Введение
Настоящее изобретение относится к внеклеточным везикулам (EV, от англ. extracellular vesicles), в частности экзосомам, композициям, содержащим данные везикулы, их применениям и способам их применения и изготовления. Настоящее изобретение также относится к применению таких везикул в терапии и генной терапии, в частности, для лечения заболеваний дыхательных путей, таких как кистозный фиброз (CF - от англ. cystic fibrosis), ХОБЛ (Хроническая обструктивная болезнь легких) и раковые заболевания легкого.
Предшествующий уровень техники
Внеклеточные везикулы представляют собой наноразмерные мембранные везикулы, инкапсулированные липидным бислоем и активно секретируемые большинством прокариотических и эукариотических клеток. Внеклеточные везикулы включают экзосомы, микровезикулы и мембранные пузырьки.
Везикулы, в общем, образуются посредством экзоцитоза из живых клеток. Экзосомы отличаются от многих других внеклеточных везикул тем, что они происходят из эндоцитозных компартментов в клетках и исключаются из клетки посредством экзоцитоза, когда мул ьти везикулярные тельца сливаются с плазматической мембраной.
Образование мультивезикулярных телец представляет собой многоэтапный процесс, который начинается с образования ранних эндосом. Данные ранние эндосомы созревают в поздние эндосомы, и на протяжении данного процесса несколько грузов, или во время деструкции в лизосомальном компартменте, или будучи упорядоченными в маленькие везикулы, подлежащие секреции, селективно заключаются внутрь эндосом посредством впячивания их мембраны и постепенно образуют множество внутрипросветных везикул. Данные эндосомы с множеством внутрипросветных везикул в настоящее время называются поздними эндосомами и затем созревают с образованием МВТ (мультивезикулярные тельца), которые в конечном итоге сливаются с плазматической мембраной с высвобождением данных везикул в виде экзосом.
Внеклеточные везикулы обычно имеют размер в интервале вплоть до примерно 1000 нм, при этом экзосомы обычно имеют размер в интервале 30-150 нм. Помимо своих мембранных молекул «подписи» везикулы, включая экзосомы, вприроде имеют внутривезикулярный груз, который может включать множество белков, РНК и микроРНКв водном растворе/суспензии.
Везикулы, в частности экзосомы, вовлечены в клеточную коммуникацию и межклеточную передачу сигнала. В результате, экзосомы и другие везикулы в последнее время начали использоваться в экспериментальных моделях заболевания для доставки лекарственных средств или желательных молекул. За счет их природного происхождения, экзосомы сохраняют явное преимущество над доставкой лекарственных средств на основе липосом как эффективное и надежное средство доставки разных терапевтических средств на основе нуклеиновых кислот, макромолекул и лекарственных средств.
Способность везикул нести белки, РНК и микроРНК в цитоплазматическое ядро делает их везикулами для удобной доставки лекарственных средств. Кроме того, наличие липидного бислоя обеспечивает структурную целостность, защищает нуклеиновые кислоты от деградации и дает возможность экзосомам выдерживать сдвиговое напряжение.
Известно, что экзосомы способны легко проникать через клеточные мембраны и также уклоняться от действия иммунной системы, частично, благодаря своему размеру. Поскольку экзосомы представляют собой нановезикулы, встречающиеся в природе, они не оказывают цитотоксического действия, которое могли бы оказывать носители лекарственных средств на основе липосом или других наночастиц, и имеют лучший период полувыведения из крови и лучшее проникание через разные биологические барьеры.
Кистозный фиброз (CF) представляет собой генетическое заболевание, при котором пациент наследует мутацию в обеих копиях гена белка регулятора трансмембранной проводимости при кистозном фиброзе (CFTR - от англ. cystic fibrosis transmembrane conductance regulator). CFTR участвует в продукции разных продуктов секреции, в частности, слизи, пищеварительных соков и пота. В то время как такие продукты секреции обычно являются негустыми, в результате мутации у пациентов с кистозным фиброзом продукты секреции являются густыми. В легких такие густые продукты секреции приводят к значительной продукции мокроты, уменьшенному клиренсу данной слизи, закупорке дыхательных путей из-за скопления слизи, постоянному кашлю, частым инфекциям в легких и затрудненному дыханию.
В то время как в настоящее время нет известных способов излечения CF, генная терапия исследуется как возможный сильнодействующий способ излечения. Разные подходы пробовались в случае генной терапии, включая липосомы и вирусные векторы. Однако обнаружено, что такие виды лечения являются неэффективными, главным образом из-за неэффективности векторов и проблем с безопасностью. Вирусные векторы могут вызывать нежелательный иммунный ответ, который препятствует повторному введению и может также вызывать потенциально канцерогенный инсерционный мутагенез за счет интеграции вирусной ДНК в геном хозяина.
Поскольку CF вызывает значимые проблемы в легких, считается, что эффективная доставка в легкие при генной терапии является требованием для эффективной терапии.
Везикулы, включая экзосомы, по-видимому, представляют собой возможные средства доставки к легкому для генной терапии и вообще других типов терапии, например, регенеративной медицины, терапии на основе белков, антител или малых молекул. Однако, существует по меньшей мере две значительные трудности. Во-первых, слизь в легком обеспечивает значимый барьер для любого средства переноса гена, достигающего клетки легких. Слизь представляет собой липкий вязкоупругий гель, который защищает от патогенов, токсинов и продуктов распада в разных точках поступления в организм, включая глаза, нос, легкие, желудочно-кишечный тракт и женские половые пути. Многие наночастицы являются сильно мукоадгезивными и оказываются эффективно захваченными в быстро высвобождающемся периферическом слое слизи, что сильно ограничивает их распространение по слизистой оболочке, а также проникновение в нижележащую ткань. Клинические испытания, тестирующие вирусные генетические векторы, включая аденовирус (AdV) и аденоассоциированный вирус (AAV - от англ. adeno-associated virus), серотип 2, и невирусные липосомные векторы для ингаляционной генной терапии, не смогли обеспечить клинически значимое положительное действие из-за неэффективного переноса генов к дыхательному эпителию, генерации иммунных реакций хозяина, инактивирующих терапию, и, в случае липосомальной генной терапии, доставка достаточных доз представляла собой еще одну общепризнанную проблему. Разные исследования показали, что захват, обеспечиваемый слизью, затрудняет доступ данных векторов к нижележащему эпителию и, таким образом, препятствует успешному переносу генов. Время удерживания данных захваченных частиц ограничивается скоростью обновления периферического слоя слизи, которая, в зависимости от органа, находится в интервале от секунд до нескольких часов. Для обеспечения эффективной доставки частиц, включая фармацевтические средства, через слизистые оболочки, такие частицы должны быть способны к быстрой диффузии через слизистый барьер, избегая адгезии слизи и избегая защиты, обусловленной мукоцил парным клиренсом. Кроме того, сразу после пересечения слизистого барьера, также необходимо пересечь барьер в виде клеточной мембраны. Он также предназначен для обеспечения барьера, предотвращающего доступ к клетке, в частности, для вирусов и чужеродных частиц.
Вирусные векторы могут быть также нейтрализованы существующими иммунными ответами или приобретенными иммунными ответами при повторном введении. Применение экзосом стволовых клеток или EV, которые являются иммунологически привилегированными, могут обходить данную проблему. Покрытие из ПЭГ (полиэтиленгликоль) может дополнительно усиливать свойства «малозаметности» экзосом стволовых клеток за счет эффективного экранирования поверхностных антигенов.
Кроме того, аэрозольная доставка терапевтически релевантных доз везикул может быть проблематичной для многих стандартных небулайзеров. Везикулы в водной наносуспензии или коллоиде могут агрегировать или образовывать агломераты. Силы, действующие между частицами, особенно в концентрированных нанофлюидах, приводят к повышенной вязкости, которая, в свою очередь, приводит к очень значимым трудностям в аэрозолизации. Кроме того, агрегация или агломерация приводит к получению везикул, не образующих истинную суспензию или коллоид, поскольку везикулы не распределяются по всей жидкости, а скапливаются или прилипают к стенкам сосуда, в котором держат наносуспензию или коллоид.
Эффективная доставка груза, в частности нуклеотидных последовательностей и белков, к клеткам дыхательного эпителия легкого остается общей проблемой для всех подходов.
В Kooijmans et, «PEGylated and targeting extracellular vesicles display enhanced cell specificity and circulation time», Journal of Controlled Release, vo. 224, 7 January 2016, стр. 77-85 описано «декорирование» внеклеточных везикул лигандами направленного действия, конъюгированными с полиэтиленгликолем, для улучшения специфичности везикул и продления их периода нахождения в системе кровообращения перед клиренсом. В данной статье описано получение внеклеточных везикул из клеток нейробластомы мыши и в ней не рассматриваются проблемы проникновения через слизь или проникновения через клеточную мембрану. В действительности в данной статье было указано на то, что ПЭГилирование негативно влияет на клеточное поглощение.
В Myung Soo Kim, «Engineered macrophage-derived exosomes for targeting paclitaxel delivery to pulmonary metastases: in vitro and in vivo evaluations*, Nanomedecine: Nanotechnology, Biology and Medicine, vol. 14, no. 1,2 October 2017, стр. 195-204 также описаны экзосомы, «декорированные» ПЭГ, конъюгированным с лигандом, специфичным к клетке/рецептору. Опять же, в данном документе не затрагивается какая-либо проблема, ассоциированная с доставкой везикул к легкому, и не упоминается пересечение слизистого барьера или клеточной мембраны. Кроме того, в данном документе описан груз - паклитаксел.
В US9901600 от имени Alexander Mitsialis описано применение экзосом для лечения и/или предупреждения заболеваний легкого. Экзосомы в качестве груза могут включать белки или нуклеиновые кислоты. Данные экзосомы без покрытия и не решают проблем слизистого барьера или барьера в виде клеточной мембраны. В то время как в данном документе предложено введение посредством инъекции или ингаляции, отсутствует информация о том, как может быть достигнута аэрозолизация экзосом, как требуется для ингаляции.
Желательно предложить средства доставки генной терапии к клеткам легкого, используя ингаляционную систему доставки. Также желательно предложить средство доставки, которое может проходить через слой слизи и проникать через клеточные мембраны, дополнительно средство доставки должно переноситься в композиции, которая может быть превращена в аэрозоль.
Цель настоящего изобретения заключается в предложении везикулы, в частности экзосомы, которая может быть превращена в аэрозоль и проходить через слой слизи с проникновением в клетки легкого. Еще одна цель изобретения заключается в предложении везикул, в частности экзосом, для применения в терапии, конкретно для терапии заболеваний легкого.
Краткое изложение сущности изобретения
Согласно изобретению предложена аэрозолизируемая композиция, содержащая внеклеточные везикулы из мезенхимальных стволовых клеток (МСК), имеющие поверхностное покрытие из гидрофильного полимера полиэтиленгликоль (ПЭГ), причем данные везикулы несут груз, содержащий одну или более из микроРНК (miR), анти-MIR, мРНК, длинной некодирующей РНК, кольцевой РНК, малой интерферирующей РНК, короткой шпилечной РНК, piwi-взаимодействующей РНК, последовательности РНК кластерных коротких палиндромных повторов, разделенных регулярными промежутками (CRISPR, от англ. clustered regularly interspaced short palindromic repeats), модификаций вышеуказанного или искусственно сконструированных последовательностей нуклеиновых кислот, белка, цитокина или липида.
Такие композиции подходящим образом содержат везикулы с поверхностным покрытием в виде коллоида или суспензии. Покрытие из гидрофильного полимера обычно приводит к получению везикул, которые могут образовывать суспензию или коллоид в водной композиции и могут проникать через слизь и поступать в клетки с доставкой терапевтического груза.
Согласно изобретению также предложены собственно везикулы и такая композиция, содержащая везикулы, например, в виде коллоида или суспензии для применения в терапии, где везикулы имеют терапевтический груз и поверхностное покрытие из гидрофильного полимера.
Такой груз может содержать микроРНК (miR), последовательность нуклеиновой кислоты, модулирующую микроРНК, такую как анти-miR/антагомир, или другие некодирующие последовательности нуклеиновых кислот, которые могут менять экспрессию генов в пораженных заболеванием клетках-мишенях (например, длинная некодирующая РНК или кольцевая РНК, которые могут действовать как губки микроРНК, таким образом, оказывая действия анти-miR; малую интерферирующую РНК или короткую шпилечную РНК, для специфичного нокдауна гена, piwi-взаимодействующую РНК для сайленсинга генов или транспозонов, участвующих в заболевании, и т.д.), мРНК, липиды, белки, цитокины или малые молекулы для доставки к клеткам-мишеням. Все из нуклеиновых кислот могут быть изменены или последовательность может быть сконструирована для модифицированных эффектов. Такие грузы могут содержать одну или более или две или более или все из следующих: микроРНК (miR), мРНК, анти-miR, другие встречающиеся в природе или искусственно полученные последовательности нуклеиновых кислот, липид, белок, цитокин и низкомолекулярное терапевтическое средство. Такие грузы могут быть введены, например, в экзосомы во время биогенеза их родительскими клетками (например, посредством сверхэкспрессии микроРНК посредством генетической модификации продуцирующего типа клеток), или терапевтическая молекула может быть введена в экзосому после ее выделения из культуральной среды (например, с использованием электро по рации, теплового шока, растворов/реагентов для переноса или других способов введения малой молекулы, белка или последовательности нуклеиновой кислоты, такой как анти-микроРНК, мРНК или микроРНК). Когда везикулы, например, экзосомы, вводят в цитоплазму клетки-мишени легкого, они могут обеспечивать лечение кистозного фиброза (CF), хронической обструктивной болезни легких (ХОБЛ), раковых заболеваний легкого, идиопатического легочного фиброза, обструктивной болезни легких, астмы, легочной гипертензии, бронхолегочной дисплазии и других патологических состояний и заболеваний легкого.
Согласно изобретению также предложена композиция, как описано выше, для применения в терапии.
Согласно изобретению также предложен способ лечения пациента, имеющего заболевание легкого, включающий:
предоставление композиции, как описано выше;
образование аэрозоля везикул композиции; и
ведение данного аэрозоля пациенту.
Подробности Изобретения
Предоставление поверхностного покрытия из гидрофильного полимера на везикулах уменьшает их склонность к агрегации или агломерации, приводя к получению коллоида или суспензии везикул с покрытием, которые могут быть превращены в аэрозоль, и также обладают способностью проходить через слизь, проникая в клетки, и, при наличии, осаждают свой дополнительный терапевтический груз, который был включен в EV, или в результате генетической модификации EV-продуцирующих клеток, или иным образом включенный экзогенный груз.
Предпочтительно везикулы представляют собой экзосомы, используемые, например, в нижеприведенных примерах. В качестве альтернативы, везикулы могут представлять собой микровезикулы, мембранные пузырьки или мембранные частицы.
Предпочтительно, везикулы, в частности экзосомы, происходят из животных клеток, более предпочтительно из клеток человека. Клетки могут представлять собой соматические клетки или стволовые клетки.
В предпочтительных воплощениях изобретения экзосомы происходят из мезенхимальных стволовых клеток, главным образом человеческих MCK (hMCK). Такие везикулы демонстрируют присущие терапевтические свойства, и доставка данных везикул в клетки-мишени, как было показано, модулирует фиброз, воспаление, регенерацию тканей и способствующие выживанию действия. Боле предпочтительно, везикулы, в частности экзосомы, происходят из hTERT (от англ. human Telomerase reverse transcriptase - обратная транскриптаза теломеразы человека) иммортализованных мезенхимальных стволовых клеток, происходящих из жировой ткани или костного мозга. МСК могут быть генетически модифицированы для сверх экспрессии конкретных miR; существуют имеющиеся в продаже конструкции со с верх экспрессией микроРНК и анти-miR (например, лентивекторы XMIRXpress от Systems Biosciences, которыми можно трансформировать клетки для сверхэкспрессии предпочтительных miR/антиR, в то время как Х-мотив направляет высокие концентрации микроРНК в экзосомы). МСК могут упаковывать данные miR в экзосомы или другие везикулы, которые могут быть собраны и очищены, используя стандартные методики для применения, как описано. Иммортализованные МСК являются идеальным источником - банком клеток, обладающим огромной способностью к стабильной сверхэкспрессии грузов в виде нуклеиновых кислот, полученных методами биоинженерии, и распространением, которое является идеальным для продукции терапевтических средств на основе везикул, в частности, экзосом, на промышленном уровне.
Везикулы, в частности экзосомы, могут быть использованы для лечения многих болезненных состояний. Везикулы с гидрофильным полимерным покрытием могут быть аэрозолизированы и, при осаждении в соответствующих областях легкого, также лучше способны проходить через слизь и неожиданно, несмотря на поверхностную модификацию, они демонстрируют сохраняющуюся способность к быстрому пересечению клеточной мембраны. В связи с этим, везикулы по изобретению особенно подходят для того, чтобы быть приспособленными для применения в лечении патологических состояний легкого, таких как кистозный фиброз (CF), хроническая обструктивная болезнь легких (ХОБЛ), рак легкого, астма, легочная гипертензия, острый респираторный дистресс-синдром (ОРДС), идиопатический легочный фиброз и другие состояния, воздействующие на легкие, но не ограничивающихся ими.
Уязвимые органы, в частности, дыхательные пути, секретируют вязкоупругую слизь для захвата и исключения чужеродного патогена и ультратонких частиц. В результате, слизь обеспечивает значительный барьер для проникновения терапевтических средств, доставляемых в легкие. В то время как низко молекулярные лекарственные средства обычно могут проникать через слизистый барьер, что приводит к тому, что аэрозольная доставка таких терапевтических средств является успешным способом доставки, слизистый барьер оказался очень проблематичным для доставки биологических молекул, таких как агенты переноса генов или векторы для применения в генной терапии.
Везикулы, в частности экзосомы, по изобретению содержат покрытие из гидрофильного полимера. Преимущественно обнаружено, что данные везикулы с покрытием обладают улучшенными свойствами образования аэрозоля. Отдельно обнаружено, что везикулы с покрытием легче проникают через слизь легкого, облегчая доступ к ткани легкого в лечении заболевания легкого или в других терапевтических применениях, при которых активные вещества доставляются в или через ткань легкого.
Обнаружено, что везикулы с покрытием, в частности экзосомы, по изобретению образуют суспензии и/или коллоиды, которые обладают пониженной вязкость, по сравнению с экзосомами без покрытия, полагают, что данный эффект является результатом стерической стабилизации экзосом в наносуспензии/коллоиде. Образование суспензии, имеющей дисперсные частицы, или коллоида облегчает образование аэрозолей с аэрозольными частицами в интервале размеров, подходящем для доставки к легким, и содержащих полезные концентрации везикул. Данные композиции полезны для доставки непосредственно везикул, например, экзосом и также полезны для доставки везикул, содержащих дополнительные компоненты для терапевтического применения, посредством доставки в, или будучи включенными в, или на везикулах. Такие экзосомы могут демонстрировать дополнительные специфичные в отношении клетки нацеливающие группировки, или присоединенные (например, посредством клик-химии), или группировки, которые были экспрессированы в результате генной инженерии. Такие нацеливающие лиганды могут усиливать поглощение экзосом в конкретных типах клеток (например, фибробластах, иммунных клетках, эндотелиальных или эпителиальных клетках).
Гидрофильные полимеры, используемые в изобретении, таким образом, приводят к получению везикул, которые демонстрируют уменьшенную агрегацию или агломерацию в растворе, образующем коллоид и суспензию. Подходящим образом, полимеры являются по существу неионными и по существу незаряженными. Один результат заключается в том, что экзосомы с покрытием по изобретению обычно имеют поверхностный заряд, который уменьшен, по сравнению с экзосомами без покрытия и имеет тенденцию к по существу нейтральному суммарному поверхностному заряду.
Поверхностное покрытие из гидрофильного полимера - полиэтиленгликоля (ПЭГ) указывает на то, что ПЭГ находится на внешней поверхности внеклеточной везикулы, таким образом, что доступна его функциональность. Это позволяет свойствам ПЭГ влиять на свойства внеклеточной везикулы, например, ее гидрофильные свойства. Ввиду этого, ПЭГ не является внутренней частью другого покрытия и не имеет какого-либо значительного покрытия на ПЭГ. ПЭГ-покрытие представляет собой самое наружное покрытие на внеклеточных везикулах. Дополнительные лиганды или другие элементы не конъюгированы на дистальных концах ПЭГ. ПЭГ образует по существу единственное покрытие на внеклеточных везикулах.
Низко молекулярный ПЭГ предпочтительно имеет молекулярную массу меньше чем 3 кДа и предпочтительно обеспечивает EV с плотным поверхностным покрытием 65% или больше, которого достаточно для по существу нейтрализации поверхностного заряда EV (а именно, от - 8 мВ до 0 мВ).
Предпочтительно поверхностное покрытие будет покрывать по меньшей мере 65%, более предпочтительно по меньшей мере 70% и наиболее предпочтительно по меньшей мере 75% поверхности везикул с обеспечением достаточной стерической стабилизации и нейтрализации поверхностного заряда с минимизацией агломерации и агрегации экзосом. В конкретных примерах, проиллюстрированных ниже, поверхностное покрытие приблизительно 70% или больше наблюдали в экзосомах с покрытием, которые образовывали аэрозоли и проникали через слизь.
Полимер поверхностного покрытия обычно имеет низкую молекулярную массу, предпочтительно меньше 5 кДа, более предпочтительно меньше 4 кДа, более предпочтительно меньше 3 кДа и наиболее предпочтительно от 2 кДа до 3 кДа. В конкретных примерах, проиллюстрированных ниже, покрытие было образовано в результате присоединения полимерных цепей приблизительно 2 кДа к поверхности везикул. В результате данные цепи не обеспечивают стерического искажения или несоответствия цепям муцина, которые могут встречаться в слизистом слое легких. Полимеры подходящим образом включены в концентрации, которая достаточна для по существу нейтрализации поверхностного заряда везикул и предотвращения агломерации или агрегации; такое плотное покрытие будет обеспечивать конформацию гидрофильных цепей в оптимальной ориентации/конформации «кисти». Требуемая концентрация полимера зависит от многих переменных, таких как длина цепи выбранного полимера, эффективность полимера, связывающегося с поверхностью везикул, и молярная концентрация и распределение частиц по размеру образца выделенных везикул. Например, полимер, инкубируемый с 100 нм экзосомами, можно использовать в концентрации приблизительно 8 мол.% или больше для обеспечения плотного покрытия и подходящей пост модификации характеристик везикул. Однако, нужно следить за тем, чтобы избежать избыточно высоких концентраций полимера, которые могут приводить к повреждению везикулярной мембраны из-за разрушающих действий, опосредованных детергентом, при более высоких концентрациях.
Везикулы по изобретению обычно имеют размер в интервале 20-1000 нм, боле подходящим образом в интервале до 300 нм. Экзосомы обычно имеют размер в интервале 30-150 нм, и микровезикулы обычно имеют размер в интервале 100-1000 нм. Обычно, экзосомы по изобретению с поверхностным покрытием имеют размер в интервале вплоть до 200 нм, предпочтительно 20-200 нм и наиболее предпочтительно 30-180 нм. Размеры могут быть измерены, используя методики динамического рассеяния света (Degiorgio, V., et al., 1979) или анализ траекторий наночастиц (NTA, от англ. па no particle tracking analysis), используемый в нижеприведенных примерах).
Как указано, предпочтительно везикулы происходят из мезенхимальной стволовой клетки, и, в связи с этим, имеют большое преимущество обладания присущими регенеративными и терапевтическими свойствами. Исследования показали, что немодифицированные МСК экзосомы и сами МСК демонстрируют антиапоптотическое, проангиогенное и противовоспалительное действие за счет множества факторов (например, микроРНК, мРНК, IncPHK, KGF (от англ. keratinocyte growth factor - фактор роста кератиноцитов), HGF (от англ. hepatocyte growth factor -фактор роста гепатоцитов), VEGF (от англ. vascular endothelial growth factor - фактор роста эндотелия сосудов), IGF-1 (от англ. insulin-like growth factor -инсулиноподобный фактор роста), TIMP3 (от англ. tissue inhibitor of metalloproteinase - тканевый ингибитор металлопротеиназ)) и за счет целого ряда механизмов (например, в результате поляризации макрофагов из провоспалительного фенотипа М1 в противовоспалительный фенотип М2, модуляции презентации антигена, модуляции высвобождения цитокинов, усиления гликолиза через АТФ-образующий ферментативный перенос и активации киназ выживания (например, ERK (от англ. extracellular signal-regulated kinase - киназа, регулируемая внеклеточными сигналами) и АКТ («АК» от англ. AKR mouse strain - линия мышей AKR, «Т» от англ. thymoma - тимома) посредством CD73), и за счет уменьшения активации комплемента с помощью CD59).
Данные полезные действия могут быть усилены посредством добавления релевантного груза к везикуле, например, экзосомам. В действительности, каким бы ни был источник везикулы, груз может обеспечивать полезное терапевтическое действие.
Везикулы, в частности экзосомы, могут также нести груз, подлежащий доставке к клеткам-мишеням. Предпочтительно, данный груз представляет собой микроРНК (miR) или анти-микроРНК (анти-miR). В качестве альтернативы, груз может представлять собой мРНК или другую эндогенную или экзогенную конструкцию нуклеиновой кислоты, сконструированную последовательность, химически модифицированную или синтетическую версию вышеуказанного, белок или фармацевтическую малую молекулу. Нуклеиновая кислота может иметь сконструированную последовательность или модифицированные пары оснований, и быть загружена в экзосомы посредством множества механизмов.
Везикулы также полезны в случае in vivo доставки в легкие CRISPR-CAS9 или других способов редактирования генов, поскольку внутриклеточная доставка к клеткам, расположенным под слизистым барьером, является проблемой, которая должна быть решена для удачного редактирования генов в случае респираторных заболеваний.
В некоторых воплощениях изобретения мРНК hTERT и каталитическая субъединица фермента hTERT могут быть включены в экзосомы в результате лентивирусной hTERT-иммортализации EV-продуцирующих стволовых клеток, данные активные группировки полезны для борьбы со старением клеток в затронутых болезнью клетках-мишенях (например, клетки-предшественники альвеолярного эпителия АЕС2), которое является признаком ХОБЛ, ИЛФ (идиопатического легочного фиброза) и все больше и больше имеет отношение к другим заболеваниям легкого. Аналогично, другой большой компонент фермента теломераза, TERC (hTR), может быть включен в EV в виде молекулы РНК. Данные грузы будут действовать в клетках-мишенях с усилением возможностей EV стволовых клеток в регенеративной медицине. Важно, что данные эффекты будут временными и обратимыми в природе, поскольку они будут действовать в цитоплазме и не будут встраиваться в геном клеток-мишеней, что является важной особенностью регулирования, поскольку никакие продолжительные воздействия не будут доступны наблюдению в клетках-мишенях или в действительности любых дочерних клетках, образованных клетками-мишенями.
МикроРНК (miR) представляют собой малые некодирующие молекулы РНК, обычно содержащие порядка 20-25 нуклеотидов, обнаруженные в растениях, животных и некоторых вирусах, которые главным образом функционируют в РНК-сайленсинге и посттранскрипционной регуляции экспрессии генов. miR функционируют посредством спаривания оснований с комплементарными последовательностями в пределах молекул мРНК и являются ключевыми регуляторами экспрессии генов. Груз может также представлять собой анти-miR или антагомиры (антаго-miR). Груз может быть подобран для применимости к состоянию, подлежащему лечению, или модуляции экспрессии генов в конкретном типе клеток-мишеней. Обычно, состояния или болезненные состояния приводят к повышающей регуляции одних miR и понижающей регуляции других miR. Когда miR подвергается понижающей регуляции, лечение с заменой данной miR может быть эффективным, в то время как в том случае, когда miR подвергается повышающей регуляции, может быть эффективным предоставление анти-miR или антагомира.
Грузы EV могут представлять собой другие некодирующие последовательности нуклеиновых кислот, которые могут менять экспрессию генов в пораженных заболеванием клетках-мишенях. Например, длинные некодирующие РНК или кольцевые РНК (эндогенные или искусственно сконструированные), которые могут действовать как «губки» или «швабры» микроРНК, таким образом, оказывая анти-miR действия; малые интерферирующие РНК или короткие шпилечные РНК, для нокдауна конкретного гена; и piwi-взаимодействующие РНК для сайленсинга гена или транспозона, имеющего отношение к заболеванию.
Когда груз представляет собой miR, и лечение предназначено для ХОБЛ, предпочтительно груз выбран из одной или более из: miR-125b-5p, miR-125b-1-3p, miR-513a-5p, miR-34c, miR-452, miR-146a, Let-7c, miR-576-3р, miR-513a-3p, miR-923, miR-937, miR-422a, miR-25, miR-99b, miR-24 и miR-187. Более предпочтительно, miR выбрана из miR-125b-5p, miR-125b-1-3p и miR-513a-5p, которые представляют новые генетические мишени для купирования заболевания. Выбранные микроРНК, hsa-miR-125b-5p, hsa-miR-125b-1-3p и hsa-miR-513a-5p, представляют собой ключевые регуляторы в ХОБЛ, нацеленные на важные гены, участвующие в ХОБЛ-патогенезе. Данные miR недостаточно экспрессируются у курильщиков с ХОБЛ, по сравнению с курильщиками без ХОБЛ, и, как известно, нацелены на и подвергают понижающей регуляции несколько важных мРНК в результате взаимодействий микроРНК-мишень (MTI - от англ. micro RNA-Target-Interact ion), которые участвуют в патогенезе заболевания.
Конкретно, микроРНК 125b-5р представляет собой подходящий груз и, как было показано, нацелена на и подвергает понижающей регуляции следующие важные гены, ассоциированные с ХОБЛ:
[1] ADAMTS4 (Aggrecanase-1) в исследованиях остеоартритических хондроцитов человека. ADAMTS4 представляет собой протеолитический фермент, участвующий в разборке внеклеточного матрикса и является наиболее сверхэкспрессируемой мРНК (увеличение в 8,91 раз) у курильщиков с ХОБЛ в сравнении с курильщиками без ХОБЛ. Кроме того, показано, что ADAMTS4 является антиангиогенным за счет связывания VEGF, понижающая регуляция ADAMTS4 может, таким образом, помогать в регенерации ангиогенеза альвеолярных компартментов; [2] SFRP5, антагонист WNT, чьи уровни, как было обнаружено, значимо выше при ХОБЛ, по сравнению со здоровыми контролями;
[3] DKK3, notch-антагонист, чья понижающая регуляция может помогать регенерации альвеол;
[4] уровень экспрессии EGFR (от англ. Epidermal Growth Factor Receptor -рецептор эпидермального фактора роста) в клетках бронхиального эпителия значимо выше у курильщиков, по сравнению со здоровыми контролями, и также выше у курильщиков с ХОБЛ, по сравнению с курильщиками без ХОБЛ. EGFR играет важную роль в регуляции продуцирования слизи и гиперплазии бокаловидных клеток в дыхательном эпителии;
[5] Рецептор для IL-6 (от англ. interleukin - интерлейкин) (IL6R), широко известный ключевой путь воспаления при ХОБЛ;
[6] ММР13, член подсемейства коллагеназ ММР (от англ. matrix metalloproteinase - матричная металлопротеиназа), который, как было показано, подвергается повышающей регуляции в альвеолярных макрофагах при ХОБЛ и альвеолярных клетках типа II. ММР13 участвует как главный медиатор патогенеза заболевания в ответ на сигаретный дым и воздействие вируса гриппа PR8 в животной модели обострений ХОБЛ;
[7] ANGPT2, антагонист ANGPT1 (ангиопоэтин). ANGPTL1 имеет пониженные уровни при запущенной ХОБЛ в сравнении со стабильной фазой/нормальными уровнями (ассоциированэ с сосудистой регрессией, уровни сильно коррелируют с потерей функции легкого);
[8] АРС (от англ. adenomatous polyposis coli - аденоматозный полипоз кишечника), регулятор сигнального пути WNT, ингибирующий, поскольку АРС является компонентом комплекса разрушения бета-катенина;
[9] МАРК14 (ингбирование Р38 является перспективным в клинических испытаниях по ХОБЛ); и
[10] Разные другие мишени, которые сильно ассоциированы с ХОБЛ: SGPL1, BMPR1B, BTG2, СЕВРА, Fas, FGFR2, FZD6, GLI1, НК2, HMGA1, HMGA2, IGF1R, IGF2, ММР2, ММР26, MUC1, SMAD4, STAT3, TNF, TNFAIP3, ТР53 и BCL2 - про- и антиапоптозные медиаторы, соответственно, а ВАК1, ВС1_2-антагонист/киллер 1, также представляет собой мишень, TP53INP1, VDR, ERBB2 и ERBB3.
Кроме того, усиленная экспрессия miR-125b-5p показала ослабление LPS-индуцированного острого повреждения легких и воспаления у мышей.
MiR-125b-1-3р является подходящим грузом и, как было показано, нацеливается на ХОБЛ-релевантные гены:
[1] Повышенный уровень экспрессии TACSTD2 (aka TROP2) в базальных клетках дыхательных путей потенциально способствует ремоделированию дыхательных путей при ХОБЛ и ремоделированию дыхательных путей за счет повышенного уровня гиперплазии базальных клеток;
[2] SGPL1, сфингозин-1-фосфат (S1P) деградирующий фермент с 4,5-кратным повышением уровня экспрессии в альвеолярных макрофагах при ХОБЛ в сравнении с контролями. Способствует дефектам альвеолярных макрофагов в фагоцитозе апоптотических клеток (эффероцитоз) при ХОБЛ;
[3] FRZB, антагонист WNT, чья понижающая регуляция, как ожидают, демонстрирует регенерирующее действие;
[4] ТР53, повышенный уровень экспрессии проапоптотического Р53 в альвеолярных клетках типа II ХОБЛ. Таким образом, ожидается, что понижающая регуляция оказывает действие, способствующее выживанию, следует отметить, что 125b-5р также нацелена на Р53;
[5] S1PR1 (адгезия клеток, иммуномодуляция);
[6] BIK, ускоряет программируемую смерть клеток;
[7] MTFP1;
[8] МАР2К7, участвующий в ответах на окружающие условия/клеточных ответах;
[9] BGLAP и [10] ITGA9.
MiR-513a-5p также представляет собой подходящий груз для везикул по изобретению, благодаря своим релевантным воздействиям на экспрессию гена ХОБЛ:
[1] CBL-miR-513a-5p, как показано, нацеливается на CBL, негативно влияя на пути фосфоинозитид-3-киназы (PI3K)-AKT и ядерного фактора-кВ (NF-кВ) через их последующие гены, тирозинкиназа селезенки (SYK) и рецептор эпидермального фактора роста (EGFR). Повышенный уровень 513а-5р, таким образом, уменьшал секрецию цитокиновых факторов - интерлейкина (IL)-10, γ-интерферона (γ-IFN), фактора некроза опухоли-α (TNF-α - от англ. tumor necrosis factor alpha) и IL-12;
[2] CB274 (лиганд программируемой смерти 1) При аде но карциномах легкого, ассоциированных с эмфизематозными буллами (ЕВ - от англ. emphysematous bullae), белок PD-L1 экспрессируется чаще, чем при аденокарциномах легкого без ЕВ. PD-L1 - иммунная контрольная точка не полностью изучена, но разрегулирована при ХОБЛ. Белок PD-L1 индуцируется в разных клетках нелимфоидной ткани, включая эпителиальные, эндотелиальные, гладкомышечные клетки, в ответ на воспалительные цитокины. Широко известные сопутствующие заболевания ХОБЛ и рак легкого делают CD274 мишенью, представляющей особый интерес;
[3] miR-513a-5p имеет много дополнительных и интересных виртуально предсказанных мишеней для исследования (TargetScan 7.2): AGER (кодирует RAGE, с повышающей регуляцией при ХОБЛ, мыши RAGE КО демонстрируют пониженный уровень воспалительных реакций, и повышающая регуляция RAGE вызывает повышенный уровень апоптоза альвеолярных клеток); ADAM9 и INHBA (уровень aka Активин А, суперсемейство TGFB, повышен при ХОБЛ и ассоциирован с ограничением воздушного потока).
Кроме того, miR, идентифицированные в данном документе как подходящие для данного изобретения, продемонстрировали функциональное MTI (слабое) для гораздо большего количества ХОБЛ-ассоциированных генов.
Рак легкого представляет собой высоко значимое и смертельное сопутствующее заболевание ХОБЛ. Повышенный риск рака легкого, демонстрируемый индивидами с ХОБЛ, не зависит от стажа курения. В приведенных ниже примерах авторы изобретения нацелены на снижение риска рака легкого посредством модуляции miR-21-5p в пораженных ХОБЛ и окружающих тканях легкого, такое снижение уровня miR-21, как считают, способно снижать риск развития рака легкого у пациентов с ХОБЛ и будет достигнуто посредством генетической модификации стволовых клеток для сверхэкспрессии последовательности анти-miR к miR-21 (hsa-miR-21-5p), которая будет загружаться в EV. Ингибитор miR-21-5р представляет собой или короткую РНК с обратнокомплементарной последовательностью микроРНК, или он может принимать вид последовательности кольцевой РНК с несколькими сайтами связывания miR-21-5р и применением концевых последовательностей интрона для управления образованием кольцевой молекулы. В качестве альтернативы, встречающаяся в природе или модифицированная длинная некодирующая последовательность РНК может быть использована для достижения уменьшения уровня miR-21.
Когда груз представляет собой miR и лечение предназначено для CF, предпочтительно груз содержит miR-17 для уменьшения гипервоспаления в легком при CF. CF представляет собой мультисистемное заболевание, но подавляющее большинство осложнений и смертельных случаев встречаются из-за заболевания, связанного с легкими. Введение противовоспалительных экзосом МСК по изобретению с грузами в виде микроРНК подходит для существенного улучшения функции легких. Кроме того, ожидают, что экзосомы, доставляемые к легким, обладают степенью системного поглощения и системным действием за счет большой площади поверхности и сосудистой системы легких.
Груз может дополнительно или в качестве альтернативы содержать мРНК. Когда лечение предназначено для CF, груз может представлять собой модифицированную мРНК CFTR. Например, модифицированная мРНК CFTR может включать мутантные или удаленные сайты связывания miR в отношении miR-101, miR-223, miR-494 и/или miR-509-3р, поскольку они подвергаются повышающей регуляции при CF и подавляют экспрессию CFTR. В качестве альтернативы, введенная мРНК CFTR может включать синтетические 3' и 5' UTR (от англ. untranslated region - нетранслируемая область), которые сконструированы таким образом, чтобы свободно экспрессироваться в большинстве типов клеток, и будут также лишены сайтов связывания miR, подвергающихся повышающей регуляции при указанном заболевании. Недавние исследования показали, что блокада сайтов-мишеней miR или сайт-направленный мутагенез в данных сайтах приводили к повышенному уровню экспрессии CFTR. Кроме того, модифицированная мРНК CFTR устойчива к in vivo внутриклеточной деградации, опосредованной микроРНК, и, вследствие этого, имеет более продолжительное время полужизни для продолжительной экспрессии CFTR in vivo. Кроме того, может достигаться усиленная эффективность, по сравнению с блокаторами сайтов-мишеней, поскольку функциональный белок CFTR будет транслироваться с кодирующей последовательности белка дикого типа, в отличие от спасения экспрессии эндогенной мутантной мРНК CFTR, которая будет вызывать клеточный стресс из-за реакций несвернутых белков.
Когда экзосомы используют в лечении рака легкого, в частности аденокарциномы, груз может представлять собой одну или более из miR-126-3p, miR-218-5p, miR-486-5p, miR-145-5p, miR-338-3p, miR-195-5p, miR-143-3p, miR-139-5p, miR-126-5p, miR-144-3p, miR-34c-5p, miR-30a-3p, let-7c-5p, miR-451a, miR-1-3p и miR-133a-3p. Дополнительно или в качестве альтернативы, груз может представлять собой одну или более из анти-miR miR-21 -5р, miR-210-3p, miR-182-5p, miR-183-5p, miR-135b-5p, miR-9-3p, miR-96-5p, miR-205-5p, miR-31-5p, miR-708-5p, miR-196b-5p, miR-375, miR-345-5p, miR-200a-3p и miR-130b-3p.
Другие подходящие грузы могут включать микроРНК или анти-miR, выбранные для лечения других заболеваний легкого, например, идиопатического легочного фиброза.
Композиции подходящим образом предложены в виде суспензий или коллоидов для доставки в легкие посредством аэрозолизации с использованием небулайзеров. Предпочтительно суспензии или коллоиды представлены на водной основе. Композиции могут также включать другие фармацевтически приемлемые вспомогательные вещества, такие как соли, поверхностно-активные вещества, стабилизаторы, консерванты, крио протекторы, буферные и рН-регулирующие средства, но не ограниченные ими.
Композиции по изобретению предназначены для образования аэрозолей, содержащих капли, которые могут быть доставлены к легкому пациента. Подходящие размеры и устройства, образующие аэрозоль, известны в данной области, и аэрозоли обычно содержат капли размером 10 мкм и меньше. Превращенные в аэрозоль капли больше 5 мкм обычно считаются слишком большими для респираторной доставки в условиях нормального дыхания. Для достижения и прилипания к бронхиальным и альвеолярным клеткам капли аэрозоля являются очень подходящими в интервале 1-5 мкм, а именно аэрозоль включает капли в данном интервале. Знакомым сданной областью будет понятно, что капли больше чем 5 мкм в диаметре могут осаждаться в желательных областях легкого, когда соответствующие ограничения накладываются на скорости инспираторного потока и скорости аэрозоля. Большие по размеру капли, будучи сферическими, несут гораздо больший объем, чем меньшие по размеру капли, например, капля диаметром 10 мкм имеет в тысячу раз больший объем (и, следовательно, емкость загрузки дозы), чем капля диаметром 1 мкм. Нацеливание осаждения аэрозоля в желательной области легкого можно эффективно контролировать посредством варьирования устройства доставки аэрозоля и параметров композиции, влияя подходящим образом на размер капель аэрозоля и скорости ингаляционного потока. Размер капли аэрозоля может быть измерен, например, посредством значений масс-медианного аэродинамического диаметра (MMAD - от англ. mass median aerodynamic diameter), которые определены как диаметр аэрозоля, при котором 50% частиц по массе больше, и 50% - меньше.
Для достижения удачной аэрозолизации обычно предпочтительно, чтобы вязкость коллоида или суспензии не была слишком высокой; данные параметры, однако, также зависят от типа и модели устройства. Предпочтительно, вязкость должна быть меньше чем 6 сП, например, меньше чем 4 сП, более предпочтительно меньше чем 3 сП и наиболее предпочтительно меньше чем 2 сП. Когда коллоид или суспензия слишком вязкие, способ аэрозолизации может приводить к пенообразованию и разбрызгиванию и, таким образом, значимо уменьшать аэрозолизацию, или если образованием аэрозоля возможно, образующиеся капли могут быть слишком маленькими для полезного осаждения в отношении предполагаемой области легкого. Равным образом, когда частицы в суспензии или коллоиде имеют тенденцию к агрегированию или агломерации, тогда агрегированные частицы не аэрозолизируются, а могут, напротив, оставаться в коллоиде или суспензии возрастающей концентрации в резервуаре небулайзера. Это, в таком случае, приводит к увеличению вязкости суспензии или коллоида и неспособности к аэрозолизации или уменьшениям в доставляемой дозе. Свободные везикулы, включая экзосомы, имеют ярко выраженную тенденцию к агрегации или агломерации, образуя вязкий коллоид или суспензию и, таким образом, не могут быть эффективно аэрозолизированы. Однако, обнаружено, что везикулы по изобретению, имеющие покрытие из гидрофильного полимера, демонстрируют значимо уменьшенную тенденцию к агрегированию и агломерации. В результате может быть образован коллоид или суспензия подходящей вязкости, и предложенные в данном документе везикулы с покрытием могут быть успешно аэрозолизированы.
Размер образованных аэрозольных капель обычно обратно пропорционален вязкости распыляемой композиции. Вязкость наноразмерных коллоидных систем, таких как композиции экзосом, резко возрастает, поскольку объемная доля экзосом возрастает. Как упомянуто ранее, стерическая стабилизация везикул по изобретению может значимо ослаблять данный эффект. Данный эффект может быть использован для содействия осаждению аэрозоля образованных аэрозолей в целевой области легкого, когда меньшие по размеру капли (с постоянной скоростью и скоростью инспираторного потока) будут осаждаться в малых дыхательных путях и периферических/дистальных областях легкого, и большие по размеру капли будут иметь более центральную картину осаждения. Например, в случае кистозного фиброза картина центрального осаждения аэрозоля является желательной, нацеливаясь на области бронхов и крупные дыхательные пути, в которых не происходит газообмен. Напротив, в случае эмфиземы, дистальные альвеолярные области и малые дыхательные пути являются главными областями, представляющими интерес, и предпочтительны распределения по размерам меньших по размеру капель.
Аэрозолизацию можно проводить, используя небулайзер. В небулайзерах используется кислород, сжатый воздух или газы и/или насадки или мощность ультразвука для того, чтобы разбить растворы, суспензии и коллоиды на мелкие капли, образуя аэрозоль, который можно вдыхать. Небулайзеры ранее конструировали таким образом, чтобы аэрозолизировать малые молекулы, и все имеющиеся в продаже типы являются успешными в этом, если лекарственное средство находится в растворе. Однако, используемый тип небулайзера более важен в случае больших по размеру биологических агентов, которые могут повреждаться небулайзерами, которые передают раствору, суспензии или коллоиду слишком много энергии и генерируют достаточно напряжения при гидродинамической нагрузке для подвергания биологических молекул усилию сдвига, что приводит к потере активности. Например, ультразвуковые небулайзеры абсолютно не подходят для доставки наносуспензий, и компрессорные небулайзеры вызывают непрерывную рециркуляцию аэрозоля, приводя к повторным сдвиговым напряжениям, которые могут повреждать биологические молекулы, такие небулайзеры также предпочтительно превращают в аэрозоль водный носитель, приводя к концентрированию композиции в резервуаре. «Мертвый объем» композиции, который не аэрозолизирован, а остается в резервуаре, обычно составляет 1 мл в компрессорных небулайзерах, и это абсолютно неприменимо и коммерчески нецелесообразно, исходя из перспективы стоимости товаров относительно передовых терапий, таких как композиции экзосом.
Из официально одобренных типов небулайзеров вибрационные меш-небулайзеры особенно подходят для доставки везикул, поскольку:
1. капли только меняют фазу с жидкой на воздушную один раз при генерации капель (в сравнении с «повторным использованием» и повторными сдвиговыми напряжениями в компрессорных небулайзерах);
2. ультразвуковые небулайзеры не подходят для доставки наносуспензий;
3. вибрационные меш-небулайзеры могут быть приспособлены для эффективной доставки маленьких объемов композиций с минимальными остаточными «мертвыми» объемами; и
4. компрессорные небулайзеры могут предпочтительно превращать в аэрозоль растворитель и концентрировать остающуюся композицию в резервуаре.
Небулайзеры на основе поверхностных акустических волн (SAW - от англ. Surface Acoustic wave), которые генерируют поверхностные акустические волны на пьезоэлектрических чипах, как было показано, аэрозолизируют суспензии малоустойчивых молекул нуклеиновой кислоты, как например, вакцины на основе плазмидной ДНК, и стволовых клеток. Высокочастотный, маломощный механизм SAW-распыления является полезным для того, чтобы избегать минимального повреждения биологических молекул, включая нуклеиновые кислоты. SAW-небулайзеры более успешны в аэрозолизации более вязких растворов и суспензий, чем многие другие типы небулайзера, что делает их идеальными для аэрозолизации суспензий и коллоидов, образованных из водной смеси экзосом, в частности экзосом, имеющих поверхностное покрытие из гидрофильного полимера. Вязкость жидкости определяет, в некоторой степени, размер капель, образующихся посредством SAW-небулайзера. Однако, данные типы небулайзеров на сегодняшний день не одобрены какими-либо регулирующими органами для применений в клинических исследованиях.
Примеры
Для того, чтобы помочь пониманию изобретения, конкретное воплощение с его вариантом теперь будет описано в качестве примера и со ссылкой на прилагаемые графические материалы, в которых:
Фиг. 1А и 1 В представляют собой графики зависимости концентрации от размера неПЭГилированных и ПЭГилированных экзосом в анализе траекторий наночастиц (NTA);
На Фиг. 2А и 2 В показано распределение дзета-потенциала ПЭГилированных и неПЭГилированных экзосом;
Фиг. 3A-3F представляют собой серию изображений просвечивающей электронной микроскопии ПЭГилированных и неПЭГилированных экзосом;
Фиг. 4A-3F представляют собой серию графиков по проточной цитометрии, показывающих мечение МСК и экзосом дальнекрасным красителем;
Фиг. 5А-5С представляют собой изображения конфокальной микроскопии обработанных и необработанных экзосом, которые поступили в эпителиальные клетки бронхов при кистозном фиброзе, сперва проникнув через слизистый слой в культуре на границе раздела воздух-жидкость;
Фиг. 6а и 6b представляют собой показания анализа траекторий наночастиц в случае dPBS (от англ. Dulbecco's Phosphate Buffered Saline - фосфатно-солевой буферный раствор Дульбекко) и 30 нм наночастиц золота (Аи) в качестве контролей;
Фиг. 7а и 7b представляют собой показания анализа траекторий наночастиц для Образца 1;
Фиг. 8а и 8b представляют собой показания анализа траекторий наночастиц для Образца 2;
Фиг. 9 представляет собой распределение согласно анализу траекторий наночастиц для Образца 3;
Фиг. 10 представляет собой распределение согласно анализу траекторий наночастиц для Образца 4;
Фиг.11 представляет собой распределение согласно анализу траекторий наночастиц для Образца 5;
Фиг. 12а и 12b представляют собой распределения согласно анализу траекторий наночастиц для Образца 6;
Фиг. 13 представляет собой распределение согласно анализу траекторий наночастиц для Образца 7;
Фиг. 14а представляет собой график, демонстрирующий показания измерения дзета-потенциала для ПЭГилированного и неПЭГилированного Образца 1;
Фиг. 14b представляет собой график распределений интенсивности дзета-потенциала для трех отдельных измерений для ПЭГилированного Образца 1;
Фиг. 15а представляет собой график, демонстрирующий показания измерения дзета-потенциала для ПЭГилированного и неПЭГилированного Образца 2;
Фиг. 15b представляет собой график, показывающий распределения интенсивности дзета-потенциала для трех отдельных измерений ПЭГилированного Образца 2;
Фиг. 16 представляет собой график, показывающий средние значения дзета-потенциала для ПЭГилированного и неПЭГилированного Образца 4;
Фиг. 17 представляет собой график, показывающий средние значения дзета-потенциала для ПЭГилированного и неПЭГилированного Образца 5;
Фиг. 18а представляет собой график, демонстрирующий показания измерения дзета-потенциала для ПЭГилированного и неПЭГилированного Образца 6;
Фиг. 18b представляет собой график распределений интенсивности дзета-потенциала для трех отдельных измерений для ПЭГилированного Образца 6;
Фиг. 19 представляет собой график, показывающий средние значения дзета-потенциала для ПЭГилированного и неПЭГилированного Образца 7;
На Фиг. 20 показана флуоресценция GFP (от англ. green fluorescent protein -зеленый флуоресцентный белок) для клеток ИМСК с трансдукцией EGFP(ot англ. enhanced green fluorescent protein - усиленный зеленый флуоресцентный белок)-CFTR; и
На Фиг. 21 показана флуоресценция GFP для клеток hMCK с трансдукцией EGFP-отрицательным контролем;
На Фиг. 22 показаны CFTR-EGFP-позитивные МСК, сфотографированные на флуоресцентном микроскопе после посева клеток на предметные стекла, фиксации и окрашивания ядер DAPI (синий) при увеличении х10;
На Фиг. 23 показаны CFTR-EGFP-позитивные МСК, сфотографированные на флуоресцентном микроскопе после посева клеток на предметные стекла, при увеличении х10;
На Фиг. 24 показаны CFTR-EGFP-позитивные МСК, сфотографированные на флуоресцентном микроскопе после посева клеток на предметные стекла, фиксации и окрашивания ядер DAPI (синий) при увеличении х20; и
На Фиг. 25 показаны CFTR-EGFP-позитивные МСК, сфотографированные на флуоресцентном микроскопе после посева клеток на предметные стекла при увеличении х20.
Пример 1
Целями данного примера было выделить экзосомы из человеческих мезенхимальных стволовых клеток (hMCK) и эффективно модифицировать их поверхность низкомолекулярным пол иэтиленгли колем, модифицированным липидом (липид-ПЭГ), для улучшения свойств экзосом в отношении аэрозолизации и проникновения через слизь.
Флуоресцентное мечение hMCK
Замороженные МСК костного мозга (Донор #163) размораживали перед тем, как их помещали непосредственно в 15 колб Т-175 (1×106 клеток в случае каждой колбы) в полные культуральные среды (ССМ, от англ. complete culture media), истощенные EV, с 10 нг/мл основного фактора роста фибробластов человека (bFGF - от англ. basic fibroblast growth factor), пересевали, когда они достигали 80-85%-ной конфлюэнтности. Клетки объединяли вместе перед тем, как их осаждали и ре суспендировал и в PBS, инкубировали в темноте с 2 мкг 1×106 клеток дальнекрасного красителя (возбуждение/испускание приблизительно 630/661 нм) (Cell Trace) в течение 20 мин при комнатной температуре (это осуществит мечение экзосом, поскольку дальнекрасный краситель метит белки цитоплазмы). После инкубации культуральные среды МСК добавляли для остановки реакции и дополнительно инкубировали в течение еще 5 мин перед осаждением клеток и ресуспендированием в EV-истощенной ССМ.
Культура hMCK для выделения экзосом
Меченные дальнекрасным красителем hMCK (Донор #163) культивировали в полной кондиционированной среде (ССМ - от англ. complete condition media), и экзосомы выделяли в соответствии с ранее опубликованными данными (Thery et al., 2006). Кратко, меченые МСК непосредственно высевали в 30 колб Т-175 (1×106 клеток в случае каждой колбы) в ССМ с 10 нг/мл основного фактора роста фибробластов человека (bFGF). Спустя 24 часа, среды из всех колб отбрасывали и заменяли свежими средами. За 3-4 суток клетки достигали 80%-ной конфлюэнтности, затем кондиционированные среды собирали из каждой колбы, и подсчитывали клетки из 5 колб, выбранных случайным образом. Кондиционированные среды из всех колб объединяли вместе перед выделением экзосом. Для выделения экзосом среды центрифугировали при 400 × g на протяжении 10 мин и 2000 × g на протяжении 30 мин для удаления обломков клеток и апоптотических телец, соответственно. После каждого вращения осадок отбрасывали, и использовали супернатант. Затем супернатант фильтровали с использованием 220 нм вакуум-фильтра для дополнительной очистки и хранили при 4°С до ультрацентрифугирования (Sorvall Discovery 100SE, Hitachi). Наконец, супернатант подвергали ультрацентрифугированию при 120000 × g в течение 75 мин для осаждения экзосом. Осадок ресуспенд провал и в PBS для промывки экзосом, крутили еще раз при той же высокой скорости, полученный осадок ре суспендировал и в 100 мкл PBS и хранили при -80°С до дальнейших экспериментов. Все стадии центрифугирования проводили при 4°С, и использовали стерильные пробирки.
Модификация экзосом после выделения посредством дистеароилфосфатидилэтаноламина-полиэтиленгликоля (ДСФЭ-ПЭГ)
Амин ДСФЭ-ПЭГ (2000) (Avanti Polar Lipids, Inc) приобретали в Sigma Aldrich (Wicklow, Ирландия). ПЭГилированные экзосомы получали на основе ранее опубликованных данных по получению ПЭГилированных липосом (Li and Huang et al., 2009). Конечный объем 300 мкл суспензии экзосом получали посредством смешивания 50 мкл концентрированной экзосомы с PBS и 37,8 мкл водного раствора ДСФЭ-ПЭГ (10 мг/мл) (конечная концентрация 1,26 мг ПЭГ в мл экзосомы). Образец затем инкубировали при 37°С в течение 1 ч и перемешивали на вибромешалке каждые 10 мин для предотвращения агрегации молекул ПЭГ или экзосом. После инкубации образец суспендировали в большем количестве PBS и, в конечном итоге, подвергали ультрацентрифугированию при 120000×g в течение 75 мин для отмывки несвязанных молекул ПЭГ. Затем осадок экзосом ресуспендировали в 50 мкл PBS и хранили при -80°С.
Анализ траекторий наночастиц (NTA)
Образцы как ПЭГилированных, так и неПЭГилированных экзосом анализировали посредством анализа траекторий наночастиц (NTA) (Malven UK) с использованием системы запуска NTA NanoSight NS 500, версия 3.2, используя оптимизированные и утвержденные протоколы (Maguire et al., 2017, Hole et al., 2013, Gerlach et al., 2017). Все образцы анализировали, используя NanoSight NS 500, оснащенный лазером 405 нм и длинноволновым пропускающим фильтром 430 нм. Все образцы хранили на сухом льду перед анализом. Разведения для NTA выполняли в буфере DPBS (Gibco), который был сертифицирован как не содержащий частиц посредством NTA сразу перед измерением. Образцы перемешивали на вибромешалке непродолжительное время перед загрузкой для обеспечения достаточного перемешивания и разрушения слабо связанных кластеров экзосом. Каждый образец разводили вручную в PBS с получением оптимальной концентрации частиц, подходящей для NTA (от 20 до 70 экзосом на поле обзора), причем каждый фактор разведения записывали в автоматически генерируемых отчетах. В общей сложности шесть х 60-секундных видео записывали для каждого образца экзосом, и порог обнаружения во время анализа выбирали с обеспечением того, чтобы анализу подвергались только четко различимые нанообъекты, для обеспечения того, чтобы артефакты были удалены.
Измерение дзета-потенциала
Дзета-потенциал экзосом измеряли, используя Litesizer 500, светорассеивающий прибор для анализа частиц (Litesizer™ 500, Anton Paar Ltd, Великобритания). Дзета-потенциал измеряли в плоскости скольжения (15°) везикулы-экзосомы, и обеспечивали, чтобы концентрация частиц в каждом разведении была достаточно высокой для обеспечения достоверных измерений посредством генерирования средней выявляемой интенсивности света выше 20 ким пульсов/с.
Анализ образца экзосом на основе проточной цитометрии (со связыванием на шариках)
Для анализа на основе проточной цитометрии экзосомы (всего 2×109 частиц), выделенные из МСК, меченных дальнекрасным красителем, инкубировали с 10 мкл альдегидных латексных шариков (0 равен 4 мкм) в PBS в течение 15 мин при комнатной температуре без вращения. Затем образец смешивали с 950 мкл PBS и инкубировали в течение ночи при 4°С без вращения. Оставшиеся сайты связывания экзосома-шарики затем блокировали добавлением 1 М глицина в PBS в течение 30 мин при комнатной температуре. Образцы шариков, покрытых экзосомами, затем центрифугировали при 1700 RCF (от англ. relative centrifugal force - относительное центробежное ускорение) в течение 5 мин, и осадок ресуспендировали в 0,5% BSA (от англ. bovine serum albumin - бычий сывороточный альбумин) в растворе PBS. Образцы промывали 3 раза аналогичным образом и ресуспендировали в конечном объеме 1 мл 0,5% BSA в PBS. Затем образцы шариков, покрытых экзосомами, или конъюгировали с РЕ(от англ. phycoerythrin - фикоэритрин)-конъюгированным мышиным антителом против человеческого антитела, для выявления CD9, CD63 и CD81, или оставляли необработанными в качестве контроля. Изотипический контроль также делали посредством инкубации шариков, покрытых экзосомами, с РЕ-конъюгированным IgG1 к мыши, изотип 1 или 2. Все образцы затем анализировали с использованием BD FACS CANTO, и данные анализировали с использованием программного обеспечения FlowJo.
Просвечивающая электронная микроскопия (ТЕМ)
Морфологическое исследование и ПЭГилированных и неПЭГилированных экзосом проводили, используя электронный микроскоп (ЕМ - от англ. electron microscope). Экзосомы фиксировали 2%-ным параформальдегидом (PFA - от англ. paraformaldehyde), промывали трижды PBS перед адсорбцией на золотых сетках для ЕМ, 200 меш, покрытых формваровой пленкой/углеродом. Затем сетки для ЕМ блокировали PBS/50 мМ глицином и PBS/5% BSA перед их обработкой первичными мышиными антителами против человеческих антител либо к CD63 (кат. №sc5275, Santa Cruz), либо TSG-101 (кат. №sc7964, Santa Cruz) (только вторичный контроль за PBS/0,1% BSA) в течение ночи и с последующей обработкой на протяжении 30 мин вторичным антителом козы против мышиного IgG, конъюгированного с золотом (кат. №G7652, Sigma). Затем сетки фиксировали 1%-ным глутаральдегидом и инкубировали в 2% фосфорновольфрамовой кислоте в PBS для добавления контраста к мембране экзосомы. Затем сетки с экзосомами промывали в PBS и давали высохнуть на воздухе перед их анализом посредством ТЕМ (от англ. transmission electron microscopy - просвечивающая электронная микроскопия) микроскопа.
Эксперименты по проникновению через слизь и клеточной интернализации с эпителиальными клетками кистозного фиброза, культивируемыми на границе раздела воздух-жидкость
Кистозный фиброз (CF) представляет собой генетическое заболевание, при котором нарушенная врожденная защита хозяина приводит к рецидивирующим тяжелым инфекциям дыхательных путей. Культуры клеток дыхательного эпителия (АЕСС - от англ. airway epithelial cell culture) на границе раздела воздух-жидкость подвергаются дифференцировке в клетки, обычно встречающиеся в эпителии бронхов (реснитчатые эпителиальные клетки и бокаловидные клетки) и продуцируют густую, вязкую дегидратированную слизь на апикальной поверхности, которая контактирует с воздухом. Модель границы раздела воздух-жидкость, таким образом, является точным повторением условий, с которыми генная терапия на основе доставки аэрозоля будет сталкиваться в фактическом легком, пораженным CF. Неожиданно, авторы изобретения обнаружили, что проникновение через слизь и клеточная интернализация достигались нанесением высоких концентраций необработанных экзосом на поверхность слизи культур ALI (от англ. air-liquid interface - граница раздела воздух-жид кость). 25 мкл обработанных экзосом и необработанных экзосом затем наносили на апикальную поверхность слизи культур ALI, причем доза обработанных экзосом приблизительно составляла половину дозы необработанных экзосом. (2,65Е плюс 0,7 экзосом на микролитр необработанного контроля и 1,22Е плюс 07 экзосом на микролитр обработанной исследуемой композиции). После 8 часов инкубации образцы 3 раза промывали PBS, фиксировали в 4% PFA и окрашивали фаллоидином (плотные контакты) и красителем Хехст (ядра).
Анализ на основе конфокальной флуоресцентной микроскопии посредством конфокального микроскопа Zeiss LSM 710. Сканирование для анализа xyz-изображений эпителиальных клеток в обработанных и необработанных образцах.
Результаты
Экзосомы, выделенные из МСК костного мозга человека, характеризовали, используя разные способы для количественной оценки их размера, для измерения их поверхностного заряда и для их идентификации в присутствии некоторых поверхностных маркеров. Как ПЭГилированные, так и неПЭГилированные экзосомы использовали для определения характеристик (за исключением проточной цитометрии). Авторы изобретения выделили экзосомы с использованием дифференциального ультрацентрифугирования, поскольку данный способ остается наиболее широко используемым стандартным способом выделения экзосом из разных биологических жидкостей или супернатантов клеточных культур (Chia et al., 2017, Chia et al., 2016, Thery et al., 2006). Анализ траекторий наночастиц (NTA) и неПЭГилированных и ПЭГилированных экзосом показал, что их средний размер составлял 143,2 плюс/минус 3,5 нм и 165,6 плюс/минус 6,0 нм, соответственно (Фиг. 1А и В). С другой стороны, когда молекулы ПЭГ, отдельно суспендированные в PBS, анализировали посредством NTA, они оказались не способны соответствовать критериям контроля качества. Это указывает на то, что число частиц, показанное в образце ПЭГилированных экзосом, не было артефактом, вызванным молекулами ПЭГ. Поскольку NTA измеряет гидродинамический диаметр частицы, измеренный диаметр обоих образцов экзосом, полученный посредством просвечивающей электронной микроскопии (ТЕМ, от англ. transmission electron microscopy), был меньше чем измеренный анализом NTA.
Как показано анализом NTA, средний размер ПЭГилированных экзосом был немного выше, чем у неПЭГилированных экзосом. Данная разница в размере обусловлена включением молекул ПЭГ в поверхность экзосом. Кроме того, распределение дзета-потенциала измеряли, используя Litesizer 500. Поверхностный заряд измеряли множество раз, и дзета-потенциал неПЭГилированных экзосом составлял примерно -16,5 мВ, и у ПЭГилированных экзосом составлял примерно -2,6 мВ (Фиг. 2А и В). На оси у показана относительная частота, которая дает вероятность конкретного заряда, записанного на протяжении конкретного периода времени, - в данном случае заряд экзосомы был записан как 1000 кадров/минута. Это явно указывает на то, что экзосомы были эффективно ПЭГилированы, и наличие молекул ПЭГ на мембране экзосом почти нейтрализовало поверхностный заряд. Кроме того, поскольку образцы экзосом требовали дополнительной стадии ультрацентрифугирования для удаления избыточных молекул ПЭГ, фактическое число частиц после выделения экзосом может быть выше, чем число, полученное после процедуры ПЭГилирования. Анализ ТЕМ экзосом, меченных иммунозолотом, подтвердил наличие CD63 и TSG-101 маркеров (Фиг. 3), и наибольшая часть экзосом была размером меньше 100 нм. На Фиг. ЗА - 3F показаны изображения просвечивающей электронной микроскопии (ТЕМ) экзосом, меченных иммунозолотом (CD63 и TSG 101) (ПЭГилированных и неПЭГилированных) (черная линия, 100 нм). Как на Фиг. ЗА, так и на Фиг. ЗВ представлены только вторичные экзосомы контрольной группы, неПЭГилированные и ПЭГилированные, соответственно; На Фиг. ЗС представлены ПЭГилированные CD63 экзосомы; на Фиг. 3D представлены неПЭГилированные CD63 экзосомы; на Фиг. ЗЕ представлены ПЭГилированные TSG 101 экзосомы; и на Фиг. 3F представлены неПЭГилированные TSG101 экзосомы. Однако, число CD63 и TSG-101-меченных ПЭГилированных экзосом было значимо меньше, чем в случае неПЭГилированных экзосом, но они не демонстрировали какого-либо наблюдаемого изменения в морфологии. Это может указывать на эффективное ПЭГилирование поверхности экзосом, которое предотвращало связывание антител с их соответствующим антигеном. Кроме того, статус мечения и МСК и экзосом дальнекрасным красителем подтверждался анализом - проточной цитометрией (Фиг. 4). На Фиг. 4А-Е показано выявление Far-Red+(дальнекрасный краситель) МСК и экзосом, конъюгированных с шариками, посредством проточной цитометрии. На Фиг. 4А-4С показаны изображения шариков, покрытых Far-Red+экзосомами, и на Фиг. 4D и 4Е показаны Far-Red+позитивные МСК (популяция, дающих сигнал выше порогового значения). На Фиг. 4А показана популяция шариков (популяция, дающая сигнал выше порогового значения), и на Фиг. 4В и 4С показана популяция, дающая сигнал выше порогового значения, для шариков, покрытых Far-Red+экзосомами.
Таким образом, авторы изобретения эффективно модифицировали поверхность экзосом, происходящих из костного мозга человека, способом простой инкубации с низкомолекулярным ДСФЭ-ПЭГ. Авторы изобретения использовали ДСФЭ-ПЭГ (а именно, ПЭГ с поперечно сшитым липидом) для данного эксперимента для включения липидного фрагмента в бислой экзосом с заякориванием полимера.
В данном примере, авторы изобретения применяли способы «последующего включения», используемые для ПЭГилирования предварительно образованных липосом (Nag et al., 2013), для заякоривания полимера на поверхности экзосом после выделения экзосом посредством ультрацентрифугирования. В данном способе авторы изобретения смешивали суспензию флуоресцентно меченных экзосом с определенной концентрацией ДСФЭ-ПЭГ для обеспечения включения ПЭГ в мембрану экзосом. В препаратах липосом образцы обычно инкубируют при более высокой температуре для достижения более быстрого и максимального включения ПЭГ. Однако, условия более высокой температуры не совместимы с получением экзосом, поэтому авторы изобретения проводили процедуры ПЭГилирования при 37°С. ПЭГилированные и неПЭГилированные экзосомы характеризовали разными способами, и результаты авторов изобретения указывают на то, что поверхность экзосом модифицирована ПЭГ в ПЭГилированных образцах, на что указывает увеличение в среднем гидродинамическом диаметре, почти нейтрализованный поверхностный заряд и уменьшенный уровень TSG 101 и CD 63-позитивных экзосом, в сравнении с неПЭГилированными экзосомами.
Анализ образцов посредством конфокальной флуоресцентной микроскопии четко показывает, что ПЭГилированные экзосомы, окрашенные флуоресцентным дальнекрасным красителем, могли поглощаться эпителиальными клетками, лежащими под слоем слизи, при дозе, составляющей менее половины дозы неПЭГилированных контрольных экзосом. Это является высоко значимым улучшением, и улучшенные эффективности будут приводить к огромному уменьшению в объемах культур стволовых клеток, требуемых для получения видов лечения на основе экзосом. Локализация экзосом в областях, окружающих ядра клетки, также является очевидной, а именно, где расположен аппарат трансляции белка клетки, то есть на рибосоме, покрывающей эндоплазматический ретикулум. Степень клеточного поглощения явно предусматривает возможность того, что меньшие по размеру вирусные векторы могут полностью задерживаться в данных слоях слизи. Следовательно, изобретением предусмотрено, что модифицированные экзосомы будут способны модулировать экспрессию генов, например, введение лишь приблизительно 100 молекул микроРНК требуется для обеспечения выявляемых изменений в клетке. Аналогично, транскрипты мРНК, образующие белок в клетке, лишь в среднем приблизительно 40 транскриптов в случае белка CFTR, что также легко достижимо при интернализации экзосом клетками-мишенями, демонстрируемой данным изобретением.
В отношении Фиг. 5, эксперименты по проникновению через слизь и клеточной интернализации демонстрируют способность внеклеточных везикул с модулированной поверхностью, в частности экзосом, проходить через слизь и поступать в апикальные клетки. Ядро клеток окрашено фиолетовым, и клеточные мембраны могут быть идентифицированы по плотным контактам, окрашенным зеленым. Ясно, что красные пятна, а именно экзосомы, проникли в клетки и объединены в кластеры вокруг ядер. Таким образом, продемонстрирована данная способность ПЭГилированных экзосом.
Фиг. 5А-5С представляют собой изображения конфокальной микроскопии культур высокодифференцированных эпителиальных клеток бронхов, пораженных кистозным фиброзом. 25 мкл экзосом наносили на поверхность слоев слизи, лежащих на поверхностях апикальных клеток, перед промывкой, фиксацией и окрашиванием образцов. Для Фиг. 5А 3,05×108 экзосом в 25 мкл PBS (экзосомы, обработанные для того, чтобы быть густо покрытыми ДСФЭ-ПЭГ2000, и с поверхностным зарядом -2,5 мВ) наносили на поверхность слоя слизи. Для Фиг. 5 В 6,63×108 экзосом в 25 мкл PBS (экзосомы представляют собой необработанные экзосомы, без модификации поверхности и поверхностным зарядом -19,5 мВ) наносили на поверхность слоя слизи. Как видно, сопоставимого проникновения через слизь и клеточной интернализации обработанных экзосом достигали с меньше чем половиной наносимых экзосом. Фиг. 5С представляет собой меченое увеличенное изображение, показывающее экзосомы в апикальных компартментах эпителиальных клеток и локализованные около ядер (эндоплазматический ретикулум и аппарат рибосомной трансляции белка).
Экзосомы со сконструированной поверхностью могут иметь полезное терапевтическое применение при доставке без какой-либо генетической модификации их содержаний, или генетическая модификация родительских клеток может оптимизировать содержания экзосом для доставки в клетки-мишени и лечения конкретных заболеваний. Планируют дополнительные исследования кинетики диффузии через слизь и клеточной интернализации, поскольку скорость доставки представляет собой важный фактор, который будет влиять на степень внутриклеточной доставки in vivo. Экзосомы с модифицированной поверхностью демонстрируют ускоренное поглощение, которое имеет значительную пользу для избегания механизмов мукоцилиарного клиренса, и это может быть продемонстрировано посредством сравнения кинетики проникновения равных «доз» экзосом с модифицированной и немодифицированной поверхностью, наносимых на поверхность слизи во множестве концентраций посредством видеоанализа на основе конфокальной микроскопии в реальном времени.
Требуемые дозы экзосом, которые нужны для доставки посредством аэрозоля, могут также быть приблизительно вычислены на основе экспериментов по проникновению через слизь in vitro. Известное число экзосом наносили на известную площадь поверхности эпителиальных клеток, покрытых слизью (60 мм2), и было продемонстрировано очень значимое поглощение эпителиальными клетками. Некоторые ключевые допущения:
1. Вязкость данной концентрации композиции экзосом и достижимый выход аэрозоля при данной вязкости:
2. Данные экзосомы с сконструированной поверхностью будут проникать относительно быстро через слой слизи, что разумным образом дает основания предположить, что концентрация будет значительным образом влиять на это через диффузию, обусловленную концентрацией, и относительно сильно разбавленные композиции экзосом использовали в пилотных исследованиях (0,00063928% (об./об.) экзосом с модифицированной поверхностью).
В следующей таблице проиллюстрирована емкость загрузки экзосомами капли аэрозоля разных размеров экзосомами, @ концентрация экзосом 1% (масс/об.):
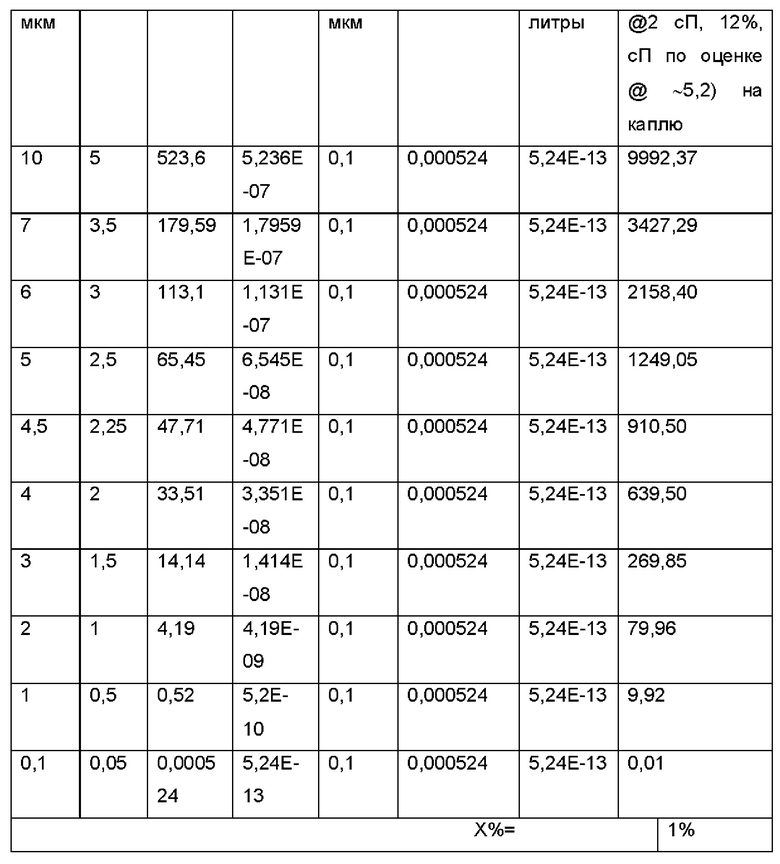
Например, в случае кистозного фиброза, целевая область легкого представляет собой общую площадь поверхности дыхательных путей от трахеи до бронхиол, которая составляет приблизительно 2,471 плюс/минус 320 см2 в легком взрослого человека. Данная площадь поверхности могла быть обработана с использованием объема осажденного аэрозоля приблизительно 70 микролитров 1% (масс/об.) композиции экзосом.
Площадь поверхности дистального легкого гораздо больше с приблизительно в 18 раз большим количеством альвеолярных клеток, чем эпителиальных клеток бронхов, и нацеливание на данную область является желательным, например, для лечения эмфиземы и идиопатического легочного фиброза. Ожидается, что уровень поглощения клетками экзосом гораздо выше в данных областях легкого за счет отсутствия слизистого барьера.
В заключение, авторы изобретения показали, что экзосомы были эффективно ПЭГилированы с использованием конъюгированного с липидом ПЭГ способом простой инкубации, и полученные экзосомы проникали через вязкую слизь CF и были интернализированы эпителиальными клетками, демонстрируя легочную платформу генной терапии на основе экзосом, или просто в качестве терапевтически полезных экзосом для лечения воспалительных заболеваний легкого.
ПЭГилирование также приводит к получению стерически стабилизированной композиции экзосом, которая обладает значительно уменьшенной вязкостью, по сравнению с неПЭГилированными композициями. Образование аэрозоля, содержащего такие экзосомы, обладающие густой гидрофильной «короной» для защиты от агломерации, вызываемой гидрофобным взаимодействием, следовательно, облегчено, что означает, что аэрозоли для доставки экзосом в приемлемой и полезной концентрации в настоящее время облегчены и доступны в более высоких концентрациях. Пэгилированные экзосомы демонстрируют улучшенное проникновение через слизь, причем данное свойство затем облегчает их прохождение через слизь легкого в ткань легкого, дополнительно усиливая эффективность пэгилированных экзосом в терапевтических применениях против заболевания легкого. Наконец, ПЭГилированные моноклональные антитела и наночастицы могут также демонстрировать усиленные «защитные» свойства, которые помогают им избежать поглощения и выведения фагоцитами, такими как альвеолярные макрофаги, которые находятся в увеличенном количестве при воспалительных заболеваниях легких.
Таким образом, согласно изобретению предложены везикулы с покрытием, например, экзосомы, которые могут быть использованы в генной терапии для CF, ХОБЛ, аденокарциномы и других патологических состояний легкого.
Пример 2
Цели данного примера заключались в том, чтобы показать, что модификация поверхности экзосом может быть эффективно достигнута, и что популяции генетически модифицированных мезенхимальных стволовых клеток, полученных посредством методов биоинженерии, сверхэкспрессирующих ген CFTr, могут быть успешно получены.
Методики культивирования клеточных линий ВТ-20 и hMCK
Клетки ВТ-20, получаемые в виде эпителиальных клеток человека ВТ-20 от АТСС® НТВ-19™, культивировали в средах DMEM (от англ. Dulbecco modified Eagle's medium - среда Иглу, модифицированная по Дульбекко) с добавлением 10% FBS (от англ. Fetal Bovine Serum - фетальная телячья сыворотка) и 100 IU/мл пенициллина/стрептомицина в качестве полной кондиционированной среды (ССМ). Использовали человеческие мезенхимальные стволовые клетки, донор №096, клетки культивировали в среде MEM Alfa Gibco с 10% FBS и 100 IU/мл пенициллина/стрептомицина и 1 нг/мл основного фактора роста фибробластов человека (bFGF).
Подпитку клеток проводили 3 дня в неделю, используя асептическую методику, которая заключалась в удалении истраченных сред из колбы для культуры посредством пипетки и добавлении в сливной сосуд с последующим восполнением колбы(колб) для культуры соответствующим объемом свежей среды (включая добавки) до верха колбы для культуры таким образом, чтобы клеточная культура (монослой) не была нарушена потоком. Затем колбу вынимали из вытяжного шкафа, наклоняя колбу таким образом, чтобы ее крышка была обращена вовнутрь, и помещали в инкубатор при 37°С с 5% CO2. Клетки рутинно исследовали с помощью светового микроскопа в отношении признаков загрязнения, таких как помутнение, клетки в суспензии, изменение цвета среды, большие комки или частицы и т.д.
Для разъединения клеток каждые 7-10 суток и перед тем, как клетки достигали 80-90% конфлюэнтности, клетки промывали 3-5 мл PBS, с таким расчетом, чтобы он доставлялся к противоположной стенке колбы таким образом, чтобы не повредить клетки с последующим добавлением 5 мл нагретого Т/Е (соответствующая концентрация, 37°С). Затем колбу аккуратно качали по своей оси в течение 30 секунд, следя за тем, чтобы все клетки были покрыты, и избыток Т/Е отливали в сливной сосуд пипеткой (оставляя в колбе приблизительно 1 мл). Клетки инкубировали с оставшимся малым количеством Т/Е при 37°С в течение 3-5 мин, при отделении клеток от дна колбы. Отделенные клетки ресуспендировали в 4-6 мл полной среды (объем зависит от ожидаемого числа клеток). Затем клеточную суспензию переносят вверх-вниз посредством пипетки, объемом 10 мл, для обеспечения гомогенности и удаляют в 15 мл фалькон. Колбы инкубировали при 37°С.
Получение сред, не содержащих EV
Среды, не содержащие EV, представляют собой необходимое условие протоколов выделения EV для очищенного выхода EV из клеток в культуре. Речь идет об исключении загрязнения изолятов EV культивируемых клеток микровезикулами снаружи от экспериментальной установки.
Получали 130-140 мл 20% FBS плюс основная среда (1:5) в проавтоклавированной стерильной стеклянной бутыли, и соответствующее количество вышеупомянутого подвергали ультрацентрифугированию при 110000 × g в течение 18 часов, при 4°С. После ультрацентрифугирования супернатант осторожно отсасывали в новую пробирку типа фалькон, объемом 50 мл, убеждаясь, что осадок не нарушен. Супернатант из фальконов, объемом 50 мл, фильтровали в только что проавтоклавированный стеклянный контейнер. К нему добавляли равную порцию основной среды для доведения до соотношения 1:1, доводя общую концентрацию FBS в вышеуказанном растворе до 10%. Это приводило к получению в результате 10% среды, не содержащей EV, подлежащей использованию для клеточной культуры для выделения EV. Кондиционированные среды хранили при 4°С вплоть до 4 недель с добавлением 100 IU/мл P/S (или других добавок) перед применением для протокола выделения EV.
Клеточная культура для выделения EV и получение кондиционированных сред, обогащенных EV
EV выделяли в соответствии с руководством MISEV2018 и дополнительными улучшенными протоколами (Thery et al., 2006). Клетки ВТ-20 доводили до достаточного количества 3-4 Т-175 см2 колбы для засева 12 × Т-175 см2 Т-колб в средах, не содержащих EV (2 × 106 клеток/колба). Клетки трипсинизировали и ресуспендировали в средах, обедненных EV, и засевали в 12 × Т-175 см2 Т-колбы в 10 мл сред, обедненных EV, на 24 часа для обеспечения того, чтобы клетки прилипли ко дну колбы. Среды затем заменяли равным количеством свежей среды, не содержащей EV (кондиционированная среда). Колбы оставляли для инкубации на 48 часов.
Спустя 48 часов, кондиционированные среды затем собирали из каждой колбы и объединяли в 50 мл фальконы, и клетки в данных колбах трипсинизировали, и подсчитанное число клеток отбирали из общего лизата клеток (соответствующее число клеток затем пересевали/замораживали/отбрасывали). Супернатант культуры сред, не содержащих EV, наливали из Т-колб в 50 мл фальконы (4 колбы на 50 мл фальконы). Затем 50 мл фильконы, содержащие супернатант, подвергали центрифугированию для удаления клеточных обломков при 300 × g в течение 10 мин, температуре -21°С. Супернатант затем отсасывали, используя 25 мл пипетки, оставляя на дне ненарушенный остаток 2-3 мл, в новые 50 мл фальконы, соответственно. Затем супернатант центрифугировали при 2000 × g в течение 10 мин при температуре -21°С. После центрифугирования супернатант собирали, оставляя осадок неживых клеток в остатке 2-3 мл на дне. Затем, используя стерильный фильтр с размером пор 0,2 мкм и шприц, верхние 25 мл супернатанта отсасывали и собирали в только что проавтоклавированные пробирки для ультрацентрифугирования. Затем отфильтрованные кондиционированные среды подвергали ультра центрифугированию при 110000 × g в течение 70 мин при 4°С на ультрацентрифуге Hitachi Micro. После ультрацентрифугирования 1 мл супернатанта отбирали из одной из пробирок в 1,5 мл пробирку и хранили при -80°С (анализ супернатанта 1). Остальную часть супернатанта отбрасывали в сливной контейнер без соскребания осадка. Осадок, содержащий EV, ресуспендировали 1 мл PBS в каждой UCT (пробирка для ультрацентрифугирования) и объединяли вместе в одну UCT. Она подвергалась ультрацентрифугированию при 110000 × g в течение 70 мин при 4°С. После ультраценрифугирования 1 мл супернатанта отбирали в 1,5 мл пробирку эппендорф из UCT и хранили при -80°С (анализ супернатанта 2). Остальную часть супернатанта отбрасывали в сливной контейнер без соскребания осадка. 85 мкл стерильного PBS отбирали пипеткой в UCT, содержащие EV, и отмеченную сторону соскабливали на протяжении примерно 5 мин с помощью 100 мкл наконечника для пипетки. После 5-8 мин соскабливания, весь ресуспендированный PBS собирали. Весь остаток, включая пузырьки, собирали в коническую пробирку для будущего ПЭГилирования. Ее подвергали импульсному центрифугированию на протяжении 10 секунд для того, чтобы избавиться от пузырьков, и супернатант смешивали в LFH с помощью пипетки. 15-20 мкл отбирали с помощью пипетки (NTA/Дзета) и 10 мкл - в пробирку для анализа белка. Все образцы выделенных EV хранили при -80°С. Все стадии проводили, используя асептические методики, и все стадии центрифугирования проводили при -4°С.
Модификация EV после выделения посредством ДСФЭ-ПЭГ 2000 (АМИН) Амин ДСФЭ-ПЭГ (2000) (Avanti Polar Lipids, Inc) приобретали у Sigma Aldrich (Wicklow, Ирландия). ПЭГилированные экзосомы получали на основе ранее опубликованных данных по получению Пэгилированных липосом (Kooijmans et al., 2016). Конечный объем соотношения 1:4 суспензии выделенных EV (50 мкл) получали посредством смешивания концентрированной суспензии EV с PBS в пластиковой или стеклянной пробирке для образцов 1, 2 и 6 и образцов 4, 5 и 7, соответственно. Для этого добавляли равное соотношение (или желательный процент) 1:1 (мкг белка:мкг белка) амина ДСФЭ-ПЭГ 2000 (10 мг/мл), которое держали для инкубации при 37°С в течение 1 ч, посредством помещения образцов EV в штатив для пробирок с регулируемой температурой сверху качающейся качалки при низких скоростях для предотвращения агрегации молекул ПЭГ или экзосом. После инкубации суспензию EV-ПЭГ вводили в 16-20 мл PBS в ультрацентрифужную пробирку. Ультра центрифугирование проводили при 110000 × g в течение 70 мин для отмывки несвязанных молекул ПЭГ. Затем осадок EV ресуспендировали в 75 мкл PBS и хранили в стеклянных контейнерах соответствующего размера с крышкой, и из данной аликвоты 20 мкл - для анализа дзета-потенциала в отдельных контейнерах. Их хранили при -80°С или -20°С для образцов 1, 2, 6 и образцов 4, 5, 7 соответственно. Анализ траекторий наночастиц (NTA)
Образцы как ПЭГилированных, так и неПЭГилированных экзосом анализировали посредством анализа траекторий наночастиц (NTA) (Malvern UK), используя систему запуска NTA NanoSight NS 500, версия 3.2, используя оптимизированные и утвержденные протоколы (Maguire et al., 2017, Hole et al., 2013, Gerlach et al., 2017). Все данные образцы анализировали, используя NanoSight NS 500, оснащенный лазером 405 нм и длинноволновым пропускающим фильтром 430 нм. Все образцы хранили на водном льду перед анализом. Разведения для NTA выполняли в D-PBS [-MgCl2,-CaCl2] (Gibco), который был сертифицирован как не содержащий частиц посредством NTA непосредственно перед измерением. Образцы пипетировали перед загрузкой для обеспечения достаточного перемешивания и разрушения слабо связанных кластеров EV. Каждый образец разводили вручную в D-PBS с получением оптимальной концентрации частиц, подходящей для NTA (от 10 до 100 частиц на поле обзора), причем каждый фактор разведения записывался в автоматически генерируемых отчетах. В общей сложности шесть × 60-секундных видео записывали для каждого образца EV, и порог обнаружения во время анализа выбирал оператор для обеспечения того, чтобы анализу подвергались только четко различимые нанообъекты, и для обеспечения того, чтобы были удалены артефакты. Поскольку NTA измеряет гидродинамический диаметр частиц, а именно, размер сольватированной частицы, которая приблизительно оценивается как сферическая, измеренные диаметры могут быть больше, чем диаметры, полученные посредством методик на основе электронной микроскопии (ЕМ). В общем, экзосомы обладают диаметрами в интервале размеров 30-150 нм. Все образцы хранили на сухом льду перед анализом. Разведения для NTA проводили в буфере DPBS (Gibco), который подтвержден как не содержащий частиц посредством NTA непосредственно перед измерением. Образцы перемешивали на вибромешалке в течение непродолжительного времени перед загрузкой для обеспечения достаточного перемешивания и разрушения слабо связанных кластеров экзосом. Каждый образец разводили вручную в PBS с получением оптимальной концентрации частиц, подходящей для NTA (от 20 до 70 экзосом на поле обзора), причем каждый фактор разведения записывали в автоматически генерируемых отчетах. В общей сложности шесть х 60-секундных видео записывали для каждого образца экзосом, и порог обнаружения во время анализа выбирали с обеспечением того, чтобы анализу подвергались только четко различимые нанообъекты, для обеспечения того, чтобы были удалены артефакты.
Измерение дзета-потенциала
Дзета-потенциал EV измеряли, используя Litesizer 500, светорассеивающий прибор для анализа частиц (Litesizer™ 500, Anton Paar Ltd, Великобритания). Дзета-потенциал измеряли посредством разведения образцов EV, ПЭГилированных/неПЭГилированных, PBS в соотношении 1:100 или 1:16 (Образец EV-PBS) в кювете Anton Paar Calliope Core Omega с использованием аппроксимации Смолуховского с фактором Дебая 1,5 со временем установления равновесия 1 минута и целевой температурой 25°С с водой в качестве дисперсионной среды, выбранными в программном обеспечении, поскольку не было варианта выбора PBS в качестве дисперсионной среды. Образцы, предназначенные для анализа дзета-потенциала, размораживали в холодильнике/контейнере и переносили в лабораторию для анализа дзета-потенциала, где образцы хранили при 4°С и вынимали при использовании для анализа, разведение проводили в исходной пробирке, содержащей ПЭГилированные или неПЭГилированные EV, непосредственно перед тем, как образец должен был быть добавлен в кювету. Кювету промывали, используя шприц, первый раз Dl-водой (от Deionized Water -деионизированная вода) и затем три раза стерильным PBS, и затем образец добавляли, используя стерильный шприц. Максимальная и минимальная емкость загрузки для каждой кюветы составляла 350 мкл и 300 мкл, соответственно. Концентрация частиц при каждом разведении была достаточно высокой для обеспечения достоверных измерений посредством генерирования максимальной интенсивности для наибольшей части распределения дзета-потенциала частиц (выбросы включены в результаты).
Лентивирусная трансдукция hMCK конструкциями гена EGFP Лентивирусные трансдукции осуществляли на hMCK донора #096. Введение EGFP-CFTR и EGFP-NEG контроля осуществляли для экспрессии белка амплифицированного CFTR в EGFP-CFTR-трансдуцированных клетках для продукции гена CFTR и мРНК инкапсулированных РНК, которые должны быть дополнительно охарактеризованы, как только достигнута жизнеспособная популяция. Трансдукции завершали, используя протокол вращения.
hMCK разделяли в обычном порядке и ресуспендировали в 5 мл полных сред. Ресуспендированные МСК осаждали посредством центрифугирования при 1000 × g в течение 4 мин, после чего супернатант полных сред отбрасывали. Клетки осторожно ресуспендировали в 5 мл основной среды (без FBS, без антибиотика), и осуществляли подсчет клеток.
1×106 (или приблизительно) клеток дополнительно разводили в приблизительно 5 мл основной среды в отдельных 50 мл пробирках фалькон для каждого лентивируса EGFP, положительного контроля и отрицательного контроля (не получает лентивирус). К каждой пробирке, экспериментальной и с положительным контролем, добавляли MOI (от англ. multiplicity of infection -множественность заражения) 3-4 (приблизительно 18 мкл) лентивируса. Отрицательный контроль не получал лентивируса, и все 50 мл пробирки типа фалькон центрифугировали в течение 90 мин при 2000 × g при 37°С после центрифугирования, отбрасывали среды и еще раз ресуспендировали осадок клеток в полных средах. Затем клетки высевали в Т-колбы соответствующего размера (Т175 или Т-75), в зависимости от числа клеток, трансдуцированных и инкубируемых в полных средах с добавлением bFGF (1 нг/мл). После 48 ч инкубации, среды меняли на полные среды с bFGF и дополняли 4 мкг/мл пуромицина для селекции трансдуцированных клеток в течение 7-14 суток, после чего у клеток меняли среду на нормальную полную среду без пуромицина. Клетки разъединяли каждые 7-10 суток после осуществления трансдукции.
Результаты
Для исходных экспериментов линию раковых клеток молочной железы человека ВТ-20 использовали за их высокую пролиферативную способность и легкость культивирования. Клетки переносили в 12 × Т-175 см2 Т-колбы. EV выделяли из вышеупомянутого, используя дифференциальное центрифугирование с последующими двумя стадиями ультрацентрифугирования в соответствии с руководством MISEV в соответствии с наиболее широко используемыми и проверенными способами, и затем разные способы осуществляли для характеристики EV, включая распределение по размерам с помощью анализа траектории наночастиц и анализ поверхностного заряда с помощью исследований дзета-потенциала с использованием прибора Litesizer™ 500. Как ПЭГилированные, так и неПЭГилированные EV характеризовали, используя установленные протоколы и методики по наиболее известному практическому опыту.
EV, выделенные из клеток ВТ-20, характеризовали, используя разные способы для количественной оценки их размера и для измерения их поверхностного заряда. Авторы изобретения выделяли экзосомы, используя дифференциальное ультрацентрифугирование, поскольку данный способ остается наиболее широко используемым стандартным способом выделения экзосом из разных биологических жидкостей или супернатантов клеточных культур (Chia et al., 2017, Thery et al., 2006). В анализе траектории наночастиц (NTA) как ПЭГилированных, так и неПЭГилированных экзосом обнаружили, что анализ NTA не является целесообразным выбором для разведенных фракций ПЭГилированных EV. ПЭГилированные EV могли подвергаться модификации поверхности за счет взаимодействия с поверхностями пластмассы в ультрацентрифжных пробирках или за счет деградации в пластиковых контейнерах, для этого авторы изобретения использовали стеклянные контейнеры для хранения фракций ПЭГилированных EV в случае образца 4, 5 и 7 и проводили анализ дзета-потенциала для характеристики данных образцов вместо анализа NTA.
Обзор культуры клеток ВТ-20
Все параметры, относящиеся к клеточной культуре, кратко представлены ниже в Таблицах 1а и 1b, в которых показаны значения подсчета количества клеток, концентрация белка EV-изолятов, процент ДСФЭ-ПЭГ:Белок в случае обработки ПЭГ, показания измерений дзета-потенциала и NTA для фракций выделенных EV. В случае показаний анализа NTA, значения для Образца 1 не являются надежными из-за низкого числа частиц на кадр (а именно, меньше 3 частиц на кадр). Это также относится к режиму NTA после ПЭГ для образцов 2 и 6. Меньше 10 частиц на кадр также отмечалось в случае режима NTA перед ПЭГ для образцов 3, 4 и 7. Однако, данные результаты удовлетворяют критериям оценки качества. Все данные представлены как число клеток/мл для подсчета количества клеток, мкл в случае объема, мВ в случае дзета-потенциала и нМ в случае типа распределения по размерам для результатов анализа NTA.
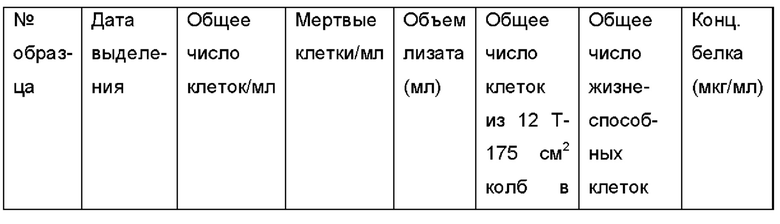


