В данной заявке заявлен приоритет патентной заявки 201911211097.Х, поданной в Управление национальной интеллектуальной собственностью Китая 2 декабря 2019 года и названной "ГЕННО-ИНЖЕНЕРНЫЕ БАКТЕРИИ ДЛЯ ПРОДУЦИРОВАНИЯ L-АРГИНИНА И СПОСОБ КОНСТРУИРОВАНИЯ И ПРИМЕНЕНИЯ ГЕННО-ИНЖЕНЕРНЫХ БАКТЕРИЙ", описание которой во всей его полноте включено в данный документ посредством ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к технической области генной инженерии и относится к генно-инженерной бактерии, способной стабильно и эффективно продуцировать L-аргинин, и к способу ее конструирования и к ее применению.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
L-аргинин представляет собой частично незаменимую основную аминокислоту или, при определенных условиях, незаменимую аминокислоту у людей и животных, и обладает важными биохимическими и физиологическими функциями. В настоящее время, L-аргинин широко используют в медицине, промышленности, питании, косметике, животноводстве и других областях, и он обладает важным экономическим и социальным значением.
Способы получения L-аргинина главным образом включают способ гидролиза белка и способ микробиологической ферментации. По сравнению со способом экстракции белкового гидролизата, способ микробиологической ферментации обладает преимуществами, которые заключаются в относительно простом процессе получения, относительно небольшом воздействии на окружающую среду и высокой чистоте продукта, и пригоден для крупномасштабного промышленного производства.
В настоящее время аргинин-продуцирующие штаммы главным образом представляют собой Corynebacterium glutamicum, имеющие проблемы, заключающиеся в длительном периоде ферментации (90 ч-120 ч) и низкой интенсивности получения в процессе получения. В то же время, на существующий процесс ферментации Corynebacterium glutamicum сильно влияет качество эксципиентов, таких как жидкий кукурузный экстракт, и производство легко меняется. Кроме того, редактирование генов Corynebacterium glutamicum затруднено, что приводит к использованию плазмидных экспрессионных векторов в существующих аргинин-продуцирующих штаммах для усиления ключевых генов, связанных с синтезом аргинина. Однако в процессе ферментации множественные копии плазмид приводят к уровню нагрузки на рост бактерии, что приводит к снижению выхода в процессе поздней ферментации. Кроме того, в процессе получения плазмидный экспрессионный вектор легко утрачивается или необходимо добавлять определенное селективное давление, что приводит к проблеме высокой стоимости в процессе промышленного производства. Многие факторы затрудняют внедрение существующих в настоящее время аргинин-продуцирующих штаммов в промышленное производство.
Поскольку существует множество механизмов обратной связи в синтетическом и метаболическом путях аргинина, и множество метаболических путей аргинина и метаболическая сеть, вовлеченная в предшественники, требующиеся для синтеза аргинина, является сложной, в исходном исследовании и разработке промышленных аргинин-продуцирующих штаммов главным образом используют традиционный способ мутагенеза, комбинированный с отбором по резистентности к структурному аналогу аргинина. Выбранные исходные штаммы представляют собой, главным образом, Brevibacterium flavum, Corynebacterium crenatum и Corynebacterium glutamicum. Стратегия исследования сосредоточена на отборе мутантов структурных аналогов аргинина для облегчения механизмов обратной регуляции в процессе синтеза аргинина и улучшения внутриклеточного накопления L-аргинина. Среди них Li Shaoping и др. отобрали штамм Corynebacterium crenatum с дефицитом по гистидину, резистентностью к сульфагуанидину, резистентностью к D-аргинину, резистентностью к гомоаргинину и резистентностью к S-метилцистеину посредством пошагового мутагенеза с NTG (N-метил-N'-нитро-N-нитрозогуанидин) (CN 201010610917.5), и после эксперимента по ферментации накопление L-аргинина составляло 32,8 г/л, когда штамм культивировали в ферментере объемом 5 л в течение 96 ч. Однако L-аргинин-продуцирующий штамм, полученный путем мутагенеза и отбора с использованием структурного аналога, трудно ввести в крупномасштабное промышленное производство из-за его плохой генетической стабильности и легкости возникновения обратных мутаций и других недостатков.
С быстрым развитием генно-инженерной технологии способ конструирования L-аргинин-продуцирующих штаммов с использованием технологии метаболической инженерии постепенно вытеснил традиционные методы селекции с мутагенезом. У Corynebacterium glutamicum отсутствуют гены, вовлеченные в деградацию аргинина; и метаболический поток внутриклеточной глюкозы, поглощенной Corynebacterium glutamicum, проходящий через гликолитический путь с получением глутаминовой кислоты, представляющей собой один из основных предшественников в синтезе аргинина, является сильным, так что Corynebacterium glutamicum представляет собой основной выбор для конструирования L-аргинин-продуцирующих штаммов. Xu Meijuan et al. (Xu M, Rao Z, Yang J, et al. J md Microbiol Biotechnol, 2012, 39(3): 495-502). Генный кластер argCJBDFRGH для синтеза L-аргинина лигировали в шаттл-экспрессионный вектор pJCtac и вводили в Corynebacterium crenatum. После 96 ч ферментации выход L-аргинина из штамма увеличивался до 45,6 г/л. Park et al. (Park S H, Kim H U, Kim T Y, et al. Nature Communications, 2014, 5:4618---) использовали Corynebacterium glutamicum в качестве исходного штамма для увеличения устойчивости глутамина к структурным аналогам L-аргинина посредством случайного мутагенеза, и системные технологии метаболической инженерии для облегчения обратного ингибирования в процессе синтеза аргинина, для увеличения поступления NADPH (восстановленного никотинамидадениндинуклеотидфосфата) в процессе синтеза и для увеличения поступление предшественников. Наконец, после 96 ч ферментации в ферментере объемом 5 л накопление L-аргинина составляло 92,5 г/л, скорость превращения составляла 0,35 г аргинина/г глюкозы, и максимальная интенсивность продуцирования составляла 0,9 г аргинина/л/ч. Вышеупомянутые L-аргинин-продуцирующие штаммы обычно имеют проблемы, заключающиеся в длительном цикле продуцирования и низкой интенсивности продуцирования. Кроме того, в процессе конструирования штамма ключевые гены в синтезе аргинина лигируют в экспрессионный вектор для увеличения транскрипционных количеств ключевых ферментов, таким образом усиливая метаболический поток в пути синтеза аргинина. Однако в процессе продуцирования экспрессионный вектор легко утрачивается, или необходимо добавлять определенное селективное давление, поэтому сложно ввести эти штаммы в промышленное производство.
Благодаря преимуществам короткого ферментационного периода, ясного генетического фона, удобной молекулярной манипуляции и стабильного процесса ферментации, Escherichia coli стала наилучшим выбором для конструирования L-аргинин-продуцирующих промышленных штаммов. Ginesy et al. (Ginesy M, Belotserkovsky J, Enman J, et al. Microbial Cell Factories, 2015, 14(1): 29.) использовали Escherichia coli в качестве исходного штамма, нокаутировали ген argR для облегчения подавления аргинина по типу обратной связи, интегрировали мутантный ген argA214 (H15Y) для облегчения подавления аргинина по типу обратной связи в ArgA, нокаутировали гену adiA, связанный с деградацией аргинина, и нокаутировали гены speC и speF, связанные с деградацией орнитина, так что больший поток углерода в виде промежуточных метаболитов направлялся в сторону L-аргинина. После 42 ч культивирования в ферментере объемом 1 л накопление L-аргинина достигало 11,64 г/л, скорость превращения была 0,44 г аргинина/г глюкозы, и интенсивность продуцирования составляла 0,29 г аргинина/л/ч. Хотя период ферментации этого штамма очевидным образом укорочен, его накопление аргинина и интенсивность продуцирования все еще не удовлетворяют требованиям промышленного производства.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В свете вышеупомянутых проблем задача настоящего изобретения заключается в том, чтобы предложить генно-инженерную бактерию, способную стабильно и эффективно продуцировать L-аргинин, и способ ее конструирования и ее применение. Сконструированная бактерия обладает хорошими перспективами для промышленного применения.
В настоящем изобретении предложены следующие технические решения:
В первом аспекте настоящего изобретения предложен генно-инженерный бактериальный штамм для продуцирования L-аргинина, который содержит гены pyrAA и pyrAB, кодирующие карбамоилфосфат-синтетазу.
В одном из воплощений для генно-инженерного бактериального штамма в качестве исходного штамма берут Escherichia coli или Corynebacterium glutamicum, такие как Е. coli W3110 или E. coli MG1655.
В одном из воплощений гены pyrAA и pyrAB интегрированы в локус гена yjiT в Е. coli.
В одном из воплощений гены pyrAA и pyrAB происходят из Bacillus subtilis, в частности, гены pyrAA и pyrAB происходят из генов, кодирующих карбамоилфосфат-синтетазу в В. subtilis А260.
В одном из воплощений генно-инженерный бактериальный штамм дополнительно содержит ген, кодирующий фермент пути биосинтеза L-аргинина, выбранный из одного или более следующих ферментов: argC, argJ, argB, argD, argF, argG, argH; ген, кодирующий фермент пути биосинтеза L-аргинина, происходит из Corynebacterium glutamicum АТСС13032; в одном из воплощений ген, кодирующий фермент пути биосинтеза L-аргинина, управляется промотором Ptrc; в одном из воплощений ген, кодирующий фермент пути биосинтеза L-аргинина, интегрирован в генный локус yghX в Е. coli.
В одном из воплощений генно-инженерный бактериальный штамм дополнительно содержит ген lysE, кодирующий транспортер аргинина (референсная последовательность NCBI (Национальный центр биотехнологической информации): WP_143758438.1), и ген транспортера происходит из Corynebacterium efficiens; в одном из воплощений ген lysE интегрирован в генный локус ilvG в Е. coli.
В одном из воплощений генно-инженерный бактериальный штамм не содержит ген деградации L-аргинина, который может быть получен путем нокаута одного или более следующих генов: гена, кодирующего аргинин-декарбоксилазу, гена, кодирующего аргинин-сукцинилтрансферазу, гена, кодирующего ацетилорнитин-деацетилазу. Ген, кодирующий аргинин-декарбоксилазу, включает по меньшей мере один из speA (NCBI-Gene ID: 12933352) и adiA (NCBI-Gene ID: 12934085); ген, кодирующий аргинин-сукцинилтрансферазу, представляет собой astA (NCBI-GeneID: 12933241); ген, кодирующий ацетилорнитин-де ацетил азу, представляет собой argE (NCBI-GeneID: 12930574). В одном из воплощений генно-инженерный бактериальный штамм представляет собой Е. coli, в котором гены speA, adiA и astA одновременно нокаутированы.
В одном из воплощений генно-инженерный бактериальный штамм содержит гены pyrAA, pyrAB, argC, argJ, argB, argD, argF, argG, argH и lysE. В одном из воплощений генно-инженерный бактериальный штамм не содержит гены speA, adiA, astA и argE.
В настоящем изобретении гены pyrAA, pyrAB, argC, argJ, argB, argD, argF, argG, argH, lysE, speA, adiA, astA и argE не ограничены генами дикого типа, но также могут представлять собой мутанты, кодирующие соответствующие белки или искусственно модифицированные гены, где соответствующие белки включают замену, делецию или вставку одного или более аминокислотных остатков в одном или более сайтах, пока белки, кодируемые этими мутантами или искусственно модифицированными генами, обладают соответствующими активностями и не имеют функциональных дефектов. Эти гены зарегистрированы в GenBank, и специалист в данной области техники может получить эти гены при помощи ПЦР (полимеразной цепной реакции). В качестве примера, ген pyrAA представляет собой NCBI-GeneID: 937368, ген pyrAB представляет собой NCBI-GeneID: 936608, ген argC представляет собой NCBI-GeneID: 1019370, ген argJ представляет собой NCBI-GeneID: 1019371, ген argB представляет собой NCBI-GeneID: 1019372, ген argD представляет собой NCBI-GeneID: 1019373, ген argF представляет собой NCBI-GeneID: 1019374, ген argG представляет собой NCBI-GeneID: 1019376, ген argH представляет собой NCBI-GeneID: 1019377, ген lysE имеет нуклеотидную последовательность, представленную в SEQ ID NO: 68 (идентификатор последовательности в NCBI: WP 143758438.1), ген speA представляет собой NCBI-GeneID: 12933352, ген adiA представляет собой NCBI-GeneID: 12934085, ген astA представляет собой NCBI-GeneID: 12933241 и ген argE представляет собой NCBI-GeneID: 12930574.
Во втором аспекте настоящего изобретения предложен способ конструирования вышеупомянутого генно-инженерного бактериального штамма, включающий следующую стадию: (1) интегрирование генов pyrAA и pyrAB в геном исходного штамма.
Например, исходный штамм представляет собой Е. coli, такой как E. coli W3110 (АТСС27325).
В одном из воплощений способ конструирования дополнительно возможно включает одну или более следующих стадий:
(2) интегрирование генов ферментов пути биосинтеза аргинина, включающих один или более из генов argC, argJ, argB, argD, argF, argG, argH; и/или интегрирование гена lysE, кодирующего транспортер аргинина;
(3) нокаутирование гена, кодирующего аргинин-декарбоксилазу, гена, кодирующего аргинин-сукцинилтрансферазу, и/или гена, кодирующего ацетилорнитин-деацетилазу; например, ген, кодирующий аргинин-декарбоксилазу, включает по меньшей мере один из генов speA и adiA; ген, кодирующий аргинин-сукцинилтрансферазу, представляет собой ген astA; ген, кодирующий ацетилорнитин-деацетилазу, представляет собой ген argE.
