tyr Tyr Ala Asp Arg Val Lys Gly
85 90 95
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe Leu Gln
100 105 110
Met Thr Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys Ser Thr
115 120 125
Gly Thr Tyr Ser Asp Leu Gln Arg Ser Ser Ser Glu Gln Ala Met Arg
130 135 140
Ser Ser Leu Ile Thr Phe Ala Tyr Trp Gly Gln Gly Thr Pro Val Thr
145 150 155 160
Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro
165 170 175
Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val
180 185 190
Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser
195 200 205
Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu
210 215 220
Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser
225 230 235 240
Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val
245 250 255
Asp Lys Lys Ile Val Pro Arg Asp Cys
260 265
<210> 114
<211> 795
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 114
caagatgagg tcacggacga ttacatcgga gacaacacca cagtggacta cactttgttc 60
gagtctttgt gctccaagaa ggacgtgcgg gaggtgcagc tggtggagtc tggtggtggt 120
ctggtcaagc ctggaggttc cctgaaactg agttgtgccg catctgggtt tacattctct 180
gactacggaa tgctgtgggt gaggcaggca ccagagaagg gcctggaatg gatcgcttat 240
atttccagcg gatctagtac tatctactat gcagacaggg tcaagggccg gttcaccatt 300
agcagagata acgccaaaaa taccctgttt ctgcagatga catcactgag gtccgaggat 360
accgctatgt attattgctc cacagggact tacagtgacc tccagaggag cagcagtgag 420
caagcgatgc gatcctctct catcacattt gcttactggg gacaggggac acccgtgacc 480
gtcagctcag ccaagaccac cccccccagc gtgtaccctc tggcccctgg ctctgccgcc 540
cagaccaaca gcatggtgac cctgggctgc ctggtgaagg gctacttccc cgagcccgtg 600
accgtgacct ggaacagcgg cagcctgagc agcggcgtgc acaccttccc cgccgtgctg 660
cagagcgacc tgtacaccct gagcagctct gtgaccgtgc ccagcagcac ctggcccagc 720
gagaccgtga catgcaacgt ggcccacccc gccagctcca ccaaggtgga caagaaaatc 780
gtgccccggg actgc 795
<210> 115
<211> 656
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 115
atgtcgtgat gactcagaat ccactgtccc tgcctgtgtc cctgggcgat caggcttcca 60
ttagctgtcg ttcctctcag tccctgatct acaacaatgg taacacctac ctgcactggt 120
atagacagaa gcccggccag tcccctaagc tgctgatcta caaagtgagt aataggttct 180
caggagtccc agaccggttt tccggcagcg gatctgggac cgatttcaca ctgaaaatct 240
ctagggtgga ggccgaagac ctgggcgtct acttttgtag tcagagcact cacgtcccct 300
tcaccttcgg cagcggaaca aaactggaaa tcaagcgcgc tgatgccgcc cctaccgtga 360
gcatcttccc ccccagcagc gagcagctga ccagcggcgg agccagcgtg gtgtgcttcc 420
tgaacaactt ctaccccaag gacatcaacg tgaagtggaa gatcgacggc agcgagcggc 480
agaacggcgt gctgaacagc tggaccgacc aggacagcaa ggactccacc tacagcatga 540
gcagcaccct gaccctgacc aaggacgagt acgagcggca caacagctac acctgcgagg 600
ccacccacaa gaccagcacc agccccatcg tgaagagctt caaccggaac gagtgc 656
<210> 116
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 116
Gly Tyr Ser Ile Thr Ser Gly Tyr Ser Trp His
1. 5 10
<210> 117
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 117
His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1. 5 10 15
<210> 118
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 118
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 119
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 119
Ser Gly Tyr Ser Trp His
1. 5
<210> 120
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 120
His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1. 5 10 15
<210> 121
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 121
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 122
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 122
Gly Tyr Ser Ile Thr Ser Gly Tyr
1. 5
<210> 123
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 123
His Ser Ser Gly Ser
1. 5
<210> 124
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 124
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 125
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 125
Gly Tyr Ser Ile Thr Ser Gly Tyr Ser
1. 5
<210> 126
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 126
Ile His Ser Ser Gly Ser Thr
1. 5
<210> 127
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 127
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr
1. 5 10
<210> 128
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 128
Asp Val Gln Leu Gln Glu Ser Gly Pro Asp Leu Val Lys Pro Ser Gln
1. 5 10 15
Ser Leu Ser Leu Thr Cys Thr Val Thr Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp His Trp Ile Arg Gln Phe Pro Gly Asn Lys Leu Glu Trp
35 40 45
Met Ala His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Ser Ile Ile Arg Asp Thr Ser Lys Asn Leu Phe Phe
65 70 75 80
Leu Gln Leu Asn Ser Val Thr Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 129
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 129
gatgtgcagc ttcaggagtc aggacctgac ctggtgaaac cttctcagtc actttcactc 60
acctgcactg tcactggcta ctccatcacc agtggttata gctggcactg gatccggcag 120
tttccaggaa acaaactgga gtggatggcc cacatccact ccagtggtag cactaactac 180
aacccatctc tcaaaagtcg catctctatc attcgagaca catccaagaa cctgttcttc 240
ctgcagttga attctgtgac tactgaggac acagccacat attactgtgc aagagggggg 300
gtacaggcct ttgcttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 130
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 130
Asp Val Gln Leu Gln Glu Ser Gly Pro Asp Leu Val Lys Pro Ser Gln
1. 5 10 15
Ser Leu Ser Leu Thr Cys Thr Val Thr Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp His Trp Ile Arg Gln Phe Pro Gly Asn Lys Leu Glu Trp
35 40 45
Met Ala His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Ser Ile Ile Arg Asp Thr Ser Lys Asn Leu Phe Phe
65 70 75 80
Leu Gln Leu Asn Ser Val Thr Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 131
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 131
gatgtgcagc ttcaggagtc aggacctgac ctggtgaaac cttctcagtc actttcactc 60
acctgcactg tcactggcta ctccatcacc agtggttata gctggcactg gatccggcag 120
tttccaggaa acaaactgga gtggatggcc cacatccact ccagtggtag cactaactac 180
aacccatctc tcaaaagtcg catctctatc attcgagaca catccaagaa cctgttcttc 240
ctgcagttga attctgtgac tactgaggac acagccacat attactgtgc aagagggggg 300
gtacaggcct ttgcttactg gggccaaggg actctggtca ctgtctctgc agctagcacc 360
aagggcccaa gtgtgtttcc cctggccccc agcagcaagt ctacttccgg cggaactgct 420
gccctgggtt gcctggtgaa ggactacttc cccgagcccg tgacagtgtc ctggaactct 480
ggggctctga cttccggcgt gcacaccttc cccgccgtgc tgcagagcag cggcctgtac 540
agcctgagca gcgtggtgac agtgccctcc agctctctgg gaacccagac ctatatctgc 600
aacgtgaacc acaagcccag caacaccaag gtggacaaga gagtggagcc caagagctgc 660
gacaagaccc acacctgccc cccctgccca gctccagaac tgctgggagg gccttccgtg 720
ttcctgttcc cccccaagcc caaggacacc ctgatgatca gcaggacccc cgaggtgacc 780
tgcgtggtgg tggacgtgtc ccacgaggac ccagaggtga agttcaactg gtacgtggac 840
ggcgtggagg tgcacaacgc caagaccaag cccagagagg agcagtacaa cagcacctac 900
agggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agaatacaag 960
tgcaaagtct ccaacaaggc cctgccagcc ccaatcgaaa agacaatcag caaggccaag 1020
ggccagccac gggagcccca ggtgtacacc ctgcccccca gccgggagga gatgaccaag 1080
aaccaggtgt ccctgacctg tctggtgaag ggcttctacc ccagcgatat cgccgtggag 1140
tgggagagca acggccagcc cgagaacaac tacaagacca cccccccagt gctggacagc 1200
gacggcagct tcttcctgta cagcaagctg accgtggaca agtccaggtg gcagcagggc 1260
aacgtgttca gctgcagcgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgagcctga gccccggcaa g 1341
<210> 132
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 132
Ser Ala Ser Ser Ser Val Ile Tyr Met His
1. 5 10
<210> 133
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 133
Asp Thr Ser Lys Leu Ala Ser
1. 5
<210> 134
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 134
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 135
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 135
Ser Ala Ser Ser Ser Val Ile Tyr Met His
1. 5 10
<210> 136
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 136
Asp Thr Ser Lys Leu Ala Ser
1. 5
<210> 137
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 137
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 138
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 138
Ser Ser Ser Val Ile Tyr
1. 5
<210> 139
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 139
Asp Thr Ser
1
<210> 140
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 140
Trp Ser Ser Asn Pro Leu
1. 5
<210> 141
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 141
Ser Ser Val Ile Tyr
1. 5
<210> 142
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 142
Asp Thr Ser
1
<210> 143
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 143
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 144
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 144
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1. 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ile Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Thr Leu Glu Leu Lys
100 105
<210> 145
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 145
caaattgtcc tcacccagtc tccagcaatc atgtctgcat ctccagggga gaaggtcacc 60
atgacctgca gtgccagttc aagtgtaatt tacatgcact ggtaccagca gaagtcaggc 120
acctccccca aaagatggat ttatgacaca tccaaactgg cttctggagt ccctgctcgc 180
ttcagtggta gtgggtctgg gacctcttac tctctcacaa tcagcagcat ggaggctgaa 240
gatgctgcca cttattactg ccagcagtgg agtagtaacc cgctcacgtt cggtgctggg 300
accacgttgg agctgaaa 318
<210> 146
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 146
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1. 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ile Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Thr Leu Glu Leu Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 147
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 147
caaattgtcc tcacccagtc tccagcaatc atgtctgcat ctccagggga gaaggtcacc 60
atgacctgca gtgccagttc aagtgtaatt tacatgcact ggtaccagca gaagtcaggc 120
acctccccca aaagatggat ttatgacaca tccaaactgg cttctggagt ccctgctcgc 180
ttcagtggta gtgggtctgg gacctcttac tctctcacaa tcagcagcat ggaggctgaa 240
gatgctgcca cttattactg ccagcagtgg agtagtaacc cgctcacgtt cggtgctggg 300
accacgttgg agctgaaacg tacggtggcc gctcccagcg tgttcatctt cccccccagc 360
gacgagcagc tgaagagtgg caccgccagc gtggtgtgcc tgctgaacaa cttctacccc 420
cgggaggcca aggtgcagtg gaaggtggac aacgccctgc agagcggcaa cagccaggag 480
agcgtcaccg agcaggacag caaggactcc acctacagcc tgagcagcac cctgaccctg 540
agcaaggccg actacgagaa gcataaggtg tacgcctgcg aggtgaccca ccagggcctg 600
tccagccccg tgaccaagag cttcaacagg ggcgagtgc 639
<210> 148
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 148
Gly Phe Thr Phe Ser Thr Tyr Ala Met Ser
1. 5 10
<210> 149
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 149
Thr Ile Ser Asp Gly Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val Lys
1. 5 10 15
Gly
<210> 150
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 150
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 151
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 151
Thr Tyr Ala Met Ser
1. 5
<210> 152
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 152
Thr Ile Ser Asp Gly Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val Lys
1. 5 10 15
Gly
<210> 153
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 153
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 154
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 154
Gly Phe Thr Phe Ser Thr Tyr
1. 5
<210> 155
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 155
Ser Asp Gly Gly Ser Tyr
1. 5
<210> 156
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 156
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 157
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 157
Gly Phe Thr Phe Ser Thr Tyr Ala
1. 5
<210> 158
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 158
Ile Ser Asp Gly Gly Ser Tyr Ser
1. 5
<210> 159
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 159
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10 15
<210> 160
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 160
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asp Gly Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Asn Leu Tyr
65 70 75 80
Leu Gln Met Ser His Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 161
<211> 366
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 161
gaagtgcagc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt acctatgcca tgtcttgggt tcgccagact 120
ccggaaaaga ggctggagtg ggtcgcaacc attagtgatg gtggtagtta ttcgtactat 180
ccagacaatg taaagggccg attcaccatc tccagagaca atgccaagaa caacctatac 240
ctgcaaatga gccatctgaa gtctgaggac acagccatgt attactgtgc aagacgaggt 300
agtaggtacg aagagtacta tgttatggac tactggggtc aaggaacctc agtcaccgtc 360
tcctca 366
<210> 162
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 162
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asp Gly Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Asn Leu Tyr
65 70 75 80
Leu Gln Met Ser His Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 163
<211> 1356
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 163
gaagtgcagc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt acctatgcca tgtcttgggt tcgccagact 120
ccggaaaaga ggctggagtg ggtcgcaacc attagtgatg gtggtagtta ttcgtactat 180
ccagacaatg taaagggccg attcaccatc tccagagaca atgccaagaa caacctatac 240
ctgcaaatga gccatctgaa gtctgaggac acagccatgt attactgtgc aagacgaggt 300
agtaggtacg aagagtacta tgttatggac tactggggtc aaggaacctc agtcaccgtc 360
tcctcagcta gcaccaaggg cccaagtgtg tttcccctgg cccccagcag caagtctact 420
tccggcggaa ctgctgccct gggttgcctg gtgaaggact acttccccga gcccgtgaca 480
gtgtcctgga actctggggc tctgacttcc ggcgtgcaca ccttccccgc cgtgctgcag 540
agcagcggcc tgtacagcct gagcagcgtg gtgacagtgc cctccagctc tctgggaacc 600
cagacctata tctgcaacgt gaaccacaag cccagcaaca ccaaggtgga caagagagtg 660
gagcccaaga gctgcgacaa gacccacacc tgccccccct gcccagctcc agaactgctg 720
ggagggcctt ccgtgttcct gttccccccc aagcccaagg acaccctgat gatcagcagg 780
acccccgagg tgacctgcgt ggtggtggac gtgtcccacg aggacccaga ggtgaagttc 840
aactggtacg tggacggcgt ggaggtgcac aacgccaaga ccaagcccag agaggagcag 900
tacaacagca cctacagggt ggtgtccgtg ctgaccgtgc tgcaccagga ctggctgaac 960
ggcaaagaat acaagtgcaa agtctccaac aaggccctgc cagccccaat cgaaaagaca 1020
atcagcaagg ccaagggcca gccacgggag ccccaggtgt acaccctgcc ccccagccgg 1080
gaggagatga ccaagaacca ggtgtccctg acctgtctgg tgaagggctt ctaccccagc 1140
gatatcgccg tggagtggga gagcaacggc cagcccgaga acaactacaa gaccaccccc 1200
ccagtgctgg acagcgacgg cagcttcttc ctgtacagca agctgaccgt ggacaagtcc 1260
aggtggcagc agggcaacgt gttcagctgc agcgtgatgc acgaggccct gcacaaccac 1320
tacacccaga agtccctgag cctgagcccc ggcaag 1356
<210> 164
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 164
Arg Ala Ser Gln Ser Ile Ser Asn Asn Leu His
1. 5 10
<210> 165
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 165
Tyr Ala Ser Gln Ser Ile Ser
1. 5
<210> 166
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 166
Gln Gln Ser Asn Ser Trp Leu Thr
1. 5
<210> 167
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 167
Arg Ala Ser Gln Ser Ile Ser Asn Asn Leu His
1. 5 10
<210> 168
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 168
Tyr Ala Ser Gln Ser Ile Ser
1. 5
<210> 169
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 169
Gln Gln Ser Asn Ser Trp Leu Thr
1. 5
<210> 170
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 170
Ser Gln Ser Ile Ser Asn Asn
1. 5
<210> 171
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 171
Tyr Ala Ser
1
<210> 172
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 172
Ser Asn Ser Trp Leu
1. 5
<210> 173
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 173
Gln Ser Ile Ser Asn Asn
1. 5
<210> 174
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 174
Tyr Ala Ser
1
<210> 175
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 175
Gln Gln Ser Asn Ser Trp Leu Thr
1. 5
<210> 176
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 176
Asp Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Thr Pro Gly
1. 5 10 15
Asp Ser Val Ser Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asn Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Ser His Glu Ser Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Thr
65 70 75 80
Glu Asp Phe Gly Met Tyr Phe Cys Gln Gln Ser Asn Ser Trp Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Gly Leu Lys
100 105
<210> 177
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 177
gatattgtgc taactcagtc tccagccacc ctgtctgtga ctccaggaga tagcgtcagt 60
ctttcctgca gggccagcca aagtattagc aacaacctac actggtatca acagaaatca 120
catgagtctc caaaacttct catcaagtat gcttcccagt ccatctctgg gatcccctcc 180
aggttcagtg gcagtggatc agggacagat ttcactctca gtatcaacag tgtggagact 240
gaagattttg gaatgtattt ctgtcaacag agtaacagct ggctcacgtt cggtgctggg 300
accaagctgg ggctgaaa 318
<210> 178
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 178
Asp Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Thr Pro Gly
1. 5 10 15
Asp Ser Val Ser Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asn Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Ser His Glu Ser Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Thr
65 70 75 80
Glu Asp Phe Gly Met Tyr Phe Cys Gln Gln Ser Asn Ser Trp Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Gly Leu Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 179
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 179
gatattgtgc taactcagtc tccagccacc ctgtctgtga ctccaggaga tagcgtcagt 60
ctttcctgca gggccagcca aagtattagc aacaacctac actggtatca acagaaatca 120
catgagtctc caaaacttct catcaagtat gcttcccagt ccatctctgg gatcccctcc 180
