Изобретение относится к биомедицинским технологиям, а конкретно к группе методов, направленных на изменение проводимости клеточных мембран для управления активностью клеток. Оно может быть использовано для фундаментальных исследований в областях нейробиологии и физиологии, а также создания медицинских технологий, направленных на компенсацию функционального дефицита клеток, тканей и органов человека, скрининга лекарственных препаратов.
Нативные и генно-инженерные белки, обладающие уникальными свойствами, находят широкое практическое применение в биомедицине, благодаря возможности контролировать использование их уникальных свойств в пространстве и времени. Среди них особое место занимают белки, которые, реагируя тем или иным образом на специфические физические и химические стимулы, запускают трансмембранный транспорт различных ионов, вызывая де- или гиперполяризацию клеточной мембраны. Таким образом, воздействуя на эти белки, можно управлять мембранным потенциалом и, следовательно, активностью возбудимых клеток таких, как, например, нервные клетки, мышечные клетки и кардиомиоциты. Экспрессия этих белков в целевых клетках путем трансгенеза широко применяется в фундаментальных научных исследованиях в областях нейробиологии и физиологии, а также служит основой для разработки новых медицинских технологий для компенсации функционального дефицита клеток, тканей и органов человека.
На момент подачи заявки из открытых источников было известно о следующих способах управления трансмембранным транспортом ионов.
Наиболее распространенным методом управления мембранным потенциалом клеток является оптогенетика. В основе метода лежит гетерологичная экспрессия в возбудимых клетках, например, в нейронах светочувствительных ионных каналов водорослей Channelrhodopsin-1 (ChR1), Channelrhodopsin-2 (ChR2) и других [Klapoetke N.C., Murata Y., Kim S.S., Pulver S.R., Birdsey-Benson A., Cho Y.K., Morimoto Т.К., Chuong A.S., Carpenter E.J., Tian Z., Wang J., Xie Y., Yan Z., Zhang Y., Chow B.Y., Surek В., Melkonian M., Jayaraman V., Constantine-Paton M., Wong G.K., Boyden E.S. Independent optical excitation of distinct neural populations. // Nature Methods. 2014. Т. 11. №3. C. 338-46] или их искусственных вариантов, полученных путем мутагенеза (US 2016096036 A1, WO 2021193731 A1, US 2019218256 A1). Эти ионные каналы при облучении их светом с соответствующей длиной волны открываются и пропускают внутрь клетки одновалентные ионы натрия. Это вызывает деполяризацию клеточной мембраны и, как следствие, генерацию потенциала действия.
Помимо активации нервных клеток, оптогенетика успешно используется для управления сокращением мышц [Vogt М., Schulz В., Wagdi A., Lebert J., van Belle G.J., Christoph J., Bruegmann Т., Patejdl R. Direct optogenetic stimulation of smooth muscle cells to control gastric contractility. // Theranostics. 2021. T. 11. №11. C. 5569-84] и сердечным ритмом [Arrenberg A.B., Stainier D.Y.R., Baier H., Huisken J. // Science. 2010. T. 330. №6006. C. 971-74; Bruegmann Т., Malan D., Hesse M., Beiert Т., Fuegemann C.J., Fleischmann B.K., Sasse P. Optogenetic control of heart muscle in vitro and in vivo. // Nature Methods. 2010. T. 7. №11. C. 897-900], а также для восстановления зрительной функции при некоторых видах слепоты у человека [Sahel J.-A., Boulanger-Scemama Е., Pagot С, Arleo A., Galluppi F., Martel J.N., Esposti S.D., Delaux A., de Saint Aubert J.-B., de Montleau C, Gutman E., Audo I., Duebel J., Picaud S., Dalkara D., Blouin L., Taiel M., Roska B. Partial recovery of visual function in a blind patient after optogenetic therapy. // Nature Medicine. 2021. T. 27. №7. C. 1223-29].
Главным недостатком оптогенетики является низкая проводимость светочувствительных каналов, из-за которой необходимо использовать свет высокой интенсивности, чтобы вызвать токи ионов, достаточные для изменения мембранного потенциала. В результате использования света высокой интенсивности происходит фототоксическое повреждение биологических тканей, что влияет на результаты научных исследований и ограничивает практическое применение оптогенентики.
Кроме того, практическое применение оптогенетики ограничено тем, что используемые светочувствительные каналы происходят из водорослей, архей и бактерий. Поэтому при их синтезе в организме человека и даже в центральной нервной системе, отделенной гематоэнцефалическим барьером, сохраняется риск конфликта с иммунитетом, который может привести к повреждению биологических тканей.
Еще одним подходом для управления активностью нервных клеток является применение генно-инженерных белков на основе G-белок зависимых рецепторов, которые мутированы таким образом, чтобы реагировать на какой-либо инертный для организма лиганд. Такой подход называют хемогенетикой. Наиболее широкое распространение получила технология DREADD (от англ. яз. designer receptor exclusively activated by designer drugs) (US 2019083652 A1, US 2021077635 A1). Она включает генно-инженерные белки на основе активирующего мускаринового ацетилхолинового рецептора М3 (hM3Dq) и ингибиторного мускаринового ацетилхоинового рецептора М4 (hM4Di). Связывание синтетического инертного лиганда клозапин-N-оксида с hM3Dq или hM4Di ведет либо к активации, либо к торможению нервных клеток соответственно. Главный недостаток хемогенетики заключается в том, что с помощью этого метода трудно контролировать активность генно-инженерных G-белок зависимых рецепторов, поскольку она зависит от локальной концентрации и кинетики распространения синтетического лиганда. Это ограничивает применение хемогенетики для управления быстрыми процессами.
Наиболее перспективной альтернативой опто- и хемогенетике является термогенетика. Эта технология эксплуатирует термочувствительные каналы надсемейства TRP (transient receptor potential channels, каналы временного рецепторного потенциала) преимущественно TRPV1 и TRPA1 [Ermakova Y.G., Lanin А.А., Fedotov I.V., Roshchin M., Kelmanson I.V., Kulik D., Bogdanova Y.A., Shokhina A.G., Bilan D.S., Staroverov D.B., Balaban P.M., Fedotov A.B., Sidorov-Biryukov D.A., Nikitin E.S., Zheltikov A.M., Belousov V. v. Thermogenetic neurostimulation with single-cell resolution. // Nature Communications. 2017. T. 8. №1. C. 15362; Roshchin M., Ermakova Y.G., Lanin A.A., Chebotarev A.S., Kelmanson I. v., Balaban P.M., Zheltikov A.M., Belousov V.V., Nikitin E.S. Thermogenetic stimulation of single neocortical pyramidal neurons transfected with TRPV1-L channels. //Neuroscience Letters. 2018. T. 687. C. 153-57., Yang Y., Pacia C.P., Ye D., Zhu L., Baek H., Yue Y., Yuan J., Miller M.J., Cui J., Culver J.P., Bruchas M.R., Chen H. Sonothermogenetics for noninvasive and cell-type specific deep brain neuromodulation. // Brain Stimulation. 2021. T. 14. №4. C. 790-800]. TRP каналы плазматической мембраны клеток относятся к семейству неселективных катионных каналов и являются сенсорами для широкого спектра физических и химических стимулов, таких как тепло, холод, механическое воздействие, рН, осмолярность и целый ряд химических соединений эндогенного и экзогенного происхождения. Активация этих каналов соответствующими стимулами вызывает изменение мембранного потенциала, запускает трансмембранный транспорт различных ионов, преимущественно кальция и натрия.