В одном из воплощений способ конструирования включает стадии:
(1) нокаутирование следующих трех генов в Е. coli: гена speA, кодирующего аргинин-декарбоксилазу, гена adiA, кодирующего аргинин-декарбоксилазу, и гена astA, кодирующего аргинин-сукцинилтрансферазу;
(2) нокаутирование гена argE, кодирующего ацетилорнитин-деацетилазу, в Е. coli, и, возможно, интегрирование гена argJ, кодирующего глутаматацетилтрансферазу в Е. coli;
(3) интегрирование следующего генного кластера, связанного с биосинтезом аргинина: argC, argJ, argB, argD, argF, argG и argH, который управляется промотором Ptrc;
(4) интегрирование генов pyrAA и pyrAB, кодирующих карбамоилфосфат-синтетазу;
(5) интегрирование гена lysE, кодирующего транспортер аргинина, в геном Е. coli. Специалисту в данной области техники может быть понятно, что
последовательность стадий (1)-(5) вышеприведенного способа конструирования по настоящему изобретению, не ограничена, и может быть осуществлена в любой последовательности, которая может быть реализована специалистами в данной области техники. Предпочтительно, стадии (1)-(5) осуществляют последовательно.
Любой способ нокаутирования гена или сайленсинга гена, известный в области техники, может быть использован для достижения вышеупомянутого нокаутирования генов, и любой способ, известный в области техники, также может быть использован для достижения интегрирования гена, такой как гомологичная рекомбинация, перекрывающаяся ПЦР, скрининг мутагенеза или редактирование гена и другие методы. Например, нокаутирование генов может быть достигнуто путем удаления конкретной области гена, так что он не обладает функцией экспрессии интересующего белка, или путем осуществления замены, делеции и вставки одного или более нуклеотидов в кодирующей области или в промоторной области путем сайт-специфической мутации и т.п., и химические реактивы также могут быть использованы для уменьшения или ликвидации транскрипции конкретного гена.
В одном из воплощений в способе конструирования используют опосредованную CRTSPR/Cas9 технологию редактирования генов для осуществления интегрирования и нокаутирования генов.
В одном из воплощений способ конструирования включает стадии конструирования рекомбинантного фрагмента и плазмиды pGRB.
В одном из воплощений стадия конструирования плазмиды pGRB включает: создание целевой последовательности, получение фрагмента ДНК, содержащего эту целевую последовательность, и рекомбинацию фрагмента ДНК, содержащего эту целевую последовательность, с линеаризованным фрагментом вектора; в конкретном воплощении целевая последовательность представляет собой 5'-NGG-3'.
В одном из воплощений в способе конструирования стадия конструирования рекомбинантного фрагмента включает конструирование рекомбинантного фрагмента для интегрирования генов или нокаутирования генов. Среди них стадия конструирования рекомбинантного фрагмента для интегрирования гена включает: использование генома исходного штамма в качестве матрицы, создание праймеров для расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч в соответствии с расположенными против хода транскрипции и по ходу транскрипции последовательностями предполагаемого сайта встраивания целевого гена, и создание праймеров в соответствии с целевым геномом для амплификации фрагмента целевого гена, и затем осуществление перекрывающейся ПЦР для получения рекомбинантного фрагмента. Стадия конструирования рекомбинантного фрагмента для нокаутирования генов включает: использование расположенных против хода транскрипции и по ходу транскрипции последовательностей нокаутируемого гена в качестве матриц, создание праймеров для расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч; соответствующую амплификацию расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч при помощи ПЦР, и затем получение рекомбинантного фрагмента посредством перекрывающейся ПЦР.
В одном из воплощений способ конструирования включает: одновременную трансформацию плазмиды pGRB и вышеупомянутого рекомбинантного фрагмента в компетентные в отношении электропорации клетки, содержащие pREDCas9, и элиминацию плазмид с получением рекомбинантного генно-инженерного бактериального штамма.
В настоящем изобретении предложено применение вышеупомянутого генно-инженерного бактериального штамма в получении L-аргинина.
В настоящем изобретении также предложен способ продуцирования L-аргинина путем использования вышеупомянутого генно-инженерный бактериального штамма, включающий: приведение в контакт вышеупомянутого генно-инженерного штамма Е. coli с ферментационной средой, и проведение ферментации с получением L-аргинина.
В соответствии с настоящим изобретением ферментация включает ферментацию во встряхиваемой колбе или ферментацию в ферментере.
В одном из воплощений количество инокулята для ферментации во встряхиваемой колбе составляет 10-15%, условия ферментации составляют 37°С, 200 об./мин на встряхивающей платформе, рН поддерживают на уровне 7,0-7,2 во время ферментации, и рН может быть скорректирован путем добавления аммиачной воды. Во время ферментации также может быть добавлен раствор глюкозы для поддержания ферментации, и масс-объемная концентрация раствора глюкозы предпочтительно составляет 60% (масс./об.). Предпочтительно, время ферментации при ферментации во встряхиваемой колбе составляет 26-30 ч. В настоящем изобретении дополнительное количество раствора глюкозы не ограничено конкретным образом, и концентрация глюкозы в ферментационном бульоне может поддерживаться на уровне менее 5 г/л, например 1-5 г/л.
В одном из воплощений ферментацию во встряхиваемой колбе осуществляют в колбе Эрленмейера объемом 500 мл для ферментации. После 26-30 ч ферментации во встряхиваемой колбе концентрация L-аргинина в ферментационном бульоне может достигать 30-32 г/л.
В одном из воплощений количество инокулята для ферментации в ферментере составляет 15-20%, температура ферментации составляет 35°С, и растворенный кислород составляет 25-35%. Во время ферментации рН контролируют таким образом, чтобы он был стабилен на уровне 7,0-7,2, и рН может быть скорректирован путем добавления аммиачной воды; когда глюкоза в среде истощается, тогда осуществляют периодическое добавление 80% (масс./об.) раствор глюкозы для поддержания концентрации глюкозы в ферментационной среде на уровне 0,1-5 г/л.
В одном из воплощений ферментацию в ферментере осуществляют в ферментере объемом 5 л для ферментации. После 50-55 ч ферментации в ферментере объемом 5 л накопление L-аргинина достигает 130-135 г/л. Скорость превращения достигает 0,48 г аргинина/г глюкозы, и интенсивность продуцирования достигает 2,5 г аргинина/л/ч.
В настоящем изобретении для ферментации может быть использована ферментационная среда для Е. coli, известная в данной области техники.
В одном из воплощений ферментационная среда для ферментации во встряхиваемой колбе состоит из: 20-40 г/л глюкозы, 1-3 г/л дрожжевого экстракта, 2-3 г/л пептона, 3-6 г/л K2HPO4, 1-2 г/л MgSO4⋅7H2O, 15-20 мг/л FeSO4⋅7H2O, 15-20 мг/л MnSO4⋅7H2O, 1-3 мг/л каждого из VB1, VB3, VB5, VB12 и VH, остальное представляет собой воду, рН 7,0-7,2.
В одном из воплощений ферментационная среда для ферментации в ферментере состоит из: 10-25 г/л глюкозы, 1-5 г/л дрожжевого экстракта, 1-5 г/л пептона, 1-5 г/л K2HPO4, 1-3 г/л MgSO4⋅7H2O, 10-30 мг/л FeSO4⋅7H2O, 10-30 мг/л MnSO4⋅H2O, 1-3 мг/л каждого из VB1, VB3, VB5, VB12 и VH, остальное представляет собой воду, рН 7,0-7,2. Благоприятные эффекты:
В настоящем изобретении в качестве исходного штамма выбрали Е. coli с коротким циклом роста, ясным метаболическим путем и удобной молекулярной манипуляцией, начиная с генной инженерии синтетического и метаболического пути L-аргинина и инженерии всей метаболической сети, проанализировали и реконструировали метаболический поток, связанный с аргинином в пути синтеза L-аргинина и полной аминокислотной метаболической сети, и, наконец, получили генно-инженерный бактериальный штамм, который обладает ясным генетическим кодом, не несет плазмиды, не претерпевает мутагенез и способен стабильно и эффективно продуцировать L-аргинин.
Штамм Е. coli, полученный в соответствии с настоящим изобретением, создает путь циркуляции L-аргинина, улучшает поток L-аргинина и поступление предшественников, уменьшает деградацию L-аргинина и способствует накоплению и транспорту L-аргинина, таким образом, эффективно увеличивая выход L-аргинина.
Продуцирующий L-аргинин генно-инженерный бактериальный штамм в соответствии с настоящим изобретением может накапливать L-аргинин в концентрации 130-135 г/л после культивирования в ферментере объемом 5 л в течение 50-55 ч. Скорость превращения может достигать 0,48 г аргинина/г глюкозы и интенсивность продуцирования может достигать 2,5 г аргинина/л/ч. По сравнению со штаммом, о котором сообщалось в Park et al. (накопление L-аргинина составляет 92,5 г/л после выращивания в ферментере объемом 5 л в течение 96 ч, скорость превращения составляет 0,35 г аргинина/г глюкозы, и максимальная интенсивность продуцирования составляет 0,9 г аргинин/л/ч), штамм в соответствии с настоящим изобретением обладает преимуществами, заключающимися в более высокой производительностью в отношении L-аргинина, без подвергания мутагенной обработке, не несет плазмидные векторы, обладает коротким циклом ферментации, ясным генетическим фоном, стабильным метаболизмом, высокой интенсивностью продуцирования, и, таким образом, обладает хорошими перспективами для промышленного применения.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 блок (а) демонстрирует карту плазмиды pREDCas9, а блок (b) демонстрирует карту плазмиды pGRB.
Фиг. 2 демонстрирует электрофореграмму конструирования и верификацию фрагмента для нокаутирования гена speA, где М: маркер ДНК 1 кб; полоса 1: расположенное против хода транскрипции гомологичное плечо; полоса 2: расположенное по ходу транскрипции гомологичное плечо; полоса 3: перекрывающийся фрагмент; полоса 4: исходный штамм (контроль); полоса 5: идентифицированный фрагмент из положительных бактерий.
Фиг. 3 демонстрирует электрофореграмму конструирования и верификации фрагмента для нокаутирования гена adiA, где М: маркер ДНК 1 кб; полоса 1: расположенное против хода транскрипции гомологичное плечо; полоса 2: расположенное по ходу транскрипции гомологичное плечо; полоса 3: перекрывающийся фрагмент; полоса 4: исходный штамм (контроль); полоса 5: идентифицированный фрагмент из положительных бактерий.
Фиг. 4 демонстрирует электрофореграмму конструирования и верификации фрагмента для нокаутирования гена astA, где М: маркер ДНК 1 кб; полоса 1: расположенное против хода транскрипции гомологичное плечо; полоса 2: расположенное по ходу транскрипции гомологичное плечо; полоса 3: перекрывающийся фрагмент; полоса 4: исходный штамм (контроль); полоса 5: идентифицированный фрагмент из положительных бактерий.
Фиг. 5 демонстрирует электрофореграмму конструирования и верификации фрагмента для интегрирования argJ, где М: маркер ДНК 1 кб; полоса 1: расположенное против хода транскрипции гомологичное плечо; полоса 2: расположенное по ходу транскрипции гомологичное плечо; полоса 3: перекрывающийся фрагмент; полоса 4: исходный штамм (контроль); полоса 5: идентифицированный фрагмент из положительных бактерий.
Фиг. 6 демонстрирует электрофореграмму конструирования и верификации фрагмента для интегрирования argC-argJ, где М: маркер ДНК 1 кб; полоса 1: расположенное против хода транскрипции гомологичное плечо; полоса 2: фрагмент argC-argJ; полоса 3: расположенное по ходу транскрипции гомологичное плечо; полоса 4: перекрывающийся фрагмент; полоса 5: исходный штамм (контроль); полоса 6: идентифицированный фрагмент из положительных бактерий.
Фиг. 7 демонстрирует электрофореграмму конструирования и верификации фрагмента для интегрирования гена argB-argD-argF, где М: маркер ДНК 1 кб; полоса 1: расположенный против хода транскрипции фрагмент argB-argD-argF-фратмет гена argB-argD-argF; полоса 2: расположенное по ходу транскрипции гомологичное плечо; полоса 3: перекрывающийся фрагмент; полоса 4: исходный штамм (контроль); полоса 5: идентифицированный фрагмент из положительных бактерий.
Фиг. 8 демонстрирует электрофореграмму конструирования и верификации фрагмента для интегрирования гена argG-argH, где М: маркер ДНК 1 кб; полоса 1: расположенное против хода транскрипции гомологичное плечо; полоса 2: фрагмент argG-argH; полоса 3: расположенное по ходу транскрипции гомологичное плечо; полоса 4: перекрывающийся фрагмент; полоса 5: исходный штамм (контроль); полоса 6: идентифицированный фрагмент из положительных бактерий.
Фиг. 9 демонстрирует электрофореграмму конструирования и верификации первого фрагмента для интегрирования pyrAA-pyrAB, где М: маркер ДНК 1 кб; полоса 1: расположенное против хода транскрипции гомологичное плечо; полоса 2: фрагмент 1-pyrAA-pyrAB; полоса 3: расположенное по ходу транскрипции гомологичное плечо; полоса 4: перекрывающийся фрагмент; полоса 5: исходный штамм (контроль); полоса 6: идентифицированный фрагмент из положительных бактерий.
Фиг. 10 демонстрирует электрофореграмму конструирования и верификации второго фрагмента для интегрирования pyrAA-pyrAB, где М: маркер ДНК 1 кб; полоса 1: расположенный против хода транскрипции фрагмент pyrAA-расположенное по ходу транскрипции гомологичное плечо pyrAA-pyrAB; полоса 2: расположенное по ходу транскрипции гомологичное плечо; полоса 3: перекрывающийся фрагмент; полоса 4: исходный штамм (контроль); полоса 5: идентифицированный фрагмент из положительных бактерий.
Фиг. 11 демонстрирует электрофореграмму конструирования и верификации фрагмента для интегрирования lysE, где М: маркер ДНК 1 кб; полоса 1: расположенное против хода транскрипции гомологичное плечо; полоса 2: фрагмент lysE; полоса 3: расположенное по ходу транскрипции гомологичное плечо; полоса 4: перекрывающийся фрагмент; полоса 5: исходный штамм (контроль); полоса 6: идентифицированный фрагмент из положительных бактерий.