Анализ NTA:
В Таблицах 2а и 2b, изложенных ниже, показаны измеренные значения для повторных снятий показаний анализа NTA в следующем виде: тип распределения частиц по размеру (нм), общая концентрация частиц и концентрация частиц в интервале 30-150 нм в NP/мл для EV-фракций, выделенных из клеток ВТ-20, культивированных в 12ХТ-175 см Т-колбах в соответствии с протоколом выделения EV (MISEV). Данные представлены как среднее и SEM (от англ. Standard error mean - Стандартная ошибка среднего). Образец 1 и все ПЭГилированные образцы обеспечивали ненадежные данные, поскольку было зарегистрировано меньше 3 частиц на кадр, что не соответствует критериям оценки качества NTA. Образцы 2 и 3 предоставляли меньше 10 частиц на кадр, хотя их измерение соответствует критериям оценки качества. Образец 7 также имеет число частиц меньше 10 на кадр при SEM, но опять же соответствует критериям оценки качества.
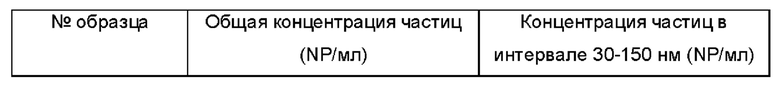
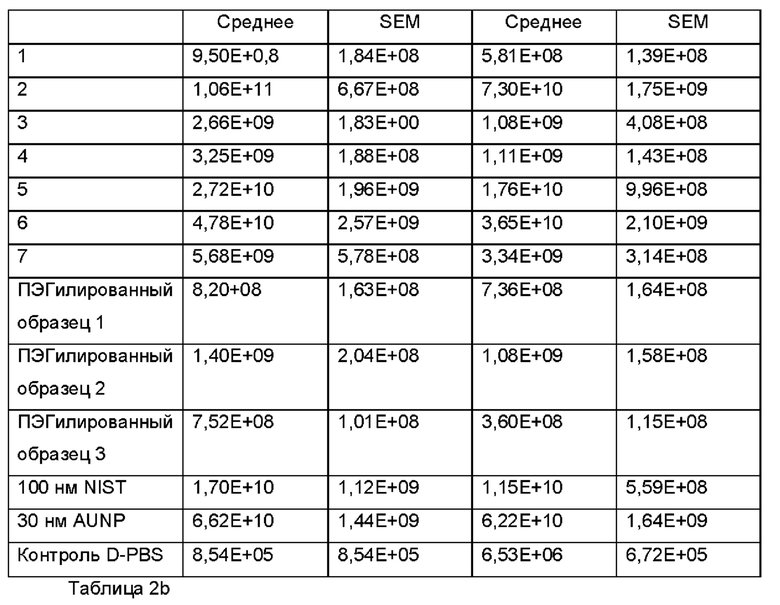
1. Калибровку системы перед каждым днем анализа проверяли, используя стандарт размера на основе полистирольных наносфер NIST (от англ. National Institute of Standards and Technology - Национальный институт стандартов и технологии), 100 нм, (100 нм NIST) и наночастицу цитрата золота, 30 нм, (30 нм AuNP).
2. Все образцы в приведенной ниже таблице соответствовали критериям оценки качества, как обсуждалось.
3. Образцы №1, ПЭГилированный образец №1, ПЭГилированный Образец №2 и ПЭГилированный образец №6 обеспечивали ненадежные результаты вследствие малого числа частиц на кадр (а именно, меньше 3 частиц на кадр, что классифицируется как «не содержащий частиц»). По этой причине, они не соответствовали критериям оценки качества.
4. Образцы №3 и №4 обеспечивали малое число частиц на кадр (а именно меньше 10). Однако, их измерения соответствовали критериям оценки качества.
5. Образцы №7 предоставляли SEM их соответствующего числа частиц на кадр, достигающего меньше 10. Однако, их измерения соответствовали критериям оценки качества, это может быть причиной незначительного рассмотрения измерения их концентраций и должно быть отражено. Контроли:
На Фиг. 6а и 6b продемонстрированы показания анализа траектории наночастиц (NTA) для dPBS и 30 нм наночастиц золота (Аи) в качестве контролей для размера частиц (нм) на оси X и распределения концентраций частиц (частицы/мл) на оси Y. dPBS демонстрирует меньше чем 1 частица на кадр, то есть раствор, не содержащий частиц, как ожидалось, и AuNP демонстрируют определенный пик со средним размером 34 плюс/минус 0,5 нм и соответствуют критерию контроля качества. В приведенной ниже Таблице 3 предложены соответствующие значения в виде среднего и SEM.
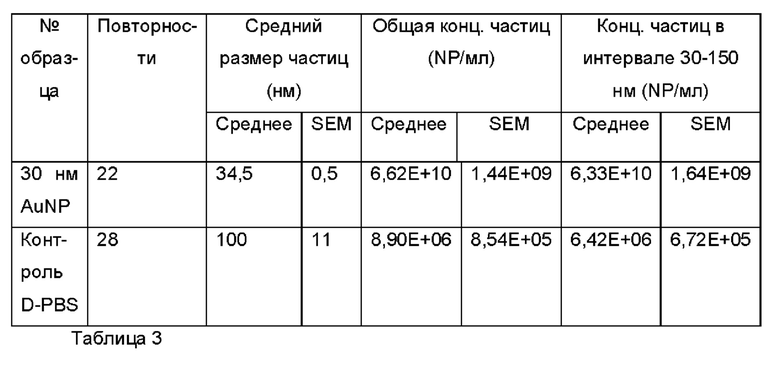
Образец 1
На Фиг. 7а и 7b показано распределение согласно анализу траектории наночастиц для образца-1. Образцы EV для размера частиц (нм) на оси X и распределений концентраций частиц (частицы/мл) на оси Y. Как неПЭГилированные, так и ПЭГилированные образцы имели менее чем 3 частицы на кадр и не соответствовали критерию контроля качества для анализа NTA, то есть раствор, не содержащий частиц. Среднее распределение по размерам и концентрация частиц изложены ниже в Таблице 4а, и общее число клеток и распределение частиц лизата исходных клеток изложены в Таблице 4b. Данные представлены, как среднее, SEM, частицы/клетка и частицы/данный объем, соответственно.
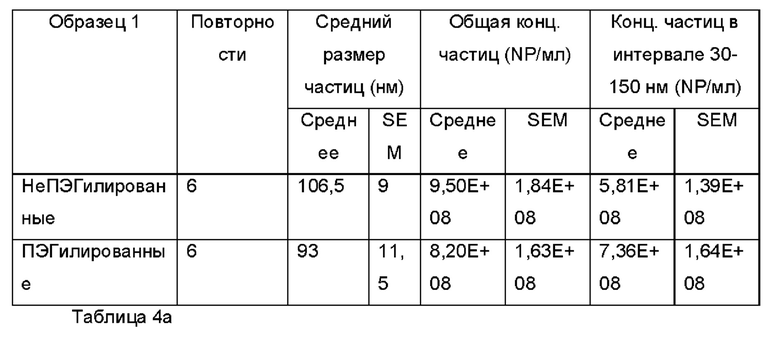