Управление работой TRP каналов при их гетерологичной экспрессии в клетках осуществляется путем незначительного нагрева, который не приводит к термическому повреждению клеточных компонентов и гибели клеток. Нагрев клеток может быть осуществлен путем их облучения инфракрасным лазером с длиной волны в диапазоне примерно 1300-1400 нм или с помощью сфокусированного ультразвука высокой интенсивности [Ermakova Y.G., Lanin А.А., Fedotov I. v., Roshchin M., Kelmanson I. v., Kulik D., Bogdanova Y.A., Shokhina A.G., Bilan D.S., Staroverov D.B., Balaban P.M., Fedotov A.B., Sidorov-Biryukov D.A., Nikitin E.S., Zheltikov A.M., Belousov V. v. Thermogenetic neurostimulation with single-cell resolution. // Nature Communications. 2017. T. 8. №1. C. 15362, Roshchin M., Ermakova Y.G., Lanin A.A., Chebotarev A.S., Kelmanson I. v., Balaban P.M., Zheltikov A.M., Belousov V. v., Nikitin E.S. Thermogenetic stimulation of single neocortical pyramidal neurons transfected with TRPV1-L channels. //Neuroscience Letters. 2018. T. 687. C. 153-57, Yang Y., Pacia C.P., Ye D., Zhu L., Baek H., Yue Y., Yuan J., Miller M.J., Cui J., Culver J.P., Bruchas M.R., Chen H. Sonothermogenetics for noninvasive and cell-type specific deep brain neuromodulation. // Brain Stimulation. 2021. T. 14. №4. C. 790-800].
Таким образом, с помощью термогенетики можно управлять активностью нервных клеток [Ermakova Y.G., Lanin А.А., Fedotov I. v., Roshchin M., Kelmanson I. v., Kulik D., Bogdanova Y.A., Shokhina A.G., Bilan D.S., Staroverov D.B., Balaban P.M., Fedotov A.B., Sidorov-Biryukov D.A., Nikitin E.S., Zheltikov A.M., Belousov V. v. Thermogenetic neurostimulation with single-cell resolution. // Nature Communications. 2017. T. 8. №1. C. 15362, Roshchin M., Ermakova Y.G., Lanin A.A., Chebotarev A.S., Kelmanson I. v., Balaban P.M., Zheltikov A.M., Belousov V. v., Nikitin E.S. Thermogenetic stimulation of single neocortical pyramidal neurons transfected with TRPV1-L channels. //Neuroscience Letters. 2018. T. 687. C. 153-57].
Одним из преимуществ термочувствительных TRP каналов, используемых в термогенетике, по сравнению со светочувствительными каналродопсинами, применяемыми в оптогенетике, является более чем на три порядка более высокая проводимость. Так, проводимость канала TRPV1 крысы для ионов натрия оценивается в 80 pS [Tominaga М., Caterina M.J., Malmberg А.В., Rosen T.A., Gilbert H., Skinner K., Raumann B.E., Basbaum A.I., Julius D. The Cloned Capsaicin Receptor Integrates Multiple Pain-Producing Stimuli. // Neuron. 1998. Т. 21. №3. С. 531-43], в то время как проводимость каналродопсина ChR2 составляет порядка 40 fS [Feldbauer K., Zimmermann D., Pintschovius V., Spitz J., Bamann C., Bamberg E. Channelrhodopsin-2 is a leaky proton pump.// Proceedings of the National Academy of Sciences. 2009. T. 106. №30. C. 12317-12322], что в 2000 раз меньше. В отличие от плохо контролируемой химической активации, которая используется в хемогенетике, быстрая активация TRP каналов облучением инфракрасным лазером или с помощью сфокусированного ультразвука высокой интенсивности не только позволяет управлять динамическими процессами, но еще и дозировать воздействие.
Известно, что для управления мембранным потенциалом с помощью термогенетики использовались только нативные TRPV1 и TRPA1 каналы различных организмов. Один из недостатков термогенетики на основе нативных TRP каналов, и в частности на основе канала TRPV1 заключается в том, что поступление ионов кальция в клетки через эти каналы может приводить перегрузу клеток кальцием, который при высоких концентрациях может оказывать цитотоксический эффект [Beider K., Rosenberg Е., Dimenshtein-Voevoda V., Sirovsky Y., Vladimirsky J., Magen H., Ostrovsky O., Shimoni A., Bromberg Z., Weiss L., Peled A., Nagler A. Blocking of Transient Receptor Potential Vanilloid 1 (TRPV1) promotes terminal mitophagy in multiple myeloma, disturbing calcium homeostasis and targeting ubiquitin pathway and bortezomib-induced unfolded protein response.// Journal of Hematology & Oncology. 2020. T. 13. №1. C. 158; Kahya M.C., Naziroglu M., 
 Modulation of Diabetes-Induced Oxidative Stress, Apoptosis, and Ca2+ Entry Through TRPM2 and TRPV1 Channels in Dorsal Root Ganglion and Hippocampus of Diabetic Rats by Melatonin and Selenium. // Molecular Neurobiology. 2017. T. 54. №3. C. 2345-2360; Olivan-Viguera A., Garcia-Otin A.L., Lozano-Gerona J., Abarca-Lachen E., Garcia-Malinis A.J., Hamilton K.L., Gilaberte Y., Pueyo E.,
Modulation of Diabetes-Induced Oxidative Stress, Apoptosis, and Ca2+ Entry Through TRPM2 and TRPV1 Channels in Dorsal Root Ganglion and Hippocampus of Diabetic Rats by Melatonin and Selenium. // Molecular Neurobiology. 2017. T. 54. №3. C. 2345-2360; Olivan-Viguera A., Garcia-Otin A.L., Lozano-Gerona J., Abarca-Lachen E., Garcia-Malinis A.J., Hamilton K.L., Gilaberte Y., Pueyo E.,  R. Pharmacological activation of TRPV4 produces immediate cell damage and induction of apoptosis in human melanoma cells and HaCaT keratinocytes. // PLOS ONE. 2018. Т. 13. №1. С.e0190307; Stueber Т., Eberhardt M.J., Caspi Y., Lev S., Binshtok A., Leffler A. Differential cytotoxicity and intracellular calcium-signalling following activation of the calcium-permeable ion channels TRPV1 and TRPA1. // Cell Calcium. 2017. T. 68. C. 34-44; Sun Z., Han J., Zhao W., Zhang Y., Wang S., Ye L., Liu Т., Zheng L. TRPV1 Activation Exacerbates Hypoxia/Reoxygenation-Induced Apoptosis in H9C2 Cells via Calcium Overload and Mitochondrial Dysfunction. // International Journal of Molecular Sciences. 2014. T. 15. №10. C. 18362-18380; Wang S., Wang S., Asgar J., Joseph J., Ro J.Y., Wei F., Campbell J.N., Chung M.-K. Ca2+ and calpain mediate capsaicin-induced ablation of axonal terminals expressing transient receptor potential vanilloid 1. // Journal of Biological Chemistry. 2017. T. 292. №20. C. 8291-8303]. Риск перегруза клеток ионами кальция при использовании термочувствительных TRP каналов в качестве инструментов термогенетики является ограничивающим фактором для данной биомедицинской технологии.