Фиг. 12 демонстрирует кривую периодической ферментации с добавлением субстрата штамма Е. coli W3110 ARG10 в ферментере объемом 5 л.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Вышеприведенные и другие характеристики и преимущества настоящего изобретения раскрыты и проиллюстрированы более подробно ниже путем описания примеров в соответствии с настоящим изобретением. Следует понимать, что следующие примеры предназначены для иллюстрации технических решений в соответствии с настоящим изобретением, а не для ограничения объема патентной охраны настоящего изобретения, определяемого формулой изобретения и ее эквивалентными решениями.
Если не указано иное, то материалы и реактивы, указанные здесь, имеются в продаже или могут быть получены специалистом в данной области техники в соответствии с предшествующим уровнем техники.
Пример 1: Конструирование генно-инженерного бактериального штамма Е. coli TRP 05
1. Способ редактирования гена
Способ редактирования гена, используемый в настоящем изобретении, приведен в литературе " Li Y, Lin Z, Huang С, et al. Metabolic engineering of Escherichia coli using CRISPR-Cas9 meditated genome editing. Metabolic engineering, 2015, 31:13-21" и карты двух плазмид, использованных в этом методе, представлены на Фиг. 1. Среди них вектор pREDCas9 несет систему элиминации гРНК экпрессионной плазмиды pGRB, систему рекомбинации Red фага "к и систему белковой экспрессии Cas9, устойчивость к спеномицину (рабочая концентрация: 100 мг/л), культивирование при 32°С; где в векторе pGRB используется pUC18 в качестве скелета и содержится промотор J23100, последовательность связывающегося с gRNA-Cas9 домена и терминирующая последовательность, устойчивость к ампициллину (рабочая концентрация: 100 мг/л), культивирование при 37°С.
Конкретные стадии данного способа:
1.1 Конструирование плазмиды pGRB
Задача конструирования плазмиды pGRB заключается в том, чтобы транскрибировать соответствующую гРНК с образованием комплекса с белком Cas9, и распознать целевой сайт целевого гена путем спаривания оснований и РАМ (мотив, смежный с протоспейсером) с получением двухцепочечного разрыва целевой ДНК. Плазмиду pGRB конструировали путем рекомбинации фрагмента ДНК, содержащего целевую последовательность с линеаризованным фрагментом вектора.
1.1.1 Создание целевой последовательности
Инструменты CRISPR RGEN использовали для конструирования целевой последовательности (РАМ: 5'-NGG-3').
1.1.2 Получение фрагмента ДНК, содержащего целевую последовательность Создавали праймер 5'-концевая последовательность линеаризованного вектора (15 п.о.) - сайт рестрикции - целевая последовательность (без последовательности РАМ)-концевая последовательность линеаризованного вектора (15 п. о)-3' и его обращенный комплементарный праймер, и фрагмент ДНК, содержащий целевую последовательность, получали путем отжига одноцепочечной ДНК. Условия реакции: предварительная денатурация при 95°С в течение 5 мин; отжиг при 30-50°С в течение 1 мин. Система отжига была следующей:

1.1.3 Получение линеаризованного вектора
Линеаризация вектора, используемого в способе амплификации посредством обратной ПЦР амплификации.
1.1.4 Реакция рекомбинации
Система рекомбинации представлена в таблице 2. Все используемые рекомбиназы представляли собой ферменты из наборов для одностадийного клонирования серий ClonExpress® П. Условия рекомбинации: 37°С, 30 мин.

1.1.5 Трансформация плазмиды
Десять мкл реакционного раствора добавляли к 100 мл химически компетентных клеток DH5a и осторожно смешивали. Полученную в результате смесь охлаждали в ледяной бане в течение 20 мин, подвергали тепловому шоку при 42°С в течение 45-90 с, немедленно охлаждали в ледяной бане в течение 2-3 мин, добавляли 900 мкл SOC, и поддерживали при 37°С в течение 1 ч. Смеси центрифугировали при 8000 об./мин в течение 2 мин, часть супернатанта удаляли, и оставшиеся 200 мкл супернатанта использовали для ресуспендирования клеток. Клетки затем распределяли на чашке, содержащей 100 мг/л ампициллина, и чашку помещали дном вверх и культивировали при 37°С в течение ночи. После выращивания единичных колоний на чашке положительные рекомбинанты идентифицировали посредством ПЦР и точечно отбирали.
1.1.6 Идентификация клонов
ПЦР-положительные колонии инокулировали в среду LB (Луриа-Бертани), содержащую 100 мг/л ампициллина для ночной культуры, и бактерии консервировали. Плазмиды экстрагировали и идентифицировали путем ферментативного расщепления.
1.2 Получение рекомбинантных фрагментов ДНК
Рекомбинантный фрагмент для нокаута состоит из расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч нокаутируемого гена (расположенное против хода транскрипции гомологичное плечо -расположенное по ходу транскрипции гомологичное плечо); рекомбинантный фрагмент для интегрирования состоит из расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч сайта интегрирования и интегрируемого фрагмента гена (расположенное против хода транскрипции гомологичное плечо-целевой ген -расположенное по ходу транскрипции гомологичное плечо). С использованием программного обеспечения для конструирования праймеров primer5 расположенные против хода транскрипции и по ходу транскрипции последовательности нокаутируемого гена или интегрируемого сайта использовали в качестве матрицы для создания праймеров для расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч (длина продукта амплификации: примерно 400-500 п.о.); интегрируемый ген использовали в качестве матрицы для создания праймеров для амплификации интегрированного гена. После амплификации, соответственно, расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч и фрагмента целевого гена при помощи ПЦР, рекомбинантный фрагмент получали посредством перекрывающейся ПЦР. Система ПЦР и способ представлены в следующей таблице 3:

Система перекрывающейся ПЦР представлена в следующей таблице 4:

Условия реакции ПЦР (фермент PrimeSTAR HS из Takara Bio): предварительная денатурация при 95°С в течение 5 мин; 30 циклов денатурации при 98°С в течение 10 с, отжиг при (Tm-3/5)°С в течение 15 с, элонгация при 72°С; и окончательная элонгация при 72°С в течение 10 мин; хранение при 4°С.
1.3 Трансформация плазмиды и рекомбинантного фрагмента ДНК
1.3.1 Трансформация pREDCas9
Плазмиду pREDCas9 подвергали электротрансформации в электропорационно-компетентные клетки W3110 посредством электротрансформации. Клетки выделяли и культивировали и затем наносили на чашку LB, содержащую спектиномицин, и культивировали при 32°С в течение ночи. Единичные колонии, выросшие на чашке с антибиотиком, подвергали ПЦР колоний с идентификацией праймеров для отбора положительных рекомбинантов.
1.3.2 Приготовление электропорационно-компетентных клеток целевого штамма, содержащих pREDCas9
Штамм выращивали при 32°С до тех пор, пока культура не достигала OD600 от 0,1 до 0,2 и затем добавляли IPTG (изопропилтиогалактозид) (до конечной концентрации 0,1 мМ). Выращивание продолжали до тех пор, пока величина OD600 не достигала от 0,6 до 0,7. Полученные клетки использовали для приготовления компетентных клеток. Цель добавления IPTG заключается в индукции экспрессии рекомбиназы в плазмиде pREDCas9. Среда и способ приготовления, требующиеся для получения компетентных клеток, относятся к обычным стандартным операциям.
1.3.3 Трансформация pGRB и рекомбинантного фрагмента ДНК
Плазмиду pGRB и рекомбинантный фрагмент ДНК подвергали одновременной электротрансформации в электропорационно-компетентные клетки, содержащие pREDCas9. После электротрансформации клетки выделяли и выращивали, и затем распределяли на чашке LB, содержащей ампициллин и спектиномицин, и культивировали при 32°С в течение ночи. Верификацию колоний при помощи ПЦР осуществляли с использованием прямого праймера для расположенного против хода транскрипции гомологичного плеча и обратного праймера для расположенного по ходу транскрипции гомологичного плеча, или с использованием специально созданных праймеров для идентификации для скрининга положительных рекомбинантов, и рекомбинанты сохраняли.
1.4 Элиминация плазмид
1.4.1 Элиминация плазмиды pGRB
Положительный рекомбинант выращивали в течение ночи в среде LB, содержащей 0,2% арабинозы, и после подходящих разведений культуру распределяли на чашке LB, содержащей спектиномицин, и культивировали при 32°С в течение ночи. Полученные рекомбинанты затем инокулировали на чашки LB, содержащие ампициллин и спектиномицин, соответственно, и единичные колонии, которые не росли на чашке, содержащей амгащиллин, но росли на чашке, содержащей спектиномицин, подвергали точечному отбору и сохраняли.
1.4.2 Элиминация плазмиды pREDCas9