Образец 2
На Фиг. 8а и 8b показано распределение согласно анализу траектории наночастиц (NTA) для примера 2. Образцы EV для размера частиц (нм) на оси х и распределений концентраций частиц (частицы/мл) на оси у. НеПЭГилированный образец демонстрирует определенный пик на уровне 110,5 плюс/минус 5 нм и соответствовал критерию качества для анализа NTA, хотя ПЭГилированные образцы имели меньше чем 3 частицы на кадр и не соответствовали критерию контроля качества для анализа NTA, то есть раствор, не содержащий частиц. Среднее распределение по размерам и концентрация частиц показаны в Таблице 5а, и общее число клеток и распределение частиц лизата исходных клеток показаны в Таблице 5b. Данные представлены, как среднее, SEM, частицы/клетка и частицы/объем образца, соответственно.
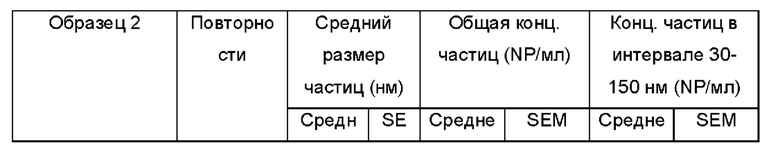
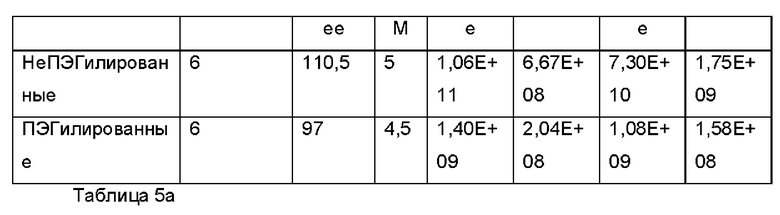

