Перекрестная ссылка на родственные заявки
Настоящая патентная заявка испрашивает приоритет согласно временной патентной заявки США № 62/280843 (подана 20 января 2016 года). Полное раскрытие приоритетной заявки включено в настоящую заявку посредством ссылки во всей полноте и для всех целей.
Предпосылки создания изобретения
Рак является одной из главных причин смерти. Это класс заболеваний, вызываемых злокачественным преобразованием здоровых клеток, которое вызвано генетическими изменениями, такими как хромосомные транслокации, мутации в генах-супрессорах опухолей, транскрипционных факторах или рецепторах факторов роста, что приводит к иммортализации клеток. Если иммортализация сочетается с чрезмерной пролиферацией, иммортализованные клетки генерируют опухоли с метастазами или без них (в случае солидных опухолей) или лейкозы и лимфомы (раковые заболевания крови). Нарушенный апоптоз или запрограммированная гибель клеток могут также способствовать злокачественному преобразованию клеток, ведущему к раку.
Семейство мембрана-ассоциированных рецепторных тирозинкиназ, состоящее из подобных рецепторной тирозинкиназе орфанных рецепторов-1 и -2 (ROR1 и ROR2), было описано как специфически связанное с конкретными типами рака (Rebagay et al. (2012) Front Oncol. 2(34)), при этом они в основном не экспрессируются в здоровой ткани, с некоторыми исключениями (Balakrishnan et al. (2016) Clin Cancer res. doi: 10.1158/1078-0432). Является или нет экспрессия ROR функционально связанной с канцерогенезом, остается неясным. Однако из-за сильной опухоль-селективной экспрессии членов ROR семейства, они являются подходящими мишенями для таргетных раковых терапий. Подобный рецепторной тирозинкиназе орфанный рецептор-1 (ROR1) представляет особый интерес как мишень для лечения рака, поскольку почти в 100% случаев он связан с хроническим лимфоцитарным лейкозом (CLL) (Cui et al. (2016) Blood 128(25), p. 2931), а также, согласно наблюдениям, экспрессируется в некоторых солидных опухолях, например, легкого и молочной железы (Balakrishnan et al. (2016) Clin Cancer res. doi: 10.1158/1078-0432). Члены ROR семейства представляют собой трансмембраннные белки типа-I, содержащие три разных внеклеточных домена, Ig, Kringle и Frizzled домен, трансмембраную область и внутриклеточную часть. Во внутриклеточной части ROR1 имеет тирозинкиназный домен, два серин/треонин-обогащенных домена и пролин-обогащенный домен. ROR были исследованы в контексте формирования эмбрионного паттерна и нейрогенеза через различные гомологи. Эти физиологические функции делятся на две части на основании требования киназного домена. Постоянно увеличивающееся число литературных источников описывает ROR1 в качестве маркера рака, например, хронического лимфоцитарного лейкоза (CLL), с которым экспрессия ROR1 связана почти в 100% случаев, некоторых острых лимфобластных лейкозов (ALL), лимфом мантийных клеток и некоторых других злокачественных новообразований. Кроме того, ROR1 играет критическую роль в прогрессировании ряда солидных опухолей, таких как нейробластома, саркома, почечноклеточная карцинома, рак молочной железы, рак легких, рак толстой кишки, рак головы и шеи, меланома и другие виды рака. Было показано, что ROR1 ингибирует апоптоз, потенцирует передачу сигналов EGFR, индуцирует эпителиально-мезенхимальный переход (EMT) и способствует образованию кавеол. Важно отметить, что ROR1 в основном можно обнаружить в эмбриональной ткани, и, как правило, он отсутствует во взрослой ткани, что делает этот белок идеальной лекарственной мишенью для лечения рака. Таким образом, ROR1 ранее был признан мишенью для разработки специфических антител к ROR1. Однако из-за высокой гомологии ROR1 между различными видами млекопитающих, который является на 100% консервативным на аминокислотном уровне между людьми и обезьянами cynomolgus, на 96,7% гомологичным между человеком и мышью и на 96,3% гомологичным между человеком и кроликом, трудно индуцировать высокоаффинные антитела против этой мишени стандартными методами, такими как иммунизация животных.
Некоторые мышиные и кроличьи антитела обсуждаются в литературе. Например, в WO 2007/051077 обсуждаются моноклональные антитела, включая гуманизированные антитела, направленные против природного ROR1, обнаруженного на лимфомах, включая CLL, мелкоклеточную лимфоцитарную лимфому, B-клеточную лимфому из клеток краевой зоны и лимфому Беркета. Способы ингибирования роста опухолевой клетки с использованием средств, которые могут представлять собой ROR1-связывающие антитела, которые ингибируют киназную активность ROR1, являются объектом изобретения, раскрытым в WO 2007/146957. В WO 2011/054007 обсуждаются способ лечения или профлактики рака, в котором внеклеточный домен ROR1 экспрессируют путем введения специфических ROR1-таргетирующтаких антител.
Кроме того, WO 2010/124188 относится к античеловеческим ROR1 антителам и, в частности, к моноклональному мышиному антителу, известному под названием 2A2, тогда как WO 2012/075158 относится к моноклональным кроличьим антителам, обозначаемым как R11 и R12. Конкретные ROR1-таргетирующие антитела также указаны в WO 2016/094873. Как в WO 2011/079902, так и в WO 2012/076066 обсуждаются биологические ингибиторы ROR1, способные индуцировать клеточную гибель, которые связываются с выбранными последовательностями внеклеточного ROR1 домена. WO 2014/031174 относится к анти-ROR1 антителам, обладающим такой же специфичностью связывания, как антитело, названное 99961. Связывающиеся эпитопы анти-ROR1 антител также обсуждаются в WO 2016/187220. В WO 2011/159847 обсуждаются конкретные конъюгаты фрагментов антител scFv, которые связываются с ROR1. В WO 2014/167022, WO 2016/055592 и WO 2016/055593 обсуждаются биспецифические ROR1-таргетирующие антитела и их применения, тогда как в WO 2015/184203 обсуждаются три-специфические связывающиеся молекулы. Более новые документы, раскрывающие гуманизированные анти-ROR1 моноклональные антитела, основаны на первоначально раскрытых мышиных или кроличьих антителах, таких как 2A2, R11, R12 или D10.
Из-за небольшого количества доступных ROR1-специфических моноклональных антител, в данной области существует потребность в лучших анти-ROR1 антителах, которые обладают более высокой аффинностью или другими функциональными свойствами, которыми не обладают известные клоны антител. Также существует потребность в дополнительных диагностических инструментах для детекции экспрессии ROR1 в связанных с ROR1 болезненных состояниях, например, методом Вестерн-блоттинга и/или иммуногистохимии (IHC). Настоящее изобретение направлено на удовлетворение этих и других потребностей.
Сущность изобретения
В одном аспекте изобретение обеспечивает новые высокоаффинные связывающиеся домены кроличьих антител, которые специфически связываются с внеклеточным доменом человеческого подобного рецепторной тирозинкиназе орфанного рецептора 1 (hROR1) и которые выбраны из чрезвычайно разнообразных фаговых библиотек неиммунизированных кроликов с использованием внеклеточных доменов человеческого ROR1 (hROR1), экспрессируемых в клетках млекопитающего, в качестве затравки. Вариабельные области кроличьих антител были выбраны путем скрининга на связывание против ECD hROR1 в качестве рекомбинантных белков, а также на основании связывания hROR1 сверхэкспрессируемого на поверхности клеток хозяина-млекопитающего. Используя эту стратегию, были идентифицированы новые антитела к hROR1 беспрецедентного качества и с выгодными функциональными свойствами. Кроме того, изобретение обеспечивает химерные полноразмерные антитела из кроличьих вариабельных доменов, слитых с доменами константной области человеческих IgG1 антител. Кроме того, изобретение обеспечивает новые высокоаффинные гуманизированные антитела, которые были образованы путем пересадки CDR кроличьих анти-ROR1 антител, раскрытых в настоящей заявке, в каркас вариабельных областей тяжелой и легкой цепей иммуноглобулинов. Такие гуманизированные антитела можно использовать для лечения заболеваний человека благодаря высокой гомологии указанных гуманизированных антител с эндогенными полностью человеческими антителами. Во втором аспекте изобретения обеспечиваются сайт-специфически конъюгированные конъюгаты антитело-лекарственное средство (ADCs) на основе химерных кроличьих-человеческих и гуманизированных анти-человеческих ROR1 (hROR1) антител с ультрасильным антрациклиновым токсином. Сайт-специфическая конъюгация достигается путем ферментативной конъюгации с использованием фермента сортазы, в частности, как раскрыто в WO2014140317, который включен в настоящую заявку в качестве ссылки. Ультрасильный антрациклиновый токсин, приводящий к анти-hROR1 ADCs с беспрецедентной активностью в различных in vitro и in vivo опухолевых моделях раскрыт в WO2016102679, который включен в настоящую заявку в качестве ссылки.
Наконец, изобретение обеспечивает химерные антигенные рецепторы (CARs) и T-клетки, сконструированные с этими CARs, т.е. так называемые CAR-T клетки, с использованием указанных анти-hROR1 связывающихся доменов, показывающих высокую эффективность in vitro.
Поэтому изобретение относится к анти-hROR1 антителам, связывающимся белкам на основе антител, фрагментам таких антител, конъюгатам антитело-лекарственное средство (ADCs) или CARs, имеющим такую же специфичность связывания в отношении hROR1, что и hROR1 специфические антитела, содержащие последовательность вариабельной области тяжелой цепи иммуноглобулина и последовательность вариабельной области легкой цепи иммуноглобулина,, соответственно показанные в (1) SEQ ID NO:1 и SEQ ID NO:14; (2) SEQ ID NO:2 и SEQ ID NO:15; (3) SEQ ID NO:3 и SEQ ID NO:16; (4) SEQ ID NO:4 и SEQ ID NO:17; (5) SEQ ID NO:5 и SEQ ID NO:18; (6) SEQ ID NO:6 и SEQ ID NO:19; (7) SEQ ID NO:7 и SEQ ID NO:20; (8) SEQ ID NO:8 и SEQ ID NO:21; (9) SEQ ID NO:9 и SEQ ID NO:22; (10) SEQ ID NO:10 и SEQ ID NO:23; (11) SEQ ID NO:11 и SEQ ID NO:24; (12) SEQ ID NO:12 и SEQ ID NO:25; (13) SEQ ID NO:13 и SEQ ID NO:26; (14) SEQ ID NO:130 и SEQ ID NO:136; (15) SEQ ID NO:131 и SEQ ID NO:137; (16) SEQ ID NO:132 и SEQ ID NO:138; (17) SEQ ID NO:133 и SEQ ID NO:139; (18) SEQ ID NO:134 и SEQ ID NO:140; или (19) SEQ ID NO:135 и SEQ ID NO:141.
Изобретение также относится к анти-hROR1 антителам, связывающимся белкам на основе антител, фрагментам таких антител, конъюгатам антитело-лекарственное средство (ADCs) или CARs, включающим последовательности CDR тяжелой цепи иммуноглобулинов и последовательности CDR легкой цепи иммуноглобулинов, которые по меньшей мере на 90% или по меньшей мере на 95% или больше чем на 95%, но меньше чем на 100% идентичны соответственно (1) SEQ ID NO:27-29 и SEQ ID NO:66-68, (2) SEQ ID NO:30-32 и SEQ ID NO:69-71, (3) SEQ ID NO:33-35 и SEQ ID NO:72-74, (4) SEQ ID NO:36-38 и SEQ ID NO:75-77, (5) SEQ ID NO:39-41 и SEQ ID NO:78-80, (6) SEQ ID NO:42-44 и SEQ ID NO:81-83, (7) SEQ ID NO:45-47 и SEQ ID NO:84-86, (8) SEQ ID NO:48-50 и SEQ ID NO:87-89, (9) SEQ ID NO:51-53 и SEQ ID NO:90-92, (10) SEQ ID NO:54-56 и SEQ ID NO:93-95, (11) SEQ ID NO:57-59 и SEQ ID NO:96-98, (12) SEQ ID NO:60-62 и SEQ ID NO:99-101, или (13) SEQ ID NO:63-65 и SEQ ID NO:102-104.
Изобретение также относится к анти-hROR1 антителам, связывающимся белкам на основе антител, фрагментам таких антител, конъюгатам антитело-лекарственное средство (ADCs) или CARs, включающим последовательности CDR тяжелой цепи иммуноглобулинов и последовательности CDR легкой цепи иммуноглобулинов, которые идентичны соответственно (1) SEQ ID NO:27-29 и SEQ ID NO:66-68, (2) SEQ ID NO:30-32 и SEQ ID NO:69-71, (3) SEQ ID NO:33-35 и SEQ ID NO:72-74, (4) SEQ ID NO:36-38 и SEQ ID NO:75-77, (5) SEQ ID NO:39-41 и SEQ ID NO:78-80, (6) SEQ ID NO:42-44 и SEQ ID NO:81-83, (7) SEQ ID NO:45-47 и SEQ ID NO:84-86, (8) SEQ ID NO:48-50 и SEQ ID NO:87-89, (9) SEQ ID NO:51-53 и SEQ ID NO:90-92, (10) SEQ ID NO:54-56 и SEQ ID NO:93-95, (11) SEQ ID NO:57-59 и SEQ ID NO:96-98, (12) SEQ ID NO:60-62 и SEQ ID NO:99-101, или (13) SEQ ID NO:63-65 и SEQ ID NO:102-104.
Изобретение также относится к анти-hROR1 антителам, связывающимся белкам на основе антител, фрагментам таких антител, конъюгатам антитело-лекарственное средство (ADCs) или CARs, включающим либо последовательность вариабельной области тяжелой цепи иммуноглобулина, либо последовательность вариабельной области легкой цепи иммуноглобулина с идентичностью по меньшей мере 90% или по меньшей мере 95% или больше чем 95%, но меньше чем 100%, на аминокислотном уровне относительно последовательности вариабельной области тяжелой цепи иммуноглобулина и последовательности вариабельной области легкой цепи иммуноглобулина, соответственно, показанной в (1) SEQ ID NO:1 и SEQ ID NO:14; (2) SEQ ID NO:2 и SEQ ID NO:15; (3) SEQ ID NO:3 и SEQ ID NO:16; (4) SEQ ID NO:4 и SEQ ID NO:17; (5) SEQ ID NO:5 и SEQ ID NO:18; (6) SEQ ID NO:6 и SEQ ID NO:19; (7) SEQ ID NO:7 и SEQ ID NO:20; (8) SEQ ID NO:8 и SEQ ID NO:21; (9) SEQ ID NO:9 и SEQ ID NO:22; (10) SEQ ID NO:10 и SEQ ID NO:23; (11) SEQ ID NO:11 и SEQ ID NO:24; (12) SEQ ID NO:12 и SEQ ID NO:25; или (13) SEQ ID NO:13 и SEQ ID NO:26.
Изобретение также относится к анти-hROR1 антителам, связывающимся белкам на основе антител, фрагментам таких антител, конъюгатам антитело-лекарственное средство (ADCs) или CARs, включающим последовательность CDR тяжелой цепи иммуноглобулина, выбранную из группы, состоящей из SEQ ID NO:27-65. Изобретение также относится к анти-hROR1 антителам, связывающимся белкам на основе антител, фрагментам таких антител, конъюгатам антитело-лекарственное средство (ADCs) или CARs, включающим последовательность CDR легкой цепи иммуноглобулина, выбранную из группы, состоящей из SEQ ID NO:66-104. В некоторых вариантах осуществления hROR1-специфические антитела, связывающиеся белки на основе антител или фрагменты таких антител, конъюгаты антитело-лекарственное средство (ADCs) или CARs содержат последовательности CDR1, CDR2 и CDR3 тяжелой цепи, которые соответственно идентичны SEQ ID NO:27-29, SEQ ID NO:30-32, SEQ ID NO:33-35, SEQ ID NO:36-38, SEQ ID NO:39-41, SEQ ID NO:42-44, SEQ ID NO:45-47, SEQ ID NO:48-50, SEQ ID NO:51-53, SEQ ID NO:54-56, SEQ ID NO:57-59, SEQ ID NO:60-62 или SEQ ID NO:63-65.
Изобретение также относится к анти-hROR1 антителам, связывающимся белкам на основе антител, фрагментам таких антител, конъюгатам антитело-лекарственное средство (ADCs) или CARs, включающий либо последовательность вариабельной области тяжелой цепи иммуноглобулина, либо последовательность вариабельной области легкой цепи иммуноглобулина, идентичную (1) SEQ ID NO:1 и SEQ ID NO:14; (2) SEQ ID NO:2 и SEQ ID NO:15; (3) SEQ ID NO:3 и SEQ ID NO:16; (4) SEQ ID NO:4 и SEQ ID NO:17; (5) SEQ ID NO:5 и SEQ ID NO:18; (6) SEQ ID NO:6 и SEQ ID NO:19; (7) SEQ ID NO:7 и SEQ ID NO:20; (8) SEQ ID NO:8 и SEQ ID NO:21; (9) SEQ ID NO:9 и SEQ ID NO:22; (10) SEQ ID NO:10 и SEQ ID NO:23; (11) SEQ ID NO:11 и SEQ ID NO:24; (12) SEQ ID NO:12 и SEQ ID NO:25; (13) SEQ ID NO:13 и SEQ ID NO:26; (14) SEQ ID NO:130 и SEQ ID NO:136; (15) SEQ ID NO:131 и SEQ ID NO:137; (16) SEQ ID NO:132 и SEQ ID NO:138; (17) SEQ ID NO:133 и SEQ ID NO:139; (18) SEQ ID NO:134 и SEQ ID NO:140; или (19) SEQ ID NO:135 и SEQ ID NO:141
Изобретение также относится к анти-hROR1 антителам, связывающимся белкам на основе антител, фрагментам таких антител, конъюгатам антитело-лекарственное средство (ADCs) или CARs, включающим анти-hROR1 специфические антитела, связывающие белки на основе антител или фрагменты таких антител, которые включают последовательность вариабельной области тяжелой цепи иммуноглобулина и последовательность вариабельной области легкой цепи иммуноглобулина, соответственно, идентичные (1) SEQ ID NO:1 и SEQ ID NO:14; (2) SEQ ID NO:2 и SEQ ID NO:15; (3) SEQ ID NO:3 и SEQ ID NO:16; (4) SEQ ID NO:4 и SEQ ID NO:17; (5) SEQ ID NO:5 и SEQ ID NO:18; (6) SEQ ID NO:6 и SEQ ID NO:19; (7) SEQ ID NO:7 и SEQ ID NO:20; (8) SEQ ID NO:8 и SEQ ID NO:21; (9) SEQ ID NO:9 и SEQ ID NO:22; (10) SEQ ID NO:10 и SEQ ID NO:23; (11) SEQ ID NO:11 и SEQ ID NO:24; (12) SEQ ID NO:12 и SEQ ID NO:25; (13) SEQ ID NO:13 и SEQ ID NO:26; (14) SEQ ID NO:130 и SEQ ID NO:136; (15) SEQ ID NO:131 и SEQ ID NO:137; (16) SEQ ID NO:132 и SEQ ID NO:138; (17) SEQ ID NO:133 и SEQ ID NO:139; (18) SEQ ID NO:134 и SEQ ID NO:140; или (19) SEQ ID NO:135 и SEQ ID NO:141.
В некоторых вариантах осуществления анти-hROR1 антитела, связывающиеся белки на основе антител, фрагменты таких антител, конъюгаты антитело-лекарственное средство (ADCs) или CARs включают последовательность CDR легкой цепи иммуноглобулина, выбранную из группы, состоящей из SEQ ID NO:66-104. Некоторые из этих молекул также включают последовательность CDR тяжелой цепи иммуноглобулина, выбранную из группы, состоящей из SEQ ID NO:27-65. Некоторые из этих молекул включают последовательности CDR1, CDR2 и CDR3 легкой цепи иммуноглобулина, которые, соответственно, идентичны SEQ ID NO:66-68, SEQ ID NO:69-71, SEQ ID NO:72-74, SEQ ID NO:75-77, SEQ ID NO:78-80, SEQ ID NO:81-83, SEQ ID NO:84-86, SEQ ID NO:87-89, SEQ ID NO:90-92, SEQ ID NO:93-95, SEQ ID NO:96-98, SEQ ID NO:99-101 или SEQ ID NO:102-104. В некоторых вариантах осуществления анти-hROR1 антитела, связывающиеся белки на основе антител, фрагменты таких антител, конъюгаты антитело-лекарственное средство (ADCs) или CARs включают последовательности CDR1, CDR2 и CDR3 тяжелой цепи иммуноглобулина и последовательности CDR1, CDR2 и CDR3 легкой цепи иммуноглобулина, соответственно показанные в (1) SEQ ID NO:27-29 и SEQ ID NO:66-68, (2) SEQ ID NO:30-32 и SEQ ID NO:69-71, (3) SEQ ID NO:33-35 и SEQ ID NO:72-74, (4) SEQ ID NO:36-38 и SEQ ID NO:75-77, (5) SEQ ID NO:39-41 и SEQ ID NO:78-80, (6) SEQ ID NO:42-44 и SEQ ID NO:81-83, (7) SEQ ID NO:45-47 и SEQ ID NO:84-86, (8) SEQ ID NO:48-50 и SEQ ID NO:87-89, (9) SEQ ID NO:51-53 и SEQ ID NO:90-92, (10) SEQ ID NO:54-56 и SEQ ID NO:93-95, (11) SEQ ID NO:57-59 и SEQ ID NO:96-98, (12) SEQ ID NO:60-62 и SEQ ID NO:99-101 или (13) SEQ ID NO:63-65 и SEQ ID NO:102-104.
Изобретение также относится к hROR1-специфическим гуманизированным антителам, связывающимся белкам на основе антител, фрагментам таких антител, конъюгатам антитело-лекарственное средство (ADCs) или CARs с гомологией по меньшей мере 90% или по меньшей мере 95% или по меньшей мере 95%, но меньше чем 100%, на аминокислотном уровне с тяжелой или легкой цепью иммуноглобулина, представленной в: (14) SEQ ID NO:130 и SEQ ID NO:136; (15) SEQ ID NO:131 и SEQ ID NO:137; (16) SEQ ID NO:132 и SEQ ID NO:138; (17) SEQ ID NO:133 и SEQ ID NO:139; (18) SEQ ID NO:134 и SEQ ID NO:140; или (19) SEQ ID NO:135 и SEQ ID NO:141.
И еще в дополнительных вариантах осуществления hROR1-специфические антитела, связывающиеся белки на основе антител или фрагменты таких антител представляют собой любое из IgAl, IgA2, IgD, IgE, IgGl, IgG2, IgG3 IgG4, или изотипы IgM, или их F(ab)2, Fv, scFv, IgGACH2, F(ab')2, scFv2CH3, Fab, VL, VH, scFv4, scFv3, scFv2, dsFv, Fv, scFv-Fc, (scFv)2 фрагменты, или неистощающие IgG, диатела или бивалентные антитела. Некоторые из молекул представляют собой IgG, выбранные из группы, состоящей из природных IgGl, IgG2, IgG3, IgG4 изотипов или синтетических IgG. Некоторые из молекул представляют собой Fab, scFv или dsFv. В некоторых вариантах осуществления hROR1-специфические антитела, связывающиеся белки на основе антител или фрагменты таких антител по изобретению конъюгированы с синтетической молекулой. Синтетическая молекула может представлять собой, например, метку, цитотоксическое средство, радиоизотоп или липосому. Цитотоксическое средство может представлять собой, например, низкомолекулярный токсин, пептидный токсин или белковый токсин. В некоторых вариантах осуществления hROR1-специфические антитела, связывающиеся белки на основе антител или фрагменты таких антител конъюгированы с трансмембранной областью и внутриклеточным сигнальным доменом T-клеточного рецептора (TCR) с образованием химерного антигенного рецептора (CAR).
Изобретение также относится к конъюгатам антитело-лекарственное средство (ADCs), включающим hROR1-специфическое гуманизированное или химерное антитело, связывающийся белок на основе антитела или фрагмент антитела с нагрузкой токсином, который осуществляет эффективный киллинг hROR1 специфических клеток. В указанных ADCs токсиновая нагрузка может быть конъюгирована не-сайт-специфическим образом с антителом, связывающимся белком на основе антитела или фрагментом антитела через боковые цепи аминокислот лизина или цистеина с использованием классических химических линкеров с малеимидной функциональной группой или других химических веществ, известных в данной области, которые могут опосредовать конъюгацию с боковыми цепями аминокислот лизина или цистеина. В указанных ADCs низкомолекулярная нагрузка также может быть конъюгирована сайт-специфически либо путем химической, либо хемо-ферментативной, либо ферментативной конъюгации, известной в данной области, такой как, например, с бифункциональными линкерами, линкерами, позволяющими осуществлять реакцию Пикте-Шпленгера по формилу-глицину с образованием фермент-модифицированных антител, посредством гликан-ремоделированных антител или посредством бактериальных ферментов трансглуаминазы или сортазы.
В некоторых соответствующих аспектах изобретение обеспечивает фармацевтические композиции или наборы, которые содержат терапевтически эффективное количество анти-hROR1 антитела, связывающегося белка на основе антитела, фрагмента антитела, конъюгата антитело-лекарственное средство (ADC), описанных в настоящей заявке, и фармацевтически приемлемый носитель. Некоторые наборы по изобретению могут дополнительно содержать один или несколько буферов для иммуноанализа. Также в изобретении обеспечиваются полинуклеотиды, кодирующие вариабельную область тяжелой цепи иммуноглобулина или легкой цепи иммуноглобулина анти-hROR1 антител, связывающихся белков на основе антител, фрагментов таких антител, конъюгатов антитело-лекарственное средство (ADCs) или CARs, раскрытых в настоящей заявке, а также векторы экспрессии, включающие такую полинуклеотидную последовательность.
В другом аспекте изобретение обеспечивает способы для киллинга или ингибирования роста клетки, экспрессирующей hROR1. Способы включают введение терапевтически эффективного количества анти-hROR1 антител, связывающегося белка на основе антител, фрагмента антитела, конъюгата антитело-лекарственное средство (ADC) или CAR по изобретению субъекту, нуждающемуся в этом, что обеспечивает киллинг или ингибирование роста клетки, экспрессирующей hROR1, у субъекта. Некоторые из этих способов специфически направлены на киллинг или ингибирование опухолевых клеток. В другом аспекте изобретение обеспечивает способы лечения заболевания или состояния, ассоциированного с повышенной экспрессией hROR1 у субъекта. Эти способы включают введение терапевтически эффективного количества анти-hROR1 антител, связывающихся белков на основе антител, фрагментов таких антител, ADCs или CARs по изобретению субъекту, страдающему заболеванием или состоянием, ассоциированным с повышенной экспрессией hROR1, что позволяет лечить заболевание или состояние, ассоциированное с повышенной экспрессией hROR1, у субъекта. Некоторые из этих терапевтических способов специфически направлены на лечение рака. Например, способы можно использовать для лечения субъектов, страдающих от различных типов рака, включая, например, CLL, ALL, лимфому из клеток мантийной зоны, нейробластому, саркому, почечноклеточную карциному, рак молочной железы, рак легкого, рак толстой кишки, рак головы и шеи и меланому.
Еще в одном аспекте изобретение обеспечивает способы детекции изменения уровня ROR1 у субъекта. Такие способы включают (a) получение биологического образца от субъекта; (b) контактирование образца с анти-hROR1 антителами, связывающимися белками на основе антител или фрагментами таких антител по изобретению; (c) определение уровня ROR1 в биологическом образце; и (d) сравнение уровня ROR1 в биологическом образце с контрольным уровнем ROR1 для определения изменился ли уровень ROR1 в биологическом образце по сравнению с контрольным уровнем ROR1. В некоторых из этих способов повышенный уровень ROR1 у субъекта по сравнению с контрольным уровнем указывает на заболевание или состояние, ассоциированное с повышенной экспрессией ROR1 у субъекта. Примеры конкретных заболеваний или состояний, подходящих для таких способов, включают, например, CLL, ALL, лимфому из клеток мантийной зоны, нейробластому, саркому, почечноклеточную карциному, рак молочной железы, рак легкого, рак толстой кишки, рак головы и шеи или меланому.
В другом соответствующем аспекте изобретение обеспечивает способы для детекции ROR1-экспрессирующей опухоли у субъекта. Способы включают (a) введение hROR1 антитела, связывающегося белка на основе антитела или фрагмента антитела по изобретению субъекту, который имеет, у которого подозревается или у которого имеется риск развития ROR1-экспрессирующей опухоли; и (b) обследование субъекта методом визуализации для определения области измененной плотности или концентрации конъюгированной метки, где плотность или концентрацию определяют относительно (i) фонового значения в проксимальной ткани или (ii) плотности или концентрации, ранее обнаруженной в той же области у субъекта, таким образом, существование области измененной плотности или концентрации конъюгированной метки является показателем присутствия ROR1-экспрессирующей опухоли у субъекта.
Природу и преимущества настоящего изобретения можно лучше понять при рассмотрении остальных частей описания и формулы изобретения.
Описание чертежей
Фиг. 1 показывает аминокислотные последовательности вариабельных областей тяжелой и легкой цепей иммуноглобулинов новых кроличьих анти-hROR1 mAbs, которые указаны. Выравнивание аминокислотных последовательностей кроличьих вариабельных доменов (Vκ, Vλ, и VH) показано с каркасными областями (FR) и определяющими комплементарность областями (CDR) с использованием нумерации Кэбата. На Фиг. 1 показаны последовательности вариабельных доменов тяжелых цепей (SEQ ID NO:1-13, соответственно) и последовательности вариабельных доменов легких цепей (SEQ ID NO:14-26, соответственно) 13 антител, обозначенных XBR1-402, ERR1-301, ERR1-306, ERR1-316, ERR1-324, ERR1-403, ERR1-409, ERR1-TOP4, ERR1-TOP15, ERR1-TOP22, ERR1-TOP40, ERR1-TOP43 и ERR1-TOP54. Как указано на Фиг. клоны XBR1-402, ERR1-301, ERR1-306, ERR1-316, ERR1-403, ERR1-409, ERR1-TOP4, ERR1-TOP15, ERR1-TOP22, ERR1-TOP43 и ERR1-TOP54 представляют собой вариабельные домены легких цепей λ иммуноглобулинов, тогда как антитела ERR1-324 и ERR1-TOP40 представляют собой вариабельные домены легких цепей κ иммуноглобулинов.
Фиг. 2 показывает активность связывания химерных кроличьих/человеческих Fabs с человеческим ROR1 (hROR1) и мышиным ROR1 (mROR1), которые были экспрессированы в виде слитых белков внеклеточного домена (ECD) hROR1 и mROR1 с человеческим Fc доменом человеческого IgG1 антитела. Связывание каждого химерного кроличьего/человеческого Fab с hROR1 и mROR1, слитыми с человеческим IgG1 Fc (hFc-hROR1 и hFc-mROR1), анализировали методом ELISA. hFc-ROR1 или hFc-mROR1 захватывались анти-человеческим IgG1 Fc антителом, иммобилизованным на планшете, и затем осуществляли инкубацию с hROR1 специфическими Fabs, включающими His-метку, используя для детекции мышиную анти-His метку. Специфичность Fabs подтверждали с использованием слитых белков внеклеточного домена (ECD) hROR2 с человеческим Fc доменом человеческого IgG1 антитела (hFc-hROR2) в качестве контроля.
Фиг. 3 показывает активность связывания химерных кроличьих/человеческих Fabs с природным человеческим ROR1 белком, экспрессируемым на клеточной поверхности мышиной preB клеточной линии 63-12 (см. Пример 1). Связывание каждого химерного кроличьего/человеческого Fab с эктопически экспрессируемым человеческим ROR1 на поверхности мышиных B-клеток предшественников (63-12) анализировали методом проточной цитометрии. ERR2-TOP35 представляет собой mAb против hROR2, который служил в качестве изотипически сходного контроля.
Фиг. 4 показывает исследования методом картирования эпитопов для химерных кроличьих/человеческих Fabs на шести разных иммобилизованных IgG1-Fc слитых белках, которые включают разные части внеклеточного домена человеческого ROR1: hFc-hROR1-Ig (включающий Иммуноглобулиновый домен hROR1), hFc-hROR1-Fr (включающий Frizzled домен hROR1), hFc-hROR1-Kr (включающий Kringle домен hROR1), hFc-hROR1-Ig-Fr (включающий Иммуноглобулиновый и Frizzled домены hROR1), hFc-hROR1-Fr-Ki (включающий Frizzled и Kringle домены hROR1) и hFc-hROR1 (включающий весь внеклеточный домен (ECD) hROR1).
Фиг. 5 показывает анализы связывания эпитопов, которые осуществляли с использованием поверхностного плазмонного резонанса. Показаны SPR сенсограммы, полученные для связывания различных Fabs с hFc-hROR1, захваченным анти-человеческим Fcγ антителом, иммобилизованным на CM5 чипе. Fabs инжектировали в разном порядке для идентификации отдельных и перекрывающихся эпитопов. Резонансная единица (RU, y ось) увеличивается, что превышало значения, найденные для ранее инжектированных Fabs, показывающих независимые эпитопы, поскольку они обеспечивают возможность одновременного связывания. Например, увеличение, обнаруженное для связывания Fab R11, превышало значения, найденные для XBR1-402 отдельно, указывая на то, что Fab R11 и XBR1-402 могут связываться одновременно с человеческим ROR1. В отличие от этого, эпитоп Fab XBR1-402 перекрывается с эпитопами ERR1-301, ERR1-403 и R12 (левая графа); эпитоп Fab ERR1-TOP43 перекрывается с эпитопом ERR1-306, XBR1-402 и ERR1-TOP40. Ось x показывает время в секундах (сек).
Фиг. 6 показывает измерения аффинности анти-hROR1 специфических Fabs к hROR1 ECD с использованием поверхностного плазмонного резонанса (SPR). (A) Показаны Biacore X100 сенсограммы, полученные для связывания каждого Fab с hFc-hROR1, захваченным анти-человеческим Fcγ антителом, иммобилизованным на CM5 чипе, после мгновенного фонового истощения. Fabs инжектировали при пяти разных концентрациях, с наивысшей концентрацией, указанной в таблице (B), одну из пяти концентраций испытывали в двух повторах. (B) Моновалентные аффинности каждого Fab показаны в таблице. Равновесную константу диссоциации (Kd) рассчитывали из koff/kon (kon - константа скорости ассоциации; koff - константа скорости диссоциации).
Фиг. 7 показывает FACS-основанное окрашивание клеток с hROR1 на различных человеческих раковых клеточных линиях анти-человеческим ROR1 антителом 2A2, как описано в Примере 9. Анализируемые клеточные линии включают 697 (человеческий острый лимфоцитарный лейкоз, ALL), Kasumi-2 (человеческий preB острый лимфоцитарный лейкоз), клеточные линии человеческого трижды отрицательного рака молочной железы MDA-MB-231, MDA-MB-468 и HS-578T, а также клеточную линию человеческого рака молочной железы T47D. За исключением T47D клеточной линии человеческого рака молочной железы, все из оцениваемых клеток являются положительными в отношении экспрессии hROR1.
Фиг. 8 показывает активность связывания выбранного химерного кроличьего/человеческого IgG1 с эндогенным hROR1, экспрессируемым на клетках рака молочной железы, измеренную методом флуоресцентного сортинга клеток (FACS). Известно, что человеческая клеточная линия рака молочной железы MDA-MB-231 экспрессирует hROR1, человеческая клеточная линия рака молочной железы T47D известна как отрицательная, что касается hROR1. В отличие от этого, T47D известна как ROR2-положительная, тогда как MDA-MB-231 известна как отрицательная, что касается экспрессии ROR-2. ERR1-Top54, ERR1-Top43, ERR1-324, XBR1-402 представляли собой выбранные анти-hROR1 специфические mAbs, XBR2-401 представлял собой hROR2 специфическое mAb, используемое в качестве контроля специфичности (A). Экспрессию эндогенных hROR1 и hROR2 на клетках рака молочной железы определяли методом проточной цитометрии с использованием коммерчески доступных козлиных анти-человеческих ROR1 и козлиных анти-человеческих ROR2 поликлональных антител (R&D Systems), соответственно, затем Alexa Fluor 647-конъюгированных AffiniPure F(ab')2 ослиных анти-козлиных IgG (H+L) поликлональных антител (Jackson ImmunoResearch Laboratories). Контрольные окрашивания осуществляли только с Alexa Fluor 647-конъюгированным AffiniPure F(ab')2 ослиным анти-козлиным IgG (H+L) поликлональным антителом. (B) Связывание химерных кроличьих/человеческих IgG1 выбранных клонов ERR1-Top54, ERR1-Top43, ERR1-324, XBR1-402 (все hROR1-специфические) и XBR2-401 (hROR2-специфический) с экспрессирующей ROR1 человеческой клеточной линией рака молочной железы MDA-MB-231 и с экспрессирующей ROR2 человеческой клеточной линией рака молочной железы T47D анализировали методом проточной цитометрии с использованием химерного кроличьего/человеческого IgG1 в качестве первичного антитела и APC-меченных козлиных анти-человеческих Fc-специфических поликлональных антител в качестве вторичного антитела.
Фиг. 9 показывает активность связывания химерных кроличьих/человеческих IgG1 XBR1-402 и ERR1-TOP43 (оба hROR1-специфические) с денатурированным hROR1 в Вестерн-блот экспериментах. hROR1, экспрессируемый на клеточной поверхности K562 клеток, или очищенный белок денатурировали и осуществляли детекцию методом Вестерн-блоттинга. Вестерн блоты содержали следующие указанные образцы: Полоса 1: K562 клетки, эктопически экспрессирующие полноразмерный hROR1. Полоса 2: нетрансфицированные K62 клетки. Полоса 3: очищенный внеклеточный домен hROR1. Полоса 4: очищенный внеклеточный домен hROR2.
Фиг. 10 показывает анализ связывания методом ELISA выбранных hROR1-специфических кроличьих-человеческих-Fc химерных антител выбранных клонов ERR1-301, XBR1-402, ERR1-306, ERR1-324, ERR1-403 и ERR1-Top43 с рекомбинантным очищенным hROR1 (панель A) и с рекомбинантным очищенным hROR2 в качестве отрицательного контроля (панель B).
Фиг. 11 схематически показывает, как сайт-специфически конъюгированные ADCs, раскрытые в настоящем изобретении, были получены. (A) схематически показывает механизм опосредованной ферментом сортазой конъюгации антитела (SMAC-технология), как раскрыто в WO2014140317. Для получения сайт-специфически конъюгированных ADCs необходимо, чтобы рекомбинантные антитела экспрессировались с C-концевым пентапептидным мотивом LPXTG (SEQ ID NO:144), которые служат в качестве сайтов распознавания для фермента сортазы A из Staphylococcus aureus (SrtA). Когда глицин-модифицированный токсиновый субстрат инкубируют с содержащим пентапептидный мотив LPXTG антителом и ферментом сортазой A, фермент сортаза A катализирует реакцию транспептидации, посредством которой глицин-модифицированный токсин заменяет C-концевой глицин LPXTG мотива и ковалентно связывается с треонином остальной LPXT (SEQ ID NO:147) последовательности. Таким образом, можно с высокой эффективностью получить ADCs, конъюгированные с токсином по C-концу. (B) показывает структуру предпочтительного токсина, PNU-159682 производного, включающего этилен-диамино (EDA) линкер, соединяющий 5x глициновый участок с карбонильной группой на C13 антрациклиновой структуры, как раскрыто в WO2016102697.
Фиг. 12 показывает эффективность для in vitro анализов киллинга клеток, осуществляемых на (панель A) иммортализованной человеческой клеточной линии рака молочной железы MDA-MB-468, с использованием известных hROR1-таргетирующих ADCs (2A2-G5-PNU, R12-G5-PNU) и нового ADC, представленного в изобретении (ERR1-Top43-G5-PNU), и на (панель B) иммортализованной человеческой клеточной линии рака молочной железы HS 578T с использованием известных hROR1-таргетирующих ADCs (2A2-G5-PNU, R12-G5-PNU) и новых ADCs, представленных в изобретении (XBR1-402-G5-PNU и ERR1-Top43-G5-PNU). CD30 таргетирующее ADC Ac10-G5-PNU использовали в качестве изотипически сходного контрольного ADC в обеих панелях.
Фиг. 13 показывает in vitro эффективность для киллинга клеток ROR1-положительной клеточной линии острого лимфоцитарного лейкоза 697 четырех анти-ROR1-PNU ADCs, включая ADCs на основе клонов антител 2A2, R11 и R12 и на основе клона XBR1-402 анти-ROR1-антитела, раскрытого в настоящей заявке. Все антитела были экспрессированы как кроличьи-человеческие (R11, R12, XBR-402) или мышиные-человеческие(2A2) химерные IgG1. (A) показывает киллинг клеток в диапазоне концентраций ADCs, сайт-специфически конъюгированных с нагрузкой токсином Gly5-EDA-PNU (сокращенно G5-PNU), с использованием каждого из ADCs. (B) показывает численные IC50 значения для киллинга клеток, рассчитанные из кривых в (A), для каждого из анти-ROR1 ADCs.
Фиг. 14 показывает in vitro киллинг клеток ROR1-положительных ALL клеточных линий Kasumi-2 (A) и 697 (B) с использованием выбранных анти-hROR1 ADCs, сайт-специфически конъюгированных с токсиновой нагрузкой Gly5-EDA-PNU (сокращенно G5-PNU). ADCs основаны на анти-ROR1 антителе клоне 2A2 и R12 и анти-ROR1 клоне XBR1-402, как показано. HER-2-специфический трастузумаб, сайт-специфически конъюгированный с Gly5-EDA-PNU токсиновой нагрузкой, использовали в качестве изотипически сходного контрольного ADC. Панели C и D показывают уровни экспрессии ROR1, измеренные на клеточной поверхности Kasumi-2 и 697, которые анализировали методом FACS с использованием антитела 2A2 в качестве первичного антитела против изотипически сходного контрольного антитела.
Фиг. 15. (A) показывает in vivo эффективность ADCs в диссеминированной мышиной модели ROR1-положительных 697 с EDA-Gly5-PNU ADCs. Мышам (группы по 8 животных) трансплантировали внутривенно 106 697 клеток человеческого ALL и обрабатывали каждое животное в течение 7 и 14 дней после этого при помощи 1 мг/кг PNU-ADC на основе антитела 2A2 и нового антитела XBR1-402 или при помощи PNU-ADC на основе HER2-специфического антитела трастузумаба в качестве отрицательного контроля. Процент выживаемости в группах мышей наносили на график в зависимости от времени. (B) показывает концентрацию ADCs в плазме, измеренную у мышей, которые получали 1 мг/кг, через 12 и 19 дней методом ELISA с использованием захвата анти-человеческим Fcg реагентом и детекции при помощи либо анти-каппа-легкой цепи детекторного антитела для концентрации антитела, либо анти-PNU детекторного антитело для концентрации ADC.