R. Pharmacological activation of TRPV4 produces immediate cell damage and induction of apoptosis in human melanoma cells and HaCaT keratinocytes. // PLOS ONE. 2018. Т. 13. №1. С.e0190307; Stueber Т., Eberhardt M.J., Caspi Y., Lev S., Binshtok A., Leffler A. Differential cytotoxicity and intracellular calcium-signalling following activation of the calcium-permeable ion channels TRPV1 and TRPA1. // Cell Calcium. 2017. T. 68. C. 34-44; Sun Z., Han J., Zhao W., Zhang Y., Wang S., Ye L., Liu Т., Zheng L. TRPV1 Activation Exacerbates Hypoxia/Reoxygenation-Induced Apoptosis in H9C2 Cells via Calcium Overload and Mitochondrial Dysfunction. // International Journal of Molecular Sciences. 2014. T. 15. №10. C. 18362-18380; Wang S., Wang S., Asgar J., Joseph J., Ro J.Y., Wei F., Campbell J.N., Chung M.-K. Ca2+ and calpain mediate capsaicin-induced ablation of axonal terminals expressing transient receptor potential vanilloid 1. // Journal of Biological Chemistry. 2017. T. 292. №20. C. 8291-8303]. Риск перегруза клеток ионами кальция при использовании термочувствительных TRP каналов в качестве инструментов термогенетики является ограничивающим фактором для данной биомедицинской технологии.
Нами поставлена задача - исключить риск повреждения и гибели возбудимых клеток в результате перегруза их ионами кальция при использовании канала TRPV1 человека в качестве термогенетического инструмента для управления трансмембранным транспортом катионов.
Технический результат, достигаемый при осуществлении изобретения, заключается в исключении повреждения и гибели возбудимых клеток за счет перегруза их ионами кальция во время термогенетического управления трансмембранным транспортом катионов с использованием канала TRPV1 человека.
Сущность изобретения заключается в следующем.
Предложена нуклеиновая кислота, кодирующая термочувствительный канал TRPV1 человека с измененной ионной селективностью, обеспечивающий деполяризацию клеточной мембраны и лишенный проводимости двухвалентных катионов кальция (TRPV1ΔCa2+), имеющий последовательность SEQ ID NO: 1.
При транскрипции и трансляции в клетках организмов предлагаемой нуклеиновой кислоты, созданной в рамках настоящего изобретении с помощью рекомбинантных технологий, образуется катионный канал TRPV1ΔCa2+ с измененной ионной селективностью, который деполяризует мембрану клеток при нагреве или действии агониста капсаицина, но при этом остается непроницаемым для двухвалентных ионов кальция.
Таким образом, при использовании TRPV1ΔCa2+ в качестве термогенетического инструмента для управления мембранным потенциалом клеток исключается возможность перегруза их ионами кальция, что в конечном счете предотвращает повреждение и гибель клеток в результате цитотоксического эффекта высоких концентраций внутриклеточного кальция.
Для решения проблемы мы предложили внести аминокислотные замены в поровую петлю канала TRPV1 человека, которая определяет ионную селективность поры канала [Liao М., Cao Е., Julius D., Cheng Y. Structure of the TRPV1 ion channel determined by electron cryo-microscopy. // Nature. 2013. T. 504. №7478. C. 107-112].
Замена с помощью рекомбинантных технологий изолейцина на валин и метионина на тирозин в канале TRPV1 человека в позициях 643 и 645, соответственно, привела к созданию варианта канала TRPV1 человека с измененной селективностью. Созданный таким образом вариант канала TRPV1 человека, названный нами TRPV1ACa2+ и определяемый аминокислотной последовательностью SEQ ID: 1, сохранил способность деполяризовать клетки при нагреве или действии агониста, но в отличие от нативного канала он стал непроницаем для двухвалентных катионов кальция.
Таким образом, настоящее изобретение представляет собой нуклеиновую кислоту, при транскрипции и трансляции которой в клетках образуется термочувствительный катионный канал TRPV1ΔCa2+ с аминокислотной последовательностью, определяемой SEQ ID NO: 1.
Также, как и нативный канал TRPV1 человека с аминокислотной последовательностью, определяемой идентификационным номером САВ66735.1 в базе данных NCBI (https://www.ncbi.nlm.nih.gov), TRPV1ΔСа2+ при его синтезе в клетках вызывает деполяризацию клеточной мембраны и поэтому может быть использован наравне с нативным TRPV1 человека для термогенетического управления мембранным потенциалом.
Однако, в отличие от нативного канала TRPV1 человека TRPV1ΔCa2+ не проводит двухвалентные катионы кальция в результате внесения с помощью рекомбинантных технологий изменений в кодирующую его нуклеотидную последовательность, которые привели к аминокислотным заменам в поровой петле, определяющей ионную селективность канала. Изолейцин в позиции 643 был заменен на валин, и метионин в позиции 645 был заменен на тирозин.