Образец 3
На Фиг. 9 показано распределение согласно анализу траектории наночастиц (NTA) для Примера 3. Образцы EV для размера частиц (нм) на оси х и распределений концентраций частиц (частицы/мл) на оси у. НеПЭГилированный образец демонстрировал среднее распределение по размерам на уровне 1,56 плюс/минус 43,5 нм, и тем не менее он соответствовал критерию качества для анализа NTA, но имел меньше чем 10 частиц на кадр для анализа NTA. Среднее распределение по размерам и концентрации частиц показаны ниже в Таблице 6а, и общее число клеток и распределение частиц лизата исходных клеток показаны в Таблице 6b. Данные представлены как среднее, SEM, частицы/клетка и частицы/объем образца, соответственно.
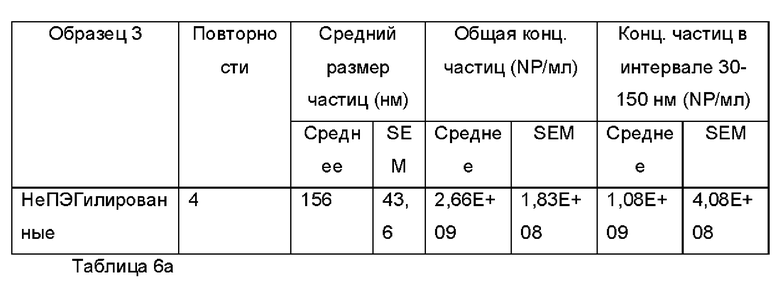
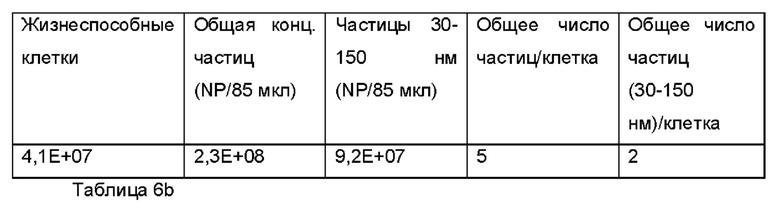
Образец 4
На Фиг. 10 показано распределение согласно анализу траектории наночастиц (NTA) для Примера 4. Образцы EV для размера частиц (нм) на оси х и распределений концентраций частиц (частицы/мл) на оси у. НеПЭГилированный образец демонстрировал среднее распределение по размерам на уровне 120 плюс/минус 26 нм и, несмотря на то, что он соответствовал критерию качества для анализа NTA, имел меньше чем 10 частиц на кадр для анализа NTA. Среднее распределение по размерам и концентрация частиц показаны ниже в Таблице 7а, и общее число клеток и распределение частиц лизата исходных клеток показаны в Таблице 7b. Данные представлены как среднее, SEM, частица/клетка и частицы/объем образца, соответственно.
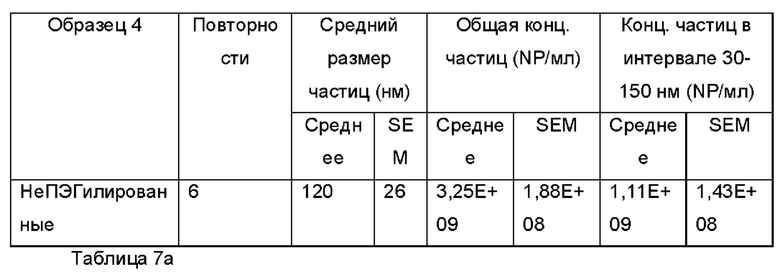
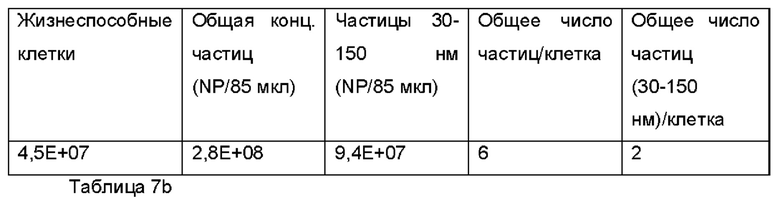
Образец 5
На Фиг. 11 показано распределение согласно анализу траектории наночастиц (NTA) для Примера 5. Образцы EV для размера частиц (нм) на оси х и распределений концентраций частиц (частицы/мл) на оси у. НеПЭГилированный образец демонстрировал среднее распределение по размерам на уровне 96 плюс/минус 3,5 нм и, несмотря на то, что он соответствовал критерию качества для анализа NTA, имел меньше чем 10 частиц на кадр для анализа NTA. Среднее распределение по размерам и концентрация частиц показаны ниже в Таблице 8а, и общее число клеток и распределение частиц лизата исходных клеток показаны в Таблице 8b. Данные представлены как среднее, SEM, частицы/клетка и частицы/объем образца, соответственно.
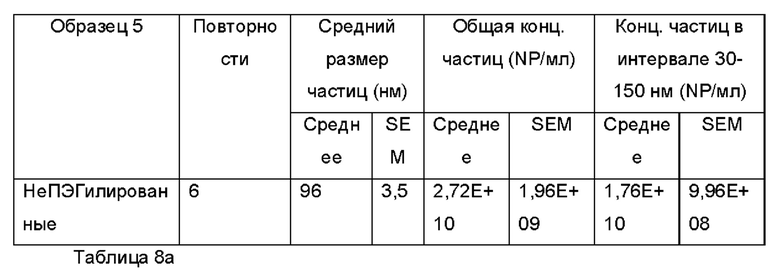