Фиг. 16. Панель (A) показывает in vitro киллинг клеток hROR1-трансфицированной мышиной клеточной линии рака молочной железы EMT6-клон14 (сокращенно EMT6-cl14) сайт-специфически конъюгированными PNU-ADCs на основе анти-ROR1 антитела R12 и нового антитела XBR1-402, которые оба были экспрессированы как химерные человеческие IgG1 антитела. Трастузумаб-G5-PNU ADC, специфический в отношении HER2, использовали в качестве изотипически сходного контрольного ADC. (B) В качестве дополнительного контроля, такой же эксперимент киллинга клеток с такими же ADCs также осуществляли на нетрансфицированных (и ROR1-отрицательных) EMT6 родительских клетках. Панель (C) показывает сравнительную экспрессию hROR1 в hROR1-трансфицированных клетках против нетрансфицированных EMT6 клеток, как определено при помощи FACS с использованием ROR1-специфического антитела 2A2.
Фиг. 17. In vitro киллинг клеток hROR1-трансфицированной мышиной клеточной линии рака молочной железы EMT6-клон14 (сокращенно EMT6-cl.14) сайт-специфически конъюгированными PNU-ADCs на основе анти-ROR1 антитела ms961 и новых антител XBR1-402 и ERR1-324, которые все были экспрессированы как химерные человеческие IgG1 антитела. Трастузумаб-G5-PNU ADC, специфический в отношении HER2, использовали в качестве изотипически сходного контрольного ADC.
Фиг. 18 (A) показывает результаты испытания эффективности in vivo с использованием ортотопической мышиной модели рака молочной железы с использованием hROR1-трансфицированной EMT6 мышиной клеточной линии рака молочной железы, которую имплантировали в жировую ткань молочной железы мышам Balb/c дикого типа. Верхняя панель показывает кривые выживаемости мышей, которых обрабатывали два раза контрольным ADC трастузумаб-G5-PNU. Средняя панель показывает кривые выживаемости мышей, которых обрабатывали PNU-ADC на основе анти-ROR1 антитела R12, и нижняя панель показывает кривые выживаемости мышей, которых обрабатывали при помощи PNU-ADC на основе нового анти-ROR1 антитела XBR1-402. Маленькие треугольники ниже x-оси показывают два вида обработки, каждая при 1 мг/кг, соответствующим ADC в день 14 и 21 после трансплантации опухолей. (B) показывает график Каплана-Мейера трех экспериментов, показанных на панели (A).
Фиг. 19 показывает in vitro стабильность XBR1-402-G5-PNU ADC в сыворотке мыши NOD SCID (панель A) и в человеческой сыворотке (панель B), которую анализировали методом ELISA с детекцией либо общего содержания антител (сплошная линия), либо интактных ADC (пунктирная линия).
Фиг. 20 показывает in vivo стабильность в плазме нового оголенного анти-ROR1 антитела XBR1-402, а также XBR1-402-G5-PNU ADC, определенную у самок CD-1 мышей. Показаны стабильность в плазме, измеренная методом ELISA, суммарных IgG, определенная с использованием реагента для детекции человеческих Fc, а также интактных ADC, определенная с использованием PNU-специфического реагента для детекции.
Фиг. 21 показывает анализ различных лизатов полученных от пациента опухолевых клеток для определения экспрессии hROR1 белка методом Вестерн-блоттинга, включая лизаты из двух контрольных клеточных линий Kasumi-2 (клеточная линия ALL человека) и A549 (клеточная линия рака легкого человека). Лизаты полученных от пациента опухолевых клеток имеют следующие обозначения и происхождение: PXF 1118: плеврамезотелиома, RXF 486: гипернефрома, PXF 541: плеврамезотелиома, SXFS 1407: нейрофибросаркома, CXF 533: аденокарцинома.
Фиг. 22 показывает эффективность сайт-специфически конъюгированного PNU-ADC, включающего XBR1-402 анти-ROR1 ADC, в различных моделях опухолей, выделенных у пациента (PDX модели), установленных у самок NMRI бестимусных мышей, которым имплантировали CXF533, RXF 486, PXF 1118, SXFS 1407 и PXF 541 опухолевый материал, полученный от пациента, по сравнению с мышами, которых обрабатывали носителем в качестве контроля.
Фиг. 23 показывает VH и VL аминокислотные последовательности клонов гуманизированного антитела, происходящих из нового анти-ROR1 антитела XBR1-402. Выравнивание аминокислотных последовательностей гуманизированных вариабельных доменов (Vλ и VH) показано с каркасными областями (FR) и определяющими комплементарность областями (CDR) с использованием нумерации Кэбата. На Фиг. показаны последовательности вариабельных доменов тяжелых цепей (SEQ ID NO:130-135, соответственно) и последовательности вариабельных доменов легких цепей (SEQ ID NO:136-141, соответственно) 6 антител, обозначенных HuXBR1-402(3), HuXBR1-402(8), HuXBR1-402(15), HuXBR1-402(17), HuXBR1-402(19) и HuXBR1-402(26).
Фиг. 24 представляет данные измерения аффинности с новыми гуманизированными клонами исходного mAb XBR1-402, включая kon и koff данные, которые указаны.
Фиг. 25 показывает (Панель A) кривые доза-ответ in vitro анализов киллинга клеток, которые осуществляли на человеческих 697 ALL раковых клетках с hROR1-таргетирующим исходным XBR1-402-G5-PNU и с ADCs на основе гуманизированных антител: huXBR1-402-3-G5-PNU, huXBR1-402-8-G5-PNU, huXBR1-402-15-G5-PNU, huXBR1-402-17-G5-PNU, huXBR1-402-19-G5-PNU и huXBR1-402-26-G5-PNU. PNU-ADC на основе HER2-таргетирующего антитела трастузумаба использовали в качестве изотипического контроля (Tras-G5-PNU). Панель B показывает количественную оценку эффективности киллинга клеток in vitro (IC50).
Фиг. 26 показывает сравнение in vitro активностей ROR1-таргетирующего XBR1-402 CAR-T и R12 CAR-T.
Фиг. 27 показывает сравнение in vitro активностей ROR1-таргетирующего XBR1-402 CAR-T с коротким и длинным спейсером.
Фиг. 28 представляет обзор анализа специфичности химерного кроличьего/человеческого IgG1 против человеческого ROR1, XBR1-402, и используемого в качестве контроля химерного кроличьего/человеческого IgG1 против человеческого ROR2, XBR2-401, с использованием Retrogenix Cell Microarray Platform.
Фиг. 29 показывает анализ специфичности химерного кроличьего/человеческого IgG1 против человеческого ROR1, XBR1-402, и используемого в качестве контроля химерного кроличьего/человеческого IgG1 против человеческого ROR2, XBR2-401, с использованием Retrogenix Cell Microarray Platform. Лучшие образцы первичного связывания из большого скрининга, включающего 4336 человеческих плазменных мембранных белков (см. Фиг. 28), объединяли на одном предметном стекле и окрашивали химерным кроличьим/человеческим IgG1 против человеческого ROR1, XBR1-402, и в качестве контролей химерным кроличьим/человеческим IgG1 против человеческого ROR2, XBR2-401, и биоподобным лекарственным препаратом ритуксимабом. ZsGreen1 сигналы слева указывают уровни экспрессии различных человеческих мембранных белков. Помимо их соответствующих антигенов (ROR1, ROR2 и CD20), испытываемые антитела в формате IgG1 также связываются с Fcγ рецепторами FCGR3B (CD16B), FCGR1A (CD64A) и FCGR2A (CD32A), как ожидалось. Окрашивание только вторичным антителом выявляет тяжелую цепь человеческого IgG3 (IGHG3), как ожидалось.
Подробное описание
I. Обзор
Изобретение основано частично на создании авторами настоящего изобретения большой библиотеки интактных химерных кроличьих/человеческих Fab и выборе связывающихся с человеческим ROR1 с помощью фагового дисплея. Подобные рецепторной тирозинкиназе орфанные рецепторы-1 и -2, ROR1 и ROR2, являются единственными членами семейства, которые определяют новое семейство рецепторных тирозинкиназ, основанное на общем структурном дизайне и некоторых функциональных сходствах. Как ROR1, так и ROR2 белки представляют собой пронизывающие мембрану один раз рецепторы типа I с внеклеточным доменом (ECD), состоящим из иммуноглобулинового домена, цистеин-обогащенного frizzled домена с и Kringle домена. После этих трех внеклеточных доменов следует трансмембранный домен, соединяющий ECD с внутриклеточной частью белка, включающей киназные домены (Rabagay et al. (2012) Frontiers Oncol. 2: 1-8). Человеческие ROR1 и ROR2 белки имеют 58% гомологию друг с другом, но каждый из ROR белков является высококонсервативным среди различных видов. Наиболее консервативным в действительности является ROR1 белок, имеющий длину 937 аминокислот, который имеет идентичность более чем 98,5% между людьми и всеми секвенированными видами отличных от человека приматов, и даже имеет гомологию 96,7 и 96,3% между человеческим и мышиным и кроличьим ROR1, соответственно (Borcherding et al. (2014) Protein Cell 5: 496-502). Поэтому была задача получения высококачественных анти-ROR1 антител путем иммунизации мыши или кролика, и существует только очень немного известных антител с приемлемой аффинностью. См., например, WO 2010/124188 (мышиное моноклональное антитело 2A2), WO 2012/075158 (кроличьи антитела R11 и R12), WO 2012/097313 (мышиное моноклональное антитело D10) и WO2014/031174 (гуманизированные варианты мышиного mAb 99961, который связывается с тем же эпитопом, что и mAb D10).
Для того чтобы не повторять генерацию анти-ROR1 антител традиционными методами иммунизации/скрининга мышей/кроликов, которые могли бы направить антитела против эпитопов наибольшей дивергенции между мышиным/кроличьим ROR1 и человеческим ROR1 (как было при идентификации mAbs 2A2, R11, R12, D10 и 99961), авторы настоящего изобретения создали очень высокосложную фаг-дисплейную наивную библиотеку Fab кроличьих антител и скринировали эту библиотеку на связывание с природным рекомбинантным ECD ROR1 млекопитающего и с экспрессируемым на клеточной поверхности человеческим ROR1 для выбора наиболее функциональных и разнообразных клонов антител, способных к взаимодействию с природным человеческим ROR1 белком. Эта стратегия была выбрана, поскольку репертуар антител, которые необходимо проанализировать, все же выделен из природных кроличьих B лимфоцитов и, таким образом, выбран для предварительно выбранных тяжелых и легких цепей иммунных антител. Однако, благодаря применяемой стратегии скрининга с вовлечением природных рекомбинантных и экспрессируемых клетками человеческих ROR1, была надежда на то, что можно будет идентифицировать hROR1 специфические антитела с хорошей способностью к развитию и функциональными качествами, и которые особенно полезны для лечения заболеваний человека, ассоциированных с экспрессией ROR1, таких как, в частности, ROR1-положительный рак.
Как результат выбранной стратегии, был идентифицирован ряд новых кроличьих высокоаффинных анти-человеческих ROR1 антител с различными CDR1, 2 и 3 клонотипами (Фиг. 1) и с высокой селективностью связывания в отношении человеческого ROR1, но не самой близкородственной ему ʺсестринской молекулыʺ, человеческого ROR2 (Фиг. 2, 3 и 10). Некоторые из hROR1-специфическтаких антител показали высокую аффинность (аффинности в одноцифровом нМ диапазоне) к hROR1 мишени (Фиг. 6). Как подробно описано в настоящей заявке, было получено тринадцать моноклональных антител (mAbs) в формате химерных кроличьих/человеческих Fab с различными последовательностями тяжелых и легких цепей. Эти mAbs были предварительно названы ʺXBR1-402ʺ, ʺERR1-301ʺ, ʺERR1-306ʺ, ʺERR1-316ʺ, ʺERR1-324ʺ, ʺERR1-403ʺ, ʺERR1-409ʺ, ʺERR1-TOP4ʺ, ʺERR1-TOP15ʺ, ʺERR1-TOP22ʺ, ʺERR1-TOP40ʺ, ʺERR1-TOP43ʺ и ʺERR1-TOP54ʺ. Все тринадцать антител связываются с очищенным человеческим ROR1, как показал анализ ELISA, и с клеточно-поверхностным человеческим ROR1, как показал анализ методом проточной цитометрии. Ни один не связывается с ROR2, который имеет близкое родство с ROR1 и имеет идентичность аминокислотной последовательности с ROR1 58%. Два mAbs (ʺERR1-306ʺ и ʺERR1-TOP22ʺ) связываются как с человеческим, так и с мышиным ROR1, тогда как остальные одиннадцать mAbs связываются только с человеческим ROR1.
Аффинность всех тринадцати mAbs определяли методом бислойной интерферометрии и поверхностного плазмонного резонанса. Кроме того, некоторые mAbs (ʺERR-301ʺ, (ʺERR-306ʺ, ʺERR-403ʺ, ʺXBR1-402ʺ, ʺERR1-324ʺ, ʺERR1-TOP43ʺ и ʺERR1-TOP54ʺ) преобразовывали в формат химерных кроличьих/человеческих IgG1, экспрессировали в клетках млекопитающего и очищали аффинной хроматографией с Белком А. В частности, клоны с наивысшей аффинностью XBR1-402 и ERR1-TOP43 при дальнейшей оценке показали самые высокие активности окрашивания при помощи FACS с клетками млекопитающего, сверхэкспрессирующими человеческий ROR1 (Фиг. 3) и природно-экспрессирующими hROR1 (Фиг. 8). Два клона с самым высоким связыванием, XBR1-402 и ERR1-TOP43, также были способны определять денатурированный ROR1 белок, как показал в Вестерн-блоттинг (Фиг. 9), что позволяет использовать эти клоны для разработки сопутствующего средства для диагностики ROR1-экспрессирующих раковых заболеваний.
Кроме того, некоторые mAbs были экспрессированы как химерные кроличьи/человеческие IgG1 с C-концевыми сортаза-распознающими метками, что делает возможной сайт-специфическую конъюгацию нагрузок с C-концами антител методом опосредованной ферментом сортазой конъюгации антител (SMAC-technologyTM), как описано по существу в WO2014140317. Эти анти-hROR1 антитела затем были сайт-специфически конъюгированы с сильнодействующим производным PNU-159682 токсина на основе антрациклина, Gly5-EDA-PNU (Фиг. 11B), для получения высокоэффективных конъюгатов антитело-лекарственное средство (ADCs), как раскрыто по существу в WO2016102679 (которая включена в настоящую заявку посредством ссылки, и текст которой включен в качестве Приложения к настоящей заявке). Эти ADCs были функционально оценены в различных in vitro и in vivo моделях опухолей против ADCs, полученных на основе известных анти-hROR1 антител. Наблюдали, что один особенно перспективный клон, названный XBR1-402, показал самую высокую активность и эффективность по сравнению с различными известными антителами (например, 2A2 (из WO 2010/124188), R11, R12 (оба из WO 2012/075158), или ms961 (из WO 2014/031174), что формирует основу гуманизированного анти-hROR1 mAb цирмтузумаба, находящегося на стадии клинических испытаний в CLL в качестве оголенного IgG1 mAb.
На основании лучших в своем классе свойств, что касается функциональности для киллинга опухолевых клеток в качестве ADC, перспективный клон XBR1-402 затем был гуманизирован, что дало несколько гуманизированных клонов с еще лучшей аффинностью к hROR1, названных ʺhuXBR1-402-3ʺ, ʺhuXBR1-402-8ʺ, ʺhuXBR1-402-15ʺ, ʺhuXBR1-402-17ʺ, ʺhuXBR1-402-19ʺ и ʺhuXBR1-402-26ʺ. Эти гуманизированные варианты ведущего клона XBR1-402 также были оценены как сайт-специфически конъюгированные PNU-ADCs, и каждый из них продемонстрировал еще более лучший киллинг опухолевых клеток в in vitro hROR1 опухолевых моделях.
Для дальнейшего исследования терапевтической пользы ROR1-таргетирующих mAbs были сконструированы CAR-T клетки на основе XBR1-402 с использованием способов, ранее описанных для известных ROR1-таргетирующих mAbs R11 и R12 (Hudecek, M., Lupo-Stanghellini, M. T., Kosasih, P. L., Sommermeyer, D., Jensen, M. C., Rader, C. and Riddell, S. R. (2013) Receptor affinity and extracellular domain modifications affect tumor recognition by ROR1-specific chimeric antigen receptor T cells. Clin. Cancer res. 19, 3153-3164). Вкратце, осуществляли лентивирусную трансдукцию ex vivo размноженных здоровых донорных CD8+ CD62L+ T клеток с использованием EF1α промотор-направляемой экспрессионной кассеты, содержащей XBR1-402 в scFv формате, с последующим коротким или длинным спейсером, трансмембранным доменом человеческого CD28, сигнальным доменом 4-1BB, сигнальным доменом CD3ζ и T2A-отделенным трансмембранным EGFR фрагментом с усеченными связывающимся с лигандом и тирозинкиназным доменами. FACS выделение EGFR+ трансдуцированных T клеток выявило устойчивое анти-ROR1 распознавание в >90% CAR-T клеток. Активность ROR1-таргетирующих XBR1-402 CAR-T с коротким спейсером испытывали против клеточных линий рака молочной железы MDA-MB-231 (ROR1+ ROR2-) и T47D (ROR1- ROR2+). В присутствии ROR1+ ROR2-, но не ROR1- ROR2+ клеток-мишеней XBR1-402 CAR-T быстро пролиферировали, массивно секретировали IFN-γ и IL-2 и эффективно убивали клетки-мишени in vitro (Фиг. 26). Примечательно, что при непосредственном сравнении было обнаружено, что XBR1-402 CAR-T имел такую же или бóльшую активность, чем у клинически исследуемого R12 CAR-T с таким же коротким спейсером и сигнальными доменами.
Кроме того, авторы настоящего изобретения предположили, что оптимальное расстояние между T-клеткой и клеткой-мишенью может достигаться путем снабжения CAR-T клеток, таргетирующих мембрана-дистальные эпитопы, более короткими спейсерами, и наоборот. На основании этого предположения, XBR1-402 CAR-T, который содержит перекрывающийся эпитоп с R12, как предполагают, более активен, когда он имеет короткий спейсер, по сравнению с тем, когда он имеет длинный спейсер. Действительно, было обнаружено, что в присутствии ROR1+ ROR2- клеток-мишеней XBR1-402 CAR-T с коротким линкером пролиферировали быстрее, чем XBR1-402 CAR-T с длинным спейсером (Фиг. 27), а также секретировали значительно больше IFN-γ и IL-2. Однако было обнаружено, что in vitro цитотоксичность является одинаково сильной (Фиг. 27).
В соответствии с этими исследованиями, настоящее изобретение обеспечивает новые моноклональные кроличьи и гуманизированные антитела и соответствующие связывающиеся белки на основе антител и фрагменты таких антител, которые специфически распознают ROR1, а также конъюгаты антитело-лекарственное средство и CARs со специфической анти-опухолевой активностью в hROR1-экспрессирующих опухолевых моделях in vitro и in vivo. Также в изобретении представлены способы использования этих средств на основе антител в терапевтических и диагностических применениях для заболеваний и состояний, ассоциированных с аномальной или повышенной экспрессией ROR1, например, рака.
Антитела и соответствующие композиции по изобретению продемонстрировали другие, к удивлению полезные свойства. Функциональная оценка новых клонов как сайт-специфически конъюгированных ADCs с сильнодействующей PNU-антрациклиновой нагрузкой с использованием различных in vitro и in vivo моделей выявила, что в особенности новый клон XBR1-402 действовал лучше, чем любое из известных антител. Например, ADCs на основе R11 имели ограниченную активность уже в in vitro опухолевых моделях (Фиг. 13). По сравнению с клонами антител 99961 (ms961), 2A2 и R12, новый XBR1-402, описанный в настоящей заявке, также неизменно действовал лучше в опухолевых моделях in vitro (Фиг. 13, 14, 17), что было даже более очевидно в опухолевых моделях in vivo (Фиг. 15 и 18). В сочетании с благоприятными свойствами XBR1-402 клона, с точки зрения его высокоспецифического распознавания ROR1 при испытании против 4336 человеческих плазменных мембранных белков (Фиг. 29) и подходящей стабильности ADC в мышиной плазме и других сыворотках, которые оценивали (Фиг. 15B, 19 & 20), ADCs на основе XBR1-402, как оказалось, имеют потенциал быть лучшими в своем классе анти-ROR1 таргетирующими продуктами с высоким потенциалом для ADC терапии.
Кроме того, гуманизированные варианты анти-ROR1 mAb XBR1-402 неожиданно показали даже еще более высокую аффинность по сравнению с исходным кроличьим клоном XBR1-402 (Фиг. 24), поскольку аффинности часто уменьшаются в процессе гуманизации не-человеческих антител (Margreitter et al. (2016) J. Mol. Recognit. 29: 266-275). Повышенная аффинность также коррелировала с улучшенной активностью этих гуманизированных mAbs при оценке на анти-опухолевую активность в качестве ADCs (Фиг. 25). Эти данные свидетельствуют о высоком потенциале оцениваемых анти-ROR1 ADC, основанных на гуманизированных XBR1-402 антителах, для лечения заболеваний человека. Этот высокий потенциал для эффективного лечения опухолей PNU-ADC на основе XBR1-402 и/или гуманизированных XBR1-402 подтверждается высокой эффективностью ADC на основе XBR1-402 в различных моделях hROR1-экспрессирующих ксенотрансплантатов, полученных от пациента (Фиг. 22).
Высокофункциональные анти-hROR1 антитела и соответствующие композиции, описанные в настоящей заявке, показали превосходные функциональные свойства для применения в качестве терапевтических средств в терапии рака человека, ассоциированного с экспрессией ROR1. Они включают различные антитела к hROR1, фрагменты антител, связывающиеся белки на основе антител, ADCs или CARs, описанные в настоящей заявке, которые обладают такими же или по существу такими же свойствами связывания, как было продемонстрировано специфическими антителами, проиллюстрированными в настоящей заявке (например, XBR1-402). Таким образом, благоприятные свойства и высокий терапевтический потенциал, продемонстрированный антителами, проиллюстрированными в настоящей заявке, могут быть распространены на гомологичные антитела, фрагменты антител, связывающиеся белки на основе антител, ADCs, CARs, которые содержат некоторые или все из последовательностей CDR вариабельных областей тяжелых и/или легких цепей, которые раскрыты в изобретении, или по существу подобные последовательности CDR вариабельных областей тяжелых и/или легких цепей, которые раскрыты в изобретении. Благоприятные свойства и/или высокий терапевтический потенциал также могут быть распространены на антитела, фрагменты антител, связывающиеся белки на основе антител, ADCs, CARs, которые содержат только одну из двух цепей иммуноглобулинов раскрытых антител (т.е. либо тяжелую, либо легкую цепь), или одну из двух цепей иммуноглобулинов (т.е. либо тяжелую, либо легкую цепь), которые гомологичны проиллюстрированным антителам.
II. Определения
Если не указано иное, все используемые технические и научные термины имеют то же значение, которое обычно понимается специалистами в области техники, к которой относится настоящее изобретение. В следующих ссылках специалист сможет найти общее определение многих терминов, используемых в настоящем изобретении: Academic Press Dictionary of Science and Technology, Morris (Ed.), Academic Press (1st ed., 1992); Oxford Dictionary of Biochemistry and Molecular Biology, Smith et al. (Eds.), Oxford University Press (revised ed., 2000); Encyclopaedic Dictionary of Chemistry, Kumar (Ed.), Anmol Publications Pvt. Ltd. (2002); Dictionary of Microbiology and Molecular Biology, Singleton et al. (Eds.), John Wiley & Sons (3rd ed., 2002); Dictionary of Chemistry, Hunt (Ed.), Routledge (1st ed., 1999); Dictionary of Pharmaceutical Medicine, Nahler (Ed.), Springer-Verlag Telos (1994); Dictionary of Organic Chemistry, Kumar and Anandand (Eds.), Anmol Publications Pvt. Ltd. (2002); and A Dictionary of Biology (Oxford Paperback Reference), Martin and Hine (Eds.), Oxford University Press (4th ed., 2000). Кроме того, следующие определения представлены, чтобы помочь читателю осуществить на практике изобретение.
Термин "антитело", также синонимично называемый "иммуноглобулины" (Ig), или "антигенсвязывающий фрагмент" относится к полипептидной цепи (целям), которая демонстрирует сильное одновалентное, двухвалентное или поливалентное связывание с данным антигеном, эпитопом или эпитопами. Если не указано иное, антитела или антигенсвязывающие фрагменты, используемые в изобретении, могут иметь последовательности, происходящие из любых видов позвоночных. Они могут быть получены с использованием любой подходящей технологии, например, гибридомной технологии, рибосомного дисплея, фагового дисплея, библиотек перетасовки генов, полусинтетических или полностью синтетических библиотек или их комбинаций. Если не указано иное, термин "антитело", используемый в настоящем изобретении, включает интактные антитела, антигенсвязывающие полипептидные фрагменты и другие сконструированные антитела, которые описаны ниже или хорошо известны в данной области (см., например, Serafini, J Nucl. Med. 34:533-6, 1993).
Интактное ʺантителоʺ типично включает по меньшей мере две тяжелые (H) цепи (около 50-70 кДа) и две легкие (L) цепи (около 25 кДа), связанные между собой дисульфидными связями. Известные гены иммуноглобулинов, кодирующие цепи антител, включают гены константных областей каппа, лямбда, альфа, гамма, дельта, эпсилон и мю, а также огромное число генов вариабельных областей иммуноглобулинов. Легкие цепи классифицируются как каппа или лямбда. Тяжелые цепи классифицируются как гамма, мю, альфа, дельта или эпсилон, которые, в свою очередь, определяют классы иммуноглобулинов, IgG, IgM, IgA, IgD и IgE, соответственно.
Каждая тяжелая цепь антитела состоит из вариабельной области тяжелой цепи (VH) и константной области тяжелой цепи. Константная область тяжелой цепи большинства IgG изотипов (подклассы) состоит из трех доменов CH1, C H2 и C H3, некоторые IgG изотипы, такие как IgM или IgE, включают четвертый домен константной области CH4, каждая легкая цепь состоит из вариабельной области легкой цепи (VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена CL. Вариабельные области тяжелой и легкой цепи содержат связывающийся домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулинов с тканями или факторами хозяина, включая различные клетки иммунной системы и первый компонент (Clq) классической системы комплемента.
VH и VL области антитела могут далее подразделяться на области гипервариабельности, также называемые определяющими комплементарность областями (CDRs), которые перемежаются с более консервативными каркасными областями (FRs). Каждая VH и VL состоит из трех CDRs и четырех FRs, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Расположение CDR и FR областей и система нумерации определены, например, Kabat et al., Sequences of Proteins of Immunological Interest, U.S. Department of Health and Human Services, U.S. Government Printing Office (1987 and 1991).
"Связывающийся белок на основе антитела", в контексте настоящей заявки, может представлять собой любой белок, который содержит по меньшей мере один происходящий из антитела VH, VL, или CH домен иммуноглобулина, в контексте других не-иммуноглобулиновых или не происходящих из антитела компонентов. Такие белки на основе антител включают, но не ограничиваются этим (i) Fc-слитые белки связывающихся белков, включая рецепторы или компоненты рецепторов со всеми или частями CH доменов иммуноглобулина, (ii) связывающиеся белки, в которых VH и или VL домены связаны с альтернативными молекулярными каркасами, или (iii) молекулы, в которых иммуноглобулиновые домены VH и/или VL и/или CH объединены и/или скомпонованы таким образом, как это обычно не наблюдается в природных антителах или фрагментах антител.
ʺАффинность связыванияʺ, как правило, выражают как равновесные константы ассоциации или диссоциации (KA или KD, соответственно), которые, в свою очередь, представляют обратные отношения констант скоростей диссоциации и ассоциации (koff и kon, соответственно). Таким образом, эквивалентные аффинности могут соответствовать разным константам скоростей, при условии, что отношение конатант скоростей остается таким же. Аффинность связывания антитела обычно выражается как KD моновалентного фрагмента (например, Fab фрагмента) антитела, при этом KD значения в одноцифровом наномолярном диапазоне или ниже (субнаномолярном или пикомолярном) считаются очень высокими и терапевтически и диагностически значимыми.
В контексте настоящей заявки, термин "специфичность связывания" относится к селективной аффинности одной молекулы относительно другой, такой как связывание антител с антигенами (или их эпитопом или антигенной детерминантой), рецепторов с лигандами и ферментов с субстратами. Таким образом, считается, что все моноклональные антитела, которые связываются с конкретной антигенной детерминантой молекулярной частицы (например, специфическим эпитопом ROR1 или ROR2), имеют одинаковую специфичность связывания в отношении этой частицы.
Термин ʺКонъюгат Антитело-Лекарственное средствоʺ, или ʺADCʺ, относится к антителу, с которым терапевтически активное вещество или активный фармацевтический ингредиент (API) ковалентно связан, так чтобы терапевтически активное вещество или активный фармацевтический ингредиент (API) могли быть нацелены на мишень, с которой связывается антитело, для проявления их фармакологической функции. Терапевтически активное вещество или активный фармацевтический ингредиент может представлять собой клеточный токсин, который способен осуществлять киллинг клеток, на которые нацелены ADCs, предпочтительно злокачественных или раковых клеток. Ковалентное присоединение терапевтически активного вещества, активного фармацевтического ингредиента или клеточного токсина можно осуществить не-сайт-специфическим образом с использованием стандартных химических линкеров, которые связывают полезные нагрузки с лизиновыми или цистеиновыми остатками, или конъюгацию предпочтительно осуществляют сайт-специфическим образом, что позволяет осуществлять полный контроль участка конъюгации и отношения лекарственного средства к антителу (DAR) в ADC, который должен быть образован.
Термин "консервативно модифицированный вариант" относится как к аминокислотным, так и к нуклеиновокислотным последовательностям. Что касается конкретных нуклеиновокислотных последовательностей, консервативно модифицированные варианты относятся к тем нуклеиновым кислотам, которые кодируют идентичные или по существу идентичные аминокислотные последовательности, или, где нуклеиновая кислота не кодирует аминокислотную последовательность - к по существу идентичным последовательностям. Из-за вырожденности генетического кода большое количество функционально идентичных нуклеиновых кислот кодирует любой заданный белок. Например, кодоны GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в каждом положении, где аланин определяется кодоном, кодон может быть изменен на любой из соответствующих описанных кодонов без изменения кодируемого полипептида. Такие варианты нуклеиновых кислот являются «молчащими вариантами», которые представляют собой одну разновидность консервативно модифицированных вариантов. Каждая нуклеиновокислотная последовательность, которая кодирует полипептид, также описывает каждый возможный молчащий вариант нуклеиновой кислоты. Специалисту будет понятно, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который обычно является единственным кодоном для метионина, и TGG, который обычно является единственным кодоном для триптофана) может быть модифицирован для получения функционально идентичной молекулы. Соответственно, каждый молчащий вариант нуклеиновой кислоты, которая кодирует полипептид, подразумевается в каждой описанной последовательности.
Что касается последовательностей полипептидов, ʺконсервативно модифицированные вариантыʺ относятся к варианту, который имеет консервативные аминокислотные замены, аминокислотные остатки, замененные другим аминокислотным остатком, имеющим боковую цепь с аналогичным зарядом. Семейства аминокислотных остатков, имеющих боковые цепи с аналогичными зарядами, определены в данной области техники. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин).
Термин ʺконтактированиеʺ имеет обычное значение и относится к совмещению двух или более веществ (например, полипептидов или фага), совмещению веществ и клеток, или совмещению двух популяций различных клеток. Контактирование может происходить in vitro, например, смешивание антитела и клетки или смешивание популяции антител с популяцией клеток в пробирке или в питательной среде. Контактирование также может происходить в клетке или in situ, например, контактирование двух полипептидов в клетке путем ко-экспрессии в клетке рекомбинантных полинуклеотидов, кодирующих два полипептида, или в клеточном лизате. Контактирование также может происходить in vivo в организме субъекта, например, путем введения средства субъекту для доставки средства к клетке-мишени.
ʺГуманизированное антителоʺ представляет собой антитело или фрагмент антитела, антиген-связывающий фрагмент или связывающийся белок на основе антитела, включающий VH или VL домены антитела с гомологией с человеческими каркасными последовательностями VH или VL антител, имеющие T20 оценку в баллах больше чем 80, как определено в соответствии с Gao et al. (2013) BMC Biotechnol. 13, pp. 55.
Термины ʺидентичныйʺ или процент ʺидентичностиʺ, в контексте двух или более нуклеиновых кислот или последовательностей полипептидов, относятся к двум или более последовательностям или субпоследовательностям, которые являются одинаковыми. Две последовательности являются "по существу идентичными", если две последовательности имеют определенный процент аминокислотных остатков или нуклеотидов, которые являются одинаковыми (т.е. 60% идентичность, опционально 65%, 70%, 75%, 80%, 85%, 90%, 95% или 99% идентичность в определенной области или, если область не определена, по всей последовательности) при сравнении и выравнивании для максимального соответствия в окне сравнения или в обозначенной области, как измерено с использованием одного из следующих алгоритмов сравнения последовательностей или путем выравнивания вручную и при визуальном наблюдении. Необязательно, идентичность существует по всей области, которая имеет по меньшей мере около 50 нуклеотидов (или 10 аминокислот) в длину, или более предпочтительно по всей области, которая имеет 100-500 или 1000 или более нуклеотидов (или 20, 50, 200 или более аминокислот) в длину.
Методы выравнивания последовательностей для сравнения хорошо известны в данной области. Оптимальное выравнивание последовательностей для сравнения можно осуществить, например, с использованием алгоритма локальной гомологии Смита и Ватермана, Adv. Appl. Math. 2:482c, 1970; алгоритма выравнивания областей гомологии Нидлмана и Вунша, J. Mol. Biol. 48:443, 1970; метода поиска сходства Пирсона и Липмана, Proc. Nat'l. Acad. Sci. USA 85:2444, 1988; путем компьютеризованного выполнения этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программ Wisconsin Genetics, Genetics Computer Group, Madison, WI); или путем выравнивания вручную и визуального наблюдения (см., например, Brent et al., Current Proticols in Molecular Biology, John Wiley & Sons, Inc. (ringbou ed., 2003)). Два примера алгоритмов, подходящих для определения процента идентичности последовательностей и сходства последовательностей, представляют собой BLAST и BLAST 2.0 алгоритмы, которые описаны в Altschul et al., Nuc. Acids Res. 25:3389-3402, 1977; и Altschul et al., J. Mol. Biol. 215:403-410, 1990, соответственно.
Термин "субъект" относится к человеку и отличным от человека животным (особенно отличным от человека млекопитающим). Термин "субъект" используется в настоящей заявке, например, в связи с терапевтическими и диагностическими способами как относящийся к субъектам людям или животным. Субъекты-животные включают, но не ограничиваются этим, животные модели, такие как модели млекопитающих состояний или расстройств, ассоциированных с повышенной экспрессией ROR1, таких как CLL, ALL, лимфома из клеток мантийной зоны, нейробластома, саркома, почечноклеточная карцинома, рак молочной железы, рак легкого, рак толстой кишки, рак головы и шеи, меланома и другие раковые заболевания. Другие конкретные примеры отличных от человека субъектов включают, например, коров, лошадей, овец, свиней, кошек, собак, мышей, крыс, кроликов, морских свинок, обезьян.
Искусственные T-клеточные рецепторы (также известные как химерные T-клеточные рецепторы, химерные иммунорецепторы, химерные антигенные рецепторы (CAR) или Т-тела) являются сконструированными рецепторами, которые передают какую-либо специфичность иммунной эффекторной клетке. Как правило, эти рецепторы используют для передачи специфичности моноклонального антитела Т-клетке; с переносом их кодирующей последовательности при помощи ретровирусных или лентивирусных векторов или транспозонов. CAR-сконструированные Т-клетки (также сокращенно CAR-T-клетки) представляют собой генно-инженерные Т-клетки, вооруженные химерными рецепторами, в которых внеклеточный участок распознавания состоит из домена распознавания, происходящего из антитела, а внутриклеточная область происходит из одного или нескольких лимфоцит-стимулирующих компонентов. Структура прототипического CAR модульная, предназначенная для размещения различных функциональных доменов и, таким образом, для обеспечения возможности выбора специфичности и контролируемой активации Т-клеток. Предпочтительной происходящей из антитела единицей распознавания является одноцепочечный вариабельный фрагмент (scFv), который объединяет специфичность и связывающиеся остатки вариабельных областей как тяжелой, так и легкой цепи моноклонального антитела. Наиболее типичные фрагменты активации лимфоцитов включают Т-клеточный костимуляторный (например, CD28) домен в тандеме с группой триггера Т-клеток (например, CD3zeta). Вооружая эффекторные лимфоциты (такие как Т-клетки и природные киллерные клетки) такими химерными рецепторами, сконструированная клетка перенаправляется с заранее определенной специфичностью к любому желаемому целевому антигену не ограничиваемым HLA образом. CAR конструкции вводят ex vivo в Т-клетки из периферических лимфоцитов данного пациента с использованием ретровирусных или лентивирусных векторов или транспозонов. После инфузии полученных CAR-сконструированных Т-клеток обратно пациенту они перемещаются, достигают своего целевого сайта и при взаимодействии с клеткой или тканью, являющейся их мишенью, происходит их активация, и они выполняют свою предопределенную эффекторную функцию. Терапевтические мишени для CAR подхода включают раковые и ВИЧ-инфицированные клетки или аутоиммунные эффекторные клетки.
Термины "лечить", "лечащий", "лечение" и "терапевтически эффективный", используемые в настоящей заявке, необязательно означают 100% или полное излечение. Скорее существуют различные степени лечения, признанные специалистами в данной области как имеющие потенциальную пользу или терапевтический эффект. В этом отношении, способ по настоящему изобретению может обеспечить любое количество любого уровня лечения. Кроме того, лечение, обеспечиваемое способом по изобретению, может включать лечение одного или нескольких состояний или симптомов заболевания, лечение которого осуществляют.
"Вектор" представляет собой репликон, такой как плазмида, фаг или космида, к которому может быть присоединен другой полинуклеотидный сегмент, чтобы обеспечить репликацию прикрепленного сегмента. Векторы, способные направлять экспрессию генов, кодирующих один или несколько полипептидов, называются «векторами экспрессии».
III. Антитела, связывающиеся белки на основе антител, фрагменты антител, конъюгаты антитело-лекарственное средство (ADCs) или CARs, специфически связывающиеся с ROR1, и их соответствующие производные.
В одном аспекте изобретение обеспечивает новые антитела, связывающиеся белки на основе антител, фрагменты таких антител, ADCs или CARs, которые специфически связываются с человеческим ROR1 с такой же специфичностью связывания, как у анти-ROR1 антитела, проиллюстрированного в настоящей заявке (Фиг. 1 и 23). Антитела по изобретению включают интактные антитела (например, IgG1 антитела, проиллюстрированные в настоящей заявке), фрагменты антител или антиген-связывающие фрагменты (например, Fab фрагменты, проиллюстрированные в настоящей заявке), связывающиеся белки на основе антител, ADCs и CARs, которые содержат антиген-связывающие части интактного антитела, которые сохраняют способность связывания родственного антигена ROR1. Примеры таких фрагментов антител включают (i) Fab фрагмент, моновалентный фрагмент, состоящий из VL, VH, CL и CH1 доменов; (ii) F(ab')2 фрагмент, бивалентный фрагмент, включающий два Fab фрагмента, связанные дисульфидным мостиком в шарнирной области; (iii) Fd фрагмент, состоящий из VH и CH1 доменов; (iv) Fv фрагмент, состоящий из VL и VH доменов одного плеча интактного антитела; (v) дисульфид-стабилизированные Fvs (dsFvs), которые имеют межцепочечную дисульфидную связь, сконструированную между структурно консервативными каркасными областями; (vi) однодоменное антитело (dAb), которое состоит из VH или VL домена (см., например, Ward et al., Nature 341:544-546, 1989); и (vii) выделенную определяющую комплементарность область (CDR) в виде линейного или циклического пептида. Примеры связывающихся белков на основе антител включают полипептиды, в которых связывающиеся домены антител объединены с другими полипептидами или полипептидными доменами, например, альтернативными молекулярными каркасами, Fc-областями, другими функциональными или связывающимися доменами других полипептидов или антител, приводя к молекулам с дополнительными свойствами связывания, например, би- или мультиспецифическим белкам или антителам. Такие полипептиды могут создавать расположение связывающих или функциональных доменов, обычно не встречающееся в природных антителах или фрагментах антител.
Антитела по изобретению также охватывают фрагменты антител (или ʺантиген-связывающие фрагментыʺ), такие как одноцепочечные антитела. Термин "одноцепочечное антитело" относится к полипептиду, включающему VH домен и VL домен в полипептидной связи, обычно связанные через спейсерный пептид, и который может включать дополнительные домены или аминокислотные последовательности на амино- и/или карбокси-концах. Например, одноцепочечное антитело может включать связывающийся сегмент для связывания с кодирующим полинуклеотидом. В качестве примера, одноцепочечный фрагмент вариабельной области (scFv) представляет собой одноцепочечное антитело. По сравнению с VL и VH доменами Fv фрагмента, которые кодируются отдельными генами, scFv имеет два домена, соединенные (например, рекомбинантными методами) синтетическим линкером. Это делает возможным их получение в виде одной белковой цепи, в которой VL и VH области спариваются с образованием моновалентных молекул.
Антитела по настоящему изобретению охватывают однодоменные антиген-связывающие единицы, которые имеют верблюжий каркас. Животные семейства верблюдовых включают верблюдов, лам и альпак. Верблюдовые продуцируют функциональные антитела, лишенные легких цепей. Вариабельный домен тяжелой цепи (VH) укладывается автономно и функционирует независимо в качестве антиген-связывающей единицы. Его связывающаяся поверхность включает только три CDRs, по сравнению с шестью CDRs в классических антиген-связывающих молекулах (Fabs) или одноцепочечных вариабельных фрагментах (scFvs). Верблюжьи антитела способны достигать аффинности связывания, которая сопоставима с аффинностью традиционных антител.