Положительный рекомбинант переносили в жидкую среду LB без антибиотиков, культивировали в течение ночи при 42°С, и после соответствующих разведений культуру распределяли на чашке LB без антибиотиков и культивировали при 37°С в течение ночи. Полученные рекомбинанты затем инокулировали на чашки LB, содержащие спектиномицин, и без антибиотиков, соответственно, единичные колонии, которые не росли на чашке со спектиномицином, но росли на чашке LB без антибиотиков, подвергали точечному отбору и сохраняли.
2. Праймеры, используемые в конструировании штамма, представлены в таблице 5:

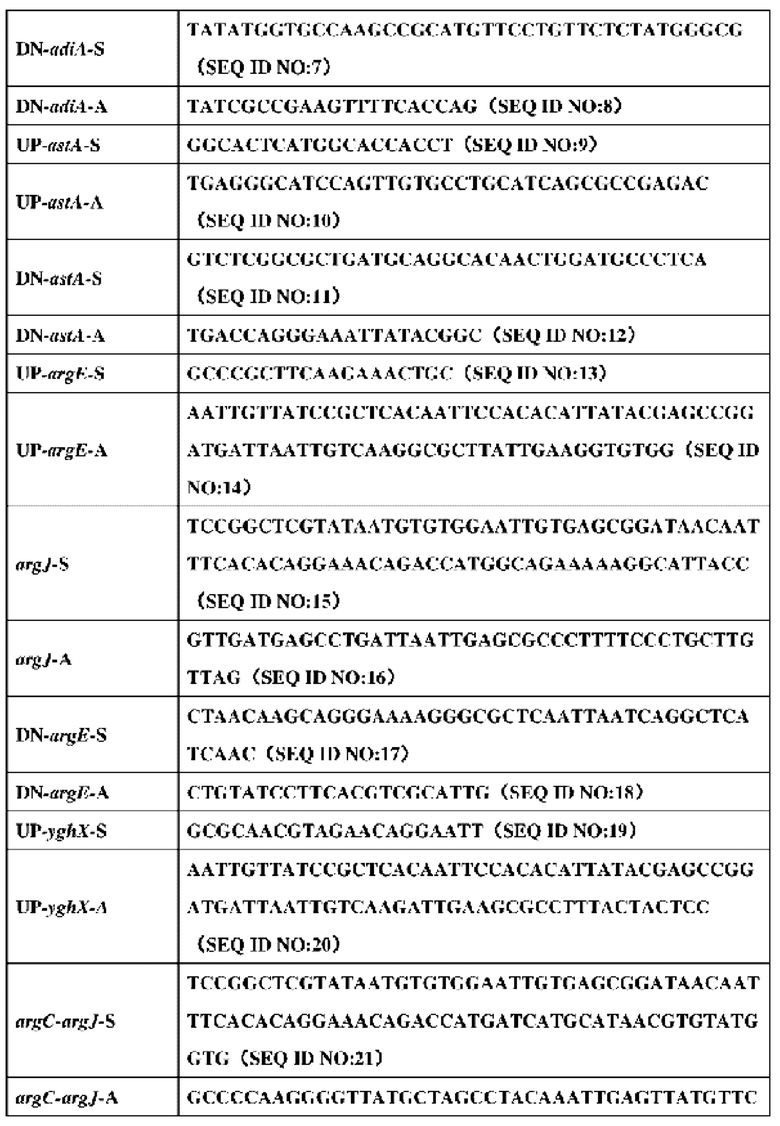
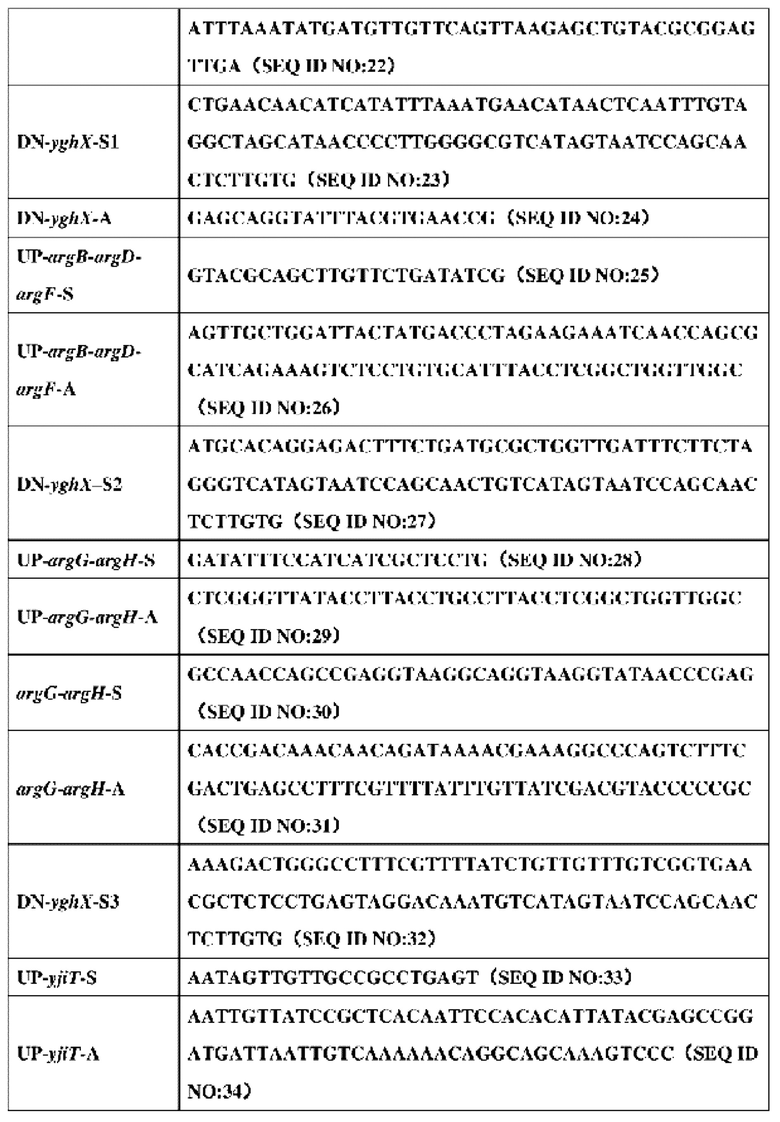
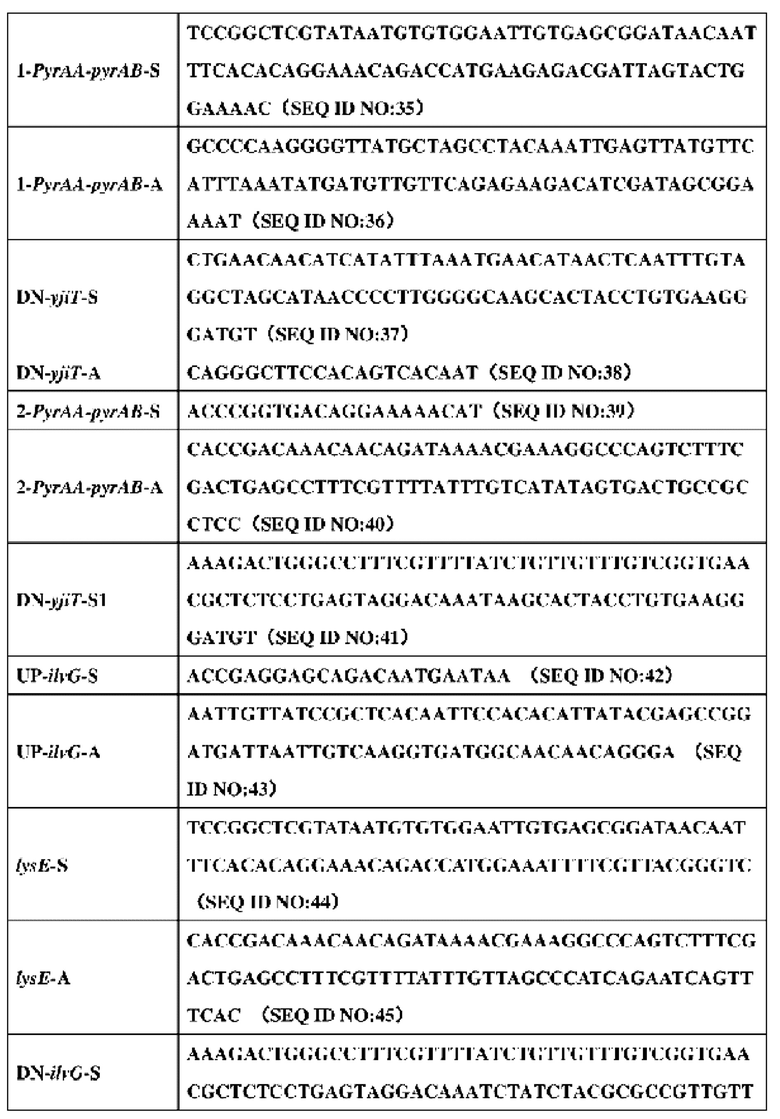
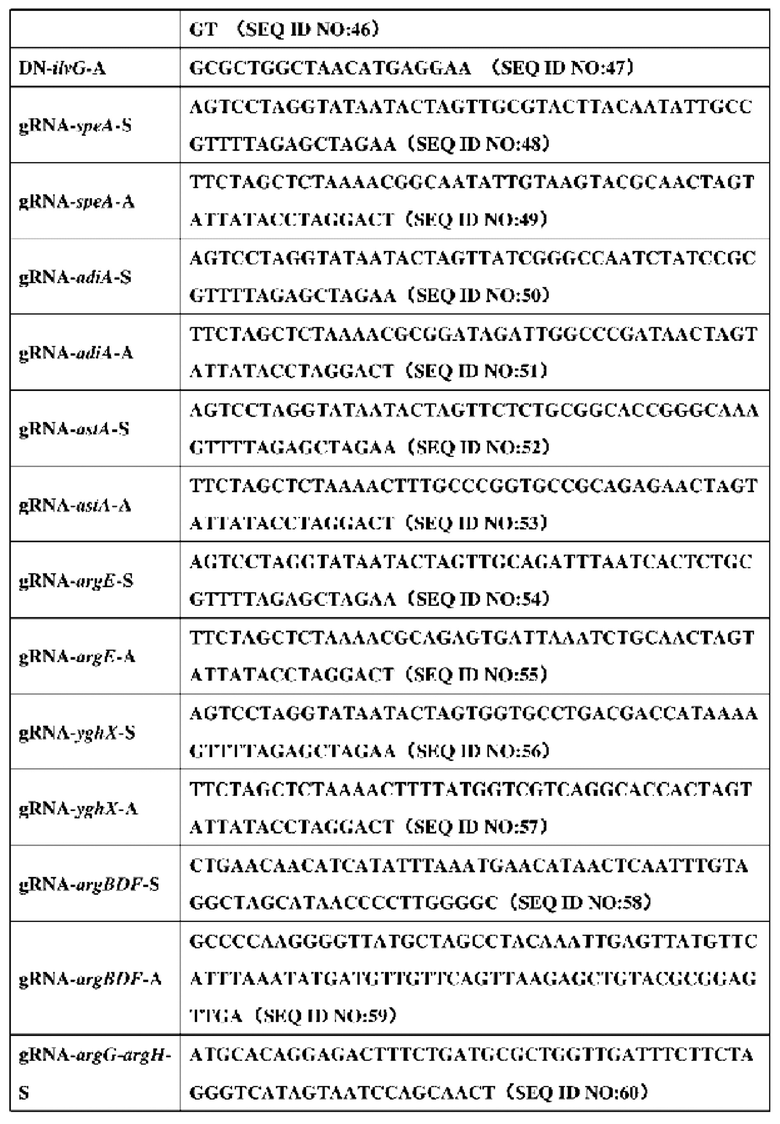

3. Конкретный процесс конструирования штамма
3.1 Нокаут трех генов speA, adiA и astA
3.1.1 Нокаут гена speA
С использованием генома Е. coli W3110 (АТСС27325) в качестве матрицы осуществляли ПЦР с праймерами для расположенного против хода транскрипции гомологичного плеча (UV-speAS, UP speA-А) и праймерами для расположенного по ходу транскрипции гомологичного плеча (DN-speA-S, DN-speA-A), созданными в соответствии с расположенными против хода транскрипции и по ходу транскрипции последовательностями его гена speA (NCBI-GeneID: 12933352), для амплификации расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч гена speA. Способ перекрывающейся ПНР применяли для слияния вышеприведенных фрагментов с получением фрагмента для нокаутирования гена speA (расположенное против хода транскрипции гомологичное плечо-расположенное по ходу транскрипции гомологичное плечо). Фрагмент ДНК, полученный путем отжига праймеров gECNA-speA-S и gRNA-speA-A, лигировали с плазмидой pGRB для конструирования рекомбинантной плазмиды pGRB-speA. Компетентные клетки Е. coli "W3110 готовили в соответствии со способами, описанными в разделах 1.3 и 1.4. Плазмиду pGRB-speA и фрагмент для нокаутирования гена speA одновременно подвергали электротрансформации в компетентные клетки, и наконец получали штамм, названный Е. coli W3110 ARG1. Электрофореграмма конструкции фрагмента для нокаутирования гена speA и ПЦР-верификация положительных бактерий представлена на Фиг. 2, где длина расположенного против хода транскрипции гомологичного плеча должна составлять 397 п.о., длина расположенного по ходу транскрипции гомологичного плеча должна составлять 468 п.о., и полная длина перекрывающегося фрагмента должна составлять 865 п.о., и для ПЦР-верификации длина амплифицированного путем ПЦР фрагмента положительных бактерий должна составлять 2752 п.о., и длина амплифицированного путем ПЦР фрагмента исходных бактерий должна составлять 865 п.о.
3.1.2 Нокаут гена adiA
С использованием генома Е. coli W3110 (АТСС27325) в качестве матрицы, осуществляли ПЦР с праймерами для расположенного против хода транскрипции гомологичного плеча (UP-adiAS, UP-adiA-A) и праймерами для расположенного по ходу транскрипции гомологичного плеча (DN-adiA-S, DN-adiA-A), созданными в соответствии с расположенными против хода транскрипции и по ходу транскрипции последовательностями его гена adiA (NCBI-GeneID: 12934085) для амплификации расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч гена adiA. Способ перекрывающейся ПЦР применяли для слияния вышеприведенных фрагментов с получением фрагмента для нокаутирования гена adiA (расположенное против хода транскрипции гомологичное плечо-расположенное по ходу транскрипции гомологичное плечо). Фрагмент ДНК, полученный путем отжига праймеров gRNA-adiA-S и gRNA-adiA-A, лигировали с плазмидой pGRB для конструирования рекомбинантной плазмиды pGRB-adiA. Компетентные клетки Е. coli W3110 ARG1 получали в соответствии со способами, описанными в разделах 1.3 и 1.4. Плазмиду pGRB-adiA и фрагмент для нокаутирования гена adiA одновременно подвергали электротрансформации в компетентные клетки, и наконец получали штамм, названный Е. coli W3110 ARG2. Электрофореграмма конструкции фрагмента для нокаутирования гена adiA и ПЦР-верификация положительных бактерий представлена на Фиг. 3, где длина расположенного против хода транскрипции гомологичного плеча должна составлять 806 п.о., длина расположенного по ходу транскрипции гомологичного плеча должна составлять 402 п.о., и полная длина перекрывающегося фрагмента должна составлять 1208 п.о., и для ПЦР-верификации длина амплифицированного путем ПЦР фрагмента положительных бактерий должна составлять 2124 п.о., и длина амплифицированного путем ПЦР фрагмента исходных бактерий должна составлять 1208 п.о.
3.1.3 Нокаут гена astA
С использованием генома Е. coli W3110 (АТСС27325) в качестве матрицы, осуществляли ПЦР с праймерами для расположенного против хода транскрипции гомологичного плеча (UP-asL4-S, UP-astA-A) и праймерами для расположенного по ходу транскрипции гомологичного плеча (DN-astA-S, DN-astA-A), созданными в соответствии с расположенными против хода транскрипции и по ходу транскрипции последовательностями его гена adiA (NCBI-GeneID: 12933241) для амплификации расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч гена astA. Способ перекрывающейся ПЦР применяли для слияния вышеприведенных фрагментов с получением фрагмента для нокаутирования гена astA (расположенное против хода транскрипции гомологичное плечо-расположенное по ходу транскрипции гомологичное плечо). Фрагмент ДНК, полученный путем отжига праймеров gRNA-ast4-S и gRNA-astA-A, лигировали с плазмидой pGRB для конструирования рекомбинантной плазмиды pGRB-astA. Компетентные клетки Е. coli W3110 ARG2 получали в соответствии со способами, описанными в разделах 1.3 и 1.4. Плазмиду pGRB-asL4 и фрагмент для нокаутирования гена astA одновременно подвергали электротрансформации в компетентные клетки, и наконец получали штамм, названный Е. coli W3110 ARG3. Электрофореграмма конструкции фрагмента для нокаутирования гена astA и ПЦР-верификация положительных бактерий представлена на Фиг. 4, где длина расположенного против хода транскрипции гомологичного плеча должна составлять 443 п.о., длина расположенного по ходу транскрипции гомологичного плеча должна составлять 523 п.о., и полная длина перекрывающегося фрагмента должна составлять 965 п.о., и для ПЦР-верификации длина амплифицированного путем ПЦР фрагмента положительных бактерий должна составлять 1869 п.о., и длина амплифицированного путем ПЦР фрагмента исходных бактерий должна составлять 965 п.о.
3.2 Нокаут гена argE в Е. coli и интегрирование гена argJ из Corynebacterium glutamicum по этому локусу
С использованием генома Е. coli W3110 (АТСС27325) в качестве матрицы, осуществляли ПЦР с праймерами для расположенного против хода транскрипции гомологичного плеча (UP-argE-S, UP-argE-A) и праймерами для расположенного по ходу транскрипции гомологичного плеча (DN-argE-S, DN-argE-A), созданными в соответствии с расположенными против хода транскрипции и по ходу транскрипции последовательностями его гена argE (NCBI-GeneID: 12930574) для амплификации расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч гена argE. С использованием генома Corynebacterium glutamicum (АТСС13032) в качестве матрицы осуществляли ПЦР с праймерами (argJ-S, argJ-A), созданными в соответствии с последовательностью его гена argJ (NCBI-GeneID: 1019371) для амплификации фрагмента argJ; промотор Ptrc конструировали в обратном праймере для расположенного против хода транскрипции гомологичного плеча и прямом праймере для гена argJ. Способ перекрывающейся ПЦР применяли для слияния вышеприведенных фрагментов с получением фрагмента для нокаутирования гена argE и интегрирования гена argJ (расположенное против хода транскрипции гомологичное плечо-Ptrc-argJ-расположенное по ходу транскрипции гомологичное плечо). Фрагмент ДНК, полученный путем отжига праймеров gRNA-argZs-S и gRNA-argE-A, лигировали с плазмидой pGRB для конструирования рекомбинантной плазмиды pGRB-argE. Компетентные клетки Е. coli W3110 ARG3 получали в соответствии со способами, описанными в разделах 1.3 и 1.4. Плазмиду pGRB-argE и фрагмент для нокаутирования гена argE и интегрирования гена argJ одновременно подвергали электротрансформации в компетентные клетки, и наконец получали штамм, названный Е. coli W3110 ARG4. Электрофореграмма конструкции фрагмента для интегрирования и ПЦР-верификация положительных бактерий во время процесса интегрирования фрагмента Ptrc-argJ представлена на Фиг. 5, где длина расположенного против хода транскрипции гомологичного плеча должна составлять 510 п.о., длина гена argJ должна составлять 1206 п.о., длина расположенного по ходу транскрипции гомологичного плеча должна составлять 668 п.о. и полная длина перекрывающегося фрагмента должна составлять 2458 п.о., и для ПЦР-верификации рекомбинантов длина амплифицированного фрагмента положительных рекомбинантов должна составлять 2458 п.о., и длина амплифицированного фрагмента исходных бактерий должна составлять 2154 п.о.