aggttcagtg gcagtggatc agggacagat ttcactctca gtatcaacag tgtggagact 240
gaagattttg gaatgtattt ctgtcaacag agtaacagct ggctcacgtt cggtgctggg 300
accaagctgg ggctgaaacg tacggtggcc gctcccagcg tgttcatctt cccccccagc 360
gacgagcagc tgaagagtgg caccgccagc gtggtgtgcc tgctgaacaa cttctacccc 420
cgggaggcca aggtgcagtg gaaggtggac aacgccctgc agagcggcaa cagccaggag 480
agcgtcaccg agcaggacag caaggactcc acctacagcc tgagcagcac cctgaccctg 540
agcaaggccg actacgagaa gcataaggtg tacgcctgcg aggtgaccca ccagggcctg 600
tccagccccg tgaccaagag cttcaacagg ggcgagtgc 639
<210> 180
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 180
Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
1. 5 10
<210> 181
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 181
Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 182
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 182
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 183
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 183
Ser Tyr Ala Met Ser
1. 5
<210> 184
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 184
Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 185
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 185
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 186
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 186
Gly Phe Thr Phe Ser Ser Tyr
1. 5
<210> 187
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 187
Ser Ser Gly Gly Ser Phe
1. 5
<210> 188
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 188
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 189
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 189
Gly Phe Thr Phe Ser Ser Tyr Ala
1. 5
<210> 190
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 190
Ile Ser Ser Gly Gly Ser Phe Thr
1. 5
<210> 191
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 191
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 192
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 192
Glu Val Met Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Ile Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val Trp Gly
100 105 110
Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 193
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 193
gaagtgatgc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt agctatgcca tgtcttggat tcgccagact 120
ccggagaaga gactggagtg ggtcgcaacc atcagtagtg gtggtagttt cacctactat 180
ccagacagtg tgaaggggcg attcaccatt tctagagaca atgtcaagaa caccctgtac 240
ctgcaaatga gcagtctgag gtctgaagac acggccatgt attactgtgc aagacgggct 300
tctacggtag taggtacgga cttcgatgtc tggggcgcag ggaccacggt caccgtctcc 360
tca 363
<210> 194
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 194
Glu Val Met Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Ile Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val Trp Gly
100 105 110
Ala Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 195
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 195
gaagtgatgc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt agctatgcca tgtcttggat tcgccagact 120
ccggagaaga gactggagtg ggtcgcaacc atcagtagtg gtggtagttt cacctactat 180
ccagacagtg tgaaggggcg attcaccatt tctagagaca atgtcaagaa caccctgtac 240
ctgcaaatga gcagtctgag gtctgaagac acggccatgt attactgtgc aagacgggct 300
tctacggtag taggtacgga cttcgatgtc tggggcgcag ggaccacggt caccgtctcc 360
tcagctagca ccaagggccc aagtgtgttt cccctggccc ccagcagcaa gtctacttcc 420
ggcggaactg ctgccctggg ttgcctggtg aaggactact tccccgagcc cgtgacagtg 480
tcctggaact ctggggctct gacttccggc gtgcacacct tccccgccgt gctgcagagc 540
agcggcctgt acagcctgag cagcgtggtg acagtgccct ccagctctct gggaacccag 600
acctatatct gcaacgtgaa ccacaagccc agcaacacca aggtggacaa gagagtggag 660
cccaagagct gcgacaagac ccacacctgc cccccctgcc cagctccaga actgctggga 720
gggccttccg tgttcctgtt cccccccaag cccaaggaca ccctgatgat cagcaggacc 780
cccgaggtga cctgcgtggt ggtggacgtg tcccacgagg acccagaggt gaagttcaac 840
tggtacgtgg acggcgtgga ggtgcacaac gccaagacca agcccagaga ggagcagtac 900
aacagcacct acagggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 960
aaagaataca agtgcaaagt ctccaacaag gccctgccag ccccaatcga aaagacaatc 1020
agcaaggcca agggccagcc acgggagccc caggtgtaca ccctgccccc cagccgggag 1080
gagatgacca agaaccaggt gtccctgacc tgtctggtga agggcttcta ccccagcgat 1140
atcgccgtgg agtgggagag caacggccag cccgagaaca actacaagac caccccccca 1200
gtgctggaca gcgacggcag cttcttcctg tacagcaagc tgaccgtgga caagtccagg 1260
tggcagcagg gcaacgtgtt cagctgcagc gtgatgcacg aggccctgca caaccactac 1320
acccagaagt ccctgagcct gagccccggc aag 1353
<210> 196
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 196
Arg Ala Ser Gln Asp Ile Gly Ser Ser Leu Asn
1. 5 10
<210> 197
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 197
Ala Thr Ser Ser Leu Asp Ser
1. 5
<210> 198
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 198
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 199
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 199
Arg Ala Ser Gln Asp Ile Gly Ser Ser Leu Asn
1. 5 10
<210> 200
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 200
Ala Thr Ser Ser Leu Asp Ser
1. 5
<210> 201
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 201
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 202
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 202
Ser Gln Asp Ile Gly Ser Ser
1. 5
<210> 203
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 203
Ala Thr Ser
1
<210> 204
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 204
Tyr Ala Ser Ser Pro Pro
1. 5
<210> 205
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 205
Gln Asp Ile Gly Ser Ser
1. 5
<210> 206
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 206
Ala Thr Ser
1
<210> 207
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 207
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 208
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 208
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1. 5 10 15
Glu Arg Val Ser Leu Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Asn Trp Leu Gln Gln Glu Pro Asp Gly Thr Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Lys Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Ser Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Val Val Tyr Tyr Cys Leu Gln Tyr Ala Ser Ser Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 209
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 209
gacatccaga tgacccagtc tccatcctcc ttatctgcct ctctgggaga aagagtcagt 60
ctcacttgtc gggcaagtca ggacattggt agtagcttaa actggcttca gcaggaacca 120
gatggaacta ttaaacgcct gatctatgcc acatccagtt tagattctgg tgtccccaaa 180
aggttcagtg gcagtaggtc tgggtcagat tattctctca ccatcagcag ccttgagtct 240
gaagattttg tagtctatta ctgtctacaa tatgctagtt cgcctccgac gttcggtgga 300
ggcaccaagc tggaaatcaa a 321
<210> 210
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 210
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1. 5 10 15
Glu Arg Val Ser Leu Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Asn Trp Leu Gln Gln Glu Pro Asp Gly Thr Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Lys Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Ser Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Val Val Tyr Tyr Cys Leu Gln Tyr Ala Ser Ser Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 211
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 211
gacatccaga tgacccagtc tccatcctcc ttatctgcct ctctgggaga aagagtcagt 60
ctcacttgtc gggcaagtca ggacattggt agtagcttaa actggcttca gcaggaacca 120
gatggaacta ttaaacgcct gatctatgcc acatccagtt tagattctgg tgtccccaaa 180
aggttcagtg gcagtaggtc tgggtcagat tattctctca ccatcagcag ccttgagtct 240
gaagattttg tagtctatta ctgtctacaa tatgctagtt cgcctccgac gttcggtgga 300
ggcaccaagc tggaaatcaa acgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 212
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 212
Gly Phe Thr Phe Ser Asn Phe Ala Met Ser
1. 5 10
<210> 213
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 213
Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 214
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 214
Arg Gly Tyr Asp Gly Val Asp Lys
1. 5
<210> 215
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 215
Ser Asn Phe Ala Met Ser
1. 5
<210> 216
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 216
Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 217
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 217
Arg Gly Tyr Asp Gly Val Asp Lys
1. 5
<210> 218
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 218
Gly Phe Thr Phe Ser Asn Phe
1. 5
<210> 219
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 219
Ser Thr Gly Gly Thr Tyr
1. 5
<210> 220
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 220
Arg Gly Tyr Asp Gly Val Asp Lys
1. 5
<210> 221
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 221
Gly Phe Thr Phe Ser Asn Phe Ala
1. 5
<210> 222
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 222
Ile Ser Thr Gly Gly Thr Tyr Thr
1. 5
<210> 223
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 223
Thr Arg Arg Gly Tyr Asp Gly Val Asp Lys
1. 5 10
<210> 224
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 224
Glu Val His Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Lys Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Thr Arg Arg Gly Tyr Asp Gly Val Asp Lys Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 225
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 225
gaagtgcatc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt aactttgcca tgtcttgggt tcgccagact 120
ccggagaaga gactggagtg ggtcgcaacc attagtactg gtggtactta cacctactat 180
ccagacagtg tgaagggtcg attcaccatc tccagagaca atgccaagaa aaccctgtac 240
ctgcaaatga gcagtctgag gtctgaggac acggccatgt attactgtac aagacggggg 300
tacgacggcg tggacaaatg gggccaaggc accactctca cagtctcctc a 351
<210> 226
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 226
Glu Val His Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Lys Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Thr Arg Arg Gly Tyr Asp Gly Val Asp Lys Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 227
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 227
gaagtgcatc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt aactttgcca tgtcttgggt tcgccagact 120
ccggagaaga gactggagtg ggtcgcaacc attagtactg gtggtactta cacctactat 180
ccagacagtg tgaagggtcg attcaccatc tccagagaca atgccaagaa aaccctgtac 240
ctgcaaatga gcagtctgag gtctgaggac acggccatgt attactgtac aagacggggg 300
tacgacggcg tggacaaatg gggccaaggc accactctca cagtctcctc agctagcacc 360
aagggcccaa gtgtgtttcc cctggccccc agcagcaagt ctacttccgg cggaactgct 420
gccctgggtt gcctggtgaa ggactacttc cccgagcccg tgacagtgtc ctggaactct 480
ggggctctga cttccggcgt gcacaccttc cccgccgtgc tgcagagcag cggcctgtac 540
agcctgagca gcgtggtgac agtgccctcc agctctctgg gaacccagac ctatatctgc 600
aacgtgaacc acaagcccag caacaccaag gtggacaaga gagtggagcc caagagctgc 660
gacaagaccc acacctgccc cccctgccca gctccagaac tgctgggagg gccttccgtg 720
ttcctgttcc cccccaagcc caaggacacc ctgatgatca gcaggacccc cgaggtgacc 780
tgcgtggtgg tggacgtgtc ccacgaggac ccagaggtga agttcaactg gtacgtggac 840
ggcgtggagg tgcacaacgc caagaccaag cccagagagg agcagtacaa cagcacctac 900
agggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agaatacaag 960
tgcaaagtct ccaacaaggc cctgccagcc ccaatcgaaa agacaatcag caaggccaag 1020
ggccagccac gggagcccca ggtgtacacc ctgcccccca gccgggagga gatgaccaag 1080
aaccaggtgt ccctgacctg tctggtgaag ggcttctacc ccagcgatat cgccgtggag 1140
tgggagagca acggccagcc cgagaacaac tacaagacca cccccccagt gctggacagc 1200
gacggcagct tcttcctgta cagcaagctg accgtggaca agtccaggtg gcagcagggc 1260
aacgtgttca gctgcagcgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgagcctga gccccggcaa g 1341
<210> 228
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 228
Lys Ser Gly Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn
1. 5 10 15
<210> 229
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 229
Leu Val Ser Lys Leu Asp Ser
1. 5
<210> 230
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 230
Trp Gln Gly Thr His Phe Pro Gln Thr
1. 5
<210> 231
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 231
Lys Ser Gly Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn
1. 5 10 15
<210> 232
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 232
Leu Val Ser Lys Leu Asp Ser
1. 5
<210> 233
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 233
Trp Gln Gly Thr His Phe Pro Gln Thr
1. 5
<210> 234
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 234
Gly Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
1. 5 10
<210> 235
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 235
Leu Val Ser
1
<210> 236
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 236
Gly Thr His Phe Pro Gln
1. 5
<210> 237
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 237
Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
1. 5 10
<210> 238
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 238
Leu Val Ser
1
<210> 239
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 239
Trp Gln Gly Thr His Phe Pro Gln Thr
1. 5
<210> 240
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 240
Asp Val Val Met Thr Gln Thr Pro Leu Thr Leu Ser Val Thr Ile Gly
1. 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Gly Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 241
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 241
gatgttgtga tgacccagac tccactcact ttgtcggtta ccattggaca accagcctcc 60
atctcttgca agtcaggtca gagcctctta gatagtgatg gaaagacata tttgaattgg 120
tttttacaga ggccaggcca gtctccaaag cgcctaatct atctggtgtc taaactggac 180
tctggagtcc ctgacaggtt cactggcagt ggatcaggga cagatttcac actgaaaatc 240
agcagagtgg aggctgagga tttgggagtt tattattgct ggcaaggtac acattttcct 300
cagacgttcg gtggaggcac caagctggaa atcaaa 336
<210> 242
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 242
Asp Val Val Met Thr Gln Thr Pro Leu Thr Leu Ser Val Thr Ile Gly
1. 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Gly Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 243
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 243
gatgttgtga tgacccagac tccactcact ttgtcggtta ccattggaca accagcctcc 60
atctcttgca agtcaggtca gagcctctta gatagtgatg gaaagacata tttgaattgg 120
tttttacaga ggccaggcca gtctccaaag cgcctaatct atctggtgtc taaactggac 180
tctggagtcc ctgacaggtt cactggcagt ggatcaggga cagatttcac actgaaaatc 240
agcagagtgg aggctgagga tttgggagtt tattattgct ggcaaggtac acattttcct 300
cagacgttcg gtggaggcac caagctggaa atcaaacgta cggtggccgc tcccagcgtg 360
ttcatcttcc cccccagcga cgagcagctg aagagtggca ccgccagcgt ggtgtgcctg 420
ctgaacaact tctacccccg ggaggccaag gtgcagtgga aggtggacaa cgccctgcag 480
agcggcaaca gccaggagag cgtcaccgag caggacagca aggactccac ctacagcctg 540
agcagcaccc tgaccctgag caaggccgac tacgagaagc ataaggtgta cgcctgcgag 600
gtgacccacc agggcctgtc cagccccgtg accaagagct tcaacagggg cgagtgc 657
<210> 244
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 244
Gly Tyr Ser Ile Thr Ser Gly Tyr Ser Trp His
1. 5 10
<210> 245
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 245
His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1. 5 10 15
<210> 246
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 246
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 247
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 247
Ser Gly Tyr Ser Trp His
1. 5
<210> 248
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 248
His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1. 5 10 15
<210> 249
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 249
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 250
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 250
Gly Tyr Ser Ile Thr Ser Gly Tyr
1. 5
<210> 251
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 251
His Ser Ser Gly Ser
1. 5
<210> 252
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 252
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 253
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 253
Gly Tyr Ser Ile Thr Ser Gly Tyr Ser
1. 5
<210> 254
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 254
Ile His Ser Ser Gly Ser Thr
1. 5
<210> 255
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 255
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr
1. 5 10
<210> 256
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 256
Asp Val Gln Leu Gln Glu Ser Gly Pro Asp Leu Val Lys Pro Ser Gln
1. 5 10 15
Ser Leu Ser Leu Thr Cys Thr Val Thr Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp His Trp Ile Arg Gln Phe Pro Gly Asn Lys Leu Glu Trp
35 40 45
Met Ala His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Ser Ile Ile Arg Asp Thr Ser Lys Asn Leu Phe Phe
65 70 75 80
Leu Gln Leu Asn Ser Val Thr Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 257
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 257
gatgtgcagc ttcaggagtc aggacctgac ctggtgaaac cttctcagtc actttcactc 60
acctgcactg tcactggcta ctccatcacc agtggttata gctggcactg gatccggcag 120
tttccaggaa acaaactgga gtggatggcc cacatccact ccagtggtag cactaactac 180
aacccatctc tcaaaagtcg catctctatc attcgagaca catccaagaa cctgttcttc 240
ctgcagttga attctgtgac tactgaggac acagccacat attactgtgc aagagggggg 300
gtacaggcct ttgcttactg gggccaaggg actctggtca ctgtctctgc a 351
<210> 258
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 258
Asp Val Gln Leu Gln Glu Ser Gly Pro Asp Leu Val Lys Pro Ser Gln
1. 5 10 15
Ser Leu Ser Leu Thr Cys Thr Val Thr Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp His Trp Ile Arg Gln Phe Pro Gly Asn Lys Leu Glu Trp