Образец 6
На Фиг. 12а и 12b показано распределение согласно анализу траектории наночастиц (NTA) для образца 5. Образцы EV для размера частиц (нм) на оси х и распределений концентраций частиц (частицы/мл) на оси у. НеПЭГилированный образец демонстрировал определенный пик на уровне 112 плюс/минус 5 нм и соответствовал критерию качества для анализа NTA, хотя ПЭГилированный образец имел меньше чем 3 частицы на кадр и не соответствовал критерию контроля качества для анализа NTA, то есть раствор, не содержащий частиц. Среднее распределение по размерам и концентрация частиц показаны ниже в Таблице 9а, и общее число клеток и распределение частиц лизата исходных клеток показаны в Таблице 9b. Данные представлены как среднее, SEM, частицы/клетка и частицы/объем образца, соответственно.
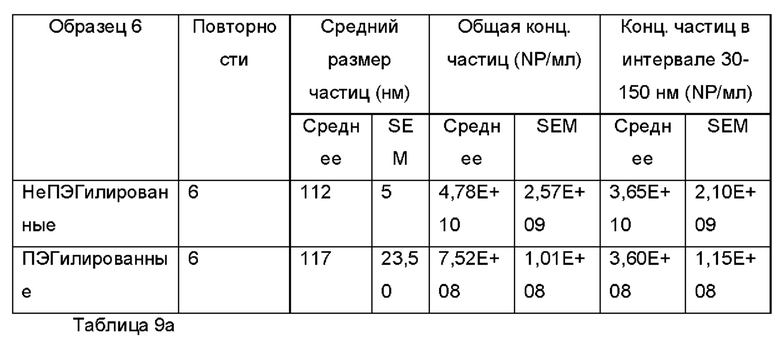
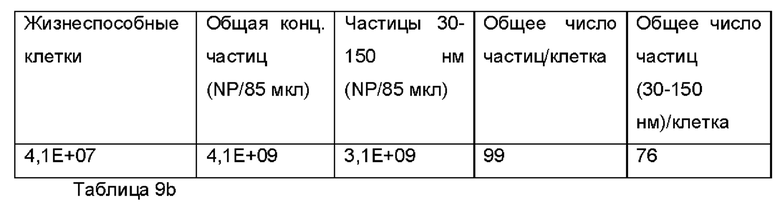
Образец 7
На Фиг. 13 показано распределение согласно анализу траектории наночастиц (NTA) для Примера 7. Образцы EV для размера частиц (нм) на оси х и распределений концентраций частиц (частицы/мл) на оси у. НеПЭГилированный образец демонстрировал среднее распределение по размерам на уровне 93,5 плюс/минус 6 нм и соответствовал критерию качества для анализа NTA, соответствуя критериям для образца очищенных EV, но имел меньше чем 10 частиц на кадр для SEM. Среднее распределение по размерам и концентрация частиц показаны ниже в Таблице 10а, и общее число клеток и распределение частиц лизата исходных клеток показаны в Таблице 10b. Данные представлены как среднее, SEM, частицы/клетка и частицы/объем образца, соответственно.

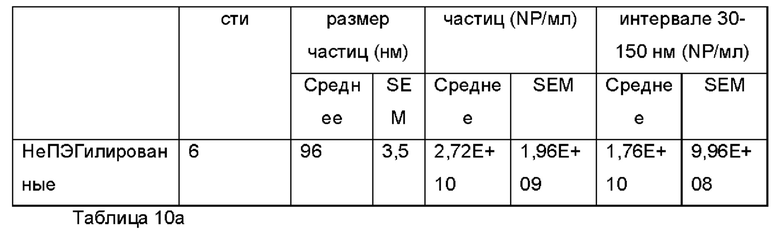
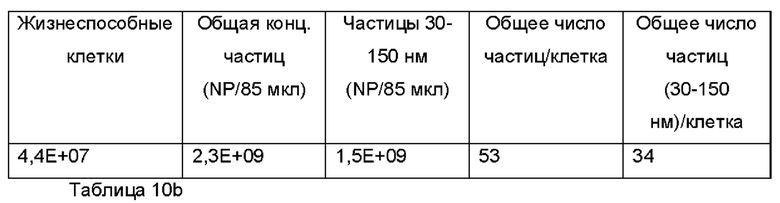
Анализ дзета-потенциала Образец 1 (ПЭГ 100%)
На Фиг. 14а и 14b показаны средние значения дзета-потенциала для образца 1 выделенных ПЭГилированных и неПЭГилированных EV (ПЭГ 100%). Фиг. 14а представляет собой график, показывающий серию считываний показаний измерений для образца выделенных ПЭГилированных EV со средним дзета-потенциалом и значением пика (моды) распределения для каждого прогона (концентрация частиц; 1:100 в PBS). На Фиг. 14b показаны распределения интенсивности дзета-потенциала для трех отдельных считываний показаний измерений для образца выделенных ПЭГилированных EV 1 (концентрация частиц: 1:100 в PBS) с относительной частотой (%) на оси у и потенциалом-дзета (мВ) на оси х. На разных кривых представлены три последовательных измерения отдельных разведений образцов из одного и того же изолята Е\/. Дзета-потенциалы определяли, используя аппроксимацию Смолуховского. Данные представлены как Среднее и SD (от англ. standard deviation - стандартное отклонение). Все данные выражены в мВ.
Образец 2 (ПЭГ 50%)
Фиг. 15а и 15b эквиваленты Фиг. 14а и 14b в отношении образца 2. Образец 4 (ПЭГ 50%)
На Фиг. 16 показаны средние значения дзета-потенциала для образца изолята ПЭГилированных и неПЭГилированных EV 4 (ПЭГ 50%). Распределения интенсивности дзета-потенциала для трех отдельных считываний показаний измерений для образца выделенных ПЭГилированных EV 4 (концентрация частиц: 1:16 в PBS) с относительной частотой (%) на оси у и дзета-потенциалом (мВ) на оси х. На разных кривых представлены три последовательных измерения одного и того же изолята ПЭГилированных EV. Ниже в Таблицах 11а и 11b показаны считывания показаний измерений одного прогона для образца изолята неПЭГилированных EV со средним дзета-потенциалом, за которыми следуют конкретные значения для того же образца после ПЭГилирования. SD и значения пика (моды) распределения для каждого прогона (концентрация частиц 1:100 в PBS). Дзета-потенциалы определяли с использованием аппроксимации Смолуховского. Данные представлены в виде Среднего и SD. Все данные выражены в мВ.

