Настоящая заявка испрашивает приоритет согласно заявке на патент Китая №201910828913.5, поданной 3 сентября 2019 г. в Национальное управление интеллектуальной собственности Китая и озаглавленной "ПРИМЕНЕНИЕ ГЕНА ТРАНСПОРТНОГО ПЕРЕНОСЧИКА, КОТОРЫЙ УЛУЧШАЕТ ЭФФЕКТИВНОСТЬ ПРОДУЦИРОВАНИЯ L-ТРИПТОФАНА У ESCHERICHIA COLI”, содержание которой включено в данный документ посредством ссылки во всей его полноте.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к метаболической инженерии и биотехнологии, в частности к гену транспортного метаболического переносчика, происходящему из Bacillus subtilis, который может эффективно улучшать эффективность продуцирования аминокислот у сконструированного продуцирующего L-триптофан штамма Escherichia coli.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Триптофан в качестве аминокислоты, которая играет важную роль в регулировании роста и развития организмов и обладает хорошими фармакологическими эффектами, такими как антидепрессивное действие и улучшение сна, играет важную роль в пищевой, медицинской, кормовой и других отраслях промышленности. Учитывая дальнейшее исследование этой аминокислоты, ее применение будет более широким, что рано или поздно обеспечит резкое увеличение рыночного спроса на триптофан. Таким образом, поиск более эффективного и дешевого способа получения триптофана постепенно привлекает внимание людей. В настоящее время способы его получения включают способ химического синтеза, способ ферментативных реакций и способ прямого микробиологического ферментирования. Способ прямого ферментирования представляет собой способ непосредственного продуцирования L-триптофана посредством ферментирования с недорогим сырьем, таким как глюкоза, в качестве субстрата. Такой способ имеет преимущества простоты манипулирования, низкой стоимости, высокой продуктивности, короткого цикла и низкого загрязнения окружающей среды. Однако исследования внутренней среды микроорганизмов и равновесных взаимоотношений между различными метаболическими путями все еще недостаточно исчерпывающи, что имеет результатом низкие выходы способов микробиологического ферментирования. Кроме того, что касается синтеза L-триптофана, то путь его синтеза является более долгим и требует большего количества предшественников. Таким образом, для того, чтобы осуществлять дальнейшее увеличение выхода, необходимо систематически совершенствовать микробную метаболическую сеть с помощью метаболической инженерии.
В настоящее время технологию метаболической инженерии широко используют для улучшения промышленных штаммов. Метаболическая инженерия использует главным образом такие технологии, как рекомбинация ДНК и CRISPR/Cas9-опосредованное редактирование генов, для оптимизации ферментов метаболического пути, непосредственно вовлеченных в синтез аминокислот, ключевых лимитирующих скорость ферментов и регуляторных факторов, не вовлеченных непосредственно в синтез аминокислот. В дополнение к этим стратегиям, усиление способности штаммов к транспорту целевого продукта также может эффективно улучшать эффективность продуцирования аминокислот.
Сообщалось о целом ряде генов, кодирующих белки, секретирующие аминокислоты, которые могут эффективно улучшать продуцирующую способность штаммов. Например, усиление экспрессии лизин-секретирующего гена lysE может улучшать потенциал продуцирования у лизин-продуцирующих бактерий рода Corynebacterium (WO9723597A2). Усиление экспрессии гена rhtB увеличивает толерантность бактерий к L-гомосерину (заявка на Европейский патент ЕР994190А2). Дополнительные копии гена rhtC увеличивают выходы L-гомосерина, L-треонина и L-лейцина (заявка на Европейский патент ЕР1013765А1). Дополнительные копии генов yahN, yeaS, yfiK и yggA увеличивают выходы L-глутамата, L-лизина, L-треонина, L-аланина и тому подобного (заявка на Европейский патент ЕР1016710А2).
Транспортный белок, наиболее часто встречающийся у продуцирующих ароматические аминокислоты бактерий, кодируется собственным геном Е. coli,yddG. Из их числа Vera Doroshenko et at. использовали Escherichia coli MG1655 в качестве исходного штамма и посредством плазмидной или геномной интеграции экспрессировали ген yddG с различными интенсивностями. В экспериментах по ферментированию в опытных пробирках накопление L-триптофана составляло 0,6 мкг/мл, а накопление L-фенилаланина составляло 30000 мкг/мл. Liu Shuangping et al. вводили ген yddG в штамм-хозяин Е. coli в форме плазмиды посредством генной инженерии. Эксперименты по ферментированию показали, что способность штамма, содержащего ген yddG, к внеклеточному транспорту L-фенилаланина была улучшена, и конечный выход L-фенилаланина достигал 62 г/л. В другом исследовании в качестве исходного штамма использовали триптофан-продуцирующий штамм SV164 (pGH5) Escherichia coli, и в клетки вводили ген yddG под контролем сильного промотора с получением штамма SV164PL-yddG (pGH5). Этот штамм подвергали культивированию со встряхиванием при 37°C в течение 48 ч в опытной пробирке 20×200 мм, и конечный выход L-триптофана составлял 4,17 г/л, что было примерно на 12% выше такового у исходного штамма.
На основании вышеизложенного можно сделать вывод, что сверхэкспрессия гена yddG может усиливать толерантность сконструированных штаммов к ароматическим аминокислотам, но мембранные белки, кодируемые этим геном, имеют большие различия в аффинности в отношении различных ароматических аминокислот, и эффект транспорта не очень хорош. Этот ген происходит из Escherichia coli, поэтому он легко подвергается саморегуляции бактериями и не может проявлять эффективную транспортную функцию. Этому гену сложно соответствовать требованиям промышленного производства.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Задача настоящего изобретения заключается в улучшении эффективности продуцирования штамма, продуцирующего L-триптофан, и в использовании этого штамма для продуцирования L-триптофана. Для того, чтобы решить указанную выше задачу, настоящее изобретение улучшает эффективность продуцирования L-триптофана штаммом-хозяином посредством введения в штамм-хозяин гена ywkB (Gene ID: 936875), кодирующего транспортер метаболитов, происходящего из Bacillus subtilis subsp. ywkB представляет собой ген, кодирующий мембранный белок, который кодирует предполагаемый транспортер метаболитов и принадлежит к семейству переносчиков потока ауксина (ТС 2.А.69), он не принимает непосредственного участия в синтезе L-триптофана, но когда этот ген вводят в штамм, продуцирующий L-триптофан, он может эффективно снижать концентрацию внутриклеточного конечного продукта и ослаблять ингибирование по типу обратной связи конечным продуктом ферментов метаболического пути, увеличивать поток углерода к пути синтеза целевой аминокислоты и в конечном счете улучшать выход продукта.
Первое техническое решение, предлагаемое в настоящем изобретении, представляет собой генетически сконструированный бактериальный штамм для продуцирования L-триптофана, где бактериальный штамм-хозяин модифицирован геном ywkB, и полученный генетически сконструированный бактериальный штамм обладает активностью продуцирования более высокого выхода L-триптофана, чем бактериальный штамм-хозяин.
В соответствии с генетически сконструированным бактериальным штаммом по настоящему изобретению, ген ywkB происходит из Bacillus subtilis, и модификация состоит в том, чтобы модифицировать псевдогенный локус уеер с помощью гена ywkB.