35 40 45
Met Ala His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Ser Ile Ile Arg Asp Thr Ser Lys Asn Leu Phe Phe
65 70 75 80
Leu Gln Leu Asn Ser Val Thr Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 259
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 259
gatgtgcagc ttcaggagtc aggacctgac ctggtgaaac cttctcagtc actttcactc 60
acctgcactg tcactggcta ctccatcacc agtggttata gctggcactg gatccggcag 120
tttccaggaa acaaactgga gtggatggcc cacatccact ccagtggtag cactaactac 180
aacccatctc tcaaaagtcg catctctatc attcgagaca catccaagaa cctgttcttc 240
ctgcagttga attctgtgac tactgaggac acagccacat attactgtgc aagagggggg 300
gtacaggcct ttgcttactg gggccaaggg actctggtca ctgtctctgc agctagcacc 360
aagggcccaa gtgtgtttcc cctggccccc agcagcaagt ctacttccgg cggaactgct 420
gccctgggtt gcctggtgaa ggactacttc ccctgtcccg tgacagtgtc ctggaactct 480
ggggctctga cttccggcgt gcacaccttc cccgccgtgc tgcagagcag cggcctgtac 540
agcctgagca gcgtggtgac agtgccctcc agctctctgg gaacccagac ctatatctgc 600
aacgtgaacc acaagcccag caacaccaag gtggacaaga gagtggagcc caagagctgc 660
gacaagaccc acacctgccc cccctgccca gctccagaac tgctgggagg gccttccgtg 720
ttcctgttcc cccccaagcc caaggacacc ctgatgatca gcaggacccc cgaggtgacc 780
tgcgtggtgg tggacgtgtc ccacgaggac ccagaggtga agttcaactg gtacgtggac 840
ggcgtggagg tgcacaacgc caagaccaag cccagagagg agcagtacaa cagcacctac 900
agggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agaatacaag 960
tgcaaagtct ccaacaaggc cctgccagcc ccaatcgaaa agacaatcag caaggccaag 1020
ggccagccac gggagcccca ggtgtacacc ctgcccccca gccgggagga gatgaccaag 1080
aaccaggtgt ccctgacctg tctggtgaag ggcttctacc cctgtgatat cgccgtggag 1140
tgggagagca acggccagcc cgagaacaac tacaagacca cccccccagt gctggacagc 1200
gacggcagct tcttcctgta cagcaagctg accgtggaca agtccaggtg gcagcagggc 1260
aacgtgttca gctgcagcgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgagcctga gccccggcaa g 1341
<210> 260
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 260
Ser Ala Ser Ser Ser Val Ile Tyr Met His
1. 5 10
<210> 261
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 261
Asp Thr Ser Lys Leu Ala Ser
1. 5
<210> 262
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 262
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 263
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 263
Ser Ala Ser Ser Ser Val Ile Tyr Met His
1. 5 10
<210> 264
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 264
Asp Thr Ser Lys Leu Ala Ser
1. 5
<210> 265
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 265
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 266
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 266
Ser Ser Ser Val Ile Tyr
1. 5
<210> 267
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 267
Asp Thr Ser
1
<210> 268
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 268
Trp Ser Ser Asn Pro Leu
1. 5
<210> 269
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 269
Ser Ser Val Ile Tyr
1. 5
<210> 270
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 270
Asp Thr Ser
1
<210> 271
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 271
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 272
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 272
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1. 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ile Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Thr Leu Glu Leu Lys
100 105
<210> 273
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 273
caaattgtcc tcacccagtc tccagcaatc atgtctgcat ctccagggga gaaggtcacc 60
atgacctgca gtgccagttc aagtgtaatt tacatgcact ggtaccagca gaagtcaggc 120
acctccccca aaagatggat ttatgacaca tccaaactgg cttctggagt ccctgctcgc 180
ttcagtggta gtgggtctgg gacctcttac tctctcacaa tcagcagcat ggaggctgaa 240
gatgctgcca cttattactg ccagcagtgg agtagtaacc cgctcacgtt cggtgctggg 300
accacgttgg agctgaaa 318
<210> 274
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 274
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1. 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ile Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Thr Leu Glu Leu Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 275
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 275
caaattgtcc tcacccagtc tccagcaatc atgtctgcat ctccagggga gaaggtcacc 60
atgacctgca gtgccagttc aagtgtaatt tacatgcact ggtaccagca gaagtcaggc 120
acctccccca aaagatggat ttatgacaca tccaaactgg cttctggagt ccctgctcgc 180
ttcagtggta gtgggtctgg gacctcttac tctctcacaa tcagcagcat ggaggctgaa 240
gatgctgcca cttattactg ccagcagtgg agtagtaacc cgctcacgtt cggtgctggg 300
accacgttgg agctgaaacg tacggtggcc gctcccagcg tgttcatctt cccccccagc 360
gacgagcagc tgaagagtgg caccgccagc gtggtgtgcc tgctgaacaa cttctacccc 420
cgggaggcca aggtgcagtg gaaggtggac aacgccctgc agagcggcaa cagccaggag 480
agcgtcaccg agcaggacag caaggactcc acctacagcc tgagcagcac cctgaccctg 540
agcaaggccg actacgagaa gcataaggtg tacgcctgcg aggtgaccca ccagggcctg 600
tccagccccg tgaccaagag cttcaacagg ggcgagtgc 639
<210> 276
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 276
Gly Tyr Ser Ile Thr Ser Gly Tyr Ser Trp His
1. 5 10
<210> 277
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 277
His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1. 5 10 15
<210> 278
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 278
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 279
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 279
Ser Gly Tyr Ser Trp His
1. 5
<210> 280
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 280
His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1. 5 10 15
<210> 281
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 281
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 282
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 282
Gly Tyr Ser Ile Thr Ser Gly Tyr
1. 5
<210> 283
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 283
His Ser Ser Gly Ser
1. 5
<210> 284
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 284
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 285
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 285
Gly Tyr Ser Ile Thr Ser Gly Tyr Ser
1. 5
<210> 286
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 286
Ile His Ser Ser Gly Ser Thr
1. 5
<210> 287
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 287
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr
1. 5 10
<210> 288
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 288
Asp Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp His Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp
35 40 45
Met Ala His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 289
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 289
gacgtgcagc tgcaggaatc tggccctggc ctggtgaaac cctcccagac cctgtccctg 60
acctgcaccg tgtccggcta ctccatcacc tccggctaca gctggcactg gatccggcag 120
caccccggca agggcctgga atggatggcc cacatccact cctccggctc caccaactac 180
aaccccagcc tgaagtccag aatcaccatc agccgggaca cctccaagaa ccagttctcc 240
ctgaagctgt cctccgtgac cgccgctgac accgccgtgt actactgtgc cagaggcggc 300
gtgcaggcct tcgcttattg gggccagggc accctggtga cagtgtcctc c 351
<210> 290
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 290
Asp Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp His Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp
35 40 45
Met Ala His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Ala Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 291
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 291
gacgtgcagc tgcaggaatc tggccctggc ctggtgaaac cctcccagac cctgtccctg 60
acctgcaccg tgtccggcta ctccatcacc tccggctaca gctggcactg gatccggcag 120
caccccggca agggcctgga atggatggcc cacatccact cctccggctc caccaactac 180
aaccccagcc tgaagtccag aatcaccatc agccgggaca cctccaagaa ccagttctcc 240
ctgaagctgt cctccgtgac cgccgctgac accgccgtgt actactgtgc cagaggcggc 300
gtgcaggcct tcgcttattg gggccagggc accctggtga cagtgtcctc cgctagcacc 360
aagggcccaa gtgtgtttcc cctggccccc agcagcaagt ctacttccgg cggaactgct 420
gccctgggtt gcctggtgaa ggactacttc ccctgtcccg tgacagtgtc ctggaactct 480
ggggctctga cttccggcgt gcacaccttc cccgccgtgc tgcagagcag cggcctgtac 540
agcctgagca gcgtggtgac agtgccctcc agctctctgg gaacccagac ctatatctgc 600
aacgtgaacc acaagcccag caacaccaag gtggacaaga gagtggagcc caagagctgc 660
gacaagaccc acacctgccc cccctgccca gctccagaac tgctgggagg gccttccgtg 720
ttcctgttcc cccccaagcc caaggacacc ctgatgatca gcaggacccc cgaggtgacc 780
tgcgtggtgg tggccgtgtc ccacgaggac ccagaggtga agttcaactg gtacgtggac 840
ggcgtggagg tgcacaacgc caagaccaag cccagagagg agcagtacaa cagcacctac 900
agggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agaatacaag 960
tgcaaagtct ccaacaaggc cctggctgcc ccaatcgaaa agacaatcag caaggccaag 1020
ggccagccac gggagcccca ggtgtacacc ctgcccccca gccgggagga gatgaccaag 1080
aaccaggtgt ccctgacctg tctggtgaag ggcttctacc cctgtgatat cgccgtggag 1140
tgggagagca acggccagcc cgagaacaac tacaagacca cccccccagt gctggacagc 1200
gacggcagct tcttcctgta cagcaagctg accgtggaca agtccaggtg gcagcagggc 1260
aacgtgttca gctgcagcgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgagcctga gccccggcaa g 1341
<210> 292
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 292
Ser Ala Ser Ser Ser Val Ile Tyr Met His
1. 5 10
<210> 293
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 293
Asp Thr Ser Lys Leu Ala Ser
1. 5
<210> 294
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 294
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 295
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 295
Ser Ala Ser Ser Ser Val Ile Tyr Met His
1. 5 10
<210> 296
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 296
Asp Thr Ser Lys Leu Ala Ser
1. 5
<210> 297
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 297
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 298
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 298
Ser Ser Ser Val Ile Tyr
1. 5
<210> 299
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 299
Asp Thr Ser
1
<210> 300
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 300
Trp Ser Ser Asn Pro Leu
1. 5
<210> 301
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 301
Ser Ser Val Ile Tyr
1. 5
<210> 302
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 302
Asp Thr Ser
1
<210> 303
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 303
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 304
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 304
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Ala Ser Pro Gly
1. 5 10 15
Glu Arg Val Thr Met Ser Cys Ser Ala Ser Ser Ser Val Ile Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Met Glu Pro Glu
65 70 75 80
Asp Ala Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 305
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 305
gagatcgtgc tgacccagtc ccctgccacc ctgtctgcta gccctggcga gcgcgtgaca 60
atgtcctgct ccgcctcctc ctccgtgatc tacatgcact ggtatcagca gaagcccggc 120
caggcccctc ggcggtggat ctacgatacc tccaagctgg cctccggcgt gcccgccaga 180
ttctccggct ctggctctgg caccgactac accctgacca tctccagcat ggaacccgag 240
gacgccgccg tgtactactg ccagcagtgg tcctccaacc ccctgacctt cggccagggc 300
accaagctgg aaatcaag 318
<210> 306
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 306
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Ala Ser Pro Gly
1. 5 10 15
Glu Arg Val Thr Met Ser Cys Ser Ala Ser Ser Ser Val Ile Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Met Glu Pro Glu
65 70 75 80
Asp Ala Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 307
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 307
gagatcgtgc tgacccagtc ccctgccacc ctgtctgcta gccctggcga gcgcgtgaca 60
atgtcctgct ccgcctcctc ctccgtgatc tacatgcact ggtatcagca gaagcccggc 120
caggcccctc ggcggtggat ctacgatacc tccaagctgg cctccggcgt gcccgccaga 180
ttctccggct ctggctctgg caccgactac accctgacca tctccagcat ggaacccgag 240
gacgccgccg tgtactactg ccagcagtgg tcctccaacc ccctgacctt cggccagggc 300
accaagctgg aaatcaagcg tacggtggcc gctcccagcg tgttcatctt cccccccagc 360
gacgagcagc tgaagagtgg caccgccagc gtggtgtgcc tgctgaacaa cttctacccc 420
cgggaggcca aggtgcagtg gaaggtggac aacgccctgc agagcggcaa cagccaggag 480
agcgtcaccg agcaggacag caaggactcc acctacagcc tgagcagcac cctgaccctg 540
agcaaggccg actacgagaa gcataaggtg tacgcctgcg aggtgaccca ccagggcctg 600
tccagccccg tgaccaagag cttcaacagg ggcgagtgc 639
<210> 308
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 308
Gly Phe Thr Phe Ser Thr Tyr Ala Met Ser
1. 5 10
<210> 309
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 309
Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val Lys
1. 5 10 15
Gly
<210> 310
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 310
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 311
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 311
Thr Tyr Ala Met Ser
1. 5
<210> 312
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 312
Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val Lys
1. 5 10 15
Gly
<210> 313
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 313
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 314
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 314
Gly Phe Thr Phe Ser Thr Tyr
1. 5
<210> 315
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 315
Ser Asp Ala Gly Ser Tyr
1. 5
<210> 316
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 316
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 317
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 317
Gly Phe Thr Phe Ser Thr Tyr Ala
1. 5
<210> 318
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 318
Ile Ser Asp Ala Gly Ser Tyr Ser
1. 5
<210> 319
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 319
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10 15
<210> 320
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 320
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asp Gly Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Asn Leu Tyr
65 70 75 80
Leu Gln Met Ser His Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 321
<211> 366
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 321
gaagtgcagc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt acctatgcca tgtcttgggt tcgccagact 120
ccggaaaaga ggctggagtg ggtcgcaacc attagtgatg gtggtagtta ttcgtactat 180
ccagacaatg taaagggccg attcaccatc tccagagaca atgccaagaa caacctatac 240
ctgcaaatga gccatctgaa gtctgaggac acagccatgt attactgtgc aagacgaggt 300
agtaggtacg aagagtacta tgttatggac tactggggtc aaggaacctc agtcaccgtc 360
tcctca 366
<210> 322
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 322
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asp Gly Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Asn Leu Tyr
65 70 75 80
Leu Gln Met Ser His Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 323
<211> 1356
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 323
gaagtgcagc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt acctatgcca tgtcttgggt tcgccagact 120
ccggaaaaga ggctggagtg ggtcgcaacc attagtgatg gtggtagtta ttcgtactat 180
ccagacaatg taaagggccg attcaccatc tccagagaca atgccaagaa caacctatac 240
ctgcaaatga gccatctgaa gtctgaggac acagccatgt attactgtgc aagacgaggt 300
agtaggtacg aagagtacta tgttatggac tactggggtc aaggaacctc agtcaccgtc 360
tcctcagcta gcaccaaggg cccaagtgtg tttcccctgg cccccagcag caagtctact 420
tccggcggaa ctgctgccct gggttgcctg gtgaaggact acttcccctg tcccgtgaca 480
gtgtcctgga actctggggc tctgacttcc ggcgtgcaca ccttccccgc cgtgctgcag 540
agcagcggcc tgtacagcct gagcagcgtg gtgacagtgc cctccagctc tctgggaacc 600
cagacctata tctgcaacgt gaaccacaag cccagcaaca ccaaggtgga caagagagtg 660
gagcccaaga gctgcgacaa gacccacacc tgccccccct gcccagctcc agaactgctg 720
ggagggcctt ccgtgttcct gttccccccc aagcccaagg acaccctgat gatcagcagg 780
acccccgagg tgacctgcgt ggtggtggac gtgtcccacg aggacccaga ggtgaagttc 840
aactggtacg tggacggcgt ggaggtgcac aacgccaaga ccaagcccag agaggagcag 900
tacaacagca cctacagggt ggtgtccgtg ctgaccgtgc tgcaccagga ctggctgaac 960
ggcaaagaat acaagtgcaa agtctccaac aaggccctgc cagccccaat cgaaaagaca 1020
atcagcaagg ccaagggcca gccacgggag ccccaggtgt acaccctgcc ccccagccgg 1080
gaggagatga ccaagaacca ggtgtccctg acctgtctgg tgaagggctt ctacccctgt 1140
gatatcgccg tggagtggga gagcaacggc cagcccgaga acaactacaa gaccaccccc 1200
ccagtgctgg acagcgacgg cagcttcttc ctgtacagca agctgaccgt ggacaagtcc 1260
aggtggcagc agggcaacgt gttcagctgc agcgtgatgc acgaggccct gcacaaccac 1320
tacacccaga agtccctgag cctgagcccc ggcaag 1356
<210> 324
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 324
Arg Ala Ser Gln Ser Ile Ser Asn Asn Leu His
1. 5 10
<210> 325
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 325
Tyr Ala Ser Gln Ser Ile Ser
1. 5
<210> 326
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 326
Gln Gln Ser Ser Ser Trp Leu Thr
1. 5
<210> 327
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 327
Arg Ala Ser Gln Ser Ile Ser Asn Asn Leu His
1. 5 10
<210> 328
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 328
Tyr Ala Ser Gln Ser Ile Ser
1. 5
<210> 329
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 329
Gln Gln Ser Ser Ser Trp Leu Thr
1. 5
<210> 330
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 330
Ser Gln Ser Ile Ser Asn Asn
1. 5
<210> 331
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 331
Tyr Ala Ser
1
<210> 332
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 332
Ser Ser Ser Trp Leu
1. 5
<210> 333
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 333