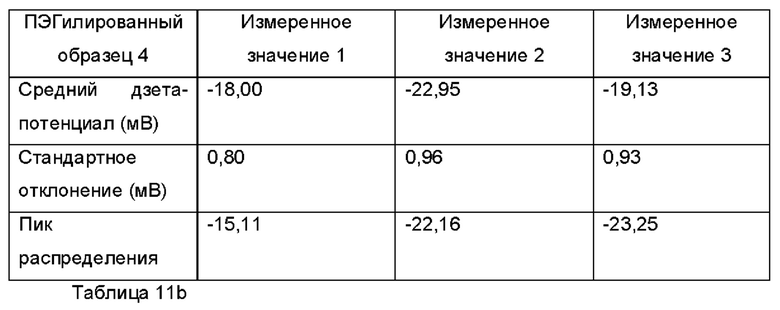
Образец 5 (ПЭГ100%)
На Фиг. 17 показаны средние значения дзета-потенциала для образца изолята ПЭГилированных и неПЭГилированных частиц 5 (ПЭГ 100%). Распределения интенсивностей дзета-потенциала для трех отдельных считываний показаний измерений для образца выделенных ПЭГилированных EV 4 (концентрация частиц: 1:16 в PBS) с относительной частотой (%) на оси-γ и дзета-потенциалом (мВ) на оси х. На разных кривых представлены три последовательных измерения одного и того же изолята ПЭГилированных EV. В представленных ниже Таблицах 12а и 12b показаны считывания показаний измерений одного прогона для образца изолята неПЭГилированных EV со средним дзета-потенциалом, за которыми следуют конкретные значения того же образца после ПЭГилирования. SD и значения пика (моды) распределения для каждого прогона (концентрация частиц 1:100 в PBS). Дзета-потенциалы определяли, используя аппроксимацию Смолуховского. Данные представлены в виде Среднего и SD. Все данные выражены в мВ.
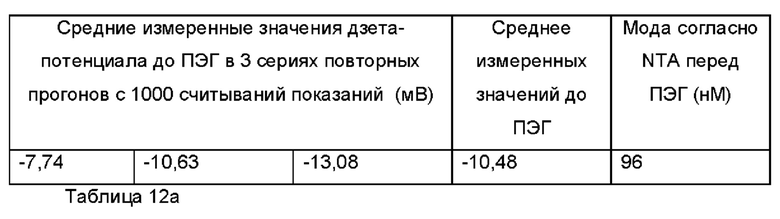
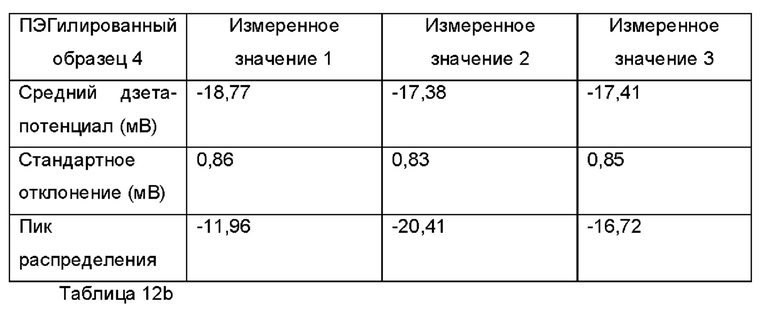
Образец 6 (ПЭГ 75%)
На Фиг. 18а и 18b показаны средние значения дзета-потенциала для образца изолята ПЭГилированных и неПЭГилированных EV 6 (ПЭГ 100%). Фиг. 18а представляет собой график, показывающий серию считываний показаний измерений для образца изолята ПЭГилированных EV со средним дзета-потенциалом и пиком (модой) распределения для каждого прогона (концентрация частиц; 1:100 в PBS). На Фиг. 18b показаны распределения интенсивности дзета-потенциала для трех отдельных считываний показаний измерений для образца изолята ПЭГилированных EV6 (концентрация частиц: 1:100 в PBS) с относительной частотой (%) на оси у и дзета- потенциалом (мВ) на оси х. Разные кривые представляют три последовательных измерения отдельных разведений образцов из одного и того же изолята Е\/. Дзета-потенциалы определяли, используя аппроксимацию Смолуховского. Данные представлены в виде Среднего и SD. Все данные выражены в мВ.
Образец 7 (ПЭГ 150%)
На Фиг. 19 показаны средние значения дзета-потенциала для образца изолята ПЭГилированных и неПЭГилированных EV 7 (ПЭГ 100%). Распределения интенсивности дзета-потенциала для трех отдельных считываний показаний измерений для образца выделенных ПЭГилированных EV 7 (концентрация частиц: 1:16 в PBS) с относительной частотой (%) на оси у и дзета-потенциалом (мВ) на оси х. Разные кривые представляют три последовательных измерения одного и того же изолята ПЭГилированных EV. Ниже в Таблицах 13а и 13b показаны считывания показаний измерений одного прогона для образца изолята неПЭГилированных EV со средним дзета-потенциалом, за которыми следуют конкретные значения того же образца после ПЭГилирования. SD и значения пика (моды) распределения для каждого прогона (концентрация частиц 1:100 в PBS). Дзета-потенциалы определяли, используя аппроксимацию Смолуховского. Данные представлены в виде среднего и SD. Все данные выражены в мВ.
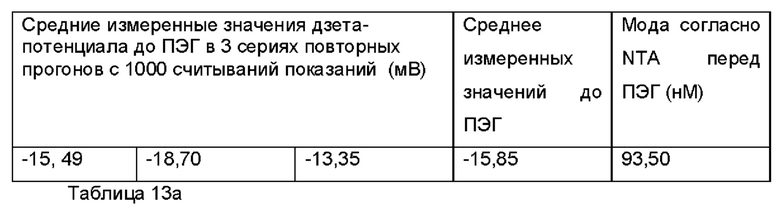
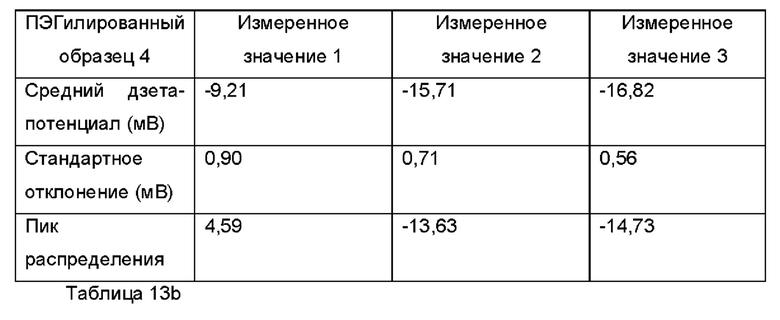
Трансдукция МСК
Трансдукцию hMCK, донор #096, выполняли, используя установленные протоколы. Лентивирусные частицы приобретали у Genecopoeia Ltd.
Авторы изобретения в настоящее время имеют 4 партии трансдуцированных клеток hMCK, перечисленных ниже, некоторые из первоначальных попыток трансдукции hMCK с MO11-2 были неуспешными и не включены.
Объемы титра лентивируса

Объемы трансдукции для трансдукции hMCK

Изображения флуоресценции показаны на Фиг. 20 и 21.
Обсуждение
В данном исследовании исследовали модификацию фракций EV, продуцированных клетками. Как показано посредством анализа NTA в ранее проведенном исследовании, средний размер ПЭГилированных экзосом был чуть больше, чем у ПЭГилированных экзосом, хотя в данных экспериментах не могли установить какой-либо конкретной взаимосвязи между различием в гидродинамическом размере ПЭГилированных EV в сравнении с неПЭГилированными EV. Это может быть обусловлено 1) включением молекул ПЭГ на поверхности EV, меняющим их поведение в суспензии; 2) вымыванием ПЭГилированных EV в результате применения пластиковых пробирок, используемых для хранения/инкубации; 3) вымыванием EV из ультрацентрифужных пробирок во время стадии промывки при ультрацентрифугировании после проведения ПЭГилирования; 4) осаждением ПЭГилированных EV в результате применения пластиковых/силиконовых пипеток. Кроме того, дзета-потенциал, измеренный, используя Litesizer, показал, что поверхностный заряд неПЭГилированных EV находился в интервале от -6,65 до -18,6 мВ для серии считываний показаний, где максимум 1000 прогонов делали посредством программного обеспечения Anton Paar до тех пор, пока не был достигнут алгоритм выполнения операций над множествами для контроля качества фаз, или среднее 1000 прогонов учитывалось автоматически прибором, тогда как в случае ПЭГилированных EV в ряде прогонов, аналогично вышеуказанному для результатов по образцам 1, 2 и 6, наблюдали максимальное среднее значение дзета-потенциала -0,40 мВдо минимум -23,6 мВ, хотя последующие считывания показаний измерения дзета-потенциала, как наблюдалось, имели заметную тенденцию, заключающуюся в значимо более низких значениях, чем исходные считывания, что свидетельствует о модификации поверхностных характеристик ПЭГилированных EV за счет приложенного напряжения и ПЭГилирования EV, эффективно приводящего поверхностный заряд EV к нейтральному значению. Это явление может в действительности помочь объяснить тот факт, что модификация поверхности экзосом обратима, что является возможным объяснением для наблюдения того, что модификация поверхности низкомолекулярным ПЭГ не препятствовала клеточному трансмембранному поглощению экзосом, как ожидали авторы изобретения.
Еще один анализ дзета-потенциала посредством повторных измерений одного и того же образца проводили в анализах с одним прогоном, хотя результаты были неубедительными, может быть из-за способности хранения или взаимодействий ПЭГ с пластмассой. Для дальнейшего уменьшения взаимодействия пластмассовых пробирок с частицами ПЭГ во время ПЭГилирования, стадию инкубации проводили посредством использования стеклянных флаконов, несмотря на то, что стадию ультрацентрифугирования с использованием пластмассовых ультрацентрифужных пробирок нельзя было обойти, также нельзя было обойти применение пластмассовых/силиконовых пипеток во время переноса ДСФЭ-ПЭГ или ПЭГилированных EV. В будущем, авторы изобретения будут модифицировать свои экспериментальные методики соответствующим образом, посредством применения стеклянных сосудов и приборов, таких как пипетки, и модификации методики очистки EV после ПЭГилирования посредством перехода на SEC хроматографию (от англ. Size-exclusion chromatography - Эксклюзионная хроматография). Также наблюдали, что ДСФЭ-ПЭГ возможно деградировал, когда проводили некоторые из данных экспериментов, хотя авторы изобретения наблюдали положительные значения в случае мод в некоторых из образцов, указывая на то, что эффективное ПЭГилирование было возможным, используя больше ПЭГ. Это является явным указанием на то, что некоторые EV были ПЭГилированы, поскольку PBS сам по себе не будет вызывать сдвиг пика распределения в сторону положительных значений, и что присутствие молекул ПЭГ на мембране экзосом почти нейтрализовало поверхностный заряд. Кроме того, поскольку образцы экзосом требовали дополнительной стадии ультрацентрифугирования для удаления избытка молекул ПЭГ, фактическое число частиц после выделения экзосом может быть выше, чем число, получаемое после процедуры ПЭГилирования.
hMCK также трансдуцировали, хотя с исходной задержкой стабильная трансдукция была достижима при примерно MOI, равной 4 или больше, используя протокол вращения. На изображениях флуоресценции, полученных от трансдуцированных клеток, показана флуоресценция GFP во всех трансдуцированных клетках, которая является результатом слитого белка CFTR-EGFP и EGFP контрольного лентивируса. Это показано на Фиг. 22-25, показаны GFP-позитивные МСК, сфотографированные на флуоресцентном микроскопе после посева клеток на предметные стекла, фиксации и окрашивания ядер DAPI (синий). Это подтверждает наличие клеток, не только автофлуоресценции. Предоставлены два увеличения, показывающие экспрессию GFP во всех клетках.
В заключение, показано, что EV могут быть ПЭГилированы для придания способностей проникновения через слизь, не оказывая влияния на внутриклеточное перемещение EV - как было предположено, возможно дополнительное исследование ПЭГилирования с использованием улучшенных и конкретных инструментов - и EV могут быть охарактеризованы - опять же это могут быть проведены дополнительные исследования для крупномасштабного производства. Характеристика EV и белков из CFTR-сверхэкспрессирующих клеток в отношении сверхэкспрессии белка CFTR с использованием анализа РНК, гена и белка дополнительно покажет эффективную трансдукцию и подтверждение концепции. Функциональный перенос экспрессии CFTR, опосредованный данными экзосомами (в сравнении с отрицательными контролями), будет оцениваться в моделях поверхности раздела воздух-жидкость эпителиальных клеток CF, поскольку данная модель воспроизводит условия, включая слизистый барьер, с которыми вдыхаемый аэрозоль столкнется, как только капли аэрозоля осядут в легком, пораженном CF. Ожидают, что данные эксперименты демонстрируют значимые улучшения, по сравнению с подходами к переносу генов, опосредованному вирусным вектором и наночастицами, современного уровня техники, а именно более высокую эффективность трансдукции, функциональный перенос белка CFTR и мРНК фактически во все клетки в культуре, и демонстрацию функциональности активных агентов, вводимых в эпителиальные клетки CF, посредством оценки транспорта хлорид-иона в экспериментах с культурами трансдуцированных клеток с использованием камеры Уссинга.
Кроме того, увеличение выделения EV с использованием технологий на основе биореактора с высокой плотностью клеток и с использованием более эффективных и коммерчески масштабируемых путей выделения EV, таких как тангенциальная поточная фильтрация и Эксклюзионная хроматография, может повышать выход EV. Применение иммортализованных стволовых клеток, которые трансдуцированы лентивирусом со сверхэкспрессией CFTR, для создания главного банка клеток, который был бы способен достоверно продуцировать сверхэкспрессируемые рекомбинантные нуклеиновые кислоты на протяжении увеличенного количества пассажей, также является крайне полезным для получения EV в промышленном масштабе. Дополнительная оптимизация протокола ПЭГилирования может нейтрализовать заряд EV, выделенных из CFTR-трансдуцированных мезенхимальных стволовых клеток, без большой потери, особенно хранение и перенос ДСФЭ-ПЭГ и ПЭГилированных образцов с использованием устройства для сохранения органического раствора.
Ссылки
Bai, L. et al. (2017) «Effects of Mesenchymal Stem Cell-Derived Exosomes on Experimental Autoimmune Uveitis*, Scientific Reports, 7(1), pp. 1-11.
Beach, A. et al. (2014) «Exosomes: An overview of biogenesis, composition and role in ovarian сапсег», Journal of Ovarian Research. Journal of Ovarian Research, 7(1), pp. 1-10.
Cheng, Y. et al. (2018) «Effect of pH, temperature and freezing-thawing on quantity changes and cellular uptake of exosomes», Protein and Cell.
Chia, D., Katsiougiannis, S., Kim, Y., Singh, R., and Wong, D. (2016). Saliva exosomesfrom pancreatic tumor-bearing mice modulate NK cell phenotype and antitumor cytotoxicity. The Faseb Journal, 31(3): 998-1010.
Chia, В., Low, Y, Wang, Q., Li, P. and Gao, Z. (2017). Advances in exosome quantification techniques. Trends in Analytical Chemistry, 0165-9936.
De Того, J. et al. (2015) «Emerging roles of exosomes in normal and pathological conditions: New insights for diagnosis and therapeutic applications*, Frontiers in Immunology, 6(MAY), pp. 1-12.
Denzer, K. et al. (2000) «Exosome: from internal vesicle of multivesicular body to intercellular signaling device*. Journal of cell science, 113 Pt 19(19), pp. 3365-74.
Deregibus, M. C. et al. (2016) «Charge-based precipitation of extracellular vesicles*, International Journal of Molecular Medicine, 38(5), pp. 1359-1366.
Degiorgio, V., Corti, M. and Giglio, M. Light Scattering in Liquids and Macromolecular Solutions. (1979). Plenum Press, New York. See Section II, p.111 on micelles, p.125 on vesicles, and p.139 on microemulsions
Gerlach, J., Maguire, C, Kruger, A., Joshi, L., Prina-Mello, A., and Griffin, M. (2017). Urinary nanovesicles captured by lectins or antibodies demonstrate variations in size and surface glycosylation profile. Nanomedicine, 12:11.
Hole, P., Sillence, K., Hannell, C, Maquire, C,, Roesslein, M., Suarez, G., Capracotta, S., Magdolenova, Z., Horev-Azaria, L., Dybowska, A., Cooke, L., Haase, A., Contal, S., Mano, S., Vennemann, A12., Sauvain, J., Staunton, K., Anguissola, S., Lunch, A., Dusinska, M., Korenstein, R., Gutleb, A., Wiemann, M., Prina-Mello, A2., Riediker, M., Wick, P. (2013). Interlaboratory comparison of size measurements on nanoparticles using nanoparticle tracking analysis (NTA). Journal of Nanoparticle Research, 15:2101
Kooijmans SAA, Fliervoet LAL, van der Meel R, Fens MHAM, Heijnen HFG, van Bergen En Henegouwen PMP, Vader P, Schiffelers RM. PEGylated and targeted extracellular vesicles display enhanced cell specificity and circulation time. J Control Release. 2016 Feb 28;224:77-85
Lai, R. C. et al. (2013) «Exosomes for drug delivery - a noval application for the mesenchymal stem cell*, Biotechnology Advances, 31(5), pp. 543-551.
Lankford, К. L. et al. (2018) «lntravenously delivered mesenchymal stem cell-derived exosomes target M2-type macrophages in the injured spinal cord», PloS ONE, 13(1), pp. 7-11.
Lin, J. et al. (2015) «Exosomes: Novel Biomarkers for Clinical Diagnosis*, The Scientific World Journal, 2015, pp. 1-8.
Lou, G. et al. (2017) «Mesenchymal stem cell-derived exosomes as a new therapeutic strategy for liver diseases*, Experimental and molecular medicine. Nature Publishing Group, 49(6), p.e346.
Li, C, Huang, Q., Zhang, G., Yang, Z., Lu, W., Zhang, R., Tian, M., Li, L. and Liang, D. (2009). Influence of anchoring ligands and particle size on the colloidal stability and in vivo bio-distribution of polyethylene glycol-coated gold na no particles in tumor-xenografted mice. Biomaterials, 30:10
Maguire, P., Parsons, M., McParland, D., Szklanna, P., Guang, M., O'connell, K., O'connor, H., McGuigan, C, Ainle, F. and McCann, A. (2017). A protocol for Improved Precision and Increased Confidence in Nanoparticle Tracking Analysis Concentration Measurements between 50 and 120 nm in Biological Fluids. Frontiers in Cardiovascular Medicine, 4:68.
Nag. O., and Awasthi, V., (2013). Surface Engineering of Liposomes for Stealth Behavior. Pharmaceutics, 5(4): 542-569.
Nag, О. K. et al. (2013) «Post-modification of preformed liposomes with novel non-phospholipid poly(ethylene glycol)-conjugated hexadecylcarbamoylmethylhexadecanoic acid for enhanced circulation persistence in vivo», International journal of pharmaceutics, 446(0), pp. 119-129.
Niu, Z. et al. (2017) «Polymer-based precipitation preserves biological activities of extracellular vesicles from an endometrial cell line», PloS ONE, 12(10), pp. 1-21.
O'Neill CP, Gilligan KE, Dwyer RM. Role of Extracellular Vesicles (Evs) in Cell Stress Response and Resistance to Cancer Therapy. Cancers (Basel). 2019 Jan 24;11(2).
Sato, Y. T. et al. (2016) «Engineering hybrid exosomes by membrane fusion with liposomes*, Scientific Reports. Nature Publishing Group, 6(February), pp. 1-11.
Stahl PD, Raposo G. Extracellular Vesicles: Exosomes and Microvesicles, Integrators of Homeostasis. Physiology (Bethesda). 2019 May 1;34(3):169-177.
Sugama, S. et al. (2017) «HHS Public Access*, pp. 39-46.
Suk, J. S. etal. (2017) «HHS Public Access*, pp. 28-51.
Thery, С, Amigorena, S., Raposo, G. and Clayton, A. (2006). Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Current Protocols in Cell Biology, 3: 3.22.
| название | год | авторы | номер документа |
|---|---|---|---|
| СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ ФИБРОЗА ТКАНЕЙ НА ОСНОВЕ КОМПОНЕНТОВ СЕКРЕТОМА МЕЗЕНХИМНЫХ СТРОМАЛЬНЫХ КЛЕТОК, СПОСОБ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ СРЕДСТВА | 2020 |
|
RU2766707C1 |
| СПОСОБ ДИАГНОСТИКИ И МОНИТОРИНГА ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ С ИСПОЛЬЗОВАНИЕМ ЭКЗОСОМ КРОВИ | 2014 |
|
RU2571507C1 |
| КОМПОЗИЦИИ, СОДЕРЖАЩИЕ КЛЕТОЧНЫЕ ВЕЗИКУЛЫ, И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2824136C2 |
| ИСПОЛЬЗОВАНИЕ БИОГЕНЕЗА МИКРОРНК В ЭКЗОСОМАХ ДЛЯ ДИАГНОСТИКИ И ЛЕЧЕНИЯ | 2014 |
|
RU2644247C2 |
| ПОЛУЧЕННЫЕ ИЗ ЛИЗАТА ТРОМБОЦИТОВ ЧЕЛОВЕКА ВНЕКЛЕТОЧНЫЕ ВЕЗИКУЛЫ ДЛЯ ПРИМЕНЕНИЯ В МЕДИЦИНЕ | 2017 |
|
RU2735139C2 |
| КОМПОЗИЦИИ ВНЕКЛЕТОЧНЫХ ВЕЗИКУЛ (EV) РАСТИТЕЛЬНОГО ПРОИСХОЖДЕНИЯ И ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2812553C2 |
| СПОСОБ ВЫДЕЛЕНИЯ МИКРОРНК ИЗ БИОЛОГИЧЕСКИХ ЖИДКОСТЕЙ | 2013 |
|
RU2548816C1 |
| ВЕЗИКУЛЫ, СОДЕРЖАЩИЕ ИНГИБИТОР PTEN, И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2800729C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ РИСКА ЗЛОКАЧЕСТВЕННОЙ ТРАНСФОРМАЦИИ КЛЕТОК ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ | 2022 |
|
RU2806518C1 |
| СПОСОБ ВЫДЕЛЕНИЯ ЭКЗОСОМ ИЗ ПЛАЗМЫ КРОВИ | 2020 |
|
RU2741776C1 |