В соответствии с генетически сконструированным бактериальным штаммом по настоящему изобретению, бактериальный штамм-хозяин представляет собой бактериальный штамм-хозяин, способный к продуцированию L-триптофана, и конкретный вид бактерий не ограничивается особым образом, при условии что он обладает способностью продуцировать L-триптофан или может получить такую способность; например, бактериальный штамм-хозяин может представлять собой штамм прокариотических бактерий, способный продуцировать L-триптофан, такой как штамм грамотрицательных бактерий, способный продуцировать L-триптофан; далее, бактериальный штамм-хозяин представляет собой штамм Escherichia coli, способный продуцировать L-триптофан; например, бактериальный штамм-хозяин представляет собой продуцирующий L-триптофан штамм Е. coli TRP 03 (этот штамм получен путем генетического модифицирования исходного штамма Escherichia coli W3110 и является тем же штаммом, что и E.coli TRP 03 (CN 108753860 А)).
В соответствии с генетически сконструированным бактериальным штаммом по настоящему изобретению, ген ywkB имеет нуклеотидную последовательность, кодирующую следующую полипептидную последовательность:
(А) белок, содержащий аминокислотную последовательность, показанную в SEQ ID NO: 2 в перечне последовательностей;
(Б) белок, который содержит аминокислотную последовательность, полученную в результате делеции, замены и/или добавления одного или более аминокислотных остатков в аминокислотной последовательности, показанной в SEQ ID NO: 2 в перечне последовательностей, и все еще обладает способностью улучшать продуцирование штаммом L-триптофана и структурных аналогов L-триптофана (таких как 5-гидрокситриптофан, и так далее);
(В) белок, который имеет по меньшей мере 85%, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 96%, 97%, 98%, 99% идентичность с аминокислотной последовательностью, показанной в SEQ ID NO: 2, происходит из Bacillus subtilis и функционирует как транспортер метаболитов.
В соответствии с генетически сконструированным бактериальным штаммом по настоящему изобретению, стадия "бактериальный штамм-хозяин модифицирован геном ywkB" относится к введению гена в бактериальный штамм-хозяин.
Способ введения кодирующего гена в бактериальный штамм-хозяин включает, но без ограничения ими, средства генной инженерии, известные специалисту в данной области техники, такие как введение целевого гена в бактериальный штамм-хозяин посредством гомологичной рекомбинации; или введение целевого гена в бактериальный штамм-хозяин посредством экспрессионных нуклеиново-кислотных конструкций и, конкретно, экспрессионная нуклеиново-кислотная конструкция может представлять собой обычную плазмиду которая известна специалисту в данной области техники, и может экспрессироваться в прокариотическом хозяине; либо целевой ген введен в хроматин хозяина посредством редактирования гена.
В соответствии с генетически сконструированным бактериальным штаммом по настоящему изобретению, где ген ywkB находится под контролем сильного промотора, этот сильный промотор представляет собой, например, промотор BBa_j23101 или промотор BBa_j23106.
Конкретно, модификация по настоящему изобретению состоит в том, чтобы использовать технологию редактирования гена Escherichia coli CRISPR/Cas9 для введения кодирующего транспортер гена ywkB в Е. coli TRP 03 и далее в том, чтобы интегрировать ген, контролируемый промотором BBa_j23106, в псевдогенный локус уеер.
Нуклеотидная последовательность гена ywkB показана в SEQ ID NO: 1.
Аминокислотная последовательность тепа ywkB показана в SEQ ID NO: 2.
Нуклеотидная последовательность промотора BBa_j23106 показана в SEQ ID NO: 3.
Второе техническое решение, предлагаемое в настоящем изобретении, представляет собой способ конструирования генетически сконструированного бактериального штамма, который продуцирует L-триптофан, где ген ywkB интегрируют в бактериальный штамм-хозяин с получением генетически сконструированного бактериального штамма, обладающего активностью продуцирования более высокого выхода L-триптофана, чем бактериальный штамм-хозяин.
Согласно способу конструирования по настоящему изобретению, стадию интегрирования осуществляют посредством модифицирования псевдогенного локуса yeep геном ywkB, например, интегрирование осуществляют посредством редактирования гена CRISPR-Cas9.
Согласно способу конструирования по настоящему изобретению, в качестве исходного штамма используют продуцирующий триптофан штамм Е. coli TRP 03, сохраняемый в лаборатории Тяньцзиньского университета науки и техники (Tianjin University of Science and Technology), и интегрируют ген ywkB, а конкретные стадии представляют собой следующее:
1) синтезирование интегрируемого генного фрагмента ywkB: конструирование фрагментов вышележащего и нижележащего гомологичного плеча псевдогена уеер, генного фрагмента ywkB и последовательности промотора BBa_j23106, где интегрируемый генный фрагмент ywkB содержит вышележащее гомологичное плечо гена уеер, нижележащее гомологичное плечо гена уеер, промоторный фрагмент ВВа_j23106 и генный фрагмент ywkB;
2) экспрессирование интегрируемого генного фрагмента ywkB в Е. coli TRP 03 с использованием технологии CRISPR/Cas9.
Согласно способу конструирования по настоящему изобретению, последовательность гена ywkB представляет собой нуклеотидную последовательность, кодирующую следующую полипептидную последовательность:
А) белок, содержащий аминокислотную последовательность, показанную в SEQ ID NO: 2 в перечне последовательностей;
Б) белок, который содержит аминокислотную последовательность, полученную в результате делеции, замены и/или добавления одного или более аминокислотных остатков в аминокислотной последовательности, показанной в SEQ ID NO: 2 в перечне последовательностей, и все еще обладает способностью улучшать продуцирование штаммом L-триптофана и структурных аналогов L-триптофана (таких как 5-гидрокситриптофан, и так далее);
(В) белок, который имеет по меньшей мере 85%, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 96%, 97%, 98%, 99% идентичности с аминокислотной последовательностью, показанной в SEQ ID NO: 2, который происходит из Bacillus subtilis и функционирует как транспортер метаболитов.
Третье техническое решение, предлагаемое в настоящем изобретении, представляет собой генетически сконструированный бактериальный штамм, полученный вышеуказанным способом конструирования генетически сконструированного бактериального штамма, который продуцирует L-триптофан.
Четвертое техническое решение, предлагаемое в настоящем изобретении, представляет собой способ продуцирования L-триптофана, который включает стадию ферментирования генетически сконструированного бактериального штамма согласно первому решению по настоящему изобретению или генетически сконструированного бактериального штамма, полученного способом согласно второму решению, с продуцированием L-триптофана.
Согласно способу продуцирования L-триптофана по настоящему изобретению, он включает следующие стадии:
1) активирование бактериального штамма;
2) приготовление посевного раствора;
3) ферментирование; например, где
на стадии (1) готовят культуру на скошенном агаре: инокулирование бактериального штамма, хранящегося при -80°C, на активированный скошенный агар методом штриха, культивирование при 37°C в течение 12 часов и однократное пересевание;
на стадии (2) готовят посевную культуру в качалочной колбе: соскабливание пятна посевного материала на скошенном агаре с помощью микробиологической петли и инокулирование в коническую колбу на 500 мл, содержащую 30 мл посевной среды, укупоривание конической колбы девятью слоями марли и культивирование при 37°C и 200 об./мин в течение 8-10 часов;
на стадии (3) готовят ферментационную культуру в качалочной колбе: инокулирование посевной культуры в концентрации 10-15% (об./об.) в коническую колбу на 500 мл, содержащую ферментационную среду (конечный объем: 30 мл), укупоривание конической колбы девятью слоями марли, культивирование при 37°C и 200 об./мин на качалке, добавление во время ферментирования аммиачной воды для поддержания рН при 7,0-7,2; добавление 60%-ного (масс./об.) раствора глюкозы для поддержания ферментации (с феноловым красным в качестве индикатора, когда цвет ферментационного бульона больше не изменяется, это расценивается как дефицит глюкозы, и добавляют 1-2 мл 60%-ного (масс./об.) раствора глюкозы для обеспечения начального значения концентрации глюкозы в ферментационном бульоне 20-40 г/л); продолжение ферментирования в течение 22-26 часов.