Различные антитела, связывающиеся белки на основе антител и фрагменты антител, описанные в настоящей заявке, можно получить путем ферментативной или химической модификации интактных антител или синтезировать de novo с использованием методов рекомбинантной ДНК или идентифицировать с использованием библиотеки фагового дисплея. Способы для генерирования этих антител, связывающихся белков на основе антител и фрагментов таких антител хорошо известны в данной области. Например, одноцепочечные антитела можно идентифицировать с использованием библиотек фагового дисплея или библиотек рибосомного дисплея, библиотек перетасовки генов (см., например, McCafferty et al., Nature 348:552-554, 1990; и патент США № 4946778). В частности, scFv антитела можно получить с использованием способов, описанных, например, в Bird et al., Science 242:423-426, 1988; и Huston et al., Proc. Natl. Acad. Sci. USA 85:5879-5883, 1988. Fv фрагменты антител можно генерировать, как описано в Skerra and Plückthun, Science 240:1038-41, 1988. Дисульфид-стабилизированные Fv фрагменты (dsFvs) можно получить с использованием способов, описанных, например, в Reiter et al., Int. J. Cancer 67:113-23, 1996. Подобным образом, однодоменные антитела (dAbs) можно получить различными способами, описанными, например, в Ward et al., Nature 341:544-546, 1989; и Cai and Garen, Proc. Natl. Acad. Sci. USA 93:6280-85, 1996. Верблюжьи однодоменные антитела можно получить с использованием способов, хорошо известных в данной области, например, Dumoulin et al., Nat. Struct. Biol. 11:500-515, 2002; Ghahroudi et al., FEBS Letters 414:521-526, 1997; и связ et al., J. Mol. Biol. 332:643-55, 2003. Другие типы антиген-связывающих фрагментов (например, Fab, F(ab')2 или Fd фрагменты) также можно легко получить обычно используемыми иммунологическими способами. См., например, Harlow & Lane, Using Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1998.
Антитела по изобретению также включают гуманизированные антитела с более высокой гомологией на аминокислотном уровне VH или VL доменов гуманизированного антитела с VH или VL доменами человеческого антитела, чем с VH или VL доменами грызунов, предпочтительно с T20 оценкой больше чем 80, как определено в Gao et al. (2013) BMC Biotechnol. 13, pp. 55.
В некоторых вариантах осуществления антитела, фрагменты антител, связывающиеся белки на основе антител, ADCs или CARs по изобретению имеют последовательности CDR1, CDR2 и CDR3 тяжелой цепи и последовательности CDR1, CDR2 и CDR3 легкой цепи, которые по существу идентичны последовательностям антител, показанным на Фиг. 1 или Фиг. 23. Последовательности CDR легкой цепи и тяжелой цепи проиллюстрированных антител все показаны на Фиг. В некоторых из этих вариантов осуществления антитела, фрагменты антител, связывающиеся белки на основе антител, ADCs или CARs имеют (1) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:27-29, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:66-68, соответственно; (2) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:30-32, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:69-71, соответственно; (3) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:33-35, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:72-74, соответственно; (4) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:36-38, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:75-77, соответственно; (5) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:39-41, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:78-80, соответственно; (6) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:42-44, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:81-83, соответственно; (7) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:45-47, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:84-86, соответственно; (8) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:48-50, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:87-89, соответственно; (9) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:51-53, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:90-92, соответственно; (10) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:54-56, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:93-95, соответственно; (11) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:57-59, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:96-98, соответственно; (12) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:60-62, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:99-101, соответственно; или (13) последовательности CDR1-3 тяжелой цепи, которые по существу идентичны SEQ ID NO:63-65, соответственно; и последовательности CDR1-3 легкой цепи, которые по существу идентичны SEQ ID NO:102-104, соответственно.
В некоторых вариантах осуществления антитела, фрагменты антител, связывающиеся белки на основе антител, ADCs или CARs по изобретению включают последовательности CDR1-CDR3 тяжелой цепи и CDR1-CDR3 легкой цепи, которые идентичны последовательностям, показанным в (1) SEQ ID NO:27-29 и SEQ ID NO:66-68 (антитело XBR1-402), (2) SEQ ID NO:30-32 и SEQ ID NO:69-71 (антитело ERR1-301), (3) SEQ ID NO:33-35 и SEQ ID NO:72-74 (антитело ERR1-306), (4) SEQ ID NO:36-38 и SEQ ID NO:75-77 (антитело ERR1-316), (5) SEQ ID NO:39-41 и SEQ ID NO:78-80 (антитело ERR1-324), (6) SEQ ID NO:42-44 и SEQ ID NO:81-83 (антитело ERR1-403), (7) SEQ ID NO:45-47 и SEQ ID NO:84-86 (антитело ERR1-409), (8) SEQ ID NO:48-50 и SEQ ID NO:87-89 (антитело ERR1-TOP4), (9) SEQ ID NO:51-53 и SEQ ID NO:90-92 (антитело ERR1-TOP15), (10) SEQ ID NO:54-56 и SEQ ID NO:93-95 (антитело ERR1-TOP22), (11) SEQ ID NO:57-59 и SEQ ID NO:96-98 (антитело ERR1-TOP40), (12) SEQ ID NO:60-62 и SEQ ID NO:99-101 (антитело ERR1-TOP43) или (13) SEQ ID NO:63-65 и SEQ ID NO:102-104 (антитело ERR1-TOP54).
В других вариантах осуществления антитела, фрагменты антител, связывающиеся белки на основе антител, ADCs или CARs по изобретению, которые специфически связываются с человеческим ROR1, содержат (a) вариабельный домен легкой цепи, имеющий последовательность, которая по существу идентична любой из SEQ ID NO:14-26 или 136-141, (b) вариабельный домен тяжелой цепи, имеющий последовательность, которая по существу идентична любой из SEQ ID NO:1-13 или 130-135, или (c) и легкую цепь (a) и тяжелую цепь (b). В некоторых вариантах осуществления антитело включает как легкую цепь (a), так и тяжелую цепь (b). В некоторых вариантах осуществления антитела, фрагменты антител, связывающиеся белки на основе антител, ADCs или CARs по изобретению содержит (a) вариабельный домен легкой цепи, имеющий идентичность по меньшей мере 90% с любой из SEQ ID NO:14-26 или 136-141, (b) вариабельный домен тяжелой цепи имеющий идентичность последовательности по меньшей мере 90% с любой из SEQ ID NO:1-13 или 130-135, или (c) и легкую цепь (a) и тяжелую цепь (b). В некоторых вариантах осуществления процент идентичности может быть по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, или по меньшей мере 99% или даже 100%. В некоторых вариантах осуществления вариабельный домен легкой цепи имеет идентичность по меньшей мере 95% с любой из SEQ ID NO:14-26 или 136-141. В некоторых вариантах осуществления вариабельный домен легкой цепи имеет 100% идентичность с любой из SEQ ID NO:14-26 или 136-141. В некоторых вариантах осуществления антитело, фрагменты антител, связывающиеся белки на основе антител, ADCs или CARs содержат вариабельный домен тяжелой цепи, имеющий идентичность по меньшей мере 90% с любой из SEQ ID NO:1-13 или 130-135. В других вариантах осуществления процент идентичности может быть по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% или даже 100%. В некоторых вариантах осуществления вариабельный домен тяжелой цепи имеет идентичность по меньшей мере 95% с любой из SEQ ID NO:1-13 или 130-135. В некоторых вариантах осуществления вариабельный домен тяжелой цепи имеет 100% идентичность с любой из SEQ ID NO:1-13 или 130-135.
В некоторых вариантах осуществления антитела, фрагменты антител, связывающиеся белки на основе антител, ADCs или CARs по изобретению могут включать любую тяжелую цепь, описанную в настоящей заявке (например, тяжелые цепи, показанные на Фиг. 1 и 23) в комбинации с любой подходящей легкой цепью, такой как цепи, проиллюстрированные в настоящей заявке. Подобным образом, антитело может включать любую их легких цепей, описанных выше (например, легкие цепи, показанные на Фиг. 1 и 23), в комбинации с любой подходящей тяжелой цепью, такой как цепи, проиллюстрированные в настоящей заявке. Например, в предпочтительных вариантах осуществления антитело включает легкую цепь, имеющую по меньшей мере 90% идентичности с SEQ ID NO:14, и тяжелую цепь, имеющую по меньшей мере 90% идентичности с SEQ ID NO:1 (антитело XBR1-402), легкую цепь, имеющую по меньшей мере 90% идентичности с SEQ ID NO:18, и тяжелую цепь, имеющую по меньшей мере 90% идентичности с SEQ ID NO:5 (антитело ERR1-324), легкую цепь, имеющую по меньшей мере 90% идентичности с SEQ ID NO:25, и тяжелую цепь, имеющую по меньшей мере 90% идентичности с SEQ ID NO:12 (антитело ERR1-TOP43), или легкую цепь, имеющую по меньшей мере 90% идентичности с SEQ ID NO:26, и тяжелую цепь, имеющую по меньшей мере 90% идентичности с SEQ ID NO:13 (антитело ERR1-TOP54). В некоторых вариантах осуществления антитело может включать последовательности легкой цепи и тяжелой цепи, соответственно показанные в (1) SEQ ID NO:14 и SEQ ID NO:1, (2) SEQ ID NO:18 и SEQ ID NO:5, (3) SEQ ID NO:25 и SEQ ID NO:12, или (4) SEQ ID NO:26 и SEQ ID NO:13. В различных вариантах осуществления процент (%) идентичности пептидных последовательностей можно рассчитать, например, как 100 x [(идентичные положения)/мин(TGA, TGB)], где TGA и TGB представляют сумму количества остатков и положений внутренних гэпов в пептидных последовательностях A и B при выравнивании, которое минимизирует TGA и TGB. См., например, Russell et al, J. Mol. Biol., 244: 332-350 (1994).
Антитело по изобретению может представлять собой любое антитело, включая полноразмерное антитело, фрагмент антитела, связывающийся белок на основе антитела, который специфически распознает или связывается с внеклеточным доменом человеческого ROR1. Например, антитело, фрагмент антитела или связывающийся белок на основе антитела может быть поликлональным, моноклональным, рекомбинантным, химерным или гуманизированным. Кроме того, антитело может быть любого изотипа, включая, без ограничения, IgA, IgD, IgE, IgG или IgM. Таким образом, например, антитело может быть любым IgA, таким как IgAl или IgA2, или любым IgG, таким как IgGl, IgG2, IgG3, IgG4 или синтетический IgG. Антитело также может представлять собой любой фрагмент антитела или связывающийся белок на основе антитела, имеющий специфичность в отношении внеклеточного домена человеческого ROR1, например, F(ab)2, Fv, scFv, IgGACH2, F(ab')2, scFv2CH3, Fab, VL, VH, scFv4, scFv3, scFv2, dsFv, Fv, scFv-Fc, (scFv)2, диатело, бивалентное, биспецифическое или мультиспецифическое антитело. Антитело может представлять собой любое модифицированное или синтетическое антитело, включая, но не ограничиваясь этим, неистощающие IgG антитела, CARs или другие Fc или Fab варианты антител.
Помимо тяжелой цепи, описанной выше, антитело, связывающиеся белки на основе антител или фрагменты таких антител по изобретению также могут включать легкую цепь, выбранную из библиотеки Fab с использованием стратегии последовательной перестановки интактных цепей. Подобным образом, помимо легкой цепи, описанной выше, антитело по изобретению также может включать тяжелую цепь, выбранную из библиотеки Fab с использованием стратегии последовательной перестановки интактных цепей. В некоторых вариантах осуществления изобретение обеспечивает антитела, связывающиеся белки на основе антител или фрагменты таких антител, которые являются консервативно модифицированными вариантами анти-ROR1 антител, проиллюстрированных в настоящей заявке. Типично, вариабельные области этих вариантов имеют аминокислотую последовательность, которая идентична одной из этих проиллюстрированных последовательностей, за исключением консервативных замен на одном или нескольких аминокислотных остатках. В некоторых вариантах осуществления антитело, фрагменты антител, связывающиеся белки на основе антител, ADCs или CARs по изобретению специфически связываются с человеческим ROR1 и содержат по меньшей мере одну CDR, имеющую последовательность, выбранную из группы, состоящей из SEQ ID NO:27-104. Изобретение также обеспечивает выделенное антитело со специфичностью в отношении ROR1, содержащее один или несколько вариантов указанных выше последовательностей CDR или по существу идентичных последовательностей CDR. Варианты последовательностей CDR в этих антителах могут включать 1, 2, или 3 замены, инсерции, делеции или их комбинации в последовательности, выбранной из группы, состоящей из SEQ ID NO:27-104. Например, рекомбинантное химерное или гуманизированное антитело (или его фрагмент) может включать одну, две, три, четыре, пять или шесть указанных выше последовательностей CDR. В некоторых вариантах осуществления, однако, рекомбинантное химерное или гуманизированное антитело (или его фрагмент) включает три последовательности CDR одной и той же легкой или тяжелой цепи, например, CDR легкой цепи, показанные в SEQ ID NO:66-68, SEQ ID NO:78-80, SEQ ID NO:99-101 или SEQ ID NO:102-104; и CDR тяжелой цепи, показанные в SEQ ID NO:27-29, SEQ ID NO:39-41, SEQ ID NO:60-62 или SEQ ID NO:63-65. В некоторых вариантах осуществления рекомбинантное химерное или гуманизированное антитело (или его фрагмент) включает шесть последовательностей CDR одного и того же антитела, например, (a) SEQ ID NO:66-68 и SEQ ID NO:27-29 (антитело XBR1-402); (b) SEQ ID NO:78-80 и SEQ ID NO:39-41(антитело ERR1-324); (c) SEQ ID NO:99-101 и SEQ ID NO:60-62 (антитело ERR1-TOP43) или (d) SEQ ID NO:102-104 и SEQ ID NO:63-65 (антитело ERR1-TOP54).
В некоторых вариантах осуществления изобретение обеспечивает антитела, связывающиеся белки на основе антител или фрагменты таких антител с авидностью к ROR1 около 10 мкМ или меньше, 5 мкМ или меньше, 2 мкМ или меньше, 1 мкМ или меньше, 500 нМ или меньше, 400 нМ или меньше, 300 нМ или меньше, или 200 нМ или меньше. В некоторых вариантах осуществления антитела, фрагменты антител, связывающиеся белки на основе антител, ADCs или CARs связываются с ROR1 с авидностью около 100 нМ или меньше, около 75 нМ или меньше, около 50 нМ или меньше, около 25 нМ или меньше, около 10 нМ или меньше, или около 5 нМ или меньше. В некоторых вариантах осуществления антитела, фрагменты антител, связывающиеся белки на основе антител, ADCs или CARs связываются с ROR1 с авидностью около 1 нМ или меньше, около 800 pM или меньше, около 700 pM или меньше, около 600 pM или меньше, около 500 pM или меньше, около 400 pM или меньше, около 300 pM или меньше, около 200 pM или меньше, или около 100 pM или меньше. Авидность можно измерить с использованием известных в данной области методов, таких как ELISA, метод бислойной интерферометрии или поверхностный плазмонный резонанс.
Антитело, связывающийся белок на основе антитела или фрагмент такого антитела по изобретению можно получить любым подходящим способом, например, с использованием любой подходящей эукариотической или не-эукариотической системы экспрессии. В некоторых вариантах осуществления антитело получают с использованием системы экспрессии млекопитающего. Некоторые специфические способы для генерирования антител, связывающихся белков на основе антител или фрагментов антител по изобретению проиллюстрирован в настоящей заявке. В некоторых вариантах осуществления антитела, связывающиеся белки на основе антител или фрагменты антител по изобретению можно получить с использованием подходящей не-эукариотической системы экспрессии, такой как бактериальная система экспрессии. Бактериальные системы экспрессии можно использовать для продукции фрагментов, таких как F(ab)2, Fv, scFv, IgGACH2, F(ab')2, scFv2CH3, Fab, VL, VH, scFv4, scFv3, scFv2, dsFv, Fv, scFv-Fc, (scFv)2, и диател. Методы изменения кодирующих последовательностей ДНК для продукции таких фрагментов известны в данной области.
Антитела, связывающиеся белки на основе антител или фрагменты антител по изобретению можно конъюгировать с синтетической молекулой с использованием любого типа подходящей конъюгации. Рекомбинантные технологии и встроенный селеноцистеин (например, как описано в Патенте США № 8916159, изданном 23 декабря 2014 года) можно использовать для конъюгации синтетической молекулы. Другие способы конъюгации могут включать ковалентное связывание с природными или полученными инженерными методами аминами боковых цепей лизинов или тиолами боковых цепей цистеинов. См., например, Wu et al., Nat. Biotechnol, 23: 1 137-1 146 (2005).
В предпочтительном варианте осуществления антитела, связывающиеся белки на основе антител или фрагменты таких антител по изобретению, конъюгированные с синтетической молекулой (конъюгат антитело-лекарственное средство с синтетической молекулой, представляющей собой токсин, называется ʺADCʺ), получают путем сайт-специфической опосредованной ферментом сортазой конъюгации антитела. Как раскрыто в WO2014140317, сортазы (также называемые транспептидазами сортазами) образуют группу прокариотических ферментов, которые модифицируют поверхностные белки, распознавая и расщепляя определенный пептидный мотив, называемый «меткой распознавания сортазой» или «сортазной меткой». Обычно данный фермент сортаза распознает одну или несколько сортаза-распознаваемых меток. Ферменты сортазы могут быть природными или могут быть получены генно-инженерными методами (Dorr et al., PNAS 2014; 111, 13343-8).
В предпочтительном варианте осуществления конъюгат получают путем сайт-специфической опосредованной ферментом сортазой конъюгации (a) антитела, связывающегося белка на основе антитела или фрагмента антитела, описанного в настоящей заявке, несущего одну или несколько сортаза-распознаваемых меток, и (b) одной или нескольких синтетических молекул, несущих глициновую или олигоглициновую метку, Gly(n).
Предпочтительно, распознаваемая сортазой метка конденсирована или конъюгирована с C-концом по меньшей мере одного субдомена антитела. Указанная распознаваемая сортазой метка предпочтительно выбрана из группы, состоящей из LPXSG (SEQ ID NO:142), LPXAG (SEQ ID NO:143), LPXTG (SEQ ID NO:144), LAXTG (SEQ ID NO:145) и NPQTG (SEQ ID NO:146).
Предпочтительно, олигоглициновая метка, Gly(n), имеет длину 1-21 глициновых остатков, где n имеет любое значение от 1 до 21), предпочтительно имеет длину 3-5 аминокислот (n имеет значение от 3 до 5, т.е. Gly(3), Gly(4) или Gly(5)).
Синтетическая молекула может представлять собой любую молекулу, такую как молекула, таргетирующая опухоль. В некоторых вариантах осуществления синтетическая молекула для конъюгации с антителом представляет собой белок (например, антитело) или РНК или ДНК аптамер.
В одном варианте осуществления антитела, связывающиеся белки на основе антител или фрагменты антител по изобретению, конъюгированные с синтетической молекулой, имеют общую формулу A - (L - P)n, в которой: A представляет собой антитело, связывающийся белок на основе антитела или фрагмент антитела, описанный в настоящей заявке, L представляет собой один или несколько линкеров, P представляет собой одну или несколько нагрузок, выбранных из группы, состоящей из метки и цитотоксического или цитостатического средства, и где n представляет собой целое число от ≥1 до ≤10.
В этом варианте осуществления линкер предпочтительно включает, или состоит из, по меньшей мере линкер, выбранный из группы, состоящей из следующих: олигопептидный линкер (включая расщепляемые и нерасщепляемые олигопептидные линкеры), гидразиновый линкер, тиомочевинный линкер, саморасщепляющийся линкер, сукцинимидил транс-4-(малеимидилметил)циклогексан-1-карбоксилатный (SMCC) линкер, малеимидный линкер, дисульфидный линкер, тиоэфирный линкер и/или малеимидный линкер.
Специалистам должно быть понятно, что другие линкеры могут быть подходящими. Такие линкеры могут быть нерасщепляемыми или могут расщепляться путем изменений pH, окислительно-восстановительного потенциала или специфических внутриклеточных ферментов. Расщепляемые олигопептидные линкеры включают расщепляемые протеазами или металлопротеазами матрикса линкеры. Должно быть понятно, что линкер может включать комбинации вышеперечисленных. Например, линкер может представлять собой валин-цитруллин PAB линкер.
В предпочтительном варианте осуществления линкер включает олигопептид с последовательностью, которая содержит пента-пептидный мотив LPXSG (SEQ ID NO:142), LPXAG (SEQ ID NO:143), LPXTG (SEQ ID NO:144), LAXTG (SEQ ID NO:145) или NPQTG (SEQ ID NO:146), где X представляет собой любую аминокислоту, с последующим олиго-глициновым участком, Gly(n), где n представляет собой целое число от ≥1 до ≤21. В предпочтительном варианте осуществления линкер конъюгируют с C-концом по меньшей мере одного субдомена антитела, связывающихся белков на основе антител или фрагментов антител.
В различных вариантах осуществления подходящие синтетические молекулы (ʺнагрузкиʺ) для конъюгации с антителом включают, например, терапевтические средства, такие как цитотоксические, цитостатические или антиангиогенные средства, радиоизотопы и липосомы. Цитотоксическое средство может представлять собой растительную, грибковую или бактериальную молекулу. В некоторых вариантах осуществления цитотоксическое средство для конъюгации с антителом по изобретению представляет собой низкомолекулярный токсин (MW< 2000 Дальтон, предпочтительно MW <1000 Дальтон), пептидный токсин или белковый токсин. Многие конкретные примеры этих токсинов хорошо известны в данной области. См., например, Dyba et al., Curr. Pharm. Des. 10:2311-34, 2004; Kuyucak et al., Future Med. Chem. 6:1645-58, 2014; Beraud et al., Inflamm. Allergy Drug Targets. 10:322-42, 2011; и Middlebrook et al., Microbiol. Rev. 48:199-221, 1984. В некоторых вариантах осуществления терапевтическое средство конъюгировано с антителом. Например, терапевтическое средство может представлять собой майтансиноид (например, майтансинол или DM1 майтансиноид), таксан, калихеамицин, семадотин, монометилауристатин (например, монометилауристатин Е или монометилауристатин F), пирролбензодиазепин (PBD) или, предпочтительно, антрациклин, более предпочтительно производное сильнодействующего антрациклина PNU-159682. Особенно предпочтительные производные сильнодействующего антрациклина PNU-159682 раскрыты в WO 0201102679 (которая включена в настоящую заявку посредством ссылки). Терапевтические средства также включают винкристин и преднизон. В различных вариантах осуществления терапевтическое средство, которое можно использовать в изобретении, может представлять собой антиметаболит (например, антифолат, такой как метотрексат, фторпиримидин, такой как 5-фторурацил, цитозин-арабинозид или аналог пурина или аденозина); интеркалирующее средство (например, антрациклин, такой как доксорубицин, неморубицин или предпочтительно производное PNU-159682), дауномицин, эпирабицин, идарубицин, митомицин-С, дактиномицин или митрамицин или другие интеркалирующие средства, такие как пирролобензодиазепин; ДНК-реакционноспособное средство, такое как калихеамицины, тианцимицины и другие ендиины; производное платины (например, цисплатин или карбоплатин); алкилирующее средство (например, азотный иприт, мелфалан, хлорамбуцил, бусульфан, циклофосфамид, ифосфамид нитрозомочевины или тиотепа); ингибитор РНК-полимеразы, такой как α-аманитин; антимитотическое средство (например, алкалоид барвинка, такой как винкристин, или таксоид, такой как паклитаксел или доцетаксел); ингибитор топоизомеразы (например, этопозид, тенипозид, амсакрин, топотекан); ингибитор клеточного цикла (например, флавопиридол); или средство, действующее на микротрубочки (например, эпотилон, тубулизин, претубулизин, аналог дискодермолида или аналог элеутеробина). Терапевтическое средство может представлять собой ингибитор протеасомы или ингибитор топоизомеразы, такой как бортезомиб, амсакрин, этопозид, этопозидфосфат, тенипозид или доксорубицин. Терапевтические радиоизотопы включают йод (131I), иттрий (90Y), лютеций (177Lu), актиний (225Ac), празеодимий, астатин (At), рений (Re), висмут (Bi или Bi) и родий (Rh). Антиангиогенные средства включают линомид, бевакузимаб, ангиостатин и разоксан.
В предпочтительном варианте осуществления синтетическая молекула токсина выбрана из PNU-159682, описанного в Quintieri et al. (2005), и его производных (см. формулу (i) ниже), майтансина, монометилауристатина MMAE и монометилауристатина MMAF. В предпочтительном варианте осуществления токсин, связанный с линкером в положении, показанном волнистой линией, имеет формулу (i), описанную в WO 2016102679 (которая включена в настоящую заявку посредством ссылки):
формула (i)
В варианте осуществления, где синтетическая молекула имеет формулу (i), предпочтительно, чтобы линкер включал алкилдиамино группу формулы NH2-(CH2)m-NH2, где m ≥1 и ≤11, предпочтительно m=2, так чтобы одна амино группа была непосредственно связана в положении, показанном волнистой линией, формулы (i) с образованием амидной связи. Также предпочтительно, чтобы вторая амино группа была связана с олигопептидным линкером, более предпочтительно с олигоглицином Gly(n), где n имеет значение ≥1 и ≤21. Наиболее предпочтительная нагрузка показана на Фиг. 11 (B).
В некоторых вариантах осуществления синтетическая молекула может быть конъюгирована с любым антителом, связывающим белком на основе антитела или фрагментом антитела. В некоторых вариантах осуществления синтетическая молекула может представлять собой метку. Метки могут быть полезны в диагностических применениях и могут включать, например, контрастные вещества. Контрастное вещество может представлять собой радиоизотопную метку, такую как йод (131I или 125I), индий (111In), технеций (99Tc), фосфор (32P), углерод (14C), тритий (3H), другой радиоизотоп (например, радиоактивный ион) или терапевтический радиоизотоп, такой как один из перечисленных выше терапевтических радиоизотопов. Кроме того, контрастные вещества могут включать рентгеноконтрастные вещества, магнитно-резонансные средства визуализации (МРТ), ультразвуковые средства визуализации и любые другие контрастные вещества, подходящие для детекции устройством, которое используют в методах визулизации организма животного. Синтетическая молекула также может представлять собой флуоресцентную метку, биологически активную ферментную метку, люминесцентную метку или хромофорную метку.
В некоторых других вариантах осуществления синтетическая молекула может представлять собой липосому, описанную в Bendas, BioDrugs, 15: 215-224, 2001. В таких вариантах осуществления антитело может быть конъюгировано с коллоидной частицей, например, липосомой, и использоваться для контролируемой доставки средства в пораженные клетки. При получении антитела, конъюгированного с липосомой, например, иммунолипосомой, средство, такое как химиотерапевтическое или другое лекарственное средство, может захватываться в липосому для доставки в клетку-мишень. В некоторых вариантах осуществления антитела, связывающие белки на основе антител или фрагменты антител по изобретению могут также иметь специфичность в отношении одного или нескольких антигенов помимо ROR1. Например, антитело по изобретению может быть сконструировано (например, в виде двухвалентного диатела или конъюгированного Fab-димера или тримера), чтобы иметь специфичность в отношении ROR1 и другого опухолевого антигена, например, антигена, ассоциированного с нейробластомой, почечноклеточной карциномой, раком молочной железы, раком желудка, раком предстательной железы, раком толстой кишки (например, аденокарциномой толстой кишки) или раком молочной железы (например, аденокарциномой молочной железы). Антитело может быть сконструировано так, чтобы иметь специфичность в отношении ROR1 и антигена, который способствует активации или нацеливанию на цитотоксические эффекторные клетки.
IV. Полинуклеотиды, векторы и клетки-хозяева для продукции антител к ROR1
Изобретение обеспечивает по существу очищенные полинуклеотиды (ДНК или РНК), которые идентичны или комплементарны последовательностям, кодирующим полипептиды, включающие сегменты или домены цепей антител, связывающихся белков на основе антител или фрагментов антител, описанных в настоящей заявке. В некоторых вариантах осуществления полинуклеотиды по изобретению кодируют последовательности доменов тяжелой цепи или легкой цепи, показанные на Фиг. 1 и 23. При экспрессии из подходящих векторов экспрессии полипептиды, кодируемые этими полинуклеотидами, способны демонстрировать способность связывания ROR1 антигена. Также, в изобретении представлены полинуклеотиды, которые кодируют по меньшей мере одну CDR область, и обычно все три CDR области из тяжелой или легкой цепи антител, описанных в настоящей заявке. Некоторые другие полинуклеотиды кодируют все или по существу все последовательности вариабельных областей тяжелой цепи и/или легкой цепи представленных антител. Например, некоторые из этих полинуклеотидов кодируют аминокислотную последовательность вариабельной области тяжелой цепи, показанную в любой из SEQ ID NO:1-13 или 130-135, и/или аминокислотную последовательность вариабельной области легкой цепи, показанную в любой из SEQ ID NO:14-26 или 136-141. Из-за вырожденности кода ряд нуклеиновокислотных последовательностей будет кодировать каждую из аминокислотных последовательностей иммуноглобулина.
Полинуклеотиды по изобретению могут кодировать только последовательности вариабельных областей представленных антител. Они также могут кодировать как вариабельную область, так и константную область антитела. Некоторые из полинуклеотидных последовательностей нуклеиновых кислот по изобретению кодируют зрелую последовательность вариабельной области тяжелой цепи, которая по существу идентична (например, по меньшей мере на 80%, 90%, 95% или 99%) зрелой последовательности вариабельной области тяжелой цепи, показанной в любой из SEQ ID NO:1-13 или 130-135. Некоторые другие полинуклеотидные последовательности кодируют зрелую последовательность вариабельной области легкой цепи, которая по существу идентична (например, по меньшей мере на 80%, 90%, 95% или 99%) зрелой последовательности вариабельной области легкой цепи, показанной в любой из SEQ ID NO:14-26 или 136-141. Некоторые из полинуклеотидных последовательностей кодируют полипептид, который включает вариабельные области тяжелой цепи или легкой цепи одного из представленных антител. Некоторые другие полинуклеотиды кодируют два полипептидных сегмента, которые по существу идентичны, соответственно, вариабельным областям тяжелой цепи или легкой цепи одного из представленных антител.
Полинуклеотидные последовательности можно получить de novo твердофазным синтезом ДНК или путем ПЦР мутагенеза существующей последовательности (например, последовательностей, описанных в примерах ниже), кодирующей представленное функциональное антитело. Прямой химический синтез нуклеиновых кислот можно осуществить способами, известными в данной области, такими как способ на основе фосфотриэфира Narang et al., Meth. Enzymol. 68:90, 1979; фосфодиэфирный способ Brown et al., Meth. Enzymol. 68:109, 1979; способ на основе диэтилфосфорамидита Beaucage et al., Tetra. Lett., 22:1859, 1981; и способ с использованием твердофазного носителя Патента США № 4458066. Введение мутаций в полинуклеотидную последовательность при помощи ПЦР можно осуществить, как описано, например, в PCR Technology: Principles and Applications for DNA Amplification, H.A. Erlich (Ed.), Freeman Press, NY, NY, 1992; PCR Протоколы: A Guide to Methods and Applications, Innis et al. (Ed.), Academic Press, San Diego, CA, 1990; Mattila et al., Nucleic Acids Res. 19:967, 1991; и Eckert et al., PCR Methods and Applications 1:17, 1991.
Также в изобретении представлены векторы экспрессии и клетки-хозяева для продукции функциональных антител, описанных в настоящей заявке. Конкретные примеры векторов на основе плазмид и транспозонов для экспрессии антител описаны в примерах ниже. Различные другие векторы экспрессии также можно использовать для экспрессии полинуклеотидов, кодирующих цепи функциональных антител или связывающиеся фрагменты. Как вирусные, так и не-вирусные векторы экспрессии можно использовать для продукции антител в клетке-хозяине млекопитающего. Не-вирусные векторы и системы включают плазмиды, эписомальные векторы, типично с экспрессионной кассетой для экспрессии белка или РНК и человеческими искусственными хромосомами (см., например, Harrington et al., Nat. Genet. 15:345, 1997). Например, не-вирусные векторы, полезные для экспрессии полинуклеотидов и полипептидов антител в клетках млекопитающих (например, человека), включают pCEP4, pREP4, pThioHis A, B & C, pcDNA3.1/His, pEBVHis A, B & C (Invitrogen, San Diego, CA), MPSV векторы и различные другие векторы, известные в данной области, для экспрессии других белков. Другие полезные не-вирусные векторы включают векторы, которые включают экспрессионные кассеты, которые могут быть мобилизованы Sleeping Beauty, PiggyBack и другими системами транспозонов. Полезные вирусные векторы включают векторы на основе лентивирусов или других ретровирусов, аденовирусов, адено-ассоциированных вирусов, вирусов герпеса, векторы на основе SV40, папилломавируса, HBP вируса Эпштейна-Барра, векторы на основе вируса коровьей оспы и вируса леса Семлики (SFV). См., Brent et al., supra; Smith, Annu. Rev. Microbiol. 49:807, 1995; и Rosenfeld et al., Cell 68:143, 1992.
Выбор вектора экспрессии зависит от предполагаемых клеток-хозяев, в которых должен экспрессироваться вектор. Обычно векторы экспрессии содержат промотор и другие регуляторные последовательности (например, энхансеры), которые функционально связаны с полинуклеотидами, кодирующими цепь или фрагмент функционального антитела. В некоторых вариантах осуществления индуцибельный промотор используется для предотвращения экспрессии вставленных последовательностей, за исключением экспрессии в индуцирующих условиях. Индуцибельные промоторы включают, например, арабинозу, lacZ, металлотионеиновый промотор или промотор генов теплового шока. Культуры трансформированных организмов могут быть размножены в неиндуцирующих условиях без изменения популяции для кодирующих последовательностей, продукты экспрессии которых лучше переносятся клетками-хозяевами. В дополнение к промоторам, также могут потребоваться или желательны другие регуляторные элементы для эффективной экспрессии цепи или фрагмента функционального антитела. Эти элементы обычно включают кодон инициации ATG и смежный сайт связывания рибосом (консенсусная последовательность Козака) или другие последовательности. Кроме того, эффективность экспрессии может быть повышена за счет включения энхансеров, соответствующих используемой клеточной системе (см., например, Scharf et al., Results Probl. Cell Differ. 20:125, 1994; и Bittner et al., Meth. Enzymol., 153:516, 1987). Например, SV40 энхансер или CMV энхансер можно использовать для повышения экспрессии в клетках-хозяевах млекопитающих.
Векторы экспрессии также могут обеспечивать положение последовательности сигнала секреции с образованием слитого белка с полипептидами, кодируемыми вставленными последовательностями функционального антитела. Более часто вставленные последовательности функционального антитела связывают с сигнальными последовательностями перед включением в вектор. Векторы, которые можно использовать для включения последовательностей, кодирующих вариабельные домены легкой и тяжелой цепи функционального антитела, иногда также кодируют константные области или их части. Такие векторы обеспечивают возможность экспрессии вариабельных областей в виде слитых белков с константными областями, таким образом, приводя к продукции интактных антител или их фрагментов. Типично, такие константные области являются человеческими, и предпочтительно из человеческих IgG1 антител.
Клетки-хозяева для экспрессии цепей функциональных антител могут быть либо прокариотическими, либо эукариотическими. В некоторых предпочтительных вариантах осуществления клетки-хозяева млекопитающих используют для экспрессии и для продукции полипептидов антител по настоящему изобретению. Например, они могут представлять собой либо гибридомную клеточную линию, экспрессирующую гены эндогенного иммуноглобулина, либо клеточную линию млекопитающего, содержащую экзогенный вектор экспрессии. Они включают любые нормальные мортальные или нормальные или аномальные иммортальные клетки животного или человека. В дополнение к клеточным линиям, приведенным в качестве примера, в данной области также известен ряд других подходящих клеточных линий хозяев, способных секретировать интактные иммуноглобулины. Они включают, например, клеточные линии СНО, различные клеточные линии HEK 293, различные клеточные линии Cos, клетки HeLa, миеломные клеточные линии, трансформированные B-клетки и гибридомы. Использование культуры клеток ткани млекопитающих для экспрессии полипептидов обсуждается в основном в, например, Winnacker, From Genes to Clones, VCH Publishers, N.Y., N.Y., 1987. Экспрессирующие векторы для клеток-хозяев млекопитающих могут включать последовательности контроля экспрессии, такие как ориджин репликации, промотор и энхансер, и необходимые сайты обработки информации, такие как сайты связывания рибосом, сайты сплайсинга РНК, сайты полиаденилирования и последовательности терминатора транскрипции. Эти векторы экспрессии обычно содержат промоторы, происходящие из генов млекопитающих или из вирусов млекопитающих. Подходящие промоторы могут быть конститутивными, специфичными для конкретного типа клеток, стадия-специфичными и/или модулируемыми или регулируемыми. Полезные промоторы включают, но не ограничиваются этим, EF1α и человеческие UbC промоторы, приведенные в качестве примеров, металлотионеиновый промотор, конститутивный аденовирусный основной поздний промотор, дексаметазон-индуцируемый MMTV промотор, SV40 промотор, MRP pol III промотор, конститутивный MPSV промотор, тетрациклин-индуцируемый CMV промотор (такой как человеческий предранний CMV промотор), конститутивный CMV промотор и комбинации промотора-энхансера, известные в данной области.
Способы введения векторов экспрессии, содержащих представляющие интерес полинуклеотидные последовательности, различаются в зависимости от типа клеток-хозяев. Например, трансформацию с использованием хлорида кальция обычно используют для прокариотических клеток, тогда как обработку фосфатом кальция или электропорацию можно использовать для других клеток-хозяев (см., в основном Sambrook et al., выше). Другие методы включают, например, электропорацию, обработку фосфатом кальция, трансформацию, опосредованную липосомами, инъекцию и микроинъекцию, баллистические методы, виросомы, иммунолипосомы, конъюгаты поликатион:нуклеиновая кислота, голую ДНК, искусственные вирионы, слияние со структурным белком вируса герпеса VP22 (Elliot and O'Hare, Cell 88: 223, 1997), агент-усиленное поглощение ДНК и трансдукцию ex vivo. Для долговременной высокопродуктивной продукции рекомбинантных белков часто требуется стабильная экспрессия. Например, клеточные линии, которые стабильно экспрессируют цепи или связывающие фрагменты антитела, могут быть получены с использованием экспрессирующих векторов по изобретению, которые содержат вирусные ориджины репликации или эндогенные экспрессирующие элементы и селектируемый маркерный ген. После введения вектора клетки можно оставить для роста на 1-2 дня в обогащенной среде до переключения их на селективные среды. Цель селективного маркера заключается в придании устойчивости к селекции, а его присутствие позволяет выращивать клетки, которые успешно экспрессируют введенные последовательности в селективных средах. Устойчивые, стабильно трансфицированные клетки могут размножаться с использованием методов культивирования тканей, подходящих для типа клеток.
Изобретение также обеспечивает эукариотические или не-эукариотические клетки (например, T лимфоциты), которые были рекомбинантно сконструированы для продукции антител, связывающихся белков на основе антител или фрагментов антител по изобретению. Эукариотические или не-эукариотические клетки можно использовать в качестве системы экспрессии для продукции антитела по изобретению. В некоторых вариантах осуществления изобретение обеспечивает нацеленные на ROR1 иммунные клетки, которые сконструированы для рекомбинантной экспрессии ROR1-специфического антитела по изобретению. Например, изобретение обеспечивает T клетку, сконструированную для экспрессии антитела по изобретению (например, scFv, scFv-Fc, или (scFv)2), которое связано с синтетической молекулой, содержащего один или несколько из следующих доменов: спейсер или шарнирная область (например, последовательность CD28 или последовательность шарнирной области-Fc IgG4), трансмембранная область (например, трансмембранный канонический домен) и внутриклеточный сигнальный домен T-клеточного рецептора (TCR), с образованием, таким образом, химерного антигенного рецептора (CAR) или T-тела. Внутриклеточные TCR сигнальные домены, которые могут быть включены в CAR (или T-тело), включают, но не ограничиваются этим, CD3ζ, FcR-γ и Syk-PT сигнальные домены, а также CD28, 4-1BB и CD134 ко-сигнальные домены. Способы для конструирования T-клеток, экспрессирующих CAR (или T-тело), известны в данной области. См., например, Marcu-Malina et al., Expert Opinion on Biological Therapy, Vol. 9, No. 5 (posted online on April 16, 2009).
V. Терапевтические и диагностические применения
ROR1 антитела, связывающиеся белки на основе антител, фрагменты антител, ADCs и CARs, раскрытые в настоящей заявке, можно использовать в различных терапевтических и диагностических применениях. ROR1 экспрессируется и участвует в развитии различных опухолей. См., например, Rebagay et al., Front Oncol. 2, 34, 2012; и Shabani et al., Expert Opin. Ther. Targets 19, 941-955, 2015. Например, ROR1 экспрессируется на поверхности опухолевых клеток в CLL, ALL, лимфоме из клеток мантийной зоны, нейробластоме, саркоме, почечноклеточной карциноме, раке молочной железы, раке легкого, раке толстой кишки, раке головы и шеи, меланоме и других раковых заболеваниях. Важно отметить, что ROR1 экспрессируется на стадии эмбриогенеза, но в основном после рождения его экспрессия прекращается. Очень немного взрослых здоровых тканей и клеток экспрессируют ROR1. Соответственно, было обнаружено, что анти-ROR1 CAR-сконструированные T клетки являются безопасными и активными у отличных от человека приматов, подтверждая ROR1 как терапевтическую мишень при раке (Berger (2015) Cancer Immunol Res. 3(2), page 2016). Таким образом, mAbs к ROR1 имеют широкую терапевтическую и диагностическую применимость при раке.
В некоторых вариантах осуществления изобретение обеспечивает способы для ингибирования клеток, которые экспрессируют ROR1 (ROR1 клетки), путем контактирования клеток с антителом, связывающимся белком на основе антитела, фрагментом антитела, ADC или CAR по изобретению. Антитело, связывающийся белок на основе антитела или фрагмент антитела может представлять собой оголенную (неконъюгированную) молекулу или антитело, связывающийся белок на основе антитела, фрагмент антитела, конъюгированный с синтетической молекулой, например, цитотоксическим, цитостатическим или антиангиогенным средством, радиоизотопом или даже с липосомой. Предпочтительно, клетки контактируют с ADC, включающим конъюгированную цитотоксическую молекулу. Способ можно использовать для ингибирования ROR1 клеток in vitro или у субъекта (т.е. in vivo). Контактированные ROR1 клеток может происходить, например, в культуре клеток или животной модели расстройства, связанного с повышенными уровнями ROR1. Такие способы полезны, например, для измерения и/или сравнительной оценки (относительно другого антитела) ингибирующей активности антитела в отношении конкретного ROR1 клеточного типа. Ингибирование ROR1 клеток может включать блокирование или снижение активности или роста ROR1 клеток. Ингибирование также может включать киллинг ROR1 клеток. Хотя такие способы не связаны или не ограничиваются каким-либо конкретным механизмом действия, ингибиторная активность может быть опосредована блокировкой ROR1-опосредованной передачи сигналов или блокировкой передачи сигналов ROR1-ассоциированного рецептора. Ингибиторная активность также может быть опосредована рекрутментом эффекторов иммунной системы, которые атакуют ROR1 клетки, например, путем активации компонентов антителозависимой клеточно-опосредованной цитотоксичности (ADCC) или систем комплемента.