Настоящее изобретение представляет собой нуклеиновую кислоту, которая кодируют полипептидную цепь с аминокислотной последовательностью, определенной SEQ ID NO: 1. Данная полипептидная цепь является одной субъединицей катионного канала TRPV1ΔCa2+ с измененной ионной селективностью. Она была получена путем внесения точечных аминокислотных замен в последовательность нативного термочувствительного канала TRPV1 человека, определяемую идентификационным номером САВ66735.1 в базе данных NCBI (https://www.ncbi.nlm.nih.gov). Для практической реализации изобретения в нуклеотидную последовательность открытой рамки считывания, кодирующей нативный канал TRPV1 человека, с использованием методов молекулярного клонирования [Green M.R and Sambrook J. Molecular cloning: a laboratory manual. 2012. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.] вносили замены таким образом, чтобы кодон в позиции 643 кодировал валин (Val), а кодон в позиции 645 кодировал тирозин (Tyr) вместо изолейцина (Ile) и метионина (Met) соответственно (Фиг. 1). На Фиг. 1 представлены выравненные аминокислотные последовательности нативного канала TRPV1 человека и генно-инженерного катионного канала TRPV1ΔСа2+ с измененной ионной селективностью, а критические замены Ile643Val и Met645Tyr выделены темным фоном. Эти замены производятся в так называемой поровой петле, которая выполняет роль фильтра ионной селективности [Liao М., Сао Е., Julius D., Cheng Y. Structure of the TRPV1 ion channel determined by electron cryo-microscopy. //Nature. 2013. T. 504. №7478. C. 107-112].
Для внесения в последовательность канала аминокислотных замен Ile643Val и Met645Tyr фрагмент открытой рамки считывания, кодирующий первые 803 аминокислоты нативного канала TRPV1 человека, был клонирован в плазмидный вектор pAL2-T.
С использованием данной конструкции в качестве матрицы была проведена полимеразная цепная реакция (ПЦР) с праймерами F1 и R1, нематричная часть 5'-конца которых вносила соответствующие нуклеотидные замены. При этом были использованы: прямой праймер F1, имеющий нуклеотидную последовательность SEQ ID NO: 2 и обратный праймер R1, имеющий нуклеотидную последовательность SEQ ID NO: 3 для внесения нуклеотидных замен в кДНК, кодирующую канал TRPV1 человека для получения канала TRPV1 человека с измененной ионной селективностью, обеспечивающего деполяризацию клеточной мембраны и лишенного проводимости двухвалентных катионов кальция (TRPV1ΔCa2+). Для аминокислотной замены Ile643Val кодон АТС был заменен на кодон GTG, для аминокислотной замены Met645Tyr кодон ATG был заменен на ТАС. Далее ПЦР-продукт был кинирован с помощью Т4 полинуклеотидкиназы. После лигирования в течение ночи при температуре 14°С лигазную смесь использовали для трансформации компетентных клеток XL 1-Blue. Успешность внесения замен была подтверждена секвенированием. По присутствующим в последовательности канала сайтам рестрикции Drain и Smal содержащий замены фрагмент был переклонирован в вектор pCAGGS. Полученный конечный вектор содержит открытую рамку считывания, имеющую нуклеотидную последовательность, кодирующую катионный канал TRPV1ΔСа2+ с измененной ионной селективностью.
Полученная нуклеиновая кислота может быть использована в дальнейшем для получения организмов трансформантов, в клетках которых продуцируется катионный канал TRPV1ΔCa2+ с измененной ионной селективностью. Встраивание нуклеиновой кислоты по настоящему изобретению в про- и эукариотические организмы может осуществляться всеми известными способами получения трансгенных организмов (все виды трансформации, трансфекции, трансдукции и рекомбинации) [Green M.R and Sambrook J. Molecular cloning: a laboratory manual. 2012. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.; Pinkert C. Transgenic animal technology: a laboratory handbook. 2002. Academic Press, Amsterdam; Boston; Joyner A.L. Gene targeting: a practical approach, Practical Approach Series. 2000. Oxford University Press, Oxford, New York],
При этом экспрессионная кассета, включающая регион инициации транскрипции, открытую рамку считывания, кодирующую катионный канал TRPV1ΔCa2+ с измененной ионной селективностью, и регион терминации транскрипции, может присутствовать в клетке-хозяине как в виде нехромосомного элемента, так и в виде элемента, встроенного в клеточный геном, исключая клетки зародышевой линии человека. В результате транскрипции и трансляции открытой рамки считывания в клетках, содержащих трансген, продуцируются полипептидные цепи одной субъединицы катионного канала TRPV1ΔCa2+ с измененной ионной селективностью. Продуцируемые субъединицы посредством межмолекулярных взаимодействий объединяются в гомотетрамерную структуру, образуя таким образом функциональный канал, способный деполяризовать клеточную мембрану в отсутствие при этом проводимости катионов кальция.
Следующий пример предлагается в качестве иллюстративного, но не ограничивающего иные применения генно-инженерного канала TRPV1ΔСа2+.
Пример.
Деполяризация клеточной мембраны без участия катионов кальция путем активации TRPV1ΔCa2+.
Для проверки того, что в результате транскрипции и трансляции созданной в настоящем изобретении нуклеиновой кислоты в клетках образуется катионный канал TRPV1ΔCa2+ с измененной ионной селективностью, методами молекулярного клонирования [Green M.R and Sambrook J. Molecular cloning: a laboratory manual. 2012. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.] была создана конструкция на базе экспрессионного вектора pCAGGS, содержащая промоторный регион (синтетический промотор CAG [Jun-ichi М., Satoshi Т., Kimi A., Fumi Т., Akira Т., Kiyoshi Т., Ken-ichi Y. Expression vector system based on the chicken β-actin promoter directs efficient production of interleukin-5. // Gene. 1989. T. 79. №2. C. 269-277], открытую рамку считывания, кодирующую TRPV1ΔCa2+ и репортерный белок tdTomato [Shaner N.C., Campbell R.E., Steinbach P.A., Giepmans B.N.G., Palmer A.E., Tsien R.Y. Improved monomelic red, orange and yellow fluorescent proteins derived from Discosoma sp.red fluorescent protein. // Nature Biotechnology. 2004. T. 22. №12. C. 1567-1572], слитые через саморасщепляющийся пептид р2А [Liu Z., Chen О., Wall J.B J., Zheng M., Zhou Y., Wang L., Ruth Vaseghi H., Qian L., Liu J. Systematic comparison of 2A peptides for cloning multi-genes in a polycistronic vector. // Scientific Reports. 2017. T. 7. №1. C. 2193], посттрансляционный элемент WPRE [Zufferey R., Donello J.E., Trono D., Hope T.J. Woodchuck Hepatitis Virus Posttranscriptional Regulatory Element Enhances Expression of Transgenes Delivered by Retroviral Vectors. // Journal of Virology. 1999. T. 73. №4. C. 2886-2892] и сигнал полиаденелирования (bGH poly (A)) [Goodwin E.C., Rottman F.M. The 3'-flanking sequence of the bovine growth hormone gene contains novel elements required for efficient and accurate polyadenylation. // The Journal of biological chemistry. 1992. T. 267. №23. C. 16330-4] (Фиг. 2).