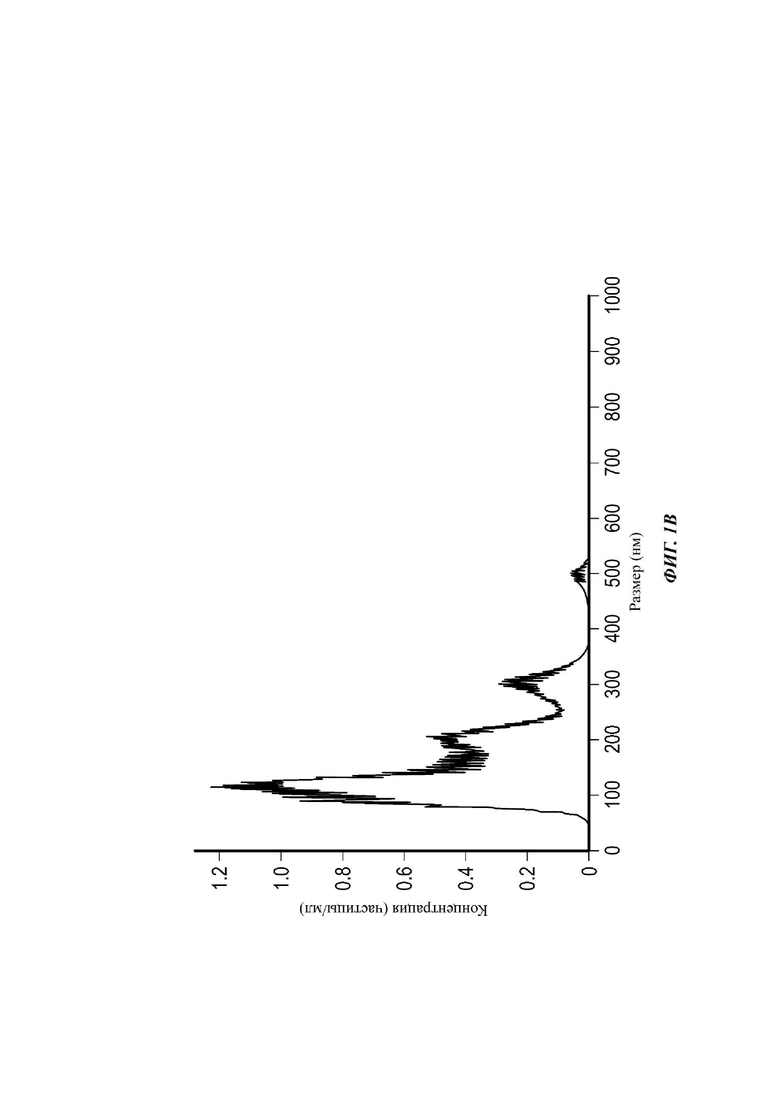
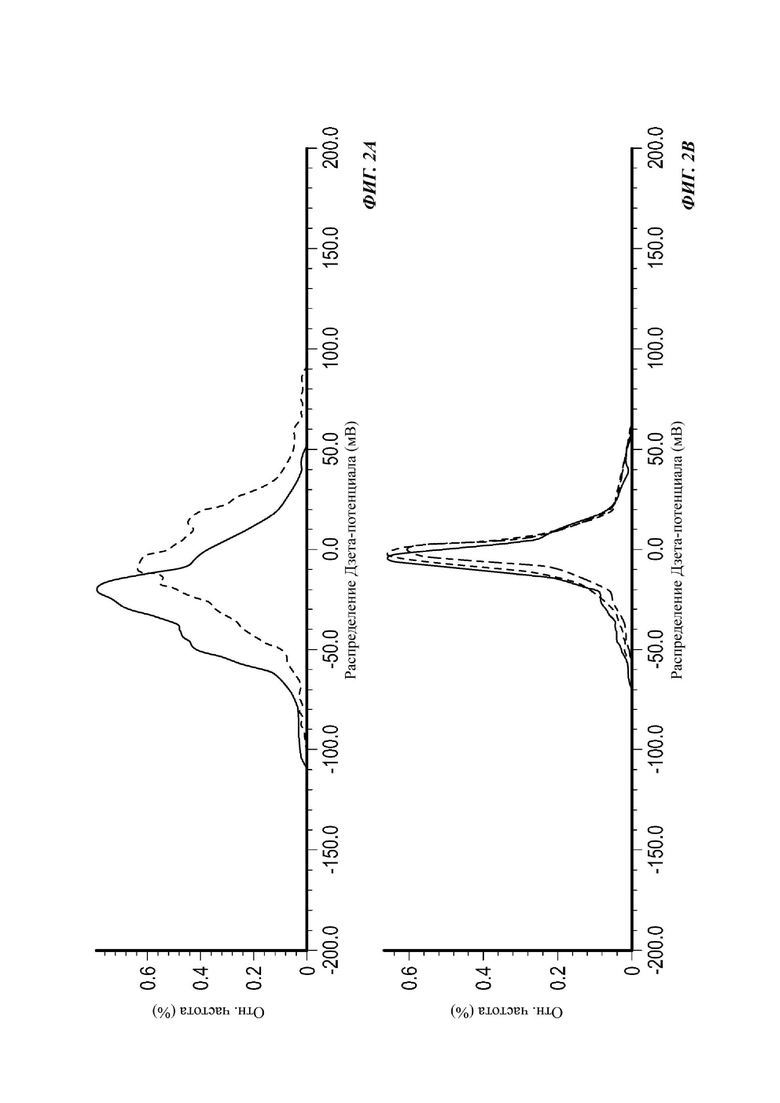
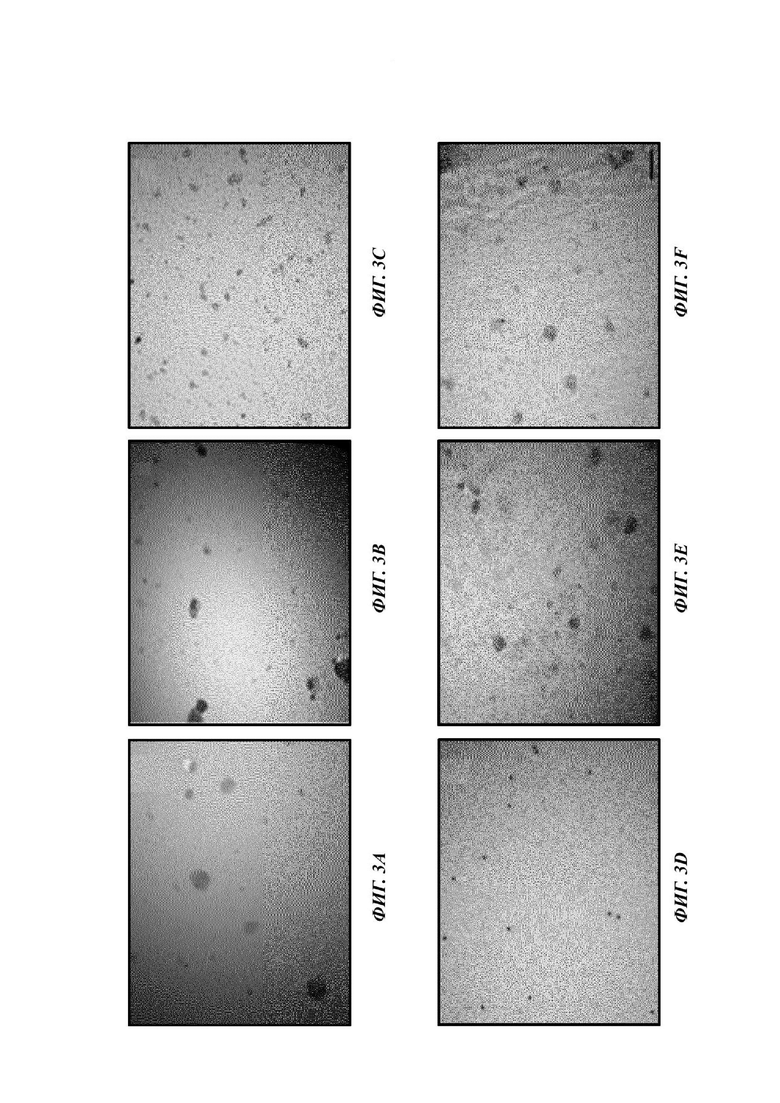
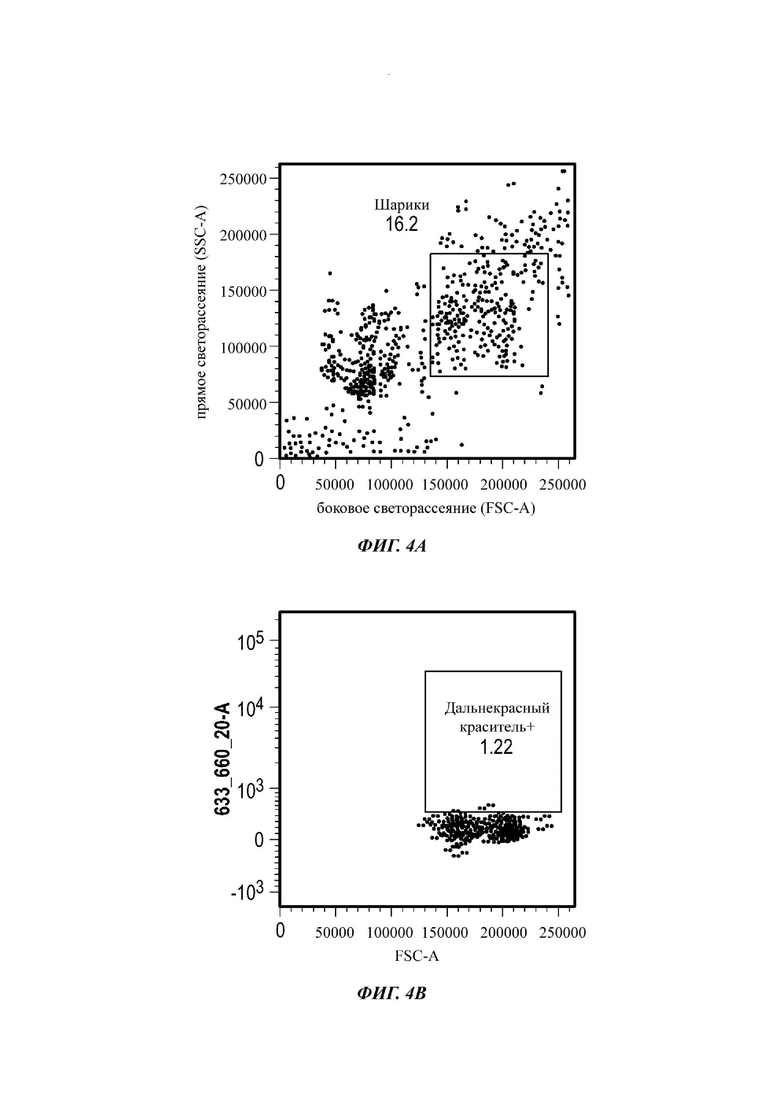
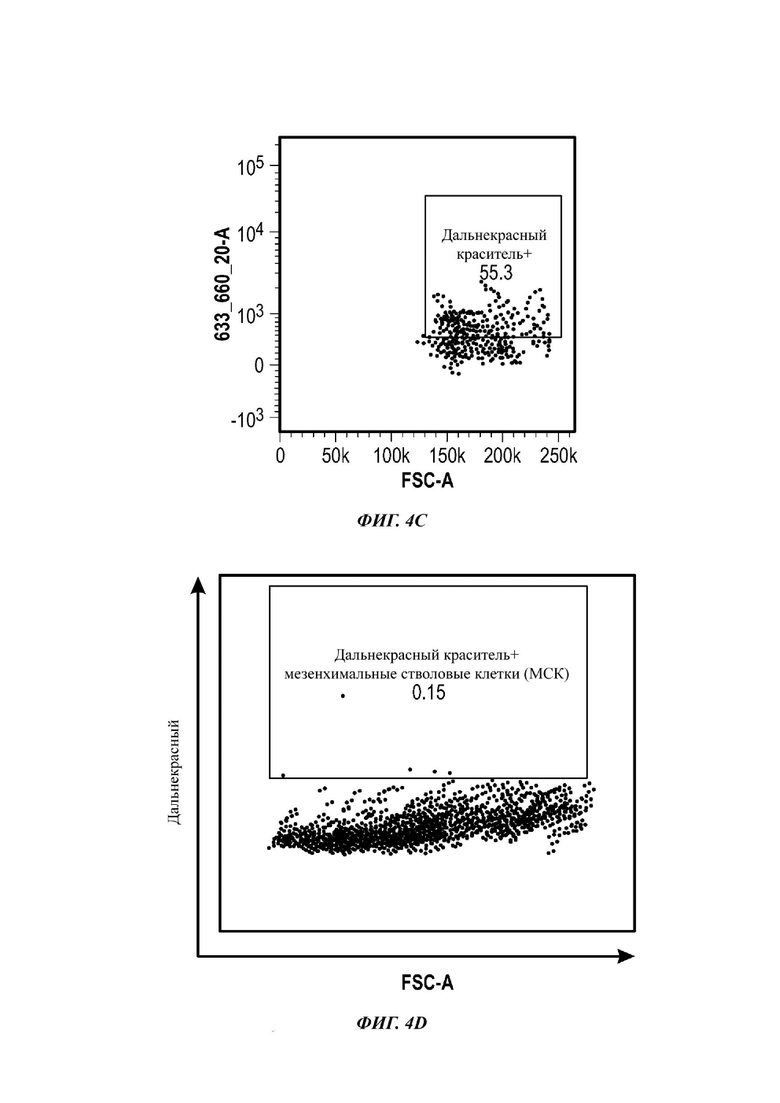
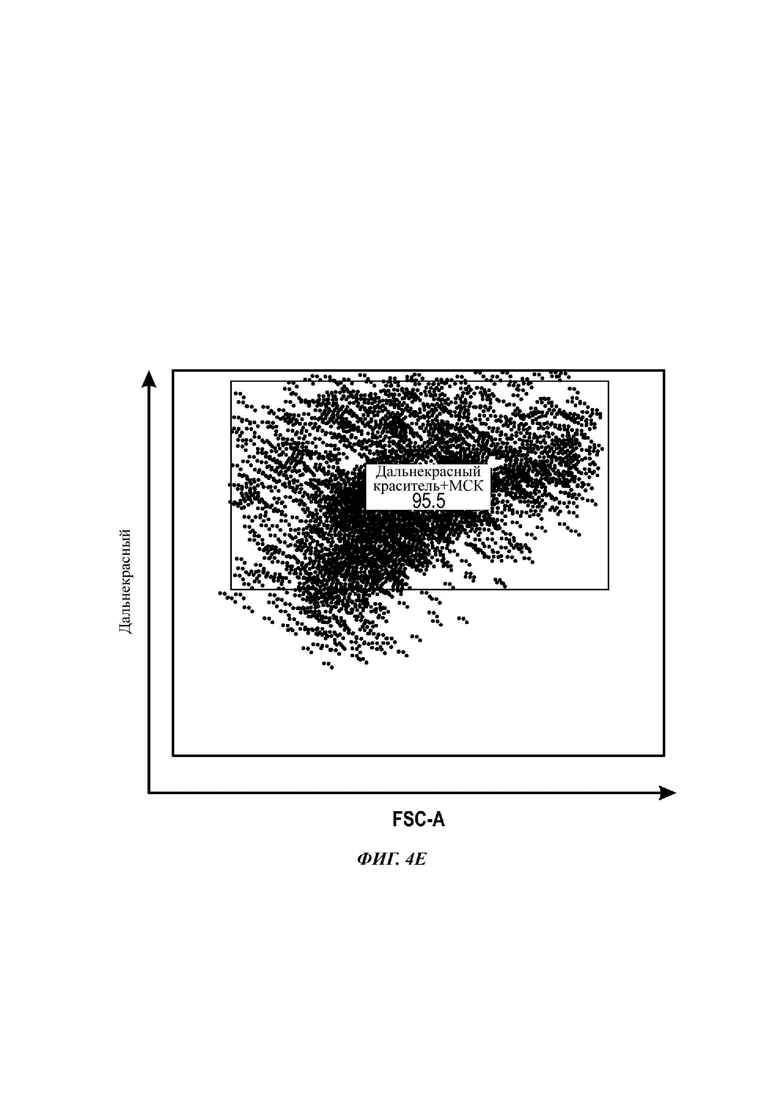

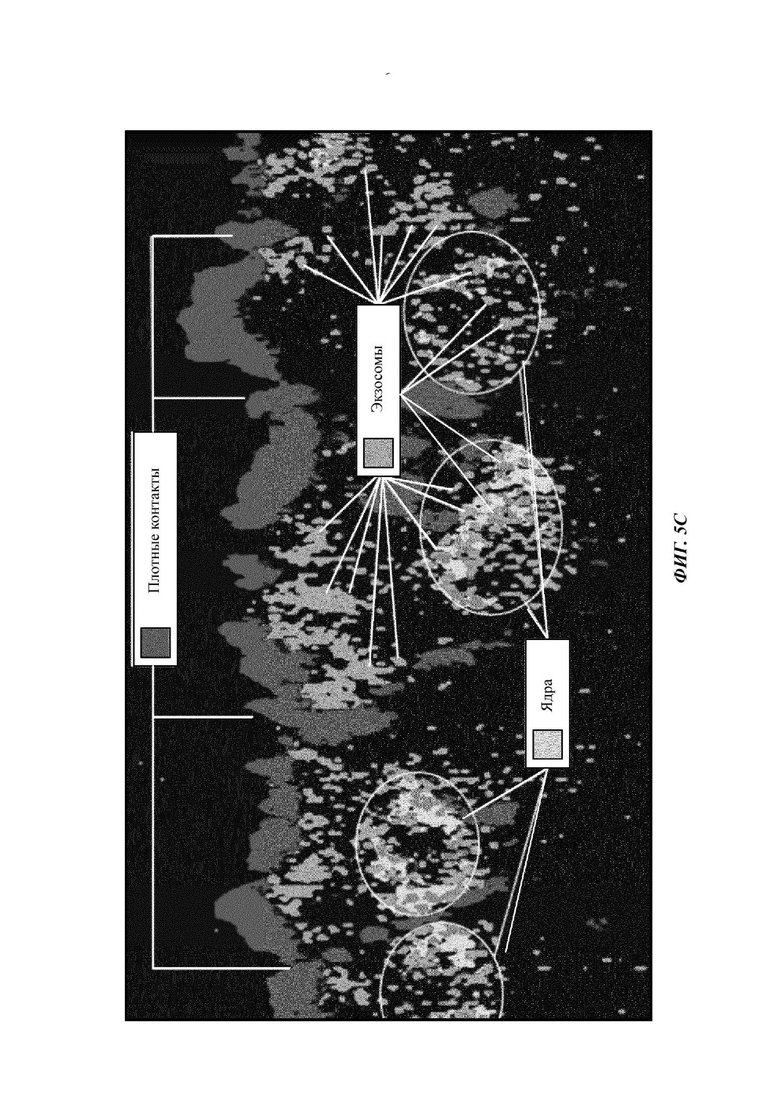

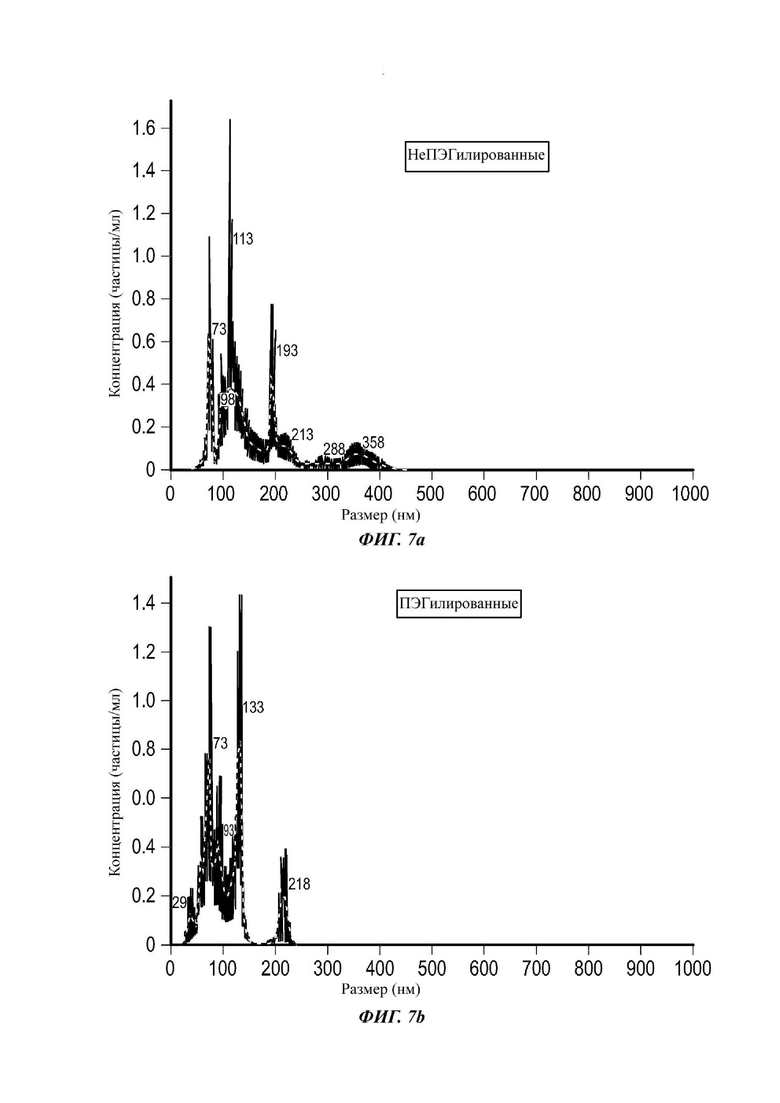

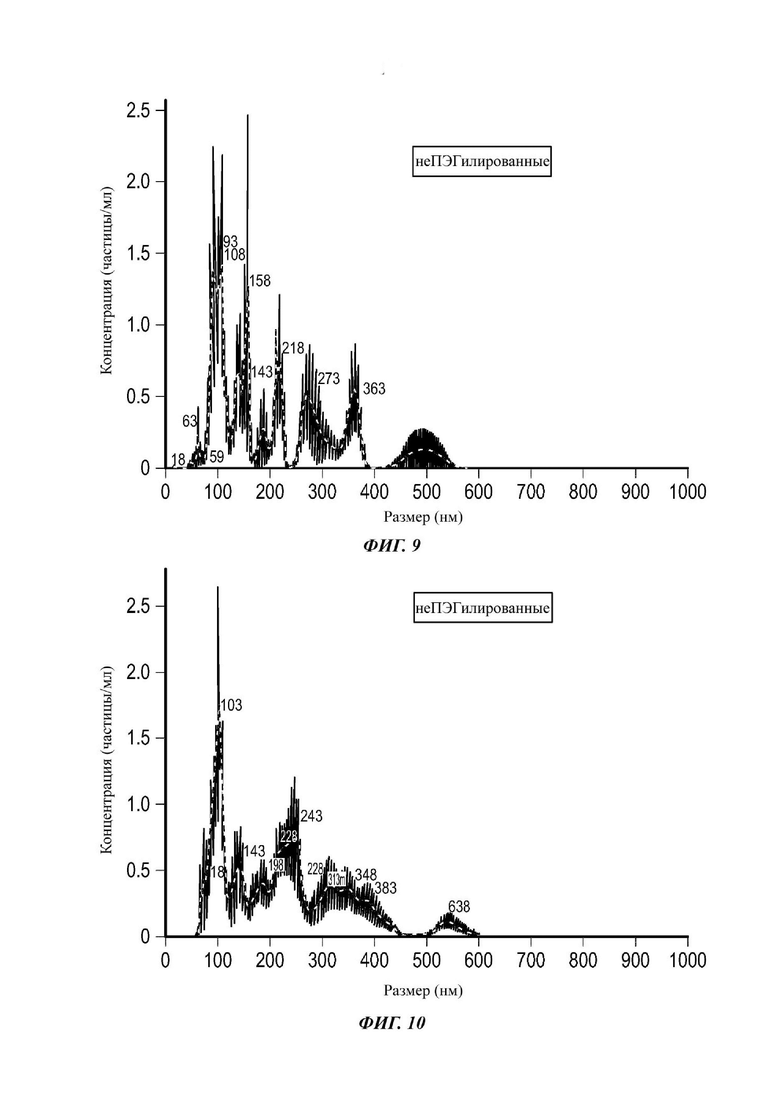
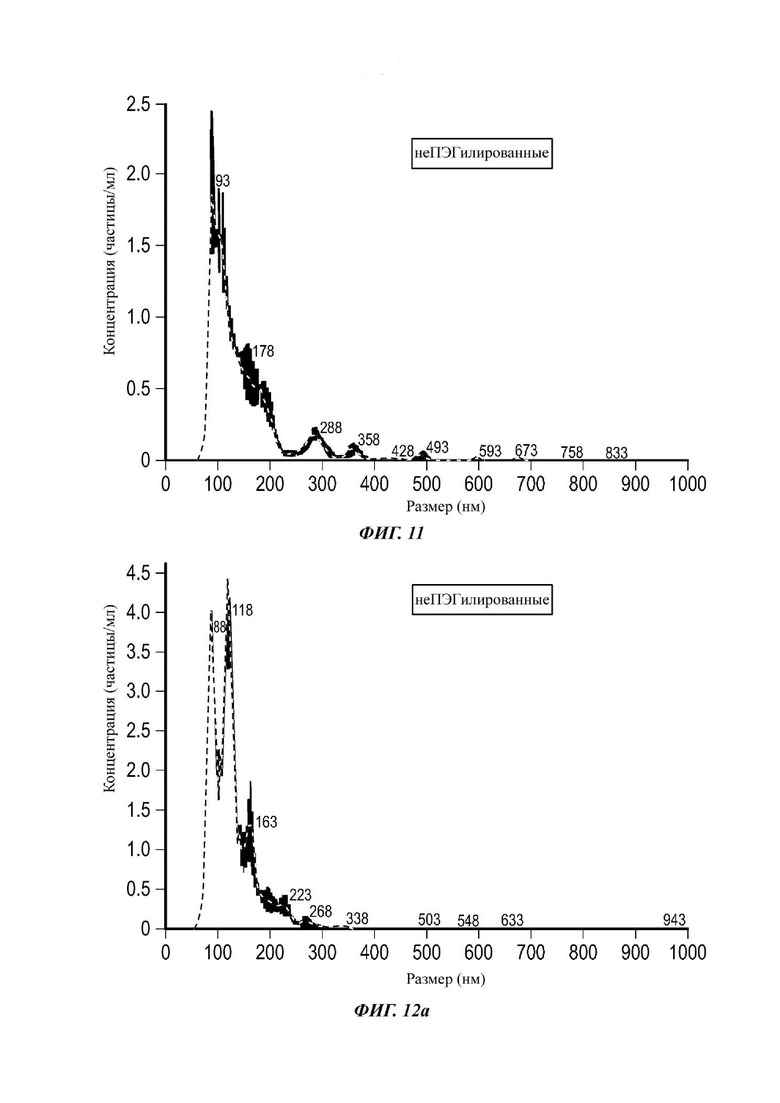
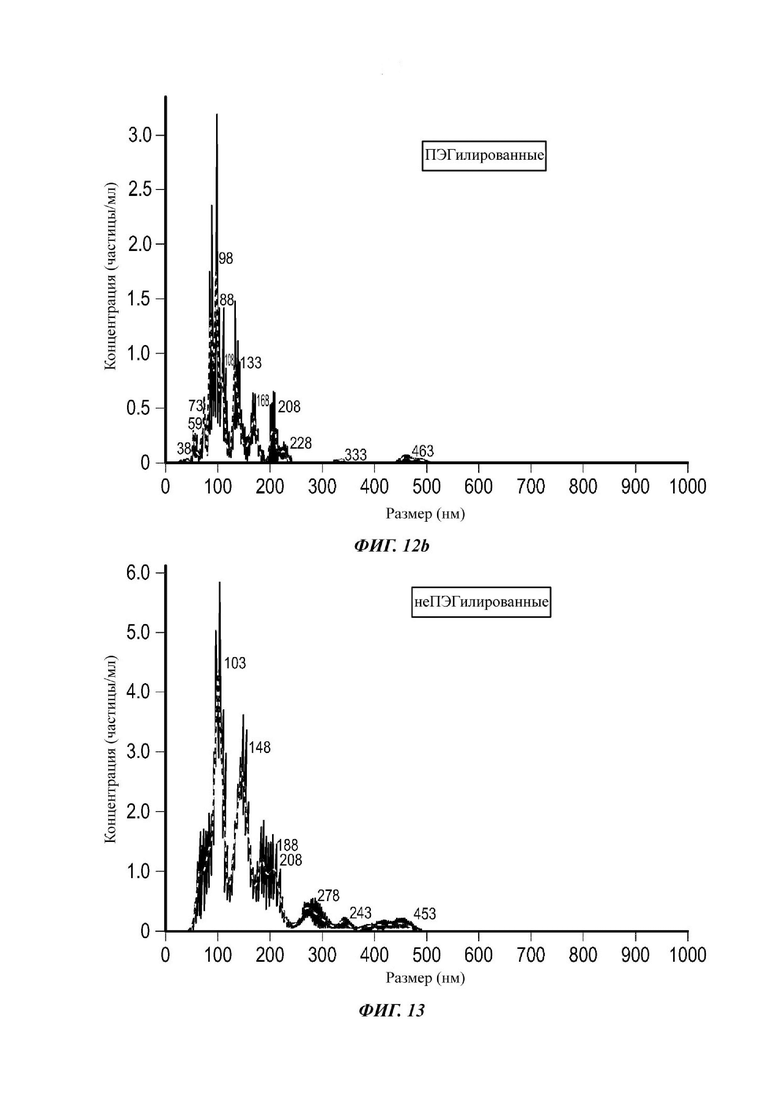