3.2 Интегрирование оперона синтеза аргинина из Corynebacterium glutamicum в локус гена yghX в Е. coli
Оператор гена синтеза аргинина из Corynebacterium glutamicum (содержащего семь генов argC, argJ, argB, argD, argF, argG и argH) успешно интегрировали в локус гена yjhX в Е. coli, и трансформацию и экспрессию этого чужеродного оперона инициировали промотором Ptrc, и наконец конструировали штамм, названный Е. coli W3110 ARG7.
Интегрирование оператора гена синтеза аргинина из Corynebacterium glutamicum разделено на три стадии.
3.2.1 Интегрирование Ptrc-argC-argJ
С использованием генома Е. coli W3110 (АТСС27325) в качестве матрицы, осуществляли ПЦР с праймерами для расположенного против хода транскрипции гомологичного плеча (UP-yghX-S, UP-yghX-A) и праймерами для расположенного по ходу транскрипции гомологичного плеча (DN-yghX-S1, DN-yghX-A), созданными в соответствии с расположенными против хода транскрипции и по ходу транскрипции последовательностями его гена yghX для амплификации расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч гена yghX. С использованием генома Corynebacterium glutamicum (АТСС13032) в качестве матрицы, осуществляли ПЦР с праймерами (argC-argJ-S, argC-argJ-A), созданными в соответствии с последовательностями его гена argC-argJ (NCBI-GeneID: 1019370, 1019371), для амплификации фрагмента argC-argJ; промотор Ptrc конструировали в обратном праймере для расположенного против хода транскрипции гомологичного плеча и прямом праймере для гена argC-argJ. Способ перекрывающейся ПЦР применяли для слияния вышеприведенных фрагментов с получением фрагмента для интегрирования генов argC-argJ (расположенное против хода транскрипции гомологичное плечо-Ptrc-argC-argJ-расположенное по ходу транскрипции гомологичное плечо). Фрагмент ДНК, содержащий целевую последовательность, получали путем отжига праймеров gRNA-yghX-S и gRNA-yghX-A, и затем лигировали с плазмидой pGRB для конструирования рекомбинантной плазмиды pGRB-yghX. Компетентные клетки Е. coli W3110 ARG4 получали в соответствии со способами, описанными в разделах 1.3 и 1.4. Плазмиду pGRB-yghX и фрагмент для интегрирования генов argC-argJ одновременно подвергали электротрансформации в компетентные клетки, и наконец получали штамм, названный Е. coli W3110 ARG5. Электрофореграмма конструкции фрагмента для интегрирования и ПЦР-верификация положительных бактерий во время процесса интегрирования фрагмента Ptrc-argC-argJ представлена на Фиг. 6, где длина расположенного против хода транскрипции гомологичного плеча должна составлять 602 п.о., длина фрагмента гена argC-argJ должна составлять 2324 п.о., длина расположенного по ходу транскрипции гомологичного плеча должна составлять 561 п.о. и полная длина перекрывающегося фрагмента должна составлять 3650 п.о., и длина амплифицированного фрагмента с использованием праймеров для идентификации должна составлять 1068 п.о., и никакие полосы не должны амплифицироваться из исходных бактерий.
3.2.2 Интегрирование argB-argD-argF
С использованием генома Corynebacterium glutamicum (АТСС13032) в качестве матрицы, осуществляли ПЦР с праймерами для расположенного против хода транскрипции гомологичного плеча (UP-argB-argD-argF-S, VP-argB-argD-argF-A), созданными в соответствии с генами argB-argD-argF (NCBI-GeneID: 1019372, 1019373, 1019374) и их расположенной против хода транскрипции последовательностью для амплификации расположенного против хода транскрипции гомологичного плеча генов argB-argD-argF. С использованием генома Е. coli W3110 (АТСС27325) в качестве матрицы, осуществляли ПЦР с праймерами для расположенного по ходу транскрипции гомологичного плеча (DN-yghX-S2, ХУН-yghX-A), созданными в соответствии с расположенной по ходу транскрипции последовательностью его гена yghX для амплификации расположенного по ходу транскрипции гомологичного плеча гена yghX. Способ перекрывающейся ПЦР применяли для слияния вышеприведенных фрагментов с получением фрагмента для интегрирования генов argB-argD-argF (argB, расположенный против хода транскрипции для интегрирования генов argB-argD-argF (argB, расположенное по ходу транскрипции гомологичное плечо). Фрагмент ДНК, содержащий целевую последовательность, получали путем отжига праймеров gRNA-argBDF-S и gRNA-argBDF- А, и затем лигировали с плазмидой pGRB для конструирования рекомбинантной плазмиды pGRB-argBDF. Компетентные клетки Е. coli W3110 ARG5 получали в соответствии со способами, описанными в разделах 1.3 и 1.4. Плазмиду pGRB-arg5DF и фрагмент для интегрирования генов argB-argD-argF одновременно подвергали электротрансформации в компетентные клетки, и наконец получали штамм, названный Е. coli W3110 ARG6. Электрофореграмма конструкции фрагмента для интегрирования и ПЦР-верификация положительных бактерий во время процесса интегрирования фрагмента argB-argD-argF представлена на Фиг. 7, где полная длина argB расположенного против хода транскрипции фратмепта-argB-argD-argF составляла 3575 п.о., длина расположенного по ходу транскрипции гомологичного плеча составляла 561 п.о., и длина перекрывающегося фрагмента составляла 4219 п.о., и длина фрагмента, амплифицированного при помощи идентифицирующих праймеров, составляла 1034 п.о., и никакие полосы не должны идентифицироваться из исходных бактерий.
3.2.3 Интегрирование argG-argH
С использованием генома Corynebacterium glutamicum (АТСС13032) в качестве матрицы, осуществляли ПЦР с праймерами для расположенного против хода транскрипции гомологичного плеча (UP-argG-argH-S, UP-argG-argH- А) и праймерами для фрагмента argG-argH (argG-argH-S, argG-argH-A), созданными в соответствии с argG-argH (NCBI-GeneID: 1019376, 1019377) и его расположенной против хода транскрипции последовательностью для амплификации расположенного против хода транскрипции гомологичного плеча генов argG-argH и фрагмента argG-argH. С использованием генома Е. coli W3110 (АТСС27325) в качестве матрицы, осуществляли ПЦР с праймерами для расположенного по ходу транскрипции гомологичного плеча (DN-yghX-S3, DN-yghX-A), созданными в соответствии с расположенным по ходу транскрипции его геном yghX для амплификации расположенного по ходу транскрипции гомологичного плеча гена yghX. Способ перекрывающейся ПЦР применяли для слияния вышеприведенных фрагментов с получением фрагмента для интегрирования генов argG-argH {argG расположенный против хода транскрипции фрагмепт-argG-argH-расположенное по ходу транскрипции гомологичное плечо). Фрагмент ДНК, содержащий целевую последовательность, получали путем отжига праймеров gRNA-argG-argH-S и gRNA-argG-argH-A, и затем лигировали с плазмидой pGRB для конструирования рекомбинантной плазмиды pGRB-argG-argH. Компетентные клетки Е. coli W3110 ARG6 получали в соответствии со способами, описанными в разделах 1.3 и 1.4. Плазмиду pGRB-argG-argH и фрагмент для интегрирования генов argG-argH одновременно подвергали электротрансформации в компетентные клетки, и, наконец, получали штамм, названный Е. coli W3110 ARG7. Электрофореграмма конструкции фрагмента для интегрирования и ПЦР-верификация положительных бактерий во время процесса интегрирования фрагмента argG-argH представлена на Фиг. 8, где полная длина расположенного против хода транскрипции фрагмента argG составляла 405 п.о., полная длина фрагмента argG-argH составляла 2826 п.о., длина расположенного по ходу транскрипции гомологичного плеча составляла 561 п.о. и длина перекрывающегося фрагмента должна составлять 3875 п.о., и длина фрагмента, амплифицированного при помощи идентифицирующих праймеров, должна составлять 1521 п.о., и никакие полосы не должны идентифицироваться из исходных бактерий.
3.3 Интегрирование генов pyrAA-pyrAB из В. subtilis в локус reuayjiT в Е. coli
В. subtilis А260 была получена из В. subtilis 168 в качестве исходного штамма путем комбинирования мутагенеза ARTP (технология плазмы при атмосферном давлении и комнатной температуре) и высокопроизводительного скрининга (этот штамм был депонирован 2 декабря 2015 года в China General Microbiological Culture Collection Center (Главный центр коллекции микробиологических культур Китая) (Address: Institute of Microbiology, Chinese Academy of Sciences, No.l West Beichen Road, Chaoyang District, Beijing, Postcode: 100101) под номером депонирования CGMCC No. 11775). Этот штамм облегчал обратную регуляцию со стороны уридиловой кислоты и аргинина в отношении карбамилфосфат-синтетазы, и путем секвенирования пиримидиново-нуклеотидного оперона гена было обнаружено, что остаток глутаминовой кислоты в положении 949 был делетирован в большой субъединице карбамилфосфата (кодируемого pyrAB) (номер публикации: CN105671007A). Гены карбамилфосфат-синтетазы (pyrAA, pyrAB) в В. subtilis А260 без обратного ингибирования аргинина интегрировали в Е. coli для улучшения поступления предшественника карбамилфосфата в процессе синтеза аргинина.
Фрагмент гена pyrAA-pyrAB длиной 4292 п.о. из В. subtilis интегрировали в Е. coli в двух сегментах, где первый сегмент составлял 2651 п.о., и второй сегмент составлял 1641 п.о.
3.3.1 Интегрирование первого сегмента Ptrc-pyrAA-pyrAB
С использованием генома Е. coli W3110 (АТСС27325) в качестве матрицы, осуществляли ПЦР с праймерами для расположенного против хода транскрипции гомологичного плеча (UP-jj/T-S, UP-yjiT-A) и праймерами для расположенного по ходу транскрипции гомологичного плеча (DN-yjiT-S, DN-yjiT-A), созданными в соответствии с расположенными против хода транскрипции и по ходу транскрипции последовательностями его гена yjiT для амплификации расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч reuayjiT. С использованием генома В. subtilis (CGMCC No. 11775) в качестве матрицы, осуществляли ПЦР с праймерами (l-pyrAA-pyrAB-S, 1-pyrAA-pyrAB-А), созданными в соответствии с геном pyrAA (NCBI-GeneID: 937368) и геном pyrAB (NCBI-GeneID: 936608) для амплификации первого сегмента фрагмента гена pyrAA-pyrAB. Промотор Pfre создавали в обратном праймере для расположенного против хода транскрипции гомологичного плеча и в прямом праймере для генов pyrAA-pyrAB. Способ перекрывающейся ПЦР применяли для слияния вышеприведенных фрагментов с получением фрагмента для интегрирования первого сегмента pyrAA-pyrAB (расположенное против хода транскрипции гомологичное плечо-Ptrc-pyrAA-pyrAB-расположенное по ходу транскрипции гомологичное плечо). Фрагмент ДНК, содержащий целевую последовательность, получали путем отжига праймеров gRNA-yjiT-S и gRNA-yjiT-A, и затем лигировали с плазмидой pGRB для конструирования рекомбинантной плазмиды pGRB-yjiT. Компетентные клетки Е. coli W3110 ARG7 получали в соответствии со способами, описанными в разделах 1.3 и 1.4. Плазмиду pGRB-yjiT и фрагмент для интегрирования первого сегмента pyrAA-pyrAB одновременно подвергали электротрансформации в компетентные клетки, и наконец получали штамм, названный Е. coli W3110 ARG8. Электрофореграмма конструкции фрагмента для интегрирования первого сегмента pyrAA-pyrAB и ПЦР-верификация положительных бактерий представлена на Фиг. 9, где длина расположенного против хода транскрипции гомологичного плеча должна составлять 316 п.о., длина первого сегмента фрагмента гена pyrAA-pyrAB должна составлять 2651 п.о., длина расположенного по ходу транскрипции гомологичного плеча должна составлять 667 п.о. и полная длина интегрированного фрагмента должна составлять 3634 п.о., и длина фрагмента, амплифицированного при помощи идентифицирующих праймеров, должна составлять 1100 п.о., и никакие полосы не должны идентифицироваться из исходных бактерий.
3.3.2 Интегрирование второго сегмента pyrAA-pyrAB
С использованием генома5. subtilis А260 (CGMCC No. 11775) в качестве матрицы, осуществляли ПЦР с праймерами для расположенного против хода транскрипции гомологичного плеча (2-pyrAA-pyrAB-S, 2-pyrAA-pyrAB-А), созданными в соответствии с о вторым сегментом pyrAA-pyrAB и его расположенной против хода транскрипции последовательностью для амплификации расположенного против хода транскрипции расположенного по ходу транскрипции гомологичного плеча (содержащего первый сегмент, расположенной против хода транскрипции последовательности pyrAA-pyrAB длиной 266 п.о., и второй последовательности pyrAA-pyrAB длиной 1641 п.о., в сумме 1907 п.о.). С использованием генома Е. coli W3110 (АТСС27325) в качестве матрицы, осуществляли ПЦР с праймерами для расположенного по ходу транскрипции гомологичного плеча (DN-yjiT-S 1, DN-yjiT-A), созданными в соответствии с расположенной по ходу транскрипции последовательностью его гена yjiT для амплификации расположенного по ходу транскрипции гомологичного плеча гена yjiT.