Настоящее изобретение обладает следующими преимуществами и полезными эффектами.
В настоящем изобретении ген ywkB, кодирующий транспортный переносчик, происходящий из Bacillus subtilis, вводят в сконструированный штамм, продуцирующий L-триптофан, что значительно улучшает выход целевого продукта. Поскольку ген происходит из Bacillus subtilis, он не будет подвергаться регулированию сконструированным штаммом и функционирует проще. Таким образом, настоящее изобретение имеет широкие перспективы применения. Сконструированный штамм, продуцирующий L-триптофан, содержащий ген ywkB, предлагаемый в настоящем изобретении, может накапливать 15,2 г/л L-триптофана после 24 часов ферментирования в качалочной колбе, что на 35% больше, чем у исходного штамма, демонстрируя потенциал для промышленного производства L-триптофана.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
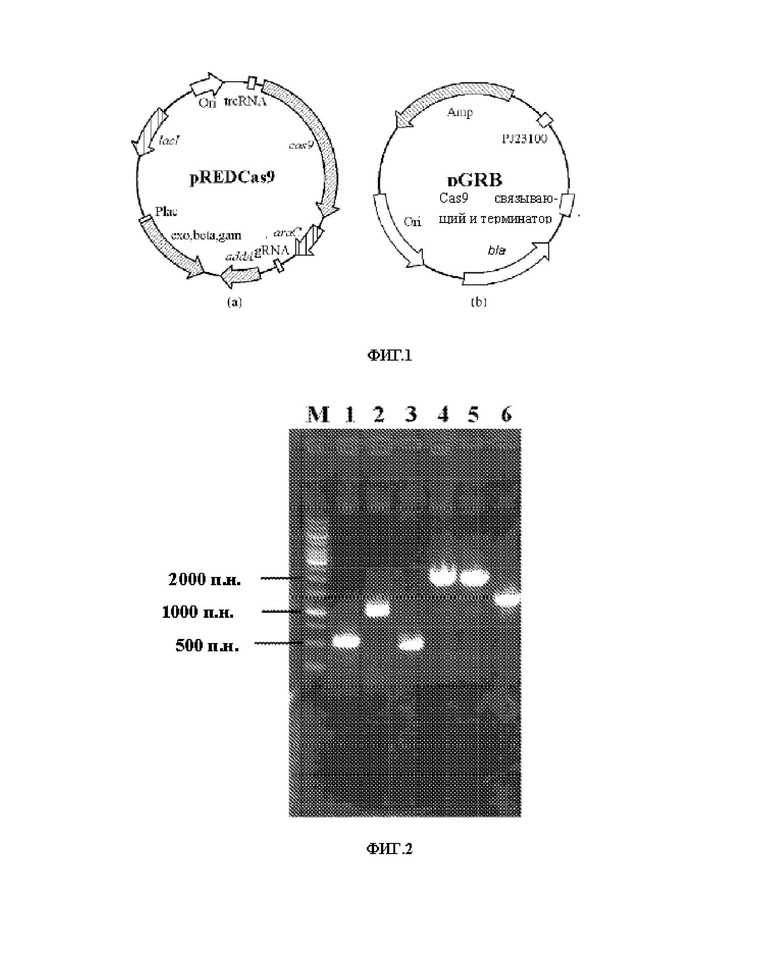
На ФИГ. 1, на панели (а) показана карта плазмиды pREDCas9, и на панели (b) показана карта плазмиды pGRB.
На ФИГ. 2 показана электрофореграмма конструкции и верификации генного фрагмента ywkB-BBa_j23106, где М: ДНК-маркер 1 т.п.н. (тысяч пар нуклеотидов); трек 1: вышележащее гомологичное плечо; трек 2: ywkB; трек 3: нижележащее гомологичное плечо; трек 4: перекрывающийся фрагмент; трек 5: фрагмент идентификации положительных бактерий; трек 6: контрольный штамм.
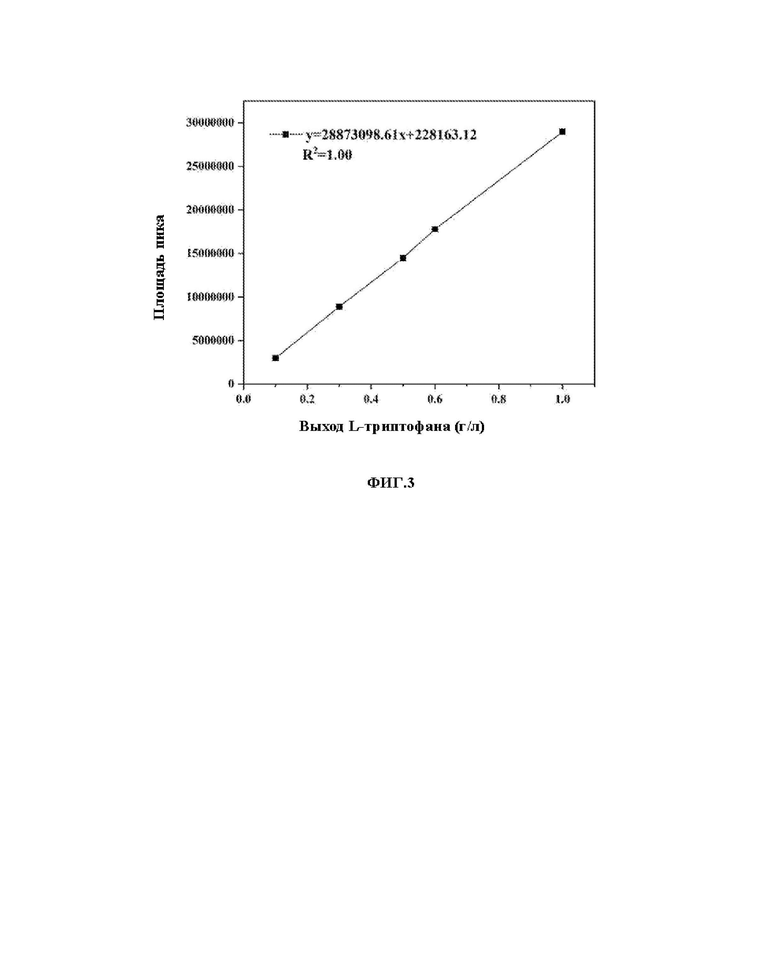
На ФИГ. 3 показана стандартная кривая для определения L-триптофана посредством ВЭЖХ (высокоэффективная жидкостная хроматография).