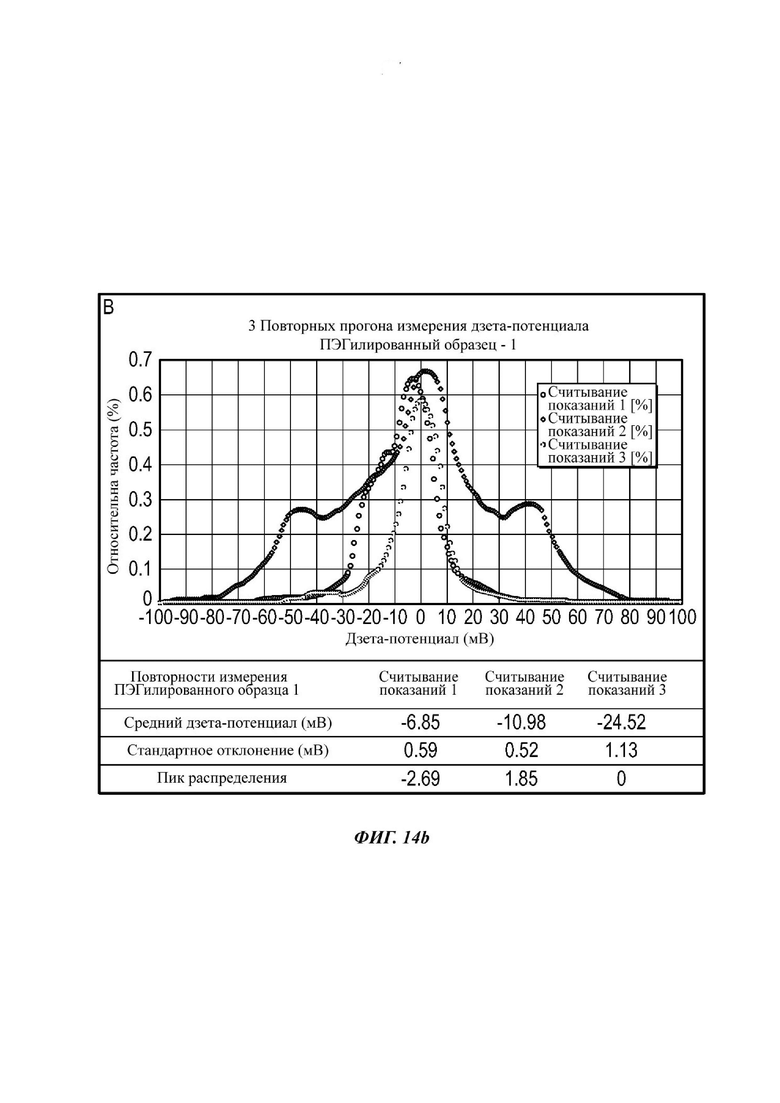
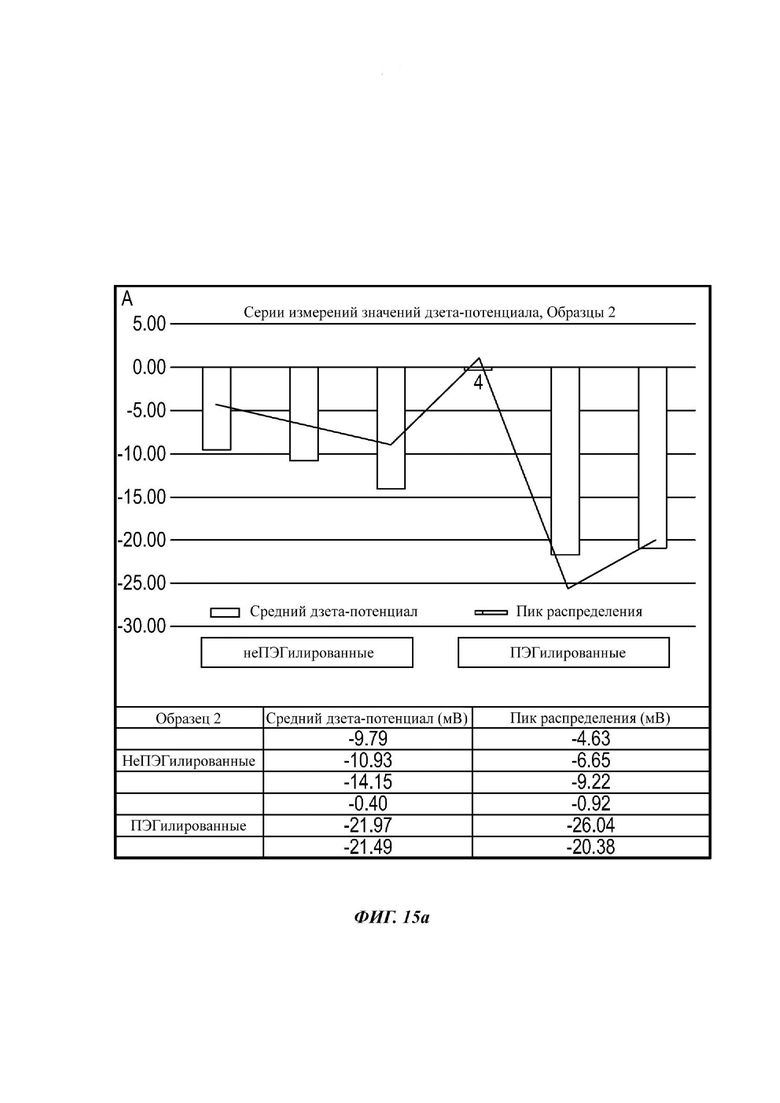
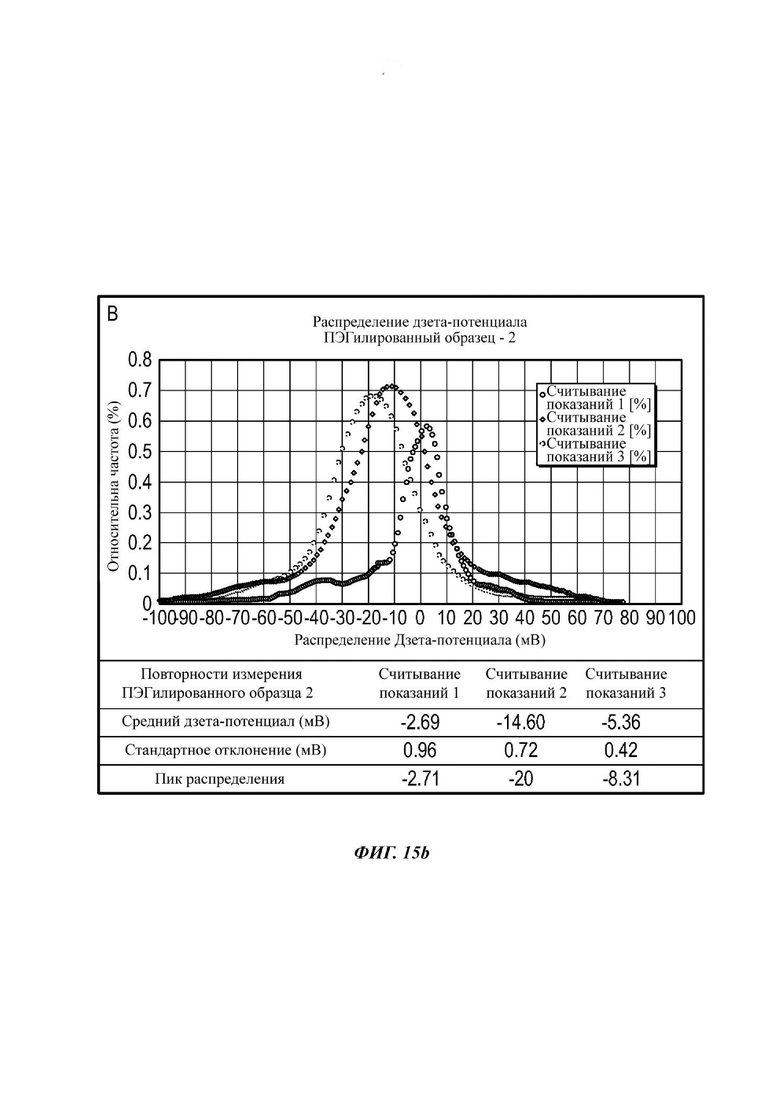
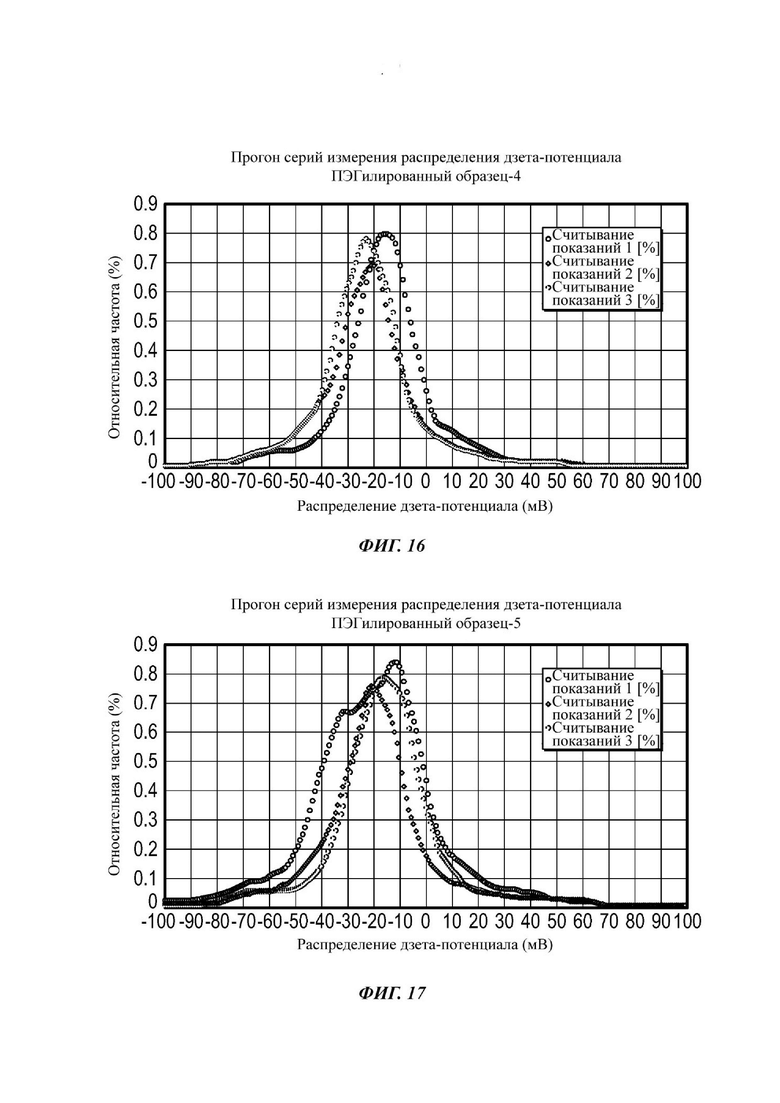
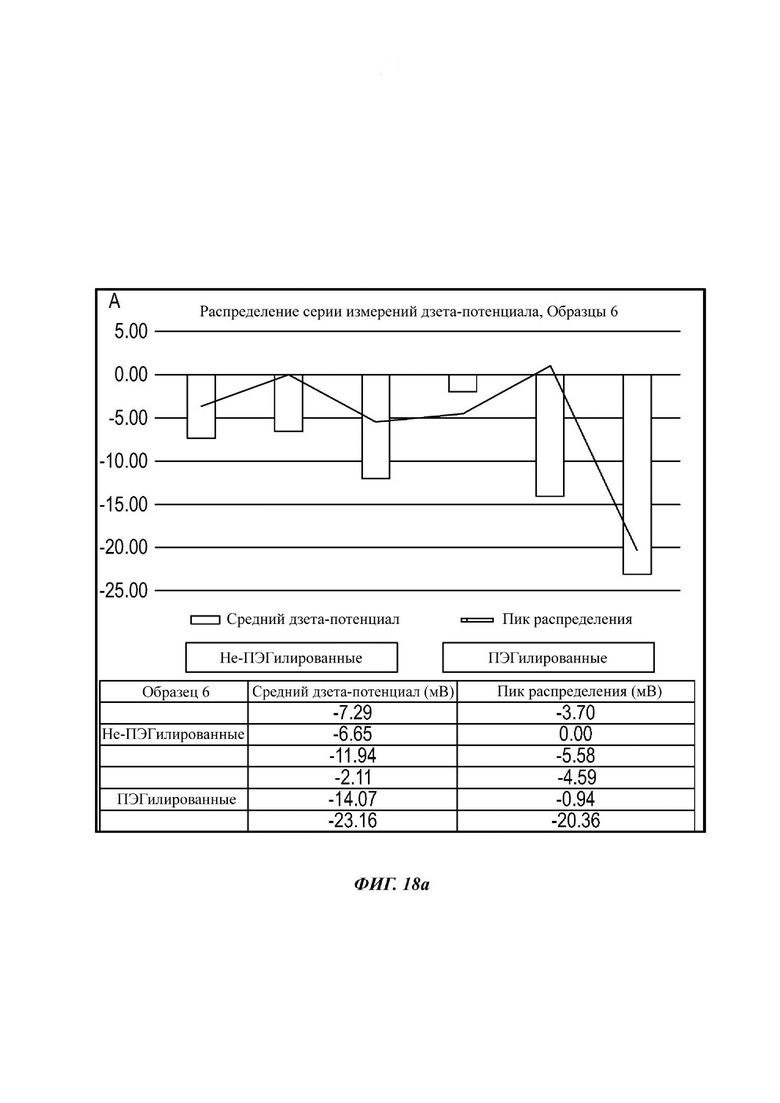

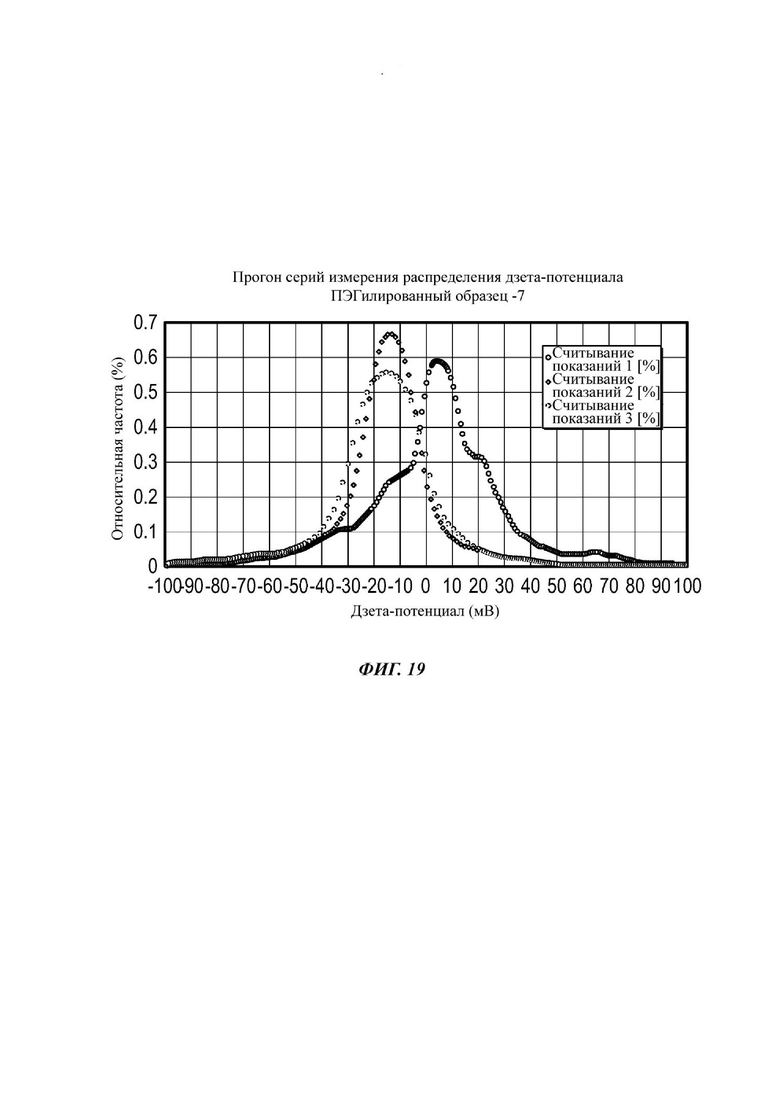



Группа изобретений относится к области фармацевтики и медицины. 1 объект представляет собой водную композицию для доставки груза к клеткам легкого, способную к аэрозолизации, содержащую экзосомы из мезенхимальных стволовых клеток (МСК), имеющие поверхностное покрытие из гидрофильного полимера полиэтиленгликоля (ПЭГ), причем экзосомы несут указанный груз, содержащий микроРНК (miR), анти-miR, мРНК, длинную некодирующую РНК, кольцевую РНК, малую интерферирующую РНК, короткую шпилечную РНК, piwi-взаимодействующую РНК, последовательность РНК кластерных коротких палиндромных повторов, разделенных регулярными промежутками (CRISPR), модификации вышеуказанного или искусственно сконструированные последовательности нуклеиновых кислот, белок, цитокин или липид, где ПЭГ имеет молекулярную массу меньше чем 5 кДа, поверхностное покрытие покрывает по меньшей мере 65% поверхности экзосом. 2 объект – способ лечения пациента, имеющего заболевание легкого, включающий предоставление композиции, образование аэрозоля экзосом композиции и введение аэрозоля пациенту. Технический результат заключается в возможности получения аэрозоля с содержанием экзосом, который проходит через слой слизи с проникновением в клетки легкого, для терапии заболеваний легкого. 2 н. и 13 з.п. ф-лы, 25 ил., 16 табл., 2 пр.
1. Водная композиция для доставки груза к клеткам легкого, способная к аэрозолизации, содержащая экзосомы из мезенхимальных стволовых клеток (МСК), имеющие поверхностное покрытие из гидрофильного полимера полиэтиленгликоля (ПЭГ), причем экзосомы несут указанный груз, содержащий микроРНК (miR), анти-miR, мРНК, длинную некодирующую РНК, кольцевую РНК, малую интерферирующую РНК, короткую шпилечную РНК, piwi-взаимодействующую РНК, последовательность РНК кластерных коротких палиндромных повторов, разделенных регулярными промежутками (CRISPR), модификации вышеуказанного или искусственно сконструированные последовательности нуклеиновых кислот, белок, цитокин или липид, где ПЭГ имеет молекулярную массу меньше чем 5 кДа, поверхностное покрытие покрывает по меньшей мере 65% поверхности экзосом.
2. Композиция по п. 1, в которой гидрофильный полимер содержит дистеароилфосфатидилэтаноламин-полиэтиленгликоль (ДСФЭ-ПЭГ).
3. Композиция по п. 1 или 2, в которой полимер, покрывающий поверхность, имеет молекулярную массу меньше 3 кДа.
4. Композиция по любому из пп. 1-3, в которой в экзосомах имеется груз, содержащий микроРНК (miR).
5. Композиция по п. 4, в которой miR выбрана из miR-125b-5p, miR-125b-1-3p, miR- 513a-5p и miR-17.
6. Композиция по любому из пп. 1-3, в которой в экзосомах имеется груз, содержащий мРНК или ее транслированный белок.
7. Композиция по п. 6, в которой мРНК представляет собой модифицированную мРНК регулятора трансмембранной проводимости при кистозном фиброзе (CFTR).
8. Композиция по любому из пп. 1-7 для применения в терапии заболевания легкого.
9. Композиция по п. 8 для применения в терапии рака.
10. Композиция по п. 8, где терапия предназначена для лечения кистозного фиброза, хронической обструктивной болезни легких (ХОБЛ), рака легкого или другого патологического состояния легкого.
11. Композиция по п. 10, где экзосомы содержат терапевтический груз, который содержит по меньшей мере одну экзогенную мРНК, белок, miR или анти-miR.
12. Композиция по любому из пп. 10, 11, где экзосомы происходят из мезенхимальных стволовых клеток, генетически модифицированных для сверхэкспрессии специфичных miR, анти-miR, мРНК или другой нуклеиновой кислоты и зрелого белка в качестве терапевтических грузов.
13. Способ лечения пациента, имеющего заболевание легкого, включающий:
предоставление композиции по любому из пп. 1-7;
образование аэрозоля экзосом данной композиции; и
введение данного аэрозоля пациенту.
14. Способ по п. 13, включающий образование аэрозоля с использованием небулайзера.
15. Способ по п. 14, включающий образование аэрозоля с использованием вибрационного меш-небулайзера.
| US 2014065240 A1, 06.03.2014 | |||
| KIM M.S | |||
| et al | |||
| Engineering macrophage-derived exosomes for targeted paclitaxel delivery to pulmonary metastases: in vitro and in vivo evaluations // Nanomedicine: Nanotechnology, Biology and Medicine | |||
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |
| - Vol | |||
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| - No | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| - P | |||
| Регулятор давления для автоматических тормозов с сжатым воздухом | 1921 |
|
SU195A1 |
| US 2015079046 A1, 19.03.2015 | |||
| WO 2017075708 A1, 11.05.2017 | |||
| WO 2015002956 | |||