Способ перекрывающейся ПЦР применяли для слияния вышеприведенных фрагментов с получением фрагмента для интегрирования второго сегмента pyrAA-pyrAB (второй сегмент pyrAA-pyrAB-расположенное по ходу транскрипции гомологичное плечо). Фрагмент ДНК, содержащий целевую последовательность, получали путем отжига праймеров gRNA-pyrAA-pyrAB-S и gRNA-pyrAA-pyrAB-A, и затем лигировали с плазмидой pGRB для конструирования рекомбинантной плазмиды pGRB-pyrAA-pyrAB. Компетентные клетки Е. coli W3110 ARG8 получали в соответствии со способами, описанными в разделах 1.3 и 1.4. Плазмиду pGRB-pyrAA-pyrAB и фрагмент для интегрирования второго сегмента pyrAA-pyrAB одновременно подвергали электротрансформации в компетентные клетки, и наконец получали штамм, названный Е. coli W3110 ARG9. Электрофореграмма конструкции интегрированного фрагмента и ПЦР-верификация положительных бактерий во время процесса интеграции второго сегмента pyrAA-pyrAB представлена на Фиг. 10, где полная длина расположенной против хода транскрипции последовательности второго сегмента pyrAA-pyrAB должна составлять 1907 п.о., длина расположенного по ходу транскрипции гомологичного плеча должна составлять 667 п.о., и полная длина перекрывающегося фрагмента должна составлять 2574 п.о., и длина фрагмента, амплифицированного при помощи идентифицирующих праймеров, должна составлять 1135 п.о., и никакие полосы не должны идентифицироваться из исходных бактерий.
3.4 Интегрирование гена lys Е из Corynebacterium efficiens в локус гена ilvG в Е. coli
С использованием генома Е. coli W3110 (АТСС27325) в качестве матрицы, осуществляли ПЦР с праймерами для расположенного против хода транскрипции гомологичного плеча (UP-ilvG-S, UP-ilvG-A) и праймерами для расположенного по ходу транскрипции гомологичного плеча (DN-ilvG-S, DN-ilvG-A), созданными в соответствии с расположенными против хода транскрипции и по ходу транскрипции последовательностями его гена ilvG для амплификации расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч гена ilvG; ПЦР осуществляли с праймерами (lysE-S, lysE-A), созданными в соответствии с последовательностью гена lysE (референсная последовательность NCBI: WP 143758438.1) (SEQ ID NO: 68) для амплификации фрагмента гена lysE. Промотор Ptrc конструировали в обратном праймере для расположенного против хода транскрипции гомологичного плеча и в прямом праймере для гена lysE. Способ перекрывающейся ПЦР применяли для слияния вышеприведенных фрагментов с получением фрагмента для интегрирования гена lysE (расположенное против хода транскрипции гомологичное плечо-Ptrc-lysE-расположенное по ходу транскрипции гомологичное плечо). Фрагмент ДНК, содержащий целевую последовательность, получали путем отжига праймеров gRNA-ilvG-S и gRNA-ilvG-A, и затем лигировали с плазмидой pGRB для конструирования рекомбинантной плазмиды pGRB-ilvG. Компетентные клетки Е. coli W3110 ARG9 получали в соответствии со способами, описанными в разделах 1.3 и 1.4. Плазмиду pGRB-ilvG и фрагмент для интегрирования гена lysE одновременно подвергали электротрансформации в компетентные клетки, и наконец получали штамм, названный Е. coli W3110 ARG10. Электрофореграмма конструкции интегрированного фрагмента Ptrc-lysE и ПЦР-верификация положительных бактерий представлена на Фиг. 11, где длина расположенного против хода транскрипции гомологичного плеча должна составлять 412 п.о., длина фрагмента гена Ptrc-lysE должна составлять 806 п.о., длина расположенного по ходу транскрипции гомологичного плеча должна составлять 481 п.о., и полная длина интегрированного фрагмента должна составлять 1699 п.о., и для ПЦР-верификации фрагмент, амплифицированный при помощи ПЦР из положительных бактерий, должен составлять 1699 п.о., и фрагмент, амплифицированный при помощи ПЦР из исходных бактерий, должен составлять 1426 п.о.
Пример 2:
Способ продуцирования аргинина путем ферментации генно-инженерного штамма Е. coli W3110 ARG10 был следующим:
(1) Ферментация во встряхиваемой колбе
культура на скошенном агаре: инокулируют бактериальный штамм, который хранили при -80°С, на активированный скошенный агар с использованием способа посева штрихами, культивируют при 37°С в течение 12 ч и осуществляют однократное пассирование;
посевная культура для ферментации во встряхиваемой колбе: соскребают кольцо посевов на скошенном агаре при помощи петли для инокуляции и инокулируют в коническую колбу объемом 500 мл, содержащую 30 мл среды для посева, коническую колбу закрывают девяти слоями марли, и культивируют при 37°С и 200 об./мин в течение 7-10 ч;
ферментация культуры во встряхиваемой колбе: инокулируют посевную культуру в концентрации 15% (об./об.) в коническую колбу объемом 500 мл, содержащую ферментационную среду (конечный объем: 30 мл), коническую колбу закрывают девяти слоями марли, культивируют при 37°С и 200 об./мин на встряхивающей платформе, во время ферментации добавляют аммиачную воду для поддержания рН на уровне 7,0-7,2; добавляют 60% (масс./об.) раствор глюкозы для поддержания ферментации; период ферментации длится в течение 26-30 ч.
Компоненты среды для скошенного агара: 1 г/л глюкозы, 10 г/л пептона, 10 г/л говяжьего экстракта, 5 г/л сухих дрожжей, 2,5 г/л NaCl, 20 г/л агара, остальное представляло собой воду, рН 7,0-7,2.
Компоненты посевной среды: 25 г/л глюкозы, 5 г/л дрожжевого экстракта, 3 г/л пептона, 1 г/л K2HPO4, 1 г/л MgSO4⋅7H2O, 10 мг/л FeSO4⋅7H2O, 10 мг/л MnSO4⋅7H2O, 1 мг/л каждого из VB1, VB3, VB5, VB12 и VH, остальное представляло собой воду, рН 7,0-7,2.
Компоненты ферментационной среды: 25 г/л глюкозы, 3 г/л дрожжевого экстракта, 2 г/л пептона, 3 г/л K2HPO4, 2 г/л MgSO4⋅7H2O, 10 мг/л FeSO4⋅7H2O, 10 мг/л MnSO4⋅7H2O, 1 мг/л каждого из VB1, VB3, VB5, VB12 и VH, остальное представляло собой воду, рН 7,0-7,2.
После 26-30 ч ферментации во встряхиваемой колбе выход L-аргинина в ферментационном бульоне штамма Е. coli W3110 ARGI0 составлял 30-32 г/л.
(2) Ферментация в ферментере
культура на активированном скошенном агаре: соскребают кольцо бактериального штамма, который хранили при -80°С, и однородно распределяют на активированном скошенном агаре, культивируют при 37°С в 12-16 ч и переносят в колбу в форме баклажана для продолжения культивирования в течение 12-16 ч;
посевная культура: необходимое количество стерилизованной воды добавляют в колбу в форме баклажана, инокулируют бактериальную суспензию в посевную среду, рН поддерживают на уровне приблизительно 7,0, температуру поддерживают на уровне 37°С и растворенный кислород поддерживают на уровне 25-35%, и культивируют клетки до достижения сухой массы клеток 5-6 г/л;
ферментационная культура: инокулируют посевную культуру в концентрации 15% в свежую ферментационную среду, начинают ферментацию, и во время ферментационного процесса поддерживают стабильное значение рН на уровне примерно 7,0, температуру на уровне 35°С и растворенный кислород на уровне 25-35%; когда глюкоза в среде истощалась, тогда 80% (масса/об.) раствор глюкозы добавляли для поддержания концентрации глюкозы в ферментационной среде на уровне 0,1-5 г/л.
Среда для скошенного агара, посевная среда и ферментационная среда были такими же, что среда для ферментации во встряхиваемой колбе.
Накопление L-аргинина достигало 130-135 г/л после культивирования в течение 50-55 ч в ферментере объемом 5 л. Скорость превращения составляла 0,48 г аргинина/г глюкозы, и интенсивность продуцирования составляла 2,5 г аргинина/л/ч. Ферментационная кривая представлена на Фиг. 12.
Воплощения настоящего изобретения описаны выше. Однако настоящее изобретение не ограничено вышеприведенными воплощениями. Любая модификация, эквивалентные замены, улучшения и т.п., осуществленные в пределах сущности и принципов настоящего изобретения, должны быть включены в объем защиты данного изобретения.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
--->
<110> NINGXIA EPPEN BIOTECH CO., LTD
<120> ГЕННО-ИНЖЕНЕРНЫЕ БАКТЕРИИ ДЛЯ ПРОДУЦИРОВАНИЯ L-АРГИНИНА И СПОСОБ
КОНСТРУИРОВАНИЯ И ПРИМЕНЕНИЯ ГЕННО-ИНЖЕНЕРНЫХ БАКТЕРИЙ
<130>
<150> 201911211097.X
<151> 2019-12-02
<160> 68
<170> SIPOSequenceListing 1.0
<210> 1
<211> 22
<212> ДНК
<213> Искусственная последовательность
<400> 1
ttaacctgtc tcaccgttct gg 22
<210> 2
<211> 40
<212> ДНК
<213> Искусственная последовательность
<400> 2
acaaacctgc ctcgaactct tccgctgacg aaggcaaacc 40
<210> 3
<211> 40
<212> ДНК
<213> Искусственная последовательность
<400> 3
ggtttgcctt cgtcagcgga agagttcgag gcaggtttgt 40
<210> 4
<211> 21
<212> ДНК
<213> Искусственная последовательность
<400> 4
catataccag atcgccgcag t 21
<210> 5
<211> 23
<212> ДНК
<213> Искусственная последовательность
<400> 5
cgagtttctc catcaagaca cct 23
<210> 6
<211> 40
<212> ДНК
<213> Искусственная последовательность
<400> 6
cgcccataga gaacaggaac atgcggcttg gcaccatata 40
<210> 7
<211> 40
<212> ДНК
<213> Искусственная последовательность
<400> 7