На фиг. 2 представлена генетическая конструкция на базе экспрессионного вектора pCAGGS для экспрессии генно-инженерного канала TRPV1ΔCa2+ с измененной ионной селективностью. Конструкция содержит синтетический промотор CAG, открытую рамку считывания, кодирующую TRPV1ΔCa2+ и репортерный белок tdTomato, слитые через саморасщепляющийся пептид р2А, посттрансляционный элемент WPRE и сигнал полиаденелирования (bGH poly (А)). Название полученного вектора - pCAGGS-CAG-TRPV1ΔCa2+-p2A-tdTomato.
Аналогичная конструкция, в которой последовательность, кодирующая катионный канал TRPV1ΔCa2+, была заменена на последовательность, кодирующую нативный канал TRPV1 человека, использовалась для сравнения электрофизиологических свойств и способности проводить катионы кальция в клетках, экспрессирующих трансгены. Полученный вектор с нативным каналом TRPV1 человека был назван pCAGGS-CAG-TRPV1-p2A-tdTomato.
Наработанные и очищенные плазмидные векторы, содержащие конструкции с последовательностями, кодирующими TRPV1ΔCa2+ и нативный TRPV1 человека, использовались для трансфекции клеток линии HEK293.
Для регистрации изменений внутриклеточной концентрации катионов кальция в ответ на температурную стимуляцию или на действие агониста TRPV1 канала капсаицина клетки дополнительно подвергались трансфекции плазмидным вектором pCS2+, содержащим последовательность флуоресцентного кальциевого сенсора GCaMP6s [Chen T.-W., Wardill T.J., Sun Y., Pulver S.R., Renninger S.L., Baohan A., Schreiter E.R., Kerr R.A., Orger M.B., Jayaraman V., Looger L.L., Svoboda K., Kim D.S. Ultrasensitive fluorescent proteins for imaging neuronal activity. // Nature. 2013. T. 499. №7458. C. 295-300], находящуюся под контролем промоторного региона CMV.
В тех клетках, в которых детектировался сигнал репортерного красного флуоресцентного белка tdTomato, проводился анализ их способности отвечать на термоактивацию или действие агониста капсаицина путем регистрации токов через клеточную мембрану или изменений внутриклеточной концентрации катионов кальция.
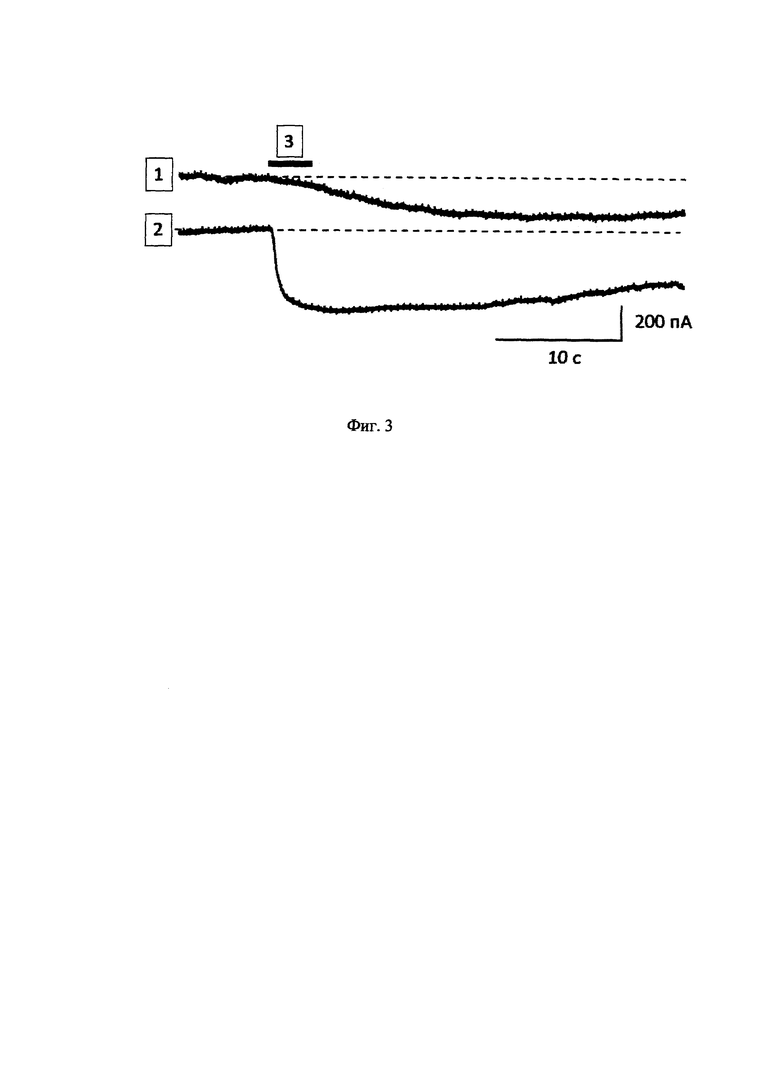
Регистрацию токов через клеточную мембрану осуществляли методом патч-кламп [The Axon Guide for Electrophysiology and Biophysics Laboratory Techniques, by R. Sherman-Gold (Editor) Axon Instruments, Inc, Part Number 2500-102 Rev A. Printed in U.S.A., 1993] на клетках линии HEK293, трансфецированных плазмидным вектором pCAGGS-CAG-TRPV1ΔCa2+-p2A-tdTomato или pCAGGS-CAG-TRPV1-p2A-tdTomato, в режиме удержания мембранного потенциала -20 мВ. На клетку, от которой производится электрофизиологическое отведение, через стеклянный микрокапилляр, подведенный непосредственно к самой клетке, подавался физиологический буферный раствор, содержащий 10 мкМ капсаицина. Капсаицин, связываясь с соответствующим сайтом связывания на канале TRPV1ΔCa2+ или на нативном канале TRPV1, вызывал возникновение токов через мембрану, направленных внутрь клетки (Фиг. 3).
На Фиг. 3 представлены изменения тока через мембрану клеток, экспрессирующих канал TRPV1ACa2+ (1) или нативный канал TRPV1 (2), на кратковременное действие 10 мкМ капсаицина (3). Активация капсаицином канала TRPV1ΔCa2+ вызывает ток, направленный внутрь клетки. Токи, направленные внутрь клетки, вызывают деполяризацию клеточной мембраны.