В некоторых близких вариантах осуществления изобретение обеспечивает способы для лечения субъекта, который имеет, у которого подозревается или у которого имеется риск развития расстройства, ассоциированного с экспрессией ROR1. Как правило, способы включают введение субъекту терапевтически эффективного количества выделенного антитела, связывающегося белка на основе антитела, фрагмента антитела, ADC или CAR по изобретению. Антитело может представлять собой любое анти-ROR1 антитело, фрагмент анти-ROR1 антитела, связывающийся белок на основе анти-ROR1 антитела по изобретению, описанного в настоящей заявке. Таким образом, антитело может быть химерным, гуманизированным, синтетическим, F(ab)2, Fv, scFv, IgGACH2, F(ab')2, scFv2CH3, Fab, VL, VH, scFv4, scFv3, scFv2, dsFv, Fv, или (scFv)2. В некоторых вариантах осуществления способ включает введение IgG, scFv, dsFv, F(ab')2, диатела или бивалентного антитела. Используемое для введения антитело, связывающийся белок на основе антитела, фрагмент антитела могут быть конъюгированы с синтетической молекулой, описанной выше, например, цитотоксическим, цитостатическим или антиангиогенным средством, терапевтическим радиоизотопом или липосомой. Примером цитотоксического средства является экзотоксин A Pseudomonas (PE38). Расстройства, которые можно лечить, включают CLL, ALL, лимфому из клеток мантийной зоны, нейробластому, саркому, почечноклеточную карциному, рак молочной железы, рак легкого, рак толстой кишки, рак головы и шеи, меланому и другие расстройства с повышенной экспрессией ROR1.
В некоторых вариантах осуществления изобретение обеспечивает способы для лечения субъекта, который имеет, у которого подозревается или у которого имеется риск развития расстройства, ассоциированного с повышенными уровнями ROR1, путем адоптивного переноса генно-инженерных T-клеток, описанных в настоящей заявке, которые экспрессируют антитело или антиген-связывающий фрагмент по изобретению в виде химерного антигенного рецептора (CAR), который селективно связывается с ROR1. Рекомбинантную технологию можно использовать для введения CAR-кодирующего генетического материала в любые подходящие Т-клетки, например, Т-клетки центральной памяти от субъекта, подлежащего лечению. Т-клетки, несущие генетический материал, могут расти (например, в присутствии цитокинов). Генно-инженерные Т-клетки переносят, как правило, путем инфузии, пациенту. Перенесенные Т-клетки по изобретению могут затем укреплять иммунный ответ против ROR1-экспрессирующих клеток у субъекта. Метод адоптивного переноса можно использовать, например, для лечения субъектов, которые имеют или у которых подозревается какое-либо раковое заболевание, связанное с ROR1, включая CLL, ALL, лимфому мантийных клеток, нейробластому, саркому, почечно-клеточную карциному, рак молочной железы, рак легкого, рак толстой кишки, рак головы и шеи, меланому и другие типы рака. В некоторых вариантах осуществления вышеуказанные способы лечения могут дополнительно включать совместное введение второго терапевтического средства для лечения расстройства, связанного с повышенным уровнем ROR1. Например, когда расстройство, подлежащее лечению, включает ROR1-экспрессирующий рак, способ может дополнительно включать совместное введение цитотоксического, цистостатического или антиангиогенного или иммуностимулирующего средства (например, антитела в качестве ингибиторов иммунных контрольных точек, например, но не ограничиваясь этим, которые связываются с PD1, PDL1, CTLA4, OX40, TIM3, GITR, LAG3 и т.п.), подходящего для лечения рака. Если рак представляет собой В-клеточное злокачественное заболевание, способ может дополнительно включать, например, совместное введение ритуксимаба, алемтузумаба, ататумумаба, окрелизумаба или химиотерапевтического режима CHOP.
В некоторых других вариантах осуществления изобретение обеспечивает способ для детекции в биологическом образце измененного уровня ROR1 (например, клеточно-поверхностного ROR1), например, относительно контроля, при помощи FACS, иммуногистохимии (IHC) или Вестерн-блоттинга. Как правило, способ включает контактирование биологического образца с антителом, связывающимся белком на основе антитела, фрагментом антитела по изобретению и определение количества антитела, которое селективно связывается с материалом (например, клетками) в образце, с определением, таким образом, уровня ROR1 в биологическом образце. Биологический образец может быть получен из культуры клеток или от испытуемого субъекта, например, из образца плазмы или ткани субъекта, который имеет, у которого подозревается или у которого имеется риск развития заболевания или состояния, ассоциированного с повышенным уровнем ROR1 у субъекта. Контрольный уровень желательно соответствует уровню ROR1, обнаруженному с использованием того же антитела в соответствующем образце(образцах) из одной или нескольких контрольных культур или от здоровых субъектов. Способы использования антитела по изобретению для определения уровней ROR1 могут включать любой иммуноанализ, такой как иммуно-(Вестерн)блоттинг, иммуноферментный анализ (ELISA), иммуногистохимия (IHC) и проточная цитометрия, например, флуоресцентно-активированный клеточный сортинг (FACS).
Можно использовать способы детекции для скрининга на присутствие расстройства, ассоциированного с повышенным ROR1. Способы включают получение образца от испытуемого субъекта, нуждающегося в скрининге, например, субъекта, который имеет, у которого подозревается или у которого имеется риск развития расстройства, ассоциированного с повышенным ROR1. Уровень ROR1 (например, количество или концентрация) в образце измеряют с использованием антитела, связывающегося белка на основе антитела, фрагмента антитела по изобретению и уровень в образце сравнивают с контрольным уровнем ROR1. Контрольный уровень представляет собой, например, средний уровень (например, количество или концентрацию) в образце(образцах) от одного или, предпочтительно, нескольких субъектов из контрольной группы, которые не имеют расстройства, ассоциированного с повышенным ROR1. Альтернативно, контрольный уровень может соответствовать уровню или среднему уровню ROR1 в одном или нескольких образцах, взятых у испытуемого субъекта в одной или нескольких более ранних точках времени, например, когда испытуемый субъект не имел или не испытывал состояние, ассоциированное с повышенным ROR1. Значительно более высокий уровень ROR1 в биологическом образце по сравнению с контрольным уровнем указывает на расстройство, ассоциированное с повышенным ROR1 у субъекта. У субъектов, таких как человек, где экспрессия ROR1 на клеточной поверхности преимущественно ограничена эмбриональным развитием, контрольный уровень ROR1 может быть нулевым или никаким. Таким образом, в некоторых вариантах осуществления способа детекции, обеспечиваемого изобретениеем, любое существенное и определяемое количество ROR1 в биологическом образце может указывать на расстройство, ассоциированное с повышенным ROR1 у субъекта.
Кроме того, способ детекции можно использовать для контроля развития расстройства, связанного с повышенным ROR1. Способ включает получение образца от субъекта, нуждающегося в скрининге, например, субъекта, у которого было диагностировано или подозревается расстройство, связанное с повышенным ROR1. Уровень ROR1 в образце измеряют с использованием антитела, связывающего белка на основе антитела, фрагмента антитела по изобретению, и уровень в образце сравнивают с контрольным уровнем, соответствующим уровню или среднему уровню ROR1 в одном или нескольких образцах, взятых у испытуемого субъекта в одной или нескольких более ранних точках времени. Уровни ROR1, которые значительно повышены или уменьшены относительно контроля, показывают, что расстройство субъекта ухудшается или улучшается, соответственно. Вышеуказанные способы детекции можно использовать для скрининга на наличие или для мониторинга прогрессирования расстройства, включающего, например, CLL, ALL, лимфому из клеток мантийной зоны, нейробластому, саркому, почечноклеточную карциному, рак молочной железы, рак легкого, рак толстой кишки, рак головы и шеи, меланому и другие раковые заболевания.
В некоторых вариантах осуществления изобретение обеспечивает способы для скрининга субъекта на изменение уровня ROR1. Как правило, способы включают введение субъекту антитела, связывающего белка на основе антитела, фрагмента антитела по изобретению, конъюгированного с меткой (например, контрастным агентом), визуализацию субъекта способом, подходящим для обнаружения метки, и определение, имеет ли область в организме субъекта измененную плотность или концентрацию метки по сравнению с фоновым уровнем метки в проксимальной ткани. Альтернативно, способы включают определение, существует ли измененная плотность или концентрация метки в области по сравнению с плотностью или концентрацией метки, ранее обнаруженной в той же области субъекта. Методы визуализации субъекта могут включать рентгенологическое исследование, рентгеновскую компьютерную томографию (CT) (например, CT-ангиографию (CTA)), магнитно-резонансную (MR) визуализацию, магнитно-резонансную ангиографию (MRA), радиоизотопный метод, ультразвук (US), оптическую визуализацию, эластографию, инфракрасную визуализацию, микроволновую визуализацию и т.п., что является подходящим для детекции метки, конъюгированной с антителом. В предпочтительном варианте осуществления субъект имеет, у него подозревается или имеется риск развития ROR1-экспрессирующей опухоли, такой как CLL, ALL, лимфома из клеток мантийной зоны, нейробластома, саркома, почечноклеточная карцинома, рак молочной железы, рак легкого, рак толстой кишки, рак головы и шеи, меланома и другие раковые заболевания, и способ используют для скрининга или детекции присутствия опухоли. В другом варианте осуществления способ можно использовать для отслеживания размера или плотности ROR1-экспрессирующей опухоли в течение времени, например, в ходе лечения.
VI. Фармацевтические композиции и комбинации
В другом аспекте изобретение обеспечивает фармацевтические композиции, которые содержат антитело, фрагмент антитела, связывающийся белок на основе антитела или ADC, описанный в настоящей заявке, и фармацевтически приемлемый носитель. Фармацевтические композиции можно получить из любого антитела, описанного в настоящей заявке. Примеры композиций включают одно или несколько из химерного антитела, имеющего SEQ ID NO:14 (легкая цепь) и/или SEQ ID NO:1 (тяжелая цепь), химерного антитела, имеющего SEQ ID NO:18 (легкая цепь) и/или SEQ ID NO:5 (тяжелая цепь), химерного антитела, имеющего SEQ ID NO:25 (легкая цепь) и/или SEQ ID NO:12 (тяжелая цепь) и химерного антитела, имеющего SEQ ID NO:26 (легкая цепь) и/или SEQ ID NO:13 (тяжелая цепь). Другие антитела, фрагменты антител, связывающиеся белки на основе антител или ADCs, подходящие для фармацевтических композиций по изобретению, включают такие, которые имеют последовательность легкой цепи, показанную в SEQ ID NO:14-26 или 136-141, и/или последовательность тяжелой цепи, показанную в SEQ ID NO:1-13 или 130-135. Другие иллюстративные композиции по изобретению могут содержать гуманизированное антитело, имеющее одну, две, три, четыре, пять или шесть CDRs, выбранных из группы, состоящей из SEQ ID NO:27-104, такие как проиллюстрированные в SEQ ID NO: 130-141. В некоторых вариантах осуществления антитело включает три последовательности CDR тех же проиллюстрированных легкой или тяжелой цепей, которые показаны на Фиг. 1. Они включают последовательности CDR1, CDR2 и CDR3 тяжелой цепи и последовательности CDR1, CDR2 и CDR3 легкой цепи, соответственно показанные в (1) SEQ ID NO:27-29 и SEQ ID NO:66-68 (антитело XBR1-402), (2) SEQ ID NO:30-32 и SEQ ID NO:69-71 (антитело ERR1-301), (3) SEQ ID NO:33-35 и SEQ ID NO:72-74 (антитело ERR1-306), (4) SEQ ID NO:36-38 и SEQ ID NO:75-77 (антитело ERR1-316), (5) SEQ ID NO:39-41 и SEQ ID NO:78-80 (антитело ERR1-324), (6) SEQ ID NO:42-44 и SEQ ID NO:81-83 (антитело ERR1-403), (7) SEQ ID NO:45-47 и SEQ ID NO:84-86 (антитело ERR1-409), (8) SEQ ID NO:48-50 и SEQ ID NO:87-89 (антитело ERR1-TOP4), (9) SEQ ID NO:51-53 и SEQ ID NO:90-92 (антитело ERR1-TOP15), (10) SEQ ID NO:54-56 и SEQ ID NO:93-95 (антитело ERR1-TOP22), (11) SEQ ID NO:57-59 и SEQ ID NO:96-98 (антитело ERR1-TOP40), (12) SEQ ID NO:60-62 и SEQ ID NO:99-101 (антитело ERR1-TOP43), или (13) SEQ ID NO:63-65 и SEQ ID NO:102-104 (антитело ERR1-TOP54). В некоторых вариантах осуществления фармацевтическая композиция включает антитело, имеющее шесть CDR последовательностей того же антитела, которое проиллюстрировано на Фиг. 1, например, (a) SEQ ID NO:66-68 и SEQ ID NO:27-29 (антитело XBR1-402); (b) SEQ ID NO:78-80 и SEQ ID NO:39-41(антитело ERR1-324); (c) SEQ ID NO:99-101 и SEQ ID NO:60-62 (антитело ERR1-TOP43) или (d) SEQ ID NO:102-104 и SEQ ID NO:63-65 (антитело ERR1-TOP54). Другие примеры фармацевтической композиции включает dsFv фрагмент, который может включать одну или несколько модификаций в аминокислотной последовательности, которые являются подходящими и понятны специалистам в данной области.
В некоторых вариантах осуществления композиции по изобретению содержат носитель для антитела, фрагмента антитела, связывающего белка на основе антитела или ADC, желательно фармацевтически приемлемый носитель. Фармацевтически приемлемый носитель может представлять собой любой подходящий фармацевтически приемлемый носитель. Это может быть один или несколько совместимых твердых или жидких наполнителей, разбавителей, других эксципиентов или инкапсулирующих веществ, которые являются подходящими для введения человеку или ветеринарному пациенту (например, физиологически приемлемый носитель или фармакологически приемлемый носитель). Термин «носитель» обозначает органический или неорганический ингредиент, природный или синтетический, с которым активный ингредиент объединяют для облегчения использования активного ингредиента, например, введения активного ингредиента субъекту. Фармацевтически приемлемый носитель можно объединить с одним или несколькими активными компонентами, например, гибридной молекулой, и друг с другом, когда в композиции присутствует более одного фармацевтически приемлемого носителя, таким образом, чтобы существенно не ухудшалась желаемая фармацевтическая эффективность. Фармацевтически приемлемые вещества обычно можно вводить субъекту, например пациенту, не вызывая при этом значительных нежелательных физиологических эффектов, таких как тошнота, головокружение, сыпь или расстройство желудка. Например, желательно, чтобы композиция, содержащая фармацевтически приемлемый носитель, не была иммуногенной при введении пациенту-человеку в терапевтических целях.
Фармацевтические композиции по изобретению могут дополнительно содержать подходящие буферные агенты, включая, например, уксусную кислоту в форме соли, лимонную кислоту в форме соли, борную кислоту в форме соли и фосфорную кислоту в форме соли. Композиции, необязательно, также могут содержать подходящие консерванты, такие как бензалконийхлорид, хлорбутанол, парабены и тимеросал. Фармацевтические композиции по изобретению могут быть представлены в стандартной лекарственной форме и могут быть получены любым подходящим способом, многие из которых хорошо известны в области фармации. Такие способы включают стадию приведения антитела по изобретению в ассоциацию с носителем, который составляет один или несколько вспомогательных ингредиентов. Как правило, композицию получают путем приведения активного агента в однородную и тесную ассоциацию с жидким носителем, тонкоизмельченным твердым носителем или обоими, и затем, если необходимо, формования продукта.
Композиция, подходящая для парентерального введения, обычно включает стерильный водный препарат композиции по изобретению, который предпочтительно является изотоническим с кровью реципиента. Этот водный препарат может быть сформулирован в соответствии с известными способами с использованием подходящих диспергирующих или смачивающих веществ и суспендирующих веществ. Стерильный препарат для инъекций также может представлять собой стерильный инъекционный раствор или суспензию в нетоксичном парентерально приемлемом разбавителе или растворителе, например, в виде раствора в 1,3-бутандиоле. Из приемлемых носителей и растворителей, которые можно использовать, можно указать воду, раствор Рингера и изотонический раствор хлорида натрия. Кроме того, стерильные, нелетучие масла обычно используют в качестве растворителя или среды для суспендирования. Для этой цели можно использовать любое мягкое нелетучее масло, такое как синтетические моно- или диглицериды. Кроме того, жирные кислоты, такие как олеиновая кислота, можно использовать для получения инъекционных препаратов. Композиции носителей, подходящие для перорального, подкожного, внутривенного, внутримышечного и т.д. введения, можно найти в Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, PA.
Получение фармацевтических композиций по изобретению их введение различными путями можно осуществлять в соответствии со способами, хорошо известными в данной области. См., Например, Remington: Science and Practice of Pharmacy, Mack Publishing Co., 20th ed., 2000; и Sustained and Controlled Release Drug delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978. Системы доставки, полезные в контексте изобретения, включают системы доставки с отсроченным высвобождением, замедленным высвобождением и пролонгированным высвобождением, так чтобы доставка композиции по изобретению происходила заранее и с достаточным временем, чтобы вызвать сенсибилизацию участка, подлежащего лечению. Композицию по изобретению можно использовать в сочетании с другими терапевтическими средствами или терапиями. Такие системы позволяют избежать повторного введения композиции согласно изобретению, тем самым повышая удобство для субъекта и лечащего врача, и могут быть особенно подходящими для некоторых композиций по изобретению.
Многие типы систем доставки доступны и известны специалистам в данной области техники. Системы доставки с подходящим высвобождением включают системы на полимерной основе, такие как поли (лактид-гликолид), сополиоксалаты, поликапролактоны, полиэфирамиды, полиортоэфиры, полигидроксимасляная кислота и полиангидриды. Микрокапсулы из вышеуказанных полимеров, содержащие лекарственные средства, описаны, например, в патенте США 5075109. Системы доставки также включают не-полимерные системы, которые представляют собой липиды, включая стеролы, такие как холестерин, сложные эфиры холестерина и жирные кислоты или нейтральные жиры, такие как моно- ди- и триглицериды; гидрогелевые системы высвобождения; силастиковые системы; системы на основе пептидов; восковые покрытия; прессованные таблетки с использованием обычных связующих и наполнителей; частично растворенные имплантаты; и т.п. Конкретные примеры включают, но не ограничиваются этим: (а) эрозионные системы, в которых активная композиция содержится в форме внутри матрицы, такие как описанные в патентах США 4452775, 4667014, 4748034 и 5239660, и (b) диффузионные системы, в которых активный компонент проникает с контролируемой скоростью из полимера, такие как описанные в патентах США 3832253 и 384480. Кроме того, могут использоваться системы доставки насосного типа, некоторые из которых адаптированы для имплантации.
Изобретение также обеспечивает наборы, подходящие для осуществления способов по изобретению. Типично, наборы содержат два или более компонентов, необходимых для осуществления терапевтических или диагностических способов по изобретению. Компоненты набора включают, но не ограничиваются этим, одно или несколько антител, связывающихся белков на основе антител, фрагментов антител или ADC по изобретению, подходящие реагенты, и/или оборудование. В некоторых вариантах осуществления наборы могут содержать антитело, связывающийся белок на основе антитела, фрагмент антитела по изобретению и буфер для иммуноанализа, подходящий для детекции ROR1 (например, методом ELISA, проточной цитометрии, магнитного сортинга или FACS). Набор может также включать один или несколько микротитровальных планшетов, стандартов, разбавителей для анализа, промывочных буферов, адгезивных крышек для планшетов, магнитных шариков, магнитов и/или инструкции для осуществления способа по изобретению с использованием набора. Набор включает антитело, связывающие белки на основе антитела, фрагменты антител по изобретению, связанные с субстратом (например, многолуночный планшет или чип), соответствующим образом упакованные и полезные для детекции ROR1. В некоторых вариантах осуществления наборы включают антитело, связывающие белки на основе антитела, фрагменты антител по изобретению, которые конъюгированы с меткой, такой как флуоресцентная метка, биологически активная ферментная метка, люминесцентная метка или хромофорная метка. Наборы могут дополнительно включать реагенты для визуализации конъюгированного антитела, связывающих белков на основе антител, фрагментов антител, например, субстрат для фермента. В некоторых вариантах осуществления наборы включают антитело, связывающие белки на основе антитела, фрагменты антител по изобретению, которые конъюгированы с контрастным агентом, и необязательно один или несколькими реагентов или частей оборудования, полезных для визуализации антитела у субъекта.
Как правило, антитело, связывающие белки на основе антитела, фрагменты антител по изобретению в наборе соответствующим образом упакованы, например, в флакон, мешочек, ампулу и/или любой контейнер, подходящий для терапевтического метода или детекции. Компоненты набора могут обеспечиваться в виде концентратов (включая лиофилизированные композиции), которые можно затем разбавить перед использованием, или они могут обеспечиваться при концентрации, подходящей для использования. Для использования антитела по изобретению in vivo единичные дозы могут обеспечиваться в стерилизованных контейнерах, содержащих желаемое количество и концентрацию компонентов.
ПРИМЕРЫ
Следующие примеры представлены для дальнейшей иллюстрации изобретения, но не для ограничения его объема. Другие варианты изобретения будут совершенно очевидны специалистам в данной области, и они охватываются прилагаемой формулой изобретения.
Пример 1. Клеточные линии
Клеточные линии: MDA-MB-231, MDA-MB-469, 697, Kasumi-2, T47D, HS578T, 63-12 и 63-12/hROR1 и 63-12/hROR2 трансфектанты, K562 и K562/hROR1 трансфектанты, EMT-6 и EMT-6/ROR1 трансфектанты культивировали в DMEM (Invitrogen; Carlsbad, CA), дополненной 10% (об./об.) термоинактивированной FBS (Thermo Scientific; Logan, UT), 100 Ед./мл пенициллина и 100 мг/мл стрептомицина (Invitrogen). HEK 293F клетки закупали у Invitrogen и поддерживали в FreeStyle среде, дополненной 1% (об./об.) термоинактивированной FBS (Thermo Scientific) для поддержания адгезивной культуры, или без FBS для суспендирования культуры, 100 Ед./мл пенициллина и 100 мг/мл стрептомицина (Invitrogen).
Клонирование полноразмерного hROR1 и hROR1 в векторы экспрессии млекопитающего: Транспозируемые векторные каркасы (pPB-Puro) собирали из модульных частей с фланкирующими сайтами рестрикции, которые были синтезированы или получены из последовательность-подтвержденных коммерчески доступных векторов и подробно описаны в Патенте WO2014013026A1. Эти исходные транспозируемые векторные каркасы модифицировали путем замены IRES-направляемой экспрессии гена резистентности к Пуромицину в исходном векторе направляемой отдельным фосфоглицераткиназным промотором (PGK) экспрессией. Это осуществляли путем замены последовательности IRES последовательностью SV40-pA, расположенной 3' от сайта множественного клонирования, с последующим введением последовательности PGK-промотора 5' от гена резистентности к Пуромицину. Открытые рамки считывания полноразмерных ROR1/2 синтезировали путем полного синтеза гена (Genscript, Piscataway) с фланкирующими сайтами рестрикции (5'NotI/3'BstBI) и затем клонировали с сайт множественного клонирования транспозируемых векторов с использованием соответствующих рестрикционных ферментов.
Конструирование клеточной линии для эктопической экспрессии hROR1 или hROR2 в 63-12 мышиной A-MuLV-трансформированной пред-B клеточной линии: Мышиную пред-B клеточную линию 63-12 мышей Abelson (Shinkai et al. (1992) Cell 68:855-67) культивировали в культуральной среде (17,7 г/л Gibco® IMDM (Life Technologies, 42200-030), 3,024 г/л NaHCO3 (Sigma-Aldrich, p.a., >99,7%), 10 мл/л 100× не являющихся незаменимыми аминокислот (Life Technologies, 11140035), 5 мг/л инсулина (Sigma-Aldrich, I-5500), 3 мл/л 10% приматона RL/UF в H2O (Sheffield Bioscience) и 1 мл/л 50 мМ 2-меркаптоэтанола (Sigma-Aldrich, M-3148) в H2O), дополненной 2% (об/об) FCS, 100 МЕ/мл пенициллина/стрептомицина/фунгизона (Amimed, 4-02F00-H), 200 мМ L-глутамина (Amimed, 5-10K00-H) и 50 мкМ 2-меркаптоэтанола (Amresco, 0482), при 37°C и 7,5% CO2. Клетки были сконструированы для сверхэкспрессии hROR1 и hROR2 путем транспозиции следующим образом: клетки центрифугировали (6 мин, 1200 об/мин, 4°C) и ресуспендировали в RPMI-1640 среде (5×106 клеток/мл). 400 мкл клеточной суспензии затем добавляли к 400 мкл RPMI, содержащей 10 мкг транспозируемого вектора pPB-PGK-Puro-ROR1 (направляющего ко-экспрессию полноразмерного ROR1 (NP_005003.2) и гена резистентности к пуромицину) или 10 мкг транспозируемого вектора pPB-PGK-Puro-ROR2 (направляющего ко-экспрессию полноразмерного ROR2 (NP_004551.2) и гена резистентности к пуромицину), вместе с 10 мкг транспозаза-содержащего вектора pCDNA3.1_hy_mPB. ДНК/63-12 клеточные смеси переносили в кюветы для электропорации (просвет 0,4 см, 165-2088, BioRad, Cressier, Switzerland) и осуществляли электропорацию с использованием Biorad Gene Pulser II с расширителем электроемкости при 300В и 950 мкФ. Затем клетки инкубировали в течение 5-10 минут при комнатной температуре. После инкубации клетки центрифугировали при 1200 об/мин в течение 6 минут (4°C), промывали один раз и затем ресуспендировали в водной культуральной среде (17,7 г/л Gibco® IMDM (Life Technologies, 42200-030), 3,024 г/л NaHCO3 (Sigma-Aldrich, p.a., >99,7%), 10 мл/л 100x не являющихся незаменимыми аминокислот (Life Technologies, 11140035), 5 мг/л инсулина (Sigma-Aldrich, I-5500), 3 мл/л 10% приматона RL/UF в H2O (Sheffield Bioscience) и 1 мл/л 50 мМ 2-меркаптоэтанола (Sigma-Aldrich, M-3148) в H2O), дополненной 2% (об/об) FCS, 100 МЕ/мл пенициллина/стрептомицина/Фунгизона (Amimed, 4-02F00-H), 200 мМ L-глутамина (Amimed, 5-10K00-H) и 50 мкМ 2-меркаптоэтанола (Amresco, 0482). После двух дней инкубации при 37°C в увлажненном инкубаторе в атмосфере 5% CO2 клеточные пулы, стабильно экспрессирующие hROR1 или hROR2, отбирали путем добавления 2 мкг/мл пуромицина (Sigma-Aldrich, P8833).
Через 4-5 дней экспрессию hROR1 или hROR2 на сконструированных клетках подтверждали методом проточной цитометрии. Вкратце, после трипсинизации 106 клеток центрифугировали в FACS пробирках; полученные клеточные осадки ресуспендировали в буфере (PBS с 2% (об./об.) FCS). В случае hROR1-сконструированных клеток, клетки затем инкубировали с 2A2 (mAb066 антитело, таргетирующее ROR1, конечная концентрация 2 мкг/мл) в течение 30 минут при 4°C, с последующим центрифугированием и промывкой. Клетки ресуспендировали, как указано выше, и инкубировали с анти-человеческим IgG антителом (Fc гамма-специфическим) PE (eBioscience, Vienna, Austria, 12-4998-82) при 1:100 разведении в темноте (30 мин, 4°C), промывали один раз в буфере и хранили на льду до FACS сортинга. Для hROR2-сконструированных 63-12 клеток следовали тому же протоколу, но с использованием EPR3779 (Abcam антитело, таргетирующее ROR2; 1:100 разведение) в качестве первичного антитела и аллофикоцианин-конъюгированного AffiniPure F(ab')2 козлиного анти-кроличьего IgG (H+L) (Jackson Immunoresearch, 111-136-144) в качестве вторичного антитела.
В случае hROR1-сконструированных 63-12 клеток, клетки подвергали сортингу в 96-луночных плоскодонных планшетах, содержащих 200 мкл дополненной культуральной среды на лунку, с использованием FACS Aria II. Планшеты инкубировали при 37°C и клоны выращивали в 6-луночных планшетах до анализа. Целевую экспрессию подтверждали методом проточной цитометрии с использованием устройства FACSCalibur (BD Biosciences) и аналитической программы FlowJo (Tree Star, Ashland, OR).
Конструирование клеточной линии для эктопической экспрессии hROR1 в мышиной клеточной линии рака молочной железы EMT-6: Мышиные клетки EMT-6 рака молочной железы (любезно предоставленные профессором доктором медицины Альфредом Циппелиусом, госпиталь университета Базеля, Швейцария) культивировали в полной DMEM (Модифицированная Дульбекко среда Игла (DMEM) с высоким содержанием глюкозы (4,5 г/л) с L-глутамином, с 10% (об./об.) фетальной телячьей сывороткой (FCS), 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-глутамина (все от Bioconcept, Allschwil, Швейцария)) при 37°C и 5% CO2. Клетки были сконструированы для сверхэкспрессии ROR1 путем транспозиции следующим образом: клетки центрифугировали (6 мин, 1200 об/мин, 4°C) и ресуспендировали в RPMI-1640 среде (5×106 клеток/мл). 400 мкл этой клеточной суспензии затем добавляли к 400 мкл RPMI, содержащей 13,3 мкг транспозируемого вектора pPB-PGK-Puro-ROR1, направляющего ко-экспрессию полноразмерного ROR1 (NP_005003.2) и гена резистентности к пуромицину, и 6,6 мкг транспозаза-содержащего вектора pCDNA3.1_hy_mPB. Смесь ДНК/EMT-6 клеток переносили в кюветы для электропорации (просвет 0,4 см, 165-2088, BioRad, Cressier, Швейцария) и осуществляли электропорацию с использованием Biorad Gene Pulser II с расширителем электроемкости при 300В и 950 мкФ. Затем клетки инкубировали в течение 5-10 минут при комнатной температуре. После инкубации клетки центрифугировали при 1200 об/мин в течение 6 мин, промывали один раз и затем ресуспендировали в полной DMEM перед инкубацией при 37°C в увлажненном инкубаторе при 5% CO2 атмосфере. Через день после электропорации клеточные пулы, стабильно экспрессирующие человеческий ROR1, отбирали путем добавления 3 мкг/мл пуромицина (Sigma-Aldrich, P8833).
Экспрессию ROR1 на выбранных EMT-6-ROR1 клетках подтверждали методом проточной цитометрии. Вкратце, после трипсинизации 106 клеток центрифугировали в FACS пробирках; полученные клеточные осадки ресуспендировали в буфере (PBS с 2% (об/об) FCS). Клетки затем инкубировали с 2A2 (mAb066); 30 мин, 4°C, конечная концентрация 2 мкг/мл) с последующим центрифугированием и промывкой. Клетки затем ресуспендировали, как указано выше, и инкубировали с анти-человеческим IgG антителом (Fc гамма-специфическим) PE (eBioscience, Vienna, Austria, 12-4998-82) при разведении 1:250 в темноте (30 мин, 4°C), промывали один раз в буфере и хранили на льду до FACS сортинга.
С использованием FACS Aria II осуществляли сортинровку по одной клетке в 96-луночный плоскодонный планшет, содержащий 200 мкл полной DMEM на лунку. Этот планшет инкубировали при 37°C и клоны выращивали в 6-луночных планшетах до анализа ROR1-экспрессии методом проточной цитометрии, как описано выше, с использованием устройства FACSCalibur (BD Biosciences) и аналитической программы FlowJo (Tree Star, Ashland, OR) для анализа.
Фиг. 16C показывает данные FACS анализа клона 14 (с высокой экспрессией ROR1) и WT (ROR1-отрицательные) EMT-6, с использованием для детекции анти-ROR1 антитела 2A2 (mAb066).
Пример 2. Получение высокосложной библиотеки кроличьих Fab и реагенты для скрининга
Конструирование, экспрессия и очистка рекомбинантных человеческих ROR1 белков: Конструирование, экспрессия, очистка и биотинилирование hFc слитых белков, содержащих различные домены человеческого ROR1 или мышиного ROR1, были описаны (Yangetal.,PloSOne6:e21018,2011). Для hROR1-AVI-6XHIS слитого белка, внеклеточный домен человеческого ROR1 (24-403) подвергали ПЦР-амплификации с праймерами pCEP4-hROR1-F и pCEP4-hROR1-Avi tag-R (необходимо отметить, что AVI метку вводили на C-конце ROR1 при помощи праймера pCEP4-hROR1-Avi tag-R), с последующей удлиняющей ПЦР с праймерами pCEP4-сигнал-F-KpnI и pCEP4-6HIS-R-XhoI для добавления сигнального пептида и 6XHIS метки к N и C концу отдельно перед клонированием в pCEP4 через KpnI/XhoI. Эту конструкцию затем транзиентно трансфицировали в HEK 293F клетки (Invitrogen) с использованием 293 фектина (Invitrogen) и белок очищали аффинной хроматографией с иммобилизованными ионами металла с использованием 1-мл HisTrap колонки (GE Healthcare), описанной в ProteinSciChapter6:Unit610,2009. Качество и количество очищенного hROR1-AVI-6XHIS анализировали при помощи SDS-PAGE и A280 поглощения, соответственно. Затем слитый белок биотинилировали с использованием набора, содержащего BirA фермент, от Avidity (Aurora, Colorado), следуя протоколу. Вкратце, 2 мг ROR1-AVI-6XHIS при 40 мкМ в 10 мМ Tris-HCl (pH 8) биотинилировали в присутстви биотина с использованием 10 мкг BirA после инкубации в течение 30 минут при 37°C, с последующей очисткой снова с использованием 1-мл HisTrap колонки (GE Healthcare), описанной выше. pCEP4-hROR1-F: 5'-atcctgtttctcgtagctgctgcaactggagcacactccgcccggggcgccgccgcccag-3' (SEQ ID NO:105); pCEP4-hROR1-Avi-tag-R: 5'-ccactcgatcttctgggcctcgaagatgtcgttcaggccctccatcttgttcttctcctt-3' (SEQ ID NO:106); pCEP4-сигнал-F-KpnI: 5' gctgggtaccggcgcgccaccatggactggacttggagaatcctgtttctcgtagctgct-3' (SEQ ID NO:107); pCEP4-6HIS-R-XhoI: 5'-gccggcctcgagtcagtgatggtgatggtggtgctcgtgccactcgatcttctgggcctc-3' (SEQ ID NO:108).
Конструирование, экспрессия и очистка рекомбинантных белков человеческого ROR1 (hROR1-His) и человеческого ROR2 (hROR2-His): hROR1-His подвергали ПЦР-амплификации с праймерами SP-hROR1_F (5' gctgggtaccggcgcgccaccatggactggacttggagaatcctgtttctcgtagctgctgcaactggagcacactccgcccggggcgcc gccgcccag 3') (SEQ ID NO:109) и hROR1-His_R (5' cggcctcgagtcagtgatggtgatggtggtgctccatcttgttcttctcctt 3') (SEQ ID NO:110) с использованием pCEP4-hFc-hROR1 (Yangetal.,PloSOne6:e21018,2011) в качестве матрицы, тогда как hROR2-His подвергали ПЦР-амплификации с праймерами SP-hROR2_F (gctgggtaccggcgcgccaccatggactggacttggagaatcctgtttctcgtagctgctgcaactggagcacactccgaagtggaggtt ctggatccg) (SEQ ID NO:111) и hROR2-His_R (cggcctcgagtcagtgatggtgatggtggtgccccatcttgctgctgtctcg) (SEQ ID NO:112) с использованием pCEP4-hFc-hROR2 в качестве матрицы. Затем их клонировали в pCEP4 (Invitrogen) отдельно через KpnI/XhoI. Эти конструкции затем отдельно и транзиентно трансфицировали в HEK 293F клетки (Invitrogen) с использованием 293фектина (Invitrogen) и соответствующие белки очищали аффинной хроматографией с иммобилизованными ионами металла с использованием 1-мл HisTrap колонки (GE Healthcare), описанной в KwongandRader, CurrProtocProteinSci Chapter 6:Unit 610, 2009. Качество и количество очищенных hROR1-His и hROR2-His анализировали при помощи SDS-PAGE и A280 поглощения, соответственно.
Получение и выбор наивных химерных кроличьих/человеческих Fab библиотек: все манипуляции с кроликами осуществлялись ветеринарами в Pocono Rabbot Farm & Laboratory (Canadensis, PA) или R & R Research (Stanwood, WA). Всего использовали девять кроликов (возраст 3-4 месяца). Пять этих кроликов были штамма New Zealand White (NZW), при этом три получены от Pocono Rabbit Farm & Laboratory (Canadensis, PA), а два получены от R & R Research (Stanwood, WA). Четыре кролика b9 дикого типа происходили из отдельной R & R Research колонии, происходящей из племенной колонии, разработанной и охарактеризованной в National Institute of Allergy and Infectious Diseases (NIAID) (McCartney-Francisetal., Proc.Natl.Acad.Sci.USA81:1794-1798,1984; и Popkovetal., J.Mol.Biol.325:325-335,2003. Селезенку и костный мозг каждого кролика собирали и обрабатывали для получения тотальной РНК и ОТ-ПЦР амплификации кроличьих Vκ, Vλ и VH кодирующих последовательностей с использованием установленных протоколов (Rader, MethodsMolBiol 525:101-128, xiv, 2009. Сборку кроличьих (rb) Vκ/человеческих (hu) Cκ/rbVH и rbVλ/huCλ/rbVH сегментов, соответственно, осуществляли в одну объединенную стадию на основе ПЦР с 3 перекрывающимися фрагментами. Необходимо отметить, что также объединяли VL от b9 кроликов с VH от NZW кроликов. Fab-кодирующие фрагменты расщепляли при помощи SfiI и лигировали с SfiI-обработанным вектором фагового дисплея pC3C (Hoferetal.,JImmunolMeth318:75-87,2007) при 16°C в течение 24 часов. Затем 15 мкг очищенных pC3C-rbVκ/hCκ/rbVH лигированных продуктов трансформировали в E. coli штамм SR320 (любезно предоставленные д-ром Сачдев С. Сидху, университет Торонто, Торонто, Онтарио, Канада) 30 отдельными электропорациями (каждая с использованием 0,5 мкг ДНК в 50 мкл электрокомпетентных клеток) и получали 7,5×109 независимых трансформантов для библиотеки κ. Для библиотеки λ, 4,8×109 независимых трансформантов были получены с использованием такой же процедуры. С использованием VCSM13 хелперного фага (Stratagene; La Jolla, CA) фагмидные библиотеки были преобразованы в библиотеки фагов, и их хранили при -80oC. Библиотеку κ и библиотеку λ фагов снова амплифицировали с использованием XL1-Blue (Stratagene) или ER2738 (Lucigen) и смешивали поровну перед четырьмя циклами пэннинга против биотинилированного hFc-hROR1 или hROR1-AVI-6HIS. В процессе пэннинга 5 мкг/мл антигена предварительно инкубировали с покрытыми стрептавидином магнитными шариками (Dynabeads MyOne Streptavidin C1; Invitrogen) при 37°C в течение 30 минут и затем связующие из библиотеки фагов захватывали в присутствии 1 мг/мл неспецифического поликлонального человеческого IgG (Thermo Scientific), когда использовали hFc-ROR1. Начиная с третьего цикла пэннинга, входящий фаг отрицательно истощался инкубацией с пустыми шариками перед селекцией против антиген-нагруженных шариков. После селекции супернатанты IPTG-индуцированных бактериальных клонов анализировали методом ELISA и методом проточной цитометрии. Посторяющиеся клоны идентифицировали методом ДНК-генотипоскопии с использованием AluI и последовательности VL и VH уникальных клонов определяли методом ДНК секвенирования (Фиг. 1).
Пример 3. Экспрессия и очистка химерных кроличьих/человеческих Fab и полноразмерных IgG1 антител
Конструирование, экспрессия и очистка химерных кроличьих/человеческих Fab и IgG1: MAb XBR1-402 в формате химерного кроличьего/человеческого Fab клонировали в E. coli экспрессирующую плазмиду pC3C-His и экспрессировали и очищали, как описано в KwongandRader, CurrProtocProteinSci Chapter 6:Unit 610, 2009. Для экспрессии mAb XBR1-402 в формате химерного кроличьего/человеческого IgG1 использовали ранее описанный вектор PIGG-R11 (Yangetal.,PloSOne6:e21018,2011). VH кодирующую последовательность Fab XBR1-402 подвергали ПЦР-амплификации с использованием праймеров 4-2_VH_F и 4-2_VH_R и клонировали через ApaI/SacI в PIGG-R11. Затем кодирующую легкую цепь последовательность XBR1-402 подвергали ПЦР-амплификации с использованием праймеров 4-2_λ_F и LEAD-B и клонировали через HindIII/XbaI в PIGG-R11 с соответствующей кодирующей тяжелую цепь последовательностью. Следует отметить, что внутренний ApaI сайт в FR4 VH-кодирующих последовательностей Fab XBR1-402 удаляли путем сайлент-мутации в праймере 4-2_VH_R. Кроме того, был изменен TAG стоп-кодон, который подавлялся в процессе селекции в E. coli штамме XL1-Blue, на CAG (глутамин), кодирующий первую аминокислоту природной VH (Фиг. 1), с использованием праймера 4-2_VH_F. Полученную PIGG-XBR1-402 плазмиду транзиентно трансфицировали в HEK 293F клетки (Invitrogen) с использованием 293фектина (Invitrogen) и белок очищали на 1-мл HiTrap колонке с рекомбинантным Белком A (GE Healthcare, Piscataway, NJ), как описано (Yangetal., PloSOne6:e21018, 2011;и YangandRader, MethodsMolBiol 901:209-232, 2012). Качество и количество очищенного IgG1 анализировали при помощи SDS-PAGE и A280 поглощения, соответственно.
