ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[1] Настоящая заявка испрашивает приоритет согласно 35 U.S.C. § 119(e) по предварительным заявкам на патент США № 62/673098, поданной 17 мая 2018 г.; № 62/574719, поданной 19 октября 2017 г.; и № 62/516656, поданной 7 июня 2017 г.; каждая из которых включена в данный документ во всей своей полноте посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ НАСТОЯЩЕЕ ИЗОБРЕТНИЕ
[2] Настоящая заявка в целом относится к композициям и способам для лечения лизосомных болезней накопления. В частности, настоящая заявка относится к целевым белковым комплексам, которые содержат заместительные ферменты, и к их применению в лечении лизосомных болезней накопления.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[3] Лизосомные болезни накопления составляют класс редких заболеваний, которые влияют на разрушение многочисленных субстратов в лизосоме. Такие субстраты включают в себя сфинголипиды, мукополисахариды, гликопротеины, гликоген и олигосахариды, которые могут накапливаться в клетках больных, что приводит к гибели клеток. Органы, поражаемые лизосомными болезнями накопления, включают в себя центральную нервную систему (CNS), периферическую нервную систему (PNS), легкие, печень, кость, скелетную и сердечную мускулатуру и высокоинтеллектуальную систему.
[4] Варианты лечения лизосомных болезней накопления включают заместительную ферментную терапию (ERT), субстрат-редуцирующую терапию, фармакологическую шаперон-опосредованную терапию, терапию с трансплантацией гемопоэтических стволовых клеток и генную терапию. Пример субстрат-редуцирующей терапии включает в себя применение миглустата или элиглустата для лечения болезнь Гоше 1 типа. Эти лекарственные средства действуют путем блокирования активности синтазы, что снижает последующее продуцирование субстрата. Терапию с использованием гемопоэтических стволовых клеток (HSCT), например, используют для облегчения и замедления негативного фенотипа центральной нервной системы у пациентов с некоторыми формами MPS. См. R.M. Boustany, «Lysosomal storage diseases--the horizon expands», 9(10) Nat. Rev. Neurol. 583-98, Oct. 2013. В таблице 1 приводятся некоторые лизосомные болезни накопления и ассоциированные с ними ферменты или другие белки.
Таблица 1. Лизосомные болезни накопления
[5] Двумя наиболее распространенными LSD являются заболевание Помпе и заболевание Фабри. Болезнь Помпе, уровень заболеваемости которой, по оценкам, составляет 1 на 10000, вызывается дефектным лизосомным ферментом альфа-глюкозидазой (GAA), который приводит к недостаточному процессингу лизосомного гликогена. Накопление лизосомного гликогена происходит преимущественно в скелетной, сердечной и печеночной тканях. Ранняя манифестация болезнь Помпе вызывает кардиомегалию, гипотонию, гепатомегалию и смерть из-за кардиореспираторной недостаточности обычно в возрасте до 2 лет. Проявление болезни Помпе во взрослом возрасте наблюдается только от двадцати до шестидесяти лет и обычно затрагивает только скелетную мускулатуру. Доступные в настоящее время средства для лечения включают MYZZME®/LUMIZYME® (алглюкозидаза альфа) от Genzyme, который представляет собой рекомбинантную альфа-глюкозидазу человека, которую получают в клетках CHO, и которую вводят посредством внутривенной инфузии.
[6] Болезнь Фабри, общий уровень заболеваемости которой, включая легкие случаи с поздним началом, по оценкам, составляет 1 на 3000, вызывается дефектным лизосомным ферментом альфа-галактозидазой A (GLA), который приводит к накоплению глоботриаозилцерамида в кровеносных сосудах и других тканях и органах. Симптомы, ассоциированные с болезнью Фабри включают в себя боль из-за повреждения нерва и/или обструкции небольших сосудов, почечную недостаточность и полное разрушение, сердечные осложнения, такие как высокое кровяное давление и кардиомиопатия, дерматологические симптомы, такие как образование ангиокератомы, ангидроз или гипергидроз, а также глазные проблемы, такие как воронковидная кератопатия, клиновидная катаракта и сосудистые нарушения конъюктивы и сетчатки. Доступные в настоящее время средства для лечения включают FABRAZYME® (агалзидаза бета) от Genzyme, который представляет собой рекомбинантную альфа-галактозидазу А человека, которую получают в клетках СНО, и которую вводят посредством внутривенной инфузии; REPLAGAL™ (агалзидаза альфа) от Shire, который представляет собой рекомбинантную альфа-галактозидазу А человека, которую получают в клетках фибробластов человека, и которую вводят посредством внутривенной инфузии; и GALAFOLD™ (мигаластат или 1-дезоксигалактоноджиримицин) от Amicus, который представляет собой низкомолекулярный шаперон для перорального введения, который обеспечивает сдвиг фолдинга альфа-галактозидазы А от аномальной к функциональной конформации.
[7] Текущие методы лечения лизосомных болезней накопления не особо оптимальны. Например, ERT, как правило, должна вводиться с высокой частотой и при высокой дозе, например, дважды в неделю и до 40 мг/кг. Также некоторые замещаемые ферменты могут быть иммунологически перекрестнореагирующими (CRIM), стимулирующими продуцирование IgG у субъекта и, таким образом, препятствующими доставке фермента в лизосому посредством манноза-6-фосфатного (M6P) рецептора. IgG может экранировать остатки M6P заместительный фермент, и комплекс антиген-IgG-антитело может попадать в клеточные лизосомы через Fc-рецептор с шунтированием тем самым заместительного фермента преимущественно в макрофаги.
[8] Доставка заместительных ферментов в соответствующие пораженные ткани также неэффективна (см. таблицу 2 и Desnick & Schuchman, «Enzyme replacement therapy for lysosomal diseases: lessons from 20 years of experience and remaining challenges», 13 Annu. Rev. Genomics Hum. Genet. 307-35, 2012). Например, пациенты, подвергшиеся длительной заместительной ферментной терапии по поводу болезни Помпе во младенческом возрасте, все равно могут страдать гиперназальной речью, остаточной мышечной слабостью, птозом, остеопенией, потерей слуха, подвергаться риску аспирации, дисфагией, сердечной аритмией и затруднением при глотании. Дозы заместительного фермента часто должны увеличиваться со временем до 40 мг/кг раз в неделю или раз в две недели.
Таблица 2. Неэффективное нацеливание ERT на ткани
[9] Эндогенный манноза-6-фосфатный рецептор (MPR) опосредует транспорт большинства рекомбинантных ферментов в лизосому. Существуют две комплементарных формы MPR: катион-независимый (CI-MPR) и катион-зависимый (CD-MPR). У нокаутов по любой из форм не секретируются лизосомные ферменты. Лизосомные гидролазы синтезируются в эндоплазматическом ретикулуме и перемещаются в сеть цис-стороны аппарата Гольджи, где они ковалентно модифицируются путем добавления манноза-6-фосфатных (M6P) групп. Образование данного маркера зависит от последовательного эффекта двух лизосомных ферментов: UDP-N-ацетилглюкозамин-l-фосфотрансферазы (G1cNac-фосфотрансфераза) и N-ацетилглюкозамин-l-фосфодиэстер-α-N-ацетил-глюкозаминидазы (экспонирующий фермент). GlcNac-фосфотрансфераза катализирует перенос G1cNAc-1-фосфатного остатка от UDP-G1cNAc в C6 положения выбранных манноз в высокоманнозных олигосахаридах гидролаз. Затем экспонирующий фермент удаляет концевой G1cNAc, экспонируя сигнал распознавания M6P. В сети транс-Гольджи сигнал M6P обеспечивает сегрегацию лизосомных гидролаз от всех других типов белков посредством селективного связывания с M6P-рецепторами. Покрытые клатрином везикулы давали отпочковывание от транс-сети Гольджи и сливались с поздними эндосомами. При низком pH в поздней эндосоме гидролазы диссоциируются от M6P-рецепторов, и пустые рецепторы возвращаются в аппарат Гольджи для следующих раундов транспорта.
[10] За исключением β-глюкоцереброзидазы, которая доставляется посредством маннозного рецептора, рекомбинантные лизосомные ферменты имеют M6P-гликозилирование и доставляются в лизосому главным образом посредством CI-MPR/IGF2R. Опосредованная гликозилированием/CI-MPR доставка заместительного фермента, однако, не достигает всех клинически релевантных тканей (таблица 2). Улучшение заместительной ферментной терапии акцентировали на улучшении доставки CI-MPR с помощью (i) усиления экспрессии на поверхности CI-MPR с использованием β2-агониста кленбутерола (Koeberl et al., «Enhanced efficacy of enzyme replacement therapy in Pompe disease through mannose-6-phosphate receptor expression in skeletal muscle», 103(2) Mol. Genet. Metab. 107-12, 2011), (ii) повышения количества остатков M6P в ферменте (Zhu et al., «Conjugation of mannose-6-phosphate-containing oligosaccharides to acid alpha-glucosidase improves the clearance of glycogen in Pompe mice», 279(48) J. Biol. Chem. 50336-41, 2004) или (iii) слияния домена IGF-II с ферментом (Maga et al., «Glycosylation-independent lysosomal targeting of acid alpha-glucosidase enhances muscle glycogen clearance in Pompe mice», 288(3) J. Biol. Chem. 1428-38, 2013).
[11] При лечении большого количества лизосомных болезней накопления с помощью заместительной ферментной терапии или генной терапии получают неудовлетворительный результат в основном из-за недостаточно эффективного нацеливания заместительного фермента на соответствующие ткань или орган, отрицательных иммунологических реакций у хозяина-реципиента и низкого значения времени полужизни в сыворотке. Существует потребность в улучшенных методах заместительной ферментной терапии, которые усилят и обеспечат лучшее биораспределение в ткани и поглощение фермента в лизосоме. Заявитель разработал улучшенную заместительную ферментную терапию в которой применяется контролируемая антителом доставка ферментов в лизосому целевых пораженных тканей, которая происходит независимо от CI-MPR.
КРАТКОЕ ОПИСАНИЕ
[12] Заявители обнаружили, что заместительные ферменты могут эффективно доставляться в клетку-мишень при связывании с доменом доставки в составе мультидоменного терапевтического белка. Мультидоменный терапевтический белок может быть доставлен в клетку, которая может находиться в условиях ex vivo или in vivo, посредством вектора для генной терапии, например, вирусного вектора, полинуклеотида без дополнительных средств, полинуклеотидного комплекса и т. д., содержащего кодирующую последовательность мультидоменного терапевтического белка.
[13] В одном аспекте настоящее изобретение предусматривает полинуклеотид, содержащий последовательность нуклеиновой кислоты, кодирующую мультидоменный терапевтический белок. В одном варианте осуществления мультидоменный терапевтический белок содержит ферментативный домен и домен доставки. В одном варианте осуществления полинуклеотид также содержит последовательность нуклеиновой кислоты вирусного вектора. В конкретном варианте осуществления полинуклеотид также содержит последовательность нуклеиновой кислоты аденоассоциированного вируса (AAV).
[14] В одном варианте осуществления ферментативный домен обладает активностью гидролазы, например гликозилазы, например гликозидазы, например альфа-глюкозидазы (GAA) или альфа-галактозидазы A (GLA). В некоторых вариантах осуществления ферментативный домен содержит аминокислотную последовательность, представленную под SEQ ID NO:1, или ее биологически активную часть. В некоторых вариантах осуществления ферментативный домен содержит аминокислотную последовательность, представленную под SEQ ID NO:78. В некоторых вариантах осуществления ферментативный домен состоит по существу из аминокислотной последовательности, представленной под SEQ ID NO:78. В некоторых вариантах осуществления ферментативный домен состоит из аминокислотной последовательности, представленной под SEQ ID NO:78. В одном варианте осуществления домен доставки представляет собой антигенсвязывающий белок, который связывается с эффектором интернализации. В одном варианте осуществления эффектор интернализации представляет собой молекулу клеточной поверхности, которая подвергается эндоцитозу и транспортируется в лизосому. В конкретном варианте осуществления эффектор интернализации представляет собой молекулу CD63. В одном варианте осуществления эффектор интернализации представляет собой молекулу ITGA7. В конкретном варианте осуществления домен доставки представляет собой антитело, фрагмент антитела или одноцепочечный вариабельный фрагмент (scFv), такой как scFv, который связывает CD63 или ITGA7 (например, фиг. 1A, панель G).
[15] В одном варианте осуществления мультидоменный терапевтический белок содержит два домена доставки (например, фиг. 1А, панель H). В одном варианте осуществления первый домен доставки связывает эффектор интернализации с обеспечением доставки фермента к соответствующей клетке-мишени или целевому субклеточному компартменту; а второй домен доставки связывает эффектор трансцитоза с обеспечением транспорта мультидоменного терапевтического белка через физиологический барьер, такой как альвеолярная мембрана, эндотелий печени, гематоэнцефалический барьер или т. п. В конкретном варианте осуществления второй домен доставки представляет собой scFv-домен к рецептору трансферрина (TfR). В конкретном варианте осуществления мультидоменный терапевтический белок содержит ферментативный домен, представляющий собой GAA, первый домен доставки, представляющий собой scFv к CD63, и второй домен доставки, представляющий собой scFv к TfR. В другом варианте осуществления мультидоменный терапевтический белок содержит ферментативный домен, представляющий собой GAA, первый домен доставки, представляющий собой scFv к ITGA7, и второй домен доставки, представляющий собой scFv к CD63.
[16] В одном аспекте настоящее изобретение предусматривает вектор для генной терапии, такой как вектор на основе AAV, полинуклеотид без дополнительных средств, полинуклеотидный комплекс и т. д., который содержит последовательность нуклеиновой кислоты, кодирующую мультидоменный терапевтический белок, содержащий ферментативный домен и домен доставки.
[17] В одном варианте осуществления ферментативный домен обладает активностью гидролазы, например гликозилазы, например гликозидазы, например альфа-глюкозидазы или альфа-галактозидазы A. В одном варианте осуществления домен доставки представляет собой антигенсвязывающий белок, который связывается с эффектором интернализации. В одном варианте осуществления эффектор интернализации представляет собой молекулу клеточной поверхности, которая подвергается эндоцитозу и транспортируется в лизосому. В конкретном варианте осуществления эффектор интернализации представляет собой молекулу CD63. В одном варианте осуществления эффектор интернализации представляет собой молекулу ITGA7. В конкретном варианте осуществления домен доставки представляет собой антитело, фрагмент антитела или одноцепочечный вариабельный фрагмент (scFv), такой как scFv, который связывает CD63 или ITGA7.
[18] В одном аспекте настоящее изобретение предусматривает мультидоменный терапевтический белой, содержащий ферментативный домен и домен доставки. В одном варианте осуществления ферментативный домен обладает активностью гидролазы, например гликозилазы, например гликозидазы, например альфа-глюкозидазы или альфа-галактозидазы A. В одном варианте осуществления домен доставки представляет собой антигенсвязывающий белок, который связывается с эффектором интернализации. В одном варианте осуществления эффектор интернализации представляет собой молекулу клеточной поверхности, которая подвергается эндоцитозу и транспортируется в лизосому. В конкретном варианте осуществления эффектор интернализации представляет собой молекулу CD63. В другом варианте осуществления эффектор интернализации представляет собой молекулу ITGA7. В конкретном варианте осуществления домен доставки представляет собой антитело, фрагмент антитела или одноцепочечный вариабельный фрагмент (scFv), такой как scFv, который связывает CD63 или ITGA7.
[19] В одном варианте осуществления мультидоменный терапевтический белок применяется для лечения пациента, нуждающегося в заместительной ферментной терапии.
[20] В одном аспекте настоящее изобретение предусматривает способ получения мультидоменного терапевтического белка, содержащего ферментативный домен и домен доставки, в клетке. В одном варианте осуществления мультидоменный терапевтический белок получают посредством обеспечения контакта клетки с вектором для генной терапии, содержащим последовательность нуклеиновой кислоты, кодирующую мультидоменный терапевтический белок. Последовательность нуклеиновой кислоты впоследствии интегрируется в геномный локус клетки, из которого последовательность нуклеиновой кислоты транскрибируется и транслируется в случае, если последовательность нуклеиновой кислоты представляет собой ДНК, или транслируется в случае, если нуклеиновая кислота представляет собой РНК, и происходит получение мультидоменного терапевтического белка. В одном варианте осуществления вектор для генной терапии представляет собой вектор, который обычно применяют для трансфекции клеток, такой как вектор на основе аденоассоциированного вируса (AAV). В одном варианте осуществления вектор для генной терапии представляет собой полинуклеотид без дополнительных средств. В одном варианте осуществления вектор для генной терапии представляет собой полинуклеотидный комплекс, например липидную наночастицу. В одном варианте осуществления геномный локус представляет собой локус «безопасной гавани», который обеспечивает высокую экспрессию мультидоменного терапевтического белка, не мешая при этом экспрессии основных генов или не стимулируя экспрессию онкогенов или других вредных генов. В одном варианте осуществления геномный локус представляет собой сайт аденоассоциированного вируса.
[21] В одном варианте осуществления клетка представляет собой клетку млекопитающего, такую как клетка человека или клетка мыши. В одном варианте осуществления клетка находится в условиях ex vivo, например клеточная линия HEK293. В другом варианте осуществления клетка находится в условиях in vivo, и вектор для генной терапии, содержащий последовательность нуклеиновой кислоты, кодирующую мультидоменный терапевтический белок, вводят субъекту, представляющему собой человека, или отличному от человека.
[22] В одном варианте осуществления ферментативный домен обладает активностью гидролазы, например гликозилазы, например гликозидазы, например альфа-глюкозидазы (GAA) или альфа-галактозидазы A (GLA). В одном варианте осуществления домен доставки представляет собой антигенсвязывающий белок, который связывается с эффектором интернализации. В одном варианте осуществления эффектор интернализации представляет собой молекулу клеточной поверхности, которая подвергается эндоцитозу и транспортируется в лизосому. В конкретном варианте осуществления эффектор интернализации представляет собой молекулу CD63. В конкретном варианте осуществления домен доставки представляет собой антитело, фрагмент антитела или одноцепочечный вариабельный фрагмент (scFv), такой как scFv, который связывает CD63 (например, SEQ ID NO:2). В некоторых вариантах осуществления мультидоменный терапевтический полипептид содержит scFv, который связывает CD63 (например, scFv, содержащий аминокислотную последовательность, представленную под SEQ ID NO:2), функционально связанный с GAA (представленной под SEQ ID NO:1) или ее биологически активной частью (представленной под SEQ ID NO:78). В некоторых вариантах осуществления мультидоменный терапевтический полипептид содержит последовательность, представленную под SEQ ID NO:10 или SEQ ID NO:79.
[23] В конкретном варианте осуществления вектор на основе AAV, содержащий полинуклеотид, кодирующий слитый белок scFv-гидролаза, вводят субъекту, представляющему собой человека, или отличному от человека. Впоследствии полинуклеотид интегрируется в геномный локус, и продуцируется кодированный слитый белок. В конкретном варианте осуществления слитый белок представляет собой слитый белок, состоящий из scFv к CD63 и GAA (например, представленный в под SEQ ID NO:10 или SEQ ID NO:79), или слитый белок, состоящий из scFv к ITGA7 и GAA, субъект, представляющий собой человека, или отличный от человека характеризуется недостатком активности эндогенной GAA, и активность GAA у субъекта эффективно восстанавливается.
[24] В одном аспекте настоящее изобретение предусматривает способ лечения пациента (представляющего собой человека или отличного от человека) с дефицитом фермента путем введения пациенту вектора для генной терапии, содержащего последовательность нуклеиновой кислоты, кодирующую мультидоменный терапевтический белок, содержащий домен доставки и ферментативный домен. В одном варианте осуществления ферментативный домен предусматривает гликозидазу, такую как GAA (например, SEQ ID NO:1) или ее биологически активная часть (например, SEQ ID NO:78) или GLA (например, UniProtKB № P06280, aa32-429, SEQ ID NO:13), и у пациента имеется болезнь Помпе или болезнь Фабри. В одном варианте осуществления домен доставки мультидоменного терапевтического белка представляет собой антигенсвязывающий белок, который связывается с эффектором интернализации, таким как CD63 или ITGA7. В одном варианте осуществления домен доставки представляет собой молекулу scFv, которая связывает CD63. В одном варианте осуществления домен доставки представляет собой молекулу scFv, которая связывает ITGA7. В другом варианте осуществления вектор для генной терапии предусматривает вектор на основе AAV, содержащий последовательность нуклеиновой кислоты, которая кодирует слитый мультидоменный терапевтический белок, содержащий фрагмент, связывающий CD63, и GAA (например, SEQ ID NO:10 или SEQ ID NO:79). В другом варианте осуществления вектор для генной терапии представляет собой вектор на основе AAV, содержащий последовательность нуклеиновой кислоты, которая кодирует слитый мультидоменный терапевтический белок, содержащий фрагмент, связывающий ITGA7, и GAA. В другом варианте осуществления вектор для генной терапии предусматривает полинуклеотид без дополнительных средств, содержащий последовательность нуклеиновой кислоты, которая кодирует слитый мультидоменный терапевтический белок, содержащий фрагмент, связывающий CD63, и GAA (например, SEQ ID NO:10 или SEQ ID NO:79). В другом варианте осуществления вектор для генной терапии предусматривает полинуклеотид без дополнительных средств, содержащий последовательность нуклеиновой кислоты, которая кодирует слитый мультидоменный терапевтический белок, содержащий фрагмент, связывающий ITGA7, и GAA. В другом варианте осуществления вектор для генной терапии содержит полинуклеотидный комплекс, содержащий последовательность нуклеиновой кислоты, которая кодирует слитый мультидоменный терапевтический белок, содержащий фрагмент, связывающий CD63, и GAA (например, SEQ ID NO:10 или SEQ ID NO:79). В другом варианте осуществления вектор для генной терапии содержит полинуклеотидный комплекс, содержащий последовательность нуклеиновой кислоты, которая кодирует слитый мультидоменный терапевтический белок, содержащий фрагмент, связывающий ITGA7, и GAA.
[25] В одном аспекте настоящее изобретение относится к композиции в соответствии с настоящим изобретением для применения в медицине. В одном варианте осуществления композиция, например фармацевтическая композиция, может содержать вектор для генной терапии, содержащий последовательность нуклеиновой кислоты, кодирующую мультидоменный терапевтический белок, содержащий домен доставки и ферментативный домен. В одном варианте осуществления домен доставки мультидоменного терапевтического белка представляет собой антигенсвязывающий белок, который связывается с эффектором интернализации, таким как CD63 или ITGA7. В одном варианте осуществления домен доставки представляет собой молекулу scFv, которая связывает CD63. В одном варианте осуществления домен доставки представляет собой молекулу scFv, которая связывает ITGA7. В другом варианте осуществления вектор для генной терапии представляет собой вектор на основе AAV, содержащий полинуклеотид, который кодирует слитый мультидоменный терапевтический белок, содержащий фрагмент, связывающий CD63, и GAA. В другом варианте осуществления вектор для генной терапии представляет собой вектор на основе AAV, содержащий полинуклеотид, который кодирует слитый мультидоменный терапевтический белок, содержащий фрагмент, связывающий ITGA7, и GAA. В некоторых вариантах осуществления полинуклеотид, например вектор для генной терапии, содержит тканеспецифический регуляторный элемент. В некоторых вариантах осуществления тканеспецифический регуляторный элемент содержит последовательность, представленную под SEQ ID NO:8, SEQ ID NO:9, или обе. В некоторых вариантах осуществления полинуклеотид, например вектор для генной терапии, содержит последовательность нуклеиновой кислоты под SEQ ID NO:11.
[26] В одном аспекте описана композиция, содержащая вектор для генной терапии, содержащий ген, кодирующий мультидоменный терапевтический белок, содержащий домен доставки и ферментативный домен, для применения в лечении пациента (представляющего собой человека или отличного от человека) с дефицитом фермента, и/или для применения с целью уменьшения накопления гликогена в ткани у субъекта, представляющего собой человека или отличного от человека. В одном варианте осуществления домен доставки мультидоменного терапевтического белка представляет собой антигенсвязывающий белок, который связывается с эффектором интернализации, таким как CD63 или ITGA7. В одном варианте осуществления домен доставки представляет собой молекулу scFv, которая связывает CD63. В одном варианте осуществления домен доставки представляет собой молекулу scFv, которая связывает ITGA7. В другом варианте осуществления вектор для генной терапии представляет собой векторы на основе AAV, содержащие полинуклеотид, который кодирует слитый мультидоменный терапевтический белок, содержащий фрагмент, связывающий CD63, и GAA. В другом варианте осуществления вектор для генной терапии представляет собой векторы на основе AAV, содержащие полинуклеотид, который кодирует слитый мультидоменный терапевтический белок, содержащий фрагмент, связывающий ITGA7, и GAA. Болезнь Помпе или болезнь Фабри, например, могут являться без ограничения клиническими показаниями, влекущими за собой дефицит фермента. В некоторых вариантах осуществления полинуклеотид, например вектор для генной терапии, содержит тканеспецифический регуляторный элемент. В некоторых вариантах осуществления тканеспецифический регуляторный элемент содержит последовательность, представленную под SEQ ID NO:8, SEQ ID NO:9, или обе. В некоторых вариантах осуществления полинуклеотид, например вектор для генной терапии, содержит последовательность нуклеиновой кислоты под SEQ ID NO:11.
[27] В одном аспекте описано применение фармацевтической композиции в соответствии с настоящим изобретением для изготовления лекарственного средства для терапевтического применения, такого как, например, лечение пациента (представляющего собой человека или отличного от человека) с дефицитом фермента, и/или уменьшение накопления гликогена в ткани у субъекта, представляющего собой человека или отличного от человека. Композиция может содержать, например, вектор для генной терапии, содержащий ген, кодирующий мультидоменный терапевтический белок, содержащий домен доставки и ферментативный домен. В одном варианте осуществления домен доставки мультидоменного терапевтического белка представляет собой антигенсвязывающий белок, который связывается с эффектором интернализации, таким как CD63 или ITGA7. В одном варианте осуществления домен доставки представляет собой молекулу scFv, которая связывает CD63. В одном варианте осуществления домен доставки представляет собой молекулу scFv, которая связывает ITGA7. В другом варианте осуществления вектор для генной терапии представляет собой векторы на основе AAV, содержащие полинуклеотид, который кодирует слитый мультидоменный терапевтический белок, содержащий фрагмент, связывающий CD63, и GAA. В другом варианте осуществления вектор для генной терапии представляет собой векторы на основе AAV, содержащие полинуклеотид, который кодирует слитый мультидоменный терапевтический белок, содержащий фрагмент, связывающий ITGA7, и GAA. Болезнь Помпе или болезнь Фабри, например, могут являться без ограничения клиническими показаниями, влекущими за собой дефицит фермента.
[28] В одном варианте осуществления мультидоменный терапевтический белок содержит ферментативный домен, представляющий собой GAA, и высокие уровни содержания GAA в сыворотке крови поддерживаются в сыворотке крови пациента в течение по меньшей мере 12 недель после введения вектора для генной терапии. В одном варианте осуществления мультидоменный терапевтический белок содержит ферментативный домен, представляющий собой GAA, и уровни содержания гликогена в сердечной, скелетной мышце и ткани печени пациента поддерживаются на уровнях, характерных для дикого типа, спустя 3 месяца после введения вектора для генной терапии. В одном варианте осуществления мультидоменный терапевтический белок содержит ферментативный домен, представляющий собой GAA, и мышечная сила пациента после лечения восстанавливается до уровней, характерных для дикого типа.
[29] В одном аспекте настоящее изобретение предусматривает способ уменьшения накопления гликогена в ткани у субъекта, представляющего собой человека или отличного от человека, путем введения вектора для генной терапии, содержащего полинуклеотид, который кодирует мультидоменный терапевтический белок. В одном варианте осуществления ткань представляет собой печень, сердце или скелетную мышцу. В одном варианте осуществления субъект, представляющий собой человека или отличный от человека, имеет болезнь Помпе. В одном варианте осуществления мультидоменный терапевтический белок содержит слитый белок, содержащий scFv к CD63 и GAA. В другом варианте осуществления мультидоменный терапевтический белок содержит слитый белок, содержащий scFv к ITGA7 и GAA.
[30] В одном аспекте в данном документе описан способ уменьшения уровня содержания перекрестнореагирующего иммунологического материала к ферменту у пациента (представляющего собой человека или отличного от человека) с дефицитом фермента, при этом способ включает введение пациенту вектора для генной терапии, содержащего ген, кодирующий только заместительный фермент, или мультидоменный терапевтический белок, содержащий домен доставки и ферментативный домен. В некоторых вариантах осуществления вектор для генной терапии представляет собой вектор на основе AAV, который может представлять собой химерный вектор на основе AAV (например, AAV2/8). В некоторых вариантах осуществления ферментативный домен предусматривает гликозидазу, такую как GAA (например, под SEQ ID NO:1) или GLA (например, UniProtKB № P06280, aa32-429, под SEQ ID NO:13), и пациент имеет болезнь Помпе или болезнь Фабри, и домен доставки представляет собой антигенсвязывающий белок, который связывается с эффектором интернализации. В некоторых вариантах осуществления вектор для генной терапии вводят в количествах, достаточных для повышения уровней содержания GAA в сыворотке крови настолько, чтобы количество перекрестнореагирующего иммунологического материала уменьшилось или перекрестнореагирующий иммунологический материал не образовывался в обнаруживаемых количествах. В некоторых вариантах осуществления вектор для генной терапии вводят в количествах, достаточных для повышения уровня содержания GAA в сыворотке крови в течение определенного периода времени. В некоторых вариантах осуществления пациента инфицируют вирусом (например, AAV), содержащим вектор для генной терапии, где необязательно вирус подвергнут псевдотипированию с целью достижения специфической нацеленности на ткань или клетку, выбранные из группы, состоящей из GALT, печени, гемопоэтической стволовой клетки, эритроцита, или их комбинацию, и/или где вектор для генной терапии содержит клеточно- или тканеспецифический энхансер и/или промотор, например, специфический для печени энхансер (например, серпина 1) и/или специфический для печени промотор (например, TTR). В одном варианте осуществления эффектор интернализации представляет собой молекулу клеточной поверхности, которая подвергается эндоцитозу и транспортируется в лизосому. В конкретном варианте осуществления эффектор интернализации представляет собой молекулу CD63. В конкретном варианте осуществления домен доставки представляет собой антитело, фрагмент антитела или одноцепочечный вариабельный фрагмент (scFv), такой как scFv, который связывает CD63 (например, SEQ ID NO:2). В некоторых вариантах осуществления способ включает введение пациенту вектора для генной терапии, описанного в данном документе, в комбинации по меньшей мере с одним иммуносупрессором, при этом вектор для генной терапии и иммуносупрессор вводят одновременно и/или последовательно. В некоторых вариантах у пациента поддерживают постоянный уровень иммуносупрессора.
[31] В одном аспекте в данном документе описан способ индуцирования толерантности к ферменту у пациента (представляющего собой человека или отличного от человека) с дефицитом фермента (т. е. индуцирования толерантности пациента к ферменту), при этом способ включает уменьшение уровня содержания перекрестнореагирующего иммунологического материала у пациента, например, введение пациенту вектора для генной терапии, содержащего ген, кодирующий только заместительный фермент, или мультидоменный терапевтический белок, содержащий домен доставки и ферментативный домен. В некоторых вариантах осуществления описан способ индуцирования толерантности к ферменту у пациента с дефицитом фермента, при этом способ включает введение пациенту вектора для генной терапии, содержащего ген, кодирующий только заместительный фермент или мультидоменный терапевтический белок, содержащий домен доставки и ферментативный домен, в течение определенного периода времени таким образом, что у пациента не появлялось увеличения обнаруживаемого перекрестнореагирующего иммунологического материала. В некоторых вариантах осуществления мультидоменный терапевтический белок содержит домен доставки и ферментативный домен. В некоторых вариантах осуществления вектор для генной терапии представляет собой вектор на основе AAV, который может представлять собой химерный вектор на основе AAV (например, AAV2/8) и/или сконструированный вектор на основе AAV (например, рекомбинантный вирусный вектор с модифицированным тропизмом, применимый для нацеливания, например, на рецептор или маркер, преимущественно или исключительно экспрессируемый клеткой или тканью, например гепатоцитом или печенью, тканью слизистой оболочки, эритроцитами, гемопоэтическими стволовыми клетками и т. д.), и где необязательно ген экспрессируется под контролем энхансера и/или промотора, специфического для клетки или ткани, например специфического для печени промотора и т. д. В некоторых вариантах осуществления ферментативный домен предусматривает гликозидазу, такую как GAA (например, под SEQ ID NO:1) или GLA (например, UniProtKB № P06280, aa32-429, под SEQ ID NO:13), и пациент имеет болезнь Помпе или болезнь Фабри, и домен доставки представляет собой антигенсвязывающий белок, который связывается с эффектором интернализации. В одном варианте осуществления эффектор интернализации представляет собой молекулу клеточной поверхности, которая подвергается эндоцитозу и транспортируется в лизосому. В конкретном варианте осуществления эффектор интернализации представляет собой молекулу CD63. В конкретном варианте осуществления домен доставки представляет собой антитело, фрагмент антитела или одноцепочечный вариабельный фрагмент (scFv), такой как scFv, который связывает CD63 (например, SEQ ID NO:2). В некоторых вариантах осуществления способ лечения пациента с дефицитом фермента включает введение пациенту рекомбинантной формы и/или изозима фермента (например, GAA у пациента с болезнью Помпе, например scFv63::GAA, GAA, оптимизированная GAA или их комбинации), где пациент является толерантным к ферменту, например, где пациент был подвергнут индуцированию толерантности к ферменту в соответствии со способом, описанным в данном документе. Соответственно, в некоторых вариантах осуществления способ введения фермента пациенту с дефицитом этого фермента включает индуцирование толерантности у пациента в отношении фермента, например, путем введения вектора для генной терапии, кодирующего фермент или мультидоменный терапевтический белок, содержащий фермент, предпочтительно в количествах, достаточных для увеличения уровня содержания фермента в сыворотке крови настолько, чтобы перекрестнореагирующий иммунологический материал находился на уровне, сопоставимом с уровнем, обнаруженным у пациента, не испытывающего дефицита фермента, при этом необязательно вектор для генной терапии специфически нацелен на печень и/или содержит специфический для печени энхансер и/или промотор. В некоторых вариантах осуществления способ дополнительно включает после введения вектора, нацеленного на ген, дополнительное введение фермента и/или его рекомбинантного варианта, включая мультидоменный терапевтический белок, содержащий фермент.
[32] В другом аспекте настоящее изобретение предусматривает антитела и их антигенсвязывающие фрагменты, которые связываются с CD63 человека. Антитела в соответствии с этим аспектом настоящего изобретения применимы, среди прочего, для специфического направления интернализации и/или лизосомного транспорта фермента, например GAA или GLA. Таким образом, этот аспект изобретения также предусматривает биспецифические антитела, их антигенсвязывающие фрагменты, которые связывают CD63 человека, и слитые конструкции антитело-белок (см., например, фиг. 1А).
[33] Иллюстративные антитела к CD63 по настоящему изобретению перечислены в таблице 10. В таблице 10 представлены идентификаторы последовательностей аминокислот и нуклеиновых кислот вариабельных областей тяжелой цепи (HCVR), вариабельных областей легкой цепи (LCVR), а также определяющих комплементарность областей тяжелой цепи (HCDR1, HCDR2 и HCDR3) и определяющих комплементарность областей легкой цепи (LCDR1, LCDR2 и LCDR3) иллюстративных антител к CD63.
[34] Настоящее изобретение предусматривает антитела или их антигенсвязывающие фрагменты, которые содержат HCVR, содержащую аминокислотную последовательность, выбранную из любой из аминокислотных последовательностей HCVR, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[35] Настоящее изобретение также предусматривает антитела или их антигенсвязывающие фрагменты, которые содержат LCVR, содержащую аминокислотную последовательность, выбранную из любой из аминокислотных последовательностей LCVR, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[36] Настоящее изобретение также предусматривает антитела или их антигенсвязывающие фрагменты, содержащие пару аминокислотных последовательностей HCVR и LCVR (HCVR/LCVR), содержащую любую из аминокислотных последовательностей HCVR, перечисленных в таблице 10, в паре с любой из аминокислотных последовательностей LCVR, перечисленных в таблице 10. В соответствии с определенными вариантами осуществления настоящее изобретение предусматривает антитела или их антигенсвязывающие фрагменты, которые содержат пару аминокислотных последовательностей HCVR/LCVR, содержащуюся в иллюстративных антителах к CD63, перечисленных в таблице 10. В определенных вариантах осуществления пара аминокислотных последовательностей HCVR/LCVR выбрана из группы, состоящей из SEQ ID NO: 14/22, SEQ ID NO: 30/38, SEQ ID NO:46/54 и SEQ ID NO: 62/70.
[37] Настоящее изобретение также предусматривает антитела или их антигенсвязывающие фрагменты, которые содержат CDR1 тяжелой цепи (HCDR1), содержащую аминокислотную последовательность, выбранную из любой из аминокислотных последовательностей HCDR1, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[38] Настоящее изобретение также предусматривает антитела или их антигенсвязывающие фрагменты, которые содержат CDR2 тяжелой цепи (HCDR2), содержащую аминокислотную последовательность, выбранную из любой из аминокислотных последовательностей HCDR2, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[39] Настоящее изобретение также предусматривает антитела или их антигенсвязывающие фрагменты, которые содержат CDR3 тяжелой цепи (HCDR3), содержащую аминокислотную последовательность, выбранную из любой из аминокислотных последовательностей HCDR3, перечисленных в таблице 11, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[40] Настоящее изобретение также предусматривает антитела или их антигенсвязывающие фрагменты, содержащие CDR1 легкой цепи (LCDR1), содержащую аминокислотную последовательность, выбранную из любой из аминокислотных последовательностей LCDR1, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[41] Настоящее изобретение также предусматривает антитела или их антигенсвязывающие фрагменты, содержащие CDR2 легкой цепи (LCDR2), содержащую аминокислотную последовательность, выбранную из любой из аминокислотных последовательностей LCDR2, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[42] Настоящее изобретение также предусматривает антитела или их антигенсвязывающие фрагменты, которые содержат CDR3 тяжелой цепи (LCDR3), содержащую аминокислотную последовательность, выбранную из любой из аминокислотных последовательностей LCDR3, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[43] Настоящее изобретение также предусматривает антитела или их антигенсвязывающие фрагменты, содержащие пару аминокислотных последовательностей HCDR3 и LCDR3 (HCDR3/LCDR3), содержащую любую из аминокислотных последовательностей HCDR3, перечисленных в таблице 10, в паре с любой из аминокислотных последовательностей LCDR3, перечисленных в таблице 10. В соответствии с определенными вариантами осуществления настоящее изобретение предусматривает антитела или их антигенсвязывающие фрагменты, которые содержат пару аминокислотных последовательностей HCDR3/LCDR3, содержащуюся в иллюстративных антителах к CD63, перечисленных в таблице 10. В определенных вариантах осуществления пара аминокислотных последовательностей HCDR3/LCDR3 выбрана из группы, состоящей из SEQ ID NO: SEQ ID NO: 20/28, SEQ ID NO: 36/44, SEQ ID NO:52/60 и SEQ ID NO: 68/76.
[44] Настоящее изобретение также предусматривает антитела или их антигенсвязывающие фрагменты, которые содержат группу из шести CDR (т. е. HCDR1-HCDR2-HCDR3-LCDR1-LCDR2-LCDR3), содержащихся в любом из иллюстративных антител к CD63, перечисленных в таблице 10. В определенных вариантах осуществления группа аминокислотных последовательностей HCDR1-HCDR2-HCDR3-LCDR1-LCDR2-LCDR3 выбран из группы, состоящей из SEQ ID NO:16-18-20-24-26-28, SEQ ID NO: 32-34-36-40-42-44, SEQ ID NO:48-50-52-56-58-60 и SEQ ID NO: 64-66-68-72-74-76.
[45] В связанном варианте осуществления настоящее изобретение предусматривает антитела или их антигенсвязывающие фрагменты, содержащие группу из шести CDR (т. е. HCDR1-HCDR2-HCDR3-LCDR1-LCDR2-LCDR3), содержащихся в паре аминокислотных последовательностей HCVR/LCVR, определенных для любого из иллюстративных антител к CD63, перечисленных в таблице 10. Например, настоящее изобретение включает антитела или их антигенсвязывающие фрагменты, содержащие группу аминокислотных последовательностей HCDR1-HCDR2-HCDR3-LCDR1-LCDR2-LCDR3, которая содержится в паре аминокислотных последовательностей HCVR/LCVR, выбранной из группы, состоящей из SEQ ID NO:14/22, SEQ ID NO: 30/38, SEQ ID NO:46/54 и SEQ ID NO: 62/70. Способы и методики идентификации CDR в аминокислотных последовательностях HCVR и LCVR хорошо известны в данной области техники и могут быть использованы для идентификации CDR в указанных аминокислотных последовательностях HCVR и/или LCVR, раскрытых в данном документе. Иллюстративные подходы, которые могут быть использованы для идентификации границ CDR, включают, например, определение по Кабат, определение по Чотиа и определение согласно AbM. В целом, определение по Кабат основано на вариабельности последовательностей, определение по Чотиа основано на положении структурных петлевых областей, и определение согласно AbM является компромиссным решением между подходами по Кабат и Чотиа. Смотри, например, Kabat, «Sequences of Proteins of Immunological Interest», National Institutes of Health, Bethesda, Md. (1991); Al-Lazikani et al., J. Mol. Biol. 273:927-948 (1997); and Martin et al., Proc. Natl. Acad. Sci. USA 86:9268-9272 (1989). Доступны также общедоступные базы данных для идентификации последовательностей CDR в антителе.