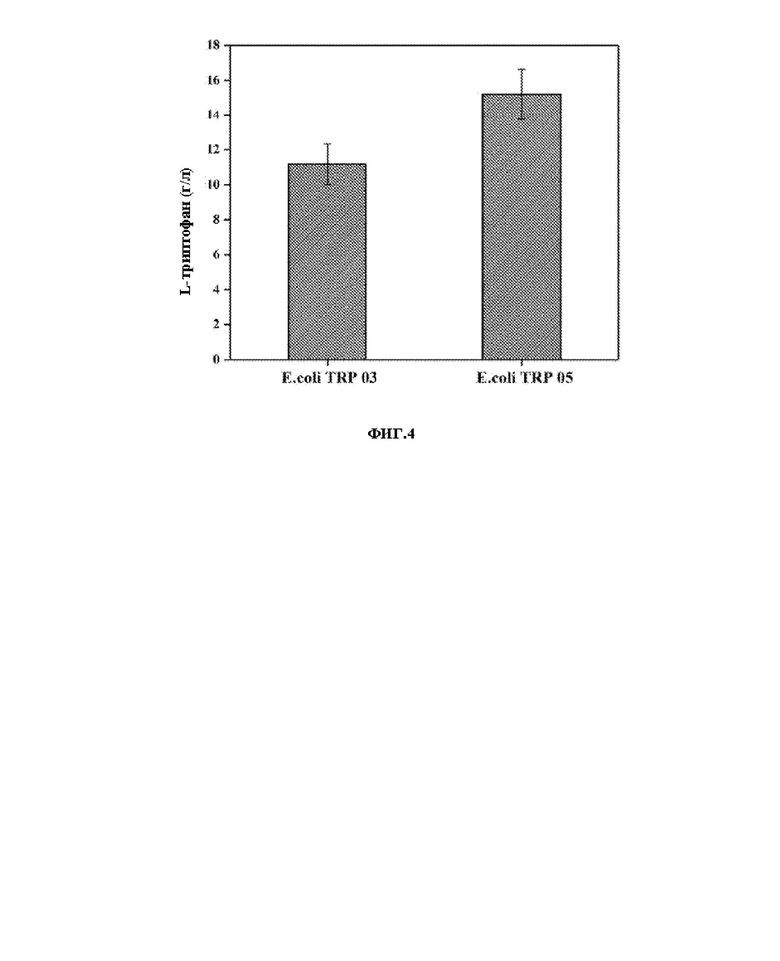
На ФИГ. 4 показаны выходы L-триптофана при ферментировании в качалочной колбе.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Пример 1: Конструирование штамма Е. coli TRP 05
1. Способ редактирования гена
Метод редактирования гена, принятый в настоящем изобретении, раскрыт в "Li Y, Lin Z, Huang С, et al. Metabolic engineering of Escherichia coli using CRISPR-Cas9 meditated genome editing. Metabolic engineering, 2015, 31:13-21", и карты двух плазмид, используемых в этом способе, показаны на ФИГ. 1. А именно, вектор pREDCas9 несет систему элиминации плазмиды pGRB для экспрессии gRNA (геномная РНК), систему Red-рекомбинации фага X и систему экспрессии белка Cas9, устойчивость к спектиномицину (рабочая концентрация: 100 мг/л), культивирование при 32°C; в векторе pGRB в качестве остова используется pUC18 и содержится промотор J23100, последовательность связывающего домена gRNA-Cas9 и последовательность терминатора, устойчивость к ампициллину (рабочая концентрация: 100 мг/л), культивирование при 37°C.
Конкретные стадии этого способа.
1.1 Конструирование плазмиды pGRB
Задача конструирования плазмиды pGRB состоит в том, чтобы транскрибировать соответствующую gRNA с образованием комплекса с белком Cas9 и распознавать целевой сайт целевого гена посредством спаривания оснований и РАМ (соседний мотив протоспейсера) для достижения двухцепочечного разрыва целевой ДНК. Плазмида pGRB была сконструирована путем рекомбинации фрагмента ДНК, содержащего целевую последовательность, с линеаризованным векторным фрагментом.
1.1.1 Конструирование целевой последовательности
Для создания целевой последовательности (РАМ: 5'-NGG-3') использовали CRISPR RGEN Tools.
1.1.2 Получение фрагмента ДНК, содержащего целевую последовательность
Конструировали праймер 5'-концевая последовательность линеаризованного вектора (15 п.н.) - сайт рестрикции целевая последовательность (без последовательности РАМ) - концевая последовательность линеаризованного вектора (15 п.н.)-3' и комплементарный обратный праймер, а фрагмент ДНК, содержащий целевую последовательность, получали посредством отжига однонитевой ДНК. Условия реакции: преденатурация при 95°C в течение 5 мин; отжиг при 30-50°C в течение 1 мин. Система отжига была следующей.
Система отжига

1.1.3 Получение линеаризованного вектора
Для линеаризации вектора использовали способ инвертированной ПЦР-амплификации.
1.1.4 Реакция рекомбинации
Система рекомбинации показана в следующей таблице. Все используемые рекомбиназы представляли собой ферменты серии ClonExpress® II One Step Cloning Kit. Условия рекомбинации: 37°C, 30 мин.
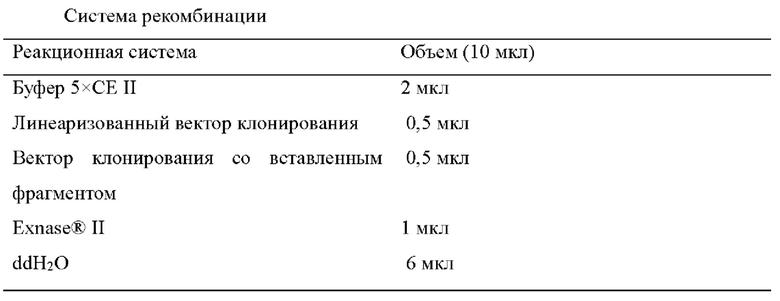
1.1.5 Трансформация плазмиды
Десять мкл реакционного раствора добавляли к 100 мл компетентных клеток DH5α и осторожно перемешивали. Полученную смесь охлаждали в ледяной бане в течение 20 мин, подвергали тепловому шоку при 42°C в течение 45-90 секунд, сразу охлаждали в ледяной бане в течение 2-3 мин, дополняли 900 мкл SOC (супероптимальная среда с катаболическим репрессором, Super Optimal broth with Catabolic repressor) и выделяли при 37°C в течение 1 часа. Смесь центрифугировали при 8000 об./мин в течение 2 мин, часть супернатанта отбрасывали и оставшиеся 200 мкл супернатанта использовали для ресуспендирования клеток. Затем клетки распределяли по чашке, содержащий 100 мг/л ампициллина, и чашку помещали перевернутой вверх дном и культивировали при 37°C в течение ночи. После того, как на чашке были выращены одиночные колонии, положительные рекомбинанты идентифицировали и отбирали посредством ПЦР методом молекулярных колоний.
1.1.6 Идентификация клонов
ПЦР-положительные колонии инокулировали в среду LB (лизогенная среда), содержащую 100 мг/л ампициллина, для культивирования в течение ночи и бактерии сохраняли. Плазмиды экстрагировали и идентифицировали с помощью ферментативного расщепления.
1.2 Получение фрагмента рекомбинантной ДНК
Рекомбинантный фрагмент для нокаута состоит из вышележащего и нижележащего гомологичных плечей гена, подлежащего нокауту (вышележащее гомологичное плечо-нижележащее гомологичное плечо); рекомбинантный фрагмент для интегрирония состоит из вышележащего и нижележащего гомологичных плечей сайта интеграции и генного фрагмента, подлежащего интегрированию (вышележащее гомологичное плечо - целевой ген - нижележащее гомологичное плечо). Используя программное обеспечение для конструирования праймеров primer5, вышележащую и нижележащую последовательности гена, подлежащего нокауту, или сайт, подлежащий интегрированию, использовали в качестве матрицы для разработки праймеров для амплификации вышележащего и нижележащего гомологичных плечей (длина продукта амплификации: около 400-500 п.н.); ген, подлежащий интегрированию, использовали в качестве матрицы для конструирования праймеров для амплификации интегрированного гена. После амплификации вышележащего и нижележащего гомологичных плечей и фрагмента целевого гена посредством ПЦР, соответственно, был получен рекомбинантный фрагмент посредством ПЦР с перекрывающимися праймерами. ДНК-полимеразы, использовавшиеся в ПЦР, приобретали в TaKaRa, включая высокоточную ДНК-полимеразу PrimeSTAR HS и ДНК-полимеразу Ex.taq для создания продуктов ПЦР с «липкими» концами. Системы и методы ПЦР показаны в следующих таблицах.