Все другие mAbs в формате химерных кроличьих/человеческих Fab клонировали в E. coli экспрессирующую плазмиду pET11a и экспрессировали и очищали, как описано (Yangetal., PloSOne6:e21018, 2011). Для экспрессии mAbs ERR1-324, ERR1-TOP43 и ERR1-TOP54 в формате химерных кроличьих/человеческих IgG1 использовали pCEP4 (Invitrogen) для клонирования тяжелой цепи и легкой цепи отдельно. Для тяжелой цепи, gBlock, содержащий последовательность, кодирующую сигнальный пептид тяжелой цепи, VH ERR2-302 (mAb к hROR2) и CH1 (1-49) человеческого IgG1, синтезировали при помощи IDT (San Jose, CA) и амплифицировали с праймерами KpnI/AscI-Сигнал и CH1-внутренний/перекрывающийся-R, и сливали с CH1 (50-88)-CH2-CH3, амплифицированным из PIGG с праймерами CH1-внутренний/перекрывающийся-F и HC-CH3-R-XhoI, путем ПЦР с перекрывающимися праймерами KpnI/AscI-Сигнал и HC-CH3-R-XhoI и затем клонировали в pCEP4 by AscI/XhoI. Следует отметить, что EheI сайт был введен в CH1 по Ala12 путем синонимичной мутации, когда синтезировали gBlock. Следовательно, эта конструкция служила в качестве вектора для клонирования тяжелых цепей других mAbs путем замены VH с использованием AscI/EheI: VH ERR1-324, ERR1-TOP43 и ERR1-TOP54 амплифицировали с прямым праймером ERR1-324 HC-F, ERR1-TOP43 HC-F и ERR1-TOP54 HC-F и обратным праймером VH-CH1-R-EheI отдельно, с последующей удлиняющей ПЦР для добавления сигнального пептида с использованием праймера KpnI/AscI-Сигнал и VH-CH1-R-EheI. Затем каждый VH встраивали в вектор при помощи AscI/EheI. Для клонирования легких цепей, в то время как лямбда легкие цепи ERR1-TOP43 и ERR1-TOP54 амплифицировали с использованием праймеров ERR1-TOP43 LC-F и ERR1-TOP54 LC-F по отдельности в комбинации с LC-R-XhoI, каппа легкие цепи ERR1-324 амплифицировали с использованием праймеров ERR1-324 KC-F и KC-R-XhoI. Затем добавляли кодирующую сигнальный пептид последовательность при помощи удлиняющей ПЦР с прямым праймером KpnI/AscI-Сигнал и обратным праймером LC-R-XhoI или KC-R-XhoI. Затем ПЦР продукты каждой легкой цепи клонировали в pCEP4 при помощи AscI/XhoI. Полученные конструкции, содержащие тяжелую цепь или легкую цепь для каждого IgG ко-трансфицировали транзиентно в HEK 293F клетки (Invitrogen) с использованием 293фектина (Invitrogen) и соответствующие белки очищали на 1-мл колонке HiTrap с рекомбинантным Белком A (GE Healthcare, Piscataway, NJ), как описано (Yangetal.,PloSOne6:e21018,2011;и YangandRader, MethodsMolBiol901:209-232,2012). Качество и количество очищенного IgG1 анализировали при помощи SDS-PAGE и A280 поглощения, соответственно.
Последовательности праймеров для клонирования последовательностей антител
Пример 4. Исследование связывающей активности антител
ELISA: для ELISA (Фиг. 2) каждую лунку 96-луночного планшета Costar 3690 (Corning, Corning, NY) покрывали 100 нг анти-человеческого IgG1 Fc в 25 мкл покровного буфера (0,1 M Na2CO3, 0,1 M NaHCO3, pH 9,6) в течение 1 часа при 37°C. После блокирования при помощи 150 мкл 3% (масс/об) BSA/TBS в течение 1 часа при 37°C осуществляли захват hFc-hROR1 или hFc-mROR1 после инкубации при 100 нг/50 мкл в течение 1 часа при 37°C. Затем 100 нг/50 мкл Fab вносили в каждую лунку при 37°C. Через 2 часа использовали 50 мкл 1:1000 разведения мышиного анти-His tag mAb, конъюгированного с пероксидазой хрена (HRP) (R&D Systems, Minneapolis, MN), в 1% (масс/об) BSA/TBS для детекции Fab. Для определения эпитопов (Фиг. 4) hFc слитые белки, содержащие различные домены hROR1, наносили непосредственно сверху, с последующей инкубацией с Fab перед детекцией с использованием мышиного анти-His tag mAb, конъюгированного с HRP. Промывку при помощи PBS повторяли и осуществляли колориметрическую детекцию с использованием 2,2'-азино-бис(3-этилбензтиазолин)-6-сульфоновой кислоты (Roche) в качестве субстрата в соответствии с инструкциями изготовителя. Поглощение измеряли при 405 нм с использованием микропланшетного ридера SpectraMax M5 (Molecular Devices; Sunnyvale, CA) и программы SoftMax Pro (Molecular Devices).
Проточная цитометрия: Клетки окрашивали с использованием стандартного метода проточной цитометрии. Вкратце, для очищенного анти-ROR1 Fab (Фиг. 3), 0,1~1 × 106 клеток окрашивали при помощи 1000 нг/100 мкл Fab на льду в течение 1 часа. После промывки два раза ледяным буфером для проточной цитометрии (PBS, содержащий 1% (об/об) BSA, 0,1% азида натрия и 1 мМ EDTA) клетки инкубировали с 1:1000 разведением мышиного анти-His tag mAb, конъюгированного с Alexa Fluor 488 (Qiagen), в 100 мкл буфера для проточной цитометрии на льду в течение 30 минут. Для очищенного анти-ROR1 IgG1 (Фиг. 8), 0,1~1 × 106 клеток блокировали при помощи 4% козьей сыворотки в течение 15 минут при комнатной температуре и затем инкубировали с 100 нг/100 мкл IgG на льду в течение 1 часа. После промывки два раза ледяным буфером для проточной цитометрии клетки окрашивали при помощи 1:500 разведения козлиных анти-человеческих IgG, Fcγ pAbs, конъюгированных с APC (Jackson ImmunoResearch), в 100 мкл буфера для проточной цитометрии на льду в течение 30 минут. Для коммерческого козлиного анти-человеческого поликлонального антитела к ROR1 (Фиг. 8), 0,1~1 × 106 клеток окрашивали при помощи 200 нг/100 мкл Ab на льду в течение 1 часа. После промывки два раза ледяным буфером для проточной цитометрии (PBS, содержащий 1% (об/об) BSA, 0,1% азида натрия и 1 мМ EDTA) клетки инкубировали с 1:1000 разведением Alexa Fluor 647-конъюгированного AffiniPure F(ab')2 ослиного антикозлиного IgG (H+L) в 100 мкл буфере для проточной цитометрии на льду в течение 30 минут. В заключение, 4',6-диамидино-2-фенилиндол (DAPI) добавляли до конечной концентрации 100 нг/мл для исключения мертвых клеток из анализа. Клетки анализировали с использованием устройства FACSCalibur (BD Biosciences) и аналитической программы FlowJo (Tree Star, Ashland, OR).
Поверхностный плазмонный резонанс: Mетод поверхностного плазмонного резонанса для измерения аффинности всех Fabs к hFc-hROR1 и для исследования методом картирования эпитопов осуществляли на устройстве Biacore X100 с использованием реагентов и программы Biacore (GE Healthcare, Piscataway, NJ). Античеловеческое IgG (Fc) антитело иммобилизовали на CM5 сенсорном чипе, следуя инструкциям, прилагаемым к набору Human Antibody Capture Kit (GE Healthcare, Piscataway, NJ). Затем hFc-hROR1 слитые белки захватывали при определенной плотности (указана на Фиг. 5 и Фиг. 6). Сенсорный чип включал пустую проточную ячейку для мгновенного фонового истощения. Для всех анализов связывания использовали 1x HBS-EP+ подвижный буфер (10 мМ HEPES, 150 мМ NaCl, 3 мМ EDTA (pH 7,4) и 0,05% (об/об) поверхностно-активного вещества P20) и скорость потока 30 мл/мин. Для измерений аффинности все Fabs инжектировали при пяти разных концентрациях (коэффициент разведения=2) и наименьшую концентрацию испытывали в двух повторах (наивысшие концентрации для каждого Fab указаны на Фиг. 6B). Сенсорный чип восстанавливали с использованием 3 MgCl2 из набора Human Antibody Capture Kit без какой-либо потери связывающей способности. Расчет констант скоростей ассоциации (kon) и диссоциации (koff) осуществляли на основании 1:1 Langmuir модели связывания. Равновесную константу диссоциации (Kd) рассчитывали из koff/kon. Для исследования методом картирования эпитопов каждый Fab получали при 500 нМ отдельно в 1x HBS-EP+ подвижном буфере и затем инжектировали в порядке, указанном на Фиг. 5.
Вестерн-блоттинг: Клетки или белки лизировали с использованием 1x буфера для образца (содержащего 1% β-меркаптоэтанола) и кипятили перед проведением электрофореза на NuPAGE Novex 4-12% Bis-Tris гелях (Invitrogen). После переноса на мембрану и блокирования 5% молоком наносили 2 мкг/мл химерного кроличьего/человеческого IgG1 XBR1-402 или ERR1-TOP43 для детекции денатурированных белков, с последующей инкубацией с 1/1000 анти-человеческим Fcγ, конъюгированным с HRP, перед проявлением с использованием реагента для детекции ECL Prime WesternDetection Reagent (GE Healthcare) (Фиг. 9).
Пример 5. Экспрессия очищенного рекомбинантного strep-меченного человеческого ROR1 и человеческого содержащего двойную strep-метку ROR2
StrepII-меченный человеческий ROR1-внеклеточный домен получали следующим образом: нуклеотидную последовательность, кодирующую внеклеточный домен человеческого ROR1 (NP_005003.2), по N-концу сливали с сигнальной последовательностью (MNFGLRLIFLVLTLKGVQC) и по C-концу сливали с последовательностью, кодирующей strepII-метку (GWSHPQFEK). Полные нуклеотидные последовательности с фланкирующими 5'NotI и 3'HindIII сайтами получали путем полного генного синтеза (GenScript, Piscataway, USA), сборки в специальный вектор экспрессии млекопитающего pEvi5 при помощи Evitria AG (Schlieren, Switzerland) и подтверждения путем ДНК секвенирования.
Экспрессию белков осуществляли в суспензия-адаптированных CHO K1 клетках при помощи Evitria AG (Schlieren, Switzerland). Супернатанты из пулов трансфицированных CHO K1 клеток собирали центрифугированием и стерильно фильтровали (0,2 мкм) перед FPLC аффинной очисткой с использованием колонок StrepTactin (IBA GmbH, Goettingen, Germany).
Рекомбинантный человеческий с двойной strep-меткой ROR2 (NP_004551.2) экспрессировали и очищали на месте в соответствии со следующим протоколом: EBNA вектор экспрессии pCB14b-ROR2-ECD-TwinStrep, направляющий экспрессию внеклеточного домена ROR2 (ECD), меченный по C-концу двойной Step-меткой, трансфицировали в HEK293T с использованием Липофектамина® LTX с реагентом PLUS™ (Thermo Fisher Scientific, 15388100). После 1-дневной инкубации (37°C, 5% CO2, среда роста: Модифицированная Дульбекко среда Игла (DMEM) с высоким содержанием глюкозы (4,5 г/л) с L-глутамином с 10% (об./об.) фетальной телячьей сывороткой (FCS), 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-глутамина (все от Bioconcept)) клетки выращивали в селективных условиях (2 мкг/мл пуромицина (Sigma-Aldrich, P8833-25 мг исходного раствора при 2 мг/мл)). Клетки расщепляли и снова выращивали (37°C, 5% CO2); после достижения конфлюентности чашки для тканевых культур покрывали 20 мкг/мл поли-L-Лизина (Sigma-Aldrich, P1524) в течение 2 часов при 37°C и промывали два раза при помощи PBS. Затем клетки трипсинизировали, промывали при помощи PBS и расщепляли 1:3 на покрытых поли-L-лизином планшетах. Снова после достижения конфлюентности клетки промывали при помощи PBS с последующей заменой среды с использованием среды для продуцирования (DMEM/F-12, Gibco/Thermo Fisher Scientific, 31330-03), дополненной 1 мкг/мл пуромицина (Sigma-Aldrich, P8833), 100 МЕ/мл пенициллина-стрептомицина-фунгизона (Bioconcept, 4-02F00-H), 161 мкг/мл N-ацетил-L-цистеина (Sigma-Aldrich, A8199) и 10 мкг/мл of L-глутатиона сокращенного (Sigma-Aldrich, G6529). Супернатант, который собирали два раза в неделю и фильтровали (0,22 мкм) для удаления клеток, хранили при 4°C до очистки. Для очистки отфильтрованный супернатант нагружали на высокопроизводительную Streptactin® Superflow® (IBA, Göttingen, Germany, 2-1238-001) колонку; очистку и элюирование осуществляли в соответствии с протоколом изготовителя на AEKTA pure (GE Healthcare). Фракции анализировали на чистоту и целостность белка при помощи SDS-PAGE. Содержащие белок фракции смешивали и подвергали буферному обмену с использованием фильтровальных установок Amicon (Millipore, Schaffhausen, Switzerland) для достижения разведения ≥1:100 в PBS и затем стерильно фильтровали с использованием фильтра с низкой удерживающей способностью (0,20 мкм, Carl Roth, Karlsruhe, Germany, PA49,1).
Пример 6. Экспрессия очищенного рекомбинантного гуманизированного анти-человеческого ROR1 и изотипических контрольных антител
Векторы экспрессии: Области, кодирующие вариабельные области антител, получали путем полного генного синтеза (GenScript) с использованием MNFGLRLIFLVLTLKGVQC в качестве лидерной последовательности и собирали вместе с человеческими IgH-γ 1 и IgL-κ или IgL-λ константными областями, соответственно, в векторе экспрессии pCB14. Этот вектор, производное эписомального вектора экспрессии млекопитающего pCEP4 (Invitrogen), содержит ориджин репликации EBV, кодирует EBV ядерный антиген (EBNA-1) для возможности внехромосомной репликации и содержит пуромициновый селективный маркер вместо исходного гена резистентности к гигромицину B.
Экспрессия и очистка на месте: Векторы экспрессии на основе pCB14 трансфицировали в HEK293T клетки с использованием Липофектамин® LTX реагента с PLUS™ реагентом (Thermo Fisher Scientific, Reinach, Switzerland, 15388100); после 1-дневной инкубации (37°C, 5% CO2, среда роста: Модифицированная Дульбекко среда Игла (DMEM) с высоким содержанием глюкозы (4,5 г/л) с L-глутамином с 10% (об./об.) фетальной телячьей сывороткой (FCS), 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-глутамина (все от Bioconcept, Allschwil, Switzerland)) клетки выращивали в селективных условиях (2 мкг/мл пуромицина (Sigma-Aldrich, Buchs SG, Switzerland, P8833-25 мг исходного раствора при 2 мг/мл)). Клетки расщепляли и снова выращивали (37°C, 5% CO2); после достижения конфлюентности чашки для тканевых культур покрывали 20 мкг/мл поли-L-Лизина (Sigma-Aldrich, P1524) в течение 2 часов при 37°C и промывали два раза при помощи PBS. Затем клетки трипсинизировали и расщепляли 1:3 на покрытых поли-L-лизином планшетах. Снова после достижения конфлюентности клетки промывали при помощи PBS, с последующей заменой среды на среду для продуцирования (DMEM/F-12, Gibco/Thermo Fisher Scientific, 31330-03), дополненную 1 мкг/мл пуромицина (Sigma, P8833), 100 МЕ/мл пенициллина-стрептомицина-фунгизона (Bioconcept), 161 мкг/мл N-ацетил-L-цистеина (Sigma-Aldrich, A8199) и 10 мкг/мл L-глутатиона сокращенного (Sigma-Aldrich, G6529). Супернатант, который собирали два раза в неделю и фильтровали (0,22 мкм) для удаления клеток, хранили при 4°C до очистки.
Для очистки отфильтрованный супернатант нагружали на PBS-уравновешенную колонку HiTrap с Белком A (GE Healthcare, Frankfurt am Main, Germany, 17-0405-01) или колонку JSR Amsphere™ с Белком A (JSR Life Sciences, Leuven, Belgium, JWT203CE) и промывали при помощи PBS; элюирование осуществляли с использованием 0,1 M глицина (pH 2,5) на AEKTA pure (GE Healthcare). Фракции сразу же нейтрализовали при помощи 1 M Tris-HCl буфера (pH 8,0) и анализировали на чистоту и целостность белка при помощи SDS-PAGE. Содержащие белок фракции смешивали и подвергали буферному обмену с использованием фильровальных установок Amicon (Millipore, Schaffhausen, Switzerland, UFC901008) для достижения разведения 1:100 в буфере, указанном в таблице 2, и затем стерильно фильтровали с использованием фильтра с низкой удерживающей способностью (0,20 мкм, Carl Roth, Karlsruhe, Germany, PA49,1).
Антитела транзиентно экспрессировали в CHO клетках способами, известными в данной области, и рекомбинантные антитела очищали стандартным способом очистки с белком A из супернатантов CHO клеток, как известно в данной области. Чистоту и целостность рекомбинантных антител анализировали при помощи SDS-PAGE.
Таблица 2 представляет антитела, используемые в последующих примерах, вместе с их конечной концентрацией и буферами.
Перечень антител, используемых в примерах
(ref.)
(HC: Тяжелая цепь,
LC: Легкая цепь)
мл)
LC: SEQ ID NO. 14
LC: G5SLPETG-Strep
LC: SEQ ID NO. 15
LC: G5SLPETG-Strep
LC: SEQ ID NO. 16
LC: G5SLPETG-Strep
LC: SEQ ID NO. 18
LC: G5SLPETG-Strep
LC: SEQ ID NO. 19
LC: G5SLPETG-Strep
LC: SEQ ID NO. 25
LC: G5SLPETG-Strep
LC: SEQ ID NO. 14
LC: G5SLPETG-Strep
(mAb188)
LC: SEQ ID NO. 18
LC: G5SLPETG-Strep
LC: SEQ ID NO. 25
LC: G5SLPETG-Strep
LC: SEQ ID NO. 14
LC: G5SLPETG-Strep
LC: SEQ ID NO. 149
LC: G5SLPETG-Strep
LC: SEQ ID NO. 151
LC: G5SLPETG-Strep
LC: SEQ ID NO. 153
LC: G5SLPETG-Strep
LC: SEQ ID NO. 155
LC: G5SLPETG-Strep
LC: SEQ ID NO. 157
LC: G5SLPETG-Strep
LC: SEQ ID NO. 159
LC: G5SLPETG-Strep
LC: SEQ ID NO. 161
LC: G5SLPETG-Strep
Ms961 соответствует анти-ROR1 антителу с тяжелой цепью в соответствии с SEQ ID NO. 1 и легкой цепью в соответствии с SEQ ID NO. 3, описанному в WO 2014/031174. 2A2 относится к анти-ROR1 антителу, описанному в WO 2010/124188, тогда как hu2A2 относится к гуманизированному 2A2, описанному в неопубликованной PCT/EP2016/076244. R11 и R12 представляют собой анти-ROR1 антитела, описанные в WO 2012/075158. Изотипические контрольные антитела трастузумаб и Ac10 таргетируют HER-2 и CD30, соответственно. Последовательности вариабельных областей тяжелой и легкой цепи моноклонального антитела брентуксимаба (клон cAc10), специфического в отношении мишени, представляющей собой человеческий CD30, были получены из патента US2008213289A1, последовательности специфического в отношении человеческого HER-2 антитела трастузумаба, содержащиеся в коммерческом антителе Герцептине (трастузумаб) или его производном ADC Kadcyla®, были получены из онлайн IMGT базы данных (VH: http://www.imgt.org/3Dstructure-DB/cgi/details.cgi?pdbcode=7637&Part= Chain&Chain=7637H & VL: http://www.imgt.org/3Dstructure-DB/cgi/details.cgi?pdbcode=7637&Part=Chain&Chain =7637L.
Пример 7. mAb ROR1 и ROR2-связывание - характеризация методом ELISA
Каждую лунку 96-луночного планшета покрывали 100 мкл 2 мкг/мл strep-меченного человеческого ROR1 (из Примера 5) в 0,1 M бикарбонатном покровном буфере (pH 9,6) и инкубировали в течение 12 часов при 4°C. Второй 96-луночный планшет получали подобным образом с меченным двойной strep-меткой человеческим ROR2 (из Примера 5).
После блокирования при помощи 150 мкл 3% (масс./об.) бычьего сывороточного альбумина (BSA)/TBS в течение 1 часа при 37°C следующие антитела добавляли в лунку в каждый планшет при концентрации 0,5 мкг/мл и осуществляли серийное разведение (коэффициент разведения 4) с использованием 1% (масс/об) BSA/TBS перед инкубацией в течение 1 часа при 37°C: ERR1-301 (mAb027), XBR1-402 (mAb031), ERR1-306 (mAb033), ERR1-324 (mAb034), ERR1-403 (mAb035) и ERR1-Top43 (mAb036). HRP-конъюгированный F(ab')2 анти-человеческий FC-гамма (Jackson Immunoresearch, 109-036-008) затем добавляли при 1:20'000 разведении, 100 мкл на лунку, и инкубировали в течение 1 часа при 37°C перед детекцией с использованием планшет-ридера Spark 10M (Tecan). Как показано на Фиг. 10, античеловеческие ROR1 антитела связываются с человеческим ROR1 (панель A) и не являются перекрестно реактивными с человеческим ROR2 (панель B).
Пример 8. Гуманизация анти-hROR1-специфического антитела XBR1-402
Гуманизированные последовательности вариабельных областей (семь hu4-2 VH вариантов и 4 hu4-2 VL варианта) конструировали при помощи Fusion Antibodies (Belfast, Ireland). Вкратце, каркасы в кроличьих вариабельных областях заменяли человеческими каркасными областями (следуя подходу CDR-прививки in-silico на основании запатентованных алгоритмов) с образованием 28 возможных пар тяжелая цепь/легкая цепь.
Экспрессию антител, представляющих все 28 возможных пар тяжелая цепь/легкая цепь осуществляли путем транзиентной трансфекции экспрессирующих конструкций на основе pCB14 в HEK-293T клетки и сбора клеточных супернатантов.
Для транзиентной экспрессии антител клетки трансфицировали в 24-луночных планшетах с использованием реагента для трансфекции Avanti Transfection Reagent I (Avanti Polar Lipids, Alabaster, USA). Для каждой лунки использовали 0,5 мкг тотальной ДНК для трансфекции, свежую питательную среду добавляли на следующий день и кондиционировали в течение 4 дней. Супернатанты стерильно фильтровали и хранили при -20°C до анализа.
Затем осуществляли скрининг на аффинность для отбора лучших связующих. Аффинность определяли с использованием устройства Biacore T200 (GE Healthcare, Buckinghamshire, UK) и данные оценивали с использованием программы Biacore Evaluation T200 V2.0. Для захвата mAbs козлиное античеловеческое Fc-гамма-специфическое IgG (Jackson ImmunoResearch, # 109-005-098) было ковалентно иммобилизовано на CM5 чипе (GE Healthcare, # BR-1005-30).
Вкратце, для захвата mAbs в HEK293T супернатантах неразбавленные супернатанты захватывали с потоком 30мкл/мин в течение 120сек. ROR1-strep разводили в подвижном буфере (HBS-EP+ pH 7,4 (10 мМ HEPES, 150 мМ NaCl, 3 мМ EDTA, 0,05% Tween 20) до 20 нМ. Ассоциацию измеряли при скорости потока 30 мкл/мин в течение 120 сек, а затем измеряли диссоциацию в течение 200 сек при скорости потока 30 мкл/мин. Уровни захвата варьировались от 31,8 RU до 59,4 RU.
Выбранные антитела экспрессировали и очищали, как в Примере 6, используя условия, указанные в Таблице 3 ниже.
Протоколы и концентрации гуманизированных mAbs, используемые в примерах
(ref.)
(HC: Тяжелая цепь, LC: Легкая цепь)
LC: SEQ ID NO. 136
LC: G5SLPETG- TwinStrep
LC: SEQ ID NO. 137
LC: G5SLPETG- TwinStrep
LC: SEQ ID NO. 138
LC: G5SLPETG- TwinStrep
LC: SEQ ID NO. 139
LC: G5SLPETG- TwinStrep
LC: SEQ ID NO. 140
LC: G5SLPETG- TwinStrep
LC: SEQ ID NO. 141
LC: G5SLPETG- TwinStrep
Пример 9. FACS окрашивание клеток для определения экспрессии hROR1
5×105 каждого типа клеток добавляли в каждую лунку 96-луночных планшетов. Планшеты центрифугировали (3 мин, 1300 об/мин) с ресуспендированием в буфере (PBS, дополненный 2% (об./об.) FCS). 2A2 (mAb066) добавляли в каждую лунку для достижения концентрации 2 мкг/мл. Планшеты затем инкубировали на льду в течение 30 минут и промывали при помощи 200 мкл буфера перед ресуспендированием в 200 мкл буфера, дополненного анти-человеческим IgG антителом (Fc гамма-специфическим) PE (eBioscience 12-4998-82) при 1:250 разведении. После 30 минут инкубации на льду и одной промывки клетки анализировали с использованием FACSCalibur устройства (BD Biosciences) и данные анализировали с использованием аналитической программы FlowJo (Tree Star, Ashland, OR).
Фиг. 7 показывает данные FACS анализа ROR1-положительных человеческих клеточных линий ALL 697 и Kasumi-2, человеческих клеточных линий трижды отрицательного рака молочной железы MDA-MB-231, MDA-MB-468 и HS-578T и ROR1-отрицательной человеческой клеточной линии рака молочной железы T47D в качестве отрицательного контроля.
Пример 10. Генерирование сайт-специфических конъюгированных ADCs с использованием SMAC-технологии
Фермент сортаза A. Рекомбинантный и аффинно очищенный фермент Сортазу A из Staphylococcus aureus продуцировали в E. coli, как раскрыто в WO2014140317A1.
Получение глицин-модифицированных токсинов. Для генерирования конъюгированных с использованием SMAC-technologyTM ADCs с пентаглицин-модифицированным PNU-159682 производным Gly5-EDA-PNU (Фиг. 11 B) получали, как раскрыто в WO2016102697. Идентичность и чистоту пентаглицин-модифицированного PNU токсина подтверждали масс-спектрометрией и ВЭЖХ. Gly5-модифицированный токсин продемонстрировал чистоту >95%, как определено по единственному пику на ВЭЖХ хроматограмме.
Сортаза-опосредованная конъюгация антител. Вышеуказанный токсин конъюгировали с анти-ROR1 антителом в соответствии с Таблицей 3 путем инкубации LPETG-меченных mAbs [10мкМ] с глицин-модифицированным токсином [200мкМ] и 3 мкМ Сортазы A в указанном буфере для конъюгации в течение 3,5 часов при 25°C. Реакцию останавливали, пропуская смесь через колонку с Белком A GraviTrap (BioRad). Связанный конъюгат элюировали с использованием 5 колоночных объемов элюирующего буфера (0,1 M глицина pH 2,5, 50 нМ NaCl), со сбором фракций 1 колоночного объема в пробирки, содержащие 25% об/об 1M HEPES pH 8 для нейтрализации кислоты. Содержащие белок фракции объединяли и формулировали в буфере для формулирования из Таблицы 7 с использованием обессоливающей колонки ZebaSpin.
ADC аналитика. DAR оценивали методом хроматографии с обращенной фазой на колонке Polymer Labs PLRP 2,1 мм × 5 см, 5 мкм при 1мл/мин/80°C с 25-мин линейным градиентом от 0,05 до 0,1% TFA/H2O и от 0,04 до 0,1% TFA/CH3CN. Образцы сначала восстанавливали путем инкубации с DTT при pH 8,0 при 37°C в течение 15 минут. DAR, определенные методом хроматографии с обращенной фазой, представлены в таблице 4 ниже.
Условия изготовления и аналитические данные ADCs, полученных в этом исследовании. DAR, отношение лекарственное средство-к-антителу. ND - не определяли.
(mAb188)
Из этих анализов можно сделать вывод, что SMAC-technologyTM конъюгация проходила при высокой эффективности, приводя к общим средним значениям DAR в диапазоне ~3,5-4,0 для комбинаций IgG-формат анти-ROR1 антитела-токсин.
Пример 11. In vitro стабильность в сыворотке ROR1-таргетирующего ADC XBR1-402-G5-PNU
In vitro стабильность в сыворотке XBR1-402-G5-PNU оценивали в анализе стабильности в сыворотке методом ELISA. Вкратце, ADC разводили до концентрации 100 мкг/мл в NOD SCID мышиной (любезно предоставлена профессором доктором медицины Альфредом Циппелиусом, госпиталь университета Базеля, Швейцария) и человеческой сыворотке (из SRK пункта взятия крови, Базель, Швейцария; 50:50 смесь мужской:женской крови, центрифугированная в течение 15 минут при 2000g с получением сыворотки) и инкубировали при 37°C. Образцы мгновенно замораживали в жидком азоте в дни 0, 3, 7 и 14 и хранили при -80°C до анализа ELISA. Для мышиной сыворотки, серийно разведенные образцы сыворотки с ADC (коэффициент разведения 3,5, от 5 до 0,0008 мкг/мл) захватывали на ELISA планшетах, покрытых 2 мкг/мл мышиного анти-PNU mAb собственной разработки (генерированного путем иммунизации мышей конъюгатом человеческий IgG-PNU и скрининга с BSA-PNU конъюгатом), для связывания ADC, или козлиным анти-человеческим Fc F(ab')2 (Jackson Immunoresearch, 109-006-098) для связывания общего IgG, и осуществляли детекцию с использованием 1:2500 разведения HRP-конъюгированного ослиного анти-человеческого IgG (Jackson Immunoresearch, 709-035-149). Для человеческой сыворотки, 2 мкг/мл рекомбинантного человеческого ROR1 (strep-меченного, Пример 5) использовали для покрытия ELISA планшетов, и 1:2500 разведение HRP-конъюгированного ослиного анти-человеческого IgG (Jackson Immunoresearch, 709-035-149) или 2 мкг/мл мышиного анти-PNU IgG (полученного на месте), затем 1:5000 разведение HRP-конъюгированного козлиного анти-мышиного Fcγ F(ab')2 (Jackson Immunoresearch, 115-036-071) использовали для детекции общего количества IgG и ADC, соответственно. Концентрации в сыворотке ADC и суммарных IgGs рассчитывали из полу-максимальных значений титрований образца путем сравнения с образцом такого же ADC известной концентрации. Как показано на Фиг. 19, XBR1-402-G5-PNU остается по существу стабильным в обеих сыворотках.
Пример 12. In vitro анализы цитотоксичности ADCs на основе анти-ROR1 антитела на EMT-6 дикого типа и ROR1-сверхэкспрессирующих EMT-6 клетках рака молочной железы
Цитотоксичность анти-ROR1 ADC XBR1-402-G5-PNU (adc262) исследовали с использованием EMT-6 дикого типа (WT) и EMT-6 клеток, сконструированных для сверхэкспрессии человеческого ROR1 (из Примера 2). Tras-G5-PNU (adc286) использовали в качестве изотипического контроля.
Для этого, 1×103 WT EMT-6 и hROR1-сверхэкспрессирующих EMT-6 клеток на лунку высевали в 96-луночные планшеты (за исключением краевых лунок, которые содержали воду) в 75 мкл DMEM, дополненной 10% по объему FCS, 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-Глутамина, и выращивали при 37°C в увлажненном инкубаторе в атмосфере 7,5% CO2. После 1-дневной инкубации каждый ADC добавляли в соответствующие лунки в количестве 25мкл 3,5-кратных серийных разведений в питательной среде (приводя к конечным концентрациям ADC от 20 мкг/мл до 0,88 нг/мл). Еще через 4 дня планшеты удаляли из инкубатора и уравновешивали до комнатной температуры. Приблизительно через 30 минут 50 мкл удаляли из каждой лунки и затем 50 мкл люминесцентного раствора CellTiter-Glo® 2.0 (Promega, G9243) добавляли в каждую лунку. После встряхивания планшетов при 450 об/мин в течение 5 минут с последующей 10-минутной инкубацией без встряхивания измеряли люминесценцию на Tecan Spark 10M планшет-ридере, с временем интегрирования 250 мсек на лунку. Кривые люминесценции против концентрации ADC (нг/мл) получали с использованием программы Graphpad Prism. IC50 значения, определенные с использованием встроенной ʺlog(ингибитор) vs. ответ -- Переменный наклон (четыре параметра)ʺ функции определения IC50 программы Prism, представлены в таблице 5.
In vitro клеточный киллинг EMT-6 клеток (WT и сконструированного клона 14) при помощи анти-ROR1 или изотипических контрольных ADC (IC50, нг/мл)
Фиг. 16 показывает кривые доза-ответ in vitro анализов киллинга клеток на WT и hROR1-сверхэкспрессирующих EMT6 клетках с использованием ADCs Таблицы 5. Как показано в Таблице выше и на Фиг. 16, XBR1-402-G5-PNU обеспечивает специфический киллинг, зависимый от статуса экспрессии ROR1.
Пример 13. In vitro анализы цитотоксичности новых и известных ADC на основе анти-ROR1 антител на ROR1-сверхэкспрессирующих EMT-6 клетках
Цитотоксичность анти-ROR1 ADC XBR1-402-G5-PNU (adc409) исследовали с использованием человеческих EMT-6 клеток, сконструированных для сверхэкспрессии человеческого ROR1 (из Примера 2), и сравнивали с анти-ROR1 ADC на основе антитела Ms961 (adc396). Tras-G5-PNU (adc394) использовали в качестве изотипического контроля.
Для этого, 1×103 hROR1-сверхэкспрессирующих EMT-6 клеток на лунку высевали в 96-луночные планшеты (за исключением краевых лунок, которые содержали воду) в 75 мкл DMEM, дополненной 10% по объему FCS, 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-Глутамина, и выращивали при 37°C в увлажненном инкубаторе в атмосфере 7,5% CO2. После 1-дневной инкубации каждый ADC добавляли в соответствующие лунки в количестве 25 мкл 3,5-кратных серийных разведений в питательной среде (приводя к конечным концентрациям ADC от 20 мкг/мл до 0,88 нг/мл). Еще через 4 дня планшеты удаляли из инкубатора и уравновешивали до комнатной температуры. Приблизительно через 30 минут 50 мкл удаляли из каждой лунки и затем в каждую лунку добавляли 50 мкл люминесцентного раствора CellTiter-Glo® 2.0 (Promega, G9243). После встряхивания планшетов при 450 об/мин в течение 5 минут с последующей 10-минутной инкубацией без встряхивания измеряли люминесценцию на Tecan Spark 10M планшет-ридере, с временем интегрирования 250 мсек на лунку. Кривые люминесценции против концентрации ADC (нг/мл) получали с использованием программы Graphpad Prism. IC50 значения, определенные с использованием встроенной ʺlog(ингибитор) vs. ответ -- Переменный наклон (четыре параметра)ʺ функции определения IC50 программы Prism, представлены в таблице 6.
In vitro клеточный киллинг сконструированных EMT-6 клеток с использованием ADC по изобретению и известных анти-ROR1 или изотипических контрольных ADCs (IC50, нг/мл)
Фиг. 17 показывает кривые доза-ответ in vitro анализов киллинга клеток на hROR1-сверхэкспрессирующих EMT6 клетках с использованием ADCs Таблицы 6. Как показано в Таблице выше и на Фиг. 17, XBR1-402-G5-PNU обеспечивает более сильный киллинг, чем сравнительный ADC на основе антитела Ms961.
Пример 14. In vitro анализы цитотоксичности новых и известных ADCs основе анти-ROR1 антител на человеческих 697 и Kasumi-2 ALL клетках
Цитотоксичность анти-ROR1 ADC XBR1-402-G5-PNU (adc262) исследовали с использованием человеческой клеточной линии 697 и сравнивали с анти-ROR1 ADCs на основе известных антител 2A2, R11 и R12 (Fig 13), или путем осуществления отдельного эксперимента с использованием снова 697 и Kasumi-2 ALL клеток по сравнению с 2A2 и R12.
Для этого, 2,5×103 697 или Kasumi-2 клеток на лунку высевали в 96-луночные планшеты (за исключением краевых лунок, которые содержали воду) в 75 мкл RPMI, дополненной 10% по объему FCS, 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-Глутамина, и выращивали при 37°C в увлажненном инкубаторе в атмосфере 7,5% CO2. После 1-дневной инкубации каждый ADC добавляли в соответствующие лунки в количестве 25 мкл 3,5-кратных серийных разведений в питательной среде (приводя к конечным концентрациям ADC от 20 мкг/мл до 0,88 нг/мл). Еще через 4 дня планшеты удаляли из инкубатора и уравновешивали до комнатной температуры. Приблизительно через 30 минут 50 мкл удаляли из каждой лунки и затем в каждую лунку добавляли 50 мкл люминесцентного раствора CellTiter-Glo® (Promega, G7570). После встряхивания планшетов при 750 об/мин в течение 5 минут с последующей 20-мин инкубацией без встряхивания измеряли люминесценцию на Tecan Infinity F200 планшет-ридере, с временем интегрирования 1 сек на лунку. Кривые люминесценции против концентрации ADC (нг/мл) получали с использованием программы Graphpad Prism. IC50 значения, определенные с использованием встроенной ʺlog(ингибитор) vs. ответ -- Переменный наклон (четыре параметра)ʺ функции определения IC50 программы Prism, представлены в таблице 7.
In vitro клеточный киллинг 697 клеток при помощи анти-ROR1 или изотипических контрольных ADCs (IC50, нг/мл)
Фиг. 13 показывает кривые доза-ответ in vitro анализов киллинга клеток на клетках 697 с использованием ADCs Таблицы 7. Как показано в Таблице выше и на Фиг. 13, ADCs по изобретению обеспечивают киллинг человеческих 697 B-клеток-предшественников лейкозных клеток, который выше, чем обеспечиваемый ADCs на основе известных анти-ROR1 антител.
Пример 15. In vitro анализы цитотоксичности ADCs на основе анти-ROR1 антител на клетках рака молочной железы MDA-MB-468 и HS 578T
Цитотоксичность анти-ROR1 ADCs 2A2-G5-PNU (adc165), XBR1-402-G5-PNU (adc135), R12-G5-PNU (adc292), ERR1-Top43-G5-PNU (adc204) и 50:50 по массе смесей ERR1-324-G5-PNU (adc202) и 2A2-G5-PNU (adc165), ERR1-324-G5-PNU (adc202) и XBR1-402-G5-PNU (adc135), ERR1-324-G5-PNU (adc202) и R12-G5-PNU (adc292) и ERR1-324-G5-PNU (adc202) и ERR1-Top43-G5-PNU (adc204) исследовали с использованием человеческих клеточных линий: MDA-MB-468, HS 578T. Ac10-G5-PNU (adc159) использовали в качестве изотипического контроля.
Для этого, следующее количество клеток на лунку высевали в 96-луночные планшеты (за исключением краевых лунок, которые содержали воду) и выращивали при 37°C в увлажненном инкубаторе в атмосфере 7,5% CO2 в питательной среде (DMEM, дополненной 10% по объему FCS, 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-Глутамина).
Посев клеток Примера 17
После 1-дневной инкубации каждый ADC добавляли в соответствующие лунки в количестве 25 мкл 3,5-кратных серийных разведений в питательной среде (приводя к конечным концентрациям ADC от 20 мкг/мл до 0,88 нг/мл). Еще через 4 дня планшеты удаляли из инкубатора и уравновешивали до комнатной температуры. Приблизительно через 30 минут 50 мкл удаляли из каждой лунки и затем в каждую лунку добавляли 50 мкл люминесцентного раствора CellTiter-Glo® 2.0 (Promega, G9423). После встряхивания планшетов при 750 об/мин в течение 5 минут с последующей 20-мин инкубацией без встряхивания измеряли люминесценцию на Tecan Infinity F200 планшет-ридере, с временем интегрирования 1 сек на лунку. Кривые люминесценции против концентрации ADC (нг/мл) получали с использованием программы Graphpad Prism. IC50 значения, определенные с использованием встроенной ʺlog(ингибитор) vs. ответ -- Переменный наклон (четыре параметра)ʺ функции определения IC50 программы Prism, представлены в таблице 9.
In vitro клеточный киллинг различных человеческий раковых клеток при помощи анти-ROR1 ADCs, а также изотипического контроля (IC50 значения представлены в нг/мл)
Фиг. 12 показывает кривые доза-ответ in vitro анализов киллинга клеток на MDA-MB-468 и HS 578T клетках с использованием ADCs Таблицы 9. Как показано в Таблице выше и на Фиг. 12, выбранные ADCs по изобретению обеспечивают киллинг некоторых человеческих раковых клеток, который выше, чем обеспечиваемый известными анти-ROR1 антителами 2A2 и R12.
Пример 16. In vitro анализы цитотоксичности ADCs на основе гуманизированных анти-ROR1 антител на 697 клетках
Цитотоксичность ADCs на основе гуманизированных анти-ROR1 антител huXBR1-402-3-G5-PNU (adc456), huXBR1-402-8-G5-PNU (adc457), huXBR1-402-15-G5-PNU (adc458), huXBR1-402-17-G5-PNU (adc459), huXBR1-402-19-G5-PNU (adc460) и huXBR1-402-26-G5-PNU (adc461) исследовали с использованием человеческих 697 клеток. Tras-G5-PNU (adc286) использовали в качестве изотипического контроля.
Для этого 5×104 697 клеток на лунку высевали в 96-луночные планшеты (за исключением краевых лунок, которые содержали воду) в 75 мкл RPMI, дополненной 10% по объему FCS, 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-Глутамина, и выращивали при 37°C в увлажненном инкубаторе в атмосфере 7,5% CO2. После 1-дневной инкубации каждый ADC добавляли в двух параллельных анализах в соответствующие лунки в количестве 25мкл 3,5-кратных серийных разведений в питательной среде (приводя к конечным концентрациям ADC от 20 мкг/мл до 0,88 нг/мл). Еще через 4 дня планшеты удаляли из инкубатора и уравновешивали до комнатной температуры. Приблизительно через 30 минут 50 мкл удаляли из каждой лунки и затем в каждую лунку добавляли 50 мкл люминесцентного раствора CellTiter-Glo® 2.0 (Promega, G9243). После встряхивания планшетов при 450 об/мин в течение 5 минут с последующей 10-минутной инкубацией без встряхивания измеряли люминесценцию на Tecan Spark 10M планшет-ридере, с временем интегрирования 250 мсек на лунку. Кривые люминесценции против концентрации ADC (нг/мл) получали с использованием программы Graphpad Prism. IC50 значения, определенные с использованием встроенной ʺlog(ингибитор) vs. ответ -- Переменный наклон (четыре параметра)ʺ функции определения IC50 программы Prism, представлены в таблице 10.