[46] Настоящее изобретение также предусматривает молекулы нуклеиновой кислоты, кодирующие антитела к CD63 или их части. Например, настоящее изобретение предусматривает молекулы нуклеиновых кислот, кодирующие любую из аминокислотных последовательностей HCVR, перечисленных в таблице 10; в определенных вариантах осуществления молекула нуклеиновой кислоты содержит полинуклеотидную последовательность, выбранную из любой из последовательностей нуклеиновых кислот HCVR, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[47] Настоящее изобретение также предусматривает молекулы нуклеиновых кислот, кодирующие любую из аминокислотных последовательностей LCVR, перечисленных в таблице 10; в определенных вариантах осуществления молекула нуклеиновой кислоты содержит полинуклеотидную последовательность, выбранную из любой из аминокислотных последовательностей LCVR, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[48] Настоящее изобретение также предусматривает молекулы нуклеиновых кислот, кодирующие любую из аминокислотных последовательностей HCDR1, перечисленных в таблице 10; в определенных вариантах осуществления молекула нуклеиновой кислоты содержит полинуклеотидную последовательность, выбранную из любой из последовательностей нуклеиновых кислот HCDR1, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[49] Настоящее изобретение также предусматривает молекулы нуклеиновых кислот, кодирующие любую из аминокислотных последовательностей HCDR2, перечисленных в таблице 10; в определенных вариантах осуществления молекула нуклеиновой кислоты содержит полинуклеотидную последовательность, выбранную из любой из последовательностей нуклеиновых кислот HCDR2, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[50] Настоящее изобретение также предусматривает молекулы нуклеиновых кислот, кодирующие любую из аминокислотных последовательностей HCDR3, перечисленных в таблице 10; в определенных вариантах осуществления молекула нуклеиновой кислоты содержит полинуклеотидную последовательность, выбранную из любой из последовательностей нуклеиновых кислот HCDR3, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[51] Настоящее изобретение также предусматривает молекулы нуклеиновых кислот, кодирующие любую из аминокислотных последовательностей LCDR1, перечисленных в таблице 10; в определенных вариантах осуществления молекула нуклеиновой кислоты содержит полинуклеотидную последовательность, выбранную из любой из последовательностей нуклеиновых кислот LCDR1, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[52] Настоящее изобретение также предусматривает молекулы нуклеиновых кислот, кодирующие любую из аминокислотных последовательностей LCDR2, перечисленных в таблице 10; в определенных вариантах осуществления молекула нуклеиновой кислоты содержит полинуклеотидную последовательность, выбранную из любой из последовательностей нуклеиновых кислот LCDR2, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[53] Настоящее изобретение также предусматривает молекулы нуклеиновых кислот, кодирующие любую из аминокислотных последовательностей LCDR3, перечисленных в таблице 10; в определенных вариантах осуществления молекула нуклеиновой кислоты содержит полинуклеотидную последовательность, выбранную из любой из последовательностей нуклеиновых кислот LCDR3, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее.
[54] Настоящее изобретение также предусматривает молекулы нуклеиновой кислоты, кодирующие HCVR, где HCVR содержит группу из трех CDR (т. е., HCDR1-HCDR2-HCDR3), при этом группа аминокислотных последовательностей HCDR1-HCDR2-HCDR3 является такой, как определено для любого из иллюстративных антител к CD63, перечисленных в таблице 10.
[55] Настоящее изобретение также предусматривает молекулы нуклеиновой кислоты, кодирующие LCVR, где LCVR содержит группу из трех CDR (т. е., LCDR1-LCDR2-LCDR3), при этом группа аминокислотных последовательностей LCDR1-LCDR2-LCDR3 является такой, как определено для любого из иллюстративных антител к CD63, перечисленных в таблице 10.
[56] Настоящее изобретение также предусматривает молекулы нуклеиновых кислот, кодирующие как HCVR, так и LCVR, где HCVR содержит аминокислотную последовательность, представляющую собой любую из аминокислотных последовательностей HCVR, перечисленных в таблице 10, и где LCVR содержит аминокислотную последовательность, представляющую собой любую из аминокислотных последовательностей LCVR, перечисленных в таблице 10. В определенных вариантах осуществления молекула нуклеиновой кислоты содержит полинуклеотидную последовательность, выбранную из любой из последовательностей нуклеиновых кислот HCVR, перечисленных в таблице 10, или по сути аналогичную ей последовательность, которая характеризуется по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности в отношении нее, и полинуклеотидную последовательность, выбранную из любой из последовательностей нуклеиновых кислот LCVR, перечисленных в таблице 10, или по сути аналогичную ей последовательность, характеризующуюся по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательностей в отношении нее. В определенных вариантах осуществления в соответствии с данным аспектом настоящего изобретения молекула нуклеиновой кислоты кодирует HCVR и LCVR, где как HCVR, так и LCVR получены из одного и того же антитела к CD63, приведенного в таблице 10.
[57] Настоящее изобретение также предусматривает рекомбинантные векторы экспрессии, способные экспрессировать полипептид, содержащий вариабельную область тяжелой или легкой цепи антитела к CD63. Например, настоящее изобретение включает рекомбинантные векторы экспрессии, содержащие любую из упомянутых выше молекул нуклеиновых кислот, т. е, молекул нуклеиновых кислот, кодирующих любую из последовательностей HCVR, LCVR и/или CDR, представленных в таблице 10. Также в объем настоящего изобретения включены клетки-хозяева, в которые были введены векторы экспрессии, а также способы получения антител или их частей путем культивирования клеток-хозяев в условиях, обеспечивающих продуцирование антител или фрагментов антител, и выделение полученных таким образом антител и фрагментов антител.
[58] В некоторых аспектах настоящее изобретение включает антитела или их антигенсвязывающие фрагменты, такие как антитела к CD63, имеющие модифицированный профиль гликозилирования. В некоторых вариантах осуществления могут быть применимыми модификация для удаления нежелательных сайтов гликозилирования, или антитело без фукозного фрагмента, который присутствует в олигосахаридной цепи, например, для усиления функции антителозависимой клеточной цитотоксичности (ADCC) (смотри Shields et al. (2002) JBC 277:26733), в случае, если цитотоксичность желательна. В других применениях модификацию галактозилирования можно осуществлять для модифицирования комплементзависимой цитотоксичности (CDC).
[59] В другом аспекте настоящее изобретение предусматривает фармацевтическую композицию, содержащую рекомбинантное антитело человека или его фрагмент, которые специфически связывают CD63, и фармацевтически приемлемый носитель. В связанном аспекте в настоящем изобретении представлена композиция, которая представляет собой комбинацию антитела к CD63 и второго терапевтического средства. В одном варианте осуществления второе терапевтическое средство представляет собой любое средство, при объединении которого с антителом к CD63 обеспечивается преимущество. Дополнительные комбинированные терапевтические препараты и комбинированные составы, включающие антитела к CD63 по настоящему изобретению, описаны в других частях данного документа.
ГРАФИЧЕСКИЕ МАТЕРИАЛЫ
[60] На фигуре 1А схематически представлены мультидоменные терапевтические белки. На панели A изображен мультидоменный терапевтический белок, содержащий биспецифическое антитело (ii) и заместительный фермент (i). На панели B изображен слитый полипептид фермент-Fc (i), ассоциирующийся со специфическим в отношении эффектора интернализации полутелом (ii) с образованием мультидоменного терапевтического белка. На панели С изображен заместительный фермент (шестиугольник), ковалентно связанный с С-концом тяжелой цепи антитела к эффектору интернализации. На панели D изображен заместительный фермент (шестиугольник), ковалентно связанный с N-концом тяжелой цепи антитела к эффектору интернализации. На панели Е изображен заместительный фермент (шестиугольник), ковалентно связанный с С-концом легкой цепи антитела к эффектору интернализации. На панели F изображен заместительный фермент (шестиугольник), ковалентно связанный с N-концом легкой цепи антитела к эффектору интернализации. На панели G изображен заместительный фермент (шестиугольник), ковалентно связанный с С-концом одноцепочечного вариабельного фрагмента (scFv), содержащего область VH (заштрихованный столбик) и область VL (незакрашенный столбик). На панели H изображен заместительный фермент (шестиугольник), ковалентно связанный с двумя доменами scFv, где первый scFv (i) служит в качестве первого домена доставки, и второй scFv (ii) служит в качестве второго домена доставки.
[61] Фигура 1В представляет собой неограничивающую иллюстративную иллюстрацию вектора для генной терапии на основе AAV, который кодирует мультидоменный терапевтический белок, представленный на панели, где scFv представляет собой scFv к CD63 человека, а заместительный фермент представляет собой GAA (например, scFv к hCD63::hGAA; см., например, аминокислотную последовательность, представленную под SEQ ID NO:10). Аминокислоты 1-119 под SEQ ID NO:10 обеспечивают аминокислотную последовательность вариабельного домена тяжелой цепи (VH) антитела H5C6; аминокислоты 120-134 под SEQ ID NO:10 обеспечивают линкерную аминокислотную последовательность между вариабельными доменами тяжелой и легкой цепей H5C6; аминокислоты 135-245 под SEQ ID NO:10 обеспечивают аминокислотную последовательность вариабельного домена легкой цепи (VL) антитела H5C6; аминокислоты 136-250 под SEQ ID NO:10 обеспечивают линкерную аминокислотную последовательность между scFv к hCD63 и GAA; и аминокислоты 251-1133 под SEQ ID NO:10 обеспечивают аминокислотную последовательность GAA. Иллюстративные последовательности 5'ITR и 3'ITR соответственно представлены под SEQ ID NO:6 и SEQ ID NO:7. Иллюстративный специфический для печени энхансер (серпина 1) представлен под SEQ ID NO:9. Иллюстративный специфический для печени промотор (TTR) представлен под SEQ ID NO:8. Дополнительные иллюстративные аминокислотные последовательности VH и VL к CD63 (и кодирующие их нуклеотидные последовательности), которые можно использовать для конструирования мультидоменного терапевтического белка, содержащего антитело к CD63 или его антигенсвязывающую часть, представлены в таблице 10.
[62] Фигура 2 представляет собой столбчатую диаграмму, изображающую количество накопленного гликогена в микрограммах на миллиграмм ткани в зависимости от доставленного фермента. На оси X изображены ткани от мыши CD63hu/hu; GAA-/- слева направо: сердце, четырехглавая мышца, икроножная мышца, диафрагма, камбаловидная мышца и длинная мышца-разгибатель пальцев (EDL). Черные ящики («1») отображают количество накопленного гликогена в мышиной модели болезни Помпе, не подвергнутой обработке. Оранжевые ящики («2») отображают количество накопленного гликогена в мышиной модели дикого типа, не подвергнутой обработке. Светло-зеленые ящики («3») отображают количество накопленного гликогена в мышиной модели болезни Помпе, которую подвергали обработке с помощью AAV-hGAA (вектор на основе аденоассоциированного вируса, содержащий ген, кодирующий GAA человека) в дозе 1010 vg. Темно-зеленые ящики («4») отображают количество накопленного гликогена в мышиной модели болезни Помпе, которую подвергали обработке с помощью AAV-hGAA в дозе 1011 vg. Светло-зеленые ящики («5») отображают количество накопленного гликогена в мышиной модели болезни Помпе, которую подвергали обработке с помощью AAV с scFv к hCD63::hGAA (вектор на основе аденоассоциированного вируса, содержащий ген, кодирующий домен scFv к CD63 человека, связанный с GAA человека) в дозе 1010 vg. Темно-синие ящики («6») отображают количество накопленного гликогена в мышиной модели болезни Помпе, которую подвергали обработке с помощью AAV с scFv к hCD63::hGAA в дозе 1011 vg.
[63] Фигура 3 представляет собой столбчатую диаграмму, изображающую средний уровень содержания гликогена (мкг/мг), измеренный в скелетной мышечной ткани каждой мыши через 3 месяца после инъекции AAV. Каждое значение измерения нанесено на график в виде зависимости от уровня воздействия GAA (т. е. уровня содержания в сыворотке крови) на каждую мышь, которую подвергали обработке с помощью конкретной конструкции на основе фермента в конкретной дозировке. Закрашенные квадраты обозначают AAV-hGAA в дозе 1010 vg. Закрашенные пирамиды обозначают AAV-hGAA в дозе 1011 vg. Закрашенные перевернутые пирамиды обозначают AAV с scFv к hCD63::hGAA в дозе 1010 vg. Закрашенные ромбы обозначают AAV с scFv к hCD63::hGAA в дозе 1011 vg.
[64] Фигура 4 представляет собой точечный график, изображающий средний уровень содержания гликогена (мкг/мг) в сердечной мышце, измеренный в ткани сердца через 3 месяца после инъекции AAV, в зависимости от уровня воздействия GAA (т. е. уровней содержания в сыворотке крови) на каждую мышь, которую подвергали обработке с помощью конкретной конструкции на основе фермента при конкретной дозировке. Закрашенные квадраты обозначают AAV-hGAA в дозе 1010 vg. Закрашенные пирамиды обозначают AAV-hGAA в дозе 1011 vg. Закрашенные перевернутые пирамиды обозначают AAV с scFv к hCD63::hGAA в дозе 1010 vg. Закрашенные ромбы обозначают AAV с scFv к hCD63::hGAA в дозе 1011 vg.
[65] Фигура 5 представляет собой точечный график, изображающий титры антител к GAA через 3 месяца после инъекции AAV в зависимости от уровня воздействия GAA (т. е. уровней содержания в сыворотке крови) на каждую мышь, которую подвергали обработке с помощью конкретной конструкции на основе фермента при конкретной дозировке. Незакрашенные квадраты обозначают AAV-hGAA в дозе 1010 vg. Незакрашенные круги обозначают AAV-hGAA в дозе 1011 vg. Незакрашенные ромбы обозначают AAV с scFv к hCD63::hGAA в дозе 1010 vg. Шестиугольники обозначают AAV с scFv к hCD63::hGAA в дозе 1011 vg.
[66] Фигура 6 представляет собой точечный график, изображающий титры антител к GAA через 3 месяца после инъекции AAV в зависимости от конструкции на основе фермента и дозы. Круги обозначают контрольных мышей, получающих пустой вектор на основе AAV. Квадраты обозначают AAV-hGAA в дозе 1010 vg. Пирамиды обозначают AAV-hGAA в дозе 1011 vg. Перевернутые пирамиды обозначают AAV с scFv к hCD63::hGAA в дозе 1010 vg. Ромбы обозначают AAV с scFv к hCD63::hGAA в дозе 1011 vg.
[67] Фигура 7А представляет собой линейный график, изображающий уровни содержания GAA в сыворотке крови (произвольные единицы «a.u.»; ось Y) в зависимости от времени в неделях после инъекции вектора для генной терапии. Квадраты обозначают AAV-hGAA в дозе 1010 vg. Пирамиды обозначают AAV-hGAA в дозе 1011 vg. Перевернутые пирамиды обозначают AAV с scFv к hCD63::hGAA в дозе 1010 vg. Ромбы обозначают AAV с scFv к hCD63::hGAA в дозе 1011 vg.
[68] Фигура 7В представляет собой гистограмму, изображающую соотношения мРНК (мРНК hGAA относительно мРНК mGAPDH) после введения конструкций на основе AAV мышам CD63 HumIn с нокаутом GAA (мыши GAA-/-, CD63hu/hu) или мышам GAA+/+, CD63hu/hu таким образом: (1) необработанный контроль (2) AAV-специфический для печени промотор-hGAA (1e10vg), (3) AAV-специфический для печени промотор-hGAA (1e11vg), (4) AAV-специфический для печени промотор-фрагмент, связывающий hCD63::hGAA (1e10vg), (5) AAV-специфический для печени промотор-фрагмент, связывающий hCD63::hGAA (1e11vg) или (6) необработанный контроль (GAA+/+, CD63hu/hu). Экспрессия GAA в печени была обнаружена при всех инъекциях конструкции на основе AAV.
[69] Фигура 7С представляет собой график, на котором сравнивается уровень содержания GAA в сыворотке крови с уровнем экспрессии РНК GAA у мышей, получающих AAV, кодирующий слитый белок (квадраты), и мышей, получающих AAV, кодирующий GAA (обе конструкции обеспечивали специфический для печени промотор (LSP) для управления экспрессией).
[70] Фигура 7D представляет собой гистограмму, изображающую гепатоциты человека Huh-7, временно трансфицированные конструкциями, управляемыми специфическим для печени промотором, кодирующими hGAA, scFv к hCD63::GAA (слитая конструкция) или контрольную несвязывающую слитую конструкцию scFv::GAA. Для обеих слитых конструкций scFv::GAA было характерным более высокое соотношение белка в секретируемом супернатанте, чем hGAA отдельно через 3 дня после трансфекции. Добавление M6P в супернатант в течение экспериментального периода для уменьшения CI-MPR-опосредованного захвата не влияло на соотношение. (* = р <0,05, n=3).
[71] Фигура 8 представляет собой флуоресцентные микрофотографии, изображающие лизосомы, окрашенные по LAMP1 в мышечных волокнах мыши, контрастно окрашенных с помощью DAPI для выявления ядер. На панелях А и А1 изображены клетки четырехглавой мышцы, полученные от необработанной мыши дикого типа (GAA+/+) и окрашенные по LAMP1 (панель А) и ядру (панель А1). На панелях B и B1 изображены клетки четырехглавой мышцы, полученные от необработанной мыши с нокаутом по GAA (GAA-/-) и окрашенные по LAMP1 (панель B) и ядру (панель B1). На панелях C и C1 изображены клетки четырехглавой мышцы, полученные от мыши GAA-/-, которую подвергали обработке с помощью конструкции на основе AAV-hGAA, и окрашенные по LAMP1 (панель C) и ядру (панель C1). На панелях D и D1 изображены клетки четырехглавой мышцы, полученные от мыши GAA-/-, которую подвергали обработке с помощью конструкции на основе AAV с scFv к hCD63::hGAA, и окрашенные по LAMP1 (панель D) и ядру (панель D1).
[72] На фигуре 9 изображены линейные графики, показывающие силу захвата и эффективность выполнения теста с вращающимся барабаном у мышей, обработанных с помощью либо AAV-LSP-hGAA, либо AAV-LSP-фрагмент, связывающий hCD63::hGAA. Измерения при ускорении вращающегося барабана (A) и измерения силы захвата передних конечностей (B) у мышей с GAA дикого типа (перевернутый треугольник), необработанного контроля (квадрат), мышей, обработанных с помощью AAV-LSP-hGAA (1e11vg/мышь) (треугольник) или мышей, обработанных с помощью AAV-LSP-фрагмент, связывающий hCD63::hGAA (1e11vg/мышь) (круг) проводились с месячными интервалами в течение 6 месяцев. Планки погрешностей представляют собой +/- SD. N=8-10 для всех групп.
[73] На фигуре 10А и фигуре 10В показано применение других мембранных белков в качестве направляющих, таких как слитые белки на основе scFv к ITGA7 (интегрин альфа-7), для направления GAA. На фигуре 10А показана активность GAA (ось у) миобластов мыши C2C12, проинкубированных в течение ночи с комплексом фрагмент, связывающий мышиный CD63-GAA, или комплексом фрагмент, связывающий мышиный ITGA7-GAA с или без 5 мМ M6P. На фигуре 10В показаны мыши с нокаутом GAA, гуманизированные по CD63 (GAA-/-; CD63hu/hu), которым вводили плазмиды, кодирующие формат scFv::GAA комплекса фрагмент, связывающий hCD63::GAA (2) или формат полноразмерный IgG4::GAA фрагмента, связывающего интегрин альфа-7 мыши (3) путем гидродинамической доставки (HDD), затем уровни содержания гликогена в ткани измеряли через 3 недели после HDD. Необработанные контрольные мыши GAA-/-; CD63hu/hu (1) и необработанные контрольные мыши дикого типа GAA+/+; CD63hu/hu (4) также тестировали на уровни содержания гликогена в тех же тканях.
[74] Фигура 11 представляет собой точечный график, изображающий уровни содержания GAA в сыворотке крови (произвольные единицы «a.u.»; ось y) спустя месяц после инъекции AAV в зависимости от в зависимости от конструкции фермента и дозы. Квадраты обозначают AAV-LSP-Δ8GAA. Пирамиды обозначают AAV-scFv к hCD63::GAA. Обе конструкции обеспечивали специфический для печени промотор (LSP) для стимуляции экспрессии. Доза представлена в виде количества вирусного генома (vg) на килограмм (кг) мыши.
[75] На фигуре 12 представлены дот-блот анализы, изображающие уровни содержания гликогена в микрограммах на миллиграмм ткани (сердце, квадрицепс, диафрагма или трицепс) в зависимости от уровня содержания GAA в сыворотке крови. Квадраты обозначают AAV-LSP-Δ8GAA. Пирамиды обозначают AAV-scFv к hCD63::GAA. Обе конструкции обеспечивали специфический для печени промотор (LSP) для стимуляции экспрессии.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[76] Настоящее изобретение не ограничено описанными конкретными вариантами осуществления, композициями, способами и условиями эксперимента, поскольку такие варианты осуществления, композиции, способы и условия могут варьировать. Терминология, используемая в данном документе, предназначена только для целей описания конкретных вариантов осуществления и не предназначена быть ограничивающей, поскольку объем настоящего изобретения будет ограничиваться только прилагаемой формулой изобретения.
[77] Хотя при практическом осуществлении или тестировании настоящего изобретения можно применять любые способы и материалы, аналогичные описанным в данном документе или эквивалентные им, в данном документе описаны лишь некоторые предпочтительные способы и материалы. Все публикации, упомянутые в данном документе, включены в данный документ посредством ссылки во всей своей полноте. Если не определено иное, все технические и научные термины, используемые в данном документе, имеют такое же значение, как это обычно понимает специалист в данной области техники, к которой принадлежит настоящее изобретение.
[78] «Заболевания, связанные с дефицитом ферментов» включают нелизосомные болезни накопления, такие как болезнь Краббе (галактозилцерамидаза), фенилкетонурия, галактоземия, болезнь кленового сиропа, митохондриальные нарушения, атаксия Фридрейха, синдром Зеллвегера, адренолейкодистрофия, болезнь Вильсона, гемохроматоз, дефицит орнитинтранскарбамилазы, метилмалоновая ацидемия, пропионовая ацидемия и лизосомные болезни накопления. Термин «лизосомные болезни накопления» включает любое нарушение, возникшее в результате дефекта функции лизосомы. На данный момент идентифицировали приблизительно 50 лизосомных болезней накопления, наиболее известные из которых включают болезнь Тея-Сакса, болезнь Гоше и болезнь Ниманна-Пика. Патогенез этих заболеваний объясняют накоплением продуктов неполного разрушения в лизосоме, обычно из-за утраты белковой функции. Лизосомные болезни накопления вызываются утратой функции или истощением вариантов в белках, чья нормальная функция заключается в разрушении или координации разрушения лизосомного содержимого. Белки, связанные с лизосомными болезнями накопления, предусматривают ферменты, рецепторы и другие трансмембранные белки (например, NPC1), посттрансляционные модифицирующие белки (например, сульфатазу), белки переноса через мембрану, а также неферментативные кофакторы и другие растворимые белки (например, GM2 ганглиозидный активатор). Таким образом, лизосомные болезни накопления охватывают больше расстройств кроме тех нарушений, которые вызваны дефективными ферментами per se, и включают в себя любое нарушение, вызванное любым молекулярным дефектом. Таким образом, используемый в данном документе термин «фермент» охватывает те другие белки, ассоциированные с лизосомными болезнями накопления.
[79] Природа молекулярного повреждения влияет на тяжесть заболевания во многих случаях, т. e. полная утрата функции, как правило, ассоциирован с пренатальным или неонатальным проявлением и предусматривает тяжелые симптомы; частичная утрата функции ассоциирована со менее выраженным (относительно) и более поздним проявлением заболевания. Как правило, требуется восстановить только небольшое процентное отношение активности, чтобы исправить метаболические дефекты в дефицитных клетках. В таблице 1 перечисляются некоторые из более распространенных лизосомных болезней накопления и ассоциированные с утратой функции при них белки. Лизосомные болезни накопления, как правило, описаны у Desnick and Schuchman, 2012.
[80] Лизосомные болезни накопления могут быть систематизированы по типу продукта, который накапливается в дефективной лизосоме. Сфинголипидоз представляет собой класс заболеваний, которые поражают метаболизм сфинголипидов, которые являются липидами, содержащими жирные кислоты, соединенные с алифатическими аминоспиртами (обзор в S. Hakomori, «Glycosphingolipids in Cellular Interaction, Differentiation, and Oncogenesis», 50 Annual Review of Biochemistry 733-764, July 1981). Накопленные продукты сфинголипидоза включают в себя ганглиозиды (например, заболевание Тея-Сакса), гликолипиды (например, заболевание Фабри) и глюкоцереброзиды (например, заболевание Гоше).
[81] Мукополисахаридоз представляет собой группу заболеваний, которые поражают метаболизм глюкозаминогликанов (GAGS или мукополисахаридов), которые представляют собой длинные неразветвленные цепи повторяющихся дисахаридов, которые помогают строить кость, хрящ, сухожилия, роговицу, кожу и соединительную ткань (обзор в Muenzer, «Early initiation of enzyme replacement therapy for the mucopolysaccharidoses», 111(2) Mol. Genet. Metab. 63-72 (Feb. 2014); Sasisekharan et al., «Glycomics approach to structure-function relationships of glycosaminoglycans», 8(1) Ann. Rev. Biomed. Eng. 181-231 (Dec. 2014)). Накопленные продукты мукополисахаридоза включают в себя гепарана сульфат, дерматана сульфат, кератина сульфат, различные формы хондроитина сульфата и гиалуроновую кислоту. Например, синдром Моркио А типа обуславливается дефектом в лизосомном ферменте галактоза-6-сульфат-сульфатаза, который приводит к накоплению кератина сульфата и хондроитин-6-сульфатав в лизосомах.
[82] Болезни накопления гликогена (также известные как гликогеноз) являются результатом неспособности клетки к метаболизму (к созданию или разрушению) гликогена. Метаболизм гликогена модерируется различными ферментами или другими белками, в том числе глюкоза-6-фосфатазой, кислой альфа-глюкозидазой, линеаризирующим гликоген ферментом, разветвляющим гликоген ферментом, мышечной гликогенфосфорилазой, печеночной гликогенфосфорилазой, мышечной фосфофруктокиназой, киназой фосфорилазы, переносчиком глюкозы, альдолазой A, бета-энолазой и гликогенсинтазой. Типичной болезнью лизосомного накопления/накопления гликогена является болезнь Помпе, при которой дефективная кислая альфа-глюкозидаза вызывает накопление гликогена в лизосомах. Симптомы включают в себя гепатомегалию, мышечную слабость, сердечную недостаточность, а в случае инфантильного варианта смерть в возрасте до 2 лет (см. DiMauro and Spiegel, «Progress and problems in muscle glycogenosis», 30(2) Acta Myol. 96-102 (Oct. 2011)).
[83] «Мультидоменный терапевтический белок» предусматривает (i) один белок, который содержит более чем один функциональный домен, (ii) белок, который содержит более чем одну полипептидную цепь, и (iii) смесь из более чем одного белка или более чем одного полипептида. Термин «полипептид», как правило, означает отдельную цепь аминокислот, соединенных вместе пептидными связями. Термин «белок» охватывает термин «полипептид», но также включает в себя более сложные структуры. То есть отдельный полипептид является белком, и белок может содержать один или более полипептидов, ассоциированных в структуру более высокого порядка. Например, гемоглобин является белком, содержащим четыре полипептида: два альфа-глобиновых полипептида и два бета-глобиновых полипептида. Миоглобин также является белком, но он содержит только один миоглобиновый полипептид.
[84] Мультидоменный терапевтический белок содержит один или более полипептидов и по меньшей мере два домена, выполняющих две функции. Один из этих доменов представляет собой «ферментативный домен», который обеспечивает замещение активности дефектного белка, ассоциированного с заболеванием, связанным с дефицитом ферментов. Другой из этих доменов представляет собой «домен доставки», который обеспечивает связывание с эффектором интернализации. Таким образом, один полипептид, который обеспечивает замещение активности фермента и способность связываться с эффектором интернализации (также известный, как белок, связывающий эффектор интернализации (активность домена доставки)), представляет собой мультидоменный терапевтический белок. Также смесь белков, где один белок обеспечивает функцию фермента, а другой белок обеспечивает активность связывания с эффектором интернализации, представляет собой мультидоменный терапевтический белок. На фигуре 1А изображены различные примеры мультидоменных терапевтических белков. В одном примере (фигура 1A, панель A) мультидоменный терапевтический белок содержит фермент (представленный шестиугольником) и биспецифическое антитело (IE-BP), которое связывает фермент (пунктирные линии) и эффектор интернализации (сплошные линии). В данном случае одно плечо биспецифического антитела нековалентно связывается с ферментом, а другое плечо нековалентно связывается с эффектором интернализации, тем самым обеспечивая интернализацию заместительного фермента в клетку или субклеточный компартмент. В другом примере (панель B) мультидоменный терапевтический белок предусматривает один белок, содержащий два полипептида, при этом один полипептид обладает функцией фермента, а другой обладает функцией домена доставки. В данном случае фермент слит с Fc-доменом иммуноглобулина или константной областью тяжелой цепи, которая ассоциируется с Fc-доменом полуантитела к ферменту, с образованием бифункционального мультидоменного терапевтического белка. Вариант осуществления, изображенный на панели B, аналогичен таковому на панели A, за исключением того, что фермент ковалентно присоединен к одному из полуантител, а не посредством взаимодействия антиген-антитело по вариабельному домену иммуноглобулина полуантитела.
[85] В других примерах мультидоменный терапевтический белок состоит из фермента, ковалентно связанного (непосредственно или опосредованно с помощью линкера) с доменом доставки. В одном варианте осуществления фермент присоединен к С-концу молекулы иммуноглобулина (например, к тяжелой цепи или, в качестве альтернативы, к легкой цепи). В других вариантах осуществления фермент присоединен к N-концу молекулы иммуноглобулина (например, к тяжелой цепи или, в качестве альтернативы, к легкой цепи). В данных примерах молекула иммуноглобулина представляет собой домен доставки. В еще одном варианте осуществления фермент присоединен к С-концу молекулы scFv, которая связывает эффектор интернализации.
[86] В одном варианте осуществления мультидоменный терапевтический белок содержит два домена доставки. В одном варианте осуществления первый домен доставки связывается с молекулой- лизосомного транспорта или другим эффектором интернализации (например, CD63). В другом варианте осуществления второй домен доставки связывается с эффектором трансцитоза для обеспечения трансклеточного транспорта мультидоменного терапевтического белка. В одном варианте осуществления эффектор трансцитоза представляет собой, среди прочего, рецептор LDL, рецептор IgA, рецептор трансферрина или неонатальный Fc-рецептор (FcRn). В конкретном варианте осуществления домен доставки, обеспечивающий трансцитоз, предусматривает молекулу, которая связывается с рецептором трансферрина, такую как, например, антитело к рецептору трансферрина или молекула scFv к рецептору трансферрина. Публикация Tuma and Hubbard, «Transcytosis: Crossing Cellular Barriers», Physiological Reviews, 83(3): 871-935 (1 июля 2003 г.) включена в данный документ посредством ссылки в отношении рецепторов клеточной поверхности, которые опосредуют трансцитоз, которые применимы при практическом осуществлении настоящего изобретения.
[87] «Ферментативный домен» или «фермент» означает любой белок, ассоциированный с этиологией или физиологическим эффектом заболевания, связанного с дефицитом ферментов. Фермент предусматривает собственно фермент, транспортный белок, рецептор или другой белок, который является дефективным и который рассматривается в роли молекулярного повреждения, вызывающего заболевание. Фермент также предусматривает любой белок, который может обеспечивать аналогичную или достаточную биохимическую или физиологическую активность, который обеспечивает замещение или обход молекулярного повреждения при заболевании. Например, «изозим» может использоваться в качестве фермента. Примеры белков, связанных с лизосомной болезнью накопления, включают белки, приведенные в таблице 1 как «вовлеченный фермент/белок», а также любой известный или открытый позже белок или другую молекулу, которые обеспечивают обход молекулярного дефекта при заболевании, связанном с дефицитом ферментов.
[88] В некоторых вариантах осуществления фермент представляет собой гидролазу, включая эстеразы, гликозилазы, гидролазы, которые действуют на эфирные связи, пептидазы, линейные амидазы, дифосфатазы, кетонгидролазы, галогеназы, фосфоамидазы, сульфогидролазы, сульфиназы, десульфиназы и т. п. В некоторых вариантах осуществления фермент представляет собой гликозилазу, включая гликозидазы и N-гликозилазы. В некоторых вариантах осуществления фермент представляет собой гликозидазу, включая альфа-амилазу, бета-амилазу, глюкан-1,4-альфа-глюкозидазу, целлюлозу, эндо-1,3(4)-бета-глюканазу, инулиназу, эндо-1,4-бета-ксиланазу, эндо-1,4-b-ксиланазу, декстраназу, хитиназу, полигалактуронидазу, лизоцим, экзо-альфа-сиалидазу, альфа-глюкозидазу, бета-глюкозидазу, альфа-галактозидазу, бета-галактозидазу, альфа-маннозидазу, бета-маннозидазу, бета-фруктофуранозидазу, альфа,альфа-трегалозу, бета-глюкуронидазу, ксилан-эндо-1,3-бета-ксилозидазу, амило-альфа-1,6-глюкозидазу, гиалуроноглюкозаминидазу, гиалуроноглюкуронидазу и т. п.
[89] В случае заболевания Помпе, при котором молекулярным дефектом является дефект активности α-глюкозидазы, ферменты включают альфа-глюкозидазу человека и «изозимы», такие как другие альфа-глюкозидазы, сконструированная рекомбинантная альфа-глюкозидаза, другие глюкозидазы, рекомбинантные глюкозидазы, любой белок, сконструированный с возможностью гидролиза концевого нередуцирующего 1-4-связанного остатка альфа-глюкозы с высвобождением одной молекулы альфа-глюкозы, любой фермент EC 3.2.1.20, природные или рекомбинантные углеводные гидролазы для гликогена или крахмалов при низком pH, а также гликозилгидролазы, такие как сахараза-изомальтаза, мальтаза-глюкоамилаза, глюкозидаза II и нейтральная альфа-глюкозидаза.
[90] Термин «интернализующий эффектор» включает белок, который способен интернализоваться в клетку, или который в противном случае участвует в ретроградном мембранном транспорте или способствует ему. В некоторых случаях интернализующий эффектор представляет собой белок, который подвергается трансцитозу; т. е. белок интернализуется с одной стороны клетки и транспортируется на другую сторону клетки (например, от апикальной к базальной). Во многих вариантах осуществления интернализующий эффекторный белок представляет собой белок, экспрессируемый на клеточной поверхности, или растворимый внеклеточный белок. Однако в настоящем изобретении также рассматриваются варианты осуществления, в которых интернализующий эффекторный белок экспрессируется во внутриклеточном компартменте, таком как эндосома, эндоплазматический ретикулум, аппарат Гольджи, лизосома и т. д. Например, белки, участвующие в ретроградном мембранном транспорте (например, пути от ранних/рециркулирующих эндосом к транс-сети Гольджи), могут служить в качестве интернализующих эффекторных белков в различных вариантах осуществления настоящего изобретения. В любом случае связывание домена доставки с интернализующим эффекторным белком приводит к тому, что весь мультидоменный терапевтический белок и любые связанные с ним молекулы (например, фермент), также интернализуются в клетку. Как объясняется ниже, интернализующие эффекторные белки включают белки, которые непосредственно интернализуются в клетку, а также белки, которые опосредованно интернализуются в клетку.
[91] Интернализующие эффекторные белки, которые непосредственно интернализуются в клетку, включают в себя связанные с мембраной молекулы по меньшей мере с одним внеклеточным доменом (например, трансмембранные белки, GPI-заякоренные белки и т. д.), которые подвергаются клеточной интернализации и предпочтительно обрабатываются посредством пути внутриклеточной деградации и/или рециркуляции. Конкретные неограничивающие примеры интернализующих эффекторных белков, которые непосредственно интернализуются в клетку, включают, например, CD63, MHC-I (например, HLA-B27), Kremen-1, Kremen-2, LRP5, LRP6, LRP8, рецептор трансферрина, LDL-рецептор, рецептор белка 1, родственного LDL, ASGR1, ASGR2, белок-2, подобный белку-предшественнику амилоида (APLP2), апелиновый рецептор (APLNR), MAL (миелиновый и лимфоцитарный белок, также известный как VIP17), IGF2R, H+ АТФазу вакуолярного типа, рецептор дифтерийного токсина, фолатный рецептор, глутаматные рецепторы, глутатионовые рецепторы, лептиновые рецепторы, скавенджер-рецепторы (например, SCARA1-5, SCARB1-3, CD36) и т. д.
[92] В определенных вариантах осуществления интернализующий эффектор представляет собой рецептор пролактина (PRLR). Было обнаружено, что PRLR представляет собой не только мишень для определенных терапевтических применений, но также является эффективным интернализующим эффекторным белком исходя из его высокой скорости интернализации и возобновления. Потенциал PRLR как интернализующего эффекторного белка, например, проиллюстрирован в WO 2015/026907, где показано, в том числе, что антитела к PRLR эффективно интернализуются PRLR-экспрессирующими клетками in vitro.
[93] В определенных вариантах осуществления эффектор интернализации представляет собой специфический для почки интернализатор, такой как CDH16 (Cadheri-16), CLDN16 (Claudn-16), KL (клото), PTH1R (рецептор паратиреоидного гормона), SLC22A13 (семейство 22 транспортеров растворенных веществ, представитель 13), SLC5A2 (котранспортер 2 натрия/глюкозы) и UMOD (уромодулин). В других определенных вариантах осуществления эффектор интернализации представляет собой специфический для мышц интернализатор, такой как BMPR1A (рецептор 1A костного морфогенетического белка), m-кадгерин, CD9, MuSK (киназа, специфическая для мышц), LGR4/GPR48 (рецептор 48, сопряженный с G-белком), холинергический рецептор (никотиновый) альфа 1, CDH15 (Cadheri-15), ITGA7 (интегрин альфа-7), CACNG1 (субъединица гамма-1 кальциевого канала L-типа), CACNAlS (субъединица альфа-15 кальциевого канала L-типа), CACNG6 (субъединица гамма-6 кальциевого канала L-типа), SCN1B (субъединица бета-1 натриевого канала), CHRNA1 (субъединица альфа рецептора ACh), CHRND (субъединица дельта рецептора ACh), LRRC14B (белок 14B, содержащий богатый лейцином повтор), дистрогликан (DAG1) и POPDC3 (белок 3, содержащий домен Popeye). В некоторых конкретных вариантах осуществления эффектор интернализации представляет собой ITGA7, CD9, CD63, ALPL2, ASGR1, ASGR2 или PRLR.
[94] В вариантах осуществления, в которых эффектор интернализации (IE) непосредственно интернализуется в клетку, домен доставки может представлять собой, например, антитело или антигенсвязывающий фрагмент антитела, которые специфически связываются с IE, или лиганд или часть лиганда, которые специфически взаимодействуют с IE. Например, если IE представляет собой Kremen-1 или Kremen-2, то домен доставки может содержать лиганд Kremen (например, DKK1) или его Kremen-связывающую часть или состоять из таких. В качестве другого примера, если IE является рецепторной молекулой, такой как ASGR1, то домен доставки может содержать лиганд, специфический в отношении рецептора (например, асиалоорозомукоида [ASOR] или бета-Ga1NAc), или его рецептор-связывающую часть или состоять из них.
[95] Интернализующие эффекторные белки, которые опосредованно интернализуются в клетку, предусматривают белки и полипептиды, которые сами по себе не интернализуются, а интернализуются в клетку после связывания или иным образом ассоциации со вторым белком или полипептидом, который непосредственно интернализуется в клетку. Белки, которые опосредованно интернализуются в клетку, предусматривают, например, растворимые лиганды, которые способны связываться с интернализующей рецепторной молекулой, экспрессируемой на клеточной поверхности. Неограничивающим примером растворимого лиганда, который (опосредованно) интернализуется в клетку благодаря его взаимодействию с интернализующей рецепторной молекулой, экспрессируемой на клеточной поверхности, является трансферрин. В вариантах осуществления, где IE представляет собой трансферрин (или другой опосредованно интернализуемый белок), связывание домена доставки с IE и взаимодействие IE с рецептором трансферрина (или другой интернализующей рецепторной молекулой, экспрессируемой на клеточной поверхности) приводит к тому, что весь домен доставки и любые молекулы, связанные с ним (например, фермент), интернализуются в клетку одновременно с интернализацией IE и его партнера по связыванию.