Система ПЦР с перекрывающимися праймерами показана в следующей таблице.

Система ПЦР методом молекулярных колоний показана в следующей таблице.

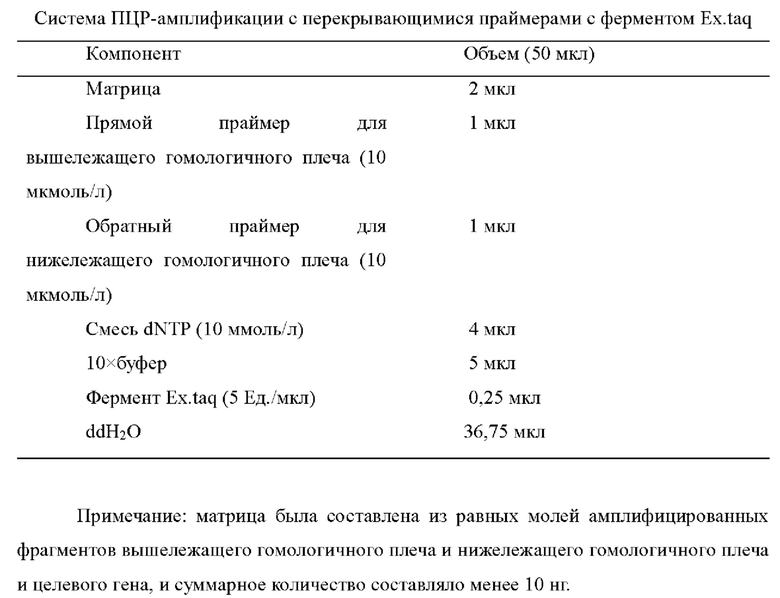
Условия реакции ПЦР: преденатурация при 95°C в течение 5 мин; 30 циклов денатурации при 98°C в течение 10 секунд, отжиг при (Tm-3/5)°C в течение 15 секунд, удлинение при 72°C (активность фермента: примерно 1 минута на т.н.); и конечное удлинение при 72°C в течение 10 мин; сохраняли при 4°C.
1.3 Трансформация плазмидой и фрагментом рекомбинантной ДНК
1.3.1 Трансформация pREDCas9
Плазмидой pREDCas9 трансформировали электропорацией компетентные для электропорации клетки W3110. Клетки выделяли, культивировали и распределяли по чашкам с LB, содержащей спектиномицин, и культивировали при 32°C в течение ночи. Одиночные колонии, выросшие на чашках с антибиотиком, подвергали ПЦР методом молекулярных колоний с помощью идентификационных праймеров для скрининга положительных рекомбинантов.
1.3.2 Получение компетентных для трансформации электропорацией клеток целевого штамма, содержащего pREDCas9
Штамм культивировали при 32°C до достижения культурой OD600 (оптическая плотность) от 0,1 до 0,2 и затем добавляли 0,1 М IPTG (изопропилтиогалактозид) (до конечной концентрации 0,1 мМ). Культивирование продолжали до достижения значения OD600 от 0,6 до 0,7. Полученные клетки использовали для получения компетентных клеток. Цель добавления IPTG состоит в том, чтобы индуцировать экспрессию рекомбиназы на плазмиде pREDCas9. Среда и способ получения, необходимые для получения компетентных клеток, относятся к общепринятым стандартным операциям.
1.3.3 Трансформация pGRB и фрагмент рекомбинантной ДНК
Плазмидой pGRB и фрагментом донорной ДНК одновременно трансформировали электропорацией компетентные для электропорации клетки, содержащие pREDCas9. После трансформации электропорацией клетки выделяли, культивировали и распределяли по чашкам с LB, содержащей ампициллин и спектиномицин, и культивировали при 32°C в течение ночи. Верификацию посредством ПЦР методом молекулярных колоний выполняли, используя прямой праймер для вышележащего гомологичного плеча и обратный праймер для нижележащего гомологичного плеча либо используя специально сконструированные праймеры для идентификации, со скринингом положительных рекомбинантов и сохраняли рекомбинантные бактерии.
1.4 Элиминация плазмиды
1.4.1 Элиминация плазмиды pGRB
Положительные рекомбинанты культивировали в течение ночи в среде LB, содержащей 0,2% арабинозы, и после надлежащего разведения их распределяли по чашкам с LB, содержащим спектиномицин, и культивировали при 32°C в течение ночи. Затем рекомбинанты инокулировали на чашки с LB, содержащие ампициллин и спектиномицин, соответственно, и одиночные колонии не росли на чашке, содержащей ампициллин, но росли на чашке, содержащей спектиномицин, отбирали и сохраняли.
1.4.2 Элиминация плазмиды pREDCas9
Положительные рекомбинанты переносили в жидкую среду LB без антибиотиков, культивировали в течение ночи при 42°C и распределяли по чашкам с LB без антибиотиков после надлежащего разведения, и культивировали при 37°C в течение ночи. Затем рекомбинанты инокулировали на чашки с LB, содержащей спектиномицин и без антибиотиков, соответственно, одиночные колонии, которые не росли на чашке со спектиномицином, но росли на чашке LB без антибиотиков, отбирали и сохраняли.
2. Конструирование модифицированного штаммаЕ. coli TRP 05, продуцирующего L-триптофан
2.1 Синтез гена ywkB
1) Используя геном Е. coli W3110 в качестве матрицы, выполняли ПЦР с праймерами для вышележащего гомологичного плеча (уеер-вверх-S, уеер-вверх-А) и праймерами для нижележащего гомологичного плеча (уеер-вниз-S, уеер-вниз-А), сконструированными по обоим концам псевдогена уеер для амплификации вышележащего и нижележащего гомологичных плечей псевдогена уеер.
2) Праймеры для ПЦР (ywkB-S, ywkB-A) конструировали в соответствии с последовательностью гена предполагаемого транспортера метаболитов ywkB Bacillus subtilis (Bacillus subtilis subsp. subtilis штамм 168), опубликованной в GENBANK, a последовательность промотора BBaj23106 конструировали в прямом праймере для гена ywkB и его фрагмент амплифицировали с HS-ферментом.
3) Амплифицированный фрагмент, полученный на стадиях (1) и (2), использовали в качестве матриц для получения интегрированного генного фрагмента BBaj23106-ywkB посредством ПЦР с перекрывающимися праймерами, и этот фрагмент состоял из вышележащего гомологичного плеча гена уеер, нижележащего гомологичного плеча гена уеер, промоторного фрагмента BBaj23106 и генного фрагмента ywkB.