In vitro клеточный киллинг 697 клеток не-гуманизированными и гуманизированными анти-ROR1 или изотипическими контрольными ADCs (IC50, нг/мл)
Фиг. 25 показывает кривые доза-ответ in vitro анализов клеточного киллинга на 697 клетках с использованием гуманизированных ADCs Таблицы 10. Как показано в Таблице выше и на Фиг. 25, гуманизированные ADCs более эффективны по сравнению с ADCs, полученными с исходными, не-гуманизированными mAb.
Пример 17. In vivo фармакокинетическое исследование анти-ROR1 XBR1-402 антитела и XBR1-402-G5-PNU ADC у мышей штамма CD-1 дикого типа
Следующие исследование осуществляли в ABPRO (Burlington, MA, USA). Самок CD-1 мышей (по меньшей мере 6-недельного возраста в начале исследования, от Taconic Biosciences, Germantown, NY, USA), которых размещали группами до 5 животных на клетку, рандомизировали по массе в две группы по 9 животных в каждой. Одна группа получала, путем внутривенного введения, одну дозу 1 мг/кг XBR1-402 (mAb202), тогда как вторая группа получала одну дозу 1 мг/кг XBR1-402-G5-PNU (adc409). Пробы крови после введения дозы собирали в соответствии с протоколом Таблицы 11 в подгруппах, состоящих из 3 животных каждая. Пробы крови брали путем прокола ланцетом подчелюстной вены (собирая приблизительно 200 мкл), за исключением сбора крови в день 21, который осуществляли путем сердечной пукнции (собирая приблизительно 600 мкл). Образцы от подгрупп объединяли для каждой точки времени. Плазму отделяли путем центрифугирования крови при 1500g в течение 10 минут и хранили в старильных криопробирках при -80°C до осуществляемого на месте анализа методом ELISA.
Используемый в фармакокинетическом исследовании протокол взятия проб крови
(количество животных)
(количество животных)
Плазму отделяли от крови центрифугированием при 1500g в течение 10 минут и переносили в стерильные криопробирки для хранения при -80°C до анализа ELISA. In vivo стабильность mAb XBR1-402 и ADC XBR1-402-G5-PNU оценивали при помощи ELISA. Серии разведений образцов ADC в сыворотке захватывали на ELISA планшетах, покрытых 2 мкг/мл мышиного анти-PNU mAb (получено на месте путем иммунизации мышей конъюгатом человеческий IgG-PNU и скрининга с BSA-PNU конъюгатом) для связывания ADC, или анти-человеческим Fc F(ab')2 (Jackson Immunoresearch) для связывания суммарных IgG, и осуществляли детекцию с использованием 1:2500 разведения HRP-конъюгированного анти-человеческого IgG F(ab')2 (Jackson Immunoresearch). Концентрации в сыворотке ADC и суммарных IgGs рассчитывали из полу-максимальных значений титрований образца путем сравнения с образцом такого же ADC известной концентрации.
Данные на Фиг. 20 показывает высокую стабильность ADC по изобретению.
Пример 18. Оценка in vivo активности ADCs в диссеминированной модели ксенотрансплантата человеческой клеточной линии острого лимфоцитарного лейкоза (ALL) 697 у NOD-SCID мышей
Следующее исследование осуществляли в Pipeline Biotech (Trige, Denmark). Эффективность XBR1-402-G5-PNU (adc288) и гуманизированного hu2A2-G5-PNU (adc287; на основании неопубликованной PCT/EP2016/076244) сравнивали в диссеминированной модели ксенотрансплантата у самок NOD-SCID мышей, которым вводили hROR1-положительные 697 ALL клетки. ADC на основе HER2-специфического антитела Трастузумаба, Tras-G5-PNU (adc286), служил в качестве отрицательного изотипического контрольного ADC.
Мышей 9-недельного возраста, каждая из которых имела массу тела по меньшей мере 20 г, инокулировали 697 опухолевыми клетками (5×106 клеток/животное, в 200 мкл PBS) в день испытания 0. Каждый ADC, сформулированный в PBS, вводили (в/в) группе из 6 мышей при 1,0 мг/кг в дни 7, 14 и 21, с введением мышиного IgG (Jackson ImmunoResearch, 015-000-003), 30 мг/кг) за 20 часов до каждого введения ADC. Любое животное, демонстрирующее клинические признаки умеренной боли, умеренного стресса, любую степень страдания или любые клинические признаки, превышающие пределы специфических для исследования гуманных конечных точек, согласно европейскому и датскому законодательству о животных в экспериментальных исследованиях, подвергалось эвтаназии. Кровь (90 мкл в EDTA пробирках) собирали из хвостовой вены каждого животного в дни 12 и 19. Плазму отделяли путем центрифугирования крови в соответствии со стандартными процедурами. Образцы из групп обработки объединяли для каждой точки времени.
Серии разведений образцов ADC в плазме захватывали на ELISA планшетах, покрытых 2 мкг/мл мышиного анти-PNU mAb (получено на месте путем иммунизации мышей конъюгатом человеческий IgG-PNU и скрининга с BSA-PNU конъюгатом) для связывания ADC, или анти-человеческим Fc F(ab')2 (Jackson Immunoresearch, 109-006-098) для связывания общего IgG, и осуществляли детекцию с использованием 1:2500 разведения HRP-конъюгированного анти-человеческого IgG F(ab')2 (Jackson Immunoresearch, 709-035-149). Концентрации в сыворотке ADC и суммарных IgG рассчитывали из полу-максимальных значений титрований образца путем сравнения с образцом такого же ADC известной концентрации.
Анализ данных осуществляли с использованием программы PRISM. Мыши, которых обрабатывали при помощи XBR1-402-G5-PNU ADC по изобретению, показали пролонгированное выживание по сравнению с теми, которых обрабатывали сравнительным ROR1-таргетирующим hu2A2-G5-PNU ADC, или с изотипическим контрольным ADC (Фиг. 15, панель A). Кроме того, концентрация XBR1-402-G5-PNU ADC остается более высокой в плазме, чем концентрация известного или изотипического контрольного ADC (Фиг. 15, панель B).
Пример 19. Оценка in vivo эффективности анти-ROR1 ADCs в ортотопических моделях рака молочной железы, установленных с hROR1-сверхэкспрессирующими EMT-6 клетками
ADC на основе нового анти-ROR1 антитела XBR1-402-G5-PNU (adc262) сравнивали с R12-G5-PNU (adc327), ADC на основе известного антитела R12, и изотипическими контрольными Tras-G5-PNU (adc286) ADCs в следующей мышиной модели в соответствии с протоколом испытания Таблицы 13.
Ортотопические модели рака молочной железы, используемые для оценки ADCs
Протоколы, используемые для оценки анти-ROR1 XBR1-402-G5-PNU ADC в ортотопических моделях рака молочной железы с ортотопически имплантированными EMT6 клетками, сверхэкспрессирующими ROR1 (клон (cl 14))
Фиг. 18 показывает объем опухоли, развившейся в ходе исследования у каждой из 6 индивидуальных мышей. Ортотопические модели рака молочной железы, установленные с клетками рака молочной железы с высокой экспрессией hROR1, отвечают существенным образом на лечение с использованием ADC по изобретению, в отличие от неотвечающей ROR1-отрицательной WT EMT-6 модели. Замечательный ответ ADC по изобретению также демонстрирует существенное улучшение по сравнению со сравнительным ADC на основе известного R12 ROR1-таргетирующего антитела.
Пример 20. Анализ экспрессии hROR1 в полученных от пациента-человека лизатах опухолей методом вестерн-блоттинга
Лизаты опухолей моделей ксенотрансплантатов (PDX), полученных от человека (Charles River, Freiburg, Germany), и лизат высоко ROR1-положительной иммортализованной клеточной линии (Kasumi-2) в качестве контроля, а также лизат иммортализованных A549 раковых клеток исследовали на экспрессию hROR1 белка вестерн-блоттингом. Для этого лизат денатурировали путем смешивания с 5x SDS-PAGE загрузочным буфером (250 мМ Tris-HCl pH 6,8, 10% SDS, 30% Глицерина, 5% μ-меркаптоэтанола) и нагревания до 99°C в течение 5 минут. После разделения при помощи SDS-PAGE белки переносили на PVDF мембраны с использованием системы переноса eblot (Genscript). Мембраны затем инкубировали при 4°C в течение ночи с коммерчески доступным поликлональным кроличьим анти-ROR1 антителом 4102 (Cell Signaling Technology, Danvers, USA), разведенным 1:200 в TBST (20 мМ Tris-HCl, 150 мМ NaCl, pH 7,6, 0,1% Tween-20), содержащем 10% лошадиную сыворотку (Amimed, Bioconcept, Switzerland). После двух промывок при помощи TBST мембраны инкубировали с HRP-конъюгированными анти-кроличьими вторичными антителами (WesternSure HRP козлиные-анти-кроличьи IgG, LiCor, Lincoln, USA) в течение 1 часа при комнатной температуре. Сигналы получали путем инкубации мембран в люминесцентном субстрате (SuperSignal West Femto (34094, Thermo Fisher) и визуализации с использованием цифрового считывающего устройства для вестерн-блоттинга (LiCor, Lincoln, USA). Детекция конститутивного гена GAPDH служила в качестве контроля загрузки.
hROR1 статус в лизатах опухолей Примера 22
Фиг. 21 показывает Вестерн-блот анализ лизатов опухолей человека, а также лизатов из иммортализованных раковых клеток.
Пример 21. Анализ in vivo эффективности ADC XBR1-402-G5-PNU в моделях ксенотрансплантатов опухолей, полученных от пациента
Следующее исследование осуществляли в Charles River (в Oncotest GmbH, Freiburg, Germany).
Таблица 15. Модели ксенотрансплантатов опухолей, полученных от пациента, использовали для оценки анти-ROR1 XBR1-402-G5-PNU ADC
Модели ксенотрансплантатов опухолей, полученных от пациента, используемые для оценки анти-ROR1 XBR1-402-G5-PNU ADC
XBR1-402-G5-PNU (adc409) ADC, ранее подтвержденный как не содержащий эндотоксина, исследовали в каждой модели в соответствии со следующим протоколом испытания:
Протоколы, используемые для оценки анти-ROR1 XBR1-402-G5-PNU ADC в моделях ксенотрансплантатов опухолей, полученных от пациента
Мышам подкожно имплантировали PDX материал (либо односторонне (PXF 111, RXF 486, CX 533), либо двусторонне (SXFS 1407, PXF 541). Мышей рандомизировали, когда опухоли достигали размера 100 см3, и обрабатывали ADC при 1мг/кг или носителем в общей сложности 3 раза. Объемы опухолей определяли путем измерения циркулем и массу тела регистрировали два раза в неделю. Мышей умерщвляли при достижении опухолевой нагрузки 2000 мм3 (односторонняя) или 1700 мм3 (двусторонняя) или при существенной потере массы тела (в целом более чем 30%, или больше чем 20% за два дня).
Фиг. 22 показывает изменение объема опухоли в процессе исследования. Опухолевые ксенотрансплантаты, установленные с полученным от пациента опухолевым материалом, имеющие высокую экспрессию hROR1, существенным образом отвечали на лечение при помощи ADC по изобретению.
Пример 22. In vitro активность XBR1-402 CAR-T
CAR-T клетки на основе XBR1-402 были сконструированы с использованием способов, ранее описанных для ROR1-таргетирующих mAbs R11 и R12 (Hudecek, M., Lupo-Stanghellini, M. T., Kosasih, P. L., Sommermeyer, D., Jensen, M. C., Rader, C. and Riddell, S. R. (2013) Receptor affinity and extracellular domain modifications affect tumor recognition by ROR1-specific chimeric antigen receptor T cells. Clin. Cancer Res. 19, 3153-3164). Осуществляли лентивирусную трансдукцию ex vivo размноженных первичных человеческих CD8+ CD62L+ T клеток при помощи XBR1-402 или R12-происходящих CARs, содержащих CD3ζ и 4-1BB сигнальные домены и короткий спейсер. Трансдуцированные T клетки очищали через tEGFR при помощи FACS и их фенотип оценивали за день до функциональных анализов. CD8+ чистота варьировалась в пределах 97% - 99%, экспрессия tEGFR варьировалась в пределах 95% - 99%. После 72 часов совместного культивирования с ROR1-положительными или ROR1-отрицательными человеческими клетками рака молочной железы CFSE-окрашенные CD8+ CD62L+ клетки анализировали методом проточной цитометрии с обнаружением мишень-зависимой пролиферации XBR1-402 и R12 CAR-T (Фиг. 26; верхняя левая панель). Концентрации IFNγ и IL2 в супернатанте, взятом после 24 часов ко-культивирования, измеряли методом ELISA (Фиг. 26; верхняя правая панель). Селективную цитотоксичность измеряли с использованием люциферазного анализа цитотоксичности после 11 часов ко-культивирования с ROR1-положительными и ROR1-отрицательными клетками (Фиг. 26; нижняя панель).
Такие же эксперименты повторяли для сравнения XBR1-402 CAR-T с коротким спейсером с XBR1-402 CAR-T с длинным спейсером. После 72 часов ко-культивирования с ROR1-положительными или ROR1-отрицательными клетками рака молочной железы человека CFSE-окрашенные CD8+ CD62L+ клетки анализировали методом проточной цитометрии, обнаружив более сильную пролиферацию XBR1-402 CAR-T с коротким спейсером (Фиг. 27; верхняя панель). Селективную цитотоксичность измеряли с использованием люциферазного анализа цитотоксичности после 11 часов ко-культивирования с ROR1-положительными и ROR1-отрицательными клетками, и селективная цитотоксичность была определена для обоих XBR1-402 CAR-T с коротким и длинным спейсером (Фиг. 27; нижняя панель).
Пример 23. Анализ специфичности XBR1-402
Фиг. 28 представляет обзор Retrogenix Cell Microarray Platform. Первичный скрининг: Очищенное химерное кроличье/человеческое IgG1 XBR1-402 и очищенное химерное кроличье/человеческое IgG1 XBR2-401 объединяли до концентрации 2 мкг/мл каждого. Пул скринировали на связывание против фиксированных HEK293 клеток/микропрепаратов, индивидуально экспрессирующих 4336 человеческих плазменных мембранных белков (13 наборов микропрепаратов; n=2 микропрепарата в каждом наборе). Эффективности всех трансфекций превышали минимальный порог. Для детекции использовали анти-человеческое IgG Fc антитело AlexaFluor647. Первичные лучшие образцы (дублированные пятна) идентифицировали путем анализа флуоресценции (AlexaFluor647 и ZsGreen1) на ImageQuant. Векторы, кодирующие все лучшие образцы, секвенировали для подтверждения их точной идентичности. Подтверждающий скрининг: Векторы, кодирующие все лучшие образцы, плюс контрольные векторы наносили в виде пятен в двух повторах на новые микропрепараты и использовали для обратной трансфекции человеческих HEK293 клеток, как ранее. Эффективности всех трансфекций превышали минимальный порог. Идентичные фиксированные микропрепараты обрабатывали каждым из двух испытываемых антител (XBR1-402 и XBR2-401) индивидуально, плюс положительными и отрицательными контролями (n=2 микропрепарата на обработку). Микропрепараты анализировали, как указано выше (Фиг. 29).
Хотя вышеприведенное изобретение было описано достаточно подробно при помощи иллюстрации и примеров для его четкого понимания, специалисту в данной области техники будет очевидно, в свете раскрытия настоящего изобретения, что возможны некоторые изменения и модификации без отступления от сущности или объема прилагаемой формулы изобретения.
Все публикации, базы данных, последовательности GenBank, патенты и патентные заявки, на которые ссылаются в настоящем описании, включены в настоящую заявку посредством ссылки, как если бы каждый ссылочный документ был специально и индивидуально указан для включения в качестве ссылки.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> НБЕ-ТЕРАПЬЮТИКС АГ
ДЗЕ СКРИППС РИСЕРЧ ИНСТИТЬЮТ
<120> КОМПОЗИЦИИ АНТИТЕЛ К ROR1 И СООТВЕТСТВУЮЩИЕ СПОСОБЫ
<130> 1713.1PC
<140>
<141> 2017-01-20
<150> 62/280,843
<151> 2016-01-20
<160> 161
<170> PatentIn version 3.5
<210> 1
<211> 117
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 1
Gln Glu Gln Gln Lys Glu Ser Gly Gly Gly Leu Phe Lys Pro Thr Asp
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Asp Ile Ser Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Asn Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Gly Ile Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Ser Arg Ser Thr Ile Thr Arg Asn Thr Asn Leu Asn Thr Val Thr Leu
65 70 75 80
Lys Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Asp His Pro Thr Tyr Gly Met Asp Leu Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 2
<211> 118
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 2
Gln Glu Gln Leu Glu Glu Ser Gly Gly Gly Leu Val Thr Pro Gly Gly
1 5 10 15
Ser Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Thr Ile Ser Thr Tyr
20 25 30
His Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Ala Gly Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Val
50 55 60
Asn Gly Arg Phe Thr Ile Ser Ser Asn Thr Thr Gln Asn Thr Val Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Thr Val Ala Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Asp His Pro Ser Tyr Gly Met Asp Leu Trp Gly Pro Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 3
<211> 125
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 3
Gln Glu Gln Leu Lys Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Lys Ala Ser Gly Phe Asp Leu Ser Asn Tyr
20 25 30
Gly Val Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asp Pro Thr Phe Asp Tyr Thr Tyr Tyr Ala Ser Trp Val
50 55 60
Asn Gly Arg Phe Ser Ile Ser Arg Glu Asn Thr Gln Asn Thr Val Ser
65 70 75 80
Leu Gln Ile Asn Ser Leu Thr Pro Ala Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Trp Val Tyr Gly Val Asp Asp Tyr Gly Asp Gly Asn Trp Leu
100 105 110
Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Ile Ser Ser
115 120 125
<210> 4
<211> 117
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 4
Gln Glu Gln Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr
1 5 10 15
Pro Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Phe Leu Ser Ser Tyr
20 25 30
Tyr Val Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Gly Thr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Ser Arg Ser Thr Ile Thr Arg Asn Thr Asn Leu Ser Thr Val Thr Leu
65 70 75 80
Lys Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Asp His Pro Asn Tyr Gly Met Asp Leu Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 5
<211> 113
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 5
Gln Ser Leu Glu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Glu Ser
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Arg Asn Gly
20 25 30
Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Ile Ile Thr Ser Ser Gly Asp Lys Tyr Tyr Ala Thr Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val Asp Leu Lys Met
65 70 75 80
Thr Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly
85 90 95
Thr Val Ser Ser Asp Ile Trp Gly Pro Gly Thr Leu Val Thr Ile Ser
100 105 110
Ser
<210> 6
<211> 115
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 6
Gln Glu Gln Leu Lys Glu Ser Gly Arg Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Lys Ala Ser Gly Phe Asp Phe Ser Gly Trp
20 25 30
Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Thr Ile Gly Thr Thr Lys Gly Arg Thr Tyr Tyr Ala Ser Trp Val
50 55 60
Asn Gly Arg Phe Thr Ile Ser Ser Asn Ala Gln Asn Thr Val Asp Leu
65 70 75 80
Gln Met Asn Ser Leu Thr Ala Ala Asp Arg Ala Thr Tyr Phe Cys Val
85 90 95
Arg Gly Ser Asp Tyr Phe Asp Leu Trp Gly Pro Gly Thr Leu Val Thr
100 105 110
Ile Ser Ser
115
<210> 7
<211> 120
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 7
Gln Glu Gln Leu Val Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Gly
1 5 10 15
Ser Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Tyr Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Thr Ile Tyr Gly Gly Ser Gly Ser Thr Trp Tyr Ala Ser Trp Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Thr Ala Val Asp Leu Lys
65 70 75 80
Ile Thr Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg
85 90 95
Ser Tyr Pro Gly Trp Thr Thr Glu Pro Tyr Phe Asp Ile Trp Gly Pro
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 8
<211> 121
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 8
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Ile Asp Leu Ser Arg Tyr Ala
20 25 30
Val Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Val Ile Ser Gly Ser Gly Thr Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly Trp
85 90 95
Ala Thr Asp Gly Ile Phe Asp Tyr Asp Asp Thr Phe Asn Leu Trp Gly
100 105 110
Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 9
<211> 115
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 9
Gln Glu Gln Leu Val Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Gly
1 5 10 15
Ser Leu Thr Leu Thr Cys Ala Ala Ser Gly Phe Ser Pro Ser Ser Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Tyr Val Asp Ser Gly Ser Ala Tyr Tyr Ala Ser Trp Ala
50 55 60
Lys Ser Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val Asp Leu
65 70 75 80
Lys Met Thr Ser Leu Thr Thr Glu Asp Thr Ala Ile Tyr Phe Cys Ala
85 90 95
Arg Asp Gly Trp Gly Ser Asp Val Trp Gly Pro Gly Thr Leu Val Thr
100 105 110
Ile Ser Ser
115
<210> 10
<211> 113
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 10
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Tyr Tyr
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Thr Ile Ser Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Val Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Met Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Asp Leu
85 90 95
Asp Tyr Asp Met Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser
100 105 110
Ser
<210> 11
<211> 117
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 11
Gln Glu Gln Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr
1 5 10 15
Pro Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Ser Asn Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Ile Ile Ser Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val Asp Leu Lys
65 70 75 80
Ile Thr Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg
85 90 95
Leu Tyr Val Ser Asp Ser Thr Ala Asn Leu Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 12
<211> 114
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 12
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Tyr Trp
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Ala Ile Tyr Gly Ser Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Asp Val
85 90 95
His Ser Thr Ala Thr Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Ile
100 105 110
Ser Ser
<210> 13
<211> 115
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 13
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Ser Asp Tyr Tyr
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Glu Gly Leu Glu Trp Ile Gly
35 40 45
Thr Ile Tyr Gly Gly Ser Gly Asn Thr Trp Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile
65 70 75 80
Thr Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Asp
85 90 95
Phe Gly Leu Ser Thr Gly Gly Leu Trp Gly Pro Gly Thr Leu Val Thr
100 105 110
Ile Ser Ser
115
<210> 14
<211> 107
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 14
Ser Tyr Glu Leu Thr Gln Leu Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Glu Gly Asn Asn Ile Gly Ser Lys Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Leu Ala Pro Gly Leu Leu Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asp Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ala Tyr Val