[96] В вариантах осуществления, в которых IE интернализуется в клетку опосредованно, домен доставки может представлять собой, например, антитело, антигенсвязывающий фрагмент антитела, или scFv, которые специфически связываются с IE, или рецептор или часть рецептора, которые специфически взаимодействуют с растворимым эффекторным белком. Например, если IE представляет собой цитокин, то домен доставки может содержать соответствующий рецептор цитокина или его лиганд-связывающую часть или состоять из таких.
[97] Примером IE является CD63, который является представителем тетраспанинового суперсемейства белков клеточной поверхности, которые пересекают клеточную мембрану четыре раза. CD63 экспрессируется практически во всех тканях и, как полагают, вовлекается в формирование и стабилизацию комплексов передачи сигнала. CD63 локализуется в клеточной мембране, лизосомной мембране и поздней эндосомальной мембране. CD63, как известно, ассоциируется с интегринами и может быть вовлечен в эпителиальный-мезенхимальный переход. См. H. Maecker et al., «The tetraspanin superfamily: molecular facilitators», 11(6) FASEB J. 428-42, May 1997, и M. Metzelaar et al., «CD63 antigen. A novel lysosomal membrane glycoprotein, cloned by a screening procedure for intracellular antigens in eukaryotic cells,” 266 J. Biol. Chem. 3239-3245, 1991.
[98] Другим примером IE является подобный предшественнику амилоида-бета (A4) белок 2 («APLP2»), повсеместно экспрессируемый представитель семейства APP (белка-предшественника амилоида). APLP2 представляет собой связанный с мембраной белок, который, как известно, взаимодействует с молекулами главного комплекса гистосовместимости (MHC) класса I (например, Kd). Он связывается с Kd на клеточной поверхности и интернализуется клатрин-зависимым образом вместе с Kd. См. Tuli et al., «Mechanism for amyloid precursor-like protein 2 enhancement of major histocompatibility complex class I molecule degradation», 284 The Journal of Biological Chemistry 34296-34307 (2009).
[99] Другим примером IE является рецептор пролактина (PRLR). Рецептор пролактина является представителем семейства цитокиновых рецепторов I типа и при связывании с лигандом и последующей димеризации активирует «тирозинкиназы Jak2, Fyn и Tec, фосфатазу SHP-2, фактор обмена гуаниновых нуклеотидов Vav и супрессор передачи сигнала SOCS», (см. Clevenger and Kline, «Prolactin receptor signal transduction», 10(10) Lupus 706-18 (2001), реферат). Рецептор пролактина подвергается эндоцитотическому рециклингу и может быть найден в лизосомных фракциях. См. Genty et al., «Endocytosis and degradation of prolactin and its receptor in Chinese hamster ovary cells stably transfected with prolactin receptor cDNA», 99(2) Mol. Cell Endocrinol. 221-8 (1994); и Ferland et al., «The effect of chloroquine on lysosomal prolactin receptors in rat liver», 115(5) Endocrinology 1842-9 (1984).
[100] Как используется в данном документе, «иммунологическая реакция» как правило, означает иммунологический ответ пациента на посторонний или «чужой» белок. Этот иммунологический ответ включает аллергическую реакцию и развитие антител, которые влияют на эффективность заместительного фермента. Некоторые пациенты не могут продуцировать какой-либо нефункционирующий белок, что, таким образом, делает заместительный фермент «чужеродным» белком. Например, повторная инъекция рекомбинантной GLA (rGLA) тем страдающим болезнью Фабри пациентам, у которых отсутствует GLA, зачастую приводит к аллергической реакции. У других пациентов продуцирование антител против rGLA, как было показано, снижает эффективность заместительного фермента в лечении заболевания. См., например, Tajima et al. («Use of a Modified α-N-Acetylgalactosaminidase (NAGA) in the Development of Enzyme Replacement Therapy for Fabry Disease», 85(5) Am. J. Hum. Genet. 569-580 (2009)), в котором обсуждается применение модифицированной NAGA в качестве «изозима» для замещения GLA. Модифицированная NAGA не обладает иммунологической перекрестной реактивностью в отношении GLA и «не реагировала с сывороткой крови от пациента с заболеванием Фабри, регулярно получающего лечение в виде рекомбинантной GLA». Id, реферат.
[101] «Иммуносупрессор» включает лекарственные средства и/или белки, которые приводят к общей иммуносупрессии, и может использоваться для предотвращения возникновения перекрестнореагирующих иммунологических материалов (CRIM) к заместительным ферментам, например GAA или GLA, соответственно, у пациентов с болезнью Помпе или Фабри. Неограничивающие примеры иммуносупрессоров включают метотрексат, микофенолата мофетил, циклофосфамид, рапамицин, алкилирующие ДНК средства, антитело к CD20, антитело к BAFF, антитело к CD3, антитело к CD4 и любую их комбинацию.
[102] Регуляторные элементы, например промоторы, которые являются специфическими для определенной ткани, например печени, усиливают экспрессию последовательностей нуклеиновых кислот, например генов, находящихся под контролем такого регуляторного элемента, в ткани, для которой данный регуляторный элемент является специфическим. Неограничивающие примеры специфических для печени регуляторных элементов, например специфических для печени промоторов, можно найти в Chuah et al. (2014) Mol. Ther. 22:1605-13.
[103] Термин «белок» означает любой аминокислотный полимер, имеющий более чем приблизительно 20 аминокислот, ковалентно связанных посредством амидных связей. Белки содержат одну или более аминокислотных полимерных цепей, общеизвестных в уровне техники как «полипептиды». Таким образом, полипептид может быть белком, а белок может содержать несколько полипептидов с образованием одной функционирующей биомолекулы. В некоторых белках могут присутствовать дисульфидные мостики (т. e. между цистеиновыми остатками с образованием цистина). Такие ковалентные связи могут находиться в одной полипептидной цепи или между двумя отдельными полипептидными цепями. Например, дисульфидные мостики необходимы для надлежащей структуры и функции инсулина, иммуноглобулинов, протамина и т.п. Последний обзор по образованию дисульфидных связей см. в Oka and Bulleid, «Forming disulfides in the endoplasmic reticulum», 1833(11) Biochim Biophys Acta 2425-9 (2013).
[104] Используемый в данном документе термин «белок» включает биотерапевтические белки, рекомбинантные белки, применяемые в исследовании или терапии, белки-ловушки и другие Fc-слитые белки, химерные белки, антитела, моноклональные антитела, антитела человека, биспецифические антитела, фрагменты антител, нанотела, химеры на основе рекомбинантных антител, слитые белки на основе scFv, цитокины, хемокины, пептидные гормоны и т. п. Белки могут быть получены с использованием рекомбинантных клеточных систем продуцирования, таких как система бакуловируса насекомых, дрожжевые системы (например, Pichia sp.), системы млекопитающих (например, клетки CHO и клетки CHO-K1, подобные производным CHO). Последний обзор обсуждения биотерапевтических белков и их продуцирования см. в Ghaderi et al., «Production platforms for biotherapeutic glycoproteins. Occurrence, impact, and challenges of non-human sialylation,” 28 Biotechnol Genet Eng Rev. 147-75 (2012).
[105] Используемый в данном документе термин «антитело» включает молекулы иммуноглобулинов, содержащие четыре полипептидные цепи, две тяжелые (H) цепи и две легкие (L) цепи, соединенные между собой дисульфидными связями. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (сокращенно обозначаемую в данном документе как HCVR или VH) и константную область тяжелой цепи. Константная область тяжелой цепи содержит три домена, CH1, CH2 и CH3. Каждая легкая цепь содержит вариабельную область легкой цепи (сокращенно обозначаемую в данном документе как LCVR или VL) и константную область легкой цепи. Константная область легкой цепи содержит один домен CL. VH- и VL-области могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), чередующиеся с более консервативными областями, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4 (CDR тяжелой цепи могут быть сокращены как HCDR1, HCDR2 и HCDR3; CDR легкой цепи могут быть сокращены как LCDR1, LCDR2 и LCDR3. Термин «высокоаффинное» антитело относится к тем антителам, которые характеризуются аффинностью связывания со своей мишенью, составляющей по меньшей мере 10-9 M, по меньшей мере 10-10 M; по меньшей мере 10-11 M или по меньшей мере 10-12 M, что измерено с помощью поверхностного плазмонного резонанса, например BIACORETM, или определения аффинности в растворе посредством ELISA. Термин «антитело» может охватывать антитела любого типа, такие как, например, моноклональные или поликлональные. Кроме того, антитело может быть любого происхождения, например, происходить от млекопитающего или не млекопитающего. В одном варианте осуществления антитело может происходить от млекопитающего или птицы. В дополнительном варианте осуществления антитело может быть человеческого происхождения и может дополнительно представлять собой человеческое моноклональное антитело.
[106] Фраза «биспецифическое антитело» включает в себя антитело, способное селективно связывать два или более эпитопов. Как правило, биспецифические антитела содержат две различные тяжелые цепи, при этом каждая тяжелая цепь специфически связывается с отдельным эпитопом - либо на двух разных молекулах (например, антигенах), либо на одной и той же молекуле (например, на одном и том же антигене). Если биспецифическое антитело способно селективно связываться с двумя разными эпитопами (первым эпитопом и вторым эпитопом), то аффинность первой тяжелой цепи по отношению к первому эпитопу, как правило, будет по меньшей мере на один, два, или три, или четыре порядка ниже аффинности первой тяжелой цепи по отношению ко второму эпитопу и наоборот. Эпитопы, распознаваемые биспецифическим антителом, могут находиться на одной и той же или на другой мишени (например, на одном и том же или на другом белке). Биспецифические антитела могут быть получены, например, путем объединения тяжелых цепей, которые распознают разные эпитопы одного и того же антигена. Например, последовательности нуклеиновой кислоты, кодирующие вариабельные последовательности тяжелой цепи, которые распознают разные эпитопы одного и того же антигена, могут быть слиты с последовательностями нуклеиновой кислоты, кодирующими разные константные области тяжелой цепи, и такие последовательности могут быть экспрессированы в клетке, которая экспрессирует легкую цепь иммуноглобулина. Типичное биспецифическое антитело имеет две тяжелые цепи, каждая из которых содержит три CDR тяжелой цепи, за которыми следует (от N-конца к C-концу) CH1-домен, шарнир, CH2-домен и CH3-домен, и легкую цепь иммуноглобулина, которая либо не обеспечивает антигенсвязывающую специфичность, но которая при этом может ассоциироваться с каждой тяжелой цепью, либо которая может ассоциироваться с каждой тяжелой цепью и которая может связываться с одним или более эпитопами, связывающимися антигенсвязывающими областями тяжелой цепи, либо которая может ассоциироваться с каждой тяжелой цепью и обеспечивать связывание либо одной, либо обеих тяжелых цепей с одним или обоими эпитопами.
[107] Фраза «тяжелая цепь» или «тяжелая цепь иммуноглобулина» включает последовательность константной области тяжелой цепи иммуноглобулина из любого организма и, если не указано иное, включает вариабельный домен тяжелой цепи. Вариабельные домены тяжелой цепи включают в себя три CDR тяжелой цепи и четыре области FR, если не указано иное. Фрагменты тяжелых цепей включают в себя CDR, CDR и FR, а также их комбинации. Типичная тяжелая цепь после вариабельного домена (от N-конца к C-концу) содержит CH1-домен, шарнир, CH2-домен и CH3-домен. Функциональный фрагмент тяжелой цепи включает в себя фрагмент, который способен специфически распознавать антиген (например, распознавать антиген с KD в микромолярном, наномолярном или пикомолярном диапазонах), который способен экспрессироваться и секретироваться клеткой и который содержит по меньшей мере одну CDR.
[108] Фраза «легкая цепь» включает последовательность константной области легкой цепи иммуноглобулина из любого организма и, если не указано иное, включает каппа и лямбда легкие цепи человека. Вариабельные (VL) домены легкой цепи, как правило, включают в себя три CDR легкой цепи и четыре каркасных (FR) области, если не указано иное. Как правило, легкая цепь полной длины включает в себя, от амино-конца к карбоксильному концу, VL домен, который включает в себя FR1-CDR1- FR2-CDR2-FR3-CDR3-FR4, и константный домен легкой цепи. Легкие цепи, которые могут быть использованы согласно настоящему изобретению, предусматривают например, те, которые не связываются селективно либо с первым, либо со вторым антигеном, селективно связываемым антигенсвязывающим белком. Подходящие легкие цепи предусматривают цепи, которые могут быть идентифицированы путем скрининга по наиболее часто используемым легким цепям в имеющихся библиотеках антител (библиотеках WET или in silico), при этом легкие цепи существенно не влияют на аффинность и/или селективность антигенсвязывающих доменов антигенсвязывающих белков. Подходящие легкие цепи включают в себя цепи, которые могут связываться с одним или обеими эпитопами, которые связываются антигенсвязывающими областями антигенсвязывающего белка.
[109] Фраза «вариабельный домен» включает аминокислотную последовательность легкой или тяжелой цепей иммуноглобулина (модифицированных, если требуется), которые содержат следующие аминокислотные области, в последовательности от N-конца к C-концу (если не указано иное): FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. «Вариабельный домен» предусматривает аминокислотную последовательность, способную сворачиваться в канонический домен (VH или VL), имеющий структуру двойных бета-складок, где бета-складки соединяются дисульфидной связью между остатками первой бета-складки и второй бета-складки.
[110] Фраза «определяющая комплементарность область» или термин «CDR» предусматривают аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты генов иммуноглобулина организма, которая в норме (т. e. у животного дикого типа) находится между двумя каркасными областями в вариабельной области легкой или тяжелой цепей молекулы иммуноглобулина (например, антитела или рецептора T-клетки). CDR может кодироваться, например, последовательностью зародышевой линии, или перегруппированной, или неперегруппированной последовательностью, и, например, наивной или зрелой B-клеткой или T-клеткой. В некоторых случая (например, для CDR3) CDR могут кодироваться двумя или более последовательностями (например, последовательностями зародышевой линии), которые не являются смежными (например, в неперегруппированной последовательности нуклеиновой кислоты), но являются смежными в B-клеточной последовательности нуклеиновой кислоты, например, как результат сплайсинга или соединения последовательностей (например, рекомбинация V-D-J с образованием CDR3 тяжелой цепи).
[111] Термин «фрагмент антитела» относится к одному или более фрагментам антитела, которые сохраняют способность специфического связывания с антигеном. Примеры связывающих фрагментов, охватываемых термином «фрагмент антитела», включают (i) Fab-фрагмент, одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) F(ab')2-фрагмент, двухвалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области; (iii) Fd-фрагмент, состоящий из доменов VH и CH1; (iv) Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела, (v) dAb-фрагмент (Ward et al. (1989) Nature 241:544-546), который состоит из домена VH, (vi) выделенную CDR и (vii) scFv, который состоит из двух доменов Fv-фрагмента, VL и VH, связанных синтетическим линкером с образованием одной белковой цепи, в которой области VL и VH спариваются с образованием одновалентных молекул. Другие формы одноцепочечных антител, таких как диатела, также охватываются термином «антитело» (см., например, Holliger et al. (1993) PNAS USA 90:6444-6448; Poljak et al. (1994) Structure 2:1121-1123).
[112] Фраза «Fc-содержащий белок» включает антитела, биспецифические антитела, иммуноадгезины и другие связывающие белки, которые содержат по меньшей мере функциональную часть областей CH2 и CH3 иммуноглобулина. «Функциональная часть» относится к CH2 и CH3 области, которая может связываться с Fc-рецептором (например, FcyR или FcRn, т. e. неонатальным Fc-рецептором) и/или которая может участвовать в активации комплемента. Если CH2 и CH3 область содержит делеции, замены и/или вставки или другие модификации, которые делают ее неспособной связываться с каким-либо Fc-рецепторам, а также неспособной активировать комплемент, то CH2 и CH3 область не является функциональной.
[113] Fc-содержащий белок может содержать модификации в доменах иммуноглобулина, в том числе модификации, которые нарушают одну или более из эффекторных функций связывающего белка (например, модификации, которые влияют на связывание FcyR, связывание FcRn и, таким образом, время полужизни и/или активность CDC). Такие модификации включают без ограничения следующие модификации и их комбинации, обозначенные на основании нумерации константной области иммуноглобулина согласно EU: 238, 239, 248, 249, 250, 252, 254, 255, 256, 258, 265, 267, 268, 269, 270, 272, 276, 278, 280, 283, 285, 286, 289, 290, 292, 293, 294, 295, 296, 297, 298, 301, 303, 305, 307, 308, 309, 311, 312, 315, 318, 320, 322, 324, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 337, 338, 339, 340, 342, 344, 356, 358, 359, 360, 361, 362, 373, 375, 376, 378, 380, 382, 383, 384, 386, 388, 389, 398, 414, 416, 419, 428, 430, 433, 434, 435, 437, 438 и 439.
[114] Например, и без ограничения, связывающий белок представляет собой Fc-содержащий белок, демонстрирующий увеличенное время полужизни в сыворотке крови (по сравнению с таким же Fc-содержащим белком без указанной модификации(-й)) и имеющий модификацию в положении 250 (например, E или Q); 250 и 428 (например, L или F); 252 (например, L/Y/F/W или T), 254 (например, S или T) и 256 (например, S/R/Q/E/D или T); или модификацию в 428 и/или 433 (например, L/R/SI/P/Q или K) и/или 434 (например, H/F или Y); или модификацию в 250 и/или 428; или модификацию в 307 или 308 (например, 308F, V308F) и 434. В другом примере модификация может предусматривать модификацию 428L (например, M428L) и 434S (например, N434S); модификацию 428L, 2591 (например, V259I) и 308F (например, V308F); модификацию 433K (например, H433K) и 434 (например, 434Y); модификацию 252, 254 и 256 (например, 252Y, 254T и 256E); модификацию 250Q и 428L (например, T250Q и M428L); модификацию 307 и/или 308 (например, 308F или 308P).
[115] Используемый в данном документе термин «антигенсвязывающий белок» относится к полипептиду или белку (один или более полипептидов, объединенных в функциональную единицу), который специфически распознает эпитоп на антигене, таком как специфический в отношении клетки антиген и/или целевой антиген в соответствии с настоящим изобретением. Антигенсвязывающий белок может быть полиспецифическим. Термин «полиспецифический» по отношению к антигенсвязывающему белку означает, что белок распознает разные эпитопы, либо на одном и том же антигене, либо на разных антигенах. В соответствии с настоящим изобретением полиспецифическая антигенсвязывающая молекула может представлять собой один многофункциональный полипептид или может представлять собой мультимерный комплекс из двух или более полипептидов, ковалентно или нековалентно связанных друг с другом. Термин «антигенсвязывающий белок» включает антитела или их фрагменты по настоящему изобретению, которые могут быть связаны или совместно экспрессированы с другой функциональной молекулой, например другим пептидом или белком. Например, антитело или его фрагмент могут быть функционально связаны (например, путем химического сочетания, генетического слияния, нековалентной ассоциации или прочего) с одним или более другими молекулярными объектами, такими как белок или его фрагмент, с получением биспецифической или полиспецифической антигенсвязывающей молекулы со второй специфичностью связывания.
[116] Используемый в данном документе термин «эпитоп» относится к части антигена, которая распознается полиспецифическим антигенсвязывающим полипептидом. Один антиген (такой как антигенный полипептид) может иметь более чем один эпитоп. Эпитопы можно определить как структурные или функциональные. Функциональные эпитопы, как правило, составляют подгруппу структуральных эпитопов и определяются как такие остатки, которые непосредственно обуславливают аффинность взаимодействия между the антигенсвязывающим полипептидом и антигеном. Также эпитопы могут быть конформационными, то есть состоять из нелинейных аминокислот. В определенных вариантах осуществления эпитопы могут включать детерминанты, которые представляют собой химически активные поверхностные структуры молекул, таких как аминокислоты, боковые цепи сахаров, фосфорильные группы или сульфонильные группы, и, в определенных вариантах осуществления могут иметь специфические характеристики трехмерных структур, и/или специфические характеристики заряда. Эпитопы, образованные непрерывными аминокислотами, как правило, сохраняются при воздействии денатурирующими растворителями, в то время как эпитопы, образованные посредством укладки в третичную структуру, утрачиваются при обработке денатурирующими растворителями.
[117] Термин «домен» относится к любой части белка или полипептида, обладающего конкретной функцией или структурой. Предпочтительно домены в соответствии с настоящим изобретением связываются со специфическими по отношению к клетке или целевыми антигенами. Домены, связывающие антиген, специфический для клетки, или целевой антиген и т. п., как используется в данном документе, включают любой имеющийся в норме, получаемый ферментативным путем, синтетический или получаемый с помощью методик генной инженерии полипептид или гликопротеин, который специфически связывает антиген.
[118] Термины «полутело» или «полуантитело», которые используются взаимозаменяемо, относятся к половине антитела, которая по существу содержит одну тяжелую цепь и одну легкую цепь. Тяжелые цепи антитела могут формировать димеры так, что тяжелая цепь одного полутела может ассоциироваться с тяжелой цепью, ассоциированной с другой молекулой (например, другим полутелом) или другим Fc-содержащим полипептидом. Два слегка отличающихся Fc-домена могут «гетеродимеризоваться», как при образовании биспецифических антител или других гетеродимеров, -тримеров, -тетрамеров и т. п. См. Vincent and Murini, «Current strategies in antibody engineering: Fc engineering and pH-dependent antigen binding, bispecific antibodies and antibody drug conjugates», 7 Biotechnol. J. 1444-1450 (20912); и Shimamoto et al., «Peptibodies: A flexible alternative format to antibodies,” 4(5) MAbs 586-91 (2012).
[119] В одном варианте осуществления вариабельный домен полутела специфически распознает эффектор интернализации, и Fc-домен полутела димеризуется с Fc-слитым белком, который содержит заместительный фермент (например, пептитело) Id, 586.
[120] Термин «одноцепочечный вариабельный фрагмент» или «scFv» включает одноцепочечный слитый полипептид, содержащий вариабельную область тяжелой цепи иммуноглобулина (VH) и вариабельную область легкой цепи иммуноглобулина (VL). В некоторых вариантах осуществления VH и VL соединены линкерной последовательностью из 10-25 аминокислот. Полипептиды ScFv могут также содержать другие аминокислотные последовательности, такие как CL- или CH1-области. Молекулы ScFv могут быть получены с помощью фагового дисплея или изготовлены путем прямого субклонирования тяжелых и легких цепей из гибридомы или В-клетки. Публикация Ahmad et al., Clinical and Developmental Immunology, volume 2012, article ID 98025 включена в данный документ посредством ссылки в отношении способов получения фрагментов scFv посредством фагового дисплея и клонирования домена антитела.
[121] Термины «альфа-глюкозидаза» (или «α-глюкозидаза»), «α-глюкозидазная активность», «GAA» и «GAA-активность» используются взаимозаменяемо и относятся к любому белку, который облегчает гидролиз 1,4-альфа-связей гликогена и крахмала в глюкозе. GAA также известна, в том числе, как EC 3.2.1.20, мальтаза, глюкоинвертаза, глюкозидосахараза, мальтаза-глюкоамилаза, альфа-глюкопиранозидаза, глюкозидоинвертаза, альфа-D-глюкозидаза, альфа-глюкозидаза гидролаза, альфа-1,4-глюкозидаза и альфа-D-глюкозидаза глюкогидролаза. GAA может быть обнаружена в лизосоме и в щеточной кайме тонкого кишечника. У пациентов, страдающих заболеванием Помпе, отсутствует функционирующая лизосомная α-глюкозидаза. См. S. Chiba, «Molecular mechanism in alpha-glucosidase and glucoamylase», 61(8) Biosci. Biotechnol. Biochem. 1233-9 (1997); и Hesselink et al., «Lysosomal dysfunction in muscle with special reference to glycogen storage disease type II», 1637(2) Biochim. Biophys. Acta. 164-70 (2003).
[122] Термины «альфа-галактозидаза A» (или «α-галактозидаза A»), «активность α-галактозидазы A», «α-галактозидаза», «активность α-галактозидазы», «GLA» и «GLA-активность» используются взаимозаменяемо и относятся к любому белку, который облегчает гидролиз терминальных α-галактозильных фрагментов из гликолипидов и гликопротеинов, а также гидролизует α-D-фукозиды. GLA также известна, в том числе, как EC 3.2.1.22, мелибиаза, α-D-галактозидаза, α-галактозидаза A, α-галактозидгалактогидролаза, α-D-галактозидгалактогидролаза. GLA является лизосомным ферментом, кодируемым связанным с Х-хромосомой геном GLA. Дефекты в GLA могут приводить к заболеванию Фабри, при котором гликолипид, известный как глоботриаозилцерамид (также известный как Gb3, GL-3 или церамидтригексозид) накапливается внутри кровеносных сосудов (т. e. выраженная васкулопатия), что приводит к боли и ухудшению в функционировании почки, сердца, кожи и/или цереброваскулярных тканей, а также в других тканях и органах. См., например, Prabakaran et al. «Mannose 6-phosphate receptor and sortilin mediated endocytosis of α-galactosidase A in kidney endothelial cells», 7(6) PLoS One e39975 pp. 1-9 (2012).
[123] В одном аспекте настоящее изобретение предусматривает способ лечения пациента (или субъекта), страдающего лизосомной болезнью накопления, путем введения пациенту «мультидоменного терапевтического белка». Мультидоменный терапевтический белок попадает в клетки пациента и доставляет в лизосомы фермент или ферментативную активность (т. e. «заместительный фермент»), которые замещают фермент (т. e «эндогенный фермент») или ферментативную активность, которые ассоциированы с LSD. В одном варианте осуществления мультидоменный терапевтический белок доставляется пациенту с помощью вектора для генной терапии, который содержит полинуклеотид, который кодирует мультидоменный терапевтический белок.
[124] Виды LSD включают сфинголипидоз, мукополисахаридоз и болезни накопления гликогена. В некоторых вариантах осуществления LSD представляет собой любую одну или более из болезни Фабри, болезни Гоше I типа, болезни Гоше II типа, болезни Гоше III типа, болезни Ниманна-Пика типа A, болезни Ниманна-Пика типа BGM1-ганглиозидоз, болезни Сандхоффа, болезни Тея-Сакса, дефицита активатора GM2, GM3-ганглиозидоза, метахроматической лейкодистрофии, дефицита активатора сфинголипида, болезни Шейе, болезни Гурлера-Шейе, болезни Гурлера, болезни Хантера, синдрома Санфилиппо A, синдрома Санфилиппо B, синдрома Санфилиппо C, синдрома Санфилиппо D, синдрома Моркио А, синдрома Моркио В, болезни Марото-Лами, болезни Слая, MPS IX и болезни Помпе. В соответствии с конкретным вариантом осуществления LSD представляет собой болезнь Фабри. В соответствии с другим вариантом осуществления LSD преставляет собой болезнь Помпе.
[125] В некоторых вариантах осуществления мультидоменный терапевтический белок содержит (a) заместительный фермент и (b) молекулярный объект, который связывается с эффектором интернализации (домен доставки). В некоторых случаях заместительный фермент представляет собой одно или более из α-галактозидазы, β-галактозидазы, α-глюкозидазы, β-глюкозидазы, активатора сапозина-С, церамидазы, сфингомиелиназы, β-гексозаминидазы, активатора GM2, GM3-синтазы, арилсульфатазы, активатора сфинголипида, α-идуронидазы, идуронидаза-2-сульфатазы, гепарин-N-сульфатазы, N-ацетил-α-глюкозаминидазы, α-глюкозамид-N-ацетилтрансферазы, N-ацетилглюкозамин-6-сульфатазы, N-ацетилгалактозамин-6-сульфат-сульфатазы, N-ацетилгалактозамин-4-сульфатазы, β-глюкуронидазы и гиалуронидазы.
[126] В некоторых случаях у пациента не может продуцироваться достаточно белка, так что заместительный фермент распознается организмом пациента как «чужой», и после введения заместительного фермента происходит иммунологическая реакция. Это не желательно. Поэтому, в некоторых вариантах осуществления заместительный фермент разрабатывают или продуцируют таким образом, чтобы избежать индуцирования иммунологической реакции у субъекта. Одним таким решением является применение «изозима» в качестве заместительного фермента. Изозим достаточно близок «своему» белку пациента, но обладает активностью заместительного фермента, достаточной для облегчения симптомов LSD.
[127] В одном конкретном варианте осуществления, при котором LSD представляет собой заболевание Помпе, и эндогенный фермент представляет собой α-глюкозидазу (GAA), изозим может представлять собой любое из кислой α-глюкозидазы, сахаразы-изомальтазы (SI), мальтазы-глюкоамилазы (MGAM), глюкозидазы II (GANAB) и нейтральной α-глюкозидазы (C GNAC). В другом конкретном варианте осуществления, при котором LSD представляет собой заболевание Фабри, и эндогенный фермент представляет собой α-галактозидазу A (GLA), изозим может представлять собой α-N- ацетилгалактозаминидазу, сконструированную с возможностью получения GLA-активности.
[128] В данном документе предусмотрены способы, отличные от применения изозима, для снижения уровня содержания перекрестнореагирующих иммунологических материалов (CRIM) к заместительному ферменту. Как показано на фигурах 5 и 6, введение мультидоменного терапевтического белка (например, посредством вектора для генной терапии), содержащего домен, связывающий интернализующий эффектор, и ферментативный домен, снижает уровень содержания CRIM к заместительному ферменту, поступающему при введении контрольного терапевтического белка (где отсутствует интернализующий эффекторный домен и содержится ферментативный домен). Таким образом, в одном варианте осуществления снижения уровня CRIM к ферменту у пациента с дефицитом фермента включает введение пациенту мультидоменного терапевтического белка (или кодирующей его нуклеиновой кислоты, например вектора для генной терапии, содержащего ген, кодирующий мультидоменный терапевтический белок), где мультидоменный терапевтический белок содержит домен доставки (например, белок, связывающий эффектор интернализации) и ферментативный домен.
[129] В мультидоменном терапевтическом белке содержится компонент, представляющий собой белок, связывающий эффектор интернализации, который обеспечивает поглощение заместительного фермента клеткой. Таким образом, в некоторых вариантах осуществления эффектор интернализации может представлять собой CD63, MHC-I, Kremen-1, Kremen-2, LRP5, LRP6, LRP8, рецептор трансферрина, LDL-рецептор, рецептор белка 1, родственного LDL, ASGR1, ASGR2, белок-2, подобный белку-предшественнику амилоида (APLP2), апелиновый рецептор (APLNR), PRLR (рецептор пролактина), MAL (миелиновый и лимфоцитарный белок, также известный, как VIP17), IGF2R, H+ АТФазу вакуолярного типа, рецептор дифтерийного токсина, фолатный рецептор, глутаматные рецепторы, глутатионовый рецептор, лептиновый рецептор, скавенджер-рецептор, SCARA1-5, SCARB1-3 и CD36. В определенных вариантах осуществления эффектор интернализации представляет собой специфический для почки интернализатор, такой как CDH16 (Cadheri-16), CLDN16 (Claudn-16), KL (клото), PTH1R (рецептор паратиреоидного гормона), SLC22A13 (семейство 22 транспортеров растворенных веществ, представитель 13), SLC5A2 (котранспортер 2 натрия/глюкозы) и UMOD (уромодулин). В других определенных вариантах осуществления эффектор интернализации представляет собой специфический для мышц интернализатор, такой как BMPR1A (рецептор 1A костного морфогенетического белка), m-кадгерин, CD9, MuSK (киназа, специфическая для мышц), LGR4/GPR48 (рецептор 48, сопряженный с G-белком), холинергический рецептор (никотиновый) альфа 1, CDH15 (Cadheri-15), ITGA7 (интегрин альфа-7), CACNG1 (субъединица гамма-1 кальциевого канала L-типа), CACNAlS (субъединица альфа-15 кальциевого канала L-типа), CACNG6 (субъединица гамма-6 кальциевого канала L-типа), SCN1B (субъединица бета-1 натриевого канала), CHRNA1 (субъединица альфа рецептора ACh), CHRND (субъединица дельта рецептора ACh), LRRC14B (белок 14B, содержащий богатый лейцином повтор), дистрогликан (DAG1) и POPDC3 (белок 3, содержащий домен Popeye). В некоторых вариантах осуществления эффектор интернализации представляет собой ITGA7, CD9, CD63, APLP2, ASGR1, ASGR2 или PRLR.
[130] В некоторых вариантах осуществления белок, связывающий эффектор интернализации, предусматривает антигенсвязывающий белок, который включает, например, молекулу слияния с рецептором, молекулу-ловушку, молекулу слияния с рецептором Fc, антитело, Fab-фрагмент, F(ab')2-фрагмент, Fd-фрагмент, Fv-фрагмент, молекулу одноцепочечного Fv (scFv), dAb-фрагмент, выделенную определяющую комплементарность область (CDR), пептид CDR3, конформационно затрудненный пептид FR3-CDR3-FR4, домен-специфическое антитело, однодоменное антитело, антитело с удаленным доменом, химерное антитело, антитело с привитой CDR, диатело, триатело, тетратело, минитело, нанотело, одновалентное нанотело, двухвалентное нанотело, иммунофармацевтическое средство на основе модульного белка малого размера (SMIP), верблюжье антитело (гомодимерное антитело на основе тяжелой цепи VHH), вариабельный домен IgNAR акулы.
[131] В одном варианте осуществления молекулярный объект, который связывается с эффектором интернализации, представляет собой антитело, фрагмент антитела или другой антигенсвязывающий белок. Например, молекулярный объект может представлять собой биспецифическое антитело, в котором одно плечо связывается с эффектором интернализации (например, ITGA7, CD9, CD63, PRLR, APLP2, ASGR1, ASGR2), а другое плечо связывается с заместительным ферментом. В данном документе мультидоменный терапевтический белок содержит биспецифическое антитело и заместительный фермент (фиг. 1A). В конкретном варианте осуществления заболевание, подлежащее лечению, представляет собой болезнь Фабри, и мультидоменный терапевтический белок содержит GLA и биспецифическое антитело, которое связывает GLA и CD63. В конкретном варианте осуществления заболевание, подлежащее лечению, представляет собой болезнь Фабри, и мультидоменный терапевтический белок содержит GLA и биспецифическое антитело, которое связывает GLA и ITGA7. В другом конкретном варианте осуществления заболевание, подлежащее лечению, представляет собой болезнь Помпе, и мультидоменный терапевтический белок содержит GAA и биспецифическое антитело, которое связывает GAA и CD63. В другом конкретном варианте осуществления заболевание, подлежащее лечению, представляет собой болезнь Помпе, и мультидоменный терапевтический белок содержит GAA и биспецифическое антитело, которое связывает GAA и ITGA7.
[132] В других вариантах осуществления молекулярный объект, который связывается с эффектором интернализации, содержит полуантитело, и заместительный фермент содержит Fc-домен (слитый полипептид фермент-Fc). В одном варианте осуществления Fc-домен слитого полипептида фермент-Fc ассоциирует с Fc-доменом полутела, специфического в отношении эффектора интернализации, с образованием мультидоменного терапевтического белка (фиг. 1B).
[133] В других вариантах осуществления заместительный фермент ковалентно связан с белком, связывающим эффектор интернализации. Вариант осуществления, представляющий собой слитый полипептид фермент-Fc:полутело, описанный в предыдущем параграфе (см. также фиг. 1B), попадает в этот класс, поскольку димер Fc может быть скреплен с помощью одного или более дисульфидных мостиков. Ковалентная связь между доменом активности фермента или полипептидом и доменом или полипептидом, связывающими эффектор интернализации, может представлять собой любой тип ковалентной связи, т. e. любую связь, которая предусматривает распределение электронов. В некоторых случаях ковалентная связь представляет собой пептидную связь между двумя аминокислотами, такую, что заместительный фермент и белок, связывающий эффектор интернализации, полностью или частично формируют непрерывную полипептидную цепь, как в белке слияния. В некоторых случаях часть заместительного фермента и белок, связывающий эффектор интернализации, связаны непосредственно. В других случаях используется линкер для прикрепления двух частей. См. Chen et al., «Fusion protein linkers: property, design and functionality», 65(10) Adv Drug Deliv Rev. 1357-69 (2013).
[134] В конкретном варианте осуществления заместительный фермент ковалентно связан с C-концом тяжелой цепи антитела к эффектору интернализации (см. фиг. 1C) или с С-концом легкой цепи (фиг. 1E). В другом конкретном варианте осуществления заместительный фермент ковалентно связан с N-концом тяжелой цепи антитела к эффектору интернализации (см. фиг. 1D) или с N-концом легкой цепи (фиг. 1F). В другом конкретном варианте осуществления фермент соединен к С-концом домена, представляющего собой scFv к эффектору интернализации (фиг. 1G).
[135] В некоторых случаях, особенно если заместительный фермент в норме не подвергается протеолитическому расщеплению в лизосоме, в такие варианты осуществления мультидоменного терапевтического белка, которые предусматривают слияние антитело-фермент, добавляют расщепляемый линкер. В некоторых вариантах осуществления между антителом и заместительным ферментом вставляют расщепляемый катепсином линкер для обеспечения удаления антитела в лизосоме с целью a) возможного способствования сохранению ферментативной активности с помощью удаления стерически крупного антитела и b) возможного увеличения времени полужизни фермента в лизосоме.
[136] В одном конкретном варианте осуществления мультидоменный терапевтический белок доставляется пациенту или клетке в составе вектора для генной терапии, который содержит полинуклеотид, который кодирует мультидоменный терапевтический белок. В одном варианте осуществления мультидоменный терапевтический белок содержит домен доставки и ферментативный домен. В конкретном варианте осуществления домен доставки связывается с интернализующим эффектором, таким как CD63, MHC-I, Kremen-1, Kremen-2, LRP5, LRP6, LRP8, рецептор трансферрина, LDL-рецептор, рецептор белка 1, родственный LDL, ASGR1, ASGR2, белок-2, подобный белку-предшественнику амилоида (APLP2), апелиновый рецептор (APLNR), MAL (миелиновый и лимфоцитарный белок (MAL)), IGF2R, H+ АТФаза вакуолярного типа, рецептор дифтерийного токсина, фолатный рецептор, глутаматные рецепторы, глутатионовый рецептор, лептиновые рецепторы, скавенджер-рецептор A1-5 (SCARA1-5), SCARB1-3 и CD36. В одном варианте осуществления домен доставки представляет собой одноцепочечный вариабельный фрагмент (scFv), который связывается с CD63 (т. е. scFv к CD63). В другом варианте осуществления домен доставки представляет собой одноцепочечный вариабельный фрагмент (scFv), который связывается с ITGA7 (т. е. scFv к ITGA7).
[137] В одном конкретном варианте осуществления ферментативный домен мультидоменного терапевтического белка предусматривает гидролазу. В конкретном варианте осуществления ферментативный домен предусматривает гидролазу, которая представляет собой гликозилазу. В более конкретном варианте осуществления ферментативный домен предусматривает гликозилазу, которая представляет собой гликозидазу. В более конкретном варианте осуществления ферментативный домен представляет собой гликозидазу, которая представляет собой альфа-глюкозидазу.
[138] В целом, в данном документе раскрыты композиции, содержащие полинуклеотиды, и их применение, например (м)РНК, ДНК и их модифицированные формы, которые кодируют мультидоменный терапевтический белок, содержащий интернализующий эффекторный домен и ферментативный домен, для лечения лизосомных болезней накопления, например, для снижения уровня гликогена и/или усиления иммунной толерантности к GAA у пациента с болезнью Помпе.
[139] Термин «полинуклеотид» включает полимер из нуклеотидов (например, РНК или ДНК), который кодирует по меньшей мере один полипептид, включая слитые полипептиды, например мультидоменный терапевтический полипептид, содержащий интернализующий эффекторный домен и ферментативный домен. Используемый в данном документе термин полинуклеотид охватывает полимеры, содержащие как модифицированные, так и немодифицированные нуклеотиды. Полинуклеотид может содержать одну или более кодирующих и некодирующих областей. Полинуклеотид может быть очищен из природных источников, получен с применением рекомбинантных систем экспрессии и необязательно очищен, химически синтезирован и т. д. Где это уместно, например, в случае химически синтезированных молекул, полинуклеотид может содержать нуклеозидные аналоги, такие как аналоги, содержащие химически модифицированные основания или сахара, модификации основной цепи и т. д. Полинуклеотидная последовательность представлена в направлении от 5' до 3', если не указано иное. В некоторых вариантах осуществления полинуклеотид представляет собой или содержит природные нуклеозиды (например, аденозин, гуанозин, цитидин, уридин); аналоги нуклеозидов (например, 2-аминоаденозин, 2-тиотимидин, инозин, пирролопиримидин, 3-метиладенозин, 5-метилцитидин, C-5-пропинилцитидин, C-5-пропинилуридин, 2-аминоаденозин, C5-бромуридин, C5-фторуридин, C5-йодуридин, C5-пропинилуридин, C5-пропинилцитидин, C5-метилцитидин, 2-аминоаденозин, 7-дезазааденозин, 7-дезазагуанозин, 8-оксоаденозин, 8-оксогуанозин, O(6)-метилгуанин и 2-тиоцитидин); химически модифицированные основания; биологически модифицированные основания (например, метилированные основания); интеркалированные основания; модифицированные сахара (например, 2'-фторрибоза, рибоза, 2'-дезоксирибоза, арабиноза и гексоза); и/или модифицированные фосфатные группы (например, фосфоротиоатные и 5'-N-фосфорамидитные связи).