2.2 Интеграция гена ywkB
1) Конструирование pGRB-yeep: плазмиду pGRB, содержащую целевую последовательность, получали в соответствии со способом, описанным в разделе 1.1, а именно, праймеры, pGRB-yeep-S и pGRB-yeep-A, конструировали в соответствии со способом согласно разделу 1.1.2 и затем фрагмент ДНК, содержащий целевую последовательность, получали посредством отжига; плазмиду pGRB линеаризировали в соответствии со способом согласно разделу 1.1.3 и затем, в соответствии со способом согласно разделу 1.1.4 получали плазмиду pGRB-yeep, содержащую целевую последовательность,
2) Получение компетентных клеток Е. coli TRP 03;
3) Получение штамма Е. coli TRP 05: в соответствии со способом, описанным в разделе 1.3, отбирали положительные клоны, затем плазмидой pREDCas9 трансформировали E.coli TRP 03 и затем получали компетентные клетки Е. coli TRP 03, содержащие плазмиду pREDCas9, трансформировали электропорацией плазмидой pGRB-yeep и фрагментами рекомбинантной ДНК, полученными на стадии (3) в разделе 2.1; после 12-16 часов культивирования на чашках осуществляли верификацию посредством ПЦР методом молекулярных колоний, положительные рекомбинанты отбирали и сохраняли; плазмиду pGRB-yeep и плазмиду pREDCas9, соответственно, элимиировали в соответствии со способом, описанным в разделе 1.4, и, наконец, штамм E.coli TRP 05 подвергали скринингу посредством ПЦР-идентификации и хранили при -80°C.
Электрофореграмма конструкта интеграционного фрагмента гена ВВа_j23106 ywkB и ПЦР-верификация положительного штамма показаны на ФИГ. 2, где длина вышележащего гомологичного плеча составляет 512 п.н., длина нижележащего гомологичного плеча составляет 513 п.н., длина фрагмента гена ywkB BBa_j23106 составляет 1040 пн., и суммарная длина фрагмента интеграции гена/wkB BBa_j23106 составляет 2065 п.н. При ПЦР-верификации длина ПЦР-амплифицированного фрагмента положительных бактерий составляла 2065 п.н., и длина ПЦР-амплифицированного фрагмента природных бактерий составляла 1396 п.н.
3. Праймеры, использованные для улучшения штамма, показаны в следующей таблице:

Пример 2: ферментация в качалочной колбе генетически сконструированной Escherichia coli с продуцированием L-триптофана
Конкретный путь использования генетически сконструированный Escherichia coli для продуцирования L-трипгофана посредством ферментирования в качалочной колбе был следующим:
культура на скошенном агаре: инокулирование бактериального штамма, хранившегося при -80°C, на активированный скошенный агар методом штриха, культивирование при 37°C в течение 12 часов и однократный пересев;
посевная культура для качалочной колбы: соскабливание пятна посевного материала на скошенном агаре с помощью микробиологической петли и инокулирование в коническую колбу на 500 мл, содержащую 30 мл посевной среды, укупоривание конической колбы девятью слоями марли и культивирование при 37°C и 200 об./мин в течение 8-10 часов;
ферментационная культура в качалочной колбе: инокулирование посевной культуры в концентрации 10-15% (об./об.) в коническую колбу на 500 мл, содержащую ферментационную среду (конечной объем: 30 мл), укупоривание конической колбы девятью слоями марли, культивирование при 37°C и 200 об./мин на качалке, добавление во время ферментации аммиачной воды для поддержания рН при 7,0-7,2; добавление 60%-ного (масс./об.) раствора глюкозы для поддержания ферментации (с феноловым красным в качестве индикатора, когда цвет ферментационного бульона больше не изменяется, это расценивается как отсутствие глюкозы, добавление 1-2 мл 60%-ного (масс./об.) раствора глюкозы для обеспечения начального значения концентрации глюкозы в ферментативном бульоне 20-40 г/л); ферментационный период продолжается 22-26 часов.
Определение концентрации L-триптофана в ферментационном бульоне: отбирали 1 мл ферментационного бульона, центрифугировали при 13000 об./мин в течение 1 мин и отбирали супернатант; собранный супернатант разбавляли (до 0,1-0,5 г/л) деионизированной водой, фильтровали через микропористый фильтр 0,22 мкм и определяли концентрацию L-триптофана посредством жидкостной хроматографии; условия хроматографии были следующими: хроматографическая колонка: колонка Kromasil С18 (250 мм × 460 мм, 5 мкм), подвижная фаза: 10%-ный раствор ацетонитрила, скорость потока: 1,0 мл/мин, температура колонки: 40°C, длина волны детектирования: 278 нм, объем вводимой пробы: 20 мкл, и время появления главного пика составляло примерно 3,5 мин; концентрацию L-триптофана в ферментационном бульоне вычисляли исходя из площади его пика в соответствии со стандартной кривой;
построение стандартной кривой L-триптофана: растворы с концентрациями L-триптофана 0,1 г/л, 0,3 г/л, 0,5 г/л, 0,6 г/л и 1 г/л подвергали вышеуказанной жидкостной хроматографии для получения площадей пиков, соответствующих концентрациям L-триптофана, и площади пиков наносили на график в зависимости от концентрации L-триптофана для получения стандартной кривой, как показано на ФИГ. 3.
Компоненты активированной среды скошенного агара: 1-3 г/л глюкозы, 5-10 г/л триптона, 5-10 г/л говяжьего экстракта, 2-5 г/л дрожжевого экстракта, 2-5 г/л NaCl, 15-30 г/л агара, остаток представляет собой воду, рН 7,0-7,2, стерилизовали в автоклаве при 121°C в течение 20 мин.
Компоненты посевной среды: 20-40 г/л глюкозы, 1-5 г/л (NH4)2SO4, 1-5 г/л KH2PO4, 0,5-2 г/л MgSO4⋅7H2O, 2-5 г/л дрожжевого экстракта, 1-3 мг/л FeSO4⋅7H2O, 1-3 мг/л MnSO4⋅H2O, 0,1-0,5 мг/л VH, 0,5-1,0 мг/л VB1, 1-3 мл/л смеси микроэлементов, 15-30 г/л фенолового красного, остаток представляет собой воду, рН 7,0-7,2, стерилизовали в автоклаве при 115°C в течение 15 мин.
Компоненты ферментационной среды: 20-40 г/л глюкозы, 2-6 г/л (NH4)2SO4, 1-5 г/л KH2PO4, 0,5-2 г/л MgSO4⋅7H2O, 1-5 г/л дрожжевого экстракта, 30-60 мг/л FeSO4⋅7H2O, 1-5 мг/л MnSO4⋅7H2O, 0,1-0,5 мг/л VH, 0,5-1,0 мг/л VB1, 1-3 мл/л смеси микроэлементов, 15-30 г/л фенолового красного, остаток представляет собой воду, рН 7,0-7,2, стерилизовали в автоклаве при 115°C в течение 15 мин.
Компоненты смеси микроэлементов: 2,5 г/л Na2MoO4⋅2H2O, 2,5 г/л AlCl3⋅6H2O, 2,5 г/л NiSO4⋅6H2O, 1,75 г/л CoCl2⋅6H2O, 10 г/л CaCl2⋅2H2O, 0,5 г/л ZnSO4⋅7H2O, 0,25 г/л CuCl2⋅2H2O, 0,125 г/л Н3ВО3.
Сконструированный штамм Е. coli TRP 05, продуцирующий L-триптофан, использовали для ферментирования в качалочных колбах, а модифицированный продуцирующий L-триптофан штамм Е. coli TRP 03, хранящийся в Лаборатории метаболической инженерии Тяньцзиньского университета науки и техники (the Metabolic Engineering Laboratory of Tianjin University of Science and Technology) использовали в качестве контроля для такой же ферментации в качалочных колбах. Результаты эксперимента показаны на ФИГ. 4. Модифицированный штамм Е. coli TRP 03, продуцирующий L-триптофан, накапливал L-триптофан вплоть до 11,2 г/л, в то время как E. coli TRP 05 накапливал L-триптофан вплоть до 15,2 г/л, что было на 35% больше, чем у контрольного штамма.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Ningxia Eppen Biotech Co., LTD; Tianjin University of Science
& Technology
<120> ПРИМЕНЕНИЕ ГЕНА ТРАНСПОРТНОГО ПЕРЕНОСЧИКА, КОТОРЫЙ УЛУЧШАЕТ
ЭФФЕКТИВНОСТЬ ПРОДУЦИРОВАНИЯ L-ТРИПТОФАНА У ESCHERICHIA COLI
<130> PUS1220487
<160> 11
<210> 1
<211> 993
<212> DNA
<213> Bacillus subtilis subsp. subtilis 168
<400> 1
TTGAGCATCT TAGATATCTT AATCCTCCTG GCGCCGATCT TCTTTGTTAT CGTGCTGGGT 60
TGGTTTGCAG GACATTTTGG AAGTTATGAT GCCAAGTCGG CAAAAGGGGT AAGTACGTTA
120
GTAACGAAAT ACGCACTTCC AGCTCACTTT ATCGCTGGTA TTTTGACAAC TTCCAGAAGT
180
GAATTTTTAT CACAAGTACC TTTAATGATT TCTTTAATTA TTGGGATTGT TGGTTTCTAT
240
ATCATCATTC TTTTGGTTTG CAGATTTATA TTCAAGTATG ATTTAACGAA CTCATCTGTA
300
TTTTCTTTGA ACTCTGCACA GCCGACATTC GCATTTATGG GTATCCCGGT ATTGGGAAGC
360
TTATTCGGAG CGAATGAAGT TGCGATTCCG ATCGCGGTCA CAGGTATCGT GGTTAACGCG
420
ATTCTTGATC CGCTCGCGAT CATTATCGCT ACTGTTGGTG AGTCTTCTAA GAAAAACGAA
480
GAGAGTGGCG ACAGCTTCTG GAAGATGACA GGAAAATCAA TCCTGCATGG TCTTTGTGAG
540
CCGCTTGCAG CTGCTCCGTT AATCAGTATG ATCTTGGTGC TGGTTTTCAA TTTCACTCTT
600
CCTGAGCTGG GTGTTAAAAT GCTTGATCAG CTTGGAAGCA CAACATCTGG TGTTGCTCTC
660
TTCGCTGTTG GTGTTACCGT TGGTATTCGT AAAATTAAAC TCAGTATGCC GGCTATCGGT
720
ATTGCGTTAC TAAAAGTTGC GGTTCAGCCT GCGTTAATGT TCCTGATTGC TCTTGCTATC
780
GGACTTCCAG CTGACCAAAC AACAAAAGCA ATCCTTCTTG TTGCATTCCC TGGTTCTGCC
840
GTTGCAGCCA TGATTGCGAC TCGTTTCGAG AAACAAGAAG AAGAAACTGC AACTGCGTTT
900
GTGGTCAGTG CGATTCTGTC ATTGATTTCA CTTCCAATCA TTATCGCGCT TACTGCGTAA
960
CAAAAAAAGC TCCCTTTAAG GGAGCTTTTT GCT
993
<210> 2
<211> 330
<212> PRT
<213> Bacillus subtilis subsp. subtilis 168
<400> 2
MSILDILILL APIFFVIVLG WFAGHFGSYD AKSAKGVSTL VTKYALPAHF IAGILTTSRS 60
EFLSQVPLMI SLIIGIVGFY IIILLVCRFI FKYDLTNSSV FSLNSAQPTF AFMGIPVLGS 120
LFGANEVAIP IAVTGIVVNA ILDPLAIIIA TVGESSKKNE ESGDSFWKMT GKSILHGLCE 180
PLAAAPLISM ILVLVFNFTL PELGVKMLDQ LGSTTSGVAL FAVGVTVGIR KIKLSMPAIG 240
IALLKVAVQP ALMFLIALAI GLPADQTTKA ILLVAFPGSA VAAMIATRFE KQEEETATAF 300
VVSAILSLIS LPIIIALTA* QKKLPLRELF A 330
<210> 3
<211> 47
<212> DNA
<213> Artificial Sequence
<400> 3
TTTACGGCTA GCTCAGTCCT AGGTATAGTG CTAGCAGGAA ACAGACC 47
<210> 4
<211> 23
<212> DNA
<213> Artificial Sequence
<400> 4
GGTCAGGAGG TAACTTATCA GCG 23
<210> 5
<211> 55
<212> DNA
<213> Artificial Sequence
<400> 5
CTGCTAGCAC TATACCTAGG ACTGAGCTAG CCGTAAAATG GCAGGGCTCC GTTTT 55
<210> 6
<211> 55
<212> DNA
<213> Artificial Sequence
<400> 6
AGGTATAGTG CTAGCAGGAA ACAGACCTTG AGCATCTTAG ATATCTTAAT CCTCC 55
<210> 7
<211> 33
<212> DNA
<213> Artificial Sequence
<400> 7
AAATCCAGTT CAGCAAAAAG CTCCCTTAAA GGG 33
<210> 8
<211> 33
<212> DNA
<213> Artificial Sequence
<400> 8
TTTTTGCTGA ACTGGATTTT CTTCTGAACC TGT 33
<210> 9
<211> 19
<212> DNA
<213> Artificial Sequence
<400> 9
ACGATGTCAG CAGCCAGCA 19
<210> 10
<211> 56
<212> DNA
<213> Artificial Sequence
<400> 10
AGTCCTAGGT ATAATACTAG TACAGAATAT TCGCGAAAAA AGTTTTAGAG CTAGAA 56
<210> 11
<211> 56
<212> DNA
<213> Artificial Sequence
<400> 11
TTCTAGCTCT AAAACTTTTT TCGCGAATAT TCTGTACTAG TATTATACCT AGGACT 56
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Генетическая модификация продуцирующих (S)-молочную кислоту термофильных бактерий | 2015 |
|
RU2671528C2 |
| ГЕННО-ИНЖЕНЕРНЫЕ БАКТЕРИИ ДЛЯ ПРОДУЦИРОВАНИЯ L-АРГИНИНА И СПОСОБ КОНСТРУИРОВАНИЯ И ПРИМЕНЕНИЯ ГЕННО-ИНЖЕНЕРНЫХ БАКТЕРИЙ | 2020 |
|
RU2820627C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ МЕТОДОМ ФЕРМЕНТАЦИИ, ШТАММ БАКТЕРИИ ESCHERICHIA COLI - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 1999 |
|
RU2207376C2 |
| МИКРООРГАНИЗМЫ ДЛЯ ПОЛУЧЕНИЯ L-ТИРОЗИНА И СПОСОБ ПОЛУЧЕНИЯ L-ТИРОЗИНА С ИХ ИСПОЛЬЗОВАНИЕМ | 2020 |
|
RU2779461C1 |
| МИКРООРГАНИЗМ, ПРОДУЦИРУЮЩИЙ ЛИЗИН, И СПОСОБ ПРОДУЦИРОВАНИЯ ЛИЗИНА | 2021 |
|
RU2797843C9 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 2006 |
|
RU2402610C2 |
| Новый экспортер L-триптофана и способ продуцирования L-триптофана с его использованием | 2019 |
|
RU2761871C1 |
| МИКРООРГАНИЗМ, СПОСОБНЫЙ ПРОДУЦИРОВАТЬ L- АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ПРИМЕНЕНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2013 |
|
RU2600875C2 |
| ШТАММЫ SPNK | 2011 |
|
RU2580015C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA | 2005 |
|
RU2311454C2 |
Изобретение относится к биотехнологии и представляет собой генетически сконструированную бактерию для продуцирования L-триптофана, где исходная бактерия модифицирована геном ywkB, и полученная генетически сконструированная бактерия обладает активностью продуцирования более высокого выхода L-триптофана, чем исходная бактерия; где модификация состоит в модифицировании псевдогенного локуса yeep геном ywkB. При осуществлении ферментирования штамма в качалочной колбе он может накапливать 15,2 г/л L-триптофана в течение 24 ч, что на 35% выше, чем у контрольного штамма. 3 н. и 10 з.п. ф-лы, 4 ил., 2 пр.
1. Генетически сконструированная бактерия для продуцирования L-триптофана, где исходная бактерия модифицирована геном ywkB, и полученная генетически сконструированная бактерия обладает активностью продуцирования более высокого выхода L-триптофана, чем исходная бактерия; где модификация состоит в модифицировании псевдогенного локуса yeep геном ywkB.
2. Генетически сконструированная бактерия по п. 1, где ген ywkB происходит из Bacillus subtilis.
3. Генетически сконструированная бактерия по пп. 1, 2, представляющая собой прокариотическую бактерию, предпочтительно грамотрицательную бактерию, более предпочтительно Escherichia coli, наиболее предпочтительно E. coli TRP 03.
4. Генетически сконструированная бактерия по п. 1, где ген ywkB имеет нуклеотидную последовательность, кодирующую следующую полипептидную последовательность:
A) белок, содержащий аминокислотную последовательность, показанную в SEQ ID NO: 2 в перечне последовательностей;
Б) белок, который содержит аминокислотную последовательность, полученную посредством делеции, замены и/или добавления одного или более аминокислотных остатков в аминокислотной последовательности, показанной в SEQ ID NO: 2 в перечне последовательностей, и при этом все еще обладает способностью улучшать продуцирование бактерией L-триптофана и структурных аналогов L-триптофана, таких как 5-гидрокситриптофан, и так далее;
В) белок, который имеет по меньшей мере 85%, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 96%, 97%, 98%, 99% идентичности с аминокислотной последовательностью, показанной в SEQ ID NO: 2, происходит из Bacillus subtilis и функционирует как транспортер метаболитов.
5. Генетически сконструированная бактерия по любому из пп. 1-4, где ген ywkB введен в исходную бактерию посредством гомологичной рекомбинации, вектора экспрессии или редактирования гена, где вектор экспрессии представляет собой предпочтительно плазмиду, например, средство редактирования гена относится к введению гена аминокислотной последовательности в хромосому исходной бактерии посредством метода CRISPR/Cas9.
6. Генетически сконструированная бактерия по любому из пп. 1-5, где ген ywkB находится под контролем сильного промотора, например, сильный промотор представляет собой промотор BBa_j23101 или промотор BBa_j23106, например, аминокислотная последовательность промотора BBa_j23106 показана в SEQ ID NO: 3.
7. Способ конструирования генетически сконструированной бактерии, которая продуцирует L-триптофан, где ген ywkB интегрируют в исходную бактерию, продуцирующую L-триптофан, посредством модифицирования псевдогенного локуса yeep геном ywkB, с получением генетически сконструированной бактерии, обладающей активностью продуцирования более высокого выхода L-триптофана, чем исходная бактерия.
8. Способ по п. 7, где исходная бактерия представляет собой E. coli TRP 03.
9. Способ по п. 7, где стадию интегрирования осуществляют посредством модифицирования псевдогенного локуса yeep геном ywkB, например, интегрирование осуществляют посредством редактирования гена с помощью CRISPR-Cas9.
10. Способ по п. 9, где редактирование гена с помощью CRISPR-Cas9 включает следующие стадии:
1) синтезирование интегрируемого генного фрагмента ywkB: конструирование фрагментов вышележащего и нижележащего гомологичного плеча псевдогена yeep, генного фрагмента ywkB и последовательности промотора BBa_j23106, где интегрируемый генный фрагмент ywkB содержит вышележащее гомологичное плечо гена yeep, нижележащее гомологичное плечо гена yeep, промоторный фрагмент BBa j23106 и генный фрагмент ywkB;
2) экспрессирование интегрируемого генного фрагмента ywkB в E. coli TRP 03 с использованием технологии CRISPR/Cas9.
11. Способ по любому из пп. 7-10, где последовательность гена ywkB представляет собой нуклеотидную последовательность, кодирующую следующую полипептидную последовательность:
A) белок, содержащий аминокислотную последовательность, показанную в SEQ ID NO: 2 в перечне последовательностей;
Б) белок, который содержит аминокислотную последовательность, полученную посредством делеции, замены и/или добавления одного или более аминокислотных остатков в аминокислотной последовательности, показанной в SEQ ID NO: 2 в перечне последовательностей, и при этом все еще обладает способностью улучшать продуцирование бактерией L-триптофана и структурных аналогов L-триптофана, таких как 5-гидрокситриптофан, и так далее;
В) белок, который имеет по меньшей мере 85%, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 96%, 97%, 98%, 99% идентичности с аминокислотной последовательностью, показанной в SEQ ID NO: 2, происходит из Bacillus subtilis и функционирует как транспортер метаболитов.
12. Способ продуцирования L-триптофана, включающий стадию ферментирования генетически сконструированной бактерии по любому из пп. 1-6 или генетически сконструированной бактерии, полученной способом по любому из пп. 7-11, с продуцированием L-триптофана.
13. Способ продуцирования L-триптофана по п. 12, включающий следующие стадии:
1) активирование бактерии;
2) приготовление посевного раствора;
3) ферментирование;
например, где на стадии (1) готовят культуру на скошенном агаре: инокулирование бактерии,
хранящегося при -80°C, на активированный скошенный агар методом штриха, культивирование при 37°C в течение 12 ч и однократное пересевание; на стадии (2) готовят посевную культуру в качалочной колбе: соскабливание пятна посевного материала на скошенном агаре с помощью микробиологической петли и инокулирование в коническую колбу на 500 мл, содержащую 30 мл посевной среды, укупоривание конической колбы девятью слоями марли и культивирование при 37°C и 200 об/мин в течение 8-10 ч; на стадии (3) готовят ферментационную культуру в качалочной колбе: инокулирование посевной культуры в концентрации 10-15% (об./об.) в коническую колбу на 500 мл, содержащую ферментационную среду с конечным объемом 30 мл, укупоривание конической колбы девятью слоями марли, культивирование при 37°C и 200 об/мин на качалке во время ферментирования, добавление аммиачной воды для поддержания pH при 7,0-7,2; добавление 60%-ного (масс./об.) раствора глюкозы для поддержания ферментации с феноловым красным в качестве индикатора, когда цвет ферментационного бульона больше не изменяется, это расценивается как отсутствие глюкозы, добавление 1-2 мл 60%-ного (масс./об.) раствора глюкозы для обеспечения начального значения концентрации глюкозы в ферментационном бульоне 20-40 г/л; период ферментирования продолжается в течение 22-26 ч.
| Микроорганизм рода Escherichia, продуцирующий L-триптофан, и способ получения L-триптофана с его использованием | 2016 |
|
RU2692645C2 |
| Микроорганизм рода Escherichia, продуцирующий L-триптофан, и способ продуцирования L-триптофана с использованием данного микроорганизма | 2015 |
|
RU2678139C2 |
| CN 108753860 B, 02.08.2019 | |||
| CN 108753860 A, 06.11.2018 | |||
| УСТРОЙСТВО ДЛЯ ПРИГОТОВЛЕНИЯ ГОРЯЧИХ НАПИТКОВ ИЗ РАСТВОРИМЫХ ИНГРЕДИЕНТОВ | 0 |
|
SU391585A1 |