85 90 95
Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly
100 105
<210> 15
<211> 109
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 15
Ser Tyr Glu Leu Thr Gln Leu Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Ser Ile Gly Ser Lys Ala Val
20 25 30
Asn Trp Tyr Gln Gln Lys Pro Gly Leu Ala Pro Gly Leu Leu Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Ala Ser
50 55 60
Asn Ser Gly Asp Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Leu Trp Asp Ser Ser Ala Val Ala
85 90 95
Tyr Val Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly
100 105
<210> 16
<211> 112
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 16
Gln Phe Val Leu Thr Gln Ser Pro Ser Val Ser Ala Ala Leu Gly Ala
1 5 10 15
Ser Ala Lys Leu Thr Cys Thr Leu Ser Ser Ala His Lys Thr Tyr Thr
20 25 30
Ile Asp Trp Tyr Gln Gln Gln Gln Gly Glu Ala Pro Arg Tyr Leu Met
35 40 45
Glu Leu Lys Ser Asp Gly Ser Tyr Thr Lys Gly Thr Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Ser Ser Gly Ala Asp Arg Tyr Leu Ile Ile Pro
65 70 75 80
Ser Val Gln Ala Asp Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Asp Tyr
85 90 95
Ser Gly Gly Tyr Val Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly
100 105 110
<210> 17
<211> 109
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 17
Ser Tyr Glu Leu Thr Gln Leu Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Ser Ile Gly Ser Lys Ala Val
20 25 30
Asn Trp Tyr Gln Gln Lys Pro Gly Leu Ala Pro Gly Leu Leu Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asp Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Leu Trp Asp Ser Ser Thr Gly Ala
85 90 95
Tyr Ala Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly
100 105
<210> 18
<211> 112
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 18
Glu Leu Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ala Ser Gln Ser Val Tyr Gly Asn
20 25 30
Asn Glu Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Arg Ala Ser Ile Leu Thr Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asn Val
65 70 75 80
Gln Arg Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Gly Tyr Val Ser
85 90 95
Gln Ser Tyr Arg Ala Ala Phe Gly Gly Gly Thr Glu Leu Glu Ile Leu
100 105 110
<210> 19
<211> 109
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 19
Ser Tyr Glu Leu Thr Gln Leu Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Ser Ile Gly Ser Lys Ala Val
20 25 30
Asn Trp Tyr Gln Gln Lys Pro Gly Leu Ala Pro Gly Leu Leu Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asp Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Leu Trp Asp Ser Ser Ala Gly Ala
85 90 95
Tyr Val Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly
100 105
<210> 20
<211> 111
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 20
Gln Phe Val Leu Thr Gln Pro Gln Ser Val Ser Gly Ser Leu Gly Gln
1 5 10 15
Thr Val Ser Ile Ser Cys Asn Arg Asp Ser Gly Asn Ile Glu Asp Tyr
20 25 30
Tyr Val His Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Thr Thr Val
35 40 45
Ile Tyr Asn Asp Asp Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Ile Asp Ser Thr Ser Asn Ser Ala Ser Leu Thr Ile Thr Gly
65 70 75 80
Leu Leu Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Leu Ser Ser Asp Ser
85 90 95
Ser Ala Tyr Val Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly
100 105 110
<210> 21
<211> 107
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 21
Ser Tyr Glu Leu Thr Gln Leu Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asp Ser Ile Glu Ser Lys Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Leu Ala Pro Gly Leu Leu Val Tyr
35 40 45
Asn Asp Asp Glu Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asp Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Leu Trp Asp Gly Ser Ala Tyr Val
85 90 95
Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly
100 105
<210> 22
<211> 109
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 22
Ser Tyr Glu Leu Thr Gln Leu Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Ser Ile Gly Ser Lys Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Leu Ala Pro Gly Leu Leu Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asp Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Leu Trp Asp Ser Ser Ala Gly Ala
85 90 95
Tyr Val Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly
100 105
<210> 23
<211> 107
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 23
Ser Tyr Glu Leu Thr Gln Leu Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Ser Ile Gly Ser Lys Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Leu Ala Pro Gly Leu Leu Ile Tyr
35 40 45
Asn Asp Asp Glu Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asp Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Leu Trp Asp Asn Asn Ala Ala Val
85 90 95
Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly
100 105
<210> 24
<211> 110
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 24
Asp Ile Val Leu Thr Gln Thr Pro Ser Ser Thr Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Gln Ser Ile Asp Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Glu Tyr Ile Leu Thr Ile Ser Gly Val Gln Arg
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Gly Tyr Pro Asn Tyr Phe
85 90 95
His Arg Thr Ala Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 25
<211> 109
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 25
Ser Tyr Glu Leu Thr Gln Leu Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ala Val
20 25 30
Asn Trp Tyr Gln Gln Lys Pro Gly Leu Ala Pro Gly Leu Leu Ile Tyr
35 40 45
Asn Asp Asp Glu Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asp Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Leu Trp Asp Ser Ser Ala Gly Ala
85 90 95
Tyr Val Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly
100 105
<210> 26
<211> 109
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 26
Ser Tyr Glu Leu Thr Gln Leu Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Ser Ile Gly Ser Lys Ala Val
20 25 30
Asn Trp Tyr Gln Gln Lys Pro Gly Leu Ala Pro Gly Leu Leu Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asp Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Leu Trp Asp Ser Ser Ala Arg Ala
85 90 95
Phe Val Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly
100 105
<210> 27
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 27
Ser Tyr Tyr Met Ser
1 5
<210> 28
<211> 16
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 28
Ala Ile Gly Ile Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys Ser
1 5 10 15
<210> 29
<211> 9
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 29
Asp His Pro Thr Tyr Gly Met Asp Leu
1 5
<210> 30
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 30
Thr Tyr His Met Ser
1 5
<210> 31
<211> 17
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 31
Ser Ile Tyr Ala Gly Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Val Asn
1 5 10 15
Gly
<210> 32
<211> 9
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 32
Asp His Pro Ser Tyr Gly Met Asp Leu
1 5
<210> 33
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 33
Asn Tyr Gly Val Ser
1 5
<210> 34
<211> 17
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 34
Tyr Ile Asp Pro Thr Phe Asp Tyr Thr Tyr Tyr Ala Ser Trp Val Asn
1 5 10 15
Gly
<210> 35
<211> 16
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 35
Trp Val Tyr Gly Val Asp Asp Tyr Gly Asp Gly Asn Trp Leu Asp Leu
1 5 10 15
<210> 36
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 36
Ser Tyr Tyr Val Ser
1 5
<210> 37
<211> 16
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 37
Ala Ile Gly Thr Gly Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Lys Ser
1 5 10 15
<210> 38
<211> 9
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 38
Asp His Pro Asn Tyr Gly Met Asp Leu
1 5
<210> 39
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 39
Arg Asn Gly Met Thr
1 5
<210> 40
<211> 16
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 40
Ile Ile Thr Ser Ser Gly Asp Lys Tyr Tyr Ala Thr Trp Ala Lys Gly
1 5 10 15
<210> 41
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 41
Gly Thr Val Ser Ser Asp Ile
1 5
<210> 42
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 42
Gly Trp Tyr Met Thr
1 5
<210> 43
<211> 17
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 43
Thr Ile Gly Thr Thr Lys Gly Arg Thr Tyr Tyr Ala Ser Trp Val Asn
1 5 10 15
Gly
<210> 44
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 44
Gly Ser Asp Tyr Phe Asp Leu
1 5
<210> 45
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 45
Ser Tyr Tyr Met Gly
1 5
<210> 46
<211> 17
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 46
Thr Ile Tyr Gly Gly Ser Gly Ser Thr Trp Tyr Ala Ser Trp Val Lys
1 5 10 15
Gly
<210> 47
<211> 13
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 47
Ser Tyr Pro Gly Trp Thr Thr Glu Pro Tyr Phe Asp Ile
1 5 10
<210> 48
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 48
Arg Tyr Ala Val Asn
1 5
<210> 49
<211> 16
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 49
Val Ile Ser Gly Ser Gly Thr Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 50
<211> 16
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 50
Gly Trp Ala Thr Asp Gly Ile Phe Asp Tyr Asp Asp Thr Phe Asn Leu
1 5 10 15
<210> 51
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 51
Ser Tyr Trp Met Ser
1 5
<210> 52
<211> 17
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 52
Ala Ile Tyr Val Asp Ser Gly Ser Ala Tyr Tyr Ala Ser Trp Ala Lys
1 5 10 15
Ser
<210> 53
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 53
Asp Gly Trp Gly Ser Asp Val
1 5
<210> 54
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 54
Ser Tyr Tyr Met Ser
1 5
<210> 55
<211> 16
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 55
Thr Ile Ser Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Val Lys Gly
1 5 10 15
<210> 56
<211> 10
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 56
Leu Tyr Val Ser Asp Ser Thr Ala Asn Leu
1 5 10
<210> 57
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 57
Asn Tyr Tyr Met Ser
1 5
<210> 58
<211> 16
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 58
Ile Ile Ser Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 59
<211> 10
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 59
Leu Tyr Val Ser Asp Ser Thr Ala Asn Leu
1 5 10
<210> 60
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 60
Ser Tyr Trp Met Ser
1 5
<210> 61
<211> 16
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 61
Ala Ile Tyr Gly Ser Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 62
<211> 9
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 62
Asp Val His Ser Thr Ala Thr Asp Leu
1 5
<210> 63
<211> 5
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 63
Asp Tyr Tyr Met Ser
1 5
<210> 64
<211> 17
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 64
Thr Ile Tyr Gly Gly Ser Gly Asn Thr Trp Tyr Ala Thr Trp Ala Lys
1 5 10 15
Gly
<210> 65
<211> 9
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 65
Asp Phe Gly Leu Ser Thr Gly Gly Leu
1 5
<210> 66
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 66
Glu Gly Asn Asn Ile Gly Ser Lys Ala Val His
1 5 10
<210> 67
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 67
Asp Asp Asp Glu Arg Pro Ser
1 5
<210> 68
<211> 9
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 68
Gln Val Trp Asp Ser Ser Ala Tyr Val
1 5
<210> 69
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 69
Gly Gly Asn Ser Ile Gly Ser Lys Ala Val Asn
1 5 10
<210> 70
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 70
Asp Asp Asp Glu Arg Pro Ser
1 5
<210> 71
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 71
Gln Leu Trp Asp Ser Ser Ala Val Ala Tyr Val
1 5 10
<210> 72
<211> 12
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 72
Thr Leu Ser Ser Ala His Lys Thr Tyr Thr Ile Asp
1 5 10
<210> 73
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 73
Leu Lys Ser Asp Gly Ser Tyr Thr Lys Gly Thr
1 5 10
<210> 74
<211> 9
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 74
Gly Thr Asp Tyr Ser Gly Gly Tyr Val
1 5
<210> 75
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 75
Gly Gly Asn Ser Ile Gly Ser Lys Ala Val Asn
1 5 10
<210> 76
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 76
Asp Asp Asp Glu Arg Pro Ser
1 5
<210> 77
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 77
Gln Leu Trp Asp Ser Ser Thr Gly Ala Tyr Ala
1 5 10
<210> 78
<211> 13
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 78
Gln Ala Ser Gln Ser Val Tyr Gly Asn Asn Glu Leu Ala
1 5 10
<210> 79
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 79
Arg Ala Ser Ile Leu Thr Ser
1 5
<210> 80
<211> 12
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 80
Leu Gly Gly Tyr Val Ser Gln Ser Tyr Arg Ala Ala
1 5 10
<210> 81
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 81
Gly Gly Asn Ser Ile Gly Ser Lys Ala Val Asn
1 5 10
<210> 82
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 82
Asp Asp Asp Glu Arg Pro Ser
1 5
<210> 83
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 83
Gln Leu Trp Asp Ser Ser Ala Gly Ala Tyr Val
1 5 10
<210> 84
<211> 13
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 84
Asn Arg Asp Ser Gly Asn Ile Glu Asp Tyr Tyr Val His
1 5 10
<210> 85
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 85
Asn Asp Asp Gln Arg Pro Ser
1 5
<210> 86
<211> 9
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 86
Leu Ser Ser Asp Ser Ser Ala Tyr Val
1 5
<210> 87
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 87
Gly Gly Asp Ser Ile Glu Ser Lys Ala Val His
1 5 10
<210> 88
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 88
Asn Asp Asp Glu Arg Pro Ser
1 5
<210> 89
<211> 9
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 89
Gln Leu Trp Asp Gly Ser Ala Tyr Val
1 5
<210> 90
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 90
Gly Gly Asn Ser Ile Gly Ser Lys Ala Val His
1 5 10
<210> 91
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 91
Asp Asp Asp Glu Arg Pro Ser
1 5
<210> 92
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 92
Gln Leu Trp Asp Ser Ser Ala Gly Ala Tyr Val
1 5 10
<210> 93
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 93
Gly Gly Asn Ser Ile Gly Ser Lys Ala Val His
1 5 10
<210> 94
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 94
Asn Asp Asp Glu Arg Pro Ser
1 5
<210> 95
<211> 9
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 95
Gln Leu Trp Asp Asn Asn Ala Ala Val
1 5
<210> 96
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 96
Gln Ala Ser Gln Ser Ile Asp Ser Tyr Leu Ala
1 5 10
<210> 97
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 97
Gly Ala Ser Asn Leu Ala Ser
1 5
<210> 98
<211> 12
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 98
Leu Gly Gly Tyr Pro Asn Tyr Phe His Arg Thr Ala
1 5 10
<210> 99
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 99
Gly Gly Asn Asn Ile Gly Ser Lys Ala Val Asn
1 5 10
<210> 100
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 100
Asn Asp Asp Glu Arg Pro Ser
1 5
<210> 101
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 101
Gln Leu Trp Asp Ser Ser Ala Gly Ala Tyr Val
1 5 10
<210> 102
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 102
Gly Gly Asn Ser Ile Gly Ser Lys Ala Val Asn
1 5 10
<210> 103
<211> 7
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 103
Asp Asp Asp Glu Arg Pro Ser
1 5
<210> 104
<211> 11
<212> БЕЛОК
<213> Oryctolagus cuniculus
<400> 104
Gln Leu Trp Asp Ser Ser Ala Arg Ala Phe Val
1 5 10
<210> 105
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 105
atcctgtttc tcgtagctgc tgcaactgga gcacactccg cccggggcgc cgccgcccag 60
<210> 106
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 106
ccactcgatc ttctgggcct cgaagatgtc gttcaggccc tccatcttgt tcttctcctt 60
<210> 107
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 107
gctgggtacc ggcgcgccac catggactgg acttggagaa tcctgtttct cgtagctgct 60
<210> 108
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 108
gccggcctcg agtcagtgat ggtgatggtg gtgctcgtgc cactcgatct tctgggcctc 60
<210> 109
<211> 99
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 109
gctgggtacc ggcgcgccac catggactgg acttggagaa tcctgtttct cgtagctgct 60
gcaactggag cacactccgc ccggggcgcc gccgcccag 99
<210> 110
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 110
cggcctcgag tcagtgatgg tgatggtggt gctccatctt gttcttctcc tt 52
<210> 111
<211> 99
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 111
gctgggtacc ggcgcgccac catggactgg acttggagaa tcctgtttct cgtagctgct 60
gcaactggag cacactccga agtggaggtt ctggatccg 99
<210> 112
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 112
cggcctcgag tcagtgatgg tgatggtggt gccccatctt gctgctgtct cg 52
<210> 113
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 113
gaggaggagc tcactctcag gagcagcaga aggagtccgg g 41
<210> 114
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 114
cgatgggccc ttggtggagg ctgaagagac ggtgacgagg gtccctggcc cccagaggtc 60
<210> 115
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 115
gagaagcttg ttgctctgga tctctggtgc ctacgggtcc tatgagctga cacagctgcc 60
<210> 116
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 116
ggccatggct ggttgggcag c 21
<210> 117
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 117
ggtaccggcg cgccaccatg gactggactt ggagaatcct gtttctcgta gctgctgcaa 60
<210> 118
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 118
gccgctggtc agggctcctg 20
<210> 119
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 119
caggagccct gaccagcggc 20
<210> 120
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 120
ggcctcgagt catttacccg gagacaggga 30
<210> 121
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 121
tttctcgtag ctgctgcaac tggagcacac tcccagtcgc tggaggagtc cggg 54
<210> 122
<211> 54
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 122
Thr Thr Thr Cys Thr Cys Gly Thr Ala Gly Cys Thr Gly Cys Thr Gly
1 5 10 15
Cys Ala Ala Cys Thr Gly Gly Ala Gly Cys Ala Cys Ala Cys Thr Cys
20 25 30
Cys Cys Ala Gly Thr Cys Gly Thr Thr Gly Gly Ala Gly Gly Ala Gly
35 40 45
Thr Cys Cys Gly Gly Gly
50
<210> 123
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 123
tttctcgtag ctgctgcaac tggagcacac tcccagtcgt tggaggagtc cggg 54
<210> 124
<211> 36
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 124
Gly Gly Ala Gly Gly Gly Cys Gly Cys Cys Ala Gly Gly Gly Gly Gly
1 5 10 15
Ala Ala Gly Ala Cys Cys Gly Ala Thr Gly Gly Gly Cys Cys Cys Thr
20 25 30
Thr Gly Gly Thr
35
<210> 125
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 125
tttctcgtag ctgctgcaac tggagcacac tcctcctatg agctgacaca gctg 54
<210> 126
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 126
tttctcgtag ctgctgcaac tggagcacac tcctcctatg agctgacaca gctg 54
<210> 127
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 127
tttctcgtag ctgctgcaac tggagcacac tccgagctcg tgctgaccca gact 54
<210> 128
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 128
ggcctcgagt tatgaacatt ctgtaggggc 30
<210> 129
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 129
ggcctcgagt taacactctc ccctgttgaa 30
<210> 130
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 130
Gln Val Gln Leu Arg Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Asp Ile Ser Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Gly Ile Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Ser Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp His Pro Thr Tyr Gly Met Asp Leu Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 131
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 131
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Asp Ile Ser Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Gly Ile Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Ser Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp His Pro Thr Tyr Gly Met Asp Leu Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 132
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 132
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Asp Ile Ser Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Gly Ile Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Ser Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp His Pro Thr Tyr Gly Met Asp Leu Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 133
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 133
Gln Val Gln Leu Arg Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Asp Ile Ser Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Gly Ile Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Ser Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp His Pro Thr Tyr Gly Met Asp Leu Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 134
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 134
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Asp Ile Ser Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Pro Pro Gly Asn Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Gly Ile Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Ser Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp His Pro Thr Tyr Gly Met Asp Leu Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 135
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 135
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Asp Ile Ser Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Pro Pro Gly Asn Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Gly Ile Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Ser Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp His Pro Thr Tyr Gly Met Asp Leu Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 136
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 136
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Glu Gly Asn Asn Ile Gly Ser Lys Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ala Tyr Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105
<210> 137
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 137
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ile Ile Thr Cys Glu Gly Asn Asn Ile Gly Ser Lys Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Leu Ser Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Val Ser Gly Thr Gln Asp Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ala Tyr Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105
<210> 138
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 138