Gln Ser Ile Ser Asn Asn
1. 5
<210> 334
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 334
Tyr Ala Ser
1
<210> 335
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 335
Gln Gln Ser Ser Ser Trp Leu Thr
1. 5
<210> 336
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 336
Asp Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Thr Pro Gly
1. 5 10 15
Asp Ser Val Ser Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asn Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Ser His Glu Ser Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Thr
65 70 75 80
Glu Asp Phe Gly Met Tyr Phe Cys Gln Gln Ser Asn Ser Trp Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Gly Leu Lys
100 105
<210> 337
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 337
gatattgtgc taactcagtc tccagccacc ctgtctgtga ctccaggaga tagcgtcagt 60
ctttcctgca gggccagcca aagtattagc aacaacctac actggtatca acagaaatca 120
catgagtctc caaaacttct catcaagtat gcttcccagt ccatctctgg gatcccctcc 180
aggttcagtg gcagtggatc agggacagat ttcactctca gtatcaacag tgtggagact 240
gaagattttg gaatgtattt ctgtcaacag agtaacagct ggctcacgtt cggtgctggg 300
accaagctgg ggctgaaa 318
<210> 338
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 338
Asp Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Thr Pro Gly
1. 5 10 15
Asp Ser Val Ser Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asn Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Ser His Glu Ser Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Thr
65 70 75 80
Glu Asp Phe Gly Met Tyr Phe Cys Gln Gln Ser Asn Ser Trp Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Gly Leu Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 339
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 339
gatattgtgc taactcagtc tccagccacc ctgtctgtga ctccaggaga tagcgtcagt 60
ctttcctgca gggccagcca aagtattagc aacaacctac actggtatca acagaaatca 120
catgagtctc caaaacttct catcaagtat gcttcccagt ccatctctgg gatcccctcc 180
aggttcagtg gcagtggatc agggacagat ttcactctca gtatcaacag tgtggagact 240
gaagattttg gaatgtattt ctgtcaacag agtaacagct ggctcacgtt cggtgctggg 300
accaagctgg ggctgaaacg tacggtggcc gctcccagcg tgttcatctt cccccccagc 360
gacgagcagc tgaagagtgg caccgccagc gtggtgtgcc tgctgaacaa cttctacccc 420
cgggaggcca aggtgcagtg gaaggtggac aacgccctgc agagcggcaa cagccaggag 480
agcgtcaccg agcaggacag caaggactcc acctacagcc tgagcagcac cctgaccctg 540
agcaaggccg actacgagaa gcataaggtg tacgcctgcg aggtgaccca ccagggcctg 600
tccagccccg tgaccaagag cttcaacagg ggcgagtgc 639
<210> 340
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 340
Gly Phe Thr Phe Ser Thr Tyr Ala Met Ser
1. 5 10
<210> 341
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 341
Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val Lys
1. 5 10 15
Gly
<210> 342
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 342
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 343
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 343
Thr Tyr Ala Met Ser
1. 5
<210> 344
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 344
Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val Lys
1. 5 10 15
Gly
<210> 345
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 345
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 346
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 346
Gly Phe Thr Phe Ser Thr Tyr
1. 5
<210> 347
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 347
Ser Asp Ala Gly Ser Tyr
1. 5
<210> 348
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 348
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 349
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 349
Gly Phe Thr Phe Ser Thr Tyr Ala
1. 5
<210> 350
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 350
Ile Ser Asp Ala Gly Ser Tyr Ser
1. 5
<210> 351
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 351
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10 15
<210> 352
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 352
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 353
<211> 366
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 353
gaggtgcagc tggtggaatc tggcggaggc ctggtgaaac ccggcggatc cctgagactg 60
tcctgcgccg cctccggctt caccttctcc acctacgcca tgtcctgggt gcggcaggct 120
cccggcaagg gcctggaatg ggtggccacc atctccgacg ccggctccta ctcctactac 180
cccgacaacg tgaagggcag attcaccatc agccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgtgc cagacggggc 300
tccagatacg aagagtacta cgtgatggac tattggggcc agggcaccac cgtgacagtg 360
tcctcc 366
<210> 354
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 354
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 355
<211> 1356
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 355
gaggtgcagc tggtggaatc tggcggaggc ctggtgaaac ccggcggatc cctgagactg 60
tcctgcgccg cctccggctt caccttctcc acctacgcca tgtcctgggt gcggcaggct 120
cccggcaagg gcctggaatg ggtggccacc atctccgacg ccggctccta ctcctactac 180
cccgacaacg tgaagggcag attcaccatc agccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgtgc cagacggggc 300
tccagatacg aagagtacta cgtgatggac tattggggcc agggcaccac cgtgacagtg 360
tcctccgcta gcaccaaggg cccaagtgtg tttcccctgg cccccagcag caagtctact 420
tccggcggaa ctgctgccct gggttgcctg gtgaaggact acttcccctg tcccgtgaca 480
gtgtcctgga actctggggc tctgacttcc ggcgtgcaca ccttccccgc cgtgctgcag 540
agcagcggcc tgtacagcct gagcagcgtg gtgacagtgc cctccagctc tctgggaacc 600
cagacctata tctgcaacgt gaaccacaag cccagcaaca ccaaggtgga caagagagtg 660
gagcccaaga gctgcgacaa gacccacacc tgccccccct gcccagctcc agaactgctg 720
ggagggcctt ccgtgttcct gttccccccc aagcccaagg acaccctgat gatcagcagg 780
acccccgagg tgacctgcgt ggtggtggcc gtgtcccacg aggacccaga ggtgaagttc 840
aactggtacg tggacggcgt ggaggtgcac aacgccaaga ccaagcccag agaggagcag 900
tacaacagca cctacagggt ggtgtccgtg ctgaccgtgc tgcaccagga ctggctgaac 960
ggcaaagaat acaagtgcaa agtctccaac aaggccctgg ctgccccaat cgaaaagaca 1020
atcagcaagg ccaagggcca gccacgggag ccccaggtgt acaccctgcc ccccagccgg 1080
gaggagatga ccaagaacca ggtgtccctg acctgtctgg tgaagggctt ctacccctgt 1140
gatatcgccg tggagtggga gagcaacggc cagcccgaga acaactacaa gaccaccccc 1200
ccagtgctgg acagcgacgg cagcttcttc ctgtacagca agctgaccgt ggacaagtcc 1260
aggtggcagc agggcaacgt gttcagctgc agcgtgatgc acgaggccct gcacaaccac 1320
tacacccaga agtccctgag cctgagcccc ggcaag 1356
<210> 356
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 356
Arg Ala Ser Gln Ser Ile Ser Asn Asn Leu His
1. 5 10
<210> 357
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 357
Tyr Ala Ser Gln Ser Ile Ser
1. 5
<210> 358
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 358
Gln Gln Ser Ser Ser Trp Leu Thr
1. 5
<210> 359
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 359
Arg Ala Ser Gln Ser Ile Ser Asn Asn Leu His
1. 5 10
<210> 360
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 360
Tyr Ala Ser Gln Ser Ile Ser
1. 5
<210> 361
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 361
Gln Gln Ser Ser Ser Trp Leu Thr
1. 5
<210> 362
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 362
Ser Gln Ser Ile Ser Asn Asn
1. 5
<210> 363
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 363
Tyr Ala Ser
1
<210> 364
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 364
Ser Ser Ser Trp Leu
1. 5
<210> 365
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 365
Gln Ser Ile Ser Asn Asn
1. 5
<210> 366
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 366
Tyr Ala Ser
1
<210> 367
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 367
Gln Gln Ser Ser Ser Trp Leu Thr
1. 5
<210> 368
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 368
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1. 5 10 15
Glu Arg Val Thr Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asn Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Ile Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Val Glu Pro
65 70 75 80
Glu Asp Phe Gly Val Tyr Phe Cys Gln Gln Ser Ser Ser Trp Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 369
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 369
gagatcgtgc tgacccagtc ccctgccacc ctgtccgtgt ctcccggcga gagagtgacc 60
ctgtcctgcc gggcctccca gtccatctcc aacaacctgc actggtatca gcagaagccc 120
ggccaggccc ctcggctgct gattaagtac gcctcccaga gcatctccgg catccctgcc 180
agattctccg gctccggcag cggcaccgac ttcaccctga ccatctccag cgtggaaccc 240
gaggacttcg gcgtgtactt ctgccagcag tcctcatcct ggctgacctt cggccagggc 300
accaagctgg aaatcaag 318
<210> 370
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 370
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1. 5 10 15
Glu Arg Val Thr Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asn Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Ile Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Val Glu Pro
65 70 75 80
Glu Asp Phe Gly Val Tyr Phe Cys Gln Gln Ser Ser Ser Trp Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 371
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 371
gagatcgtgc tgacccagtc ccctgccacc ctgtccgtgt ctcccggcga gagagtgacc 60
ctgtcctgcc gggcctccca gtccatctcc aacaacctgc actggtatca gcagaagccc 120
ggccaggccc ctcggctgct gattaagtac gcctcccaga gcatctccgg catccctgcc 180
agattctccg gctccggcag cggcaccgac ttcaccctga ccatctccag cgtggaaccc 240
gaggacttcg gcgtgtactt ctgccagcag tcctcatcct ggctgacctt cggccagggc 300
accaagctgg aaatcaagcg tacggtggcc gctcccagcg tgttcatctt cccccccagc 360
gacgagcagc tgaagagtgg caccgccagc gtggtgtgcc tgctgaacaa cttctacccc 420
cgggaggcca aggtgcagtg gaaggtggac aacgccctgc agagcggcaa cagccaggag 480
agcgtcaccg agcaggacag caaggactcc acctacagcc tgagcagcac cctgaccctg 540
agcaaggccg actacgagaa gcataaggtg tacgcctgcg aggtgaccca ccagggcctg 600
tccagccccg tgaccaagag cttcaacagg ggcgagtgc 639
<210> 372
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 372
Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
1. 5 10
<210> 373
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 373
Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 374
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 374
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 375
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 375
Ser Tyr Ala Met Ser
1. 5
<210> 376
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 376
Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 377
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 377
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 378
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 378
Gly Phe Thr Phe Ser Ser Tyr
1. 5
<210> 379
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 379
Ser Ser Gly Gly Ser Phe
1. 5
<210> 380
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 380
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 381
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 381
Gly Phe Thr Phe Ser Ser Tyr Ala
1. 5
<210> 382
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 382
Ile Ser Ser Gly Gly Ser Phe Thr
1. 5
<210> 383
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 383
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 384
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 384
Glu Val Met Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Ile Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val Trp Gly
100 105 110
Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 385
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 385
gaagtgatgc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt agctatgcca tgtcttggat tcgccagact 120
ccggagaaga gactggagtg ggtcgcaacc atcagtagtg gtggtagttt cacctactat 180
ccagacagtg tgaaggggcg attcaccatt tctagagaca atgtcaagaa caccctgtac 240
ctgcaaatga gcagtctgag gtctgaagac acggccatgt attactgtgc aagacgggct 300
tctacggtag taggtacgga cttcgatgtc tggggcgcag ggaccacggt caccgtctcc 360
tca 363
<210> 386
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 386
Glu Val Met Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Ile Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val Trp Gly
100 105 110
Ala Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 387
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 387
gaagtgatgc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt agctatgcca tgtcttggat tcgccagact 120
ccggagaaga gactggagtg ggtcgcaacc atcagtagtg gtggtagttt cacctactat 180
ccagacagtg tgaaggggcg attcaccatt tctagagaca atgtcaagaa caccctgtac 240
ctgcaaatga gcagtctgag gtctgaagac acggccatgt attactgtgc aagacgggct 300
tctacggtag taggtacgga cttcgatgtc tggggcgcag ggaccacggt caccgtctcc 360
tcagctagca ccaagggccc aagtgtgttt cccctggccc ccagcagcaa gtctacttcc 420
ggcggaactg ctgccctggg ttgcctggtg aaggactact tcccctgtcc cgtgacagtg 480
tcctggaact ctggggctct gacttccggc gtgcacacct tccccgccgt gctgcagagc 540
agcggcctgt acagcctgag cagcgtggtg acagtgccct ccagctctct gggaacccag 600
acctatatct gcaacgtgaa ccacaagccc agcaacacca aggtggacaa gagagtggag 660
cccaagagct gcgacaagac ccacacctgc cccccctgcc cagctccaga actgctggga 720
gggccttccg tgttcctgtt cccccccaag cccaaggaca ccctgatgat cagcaggacc 780
cccgaggtga cctgcgtggt ggtggacgtg tcccacgagg acccagaggt gaagttcaac 840
tggtacgtgg acggcgtgga ggtgcacaac gccaagacca agcccagaga ggagcagtac 900
aacagcacct acagggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 960
aaagaataca agtgcaaagt ctccaacaag gccctgccag ccccaatcga aaagacaatc 1020
agcaaggcca agggccagcc acgggagccc caggtgtaca ccctgccccc cagccgggag 1080
gagatgacca agaaccaggt gtccctgacc tgtctggtga agggcttcta cccctgtgat 1140
atcgccgtgg agtgggagag caacggccag cccgagaaca actacaagac caccccccca 1200
gtgctggaca gcgacggcag cttcttcctg tacagcaagc tgaccgtgga caagtccagg 1260
tggcagcagg gcaacgtgtt cagctgcagc gtgatgcacg aggccctgca caaccactac 1320
acccagaagt ccctgagcct gagccccggc aag 1353
<210> 388
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 388
Arg Ala Ser Gln Asp Ile Gly Ser Ser Leu Asn
1. 5 10
<210> 389
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 389
Ala Thr Ser Ser Leu Asp Ser
1. 5
<210> 390
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 390
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 391
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 391
Arg Ala Ser Gln Asp Ile Gly Ser Ser Leu Asn
1. 5 10
<210> 392
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 392
Ala Thr Ser Ser Leu Asp Ser
1. 5
<210> 393
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 393
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 394
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 394
Ser Gln Asp Ile Gly Ser Ser
1. 5
<210> 395
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 395
Ala Thr Ser
1
<210> 396
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 396
Tyr Ala Ser Ser Pro Pro
1. 5
<210> 397
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 397
Gln Asp Ile Gly Ser Ser
1. 5
<210> 398
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 398
Ala Thr Ser
1
<210> 399
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 399
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 400
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 400
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1. 5 10 15
Glu Arg Val Ser Leu Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Asn Trp Leu Gln Gln Glu Pro Asp Gly Thr Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Lys Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Ser Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Val Val Tyr Tyr Cys Leu Gln Tyr Ala Ser Ser Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 401
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 401
gacatccaga tgacccagtc tccatcctcc ttatctgcct ctctgggaga aagagtcagt 60
ctcacttgtc gggcaagtca ggacattggt agtagcttaa actggcttca gcaggaacca 120
gatggaacta ttaaacgcct gatctatgcc acatccagtt tagattctgg tgtccccaaa 180
aggttcagtg gcagtaggtc tgggtcagat tattctctca ccatcagcag ccttgagtct 240
gaagattttg tagtctatta ctgtctacaa tatgctagtt cgcctccgac gttcggtgga 300
ggcaccaagc tggaaatcaa a 321
<210> 402
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 402
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1. 5 10 15
Glu Arg Val Ser Leu Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Asn Trp Leu Gln Gln Glu Pro Asp Gly Thr Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Lys Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Ser Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Val Val Tyr Tyr Cys Leu Gln Tyr Ala Ser Ser Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 403
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 403
gacatccaga tgacccagtc tccatcctcc ttatctgcct ctctgggaga aagagtcagt 60
ctcacttgtc gggcaagtca ggacattggt agtagcttaa actggcttca gcaggaacca 120
gatggaacta ttaaacgcct gatctatgcc acatccagtt tagattctgg tgtccccaaa 180
aggttcagtg gcagtaggtc tgggtcagat tattctctca ccatcagcag ccttgagtct 240
gaagattttg tagtctatta ctgtctacaa tatgctagtt cgcctccgac gttcggtgga 300
ggcaccaagc tggaaatcaa acgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 404
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 404
Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
1. 5 10
<210> 405
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 405
Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 406
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 406
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 407
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 407
Ser Tyr Ala Met Ser
1. 5
<210> 408
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 408
Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 409
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 409
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 410
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 410
Gly Phe Thr Phe Ser Ser Tyr
1. 5
<210> 411
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 411
Ser Ser Gly Gly Ser Phe
1. 5
<210> 412
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 412
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 413
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 413
Gly Phe Thr Phe Ser Ser Tyr Ala
1. 5
<210> 414
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 414
Ile Ser Ser Gly Gly Ser Phe Thr
1. 5
<210> 415
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 415
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 416
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 416
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 417
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 417
gaggtgcagc tggtggaatc tggcggaggc ctggtgaaac ccggcggatc cctgagactg 60
tcctgcgccg cctccggctt caccttctcc agctacgcca tgtcctggat ccggcaggct 120
cccggcaagg gcctggaatg ggtggccacc atctcctccg gcggcagctt cacctactac 180
cccgacagcg tgaagggcag attcaccatc agccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgtgc cagacgggcc 300
tccaccgtcg tgggaaccga cttcgatgtg tggggccagg gcaccaccgt gacagtgtcc 360
tcc 363
<210> 418
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 418
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 419
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 419
gaggtgcagc tggtggaatc tggcggaggc ctggtgaaac ccggcggatc cctgagactg 60
tcctgcgccg cctccggctt caccttctcc agctacgcca tgtcctggat ccggcaggct 120
cccggcaagg gcctggaatg ggtggccacc atctcctccg gcggcagctt cacctactac 180
cccgacagcg tgaagggcag attcaccatc agccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgtgc cagacgggcc 300
tccaccgtcg tgggaaccga cttcgatgtg tggggccagg gcaccaccgt gacagtgtcc 360
tccgctagca ccaagggccc aagtgtgttt cccctggccc ccagcagcaa gtctacttcc 420
ggcggaactg ctgccctggg ttgcctggtg aaggactact tcccctgtcc cgtgacagtg 480
tcctggaact ctggggctct gacttccggc gtgcacacct tccccgccgt gctgcagagc 540
agcggcctgt acagcctgag cagcgtggtg acagtgccct ccagctctct gggaacccag 600
acctatatct gcaacgtgaa ccacaagccc agcaacacca aggtggacaa gagagtggag 660
cccaagagct gcgacaagac ccacacctgc cccccctgcc cagctccaga actgctggga 720
gggccttccg tgttcctgtt cccccccaag cccaaggaca ccctgatgat cagcaggacc 780
cccgaggtga cctgcgtggt ggtggccgtg tcccacgagg acccagaggt gaagttcaac 840
tggtacgtgg acggcgtgga ggtgcacaac gccaagacca agcccagaga ggagcagtac 900
aacagcacct acagggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 960
aaagaataca agtgcaaagt ctccaacaag gccctggctg ccccaatcga aaagacaatc 1020
agcaaggcca agggccagcc acgggagccc caggtgtaca ccctgccccc cagccgggag 1080
gagatgacca agaaccaggt gtccctgacc tgtctggtga agggcttcta cccctgtgat 1140
atcgccgtgg agtgggagag caacggccag cccgagaaca actacaagac caccccccca 1200
gtgctggaca gcgacggcag cttcttcctg tacagcaagc tgaccgtgga caagtccagg 1260
tggcagcagg gcaacgtgtt cagctgcagc gtgatgcacg aggccctgca caaccactac 1320
acccagaagt ccctgagcct gagccccggc aag 1353
<210> 420
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 420
Arg Ala Ser Gln Asp Ile Gly Ser Ser Leu Asn
1. 5 10
<210> 421
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 421
Ala Thr Ser Ser Leu Asp Ser
1. 5
<210> 422
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 422
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 423
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 423
Arg Ala Ser Gln Asp Ile Gly Ser Ser Leu Asn
1. 5 10
<210> 424
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 424
Ala Thr Ser Ser Leu Asp Ser
1. 5
<210> 425
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 425
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 426
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 426
Ser Gln Asp Ile Gly Ser Ser
1. 5
<210> 427
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 427
Ala Thr Ser
1
<210> 428
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 428
Tyr Ala Ser Ser Pro Pro
1. 5
<210> 429
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 429
Gln Asp Ile Gly Ser Ser
1. 5
<210> 430
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 430
Ala Thr Ser
1
<210> 431
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 431
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 432
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 432
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Leu Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Asn Trp Leu Gln Gln Lys Pro Gly Lys Ala Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Val Val Tyr Tyr Cys Leu Gln Tyr Ala Ser Ser Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 433
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 433
gacatccaga tgacccagtc cccctccagc ctgtccgcct ccgtgggcga tagagtgacc 60
ctgacctgcc gggcctccca ggacatcggc tcctccctga actggctgca gcagaagccc 120
ggcaaggcca tcaagcggct gatctacgcc acctcctccc tggactccgg cgtgccctcc 180
cggttctccg gctctagatc cggcaccgac tacaccctga ccatctccag cctgcagccc 240
gaggacttcg tggtgtacta ctgcctgcag tacgcctcca gcccccccac ctttggcgga 300
ggcaccaagc tggaaatcaa g 321
<210> 434
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 434
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Leu Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Asn Trp Leu Gln Gln Lys Pro Gly Lys Ala Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Val Val Tyr Tyr Cys Leu Gln Tyr Ala Ser Ser Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 435
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 435
gacatccaga tgacccagtc cccctccagc ctgtccgcct ccgtgggcga tagagtgacc 60
ctgacctgcc gggcctccca ggacatcggc tcctccctga actggctgca gcagaagccc 120
ggcaaggcca tcaagcggct gatctacgcc acctcctccc tggactccgg cgtgccctcc 180
cggttctccg gctctagatc cggcaccgac tacaccctga ccatctccag cctgcagccc 240
gaggacttcg tggtgtacta ctgcctgcag tacgcctcca gcccccccac ctttggcgga 300
ggcaccaagc tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 436
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 436
Gly Phe Thr Phe Ser Asn Phe Ala Met Ser
1. 5 10
<210> 437
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 437
Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 438
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 438
Arg Gly Tyr Asp Gly Val Asp Lys
1. 5
<210> 439
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 439
Ser Asn Phe Ala Met Ser
1. 5
<210> 440
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 440
Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 441
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 441
Arg Gly Tyr Asp Gly Val Asp Lys
1. 5
<210> 442
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 442
Gly Phe Thr Phe Ser Asn Phe
1. 5
<210> 443
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 443
Ser Thr Gly Gly Thr Tyr
1. 5
<210> 444
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 444
Arg Gly Tyr Asp Gly Val Asp Lys
1. 5
<210> 445
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 445
Gly Phe Thr Phe Ser Asn Phe Ala
1. 5
<210> 446
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 446
Ile Ser Thr Gly Gly Thr Tyr Thr
1. 5
<210> 447
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 447
Thr Arg Arg Gly Tyr Asp Gly Val Asp Lys
1. 5 10
<210> 448
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 448
Glu Val His Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Lys Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Thr Arg Arg Gly Tyr Asp Gly Val Asp Lys Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 449
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 449
gaagtgcatc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt aactttgcca tgtcttgggt tcgccagact 120
ccggagaaga gactggagtg ggtcgcaacc attagtactg gtggtactta cacctactat 180
ccagacagtg tgaagggtcg attcaccatc tccagagaca atgccaagaa aaccctgtac 240
ctgcaaatga gcagtctgag gtctgaggac acggccatgt attactgtac aagacggggg 300
tacgacggcg tggacaaatg gggccaaggc accactctca cagtctcctc a 351
<210> 450
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 450
Glu Val His Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Lys Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Thr Arg Arg Gly Tyr Asp Gly Val Asp Lys Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 451
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 451
gaagtgcatc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt aactttgcca tgtcttgggt tcgccagact 120
ccggagaaga gactggagtg ggtcgcaacc attagtactg gtggtactta cacctactat 180
ccagacagtg tgaagggtcg attcaccatc tccagagaca atgccaagaa aaccctgtac 240
ctgcaaatga gcagtctgag gtctgaggac acggccatgt attactgtac aagacggggg 300
tacgacggcg tggacaaatg gggccaaggc accactctca cagtctcctc agctagcacc 360
aagggcccaa gtgtgtttcc cctggccccc agcagcaagt ctacttccgg cggaactgct 420
gccctgggtt gcctggtgaa ggactacttc ccctgtcccg tgacagtgtc ctggaactct 480
ggggctctga cttccggcgt gcacaccttc cccgccgtgc tgcagagcag cggcctgtac 540
agcctgagca gcgtggtgac agtgccctcc agctctctgg gaacccagac ctatatctgc 600
aacgtgaacc acaagcccag caacaccaag gtggacaaga gagtggagcc caagagctgc 660
gacaagaccc acacctgccc cccctgccca gctccagaac tgctgggagg gccttccgtg 720
ttcctgttcc cccccaagcc caaggacacc ctgatgatca gcaggacccc cgaggtgacc 780
tgcgtggtgg tggacgtgtc ccacgaggac ccagaggtga agttcaactg gtacgtggac 840
ggcgtggagg tgcacaacgc caagaccaag cccagagagg agcagtacaa cagcacctac 900
agggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agaatacaag 960
tgcaaagtct ccaacaaggc cctgccagcc ccaatcgaaa agacaatcag caaggccaag 1020
ggccagccac gggagcccca ggtgtacacc ctgcccccca gccgggagga gatgaccaag 1080
aaccaggtgt ccctgacctg tctggtgaag ggcttctacc cctgtgatat cgccgtggag 1140
tgggagagca acggccagcc cgagaacaac tacaagacca cccccccagt gctggacagc 1200
gacggcagct tcttcctgta cagcaagctg accgtggaca agtccaggtg gcagcagggc 1260
aacgtgttca gctgcagcgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgagcctga gccccggcaa g 1341
<210> 452
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 452
Lys Ser Gly Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn
1. 5 10 15
<210> 453
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 453
Leu Val Ser Lys Leu Asp Ser
1. 5
<210> 454
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 454
Trp Gln Gly Thr His Phe Pro Gln Thr
1. 5
<210> 455
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 455
Lys Ser Gly Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn
1. 5 10 15
<210> 456
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 456
Leu Val Ser Lys Leu Asp Ser
1. 5
<210> 457
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 457
Trp Gln Gly Thr His Phe Pro Gln Thr
1. 5
<210> 458
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 458
Gly Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
1. 5 10
<210> 459
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 459
Leu Val Ser
1
<210> 460
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 460
Gly Thr His Phe Pro Gln
1. 5
<210> 461
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 461
Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
1. 5 10
<210> 462
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 462
Leu Val Ser
1
<210> 463
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 463
Trp Gln Gly Thr His Phe Pro Gln Thr
1. 5
<210> 464
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 464
Asp Val Val Met Thr Gln Thr Pro Leu Thr Leu Ser Val Thr Ile Gly
1. 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Gly Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 465
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 465
gatgttgtga tgacccagac tccactcact ttgtcggtta ccattggaca accagcctcc 60
atctcttgca agtcaggtca gagcctctta gatagtgatg gaaagacata tttgaattgg 120
tttttacaga ggccaggcca gtctccaaag cgcctaatct atctggtgtc taaactggac 180
tctggagtcc ctgacaggtt cactggcagt ggatcaggga cagatttcac actgaaaatc 240
agcagagtgg aggctgagga tttgggagtt tattattgct ggcaaggtac acattttcct 300
cagacgttcg gtggaggcac caagctggaa atcaaa 336
<210> 466
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 466
Asp Val Val Met Thr Gln Thr Pro Leu Thr Leu Ser Val Thr Ile Gly
1. 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Gly Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 467
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 467
gatgttgtga tgacccagac tccactcact ttgtcggtta ccattggaca accagcctcc 60
atctcttgca agtcaggtca gagcctctta gatagtgatg gaaagacata tttgaattgg 120
tttttacaga ggccaggcca gtctccaaag cgcctaatct atctggtgtc taaactggac 180
tctggagtcc ctgacaggtt cactggcagt ggatcaggga cagatttcac actgaaaatc 240
agcagagtgg aggctgagga tttgggagtt tattattgct ggcaaggtac acattttcct 300
cagacgttcg gtggaggcac caagctggaa atcaaacgta cggtggccgc tcccagcgtg 360
ttcatcttcc cccccagcga cgagcagctg aagagtggca ccgccagcgt ggtgtgcctg 420
ctgaacaact tctacccccg ggaggccaag gtgcagtgga aggtggacaa cgccctgcag 480
agcggcaaca gccaggagag cgtcaccgag caggacagca aggactccac ctacagcctg 540
agcagcaccc tgaccctgag caaggccgac tacgagaagc ataaggtgta cgcctgcgag 600
gtgacccacc agggcctgtc cagccccgtg accaagagct tcaacagggg cgagtgc 657
<210> 468
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 468
Gly Phe Thr Phe Ser Asn Phe Ala Met Ser
1. 5 10
<210> 469
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 469
Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 470
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 470
Arg Gly Tyr Asp Gly Val Asp Lys
1. 5
<210> 471
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 471
Ser Asn Phe Ala Met Ser
1. 5
<210> 472
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 472
Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 473
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 473
Arg Gly Tyr Asp Gly Val Asp Lys
1. 5
<210> 474
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 474
Gly Phe Thr Phe Ser Asn Phe
1. 5
<210> 475
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 475
Ser Thr Gly Gly Thr Tyr
1. 5
<210> 476
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 476
Arg Gly Tyr Asp Gly Val Asp Lys
1. 5
<210> 477
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 477
Gly Phe Thr Phe Ser Asn Phe Ala
1. 5
<210> 478
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 478
Ile Ser Thr Gly Gly Thr Tyr Thr
1. 5
<210> 479
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 479
Thr Arg Arg Gly Tyr Asp Gly Val Asp Lys
1. 5 10
<210> 480
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 480
Glu Val His Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Lys Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Thr Arg Arg Gly Tyr Asp Gly Val Asp Lys Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 481
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 481
gaagtgcatc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt aactttgcca tgtcttgggt tcgccagact 120
ccggagaaga gactggagtg ggtcgcaacc attagtactg gtggtactta cacctactat 180
ccagacagtg tgaagggtcg attcaccatc tccagagaca atgccaagaa aaccctgtac 240
ctgcaaatga gcagtctgag gtctgaggac acggccatgt attactgtac aagacggggg 300
tacgacggcg tggacaaatg gggccaaggc accactctca cagtctcctc a 351
<210> 482
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 482
Glu Val His Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Lys Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Thr Arg Arg Gly Tyr Asp Gly Val Asp Lys Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Ala Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 483
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 483
gaagtgcatc tggtggagtc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt aactttgcca tgtcttgggt tcgccagact 120
ccggagaaga gactggagtg ggtcgcaacc attagtactg gtggtactta cacctactat 180
ccagacagtg tgaagggtcg attcaccatc tccagagaca atgccaagaa aaccctgtac 240
ctgcaaatga gcagtctgag gtctgaggac acggccatgt attactgtac aagacggggg 300
tacgacggcg tggacaaatg gggccaaggc accactctca cagtctcctc agctagcacc 360
aagggcccaa gtgtgtttcc cctggccccc agcagcaagt ctacttccgg cggaactgct 420
gccctgggtt gcctggtgaa ggactacttc ccctgtcccg tgacagtgtc ctggaactct 480
ggggctctga cttccggcgt gcacaccttc cccgccgtgc tgcagagcag cggcctgtac 540
agcctgagca gcgtggtgac agtgccctcc agctctctgg gaacccagac ctatatctgc 600
aacgtgaacc acaagcccag caacaccaag gtggacaaga gagtggagcc caagagctgc 660
gacaagaccc acacctgccc cccctgccca gctccagaac tgctgggagg gccttccgtg 720
ttcctgttcc cccccaagcc caaggacacc ctgatgatca gcaggacccc cgaggtgacc 780
tgcgtggtgg tggccgtgtc ccacgaggac ccagaggtga agttcaactg gtacgtggac 840
ggcgtggagg tgcacaacgc caagaccaag cccagagagg agcagtacaa cagcacctac 900
agggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agaatacaag 960
tgcaaagtct ccaacaaggc cctggctgcc ccaatcgaaa agacaatcag caaggccaag 1020
ggccagccac gggagcccca ggtgtacacc ctgcccccca gccgggagga gatgaccaag 1080
aaccaggtgt ccctgacctg tctggtgaag ggcttctacc cctgtgatat cgccgtggag 1140
tgggagagca acggccagcc cgagaacaac tacaagacca cccccccagt gctggacagc 1200
gacggcagct tcttcctgta cagcaagctg accgtggaca agtccaggtg gcagcagggc 1260
aacgtgttca gctgcagcgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgagcctga gccccggcaa g 1341
<210> 484
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 484
Lys Ser Gly Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn
1. 5 10 15
<210> 485
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 485
Leu Val Ser Lys Leu Asp Ser
1. 5
<210> 486