[140] В некоторых вариантах осуществления полинуклеотид содержит один или более нестандартных нуклеотидных остатков. Нестандартные нуклеотидные остатки могут включать, например, 5-метилцитидин («5mC»), псевдоуридин («.psi.U»), и/или 2-тиоуридин («2sU»). См., например, патент США № 8278036 или WO2011012316, каждый из которых включен в данный документ посредством ссылки во всей своей полноте в связи с обсуждением таких остатков и их включения в полинуклеотид. Присутствие нестандартных нуклеотидных остатков может сделать полинуклеотид более стабильным и/или менее иммуногенным, чем контрольный полинуклеотид с той же последовательностью, но содержащий только стандартные остатки. В дополнительных вариантах осуществления полинуклеотид может содержать один или более нестандартных нуклеотидных остатков, выбранных из изоцитозина, псевдоизоцитозина, 5-бромурацила, 5-пропинилурацила, 6-аминопурина, 2-аминопурина, инозина, диаминопурина и 2-хлор-6-аминопурина, цитозинов. а также комбинации этих модификаций и других модификаций нуклеиновых оснований. Определенные варианты осуществления могут, кроме того, включать дополнительные модификации фуранозного кольца или нуклеинового основания. Дополнительные модификации могут включать, например, модификации или замены сахара (например, одну или более из модификации 2'-O-алкила, блокированной нуклеиновой кислоты (LNA)). В некоторых вариантах осуществления полинуклеотид может образовывать комплекс или быть гибридизованным с дополнительными полинуклеотидами и/или пептидо-полинуклеотидами (PNA). В вариантах осуществления, где модификация сахара представляет собой 2'-O-алкил-модификацию, такая модификация может включать без ограничения 2'-дезокси-2'-фтор-модификацию, 2'-O-метил-модификацию, 2'-О-метоксиэтил-модификацию и 2'-дезокси-модификацию. В определенных вариантах осуществления любая из этих модификаций может присутствовать в 0-100% нуклеотидов, например, в более чем 0%, 1%, 10%, 25%, 50%, 75%, 85%, 90%, 95% или в 100% нуклеотидов, входящих в состав, по отдельности или в комбинации. В некоторых вариантах осуществления полинуклеотид включает молекулы информационной РНК (мРНК), которые могут быть или могут не быть модифицированными, например, которые могут содержать или могут не содержать модифицированы нуклеотид, с помощью хорошо известных способов с целью повышения их стабильности и/или снижения их иммуногенности. В некоторых вариантах осуществления полинуклеотид включает молекулы ДНК, которые могут, которые могут быть или могут не быть модифицированы, например, которые могут содержать или могут не содержать модифицированный нуклеотид, с помощью хорошо известных способов с целью повышения их стабильности и/или снижения их иммуногенности.
[141] В некоторых вариантах осуществления полинуклеотид также включает «последовательность нуклеиновой кислоты, нацеливающуюся на локус». Последовательность, нацеливающаяся на локус, обеспечивает возможность интеграции полинуклеотида, кодирующего мультидоменный терапевтический белок, в геном клетки-хозяина реципиента. В некоторых вариантах осуществления последовательность, нацеливающаяся на локус, включает фланкирующие гомологичные плечи для обеспечения гомологичной рекомбинации. В некоторых вариантах осуществления последовательность, нацеливающаяся на локус, включает последовательности направляющей РНК и фермент Cas типа II для обеспечения интеграции (т. е. способ на основе CRISPR-Cas9). В некоторых вариантах осуществления последовательность, нацеливающаяся на локус, включает направляющие последовательности распознавания нуклеазы с «цинковыми пальцами» (ZFN) для обеспечения интеграции. В некоторых вариантах осуществления последовательность, нацеливающаяся на локус, включает последовательности распознавания эффекторной нуклеазы, подобной активаторам транскрипции (TALEN), для обеспечения интеграции. В других вариантах осуществления последовательность, нацеливающаяся на локус, включает код «один остаток-один нуклеотид», используемый нуклеазами, полученными на основе BuD, для обеспечения интеграции.
[142] В некоторых вариантах осуществления геномный локус, в который интегрируется полинуклеотид, кодирующий мультидоменный терапевтический белок, представляет собой локус «безопасной гавани». В одном варианте осуществления локус «безопасной гавани» обеспечивает высокую экспрессию мультидоменного терапевтического белка, не мешая при этом экспрессии основных генов или не стимулируя экспрессию онкогенов или других вредных генов. В одном варианте осуществления геномный локус находится внутри или поблизости локуса экспрессируемого в печени альбумина (Alb), локуса EESYR, локуса SARS, положения 188083272 на хромосоме 1 человека или его ортолога у млекопитающего, отличного от человека, положения 3046320 на хромосоме 10 человека или его ортолога у млекопитающего, отличного от человека, положения 67328980 на хромосоме 17 человека или его ортолога у млекопитающего, отличного от человека, сайта аденоассоциированного вируса 1 (AAVS1) на хромосоме, в норме имеющегося сайта интеграции вируса AAV на хромосоме 19 человека или его ортолога у млекопитающего, отличного от человека, гена рецептора хемокина 5 (CCR5), гена рецептора хемокина, кодирующего корецептор HIV-1, или локуса Rosa26 у мыши или его ортолога у млекопитающего, отличного от мыши. В одном варианте осуществления геномный локус представляет собой сайт аденоассоциированного вируса. В одном варианте осуществления геномный локус для интеграции выбирают в соответствии со способом согласно публикации Papapetrou и Schambach, J. Molecular Therapy, vol. 24 (4):678-684, April 2016, которая включена в данный документ посредством ссылки в отношении поэтапного выбора геномного локуса «безопасной гавани» для интеграции вектора для генной терапии; см. также публикацию Barzel et al. Nature, vol. 517: 360-364, полностью включенную в данный документ посредством ссылки в отношении нацеливания гена без промотора в локус экспрессируемого в печени альбумина (Alb).
[143] В некоторых вариантах осуществления полинуклеотид, например ДНК, также содержит промотор, функционально связанный с последовательностью нуклеиновой кислоты, кодирующей мультидоменный терапевтический белок. В конкретном варианте осуществления промотор представляет собой тканеспецифический промотор, который управляет экспрессией гена в конкретной ткани. В одном варианте осуществления тканеспецифический промотор представляет собой специфический для печени энхансер/промотор, полученный из serpina1 (например, SEQ ID NO:9), и/или представляет собой промотор TTR (SEQ ID NO:8). В других вариантах осуществления промотор представляет собой промотор CMV. В других вариантах осуществления промотор представляет собой промотор убиквитина С.
[144] В одном варианте осуществления «вектор для генной терапии», кодирующий мультидоменный терапевтический белок, представляет собой любой вектор, способный доставлять полинуклеотид, кодирующий мультидоменный терапевтический белок, хозяину, например, пациенту. В некоторых вариантах осуществления вектор для генной терапии нацелен на конкретную клетку-хозяина или орган, например, для локальной доставки, например тканеспецифической доставки. Как правило, в случае локальной доставки требуется, чтобы белок (например, мультидоменный терапевтический белок), кодируемый мРНК, транслировался и экспрессировался главным образом в органе и/или органом, например, в печени, где в результате этого образовывалось бы депо, например, депо в печени для продуцирования (и секреции) белка. В некоторых вариантах осуществления вектор для генной терапии доставляет полинуклеотид мультидоменного терапевтического белка в печень пациента с образованием депо в печени. См., например, публикацию DeRosa et al. Gene Therapy, vol. 10:699-707, полностью включенную в настоящий документ посредством ссылки. В некоторых вариантах осуществления вектор для генной терапии доставляет полинуклеотид, кодирующий мультидоменный терапевтический белок, в мышечную ткань пациента. В некоторых вариантах осуществления вектор для генной терапии доставляет полинуклеотид, кодирующий мультидоменный терапевтический белок, в мозг пациента.
[145] Любой природный или сконструированный вектор для генной терапии доставки, известный в настоящее время или который будет разработан в будущем, может быть использован в осуществлении настоящего изобретения. В некоторых вариантах осуществления вектор для генной терапии представляет собой вирусный вектор, например, содержит вирус, вирусный капсид, вирусный геном и т. д. В некоторых вариантах осуществления вектор для генной терапии представляет собой полинуклеотид без дополнительных средств, например эписому. В некоторых вариантах осуществления вектор для генной терапии включает полинуклеотидный комплекс. Иллюстративные неограничивающие полинуклеотидные комплексы для применения в качестве вектора для генной терапии включают липоплексы, полимерсомы, полипексы, дендримеры, неорганические наночастицы (например, покрытые полинуклеотидом золото, диоксид кремния, оксид железа, фосфат кальция и т. д.). В некоторых вариантах осуществления вектор для генной терапии, описанный в данном документе, содержит комбинацию вирусного вектора, полинуклеотидов без дополнительных средств и полинуклеотидных комплексов.
[146] В одном варианте осуществления вектор для генной терапии представляет собой вирус, включая ретровирус, аденовирус, вирус простого герпеса, поксвирус, вирус осповакцины, лентивирус или аденоассоциированный вирус. В одном варианте осуществления вектор для генной терапии представляет собой аденоассоциированный вирус (AAV), включая серотипы AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10 и AAV11 или их сконструированные варианты или варианты, возникшие в результате естественного отбора.
[147] В одном варианте осуществления полинуклеотид также содержит последовательность нуклеиновой кислоты аденоассоциированного вируса (AAV). В одном варианте осуществления вектор для генной терапии представляет собой химерный аденоассоциированный вирус, содержащий генетические элементы из двух или более серотипов. Например, вектор на основе AAV с генами rep из AAV1 и генами cap из AAV2 (обозначенные как AAV1/2 или AAV RC1/2) можно использовать в качестве вектора для генной терапии для доставки полинуклеотида мультидоменного терапевтического белка в клетку или клетку нуждающегося пациента. В одном варианте осуществления вектор для генной терапии представляет собой AAV1/2, AAV1/3, AAV1/4, AAV1/5, AAV1/6, AAV1/7, AAV1/8, AAV1/9, AAV1/10, AAV1/11, AAV2/1, AAV2/3, AAV2/4, AAV2/5, AAV2/6, AAV2/7, AAV2/8, AAV2/9, AAV2/10, AAV2/11, AAV3/1, AAV3/2, AAV3/4, AAV3/5, AAV3/6, AAV3/7, AAV3/8, AAV3/9, AAV3/10, AAV3/10, AAV4/1, AAV4/2, AAV4/3, AAV4/5, AAV4/6, AAV4/7, AAV4/8, AAV4/9, AAV4/10, AAV4/11, AAV5/1, AAV5/2, AAV5/3, AAV5/4, AAV5/6, AAV5/7, AAV5/8, AAV5/9, AAV5/10, AAV5/11, AAV6/1, AAV6/2, AAV6/3, AAV6/4, AAV6/5, AAV6/7, AAV6/8, AAV6/9, AAV6/10, AAV6/10, AAV7/1, AAV7/2, AAV7/3, AAV7/4, AAV7/5, AAV7/6, AAV7/8, AAV7/9, AAV7/10, AAV7/11, AAV8/1, AAV8/2, AAV8/3, AAV8/4, AAV8/5, AAV8/6, AAV8/7, AAV8/9, AAV8/10, AAV8/11, AAV9/1, AAV9/2, AAV9/3, AAV9/4, AAV9/5, AAV9/6, AAV9/7, AAV9/8, AAV9/10, AAV9/11, AAV10/1, AAV10/2, AAV10/3, AAV10/4, AAV10/5, AAV10/6, AAV10/7, AAV10/8, AAV10/9, AAV10/11, AAV11/1, AAV11/2, AAV11/3, AAV11/4, AAV11/5, AAV11/6, AAV11/7, AAV11/8, AAV11/9, AAV11/10, химерный вирион или их производные. Публикация Gao et al., «Novel adeno-associated viruses from rhesus monkeys as vectors for human gene therapy», PNAS 99(18): 11854-11859, Sep. 3, 2002, включена в данный документ посредством ссылки в отношении векторов на основе AAV и химерных вирионов, применимых в качестве векторов для генной терапии, а также их конструирования и применения.
[148] В более конкретном варианте осуществления вектор для генной терапии представляет собой химерный вектор на основе AAV с последовательностью гена rep серотипа 2 и последовательностью cap серотипа 8 («AAV2/8» или «AAV RC2/8»).
[149] В некоторых вариантах осуществления вектор для генной терапии представляет собой вирусный вектор, который был подвергнут псевдотипированию (например, сконструирован) для нацеливания на конкретную клетку, например гепатоцит. Многие из достижений в направленной генной терапии с применением вирусных векторов можно кратко описать как нерекомбинаторную (отличную от генетической) или рекомбинаторную (генетическую) модификацию вирусного вектора, которая приводит к псевдотипированию, расширению тропизма и/или изменению мишени тропизма естественного вирусного вектора. (Обзор представлен в публикации Nicklin and Baker (2002) Curr. Gene Ther. 2:273-93; Verheiji and Rottier (2012) Advances Virol 2012:1-15). В подходах, отличных от генетических, обычно используют адаптер, который распознает как поверхностный белок вируса дикого типа (немодифицированный), так и клетку-мишень. Растворимые псевдорецепторы (для вируса дикого типа), полимеры, такие как полиэтиленгликоль, и антитела или их части, применяли в качестве вирус-связывающего домена адаптеров, тогда как для связывающего клетку домена адаптеров, описанных выше, применяли природные лиганды, представляющие собой пептиды или витамины, а также антитела и их части. Например, замена мишени вирусного вектора на клетку-мишень может быть выполнена после связывания комплекса вектор:адаптер с белком, экспрессируемым на поверхности клетки-мишени, например белком клеточной поверхности. Такой подход применяли для AAV (Bartlett et al. (1999) Nat. Biotechnol. 74: 2777-2785), аденовирусов (Hemminki et al. (2001) Cancer Res. 61: 6377-81; van Beusechem et al. (2003) Gene Therapy 10:1982-1991; Einfeld, et al. (2001) J. Virol. 75:11284-91; Glasgow et al. (2009) PLOS One 4:e8355), герпесвирусов (Nakano et al. (2005) Mol. Ther. 11:617-24) и парамиксовирусов (Bian et al. (2005) Cancer Gene Ther. 12:295-303; Bian et al. (2005) Int. J. Oncol. 29:1359-69), коронавирусов (Haijema et al. (2003) J. Virol. 77:4528-4538; Wurdinger et al. (2005) Gene Therapy 12:1394-1404).
[150] Более популярным подходом являлась рекомбинаторная генетическая модификация вирусных капсидных белков и, таким образом, поверхности вирусного капсида. При непрямых рекомбинаторных подходах вирусный капсид модифицируется с помощью гетерологичного «каркаса», который затем связывается с адаптером. Адаптер связывается с каркасом и клеткой-мишенью. (Arnold et al. (2006) Mol. Ther. 5:125-132; Ponnazhagen et al. (2002) J. Virol. 76:12900-907; см. также WO 97/05266). Каркасы, такие как (1) Fc-связывающие молекулы (например, Fc-рецепторы, белок A и т. д.), которые связываются с Fc антител-адаптеров, (2) (стрепт)авидин, который связывается с биотинилированными адаптерами, (3) биотин, который связывается с адаптерами, слитыми с (стрепт)авидином, и (4) пары связывания белок:белок, которые образуют изометрические пептидные связи, такие как SpyCatcher, который связывает адаптер SpyTagged, были включены в Ad (Pereboeva et al. (2007) Gene Therapy 14: 627-637; Park et al. (2008) Biochemical and Biophysical Research Communications 366: 769-774; Henning et al. (2002) Human Gene Therapy 13:1427-1439; Banerjee et al. (2011) Bioorganic and Medicinal Chemistry Letters 21:4985-4988), AAV (Gigout et al. (2005) Molecular Therapy 11:856-865; Stachler et al. (2008) Molecular Therapy 16:1467-1473), и тогавирусы (Quetglas et al. (2010) Virus Research 153:179-196; Ohno et al. (1997) Nature Biotechnology 15:763-767; Klimstra et al. (2005) Virology 338:9-21).
[151] При прямом рекомбинаторном подходе к нацеливанию нацеливающий лиганд непосредственно вставляется в вирусный капсид или соединяется с ним, то есть белковые вирусные капсиды модифицируются для экспрессии гетерологичного лиганда. Лиганд, кроме того, перенаправляет, например, связывает рецептор или маркер, преимущественно или исключительно экспрессируемый на клетке-мишени. (Stachler et al. (2006) Gene Ther. 13:926-931; White et al. (2004) Circulation 109:513-519.). Прямые рекомбинаторные подходы применяли для AAV (Park et al., (2007) Frontiers in Bioscience 13:2653-59; Girod et al. (1999) Nature Medicine 5:1052-56; Grifman et al. (2001) Molecular Therapy 3:964-75; Shi et al. (2001) Human Gene Therapy 12:1697-1711; Shi and Bartlett (2003) Molecular Therapy 7:515-525), ретровируса (Dalba et al. Current Gene Therapy 5:655-667; Tai and Kasahara (2008) Frontiers in Bioscience 13:3083-3095; Russell and Cosset (1999) Journal of Gene Medicine 1:300-311; Erlwein et al. (2002) Virology 302:333-341; Chadwick et al. (1999) Journal of Molecular Biology 285:485-494; Pizzato et al. (2001) Gene Therapy 8:1088-1096), поксвируса (Guse et al. (2011) Expert Opinion on Biological Therapy 11:595-608; Galmiche et al. (1997) Journal of General Virology 78:3019-3027; Paul et al. (2007) Viral Immunology 20:664-671), парамиксовируса (Nakamura and Russell (2004) Expert Opinion on Biological Therapy 4:1685-1692; Hammond et al. (2001) Journal of Virology 75:2087-2096; Galanis (2010) Clinical Pharmacology and Therapeutics 88:620-625; Blechacz and Russell (2008) Current Gene Therapy 8:162-175; Russell and Peng (2009) Current Topics in Microbiology and Immunology 330:213-241) и герпесвируса (Shah and Breakefield (2006) Current Gene Therapy 6:361-370; Campadelli-Fiume et al. (2011) Reviews in Medical Virology 21:213-226).
[152] В некоторых вариантах осуществления вектор для генной терапии, описанный в данном документе, подвергается псевдотипированию в отношении тех тканей, которые особенно подходят для генерации регуляторного ответа, например толерантности, например, к заместительному ферменту. Такие ткани включают без ограничения слизистую ткань, например кишечно-ассоциированную лимфоидную ткань (GALT), гемопоэтические стволовые клетки и печень. В некоторых вариантах осуществления вектор для генной терапии или ген, кодирующий мультидоменный терапевтический белок, описанный в данном документе, экспрессируются под контролем промоторов, специфических для этих тканей, например специфического для печени промотора.
[153] В некоторых вариантах осуществления вектор для генной терапии, описанный в данном документе, предусматривает полинуклеотид без дополнительных средств. Например, в некоторых вариантах осуществления полинуклеотид, кодирующий мультидоменный терапевтический полипептид, можно вводить, например, внутримышечно, непосредственно в орган для образования депо, внутривенно и т. д. Дополнительные хорошо известные способы улучшенной доставки полинуклеотидов без дополнительных средств включают без ограничения электропорацию, сонопорацию, применение генной пушки для «стрельбы» частицами золота, покрытыми полинуклеотидами, магнитофекцию и гидродинамическую доставку.
[154] В некоторых вариантах осуществления вектор для генной терапии, описанный в данном документе, содержит полинуклеотидные комплексы, такие как без ограничения наночастицы (например, самоорганизующиеся наночастицы полинуклеотидов, самоорганизующиеся наночастицы на основе полимеров, неорганические наночастицы, липидные наночастицы, полупроводниковые/металлические наночастицы), гели и гидрогели, полинуклеотидные комплексы с катионами и анионами, микрочастицы и любые их комбинации.
[155] В некоторых вариантах осуществления полинуклеотиды, раскрытые в данном документе, могут быть составлены в виде самоорганизующихся наночастиц. В качестве неограничивающего примера, полинуклеотиды могут применяться для получения наночастиц, которые могут применяться в системе доставки для полинуклеотидов (см., например, международную публикацию № WO2012125987; которая включена в данный документ посредством ссылки во всей своей полноте). В некоторых вариантах осуществления полинуклеотидные самоорганизующиеся наночастицы могут содержать ядро из полинуклеотидов, раскрытых в данном документе, и полимерную оболочку. Полимерная оболочка может представлять собой любой из полимеров, описанных в данном документе, и они известны в данной области техники. В дополнительном варианте осуществления полимерная оболочка может применяться для защиты полинуклеотидов в ядре.
[156] В некоторых вариантах осуществления эти самоорганизующиеся наночастицы могут представлять собой микрогубки, образованные из длинных полимеров, состоящих из полинуклеотидных «шпилек», которые формируются в кристаллические «плиссированные» листы перед самоорганизацией в микрогубки. Эти микрогубки представляют собой плотно упакованные губчатые микрочастицы, которые могут функционировать как эффективный носитель и могут доставлять груз в клетку. Диаметр микрогубок может составлять от 1 до 300 нм. Микрогубки можно объединять в комплексы с другими веществами, известными в данной области техники, для образования более крупных микрогубок. В качестве неограничивающего примера, микрогубку можно объединять в комплекс со средством для формирования наружного слоя, способствующего клеточному поглощению, таким как поликатион полиэтиленим (PEI). Этот комплекс может образовывать частицы диаметром 250 нм, которые могут оставаться стабильными при высоких температурах (150°C.) (публикация Grabow and Jaegar, Nature Materials 2012, 11: 269-269; которая включена в данный документ посредством ссылки во всей своей полноте). Кроме того, эти микрогубки могут обладать способностью проявлять исключительную степень защиты от разложения рибонуклеазами. В другом варианте осуществления самоорганизующиеся наночастицы на основе полимера, такие как без ограничения микрогубки, могут быть полностью программируемыми наночастицами. Геометрию, размер и стехиометрию наночастиц можно точно контролировать для создания оптимальной наночастицы для доставки груза, такого как без ограничения полинуклеотиды.
[157] В некоторых вариантах осуществления полинуклеотиды могут быть составлены в виде неорганических наночастиц (патент США № 8257745, включенный в данный документ посредством ссылки во всей своей полноте). Неорганические наночастицы могут включать без ограничения глинистые вещества, которые набухают в воде. В качестве неограничивающего примера, неорганическая наночастица может включать синтетические смектитовые глины, которые изготовлены из простых силикатов (см., например, патенты США № 5585108 и 8257745, каждый из которых включен в данный документ посредством ссылки во всей их полноте).
[158] В некоторых вариантах осуществления полинуклеотид может быть составлен в виде диспергируемой в воде наночастицы, содержащей полупроводниковый или металлический материал (публикация патента США № 20120228565; включенная в данный документ посредством ссылки во всей своей полноте) или составлен в виде магнитной наночастицы (публикации патента США № 20120265001 и 20120283503; каждая из которых включена в данный документ посредством ссылки во всей своей полноте). Диспергируемые в воде наночастицы могут представлять собой гидрофобные наночастицы или гидрофильные наночастицы.
[159] В некоторых вариантах осуществления полинуклеотиды, раскрытые в данном документе, могут быть инкапсулированы в любой гидрогель, известный в данной области техники, который может образовывать гель при инъекции субъекту. Гидрогели представляют собой сеть полимерных цепей, которые являются гидрофильными, и иногда находятся в виде коллоидного геля, в котором вода является дисперсионной средой. Гидрогели являются (они могут содержать более 99% воды) природными или синтетическими полимерами с высокой абсорбционной способностью. Гидрогели также обладают степенью гибкости, очень близкой к такой в живых тканях, благодаря значительному содержанию в них воды. Описанный в данном документе гидрогель может применяться для инкапсулирования липидных наночастиц, которые являются биосовместимыми, биоразлагаемыми и/или пористыми.
[160] В качестве неограничивающего примера гидрогель может представлять собой гидрогель, функционализированный с помощью аптамера. Гидрогель, функционализированный с помощью аптамера, может быть запрограммирован на высвобождение одного или более полинуклеотидов с применением гибридизации полинуклеотидов. (Публикация Battig et al., J. Am. Chem. Society. 2012 134:12410-12413; которая включена в данный документ посредством ссылки во всей своей полноте). В некотором варианте осуществления полинуклеотид может быть инкапсулирован в липидную наночастицу, и затем липидная наночастица может быть инкапсулирована в гидрогель.
[161] В некоторых вариантах осуществления полинуклеотиды, раскрытые в данном документе, могут быть инкапсулированы в фибриновый гель, фибриновый гидрогель или фибриновый клей. В другом варианте осуществления полинуклеотиды могут быть составлены в виде липидной наночастицы или быстро удаляемой липидной наночастицы перед инкапсулированием в фибриновый гель, фибриновый гидрогель или фибриновый клей. В еще одном варианте осуществления полинуклеотиды могут быть составлены в виде липоплекса перед инкапсулированием в фибриновый гель, гидрогель или фибриновый клей. Фибриновые гели, гидрогели и клеи содержат два компонента: раствор фибриногена и раствор тромбина с высоким содержанием кальция (см., например, публикации Spicer and Mikos, Journal of Controlled Release 2010. 148: 49-55; Kidd et al. Journal of Controlled Release 2012. 157:80-85; каждая из которых включена в данный документ посредством ссылки во всей своей полноте). Концентрация компонентов фибринового геля, гидрогеля и/или клея может быть изменена для изменения характеристик, размера ячеек сети и/или характеристик разложения геля, гидрогеля и/или клея, таких как без ограничения изменение характеристик высвобождения фибринового геля, гидрогеля и/или клея. (См., например, публикации Spicer and Mikos, Journal of Controlled Release 2010. 148: 49-55; Kidd et al. Journal of Controlled Release 2012. 157:80-85; Catelas et al. Tissue Engineering 2008. 14:119-128; каждая из которых включена в данный документ посредством ссылки во всей своей полноте). Этот признак может быть полезным при использовании для доставки полинуклеотида, раскрытого в данном документе. (См., например, публикации Kidd et al. Journal of Controlled Release 2012. 157:80-85; Catelas et al. Tissue Engineering 2008. 14:119-128; каждая из которых включена в данный документ посредством ссылки во всей своей полноте).
[162] В некоторых вариантах осуществления полинуклеотид, раскрытый в данном документе, может содержать катионы или анионы. В одном варианте осуществления составы содержат катионы металлов, такие как без ограничения Zn2+, Ca2+, Cu2+, Mg+ и их комбинации. В качестве неограничивающего примера, составы могут содержать полимеры и полинуклеотид, образующие комплекс с катионом металла (см., например, патенты США №№ 6265389 и 6555525, каждый из которых включен в данный документ посредством ссылки во всей своей полноте).
[163] В некоторых вариантах осуществления полинуклеотид может быть составлен в виде наночастиц и/или микрочастиц. Эти наночастицы и/или микрочастицы могут быть отлиты в форму любого размера и химического состава. В качестве примера, наночастицы и/или микрочастицы могут быть получены с применением технологии PRINT® от LIQUIDA TECHNOLOGIES.RTM. (Моррисвилль, Северная Каролина, США) (см., например, международную публикацию № WO2007024323; которая включена в данный документ посредством ссылки во всей своей полноте).
[164] В некоторых вариантах осуществления полинуклеотиды, раскрытые в данном документе, могут быть составлены в виде наночастиц NanoJacket и NanoLiposome от Keystone Nano (Стейт Колледж, Пенсильвания, США). Наночастицы NanoJacket изготовлены из соединений, которые в норме содержатся в организме, включая кальций, фосфат, и могут также включать небольшое количество силикатов. Размер наночастиц NanoJacket может составлять от 5 до 50 нм, и их можно применять для доставки гидрофильных и гидрофобных соединений, таких как без ограничения полинуклеотиды, первичные конструкции и/или полинуклеотид. Наночастицы NanoLiposome получают из липидов, таких как без ограничения липиды, которые в норме содержатся организме. Размер наночастиц NanoLiposome может составлять 60-80 нм и их можно применять для доставки гидрофильных и гидрофобных соединений, таких как без ограничения полинуклеотиды, первичные конструкции и/или полинуклеотиды. В одном аспекте полинуклеотиды, раскрытые в данном документе, составлены в виде наночастицы NanoLiposome, такой как без ограничения Ceramide NanoLiposome.
[165] В одном варианте осуществления мультидоменный терапевтический белок представляет собой слитый белок, содержащий scFv к CD63 и GAA, или слитый белок, содержащий scFv к ITGA7 и GAA. Введение слитого белка, содержащего scFv к CD63 и GAA, или слитого белка, содержащего scFv к ITGA7 и GAA, посредством AAV-доставки обеспечивает долгосрочное стабильное продуцирование GAA в сыворотке крови пациента после введения вектора для генной терапии, содержащего мультидоменный терапевтический белок. В одном варианте осуществления уровень содержания GAA в сыворотке крови пациента-реципиента в ≥ 1,5 раза-100 раз, в ≥ 1,5 раза-10 раз, в ≥ 2,5 раза, в 2,5 раза-3 раза, в 2,5 раза, в 2,6 раза, в 2,7 раза, в 2,8 раза, в 2,9 раза, в 3,0 раза, в 3,1 раза, в 3,2 раза, в 3,3 раза, в 3,4 раза, в 3,5 раза, в 3,6 раза, в 3,7 раза, в 3,8 раза, в 3,9 раза, в 4 раза, в 5 раз, в 6 раз, в 7 раз, в 8 раз, в 9 раз или в 10 раз выше, чем уровни содержания в сыворотке крови пациента, получающего GAA, не соединенный с доменом доставки, спустя 1 месяц, 3 месяца, 4 месяца, 5 месяцев или 6 месяцев после введения вектора для генной терапии, содержащего мультидоменный терапевтический белок.
[166] В одном варианте осуществления введение слитого белка, содержащего scFv к CD63 и GAA, или слитого белка, содержащего scFv к ITGA7 и GAA, посредством AAV-доставки обеспечивает долговременное стабильное снижение уровней содержания накопленного гликогена у пациентов с болезнью Помпе. В одном варианте осуществления уровни содержания гликогена в сердечной, скелетной мышце и ткани печени у пациента снижаются до уровней, характерных для дикого типа (без заболевания). В одном варианте осуществления уровни содержания гликогена в сердечной, скелетной мышце и ткани печени у пациента поддерживаются на уровнях, характерных для дикого типа, спустя 1 месяц, 2 месяца, 3 месяца, 4 месяца, 5 месяцев или 6 месяцев после введения вектора для генной терапии, содержащего мультидоменный терапевтический белок.
[167] В одном варианте осуществления введение слитого белка, содержащего scFv к CD63 и GAA, или слитого белка, содержащего scFv к ITGA7 и GAA, посредством AAV-доставки обеспечивает долгосрочное восстановление мышечной силы у пациентов с болезнью Помпе. В одном варианте осуществления сила пациента, измеренная по силе захвата, восстанавливается до нормы (то есть нормального уровня, характерного для состояния без заболевания) через 1 месяц, 2 месяца, 3 месяца, 4 месяца, 5 месяцев или 6 месяцев после введения вектора для генной терапии, содержащего мультидоменный терапевтический белок.
[168] В другом аспекте настоящее изобретение предусматривает композицию, обладающую ферментативной активностью и содержащую антигенсвязывающий белок, при этом фермент ассоциирован с заболеванием, связанным с дефицитом ферментов (LSD), и белком, связывающим эффектор интернализации. Ферменты (включающие белки, которые не являются каталитическими per se), ассоциированные с лизосомными болезнями накопления, включают, например, любые возможные гидролазы, α-галактозидазу, β-галактозидазу, α-глюкозидазу, β-глюкозидазу, активатор сапозина-С, церамидазу, сфингомиелиназу, β-гексозаминидазу, активатор GM2, GM3-синтазу, арилсульфатазу, активатор сфинголипида, α-идуронидазу, идуронидаза-2-сульфатазу, гепарин-N-сульфатазу, N-ацетил-α-глюкозаминидазу, α-глюкозамид-N-ацетилтрансферазу, N-ацетилглюкозамин-6-сульфатазу, N-ацетилгалактозамин-6-сульфат-сульфатазу, N-ацетилгалактозамин-4-сульфатазу, β-глюкуронидазу, гиалуронидазу и т. п.
[169] Белки, связывающие эффектор интернализации, например, включают молекулу слияния с рецептором, молекулу-ловушку, молекулу слияния с рецептором Fc, антитело, Fab-фрагмент, F(ab')2-фрагмент, Fd-фрагмент, Fv-фрагмент, молекулу одноцепочечного Fv (scFv), dAb-фрагмент, выделенную определяющую комплементарность область (CDR), пептид CDR3, конформационно затрудненный пептид FR3-CDR3-FR4, домен-специфическое антитело, однодоменное антитело, антитело с удаленным доменом, химерное антитело, антитело с привитой CDR, диатело, триатело, тетратело, минитело, нанотело, одновалентное нанотело, двухвалентное нанотело, иммунофармацевтическое средство на основе модульного белка малого размера (SMIP), верблюжье антитело (гомодимерное антитело на основе тяжелой цепи VHH), вариабельный домен IgNAR акулы, другие антигенсвязывающие белки и т. п.
[170] Эффекторы интернализации включают, например, CD63, MHC-I, Kremen-1, Kremen-2, LRP5, LRP6, LRP8, рецептор трансферрина, LDL-рецептор, рецептор белка 1, родственного LDL, ASGR1, ASGR2, белок-2, подобный белку-предшественнику амилоида (APLP2), апелиновый рецептор (APLNR), PRLR (рецептор пролактина), MAL (миелиновый и лимфоцитарный белок, также известный, как VIP17), IGF2R, H+ АТФазу вакуолярного типа, рецептор дифтерийного токсина, фолатный рецептор, глутаматные рецепторы, глутатионовый рецептор, лептиновый рецептор, скавенджер-рецептор, SCARA1-5, SCARB1-3 и CD36. В определенных вариантах осуществления эффектор интернализации представляет собой специфический для почки интернализатор, такой как CDH16 (Cadheri-16), CLDN16 (Claudn-16), KL (клото), PTH1R (рецептор паратиреоидного гормона), SLC22A13 (семейство 22 транспортеров растворенных веществ, представитель 13), SLC5A2 (котранспортер 2 натрия/глюкозы) и UMOD (уромодулин). В других определенных вариантах осуществления эффектор интернализации представляет собой специфический для мышц интернализатор, такой как BMPR1A (рецептор 1A костного морфогенетического белка), m-кадгерин, CD9, MuSK (киназа, специфическая для мышц), LGR4/GPR48 (рецептор 48, сопряженный с G-белком), холинергический рецептор (никотиновый) альфа 1, CDH15 (Cadheri-15), ITGA7 (интегрин альфа-7), CACNG1 (субъединица гамма-1 кальциевого канала L-типа), CACNAlS (субъединица альфа-15 кальциевого канала L-типа), CACNG6 (субъединица гамма-6 кальциевого канала L-типа), SCN1B (субъединица бета-1 натриевого канала), CHRNA1 (субъединица альфа рецептора ACh), CHRND (субъединица дельта рецептора ACh), LRRC14B (белок 14B, содержащий богатый лейцином повтор), дистрогликан (DAG1) и POPDC3 (белок 3, содержащий домен Popeye). В некоторых вариантах осуществления эффектор интернализации представляет собой ITGA7, CD9, CD63, ALPL2, ASGR1, ASGR2 или PRLR.
[171] В некоторых вариантах осуществления фермент ковалентно связан (т. e. электроны распределяются между атомами) с антигенсвязывающим белком. В одном конкретном варианте осуществления белок, связывающий эффектор интернализации, состоит из полутела или содержит таковое; фермент слит с Fc-слитым доменом (например, на C-конце); и Fc-домен, который ковалентно связан с ферментом, ассоциируется с Fc-доменом антигенсвязывающего белка так, что ассоциация предусматривает один или более дисульфидных мостиков. Данный конкретный вариант осуществления схематически изображен на фиг. 1A, панель B.
[172] В другом конкретном варианте осуществления белок, связывающий эффектор интернализации (домен доставки), состоит из антитела или фрагмента антитела или содержит таковые, и фермент ковалентно связан с антителом или фрагментом антитела. В конкретном варианте осуществления домен доставки представляет собой антитело, и фермент ковалентно связан (непосредственно с помощью пептидной связи или опосредованно с помощью линкера) с С-концом тяжелой цепи или легкой цепи антитела (фиг. 1А, панели С или Е соответственно). В другом конкретном варианте осуществления домен доставки представляет собой антитело, и фермент ковалентно связан (непосредственно с помощью пептидной связи или опосредованно с помощью линкера) с N-концом тяжелой цепи или легкой цепи антитела (фиг. 1А, панели D или F соответственно).
[173] В некоторых вариантах осуществления фермент и домен доставки не связаны ковалентно, а объединены в смесь. Домен доставки и фермент могут связываться посредством нековалентных сил с образованием комплекса. Например, в одном конкретном варианте осуществления домен доставки представляет собой биспецифическое антитело, в котором одно плечо антитела связывается с эффектором интернализации, а другое плечо связывается с ферментом. Данный вариант осуществления схематически изображен на фиг. 1A, панель A.
[174] В некоторых вариантах осуществления фермент представляет собой GAA или обладает активностью GAA (например, изозим с активностью GAA), и эффектор интернализации представляет собой ITGA7, CDH15, CD9, CD63, APLP2, ASGR1, ASGR2 или PRLR. В конкретном варианте осуществления фермент представляет собой GAA или обладает активностью GAA, домен интернализации представляет собой CD63, и домен доставки представляет собой биспецифическое антитело со специфичностью к CD63 и GAA. В конкретном варианте осуществления фермент представляет собой GAA или обладает активностью GAA, домен интернализации представляет собой ITGA7, и домен доставки представляет собой биспецифическое антитело со специфичностью к ITGA7 и GAA.
[175] В некоторых вариантах осуществления фермент представляет собой GLA или обладает активностью GLA (например, изозим с активностью GAA), и эффектор интернализации представляет собой ITGA7, CD9, CD63, APLP2, ASGR1, ASGR2 или PRLR. В конкретном варианте осуществления фермент представляет собой GLA или обладает активностью GLA, домен интернализации представляет собой CD63, и домен доставки представляет собой биспецифическое антитело со специфичностью к CD63 и GLA. В конкретном варианте осуществления фермент представляет собой GLA или обладает активностью GLA, домен интернализации представляет собой ITGA7, и домен доставки представляет собой биспецифическое антитело со специфичностью к ITGA7 и GLA.
Фармацевтические композиции и их введение
[176] Фармацевтические составы могут дополнительно содержать фармацевтически приемлемое вспомогательное вещество, которой, как используется в данном документе, включает любые возможные растворители, дисперсионные среды, разбавители или другие жидкие среды-носители, средства для диспергирования или суспендирования, поверхностно-активные вещества, изотонические средства, загущающие или эмульгирующие средства, консерванты, твердые связующие вещества, смазывающие вещества и т. п., подходящие для конкретной необходимой лекарственной формы. В публикации Remington's The Science and Practice of Pharmacy, 21.sup.st Edition, A. R. Gennaro (Lippincott, Williams & Wilkins, Baltimore, Md., 2006; которая включена в данный документ посредством ссылки во всей своей полноте), раскрыты различные вспомогательные вещества, используемые при составлении фармацевтических композиций, и известные методики для их получения. За исключением случаев, когда какая-либо обычная среда на основе вспомогательного вещества несовместима с веществом или его производными, например, из-за какого-либо нежелательного биологического эффекта или из-за другого взаимодействия вредным образом с любым другим компонентом (компонентами) фармацевтической композиции, предполагается, что ее использование находится в пределах объема настоящего изобретения.
[177] В некоторых вариантах осуществления фармацевтически приемлемое вспомогательное вещество является чистым, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или по меньшей мере на 100%. В некоторых вариантах осуществления вспомогательное вещество одобрено для применения у людей и для ветеринарного применения. В некоторых вариантах осуществления вспомогательное вещество одобрено Управлением по контролю качества продуктов и лекарственных препаратов США. В некоторых вариантах вспомогательное вещество характеризуется фармацевтической степенью чистоты. В некоторых вариантах осуществления вспомогательное вещество соответствует стандартам Фармакопеи США (USP), Европейской Фармакопеи (EP), Британской Фармакопеи и/или Международной Фармакопеи.
[178] Фармацевтически приемлемые вспомогательные вещества, применяемые в производстве фармацевтических композиций, включают без ограничения инертные разбавители, диспергирующие и/или гранулирующие средства, поверхностно-активные вещества и/или эмульгаторы, дезинтегрирующие средства, связующие средства, консерванты, буферные средства, смазывающие средства и/или масла. Такие вспомогательные вещества могут быть необязательно включены в фармацевтические композиции.