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Glu Gly Asn Asn Ile Gly Ser Lys Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ala Tyr Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105
<210> 139
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 139
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Glu Gly Asn Asn Ile Gly Ser Lys Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ala Tyr Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105
<210> 140
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 140
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Glu Gly Asn Asn Ile Gly Ser Lys Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ala Tyr Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105
<210> 141
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 141
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Glu Gly Asn Asn Ile Gly Ser Lys Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Leu Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ala Tyr Val
85 90 95
Phe Gly Gly Gly Thr Gln Leu Thr Val Leu Gly
100 105
<210> 142
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<220>
<221> MISC_FEATURE
<222> (3)..(3)
<223> X is any amino acid residue
<400> 142
Leu Pro Xaa Ser Gly
1 5
<210> 143
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<220>
<221> MISC_FEATURE
<222> (3)..(3)
<223> X is any amino acid residue
<400> 143
Leu Pro Xaa Ala Gly
1 5
<210> 144
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<220>
<221> MISC_FEATURE
<222> (3)..(3)
<223> X is any amino acid residue
<400> 144
Leu Pro Xaa Thr Gly
1 5
<210> 145
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<220>
<221> MISC_FEATURE
<222> (3)..(3)
<223> X is any amino acid residue
<400> 145
Leu Ala Xaa Thr Gly
1 5
<210> 146
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 146
Asn Pro Gln Thr Gly
1 5
<210> 147
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<220>
<221> MISC_FEATURE
<222> (3)..(3)
<223> X is any amino acid residue
<400> 147
Leu Pro Xaa Thr
1
<210> 148
<211> 460
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 148
Glu Ile Gln Leu Gln Gln Ser Gly Pro Val Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Thr Ala Tyr
20 25 30
Asn Ile His Trp Val Arg Gln Ser His Gly Lys Arg Leu Glu Trp Ile
35 40 45
Gly Ser Phe Asp Pro Tyr Asp Gly Gly Ser Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Thr Thr Ala Tyr
65 70 75 80
Met His Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Trp Tyr Tyr Phe Asp Tyr Trp Gly His Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Leu Pro
435 440 445
Glu Thr Gly Gly Trp Ser His Pro Gln Phe Glu Lys
450 455 460
<210> 149
<211> 233
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 149
Asp Val Gln Ile Thr Gln Ser Pro Ser Tyr Leu Ala Ala Ser Pro Gly
1 5 10 15
Glu Thr Ile Thr Ile Asn Cys Arg Ala Ser Lys Ser Ile Ser Lys Tyr
20 25 30
Leu Ala Trp Tyr Gln Glu Lys Pro Gly Lys Thr Asn Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Ser Thr Leu Gln Ser Gly Ile Pro Ser Arg Phe Arg Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Met Tyr Tyr Cys Gln Gln His Asp Glu Ser Pro Tyr
85 90 95
Thr Phe Gly Glu Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Leu Pro Glu Thr Gly
210 215 220
Gly Trp Ser His Pro Gln Phe Glu Lys
225 230
<210> 150
<211> 462
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 150
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Glu Met His Trp Val Ile Gln Thr Pro Val His Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Ile Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Gly Tyr Tyr Asp Tyr Asp Ser Phe Thr Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
Leu Pro Glu Thr Gly Gly Trp Ser His Pro Gln Phe Glu Lys
450 455 460
<210> 151
<211> 233
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 151
Asp Ile Val Met Thr Gln Ser Gln Lys Ile Met Ser Thr Thr Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asn Val Asp Ala Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Met Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Asp Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Leu Pro Glu Thr Gly
210 215 220
Gly Trp Ser His Pro Gln Phe Glu Lys
225 230
<210> 152
<211> 462
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 152
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Leu
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Gly Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Thr Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Gly Tyr Tyr Asp Tyr Asp Ser Phe Thr Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Ser Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
Leu Pro Glu Thr Gly Gly Trp Ser His Pro Gln Phe Glu Lys
450 455 460
<210> 153
<211> 233
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 153
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asn Val Asp Ala Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Val Ala Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Asp Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Leu Pro Glu Thr Gly
210 215 220
Gly Trp Ser His Pro Gln Phe Glu Lys
225 230
<210> 154
<211> 460
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 154
Gln Ser Val Lys Glu Ser Glu Gly Asp Leu Val Thr Pro Ala Gly Asn
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Ser Asp Ile Asn Asp Tyr Pro
20 25 30
Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Phe Ile Asn Ser Gly Gly Ser Thr Trp Tyr Ala Ser Trp Val Lys Gly
50 55 60
Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr Val Asp Leu Lys Met Thr
65 70 75 80
Ser Leu Thr Thr Asp Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly Tyr
85 90 95
Ser Thr Tyr Tyr Gly Asp Phe Asn Ile Trp Gly Pro Gly Thr Leu Val
100 105 110
Thr Ile Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Leu Pro
435 440 445
Glu Thr Gly Gly Trp Ser His Pro Gln Phe Glu Lys
450 455 460
<210> 155
<211> 236
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 155
Glu Leu Val Met Thr Gln Thr Pro Ser Ser Thr Ser Gly Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ala Ser Gln Ser Ile Asp Ser Asn
20 25 30
Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Thr Leu Leu Ile
35 40 45
Tyr Arg Ala Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Glu Tyr Thr Leu Thr Ile Ser Gly Val Gln Arg
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Gly Val Gly Asn Val Ser
85 90 95
Tyr Arg Thr Ser Phe Gly Gly Gly Thr Glu Val Val Val Lys Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Leu Pro
210 215 220
Glu Thr Gly Gly Trp Ser His Pro Gln Phe Glu Lys
225 230 235
<210> 156
<211> 465
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 156
Gln Glu Gln Leu Val Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Gly
1 5 10 15
Ser Leu Thr Leu Ser Cys Lys Ala Ser Gly Phe Asp Phe Ser Ala Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Thr Ile Tyr Pro Ser Ser Gly Lys Thr Tyr Tyr Ala Thr Trp Val
50 55 60
Asn Gly Arg Phe Thr Ile Ser Ser Asp Asn Ala Gln Asn Thr Val Asp
65 70 75 80
Leu Gln Met Asn Ser Leu Thr Ala Ala Asp Arg Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Asp Ser Tyr Ala Asp Asp Gly Ala Leu Phe Asn Ile Trp Gly
100 105 110
Pro Gly Thr Leu Val Thr Ile Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys Leu Pro Glu Thr Gly Gly Trp Ser His Pro Gln Phe Glu
450 455 460
Lys
465
<210> 157
<211> 236
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 157
Glu Leu Val Leu Thr Gln Ser Pro Ser Val Ser Ala Ala Leu Gly Ser
1 5 10 15
Pro Ala Lys Ile Thr Cys Thr Leu Ser Ser Ala His Lys Thr Asp Thr
20 25 30
Ile Asp Trp Tyr Gln Gln Leu Gln Gly Glu Ala Pro Arg Tyr Leu Met
35 40 45
Gln Val Gln Ser Asp Gly Ser Tyr Thr Lys Arg Pro Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Ser Ser Gly Ala Asp Arg Tyr Leu Ile Ile Pro
65 70 75 80
Ser Val Gln Ala Asp Asp Glu Ala Asp Tyr Tyr Cys Gly Ala Asp Tyr
85 90 95
Ile Gly Gly Tyr Val Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly
100 105 110
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
115 120 125
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
130 135 140
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
145 150 155 160
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
165 170 175
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
180 185 190
His Lys Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
195 200 205
Lys Thr Val Ala Pro Thr Glu Cys Ser Gly Gly Gly Gly Ser Leu Pro
210 215 220
Glu Thr Gly Gly Trp Ser His Pro Gln Phe Glu Lys
225 230 235
<210> 158
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 158
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 159
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 159
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 160
<211> 461
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 160
Gln Ile Gln Leu Gln Gln Ser Gly Pro Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Ile Thr Trp Val Lys Gln Lys Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Ser Gly Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Thr Ser Ser Ser Thr Ala Phe
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Asn Tyr Gly Asn Tyr Trp Phe Ala Tyr Trp Gly Gln Gly Thr Gln
100 105 110
Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Leu
435 440 445
Pro Glu Thr Gly Gly Trp Ser His Pro Gln Phe Glu Lys
450 455 460
<210> 161
<211> 237
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 161
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Val Leu Ile Tyr Ala Ala Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys Gly Gly Gly Gly Ser Leu
210 215 220
Pro Glu Thr Gly Gly Trp Ser His Pro Gln Phe Glu Lys
225 230 235
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-ROR1/АНТИ-CD3 БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2831608C2 |
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С ROR2 | 2018 |
|
RU2784586C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2837770C2 |
| БЕЛКИ УПРАВЛЕНИЯ, НАВИГАЦИИ И КОНТРОЛЯ И СПОСОБ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2019 |
|
RU2811457C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ НАПРАВЛЯЮЩИХ И НАВИГАЦИОННЫХ КОНТРОЛЬНЫХ БЕЛКОВ | 2019 |
|
RU2824896C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ ЛЕЧЕНИЯ РАКА | 2017 |
|
RU2826270C2 |
| CD20 ТЕРАПИЯ, CD22 ТЕРАПИЯ И КОМБИНИРОВАННАЯ ТЕРАПИЯ КЛЕТКАМИ, ЭКСПРЕССИРУЮЩИМИ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) K CD19 | 2016 |
|
RU2752918C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2019 |
|
RU2810092C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| ЛИНКЕРЫ НА ОСНОВЕ СУЛЬФОМАЛЕИМИДА И СООТВЕТСТВУЮЩИЕ КОНЪЮГАТЫ | 2019 |
|
RU2815199C2 |
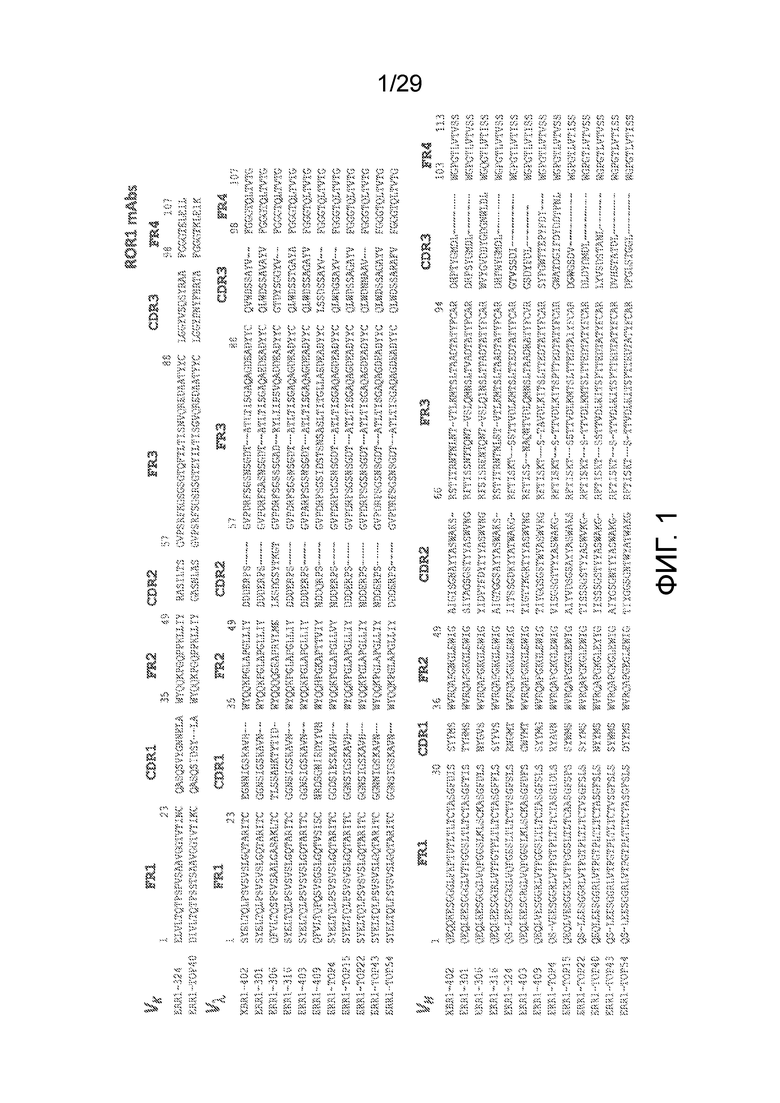

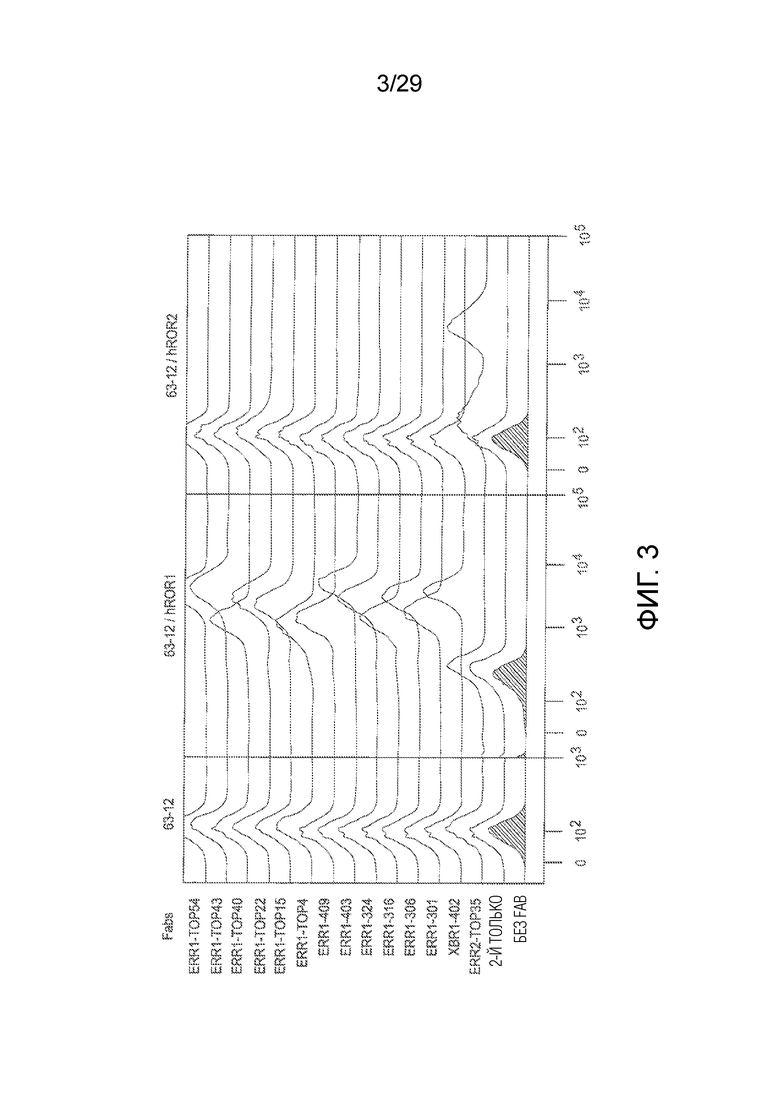
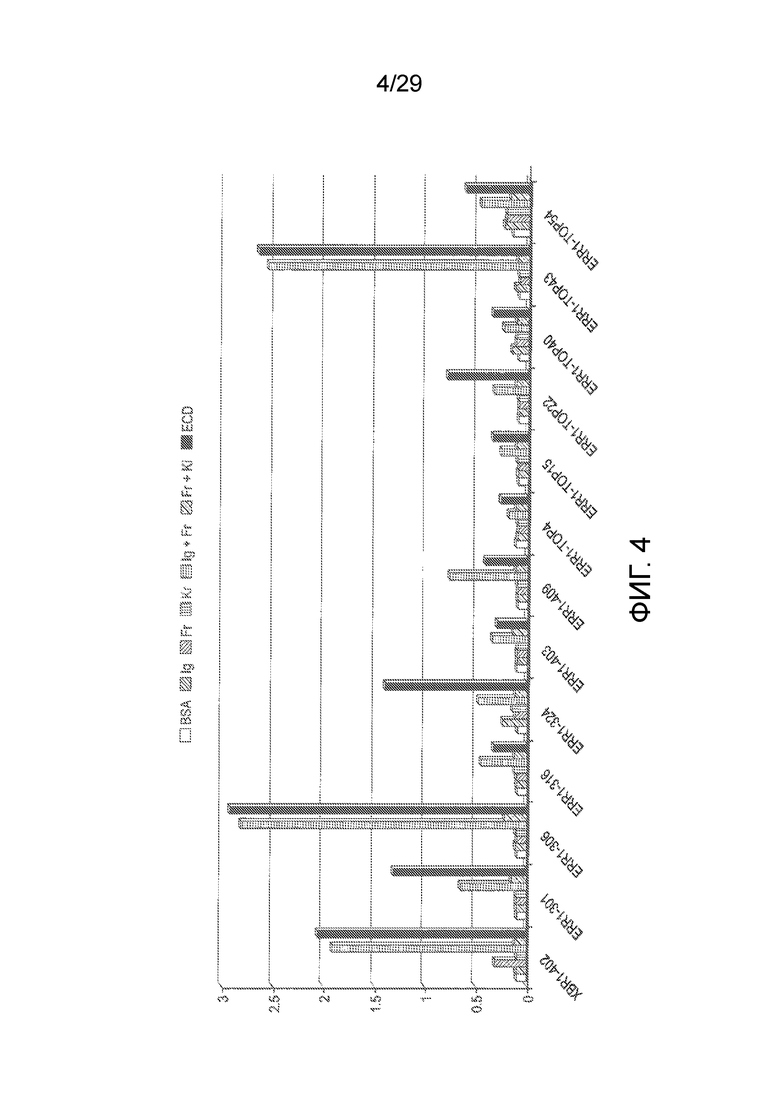
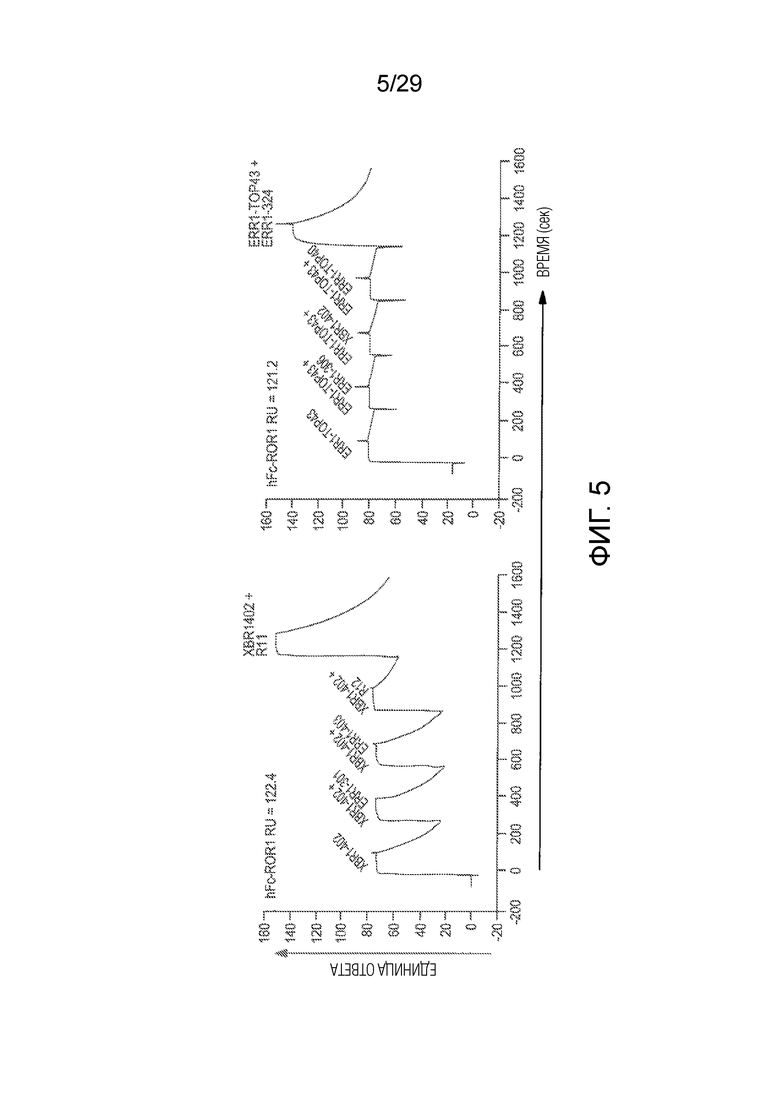
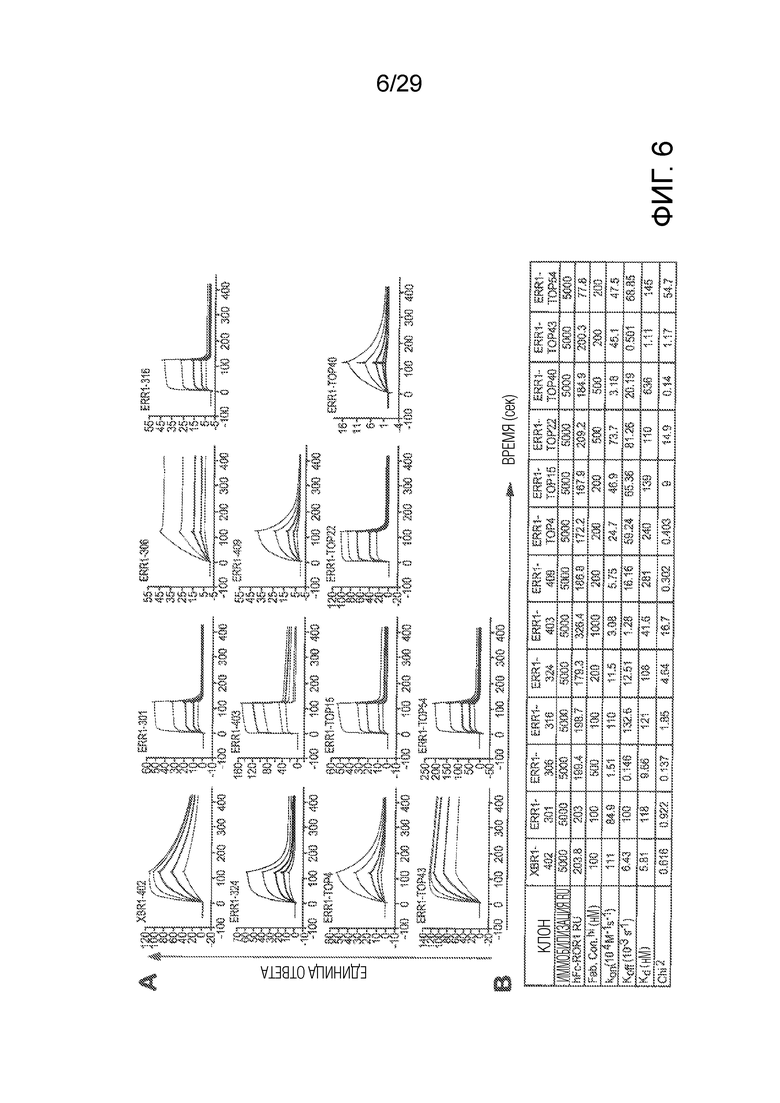

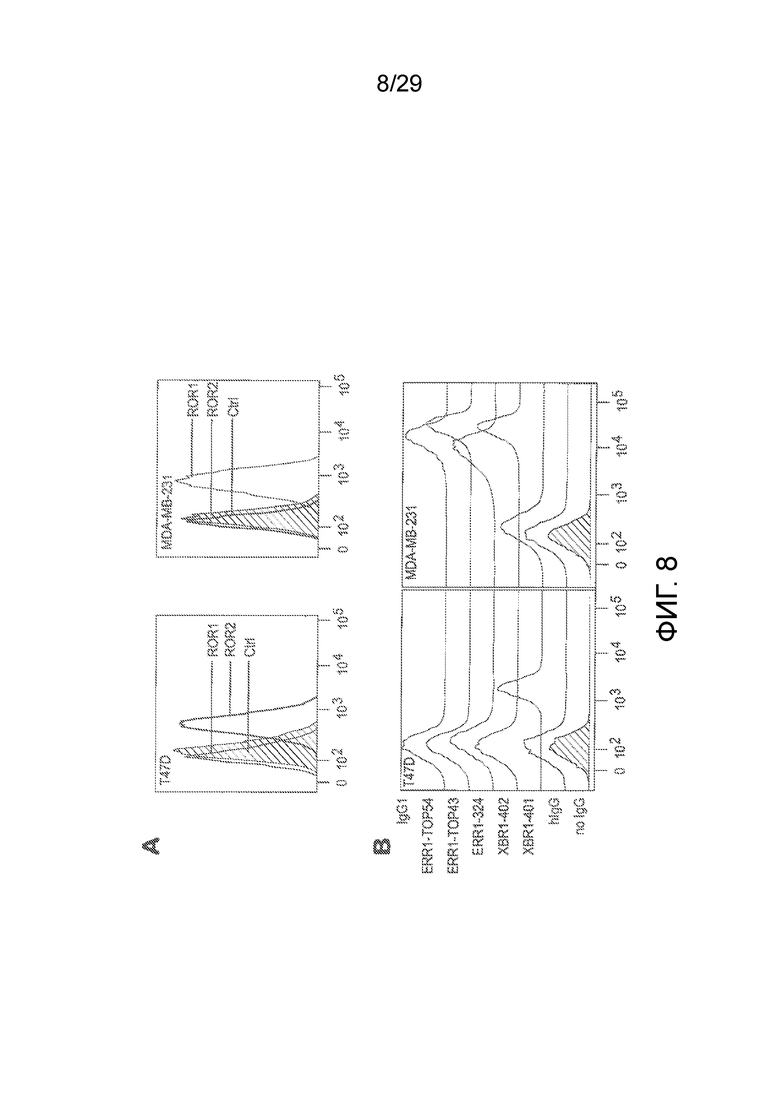
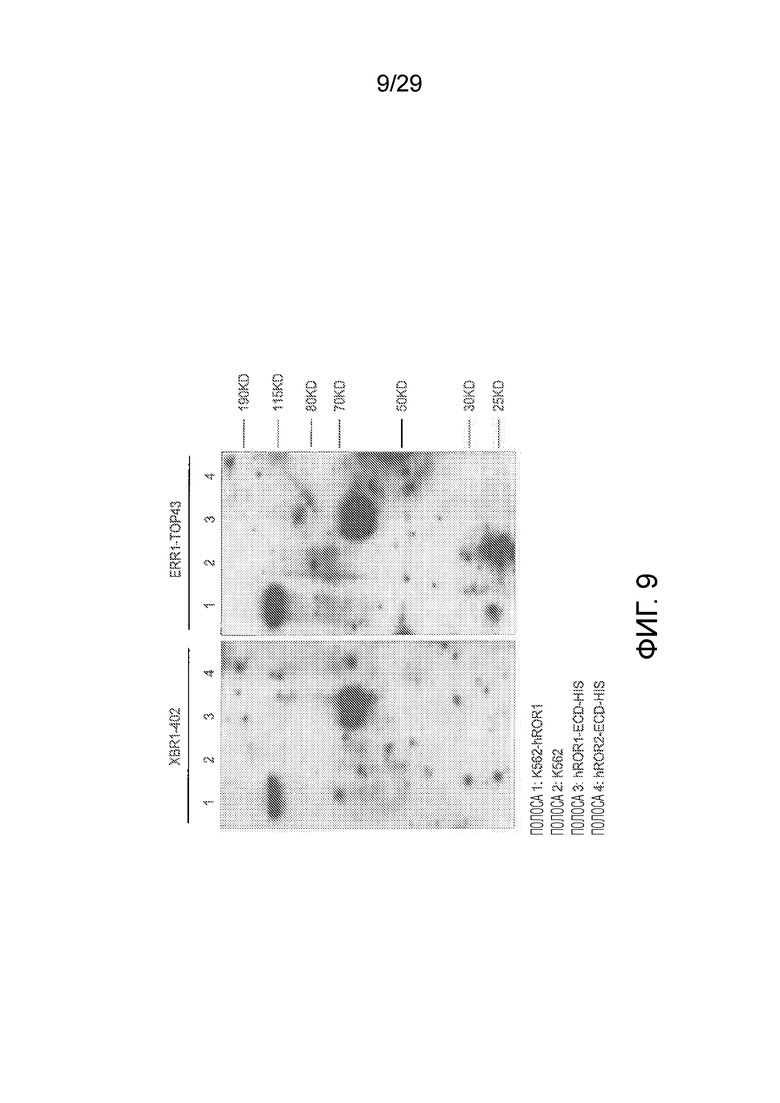

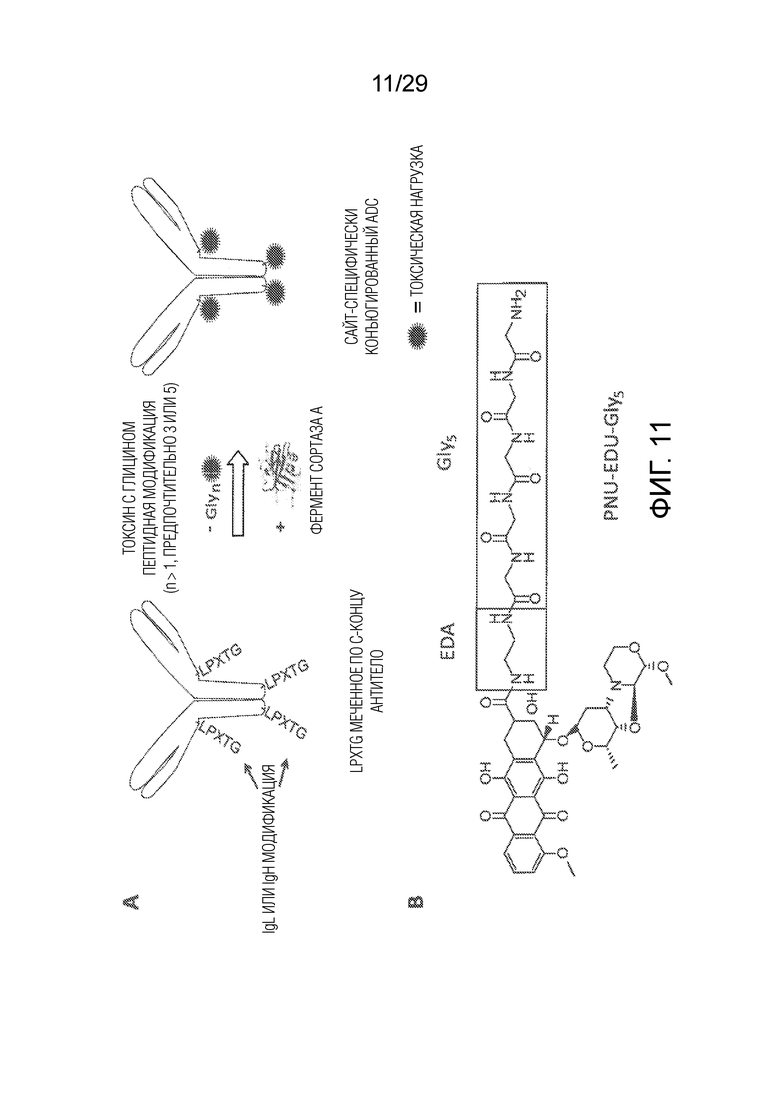
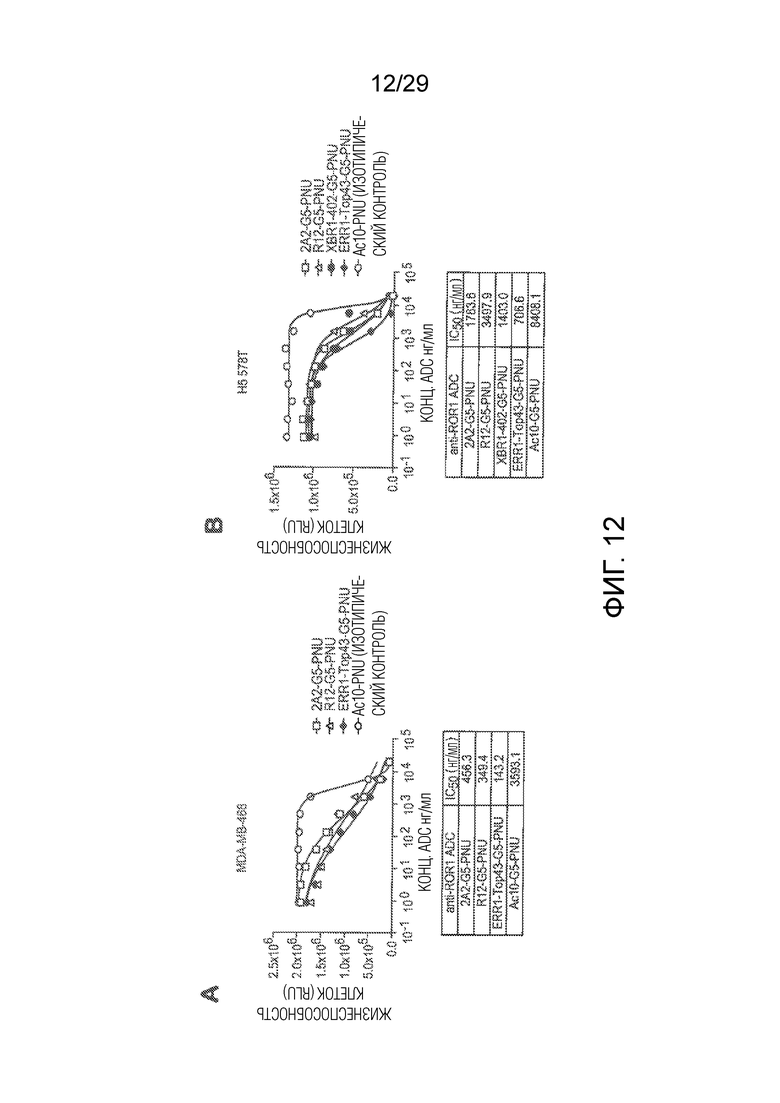
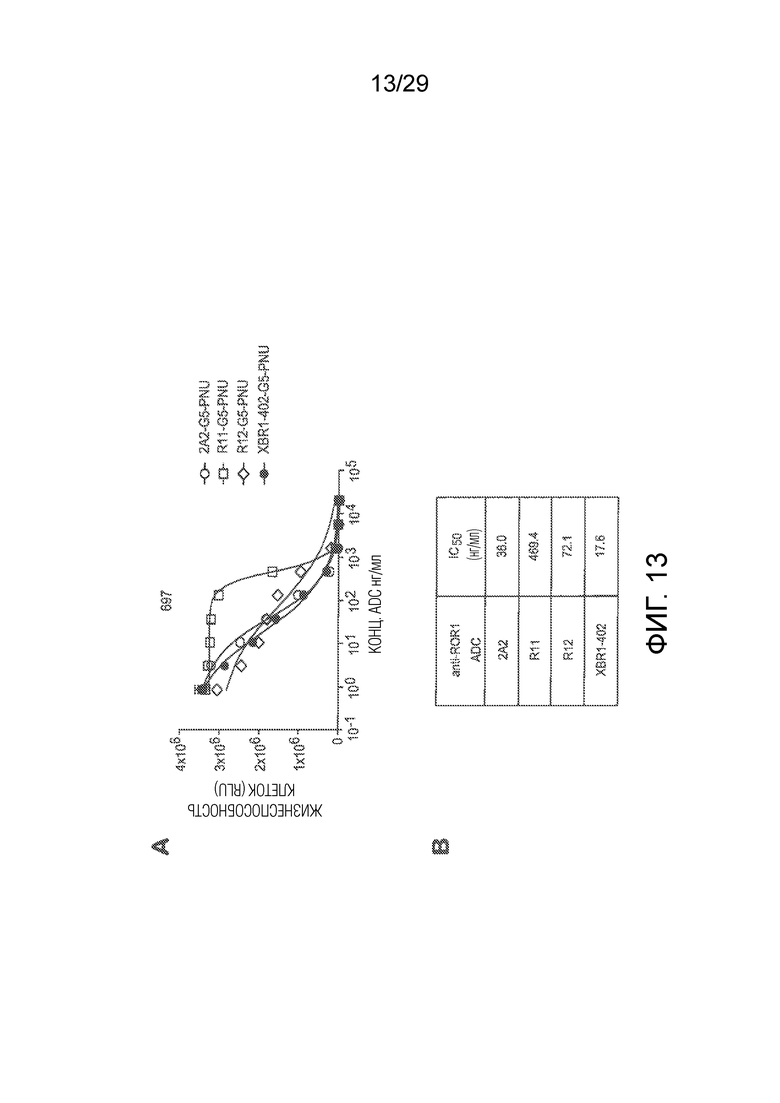
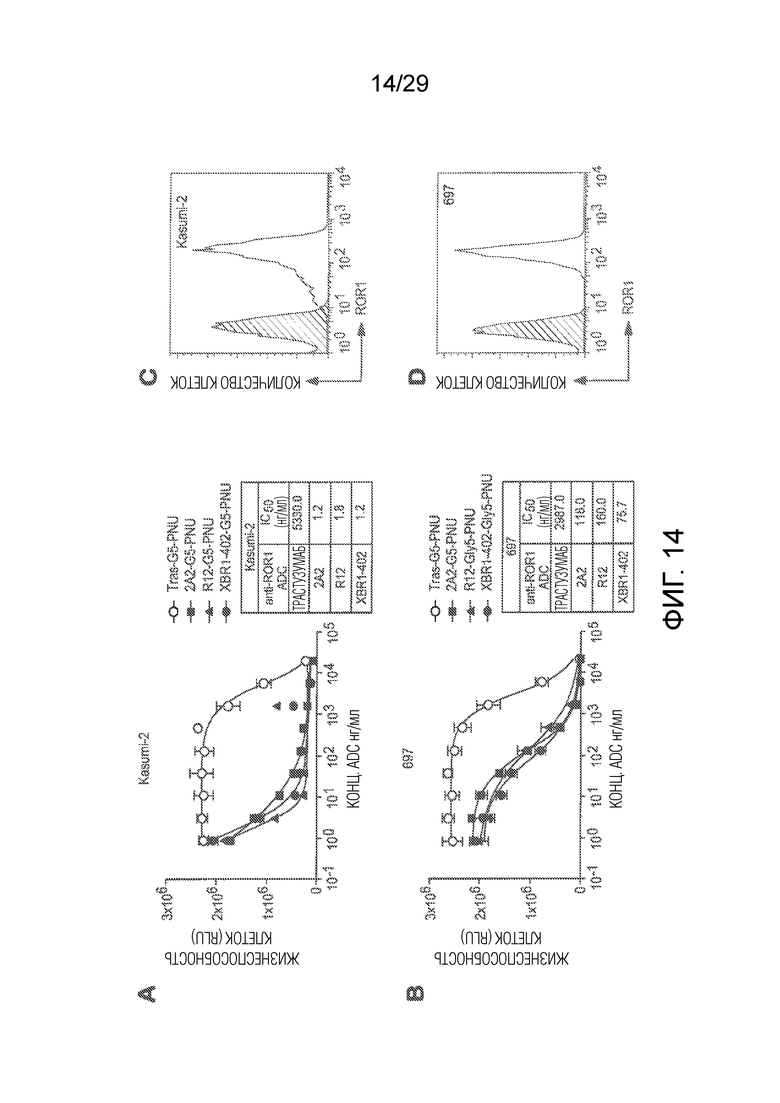
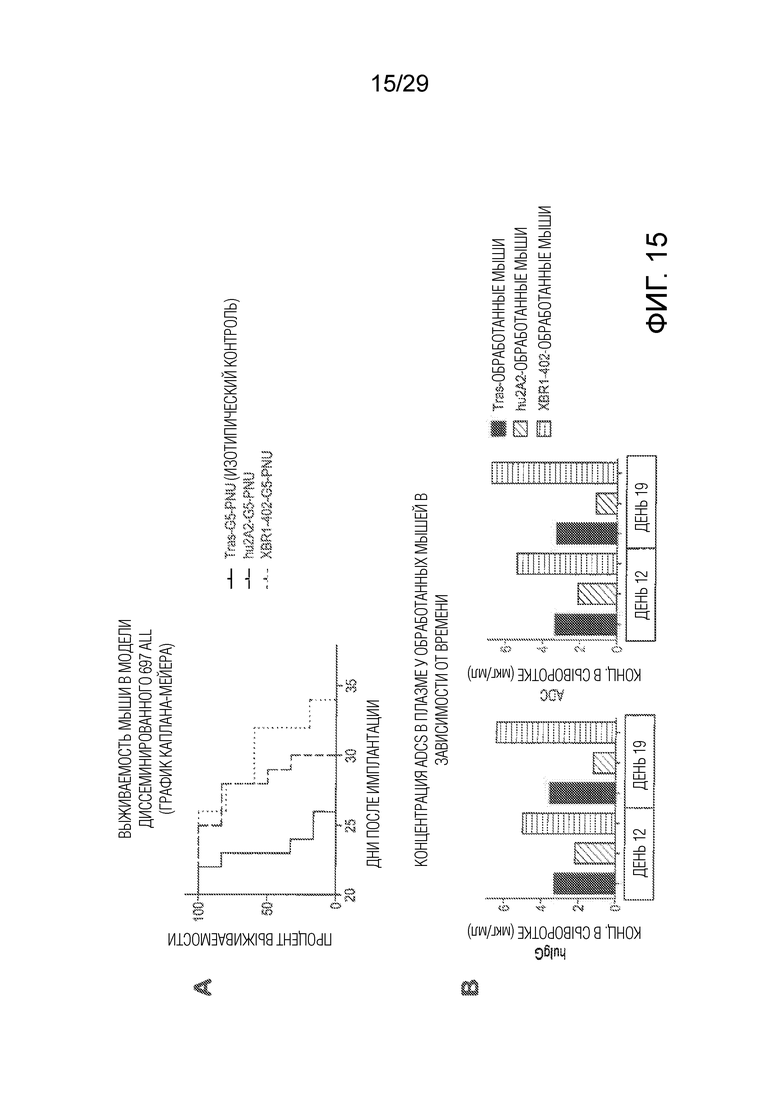
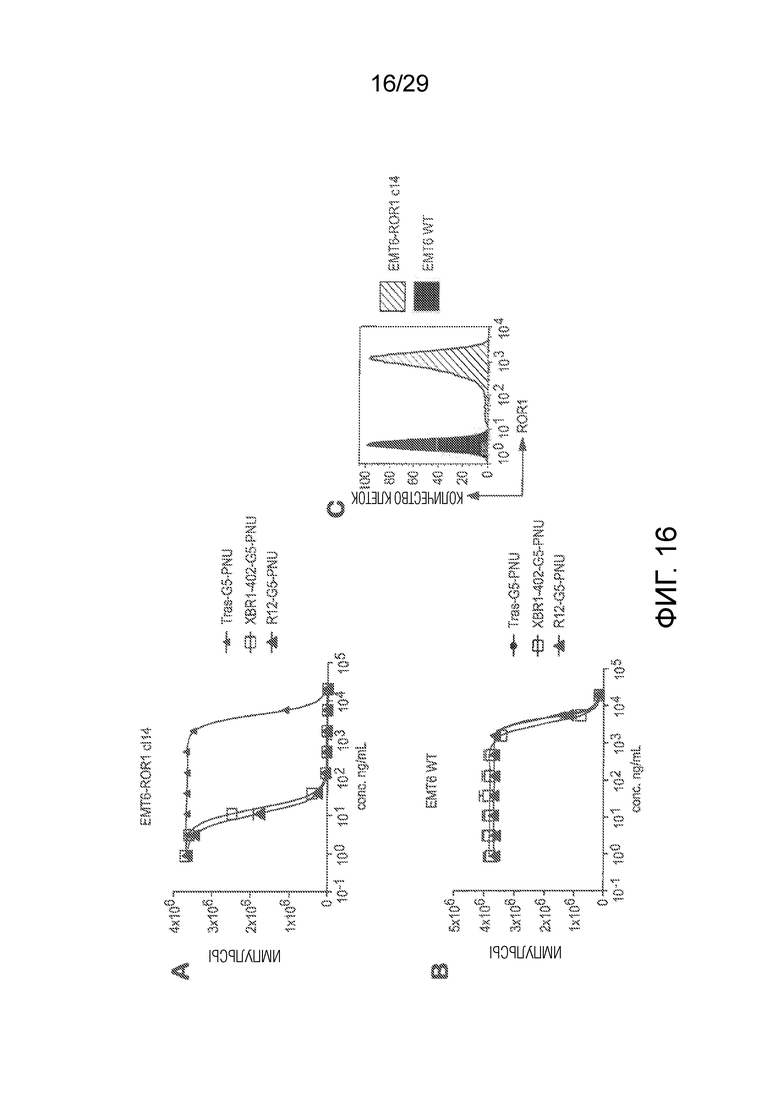
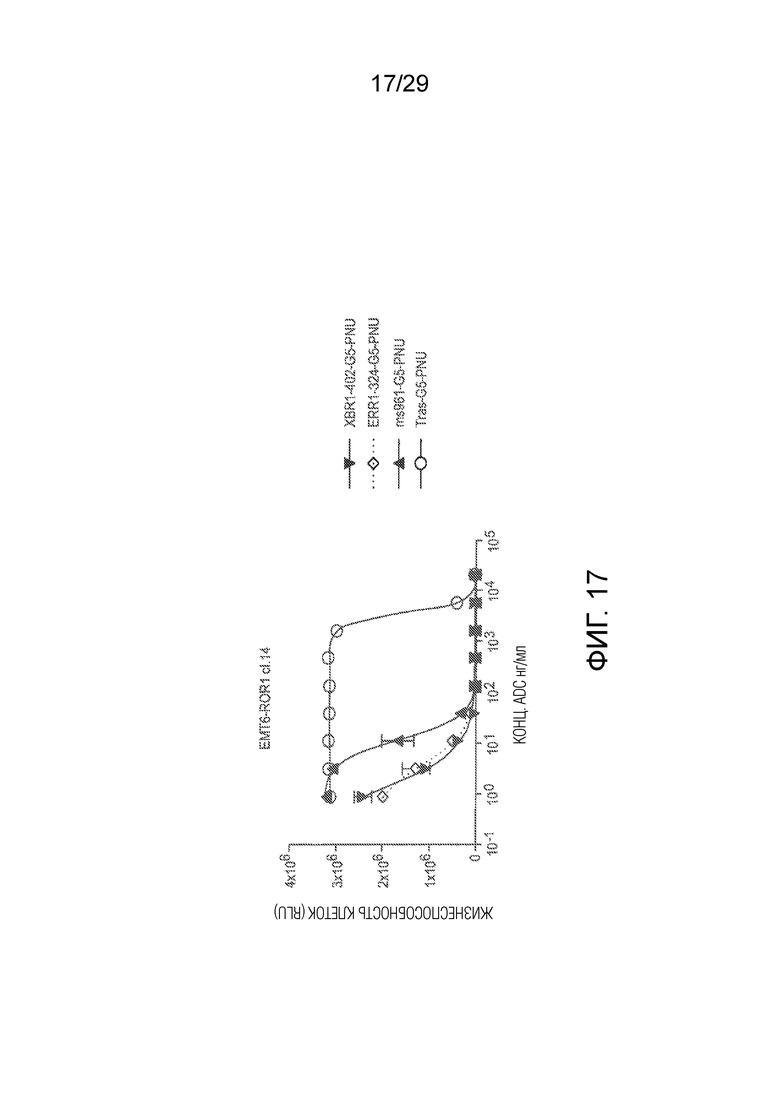
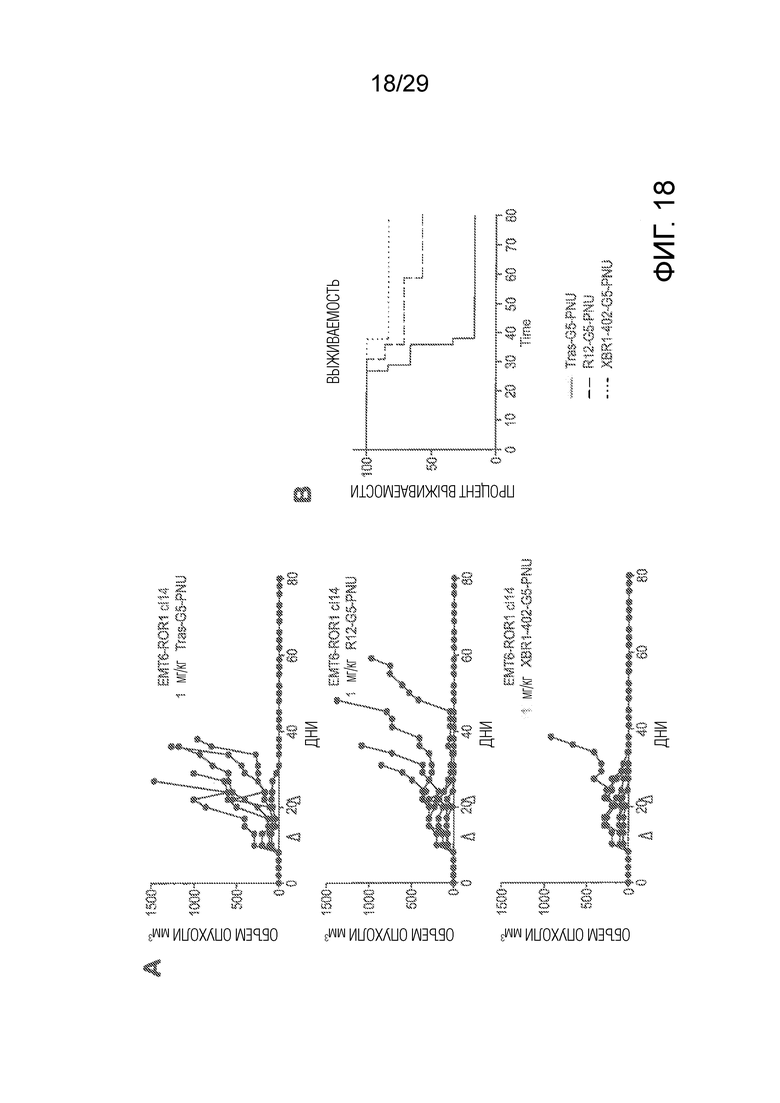
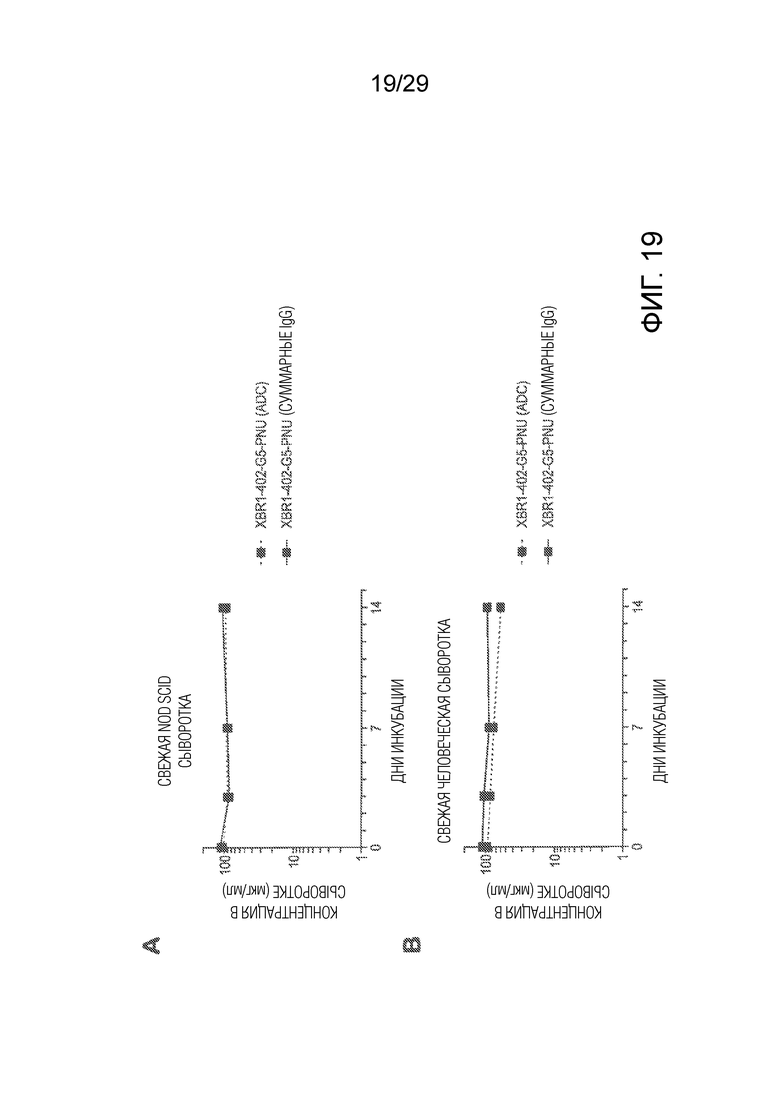
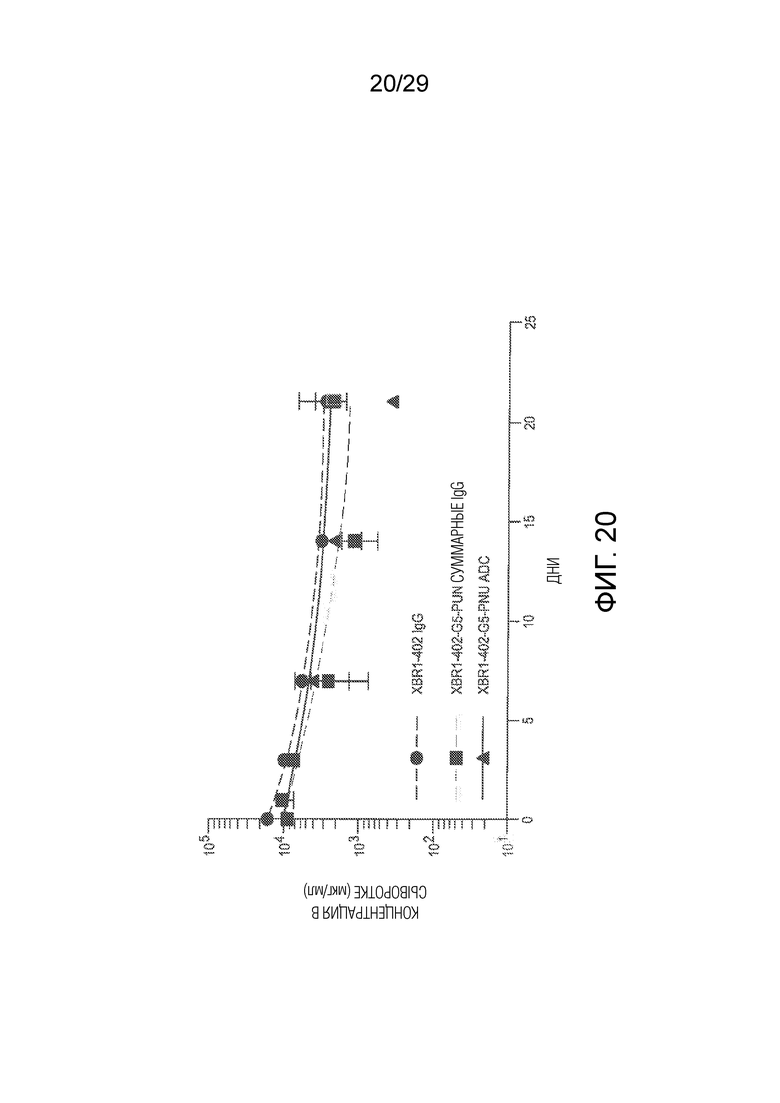
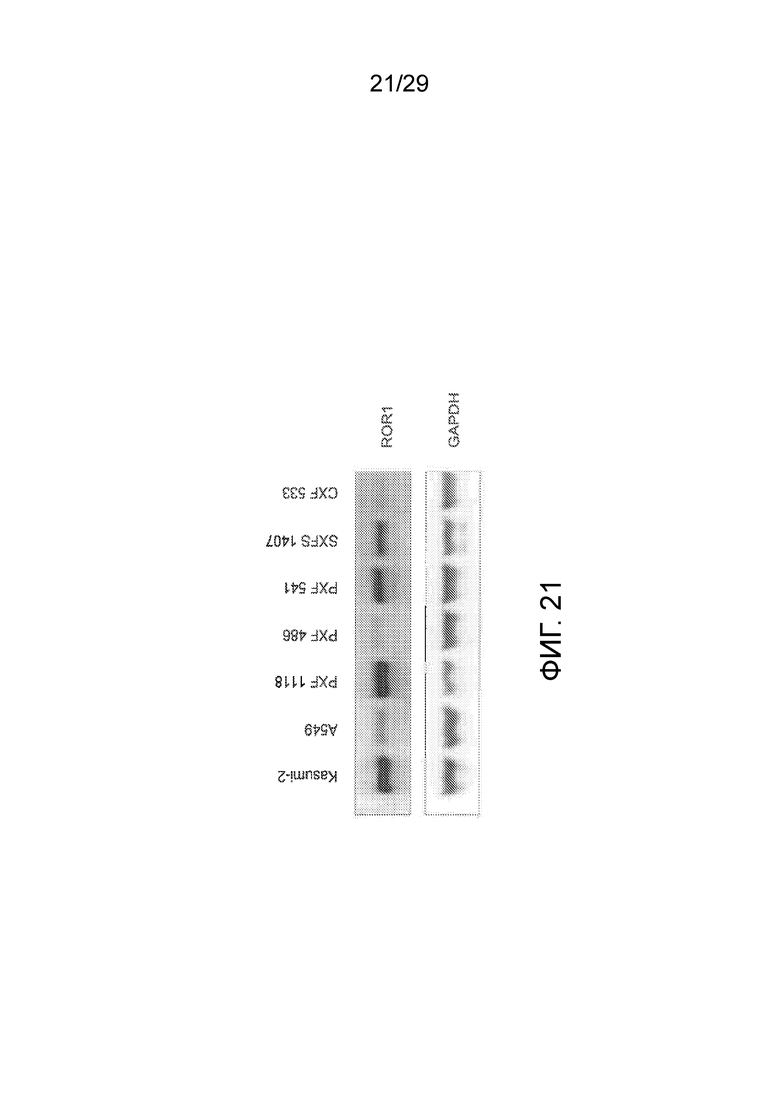
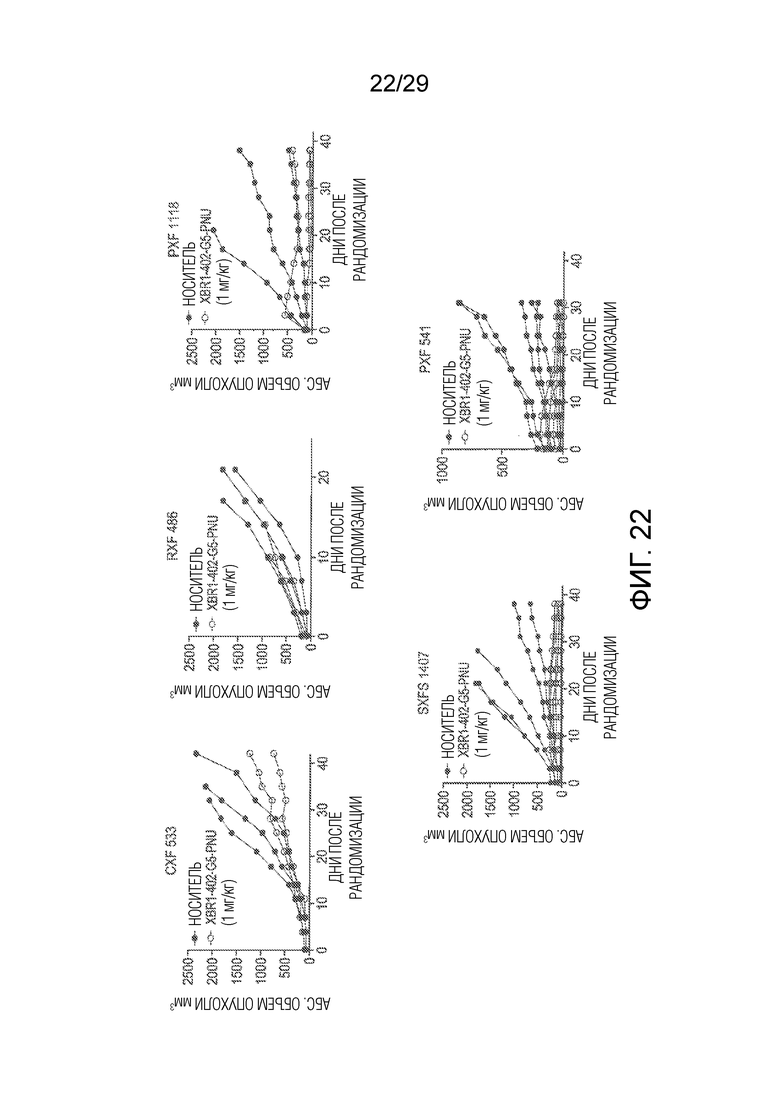

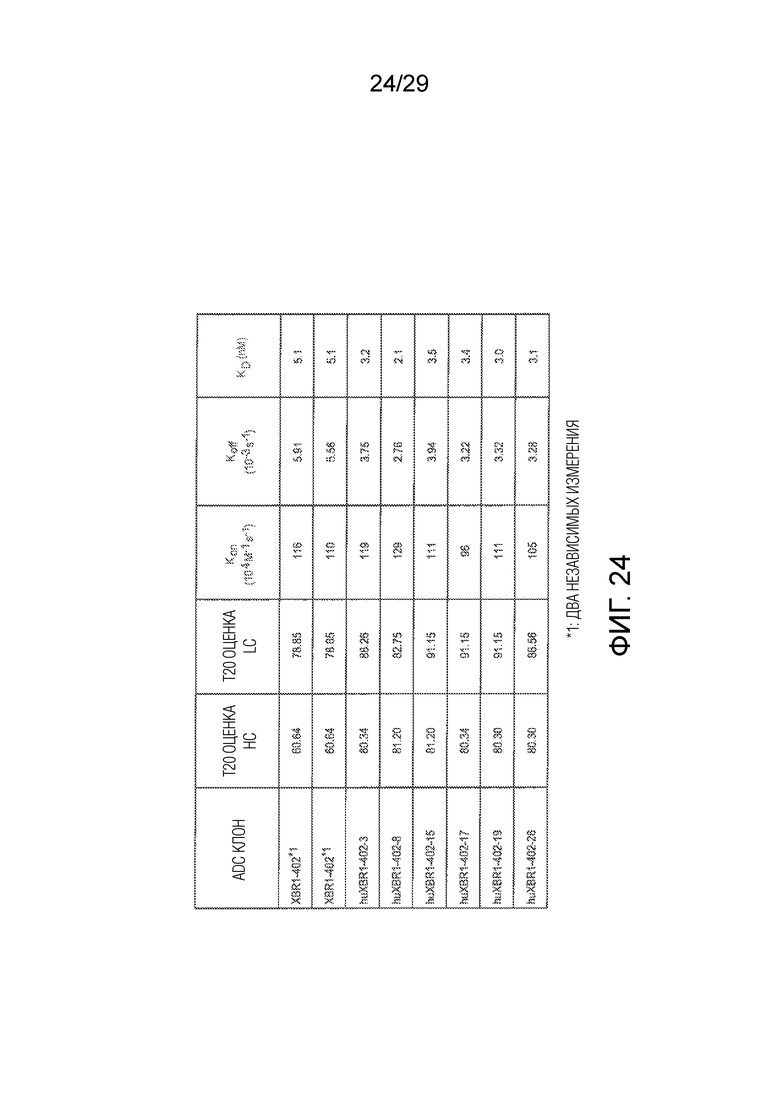
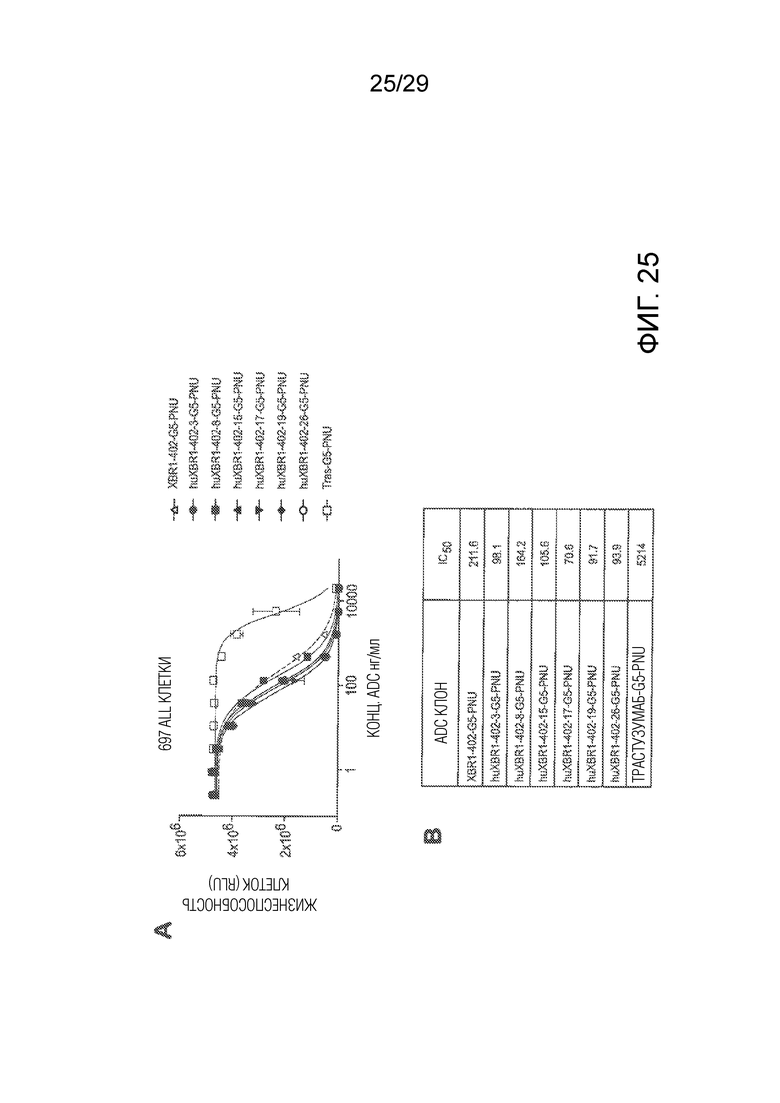
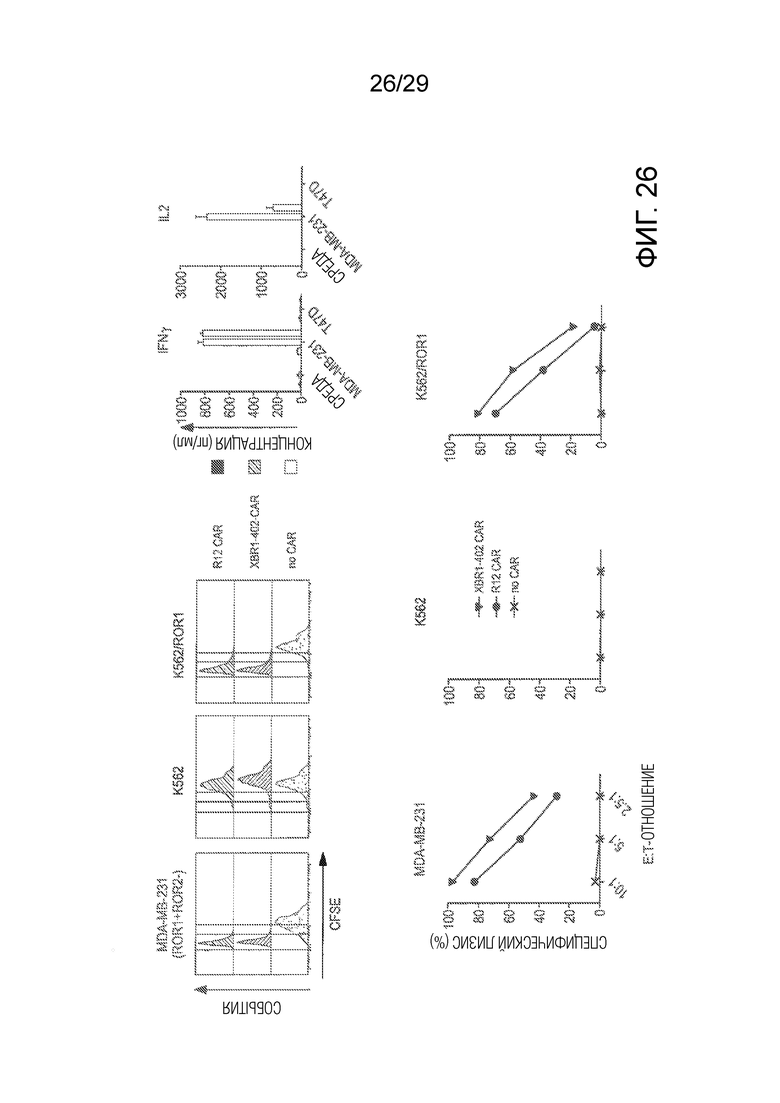
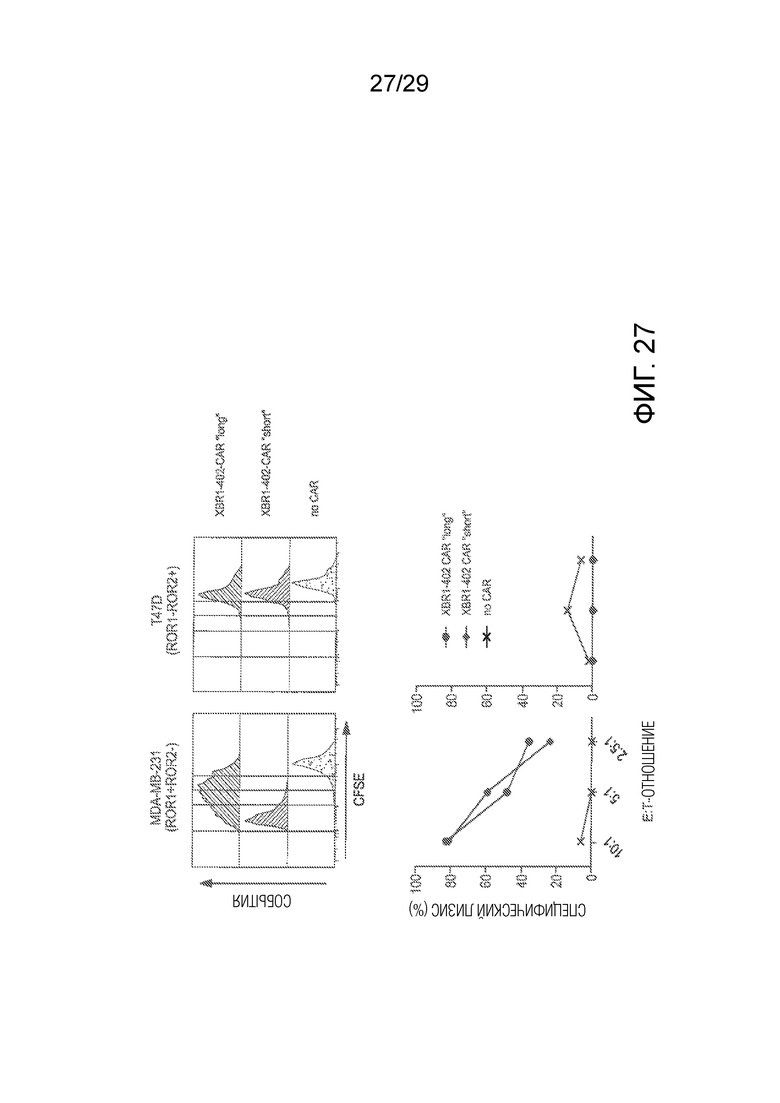
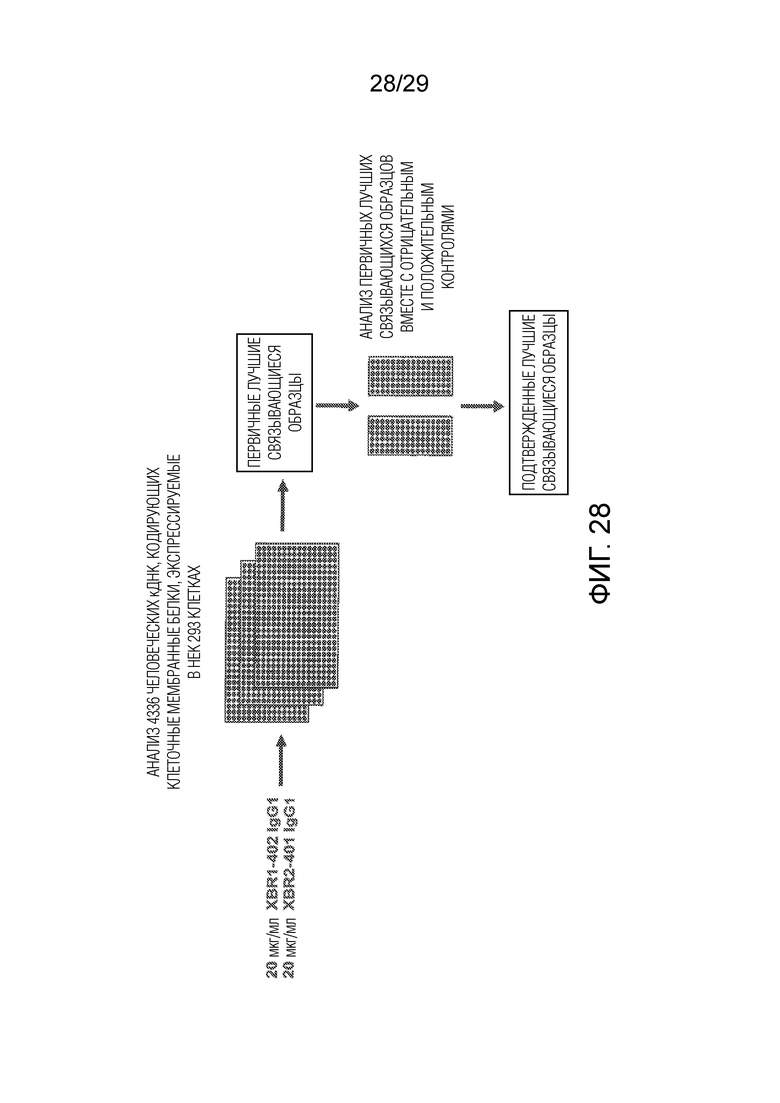

Группа изобретений относится к иммунологии и терапии заболеваний или состояний, ассоциированных с повышенной экспрессией человеческого ROR1. Предложены антитела, которые специфически распознают человеческий ROR1, конъюгаты антитело-лекарственное средство (ADCs), химерные антигенные рецепторы (CARs), композиции на их основе и их применения. Антитело или антигенсвязывающий фрагмент антитела включает последовательности CDRs тяжелой и легкой цепей, показанные в (1) SEQ ID NO:27-29 и SEQ ID NO:66-68; (2) SEQ ID NO:30-32 и SEQ ID NO:69-71; (3) SEQ ID NO:33-35 и SEQ ID NO:72-74; (4) SEQ ID NO:36-38 и SEQ ID NO:75-77; (5) SEQ ID NO:39-41 и SEQ ID NO:78-80; (6) SEQ ID NO:42-44 и SEQ ID NO:81-83; (7) SEQ ID NO:45-47 и SEQ ID NO:84-86; (8) SEQ ID NO:48-50 и SEQ ID NO:87-89; (9) SEQ ID NO:51-53 и SEQ ID NO:90-92; (10) SEQ ID NO:54-56 и SEQ ID NO:93-95; (11) SEQ ID NO:57-59 и SEQ ID NO:96-98; (12) SEQ ID NO:60-62 и SEQ ID NO:99-101; или (13) SEQ ID NO:63-65 и SEQ ID NO:102-104. Предложенные анти-ROR1 антитела обладают высокой аффинностью. CARs и CAR-T клетки, сконструированные с использованием указанных анти-hROR1 связывающихся доменов, показывают высокую эффективность in vitro. ADCs, включающие hROR1-специфическое антитело, осуществляют эффективный киллинг hROR1 специфических клеток. Изобретения позволяют проводить эффективную терапию рака человека, ассоциированного с экспрессией ROR1. 14 н. и 23 з.п. ф-лы, 29 ил., 16 табл., 23 пр.
1. Антитело или антигенсвязывающий фрагмент антитела, который специфически связывается с внеклеточным доменом человеческого подобного рецепторной тирозинкиназе орфанного рецептора 1 (ROR1), где указанное антитело или фрагмент антитела включает последовательности CDR1, CDR2 и CDR3 тяжелой цепи и последовательности CDR1, CDR2 и CDR3 легкой цепи, соответственно показанные в
(1) SEQ ID NO:27-29 и SEQ ID NO:66-68;
(2) SEQ ID NO:30-32 и SEQ ID NO:69-71;
(3) SEQ ID NO:33-35 и SEQ ID NO:72-74;
(4) SEQ ID NO:36-38 и SEQ ID NO:75-77;
(5) SEQ ID NO:39-41 и SEQ ID NO:78-80;
(6) SEQ ID NO:42-44 и SEQ ID NO:81-83;
(7) SEQ ID NO:45-47 и SEQ ID NO:84-86;
(8) SEQ ID NO:48-50 и SEQ ID NO:87-89;
(9) SEQ ID NO:51-53 и SEQ ID NO:90-92;
(10) SEQ ID NO:54-56 и SEQ ID NO:93-95;
(11) SEQ ID NO:57-59 и SEQ ID NO:96-98;
(12) SEQ ID NO:60-62 и SEQ ID NO:99-101; или
(13) SEQ ID NO:63-65 и SEQ ID NO:102-104.
2. Антитело или антигенсвязывающий фрагмент антитела по п.1, включающее последовательности CDR1, CDR2 и CDR3 тяжелой цепи и последовательности CDR1, CDR2 и CDR3 легкой цепи, соответственно показанные в SEQ ID NO:27-29 и SEQ ID NO:66-68.
3. Антитело или антигенсвязывающий фрагмент антитела по п.1, включающее последовательности вариабельной области тяжелой цепи и последовательности вариабельной области легкой цепи, соответственно показанные в:
(1) SEQ ID NO:1 и SEQ ID NO:14;
(2) SEQ ID NO:130 и SEQ ID NO:136;
(3) SEQ ID NO:131 и SEQ ID NO:137;
(4) SEQ ID NO:132 и SEQ ID NO:138;
(5) SEQ ID NO:133 и SEQ ID NO:139;
(6) SEQ ID NO:134 и SEQ ID NO:140; или
(7) SEQ ID NO:135 и SEQ ID NO:141.
4. Антитело или антигенсвязывающий фрагмент антитела по п. 3, включающий последовательность вариабельной области тяжелой цепи и последовательность вариабельной области легкой цепи соответственно, показанные в SEQ ID NO:133 и SEQ ID NO:139.
5. Антитело или антигенсвязывающий фрагмент антитела по п. 1, которое выбрано из группы, состоящей из IgAl, IgA2, IgD, IgE, IgGl, IgG2, IgG3, IgG4, синтетическое IgG, IgM, F(ab)2, Fv, scFv, IgGACH2, F(ab')2, scFv2CH3, Fab, scFv4, scFv3, scFv2, dsFv, Fv, scFv-Fc, (scFv)2, неистощающее IgG и бивалентного антитела.
6. Конъюгат антитело-лекарственное средство (ADC) для лечения рака, ассоциированного с повышенной экспрессией ROR1, включающий антитело или антигенсвязывающий фрагмент антитела по п. 1 и по меньшей мере одно цитотоксическое средство.
7. Конъюгат антитело-лекарственное средство по п. 6, где цитотоксическое средство представляет собой низкомолекулярный токсин, пептидный токсин или белковый токсин.
8. Конъюгат антитело-лекарственное средство по п. 6, где антитело или антигенсвязывающий фрагмент антитела конъюгирован с цитотоксическим средством через опосредованную ферментом сортазой конъюгацию антитела (SMAC).
9. Конъюгат антитело-лекарственное средство по п. 6, где цитотоксическое средство представляет собой производное антрациклинового (PNU) токсина Gly(n)-EDA-PNU, где n представляет собой любое число от 1 до 21.
10. Конъюгат антитело-лекарственное средство по п. 6, где антитело или антигенсвязывающий фрагмент антитела является химерным или гуманизированным.
11. Конъюгат антитело-лекарственное средство по п. 6, где антитело или антигенсвязывающий фрагмент антитела включает последовательности CDR1, CDR2 и CDR3 тяжелой цепи и последовательности CDR1, CDR2 и CDR3 легкой цепи, соответственно показанные в SEQ ID NO:27-29 и SEQ ID NO:66-68.
12. Конъюгат антитело-лекарственное средство по п. 6, где антитело или антигенсвязывающий фрагмент антитела включает последовательность вариабельной области тяжелой цепи и последовательность вариабельной области легкой цепи, соответственно показанные в
(1) SEQ ID NO:1 и SEQ ID NO:14;
(2) SEQ ID NO:130 и SEQ ID NO:136;
(3) SEQ ID NO:131 и SEQ ID NO:137;
(4) SEQ ID NO:132 и SEQ ID NO:138;
(5) SEQ ID NO:133 и SEQ ID NO:139;
(6) SEQ ID NO:134 и SEQ ID NO:140; или
(7) SEQ ID NO:135 и SEQ ID NO:141.
13. Химерный антигенный рецептор (CAR) для лечения рака, ассоциированного с повышенной экспрессией ROR1, включающий антитело или антигенсвязывающий фрагмент антитела по п. 1, который слит с трансмембранной областью и внутриклеточным сигнальным доменом T-клеточного рецептора (TCR).
14. Химерный антигенный рецептор по п. 13, где антитело или антигенсвязывающий фрагмент антитела является химерным или гуманизированным.
15. Химерный антигенный рецептор по п. 13, где антитело или антигенсвязывающий фрагмент антитела включает последовательности CDR1, CDR2 и CDR3 тяжелой цепи и последовательности CDR1, CDR2 и CDR3 легкой цепи, соответственно показанные в SEQ ID NO:27-29 и SEQ ID NO:66-68.
16. Химерный антигенный рецептор по п. 13, где антитело или антигенсвязывающий фрагмент антитела включает последовательность вариабельной области тяжелой цепи и последовательность вариабельной области легкой цепи, соответственно показанные в
(1) SEQ ID NO:1 и SEQ ID NO:14;
(2) SEQ ID NO:130 и SEQ ID NO:136;
(3) SEQ ID NO:131 и SEQ ID NO:137;
(4) SEQ ID NO:132 и SEQ ID NO:138;
(5) SEQ ID NO:133 и SEQ ID NO:139;
(6) SEQ ID NO:134 и SEQ ID NO:140; или
(7) SEQ ID NO:135 и SEQ ID NO:141.
17. Фармацевтическая композиция для лечения рака, ассоциированного с повышенной экспрессией ROR1, включающая (1) терапевтически эффективное количество (a) антитела или антигенсвязывающего фрагмента антитела по п. 1 или (b) конъюгата антитело-лекарственное средство (ADC) по п. 6 и (2) фармацевтически приемлемый носитель.
18. Фармацевтическая композиция по п. 17, в которой рак выбран из хронического лимфоцитарного лейкоза (CLL), острого лимфобластного лейкоза (ALL), лимфомы из клеток мантийной зоны, нейробластомы, саркомы, почечноклеточной карциномы, рака молочной железы, рака легкого, рака толстой кишки, рака головы и шеи и меланомы.
19. Набор для лечения рака, ассоциированного с повышенной экспрессией ROR1, включающий (1) антитело или антигенсвязывающий фрагмент антитела по п. 1 или конъюгат антитело-лекарственное средство (ADC) по п. 6; (2) подходящие реагенты, и/или оборудование.
20. Полинуклеотид, кодирующий вариабельную область тяжелой цепи и легкой цепи антитела или антигенсвязывающего фрагмента антитела по п. 1.
21. Вектор, включающий полинуклеотид по п. 20, для экспрессии антитела или антигенсвязывающего фрагмента антитела по п.1.
22. Способ лизиса или ингибирования роста клетки, сверхэкспрессирующей ROR1, у субъекта, включающий введение эффективного количества фармацевтической композиции по п. 17 субъекту, нуждающемуся в этом, осуществляя таким образом лизис или ингибирование роста клетки, экспрессирующей ROR1, у субъекта.
23. Способ по п. 22, где клетка представляет собой опухолевую клетку.
24. Способ по п. 22, где фармацевтическая композиция включает конъюгат антитело-лекарственное средство (ADC), включающий антитело, связывающийся белок на основе антитела или фрагмент антитела по п. 1, который конъюгирован с цитотоксическим средством.
25. Способ по п. 24, где цитотоксическое средство представляет собой низкомолекулярный токсин, пептидный токсин или белковый токсин.
26. Способ лечения заболевания или состояния, ассоциированного с повышенной экспрессией ROR1, у субъекта, включающий введение эффективного количества фармацевтической композиции по п. 17 субъекту, имеющему заболевание или состояние, ассоциированное с повышенной экспрессией ROR1, осуществляя таким образом лечение заболевания или состояния, ассоциированного с повышенной экспрессией ROR1, у субъекта.
27. Способ по п. 26, где заболевание или состояние представляет собой рак.
28. Способ по п. 26, где заболевание или состояние выбрано из группы, состоящей из хронического лимфоцитарного лейкоза (CLL), острого лимфобластного лейкоза (ALL), лимфомы из клеток мантийной зоны, нейробластомы, саркомы, почечноклеточной карциномы, рака молочной железы, рака легкого, рака толстой кишки, рака головы и шеи и меланомы.
29. Способ по п. 26, где фармацевтическая композиция включает конъюгат антитело-лекарственное средство (ADC), включающий антитело, связывающийся белок на основе антитела или фрагмент антитела по п. 1, который конъюгирован с цитотоксическим средством.
30. Способ по п. 29, где цитотоксическое средство представляет собой низкомолекулярный токсин, пептидный токсин или белковый токсин.
31. Способ по п. 26, где антитело или фрагмент антитела слит с трансмембранной областью и внутриклеточным сигнальным доменом T-клеточного рецептора (TCR) с образованием химерного антигенного рецептора (CAR).
32. Применение антитела или антигенсвязывающего фрагмента антитела по любому из пп.1-5 в лечении рака, ассоциированного с повышенной экспрессией ROR1.
33. Применение ADC по любому из пп. 6-12 в лечении рака, ассоциированного с повышенной экспрессией ROR1.
34. Применение CAR по любому из пп. 13-16 для получения T-клеток, сконструированных с CAR, для лечения рака, ассоциированного с повышенной экспрессией ROR1.
35. Применение фармацевтической композиции по п. 17 или 18 в лечении рака.
36. Применение набора по п. 19 в лечении рака, ассоциированного с повышенной экспрессией ROR1.
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| Плавучий мост | 1921 |
|
SU961A1 |
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| Прибор для запора стрелок | 1921 |
|
SU167A1 |