tatatggtgc caagccgcat gttcctgttc tctatgggcg 40
<210> 8
<211> 21
<212> ДНК
<213> Искусственная последовательность
<400> 8
tatcgccgaa gttttcacca g 21
<210> 9
<211> 19
<212> ДНК
<213> Искусственная последовательность
<400> 9
ggcactcatg gcaccacct 19
<210> 10
<211> 37
<212> ДНК
<213> Искусственная последовательность
<400> 10
tgagggcatc cagttgtgcc tgcatcagcg ccgagac 37
<210> 11
<211> 37
<212> ДНК
<213> Искусственная последовательность
<400> 11
gtctcggcgc tgatgcaggc acaactggat gccctca 37
<210> 12
<211> 21
<212> ДНК
<213> Искусственная последовательность
<400> 12
tgaccaggga aattatacgg c 21
<210> 13
<211> 19
<212> ДНК
<213> Искусственная последовательность
<400> 13
gcccgcttca agaaactgc 19
<210> 14
<211> 76
<212> ДНК
<213> Искусственная последовательность
<400> 14
aattgttatc cgctcacaat tccacacatt atacgagccg gatgattaat tgtcaaggcg 60
cttattgaag gtgtgg 76
<210> 15
<211> 81
<212> ДНК
<213> Искусственная последовательность
<400> 15
tccggctcgt ataatgtgtg gaattgtgag cggataacaa tttcacacag gaaacagacc 60
atggcagaaa aaggcattac c 81
<210> 16
<211> 45
<212> ДНК
<213> Искусственная последовательность
<400> 16
gttgatgagc ctgattaatt gagcgccctt ttccctgctt gttag 45
<210> 17
<211> 45
<212> ДНК
<213> Искусственная последовательность
<400> 17
ctaacaagca gggaaaaggg cgctcaatta atcaggctca tcaac 45
<210> 18
<211> 22
<212> ДНК
<213> Искусственная последовательность
<400> 18
ctgtatcctt cacgtcgcat tg 22
<210> 19
<211> 21
<212> ДНК
<213> Искусственная последовательность
<400> 19
gcgcaacgta gaacaggaat t 21
<210> 20
<211> 79
<212> ДНК
<213> Искусственная последовательность
<400> 20
aattgttatc cgctcacaat tccacacatt atacgagccg gatgattaat tgtcaagatt 60
gaagcgcctt tactactcc 79
<210> 21
<211> 85
<212> ДНК
<213> Искусственная последовательность
<400> 21
tccggctcgt ataatgtgtg gaattgtgag cggataacaa tttcacacag gaaacagacc 60
atgatcatgc ataacgtgta tggtg 85
<210> 22
<211> 86
<212> ДНК
<213> Искусственная последовательность
<400> 22
gccccaaggg gttatgctag cctacaaatt gagttatgtt catttaaata tgatgttgtt 60
cagttaagag ctgtacgcgg agttga 86
<210> 23
<211> 89
<212> ДНК
<213> Искусственная последовательность
<400> 23
ctgaacaaca tcatatttaa atgaacataa ctcaatttgt aggctagcat aaccccttgg 60
ggcgtcatag taatccagca actcttgtg 89
<210> 24
<211> 22
<212> ДНК
<213> Искусственная последовательность
<400> 24
gagcaggtat ttacgtgaac cg 22
<210> 25
<211> 23
<212> ДНК
<213> Искусственная последовательность
<400> 25
gtacgcagct tgttctgata tcg 23
<210> 26
<211> 81
<212> ДНК
<213> Искусственная последовательность
<400> 26
agttgctgga ttactatgac cctagaagaa atcaaccagc gcatcagaaa gtctcctgtg 60
catttacctc ggctggttgg c 81
<210> 27
<211> 89
<212> ДНК
<213> Искусственная последовательность
<400> 27
atgcacagga gactttctga tgcgctggtt gatttcttct agggtcatag taatccagca 60
actgtcatag taatccagca actcttgtg 89
<210> 28
<211> 22
<212> ДНК
<213> Искусственная последовательность
<400> 28
gatatttcca tcatcgctcc tg 22
<210> 29
<211> 40
<212> ДНК
<213> Искусственная последовательность
<400> 29
ctcgggttat accttacctg ccttacctcg gctggttggc 40
<210> 30
<211> 40
<212> ДНК
<213> Искусственная последовательность
<400> 30
gccaaccagc cgaggtaagg caggtaaggt ataacccgag 40
<210> 31
<211> 81
<212> ДНК
<213> Искусственная последовательность
<400> 31
caccgacaaa caacagataa aacgaaaggc ccagtctttc gactgagcct ttcgttttat 60
ttgttatcga cgtacccccg c 81
<210> 32
<211> 89
<212> ДНК
<213> Искусственная последовательность
<400> 32
aaagactggg cctttcgttt tatctgttgt ttgtcggtga acgctctcct gagtaggaca 60
aatgtcatag taatccagca actcttgtg 89
<210> 33
<211> 21
<212> ДНК
<213> Искусственная последовательность
<400> 33
aatagttgtt gccgcctgag t 21
<210> 34
<211> 76
<212> ДНК
<213> Искусственная последовательность
<400> 34
aattgttatc cgctcacaat tccacacatt atacgagccg gatgattaat tgtcaaaaaa 60
caggcagcaa agtccc 76
<210> 35
<211> 87
<212> ДНК
<213> Искусственная последовательность
<400> 35
tccggctcgt ataatgtgtg gaattgtgag cggataacaa tttcacacag gaaacagacc 60
atgaagagac gattagtact ggaaaac 87
<210> 36
<211> 86
<212> ДНК
<213> Искусственная последовательность
<400> 36
gccccaaggg gttatgctag cctacaaatt gagttatgtt catttaaata tgatgttgtt 60
cagagaagac atcgatagcg gaaaat 86
<210> 37
<211> 86
<212> ДНК
<213> Искусственная последовательность
<400> 37
ctgaacaaca tcatatttaa atgaacataa ctcaatttgt aggctagcat aaccccttgg 60
ggcaagcact acctgtgaag ggatgt 86
<210> 38
<211> 21
<212> ДНК
<213> Искусственная последовательность
<400> 38
cagggcttcc acagtcacaa t 21
<210> 39
<211> 21
<212> ДНК
<213> Искусственная последовательность
<400> 39
acccggtgac aggaaaaaca t 21
<210> 40
<211> 85
<212> ДНК
<213> Искусственная последовательность
<400> 40
caccgacaaa caacagataa aacgaaaggc ccagtctttc gactgagcct ttcgttttat 60
ttgtcatata gtgactgccg cctcc 85
<210> 41
<211> 86
<212> ДНК
<213> Искусственная последовательность
<400> 41
aaagactggg cctttcgttt tatctgttgt ttgtcggtga acgctctcct gagtaggaca 60
aataagcact acctgtgaag ggatgt 86
<210> 42
<211> 23
<212> ДНК
<213> Искусственная последовательность
<400> 42
accgaggagc agacaatgaa taa 23
<210> 43
<211> 76
<212> ДНК
<213> Искусственная последовательность
<400> 43
aattgttatc cgctcacaat tccacacatt atacgagccg gatgattaat tgtcaaggtg 60
atggcaacaa caggga 76
<210> 44
<211> 82
<212> ДНК
<213> Искусственная последовательность
<400> 44
tccggctcgt ataatgtgtg gaattgtgag cggataacaa tttcacacag gaaacagacc 60
atggaaattt tcgttacggg tc 82
<210> 45
<211> 87
<212> ДНК
<213> Искусственная последовательность
<400> 45
caccgacaaa caacagataa aacgaaaggc ccagtctttc gactgagcct ttcgttttat 60
ttgttagccc atcagaatca gtttcac 87
<210> 46
<211> 84
<212> ДНК
<213> Искусственная последовательность
<400> 46
aaagactggg cctttcgttt tatctgttgt ttgtcggtga acgctctcct gagtaggaca 60
aatctatcta cgcgccgttg ttgt 84
<210> 47
<211> 20
<212> ДНК
<213> Искусственная последовательность
<400> 47
gcgctggcta acatgaggaa 20
<210> 48
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 48
agtcctaggt ataatactag ttgcgtactt acaatattgc cgttttagag ctagaa 56
<210> 49
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 49
ttctagctct aaaacggcaa tattgtaagt acgcaactag tattatacct aggact 56
<210> 50
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 50
agtcctaggt ataatactag ttatcgggcc aatctatccg cgttttagag ctagaa 56
<210> 51
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 51
ttctagctct aaaacgcgga tagattggcc cgataactag tattatacct aggact 56
<210> 52
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 52
agtcctaggt ataatactag ttctctgcgg caccgggcaa agttttagag ctagaa 56
<210> 53
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 53
ttctagctct aaaactttgc ccggtgccgc agagaactag tattatacct aggact 56
<210> 54
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 54
agtcctaggt ataatactag ttgcagattt aatcactctg cgttttagag ctagaa 56
<210> 55
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 55
ttctagctct aaaacgcaga gtgattaaat ctgcaactag tattatacct aggact 56
<210> 56
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 56
agtcctaggt ataatactag tggtgcctga cgaccataaa agttttagag ctagaa 56
<210> 57
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 57
ttctagctct aaaactttta tggtcgtcag gcaccactag tattatacct aggact 56
<210> 58
<211> 63
<212> ДНК
<213> Искусственная последовательность
<400> 58
ctgaacaaca tcatatttaa atgaacataa ctcaatttgt aggctagcat aaccccttgg 60
ggc 63
<210> 59
<211> 86
<212> ДНК
<213> Искусственная последовательность
<400> 59
gccccaaggg gttatgctag cctacaaatt gagttatgtt catttaaata tgatgttgtt 60
cagttaagag ctgtacgcgg agttga 86
<210> 60
<211> 63
<212> ДНК
<213> Искусственная последовательность
<400> 60
atgcacagga gactttctga tgcgctggtt gatttcttct agggtcatag taatccagca 60
act 63
<210> 61
<211> 63
<212> ДНК
<213> Искусственная последовательность
<400> 61
agttgctgga ttactatgac cctagaagaa atcaaccagc gcatcagaaa gtctcctgtg 60
cat 63
<210> 62
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 62
agtcctaggt ataatactag tagggattat gaacggcaat ggttttagag ctagaa 56
<210> 63
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 63
ttctagctct aaaaccattg ccgttcataa tccctactag tattatacct aggact 56
<210> 64
<211> 63
<212> ДНК
<213> Искусственная последовательность
<400> 64
ctgaacaaca tcatatttaa atgaacataa ctcaatttgt aggctagcat aaccccttgg 60
ggc 63
<210> 65
<211> 63
<212> ДНК
<213> Искусственная последовательность
<400> 65
gccccaaggg gttatgctag cctacaaatt gagttatgtt catttaaata tgatgttgtt 60
cag 63
<210> 66
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 66
agtcctaggt ataatactag tggaagagtt gccgcgcatc agttttagag ctagaa 56
<210> 67
<211> 56
<212> ДНК
<213> Искусственная последовательность
<400> 67
ttctagctct aaaactgatg cgcggcaact cttccactag tattatacct aggact 56
<210> 68
<211> 687
<212> ДНК
<213> Искусственная последовательность
<400> 68
atggaaattt tcgttacggg tctgctgctg ggtgccagtc tgctgctggc catcggtccg 60
cagaacgtgc tcgtgatcaa acaaggcatc aagcgcgaag gtatcaccgc ggttatcatc 120
gtgtgtctgc tgagcgacgt tgtgctgttc acgctgggca cgctgggcgt tggtctgatc 180
agcgataccg cgccgatcat tctggatatt ctgcgctggt gcggtatcgc ctatctgctg 240
tggtttgcgg ttatggccgc gcgtgatgcg ctgcgtgcgc gtaccgaagt gacgttcgtt 300
gaacacagcg aaccagttgc cgcggccagt gcgagtggtg gtggtgttac gaccaaacag 360
cgcccacgtc tgcgtatcac gagcggtacc cgccaagttt gggttcgccc gatgctgatg 420
gcgatcgttc tgacgtggct gaatccgaac gcgtatctgg atgcgttcgt tttcatcggc 480
ggtgttggcg cgcagtatgg cgaaaccggt cgctggattt ttgcggcggg tgcgtttgcc 540
gcgagtctgg tttggtttcc gctggttggt tatggtgccg ccgcgctgag tcgtccactg 600
agtagcccac gcgtgtggcg ctggatcaat atcggcgttg ccgtggttct gaccggtctg 660
gcggtgaaac tgattctgat gggctaa 687
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРИМЕНЕНИЕ ГЕНА ТРАНСПОРТНОГО ПЕРЕНОСЧИКА, КОТОРЫЙ УЛУЧШАЕТ ЭФФЕКТИВНОСТЬ ПРОДУЦИРОВАНИЯ L-ТРИПТОФАНА У ESCHERICHIA COLI | 2019 |
|
RU2806731C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АРГИНИНА И L-ЦИТРУЛЛИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE, В КОТОРОЙ ОСЛАБЛЕНА ЭКСПРЕССИЯ ГЕНА pepA | 2010 |
|
RU2471870C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2007 |
|
RU2364628C2 |
| Новый экспортер L-триптофана и способ продуцирования L-триптофана с его использованием | 2019 |
|
RU2761871C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ НА ОСНОВЕ ESCHERICHIA COLI, СПОСОБ ЕГО КОНСТРУИРОВАНИЯ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2813283C2 |
| Способ получения L-лизина модифицированием гена аконитазы и/или его регуляторных элементов | 2014 |
|
RU2792116C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pF646, КОДИРУЮЩАЯ ГИБРИДНЫЙ ПОЛИПЕПТИД, СОДЕРЖАЩИЙ ПРОИНСУЛИН АСПАРТ, И ШТАММ БАКТЕРИЙ Escherichia coli - ПРОДУЦЕНТ ГИБРИДНОГО ПОЛИПЕПТИДА, СОДЕРЖАЩЕГО ПРОИНСУЛИН АСПАРТ | 2019 |
|
RU2729353C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДЛЯ ПРОДУЦИРОВАНИЯ L-АМИНОКИСЛОТЫ, СПОСОБ ЕГО КОНСТРУИРОВАНИЯ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2832588C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ С МОДИФИЦИРОВАННЫМ ГЕНОМ BBD29_11265 ДЛЯ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ, А ТАКЖЕ СПОСОБ ЕГО КОНСТРУИРОВАНИЯ И ПРИМЕНЕНИЯ | 2021 |
|
RU2841248C1 |
| Микроорганизм для продуцирования путресцина и способ получения путресцина с его применением | 2021 |
|
RU2817284C1 |
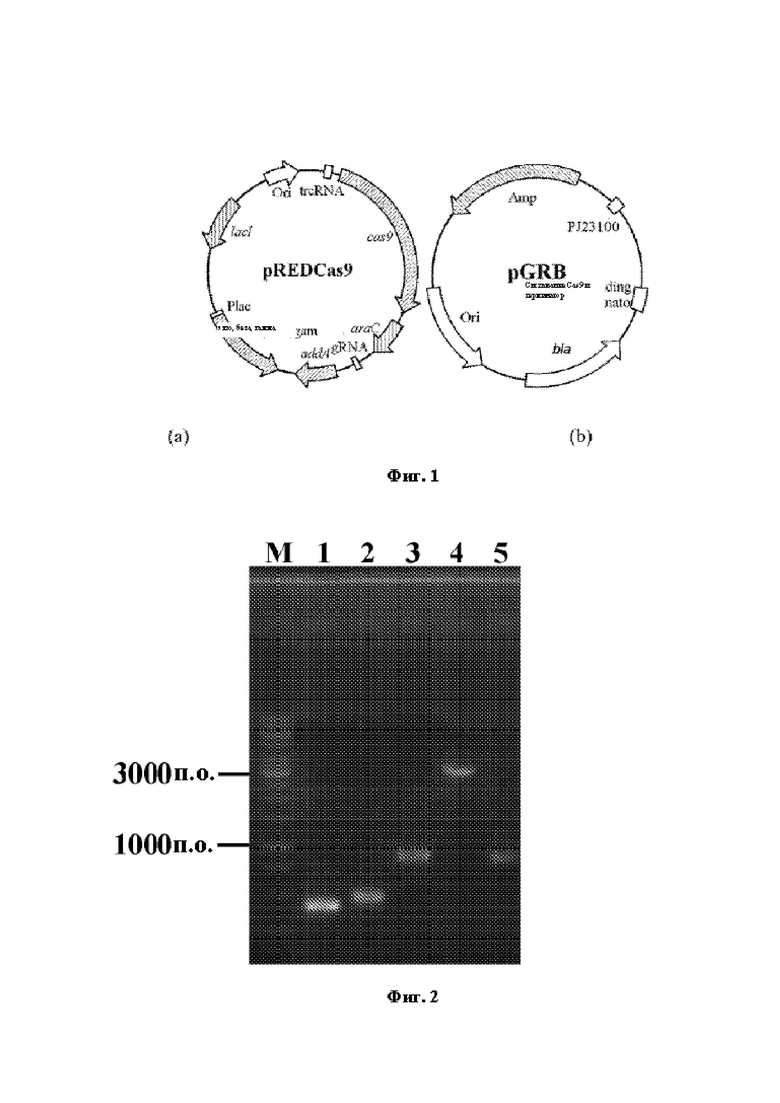


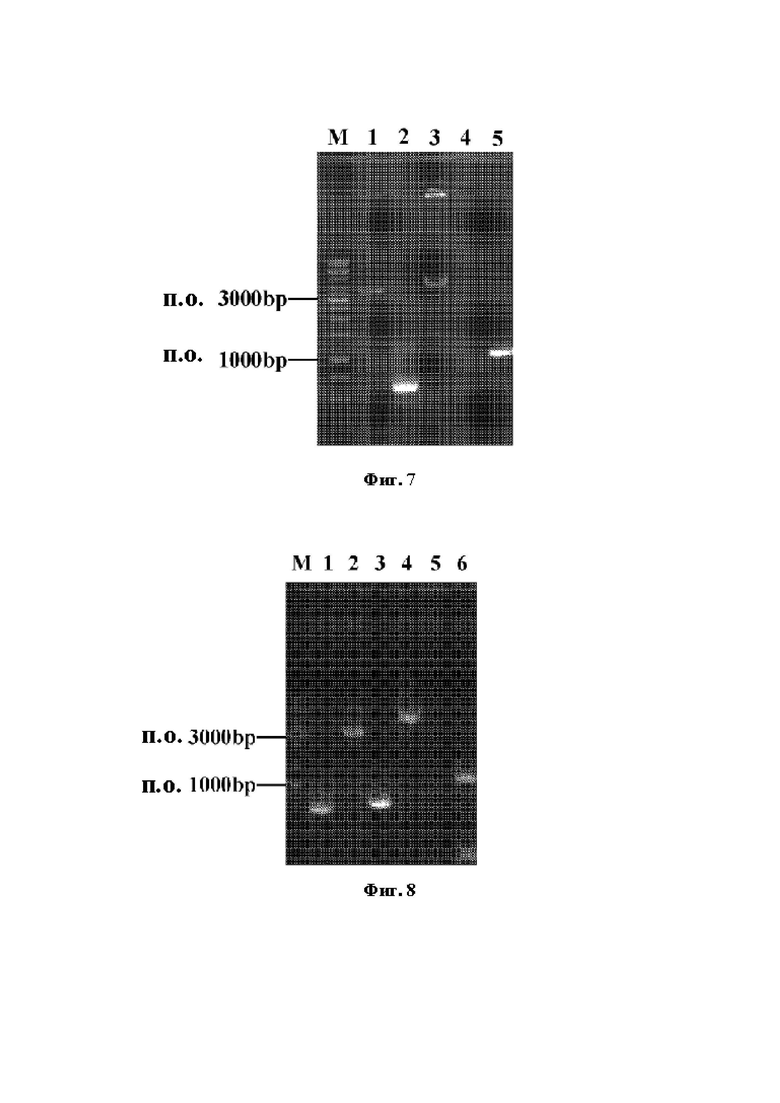
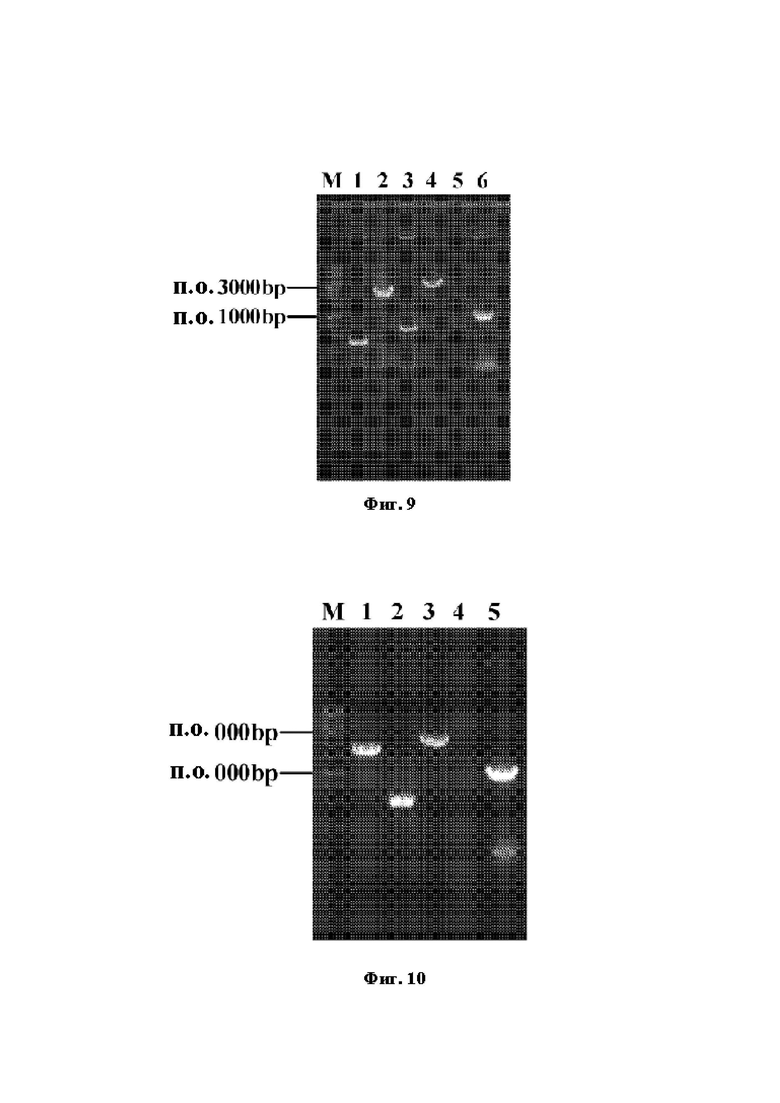
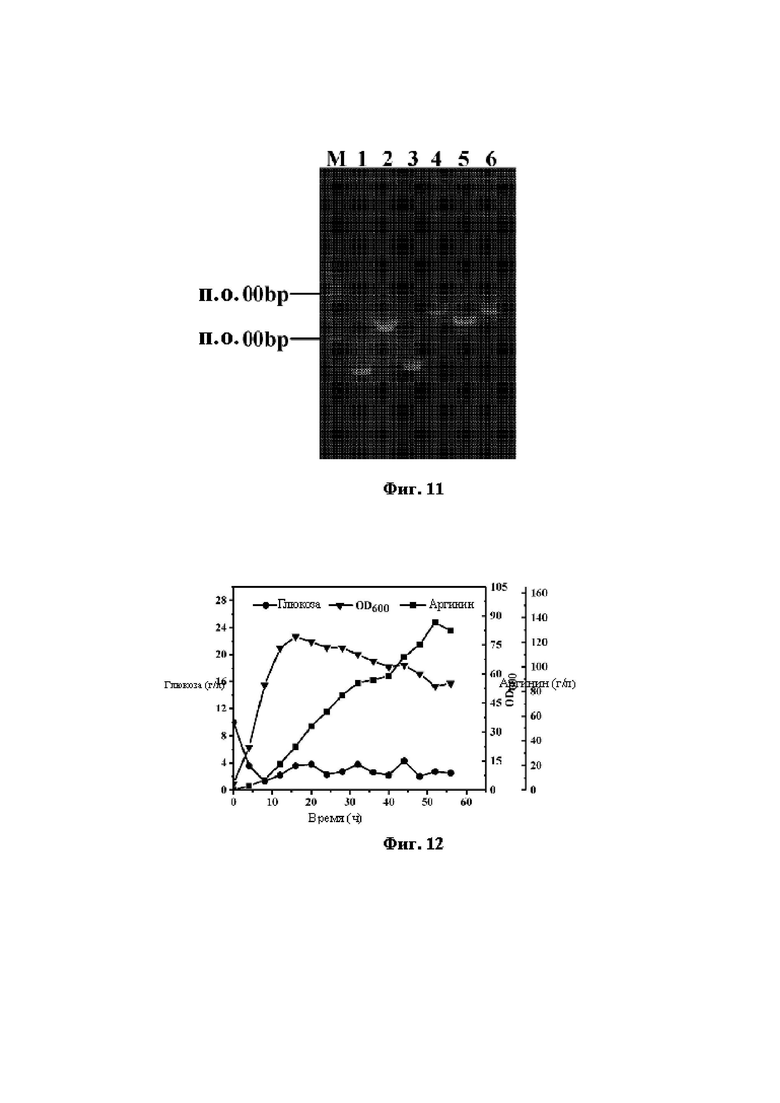
Изобретение относится к биотехнологии и представляет собой генно-инженерную бактерию для продуцирования L-аргинина, которая содержит два гена pyrAA и pyrAB, кодирующие карбамоилфосфат-синтетазу, и где гены pyrAA и pyrAB происходят из Bacillus subtilis; которая дополнительно содержит ген, кодирующий фермент пути биосинтеза L-аргинина, выбранный из одного или более из следующих ферментов: argC, argJ, argB, argD, argF, argG, argH; которая дополнительно содержит ген lysE, кодирующий транспортер аргинина; и для которой в качестве исходной бактерии берут Escherichia coli с одновременным нокаутом генов speA, adiA, astA и argE. Изобретение позволяет получать L-аргинин с высокой степенью эффективности. 3 н. и 12 з.п. ф-лы, 12 ил., 5 табл., 2 пр.
1. Генно-инженерная бактерия для продуцирования L-аргинина, которая содержит два гена pyrAA и pyrAB, кодирующие карбамоилфосфат-синтетазу, и где гены pyrAA и pyrAB происходят из Bacillus subtilis; которая дополнительно содержит ген, кодирующий фермент пути биосинтеза L-аргинина, выбранный из одного или более из следующих ферментов: argC, argJ, argB, argD, argF, argG, argH; которая дополнительно содержит ген lysE, кодирующий транспортер аргинина; и для которой в качестве исходной бактерии берут Escherichia coli с одновременным нокаутом генов speA, adiA, astA и argE.
2. Генно-инженерная бактерия по п. 1, где Escherichia coli выбрана из E. Coli W3110 или E. coli MG1655.
3. Генно-инженерная бактерия по п. 1 или 2, где гены pyrAA и pyrAB интегрированы в локус гена yjiT в E. coli.
4. Генно-инженерная бактерия по п. 1 или 2, где ген, кодирующий фермент пути биосинтеза L-аргинина, управляется промотором Ptrc.
5. Генно-инженерная бактерия по п. 1 или 2, где ген, кодирующий фермент пути биосинтеза L-аргинина, интегрирован в локус гена yghX в E. coli.
6. Генно-инженерная бактерия по п. 1 или 2, где ген lysE интегрирован в локус гена ilvG в E. coli; предпочтительно, ген lysE имеет нуклеотидную последовательность, представленную в SEQ ID NO: 68.
7. Способ конструирования генно-инженерной бактерии, включающий следующие стадии:
(1) интегрирование генов pyrAA и pyrAB в геном исходной бактерии, где в качестве исходной бактерии для указанной генно-инженерной бактерии берут Escherichia coli и гены pyrAA и pyrAB происходят из Bacillus subtilis;
(2) интегрирование генов ферментов пути биосинтеза аргинина, включающих гены argC, argJ, argB, argD, argF, argG и argH; интегрирование гена lysE, кодирующего транспортер аргинина; (3) нокаутирование генов speA и adiA, кодирующих аргинин-декарбоксилазу, гена, кодирующего аргинин-сукцинилтрансферазу astA, и гена, кодирующего ацетилорнитин-деацетилазу argE.
8. Способ конструирования по п. 7, который включает стадии:
(1) нокаутирования следующих трех генов в E. coli: гена speA, кодирующего аргинин-декарбоксилазу, гена adiA, кодирующего аргинин-декарбоксилазу, и гена astA, кодирующего аргинин-сукцинилтрансферазу;
(2) нокаутирование гена argE, кодирующего ацетилорнитин-деацетилазу, в E.coli, и интегрирование гена argJ, кодирующего глутаматацетилтрансферазу, в E. coli;
(3) интегрирование следующего генного кластера, связанного с биосинтезом аргинина: argC, argJ, argB, argD, argF, argG и argH;
(4) интегрирование генов pyrAA и pyrAB, кодирующих карбамоилфосфат-синтетазу;
(5) интегрирование гена lysE, кодирующего транспортер аргинина, в геном E. coli.
9. Способ конструирования по п. 7 или 8, включающий применение опосредованной CRISPR/Cas9 технологии редактирования гена для осуществления интегрирования и нокаутирования гена.
10. Способ конструирования по п. 7 или 8, включающий стадии конструирования рекомбинантного фрагмента и плазмиды pGRB.
11. Способ конструирования по п. 10, где стадия конструирования плазмиды pGRB включает: создание целевой последовательности, получение фрагмента ДНК, содержащего целевую последовательность, и рекомбинацию фрагмента ДНК, содержащего целевую последовательность, с линеаризованным фрагментом вектора;
предпочтительно, в данном способе конструирования стадия конструирования рекомбинантного фрагмента включает конструирование рекомбинантного фрагмента для интегрирования гена или для нокаутирования генов.
12. Способ конструирования по п. 11, где стадия конструирования рекомбинантного фрагмента для интегрирования гена включает: использование генома исходной бактерии в качестве матрицы, создание праймеров для расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч в соответствии с расположенными против хода транскрипции и по ходу транскрипции последовательностями предполагаемого сайта встраивания целевого гена, и создание праймеров в соответствии с целевым геномом для амплификации фрагмента целевого гена, и затем осуществление перекрывающейся ПЦР (полимеразная цепная реакция) с получением рекомбинантного фрагмента; предпочтительно, стадия конструирования рекомбинантного фрагмента для нокаутирования генов включает: использование расположенных против хода транскрипции и по ходу транскрипции последовательностей нокаутируемого гена в качестве матриц, создание праймеров для расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч; соответственно амплификацию расположенных против хода транскрипции и по ходу транскрипции гомологичных плеч посредством ПЦР, и затем получение рекомбинантного фрагмента при помощи перекрывающейся ПЦР.
13. Способ конструирования по п. 12, где данный способ конструирования включает: одновременную трансформацию плазмиды pGRB и вышеупомянутого рекомбинантного фрагмента в электропорационно-компетентные клетки, содержащие pREDCas9, и элиминацию плазмид с получением рекомбинантной генно-инженерной бактерии.
14. Способ продуцирования L-аргинина путем ферментации генно-инженерной бактерии по любому из пп. 1-6, включающий: приведение вышеупомянутой генно-инженерной бактерии в контакт с ферментационной средой и осуществление ферментации с получением L-аргинина.
15. Способ по п. 14, где ферментация включает ферментацию во встряхиваемой колбе или ферментацию в ферментере; предпочтительно, накопление L-аргинина достигает 130-135 г/л, скорость превращения достигает 0,48 г аргинина/г глюкозы, и интенсивность продуцирования
достигает 2,5 г аргинина/л/ч.
| Валковая мельница | 1984 |
|
SU1204255A1 |
| ВС^'СОЮЗНДЯ П5ТП1ГнП„ТГ'Гтг--;--'г | 0 |
|
SU389434A1 |
| subtilis str | |||
| Приспособление, заменяющее сигнальную веревку | 1921 |
|
SU168A1 |
| 0 |
|
SU389435A1 | |