[179] Иллюстративные разбавители включают без ограничения карбонат кальция, карбонат натрия, фосфат кальция, дикальция фосфат, сульфат кальция, гидрофосфат кальция, фосфат натрия, лактозу, сахарозу, целлюлозу, микрокристаллическую целлюлозу, каолин, маннит, сорбит, инозит, хлорид натрия, сухой крахмал, кукурузный крахмал, сахарную пудру и т. д. и/или их комбинации.
[180] Иллюстративные гранулирующие и/или диспергирующие средства включают без ограничения картофельный крахмал, кукурузный крахмал, крахмал тапиоки, натрия крахмала гликолят, глины, альгиновую кислоту, гуаровую камедь, мякоть цитрусовых, агар, бентонит, целлюлозу и продукты переработки древесины, натуральную губку, катионообменные смолы, карбонат кальция, силикаты, карбонат натрия, сшитый поли(винилпирролидон) (кросповидон), карбоксиметилкрахмал натрия (крахмалгликолят натрия), карбоксиметилцеллюлоза, сшитая карбоксиметилцеллюлоза натрия (кроскармеллоза), метилцеллюлоза, предварительно желатинизированный крахмал (крахмал 1500), микрокристаллический крахмал, нерастворимый в воде крахмал, карбоксиметилцеллюлоза кальция, алюмосиликат магния (VEEGUM®), лаурилсульфат натрия, соединения четвертичного аммония и т. д. и/или их комбинации.
[181] Иллюстративные поверхностно-активные вещества и/или эмульгаторы включают без ограничения природные эмульгаторы (например, акацию, агар, альгиновую кислоту, альгинат натрия, трагакант, хондрукс, холестерин, ксантан, пектин, желатин, яичный желток, казеин, ланолин, холестерин, воск и лецитин), коллоидные глины (например, бентонит [силикат алюминия] и VEEGUM® [алюмосиликат магния]), производные аминокислот с длинной цепью, высокомолекулярные спирты (например, стеариловый спирт, цетиловый спирт, олеиловый спирт, моностеарат триацетина, дистеарат этиленгликоля, глицерилмоностеарат и моностеарат пропиленгликоля, поливиниловый спирт), карбомеры (например, карбоксиполиметилен, полиакриловая кислота, полимер акриловой кислоты и карбоксивиниловый полимер), каррагенан, производные целлюлозы (например, карбоксиметилцеллюлоза натрия, порошкообразная целлюлоза, гидроксиметилцеллюлоза, гидроксипропилцеллюлоза, гидроксипропилметилцеллюлоза, метилцеллюлоза), сложные эфиры сорбитана и жирных кислот (например, полиоксиэтиленсорбитанмонолаурат [TWEEN® 20], полиоксиэтиленсорбитан [TWEEN® 60], полиоксиэтиленсорбитанмоноолеат [TWEEN® 80], сорбитанмонопальмитат [SPAN® 40], сорбитанмоностеарат [SPAN® 60], сорбитантристеарат [SPAN® 65], глицерилмоноолеат, сорбитанмоноолеат [SPAN® 80]) сложные эфиры полиоксиэтилена (например, полиоксиэтиленмоностеарат [MYRJ® 45], полиоксиэтиленовое гидрогенизированное касторовое масло, полиэтоксилированное касторовое масло, полиоксиметиленстеарат и SOLUTOL®), сложные эфиры сахарозы и жирных кислот, сложные эфиры жирных кислот и полиэтиленгликоля (например, CREMOPHOR®), простые эфиры полиоксиэтилена (например, полиоксиэтиленовый эфир лаурилового спирта [BRIJ® 30]), поли(винилпирролидон), монолаурат диэтиленгликоля, олеат триэтаноламина, олеат натрия, олеат калия, этилолеат, олеиновая кислота, этиллаурат, лаурилсульфат натрия, PLUORINC® F 68, POLOXAMER® 188, бромид цетримония, хлорид цетилпиридиния, хлорид бензалкония, докузат натрия и т. д. и/или их комбинации.
[182] Иллюстративные связывающие средства включают без ограничения крахмал (например, кукурузный крахмал и крахмальную пасту); желатин; сахара (например, сахарозу, глюкозу, декстрозу, декстрин, мелассу, лактозу, лактит, маннит); природные и синтетические камеди (например, гуммиарабик, альгинат натрия, экстракт ирландского мха, камедь «панвар», камедь гхатти, слизь из оболочек семян подорожника блошного, карбоксиметилцеллюлоза, метилцеллюлоза, этилцеллюлоза, гидроксиэтилцеллюлоза, гидроксипропилцеллюлоза, гидроксипропилметилцеллюлоза, микрокристаллическая целлюлоза, ацетилцеллюлоза, поливинилпирролидон, алюмосиликат магния (Veegum®) и арабогалактан лиственницы); альгинаты; полиэтиленоксид; полиэтиленгликоль; неорганические соли кальция; кремниевая кислота; полиметакрилаты; воски; вода; спирт; и т. д.; и их комбинации.
[183] Иллюстративные консерванты могут включать без ограничения антиоксиданты, хелатирующие средства, антимикробные консерванты, противогрибковые консерванты, спиртовые консерванты, кислотные консерванты и/или другие консерванты. Иллюстративные антиоксиданты включают без ограничения альфа-токоферол, аскорбиновую кислоту, аскорбилпальмитат, бутилированный гидроксианизол, бутилированный гидрокситолуол, монотиоглицерин, метабисульфит калия, пропионовую кислоту, пропилгаллат, аскорбат натрия, бисульфат натрия, метабисульфит натрия и/или сульфит натрия. Иллюстративные хелатирующие средства включают этилендиаминтетрауксусную кислоту (EDTA), моногидрат лимонной кислоты, динатрия эдетат, дикалия эдетат, эдетовую кислоту, фумаровую кислоту, яблочную кислоту, фосфорную кислоту, натрия эдетат, винную кислоту и/или тринатрия эдетат. Иллюстративные антимикробные консерванты включают без ограничения хлорид бензалкония, хлорид бензетония, бензиловый спирт, бронопол, цетримид, хлорид цетилпиридиния, хлоргексидин, хлорбутанол, хлоркрезол, хлороксиленол, крезол, этиловый спирт, глицерин, гексетидин, имидомочевину, фенол, феноксиэтанол, фенилэтиловый спирт, фенилмеркуриевый нитрат, пропиленгликоль и/или тимеросал. Иллюстративные противогрибковые консерванты включают без ограничения бутилпарабен, метилпарабен, этилпарабен, пропилпарабен, бензойную кислоту, гидроксибензойную кислоту, бензоат калия, сорбат калия, бензоат натрия, пропионат натрия и/или сорбиновую кислоту. Иллюстративные спиртовые консерванты включают без ограничения этанол, полиэтиленгликоль, фенол, фенольные соединения, бисфенол, хлорбутанол, гидроксибензоат и/или фенилэтиловый спирт. Иллюстративные кислотные консерванты включают без ограничения витамин А, витамин С, витамин Е, бета-каротин, лимонную кислоту, уксусную кислоту, дегидроуксусную кислоту, аскорбиновую кислоту, сорбиновую кислоту и/или фитиновую кислоту. Другие консерванты включают без ограничения токоферол, токоферола ацетат, детероксим мезилат, цетримид, бутилированный гидроксианизол (BHA), бутилированный гидрокситолуол (BHT), этилендиамин, лаурилсульфат натрия (SLS), лауретсульфат натрия (SLES), бисульфит натрия, метабисульфит натрия, сульфит калия, метабисульфит калия, GLYDANT PLUS®, PHENONIP®, метилпарабен, GERMALL® 115, GERMABEN® II, NEOLONE.TM., KATHON.TM. и/или EUXYL®.
[184] Иллюстративные буферные средства включают без ограничения цитратные буферные растворы, ацетатные буферные растворы, фосфатные буферные растворы, хлорид аммония, карбонат кальция, хлорид кальция, цитрат кальция, глубионат кальция, глюцептат кальция, глюконат кальция, D-глюконовая кислота, глицерофосфат кальция, лактат кальция, пропановая кислота, левулинат кальция, пентановая кислота, двухосновный фосфат кальция, фосфорная кислота, трехосновный фосфат кальция, гидроксид-фосфат кальция, ацетат калия, хлорид калия, глюконат калия, смеси на основе калия, двухосновный фосфат калия, одноосновный фосфат калия, смеси на основе фосфата калия, ацетат натрия, бикарбонат натрия, хлорид натрия, цитрат натрия, лактат натрия, двухосновный фосфат натрия, одноосновный фосфат натрия, смеси на основе фосфата натрия, трометамин, гидроксид магния, гидроксид алюминия, альгиновая кислота, апирогенная вода, изотонический солевой раствор, раствор Рингера, этиловый спирт и т. д. и/или их комбинации.
[185] Иллюстративные смазывающие средства включают без ограничения стеарат магния, стеарат кальция, стеариновую кислоту, диоксид кремния, тальк, солод, глицерилбегенат, гидрогенизированные растительные масла, полиэтиленгликоль, бензоат натрия, ацетат натрия, хлорид натрия, лейцин, лаурилсульфат магния, лаурилсульфат натрия и т. д. и их комбинации.
[186] Иллюстративные масла включают без ограничения масла/жиры миндаля, косточки абрикоса, авокадо, бабассу, бергамота, семян черной смородины, бурачника, можжевельника, ромашки, канола, тмина, карнаубы, касторовое, корицы, масло какао, кокосовое, печени трески, кофе, кукурузы, семян хлопчатника, эму, эвкалипта, примулы вечерней, рыбы, семян льна, гераниоловое, тыквы, виноградных косточек, фундука, иссопа, изопропилмиристат, жожобы, плодов лакового дерева, лавандиновое, лавандовое, лимона, лицеа кубебы, макадамского ореха, мальвы, семян манго, семян пенника лугового, норки, мускатного ореха, оливковое, апельсина, атлантического большеголова, пальмовое, пальмоядровое, косточки персика, арахисовое, маковое, семян тыквы, рапсовое, рисовых отрубей, розмарина, сафлоровое, сандаловое, камелии горной, пряное, облепиховое, кунжутное, масло ши, кремниевое, соевое, подсолнечное, чайного дерева, расторопши, камелии, ветиверовое, грецкого ореха и зародышей пшеницы. Иллюстративные масла включают без ограничения бутилстеарат, каприловый триглицерид, каприновый триглицерид, циклометикон, диэтилсебацинат, диметикон 360, изопропилмиристат, минеральное масло, октилдодеканол, олеиловый спирт, силиконовое масло и/или их комбинации.
[187] Вспомогательные вещества, такие как масло какао и воски для суппозиториев, красящие средства, покрывающие средства, подсластители, вкусовые добавки и/или ароматизирующие добавки, могут присутствовать в композиции, по усмотрению составителя.
[188] Доставка
[189] Настоящее изобретение охватывает доставку вектора для генной терапии (например, полинуклеотидов) любым подходящим путем, принимая во внимание вероятные достижения в области доставки лекарственных средств. Доставка может быть без дополнительных средств или в составе.
[190] Доставка без дополнительных средств
[191] Полинуклеотиды по настоящему изобретению могут быть доставлены к клетке без применения дополнительных веществ. Используемый в данном документе термин «без дополнительных средств» относится к доставке полинуклеотидов без средств, которые способствуют трансфекции. Например, полинуклеотиды, доставленные в клетку, могут не содержать модификаций. Полинуклеотиды без дополнительных средств могут быть доставлены в клетку с применением способов введения, известных в данной области техники и описанных в данном документе.
[192] Доставка в составе
[193] Полинуклеотиды могут быть составлены с применением способов, описанных в данном документе. Составы могут содержать полинуклеотиды и могут дополнительно содержать без ограничения средства для проникновения в клетку, фармацевтически приемлемый носитель, средство доставки, биоразрушаемый или биосовместимый полимер, растворитель и депо для доставки с замедленным высвобождением. мРНК в виде составленных полинуклеотидов может быть доставлена в клетку с применением способов введения, известных в данной области техники и описанных в данном документе.
[194] Введение
[195] Полинуклеотиды по настоящему изобретению можно вводить с помощью любого пути, который обеспечивает терапевтически эффективный результат. К этим путям относятся без ограничения: энтеральное, гастроэнтеральное, эпидуральное, пероральное, трансдермальное, эпидуральное (перидуральное), интрацеребральное (в головной мозг), интрацеребровентрикулярное (в желудочки головного мозга), накожное (нанесение на кожу), внутрикожное (собственно в кожу), подкожное (под кожу), назальное введение (через нос), внутривенная (в вену), внутриартериальная (в артерию), внутримышечная (в мышцу), внутрисердечная (в сердце), внутрикостная инфузия (в костный мозг), интратекальная (в позвоночный канал), внутрибрюшинная (инфузия или инъекция в брюшину), внутрипузырная инфузия, интравитреальная (в глаз), интракавернозная инъекция (в основание полового члена), интравагинальное введение, внутриматочное, экстраамниотическое введение, трансдермальное (диффузия через неповрежденную кожу для системного распределения), трансмукозальное (диффузия через слизистую оболочку), инсуффляция (вдувание), сублингвальное, сублабиальное, клизма, глазные капли (на конъюнктиву) или ушные капли. В конкретных вариантах осуществления композиции можно вводить таким образом, чтобы они могли проникать через гематоэнцефалический барьер, сосудистый барьер или другой эпителиальный барьер. Неограничивающие пути введения полинуклеотидов, первичных конструкций или мРНК по настоящему изобретению описаны ниже.
[196] Парентеральное и инъекционное введение
[197] Жидкие лекарственные формы для парентерального введения включают без ограничения фармацевтически приемлемые эмульсии, микроэмульсии, растворы, суспензии, сиропы и/или настойки. В дополнение к действующим веществам жидкие лекарственные формы могут содержать инертные разбавители, обычно используемые в данной области техники, такие как, например, вода или другие растворители, солюбилизирующие средства и эмульгаторы, такие как этиловый спирт, изопропиловый спирт, этилкарбонат, этилацетат, бензиловый спирт, бензилбензоат, пропиленгликоль, 1,3-бутиленгликоль, диметилформамид, масла (в частности, хлопковое, арахисовое, кукурузное, зародышей пшеницы, оливковое, касторовое и кунжутное масла), глицерин, тетрагидрофурфуриловый спирт, полиэтиленгликоли и сложные эфиры жирных кислот и сорбитана и их смеси. Помимо инертных разбавителей пероральные композиции могут содержать вспомогательные вещества, такие как смачивающие средства, эмульгирующие и суспендирующие средства, подсластители, вкусовые добавки и/или ароматизирующие средства. В определенных вариантах осуществления для парентерального введения композиции смешивают с солюбилизирующими средствами, такими как CREMOPHOR®, спирты, масла, модифицированные масла, гликоли, полисорбаты, циклодекстрины, полимеры и/или их комбинации.
[198] Инъекционные лекарственные препараты, например, стерильные инъекционные водные или масляные суспензии, могут быть составлены в соответствии с известным уровнем техники с применением подходящих диспергирующих средств, смачивающих средств и/или суспендирующих средств. Стерильные инъекционные препараты могут представлять собой стерильные инъекционные растворы, суспензии и/или эмульсии в нетоксичных приемлемых для парентерального введения разбавителях и/или растворителях, например, как раствор в 1,3-бутандиоле. Среди приемлемых сред-носителей и растворителей, которые можно применять, представлены вода, раствор Рингера, U.S.P. и изотонический раствор хлорида натрия. Стерильные нелетучие масла обычно используются в качестве растворителя или суспендирующей среды. Для этой цели можно использовать любое щадящее нелетучее масло, включая синтетические моно- или диглицериды. Жирные кислоты, такие как олеиновая кислота, могут применяться при получении инъекционных препаратов.
[199] Инъекционные составы могут быть стерилизованы, например, путем фильтрования через задерживающий бактерии фильтр и/или путем включения стерилизующих средств в форме стерильных твердых композиций, которые могут быть растворены или диспергированы в стерильной воде или другой стерильной инъекционной среде перед применением.
[200] Чтобы продлить эффект действующего вещества, часто желательно замедлить абсорбцию действующего вещества при подкожной или внутримышечной инъекции. Это может быть достигнуто путем применения жидкой суспензии кристаллического или аморфного материала с плохой растворимостью в воде. Скорость абсорбции лекарственного средства затем зависит от скорости его растворения, которая, в свою очередь, может зависеть от размера кристаллов и кристаллической формы. В качестве альтернативы, отсроченная абсорбция парентерально вводимой лекарственной формы достигается путем растворения или суспендирования лекарственного средства в масляной среде-носителе. Инъекционные депо-формы получают путем формирования микроинкапсулированных матриц лекарственного средства в биоразлагаемых полимерах, таких как полилактид-полигликолид. В зависимости от соотношения лекарственного средства и полимера и природы конкретного применяемого полимера можно контролировать скорость высвобождения лекарственного средства. Примеры других биоразлагаемых полимеров включают сложные поли(ортоэфиры) и поли(ангидриды). Составы, представляющие собой инъекционные депо, получают путем захватывания лекарственного средства в липосомы или микроэмульсии, которые совместимы с тканями организма.
[201] Введение депо
[202] Как описано в данном документе, в некоторых вариантах осуществления композиция составлена в виде депо для пролонгированного высвобождения. Как правило, мишенью для введения является конкретный орган или ткань («ткань-мишень»).
[203] В некоторых аспектах изобретения полинуклеотиды пространственно удерживаются внутри или вблизи ткани-мишени. Предложен способ обеспечения ткани-мишени субъекта-млекопитающего композицией путем приведения ткани-мишени (которая содержит одну или более клеток-мишеней) в контакт с композицией в условиях, при которых композиция, в частности компонент(ы) нуклеиновой кислоты композиции, по сути удерживается в ткани-мишени, что означает, что по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 85, 90, 95, 96, 97, 98, 99, 99,9, 99,99 или более чем 99,99% композиции удерживается в ткани-мишени. Преимущественно уровень удержания определяют путем измерения количества нуклеиновой кислоты, присутствующей в композиции, которая попадает в одну или более клеток-мишеней. Например, по меньшей мере 1, 5, 10, 20, 30, 40, 50, 60, 70, 80, 85, 90, 95, 96, 97, 98, 99, 99,9, 99,99 или более 99,99% нуклеиновых кислоты, вводимых субъекту, присутствуют в клетке в период времени после введения. Например, внутримышечную инъекцию субъекту-млекопитающему выполняют с применением водной композиции, содержащей полинуклеотид и реагент для трансфекции, и удержание композиции определяют путем измерения количества рибонуклеиновой кислоты, содержащейся в мышечных клетках.
[204] Аспекты настоящего изобретения направлены на способы обеспечения ткани-мишени субъекта-млекопитающего композицией путем приведения ткани-мишени (содержащей одну или более клеток-мишеней) в контакт с композицией в условиях, при которых композиция по сути удерживается в ткани-мишени. Композиция содержит эффективное количество полинуклеотида, такое что полипептид, представляющий интерес, продуцируется по меньшей мере в одной клетке-мишени. Композиции, как правило, содержат средство для проникновения в клетку, хотя также рассматривается нуклеиновая кислота без дополнительных средств (такая как нуклеиновые кислоты без средства для проникновения в клетку или другого средства), и фармацевтически приемлемый носитель.
[205] В некоторых обстоятельствах необходимо увеличение количества белка, продуцируемого клетками в ткани. Предпочтительно, это увеличение продуцирования белка пространственно ограничено клетками в ткани-мишени. Таким образом, предоставлены способы увеличения продуцирования белка, представляющего интерес, в ткани субъекта-млекопитающего. Предусматривается композиция, которая содержит полинуклеотиды, отличающаяся тем, что для единицы количества композиции установлено, что она обеспечивает продуцирование полипептида, представляющего интерес, в значительном проценте клеток, содержащихся в предварительно определенном объеме ткани-мишени.
[206] В некоторых вариантах осуществления композиция содержит несколько различных полинуклеотидов, где один или более полинуклеотидов кодируют полипептид, представляющий интерес. Необязательно композиция также содержит средство для проникновения в клетку, чтобы способствовать внутриклеточной доставке композиции. Определяют дозу композиции, необходимую для обеспечения продуцирования полипептида, представляющего интерес, в значительном проценте клеток, содержащихся в предварительно определенном объеме ткани-мишени (как правило, без индуцирования значительного уровня продуцирования полипептида, представляющего интерес, в ткани, расположенной рядом с предварительно определенным объемом или дистально от ткани-мишени). После этого определения определенную дозу вводят непосредственно в ткань субъекта-млекопитающего.
[207] В одном варианте осуществления настоящее изобретение предусматривает доставку полинуклеотидов с помощью более чем одной инъекции или с помощью инъекций в соответствии с методом дробления дозы.
[208] В одном варианте осуществления настоящее изобретение может удерживаться вблизи ткани-мишени с применением небольшого одноразового резервуара с лекарственным средством, пластыря-насоса или осмотического насоса. Неограничивающие примеры пластырей-насосов включают насосы, производимые и/или распространяемые BD® (Франклин Лейкс, Нью-Джерси, США), Insula Corporation (Бедфорд, Массачусетс, США), SteadyMed Therapeutics (Сан-Франциско, Калифорния, США), Medtronic (Миннеаполис, Миннесота, США) (например, MiniMed), UniLife (Йорк, Пенсильвания, США), Valeritas (Бриджуотер, Нью-Джерси, США) и SpringLeaf Therapeutics (Бостон, Массачусетс, США). Неограничивающий пример осмотического насоса включает насос, производимый DURECT® (Купертино, Калифорния, США) (например, DUROS® и ALZET®).
[209] Режим дозирования
[210] Настоящее изобретение предусматривает способы, включающие введение вектора для генной терапии, содержащего полинуклеотид, кодирующий мультидоменный терапевтический полипептид, и необязательно введение впоследствии мультидоменного терапевтического полипептида субъекту, нуждающемуся в этом. В некоторых вариантах осуществления способ включает введение вектора для генной терапии, содержащего полинуклеотид, кодирующий мультидоменный терапевтический полипептид, в терапевтически эффективном количестве пациенту, нуждающемуся в этом, при этом терапевтически эффективное количество является достаточным для устранить необходимость в последующем введении мультидоменного терапевтического полипептида. Соответственно, в некоторых вариантах осуществления способ лечения пациента с дефицитом фермента, нуждающегося в этом, например снижение уровней содержания гликогена и/или снижение CRIM к GAA у пациента с болезнью Помпе, включает введение пациенту вектора для генной терапии, содержащего полинуклеотид, кодирующий мультидоменный терапевтический белок, содержащий заместительный фермент, например слитый белок, содержащий scFv к CD63::GAA, например, мультидоменный терапевтический белок, содержащий последовательность, представленную под SEQ ID NO:11, в терапевтически эффективном количестве, где терапевтически эффективное количество устраняет необходимость в последующем введении пациенту заместительного фермента, например GAA или его производных. В некоторых вариантах осуществления способ лечения пациента с дефицитом фермента и который нуждается в этом, например снижение уровня содержания гликогена и/или снижение CRIM к GAA у пациента с болезнью Помпе, включает введение пациенту вектора для генной терапии, содержащего полинуклеотид, кодирующий мультидоменный терапевтический белок, содержащий заместительный фермент, например слитый белок scFv к CD63::GAA, например мультидоменный терапевтический белок, содержащий последовательность, представленную под SEQ ID NO:11, в терапевтически эффективном количестве, и дополнительно включает введение пациенту терапевтически эффективного количества заместительного фермента. Нуклеиновые кислоты, белки или комплексы или фармацевтические композиции, композиции для визуализации, диагностические или профилактические композиции на их основе могут быть введены субъекту с применением любого количества и любого пути введения, эффективного для предотвращения, лечения, диагностики или визуализации заболевания, нарушения и/или состояния (например, заболевания, нарушения и/или состояния, связанного с дефицитом кратковременной памяти). Точное требуемое количество будет варьироваться от субъекта к субъекту, в зависимости от вида, возраста и общего состояния субъекта, тяжести заболевания, конкретной композиции, способа ее введения, способа ее действия и т. п. Композиции в соответствии с настоящим изобретением обычно составляют в виде дозированной лекарственной формы для простоты введения и однородности дозирования. Понятно, однако, что общее суточное применение композиций по настоящему изобретению может определяться лечащим врачом исходя из результатов медицинской оценки. Конкретный терапевтически эффективный, профилактически эффективный или подходящий для визуализации уровень дозы для любого конкретного пациента будет зависеть от множества факторов, включая нарушение, подлежащее лечению, и тяжесть нарушения; активность конкретного применяемого соединения; конкретную применяемую композицию; возраст, массу тела, общее состояние здоровья, пол и режим питания пациента; время введения, способ введения и скорость выведения конкретного применяемого соединения; продолжительность лечения; лекарственные средства, применяемые в комбинации или одновременно с конкретным применяемым соединением; и подобные факторы, хорошо известные в медицине.
[211] Следующие примеры представлены с целью дальнейшей иллюстрации способов согласно настоящему изобретению. Данные примеры являются исключительно иллюстративными и не предназначены для ограничения объема настоящего изобретения каким-либо образом.
ПРИМЕРЫ
Пример 1. Конструирование полинуклеотида ScFv к hCD63::GAA и вектора для генной терапии
[212] Вирусы AAV2/8, кодирующие экспрессию GAA человека (hGAA; SEQ ID NO:1; последовательность нуклеиновой кислоты, представленная под SEQ ID NO:12) или одноцепочечный вариабельный фрагмент к CD63 человека (ScFv), слитый на его С-конце с GAA человека (scFv к hCD63-hGAA; SEQ ID NO:10; нуклеиновая кислота, представленная под SEQ ID NO: 11), генерировали с применением стандартного протокола тройной трансфекции (Gray et al. 2011; см. также «Production of recombinant adeno-associated viral vectors and use in vitro and in vivo administration», Current Protocols in Neuroscience, John Wiley & Sons, New York (1999), pp. 4.17.1-4.17.25, Vol 1). Для продуцирования 1×107 клеток HEK293 высевали на чашки диаметром 15 см. На следующий день клетки трансфицировали с помощью (А) либо 8 мкг контрольного вектора pAAV, содержащего специфический для печени энхансер серпина 1 (SEQ ID NO:9) и кодирующего TTR-управляемую GAA человека, либо тестируемого pAAV, содержащего специфический для печени энхансер серпина 1 (SEQ ID NO:9) и кодирующего TCD-управляемый scFv к hCD63-hGAA (см. фигуру 1B) и (B) вектора, полученного из pAAV RC2/8, (Gao, 2002) и 16 мкг pHelper (Agilent, № по кат. 240074), с применением PEIpro (Polyplus transfection, Нью-Йорк, Нью-Йорк, США № по кат. 115-100)-опосредованной трансфекции, в соотношении 1:1 (1 мкл PEIpro: 1 мкг ДНК). Через семьдесят два часа после трансфекции клетки собирали и лизировали в буфере, содержащем 20 мМ Tris-HCl, 1 мМ MgCl2, 2,5 мМ KCl, 100 мМ NaCl, с применением стандартного метода замораживания-оттаивания. Затем к образцам добавляли бензоназу (Sigma, № по кат. E1014-25KU) в конечной концентрации 0,5 ед./мкл, и затем инкубировали при 37°С в течение 60 минут. Затем вирусы очищали с применением ультрацентрифугирования в градиенте йодиксанола, как описано в (Zolotukhin et al., 1999, Gene Ther 1999;6:973-985), и затем титровали с помощью qPCR.
[213] Образцы AAV обрабатывали с помощью DNaseI (Thermofisher Scientific, № по кат. EN0525) при 37°C в течение одного часа и лизировали с применением экстракта ДНК All Reagents (Thermofisher Scientific № по кат. 4403319). Инкапсидированные вирусные геномы определяли количественно с применением системы ПЦР в режиме реального времени QuantStudio 3 (Thermofisher Scientific) с применением праймеров, направленных на ITR AAV2. Последовательности праймеров к ITR AAV2 представляют собой 5′-GGAACCCCTAGTGATGGAGTT-3′ (прямой праймер к ITR; SEQ ID NO:3) и 5′-CGGCCTCAGTGAGCGA-3′ (обратный праймер к ITR; SEQ ID NO:4) (Aurnhammer et al., 2012), с помощью которых получают последовательность левого внутреннего инвертированного повтора (ITR) AAV (SEQ ID NO:6) и правого внутреннего инвертированного повтора (ITR) AAV (SEQ ID NO:7), соответственно. Последовательность зонда ITR AAV2 представляет собой 5'-6-FAM-CACTCCCTCTCTGCGCGCTCG-TAMRA-3' (SEQ ID NO:5) (Aurnhammer C., Haase M., Muether N., et al., 2012, Hum. Gene Ther. Methods 23, 18-28). После этапа активации при 95°С, длившегося 10 минут, проводили цикл двухстадийной ПЦР при 95°С в течение 15 секунд и при 60°С в течение 30 секунд, всего 40 циклов. Для qPCR применяли TaqMan Universal PCR Master Mix (Thermofisher Scientific, № по кат. 4304437). ДНК-плазмиду (Agilent, № по кат. 240074) применяли в качестве стандарта для определения абсолютных титров.
[214] Для антител к CD63 человека и слитых молекул на их основе применяли вариабельные домены мышиного H5C6 к CD63 человека (аминокислоты 1-119 из SEQ ID NO:10 обеспечивают аминокислотную последовательность вариабельного домена тяжелой цепи (VH) антитела H5C6, а аминокислоты 135-245 из SEQ ID NO:10 обеспечивают аминокислотную последовательность вариабельного домена легкой цепи (VL) антитела H5C6). Используемый в данном документе scFv к hCD63 (SEQ ID NO:2) получали из клона H5C6, который представляет собой легкую цепь антитела каппа из моноклонального мышиного IgG1 к hCD63, (H5C6 внесли в Банк гибридом для исследований развития (Developmental Studies Hybridoma Bank) Университета Айовы к августу, J.T. / Hildreth, J.E.K. (DSHB Hybridoma Product H5C6; DSHB № по кат. h5c6, RRID:AB_528158). ScFv-версии антител клонировали с вариабельными доменами в порядке тяжелая цепь-легкая цепь с глицин-сериновым линкером между ними (5'-VH-Gly-Ser-VL-3').
Пример 2. Содержание гликогена в мышиной модели болезни Помпе после введения AAV
[215] Чтобы определить эффект доставленного с помощью AAV слитого белка, содержащего scFv к hCD63 и GAA, по сравнению с доставленной с помощью AAV GAA в соответствующей модели накопления гликогена in vivo, оба средства терапии доставляли в мышиную модель болезни Помпе, где мыши были гомозиготными по делеции гена GAA мыши и были гомозиготными по экспрессии CD63 человека вместо CD63 мыши с фоновым генотипом штамма, составляющим 75% C57BL/6; 25% 129SvJ. Эти мыши в данном документе упоминаются как мыши CD63 HumIn с нокаутом GAA или, в качестве альтернативы, CD63hu/hu; мыши GAA-/-.
[216] Для эксперимента 2-месячным мышам CD63 HumIn с нокаутом GAA вводили посредством инъекции в хвостовую вену либо вирус AAV2/8, содержащий геном либо со специфическим для печени TTR-промотором, управляющим GAA человека (AAV-hGAA; описанный в примере 1), либо со специфическим для печени TTR-промотором, управляющим scFv к CD63 человека, слитым на своем С-конце с GAA человека (AAV-scFv к hCD63-hGAA; описанный в примере 1). Оба вируса AAV2/8 доставляли в одной из двух доз, 1e10 vg/мышь или 1e11 vg/мышь. В качестве контроля в анализ были включены необработанные мыши CD63 HumIn с нокаутом GAA и необработанные мыши CD63 HumIn с интактным геном GAA. Мышей содержали в течение 3 месяцев после обработки и периодически (ежемесячно) брали у них кровь в течение этого периода для измерений уровней содержания GAA и антител к GAA в сыворотке крови. Через 3 месяца всех мышей умерщвляли и собирали отдельные ткани для измерений уровней гликогена, окрашивания с помощью PAS-H, количественного определения центрально расположенных ядер, измерения уровня пролиферации лизосом и измерения уровня экспрессии LC3b. Протокол дозирования и обработки для групп мышей в ходе эксперимента показан в таблице 3.
Таблица 3. Протокол дозирования и обработки для групп мышей в ходе эксперимента
[217] Результаты также представлены на фигуре 2, из которой видно, что scFv к hCD63::GAA понижает уровень содержания гликогена в скелетной мышце до уровней, характерных для дикого типа, в отличие от GAA отдельно. Обработка с помощью мультидоменного терапевтического белка, представляющего собой двухдоменный scFv к hCD63::GAA, привела к гораздо большему снижению содержания накопленного гликогена по сравнению с обработкой с помощью однодоменного заместительного фермента GAA. Из графика зависимости уровней содержания гликогена в квадрицепсе (фигура 3) или уровней содержания гликогена в сердце (фигура 4) от общей экспрессии GAA или scFv-GAA в сыворотке крови в течение трех месяцев для отдельных мышей видно, что слитый белок scFv к hCD63::GAA удаляет больше гликогена, чем фермент GAA отдельно, даже при аналогичных уровнях содержания в сыворотке крови (фигуры 3 и 4).
Пример 3. Иммунный ответ на GAA
[218] Для измерения уровней содержания антител к GAA человека в сыворотке крови во всех группах обработки отделение сыворотки от крови, собранной во время терминального кровотечения, осуществляли с применением пробирок для отделения сыворотки (BD Biosciences, № по кат. 365967) в соответствии с инструкциями производителя. Отдельно 96-луночные планшеты с высокой эффективностью связывания белков (ThermoFisher, № по кат. 15041) покрывали с помощью 20 мкг hGAA (R&D Systems, № по кат. 8329-GH-025), разведенного в PBS в течение ночи. Планшеты промывали с помощью PBS + 0,05% Tween (PBS-T) 3 раза. Планшеты блокировали с помощью 0,5% BSA в PBS-T, и серийные разведения мышиной сыворотки в диапазоне от 1:300 до 1:5,1e7 добавляли в планшет на ночь. Общее количество антител к IgG мыши (подклассы 1 + 2a + 2b + 3) измеряли с применением конъюгированного с HRP козьего антитела к IgG мыши (Jackson Immuno Research, № по кат. 115-035-164) и набора субстратов BD Opt EIA. Колориметрические реакции останавливали с применением 1 н. фосфорной кислоты. Затем измеряли поглощение при 450 нм на планшет-ридере Spectramax i3 (Molecular Devices). Кривые разведений аппроксимировали по сигмоидальным кривым, и из кривых рассчитывали титры. Титры, выраженные в виде среднего общего титра IgG +/- SD, показаны в таблице 4.
[219] Как показано в таблице 4, мыши, которые не подвергались обработке, показывали средний фоновый титр со средними уровнями 1,1E+03. Мыши, которых обрабатывали с помощью низкой дозы вируса (1e10 vg/мышь), либо AAV-scFv к hCD63-hGAA, либо AAV-hGAA, демонстрировали высокие титры, тогда как у мышей, обработанных высокой дозой (1e11 vg/мышь), титры были ниже. Мыши, обработанные с помощью 1e11vg AAV-scFv к hCD63-hGAA, которые показывали наиболее высокие уровни GAA в сыворотке крови, имели титры, находящиеся в диапазоне таковых у необработанных мышей.
Таблица 4. Уровни содержания антител к GAA в сыворотке крови
(1e10 vg)
(1e11 vg)
(1e10 vg)
(1e11 vg)
[220] Более высокие уровни содержания GAA или scFv к hCD63::GAA после введения AAV соответствуют более низким титрам антител к GAA. Сыворотку мышей с нокаутом по GAA, обработанных высокими или низкими титрами AAV-scFv к hCD63::GAA или AAV-GAA, оценивали в отношении содержания антител к GAA в течение трех месяцев после инъекции. На фигуре 5 изображена зависимость сывороточных титров антител к GAA от уровня воздействия GAA (т. е. общая экспрессия GAA или scFv-GAA в сыворотке в течение 3 месяцев) для отдельных мышей, что демонстрирует отрицательную корреляцию между титром антител и уровнем воздействия GAA на сыворотку, демонстрируя, что мыши с высоким уровнем воздействия GAA были подвергнуты индуцированию толерантности к GAA. Аналогично, фигура 6, на которой представлены титры антител к GAA для различных групп, инфицированных AAV, кодирующим GAA или белок scFv к hCD63::GAA, демонстрирует, что более высокие дозы конструкции приводили к более низким титрам антител к GAA.
Пример 4. Содержание GAA в сыворотке
[221] Чтобы измерить уровни содержания GAA человека в сыворотке крови в течение эксперимента, образцы отбирали в определенные моменты каждый месяц, вызывая хвостовое кровотечение. Сыворотку отделяли от крови, используя пробирки для отделения сыворотки (BD Biosciences, № по кат. 365967) в соответствии с инструкциями производителя. Затем 1 мкл выделенной сыворотки загружали в 4-20% предварительно залитый гель Novex wedgewell, прогоняли при 220 В в течение 45 минут и переносили на нитроцеллюлозную мембрану при 200 мА в течение 1 часа, применяя стандартные процедуры. Затем нитроцеллюлозную мембрану изучали с помощью первичного антитела к GAA (Abcam, № ab137068), используемого в разведении 1:2000, и антитела к GAPDH (Abcam, № AB9484), используемого в разведении 1:1000, в 12 мл и инкубировали в течение ночи при 4°С. После первичной инкубации с антителами мембрану трижды промывали с помощью 1 × TBST, по 5 минут на каждую промывку. Затем к мембране добавляли вторичные антитела к IgG кролика (LiCor, 926-32211) и к IgG мыши (LiCor, 925-68070) (LiCor, Линкольн, Небраска) в разведении 1:15000 в 12 мл и инкубировали в течение 1 часа при комнатной температуре. После инкубации с вторичными антителами мембрану дважды промывали с помощью 1 х TBST, по 5 минут на каждую промывку, и один раз с помощью 1 х TBS в течение 5 минут. Затем мембрану визуализировали и осуществляли количественное определение с применением прибора LiCor Odyssey (LI-COR Biotechnology). Уровни содержания GAA в сыворотке крови, выраженные как среднее +/- стандартное отклонение (SD) в произвольных единицах, показаны в таблице 5.
[222] Как показано в таблице 5, мыши CD63 HumIn с нокаутом GAA, обработанные высокой дозой (1011 vg/мышь) исследуемых AAV-scFv к hCD63-hGAA или AAV-hGAA, продемонстрировали устойчивые уровни содержания GAA в сыворотке крови в течение эксперимента, причем уровни содержания GAA в сыворотке крови были несколько выше у мышей, обработанных с помощью AAV-scFv к hCD63-hGAA, чем у мышей, обработанных с помощью AAV-hGAA. У мышей, обработанных низкой дозой (1010 vg/мышь) либо AAV-scFv к hCD63-hGAA, либо AAV-hGAA, уровни GAA снижались в течение эксперимента, приближаясь к пренебрежительно малым уровням у некоторых мышей к моменту времени на 12 неделе.
Таблица 5. Уровни содержания GAA в сыворотке
[223] Экспрессия GAA или scFv к hCD63::GAA сохранялась в течение времени у мышей, получавших высокую дозу AAV (1011 vg/мышь), но снижалась у мышей, получавших более низкую дозу (1010 vg/мышь). На фигуре 7А представлен график изменения уровней содержания GAA в сыворотке крови, измеренных с помощью вестерн-блоттинга, с течением времени для различных групп, инфицированных AAV, кодирующим GAA или scFv к hCD63, слитый с GAA. В случае слитого белка (scFv::GAA) демонстрировали неизменно более высокие уровни (например, в 2,5-3 раза) содержания GAA в сыворотке, чем в случае фермента GAA без домена доставки (фигура 7A).
[224] Количественная оценка экспрессии с помощью ПЦР в режиме реального времени в лизатах печени, сердца и квадрицепса спустя 3 месяца после инъекции показана на фиг. 7В. Экспрессия в печени была обнаружена при всех инъекциях конструкции на основе AAV, причем наивысшие уровни были обнаружены при инъекциях 1e11vg/мышь как для AAV-hGAA, так и для AAV-фрагмент, связывающий hCD63::hGAA (оба управляются специфическим для печени промотором, LSP). Провели также сравнение уровня содержания GAA в сыворотке крови с уровнем экспрессии РНК GAA (фиг. 7C), и результаты показывают, что мыши, получающие AAV, кодирующий слитый белок, демонстрировали более низкую экспрессию РНК GAA, локализующейся в печени, через 3 месяца, однако уровни содержания GAA в сыворотке крови были высокими у этой конкретной мыши. В случае инъекций AAV-LSP-hGAA не показали высокого уровня содержания GAA в сыворотке крови при низких уровнях содержания РНК в печени. См. фиг. 7C. Эти данные свидетельствуют о том, что AAV, кодирующий белок слияния (экспрессия управляется специфическим для печени промотором), обеспечивает улучшенный профиль секреции GAA.
[225] Также в гепатоцитах Huh-7 наблюдали более высокое отношение секретируемого к внутриклеточному антителу:hGAA по сравнению с введением hGAA отдельно. В одном эксперименте гепатоциты Huh-7 человека были временно трансфицированы конструкциями, управляемыми специфическим для печени промотором, которые кодировали hGAA, слитый белок scFv к hCD63::GAA или контрольный слитый белок, содержащий несвязывающий scFv и GAA. Обе слитые конструкции scFv::GAA характеризовались более высоким соотношением белка в секретируемом супернатанте, чем в случае hGAA отдельно, через 3 дня после трансфекции (статистически значимо для p <0,05, n=3). Добавление M6P в супернатант в течение экспериментального периода для уменьшения CI-MPR-опосредованного захвата не влияло на соотношение.
Пример 5. Измерение уровня содержания гликогена в ткани и гистологическая характеристика мышечной ткани
[226] Измерение уровня содержания гликогена в ткани. Для измерения содержания гликогена в отдельных тканях - ткани сердца, квадрицепса, икроножной мышцы, диафрагмы, камбаловидной мышцы и EDL, - ткани вырезали у мышей из всех групп сразу же после асфиксии при помощи CO2, а затем мгновенно замораживали в жидком азоте и хранились при -80°C. ~50 мг каждой ткани лизировали на настольном гомогенизаторе с шариками из нержавеющей стали в дистиллированной воде в соотношении от 1 мг до 25 мкл воды для измерений уровня содержания гликогена. Лизаты гликогена для анализа нагревали при 105°С в течение 15 минут и центрифугировали при 21000 x g для удаления дебриса. Измерения уровня гликогена проводили с применением набора для анализа гликогена (Sigma-Aldrich, № MAK016) в соответствии с инструкциями производителя для флуорометрических анализов. Флуоресценцию каждого образца измеряли при длине волны возбуждения 535 нм и длине волны излучения 587 нм на флуоресцентном планшет-ридере (Molecular Devices, Spectramax i3). Рассчитанное количество гликогена рассчитывали с применением следующей формулы, предоставленной производителем. Рассчитанное количество гликогена из каждой ткани в каждой группе обработки затем усредняли и полученные значения выражены в виде среднего +/- стандартное отклонение (SD) в таблице 6.
[227] Как показано в таблице 6, потеря Gaa вызывает значительное повышение средних уровней содержания гликогена во всех тканях, в отношении которых осуществляли измерения, по сравнению с мышами с GAA WT. Обработка с помощью AAV-scFv к hCD63-hGAA при 1011 vg/мышь снижала уровень содержания гликогена до уровней, характерных для WT, или почти до уровней, характерных для WT, во всех протестированных тканях, в отличие от обработки с помощью AAV-GAA, которая обеспечивала лишь частичное снижение содержания накопленного гликогена. Низкие дозы любого вируса также снижают уровень содержания гликогена, но в меньшей степени, чем высокие дозы. Доза 1010 vg/мышь AAV-scFv к hCD63-hGAA снижала уровни содержания гликогена аналогично дозе 1011 vg/мышь AAV-GAA.
Таблица 6. Средний уровень содержания гликогена +/- SD, измеренный в сердце, квадрицепсе, икроножной мышце, диафрагме, камбаловидной мышце и EDL
(1010 vg)
(1011 vg)
(1010 vg)
(1011 vg)
[228] Сбор квадрицепса для гистопатологического исследования и количественного определения. Образцы ткани квадрицепса от мышей из каждой группы, кроме группы, которую обрабатывали с помощью низкой дозы (1e10 vg/мышь), либо мгновенно замораживали в жидком азоте сразу после вырезания и хранили при -80°C для количественного определения экспрессии LC3b, либо помещали в блоки, содержащие среду O.C.T (Tissue-Tek, №4583).
[229] Образцы тканей в среде O.C.T отправляли в Histoserv, Inc (Джермантуан, Мэриленд, США) для приготовления срезов и окрашивания с помощью реактива Шиффа (PAS) для обнаружения полисахаридов. Дополнительные срезы были получены и возвращены для окрашивания с целью определения центрально расположенных ядер и уровня пролиферации лизосом.
[230] Окрашивание с помощью PAS. Окрашенные с помощью PAS срезы визуализировали с применением сканера для препаратов Leica при увеличении 20x. Полученные изображения от мышей, являющихся репрезентативными для каждой группы обработки, показаны на фиг. 8.
[231] Как показано на фиг. 8, CD63 HumIn с нокаутом GAA, которых обработали с помощью AAV-scFv к hCD63-hGAA, через 3 месяца демонстрировали заметное снижение окрашивания по сравнению как с мышами CD63 HumIn с нокаутом GAA без обработки, так и с мышами CD63 HumIn GAA, обработанными с помощью AAV-hGAA, которые показали высокий уровень окрашивания с помощью PAS. Это дополнительно указывает на то, что обработка с помощью AAV-scFv к hCD63-hGAA может снизить уровень аккумуляции полисахаридов у мышей CD63 HumIn с нокаутом GAA и может действовать таким образом одинаково на все мышечные волокна.
[232] Количественное определение центральных ядер и пролиферации лизосом. Неокрашенные срезы от Histoserv извлекали из морозильной камеры и затем фиксировали с помощью 4% параформальдегида в PBS в течение 15 минут в камере для окрашивания. Фиксированные препараты затем дважды промывали по 5 минут в PBS и затем инкубировали с блокирующим буфером (eBiosciences, 00-4953-54) в течение 1 часа при комнатной температуре. Затем препараты либо окрашивали с помощью крысиного антитела к Lamp-1(Abcam, №AB25245) в разведении 1:50 в блокирующем буфере, с помощью кроличьего антитела к ламинину (Sigma, №L9393) в разведении 1:1000 в блокирующем буфере или с помощью блокирующего буфера без добавления антител в камере для окрашивания с высокой влажностью, а затем переносили в 4°C для инкубации в течение ночи. На следующий день препараты затем дважды промывали по 5 минут в PBS и затем окрашивали либо козьим суперклональным вторичным антителом к IgG кролика (H+L), конъюгированным с Alexa Fluor 647 (Life Tech Thermo, №A27040), либо козьим перекрестно-адсорбированным вторичным антитело к IgG крысы (H+L), конъюгированным с Alexa Fluor 555 (Life Tech Thermo, №A21434) в камере для окрашивания, затем оставляли для инкубации в течение 1 часа при комнатной температуре. Затем окрашенные препараты дважды промывали по 5 минут в PBS, после чего их заливали средой Fluoromount-G с DAPI (Life Tech Thermo, №00-4959-52) и визуализировали с помощью прибора Zeiss LSM710 (Carl Zeiss Microscopy GmbH). Количество ядер, расположенных по центру, количественно определяли с применением программного обеспечения Halo (Indica Labs, Нью-Мексико, США), и это количество выражено в виде процента волокон, для которых показано центральное расположение ядра, +/- стандартное отклонение и показано в таблице 7. Пролиферация лизосом изображена на фигуре 8.
Таблица 7. Количественное определение центрально расположенных ядер
[233] Количественное определение уровня экспрессии LC3b. Для количественного определения уровня экспрессии LC3b образцы, подвергнутые мгновенному замораживанию, размораживали, гомогенизировали и затем лизировали в буфере RIPA при соотношении 1 мг ткани к 25 мкл буфера RIPA (150 мМ NaCl, 1,0% IGEPAL® CA-630, 0,5% дезоксихолата натрия, 0,1% SDS, 50 мМ Tris, рН 8,0, Sigma Aldrich, R0278) с помощью соударения с шариками в течение 45 секунд (MP Biomedical). Лизаты очищали от нерастворимого материала путем центрифугирования при 21000 x g, а затем 300 мкг лизата в буфере RIPA загружали в 4-20% предварительно залитый гель Novex wedgewell, переносили на нитроцеллюлозную мембрану и анализировали с помощью вестерн-блоттинга, используя протокол, аналогичный ранее описанному, для анализа уровней содержания GAA в сыворотке крови, заменяя первичное антитело к GAA на использование первичного антитела, которое распознает мышиные LC3b-I и LC3b-II (Sigma, №L7543). Затем мембрану визуализировали и осуществляли количественное определение с применением прибора LiCor Odyssey (LI-COR Biotechnology). Уровни LC3b-I и LC3b-II, выраженные в виде (среднее +/- стандартное отклонение) в произвольных единицах, показаны в таблице 8.
[234] Как показано в таблице 8, наблюдалось значительное увеличение как средних уровней LC3b-I, так и LC3b-II у мышей с дефицитом GAA, по сравнению с мышами CD63 HumIn с GAA WT. Обработка с помощью AAV-scFv к hCD63-hGAA снизила средние уровни LC3b-I и LC3b-II в CD63 HumIn с нокаутом GAA до уровней, характерных для WT, или почти до уровней, характерных для WT. CD63 HumIn с нокаутом GAA, обработанные с помощью AAV-hGAA, демонстрировали слегка пониженные средние уровни LC3b-I и LC3b-II по сравнению с мышами CD63 HumIn с нокаутом GAA, но это снижение было не таким выраженным, как при обработке с помощью AAV-scFv к hCD63-hGAA.
Таблица 8. Уровни LC3b-I и LC3b-II в квадрицепсах мышей
Пример 6. Обработка с помощью AAV, содержащего фрагмент, связывающий hCD63::GAA, приводит к значительным улучшениям в тестах мышечной силы и координации
[235] Сила захвата и эффективность выполнения теста с вращающимся барабаном у мышей, обработанных (см. выше) с помощью либо AAV-LSP-hGAA, либо AAV-LSP-фрагмент, связывающий hCD63::hGAA. Измерения при ускорении вращающегося барабана (фиг. 9A) и измерения силы захвата передних конечностей (фиг. 9B) у мышей с GAA дикого типа, необработанного контроля, при обработке с помощью AAV-LSP-hGAA (1e11vg/мышь) или AAV-LSP-фрагмент, связывающий hCD63::hGAA (1e11vg/мышь) проводились с месячными интервалами в течение 6 месяцев. Планки погрешностей представляют собой +/- SD. N=8-10 для всех групп.
Пример 7. Другие мембранные белки в качестве «направляющих», которые направляют GAA в ткани
[236] Протестировали другие мембранные белки, такие как слитые белки на основе фрагмента, связывающего ITGA7 (интегрин альфа-7) в отношении направления GAA в ткани для замещения GAA у мышей с дефицитом ферментов. Мышиные миобласты C2C12 инкубировали в течение ночи с комплексом фрагмента, связывающего mCD63, и GAA, или комплексом фрагмента, связывающего ITGA7, и GAA, с или без 5 мМ M6P. Динамику содержания активного фермента GAA в лизатах миобластов выявляли в случае обоих слитых белков (фиг. 10А). В дальнейших экспериментах мышам с нокаутом GAA, гуманизированным по CD63 (GAA-/-; CD63hu/hu), вводили плазмиды, кодирующие фрагмент, связывающий hCD63::GAA в формате scFv::GAA или фрагмент, связывающий интегрин альфа-7, в формате полноразмерный IgG4::GAA путем гидродинамической доставки (HDD), и мышей умерщвляли через 3 недели после HDD. Уровни содержания гликогена в тканях измеряли в сердце, квадрицепсе, икроножной мышце и диафрагме. Необработанных контрольных мышей GAA-/- xCD63hu/hu и необработанных контрольных мышей дикого типа GAA+/+; CD63hu/hu (4) также тестировали в тех же условиях. Уровни содержания гликогена были на очень низком уровне как у мышей, обработанных с помощью комплекса фрагмент, связывающий hCD63::GAA, так и у мышей, обработанных с помощью комплекса фрагмент, связывающий ITGA7::GAA, как у мышей дикого типа. См. фигуру 10B.
Пример 8. При сопоставимых уровнях содержания в сыворотке крови обработка с помощью AAV-фрагмент, связывающий CD63::GAA является более эффективной, чем конструкция AAV с оптимизированным GAA
[237] Мыши CD63 HumIn с нокаутом GAA (GAA-/- x CD63hu/hu), инфицированные с помощью AAV, содержащих специфический для печени энхансер (серпина 1; SEQ ID NO:9) и специфический для печени промотор (LSP; TTR; SEQ ID NO:8), управляющие экспрессией мультидоменного терапевтического средства на основе комплекса фрагмент, связывающий hCD63::GAA (SEQ ID NO:10), в котором используется сигнальный пептид химотрипсиноген B2 (SP7) и содержатся аминокислоты 36-952 GAA человека (Δ8GAA; SEQ ID NO:78), показали значительный прирост в тестах мышечной силы и координации. Вводили три разные дозы каждого вируса: 5e11vg/кг, 2e12vg/кг и 4e12vg/кг. Сыворотку собирали путем вызывания подчелюстного кровотечения на регулярной основе. Через месяц после инфицирования с помощью AAV мышей умерщвляли. Образцы ткани сердечной и скелетной мышцы собирали и мгновенно замораживали в жидком азоте и поддерживали при -80°C для хранения. Уровень содержания гликогена в тканях измеряли путем гомогенизации тканей с помощью соударения с шариками в дистиллированной воде. Образцы кипятили и центрифугировали, а супернатанты использовали в коммерческом наборе для анализа гликогена. В случае сыворотки количественное определение осуществляли с применением вестерн-блоттинга с антителом к GAA человека, как описано в предыдущих примерах. Для каждой мыши строили график зависимости уровня содержания гликогена в каждой ткани и уровня содержания конструкции в сыворотке крови через 1 месяц. Для определения EC50 для двух средств обработки в каждой ткани применяли аппроксимацию кривой по 4 параметрам.
[238] Инфицирование с помощью AAV, содержащих специфический для печени промотор (LSP), кодирующий либо комплекс фрагмент, связывающий hCD63::GAA, либо sp7-Δ8GAA, обеспечивало сопоставимые уровни содержания GAA в сыворотке крови при каждой инфицирующей дозе. Фигура 11. Тем не менее, в каждой анализируемой мышечной ткани при применении комплекса фрагмент, связывающий hCD63::GAA, в отличии от sp7-Δ8GAA, наблюдалось снижение EC50 в ~2,2 раза, демонстрируя, что при эквивалентных уровнях содержания в сыворотке крови комплекс фрагмент, связывающий CD63::GAA более эффективно удаляет гликоген, чем модифицированная конструкция для экспрессии GAA, которая не слита с антителом. См. фигуру 12 и таблицу 9.
Таблица 9. EC50 (95% доверительный интервал) для AAV-фрагмент, связывающий hCD63::GAA и AAV-sp7-Δ8GAA в сердце, диафрагме, квадрицепсе и трицепсе
Пример 9. Иллюстративные антитела к CD63
[239] Получение антител к CD63 человека
[240] Антитела к CD63 человека были получены путем иммунизации мыши (например, сконструированной мыши, содержащей ДНК, кодирующую вариабельные области тяжелой цепи и легкой каппа-цепи иммуноглобулина человека) с помощью CD63 человека.
[241] После иммунизации спленоциты собирали у каждой мыши и (1) проводили слияние с клетками миеломы мыши для сохранения их жизнеспособности и формирования клеток гибридомы и проводили скрининг на специфичность к CD63 человека или (2) отсортировали В-клетки (как описано в US 2007/0280945A1) с помощью фрагмента CD63 человека в качестве сортирующего реагента, который связывает реактивные антитела и обеспечивает их идентификацию (антиген-позитивные В-клетки).
[242] Химерные антитела к CD63 человека были первоначально выделены с вариабельной областью человека и константной областью мыши с применением, например, технологии VELOCIMMUNE, как описано в патенте США № 7105348; патенте США № 8642835; и патенте США 9622459, каждый из которых включен в данный документ посредством ссылки.
[243] В некоторых антителах в целях тестирования константные области мыши заменяли необходимой константной областью человека, например СН человека дикого типа или модифицированным СН человека (например, изотипов IgG1, IgG2 или IgG4), и константной областью легкой цепи (CL) для получения полностью человеческого фрагмента, связывающего hCD63, включая полностью человеческое биспецифическое антитело, предусматривающее антитело к hCD63 или его антигенсвязывающую часть. Хотя выбранная константная область может варьировать в зависимости от конкретного применения, свойства, представляющие собой высокую аффинность связывания с антигеном или специфичность к мишеням, характерны для вариабельной области.
[244] Некоторые биологические свойства иллюстративных биспецифических антител, содержащих связывающее плечо антитела к CD63 человека, полученных в соответствии со способами из данного примера, подробно описаны в представленных ниже примерах.
[245] Последовательности аминокислот и нуклеиновых кислот вариабельных областей тяжелой и легкой цепей антител к CD63
[246] В таблице 10 приведены идентификаторы кодирующей последовательности нуклеиновой кислоты (NA), а в скобках представлена сама аминокислотная (AA) последовательность вариабельной области тяжелой или легкой цепи (HCVR или LCVR соответственно) или CDR тяжелой или легкой цепи (HCDR и LCDR соответственно) выбранных антител к CD63, которые применяли для получения мультидоменных терапевтических белков, содержащих фрагмент, связывающий CD63::GAA, описанных в данном документе.
Таблица 10. Идентификаторы последовательности фрагмента, связывающего CD63
NA
(AA)
NA
(AA)
NA
(AA)
NA
(AA)
NA
(AA)
NA
(AA)
NA
(AA)
NA
(AA)
(14)
(16)
(18)
(20)
(22)
(24)
(26)
(28)
(30)
(32)
(34)
(36)
(38)
(40)
(42)
(44)
(46)
(48)
(50)
(52)
(54)
(56)
(58)
(60)
(62)
(64)
(66)
(68)
(70)
(72)
(74)
(76)
[247] Связывание с помощью родительских антител к CD63 человека
[248] Относительный уровень связывания с клеточной поверхностью в случае связывания антител к CD63 с клетками, экспрессирующими CD63 человека, оценивали с помощью проточной цитометрии с применением CD63-положительных клеток HEK293 (ATCC, № по кат. CRL-1573), которые эндогенно экспрессируют CD63 человека, и CD63-отрицательных клеток с нокаутом HEK293/CD63. Для анализа клетки высевали в PBS без кальция и магния (VWR, № по кат. 45000-446), содержащий 2% FBS (Saradigm № по кат. 1500-500) (окрашивающий буфер) в 96-луночные планшеты с V-образным дном (Axygen Scientific, № по кат. P-96-450-VCS). Затем клетки инкубировали с антителами к CD63 или антителами изотипического контроля в концентрациях от 100 нМ до 1,7 пМ в течение 30 минут на льду. Лунки, не содержащие антител, применяли в качестве контроля. Клетки HEK293/клетки с нокаутом CD63 окрашивались только с помощью наиболее высокой концентрации (100 нМ) антител. Затем клетки один раз промывали с помощью окрашивающего буфера и инкубировали с вторичным антителом к Fc мыши, конъюгированным с PE (Jackson ImmunoResearch, № по кат. 115-115-164) при 100 нМ в течение 30 минут при 4°C. Затем клетки промывали и фиксировали с применением 50% раствора Cytofix (BD Biosciences, № по кат. 554655), разведенного в PBS. Образцы загружали в проточный цитометр Intellicyte Hypercyt, и результаты анализировали в программном обеспечении ForeCyt (Intellicyte) для расчета средней интенсивности флуоресценции (MFI). Измеренные значения анализировали с применением четырех-параметрического логистического уравнения по 12-точечной кривой ответа с применением GraphPad Prism, и были получены результирующие значения EC50 (таблица 11). Отношение сигнал/шум (S/N) определяли путем расчета отношения MFI антител к CD63 или контрольных антител к MFI лунок, не содержащих антител (таблица 11).
[249] Как показано в таблице 11, три антитела из антител к CD63 по настоящему изобретению продемонстрировали связывание с клетками HEK293 со значениями S/N в диапазоне от 21,0 до 31,6 и значениями EC50 в диапазоне от 0,5 нМ до 1,9 нМ. Несвязывающиеся контроли не демонстрировали связывания с клетками HEK293 (S/N ≤ 1,5). Как антитела к CD63, так и антитела изотипического контроля, демонстрировали слабое или незначительное связывание с клетками HEK293/клетками с нокаутом CD63 (S/N ≤ 4,4).
Таблица 11. Связывание антител к CD63 с клетками HEK293 и с клетками HEK293 с нокаутом CD63 по данным проточной цитометрии
(нМ)
(S/N)
ND = не определено
[250] Способность моноклональных антител к CD63 по настоящему изобретению связываться с клетками, экспрессирующими CD63 человека, также определяли с применением анализа с обнаружением на основе электрохемилюминесценции (ECL).
[251] Для получения сверхэкспрессирующих клеток мышиные эмбриональные фибробластные клетки NIH3T3 (ATCC, № по кат. CRL-1658) трансфицировали с образованием линии клеток «NIH3T3/hCD63», которая стабильно экспрессирует CD63 человека (hCD63; аминокислоты M1-M238 с учетным номером NP_001771; SEQ ID №:77). Были проанализированы уровни экспрессии CD63 человека в эндогенно экспрессирующих клетках, человеческой линии клеток аденокарциномы простаты, чувствительной к андрогенам, LNCAP (ATCC, № по кат. CRL-1740), и линии клеток первичной глиобластомы человека, U87MG (ATCC, № по кат. HTB-14), с помощью Quantum™ Alexa Fluor® 647 MESF (Bangs Laboratories, № по кат. 647B) и антитела к IgG мыши Simply Cellular® (Bangs Laboratories Inc, № по кат. 815) в соответствии с инструкциями производителя. Было установлено, что клетки LNCAP содержат меньшее количество копий CD63 человека, чем клетки U87MG. Нетрансфицированные клетки NIH3T3, у которых нет детектируемой путем сортировки флуоресцентно-активированных клеток (FACS) экспрессии CD63 человека, были включены в качестве отрицательного контроля.
[252] Вкратце, клеточные линии промывали один раз в буфере PBS без Ca2+/Mg2+ и инкубировали в течение 10 минут при 37°C с раствором для диссоциации клеток, не содержащим ферментов (Millipore, № по кат. S-004-C), чтобы отделить клетки. Затем клетки один раз промывали с помощью PBS с Ca 2+/Mg2+ и подсчитывали с помощью счетчика клеток Cellometer™ Auto T4 (Nexcelom Bioscience, LLC). Приблизительно 2,0×104 клеток NIH3T3/hCD63, LNCAP, U87MG или NIH3T3 высевали отдельно на 96-луночные планшеты с угольными электродами (Meso Scale Discovery, № по кат. L15XB-6) и затем инкубировали в течение одного часа при 37°C. Сайты неспецифического связывания блокировали с помощью 2% BSA (вес/объем) в PBS с Ca2+/Mg2+ в течение одного часа при комнатной температуре (к. т.). Растворы, содержащие антитела к CD63 или антитела изотипического контроля в диапазоне концентраций (от 1,7 пМ до 100 нМ) в 0,5% BSA (вес/объем) в PBS с Ca2+/Mg2+, а также отдельно контрольный буфер добавляли затем в двух повторностях к связанным с планшетами клеткам NIH3T3/hCD63, LNCAP, U87MG или NIH3T3 и инкубировали в течение одного часа при к. т. Затем планшеты промывали для удаления несвязанных антител с применением устройства для промывания планшетов AquaMax2000 с головкой для отмывания клеток (MDS Analytical Technologies). Связанные с планшетами антитела определяли с применением 1 μг/мл либо козьего поликлонального антитела к IgG человека, специфического к фрагменту Fcγ, конъюгированного с SULFO-TAG™(Jackson Immunoresearch, № по кат. 109-005-098), либо козьего поликлонального антитела к IgG мыши, специфического к фрагменту Fcγ (Jackson Immunoresearch, № по кат. 115-005-164), конъюгированного с SULFO-TAG™, в течение одного часа при к. т. Планшеты промывали и затем инкубировали с буфером для считывания (MSD, № по кат. R92TD-2) в соответствии с инструкциями производителя. Люминесцентные сигналы измеряли с применением устройства для визуализации SECTOR (MSD). Интенсивность люминесценции, измеренную в относительных световых единицах (RLU), регистрировали, чтобы установить интенсивность связывания каждого антитела в диапазоне тестируемых концентраций. Соотношение сигнала, обнаруженного при связывании 3,7 нМ антитела с клетками, экспрессирующими CD63 человека, и такового для той же концентрации антитела, связывающегося с отрицательными клетками, было принято в качестве показателя специфичности связывания с CD63. Антитела с соотношением значений связывания более 3 классифицировали как специфические связывающие средства, а антитела с соотношением значений связывания, меньшим или равным 3, классифицировали как несвязывающие и отмечали как NB в таблице 12. Кроме того, сигналы прямого связывания (в RLU) анализировали в зависимости от концентрации антитела, и данные аппроксимировали по сигмоидальной (четырех-параметрической логистической) модели доза-ответ с применением программного обеспечения GraphPad Prism™. Значение ЕС50 для связывания с клетками, экспрессирующими CD63 человека, определенное как концентрация антитела, при которой обнаруживается 50% максимального сигнала связывания, было определено для установления активности каждого антитела и представлено в таблице 12 только для конкретных связывающих средств.
[253] Как показано в таблице 12, четыре антитела к CD63, полученные, как описано в примере 9, и Ab-компаратор (компаратор 1), специфически связывались с CD63 человека, экспрессируемым на сконструированных клетках NIH3T3/hCD63, а также эндогенно экспрессируемым на линиях клеток LNCAP и U87MG. Четыре антитела к CD63, описанные в примере 9, связывались с клетками NIH3T3/hCD63 со значениями EC50 в диапазоне от 280 пМ до 970 пМ и соотношением значений связывания относительно отрицательной линии клеток в диапазоне от 91 до 281 раз. Четыре антитела к CD63 по настоящему изобретению связывались с клетками U87MG со значениями ЕС50 в диапазоне от 500 пМ до 1,4 нМ и соотношением значений связывания относительно отрицательной линии клеток в диапазоне от 52 до 272 раз. Четыре антитела к CD63, полученные, как описано в примере 9, связывались с клетками LNCAP со значениями EC50 в диапазоне от 210 пМ до 1,7 нМ и соотношением значений связывания относительно отрицательной линии клеток в диапазоне от 7 до 20 раз. Более низкие отношения значений связывания для клеток LNCAP согласуются с более низким числом копий CD63 на этих клетках по сравнению с клетками U87MG. Как и ожидалось, антитела изотипического контроля являлись несвязывающими и для них соотношение значений связывания с клетками составляло 3 или меньше.
Таблица 12. Связывание антител к CD63 с клетками, экспрессирующими CD63 человека, измеренное с помощью электрохемилюминесцентного обнаружения
NB - несвязывающий; антитела с соотношением значений связывания меньшим или равным 3 были классифицированы как несвязывающие.
[254] Цитотоксичность, опосредованная родительскими антителами к CD63 человека
[255] Чтобы оценить способность антитела к CD63, описанного в данном документе, интернализоваться в экспрессирующих CD63 клетках, был проведен непрямой анализ цитотоксичности in vitro. Человеческие CD63-позитивные клетки T47D (ATCC, № по кат. HTB-133) и человеческие CD63-отрицательные клетки NIH3T3 (ATCC, № по кат. CRL-1658) соответственно высевали в покрытые с помощью PDL 96-луночные планшеты (BD Biocoat, № по кат. 356461) при либо 6000 клеток на лунку в RPMI (Irvine Scientific, № по кат. 9160), содержащем 10% FBS (ATCC, № по кат. 30-2020), пенициллин/стрептомицин/L-глутамин (Gibco, № по кат. 10378-016), 50 мкМ бета-меркаптоэтанола ( Sigma, № по кат. M7522) (питательная среда), пируват натрия 100 мМ (Millipore, № по кат. TMS-005-C), HEPES 1M (Irvine Scientific, № по кат. 9319) и бычий инсулин 10 мкг/мл (Gemini BioProducts, № по кат. 700-912P), либо 2000 клеток на лунку в DME с высоким содержанием глюкозы (Irvine Scientific, № по кат. 9033), 10% телячьей сыворотки (Hyclone, № по кат. SH30072.03), а также пенициллин/стрептомицин/L-глутамин (Gibco, № по кат. 10378-016), и выращивали в течение ночи при 37°С в 5% СО2. Для получения кривых жизнеспособности клеток клетки инкубировали в течение 5 минут при 37°С с серийно разведенным антителом к CD63 (H2M12450N) или несвязывающим антителом изотипического контроля в концентрациях от 3,0 пМ до 2,2 нМ. Вторичное антитело, представляющее собой Fab к mFc, конъюгированное с цитотоксической нагрузкой MMAF (Moradec, № по кат. AM-201AF-50), затем добавляли при 20 нМ в каждую лунку. Отдельно среда служила отрицательным контролем, и 33 мкМ дигитонина (Promega, № по кат. G9441) применяли для определения максимальной цитотоксичности. После 72-часовой инкубации жизнеспособность клеток измеряли с применением набора Cell Counting Kit-8 (Dojindo, № по кат. CK04) в соответствии с протоколами производителя с продолжительностью инкубации в диапазоне 1-3 часа. Поглощение при 450 нм (OD450) измеряли на ридере Envision (PerkinElmer). Для всех лунок с обработанными дигитонином клетками вычитали значения фоновых уровней OD450, и жизнеспособность выражали в виде процента от необработанных контролей (% жизнеспособности). Значения IC50 определяли из четырех-параметрического логистического уравнения по 8-точечной кривой ответа (GraphPad Prism). Все значения IC50 выражены в концентрации в нМ, и представлены значения минимального % жизнеспособных клеток, остающихся после обработки.
[256] Как показано в таблице 13, антитело к CD63, H2M12450N, снижало жизнеспособность T47D до 21% со значением IC50, составляющим 0,24 нМ, тогда как изотипический контроль снижал жизнеспособность только до 64%. Антитела практически не влияли на жизнеспособность линии клеток NIH3T3.
Таблица 13. Уровень интернализации антител к CD63, измеренный с помощью непрямого анализа цитотоксичности в клетках T47D и NIH3T3
(нМ)
IC50
(нМ)
IC50
ND = не определено
[257] Кинетика связывания Biacore моноклональных антител к CD63, связывающихся с реагентами петли CD63 (EC2), измеренная при 25°C и 37°C
[258] Константы равновесной диссоциации (KD) для различных реагентов петли CD63, связывающихся с очищенными моноклональными антителами к CD63, определяли с применением биосенсора Biacore T200 или биосенсора Biacore 2000 на основе поверхностного плазмонного резонанса в реальном времени. Все исследования связывания проводили в подвижном буфере, состоящем из 10 мМ HEPES, 150 мМ NaCl, 3 мМ EDTA и 0,05% об./об. поверхностно-активного вещества Tween-20, pH 7,4 (HBS-ET), при 25°C и 37°C. Поверхность сенсорного чипа Biacore CM5 была сначала дериватизировали путем иммобилизации по амино-группе либо с кроличьим специфическим поликлональным антителом к Fc мыши (GE Healthcare, № по кат. BR100838), либо с набором антител к Fab человека (GE Healthcare, № по кат. 28958325) для захвата моноклональных антител к CD63. Исследования связывания проводили либо на рекомбинантной внеклеточной петле 2 CD63 человека, экспрессированной с С-концевым Myc-Myc-гексагистидином (EC-петля 2 hCD63-MMH; SEQ ID NO:80), либо на рекомбинантной внеклеточной петле 2 CD63 человека, экспрессированной с С-концевой меткой, представляющей собой Fc человека (EC-петля 2 hCD63-hFc; SEQ ID NO:81). Различные концентрации EC-петли 2 hCD63-MMH или EC-петли 2 hCD63-hFc (обе тестировали при концентрации 50нM-12,5 нМ в 4-кратном разведении, либо при концентрации 90 нМ-0,37 нМ в 3-кратных серийных разведениях) сначала получили в подвижном буфере HBS-ET и наносили на поверхность с моноклональным антителом к CD63, захваченным антителом к Fc мыши, на 4 минуты при скорости потока 35 мкл/минуту или 50 мкл/минуту, в то время как мониторинг диссоциации реагента CD63, связанного с моноклональным антителом, осуществляли в течение 8 или 10 минут в подвижном буфере HBS-ET. Скорость ассоциации (ka) и скорость диссоциации (kd) определяли путем аппроксимации сенсограмм связывания в режиме реального времени по модели связывания 1:1 с ограничением массопереноса с применением программного обеспечения для подбора кривой Scrubber 2.0c. Равновесную константу диссоциации связывания (KD) и значения периодов полудиссоциации (t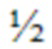 ) рассчитывали исходя из кинетических скоростей следующим образом:
) рассчитывали исходя из кинетических скоростей следующим образом:
KD (M) =  , и t½ (мин.) =
, и t½ (мин.) = 
[259] Кинетические параметры связывания для связывания белка, представляющего собой EC-петлю 2 CD63 человека, с различными моноклональными антителами к CD63 по настоящему изобретению при 25°C и 37°C показаны в таблицах 14-17.
[260] При 25°С все моноклональные антитела к CD63 по настоящему изобретению связывались с EC-петлей 2 hCD63-MMH со значениями KD в диапазоне от 530 пМ до 11,0 нМ, как показано в таблице 14. При 37°С все моноклональные антитела к CD63 по настоящему изобретению связывались с EC-петлей 2 hCD63-MMH со значениями KD в диапазоне от 1,15 нМ до 56,4 нМ, как показано в таблице 15.
[261] При 25°С все моноклональные антитела к CD63 по настоящему изобретению связывались с EC-петлей 2 hCD63-hFc со значениями KD в диапазоне от 150 пМ до 753 пМ, как показано в таблице 16. При 37°С все моноклональные антитела к CD63 по настоящему изобретению связывались с EC-петлей 2 hCD63-hFc со значениями KD в диапазоне от 119 пМ до 3,38 нМ, как показано в таблице 17.
Таблица 14. Кинетические параметры связывания для связывания EC-петли 2 hCD63-MMH человека с моноклональными антителами к CD63 при 25°C
(M-1с-1)
(1/с)
(M)
(мин.)
Таблица 15. Кинетические параметры связывания для связывания EC-петли 2 hCD63-MMH человека с моноклональными антителами к CD63 при 37°C
(M-1с-1)
(1/с)
(M)
(мин.)
Таблица 16. Кинетические параметры связывания для связывания EC-петли 2 hCD63-hFC человека с моноклональными антителами к CD63 при 25°C
(M-1с-1)
(1/с)
(M)
(мин.)
Таблица 17. Кинетические параметры связывания для связывания EC-петли 2 hCD63-hFC человека с моноклональными антителами к CD63 при 37°C
(M-1с-1)
(1/с)
(M)
(мин.)
[262] Получение биспецифических антител для определения уровня интернализации биспецифического комплекса, содержащего связывающее плечо к CD63
[263] Чтобы оценить способность антител к CD63, получаемых, как описано в этом примере, интернализоваться в виде части биспецифической антигенсвязывающей молекулы, антитела реконструировали в виде биспецифических форматов, где одно связывающее плечо было представлено парой VH/VL антитела к CD63 (см. таблица 10 родительские антитела), а другое было представлено посторонним связывающим плечом. Использовали стандартные способы получения биспецифических антител, и иллюстративные способы описаны, например, в публикации заявки США № 2010/0331527 и патенте США № US5731168, каждый из которых включен в настоящий документ посредством ссылки. Биспецифические антитела тестировали на их способность к интернализации с применением клеток, экспрессирующих CD63 человека. Для анализа клетки HEK293, которые эндогенно экспрессируют CD63 человека, высевали с плотностью 10000 клеток/лунка в DMEM, содержащем 10% FBS и пенициллин-стрептомицин/L-глутамин (Gibco, № по кат. 10378016), в черные 96-луночные планшеты с прозрачным дном, покрытые поли-D-лизином (Greiner, № по кат. 655946). Два дня спустя среду заменяли свежей средой, содержащей биспецифические антитела к CD63 и антитело отрицательного контроля в серии 2-кратных разведений, начинающихся с 10 мкг/мл до 0,157 мкг/мл, вместе с контролем, представляющим собой отдельно среду. Затем клетки инкубировали при 37°С в течение 3 часов, чтобы обеспечить интернализацию антител. После инкубации клетки промывали с помощью PBS, фиксировали в 4% параформальдегиде (Thermo Scientific, № по кат. 28908) в течение 20 минут при комнатной температуре и затем пермеабилизировали с помощью 0,2% Triton X-100 (Spectrum Chemical, № по кат. TR135) в 5% нормальной козьей сыворотке (NGS) (Gibco, № по кат. PCN5000) в течение 20 минут при комнатной температуре. Затем клетки инкубировали либо с 2 мкг/мл Fab осла к IgG мыши с Alexa Fluor-647 (производство, № по кат. 115-606-006), либо с 2 мкг/мл Fab козы к IgG человека с Alexa Fluor-647 (Jackson ImmunoResearch, № по кат. 115-606-006) в 5% NGS в течение 1 часа при комнатной температуре. Раствор вторичного антитела удаляли, затем клетки промывали с помощью PBS и затем добавляли свежий PBS, содержащий 2 капли/мл NucBlue (Invitrogen, № по кат. R37605), для окрашивания ядер живых клеток. Интернализацию антител и ядра визуализировали при увеличении 40x с помощью системы для одновременной многопараметрической визуализации ImageXpress (Molecular Devices), а количественное определение интернализации антител осуществляли с применением программного модуля MetaXpress Transfluor Application Module (Molecular Devices). Уровень интернализации антитела представлен в виде точечно интегрированной интенсивности на клетку ± стандартное отклонение (SD).
[264] Как показано в таблице 18, все биспецифические антитела, содержащее одно плечо, связывающееся с CD63 человека (полученное из H1M12451, H2M12450 или H2M12395), и постороннее несвязывающее плечо, продемонстрировали эффективную интернализацию в клетки HEK293. Биспецифическое антитело, содержащее одно связывающее плечо mAb12450, продемонстрировало более высокую степень интернализации, чем другие протестированные биспецифические антитела.
Таблица 18. Уровень интернализации биспецифических антител к CD63 клетками HEK293
би-специфическое
би-специфическое
би-
специфическое
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Регенерон Фармасьютикалс, Инк.
Байк, Эндрю
Сигнэр, Кэтрин
Шенхерр, Кристофер
Кирацус, Христос
Ванг, Ченг
<120> КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИНТЕРНАЛИЗАЦИИ ФЕРМЕНТОВ
<130> 009108.361WO1/10361WO01
<150> 62/673,098
<151> 2018-05-17
<150> 62/574,719
<151> 2017-10-19
<150> 62/516,656
<151> 2017-06-07
<160> 81
<170> PatentIn версия 3.5
<210> 1
<211> 952
<212> Белок
<213> Homo sapiens
<400> 1
Met Gly Val Arg His Pro Pro Cys Ser His Arg Leu Leu Ala Val Cys
1 5 10 15
Ala Leu Val Ser Leu Ala Thr Ala Ala Leu Leu Gly His Ile Leu Leu
20 25 30
His Asp Phe Leu Leu Val Pro Arg Glu Leu Ser Gly Ser Ser Pro Val
35 40 45
Leu Glu Glu Thr His Pro Ala His Gln Gln Gly Ala Ser Arg Pro Gly
50 55 60
Pro Arg Asp Ala Gln Ala His Pro Gly Arg Pro Arg Ala Val Pro Thr
65 70 75 80
Gln Cys Asp Val Pro Pro Asn Ser Arg Phe Asp Cys Ala Pro Asp Lys
85 90 95
Ala Ile Thr Gln Glu Gln Cys Glu Ala Arg Gly Cys Cys Tyr Ile Pro
100 105 110
Ala Lys Gln Gly Leu Gln Gly Ala Gln Met Gly Gln Pro Trp Cys Phe
115 120 125
Phe Pro Pro Ser Tyr Pro Ser Tyr Lys Leu Glu Asn Leu Ser Ser Ser
130 135 140
Glu Met Gly Tyr Thr Ala Thr Leu Thr Arg Thr Thr Pro Thr Phe Phe
145 150 155 160
Pro Lys Asp Ile Leu Thr Leu Arg Leu Asp Val Met Met Glu Thr Glu
165 170 175
Asn Arg Leu His Phe Thr Ile Lys Asp Pro Ala Asn Arg Arg Tyr Glu
180 185 190
Val Pro Leu Glu Thr Pro His Val His Ser Arg Ala Pro Ser Pro Leu
195 200 205
Tyr Ser Val Glu Phe Ser Glu Glu Pro Phe Gly Val Ile Val Arg Arg
210 215 220
Gln Leu Asp Gly Arg Val Leu Leu Asn Thr Thr Val Ala Pro Leu Phe
225 230 235 240
Phe Ala Asp Gln Phe Leu Gln Leu Ser Thr Ser Leu Pro Ser Gln Tyr
245 250 255
Ile Thr Gly Leu Ala Glu His Leu Ser Pro Leu Met Leu Ser Thr Ser
260 265 270
Trp Thr Arg Ile Thr Leu Trp Asn Arg Asp Leu Ala Pro Thr Pro Gly
275 280 285
Ala Asn Leu Tyr Gly Ser His Pro Phe Tyr Leu Ala Leu Glu Asp Gly
290 295 300
Gly Ser Ala His Gly Val Phe Leu Leu Asn Ser Asn Ala Met Asp Val
305 310 315 320
Val Leu Gln Pro Ser Pro Ala Leu Ser Trp Arg Ser Thr Gly Gly Ile
325 330 335
Leu Asp Val Tyr Ile Phe Leu Gly Pro Glu Pro Lys Ser Val Val Gln
340 345 350
Gln Tyr Leu Asp Val Val Gly Tyr Pro Phe Met Pro Pro Tyr Trp Gly
355 360 365
Leu Gly Phe His Leu Cys Arg Trp Gly Tyr Ser Ser Thr Ala Ile Thr
370 375 380
Arg Gln Val Val Glu Asn Met Thr Arg Ala His Phe Pro Leu Asp Val
385 390 395 400
Gln Trp Asn Asp Leu Asp Tyr Met Asp Ser Arg Arg Asp Phe Thr Phe
405 410 415
Asn Lys Asp Gly Phe Arg Asp Phe Pro Ala Met Val Gln Glu Leu His
420 425 430
Gln Gly Gly Arg Arg Tyr Met Met Ile Val Asp Pro Ala Ile Ser Ser
435 440 445
Ser Gly Pro Ala Gly Ser Tyr Arg Pro Tyr Asp Glu Gly Leu Arg Arg
450 455 460
Gly Val Phe Ile Thr Asn Glu Thr Gly Gln Pro Leu Ile Gly Lys Val
465 470 475 480
Trp Pro Gly Ser Thr Ala Phe Pro Asp Phe Thr Asn Pro Thr Ala Leu
485 490 495
Ala Trp Trp Glu Asp Met Val Ala Glu Phe His Asp Gln Val Pro Phe
500 505 510
Asp Gly Met Trp Ile Asp Met Asn Glu Pro Ser Asn Phe Ile Arg Gly
515 520 525
Ser Glu Asp Gly Cys Pro Asn Asn Glu Leu Glu Asn Pro Pro Tyr Val
530 535 540
Pro Gly Val Val Gly Gly Thr Leu Gln Ala Ala Thr Ile Cys Ala Ser
545 550 555 560
Ser His Gln Phe Leu Ser Thr His Tyr Asn Leu His Asn Leu Tyr Gly
565 570 575
Leu Thr Glu Ala Ile Ala Ser His Arg Ala Leu Val Lys Ala Arg Gly
580 585 590
Thr Arg Pro Phe Val Ile Ser Arg Ser Thr Phe Ala Gly His Gly Arg
595 600 605
Tyr Ala Gly His Trp Thr Gly Asp Val Trp Ser Ser Trp Glu Gln Leu
610 615 620
Ala Ser Ser Val Pro Glu Ile Leu Gln Phe Asn Leu Leu Gly Val Pro
625 630 635 640
Leu Val Gly Ala Asp Val Cys Gly Phe Leu Gly Asn Thr Ser Glu Glu
645 650 655
Leu Cys Val Arg Trp Thr Gln Leu Gly Ala Phe Tyr Pro Phe Met Arg
660 665 670
Asn His Asn Ser Leu Leu Ser Leu Pro Gln Glu Pro Tyr Ser Phe Ser
675 680 685
Glu Pro Ala Gln Gln Ala Met Arg Lys Ala Leu Thr Leu Arg Tyr Ala
690 695 700
Leu Leu Pro His Leu Tyr Thr Leu Phe His Gln Ala His Val Ala Gly
705 710 715 720
Glu Thr Val Ala Arg Pro Leu Phe Leu Glu Phe Pro Lys Asp Ser Ser
725 730 735
Thr Trp Thr Val Asp His Gln Leu Leu Trp Gly Glu Ala Leu Leu Ile
740 745 750
Thr Pro Val Leu Gln Ala Gly Lys Ala Glu Val Thr Gly Tyr Phe Pro
755 760 765
Leu Gly Thr Trp Tyr Asp Leu Gln Thr Val Pro Val Glu Ala Leu Gly
770 775 780
Ser Leu Pro Pro Pro Pro Ala Ala Pro Arg Glu Pro Ala Ile His Ser
785 790 795 800
Glu Gly Gln Trp Val Thr Leu Pro Ala Pro Leu Asp Thr Ile Asn Val
805 810 815
His Leu Arg Ala Gly Tyr Ile Ile Pro Leu Gln Gly Pro Gly Leu Thr
820 825 830
Thr Thr Glu Ser Arg Gln Gln Pro Met Ala Leu Ala Val Ala Leu Thr
835 840 845
Lys Gly Gly Glu Ala Arg Gly Glu Leu Phe Trp Asp Asp Gly Glu Ser
850 855 860
Leu Glu Val Leu Glu Arg Gly Ala Tyr Thr Gln Val Ile Phe Leu Ala
865 870 875 880
Arg Asn Asn Thr Ile Val Asn Glu Leu Val Arg Val Thr Ser Glu Gly
885 890 895
Ala Gly Leu Gln Leu Gln Lys Val Thr Val Leu Gly Val Ala Thr Ala
900 905 910
Pro Gln Gln Val Leu Ser Asn Gly Val Pro Val Ser Asn Phe Thr Tyr
915 920 925
Ser Pro Asp Thr Lys Val Leu Asp Ile Cys Val Ser Leu Leu Met Gly
930 935 940
Glu Gln Phe Leu Val Ser Trp Cys
945 950
<210> 2
<211> 245
<212> Белок
<213> Искуственная
<220>
<223> синтетическая
<400> 2
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Ser Gly Gly Ser Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Lys Gly Gln Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Arg Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Glu Asp Tyr Asp Gly Arg Leu Thr Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Ile Ser Ala Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Asp Ile Val Leu Thr Gln Ser Pro Ala Ser
130 135 140
Leu Ala Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser
145 150 155 160
Lys Ser Val Ser Thr Ser Gly Tyr Ser Tyr Met Asn Trp Tyr Gln Gln
165 170 175
Lys Pro Gly Gln Pro Pro Lys Val Leu Ile Tyr Leu Ala Ser Lys Leu
180 185 190
Glu Ser Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
195 200 205
Phe Thr Leu Asn Ile His Pro Val Glu Glu Glu Asp Ala Ala Thr Tyr
210 215 220
Tyr Cys Gln His Ser Arg Glu Leu Pro Tyr Thr Phe Gly Gly Gly Thr
225 230 235 240
Lys Leu Glu Ile Lys
245
<210> 3
<211> 21
<212> ДНК
<213> Искуственная
<220>
<223> синтетическая
<400> 3
ggaaccccta gtgatggagt t
21
<210> 4
<211> 16
<212> ДНК
<213> Искуственная
<220>
<223> синтетическая
<400> 4
cggcctcagt gagcga
16
<210> 5
<211> 21
<212> ДНК
<213> Искуственная
<220>
<223> синтетическая
<400> 5
cactccctct ctgcgcgctc g
21
<210> 6
<211> 141
<212> ДНК
<213> Искуственная
<220>
<223> синтетическая
<400> 6
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc
60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca
120
actccatcac taggggttcc t
141
<210> 7
<211> 141
<212> ДНК
<213> Искуственная
<220>
<223> синтетическая
<400> 7
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg
60
ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc
120
gagcgcgcag ctgcctgcag g
141
<210> 8
<211> 224
<212> ДНК
<213> Искуственная
<220>
<223> синтетическая
<400> 8
gtctgtctgc acatttcgta gagcgagtgt tccgatactc taatctccct aggcaaggtt
60
catatttgtg taggttactt attctccttt tgttgactaa gtcaataatc agaatcagca
120
ggtttggagt cagcttggca gggatcagca gcctgggttg gaaggagggg gtataaaagc
180
cccttcacca ggagaagccg tcacacagat ccacaagctc ctga
224
<210> 9
<211> 72
<212> ДНК
<213> Искуственная
<220>
<223> синтетическая
<400> 9
gggggaggct gctggtgaat attaaccaag gtcaccccag ttatcggagg agcaaacagg
60
ggctaagtcc ac
72
<210> 10
<211> 1133
<212> Белок
<213> Искуственная
<220>
<223> синтетическая
<400> 10
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Ser Gly Gly Ser Thr Tyr Tyr Ser Asp Thr Val
50 55 60
Lys Gly Gln Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Arg Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Glu Asp Tyr Asp Gly Arg Leu Thr Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Ile Ser Ala Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Asp Ile Val Leu Thr Gln Ser Pro Ala Ser
130 135 140
Leu Ala Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser
145 150 155 160
Lys Ser Val Ser Thr Ser Gly Tyr Ser Tyr Met Asn Trp Tyr Gln Gln
165 170 175
Lys Pro Gly Gln Pro Pro Lys Val Leu Ile Tyr Leu Ala Ser Lys Leu
180 185 190
Glu Ser Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
195 200 205
Phe Thr Leu Asn Ile His Pro Val Glu Glu Glu Asp Ala Ala Thr Tyr
210 215 220
Tyr Cys Gln His Ser Arg Glu Leu Pro Tyr Thr Phe Gly Gly Gly Thr
225 230 235 240
Lys Leu Glu Ile Lys Gly Gly Gly Gly Ser Ala His Pro Gly Arg Pro
245 250 255
Arg Ala Val Pro Thr Gln Cys Asp Val Pro Pro Asn Ser Arg Phe Asp
260 265 270
Cys Ala Pro Asp Lys Ala Ile Thr Gln Glu Gln Cys Glu Ala Arg Gly
275 280 285
Cys Cys Tyr Ile Pro Ala Lys Gln Gly Leu Gln Gly Ala Gln Met Gly
290 295 300
Gln Pro Trp Cys Phe Phe Pro Pro Ser Tyr Pro Ser Tyr Lys Leu Glu
305 310 315 320
Asn Leu Ser Ser Ser Glu Met Gly Tyr Thr Ala Thr Leu Thr Arg Thr
325 330 335
Thr Pro Thr Phe Phe Pro Lys Asp Ile Leu Thr Leu Arg Leu Asp Val
340 345 350
Met Met Glu Thr Glu Asn Arg Leu His Phe Thr Ile Lys Asp Pro Ala
355 360 365
Asn Arg Arg Tyr Glu Val Pro Leu Glu Thr Pro His Val His Ser Arg
370 375 380
Ala Pro Ser Pro Leu Tyr Ser Val Glu Phe Ser Glu Glu Pro Phe Gly
385 390 395 400
Val Ile Val Arg Arg Gln Leu Asp Gly Arg Val Leu Leu Asn Thr Thr
405 410 415
Val Ala Pro Leu Phe Phe Ala Asp Gln Phe Leu Gln Leu Ser Thr Ser
420 425 430
Leu Pro Ser Gln Tyr Ile Thr Gly Leu Ala Glu His Leu Ser Pro Leu
435 440 445
Met Leu Ser Thr Ser Trp Thr Arg Ile Thr Leu Trp Asn Arg Asp Leu
450 455 460
Ala Pro Thr Pro Gly Ala Asn Leu Tyr Gly Ser His Pro Phe Tyr Leu
465 470 475 480
Ala Leu Glu Asp Gly Gly Ser Ala His Gly Val Phe Leu Leu Asn Ser
485 490 495
Asn Ala Met Asp Val Val Leu Gln Pro Ser Pro Ala Leu Ser Trp Arg
500 505 510
Ser Thr Gly Gly Ile Leu Asp Val Tyr Ile Phe Leu Gly Pro Glu Pro
515 520 525
Lys Ser Val Val Gln Gln Tyr Leu Asp Val Val Gly Tyr Pro Phe Met
530 535 540
Pro Pro Tyr Trp Gly Leu Gly Phe His Leu Cys Arg Trp Gly Tyr Ser
545 550 555 560
Ser Thr Ala Ile Thr Arg Gln Val Val Glu Asn Met Thr Arg Ala His
565 570 575
Phe Pro Leu Asp Val Gln Trp Asn Asp Leu Asp Tyr Met Asp Ser Arg
580 585 590
Arg Asp Phe Thr Phe Asn Lys Asp Gly Phe Arg Asp Phe Pro Ala Met
595 600 605
Val Gln Glu Leu His Gln Gly Gly Arg Arg Tyr Met Met Ile Val Asp
610 615 620
Pro Ala Ile Ser Ser Ser Gly Pro Ala Gly Ser Tyr Arg Pro Tyr Asp
625 630 635 640
Glu Gly Leu Arg Arg Gly Val Phe Ile Thr Asn Glu Thr Gly Gln Pro
645 650 655
Leu Ile Gly Lys Val Trp Pro Gly Ser Thr Ala Phe Pro Asp Phe Thr
660 665 670
Asn Pro Thr Ala Leu Ala Trp Trp Glu Asp Met Val Ala Glu Phe His
675 680 685
Asp Gln Val Pro Phe Asp Gly Met Trp Ile Asp Met Asn Glu Pro Ser
690 695 700
Asn Phe Ile Arg Gly Ser Glu Asp Gly Cys Pro Asn Asn Glu Leu Glu
705 710 715 720
Asn Pro Pro Tyr Val Pro Gly Val Val Gly Gly Thr Leu Gln Ala Ala
725 730 735
Thr Ile Cys Ala Ser Ser His Gln Phe Leu Ser Thr His Tyr Asn Leu
740 745 750
His Asn Leu Tyr Gly Leu Thr Glu Ala Ile Ala Ser His Arg Ala Leu
755 760 765
Val Lys Ala Arg Gly Thr Arg Pro Phe Val Ile Ser Arg Ser Thr Phe
770 775 780
Ala Gly His Gly Arg Tyr Ala Gly His Trp Thr Gly Asp Val Trp Ser
785 790 795 800
Ser Trp Glu Gln Leu Ala Ser Ser Val Pro Glu Ile Leu Gln Phe Asn
805 810 815
Leu Leu Gly Val Pro Leu Val Gly Ala Asp Val Cys Gly Phe Leu Gly
820 825 830
Asn Thr Ser Glu Glu Leu Cys Val Arg Trp Thr Gln Leu Gly Ala Phe
835 840 845
Tyr Pro Phe Met Arg Asn His Asn Ser Leu Leu Ser Leu Pro Gln Glu
850 855 860
Pro Tyr Ser Phe Ser Glu Pro Ala Gln Gln Ala Met Arg Lys Ala Leu
865 870 875 880
Thr Leu Arg Tyr Ala Leu Leu Pro His Leu Tyr Thr Leu Phe His Gln
885 890 895
Ala His Val Ala Gly Glu Thr Val Ala Arg Pro Leu Phe Leu Glu Phe
900 905 910
Pro Lys Asp Ser Ser Thr Trp Thr Val Asp His Gln Leu Leu Trp Gly
915 920 925
Glu Ala Leu Leu Ile Thr Pro Val Leu Gln Ala Gly Lys Ala Glu Val
930 935 940
Thr Gly Tyr Phe Pro Leu Gly Thr Trp Tyr Asp Leu Gln Thr Val Pro
945 950 955 960
Val Glu Ala Leu Gly Ser Leu Pro Pro Pro Pro Ala Ala Pro Arg Glu
965 970 975
Pro Ala Ile His Ser Glu Gly Gln Trp Val Thr Leu Pro Ala Pro Leu
980 985 990
Asp Thr Ile Asn Val His Leu Arg Ala Gly Tyr Ile Ile Pro Leu Gln
995 1000 1005
Gly Pro Gly Leu Thr Thr Thr Glu Ser Arg Gln Gln Pro Met Ala
1010 1015 1020
Leu Ala Val Ala Leu Thr Lys Gly Gly Glu Ala Arg Gly Glu Leu
1025 1030 1035
Phe Trp Asp Asp Gly Glu Ser Leu Glu Val Leu Glu Arg Gly Ala
1040 1045 1050
Tyr Thr Gln Val Ile Phe Leu Ala Arg Asn Asn Thr Ile Val Asn
1055 1060 1065
Glu Leu Val Arg Val Thr Ser Glu Gly Ala Gly Leu Gln Leu Gln
1070 1075 1080
Lys Val Thr Val Leu Gly Val Ala Thr Ala Pro Gln Gln Val Leu
1085 1090 1095
Ser Asn Gly Val Pro Val Ser Asn Phe Thr Tyr Ser Pro Asp Thr
1100 1105 1110
Lys Val Leu Asp Ile Cys Val Ser Leu Leu Met Gly Glu Gln Phe
1115 1120 1125
Leu Val Ser Trp Cys
1130
<210> 11
<211> 3489
<212> ДНК
<213> Искуственная
<220>
<223> синтетическая
<400> 11
atgcacagac ctagacgtcg tggaactcgt ccacctccac tggcactgct cgctgctctc
60
ctcctggctg cacgtggtgc tgatgcagaa gtgaagctgg tggagtctgg gggaggctta
120
gtgcagcctg gagggtccct gaaactctcc tgtgcaacct ctggattcac tttcagtgac
180
tattacatgt cttgggttcg ccagactcca gagaagaggc tggagtgggt cgcatatatt
240
agtagtagtg gtggtagcac ctattattca gacactgtaa agggccaatt caccatctcc
300
agagacaatg ccaagaacac cctgtacctg caaatgagcc gtctgaagtc tgaggacaca
360
gccatgtatt actgtgcaag acgagaagat tacgacggaa gacttactta ctggggccaa
420
gggactctgg tcaccatctc tgcaggagga agtggtggag gcgggtcagg aggtggcggg
480
agcggcggtg acattgtgct gacacagtct cctgcttcct tagctgtatc tctggggcag
540
agggccacca tctcctgcag ggccagcaaa agtgtcagta catctggtta tagttatatg
600
aactggtacc aacagaaacc aggacagcca cccaaagtcc tcatctatct tgcatccaaa
660
ctagaatctg gggtccctgc caggttcagt ggcagtgggt cagggacaga cttcaccctc
720
aacatccatc ctgtggagga ggaggatgct gcaacctatt actgtcagca cagtagggag
780
cttccgtaca cgttcggagg ggggaccaaa ctggaaataa aaggtggtgg cggttcagca
840
caccccggcc gtcccagagc agtgcccaca cagtgcgacg tcccccccaa cagccgcttc
900
gattgcgccc ctgacaaggc catcacccag gaacagtgcg aggcccgcgg ctgttgctac
960
atccctgcaa agcaggggct gcagggagcc cagatggggc agccctggtg cttcttccca
1020
cccagctacc ccagctacaa gctggagaac ctgagctcct ctgaaatggg ctacacggcc
1080
accctgaccc gtaccacccc caccttcttc cccaaggaca tcctgaccct gcggctggac
1140
gtgatgatgg agactgagaa ccgcctccac ttcacgatca aagatccagc taacaggcgc
1200
tacgaggtgc ccttggagac cccgcatgtc cacagccggg caccgtcccc actctacagc
1260
gtggagttct ccgaggagcc cttcggggtg atcgtgcgcc ggcagctgga cggccgcgtg
1320
ctgctgaaca cgacggtggc gcccctgttc tttgcggacc agttccttca gctgtccacc
1380
tcgctgccct cgcagtatat cacaggcctc gccgagcacc tcagtcccct gatgctcagc
1440
accagctgga ccaggatcac cctgtggaac cgggaccttg cgcccacgcc cggtgcgaac
1500
ctctacgggt ctcacccttt ctacctggcg ctggaggacg gcgggtcggc acacggggtg
1560
ttcctgctaa acagcaatgc catggatgtg gtcctgcagc cgagccctgc ccttagctgg
1620
aggtcgacag gtgggatcct ggatgtctac atcttcctgg gcccagagcc caagagcgtg
1680
gtgcagcagt acctggacgt tgtgggatac ccgttcatgc cgccatactg gggcctgggc
1740
ttccacctgt gccgctgggg ctactcctcc accgctatca cccgccaggt ggtggagaac
1800
atgaccaggg cccacttccc cctggacgtc cagtggaacg acctggacta catggactcc
1860
cggagggact tcacgttcaa caaggatggc ttccgggact tcccggccat ggtgcaggag
1920
ctgcaccagg gcggccggcg ctacatgatg atcgtggatc ctgccatcag cagctcgggc
1980
cctgccggga gctacaggcc ctacgacgag ggtctgcgga ggggggtttt catcaccaac
2040
gagaccggcc agccgctgat tgggaaggta tggcccgggt ccactgcctt ccccgacttc
2100
accaacccca cagccctggc ctggtgggag gacatggtgg ctgagttcca tgaccaggtg
2160
cccttcgacg gcatgtggat tgacatgaac gagccttcca acttcatcag gggctctgag
2220
gacggctgcc ccaacaatga gctggagaac ccaccctacg tgcctggggt ggttgggggg
2280
accctccagg cggccaccat ctgtgcctcc agccaccagt ttctctccac acactacaac
2340
ctgcacaacc tctacggcct gaccgaagcc atcgcctccc acagggcgct ggtgaaggct
2400
cgggggacac gcccatttgt gatctcccgc tcgacctttg ctggccacgg ccgatacgcc
2460
ggccactgga cgggggacgt gtggagctcc tgggagcagc tcgcctcctc cgtgccagaa
2520
atcctgcagt ttaacctgct gggggtgcct ctggtcgggg ccgacgtctg cggcttcctg
2580
ggcaacacct cagaggagct gtgtgtgcgc tggacccagc tgggggcctt ctaccccttc
2640
atgcggaacc acaacagcct gctcagtctg ccccaggagc cgtacagctt cagcgagccg
2700
gcccagcagg ccatgaggaa ggccctcacc ctgcgctacg cactcctccc ccacctctac
2760
acactgttcc accaggccca cgtcgcgggg gagaccgtgg cccggcccct cttcctggag
2820
ttccccaagg actctagcac ctggactgtg gaccaccagc tcctgtgggg ggaggccctg
2880
ctcatcaccc cagtgctcca ggccgggaag gccgaagtga ctggctactt ccccttgggc
2940
acatggtacg acctgcagac ggtgccagta gaggcccttg gcagcctccc acccccacct
3000
gcagctcccc gtgagccagc catccacagc gaggggcagt gggtgacgct gccggccccc
3060
ctggacacca tcaacgtcca cctccgggct gggtacatca tccccctgca gggccctggc
3120
ctcacaacca cagagtcccg ccagcagccc atggccctgg ctgtggccct gaccaagggt
3180
ggggaggccc gaggggagct gttctgggac gatggagaga gcctggaagt gctggagcga
3240
ggggcctaca cacaggtcat cttcctggcc aggaataaca cgatcgtgaa tgagctggta
3300
cgtgtgacca gtgagggagc tggcctgcag ctgcagaagg tgactgtcct gggcgtggcc
3360
acggcgcccc agcaggtcct ctccaacggt gtccctgtct ccaacttcac ctacagcccc
3420
gacaccaagg tcctggacat ctgtgtctcg ctgttgatgg gagagcagtt tctcgtcagc
3480
tggtgttag
3489
<210> 12
<211> 2859
<212> ДНК
<213> Homo sapiens
<400> 12
atgggagtga ggcacccgcc ctgctcccac cggctcctgg ccgtctgcgc cctcgtgtcc
60
ttggcaaccg ctgcactcct ggggcacatc ctactccatg atttcctgct ggttccccga
120
gagctgagtg gctcctcccc agtcctggag gagactcacc cagctcacca gcagggagcc
180
agcagaccag ggccccggga tgcccaggca caccccggcc gtcccagagc agtgcccaca
240
cagtgcgacg tcccccccaa cagccgcttc gattgcgccc ctgacaaggc catcacccag
300
gaacagtgcg aggcccgcgg ctgttgctac atccctgcaa agcaggggct gcagggagcc
360
cagatggggc agccctggtg cttcttccca cccagctacc ccagctacaa gctggagaac
420
ctgagctcct ctgaaatggg ctacacggcc accctgaccc gtaccacccc caccttcttc
480
cccaaggaca tcctgaccct gcggctggac gtgatgatgg agactgagaa ccgcctccac
540
ttcacgatca aagatccagc taacaggcgc tacgaggtgc ccttggagac cccgcatgtc
600
cacagccggg caccgtcccc actctacagc gtggagttct ccgaggagcc cttcggggtg
660
atcgtgcgcc ggcagctgga cggccgcgtg ctgctgaaca cgacggtggc gcccctgttc
720
tttgcggacc agttccttca gctgtccacc tcgctgccct cgcagtatat cacaggcctc
780
gccgagcacc tcagtcccct gatgctcagc accagctgga ccaggatcac cctgtggaac
840
cgggaccttg cgcccacgcc cggtgcgaac ctctacgggt ctcacccttt ctacctggcg
900
ctggaggacg gcgggtcggc acacggggtg ttcctgctaa acagcaatgc catggatgtg
960
gtcctgcagc cgagccctgc ccttagctgg aggtcgacag gtgggatcct ggatgtctac
1020
atcttcctgg gcccagagcc caagagcgtg gtgcagcagt acctggacgt tgtgggatac
1080
ccgttcatgc cgccatactg gggcctgggc ttccacctgt gccgctgggg ctactcctcc
1140
accgctatca cccgccaggt ggtggagaac atgaccaggg cccacttccc cctggacgtc
1200
cagtggaacg acctggacta catggactcc cggagggact tcacgttcaa caaggatggc
1260
ttccgggact tcccggccat ggtgcaggag ctgcaccagg gcggccggcg ctacatgatg
1320
atcgtggatc ctgccatcag cagctcgggc cctgccggga gctacaggcc ctacgacgag
1380
ggtctgcgga ggggggtttt catcaccaac gagaccggcc agccgctgat tgggaaggta
1440
tggcccgggt ccactgcctt ccccgacttc accaacccca cagccctggc ctggtgggag
1500
gacatggtgg ctgagttcca tgaccaggtg cccttcgacg gcatgtggat tgacatgaac
1560
gagccttcca acttcatcag gggctctgag gacggctgcc ccaacaatga gctggagaac
1620
ccaccctacg tgcctggggt ggttgggggg accctccagg cggccaccat ctgtgcctcc
1680
agccaccagt ttctctccac acactacaac ctgcacaacc tctacggcct gaccgaagcc
1740
atcgcctccc acagggcgct ggtgaaggct cgggggacac gcccatttgt gatctcccgc
1800
tcgacctttg ctggccacgg ccgatacgcc ggccactgga cgggggacgt gtggagctcc
1860
tgggagcagc tcgcctcctc cgtgccagaa atcctgcagt ttaacctgct gggggtgcct
1920
ctggtcgggg ccgacgtctg cggcttcctg ggcaacacct cagaggagct gtgtgtgcgc
1980
tggacccagc tgggggcctt ctaccccttc atgcggaacc acaacagcct gctcagtctg
2040
ccccaggagc cgtacagctt cagcgagccg gcccagcagg ccatgaggaa ggccctcacc
2100
ctgcgctacg cactcctccc ccacctctac acactgttcc accaggccca cgtcgcgggg
2160
gagaccgtgg cccggcccct cttcctggag ttccccaagg actctagcac ctggactgtg
2220
gaccaccagc tcctgtgggg ggaggccctg ctcatcaccc cagtgctcca ggccgggaag
2280
gccgaagtga ctggctactt ccccttgggc acatggtacg acctgcagac ggtgccagta
2340
gaggcccttg gcagcctccc acccccacct gcagctcccc gtgagccagc catccacagc
2400
gaggggcagt gggtgacgct gccggccccc ctggacacca tcaacgtcca cctccgggct
2460
gggtacatca tccccctgca gggccctggc ctcacaacca cagagtcccg ccagcagccc
2520
atggccctgg ctgtggccct gaccaagggt ggggaggccc gaggggagct gttctgggac
2580
gatggagaga gcctggaagt gctggagcga ggggcctaca cacaggtcat cttcctggcc
2640
aggaataaca cgatcgtgaa tgagctggta cgtgtgacca gtgagggagc tggcctgcag
2700
ctgcagaagg tgactgtcct gggcgtggcc acggcgcccc agcaggtcct ctccaacggt
2760
gtccctgtct ccaacttcac ctacagcccc gacaccaagg tcctggacat ctgtgtctcg
2820
ctgttgatgg gagagcagtt tctcgtcagc tggtgttag
2859
<210> 13
<211> 354
<212> ДНК
<213> Homo sapiens
<400> 13
caggtgcagc tgcaggagtc gggccccgga ctgatgaagc cttcggagac cctgtccctc
60
acctgcactg tctctggtgg ctccttcagc agttactatt ggaactggat ccggcagtcc
120
ccagggaagg gactggagtg gattgggtat atccgttata gtggggacac caactacaag
180
ccctccctca agagtcgatt caccatatca attgacacgt ccaagaacct tttctccctg
240
aggctgaaat ctgtgaccgc tgcggacacg gccgtgtatt actgtgcgag gatgggactg
300
gggagtgatg cttttgatat ctggggccaa gggacaatgg tcaccgtctc ttca
354
<210> 14
<211> 118
<212> Белок
<213> Homo sapiens
<400> 14
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Met Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Phe Ser Ser Tyr
20 25 30
Tyr Trp Asn Trp Ile Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Arg Tyr Ser Gly Asp Thr Asn Tyr Lys Pro Ser Leu Lys
50 55 60
Ser Arg Phe Thr Ile Ser Ile Asp Thr Ser Lys Asn Leu Phe Ser Leu
65 70 75 80
Arg Leu Lys Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Met Gly Leu Gly Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser
115
<210> 15
<211> 24
<212> ДНК
<213> Homo sapiens
<400> 15
ggtggctcct tcagcagtta ctat
24
<210> 16
<211> 8
<212> Белок
<213> Homo sapiens
<400> 16
Gly Gly Ser Phe Ser Ser Tyr Tyr
1 5
<210> 17
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 17
atccgttata gtggggacac c
21
<210> 18
<211> 7
<212> Белок
<213> Homo sapiens
<400> 18
Ile Arg Tyr Ser Gly Asp Thr
1 5
<210> 19
<211> 36
<212> ДНК
<213> Homo sapiens
<400> 19
gcgaggatgg gactggggag tgatgctttt gatatc
36
<210> 20
<211> 12
<212> Белок
<213> Homo sapiens
<400> 20
Ala Arg Met Gly Leu Gly Ser Asp Ala Phe Asp Ile
1 5 10
<210> 21
<211> 324
<212> ДНК
<213> Homo sapiens
<400> 21
gaaattgtgt tgacgcagtc tccaggcacc ctgtctttgt ctccagggga aagagccacc
60
ctctcctgca gggccagtca gagtgttaac aacaattatt tagcctggta ccagcagaaa
120
cctggccagg ctcccaggct cctcatctat ggtgtattca acagggccac taacatccca
180
gacaggttca gtggcagtgg gtctgggaca gatttcactc tcaccatcag cagactggag
240
cctgaagatt ttgcagtgta ttactgtcag cagtatggta gttcaccttg gacgttcggc
300
caagggacca aggtggaaat caaa
324
<210> 22
<211> 108
<212> Белок
<213> Homo sapiens
<400> 22
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Asn Asn
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Val Phe Asn Arg Ala Thr Asn Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 23
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 23
cagagtgtta acaacaatta t
21
<210> 24
<211> 7
<212> Белок
<213> Homo sapiens
<400> 24
Gln Ser Val Asn Asn Asn Tyr
1 5
<210> 25
<211> 9
<212> ДНК
<213> Homo sapiens
<400> 25
ggtgtattc
9
<210> 26
<211> 3
<212> Белок
<213> Homo sapiens
<400> 26
Gly Val Phe
1
<210> 27
<211> 27
<212> ДНК
<213> Homo sapiens
<400> 27
cagcagtatg gtagttcacc ttggacg
27
<210> 28
<211> 9
<212> Белок
<213> Homo sapiens
<400> 28
Gln Gln Tyr Gly Ser Ser Pro Trp Thr
1 5
<210> 29
<211> 354
<212> ДНК
<213> Homo sapiens
<400> 29
caggtgcagc tgcaggagtc gggcccaaga ctggtgaagc cttcggagac cctgtccctc
60
acctgcattg tctctggtgg ctccatcagt aatttctact ggaactggat ccggcagtcc
120
ccagggaagg gactggaatg gattggatat ttcttttaca ctgggactat cgactacaac
180
ccctccctca agagtcgagt caccatatca ctggacacgt ccaagaacca gttctccctg
240
aacctgcgtc ttctgaccgc cgcagacgcg gccgtttatt attgtgcgag gatggggctg
300
ggggctaatg cttttgacat ctggggccac gggacaatgg tcaccgtctc ttca
354
<210> 30
<211> 118
<212> Белок
<213> Homo sapiens
<400> 30
Gln Val Gln Leu Gln Glu Ser Gly Pro Arg Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ile Val Ser Gly Gly Ser Ile Ser Asn Phe
20 25 30
Tyr Trp Asn Trp Ile Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Phe Phe Tyr Thr Gly Thr Ile Asp Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Leu Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Asn Leu Arg Leu Leu Thr Ala Ala Asp Ala Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Met Gly Leu Gly Ala Asn Ala Phe Asp Ile Trp Gly His Gly Thr
100 105 110
Met Val Thr Val Ser Ser
115
<210> 31
<211> 24
<212> ДНК
<213> Homo sapiens
<400> 31
ggtggctcca tcagtaattt ctac
24
<210> 32
<211> 8
<212> Белок
<213> Homo sapiens
<400> 32
Gly Gly Ser Ile Ser Asn Phe Tyr
1 5
<210> 33
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 33
ttcttttaca ctgggactat c
21
<210> 34
<211> 7
<212> Белок
<213> Homo sapiens
<400> 34
Phe Phe Tyr Thr Gly Thr Ile
1 5
<210> 35
<211> 36
<212> ДНК
<213> Homo sapiens
<400> 35
gcgaggatgg ggctgggggc taatgctttt gacatc
36
<210> 36
<211> 12
<212> Белок
<213> Homo sapiens
<400> 36
Ala Arg Met Gly Leu Gly Ala Asn Ala Phe Asp Ile
1 5 10
<210> 37
<211> 324
<212> ДНК
<213> Homo sapiens
<400> 37
gaaattgtgt tgacgcagtc tccaggcacc ctgtctttgt ctcctggaga aagagccacc
60
ctctcctgca gggccagtca gcatgttagc agcaactact tagcctggta ccagcagaaa
120
cctggccagg ctcccaggct cctcatctat ggtggatcca gcagggccac tggcatccca
180
gacaggttca gtggcagtgg gtctgggaca gacttcactc tcaccatcag cagactggag
240
cctgcagatt ttgcagtgtt ttactgtcag cagtatggta actcaccttg gacgttcggc
300
caagggacca aggtggaaat gaaa
324
<210> 38
<211> 108
<212> Белок
<213> Homo sapiens
<400> 38
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln His Val Ser Ser Asn
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Gly Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Ala Asp Phe Ala Val Phe Tyr Cys Gln Gln Tyr Gly Asn Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Met Lys
100 105
<210> 39
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 39
cagcatgtta gcagcaacta c
21
<210> 40
<211> 7
<212> Белок
<213> Homo sapiens
<400> 40
Gln His Val Ser Ser Asn Tyr
1 5
<210> 41
<211> 9
<212> ДНК
<213> Homo sapiens
<400> 41
ggtggatcc
9
<210> 42
<211> 3
<212> Белок
<213> Homo sapiens
<400> 42
Gly Gly Ser
1
<210> 43
<211> 27
<212> ДНК
<213> Homo sapiens
<400> 43
cagcagtatg gtaactcacc ttggacg
27
<210> 44
<211> 9
<212> Белок
<213> Homo sapiens
<400> 44
Gln Gln Tyr Gly Asn Ser Pro Trp Thr
1 5
<210> 45
<211> 351
<212> ДНК
<213> Homo sapiens
<400> 45
caggtgcaac tacaggagtc gggcccaaag gtggtgaagc cttcggagac cctgtccctc
60
acctgcactg tctctggtgg ctccatcagt agttactact ggaattggat ccgccagtcc
120
ccagggaagg gactggagtg gattggatat accaaaagag ggtataccga ctacaacccc
180
tccctcagga gtcgcgtcac tatatcagaa gacacgtcca agaaccagtt ctccctgagg
240
atcagctctg tgaccgccgc agacacggcc gtatattact gtgcacaaat ggggtgggga
300
tcccatgctt ttgacatgtg gggccaaggg acaatggtcg ccgtctcttc a
351
<210> 46
<211> 117
<212> Белок
<213> Homo sapiens
<400> 46
Gln Val Gln Leu Gln Glu Ser Gly Pro Lys Val Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Tyr
20 25 30
Tyr Trp Asn Trp Ile Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Thr Lys Arg Gly Tyr Thr Asp Tyr Asn Pro Ser Leu Arg Ser
50 55 60
Arg Val Thr Ile Ser Glu Asp Thr Ser Lys Asn Gln Phe Ser Leu Arg
65 70 75 80
Ile Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala Gln
85 90 95
Met Gly Trp Gly Ser His Ala Phe Asp Met Trp Gly Gln Gly Thr Met
100 105 110
Val Ala Val Ser Ser
115
<210> 47
<211> 24
<212> ДНК
<213> Homo sapiens
<400> 47
ggtggctcca tcagtagtta ctac
24
<210> 48
<211> 8
<212> Белок
<213> Homo sapiens
<400> 48
Gly Gly Ser Ile Ser Ser Tyr Tyr
1 5
<210> 49
<211> 18
<212> ДНК
<213> Homo sapiens
<400> 49
accaaaagag ggtatacc
18
<210> 50
<211> 6
<212> Белок
<213> Homo sapiens
<400> 50
Thr Lys Arg Gly Tyr Thr
1 5
<210> 51
<211> 36
<212> ДНК
<213> Homo sapiens
<400> 51
gcacaaatgg ggtggggatc ccatgctttt gacatg
36
<210> 52
<211> 12
<212> Белок
<213> Homo sapiens
<400> 52
Ala Gln Met Gly Trp Gly Ser His Ala Phe Asp Met
1 5 10
<210> 53
<211> 324
<212> ДНК
<213> Homo sapiens
<400> 53
gaaattgtgt tgacgcagtc tccaggcacc ctgtctttgt ctccagggga aagagccacc
60
ctctcctgca gggccagtca gagtgttagc agcagctact tagcctggta ccagcagaaa
120
cctggccagg ctcccaggct cctcatctat ggtgcatcca gcagggccac tggcatccca
180
gacaggttca gtggcagtgg gtctgggaca gacttcactc tcaccatcag cagactggag
240
cctgaagatt ttgcagtgta ttactgtcag cagtatggta gctcaccttg gacgttcggc
300
caagggacca aggtggaaat caaa
324
<210> 54
<211> 108
<212> Белок
<213> Homo sapiens
<400> 54
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 55
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 55
cagagtgtta gcagcagcta c
21
<210> 56
<211> 7
<212> Белок
<213> Homo sapiens
<400> 56
Gln Ser Val Ser Ser Ser Tyr
1 5
<210> 57
<211> 9
<212> ДНК
<213> Homo sapiens
<400> 57
ggtgcatcc
9
<210> 58
<211> 3
<212> Белок
<213> Homo sapiens
<400> 58
Gly Ala Ser
1
<210> 59
<211> 27
<212> ДНК
<213> Homo sapiens
<400> 59
cagcagtatg gtagctcacc ttggacg
27
<210> 60
<211> 9
<212> Белок
<213> Homo sapiens
<400> 60
Gln Gln Tyr Gly Ser Ser Pro Trp Thr
1 5
<210> 61
<211> 351
<212> ДНК
<213> Homo sapiens
<400> 61
caggtgcaac tacaggagtc gggcccaaag gtggtgaagc cttcggagac cctgtccctc
60
acctgcactg tctctggtgg ctccatcagt agttactact ggaattggat ccgccagtcc
120
ccagggaagg gactggagtg gattggatat accaaaagag ggtataccga ctacaacccc
180
tccctcagga gtcgcgtcac tatatcagaa gacacgtcca agaaccagtt ctccctgagg
240
atcagctctg tgaccgccgc agacacggcc gtatattact gtgcacaaat ggggtgggga
300
tcccatgctt ttgacatgtg gggccaaggg acaatggtcg ccgtctcttc a
351
<210> 62
<211> 117
<212> Белок
<213> Homo sapiens
<400> 62
Gln Val Gln Leu Gln Glu Ser Gly Pro Lys Val Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Tyr
20 25 30
Tyr Trp Asn Trp Ile Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Thr Lys Arg Gly Tyr Thr Asp Tyr Asn Pro Ser Leu Arg Ser
50 55 60
Arg Val Thr Ile Ser Glu Asp Thr Ser Lys Asn Gln Phe Ser Leu Arg
65 70 75 80
Ile Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala Gln
85 90 95
Met Gly Trp Gly Ser His Ala Phe Asp Met Trp Gly Gln Gly Thr Met
100 105 110
Val Ala Val Ser Ser
115
<210> 63
<211> 24
<212> ДНК
<213> Homo sapiens
<400> 63
ggtggctcca tcagtagtta ctac
24
<210> 64
<211> 8
<212> Белок
<213> Homo sapiens
<400> 64
Gly Gly Ser Ile Ser Ser Tyr Tyr
1 5
<210> 65
<211> 18
<212> ДНК
<213> Homo sapiens
<400> 65
accaaaagag ggtatacc
18
<210> 66
<211> 6
<212> Белок
<213> Homo sapiens
<400> 66
Thr Lys Arg Gly Tyr Thr
1 5
<210> 67
<211> 36
<212> ДНК
<213> Homo sapiens
<400> 67
gcacaaatgg ggtggggatc ccatgctttt gacatg
36
<210> 68
<211> 12
<212> Белок
<213> Homo sapiens
<400> 68
Ala Gln Met Gly Trp Gly Ser His Ala Phe Asp Met
1 5 10
<210> 69
<211> 324
<212> ДНК
<213> Homo sapiens
<400> 69
gaaattgtgt tgacgcagtc tccaggcacc ctgtctttgt ctccagggga aagagccacc
60
ctctcctgca gggccagtca gagtgttaat agtaggtact tagcctggta ccagcagaaa
120
cctggccagg ctcccaggct cctcatctat ggtgcatcca gcagggccac tggcatccca
180
gacaggtgca gtggcagtgg gtccgggaca gacttcactc tcaccatcag cagactggag
240
cctgaagatt ttgcagtgta ttactgtcag cagtatggta gctcaccttg gacgttcggc
300
caggggacca aggtggaaat caaa
324
<210> 70
<211> 108
<212> Белок
<213> Homo sapiens
<400> 70
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Arg
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Cys Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 71
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 71
cagagtgtta atagtaggta c
21
<210> 72
<211> 7
<212> Белок
<213> Homo sapiens
<400> 72
Gln Ser Val Asn Ser Arg Tyr
1 5
<210> 73
<211> 9
<212> ДНК
<213> Homo sapiens
<400> 73
ggtgcatcc
9
<210> 74
<211> 3
<212> Белок
<213> Homo sapiens
<400> 74
Gly Ala Ser
1
<210> 75
<211> 27
<212> ДНК
<213> Homo sapiens
<400> 75
cagcagtatg gtagctcacc ttggacg
27
<210> 76
<211> 9
<212> Белок
<213> Homo sapiens
<400> 76
Gln Gln Tyr Gly Ser Ser Pro Trp Thr
1 5
<210> 77
<211> 238
<212> Белок
<213> Homo sapiens
<400> 77
Met Ala Val Glu Gly Gly Met Lys Cys Val Lys Phe Leu Leu Tyr Val
1 5 10 15
Leu Leu Leu Ala Phe Cys Ala Cys Ala Val Gly Leu Ile Ala Val Gly
20 25 30
Val Gly Ala Gln Leu Val Leu Ser Gln Thr Ile Ile Gln Gly Ala Thr
35 40 45
Pro Gly Ser Leu Leu Pro Val Val Ile Ile Ala Val Gly Val Phe Leu
50 55 60
Phe Leu Val Ala Phe Val Gly Cys Cys Gly Ala Cys Lys Glu Asn Tyr
65 70 75 80
Cys Leu Met Ile Thr Phe Ala Ile Phe Leu Ser Leu Ile Met Leu Val
85 90 95
Glu Val Ala Ala Ala Ile Ala Gly Tyr Val Phe Arg Asp Lys Val Met
100 105 110
Ser Glu Phe Asn Asn Asn Phe Arg Gln Gln Met Glu Asn Tyr Pro Lys
115 120 125
Asn Asn His Thr Ala Ser Ile Leu Asp Arg Met Gln Ala Asp Phe Lys
130 135 140
Cys Cys Gly Ala Ala Asn Tyr Thr Asp Trp Glu Lys Ile Pro Ser Met
145 150 155 160
Ser Lys Asn Arg Val Pro Asp Ser Cys Cys Ile Asn Val Thr Val Gly
165 170 175
Cys Gly Ile Asn Phe Asn Glu Lys Ala Ile His Lys Glu Gly Cys Val
180 185 190
Glu Lys Ile Gly Gly Trp Leu Arg Lys Asn Val Leu Val Val Ala Ala
195 200 205
Ala Ala Leu Gly Ile Ala Phe Val Glu Val Leu Gly Ile Val Phe Ala
210 215 220
Cys Cys Leu Val Lys Ser Ile Arg Ser Gly Tyr Glu Val Met
225 230 235
<210> 78
<211> 917
<212> Белок
<213> Homo sapiens
<400> 78
Leu Leu Val Pro Arg Glu Leu Ser Gly Ser Ser Pro Val Leu Glu Glu
1 5 10 15
Thr His Pro Ala His Gln Gln Gly Ala Ser Arg Pro Gly Pro Arg Asp
20 25 30
Ala Gln Ala His Pro Gly Arg Pro Arg Ala Val Pro Thr Gln Cys Asp
35 40 45
Val Pro Pro Asn Ser Arg Phe Asp Cys Ala Pro Asp Lys Ala Ile Thr
50 55 60
Gln Glu Gln Cys Glu Ala Arg Gly Cys Cys Tyr Ile Pro Ala Lys Gln
65 70 75 80
Gly Leu Gln Gly Ala Gln Met Gly Gln Pro Trp Cys Phe Phe Pro Pro
85 90 95
Ser Tyr Pro Ser Tyr Lys Leu Glu Asn Leu Ser Ser Ser Glu Met Gly
100 105 110
Tyr Thr Ala Thr Leu Thr Arg Thr Thr Pro Thr Phe Phe Pro Lys Asp
115 120 125
Ile Leu Thr Leu Arg Leu Asp Val Met Met Glu Thr Glu Asn Arg Leu
130 135 140
His Phe Thr Ile Lys Asp Pro Ala Asn Arg Arg Tyr Glu Val Pro Leu
145 150 155 160
Glu Thr Pro His Val His Ser Arg Ala Pro Ser Pro Leu Tyr Ser Val
165 170 175
Glu Phe Ser Glu Glu Pro Phe Gly Val Ile Val Arg Arg Gln Leu Asp
180 185 190
Gly Arg Val Leu Leu Asn Thr Thr Val Ala Pro Leu Phe Phe Ala Asp
195 200 205
Gln Phe Leu Gln Leu Ser Thr Ser Leu Pro Ser Gln Tyr Ile Thr Gly
210 215 220
Leu Ala Glu His Leu Ser Pro Leu Met Leu Ser Thr Ser Trp Thr Arg
225 230 235 240
Ile Thr Leu Trp Asn Arg Asp Leu Ala Pro Thr Pro Gly Ala Asn Leu
245 250 255
Tyr Gly Ser His Pro Phe Tyr Leu Ala Leu Glu Asp Gly Gly Ser Ala
260 265 270
His Gly Val Phe Leu Leu Asn Ser Asn Ala Met Asp Val Val Leu Gln
275 280 285
Pro Ser Pro Ala Leu Ser Trp Arg Ser Thr Gly Gly Ile Leu Asp Val
290 295 300
Tyr Ile Phe Leu Gly Pro Glu Pro Lys Ser Val Val Gln Gln Tyr Leu
305 310 315 320
Asp Val Val Gly Tyr Pro Phe Met Pro Pro Tyr Trp Gly Leu Gly Phe
325 330 335
His Leu Cys Arg Trp Gly Tyr Ser Ser Thr Ala Ile Thr Arg Gln Val
340 345 350
Val Glu Asn Met Thr Arg Ala His Phe Pro Leu Asp Val Gln Trp Asn
355 360 365
Asp Leu Asp Tyr Met Asp Ser Arg Arg Asp Phe Thr Phe Asn Lys Asp
370 375 380
Gly Phe Arg Asp Phe Pro Ala Met Val Gln Glu Leu His Gln Gly Gly
385 390 395 400
Arg Arg Tyr Met Met Ile Val Asp Pro Ala Ile Ser Ser Ser Gly Pro
405 410 415
Ala Gly Ser Tyr Arg Pro Tyr Asp Glu Gly Leu Arg Arg Gly Val Phe
420 425 430
Ile Thr Asn Glu Thr Gly Gln Pro Leu Ile Gly Lys Val Trp Pro Gly
435 440 445
Ser Thr Ala Phe Pro Asp Phe Thr Asn Pro Thr Ala Leu Ala Trp Trp
450 455 460
Glu Asp Met Val Ala Glu Phe His Asp Gln Val Pro Phe Asp Gly Met
465 470 475 480
Trp Ile Asp Met Asn Glu Pro Ser Asn Phe Ile Arg Gly Ser Glu Asp
485 490 495
Gly Cys Pro Asn Asn Glu Leu Glu Asn Pro Pro Tyr Val Pro Gly Val
500 505 510
Val Gly Gly Thr Leu Gln Ala Ala Thr Ile Cys Ala Ser Ser His Gln
515 520 525
Phe Leu Ser Thr His Tyr Asn Leu His Asn Leu Tyr Gly Leu Thr Glu
530 535 540
Ala Ile Ala Ser His Arg Ala Leu Val Lys Ala Arg Gly Thr Arg Pro
545 550 555 560
Phe Val Ile Ser Arg Ser Thr Phe Ala Gly His Gly Arg Tyr Ala Gly
565 570 575
His Trp Thr Gly Asp Val Trp Ser Ser Trp Glu Gln Leu Ala Ser Ser
580 585 590
Val Pro Glu Ile Leu Gln Phe Asn Leu Leu Gly Val Pro Leu Val Gly
595 600 605
Ala Asp Val Cys Gly Phe Leu Gly Asn Thr Ser Glu Glu Leu Cys Val
610 615 620
Arg Trp Thr Gln Leu Gly Ala Phe Tyr Pro Phe Met Arg Asn His Asn
625 630 635 640
Ser Leu Leu Ser Leu Pro Gln Glu Pro Tyr Ser Phe Ser Glu Pro Ala
645 650 655
Gln Gln Ala Met Arg Lys Ala Leu Thr Leu Arg Tyr Ala Leu Leu Pro
660 665 670
His Leu Tyr Thr Leu Phe His Gln Ala His Val Ala Gly Glu Thr Val
675 680 685
Ala Arg Pro Leu Phe Leu Glu Phe Pro Lys Asp Ser Ser Thr Trp Thr
690 695 700
Val Asp His Gln Leu Leu Trp Gly Glu Ala Leu Leu Ile Thr Pro Val
705 710 715 720
Leu Gln Ala Gly Lys Ala Glu Val Thr Gly Tyr Phe Pro Leu Gly Thr
725 730 735
Trp Tyr Asp Leu Gln Thr Val Pro Val Glu Ala Leu Gly Ser Leu Pro
740 745 750
Pro Pro Pro Ala Ala Pro Arg Glu Pro Ala Ile His Ser Glu Gly Gln
755 760 765
Trp Val Thr Leu Pro Ala Pro Leu Asp Thr Ile Asn Val His Leu Arg
770 775 780
Ala Gly Tyr Ile Ile Pro Leu Gln Gly Pro Gly Leu Thr Thr Thr Glu
785 790 795 800
Ser Arg Gln Gln Pro Met Ala Leu Ala Val Ala Leu Thr Lys Gly Gly
805 810 815
Glu Ala Arg Gly Glu Leu Phe Trp Asp Asp Gly Glu Ser Leu Glu Val
820 825 830
Leu Glu Arg Gly Ala Tyr Thr Gln Val Ile Phe Leu Ala Arg Asn Asn
835 840 845
Thr Ile Val Asn Glu Leu Val Arg Val Thr Ser Glu Gly Ala Gly Leu
850 855 860
Gln Leu Gln Lys Val Thr Val Leu Gly Val Ala Thr Ala Pro Gln Gln
865 870 875 880
Val Leu Ser Asn Gly Val Pro Val Ser Asn Phe Thr Tyr Ser Pro Asp
885 890 895
Thr Lys Val Leu Asp Ile Cys Val Ser Leu Leu Met Gly Glu Gln Phe
900 905 910
Leu Val Ser Trp Cys
915
<210> 79
<211> 1128
<212> Белок
<213> Искуственная последовательность
<220>
<223> scFv_H4H123450(VH-linker-VL)_linker_GAA
<400> 79
Gln Val Gln Leu Gln Glu Ser Gly Pro Lys Val Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Tyr
20 25 30
Tyr Trp Asn Trp Ile Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Thr Lys Arg Gly Tyr Thr Asp Tyr Asn Pro Ser Leu Arg Ser
50 55 60
Arg Val Thr Ile Ser Glu Asp Thr Ser Lys Asn Gln Phe Ser Leu Arg
65 70 75 80
Ile Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala Gln
85 90 95
Met Gly Trp Gly Ser His Ala Phe Asp Met Trp Gly Gln Gly Thr Met
100 105 110
Val Ala Val Ser Ser Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser
130 135 140
Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser
145 150 155 160
Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala
165 170 175
Pro Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro
180 185 190
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
195 200 205
Ser Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr
210 215 220
Gly Ser Ser Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
225 230 235 240
Gly Gly Gly Gly Ser Ala His Pro Gly Arg Pro Arg Ala Val Pro Thr
245 250 255
Gln Cys Asp Val Pro Pro Asn Ser Arg Phe Asp Cys Ala Pro Asp Lys
260 265 270
Ala Ile Thr Gln Glu Gln Cys Glu Ala Arg Gly Cys Cys Tyr Ile Pro
275 280 285
Ala Lys Gln Gly Leu Gln Gly Ala Gln Met Gly Gln Pro Trp Cys Phe
290 295 300
Phe Pro Pro Ser Tyr Pro Ser Tyr Lys Leu Glu Asn Leu Ser Ser Ser
305 310 315 320
Glu Met Gly Tyr Thr Ala Thr Leu Thr Arg Thr Thr Pro Thr Phe Phe
325 330 335
Pro Lys Asp Ile Leu Thr Leu Arg Leu Asp Val Met Met Glu Thr Glu
340 345 350
Asn Arg Leu His Phe Thr Ile Lys Asp Pro Ala Asn Arg Arg Tyr Glu
355 360 365
Val Pro Leu Glu Thr Pro His Val His Ser Arg Ala Pro Ser Pro Leu
370 375 380
Tyr Ser Val Glu Phe Ser Glu Glu Pro Phe Gly Val Ile Val Arg Arg
385 390 395 400
Gln Leu Asp Gly Arg Val Leu Leu Asn Thr Thr Val Ala Pro Leu Phe
405 410 415
Phe Ala Asp Gln Phe Leu Gln Leu Ser Thr Ser Leu Pro Ser Gln Tyr
420 425 430
Ile Thr Gly Leu Ala Glu His Leu Ser Pro Leu Met Leu Ser Thr Ser
435 440 445
Trp Thr Arg Ile Thr Leu Trp Asn Arg Asp Leu Ala Pro Thr Pro Gly
450 455 460
Ala Asn Leu Tyr Gly Ser His Pro Phe Tyr Leu Ala Leu Glu Asp Gly
465 470 475 480
Gly Ser Ala His Gly Val Phe Leu Leu Asn Ser Asn Ala Met Asp Val
485 490 495
Val Leu Gln Pro Ser Pro Ala Leu Ser Trp Arg Ser Thr Gly Gly Ile
500 505 510
Leu Asp Val Tyr Ile Phe Leu Gly Pro Glu Pro Lys Ser Val Val Gln
515 520 525
Gln Tyr Leu Asp Val Val Gly Tyr Pro Phe Met Pro Pro Tyr Trp Gly
530 535 540
Leu Gly Phe His Leu Cys Arg Trp Gly Tyr Ser Ser Thr Ala Ile Thr
545 550 555 560
Arg Gln Val Val Glu Asn Met Thr Arg Ala His Phe Pro Leu Asp Val
565 570 575
Gln Trp Asn Asp Leu Asp Tyr Met Asp Ser Arg Arg Asp Phe Thr Phe
580 585 590
Asn Lys Asp Gly Phe Arg Asp Phe Pro Ala Met Val Gln Glu Leu His
595 600 605
Gln Gly Gly Arg Arg Tyr Met Met Ile Val Asp Pro Ala Ile Ser Ser
610 615 620
Ser Gly Pro Ala Gly Ser Tyr Arg Pro Tyr Asp Glu Gly Leu Arg Arg
625 630 635 640
Gly Val Phe Ile Thr Asn Glu Thr Gly Gln Pro Leu Ile Gly Lys Val
645 650 655
Trp Pro Gly Ser Thr Ala Phe Pro Asp Phe Thr Asn Pro Thr Ala Leu
660 665 670
Ala Trp Trp Glu Asp Met Val Ala Glu Phe His Asp Gln Val Pro Phe
675 680 685
Asp Gly Met Trp Ile Asp Met Asn Glu Pro Ser Asn Phe Ile Arg Gly
690 695 700
Ser Glu Asp Gly Cys Pro Asn Asn Glu Leu Glu Asn Pro Pro Tyr Val
705 710 715 720
Pro Gly Val Val Gly Gly Thr Leu Gln Ala Ala Thr Ile Cys Ala Ser
725 730 735
Ser His Gln Phe Leu Ser Thr His Tyr Asn Leu His Asn Leu Tyr Gly
740 745 750
Leu Thr Glu Ala Ile Ala Ser His Arg Ala Leu Val Lys Ala Arg Gly
755 760 765
Thr Arg Pro Phe Val Ile Ser Arg Ser Thr Phe Ala Gly His Gly Arg
770 775 780
Tyr Ala Gly His Trp Thr Gly Asp Val Trp Ser Ser Trp Glu Gln Leu
785 790 795 800
Ala Ser Ser Val Pro Glu Ile Leu Gln Phe Asn Leu Leu Gly Val Pro
805 810 815
Leu Val Gly Ala Asp Val Cys Gly Phe Leu Gly Asn Thr Ser Glu Glu
820 825 830
Leu Cys Val Arg Trp Thr Gln Leu Gly Ala Phe Tyr Pro Phe Met Arg
835 840 845
Asn His Asn Ser Leu Leu Ser Leu Pro Gln Glu Pro Tyr Ser Phe Ser
850 855 860
Glu Pro Ala Gln Gln Ala Met Arg Lys Ala Leu Thr Leu Arg Tyr Ala
865 870 875 880
Leu Leu Pro His Leu Tyr Thr Leu Phe His Gln Ala His Val Ala Gly
885 890 895
Glu Thr Val Ala Arg Pro Leu Phe Leu Glu Phe Pro Lys Asp Ser Ser
900 905 910
Thr Trp Thr Val Asp His Gln Leu Leu Trp Gly Glu Ala Leu Leu Ile
915 920 925
Thr Pro Val Leu Gln Ala Gly Lys Ala Glu Val Thr Gly Tyr Phe Pro
930 935 940
Leu Gly Thr Trp Tyr Asp Leu Gln Thr Val Pro Val Glu Ala Leu Gly
945 950 955 960
Ser Leu Pro Pro Pro Pro Ala Ala Pro Arg Glu Pro Ala Ile His Ser
965 970 975
Glu Gly Gln Trp Val Thr Leu Pro Ala Pro Leu Asp Thr Ile Asn Val
980 985 990
His Leu Arg Ala Gly Tyr Ile Ile Pro Leu Gln Gly Pro Gly Leu Thr
995 1000 1005
Thr Thr Glu Ser Arg Gln Gln Pro Met Ala Leu Ala Val Ala Leu
1010 1015 1020
Thr Lys Gly Gly Glu Ala Arg Gly Glu Leu Phe Trp Asp Asp Gly
1025 1030 1035
Glu Ser Leu Glu Val Leu Glu Arg Gly Ala Tyr Thr Gln Val Ile
1040 1045 1050
Phe Leu Ala Arg Asn Asn Thr Ile Val Asn Glu Leu Val Arg Val
1055 1060 1065
Thr Ser Glu Gly Ala Gly Leu Gln Leu Gln Lys Val Thr Val Leu
1070 1075 1080
Gly Val Ala Thr Ala Pro Gln Gln Val Leu Ser Asn Gly Val Pro
1085 1090 1095
Val Ser Asn Phe Thr Tyr Ser Pro Asp Thr Lys Val Leu Asp Ile
1100 1105 1110
Cys Val Ser Leu Leu Met Gly Glu Gln Phe Leu Val Ser Trp Cys
1115 1120 1125
<210> 80
<211> 125
<212> Белок
<213> Искуственная последовательность
<220>
<223> EC-петля 2 CD63 MMH
<400> 80
Arg Asp Lys Val Met Ser Glu Phe Asn Asn Asn Phe Arg Gln Gln Met
1 5 10 15
Glu Asn Tyr Pro Lys Asn Asn His Thr Ala Ser Ile Leu Asp Arg Met
20 25 30
Gln Ala Asp Phe Lys Cys Cys Gly Ala Ala Asn Tyr Thr Asp Trp Glu
35 40 45
Lys Ile Pro Ser Met Ser Lys Asn Arg Val Pro Asp Ser Cys Cys Ile
50 55 60
Asn Val Thr Val Gly Cys Gly Ile Asn Phe Asn Glu Lys Ala Ile His
65 70 75 80
Lys Glu Gly Cys Val Glu Lys Ile Gly Gly Trp Leu Arg Lys Asn Val
85 90 95
Leu Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Gly Gly Glu Gln Lys
100 105 110
Leu Ile Ser Glu Glu Asp Leu His His His His His His
115 120 125
<210> 81
<211> 324
<212> Белок
<213> Искуственная последовательность
<220>
<223> EC-петля 2 CD63 hFC
<400> 81
Arg Asp Lys Val Met Ser Glu Phe Asn Asn Asn Phe Arg Gln Gln Met
1 5 10 15
Glu Asn Tyr Pro Lys Asn Asn His Thr Ala Ser Ile Leu Asp Arg Met
20 25 30
Gln Ala Asp Phe Lys Cys Cys Gly Ala Ala Asn Tyr Thr Asp Trp Glu
35 40 45
Lys Ile Pro Ser Met Ser Lys Asn Arg Val Pro Asp Ser Cys Cys Ile
50 55 60
Asn Val Thr Val Gly Cys Gly Ile Asn Phe Asn Glu Lys Ala Ile His
65 70 75 80
Lys Glu Gly Cys Val Glu Lys Ile Gly Gly Trp Leu Arg Lys Asn Val
85 90 95
Leu Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
100 105 110
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
115 120 125
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
130 135 140
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
145 150 155 160
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
165 170 175
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
180 185 190
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
195 200 205
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
210 215 220
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
225 230 235 240
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
245 250 255
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
260 265 270
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
275 280 285
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
290 295 300
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
305 310 315 320
Ser Pro Gly Lys
<---

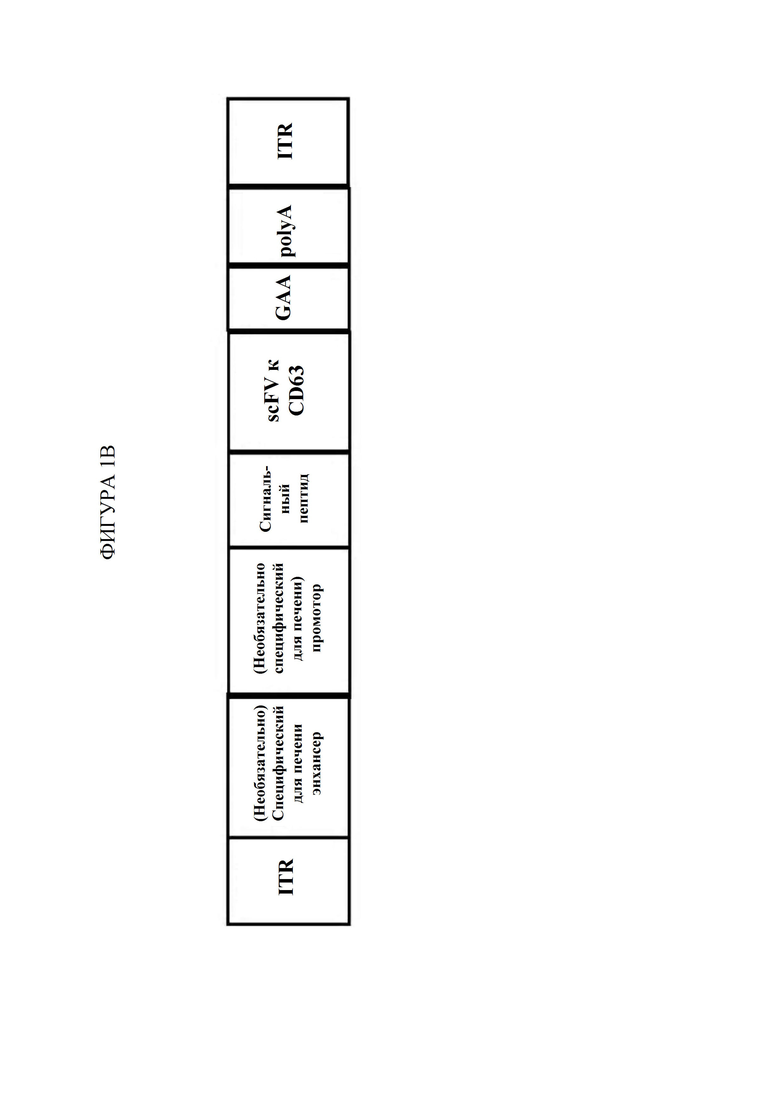
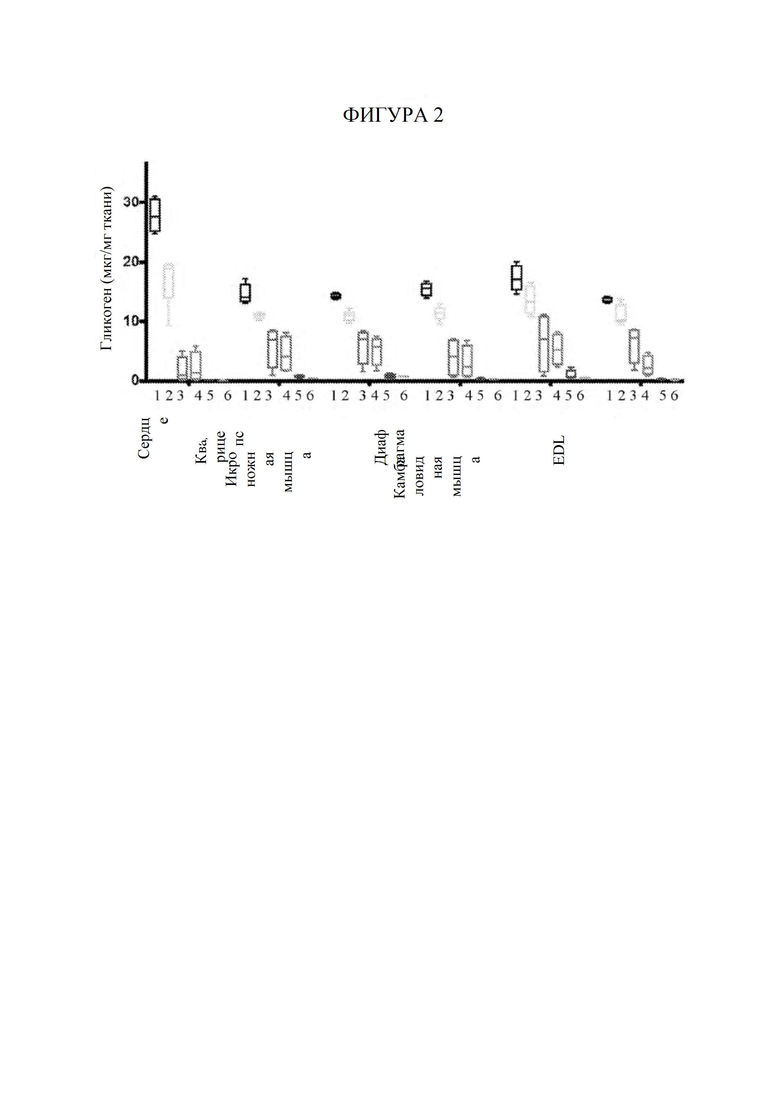

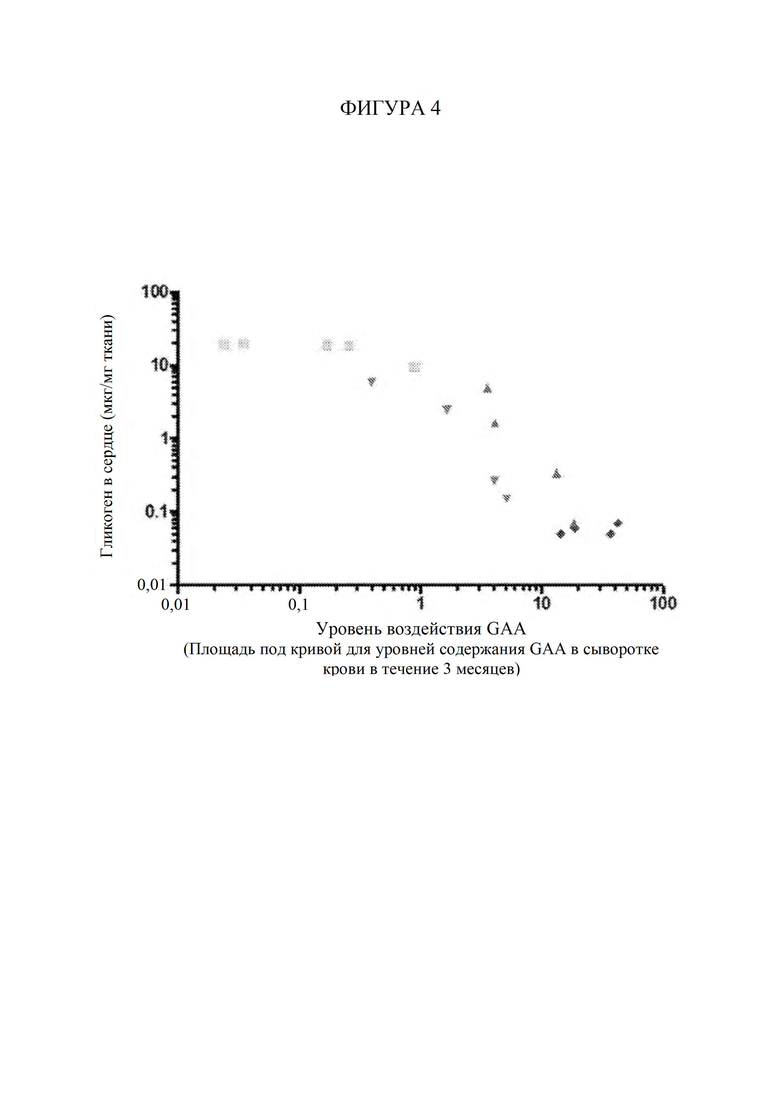
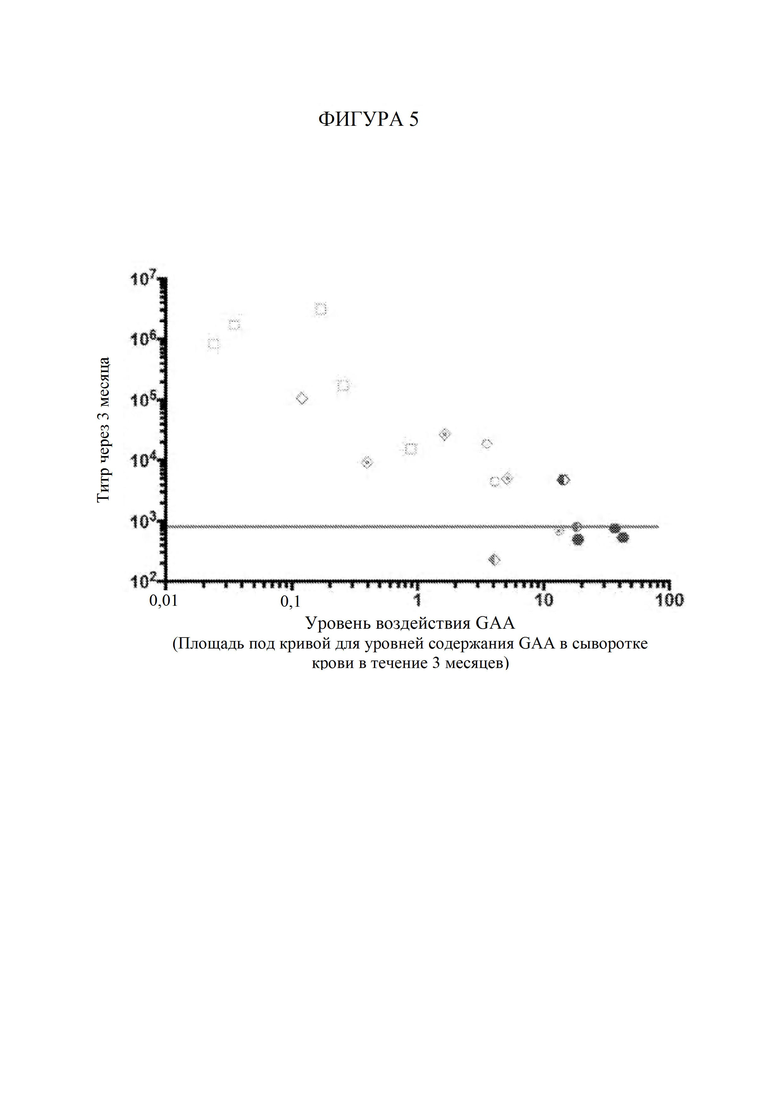

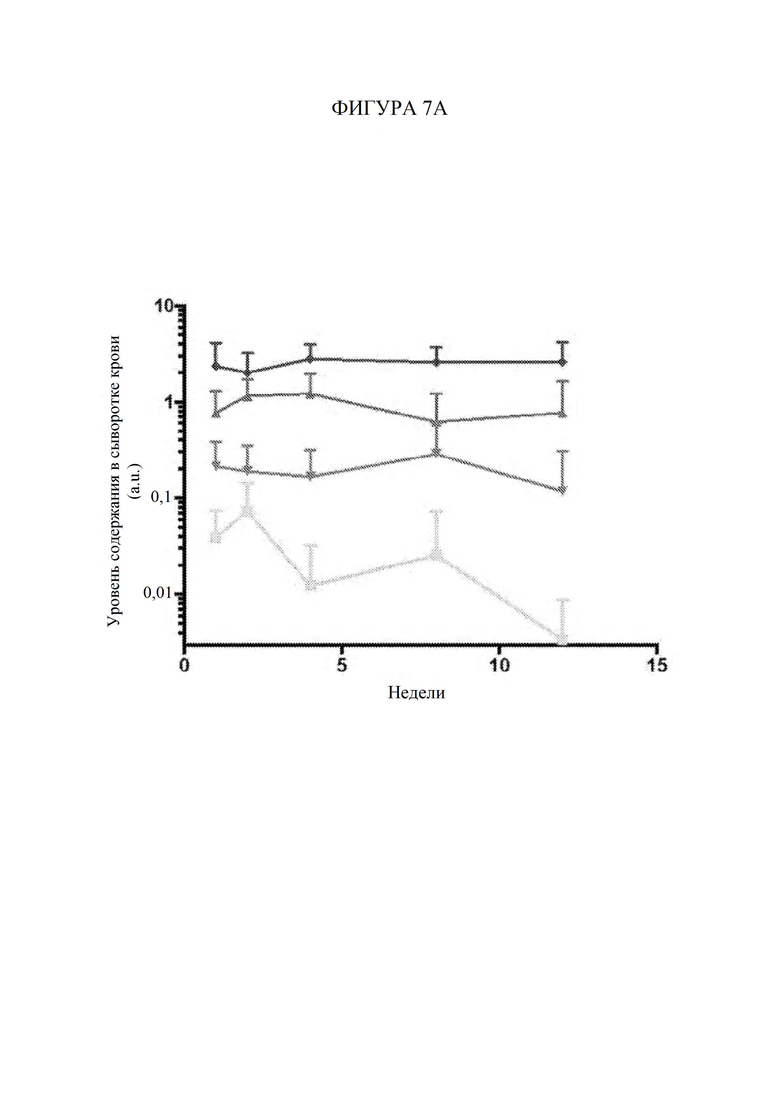










Изобретение относится к области биотехнологии. Раскрыта композиция для применения в лечении лизосомных болезней накопления, содержащая полинуклеотид, кодирующий мультидоменный терапевтический белок, содержащий домен доставки и ферментативный домен, и фармацевтически приемлемый носитель. В одном варианте реализации домен доставки связывается с эффектором интернализации, эффектор интернализации представляет собой CD63. Изобретение расширяет арсенал средств для лечения лизосомных болезней накопления. 14 з.п. ф-лы, 17 ил., 18 табл., 9 пр.
1. Композиция для применения в лечении лизосомных болезней накопления, содержащая полинуклеотид, кодирующий мультидоменный терапевтический белок, содержащий домен доставки и ферментативный домен, и фармацевтически приемлемый носитель,
где домен доставки связывается с эффектором интернализации;
где эффектор интернализации представляет собой CD63; и
где домен доставки содержит:
аминокислотную последовательность под SEQ ID NO:2 или
аминокислотную последовательность, выбранную из группы, состоящей из
аминокислотной последовательности HCVR, выбранной из SEQ ID NOs:14, 30, 46 и 62, аминокислотной последовательности LCVR, выбранной из SEQ ID NOs:22, 38, 54 и 70, аминокислотной последовательности HCDR1, выбранной из SEQ ID NOs:16, 32, 48 и 64, аминокислотной последовательности HCDR2, выбранной из SEQ ID NOs:18, 34, 50 и 66, аминокислотной последовательности HCDR3, выбранной из SEQ ID NOs:20, 36, 52 и 68, аминокислотной последовательности LCDR1, выбранной из SEQ ID NOs:24, 40, 56 и 72, аминокислотной последовательности LCDR2, выбранной из SEQ ID NOs:26, 42, 58 и 74, аминокислотной последовательности LCDR3, выбранной из SEQ ID NOs:28, 44, 60 и 76, пары из аминокислот HCVR/LCVR, представленной под SEQ ID NO: 14/22, SEQ ID NO: 30/38,
SEQ ID NO:46/54 или SEQ ID NO:62/70, где необязательно пара из аминокислот HCVR/LCVR содержит от N-конца до C-конца: HCVR, необязательный линкер и LCVR, и/или где необязательно ферментативный домен присоединен к С-концу LCVR, и любую их комбинацию.
2. Композиция по п. 1, где ферментативный домен присоединен к С-концу LCVR посредством линкера.
3. Композиция по п. 1 или 2, где домен доставки представляет собой антигенсвязывающий белок.
4. Композиция по пп. 1-3, где домен доставки представляет собой одноцепочечный вариабельный фрагмент (scFv).
5. Композиция по любому из пп. 1-4, где ферментативный домен предусматривает гидролазу.
6. Композиция по любому из пп. 1-5, где ферментативный домен предусматривает гликозилазу.
7. Композиция по любому из пп. 1-6, где ферментативный домен предусматривает гликозидазу.
8. Композиция по любому из пп. 1-7, где ферментативный домен предусматривает альфа-глюкозидазу.
9. Композиция по любому из пп. 1-8, где ферментативный домен содержит аминокислотную последовательность под SEQ ID NO:1, SEQ ID NO:13, SEQ ID NO:78 или ее фрагмент.
10. Композиция по любому из пп. 1-9, где полинуклеотид дополнительно содержит последовательность нуклеиновой кислоты вируса.
11. Композиция по любому из пп. 1-10, где полинуклеотид дополнительно содержит последовательность нуклеиновой кислоты вируса, где последовательность нуклеиновой кислоты вируса представляет собой последовательность нуклеиновой кислоты аденоассоциированного вируса (AAV).
12. Композиция по любому из пп. 1-11, где полинуклеотид дополнительно содержит последовательность нуклеиновой кислоты вируса и необязательно последовательность нуклеиновой кислоты, нацеливающуюся на локус, при этом последовательность нуклеиновой кислоты вируса представляет собой последовательность нуклеиновой кислоты аденоассоциированного вируса (AAV), и где последовательность нуклеиновой кислоты AAV содержит последовательность внутреннего концевого повтора и необязательно тканеспецифический регуляторный элемент, такой как специфический для печени промотор.
13. Композиция по любому из пп. 1-12, где полинуклеотид дополнительно содержит последовательность нуклеиновой кислоты вируса и необязательно последовательность нуклеиновой кислоты, нацеливающуюся на локус, где последовательность нуклеиновой кислоты вируса представляет собой последовательность нуклеиновой кислоты аденоассоциированного вируса (AAV), содержащую последовательность внутреннего концевого повтора, которая содержит SEQ ID NO:6, SEQ ID NO:7 или обе, и необязательно тканеспецифический регуляторный элемент, такой как специфический для печени промотор.
14. Композиция по любому из пп. 1-13, где полинуклеотид дополнительно содержит тканеспецифический регуляторный элемент, содержащий последовательность, представленную под SEQ ID NO:8, SEQ ID NO:9 или обе.
15. Композиция по любому из пп. 1-14, где полинуклеотид содержит последовательность нуклеиновой кислоты под SEQ ID NO:11 и/или кодирует аминокислотную последовательность, представленную под SEQ ID NO:10 или SEQ ID NO:78.
| WO 2017100467 A4, 21.09.2017 | |||
| WO 2013014405 A2, 31.01.2013 | |||
| ЛЕЧЕНИЕ α-ГАЛАКТОЗИДАЗНОЙ А НЕДОСТАТОЧНОСТИ | 2000 |
|
RU2248213C2 |
| Boado, R | |||
| J | |||
| et al | |||
| IgG-Enzyme Fusion Protein: Pharmacokinetics and Anti-Drug Antibody Response in Rhesus Monkeys | |||
| Bioconjugate Chemistry, 2012, 24 (1), 97-104 | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |