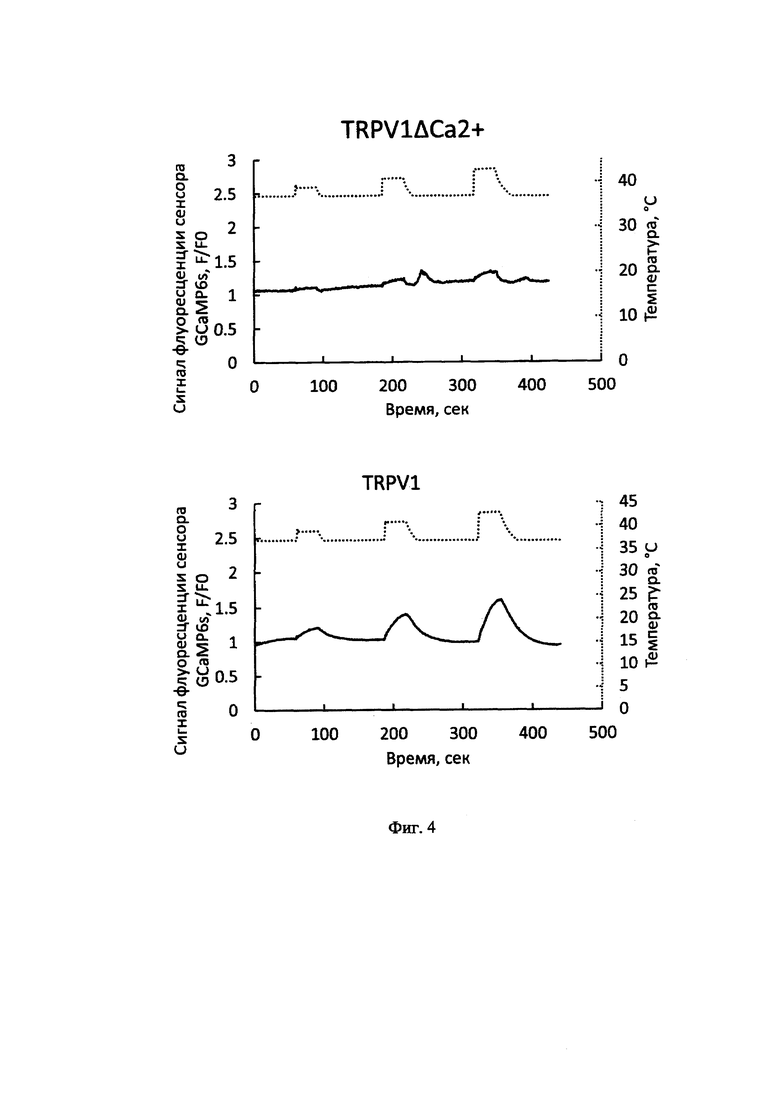
Кальциевый имиджинг (визуализация изменений концентрации внутриклеточного кальция) на клетках, трансфецированных плазмидными векторами, проводился с использованием эпифлуоресцентного микроскопа. Флуоресцентный кальциевый сенсор GCaMP6s возбуждался светом диодного осветителя с длиной волны 470 нм. Регистрацию сигнала флуоресценции осуществляли с помощью цифровой камеры в режиме периодической записи изображений с интервалом 1 сек. С помощью программных средств определяли среднее значение сигнала от каждой клетки, экспрессирующей либо канал TRPV1ΔCa2+ (1), либо нативный канал TRPV1 человека. Сигнал далее усреднялся по более чем ста клеткам и нормализовался на среднее значение интенсивности флуоресценции в первые 60 циклов записи (первые 60 сек каждой съемки). Далее строились кривые зависимости нормализованного сигнала от времени. Термоактивацию канала TRPV1ΔСа2+ или нативного канала TRPV1 человека осуществляли путем последовательного нагрева окружающего клетки физиологического буферного раствора с 37°С до 39, 41 и 43°С на 30 сек с интервалом между нагревами в 90 сек. Анализ сигнала в клетках, экспрессирующих канал TRPV1ΔСа2+, не выявил изменений концентрации внутриклеточного кальция при нагревах в отличие от клеток, экспрессирующих нативный канал TRPV1 человека, в которых эти изменения были (Фиг. 4).
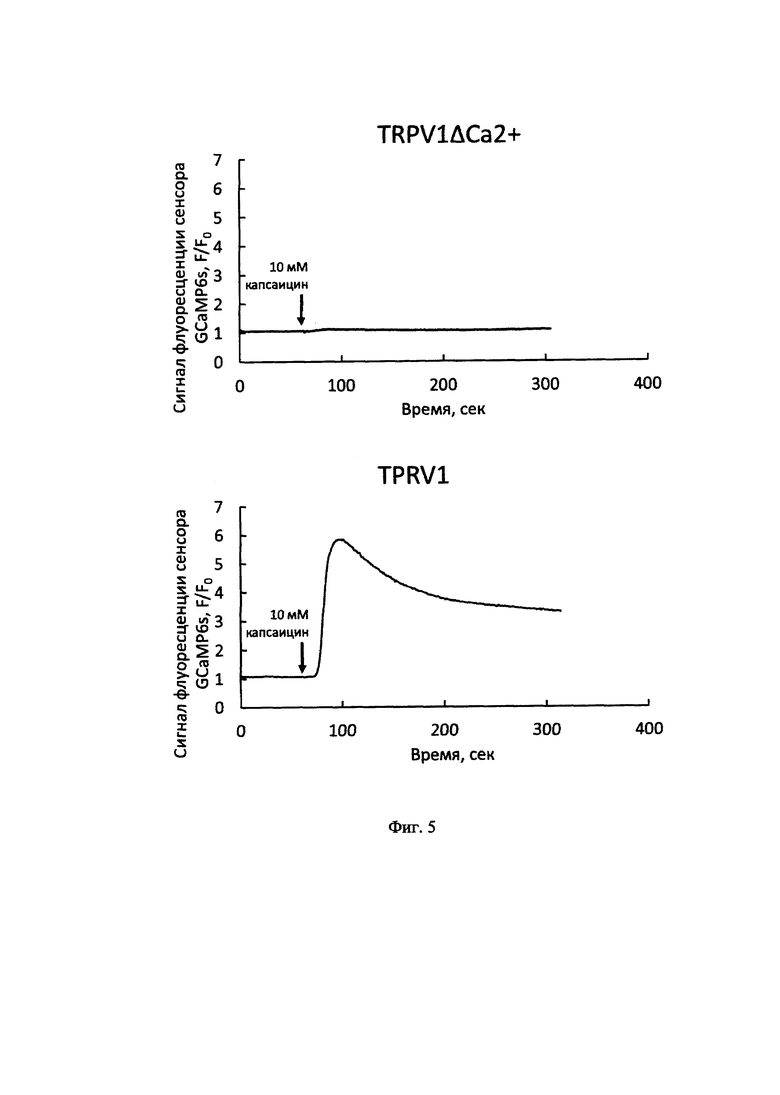
На Фиг. 4 представлены усредненные ответы клеток на нагревы (сплошная линия, шкала слева) и температура окружающего физиологического буфера (пунктирная линия, шкала слева). Аналогичным образом добавление капсицина (конечная концентрация 10 мкМ) к клеткам, экспрессирующим канал TRPV1ΔСа2+, не вызывала изменений концентрации внутриклеточного кальция, тогда клетки с нативным каналом TRPV1 человека демонстрировали увеличение сигнала флуоресценции кальциевого сенсора в ответ на добавление капсаицина в той же концентрации (Фиг. 5).
На фиг. 5 представлены усредненные ответы клеток (изменение интенсивности сигнала флуоресцентного кальциевого сенсора GCaMP6s) на добавление 10 мкМ капсаицина.
Суммарно, полученные данные подтверждают, что канал TRPV1ΔСа2+ деполяризует клеточную мембрану в отсутствие проводимости двухвалентных катионов кальция.
Настоящее изобретение не предназначено для модификации генетической целостности клеток зародышевой линии человека, не предполагает использование человеческих эмбрионов.
--->
Перечень последовательностей
<110> Федеральное государственное автономное образовательное учреждение
высшего образования «Российский национальный исследовательский медицинский
университет имени Н.И. Пирогова» Министерства здравоохранения Российской
Федерации (Federalnoe gosudarstvennoe autonomnoe obrazovatelnoe uchrezhdenie
vysshego obrazovaniya «Rossijskij natsionalnyj issledovatelskij meditsinskij
universitet imeni N.I. Pirogova» Ministerstva zdravookhraneniya Rossijskoj
Federatsii)
<120> Нуклеиновая кислота, кодирующая термочувствительный канал TRPV1 человека
с измененной ионной селективностью для управления мембранным потенциалом клеток
<160> 3
<210> SEQ ID NO: 1
<211> 839
<212> ПРТ
<213> Искусственная
<220>
<221> Мутаген
<222> (643), (645)
<223> Термочувствительный канал TRPV1 человека с измененной ионной
селективностью, обеспечивающий деполяризацию клеточной мембраны и лишенный
проводимости двухвалентных катионов кальция (TRPV1ΔCa2+)
<400> 1
Met Lys Lys Trp Ser Ser Thr Asp Leu Gly Ala Ala Ala Asp Pro Leu
1 5 10 15
Gln Lys Asp Thr Cys Pro Asp Pro Leu Asp Gly Asp Pro Asn Ser Arg
20 25 30
Pro Pro Pro Ala Lys Pro Gln Leu Ser Thr Ala Lys Ser Arg Thr Arg
35 40 45
Leu Phe Gly Lys Gly Asp Ser Glu Glu Ala Phe Pro Val Asp Cys Pro
50 55 60
His Glu Glu Gly Glu Leu Asp Ser Cys Pro Thr Ile Thr Val Ser Pro
65 70 75 80
Val Ile Thr Ile Gln Arg Pro Gly Asp Gly Pro Thr Gly Ala Arg Leu
85 90 95
Leu Ser Gln Asp Ser Val Ala Ala Ser Thr Glu Lys Thr Leu Arg Leu
100 105 110
Tyr Asp Arg Arg Ser Ile Phe Glu Ala Val Ala Gln Asn Asn Cys Gln
115 120 125
Asp Leu Glu Ser Leu Leu Leu Phe Leu Gln Lys Ser Lys Lys His Leu
130 135 140
Thr Asp Asn Glu Phe Lys Asp Pro Glu Thr Gly Lys Thr Cys Leu Leu
145 150 155 160
Lys Ala Met Leu Asn Leu His Asp Gly Gln Asn Thr Thr Ile Pro Leu
165 170 175
Leu Leu Glu Ile Ala Arg Gln Thr Asp Ser Leu Lys Glu Leu Val Asn
180 185 190
Ala Ser Tyr Thr Asp Ser Tyr Tyr Lys Gly Gln Thr Ala Leu His Ile
195 200 205
Ala Ile Glu Arg Arg Asn Met Ala Leu Val Thr Leu Leu Val Glu Asn
210 215 220
Gly Ala Asp Val Gln Ala Ala Ala His Gly Asp Phe Phe Lys Lys Thr
225 230 235 240
Lys Gly Arg Pro Gly Phe Tyr Phe Gly Glu Leu Pro Leu Ser Leu Ala
245 250 255
Ala Cys Thr Asn Gln Leu Gly Ile Val Lys Phe Leu Leu Gln Asn Ser
260 265 270
Trp Gln Thr Ala Asp Ile Ser Ala Arg Asp Ser Val Gly Asn Thr Val
275 280 285
Leu His Ala Leu Val Glu Val Ala Asp Asn Thr Ala Asp Asn Thr Lys
290 295 300
Phe Val Thr Ser Met Tyr Asn Glu Ile Leu Ile Leu Gly Ala Lys Leu
305 310 315 320
His Pro Thr Leu Lys Leu Glu Glu Leu Thr Asn Lys Lys Gly Met Thr
325 330 335
Pro Leu Ala Leu Ala Ala Gly Thr Gly Lys Ile Gly Val Leu Ala Tyr
340 345 350
Ile Leu Gln Arg Glu Ile Gln Glu Pro Glu Cys Arg His Leu Ser Arg
355 360 365
Lys Phe Thr Glu Trp Ala Tyr Gly Pro Val His Ser Ser Leu Tyr Asp
370 375 380
Leu Ser Cys Ile Asp Thr Cys Glu Lys Asn Ser Val Leu Glu Val Ile
385 390 395 400
Ala Tyr Ser Ser Ser Glu Thr Pro Asn Arg His Asp Met Leu Leu Val
405 410 415
Glu Pro Leu Asn Arg Leu Leu Gln Asp Lys Trp Asp Arg Phe Val Lys
420 425 430
Arg Ile Phe Tyr Phe Asn Phe Leu Val Tyr Cys Leu Tyr Met Ile Ile
435 440 445
Phe Thr Met Ala Ala Tyr Tyr Arg Pro Val Asp Gly Leu Pro Pro Phe
450 455 460
Lys Met Glu Lys Ile Gly Asp Tyr Phe Arg Val Thr Gly Glu Ile Leu
465 470 475 480
Ser Val Leu Gly Gly Val Tyr Phe Phe Phe Arg Gly Ile Gln Tyr Phe
485 490 495
Leu Gln Arg Arg Pro Ser Met Lys Thr Leu Phe Val Asp Ser Tyr Ser
500 505 510
Glu Met Leu Phe Phe Leu Gln Ser Leu Phe Met Leu Ala Thr Val Val
515 520 525
Leu Tyr Phe Ser His Leu Lys Glu Tyr Val Ala Ser Met Val Phe Ser
530 535 540
Leu Ala Leu Gly Trp Thr Asn Met Leu Tyr Tyr Thr Arg Gly Phe Gln
545 550 555 560
Gln Met Gly Ile Tyr Ala Val Met Ile Glu Lys Met Ile Leu Arg Asp
565 570 575
Leu Cys Arg Phe Met Phe Val Tyr Ile Val Phe Leu Phe Gly Phe Ser
580 585 590
Thr Ala Val Val Thr Leu Ile Glu Asp Gly Lys Asn Asp Ser Leu Pro
595 600 605
Ser Glu Ser Thr Ser His Arg Trp Arg Gly Pro Ala Cys Arg Pro Pro
610 615 620
Asp Ser Ser Tyr Asn Ser Leu Tyr Ser Thr Cys Leu Glu Leu Phe Lys
625 630 635 640
Phe Thr Val Gly Tyr Gly Asp Leu Glu Phe Thr Glu Asn Tyr Asp Phe
645 650 655
Lys Ala Val Phe Ile Ile Leu Leu Leu Ala Tyr Val Ile Leu Thr Tyr
660 665 670
Ile Leu Leu Leu Asn Met Leu Ile Ala Leu Met Gly Glu Thr Val Asn
675 680 685
Lys Ile Ala Gln Glu Ser Lys Asn Ile Trp Lys Leu Gln Arg Ala Ile
690 695 700
Thr Ile Leu Asp Thr Glu Lys Ser Phe Leu Lys Cys Met Arg Lys Ala
705 710 715 720
Phe Arg Ser Gly Lys Leu Leu Gln Val Gly Tyr Thr Pro Asp Gly Lys
725 730 735
Asp Asp Tyr Arg Trp Cys Phe Arg Val Asp Glu Val Asn Trp Thr Thr
740 745 750
Trp Asn Thr Asn Val Gly Ile Ile Asn Glu Asp Pro Gly Asn Cys Glu
755 760 765
Gly Val Lys Arg Thr Leu Ser Phe Ser Leu Arg Ser Ser Arg Val Ser
770 775 780
Gly Arg His Trp Lys Asn Phe Ala Leu Val Pro Leu Leu Arg Glu Ala
785 790 795 800
Ser Ala Arg Asp Arg Gln Ser Ala Gln Pro Glu Glu Val Tyr Leu Arg
805 810 815
Gln Phe Ser Gly Ser Leu Lys Pro Glu Asp Ala Glu Val Phe Lys Ser
820 825 830
Pro Ala Ala Ser Gly Glu Lys
835
<210> SEQ ID NO: 2
<211> 22
<212> ДНК
<213> Искусственная
<220>
<223> Прямой праймер F1 для внесения нуклеотидных замен в кДНК, кодирующую
канал TRPV1 человека для получения канала TRPV1 человека с измененной ионной
селективностью, обеспечивающего деполяризацию клеточной мембраны и лишенного
проводимости двухвалентных катионов кальция (TRPV1ΔCa2+)
<400> 2
tacggcgacc tggagttcac tg 22
<210> SEQ ID NO: 3
<211> 24
<212> ДНК
<213> Искусственная
<220>
<223> Обратный праймер R1 для внесения нуклеотидных замен в кДНК, кодирующую
канал TRPV1 человека для получения канала TRPV1 человека с измененной ионной
селективностью, обеспечивающего деполяризацию клеточной мембраны и лишенного
проводимости двухвалентных катионов кальция (TRPV1ΔCa2+)
<400> 3
gcccacggtg aacttgaaca gctc 24
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ УПРАВЛЕНИЯ РИТМОМ СЕРДЦА И СОКРАЩЕНИЕМ ОТДЕЛЬНЫХ КАРДИОМИОЦИТОВ ПРИ ПОМОЩИ ТЕРМОГЕНЕТИКИ | 2022 |
|
RU2802995C1 |
| НОВЫЕ СПОСОБЫ СЕЛЕКЦИИ ЭПИТОПОВ | 2016 |
|
RU2764992C2 |
| СОДЕРЖАЩИЙ ЛЕКАРСТВЕННОЕ СРЕДСТВО ПЕРЕНОСЧИК В КЛЕТКУ ДЛЯ ФОРМИРОВАНИЯ ИММУННОГО КОМПЛЕКСА | 2012 |
|
RU2739792C1 |
| МОДИФИЦИРОВАННАЯ J-ЦЕПЬ | 2015 |
|
RU2761118C2 |
| HPV-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2018 |
|
RU2804664C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ИНДУЦИРУЮЩАЯ ИММУННЫЙ ОТВЕТ НА АНТИГЕН-МИШЕНЬ | 2012 |
|
RU2722829C2 |
| ВЫСОКОГЛИКОЗИЛИРОВАННЫЙ СЛИТЫЙ БЕЛОК НА ОСНОВЕ ФАКТОРА СВЕРТЫВАНИЯ КРОВИ ЧЕЛОВЕКА VIII, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ | 2016 |
|
RU2722374C1 |
| НЕВИРУСНЫЙ ИММУНОТАРГЕТИНГ | 2020 |
|
RU2833379C2 |
| Антитела против белка р17 ВИЧ-1 субтипа А | 2019 |
|
RU2727673C1 |
| СУБЪЕДИНИЧНАЯ ВАКЦИНА ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРЕДОТВРАЩЕНИЯ ИНФЕКЦИИ ДЫХАТЕЛЬНЫХ ПУТЕЙ | 2020 |
|
RU2811991C2 |

Изобретение относится к биомедицинским технологиям, а конкретно к группе методов, направленных на изменение проводимости клеточных мембран для управления активностью клеток. Предложена нуклеиновая кислота, кодирующая термочувствительный канал TRPV1 человека с измененной ионной селективностью (TRPV1ΔCa2+), обеспечивающий деполяризацию клеточной мембраны и лишенный проводимости двухвалентных катионов кальция, имеющий последовательность SEQ ID NO: 01. Технический результат, достигаемый при осуществлении изобретения, заключается в исключении повреждения и гибели возбудимых клеток за счет перегруза их ионами кальция во время термогенетического управления трансмембранным транспортом катионов с использованием канала TRPV1 человека. 5 ил., 1 пр.
Нуклеиновая кислота, кодирующая термочувствительный канал TRPV1 человека с измененной ионной селективностью, обеспечивающий деполяризацию клеточной мембраны и лишенный проводимости двухвалентных катионов кальция, имеющий последовательность SEQ ID NO: 1.
| US 0010696728 B2, 30.06.2020 | |||
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ГИПЕРАЛГЕЗИИ | 2007 |
|
RU2430750C2 |
| RU 2022100590 A, 10.03.2022 | |||
| Lina Duo, Ting Wu и др., Gain of Function of Ion Channel TRPV1 Exacerbates Experimental Colitis by Promoting Dendritic Cell Activation, Molecular Therapy: Nucleic Acids Vol | |||
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |