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 486
Trp Gln Gly Thr His Phe Pro Gln Thr
1. 5
<210> 487
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 487
Lys Ser Gly Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn
1. 5 10 15
<210> 488
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 488
Leu Val Ser Lys Leu Asp Ser
1. 5
<210> 489
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 489
Trp Gln Gly Thr His Phe Pro Gln Thr
1. 5
<210> 490
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 490
Gly Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
1. 5 10
<210> 491
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 491
Leu Val Ser
1
<210> 492
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 492
Gly Thr His Phe Pro Gln
1. 5
<210> 493
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 493
Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
1. 5 10
<210> 494
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 494
Leu Val Ser
1
<210> 495
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 495
Trp Gln Gly Thr His Phe Pro Gln Thr
1. 5
<210> 496
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 496
Asp Val Val Met Thr Gln Thr Pro Leu Thr Leu Ser Val Thr Ile Gly
1. 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Gly Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 497
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 497
gatgttgtga tgacccagac tccactcact ttgtcggtta ccattggaca accagcctcc 60
atctcttgca agtcaggtca gagcctctta gatagtgatg gaaagacata tttgaattgg 120
tttttacaga ggccaggcca gtctccaaag cgcctaatct atctggtgtc taaactggac 180
tctggagtcc ctgacaggtt cactggcagt ggatcaggga cagatttcac actgaaaatc 240
agcagagtgg aggctgagga tttgggagtt tattattgct ggcaaggtac acattttcct 300
cagacgttcg gtggaggcac caagctggaa atcaaa 336
<210> 498
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 498
Asp Val Val Met Thr Gln Thr Pro Leu Thr Leu Ser Val Thr Ile Gly
1. 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Gly Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Phe Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 499
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 499
gatgttgtga tgacccagac tccactcact ttgtcggtta ccattggaca accagcctcc 60
atctcttgca agtcaggtca gagcctctta gatagtgatg gaaagacata tttgaattgg 120
tttttacaga ggccaggcca gtctccaaag cgcctaatct atctggtgtc taaactggac 180
tctggagtcc ctgacaggtt cactggcagt ggatcaggga cagatttcac actgaaaatc 240
agcagagtgg aggctgagga tttgggagtt tattattgct ggcaaggtac acattttcct 300
cagacgttcg gtggaggcac caagctggaa atcaaacgta cggtggccgc tcccagcgtg 360
ttcatcttcc cccccagcga cgagcagctg aagagtggca ccgccagcgt ggtgtgcctg 420
ctgaacaact tctacccccg ggaggccaag gtgcagtgga aggtggacaa cgccctgcag 480
agcggcaaca gccaggagag cgtcaccgag caggacagca aggactccac ctacagcctg 540
agcagcaccc tgaccctgag caaggccgac tacgagaagc ataaggtgta cgcctgcgag 600
gtgacccacc agggcctgtc cagccccgtg accaagagct tcaacagggg cgagtgc 657
<210> 500
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 500
Gly Tyr Ser Ile Thr Ser Gly Tyr Ser Trp His
1. 5 10
<210> 501
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 501
His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1. 5 10 15
<210> 502
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 502
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 503
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 503
Ser Gly Tyr Ser Trp His
1. 5
<210> 504
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 504
His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1. 5 10 15
<210> 505
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 505
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 506
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 506
Gly Tyr Ser Ile Thr Ser Gly Tyr
1. 5
<210> 507
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 507
His Ser Ser Gly Ser
1. 5
<210> 508
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 508
Gly Gly Val Gln Ala Phe Ala Tyr
1. 5
<210> 509
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 509
Gly Tyr Ser Ile Thr Ser Gly Tyr Ser
1. 5
<210> 510
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 510
Ile His Ser Ser Gly Ser Thr
1. 5
<210> 511
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 511
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr
1. 5 10
<210> 512
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 512
Asp Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp His Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp
35 40 45
Met Ala His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 513
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 513
gacgtgcagc tgcaggaatc tggccctggc ctggtgaaac cctcccagac cctgtccctg 60
acctgcaccg tgtccggcta ctccatcacc tccggctaca gctggcactg gatccggcag 120
caccccggca agggcctgga atggatggcc cacatccact cctccggctc caccaactac 180
aaccccagcc tgaagtccag aatcaccatc agccgggaca cctccaagaa ccagttctcc 240
ctgaagctgt cctccgtgac cgccgctgac accgccgtgt actactgtgc cagaggcggc 300
gtgcaggcct tcgcttattg gggccagggc accctggtga cagtgtcctc c 351
<210> 514
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 514
Asp Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Ser Trp His Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp
35 40 45
Met Ala His Ile His Ser Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Val Gln Ala Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Ala Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 515
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 515
gacgtgcagc tgcaggaatc tggccctggc ctggtgaaac cctcccagac cctgtccctg 60
acctgcaccg tgtccggcta ctccatcacc tccggctaca gctggcactg gatccggcag 120
caccccggca agggcctgga atggatggcc cacatccact cctccggctc caccaactac 180
aaccccagcc tgaagtccag aatcaccatc agccgggaca cctccaagaa ccagttctcc 240
ctgaagctgt cctccgtgac cgccgctgac accgccgtgt actactgtgc cagaggcggc 300
gtgcaggcct tcgcttattg gggccagggc accctggtga cagtgtcctc cgctagcacc 360
aagggcccaa gtgtgtttcc cctggccccc agcagcaagt ctacttccgg cggaactgct 420
gccctgggtt gcctggtgaa ggactacttc cccgagcccg tgacagtgtc ctggaactct 480
ggggctctga cttccggcgt gcacaccttc cccgccgtgc tgcagagcag cggcctgtac 540
agcctgagca gcgtggtgac agtgccctcc agctctctgg gaacccagac ctatatctgc 600
aacgtgaacc acaagcccag caacaccaag gtggacaaga gagtggagcc caagagctgc 660
gacaagaccc acacctgccc cccctgccca gctccagaac tgctgggagg gccttccgtg 720
ttcctgttcc cccccaagcc caaggacacc ctgatgatca gcaggacccc cgaggtgacc 780
tgcgtggtgg tggccgtgtc ccacgaggac ccagaggtga agttcaactg gtacgtggac 840
ggcgtggagg tgcacaacgc caagaccaag cccagagagg agcagtacaa cagcacctac 900
agggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agaatacaag 960
tgcaaagtct ccaacaaggc cctggctgcc ccaatcgaaa agacaatcag caaggccaag 1020
ggccagccac gggagcccca ggtgtacacc ctgcccccca gccgggagga gatgaccaag 1080
aaccaggtgt ccctgacctg tctggtgaag ggcttctacc ccagcgatat cgccgtggag 1140
tgggagagca acggccagcc cgagaacaac tacaagacca cccccccagt gctggacagc 1200
gacggcagct tcttcctgta cagcaagctg accgtggaca agtccaggtg gcagcagggc 1260
aacgtgttca gctgcagcgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgagcctga gccccggcaa g 1341
<210> 516
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 516
Ser Ala Ser Ser Ser Val Ile Tyr Met His
1. 5 10
<210> 517
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 517
Asp Thr Ser Lys Leu Ala Ser
1. 5
<210> 518
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 518
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 519
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 519
Ser Ala Ser Ser Ser Val Ile Tyr Met His
1. 5 10
<210> 520
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 520
Asp Thr Ser Lys Leu Ala Ser
1. 5
<210> 521
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 521
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 522
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 522
Ser Ser Ser Val Ile Tyr
1. 5
<210> 523
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 523
Asp Thr Ser
1
<210> 524
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 524
Trp Ser Ser Asn Pro Leu
1. 5
<210> 525
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 525
Ser Ser Val Ile Tyr
1. 5
<210> 526
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 526
Asp Thr Ser
1
<210> 527
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 527
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 528
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 528
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Ala Ser Pro Gly
1. 5 10 15
Glu Arg Val Thr Met Ser Cys Ser Ala Ser Ser Ser Val Ile Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Met Glu Pro Glu
65 70 75 80
Asp Ala Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 529
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 529
gagatcgtgc tgacccagtc ccctgccacc ctgtctgcta gccctggcga gcgcgtgaca 60
atgtcctgct ccgcctcctc ctccgtgatc tacatgcact ggtatcagca gaagcccggc 120
caggcccctc ggcggtggat ctacgatacc tccaagctgg cctccggcgt gcccgccaga 180
ttctccggct ctggctctgg caccgactac accctgacca tctccagcat ggaacccgag 240
gacgccgccg tgtactactg ccagcagtgg tcctccaacc ccctgacctt cggccagggc 300
accaagctgg aaatcaag 318
<210> 530
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 530
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Ala Ser Pro Gly
1. 5 10 15
Glu Arg Val Thr Met Ser Cys Ser Ala Ser Ser Ser Val Ile Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Met Glu Pro Glu
65 70 75 80
Asp Ala Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 531
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 531
gagatcgtgc tgacccagtc ccctgccacc ctgtctgcta gccctggcga gcgcgtgaca 60
atgtcctgct ccgcctcctc ctccgtgatc tacatgcact ggtatcagca gaagcccggc 120
caggcccctc ggcggtggat ctacgatacc tccaagctgg cctccggcgt gcccgccaga 180
ttctccggct ctggctctgg caccgactac accctgacca tctccagcat ggaacccgag 240
gacgccgccg tgtactactg ccagcagtgg tcctccaacc ccctgacctt cggccagggc 300
accaagctgg aaatcaagcg tacggtggcc gctcccagcg tgttcatctt cccccccagc 360
gacgagcagc tgaagagtgg caccgccagc gtggtgtgcc tgctgaacaa cttctacccc 420
cgggaggcca aggtgcagtg gaaggtggac aacgccctgc agagcggcaa cagccaggag 480
agcgtcaccg agcaggacag caaggactcc acctacagcc tgagcagcac cctgaccctg 540
agcaaggccg actacgagaa gcataaggtg tacgcctgcg aggtgaccca ccagggcctg 600
tccagccccg tgaccaagag cttcaacagg ggcgagtgc 639
<210> 532
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 532
Gly Phe Thr Phe Ser Thr Tyr Ala Met Ser
1. 5 10
<210> 533
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 533
Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val Lys
1. 5 10 15
Gly
<210> 534
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 534
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 535
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 535
Thr Tyr Ala Met Ser
1. 5
<210> 536
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 536
Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val Lys
1. 5 10 15
Gly
<210> 537
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 537
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 538
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 538
Gly Phe Thr Phe Ser Thr Tyr
1. 5
<210> 539
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 539
Ser Asp Ala Gly Ser Tyr
1. 5
<210> 540
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 540
Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10
<210> 541
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 541
Gly Phe Thr Phe Ser Thr Tyr Ala
1. 5
<210> 542
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 542
Ile Ser Asp Ala Gly Ser Tyr Ser
1. 5
<210> 543
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 543
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr
1. 5 10 15
<210> 544
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 544
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 545
<211> 366
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 545
gaggtgcagc tggtggaatc tggcggaggc ctggtgaaac ccggcggatc cctgagactg 60
tcctgcgccg cctccggctt caccttctcc acctacgcca tgtcctgggt gcggcaggct 120
cccggcaagg gcctggaatg ggtggccacc atctccgacg ccggctccta ctcctactac 180
cccgacaacg tgaagggcag attcaccatc agccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgtgc cagacggggc 300
tccagatacg aagagtacta cgtgatggac tattggggcc agggcaccac cgtgacagtg 360
tcctcc 366
<210> 546
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 546
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Asp Ala Gly Ser Tyr Ser Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Ser Arg Tyr Glu Glu Tyr Tyr Val Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 547
<211> 1356
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 547
gaggtgcagc tggtggaatc tggcggaggc ctggtgaaac ccggcggatc cctgagactg 60
tcctgcgccg cctccggctt caccttctcc acctacgcca tgtcctgggt gcggcaggct 120
cccggcaagg gcctggaatg ggtggccacc atctccgacg ccggctccta ctcctactac 180
cccgacaacg tgaagggcag attcaccatc agccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgtgc cagacggggc 300
tccagatacg aagagtacta cgtgatggac tattggggcc agggcaccac cgtgacagtg 360
tcctccgcta gcaccaaggg cccaagtgtg tttcccctgg cccccagcag caagtctact 420
tccggcggaa ctgctgccct gggttgcctg gtgaaggact acttccccga gcccgtgaca 480
gtgtcctgga actctggggc tctgacttcc ggcgtgcaca ccttccccgc cgtgctgcag 540
agcagcggcc tgtacagcct gagcagcgtg gtgacagtgc cctccagctc tctgggaacc 600
cagacctata tctgcaacgt gaaccacaag cccagcaaca ccaaggtgga caagagagtg 660
gagcccaaga gctgcgacaa gacccacacc tgccccccct gcccagctcc agaactgctg 720
ggagggcctt ccgtgttcct gttccccccc aagcccaagg acaccctgat gatcagcagg 780
acccccgagg tgacctgcgt ggtggtggcc gtgtcccacg aggacccaga ggtgaagttc 840
aactggtacg tggacggcgt ggaggtgcac aacgccaaga ccaagcccag agaggagcag 900
tacaacagca cctacagggt ggtgtccgtg ctgaccgtgc tgcaccagga ctggctgaac 960
ggcaaagaat acaagtgcaa agtctccaac aaggccctgg ctgccccaat cgaaaagaca 1020
atcagcaagg ccaagggcca gccacgggag ccccaggtgt acaccctgcc ccccagccgg 1080
gaggagatga ccaagaacca ggtgtccctg acctgtctgg tgaagggctt ctaccccagc 1140
gatatcgccg tggagtggga gagcaacggc cagcccgaga acaactacaa gaccaccccc 1200
ccagtgctgg acagcgacgg cagcttcttc ctgtacagca agctgaccgt ggacaagtcc 1260
aggtggcagc agggcaacgt gttcagctgc agcgtgatgc acgaggccct gcacaaccac 1320
tacacccaga agtccctgag cctgagcccc ggcaag 1356
<210> 548
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 548
Arg Ala Ser Gln Ser Ile Ser Asn Asn Leu His
1. 5 10
<210> 549
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 549
Tyr Ala Ser Gln Ser Ile Ser
1. 5
<210> 550
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 550
Gln Gln Ser Ser Ser Trp Leu Thr
1. 5
<210> 551
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 551
Arg Ala Ser Gln Ser Ile Ser Asn Asn Leu His
1. 5 10
<210> 552
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 552
Tyr Ala Ser Gln Ser Ile Ser
1. 5
<210> 553
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 553
Gln Gln Ser Ser Ser Trp Leu Thr
1. 5
<210> 554
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 554
Ser Gln Ser Ile Ser Asn Asn
1. 5
<210> 555
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 555
Tyr Ala Ser
1
<210> 556
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 556
Ser Ser Ser Trp Leu
1. 5
<210> 557
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 557
Gln Ser Ile Ser Asn Asn
1. 5
<210> 558
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 558
Tyr Ala Ser
1
<210> 559
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 559
Gln Gln Ser Ser Ser Trp Leu Thr
1. 5
<210> 560
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 560
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1. 5 10 15
Glu Arg Val Thr Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asn Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Ile Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Val Glu Pro
65 70 75 80
Glu Asp Phe Gly Val Tyr Phe Cys Gln Gln Ser Ser Ser Trp Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 561
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 561
gagatcgtgc tgacccagtc ccctgccacc ctgtccgtgt ctcccggcga gagagtgacc 60
ctgtcctgcc gggcctccca gtccatctcc aacaacctgc actggtatca gcagaagccc 120
ggccaggccc ctcggctgct gattaagtac gcctcccaga gcatctccgg catccctgcc 180
agattctccg gctccggcag cggcaccgac ttcaccctga ccatctccag cgtggaaccc 240
gaggacttcg gcgtgtactt ctgccagcag tcctcatcct ggctgacctt cggccagggc 300
accaagctgg aaatcaag 318
<210> 562
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 562
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1. 5 10 15
Glu Arg Val Thr Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asn Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Ile Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Val Glu Pro
65 70 75 80
Glu Asp Phe Gly Val Tyr Phe Cys Gln Gln Ser Ser Ser Trp Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 563
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 563
gagatcgtgc tgacccagtc ccctgccacc ctgtccgtgt ctcccggcga gagagtgacc 60
ctgtcctgcc gggcctccca gtccatctcc aacaacctgc actggtatca gcagaagccc 120
ggccaggccc ctcggctgct gattaagtac gcctcccaga gcatctccgg catccctgcc 180
agattctccg gctccggcag cggcaccgac ttcaccctga ccatctccag cgtggaaccc 240
gaggacttcg gcgtgtactt ctgccagcag tcctcatcct ggctgacctt cggccagggc 300
accaagctgg aaatcaagcg tacggtggcc gctcccagcg tgttcatctt cccccccagc 360
gacgagcagc tgaagagtgg caccgccagc gtggtgtgcc tgctgaacaa cttctacccc 420
cgggaggcca aggtgcagtg gaaggtggac aacgccctgc agagcggcaa cagccaggag 480
agcgtcaccg agcaggacag caaggactcc acctacagcc tgagcagcac cctgaccctg 540
agcaaggccg actacgagaa gcataaggtg tacgcctgcg aggtgaccca ccagggcctg 600
tccagccccg tgaccaagag cttcaacagg ggcgagtgc 639
<210> 564
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 564
Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
1. 5 10
<210> 565
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 565
Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 566
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 566
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 567
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 567
Ser Tyr Ala Met Ser
1. 5
<210> 568
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 568
Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 569
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 569
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 570
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 570
Gly Phe Thr Phe Ser Ser Tyr
1. 5
<210> 571
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 571
Ser Ser Gly Gly Ser Phe
1. 5
<210> 572
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 572
Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 573
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 573
Gly Phe Thr Phe Ser Ser Tyr Ala
1. 5
<210> 574
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 574
Ile Ser Ser Gly Gly Ser Phe Thr
1. 5
<210> 575
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 575
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val
1. 5 10
<210> 576
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 576
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 577
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 577
gaggtgcagc tggtggaatc tggcggaggc ctggtgaaac ccggcggatc cctgagactg 60
tcctgcgccg cctccggctt caccttctcc agctacgcca tgtcctggat ccggcaggct 120
cccggcaagg gcctggaatg ggtggccacc atctcctccg gcggcagctt cacctactac 180
cccgacagcg tgaagggcag attcaccatc agccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgtgc cagacgggcc 300
tccaccgtcg tgggaaccga cttcgatgtg tggggccagg gcaccaccgt gacagtgtcc 360
tcc 363
<210> 578
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 578
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Ser Thr Val Val Gly Thr Asp Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 579
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 579
gaggtgcagc tggtggaatc tggcggaggc ctggtgaaac ccggcggatc cctgagactg 60
tcctgcgccg cctccggctt caccttctcc agctacgcca tgtcctggat ccggcaggct 120
cccggcaagg gcctggaatg ggtggccacc atctcctccg gcggcagctt cacctactac 180
cccgacagcg tgaagggcag attcaccatc agccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgtgc cagacgggcc 300
tccaccgtcg tgggaaccga cttcgatgtg tggggccagg gcaccaccgt gacagtgtcc 360
tccgctagca ccaagggccc aagtgtgttt cccctggccc ccagcagcaa gtctacttcc 420
ggcggaactg ctgccctggg ttgcctggtg aaggactact tccccgagcc cgtgacagtg 480
tcctggaact ctggggctct gacttccggc gtgcacacct tccccgccgt gctgcagagc 540
agcggcctgt acagcctgag cagcgtggtg acagtgccct ccagctctct gggaacccag 600
acctatatct gcaacgtgaa ccacaagccc agcaacacca aggtggacaa gagagtggag 660
cccaagagct gcgacaagac ccacacctgc cccccctgcc cagctccaga actgctggga 720
gggccttccg tgttcctgtt cccccccaag cccaaggaca ccctgatgat cagcaggacc 780
cccgaggtga cctgcgtggt ggtggccgtg tcccacgagg acccagaggt gaagttcaac 840
tggtacgtgg acggcgtgga ggtgcacaac gccaagacca agcccagaga ggagcagtac 900
aacagcacct acagggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 960
aaagaataca agtgcaaagt ctccaacaag gccctggctg ccccaatcga aaagacaatc 1020
agcaaggcca agggccagcc acgggagccc caggtgtaca ccctgccccc cagccgggag 1080
gagatgacca agaaccaggt gtccctgacc tgtctggtga agggcttcta ccccagcgat 1140
atcgccgtgg agtgggagag caacggccag cccgagaaca actacaagac caccccccca 1200
gtgctggaca gcgacggcag cttcttcctg tacagcaagc tgaccgtgga caagtccagg 1260
tggcagcagg gcaacgtgtt cagctgcagc gtgatgcacg aggccctgca caaccactac 1320
acccagaagt ccctgagcct gagccccggc aag 1353
<210> 580
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 580
Arg Ala Ser Gln Asp Ile Gly Ser Ser Leu Asn
1. 5 10
<210> 581
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 581
Ala Thr Ser Ser Leu Asp Ser
1. 5
<210> 582
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 582
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 583
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 583
Arg Ala Ser Gln Asp Ile Gly Ser Ser Leu Asn
1. 5 10
<210> 584
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 584
Ala Thr Ser Ser Leu Asp Ser
1. 5
<210> 585
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 585
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 586
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 586
Ser Gln Asp Ile Gly Ser Ser
1. 5
<210> 587
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 587
Ala Thr Ser
1
<210> 588
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 588
Tyr Ala Ser Ser Pro Pro
1. 5
<210> 589
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 589
Gln Asp Ile Gly Ser Ser
1. 5
<210> 590
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 590
Ala Thr Ser
1
<210> 591
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 591
Leu Gln Tyr Ala Ser Ser Pro Pro Thr
1. 5
<210> 592
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 592
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Leu Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Asn Trp Leu Gln Gln Lys Pro Gly Lys Ala Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Val Val Tyr Tyr Cys Leu Gln Tyr Ala Ser Ser Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 593
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 593
gacatccaga tgacccagtc cccctccagc ctgtccgcct ccgtgggcga tagagtgacc 60
ctgacctgcc gggcctccca ggacatcggc tcctccctga actggctgca gcagaagccc 120
ggcaaggcca tcaagcggct gatctacgcc acctcctccc tggactccgg cgtgccctcc 180
cggttctccg gctctagatc cggcaccgac tacaccctga ccatctccag cctgcagccc 240
gaggacttcg tggtgtacta ctgcctgcag tacgcctcca gcccccccac ctttggcgga 300
ggcaccaagc tggaaatcaa g 321
<210> 594
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 594
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Leu Thr Cys Arg Ala Ser Gln Asp Ile Gly Ser Ser
20 25 30
Leu Asn Trp Leu Gln Gln Lys Pro Gly Lys Ala Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Val Val Tyr Tyr Cys Leu Gln Tyr Ala Ser Ser Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 595
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 595
gacatccaga tgacccagtc cccctccagc ctgtccgcct ccgtgggcga tagagtgacc 60
ctgacctgcc gggcctccca ggacatcggc tcctccctga actggctgca gcagaagccc 120
ggcaaggcca tcaagcggct gatctacgcc acctcctccc tggactccgg cgtgccctcc 180
cggttctccg gctctagatc cggcaccgac tacaccctga ccatctccag cctgcagccc 240
gaggacttcg tggtgtacta ctgcctgcag tacgcctcca gcccccccac ctttggcgga 300
ggcaccaagc tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc 360
agcgacgagc agctgaagag tggcaccgcc agcgtggtgt gcctgctgaa caacttctac 420
ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcataag gtgtacgcct gcgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 596
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 596
Gly Phe Thr Phe Ser Asn Phe Ala Met Ser
1. 5 10
<210> 597
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 597
Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 598
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 598
Arg Gly Tyr Ser Gly Val Asp Lys
1. 5
<210> 599
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 599
Ser Asn Phe Ala Met Ser
1. 5
<210> 600
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 600
Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val Lys
1. 5 10 15
Gly
<210> 601
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 601
Arg Gly Tyr Ser Gly Val Asp Lys
1. 5
<210> 602
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 602
Gly Phe Thr Phe Ser Asn Phe
1. 5
<210> 603
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 603
Ser Thr Gly Gly Thr Tyr
1. 5
<210> 604
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 604
Arg Gly Tyr Ser Gly Val Asp Lys
1. 5
<210> 605
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 605
Gly Phe Thr Phe Ser Asn Phe Ala
1. 5
<210> 606
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 606
Ile Ser Thr Gly Gly Thr Tyr Thr
1. 5
<210> 607
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 607
Thr Arg Arg Gly Tyr Ser Gly Val Asp Lys
1. 5 10
<210> 608
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 608
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Tyr Ser Gly Val Asp Lys Trp Gly Gln Gly Thr Thr
100 105 110
Val Thr Val Ser Ser
115
<210> 609
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 609
gaggtgcagc tggtggaatc tggcggaggc ctggtgaaac ccggcggatc cctgagactg 60
tcctgcgccg cctccggctt caccttctcc aacttcgcca tgtcctgggt gcggcaggct 120
cccggcaagg gcctggaatg ggtgtccacc atctccaccg gcggcaccta cacctactac 180
cccgacagcg tgaagggcag attcaccatc agccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgtgc cagacggggc 300
tactcaggcg tggacaaatg gggccagggc accaccgtga cagtgtcctc c 351
<210> 610
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 610
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Ser Thr Gly Gly Thr Tyr Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Tyr Ser Gly Val Asp Lys Trp Gly Gln Gly Thr Thr
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Cys Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Ala Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Cys Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 611
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 611
gaggtgcagc tggtggaatc tggcggaggc ctggtgaaac ccggcggatc cctgagactg 60
tcctgcgccg cctccggctt caccttctcc aacttcgcca tgtcctgggt gcggcaggct 120
cccggcaagg gcctggaatg ggtgtccacc atctccaccg gcggcaccta cacctactac 180
cccgacagcg tgaagggcag attcaccatc agccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac accgccgtgt actactgtgc cagacggggc 300
tactcaggcg tggacaaatg gggccagggc accaccgtga cagtgtcctc cgctagcacc 360
aagggcccaa gtgtgtttcc cctggccccc agcagcaagt ctacttccgg cggaactgct 420
gccctgggtt gcctggtgaa ggactacttc ccctgtcccg tgacagtgtc ctggaactct 480
ggggctctga cttccggcgt gcacaccttc cccgccgtgc tgcagagcag cggcctgtac 540
agcctgagca gcgtggtgac agtgccctcc agctctctgg gaacccagac ctatatctgc 600
aacgtgaacc acaagcccag caacaccaag gtggacaaga gagtggagcc caagagctgc 660
gacaagaccc acacctgccc cccctgccca gctccagaac tgctgggagg gccttccgtg 720
ttcctgttcc cccccaagcc caaggacacc ctgatgatca gcaggacccc cgaggtgacc 780
tgcgtggtgg tggccgtgtc ccacgaggac ccagaggtga agttcaactg gtacgtggac 840
ggcgtggagg tgcacaacgc caagaccaag cccagagagg agcagtacaa cagcacctac 900
agggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agaatacaag 960
tgcaaagtct ccaacaaggc cctggctgcc ccaatcgaaa agacaatcag caaggccaag 1020
ggccagccac gggagcccca ggtgtacacc ctgcccccca gccgggagga gatgaccaag 1080
aaccaggtgt ccctgacctg tctggtgaag ggcttctacc cctgtgatat cgccgtggag 1140
tgggagagca acggccagcc cgagaacaac tacaagacca cccccccagt gctggacagc 1200
gacggcagct tcttcctgta cagcaagctg accgtggaca agtccaggtg gcagcagggc 1260
aacgtgttca gctgcagcgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgagcctga gccccggcaa g 1341
<210> 612
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 612
Lys Ser Gly Gln Ser Leu Leu Asp Ser Thr Gly Lys Thr Tyr Leu Asn
1. 5 10 15
<210> 613
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 613
Leu Val Ser Lys Leu Asp Ser
1. 5
<210> 614
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 614
Trp Gln Gly Thr His Phe Pro Gln Thr
1. 5
<210> 615
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 615
Lys Ser Gly Gln Ser Leu Leu Asp Ser Thr Gly Lys Thr Tyr Leu Asn
1. 5 10 15
<210> 616
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 616
Leu Val Ser Lys Leu Asp Ser
1. 5
<210> 617
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 617
Trp Gln Gly Thr His Phe Pro Gln Thr
1. 5
<210> 618
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 618
Gly Gln Ser Leu Leu Asp Ser Thr Gly Lys Thr Tyr
1. 5 10
<210> 619
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 619
Leu Val Ser
1
<210> 620
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 620
Gly Thr His Phe Pro Gln
1. 5
<210> 621
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 621
Gln Ser Leu Leu Asp Ser Thr Gly Lys Thr Tyr
1. 5 10
<210> 622
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 622
Leu Val Ser
1
<210> 623
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 623
Trp Gln Gly Thr His Phe Pro Gln Thr
1. 5
<210> 624
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 624
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1. 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Gly Gln Ser Leu Leu Asp Ser
20 25 30
Thr Gly Lys Thr Tyr Leu Asn Trp Phe Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 625
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 625
gacgtggtga tgacccagtc ccccctgtcc ctgcctgtga ccctgggcca gcctgcctcc 60
atctcctgca agtccggcca gtccctgctg gactccactg gcaagaccta cctgaactgg 120
ttcctgcagc ggcctggcca gtcccctcgg cggctgatct acctggtgtc caagctggac 180
agcggcgtgc ccgacagatt ctccggctct ggctccggca ccgacttcac cctgaagatc 240
tcccgggtgg aagccgagga cgtgggcgtg tactactgct ggcagggcac ccacttcccc 300
cagaccttcg gcggaggcac caagctggaa atcaag 336
<210> 626
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полипептид"
<400> 626
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1. 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Gly Gln Ser Leu Leu Asp Ser
20 25 30
Thr Gly Lys Thr Tyr Leu Asn Trp Phe Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 627
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
полинуклеотид"
<400> 627
gacgtggtga tgacccagtc ccccctgtcc ctgcctgtga ccctgggcca gcctgcctcc 60
atctcctgca agtccggcca gtccctgctg gactccactg gcaagaccta cctgaactgg 120
ttcctgcagc ggcctggcca gtcccctcgg cggctgatct acctggtgtc caagctggac 180
agcggcgtgc ccgacagatt ctccggctct ggctccggca ccgacttcac cctgaagatc 240
tcccgggtgg aagccgagga cgtgggcgtg tactactgct ggcagggcac ccacttcccc 300
cagaccttcg gcggaggcac caagctggaa atcaagcgta cggtggccgc tcccagcgtg 360
ttcatcttcc cccccagcga cgagcagctg aagagtggca ccgccagcgt ggtgtgcctg 420
ctgaacaact tctacccccg ggaggccaag gtgcagtgga aggtggacaa cgccctgcag 480
agcggcaaca gccaggagag cgtcaccgag caggacagca aggactccac ctacagcctg 540
agcagcaccc tgaccctgag caaggccgac tacgagaagc ataaggtgta cgcctgcgag 600
gtgacccacc agggcctgtc cagccccgtg accaagagct tcaacagggg cgagtgc 657
<210> 628
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: Синтетическая
6xHis метка"
<400> 628
His His His His His His
1. 5
<210> 629
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
пептид"
<400> 629
Tyr Gly Arg Lys Lys Arg Arg Gln Arg Arg Arg Leu Tyr Arg Ser Pro
1. 5 10 15
Ala Met Pro Glu Asn Leu
20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТ ОПУХОЛЕВУЮ ТКАНЬ, ДЛЯ ДИАГНОСТИКИ И ЛЕЧЕНИЯ | 2020 |
|
RU2806915C2 |
| АМАТОКСИНОВЫЕ КОНЪЮГАТЫ АНТИТЕЛА С ЛЕКАРСТВЕННЫМ СРЕДСТВОМ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2826004C2 |
| АНТИТЕЛА К ТАУ И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2787779C2 |
| АНТИТЕЛО, СПОСОБНОЕ СВЯЗЫВАТЬСЯ С ТИМИЧЕСКИМ СТРОМАЛЬНЫМ ЛИМФОПОЭТИНОМ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2825304C2 |
| СТАБИЛЬНЫЕ СОСТАВЫ АНТИТЕЛ ПРОТИВ TIGIT, ОТДЕЛЬНО И В КОМБИНАЦИИ С АНТИТЕЛАМИ ПРОТИВ РЕЦЕПТОРА 1 ПРОГРАММИРУЕМОЙ СМЕРТИ (PD-1), И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2820576C2 |
| АНТИТЕЛА, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ CD70, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2801093C2 |
| АНТИТЕЛА К ENTPD2, ВИДЫ КОМБИНИРОВАННОЙ ТЕРАПИИ И СПОСОБЫ ПРИМЕНЕНИЯ АНТИТЕЛ И ВИДОВ КОМБИНИРОВАННОЙ ТЕРАПИИ | 2019 |
|
RU2790991C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛЕННЫЕ НА АНТИГЕН СОЗРЕВАНИЯ В-КЛЕТОК, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2829097C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ И СВЯЗЫВАЮЩИЕ АГЕНТЫ, НАЦЕЛЕННЫЕ НА DLL3 | 2020 |
|
RU2822366C2 |
| БИСПЕЦИФИЧЕСКИЙ БЕЛОК | 2019 |
|
RU2784486C1 |
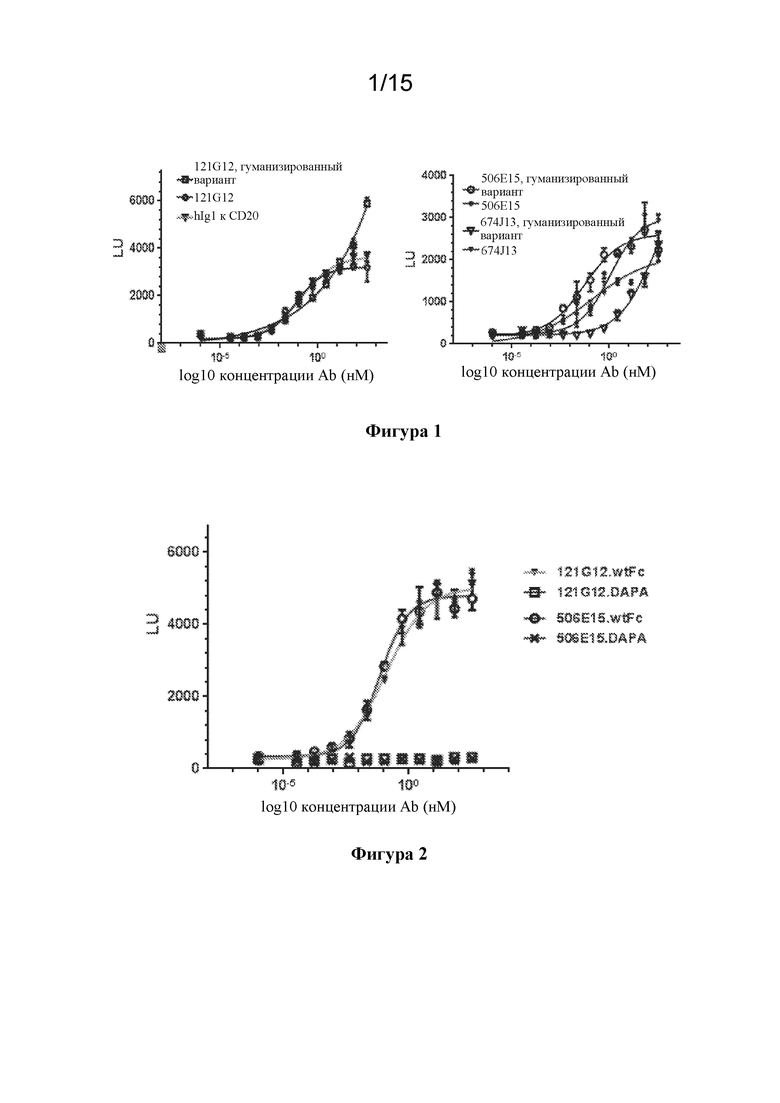
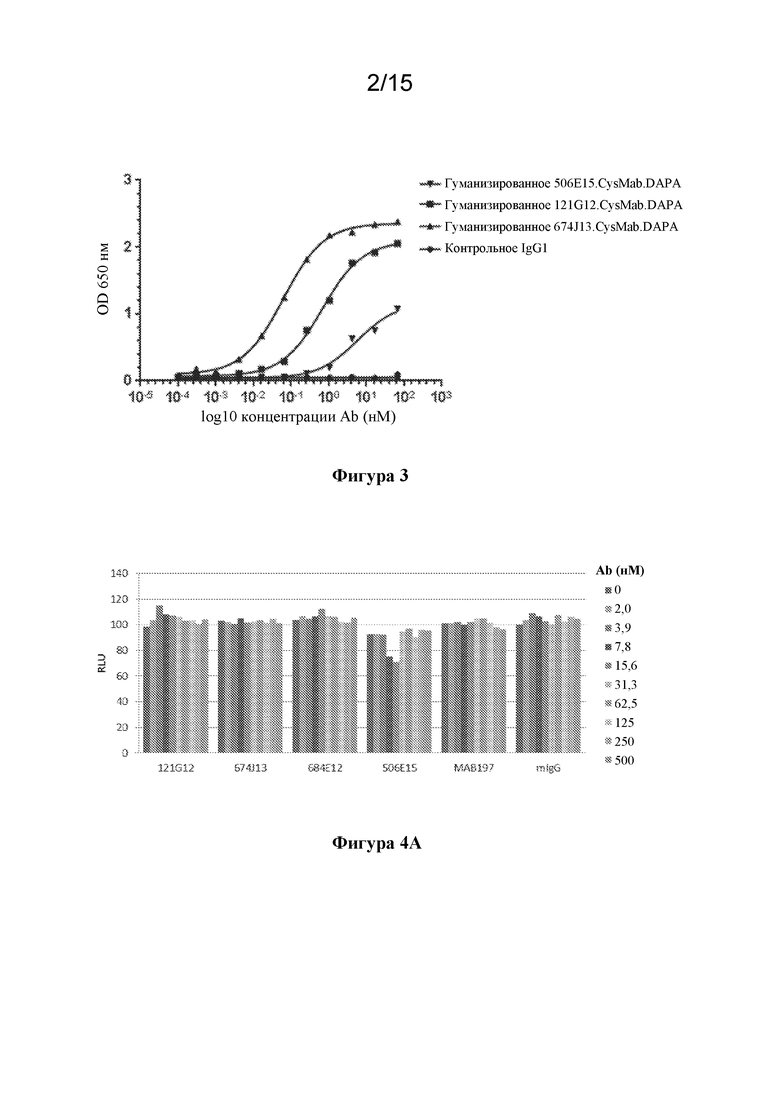
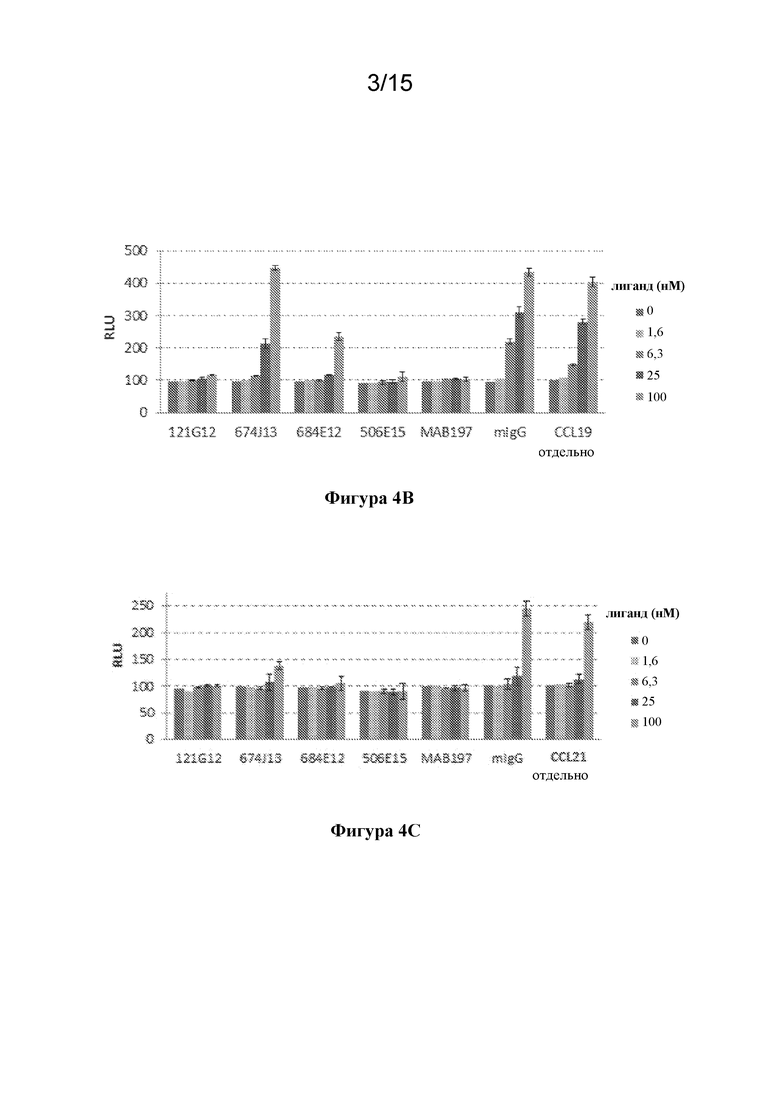
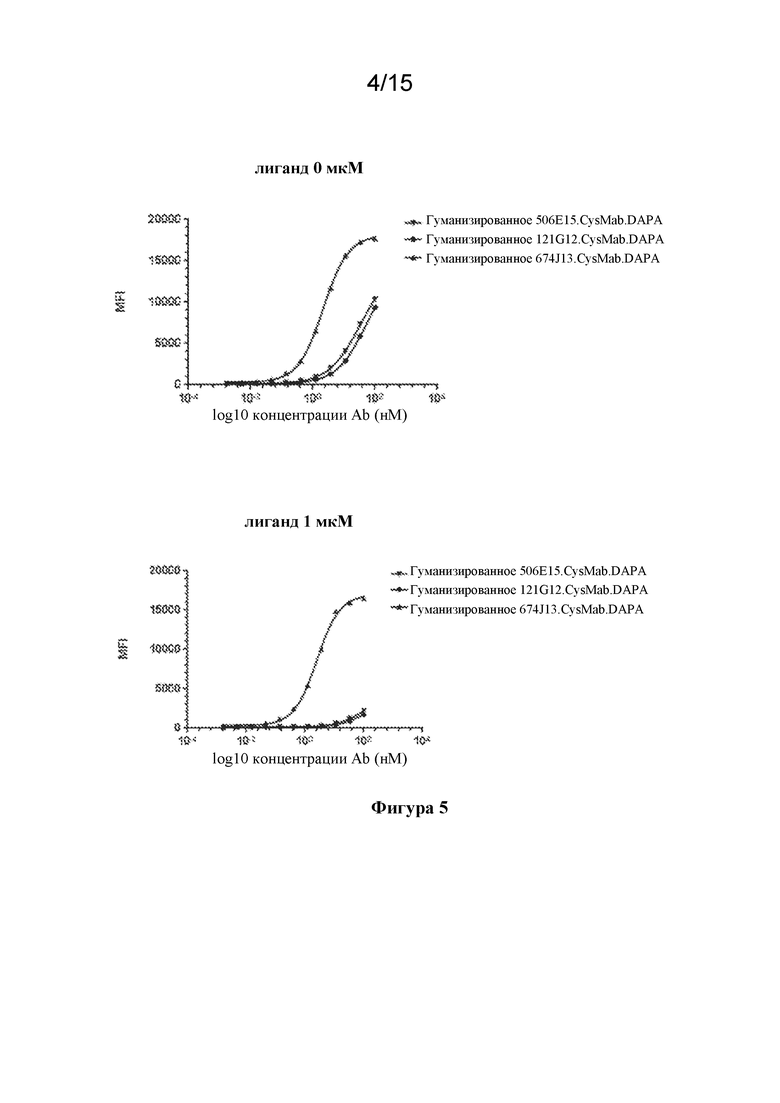
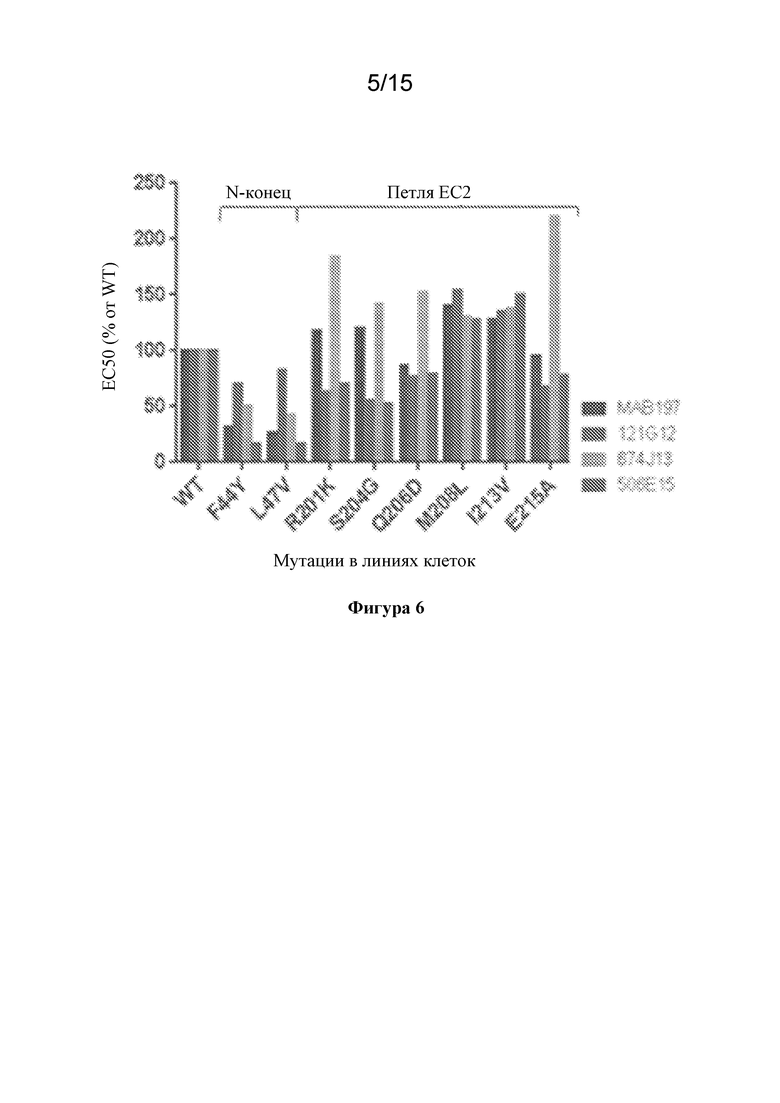
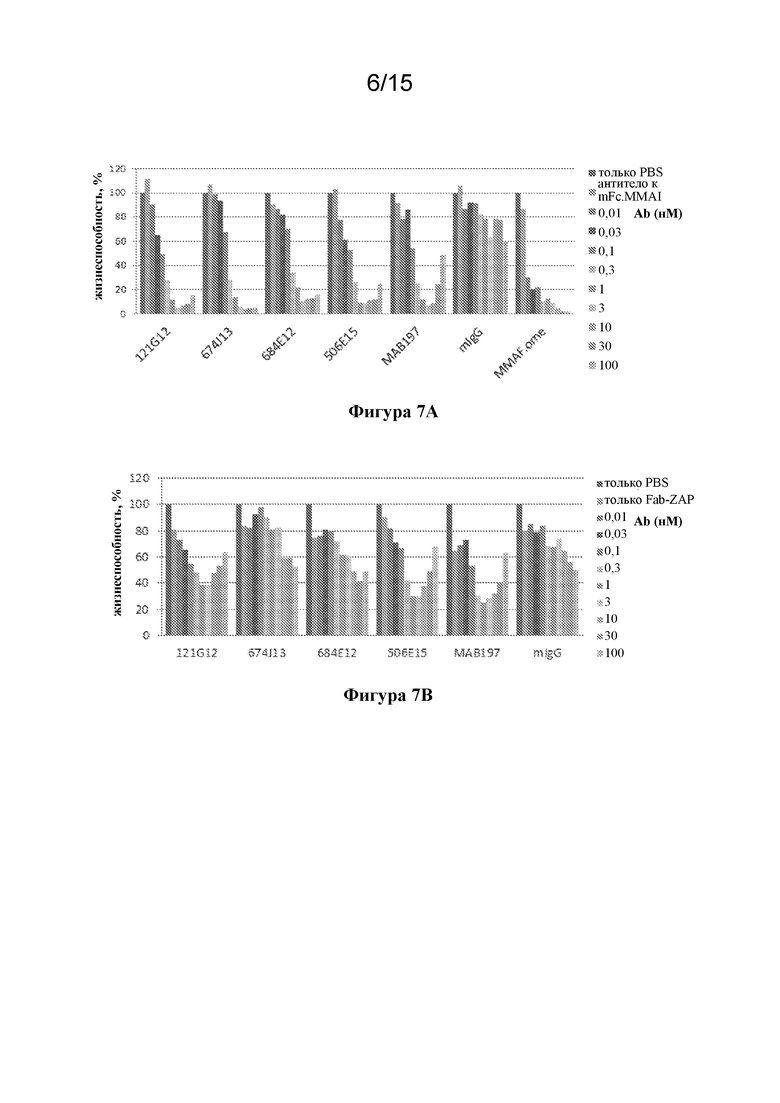
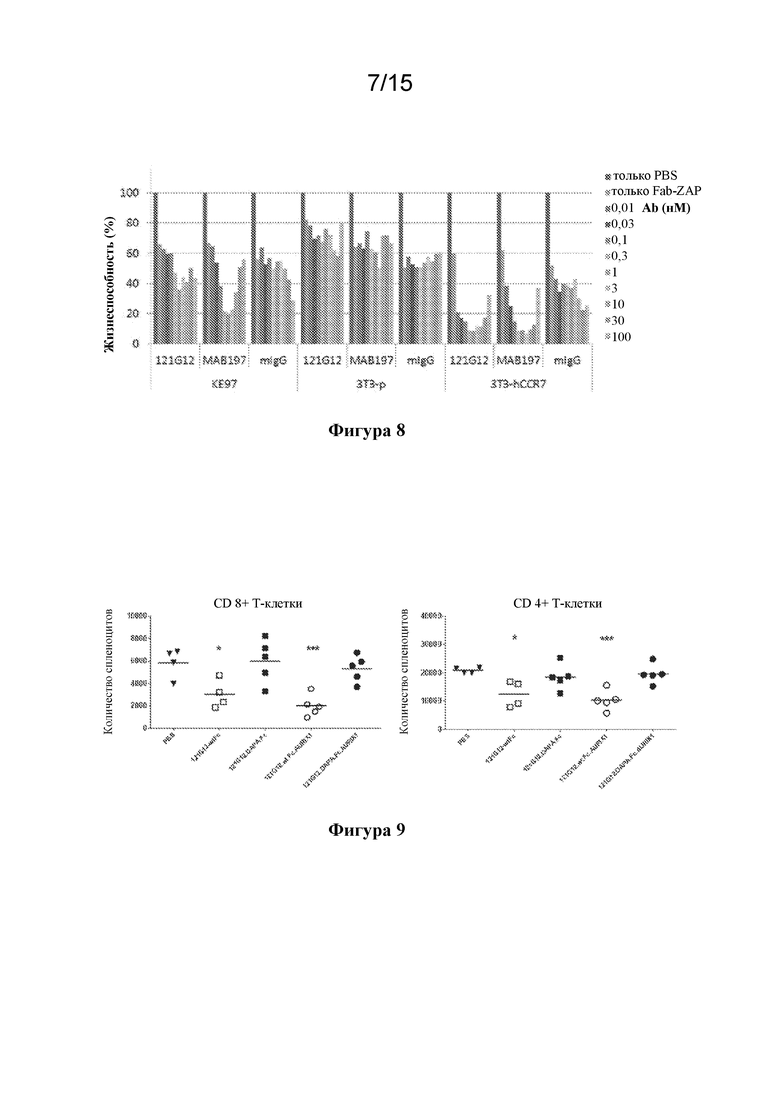
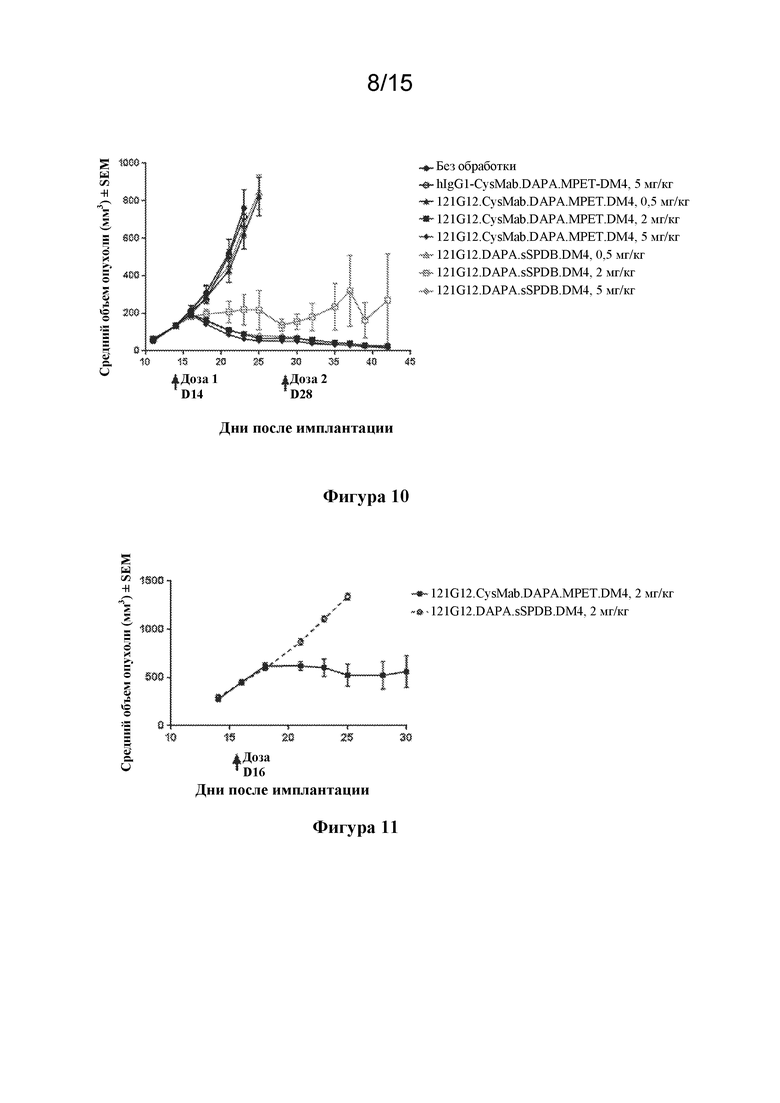
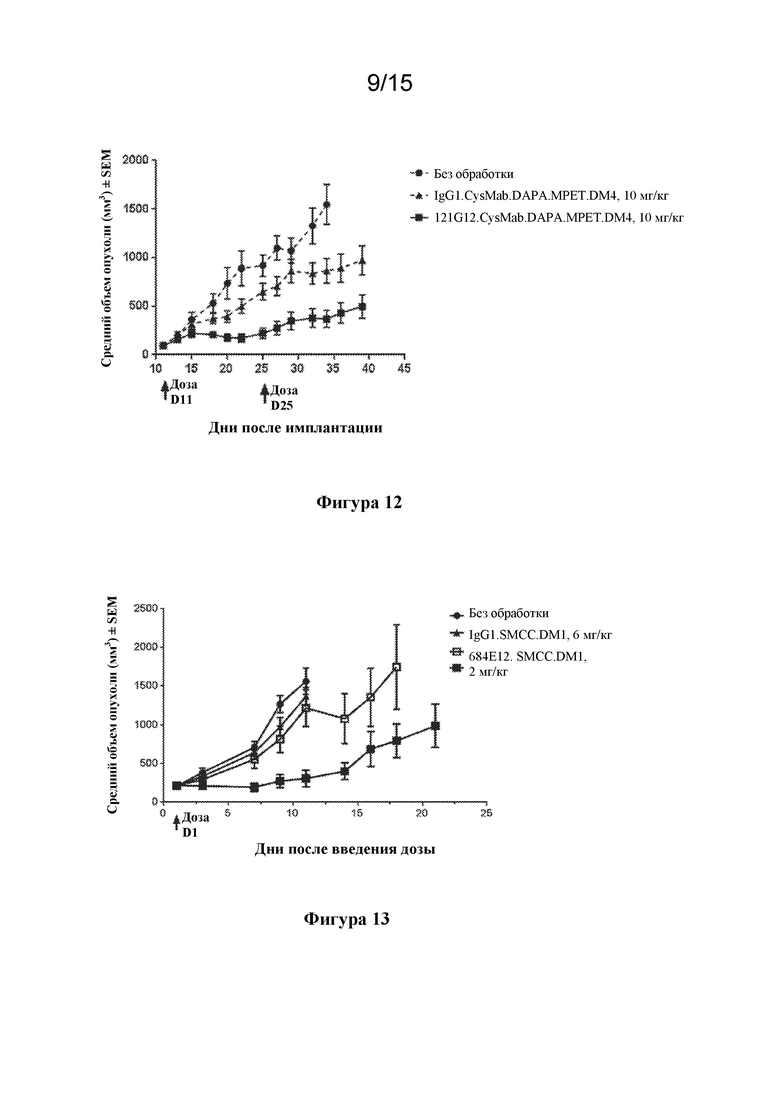

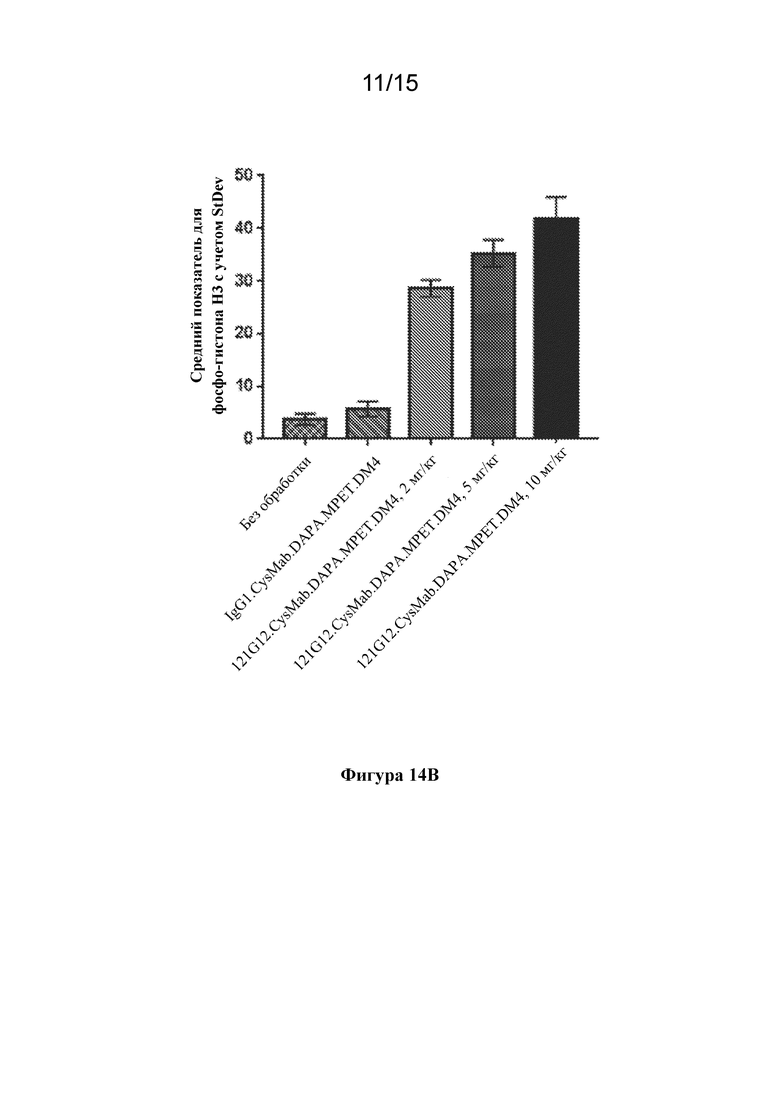

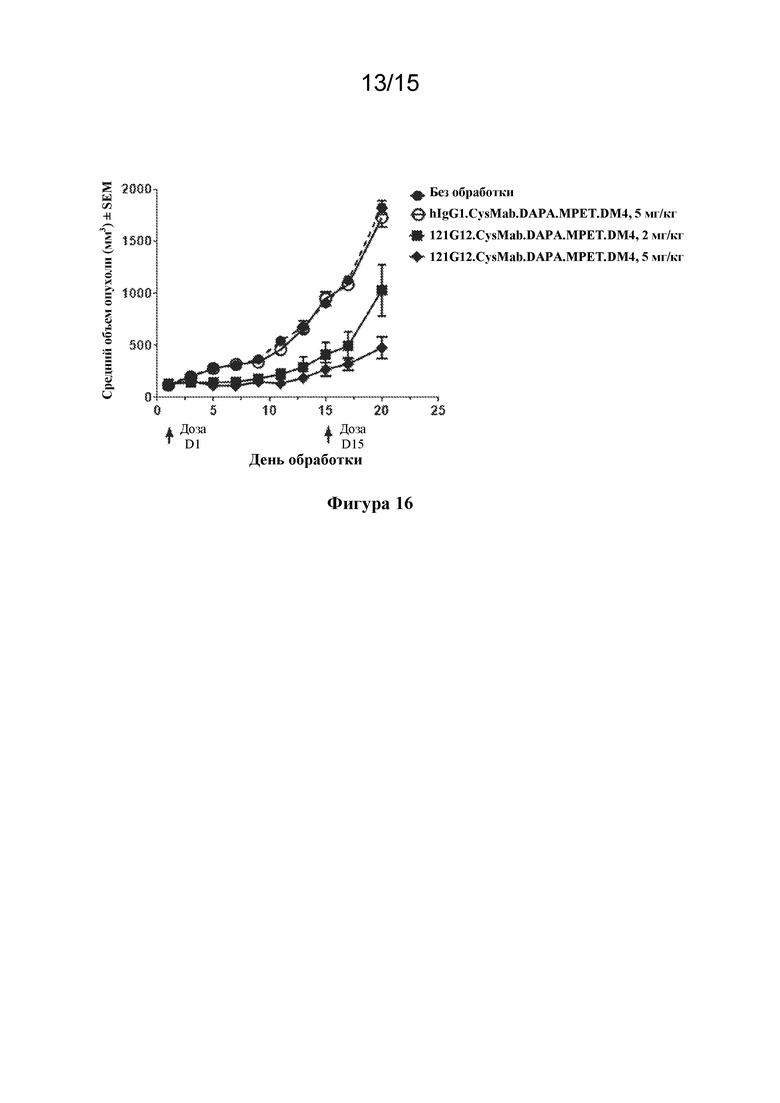
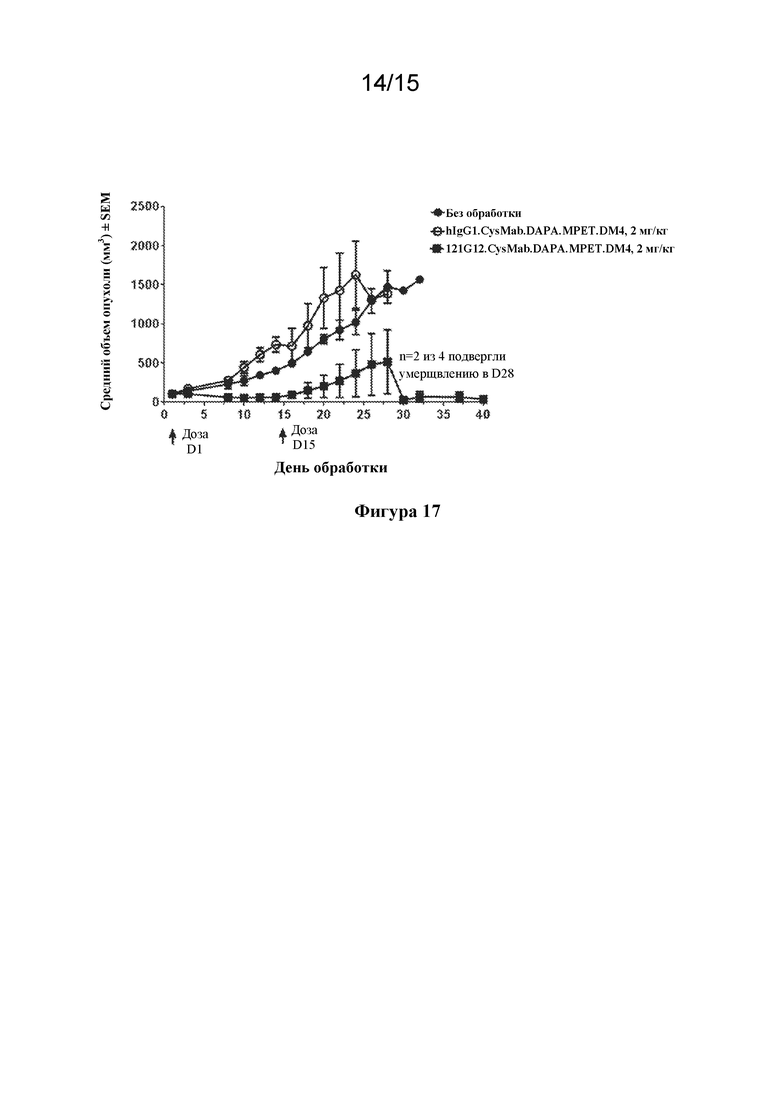
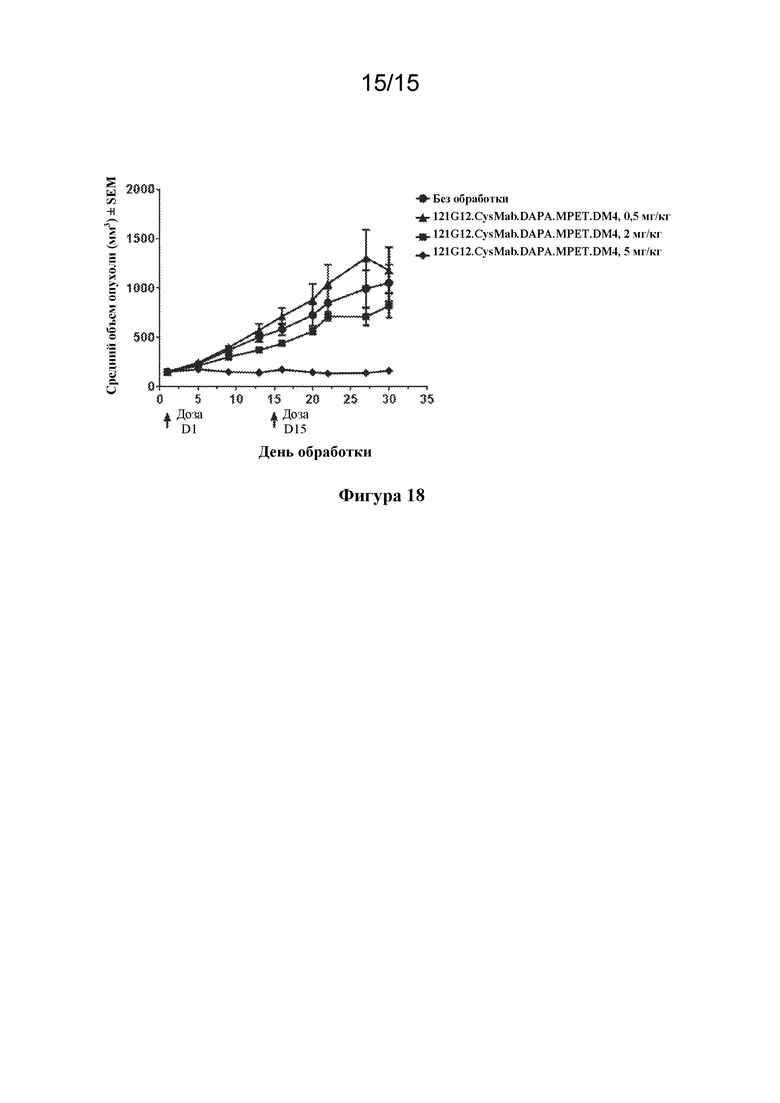
Изобретение относится к области биотехнологии. Предложены антитела к CCR7, их антигенсвязывающие фрагменты и конъюгаты антитела и лекарственного средства на основе указанных антител или антигенсвязывающих фрагментов. Также изобретение относится к фармацевтическим композициям и способам лечения или предупреждения рака с применением указанных антител, антигенсвязывающих фрагментов и конъюгатов. Кроме того, раскрыты нуклеиновые кислоты, векторы экспрессии и клетки-хозяева для получения антител. Изобретение может быть использовано в терапии рака при отсутствии значительного истощения нормальных гемопоэтических клеток, экспрессирующих CCR7. 10 н. и 30 з.п. ф-лы, 22 ил., 27 табл., 21 пр.
1. Антитело или его антигенсвязывающий фрагмент, которые связываются с белком CCR7 человека, где антитело или его антигенсвязывающий фрагмент характеризуются сниженной или незначительной эффекторной функцией по сравнению с антителом дикого типа того же изотипа, содержащие:
a. вариабельную область тяжелой цепи, которая содержит HCDR1 (определяющая комплементарность область 1 тяжелой цепи) под SEQ ID NO:1, HCDR2 (определяющая комплементарность область 2 тяжелой цепи) под SEQ ID NO:2 и HCDR3 (определяющая комплементарность область 3 тяжелой цепи) под SEQ ID NO:3; и вариабельную область легкой цепи, которая содержит LCDR1 (определяющая комплементарность область 1 легкой цепи) под SEQ ID NO:17, LCDR2 (определяющая комплементарность область 2 легкой цепи) под SEQ ID NO:18 и LCDR3 (определяющая комплементарность область 3 легкой цепи) под SEQ ID NO:19;
b. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:4, HCDR2 под SEQ ID NO:5 и HCDR3 под SEQ ID NO:6; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:20, LCDR2 под SEQ ID NO:21 и LCDR3 под SEQ ID NO:22;
c. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:7, HCDR2 под SEQ ID NO:8 и HCDR3 под SEQ ID NO:9; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:23, LCDR2 под SEQ ID NO:24 и LCDR3 под SEQ ID NO:25;
d. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:10, HCDR2 под SEQ ID NO:11 и HCDR3 под SEQ ID NO:12; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:26, LCDR2 под SEQ ID NO:27 и LCDR3 под SEQ ID NO:28;
e. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:33, HCDR2 под SEQ ID NO:34 и HCDR3 под SEQ ID NO:35; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:49, LCDR2 под SEQ ID NO:50 и LCDR3 под SEQ ID NO:51;
f. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:36, HCDR2 под SEQ ID NO:37 и HCDR3 под SEQ ID NO:38; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:52, LCDR2 под SEQ ID NO:53 и LCDR3 под SEQ ID NO:54;
g. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:39, HCDR2 под SEQ ID NO:40 и HCDR3 под SEQ ID NO:41; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:55, LCDR2 под SEQ ID NO:56 и LCDR3 под SEQ ID NO:57;
h. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:42, HCDR2 под SEQ ID NO:43 и HCDR3 под SEQ ID NO:44; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:58, LCDR2 под SEQ ID NO:59 и LCDR3 под SEQ ID NO:60;
i. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:65, HCDR2 под SEQ ID NO:66 и HCDR3 под SEQ ID NO:67; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:81, LCDR2 под SEQ ID NO:82 и LCDR3 под SEQ ID NO:83;
j. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:68, HCDR2 под SEQ ID NO:69 и HCDR3 под SEQ ID NO:70; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:84, LCDR2 под SEQ ID NO:85 и LCDR3 под SEQ ID NO:86;
k. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:71, HCDR2 под SEQ ID NO:72 и HCDR3 под SEQ ID NO:73; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:87, LCDR2 под SEQ ID NO:88 и LCDR3 под SEQ ID NO:89;
l. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:74, HCDR2 под SEQ ID NO:75 и HCDR3 под SEQ ID NO:76; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:90, LCDR2 под SEQ ID NO:91 и LCDR3 под SEQ ID NO:92;
m. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:596, HCDR2 под SEQ ID NO:597 и HCDR3 под SEQ ID NO:598; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:612, LCDR2 под SEQ ID NO:613 и LCDR3 под SEQ ID NO:614;
n. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:599, HCDR2 под SEQ ID NO:600 и HCDR3 под SEQ ID NO:601; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:615, LCDR2 под SEQ ID NO:616 и LCDR3 под SEQ ID NO:617;
o. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:602, HCDR2 под SEQ ID NO:603 и HCDR3 под SEQ ID NO:604; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:618, LCDR2 под SEQ ID NO:619 и LCDR3 под SEQ ID NO:620; или
p. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:605, HCDR2 под SEQ ID NO:606 и HCDR3 под SEQ ID NO:607; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:621, LCDR2 под SEQ ID NO:622 и LCDR3 под SEQ ID NO:623.
2. Антитело по п. 1, где антитело или его антигенсвязывающий фрагмент характеризуются сниженным уровнем или незначительным уровнем активности антителозависимой клеточноопосредованной цитотоксичности (ADCC).
3. Антитело по п. 1 или 2, где антитело или его антигенсвязывающий фрагмент содержат "молчащую" область Fc.
4. Антитело по п. 3, где антитело или его антигенсвязывающий фрагмент содержат мутацию в области Fc, выбранную из: D265A; P329A; P329G; N297A; D265A и P329A; D265A и N297A; L234 и L235A; P329A, L234A и L235A; а также P329G, L234A и L235A.
5. Антитело по любому из пп. 1-4, где антитело или его антигенсвязывающий фрагмент характеризуются незначительной активностью цитолиза клеток.
6. Антитело по любому из пп. 1-5, где антитело или его антигенсвязывающий фрагмент связываются с большей аффинностью с клетками, экспрессирующими более высокие уровни CCR7, чем с клетками, экспрессирующими более низкие уровни CCR7.
7. Антитело по п. 6, где антитело или его антигенсвязывающий фрагмент связываются с большей аффинностью с раковыми клетками, которые экспрессируют более высокие уровни CCR7, чем с нормальными клетками, которые экспрессируют более низкий уровень CCR7.
8. Антитело по любому из пп. 1-7, где антитело или его антигенсвязывающий фрагмент в незначительной степени истощают нормальные гемопоэтические клетки, которые экспрессируют CCR7.
9. Антитело или его антигенсвязывающий фрагмент, которые связывают CCR7, содержащие:
a. вариабельную область тяжелой цепи, которая содержит HCDR1 (определяющая комплементарность область 1 тяжелой цепи) под SEQ ID NO:1, HCDR2 (определяющая комплементарность область 2 тяжелой цепи) под SEQ ID NO:2 и HCDR3 (определяющая комплементарность область 3 тяжелой цепи) под SEQ ID NO:3; и вариабельную область легкой цепи, которая содержит LCDR1 (определяющая комплементарность область 1 легкой цепи) под SEQ ID NO:17, LCDR2 (определяющая комплементарность область 2 легкой цепи) под SEQ ID NO:18 и LCDR3 (определяющая комплементарность область 3 легкой цепи) под SEQ ID NO:19;
b. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:4, HCDR2 под SEQ ID NO:5 и HCDR3 под SEQ ID NO:6; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:20, LCDR2 под SEQ ID NO:21 и LCDR3 под SEQ ID NO:22;
c. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:7, HCDR2 под SEQ ID NO:8 и HCDR3 под SEQ ID NO:9; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:23, LCDR2 под SEQ ID NO:24 и LCDR3 под SEQ ID NO:25;
d. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:10, HCDR2 под SEQ ID NO:11 и HCDR3 под SEQ ID NO:12; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:26, LCDR2 под SEQ ID NO:27 и LCDR3 под SEQ ID NO:28;
e. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:33, HCDR2 под SEQ ID NO:34 и HCDR3 под SEQ ID NO:35; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:49, LCDR2 под SEQ ID NO:50 и LCDR3 под SEQ ID NO:51;
f. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:36, HCDR2 под SEQ ID NO:37 и HCDR3 под SEQ ID NO:38; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:52, LCDR2 под SEQ ID NO:53 и LCDR3 под SEQ ID NO:54;
g. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:39, HCDR2 под SEQ ID NO:40 и HCDR3 под SEQ ID NO:41; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:55, LCDR2 под SEQ ID NO:56 и LCDR3 под SEQ ID NO:57;
h. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:42, HCDR2 под SEQ ID NO:43 и HCDR3 под SEQ ID NO:44; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:58, LCDR2 под SEQ ID NO:59 и LCDR3 под SEQ ID NO:60;
i. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:65, HCDR2 под SEQ ID NO:66 и HCDR3 под SEQ ID NO:67; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:81, LCDR2 под SEQ ID NO:82 и LCDR3 под SEQ ID NO:83;
j. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:68, HCDR2 под SEQ ID NO:69 и HCDR3 под SEQ ID NO:70; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:84, LCDR2 под SEQ ID NO:85 и LCDR3 под SEQ ID NO:86;
k. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:71, HCDR2 под SEQ ID NO:72 и HCDR3 под SEQ ID NO:73; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:87, LCDR2 под SEQ ID NO:88 и LCDR3 под SEQ ID NO:89;
l. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:74, HCDR2 под SEQ ID NO:75 и HCDR3 под SEQ ID NO:76; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:90, LCDR2 под SEQ ID NO:91 и LCDR3 под SEQ ID NO:92;
m. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:596, HCDR2 под SEQ ID NO:597 и HCDR3 под SEQ ID NO:598; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:612, LCDR2 под SEQ ID NO:613 и LCDR3 под SEQ ID NO:614;
n. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:599, HCDR2 под SEQ ID NO:600 и HCDR3 под SEQ ID NO:601; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:615, LCDR2 под SEQ ID NO:616 и LCDR3 под SEQ ID NO:617;
o. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:602, HCDR2 под SEQ ID NO:603 и HCDR3 под SEQ ID NO:604; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:618, LCDR2 под SEQ ID NO:619 и LCDR3 под SEQ ID NO:620; или
p. вариабельную область тяжелой цепи, которая содержит HCDR1 под SEQ ID NO:605, HCDR2 под SEQ ID NO:606 и HCDR3 под SEQ ID NO:607; и вариабельную область легкой цепи, которая содержит LCDR1 под SEQ ID NO:621, LCDR2 под SEQ ID NO:622 и LCDR3 под SEQ ID NO:623.
10. Антитело или его антигенсвязывающий фрагмент, которые связывают CCR7, по п. 9, содержащие:
a. вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:13, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:29;
b. вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:45, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:61;
c. вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:77, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:93; или
d. вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность под SEQ ID NO:608, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность под SEQ ID NO:624.
11. Антитело или его антигенсвязывающий фрагмент, которые связывают CCR7, по п. 9, содержащие:
a. тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:15, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:31;
b. тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:47, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:63;
c. тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:79, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:95; или
d. тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:610, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:626.
12. Антитело по любому из пп. 1-11, где антитело или его антигенсвязывающий фрагмент содержат одну или несколько цистеиновых замен.
13. Антитело по п. 12, где антитело или его антигенсвязывающий фрагмент содержат одну или несколько цистеиновых замен, выбранных из S152C, S375C, или обеих S152C и S375C, в тяжелой цепи антитела или его антигенсвязывающего фрагмента, где нумерация положения соответствует системе EU.
14. Антитело по любому из пп. 1-13, где указанное антитело представляет собой моноклональное антитело.
15. Конъюгат антитела и лекарственного средства для применения в лечении рака, предусматривающий формулу:
Ab-(L-(D)m)n,
или ее фармацевтически приемлемую соль; где
Ab представляет собой антитело или его антигенсвязывающий антигенсвязывающий фрагмент по любому из пп. 1-14;
L представляет собой линкер;
D представляет собой фрагмент, представляющий собой лекарственное средство;
m составляет целое число от 1 до 8; и
n составляет целое число от 1 до 12.
16. Конъюгат антитела и лекарственного средства по п. 15, где указанный m составляет 1.
17. Конъюгат антитела и лекарственного средства по любому из пп. 15 или 16, где указанный n составляет от приблизительно 3 до приблизительно 4.
18. Конъюгат антитела и лекарственного средства по любому из пп. 15-17, где указанный линкер выбран из группы, состоящей из расщепляемого линкера, нерасщепляемого линкера, гидрофильного линкера, предварительно заряженного линкера и линкера на основе дикарбоновой кислоты.
19. Конъюгат антитела и лекарственного средства по п. 18, где линкер получен из сшивающего реагента, выбранного из группы, состоящей из N-сукцинимидил-3-(2-пиридилдитио)пропионата (SPDP), N-сукцинимидил-4-(2-пиридилдитио)пентаноата (SPP), N-сукцинимидил-4-(2-пиридилдитио)бутаноата (SPDB), N-сукцинимидил-4-(2-пиридилдитио)-2-сульфо-бутаноата (сульфо-SPDB), N-сукцинимидилйодацетата (SIA), N-сукцинимидил-(4-йодацетил)аминобензоата (SIAB), малеимид-PEG-NHS, N-сукцинимидил-4-(малеимидометил)циклогексанкарбоксилата (SMCC), N-сульфосукцинимидил-4-(малеимидометил)циклогексанкарбоксилата (сульфо-SMCC) и 2,5-диоксопирролидин-1-ил-17-(2,5-диоксо-2,5-дигидро-1H-пиррол-1-ил)-5,8,11,14-тетраоксо-4,7,10,13-тетраазагептадекан-1-оата (CX1-1).
20. Конъюгат антитела и лекарственного средства по п. 18, где указанный линкер имеет следующую формулу (IIA):
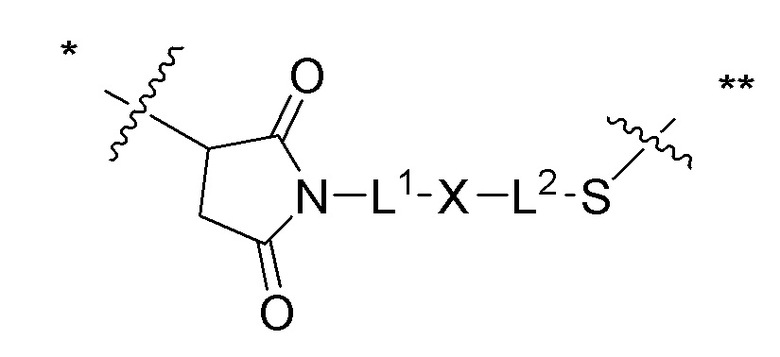 (IIA);
(IIA);
где * соединен с тиольной функциональной группой на антителе, и ** соединен с тиольной функциональной группой фрагмента, представляющего собой лекарственное средство; и где:
L1 представляет собой C1-6алкилен, при этом одна из метиленовых групп может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11;
X представляет собой -C(O)-NH-, -NHC(O)- или триазол; а алкилен является линейным или разветвленным.
21. Конъюгат антитела и лекарственного средства по п. 20, где указанный линкер имеет следующую формулу:
где y составляет от 1 до 11; * соединен с тиольной функциональной группой на антителе, и ** соединен с тиольной функциональной группой фрагмента, представляющего собой лекарственное средство.
22. Конъюгат антитела и лекарственного средства по любому из пп. 15-21, где указанный фрагмент, представляющий собой лекарственное средство, выбран из группы, состоящей из ингибитора V-АТФазы, проапоптического средства, ингибитора Bcl2, ингибитора MCL1, ингибитора HSP90, ингибитора IAP, ингибитора mTor, стабилизатора микротрубочек, дестабилизатора микротрубочек, ауристатина, доластатина, майтанзиноида, MetAP (метионинаминопептидазы), ауристатина, аманитина, пирролобензодиазепина, ингибитора РНК-полимеразы, ингибитора ядерного экспорта белков CRM1, ингибитора DPPIV, ингибиторов протеасомы, ингибиторов реакций переноса фосфорила в митохондриях, ингибитора синтеза белка, ингибитора киназ, ингибитора CDK2, ингибитора CDK9, ингибитора кинезина, ингибитора HDAC, ДНК-повреждающего средства, ДНК-алкилирующего средства, ДНК-интеркалятора, средства, связывающего малую бороздку ДНК, и ингибитора DHFR.
23. Конъюгат антитела и лекарственного средства по п. 22, где цитотоксическое средство представляет собой майтанзиноид.
24. Конъюгат антитела и лекарственного средства по п. 23, где майтанзиноид представляет собой N(2')-деацетил-N(2')-(3-меркапто-l-оксопропил)-майтанзин (DM1), N(2')-деацетил-N(2')-(4-меркапто-1-оксопентил)-майтанзин (DM3) или N(2')-деацетил-N2-(4-меркапто-4-метил-1-оксопентил)-майтанзин (DM4).
25. Конъюгат антитела и лекарственного средства по любому из пп. 15-24, имеющий следующую формулу (VIII):
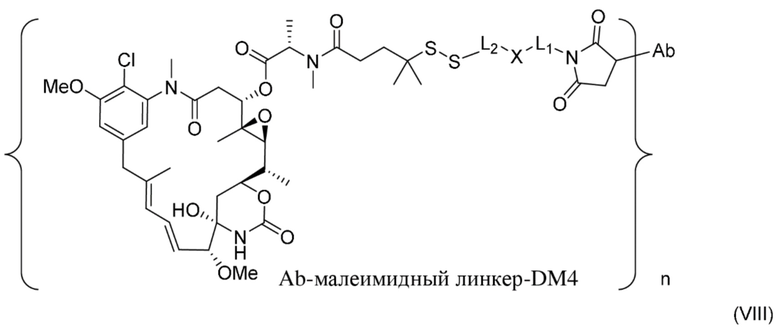
где L1 представляет собой C1-6алкилен, при этом одна метиленовая группа может быть заменена на кислород;
L2 представляет собой C1-6алкилен или представляет собой -(CH2CH2O)y-CH2-CH2-, где y составляет от 1 до 11;
X представляет собой -C(O)-NH-, -NHC(O)- или триазол; и алкилен является линейным или разветвленным; и где n составляет от приблизительно 3 до приблизительно 4; или ее фармацевтически приемлемую соль.
26. Конъюгат антитела и лекарственного средства для применения в лечении рака, имеющий следующую формулу:
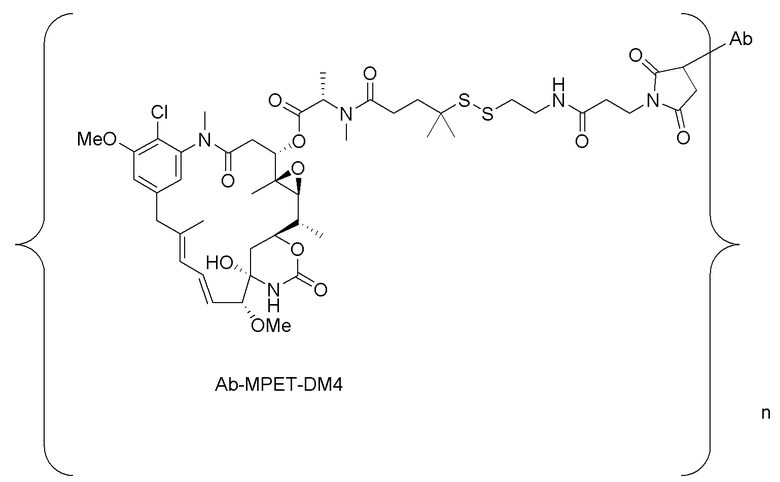
где n составляет от приблизительно 3 до приблизительно 4, и Ab представляет собой антитело, содержащее тяжелую цепь, содержащую аминокислотную последовательность под SEQ ID NO:47, и легкую цепь, содержащую аминокислотную последовательность под SEQ ID NO:63; или ее фармацевтически приемлемую соль.
27. Фармацевтическая композиция для применения в лечении рака, содержащая конъюгат антитела и лекарственного средства по любому из пп. 15-26 и фармацевтически приемлемый носитель.
28. Способ лечения или предупреждения рака у пациента, нуждающегося в этом, предусматривающий введение указанному пациенту конъюгата антитела и лекарственного средства по любому из пп. 15-26 или фармацевтической композиции по п. 27, где рак экспрессирует CCR7.
29. Способ по п. 28, где конъюгат антитела и лекарственного средства или фармацевтическую композицию вводят пациенту в комбинации с одним или несколькими дополнительными терапевтическими соединениями.
30. Способ по п. 29, где одно или несколько дополнительных терапевтических соединений выбраны из стандартного химиотерапевтического средства, костимулирующей молекулы или ингибитора контрольных точек.
31. Способ по п. 30, где костимулирующая молекула выбрана из агониста OX40, CD2, CD27, CDS, ICAM-1, LFA-1 (CD11a/CD18), ICOS (CD278), 4-1BB (CD137), GITR, CD30, CD40, BAFFR, HVEM, CD7, LIGHT, NKG2C, SLAMF7, NKp80, CD160, B7-H3, STING или лиганда CD83.
32. Способ по п. 30, где ингибитор контрольных точек выбран из ингибитора PD-1, PD-L1, PD-L2, CTLA4, TIM3, LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2B4 и/или TGFR-бета.
33. Конъюгат антитела и лекарственного средства по любому из пп. 15-26 или фармацевтическая композиция по п. 27 для применения в качестве лекарственного препарата.
34. Конъюгат антитела и лекарственного средства по любому из пп. 15-26 или фармацевтическая композиция по п. 27 для применения в лечении или предупреждении рака, экспрессирующего CCR7, у пациента, нуждающегося в этом.
35. Применение антитела или его антигенсвязывающего фрагмента по любому из пп. 1-14, или конъюгата антитела и лекарственного средства по любому из пп. 15-26, или фармацевтической композиции по п. 27 при изготовлении лекарственного препарата для применения в лечении рака.
36. Способ по любому из пп. 28-32 или конъюгат антитела и лекарственного средства по п. 34, где рак выбран из группы, состоящей из хронического лимфоцитарного лейкоза (CLL), периферических T-клеточных лимфом (PTCL), таких как T-клеточный лейкоз/лимфома взрослых (ATLL) и анапластическая крупноклеточная лимфома (ALCL), неходжкинской лимфомы (NHL), такой как лимфома из клеток мантийной зоны (MCL), лимфома Беркитта и диффузная В-крупноклеточная лимфома (DLBCL), карциномы желудка, немелкоклеточного рака легкого, мелкоклеточного рака легкого, рака головы и шеи, носоглоточной карциномы (NPC), рака пищевода, колоректальной карциномы, рака поджелудочной железы, рака щитовидной железы, рака молочной железы, почечноклеточного рака и рака шейки матки.
37. Способ по любому из пп. 28-32 или конъюгат антитела и лекарственного средства по п. 34, где рак выбран из группы, состоящей из хронического лимфоцитарного лейкоза (CLL), периферических T-клеточных лимфом (PTCL), таких как T-клеточный лейкоз/лимфома взрослых (ATLL) и анапластическая крупноклеточная лимфома (ALCL), неходжкинской лимфомы (NHL), такой как лимфома из клеток мантийной зоны (MCL), лимфома Беркитта и диффузная В-крупноклеточная лимфома (DLBCL), и немелкоклеточного рака легкого.
38. Нуклеиновая кислота, которая кодирует антитело или антигенсвязывающий фрагмент по любому из пп. 1-14.
39. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 38.
40. Клетка-хозяин, содержащая вектор по п. 39 или нуклеиновую кислоту по п. 38, при этом указанная клетка-хозяин предназначена для получения антитела по любому из пп. 1-14.
| РОТОРНЫЙ ГАЗОТУРБИННЫЙ ДВИГАТЕЛЬ | 2016 |
|
RU2623592C1 |
| WO 2013184200 A1, 12.12.2013 | |||
| WO 2014150937 A1, 25.09.2014 | |||
| АНТИТЕЛА ПРОТИВ РЕЦЕПТОРА CCR7 ДЛЯ ЛЕЧЕНИЯ РАКА | 2006 |
|
RU2404808C2 |
| ALFONSO-PEREZ M | |||
| et al., Anti-CCR7 monoclonal antibodies as a novel tool for the treatment of chronic lymphocyte leukemia, JOURNAL OF LEUKOCYTE BIOLOGY, 2006, Volume 79, Issue 6, pp.1157-1165. | |||

